interim report ir-06-034 mothers determine offspring size...
TRANSCRIPT

International Institute for Applied Systems Analysis Schlossplatz 1 A-2361 Laxenburg, Austria
Tel: +43 2236 807 342Fax: +43 2236 71313
E-mail: [email protected]: www.iiasa.ac.at
Interim Reports on work of the International Institute for Applied Systems Analysis receive onlylimited review. Views or opinions expressed herein do not necessarily represent those of theInstitute, its National Member Organizations, or other organizations supporting the work.
Interim Report IR-06-034
Mothers determine offspring size in response to own juvenile growth conditions Barbara Taborsky ([email protected])
Approved by Ulf Dieckmann Program Leader, Evolution and Ecology Program
September 2006

IIASA STUDIES IN ADAPTIVE DYNAMICS NO. 116
EEP
The Evolution and Ecology Program at IIASA fosters the devel-opment of new mathematical and conceptual techniques for un-derstanding the evolution of complex adaptive systems.Focusing on these long-term implications of adaptive processesin systems of limited growth, the Evolution and Ecology Programbrings together scientists and institutions from around the worldwith IIASA acting as the central node.Scientific progress within the network is collected in the IIASAStudies in Adaptive Dynamics series.
No. 1 Metz JAJ, Geritz SAH, Meszéna G, Jacobs FJA, vanHeerwaarden JS: Adaptive Dynamics: A Geometrical Studyof the Consequences of Nearly Faithful Reproduction. IIASAWorking Paper WP-95-099 (1995). van Strien SJ, VerduynLunel SM (eds): Stochastic and Spatial Structures of Dynami-cal Systems, Proceedings of the Royal Dutch Academy of Sci-ence (KNAW Verhandelingen), North Holland, Amsterdam,pp. 183-231 (1996).
No. 2 Dieckmann U, Law R: The Dynamical Theory of Co-evolution: A Derivation from Stochastic Ecological Processes.IIASA Working Paper WP-96-001 (1996). Journal of Mathe-matical Biology 34:579-612 (1996).
No. 3 Dieckmann U, Marrow P, Law R: Evolutionary Cy-cling of Predator-PreyInteractions: Population Dynamicsandthe Red Queen. IIASA Preprint (1995). Journal of Theoreti-cal Biology 176:91-102 (1995).
No. 4 Marrow P, Dieckmann U, Law R: Evolutionary Dy-namics of Predator-Prey Systems: An Ecological Perspective.IIASA Working Paper WP-96-002 (1996). Journal of Mathe-matical Biology 34:556-578 (1996).
No. 5 Law R, Marrow P, Dieckmann U: On Evolution underAsymmetric Competition. IIASA Working Paper WP-96-003(1996). Evolutionary Ecology 11:485-501 (1997).
No. 6 Metz JAJ, Mylius SD, Diekmann O: When Does Evo-lution Optimize? On the Relation Between Types of DensityDependence and Evolutionarily Stable Life History Parame-ters. IIASA Working Paper WP-96-004 (1996).
No. 7 Ferrière R, Gatto M: Lyapunov Exponents and theMathematics of Invasion in Oscillatory or Chaotic Popula-tions. Theoretical Population Biology 48:126-171 (1995).
No. 8 Ferrière R, Fox GA: Chaos and Evolution. IIASAPreprint (1996). Trends in Ecology and Evolution 10:480-485 (1995).
No. 9 Ferrière R, Michod RE: The Evolution of Cooperationin Spatially Heterogeneous Populations. IIASA Working Pa-per WP-96-029 (1996). The American Naturalist 147:692-717 (1996).
No. 10 van Dooren TJM, Metz JAJ: Delayed Maturation inTemporally Structured Populations with Non-Equilibrium Dy-namics. IIASA Working Paper WP-96-070 (1996). Journalof Evolutionary Biology 11:41-62 (1998).
No. 11 Geritz SAH, Metz JAJ, Kisdi É, Meszéna G: The Dy-namics of Adaptation and Evolutionary Branching. IIASAWorking Paper WP-96-077 (1996). Physical Review Letters78:2024-2027 (1997).
No. 12 Geritz SAH, Kisdi É, Meszéna G, Metz JAJ: Evo-lutionary Singular Strategies and the Adaptive Growth andBranching of the Evolutionary Tree. IIASA Working PaperWP-96-114 (1996). Evolutionary Ecology 12:35-57 (1998).
No. 13 Heino M, Metz JAJ, Kaitala V: Evolution of MixedMaturation Strategies in Semelparous Life-Histories: TheCrucial Role of Dimensionality of Feedback Environment.IIASA Working Paper WP-96-126 (1996). Philosophi-cal Transactions of the Royal Society of London Series B352:1647-1655 (1997).
No. 14 Dieckmann U: Can Adaptive Dynamics Invade?IIASA Working Paper WP-96-152 (1996). Trends in Ecol-ogy and Evolution 12:128-131 (1997).
No. 15 Meszéna G, Czibula I, Geritz SAH: Adaptive Dynam-ics in a 2-Patch Environment: A Simple Model for Allopatricand Parapatric Speciation. IIASA Interim Report IR-97-001(1997). Journal of Biological Systems 5:265-284 (1997).
No. 16 Heino M, Metz JAJ, Kaitala V: The Enigma ofFrequency-Dependent Selection. IIASA Interim Report IR-97-061 (1997). Trends in Ecology and Evolution 13:367-370(1998).
No. 17 Heino M: Management of Evolving Fish Stocks.IIASA Interim Report IR-97-062 (1997). Canadian Journalof Fisheries and Aquatic Sciences 55:1971-1982 (1998).
No. 18 Heino M: Evolution of Mixed Reproductive Strategiesin Simple Life-History Models. IIASA Interim Report IR-97-063 (1997).
No. 19 Geritz SAH, van der Meijden E, Metz JAJ: Evolution-ary Dynamics of Seed Size and Seedling Competitive Ability.IIASA Interim Report IR-97-071 (1997). Theoretical Popu-lation Biology 55:324-343 (1999).
No. 20 Galis F, Metz JAJ: Why Are There So Many CichlidSpecies? On the Interplay of Speciation and Adaptive Radi-ation. IIASA Interim Report IR-97-072 (1997). Trends inEcology and Evolution 13:1-2 (1998).

No. 21 Boerlijst MC, Nowak MA, Sigmund K: Equal Payfor all Prisoners/ The Logic of Contrition. IIASA InterimReport IR-97-073 (1997). American Mathematical SocietyMonthly 104:303-307 (1997). Journal of Theoretical Biology185:281-293 (1997).
No. 22 Law R, Dieckmann U: Symbiosis Without Mutualismand the Merger of Lineages in Evolution. IIASA Interim Re-port IR-97-074 (1997). Proceedings of the Royal Society ofLondon Series B 265:1245-1253 (1998).
No. 23 Klinkhamer PGL, de Jong TJ, Metz JAJ: Sex and Sizein Cosexual Plants. IIASA Interim Report IR-97-078 (1997).Trends in Ecology and Evolution 12:260-265 (1997).
No. 24 Fontana W, Schuster P: Shaping Space: The Possi-ble and the Attainable in RNA Genotype-Phenotype Mapping.IIASA Interim Report IR-98-004 (1998). Journal of Theoret-ical Biology 194:491-515 (1998).
No. 25 Kisdi É, Geritz SAH: Adaptive Dynamics in AlleleSpace: Evolution of Genetic Polymorphism by Small Muta-tions in a HeterogeneousEnvironment. IIASA Interim ReportIR-98-038 (1998). Evolution 53:993-1008 (1999).
No. 26 Fontana W, Schuster P: Continuity in Evolution: Onthe Nature of Transitions. IIASA Interim Report IR-98-039(1998). Science 280:1451-1455 (1998).
No. 27 Nowak MA, Sigmund K: Evolution of Indirect Reci-procity by Image Scoring/ The Dynamics of Indirect Reci-procity. IIASA Interim Report IR-98-040 (1998). Nature393:573-577 (1998). Journal of Theoretical Biology 194:561-574 (1998).
No. 28 Kisdi É: Evolutionary Branching Under AsymmetricCompetition. IIASA Interim Report IR-98-045 (1998). Jour-nal of Theoretical Biology 197:149-162 (1999).
No. 29 Berger U: Best ResponseAdaptation for Role Games.IIASA Interim Report IR-98-086 (1998).
No. 30 van Dooren TJM: The Evolutionary Ecology ofDominance-Recessivity. IIASA Interim Report IR-98-096(1998). Journal of Theoretical Biology 198:519-532 (1999).
No. 31 Dieckmann U, O’Hara B, Weisser W: The Evolution-ary Ecology of Dispersal. IIASA Interim Report IR-98-108(1998). Trends in Ecology and Evolution 14:88-90 (1999).
No. 32 Sigmund K: Complex Adaptive Systems and the Evo-lution of Reciprocation. IIASA Interim Report IR-98-100(1998). Ecosystems 1:444-448 (1998).
No. 33 Posch M, Pichler A, Sigmund K: The Efficiency ofAdapting Aspiration Levels. IIASA Interim Report IR-98-103 (1998). Proceedings of the Royal Society London SeriesB 266:1427-1435 (1999).
No. 34 Mathias A, Kisdi É: Evolutionary Branching and Co-existence of Germination Strategies. IIASA Interim ReportIR-99-014 (1999).
No. 35 Dieckmann U, Doebeli M: On the Origin of Speciesby Sympatric Speciation. IIASA Interim Report IR-99-013(1999). Nature 400:354-357 (1999).
No. 36 Metz JAJ, Gyllenberg M: How Should We Define Fit-ness in Structured Metapopulation Models? Including an Ap-plication to the Calculation of Evolutionarily Stable DispersalStrategies. IIASA Interim Report IR-99-019 (1999). Pro-ceedings of the Royal Society of London Series B 268:499-508 (2001).
No. 37 Gyllenberg M, Metz JAJ: On Fitness in StructuredMetapopulations. IIASA Interim Report IR-99-037 (1999).Journal of Mathematical Biology 43:545-560 (2001).
No. 38 Meszéna G, Metz JAJ: Species Diversity and Popula-tion Regulation: The Importance of Environmental FeedbackDimensionality. IIASA Interim Report IR-99-045 (1999).
No. 39 Kisdi É, Geritz SAH: Evolutionary Branching andSympatric Speciation in Diploid Populations. IIASA InterimReport IR-99-048 (1999).
No. 40 Ylikarjula J, Heino M, Dieckmann U: Ecology andAdaptation of Stunted Growth in Fish. IIASA Interim ReportIR-99-050 (1999). Evolutionary Ecology 13:433-453 (1999).
No. 41 Nowak MA, Sigmund K: Games on Grids. IIASAInterim Report IR-99-038 (1999). Dieckmann U, Law R,Metz JAJ (eds): The Geometry of Ecological Interactions:Simplifying Spatial Complexity, Cambridge University Press,Cambridge, UK, pp. 135-150 (2000).
No. 42 Ferrière R, Michod RE: Wave Patterns in SpatialGames and the Evolution of Cooperation. IIASA InterimReport IR-99-041 (1999). Dieckmann U, Law R, Metz JAJ(eds): The Geometry of Ecological Interactions: SimplifyingSpatial Complexity, Cambridge University Press, Cambridge,UK, pp. 318-332 (2000).
No. 43 Kisdi É, Jacobs FJA, Geritz SAH: Red Queen Evo-lution by Cycles of Evolutionary Branching and Extinction.IIASA Interim Report IR-00-030 (2000). Selection 2:161-176 (2001).
No. 44 MeszénaG, Kisdi É, DieckmannU, Geritz SAH, MetzJAJ: Evolutionary Optimisation Models and Matrix Games inthe Unified Perspectiveof Adaptive Dynamics. IIASA InterimReport IR-00-039 (2000). Selection 2:193-210 (2001).
No. 45 Parvinen K, Dieckmann U, Gyllenberg M, Metz JAJ:Evolution of Dispersal in Metapopulations with Local DensityDependence and Demographic Stochasticity. IIASA InterimReport IR-00-035 (2000). Journal of Evolutionary Biology16:143-153 (2003).
No. 46 Doebeli M, Dieckmann U: Evolutionary Branch-ing and Sympatric Speciation Caused by Different Types ofEcological Interactions. IIASA Interim Report IR-00-040(2000). The American Naturalist 156:S77-S101 (2000).
No. 47 Heino M, Hanski I: Evolution of Migration Rate ina Spatially Realistic Metapopulation Model. IIASA InterimReport IR-00-044 (2000). The American Naturalist 157:495-511 (2001).
No. 48 Gyllenberg M, Parvinen K, Dieckmann U: Evolution-ary Suicide and Evolution of Dispersal in StructuredMetapop-ulations. IIASA Interim Report IR-00-056 (2000). Journalof Mathematical Biology 45:79-105 (2002).
No. 49 van Dooren TJM: The Evolutionary Dynamics of Di-rect Phenotypic Overdominance: Emergence Possible, LossProbable. IIASA Interim Report IR-00-048 (2000). Evolu-tion 54:1899-1914 (2000).
No. 50 Nowak MA, Page KM, Sigmund K: Fairness VersusReason in the Ultimatum Game. IIASA Interim Report IR-00-57 (2000). Science 289:1773-1775 (2000).
No. 51 de Feo O, Ferrière R: Bifurcation Analysis of Pop-ulation Invasion: On-Off Intermittency and Basin Riddling.IIASA Interim Report IR-00-074 (2000). International Jour-nal of Bifurcation and Chaos 10:443-452 (2000).

No. 52 Heino M, Laaka-Lindberg S: Clonal Dynamics andEvolution of Dormancy in the Leafy Hepatic Lophozia Sil-vicola. IIASA Interim Report IR-01-018 (2001). Oikos94:525-532 (2001).
No. 53 Sigmund K, Hauert C, Nowak MA: Reward and Pun-ishment in Minigames. IIASA Interim Report IR-01-031(2001). Proceedings of the National Academy of Sciencesof the USA 98:10757-10762 (2001).
No. 54 Hauert C, De Monte S, Sigmund K, Hofbauer J: Os-cillations in Optional Public Good Games. IIASA InterimReport IR-01-036 (2001).
No. 55 Ferrière R, Le Galliard J: Invasion Fitness and Adap-tive Dynamics in Spatial Population Models. IIASA InterimReport IR-01-043 (2001). Clobert J, Dhondt A, Danchin E,Nichols J (eds): Dispersal, Oxford University Press, pp. 57-79(2001).
No. 56 de Mazancourt C, Loreau M, Dieckmann U: Can theEvolution of Plant Defense Lead to Plant-Herbivore Mutual-ism. IIASA Interim Report IR-01-053 (2001). The AmericanNaturalist 158:109-123 (2001).
No. 57 Claessen D, Dieckmann U: Ontogenetic Niche Shiftsand Evolutionary Branching in Size-Structured Populations.IIASA Interim Report IR-01-056 (2001). Evolutionary Ecol-ogy Research 4:189-217 (2002).
No. 58 Brandt H: Correlation Analysis of Fitness Land-scapes. IIASA Interim Report IR-01-058 (2001).
No. 59 Dieckmann U: Adaptive Dynamics of Pathogen-HostInteracations. IIASA Interim Report IR-02-007 (2002).Dieckmann U, Metz JAJ, Sabelis MW, Sigmund K (eds):Adaptive Dynamics of Infectious Diseases: In Pursuit of Viru-lence Management, Cambridge University Press, Cambridge,UK, pp. 39-59 (2002).
No. 60 Nowak MA, Sigmund K: Super- and Coinfection:The Two Extremes. IIASA Interim Report IR-02-008 (2002).Dieckmann U, Metz JAJ, Sabelis MW, Sigmund K (eds):Adaptive Dynamics of Infectious Diseases: In Pursuit of Viru-lence Management, Cambridge University Press, Cambridge,UK, pp. 124-137 (2002).
No. 61 Sabelis MW, Metz JAJ: Taking Stock: Relating The-ory to Experiment. IIASA Interim Report IR-02-009 (2002).Dieckmann U, Metz JAJ, Sabelis MW, Sigmund K (eds):Adaptive Dynamics of Infectious Diseases: In Pursuit of Viru-lence Management, Cambridge University Press, Cambridge,UK, pp. 379-398 (2002).
No. 62 Cheptou P, Dieckmann U: The Evolution of Self-Fertilization in Density-Regulated Populations . IIASA In-terim Report IR-02-024 (2002). Proceedings of the RoyalSociety of London Series B 269:1177-1186 (2002).
No. 63 Bürger R: Additive Genetic Variation Under Intraspe-cific Competition and Stabilizing Selection: A Two-LocusStudy. IIASA Interim Report IR-02-013 (2002). Theoret-ical Population Biology 61:197-213 (2002).
No. 64 Hauert C, De Monte S, Hofbauer J, Sigmund K: Vol-unteering as Red Queen Mechanism for Co-operation in Pub-lic Goods Games. IIASA Interim Report IR-02-041 (2002).Science 296:1129-1132 (2002).
No. 65 Dercole F, Ferrière R, Rinaldi S: Ecological Bistabil-ity and Evolutionary Reversals under Asymmetrical Competi-tion. IIASA Interim Report IR-02-053 (2002). Evolution56:1081-1090 (2002).
No. 66 Dercole F, Rinaldi S: Evolution of CannibalisticTraits: Scenarios Derived from Adaptive Dynamics. IIASAInterim Report IR-02-054 (2002). Theoretical Population Bi-ology 62:365-374 (2002).
No. 67 Bürger R, Gimelfarb A: Fluctuating Environmentsand the Role of Mutation in Maintaining Quantitative GeneticVariation. IIASA Interim Report IR-02-058 (2002). Geneti-cal Research 80:31-46 (2002).
No. 68 Bürger R: On a Genetic Model of Intraspecific Com-petition and Stabilizing Selection. IIASA Interim Report IR-02-062 (2002). Amer. Natur. 160:661-682 (2002).
No. 69 Doebeli M, Dieckmann U: Speciation Along Environ-mental Gradients. IIASA Interim Report IR-02-079 (2002).Nature 421:259-264 (2003).
No. 70 Dercole F, Irisson J, Rinaldi S: Bifurcation Analysis ofa Prey-Predator Coevolution Model. IIASA Interim ReportIR-02-078 (2002). SIAM Journal on Applied Mathematics63:1378-1391 (2003).
No. 71 Le Galliard J, Ferrière R, DieckmannU: The AdaptiveDynamics of Altruism in Spatially HeterogeneousPopulations.IIASA Interim Report IR-03-006 (2003). Evolution 57:1-17(2003).
No. 72 Taborsky B, Dieckmann U, Heino M: Unex-pected Discontinuities in Life-History Evolution under Size-Dependent Mortality. IIASA Interim Report IR-03-004(2003). Proceedings of the Royal Society of London Series B270:713-721 (2003).
No. 73 Gardmark A, Dieckmann U, Lundberg P: Life-History Evolution in Harvested Populations: The Role of Nat-ural Predation. IIASA Interim Report IR-03-008 (2003).Evolutionary Ecology Research 5:239-257 (2003).
No. 74 Mizera F, Meszéna G: Spatial Niche Packing, Char-acter Displacement and Adaptive Speciation Along an En-vironmental Gradient. IIASA Interim Report IR-03-062(2003). Evolutionary Ecology Research 5:363-382 (2003).
No. 75 Dercole F: Remarks on Branching-Extinction Evolu-tionary Cycles. IIASA Interim Report IR-03-077 (2003).Journal of Mathematical Biology 47:569-580 (2003).
No. 76 Hofbauer J, Sigmund K: Evolutionary Game Dynam-ics. IIASA Interim Report IR-03-078 (2003). Bulletin of theAmerican Mathematical Society 40:479-519 (2003).
No. 77 Ernande B, Dieckmann U, Heino M: AdaptiveChanges in Harvested Populations: Plasticity and Evolutionof Age and Size at Maturation. IIASA Interim Report IR-03-058 (2003). Proceedings of the Royal Society of LondonSeries B-Biological Sciences 271:415-423 (2004).
No. 78 Hanski I, Heino M: Metapopulation-Level Adaptationof Insect Host Plant Preference and Extinction-ColonizationDynamics in Heterogeneous Landscapes. IIASA InterimReport IR-03-028 (2003). Theoretical Population Biology63:309-338 (2003).
No. 79 van Doorn G, Dieckmann U, Weissing FJ: SympatricSpeciation by Sexual Selection: A Critical Re-Evaluation.IIASA Interim Report IR-04-003 (2004). American Natu-ralist 163:709-725 (2004).
No. 80 Egas M, Dieckmann U, Sabelis MW: Evolution Re-stricts the Coexistence of Specialists and Generalists - theRole of Trade-off Structure. IIASA Interim Report IR-04-004(2004). American Naturalist 163:518-531 (2004).

No. 81 Ernande B, Dieckmann U: The Evolution of Pheno-typic Plasticity in Spatially StructuredEnvironments: Implica-tions of Intraspecific Competition, Plasticity Costs, and Envi-ronmental Characteristics. IIASA Interim Report IR-04-006(2004). Journal of Evolutionary Biology 17:613-628 (2004).
No. 82 Cressman R, Hofbauer J: Measure Dynamics on aOne-Dimensional Continuous Trait Space: Theoretical Foun-dations for Adaptive Dynamics. IIASA Interim Report IR-04-016 (2004).
No. 83 Cressman R: Dynamic Stability of the ReplicatorEquation with Continuous Strategy Space. IIASA InterimReport IR-04-017 (2004).
No. 84 Ravigné V, Olivieri I, Dieckmann U: Implications ofHabitat Choice for Protected Polymorphisms. IIASA InterimReport IR-04-005 (2004). Evolutionary Ecology Research6:125-145 (2004).
No. 85 Nowak MA, Sigmund K: Evolutionary Dynamics ofBiological Games. IIASA Interim Report IR-04-013 (2004).Science 303:793-799 (2004).
No. 86 Vukics A, Asbóth J, Meszéna G: Speciation in Mul-tidimensional Evolutionary Space. IIASA Interim ReportIR-04-028 (2004). Physical Review 68:041-903 (2003).
No. 87 de Mazancourt C, Dieckmann U: Trade-off Geome-tries and Frequency-dependent Selection. IIASA Interim Re-port IR-04-039 (2004). American Naturalist 164:765-778(2004).
No. 88 Cadet CR, Metz JAJ, Klinkhamer PGL: Size and theNot-So-Single Sex: disentangling the effects of size on sex al-location. IIASA Interim Report IR-04-084 (2004). Ameri-can Naturalist 164:779-792 (2004).
No. 89 Rueffler C, van Dooren TJM, Metz JAJ: AdaptiveWalks on Changing Landscapes: Levins’ Approach Extended.IIASA Interim Report IR-04-083 (2004). Theoretical Popu-lation Biology 65:165-178 (2004).
No. 90 de Mazancourt C, Loreau M, Dieckmann U: Under-standing Mutualism When There is Adaptation to the Partner.IIASA Interim Report IR-05-016 (2005). Journal of Ecology93:305-314 (2005).
No. 91 Dieckmann U, Doebeli M: Pluralism in EvolutionaryTheory. IIASA Interim Report IR-05-017 (2005). Journal ofEvolutionary Biology 18:1209-1213 (2005).
No. 92 Doebeli M, Dieckmann U, Metz JAJ, Tautz D: WhatWe Have Also Learned: Adaptive Speciation is TheoreticallyPlausible. IIASA Interim Report IR-05-018 (2005). Evolu-tion 59:691-695 (2005).
No. 93 Egas M, Sabelis MW, Dieckmann U: Evolution ofSpecialization and Ecological Character Displacement ofHerbivoresAlong a Gradient of Plant Quality. IIASA InterimReport IR-05-019 (2005). Evolution 59:507-520 (2005).
No. 94 Le Galliard J, Ferrière R, Dieckmann U: AdaptiveEvolution of Social Traits: Origin, Trajectories, and Corre-lations of Altruism and Mobility. IIASA Interim Report IR-05-020 (2005). American Naturalist 165:206-224 (2005).
No. 95 Doebeli M, Dieckmann U: Adaptive Dynamics asa Mathematical Tool for Studying the Ecology of SpeciationProcesses. IIASA Interim Report IR-05-022 (2005). Journalof Evolutionary Biology 18:1194-1200 (2005).
No. 96 Brandt H, Sigmund K: The Logic of Reprobation: As-sessment and Action Rules for Indirect Reciprocity. IIASAInterim Report IR-04-085 (2004). Journal of Theoretical Bi-ology 231:475-486 (2004).
No. 97 Hauert C, Haiden N, Sigmund K: The Dynamics ofPublic Goods. IIASA Interim Report IR-04-086 (2004). Dis-crete and Continuous Dynamical Systems - Series B 4:575-587 (2004).
No. 98 Meszéna G, Gyllenberg M, Jacobs FJA, Metz JAJ:Link Between Population Dynamics and Dynamics of Dar-winian Evolution. IIASA Interim Report IR-05-026 (2005).Physical Review Letters 95:Article 078105 (2005).
No. 99 Meszéna G: Adaptive Dynamics: The Continuity Ar-gument. IIASA Interim Report IR-05-032 (2005).
No. 100 Brännström NA, Dieckmann U: Evolutionary Dy-namics of Altruism and Cheating Among Social Amoebas.IIASA Interim Report IR-05-039 (2005). Proceedings of theRoyal Society London Series B 272:1609-1616 (2005).
No. 101 Meszéna G, Gyllenberg M, Pasztor L, Metz JAJ:Competitive Exclusion and Limiting Similarity: A UnifiedTheory. IIASA Interim Report IR-05-040 (2005).
No. 102 Szabo P, Meszéna G: Limiting Similarity Revisited.IIASA Interim Report IR-05-050 (2005).
No. 103 Krakauer DC, Sasaki A: The Greater than Two-FoldCost of Integration for Retroviruses. IIASA Interim ReportIR-05-069 (2005).
No. 104 Metz JAJ: Eight Personal Rules for Doing Science.IIASA Interim Report IR-05-073 (2005). Journal of Evolu-tionary Biology 18:1178-1181 (2005).
No. 105 Beltman JB, Metz JAJ: Speciation: More LikelyThrough a Genetic or Through a Learned Habitat Preference?IIASA Interim Report IR-05-072 (2005). Proceedings of theRoyal Society of London Series B 272:1455-1463 (2005).
No. 106 Durinx M, Metz JAJ: Multi-type Branching Pro-cesses and Adaptive Dynamics of Structured Populations.IIASA Interim Report IR-05-074 (2005). Haccou P, JagerP, Vatutin V (eds): Branching Processes: Variation, Growthand Extinction of Populations, Cambridge University Press,Cambridge, UK, pp. 266-278 (2005).
No. 107 Brandt H, Sigmund K: The Good, the Bad andthe Discriminator - Errors in Direct and Indirect Reciprocity.IIASA Interim Report IR-05-070 (2005). Journal of Theoret-ical Biology 239:183-194 (2006).
No. 108 Brandt H, Hauert C, Sigmund K: Punishing and Ab-staining for Public Goods. IIASA Interim Report IR-05-071(2005). Proceedings of the National Academy of Sciences ofthe United States of America 103:495-497 (2006).
No. 109 Ohtsuki A, Sasaki A: Epidemiology and Disease-Control Under Gene-for-Gene Plant-Pathogen Interaction.IIASA Interim Report IR-05-068 (2005).
No. 110 Brandt H, Sigmund K: Indirect Reciprocity, Image-Scoring, and Moral Hazard. IIASA Interim Report IR-05-078 (2005). Proceedings of the National Academy of Sci-ences of the United States of America 102:2666-2670 (2005).
No. 111 Nowak MA, Sigmund K: Evolution of Indirect Reci-procity. IIASA Interim Report IR-05-079 (2005). Nature437:1292-1298 (2005).
No. 112 Kamo M, Sasaki A: Evolution Towards Multi-YearPeriodicity in Epidemics. IIASA Interim Report IR-05-080(2005). Ecology Letters 8:378-385 (2005).

No. 113 Dercole F, Ferrière R, Gragnani A, Rinaldi S: Co-evolution of Slow-fast Populations: EvolutionarySliding, Evo-lutionoary Pseudo-equilibria, and Complex Red Queen Dy-namics. IIASA Interim Report IR-06-006 (2006). Proceed-ings of the Royal Society B-Biological Sciences 273:983-990(2006).
No. 114 Dercole F: Border Collision Bifurcations in the Evo-lution of Mutualistic Interactions. IIASA Interim ReportIR-05-083 (2005). International Journal of Bifurcation and
Chaos 15:2179-2190 (2005).
No. 115 Taborsky B: The influence of juvenile and adult en-vironments on life-history trajectories. IIASA Interim Re-port IR-06-033 (2006). Proceedings of the Royal SocietyB-Biological Sciences 273 :741-750 (2006).
No. 116 Taborsky B: Mothers determine offspring size in re-sponse to own juvenile growth conditions. IIASA InterimReport IR-06-034 (2006).
Issues of the IIASA Studies in Adaptive Dynamics series can be obtained at www.iiasa.ac.at/Research/EEP/Series.html or bywriting to [email protected].

Contents
Summary........................................................................................................................... 2
Introduction ...................................................................................................................... 2
Materials and Methods ..................................................................................................... 3
Results .............................................................................................................................. 5
Discussion......................................................................................................................... 6
Acknowledgements .......................................................................................................... 8
References ........................................................................................................................ 8
Figures ............................................................................................................................ 10
Electronic Appendix A .................................................................................................... 1
Study species ................................................................................................................ 1
Expermental fish and housing ...................................................................................... 1
Transition between juvenile and adult treatment.......................................................... 1
Food source for young during adult treatment ............................................................. 2
Field survey .................................................................................................................. 2
References .................................................................................................................... 3

Mothers determine offspring size in response to own juvenile growth
conditions
Barbara Taborsky1,2
1Behavioural Ecology, University of Bern, Wohlenstrasse 50A. CH-3032 Hinterkappelen,
Switzerland,
2EEP, International Institute for Applied Systems Analysis (IIASA), A-2361 Laxenburg, Austria
Running title: Juvenile growth of mothers determines offspring size
Correspondence:
Behavioural Ecology, University of Bern, Wohlenstrasse 50A, CH-3032 Hinterkappelen, Switzerland e-mail: [email protected]

2
Through non-genetic maternal effects, mothers can tailor offspring phenotype to the
environment in which young will grow up. If juvenile and adult ecologies differ, the
conditions mothers had experienced as juveniles may better predict their offspring’s
environment than the adult environment of mothers. In this case maternal decisions about
investment in offspring quality should be determined already during the juvenile phase of
mothers. I tested this hypothesis by manipulating juvenile and adult maternal
environments independently in a cichlid fish. Females raised in a poor environment
produced larger young than females raised without food limitations, irrespective of the
feeding conditions experienced during adulthood. This maternal boost was due to a higher
investment in eggs, and to faster larval growth. Apparently mothers prepare their
offspring for similar environmental conditions as they had encountered as juveniles. This
explanation is supported by the distribution of these fish under natural conditions.
Juveniles live in a different and much narrower range of habitats than adults. Therefore,
the habitat mothers experienced as juveniles will allow them to predict their offspring's
environment better than the conditions in the adult home range.
INTRODUCTION
Non-genetic maternal effects are widespread and strongly influence the fitness of mothers and
offspring (reviewed in Mousseau & Fox 1998, Lindström 1999, Lummaa & Clutton-Brock
2002). Egg size (Bernardo 1996, Mousseau & Fox 1998) and quality (e.g. Schwabl et al. 1997,
Blount et al. 2002) are important maternal effects, which can determine the entire life histories of
offspring (Lindström 1999). Maternal effects may depend on maternal condition (e.g. Blount et
al. 2002) and on the conditions in the current environment (Mousseau & Fox 1998, Lindström
1999) or during preceding breeding events (Reznick & Yang 1993).

3
While this appears to be adequate in some cases, in many animals successive life stages use
different habitats (reviewed in Werner & Gilliam 1984) or they use resources differently within
the same habitat (e.g. Lind & Welsh 1994). If juvenile and adult ecologies are uncorrelated, cues
from the current conditions are a poor predictor of the environment offspring are likely to
encounter (Bernardo 1996). In such cases the conditions a female encountered as juvenile may
allow her to predict her offspring's environment with a much higher precision. Consequently,
maternal investment decisions should be determined by a mother's early development (sensu
Lindström 1999), but should be independent of the environmental conditions in which young are
produced.
I tested this hypothesis experimentally with the African cichlid Simochromis pleurospilus by
independently varying the resource availability of juveniles and adult females. In the field, I
tested the prediction that the habitat use of adult females and juveniles differs, so that the
environmental conditions experienced these stages are rather independent of each other.
MATERIALS AND METHODS
I raised 120 fish (among them 55 females) of the maternally mouthbrooding cichlid Simochromis
pleurospilus, each in a separate tank, exposing aliquot numbers to high-food (H) and low-food (L)
conditions (juvenile treatment, J) (see electronic Appendix A and Taborsky (2005) for details on
experimental set-up). Fish were fed six days a week standardized agarose gel cubes containing an amount
of Tetramin® flake food corresponding to 12% or 4% of mean body weight plus 5% Spirulina algae. I
adjusted the food amounts to increasing body weight every 14 days. The juvenile treatment took on
average 265 days and covered the period between independence of young (mean age 29 days) and the end
of the first breeding attempt. Afterwards, half of all reproductively active females (n=23) were switched
to the opposite treatment, while the remaining 23 females stayed on the original ration (adult treatment,
A). Siblings were assigned alternately to high or low food ration as juveniles, and to switched or original
ration during the adult treatment to achieve equal brood splitting. The resulting treatment groups were

4
denoted as HH (n=13), HL (n=10), LH (n=11) and LL (n=12). For each successive brood a female was
mated with a different male (Taborsky (2005) gives details about schedules of male presence in female
tanks). About 50% of females never raised a brood successfully despite spawning several clutches.
Females remained in the adult treatment until they stopped to produce further clutches (mean
reproductive lifespan in experiment: 264.5d; Taborsky 2005).
Brood care of S. pleurospilus consists of two phases, two weeks of continuous incubation when young
use up yolk reserves ('1st incubation phase') and two weeks when young are periodically released from the
mouth for feeding ('2nd incubation phase'). Total length (TL; nearest 0.5mm) and weight (W; nearest
0.01g) of females and young (TL: nearest 0.1mm, W: nearest 0.0001g) and weight of females and young
were measured at the end of both incubation phases. I calculated Fulton's condition factor K as
K=100×W/TL3 , and specific growth rates of young (SGR) as ln(TL2/TL1)/t, where TL1, TL2 are lengths at
the beginning and end of the second incubation phase and t is the duration of this phase. As SGR depends
on absolute size, it was corrected for TL1. The experiment lasted from November 2001 to May 2004. One
clutch each was collected of all females that spawned during the last six months of this period (3HH,
4HL, 4LH, and 4LL females) to measure egg mass. Clutches could not be collected earlier, as this would
have interfered with the measurement of reproductive rates of females (see Taborsky 2005). Mean clutch
mass was calculated from individual egg dry mass measured to the nearest 0.0001g.
I calculated two-way analyses of variance with juvenile treatment (J) and adult treatment (A) as
independent factors and individual females as independent units of analysis (using female means of clutch
means). Female size and body condition was not related to size or weight of young after the 1st and 2nd
incubation phase, and was therefore not included as a covariate in the ANOVAs.
I determined the size-frequency distribution of juveniles and females across the natural depth range of S.
pleurospilus by transect counts along a 150-m stretch of pebble shore in Mbete Bay near Mpulungu,
Zambia (see electronic Appendix A for details on survey methods). The mean distributions of four study
years are presented here. In the lab experiment, females started to reproduce at a mean size of 5.7cm TL

5
(Taborsky 2005). Therefore, I considered individuals in size classes of ≤ 4.5-5.4cm as 'juveniles' and
individuals above 5.5cm as 'adults'.
RESULTS
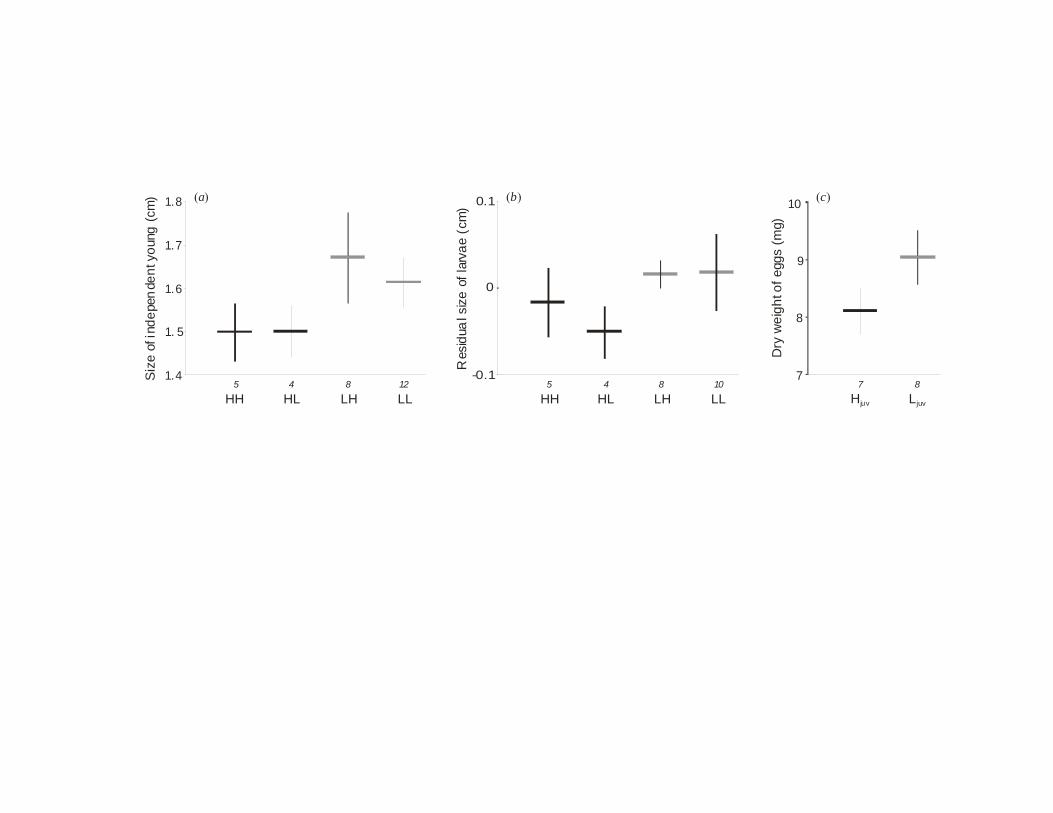
When mothers had been raised with little food, offspring were on average 1.4mm larger at the
end of brood care than when mothers had been well fed, irrespective of the ration mothers
received during adulthood (Figure 1a; Table 1). The larger size of young was not explained by
extended brood care, as the results did not change when controlling for the age of young (Table
1). Rather young of poorly-reared mothers grew faster during the second incubation phase
(independent of their size at the beginning of this phase; Table 1). The condition factor K of
young was not affected by treatment (Table 1), showing that faster growth was not compromised
by a slower increase of body mass.
Young of poorly-raised mothers were already larger for their age after the first incubation phase
(Figure 1b, Table 1), when larvae depend entirely on yolk reserves. Accordingly, females raised
with little food laid eggs with a higher dry mass (ANOVA, F1,13=8.74, p=0.011; Figure 1c). Egg
mass was not related to female length (regression analysis: R2=0.02, p=0.64, d.f.=1,13) or body
condition (R2=0.04, p=0.52, d.f.=1,12).
In their natural habitat, the majority of juveniles (<5.5cm) stayed at a depth of 0.5m and were
rarely found below 1m (median depth: 0.5m, inter quartile range (iqr) 0.5-1; Figure 2a,b). In
contrast, fish of adult size occurred predominantly in deeper water (median: 1.5m, iqr 1.0-3.5;
Figure 2a,b). Juvenile and adult habitat use differed with respect to depth distribution (Chi-
square test, d.f.=5, χ2=36.94, p<0.001; frequencies ≥3m were pooled because of low expected
frequencies in these depths) and central tendency of depth distribution (Median test, d.f.=1,
χ2=11.16, p<0.001).

6
Above 0.5m water depth, only small specimen (1.5–4.4cm were found). As they could not be
counted by transect swimming, these numbers are not included in the results (separate data set of
shallow-water counts given in electronic Appendix A).
DISCUSSION
At the end of broodcare, young of S. pleurospilus mothers raised on a poor diet were on average
8.8% longer than young of mothers raised without food limitation. This size difference is likely
to affect offspring fitness, as even a size advantage of 3.3% can significantly increase the
survival chances of fish larvae (McCormick & Hoey 2004). This is the first experimental
evidence showing that maternal effects induced by pre-reproductive conditions can be similarly
expressed in markedly different reproductive environments, while not being influenced by the
conditions in which females produce their young. Moreover, the reported size differences of
young were based on up to four successful broods spread over female reproductive life span,
suggesting that such maternal effects can persist during the entire adulthood even in iteroparous,
long-lived animals.
Poorly-raised females paid costs for producing larger young by having smaller clutches, while
there was no indication for such a trade-off in females raised on the high-food diet (Taborsky
2005). If females raised on a poor diet had resulted in poorer quality adults than females raised
on a rich diet, the former may have been more affected by the costs of reproduction than the
latter.
S. pleurospilus females raised with little food apparently used two different mechanisms to boost
the size of young. (i) They produced eggs with a higher energy content as indicated by higher
egg dry mass. (ii) Young grew faster during the second incubation phase, when most yolk is used

7
up and they use already external food sources. The mechanism responsible for faster growth
needs still to be clarified.
In several fish, insects and amphibians larger offspring have survival advantages under adverse
growth conditions, while under benign conditions smaller young do equally well (Hutchings
1991, Mousseau & Fox 1998, Einum & Fleming 1999) or better (Kaplan 1992, Rotem et al.
2003). Therefore, the experimental results strongly suggest that S. pleurospilus females prepared
their young for similar environmental conditions as they had encountered themselves as
juveniles. In humans, it has been hypothesized that predicting the quality of the offspring
environment incorrectly may have detrimental consequences for offspring health and survival
(Bateson et al. 2004). The conditions mothers experienced as juveniles may predict their
offspring's environment better than current environmental cues, if (i) environmental conditions
for adults and juveniles vary independently, and (ii) juvenile conditions of successive
generations are similar. When juveniles and adults have entirely different ecologies like in
metamorphosing animals or anadromous fish, some species appear indeed to adjust offspring
phenotype to their juvenile environment (Jonsson et al. 1996, Rotem et al. 2003) while others do
not (Fox et al. 1995).
In contrast, juvenile and adult S. pleurospilus co-occur along the rocky shores of Lake
Tanganyika use the same major food source, filamentous turf algae. However juveniles mainly
live in shallow water using only a narrow range of water depths, while after maturity females
mainly use deeper habitats for breeding (Figure 2a). The juvenile habitat has a high productivity
of turf algae, while productivity varies by at least two orders of magnitude over the depth range
inhabited by adults (Taborsky 1999). Because of this large variation the ability to predict food
availability for offspring reliably is limited if females use cues from their ambient environment.

8
A much better estimate can be achieved if females use the growth conditions experienced during
their own juvenile phase.
As differential habitat use of juveniles and adult is very widespread in animals, parents will often
not be able to predict the conditions for the early life stages of offspring by using cues from their
current environment. Therefore I predict that it is a common parental strategy to base decisions
about the investment in individual offspring more strongly on the own early environment than on
present conditions. When studying the origin and function of parental effects, it is hence
inevitable to incorporate the influence on parents of the environment experienced during early
development.
I am indebted to D. Heg, I. Hamilton and M. Taborsky for helpful comments on the manuscript, to S.
Aschwanden, Z. Bachar, D. Bonfils, R. Eggler C. Grüter, N. Hirt, S. Immler, S. Lehner, M. Massironi, G.
Pachler, T. Reichlin, R. Schürch and P. Vonlanthen for assistance with data collection and analysis and to
the Austrian Science Fund (FWF grant P14327-B06) and the Forschungsstiftung, University of Bern
(48/2003) for financial support.
Bateson,P., Barker,D., Clutton-Brock,T., Deb,D., D'Udine,B., Foley,R.A., Gluckman,P., Godfrey,K., Kirkwood,T., Lahr,M.M., McNamara,J., Metcalfe,N.B., Monaghan,P., Spencer,H.G. & Sultan,S.E. 2004 Developmental plasticity and human health. Nature 430, 419-421.
Bernardo,J. 1996 Maternal effects in animal ecology. Am. Zool. 36, 83-105.
Blount,J.D., Surai,P.F., Nager,R.G., Houston,D.C., Moller,A.P., Trewby,M.L. & Kennedy,M.W. 2002 Carotenoids and egg quality in the lesser black-backed gull Larus fuscus: a supplemental feeding study of maternal effects. Proc. Roy. Soc. Lond. Ser. B 269, 29-36.
Einum,S. & Fleming,I.A. 1999 Maternal effects of egg size in brown trout (Salmo trutta): norms of reaction to environmental quality. Proc. Roy. Soc. Lond. Ser. B 266, 2095-2100.
Fox,C.W., Waddell,K.J. & Mousseau,T.A. 1995 Parental host plant affects offspring life histories in a seed beetle. Ecology 76, 402-411.
Hutchings,J.A. 1991 Fitness consequences of variation in egg size and food abundance in Brook Trout Salvelinus fontinalis. Evolution 45, 1162-1168.

9
Jonsson,N., Jonsson,B. & Fleming,I.A. 1996 Does early growth cause a phenotypically plastic response in egg production of Atlantic Salmon? Func. Ecol. 10, 89-96.
Kaplan,R.H. 1992 Greater maternal investment can decrease offspring survival in the frog Bombina orientalis. Ecology 73, 280-288.
Lind,A.J. & Welsh,H.H. 1994 Ontogenetic changes in foraging behaviour and habitat use by the Oregon garter snake Thamnophis atratus hydrophilus. Anim. Behav. 48, 1261-1273.
Lindström,J. 1999 Early development and fitness in birds and mammals. Trends Ecol. Evol. 14, 343-348.
Lummaa,V. & Clutton-Brock,T. 2002 Early development, survival and reproduction in humans. Trends Ecol. Evol. 17, 141-147.
McCormick,M.I. & Hoey,A.S. 2004 Larval growth history determines juvenile growth and survival in a tropical marine fish. Oikos 106, 225-242.
Mousseau,T.A. & Fox,C.W. 1998 The adaptive significance of maternal effects. Trends Ecol. Evol. 13, 403-407.
Reznick,D. & Yang,A.P. 1993 The influence of fluctuating resources on life-history patterns of allocation and plasticity in female guppies. Ecology 74, 2011-2019.
Rotem,K., Agrawal,A.A. & Kott,L. 2003 Parental effects in Pieris rapae in response to variation in food quality: adaptive plasticity across generations? Ecol. Entomol. 28, 211-218.
Schwabl,H., Mock,D.W. & Gieg,J.A. 1997 A hormonal mechanism for parental favouritism. Nature 386, 231.
Taborsky,B. 1999 Size-dependent distribution in littoral fish: optimization or competitive exclusion? In: Behaviour and Conservation of Littoral Fishes (Ed. by V.C. Almada, R.F. Oliveira & E.J. Goncalves), pp. 351-376. Lisboa, ISPA.
Taborsky,B. 2005 The influence of juvenile and adult environments on life history trajectories. Proc. Royal. Soc. Lond B, doi:10.1098/rspb.2005.3347
Werner,E.E. & Gilliam,J.F. 1984 The ontogenetic niche and species interactions in size structured populations. Ann. Rev. Ecol. Syst. 15, 393-425

10
Figures
Figure 1. Total length of offspring at the end of (a) 2nd incubation phase, (b) 1st incubation phase
(residuals of the model with offspring age as covariate are shown) and (c) egg dry mass of
females (means ±s.e. of brood means).
Figure 2. (a) Size-depth distribution of S. pleurospilus (mean of four study seasons); grey shades
indicate frequencies of fish recorded for each size class and depth. (b) Medians and quartiles for
the depth distributions of 1-cm size classes. Stippled lines indicate size at maturity.
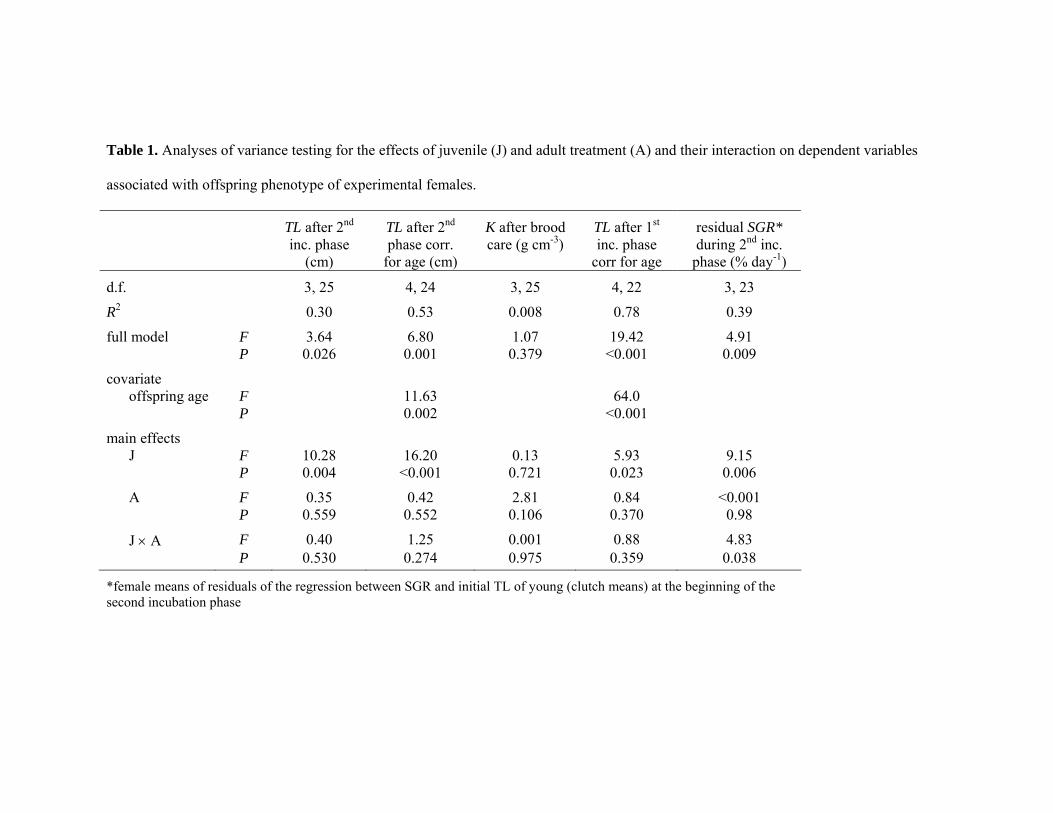
Table 1. Analyses of variance testing for the effects of juvenile (J) and adult treatment (A) and their interaction on dependent variables
associated with offspring phenotype of experimental females.
TL after 2nd inc. phase
(cm)
TL after 2nd phase corr.
for age (cm)
K after brood care (g cm-3)
TL after 1st inc. phase
corr for age
residual SGR* during 2nd inc.
phase (% day-1)
d.f. 3, 25 4, 24 3, 25 4, 22 3, 23
R2 0.30 0.53 0.008 0.78 0.39
full model F 3.64 6.80 1.07 19.42 4.91 P 0.026 0.001 0.379 <0.001 0.009
covariate offspring age F 11.63 64.0
P 0.002 <0.001
main effects J F 10.28 16.20 0.13 5.93 9.15
P 0.004 <0.001 0.721 0.023 0.006
A F 0.35 0.42 2.81 0.84 <0.001 P 0.559 0.552 0.106 0.370 0.98
J × A F 0.40 1.25 0.001 0.88 4.83 P 0.530 0.274 0.975 0.359 0.038
*female means of residuals of the regression between SGR and initial TL of young (clutch means) at the beginning of the second incubation phase
LjuvHjuv
7
8
9
10
Dry
wei
ght o
f egg
s (m
g)
( )c
7 8
LLLHHLHH
1. 5
Size
of i
ndep
ende
nt y
oung
(cm
) 1.8
1.7
1.6
1.4
( )a
5 4 8 12
Res
idua
l size
of l
arva
e (c
m) 0.1
0
-0.1
( )b
LLLHHLHH5 4 8 10

Size class (cm)
Size class (cm)
3
30
4
4
5
5
6
6
7
7
8
8
9
9
10
10
11
11
Dep
th (m
)D
epth
(m)
1
2
2
3
4
4
5
6
6
0.5 1.0 1.5 2.0 2.5 3.0
( )b
( )a

1
Electronic Appendix A
Mothers determine offspring size in response to own juvenile growth conditions
Barbara Taborsky1,2
1Behavioural Ecology, University of Bern, Wohlenstrasse 50A. CH-3032 Hinterkappelen, Switzerland,
2ADN, International Institute for Applied Systems Analysis (IIASA), A-2361 Laxenburg, Austria
Study species
In Simochromis pleurospilus Nelissen (Subfamily: Tropheini), a mouthbrooding cichlid endemic to Lake
Tanganyika, females produce clutches year-round and care for the young alone. Brood care consists of two
phases; two weeks of continuous incubation when young only use yolk reserves are followed by another one
to two weeks when young are periodically released from the mouth for feeding on the surrounding substrate.
Afterwards, young are independent.
Experimental fish and housing
The experimental fish were recruited from 14 broods, bred from closely related parents (siblings and half-
siblings) to minimize genetic variability. I raised 120 young S. pleurospilus, each in a separate 20-litre tank.
Each 20-litre tank contained an internal biological filter and a clay flower pot half (10 cm diameter), both
used as shelters by the fish, and a layer of river sand. Water temperature was kept at 27°C by controlled
heating of the room temperature. The light:dark regime was set to 13h:11h including 10 min periods of
dimmed light in the mornings and evenings to simulate natural light conditions at Lake Tanganyika. The
sexes of the experimental fish could only be determined from an age of 5 to 6 month, when there were 55
females in the experimental population. Of these, 46 females (23 high-food and 23 low-food females)
produced clutches of eggs during the experiment. These females were used in both the juvenile and the adult
treatment phases. Of the 46 females, 23 females produced 'successful broods', i.e. they incubated and raised
their young. The probability of successful incubation was not influenced by the food treatment (Taborksy
2005), and it is unknown why the remaining females always abandoned or swallowed their eggs.
Transition between juvenile and adult treatment
I switched the females from the juvenile to the adult treatment after their first breeding attempt. Most first-
spawning females (37 of 46) interrupted incubation after 3.1 days (mean± 0.29 SE) and swallowed the eggs, a
phenomenon generally observed in mouthbrooding cichlids. Only 9 females raised their first clutch. I wanted
to ensure that all females were in the same developmental stage at the switch. Therefore, the day after the
first breeding attempt was chosen as starting point for the adult treatment as first spawning is the first

2
unequivocal sign of maturity in these fish. Hence, the juvenile treatment period covered the entire phase of
'early development' (the time from birth to developmental maturity, Lindström 1999). Additionally it
included a short period of ovary maturation before first spawning (2-3 weeks in Tropheini, Yanagisawa &
Nishida 1991, B. Taborsky unpub. data, i.e., 6-7% of the juvenile treatment period), and in 9 females also
incubation.
Of the 9 females that did raise the first clutch, six were switched to the opposite food level after
incubation, according to the experimental protocol. As the first young of these 6 females were produced still
under the previous (juvenile) food conditions, data of these clutches were analysed together with data of the
respective non-switched groups whenever all four treatment groups were compared.
The majority (83%) of the 54 successful broods (produced by 23 females, mean: 1.9 broods ±0.18 SE
per female, range 1-4), were produced during the adult treatment. Therefore, the timing of the treatment
switch closely matched the divide between juvenile and adult stages of the experimental females.
Food source for young during adult treatment
Overall, clutch size increased with female size, while the latter was influenced by the food treatment
(Taborsky 2005). In principle, it would be possible that the differences of growth rates during the second
incubation phase have been caused by differential levels of density-dependent competition for the food
rations provided daily. However, this possibility is unlikely because (i) larvae fed mainly on detritus and
algae, which were plentiful in all tanks, and were thereby largely independent of the daily food rations and
(ii) growth rates did not correlate with clutch size within treatments (except for the LH group, r=-0.55,
P=0.034, n=15 individual broods).
Field survey
Data were collected along a 150m stretch of coastline in Mbete Bay at the southern tip of Lake Tanganyika
about 1 km west of Mpulungu, Zambia. Data include counts of four study periods (25 Dec 96 to 26 Jan 97, 28
Jan to 2 Mar 98, 1 Mar to 10 Mar 2002, 1 Mar 20 Mar 2003).
Before starting with the actual data collection each observer did several training trials allow for a direct
comparison of count data collected by different observers. During these training trials, the observer first
estimated the size of a randomly chosen focal individual, then he caught it and measured its total length to
verify his estimate. At least 5 successive estimates had to be done correctly before an observer started to
collect the data. This condition was usually fulfilled after very few training trials.
When conducting a transect count, the observer swam in parallel to the shoreline in constant water
depth either by snorkelling or by SCUBA diving. Transects were conducted at each half depth meter between
0.5 and 6 m depth (12 transects per study period). Every 10 m along a transect the observer stopped,
remained motionless for 2 min, counted all S. pleurospilus by once scanning the substrate and the water

3
column in a half-circle with a radius of 2 m. He then noted the size of each individual, which was estimated
to the nearest 1.0 cm resulting size classes from 2.5-3.4 cm to 10.5-11.4 cm for females and juveniles. Male
counts are not presented in this paper, as only the habitat use of females and juveniles is relevant for the focus
of this study. Adult males were easily distinguished from juveniles and females by their bright colour pattern.
For further analysis the counts of all 15 positions of a transect were summed. Whenever a transect was
repeated (all transects of 2002 and some transects in 1998), the counts of the repeated transects were
averaged by taking the arithmetic mean.
The total numbers of juvenile fish detected during the transect counts was relatively low compared to
the number of adults. This is because a large number of juveniles occurred above 0.5 m of depth, where the
counting method described above is not feasible. To account for this problem, two additional counts were
done in 2002 by walking on land along 100 m of shoreline in front of the study site (shore transects). For
these transects fish were counted every 5 m on an area of a half-circle with 1 m radius. An additional size
class (1.5-2.4 cm) was found during these counts. The numbers of fish in count 1 and 2, respectively, were 27
and 22 in size class 1.5 to 2.4 cm, 16 and 25 in class 2.5-3.4 cm and 7 and 9 in class 3.5-4.4 cm. No S.
pleurospilus larger than 4.4 cm were found during these shore transects. As the counting method used in the
shore transects is not directly comparable with the transects done by swimming, the results of these two
additional transects were not included in the data presented in the 'Results' section.
References
Lindström, J. (1999). Early development and fitness in birds and mammals. Trends Ecol. Evol. 14, 343-348. Taborsky, B. (2005). The influence of juvenile and adult environments on life history trajectories. Proc.
Royal. Soc. Lond B, doi: 10.1098/rspb.2005.3347 Yanagisawa, Y. & Nishida, M. (1991). The social and mating system of the maternal mouthbrooder Tropheus
moorii (Cichlidae) in Lake Tanganyika. Jap. J. Ichthyol. 38, 271-282.