introduction to biological psychiatrypsych.lf1.cuni.cz/bpen/bpen1.pdf · 3 introduction biological...
TRANSCRIPT

INTRODUCTION TO BIOLOGICAL PSYCHIATRY
(http://www1.lf1.cuni.cz/~zfisar/bpen/default.htm)
RNDr. Zdeněk Fišar, CSc. Department of Psychiatry
1st Faculty of Medicine, Charles University, Prague
E-mail: [email protected]
This work was supported by FRVŠ 2448/2002 grant.

Zdeněk Fišar: Introduction to Biological Psychiatry
2
CONTENTS INTRODUCTION ......................................................................................................................................................... 3
1. NEUROBIOLOGY ........................................................................................................................................... 4 1.1. NEURON...................................................................................................................................................... 4 1.2. GLIA ........................................................................................................................................................... 6 1.3. PLASMA MEMBRANE ................................................................................................................................... 8 1.4. ACTION POTENTIAL................................................................................................................................... 11 1.5. ION CHANNELS.......................................................................................................................................... 13 1.6. SYNAPSE................................................................................................................................................... 15 1.7. CHEMICAL SYNAPSE ................................................................................................................................. 18
2. NEUROCHEMISTRY.................................................................................................................................... 22 2.1. CRITERIA TO IDENTIFY NEUROTRANSMITTERS .......................................................................................... 22 2.2. CLASSICAL NEUROTRANSMITTERS ............................................................................................................ 23 2.3. NEUROPEPTIDES ....................................................................................................................................... 27 2.4. MEMBRANE TRANSPORTERS ..................................................................................................................... 28 2.5. GROWTH FACTORS.................................................................................................................................... 30 2.6. MEMBRANE RECEPTORS............................................................................................................................ 31 2.7. RECEPTOR CLASSIFICATION ...................................................................................................................... 33 2.8. RECEPTORS COUPLED TO ION CHANNELS .................................................................................................. 33 2.9. RECEPTORS ASSOCIATED WITH G PROTEINS.............................................................................................. 35 2.10. RECEPTORS WITH INTRINSIC GUANYLYL CYCLASE ACTIVITY .................................................................... 35 2.11. RECEPTOR TYPES ...................................................................................................................................... 40 2.12. FEEDBACK AND CROSSCONNECTION ......................................................................................................... 45
3. PSYCHOPHARMACOLOGY....................................................................................................................... 49 3.1. ANTIPSYCHOTICS...................................................................................................................................... 51 3.2. ANTIDEPRESSANTS ................................................................................................................................... 53 3.3. MECHANISM OF ACTION OF ANTIDEPRESSANTS......................................................................................... 56
4. SCHIZOPHRENIA ......................................................................................................................................... 59 4.1. ENVIRONMENTAL MODELS........................................................................................................................ 60 4.2. GENETIC MODELS ..................................................................................................................................... 61 4.3. NEURODEVELOPMENTAL MODELS ............................................................................................................ 62 4.4. DELAYED ONSET OF SYMPTOMS................................................................................................................ 63 4.5. NEURODEGENERATIVE HYPOTHESIS ......................................................................................................... 63 4.6. BIOCHEMICAL BASIS OF SCHIZOPHRENIA .................................................................................................. 64
5. AFFECTIVE DISORDERS............................................................................................................................ 68 5.1. NEUROENDOCRINE HYPOTHESES............................................................................................................... 68 5.2. NEUROCHEMICAL HYPOTHESES ................................................................................................................ 70 5.3. NEUROTRANSMITTER HYPOTHESES........................................................................................................... 71 5.4. RECEPTOR HYPOTHESES............................................................................................................................ 73 5.5. POSTRECEPTOR HYPOTHESES .................................................................................................................... 76 5.6. MOLECULAR AND CELLULAR THEORY OF DEPRESSION.............................................................................. 77 5.7. MOLECULAR MECHANISM OF ACTION OF ANTIDEPRESSANT TREATMENTS ................................................ 77
6. LABORATORY SURVEY IN PSYCHIATRY ............................................................................................ 79 REFERENCES....................................................................................................................................................... 83

3
Introduction
Biological psychiatry occupies itself by mental disorders from biological, chemical and
physical point of view. The base of biological psychiatry is the assumption that human
mind is connected with human body so that mental disorders (processes) are
accompanied with biochemical changes, which can be measured. The second issue of
biological psychiatry is causality of all natural processes, including human mind. This
causality is very complexive and facts described by genetics, neuroendocrinology,
immunology, biochemistry, physics etc. are included.
Biological psychiatry studies disorders in human mind from the neurochemical,
neuroendocrine and genetic point of view mainly. It is postulated that changes in brain
signal transmission are essential in development of mental disorders.
The purpose of this teaching material is to describe cellular and molecular properties of
neurons, mechanisms of psychotropic drugs action and biological basis of mental
disorders. Familiarity with biochemistry and physiology is supposed.
Apportionable parts of this teaching material are dedicated to essence of neurobiology
and neurophysiology (neurons, synapses), neurochemistry (neurotransmitters,
neurotrophins and receptors), psychopharmacology and biological hypothesis of
schizophrenia or affective disorders.

Zdeněk Fišar: Introduction to Biological Psychiatry
4
1. NEUROBIOLOGY
The brain consists of neurons and glia. There are many different kinds of neurons and
several classes of glial cells. The glia outnumber the neurons ten times or more.
1.1. Neuron The neurons are the brain cells that are responsible for intracellular and intercellular
signalling. Neurons (Fig. 1.1) share many features in common with other cells, but they
are distinguished by their highly asymmetric shapes and by existence of dendrite
(whose function is reception of signals from the other neurons) and axon (which is
specialized for intracellular transmission of action potential from cell body to synapses).
Action potential is large and rapidly reversible fluctuation in the membrane potential,
that propagate along the axon. At the end of axon there are many nerve endings (or
synaptic terminals, presynaptic parts, synaptic buttons, knobs). Nerve ending form an
integral parts of synapse. Synapse mediates the signal transmission from one neuron
to another. The dendritic tree is in continual flux and revises its synaptic connections in
the course of life.
Cytoskeleton is important part of neuron. Cytoskeleton is heterogeneous network of
filamentous structures: major components are microfilaments, neurofilaments and
microtubules. The neuronal cytoskeleton is essential for establishing this cell shape
and for axonal transport (for moving vesicles and other organelles to regions remote
from the neuronal cell body). Protein called molecular motors makes use of the energy
released by hydrolysis of ATP to drive axonal transport.

1. NEUROBIOLOGY
5
Figure 1.1. Neurone
dendritic tree dendritic spines
mitochondria
Golgi complex
ribosome
lysosomes
microtubules
axon hillock
initial segment
myelin sheath
node of Ranvier
internodium
synaptic vesicles
plasma membrane
nucleus rough endoplasmic reticulum (Nissl substance) axon telodendrium synaptic terminals
neuronal cell body (soma)

Zdeněk Fišar: Introduction to Biological Psychiatry
6
1.2. Glia Glial cells (Fig. 1.2) are:
astrocytes
oligodendrocytes (forming the myelin sheath)
microglia
Glia are known to participate in many neuronal functions (Table 1.1). Myelination is
one role of glia that is well understood. The myelin sheath surrounds many, but not all,
axons in nervous system. It is formed by oligodendrocytes on central nervous system
axons and by Schwann cells in the peripheral nervous system. On Figure 1.3 you can
see cross section with an electron microscope - axon is surrounded by concentric
circles of glial membrane. A single Schwann cell may occupy up to about 1 mm of the
length of axon. There are gaps between adjacent Schwann cells of several
micrometers, known as nodes of Ranvier. Region between the nodes of Ranvier is
known as internode.
Table 1.1. Functions of glia
1. act as electrical insulator between neurons
2. participate in the uptake and metabolism of the neurotransmitters
3. act as a scaffolding for neuronal migration and axon outgrowth
4. take up and buffer ions from the extracellular environment
5. act as scavengers to remove debris produced by dying neurons
6. segregate groups of neurons one from another
7. provide structural support for neurons
8. play a role in information handling and memory storage

1. NEUROBIOLOGY
7
Figure 1.2. Glial cells
Figure 1.3. Myelin sheath
axon glial cell
glia
axon
paranodium nodium paranodium
node of Ranvier
astrocyte oligodendrocyte microglia

Zdeněk Fišar: Introduction to Biological Psychiatry
8
1.3. Plasma membrane All cells, including neurons and glia, are enclosed by a plasma membrane.
Plasma membrane acts as:
A barrier preventing the content of the cell from mixing with those of the
extracellular space
Effective electrical insulator
Regulator of transport of charged and no charged molecules in and out of the cell
Plasma membranes consist of (Figure 1.4):
Lipid bilayer (phospholipids, glycolipids, sphingomyelin, cholesterol)
Integral and peripheral proteins (ion channels, receptors, enzymes, transporters)
Main properties of lipid bilayer are included in model of fluid mosaic:
1. Heterogeneity in the plane horizontal and vertical
2. Fluid state of most lipids at physiological conditions
3. Translation and rotation movement of membrane molecules
Dynamic properties of biological membranes enable movement and mutual interactions
of membrane proteins (Figure 1.5), so membrane lipids can participate on signal
transduction.

1. NEUROBIOLOGY
9
Figure 1.4. Model of plasma membrane
peripheral glycoproteins glycolipids proteins
cholesterol integral proteins ion α-helix phospholipids channels

Zdeněk Fišar: Introduction to Biological Psychiatry
10
Figure 1.5. Lipid bilayer and function of membrane proteins
(See: Shinitzky M.: Membrane fluidity and receptor function. In: Membrane Fluidity, M. Kates, L.A. Manson, eds., Plenum Publ. Corp. 1984, pp. 585-601.)
movement movement perpendicular in the plane to membrane of membrane
lipid bilayer in fluid state
lipid bilayer in gel state

1. NEUROBIOLOGY
11
1.4. Action potential All living cell, including neurons, exhibit a voltage difference across plasma membrane
known as membrane potential. In neurons the resting potential is usually in the range
-40 to -90 mV. Information is carried from one part of the neuron to another in the form
of active response of membrane termed as action potential. Action potential is a large
and rapidly reversible fluctuation in the membrane potential, that propagate along the
axon. Generation and propagation of action potentials are enabled by membrane ion
channels.
Specific properties of axonal membranes allowing signal transmission in the form of
action potentials:
1. There is a threshold for generation of action potential; i.e. small random
fluctuations in the membrane potential are not interpreted as useful information.
2. The all-or-none law ensures full size of action potential.
3. There is frequency coding of the strength and latency of the initial stimulus.
4. Properties of the axonal membrane allow passive spread of the action potential
depolarization along the axon, which is essential in propagation of active
response. Saltatory conduction, i.e. jumping of action potentials along
myelinated axons, is enabled by passive spread of depolarizing stimulus and by
accumulation of ion channels in the nodes of Ranvier.
When the depolarization exceeds threshold the active response of the membrane
occurs and a large change in membrane potential is observed several milliseconds in
duration (Figure 1.6). The amplitude of the action potential is independent on stimulus
intensity. The membrane potential depolarises very rapidly, and then there is a slightly
less rapid return to the resting level. It is impossible to evoke another action potential
immediately after firing of an action potential; this absolute refractory period is followed
by relative refractory period, during which initial depolarization must be larger to evoke
action potential.
Amplitude of action potential carries little information about stimulus that triggered them;
in fact information about stimulus strength and latency can be coded in the frequency of
action potential firing (Figure 1.7).

Zdeněk Fišar: Introduction to Biological Psychiatry
12
Figure 1.6. Action potential
Figure 1.7. Frequency coding
(See: Levitan I.B. a Kaczmarek L.K.: The Neuron. Oxford Univ. Press, New York, Oxford, 1997; Ganong W.F.: Přehled lékařské fyziologie. Nakl. a vyd. H&H, 1995.)
equilibrium potential for Na+
40
∆ψ (mV) 20
0
- 20
- 40 threshold
- 60 resting potential
- 80 equilibrium potential for K+
1 2 4 8 16 32 depolarizing time (ms) stimulus
following following latency depolarization hyperpolarization
depolarizing stimulus
small short longer long large+long depolarization stimulus (mV)
+20
0
response ∆ψ (mV) +30
threshold resting potential
time (ms)

1. NEUROBIOLOGY
13
1.5. Ion channels Ion channels underlie electrical signalling in neurons. It is the sum of the various
currents flowing at any point that determines the neuron's membrane potential (Figure
1.8). The activities of the sodium and potassium channels responsible for axonal action
potentials are dependent on voltage.
Sequence of events during the nerve impulse in a large unmyelinated axon (Figure 1.9):
1. Membrane at rest
2. Initial stimulus → sodium channel opens → Na+ enters axon → membrane
depolarisation
3. At peak of impulse → sodium channel inactivates, potassium channel opens →
K+ leaves axon → refractory period
4. After refractory period → sodium channel inactivation removed → both channels
closed
It is now evident that there is a considerable diversity of ion currents in axons, cell
bodies and dendrites. Voltage clamp and patch clamp techniques make possible
revealing of heterogeneity of voltage dependent sodium, calcium, and potassium
channels.

Zdeněk Fišar: Introduction to Biological Psychiatry
14
Figure 1.8. Membrane permeability for Na+ and K+ during action potential
(see Voet D., Voetová J.G.: Biochemie. Victoria Publ., Praha, 1995.)
Figure 1.9. Ion flow during action potential
(See: Atwood H.L., MacKay W.A.: Essentials of Neurochemistry. B.C. Decker Inc., Toronto, Philadelphia, 1989.)
30
20
10
0
1 2 3 time (ms)
20
0
- 20
- 40
- 60
Ion
perm
eabi
lity
(mS/
cm2 )
∆ψ (m
V)
action potential
permeability for Na+
permeability for K+
Na+ channel K+ channel
resting state
depolarization
repolarization
hyperpolarization
beginning of depolarizing stimulus rapid opening of Na+ channels; slow opening of K+ channels K+ channels open; Na+ channels inactivated (refractory period) slow closing of K+ channels; removal of inactivation and closed Na+ channels

1. NEUROBIOLOGY
15
1.6. Synapse Neurons communicate with one another by direct electrical coupling or by the secretion
of neurotransmitters (Table 1.2).
Table 1.2. Intercellular communication
1. direct transfer of molecules and ions from the cytoplasm of one cell into that of
another (gap junction; electrical synapse)
2. the release of a chemical that diffuses to, and acts on, another cell (secretion;
chemical synapse)
3. direct physical contact of cell surfaces
Electrical coupling is mediated by connexins, i.e. proteins that form pores linking the
cytoplasm of adjacent cells, so ions and small molecules can diffuse through pores from
one cell to another.
Neurosecretion is a more complex process. Stimulation of cells is followed by an
elevation of intracellular calcium, which is important intracellular messenger. Increased
presynaptic Ca2+ levels allow the vesicles to fuse with the plasma membrane and to
release their contents in the synaptic cleft.
Synapses are specialized structures for signal transduction from one neuron to other.
Chemical synapses are studied in the biological psychiatry. Morphology of chemical
synapse is shown on Figure 1.10. Different kinds of synapses are shown on Figure
1.11.

Zdeněk Fišar: Introduction to Biological Psychiatry
16
Figure 1.10. Morphology of chemical synapse
(See: Němeček S. a kol.: Neurobiologie, Avicenum, Praha 1972)
axon
microtubules mitochondria synaptic vesicles
presynaptic membrane
synaptic cleft postsynaptic membrane
postsynaptic cell
glia presynaptic cell

1. NEUROBIOLOGY
17
Figure 1.11. Synapses
(See: Němeček S. a kol.: Neurobiologie, Avicenum, Praha 1972; modified.)
axo-dendritic synapse axo-somatic synapse axo-axonic synapse button from node of Ranvier axo-axonic synapse
reverse collaterale reverse collaterale presynaptic elements en passant synaptic knob (terminal button)

Zdeněk Fišar: Introduction to Biological Psychiatry
18
1.7. Chemical synapse The main interest of biological psychiatry is knowledge of changes in signal transduction
through chemical synapse (Figure 1.12) in the mental disorder and during its treatment.
It is known that many psychotropic drugs act on the level of chemical synapse; it is on
the level of changes of availability of neurotransmitters, regulation of receptors and
adaptation of post receptor events. Therefore this teaching material is dedicated to
molecular mechanisms of nerve signal transduction.
Table 1.3. Basic steps in signal transduction
1. Depolarisation of presynaptic membrane leads to opening of voltage gated
calcium channels.
2. Ca2+ enters to presynaptic part and activates many enzymes.
3. Neurotransmitters are released from synaptic vesicles by exocytosis to synaptic
cleft. They diffuse to postsynaptic and presynaptic membrane receptors and
specifically bind to them.
4. Receptors are activated and either ion channels are opened or closed, or
activities of intracellular enzymes are changed. These changes lead to
physiological effect of signal transduction.
Release of neurotransmitters is closely linked to the entry of calcium. With repetitive
stimulation, phenomenon as facilitation, potentiation and depression of neurotransmitter
release may occur.
Following neurotransmitter release postsynaptic or presynaptic membrane receptors are
activated. In inhibition synapse efflux of potassium ions causes hyperpolarization of
postsynaptic membrane; in excitation synapse influx of sodium ions causes
depolarization of postsynaptic membrane (Figure 1.13).
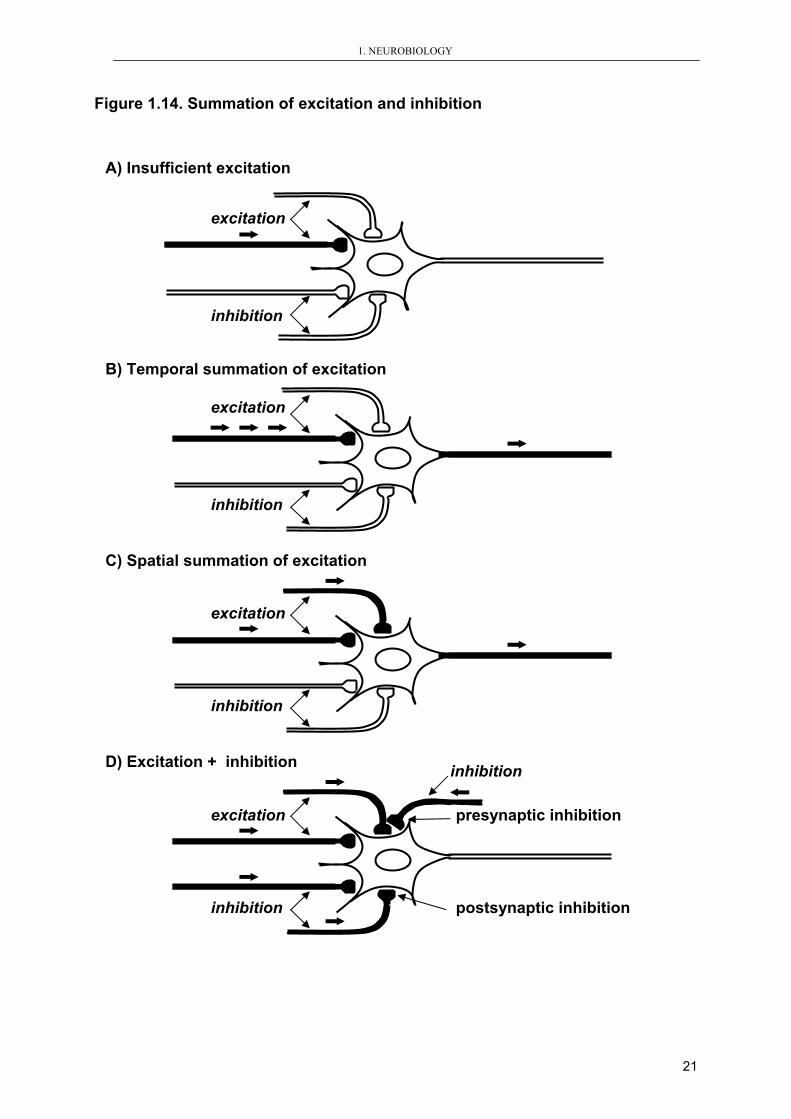
Temporal and/or spatial summation of synaptic potentials is necessary to the evocation
of desirable physiological effect since the amount of neurotransmitters released from
one vesicle is generally insufficient (Figure 1.14).

1. NEUROBIOLOGY
19
Figure 1.12. Chemical synapse - signal transduction
PRESYNAPTIC PART action potential
- Ca2+ entry through voltage-gated channel
- Ca2+-catalysed reaction → exocytosis
- Inactivation of intracellular Ca2+
- Diffusion of neurotransmitter
- Reaction with receptors of postsynaptic cell and changes in membrane permeability
Ca2+

Zdeněk Fišar: Introduction to Biological Psychiatry
20
Figure 1.13. Excitation and inhibition postsynaptic potentials
(See: Netter F.H.: Physiology and Functional Neuroanatomy. Nervous System I. CIBA-GEIGY Ltd., Basle, 1989.)
Na+ K+ Cl- K+
0 4 8 12 16time (ms)
∆ψ (mV)
-65
-70
resulted ion current
excitation postsynaptic potential
time (ms) 0 4 8 12 16
-70
-65
∆ψ (mV)
resulted ion current
inhibition postsynaptic potential
synaptic vesicles
synaptic cleft (released transmitters)
postsynaptic membrane
excitation synapse inhibition synapse
1. NEUROBIOLOGY
21
Figure 1.14. Summation of excitation and inhibition
excitation
inhibition
B) Temporal summation of excitation
excitation
inhibition
A) Insufficient excitation
excitation
inhibition
C) Spatial summation of excitation
excitation presynaptic inhibition
inhibition postsynaptic inhibition
inhibition D) Excitation + inhibition

Zdeněk Fišar: Introduction to Biological Psychiatry
22
2. NEUROCHEMISTRY
2.1. Criteria to identify neurotransmitters A multitude of chemicals called neurotransmitters mediate intercellular communication
in the nervous system. Although they exhibit great diversity in many of their properties,
all are stored in vesicles in nerve terminals and are released to the extracellular space
via process requiring calcium ions. Their action is terminated by reuptake into
presynaptic terminal or glial cells by specific transporter proteins or by catabolism in
synaptic cleft or in presynaptic terminal. Criteria to identify neurotransmitters are shown
in the Table 2.1.
Table 2.1. Criteria to identify neurotransmitter
1. presence in presynaptic nerve terminal
2. synthesis by presynaptic neuron
3. releasing on stimulation (membrane depolarisation)
4. producing rapid-onset and rapidly reversible responses in the target cell
5. existence of specific receptor
From the chemical point of view are neurotransmitters monoamines, amino acids and
peptides. There are two main groups of neurotransmitters:
1. classical neurotransmitters
2. neuropeptides

2. NEUROCHEMISTRY
23
2.2. Classical neurotransmitters All classical neurotransmitters (Table 2.2) are synthesized in nerve terminals.
The first molecule to be implicated as neurotransmitter was acetylcholine (Ach). It was
demonstrated that acetylcholine is the transmitter at neuromuscular synapses, as well
as at a variety of neuron-neuron synapses. Ach is synthesized from choline and
acetyl-coenzyme A in the nerve endings; reaction is catalyzed by the enzyme choline
acetyltransferase (Figure 2.1). ACh is rapidly degraded in synaptic cleft by the enzyme
acetylcholinesterase to choline and acetate.
Catecholamines (dopamine, norepinephrine and epinephrine) are synthesized from
tyrosine (Figure 2.2). Indolamines (serotonin and tryptamine) are synthesized from
tryptophan (Figure 2.3.). Action of catecholamines and indolamines on target cells is
terminated much more slowly than those of acetylcholine; they are removed from
synaptic cleft by reuptake. The major enzymes involved in the catabolism of
catecholamines are monoamine oxidase (MAO) or catechol-O-methyltransferase
(COMT). Serotonin is metabolized by MAO.
There are evidence that dysfunction in brain catecholamine or indolamine pathways
contribute to affective disorders and schizophrenia. Some antidepressant prolongs
monoamine neurotransmitters action by inhibiting their high affinity reuptake system.
Catecholamine theory of psychotic illness focuses on dysfunction at dopaminergic
synapses.
There are three major amino acid neurotransmitters in the nervous system: γ-amino
butyric acid (GABA), glycine and glutamic acid. GABA and glycine are inhibitory
neurotransmitters; glutamate and aspartate are excitatory neurotransmitters.
Specific properties have nitric oxide, which does not interact with membrane receptors
but diffuse to target intracellular receptor.

Zdeněk Fišar: Introduction to Biological Psychiatry
24
Table 2.2. Selected classical neurotransmitters
System Transmitter Precursor
cholinergic acetylcholine choline+acetylcoenzym A
GABA glucose → glutamate
aspartic acid glucose+glutamine; glutamate
glutamic acid glucose+glutamine; aspartate
glycine serine
aminoacidergic
homocysteine cysteine→cystine
monoaminergic
dopamine tyrosine→DOPA→dopamine
norepinephrine →norepinephrine→epinephrine
catecholamines
epinephrine
tryptamine indolamines
serotonin tryptophan→5-hydroxytryptophan
histamine histidine others, related to aa
taurine cysteine→cysteamine
adenosine
ADP
AMP
purinergic
ATP

2. NEUROCHEMISTRY
25
Figure 2.1. Synthesis and release of acetylcholine AXON axoplasmic transport
of choline acetyltransferase (synthesized in soma)
NERVE TERMINAL glucose uptake choline uptake glucose metabolism mitochondrial enzymes acetyl-CoA + choline choline acetyltransferase acetylcholine + CoA vesicles free acetylcholine (cytoplasmic) SYNAPTIC released acetylcholine CLEFT
hydrolysis by acetylcholinesterase choline + acetate

Zdeněk Fišar: Introduction to Biological Psychiatry
26
Figure 2.2. Catecholamine biosynthesis
L-tyrosine
tyrosine hydroxylase L-DOPA (3,4-dihydroxyphenylalanine)
DOPA decarboxylase dopamine (DA)
dopamine-β-hydroxylase norepinephrine (NE; also called noradrenaline or NA)
phenylethanolamine N-methyltransferase epinephrine (adrenaline)
Figure 2.3. Serotonin biosynthesis tryptophan
tryptophan hydroxylase 5-hydroxytryptophan
decarboxylase 5-hydroxytryptamine (5-HT, serotonin)

2. NEUROCHEMISTRY
27
2.3. Neuropeptides It was identified a great number of neuropeptides (Table 2.3) and new neuropeptides
are discovered continually. β-endorphin is important in psychiatry, since it has a role in
pain and in stress. All neuropeptides are synthesized in the cell body in following steps:
1. transcription of pre-propeptide gene and creation of pre-propeptide RNA; 2.
translation into pre-propeptide, which enters the endoplasmic reticulum; 3. forming of
the propeptide, which is direct precursor of neuropeptide; 4. propeptide enters vesicles,
where it is converted into the neuropeptide. The action of neuropeptides in the synaptic
cleft is terminated by peptidases; there is no reuptake for neuropeptides. Neuropeptides
are co-transmitters usually and their transduction mechanism is coupled with G
proteins.
Table 2.3. Selected bioactive peptides
Peptide Group substance P, substance K (tachykinins) neurotensin cholecystokinin (CCK) gastrin bombesin
brain and gastrointestinal peptides
galanin neuromedin K neuropeptideY (NPY) peptide YY (PYY)
neuronal
cortikotropin releasing hormone (CRH) growth hormone releasing hormone (GHRH) gonadotropin releasing hormone (GnRH) somatostatin thyrotropin releasing hormone (TRH)
hypothalamic releasing factors
adrenocorticotropic hormone (ACTH) growth hormone (GH) prolactin (PRL) lutenizing hormone (LH) thyrotropin (TSH)
pituitary hormones
oxytocin vasopressin neurohypophyseal peptides
atrial natriuretic peptide (ANF) vasoactive intestinal peptide (VIP) neuronal and endocrine
enkephalines (met-, leu-) dynorphin β-endorphin
opiate peptides

Zdeněk Fišar: Introduction to Biological Psychiatry
28
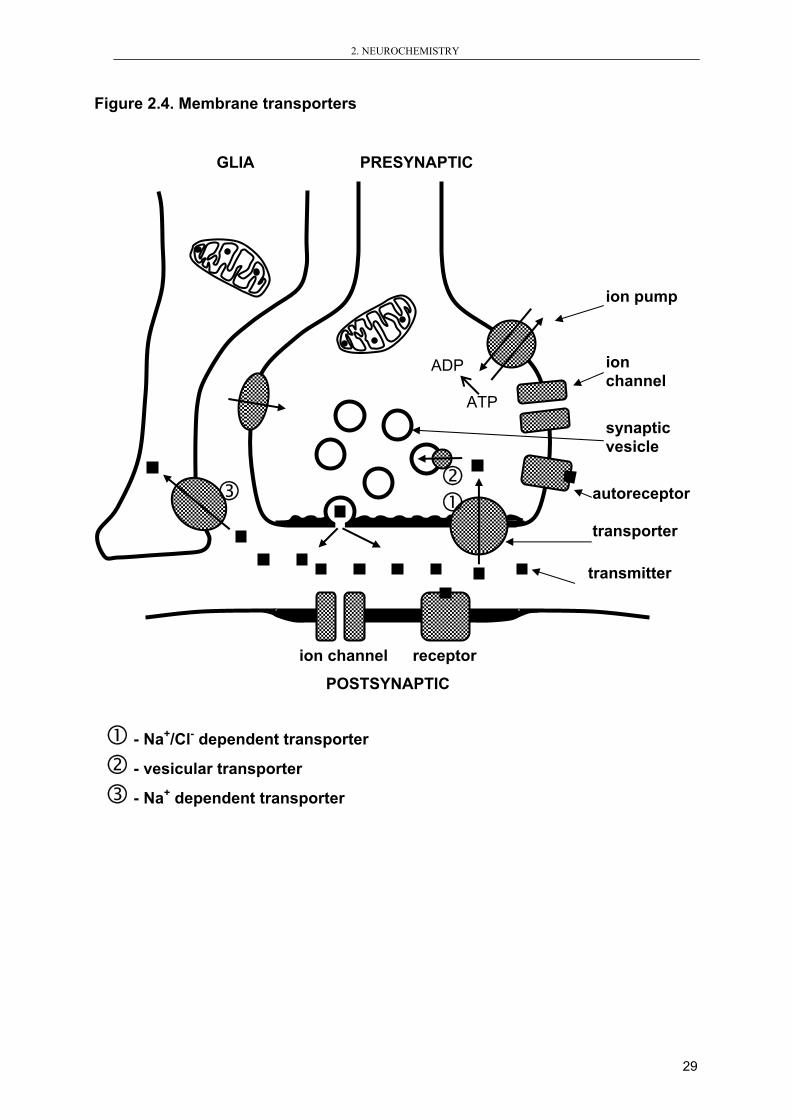
2.4. Membrane transporters Neurotransmitters are removed from synaptic cleft by enzymatic degradation or by
active transport to presynaptic button or to surrounding glia cells. The main way of
acetylcholine removing from synaptic cleft is metabolism by acetylcholinesterase. But
for majority of neurotransmitters speculated in biochemical hypothesis of mental
disorders there are specific membrane transporters (Figure 2.4).
The main classes of membrane transporters:
1. Na+/Cl- dependent transporters are embedded in the presynaptic membrane
mainly; they transport serotonin, norepinephrine or dopamine
2. Vesicular transporters carries neurotransmitters into synaptic vesicles (it is
important to prevent neurotransmitter degradation)
3. Na+ dependent transporters are localized in the membrane of glia cells; they
transport GABA, glutamate or aspartate
2. NEUROCHEMISTRY
29
Figure 2.4. Membrane transporters
GLIA PRESYNAPTIC
ion channel receptor
POSTSYNAPTIC
- Na+/Cl- dependent transporter
- vesicular transporter
- Na+ dependent transporter
ion pump
ion channel
synaptic vesicle
autoreceptor transporter transmitter
ADP ATP

Zdeněk Fišar: Introduction to Biological Psychiatry
30
2.5. Growth factors Growth factors are proteins that stimulate cellular proliferation and promote cellular
survival. They are essential for nervous system development and function. Different
cells including neurons and glia produce growth factors. They are released from cell and
after interaction with membrane receptors changes in activity of intracellular enzymes
occur. So, mechanism of their action is similar to action of neurotransmitters, but they
are not released in response to membrane depolarization and changes intracellular
calcium levels.
There are 6 major classes of growth factors, which act within nervous system (Table
2.4.). From the psychiatry point of view neurotrophins are most important.
Neurotrophins support the survival and phenotypic specificity of subsets of neurons.
Brain-derived neurotrophic factor (BDNF) has a role in response on stress and in
action of antidepressants.
Table 2.4. Growth factors in the nervous system
Neurotrophins
Nerve growth factor (NGF)
Brain-derived neurotrophic factor (BDNF) Neurotrophin 3 (NT3)
Neurotrophin 4/5 (NT4/5)
Neurokines Ciliary neurotrophic factor (CNTF)
Leukemia inhibitory factor (LIF)
Interleukin 6 (IL-6)
Cardiotrophin 1 (CT-1)
Fibroblast growth factors FGF-1
FGF-2
Transforming growth factor β superfamily Transforming growth factors β (TGFβ)
Bone morphogenetic factors (BMPs)
Glial-derived neurotrophic factor (GDNF)
Neurturin
Epidermal growth factor superfamily Epidermal growth factor (EGF)
Transforming growth factor α (TGFα)
Neuregilins
Other growth factors Platelet-derived growth factor (PDGF)
Insulin-like growth factor I (IGF-I)

2. NEUROCHEMISTRY
31
2.6. Membrane receptors Receptor is macromolecule specialized on transmission of information. It is defined as
binding site with functional relationships. A binding site is not necessarily a physiological
receptor, since neurotransmitters must bind to uptake systems and metabolic enzymes
also. To determine whether a binding site is real receptor pharmacology of binding site
and biological response must coincide.
Receptor complex includes:
1. Specific binding site
2. Transduction element
3. Effector system (2nd messengers)
Effector system includes enzymes adenylyl cyclase or phospholipase, which generate
2nd messengers, such as cyclic adenosine monophosphate (cAMP), cyclic guanosine
monophosphate (cGMP), inositoltriphosphate (IP3), diacylglycerol (DG), calcium (Ca2+);
second messengers activate protein kinases A, G, C or CaM-II.
Receptors are able to adapt their properties to increased or decreased activation (Table
2.5, Figure 2.5). Changes in the number of receptors are known mechanism of their
adaptation. But response to receptor activation can be altered at unchanged density of
receptors too. Regulation of properties of receptors consists of decreased or increased
activity of post receptor events, which results in decreased or increased final
physiological response to receptor stimulation (Table 2.6).
Table 2.5. Regulation of receptors
Number of receptors (down-regulation, up-regulation)
Properties of receptors (desensitisation, hypersensitivity)
Table 2.6. Mechanisms of receptor desensitisation
Interactions of subunits of active molecules (by phosphorylation, ribosylation, changes in membrane lipid composition, etc.) Production of messengers - inhibitors of receptors
Production of receptor clusters (by cytoskeleton)
Internalisation of receptors (by endocytosis)

Zdeněk Fišar: Introduction to Biological Psychiatry
32
Figure 2.5. Adaptation of receptors
reactive hyperfunction (upregulation)
balance
secondary hyperfunction
adaptive hypofunction (desensitization)
secondary hypofunction
RECEPTOR
NEUROTRANSMITTER

2. NEUROCHEMISTRY
33
2.7. Receptor classification There are many types of receptors and they can be classified by different criteria, for
example according to their pharmacological properties (it is according to activating
neurotransmitter or other agonist) or according to effectors system, which is connected
with their function (Table 2.7).
Table 2.7. Classification of receptors by their effector system
1. Receptor coupled directly to the ion channel
2. Receptor associated with G proteins
3. Receptor with intrinsic guanylyl cyclase activity
4. Receptor with intrinsic tyrosine kinase activity
2.8. Receptors coupled to ion channels Direct coupling of the neurotransmitter receptor to the ion channel whose activity it
regulates is the simplest and the most rapid way of signal transduction. An example of
receptor with internal ion channel is GABAA receptor, nicotinic acetylcholine receptors,
or ionotropic glutamate receptors.
Nicotinic acetylcholine receptor complex is one of ligand-gated ion channels; it
contains both the acetylcholine binding site and the ion channel that is activated by
acetylcholine binding. Following receptor activation, nicotinic acetylcholine receptors as
well as ionotropic glutamate receptors increase membrane permeability for cations
Na+, K+ or Ca2+; they are excitatory receptors.
Receptor for γ-amino butyric acid includes chlorine (Cl-) channel, which is opened in
response to binding of GABA (Figure 2.6). Chlorine inputs into cell and
hyperpolarization of membrane occurs; so it is inhibitory receptor. There are many
modulation sites on GABAA receptor, for example benzodiazepines positively modulate
activity of this receptor.

Zdeněk Fišar: Introduction to Biological Psychiatry
34
Figure 2.6. GABAA receptor
Pi
Cl-
barbiturates GABA, bicuculine neurotransmitter binding site neurosteroids benzodiazepines
allosteric modulatory sites picrotoxin phosphorylation pore ~0.5 nm
receptor formed by 5 subunits

2. NEUROCHEMISTRY
35
2.9. Receptors associated with G proteins It is assumed that signal transduction mediated by receptors associated with guanyl
nucleotide-binding proteins (or G proteins) and 2nd messenger systems are altered
at mental disorders and during treatment with psychotropic drugs. Scheme for
hypothesis of second messengers in signal transduction (Figure 2.7) show that after
activation of receptor by first messenger G proteins are activated and activated G
proteins activate effectors enzymes, such as adenylyl cyclase or phospholipase C, and
second messengers are produced. Second messenger activate protein kinase of type
A, C or CaM-II, which catalyses phosphorylation of cell proteins and physiological
response to receptor activation arises. Transcription factors can be phosphorylated too,
and phosphorylated transcription factors serve as third messengers, which activate
gene expression.
According to type of 2nd messenger used there are two main pathways of signal
transduction: 1. adenylyl cyclase system; 2. phosphoinositide system.
In adenylyl cyclase system (Figure 2.8) activated receptor activates G protein, which
activates adenylyl cyclase and cyclic adenosine monophosphate (cAMP) is produced as
2nd messenger. cAMP activates protein kinase A, which phosphorylates cellular proteins
(receptors, ion channels, enzymes, transcription factors, etc.).
In phosphoinositide system (Figure 2.9) activated receptor activates G proteins,
which activate phospholipase C, and inositoltriphosphate (IP3) and diacylglycerol (DG)
are produced as 2nd messengers. DG activates protein kinase C, IP3 activate releasing
of Ca2+ from intracellular pool and Ca2+ activate many enzymes including calmoduline
and protein kinase dependent on Ca2+ and calmoduline.
2.10. Receptors with intrinsic guanylyl cyclase activity Cyclic guanosine monophosphate (cGMP) is produced as second messenger after
activation of membrane bound or soluble form of guanylyl cyclase system (Figure
2.10). cGMP can activate protein kinase, activate or inhibit several forms of
phosphodiesterase, open up cation channels (in retina), etc.

Zdeněk Fišar: Introduction to Biological Psychiatry
36
Figure 2.7. Scheme for hypothesis of second messengers
receptors
G proteins
effector proteins
second messengers
protein kinase
biological response
gene expression etc.
first messengers
third messengers

2. NEUROCHEMISTRY
37
Figure 2.8. Signal transduction: Adenylyl cyclase system
Gs/i/o/x – G proteins, CaM – calmodulin, GTP – guanosine triphosphate, ATP – adenosine triphosphate, ADP – adenosine diphosphate, cAMP – cyclic adenosine monophosphate, 5´-AMP - 5´-adenosine monophosphate, PKA – protein kinase A, PKC – protein kinase C
free Ca2+ PKA, PKC Ca2+/CaM
5´-AMP
fosfodiesterase protein kinase A
γ γ β β Gs Gi/o/x αs-GTP αi/o/x-GTP Ca2+
activated adenylyl cyclase activated Ca2+- receptor receptor channel forskolin
ATP
cAMP + ADP
activated protein
ADP + Pi
ATP

Zdeněk Fišar: Introduction to Biological Psychiatry
38
Figure 2.9. Signal transduction: Phosphoinositide system
PI-PLC - phospholipase C specific for phosphoinositides, PIP2 - phosphatidylinositol-4,5-biphosphate, IP3 - inositol-1,4,5-triphosphate; DG - diacylglycerol; PKC – protein kinase C
γ γ Pi Pi β β
Gq αq PKC Gi/o PI-PLCγ αi/o
IP3 protein phosphorylation
Ca2+-dependent Ca2+
processes IP3 receptor
endoplasmic reticulum
receptor receptor receptor Ca2+- with intrinsic associated with s Gq associated with Gi/o channel tyrosin kinase PI-PLCβ1 DG PI-PLCβ2
PIP2 PIP2

2. NEUROCHEMISTRY
39
Figure 2.10. Signal transduction: Guanylyl cyclase system
GC – guanylyl cyclase, CaM – calmodulin, NO – nitric oxide, nNOS – nitric oxide synthase, ATP – adenosine triphosphate, cGMP – cyclic guanosine monophosphate
GC-A GC-B GC-C Ca2+-channel
Ca2+
Ca2+/CaM
nNOS
NO cGMP
biological response
ATP
catalytic domain
soluble GC
membrane bound GC
intracellular stores of Ca2+

Zdeněk Fišar: Introduction to Biological Psychiatry
40
2.11. Receptor types Basic characteristics of receptors which can be used to their classification are shown in
Table 2.8. Primary types and selected subtypes of receptors (according to the
international agreement) are summarized in Tables 2.9, 2.10, 2.11, 2.12 and 2.13.
Table 2.8. Basic characteristics of receptors
Name
Endogenous ligands and their efficiency
Selective agonists
Selective antagonists
Blockers of ion channels
Selective radioligands
cGMP (internal guanylyl cyclase)
Gi/o (K+↑, Ca2+↓, cAMP↓)
Gq/11 (IP3, DG)
Gs (cAMP↑)
Effectors
internal ion channel (Na+/K+/Ca2+/Cl-)
Gen
Structure

2. NEUROCHEMISTRY
41
Table 2.9. Type of receptors
System Type
acetylcholine nicotinic receptors acetylcholinergic
acetylcholine muscarinic receptors
α1-adrenoceptors
α2-adrenoceptors
β-adrenoceptors
dopamine receptors
monoaminergic
serotonin receptor
GABA receptors
glutamate ionotropic receptors
glutamate metabotropic receptors
glycine receptors
aminoacidergic
histamine receptors
opioid receptors peptidergic
other peptide receptors
adenosine receptors (P1 purinoceptors) purinergic
P2 purinoceptors

Zdeněk Fišar: Introduction to Biological Psychiatry
42
Table 2.10. Subtypes of monoamine receptors
RECEPTORS Subtype Transducer Structure (aa/TM)α1A Gq/11 ↑IP3/DAG 466/7 α1B Gq/11 ↑IP3/DAG 519/7
α1-adrenoceptors
α1D Gq/11 ↑IP3/DAG 572/7 α2A Gi/o cAMP 450/7 α2B Gi/o cAMP 450/7 α2C Gi/o cAMP 461/7
α2-adrenoceptors
α2D Gi/o cAMP 450/7 β1 Gs ↑cAMP 477/7 β2 Gs ↑cAMP 413/7
β-adrenoceptors
β3 Gs, Gi/o ↑cAMP 408/7 D1 Gs ↑cAMP 446/7 D2 Gi
Gq/11 cAMP ↑IP3/DAG, ↑K+, ↓Ca2+
443/7
D3 Gi cAMP 400/7 D4 Gi cAMP, ↑K+ 386/7
dopamine
D5 Gs ↑cAMP 477/7 5-HT1A Gi/o cAMP 421/7 5-HT1B Gi/o cAMP 390/7 5-HT1D Gi/o cAMP 377/7 5-ht1E Gi/o cAMP 365/7 5-ht1F Gi/o cAMP 366/7 5-HT2A Gq/11 ↑IP3/DAG 471/7 5-HT2B Gq/11 ↑IP3/DAG 481/7 5-HT2C Gq/11 ↑IP3/DAG 458/7 5-HT3 internal cationic channel 478 5-HT4 Gs ↑cAMP 387/7 5-ht5A ? 357/7 5-ht5B ? 370/7 5-ht6 Gs ↑cAMP 440/7
5-hydroxytryptamine
5-HT7 Gs ↑cAMP 445/7

2. NEUROCHEMISTRY
43
Table 2.11. Subtypes of aminoacidergic receptors
RECEPTORS Subtypes Transducer Structure (aa/TM)
GABAA int. Cl- more subunits GABA
GABAB Gi/o, Gs cAMP, ↑cAMP,
↑K+(G), ↑Ca2+(G)
961,941/7
NMDA int. Na+/K+/Ca2+ more subunits
AMPA int. Na+/K+/Ca2+ more subunits
excitatory aa
(ionotropic)
kainate int. Na+/K+/Ca2+ more subunits
mglu1 Gq/11 ↑IP3/DAG 1194/7
mglu2 Gi/o cAMP 872/7
mglu3 Gi/o cAMP 877/7
mglu4 Gi/o cAMP 912/7
mglu5 Gq/11 ↑IP3/DAG 1212/7
mglu6 Gi/o cAMP 877/7
mglu7 Gi/o cAMP 915/7
excitatory aa
(metabotropic)
mglu8 Gi/o cAMP 908/7
glycine glycine int. Cl- pentamer
H1 Gq/11 ↑IP3/DAG 487/7
H2 Gs ↑cAMP 359/7
histamine
H3 Gi/o cAMP 445/7

Zdeněk Fišar: Introduction to Biological Psychiatry
44
Table 2.12. Subtypes of acetylcholine receptors
RECEPTORS Subtypes Transducer Structure (aa/TM)
M1 Gq/11, NO ↑IP3/DAG 460/7
M2 Gi/o cAMP,↑K+(G) 466/7
M3 Gq/11, NO ↑IP3/DAG 590/7
M4 Gi/o cAMP,↑K+(G) 479/7
muscarinic
M5 Gq/11, NO ↑IP3/DAG 532/7
α1 int. Na+/K+/Ca2+ pentamer
α2 int. Na+/K+/Ca2+ pentamer
α3 int. Na+/K+/Ca2+ pentamer
α4 int. Na+/K+/Ca2+ pentamer
α6 int. Na+/K+/Ca2+ pentamer
α7 int. Na+/K+/Ca2+ pentamer
α8 int. Na+/K+/Ca2+ pentamer
nicotinic
α9 int. Na+/K+/Ca2+ pentamer
Table 2.13. Subtypes of opioid and related peptidergic receptors
RECEPTORS Subtypes Transducer Structure (aa/TM)
mu opioid peptidergic
(µ)
Gi/
o
cAMP,
↑K+(G),↓Ca2+(G)
400/7
delta opioid
peptidergic (δ )
Gi/
o
cAMP,
↑K+(G),↓Ca2+(G)
372/7
kappa opioid
peptidergic (κ)
Gi/
o
cAMP,
↑K+(G),↓Ca2+(G)
380/7
opioid and
similar receptors
N/OFQ receptor Gi/
o
cAMP,
↑K+(G),↓Ca2+(G)
370/7

2. NEUROCHEMISTRY
45
2.12. Feedback and crossconnection There are many neurotransmitters, many receptors, less G proteins and a few effector
systems. But there are many feedbacks at the cellular level and many cross-reactions at
the intracellular level. That results in divergence or convergence of signals.
Example of negative feedback in synapse (Figure 2.11):
Norepinephrine released from noradrenergic synapse can diffuse to adjacent
acetylcholinergic synapse and can activate presynaptic α2 receptors. Because α2
receptors are inhibitory receptors, activation of noradrenergic synapse can lead to
inhibition of acetylcholinergic synapse. There is great number of similar feedbacks.
Example of intracellular cross connection (Figure 2.12):
Activation of serotonin 2 receptor leads to production of diacylglycerol (DG) as 2nd
messenger and DG activates protein kinase C (PKC). PKC can phosphorylate
adenylyl cyclase and positive regulation of transduction system connected with
adrenoreceptors occurs. PKC can affect noradrenergic signal transduction through
direct phosphorylation of receptors or G proteins also.
General scheme of signal transduction implying feedbacks is shown on Figure 2.13.

Zdeněk Fišar: Introduction to Biological Psychiatry
46
Figure 2.11. Feedback to transmitter-releasing
(See: Atwood H.L., MacKay W.A.: Essentials of Neurochemistry. B.C. Decker Inc., Toronto, Philadelphia, 1989.)
-
-
α2-adrenoceptor (autoinhibition)
NE
NE
α1,2-adrenoceptors
+ -
muscarinic receptor (M2,M4) (autoinhibition)
muscarinic or nicotinic receptors
ACh
ACh
+ -
-
α2
muscarinic receptor
ACh
ACh
muscarinic receptor
NE
NE
α1-adrenoceptor
+ -
- -
-
adenosine receptor (autoinhibition)
P2 purinoceptor
+ -
ATP
ATP

2. NEUROCHEMISTRY
47
Figure 2.12. Crossconnection of transducing systems on postreceptor level
AR – adrenoceptor, G – G protein, PI-PLC – phosphoinositide specific phospholipase C, IP3 – inositoltriphosphate, DG – diacylglycerol, CaM – calmodulin, AC – adenylyl cyclase, PKC – protein kinase C
PKC
AC
5-HT2 β-AR α2-AR
Ca2+
Ca2+/CaM
IP3 + DAG
PIP2
Gq/11 Gs Gi/o
PI-PLC
-
+
+
-

Zdeněk Fišar: Introduction to Biological Psychiatry
48
normal
Pi
++ -
+
+
excitation inhibition
GαGTP GαGTP
IP3, DG, Ca2+, cAMP, FFA
PLC, PLD, PLA2, AC
gene expression
PKC, PKA, PKCaM
PP
neurotransmitters membrane receptors G proteins effector enzymes second messengers protein kinases and gene expression phosphoprotein phosphatases
+-+-
Pi Pi Pi
Figure 2.13. Scheme of signal transduction (implying feedback)
G – G protein, GTP – guanosine triphosphate, PLC - phospholipase C, PLD - phospholipase D, PLA2 - phospholipase A2, AC – adenylyl cyclase, IP3 – inositoltriphosphate, DG – diacylglycerol, cAMP – cyclic adenosine monophosphate, FFA – free fatty acids, PKC – protein kinase C, PKA – protein kinase A, PKCaM – Ca2+ and calmodulin dependent protein kinase, PP – phosphoprotein phosphatase

3. PSYCHOPHARMACOLOGY
49
3. Psychopharmacology Psychopharmacology is very extensive branch of medicine. This chapter will emphasize
basic pharmacological concepts of action of antipsychotics and antidepressants only.
The reader should consult standard reference sources for more information from
psychopharmacology.
Psychotropic drugs are amphiphilic molecules frequently; i.e. they are soluble both in
the water phase and in the lipid bilayer. Amphiphilic drugs rapidly permeate through
plasma membrane and/or accumulate in hydrophobic interior of lipid bilayer (Figure
3.1), so, interactions are possible both with membrane macromolecules and with
cytoplasmatic or nuclear molecules.
Central nervous system drugs act primary as agonists or antagonists of
neurotransmitter receptors, inhibitors of regulatory enzymes or blockers of stimulators of
neurotransmitter membrane transporters (Table 3.1). Classification psychotropic drugs
according to effects on mental functions are shown in Table 3.2.
Figure 3.1. Interaction of amphiphilic drugs with membrane
out
in
membrane

Zdeněk Fišar: Introduction to Biological Psychiatry
50
Table 3.1. Potential action of psychotropics
1. synthesis and storage of neurotransmitter
2. releasing of neurotransmitter
3. receptor-neurotransmitter interactions (blockade of receptors)
4. catabolism of neurotransmitter
5. reuptake of neurotransmitter
6. transduction element (G protein)
7. effector system
Table 3.2. Classification psychotropics according to effects on mental functions
parameter effect group of psychotropics
examples
positive psychostimulant drugs
amphetamine, amphetaminile, ephedrine, fenmetrazine, mazindole, mezokarb, pemoline, methylphenidate
watchfulness (vigility)
negative hypnotic drugs barbital, amobarbital, hexobarbital; glutethimid, metachalone, clomethiazol; nitrazepam, flunitrazepam, triazolam; zopiclone, zolpidem
antidepressants imipramine, desipramine, amitriptyline, nortriptyline, clomipramine, maprotiline, citalopram, fluoxetine, fluvoxamine, mianserine, buspirone, moclobemide, Li+
positive
anxiolytics guaifenezine, meprobamate; diazepam, alprazolam, oxazepam; neuroleptics and antidepressants
affectivity
negative dysphoric drugs reserpine, clonidine, α-methylDOPA positive neuroleptics,
atypical antipsychotics
thioridazine, chlorpromazine, chlorprotixene, levopromazine, haloperidol, perfenazine, clozapine, amisulpride, risperidone, sertindole, quetiapine, olanzapine
psychic integrations
negative hallucinogenic agents
lysergid, cocaine, amphetamines, ketamine, hashish, marihuana, phencyclidine, mescaline
positive nootropics piracetam, pyritinol, meclophenoxate memory negative amnestic drugs anticholinergics
(See: Švestka J. a kol.: Psychofarmaka v klinické praxi. GRADA Publishing, 1995; Hynie S.: Psychofarmakologie v praxi. Galén, 1995.)

3. PSYCHOPHARMACOLOGY
51
3.1. Antipsychotics Antipsychotics are divided into two groups (Table 3.3):
1. Conventional antipsychotics (basal or incisive)
2. Atypical antipsychotics
Incisive antipsychotics have high affinity to D2 receptors and low affinity to other
types of receptors. They affect positive symptoms of schizophrenia mainly. They bring
about extrapyramidal symptoms (EPS).
Basal antipsychotics have lower affinity to D2 receptors and higher affinity to other
receptor types beside incisive antipsychotics. They affect positive and affective
symptoms of schizophrenia. They have sedative effects, they bring about
anticholinergic, antihistaminic and cardiovascular side effect, and extrapyramidal
symptoms are less frequent.
Atypical antipsychotics (serotonin-dopamine antagonists) are antagonists of D2 and
serotonin 2A receptors, but they can affect many other types of receptors (Table 3.5).
They are much more efficient in treatment of negative symptoms of schizophrenia in
comparison with conventional antipsychotics. Atypical antipsychotics have lower side
effects (lower EPS or tardive dyskinesis)
Table 3.3. Classification of antipsychotics
group examples
basal (sedative)
antipsychotics
chlorpromazine, chlorprotixene, clopenthixole,
levopromazine, periciazine, thioridazine
conventional antipsychotics
(classical neuroleptics) incisive
antipsychotics
droperidole, flupentixol, fluphenazine,
fluspirilene, haloperidol, melperone,
oxyprothepine, penfluridol, perphenazine,
pimozide, prochlorperazine, trifluoperazine
atypical antipsychotics
(antipsychotics of 2nd generation)
amisulpiride, clozapine, olanzapine,
quetiapine, risperidone, sertindole, sulpiride
(See: Švestka J.: Nová (atypická) antipsychotika 2. generace. Remedia 9, 366-385, 1999.)
Table 3.4. Mechanisms of action of antipsychotics

Zdeněk Fišar: Introduction to Biological Psychiatry
52
conventional antipsychotics • D2 receptor blockade of postsynaptic in
the mesolimbic pathway
atypical antipsychotics • D2 receptor blockade of postsynaptic in the
mesolimbic pathway to reduce positive symptoms;
• enhanced dopamine release and 5-HT2A receptor
blockade in the mesocortical pathway to reduce
negative symptoms;
• other receptor-binding properties may contribute to
efficacy in treating cognitive symptoms, aggressive
symptoms and depression in schizophrenia
Table 3.5. Receptor systems affected by atypical antipsychotics
risperidone D2, 5-HT2A, 5-HT7, α1, α2
sertindole D2, 5-HT2A, 5-HT2C, 5-HT6, 5-HT7, D3, α1
ziprasidone D2, 5-HT2A, 5-HT1A, 5-HT1D, 5-HT2C, 5-HT7, D3, α1, NRI, SRI
loxapine D2, 5-HT2A, 5-HT6, 5-HT7, D1, D4, α1, M1, H1, NRI
zotepine D2, 5-HT2A, 5-HT2C, 5-HT6, 5-HT7, D1, D3, D4, α1, H1, NRI
clozapine D2, 5-HT2A, 5-HT1A, 5-HT2C, 5-HT3, 5-HT6, 5-HT7, D1, D3, D4, α1, α2, M1, H1
olanzapine D2, 5-HT2A, 5-HT2C, 5-HT3, 5-HT6, D1, D3, D4, D5, α1, M1-5, H1
quetiapine D2, 5-HT2A, 5-HT6, 5-HT7, α1, α2, H1
NRI - norepinephrine reuptake inhibitor, SRI – serotonin reuptake inhibitor
(See: Stahl S.M.: Psychopharmacology of Antipsychotics, Martin Dunitz, London, 1999.)

3. PSYCHOPHARMACOLOGY
53
3.2. Antidepressants Molecular mechanisms of action of antidepressants are much more diverse than that of
antipsychotics. Classification of antidepressants based on their acute pharmacological
actions is shown in Table 3.6. Antidepressants are amphiphilic molecules; so, they easy
permeate through the cell membrane and may affect molecules on the outer and inner
membrane surface, cytoplasmic elements and nuclear molecules (Figure 3.2).
Table 3.6. Classification of antidepressants based on acute pharmacological actions
inhibitors of neurotransmitter
catabolism
monoamine oxidase inhibitors (IMAO)
reuptake inhibitors serotonin reuptake inhibitors (SRI)
norepinephrine reuptake inhibitors (NRI)
selective SRI (SSRI)
selective NRI (SNRI)
serotonin/norepinephrine (dual) inhibitors (SNRI)
norepinephrine and dopamine reuptake inhibitors (NDRI)
serotonin 2A antagonist/reuptake inhibitors (SARI)
agonists of receptors 5-HT1A
antagonists of receptors α2-AR, 5-HT2
inhibitors or stimulators of
other components of signal
transduction
G proteins, adenylyl cyclase (AC), phospholipase (PL),
protein kinase (PK), phosphatase, ATPase, phospholipid
dependent proteins, transcription factors, 2nd a 3rd
messengers
Therapeutic response is observed after a few weeks of antidepressant treatment; so,
many adaptive changes in cellular functions occur. The neurotransmitter receptor
hypothesis of antidepressant action explains the ultimate mechanism of their
therapeutic action by receptor sensitivity changes. Currently, there is focus on the gene
expression that is activated by antidepressants (see Chapter 5).

Zdeněk Fišar: Introduction to Biological Psychiatry
54
Figure 3.2. Potential mechanisms of antidepressants action
transporter COMT
transmitter synthesis metabolic degradation transport to vesicles releasing to synaptic cleft reuptake interaction with receptors interaction with transducers
interaction with presynaptic autoreceptors and heteroreceptors metabolic degradation of neurotransmitter
MAO
ATPase synaptic vesicles
PLC AC
ATP cAMP DG IP3
PIP2
ATPase ion-channel receptor
nucleus
protein kinase
Ca2+
transcription factors
A C CaM

3. PSYCHOPHARMACOLOGY
55
Table 3.7. Examples of antidepressants
group subgroup examples
activating desipramine, nortriptyline, protriptyline, dosulepine
tricyclic (thymoleptics)
sedative, anxiolytic imipramine, amitriptyline, trimipramine, clomipramine
II. generation - activating viloxazine, buspirone, amineptine, maprotiline
II. generation - sedative, anxiolytic
mianserine, trazodone, nefazodone, pirlindol
III. generation - SSRI citalopram, fluvoxamine, fluoxetine, sertraline, paroxetine
II. a III. generation
enhancing 5-HT uptake tianeptine
nonselective irreversible phenelzine, tranylcypromine, isocarboxazid
selective MAOI-B irreversible deprenyl
selective MAOI-B reversibile
selective MAOI-A irreversible
MAO inhibitors
selective MAOI-A reversible moclobemide, brofaromine, toloxatone, amiflamine
thymoprophylactics lithium, carbamazepine, sodium valproate, valpromide
(See: Švestka J. a kol.: Psychofarmaka v klinické praxi. GRADA Publishing, 1995.)

Zdeněk Fišar: Introduction to Biological Psychiatry
56
3.3. Mechanism of action of antidepressants Selective serotonin reuptake inhibitors (SSRI) are the most frequently used
antidepressants. Their mechanism of action on serotonergic neuron in a depressed
patient is shown on Figure 3.3.
Before treatment (Figure 3.3 A):
There is relative deficiency of 5-HT in serotonin neurone in a depressed patient.
Number of serotonin receptors is up-regulated, including presynaptic
autoreceptors as well as postsynaptic receptors.
Releasing of serotonin from synaptic knob can be affected: 1. positively by
activity of serotonin transporter or by tryptophan (serotonin precursor) availability;
2. negatively by activation both presynaptic inhibitory receptors, 5-HT1B or α2-AR,
and somatodendritic receptors, 5-HT1A.
After acute administration of SSRI (Figure 3.3 B):
A considerable part of serotonin transporters is blocked and serotonin remains
for a longer time in extracellular space. This causes serotonin to increase in the
somatodendritic area mainly.
Negative feedback mediated by inhibitory presynaptic and somatodendritic
receptors is increased and both frequency of firing of action potentials and
amount of serotonin released from presynaptic button is decreased.
After chronic treatment by SSRI (Figure 3.3 C):
The increased 5-HT at the inhibitory somatodendritic receptors causes them to
down-regulate and/or desensitise. It results in increase of frequency of firing of
action potentials and in increase of the amount of serotonin released to synaptic
cleft. The marked increase of serotonin release in the axon terminal is delayed as
compared with the processes after acute administration of SSRI. This delay may
explain why therapeutic action of antidepressants is not immediate.
The increased 5-HT at the axon terminal causes down-regulation and/or
desensitization of postsynaptic and presynaptic receptors. This desensitization
may mediate the reduction of side effects of SSRI.
Mechanism of action of α2-adrenoceptor blockers is shown on Figure 3.4; mechanism
of action of reversible inhibitors of monoamine oxidase A (RIMA) is on Figure 3.5.

3. PSYCHOPHARMACOLOGY
57
5-HT1A 5-HT1B 5-HT2A 5-HT2C 5-HT3 5-HT4
...
5-HT1A TRP
5-HT
MAO
5-HIAA
SERT
A) before treatment α2-AR 5-HT1B
C) chronic treatment by SSRI α2-AR
TRP
5-HT
MAO
5-HIAA
SERT
5-HT1A
5-HT1A TRP
5-HT
MAO
5-HIAA
SERT
B) acute administration of SSRI α2-AR 5-HT1B
Figure 3.3. Mechanism of action of selective serotonin reuptake inhibitors (SSRI)

Zdeněk Fišar: Introduction to Biological Psychiatry
58
Figure 3.4. Mechanism of action of α2-adrenoceptor blockers
Figure 3.5. Mechanism of action of reversible inhibitors of MAO-A (RIMA)
presynaptic α2-AR α2-AR α2-AR
NORMAL DEPRESSION ↑ density and sensitivity of
presynaptic α2-AR ↓ NE or 5-HT transmission
AFTER TREATMENT α2-AR blockade
↑ NE or 5-HT transmission
NE or 5-HT
NE or 5-HT neurone
serotonin dopamine phenylethylaminenorepinephrine tyramine benzylamine epinephrine tryptamine methylhistamine
TYR
AM
INE
RIM
A
MAO-BMAO-A
TYR
AM
INE
TYR
AM
INE
TYR
AM
INE
TYR
AM
INE
TYR
AM
INE
RIM
A
MAO-BMAO-A
TYR
AM
INE
TYR
AM
INE
TYR
AM
INE
TYR
AM
INE

4. SCHIZOPHRENIA
59
4. Schizophrenia
Schizophrenia is specific human disease. It is group of diseases characterized by
delusions, hallucinations, disorganized speech, grossly disorganized or catatonic
behaviour, of negative symptoms (Table 4.1). Symptoms of schizophrenia can be
subcategorized into:
1. Positive symptoms
2. Negative symptoms
3. Cognitive symptoms
4. Aggressive/hostile symptoms
5. Depressive/anxious symptoms
In this chapter, environmental, genetic, neurodevelopmental and biochemical
hypotheses of schizophrenia are presented. Classification, clinical description and
criteria for diagnosis of schizophrenia are not described.
For practical purposes the descriptive psychopathology of schizophrenia can be treated
in three sections:
1. Purely positive symptoms (hallucinations and other abnormal experiences,
delusions and catatonia)
2. What used to be referred to as psychological deficit, purely negative symptoms –
impaired attention, intelligence, memory, perception and will.
3. Traditional psychopathological groupings containing positive and negative
symptoms (mixed) – thought disorder and disturbances of emotions.
Table 4.1. Positive and negative symptoms of schizophrenia
negative positive
alogia hallucination
affective flattening delusions
avolition – apathy bizarre behaviour
anhedonia – asociality positive formal thought disorder
attentional impairment
Biological models of schizophrenia can be divided into three related classes:

Zdeněk Fišar: Introduction to Biological Psychiatry
60
Environmental models
Genetic models
Neurodevelopmental models
4.1. Environmental models Environmental models suppose that either stress or physical factors are the main
evoking phenomenon in schizophrenia (Table 4.2).
Table 4.2. Environmental models of schizophrenia
Model of “evocative influence of complex social demands”:
There are four criteria for schizophrenia-evoking stress:
1. A situation demanding action or decision
2. Complexity or ambiguity of the information supplied to deal with the task
3. Unless resolved, the situation demanding action or decision persist
4. The subject has no “escape route” available
Most stresses of this nature will be non-pathogenic; a schizophrenia-evoking effect
occurs only in conjunction with a specific genetic liability.
Non-psychosocial environmental model:
An unspecified number of varieties of physical factors will be sufficient to produce the
specific cerebral lesions and dysfunction thought to be characteristic of schizophrenia,
regardless of the presence or absence of a genetic susceptibility (see
“Neurodevelopmental models” below). (See: Jablensky A.: Schizophrenia: the epidemiological horizon. In: Schizophrenia, Hirsch S.R. and Weinberger D.R., eds., Blackwell Science, pp. 206-252, 1995.)

4. SCHIZOPHRENIA
61
4.2. Genetic models Evidence for a genetic contribution to schizophrenia comes from twin and familiar
studies. The genetic factors in schizophrenia have specificity as they do not increase
the risk for major affective disorders or delusional disorder. Clearly, schizophrenia is
clinically or phenotypically heterogeneous, but whether this variety is paralleled by
etiological heterogeneity or to what extent is problematic. Genetic models are
summarized in Table 4.3. Published data indicates that monogenic models could be
rejected and multifactorial threshold model or mixed models are favoured.
Table 4.3 Genetic models
Distinct heterogeneity model:
Schizophrenia is a collection of several separate diseases, each associated with single
major locus (SML) that may be inherited either dominantly or recessively. In addition,
there are sporadic, environmentally caused cases.
Monogenic models (single major locus models):
Schizophrenia might be a single-gene dominant disorder with highly variable
expression or reduced penetrance of the trait; i.e. all cases of schizophrenia share the
same single major locus (SML).
Multifactorial-polygenic threshold model:
Schizophrenia is the result of a combined effect of multiple genes interacting with
variety of environmental factors; i.e. several or many genes, each of small effect,
combine additively with the effects of non-inherited factors. The liability to
schizophrenia is linked to one end of the distribution of a continuous trait, and there
may be a threshold for the clinical expression of the disease.
A mixed or combined model:
The model includes the elements of some, or all, of the above three. (See: Jablensky A.: Schizophrenia: the epidemiological horizon. In: Schizophrenia, Hirsch S.R. and Weinberger D.R., eds., Blackwell Science, pp. 206-252, 1995; Asherson P., Mant R., McGuffin P.: Genetics and schizophrenia. In: Schizophrenia, Hirsch S.R. and Weinberger D.R., eds., Blackwell Science, pp. 253-274, 1995.)

Zdeněk Fišar: Introduction to Biological Psychiatry
62
4.3. Neurodevelopmental models The leading hypothesis for the aetiology of schizophrenia is related to disturbance in
normal brain development. The principal assumption is that normal brain development
is disrupted in specific ways at critical periods and the resulting lesion produces the
symptoms of schizophrenia only through interaction with the normal maturation
processes in the brain, which occur in late adolescence or early adulthood.
Neurodevelopmental hypothesis states that:
A substantial group of patients, who receive diagnosis of schizophrenia in adult life,
have experienced a disturbance of the orderly development of the brain decades before
the symptomatic phase of the illness.
The neurodevelopmental model therefore directs attention to both genetic and no
genetic risk factors that may have impacted on the developing brain during prenatal
and perinatal life; pregnancy and birth complications (PBCs) are considered in
psychiatry:
viral infections in utero
gluten sensitivity
brain malformations
obstetric complications
Genetic contribution to schizophrenia development consists in wrong genetic program
for the normal formation of synapses and migration of neurons in the developing brain.
These risk factors may have the final common effect on nerve growth factors reduction
resulting in structural abnormalities, selection of wrong neurons to survive in the fetal
brain, neuron migration to the wrong places, neuron inervation of wrong targets or mix-
up of the nurturing signals.

4. SCHIZOPHRENIA
63
4.4. Delayed onset of symptoms We can propound the question, why is the illness manifestation delayed typically for
about two decades after birth? Delayed onset of symptoms of schizophrenia (in early
adult) can be explained by different mechanisms:
1. There is an additional pathological process occurring around the time of onset of
the clinical symptoms
2. An interaction between a static developmental defect and normal developmental
events that occur in early adulthood is necessary
It is supposed that onset of schizophrenia can be initiated by wrong organization,
elimination and restructuring of synapses during adolescence, which may be or not
secondary to the maldevelopment in utero.
4.5. Neurodegenerative hypothesis Recent post mortem studies on schizophrenics described a multitude of morphological
changes in different brain structures. The most often reported structural alterations are
in the limbic system, mainly in hippocampal formation and parahippocampal and
cingulate gyri. However, other structures such as the thalamus, frontal and temporal
cortex and basal ganglia seem to be affected too.
Neurodegenerative hypothesis of schizophrenia suggest an ongoing
neurodegenerative processes with loss of neuronal function during the course of the
disease. Excitotoxic hypothesis proposes that neurons degenerate as a consequence of
excessive glutamatergic neurotransmission.
Combined neurodevelopmental/neurodegenerative hypothesis suggest that
schizophrenia may be a neurodegenerative process superimposed on a
neurodevelopmental abnormality.

Zdeněk Fišar: Introduction to Biological Psychiatry
64
4.6. Biochemical basis of schizophrenia The biochemical hypotheses of schizophrenia are orientated towards the role of
neurotransmitters and their receptors; dopamine, serotonin, glutamate, GABA,
norepinephrine and so on are considered (Table 4.4). Dopamine plays a key role in
biochemical hypotheses of schizophrenia. Dopamine hypothesis of schizophrenia was
formulated almost 40 years ago by Randrup and Munkvad (1965) and plays a
prominent role in schizophrenia research hitherto. Basis or motives for dopamine
hypothesis are summarized in Table 4.5.
According to the classical dopamine hypothesis of schizophrenia, psychotic
symptoms are related to dopaminergic hyperactivity in the brain. Hyperactivity of
dopaminergic systems during schizophrenia is result of increased sensitivity and density
of dopamine D2 receptors. This increased activity can be localized in specific brain
regions.
Table 4.4. Biochemical hypotheses of schizophrenia
classical dopamine GABAergic
norepinephrine glutamatergic
serotonin peptidergic
monoaminoxidase membrane
revisited dopamine transmethylation
Table 4.5. Basis of classical dopamine hypothesis of schizophrenia
Dopamine-releasing drugs (amphetamine, mescaline, diethyl amide of lysergic acid -
LSD) can induce state closely resembling paranoid schizophrenia.
Conventional neuroleptic drugs, that are effective in the treatment of schizophrenia,
have in common the ability to inhibit the dopaminergic system by blocking action of
dopamine in the brain.
Neuroleptics raise dopamine turnover as a result of blockade of postsynaptic dopamine
receptors or as a result of desensitisation of inhibitory dopamine autoreceptors localized
on cell bodies.

4. SCHIZOPHRENIA
65
Revised dopamine hypothesis of schizophrenia postulate that a reduced striatal
inhibition on the thalamus, caused by either an increased dopaminergic or a reduced
glutamatergic tone, should lead to an increase in arousal and psychomotor activity and
to an increased sensory input transmitted to the cortex. If a certain threshold is
exceeded the integrative capacity of the cortex will become insufficient and this will lead
to positive symptoms of schizophrenia. An excessive dopaminergic function may also
lead to a disintegration of motor functions. This inhibitory function of the striatum
appears to be exerted via the indirect pathways (Figure 4.1 A). The direct pathways
(Figure 4.1 B) should be able to mediate the excitatory glutamatergic input from the
cortex to the thalamus; the dopamine input in the direct pathways appears to be
excitatory, and dopamine should thus be behaviourally stimulating also via the direct
pathways.
Hypothetical scheme of interactions, leading to psychotogenic responses, is shown in
Figure 4.2.

Zdeněk Fišar: Introduction to Biological Psychiatry
66
Figure 4.1. Excitatory and inhibitory influence of dopamine on the direct and
indirect pathways
Glu – glutamate, DA – dopamine, GABA - γ-amino butyric acid (See: Carlsson A.: The dopamine theory revisited. In: Schizophrenia, Hirsch S.R. and Weinberger D.R., eds., Blackwell Science, pp. 379-400, 1995.)
+ excitatory action - inhibitory action
cerebral cortex
striatum
substantia nigra, area ventralis tegmenti
thalamus sensory input
behaviour
Glu + Glu +
DA -
3 GABA
-
cerebral cortex
striatum
substantia nigra, area ventralis tegmenti
thalamus sensory input
behaviour
Glu + Glu +
DA +
2 GABA
+
A) „indirect“ pathways (negative feedback)
B) „direct“ pathways (positive feedback)

4. SCHIZOPHRENIA
67
cortex
GluGlu
substantia nigra, area ventralis tegmenti
DA
striatum
ACh
GABA
nucleus raphe
5-HT
locus coeruleus
NE
GABA
LSD (5-HT2 agonist) muscimol (GABAA agonist)
PCP (NMDA antagonist)
atropine (M1 antagonist)
PCP (NMDA antag.)
LSD
(5-HT2 agon.)
amphetamine (DA releaser)
Figure 4.2. Potential psychogenic pathways and sites of action of psychotogenic
and antipsychotic agents
LSD – diethyl amid of lysergic acid, PCP – phencyclidine, 5-HT – serotonin, Glu – glutamate, DA – dopamine, GABA – γ-amino butyric acid, ACh – acetylcholine, NE – norepinephrine, NMDA – N-methyl-D-aspartate (See: Carlsson A.: The dopamine theory revisited. In: Schizophrenia, Hirsch S.R. and Weinberger D.R., eds., Blackwell Science, pp. 379-400, 1995.)

Zdeněk Fišar: Introduction to Biological Psychiatry
68
5. Affective disorders
Mood disorders are characterized by depression, mania, or both. In this chapter,
biochemical hypothesis of affective disorders are presented. Classification, clinical
description and criteria for diagnosis of disorders of mood are not described.
Depression and mania are thought to be heterogeneous illnesses that can result from
dysfunction of several neurotransmitter or metabolic systems. Approaches of biological
psychiatry to the affective disorders are summarized in Table 5.1.
Table 5.1 Biological psychiatry and affective disorders
genetics vulnerability to mental disorders stress increased sensitivity
BIOLOGY
chronobiology desynchronisation of biological rhythms
neurotransmitters availability, metabolism receptors number, affinity, sensitivity
NEUROCHEMISTRY
postreceptor processes G proteins, 2nd messengers, phosphorylation, transcription
hypothalamic-pituitary-adrenocortical system
increased activity during depression
IMMUNONEURO- ENDOCRINOLOGY
immune function different changes during depression
Theoretical and clinical studies have provided evidence that the monoamine
neurotransmitter systems are involved in the treatment of affective disorders. These
studies have led to a series of hypotheses concerning the mechanism of the action of
antidepressant treatments, as well as pathophysiology of depression, that have focused
on alterations in brain levels of serotonin and norepinephrine or their receptors.
Studies at the level of neurotransmitters and receptors have not generated a compelling
model of antidepressant action or the pathophysiology of depression. For example,
common action of antidepressant treatments at the level of monoamines or their
receptors was not identified so far. It is possible that there is more than a single
mechanism by which antidepressant treatments exert their therapeutic actions.
An updated hypothesis suggests that postreceptor intracellular targets mediate the long-
term, therapeutic action of antidepressant treatments.
5.1. Neuroendocrine hypotheses

5. AFFECTIVE DISORDERS
69
Changes in the activity of the hypothalamic-pituitary-adrenal (HPA) axis or
hypothalamic-pituitary-thyroid (HPT) axis were observed during depression. An
increased function of the HPA axis was described in more than 50% of depressed
patients. Abnormalities of the HPA axis in patients with depression are summarized in
Table 5.2.
The dexamethasone suppression test (DST) is used to determine the sensitivity of
the HPA axis to negative feedback. 1 mg of dexamethasone is usually administered at
23:00 h and plasma cortisol is monitored following morning. In healthy subjects
dexamethasone suppresses plasma cortisol. DST was suggested as test for diagnosis
of major depressive disorder, since increased function of HPA axis can be expressed in
reduced decrease of ACTH and cortisol levels after dexamethasone administration. But
it was demonstrated that DST test is unsufficiently specific.
Table 5.2. Abnormalities of the hypothalamic-pituitary-adrenal (HPA) axis in patients
with depression
cortisol hypersecretion
increased urinary free cortisol
increased CSF corticotrophin releasing factor
increased circulating ACTH
abnormal circadian rhythms of cortisol
abnormal dexamethasone suppression
decreased glucocorticoid receptor sensitivity
decreased release of ACTH to CRF
increased adrenal gland size CSF – cerebrospinal fluid, ACTH – adrenocorticotrophic hormone, CRF – corticotrophin-releasing factor (See: Trimble M.R.: Biological Psychiatry. 2nd ed., John Wiley & Sons, New York, 1996.)

Zdeněk Fišar: Introduction to Biological Psychiatry
70
Neuroendocrine hypothesis of affective disorders were formulated on the base of
abnormalities in HPA axis function and disturbed glucocorticoid feedback in depression
(Table 5.3).
Table 5.3. Neuroendocrine hypotheses of affective disorders
Endocrine abnormalities associated with depression are results of increased releasing
of CRF both in hypothalamus and out of them.
Disturbed glucocorticoid feedback in depression is caused by lower density of
corticosteroid receptors in hippocampus and hypothalamus during depression (i.e. by
disturbed regulation of gene expression of these receptors).
5.2. Neurochemical hypotheses There are evidences that signal transmission through chemical synapse is disturbed in
the affective disorders. Changes in neurotransmitters, membrane receptors and
postreceptor pathways are studied during mental illness and its treatment. The
neurochemical parameters studied in affective disorders are summarized in Table 5.4.
Table 5.4. Neurochemical parameters studied in affective disorders
availability neurotransmitters
metabolism
number or density
affinity
receptors
sensitivity
number and activity of G proteins
effector enzymes
2nd messengers systems
protein kinases
phosphatases
postreceptor systems
transcription factors

5. AFFECTIVE DISORDERS
71
5.3. Neurotransmitter hypotheses Discovery of the tricyclic antidepressant drugs supported hypothesis about significant
role for the biogenic amine, particularly NE and 5-HT in the ethiopathogenesis of
affective disorders. Reduced activity of the serotonergic and noradrenergic systems has
been reported in subgroups of patients with major depression, but this has not been
observed in all depressed patients.
Initial neurotransmitter hypotheses were supported by effect of tricyclic antidepressants
on reuptake of NE and 5-HT, effect of monoamine oxidase inhibitors (MAOI) on
catabolism of monoamine neurotransmitters and effect of reserpine on vesicular
transport of neurotransmitters (Table 5.5). Reuptake inhibition results in an enrichment
of the NE and 5-HT in the synaptic cleft. Similar effect can be achieved with MAO
inhibitors. For many years this effect was considered to be the precondition for
antidepressant activity. The first major theory about the biological ethiology of
depression was monoamine hypothesis supposing that:
Depression was due to a deficiency of monoamine neurotransmitters, norepinephrine
and serotonin. MAOI act as antidepressants by blocking of enzyme MAO, thus allowing
presynaptic accumulation of monoamine neurotransmitters. Tricyclic antidepressants
act as antidepressants by blocking membrane transporters ensuring reuptake of 5-HT
or NE, thus causing increased extracellular neurotransmitter concentrations.
Table 5.5. Data for neurotransmitter hypothesis
1. Tricyclic antidepressants through blockade of neurotransmitter reuptake increase
neurotransmission at noradrenergic synapses
2. MAOIs increase availability of monoamine neurotransmitters in synaptic cleft
3. Depressive symptoms are observed after treatment by reserpine, which depletes
biogenic amines in synapse

Zdeněk Fišar: Introduction to Biological Psychiatry
72
In addition to the role of noradrenergic and serotonergic systems in depression,
cholinergic and dopaminergic and others systems were considered, so, many
neurotransmitter hypotheses were formulated (Table 5.6).
Table 5.6. Neurotransmitter hypothesis of affective disorders
catecholamine hypothesis
indolamine hypothesis
cholinergic-adrenergic balance hypothesis
„permissive“ hypothesis
dopamine hypothesis
hypothesis of biogenic amine
monoamine hypothesis
Permissive biogenic amine hypothesis persists as part of more recent hypothesis:
A deficit in central indolaminergic transmission permits affective disorder, but is
insufficient for its cause; changes in central catecholaminergic transmission, when they
occur in the context of a deficit in indoleaminergic transmission, act as a proximate
cause for affective disorders and determine their quality, catecholaminergic
transmission being elevated in mania and diminished in depression.
However, neither the catecholamine nor the serotonin hypothesis could be confirmed in
depressive patients. Clinically, the onset of the antidepressant action differs from the
biochemical effects; the uptake inhibition occurs suddenly, whereas the therapeutic
effect needs two or three weeks of pharmacotherapy. Furthermore, depletion of 5-HT or
NE in healthy individuals does not induce clinically significant depressive
symptomatology. These are reasons why receptor and postreceptor events are
observed during affective disorders and their treatment at present time.

5. AFFECTIVE DISORDERS
73
5.4. Receptor hypotheses The neurotransmitter receptor hypothesis posits that disturbance occurs in function of
receptors for the key monoamine neurotransmitters. Such wrong receptor function may
be caused by depletion of monoamine neurotransmitters, by abnormalities in the
receptor, or by problems with signal transduction on postreceptor level.
The common final result of chronic treatment by majority of antidepressants is the
down-regulation or up-regulation of postsynaptic or presynaptic receptors (Table 5.7).
The delay of clinical response corresponds with these receptor alterations, hence many
receptor hypotheses of affective disorders were formulated and tested.
Table 5.7. Effect of depression and antidepressant treatment on receptor sensitivity
system receptor system treatment by antidepressants
depression
β1-AR (postsynaptic, exc.) ↓ ↑
α2-AR (presynaptic, inh.) ↓ ↑
α2-AR (postsynaptic, inh.) ↑ ↓
adrenergic
α1-AR (postsynaptic, exc.) ↑
5-HT2 (exc.) ↓ ↑
5-HT1A (somatodendritic autoreceptors, inh.)
↓ ↓?
5-HT1A (postsynaptic, inh.) ↑ ↓
serotonergic
5-HT1B (terminal autoreceptors, inh.) ↓
ACh ↓ ↑
GABA ↑ ↓
DA ↑ ↓
other
corticosteroid receptors ↑ ↓
ACh - acetylcholine, GABA - γ-amino butyric acid, DA - dopamine, AR – adrenoceptors. ↑ - increased, ↓ - decreased

Zdeněk Fišar: Introduction to Biological Psychiatry
74
When activated by catecholamines, the presynaptic α2-adrenergic receptors can inhibit
NE or 5-HT release. Some antidepressants reduce sensitivity of brain
α2-adrenoreceptors. Hypothesis about role of presynaptic α2-AR was formulated:
There is increased density of presynaptic α2-adrenergic receptors in high affinity state in
depression.
Several lines of evidence indicate that an enhancement of 5-HT neurotransmission
might underlie the therapeutic response to different types of antidepressant treatment,
so, general serotonin receptor hypothesis was suggested:
Depression is connected with these abnormalities in serotonin receptors:
5-HT2 up-regulation
5-HT1A desensitisation
Abnormal signal transduction following 5-HT binding to receptor
Very elegant but not confirmed was receptor catecholamine hypothesis:
Supersensitivity of catecholamine receptors in the presence of low levels of serotonin is
the biochemical basis of depression.
The common result of chronic treatment by majority of antidepressants is the down
regulation of β-adrenergic receptors, which can be modulated by interaction with the 5-
HT system, DA system, neuropeptides and hormones. Classical norepinephrine
receptor hypothesis of affective disorders was formulated (Figure 5.1):
There is increased density of postsynaptic β-AR in depression (due to decreased NE
release, disturbed interactions of noradrenergic, serotonergic and dopaminergic
systems, etc.). Long-term antidepressant treatment causes down regulation of β1-AR
(by inhibition of NE reuptake, stimulation or blockade of receptors, regulation through
serotonergic or dopaminergic systems, etc.). Transient increase of neurotransmitter
availability can cause fault to mania.
Apparently paradoxical increase of intracellular cAMP levels were observed at
decreased density of β-AR, so increased cAMP system activity seems to be
fundamental in therapeutic action of antidepressants. So, postreceptor hypotheses of
affective disorders are tested at present time.

5. AFFECTIVE DISORDERS
75
Figure 5.1. Classical noradrenergic receptor hypothesis
(See: Lopez D. et al.: The Essential Brain. Current Topics Sci. Med. Merck 1991.)
5-HT
NE
DA
5-HT
NE
DA
DA 5-HT NE NE - receptors
NE reuptake
pre- postsynaptic
DEPRESSION
5-HT
NE
DA
MANIA
DA
NE
5-HT

Zdeněk Fišar: Introduction to Biological Psychiatry
76
5.5. Postreceptor hypotheses The introduction of new, more selective antidepressants led to new reflection upon the
mechanism of their action. Most promising are recent investigations of the second
messenger systems, the adenylyl cyclase system, the phosphatidylinositol system,
G proteins, transcription factors etc. Modulation of receptor sensitivity, either
down-regulation or up-regulation, is most probably dependent on the second and third
messenger systems. Selected postreceptor hypotheses are shown in Table 5.8.
Table 5.8. Postreceptor hypotheses
Second messenger dysbalance hypothesis:
Affective disorders arise from the dysbalance of the two major intraneuronal signal-
amplification systems, with depression resulting from a hypofunction of the AC-cAMP
kinases pathway together with dominance of the PLC-CaM/C kinases system, and
mania resulting from the converse.
G protein hypothesis:
They are changes either density or function of subunits of G proteins during depression.
(e.g. increase of Gs protein)
Molecular model of affective disorders:
Clinical heterogeneity observed at depressed patients can be explained by changes in
signal transduction pathways which regulate two or more different neurotransmitter
systems and which affect the neuron activity. G proteins, phosphatases and
transcription factors are studied mainly.
Molecular and cellular theory of depression:
Transcription factor, cAMP response element-binding protein (CREB), is one
intracellular target of long-term antidepressant treatment and brain-derived neurotrophic
factor (BDNF) is one target gene of CREB. Chronic stress leads to decrease in
expression of BDNF in hippocampus. Long-term increase in levels of glucocorticoids,
ischemia, neurotoxins, hypoglycaemia etc. decreases neuron survival. Long-term
antidepressant treatment leads to increase in expression of BDNF and his receptor trkB
through elevated function of serotonin and norepinephrine systems.

5. AFFECTIVE DISORDERS
77
5.6. Molecular and cellular theory of depression Recent studies have begun to characterize the action of stress and antidepressant
treatments on signal transduction at postreceptor level. Long-term antidepressant
treatments result in the sustained activation of the cAMP system in specific brain
regions, including the increased function and expression of the transcription factor
cAMP response element-binding protein (CREB). The activated cAMP system leads to
the regulation of specific target genes, including the increased expression of brain-
derived neurotrophic factor (BDNF) in hippocampus and cortex.
It was found that stress can decrease the expression of BDNF and lead to atrophy of
the same populations of stress-vulnerable hippocampal neurons. Decreased size and
impaired function of these neurons may be involved in depression; this possibility is
supported by clinical imaging studies, which demonstrate a decreased volume of certain
brain structures. These findings constitute the framework for a molecular and cellular
hypothesis of depression (Table 5.8), which assume that stress-induced vulnerability
and therapeutic action of antidepressant treatments occur via intracellular mechanisms
that decrease or increase, respectively, neurotrophic factors necessary for the survival
and function of particular neurons. This hypothesis also explains how stress and other
types of neuronal insult can lead to depression in vulnerable individuals.
5.7. Molecular mechanism of action of antidepressant treatments A model for the molecular mechanism of action of antidepressant treatments is shown
on Figure 5.2. Antidepressant treatment causes inhibition of serotonin and
norepinephrine reuptake or breakdown. Short-term antidepressant treatment increase
extracellular levels of serotonin and norepinephrine. Long-term treatment leads to
decrease in the function and expression of serotonin and norepinephrine receptors, to
increase in the cAMP signal transduction and to increase in expression of CREB.
Increased activity of the cAMP signal transduction cascade indicates that the functional
output of 5-HT and NE are up-regulated, even though levels of certain 5-HT and NE
receptors are down-regulated. Expression of BDNF and its receptor trkB is also
increased by long-term antidepressant treatment, so increased neuronal survival,
function, and remodelling of synaptic architecture are provided.

Zdeněk Fišar: Introduction to Biological Psychiatry
78
Figure 5.2. A model for the molecular mechanism of action of antidepressant
treatments
Antidepressant treatments (AT)
Inhibition of serotonin and norepinephrine reuptake or breakdown.
Short-Term AT: Increase in levels of serotonin or norepinephrine.
Long-Term AT: Decrease in the function and expression of 5-HT and NE
receptors.
Increase in the cAMP signal transduction pathway (increased
levels of adenylyl cyclase and PKA and translocation of PKA to
the cell nucleus).
Increase in expression of the transcription factor CREB (cAMP
response element-binding protein); it is suggested that CREB is
a common postreceptor target for antidepressants.
(See: Duman et al.: A Molecular and Cellular Theory of Depression. Arch. Gen. Psychiatry, 1997; 54: 597-606.)

6. LABORATORY SURVEY IN PSYCHIATRY
79
6. Laboratory survey in psychiatry
Biochemical, immunological, genetic, electrophysiological and neuroendocrine
parameters are studied in mental disorders. Laboratory survey methods in psychiatry
coincide with internal and neurological methods:
Classic and special biochemical and neuroendocrine tests
Immunological tests
Electrocardiography (ECG)
Electroencephalography (EEG)
Computed tomography (CT)
Nuclear magnetic resonance (NMR)
Phallopletysmography

Zdeněk Fišar: Introduction to Biological Psychiatry
80
Table 6.1. Classic and special biochemical tests
Test Indication
serum cholesterol (3,7-6,5 mmol/l) and lipemia (5-
8 g/l)
brain disease at atherosclerosis
cholesterolemia, TSH, T3, T4, blood pressure,
mineralogram (calcemia, phosphatemia)
thyroid disorder,
hyperparathyreosis or
hypothyroidism can be an
undesirable side effect of lithium
therapy
hepatic tests: bilirubin (total < 17mmol/l),
cholesterol, aminotranspherase (AST, ALT, TZR,
TVR), alkaline phosphatase
before pharmacotherapy and in
alcoholics
glycaemia diabetes mellitus
blood picture during pharmacotherapy
determination of metabolites of psychotropics in
urine or in blood
control or toxicology
lithemia (0,4-1,2 mmol/l), function of thyroid and
kidney (serum creatinine, urea), pH of urine,
molality, clearance, serum mineralogram (Na, K)
during lithiotherapy
determination of neurotransmitter metabolites, e.g.
homovanilic acid (HVA, dopamine metabolite),
hydroxyindolacetic acid (HIAA, serotonin
metabolite), methoxyhydroxyphenylglycole
(MHPG, norepinephrine metabolite)
research
neurotransmitter receptors and transporters research
cerebrospinal fluid: pH, tension, elements,
abundance of globulins (by electrophoresis)
diagnosis of progressive
paralysis, …
neuroendocrinne stimulative or suppressive tests:
dexamethasone suppressive test (DST), TRH test,
fenfluramine test
depressive disorders
prolactin determination increased during treatment with
neuroleptics

6. LABORATORY SURVEY IN PSYCHIATRY
81
Table 6.2. Recommended laboratory survey in selected psychiatric cases Alcoholism and drug dependence Atypical psychosis
blood picture + differential blood picture + differential serum electrolytes, glycaemia, S-urea, creatinine serum electrolytes, glycaemia, S-urea,
creatinine hepatic tests: GGT, AST, ALT, AF hepatic tests: GGT, AST, ALT, AF total protein, S-albumin glycaemia prothrombine time nephric function (N-urea, S-creatinine) serum Ca, P serum Ca, P lithic acid thyroidal tests toxicological survey of urine cortisolemia alcoholemia screening on syphilis (RRR, BWR) and Lyme
Borreliosis screening on syphilis (RRR, BWR) think on hypovitaminosis B12 ECG HIV test at risk person thorax Rtg urine + deposit urine + deposit toxicological survey of urine ordure uropophyrine and porphobilinogene in urine S-ammonia thorax Rtg cranium Rtg or CT ECG HIV survey EEG or CT
Anorexia Dementia blood picture + differential blood picture + differential FW serum electrolytes, glycaemia, S-urea,
creatinine hepatic tests hepatic tests: GGT, AST, ALT, AF thorax Rtg thyroidal tests ECG screening on syphilis (RRR, BWR) serum minerals and electrolytes think on hypovitaminosis B12 total protein urine + deposit urine + deposit HIV tests special survey: cortisolemia, thyroid tests, serum prolactin, nephric function, EEG, cranium Rtg or CT, special test on laxative
cranium Rtg
Bulimia ECG glycaemia EEG or CT serum Ca, P Malign neuroleptic syndrome ECG all basic blood and urine tests toxicological survey of urine S-creatine kinase special test on laxative myoglobin in urine lumbal puncture to rule out CNS infection ECG thorax Rtg

Zdeněk Fišar: Introduction to Biological Psychiatry
82
Table 6.3. Immunological tests
Test Indication
venereal disease research laboratories
(VDRL) Test
syphilis
rapid reagine reaction (RRR) syphilis
antibodies against Borrelia Burgdorferi Lyme Borreliosis
complement fixative test brucellosis
antibodies against HIV HIV infection
hepatic tests (ALT, AST) hepatitis
specific test on antigen EBV and
antibodies
Epstein-Barr virus (EBV, infectious
mononucleosis) and Cytomegalovirus
(CMV, seronegative mononucleosis), …
antinuclear antibodies and lupus
erytematosus cell preparation
systemic lupus erytematodes

83
REFERENCES (RECOMMENDED LITERATURE)
Atwood H.L., MacKay W.A.: Essentials of Neurochemistry. B.C.Decker Inc., Toronto-Philadelphia, 1989.
Basic Neurochemistry: Molecular, Cellular and Medical Aspects, 6th Ed., edited by G.J.Siegel et
al., Lippincott-Raven, Philadelphia, New York, 1999. Caron M.G.: TiPS Receptor and Ion Channel Nomenclature Supplement, 1996.
Duman et al.: A Molecular and Cellular Theory of Depression. Arch. Gen. Psychiatry, 1997; 54: 597-606.
Fišar Z.: Biochemické hypotézy afektivních poruch, Galén, Praha 1998.
Fišar Z., Jirák R.: Vybrané kapitoly z biologické psychiatrie, GRADA, Praha, 2001.
Ganong W.F.: Přehled lékařské fyziologie. Nakl. a vyd. H&H, 1995.
Höschl C.: Psychiatrie pro praktické lékaře, H&H, Jinočany, 1996
Höschl C., Libiger J., Švestka J. et al.: Psychiatrie. Tigis, 2002.
Hynie S.: Psychofarmakologie v praxi. Galén, Praha, 1995.
Kotyk A., Janáček K.: Membrane Transport. Academia, Praha, 1977.
Levitan I.B., Kaczmarek L.K.: The Neuron. Cell and Molecular Biology. Second Edition, Oxford
Univ. Press, New York, Oxford, 1997. Němeček S., Lodin Z., Wolf J., Vyskočil F.: Neurobiologie, Avicenum, zdrav. nakl., Praha, 1972.
Raboch J., Zvolský P. et al.: Psychiatrie. Galén, Karolinum, 2001.
Randrup A., Munkvad I.: Special antagonism of amphetamine-induced abnormal behaviour. Inhibition of
stereotyped activity with increase of some normal activities. Psychopharmacologia, 7: 416-422, 1965.
Schizophrenia, edited by S.R. Hirsch and D.R. Weinberger, Blackwell Science, 1995. Shinitzky M.: Membrane fluidity and receptor function. In: Membrane Fluidity (M. Kates, L.A. Manson,
eds.), Plenum Publ. Corp., pp. 585-601, 1984.
Stahl S.M.: Psychopharmacology of Antipsychotics, Martin Dunitz, London, 1999.
Stahl S.M.: Essentials Psychopharmacology, Cambridge Univ. Press, cambridge, 2000.
Švestka J. a kol.: Psychofarmaka v klinické praxi. Grada, Praha, 1995
Švestka J.: Nová (atypická) antipsychotika 2. generace. Remedia 9, 366-385, 1999.
The Essential Brain. Current Topics in Science and Medicine. Merck, 1991.
The RBI Handbook of Receptor Classification and Signal Transduction, 3rd Ed., edited by K.J.Watling,
RBI, Natick, 1998.
TiPS Receptor and Ion Channel Nomenclature Supplement 2000, 11th Ed., compiled by S.P.H.Alexander
and J.A.Peters.
Trimble M.R.: Biological Psychiatry. 2nd ed., John Wiley & Sons, New York, 1996. Vinař O.: Psychofarmaka. Minimum pro praxi. Triton, Praha, 1998.
Voyet D., Voyetová J.G.: Biochemie. Victoria Publishing a.s., Praha, 1995.
Zvolský P.: Rozvoj genetiky v psychiatrii. Avicenum, Praha, 1990.