initial experience with volumetric 68ga-psma i&t pet/ct for … · initial experience with...
TRANSCRIPT

AEM4123-14, revised Tropical R. solanacearum outcompetes R3bv2 strains
1
1 2 Tropical strains of Ralstonia solanacearum outcompete Race 3 biovar 2 strains at lowland 3
tropical temperatures 4 5 6 7 Alejandra I. Huerta1, Annett Milling2, and Caitilyn Allen1* 8 9 10 1Department of Plant Pathology, University of Wisconsin-Madison, Madison 53706 11 2Current address: R. J. Reynolds, Inc., Winston-Salem NC 12 13 Revised version of AEM4123-14 14 15 Running Head: Tropical R. solanacearum outcompetes R3bv2 strains 16 17 18 19 20 Keywords: bacterial wilt, brown rot, intra-strain competition, rhizosphere, competitive index, 21 plant pathogen, bacteriocin 22 23 24 *Corresponding author. Tel. 608-262-9578; email: [email protected] 25 26
AEM Accepted Manuscript Posted Online 13 March 2015Appl. Environ. Microbiol. doi:10.1128/AEM.04123-14Copyright © 2015, American Society for Microbiology. All Rights Reserved.
on June 5, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from

AEM4123-14, revised Tropical R. solanacearum outcompetes R3bv2 strains
2
Abstract 27 28 Bacterial wilt, caused by members of the heterogenous Ralstonia solanacearum species 29 complex, is an economically important vascular disease affecting many crops. Human 30 activity has widely disseminated R. solanacearum strains, increasing their global 31 agricultural impact. However, tropical highland Race 3 biovar 2 (R3bv2) strains do not 32 cause disease in tropical lowlands, even though they are virulent at warm temperatures. 33 We tested the hypothesis that differences in temperature adaptation and competitive 34 fitness explain the uneven geographical distribution of R. solanacearum strains. Using three 35 phylogenetically and ecologically distinct strains, we measured competitive fitness at two 36 temperatures following paired-strain inoculations of their shared host, tomato. Lowland 37 tropical strain GMI1000 was only weakly virulent on tomato under temperate conditions 38 (24°C day/19°C night) but highland tropical R3bv2 strain UW551 and U.S. warm-temperate 39 strain K60 were highly virulent at both 24/19°C and 28°C. Strain K60 was significantly 40 more competitive than both GMI1000 and UW551 in tomato rhizospheres and stems at 41 28°C, and GMI1000 also outcompeted UW551 at 28°C. The results were reversed at cooler 42 temperatures, where highland strain UW551 generally outcompeted GMI1000 and K60 in 43 planta. The superior competitive index of UW551 at 24/19°C suggests that adaptation to 44 cool temperatures could explain why only R3bv2 strains threaten highland agriculture. 45 Strains K60 and GMI1000 each produced different bacteriocins that inhibited growth of 46 UW551 in culture. Such interstrain inhibition could explain why R3bv2 strains do not cause 47 disease in tropical lowlands. 48 49 50
on June 5, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from

AEM4123-14, revised Tropical R. solanacearum outcompetes R3bv2 strains
3
INTRODUCTION 51 52 Ralstonia solanacearum, which causes bacterial wilt disease of many crops, is found on six 53 continents and its hosts include plants in over 50 dicot and monocot families. The 54 pathogen forms a heterogenous species complex that contains thousands of distinct strains 55 (1, 2). No single strain can attack all known hosts, but most members of the species 56 complex can wilt tomato. The best way to manage bacterial wilt is by planting resistant 57 crop varieties, but the high variability of the pathogen complicates breeding efforts because 58 wilt resistance is often strain-specific (3, 4). Sequencing over 20 diverse strains revealed 59 that R. solanacearum has a fluid genome with evidence of extensive horizontal gene 60 transfer (5-7). However, the specific mechanisms by which this pathogen has adapted to 61 diverse environments and hosts are unknown. For example, most strains of R. 62 solanacearum are tropical or subtropical but one highly adapted group, known historically 63 and for regulatory purposes as Race 3 biovar 2 (R3bv2), causes economically important 64 wilt of both potato and tomato in cool highland tropics (8). 65 66 R. solanacearum strains fall into four genetically distinct phylotypes that correspond to 67 their geographic origin: phylotype I (Asia), II (the Americas), III (Africa), and IV (Indonesia) 68 (1) However, because of human activity, multiple phylotypes are now commonly present in 69 a single region. Recent surveys found strains from two or three phylotypes in Guatemala, 70 Florida, Reunion Island, and Cameroon (9-12). Despite this homogenization, highland 71 tropical R3bv2 strains and warm-temperate or tropical strains are not known to cause 72 disease in the same plants or even in the same fields. Field surveys have not found tropical 73 lowland or warm-temperate strains in the cool temperate tropical highlands where R3bv2 74 is endemic (10, 11). Conversely, R3bv2 has not been isolated from wilting plants in tropical 75 lowland regions, even though these strains frequently enter tropical zones via infected 76 plant material and contaminated surface water (13). 77 78 An obvious explanation for the failure of R3bv2 strains to proliferate in the lowland tropics 79 is that they cannot grow or cause disease effectively at high temperatures. However, we 80 previously found no difference in either growth rate or virulence between R3bv2 strain 81
on June 5, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from

AEM4123-14, revised Tropical R. solanacearum outcompetes R3bv2 strains
4
UW551 and a tropical or a subtropical strain at the tropical temperature of 28°C (14, 15). 82 In contrast, the tropical strains were significantly less virulent than R3bv2 at cool 83 temperatures. It appears that UW551 has adapted to growth and virulence at cool 84 temperatures without losing its ability to thrive under warmer conditions. 85 86 These findings raise several questions. If R3bv2 strains are highly virulent at tropical 87 temperatures, why don’t they cause disease in the lowland tropics? This is especially 88 puzzling since R3bv2 strains can overcome the bacterial wilt resistance in most 89 commercial tomato varieties, which should give them a competitive advantage over the 90 lowland strains. Why aren’t strains from multiple phylotypes isolated from the same host 91 plant in regions where they co-occur? How do temperature and intraspecific competition 92 affect the ability of co-occurring R. solanacearum strains to cause disease? 93 94 We hypothesized that R3bv2 is not found in warm lowland sites because of differences in 95 competitive fitness among R. solanacearum strains in those environments. We predicted 96 that lowland tropical R. solanacearum strains have a competitive advantage over tropical 97 highland strains in a warm environment, while highland strains are more competitive at 98 cooler temperatures. Such differences would lead to niche exclusion of non-adapted strains 99 and could explain the observed differences in geographic distribution of strains. We tested 100 this hypothesis with paired-strain competition experiments at two temperatures in planta 101 and in culture. GMI1000 was isolated from tomato in the lowland tropics (16). K60, the R. 102 solanacearum type strain, was isolated from tomato in warm-temperate North Carolina 103 USA (17, 18). UW551 is a typical R3bv2 strain originally isolated from geraniums grown in 104 the cool highlands of Kenya (19). It is highly virulent on both potato and tomato plants at 105 temperatures ranging from 18°C to 28°C (15, 20). 106 107 Under cool temperate conditions, highland tropical strain UW551 was most competitive in 108 rhizospheres of susceptible tomato plants. At a tropical lowland temperature, however, the 109 warm-temperate and tropical lowland strains outcompeted UW551 in both rhizospheres 110 and stems. These more competitive strains secreted proteins that specifically inhibited 111 growth of UW551. 112
on June 5, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from

AEM4123-14, revised Tropical R. solanacearum outcompetes R3bv2 strains
5
113 MATERIALS AND METHODS 114 115 Bacterial strains and culture conditions. The bacterial strains used in this work are 116 listed in Table 1, along with their relevant characteristics. R. solanacearum strains were 117 cultured on casamino acid-peptone-glucose broth medium (CPG) (21) or on CPG plates 118 containing TZC (22) at 28°C, unless otherwise stated. Boucher’s Minimal Medium (BMM) 119 was used where defined medium was required (23). Pectobacterium carotovorum, 120 Xanthomonas campestris pv. vesicatoria, X. campestris pv. campestris, Agrobacterium 121 tumefaciens, Escherichia coli, and Bacillus subtilis, were grown on Luria-Bertani (LB) (24) 122 medium at 37°C or 28°C. Pseudomonas fluorescens and P. aureofaciens were grown at 28°C 123 on King’s B Medium (25). When necessary, antibiotics were added to the medium at the 124 following concentrations: rifampin, 25 mg/L and tetracycline, 15mg/L. Stock inoculum for 125 all strains was prepared from 24 h cultures grown with shaking at 225 rpm at 28°C or 37°C 126 according to the optimal growth temperature of the strain. Bacteria were pelleted and 127 washed with sterile Milli-Q (SMQ) water, re-pelleted, and re-suspended in SMQ water to an 128 O.D.600 of 0.2 and further diluted to the desired final cell density. 129 130 Growth Curves. Growth rates of the four antibiotic-resistant R. solanaceraum strains used 131 in this study, GMI1000-RifR, K60-TetR, K60-RifR, and UW551-TetR,were measured and 132 compared to their wild type (WT) parents, GMI1000, K60 and UW551. Bacterial growth 133 was measured by optical density OD600 using a Synergy/HT microplate reader (Bio Tek 134 Instruments, Winooski, Vermont). Two hundred microliters of a bacterial culture at OD600 135 =0.01 were aliquoted into a 96-well sterile microplate in triplicate and rapid shaking. 136 Optical density was measured every hour for 94 h. Cultures were grown at a constant 28°C, 137 simulating a tropical lowland environment, or at cycling temperatures of 24°C for 14 h and 138 19°C for 10 h (24/19°C), simulating a tropical highland environment. 139 140 CPG broth competition assay. The competitive fitness of strains GMI1000, K60 and 141 UW551 was measured in liquid media by inoculating 50ml of CPG in 125ml Erlenmeyer 142 flasks with a 1:1 bacterial suspension of either GMI1000-RifR and UW551-TetR; GMI1000-143
on June 5, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from

AEM4123-14, revised Tropical R. solanacearum outcompetes R3bv2 strains
6
RifR and K60-TetR; or UW551-TetR and K60-RifR at a final concentration of ~2.0x105 144 CFU/ml of media (1x105 CFU/ml of each competing strain). Cultures were shaken at 225 145 rpm and bacterial populations were measured every 12 h for four days by serial dilution 146 and plating onto selective media. Plates were incubated at 28°C and colonies were counted 147 after 48 h. All three paired strain combinations were grown in triplicate at temperatures 148 simulating tropical lowland and tropical highland conditions as indicated above. 149 150 Sampling bacterial populations in rhizospheres and stems. Unwounded tomato plants 151 (bacterial wilt-susceptible cv. Bonny Best) were inoculated via a naturalistic soil soak as 152 previously described (26). Briefly, 50ml of a bacterial suspension containing a 1:1 mixture 153 of two competing R. solanacearum strains, each at a concentration of 2x107 CFU/ml, was 154 poured over the soil of 16-day-old unwounded tomato plants. Competing strain 155 combinations were as described above and each pairwise combination was performed on 156 at least 45 plants at both temperature conditions. At first sign of wilt symptoms (1-25% of 157 the leaves displaying wilt symptoms), bacterial population sizes were quantified in the 158 stem and rhizosphere of each tomato plant. Rhizospheres were sampled by excising the 159 entire root system from each plant. These were shaken to remove loosely adhering soil, 160 weighed, placed in a 50 ml conical test tube, and vortexed in 10 volumes sterile water for 1 161 min.. The supernatant was serially diluted and plated onto TZC plates containing the 162 relevant selective antibiotic. Bacterial population sizes in tomato stems were measured by 163 grinding a ~100 mg transverse slice of tomato stem harvested at the height of the first leaf 164 petiole in 1ml sterile water. The supernatant was serially diluted and plated onto selective 165 media as above. Colonies were counted after 48 h incubation at 28˚C. R. solanacearum 166 populations in each sample were normalized to log CFU/g of plant tissue +1. 167 168 In planta fitness of individual strains. We determined the fitness of each individual 169 strain in the absence of competition in tomato plant rhizospheres and stems at both 170 tropical and cool temperatures by inoculating tomato plants with one of the three 171 competing strains as described above. Bacterial population sizes in the rhizospheres and 172 stems were measured as described above. Each treatment included ten plants. 173 174
on June 5, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from

AEM4123-14, revised Tropical R. solanacearum outcompetes R3bv2 strains
7
Competitive indices. Competitive index (CI) values were calculated as previously 175 described (27). Briefly, the proportion of the two bacterial populations recovered in a 176 paired strain competition assay was divided by the proportion of the same two bacterial 177 populations in the initial inoculum for that assay. The CI value was calculated individually 178 for each rhizosphere and stem sample and for all three paired strain combinations. A CI 179 value of 1 indicates no competitive advantage of one strain over another. A CI value greater 180 than one indicates an advantage of the strain in the numerator to that in the denominator 181 and a CI value less than one indicates a competitive disadvantage for the strain in the 182 numerator as compared to the strain in the denominator. 183 184 Inhibition Activity Assays. A qualitative assay for antimicrobial activity was adapted from 185 a previously described overlay plate growth inhibition assay (28). R. solanacearum strains 186 were tested for their ability to inhibit growth of other strains from the same species and 187 selected non-R. solanaceraum strains. The overlay consisted of molten media inoculated 188 with a target bacterial strain to a final concentration of 2x105 CFU/ml. This inoculated 189 overlay was vigorously vortexed for 30 s and 20 ml aliquots were quickly poured onto Petri 190 plates containing 20 mL of the solid medium on which the target strain was inoculated. 191 After the agar solidified, 3-mm wells were created in each plate with a sterile #1 cork 192 borer. Each well was then filled with 25 μl of culture supernatant from a test strain. 193 Supernatants were prepared by centrifuging an overnight broth culture of the test strain at 194 6000 x g for 10 min and passing the supernatant through a sterile 0.2 µm cellulose acetate 195 syringe filter. The overlay inhibition plates were incubated at either 24/19°C or 28°C. 196 Plates were photographed at 48 h post inoculation. Zones of inhibition were measured 197 using ImageJ (http://imagej.nih.gov). Each assay was replicated at least three times with 198 two technical replications for each bioreplicate. 199 200 Characterization of inhibitory activity in culture supernatants. Supernatants from R. 201 solanacearum strains GMI1000, UW551 and K60 were obtained from 50 ml overnight 202 cultures grown in CPG medium as described above. Supernatants were passed through a 203 10kDa Amicon centrifugal filter (Millipore Corporation, Billerica, MA) following the 204
on June 5, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from

AEM4123-14, revised Tropical R. solanacearum outcompetes R3bv2 strains
8
manufacturer’s instructions. Filtrate (<10kDa) and concentrate (>10kDa) were each 205 collected and used immediately. Fifty micoliters of both filtrate and concentrate were heat 206 treated for 10 min at 65°C. Proteinase K was added to supernatant filtrate or concentrate to 207 a final concentration of 50 μg/ml and incubated at 37°C for 1 h. Untreated supernatant 208 from each of the three strains was used as control. Overlay plate inhibition activity assays 209 were then performed as described above. 210 211 Statistical Analysis. One-way ANOVA was used to analyze data from in planta fitness 212 assays of individual strains, inhibition activity assays and growth curves. Data describing 213 bacterial population sizes recovered from paired-strain combination in plants were 214 analyzed using a Wilcoxon signed-rank test (29). Statistical analyses were carried out using 215 JMP, Version 9 (SAS Institute Inc., Cary, NC). 216 217 218 RESULTS 219 220 We assessed the competitive interactions among three phylogenetically and ecologically 221 distinct R. solanacearum strains in the contexts of varying temperature and a shared plant 222 host, tomato. We first measured the ability of each strain to multiply in culture and in 223 planta in the absence of competition, and then in paired-strain competition assays in 224 culture and in planta. Because temperature can affect competitive interactions, each 225 experiment was conducted at both a typical lowland tropical temperature of 28°C, and a 226 highland tropical regime of 24°C day/19°C night. 227 228 Antibiotic resistance markers did not decrease strain fitness. Each strain was marked 229 with antibiotic resistance genes to allow quantification of the strains in planta and in 230 paired-strain competition studies. There were no significant differences between growth 231 rates of the antibiotic-resistant strain and its wild-type parent over a 144 h period in 232 minimal broth at either 24/19°C or 28°C, indicating that the antibiotic resistance genes did 233 not cause a measurable fitness loss (data not shown). R. solanacearum strains carrying the 234
on June 5, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from

AEM4123-14, revised Tropical R. solanacearum outcompetes R3bv2 strains
9
spontaneous rifampin resistance used here retain wild-type bacterial wilt virulence on 235 tomato (30, 31). 236 237 In culture, UW551 grew better than GMI1000 and K60 at cool temperatures, but lost 238 its advantage in tropical conditions. To assess the effect of temperature in the absence of 239 competition, we compared growth curves of all three strains in rich CPG broth at either 240 24/19°C or 28°C. As expected, at cool temperatures R3bv2 strain UW551 grew faster than 241 tropical lowland strain GMI1000 or warm-temperate strain K60 (p <.0001, repeated 242 measures ANOVA, Fig. 1). K60 and GMI1000 had similar growth rates (p =.0604, t-test). 243 Under tropical conditions UW551 lost its advantage, with a growth curve indistinguishable 244 from that of K60 at 28°C (p = 0.0667, Fig. 1B). Interestingly, at 28°C GMI1000 grew more 245 slowly than UW551 and K60 (p <.0001, repeated measures ANOVA). 246 247 Without competition, all three strains grew similarly in planta at both temperatures. 248 R. solanacearum strains generally invade host plant roots from the soil, then colonize and 249 spread through the xylem elements of the plant’s vascular system (7, 32). We therefore 250 focused on two plant-associated environments where competing pathogen strains interact: 251 the rhizosphere, known to be a microbially diverse and highly competitive niche (33, 34), 252 and the mid-stem, where R. solanacearum strains can reach very high population sizes and 253 interact with a relatively small number of other xylem-inhabiting microbes (35-37). Any 254 growth differences between strains in planta could result from either competitive 255 interactions between strains or from innate differences in ability to colonize tomato 256 rhizospheres and stems. To separate these traits, we first determined the reproductive 257 success of each strain growing alone in plants following a naturalistic soil-soak inoculation 258 and incubation in either warm or cool conditions. Bacteria were isolated from 259 rhizospheres and stems of tomato plants when wilt symptoms first appeared. The three 260 strains reached indistinguishable population sizes of around 107 CFU/g in rhizospheres of 261 tomato plants incubated at 24/19°C (Fig 2, p = 0.8775). The stems of these same plants 262 contained over 108 CFU/g of each strain, with UW551 colonizing stems slightly better than 263 the other two strains in the cool condition (Fig. 2). At 28°C, the three strains reached 264
on June 5, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from

AEM4123-14, revised Tropical R. solanacearum outcompetes R3bv2 strains
10
comparable cell densities of around 2.5 x 108 CFU/g in tomato rhizospheres (p = 0.8739, 265 one-way ANOVA) and 2.6 x 109 CFU/g in stems (p = 0.4836, one-way ANOVA). 266 267 Strain competitive fitness differed in culture, with warm temperatures favoring K60 268 and cooler temperatures favoring R3bv2 strain UW551. To determine the effect of 269 temperature on bacterial strain interactions in the absence of a plant host, we conducted 270 paired competition assays in vitro under both temperature regimes. Bacterial populations 271 were enumerated after 48 h growth in CPG rich broth inoculated with equal numbers of 272 each competing strain. At the cooler temperatures in culture, GMI1000 was outcompeted 273 by both K60 (competitive index value (CI) of 2.65,) and by UW551 (CI = 6.17) (Fig 3, Table 274 2). There was no significant difference in competitiveness between UW551 and K60 at cool 275 temperatures, although the mean population size of K60 was slightly larger than that of 276 UW551 (1.08 x 106 CFU/ml and 7.06 x 105 CFU/ml, respectively) (Table 2). At 28°C, warm-277 temperate strain K60 easily outcompeted both tropical strain GMI1000 (CI=6.37 x 104) and 278 R3bv2 strain UW551 (CI=8.57 x 104). UW551 was outcompeted to a lesser degree at 28°C 279 by tropical strain GMI1000 (CI = 1.74x10-1, Fig. 3, Table 2). Overall, temperate strain K60 280 dominated in culture, outcompeting both other strains at 28°C and GMI1000 at 24°/19°C, 281 while matching UW551 at the cooler temperatures. 282 283 In tomato plants, tropical highland strain UW551 outcompeted other strains under 284 cool conditions. Having established the effects of temperature on competitive interactions 285 between R. solanacearum, we added a biologically relevant context for these interactions in 286 the form of the plant environment. Unwounded tomato plants were inoculated with a 1:1 287 mixture of each pathogen strain pair, and the population size of each strain was measured 288 in rhizospheres and stems when symptoms first appeared (approximately 4-8 days post-289 inoculation). At a cool 24°/19°C, UW551 significantly outcompeted both GMI1000 and K60 290 in tomato rhizospheres, but K60 and GMI1000 reached similar population sizes when they 291 competed under these conditions (Fig. 4, Table 3). Once the bacteria invaded plant stems, 292 UW551 outcompeted K60 to an even greater degree, with a CI of 4.66 x 106. Results of the 293 other competitions in stems at 24°/19°C were more complex. GMI1000 slightly 294
on June 5, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from

AEM4123-14, revised Tropical R. solanacearum outcompetes R3bv2 strains
11
outcompeted UW551 (CI=12) but it was strongly outcompeted by warm-temperate strain 295 K60 (CI=6.84 x 109) (Fig. 5, Table 3).. 296 297 In planta, warm-temperate strain K60 outcompeted the other strains at 28°C. When 298 strains were co-inoculated onto tomato plants growing in a lowland tropical environment, 299 warm-temperate strain K60 outcompeted tropical highland strain UW551 and tropical 300 lowland strain GMI1000 in both rhizospheres and stems (Fig. 4 and 5). With CI values 301 greater than 108 in the stems of infected tomato plants, K60 was a much stronger 302 competitor when strains shared this specialized niche (Table 3). Tropical lowland strain 303 GMI1000 also outcompeted UW551 in both tomato rhizospheres and stems at 28°C (Fig. 4 304 and 5, Table 3). Together these data suggest that tropical highland strain UW551 is not 305 well adapted to compete with warm-temperate and tropical lowland strains in plants 306 growing at warm temperatures (summarized in Fig. 6). 307 308 Strains K60 and GMI1000 secrete intra-species growth inhibitors. The most direct 309 explanation for the observed differences in competitive fitness between R. solanacearum 310 strains is interstrain chemical inhibition. To test this hypothesis, we measured the ability of 311 each strain to inhibit growth of the others in vitro using an assay wherein cell-free 312 supernatants of one strain were placed in a well on an agar plate overlaid with a lawn of 313 the putative target strain. These experiments revealed that both warm-temperate strain 314 K60 and tropical lowland strain GMI1000 produced diffusible factor(s) that limited growth 315 of highland tropical strain UW551, resulting in a clear zone of inhibition around the well 316 (Fig. 7). GMI1000 supernatants slightly inhibited growth of K60, while K60 supernatants 317 had a noticeably greater effect on GMI1000, indicating that K60 and GMI1000 produce 318 different inhibitors. In contrast, UW551 did not inhibit either K60 or GMI1000 (Fig. 7). No 319 strain was inhibited by its own supernatant. Temperature was not a factor in inhibitor 320 production or response, since similar zones of inhibition were observed when cultures 321 were grown and assayed at 24/19°C (data not shown). 322 323 To determine if these inhibitors affected strains outside the R. solanacearum species 324 complex, we used the plate overlay assay to measure the effects of supernatants from K60, 325
on June 5, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from

AEM4123-14, revised Tropical R. solanacearum outcompetes R3bv2 strains
12
UW551, and GMI1000 on growth of six plant-associated bacterial species, as well as E. coli. 326 None of the R. solanacearum culture supernatants noticeably affected growth of 327 Pectobacterium carotovorum subsp. carotovorum, Xanthomonas campestris pv. vesicatoria, 328 X. campestris pv. campestris, Agrobacterium tumefaciens, Bacillus subtilis, Pseudomonas 329 fluorescens, P. aureofaciens, or E. coli (data not shown). This suggests that R. solanacearum 330 strains produce narrowly targeted intra-specific inhibitors. 331 332 Preliminary characterization of inhibitors. We further characterized the compound(s) 333 responsible for the zones of growth inhibition. Cell-free culture supernatants from K60 334 and GMI1000 completely lost their ability to inhibit growth of other R. solanacearum 335 strains after 10 min at 65°C or 1 h incubation with 50 ug/ml Proteinase K at 37°C. When 336 cell-free culture supernatants were passed through a 10 kDa centrifugal concentrator, the 337 eluates had no detectable activity, but the concentrated liquid on the membrane retained 338 inhibitory activity (Table 4). Thus, the inhibitory activity is proteinaceaous and larger than 339 10 kDa. These results suggest that K60 and GMI1000 inhibit growth of UW551 with 340 bacteriocins, commonly defined as antibacterial proteins that affect only closely related 341 strains (38-40). 342 343 344 DISCUSSION 345 346 To our knowledge, only R3bv2 strains of R. solanacearum cause crop losses in cool-347 temperate tropical highlands (41). Based on extensive field observations, Thurston 348 speculated that R3bv2 strains are specifically adapted to lower temperatures (42). Some 349 laboratory studies show that R3bv2 strains grow better at cool temperatures in culture 350 than tropical strains, but growth in culture at a given temperature does not always 351 correlate with ability to cause disease (14, 41, 43). Under conditions tested here, R3bv2 352 strain UW551 reached modestly higher cell densities in culture than K60 or GMI1000 at the 353 cool temperature of 24°/19°C. 354 355
on June 5, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from

AEM4123-14, revised Tropical R. solanacearum outcompetes R3bv2 strains
13
The cold adaptation phenotype of R3bv2 strains is more than a physiological capacity to 356 grow at cooler temperatures, because it is specifically expressed during interaction with 357 host plants. For example, at the typical potato storage temperature of 4°C, UW551 survives 358 in potato tubers two to four months longer than K60 or GMI1000 even though in 4°C water 359 K60 survives better than UW551 (14). Similarly, R3bv2 strains are dramatically more 360 virulent on tomato than lowland tropical strains at cool temperatures (14, 15). Other 361 controlled inoculation studies have reported that R3bv2 strains can wilt tomato and potato 362 plants at temperatures as low as 16°C, while warm-temperate and tropical strains like K60 363 and GMI1000 cause little or no disease below 20°C (22, 43, 44). We found that at 24°/19°C, 364 UW551 colonized tomato stems better than the other two strains in the absence of 365 competition. Thus, a substantial body of research supports the idea that R3bv2 strains are 366 better adapted to survive in planta and cause disease at cool temperatures than other R. 367 solanacearum strains. It should be noted however that cold adaptation is not entirely 368 unique to R3bv2 strains, since under laboratory conditions phylotype II sequevar 4 strains 369 isolated from Pothos and Anthurium could wilt tomato and potato plants at 18°C (45). 370 371 Although differential temperature adaptation appears to explain the absence of lowland 372 tropical R. solanacearum strains from cool regions, it does not explain why R3bv2 strains 373 are not a problem in warmer cropping systems. In the laboratory at 28 to 30°C, there is 374 little difference between R3bv2 and tropical strains with respect to either growth in culture 375 or virulence on plants (14, 15). Consistently with these previous studies, R3bv2 strain 376 UW551 grew as well as warm-temperate strain K60 and tropical strain GMI1000 at 28°C 377 under our conditions. Accumulated experimental data thus demonstrate that R3bv2 strains 378 are capable of infecting, colonizing, and wilting plants at warm temperatures in the absence 379 of competition. 380 381 No obvious physical barrier confines R3bv2 strains to the tropical highlands. These 382 widespread bacteria contaminate surface waterways in many tropical highlands, including 383 rivers used to irrigate downstream lowlands (11). Such rivers transport R3bv2 cells from 384 highlands to lowland fields where they likely encounter potential hosts, such as tomato. 385 However, R3bv2 has not been identified from tropical lowland sites. Our goal was to 386
on June 5, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from

AEM4123-14, revised Tropical R. solanacearum outcompetes R3bv2 strains
14
understand why R3bv2 strains are not found in the warm lowland tropics despite their 387 demonstrated biological ability to thrive and cause disease in hot conditions. One 388 explanation for this absence is that R3bv2 strains are directly outcompeted by closely 389 related lowland R. solanacearum strains that occupy the same niche in host plant xylem 390 tissue. We explored this hypothesis by co-inoculating tomato plants with pairs of 391 ecologically distinct R. solanacearum strains under controlled conditions. 392 393 We first measured the independent growth of the three bacterial strains on and in tomato 394 plants at warm and cool temperatures. Although highland tropical strain UW551 reached a 395 slightly higher population size in tomato stems at 24/19°C, all strains colonized the plant 396 habitats equally well in the other three conditions. These results indicated that any 397 observed differences in population size following co-inoculation resulted from competitive 398 interactions between bacterial strains. 399 400 Following paired-strain inoculation of tomato plants, we determined the relative 401 competitive indices (CIs) of each strain in both the rhizospheres and the mid-stems. 402 Although there was little or no difference between the three strains in their ability to 403 colonize tomato without competition at either temperature, highland tropical strain 404 UW551 generally outcompeted K60 and GMI1000 in planta at cool temperatures, while 405 warm-temperate strain K60 was the best competitor under tropical lowland conditions. 406 These results demonstrate that in vitro experiments cannot accurately predict competitive 407 interactions among strains in plants. Whereas K60 and UW551 grew to equal population 408 sizes in paired-strain competition in culture at 24°/19°C, at that temperature UW551 409 outcompeted K60 by three and six orders of magnitude in tomato rhizospheres and stems, 410 respectively. The host plant environment is thus a critically important factor mediating 411 interactions between R. solanacearum strains. 412 413 Although strain CI values varied among plants within assays (Supplemental Fig. 2 and 3), 414 the mean CI for a given strain pair was generally much higher in stems than in 415 rhizospheres. For example, K60 outcompeted UW551 by 108-fold in tomato rhizospheres 416 at 28°C, but it had a 4.89 x 108-fold advantage in tomato midstems. These large differences 417
on June 5, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from

AEM4123-14, revised Tropical R. solanacearum outcompetes R3bv2 strains
15
may reflect the fact that the inoculum containing equal numbers of each competing strain 418 was applied directly to the soil, so at the beginning of the assay the rhizosphere contained 419 comparable populations of both strains. However, bacteria in the plant midstem have 420 survived intense successive selective pressures that would magnify any minor advantage of 421 a strain. These pressures include persisting in the pathogen’s microbially diverse infection 422 court on the root surface, invading the plant by traversing the root cortex and entering the 423 developing protoxylem, overcoming plant defenses, multiplying in the relatively nutrient-424 poor xylem fluid, and finally spreading up into aboveground stems. During these processes 425 the co-inoculated strains are in direct competition for optimal colonization sites and 426 limited nutrients. Moreover, they are confined together in xylem vessels; this close physical 427 proximity would strongly favor bacteria that could chemically inhibit other strains. 428 429 We explored the possibility of such a direct inhibitor using paired-strain growth assays in 430 culture. Indeed, both GMI1000 and K60 out-competed R3bv2 strain UW551 in rich 431 medium. These in vitro CI differences suggested that the tropical lowland strains’ higher 432 competitive fitness could result from chemical inhibition that acts even in the absence of a 433 plant host. This would not be surprising because microbes often use antibiosis against 434 competitors (46). As described above, the biology of R. solanacearum would make chemical 435 inhibition an effective competitive strategy, especially within xylem vessels. Niche theory, 436 increasingly applied to microbial ecology, predicts that antibiosis would be particularly 437 important in competition with a closely related strain (47, 48). Different species that 438 occupy the same habitat can functionally partition resources to avoid competition, but 439 members of a single species that exploit the same niche must compete directly for 440 resources (49). 441 442 We found that strains K60 and GMI1000 each produced distinct proteins that inhibited 443 growth of UW551 on plates. These proteins had no effect on seven other bacterial species, 444 indicating that they are putative bacteriocins, proteins that specifically target conspecific or 445 closely related strains (28, 50). R. solanacearum has long been known to produce 446 bacteriocins, but their biological role(s) were unknown (38, 51-53). Many plant-associated 447 bacteria make bacteriocins, such as agrocin, produced by Agrobacterium radiobacter strain 448
on June 5, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from

AEM4123-14, revised Tropical R. solanacearum outcompetes R3bv2 strains
16
K84. This bacteriocin excludes the crown gall pathogen A. tumefaciens from the infection 449 court so effectively that A. radiobacter is widely used for biocontrol (54). 450 451 Interestingly, both the lowland strains studied here could inhibit highland tropical strain 452 UW551. This observation is consistent with the latitudinal diversity gradient hypothesis, 453 which postulates that adaptation to biotic competition drives increasing diversity in 454 tropical zones, while adaptation to environmental stresses is a more important limiting 455 factor for organisms living in colder habitats closer to the poles or at higher elevation (55). 456 This geographic diversity gradient has been observed within species (56) and in microbial 457 populations (57). It is possible that by adapting to cooler temperatures in the tropical 458 highlands, R3bv2 strains escaped the interstrain competition experienced by lowland 459 tropical R. solanacearum strains and as a result they were no longer under selection 460 pressure to maintain bacteriocin production and immunity. 461 462 We identified a consistent correlation between the ability of strains K60 and GMI1000 to 463 outcompete and exclude UW551 in planta and production by these strains of bacteriocin(s) 464 that specifically inhibited UW551 growth. Bacteriocin production could explain why 465 GMI1000 outcompeted UW551 in tomato stems at 24/19°C even though UW551 466 outcompeted GMI1000 in the rhizospheres of these same plants and GMI1000 is not as 467 virulent as UW551 at this temperature. To directly determine if these bacteriocins 468 contribute to in planta competitive fitness of R. solanacearum strains, it will be necessary to 469 identify and mutate the gene(s) encoding bacteriocin production and see if the resulting 470 mutant strain has reduced competitive fitness. 471 472 Our results suggest that interstrain competition mediated by bacteriocins can explain the 473 geographical distribution of R3bv2 strains. However, in the field other factors may also 474 contribute to this phenomenon. For example, R3bv2 may be a generally poor competitor in 475 lowland soils relative to tropical R. solanacearum strains. It would be interesting to 476 compare survival of R3bv2 and tropical strains in the natural microbiome of plant roots in 477 live tropical soils. Alternatively, tomato plants growing at 24/19°C and 28°C may vary in 478
on June 5, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from

AEM4123-14, revised Tropical R. solanacearum outcompetes R3bv2 strains
17
antimicrobial defenses or other physiological attributes that differentially affect the 479 bacterial strains studied here. 480 481 The bacteriocins described here may be practically useful for management of potato brown 482 rot and southern wilt of geranium. Because R. solanacearum R3bv2 causes both diseases, it 483 inflicts both direct yield losses and indirect losses associated with the fact that R3bv2 is a 484 quarantine pathogen in Europe and North America, and a highly regulated Select Agent 485 pathogen in the U.S. We identified at least two different bacteriocins that inhibit UW551, 486 which is typical of the nearly clonal group of R3bv2 strains that have been globally 487 distributed along with the potato (29). Transgenic potato or geranium plants that express 488 these bacteriocins might have increased resistance to infection by R3bv2 strains of R. 489 solanacearum. 490 491 492 ACKNOWLEDGEMENTS 493 494 This study was supported by a National Science Foundation Predoctoral Fellowship to 495 A.I.H, a USDA-ARS Floral and Nursery Crops Research Initiative grant, a USDA-Hatch 496 project WIS01776 and by the University of Wisconsin-Madison College of Agricultural and 497 Life Sciences. The authors gratefully acknowledge Amy Charkowski, Jeri Barak and 498 Jonathan M. Jacobs for insightful discussions and Nick Keuler for statistical advice. We also 499 thank Nicole Bacheller, Jordan Weibel, and Manav Khanna for technical assistance. 500 501
on June 5, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from

AEM4123-14, revised Tropical R. solanacearum outcompetes R3bv2 strains
18
REFERENCES 502 503 1. Fegan M, Prior P. 2005. How complex is the "Ralstonia solanacearum species 504 complex"?, p 449-461. In Allen C, Prior P, Hayward AC (ed), Bacterial Wilt Disease 505 and the Ralstonia solanacearum species complex. APS Press, St. Paul. 506 2. Hayward AC. 1994. Systematics and phylogeny of Pseudomonas solanacearum and 507 related bacteria, p 123-135. In Hayward AC, Hartman GL (ed), Bacterial Wilt: The 508 Disease and its Causative Agent, Pseudomonas solanacearum CAB International, 509 Wallingford. 510 3. Liao B, Shan ZH, Duan NX, Tan YJ, Lei Y, Li D. 1998. Relationship between latent 511 infection and groundnut bacterial wilt resistance, p 294-299. In Prior P, Allen C, 512 Elphinstone J (ed), Bacterial Wilt Disease: Molecular and Ecological Aspects. 513 Springer Verlag, Berlin. 514 4. Hanson PM, Wang J, Licardo O, Haudin, Mah S, Hartman GL, Lin Y, Chen J. 1996. 515 Variable reaction ot tomato lines to bacterial wilt evaluated at several locations in 516 southeast Asia. Hort Sci 31:143-146. 517 5. Remenant B, Coupat-Goutaland B, Guidot A, Cellier G, Wicker E, Allen C, Fegan 518
M, Pruvost O, Elbaz M, Calteau A, Salvignol G, Mornico D, Mangenot S, Barbe V, 519 Médigue C, Prior P. 2010. Genomes of three tomato pathogens within the Ralstonia 520 solanacearum species complex reveal significant evolutionary divergence. BMC 521 Genomics 11:379. 522 6. Remenant B, Cambiaire J-CD, Cellier G, Barbe V, Medigue C, Jacobs JM, Fegan M, 523 Allen C, Prior P. 2011. Phylotype IV strains of Ralstonia solanacearum, R. syzygii 524 and the Blood Disease Bacterium form a single genomic species despite their 525 divergent life-styles PLoS One 6:e24356. 526 7. Genin S, Denny TP. 2012. Pathogenomics of the Ralstonia solanacearum species 527 complex. Annu Rev Phytopathol 50:67-89. 528 8. Allen C, Kelman A, French ER. 2001. Brown rot of potatoes, p 11-13. In Stevenson 529 WR, Loria R, Franc GD, Weingartner DP (ed), Compendium of Potato Diseases, 2nd 530 ed. APS Press, St. Paul, MN. 531 9. Hong J, Momol M, Jones J, Ji P, Olson S, Allen C, Perez A, Pradhanang P, Guven K. 532 2008. Detection of Ralstonia solanacearum in irrigation ponds and aquatic weeds 533 associated with ponds in North Florida. Plant Dis 92:1674-1682. 534 10. Toukam G, Cellier G, Wicker E, Guilbaud C, Kahane R, Allen C, Prior P. 2009. 535 Broad diversity of Ralstonia solanacearum strains in Cameroon. Plant Dis 93:1123-536 1130. 537 11. Sanchez-Perez A, Mejia L, Fegan M, Allen C. 2008. Diversity and distribution of 538 Ralstonia solanacearum strains in Guatemala and rare occurence of tomato fruit 539 infection. Plant Pathol 57:320-331. 540 12. Cellier G, Prior P. 2010. Deciphering phenotypic diversity of Ralstonia 541 solanacearum strains pathogenic to potato. Phytopathology 11:1250-1261. 542 13. Elphinstone JG. 2005. The current bacterial wilt situation: A global overview, p 9-543 28. In Allen C, Prior P, Hayward AC (ed), Bacterial Wilt: The Disease and the 544 Ralstonia solanacearum Species Complex. APS Press, St Paul. 545
on June 5, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from

AEM4123-14, revised Tropical R. solanacearum outcompetes R3bv2 strains
19
14. Milling A, Meng F, Denny TP, Allen C. 2009. Interactions with hosts at cool 546 temperatures, not cold tolerance, explain the unique epidemiology of Ralstonia 547 solanacearum Race 3 biovar 2. Phytopathology 99:1127-1134. 548 15. Swanson J, Yao J, Tans-Kersten J, Allen C. 2005. Behavior of Ralstonia 549 solanacearum Race 3 biovar 2 during latent and active infection of geranium. 550 Phytopathology 95:136-146. 551 16. Salanoubat M, Genin S, Artiguenave F, Gouzy J, Mangenot S, Arlat M, Billault A, 552 Brottier P, Camus JC, Cattolico L, Chandler M, Choisne N, Claudel-Renard C, 553 Cunnac S, Demange N, Gaspin C, Lavie M, Moisan A, Robert C, Saurin W, Schiex 554 T, Siguier P, Thebault P, Whalen M, Wincker P, Levy M, Weissenbach J, Boucher 555 CA. 2002. Genome sequence of the plant pathogen Ralstonia solanacearum. Nature 556 415:497-502. 557 17. Kelman A. 1953. The bacterial wilt caused by Pseudomonas solanacearum. North 558 Carolina Agricultural Experiment Station, Technical Bull No. 99. 559 18. Remenant B, Babujee L, Lajus A, Médigue C, Prior P, Allen C. 2012. Sequencing of 560 K60, type strain of the major plant pathogen Ralstonia solanacearum. J Bacteriol 561 194:2742-2743. 562 19. Gabriel DW, Allen C, Schell M, Denny TP, Greenberg JT, Duan YP, Flores-Cruz Z, 563 Huang Q, Clifford JM, Presting G, González ET, Reddy J, Elphinstone J, Swanson 564 J, Yao J, Mulholland V, Liu L, Farmerie W, Patnaikuni M, Balogh B, Norman D, 565 Alvarez A, Castillo JA, Jones J, Saddler G, Walunas T, Zhukov A, Mikhailova N. 566 2006. Identification of open reading frames unique to a select agent: Ralstonia 567 solanacearum Race 3 Biovar 2. Mol Plant-Microbe Interact 19:69-79. 568 20. Williamson L, Kazuhiro N, Hudelson B, Allen C. 2002. Ralstonia solanacearum 569 Race 3, Biovar 2 strains isolated from geranium are pathogenic on potato. Plant Dis 570 86:987-991. 571 21. Hendrick CA, Sequeira L. 1984. Lipopolysaccharide-defective mutants of the wilt 572 pathogen Pseudomonas solanacearum. Appl Environ Microbiol 48:94-101. 573 22. Kelman A. 1954. The relationship of pathogenicity of Pseudomonas solanacearum to 574 colony appearance in tetrazolium medium. Phytopathology 44:693-695. 575 23. Boucher C, Barberis P, Trigalet A, Demery D. 1985. Transposon mutagenesis of 576 Pseudomonas solanacearum: isolation of Tn5-induced avirulent mutants. J Gen 577 Microbiol 131:2449-2457. 578 24. Miller JH. 1992. A short course in bacterial genetics: a laboratory manual and 579 handbook for Escherichia coli and related bacteria. Cold Spring Harbor Laboratory 580 Press, Cold Spring Harbor, NY. 581 25. Schaad NW, Jones JB, Chun W. 2001. Laboratory guide for the identification of 582 plant pathogenic bacteria, 3rd ed. APS Press, St Paul. 583 26. Tans-Kersten J, Guan Y, Allen C. 1998. Ralstonia solanacearum pectin 584 methylesterase is required for growth on methylated pectin but not for bacterial 585 wilt virulence. Appl Environ Microbiol 64:4918-4923. 586 27. Monk I, Casey P, Cronin M, Gahan C, Hill C. 2008. Development of multiple strain 587 competitive index assays for Listeria monocytogenes using pIMC; a new site-specific 588 integrative vector. BMC Microbiology 8:96. 589
on June 5, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from

AEM4123-14, revised Tropical R. solanacearum outcompetes R3bv2 strains
20
28. Sisto A, Cipriani MG, Morea M, Lonigro SL, Valerio F, Lavermicocca P. 2010. An 590 Rhs-like genetic element is involved in bacteriocin production by Pseudomonas 591 savastanoi pv. savastanoi. Anton Leeuw Int J G 98:505-517. 592 29. Wilcoxon F. 1945. Individual comparisons by ranking methods. Biometrics Bull 593 1:80-83. 594 30. Yao J, Allen C. 2006. Chemotaxis is required for virulence and competitive fitness of 595 the bacterial wilt pathogen Ralstonia solanacearum. J Bacteriol 188:3697-3708. 596 31. Scherf JM, Milling A, Allen C. 2010. Moderate temperature fluctuations rapidly 597 reduce viability of Ralstonia solanacearum Race 3 biovar 2 in infected geranium, 598 tomato, and potato. Appl Environ Microbiol 76:7061-7067. 599 32. Valls M, Genin S, Boucher C. 2006. Integrated regulation of the type III secretion 600 system and other virulence determinants in Ralstonia solanacearum. PLoS Pathog 601 2:798-807. 602 33. Mendes R, Kruijt M, de Bruijn I, Dekkers E, van der Voort M, Schneider JHM, 603 Piceno YM, DeSantis TZ, Andersen GL, Bakker PAHM, Raaijmakers JM. 2011. 604 Deciphering the rhizosphere microbiome for disease-suppressive bacteria. Science 605 332:1097-1100. 606 34. Berendsen RL, Pieterse CMJ, Bakker PAHM. 2012. The rhizosphere microbiome 607 and plant health. Trends in Plant Sci 17:478-486. 608 35. Cruz APZ, Ferreira V, Pianzzola MJ, Siri MI, Coll NS, Valls M. 2013. A novel, 609 sensitive method to evaluate potato germplasm for bacterial wilt resistance using a 610 luminescent Ralstonia solanacearum reporter strain. Mol Plant-Microbe Interact 611 27:277-285. 612 36. Grimault V, Anais, G. and Prior, P. 1994. Distribution of Pseudomonas 613 solanacearum in the stem tissues of tomato plants with different levels of resistance 614 to bacterial wilt. Plant Pathol 43:663-668. 615 37. Yadeta KA, J. Thomma BPH. 2013. The xylem as battleground for plant hosts and 616 vascular wilt pathogens. Front in Plant Sci 4:97. 617 38. Cuppels DA, Hanson RS, Kelman A. 1978. Isolation and characterization of a 618 bacteriocin produced by Pseudomonas solanacearum. J Gen Microbiol 109:295-303. 619 39. Riley MA, Wertz JE. 2002. Bacteriocins: evolution, ecology, and application. Annu 620 Rev Microbiol 56:117-137. 621 40. Riley M. 2011. Bacteriocin-mediated competitive interactions of bacterial 622 populations and communities, p 13-26. In Drider D, Rebuffat S (ed), Prokaryotic 623 Antimicrobial Peptides: From Genes to Aplications. Springer Science-Business LLC. 624 41. Hayward AC. 1991. Biology and epidemiology of bacterial wilt caused by 625 Pseudomonas solanacearum. Annu Rev Phytopathol 29:65-87. 626 42. Thurston HD. 1963. Bacterial wilt of potatoes in Colombia. Am Potato J 40:381-390. 627 43. Ciampi L, Sequeira, L. and French, E.R. 1980. Latent infection of potato tubers by 628 Pseudomonas solanacearum. Am Potato J 57:377-386. 629 44. Ciampi L, Sequeira L. 1980. Influence of temperature on virulence of Race 3 strains 630 of Pseudomonas solanacearum. Am Potato J 57:307-317. 631 45. Bocsanczy AM, Achenbach UCM, Mangravita-Novo A, Yuen J, Norman DJ. 2012. 632 Comparative effect of low temperature on virulence and twitching motility of 633 Ralstonia solanacearum strains present in Florida. Phytopathology 102:185-194. 634
on June 5, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from

AEM4123-14, revised Tropical R. solanacearum outcompetes R3bv2 strains
21
46. Bardgett RD, van der Putten WH. 2014. Belowground biodiversity and ecosystem 635 functioning. Nature 515:505-511. 636 47. Horner-Devine MC, Bohannan BJM. 2006. Phylogenetic clustering and 637 overdispersion in bacterial communities. Ecology 87:S100-S108. 638 48. Lennon JT, Aanderud ZT, Lehmkuhl BK, Schoolmaster DR. 2012. Mapping the 639 niche space of soil microorganisms using taxonomy and traits. Ecology 93:1867-640 1879. 641 49. Levine JM, HilleRisLambers J. 2009. The importance of niches for the maintenance 642 of species diversity. Nature 461:254-257. 643 50. Holtsmark I, Eijsink VGH, Brurberg MB. 2008. Bacteriocins from plant pathogenic 644 bacteria. FEMS Microbiol Lett 280:1-7. 645 51. Chen WY, Echandi E. 1984. Effects of avirulent bacteriocin producing strains of 646 Pseudomonas solanacearum on the control of bacterial wilt of tobacco. Plant Pathol 647 33:245-253. 648 52. Arwiyanto T, Goto M, Takikawa Y. 1993. Characteristics of bacteriocins produced 649 by Pseudomonas solanacearum. Ann Phytopath Soc Jpn 59:114-122. 650 53. Okabe N. 1954. Studies on Pseudomonas solanacearum. V. Antagonism among the 651 strains of P. solanacearum. Rep Fac Agric Shizuoka Univ 4:37-40. 652 54. Kim J-G, Park BK, Kim S-U, Choi D, Nahm BH, Moon JS, Reader JS, Farrand SK, 653 Hwang I. 2006. Bases of biocontrol: Sequence predicts synthesis and mode of action 654 of agrocin 84, the Trojan Horse antibiotic that controls crown gall. Proc Natl Acad 655 Sci USA 103:8846-8851. 656 55. Schemske DW, Mittelbach GG, Cornell HV, Sobel JM, Roy K. 2009. Is there a 657 latitudinal gradient in the importance of biotic interactions? Annu Rev Ecol Syst 658 40:245-269. 659 56. Araújo MS, Costa-Pereira R. 2013. Latitudinal gradients in intraspecific ecological 660 diversity. Biol Lett 9. 661 57. Fuhrman JA, Steele JA, Hewson I, Schwalbach MS, Brown MV, Green JL, Brown 662 JH. 2008. A latitudinal diversity gradient in planktonic marine bacteria. Proc Natl 663 Acad Sci USA 105:7774-7778. 664 58. Colburn-Clifford JM, Scherf JM, Allen C. 2010. Ralstonia solanacearum Dps 665 contributes to oxidative stress tolerance and to colonization of and virulence on 666 tomato plants. Appl Environ Microbiol 76:7392-7399. 667 59. Yap M-N, Barak JD, Charkowski AO. 2004. Genomic diversity of Erwinia 668 carotovora subsp. carotovora and its correlation with virulence. Appl Environ 669 Microbiol 70:3013-3023. 670 60. Hanahan D. 1983. Studies on transformation of Escherichia coli with plasmids. J Mol 671 Biol 166:557-580. 672
673 674
on June 5, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from

AEM4123-14, revised Tropical R. solanacearum outcompetes R3bv2 strains
22
Table 1. Bacterial strains used in this study, with relevant characteristics 675 Strain Phylotype /
Sequevara
Race / Biovarb
Geographic Origin
Isolation Host
Reference
Ralstonia solanacearum
(alternate name)
K60 (UW25) IIA/7 1/1 North
Carolina, USA
Tomato (22)
K60-RifR IIA/7 1/1 North
Carolina, USA
Tomato (30)
K60-TetR IIA/7 1/1 North
Carolina, USA
Tomato (30)
UW551 IIB/I 3/2 Kenya Geraniu
m
(20)
UW551-TetR IIB/I 3/2 Kenya Geraniu
m
(58)
GMI1000 (UW643) I/18 1/3 French
Guyana
Tomato (23)
GMI1000-RifR I/18 1/3 French
Guyana
Tomato (14)
Pectobacterium
carotovorum subsp.
carotovorum WPP14
n/a n/a Wisconsin,
USA
Potato (59)
Xanthomonas campestris
pv. vesicatoria
n/a n/a Allen lab
collection
Xanthomonas campestris
pv. campestris
n/a n/a Allen lab
collection
Agrobacterium
tumefaciens n/a n/a Allen lab
collection
Escherichia coli (DH5α) n/a n/a (60)
Bacillus subtilis
n/a n/a Allen lab
collection
Pseudomonas
fluorescens
n/a n/a Allen lab
collection
Pseudomonas
aureofaciens
n/a n/a Allen lab
collection
676
on June 5, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from

AEM4123-14, revised Tropical R. solanacearum outcompetes R3bv2 strains
23
aPhylotype corresponds to strain phylogeny and geographic origin; sequevar clusters strains by phylogenetic 677 analysis of partial endoglucanase gene sequence. 678 679 bRace is a historic designation with little predictive value, based loosely on host range. Strains are divided into 680 biovars based on ability to oxidize a panel of disaccharides and hexose alcohols. 681 682
on June 5, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from

AEM4123-14, revised Tropical R. solanacearum outcompetes R3bv2 strains
24
Table 2. Results of paired-strain competitions in culture with statistical analysis 683 684 Competing Strains Competing Strains Competing Strains
GMI1000a K60b
GMI1000a UW551b K60b UW551a
CPG at 24/19°C
Mean Population
Size (CFU/g)
8.44x104 2.24x105
1.62x105 1.00x106
1.08x106 7.06x105
Competitive Indexc 2.65 6.17 1.53 Fd 11.847 19.0799 0.0676
p-valuee 0.0034 0.0005 0.7982
CPG at 28°C
Mean Population
Size (CFU/g)
1.94x104 1.24x109
2.37x107 4.15x106
2.44x104 2.09x109
Competitive Indexc 6.37x104 1.74x10-1 8.57x104 Fd 3340 5.355 2997
p-valuee <.0001 0.0343 <.0001 685 a This strain is the reference strain for competitive index calculations (denominator). 686 b This strain is the competing strain for competitive index calculations (numerator). 687 c Competitive index values represent the proportional competitive advantage of the competing strain over the 688 reference strain. A competitive index of 1 indicates no competitive advantage. 689 d F is the critical value associated with one-way ANOVA statistical analysis. 690 e p-values indicate the level of difference in bacterial population sizes between competing strains in each paired-691 strain combination in liquid broth medium as determined by one-way ANOVA. 692 693 694 695
on June 5, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from

AEM4123-14, revised Tropical R. solanacearum outcompetes R3bv2 strains
25
Table 3. Results of paired-strain competitions in planta with statistical analysis 696 697 Competing Strains Competing Strains Competing Strains
GMI1000a K60b
GMI1000a UW551b K60a UW551b
Rhizosphere at 24/19°C
Median Population
Size (CFU/g)
4.66x106 3.70x107
1.27 x106 1.20x107
1.08x105 8.97x107
Competitive Indexc 7.81x103 10 6.5x10-3 Zd 0.703 3.39 5.85
p-valuee .4820 <.0007 <.0001
Rhizosphere at 28°C
Median Population
Size (CFU/g)
6.42x107 2.59x105
1.20x107 5.25x105
4.59x107 2.58x105
Competitive Indexc -243 -5x10-5 -108 Zd 5.93 3.24 7.96
p-valuee <.0001 .0012 <.0001 Stem at 24/19°C
Median Population
Size (CFU/g)
4.66x106 5.33x109
1.98x108 1.40x106
1 2.99x108
Competitive Indexc 6.84x10-9 -12 4.66x106 Zd 4.68 0.114 4.18
p-valuee <.0001 .909 <.0001
Stem at 28°C
Median Population
Size (CFU/g)
1 1.90x109
2.31x108 1
6.41x108 1
Competitive Indexc 1.39x109 -5.0x10-5 -4.89x108 Zd 6.73 4.67 8.23
p-valuee <.0001 <.0001 <.0001 a This strain is the reference strain for competitive index calculations (denominator). 698 b This strain is the competing strain for competitive index calculations (numerator). 699 c Competitive index values represent the proportional competitive advantage of the competing strain over the 700 reference strain. A competitive index of 1 indicates no competitive advantage. 701 d Z is the critical value associated with the Wilcoxon-signed rank test for each paired-strain competition. 702 ep-values indicate the level of difference in bacterial population sizes between competing strains in each paired-703 strain combination in plants as determined by Wilcoxon signed-rank test. 704 705
on June 5, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from
AEM4123-14, revised Tropical R. solanacearum outcompetes R3bv2 strains
26
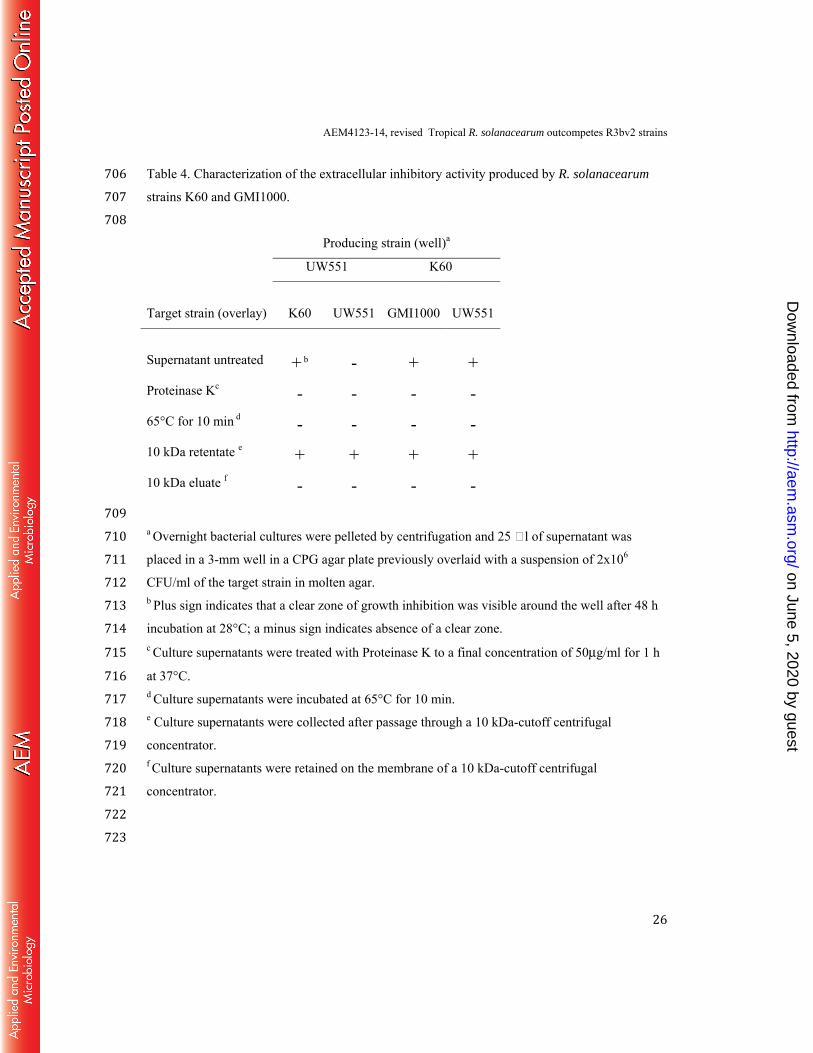
Table 4. Characterization of the extracellular inhibitory activity produced by R. solanacearum 706 strains K60 and GMI1000. 707 708 Producing strain (well)a
UW551 K60
Target strain (overlay)
K60
UW551
GMI1000
UW551
Supernatant untreated
+ b
-
+
+ Proteinase Kc - - - - 65°C for 10 min d - - - - 10 kDa retentate e + + + + 10 kDa eluate f - - - - 709 a Overnight bacterial cultures were pelleted by centrifugation and 25 l of supernatant was 710 placed in a 3-mm well in a CPG agar plate previously overlaid with a suspension of 2x106 711 CFU/ml of the target strain in molten agar. 712 b Plus sign indicates that a clear zone of growth inhibition was visible around the well after 48 h 713 incubation at 28°C; a minus sign indicates absence of a clear zone. 714 c Culture supernatants were treated with Proteinase K to a final concentration of 50μg/ml for 1 h 715 at 37°C. 716 d Culture supernatants were incubated at 65°C for 10 min. 717 e Culture supernatants were collected after passage through a 10 kDa-cutoff centrifugal 718 concentrator. 719 f Culture supernatants were retained on the membrane of a 10 kDa-cutoff centrifugal 720 concentrator. 721 722 723
on June 5, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from

AEM4123-14, revised Tropical R. solanacearum outcompetes R3bv2 strains
27
FIGURE LEGENDS 724 725 Fig. 1. Growth of ecologically distinct R. solanacearum strains in culture without 726 competition. Rich CPG broth was inoculated with strain GMI1000, K60, or UW551 to a final 727 OD600 of 0.01 and cultured with shaking for 36 h in a 96 well plate at either 24/19°C (top) or 728 28°C (bottom). The OD600 of the samples was determined every 30 min by a Biotek plate reader. 729 At 28°C tropical lowland strain GMI1000 grew significantly more slowly than UW551 and K60 730 (P < .0008, One-way ANOVA). At the cooler temperatures strain UW551 grew more quickly 731 and reached a higher final optical density than either K60 or GMI1000 (p < 0.0001, One-way 732 ANOVA ). Data shown represent the means of three technical replicates in a representative 733 experiment. 734 735 Fig. 2. Growth of ecologically distinct R. solanacearum strains in tomato stems and 736 rhizospheres under cool and warm conditions without competition. Sixteen-day-old 737 unwounded tomato plants were inoculated via a naturalistic soil soak with a bacterial suspension 738 of R. solanacearum strain GMI1000, K60 or UW551 at a concentration of 6.0 x 106 CFU/g of 739 soil. At the first sign of wilt symptoms, bacterial population sizes in rhizospheres and stems were 740 determined by serial dilution plating. There were no significant differences between group 741 mean population sizes in tomato rhizospheres at 24/19°C (top) and 28°(bottom) and in tomato 742 stems at 28°C, as determined by one-way ANOVA. In tomato stems at 19°/24°C, strain UW551 743 populations were significantly higher than those of GMI1000 and K60, as indicated by an 744 asterisk (F=4.34, P =.02, n=10). 745 746 Fig. 3. Competitive fitness of ecologically distinct R. solanacearum strains growing in co-747 inoculated culture at cool and warm temperatures. 50 ml of CPG broth was inoculated with 748 a 1:1 bacterial suspension containing 3 x 105 cells each of strain GMI1000 and K60; GMI1000 749 and UW551; or K60 and UW551. Population size of each strain was determined by serial 750 dilution plating after 48 h shaking incubation at either 24/19°C (top) or 28°C (bottom). Results 751 shown are the means of three biological replicates. Asterisks indicate different mean population 752 sizes (P < 0.01); for detailed statistical analysis see Table 2. 753 754
on June 5, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from

AEM4123-14, revised Tropical R. solanacearum outcompetes R3bv2 strains
28
Fig. 4. Competitive fitness of R. solanacearum strains in the rhizosphere at two 755 temperatures. Box-whisker plots show the distribution of bacterial population sizes recovered 756 from the rhizospheres of 16-day-old tomato plants soil-soak inoculated with a 1:1 mixture of two 757 different R. solanacearum strains as indicated and grown at either 19°/24°C (top) or 28°C 758 (bottom). At first wilt symptoms, the population size of each strain was determined by serial 759 dilution plating of rhizosphere soil. The black line in the middle of each box represents the 760 median, and dashed lines highlight the relative trends in population size for each paired-strain 761 competition. The boxes represent the interquantile range (from 25-75%) of the results and each 762 dot represents the bacterial population size in an individual plant (n = 50). See Table 3 for 763 detailed statistical analyses. 764 765 Fig. 5. Competitive fitness of R. solanacearum strains in plant stems at two temperatures. 766 Box-whisker plots show the distribution of bacterial population sizes recovered from the stems 767 of 16-day-old tomato plants soil-soak inoculated with a 1:1 mixture of two different R. 768 solanacearum strains as indicated and grown at either 19°/24°C (top) or 28°C (bottom). At first 769 wilt symptoms, the population size of each strain was determined by serial dilution plating of 770 ground mid-stem tissue. The black line in the middle of each box represents the median, and 771 dashed lines highlight the relative trends in population size for each paired-strain competition. 772 The boxes represent the interquantile range (from 25-75%) of the results and each dot represents 773 the bacterial population size in an individual plant (n = 50). See Table 3 for detailed statistical 774 analyses. 775 776 Fig. 6. Summary of the relative competitive fitness of three ecologically distinct R. 777 solanacearum strains in tomato plants following paired-strain inoculations. The competition 778 outcomes are shown for each strain pair in rhizospheres (A, C) and in tomato mid-stems (B, D) 779 at temperate (A, B, in blue) and tropical (C, D, in red) conditions. The colored shading and 780 connecting shapes represent the relative competitive success of each strain pair in the given 781 environment. For example, in tomato rhizospheres at 24/19°C, tropical highland R3bv2 strain 782 UW551 outcompeted both tropical lowland strain GMI1000 and warm-temperate strain K60, as 783 indicated by the spear points, while GMI1000 and K60, connected by a rectangle, had similar 784 fitness. 785
on June 5, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from

AEM4123-14, revised Tropical R. solanacearum outcompetes R3bv2 strains
29
786 Fig. 7. Cell-free culture supernatants from R. solanacearum strains inhibited growth of 787 other R. solanacearum strains in culture. Agar plates were overlaid with a suspension of the 788 target strain in molten agar. Wells in the plate were filled with 25 μl of cell-free supernatant from 789 an overnight culture of the producing strain. Plates were photographed after 48 h and the 790 diameter of the growth inhibition zone was measured; representative images of inhibition zones 791 are shown under the corresponding bar. Each bar represents the mean inhibition zone diameter 792 from 6 replicates. Results shown are from cultures grown and plates incubated at 28°C; similar 793 results were observed when cultures were grown and plates were incubated at 24/19°C. Asterisks 794 indicate that inhibition zones were different from empty well control (P<0.0001, as determined 795 by one-way ANOVA. UW551 was inhibited by producing strains GMI000 and K60 (F=435); 796 K60 was inhibited by producing strain GMI1000 (F = 471) and K60 was inhibited by producing 797 strain GMI1000 (F=397). 798
on June 5, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from

0.0
0.2
0.4
0.6
0.8
K60 UW551 GMI1000
24/19°C
0 6 12 18 24 30 360.0
0.2
0.4
0.6
0.8
Time (hrs)
28°C
Cell
Density
(O
D6
00)
on June 5, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from

0
2
4
6
8
10
GMI1000K60 UW551
24/19°C
*
Rhizosphere Stem0
2
4
6
8
10
28°C
Log(CFU/g)
on June 5, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from

0
2
4
6
8
10K60 UW5551 GMI1000
24/19°C
**
0
2
4
6
8
10
28°C
** **
*
GMI1000 K60 GMI1000 UW551 K60 UW551
Log(C
FU
/g)
on June 5, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from

GMI1000 K60 GMI1000 UW551 K60 UW551
Strain
L
og
(C
FU
/g)
24
/19
°C 2
8°C
on June 5, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from



0.0
0.5
1.0
Target Strain (overlay)
Zone o
f In
hib
itio
n (
cm
)
GMI1000 K60 UW551
Producing Strain (well)
GMI1000 K60 UW551
*
**
*
on June 5, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from