interleukin 17, a t-cell-derived cytokine, promotes tumorigenicity
TRANSCRIPT

[CANCER RESEARCH 59, 3698–3704, August 1, 1999]
Interleukin 17, a T-cell-derived Cytokine, Promotes Tumorigenicity of HumanCervical Tumors in Nude Mice1
Eric Tartour, 2 Francois Fossiez, Isabelle Joyeux, Annie Galinha, Alain Gey, Emmanuel Claret, Xavier Sastre-Garau,Jerome Couturier, Veronique Mosseri, Virginie Vives, Jacques Banchereau, Wolf H. Fridman, John Wijdenes,Serge Lebecque, and Catherine Sautes-FridmanUnite d’Immunologie Clinique, INSERM U 225 and Universite Pierre et Marie Curie [E. T., I. J., A. Ga., A. Ge., V. V., W. H. F., C. S-F.], Departement de Pathologie [X. S-G.,J. C.], and Unite de Biostatistique [V. M.], Institut Curie, 75248 Paris, Cedex 05, France; Schering Plough, 69571 Dardilly, France [F. F., J. B., S. L.]; and Diaclone, BP1985,F25020 Besancon, France [E. C., J. W.]
ABSTRACT
Interleukin (IL) 17 is a proinflammatory cytokine secreted mainly byactivated human memory CD4 T cells that induces IL-6, IL-8, and nitricoxide. Because IL-6 and IL-8 have been implicated in the pathogenesis ofcervical cancer, we investigated the action of IL-17 on human cervicaltumor cell lines in vitro and in vivo. We showed that in vitro, IL-17increases IL-6 and IL-8 secretion by cervical carcinoma cell lines at bothprotein and mRNA levels. No direct effect of IL-17 onin vitro proliferationof cervical tumor cell lines could be demonstrated. However, two cervicalcell lines transfected with a cDNA encoding IL-17 exhibited a significantincrease in tumor size as compared to the parent tumor when trans-planted in nude mice. This enhanced tumor growth elicited by IL-17 wasassociated with increased expression of IL-6 and macrophage recruitmentat the tumor site. A potential role of IL-17 in modulation of the humancervical tumor phenotype was also supported by its expression on thecervical tumor in patients with CD4 infiltration. IL-17 therefore behaveslike a T-cell-specific cytokine with paradoxical tumor-promoting activity.This may partially explain previous reports concerning the deleteriouseffect of CD4 T cells in cancer.
INTRODUCTION
IL-17,3 previously termed CTL-associated antigen 8 (CTLA-8), isa Mr 17,000 cytokine expressed mainly by activated human memoryCD4 T cells (1–3). Its amino acid sequence shares remarkable ho-mology with that of the thirteenth open reading frame ofherpesvirussaimiri, a g herpes virus causing T-cell lymphoma in monkeys andrabbits (4).In vitro, this virus could immortalize human CD4 and CD8T cells via an unknown mechanism that confers an IL-2-based auto-crine growth after activation by CD2 upon these cells (5, 6). The roleof IL-17 in the proliferation of T cells is a matter of discussion. Inmice, IL-17 plus suboptimal concentrations of phytohemagglutininresulted in a modest enhancement of T-cell growth (7). No similargrowth factor activity was observed in the human system (2). IL-17 isconsidered to be a proinflammatory cytokine because it has beenshown to increase the production of IL-6 and IL-8 in macrophages,fibroblasts, keratinocytes, and synovial cells (2, 7–10) and of nitricoxide in human osteoarthritis cartilage via activation of mitogen-activated protein kinase and nuclear factorkB (11, 12). The release ofIL-10, IL-12, IL-1R antagonist, and stromelysin was also stimulatedby rhIL-17 in human activated macrophages (8).
Interestingly,in vitro, IL-6 behaves as a growth factor for manytumor cell lines derived from myeloma, lymphoma, Kaposi’s sar-coma, melanoma, and ovarian and renal or bladder cell carcinoma(13). In mice, IL-6-transfected tumors often exhibited increased tu-morigenicity (14–16). In myeloma patients, anti-IL-6 mAb adminis-tration transiently inhibited myeloma cell proliferation (17). IL-8, amember of the CXC family of chemotactic cytokines, also stimulatesthe proliferation of tumor cells because IL-8 is an autocrine growthfactor for human melanoma (18). Its expression by tumor cells isdirectly correlated with their metastatic potential in nude mice (19).Therefore, if the activity of IL-17 is mediated via these proinflamma-tory cytokines, a potential role for IL-17 in tumor cell proliferationmay be hypothesized.
Cervical cancer is associated with HPV infection, but additionalfactors must contribute to its pathogenesis because only a minority ofHPV infections result in persistent lesions or progress to malignancy(20). Various arguments suggest that IL-6 may be involved in thepathogenesis and development of cervical cancers. IL-6 stimulates thegrowth of both normal cervical cells and HPV-immortalized andcervical carcinoma-derived cell lines (21, 22).In vitro, cervical car-cinoma cells also secrete higher levels of IL-6 and IL-8 than HPV-infected and normal cervical epithelial cells (23). Finally, an increasedexpression of IL-6 mRNA was demonstrated in biopsies derived frominvasive cervical carcinoma compared to biopsies derived from cer-vical intraepithelial neoplasia or normal cervix (24). The role of IL-17in the up-regulation of IL-6 and IL-8 expression therefore promptedus to investigate the action of this cytokine on thein vitro andin vivoproliferation of human cervical tumors.
MATERIALS AND METHODS
Tumor Cell Lines. The HeLa cervical tumor cell line was obtained fromthe American Type Culture Collection (Manassas, VA), and the IC1 cervicalcarcinoma cell line was provided by Dr J. Couturier (Institut Curie, Paris,France; Ref. 25). The human melanoma cell lines WM793, 1341D, MZ2, andHT144 have been described previously (26). These cell lines were grown inRPMI 1640 (Biowhittaker, Walkersville, MD) supplemented with 10% FCS,100 units/ml penicillin-streptomycin, 5% sodium pyruvate, and 0.01% mer-captoethanol (all from Sigma Chemical Co., St. Louis, MO).
Transfection of HeLa and IC1 Cells. A cDNA encoding hIL-17 wasinserted into an expression vector under the control of the SRa promoter inpBR322 into which a neomycin resistance gene was introduced (NT-Neo; Ref.27). NT-Neo containing or lacking the 640-bp IL-17 cDNA was linearizedwith ScaI restriction enzyme and stably transfected by electroporation intoHeLa and IC1 cells. Electroporation was performed with a Bio-Rad GenePulser at a voltage of 260 V with a capacitance of 960mF. At 72 h afterelectroporation, transfectants were selected by culture in RPMI 1640 (LifeTechnologies, Inc.) supplemented with 1 mg/ml G418 (Geneticin; Life Tech-nologies, Inc., Paisley, United Kingdom). G418-resistant clones were ex-panded in selection medium and tested for IL-17 expression.
Recombinant Proteins and Antibodies.rhIL-17 was purified from thesupernatant of IL-17-transfected NSO cells, as described previously (2)
Neutralizing anti-IL-17 mAb 5 as well as anti-IL-17 mAbs 16 and 25 wereproduced in ascites and purified by anion-exchange chromatography (2).
Received 2/18/99; accepted 6/3/99.The costs of publication of this article were defrayed in part by the payment of page
charges. This article must therefore be hereby markedadvertisementin accordance with18 U.S.C. Section 1734 solely to indicate this fact.
1 Supported by grants from Comite de Paris de la Ligue Nationale de Lutte contre leCancer (France) and the Indo-French Center for the Promotion of Advanced Research andby the Institut Curie and the Institut National de la Sante et de la Recherche Medicale.
2 To whom requests for reprints should be addressed, at Unite d’Immunologie Clin-ique, Institut Curie, 26 Rue d’Ulm, 75248 Paris, Cedex 05, France. Phone: 33-1-44-32-42-18. Fax: 33-1-40-51-04-20; E-mail: [email protected].
3 The abbreviations used are: IL, interleukin; rhIL, recombinant human IL; HPV,human papillomavirus; hIL, human IL; mAb, monoclonal antibody; RT-PCR, reversetranscription-PCR; MTT, 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide;TBS, Tris-buffered saline; MCP-1, monocyte chemoattractant protein 1.
3698
Research. on November 22, 2018. © 1999 American Association for Cancercancerres.aacrjournals.org Downloaded from
Assays.hIL-6 and hIL-8 were assayed using ELISA kits purchased fromImmunotech (Marseille, France) and Medgenix (Brussels, Belgium), respec-tively.
An ELISA was used to measure hIL-17 concentrations in culture superna-tants. Briefly, 96 break-away, flat-bottomed-well (Nunc) microtiter plates werecoated with 50ml of anti-IL-17 mAb 25 (10mg/ml) diluted in carbonate buffer[0.1 M Na2CO3/NaHCO3 (pH 9.6)] overnight at 4°C. The plate was thensaturated with 200ml of PBS-1% BSA for 1 h at room temperature. Afterwashes with PBS-0.05% Tween 20 (Merck, Schuchardt, Germany), 50ml ofrhIL-17 or samples diluted in PBS-1% BSA were added and incubated for 3 hat 37°C. After washes, the plates were incubated with 50ml of peroxidase-labeled anti-IL-17 mAb 16 (5mg/ml) diluted in PBS-1% BSA for 2 h at 37°C.The reaction was revealed by the addition of the peroxidase substrate(o-phenylenediaminedihydrochloride), and the optical density was read at awavelength of 492 nm. The lowest concentration of hIL-17 detected was 0.05ng/ml.
RT-PCR Amplification. RT-PCR was performed as described previously(28). The following oligonucleotide primers were used: (a) humanb-actinsense, TCGTCGACAACGGCTCCGGCATGTGC; (b) humanb-actin anti-sense, TTCTGCAGGGAGGAGCTGGAAGCAGC; (c) hIL-6 sense, AC-GAATTCACAAACAAATTCGGTACA; (d) hIL-6 antisense, CATCTAGAT-TCTTTGCCTTTTTCTGC; (e) hIL-8 sense, TTCTGCAGCTCTGTGTGA-AGG; (f) hIL-8 antisense, GAAGAGGGCTGAGAATTCAT; (g) hIL-17sense, ACTCCTGGGAAGACCTCATTG; (h) hIL-17 antisense, GGCCA-CATGGTGGACAATCG; (i) human CD4 sense, GGAGTCCCTTTTAGGC-ACTTGC; (j) human CD4 antisense, GAACTCCACCTGTTCCCCCTC; (k)human CD8 sense, CTCCTCCTGGCCGCGCAGCTG; (l) human CD8 anti-sense, GCCGGGCTCTCCTCCGCCG; (m) murine IL-6 sense, TGGAGTCA-CAGAAGGAGTGGCTAAG; (n) murine IL-6 antisense, TCTGACCACAGT-GAGGAATGTCCAC; (o) murine hypoxanthine phosphoribosyltransferasesense, GTTGGATACAGGCCAGACTTTGTTG; and (p) murine hypoxan-thine phosphoribosyltransferase antisense, GATTCAACTTGCGCTCATCT-TAGGC.
In Vitro Growth Assay. Cells were plated into flat-bottomed 96-wellplates at a density of 104 cells/well. The cells were cultured for 3 days, andtheir proliferation was determined by a MTT assay (29). Briefly, 20ml of MTT(Sigma) at a concentration of 5 mg/ml were added to each well and incubatedfor 4 h in thedark. The formazan grain was then dissolved in DMSO, and theabsorbance at 570 nm was read using an ELISA plate reader. A standard curvebetween the absorbance of the MTT test and the number of cells was deter-mined for each cell line. The conversion of MTT to formazan was directlycorrelated with the number of viable cells.
Tumor Growth in Nude Mice. Male 8-week-old athymicnu/numice (IffaCredo, L’Arbresle, France) were used for the experiments described here.Human cervical tumors were injected into mice by subdermal inoculation of
106 cells. A total of 8–10 mice/group were used per experiment. Tumorvolume (in mm3) was estimated from the length (a) and width (b) of the tumorby using the following formula: volume5 ab2/2. Biopsies were snap-frozen inliquid nitrogen and stored at270°C for RNA extraction.
Immunocytochemistry. Cryostat sections (5mM) of tumor xenograftswere fixed in acetone at 4°C for 5 min. After washes in TBS, they wereincubated with biotinylated rat mAb against Mac1 (M1/70 hybridoma) or withisotype-matched control biotinylated rat mAb (PharMingen, San Diego, CA).After washes in TBS, slides were incubated with alkaline phosphatase-conju-gated streptavidin (DAKO, Trappes, France), and enzymatic activity wasrevealed with Fast Red reagent (DAKO) associated with 1 mM of levamisole,a known inhibitor of endogenous alkaline phosphatase. Sections were coun-terstained with Mayer’s Hematoxylin.
Statistical Analysis. The in vivo comparison of tumor size between thevarious groups of mice was analyzed by the Mann-Whitney test.
RESULTS
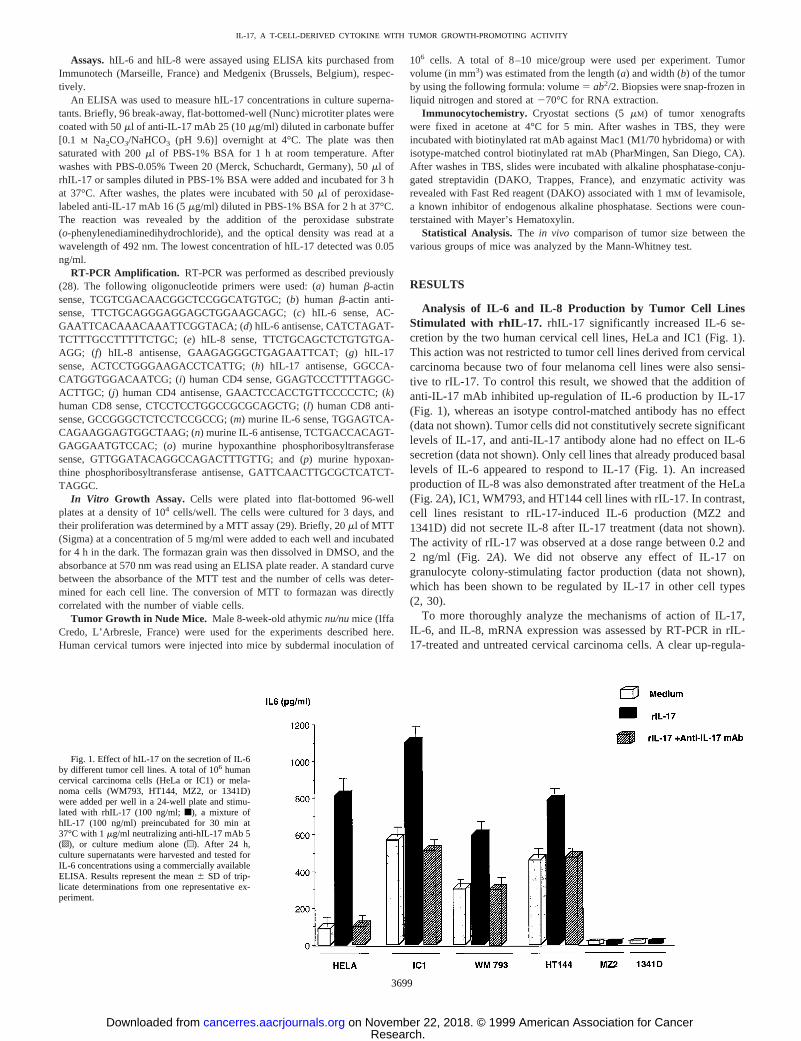
Analysis of IL-6 and IL-8 Production by Tumor Cell LinesStimulated with rhIL-17. rhIL-17 significantly increased IL-6 se-cretion by the two human cervical cell lines, HeLa and IC1 (Fig. 1).This action was not restricted to tumor cell lines derived from cervicalcarcinoma because two of four melanoma cell lines were also sensi-tive to rIL-17. To control this result, we showed that the addition ofanti-IL-17 mAb inhibited up-regulation of IL-6 production by IL-17(Fig. 1), whereas an isotype control-matched antibody has no effect(data not shown). Tumor cells did not constitutively secrete significantlevels of IL-17, and anti-IL-17 antibody alone had no effect on IL-6secretion (data not shown). Only cell lines that already produced basallevels of IL-6 appeared to respond to IL-17 (Fig. 1). An increasedproduction of IL-8 was also demonstrated after treatment of the HeLa(Fig. 2A), IC1, WM793, and HT144 cell lines with rIL-17. In contrast,cell lines resistant to rIL-17-induced IL-6 production (MZ2 and1341D) did not secrete IL-8 after IL-17 treatment (data not shown).The activity of rIL-17 was observed at a dose range between 0.2 and2 ng/ml (Fig. 2A). We did not observe any effect of IL-17 ongranulocyte colony-stimulating factor production (data not shown),which has been shown to be regulated by IL-17 in other cell types(2, 30).
To more thoroughly analyze the mechanisms of action of IL-17,IL-6, and IL-8, mRNA expression was assessed by RT-PCR in rIL-17-treated and untreated cervical carcinoma cells. A clear up-regula-
Fig. 1. Effect of hIL-17 on the secretion of IL-6by different tumor cell lines. A total of 106 humancervical carcinoma cells (HeLa or IC1) or mela-noma cells (WM793, HT144, MZ2, or 1341D)were added per well in a 24-well plate and stimu-lated with rhIL-17 (100 ng/ml;f), a mixture ofhIL-17 (100 ng/ml) preincubated for 30 min at37°C with 1mg/ml neutralizing anti-hIL-17 mAb 5(o), or culture medium alone (u). After 24 h,culture supernatants were harvested and tested forIL-6 concentrations using a commercially availableELISA. Results represent the mean6 SD of trip-licate determinations from one representative ex-periment.
3699
IL-17, A T-CELL-DERIVED CYTOKINE WITH TUMOR GROWTH-PROMOTING ACTIVITY
Research. on November 22, 2018. © 1999 American Association for Cancercancerres.aacrjournals.org Downloaded from
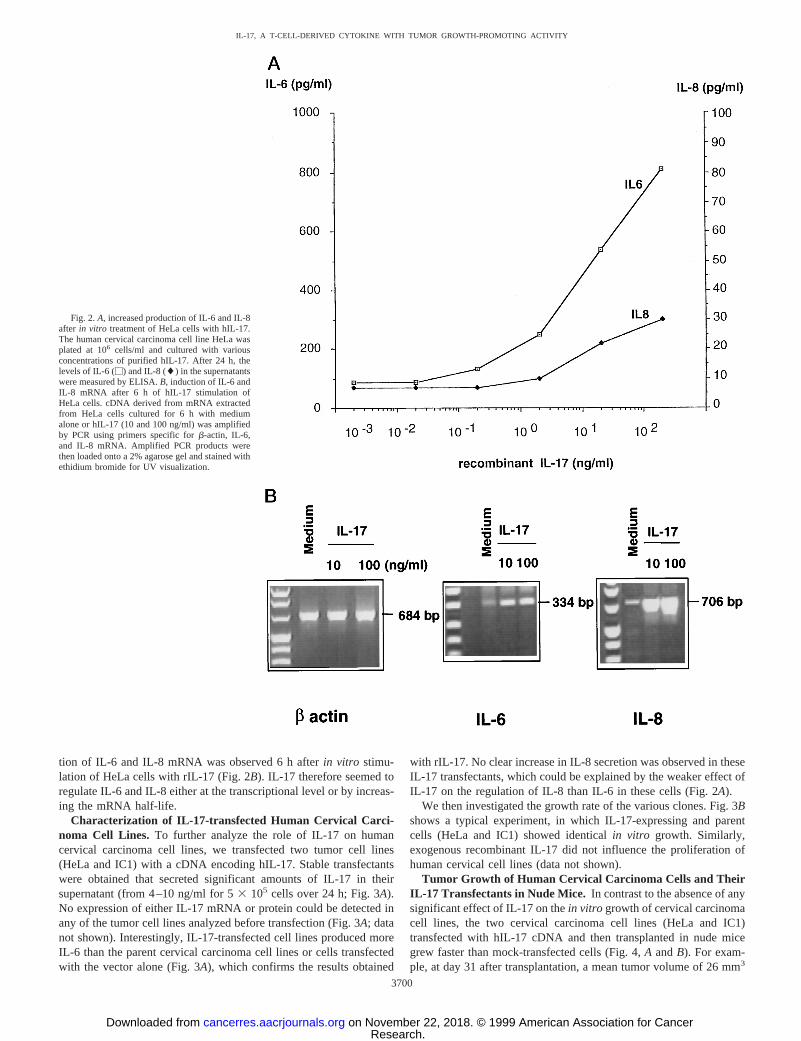
tion of IL-6 and IL-8 mRNA was observed 6 h afterin vitro stimu-lation of HeLa cells with rIL-17 (Fig. 2B). IL-17 therefore seemed toregulate IL-6 and IL-8 either at the transcriptional level or by increas-ing the mRNA half-life.
Characterization of IL-17-transfected Human Cervical Carci-noma Cell Lines. To further analyze the role of IL-17 on humancervical carcinoma cell lines, we transfected two tumor cell lines(HeLa and IC1) with a cDNA encoding hIL-17. Stable transfectantswere obtained that secreted significant amounts of IL-17 in theirsupernatant (from 4–10 ng/ml for 53 105 cells over 24 h; Fig. 3A).No expression of either IL-17 mRNA or protein could be detected inany of the tumor cell lines analyzed before transfection (Fig. 3A; datanot shown). Interestingly, IL-17-transfected cell lines produced moreIL-6 than the parent cervical carcinoma cell lines or cells transfectedwith the vector alone (Fig. 3A), which confirms the results obtained
with rIL-17. No clear increase in IL-8 secretion was observed in theseIL-17 transfectants, which could be explained by the weaker effect ofIL-17 on the regulation of IL-8 than IL-6 in these cells (Fig. 2A).
We then investigated the growth rate of the various clones. Fig. 3Bshows a typical experiment, in which IL-17-expressing and parentcells (HeLa and IC1) showed identicalin vitro growth. Similarly,exogenous recombinant IL-17 did not influence the proliferation ofhuman cervical cell lines (data not shown).
Tumor Growth of Human Cervical Carcinoma Cells and TheirIL-17 Transfectants in Nude Mice. In contrast to the absence of anysignificant effect of IL-17 on thein vitro growth of cervical carcinomacell lines, the two cervical carcinoma cell lines (HeLa and IC1)transfected with hIL-17 cDNA and then transplanted in nude micegrew faster than mock-transfected cells (Fig. 4,A andB). For exam-ple, at day 31 after transplantation, a mean tumor volume of 26 mm3
Fig. 2.A, increased production of IL-6 and IL-8after in vitro treatment of HeLa cells with hIL-17.The human cervical carcinoma cell line HeLa wasplated at 106 cells/ml and cultured with variousconcentrations of purified hIL-17. After 24 h, thelevels of IL-6 (M) and IL-8 (l) in the supernatantswere measured by ELISA.B, induction of IL-6 andIL-8 mRNA after 6 h of hIL-17 stimulation ofHeLa cells. cDNA derived from mRNA extractedfrom HeLa cells cultured for 6 h with mediumalone or hIL-17 (10 and 100 ng/ml) was amplifiedby PCR using primers specific forb-actin, IL-6,and IL-8 mRNA. Amplified PCR products werethen loaded onto a 2% agarose gel and stained withethidium bromide for UV visualization.
3700
IL-17, A T-CELL-DERIVED CYTOKINE WITH TUMOR GROWTH-PROMOTING ACTIVITY
Research. on November 22, 2018. © 1999 American Association for Cancercancerres.aacrjournals.org Downloaded from
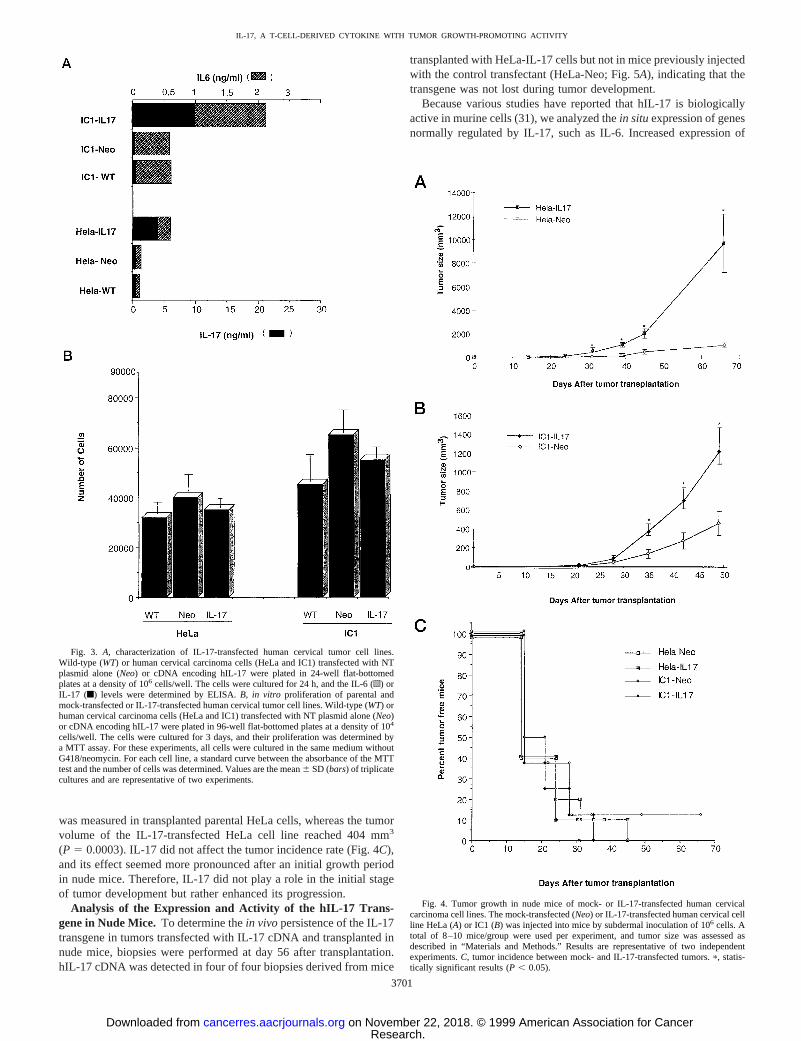
was measured in transplanted parental HeLa cells, whereas the tumorvolume of the IL-17-transfected HeLa cell line reached 404 mm3
(P 5 0.0003). IL-17 did not affect the tumor incidence rate (Fig. 4C),and its effect seemed more pronounced after an initial growth periodin nude mice. Therefore, IL-17 did not play a role in the initial stageof tumor development but rather enhanced its progression.
Analysis of the Expression and Activity of the hIL-17 Trans-gene in Nude Mice.To determine thein vivopersistence of the IL-17transgene in tumors transfected with IL-17 cDNA and transplanted innude mice, biopsies were performed at day 56 after transplantation.hIL-17 cDNA was detected in four of four biopsies derived from mice
transplanted with HeLa-IL-17 cells but not in mice previously injectedwith the control transfectant (HeLa-Neo; Fig. 5A), indicating that thetransgene was not lost during tumor development.
Because various studies have reported that hIL-17 is biologicallyactive in murine cells (31), we analyzed thein situexpression of genesnormally regulated by IL-17, such as IL-6. Increased expression of
Fig. 3. A, characterization of IL-17-transfected human cervical tumor cell lines.Wild-type (WT) or human cervical carcinoma cells (HeLa and IC1) transfected with NTplasmid alone (Neo) or cDNA encoding hIL-17 were plated in 24-well flat-bottomedplates at a density of 106 cells/well. The cells were cultured for 24 h, and the IL-6 (o) orIL-17 (f) levels were determined by ELISA.B, in vitro proliferation of parental andmock-transfected or IL-17-transfected human cervical tumor cell lines. Wild-type (WT) orhuman cervical carcinoma cells (HeLa and IC1) transfected with NT plasmid alone (Neo)or cDNA encoding hIL-17 were plated in 96-well flat-bottomed plates at a density of 104
cells/well. The cells were cultured for 3 days, and their proliferation was determined bya MTT assay. For these experiments, all cells were cultured in the same medium withoutG418/neomycin. For each cell line, a standard curve between the absorbance of the MTTtest and the number of cells was determined. Values are the mean6 SD (bars) of triplicatecultures and are representative of two experiments.
Fig. 4. Tumor growth in nude mice of mock- or IL-17-transfected human cervicalcarcinoma cell lines. The mock-transfected (Neo) or IL-17-transfected human cervical cellline HeLa (A) or IC1 (B) was injected into mice by subdermal inoculation of 106 cells. Atotal of 8–10 mice/group were used per experiment, and tumor size was assessed asdescribed in “Materials and Methods.” Results are representative of two independentexperiments.C, tumor incidence between mock- and IL-17-transfected tumors.p, statis-tically significant results (P , 0.05).
3701
IL-17, A T-CELL-DERIVED CYTOKINE WITH TUMOR GROWTH-PROMOTING ACTIVITY
Research. on November 22, 2018. © 1999 American Association for Cancercancerres.aacrjournals.org Downloaded from
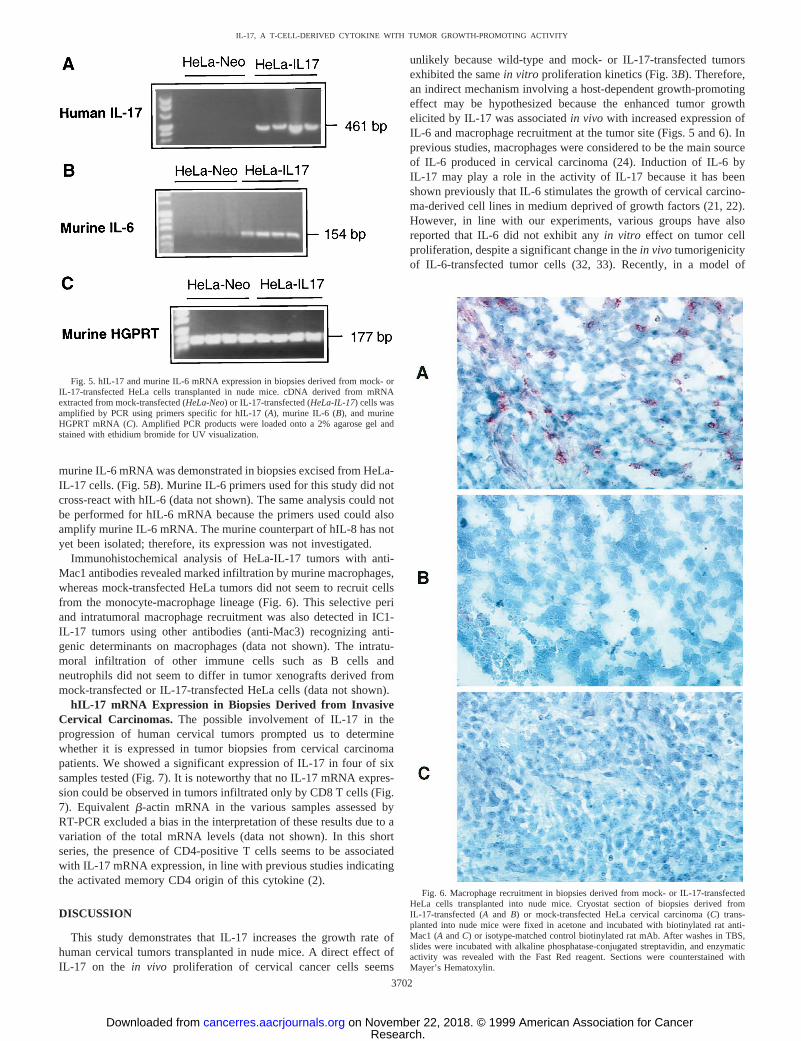
murine IL-6 mRNA was demonstrated in biopsies excised from HeLa-IL-17 cells. (Fig. 5B). Murine IL-6 primers used for this study did notcross-react with hIL-6 (data not shown). The same analysis could notbe performed for hIL-6 mRNA because the primers used could alsoamplify murine IL-6 mRNA. The murine counterpart of hIL-8 has notyet been isolated; therefore, its expression was not investigated.
Immunohistochemical analysis of HeLa-IL-17 tumors with anti-Mac1 antibodies revealed marked infiltration by murine macrophages,whereas mock-transfected HeLa tumors did not seem to recruit cellsfrom the monocyte-macrophage lineage (Fig. 6). This selective periand intratumoral macrophage recruitment was also detected in IC1-IL-17 tumors using other antibodies (anti-Mac3) recognizing anti-genic determinants on macrophages (data not shown). The intratu-moral infiltration of other immune cells such as B cells andneutrophils did not seem to differ in tumor xenografts derived frommock-transfected or IL-17-transfected HeLa cells (data not shown).
hIL-17 mRNA Expression in Biopsies Derived from InvasiveCervical Carcinomas. The possible involvement of IL-17 in theprogression of human cervical tumors prompted us to determinewhether it is expressed in tumor biopsies from cervical carcinomapatients. We showed a significant expression of IL-17 in four of sixsamples tested (Fig. 7). It is noteworthy that no IL-17 mRNA expres-sion could be observed in tumors infiltrated only by CD8 T cells (Fig.7). Equivalentb-actin mRNA in the various samples assessed byRT-PCR excluded a bias in the interpretation of these results due to avariation of the total mRNA levels (data not shown). In this shortseries, the presence of CD4-positive T cells seems to be associatedwith IL-17 mRNA expression, in line with previous studies indicatingthe activated memory CD4 origin of this cytokine (2).
DISCUSSION
This study demonstrates that IL-17 increases the growth rate ofhuman cervical tumors transplanted in nude mice. A direct effect ofIL-17 on the in vivo proliferation of cervical cancer cells seems
unlikely because wild-type and mock- or IL-17-transfected tumorsexhibited the samein vitro proliferation kinetics (Fig. 3B). Therefore,an indirect mechanism involving a host-dependent growth-promotingeffect may be hypothesized because the enhanced tumor growthelicited by IL-17 was associatedin vivo with increased expression ofIL-6 and macrophage recruitment at the tumor site (Figs. 5 and 6). Inprevious studies, macrophages were considered to be the main sourceof IL-6 produced in cervical carcinoma (24). Induction of IL-6 byIL-17 may play a role in the activity of IL-17 because it has beenshown previously that IL-6 stimulates the growth of cervical carcino-ma-derived cell lines in medium deprived of growth factors (21, 22).However, in line with our experiments, various groups have alsoreported that IL-6 did not exhibit anyin vitro effect on tumor cellproliferation, despite a significant change in thein vivo tumorigenicityof IL-6-transfected tumor cells (32, 33). Recently, in a model of
Fig. 5. hIL-17 and murine IL-6 mRNA expression in biopsies derived from mock- orIL-17-transfected HeLa cells transplanted in nude mice. cDNA derived from mRNAextracted from mock-transfected (HeLa-Neo) or IL-17-transfected (HeLa-IL-17) cells wasamplified by PCR using primers specific for hIL-17 (A), murine IL-6 (B), and murineHGPRT mRNA (C). Amplified PCR products were loaded onto a 2% agarose gel andstained with ethidium bromide for UV visualization.
Fig. 6. Macrophage recruitment in biopsies derived from mock- or IL-17-transfectedHeLa cells transplanted into nude mice. Cryostat section of biopsies derived fromIL-17-transfected (Aand B) or mock-transfected HeLa cervical carcinoma (C) trans-planted into nude mice were fixed in acetone and incubated with biotinylated rat anti-Mac1 (AandC) or isotype-matched control biotinylated rat mAb. After washes in TBS,slides were incubated with alkaline phosphatase-conjugated streptavidin, and enzymaticactivity was revealed with the Fast Red reagent. Sections were counterstained withMayer’s Hematoxylin.
3702
IL-17, A T-CELL-DERIVED CYTOKINE WITH TUMOR GROWTH-PROMOTING ACTIVITY
Research. on November 22, 2018. © 1999 American Association for Cancercancerres.aacrjournals.org Downloaded from

human melanoma transplanted in nude mice, Luet al. (34) demon-strated that IL-6 could indeed function as anin vivoparacrine growth-stimulating factor for advanced-stage human melanoma cells.
A tumor-promoting activity of macrophages acting on the forma-tion of tumor stroma has also been described. For example, tumorgrowth was markedly impaired in theop/opmouse, which possessesa profound macrophage deficiency compared with normal littermates.Treatment of tumor-bearingop/op mice with recombinant colony-stimulating factor 1 corrected this impairment (35). Moreover, in nudemice, the reduced tumorigenicity of IL-10-transfected Chinese ham-ster ovary cells compared to parent cells was found to be associatedwith inhibition of macrophage recruitment at the tumor site (36). Themechanisms that may explain the selective tumor homing of macro-phages in IL-17-producing cervical carcinoma remain to be eluci-dated. It has been shown that IL-17 induces MCP-1, a known selectivemacrophage chemoattractant, in epithelial cells (37). However, wefound no difference in the expression of JE and MCP-5 mRNA, themurine counterpart of MCP-1, in biopsies derived from mock- orIL-17-transfected cervical carcinoma cell lines transplanted in nudemice. In addition, IL-17 did not induce human MCP-1 in humancervical cell lines (data not shown).
Although it has been shown that IL-17 stimulated granulopoiesisand elicited a rise in peripheral neutrophil counts after s.c. adminis-tration (2, 38), we could not demonstrate any significant neutrophilinfiltrate in established cervical tumor xenografts.
Of course, only neutralization of macrophages and/or IL-6 activityin vivo would constitute direct evidence of the role of these factors asmediators of IL-17 activity on tumor enhancement. Unfortunately, inthree independent experiments, the unexpected inhibitory effects ofcontrol immunoglobulins administered to tumor-bearing nude miceprevented us from drawing any definitive conclusions about the roleof anti-IL-6 or anti-Mac1 antibodies in this model.
Because IL-17 is secreted mainly by CD4-positive T cells in man[Ref. 2; its source in mice has not been thoroughly investigated (3,31)], it may seem paradoxical that immune cells secrete tumor-promoting factors. However, other studies have also reported that Tlymphocytes could secrete basic fibroblast growth factor or vascularendothelial growth factor, which stimulate tumor angiogenesis (39,40), or heparin binding epidermal growth factor, which is consideredto be a tumor survival factor (40, 41).
In mice, although CD4 and CD8 T cells were often both requiredfor induction of immune control on tumor growth (42–44), variousmodels have clearly implicated the CD4 T-cell population as cells thatmay enhance tumor development (45). A spontaneously arising B-celllymphoma in SJL/J mice has been shown to depend on host CD4 Tcells for its proliferation and growth (46). Treatment with anti-CD4mAb before or after inoculation of a lethal dose of lymphoma cellsinhibited tumor growth and allowed mice to survive (47, 48). Variousimmunotherapy protocols in mice, including systemic administrationof IL-2 or IL-12, were greatly improved after elimination of CD4 Tcells (49–51). Similarly, depletion of CD4 T cells increased the
efficiency of T-cell-mediated immunity after peptide vaccination in amodel of established micrometastases from lung carcinoma (52).
In man, several tumors seem to be sensitive to the enhancing effectsof CD4 T cells. Nakamuraet al. (53) characterized Kaposi’s sarcomacell lines that were dependent on growth factors released by CD4 Tcells. Umetsuet al. (54) also identified a group of follicular B-celllymphomas highly infiltrated with CD4 T cells that play an essentialrole in sustaining rather than inhibiting tumor cell growthin vivo.These examples may explain some reports in which a high number oftumor-infiltrating lymphocytes in certain cancers is considered to be apoor prognostic marker (55).
Finally the expression of IL-17 in biopsies derived from cervicalcancer patients (Fig. 7), together with the fact that the amount of IL-17secreted by activated human T cells (2) was in the same range as thatproduced by the stably transfected cervical carcinoma cell lines,suggests that our findings may have some clinical relevance.
Future studies will be designed to assess the clinical prognosticvalue of this factor.
ACKNOWLEDGMENTS
We thank John Abrams (DNAX) for providing us with some antibodiesagainst IL-17.
REFERENCES
1. Rouvier, E., Luciani, M. F., Mattei, M. G., Denizot, F., and Golstein, P. CTLA-8,cloned from an activated T cell, bearing AU-rich messenger RNA instability se-quences, and homologous to aherpesvirus saimirigene. J. Immunol.,150: 5445–5556, 1993.
2. Fossiez, F., Djossou, O., Chomarat, P., Flores-Romo, L., Ait-Yahia, S., Maat, C., Pin,J. J., Garrone, P., Garcia, E., Saeland, S., Blanchard, D., Gaillard, C., Das Mahapatra,B., Rouvier, E., Golstein, P., Banchereau, J., and Lebecque, S. T cell interleukin-17induces stromal cells to produce proinflammatory and hematopoietic cytokines. J.Exp. Med.,183: 2593–2603, 1996.
3. Spriggs, M. K. Interleukin-17 and its receptor. J. Clin. Immunol.,17: 366–369, 1997.4. Fleckenstein, B., and Desrosiers, R. C.Herpesvirus saimiriandHerpesvirus ateles.
In: I. B. Roizman (ed.), The Herpesviruses, pp 253–332. New York: Plenum Pub-lishing Press, 1992.
5. Mittrucker, H. W., Muller-Fleckenstein, I., Fleckenstein, B., and Fleischer, B. CD2-mediated autocrine growth ofHerpesvirus saimiri-transformed human T lympho-cytes. J. Exp. Med.,176: 909–913, 1992.
6. Biesinger, B., Muller-Fleckenstein, I., Simmer, B., Lang, G., Wittmann, S., Platzer,E., Desrosiers, R. C., and Fleckenstein, B. Stable growth transformation of human Tlymphocytes byHerpesvirus saimiri. Proc. Natl. Acad. Sci. USA,89: 3116–3119,1992.
7. Yao, Z., Fanslow, W. C., Seldin, M. F., Rousseau, A. M., Painter, S. L., Comeau,M. R., Cohen, J. I., and Spriggs, M. K.Herpesvirus saimiriencodes a new cytokine,IL-17, which binds to a novel cytokine receptor. Immunity,3: 811–821, 1995.
8. Jovanovic, D. V., Di Battista, J. A., Martel-Pelletier, J., Jolicoeur, F. C., He, Y.,Mineau, F., and Pelletier, J. P. IL-17 stimulates the production and expression ofproinflammatory cytokines, IL1-b and TNF-a, by human macrophages. J. Immunol.,160: 3513–3521, 1998.
9. Teunissen, M. B., Koomen, C. W., de Waal Malefyt, R., Wierenga, E. A., and Bos,J. D. Interleukin-17 and interferon-g synergize in the enhancement of proinflamma-tory cytokine production by human keratinocytes. J. Investig. Dermatol.,111: 645–649, 1998.
10. Chabaud, M., Fossiez, F., Taupin, J. L., and Miossec, P. Enhancing effect of IL-17 onIL-1-induced IL-6 and leukemia inhibitory factor production by rheumatoid arthritissynoviocytes and its regulation by Th2 cytokines. J. Immunol.,161: 409–414, 1998.
Fig. 7. IL-17 mRNA expression in biopsiesderived from invasive cervical carcinomas. SixcDNAs derived from mRNA extracted from sixprimary invasive cervical carcinomas were ampli-fied by PCR using oligonucleotide primers specificfor hIL-17, CD4, and CD8. Amplified PCR prod-ucts were loaded onto a 2% agarose gel and stainedwith ethidium bromide for UV visualization.
3703
IL-17, A T-CELL-DERIVED CYTOKINE WITH TUMOR GROWTH-PROMOTING ACTIVITY
Research. on November 22, 2018. © 1999 American Association for Cancercancerres.aacrjournals.org Downloaded from

11. Shalom-Barak, T., Quach, J., and Lotz, M. Interleukin-17-induced gene expression inarticular chondrocytes is associated with activation of mitogen-activated proteinkinases and NF-kB. J. Biol. Chem.,273: 27467–27473, 1998.
12. Attur, M. G., Patel, R. N., Abramson, S. B., and Amin, A. R. Interleukin-17up-regulation of nitric oxide production in human osteoarthritis cartilage. ArthritisRheum.,40: 1050–1053, 1997.
13. Fridman, W. H., and Tartour, E. Cytokines and cell regulation.In: Molecular Aspectsof Medicine, pp. 3–90. Oxford, United Kingdom: Elsevier Science, 1997.
14. Vink, A., Coulie, P., Warnier, G., Renauld, J. C., Stevens, M., Donckers, D., and VanSnick, J. Mouse plasmacytoma growthin vivo: enhancement by interleukin 6 (IL-6)and inhibition by antibodies directed against IL-6 or its receptor. J. Exp. Med.,172:997–1000, 1990.
15. Scala, G., Quinto, I., Ruocco, M. R., Arcucci, A., Mallardo, M., Caretto, P., Forni, G.,and Venuta, S. Expression of an exogenous interleukin 6 gene in human Epstein Barrvirus B cells confers growth advantage andin vivo tumorigenicity. J. Exp. Med.,172:61–68, 1990.
16. Durandy, A., Emilie, D., Peuchmaur, M., Forveille, M., Clement, C., Wijdenes, J., andFischer, A. Role of IL-6 in promoting growth of human EBV-induced B-cell tumors insevere combined immunodeficient mice. J. Immunol.,152: 5361–5367, 1994.
17. Bataille, R., Barlogie, B., Lu, Z. Y., Rossi, J. F., Lavabre-Bertrand, T., Beck, T.,Wijdenes, J., Brochier, J., and Klein, B. Biologic effects of anti-interleukin-6 murinemonoclonal antibody in advanced multiple myeloma. Blood,86: 685–691, 1995.
18. Schadendorf, D., Moller, A., Algermissen, B., Worm, M., Sticherling, M., andCzarnetzki, B. M. IL-8 produced by human malignant melanoma cellsin vitro is anessential autocrine growth factor. J. Immunol.,151: 2667–2675, 1993.
19. Singh, R. K., Gutman, M., Radinsky, R., Bucana, C. D., and Fidler, I. J. Expressionof interleukin 8 correlates with the metastatic potential of human melanoma cells innude mice. Cancer Res.,54: 3242–3247, 1994.
20. Schiffmank, M. H., and Brinton, L. A. The epidemiology of cervical carcinogenesis.Cancer (Phila.),76 (Suppl. 10):1888–1901, 1995.
21. Iglesias, M., Plowman, G. D., and Woodworth, C. D. Interleukin-6 and interleukin-6soluble receptor regulate proliferation of normal, human papillomavirus-immortalized,and carcinoma-derived cervical cellsin vitro. Am. J. Pathol.,146: 944–952, 1995.
22. Eustace, D., Han, X., Gooding, R., Rowbottom, A., Riches, P., and Heyderman, E.Interleukin-6 (IL-6) functions as an autocrine growth factor in cervical carcinomasinvitro. Gynecol. Oncol.,50: 15–19, 1993.
23. Woodworth, C. D., and Simpson, S. Comparative lymphokine secretion by culturednormal human cervical keratinocytes, papillomavirus-immortalized, and carcinomacell lines. Am. J. Pathol.,142: 1544–1555, 1993.
24. Tartour, E., Gey, A., Sastre-Garau, X., Pannetier, C., Mosseri, V., Kourilsky, P., andFridman, W. H. Analysis of interleukin 6 gene expression in cervical neoplasia usinga quantitative polymerase chain reaction assay: evidence for enhanced interleukin 6gene expression in invasive carcinoma. Cancer Res.,54: 6243–6248, 1994.
25. Couturier, J., Sastre-Garau, X., Schneider-Maunoury, S., Labib, A., and Orth, G.Integration of papillomavirus DNA nearmyc genes in genital carcinomas and itsconsequences for proto-oncogene expression. J. Virol.,65: 4534–4538, 1991.
26. Montero, F., Brailly, H., Sautes, C., Joyeux, I., Dorval, T., Mosseri, V., Yasukawa,K., Wijdenes, J., Adler, A., Gorin, I., Fridman, W. H., and Tartour, E. Characteriza-tion of soluble gp130 released by melanoma cell lines: a polyvalent antagonist ofcytokines from IL6 family. Clin. Cancer Res.,3: 1443–1451, 1997.
27. Daeron, M., Latour, S., Malbec, O., Espinosa, E., Pina, P., Pasmans, S., and Fridman,W. H. The same tyrosine-based inhibition motif, in the intracytoplasmic domain of Fcg RIIB, regulates negatively BCR-, TCR-, and FcR-dependent cell activation. Im-munity, 3: 635–646, 1995.
28. Tartour, E., Gey, A., Sastre-Garau, X., Lombard Surin, I., Mosseri, V., and Fridman,W. H. Prognostic value of intratumoral interferong messenger RNA expression ininvasive cervical carcinoma. J. Natl. Cancer Inst.,4: 287–294, 1998.
29. Hansen, M. B., Nielsen, S. E., and Berg, K. Re-examination and further developmentof a precise and rapid dye method for measuring cell growth/cell kill. J. Immunol.Methods,119: 203–210, 1989.
30. Cai, X. Y., Gommoll, C. P., Jr., Justice, L., Narula, S. K., and Fine, J. S. Regulationof granulocyte colony-stimulating factor gene expression by interleukin-17. Immunol.Lett., 62: 51–58, 1998.
31. Kennedy, J., Rossi, D. L., Zurawski, S. M., Vega, F., Jr., Kastelein, R. A., Wagner,J. L., Hannum, C. H., and Zlotnik, A. Mouse IL-17: a cytokine preferentiallyexpressed bya b TCR 1 CD4-CD8-T cells. J. Interferon Cytokine Res.,16:611–617, 1996.
32. Sun, W. H., Kreisle, R. A., Phillips, A. W., and Ershler, W. B.In vivo and in vitrocharacteristics of interleukin 6-transfected B16 melanoma cells. Cancer Res.,52:5412–5415, 1992.
33. Mullen, C. A., Coale, M. M., Levy, A. T., Stetler-Stevenson, W. G., Liotta, L. A.,Brandt, S., and Blaese, R. M. Fibrosarcoma cells transduced with the IL-6 geneexhibited reduced tumorigenicity, increased immunogenicity, and decreased meta-static potential. Cancer Res.,52: 6020–6024, 1992.
34. Lu, C., Sheehan, C., Rak, J. W., Chambers, C. A., Hozumi, N., and Kerbel, R. S.Endogenous interleukin 6 can function as anin vivo growth-stimulatory factor foradvanced-stage human melanoma cells. Clin. Cancer Res.,2: 1417–1425, 1996.
35. Nowicki, A., Szenajch, J., Ostrowska, G., Wojtowicz, A., Wojtowicz, K., Krusze-wski, A. A., Maruszynski, M., Aukerman, S. L., and Wiktor-Jedrzejczak, W. Im-paired tumor growth in colony-stimulating factor 1 (CSF-1)-deficient, macrophage-deficient op/op mouse: evidence for a role of CSF-1-dependent macrophages information of tumor stroma. Int. J. Cancer,65: 112–119, 1996.
36. Richter, G., Kruger-Krasagakes, S., Hein, G., Huls, C., Schmitt, E., Diamantstein, T.,and Blankenstein, T. Interleukin 10 transfected into Chinese hamster ovary cellsprevents tumor growth and macrophage infiltration. Cancer Res.,53: 4134–4137,1993.
37. Van Kooten, C., Boonstra, J. G., Paape, M. E., Fossiez, F., Banchereau, J., Lebecque,S., Bruijn, J. A., De Fijter, J. W., Van Es, L. A., and Daha, M. R. Interleukin-17activates human renal epithelial cellsin vitro and is expressed during renal allograftrejection.J. Am. Soc. Nephrol.,9: 1526–1534, 1998.
38. Schwarzenberger, P., La Russa, V., Miller, A., Ye, P., Huang, W., Zieske, A., Nelson,S., Bagby, G. J., Stoltz, D., Mynatt, R. L., Spriggs, M., and Kolls, J. K. IL-17stimulates granulopoiesis in mice: use of an alternate, novel gene therapy-derivedmethod forin vivo evaluation of cytokines. J. Immunol.,161: 6383–6389, 1998.
39. Freeman, M. R., Schneck, F. X., Gagnon, M. L., Corless, C., Soker, S., Niknejad, K.,Peoples, G. E., and Klagsbrun, M. Peripheral blood T lymphocytes and lymphocytesinfiltrating human cancers express vascular endothelial growth factor: a potential rolefor T cells in angiogenesis. Cancer Res.,55: 4140–4145, 1995.
40. Blotnick, S., Peoples, G. E., Freeman, M. R., Eberlein, T. J., and Klagsbrun, M. T. Tlymphocytes synthesize and export heparin-binding epidermal growth factor-likegrowth factor and basic fibroblast growth factor, mitogens for vascular cells andfibroblasts: differential production and release by CD41 and CD81T cells. Proc.Natl. Acad. Sci. USA,91: 2890–2894, 1994.
41. Miyoshi, E., Higashiyama, S., Nakagawa, T., Hayashi, N., and Taniguchi, N. Mem-brane-anchored heparin-binding epidermal growth factor-like growth factor acts as atumor survival factor in a hepatoma cell line. J. Biol. Chem.,272: 14349–14355,1997.
42. Kaido, T. J., Maury, C., and Gresser, I. Host CD41 T lymphocytes are required forthe synergistic action of interferon-a/b and adoptively transferred immune cells in theinhibition of visceral ESb metastases. Cancer Res.,55: 6133–6139, 1995.
43. Hock, H., Dorsch, M., Diamantstein, T., and Blankenstein, T. Interleukin 7 inducesCD41 T cell-dependent tumor rejection. J. Exp. Med.,174: 1291–1298, 1991.
44. Flamand, V., Biernaux, C., Van Mechelen, M., Sornasse, T., Urbain, J., Leo, O., andMoser, M. Immune surveillance: both CD31CD41 and CD31 CD81 T cellscontrol in vivo growth of P815 mastocytoma. Int. J. Cancer,45: 757–762, 1990.
45. North, R. J. Down-regulation of the antitumor immune response. Adv. Cancer Res.,45: 1–43, 1985.
46. Katz, I. R., Chapman-Alexander, J., Jacobson, E. B., Lerman, S. P., and Thorbecke,G. J. Growth of SJL/J-derived transplantable reticulum cell sarcoma as related to itsability to induce T-cell proliferation in the host. III. Studies on thymectomized andcongenitally athymic SJL mice. Cell. Immunol.,65: 84–92, 1981.
47. Ohnishi, K., and Bonavida, B. Regulation of Ia1 reticulum cell sarcoma (RCS)growth in syngeneic SJL/J mice. I. Inhibition of tumor growth by passive adminis-tration of L3T4 monoclonal antibody before or after tumor inoculation. J. Immunol.,138: 4524–4529, 1987.
48. Ohnishi, K., and Bonavida, B. Regulation of B-cell lymphoma growth in syngeneic SJL/Jmice. Establishment of tumor dormancy following administration of anti-CD4 mono-clonal antibody into tumor-bearing mice. Leukemia (Baltimore),7: 1801–1806, 1993.
49. Rakhmilevich, A. L., and North, R. J. Elimination of CD41 T cells in mice bearingan advanced sarcoma augments the antitumor action of interleukin-2. Cancer Immu-nol. Immunother.,38: 107–112, 1994.
50. Colombo, M. P., Vagliani, M., Spreafico, F., Parenza, M., Chiodoni, C., Melani, C.,and Stoppacciaro, A. Amount of interleukin 12 available at the tumor site is criticalfor tumor regression. Cancer Res.,56: 2531–2534, 1996.
51. Martinotti, A., Stoppacciaro, A., Vagliani, M., Melani, C., Spreafico, F., Wysocka,M., Parmiani, G., Trinchieri, G., and Colombo, M. P. CD4 T cells inhibitin vivo theCD8-mediated immune response against murine colon carcinoma cells transducedwith interleukin-12 genes. Eur. J. Immunol.,25: 137–146, 1995.
52. Mandelboim, O., Vadai, E., Fridkin, M., Katz-Hillel, A., Feldman, M., Berke, G., andEisenbach, L. Regression of established murine carcinoma metastases followingvaccination with tumour-associated antigen peptides. Nat. Med.,1: 1179–1183, 1995.
53. Nakamura, S., Salahuddin, S. Z., Biberfeld, P., Ensoli, B., Markham, P. D., Wong-Staal, F., and Gallo, R. C. Kaposi’s sarcoma cells: long-term culture with growthfactor from retrovirus-infected CD41 T cells. Science (Washington DC),242:426–430, 1988.
54. Umetsu, D. T., Esserman, L., Donlon, T. A., DeKruyff, R. H., and Levy, R. Inductionof proliferation of human follicular (B type) lymphoma cells by cognate interactionwith CD41 T cell clones. J. Immunol.,144: 2550–2557, 1990.
55. Rosen, P. R., Groshen, S., Saigo, P. E., Kinne, D. W., and Hellman, S. A long-termfollow-up study of survival in stage I (T1N0M0) and stage II (T1N1M0) breastcarcinoma. J. Clin. Oncol.,7: 355–366, 1989.
3704
IL-17, A T-CELL-DERIVED CYTOKINE WITH TUMOR GROWTH-PROMOTING ACTIVITY
Research. on November 22, 2018. © 1999 American Association for Cancercancerres.aacrjournals.org Downloaded from

1999;59:3698-3704. Cancer Res Eric Tartour, François Fossiez, Isabelle Joyeux, et al. Tumorigenicity of Human Cervical Tumors in Nude MiceInterleukin 17, a T-cell-derived Cytokine, Promotes
Updated version
http://cancerres.aacrjournals.org/content/59/15/3698
Access the most recent version of this article at:
Cited articles
http://cancerres.aacrjournals.org/content/59/15/3698.full#ref-list-1
This article cites 52 articles, 32 of which you can access for free at:
Citing articles
http://cancerres.aacrjournals.org/content/59/15/3698.full#related-urls
This article has been cited by 29 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cancerres.aacrjournals.org/content/59/15/3698To request permission to re-use all or part of this article, use this link
Research. on November 22, 2018. © 1999 American Association for Cancercancerres.aacrjournals.org Downloaded from