integrated effect of sunflower residues and...
TRANSCRIPT

Republic of Iraq
Ministry of Higher Education
& Scientific Research
University of Baghdad
College of Science
Integrated effect of sunflower residues and
chevalier on weeds of wheat and growth of
mycorrhiza.
A Thesis Submitted to the Council of the College of Science,
Department of Biology, University of Baghdad as a Partial
Fulfillment of the Requirements for the Degree of Doctor of
Philosophy (Ph.D.)
in Plant Physiology, Botany.
By
Srraaۥ Nsayef Muslim Al-Eqaili
B. Sc. Microbiology/ College of Science/ University of
Mustansiriya 2006
M.Sc. Botany/ College of Science/ University of Mustansiriya
2009.
Supervised By
Ibrahim S. Alsaadawi Dr. Hadi M. Aboud Prof. Dr.
2014 A. H. 1435 A. D.

الرحيـمبســم هللا الرحمـن
يسجد له من في السماوات ومن في األرض ا﴿ لم تر أن الل والقمر والنجوم والجبـال والشجر والدواب وكثير والشمس
فما له وكثير من النـاس حق عليه العذاب ومن يهن الل يفعل ما يشاء ﴾ من مكرم إن الل
صـدق هللا الـعـظيم (81سورة الحج ) أية

This is to certify that the thesis titled: " Integrated effect of
sunflower residues and chevalier on weeds of wheat and growth of
mycorrhiza ".
Submitted by: Srraaۥ Nsayef Muslim Al-Eqaili
Department: Biology
College: Science
has been linguistically corrected and its language, in its present form, is
acceptable.
Name: Professor Shatha K. AL-Saadi
Address: Department of English, College of Education for Women,
University of Baghdad
Signature:

SUPERVISOR CERTIFICATION
We certify that thesis entitled '' Integrated effect of sunflower
residues and chevalier on weeds of wheat and growth of mycorrhiza
'' is prepared by " Srraaۥ Nsayef Muslim Al-Eqaili " under our
supervision at the College of Science, University of Baghdad in partial
fulfillment of requirements for the degree of Doctor of Philosophy in
Biology.
Signature:
Name : Dr. Ibrahim S. Alsaadawi
Title : Professor
Address : College of Science,
Department of Biology, University of
Baghdad.
Date: / / 2014
Signature:
Name : Dr. Hadi M. Aboud
Title : Chief Scientific Researchers
Address: Agricultural Research Center,
Ministry of Science and Technology.
Date: / / 2014
In view of the available recommendation, I forward this thesis for debate
by the examination committee.
Signature:
Dr. Sabah N. Alwachi
Title: Professor
Head Department of Biology
Date: / / 2014

Committee Certification We, the Examining Committee, certify that we have read this thesis in
titled '' Integrated effect of sunflower residues and chevalier on weeds
of wheat and growth of mycorrhiza '' and have examined the student
"Srraa Nsayef Muslim Al-Eqaili" in its contents and that in our opinion; it is
accepted as a thesis for awarding the Degree of Doctor of Philosophy in
Biology.
Dr. Abd oun H. Alwan Dr. Abdullah I. Shaheed
Professor Professor
Member Member
/ / 2014 / / 2014
Dr. Risan K. Shatti Dr. Ayyad W. AL-Shahwany
Professor Assist. Professor
Member Member
/ / 2014 / / 2014
Dr. Muna H. Al Jubori
Professor
Chairman
/ /2014
Dr. Ibrahim S. Alsaadawi Dr. Hadi M. Aboud
Professor Chief Scientific Researchers
(Advisor) (Advisor)
/ /2014 / / 2014
Approved for the College of committee of graduate studies
Assist. Professor Dr. Mohammad A. Atiya
Dean
College of Science, Baghdad University
/ / 2014

Acknowledgment
First of all, I would like to thank the grace of God for completing
this work at this final shape. All respects are for His Holy Prophet
Mohammad (Peace be upon him and his family), for enlightening our
conscience with the essence of faith in Allah.
My deepest thanks and gratitude to my supervisor, Dr. Ibrahim
Alsaadawi and Dr. Hadi M. Aboud for supervision, scientific guidance,
helpful discussion, generosity, providing the possible laboratory
materials and support during all the period of the research.
Many thanks to the Head of the Department of Biology, College of
Science, University of Baghdad, also I am grateful to Dr. Hind Hussain
for her help and moral support. Appreciation is extended to Dean of
College of Science, University of Baghdad.
Deep thanks to Dr. Nabil R. Al-bedairy from University of
Wasit who enable me, with their endless support, and encouragement to
complete this thesis , also to Mrs. Falah Abdul Hassan from Dept. of
Food analysis.
Finally, I would like to express my sincere thanks and gratitude to all my
best friends (Hamed, Ali, Ala, laith, Tamara, Arwa) who enable me,
with their endless support, and encouragement to complete this thesis.

Dedication
I dedicate the fruit of my efforts
To those who taught us letters of gold and
words of jewel of the utmost and sweetest
sentences in the whole knowledge… My supervisors
To the Spring that never stops giving, who
weaves my happiness with strings from her
merciful heart… The pure soul of my lovely mother
To whose love flows in my veins, and my heart always
remembers them… My brothers and sisters
Srraۥa

Abstract
A set of experiments was performed to test the allelopathic
potential of sunflower residues alone or in combination with reduced rate
(50% of recommended dose) of chevalier herbicide on weeds, wheat crop
and growth of mycorrhiza associated with wheat roots. The field
experiment was conducted during 2012-2013 season at the Research
Farm of Biology Department, College of Science, Baghdad University by
using the randomized complete block design (RCBD) with 4 replications
to test the effect of sunflower residues at 3 and 6 t ha-1
alone or in
combination with reduced dose of Chevalier (150 g ha-1
) on weed and
wheat crop. Weedy check and label rate of chevalier were also included
for comparison. Each treatment was replicated four times. Total
phenolics in field soil amended with sunflower residues at 6 t ha-1
was
determined during different periods after sowing. Mycorrhizal sporulation
was determined during different growth stages of wheat and colonization
rate and intensity were determined at flowering stage of wheat.
Result showed that incorporation of sunflower residues at 3 t ha-1
reduced weed density by 88 and 97% of control after 90 and 120 days
after sowing (DAS), respectively. The reduction was increased when
sunflower residues were incorporated at 6 t ha-1
and reached to 89 and
78% of the control after 90 and 120 DAS, respectively. However, the
suppression of weed population and dry weight biomass was further
improved when the plots were treated with 50% of labeled rate of
herbicide and amended with sunflower residues. Integration of reduced
herbicide rate and sunflower residues at 3 t ha-1
and 6 t ha-1
resulted in
more weed suppression than sole application of the respected sunflower
residues.

The results also revealed that weed suppression was directly
translated into yield of wheat. Application of Chevalier herbicide at 50%
rate in plots amended with sunflower residue at rates of 3 t ha-1
resulted in
similar biological and grain yields, number of spikes per plant, number of
grains per spike and harvest index was achieved by the label herbicide
rate treatment.
Chemical analyses indicated that total phenolics started to
increase at 14 and 28 days of decomposition and declined thereafter until
vanished 6 weeks of decomposition. Biological activity test of field soil
revealed that suppression of Malva rotundifolia weed was highly
correlated with total phenolics of soil suggesting that high weed
suppression was mainly due to high activity of phenolics.
With respect to mycorrhizal studies, the number of spores in field
soil amended with sunflower residues was significantly increased at 2, 4
and 6 weeks of residue decomposition compared to the control treatment
(without sunflower residue). At flowering stage, it was found that
chevalier at reduced (50% of the label rate) rate applied to plots amended
with sunflower residue at 3 t ha-1
scored spore number significantly lower
than that of the control treatment, but when the reduced dose was applied
to plots amended with higher residues rate, the number of spores was
significantly increased over the control. Sunflower residues incorporated
in to field soil at rates of 3 t ha-1
increased rate and intensity of
colonization by 49% and 44% of control, respectively. Application of
reduced dose of herbicide on plants grown in plots amended with
sunflower residues significantly increased rate of colonization compared
to the control.

List of Contents Item
No. Item Page No.
Abstract I
List of contents Ш
List of tables VII
List of figures X
Chapter one
Literatures review
Introduction 1
1 Literatures review 4
1.1. Allelopathy of crop on weeds 4
1.1.1 Effect of allelopathic plant extracts 5
1.1.2. Effects of allelopathic crop residues 11
1.1.3. Combined effect of allelopathic extract with
herbicides
19
1.1.4. Combined effect of allelopathic residues with
herbicides
26
1.2 Effect of phenolic compounds on arbuscular
mycorrhiza
29
Chapter two
Material and methods
2 Material and methods 35
2.1 Field preparations 35
2.1.1. Site selection 35
2.1.2. Seeds and herbicides sources 35
2.1.3. Soil Sampling and Analyses 35
2.1.4. Preparation of sunflower residues 37
2.2. Field trial 37
2.3 Weed measurements 38

2.3.1. Weed density (plant m‾2) 38
2.3.2. Weed biomass (g m‾2) 39
2.4. Wheat crop measurements 39
2.4.1. Dry weight of plant through different stages of
crop growth (g/m2)
39
2.4.2. Crop Growth Rate CGR (g/m2/day) 39
2.4.3. Plant height 40
2.4.4. Number of spikes/m2 40
2.4.5. Number of grains per spike 40
2.4.6. 1000-grain weight (g) 40
2.4.7. Total wheat biomass (t ha1ـ) 40
2.4.8. Grain yield (t haـ 1) 40
2.4.9. Biological yield 41
2.4.10. Harvest index (%) 41
2.5. Mycorrhizal studies 41
2.5.1. Spore extraction 41
2.5.2. Isolation and identification of mycorrhiza spores 42
2.5.3. Preparation of Mycorrhiza inoculums 42
2.5.4. Spore counting 42
2.5.5. Mycorrhizal colonization rate (%) 43
2.6. Determination of total phenolics 44
2.7. Bioassay of soil amended with sunflower
residues
45
2.8. statistical analyzsis 45
Chapter three
Results
3 Results 46
3.1. Weed parameter 46

3.1.1. Effect of different rates of chevalier 15 WG
herbicide and sunflower residues cv. Asgrow on
total weed density in wheat field
46
3.1.2. Effect of different rates of chevalier 15 WG
herbicide and sunflower residues cv. Asgrow on
dry weight biomass of weeds in wheat.
49
3.2. Crop parameters 49
3.2.1 Effect of different rates of chevalier 15 WG
herbicide and sunflower residues cv. Asgrow on
dry weight of wheat during different growth
stages.
49
3.2.2 Effect of different rates of chevalier 15 WG
herbicide and sunflower residues cv. Asgrow on
crop growth rate of wheat crop
52
3.2.3. Grain yield (t ha-1
) 52
3.2.4. Effect of different rates of chevalier 15 WG
herbicide and sunflower residues cv. Asgrow on
yield components of wheat
55
3.2.4.1. Effect on number of spikes per m2 55
3.2.4.2. Effect on grains per spike 55
3.2.4.3. Effect on 1000-grain weight (g) 56
3.2.5. Biological yield (t ha-1
) 59
3.2.6. Biomass (t ha-1
) 59
3.2.7. Harvest index % 61
3.2.8. Effect of different rates of chevalier 15 WG
herbicide and sunflower residues cv. Asgrow on
plant height of wheat
62
3.3. Determination of Total phenolics in field soil 63
3.4. Weed bioassay 64
3.4.1. Seed germination of malva rotundifolia 64
3.4.2. Total dry weight of Malva rotundifolia 65

3.5. Mycorrhizal studies 66
3.5.1. Isolation and identification of mycorrhiza from
field soil of wheat crop
66
3.5.2. Effect of different periods of decomposition of
sunflower residues in field soil before chevalier
application on sporulation
67
3.5.3. Effect of different rates of chevalier 15 WG
herbicide and sunflower residues cv. Asgrow on
sporulation at flowering stage
68
3.5.4. Effect of different rates of chevalier 15 WG
herbicide and sunflower residues cv. Asgrow on
colonization rate of Glomus mosseae
68
3.5.5. Effect of different rates of chevalier 15 WG
herbicide and sunflower residues cv. Asgrow on
colonization intensity of Glomus mosseae
71
4 Chapter four-Discussion 72
5 Conclusions and Recommendations 79
6 References 81
7 Appendices 103
8 Abstract in Arabic

List of tables
No. Title Page
1
Some Physical and chemical properties of field
soil before and after sunflower residue
incorporation.
36
2 Weed species grown in the test wheat field. 47
3
Effect of different rates of chevalier 15 WG
herbicide and sunflower residues cv. Asgrow on
weeds population grown in wheat .
48
4
Effect of different rates of chevalier 15 WG
herbicide and sunflower residues cv. Asgrow on
dry weight biomass of weeds grown in wheat.
50
5
Effect of different rates of chevalier 15 WG
herbicide and sunflower residues cv. Asgrow
on grain yield of wheat crop.
54
6
Effect of different rates of chevalier 15 WG
herbicide and sunflower residues cv. Asgrow
on number of spikes of wheat.
56
7
Effect of different rates of chevalier 15 WG
herbicide and sunflower residues cv. Asgrow
on number of grains per spike of wheat.
57
8
Effect of different rates of chevalier 15 WG
herbicide and sunflower residues cv. Asgrow
on weight of 1000 grains of wheat.
58
9
Effect of different rates of chevalier 15 WG
herbicide and sunflower residues cv. Asgrow
on biological yield of wheat crop.
60
10
Effect of different rates of chevalier 15 WG
herbicide and sunflower residues cv. Asgrow on
biomass of wheat.
60
11
Effect of different rates of chevalier 15
WG herbicide and sunflower residues
cv. Asgrow on number of spores of
Glomus mosseae associated with wheat
roots.
69

12
Effect of different rates of chevalier 15 WG
herbicide and sunflower residues cv. Asgrow
on colonization rate of Glomus mosseae
associated with wheat roots.
70
13
Effect of different rates of chevalier 15 WG
herbicide and sunflower residues cv. Asgrow
on colonization intensity Glomus mosseae
associated with wheat roots.
71

List of figure
No. Title Page
1
Effect of different rates of chevalier 15 WG
herbicide and sunflower residues cv. Asgrow on
dry weight of wheat.
51
2
Effect of different rates of chevalier 15 WG
herbicide and sunflower residues cv. Asgrow on
crop growth rate of wheat.
53
3
Effect of different rates of chevalier15 WG
herbicide and sunflower residues cv. Asgrow
on Harvest index of wheat.
61
4
Effect of different rates of chevalier 15 WG
herbicide and sunflower residues cv. Asgrow
on plant height of wheat.
62
5
Total phenolics released in field soil amended
with sunflower residues at 6 t ha-1
during
different decomposition periods.
63
6
Seed germination of Malva rotundifolia in field
soil amended with sunflower residues at 6 t ha-1
during different decomposition periods.
64
7
Total dry weight of Malva rotundifolia in field
soil amended with sunflower residues at 6 t ha-1
during different decomposition periods.
65
8 spore of Glomus mosseae 66
9 Sporocarp 66
10
Sporulation of Glomus mosseae in field soil
amended with sunflower residues at 6 t ha-1
during different decomposition periods.
67

Introduction

Introduction
Weeds represent a global agronomic problem that reduces the
productivity of cultivated crops. Weeds compete with cultivated crops for
the available moisture, nutrients and light. Consequently, weeds
significantly reduce either crop yield or quality. Control of weeds is
essential to maintaining the production of economic crops. Currently,
Agriculture worldwide is using about 3 million tons of herbicides per
year, and herbicide-resistant weeds have become more prolific, which has
further expanded the use of herbicides (Shibayama, 2001). The overuse of
agrochemicals has caused environmental pollution, weed tolerance and
human health concerns. To solve these problems, it is necessary to
develop sustainable weed management systems that may reduce herbicide
dependency, inexpensive, easy to use and helpful in maintaining the
ecosystem stability (Khanh et al., 2013).
Allelopathy has been accepted as environment-friendly
phenomenon, which may prove to be a useful means for weed
management and thereby increase crop yields (Putnam et al., 1983). It
involves direct or indirect effects of one plant on another through the
production of secondary chemical compounds that escape into the
environment (Rice, 1984). Chemicals with allelopathic potential are
present in virtually all plant parts or tissue. These allelochemicals offer
great potential as natural pesticides and can be used for weed control
directly or their chemistry could be used to develop new herbicides
(Putnam et al, 1990). However, in most cases, allelopathic extracts or
crop residues provide limited weed suppression, and most often
reductions in weed growth are not comparable to those observed with
labeled herbicides. Therefore, other methods to increase the efficacy of
allelopathic extracts may be critical to enhance weed suppression while at
the same time reducing our reliance on herbicides. Substantial scope to

reduce the herbicide rate has been observed by various investigators when
herbicides are applied together with aqueous sorghum extracts. The
results of the research conducted in recent years has revealed that one-
third or half dose of herbicides combined with sunflower extracts provide
similar weed control to the labeled (full) dose of herbicides in different
field crops (Cheema et al., 2003a, b, c; Iqbal et al., 2009). Although
successful results have been obtained from sorghum extracts applied with
low herbicide rates, additional work in other soil types and locations
should be performed. In addition, to employ this technology on large
scale operations, large volumes of sprays are required for field
application, and therefore appropriate concentrations for each crop should
be determined.
Due to these limitations, an alternative practical and feasible
approach has been developed for weed management, where low doses of
herbicides are applied along with the residues of allelopathic crops. The
residues of allelopathic crops were left after harvest to dry then
incorporated into field soil during tilling process (Alsaadawi et al.,
2013). By using this approach with sunflower and sorghum, it was found
that a combination of herbicide with sunflower or sorghum residues
resulted in a significant reduction in weed number and biomass as was
achieved by the label dose of herbicide )Alsaadawi et al., 2011; Lahmod,
2012). However, the effects of this combination on beneficial
microorganisms such as mycorrhiza, rhizobia and others could not be
excluded.
Customarily, farmers in Iraq cultivate wheat after sunflower harvest
during October. Several types of herbicides are generally used to control
weeds of wheat. Therefore, it was contemplated in the present studies to:

1. Evaluate the effect of allelopathic residues alone or in
combination with lower (one half of label dose) chevalier dose
on weed population and growth.
2. Test the effect of the above aforementioned combinations on
yield and yield components of wheat crop.
3. To test if the residues of sunflower and reduced chevalier
herbicide alone or in combination with each affect number of
spores, colonization rate and intensity of mycorrhiza associated
with at roots of wheat.

Chapter One
Literature Review

CHAPTER ONE
1. Literatures review
1.1. Allelopathy of crop on weeds
Weed management effects has been a serious problem for
many world countries. Worldwide, a 10% loss of agricultural
products can be attributed to competitive and allelopathic effects
of weeds, despite their intensive control (yarnia, 2011). Potential
yield reductions caused by uncontrolled weed growth throughout
growing season have been estimated to be 45 to 95%, depending
on crop species, weed species, weed densities and ecological
factors (Ampong-Nyarko and De Datta, 1991). Therefore, weed
management is a key element of most agricultural systems.
Although there are various methods to weed control including
cultural, mechanical and biological methods, nevertheless,
application of herbicides has been a major chemical factor
enabling the intensification of agriculture in past decades
(Abdollahi and Ghadiri, 2004). Indeed, three million tones of
herbicides per year are used in most agricultural systems
(Stephenson, 2000). There has been increasing herbicide
resistance in weeds and widespread concern about adverse
environmental effects from herbicide use (Stephenson, 2000). For
this reason, searching of alternative method becomes viable option
to reduce the rely on herbicides. In recent years allelopathy may
provide an alternative to minimize the risk of herbicides towards
agroecosystems.
Several strategies in which allelopathy can play a beneficial
role in cropping systems have been adopted including mixed
cropping, multiple cropping, cover cropping and mulches,

incorporation of crop residues, crop rotations, minimum and no–
tillage systems and using of allelopathic water extracts. Of all
these strategies, utilization of crop residues is the most successful
one (An et al., 1996). In this section, The application of water
extract and residues of allelopathic crops alone or in combination
with low recommended dose of herbicides on weeds and crops are
reviewed.
1.1.1. Effect of allelopathic plant extracts
In the earlier research work on allelopathy, the biological
activities of extract was performed to examine if the allelopathic
crop has phytotoxins which may be responsible for the
allelopathic activity of the tested plant ( Rice , 1984). Later, the
use of allelopathic crops extract has been developed as one of the
strategies used for weed control (Khaliq et al., 2002).
The effect of allelopathic crops extract on weeds has been
also studied under field conditions. Javid et al. (2006) evaluated
the allelopathic potential of aqueous root and shoot extracts of
sunflower, sorghum and rice (Oryza sativa L.) on germination and
growth of the noxious alien weed Parthenium hysterophorus L.
using petri dishes technique . Aqueous root and shoot extracts (5,
10, 15, 20 and 25% (w/v) ) of fresh plant materials of the tested
crops insignificantly affected shoot length and seedling biomass
while germination and root length were significantly reduced by
extracts of all the test crops. In a foliar spray bioassay, when
aqueous shoot extracts at 50 and 100% w/v (on a fresh weight
basis) of sunflower and sorghum were applied on 10 day old
Parthenium plants, the root biomass of Parthenium plants was
significantly suppressed. Both concentrations of sorghum extracts

significantly reduced shoot biomass, but sunflower extract was
inhibitory by the lower concentration only.
Sunflower leaf extracts successfully killed Chenopodium
album and Rumex dentatus weeds in wheat crop and thereby
increased wheat yield significantly (Anjam and Bajwa, 2007).
They suggested that the considerable allelopathic potential of
sunflower could be used as natural herbicide.
Cheema et al. (2008) conducted a series of field
experiments to test allelopathic effects of this crop on weed
control and yield of wheat. They found that 10% w/v water
leachate of aerial parts of sorghum (also called sorgaab) applied at
30 and 60 days after sowing can reduce weed biomass by as much
as 49% with concomitant increase of wheat yield over 20%
compared to the control. They concluded that sorgaab has a great
potential in increasing weed control and grain yield of wheat.
Application of this method of weed management has enormous
economic and environmental benefits in wheat cultivation.
Naseem et al. (2009) studied the allelopathic influence of
sunflower plant water extract (1:10 w/v) on weeds in wheat crop
under field conditions. Results indicated that the highest inhibitory
effects on germination of Phalaris minor was achieved by pre-
emergence application, whereas wheat remained unaffected by
this treatment. Application of water extract at pre-emergence + 25
DAS, 25 + 35 DAS and pre-emergence + 25 + 35 DAS
suppressed growth of Phalaris minor Retz., Chenopodium album
L., Coronopus didymus L. and Avena fatua L.. The inhibitory
effects were species specific and increased with increasing the
water extract application frequency. All the treatments except pre-

emergence + 25 + 35 DAS increased the wheat yield significantly
over control.
Hozayn et al. (2011) conducted field experiment to determine the
response of lentil and its weeds to foliar application of different
concentrations of sorghum or sunflower water extracts. Both sorghum
and sunflower extracts significantly reduced the weeds dry weight by
65.62% and 63.56% and thus increased the lentil dry weight by 67.66%
and 64.41% over the control and the lentil seed yield by 61.34% and
56.18% of control, respectively.
Nikneshan et al. (2011) evaluated the allelopathic potential
of different concentrations (25, 50 and 100% w/v) of sunflower
cultivars on several crops and associated weeds. Results showed
that Amaranthus retroflexus was the most sensitive to sunflower
allelopathy and Portulaca oleracea was the most resistant. As
extract concentration increased from 25 to 100%, the inhibitory
effect on germination indices increased, while with 25% extract
concentration was observed to have stimulatory effects on wheat
and P. oleracea germination. The Megasun sunflower cultivar had
the most effect and Hysun 36 had the least effect on the target
plants. Megasun extract at 100% concentration suppressed
selected weeds by 80%. The results also indicated that the
allelopathic properties of some sunflower cultivars can affect
noxious weed species such as Hordeum spontaneum and Lolium
rigidum in wheat and A. retroflexus in safflower crop.
Awan et al. (2012) tested the allelopathic effect of different
combinations of water extracts of sorghum, sunflower and
brassica on weeds in wheat crop under field conditions. Extracts
were prepared by soaking chopped dry sorghum, sunflower and
brassica herbage separately in water for one day in the proportion

of 1:10 (w/v). The treatments were control, hand weeding, foliar
application of sorghum, sunflower and brassica individually and
combined as sorghum + sunflower, sorghum + brassica, sunflower
+ brassica and sorghum + sunflower + brassica. Results revealed
that the highest weed density and biomass suppression was
achieved by hand weeding. Among the foliar application of
extracts, the highest weed control was recorded with the combined
application of concentrated aqueous extracts of sorghum,
sunflower and brassica at 45 and 75 days after sowing followed by
sorghum + sunflower extracts. The highest increase of yield and
yield components of wheat was recorded by manual weeding and
the lowest by control. Among extracts application treatments, the
highest increase in wheat biomass and yield attributes was found
by combined application of sorghum, sunflower and brassica
extracts compared to rest of the treatments.
Khan and Khan (2012) conducted a field experiment
during spring of 2010 to test pre and post application of plants
extract for weed control and their impact on grain yield of wheat.
Water extracts of sorghum, sunflower, Parthenium, (Phragmites
australis L., Johnson grass (Sorghum halepense) and rice extract,
herbicide (Logran Extra 64 WG), and a weedy check. They found
that pre-emergence application of plant water extracts proved to
be superior than post-emergence application in controlling weeds.
Pre-emergence application of P. australis and sunflower gave 68
and 65% reduction in weed control, respectively. Sorghum gave
maximum grain yield 5015 kg ha-1
in comparison to weedy check
2700.6 kg ha-1
. The instant results suggest that P. australis, H.
annuus and sorghum extracts could be successfully incorporated
in weed management approaches in wheat.

The allelopathic effect of sunflower extract on weeds was also
studied under laboratory conditions (Ghafar et al., 2000; Batlang et al.,
2007; Mubeen et al., 2012; Azadbackht et al., 2013; Khaliq et al.,
2013). In most of these studies, suppression of test weeds and crops has
been observed, particularly at the higher concentrations of the extracts.
1.1.2. Effects of allelopathic crop residues
Allelopathic crops residue decomposition produces a variety
of phytotoxins in the soil causing adverse effects on other plants
(Nelson, 1996), and have the potential to sustain a chemical as
well as physical effect on the growth and development of
subsequent crops and weeds (Reddy, 2001). Allelopathic crop
residues can be exploited for weed suppression, and can thus be
helpful in reducing reliance on herbicides (Weston, 1996). In this
section, the potential role of allelopathic crop residues in weed
management is briefly reviewed.
Putnam and DeFrank in 1983 found that the populations of
Portulaca oleracea L. and Digitaria ischaemum L. (Schreb) were
reduced by 70% and 90% by residues of sorghum, respectively. In
general, the larger-seeded vegetables, particularly legumes grew
normally or were sometimes stimulated by the residues, whereas
several species of smaller-seeded vegetables were severely
injured. Glasshouse experiments with two soils confirmed both
weed-suppressing and crop-stimulating effects of sorghum
residues.
In Pakistan, Cheema and Khaliq (2000) found in a field experiment
that incorporation of whole sorghum plant or its various parts alone or
mixed with each other suppressed total weed dry weight from 26-56%
and increased wheat yield from 6-17% over weedy check.

Srisa-ard (2007) investigated the effects of residues of sunflower,
maize (Zea mays L.) and soybean (Glycine max L) on total dry weight,
top dry weight, plant height, root dry weight and seed yield of sunflower
plants using Koral soil series (Oxic paleustults) during the rainy season
(July-october) of the 2001. Whilst crop residues of roots, top growth and
roots top growth were used as subplots. The results showed that crop
residues derived from roots of both sunflower and soybean plants
significantly inhibited plant height, root dry weight, top growth dry
weight and total dry weight plant of the sunflower plants compared to
those derived from top growth of both crops alone (sunflower and
soybean). Maize plant residues had no significant inhibitory effect on
growth of subsequent sunflower crop.
In Iraq, several hundreds of sorghum cultivars were introduced and
cultivated to select the most promising genotypes in terms of production,
weed competition, and fitness to local environment. Field observation by
Alsaadawi et al. (2007) revealed that growth and population of
companion weeds were variable among stands of selected genotypes.
Additional work indicated that residues of all test cultivars significantly
inhibited the growth of Lolium temulentum weed. The phytotoxicity of
residues differed among the test genotypes. Experiment conducted in
afield infested with Lolium rigidum, L. temulentum, Malva pariflora,
Carthumus oxycantha, Chenopodium album, Beta vulgaris, Trifolium
repense and Plantago ovate revealed that the aboveground biomass and
number of all weeds were reduced by the residues of test sorghum
cultivars incorporated into field soil at rates of 3 and 6 g/ kg soil.
However, the response varied among the weed species. Residues of
cultivars Giza 15, Giza 115 and Enkath provided 67, 59, and 63%
reduction in average weed number and 58, 66, and 58% reduction in

average weed biomass, respectively. Residues of Rabeh cultivars
inhibited average weed numbers and average weed biomass by 41 and
52%, respectively. Weeds numbers were significantly decreased with
increasing rate of residues of the stronger allelopathic cultivars in soil.
The phytotoxicity of the residues started after 1 week of decomposition
and persisted for 8 weeks at low rate of residues and for 10 weeks at the
higher rate of residues. The reduction was proportional to the amount of
residues in soil during the first 6 weeks of decomposition. Giza 15, Giza
115, and Enkath showed greater phytotoxicity than Rabeh cultivar at all
decomposition periods.
Ashrafi et al. (2008) conducted greenhouse and laboratory
experiments to determine the effects of sunflower residues on
germination and seedling growth of wild barley (Hordeum spontaneum
Koch.). They showed that incorporation of fresh roots of sunflower alone
or in combination with shoots significantly reduced wild barley
germination, plant height and weight as compared with control (no
residue). Laboratory experiment revealed that the degree of toxicity of
different sunflower plant parts showed the following order of inhibition:
leaves > flowers > mixture of all plant parts > stems > roots.
Khaliq et al. (2010) conducted field experiment to test the
weed suppression potential of mixture of allelopathic crop
residues and their effects on maize yield. Residues of sorghum,
sunflower, rice and brassica (Brassica campestris L.) in various
combinations were incorporated in soil at 5 and 7.5 t ha-1
.
Results revealed that a combination of sorghum + sunflower +
brassica residues at 7.5 t ha-1
provided > 90% suppression in
density and dry weight of horse purslane (Trianthema
potrtulacastrum L.) and purple nutsedge (Cyperus rotundus L.) as
compared with control (no residue). This treatment also accounted

for maximum maize grain yield and net benefits. They concluded
that for better management of natural resources and to decrease
environmental pollution, soil incorporation of crop residues may
serve as an important weed management tool in maize fields.
Matloob et al. (2010) evaluated allelopathic potential of different
crop residues viz. sorghum, sunflower, brassica (applied alone or in
combination) for the purple nutsedge suppression in a pot study. Chopped
residues were incorporated at 12 t ha-1
(6 g kg-1
of the soil) into the soil
and a weedy check was also maintained. There were six tubers of purple
nutsedge in each soil filled pot. Soil incorporation of all the residues
substantially delayed the tuber sprouting. Nonetheless combinations of
residues showed more effective in purple nutsedge suppression than sole
application of either of them. Sorghum and brassica residues, when
applied in combination did not allow any tuber to sprout. There was
substantial suppression in final germination by 41- 45% from sole
application and 27-100% from combination of crop residues. These
residues exerted a pronounced negative influence on the shoot and root
length by 21-100 and 17- 100%, respectively. Likewise, there was 50-
100% and 47-100% suppression in shoot and root dry weights,
respectively. Hence, soil incorporation of the test allelopathic crop
residues may be employed in the integrated approach for purple nutsedge
management.
Khaliq et al.(2011) evaluated the allelopathic potential of different
of residues of sorghum, sunflower and brassica on rice crop and jungle
rice weed (Echinochloa coloma [L.] Link). Soil incorporation of residues
substantially delayed germination of jungle rice. The time to start
germination, time to 50% emergence, mean emergence time, emergence
index and final germination percentage were all depressed by residue

incorporation. Final germination of rice and jungle rice dropped by 11 to
15% and 11 to 27% with residue application alone and by 18 to 22% and
8 to 34% with a combination of crop residues, respectively. Residues
were more suppressive to germination dynamics of jungle rice than rice.
Crop residues exerted a pronounced negative influence on the shoot (25
to 100% and 14 to 44%) and root lengths (22 to 100% and 10 to 43%) of
rice and jungle rice, respectively. Shoot and root dry weight of both rice
and jungle rice were also decreased significantly. An appreciable
quantity of phenolics was recorded in soil amended with sorghum +
sunflower + brassica residues. They suggested that the time of residue
application for jungle rice suppression and rice seeding time need to be
adjusted to minimize rice crop damage.
Alsaadawi et al. (2011b) evaluated eight genotypes of sunflower for
their allelopathic potential against weeds and wheat crop which is
customarily grown after sunflower in crop rotation in Iraq. They are
revealed that all sunflower genotypes significantly inhibited total number
and biomass of companion weeds and the magnitude of inhibition is
genotype dependent. Of the 8 genotypes tested Sin- Altheeb and Coupan,
were the most allelopathic potential cultivars with the reduction in total
weed number by 47.25 and 86.81 % of control and weed biomass by
74.23 and 80.79% of the control respectively. Euroflor and Shumoos
were the least allelopathic potential genotypes with an inhibition in total
weed number by 21.50 and 9.59% and weed biomass by 42.28 and
33.67% of the control respectively. Subsequent field experiment indicated
that the residues of sunflower incorporated into the field soil significantly
inhibited total number of weeds grown in wheat field by 24.51-74.52% of
control at low residues rate (3g kg-1
soil) and by 49.05-75.47% at the
high residues rate (7g kg-1
soil). Weeds biomass significantly reduced

with a range of 12.27-64.52% at low residue rate and 40.33-66.75% at
high residue rate. However, sunflower genotypes Sin Altheeb and
Coupan appeared to be more inhibitory to total weeds number and
biomass and significantly increased yield of wheat compared to the least
allelopathic potential genotypes (Euroflor and Shumoos genotypes).
Chromatographic analyses by HPLC revealed the presence of 13
phytotoxins in the residues of the tested sunflower genotypes. All the
isolated compounds appeared to have different retention times and were
identified as phenolic compounds with the exception of terpinol which is
known as terpenoid derivative. The total concentration of phytotoxins
appeared to be dramatically increased in the most allelopathic potential
genotypes compared to the least allelopathic potential genotypes.
Alsaadawi et al. (2014) tested if the variation in weed
population and biomass between the stands of Enkath and Rabeh
sorghum cultivars, which was observed in the field, was due to
differences in their allelopathic potential. Field experiment
revealed that Enkath cultivar significantly suppressed weed
density and dry weight biomass over Rabeh cultivar by 34 and
29% of control after 35 DAS, and 22 and 24% after 65 DAS. Stair
case experiment indicated that root exudates of Enkath cultivar
showed more suppression to weeds than Rabeh giving additional
evidence for the superiority of Enkath cultivar in its allelopathic
weed suppression. Chemical analysis revealed that sorgoleone and
several phenolic acids were present in higher concentrations in
root exudates of Enkath compared to Rabeh. Thus it appears that
the variation in weed suppressive ability between the test sorghum
cultivars was attributed to their differences in allelopathic
potential of root exudates and the phytotoxicity of root exudates
was not restricted to sorgoleone alone but with phenolic acids.

The results recommend screening more sorghum cultivars in order
to offer a potential source of allelopathic germplasm that could be
manipulated to enhance weed suppression in an effective and
environmentally sustainable approach.
1.1.3. Combined effect of allelopathic extract with
herbicides
As in sections 1.1.1 and 1.1.2 the allelopathic extracts or crop
residues provide limited weed suppression, and most often reductions in
weed growth are not comparable to those observed with labeled
herbicides. Therefore, other methods to increase the efficacy of
allelopathic extracts may be critical to enhance weed suppression while at
the same time reducing our reliance on herbicides. Furthermore it has
been reported that allelochemicals and herbicides work in concert to
inhibit the growth of weeds, e.g. adequate allelopathic crop plus reduced
input of herbicide give adequate weed control (Einhellig and Leather,
1988; Einhellig, 1995, 1996; Streibig et al., 1999). In this section, the use
of integration of allelopathic extracts and lower dose of herbicide is
reviewed.
Mahmood et al. (2009) indicated that combined application of
sorghum water extract (sorgaab) and sunflower water extract (sunfaab)
each @ 15 L ha-1
with 1/3rd
of the label dose of phenoxoprop-p-ethyl
showed maximum weed suppression at 45 and 65 days of sowing of
wheat. Bromoxinil + MCPA alone did not show effective weed control at
lower rates. Sorgaab and sunfaab 15 L ha-1
each in combination with 1/3rd
dose of phenoxoprop p-ethyl resulted in maximum grain yield and net
benefits, but bromixinil + MCPA alone was effective only at higher rates.
Increase in yield was only due to effective weed control and
improvements in yield components of wheat crop.

Khan et al. (2009) conducted a field trial to investigate the
response of sunflower and its weeds to sorghum water extract
(sorgaab) in combination with reduced doses of Dual gold
herbicide. Results revealed that Sorgaab @ 15 L ha-1
+ 1/3rd
dose
of Dual gold reduced Chenopodium album density by 92% at 70
DAS. Sorgaab alone @ 15 L ha-1
, full dose reduced Chenopodium
album density by 81% at 70 DAS. Sorgaab + 1/3rd
dose of Dual
gold reduced Coronopus didymus density by 68%. While Sorgaab
alone @ 15 L ha-1
, decreased C. didymus density by 36%. Sorgaab
+1/3rd
dose of Dual gold, decreased total weed density by 89%,
over control at 70 DAS. While Sorgaab alone @ 15 L ha-1
reduced
weed density by 76% at 70 DAS. Total dry weed biomass
suppressed by sorgaab + Dual gold, 1/3 rd
dose was 92% at 70
DAS. While sorgaab alone @ 15 L ha-1
reduced total dry weed
biomass by 85% at 70 DAS. It was also observed during this study
that plant height, head diameter, stem diameter, number of
achenes per head, 1000-achene weight and achene yield were
significantly affected by the combination of sorgaab 15 L ha-1
, full
dose and 1/3rd
dose of Dual gold herbicide.
Cheema et al. (2010) conducted a field trial to determine the
effect of allelopathic water extracts (sorghum + sunflower, and rice husk)
alone or in combination with lower rates of penoxsulam (Ryzelan). The
mixture of sorghum + sunflower @ 18 L ha-1
suppressed the density of
different weeds (up to 50%) and dry weight (up to 49%) while rice husk
@ 18 L ha-1
suppressed the density (up to 46%) and dry weight of weeds
(up to 49%). The water extracts with half dose of herbicide were effective
in controlling weeds and their effectiveness was equal to full dose of
herbicide at 20 and 40 DAS, while at 60 DAS, they provided good
control of weeds but not equal to the full dose of herbicide. The mixtures

of water extracts with 1/3rd
dose of herbicide were less effective in
reducing the weed density and dry weight of weeds as compared to
mixture of water extracts with half dose of herbicide. However, full dose
of penoxsulam was the most effective treatment in reducing the weed
density and dry weight of the weeds.
Jabran et al. (2010) found that when water extracts of sorghum
sunflower, mustard and rice each at 15 L ha-1
were tank mixed with 0.4
and 0.6 kg active ingredient (a.i.) ha-1
pendimethalin and sprayed
immediately after sowing of canola (Brassica napus L.), density, fresh
weight and dry weight of weeds were lower than that of control in all the
treatments; however the performance of various combinations of
allelopathic crop water extracts and lower pendimathalin rates was better
than the standard dose of herbicide particularly in case of purple
nutsedge. All yield parameters including number of branches per plant,
number of pods per plant, numbers of seeds per pod and 1000-seed
weight were higher, when combinations of allelopathic extracts were used
with lower herbicide rates.
Razzaq et al. (2010) tested the allelopathic crop water extracts
with reduced herbicide doses for weed management in wheat. Sorghum
and sunflower water extracts combinations each at 18 L ha-1
with reduced
doses of 70% of mesosulfuron + idosulfuron (Atlantis 3.6 WG),
mesosulfuron + idosulfuron (Atlantis 12 EC), metribuzin + fenoxaprop
(Bullet 38 SC), bensulfuron + isoproturon (Cleaner 70 WP) and
metribuzin (Sencor 70 WP) were compared with their label doses which
were sprayed alone 30 days after sowing for weed control in wheat.
Combination of sorghum + sunflower each at 18 L ha-1
with reduced dose
of metribuzin + fenoxaprop significantly reduced dry weed biomass by
92% compared to label dose of mesosulfuron + idosulfuron (93%).
Treatment combination of sorghum + sunflower each at 18 L ha-1
with

reduced dose of metribuzin + fenoxaprop by 70% produced maximum
(2.82 t ha-1
) grain yield with 34 % increase over control and it was
significantly higher by 17% than its label dose .
Rehman et al. (2010) revealed that combined application of
mixture of allelopathic crops water extracts of sorghum, sunflower and
rice with ½ of the recommended dose of pre-emergence herbicides
butachlor (600 g a.i. ha-1
), pretilachlor (313 g a.i. ha-1
) and
ethoxysulfuronethyl (15 g a.i. ha-1
) reduced barnyard grass (Echinochloa
crusgalli L.), flat sedge (Carex spicata L.) and crowfoot grass
(Dactyloctenium aegyptium L.) density by 75, 67 and 74% and their dry
weight by 66, 71 and 76%, respectively, while 1/3rd
of the recommended
dose of butachlor (400 g a.i. ha-1
), pretilachlor (208 g a.i. ha-1
) and
ethoxysulfuronethyl (10 g a.i. ha-1
) in combination with mixture of
allelopathic water extracts reduced the density and dry weight of these
weeds by 68, 60 and 67% and 63, 67 and 72%, respectively. Application
of water extracts mixture with ½ of the label rates of pre-emergence
herbicides improved rice grain yield by 61, 59 and 41%, respectively,
while mixture of allelopathic extracts alone enhanced rice grain yield by
29% over control.
Iqbal et al. (2010) carried out afield trial to investigate the
response of wheat crop and its weeds to various crop water
extracts sorghum, brassica and sunflower in combination with
reduced rates of herbicide (Bromoxynil + MCPA 20 + 20 EC).
The data recorded at 70 DAS (days after sowing) showed that
water extracts @ 18 L ha-1
combined with Bromoxynil + MCPA
50 g a.i ha-1
inhibited total weeds density by 88%, total weeds
fresh weight by 90% and total weeds dry biomass by 95% and
increased grain yield by 35% over control.

Razzaq et al. (2012) indicated that spraying of sorghum + sunflower
extract each at 18 L ha-1
combined with 70% reduced dose of
mesosulfuron + idosulfuron or metribuzin + phenoxaprop or
mesosulfuron + idosulfuron reduced total weed dry weight by more than
90% over the control. On the other hand, sorghum and sunflower water
extracts each at 18 L ha-1
combined with metribuzin + phenoxaprop
produced a maximum number of productive tillers, spikelet's per spike,
number of grains per spike, biological yield and grain yield.
Khan et al. (2012) investigated the possible effects of allelopathic
plant water extracts of sorghum, brassica, sunflower and mulberry in
combination with reduced doses of atrazine for weed control in maize.
The four levels (full, ½,⅓ and ¼ dose) of atrazine showed 65-81%
suppression of weeds density and weeds dry weight over control (weedy
check), while allelopathic plant water extracts showed 70-75%
suppression of weeds density and dry weight when used in combination
with half and 1/3rd
dose of atrazine over control. Nonetheless, 49%, 36%
and 31% more grain yield was obtained when full dose (alone) and half
and ⅓rd
dose of atrazine in combination of allelopathic plant water extract
was applied, respectively.

1.1.4. Combined effect of allelopathic residues with
herbicides
Although successful results have been obtained from crop extracts
applied with low herbicide rates, additional work in other soil types and
locations should be performed. To employ this technology, large volumes
of sprays are required for field application, and therefore appropriate
concentrations for each crop should be determined for large scale field
operations. Due to these limitations, an alternative practical and feasible
approach has been developed where the residues of allelopathic crops
including sorghum have been left to dry under field conditions and then
promptly incorporated into production sites for weed management (Al-
Bedairy, 2011). In this section, a review will be made to include all the
activities which have been done on this approach.
Alsaadawi et al. (2011a) conducted field trial with the aim of
utilizing allelopathic crop residues to reduce the use of synthetic
herbicides in broad bean (Vicia faba) fields. They found that a
combination of trifluralin herbicide with sunflower residue had the least
total count and biomass of weeds in broad bean, which was even better
than herbicide used alone. Integration of recommended dose of trifluralin
with sunflower residue at 1400 g m-2
produced maximum (987.5 g m-2
)
aboveground biomass of broad bean, which was 74 and 36% higher than
control and recommended herbicide dose applied alone, respectively.
Combination of herbicide and sunflower residue appeared to better
enhance pod number and yield per unit area than herbicide alone.
Application of 50% dose of trifluralin in plots amended with sunflower
residue resulted in similar yield advantage as was noticed with 100%
herbicide dose. Chromatographic analysis revealed the presence of
several phenolic compounds in the soil containing sunflower

residues and none of these appeared in soil without sunflower
residues. Concentration of total phenolic compounds appeared to be
increased at two weeks of decomposition, reached its maximum at
the 4th
week of decomposition and started to decline thereafter until
vanished at the 8th week of decomposition. Weeds population started to
increase after 6 weeks of residues decomposition when the phytotoxins
concentration was sharply reduced in the soil.
Alsaadawi and Al-Temimi (2011) evaluated the allelopathic
potential of sunflower residues alone and in combination with
subrecommended doses of 2,4-D and Topic herbicides against weeds of
barley crop. Results revealed that combination of recommended dose of
herbicides with sunflower residues at 1400 g m-2
produced minimum
above-ground biomass (122 g m-2
) and weeds number (155.5 weeds per
m2 ), which were 35 and 50% less than recommended herbicide dose
applied alone, respectively. Meanwhile, integration of herbicides and
sunflower residue appeared superior in enhancing yield per unit area than
herbicide alone. Application of 50% dose of herbicides on plants growing
in plots containing sunflower residues at 1400 g m-2
resulted in similar
yield advantage as was noticed with 100% herbicide dose.
Alsaadawi et al. (2013) conducted two year field study to explore
the response of broad bean crop and its weeds to soil incorporated
allelopathic sorghum residues in combination with lower rate of a pre-
plant herbicide (trifluralin). Results indicated that plots treated with 50%
of label rate of herbicide and amended with sorghum residues recorded
least weed density and dry biomass and this suppression was much
greater than the residue treatments alone. Application of herbicide at 50%
rate in plots amended with sorghum residue resulted in similar yield as
with the 100% herbicide rate treatment. Chromatographic analysis of

sorghum amended field soil revealed the presence of several potent
allelopathic compounds of phenolics in nature. Periodic data revealed that
maximum quantities of these phytotoxins were coincided with the period
in which maximum suppressive activity against weeds was noticed under
field condition, which explain the activity of phytotoxins on weed
suppression. They suggested that integration of sorghum residues with a
lower herbicide rate can furnish adequate weed suppression without
compromising yield, which could be used as a feasible and
environmentally sound weed management approach in broad bean fields.
Lahmod and Alsaadawi (2014) conducted A two-year field trial to
test the response of wheat crop and its weeds to different rates of
sorghum residues alone or in combination with 50% of the label rate of
Chevalier herbicide. Results showed that all treatments significantly
reduced weed population and dry weight of weeds in comparison to
weedy check treatments during both years of the study. However, plots
treated with 50% of label rate of herbicide and amended with sorghum
residues recorded least weed density and dry biomass and this
suppression was much greater than the residue treatments alone.
Application of chevalier herbicide at 50% rate in plots amended with
sorghum residue at rates 3.50 and resulted in similar yield as with the
100% herbicide rate treatment. The increase in yield apparently due to
increase number of grains and weight of grains over control. They
concluded that integration of sorghum residues with a lower herbicide
rate can furnish adequate weed suppression without compromising yield.

1.2. Effect of phenolic compounds on arbuscular mycorrhiza
Cooper (2004) mentioned that some phenolic compounds are
important signal molecules for the development and growth of both plants
and microbes. During the arbuscular mycorrhiza fungi (AMF)
colonization. The effects of phenolic compounds on spore germination
and the subsequent colonization have been intensively investigated
(Becard et al., 1992; Gianinazzi and Gianinazzi-pearson, 1992). Factors
inducing the germination of AMF spores and the following a symbiotic
growth of germ tube have been determined including soil-derived stimuli
(e.g. CO2, ethylene) and plant-derived signals (e.g. phenolic compounds)
(Nair et al., 1991). Nagahashi et al. (1996) compared the phenolic
constituents in cell walls and cytoplasm's of host (carrot) and non-host
(sugar beet) of AMF, and observed some phenolic acids unique to host
while others unique to non host. In further bioassay, they demonstrated
that phenolic constituents in host roots were not always stimulatory and
those in non host roots were not always inhibitory to the growth of AMF
(Douds et al., 1996).
Some plant phenolic compounds have been found to be potential
candidates as signals during mycorrhizal formation. Some reports show
that exogenous application of flavonoids exerts a positive effect on
hyphal growth during symbiosis (Gianinazzi-Pearson, 1989; Tsai and
Phillips, 1991; Poulin et al., 1997). These effects range from increased
spore germination to enhanced hyphal growth, hyphal branching and
formation of secondary spores. It has been shown that the arbuscular
mycorrhizal formation alters the flavonoid profile of root extracts through
modifications to the expression of genes involved in phenylpropanoid,
flavonoid and isoflavonoid metabolism (Harrison and Dixon, 1993;1994).
Flavonoids are specific to mycorrhizal symbiosis formation, a single
flavonoid might exert a positive, negative or neutral effect on different

fungi (Siqueira et al., 1991a; Poulin et al., 1997). This may be explained
by specific effects of each flavonoid, as recently reported (Scervino et al.,
2005). Therefore it is likely that the role of flavonoids is limited to a
stimulatory effect on AM fungal growth (Silva-Junior and Siqueira,
1998).
Among a large variety of phenolic compounds, those stimulatory or
inhibitory to AMF spore germination have been described. Chabot et al.
1992 reported that at 2% CO2 conditions, flavonols (Kaempferol,
quercetin and morin) with at least one hydroxyl group on the B ring
improved spore germination and hyphal growth of G. margarita, while
biochanin A, genistein, hesperetin, galangin and chrysin inhibited the
hyphal growth. The latter two compounds possess no hydroxyl group on
the B ring. Buee et al. (2000) have reported a root factor (one or several
molecules) that stimulated the nuclear division and thus improved the
hyphal branching of G. gigantean germ tube. They demonstrate the
presence of this active factor in root exudates of all mycotrophic plant
species tested (eight species) but not in those of nonhost plant species
(four species), and further they hypothesized that this lipophilic
compound was not a flavonoid or a compound synthesized via the
flavonoid pathway. This hypothesis is consistent with the results of
Becard et al. (1995) who stated that root metabolites in addition to
flavonoids may stimulate AM fungal growth. Recently, Akiyama et al.
(2005) isolated a branching factor from the root exudates of Lotus
japanicus and further identified it as sesquiterpene. They found that
natural strigolactones, 5-deoxy-strigol, sorgolactone, strigol sythetic and
strigolactone analog GR24 induced extensive hyphal branching in
germinating spores of Gigaspora margarita at concentration as low as 3-
10 pg per disk.

Several workers have observed considerable increases in phenolic
compounds in the host as a result of AMF inoculation (Ling-lee and
Chilvers 1977; Selvaray and Subramanian, 1990). Giovannetti et
al.(1996) speculated that only the perception of the right chemical signals,
coming from the roots of host plants, promotes differential
morphogenesis of AMF hyphae in the rhizosphere and that root cells
penetration by the fungus depends on the host genome. Therefore,
phenolic compounds could enhance the initial stages of AMF
establishment, but root penetration and AMF development are likely
regulated by the host plant and subsequent interactions with the fungal
partner.
The effects of the application of 0.25 or 1.0 mM p-coumaric acid,
p-hydroxybenzoic acid, or quercetin on growth and colonization of clover
(Trifolium repens L. cv. Ladino) and sorghum (Sorghum bicolor L.) roots
by the arbuscular mycorrhizal (AM) fungus Glomus intraradices were
studied (Schenck and Smith, 1982). In general, soil application of these
compounds at 0.25 mM stimulated plant growth and AMF colonization,
whereas at 1.0 mM these phenolics were inhibitory to both growth and
colonization. Such effects were noted for both clover and sorghum. The
results suggest that phenolic compounds, commonly found in many soils,
influence the establishment of AMF symbioses, and these compounds
may have immediate effects on host growth. They indicated that studies
involving these chemicals and their effects on mycorrhizal associations
may provide new insights concerning the importance of the AMF
symbiosis in agricultural systems. These phenolic compounds may be
used as potential soil amendments to enhance AM fungal colonization,
and thus, exploit indigenous populations of AM fungi (Fries et al., 1997).
In another study, Martin et al, (2002) also mentioned that among
the various phenolic compounds, flavonoids are the most intensively

studied in AMF colonization. They revealed that the effects of flavonoids
on the AMF colonization are stimulatory, null and even inhibitory,
depending on the molecular species and concentration.
Siqueira et al. (1991a) studied the effects of flavonoid compounds
on VA (AM) mycorrhiza root colonization and growth of white clover
(Trifolium repens L.) plants under growth chamber conditions. The
isoflavonoids, formononetin and biochanin A, previously identified from
clover roots, stimulated colonization and growth of clover, while several
other flavonoid compounds were inactive when tested at concentrations
of 5 mg 1−1
. The flavone, chrysin, when applied at concentrations higher
than those tested for formononetin and biochanin A, also increased root
colonization and plant growth. The stimulatory effects of the
isoflavonoids on plant growth were mediated by VA mycorrhizal fungi
and were dependent on concentration, period of growth and soil spore
density. Maximum responses were found when 5 mg 1−1
solutions were
applied to soil containing 2 to 4 VA mycorrhiza spores g−1
of soil. These
results may provide insights on the molecular mechanisms of host-fungus
interaction and for the development of technology to exploit the potential
of the indigenous VA mycorrhizal fungi in field soil. They also suggested
that the stimulatory effects of these compounds on colonization were
mediated by concentration, because flavones and chrysin increased root
colonization and plant growth at the concentration above 5 mg l-1
. Caffeic
acid showed a stimulatory effect on hyphal mycorrhizal colonization at
5×10-5
M, but it produced an inhibitory effect at 5×10-4
and 5×10-3
M.
(Siqueira et al. 1991b).
The effects of an aqueous extract of Artemisia princeps var.
orientalis and two phenolic compounds on mycorrhizal colonization and
plant growth have been investigated (Yun and Choi., 2002). Under
greenhouse studies, they showed that the inhibitory effect of the extract

on mycorrhizal colonization and plant growth increased in proportion to
the concentration of the extract. When the mycorrhizal test plants were
treated with an increasing concentration of phenolic compounds, the
mycorrhizal colonization in roots of the test plant and the plant growth
were decreased. There were strong indications that mycorrhizal fungi
mitigated the inhibitory influence of shoot extract of A. princeps var.
orientalis and phenolic compounds.
In contrast, the reduced colonization due to the application of
phenolic compounds has also been reported. Olive mill residue normally
contains phenolic compounds of 15 mg kg-1
. When AMF inoculation was
done 4 weeks prior to the application of olive mill residue, no effect was
observed on the colonization, whereas the application of AMF
inoculation significantly decreased the colonization (Martin et al., 2002).
This case indicates that phenolic compounds probably exert inhibitory
effects on colonization by suppressing spore germination.
AMF colonization is the function of many factors (spore
germination, growth and branching of germ tube, root exudation, root
signaling). Recent experiments showed that phenolic compounds may
also act as a signaling molecules. With exogenous flavonoids, apigenin
and daidzein. Dong and Zhao (2004) successfully induced the
colonization of non-mycorrhizal plant Brassica juncea Cross by G.
intraradices or G.mosseae. The alkaline phosphatase activity in colonized
roots was observed and new spore were also produced 13 weeks after
treatment. However, they found the effect of flavonoids were
concentration dependent,with 150 nmol l-1
better than 15 or 1500 nmol l-1
.
Vierheilig et al.(2000) indicated that although some phenolic
compounds are stimulatory to AMF colonization, but recent researches
showed that AMF infection inhibited the root colonization in barley
plants by AMF due to effect of phenolic compounds.

Afzal et al. (2000) have reported that the application of aqueous
extracts of Imperata cylindrica, an allelopathic grass, markedly reduced
AMF colocization in Vigna radiate. In a review article by Javaid (2007),
the two allelopathic grasses namely, I. cylindrica and Dichanthium
annulatum have been shown to adversely affect the AMF colonization
extent in the associated weed species. This reduction in AMF
colonization under the allelopathic effects is mediated by an array of
biochemicals released into the soil by plants. These may include phenolic
substances, flavonoids and many more (Siqueira et al. 1991b; Lynn and
Chang, 1990). These phenolic compounds are produced as plant exudates
or as a result of decomposition of plant debris. These biochemicals are
signal transduction molecules and are liable to produce both stimulatory
and inhibitory effects depending upon the concentration (Becard et al.
1992).

Chapter Two
Materials
and
Methods
CHAPTER TWO

2. Materials and Methods
2.1. Field preparations
2.1.1. Site selection
The proposed study was conducted at Research Farm of Biology
Department, College of Science, Baghdad University, Baghdad, Iraq. The
soil of experimental site was calcareous silt clay loam.
2.1.2. Seeds and herbicides sources
Seeds of sunflower cv. Asgrow and grains of wheat cv. Abu Ghraib
were obtained from Department of Crop Sciences, College of
Agriculture, Baghdad University. Chevalier WG herbicide
(Mesosulfuron + Iodosulfuron) which belongs to a group of Sulfonyl-
urea. The herbicide is a product of Bayr Crop Science Company.
2.1.3. Soil Sampling and Analyses
Soil samples were taken at flowering stage from control and treated
(plots amended with sunflower residue at 6 t ha-1
) plots from the wheat
field . Two samples were taken randomly from each plot, 10-15 cm deep.
The samples were mixed, air- dried, sieved through a sieve with 2 mm
openings to remove large rock and plant debris, and pulverized. The
small roots and stones were picked out. Soil texture, organic matter and
electrical conductivity (Ec) , pH , inorganic nutrients NH4, NO3, P, N, Mg
and K were carried out in Ministry of Science and Technology. The
physical and chemical characteristics of the soil were listed .

Table 1. Some Physical and chemical properties of field soil before and
after sunflower residue incorporation.
Values ParM Mmeterss
Parameter Plots amended with
sunflower residues
Control Plots
87.52 88.22 % Sand
68.88 62.73 % Clay
65.83 69.88 % Silt
Clay loam Clay loam Soil texture
5.9 5.3
pH**
8.63 9.92
Electrical
conductivity(dS.m-1
) 1.70 3.53 % Organic carbon
50.34 93.68 NH4+-N ( ppm)
62.53 73.9 NO3--N (ppm)
33.38 4.54 P (ppm)
833.3 883.92 Na (ppm)
353.88 387.33 Mg++
(ppm)
70.12 93.39
K+(ppm)
Each value was an average of 4 replicates. **
In saturated past extract at 25 C

2.1.4. Preparation of sunflower residues
To prepare residues of sunflower plants, field plot (7×2 m) were
tilled twice at the beginning of July 2012. Seeds of sunflower was sown
in 75 cm spaced crop rows with distances of 25 cm between seeds. Plot
(12×2 m) without crop was used as a control. Fertilizers were nitrogen as
urea (46% N) at 240 kg ha-1
and phosphorus as triple super phosphate
(46% P2O5) at 240 kg ha-1
. The whole phosphorus and half of the nitrogen
were applied at planting while the remaining half of the nitrogen was
applied after two months (AL-bedairy, 2011). Irrigation was applied as
recommended for this crop. At physiological maturity, the heads were
removed then 50% of the half sunflower plant parts while, the other half
were left for drying for several days. These plot areas were then tilled
twice at mid November 2012 by using a disc plough to incorporate
sunflower residues in to the soil. Residue free treatment was maintained
as control on an intentionally un-cropped area of the same field. Field
measurements revealed that the residues of sunflower incorporated in
field soil before and after removal half of sunflower plant parts
correspond to sunflower residue rate of about 6 and 3 t ha-1
.
2.2. Field trial
The plots that received sunflower residues at 0, 3 and 6 t ha-1
of
previous experiment were subdivided into plots measuring 2×1.5 m at the
middle of November 2012. Fertilizers Nitrogen as urea (46% N) and
phosphorus as triple super phosphate (46% P2O5) were applied to these
plots as recommended for wheat crop (Cheema and Khaliq, 2000). Each
field plot was treated on lines with 250 g of Arbuscular mycorrhizal
(Glomus mosseae) inoculum (spores, hyphea and roots of the host) before

sown wheat. Grains of wheat cv. Abu-Grab were manually sown in all
plots in 20 cm apart crop rows at seeding rate of 120 kg ha-1
. All plots
received equal irrigation water during the entire course of study. The
experiment consists of the following treatments:
a. Control (un cultivated sunflower field)
b. Residues at 3 t ha-1
c. Residues at 3 t ha-1
+ 50% of label rate of chevalier 15 WG
d. Residues at 6 t ha-1
e. Residues at 6 t ha-1
+ 50% of label rate of chevalier 15 WG
f. Label rate of chevalier 15 WG (300 g/ha)
Chevalier 15 WG herbicide was sprayed on weeds of their
respective plots after 45 days from sowing of wheat, using hand sprayer a
Knapsack hand sprayer fitted with T-Jet nozzle at a pressure of 207 k Pa.
The experiment was conducted in split plot design with four replications
for each treatment. The herbicide rates were kept in the subplot while
sunflower residue rates were assigned as main plot. All plots received
equal irrigated water during the growing season of the crop.
2.3. Weed measurements:
2.3.1. Weed density (plant m‾2)
A quadrate measuring 0.5 × 0.5 m was randomly placed in
respective plots at 90 and 120 days after sowing to record weed density.
their average was calculated and converted to weed density per meter
square.

2.3.2. Weed biomass (g m‾2)
The weeds were counted at 120 days after sowing , cut from ground
surface, stored in polythene bags and then brought to laboratory for
recording their biomass. Total dry weight of weeds was determined after
oven-drying at 70˚C for 3 days until constant weight was achieved. The
weight was measured using an electric balance, averaged and calculated
on square meter basis.
2.4. Wheat crop measurements
2.4.1. Dry weight of plant through different stages of crop growth
(g/m2)
An area 20 cm was selected randomly from each plot at 27 ,44, 75,
and 103 day after sowing, cut from ground surface, stored in polythene
bags and then brought to laboratory for recording their dry biomass. The
dry biomass of plants was determined after oven-drying at 70˚C for 3
days until constant weight was achieved. The weight was measured using
an electrical balance, averaged and calculated on square meter basis.
2.4.2. Crop Growth Rate CGR (g/m2/day)
The following formulae proposed by Hunt (1982) was used to
calculate crop growth.
.1
12
12
TT
WW
ACGR
Where: W2 and W1 were the dry weight of plant and T2 and
T1 were the time of sampling.
2.4.3. Plant height
At harvesting, ten plants were selected randomly from each plot
and their height was measured from ground surface to the top of the plant
with the help of meter rod and the average height was calculated in cm.

2.4.4. Number of spikes/m2
An area of 0.5 × 0.5 m were selected at random in each plot to count
number of spikes at maturity and converted to number of spikes/m2
2.4.5. Number of grains per spike
Twenty spikes were selected randomly from each experimental
unit and threshed individually. The grains were counted and average
number of grains per spike was calculated.
2.4.6.1000-grain weight (g)
Two samples, each of 1000-grain, were taken from the produce of
each plot. These samples were weighed on an electric balance and
average 1000-grain weight was calculated.
2.4.7. Total wheat biomass (t ha1ـ)
The crop was harvested, tied into bundles and allowed to sundry for
a week in respective plant. Total wheat biomass of the sun dried samples
was recorded for each treatment. The total biomass yield per plot was
converted to tones per hectare (t haـ 1).
2.4.8. Grain yield (t haـ 1)
The harvested and sun dried crop was threshed manually. The grains
weight for each treatment was recorded in gram and later expressed in
tones per hectare (t haـ 1).
2.4.9. Biological yield
Biological yield was calculated by summation of straw and grain
weights.
2.4.10. Harvest index
Harvest index for each treatment was calculated by using the
following formula (Donald and Hamblin, 3454 ).
Harvest index = (Grain yield / Total biomass yield) ×100

2.5. Mycorrhizal studies
2.5.1. Spore extraction
Soil samples were collected from each plot, air dried and sieved
through a 2 mm openings sieve to remove large debris. A sub sample
(100 g) was taken from each sample and placed in a 500 ml beaker
containing 200 ml 0.08 M sodium hexametaphosphate solution to break
up clay clumps. The suspension was agitated for 5 minutes and left to
settle for 15 seconds (Smith and Dickson, 1997). The supernatant was
decanted through a nest of sieves with reducing mesh sizes from 500 μm,
250 μm, 125 μm to 45 μm. This step was repeated with water twice and
the debris from the 45 μm was discarded. The debris on the remaining
sieves, containing the AM spores was washed and placed in 40 ml
centrifuge tubes for purification. The spore suspension was centrifuged at
3000 rpm for 5 minutes, after which the supernatant was discarded. The
pellet was re-suspended in 60% sucrose solution and centrifuged for
another 5 minutes. The supernatant containing AM fungal spores was
decanted into a 45 μm sieve and washed with water to remove sucrose on
the spores (Smith and Dickson, 1997).
2.5.2. Isolation and identification of mycorrhiza spores
Some spores were mounted in small glass capsules containing water
and a drop of chloroform for identification. The identified according to
Schenck and Smith, year 1982.
2.5.3. Preparation of Mycorrhiza inoculums
Loamy soil was brought from field, sterilized in autoclave for 0.5 hr
period for two days and packed in 10 plastic pots of 1 kg capacity.
Spores of mycorrhiza Glomus mosseae were extracted by procedure
outlined by Smith and Dickson (1997). The extract was mixed with the

upper part of pot soil . Ten seeds of sorghum and 10 of millet were sown
separately and irrigated with appropriate amount of water. Three months
after planting, the above ground of plants were cut. The soil plus roots of
both plant species were taken, mixed thoroughly and used for inoculation
process (Matloob, 2012).
2.5.4. Spore counting
Based on previous field study, total phenolics was increased after
two weeks of residue decomposition and reach maximum accumulation
in December 15, 2012 of residues decomposition and declined drastically
thereafter. To test if the mycorrhizal population is correlated with the
phenolics profile during this period, spore counting was conducted by
taking soil samples from rhizosphere of wheat plants growing in plots
containing 6 t ha-1
and in plots without sunflower residues (Control) at 1,
14, 28 and 42 days after sowing (DAS).
Spore counting was also made at the end of wheat crop maturity to
determine the possible effect of test treatments on mycorrhizal population
and growth. Soil samples from rhizosphere of wheat plants growing in
plots of all treatments were taken and used for counting number of spores
using microscopic slide.
2.5.5. Mycorrhizal colonization rate (%)
Wheat plant roots were taken from the field at the end of crop
maturity for determining the colonization rate and colonization intensity.
Fresh roots were carefully washed and cut into 1-3 cm pieces. The pieces
were immersed with 1% KOH solution and incubated at 70°C for 20
minutes to remove the cytoplasm. The KOH solution was discarded and
the roots were rinsed well with distilled water. Roots were covered with a
freshly prepared alkaline H2O2 solution for 10 minutes. The bleaching

solution was discarded and the roots were rinsed with water. Roots were
acidified in a 0.1 M HCl solution overnight to ensure adequate binding of
stain to fungal structures. The HCl solution was discarded and roots were
covered with Lacto glycerol Trypan Blue (0.05%) stain and incubated for
45 minutes at 90°C. The stain was poured off and roots were covered
with lacto glycerol destain. Roots were allowed to destain overnight
before microscopic examination (Smith and Dickson, 1997). Finally,
roots were mounted on microscopic slides and using a compound
microscope for examined. The percentage root colonization was
calculated by the following equation using a modified Line Intersect
Method (Mc Gonigle et al., 1990) (Appendix 1,2,3,4).
% Colonization = (Total number of AM positive segments / Total number
of segment studied) × 100
Mycorrhizal intensity was calculated based on the following rate
index.
0 = the root fragment was not mycorrhizaled
1 = 1-25 of the root fragment was mycorrhizaled
2 = 26-50 of the root fragment was mycorrhizaled
3 = 51-75 of the root fragment was mycorrhizaled
4 = 76-100 of the root fragment was mycorrhizaled
Mycorrhizal intensity (MI) was calculated according to the
following equation:
MI = Total (number of fragments × their rate index) / number of observed
fragments × highest rate
2.6. Determination of total phenolics Folin-Denis was used for total phenol analysis (A.O.A.C., 1990) and
ferulic acid was used as standard since it is an allelopathic agent present
in sunflower plant (Haslam, 1988). Soil samples were taken from soil of

plots amended with 6 t ha-1
and 0 t ha-1
(control) at a depth of 30 cm at 1,
14, 28 and 42 days after sowing (DAS). The soils were mixed thoroughly
and allowed to dry at room temperature for 3 days. Samples of 250 g dry
soil were extracted separately in 250 ml of distilled water by shaking for
24 hrs at 200 rpm (Ben- Hammouda et al.,1995). Soil suspensions were
filtered through Whatman No. 2 filter paper under vacuum. Folin-Denis
(0.5 ml) and Na2CO3 (one ml) were added to one ml of soil water extract
and left to stand for 30 minutes. Absorbance was determined at 750 nm
on aspectrophotometer (Blum et al.,1991). The total phenolic content was
obtained by standard curve using different concentrations of ferulic acid
(Appendix 5,6,7).
2.7. Bioassay of soil amended with sunflower residues
Soil samples were taken from soil of plots amended with 6 t ha-1
and 0 t ha-1
(control) at a depth of 30 cm at 1, 14, 28 and 42 days after
sowing (DAS). The soils were mixed thoroughly and allowed to dry at
room temperature for 3 days. Samples of 200 g dry soil packed in plastic
pots of 250 g capacity. Ten seeds of Malva rotundifolia L. was sown in
each pots. The pots were arranged in randomized complete blocks in
three replications in plastic house at the beginning of December. The pots
were irrigated with appropriate amount of water whenever needed. Seed
germination was recorded 15 days after sowing, after which time, the
seedlings were thinned to 3 per pot and allowed to grow for additional
two weeks. The plants were pulled from the pots using running water.
Total dry weight of M. rotundifolia L was determined after oven-drying
the plants at 70˚C for 3 days. The weight was measured using an
electrical balance.

2.8. statistical analysis
The collected data were statistically analyzed using analysis
of variance (ANOVA) by GENSTAT computer software package.
Differences among treatment averages were compared using Least
Significant Difference (LSD) ≤ 0.05 probability level (Steel et al.,
1980).

Chapter Three
Results

CHAPTER THREE
RESULTS
3.1. Weed parameter
3.1.1. Effect of different rates of chevalier 15 WG herbicide and
sunflower residues cv. Asgrow on total weed density in wheat
field
Field observations showed that 70% of weeds species
grown in the wheat field were broad leaf viz. Beta vulgaris L.,
Ammi majus L., Malva rotundifolia L., Ducus carota L.,
Carthamus oxyacanthus M.B., Spergularia salina L., and the
remain was grass weeds, namely Avena fatua L., Lolium
temulentum L. and Phalaris minor L. (Table 2).
The result presented in table 3 revealed that soil
incorporation of sunflower residues at 3 t ha-1
significantly
reduced total weed density by 88 and 70% of control after 90 and
120 days after sowing (DAS), respectively. Similarly, weed
population was significantly increased by 89 and 78% over
control by the residue rate to 6 t ha-1
after 90 and 120 DAS,
respectively. The suppression of weed density was increased with
increasing rate of residues to 6 t ha-1
at 120 DAS only. However,
the inhibition of weed population by the residues rates was
significantly averted when the residues combined with the
herbicide. Foliar application of 50% of label rate of chevalier to
plots amended with sunflower residue at 3 and 6 t ha-1
significantly suppressed weed density more than the respective
residue rates when applied alone after 90 and 120 DAS.
Application of label rate of chevalier recorded the highest
reduction in weed density compared to all treatments.

Table 2. Weed species grown in the test wheat field.
Common name
Scientific name Family
Broad leaf - weeds
Common beet Beta vulgaris L.
Chenopodiaceae
Large bull wort Ammi majus L.
Umberlliferae
Mallow Malva rotundifolia L.
Malvaceae
Wild carrot Ducus carota L.
Umberlliferae
Wild safflower Carthamus oxyacanthus
Asteraceae
Salt sandsparry Spergularia salina L.
Caryophllaceae
Narrow- leaf weeds
Wild oat Avena fatua L.
Poaceae
Rye grass Lolium temulentum L.
Poaceae
Canary grass Phalaris minor L.
Poaceae

Table 3. Effect of different rates of chevalier 15 WG herbicide and sunflower residues cv. Asgrow on weeds population
grown in wheat .
Treatments
Population density
(Plants per m2)
at 90 DAS*
% of control
Population density
(Plants per m2)
at 120 DAS
of control %
Weedy check (Control) 480 ---- 184 ---
Residues at 3 t ha-1
60 12.5 55 29.9
Residues at 3 t ha-1
+ 50% of label
rate of chevalier15 WG 33 6.9 36
19.6
Residues at 6 t ha-1
53 11.0 40 21.7
Residues at 6 t ha-1
+ 50% of label
rate of chevalier15 WG 29 6.0 26 14.1
Chevalier15 WG (Label rate) 11 2.3 0 0
LSD ≤ 0.05 5.5 --- 10.9 ---
* DAS= day after sowing

3.1.2. Effect of different rates of chevalier 15 WG herbicide and
sunflower residues cv. Asgrow on dry weight biomass of
weeds in wheat
The results presented in table 4 revealed that average weed biomass
was significantly reduced by the residues of sunflower incorporated in
field soil. The reduction increased from 87% of control at 3 t ha-1
sunflower residue to 88% of control at 6 t ha-1
sunflower residue.
The interaction of herbicides and sunflower residues showed
significant effect on weed biomass. When chevalier applied at reduced
50% of label rate to plots amended with sunflower residues at 3 and 6 t
ha-1
scored weed biomass (94 and 96% of control respectively) more than
the residue rates when applied alone. However, chevalier application
(label dose) recorded maximum reduction (100%) in weed biomass.
3.2. Crop parameters
3.2.1. Effect of different rates of chevalier 15 WG herbicide and
sunflower residues cv. Asgrow on dry weight of wheat during
different growth stages
Result presented in figure 1 revealed the soil incorporation of
sunflower residue at 3 and 6 t ha-1
significantly inhibited dry
weight of wheat through the first two months after sowing and
the inhibition increased with increasing sunflower residues. At 75
DAS, no significant effect was appeared on dry weight of wheat
by application of sunflower residues alone. However, plots treated
with 50% of label rate of chevalier herbicide and amended with
sunflower residue at 3 t ha-1
recorded more dry weight (921.72
and 2023.2) than sole application of the 3 t ha-1
residue rate at 75
and 103 DAS, respectively. Similarly, plots treated with 50% of
label rate of chevalier and amended with sunflower residue at 6 t

ha-1
posed more dry weight (617.92 and 1884) than the 6 t ha-1
residue rate applied alone after 75 and 103 DAS, respectively.
Application of label rate of chevalier recorded maximum increase
in dry weight of wheat compared to all treatment at 75 and 103
DAS.
Table 4. Effect of different rates of chevalier 15 WG herbicide and
sunflower residues cv. Asgrow on dry weight biomass of
weeds grown in wheat.
Treatments dry weight biomass
(g per m2)
% of control
Weedy check (Control) 149.62 -----
Residues at 3 t ha-1
19.76 13.2
Residues at 3 t ha-1
+ 50% of
label rate of chevalier15 WG 9.26 6.2
Residues at 6 t ha-1
17.37 11.6
Residues at 6 t ha-1
+ 50% of
label rate of chevalier15 WG 6.46 4.3
Chevalier15 WG (Label rate) 0.00 -----
LSD ≤ 0.05 3.5 -----

Figure 1. Effect of different rates of chevalier 15 WG herbicide and
sunflower residues cv. Asgrow on dry weight of wheat.
T1. Control (uncultivated sunflower field), T2. Residues at 3 t ha-1
, T3. Residues at 3
t ha-1
+ 50% of label rate of chevalier WG, T4. Residues at 6 t ha-1, T5. Residues at 6 t
ha-1
+ 50% of label rate of chevalier15 WG, T6. Chevalier WG (Label rate).
0
500
1000
1500
2000
2500
27 44 75 103
Days after sowing
Dry
we
igh
t (g
/m2)
T1 T2 T3 T4 T5 T6
LSD≤ 0.05 2.43 8.93 77.5 87.7

3.2.2. Effect of different rates of chevalier 15 WG herbicide
and sunflower residues cv. Asgrow on crop growth rate
of wheat crop
The result presented in figure 2 revealed that soil
incorporation of sunflower residue at 3 and 6 t ha-1
showed
significant effect on crop growth rate of wheat during first 75
days after sown. Crop growth rate was decreased by 31 and 48%
of control by application of sunflower residue at 3 and 6 t ha-1
at
27 DAS. While the effect continue until 44 DAS, plots treated
with 50% of label rate of chevalier herbicide and amended with
sunflower residue at 3 and 6 t ha-1
more increase of crop growth
rate than the respective residue rate applied alone and control
treatments. Maximum crop growth rate was recorded by the
reduced rate of herbicide and sunflower residue rate at 6 t ha-1
after 75 DAS followed by a decrease in order by T6> T3> T2 >T1
respectively.
3.2.3. Grain yield (t ha-1
)
The result in table 5 revealed that the grain yield appeared to be
significantly affected by herbicide and sunflower residue treatments. The
grain yield was increased by 58 and 37% of the control when sunflower
residues were incorporated into the field soil at 3 and 6 t ha-1
,
respectively. However, plots treated with 50% of label rate of chevalier
herbicide and amended with sunflower residue at 3 t ha-1
recorded higher
yield than the respective residue rate applied alone and the label rate of
herbicide treatments. Similar trend of increase was observed with the
higher residue rate when combined with the reduced dose of herbicide.
Plots treated with 50% of label rate of chevalier herbicide and amended
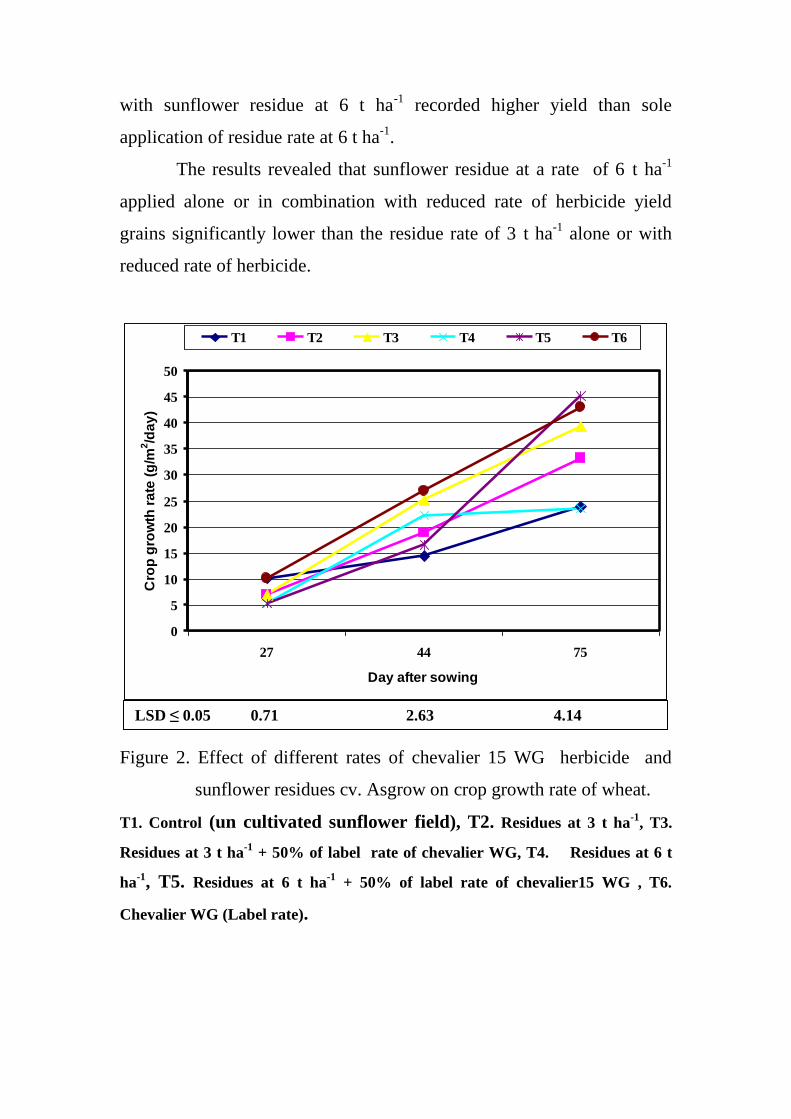
with sunflower residue at 6 t ha-1
recorded higher yield than sole
application of residue rate at 6 t ha-1
.
The results revealed that sunflower residue at a rate of 6 t ha-1
applied alone or in combination with reduced rate of herbicide yield
grains significantly lower than the residue rate of 3 t ha-1
alone or with
reduced rate of herbicide.
Figure 2. Effect of different rates of chevalier 15 WG herbicide and
sunflower residues cv. Asgrow on crop growth rate of wheat.
T1. Control (un cultivated sunflower field), T2. Residues at 3 t ha-1
, T3.
Residues at 3 t ha-1
+ 50% of label rate of chevalier WG, T4. Residues at 6 t
ha-1, T5. Residues at 6 t ha
-1 + 50% of label rate of chevalier15 WG , T6.
Chevalier WG (Label rate).
0
5
10
15
20
25
30
35
40
45
50
27 44 75
Day after sowing
Cro
p g
row
th r
ate
(g
/m2/d
ay
)
T1 T2 T3 T4 T5 T6
LSD ≤ 0.05 0.71 2.63 4.14
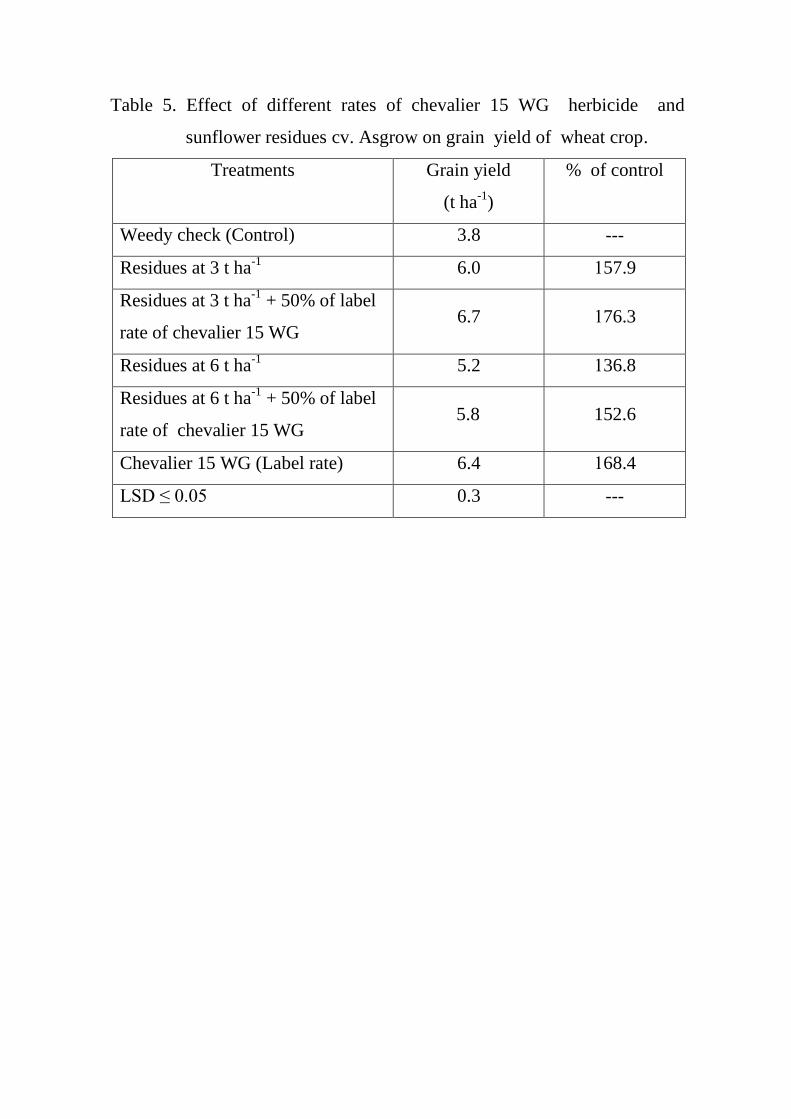
Table 5. Effect of different rates of chevalier 15 WG herbicide and
sunflower residues cv. Asgrow on grain yield of wheat crop.
Treatments Grain yield
(t ha-1
)
% of control
Weedy check (Control) 3.8 ---
Residues at 3 t ha-1
6.0 57.93
Residues at 3 t ha-1
+ 50% of label
rate of chevalier 15 WG 6.7 76.33
Residues at 6 t ha-1
5.2 36.83
Residues at 6 t ha-1
+ 50% of label
rate of chevalier 15 WG 5.8 152.6
Chevalier 15 WG (Label rate) 6.4 68.43
LSD ≤ 0.05 0.3 ---

3.2.4. Effect of different rates of chevalier 15 WG herbicide and
sunflower residues cv. Asgrow on yield components of wheat
3.2.4.1. Effect on number of spikes per m2
All treatments significantly affected number of spikes
compared to the control (Table 6). Plots amended with sunflower
residues at 3 and 6 t ha-1
increased number of spikes by 43 and
37% over control, respectively. Plots treated with 50% of label
rate of chevalier herbicide and amended with sunflower residue at
3 t ha-1
recorded significantly more number of spikes than plots
treated with 3 t ha-1
residues alone. However, combination of 50%
of label rate of herbicide with sunflower residue at 6 t ha-1
did not
significantly increase the number of spikes over 6 t ha-1
residues
treatment alone. Maximum increase was recorded in plots treated
with label rate of chevalier herbicide.
3.2.4.2. Effect on grains per spike
The average number of grains per spike was significantly
increased by 16 and 23% over control by incorporation of
sunflower residues at 3 and 6 t ha-1
, respectively (Table 7 ).
Herbicide application at reduced (50%) dose to plots amended
with test sunflower residues scored decrease number of grains per
spike compared to this sunflower residues when applied alone and
control (without sunflower residue). Meanwhile, Reduced
herbicide (50 %) dose in combination with 3 t ha-1
sunflower
residue rate scored similar number of grains per spike as was
observed under sole application of the label dose.

3.2.4.3. Effect on 1000-grain weight (g)
All test treatments significantly affected 1000-grain
weight compared to the control (Table 8). Incorporation of
sunflower residues in the field soil at rates of 3 and 6 t ha-1
significantly reduced 1000-grain weight by 8 and 7 over control
respectively; however, this increment was further hastened to
become greater than the label rate of herbicide treatment when
reduced dose (50%) of chevalier was applied to plots amended
with sunflower residues at 3 and 6 t ha-1
.
Table 6. Effect of different rates of chevalier 15 WG herbicide and
sunflower residues cv. Asgrow on number of spikes of wheat.
Treatments Number of spikes per m2 % of control
Weedy check (Control) 368 ---
Residues at 3 t ha-1
526 142.9
Residues at 3 t ha-1
+ 50% of
label rate of chevalier15 WG 577 156.8
Residues at 6 t ha-1
504 137.0
Residues at 6 t ha-1
+ 50% of
label rate of chevalier15 WG 537 145.9
Chevalier15 WG (Label rate) 634 172.3
LSD ≤ 0.05 29.05 ---
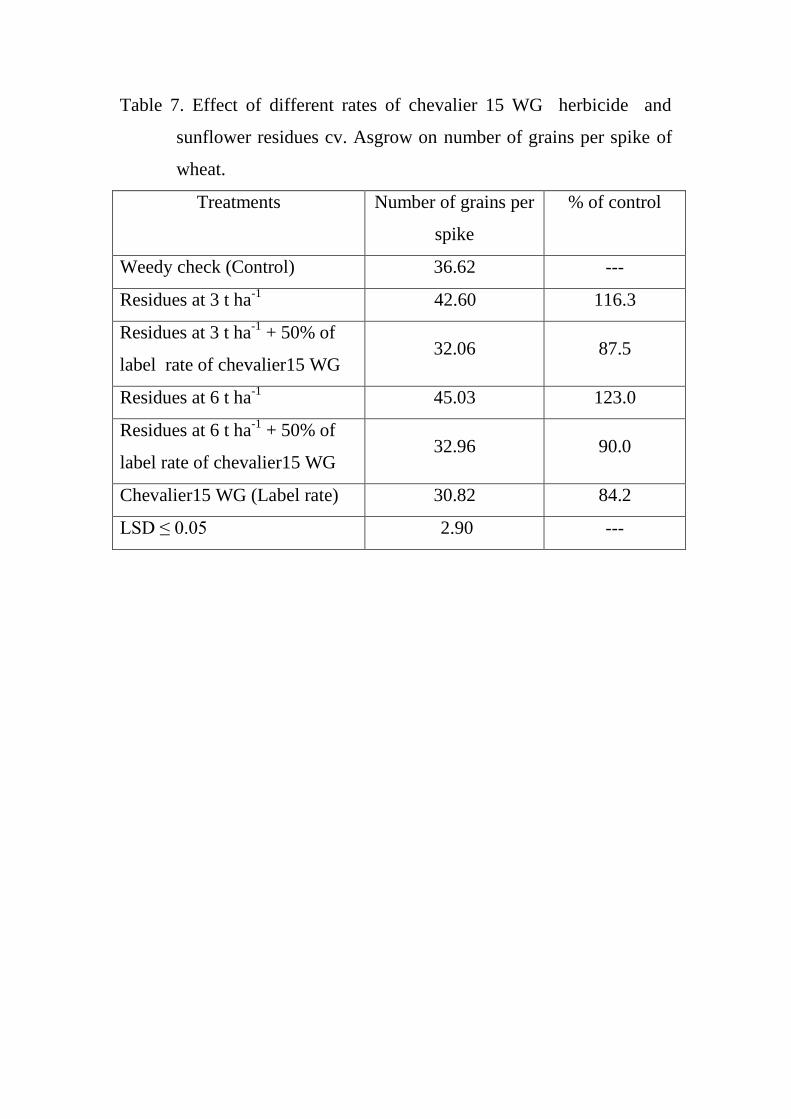
Table 7. Effect of different rates of chevalier 15 WG herbicide and
sunflower residues cv. Asgrow on number of grains per spike of
wheat.
Treatments Number of grains per
spike
% of control
Weedy check (Control) 36.62 ---
Residues at 3 t ha-1
42.60 116.3
Residues at 3 t ha-1
+ 50% of
label rate of chevalier15 WG 32.06 87.5
Residues at 6 t ha-1
45.03 123.0
Residues at 6 t ha-1
+ 50% of
label rate of chevalier15 WG 32.96 90.0
Chevalier15 WG (Label rate) 30.82 84.2
LSD ≤ 0.05 2.90 ---

Table 8. Effect of different rates of chevalier 15 WG herbicide and
sunflower residues cv. Asgrow on weight of 1000 grains of
wheat.
Treatments Weight of 1000
grains (g)
% of control
Weedy check (Control) 33.72 ---
Residues at 3 t ha-1
36.46 108.1
Residues at 3 t ha-1
+ 50% of label
rate of chevalier15 WG 39.30 116.5
Residues at 6 t ha-1
36.06 106.9
Residues at 6 t ha-1
+ 50% of label
rate of chevalier15 WG 37.77 112.0
Chevalier15 WG (Label rate) 37.30 110.6
LSD ≤ 0.05 1.47 ---

3.2.5. Biological yield (t ha-1
)
The results in table 9 revealed that the biological yield was
significantly affected by herbicide and sunflower residues
treatments compared to control. The biological yield was
increased by 36 and 22% of the control when sunflower residues
were incorporated in to the field soil at 3 and 6 t ha-1
, respectively.
However, these increments were significantly (56% of control) or
appreciably (29% of control) improved when the reduced dose of
herbicide was applied to plots amended with sunflower residue at
3 and 6 t ha-1
, respectively. Apparently, the residue rate 6 t ha-1
alone or in combination with the reduced dose of chevalier
herbicide recorded biological yield lower than the respective
treatments of 3 t ha-1
and the label rate of herbicide.
3.2.5. Biomass (t ha-1
)
The results in table 10 revealed that the biomass was
significantly affected by herbicide and sunflower residues
treatments compared to control. The biomass was increased by 29
and 20% of the control when sunflower residues were
incorporated in to the field soil at 3 and 6 t ha-1
, respectively.
However, these increments were significantly improved when the
reduced dose of herbicide was applied to plots amended with
sunflower residue at 3 and 6 t ha-1
, respectively. Apparently, the
residue rate 6 t ha-1
alone or in combination with the reduced dose
of chevalier herbicide recorded biomass lower than the respective
treatments of 3 t ha-1
and the label rate of herbicide.
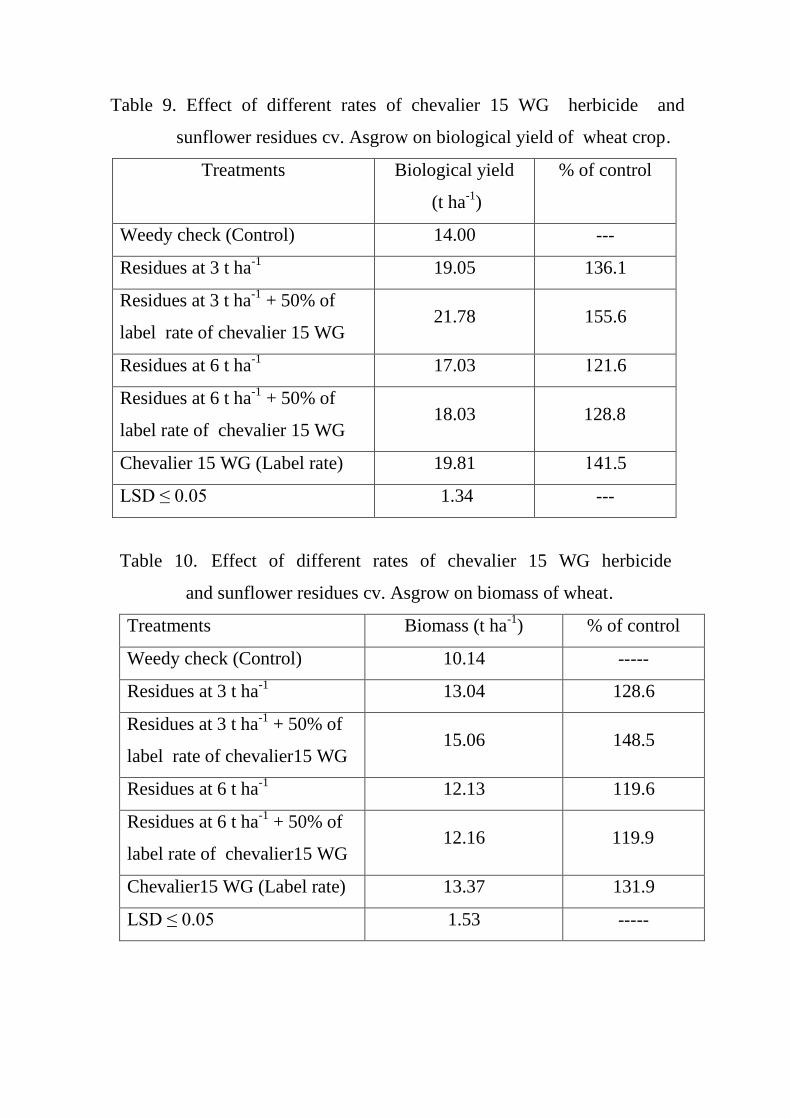
Table 9. Effect of different rates of chevalier 15 WG herbicide and
sunflower residues cv. Asgrow on biological yield of wheat crop.
Treatments Biological yield
(t ha-1
)
% of control
Weedy check (Control) 14.00 ---
Residues at 3 t ha-1
19.05 136.1
Residues at 3 t ha-1
+ 50% of
label rate of chevalier 15 WG 21.78 155.6
Residues at 6 t ha-1
17.03 21.63
Residues at 6 t ha-1
+ 50% of
label rate of chevalier 15 WG 18.03 128.8
Chevalier 15 WG (Label rate) 19.81 41.53
LSD ≤ 0.05 1.34 ---
Table 10. Effect of different rates of chevalier 15 WG herbicide
and sunflower residues cv. Asgrow on biomass of wheat.
Treatments Biomass (t ha-1
) % of control
Weedy check (Control) 10.14 -----
Residues at 3 t ha-1
13.04 128.6
Residues at 3 t ha-1
+ 50% of
label rate of chevalier15 WG 15.06 148.5
Residues at 6 t ha-1
12.13 119.6
Residues at 6 t ha-1
+ 50% of
label rate of chevalier15 WG 12.16 119.9
Chevalier15 WG (Label rate) 13.37 131.9
LSD ≤ 0.05 1.53 -----

3.2.7. Harvest index
All treatments enhanced the harvest index over the control
(Figure 3). Application of sunflower residue at 3 and 6 t ha-1
significantly increased harvest index by 14 and 12% over the
control. Maximum harvest index (32.32) was recorded by
treatment of residues at 6 t ha-1
+ 50% of label rate of chevalier15
WG, which is statistically at par with treatment of Label rate of
Chevalier 15 WG.
Figure 3. Effect of different rates of chevalier15 WG herbicide
and sunflower residues cv. Asgrow on Harvest index
of wheat.
T1. Control (un cultivated sunflower field), T2. Residues at 3 t ha-1
, T3.
Residues at 3 t ha-1
+ 50% of label rate of chevalier WG, T4. Residues
at 6 t ha-1
, T5. Residues at 6 t ha-1
+ 50% of label rate of chevalier15 WG
, T6. Chevalier WG (Label rate).
0
5
10
15
20
25
30
35
T1 T2 T3 T4 T5 T6 LSD ≤ 0.05
Treatments
He
rve
st
ind
ex

3.2.8. Effect of different rates of chevalier 15 WG herbicide and
sunflower residues cv. Asgrow on plant height of wheat
Plant height was significantly affected by the rates of
herbicide and sunflower residues and their interaction (Figure 4).
Application of sunflower residues at 3 and 6 t ha-1
significantly
reduced plant height by 39 and 6% of control respectively.
However, this reduction was further increased (4 and 8% of
control) when plots treated with 50% of label rate of chevalier
herbicide and amended with sunflower residues at 3 and 6 t ha-1
respectively. Maximum plant height (94.23 cm) was recorded by
control treatment and 3 t ha-1
treatments.
Figure 4. Effect of different rates of chevalier 15 WG herbicide and
sunflower residues cv. Asgrow on plant height of wheat.
T1. Control (un cultivated sunflower field), T2. Residues at 3 t ha-1
, T3. Residues
at 3 t ha-1
+ 50% of label rate of chevalier WG, T4. Residues at 6 t ha-1
, T5.
Residues at 6 t ha-1
+ 50% of label rate of chevalier15 WG , T6. Chevalier WG
(Label rate).
0
10
20
30
40
50
60
70
80
90
100
pla
nt
hei
gh
er (
cm)
T1 T2 T3 T4 T5 T6 LSD ≤
0.05
Treatments

3.3. Determination of Total phenolics in field soil
Total phenolics in field soil significantly increased after
incorporation of sunflower residues and reached their peak at 4
weeks of residues decomposition, then decreased significantly at 6
weeks and vanished at 6 weeks (Figure 5).
Figure 5. Total phenolics released in field soil amended with sunflower
residues at 6 t ha-1
during different decomposition periods.
0
0.2
0.4
0.6
0.8
1
1.2
1.4
1.6
1.8
2
1 14 28 42 L.S.D ≤ 0.05
Decomposition periods (days)
To
tal p
he
no
lic
s (
mg
/g d
ry
so
il)
Sunflower residues rate of 6 t.ha-¹ control (without sunflower residue) L.S.D ≤ 0.05

3.4. Weed bioassay
3.4.1. Seed germination of malva rotundifolia
Seed germination of malva rotundifolia was significantly
reduced after incorporation of sunflower residues in to the soil and
reached its maximum reduction at 4 weeks of residues
decomposition, then decreased significantly at 6 weeks (Figure 6).
Figure 6. Seed germination of Malva rotundifolia in field soil
amended with sunflower residues at 6 t ha-1
during
different decomposition periods.
0
0.2
0.4
0.6
0.8
1
1.2
1 14 28 42 L.S.D ≤ 0.05
Decomposition periods (days)
Seed
germ
ina
tio
n %
Sunflower residues rate of 6 t.ha-¹ control (without sunflower residue) L.S.D ≤ 0.05

3.4.2. Total dry weight of Malva rotundifolia
Total dry weight of malva rotundifolia in pots containing
field soil significantly reduced after incorporation of sunflower
residues and reached the maximum reduction at 4 weeks of
residues decomposition and gradually increased at the end of the
6th
week (Figure 7).
Figure 7. Total dry weight of Malva rotundifolia in field soil amended
with sunflower residues at 6 t ha-1
during different
decomposition periods.
0
0.02
0.04
0.06
0.08
0.1
0.12
0.14
0.16
0.18
0.2
1 14 28 42 L.S.D ≤ 0.05
Decompsition periods (days)
Total
dry w
eig
ht
Sunflower residues rate of 6 t.ha-¹ control (without sunflower residue) L.S.D ≤ 0.05

3.5. Mycorrhizal studies
3.5.1. Isolation and identification of mycorrhiza from field soil of
wheat crop
Microscopic examination of the collected samples from
wheat field revealed that the dominant (90 % of the population)
species of mycorrhiza is Glomus mosseae. The species is
characterized by the following characters: spores are rarely filled
with hyphae, sporocarp containing 1-10 spores (sometimes),
diameter of subtending hyphae at widest part 18-50 µm, outer
surface of inner wall is not ornamented; thin, hyaline outer wall
may not be obvios. Spores are more than 100 µm in size,
subtending hyphae is generally funnel-shaped with cup-shaped
septum (Figure 8, 9).
Figure 8. spore of Glomus mosseae Figure 9. Sporocarp
X40 X4
0
hyphea
spore

3.5.2. Effect of different periods of decomposition of sunflower
residues in field soil before chevalier application on sporulation
Figure 10 showed the sporulation of Glomus mosseae in field
soil amended with sunflower residues at different periods of
decomposition and before the time of chevalier herbicide
application. The results revealed that incorporation of the residues
significantly increased spores number at 2, 4 and 6 weeks of
residue decomposition compared to control treatment (without
sunflower residue).
Figure 10. Sporulation of Glomus mosseae in field soil amended with
sunflower residues at 6 t ha-1
during different decomposition
periods.
0
50
100
150
200
250
300
1 14 28 42 L.S.D ≤ 0.05
Decomposition periods (days)
Sp
oru
lati
on
(sp
ore/1
00
dry s
oil
)
Sunflower residues rate of 6 t.ha-¹ control (without sunflower residue) L.S.D ≤ 0.05

3.5.3. Effect of different rates of chevalier 15 WG herbicide and
sunflower residues cv. Asgrow on sporulation at flowering
stage
The results presented in table 11 exhibited that sunflower residues
incorporated in field soil had a significant effect on number of spores.
Field soil amended with sunflower residues at 3 and 6 t ha-1
increased
number of spores by 16 and 23% over control. However, the number of
spores was decreased significantly when chevalier was applied at the
label dose. Chevalier application at reduced (50% of the label rate) rate
applied to plots amended with sunflower residue at 3 t ha-1
scored spore
number significantly lower than control treatment.
3.5.4. Effect of different rates of chevalier 15 WG herbicide and
sunflower residues cv. Asgrow on colonization rate of Glomus
mosseae
The results showed that colonization intensity was
significantly affected by herbicide and sunflower residues
application (Table 12). Sunflower residues incorporated into field
soil at rates of 3 and 6 t ha-1
significantly increased colonization
rate by 94 and 113% over control by 94 and 113 over the label
rate of herbicide, respectively. However, combination of reduced
dose of herbicide and the test rates of sunflower residues
significantly lowered colonization rate compared to sole
application of sunflower residue rates. Application of chevalier
did not affect rate and intensity of colonization significantly in
comparison to the control.

Table 11. Effect of different rates of chevalier 15 WG herbicide and
sunflower residues cv. Asgrow on number of spores of
Glomus mosseae associated with wheat roots.
Treatments Number of spores per
100 g dry soil
% of control
Weedy check (Control) 363.5 ---
Residues at 3 t ha-1
420.5 115.7
Residues at 3 t ha-1
+ 50% of
label rate of chevalier15 WG
299.0 82.3
Residues at 6 t ha-1
448.8 123.5
Residues at 6 t ha-1
+ 50% of
label rate of chevalier15 WG
341.2 93.9
Chevalier15 WG (Label rate) 241.5 66.4
LSD ≤ 0.05 27.9 ---

Table 12. Effect of different rates of chevalier 15 WG herbicide and
sunflower residues cv. Asgrow on colonization rate of Glomus
mosseae associated with wheat roots.
Treatments Colonization rate (%) % of control
Weedy check (Control) 40.00 ---
Residues at 3 t ha-1
77.50 193.8
Residues at 3 t ha-1
+ 50% of
label rate of chevalier15 WG
45.00 112.5
Residues at 6 t ha-1
85.00 212.5
Residues at 6 t ha-1
+ 50% of
label rate of chevalier15 WG
50.00 125.0
Chevalier15 WG (Label rate) 40.00 100.0
LSD ≤ 0.05 7.41 ---
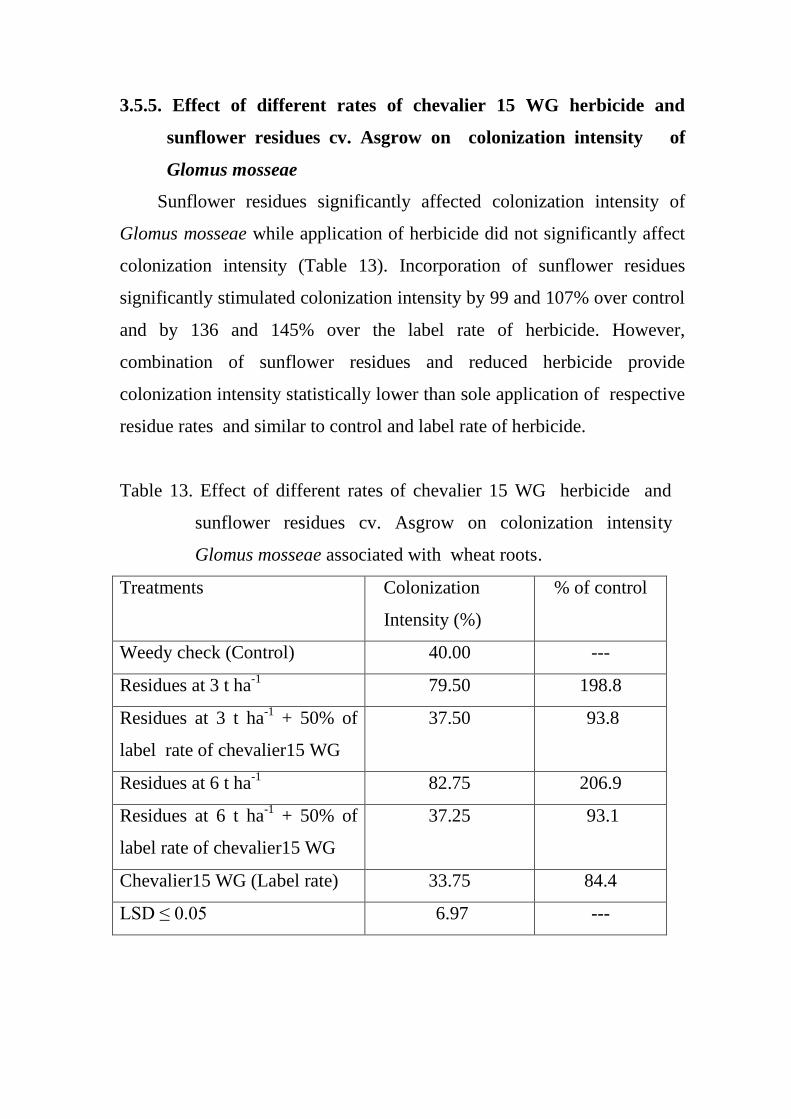
3.5.5. Effect of different rates of chevalier 15 WG herbicide and
sunflower residues cv. Asgrow on colonization intensity of
Glomus mosseae
Sunflower residues significantly affected colonization intensity of
Glomus mosseae while application of herbicide did not significantly affect
colonization intensity (Table 13). Incorporation of sunflower residues
significantly stimulated colonization intensity by 99 and 107% over control
and by 136 and 145% over the label rate of herbicide. However,
combination of sunflower residues and reduced herbicide provide
colonization intensity statistically lower than sole application of respective
residue rates and similar to control and label rate of herbicide.
Table 13. Effect of different rates of chevalier 15 WG herbicide and
sunflower residues cv. Asgrow on colonization intensity
Glomus mosseae associated with wheat roots.
Treatments Colonization
Intensity (%)
% of control
Weedy check (Control) 40.00 ---
Residues at 3 t ha-1
79.50 198.8
Residues at 3 t ha-1
+ 50% of
label rate of chevalier15 WG
37.50 93.8
Residues at 6 t ha-1
82.75 206.9
Residues at 6 t ha-1
+ 50% of
label rate of chevalier15 WG
37.25 93.1
Chevalier15 WG (Label rate) 33.75 84.4
LSD ≤ 0.05 6.97 ---

Chapter Four
Discussion

CHAPTER FOUR
Discussion Allelopathy has been reported to offer a significant role in weed
control (Sodaeizadeh and Hosseini, 2012). Several non herbicidal weed
control strategies in which Allelopathy is involved has been explored such
as rotational crop, cover crop, smother crop, intercropping, crop mixtures,
water extract and use of allelopathic crop residues as mulch or incorporated
in field soil (Mohammadi, 2013) . Of the all of these strategies, use of
allelopathic plant residues is the most successful, effective and readily
available (Kelton et al., 2012). Nevertheless, the efficacy of crop residues
is not comparing with the efficacy of the herbicides. Hence the present
study was focused to test if we can improve the efficacy of sunflower
residues by integrated it with the lower doses of herbicide.
In present study, the inhibitory effect of sunflower residues
incorporated into the field soil suggests that the residues contain phytotoxic
materials released into the rhizosphere by dissolving in irrigation water
and/or by the action of soil microorganisms and affect the receiver plants.
Several investigators indicated that the residues of allelopathic plants are
the main source of allelopathic compounds in natural and agricultural
ecosystems (Singh et al., 2003; Batish et al., 2001; Alsaadawi and Dayan,
2009). The increase in phytotoxicity against weed density and biomass due
to increase in rate of residues in soil may be attributed to the increase of the
concentration of allelopathic compounds released from the residues into the
field soil. Similar results were reported in sunflower and other plant species
by several researchers (Alsaadawi et al., 1986b; Alsaadawi et al., 1998;
Sarbout, 2010). However, this trend of inhibition was not taken place with
wheat crop. The lower dose (3 t ha-1
) was found to have more stimulatory
effect on growth and yield than the higher dose (6 t ha-1
). This suggests that
the response of plant to phytotoxins is a concentration and species

dependent. The differential response of plant species to phytotoxins was
reported by different investigators. Weston (1996) mentioned that the large
seed plants are more tolerant to phytotoxins than the small seeds plants
such as weed. An et al, (1996) reported that allelopathic residues added to
the soil should be regulated to avoid the negative impact of the higher
amount of allelopathic residues by increasing seeds rate and/or remove
some of the allelopathic residues.
The presence and release of phytotoxins from the residues
incorporated into soil is further confirmed by the experiment of phenolics
determination in soil. Phenolics appeared to be released after incorporation
in soil, reached maximum beak at 4th
week of decomposition and then
declined until vanished at the end of 6th week (Figure 5). No attempt was
made to isolate and identify the phenolic acids in the decomposed
sunflower residues in soil; however, several investigators (Al-Temimi,
2010 ; Sarbout, 2010) were able to isolate and identify phenolic acids,
namely Chlorogenic acid, isolchlogenic acid, caffeic acid, gallic acid,
syrinigic acid, hydroxy benzoic acid, p- coumaric acid, ferullic acid,
vanillic acid and Catechol from the soil containing sunflower residues
and they reached their maximum beak at 4th week and vanished at 2
nd
month from incorporation of residues in to the field soil. These phytotoxins
are reported to have inhibitory effects on several metabolic processes such
as inhibition of chlorophyll biosynthesis (Alsaadawi et al., 1986a ; Weir et
al., 2004), ions uptake (Olmsted and Rice, 1970; Alsaadawi et al., 1986a;
Lehman and Blum, 1999), photosynthesis (Hejl et al., 1993), inhibition of
activity of plasma H+-ATPase which leads to decreased ions and water
absorption by guard cells of leaves and causing close of stomata (Hejl and
Koster, 2004), inhibition of photosystem II and thus decreases the
production of ATP and NADPH2 required for CO2 fixation in dark reaction
(Barkosky et al., 2000) , inhibition of oxidation phosphorylation (Koeppe

and Miller, 1974), inhibition of activity of several enzymes involved in
essential metabolic processes (Politycka and Gmerek, 2008), interfering
with hormones metabolism in plant. Inhibition of stomata opening (Rai et
al., 2003). Also phenolic acids are reported to reduce the number of
mitochondria and disrupt the membranes surrounding nuclei, mitochondria
and dictyosomes (Lorber and Muller, 1976).
Field observations indicated that low percentage of weeds emerged
at the beginning of sowing time and continued for about 6 weeks after
emergence then started to increase rapidly. Chemical analysis revealed that
phenolic acids were highly increased until reached maximum at 4 weeks
from sowing time then started to decline until vanished at 6 weeks from
sowing time. Thus, it appeared that the increase in concentration of total
phenolic acids in soil was parallel with high suppressive activity of weeds
in the field and vice versa. These results suggested that phenolics are the
main cause of the suppressive activity of weeds. Another evidence of
phenolic responsibility for the weed inhibition is the results of weed
bioassay experiment. The inhibition of seed germination and seedling
growth of Malva rotundifolia weed was highly correlated with the
increase concentration of total phenolics in field soil (r = 0.71 and 0.94
between phenolics concentration , seed germination and seedling growth,
respectively). At the end of 2nd
month from sowing, weeds started to
emerge and grow rapidly; however at that time, the crop plants became
large and highly competitive to weeds. These observations were also
reported when the residues of sorghum were added in broad bean field AL-
bedairy, 2011).
The potential inhibition of incorporated sunflower residues against
weeds population suggested that sunflower residues provide a significant
inhibitory effect. Maximum inhibition (78%) was achieved by incorporated
sunflower residues at 6 t ha-1
and this inhibition did not match with the

efficacy of the test herbicide. However, when this amount of residues
integrated with reduced dose of chevalier herbicide, a great reduction of
weed population (86%) and weed biomass (96%) was achieved and this
reduction can be comparable with efficacy of standard herbicides. This
result confirmed the previous hypothesis proposed by Bhowmik and
Inderjit (2003) that herbicides applied in combination with allelopathic
conditions could have a complementary interaction, and may help to
minimize herbicide usage for weed management in field crops. It seems
that a reduced level of herbicide may be feasible for providing satisfactory
weed control when it works simultaneously with allelopathic conditions.
The combination of allelopathy and herbicides has been suggested by
several scientists for a long time and very interesting results have been
obtained (Cheema et al., 2003a; Cheema et al., 2003c ; Iqbal and Cheema,
2008). However most of the work was done on water extract and no
experiments were conducted on the use of residues of allelopathic crops
including sunflower with herbicides. Our method is easy to conduct in the
field and the added residues would improve the soil physical properties and
nutritional status in addition to allelopathic potential against weeds.
The improvement in grains and biological yields by sunflower
residues alone or in combination with reduced dose of chevalier herbicide
seems an outcome of reduced weed-crop competition for any of the growth
factors which might have contributed to higher yields. It appeared that
weed suppression was directly translocated into yield so that significant
yield improvement over weedy check was realized by all treatments and
their various combinations. Improvement in crop yield and related
components by integrating allelopathy with reduced herbicide dose is in
line with the previous findings of Alsaadawi et al. 2011a and Khaliq et al.
(2012a,2012b).

Analysis of yield components indicated that the increase in number
of spikes per plant was the only one responsible for the increase in yield of
wheat crop. Similar result was recorded when the effect of sorghum residue
was tested alone or in combination with reduced dose of chevalier on wheat
yield and yield components (AL-bedairy, 2011).
The increase in the yield of wheat due to sunflower residue and
herbicide treatments is parallel to significant reduction in weeds population
and biomass. The higher residue concentration alone or in combination
with reduced dose of herbicide increased yield of wheat over control but
inhibited the yield when compared to the label rate of herbicide. This
suggests that the inhibition was due to the phytotoxicity of the higher
concentration of sunflower residues. The lowest level of residue (3 t ha-1
)
apparently was sufficient, when combined with reduced dose of chevalier,
to reduce weeds population and biomass (80 and 94%) over control and
provided yield greater than the label rate of the herbicide. Similar trend of
effect was reported when residues of sorghum and sunflower were applied
to different types of crops treated with lower rates of pre and post
emergence herbicides (Alsaadawi et al, 2011a; Alsaadawi and AI-Temimi,
2011; AL-bedairy, 2011).
The increase of harvest index due to residues application alone or in
combination with the reduced rate of herbicide suggests that much of
nutrients provided from weed suppression is translocated towards the
production of grains rather than to vegetative structures of wheat.
The present study revealed that the combined effect of crop residues
and reduced chevalier herbicide not only directly affect wheat crop but also
indirectly through their positive impact on sporulation, colonization and
hyphal growth of Glomus mosseae. In our study, the better sporulation was
observed during the period of phytotoxins relase from sunflower residues
in to soil (during the first 6 weeks from sowing). This suggests that the

phenolics pose stimulatory or at least did not interfere with the growth of
Glomus mosseae fungi. Reports on the effect of phenolic acids on AM
fungi are controversial. Siqueira et al. (1992) indicated that
allelochemicals specially phenolics stimulated mycorrhizal colonization,
while others found that mycorrhizal colonization was suppressed by
phenolic acids released from allelopathic plants (Javaid, 2008; Riaz et al.,
2007; Wacker et al., 1990). The inhibition or stimulation of AM spore
germination and hyphal growth and branching was mainly depended on
concentration and type of allelochemicals present in soil rhizosphere
(Batish et al., 2007).
The inhibition of AM sporulation and colonization intensity by the
label dose of chevalier is coincided with general trends of the effect of most
herbicides on mycorrhiza (Abd-Alla et al., 2000). However, reduced dose
of chevalier in combination with higher rate of sunflower residues scored
sporulation and colonization intensity similar to that of control treatment. It
is possible that the residues may mitigate the effect of chevalier herbicide
when applied in combination.
The results of this study led to the conclusion that the sunflower
residues amended in field soil provided a good medium for growing
Glomus mosseae fungi and the allelochemicals released from the residues
did not interfere with the test growth parameters of Glomus mosseae.

Conclusions
and
Recommendations
Conclusions and Recommendations

Conclusions
1. Sunflower residue showed allelopathic inhibition on weeds grown
wheat field. However the efficacy of the residue is not comparable
with the herbicide.
2. Combination of sunflower residues and reduced rate (50% of the
labeled rate) of herbicides appeared to be more effective in
controlling weeds population and growth than sole application of
residues.
3. Integration of sunflower residues at 3 t ha-1
with reduced rate of
herbicide provided grain and biomass yields of wheat similar to
that achieved by labeled rate of herbicide.
4. Phenolic compounds in sunflower residues appeared to be released
in higher amount into the soil after two weeks of decomposition
and continued until 6 weeks then vanished. Weed suppression was
coincided with the higher concentration of phenolics in soil.
5. Sunflower residues amended in field soil was found to provide a
good medium for growing Glomus mosseae fungus and the
allelochemicals released from the residues did not interfere with
the test growth parameters of this fungus.

Recommendations
According to the results of this study, the following points are
recommended:
1. The allelopathic phenomenon can be integrated with lower rate of
herbicide as a useful agricultural practice for weed management in
order to reduce dependence on herbicides and achieve
agroecosystems sustainability.
2. Further investigations into the allelopathic potential of other crop
residues along with different herbicides under varying
environmental conditions need to be conducted to provide data
base on utilization of this practice in weed and crop management.
3. Redesigning of crop rotations based on allelopathic phenomenon
needs to be considered to achieve sustainability and reduce the
pesticide input in agroecosystems and thereby reducing the
environmental pollution.
4. The possible effect of this practice on beneficial soil microflora
other than mycorrhiza such as nitrogen fixing and nitrifying
bacteria needs to be investigated to obtain a comprehensive picture
on the importance of this practice in agricultural production.

References
References

A.O.A.C. (1990). Official methods of analysis of the association of
official analytical chemists. Tannin. 15th
ed. Washington,
D.C.p. 746.
Abd-Alla, M. H., Omar, S. A. and Karanxha, S. (2000). The impact of
pesticides on arbuscular mycorrhizal and nitrogen-fixing
symbioses in legumes. Applied Soil Ecology 14:191-200.
Abdollahi, F. and Ghadiri, H. (2004). Effect of separate and
combined application of herbicides on weed control and
yield of sugar beet. Weed Technology 18: 968-976.
Afzal, B., Bajwa, R. and Javaid, A. (2000). Allelopathy and VA
mycorrhiza VII. Cultivation of Vigna radiata and
Phaseolus vulgaris under allelopathic stress caused by
Imperata cylindrica. Pakistan Journal of Biological Sciences 3:
485-487.
AL-bedairy, N. R., ALsaadawi, I. S., and Shati, R. K. (2011).
Effect of combination of Sorghum bicolor L. (Moench)
residues and Trifluralin herbicide on broad bean and its
weeds. Iraqi Journal of agriculture 94–102.
Alsaadawi, I. S., Al-Hadithy, S. M. and Arif, M. B. (1986a). Effect of
three phenolic acids on chlorophyll content and ions uptake in
cowpea seedlings. Journal of Chemical Ecology 12: 221-227.
Alsaadawi, I. S., Al-Uqaili, J. K., Al-Rubeaa, A. J. and Al-Hadithy,
S. M. (1986b). Allelopathic suppression of weed and
nitrification by selected cultivars of Sorghum bicolor L.
(Moench). Journal of Chemical Ecology 12:209–219.
Alsaadawi, I. S., Zwain, K. H. Y. and Shahata, H. A. (1998). Allelopathic
inhibition of growth of rice by wheat residues. Allelopathy
Journal 5: 163-169.

Alsaadawi, I. S., Al-Ekeelie, M. H. S. and Al-Hamzawi, M. K.
(2007). Defferential allelopathic potential of grain sorghum
genotypes to weed. Allelopathy Journal 19:153–160.
Alsaadawi, A. S. and Dayan, F. E. (2009). Potentials and prospects of
sorghum allelopathy in agroecosystems. Allelopathy Journal 24
:255-270.
Alsaadawi, I. S. and AI-Temimi, A. A.(2011). Use of Sunflower
residues in combination with Sub-recommended dose of
herbicides for weeds control in barley field. Academic
Journal 12:83-93.
Alsaadawi, I. S., Khaliq, A., Al-Temimi, A. O. and Matloob, A. (2011a).
Integration of sunflower (Helianthus annuus L.) residues with a
pre-plant herbicide enhances weed suppression in broad bean
(Vicia faba L.) fields. Journal Planta Daninha 29: 849-859 .
Alsaadawi, I. S., Sarbout, A. K. and AI-Shamma, L. M. (2011b).
Differential allelopathic potential of sunflower(Helianthus annuus
L.) genotypes on weeds and wheat (Triticum aestivum L.) crop.
Agronomy and Soil Science Journal 58:1139-1148.
Alsaadawi, I. S., Khaliq, A., Lahmod, N. R. and Matloob, A.
(2013). Weed management in broad bean (Vicia faba L.)
through allelopathic Sorghum bicolor (L.) Moench residues
and reduced rate of a pre-plant herbicide. Allelopathy
Journal 32: 203-212.
Alsaadawi, I. S., Alkhateeb, T. A., Hadwan, H.A. and Lahmood,
N. R. (2014). A chemical basis for differential allelopathic
potential of root exudates of Sorghum bicolor L. (Moench)
cultivars on companion weeds. Submitted for publication in
Iraqi Journal of agriculture.

Al-Temimi, A. O. (2010). Effect of interaction of sunflower residues and
herbicides on weeds and barley crop. M.Sc thesis, Biology
Department, College of Science , University of Baghdad, Iraq.
Ampong, N. K. and De Datta, S. K. (1991). A handbook for weed
control in rice. Manila (Philippines) : International Rice
Research Institute, pp 113.
An, M., Johnson, I. R. and Lovitt, J. V. (1996). Mathematical Modeling of
Allelopathy. III. A Model for Curve-Fitting Allelochemical Dose
Responses. Allelopathy Journal 3: 33-42.
Anjum, T. and Bajwa, R. (2007). Field appraisal of herbicide
potential of sunflower leaf extract against Rumex
dentatus. Field Crops Research 100:139-142.
Ashrafi, Z. Y., Sadeghi, S., Mashhadi, H. R. and Hassan, M. A.
(2008). Allelopathic Effects of sunflower (Helianthus
annuus L.) on germination and growth of wild Barley
(Hordeum spontaneum L.). Journal of Agricultural
Technology 4: 219-229.
Awan, F. K., Rasheed, M., Ashraf, M. and Khurshid, M. Y.
(2012). Efficacy of brassica , sorghum and sunflower
aqueous extracts to control wheat weeds under rainfed
conditions of pathway, Pakistan. The Journal of Animal
and Plant Sciences 22: 715-721.
Azadbakht, A., Mahmoodi, S., Mahmoodi, R., Amraei, B. and
Nasrollahi, H.(2013). Evaluation the allelopathic effects of
aerial and underground extract of sunflower (Helianthus
annuus L.) on germination characteristics and seedling
growth of Hoary cress (Cardaria draba). Annals of
Biological Research 4:188-195.

Barkosky, R. R., Einhellig F. A. and Butler J. L. (2000). Caffeic acid-
induced changes in plant-water relationships and photosynthesis in
leafy spurge Euphorbiaesula. Journal of Chemical Ecology 26:
2095-2109.
Batish, D. R., Singh, H. P., Kholi, R. K. and Kaur, S. (2001). Crop
allelopathy and its role in ecological agriculture. Journal of Crop
Production 4:121-161.
Batish, D. R., Lavanya, K., Singh, H. P. and Kohli, R. K. (2007). Phenolic
allelochemicals released by Chenopodium murale affect the
growth, nodulation and macromolecule content in chickpea and
pea. Plant Growth Regulations 51: 119–128.
Batlang, U. and Shushu, D. D.(2007). Allelopathic activity of
sunflower (Helianthus annuus L.) on growth and
Nodulation of Bambara groundnut (Vigna subterranean L.
Verdc.). Journal of Agronomy 6:541-547.
Becard, G., Douds, D. D. and Pfeffer, P. E. (1992). Extensive in
vitro hyphal growth of vesicular- arbuscular mycorrhizal
fungi in the presence of CO2 and flavonols. Applied and
Environmental Microbiology 58: 821-825.
Becard, G., Taylor, L. P., Douds, D. D., Pfeffer, P. E. and Doner, L.
W.(1995). Flavonoids are not necessary plant signal compounds in
arbuscular mycorrhizal symbioses. Molecular Plant-Microbe
Interactions 8:252-258.
Ben-Hammouda, M., Robert, J. K., Harry, C. M. and Sarwar, M.
(1995). A Chemical basis for differential allelopathic
potential of sorghum hybrids on wheat. Journal of
Chemical Ecology 21:775-786.

Bhatti, M. Q. L., Cheema, Z. A. and Mehmood, T. (2000). Efficacy of sorgaab
as natural weed inhibitor in ray. Pakistan Journal of Biological
Sciences 3:1128-1130.
Bhowmik, P. and Inderjit, C. (2003). Challenges and opportunities in implementing
allelopathy for natural weed management. Journal of Crop Protection 22 :
661-671.
Blum, U., Wentworth, T. R., Klein, A. D., King, L. D., Gerig, T.
M. and Lyu, S. W. (1991). Phenolic acid content of soils
from wheat-no till, wheat-conventional till and fallow-
conventional till soybean cropping systems. Journal of
Chemical Ecology 17:1045-1067.
Buée, M., Rossignol, M., Jauneau, A., Ranjeva, R. and Bécard, G. (2000).
The pre-symbiotic growth of arbuscular mycorrhizal fungi is induced
by a branching factor partially purified from plant root exudates.
Molecular Plant Microbe Interaction 13:693-698.
Chabot, S., Bécard, G., and Piché, Y. (1992). Life cycle of Glomus
intraradix in root organ culture. Mycologia 84: 315–321.
Cheema, Z. A. and Khaliq, A. (2000). Use of sorghum allelopathic
properties to control weeds in irrigated wheat and semi-arid
region of Punjab. Agriculture Ecosystems and
Environment Journal 79:105-112.
Cheema, Z. A., Farid, M. S. and Khaliq, A. (2003a). Efficacy of
concentrated sorgaab with low rates of atrazine for weed control in
maize. Journal of Animal and plant S c i e n c e s 13:48-51.
Cheema, Z. A., Khaliq, A. and Farooq, R. (2003b). Effect of
concentrated sorgaab alone and in combination with
herbicides and surfactant in wheat. The Journal of Animal
& Plant Sciences 13: 10-13

Cheema, Z. A., Khaliq, A. and Hussain, R. (2003c). Reducing herbicide
rate in combination with allelopathic sorgaab for weed control in
cotton. International Journal of Agriculture and Biology 5: 4–6.
Cheema, Z. A., Khaliq, A. and Saeed. S. (2004). Weed control in maize
(Zea mays L.) through sorghum allelopathy. Journal of Sustainable
Agriculture 23:73-86.
Cheema, Z. A, Khaliq, A. and Farooq, M. (2008). Sorghum
allelopathy for weed management in wheat. Allelopathy
in Sustainable Agriculture and Forestry 3:255-270.
Cheema, Z. A., Muhammad, Z., Ahmad, R. and Murtaza, G.
(2010). Application of allelopathic water extracts for
suppressing the rice weeds. Crop Environment 1: 1-5.
Cooper, J. E.(2004). Dominant role of flavonoids among signal
molecules involved in the formation of legume rhizobia
symbiosis. Polyphenols communications 87.
Donald , C. M., and Hamblin, J . (1979) . The biological yield and
harves index of cereals as agronomic and plant breeding
criteria - Adv . Agronomy Journal 28 : 361 - 405 .
Dong, C. J. and Zhao, B. (2004). Arbuscular mycorrhizal formation of
crucifer leaf mustard induced by flavonoids apigenin and daidzein.
Chinese Science Bulletin 49:1254-1261.
Douds, D. D., Nagahashi, G. and Abney, G. D. (1996). The
differential effects of cell wall-associated phenolics, cell
walls, and cytosolic phenolics of host and non-host roots on
the growth of two species of AM fungi. New Phytologist
133:289–294.
Einhellig, F. A. and Leather, G. R. (1988). Potentials for exploiting
allelopathy to enhance crop production. Journal of
Chemical Ecology 14:1829-1844.

Einhellig, F. A. (1995), Allelopathy: Current status and future goals. In:
Allelopathy: Organisms, Processes and Applications, Inderjit,
Dakshini, K.M.M. and Einhellig, F.A, Eds.; Am. Chem. Soc.
Washington DC, USA, pp. 01-24.
Einhellig, F. A. (1996). Interactions involving allelopathy in
cropping systems. Agronomy Journal 88:886-893.
Fries. L. M., Pacovsky, R. S., Safir, G. R. and Siqueira, J. O.
(1997). Plant growth and arbuscular mycorrhizal fungal
colonization affected by exogenously applied phenolic
compounds. Journal Of Chemical Ecology 23:755–1767.
Ghafar, A., Saleem, B. and Qureshi, M. J. (2000). Allelopathic
effects of sunflower (Helianthus annuus L.) on germination
and seedling growth of wheat (Triticum aestivum L.).
Pakistan Journal of Biological Sciences 3: 1301–1302.
Gianinazzi, S. and Gianinazzi-Pearson, V.(1992). Cytology, histochemistry
and immunocytochemistry as tools for studying structure and
function in endomycorrhiza. Methods in Microbiology 24: 109–139.
Gianinazzi-Pearson, V., Branzanti, B. and Gianinazzi, S. (1989) In vitro
enhancement of spore germination and early hyphal growth of a
vesicular-arbuscular mycorrhizal fungus by root exudates and plant
flavonoids. Symbiosis 7: 243-255.
Giovannettim, M., Sbranam, C., Citernesi, A. S. and Avio, L. (1996). Analysis
of factors involved in fungal recognition responses to host derived
signals by arbuscular mycorrhizal fungi. New Phytologist
Journal 133:65–71.
Harrison, M. J. and Dixon, R.A. (1993). Isoflavonoid accumulation
and expression of defense gene transcripts during the
establishment of vesicular–arbuscular mycorrhizal
associations in roots of Medicago truncatula. Molecular
Plant–Microbe Interactions 6: 643–654.

Harrison, M. and Dixon, R.(1994). Spatial patterns of expression of
flavonoid/isoflavonoid pathway genes during interactions between roots of
Medicago truncatula and the mycorrhizal fungus Glomus
versiforme. The Plant Journal 6:9–20.
Haslam, E. (1988). Plant polyphenols (syn. Vegetable tannins) and
chemical defense- are appraisal. Journal Chemical Ecological
14:1789-1805.
Hejl, A. M., Einhellig, F. A. and Rasmussen, J. A. (1993) Effects of
juglone on growth, photosynthesis, and respiration. Journal
Chemical Ecology 19: 559-568.
Hejl, A. M. and Koster, K. L. (2004) Juglone disrupts root plasma
membrane H (+)ATPase activity and impairs water uptake, root
respiration, and growth in soybean (Glycine max) and corn (Zea
mays). Journal Chemical Ecology 30: 453-471.
Hozayn, M., Abd El-Lateef, E. M., Sharar, F. M. and Abd El-
Monem, A. A.(2011). Potential uses of sorghum and
sunflower residues for weed control and to improve lentil
yields. Allelopathy Journal 27 :15-22.
Hunt, R. (1982). Plant Growth Curves: The Functional Approach
to Plant Growth analysis. London, Edward Arnold. PP:
248.
Iqbal, J. and Cheema, A. (2008). Purple nutsedge ( Cyperus rotundus L.)
management in cotton with combined application of sorgaab and
s- metolachlor. Pakistan Journal of Botany 40: 2383-2391.
Iqbal, J., Cheema, Z. A. and Mushtaq, M. N. (2009). Allelopathic
crop water extracts reduce the herbicide dose for weed
control in cotton (Gossypium hirsutum). International
Journal of Agriculture and Biology 11:360–366.

Iqbal, J., Karim, F. and Hussain, S. (2010). Response of wheat
sponse of wheat crop (Triticum aestivum L.) and Its weeds
to allelopathic crop water extracts in combination with
reduced herbicide rates. Pakistan Journal of Agricultural
Sciences 47: 309-316.
Jabran, K., Cheema, Z. A., Farooq, M. and Hussain, M. (2010).
Lower doses of pendimethalin mixed with allelopathic
crop water extracts for weed management in canola
(Brassica napus). International Journal of Agriculture and
Biology 2:335–340.
Javaid, A., Shafique, S., Bajwa, R. and Shafique, S. (2006).
Biological control of noxious alien weed Parthenium
hysterophorus L. in Pakistan. International Journal of
Biology and Biotechnology 3: 721-724.
Javaid, A. (2007). Allelopathic interactions in mycorrhizal
associations. Allelopathy Journal 20: 29-42.
Javaid, A. (2008). Allelopathy in mycorrhizal symbiosis in the Poaceae
family. Allelopathy Journal 21: 207-218.
Kelton, J., Price, A. J. and Mosjidis, J. (2012). Allelopathic Weed
Suppression Through the Use of Cover Crops, Weed Control, Dr.
Andrew Price (Ed.), ISBN: 978-953-51-0159-8, InTech, DOI:
10.5772/34196. Available from:
http://www.intechopen.com/books/weed-control/allelopathic
weed-suppression-through-the-use-of-cover-crops.
Khaliq, A., Aslam, Z. and Cheema, Z. A. (2002). Efficacy of
different weeds management strategies in mungbean
(Vigna radiata L.). International Journal of Agriculture
and Biology 4: 237–239.

Khaliq, A., Matloob, A., Irshad, M. S., Tanveer, A. and Zamir, M.
S. (2010). Organic weed management in maize (Zea mays
L.) through integration of allelopathic crop residues.
Pakistan Journal of Weed Science Research 16: 409-420.
Khaliq, A., Matloob, A., Cheema, Z. A. and Farooq, M. (2011).
Allelopathic activity of crop residue incorporation alone or
mixed against rice and its associated grass weed jungle rice
(Echinochloa colona L. Link).Chilean Journal of
Agricultural Research 71:418-423.
Khaliq, A., Matloob, A. and Riaz, Y. (2012a). Bio-economic and
qualitative impact of reduced herbicide usage in direct seeded fine
rice through multipurpose tree water extracts. Chilean Journal of
Agricultural Research 72:350-357.
Khaliq, A., Matloob, A., Tanweer, A. and Khan, M. B. (2012b). Naturally
occurring phytotoxins in allelopathic plants helps reduce herbicide
dose in wheat. Natural Products Research 26: 1156-1160.
Khaliq, A., Matloob, A. Khan, M. B. and Tanveer, A. (2013).
Differential suppression of rice weeds by allelopathic plant
aqueous extracts . Journal Planta Daninha 31: 21-28.
Khan, E. A. Shah, S. H. and Sadozai, G. U.(2009). Reduced
herbicide doses in combination with allelopathic sorghum
water for weed control in sunflower (Helianthus annuus
L.). Pakistan Journal of Weed Science Research 15: 145-
154.
Khan, M. B., Ahmad, M., Hussain, M., Jabran, K., Farooq, S. and
Waqas-Ul-Haq, M. (2012). Allelopathic plant water
extracts tank mixed with reduced doses of atrazine
efficiently control Trianthema portulacastrum L. in Zea

mays L. The Journal of Animal & Plant Sciences 22:339-
346.
Khan, R. and Khan, M. A.(2012). Weed control efficiency of
bioherbicides and their impact on grain yield of wheat
(Triticum aestivum L.). European Journal of Applied
Sciences 4: 216-219.
Khanh, T. D., Linh, L. H., Linh, T. H., Quan, N. T., Cuong, D. M.,
Hien,V. T. T., Ham, L. H., Trung, K. H., and Xuan, T. D.,
(2013). Integration of Allelopathy to Control Weeds in
Rice. Ch. 4:75-99. In Herbicides - Current Research and
Case Studies in Use. dx.doi.org/10.5772/56035.
Koeppe, D. E. and Miller, R. J. (1974) Kaempferol inhibition of corn
mitochondrial phosphorylation. Plant Physiology 54: 374-378.
Lahmod, N. R. (2012). Allelopathic effects of sorghum (Sorghum bicolor
L.) Moench on companion weeds and subsequence crop. Ph. D
thesis , Field Crop Sciences College of Agricultural, University
of Baghdad, Iraq.
Lahmod, N. R. and Alsaadawi, I. S. (2014). Evaluation of Sorghum bicolor
L. (Moench) residues alone and in combination with reduced dose
of post-emergence herbicide for weed control in wheat. 1st Africa-
International Allelopathy Congress, Sousse, Tunisia. February 6-
9.
Lehman, M. E. and Blum, U. (1999). Evaluation of ferulic acid uptake as
a measurement of allelochemical dose: effective concentration.
Journal Chemical Ecology 25:2585–2600.
Ling-Lee, M., Chilvers, G. A. and Ashford, A. E. (1977). A histochemical
study of phenolic materials in mycorrhizal and uninfected roots
of Eucalyptus fastigata Dean and Maid. New Phytologist
Journal 78:313–328.

Lorber, P. and Muller, W. H. (1976). Volatile growth inhibitors produced
by Salvia leucophylla: effects on seedling root tip ultra structure.
American Journal of Botany 63, 196-200.
Lynn, D. G. and Chang, M.(1990). Phenolic signals in cohabitation:
implications for plant development. Annual Review of Plant
Physiology and Plant Molecular Biology 41:497-526.
Mahmood, K., Muhammad, A., Khan, B., Hussain, M. and
Gorchani, M. A. (2009). Weed management in wheat field
(Triticum aestivum) using allelopathic crop water extracts.
International Journal of Agriculture and Biology 11:751–
755.
Martin, T., Sampedro, I., Garcia-Romera, I., Garcia-Garrido, J. M. and
Ocampo, J. A.(2002). Arbuscular mycorrhizal colonization and
growth of soybean (Glycine max) and lettuce (Lactuca sativa) and
phytotoxic effect of olive mill residues. Soil Biology and
Biochemistry 34:1769-1775.
Matloob, A., Khaliq, A., Farooq, M. and Cheema, Z. A.(2010).
Quantification of allelopathic potential of different crop
residues for the purple nutsedge suppression. Pakistan
Journal of Weed Science Research 16: 1-12.
Matloob, A. A. (2012). Determination of the causeses of bean foot
and root rot disease and evaluation of the efficacy of some
biocontrol agents in their control. Ph. D thesis, Agriculture
Sciences College of Agricultural, University of Baghdad,
Iraq.
Mc Gonigle, T. P., Miller, M. H., Evans, D. G., Fairchild, G. L.
and Swan, J. A. (1990). A new method which gives an
objective measure of colonisation of roots by vesicular-

arbuscular mycorrhizal fungi. New Phytologist 115: 495-
501.
Mohammadi, G. R. (2013). Alternative weed control methods: A Review.
http://dx.doi.org/10.5772/54164.
Mubeen, K., Nadeem, M. A., Tanveer, A. and Zahir, Z. A. (2012).
Allelopathic effect of Sorghum and Sunflower water
extracts on germination and seedling growth of Rice
(Oryza sativa L.) and thee weed species. The Journal of
Animal & Plant Sciences 22: 738-746.
Nagahashi, G., Douds, D. D. and Abney, G. D. (1996). Phosphorus
amendment inhibits hyphal branching of the VAM fungus
Gigaspora margarita directly and indirectly through its
effect on root exudation. Mycorrhiza 6: 403–408.
Nair, M .G. Safir, G. R. and Siqueira, J. O.(1991). Isolation and identification
of vesicular-arbuscular mycorrhiza stimulatory compounds from
Clover (Trifolium repens) roots. Applied and Environment
microbiology 57:434-439.
Naseem, M., Aslam, M., Ansar, M. and Azhar, M. (2009).
Allelopathic effects of sunflower water extract on weed
control and wheat productivity . Pakistan Journal of Weed
Science Research 15: 107-116.
Nelson, C. J. (1996). Allelopathy in cropping systems. Agronomy
Journal 88 : 991-996.
Nikneshan, P., Karimmojeni, H., Moghanibashi, M. and Hosseini,
N. A. S. (2011). Allelopathic potential of sunflower on
weed management in safflower and wheat. Australian
Journal of Crop Science 5: 1434-1440.

Olmsted, C. E. and Rice, E. L. (1970). Relative effects of known plant
inhibitors on species from first two stages of old field succession
.Southwestern Naturalist 15:165-173.
Politycka, B. and Gmerek, J. (2008). Effects of ferulic and p-coumaric
acids on the activity of hydrolytic enzymes and the growth of
radicles in germinating seeds of cucumber and pea. Allelopathy
Journal 21: 227-238.
Poulin, M. J., Simard, J., Catford, J. G., Labrie, F. and Piche, Y. (1997).
Response of symbiotic endomycorrhizal fungi to estrogen and
antiestrogens. Molecular Plant-Microbe Interaction 10:481–487.
Putnam, A. R. and DeFrank, J. (1983). Use of phytotoxic plant
residues for selective weed control. Crop Protection 2:
173-181.
Putnam, A. R., Nair, M. G. and Barnes, J. B.(1990). Allelopathy:
a viable weed control strategy. In: New directions in
biological control. Alan R. Liss Inc. pp. 317-322.
Putnam, A. R., DeFrank, J. and Baornes, J. P. (1983).
Exploitation of allelopathy for weed control in annual and
perennial cropping system. Journal of Chemical Ecology.
9:1001-1010.
Rai, V. K., Gupta, S. C. and Singh, B. (2003). Volatile monoterpenes
from Prinsepia utilis L. leaves inhibit stomata opening in Vicia
faba L. Journal of Plant Biology 46: 121-124.
Razzaq, A., Cheema, Z. A., Jabran, K., Farooq, M., Khaliq, A.,
Haider, G. and Basra, S. M. A. (2010). Weed management
in wheat through combination of allelopathic water extract
with reduced doses of herbicides. Pakistan Journal of
Weed Sciences Research 16: 247-256.

Razzaq, A., Cheema, Z. A., Jabran, K., Hussain, M., Farooq, M.
and Zafar, M. (2012). Reduced herbicide doses used
together with allelopathic Sorghum and Sunflower water
extracts for weed control in Wheat. Journal of Plant
Protection Research 52: 281–285.
Reddy, K. N. (2001). Effect of cereal and legume cover crop residues on
weeds, yield and net return in soybean (Glycine max). Weed
Technology 15:660-668.
Rehman, A., Cheema, Z. A., Khaliq, A., Arshad, M. and Mohsan,
S. (2010). Application of sorghum, sunflower and rice
water extract combinations helps in reducing herbicide
dose for weed management in rice. International Journal of
Agriculture and Biology 12: 901–906.
Riaz, T., Khan, S. N. and Javaid, A. (2007). Effects of incorporation of
allelopathic plants leaf residues on mycorrhizal colonization and
Gladiolus diseases. Allelopathy Journal 20: 61-70.
Rice, E. L. (1984). Allelopathy. 2nd
Ed. Academic Press, Orlando,
Florida, USA.
Roth, C. M., James, P. S. and Gary, M. P. (2000). Allelopathy of
sorghum on wheat under several tillage systems.
Agronomy Journal 92:855-860.
Sarbout, A. K. (2010). Study the allelopathic potential of sunflower
cultivars on weeds and wheat. M.Sc thesis, Biology Department,
College of Science , University of Baghdad, Iraq.
Scervino, J. M., Ponce, M. A., Erra-Bassells, R., Vierheilig, H., Ocampo,
J. A. and Godeas, A. (2005). Flavonoids exhibit fungal species
and genus specific effects on the presymbiotic growth
of Gigaspora and Glomus. Mycological Research 109:789–794.

Schenck, N. C. and Smith, G. S. (1982). Additional new and unreported
species of mycorrhizal fungi (Endogonaceae) from Florida,
Mycologia, 74-77. In: powell, C. L. and Bagyaraj, D. J. (1984).VA
Mycorrhiza. CRC press, Inc. Boca Raton, Florida, pp 234.
Selvaraj, J. T. and Subramanian, G.(1990). Phenols and lipids in
mycorrhizal and non-mycorrhizal roots of Sesamum
indicum. Current Science 59:471–473.
Shibayama, H. (2001). Weeds and weed management in rice
production in Japan. Weed Biology and Management 1:
53–60.
Silva-Junior, P. and Siqueira, J. O. (1998). Colonização
micorrízicae crescimento da soja com diferentes fungos e
aplicação do isoflavonóide formononetina. Pesquisa
Agropecuária Brasileira. 33:953–959.
Singh, H. P., Batish, D. R. and Kohli, R. K. (2003). Allelopathic
interaction and allelochemicals: new possibilities for sustainable
weeds management. Critical Reviews in Plant Sciences 22: 239–
311.
Siqueira, J. O., Safir, G. R. and Nair, M. G. (1991a). Simulation of
VAM formation and growth of white clover by flavonoid
compound. New Physiologist Journal 118:87-93.
Siqueira, J. O., Safir, G. R. and Nair, M.G. (1991b). significance of
phenolic compounds in plant‐soil‐microbial systems.
Critical Reviews in Plant Sciences 10:63-121.
Siqueira, J. O., Brown, D. G., Safir, G. R. and Nair, M. G. (1992). Field
application of the VAM stimulating isoflavonoid formononetin on
corn and soybean in Brazil. In: International Symposium of

Management of Mycorrhizae. Perth, University of Western
Australia. pp. 132.
Smith, F. A. and Dickson, S. E. (1997). Tansley review no. 96.
Structural diversity in (vesicular)-arbuscular mycorrhizal
symbioses. New Phytologist 137: 373-388.
Sodaeizadeh1, H. and Hosseini, Z. (2012). Allelopathy an
environmentally friendly method for weed control. International
Conference on Applied Life Sciences. Turkey, September 10-12.
Srisa-ard, K. (2007). Effect of crop residues of sunflower
(Helianthus annuus), Maize (Zea mays L.) and Soybean
(Glycine max) on growth and seed yields of Sunflower.
Pakistan Journal of Biological Sciences 10:1282-1287.
Steel, R. G. and Torrie, H. (1920). Principles and Procedures of
Statistics. Mc grow. Hill Book Company, Inc. New York.
Stephenson, G. R. (2000). Herbicide use and world food
production: Risk and Benefits. Abstract of 3rd Int. Weed
Science Congress. Foz Do Iguassu, Brazil, pp 240.
Streibig, J.C., Dayan, F. E., Rimando, A. M. and Duke, S. O. (1999). Joint
action of natural and synthetic photosystem II inhibitors. Journal
of Pesticide Science 55:137–146.
Tsai, S. M. and Phillips, D. A. (1991). Flavonoids released
naturally from alfalfa promote development of symbiotic
Glomus spores in vitro. Applied Environmental
Microbiology 57:85–88.
Vierheilig, H., Gagnon, H., Strack, D. and Maier, W. (2000).
Accumulation of cyclohexenone derivatives in barley,
wheat and maize roots in response to inoculation with

different arbuscular mycorrhizal fungi. Mycorrhiza 9:291–
293.
Wacker, T. L., Safir, G. R. and Stephenson, S. N. (1990). Evidence for
succession of mycorrhizal fungi in Michigan asparagus fields.
Acta Horticulturae 271: 273-279.
Weir, T. L., Sang-Wook, P. and Vivanco, J. M. (2004). Biological and
physiological mechanisms mediated by allelochemicals. Current
in Plant Biology 7: 472-479.
Weston, L. A. (1996). Utilization of allelopathy for weed management in
agroecosystems. Agronomy Journal 88: 860-866.
Yarnia, M. (2011). Effect of Chenopodium Album Allelopathic
Extract and Salcylic Acid Priming Under Seed Aging
Condition on Germination and Seedling Growth of
Foeniculum vulgare Mill. Journal of Applied
Environmental and Biological Sciences 1:602-607.
Yun, K. W. and Choi, S. K.(2002). Mycorrhizal colonization and
plant growth affected by aqueous extract of artemisia
princeps var. orientalis and two phenolic compounds.
Journal of Chemical Ecology 28:353-362.

Appendices
Appendices

Appendix 1.
Preparation of Alkaline Peroxide H2O2
3 ml NH4OH (Ammonia) + 30 ml 10% H2O2 + 567 ml distilled
water
Appendix 2.
Preparation of 0.1 M HCl (32% MW 36.46)
22.79 ml HCl
2 L Distilled water
Appendix 3.
Preparation of Lacto glycerol Trypan blue stain
520 ml lactic acid
480 ml Glycerol
640 ml distilled water
Mix the above compounds in a ratio of 13:12:16, respectively.
Then add 0.82 g Trypan blue to the mixture.
Appendix 4.
Preparation of Lacto glycerol Destain
520 ml lactic acid
480 ml Glycerol
640 ml distilled water
Mix the above compounds in a ratio 13:12:16, respectively.
Appendix 5. preparation of Folin-Denis Reagent

Amixture of 10g Na2WO4, 2 g phosphomolybdic acid, and 5 ml H3PO4 in
75 ml distilled water was refluxed for 2 h, cooled, and diluted to 100 ml
with distilled water.
Appendix 6. preparation of Sodium carbonate-saturated
anhydrous
Na2CO3 (4 g) was added to 10 ml distilled water and dissolved for 1 h at
70-80˚C, cooled, overnight, and filtered through glass wool.
Appendix 7. Ferulic acid standard solution
Ferulic acid (10 mg) was dissolved in 100 ml distilled water.
Aliquots of 0, 0.2, 0.4, 0.6, 0.8, and 1 ml of the standard ferulic
acid solution were dispensed into tubes containing 0.5 ml Folin-
Denis reagent and 1 ml saturated Na2CO3 solution. The standareds
were diluted to 10 ml with distilled weter and quickly shaken. A
bsorbance was determined after 30 min at 750 nm (Blum et
al.,1991) on a spectrophotometer (Pharmacia Ultra
spectrophotometer III).
Appendix 8. Effect of different rates of chevalier 15 WG herbicide
and sunflower residues cv. Asgrow on dry weight of wheat.

T1. Control (uncultivated sunflower field), T2. Residues at 3 t ha-1
, T3. Residues at 3
t ha-1
+ 50% of label rate of chevalier WG, T4. Residues at 6 t ha-1, T5. Residues at 6 t
ha-1
+ 50% of label rate of chevalier15 WG, T6. Chevalier WG (Label rate).
Appendix 9. Local, common and scientific names of plant species
appeared in the thesis
0
500
1000
1500
2000
2500
Days after sowing
Dry
weig
ht
(g/m
2)
T1 T2 T3 T4 T5 T6
T1 23.92 196.88 644.2 1311.6
T2 16.84 136.4 721.8 1654
T3 16.84 136.4 921.72 2023.2
T4 14.84 104.08 796.04 1457.2
T5 14.84 104.08 617.92 1884
T6 23.92 196.88 1033.56 2240.4
27 44 75 103
Local Arabic
name
Common name Scientific name
Acacia bark السنط النيلي
Acacia nilotica L.
.Alfalfa Medicago sativa L جت
Annual ryegrass حنيطة
Lolium rigidum L.
.Barley Hordeum vulgare L شعير
Barnyard grass دنان
Echinochloa crus-galli L.
Beet root wild سليجة
Beta vulgaris L.
Blady grass الحلفة
Imperata cylindrical L.
.Blond Plantago Plantago ovate Forssk لسان الحمل
.Broad bean Vicia faba L الباقالء
Broad leaved دنان
platran
Plantogo major L.
.Canola Brassica napus L سلجم
.Canary grass Phalaris minor Retz ابو دميم
.Celery cabbage Brassica campestris L سلجم صيني
.Cheese weed Malva pariflora L خباز
Clover البرسيم
Trifolium repens L.
.Cotton Gossypium hirsutum L القطن
.Belmude grass Cynodon dartylonl L الثيل المصري
.Darnel ryegrass Lolium temulentum L رويطة
Eucalyptus Eucalyptus camaldulensis يوكالبتوس
Dehnh.
.Flat sedge Carex spicata L البردي المفلطح

.Horse purslane Trianthema potrtulacastrum L خف الحصان
Johnson grass سفرندا
Sorghum halepense L.
Jungle rice دهنان
Echinochloa colona L.
Lamb's quarters رغيلة
Chenopodium album L.
Lentil Lens culinaris Medikus العدس
-Lesser swine الرشاد البري
cress
Coronopus didymus L.
الصفراءالذرة Maize Zea mays L.
Hairy-node bear زمزوم
grass
Dichanthium annulatum
Forssk.
Mulberry Morus Moraceae توت االبيض
Morus alba L.
.Mung bean Vigna radiate L الماش
Neem النيم
Azadirachta indica A.Juss
.Olive mill Olea europaea L زيتون
Pearl millet دخن
Pennisetum glaucum L.
.pigweed Portulaca oleracea L بربين بري
purple nutsedge سعد
Cyperus rotundus L.
.Common reed Phragmites australis L قصب البري
.rye Secale cereal L شيلم
red-root عرف الديك
amaranth
Amaranthus retroflexus L.
Rice الرز
Oryza sativa L.
.Safflower Carthamus tinctorius L العصفر

sorghum Sorghum bicolor L. (Moench) الذرة البيضاء
Soybean فول الصويا
Glycine max L.
Sunflower زهرة الشمس
Helianthus annuus L.
toothed dock ضرس العجوز
Rumex dentatus L.
.White top Weed Parthenium hysterophorus L االقحوان
.wheat Triticum aestivum L الحنطة
.wild oat Avena fatua L شوفان البري
الخالصة

والمختبريذذة الختبذذار الجالذذد االليلوبذذااي لمخلفذذات مجموعذذة مذذن التجذذارل الحقليذذةنفذذذت
فذي ماافحذة اغدغذال وحاصذل زهرة الشذمس بمفردهذا او مذر جرعذة منخفنذة مذن مبيذد الشذيفالير
الموسذم قليذة فذي الحتجربذة . فقذد نفذذت النمو الماياورايزا المتعايشذة مذر جذذور الحنطذة والحنطة
، اذ كليذة العلذوم، جامعذة بدذداد ،م الحيذاة في حقل تجارل قسم علو 8336-8338لعام الزراعي ل
طذذن هذذـ 8و 6 المضذذافة بمعذذدل Asgrowمخلفذذات زهذذرة الشذذمس صذذنف سذذتعملتا-3
بصذذورة
غم. هـ 373) مر نصف الاميةاو منفردة -3
( الموصى بالا من مبيد الشيفالير في ماافحة اغدغال
ومعاملذة مبيذد وحاصل الحنطذة. كمذا تضذمنت الدراسذة معاملذة مقارنذة بذدون ماافحذة ) مدغلذة(
ةأربعذوب RCBDتصذميم القطاعذات العشذوا ية الااملذة سذتعملللمقارنذة. ابالجرعة الموصى بالا
وحذذدد المحتذذوك الالذذي للفينذذوالت فذذي تربذذة الحقذذل المخلوطذذة بمخلفذذات زهذذرة الشذذمس .ماذذررات
هذذـطذذن 8بتركيذذز -3
المذذاياورايزا كمذذا تذذم عذذزل وتشذذخي مختلفذذة مذذن الزراعذذة. مذذدد خذذالل
و وحسذال عذدد السذبورات خذالل المراحذل اغولذى مذن نمذو الحنطذة المتعايشة مر جذور الحنطذة
يذد نسذبة ااصذابة وشذدتالا فذي مرحلذة تزهيذر نبذات الحنطذة فذي وعنذد مرحلذة التزهيذر. وتذم تحد
الحقل.
هطذذن 6تربذذة الحقذذل بمعذذدل ضذذافة الذذى مخلفذذات زهذذرة الشذذمس الم أنأظالذذرت النتذذا
يوما من الزراعة علذى 383و 43% عن المقارنة بعد 45% و 22اختزلت كثافة اغدغال بنسبة
ه طن 8وازداد هذا االختزال عند زيادة المخلفات المخلوطة إلى تابرالت-3
% 52و 24ليصل إلى
االختذزال ازداد بشذال كبيذر أال أن . تذابريوما مذن الزراعذة علذى الت 383و 43عن المقارنة بعد
إضافة نصذف الجرعذة وادت .نصف كمية مبيد الشيفالير مر مخلفات زهرة الشمس عملتاست عند
هطن 6مر مخلفات زهرة الشمس بتركيز من المبيد-3
مذن مذا حققتذ لألدغذال اعلذي اختذزالإلذى
ها.مفردب النسبة نفسالا من المخلفات إضافة معاملة
علذذى قذذد انعاذذس تذذاايرب ايجابيذذاالنتذذا إن االنخفذذا فذذي كثافذذة اغدغذذال لقذذد أظالذذرت
بينت النتا إن إضافة نصف الجرعة من المبيد مر مخلفذات زهذرة الشذمس بتركيذز فقد الحاصل.
هطن 6-3
لحبول بالسنبلة ودليذل للسنابل وعدد لحبول وعدد وحاصال لل ابيولوجي حاصالاعطت
من مبيد الشيفالير. الموصى بالامعاملة الجرعة مشاب لما اعطت لحصاد ل
يومذذا 39بعذذد الالذذي للفينذذوالت قذذد ازداد المحتذذوك وأظالذذرت نتذذا التحليذذل الايميذذا ي إن
أسابير مذن 8بعد ىهبط إلى إن تالش بعدهايوما من التحلل 82صل إلى اعلي مستوك ل بعد وو
محتذوك الالذي للفينذوالت فذي المذر بصورة معنويذة عاليذة تثبيط دغل الخباز وقد ارتبط التحلل.
فينوالت.للباز كان نتيجة الفعالية العالية لخاتثبيط دغل ان يشير إلى، التربة وهذا

رات في تربة الحقل المضاف إلي مخلفات زهرة وسبما نتا دراسة الماياورايزا، إن عدد االأ
أسذذابير مذذن تحلذذل مخلفذذات زهذذرة الشذذمس مقارنذذة بمعاملذذة 8 و 9 و 8 الشذذمس ازداد معنويذذا عنذذد
إما عند إضافة نصف الجرعة من المبيد مر مخلفات زهذرة المقارنة )بدون مخلفات زهرة الشمس(.
هطن 6الشمس بتركيز -3
فقد انخفض معنويا عدد السبورات أكثر من معاملة المقارنذة، إال إنذ عنذد
هطن 8إضافة نصف الجرعة من المبيد مر مخلفات زهرة الشمس بتركيز -3
عذدد السذبورات ازداد
6. إما عند إضافة مخلفات زهرة الشمس إلذى تربذة بتركيذز ارنةلتصل لما هو علي في المعاملة المق
هطن -3
وقد سجلت ألواح % عن المقارنة.44 و 49 الماياورايزا بنسبةبزادت نسبة وشدة ااصابة
هطذن 8و 6فيالا المخلفات إلى تربة الحقذل بنسذبة تالمعاملة التي أضيف-3
مذر نصذف الجرعذة مذن
ااصابة مقارنة بالسيطرة.المبيد زيادة معنوية في نسبة

جمهورية العراق
وزارة التعليم العالي والبحث العلمي
جامعة بغداد / كلية العلوم
قسم علوم الحياة
تأثير التكامل بين مخلفات زهرة الشمس و مبيد الشيفالير في
المايكورايزا ونمو مكافحة أدغال محصول الحنطة
جامعة بغداد -قسم علوم الحياة –كلية العلوم مجلس مقدمة إلى اطروحة
نباتفسلجة ال/علوم الحياة الدكتوراهوهي جزء من متطلبات نيل درجة
من قبل
ياء نصيف مسلم العكيلسر
6002 المستنصرية جامعةال/ كلية العلوم / بكلوريوس علوم الحياة
6009 الجامعة المستنصرية/ كلية العلوم / نباتماجستير
بإشراف
د. إبراهيم شعبان السعداوي . أ
هادي مهدي عبود د.
م 4153 هـ 5341