interference contrast and phase contrast microscopy of sporulation
TRANSCRIPT

JOURNAL OF BACTERIOLOGY, Nov. 1968, p. 1811-1817Copyright © 1968 American Society for Microbiology
Vol. 96, No. 5Printed in U.S.A.
Interference Contrast and Phase Contrast Microscopyof Sporulation and Germination of
Bacillus megateriumANTHONY D. HITCHINS, ARNOLD J. KAHN, AND RALPH A. SLEPECKY
Biological Research Laboratories, Syracuse University, Syracuse, New York 13210
Received for publication 30 August 1968
The techniques of Nomarski interference contrast microscopy and phase-contrastmicroscopy were compared for their utility in monitoring sporulation and germina-tion in Bacillus megaterium. The Nomarski technique permitted rapid and easydelineation of septation and engulfment during sporulation, whereas with phasecontrast microscopy these stages were not detected at all. The later stages of sporu-lation were easily seen by either technique. Thus, of the seven stages of sporulationas recognized by the electron microscopy of thin sections, five can now be rou-tinely detected quantitatively by optical microscopy: septation (stage II), engulf-ment (stage III), phase-dark forespore (corresponding to cortex formation, stageIV), phase-bright spore in a sporangium (corresponding to coat formation, stageV), and the free spore (stage VII). This means that now only stage I (axial filament)and stage VI (maturation of the refractile spore) require electron microscopy forroutine detection. There was no advantage in using Nomarski optics for germina-tion studies.
Bacterial sporulation has been divided intoseven distinct stages based on the examination ofstained preparations in the bright field micro-scope, of living cells with the phase-contrastmicroscope, and of shadowed whole cells or thinsections of cells with the electron microscope.These stages are: I, preseptation or axial chroma-tin; II, septation; III, protoplast envelopment;IV, cortex formation; V, coat formation; VI,maturation; and VII, the free spore (see reviewby Murrell, reference 9). All seven stages can bereadily distinguished in thin sections with theelectron microscope; however, this techniquedoes not allow for easy quantitative monitoringof the course of sporulation or the determinationof the degree of synchrony of the sporulationprocess. The usual practice for the assessment ofthe degree of development is to follow the processby phase-contrast microscopy, but stages I toIII cannot be seen by this method. By phase-contrast microscopy, stage IV appears as aphase-dark cell containing a still darker body,usually referred to as the forespore or prespore,whereas stage V appears as a phase-dark cellcontaining the partly or fully refractile spore.Stage VI, maturation, cannot be distinguishedfrom stage V with phase contrast microscopy.Stage VII, the free spore, is of course easily
detected with phase contrast microscopy. Thus,phase microscopy can only be used to assessrelatively late periods of sporogenesis.
In this investigation, we examined Bacillusmegaterium cells during sporulation and germina-tion by Nomarski interference contrast optics (1,7, 10). The same cells were then examined withdark phase contrast. Unlike phase contrast, theNomarski technique permitted rapid and easydelineation of stages II and III cells.
MATERIALS AND METHODS
Organism. The organism used was B. megateriumATCC 19213.
Culture techniiques. For studies on the growth andsporulation of B. megaterium, a portion of a standardspore suspension (6) was heated at 70 C for 0.5 hr,inoculated into a defined sucrose salts medium (SS)(14) containing the germinants L-alanine and inosine(each 100 pg/ml; NBC), incubated at 30 C with shak-ing to a turbidity of 70 Klett units (about 2 X 108cells/ml), and measured in a Klett-Summerson photo-electric colorimeter with a no. 54 filter. The cells werethen harvested by centrifugation in a Sorvall Super-speed refrigerated centrifuge at 2,000 X g for 15 minat 6 C. This cell suspension was placed into 50 ml ofSS medium in a Klett sidearm flask and was incubatedat 30 C on a New Brunswick rotary shaker (160 rev/min). For the germination studies, stock spores wereheated at 70 C for 0.5 hr, inoculated into Nutrient
1811
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
01
Dec
embe
r 20
21 b
y 22
0.88
.11.
243.

HITCHINS, KAHN, AND SLEPECKY
Broth (Difco), and incubated as described for thesporulation studies.We adopted the convention (9, 16) of arbitrarily
designating the commencement of sporulation as thepoint at which exponential growth ends (To), corre-sponding to the axial chromatin or preseptation stage(stage I) as determined by exaniination of thin sectionsin the electron microscope. Stages I through VI areusually completed in 6 to 8 hr (T6 to T8) and representseveral generation times in terms of exponentialgrowth division.
Microscopy and photography. Samples (2 ml) ofcultures were removed from the Klett flasks at inter-vals of 0.5 or 1 hr and were stored in a freezer untilexamined. Cells could be held at 0 C for 1 week with-out damage. In preparing wet mounts for microscopicexamination, the suspension was spread as thin aspossible on a glass slide in order to insure optimalcontrast and the glass coverslip was paraffin-sealed tominimize movement of the cells. Observations weremade with a Zeiss photomicroscope equipped withphase contrast and Nomarski interference contrastoptics. All photographs were taken through oil im-mersion objectives (phase contrast, neofluor, NA, 1.3;interference contrast, Planachromat, NA, 1.25) onPlus X 35-mm film. Contrast and resolution wereenhanced by placing immersion oil between the con-denser and the slide and by using a green filter. Finalmicroscope magnification was X 1,250.
Enumeration of cell types. For quantitative meas-urement of cell types in the population at a given timeduring sporulation, 200 cells were counted per sampleby use of either Nomarski or phase contrast optics.
RESULTSGeneral comparison of phase contrast with in-
terference contrast. Figures 1 to 4 show theappearance of B. megaterium during its vegetativecell-spore-vegetative cell developmental cycle, asviewed with phase contrast and Nomarski inter-ference contrast optics. In general, the cells seenin the Nomarski system (i) lacked the light halocharacteristic of the phase-contrast system; (ii)appeared larger; (iii) presented an optically flatappearance; (iv) showed a "shadow effect"reminiscent of shadowed preparations seen in theelectron microscope; and, most importantly, (v)showed more detail, particularly in the earlystages of sporulation. With regard to the lastpoint, the division septa and the sporulation septawere especially well defined. In addition to somegranular objects visible in phase contrast, otherobjects, which are not seen under phase opticswere visible.
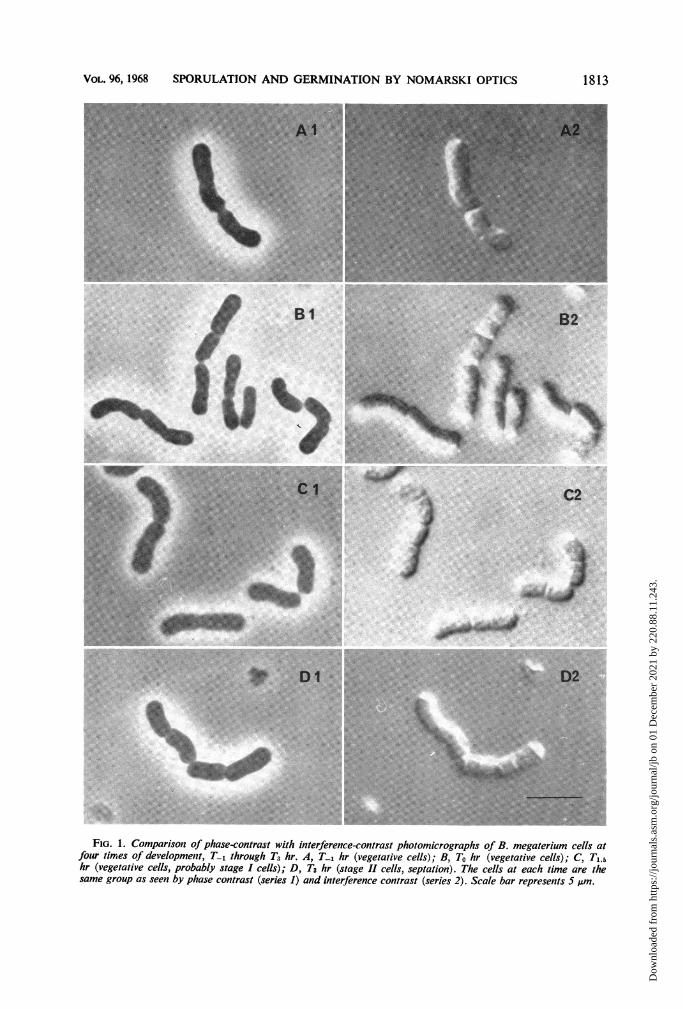
Sporulation viewed under phase contrast. Ingeneral, the phase-contrast pictures of the courseof sporulation in B. megaterium (Fig. 1, Al, BI,Cl, and Dl; Fig. 2, Al, Bi, Cl, and Dl) pre-sented essentially the same details as describedfor this method with other sporulating bacteria(4, 18). In the early stages (Fig. 1), except for theappearance of some phase-bright granules, par-
ticularly at T-1 and To (Fig. lAl, B1), the cellshad the typical featureless phase-dark appearanceof vegetative cells. The granules, which wererelatively scarce, were probably poly-j3-hydroxy-butyric acid (PHB) granules; in a previous studywith the same organism and cultural conditions,the granules were shown to be PHB and werealso rare at this stage (15).
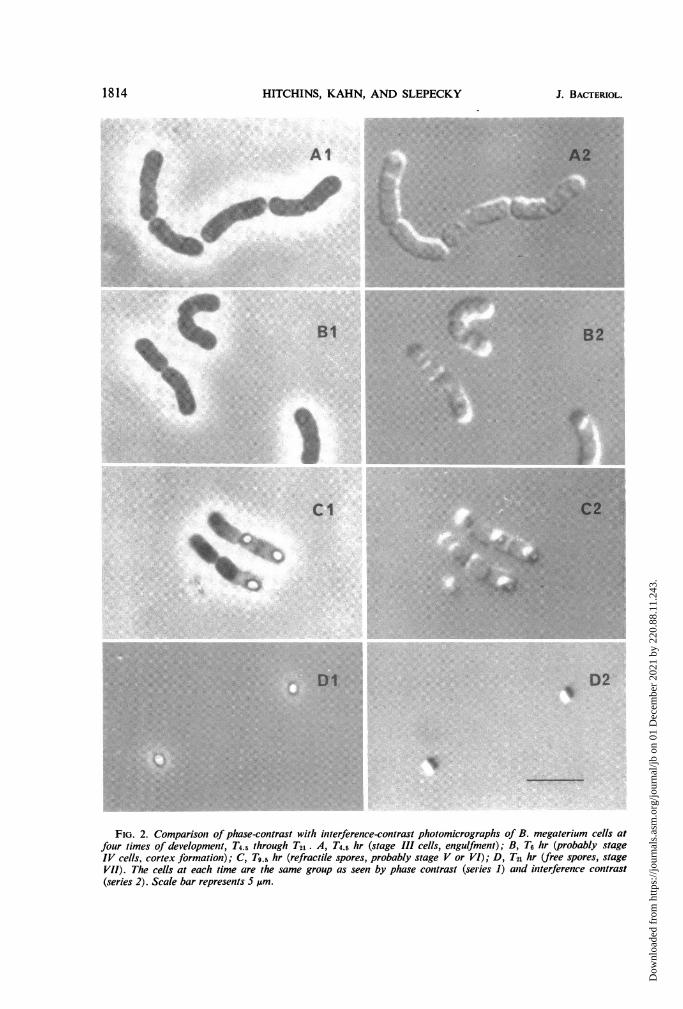
In the later stages of sporulation (Fig. 2), thephase-contrast pictures revealed the featureswhich have been routinely observed and havebeen extremely useful in assessing the laterstages. Phase-dark bodies at the poles were seenstarting at T4.5 (Fig. 2, Al). These bodies ex-hibited a slight refractility at T6 (Fig. 2, Bl), andcomparable studies have defined them as fore-spores (6). At T0.5 (Fig. 2, Cl), refractile sporeswere present in the partially lysed sporangia.Prolonged incubation to T21 resulted in completelysis with the release of refractile mature spores(Fig. 2, Dl).
Sporulation viewed under interference contrastand compared with phase contrast. Interferencecontrast provided much greater detail thanphase contrast, particularly in the early stages ofsporulation. Whereas the cytoplasm did notappear very granular under phase contrast in theearly stages (Fig. 1, Al, B1, Cl), cells examinedby interference contrast appeared much moregranulated (Fig. 1, A2, B2, C2); this additionalgranulation did not appear to correspond withthe phase-bright granules. The division septawere more clearly seen by interference contrast,and, unlike with phase contrast, it appeared thatdifferent stages of division septum formationcould be discerned. For example, the early stagesof division septation can clearly be seen by inter-ference contrast (the lower cell of Fig. 1, A2),whereas only an indentation of the cell outline isvisible by phase contrast (Fig. 1, Al).The most important and striking difference
between the two methods of microscopy wasrevealed at about T3. Interference contrastmicroscopy clearly showed the presence of septaat the poles of the cells (Fig. 1, D2), whereasthis could not be easily discerned in most cellsby phase contrast (Fig. 1, D1). These septa werejudged to be spore septa on the basis of thefollowing criteria: the time of their appearancein the culture, subsequent events, the fact thatthey could also be detected by staining withcrystal violet (3), and their presence, as revealedby examination of thin sections in the electronmicroscope, in samples taken at equivalenttimes with the same organism under similar cul-tural conditions (2). However, the prime reasonfor considering them as spore septa was theasymmetric positioning of these septa in the cell.By T4 .5, further stages of development were
1812 J. BAcTERioL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
01
Dec
embe
r 20
21 b
y 22
0.88
.11.
243.
VOL. 96, 1968 SPORULATION AND GERMINATION BY NOMARSKI OPTICS
^ ~B1
_-~~~~~~~~~~~~~~~~....
B2
.. .....
i.l.....I ..
C2
r,NILt'":L
4.~-.ls4
AL~ ~ ~ -
".. .........
.~~~ ~~~~~~~~~~~~~~
.7si.
FIG. 1. Comparisonz of phase-contrast with interference-contrast photomicrographs of B. megaterium cells atfouir times of development, T-1 through T3 hr. A, T-1 hr (vegetative cells); B, To hr (vegetative cells); C, Ti.5hr (vegetative cells, probably stage I cells); D, T3 hr (stage II cells, septation). The cells at each time are thesame group as seen by phase contrast (series 1) and interference contrast (series 2). Scale bar represents 5 pAm.
1813
...............
..............
....................
........................
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
01
Dec
embe
r 20
21 b
y 22
0.88
.11.
243.
HITCHINS, KAHN, AND SLEPECKY
Al
BI
C1
.... .. .. ..
*: :. .:*: ..
A2...T : .. ... ,: .: .
_ $' Ri ';*t i;00 '" ,0d''.' . ';.:f 00:*Gi . .: '' '.: .' 7 : :: : :: . . . . : ::% %. . ,' :*--x Msz. .: ' . : .|*hS;W . :
:.
B2
I.f.
i.~~~~~~~~~ .:
... ...............
DlDIt
D2
si
FIG. 2. Comparison of phase-contrast with interference-contrast photomicrographs of B. megaterium cells atfour times of development, T4.5 through T21. A, T4.5 hr (stage Ill cells, engulfment); B, T6 hr (probably stageIV cells, cortex formation); C, T9.s hr (refractile spores, probably stage V or VI); D, T2n hr (free spores, stageVil). The cells at each time are the same group as seen by phase contrast (series 1) and interference contrast(series 2). Scale bar represents 5 pm.
1814 J. BACTERIOL.
.. ...... :.:.... ...I... :.I"
.. ..
i. .,
?-x..:" 'iU
!-. Alf.V.t :.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
01
Dec
embe
r 20
21 b
y 22
0.88
.11.
243.
VOL. 96, 1968 SPORULATION AND GERMINATION BY NOMARSKI OPTICS
becoming visible in phase contrast (Fig. 2, Al);again, however, these stages were more clearlyseen with Nomarski optics (Fig. 2, A2). Theshape of the cells and the time of their appear-ance suggested that these cells were at stage III,the protoplast envelopment stage.By T6 and later, the stages of sporulation
could be as readily distinguished in phase con-trast (Fig. 2, Bi, Cl, DI) as in interferencecontrast (Fig. 2, B2, C2, D2). The developmentof refractility under phase contrast was paral-leled by an increase in the characteristic strong"shadowing effect" of spores with Nomarskioptics.
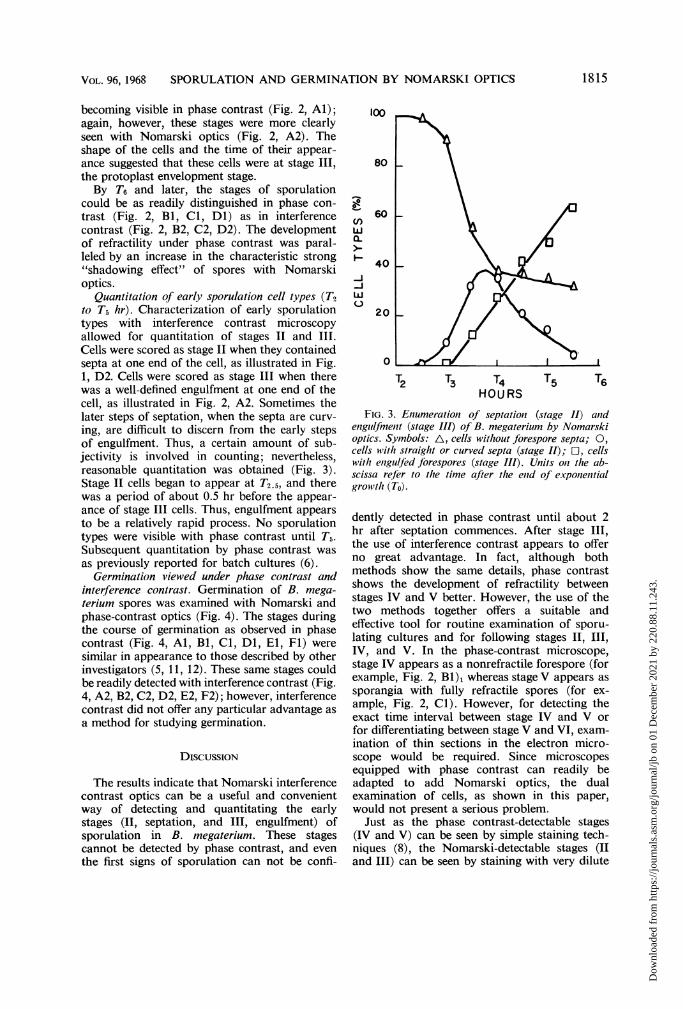
Quantitation of early sporulation cell types (T2to T5 hr). Characterization of early sporulationtypes with interference contrast microscopyallowed for quantitation of stages II and III.Cells were scored as stage II when they containedsepta at one end of the cell, as illustrated in Fig.1, D2. Cells were scored as stage III when therewas a well-defined engulfment at one end of thecell, as illustrated in Fig. 2, A2. Sometimes thelater steps of septation, when the septa are curv-ing, are difficult to discern from the early stepsof engulfment. Thus, a certain amount of sub-jectivity is involved in counting; nevertheless,reasonable quantitation was obtained (Fig. 3).Stage II cells began to appear at T2.5, and therewas a period of about 0.5 hr before the appear-ance of stage III cells. Thus, engulfment appearsto be a relatively rapid process. No sporulationtypes were visible with phase contrast until T5.Subsequent quantitation by phase contrast wasas previously reported for batch cultures (6).
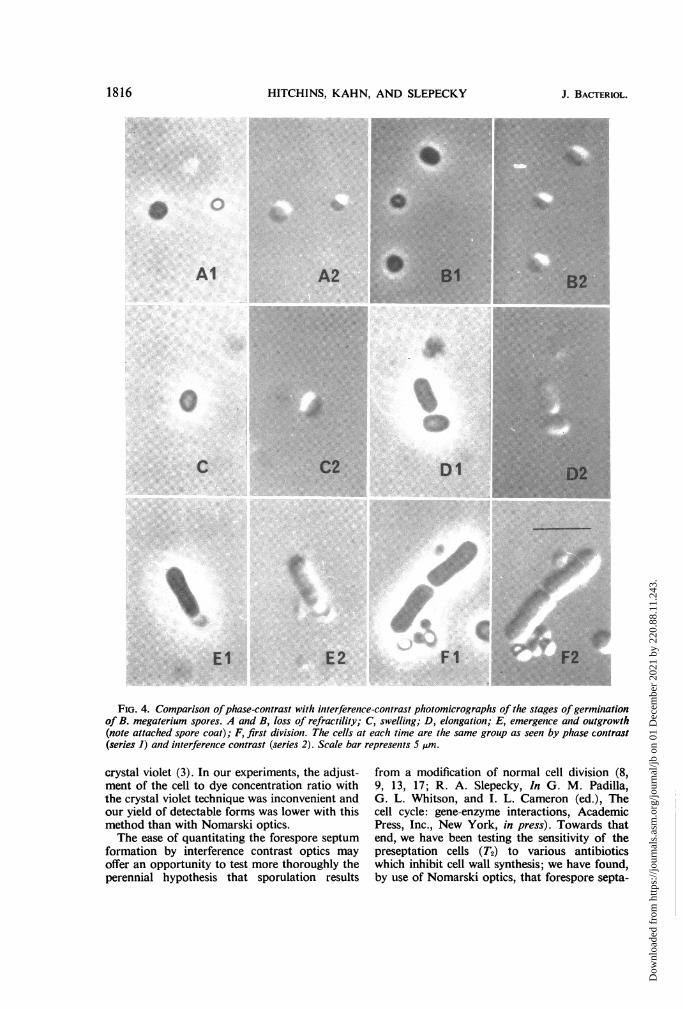
Germination viewed under phase contrast andinterference contrast. Germination of B. mega-terium spores was examined with Nomarski andphase-contrast optics (Fig. 4). The stages duringthe course of germination as observed in phasecontrast (Fig. 4, Al, Bi, Cl, Dl, El, Fl) weresimilar in appearance to those described by otherinvestigators (5, 11, 12). These same stages couldbe readily detected with interference contrast (Fig.4, A2, B2, C2, D2, E2, F2); however, interferencecontrast did not offer any particular advantage asa method for studying germination.
DISCUSSION
The results indicate that Nomarski interferencecontrast optics can be a useful and convenientway of detecting and quantitating the earlystages (II, septation, and III, engulfment) ofsporulation in B. megaterium. These stagescannot be detected by phase contrast, and eventhe first signs of sporulation can not be confi-
100
80
n(I)wa-
-J-Jw
60
40
20
0o1 rt I I I
T2 T3 T4 T5 T6HOURS
FIG. 3. Enumeration of septation2 (stage 11) aiidenguilfmenit (stage III) of B. megaterium by Nomarskioptics. Symbols: A, cells without forespore septa; 0,cells with straight or curved septa (stage II); 0, cellswith enzgulfed forespores (stage III). Units oni the ab-scissa refer to the time after the enid of exponentialgrowvth (To).
dently detected in phase contrast until about 2hr after septation commences. After stage III,the use of interference contrast appears to offerno great advantage. In fact, although bothmethods show the same details, phase contrastshows the development of refractility betweenstages IV and V better. However, the use of thetwo methods together offers a suitable andeffective tool for routine examination of sporu-lating cultures and for following stages II, III,IV, and V. In the phase-contrast microscope,stage IV appears as a nonrefractile forespore (forexample, Fig. 2, Bl), whereas stage V appears assporangia with fully refractile spores (for ex-ample, Fig. 2, Cl). However, for detecting theexact time interval between stage IV and V orfor differentiating between stage V and VI, exam-ination of thin sections in the electron micro-scope would be required. Since microscopesequipped with phase contrast can readily beadapted to add Nomarski optics, the dualexamination of cells, as shown in this paper,would not present a serious problem.
Just as the phase contrast-detectable stages(IV and V) can be seen by simple staining tech-niques (8), the Nomarski-detectable stages (IIand III) can be seen by staining with very dilute
1815
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
01
Dec
embe
r 20
21 b
y 22
0.88
.11.
243.
HITCHINS, KAHN, AND SLEPECKY J. BACTERI
SM.P...
S X
.B2A2 .....B.
,C..... ... . .D
El E2 FlI F2.
FiG,. 4. Comparison ofphase-contrast with interferenice-contrast photomicrographs of the stages of germiniationof B. megaterium spores. A and B, loss of refractility; C, swelling; D, elongation; E, emergence and outgrowth
(note attached spore coat); F, first division. The cells at each time are the same group as seen by phase contrast
(series 1) and initerferenice contrast (series 2). Scale bar represenits 5 pAm.
crystal violet (3). In our experiments, the adjust-ment of the cell to dye concentration ratio withthe crystal violet technique was inconvenient andour yield of detectable forms was lower with thismethod than with Nomarski optics.The ease of quantitating the forespore septum
formation by interference contrast optics mayoffer an opportunity to test more thoroughly theperennial hypothesis that sporulation results
from a modification of normal cell division (8,9, 13, 17; R. A. Slepecky, In G. M. Padilla,G. L. Whitson, and I. L. Cameron (ed.), Thecell cycle: gene-enzyme interactions, AcademicPress, Inc., New York, in press). Towards thatend, we have been testing the sensitivity of thepreseptation cells (T2) to various antibioticswhich inhibit cell wall synthesis; we have found,by use of Nomarski optics, that forespore septa-
1816 IOL.
SM2 2
Al
C
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
01
Dec
embe
r 20
21 b
y 22
0.88
.11.
243.

VOL. 96, 1968 SPORULATION AND GERMINATION BY NOMARSKI OPTICS
tion is inhibited (Hitchins and Slepecky, unpub-lished data).To the best of our knowledge, this is the first
investigation in which Nomarski interferencecontrast optics have been used to study bacteria.This technique may be applicable to the study ofother differentiating bacterial systems.
ACKNOWLEDGMENTS
This investigation was supported by NationalScience Foundation grants GB 7275 (to Arnold Kahn)and GB 6433 (to Ralph Slepecky).
LITERATURE CITED
1. Bajer, A., and R. D. Allen. 1966. Structure andorganization of the living mitotic spindle ofHaematithus endosperm. Science 151:572-274.
2. Ellar, D. J., D. G. Lundgren, and R. A. Slepecky.1967. Fine structure of Bacillus megateriurnduring synchronous growth. J. Bacteriol. 94:1189-1205.
3. Gordon, R. A., and W. G. Murrell. 1967. Simplemethod of detecting spore septum formationand synchrony of sporulation. J. Bacteriol. 93:495-496.
4. Hashimoto, T., S. H. Black, and P. Gerhardt.1960. Development of fine structure, thermo-stability, and dipicolinate during sporogenesisin a bacillus. Can. J. Microbiol. 6:203-212.
5. Hitchins, A. D., G. W. Gould, and A. Hurst.1963. The swelling of bacterial spores duringgermination and outgrowth. J. Gen. Microbiol.30:445-453.
6. Imanaka, H., J. R. Gillis, and R. A. Slepecky.1967. Synchronous growth and sporulation ofBacillus megaterium. J. Bacteriol. 93:1624-1630.
7. Maguire, M. P. 1968. Nomarski interference con-trast resolution of subchromatid structure.Proc. Natl. Acad. Sci. U.S. 60:533-536.
8. Murrell, W. G. 1961. Spore formation and germi-
nation as a microbial reaction to environment.Symp. Soc. Gen. Microbiol. 11:100-150.
9. Murrell, W. G. 1967. The biochemistry of thebacterial endospore. Advan. Microbiol.Physiol. 1:133-251.
10. Nomarski, G. 1955. Microinterferonmetre dif-ferentiel a ondes polarisees. J. Phys. Radium16:9s-1 3s.
11. Powell, E. 0. 1957. The appearance of bacterialspores under phase-contrast illumination. J.Appl. Bacteriol. 3:342-348.
12. Pulvertaft, R. J. V., and J. A. Haynes. 1951.Adenosine and spore germination; phase-contrast studies. J. Gen. Microbiol. 5:657-663.
13. Robinow, C. F. 1960. Morphology of bacterialspores, their development and germination, p.207-248. In I. C. Gunsalus and R. Y. Stanier(ed.), The Bacteria, vol. 1. Academic Press,Inc., New York.
14. Slepecky, R., and J. W. Foster. 1959. Alterationsin metal content of spores of Bacillus mega-rerium and the effect of some spore properties.J. Bacteriol. 78:117-123.
15. Slepecky, R. A., and J. H. Law. 1961. Synthesisand degradation of poly-,-hydroxybutyric acidin connection with sporulation of Bacillusn7egaterium. J. Bacteriol. 82:37-42.
16. Szulmajster, J., and P. Schaeffer. 1961. Augmen-tation de l'activite DPNH-oxydasique au coursde la sporulation de Bacillus subtilis. Compt.Rend. 252:220-222.
17. Vinter, V. 1963. Spores of microorganisms.Chloramphenicol-sensitive and penicillin-re-sistant incorporation of '4C-diaminopimelicacid into sporulating cells of Bacillus cereus.Experientia 19:307-310.
18. Young, E. I., and P. C. Fitz-James. 1959. Chemi-cal and morphological studies of bacterial sporeformation. I. The formation of spores inBacillus cereus. J. Biophys. Biochem. Cytol.6:467-482.
1817
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
01
Dec
embe
r 20
21 b
y 22
0.88
.11.
243.