investigating enteric coccidiosis in the endangered black
TRANSCRIPT

Investigating Enteric Coccidiosis in the Black-footed (Mustela nigripes) and
Domestic Ferret (Mustela putorius furo)
by
Adriana R Pastor
A Thesis
presented to
The University of Guelph
In partial fulfilment of requirements
for the degree of
Doctor of Veterinary Science
in
Zoological Medicine and Pathology
Guelph Ontario Canada
copy Adriana R Pastor November 2017
ABSTRACT INVESTIGATING ENTERIC COCCIDIOSIS IN THE BLACK-FOOTED (MUSTELA NIGRIPES) AND
DOMESTIC FERRET (MUSTELA PUTORIUS FURO)
Adriana R Pastor Advisors
University of Guelph 2017 Dr D A Smith
Dr J R Barta
Enteric coccidiosis is a major cause of death in both juvenile and adult black-footed ferrets (BFF
Mustela nigripes) in captive breeding programs that reduces the availability of animals for release to their
former North American range Coccidiosis is poorly understood in BFF but in vivo experimental infection
in this endangered host is untenable The goal of this research was to better characterize the etiologic
agents and natural history of enteric coccidiosis in BFF and to evaluate the domestic ferret (DF Mustela
putorius furo) as a model for experimental infection
Morphometric and molecular characterization of coccidia from BFF and DF was undertaken
Only Eimeria ictidea was identified in juvenile and adult BFF from 1999-2016 at the Toronto Zoo and
from BFF at the Louisville Zoo in 2016 Eimeria furonis and Isospora (=Cystoisospora) laidlawi were
identified in DF fecal and necropsy samples from Canadian and European diagnostic laboratories during
2008-2017 Molecular characterization of these parasites included generation of complete mitochondrial
genomes and nuclear 18S rDNA sequences for Eimeria ictidea and Eimeria furonis from BFF and DF
respectively Partial sequences were obtained from the same genetic targets from I (=C) laidlawi from
DF DNA isolation from formalin fixed paraffin embedded tissues and PCR amplicon sequencing
permitted identification of coccidia in BFF and DF tissues dating from 1999 to present
Retrospective and prospective analyses of medical and pathology records supplemented with
parasitological evaluation of repeated fecal samples was performed to determine the natural history of
coccidiosis in captive BFF Clinical signs and histopathologic changes associated with infection in BFF
were as described previously in the published literature Average yearly coccidia associated mortality
rates were 053 in adults and 195 in juveniles
Domestic ferrets were confirmed as experimental hosts of E ictidea isolated from BFF Seven of
10 juvenile DF inoculated with oocysts from a BFF developed patent infections and mild clinical disease
was observed in six of these seven Infection was confirmed via morphometric molecular and histologic
examination of samples from infected DF While much is still unknown about enteric coccidiosis in BFF
domestic ferrets provide a promising model for further investigation of this disease
iv
DEDICATION
For my mother Anna Pastorhellip
v
ACKNOWLEDGEMENTS
Itrsquos hard to believe that my residency and thesis have been completed and I have a lot of people
to thank for that
Dale and Graham the two people I wanted to be when I grew up thank you for your mentorship
for many years even before this program I know that you werenrsquot convinced that this project was DVSc
worthy when I first proposed it but Irsquom hoping that the results have changed your mind
I am sincerely grateful to all the members of my advisory committee Dale Smith John Barta and
Simon Hollamby for their insight support and interest in this project Dale you have been an exceptional
advisor I donrsquot know that I will ever get to your level but thank you for showing me that being a great
clinical zoo vet and pathologist are not mutually exclusive John thank you for spontaneously agreeing to
be my advisor when I came to you with this project proposal in my first semester for your energy and
enthusiasm and for supporting my widening interest in parasitology research
My heartfelt appreciation for the Toronto Zoo WHC veterinarians past and present Chris
Dutton Pauline Delnatte Simon Hollamby and Graham Crawshaw I have learned so much from all of
you that I will take forward into my future endeavours I appreciate the extra time you put in including
comps study sessions after-hours tecircte-agrave-tecirctes and the fact that your doors were open when I needed it
For the Toronto Zoo vet techs extraordinaire Michelle Lovering Cassia Devison Dawn
Mihailovic and Tasha Long ndash you have been indispensable during this program and there are not enough
words to express my gratitude
I would especially like to thank all the Wildlife Health Center staff (2013-2016) Mark Bongelli
Charles Guthrie Christine McKenzie Brian Telford Rick Vos Gerri Mintha Margaret Kolakowski
Andrew Lentini Rebecca Clark Lydia Attard Nigel Parr Paula Roberts Andrea Dada Mindy Waisglass
and Julie Digiandomenico for three very memorable years It is all of you that make the WHC such an
amazing place to be Irsquom not sure I have laughed so hard or so often as I did in that lunchroom and I hope
our paths will cross again
vi
I donrsquot think that I can truly express how thankful I am to Pathobiology laboratory technicians
Julie Cobean and Julia Whale Without your assistance patient teaching and friendship I would probably
still be screening fecal samples years from now and scratching my head as to how our lab protocols
actually work It is people like you who make sure graduate students become successful doctorates and I
canrsquot imagine Pathobio without you both in it
I would also like to thank my labmates in the Barta lab mdash Mian Hafeez Evelyn Rejman Rachel
Imai Perryn Kruth Ryan Snyder and Mosun Ogedengbe A special thank you goes to Alex Leveille
without whom my many adventures in parasitology research from coccidia to Babesia would not have
been as successful
To all the students who helped with ferret fecal sample processing data compilation and
necropsies Nathalie Ferriman Janessa Price Thisuri Eagalle Sarah Brisson thank you so much for your
hard work and excitement about my projecthellip even when it was very smelly
So many thanks to the amazing staff of Central Animal Facility - Linda Groocock Vicky Carson
Tony Cengija and Mary Fowler for the daily care and enrichment of my experimental ferrets Your
excitement about working with our ferrets and your assistance with all parts of the process helped made
this project a success
To Adriana Nielsen who was not only my better half but the other fifty percent of my brain for
several years It is your friendship fortitude and our endless phone conversations that got me through the
never-ending Toronto-Guelph commute and this program
To all the ldquoscope roomrdquo pathology co-residents past and present - thank you for being wonderful
friends and colleagues It is indeed rare to find so many amazing people in one place and I know this
program and my sanity would not have been the same without you
To the anatomic pathology faculty and senior graduate students - thank you for all the time
teaching and guidance you provided during my program While I canrsquot say that I have become an amazing
pathologist I can say that because of your mentorship I am a better diagnostician and the type of clinician
who asks better questions takes better samples and understands that you canrsquot ldquojust make a PCR for thatrdquo
vii
A special thank you to Tony van Dreumel who came out of retirement for a semester to try to teach the
Adrianas zoo pathology screening cases with you was always a pleasure
To all the lovely Histo Ladies PM room staff and the other AHL staff who helped me with
Toronto Zoo and HSC pathology cases along the way - I donrsquot think the anatomic path students could
survive without you Thank you for always smiling assisting and accommodating me even when I made
near-impossible processing requests during my weekly Guelph visits
I would also like to acknowledge and sincerely thank all the individuals who helped with resource
and sample acquisition for this project A special mention for those who went above and beyond because
of their interest in this project Don Duszynski who was instrumental in acquiring and then providing a
translator for many of the original mustelid Eimeria descriptions and Majda Globokar Nikola Pantchev
and Donald Martin who supplied my domestic ferret fecal samples and historical data
A shout-out to Julie Swenson Gary West and the Phoenix Zoo BFF team who fostered my love
of this endangered species and helped develop the idea for this project
As always I continue to go out into the world and pursue my dreams with the knowledge that I
have the support of my incredible family long-time friends and my partner Keith Morris I am so lucky
that my residency brought me home and that it afforded us all more time spent together For my aunt
Veronica Lacey who has never failed to believe in my potential and always pushed me to become an
academic ndash yoursquoll never get that PhD from me but I think this is pretty close Finally for my mother
Anna Pastor who never lived to see my greatest achievements but had absolute faith that I could reach
any goal I worked towardshellip this is for you
Finally none of this would have been possible without the generous support of the Toronto Zoo
Residency program and funding through the Barta Laboratory University of Guelph
Adriana Pastor
Toronto August 2017
viii
DECLARATION OF WORK PERFORMED
I declare that all the work reported in this thesis was performed by myself with the following
exceptions
Fecal samples were collected by personnel at the Toronto Zoo Louisville Zoo and participating
diagnostic laboratories
Fecal oocyst per gram counts (routine salt flotation and McMaster counts) were performed by
myself Julie Cobean Julia Whale Evelin Rejman Sarah Brisson Adriana Rodriguez and Perryn Kruth
Whole mitochondrial genome PCR and sequencing was performed by me in conjunction with
Julia Whale and Dr Mian Hafeez
Sequencing of PCR samples was performed at the University of Guelph Laboratory Services
(Guelph Ontario Canada) and results were obtained electronically
ix
TABLE OF CONTENTS
ABSTRACT ii
DEDICATION iv
ACKNOWLEDGEMENTS v
DECLARATION OF WORK PERFORMED viii
TABLE OF CONTENTS ix
LIST OF TABLES xiii
LIST OF FIGURES xiv
LIST OF APPENDICES xv
ABBREVIATIONS xvi
CHAPTER 1 LITERATURE REVIEW 1
11 INTRODUCTION 1
12 APICOMPLEXA 1
121 Brief introduction to apicomplexan pathogens 1
122 Life cycles of the Eimeria and Isospora species implicated in enteric coccidiosis 3
123 Methods of characterization 5
13 RECLASSIFICATION OF MAMMALIAN ISOSPORA 8
14 EIMERIID SPECIES CHARACTERIZED IN MUSTELIDS 8
141 The family Mustelidae 8
142 Eimeriid coccidia described from mustelids 9
143 Eimeriid coccidia described from domestic ferrets 16
144 Molecular characterization 19
145 Clinical signs of disease in domestic ferrets 21
146 Gross necropsy and histologic findings 21
15 INTRODUCTION TO ENTERIC COCCIDIOSIS IN THE BLACK-FOOTED FERRET
25
151 Natural history and conservation of the black-footed ferret in North America 25
152 Coccidia identified from black-footed ferrets 26
153 Morbidity mortality and clinical signs associated with enteric coccidiosis in black-footed
ferrets 28
16 TREATMENT PREVENTION AND CONTROL OF INFECTION BY EIMERIA SPP 29
161 Current recommendations for treatment of eimeriid coccidia in carnivores 29
x
162 Current recommendations for anticoccidial treatment and prophylaxis in domestic and
black-footed ferrets 30
17 VACCINES AGAINST COCCIDIA 32
171 Theory 32
172 Species successes in anticoccidial vaccination 34
18 RESEARCH GOALS AND OBJECTIVES 36
181 Objectives 36
182 Hypotheses 36
183 Applications 36
CHAPTER 2 MOLECULAR CHARACTERIZATION OF ENTERIC COCCIDIA FROM DOMESTIC
FERRETS (MUSTELA PUTORIUS FURO) 38
21 INTRODUCTION 39
22 MATERIALS amp METHODS 43
221 Fecal samples 43
222 Formalin fixed intestinal tissues 44
223 Molecular characterization 44
224 Phylogenetic analysis 46
23 RESULTS 47
231 Fresh fecal samples 47
232 Formalin fixed samples 48
233 Molecular characterization 49
234 Phylogenetic analysis 50
24 DISCUSSION 50
CHAPTER 3 MORPHOLOGICAL AND MOLECULAR CHARACTERIZATION OF ENTERIC
COCCIDIA ISOLATED FROM BLACK-FOOTED FERRETS (MUSTELA NIGRIPES) 60
31 INTRODUCTION 60
32 MATERIALS AND METHODS 64
321 Fecal samples 64
322 Formalin fixed intestinal tissues 65
323 Molecular characterization 66
33 RESULTS 66
331 Morphometric characterization 67
332 Molecular characterization 68
34 DISCUSSION 69
xi
CHAPTER 4 NATURAL HISTORY OF ENTERIC COCCIDIOSIS IN THE BLACK-FOOTED
FERRET (MUSTELA NIGRIPES) 78
41 INTRODUCTION 78
42 MATERIALS AND METHODS 80
421 Toronto Zoo BFF breeding program 80
422 Fecal oocyst evaluation 81
423 Retrospective review of pathology records 82
424 Prospective modified necropsy protocol 82
425 Retrospective medical history review 83
43 RESULTS 83
431 Fecal oocyst evaluation and retrospective medical history review 83
432 Pathology 86
433 Morbidity and mortality 88
44 DISCUSSION 88
CHAPTER 5 EVALUATING THE DOMESTIC FERRET (MUSTELA PUTORIUS FURO) AS AN
EXPERIMENTAL MODEL FOR ENTERIC COCCIDIOSIS IN THE BLACK-FOOTED FERRET
(MUSTELA NIGRIPES) 104
51 INTRODUCTION 104
52 MATERIALS AND METHODS 106
521 Animal care 106
522 Oocyst preparation 107
523 Experimental infections 108
524 Animal welfare 109
525 Hematology 110
526 Morphologic and molecular characterization 110
527 Necropsy protocol 111
53 RESULTS 111
531 Oocyst shedding 112
532 Morphologic and molecular characterization 113
533 Clinical signs 113
534 Hematology 113
535 Necropsy 114
54 DISCUSSION 115
xii
CHAPTER 6 WHOLE MITOCHONDRIAL GENOME SEQUENCES OF TWO EIMERIA SPECIES
ISOLATED FROM DOMESTIC (MUSTELA PUTORIUS FURO) AND BLACK- FOOTED FERRETS
(MUSTELA NIGRIPES) 129
61 INTRODUCTION 129
62 MATERIALS amp METHODS 130
621 Parasites 130
622 DNA isolation from coccidia in feces 131
623 Whole genome sequencing 131
624 Phylogenetic analysis 132
63 RESULTS 133
64 DISCUSSION 134
CHAPTER 7 CONCLUSIONS AND FUTURE DIRECTIONS 145
REFERENCES 148
APPENDICES 157
xiii
LIST OF TABLES
Table 11 Morphometrics of Eimeria and Isospora (=Cystoisospora) species affecting mustelids 10
Table 21 Amplification primers for nuclear 18S rDNA and mitochondrial COI loci used in the
identification of enteric coccidia from domestic ferrets 55
Table 22 Summary of fecal samples from domestic ferrets submitted to two diagnostic laboratories
from 2008-2015 56
Table 23 Morphologic and molecular identification of coccidia from domestic ferrets 57
Table 31 Amplification primers for nuclear 18S rDNA mitochondrial COI and COIII loci used in the
identification of coccidia from black-footed ferrets 73
Table 32 Morphologic and molecular characterization of coccidia from fecal and FFPE necropsy
samples from black-footed ferrets 76
Table 33 Morphometric characterization of Eimeria ictidea oocysts from black-footed ferrets 77
Table 41 Eimeria ictidea shedding in black-footed ferret dam and kit family groups - 2014-2016 97
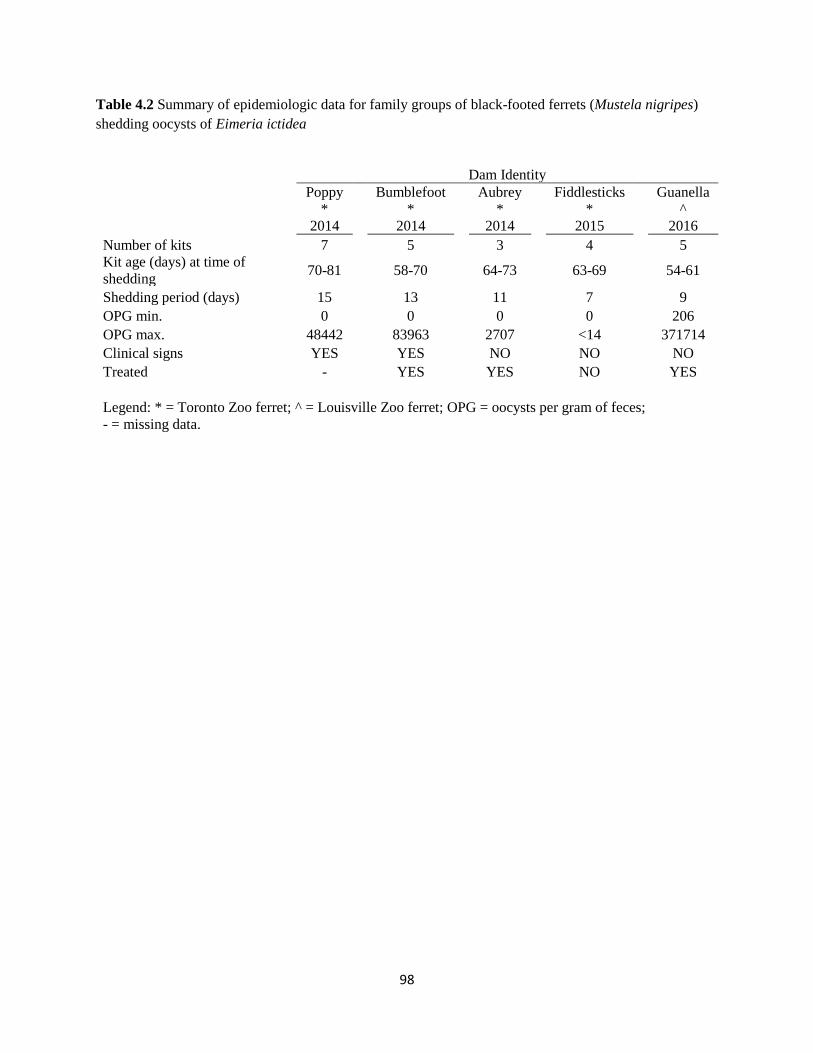
Table 42 Epidemiologic data for family groups of black-footed ferrets shedding Eimeria ictidea 98
Table 43 Shedding of Eimeria ictidea in adult black-footed ferrets - 2015-2016 99
Table 44 Epidemiologic data for adult black-footed ferrets shedding Eimeria ictidea 100
Table 45 Histologic findings from black-footed ferrets with enteric coccidiosis 101
Table 46 Incidence of coccidial infections in black-footed ferrets at the Cheyenne Mountain Zoo 102
Table 47 Yearly mortality associated with coccidiosis in black-footed ferrets at the Toronto Zoo 103
Table 51 Prepatent period and oocyst shedding of Eimeria ictidea in experimentally infected
domestic ferrets 126
Table 52 Results of oral inoculation of domestic ferrets with oocysts of Eimeria ictidea 127
Table 53 Distribution of coccidial life stages in intestinal tract of domestic ferrets orally
inoculated with oocysts of Eimeria ictidea 128
Table 61 PCR primers used to sequence the mitochondrial genome of Eimeria furonis 136
Table 62 PCR primers used to sequence the mitochondrial genome of Eimeria ictidea 137
Table 63 Coding regions in the mitochondrial genome of Eimeria furonis from a domestic ferret 138
Table 64 Coding regions in the mitochondrial genome of Eimeria ictidea from a black-footed ferret 139
Table 65 Pairwise comparison of coding regions in the mitochondrial genomes of Eimeria furonis
and Eimeria ictidea 140
xiv
LIST OF FIGURES
Figure 11 Phylogeny of the Apicomplexa 2
Figure 12 Classical life cycle of coccidian parasites 4
Figure 13 Morphologic characteristics used for identification of eimeriid oocysts 6
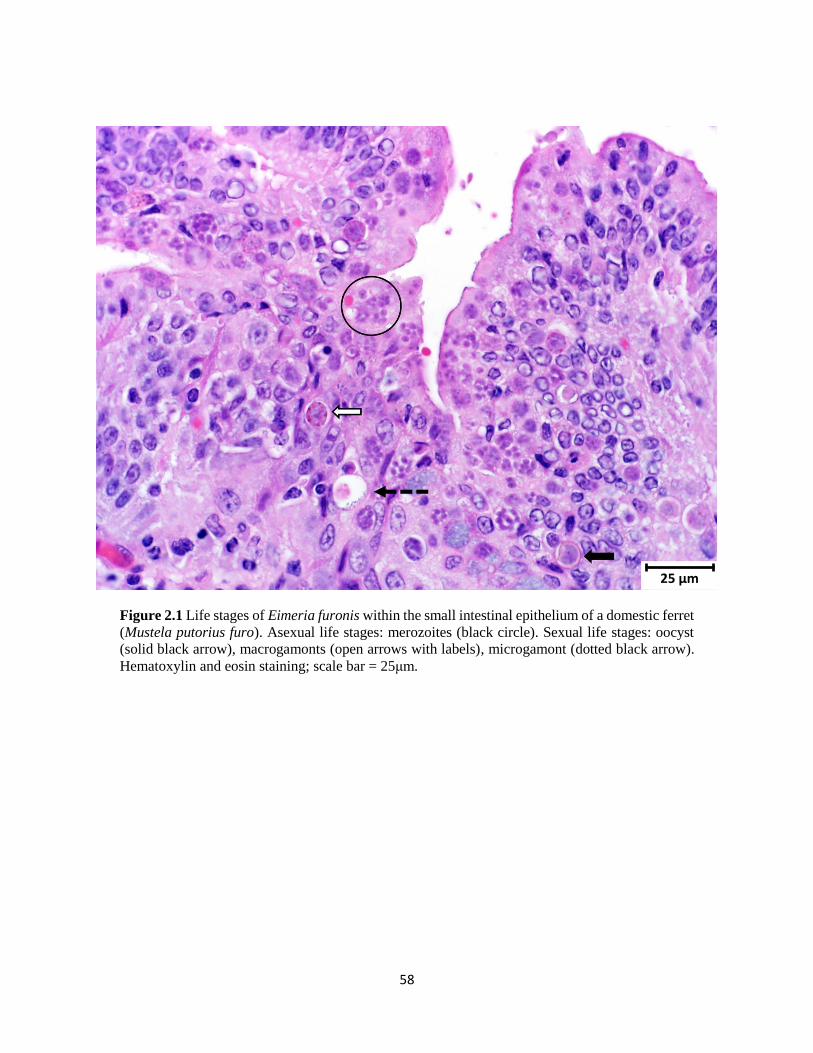
Figure 21 Life stages of Eimeria furonis within the small intestine of a domestic ferret 58
Figure 22 Phylogenetic relationships of coccidia (Eimeria ictidea Eimeria furonis and Isospora
(=Cystoisospora) laidlawi) from domestic or black-footed ferrets 59
Figure 31 Nuclear and mitochondrial genetic loci targeted by primers listed in Table 31 73
Figure 32 Morphometrics of Eimeria ictidea from a black-footed ferret (Mustela nigripes) 74
Figure 33 Nuclear 18S rDNA sequences of Eimeria ictidea to newly generated (see Chapter 2) and
published sequences of Eimeria furonis 75
Figure 34 Mitochondrial cytochrome c oxidase subunit I sequences of Eimeria ictidea to sequences
from other eimeriid parasites of carnivores 75
Figure 41 Oocyst per gram counts and shedding period of Eimeria ictidea from black-footed ferret
family groups from 2014-2016 95
Figure 42 Sexual life stages of Eimeria ictidea in the small intestine of a black-footed ferret 96
Figure 51 Exogenous life stages of Eimeria ictidea 123
Figure 52 Endogenous life stages of Eimeria ictidea within the small intestine of an experimentally
infected domestic ferret 124
Figure 53 Distribution of sexual and asexual life stages of Eimeria ictidea along the intestinal tract
of experimentally infected domestic ferrets 125
Figure 61 Map of the mitochondrial genome of Eimeria furonis 141
Figure 62 Map of the mitochondrial genome of Eimeria ictidea 142
Figure 63 Comparison of the mitochondrial genomes of Eimeria furonis and Eimeria ictidea 143
Figure 64 Phylogenetic relationships of coccidia from domestic and black-footed ferrets based on
complete mitochondrial genome sequences 144
xv
LIST OF APPENDICES
Appendix 1 Shedding of oocysts of Eimeria ictidea in black-footed ferret (Mustela nigripes) dam and
kit family groups from 2014-2016 158
Appendix 2a Hematology values for domestic ferrets (Mustela putorius furo) from 49-51 days of
age prior to experimental inoculation 161
Appendix 2b Serum biochemistry values for domestic ferrets (Mustela putorius furo) from
49-51 days of age prior to experimental inoculation 162
Appendix 3a Hematology values for domestic ferrets (Mustela putorius furo) inoculated orally
with Eimeria ictidea 163
Appendix 3b Serum biochemistry values for domestic ferrets (Mustela putorius furo) inoculated
orally with Eimeria ictidea 164
Appendix 4 Domestic ferret (Mustela putorius furo) weekly monitoring sheet 165
Appendix 5 Domestic ferret (Mustela putorius furo) 24 hour intensive monitoring sheet 167
Appendix 6 Domestic ferret (Mustela putorius furo) infection trial standard operating procedures 172
xvi
ABBREVIATIONS
ATP Adenosine triphosphate
BFF Black-footed ferret(s)
BI Bayesian inference
bp Base pair
CAPC Companion Animal Parasitology Council
CDS Coding DNA sequence
CITES Convention on International Trade in Endangered Species of Wild Fauna and Flora
COI Cytochrome c oxidase subunit 1
COIII Cytochrome c oxidase subunit 3
CytB Cytochrome b
DF Domestic ferret(s)
DNA Deoxyribonucleic acid
FFPE Formalin-fixed paraffin embedded tissue
IUCN International Union on the Conservation of Nature
L Length
LSU Large subunit
mt Mitochondrial
NaOH Sodium hydroxide
nu Nuclear
OPG Oocyst per gram count
PCR Polymerase chain reaction
rDNA Ribosomal DNA
SI Shape index
SND Single nucleotide difference
SOP Standard operating procedure
sp spp Species (singular plural)
SSP Species Survival Plan
SSU Small subunit
TMS Trimethoprim sulfadimethoxine
USFWS United States Fish and Wildlife Service
W Width
1
CHAPTER 1 LITERATURE REVIEW
11 INTRODUCTION
Black-footed ferrets (Mustela nigripes) are one of three wild ferret species worldwide Although
formerly distributed throughout the North American prairies black-footed ferrets (BFF) had been
extirpated from the majority of their range by the 1970s and were declared extinct in the wild in 1987
Since 1986 a multi-institutional effort has been breeding this species in captivity with reintroduction back
into the wild at select sites within Canada the USA and Mexico
Coccidial enteritis is a major cause of death in young captive black-footed ferrets (Bronson et al
2007) but coccidiosis can affect all age classes (personal observation) As a result fewer captive-bred
ferrets may be reared successfully for release to the wild The significance of coccidiosis in wild ferrets is
unknown Consequently the prevention and control of coccidial outbreaks is an important part of black-
footed ferret captive breeding programs and management This research is intended to improve the in situ
and ex situ health of the black-footed ferret through the provision of a better understanding of the
pathogenesis of enteric coccidiosis in this species and to pave the way for the investigation of novel
methods for disease treatment and control
12 APICOMPLEXA
121 Brief introduction to apicomplexan pathogens
The phylum Apicomplexa comprises a large number of eukaryotic intracellular parasitic
organisms many of which are of importance to human and veterinary medicine As indicated by their
name these parasites are characterized by the presence of an apical complex at the anterior aspect of the
infective stage of the life-cycle (Tenter et al 2002) The taxonomic classifications of members of the
Apicomplexa continue to be in a state of flux (reviewed by Adl et al 2005 Cavalier-Smith 2014 Tenter
et al 2002) For this reason a more simplified taxonomic structure has been used in this review (see
2
Figure 11) The subclass Coccidia is a speciose group within the Apicomplexa with most genera falling
into one of two major coccidian suborders within the Eucoccidiorida To date greater than 2000 species
of coccidia have been named (Duszynski Upton amp Couch nd Upton 2000) The adeleid coccidia
(suborder Adeleorina) include monoxenous (single host) and heteroxenous (multiple hosts) parasites in
genera such as Adelea Haemogregarina Hepatozoon and Karyolysus The eimeriorinid coccidia
(suborder Eimeriorina) include the typical intestinal coccidia such as Eimeria Isospora and Cyclospora
species belonging to the family Eimeriidae as well as tissue (cyst forming) coccidia such as
Cystoisospora Besnoitia Toxoplasma and Sarcocystis species that belong to the family Sarcocystidae
(Cox 1994)
Figure 11 Phylogeny of the Apicomplexa Numbers on branches and thickness indicate diversity
(ie named species) Taxonomic groupings demonstrated by the phylogenetic tree (1) subclass
Coccidia (2) suborder Adeleorina (3) suborder Eimeriorina (4) family Eimeriidae and (5) family
Sarcocystidae Adapted from Šlapeta J Morin-Adeline V (2011) Apicomplexa Levine 1970
Sporozoa Leucart 1879 httptolweborgApicomplexa2446 in The Tree of Life Web Project
httptolweborg
2
1
3
4
5
3
122 Life cycles of the Eimeria and Isospora species implicated in enteric coccidiosis
The life cycle of Eimeria species is considered the classical coccidian life cycle which is
typically completed in one host (monoxenous) with many Eimeria species parasitizing only a single host
species (stenoxenous) (Figure 12) The life cycle has two main phases of development one that takes
place within the host (endogenous) and the other that takes places outside of the host (exogenous)
Classically the endogenous stages of the Eimeria life cycle take place within the intestinal epithelium
however some Eimeria species undergo extraintestinal endogenous development such as Eimeria stiedae
in rabbits which replicates within the epithelium of the biliary tree During the exogenous phase of the
life cycle unsporulated oocysts that are shed in the feces of the host sporulate within the environment
resulting in the formation of four sporocysts within each oocyst (tetrasporocystic) Each sporocyst
contains two sporozoites (dizoic) Sporulation is affected by three main factors temperature moisture and
aerobic conditions (Fayer 1980)
Once ingested by the host the wall of the sporulated oocyst is broken to release sporocysts from
which the sporozoites (infective stage) excyst The freed sporozoites penetrate the intestinal epithelial
cells and undergo multiple mitotic divisions to form a single multinucleate meront The meront then
undergoes simultaneous cytokinesis to form first generation merozoites which leave the host cell to infect
new cells and undergo further asexual replications The undifferentiated uninucleate tissue stage of the
parasite within the intestinal epithelial cell is called a trophozoite The number of cycles of asexual
replication (merogony) is predetermined after which the last generation of merozoites penetrate host cells
and undergo sexual differentiation into male and female gamonts (gametogony) Each microgamont
(male) undergoes simultaneous fission to produce numerous motile microgametes each macrogamont
(female) develops into a single mature macrogamete Fertilization of a macrogamete by a motile
microgamete results in formation of a zygote that is rapidly enclosed in a thick wall to form an
unsporulated oocyst Oocysts are shed with the hostrsquos feces into the environment where they are
protected from desiccation and chemical disinfection by the oocyst wall Traditionally Eimeria species
4
have been differentiated based on the host species or host genus affected the site of endogenous life cycle
development and the microscopic cellular characteristics of the different life stages Interestingly
experimental cross infection of Eimeria species from their natural host to a novel host of a taxonomically
similar species has been successful in some cases (De Vos 1970 Levine and Ivens 1970 Haberkorn
1971) challenging the notion that Eimeria are truly stenoxenous parasites
Figure 12 Classical life cycle of coccidian parasites This apicomplexan life cycle includes both
sexual and asexual development The three processes in the life cycle are merogony (asexual
replication A-D) followed by gametogony (formation of gametes E-H) within the digestive tract
of the host with release of unsporulated oocysts (I) Exogenous sporogony (I-L) results in the
production of infective sporulated oocysts (L) Adapted from Barta 2001 with permission of the
author
The life cycle of Isospora spp is similar to that of species in the genus Eimeria (see Figure 12)
but the number of sporocysts and sporozoites differ sporulated oocysts contain two sporocysts (disporic)
5
each of which contains four sporozoites (tetrazoic) These characteristics are not unique to Isospora spp
because diasporic tetrazoic sporulated oocysts are also found in the genera Besnoitia Frenkelia
Hammondia Sarcocystis and Toxoplasma However the sporocysts in the latter parasites are
morphologically distinct in that they lack Stieda bodies
123 Methods of characterization
1231 Morphological features
Historically eimeriid coccidia have been classified based on the cellular morphology of the
different life stages (particularly the morphometrics of sporulated oocysts) where these stages occur in
the host and apparent host specificity (frequently assumed and not tested experimentally) The
morphological features and dimensions of oocysts and their components are important diagnostic features
because of the availability of these stages in clinical specimens these characteristics can include size
(length [L] width [W] shape index [SI=LW]) number of sporocysts wall morphology
presenceabsence of a micropyle micropyle cap residual body or polar granules for oocysts size number
of sporozoites wall morphology presenceabsence of Stieda body subStieda body paraStieda body or
residual body for sporocysts and presenceabsence of refractile bodies for sporozoites (see Figure 13)
Pertinent life cycle information includes type of life cycle (monoxenous versus heteroxenous) tissue
sites of merogony and gametogony (intestinal versus extraintestinal) and the presence or absence of
extraintestinal hypobiotic stages (eg dormozoites or hypnozoites) Further information used to
characterize coccidia that form tissue cysts generally includes details on life stages in the definitive and
intermediate hosts location and morphology of tissue cysts route(s) of transmission among host species
and morphologic descriptions of merozoites (eg tachyzoites or bradyzoites) in tissue culture
6
Figure 13 Morphologic characteristics used for identification of eimeriid oocysts 1) Oocyst in cross
section ol - oocyst length or - oocyst residual body ow - oocyst width pg - polar granule row -rough
outer wall 2) The top of a hypothetical oocyst mcd - depth of the micropyle cap mcw - width of the
micropyle cap mw - width of the micropyle sow - smooth outer wall 3) Sporocyst in cross section
psb - paraStieda body sb - Stieda body sl - sporocyst length sp - sporozoite sr - sporocyst residual
body srb - sporozoite refractile body ssb - subStieda body sw - sporocyst width From Duszynski D
Wilber PG (1997) A guideline for the preparation of species descriptions in the Eimeriidae Journal of
Parasitology 83(2)333-336 reproduced with permission of Allen Press Publishing Services
1232 Molecular characterization (genetic loci and methods)
More recently molecular techniques have been used to infer phylogenetic or evolutionary
relationships among coccidia and to reclassify taxonomic assignments to better reflect the evolutionary
history of these parasites Molecular data can be more informative than phenotypic data because recent
evolutionary divergence among coccidia is unlikely to be reflected in morphologic differences but may
be detectable using molecular data The principle behind the use of molecular sequencing to describe
evolutionary relationships is that nucleotide sequences like morphological features diverge over time
under selective pressure however nucleotide sequences evolve at a more regular rate than do
morphologic characteristics Phenotypic data is thus less likely to detect recent evolutionary divergence
Sequences that are more similar are inferred to be more closely related and to have diverged more
recently (Cox 1994) Molecular characterization can be performed using DNA RNA or protein
sequences Most of the early molecular phylogenetic analyses of coccidia performed used ribosomal RNA
sequences usually by PCR amplification of ribosomal DNA (rDNA) in the nuclear genome of the
7
parasites Ribosomes contain both small and large RNA subunits in eukaryotes the large ribosomal RNA
consists of two forms 5S and 28S while the small ribosomal RNA exists only as 18S Sequences from
several genetic loci have been used for characterization of parasites most commonly 18S rDNA 28S
rDNA and ribosomal internal transcribed spacer regions (ITS) from the nuclear genome and more
recently mitochondrial cytochrome c oxidase subunits I (COI) and III (COIII) however sequencing of
nuclear 18S rDNA (nu 18S rDNA) has been the most prevalent in the literature by far Early attempts to
use 5S RNA sequences formed unlikely phylogenies and too few 28S ribosomal DNA sequences have
been obtained to make this locus useful (Cox 1994 Tenter et al 2002) The disadvantage of nu 18S
rDNA is that it is comparatively poor at distinguishing among closely related eimeriid coccidial species
because of its conserved nature but for that reason the nu18S rDNA locus is useful for inferring
relationships among species with greater evolutionary divergence Although only exploited recently
because of the paucity of suitable PCR primers the mitochondrial COI locus appears to be more useful
for distinguishing closely related eimeriid coccidia (Ogedengbe Hanner amp Barta 2011) but COI
sequences are less useful for inferring more ancient relationships between highly divergent coccidial
species Consequently the combined use of nu 18S rDNA and mitochondrial COI sequencing has been
recommended for improved species description and phylogenetic analysis (El-Sherry et al 2013)
Molecular characterization has also been used for diagnostic purposes and is well-suited to the
identification of coccidia when information on host specificity parasite life cycle and life stages is not
available as the molecular (genetic) data is the same for a given parasite during each of its life cycle
stages This information can be particularly useful in identifying the relationship between different life
stages of heteroxenous parasites collected from different hosts (intermediate definitive) Furthermore for
previously unidentified coccidia or those for which limited information is available molecular
characterization could be used to predict likely definitive hosts or parasite life cycle traits based on
phylogenetic relationships to other known species
8
13 RECLASSIFICATION OF MAMMALIAN ISOSPORA
Recommendations have been made to reclassify the avian and mammalian Isospora into two
separate genera based on life cycle molecular phylogenetic studies and morphologic description of
sporulated oocysts (Frenkel 1977 Barta et al 2005) Due to their classical coccidian life cycle presence
of Stieda bodies within sporocysts and close phylogenetic association with Eimeria species the avian
Atoxoplasma and Isospora have been retained in the genus Isospora (see Barta et al 2005) Conversely
the presence of tissue life cycle stages lack of Stieda bodies within sporocysts and close phylogenetic
association with other genera within the family Sarcocystidae have required many mammalian Isospora
to be reclassified as members of the genus Cystoisospora Frenkel 1977 (Frenkel 1977 Barta et al 2005)
Consequently for the remainder of this thesis Isospora species from mustelids will be referred to as
Isospora (=Cystoisospora) to reflect their probable generic association
14 EIMERIID SPECIES CHARACTERIZED IN MUSTELIDS
141 The family Mustelidae
The family Mustelidae within the order Carnivora comprises a group of approximately 59
carnivorous mammalian species within 22 genera Native mustelids are found in terrestrial and aquatic
environments on almost every continent with the exception of Australia and Antarctica The Mustelidae
are classically divided into two subfamilies as defined by Wozencraft (2005) 1) Mustelinae (weasels
mink ferrets marten wolverine) the larger subfamily including the following genera Arctonyx Eira
Galictis Gulo Ictonyx Lyncodon Martes Meles Mellivora Melogale Mustela Neovison Poecilogale
Taxidea and Vormela and 2) Lutrinae (otters) including seven genera Aonyx Enhydra Hydrictis
Lontra Lutra Lutrogale and Pteronura More recently molecular data suggest the Mustelidae should be
separated into eight subfamilies although this is not universally accepted (Koepfli et al 2008 Lariviegravere
and Jennings 2009 Yu et al 2011)
9
142 Eimeriid coccidia described from mustelids
Ten named Eimeria species and twelve named Isospora (=Cystoisospora) species have been
described in the Mustelidae and are summarized in Table 11 This table includes information on host
range life cycle and detailed morphologic data used to identify and classify the individual parasites Two
coccidial parasites isolated from the Libyan striped weasel (Ictonyx libyca) and the European polecat
(Mustela putorius) initially ascribed to the genus Isospora Isospora zorillae and Isospora putori
respectively have since been reclassified as Sarcocystis spp (see footnote to Table 2 of Yi-Fan et al
2012)
10
Table 11 Morphologic characteristics of Eimeria and Isospora (=Cystoisospora) species affecting mustelids
Coccidial species Host genus and
species
Life cycle
Location
Oocyst shape and
size
Oocyst
description
Sporocyst
description
Sporozoite
description
References
Cytoisospora
eversmanni
Mustela
eversmanii
(Steppe polecat)
Mustela
putorius
(European
polecat)
Homoxenous L185 (16ndash20)
W 148 (16ndash12)
LW 13 (11ndash16)
M absent
PG absent
OR absent
L 115
(10ndash135)
W 98
(9ndash11)
LW 12
(11ndash15)
SB absent
SR present
SRB present Yi-Fan et al 2012
Svanbaev 1956
Nukerbaeva amp
Svanbaev 1973
1977
Cystoisospora
pavlovskyi
Mustela
eversmanii
Mustela
putorius
Homoxenous L 322 (29ndash36)
W 273 (265ndash285)
LW 12 (11ndash14)
M absent
PG absent
OR absent
L 195
(18ndash21)
W 144
(12ndash15)
LW 14
(12ndash15)
SB absent
SR present
SRB present Yi-Fan et al 2012
Svanbaev 1956
Nukerbaeva amp
Svanbaev 1973
1977
Eimeria
baskanica^
Mustela
erminae
(ermine)
Homoxenous Oval with tapered
ends
L 112-126
W 84-98
M absent
PG absent
OR present
SR absent Bean shaped Nukerbaeva amp
Svanbaev 1977
Eimeria furonis Mustela
putorius
Mustela
putorius furo
(dom ferret)
Mustela
nigripes (BFF)
Mustela vison
(mink)
Homoxenous
Small intestine
rectum (H 1927)
Jejunumileum (BP
1993)
Spherical ndash
subspherical
L 11-14
W 10-13
OW 2 layers
M absent
PG absent
OR absent
Spindloid
L 8-9
W 4
SB present
SR present
Vermiform Blankenship-Paris
et al 1993
Hoare 1927 1935b
Jolley et al 1994
Nukerbaeva amp
Svanbaev
19731977
Williams et al 1988
1992 1996
Eimeria hiepei Mustela vison Homoxenous
Bile duct
Spherical
L 13-17
W 13-17
OW 2 layers
(outer
smooth)
M absent
PG absent
OR absent
L 6
W 4
SB absent
SR absent
Banana shaped Davis et al 1953
Grafner et al 1967
11
Coccidial species Host genus and
species
Life cycle
Location
Oocyst shape and
size
Oocyst
description
Sporocyst
description
Sporozoite
description
References
Eimeria ictidea Mustela
eversmanni
Mustela
nigripes
Mustela
putorius
Mustela
putorius furo
Homoxenous
Small intestine
Ovoid ndashellipsoid
L 13-27
W 13-21
OW 2 layers
M present
PG absent
OR absent
Ovoid
(irregular)
L 115
W65
SB present
SR present
- Hoare 1927 1935a
1935b
Jolley et al 1994
Litvenkova 1969
Svanbaev 1956
Tinar 1985
Williams et al 1988
1992
Eimeria irara Eira barbara
(tayra)
Homoxenous
Feces
Ovoid
L 21-25
W 18-20
OW outer
layer smooth
M absent
PG absent
OR absent
Ellipsoid
L 10-12
W 65
SB present
SR present
Elongate (one
end broader than
the other)
Carini amp da
Fonseca 1938
Eimeria melis Meles meles
(European
badger)
Homoxenous Ellipsoid
L 20plusmn018
W 157plusmn002
LW128plusmn0017
(112-15)
OW 2 layers
(outer
smooth)
M absent
PG present
OR present
Ovoid
L
119plusmn0018
W 65plusmn008
LW 183
(155-24)
SB present
L 90plusmn005
W 324plusmn0025
SRB present
Anwar et al 2000
Kotlan amp Pospesch
1933
Eimeria mustelae Mustela vison
Mustela nivalis
(snow weasel)
Homoxenous
Duodenumileum
Spherical or
Ellipsoid
L 18-26
W 14-24
OW 2 layers
M absent
PG present
OR absent
Ovoid
L 8
W 5
SB present
SR present
Broad at one
end and tapered
at other
L 7
W 3
Glebezdin 1978
Iwanoff-Gobzem
1934
Levine 1948
Musaev amp Veisov
1983
Tinar 1985
Eimeria sablii Martes zibellina
(sable)
Homoxenous
Gut
Spherical or
subspherical
L 112-126
W 112
OW 2 layers
M absent
OR absent
Ovoid
L 56
W 42
SR present
Elongate Nukerbaeva 1981
Eimeria sibirica Martes zibellina Homoxenous Ovoid
L avg 216
W avg 180
LW 1076
OW 2 layers
M absent
PG absent
OR absent
Ovoid
L 96-112
W 56-72
SR absent
Elongate Nukerbaeva 1981
Yakimoff amp
Gousseff 1934
Yakimoff amp
Terwinsky 1930
1931
12
Coccidial species Host genus and
species
Life cycle
Location
Oocyst shape and
size
Oocyst
description
Sporocyst
description
Sporozoite
description
References
Eimeria vison
(Eimeria
mustelae)
Mustela
putorius
Mustela
putorius furo
Mustela vison
Homoxenous
Small intestine
+- large intestine
Ovoid
L 17-22
W 9-18
OW 2 layers
M absent
OR
sometimes
present
Ovoid or
Piriform
L 10
W 55
SB absent
SR present
Curved or Club
shaped
L 9
W 25
Foreyt amp Todd 1976
Foreyt et al 1977
Kingscote 1934
1935
Levine 1948
McTaggart 1960
Nukerbaeva amp
Svanbaev
19731977
Tinar 1985
Umurzakov amp
Nukerbaeva 1985
Wolter 1961
Zimmermann 1959
Isospora africana Ictonyx libyca
(Libyan striped
weasel)
Homoxenous
Feces
Spherical
L 25-27
W 25-27
OW 2 layers
(outer
smooth)
M absent
PG absent
OR absent
Ovoid
L 15-17
W 10-12
SB absent
SR present
Elongate
L 135
W 3
Prasad 1961
Isospora altaica Mustela altaica
(mountain
weasel)
Homoxenous
Gut
Oval or spherical
L 280-336
W 252-280
LW 121 (111-
124)
OW 2 layers
M absent
PG absent
OR absent
Ovoid or
spherical
L 140-168
W 111-168
SR present
Svanbaev amp
Rachmatullina
1971
Isospora goussevi Mustela nivalis Homoxenous
Large intestine
Ovoid
L 224 (220-250)
W 174 (160-190)
LW 135 (133-
137)
OW 1 layer
PG present
OR present
Ovoid
L 120
(100-130)
W 70 (60-
80)
SB present
SR present
Elongate Musaev amp Veisov
1983
13
Coccidial species Host genus and
species
Life cycle
Location
Oocyst shape and
size
Oocyst
description
Sporocyst
description
Sporozoite
description
References
Isospora
hoogstraali
Ictonyx libyca Homoxenous
Feces
Ellipsoid
L 37-41
W 32-34
OW 2 layers
(outer
smooth)
M absent
PG some
OR absent
Ovoid
L 19-21
W 13-15
SB absent
SR present
Club-shaped
L 18-19
W 4-6
Prasad 1961
Isospora laidlawi Mustela
putorius
Mustela
putorius furo
Mustela vison
Homoxenous
Feces
Intestinal contents
Ovoid L
320-368
W 272-304
OW 2 layers
M absent
PG absent
OR absent
Ellipsoid
L 208
W 144
SB absent
SR present
Sausage shaped Foreyt et al 1977
Hoare 1927
Levine 1948
McTaggart 1960
Nukerbaeva amp
Svanbaev 1973
1974 1977
Tinar 1985
Isospora lutrae Lutra lutra
(European
otter)
Lutra
canadensis
(North
American river
otter)
Homoxenous Spherical
L 312 (275-32)
W 296 (28-31)
LW 104
(10-112)
OW 2 layers
(outer
smooth)
M absent
PG absent
OR absent
Ellipsoid
L 182 (17-
19)
W 144 (14-
16)
LW128
(12-14)
Sb absent
sSB absent
SR present
Spindle- shaped
L 124
W 25
SRB present
Torres et al 2000
Hoover et al 1985
Isospora
martessii
Martes zibellina Homoxenous
Gut
Ovoid short oval or
spherical
L 252 ndash 280 196
168
W 168 ndash 224 168
168
OW 2 layers
M absent
OR absent
Ovoid
L 112-168
W 84-112
SR present
Elongate Nukerbaeva 1981
Isospora melis Meles meles Homoxenous Ovoid
L 328plusmn034
W 269plusmn019
LW122 (110-
157)
OW 2 layers
(outer
smooth)
M absent
PG absent
OR absent
Ellipsoid
L
215plusmn0166
W 14plusmn012
LW 155
(133-185)
SR absent
Round at one
end other end
tapered
L 142plusmn116
W 40plusmn017
SRB absent
Anwar et al 2000
Glebezdin 1978
Kotlan amp Pospesch
1933
Pelleacuterdy 1955
14
Coccidial species Host genus and
species
Life cycle
Location
Oocyst shape and
size
Oocyst
description
Sporocyst
description
Sporozoite
description
References
Isospora
mustelae (nomen
nudum)
Martes martes Ovoid L
7 W
225
M present - - Galli-Valerio 1932
Isospora nivalis Mustela nivalis Homoxenous
Large intestine
Ovoid
L 206 (200-230)
W 184 (180-210)
LW 11 (109-111)
OW 1 layer
PG absent
OR absent
Ovoid
L 125
(120-130)
W 80 (70-
90)
SR present
Lemon or pear
shaped
Musaev amp Veisov
1983
Unnamed
ldquoCoccidiardquo^
Mustela
nigripes
Urinary bladder - - - - Jolley et al 1994
Unnamed
ldquoCoccidiardquo^
Mustela
nigripes
Trachea bronchus
bronchial glands
- - - - Jolley et al 1994
Unnamed
Eimeria sp^
Mustela
nigripes
Feces
intestinal contents
Ovoid
L 350-386
W 212-232
- - - Jolley et al 1994
Williams et al
1992
Unnamed
Eimeria sp^
Mustela
putorius furo
Small intestine - - - - Blankenship-Paris
et al 1993
Unnamed
Eimeria sp^
Mustela nivalis Homoxenous
Large intestine
Ovoid-ellipsoid L
2031 (1712-2162)
W 148 (1225-
1681)
LW 136 (121-16-
)
OW 1 layer
PG absent
OR absent
Ovoid or
pear-shaped
L 60-100
W 40-80
SR present
Elongate
L 50-90
W 30-70
Musaev amp Veisov
1983
Unnamed
Eimeria sp^
Martes martes
(marten)
Homoxenous Ovoid
L avg 216
W avg 180
LW 1076
OR absent 4 sporocysts
SR present
L 126
W 60
Yakimoff and
Gousseff 1934
Unnamed
Isospora sp^
Mustela
putorius furo
Feces - - - - Bell 1994
Unnamed
Isospora sp^
Mustela
putorius furo
Feces - - - - Bell 1994
Legend L = length W = width LW = length-width ratio avg = average OW = oocyst wall PG = polar granules M = micropyle SB = Stieda body sSB =
subStieda body OR = oocyst residuum SR = sporocyst residuum SRB = sporozoite refractile body ^ = species inquirendae - = no information provided by
author(s) = information obtained from secondary sources (primary reference could not be obtained) All measurements are in micrometers Bolded references
15
are those from which morphometric data were assembled Remaining references indicate other authors who have identified that parasite species in the same or
similar host
16
143 Eimeriid coccidia described from domestic ferrets
Three species of coccidia were originally described from 50 domestic ferrets (Mustela putorius
furo) Eimeria ictidea Eimeria furonis and Isospora (= Cystoisospora) laidlawi (Hoare 1927) All three
species were detected in feces from domestic ferrets at a research facility undergoing an outbreak of
canine distemper Sick ferrets appeared more frequently infected than healthy ones As per Hoare (1927)
none of the ferrets appeared to display clinical signs associated with protozoal infection For each
parasite the author described morphology of sporulated oocysts isolated from feces and sporulation time
(exogenous life stages) The pre-patent period (minimum duration of endogenous development) in an
inoculated naiumlve ferret was described only for E furonis and E ictidea due to insufficient sample size of
I (=C) laidlawi oocysts for an experimental infection trial Sporulation of oocysts occurred within 5-6
days for E furonis 3 days for E ictidea and 4 days for I (=C) laidlawi The sporulated oocysts of E
furonis were spherical with a double outer wall with a thin colourless outer layer and thick yellowish
inner layer no micropyle or residual body and measured on average 128 times 120 microm (length [L] 112-
144 width [W] 104-128 shape index [SI] 107) Unsporulated oocysts contained a zygote with a
diameter of 96 microm Sporocysts were spindle-shaped with one end constrictedblunted contained a
residual body and on average measured 8-88 times 4 microm Sporozoites were vermiform with one end
narrower than the other arranged head to tail and had a central nucleus a clear vacuole was identified in
some at the broad end The sporulated oocysts of E ictidea were oval or elliptical with a double outer
wall with a thin colourless outer layer and thick yellowish inner layer no micropyle or residual body
and measured on average 236 times 175 microm (L 184-272 W 128-208 shape index 135) The zygote in
unsporulated oocysts was elongate with a diameter of 15 times 12 microm when originally passed in feces but
became more spherical with time Sporocysts were irregularly oval with one end broad and the other
more constricted contained a residual body and on average measured 115 times 65 microm Sporozoites were
vermiform with one end narrower than the other arranged head to tail and had a central nucleus and a
clear vacuole at the broad end The sporulated oocysts of Isospora (=Cystoisospora) laidlawi were ovoid
with a double outer wall with a thin colourless outer layer and thick yellowish inner layer no micropyle
17
or residual body and measured on average 34 times 29 microm (L 320-368 W 272-304) Unsporulated
oocysts contained a spherical zygote with a diameter of 236 microm Two sporocysts were identified each
containing 4 sporozoites and no Stieda body sporocysts were elliptical contained a residual body and on
average measured on 208 times 144 microm Sporozoites were sausage shaped with one end slightly pointed
and had a central nucleus and a clear vacuole identified at the pointed end Sporozoites were arranged
with pointed ends all at the same pole of the sporocyst The pre-patent periods described for E furonis
and E ictidea were 6 days and 7 days respectively (Hoare 1927)
Since Hoarersquos initial description (Hoare 1927 Hoare 1935) multiple single case reports and
outbreaks of severe clinical disease associated with intestinal coccidiosis have been reported in domestic
ferrets Blankenship-Paris et al (1993) described a single case of a four-month-old domestic ferret that
presented depressed in thin body condition dehydrated and with pasty dark feces on the perineum This
ferret had been housed with its dam and another sibling neither dam nor sibling showed clinical signs of
enteric disease and both had negative fecal examination results on repeated evaluation Routine fecal
examination of the rest of the colony and necropsies on eight other ferrets in the colony revealed no
evidence of coccidial infection Enteric coccidiosis was determined to be the cause of disease in the four-
month-old ferret based on necropsy findings but the coccidia could not be speciated because diagnosis
was made on histologic findings only
Sledge et al ( 2011) described three separate outbreaks of severe enteric coccidiosis in domestic
ferrets from one ferret rescue centre (group 1) and two shelters (groups 2 and 3) all affected by the same
Eimeria sp The morphologic characteristics of sporulated oocysts were only described for group 1 no
coccidial oocysts were detected on direct smear or fecal flotation of diarrheic samples submitted from
groups 2 and 3 Oocysts were identified as spherical measuring 12-13 microm in diameter with four
sporocysts each containing two sporozoites Oocyst morphometrics histopathologic findings and nu 18S
rDNA partial sequences from all three groups were used collectively to confirm the coccidial species
identify in each outbreak as E furonis
18
Two cases of biliary coccidiosis with E furonis have been reported in domestic ferrets The first
was in a nine-week-old male ferret from a research facility (Williams Chimes amp Gardiner 1996) The
ferret presented with signs of hepatic disease and was negative for coccidia on fecal flotation and direct
smears Endogenous coccidial life stages were described from the gall bladder and liver on histologic
examination In tissue section the oocysts were oval to spherical and measured 125 times 120 microm Meronts
measured 108-130 times 89-93 microm and contained up to 16 merozoites The merozoites exhibited a double-
layered pellicle prominent conoid few rhoptries and many micronemes anterior to the nucleus Based on
the morphologic description of the life stages in this case the coccidia were identified by the authors as
an Eimeria species most likely E furonis Kaye et al (2015) described a second case of biliary
coccidiosis in an 18-month-old female pet domestic ferret with concurrent pure red cell aplasia In this
case all endogenous coccidial life stages were observed on histologic examination of the epithelium of
the extrahepatic biliary tree The oocysts were ovoid and measured 12 times 13 microm Meronts measured 12 times
15 microm and contained up to 16 merozoites each measuring 2 times 5 microm Based on the morphologic
description of the life stages in this case and nu 18S rDNA sequences the pathogen was also determined
to be E furonis Biliary coccidiosis has also been identified in mink (Mustela vison) with the etiologic
agent identified as Eimeria hiepei (Davis Chow amp Gorham 1953 Grafner Graubmann amp Dobbriner
1967)
Oocysts from Cystoisospora ohioensis have been reported from fecal samples collected from
healthy domestic ferret kits in a large American ferret breeding operation that were raised on the same
premise as juvenile domestic dogs (Patterson amp Fox 2007) The method of identification of this parasite
was not described by Patterson amp Fox A second similar institution reported the presence of a
Cystoisospora species also thought to be C ohioensis in routine fecal examination of their ferret colony
(Dr Bambi Jasmin personal communication) Coccidial identification in this case was performed by the
Animal Health Diagnostic Center at Cornell University The significance of these findings is unknown as
no clinical signs or histologic lesions have been described in domestic ferrets associated with shedding of
19
oocysts and the definitive host for C ohioensis is the domestic dog It is most likely that fecal
identification of C ohioensis represents a pseudoparasite in both of these cases or perhaps an
undescribed Cystoisospora sp that is morphologically indistinguishable from C ohioensis
It is difficult to estimate the prevalence of enteric coccidia within the North American domestic
ferret population Fecal samples submitted to university or large veterinary diagnostic laboratories from
domestic ferrets in Canada are uncommon and samples positive for coccidia appear infrequently (Dr
Donald Martin personal communication) Data from Idexx Vet Med Lab in Ludwigsburg Germany was
compiled to review the prevalence of coccidia and Giardia within fecal samples from domestic ferrets
(Pantchev et al 2011) The authors reported that of 284 fecal samples submitted from 2002-2004 18
(63) had detectable coccidial oocysts on fecal flotation Oocysts were identified based on morphologic
characteristics as E ictidea E furonis I (=C) laidlawi and another unidentified Isospora species
Comparative data from the same laboratory from 2009-2010 included sample submissions from 253
ferrets 21 (83) of which were positive for coccidial oocysts on fecal flotation Nine of the samples
were positive for E furonis three were positive with both E furonis and I(=C) laidlawi present eight
were positive only for I(=C) laidlawi and one sample contained both E furonis and E ictidea
identification in all cases was based on morphologic characteristics No statistically significant difference
in the occurrence of coccidial oocysts was detected when data from the two periods were compared
(Fisherrsquos exact test P=041) (Pantchev et al 2011)
144 Molecular characterization
Molecular characterization of Eimeria furonis was first performed by Abe et al (2008) using
oocysts purified from the feces of a single domestic ferret with clinical signs of coccidial enteritis Small
subunit ribosomal DNA (nu 18S rDNA) primers CYC1FE (5ʹ-TAC CCA ATG AAA ACA GTT T-3prime) and
CYC4RB (5prime-CGT CTT CAA ACC CCC TAC TG-3prime) were used to amplify a 347 base pair (bp) fragment
of nu 18S rDNA These primers were initially developed for molecular identification of Cyclospora
species but have since been shown to amplify nu 18S rDNA from several Eimeria species (Matsubayashi
20
et al 2005) The amplicon was sequenced (GenBank AB329724) and compared with previously
published partial nu 18S rDNA sequences from 40 Eimeria two Isospora and four Cyclospora species
The resulting phylogram grouped E furonis with E alabamensis (cattle) and E meleagrimitis (turkey) In
the same study the microscopic morphology of the oocysts was used to identify this coccidial species as
E furonis by comparison with published descriptions of E furonis E ictidea and E heipei by Hoare
(1927) Hoare (1935) and Grafner Graubmann amp Dobbriner (1967) respectively
Nuclear 18S rDNA was also used by Sledge et al (2011) for molecular identification of the
eimeriid coccidia implicated in the three distinct outbreaks of enteric disease in domestic ferrets As
described above initial identification and speciation of the coccidia was performed using morphologic
characteristics of the sporulated oocysts collected from feces in one of the three outbreaks being
investigated the oocysts were identified as E furonis Histologic sections of formalin fixed intestinal
segments from ferrets from each of the three outbreaks contained multiple coccidial life stages DNA was
then isolated from stored formalin-fixed tissues for further genetic analysis Using the partial nu 18S
rDNA gene sequence reported by Abe et al (2008) (GenBank AB329724) the following PCR primers
were created 5ʹ-ACA ATT GGA GGG CAA GTC TG-3ʹ and 5ʹ-GGCGAC AAG CCT GCT TGA AAC-
3ʹ PCR amplification produced a 247 bp amplicon from each of the three groups Analysis and
sequencing of amplicons from all three groups showed 100 homology to nucleic acid sequences
previously reported by Abe et al (2008) for the gene encoding E furonis nu 18S rDNA
Coccidia were identified within hepatobiliary lesions in a domestic ferret receiving
immunosuppressive therapy for red cell aplasia (Kaye et al 2015) DNA was extracted from frozen liver
and a 247 bp fragment of the nu 18S rDNA was amplified using the primers previously described by
Sledge et al (2011) and sequenced Kaye et al (2015) reported that the DNA sequence of the amplicon
was 100 homologous to the published nu 18S rDNA of E furonis and 95 homologous to the nu 18S
rDNA of E myoxi (rodent) E alabamensis (cattle) and I robini (avian)
21
145 Clinical signs of disease in domestic ferrets
Hoare (1927 1935b) in his initial descriptions of enteric coccidiosis in domestic ferrets
observed that clinical signs of intestinal disease were not evident The recent literature supports the
finding of subclinical disease but also describes signs ranging from mild transient diarrhea in young or
stressed animals to more severe disease with dehydration lethargy depression weight lossemaciation
inappetence and death (Blankenship-Paris et al 1993 Powers 2009 Sledge et al 2011 Hoefer et al
2012 Patterson et al 2014) Rectal prolapse has also been reported in ferrets with enteric coccidiosis
(Hillyer 1992 Hoefer et al 2012) In one study co-infection with coccidia and Lawsonia intracellularis
(Desulfovibrio sp) was diagnosed in 4 of 19 ferrets with proliferative bowel disease (Li et al 1996)
These ferrets presented with variable clinical signs including diarrhea lethargy anorexia weight loss
dehydration and emaciation
In the two reports of biliary coccidiosis clinical signs conformed to those expected with
hepatobiliary disease Williams et al (1996) described their case to have presented with emaciation poor
appetite abdominal distension and icterus Kaye et al (2015) described a one week history of lethargy
inappetence and icterus with serum biochemistry results consistent with cholestasis later clinical signs in
this case included melena anemia and cachexia
146 Gross necropsy and histologic findings
The pathology of enteric coccidiosis in domestic ferrets was described by Hoare (1927 1935b)
Two healthy domestic ferrets were experimentally inoculated one each with large numbers of mature
oocysts of either E furonis or E ictidea that were isolated during his initial work The inoculated ferrets
were killed humanely for histologic examination of intestinal sections at the time of first detection of fecal
oocyst shedding no clinical signs of coccidiosis were detected in these ferrets prior to death Infection
with E furonis resulted in invasion of the epithelium of the small intestine and rectum Within the small
intestine the parasites were concentrated in the tips of the villi but could be found to the level of the
22
opening of the crypts of Lieberkuumlhn In rectal sections life stages were limited to the epithelial ridges
between the openings of the glands of Lieberkuumlhn Organisms were located within the apical portion of
the epithelial cells and intensely infected regions exhibited multiple parasites within a single host cell
Both asexual and sexual life stages were present within the same sections Hoare (1927) described similar
histopathologic changes in naturally infected ferrets but the proportion of asexual versus sexual life
stages differed In natural infections sexual life stages were more numerous whereas in experimental
infections asexual life stages predominated these findings would be expected to correlate with the stage
of infection at which ferrets died or were humanely killed for tissue collection and would not be
reflective of differences between natural and experimental infection with this parasite Hoare also
described the morphology of the different endogenous stages including trophozoite (3-4 microm) merozoite
(stumpy sausage shaped L 3-4 microm W 2 microm) macrogamete (spherical 8 microm diameter with darkly
staining globular inclusions of reserve material) and microgamete (described as similar to those of other
Eimeria species) Two types of merogony are described from histologic sections the first with stumpy
merozoites as described above and the second with merozoites with elongated curved bodies and a
compact polar nucleus measuring 60 times 13 microm This second merogonic generation was observed almost
exclusively in the naturally infected ferrets and was associated with initiation of sexual differentiation and
reproduction
The pathology of experimental and non-experimental infection with E ictidea in domestic ferrets
was also described by Hoare (1927 1935b) Parasitic invasion of the epithelium was noted only in the
small intestine with patchy distribution of the parasite life stages throughout affected sections Within the
small intestinal villi the parasites were again concentrated in the tips of the villi with infected epithelial
cells never containing more than one parasite As each intracellular parasite grew it filled the entire host
cell displacing the nucleus to the base of the cell Predominantly sexual life stages were detected in tissue
sections with few asexual generations observed Interestingly the parasites were arranged into age
groups with forms of the same life stage grouped together within the affected epithelial sections this is in
23
contrast to E furonis where life stages of different maturities were found together in affected sections
Hoare described the morphology of the different endogenous stages of E ictidea including merozoites
(free within the lumen elongated vermiform with one pointed end and a nucleus located at the rounded
end 11 microm times 1 microm within the epithelium shortened and rounded 3-4 microm diameter) macrogametes
(elongated 20 times 7 microm occupying the entire host cell with darkly staining globular inclusions of reserve
material) and mature microgamonts (morphologically similar to those of other Eimeria species but larger
than those of E furonis) Of note a tissue reaction was observed specifically in association with more
developed life stages of E ictidea (eg mature meronts mature gamonts unsporulated oocysts) which
was not observed when cells contained earlier stages of development (eg trophozoites immature
gamonts) This tissue reaction was described by Hoare (1935a 1935b) as the development of an annular
constriction of the apical portion of the villus separating infected epithelial cells from unaffected cells
The constriction involved the epithelium but could also extend inwards into the core of the villus These
changes were associated with congestion of capillaries and extravasation of red blood cells within the
constricted segment and in some sections villar tip necrosis
In their case report of one domestic ferret Blankenship-Paris et al (1993) described the gross
pathologic lesions associated with intestinal coccidiosis in this case there was diffuse dilation and
reddening of the small intestine which was empty and the colon contained dark watery material
Histologic lesions were confined to the ileum and jejunum The jejunum exhibited thickening of the villi
with a crypt to villus ratio of 15 mild granulomatous inflammation in the lamina propria and large
numbers of coccidial meronts gamonts and oocysts within the enterocytes of the villar tips
The gross lesions described by Sledge et al (2011) from 20 domestic ferrets are as follows thin
body condition with moderate to marked dehydration perineal staining with diarrhea moderate dilation
of the small and large intestines and the presence of pasty tan to tarry black digesta within the distal small
intestine and colon Other findings in one to a small number of ferrets included enlarged pale tan livers
splenomegaly with dark red colouration and multiple superficial gastric or duodenal ulcers The
24
histologic lesions from 10 ferrets included moderate blunting and occasional fusion of jejunal and ileal
villi focal attenuation and erosion of the epithelium of the villar tips with exudation of fibrin neutrophils
and blood into the intestinal lumen in regions with severe erosion Intact epithelial cells at the villus tips
and rarely sloughed epithelial cells in the intestinal lumen contained numerous intracytoplasmic coccidia
representing a range of asexual and sexual life stages (meronts macrogamonts microgamonts and
oocysts) The subjacent lamina propria of the small intestine and of the large intestine exhibited moderate
lymphoplasmacytic infiltration with occasional neutrophils and congestion of blood vessels Marked
mucosal hemorrhage was identified in the most severely affected sections
Marked gross and histopathologic hepatobiliary lesions were described in a single ferret by
Williams et al (1996) On gross necropsy the liver was pale and enlarged with dilated firm bile ducts
and thickening of the gall bladder wall Similar gross necropsy findings were described by Kaye et al
(2015) marked dilation and mural thickening of the entire biliary tree (including gall bladder intrahepatic
and extrahepatic bile ducts) On histopathology Williams et al (1996) noted that the marked thickening
of the gallbladder wall was a result of cystic proliferation of mucosal glands which were separated by
tracts of fibrous connective tissue and marked granulomatous inflammation Liver sections exhibited
marked biliary hyperplasia marked periductular fibrosis and moderate periportal lymphoplasmacytic
cuffing There was multifocal papillary proliferation of bile duct epithelium and dilation of the bile ducts
and within the ductular lumens there were moderate numbers of lymphocytes and plasma cells small
numbers of degenerate neutrophils sloughed epithelial cells and debris All endogenous coccidial life
stages were present within the gall bladder and biliary epithelium with meronts visible in 20 of the
intact epithelial cells of the biliary tree and gallbladder and oocysts free within the lumen of the
intrahepatic bile ducts Similar lesions were present in the case described by Kaye et al (2015) and as
well as in juvenile and adult farmed mink (Mustela vison) with hepatobiliary coccidiosis (Davis Chow amp
Gorham 1953)
25
15 INTRODUCTION TO ENTERIC COCCIDIOSIS IN THE BLACK-FOOTED FERRET
151 Natural history and conservation of the black-footed ferret in North America
Black-footed ferrets are one of only three wild ferret species worldwide the other species are the
European polecat (Mustela putorius) and the Siberian polecat or steppe polecat (Mustela eversmanii)
They are the only native North American ferret species and the most endangered North American
carnivore They are nocturnal carnivores whose diet and lifestyle are highly dependent on local prairie
dog (Cynomys sp) populations Prairie dogs comprise almost exclusively the diet for the BFF who also
use the complex burrow systems made by prairie dogs to escape their predators and raise their young
(Santymire et al 2014 USFWS BFF Recovery Program 2017)
While formerly distributed throughout the North America prairie ecosystem BFF were
considered extinct by the late 1950s In 1964 a single population was discovered in Mellette County
South Dakota Progressive decline of this population in subsequent years resulted in the decision by
United States Fish and Wildlife Service (USFWS) to initiate a captive breeding program for the species
From 1971-1973 four females and five males were captured for this purpose Despite successful breeding
no kits survived and the last adult ferret in this captive colony died in 1979 at that time BFF were again
presumed extinct in the wild based on annual surveys of the initial capture site In 1981 a dead BFF was
discovered by a ranch dog outside of Meeteetse Wyoming allowing wildlife biologists to identify
another colony of BFF This colony flourished until 1985 when an outbreak of canine distemper in the
BFF population and an outbreak of sylvatic plague in the local prairie dog population resulted in sharp
population declines From 1985 through 1987 all 24 of the remaining BFF were trapped and brought into
captivity to re-initiate the captive breeding program Six ferrets in this initial group died of canine
distemper while in captivity and of the remaining 18 survivors 7 bred successfully to create the founding
population of the current captive breeding population Today this captive breeding population consists of
approximately 300 BFF distributed among multiple institutions (Santymire et al 2014)
26
Since 1986 this multi-institutional effort has been breeding BFF in captivity with reintroductions
back into the wild in 28 selected locations in Canada the USA and Mexico Currently six facilities
participate in the BFF Species Survival Plan (SSP) the Toronto Zoo USFWS National Black-footed
Ferret Conservation Center National Zoorsquos Smithsonian Conservation Biology Institute Louisville
Zoological Garden Cheyenne Mountain Zoo and the Phoenix Zoo (Black-footed Ferret Recovery
Implementation Team 2011) As of 2011 over 8000 BFF kits had been produced in captive breeding
facilities (Black-footed Ferret Recovery Implementation Team 2011)
Multiple infectious diseases pose a significant risk to the captive breeding and post-release
survival of BFF including canine distemper and sylvatic plague Coccidiosis is recognized as a cause of
significant juvenile morbidity and mortality in captive breeding programs and can result in significant
population losses (Bronson et al 2007 Santymire et al 2014 USFWS BFF Recovery Program 2017)
152 Coccidia identified from black-footed ferrets
Eimeria ictidea and Eimeria furonis have been identified in black-footed ferrets based on
morphologic criteria (Jolley et al 1994) Jolley et al examined fecal samples from six captive BFF during
a distemper outbreak as well as samples from wild BFF They described one medium-sized ovoid
tetrasporic dizoic oocyst with a double wall presence of a polar body and lacking both an oocyst residual
body and micropyle The oocysts measured 232 microm (range 182-274) by 155microm (range 130-162) with
a SI of 150 The sporocysts were elongate with the presence of both sporocyst residuum and a Stieda
body Sporozoites contained prominent refractile bodies at the posterior end and were aligned anterior to
posterior within sporocysts These oocysts were shed by all six captive ferrets On histopathology of
intestinal sections merogony and gametogony were observed within the villar epithelium throughout the
small intestine but were concentrated in the jejunum Two morphologically distinct meronts were
detected in these sections one at the villar tips which was larger and lacking in undifferentiated mass
and the other at the base of the villi or rarely in the intestinal crypts Gametogony was predominantly
27
observed at the villar tips and was noted throughout the small intestine These organisms were considered
consistent with Eimeria ictidea based on descriptions by Hoare (1927) from domestic ferrets
A second small spherical to subspherical tetrasporic dizoic oocyst was documented that had a
pink double wall a granular residual body and lacked both oocyst polar body and micropyle This
smaller oocyst measured 126plusmn12 microm (108-152) by 119plusmn09 microm (101-129) with a SI of 106 The
sporocysts were elongate with the presence of a Stieda body and sporozoites contained refractile bodies
Similar to the larger Eimeria species described above merogony and gametogony were observed within
the villar epithelium throughout the small intestine with endogenous developmental stages most
numerous in the jejunum The meronts were small with 16 or fewer merozoites Micro- and
macrogamonts were observed clustered within the apical third of the villar epithelium as were meronts
Jolley et al (1994) determined these small spherical oocysts to be consistent with Eimeria furonis as
described by Hoare (1927) from domestic ferrets
Jolley et al (1994) described a third type of coccidial oocyst occasionally detected in small
numbers within the BFF fecal samples the authors did not state whether this third type of oocyst was
recovered from wild or captive BFF The oocysts measured 370plusmn13 microm (350-386) by 223plusmn23 microm
(212-232) with a SI of 106 Attempts to sporulate collected oocysts were largely unsuccessful and
corresponding endogenous stages were not identified on histopathologic examination of necropsied
ferrets precluding further morphologic identification of the parasite It should be noted that coccidial
oocysts with similar measurements had not been detected in wild or captive prey species available for
ingestion by BFF (Jolley et al 1994)
Previous to this report coccidial oocysts had been isolated from the feces of BFF in two captive
populations (Carpenter amp Hillman 1979 Williams et al 1988) The abstract by Carpenter amp Hillman
(1979) did not describe the oocysts whereas Williams et al (1988) stated that two Eimeria species (one
with larger oocysts and one with smaller oocysts) were identified within the fecal samples but they were
28
not identified further Interestingly Williams et al reported both Eimeria species to be shed in the feces
of all ferrets concurrently affected by distemper and by approximately 30 of the clinically healthy
ferrets at the time of investigation
Non-enteric coccidia have been reported from captive BFF in one facility by two authors (Jolley
et al 1994 Williams et al 1988) Both reports presumably describing the same case(s) noted the
presence of endogenous coccidial life stages in histologic sections of respiratory tissue and merozoites of
an unidentified coccidium in impression smears of the urinary bladder from BFF diagnosed with canine
distemper Meronts were observed within the epithelium of the trachea a large bronchus and associated
bronchial glands Jolley et al (1994) described the lesions as occurring in the same ferret whereas in the
earlier report by Williams et al (1988) they are described as occurring in two different ferrets There have
been no subsequent published reports of systemic coccidiosis in black-footed ferrets and no cases have
been identified within the pathology database of the Toronto Zoo captive BFF population or by the
current SSP pathologist (Dr Michael M Garner personal communication)
There is a significant information gap regarding the pre-patent periods and pathogenicity of both
identified Eimeria species in BFF and studies to further characterize the eimeriid coccidia of the BFF are
lacking
153 Morbidity mortality and clinical signs associated with enteric coccidiosis in black-footed ferrets
The clinical signs of enteric coccidiosis in black-footed ferrets include mucoid to hemorrhagic
diarrhea abdominal discomfort lethargy appetite loss vomiting and dehydration In some cases sudden
death precedes the development of diarrhea Both adult and juvenile BFF are affected by the disease
which causes significant morbidity and mortality in captive populations (Bronson et al 2007) One
retrospective study of the captive BFF population at the Smithsonian National Zoological Park
determined that the most common cause of death in juvenile BFF (aged 30 days ndash 11 months) was
gastrointestinal pathology (524 of juvenile deaths) with 636 of these cases caused by enteric
29
coccidiosis (Bronson et al 2007) Despite the significance of this disease to the captive population its
effect on morbidity and mortality in wild BFF populations is unknown To the authorrsquos knowledge no
routine surveys of fecal parasites have been conducted on wild-born or captive released BFF during
yearly spotlighting events at ferret release sites However samples may be collected opportunistically if
fecal material is identified within the traps used to catch wild BFF during yearly surveys at release sites
Where fecal samples have been analyzed a 13 prevalence of coccidiosis has been identified in wild
born BFF (Dr Rachel Santymire personal communication) Fecal samples have been collected from BFF
at four release sites within the USA Wind Cave National Park (South Dakota) Badlands (South Dakota)
Conata Basin (South Dakota) and Aubrey Valley (Arizona) and positive samples were identified only at
the first site (Dr Rachel Santymire personal communication) Although radio-telemetry has been used at
some release sites to determine sources of mortality and factors involved in survival its use is not
widespread Furthermore the nocturnal and fossorial lifestyle of the BFF is a significant impediment to
the surveillance and monitoring of disease in this species
16 TREATMENT PREVENTION AND CONTROL OF INFECTION BY EIMERIA SPP
161 Current recommendations for treatment of eimeriid coccidia in carnivores
Described anticoccidial therapies for carnivores come from research in domestic cats and dogs
infected by Cystoisospora species these tissue coccidia (family Sarcocystidae) are only distantly related
to the Eimeria species infecting the BFF and other ferrets Current therapeutic recommendations by the
Companion Animal Parasite Council (CAPC 2013) for treatment of described Cystoisospora species
isolated from cats and dogs include the following amprolium (300-400 mg daily for 5 days in dogs 110-
200 mg daily for 7-12 days in dogs 60-100 mgkg daily for 7 days in cats) amproliumsulfadimethoxine
(150 mgkg amprolium and 25 mgkg sulfadimethoxine daily for 14 days in dogs) diclazuril (25 mgkg
for one dose in cats) furazolidone (8-20 mgkg 1-2 times daily for 5 days in dogs and cats) ponazuril (20
mgkg daily for 1-3 days in dogs and cats) quinacrine (10 mgkg daily for 5 days in cats)
30
sulfadimethoxine (50-60 mgkg daily for 5-20 days in dogs and cats) sulfadimethoxineormetoprim (55
mgkg sulfadimethoxine and 11 mgkg ormetoprim daily for 7-23 days in dogs) sulfaguanidine (150 or
200 mgkg daily for 6 days or 100-200 mgkg every 8 hours for 5 days in dogs and cats) toltrazuril (10-
30 mgkg daily for 1-3 days in dogs) trimethoprimsulfonamide (30-60 mgkg trimethoprim daily for 6
days if gt4kg 15-30 mgkg trimethoprim daily for 6 days if lt4kg) (CAPC 2013) Notably the use of all
drugs listed by the CAPC is considered off-label with the exception of sulfadimethoxine
162 Current recommendations for anticoccidial treatment and prophylaxis in domestic and black-
footed ferrets
1621 Domestic ferrets
Recommended daily oral treatment regimens for enteric coccidiosis in domestic ferrets include
amprolium (19 mgkg once daily 05 mgkg) decoquinate (05 mgkg) sulfadimethoxine (300 mgkg in
drinking water) or sulfadiazine-trimethoprim (30 mgkg once daily) all administered for a minimum of
two weeks (Bell 1994 Patterson amp Fox 2007 Patterson et al 2014) Both the aforementioned
coccidiostats amprolium and decoquinate are sold in large formats and are ideal for use in larger
operations such as breeding facilities research facilities or rescue centers Other anticoccidial therapies
used in domestic ferrets include toltrazuril (20 mgkg) and ponazuril (30-50 mgkg) once daily It should
be noted that all anticoccidial therapy used in domestic ferrets is considered off-label drug use
Multiple follow up fecal examinations should be performed after the treatment regimen is
complete and large groups may need to be treated multiple times Routine cage cleaning is also important
to decrease the environmental oocyst burden and prevent re-infection and in the case of coccidial
outbreaks ferrets should be transferred to clean cages multiple times during the course of anticoccidial
therapy Disinfectants such as bleach or quaternary ammonium compounds or dry heat should be used
for effective environmental decontamination (Patterson et al 2014)
31
1622 Species Survival Plan recommendations for black-footed ferrets
Treatment and prophylaxis of enteric coccidiosis with oral sulfadimethoxine was previously
recommended by the BFF Species Survival Plan (SSP) However due to a suspicion of decreasing
efficacy of treatment ponazuril has been recommended recently for treatment Due to the perceived
exquisite sensitivity of BFF to enteric coccidia the current SSP recommendation for treatment is oral
ponazuril at 30 mgkg once if ferrets are to be transported anesthetized stressed or are otherwise
suffering from another illness or injury (even in the absence of clinical signs or fecal shedding) The same
single oral dose of 30 mgkg is also recommended for kits at weaning (30-35 days of age) post weaning
(40-45 days of age) and prior to anesthesia for initial examination and vaccines (50-60 days of age)
Large crowded or otherwise stressed litters should be administered 30 mgkg orally once every 7-10 days
during the period of stress For treatment of coccidial diarrhea diagnosed by fecal examination 30 mgkg
orally once every 7 days for two doses or 50 mgkg orally once daily for 3 days in food (repeated in 7
days) is recommended In BFF with clinical signs of dehydration administration of subcutaneous or
intravenous fluid therapy has been performed Additional therapy with other antibiotics is sometimes
provided in cases with severe clinical signs or where secondary or primary bacterial enteritis is suspected
There is no pharmacokinetic or pharmacodynamic information available for the use of
anticoccidial drugs in BFF or other Mustelidae and thus it is unknown whether the current dose or
frequency of administration is truly appropriate for treatment of coccidiosis In 2 to 3-month-old piglets
administered a single dose of ponazuril orally at 5 mgkg peak serum concentration occurred at 42 hours
(36-48 hr) and elimination half-life was ~56 days (Zou et al 2014) In llamas administered ponazuril as
a single dose of 20 mgkg orally peak serum concentration occurred at 84 hours and elimination half-life
was ~56 days (Prado et al 2011) In domestic cows administered ponazuril as a single 5 mgkg dose
orally peak serum concentration occurred at 48 hours and elimination half-life was 58 hours (Dirikolu et
al 2009) The relevance of serum drug concentrations for treating an intestinal infection that lacks
extraintestinal life stages is likely minimal because the highest drug dose will reach the site of concern
(intestines) and systemic distribution is not required
32
Furthermore no safety or efficacy studies have been performed in any ferret species to validate
the current uses of either sulfadimethoxine or ponazuril for treatment nor have the current recommended
treatment lengths been validated However anecdotal information based on current usage would indicate
that they are safe at the current dosages and frequencies of administration as no adverse effects have been
reported A recent efficacy study in shelter dogs and cats showed that oral ponazuril (50 mgkg)
administered once daily for 3 days was effective for treatment of infection with Cystoisospora as
determined by a reduction in or cessation of fecal oocyst shedding at 4 and 8 days post treatment
Treatment efficacy in this study was inversely correlated to fecal oocyst counts at the initiation of
treatment (Litster et al 2014) Interestingly efficacy of this dose compared to the other two treatment
groups (single 50 mgkg or 20 mgkg oral dose) did not seem to differ but no statistical analysis was
performed Given the ubiquitous use of ponazuril in captive breeding facilities and concerns regarding
resistance of coccidia species to sulfadimethoxine therapy information on minimum effective doses and
dose regimes would be necessary to inform appropriate future SSP treatment and management plans and
to minimize development of drug resistance
17 VACCINES AGAINST COCCIDIA
171 Theory
The development of resistance of protozoal parasites to chemotherapeutic agents has resulted in a
shift towards the development of vaccines for the protection of domestic livestock Immunity to enteric
coccidiosis in avian and mammalian species involves both humoral and cell mediated responses Eimeria
spp infection in sheep rats poultry and other species generally results in a protective immune response
against subsequent re-infections (Catchpole et al 1993 Shi et al 2000) Interestingly this is not the case
for some host parasite interactions for example a recent report indicated that primary infection with E
ninakohlyakimovae in goat kids did not provide protective immunity against subsequent challenge with
the same parasite (Ruiz et al 2013)
33
Vaccines can be divided into four general categories live vaccines inactivatedkilled vaccines
subunit vaccines and recombinant vaccines Live vaccines are orally administered using small numbers of
infectious oocysts or oocysts from strains with low pathogenicity and result in patent but ideally sub-
clinical infections in the host that will elicit a protective immune response Such live vaccines can be
produced using attenuated forms of the pathogen of interest for example in chickens using ldquoprecociousrdquo
strains of Eimeria spp These precocious strains undergo a reduced number of merogonic replications
within the host cells and thus fewer oocysts are shed in the feces of vaccinated animals This reduction in
endogenous merogonic cycles reduces the amount of damage to the intestinal epithelium as well as
reducing the number of oocysts contaminating the environment
Another strategy has been to use live parasites with truncated life cycles An example of this is
the Toxoplasma gondii vaccine developed to prevent abortion in sheep This parasite was passaged
multiple times through a mouse host resulting in an inability to produce tissue cysts (Meeusen et al
2007) This is desirable as the cyst stage of this parasite normally inhibited by the immune system can be
reactivated during periods of stress or immunocompromise The potential drawbacks of live vaccines
include 1) the ability to produce and isolate adequate numbers of coccidial oocysts to meet vaccine
production requirements 2) the potential development of clinical disease in the host as a result of
inoculation 3) the need for all susceptible individuals to receive the vaccine simultaneously to prevent
fecal-oral inoculation of unvaccinated animals with high doses of the infective agent likely to be present
in a shared environment through fecal shedding
Inactivated vaccines are produced when the microbe of interest is killed via application of heat
radiation or chemical treatment prior to inoculation into the host species While safer because they cannot
induce disease in the inoculated patient inactivated vaccines stimulate a reduced immune response
compared with live vaccines and are consequently less effective Subunit vaccines contain single or
multiple antigens of importance in initiating the host immune response rather than the entire pathogen of
concern Subunit vaccines cannot induce disease in the immunized host but are more difficult to produce
34
because they require a detailed understanding of host immune response to infection Recombinant
vaccines involve the genetic modification of a vector (virus or bacteria) one capable of infecting the host
of interest to contain DNA of the pathogen of interest These vectors induce an immune response in the
vaccinated host but as with subunit vaccines cannot induce disease However recombinant vaccines are
again difficult to produce because they require an in depth understanding of the life cycle stages genes
and antigens targeted by the host immune response to infection There are currently no recombinant
vaccines marketed in Canada for use in veterinary medicine against protozoal disease
Creation of effective vaccines against protozoal parasites is complicated by parasite antigenic
diversity during the different life cycle stages and among protozoal species and strains of the same species
(Meeusen et al 2007) Although most parasites induce some level of immunity in their host species the
immunological response to different parasite life stages and species has been poorly characterized for
most coccidia Furthermore many parasites have developed mechanisms to evade host immune responses
or to continue survive and replicate in and transmission by previously infected hosts Our limited
understanding of the immune responses against coccidial antigens has restricted commercial vaccine
production to live or attenuated vaccines (Meeusen et al 2007)
A notable disadvantage of anticoccidial vaccines is that they need to be developed for each
coccidial species of interest because of the species-specific nature of the immune responses this is a
considerable limitation compared with anticoccidial drugs that can have a much wider spectrum of action
(Vermeulen 2005) While the requirement for mass production of vaccine is a limiting factor for vaccines
developed for the agricultural industry this drawback would be less important for production of a vaccine
to be used in an endangered species
172 Species successes in anticoccidial vaccination
The first successful immunization against coccidiosis was reported in 1918 in dogs (Hall amp
Wigdor) In this report a dog that had previously recovered from coccidial infection with Diplospora
35
bigemina was fed three increasing doses of live non-attenuated coccidial culture (at 14 32 and 48 days
post recovery from primary infection) which resulted in no development of clinical signs and no oocyst
shedding for 11mdash18 days after each challenge Subsequently immunization of dogs and cats against
coccidia with protection lasting up to seven months was reported by Andrews (1926) Immunization of
albino rats to eimeriid infection after administration of three or more sublethal doses of Eimeria
nieschulzi via gastric intubation was reported by Morehouse (1938) further experiments showed that
sporozoites did not enter the host intestinal epithelium in immunized rats given a challenge dose
(Morehouse 1938) Similar findings were reported in chickens immunized against Eimeria tenella that
had 50 fewer intra-epithelial sporozoites following challenge compared to naiumlve birds (Augustine and
Danforth 1986) Conversely chickens previously inoculated with Eimeria acervulina exhibited more
intracellular sporozoites after challenge than naiumlve birds but sporozoites were not observed to develop in
previously immunized birds (Augustine and Danforth 1986) These findings provide further evidence that
the immune response to Eimeria spp may differ among host species
Vaccination against Eimeria species has been most successful and is most widely used in the
poultry industry particularly in breeder and layer flocks Almost all vaccines marketed for poultry are
live vaccines (attenuated and non-attenuated) Vaccination against other apicomplexan parasites in
domestic mammals has also been achieved but has been generally less effective for disease prevention
and is less widely available Marketed killed and inactivated (attenuated) vaccines include those
containing killed tachyzoites of Neospora caninum for cattle (Neoguard Merck Animal Health) and
chemically inactivated merozoites of Sarcocystis neurona for horses (EPM Vaccine Fort Dodge ndash no
longer in production) A subunit vaccine for Babesia canis in dogs uses cultured antigen (Pirodog
Merial) Available live vaccines include a vaccine against Toxoplasma gondii in sheep (Ovilis Toxovax
Intervet) that uses an attenuated temperature sensitive strain (S48)
36
18 RESEARCH GOALS AND OBJECTIVES
181 Objectives
a) To determine and characterize (morphologically and molecularly) the enteric coccidial species
currently affecting the black-footed ferret population
b) To describe the natural history of enteric coccidiosis in captive black-footed ferrets including
pre-patent period shedding frequency and burdens and morbidity and mortality rates
c) To compare molecular morphologic and life history characteristics of enteric coccidial species
identified in domestic ferrets to those in black-footed ferrets
d) To validate domestic ferrets as an experimental model for intestinal coccidiosis in the black-
footed ferret
182 Hypotheses
a) Multiple Eimeria species will be isolated from the black-footed ferret population
b) The Eimeria species identified from black-footed ferrets will be the same as those previously
described in domestic ferrets
c) A single pathogenic Eimeria species will be implicated in recorded outbreaks of clinical
coccidiosis during the period of study
d) Domestic ferrets can act as an experimental model of intestinal coccidiosis for black-footed
ferrets
183 Applications
The goal of this project is to better characterize the enteric coccidia of the endangered black-
footed ferret in order to set the stage for improved disease prevention and treatment To the authorrsquos
knowledge this project is the first attempt to isolate and perform molecular characterization of the
coccidial species endemic in the black-footed ferret population This information will be used to compare
these species to known coccidia from domestic ferrets and other related mammals As experimental work
37
cannot be carried out on enteric coccidiosis in the BFF due to its endangered status if the domestic ferret
can be validated as an experimental model studies of the patterns of anticoccidial resistance and
development of immunity against Eimeria spp can be undertaken in vivo The ultimate goal would be the
development of an autogenous vaccine used to improve survival of ferret kits and reduce morbidity and
mortality associated with coccidiosis in BFF captive breeding programs Based on clinical experience
stressful life events such a breeding weaning and transfer between institutions appear to increase the risk
of coccidial outbreaks in adult BFF As such vaccination could assist in reducing disease outbreaks in
BFF associated with various management activities There is no data on the significance of coccidiosis in
wild populations and limited means of disease surveillance following release vaccination during captive-
rearing or pre-release conditioning of BFF would be an ideal method of reducing the potential effects of
this disease in released and free-living BFF Increasing the numbers of ferrets being released to the wild
and releasing ferrets immune to the subsequent threat of coccidiosis would support the goals of the
conservation initiative for the black-footed ferret
38
CHAPTER 2 MOLECULAR CHARACTERIZATION OF ENTERIC COCCIDIA FROM
DOMESTIC FERRETS (MUSTELA PUTORIUS FURO)
This chapter has been submitted for publication as
Adriana R Pastor Dale A Smith and John R Barta (2017) Molecular Characterization of Enteric
Coccidia from Domestic Ferrets (Mustela putorius furo) Vet Parasitol Regional Studies and Reports (In
review)
ABSTRACT
Combined morphometric and molecular characterization of coccidia that infect domestic ferrets
(Mustela putorius furo) was completed to improve the diagnostic specificity of enteric lsquococcidiosisrsquo in
this host Coccidia positive fecal samples (n=11) and formalin fixed paraffin embedded intestinal tissues
(n=3) from domestic ferrets were collected from diagnostic laboratories in Canada and Europe An
average of 35 and 13 domestic ferret fecal samples per year were coccidia-positive when tested by
Canadian and European diagnostic laboratories respectively during the period 2008-2015 Oocyst
morphometrics and sequence genotyping at two loci (nuclear 18S rDNA [nu 18S rDNA] and
mitochondrial cytochrome c oxidase subunit I [mt COI]) were conducted on all samples The first nu 18S
rDNA and mt COI sequences for Isospora (=Cystoisospora) laidlawi and the first mt COI sequence for
Eimeria furonis were generated during this study Phylogenetic analysis of the mitochondrial COI
sequences demonstrated that E furonis was most closely related to E cf ictidea isolated from a black-
footed ferret (Mustela nigripes) and that I (=C) laidlawi was closely related to C canis and C felis The
identifications provided by diagnostic laboratories of the specific parasite species present in a sample
showed poor agreement with their identifications based on genotyping obtained in this study Molecular
techniques appear to be essential for accurate determination of coccidial species responsible for individual
and group outbreaks of coccidiosis and for further understanding of eimeriid host-parasite relationships
Key words coccidia Cystoisospora laidlawi domestic ferret Eimeria furonis Eimeria ictidea Mustela
putorius furo
39
21 INTRODUCTION
Coccidia are host-specific parasites of the phylum Apicomplexa with greater than 2000 species
named to date (Duszynski et al 2000 Upton 2000) The eimeriorinid coccidia (suborder Eimeriorina)
include typical intestinal coccidia such as Eimeria Isospora and Cyclospora species belonging to the
family Eimeriidae as well as tissue (cyst-forming) coccidia such as Cystoisospora Besnoitia Toxoplasma
and Sarcocystis species that belong to the family Sarcocystidae (see Cox 1994)
Enteric coccidia affect both domestic ferrets (Mustela putorius furo) and their wild counterparts
In his initial descriptions of enteric coccidiosis in domestic ferrets Hoare (1927 1935b) did not observe
clinical signs of intestinal disease associated with infection More recently it has been recognized that
enteric coccidiosis can result in clinical signs ranging from mild transient diarrhea to more severe disease
with dehydration lethargy depression weight lossemaciation inappetence and death (Blankenship-Paris
et al 1993 Powers 2009 Sledge et al 2011 Hoefer et al 2012 Patterson et al 2014) Rectal prolapse
has also been reported in ferrets with enteric coccidiosis (Hillyer 1992 Hoefer et al 2012) Disease
appears to be most common in young or stressed animals In one study co-infection with coccidia and
Lawsonia intracellularis (Desulfovibrio sp) was diagnosed in 4 of 19 ferrets with proliferative bowel
disease (Li et al 1996) these ferrets presented with clinical signs including diarrhea lethargy anorexia
weight loss dehydration and emaciation Two cases of biliary coccidiosis have also been reported in
domestic ferrets infection was associated with biliary epithelial hyperplasia cholecystitis and
cholangiohepatitis (Williams et al 1996 Kaye et al 2015)
Three species of coccidia affecting domestic ferrets were originally described and named by
Hoare (1927) Eimeria ictidea Eimeria furonis and Isospora (=Cystoisospora) laidlawi The three
species were detected in feces from 50 domestic ferrets involved in an outbreak of canine distemper at a
research facility For each parasite the author described the morphology of sporulated oocysts isolated
from feces as well as sporulation time (exogenous life stages) All subsequent reports of morphologic
diagnoses of these coccidia have been based on Hoarersquos original descriptions The pre-patent period
40
(minimum duration of endogenous development) in inoculated naiumlve ferrets was described for E furonis
and E ictidea as 6 and 7 days respectively (Hoare 1935b) The pre-patent period for I (=C) laidlawi
was not determined because the number of oocysts available was insufficient for an experimental
infection trial
Hoare (1927) described the sporulated oocysts of E furonis as follows spherical double outer
wall with a thin colourless outer layer and a thick yellowish inner layer no micropyle or residual body
and measuring on average 128 times 120 microm (length [L] 112-144 width [W] 104-128 shape index [SI]
107) Sporocysts were spindle-shaped with one end constrictedblunted contained a residual body and
on average measured 8-88 times 4 microm Sporozoites were vermiform with one end narrower than the other
arranged head to tail and each had a central nucleus a clear vacuole was identified in some sporozoites at
their broad posterior end
The sporulated oocysts of E ictidea were described as follows oval or elliptical with a double
outer wall with a thin colourless outer layer and a thick yellowish inner layer no micropyle or residual
body and measuring on average 236 times 175 microm (L 184-272 W 128-208 SI 135) Sporocysts were
irregularly oval with one end broad and the other more constricted contained a residual body and
measured 115 times 65 microm on average Sporozoites were vermiform with one end narrower than the other
arranged head to tail in the sporocysts and had a central nucleus and a clear vacuole at their broad
posterior end
The sporulated oocysts of I (=C) laidlawi were ovoid with a double outer wall with a thin
colourless outer layer and a thick yellowish inner layer had no micropyle or residual body and measured
on average 34 times 29 microm (L 320-368 W 272-304) A SI of 117 can be calculated from the original
mean dimensions Two sporocysts were identified each containing four sporozoites and no Stieda body
sporocysts were elliptical contained a residual body and measured 208 times 144 microm on average
Sporozoites were sausage shaped with one end slightly pointed and had a central nucleus and a clear
41
vacuole identified at the pointed end Sporozoites were arranged with pointed ends all at the same pole of
the sporocyst
Oocysts identified as Cystoisospora ohioensis have been reported from fecal samples collected
from healthy domestic ferret kits in a large American ferret breeding operation that also housed juvenile
domestic dogs (Patterson and Fox 2007) The method of identification of this parasite was not described
A second similar institution reported the presence of Cystoisospora (=Isospora) species also thought to
be C ohioensis in routine fecal examination of their colony (Dr Bambi Jasmin personal
communication) Identification in this case was performed by the Animal Health Diagnostic Center at
Cornell University and was based on morphometrics using light microscopy The significance of these
findings is unknown but no clinical signs or histologic lesions were described in the ferrets shedding
these oocysts The definitive hosts for C ohioensis are canids including the domestic dog
More recently molecular techniques have been used for the more precise identification of
coccidia Nucleotide sequences like morphological features diverge over time under selective pressure
however recent evolutionary divergence among coccidia is more likely to be reflected in molecular as
compared to morphologic differences Thus nucleotide sequences that are more similar are inferred to be
more closely related and to have diverged more recently (Cox 1994)
Molecular characterization of ferret coccidia has only been performed for one species Eimeria
furonis Abe et al (2008) extracted DNA from oocysts from the feces of a single domestic ferret with
clinical signs resulting from coccidial enteritis Using primers initially developed for molecular
identification of Cyclospora species (see Matsubayashi et al 2005) small subunit ribosomal DNA (nu
18S rDNA) primers CYC1FE (5ʹ-TAC CCA ATG AAA ACA GTT T-3prime) and CYC4RB (5prime-CGT CTT
CAA ACC CCC TAC TG-3prime) were used to amplify a 347 base pair fragment of nu 18S rDNA The
amplicon was sequenced (GenBank AB329724) and compared with previously published partial nu 18S
rDNA sequences from 40 Eimeria two Isospora and four Cyclospora species The resulting phylogram
42
grouped E furonis with E alabamensis (cattle) and E meleagrimitis (turkey) In the same study the
microscopic morphology of the oocysts was used to identify this coccidial species as Eimeria furonis by
comparison with Hoarersquos (1927 1935b) published descriptions of Eimeria furonis and Eimeria ictidea
Sledge et al (2011) also used nu 18S rDNA to identify Eimeria furonis as the cause of three
distinct outbreaks of enteric disease in domestic ferrets Initial identification was performed using
morphometrics of sporulated oocysts collected from feces in one of the three outbreaks being
investigated Formalin fixed paraffin embedded intestinal segments from ferrets from each of the
outbreaks contained multiple coccidial life stages when examined by light microscopy PCR amplification
of a 247 base pair (bp) amplicon of the nu 18S rDNA was generated from DNA isolated from stored
formalin-fixed tissues for further genetic analysis Analysis and sequencing of amplicons from all three
groups showed 100 identity to sequences previously reported by Abe et al (2008) for the gene encoding
E furonis nu 18S
In 2015 Kaye et al identified coccidia within hepatobiliary lesions in a domestic ferret receiving
immunosuppressive therapy for red cell aplasia DNA was extracted from frozen liver and a fragment of
the nu 18S rDNA was amplified using the primers previously described by Sledge et al(2011) The
authors reported that the DNA sequence of the amplicon had 100 identity to the published nu 18S
rDNA sequence of E furonis and 95 identity to the nu 18S rDNA of E myoxi (rodent) E alabamensis
(cattle) and Isospora robini (avian) (Kaye et al 2015)
It is difficult to estimate the current prevalence of enteric coccidia within the North American
domestic ferret population and no studies have been conducted to do so Fecal samples submitted to
veterinary diagnostic laboratories from domestic ferrets in Canada are uncommon and samples positive
for coccidia appear infrequently (Dr Donald Martin personal communication) Conversely in Europe
the prevalence of coccidia within the domestic ferret population appears to be higher based on
submissions to a large veterinary diagnostic laboratory in Germany Data from Idexx Vet Med Lab in
Ludwigsburg Germany was compiled to review the prevalence of coccidia and Giardia within fecal
samples from domestic ferrets (Pantchev et al 2011) The authors reported that of 284 fecal samples
43
submitted from 2002-2004 18 (63) had detectable coccidial oocysts on fecal flotation Based on
morphologic characteristics oocysts were identified as E ictidea E furonis I laidlawi (herein referred
to as I (=C) laidlawi as noted above) and another unidentified Isospora species Comparative data from
the same laboratory from 2009-2010 included sample submissions from 253 ferrets 21 (83) of which
were positive for coccidial oocysts on fecal flotation Nine of the samples were identified as containing E
furonis three contained both E furonis and I (=C) laidlawi eight contained only I (=C) laidlawi and
one sample contained both E furonis and E ictidea identification in all cases was again based on oocyst
morphometrics No statistically significant difference in the occurrence of coccidial oocysts was detected
when data from the two periods were compared (Fisherrsquos exact test P=041) (Pantchev et al 2011)
The purpose of the present study was to perform a more detailed molecular characterization of the
coccidial species isolated from domestic ferrets to estimate prevalence of the different coccidial species
within the Canadian domestic ferret population and to associate morphologic and molecular
characteristics of a greater range of enteric coccidial species in order to improve diagnostic accuracy
22 MATERIALS amp METHODS
221 Fecal samples
Multiple diagnostic laboratories within Ontario Canada1 and a major European diagnostic
laboratory 2 were solicited for fecal samples from domestic ferrets shedding coccidial oocysts Fecal
samples were diagnosed positive for coccidia based on fecal flotation and light microscopic identification
of Eimeria or Cystoisospora species Eleven samples were collected during the study period (from 2014-
2017) and preserved in potassium dichromate (25 wv) eight from Europe and three from Canada
Centrifugal flotation with saturated salt solution (Ryley et al 1976) was used to isolate oocysts from fecal
samples for genomic DNA extraction Genomic DNA extraction and purification were performed using a
1 Animal Health Laboratory Guelph ON Antech Diagnostics Canada Ltd Mississauga ON IDEXX Canada
Markham ON 2 Vet Med Labor GmbH Division of IDEXX Laboratories Ludwigsburg Germany
44
QIAamp DNA Mini Kit (Qiagen Hilden Germany) according to manufacturerrsquos instructions After
addition of DNAzol to the samples (Qiagen Hilden Germany) samples were vortexed using 05 mm
glass beads (Biospec Products Inc Bartlesville OK USA) prior to extraction in order to fracture the
oocyst walls and release the sporocysts Concentrations of the resultant DNA were estimating using a
Nanodrop 2000 spectrophotometer (NanoDrop Products Wilmington DE USA) and stored at 4 degC for
immediate use or minus20 degC for later use
For each laboratory the number of domestic ferret fecal sample submissions numbers diagnosed
positive for coccidial oocysts and number of each coccidial species identified in positive samples were
tabulated for each of the years 2008-2015
222 Formalin fixed intestinal tissues
Major diagnostic pathology services across Canada3 were contacted to identify cases of enteric
coccidiosis identified on necropsy of domestic ferrets Cases were considered positive based on the
presence of asexual or sexual life stages of the parasites in intestinal sections The histologic sections on
each positive case were reviewed re-described and organisms measured (AP DAS) Gross necropsy
reports for all cases were also reviewed to identify any clinical correlates associated with enteric
coccidiosis DNA was extracted from ten 5-6 microm scrolls of formalin fixed paraffin embedded tissue
(FFPE) using the QIAamp DNA FFPE Tissue Kit (Qiagen) as per manufacturer instructions
223 Molecular characterization
Regions from the nu 18S rDNA and mitochondrial cytochrome c oxidase subunit I (mt COI)
DNA were amplified by polymerase chain reaction (PCR) from each sample using the primers listed in
Table 21 PCR amplification was performed for all samples in a volume of 25 microl containing ~100 ng of
3 Animal Health Centre Abbotsford BC Animal Health Laboratory Guelph ON Faculteacute de meacutedecine veacuteteacuterinaire
Universiteacute de Montreacuteal Saint-Hyacinthe QC Histovet Surgical Pathology Guelph ON IDEXX Canada
Markham ON Prairie Diagnostic Services Inc Saskatoon SK
45
genomic DNA 1times PCR buffer 15 mM MgCl2 02 mM deoxyribonucleotide triphosphates (dNTPs) 400
nM of each primer and 1 U of Invitrogen Platinum Taq DNA Polymerase (Thermo Fisher Scientific
Toronto ON Canada) Reactions were performed on a Bio-Rad T100 PCR thermal cycler (Bio-Rad
Laboratories Singapore) Samples were denatured and Taq polymerase activated at 95 degC for 3 min then
subjected to 35 cycles of 94 degC for 30s anneal at 50-62 degC (see Table 21 for specific anneal conditions
for the various primer pairs) for 30s and extension at 72 degC for 30-75s (see Table 21) followed by a
final extension at 72 degC for 7 min Suitable DNA (ie genomic DNA from an Eimeria or Sarcocystis sp)
was included in the PCR reactions to act as a positive control for the reaction chemistry All amplification
products were subjected to electrophoretic separation using 15 submarine agarose gel stained with
ethidium bromide and visualized on an ultraviolet transilluminator (Spectronics Corporation New York
NY USA) DNA band size was determined by comparison with a 1 kb DNA ladder (GeneRuler 1kb Plus
DNA ladder Thermo Fisher Scientific Waltham MA USA) Bands were excised with a new sterile
scalpel blade and PCR products were purified from the gel using a QIAquick Gel Extraction Kit (Qiagen)
PCR products were cycle sequenced using an ABI Prism 7000 Sequence Detection System (Applied
Biosystems Inc Foster City CA USA) by the Molecular Biology Unit of the Laboratory Services
Division University of Guelph (Guelph ON Canada) using the amplification primers to obtain
sequences in both directions The resulting chromatograms were aligned and analyzed with Geneious Ver
818 or later (Biomatters Limited Auckland New Zealand) and high quality consensus sequences
generated The resulting consensus sequences were searched from within Geneious against publically
available sequences on the BLAST server (blastncbinlmnihgovBlastcgi) using the blastn search
algorithm against the nrnt database (GenBank+EMBL+DDBJ+RefSeq ndash AA or DNA) Resultant new
nucleotide sequences were submitted to GenBank
46
224 Phylogenetic analysis
To determine the phylogenetic affinities of the newly obtained sequences with sequences from
related apicomplexan taxa representative nu 18S rDNA and mt COI sequences were downloaded from
GenBank with special reference to sequences from parasites that infect members of the order Carnivora
Nuclear 18S and mt COI sequences were aligned independently using MAFTT v7017 (Katoh et
al 2002) executed from within Geneious and then concatenated into a combined nu18S rDNA mt COI
dataset Multiple sequences from a single parasite were used to generate consensus sequences for each
locus as described by Ogedengbe et al (2017) Aligned sequences were trimmed to the length of the
largest newly generated nu 18S sequence Phylogenetic trees were generated using Bayesian Inference
(BI) using MrBayes Ver 326 (Huelsenbeck and Ronquist 2001) executed from within Geneious the
combined nu 18S and mt COI alignment was partitioned to permit locus-appropriate substitution models
to be applied to each partition For the nu 18S sequence partition the general time reversible (GTR)
substitution model (nst=6) with gamma rate variation (ie a GTR+G+I model) was applied For the mt
COI sequence partition the codon (M1) substitution model (using translation table 4 [ie lsquometmtrsquo]) was
used instead of the GTR with the remaining parameters remaining the same
The resulting tree was rooted using a pair of adeleid coccidia (Hepatozoon spp) as the taxonomic
outgroup All BI analyses were run for a chain length of 1000000 with tree sampling every 1000
following a burn-in of 100000 with default settings of 4 heated chains and heated chain temp of 02
47
23 RESULTS
231 Fresh fecal samples
From 2008-2015 inclusive the Canadian diagnostic parasitology laboratory4 received an average
of 1206 (range 81-160) domestic ferret fecal samples yearly the European parasitology laboratory5
received a yearly average of 230 samples (range 213-270) The number of fecal samples diagnosed as
positive for coccidial oocysts per year on fecal flotation during this time averaged 35 (range 0-8) and
130 (range 6-20) for the Canadian and European laboratories respectively The diagnosing laboratories
used oocyst morphometrics to identify the species of coccidia present Almost all coccidia-positive
submissions to the Canadian laboratory were identified as containing an I (=C) species based on light
microscopy Coccidia in only three samples from the Canadian laboratory were identified as E furonis
one in each of 2010 2012 and 2014 E ictidea was not identified in any samples submitted to the
Canadian laboratory Approximately equal numbers of coccidia-positive samples from the European
laboratory were identified as E furonis and I (=C) laidlawi each year Only two samples from the
European laboratory contained oocysts that were identified as Eimeria ictidea using morphometrics one
from each of 2011 and 2013 Laboratory submissions to both laboratories are summarized in Table 22
Twelve fecal samples preserved in potassium dichromate were received for analysis by the
authors Eleven samples had previously been identified as containing a single coccidial species five
containing E furonis two containing E ictidea and four containing I (=C) laidlawi A final sample had
been identified as containing a mix of E furonis and Cystoisospora canis Results of microscopic and
molecular characterization of these samples are summarized in Table 23
4 IDEXX Canada Markham ON
5 Vet Med Labor GmbH Division of IDEXX Laboratories Ludwigsburg Germany
48
232 Formalin fixed samples
Only three cases of coccidiosis were identified in domestic ferrets within the databases of the five
diagnostic laboratories that participated in the retrospective study Histologic sections of intestine were
received from these three cases which originated in Ontario6 and Quebec7 The Quebec sample (P2010-I)
was collected in 2010 and the Ontario samples (93-40404 and 17-008571) in 1993 and 2017 respectively
On gross necropsy the small intestinal contents of case P2010-I were described as pasty mucoid
yellow-brown feces with some blood For case 93-40404 the small intestines were described as empty
but melena was present within the terminal portion of the large intestine Scant intestinal contents and
dark brown fecal material in the colon were described in case 17-008571
In all cases endogenous developmental stages of coccidia were visible in histological sections
(Figure 21 is exemplary of the findings from one case) Hematoxylin and eosin stained sections from
P2010-I contained two affected regions of small intestine The intestinal mucosa of the first region
contained numerous asexual life stages and moderate numbers of sexual life stages as well as a small
number of oocysts free within the lumen The second section contained tissues that were poorly
preserved nonetheless 0-4 oocysts per 400times field were identifiable within the intestinal lumen Two
regions of affected small intestine were identified from 93-40404 after screening of all submitted sections
both contained low numbers of sexual and asexual endogenous stages Within one region there were small
numbers of meronts within the intestinal mucosa and lamina propria The second region had small
numbers of oocysts within cells of the epithelium and lamina propria as well as free within the intestinal
lumen In case 17-008571 multiple sections of jejunum contained numerous coccidian meronts gamonts
6 Animal Health Laboratory Guelph ON
7 Faculteacute de meacutedecine veacuteteacuterinaire Universiteacute de Montreacuteal Saint-Hyacinthe QC
49
and oocysts within intestinal villi within the ileum scattered epithelial cells also contained these various
life stages
Average length and width of oocysts were measured from slide sections for all cases For P2010-
I oocyst average length and width were determined from seven oocysts to be 94 microm (range 85-105) and
75 microm (range 69-84) respectively with a SI of 125 (range112-140) Average length and width of
oocysts measured from 93-40404 were determined from 5 oocysts to be 2814 microm (range 229-341) and
233 microm (range 180-308) respectively with a SI of 123 (range 110-146) For the third case 17-
008571 only 2 oocysts were identified and average length and width of oocysts measured 982 microm (range
973-992) and 845 microm (range 821-870) respectively with a SI of 116 (range 114-118)
233 Molecular characterization
DNA was successfully extracted from all twelve fecal samples and two cases with formalin fixed
tissue samples Attempts at amplification of DNA extracted from sample 93-40404 using the primer pairs
listed in Table 21 were unsuccessful Molecular identification results and GenBank accession numbers
for the remaining samples are summarized in Table 23 Both the nu 18S rDNA and mt COI sequences
from I (=C) laidlawi were unique when compared with available sequences from other Cystoisospora
species within the public databases However sequences from I (=C) laidlawi were most similar to
sequences from C canis and C felis and somewhat more divergent from sequences from members of the
C ohioensis species complex Two apparent genotypes of E furonis were identified based on nu 18S and
mt COI sequencing results Genotype 1 represented by EU sample 9014 had 100 identity to previously
published sequences of the nu 18S locus from two isolates from Japan (GenBank AB239130 and
AB329724) Genotype 2 represented by EU sample 907 and Canadian sample 17-008571 had 994
identity at the nu 18S locus (3 single nucleotide differences [SNDs] over 561 base pair region [bp]) to the
three sequences above belonging to E furonis genotype 1 Pairwise alignment of mt COI sequences from
both genotypes identified only 2 SNDs (996 pairwise identity over 513 bp region) Partial mt COI
50
sequences of E furonis from both genotypes were only distantly related (941 pairwise identity 30
SNDs over 513 bp and 905 pairwise identity 49 SNDs over 513 bp respectively) to publicly available
sequences from Eimeria ictidea from the black-footed ferret (Mustela nigripes) (GenBank KT203399)
and Eimeria mephitidis from the striped skunk (Mephitis mephitis) (GenBank KT203398) the only other
Eimeria species infecting members of the Carnivora for which sequence was available
234 Phylogenetic analysis
A phylogenetic reconstruction based on concatenated partial nu 18S rDNA and mt COI sequences
of E furonis I (=C) laidlawi and related coccidia is illustrated in Figure 22 The combined 18SCOI-
based tree demonstrates that the two Eimeria species from ferrets form a well-supported monophyletic
group that branches among a collection of other eimeriid coccidia that infect mammals The sarcocystid
parasite of the domestic ferret I (=C) laidlawi was found to group as the sister taxon to C canis that
together formed a monophyletic group with the closely related C felis all three of these closely related
Cystoisopora species possess comparatively large egg-shaped oocysts that are similar morphologically
24 DISCUSSION
The present work has generated the first nu 18S rDNA and mt COI sequences for Cystoisospora
laidlawi and the first mt COI sequence for Eimeria furonis both isolated from the domestic ferret In this
study histologic presence of organisms and microscopic identification of oocysts shed in feces have been
correlated with published and novel nu 18S and mt COI sequences
Eimeria ictidea was not identified in any Canadian sample and this coccidium was reported in
only 2 of 1840 fecal samples submitted from across the European Union (EU) to IDEXX Germany during
2008-2015 suggesting that E ictidea is not a frequent cause of enteric coccidiosis in domestic ferrets in
Canada or the EU
51
During the study period (2008-2015) almost twice as many domestic ferret fecal submissions
were made to the European as compared to the Canadian diagnostic laboratory however the prevalence
of coccidia-positive samples was similar The methodology used in this report cannot be used to
determine the actual prevalence of enteric coccidial infection (coccidiasis) or disease (coccidiosis) within
the domestic ferret population Fecal samples may be submitted to laboratories either as a result of
investigation into enteric disease or as part of a routine health examination Thus without historic
information accompanying each sample one can simply identify the proportion of positive samples and
compare the frequency of the finding of different coccidial species Prospective surveys of fecal samples
from healthy and sick domestic ferrets with greater sample size would be necessary to determine the true
prevalence of these parasites within the population and to infer their clinical significance
Comparatively few mitochondrial COI sequences have been generated for apicomplexan parasites
compared with other genetic loci the majority of published sequences obtained from Apicomplexa are
from nu 18S The disadvantage of using nu 18S rDNA sequences for parasite identification is that they
are poor at distinguishing among closely related eimeriid coccidia due to the highly conserved nature of
the nuclear ribosomal RNA locus In contrast mt COI sequences appear to be more useful for
distinguishing closely related coccidian species (Ogedengbe et al 2011) but are less useful than nu 18S
rDNA sequences for inferring more ancient relationships among more distantly related coccidia
Consequently the combined use of nu 18S rDNA and mt COI sequencing has been recommended for
improved species description and phylogenetic analysis (El-Sherry et al 2013) For these reasons both nu
18S and mt COI sequences were analysed in the present study
Despite adequate quantities of DNA extracted from the Ontario laboratory sample (93-40404)
successful amplification did not result with any primer pair (Table 21) Potential reasons for this include
degradation of formalin-fixed DNA into fragments too small for amplification with the desired primers
perhaps as a result of extended length of time in formalin prior to paraffin embedding or length of time
stored as FFPE tissue (23 years) or insufficient parasite DNA within the paraffin scrolls The primer pairs
52
used appear to be useful for most eimeriid coccidia (Ogedengbe 2015) and successfully amplified both
Eimeria species from DNA isolated from oocysts so it is unlikely that failure to amplify DNA from this
sample resulted from an inability of the primers used to recognize the parasite seen on section
Two genotypes of E furonis were identified in this study Genotype 1 was identified only from
samples originating from domestic ferrets in Europe but exhibited 100 identity based on nu 18S
sequencing with previously published sequences from both Japan and the USA Genotype 2 was
identified from samples originating from domestic ferrets in both Canada and Europe The small number
of single nucleotide differences between the two genotypes at two genetic loci in different genomes are
consistent with intraspecific variation (ie strain variation)
As might have been expected because of their morphological and host similarities nu 18S and mt
COI sequences of E furonis were determined to be most similar to an Eimeria species (E ictidea)
previously isolated from black-footed ferrets (Mustela nigripes) these eimeriid coccidia formed a
monophyletic group that was distinct from other eimeriid coccidia infecting mammals in the phylogenetic
analyses based on combined nu 18S rDNA and mt COI sequences Similarly the nu 18S rDNA and mt
COI sequences of I (=C) laidlawi are most similar to sequences from two other Cystoisospora species of
carnivores (C canis and C felis) that both have large egg-shaped oocysts comparable to those of I (=C)
laidlawi Both morphometrics and genotyping support the close relationships among these three
sarcocystid coccidia of carnivores These molecular data confirm that transfer of Isospora laidlawi to the
genus Cystoisospora by Barta et al (2005) is warranted
The previous light microscopic identifications of coccidial species in 3 of the 11 fecal samples
were not in agreement with the molecular findings These results were not surprising because light
microscopy has been shown to be an insensitive tool for distinguishing among apicomplexan parasites at
both the genus and species level Furthermore re-evaluation of these samples by the authors revealed that
many of the samples that were identified incorrectly based on morphometrics contained primarily
53
unsporulated oocysts making accurate identification based on microscopic appearance highly
challenging These findings further underscore the importance of molecular methods in accurate parasite
identification In the absence of molecular tools accurate measurement of oocyst size shape and
determination of SI can be useful for differentiating among species of Eimeria and Cystoisospora
however this can only be performed accurately on sporulated oocysts from feces Interestingly the size
and shape indices of oocysts of E furonis measured in histologic sections did not match those previously
described by Hoare (1927) for the same oocysts in feces despite molecular confirmation of identity
Thus measurements of oocysts in histologic sections are not recommended for use in coccidial
identification
Our observations highlight the utility of molecular methods for identifying enteric coccidia
infecting domestic ferrets and suggest that diagnoses based on morphological methods should perhaps be
limited to broad determinations of disease etiology (ie lsquococcidiosisrsquo or lsquococcidiasisrsquo) Using molecular
techniques we were able to differentiate morphologically similar coccidial species isolated from the feces
of domestic ferrets and specifically identify parasites seen in histological sections of ferret intestine
Molecular techniques thus appear to be essential for determining the coccidial species responsible for
individual and group outbreaks of coccidiosis and for further understanding of eimeriid host-parasite
relationships
ACKNOWLEDGEMENTS
Many thanks to Julia Whale and Alex Leveille for their assistance and encouragement during the course
of this project The authors would like to recognize the contributions of Dr Donald Martin (IDEXX
Canada) and Drs Nikola Pantchev and Majda Globokar (IDEXX Germany) for the contributions of data
and samples to this project The authors would also like to recognize the Laboratoire de Pathologie
(Service de diagnostic Faculteacute de meacutedecine veacuteteacuterinaire St Hyacinthe Quebec) and the Animal Health
Laboratory (Guelph Canada) for contributions of samples and data to this project Finally this project
was made possible through funding by the Toronto Zoo Residency Research Fund to DASAP and partial
54
funding from a Natural Sciences and Engineering Research Council of Canada (NSERC) Discovery Grant
(400566) to JRB
55
Table 21 Amplification primers for nuclear 18S rDNA and mitochondrial COI loci anneal temperatures (Ta) extension times and expected PCR
product sizes used in the identification of enteric coccidia from domestic ferrets (Mustela putorious furo)
Gene Target Primer Pairs Primer Sequence (5ʹ-3ʹ) Size (bp) Ta (degC) Anneal (sec) Reference
nu 18S rDNA CYC1FE TACCCAATGAAAACAGTTT 560 52 45 Matsubayashi et al (2005)
CYC4RB CGTCTTCAAACCCCCTACTG Matsubayashi et al (2005)
Cocci_18S_595F CCGCGGTAATTCCAGCTCCAAT 216 62 30 Present study
Cocci_18S_847R GCTGMAGTATTCAGGGCGACAA Present study
Lank_18S_224F TCATAGTAACCGAACGGATC 1080 54 60 Ogedengbe (2015)
Api_SSU_2733R CGGAATTAACCAGACAAATC Mathew et al (2000)
mt COI COI_10F GGWDSWGGWRYWGGWTGGAC 500 52 30 Ogedengbe et al (2011)
COI_500R CATRTGRTGDGCCCAWAC Ogedengbe et al (2011)
COI 272F CAATTCTAYGATGCCGCWTT 222 52 30 Present study
COI_500R CATRTGRTGDGCCCAWAC Ogedengbe et al (2011)
Sdae-COI_260F GATCTTTATGTTYTTRATGCC 890 50 75 Ogedengbe (2015)
Sdae-COI_1147R CATTACCCATAACYACACC Ogedengbe (2015)
56
Table 22 Summary of fecal samples from domestic ferrets (Mustela putorius furo) submitted to two diagnostic laboratories from 2008-2015
No fecal samples positive for coccidia
No samples submitted
(percentage positive)
No samples positive for
Cystoisospora sp
No samples positive for
Eimeria furonis
No samples positive for
Eimeria ictidea
Year Canada Europe Canada Europe Canada Europe Canada Europe
2008 3140 (21) 6214 (28) 3 2 0 4 0 0
2009 2160 (12) 14214 (65) 2 9 0 5 0 0
2010 8127 (63) 20213 (94) 7 10 1 10 0 0
2011 0114 (0) 17215 (79) 0 9 0 7 0 1
2012 3108 (28) 10231 (43) 2 4 1 6 0 0
2013 281 (25) 16270 (59) 2 13 0 2 0 1
2014 6127 (47) 12234 (51) 5 6 1 6 0 0
2015 4108 (37) 9249 (36) 4 3 0 6 0 0
Total 28 (29) 104 (56) 25 56 3 46 0 2
Average
year 35 130 31 70 04 58 00 03
Legend Numbers in brackets refer to the percent of the total number of fecal samples submitted
57
Table 23 Morphologic and molecular identification of coccidia from domestic ferrets (Mustela putorius furo)
Sample ID Source
External Lab Morphologic
Diagnosis Morphologic Diagnosis (ARP) Molecular Diagnosis
mt COI GenBank
Accession
nu 18S rDNA
GenBank Accession
93-40404 FFPE enteric coccidia Histologic sample - - -
P2010-I FFPE enteric coccidia Histologic sample E furonis Identical to MF774036 Identical to MF774678
17-008571 FFPE NP Histologic sample E furonis Same as MF774036 Same as MF774678
17-008571 feces NP E furonis E furonis MF774036 MF774678
907 feces E furonis E furonis E furonis MF774035 MF774679
938 feces I (=C) laidlawi no oocysts visualized I (=C) laidlawi MF774037 MF774677
952-A feces E ictidea Cystoisospora sp I (=C) laidlawi Identical to MF774037 Identical to MF774677
9958 feces E furonis no oocysts visualized E furonis Identical to MF774034 Identical to MF774680
9011 feces E furonis E furonis E furonis Identical to MF774035 Identical to MF774679
9014 feces E furonis E furonis E furonis MF774034 MF774680
9017 feces I (=C) laidlawi Cystoisospora sp I (=C) laidlawi Same as MF774037 Same as MF774677
9040 feces I (=C) laidlawi no oocysts visualized I (=C) laidlawi Same as MF774037 Same as MF774677
912-260 feces I (=C) laidlawi Cystoisospora sp I (=C) laidlawi Same as MF774037 Same as MF774677
CAN-2016-1 feces C canis + E furonis Cystoisospora sp I (=C) laidlawi MF774038 MF774676
Legend FFPE = formalin fixed paraffin embedded intestinal sections - = unsuccessful = morphologic diagnosis performed by JRB same as = 100 sequence
identity with listed GenBank entry over entire sequence length identical to = 100 sequence identity but shorter sequence than listed GenBank entry
58
Figure 21 Life stages of Eimeria furonis within the small intestinal epithelium of a domestic ferret
(Mustela putorius furo) Asexual life stages merozoites (black circle) Sexual life stages oocyst
(solid black arrow) macrogamonts (open arrows with labels) microgamont (dotted black arrow)
Hematoxylin and eosin staining scale bar = 25μm
25 microm
59
Figure 22 Phylogenetic relationships of coccidia (Eimeria ictidea Eimeria furonis and Isospora
(=Cystoisospora) laidlawi) from domestic (Mustela putorius furo) or black-footed (Mustela
nigripes) ferrets based on partial nuclear 18S rDNA and mitochondrial COI sequences of these
parasites and related apicomplexan parasites A summary of the sources of the molecular data for
the remaining taxa included in this phylogenetic analysis are found in Supplementary Table 1 of
Ogedengbe et al (2017) Bayesian support is indicated for each node horizontal distance is
proportional to hypothesized evolutionary change (scale indicates sequence divergence of 10)
60
CHAPTER 3 MORPHOLOGICAL AND MOLECULAR CHARACTERIZATION OF
ENTERIC COCCIDIA ISOLATED FROM BLACK-FOOTED FERRETS (MUSTELA
NIGRIPES)
ABSTRACT
Black-footed ferrets (BFF Mustela nigripes) are the only ferret species native to North America
and have been identified as endangered since 1967 Starting in 1986 a multi-institutional effort has been
breeding this species in captivity with successful reintroductions back into the wild Coccidiosis is
recognized as a cause of significant juvenile morbidity and mortality in captive breeding programs and
can result in significant population losses Little is known about the etiology of enteric coccidiosis in
BFF Coccidia positive fecal samples (n=12) and formalin fixed paraffin embedded intestinal tissues
(n=11) were obtained from BFF in the Toronto Zoo and Louisville Zoo Species Survival Plan (SSP)
populations Oocyst morphometrics and sequence genotyping at three loci (nuclear 18S rDNA
mitochondrial cytochrome c oxidase subunit I and mitochondrial cytochrome c oxidase subunit III) were
conducted Results suggest that the same Eimeria species E ictidea was the cause of enteric coccidiosis
in both SSP populations in both juvenile and adult age classes Wider research is indicated to determine
whether these findings are representative of the larger captive and wild BFF populations
31 INTRODUCTION
Black-footed ferrets (BFF) are one of only three wild ferret species worldwide the other two
being the European polecat (Mustela putorius) and the Siberian polecat or steppe polecat (Mustela
eversmanii) The BFF the only native North American ferret species was formerly distributed throughout
the North America prairie ecosystem but were considered extinct by the late 1950s In 1964 a single
population was discovered in Mellette County South Dakota Progressive decline of this population in
subsequent years resulted in the decision by United States Fish and Wildlife Service to initiate a captive
breeding program for the species From 1971-1973 four females and five males were captured for this
61
purpose Despite successful breeding no kits survived and the last adult ferret in this captive colony died
in 1979 BFF were again presumed extinct in the wild based on annual surveys of the initial capture site
In 1981 a dead BFF was discovered by a ranch dog outside of Meeteetse Wyoming allowing wildlife
biologists to identify another colony of BFF This colony flourished until 1985 when an outbreak of
canine distemper in this wild BFF population and an outbreak of sylvatic plague in the local prairie dog
population resulted in sharp population declines From 1985 through 1987 all 24 of the remaining BFF
were trapped and brought into captivity to re-initiate the captive breeding program Six ferrets in this
initial group died of canine distemper while in captivity and seven of the remaining eighteen survivors
are the founding population of the current captive breeding population Today this captive breeding
population consists of approximately 300 BFF distributed among multiple institutions (Santymire et al
2014)
Since 1986 a multi-institutional effort has been breeding BFF in captivity with reintroductions
back into the wild in selected locations in Canada the USA and Mexico Currently six facilities
participate in the BFF Species Survival Plan (SSP) the Toronto Zoo United States Fish and Wildlife
Services National Black-footed Ferret Conservation Center National Zoorsquos Smithsonian Conservation
Biology Institute Louisville Zoo Cheyenne Mountain Zoo and the Phoenix Zoo (Black-footed Ferret
Recovery Implementation Team 2011) In order to provide the best genetic matches BFF are transferred
among the six institutions for breeding Approximately 300-400 kits are produced annually between the
six SSP facilities with ~200 of these kits allocated for release to the wild yearly (Santymire et al 2014)
As of 2011 over 8000 BFF kits had been produced in captive breeding facilities (Black-footed Ferret
Recovery Implementation Team 2011)
Multiple infectious diseases pose a significant risk to the captive breeding and post-release
survival of BFF including canine distemper and sylvatic plague (Santymire et al 2014 USFWS BFF
Recovery Program 2017) Coccidiosis is recognized as a cause of significant juvenile morbidity and
mortality in captive breeding programs and can result in significant population losses (Bronson et al
62
2007 Santymire et al 2014 USFWS BFF Recovery Program 2017) While the effects of the disease on
the wild population are not clear a prevalence of approximately 13 has been reported based on fecal
samples collected from wild BFF born at release sites (Dr R Santymire personal commication)
Coccidia are eukaryotic host-specific parasites of the phylum Apicomplexa affecting numerous
hosts within a wide taxonomic range Two species of coccidia Eimeria ictidea Hoare 1927 and Eimeria
furonis Hoare 1927 have been identified in black-footed ferrets based on morphometrics (Jolley et al
1994) Jolley et al examined fecal samples from six captive BFF during a distemper outbreak as well as
samples from wild BFF They described one medium-sized ovoid eimeriid oocyst with a double wall
presence of a polar body and lacking both an oocyst residual body and micropyle Oocysts of this Eimeria
species (sp) measured 232 times 155 microm (range 182-274 times 130-162) with a shape index (SI) of 150 The
sporocysts were elongate with the presence of both sporocyst residuum and a Stieda body Sporozoites
contained prominent refractile bodies at the posterior end and were aligned anterior to posterior within
sporocysts These oocysts shed by all six captive ferrets were considered consistent with Eimeria ictidea
based on descriptions by Hoare (1927) On histopathologic examination of intestinal sections parasites
undergoing merogony and gamogony were observed within the villar epithelium throughout the small
intestine but were concentrated in the jejunum (Hoare 1935b) parasite life stages were not described
from other tissuesorgans
A second small spherical to subspherical eimeriid oocyst was also documented in the captive
ferrets by Jolley et al (1994) this second species had a pink double oocyst wall a granular residual body
and lacked both oocyst polar body and micropyle This smaller species measured 126 times 119 microm (range
108-152 times 101-129) with a SI of 106 The sporocysts were elongate and possessed a Stieda body and
sporozoites contained refractile bodies Similar to the larger Eimeria sp described above merogonic and
gamogonic stages were observed within the villar epithelium throughout the small intestine but were
most numerous in the jejunum Jolley et al (1994) concluded these small spherical oocysts were
consistent with E furonis described by Hoare (1927) from domestic ferrets
63
Jolley et al (1994) described a third type of coccidial oocyst occasionally detected in small
numbers within BFF fecal samples however the authors did not state whether this third oocyst
morphotype was recovered from wild or captive animals The oocysts measured 370 times 223 microm (range
350-386 times 212-232) with a SI of 106 Attempts to sporulate collected oocysts were unsuccessful and
corresponding endogenous stages were not identified on histopathologic examination of necropsied
ferrets precluding further morphologic identification of the parasite It should be noted that coccidial
oocysts with similar measurements had not been detected in wild or captive prey species available for
ingestion by BFF making it unlikely that this coccidial species would have been a pseudoparasite (Jolley
et al 1994)
Previous to this report by Jolley coccidial oocysts had been isolated from the feces of BFF in two
captive populations (Carpenter amp Hillman 1979 Williams et al 1988) The abstract by Carpenter amp
Hillman (1979) did not describe the oocysts whereas Williams et al (1988) stated that two Eimeria sp
(one with larger oocysts and one with smaller oocysts) were observed within the fecal samples but they
were not identified further Interestingly Williams et al reported both Eimeria sp to be shed in the feces
of all ferrets concurrently affected by distemper and by approximately 30 of the clinically healthy
ferrets at the time of investigation
Non-enteric coccidia have also been reported by two authors from captive BFF at one facility
(Jolley et al 1994 Williams et al 1988) Both reports which presumably described the same case(s)
noted the presence of endogenous coccidial life stages in histologic sections of respiratory tissue and
merozoites of an unidentified coccidium in impression smears of the urinary bladder from BFF diagnosed
with canine distemper Meronts were observed within the epithelium of the trachea a large bronchus and
associated bronchial glands In the later report Jolley et al (1994) described the lesions as occurring in the
same ferret whereas in the earlier report by Williams et al (1988) they are described as occurring in two
different ferrets Paraffin blocks containing formalin fixed tissues from these cases have since been
discarded precluding further attempts at parasite identification with molecular methods Subsequent to
64
these reports further cases of systemic coccidiosis in BFF have neither been published nor identified
within the pathology database of the Toronto Zoo captive BFF population nor by the current SSP
pathologist (Dr Michael M Garner personal communication)
Previous characterization of coccidia from black-footed ferrets has been based on host species
affected tissues in the host and morphometric characterization of life stages in histologic sections and
oocyst characteristics using light microscopy It is known that morphologically similar Eimeria species
are not necessarily conspecific and may vary in host specificity and pathogenicity Molecular
characterization is thus required to accurately identify coccidia to the species level No molecular
characterization of coccidian parasites from black-footed ferrets has been performed to date
There is a significant information gap regarding which parasite species are implicated in
morbidity and mortality events associated with enteric coccidiosis in BFF and whether different coccidia
are associated with this disease in adult versus juvenile age classes or in different SSP institutions
Studies to further characterize the eimeriid coccidia of the BFF are warranted to improve the management
of this disease in the captive population The objectives of this research were to morphologically and
molecularly characterize coccidia associated with enteric disease in BFF at the Toronto Zoo and in other
SSP facilities
32 MATERIALS AND METHODS
321 Fecal samples
Twelve fecal samples were collected during the study period (from 2014-2016) and preserved in
potassium dichromate (25 wv aqueous) seven from the Toronto Zoo and five from the Louisville Zoo
Centrifugal flotation with saturated salt solution (Ryley et al 1976) was used to isolate and concentrate
oocysts from fecal samples for light microscopic examination and genomic DNA extraction
One to two drops of the supernatant from the centrifugal flotation were placed directly on a slide
and beneath a coverslip The morphology and dimensions of sporulated oocysts were documented using a
65
Provis AX70 photomicroscope (Olympus Canada Richmond Hill ON Canada) fitted with a digital
imaging device (Infinity3-1C Lumenera Corporation Ottawa ON Canada) controlled using iSolution
Lite image analysis software (Hoskin Scientific Burlington ON Canada) operated at a total
magnification of 1000times Morphologic features noted for each oocyst included oocyst wall morphology
number of sporocysts presence or absence of a micropyle micropyle cap residual body and polar
granules For sporocysts size number of sporozoites per sporocyst and presence or absence of Stieda
body and sporocyst residuum were noted Alignment of sporozoites within the sporocyst and
presenceabsence of refractile bodies within sporozoites were also described The sporulated oocyst
length and width measurements (in microm) were then used to calculate the SI for each measured oocyst
Morphologic and morphometric features were compared to previously published values for E furonis and
E ictidea from domestic and black-footed ferrets
Genomic DNA extraction and purification were performed using a QIAamp DNA Mini Kit
(Qiagen Hilden Germany) according to manufacturerrsquos instructions as described in Chapter 2 (Materials
amp Methods)
322 Formalin fixed intestinal tissues
The pathology records of the Toronto Zoo were searched from 1993-2016 for cases of BFF
diagnosed with enteric coccidiosis on histopathology For each case slides of histologic sections from all
submitted intestinal segments were reviewed to confirm the presence of sexual andor asexual life stages
within the intestinal epithelium Scrolls (5-6 microm) were cut from the paraffin blocks containing affected
intestinal sections and DNA extracted from the formalin fixed paraffin-embedded tissue (FFPE) using
the QIAamp DNA FFPE Tissue Kit (Qiagen Toronto Ontario) as per manufacturerrsquos instructions
66
323 Molecular characterization
Molecular characterization of coccidial isolates was performed on oocysts purified from fresh
fecal samples (isolated as described above) that were collected from juvenile and adult ferrets from
2014-2016 and DNA extracted from FFPE samples of BFF intestine containing parasite life stages
Regions from the nuclear 18S (SSU) rDNA (nu 18S rDNA) mitochondrial cytochrome c oxidase
subunit I (mt COI) DNA and mitochondrial cytochrome c oxidase subunit III (mt COIII) DNA were
amplified by polymerase chain reaction (PCR) from each sample using the primers listed in Table 31 and
methodology described in the Materials amp Methods section of Chapter 2 Table 31 also contains the
specific anneal conditions used for the various primer pairs Genomic DNA from an Eimeria species of
poultry was included in the PCR reactions to act as a positive control for the reaction chemistry A
representative selection of the newly generated nucleotide sequences resulting from the above were
submitted to GenBank
DNA obtained from oocysts collected from fecal samples during the first year of the study (2014)
was used to generate a complete mitochondrial genome (see Chapter 6 for details) using primer pairs and
sequencing primers summarized in Table 31 All subsequent samples collected in 2015 and 2016 had
shorter mt COI and mt COIII sequences obtained to permit genotyping of all collected oocysts at these
two loci The location of each primer in the nu 18S mt COI and mt COIII genetic locus is illustrated in
Figure 31
33 RESULTS
From 2014-2016 coccidia-positive fecal samples were obtained from twelve BFF ferretsferret
groups from the Toronto Zoo and Louisville Zoo SSP populations (see Table 32) Nine samples were from
single housed adults between the ages of 1-5 years (63 MaleFemale) Two samples were from mixed
groups one pooled fecal sample from four adults (FERA-1 13 MF) and one fecal sample from a family
67
group consisting of a dam and five kits (23 MF) One fecal sample was collected from a juvenile male
ferret at the time of necropsy
Eleven BFF with enteric coccidiosis were identified in the Toronto Zoo necropsy reports from
1998-2016 and all were confirmed by histological re-evaluation (Table 32) Both juvenile (n=9 36 MF)
and adult ferrets (n=2 20 MF) were represented
331 Morphometric characterization
Twelve coccidia-positive fecal samples were identified from adult and juvenile BFF from 2014-
2016 by on site laboratories at either the Toronto Zoo or the Louisville Zoo Fecal flotation and light
microscopic re-examination of the samples identified coccidial oocysts in 10 of these 12 samples
Morphometric characterization was performed on six samples in which there was adequate
quantity and quality of sporulated oocysts for examination These included three samples from single-
housed adults one from a juvenile at the time of necropsy one of pooled feces from a group of adult
ferrets and one of pooled feces from a family group (dam and kits) Two of the three samples from
single-housed adults were from the same ferret on different dates in 2016 the dates of collection were
separated by a period in which shedding of oocysts was not identified on routine repeated fecal
examinations Oocysts were elliptical with a colourless double wall and contained four sporocysts each
with two sporozoites Sporocysts were ovoid and both Stieda body and residual body were present
Sporozoites exhibited an anterior to posterior alignment within the sporocysts and refractile bodies were
identified (Figure 32) Results for length width and shape index of sporulated oocysts including range
and average values are summarized in Table 33 and Figure 32 The average measurements based on the
results of all 148 oocysts measured were length 2398 microm (1859-3057) width 1855 microm (1373-2383)
and shape index 130 (101-160)
The same measurements were performed on 59 sporocysts from a single ferret (Noodle) and
results are as follows average length 1280 microm (898-1480) average width 738 microm (505-1028) and
average SI 176 (124-247) In one sample (Mohawk-2) sporozoites were visible free on the slide
68
Measurement of three of these provided an average length of 1068 microm (1044-1117) and an average
width of 341 microm (316-393)
332 Molecular characterization
Molecular characterization was successfully performed on oocysts from seven of 10 fecal
samples containing coccidial oocysts and FFPE tissue from nine of the 11 necropsy cases (see Table 32)
Attempts at amplification of DNA extracted from necropsy samples Z228-98 and Z137-14 using the
primer pairs listed in Table 31 were unsuccessful Similarly attempts at PCR and sequencing of DNA
extracted from fecal oocysts from two Toronto Zoo BFF Jenna and Ruckus were unsuccessful
Molecular identification results for the remaining samples are summarized in Table 32
Only one Eimeria species E ictidea was identified in all enteric coccidiosis cases diagnosed at
necropsy in both juvenile and adult BFF at the Toronto Zoo from 1998-2014 This same species was
identified in all Toronto and Louisville Zoo BFF fecal samples that were sequenced successfully (n=8)
with the exception of a single case from Louisville This Louisville ferret was identified as having a
rodent pseudoparasite (Eimeria species) in the submitted fecal sample the eimeriid pseudoparasite had
986 sequence identity at the mt COI locus to the murine coccidium Eimeria falciformis All sequences
generated for E ictidea exhibited 100 sequence identity at the mt COI and COIII loci
Novel nu 18S rDNA mt COI and mt COIII sequences were generated for E ictidea from both
geographic locations and deposited in GenBank (Accessions MF860826 MF860827 MF860823
MF860825 MF860822 MF860824) Sequences were compared to those previously published for related
eimeriid coccidia The nu 18S rDNA sequence from Eimeria ictidea isolated from the Toronto Zoo BFF
had 9736 identity (14 single nucleotide differences) to the previously published sequences from
isolates of E furonis from domestic ferrets (Mustela putorius furo) in Japan (GenBank AB239130 and
AB329724) and newly generated sequences from Canadian and European isolates (GenBank MF774678-
MF774680 see Chapter 2 and Figure 33) In contrast nu 18S rDNA sequence of E furonis from
domestic ferrets (see Chapter 2) showed 9953 to 100 identity (0 to 3 SND) to the Japanese
69
sequences Comparison of newly generated partial sequences of the mt COI region from E ictidea from
BFF to isolates of E furonis (GenBank MF774034-MF774036) from DF and E mephitidis (GenBank
KT203398) from the striped skunk (Mephitis mephitis) the only carnivore Eimeria sp for which a mt
COI sequence was previously available reveals only 9415 and 9084 sequence identity respectively
with these other Eimeria spp of carnivores (Figure 34)
34 DISCUSSION
This work presents the first nu 18S rDNA mt COI and mt COIII sequences (nu 18S rDNA -
MF860826 MF860827 mt COI - MF860823 MF860825 mt COIII - MF860822 MF860824) generated
from an intestinal eimeriid parasite of the BFF referred to here as E ictidea collected from multiple BFF
of different ages from two separate captive populations (Toronto Zoo Toronto Ontario Canada and
Louisville Zoo Louisville Kentucky USA)
The morphometric description of coccidial oocysts from BFF in this work are consistent with
previous descriptions of E ictidea from mustelids including BFF the Steppe polecat the European
polecat and domestic ferrets (Hoare 1927 Svanbaev 1956 Jolley et al 1994) Thus I propose the name
E ictidea for the enteric coccidium described from BFF reflecting the similarity in morphology host
species and location of infection in intestinal tissues yet recognizing the absence of species identification
by molecular techniques Molecular characterization of parasites that agree with the description of E
ictidea morphologically from various mustelid host species would allow not only for determination of
whether the parasites are conspecific but would also provide insight into the potential for cross-
transmission among related mustelid hosts
DNA extraction from FFPE samples allowed successful PCR and sequencing of small DNA
fragments (220 bp) in nine of the eleven cases in which the technique was attempted Age of the samples
did not appear to be the major factor associated with successful extraction of good quality DNA the two
samples for which it was unsuccessful were the most recent (2014) and oldest (1998) casesConsequently
it may be possible to use banked FFPE tissues from historic necropsy cases from other SSP institutions
70
and necropsies of wild-born or re-introduced ferrets to determine the identity of the coccidial species
underlying disease in these cases and to better characterize the disease in the greater BFF captive and wild
populations Williams et al (1988) were contacted regarding their historic FFPE samples but formalin
blocks were no longer available for these cases and thus comparisons could not be made Banked FFPE
samples were requested from other SSP institutions however the Convention on International Trade in
Endangered Species of Wild Fauna and Flora (CITES) restrictions on the international transport of DNA
from endangered species did not allow for sample acquisition during the period in which this research was
conducted
Evaluation of FFPE samples from the Toronto Zoo indicate that the same Eimeria species has
been implicated in deaths associated with enteric coccidiosis from 1999-2014 as well as episodes of
clinical disease in ferrets in the Toronto Zoo population from 2014-2016 Samples from coccidia-positive
BFF at the Louisville Zoo in 2016 also contained the same Eimeria species Finding the same parasite at
multiple SSP locations was expected because BFF are transferred among institutions on a yearly basis for
breeding and potential release Consequently these parasites have repeated opportunities to move
between institutions in infected hosts or on contaminated cage materials to become established at a new
location Furthermore the stress of transport and transfer to a new environment may precipitate shedding
of endemic coccidia and increase the risk of a coccidial outbreak this concern is reflected in the SSP
recommendations for prophylactic treatment of all BFF with anti-coccidial medication prior to shipment
(USFWS BFF Recovery Program 2017)
A single BFF from the Louisville SSP population not showing clinical signs consistent with
coccidiosis was identified as having a rodent Eimeria species in the submitted fecal sample
Morphometric characterization of oocysts in this sample was not performed due to the paucity of visible
oocysts however examination at 100times suggested that the oocysts in the sample were ovoid in shape and
of comparable size to oocysts identified in other BFF samples The finding of a rodent Eimeria in a BFF
fecal sample is not unexpected as whole rodents comprise a significant part of the captive BFF diet The
oocysts shed by the BFF were most likely acquired through ingestion of an infected prey item and thus
71
most likely represent pseudoparasitism Molecular characterization was however required to
differentiate this from a case of true enteric coccidiasis
Reports from the first captive BFF population derived from South Dakota indicate the presence
of an unidentified species of enteric coccidium (Carpenter and Hillman 1979) in this group before its
demise in 1979 No reports containing morphometric descriptions of the coccidia from this group were
found on literature review and all parasites of this group have been lost with their hosts All subsequent
reports on enteric coccidiosis in BFF are from ferrets derived from the second founder group from
Wyoming in the 1980s The frequent transfer of ferrets among SSP institutions within the captive
breeding program and to different release sites within North America would be expected to result in the
same Eimeria species being found in all populations The exception to this would be the potential for
cross-transfer of other eimeriid parasites to wild BFF from sympatric mustelid species such as the long-
tailed weasel (Mustela frenata) Jolley et al described two other species of enteric coccidia from this
second captive population in 1994 the first was similar to E furonis of domestic ferrets and the second a
large coccidian parasite of unknown genus Neither of these parasites was identified in the Toronto and
Louisville Zoo populations during the course of this study In order to determine whether these parasites
persist within the present-day BFF populations and their impact on this species more detailed
examination of coccidia-positive fecal samples from captive and wild BFF populations is recommended
Furthermore the molecular identification of enteric coccidia from historic and future necropsy samples of
wild and captive BFF could aid in determining the presence of and contribution to mortality events by
these additional coccidia species
ACKNOWLEDGEMENTS
The authors would like to recognize the Wildlife Health Centre staff at the Toronto Zoo for their
assistance with the collection of fecal samples from the BFF from 2014-2016 The authors would also like
to recognize the Louisville Zoo for their contribution of samples to this project Finally this project was
made possible through funding by the Toronto Zoo Residency Research Fund to DASAP and partial
72
funding from a Natural Sciences and Engineering Research Council of Canada (NSERC) Discovery
Grant (400566) to JRB
73
Table 31 Amplification primers used to sequence the nuclear 18S rDNA mitochondrial COI and COIII loci of Eimeria ictidea originating from
fecal and formalin-fixed paraffin embedded tissue samples from black-footed ferrets (Mustela nigripes) including anneal temeratures (Ta)
extension times and expected PCR product sizes
Figure 31 Nuclear and mitochondrial genetic loci targeted by primers listed in Table 31 and used to characterize Eimeria ictidea originating
from black-footed ferrets (Mustela nigripes)
Gene Target Primer Pairs Primer Sequence (5ʹ-3ʹ) Size (bp) Ta (degC) Anneal (sec) Reference
18SrDNA Sarco_18S_123F TATCAGCTTTCGACGGTAGTGTATT 1480 60 30 Ogedengbe et al (2016)
ERIB10_REV CTTCCGCAGGTTCACCTACGG
mt COI T_Eim_COI_272F CAATTCTAYGATGCCGCWTT 220 52 30 Chapter 2 (Table 21)
COX1-500R CATRTGRTGDGCCCAWAC Ogedengbe et al (2011)
COI-400F GGDTCAGGTRTTGGTTGGAC 800 52 60 El-Sherry et al (2013)
COI-1202R CAAKRAYHGCACCAAGAGATA El-Sherry et al (2013)
mt COIII WG-MT_4140F AGAAAACCTAAAATCATCATGT 1000 52 60 Ogedengbe et al (2015)
Eimeriid_CO3_799R AAGTGAGTTCGCATGTTTAC Ogedengbe et al (2015)
74
Figure 32 A+B) Features and cellular contents of Eimeria ictidea from a black-footed ferret
(Mustela nigripes) Legend Oocyst dotted thin white arrow = polar granule Sporocyst dotted thin
black arrow = Stieda body thick white arrow= sporozoite refractile body thick black arrow=
residuum scale bars as indicated C) Shape index length and width measurements of sporulated
oocysts of Eimeria ictidea from black-footed ferrets (Mustela nigripes) Legend times indicates the
mean Dotted oval indicates one standard deviation around the mean
10 microm 5 microm
75
Figure 33 Comparison of nuclear 18S rDNA sequence alignment of Eimeria ictidea from two black-footed ferrets (Mustela nigripes) to
newly generated (see Chapter 2) and published sequences of Eimeria furonis from domestic ferrets (Mustela putorius furo)
Figure 34 Comparison of mitochondrial cytochrome c oxidase subunit I sequence alignment of Eimeria ictidea from two black-footed
ferrets (Mustela nigripes) to sequences from other eimeriid parasites of carnivores
Identity
Eimeria mephitidis ndash KT2033981
Eimeria ictidea ndash MF860823 (Guanella ndash LZ)
Eimeria ictidea ndash MF860825 (Mystery ndash MTZ)
Eimeria furonis ndash MF774034 (Type 1)
Eimeria furonis ndash MF774035 (Type 2)
Identity
Eimeria ictidea ndash MF860827 (Guanella ndash LZ) Eimeria ictidea ndash MF860826 (Mystery ndash MTZ)
Eimeria furonis ndash MF774680 (Type 1) Eimeria furonis ndash AB3297241 (Type 1) Eimeria furonis ndash AB2391302 (Type 1)
Eimeria furonis ndash MF774678 (Type 2) Eimeria furonis ndash MF774679 (Type 2)
76
Table 32 Morphologic and molecular characterization of coccidia from fecal and FFPE necropsy samples from black-footed ferrets (Mustela
nigripes)
Sample ID Sample Source Year Age (years) Sex Source Microscopic Description^ Molecular Diagnosis
Z228-98 Toronto Zoo 1998 7 M FFPE rare asexual stages -
Z143-99 Toronto Zoo 1999 1 M FFPE sexual and asexual stages E ictidea
Z106-02 Toronto Zoo 2002 008 F FFPE rare asexual stages E ictidea
Z108-03 Toronto Zoo 2003 008 F FFPE sexual and asexual stages E ictidea
Z109-03 Toronto Zoo 2003 008 F FFPE sexual and asexual stages E ictidea
Z124-12 Toronto Zoo 2012 021 F FFPE sexual and asexual stages E ictidea
Z113-13 Toronto Zoo 2013 017 F FFPE sexual and asexual stages E ictidea
Z117-13 Toronto Zoo 2013 025 M FFPE sexual and asexual stages E ictidea
Z118-13 Toronto Zoo 2013 025 M FFPE sexual and asexual stages E ictidea
Z119-13 Toronto Zoo 2013 025 F FFPE sexual and asexual stages E ictidea
Z137-14 Toronto Zoo 2014 021 M FFPE
fresh feces
sexual and asexual stages
POS E cf ictidea
-
E ictidea
FERA_1 Toronto Zoo 2014 gt 1 MF fresh feces POS E cf ictidea E ictidea
Noodle Toronto Zoo 2015 1 M fresh feces POS E cf ictidea E ictidea
Ruckus Toronto Zoo 2016 3 F fresh feces POS -
Mystery Toronto Zoo 2016 1 M fresh feces POS E ictidea
Mohawk Toronto Zoo 2016 1 M fresh feces POS E cf ictidea E ictidea
Jenna Toronto Zoo 2016 1 F fresh feces POS -
Thrope Louisville Zoo 2016 3 M fresh feces NEG -
FloJean Louisville Zoo 2016 2 F fresh feces NEG -
Rigatoni Louisville Zoo 2016 5 M fresh feces POS rodent Eimeria
Guanella +kits Louisville Zoo 2016 2 015 F kits 2M3F fresh feces POS E cf ictidea E ictidea
Clive Louisville Zoo 2016 1 M fresh feces POS E ictidea
Legend FFPE= formalin fixed paraffin embedded Sex MF = samples from family groups containing both sexes - = PCR and sequencing unsuccessful ^ = life stages identified on
histologic section
= mitochondrial COI andor COIII sequencing results Presence (POS) or absence (NEG) of oocysts and morphological identification of oocysts based on measurements when
77
Table 33 Morphometric (length width shape index) characterization of Eimeria ictidea oocysts from fecal samples from black-footed ferrets
(Mustela nigripes)
Sample ID FERA - 1 ^ Z137-14 Noodle Mohawk -1 Mohawk - 2 Guanella Total
Number of oocysts 12 36 32 10 21 37 148
Length (microm) 2333 (2055-2583) 2456 (2111-2848) 2505 (2079-3008) 2779 (2590-3060) 2493 (2036-2822) 2139 (1859-2372) 2398 (1859-3057)
Width (microm) 1676 (1373-2180) 1835 (1643-2232) 1975 (1509-2360) 2253 (2092-2383) 1803 (1549-2017) 1751 (1610-1888) 1855 (1373-2383)
Shape index 135 (103-160) 134 (113-156) 127 (105-155) 124 (113-138) 139 (114-154) 122 (101-145) 130 (101-160)
Legend ^= mixed adult group = dam and kit group
78
CHAPTER 4 NATURAL HISTORY OF ENTERIC COCCIDIOSIS IN THE BLACK-
FOOTED FERRET (MUSTELA NIGRIPES)
ABSTRACT
Black-footed ferrets (BFF Mustela nigripes) the only native North American ferret species are
endangered throughout their former geographic range An intensive captive breeding program produces
animals to supplement re-established wild populations Coccidial enteritis is a major cause of morbidity in
young captive ferrets but the disease also affects adults Limited information is available on the
pathogenesis of intestinal coccidiosis in captive BFF and characterization of the natural history of the
disease for improved prevention and management is imperative The objectives of this research were to
determine morbidity and mortality rates in the Toronto Zoo captive BFF population as well as
characterizing the natural history of the disease in this species through evaluation of shedding patterns
body tissues affected pre-patent period and periods of enhanced host susceptibility to infection
Coccidia-associated mortality in BFF at the Toronto Zoo from 1997-2016 averaged 053 yearly in
adults (range 0-526) and 195 in juveniles (range 0-1667) Clinical signs and histologic lesions in
Toronto Zoo BFF were similar to those described in previous publications A seasonal influence on
oocyst shedding was identified in adult BFF and ferrets appeared to maintain persistent infection with E
ictidea shedding coccidia in multiple years A larger multi-institutional study is required to better
elucidate the natural history of enteric coccidiosis in this species
41 INTRODUCTION
Black-footed ferrets (BFF Mustela nigripes) are the only native North American ferret species
and are endangered throughout their former geographic range When the last remaining truly wild
population underwent serious decline as a result of disease the decision was made by the by United States
Fish and Wildlife Service to capture the remaining 24 animals and establish a captive breeding program
this occurred between 1985 and 1987 Only seven of the captured ferrets bred successfully and are the
79
founders of the current North American BFF population (USFWS BFF Recovery Program 2017) The
captive population which now numbers approximately 300 individuals is distributed among and managed
by six collaborating facilities these include the Toronto Zoo United States Fish and Wildlife Services
National Black-Footed Ferret Conservation Center National Zoorsquos Smithsonian Conservation Biology
Institute Louisville Zoo Cheyenne Mountain Zoo and the Phoenix Zoo (Black-footed Ferret Recovery
Implementation Team 2011 Santymire et al 2014) Since 1991 BFF have been released into
reintroduced into the wild at multiple sites within their former range and over 8000 BFF kits had been
produced in captive breeding facilities as of 2011(Black-footed Ferret Recovery Implementation Team
2011) Twenty-eight BFF reintroduction sites currently exist throughout North America however there
continues to be a need to support wild populations as only a four of the re-established groups are truly
self-sustaining
Enteric coccidiosis is recognized as a cause of significant morbidity and mortality in captive
breeding programs affecting both juvenile and adult animals (Bronson et al 2007 USFWS BFF
Recovery Program 2017) Two Eimeria species Eimeria ictidea and Eimeria furonis have been
identified from cases of entric coccidiosis in BFF (Jolley et al 1994) Jolley et al examined fecal samples
from both wild and captive BFF and provided detailed morphologic descriptions of the oocysts of both
Eimeria spp as well as descriptions of the intestinal pathology associated with infection Asexual and
sexual life stages of both of the aforementioned Eimeria spp were identified on histologic section within
the villar epithelium throughout the small intestine but were concentrated in the jejunum Intestinal
sections from BFF infected with E ictidea exhibited two morphologically distinct meronts one at the
villar tips which was larger and lacking in undifferentiated mass and the other at the base of the villi or
rarely in the intestinal crypts gamogony was predominantly observed at the villar tips and was noted
throughout the small intestine
80
Extraintestinal coccidia have also been reported from captive BFF at one facility (Jolley et al
1994 Williams et al 1988) The authors identified the presence of endogenous coccidial life stages in
histologic sections of respiratory tissue and in impression smears of the urinary bladder from BFF
diagnosed with canine distemper No subsequent reports of systemic coccidiosis in BFF have been
published or identified within the pathology database of the Toronto Zoo captive BFF population or by
the current SSP pathologist (Dr Michael M Garner personal communication)
Recent investigations into the etiologic agents of enteric coccidiosis in BFF at the Toronto Zoo
have identified a single Eimeria species associated with all cases of enteric coccidiosis and associated
mortality in juvenile and adult BFF from 1999-2016 Furthermore this pathogen was identified in fecal
samples based on morphologic and molecular characterization from adult and juvenile BFF in another
zoological collection (Louisville Zoo Kentucky USA) (see Chapter 3) This coccidium is
morphologically consistent with Hoarersquos original description of E ictidea (1927) and is referred to
henceforth as Eimeria ictidea
There is a significant information gap regarding the pathogenicity of E ictidea in BFF The
objectives of this research were to determine morbidity and mortality rates in the Toronto Zoo and
additional captive BFF SSP populations as well as characterizing the natural history of the disease in this
species through evaluation of shedding patterns body tissues affected pre-patent period and periods of
enhanced host susceptibility to infection
42 MATERIALS AND METHODS
421 Toronto Zoo BFF breeding program
At the Toronto Zoo black-footed ferret breeding program all adult ferrets are housed
individually with the exception of dams and kits After the birth of the kits dams are housed with their
offspring from whelp date until removal at approximately 4-6 months of age Routine monthly fecal
81
examinations (direct examination and flotation) are performed in house for all ferrets in the breeding
program based on SSP recommendations to evaluate for the presence of coccidia
422 Fecal oocyst evaluation
Family groups
From 2014-2016 daily fecal examination for coccidial oocysts was initiated for all group-housed
dams and kits In 2014 fecal samples were collected daily from all dams and kits from weaning (30 days
after whelping) to 72 days post whelping Based on 2014 data in 2015 this surveillance was extended
from weaning (35 days post whelping) to 135 days of age Furthermore fecal samples were collected
from the dam for an additional 14 days after removal of kits In 2016 no fecal samples were collected
from dam and kit groups at the Toronto Zoo but samples were submitted from one group of dam and kits
from another SSP population at the Louisville Zoo (Kentucky USA)
Adults
From 2015-2016 daily fecal samples were also collected from all adult ferrets identified as
shedding coccidial oocysts on their monthly routine fecal examination and from clinically ill BFF
Samples were collected for 10-14 days after initial positive sample identification In 2016 fecal samples
were also submitted from four coccidia-positive adult ferrets from the Louisville Zoo population samples
were collected for 7 days post initial identification of shedding
Individual fecal samples were analyzed via flotation using the McMaster method followed by
routine flotation in saturated salt solution (Dryden et al 2005) to determine the presence or absence of
oocysts and oocyst burden (oocysts per gram of feces OPG) Temporal trends in oocyst shedding were
monitored Coccidia-positive ferrets were evaluated visually on a daily basis for presence of clinical signs
consistent with infection Infected juvenile ferrets and adult ferrets were treated with oral ponazuril or
toltrazuril regardless of the presence of clinical signs as per the black-footed ferret SSP
recommendations Based on these recommendations ponazuril is typically administered orally at 30-50
82
mgkg once daily for 3-7 days until clinical signs have resolved or oocyst shedding has been significantly
reduced (USFWS BFF Recovery Program 2017)
423 Retrospective review of pathology records
The pathology records of the Toronto Zoo were searched from 1993-2016 for cases of BFF
diagnosed with enteric coccidiosis on histopathology For each case gross necropsy reports were
reviewed and slides of histologic sections from all submitted intestinal segments re-examined to confirm
the presence of sexual andor asexual life stages within the intestinal epithelium and describe the
histologic lesions associated with presence of the parasite life stages
424 Prospective modified necropsy protocol
During the study period 2014-2016 necropsy protocols for all BFF were modified to improve
detection of coccidial life stages and better to determine which portions of the intestinal tract were
affected The entire length of the intestine from duodenum to anus was measured and intestinal contents
were flushed with 12 mL of sterile saline into a sterile container Intestinal contents were preserved in
25 potassium dichromate solution (mixed 11 with intestinal contents vv) for molecular diagnostics
Paired 2-cm long intestinal samples were collected from all sections of small and large bowel duodenum
(1) jejunum (6) ileum (1) and colon (2) The eight small intestinal samples were collected at equal
distances from the pyloric sphincter to the beginning of the colon and the distance from the pylorus noted
for each Colon samples were taken at 25 and 75 of the length of the colon One sample from each
pair was preserved in Serra solution (100 ethanol (60 vv) 37 formaldehyde (30 vv) glacial
acetic acid (10 vv) and the second sample was frozen Representative tissues from all internal organs
as well as additional intestinal samples skin muscle and brain were also collected and preserved in 10
buffered formalin Histopathologic examination was performed on all tissues collected
83
425 Retrospective medical history review
Medical histories of all BFF held by the Toronto Zoo since the initiation of the SSP program were
reviewed for data on frequency of occurrence of shedding of coccidial oocysts in adults and juveniles as
well as any association of shedding with clinical signs and administration of anticoccidial treatment Data
was tabulated yearly for adult and juvenile ferrets to determine annual morbidity and mortality rates
associated with enteric coccidiosis Medical records and pathology reports were solicited from the other
SSP institutions to determine comparative morbidity and mortality rates associated with enteric
coccidiosis in BFF at other facilities Both morbidity and mortality rates were calculated as
incidenceattack rates with yearly adult population size or number of family groups (dam and kits) as the
denominator for morbidity rates and number of yearly deaths in each age class as the denominator for
mortality rates
43 RESULTS
431 Fecal oocyst evaluation and retrospective medical history review
Family groups
Fecal samples were collected from seven groups of dams and kits housed together at the Toronto
and the Louisville Zoos from 2014-2016 All data from first to last day of collection for all family groups
is listed in Appendix 1 selected pertinent data for each group is presented in Table 41 Five groups of
dams and kits were sampled in 2014 and one group in each of 2015 and 2016 Shedding occurred no
earlier than 55 days of kit-age in any of the groups and was identified from 55-81 days of age (Table 41
Table 42 Figure 41)
In 2014 fecal oocyst shedding was identified in three of the five surveyed groups In two of the
three groups (dams Poppy and Bumblefoot) changes to fecal colour and consistency were identified
concurrently with periods of oocyst shedding both groups shed higher numbers of oocysts than the other
dam and kit groups in 2014 and 2015 Both Poppy and Bumblefoot had had litters in the previous one and
84
two years prior to this study respectively based on medical record review these dams and their litters
were also diagnosed as shedding coccidial oocysts that were too numerous to count on direct exam and
fecal flotation Clinical signs in the previous years included dark tarry hemorrhagic or soft mucoid feces
and reduced appetite both groups received treatment with toltrazuril (Baycox Coccidiocide Solution
25 Bayer Inc Mississauga Canada) and trimethoprim sulfamethoxazole (Novo-Trimel Teva Canada
Ltd Scarborough Canada) (TMS) One of four kits from Poppyrsquos 2013 litter (Z113-13) died of enteric
coccidiosis three days after the group was diagnosed as shedding coccidial oocysts and the initiation of
treatment with TMS
In 2015 low grade fecal oocyst shedding (lt14 oocysts per gram of feces) without associated
clinical signs was noted in the Fiddlesticks group on three days during a seven day period from 63-69
days of kit-age and again for a single day at 128 days of kit-age The dam had been diagnosed and treated
for enteric coccidiosis in 2014 at which time she exhibited clinical signs of loose green feces to
hemorrhagic diarrhea lethargy and dehydration In 2016 she was diagnosed as shedding low numbers of
coccidia exhibited no clinical signs and did not receive treatment prior to resolution of shedding
In 2016 Guanella and kits shed oocysts over a nine day period and daily fecal oocyst shedding
ranged from 206 ndash 371714 OPG Combined treatment with ponazuril (first four days of shedding)
sulfadimethoxine injectable (first two days of shedding additional product information not available)
amoxicillin oral (first two days of shedding additional product information not available) penicillin
injectable (first two days of shedding additional product information not available) and subcutaneous
fluids (first two days of shedding additional product information not available) was administered to this
group Previous medical history was not available for this female for review
In 2014 and 2015 fecal oocyst shedding in all groups in the Toronto Zoo population started in the
three week period from the last week of July to mid-August In 2016 shedding was first identified in the
Louisville Zoo group in mid-July
85
Adults
Seven single-housed adult BFF (52 MF) were detected to have shed coccidia during the study
period (Table 43) Shedding periods lasted from 2-10 days and oocyst per gram counts ranged from 104
ndash 554274 (Table 44) Clinical signs were identified in four of the seven ferrets and consisted of
lossreduction of appetite (n=2) weight loss (n=1) lethargy (n=1) blood in feces (n=1) loose or runny
feces (n=3) soft mucoid feces (n=1) green colour of feces (n=2) Five of the seven adults received
treatment after detection of oocyst shedding two of which received treatment in the absence of clinical
signs Treatment consisted of oral toltrazuril in four cases toltrazuril in combination with trimethoprim
sulfamethoxazole in one case (Mohawk-A) and ponazuril and sulfadimethoxine (manufacturerrsquos
information not available) in one case (Clive) (Table 43)
Three of the adults in this study Mohawk Mystery and Jenna shed oocysts during multiple
different periods in 2016 Mohawk shed oocysts in May July and September of 2016 data from the first
two periods are reported in Tables 43 and 44 Mystery shed oocysts in June and July of 2016 Clinical
signs were observed only during the first shedding period and included poor appetite and hemorrhagic to
soft mucoid feces Jenna shed oocysts in July September and November of 2016 and again in February
and May of 2017 Although clinical signs were not detected in association with the initial period of
shedding in July 2016 (see Table 44) depressed mentation and hemorrhagic mucoid feces were identified
in the subsequent two shedding periods In both Mohawk and Jenna oocysts were not detected in feces on
multiple recheck and routine monthly fecal examination between shedding periods
Ruckus the fourth ferret shed low numbers of oocysts for two days in 2016 while housed alone
and had been reported to have shed oocysts during a 30 day period in 2014 while housed in a family
group with her kits Diarrhea to soft mucoid feces and loss of appetite were reported in 2014 Clinical
signs in 2016 consisted of small amounts of runny feces for a two day period two days after oocyst
shedding was no longer detected Treatment consisted of toltrazuril and TMS in 2014 and toltrazuril
again in 2016 Similarly Noodle shed low numbers of oocysts in July 2015 and had been identified as
shedding low numbers of oocysts during September of the previous year
86
The final ferret Rigatoni shed oocysts in feces in low numbers (54-732 OPG) over 5 days All
positive samples were pooled for molecular diagnostics and sequencing results showed 990 identity (7
single nucleotide differences SNDs) at the mt COI locus with a pair of rodent Eimeria species (ie
GenBank HM771682 JQ993704) (see Chapter 3) This ferret did not show clinical signs associated with
shedding and was not treated Previous medical history was not available for Clive the single adult ferret
from the Louisville Zoo
Based on sample collection dates and medical record review for the adult BFF for 2014-2016
shedding occurred during spring summer and fall with three ferrets shedding in May (Jenna Ruckus and
Mohawk) one ferret shedding in June (Mystery) three ferrets shedding in July (Mohawk Jenna Noodle)
one ferret shedding in August (Clive) two ferrets shedding in September (Noodle Jenna) and one ferret
shedding in November (Jenna)
432 Pathology
Eleven BFF with enteric coccidiosis were identified in the Toronto Zoo necropsy reports from
1993-2016 (Table 45) Cases were identified from 1998-2014 inclusive and all were confirmed by
histological re-evaluation (Chapter 3 Table 32) Both juvenile (n=9 36 MF) and adult ferrets (n=2 20
MF) were represented
Gross Pathology
Gross necropsy findings were similar across the 11 cases and included mucoid to fluid luminal
contents (n=7 636) beige to white pasty coating of the mucosal surface of the small intestine (n=6
545) and colon (n=4 364) gaseous dilation of intestinal segments (n=3 272) segmental enteritis
and hemorrhage (n=1 91) In one case Z228-98 no gross lesions were identified within the intestines
Impression smears of luminal contents or scrapings of intestinal mucosa were performed in four cases and
coccidia were identified in all four
Histopathology
87
Both sexual and asexual life stages were identified within the small intestinal segments in all
cases except Z228-98 and Z106-02 in which only rare asexual parasite life stages were identified (Table
45 Figure 42) Other histologic lesions seen in intestinal segments containing coccidia included
lymphoplasmacytic inflammation of the lamina propria (n=4) neutrophilic infiltration of the lamina
propria (n=2) villar necrosis (n=2) villar atrophy or blunting (n=3) and thrombi within the villar tips
(n=1)
Additional necropsy diagnoses included cholangiolar hyperplasia multiple hepatobiliary cysts
with suppuration renal adenocarcinoma apocrine gland adenocarcinoma (Z228-98) concurrent
clostridial enteritis (Z143-99) presumptive Salmonella sp septicemia (Z106-02) interstitial pneumonitis
(Z108-03) myocardial mineralization interstitial pneumonia and nephritis periportal hepatitis and
bacteremia (Z124-12) and suppurative esophagitis (Z137-14)
From 2014-2016 three black-footed ferrets were necropsied using the detailed protocol described
above Only one of the three cases Z137-14 was diagnosed with enteric coccidiosis based on
histopathology Two duodenal five jejunal two ileal and two colonic sections were collected at measured
lengths from the pylorus Parasite life stages were identified within the mucosal epithelium of all
intestinal segments extending from the distal duodenum (10-12 cm aboral to pylorus) through to the distal
colon (157-159 cm aboral from pylorus) The distal duodenal section contained asexual life stages only
with a single focus of epithelial cells containing meronts Sexual life stages (microgamonts
macrogamonts unsporulated oocysts) were identified within villar epithelial cells in all remaining
sections of the small intestine with numerous oocysts in the bowel lumen Mild lymphoplasmacytic
inflammation of the lamina propria was associated with the jejunal and ileal lesions and blunting of the
villi was identified within one jejunal segment The colonic sections contained small to moderate numbers
of sexual life stages identified within both superficial and deep crypt epithelium with occasional life
stages identified near the germinal cells Large numbers of oocysts and bacteria were identified within the
88
lumen of these colonic sections and both sections contained abscesses within the crypts The proximal
duodenum (0-2 cm) was the only section of the intestines not containing parasitic life stages
433 Morbidity and mortality
Annual morbidity rates for enteric coccidiosis at the Cheyenne Mountain Zoo and mortality rates
from enteric coccidiosis in Toronto Zoo BFF are summarized in Tables 46 and 47 During 2003-2016
yearly incidence of coccidiosis in adult BFF at the Cheyenne Mountain Zoo averaged 69 (range 0-
421) For family groups consisting of juvenile ferrets housed with their dams yearly incidence of
enteric coccidiosis averaged 115 (range 0-600)
From 1997-2016 coccidia-associated mortality in adult BFF at the Toronto Zoo averaged 053
yearly (range 0-526) with an average total mortality rate of 141 per year (range 0-526) For
juvenile ferrets (under 1 year of age) during the same period coccidia-associated mortality accounted for
an average of 133 of deaths yearly (range 0-100) with an overall average mortality rate of 170 per
year (range 0-3404) from all causes
Multiple additional SSP institutions provided partial medical and pathology data sets for use in
this study which were not sufficiently detailed to permit computation of morbidity and mortality rates for
those populations
44 DISCUSSION
The work described here supports previous clinical findings regarding the impact through both
morbidity and mortality associated with enteric coccidiosis in BFF No previous studies have been
undertaken to determine morbidity and mortality rates associated with enteric coccidiosis across BFF SSP
institutions
89
In a retrospective mortality study of captive BFF from 1984-2004 at the Smithsonianrsquos National
Zoological Park Bronson et al (2007) reported that gastrointestinal disease was the most common cause
of death in juvenile BFF (524) with 333 of juvenile mortality cases in the study caused by enteric
coccidiosis While the data is not directly comparable the findings reported here also reflect enteric
coccidiosis as a common cause of death in juveniles with increased mortality associated with the disease
compared with adult counterparts All Toronto Zoo mortalities in both juvenile and adult age classes for
which necropsy tissues were available have been attributed to infection with a single coccidia species
Eimeria ictidea (see Chapter 3)
Multiple SSP institutions provided partial medical and pathology data sets for use in this study
which were not sufficiently detailed to permit computation of morbidity and mortality rates for those
populations In future it would be useful to determine whether morbidity and mortality rates associated
with enteric coccidiosis vary among SSP institutions as this may allow for improved identification of
host parasite and environmental factors that increase risk
Clinical signs reported here are consistent with those described from both BFF and domestic
ferrets with enteric coccidiosis (Sledge et al 2011 Santymire et al 2014 USFWS BFF Recovery
Program 2017) Changes to fecal colour and consistency were the most common abnormalities identified
at the time of first detection of oocyst shedding While clinical signs in the cases described here do not
always correlate directly with the quantity of oocysts shed individuals shedding higher number of oocysts
showed clinical signs more frequently than those shedding lower numbers
In this study oocyst shedding from single-housed adult BFF ranged from 104 ndash 554274 oocysts
per gram Daily fecal samples produced from individual adult ferrets range in size from approximately 1-
16 grams In light of these findings during peak shedding from ~5times105 to 9times106 oocysts can be shed into
the environment in one day providing a massive infective dose The large numbers of oocysts shed
90
combined with confinement in a small enclosure space and hardiness of Eimeria oocysts in the
environment would be expected to markedly increase risk of infection in captive BFF
Oocyst shedding from family groups ranged from 0 ndash 371714 OPG The wide variability seen in
OPG counts between days as seen in Figure 41 and Table 41 may be accounted for by the staggered
initiation and resolution of shedding by different ferrets It is most likely that the source of infection in
these family groups is shedding by the dams some of which were identified to shed in multiple years
although environmental contamination cannot be excluded
Shedding in the adult ferrets was clustered during particular time periods specifically May July
and September If shedding is associated with stress and immunosuppression activities such as breeding
whelping electro-ejaculation of male ferrets and shipmenttransfer could act as stressors Whelping
which occurs primarily in May and June could also act as a stressor to other ferrets in the facility either
through social cues or as there would be associated changes in husbandry protocols The September
cluster could be associated with the transfer of ferrets between institutions kits are pulled during this time
and adult ferrets are moved among institutions thus changing the population dynamics of each SSP site A
large cluster of shedding was recorded in July With the exception of weaning of kits no other major
stressors are expected to occur during this time thus the increase in shedding by adults in this group
cannot be easily explained Interestingly shedding was not identified in ferrets from March through April
which is the typical breeding season and when ferrets are introduced for breeding a presumably stressful
time Shedding was not noted from December through April which could reflect reduced environmental
burdens due to low humidity levels as would be expected in a Nordic climate during the winter (which
would kill oocysts and thus block transmission) or may be consistent with reduced stress during this
period The results from single-housed adults are in contrast to the dam and juvenile ferret groups in
which oocyst shedding appeared to be correlated to a period of 55-81 days of kit age These results are
consistent with reports from other facilities of increased incidence of shedding by kits after 70 days of age
(USFWS BFF Recovery Program 2017)
91
Retrospective and prospective review of histologic sections of intestines from affected BFF at the
Toronto Zoo showed the presence parasitic life stages in epithelial cells of both the small and large
intestines Neither Hoare (1935b) nor Jolley et al (1994) mentioned the presence of parasites in the large
intestine of experimentally infected domestic ferrets or naturally infected BFF respectively In the study
described here asexual and sexual life stages were identified within the epithelial cells of the small
intestinal villi from base to tip and were most numerous in jejunum This matches the description by
Jolley et al (1994) however Hoare (1935a b) found E ictidea to be present primarily in the villar tips
Jolley et al (1994) also described two morphologically distinct meronts of E ictidea within the small
intestinal sections one at the villar tip that was larger and lacking in undifferentiated mass and the other
at the base of the villi or in the intestinal crypts these findings were not echoed in this study as
merogonic stages were identified throughout the intestinal epithelium from villus to the base of the crypt
and no visual differences between meronts in any location were identified Hoare (1935a b) also
described a resulting annular constriction of the villus separating the affected and non-affected segments
this constriction was neither seen in the cases described here nor mentioned by Jolley et al (1994)
Whether these differences result from E ictidea from BFF and E ictidea from domestic ferrets being
different parasites or from differences in tissue tropism of a single parasite in two different hosts cannot
be ascertained from the available information
Histologic lesions such as necrosis hemorrhage villar atrophy and inflammation associated with
the presence of parasitic life stages were rare These changes are normally elicited by the host immune
system (inflammation) and the parasite (cellular rupture to release life stages resulting in hemorrhage and
necrosis) in response to infection In light of the fact that acute death occurred in a number of these ferrets
(Z113-13 Z117-13 Z118-13 Z119-13 Z137-14) in the absence of secondary disease processes and
with the intestinal epithelium intact but containing myriad parasitic life stages an alternative mechanism
for mortality associated with the infection must be proposed It is possible that these parasites elaborate
exotoxins during their life cycle and when at high density result in sudden death of the host with minimal
92
tissue changes The presence of parasitic life stages occupying the majority of both small and large
intestinal epithelial cells could also potentially impair fluid and protein movement in and out of the
mucosa however clinical signs associated with malabsorptive diarrhea were not identified in any of these
cases The presence of bacteria within the blood or other organ tissues was not identified in any cases and
consequently sepsis is unlikely to be the cause of death
Black-footed ferrets appear to maintain persistent infection with E ictidea Adult BFF in the
Toronto Zoo population shed coccidia in multiple years and in two adult ferrets multiple times in the
same year While the coccidia seen in all cases were not confirmed as E ictidea using molecular
techniques morphologic similarities and a lack of additional Eimeria spp identified on molecular work
undertaken suggest that only one species of parasite is and has been present in the collection Two dams
that had been identified as infected based on routine fecal screening in previous years presumably acted as
the source of infection to their litters of kits in multiple years While continued environmental
contamination cannot be ruled out these findings imply a failure of the immune response of the BFF to
clear infection with E ictidea or even to protect against sufficient replication of organisms to result in
clinical disease
Based on clinical experience and review of the literature BFF appear to be much more sensitive
to infection with E ictidea compared with their domestic counterparts In domestic ferrets subclinical
shedding of oocysts appears to be the most common with rare reports in the literature of overt disease
and that only in juveniles (Blankenship-Paris et al 1993 Abe et al 2008) However a single report exists
of three separate clinical outbreaks of Eimeria furonis infection in domestic ferrets under intensive
management with increased morbidity and mortality affecting all ages classes (Sledge et al 2011) The
role of genetics in the apparent increased susceptibility of BFF to enteric coccidiosis is unknown but the
current captive BFF population is derived from seven founders and inbreeding depression or familial
genetic susceptibility may play a role in their increased susceptibility to disease caused by E ictidea
93
Black-footed ferrets diagnosed with enteric coccidiosis during the course of the study were
treated with either ponazuril or toltrazuril sulfonamide drugs or often a combination of the two groups
of therapeutic agents Toltrazuril and ponazuril are triazine coccidiocides with proven efficacy against
both asexual and sexual life stages of mammalian and avian Eimeria spp (Mehlhorn and Aspock 2008)
The sulfonamides are antimicrobial drugs that exhibit coccidiostatic or coccidiocidal effects depending on
dose they act by blocking folate synthesis and have effects on first and second generation meronts
(asexual life stages) as well as potentially acting on sexual life stages (Mehlhorn and Aspock 2008)
Based on the limited data available from this study and the fact that treatment was initiated in almost all
adult BFF and family groups at the time of oocyst detection regardless of the presence of clinical disease
the effects of treatment on duration of clinical signs cannot be effectively evaluated It appears
subjectively that adult ferrets treated with toltrazuril and in one case a combination of toltrazuril and
TMS showed reduction in oocyst shedding after 3-5 days of oral anti-coccidial therapy (see Table 43)
The effects of treatment with either sulfonamides or triazines would be expected to reduce oocyst
shedding consequently the duration and amount of oocyst shedding reported in this study may not
accurately characterize the natural course of disease
Perceived resistance to sulfa drugs has been reported from multiple SSP facilities In light of this
and their potential negative effects on ferret reproduction (eg prevention of embryo implantation in the
uterus and impairment of sperm development) sulfonamides are no longer recommended by the SSP for
treatment of coccidia in this species (USFWS BFF Recovery Program 2017) The frequent and
widespread use of triazines in the management of enteric coccidiosis in BFF presents a risk for
development of resistance to this drug class in the future Neither pharmacokinetic (PK) nor
pharmacodynamic (PD) studies have been published to validate the dose and frequency of dosage in
either class of drugs in BFF and consequently it is unclear whether this perceived failure of some ferrets
to respond to treatment is based on true resistance versus inappropriate dosing The only work evaluating
ponazuril in BFF evaluated serum levels of ponazuril after a single oral dose of 50 mgkg and reported
94
therapeutic levels for 10 days after administration (USFWS BFF Recovery Program 2017) No
information was provided on number or age of ferrets that participated in the study or on how the
determination of what were therapeutic levels was made Furthermore as life cycles of the coccidia
affecting BFF are limited to the gastrointestinal tract and do not exhibit tissue stages the validity and
usefulness of assessing blood levels of ponazuril in determining appropriate dosage and dose schedules is
questionable Further work to determine the PK and PD of triazines in ferrets is warranted to provide safe
and efficacious treatment and to reduce the risk of development of resistance Furthermore the creation
and validation of a model for enteric coccidiosis in a related species would allow for in vivo studies of
drug resistance
95
Figure 41 Oocyst per gram counts and shedding period of Eimeria ictidea from black-footed ferret
(Mustela nigripes) family groups from 2014-2016
0
50000
100000
150000
200000
250000
300000
350000
400000
29
33
37
41
45
49
53
57
61
65
69
73
77
81
85
89
93
97
10
1
10
5
10
9
11
3
11
7
12
1
12
5
12
9
13
3
13
7
14
1
14
5
14
9
OP
G
Age of Kits
Poppy Bumblefoot Calico Aubrey Ruckus Fiddlesticks Guanella
96
Figure 42 Small intestinal epithelium of a black-footed ferret (Mustela nigripes) containing sexual
life stages of Eimeria ictidea Legend Solid black arrow = oocyst Hatched arrow = macrogamont
Outlined arrow = microgamont Hematoxylin and eosin staining scale bar = 25 microm
25 microm
97
Table 41 Shedding of oocysts of Eimeria ictidea in black-footed ferret (Mustela nigripes) dam and kit
family groups from 2014-2016
Collection Year 2014 2014 2014 2014 2014 2015 2016
Age of kits (days) Poppy Bumblefoot Calico Aubrey Ruckus Fiddlesticks Guanella^
29 - - - 0 - - -
30 - 0 - 0 - - -
- - - - -
34 0 0 - 0 0 - -
35 0 0 - 0 0 0 -
- -
46 0 0 - 0 0 0 -
47 0 0 - 0 0 0 -
48 0 lt14 - 0 0 0 -
- -
53 0 0 - 0 0 0 +
54 0 0 - 0 0 0 +
55 0 0 - 0 0 0 3717146
56 0 0 - 0 - 0 1084436
57 0 0 - 0 - 0 64133
58 0 324 - 0 0 0 20654
59 0 0 - 0 0 0 0
60 0 446688 - 0 0 0 0
61 0 934828 - 0 0 0 41111
62 0 530469 - 0 0 0
63 0 1617131 - 0 0 lt14
64 0 8396357 - 114 0 0
65 0 234876 - 5368 0 lt14
66 0 374625 - 9455 0 0
67 lt14 2311575 - 39579 0 -
68 0 603563 - 125051 0 0
69 - 4103036 - 27067 0 lt14
70 2470327 7759974 - 0 0 0
71 1073085 - 1784 0 0
72 3430966 - 0 0 0
73 4459536 - 7326 0 0
74 4842212 - 2 0 0
75 3627307 - 0 0 0
76 5801885 - 0 0 0
77 5994 0 0 0 0
78 589835 0 0 0
79 24815 0 0 0
80 144016 0 0 0
81 97862 0 0 0
82 0 0 0
127 0 0
128 0 2843
136 0 0
150 0
Numbers of Oocysts Shed (oocysts per gram of feces)
Dam Identity
Legend lt 14 = oocyst positive samples with less than 14 oocyst per gram of feces - = no sample recorded for this date underline = last
sampling date + = coccidia present but OPG count not performed = Toronto Zoo ferret ^ = Louisville Zoo ferret thick outer border =
days treatment was received = range of sequential dates between previous and subsequent number during which OPG counts were
performed and samples contained 0 oocysts
98
Table 42 Summary of epidemiologic data for family groups of black-footed ferrets (Mustela nigripes)
shedding oocysts of Eimeria ictidea
Dam Identity
Poppy
2014
Bumblefoot
2014
Aubrey
2014
Fiddlesticks
2015
Guanella
^
2016
Number of kits 7 5 3 4 5
Kit age (days) at time of
shedding 70-81 58-70 64-73 63-69 54-61
Shedding period (days) 15 13 11 7 9
OPG min 0 0 0 0 206
OPG max 48442 83963 2707 lt14 371714
Clinical signs YES YES NO NO NO
Treated - YES YES NO YES
Legend = Toronto Zoo ferret ^ = Louisville Zoo ferret OPG = oocysts per gram of feces
- = missing data
Table 43 Shedding of oocysts of Eimeria ictidea in single-housed adult black-footed ferrets (Mustela nigripes) 2015-2016
Numbers of Oocysts Shed (oocysts per gram of feces)
Collection
Year 2015 2016 2016 2016 2016 2016 2016
Ferret Identity Noodle Ruckus Mohawk-A Mohawk-B Mystery Jenna Clive^
Age (years) 1 3 1 1 1 1 1
1084058 0 + + + + +
+ 0 42307650 6286676 + 183150 554274
857808 16650 12805238 7777929 + 215710 377920
1604894 16650 309690 139860 + 0 25808
377042 0 599400 119880 + - 37294
554445 0 34688 385579 117920 0 5363
26640 0 16650 0 0 0 7500
0 10406 0 0 0 1090
0 20813 0 0 0
- 0 1761 0
0 0 0 0
- 0
0
0
Legend lt 1 = oocyst positive samples with less than 1 oocyst per gram of feces underline = last sampling date + = coccidia present but OPG
count not performed = Toronto Zoo ferret ^ = Louisville Zoo ferret thick outer border = days treatment was received
100
Table 44 Summary of epidemiologic data for single housed adult black-footed ferrets (Mustela nigripes) shedding oocysts of Eimeria ictidea
Noodle Ruckus Mohawk-A Mohawk-B Mystery Jenna Clive^
Sex M F M M M F M
Age (years) 1 3 1 1 1 1 1
Shedding period (days) 7 2 9 6 10 4 8
OPG min 266 166 104 1199 1761 1831 1090
OPG max 10840 166 423076 77779 - 2157 554274
Clinical signs YES YES NO NO YES NO YES
Treated NO YES YES YES YES YES YES
Legend = Toronto Zoo ferret ^ = Louisville Zoo ferret M = male F= female OPG = oocysts per gram of feces - = missing data
NOTE Mohawk-A and Mohawk-B refer to two separate episodes of oocyst shedding by the same ferret
101
Table 45 Histologic findings from necropsies of black-footed ferrets (Mustela nigripes) with enteric coccidiosis
Number of Sections Affected
Ferret
ID Year
Age
(years) Sex Coccidia in Intestinal Sections Small Intestinea Large Intestinea
Z228-98 1998 7 M rare asexual stages S - 01 A - 11 S - 01 A - 01
Z143-99 1999 1 M sexual and asexual stages S - 24 A - 24 S - 03 A - 03
Z106-02 2002 008 F rare sexual stages S - 14 A - 04 S - 01 A - 01
Z108-03 2003 008 F sexual and asexual stages S - 12 A - 12 S - 01 A - 01
Z109-03 2003 008 F sexual and asexual stages S - 23 A - 23 S - 11 A - 11
Z124-12 2012 021 F sexual and asexual stages S - 24 A - 04 none
Z113-13 2013 017 F sexual and asexual stages S - 77 A - 07 none
Z117-13 2013 025 M sexual and asexual stages S - 56 A - 56 S - 11 A - 01
Z118-13 2013 025 M sexual and asexual stages S - 56 A - 46 S - 11 A - 01
Z119-13 2013 025 F sexual and asexual stages S - 45 A - 35 none
Z137-14 2014 021 M sexual and asexual stages S - 1011 A - 911 S - 22 A - 02
Legend a= xn where x is number of sections containing sexual or asexual lifestages n is the number of sections examined S = sexual life
stages A= asexual life stages
102
Table 46 Yearly incidence of coccidial infection in black-footed ferrets (Mustela nigripes) at the
Cheyenne Mountain Zoo
Cheyenne Mountain Zoo
Year Adult Family
2003 116 (625) -
2004 819 (4211) -
2005 021 (000) 14 (2500)
2006 021 (000) 07 (000)
2007 023 (000) 08 (000)
2008 224 (833) 14 (2500)
2009 025 (000) 06 (000)
2010 326 (1154) 07 (000)
2011 125 (400) 09 (000)
2012 125 (400) 08 (000)
2013 028 (000) 05 (000)
2014 430 (1333) 09 (000)
2015 035 (000) 35 (6000)
2016 - 27 (2857)
Mean annual () 689 1155
Legend - = missing data xn= where x is the number of ferrets shedding coccidial oocysts and n is the
total number of adult ferrets or family groups in a given year () = incidence expressed as a percentage
103
Table 47 Yearly mortality rate and incidence of mortality associated with coccidial infection in
black-footed ferrets (Mustela nigripes) at the Toronto Zoo
Total Mortality
Coccidia Other Causes
Year Kit Adult Kit Adult
1997 015 (000) 023 (000) 315 (2000) 423 (1739) 1998 038 (000) 119 (526) 838 (2105) 919 (4734) 1999 047 (000) 119 (526) 1647 (3404) 119 (526) 2000 034 (000) 015 (000) 434 (1176) 315 (2000) 2001 032 (000) 016 (000) 532 (1563) 116 (625) 2002 150 (200) 020 (000) 450 (800) 220 (1000) 2003 227 (741) 018 (000) 327 (1111) 118 (555) 2004 020 (000) 016 (000) 620 (3000) 216 (1250) 2005 016 (000) 015 (000) 416 (2500) 215 (1333) 2006 030 (000) 016 (000) 230 (667) 016 (000) 2007 019 (000) 015 (000) 419 (2105) 215 (1333) 2008 034 (000) 016 (000) 1134 (3235) 316 (1875) 2009 017 (000) 016 (000) 017 (000) 116 (625) 2010 017 (000) 016 (000) 317 (1765) 316 (1875) 2011 011 (000) 016 (000) 111 (909) 216 (1250) 2012 111 (909) 017 (000) 111 (909) 317 (1765) 2013 424 (1667) 017 (000) 424 (1667) 317 (1765) 2014 126 (384) 017 (000) 326 (1154) 217 (1176) 2015 04 (000) 017 (000) 04 (000) 217 (1176) 2016 011 (000) 017 (000) 211 (1818) 117 (588)
Mean annual () 195 053 1594 1359
Legend xn= where x is the number of ferrets that died with coccidial infection or of other
causes and n is the total number of adult ferrets or kits in a given year () = incidence expressed
as a percentage
104
CHAPTER 5 EVALUATING THE DOMESTIC FERRET (MUSTELA PUTORIUS FURO) AS
AN EXPERIMENTAL MODEL FOR ENTERIC COCCIDIOSIS IN THE BLACK-FOOTED
FERRET (MUSTELA NIGRIPES)
ABSTRACT
The purpose of this study was to determine whether the domestic ferret (Mustela putorius furo) is
susceptible to an isolate of Eimeria ictidea originating from black-footed ferrets (BFF Mustela nigripes)
and thus could act as a suitable experimental model in which to investigate the pathogenesis and
management of this disease A pilot study was performed with 10 male intact juvenile domestic ferrets
Ferrets were administered an oral inoculum containing either a high dose (1 times 106 oocysts) moderate
dose (5 times 104 oocysts) or saline control and observed for shedding of oocysts and development of
clinical signs Seven of ten ferrets developed patent infection all of which had received the high dose
inoculum The prepatent period was 7-9 days and duration of shedding varied from 1-7 days Clinical
signs were identified in six of the seven infected ferrets and were consistent with those previously
described for enteric coccidiosis in domestic and BFF Parasite life stages were identified within the
intestines of four of the seven ferrets with patent infection and were limited to the distal jejunum and
ileum The demonstrated ability to produce patent infections in domestic ferrets following oral inoculation
of a high dose of E ictidea (1 times 106 oocysts) isolated from BFF provides an avenue for future
experimental investigations into the control and treatment of enteric coccidiosis in this endangered
species
51 INTRODUCTION
Black-footed ferrets (BFF Mustela nigripes) are one of only three ferret species worldwide
While formerly distributed throughout the North American prairies black-footed ferrets were declared
extinct in the wild in the 1980s Since 1986 a multi-institutional consortium has been breeding this
species in captivity with reintroductions back into the wild within their historic range in selected locations
in Canada the USA and Mexico Introduced colonies of BFF are present in Arizona Colorado Kansas
105
Montana New Mexico South Dakota Utah Wyoming and Chihuahua (Mexico) Reintroduction
attempts in Saskatchewan Canada have been unsuccessful to date
Multiple infectious diseases pose a significant risk to the captive breeding and post-release
survival of BFF including distemper and sylvatic plague (Santymire et al 2014 USFWS BFF Recovery
Program 2017) Coccidiosis is a recognized cause of juvenile and adult morbidity and mortality in captive
breeding programs and can result in significant losses (Bronson et al 2007 Santymire et al 2014
USFWS BFF Recovery Program 2017) The effect of the disease on wild populations is unknown
Clinical signs of coccidiosis include mucoid to hemorrhagic diarrhea abdominal discomfort lethargy
appetite loss vomiting and dehydration Recent investigations into diseases affecting BFF at the Toronto
Zoo have identified a single Eimeria species E ictidea associated with all cases of enteric coccidiosis in
juvenile and adult BFF from 2014-2016 (see Chapter 3) This same Eimeria species was identified
retrospectively as the cause of juvenile and adult mortalities in previous years (1999 through 2014
inclusive) (Chapters 3 and 4) Furthermore this pathogen was identified in fecal samples based on
morphologic and molecular characterization from adult and juvenile BFF in another zoological collection
(Louisville Zoo Kentucky USA) (see Chapter 3)
Enteric coccidiosis also occurs in domestic ferrets (Mustela putorius furo) with three
morphologically distinct species of coccidia Eimeria ictidea Eimeria furonis and Isospora (=
Cystoisospora) laidlawi Both of the aforementioned Eimeria species have been identified in black-footed
ferrets based on morphologic criteria but molecular characterization was needed to confirm whether the
same species of parasite infects both ferret species (see Chapter 2 and 3) To this end nuclear and
mitochondrial sequences for E furonis and for I=(C) laidlawi were generated (Chapter 2) expanding the
existing limited sequence data from the nuclear 18S rRNA locus of Eimeria furonis Molecular
characterization of E ictidea from domestic ferrets was not possible because samples containing this
parasite were not available for study consequently it is unclear whether the same coccidium affects both
domestic and black-footed ferrets
106
There is no published information describing the pre-patent periods and pathogenicity of enteric
coccidia in BFF and given the conservation status of the BFF experimental work cannot be undertaken
in the natural host The purpose of this study was to determine whether the domestic ferret is susceptible
to E ictidea isolated from BFF if susceptible the domestic ferret could act as a suitable experimental
model in which to investigate the pathogenesis prevention and treatment of coccidiosis caused by E
ictidea
52 MATERIALS AND METHODS
521 Animal care
Ten juvenile male intact ferrets of 48 (n=6) or 50 (n=4) days of age were obtained from a
commercial source (Marshall BioResources North Rose New York USA) and were housed in the
University of Guelph Central Animal Facility Isolation Facility Ferret weights on arrival ranged from
334-475 g (average= 3928 g) All ferrets were housed individually in wire bottom cages of 813 times 1117
times 457 cm size and were divided in equal numbers between two non-adjoining rooms They received ad
libitum access to Envigo Teklad Certified Global Ferret Diet (Madison Wisconsin USA) and water
changed daily Room temperature was maintained at 18-21 degC and a 16 hour light 8 hour dark
photoperiod was provided All personnel working with the ferrets were required to wear personal
protective equipment including disposable facemasks gloves gowns and bouffant caps Shoes were
provided for use in each room This study was carried out in accordance with the recommendations in the
Canadian Council on Animal Care guidelines The protocol was approved by the Animal Care Committee
of the University of Guelph (Animal Use Protocol 3289) and by both the Animal Welfare Committee
and Animal Care and Research Committee of the Toronto Zoo
An initial physical examination and blood collection were performed on each ferret by the
principal investigator (ARP) one day after arrival to assess health status prior to enrollment in the study
Ferrets were mask induced with isoflurane (Isoflurane USP Fresenius Kabi Richmond Hill Ontario) in
107
oxygen placed on a heat disc (SnuggleSafe Lenric C21 Ltd Littlehampton United Kingdom) weighed
examined and blood was collected from the jugular vein for routine CBC and biochemical profiles
All ferrets subsequently underwent an acclimation period of two weeks During this time fecal
samples were collected daily from each ferret and examined for the presence of coccidial oocysts using a
standard salt flotation technique (Dryden et al 2005) to ensure that all individuals were free of coccidia
prior to initiation of experimental work Any ferret positive for coccidia was to be removed from the
study
522 Oocyst preparation
Oocysts used for inoculation originated from fecal samples from two naturally infected BFF These
samples were stored in potassium dichromate for four weeks prior to oocyst purification and use in this
infection trial Stored fecal samples were mixed with distilled water and passed through a small sieve to
remove debris The strained contents were transferred to a 50 mL conical vial and topped up to 50 mL
with additional distilled water Samples were centrifuged (Sorvall ST40R Centrifuge Thermo Scientific)
at 2800 rpm (1315 timesG) for 10 minutes at 12 degC A drop of supernatant was evaluated microscopically at
100times for the presence of oocysts If oocysts were observed the supernatant was poured off into a second
50 mL conical tube and again topped up to 50 mL with distilled water and re-centrifuged under the same
conditions Otherwise the supernatant was discarded The pellets from both the first and second tubes
were combined with saturated salt solution at a 14 ratio by volume Oocysts were floated in the salt
solution by centrifugation at 1500 rpm (377 timesG) for 10 minutes at 12 degC The top 5 mL of supernatant
were collected and transferred to a clean 50 mL conical tube topped up to 40 mL with distilled water and
washed via centrifugation at 2800 rpm (1315 timesG) for 10 minutes After the wash step the supernatant
was again checked for presence of oocysts and discarded if no oocysts were observed The pellet was
collected and the presence of oocysts confirmed by examination of a drop placed on a clean glass slide at
100times Once verified the contents of the pellet of concentrated oocysts was placed in a 250 mL storage
container and mixed with approximately 200 mL of sterile saline (09 sodium chloride Hospira
108
Montreal Quebec) prior to storage for two to four weeks at 4 degC until inoculation Prior to inoculation a
McMaster count was performed to determine the number of oocysts per mL in order to determine
appropriate volume of inoculum
523 Experimental infections
Part 1
Five ferrets were randomly assigned to each of the control and infection groups After the
acclimation period on day 0 four ferrets in the infection group were inoculated orally with a high dose
oocyst suspension (1 times 106 oocysts in 025 mL of saline) mixed into 1 mL of FerreTone Skin amp Coat
Supplement (United Pet Group Inc Blacksburg Virginia USA) a fifth ferret was inoculated with a
moderate dose oocyst suspension (5 times 104 oocysts in 025 mL of saline) in the same volume of FerreTone
Ferrets in the control group were inoculated with a placebo (025 mL of saline) in 1 mL of FerreTone
Inoculation was performed by offering the oocyst suspension or placebo to the ferrets in a plastic
container
Fecal samples were collected daily from each inoculated ferret for 14 days post-inoculation
Samples were analyzed via fecal flotation using the McMaster method followed by routine flotation in
saturated salt solution (Dryden et al 2005) to determine the presence or absence of oocysts and oocyst
burden (oocysts per gram of feces [OPG])Temporal trends in oocyst shedding were monitored Ferrets
were evaluated visually twice daily for the presence of clinical signs of coccidial disease The first of
every two ferrets identified to shed oocysts was to be humanely killed at the time of peak oocyst shedding
(ie the first day that fecal oocyst counts remained static or declined) and necropsied to confirm the
presence of and describe parasitic replication in the intestinal mucosa Any remaining animals that shed
oocysts were to be monitored throughout the 14 day period following inoculation in order to determine
the duration and intensity of oocyst shedding for these individuals the total number oocysts shed during
patency was determined
109
Part 2
All ferrets from the infection group that did not shed oocysts during Part 1 (n=4) and all but one
ferret from the previous control group (n=4) were orally inoculated with the high dose oocyst suspension
(1 times 106 in 1 ml of saline) mixed with an equal volume of FerreTone Consequently between phases 1
and 2 all but one ferret were inoculated at least once with the BFF coccidia in order to increase
experimental animal numbers and determine if ferret age played a role in susceptibility to infection
One ferret from the previous control group was inoculated with a lower dose (2 times 105 oocysts in
075 mL of 09 saline mixed with 1 mL FerreTone) of oocysts that had been collected from the single
domestic ferret that shed in Part 1 oocysts were purified as described above for the initial inocula Fecal
collection and analysis were performed as previously As in Part 1 one in every two ferrets sequentially
identified to be shedding oocysts in feces was killed humanely at the time of peak shedding and a
complete necropsy examination performed The remainder of the ferrets observed to be shedding were
monitored for the full 14 days of the trial after which they were killed humanely and necropsied and total
number of oocysts shed during patency was determined All ferrets that did not shed coccidial oocysts
during the infection trial were rehomed at the end of the trial
524 Animal welfare
Ferrets were evaluated twice daily for development of clinical signs of coccidial disease and any
animal showing clinical disease was to be treated as determined by a veterinarian with supportive care
including fluid therapy A grading system for clinical signs including intervention points and removal
criteria was created for use during daily evaluation (see Appendices 3 and 4) Animals whose clinical
signs could not be ameliorated without the use of specific anticoccidial therapy were to be euthanized
Should the inoculation in Part 1 have resulted in clinical disease that required extensive treatment andor
necessitated euthanasia a lower number of oocysts would be used for subsequent inoculation in Part 2
Ferrets to be euthanized were anesthetized by mask induction with isoflurane in oxygen a 1 mL blood
110
sample was collected from the cranial vena cava and then an intracardiac dose of potassium chloride (2
mEqkg) was administered
525 Hematology
Blood was collected from all ferrets under isoflurane anesthesia at the time of pre-trial health
examination and again at the time of humane killing Blood was collected via jugular venipuncture
initially due to the small size of the ferrets at arrival and then by cranial vena cava venipuncture or
cardiocentesis prior to euthanasia Complete blood count and serum biochemistries were performed by the
Animal Health Laboratory of the University of Guelph Guelph Ontario
526 Morphologic and molecular characterization
Morphologic and molecular characterization of oocysts shed by the domestic ferrets during the
course of the infection trial was performed to ensure that the ferrets were shedding the same species of
Eimeria with which they were inoculated Oocysts were concentrated from positive fecal samples as
described above A drop of concentrated oocyst solution was viewed photographed and measured at
400times and 600times for comparison with previously determined morphometrics of Eimeria ictidea oocysts
(Chapter 3)
Regions from the mitochondrial cytochrome c oxidase subunit I and III (mt COI and mt COIII)
DNA were amplified by polymerase chain reaction (PCR) from each sample using primer pairs
400F1202R and -172F799R respectively For all PCR reactions samples were denatured at 95 degC for 5
min then subjected to 35 cycles of 94 degC for 30s anneal at 52degC for 30s and extension at 72 degC for 60s
followed by a final extension at 72 degC for 7 min PCR gel electrophoresis and sequencing methods used
were as described in the Materials amp Methods section of Chapter 2
The resulting consensus sequences were searched from within Geneious against previous
sequences for E ictidea produced by the authors and against publically available sequences on the
111
BLAST server (blastncbinlmnihgovBlastcgi) using the blastn search algorithm against the nrnt
database (GenBank+EMBL+DDBJ+RefSeq ndash AA or DNA)
527 Necropsy protocol
All humanely killed ferrets underwent a complete necropsy (Appendix 6) using the modified
protocol described in Materials amp Methods section 424 of Chapter 4
53 RESULTS
Initial physical examination was unremarkable with the exception of mild to moderate bilateral
ceruminous discharge within the external ear canal of all ferrets Complete blood count and serum
biochemistry results for all ferrets were within normal reference intervals for juvenile domestic ferrets
(Appendices 2a and b) (Fox 2014) Six days after arrival a single ferret (103) in the control group
developed mild upper respiratory signs consisting of sneezing and clear nasal and ocular discharge these
clinical signs were associated with mild dehydration and decreased food and water consumption The
ferret was treated with subcutaneous fluid therapy (10 mL Plasmalyte-A subcutaneous Baxter Alliston
Ontario) heat and supportive care and all clinical signs resolved within three days This ferret was
deemed healthy to participate in the remainder of the clinical trial A second ferret (105) in the control
group developed unilateral purulent ocular discharge 14 days after arrival one day prior to placebo
inoculation The ferret was treated topically twice daily for five days with Isathal ophthalmic gel (fusidic
acid 10 mgg Dechra Veterinary Products Inc Point-Claire Quebec) and the discharge resolved but
reoccurred within 2 days of treatment cessation Ocular examination showed mild conjunctivitis but no
evidence of corneal lesions and fluorescein staining did not indicate the presence of corneal ulceration
The ferret was treated for an additional six days with tobramycin ophthalmic solution (3 mgmL Sandoz
Tobramycin 03 Boucherville Quebec) after which clinical signs resolved completely No coccidial
oocysts were shed in feces from any of the ferrets during the two week acclimation period
112
531 Oocyst shedding
All ferrets readily ingested the inoculum with either placebo or concentrated oocysts In Part 1
one ferret (203) in the infection group which had received the high dose (1 times 106 oocysts) shed oocysts
on day 8 and day 9 after inoculation (Tables 51 52) This ferret was 71 days of age at the time shedding
was initially identified The ferret was killed humanely 11 days post inoculation later than had been
outlined in the protocol as processing of fecal samples had been delayed by two days resulting in late
detection of oocyst shedding in this individual Oocysts were not identified in the feces of the three
remaining ferrets that received the high inoculation dose the single ferret that received the lower dose (5
times 104 oocysts) or in the ferrets within the control group
In Part 2 six of eight ferrets inoculated with the high dose (1 times 106 oocysts) shed oocysts during
the 14 day observation period (Tables 51 52) Four of these ferrets were from the previous control
group One of the ferrets previously inoculated with the high dose inoculum in Phase 1 that had not shed
oocysts did shed oocysts after being inoculated a second time with the same dose during Phase 2 The
ferret that had previously received the low dose (5 times 104 oocysts) of oocysts in Phase 1 also shed after
inoculation with the high dose in Phase 2 Three ferrets did not shed oocysts after high dose inoculation in
Phase 2 one of these had been part of the previous control group the other two had received the high
dose inoculation previously in Phase 1
The pre-patent period ranged from 7-9 days (Table 51) with equivalent numbers of ferrets
commencing shedding on each of days 7 through 9 All six ferrets were between 91 and 93 days of age at
the time shedding was initially identified Oocyst per gram counts and shedding trends for all individuals
are shown in Table 51 Total oocyst shedding during patency was lt14 oocysts 8904 oocysts and
172291 oocysts for ferrets 201 104 and 105 respectively The two ferrets for which the prepatent period
was 9 days only shed oocysts for one day and in low numbers Oocysts were not identified in the feces of
the three remaining ferrets two of which received the high inoculation dose (1 times 106 oocysts) and the
third that received the lower inoculation dose (2 times 105 oocysts) (Table 52)
113
532 Morphologic and molecular characterization
In all seven ferrets that shed oocysts the morphologic features and measurements (length width
shape index) of the shed oocysts were consistent with those of the E ictidea administered in the inoculum
(Figure 51) Molecular confirmation of the identity of the oocysts shed was successful in 3 out of the 7
ferrets (102 103 203) samples from the four remaining ferrets did not contain adequate quantity or
quality of DNA for confirmation
533 Clinical signs
Clinical signs associated with patent infection were identified in 6 of 7 ferrets (Table 52) These
signs included weight loss (n=5) diarrhea (n=1) mucoid soft feces (n=2) feces containing blood (n=2)
and malodorous feces (n=1) Appetite reduction was noted in two ferrets from the infection group in Part
1 between 6-8 days post infection however no oocyst shedding was detected from either ferret during
this time
534 Hematology
CBC and serum biochemistry values from ferrets collected during pre-trial health screening are
displayed in Appendices 1a and 1b Values obtained for ferrets euthanized during or after the
experimental trial are displayed in Appendices 3a and 3b Minor variances from reference range values
for CBC and serum biochemistry were identified in six of the seven ferrets with patent infection In all six
ferrets for which a complete serum biochemistry was obtained immediately prior to death creatinine
kinase (CK) values were elevated (see Appendix 3b) Ferret 103 exhibited a mild hypoalbuminemia (20
ref 24-40 gL) on ante-mortem serum biochemistry (see Appendix 3b)
114
535 Necropsy
No gross or histopathologic lesions were present and coccidia could not be identified in sections
of intestine from the single ferret (203) humanely killed in Part 1 For the ferrets humanely killed in Part
2 no evidence of diarrhea hematochezia or mucoid fecal material was identified grossly Ferret 105
killed at the termination of the experiment but still shedding low numbers of oocysts in its feces
exhibited a 7 cm region of congested mucosa within the distal jejunum
Coccidial life stages were identified in small intestinal sections from four of the seven ferrets that
were identified to shed oocysts at some point prior to necropsy (Figure 52 Tables 52 and 53) Affected
sections included jejunum in all four animals as well as ileum in one and were collected from 114 to 218
cm aboral from the pylorus (see Figure 53) Coccidia were not identified in sections of duodenum
proximal jejunum or large intestine however oocysts were identified within fecal material in the lumen
of the large intestine from one ferret (103) Of the 11-19 sections of intestine examined for each ferret the
number of sections containing parasites ranged from one to eight 1 section in ferret 102 2 sections in
ferret 201 4 sections in ferret 105 8 sections in ferret 103 A mix of sexual and asexual life stages was
observed within the enterocytes in small intestinal sections from ferret 103 the remainder of the ferrets
showed either asexual (102 201) or sexual (105) life stages in affected segments
Pathologic changes and additional histologic findings in small intestinal sections of ferrets with
enteric coccidia included rare regions of blunting of the villi and sloughing of the epithelium associated
with hemorrhage and inflammation The primary lesions identified were subjectively increased numbers
of eosinophils lymphocytes and plasma cells within the lamina propria of the small intestine and
similarly increased neutrophils lymphocytes and plasma cells within the lamina propria of the large
intestine Neutrophils were rarely present in intestinal crypts and glands
Other gross necropsy findings included two ferrets with renal cortico-medullary cysts and one
ferret with mild thickening of the esophageal mucosa midway along the esophageal length No histologic
changes were identified within a sample of esophagus taken from this region
115
54 DISCUSSION
The findings of this study show that domestic ferrets are susceptible to infection with the enteric
coccidium Eimeria ictidea isolated from black-footed ferrets Both morphometric and molecular
diagnostic methods were used to confirm that ferrets were shedding oocysts of the same species with
which they were inoculated Molecular characterization was successful in three of the seven ferrets that
developed patent infections and as no other coccidial species was identified during pre-trial observation
morphometry was considered to be confirmatory in the remaining four animals
We have referred to the eimeriid coccidium affecting BFF and used in this experimental trial as
E ictidea based on morphometric similarity of their oocysts with those of E ictidea as described from
domestic ferrets (see Chapter 3) There is limited published information on infection of domestic ferrets
with E ictidea outside of Hoarersquos original descriptions (1927 1935a b) which form the basis for all
subsequent identifications of E ictidea in domestic ferrets and in BFF Attempts to obtain exemplars of E
ictidea from domestic ferrets to characterize using molecular techniques were unsuccessful (Chapter 2)
Multiple diagnostic laboratories in Canada and Europe were solicited for coccidia-positive fecal samples
from domestic ferrets but no samples of E ictidea were received over a 4 year period (2014-2017)
Eimeria ictidea was identified based on microscopic examination in only two samples submitted to a
European diagnostic laboratory from 2008-2015 It is unproven whether the E ictidea described from
domestic ferrets and the E ictidea identified from black-footed ferrets and used in this experimental
work are the same or are simply morphologically indistinguishable Eimeria species However the
consistency in morphology host genus and location of infection within the intestinal tissues combined
with the successful cross-transmission of this parasite to domestic ferrets described in the present study
suggests they are likely conspecific
116
The pre-patent period (minimum duration of endogenous development) for infection with E
ictidea in the inoculated domestic ferrets ranged from 7-9 days (see Table 51) the pre-patent period for
this parasite in the BFF the natural host for this coccidium is unknown Hoare experimentally infected
naiumlve domestic ferrets with E ictidea derived from naturally occurring infection in this species (Hoare
1935b) The inoculated ferrets shed oocysts after a pre-patent period of 7 days consistent with the 7-9
days seen in the work described here with E ictidea
Shedding of oocysts was identified over a period of 1-7 days (see Table 51) and intensity ranged
from less than 14 up to 15624 OPG These results may be skewed with erroneously low duration of
shedding and number of oocysts shed as three of the seven ferrets were humanely killed at or prior to the
expected peak of oocyst shedding for tissue collection and histologic examination in order to increase the
probability of identifying parasite life stages within the intestinal sections Shedding periods were similar
to those identified in adult single-housed BFF which ranged from 2-9 days however oocyst per gram
counts from the domestic ferrets were consistently lower than OPG counts from BFF (104 - 554274
OPG) infected with the same parasite (see Chapter 4) Furthermore the total number of oocysts shed by
individual domestic ferrets (14 - 172291 oocysts) during patency was reduced compared to BFF despite
similar length of shedding period (see Chapter 4) The domestic ferrets in this study were naiumlve individual
juveniles whereas the BFF were adults 1-5 years of age some of which were showing clinical signs at
the time of oocyst shedding The relative influences of age species and previous exposure to the parasite
on our observations are unknown
Two different fecal flotation methods were used on all samples to increase the probability of
oocyst detection The McMaster method was used to provide accurate OPG counts for quantification of
oocyst shedding however this method had a minimum detection limit of ~13 oocysts per gram (1333
OPG calculated) because it is based on dilution of the initial sample with flotation media (saturated salt)
In samples with few oocysts oocysts may be missed or to be present in numbers below this detection
limit Routine salt flotation is in contrast performed using the entire sample allowing for detection of
117
small numbers of oocysts Consequently in cases where oocyst per gram counts were low shedding was
identified on routine salt flotation but not by the McMaster method and recorded as lsquopositiversquo but below
the detection limit of the enumeration method
Subclinical to clinical disease occurred in six of the seven ferrets that developed patent infection
with weight loss being the most frequent clinical sign Other clinical signs were typical of enteric
coccidiosis including diarrhea hematochezia and mucoid andor soft feces These clinical signs are
similar to those previously described for black-footed ferrets infected with this parasite (USFWS BFF
Recovery Program 2017 Chapter 4) and for domestic ferrets with enteric coccidiosis (Sledge et al
2011) Interestingly development of clinical disease was not described in the naiumlve domestic ferret
inoculated by Hoare (1935) with E ictidea derived from naturally occurring infection Based on review of
the literature severe clinical disease resulting from intestinal coccidiosis is rare in domestic ferrets
Black-footed ferrets however appear more susceptible to disease development and more frequently show
significant clinical signs No domestic ferret required treatment for clinical coccidiosis during the course
of this study It is possible that the more pronounced clinical signs associated with enteric coccidiosis in
BFF may result from the limited genetic diversity in a population derived from so few individuals and
increased susceptibility of BFF to other diseases such as sylvatic plague have been described in
comparison with their domestic counterparts (Williams et al 1994) Although unlikely recent acquisition
of E ictidea from a related host species such as the domestic ferret could have resulted in increased
pathogenicity and severity of clinical disease from infection with this parasite in BFF
Minor variances from reference range values for CBC and serum biochemistry were identified in
six of the seven ferrets with patent infection but only in one case (ferret 103) did this appear to be
correlated with infectiondisease In this animal a mild hypoalbuminemia (20 ref 24-40 gL) was noted
(Appendix 3b) On histopathology large regions of the small intestine contained parasite life stages
however inflammation lysis of epithelial cells and necrosis of affected areas that could be expected to
result in protein loss into the intestinal lumen were not identified
118
Creatine kinase values were elevated in all six ferrets for which a complete serum biochemistry
was obtained immediately prior to death these findings are consistent with release from CK rich tissues
such a skeletal muscle during venipuncture and manual restraint
The pathologic lesions identified within the intestinal sections of ferrets euthanized at the time of
oocyst shedding were similar to those identified in affected BFF but in most cases were less locally
extensive or widespread throughout the small intestine than those observed in necropsy cases of BFF (see
Chapter 4) Coccidia were seen in the histologic sections of four ferrets all three ferrets that were actively
shedding oocysts at the time of necropsy (102 103 105) and one of four in which necropsies were
performed after oocyst shedding had ceased (201)
Although the primary objective of the examination of histologic sections from affected ferrets
was to identify coccidial life stages attempts were made to describe the pathologic changes associated
with the presence of the parasite Despite necropsies being performed almost immediately after death and
the use of Serra solution fixative to improve parasite and tissue preservation the villi and villar
epithelium of the trimmed sections were frequently distorted or absent and consequently accurate
commentary on these areas was precluded and was made only on visible components of the lamina
propria and crypts or glands A deliberate decision was made not to kill and collect samples from the
saline inoculated control ferrets after Phase 1 and those not shedding oocysts during Phase 1 or 2 thus no
age-matched intestinal sections were available for comparison It is difficult to comment on the
significance of the inflammatory cells observed in the lamina propria of the small and large intestinal
segments or the proliferative rate of the crypt epithelium The only changes identified which may be
considered significant are the presence of neutrophils within the crypts and glands of small and large
intestinal sections respectively but these lesions were rare and not associated with the presence of
parasitic life stages
119
While no parasitic life stages were identified in intestinal sections from ferrets 104 203 and 205
one of the three ferrets ferret 104 exhibited lymphoplasmacytic inflammation and blunting of jejunal
villi in one section (36-38 cm aboral from pylorus) These findings might be expected as the most
extensive histologic lesions would occur associated with lysis of the intestinal epithelial cells as oocysts
were shed into the feces after which new intestinal epithelial cells would re-cover the denuded villar
surface Thus for those cases in which histologic examination was performed after shedding had ceased
presence of the protozoal life stages in the intestines would be expected to be significantly reduced or
absent
Hoare (1935a b) described a particular reaction to the presence of parasitic life stages of Eimeria
ictidea in the small intestine of domestic ferrets in which only the villar tips were affected and there was
resultant annular constriction of the villus separating the affected and non-affected segments These
particular changes were not identified in any of the ferrets in this study and had not been noted
retrospectively in naturally infected BFF (Chapter 4)
While patent infection and intestinal disease could be experimentally created in domestic ferrets
without equivalent experimental work in BFF it is difficult to fully compare the susceptibility to infection
and to the development of disease between the two species Eimeria species tend to be host specific thus
if E ictidea from BFF is not conspecific with E ictidea in domestic ferrets it might be expected that the
domestic ferret would be less susceptible to infection and the development of disease than is the BFF
Even if the two parasites are identical natural passaging through the BFF may alter affinity for the
domestic ferret
It appears that the infectious dose of oocysts of E ictidea derived from BFF required to initiate a
patent infection in domestic ferrets is high The ferrets that developed patent infections were administered
an inoculum containing 1 times 106 sporulated oocysts and even with this extremely high inoculating dose
only a proportion of inoculated ferrets became infected Neither the ferret that received the low dose
120
inoculum nor the ferret that received the passaged oocysts from Part 1 of the study shed oocysts during
the 14 day period post inoculation The latter finding was unexpected as fresh passaged oocysts would be
expected to contain larger proportions of viable oocysts and be comparatively more infective than oocysts
that had been stored for 2-4 weeks prior to inoculation
The only publication describing oral inoculation of Eimeria species in domestic ferrets (Hoare
1935b) did not quantify the number of oocysts administered A study performed in 16 farmed juvenile
mink (Mustela vison) administered 2000 sporulated oocysts of each of three coccidial species (I
laidlawi E vison and an unknown Eimeria species) resulting in patent infection with one of the three
species (Foreyt et al 1977) as determined by the presence of oocysts on fecal examination The authors
did not reveal which type of oocysts resulted in the infection
The number of oocysts required to result in infection in BFF is unknown Based on the
authorsrsquo observations of over 100000 oocysts per gram of feces being shed by black-footed ferrets into
cages of lt1m2 floor space (see Chapter 4) we estimated that under normal caged conditions animals
would likely ingest thousands of oocysts over a short period of time This was in part why a large number
of oocysts (up to 1000000 as available from our store of viable oocysts) was administered to each
domestic ferret in order to increase the probability that infection and shedding would result Furthermore
for the parasite to persist within the ferret population the total number of oocysts shed into the
environment would have to be several times higher than the infective dose required to generate a patent
infection otherwise the parasite would be expected to die out If E ictidea of domestic ferrets and E
ictidea of BFF are conspecific the difference in oocyst shedding between the host species during patent
infection could potentially explain the low prevalence of E ictidea reported from the domestic ferret
population (as seen in Chapter 2)
Both humoral and cell mediated immunity are involved in the immune response to coccidia The
role of maternal derived antibodies in combatting protozoal infection in mustelids has not been studied
121
but in carnivores maternal antibodies to viruses can last up to 16 weeks (Chappuis 1998) In poultry
maternal Eimeria-specific IgG is transferred via the egg yolk to offspring In one study breeding hens
were infected with a single species of Eimeria 28-39 days prior to lay Their hatched chicks were
challenged by inoculation with oocysts of the same and a related Eimeria species and showed reduced
oocyst shedding compared to age matched controls indicative of passive transfer of immunity (Smith et
al 1994) It is possible then that the presence or absence of maternal antibodies may be a factor in the
age at which ferrets are susceptible to coccidial infection and the development of disease The facility
from which the domestic ferrets were acquired has not previously detected Eimeria species on routine
fecal screening (Dr Bambi Jasmin personal communication) consequently it is unlikely that they would
have received maternal immunity to or been exposed to this parasite and thus can be considered to be
naive
Despite a theoretical lack of maternal immunity there did appear to be an effect of age on
susceptibility to infection In Part 1 when the ferrets were approximately 70 days of age only 1 of 4
ferrets inoculated with the high dose of oocysts developed patent infection In comparison in Part 2
when the ferrets were 91-93 days of age 3 of 4 ferrets that had been in the saline control group for Part 1
developed patent infections as did 2 of 4 ferrets that had been inoculated in Part 1 but had not shed
oocysts One of these previously inoculated ferrets was the individual that had received the lower dose of
oocysts Thus it appears that patent infection could be produced more easily in the older ferret kits
However the two ferrets that had previously been inoculated exhibited the shortest shedding periods (1
day) and lowest oocyst per gram counts and parasite life stages in these cases were rare (201) to absent
(205) on histologic examination of numerous sections of intestine It is thus possible that the primary
inoculation resulted in abbreviated infections or infections in which so few oocysts were shed that
infection was not detected Our observation of endogenous stages in the intestinal tissues of some of the
ferrets following cessation of oocyst shedding suggests that the pre-patent period and duration of patency
may vary considerably from animal to animal consequently it is possible that some of the kits would
122
have ultimately shed a few oocysts from the primary inoculum if followed beyond 14 days post-
inoculation Whether through an aborted or undetected infection previous exposure of these kits to the
parasite probably generated partial immunity against E ictidea and therefore the intensity of infection
upon challenge in previously exposed domestic ferret kits was reduced
The authors acknowledge the limitations of this initial pilot study however it was proven that
patent infection with E ictidea isolated from black-footed ferrets could be generated in a novel host the
domestic ferret Further studies will be required to investigate the effect of age on susceptibility to
infection as well as the possibility of development of immunity after exposure and its role in reducing
parasite replication and disease in subsequent infection With so few BFF in existence the use of BFF for
in vivo infection trials cannot be contemplated consequently refining the domestic ferret infection model
will be essential for carrying out research specifically intended to help manage coccidiosis in the
endangered black-footed ferret
Figure 51 Exogenous life stages of Eimeria ictidea shed from a domestic ferret (Mustela putorius
furo) experimentally inoculated with oocysts originating from black-footed ferrets (Mustela
nigripes) Unsporulated oocyst (solid black arrow) Sporulated oocyst (solid white arrow) Bright
field microscopy scale bar = 25 microm
25 microm
124
Figure 52 Life stages of Eimeria ictidea within the small intestinal epithelium of an experimentally
infected domestic ferret (Mustela putorius furo) A) Sexual life stages (micro- and macrogamonts
- white arrows) crowding the villar enterocytes all stages are found between the nucleus and
luminal surface of infected enterocytes Hematoxylin and eosin staining scale bar = 25microm B) At
higher magnification meronts (black arrows) and gamonts (open arrows) are crowded between the
enterocyte nuclei and brush border Hematoxylin and eosin staining scale bar = 25microm
25 microm
25 microm
125
Figure 53 Presence and location of sexual and asexual life stages of Eimeria ictidea within the
intestinal epithelium of domestic ferrets (Mustela putorius furo) (n=7) that developed patent
infection after experimental inoculation with oocysts originating from black-footed ferrets (Mustela
nigripes) Legend duod = duodenum jej= jejunum Sequential numbers for jejunal and colonic
sections represent the order aboral from the pylorus from which the samples were collected SI =
additional section(s) of small intestine whose aboral sequence was not recorded
0
1
2
3
duod jej 1 jej 2 jej 3 jej 4 jej 5 ileum colon 1 colon 2 SI
Nu
mb
er
of
Ferr
ets
Aff
ect
ed
Intestinal Section Containing Life Stages of Eimeria ictidea
Asexual life stages Sexual life stages
126
Table 51 Prepatent period and oocyst shedding patterns in domestic ferrets (Mustela putorius furo)
experimentally inoculated with 1 times 106 oocysts of Eimeria ictidea originating from black-footed ferret
(Mustela nigripes) that developed patent infections
Oocysts shed per gram of feces
Ferret Identity
Day post inoculation 102 103 104 105 201 203 205
1 0 0 0 0 0 0 0
2 0 0 0 0 0 0 0
3 0 0 0 0 0 0 0
4 0 0 0 0 0 0 0
5 0 0 0 0 0 0 0
6 0 0 0 0 0 0 0
7 lt 14 1807 0 0 0 0 0
8 11053 139 7091 0 156238 0
9 463 11733 lt 14 203 lt 14
10 578 7549 0 0 0
11 lt 14 0 0 0
12 0 lt 14 0
13 0 lt 14 0
14 0 lt 14 0
Legend ferret euthanized as of this date lt 14 = oocyst positive samples with less than 14 oocysts per
gram of feces
127
Table 52 Results of oral inoculation of domestic ferrets (Mustela putorius furo) with oocysts of Eimeria ictidea originating from black-footed
ferrets (Mustela nigripes)
Ferret
Identity
Results Part 1 Results Part 2
Inoculum Oocyst
Shedding
Presence
of clinical
disease
Coccidia
present in
sections
Inoculum Oocyst
Shedding
Presence
of clinical
disease
Coccidia
present in
sections
101 Saline N N - 2 times 105 oocystsa N N -
102 Saline N N - 1 times 106 oocysts Y N Y^
103 Saline N N - 1 times 106 oocysts Y Y Y^
104 Saline N N - 1 times 106 oocysts Y Y Nyen
105 Saline N N - 1 times 106 oocysts Y Y Yyen
201 1 times 106 oocysts N N - 1 times 106 oocysts Y Y Yyen
202 1 times 106 oocysts N N - 1 times 106 oocysts N N -
203 1 times 106 oocysts Y Y Nyen
204 1 times 106 oocysts N N - 1 times 106 oocysts N N -
205 5 times 104 oocysts N N - 1 times 106 oocysts Y Y N^
Legend includes any of the following weight loss diarrhea mucoid feces malodorous feces inappetence N = no Y= yes - = necropsy not
performed a = oocysts collected after passage through ferret 203 ^ = shedding oocysts at time of necropsy yen = not shedding oocysts at time of
necropsy
128
Table 53 Distribution of coccidial life stages in domestic ferrets (Mustela putorius furo) orally inoculated with oocysts of Eimeria ictidea
originating from black-footed ferrets (Mustela nigripes)
Ferret Identity
Intestinal level 102
103
104 105
201
203
205
Duodenum N N N N N N N
Jejunum 1 N N N N N N N
Jejunum 2 N N N N N N N
Jejunum 3 N S A N N N N N
Jejunum 4 N S A N N N N N
Jejunum 5 N S A N S A N N
Ileum N S A N N N N N
Colon 1 N Na N N N N N
Colon 2 N Na N N N N N
Unmeasured small intestinebc
S - 05
A - 15
S - 45
A - 25
S - 05
A - 05
S - 26
A - 06
S - 06
A - 06
S - 06
A - 06
S - 08
A - 08
Unmeasured large intestinebc
S - 01
A - 01
S - 01
A - 01
S - 01
A - 01
S - 02
A - 02
S - 01
A - 01 none
S - 01
A - 01
Legend N = no parasite life stages S = sexual life stages present A = asexual life stages present a= oocysts present in feces b=
additional sections of intestine for which the location measured from the pylorus was not obtained c= xn where x is number of
sections containing sexual or asexual lifestages n is the number of sections examined
129
CHAPTER 6 WHOLE MITOCHONDRIAL GENOME SEQUENCES OF TWO EIMERIA
SPECIES ISOLATED FROM DOMESTIC (MUSTELA PUTORIUS FURO) AND BLACK-
FOOTED FERRETS (MUSTELA NIGRIPES)
ABSTRACT
The complete mitochondrial (mt) genomes of Eimeria furonis and Eimeria ictidea (Eimeriidae
Coccidia Apicomplexa) originating from single fecal samples from a domestic (Mustela putorius furo)
and a black-footed ferret (Mustela nigripes) respectively were sequenced Both mt genomes were
circular-mapping with lengths of 6165 base pairs (Eimeria furonis - GenBank MF795598) and 6182
base pairs (Eimeria ictidea - GenBank KT203399) Genome organization and gene contents were
comparable with those of other publically available mt genomes from a variety of Eimeria species and
related coccidia there were three complete coding DNA sequence regions encoding cytochrome c
oxidase subunit I cytochrome c oxidase subunit III and cytochrome B and 33 regions encoding
fragmented rDNA Alignment of these mt genome sequences demonstrates a relatively high (945 340
single nucleotide differences [SNDs]) pairwise sequence identity between these Eimeria spp infecting
ferrets the majority of the SNDs resulted in synonymous codon changes with no changes to their protein
products Alignment of the newly sequenced mt genomes demonstrates and phylogenetic reconstructions
support the monophyly of these Eimeria spp of ferrets with another Eimeria sp of carnivores as the
sister taxon to this clade
61 INTRODUCTION
Coccidia are protozoal eukaryotic host-specific parasites of the phylum Apicomplexa and can
be divided into two major taxonomic suborders the eimerioirinid and adeleid coccidia The eimeriorinid
coccidia include both the typical intestinal coccidia (eg Eimeria Isospora Cyclospora) species
belonging to the family Eimeriidae as well as tissue or cyst-forming coccidia (eg Cystoisospora
130
Besnoitia Toxoplasma Sarcocystis) of the family Sarcocystidae (see Cox 1994) Ten species of
Eimeria and twelve species of Isospora (=Cystoisospora) have been described in the Mustelidae
(see Chapter 1 Table 11) Eimeria furonis has been reported in the European polecat (Mustela
putorius) domestic ferret (DF Mustela putorius furo) black-footed ferret (BFF Mustela nigripes) and
mink (Mustela vison) (Hoare 1927 Nukerbaeva and Svanbaev 1973 Jolley et al 1994) Eimeria ictidea
has been reported in the Steppe polecat (Mustela eversmanii) as well as the European polecat domestic
ferret and black-footed ferret (Hoare 1927 Svanbaev 1956 Jolley et al 1994) These reports are based on
the morphometric characteristics of oocysts identified in the feces of the aforementioned host species
without the use of molecular techniques to confirm specific parasite identities
Recently sequences of the mitochondrial cytochrome c oxidase subunit I gene (mt COI) and
nuclear small subunit ribosomal DNA (nu 18S rDNA) of E furonis originating from a domestic ferret (nu
18S rDNA GenBank MF774678-MF774680 mt COI GenBank MF774034-MF774036) and E ictidea
originating from a black-footed ferret (nu 18S rDNA GenBank MF860826-MF860827 mt COI
GenBank MF860823 MF860825) were generated (see Chapters 2 and 3) The parasite originating from
the black-footed ferret was identified as E ictidea based on morphologic similarity to the original
descriptions of E ictidea from domestic ferrets however sequence-based genotyping of E ictidea from
domestic ferrets has not been completed and consequently it has not been demonstrated unequivocally
that the two parasites are conspecific In the present work the complete mitochondrial genomes of E
furonis from the DF and E ictidea from the BFF are described and compared with the mitochondrial
genomes of related coccidia
62 MATERIALS amp METHODS
621 Parasites
Two isolates of morphologically distinct Eimeria species were used in this study Isolate one
identified morphologically and by nu 18S rDNA and mt COI sequences as Eimeria furonis was obtained
131
from a fecal sample from a DF that was submitted for routine ova and parasite examination to a European
diagnostic laboratory8 Isolate two identified morphologically and by nu 18S rDNA and mt COI
sequences as Eimeria ictidea was obtained from a fecal sample from a BFF and was collected during
routine cage cleaning in a captive breeding facility (see Chapters 3 and 4) Fecal collection techniques for
the BFF were reviewed and approved by both the Animal Welfare Committee and the Animal Care and
Research Committee of the Toronto Zoo
622 DNA isolation from coccidia in feces
Genomic DNA was isolated from fecal derived coccidial oocysts as described section 221 of the
Chapter 2 Materials amp Methods Parasite DNA concentration was estimated using a Nanodrop 2000
spectrophotometer (NanoDrop Products Wilmington DE USA) and DNA was stored at 4 degC for
immediate use or minus20 degC for later use
623 Whole genome sequencing
Mitochondrial whole genome amplification for both Eimeria species was initiated using sets of
mt-specific primers that generated overlapping polymerase chain reaction (PCR) fragments (Tables 61
and 62) PCR amplification was performed for all samples in a volume of 25 microl containing ~100 ng of
genomic DNA 1times PCR buffer 3 mM MgCl2 06 mM deoxyribonucleotide triphosphates (dNTPs) 500
nM of each primer and 4 U of Invitrogen Platinum Taq DNA Polymerase (Thermo Fisher Scientific
Toronto ON Canada) PCR reactions were performed on a Bio-Rad T100 PCR thermal cycler (Bio-Rad
Laboratories Singapore) using settings as described previously in the Materials amp Methods section of
Chapter 2 Table 61 details the specific anneal conditions used for the various primer pairs Genomic
DNA from either Eimeria maxima or Eimeria tenella acted as a positive control for the reaction
chemistry Gel electrophoresis purification and sequencing of the PCR amplification products were
8 Vet Med Labor GmbH Division of IDEXX Laboratories Ludwigsburg Germany
132
performed as described in Chapter 2 The resulting chromatograms were aligned and analyzed with
Geneious Ver 818 or later (Biomatters Limited Auckland New Zealand) and high quality consensus
sequences generated The completed mt genome sequences were annotated by comparison with
previously annotated mt genomes from other Eimeria species (eg Eimeria innocua - KR1082961) and
the annotated mt genomes deposited in GenBank
624 Phylogenetic analysis
To determine the phylogenetic affinities of the newly obtained sequences with sequences from
related apicomplexan taxa representative whole mt genome sequences from eimeriid coccidia were
downloaded from GenBank A complete mt genome sequence from an unnamed Choleoeimeria sp was
used to root the ingroup taxa several small genomic rearrangements in the Choleoeimeria sp sequence
required some rearrangement of the genome sequence to unambiguously align homologous regions across
the complete mt genomes
Whole genome sequences were aligned using MAFTT v7017 (Katoh et al 2002) executed from
within Geneious the resulting alignment was examined by eye to adjust start and stop codon positions in
aligned coding DNA sequence [CDS] regions (ie mt COI mitochondrial cytochrome c oxidase subunit
III gene [mtCOIII] mitochondrial cytochrome b gene [CytB]) Phylogenetic trees were generated using
Bayesian Inference (BI) using MrBayes Ver 326 (Huelsenbeck and Ronquist 2001) executed from
within Geneious The aligned complete mt genomes were partitioned into coding (ie CDS) and non-
coding regions so that region-specific models of nucleotide substitution could be applied Characters in
the non-coding region were analysed with the general time reversible (GTR) model (Tavareacute 1986) with
the following parameters nucmodel=4by4 nst=6 rates=invgamma (ie GTR+I+G) Characters in the
coding regions were analysed using the codon nucleotide model (ie lset nucmodel=codon rates=gamma
ngammacat=4) using metazoan mitochondrial translation (ie code=metmt)
133
All BI analyses were run for a chain length of 1000000 with tree sampling every 1000
following a burn-in of 100000 with default settings of 4 heated chains and heated chain temp of 02
63 RESULTS
The whole mt genome sequences of the single isolates of E furonis and E ictidea were
respectively 6165 base pairs (bp) (Figure 61 GenBank MF795598) and 6182 bp (Figure 62 GenBank
KT203399) Content and organization of both mt genomes consisted of three protein-coding genes (mt
COI mt COIII and CytB) interspersed with large and small subunit ribosomal DNA (rDNA) fragments
Details of the various CDS and rDNA fragments are summarized in Table 63 (for E furonis) and Table
64 (for E ictidea)
Pairwise alignment of the mt genome sequences from E furonis and E ictidea demonstrated a
relatively high pairwise sequence identity (946 333 single nucleotide differences [SNDs]) between
these two parasites The bulk of the SNDs (676 225333) were clustered within the three CDS regions
that encode CytB mt COI and mt COIII (see Figure 63 and Table 65) However the majority of these
SNDs (826 186225) were synonymous codon changes that resulted in no changes to the protein
products Only 41 SNDs were involved in 34 amino acid changes distributed among the three CDS
The 33 rDNA fragments comprised 2108 and 2109 bp respectively of the mt genomes of
E furonis and E ictidea Pairwise comparison of these rDNA fragments demonstrated high (986 30
SNDs) sequence identity between the two parasites The remaining 778 and 794 bp respectively of the
mt genomes of E furonis and E ictidea were intergenic stretches between the various rDNA and CDS
regions these intergenic regions were more variable that other regions of the genomes with 78 SNDs
(almost 10 sequence divergence) Additionally all indels were restricted to these variable intergenic
regions
The BI phylogeny generated from aligned complete mt genomes (Figure 64) supported the close
relationship between E furonis and E ictidea within a clade of Eimeria species that include the only three
sequences available for Eimeria spp of carnivores Eimeria mephitidis from the striped skunk (Mephitis
134
mephitis Family Mephitidae) was the sister taxon to the two Eimeria species of ferrets (Family
Mustelidae)
64 DISCUSSION
This work generated the first complete mt genomes from coccidia that infect domestic and black-
footed ferrets (Carnivora Mustelidae) Eimeria mephitidis from the striped skunk Mephitis mephitis
(Carnivora Mephitidae) is the only other Eimeria species from a carnivore for which a complete mt
genome has been reported
Comparatively few eimeriid coccidia only 26 Eimeria species have been described from
carnivores there are 14 named species from the mustelids four from the procyonids four from the ursids
three from the herpestids and one from the viverrids (Duszynski et al 2000) The majority of coccidia
that infect the digestive tract of carnivores belong to the family Sarcocystidae including monoxenous or
facultatively heteroxenous Cystoisospora species or heteroxenous parasites in the genera Sarcocystis
Hammondia and Neospora So far as is known none of the parasites in the Sarcocystidae possess typical
apicomplexan mt genomes with 3 complete CDS and many rDNA fragments (Ogedengbe 2015)
The mt genomes from the two Eimeria sp of mustelid origin demonstrate the same structural
organization (ie the order and number of CDS and rDNA fragments) and circular mapping as the mt
genomes from other Eimeria spp and other closely related eimeriid coccidia such as Isospora
Cyclospora and Lankesterella species Despite the ability of the eimeriid sequences to be mapped
circularly the physical form of Eimeria spp mt genomes may be a linear concatemer of multiple genome
copies as demonstrated for Eimeria tenella (Hikosaka et al 2011) As in the mt genomes of other
eimeriid coccidia (Ogedengbe et al 2013 2014) the CDS for mt COIII demonstrated the highest
sequence divergence between E furonis and E ictidea the mt COI CDS was somewhat more conserved
and CytB CDS demonstrated the fewest SNDs
As expected based on limited sequence divergence between E furonis and E ictidea a BI
phylogenetic analysis using aligned complete mt genome sequences generated a tree that placed these
135
two Eimeria species that infect mustelids within a well-supported monophyletic group The sister taxon
for these ferret parasites was the only other Eimeria species from carnivores for which a complete mt
genome is available E mephitidis which infects hosts belonging to a different family of carnivores
Eimeriid parasites that infect closely-related definitive hosts are commonly found in a single or limited
number of clades based on mitochondrial and nuclear genetic loci (Ogedengbe et al in press)
Sequencing of the mt genomes and at least one nuclear genetic locus (ie nu 18S rDNA) from additional
Eimeria species infecting carnivores will be required to determine if all carnivore-specific Eimeria
species share a common ancestor
136
Table 61 PCR primer pairs and resulting fragments used for sequencing the mitochondrial genome sequence of an isolate of Eimeria furonis
originating from a fecal sample from a domestic ferret (Mustela putorius furo)
Fragment Primer names Primer sequences (5ʹ-3ʹ) Size (bp) Anneal Temp References
1 WG-MT_4140F AGAAAACCTAAAATCATCATGT 1000 52 Ogedengbe et al (2015)
Eim_CO3_799R AAGTGAGTTCGCATGTTTAC Ogedengbe et al (2015)
2 Eim_COI_19F ACTGCYGCAAACCATAARGAA 1700 60 Present study
Api_LSUG_UNI_R AGATAGGGAACAAACTGYCTCAA Present study
3 WG_MT_5416F GGTCCAGATAAGCGATCTCATG 3400 53 Ogedengbe et al (2013)
Eim_COI_1436R CACATTGTGTTCARATAAGTTA Present study
4 WG-MT_6219F GCATCCATCTACAGCTGCGG 500 55 Ogedengbe et al (2013)
WG-MT_344R GTAGGAATCTRAATTCCCAACC Ogedengbe et al (2013)
5 Api_LSUE UNI_F AGGTGCTCAGGGTCTTACCG 500 55 Present study
WG_MT_63R CTGGTATGGATGGATAACACT Ogedengbe et al (2015)
6 Lank_COB-30F CCAGGCCAACTGAACTCGTT 1300 55 Present study
q_Eim_COI_221R GGCATAACTACAAAGAARATCATA Present study
7 Cocci_MT_WG_F TACACCTAGCCAACACGAT 1600 55 Ogedengbe et al (2014)
q_Eim_COI_221R GGCATAACTACAAAGAARATCATA Present study
137
Table 62 PCR primer pairs and resulting fragments used for sequencing the mitochondrial genome sequence of an isolate of Eimeria ictidea
originating from a fecal sample from a black-footed ferret (Mustela nigripes)
Fragment Primer names Primer sequences (5ʹ-3ʹ) Size (bp) Anneal Temp References
1 WG_MT_63R CTGGTATGGATGGATAACACT 2180 52 Ogedengbe et al (2015)
WG-MT_4140F AGAAAACCTAAAATCATCATGT Ogedengbe et al (2015)
2 Cocci_MT_WG_F TACACCTAGCCAACACGAT 1551 58 Ogedengbe et al (2014)
q_Eim_COI_221R GGCATAACTACAAAGAARATCATA Present study
3 WG-MT_3658F CTGGCGAGAAGGGAAGTGTG 1329 55 Ogedengbe et al (2013)
Eim_CO3_799R AAGTGAGTTCGCATGTTTAC Ogedengbe et al (2015)
4 Lank_COB-30F CCAGGCCAACTGAACTCGTT 3899 57 Present study
WG_MT_4072R GGTTGTTTCCATCTCGACTC Ogedengbe et al (2013)
138
Table 63 Coding regions within the mitochondrial genome of the eimeriid parasite Eimeria furonis from a domestic ferret (Mustela putorius faro)
139
Table 64 Coding regions of the mitochondrial genome of the eimeriid parasite Eimeria ictidea originating from a black-footed ferret (Mustela
nigripes)
Table 64 Features associated with the protein-coding regions of the mitochondrial genome of the eimeriid parasite Eimeria cf ictidea originating from a black-footed ferret (Mustela nigripes )
Protein coding regions (CDS) Sequence size (bp) Start position (bp) Stop position (bp) Direction Translation start codon Translation stop codon
Cytochrome c oxidase subunit I (COI) 1443 1343 2785 Forward ATG TAA
Cytochrome c oxidase subunit III (COIII) 756 4333 5088 Forward TTA TAA
Cytochrome b (CytB) 1080 226 1305 Forward ATG TAA
Ribosomal DNA fragments (rDNA) Product
SSUrRNA 46 17 62 forward RNA9 SSU8
SSUrRNA 77 104 180 forward SSUA SSU4 (partial)
SSUrRNA 35 181 215 forward RNA23t
LSUrRNA 20 2790 2809 forward RNA20 (partial) LSU
LSUrRNA 112 2835 2946 forward LSUF LSU11
LSUrRNA 106 2947 3052 forward LSUG LSU12
LSUrRNA 25 3087 3063 reverse LSU
SSUrRNA 39 3129 3167 forward RNA14 SSU1
LSUrRNA 16 3188 3173 reverse LSUC LSU4
SSUrRNA 33 3211 3243 forward SSU
SSUrRNA 61 3386 3326 reverse SSUF SSU12
LSUrRNA 74 3387 3460 forward RNA10 LSU13 (partial)
LSUrRNA 49 3476 3524 forward RNA11 LSU5
SSUrRNA 65 3533 3597 forward SSUD SSU10
SSUrRNA 40 3631 3670 forward RNA17 SSU3
SSUrRNA 30 3677 3706 forward RNA15 SSU
LSUrRNA 30 3711 3740 forward RNA13 LSU10
LSUrRNA 56 3749 3804 forward RNA6 LSU15
LSUrRNA 79 3899 3821 reverse LSUD LSU8
LSUrRNA 24 3931 3908 reverse RNA16 (partial)
SSUrRNA 92 4036 3945 reverse RNA8 SSU5
LSUrRNA 66 4054 4119 forward RNA2 LSU2
LSUrRNA 177 4316 4140 reverse LSUA LSU1
SSUrRNA 30 5125 5096 reverse RNA19 SSU7
LSUrRNA 88 5154 5241 forward RNA1 LSU6
LSUrRNA 26 5342 5317 reverse LSUB LSU3
LSUrRNA 73 5447 5375 reverse RNA3 LSU7
LSUrRNA 20 5479 5460 reverse RNA18 LSU14
SSUrRNA 119 5615 5497 reverse SSUB SSU6
LSUrRNA 80 5699 5620 reverse RNA7
LSUrRNA 188 5897 5710 reverse LSUE LSU9
SSUrRNA 33 6033 6001 reverse SSUE SSU11 (partial)
SSUrRNA 99 6156 6058 reverse RNA5SSU9
140
Table 65 Pairwise comparison of coding DNA and concatenated rDNA fragment sequences between the
mitochondrial genomes of Eimeria furonis originating from a domestic ferret (Mustela putorius furo) and
Eimeria ictidea originating from a black-footed ferret (Mustela nigripes)
Total length
(nucleotides)
Nucleotide
identity
Total amino
acids
Amino acid
identity
COI CDS 1443 934 (95) 480 975 (12)
COIII CDS 756 899 (76) 251 932 (17)
CytB CDS 1080 950 (54) 359 986 (5)
rDNA fragments 2109 985 (32) na na
Legend Numbers in brackets indicate the number of single nucleotide differences na = not
applicable
141
Figure 61 Circular mapping and organization of the mitochondrial genome content of Eimeria
furonis showing three protein-coding genes (COI COIII and CytB) interspersed with large and
small subunit rRNA fragments
142
Figure 62 Circular mapping and organization of the mitochondrial genome content of Eimeria
ictidea showing three protein-coding genes (COI COIII and CytB) interspersed with large and
small subunit rRNA fragments
143
Figure 63 Pairwise comparison of coding DNA and concatenated rDNA fragment sequences of
the mitochondrial genomes of Eimeria furonis and Eimeria ictidea Legend Yellow bands
correspond to coding DNA fragments Red arrows correspond to rDNA fragments Arrows of both
colours indicate the forward or reverse direction of these segments
144
Figure 64 Phylogenetic relationships of coccidia (Eimeria furonis and Eimeria ictidea) from
domestic (Mustela putorius furo) and black-footed ferrets (Mustela nigripes) based on the complete
mitochondrial genome sequences these Eimeria species and a selection of related apicomplexan
parasites
145
CHAPTER 7 CONCLUSIONS AND FUTURE DIRECTIONS
The primary objective of this project was to better characterize the enteric coccidia of the
endangered black-footed ferret (BFF) in order to set the stage for improved disease prevention
management and treatment To the authorrsquos knowledge this work was the first attempt to isolate and
perform molecular characterization of the coccidial species endemic in the black-footed ferret population
and to characterize the natural history of the disease in this host
Two Eimeria species Eimeria ictidea and Eimeria furonis have been described from BFF and
domestic ferrets A single Eimeria species morphologically resembling E ictidea of the domestic ferret
and referred to in this work as E ictidea was identified from all historic and active cases of enteric
coccidiosis in the Toronto Zoo BFF population The same species was identified in both BFF family
groups and single-housed adults that shed coccidia during the study period (2014-2016) as well as from
historic necropsy samples of juvenile and adult Toronto Zoo BFF from 1999-2014 Furthermore this
same species was identified from both family groups and single-housed adults shedding coccidia at an
additional BFF Species Survival Plan institution the Louisville Zoo in 2016 These findings are contrary
to previous published reports that indicated multiple coccidia species were affecting captive and wild
BFF In order to determine whether these additional parasites continue to exist within the present-day
BFF population and their impact on this species further examination of coccidia-positive fecal samples
from captive and wild BFF populations is recommended Moreover determination of which parasite
species impact BFF morbidity and mortality would allow for development of targeted therapies for
disease management
Coccidia-positive fecal samples and necropsy samples from domestic ferrets were collected from
multiple diagnostic laboratories in Canada and Europe for comparison with results from BFF Eimeria
furonis and Isospora (=Cystoisospora) laidlawi were identified from fecal parasitology reports from
2008-2015 and in fecal samples obtained prospectively from 2014-2016 from domestic ferrets submitted
to Canadian and European laboratories E furonis was also identified in necropsy samples from 2010 and
146
2017 from two Canadian diagnostic laboratories No samples containing E ictidea were submitted to the
Canadian laboratories however samples containing E ictidea were identified twice by the European
diagnostic laboratory with one sample in 2011 and a second in 2013 Consequently it appears that E
ictidea is rarely identified from domestic ferrets Since no samples of E ictidea were acquired from
domestic ferrets during the study period it remains undetermined whether E ictidea from domestic
ferrets and E ictidea of BFF are conspecific Future molecular characterization of E ictidea isolated from
various mustelid host species would allow not only for determination of whether the parasites are
conspecific but would also provide insight into the potential for cross-transmission of parasites between
related mustelid hosts
The identifications provided by diagnostic laboratories of the specific parasite species present in
the domestic ferret fecal samples showed poor agreement with their identifications based on genotyping
obtained in this study In the authorrsquos opinion molecular techniques are essential tools for determining
the specific coccidial species responsible for individual and group outbreaks of coccidiosis and for
further understanding of the eimeriid host-parasite relationships To this end the nu 18S rDNA region and
whole mitochondrial (mt) genomes of E ictidea and E furonis were sequenced these mt whole genome
sequences are the first for the Eimeria species of mustelids to be entered into GenBank These sequences
may provide suitable targets for the development of highly discriminatory PCR-based methods of
identification that could be applied to fecal tissue or even formalin-fixed paraffin-embedded samples
(see Chapters 2 and 3) Methodological improvements such as less expensive next generation sequencing
methods that could be applied to diagnostics will depend on the availability of high quality reference
sequences such as generated herein Furthermore evaluation of the evolution of the eimeriid coccidia
using phylogenetic analysis based on additional whole mitochondrial genome sequences may allow for
more accurate determinations of relationships between parasite species and timing of their divergence
from common ancestors
One of the goals of this research was to investigate the possibility of using the domestic ferret as
an experimental model to study enteric coccidiosis in the BFF Experimental work cannot be carried out
147
in the BFF due to its endangered status We showed that the domestic ferret is susceptible to infection
with Eimeria ictidea originating from BFF Infected domestic ferrets showed similar clinical signs and
pathologic lesions to BFF strengthening the possibility of their serving as a suitable model Future
investigations would include studies of the domestic ferretrsquos immune response to enteric coccidial
infection in vivo drug trials including pharmacokinetic pharmacodynamic and efficacy studies of
anticoccidial medications investigations into patterns of drug resistance in coccidial species of ferrets
and oral vaccine development
Ultimately the goal of research on coccidiosis in BFF may be the development of an autogenous
vaccine to improve the survival of ferret kits and reduce coccidiosis-related morbidity and mortality in
BFF captive breeding programs It may be possible to use the domestic ferret to select for E ictidea that
are more highly adapted to this host perhaps with a commensurate loss of virulence to its original host If
this were to be the case domestic ferret-derived coccidia could be used in BFF as vaccine organisms
Alternately methods such as use of a bioshuttle (live vaccination with coccidia followed by anticoccidial
treatment to limit pathogenicity) might be useful for generating long-lasting immunological protection in
BFF against E ictidea Improving immunity through vaccination could assist in reducing coccidial
shedding and disease in BFF associated with stressful life events such a breeding weaning and transfer
between institutions There is minimal available data on the significance of coccidiosis in wild
populations of BFF and limited means of disease surveillance post release however it is logical to
assume that the stresses associated with release to the wild might result in clinical disease as happens with
stressed BFF in captivity Reducing morbidity and mortality associated with coccidiosis in BFF could
result in increased numbers of ferrets being released to the wild and higher survival of released ferrets
increasing the number of BFF in the wild supports the goals of the conservation initiative for the black-
footed ferret and will support the longer-term survival and recovery of this species
148
REFERENCES
Abe N Tanoue T Ohta G Iseki M (2008) First record of Eimeria furonis infection in a ferret Japan with
notes on the usefulness of partial small subunit ribosomal RNA gene sequencing analysis for
discriminating among Eimeria species Parasitol Res 103967ndash70 doi 101007s00436-008-1037-x
Adl SM Simpson AGB Farmer MA et al (2005) The new higher level classification of eukaryotes with
emphasis on the taxonomy of protists J Eukaryot Microbiol 52399ndash451 doi 101111j1550-
7408200500053x
Andrews JM (1926) Coccidiosis in mammals Am J Hyg 6784ndash798
Augustine PC Danforth HD (1986) A study of the dynamics of the invasion of immunized birds by
Eimeria sporozoites Avian Dis 30347ndash351
Barta JR Schrenzel MD Carreno R Rideout BA (2005) The genus Atoxoplasma (Garnham 1950) as a
junior objective synonym of the genus Isospora (Schneider 1881) species infecting birds and
resurrection of Cystoisospora (Frenkel 1977) as the correct genus for Isospora species infecting
mammals J Parasitol 91726ndash727 doi 101645GE-33411
Bell JA (1994) Parasites of Domesticated Pet Ferrets Compend Contin Educ Pract Vet 16617ndash620
Black-footed Ferret Recovery Implementation Team (2011) Captive Breeding
httpwwwblackfootedferretorgcaptive-breeding Accessed 8 Jan 2014
Blankenship-Paris TL Chang J Bagnell CR (1993) Enteric coccidiosis in a ferret Lab Anim Sci 43361ndash
363
Bronson E Bush M Viner T et al (2007) Mortality of captive black-footed ferrets (Mustela nigripes) at
Smithsonianrsquos National Zoological Park 1989 ndash 2004 J Zoo Wildl Med 38169ndash176
149
CAPC (2013) Current Advice on Parasite Control Intestinal Parasites - Coccidia
httpwwwcapcvetorgcapc-recommendationscoccidia Accessed 6 Jun 2015
Carpenter JW Hillman CN (1979) Husbandry reproduction and veterinary care of captive ferrets In
1978 Proceedings of the Annual Meeting of the American Association of Zoo Veterinarians
Washington DC pp 36ndash47
Catchpole J Norton CC Gregory MW (1993) Immunisation of lambs against coccidosis Vet Rec
13256ndash59
Cavalier-Smith T (2014) Gregarine site-heterogeneous 18S rDNA trees revision of gregarine higher
classification and the evolutionary diversification of Sporozoa Eur J Protistol 50472ndash495 doi
101016jejop201407002
Chappuis G (1998) Neonatal immunity and immunisation in early age lessons from veterinary medicine
Vaccine 161468ndash1472
Cox FE (1994) The evolutionary expansion of the Sporozoa Int J Parasitol 241301ndash1316
Davis CL Chow TL Gorham JR (1953) Hepatic coccidiosis in mink Vet Med 48371ndash375
De Vos AJ (1970) Studies on the host range of ltigtEimeria chinchillaeltIgt de Vos and Van Der
Westhuizen 1968 Onderstepoort J Vet Res 3729ndash36
Dirikolu L Yohn R Garrett EF et al (2009) Detection quantifications and pharmacokinetics of
toltrazuril sulfone (Ponazuril) in cattle J Vet Pharmacol Ther 32280ndash288
Dryden MW Payne PA Ridley R Smith V (2005) Comparison of common fecal flotation techniques for
the recovery of parasite eggs and oocysts Vet Ther 615ndash28
Duszynski D Wilber PG (1997) A Guideline for the Preparation of Species Descriptions in the
Eimeriidae J Parasitol 83333ndash336
150
Duszynski DW Couch L Upton SJ (2000) The Coccidia of the World
httpbiologyunmeducoccidiacarniv2html Accessed 1 Sep 2017
El-Sherry S Ogedengbe ME Hafeez MA Barta JR (2013) Divergent nuclear 18S rDNA paralogs in a
turkey coccidium Eimeria meleagrimitis complicate molecular systematics and identification Int J
Parasitol 43679ndash685 doi 101016jijpara201303005
Evans HE An NQ (2014) Anatomy of the ferret In Fox JG Marini RP (eds) Biology and Diseases of the
Ferret 3rd edn Wiley Blackwell pp 23ndash67
Fayer R (1980) Epidemiology of Protozoan Infections The Coccidia Vet Parasitol 675ndash103
Foreyt WJ Todd AC Hartsough GR (1977) Anticoccidial Activity of Eight Compounds in Domestic
Mink AJVR 38391ndash394
Fox JG (2014) Normal Clinical and Biological Parameters In Fox JG Marini RP (eds) Biology and
Diseases of the Ferret 3rd edn John Wiley amp Sons Inc pp 157ndash185
Frenkel JK (1977) Besnoitia wallacei of Cats and Rodents With a Reclassification of Other Cyst-
Forming Isosporoid Coccidia J Parasitol 63611ndash628
Grafner G Graubmann HD Dobbriner W (1967) Leberkokzidiose beim Nerz (Lutreola vison Schreb)
hervorgerufen durch eine neue Kokzidienart Eimeria hiepei n sp Monatshefte fur
Veterinearmedizin 22696ndash700
Haberkorn A (1971) Zur Wirtsspezifitat yon Eimeria contorta nsp (Sporozoa Eimeriidae) Z
Parasitenkd 37303ndash314
Hall MC Wigdor M (1918) Canine coccidiosis with a note regarding other protozoan parasites from the
dog J Am Vet Med Assn 5364ndash76
Hikosaka K Nakai Y Watanabe YI et al (2011) Concatenated mitochondrial DNA of the coccidian
151
parasite Eimeria tenella Mitochondrion 11273ndash278
Hillyer E V (1992) Gastrointestinal diseases of ferrets (Mustela putorius furo) J Small Anim Med 244ndash
45
Hoare CA (1927) On the coccidia of the ferret Ann Trop Med Parasitol 21313ndash320
Hoare CA (1935a) A histopathological reaction of a special type on the part of the intestinal villi in ferret
coccidiosis Trans R Soc Trop Med Hyg 292
Hoare CA (1935b) The endogenous development of the coccidia of the ferret and the histopathological
reaction of the infected intestinal villi Ann Trop Med Parasitol 29111ndash122
Hoefer HL Fox JG Bell JA (2012) Gastrointestinal Diseases In Quesenberry KE Carpenter JW (eds)
Ferrets Rabbits and Rodents Clinical Medicine and Surgery 3rd edn Elsevier Saunders St Louis
Missouri pp 27ndash45
Huelsenbeck JP Ronquist F (2001) MrBayes Bayesian inference of phylogenetic trees Bioinformatics
17754ndash755 doi 101093bioinformatics178754
Jolley WR Kingston N Williams ES Lynn C (1994) Coccidia Giardia sp and a Physalopteran
Nematode Parasite from Black-footed Ferrets (Mustela nigripes) in Wyoming J Helminthol Soc
Washingt 6189ndash94
Katoh K Misawa K Kuma K Miyata T (2002) MAFFT a novel method for rapid multiple sequence
alignment based on fast Fourier transform Nucleic Acids Res 303059ndash3066 doi
101093nargkf436
Kaye SW Ossiboff RJ Noonan B et al (2015) Biliary coccidiosis associated with immunosuppressive
treatment of pure red cell aplasia in an adult ferret (Mustela putoris furo) J Exot Pet Med doi
101053jjepm201504012
152
Koepfli KP Deere K Slater GJ et al (2008) Multigene phylogeny of the Mustelidae resolving
relationships tempo and biogeographic history of a mammalian adaptive radiation BMC Biol 610
Lariviegravere S Jennings AP (2009) Family Mustelidae (weasels and relatives) In Wilson DE Mittermeier
RA (eds) Handbook of the Mammals of the World Volume 1 Carnivores Lynx Edicions
Barcelona pp 564ndash656
Levine ND Ivens V (1970) The coccidian parasites (Protozoa Sporozoa) of ruminants
Li X Pang J Fox JG (1996) Coinfection with intracellular Desulfovibrio species and coccidia in ferrets
with proliferative bowel disease Lab Anim Sci 46569ndash571
Litster AL Nichols J Hall K et al (2014) Use of ponazuril paste to treat coccidiosis in shelter-housed
cats and dogs Vet Parasitol 202319ndash325 doi 101016jvetpar201403003
Matsubayashi M Takami K Abe N et al (2005) Molecular characterization of crane coccidia Eimeria
gruis and E reichenowi found in feces of migratory cranes Parasitol Res 9780ndash83
Meeusen ENT Walker J Peters A et al (2007) Current status of veterinary vaccines Clin Microbiol Rev
20489ndash510 doi 101128CMR00005-07
Mehlhorn H Aspock H (eds) (2008) Coccidial Drugs In Encyclopedia of Parasitology 3rd edn Springer
Berlin Heidelberg New York pp 269ndash286
Morehouse NF (1938) The Reaction of the Immune Intestinal Epithelium of the Rat to Reingection wiht
Eimeria nieschulzi J Parasitol 24311ndash317
Nukerbaeva KK Svanbaev SK (1973) Coccidia of fur bearing mammals in Kazakhstan Vestn Selrsquoskokh
Nauk Kazakh 1250ndash54
Ogedengbe JD Hanner RH Barta JR (2011) DNA barcoding identifies Eimeria species and contributes to
the phylogenetics of coccidian parasites (Eimeriorina Apicomplexa Alveolata) Int J Parasitol
153
41843ndash850
Ogedengbe ME (2015) DNA Barcoding of Apicomplexa Mitochondrial Evolution across the Phylum
University of Guelph
Ogedengbe ME El-Sherry S Ogedengbe JD et al Whatrsquos in a name Phylogenies based on combined
mitochondrial and nuclear sequences conflict with morphologically defined genera in the eimeriid
coccidia (Apicomplexa)
Ogedengbe ME El-Sherry S Whale J Barta JR (2014) Complete mitochondrial genome sequences from
five Eimeria species (Apicomplexa Coccidia Eimeriidae) infecting domestic turkeys Parasit
Vectors 7335 doi 1011861756-3305-7-335
Ogedengbe ME Hafeez MA Barta JR (2013) Sequencing the complete mitochondrial genome of Eimeria
mitis strain USDA 50 (Apicomplexa Eimeriidae) suggests conserved start positions for mtCOI- and
mtCOIII-coding regions Parasitol Res 1124129ndash4136 doi 101007s00436-013-3604-z
Pantchev N Gassmann D Globokar-Vrhovec M (2011) Increasing numbers of Giardia (but not
coccidian) infections in ferrets 2002 to 2010 Vet Rec 168519 doi 101136vrd2962
Patterson M Fox JG (2007) Parasites of Ferrets In Baker DG (ed) Flynnrsquos Parasites of Laboratory
Animals 2nd edn Wiley Blackwell pp 501ndash508
Patterson MM Fox JG Eberhard ML (2014) Parasitic Diseases In Fox JG Marini RP (eds) Biology and
Diseases of the Ferret 3rd edn Wiley Blackwell pp 553ndash572
Powers L V (2009) Bacterial and parasitic diseases of ferrets Vet Clin North Am - Exot Anim Pract
12531ndash561 doi 101016jcvex200906001
Prado ME Ryman JT Boileau MJ et al (2011) Pharmacokinetics of ponazuril after oral administration to
healthy llamas (Llama glama) Am J Vet Res 721386ndash9
154
Ruiz A Muntildeoz MC Molina JM et al (2013) Primary infection of goats with Eimeria ninakohlyakimovae
does not provide protective immunity against high challenge infections Small Rumin Res 113258ndash
266 doi 101016jsmallrumres201301006
Ryley J Meade R Hazelhurst J Robinson T (1976) Methods in coccidiosis research separation of
oocysts from faeces Parasitology 73311ndash326
Santymire R Branvold-Faber H Marinari PE (2014) Recovery of the Black-Footed Ferret In Fox JG
Marini RP (eds) Biology and Diseases of the Ferret 3rd edn Wiley Blackwell pp 219ndash231
Shi MQ Huther S Burkhardt E Zahner H (2000) Immunity in rats against Eimeria separata oocyst
excretion effects on endogenous stages and local tissue response after primary and challenge
infections Parasitol Res 86891ndash898
Sledge DG Bolin SR Lim A et al (2011) Outbreaks of severe enteric disease associated with Eimeria
furonis infection in ferrets (Mustela putorius furo) of 3 densely populated groups J Am Vet Med
Assoc 2391584ndash1588 doi 102460javma239121584
Smith NC Wallach M Petracca M et al (1994) Maternal transfer of antibodies induced by infection with
Eimeria maxima partially protects chickens against challenge with Eimeria tenella Parasitology
109551ndash557
Svanbaev SK (1956) Materials on the fauna of coccidia of wild mammals in western Kazakhstan Tr
Instituta Zool Akad Nauk Kazachskoi SSR 5180ndash191
Tavareacute S (1986) Some probabilistic and statistical problems in the analysis of DNA sequences Am Math
Soc Lect Math Life Sci 1757ndash86 doi citeulike-article-id4801403
Tenter AM Barta JR Beveridge I et al (2002) The conceptual basis for a new classification of the
coccidia Int J Parasitol 32595ndash616
155
Upton SJ (2000) Suborder Eimeriorina Leacuteger 1911 In Lee JJ Leedale GF Bradbury P (eds) An
Illustrated Guide to the Protozoa vol 1 2nd edn Allen Press Lawrence Kansas pp 318ndash339
USFWS BFF Recovery Program (2017) Black-footed Ferret Managed Care Operations Manual
(BFFMCOM)
Vermeulen AN (2005) Vaccination against coccidial parasites The method of choice In Proceeding of
the 9th International Coccidiosis Conference
Williams BH Chimes MJ Gardiner CH (1996) Biliary coccidiosis in a ferret (Mustela putorius furo) Vet
Pathol 33437ndash439 doi 101177030098589603300412
Williams ES Mills K Kwiatkowski DR et al (1994) Plague in a Black-footed (Mustela nigripes)
presence J Wildl Dis 30581ndash585
Williams ES Thome ET Appel MJG Belitsky DW (1988) Canine Distemper in Black-Footed (Mustela
nigripes) from Wyoming J Wildl Dis 24385ndash398
Wozencraft WC (2005) Order Carnivora In Wilson DE Reeder DM (eds) Mammal Species of the
World A Taxonomic and Geographic Reference 3rd edn Johnrsquos Hopkins University Press
Baltimore Maryland p 2142
Yi-Fan C Le Y Yin D et al (2012) Emendation of 2 Isospora Species (Apicomplexa Eimeriidae)
Infecting the Steppe Polecat Mustela eversmanii Lesson 1827 in China to the Genus
Cystoisospora (Apicomplexa Sarcocystidae) Comp Parasitol 79147ndash152 doi 10165445311
Yu L Peng D Liu J et al (2011) On the phylogeny of Mustelidae subfamilies analysis of seventeen
nuclear non-coding loci and mitochondrial complete genomes BMC Evol Biol 1192 doi
1011861471-2148-11-92
Zou M Guo G Zhao Y Zhang Q (2014) Detection quantifications and pharmacokinetics of ponazuril in
156
healthy swine J Vet Pharmacol Ther 37598ndash602 doi 101111jvp12126
157
APPENDICES
158
Appendix 1 Shedding of oocysts of Eimeria ictidea in black-footed ferret (Mustela nigripes) dam and kit
family groups from 2014-2016
Collection Year 2014 2014 2014 2014 2014 2015 2016
Age of kits (days) Poppy Bumblefoot Calico Aubrey Ruckus Fiddlesticks Guanella^
29 - - - 0 - - -
30 - 0 - 0 - - -
31 - 0 - 0 - - -
32 - 0 - 0 - - -
33 - 0 - 0 - - -
34 0 0 - 0 0 - -
35 0 0 - 0 0 0 -
36 0 0 - 0 0 0 -
37 - 0 - 0 - 0 -
38 - 0 - 0 - 0 -
39 - 0 - 0 - 0 -
40 0 0 - 0 0 0 -
41 - 0 - 0 - 0 -
42 0 0 - 0 0 - -
43 0 0 - 0 0 0 -
44 0 0 - - 0 0 -
45 0 0 - 0 0 - -
46 0 0 - 0 0 0 -
47 0 0 - 0 0 0 -
48 0 lt14 - 0 0 0 -
49 0 0 - 0 0 0 -
50 0 0 - - 0 0 -
51 0 0 - 0 0 0 -
52 0 0 - 0 0 0 -
53 0 0 - 0 0 0 +
54 0 0 - 0 0 0 +
55 0 0 - 0 0 0 3717146
56 0 0 - 0 - 0 1084436
57 0 0 - 0 - 0 64133
58 0 324 - 0 0 0 20654
59 0 0 - 0 0 0 0
60 0 446688 - 0 0 0 0
61 0 934828 - 0 0 0 41111
62 0 530469 - 0 0 0
63 0 1617131 - 0 0 lt14
64 0 8396357 - 114 0 0
65 0 234876 - 5368 0 lt14
66 0 374625 - 9455 0 0
67 lt14 2311575 - 39579 0 -
68 0 603563 - 125051 0 0
69 - 4103036 - 27067 0 lt14
70 2470327 7759974 - 0 0 0
71 1073085 - 1784 0 0
72 3430966 - 0 0 0
73 4459536 - 7326 0 0
74 4842212 - 2 0 0
75 3627307 - 0 0 0
76 5801885 - 0 0 0
77 5994 0 0 0 0
78 589835 0 0 0
Numbers of Oocysts Shed (oocysts per gram of feces)
Dam Identity
159
Appendix 1 continued Shedding of oocysts of Eimeria ictidea in black-footed ferret (Mustela nigripes)
dam and kit family groups from 2014-2016
Collection Year 2014 2014 2014 2014 2014 2015 2016
Age of kits (days) Poppy Bumblefoot Calico Aubrey Ruckus Fiddlesticks Guanella^
79 24815 0 0 0
80 144016 0 0 0
81 97862 0 0 0
82 0 0 0
83 0 0 0
84 0 0 0
85 0 0 0
86 0 0 0
87 0 0 0
88 0 0 0
89 0 0 0
90 0 0 0
91 0 0 0
92 0 - -
93 0 - 0
94 0 - 0
95 0 0 -
96 0 - 0
97 0 0 -
98 0 0 0
99 0 0 0
100 0 0 0
101 0 0 0
102 0 0 0
103 0 0 0
104 0 0 0
105 0 0 -
106 0 0 0
107 0 0 0
108 - 0 0
109 0 0 -
110 0 0 0
111 0 0 -
112 0 0 0
113 0 0 -
114 0 0 -
115 0 0 -
116 0 0 -
117 0 -
118 0 -
119 0 0
120 0 0
121 0 0
122 0 -
Numbers of Oocysts Shed (oocysts per gram of feces)
Dam Identity
160
Appendix 1 continued Shedding of oocysts of Eimeria ictidea in black-footed ferret (Mustela nigripes)
dam and kit family groups from 2014-2016
Collection Year 2014 2014 2014 2014 2014 2015 2016
Age of kits (days) Poppy Bumblefoot Calico Aubrey Ruckus Fiddlesticks Guanella^
123 0 0
124 0 -
125 0 -
126 0 0
127 0 0
128 0 2843
129 0 0
130 0 -
131 0 0
132 0 0
133 0 0
134 0 0
135 0 0
136 0 0
137 0
138 0
139 0
140 0
141 0
142 0
143 0
144 0
145 0
146 -
147 0
148 0
149 0
150 0
Legend lt 14 = oocyst positive samples with less than 14 oocyst per gram of feces - = no sample recorded for this date
underline = last sampling date + = coccidia present but OPG count not performed = Toronto Zoo ferret ^ = Louisville Zoo
ferret thick outer border = days treatment was received
Numbers of Oocysts Shed (oocysts per gram of feces)
Dam Identity
161
Appendix 2a Hematology values for domestic ferrets (Mustela putorius furo) from 49-51 days of age
prior to experimental inoculation
Test Reference
Intervala
Reference
Intervalb
101 102 103 104 105 201 202 203 204 205
Age (days) adult 70 50 50 50 50 50 48 48 48 50 48
WBC (x 109L) 27-112 53-120 86 64 77 97 76 48 8 72 104 76
RBC (x 1012
L) 50-108 55ndash74 66 58 45 5 48 47 67 5 5 42
Hb (gL) 87-177 104ndash136 121 106 90 58 94 91 122 96 98 80
HCT (LL) 04 - 051 029ndash037 037 033 027 030 029 027 038 030 030 024
MCV (fL) 44-52 478ndash548 55 56 60 60 60 58 57 59 61 58
MCH (pg) 15-18 175ndash191 18 18 20 12 20 20 18 19 20 18
MCHC (gL) 325-362 347ndash370 331 327 328 196 325 337 321 327 324 331
RDW () 12-16 - 134 127 139 139 133 131 122 136 131 127
Platelets (x 109L) 54-695 629ndash775 777 329 529 480 640 402 534 336 527 363
MPV (fL) 5-10 - 78 78 96 75 66 74 76 81 74 82
TS Protein (gL) 49-76 - 54 51 - - - - - - - -
Seg Neuts (x 109L) 1-8 15ndash48 292 141 216 281 205 187 152 151 354 251
Lymphocytes (x 109L) 1-63 28ndash63 525 416 516 64 509 254 608 468 645 456
Monocytes (x 109L) 0-09 01ndash05 026 07 031 039 038 034 024 094 031 038
Eosinophils (x 109L) 0-13 01ndash06 009 013 008 01 008 005 016 007 010 015
Basophils (x 109L) 0-02 0 009 0 0 0 0 0 0 0 0 0
Polychromasia 2-5 - 5-10 2-5 10-15 10-15 10-15 10-15 2-5 10-15 10-15 5-10
Anisocytosis Occ 1+ 1+ 1+ 1+ 1+ 1+ 1+
HJ bodies rare rare rare rare rare
crenation Occ
poikilocytosis Occ Occ Occ
shift platelets Occ Occ
hemolysis Neg Neg Neg Neg Neg Neg Neg Neg Neg Neg Neg
lipemia mild mild mild mild mild Neg Neg Neg mild mild
Ferret Identity
Legend - = reference value unavailable bolded = outside reference range a = adult ferret reference ranges from Animal Health Laboratory
(University of Guelph Guelph Canada) b
= reference ranges for 10-week old ferrets (Fox JG 2014) Neg = negative Occ = occasional
162
Appendix 2b Serum biochemistry values for domestic ferrets (Mustela putorius furo) from 49-51 days of
age prior to experimental inoculation
Test Reference
Intervala
Reference
Intervalb
101 102 103 104 105 201 202 203 204 205
Age (days) adult 70 50 50 50 50 50 48 48 48 50 48
Calcium (mmolL) 185-242 253-302 239 233 241 244 24 221 242 234 253 242
Phosphorus (mmolL) 112-256 278-475 239 267 29 264 31 218 286 284 315 303
Magnesium (mmolL) 08-139 - 08 08 07 08 08 06 08 07 08 08
Sodium (mmolL) 147-159 146-154 149 149 149 149 148 144 153 148 152 152
Potassium (mmolL) 37-57 47-83 44 46 42 46 48 45 47 44 47 46
Chloride (mmolL) 111-129 115-121 110 112 115 113 113 110 119 117 117 120
Carbon dioxide (mmolL) 17-29 13-27 18 18 18 17 17 16 15 15 19 17
Anion gap (mmolL) 6 - 23 - 25 24 20 24 23 23 24 20 21 20
NaK ratio - - 34 32 35 32 31 32 33 34 32 33
Total protein (gL) 51-75 44-56 49 46 44 47 44 41 52 45 49 43
Albumin (gL) 24-40 26-32 29 26 28 28 28 25 28 27 29 25
Globulin (gL) 19-41 17-24 20 20 16 19 16 16 24 18 20 18
AG ratio 053-167 13ndash12 145 130 175 147 175 156 117 15 145 139
Blood urea nitrogen (mmolL) 45-153 71-139 83 89 107 11 136 73 119 105 118 135
Creatinine (umolL) 8-67 53-124 50 58 68 59 50 7 68 41 62 92
Glucose (mmolL) 32-91 64-138 47 54 54 42 55 53 57 56 52 59
Cholesterol (mmolL) 294-894 619-860 413 386 375 393 344 349 227 357 373 346
Total bilirubin (umolL) 2 to 7 - 1 L 0 L 0 0 0 1 0 0 0 0
Conjugated biilrubin (umolL) - 0-10 1 0 0 1 1 1 0 0 1 0
Free bilirubin (umolL) 0-2 0-15 0 0 0 0 0 0 0 0 0 0
ALKP (UL) 13-237 117ndash277 180 169 172 215 175 168 241 184 177 179
GGT (UL) 0-40 2ndash20 1 1 6 10 4 0 1 1 5 9
AST (UL) - 63ndash152 61 58 48 61 64 59 93 61 58 69
ALT (UL) 39-196 95ndash544 95 105 89 105 106 82 234 137 115 156
CK (UL) 74-294 - 513 330 496 560 530 492 793 539 479 724
Amylase (UL) - - 23 28 35 35 29 28 29 24 36 23
Lipase (UL) - - 65 63 60 64 62 56 67 69 60 68
Calculated osmo (mmolL) - - 298 300 301 301 303 289 311 300 308 311
Ferret Identity
Legend - = reference value unavailable bolded = outside reference range a = adult ferret reference ranges from Animal Health Laboratory
(University of Guelph Guelph Canada) b = reference ranges for 10-week old ferrets (Fox JG 2014)
163
Appendix 3a Hematology values for domestic ferrets (Mustela putorius furo) inoculated orally with
Eimeria ictidea
Test Reference
Intervala
Reference
Intervalb
102 103 104 105 201 203 205
Age (days) adult 98-112 92 91 99 99 97 74 92
Days post inocculation 8 7 15 15 15 11 10
WBC (x 109L) 27-112 52ndash150 94 124 81 85 88 85 142
RBC (x 1012
L) 50-108 62ndash92 71 62 73 60 65 63 65
Hb (gL) 87-177 127ndash159 122 110 122 98 103 114 102
HCT (LL) 04-051 030ndash043 037 033 037 031 032 035 032
MCV (fL) 44-52 50ndash54 53 53 52 51 50 55 49
MCH (pg) 15-18 16-21 17 18 17 16 16 18 16
MCHC (gL) 325-362 351ndash426 328 332 325 319 320 326 317
RDW () 12-16 - 122 126 129 129 145 127 145
Platelets (x 109L) 54-695 376ndash610 524 413 445 117 303 42 429
MPV (fL) 5-10 - 8 78 72 66 67 14 8
TS Protein (gL) 49-76 - 66 63 58 66 55 59 59
Seg Neuts (x 109L) 1-8 21ndash62 160 446 203 170 211 170 554
Lymphocytes (x 109L) 1-63 16ndash79 761 657 551 646 59 646 682
Monocytes (x 109L) 0-09 01ndash02 019 087 024 026 044 026 085
Eosinophils (x 109L) 0-13 03ndash09 0 037 032 009 026 009 099
Basophils (x 109L) 0-02 0 0 012 0 0 009 0 0
Polychromasia 2-5 - 1-3 0-2 0-2 1-3 1-3 2-5 5-10
Anisocytosis Occ Occ Occ
HJ bodies rare
crenation
rouleaux
poikilocytosis
shift platelets Occ Occ
hemolysis Neg
Ferret Identity
Legend - = reference value unavailable bolded = outside reference range a = adult ferret reference ranges from Animal Health
Laboratory (University of Guelph Guelph Canada) b
= reference ranges for 10-week old ferrets (Fox JG 2014) Neg = negative
Occ = occasional = many platelet clumps
164
Appendix 3b Serum biochemistry values for domestic ferrets (Mustela putorius furo) inoculated orally
with Eimeria ictidea
Test Reference
Intervala
Reference
Intervalb
102 103 104 105 201 203 205
Age (days) adult 98-112 92 91 99 99 97 74 92
Days post inocculation 8 7 15 15 15 11 10
Calcium (mmolL) 185-242 245-268 243 222 238 233 232 236 240
Phosphorus (mmolL) 112-256 200-323 292 223 248 298 268 288 306
Magnesium (mmolL) 08-139 - 06 08 08 08 09 08
Sodium (mmolL) 147-159 148-155 147 150 150 148 148 150
Potassium (mmolL) 37-57 45-55 42 41 51 41 50 44
Chloride (mmolL) 111-129 114-124 114 113 118 112 111 115
Carbon dioxide (mmolL) 17-29 16-24 16 21 19 20 12 18
Anion gap (mmolL) 6 - 23 - 21 20 18 20 30 21
NaK ratio - - 35 37 29 36 30 34
Total protein (gL) 51-75 49-64 52 55 55 49 56 57
Albumin (gL) 24-40 30-36 28 20 27 27 26 29 27
Globulin (gL) 19-41 19-30 32 28 28 23 27 30
AG ratio 053-167 11-17 063 096 096 113 107 090
Blood urea nitrogen (mmolL) 45-153 50-150 92 87 119 92 110 98 149
Creatinine (umolL) 8-67 706-1414 56 26 49 40 50 46 79
Glucose (mmolL) 32-91 688-943 52 59 64 71 58 17 66
Cholesterol (mmolL) 294-894 440-640 453 429 262 475 370 285
Total bilirubin (umolL) 2 to 7 - 0 0 0 0 1 1
Conjugated biilrubin (umolL)- - 1 0 0 0 0 0
Free bilirubin (umolL) 0-2 - 0 0 0 0 1 1
ALKP (UL) 13-237 41-181 124 120 213 120 146 170 196
GGT (UL) 0-40 1-2 2 1 3 0 0 2
AST (UL) - 47-128 48 95 104 66 100 100
ALT (UL) 39-196 78-279 133 110 140 203 158 183 281
CK (UL) 74-294 - 382 765 1190 578 680 930
Amylase (UL) - - 24 37 31 28 33 35
Lipase (UL) - - 60 65 72 72 86 79
Calculated osmo (mmolL) - - 296 305 305 300 296 309
Ferret Identity
Legend - = reference value unavailable bolded = outside reference range a = adult ferret reference ranges from Animal Health
Laboratory (University of Guelph Guelph Canada) b
= reference ranges for 10-week old ferrets (Fox JG 2014)
165
Appendix 4 Domestic ferret (Mustela putorius furo) weekly monitoring sheet
Mon
8 AM
Mon
4 PM
Tues
8 AM
Tues
4 PM
Wed
8 AM
Wed
4 PM
Thurs
8 AM
Thurs
8 PM
Fri
8 AM
Fri
4 PM
Sat
8 AM
Sat
4 PM
Sun
8AM
Sun
4 PM
Mentation
Weight (g)
Respiratory
Rate
Vomit
(+ ++ +++)
Diarrhea
(+ ++ +++)
Urination
(+ ++ +++)
Defecation
(+ ++ +++)
Food
offered
Food
remaining
Water remaining
(ml)
Treatments
Other
observations
Initials of
observer
166
Animal ID ________________________________________ Week ______________
Monitoring Criteria
Mentation BAR (bright alert responsive) QAR (quiet alert responsive) depressed lethargic quiet
Weight measured in grams to be performed once weekly (pre-inoculation) and once daily (post-inoculation)
Respiratory rate measured in breaths per minute
Vomit + (small amount) ++ (moderate amount) +++ (large amount)
Diarrhea + (small amount) ++ (moderate amount) +++ (large amount)
Urination + (small amount) ++ (normal amount) +++ (large amount)
Defecation + (one pile) ++ (two piles) +++ (3+ piles)
Food offered Y (yes) N (no)
Food remaining A (all) P (partial) N (none)
Please note any additional observations in the appropriate section
Monitoring Times
Pre-inoculation ndash ONCE daily at 8am during cage cleaningfeeding
Post-inoculation ndash TWICE daily at 8am and 4pm
Critical patients (as determined by veterinary examination) ndash 3-6 times daily (based on veterinarian recommendation) ndash switch to 24
hour care sheet
Intervention Points
If any ferret develops vomiting diarrhea goes off food or exhibits gt 3 weight loss both the PI and graduate student listed below are
to be notified and the animal will be assessed to determine further treatment plans
Removal Criteria
If an individual ferret develops severe gastrointestinal or systemic disease during the course of the study as characterized by abnormal
mentation marked dehydration 10 weight loss within 24-48 hours profuse diarrhea unresponsive to treatment anorexia of 2 or
more days or frequent vomiting unresponsive to treatment the individual will be removed from the study and humanely killed
167
Appendix 5 Domestic ferret (Mustela putorius furo) 24 hour intensive monitoring sheet
Animal ID ___________________________________________ Date___________________
0700 800 900 1000 1100 1200 1300 1400 1500 1600 1700 1800
Mentation
Weight (g)
Heart Rate
Respiratory Rate
Temperature (C)
Water offered
(mL)
Water consumed
(mL)
Food offered
(type and
amount in
grams)
Food consumed
(type and
amount in
grams)
Presence of urine
Presence of feces
168
Character of
feces
Presence of
vomit
Character of
vomit
Medications to
be administered
Other
observations
169
1900 2000 2100 2200 2300 2400 100 200 300 400 500 600
Mentation
Weight (g)
Heart Rate
Respiratory Rate
Temperature (C)
Water offered
(mL)
Water consumed
(mL)
Food offered
(type and amount
in grams)
Food consumed
(type and amount
in grams)
Presence of urine
Presence of feces
Character of
feces
Presence of vomit
Character of
vomit
170
Medications to be
administered
Other
observations
Monitoring Criteria
Mentation BAR (bright alert responsive) QAR (quiet alert responsive) depressed lethargic quiet
Weight measured in grams to be performed once weekly (pre-inocculation) and once daily (post-inocculation)
Respiratory rate measured in breaths per minute
Vomit + (small amount) ++ (moderate amount) +++ (large amount)
Diarrhea + (small amount) ++ (moderate amount) +++ (large amount)
Urination + (small amount) ++ (normal amount) +++ (large amount)
Defecation + (one pile) ++ (two piles) +++ (3+ piles)
171
Food offered Y (yes) N (no)
Food remaining A (all) P (partial) N (none)
Please note any additional observations in the appropriate section
Monitoring Times
Critical patients (as determined by veterinary examination) ndash 3-6 times daily (based on veterinarian recommendation) ndash switch to 24
hour care sheet
Intervention Points
If any ferret develops vomiting diarrhea goes off food or exhibits gt 3 weight loss both the PI and graduate student listed below are
to be notified and the animal will be assessed to determine further treatment plans
Removal Criteria
If an individual ferret develops severe gastrointestinal or systemic disease during the course of the study as characterized by abnormal
mentation marked dehydration 10 weight loss within 24-48 hours profuse diarrhea unresponsive to treatment anorexia of 2 or
more days or frequent vomiting unresponsive to treatment the individual will be removed from the study and humanely killed
172
Appendix 6 Domestic ferret (Mustela putorius furo) infection trial standard operating procedures
Daily Fecal Collection
1 Collection of all feces passed in a 24 hour period will be performed once daily for all ferrets from
July 13 to July 24 inclusive
2 CAF Isolation staff will collect all fecal material present at the time of daily cage cleaning and
place in individual pre-labelled plastic bags (one per cage)
3 The amount of non-fecal matter (bedding etc) collected should be as minimal as possible
4 CAF isolation staff will record fecal character observations daily for each sample by ticking the
appropriate box on the baggie label
5 Isolation staff will place samples in the necropsy cooler (4 degC) while awaiting collection by
summer student
6 Monday to Friday fecal samples will be collected by A Rodriguez and brought for processing to
the Barta Lab in Pathobiology Samples collected Saturday and Sunday will be put on hold in a
refrigerator (4degC) until pick up on Monday
Example Bag label
Ferret ID Date
Weight of Feces
Fecal Character Normal
Soft
Liquid
Bloody
Abnormal odour
Physical Examination and Health Assessments
1 Manual restraint by hand or in towel by CAF Isolation staff and project personnel If required
examination +- blood collection may be performed under general anesthesia with isoflurane
2 Physical examination body weight HR RR temperature to be performed by PIs (Adriana Pastor
and Dale Smith)
3 Blood collection (~1mL per ferret) is to be performed from the jugular vein if under manual
restraint or the cranial vena cava under anesthesia by PIs using a 25g needle and 1cc syringe
Blood will be collected into small heparinized tubes and submitted to AHL for
CBCBiochemistry
4 All physical examination findings will be recorded on the Exam Sheet
5 Any minor wounds will be treated as appropriate (to be determined by PIs)
Inoculation of Ferrets with Coccidia
1 All ferrets to be inoculated will have been confirmed negative on daily fecals for two weeks
173
2 Brief physical examination by PIs to confirm that the ferrets are healthy to continue in study
3 The concentrated oocyst solution in sterile saline (up to a volume of 1mL) will be combined with
up to 1 mL of FerreTone (or another highly palatable substance if more preferred by the ferrets)
and ferrets will be allowed to consume the mixture ad lib while being monitored
4 Control ferrets will be administered saline only (equal mL to oocyst solution) with 1 mL of
FerreTone (or other substance as used for experimental group)
5 If ferrets refuse to consume the mixture oral inoculation via syringe of concentrated oocyst
solution will be performed by PIs under manual restraint
6 If ferrets are resistant to manual restraint for oral inoculation then inoculation will be performed
by PIs under general anesthesia via gastric tube (8 Fr red rubber)
Euthanasia Protocol
1 Ferrets to be euthanized will be masked down with isoflurane under manual restraint (or in an
anesthetic chamber as judged most appropriate by the PI)
2 Once anesthetized the ferret will be weighed and any blood fecal or other samples will be
collected as required (as determined by PIs)
3 Once an adequate plane of anesthesia is obtained (as determined by PIs) potassium chloride at a
dose of 2 mEq K+kg will be administered either via vena caval puncture or cardiac puncture to
induce cardiac arrest
4 Presenceabsence of respirations heartbeat corneal reflex will be used to assess death
Necropsy Protocol
1 Measure the ferret from nose to tail base (body length)
2 Perform standard necropsy but start with gastrointestinal tract first then thoracic and abdominal
viscera
3 Gently flush the entire contents of the intestine with 12 mL saline into a sterile urine cup
Potassium dichromate should be added in a 11 volumevolume ratio to the same container and
mixed with the combined intestinal contents and saline Label the container with the ferretrsquos ID
ferret group ID date and place on appropriate shelf in the refrigerator (4 degC) in the Barta lab
4 Measure the length of the gut from duodenum to anus
5 Collect paired sections of intestine for histological sectioning and frozen These sections should
be collected along the entire length of the gut from duodenum to rectum (see below for GI length
calculations)
6 For histological sections cut a 2 cm long section of bowel open completely on one side place on
a pre-cut section of box-board and staple both ends to the board to create a flat section Place all
gastrointestinal sections (attached to the board) in Serra fixative solution (100 ethanol (60
VV) 37 formaldehyde (30 VV) glacial acetic acid (10 VV)) for fixation and trim in
cross (transverse) sections
7 Adjacent to each sample removed for histopathology remove another 2 cm section intact and
place in a labelled Whirl-pack for freezing
8 Box-board and pre-labelled Whirl-packs should be labelled in pencil and sharpie respectively
with the following information Animal ID Zoo pm number section of gut (information should
include the region of the gut - jejunum colon etc and the length from the pylorus to the section)
174
Ferret gastrointestinal length calculations (from Evans amp An 2014)
Adult ferret body length 36-41 cm
NB In domestic ferrets there is ~51 ratio of small intestine to body length
Adult domestic ferret GI lengths
Small intestine ndash 182-198 cm
Large intestine - ~10 cm long 06 cm diameter (colon ndash 7 cm rectum ndash 2 cm anus ndash 1 cm)
Based on the above information
Small intestine Six sections of the small intestine from duodenum to ileum will be collected each ~ 25-40
cm apart depending on the size of the ferret
Large intestine Collect two sections of colon (4 cm each) at 25 and 75 of the length of the colon from
the junction of the small and large intestine to the rectum Smaller ferrets may allow only on section
based on colon length
Laboratory SOPs
General notes on processing fecal samples
1 Samples will be collected from the isolation facility necropsy room cooler daily from Monday-
Friday for the entire 6 weeks of the project
2 Upon transfer to the lab each baggie will be weighed after filling to determine the amount of
feces (grams) in each bag and that number entered onto the label on the bag
3 All information regarding fecal processing will be entered onto the spreadsheet for the individual
ferret including ID weight of feces feces character date of sample collection date of sample
processing fecal flotation method presence or absence of oocysts oocyst quantification
Fecal Processing - Weeks 1+2 (acclimation period)
1 After weighing transfer the contents of one baggie to a small sieve on top of a paper cup
2 Fill a single glass container (for fecal flotation) with saturated salt solution
3 Slowly pour small amounts of the solution over the feces mix and strain liquid contents in the paper
cup using a tongue depressor
4 Dispose of the remaining fecal matter in the sieve
5 Pour the contents of the paper cup back into the glass container and cover slip for 5-7 minutes
6 Place coverslip on a clean glass slide and viewscan under microscope at 10x power for presence of
oocysts
7 Record all findings on the provided spreadsheet
8 Notify A Pastor if any oocysts are detected If detected contents of the slide should be flushed back
into the glass container with distilled water and contents of glass container should be transferred to a
50 mL plastic conical tube Clean to remove salt as per Barta lab SOP and mix with potassium
dichromate equal parts by volume to amount of oocysts in diH2O Place a checkmark on the lid of the
50 mL conical vial containing the remainder of the feces and store in the lab fridge at 4 degC
175
Fecal Processing - Weeks 3-7 (infection trial)
1 After weighing transfer the entire contents of one baggie to a small sieve on top of a paper cup
2 Slowly pour small amounts of distilled water over the feces (enough to wet) mix and strain liquid
contents in the paper cup using a tongue depressor until feces appear almost dry
3 Dispose of the remaining fecal matter in the sieve
4 Pour the contents of the paper cup into a 50 mL conical tube If samples are not going to be counted
on the same day then mix potassium dichromate (25 wv) 11 by volume with sieved fluid from
the paper cup and store at 4 degC (refrigerator) until ready to perform OPG counts
If proceeding with the count the same day
5 Pipette and place one drop of the sieved fecal fluid on a slide to determine approximate oocyst
concentration If oocysts rare to none ndash dilute 12 during step 6 if moderate numbers ndash dilute 19 if
too numerous to count ndash dilute 199 (or perform serial dilutions of 10x from initial 19 dilution)
6 Transfer x mL of the mixture from the 50 mL conical tube into a clean 15 mL conical tube and mix
with appropriate amount of saturated salt solution for desired dilution
7 Fill both sides of the McMaster counter chamber and count the number of oocysts per side (total for
one side= total number of oocysts from all 6 sections of the chamber )
8 Average the total counts from both sides
9 Use the following calculation to determine the oocyst per gram count
OocystsmL = oocysts counted times 666 times dilution (ie 3 if dilution 12)
OPG= oocystmL times total volume recorded at end of step 4
10 Record all findings on the provided spreadsheet
11 Notify A Pastor if any oocysts are detected If detected place a checkmark on the lid of the 50 mL
conical vial containing the remainder of the feces and store in the lab fridge at 4 degC
12 For all samples for which no oocysts are detected during steps 6-8 follow up with routine salt
flotation of the remainder of the fecal sample from step 6 (use instructions for fecal processing from
weeks 1-2)

ABSTRACT INVESTIGATING ENTERIC COCCIDIOSIS IN THE BLACK-FOOTED (MUSTELA NIGRIPES) AND
DOMESTIC FERRET (MUSTELA PUTORIUS FURO)
Adriana R Pastor Advisors
University of Guelph 2017 Dr D A Smith
Dr J R Barta
Enteric coccidiosis is a major cause of death in both juvenile and adult black-footed ferrets (BFF
Mustela nigripes) in captive breeding programs that reduces the availability of animals for release to their
former North American range Coccidiosis is poorly understood in BFF but in vivo experimental infection
in this endangered host is untenable The goal of this research was to better characterize the etiologic
agents and natural history of enteric coccidiosis in BFF and to evaluate the domestic ferret (DF Mustela
putorius furo) as a model for experimental infection
Morphometric and molecular characterization of coccidia from BFF and DF was undertaken
Only Eimeria ictidea was identified in juvenile and adult BFF from 1999-2016 at the Toronto Zoo and
from BFF at the Louisville Zoo in 2016 Eimeria furonis and Isospora (=Cystoisospora) laidlawi were
identified in DF fecal and necropsy samples from Canadian and European diagnostic laboratories during
2008-2017 Molecular characterization of these parasites included generation of complete mitochondrial
genomes and nuclear 18S rDNA sequences for Eimeria ictidea and Eimeria furonis from BFF and DF
respectively Partial sequences were obtained from the same genetic targets from I (=C) laidlawi from
DF DNA isolation from formalin fixed paraffin embedded tissues and PCR amplicon sequencing
permitted identification of coccidia in BFF and DF tissues dating from 1999 to present
Retrospective and prospective analyses of medical and pathology records supplemented with
parasitological evaluation of repeated fecal samples was performed to determine the natural history of
coccidiosis in captive BFF Clinical signs and histopathologic changes associated with infection in BFF
were as described previously in the published literature Average yearly coccidia associated mortality
rates were 053 in adults and 195 in juveniles
Domestic ferrets were confirmed as experimental hosts of E ictidea isolated from BFF Seven of
10 juvenile DF inoculated with oocysts from a BFF developed patent infections and mild clinical disease
was observed in six of these seven Infection was confirmed via morphometric molecular and histologic
examination of samples from infected DF While much is still unknown about enteric coccidiosis in BFF
domestic ferrets provide a promising model for further investigation of this disease
iv
DEDICATION
For my mother Anna Pastorhellip
v
ACKNOWLEDGEMENTS
Itrsquos hard to believe that my residency and thesis have been completed and I have a lot of people
to thank for that
Dale and Graham the two people I wanted to be when I grew up thank you for your mentorship
for many years even before this program I know that you werenrsquot convinced that this project was DVSc
worthy when I first proposed it but Irsquom hoping that the results have changed your mind
I am sincerely grateful to all the members of my advisory committee Dale Smith John Barta and
Simon Hollamby for their insight support and interest in this project Dale you have been an exceptional
advisor I donrsquot know that I will ever get to your level but thank you for showing me that being a great
clinical zoo vet and pathologist are not mutually exclusive John thank you for spontaneously agreeing to
be my advisor when I came to you with this project proposal in my first semester for your energy and
enthusiasm and for supporting my widening interest in parasitology research
My heartfelt appreciation for the Toronto Zoo WHC veterinarians past and present Chris
Dutton Pauline Delnatte Simon Hollamby and Graham Crawshaw I have learned so much from all of
you that I will take forward into my future endeavours I appreciate the extra time you put in including
comps study sessions after-hours tecircte-agrave-tecirctes and the fact that your doors were open when I needed it
For the Toronto Zoo vet techs extraordinaire Michelle Lovering Cassia Devison Dawn
Mihailovic and Tasha Long ndash you have been indispensable during this program and there are not enough
words to express my gratitude
I would especially like to thank all the Wildlife Health Center staff (2013-2016) Mark Bongelli
Charles Guthrie Christine McKenzie Brian Telford Rick Vos Gerri Mintha Margaret Kolakowski
Andrew Lentini Rebecca Clark Lydia Attard Nigel Parr Paula Roberts Andrea Dada Mindy Waisglass
and Julie Digiandomenico for three very memorable years It is all of you that make the WHC such an
amazing place to be Irsquom not sure I have laughed so hard or so often as I did in that lunchroom and I hope
our paths will cross again
vi
I donrsquot think that I can truly express how thankful I am to Pathobiology laboratory technicians
Julie Cobean and Julia Whale Without your assistance patient teaching and friendship I would probably
still be screening fecal samples years from now and scratching my head as to how our lab protocols
actually work It is people like you who make sure graduate students become successful doctorates and I
canrsquot imagine Pathobio without you both in it
I would also like to thank my labmates in the Barta lab mdash Mian Hafeez Evelyn Rejman Rachel
Imai Perryn Kruth Ryan Snyder and Mosun Ogedengbe A special thank you goes to Alex Leveille
without whom my many adventures in parasitology research from coccidia to Babesia would not have
been as successful
To all the students who helped with ferret fecal sample processing data compilation and
necropsies Nathalie Ferriman Janessa Price Thisuri Eagalle Sarah Brisson thank you so much for your
hard work and excitement about my projecthellip even when it was very smelly
So many thanks to the amazing staff of Central Animal Facility - Linda Groocock Vicky Carson
Tony Cengija and Mary Fowler for the daily care and enrichment of my experimental ferrets Your
excitement about working with our ferrets and your assistance with all parts of the process helped made
this project a success
To Adriana Nielsen who was not only my better half but the other fifty percent of my brain for
several years It is your friendship fortitude and our endless phone conversations that got me through the
never-ending Toronto-Guelph commute and this program
To all the ldquoscope roomrdquo pathology co-residents past and present - thank you for being wonderful
friends and colleagues It is indeed rare to find so many amazing people in one place and I know this
program and my sanity would not have been the same without you
To the anatomic pathology faculty and senior graduate students - thank you for all the time
teaching and guidance you provided during my program While I canrsquot say that I have become an amazing
pathologist I can say that because of your mentorship I am a better diagnostician and the type of clinician
who asks better questions takes better samples and understands that you canrsquot ldquojust make a PCR for thatrdquo
vii
A special thank you to Tony van Dreumel who came out of retirement for a semester to try to teach the
Adrianas zoo pathology screening cases with you was always a pleasure
To all the lovely Histo Ladies PM room staff and the other AHL staff who helped me with
Toronto Zoo and HSC pathology cases along the way - I donrsquot think the anatomic path students could
survive without you Thank you for always smiling assisting and accommodating me even when I made
near-impossible processing requests during my weekly Guelph visits
I would also like to acknowledge and sincerely thank all the individuals who helped with resource
and sample acquisition for this project A special mention for those who went above and beyond because
of their interest in this project Don Duszynski who was instrumental in acquiring and then providing a
translator for many of the original mustelid Eimeria descriptions and Majda Globokar Nikola Pantchev
and Donald Martin who supplied my domestic ferret fecal samples and historical data
A shout-out to Julie Swenson Gary West and the Phoenix Zoo BFF team who fostered my love
of this endangered species and helped develop the idea for this project
As always I continue to go out into the world and pursue my dreams with the knowledge that I
have the support of my incredible family long-time friends and my partner Keith Morris I am so lucky
that my residency brought me home and that it afforded us all more time spent together For my aunt
Veronica Lacey who has never failed to believe in my potential and always pushed me to become an
academic ndash yoursquoll never get that PhD from me but I think this is pretty close Finally for my mother
Anna Pastor who never lived to see my greatest achievements but had absolute faith that I could reach
any goal I worked towardshellip this is for you
Finally none of this would have been possible without the generous support of the Toronto Zoo
Residency program and funding through the Barta Laboratory University of Guelph
Adriana Pastor
Toronto August 2017
viii
DECLARATION OF WORK PERFORMED
I declare that all the work reported in this thesis was performed by myself with the following
exceptions
Fecal samples were collected by personnel at the Toronto Zoo Louisville Zoo and participating
diagnostic laboratories
Fecal oocyst per gram counts (routine salt flotation and McMaster counts) were performed by
myself Julie Cobean Julia Whale Evelin Rejman Sarah Brisson Adriana Rodriguez and Perryn Kruth
Whole mitochondrial genome PCR and sequencing was performed by me in conjunction with
Julia Whale and Dr Mian Hafeez
Sequencing of PCR samples was performed at the University of Guelph Laboratory Services
(Guelph Ontario Canada) and results were obtained electronically
ix
TABLE OF CONTENTS
ABSTRACT ii
DEDICATION iv
ACKNOWLEDGEMENTS v
DECLARATION OF WORK PERFORMED viii
TABLE OF CONTENTS ix
LIST OF TABLES xiii
LIST OF FIGURES xiv
LIST OF APPENDICES xv
ABBREVIATIONS xvi
CHAPTER 1 LITERATURE REVIEW 1
11 INTRODUCTION 1
12 APICOMPLEXA 1
121 Brief introduction to apicomplexan pathogens 1
122 Life cycles of the Eimeria and Isospora species implicated in enteric coccidiosis 3
123 Methods of characterization 5
13 RECLASSIFICATION OF MAMMALIAN ISOSPORA 8
14 EIMERIID SPECIES CHARACTERIZED IN MUSTELIDS 8
141 The family Mustelidae 8
142 Eimeriid coccidia described from mustelids 9
143 Eimeriid coccidia described from domestic ferrets 16
144 Molecular characterization 19
145 Clinical signs of disease in domestic ferrets 21
146 Gross necropsy and histologic findings 21
15 INTRODUCTION TO ENTERIC COCCIDIOSIS IN THE BLACK-FOOTED FERRET
25
151 Natural history and conservation of the black-footed ferret in North America 25
152 Coccidia identified from black-footed ferrets 26
153 Morbidity mortality and clinical signs associated with enteric coccidiosis in black-footed
ferrets 28
16 TREATMENT PREVENTION AND CONTROL OF INFECTION BY EIMERIA SPP 29
161 Current recommendations for treatment of eimeriid coccidia in carnivores 29
x
162 Current recommendations for anticoccidial treatment and prophylaxis in domestic and
black-footed ferrets 30
17 VACCINES AGAINST COCCIDIA 32
171 Theory 32
172 Species successes in anticoccidial vaccination 34
18 RESEARCH GOALS AND OBJECTIVES 36
181 Objectives 36
182 Hypotheses 36
183 Applications 36
CHAPTER 2 MOLECULAR CHARACTERIZATION OF ENTERIC COCCIDIA FROM DOMESTIC
FERRETS (MUSTELA PUTORIUS FURO) 38
21 INTRODUCTION 39
22 MATERIALS amp METHODS 43
221 Fecal samples 43
222 Formalin fixed intestinal tissues 44
223 Molecular characterization 44
224 Phylogenetic analysis 46
23 RESULTS 47
231 Fresh fecal samples 47
232 Formalin fixed samples 48
233 Molecular characterization 49
234 Phylogenetic analysis 50
24 DISCUSSION 50
CHAPTER 3 MORPHOLOGICAL AND MOLECULAR CHARACTERIZATION OF ENTERIC
COCCIDIA ISOLATED FROM BLACK-FOOTED FERRETS (MUSTELA NIGRIPES) 60
31 INTRODUCTION 60
32 MATERIALS AND METHODS 64
321 Fecal samples 64
322 Formalin fixed intestinal tissues 65
323 Molecular characterization 66
33 RESULTS 66
331 Morphometric characterization 67
332 Molecular characterization 68
34 DISCUSSION 69
xi
CHAPTER 4 NATURAL HISTORY OF ENTERIC COCCIDIOSIS IN THE BLACK-FOOTED
FERRET (MUSTELA NIGRIPES) 78
41 INTRODUCTION 78
42 MATERIALS AND METHODS 80
421 Toronto Zoo BFF breeding program 80
422 Fecal oocyst evaluation 81
423 Retrospective review of pathology records 82
424 Prospective modified necropsy protocol 82
425 Retrospective medical history review 83
43 RESULTS 83
431 Fecal oocyst evaluation and retrospective medical history review 83
432 Pathology 86
433 Morbidity and mortality 88
44 DISCUSSION 88
CHAPTER 5 EVALUATING THE DOMESTIC FERRET (MUSTELA PUTORIUS FURO) AS AN
EXPERIMENTAL MODEL FOR ENTERIC COCCIDIOSIS IN THE BLACK-FOOTED FERRET
(MUSTELA NIGRIPES) 104
51 INTRODUCTION 104
52 MATERIALS AND METHODS 106
521 Animal care 106
522 Oocyst preparation 107
523 Experimental infections 108
524 Animal welfare 109
525 Hematology 110
526 Morphologic and molecular characterization 110
527 Necropsy protocol 111
53 RESULTS 111
531 Oocyst shedding 112
532 Morphologic and molecular characterization 113
533 Clinical signs 113
534 Hematology 113
535 Necropsy 114
54 DISCUSSION 115
xii
CHAPTER 6 WHOLE MITOCHONDRIAL GENOME SEQUENCES OF TWO EIMERIA SPECIES
ISOLATED FROM DOMESTIC (MUSTELA PUTORIUS FURO) AND BLACK- FOOTED FERRETS
(MUSTELA NIGRIPES) 129
61 INTRODUCTION 129
62 MATERIALS amp METHODS 130
621 Parasites 130
622 DNA isolation from coccidia in feces 131
623 Whole genome sequencing 131
624 Phylogenetic analysis 132
63 RESULTS 133
64 DISCUSSION 134
CHAPTER 7 CONCLUSIONS AND FUTURE DIRECTIONS 145
REFERENCES 148
APPENDICES 157
xiii
LIST OF TABLES
Table 11 Morphometrics of Eimeria and Isospora (=Cystoisospora) species affecting mustelids 10
Table 21 Amplification primers for nuclear 18S rDNA and mitochondrial COI loci used in the
identification of enteric coccidia from domestic ferrets 55
Table 22 Summary of fecal samples from domestic ferrets submitted to two diagnostic laboratories
from 2008-2015 56
Table 23 Morphologic and molecular identification of coccidia from domestic ferrets 57
Table 31 Amplification primers for nuclear 18S rDNA mitochondrial COI and COIII loci used in the
identification of coccidia from black-footed ferrets 73
Table 32 Morphologic and molecular characterization of coccidia from fecal and FFPE necropsy
samples from black-footed ferrets 76
Table 33 Morphometric characterization of Eimeria ictidea oocysts from black-footed ferrets 77
Table 41 Eimeria ictidea shedding in black-footed ferret dam and kit family groups - 2014-2016 97
Table 42 Epidemiologic data for family groups of black-footed ferrets shedding Eimeria ictidea 98
Table 43 Shedding of Eimeria ictidea in adult black-footed ferrets - 2015-2016 99
Table 44 Epidemiologic data for adult black-footed ferrets shedding Eimeria ictidea 100
Table 45 Histologic findings from black-footed ferrets with enteric coccidiosis 101
Table 46 Incidence of coccidial infections in black-footed ferrets at the Cheyenne Mountain Zoo 102
Table 47 Yearly mortality associated with coccidiosis in black-footed ferrets at the Toronto Zoo 103
Table 51 Prepatent period and oocyst shedding of Eimeria ictidea in experimentally infected
domestic ferrets 126
Table 52 Results of oral inoculation of domestic ferrets with oocysts of Eimeria ictidea 127
Table 53 Distribution of coccidial life stages in intestinal tract of domestic ferrets orally
inoculated with oocysts of Eimeria ictidea 128
Table 61 PCR primers used to sequence the mitochondrial genome of Eimeria furonis 136
Table 62 PCR primers used to sequence the mitochondrial genome of Eimeria ictidea 137
Table 63 Coding regions in the mitochondrial genome of Eimeria furonis from a domestic ferret 138
Table 64 Coding regions in the mitochondrial genome of Eimeria ictidea from a black-footed ferret 139
Table 65 Pairwise comparison of coding regions in the mitochondrial genomes of Eimeria furonis
and Eimeria ictidea 140
xiv
LIST OF FIGURES
Figure 11 Phylogeny of the Apicomplexa 2
Figure 12 Classical life cycle of coccidian parasites 4
Figure 13 Morphologic characteristics used for identification of eimeriid oocysts 6
Figure 21 Life stages of Eimeria furonis within the small intestine of a domestic ferret 58
Figure 22 Phylogenetic relationships of coccidia (Eimeria ictidea Eimeria furonis and Isospora
(=Cystoisospora) laidlawi) from domestic or black-footed ferrets 59
Figure 31 Nuclear and mitochondrial genetic loci targeted by primers listed in Table 31 73
Figure 32 Morphometrics of Eimeria ictidea from a black-footed ferret (Mustela nigripes) 74
Figure 33 Nuclear 18S rDNA sequences of Eimeria ictidea to newly generated (see Chapter 2) and
published sequences of Eimeria furonis 75
Figure 34 Mitochondrial cytochrome c oxidase subunit I sequences of Eimeria ictidea to sequences
from other eimeriid parasites of carnivores 75
Figure 41 Oocyst per gram counts and shedding period of Eimeria ictidea from black-footed ferret
family groups from 2014-2016 95
Figure 42 Sexual life stages of Eimeria ictidea in the small intestine of a black-footed ferret 96
Figure 51 Exogenous life stages of Eimeria ictidea 123
Figure 52 Endogenous life stages of Eimeria ictidea within the small intestine of an experimentally
infected domestic ferret 124
Figure 53 Distribution of sexual and asexual life stages of Eimeria ictidea along the intestinal tract
of experimentally infected domestic ferrets 125
Figure 61 Map of the mitochondrial genome of Eimeria furonis 141
Figure 62 Map of the mitochondrial genome of Eimeria ictidea 142
Figure 63 Comparison of the mitochondrial genomes of Eimeria furonis and Eimeria ictidea 143
Figure 64 Phylogenetic relationships of coccidia from domestic and black-footed ferrets based on
complete mitochondrial genome sequences 144
xv
LIST OF APPENDICES
Appendix 1 Shedding of oocysts of Eimeria ictidea in black-footed ferret (Mustela nigripes) dam and
kit family groups from 2014-2016 158
Appendix 2a Hematology values for domestic ferrets (Mustela putorius furo) from 49-51 days of
age prior to experimental inoculation 161
Appendix 2b Serum biochemistry values for domestic ferrets (Mustela putorius furo) from
49-51 days of age prior to experimental inoculation 162
Appendix 3a Hematology values for domestic ferrets (Mustela putorius furo) inoculated orally
with Eimeria ictidea 163
Appendix 3b Serum biochemistry values for domestic ferrets (Mustela putorius furo) inoculated
orally with Eimeria ictidea 164
Appendix 4 Domestic ferret (Mustela putorius furo) weekly monitoring sheet 165
Appendix 5 Domestic ferret (Mustela putorius furo) 24 hour intensive monitoring sheet 167
Appendix 6 Domestic ferret (Mustela putorius furo) infection trial standard operating procedures 172
xvi
ABBREVIATIONS
ATP Adenosine triphosphate
BFF Black-footed ferret(s)
BI Bayesian inference
bp Base pair
CAPC Companion Animal Parasitology Council
CDS Coding DNA sequence
CITES Convention on International Trade in Endangered Species of Wild Fauna and Flora
COI Cytochrome c oxidase subunit 1
COIII Cytochrome c oxidase subunit 3
CytB Cytochrome b
DF Domestic ferret(s)
DNA Deoxyribonucleic acid
FFPE Formalin-fixed paraffin embedded tissue
IUCN International Union on the Conservation of Nature
L Length
LSU Large subunit
mt Mitochondrial
NaOH Sodium hydroxide
nu Nuclear
OPG Oocyst per gram count
PCR Polymerase chain reaction
rDNA Ribosomal DNA
SI Shape index
SND Single nucleotide difference
SOP Standard operating procedure
sp spp Species (singular plural)
SSP Species Survival Plan
SSU Small subunit
TMS Trimethoprim sulfadimethoxine
USFWS United States Fish and Wildlife Service
W Width
1
CHAPTER 1 LITERATURE REVIEW
11 INTRODUCTION
Black-footed ferrets (Mustela nigripes) are one of three wild ferret species worldwide Although
formerly distributed throughout the North American prairies black-footed ferrets (BFF) had been
extirpated from the majority of their range by the 1970s and were declared extinct in the wild in 1987
Since 1986 a multi-institutional effort has been breeding this species in captivity with reintroduction back
into the wild at select sites within Canada the USA and Mexico
Coccidial enteritis is a major cause of death in young captive black-footed ferrets (Bronson et al
2007) but coccidiosis can affect all age classes (personal observation) As a result fewer captive-bred
ferrets may be reared successfully for release to the wild The significance of coccidiosis in wild ferrets is
unknown Consequently the prevention and control of coccidial outbreaks is an important part of black-
footed ferret captive breeding programs and management This research is intended to improve the in situ
and ex situ health of the black-footed ferret through the provision of a better understanding of the
pathogenesis of enteric coccidiosis in this species and to pave the way for the investigation of novel
methods for disease treatment and control
12 APICOMPLEXA
121 Brief introduction to apicomplexan pathogens
The phylum Apicomplexa comprises a large number of eukaryotic intracellular parasitic
organisms many of which are of importance to human and veterinary medicine As indicated by their
name these parasites are characterized by the presence of an apical complex at the anterior aspect of the
infective stage of the life-cycle (Tenter et al 2002) The taxonomic classifications of members of the
Apicomplexa continue to be in a state of flux (reviewed by Adl et al 2005 Cavalier-Smith 2014 Tenter
et al 2002) For this reason a more simplified taxonomic structure has been used in this review (see
2
Figure 11) The subclass Coccidia is a speciose group within the Apicomplexa with most genera falling
into one of two major coccidian suborders within the Eucoccidiorida To date greater than 2000 species
of coccidia have been named (Duszynski Upton amp Couch nd Upton 2000) The adeleid coccidia
(suborder Adeleorina) include monoxenous (single host) and heteroxenous (multiple hosts) parasites in
genera such as Adelea Haemogregarina Hepatozoon and Karyolysus The eimeriorinid coccidia
(suborder Eimeriorina) include the typical intestinal coccidia such as Eimeria Isospora and Cyclospora
species belonging to the family Eimeriidae as well as tissue (cyst forming) coccidia such as
Cystoisospora Besnoitia Toxoplasma and Sarcocystis species that belong to the family Sarcocystidae
(Cox 1994)
Figure 11 Phylogeny of the Apicomplexa Numbers on branches and thickness indicate diversity
(ie named species) Taxonomic groupings demonstrated by the phylogenetic tree (1) subclass
Coccidia (2) suborder Adeleorina (3) suborder Eimeriorina (4) family Eimeriidae and (5) family
Sarcocystidae Adapted from Šlapeta J Morin-Adeline V (2011) Apicomplexa Levine 1970
Sporozoa Leucart 1879 httptolweborgApicomplexa2446 in The Tree of Life Web Project
httptolweborg
2
1
3
4
5
3
122 Life cycles of the Eimeria and Isospora species implicated in enteric coccidiosis
The life cycle of Eimeria species is considered the classical coccidian life cycle which is
typically completed in one host (monoxenous) with many Eimeria species parasitizing only a single host
species (stenoxenous) (Figure 12) The life cycle has two main phases of development one that takes
place within the host (endogenous) and the other that takes places outside of the host (exogenous)
Classically the endogenous stages of the Eimeria life cycle take place within the intestinal epithelium
however some Eimeria species undergo extraintestinal endogenous development such as Eimeria stiedae
in rabbits which replicates within the epithelium of the biliary tree During the exogenous phase of the
life cycle unsporulated oocysts that are shed in the feces of the host sporulate within the environment
resulting in the formation of four sporocysts within each oocyst (tetrasporocystic) Each sporocyst
contains two sporozoites (dizoic) Sporulation is affected by three main factors temperature moisture and
aerobic conditions (Fayer 1980)
Once ingested by the host the wall of the sporulated oocyst is broken to release sporocysts from
which the sporozoites (infective stage) excyst The freed sporozoites penetrate the intestinal epithelial
cells and undergo multiple mitotic divisions to form a single multinucleate meront The meront then
undergoes simultaneous cytokinesis to form first generation merozoites which leave the host cell to infect
new cells and undergo further asexual replications The undifferentiated uninucleate tissue stage of the
parasite within the intestinal epithelial cell is called a trophozoite The number of cycles of asexual
replication (merogony) is predetermined after which the last generation of merozoites penetrate host cells
and undergo sexual differentiation into male and female gamonts (gametogony) Each microgamont
(male) undergoes simultaneous fission to produce numerous motile microgametes each macrogamont
(female) develops into a single mature macrogamete Fertilization of a macrogamete by a motile
microgamete results in formation of a zygote that is rapidly enclosed in a thick wall to form an
unsporulated oocyst Oocysts are shed with the hostrsquos feces into the environment where they are
protected from desiccation and chemical disinfection by the oocyst wall Traditionally Eimeria species
4
have been differentiated based on the host species or host genus affected the site of endogenous life cycle
development and the microscopic cellular characteristics of the different life stages Interestingly
experimental cross infection of Eimeria species from their natural host to a novel host of a taxonomically
similar species has been successful in some cases (De Vos 1970 Levine and Ivens 1970 Haberkorn
1971) challenging the notion that Eimeria are truly stenoxenous parasites
Figure 12 Classical life cycle of coccidian parasites This apicomplexan life cycle includes both
sexual and asexual development The three processes in the life cycle are merogony (asexual
replication A-D) followed by gametogony (formation of gametes E-H) within the digestive tract
of the host with release of unsporulated oocysts (I) Exogenous sporogony (I-L) results in the
production of infective sporulated oocysts (L) Adapted from Barta 2001 with permission of the
author
The life cycle of Isospora spp is similar to that of species in the genus Eimeria (see Figure 12)
but the number of sporocysts and sporozoites differ sporulated oocysts contain two sporocysts (disporic)
5
each of which contains four sporozoites (tetrazoic) These characteristics are not unique to Isospora spp
because diasporic tetrazoic sporulated oocysts are also found in the genera Besnoitia Frenkelia
Hammondia Sarcocystis and Toxoplasma However the sporocysts in the latter parasites are
morphologically distinct in that they lack Stieda bodies
123 Methods of characterization
1231 Morphological features
Historically eimeriid coccidia have been classified based on the cellular morphology of the
different life stages (particularly the morphometrics of sporulated oocysts) where these stages occur in
the host and apparent host specificity (frequently assumed and not tested experimentally) The
morphological features and dimensions of oocysts and their components are important diagnostic features
because of the availability of these stages in clinical specimens these characteristics can include size
(length [L] width [W] shape index [SI=LW]) number of sporocysts wall morphology
presenceabsence of a micropyle micropyle cap residual body or polar granules for oocysts size number
of sporozoites wall morphology presenceabsence of Stieda body subStieda body paraStieda body or
residual body for sporocysts and presenceabsence of refractile bodies for sporozoites (see Figure 13)
Pertinent life cycle information includes type of life cycle (monoxenous versus heteroxenous) tissue
sites of merogony and gametogony (intestinal versus extraintestinal) and the presence or absence of
extraintestinal hypobiotic stages (eg dormozoites or hypnozoites) Further information used to
characterize coccidia that form tissue cysts generally includes details on life stages in the definitive and
intermediate hosts location and morphology of tissue cysts route(s) of transmission among host species
and morphologic descriptions of merozoites (eg tachyzoites or bradyzoites) in tissue culture
6
Figure 13 Morphologic characteristics used for identification of eimeriid oocysts 1) Oocyst in cross
section ol - oocyst length or - oocyst residual body ow - oocyst width pg - polar granule row -rough
outer wall 2) The top of a hypothetical oocyst mcd - depth of the micropyle cap mcw - width of the
micropyle cap mw - width of the micropyle sow - smooth outer wall 3) Sporocyst in cross section
psb - paraStieda body sb - Stieda body sl - sporocyst length sp - sporozoite sr - sporocyst residual
body srb - sporozoite refractile body ssb - subStieda body sw - sporocyst width From Duszynski D
Wilber PG (1997) A guideline for the preparation of species descriptions in the Eimeriidae Journal of
Parasitology 83(2)333-336 reproduced with permission of Allen Press Publishing Services
1232 Molecular characterization (genetic loci and methods)
More recently molecular techniques have been used to infer phylogenetic or evolutionary
relationships among coccidia and to reclassify taxonomic assignments to better reflect the evolutionary
history of these parasites Molecular data can be more informative than phenotypic data because recent
evolutionary divergence among coccidia is unlikely to be reflected in morphologic differences but may
be detectable using molecular data The principle behind the use of molecular sequencing to describe
evolutionary relationships is that nucleotide sequences like morphological features diverge over time
under selective pressure however nucleotide sequences evolve at a more regular rate than do
morphologic characteristics Phenotypic data is thus less likely to detect recent evolutionary divergence
Sequences that are more similar are inferred to be more closely related and to have diverged more
recently (Cox 1994) Molecular characterization can be performed using DNA RNA or protein
sequences Most of the early molecular phylogenetic analyses of coccidia performed used ribosomal RNA
sequences usually by PCR amplification of ribosomal DNA (rDNA) in the nuclear genome of the
7
parasites Ribosomes contain both small and large RNA subunits in eukaryotes the large ribosomal RNA
consists of two forms 5S and 28S while the small ribosomal RNA exists only as 18S Sequences from
several genetic loci have been used for characterization of parasites most commonly 18S rDNA 28S
rDNA and ribosomal internal transcribed spacer regions (ITS) from the nuclear genome and more
recently mitochondrial cytochrome c oxidase subunits I (COI) and III (COIII) however sequencing of
nuclear 18S rDNA (nu 18S rDNA) has been the most prevalent in the literature by far Early attempts to
use 5S RNA sequences formed unlikely phylogenies and too few 28S ribosomal DNA sequences have
been obtained to make this locus useful (Cox 1994 Tenter et al 2002) The disadvantage of nu 18S
rDNA is that it is comparatively poor at distinguishing among closely related eimeriid coccidial species
because of its conserved nature but for that reason the nu18S rDNA locus is useful for inferring
relationships among species with greater evolutionary divergence Although only exploited recently
because of the paucity of suitable PCR primers the mitochondrial COI locus appears to be more useful
for distinguishing closely related eimeriid coccidia (Ogedengbe Hanner amp Barta 2011) but COI
sequences are less useful for inferring more ancient relationships between highly divergent coccidial
species Consequently the combined use of nu 18S rDNA and mitochondrial COI sequencing has been
recommended for improved species description and phylogenetic analysis (El-Sherry et al 2013)
Molecular characterization has also been used for diagnostic purposes and is well-suited to the
identification of coccidia when information on host specificity parasite life cycle and life stages is not
available as the molecular (genetic) data is the same for a given parasite during each of its life cycle
stages This information can be particularly useful in identifying the relationship between different life
stages of heteroxenous parasites collected from different hosts (intermediate definitive) Furthermore for
previously unidentified coccidia or those for which limited information is available molecular
characterization could be used to predict likely definitive hosts or parasite life cycle traits based on
phylogenetic relationships to other known species
8
13 RECLASSIFICATION OF MAMMALIAN ISOSPORA
Recommendations have been made to reclassify the avian and mammalian Isospora into two
separate genera based on life cycle molecular phylogenetic studies and morphologic description of
sporulated oocysts (Frenkel 1977 Barta et al 2005) Due to their classical coccidian life cycle presence
of Stieda bodies within sporocysts and close phylogenetic association with Eimeria species the avian
Atoxoplasma and Isospora have been retained in the genus Isospora (see Barta et al 2005) Conversely
the presence of tissue life cycle stages lack of Stieda bodies within sporocysts and close phylogenetic
association with other genera within the family Sarcocystidae have required many mammalian Isospora
to be reclassified as members of the genus Cystoisospora Frenkel 1977 (Frenkel 1977 Barta et al 2005)
Consequently for the remainder of this thesis Isospora species from mustelids will be referred to as
Isospora (=Cystoisospora) to reflect their probable generic association
14 EIMERIID SPECIES CHARACTERIZED IN MUSTELIDS
141 The family Mustelidae
The family Mustelidae within the order Carnivora comprises a group of approximately 59
carnivorous mammalian species within 22 genera Native mustelids are found in terrestrial and aquatic
environments on almost every continent with the exception of Australia and Antarctica The Mustelidae
are classically divided into two subfamilies as defined by Wozencraft (2005) 1) Mustelinae (weasels
mink ferrets marten wolverine) the larger subfamily including the following genera Arctonyx Eira
Galictis Gulo Ictonyx Lyncodon Martes Meles Mellivora Melogale Mustela Neovison Poecilogale
Taxidea and Vormela and 2) Lutrinae (otters) including seven genera Aonyx Enhydra Hydrictis
Lontra Lutra Lutrogale and Pteronura More recently molecular data suggest the Mustelidae should be
separated into eight subfamilies although this is not universally accepted (Koepfli et al 2008 Lariviegravere
and Jennings 2009 Yu et al 2011)
9
142 Eimeriid coccidia described from mustelids
Ten named Eimeria species and twelve named Isospora (=Cystoisospora) species have been
described in the Mustelidae and are summarized in Table 11 This table includes information on host
range life cycle and detailed morphologic data used to identify and classify the individual parasites Two
coccidial parasites isolated from the Libyan striped weasel (Ictonyx libyca) and the European polecat
(Mustela putorius) initially ascribed to the genus Isospora Isospora zorillae and Isospora putori
respectively have since been reclassified as Sarcocystis spp (see footnote to Table 2 of Yi-Fan et al
2012)
10
Table 11 Morphologic characteristics of Eimeria and Isospora (=Cystoisospora) species affecting mustelids
Coccidial species Host genus and
species
Life cycle
Location
Oocyst shape and
size
Oocyst
description
Sporocyst
description
Sporozoite
description
References
Cytoisospora
eversmanni
Mustela
eversmanii
(Steppe polecat)
Mustela
putorius
(European
polecat)
Homoxenous L185 (16ndash20)
W 148 (16ndash12)
LW 13 (11ndash16)
M absent
PG absent
OR absent
L 115
(10ndash135)
W 98
(9ndash11)
LW 12
(11ndash15)
SB absent
SR present
SRB present Yi-Fan et al 2012
Svanbaev 1956
Nukerbaeva amp
Svanbaev 1973
1977
Cystoisospora
pavlovskyi
Mustela
eversmanii
Mustela
putorius
Homoxenous L 322 (29ndash36)
W 273 (265ndash285)
LW 12 (11ndash14)
M absent
PG absent
OR absent
L 195
(18ndash21)
W 144
(12ndash15)
LW 14
(12ndash15)
SB absent
SR present
SRB present Yi-Fan et al 2012
Svanbaev 1956
Nukerbaeva amp
Svanbaev 1973
1977
Eimeria
baskanica^
Mustela
erminae
(ermine)
Homoxenous Oval with tapered
ends
L 112-126
W 84-98
M absent
PG absent
OR present
SR absent Bean shaped Nukerbaeva amp
Svanbaev 1977
Eimeria furonis Mustela
putorius
Mustela
putorius furo
(dom ferret)
Mustela
nigripes (BFF)
Mustela vison
(mink)
Homoxenous
Small intestine
rectum (H 1927)
Jejunumileum (BP
1993)
Spherical ndash
subspherical
L 11-14
W 10-13
OW 2 layers
M absent
PG absent
OR absent
Spindloid
L 8-9
W 4
SB present
SR present
Vermiform Blankenship-Paris
et al 1993
Hoare 1927 1935b
Jolley et al 1994
Nukerbaeva amp
Svanbaev
19731977
Williams et al 1988
1992 1996
Eimeria hiepei Mustela vison Homoxenous
Bile duct
Spherical
L 13-17
W 13-17
OW 2 layers
(outer
smooth)
M absent
PG absent
OR absent
L 6
W 4
SB absent
SR absent
Banana shaped Davis et al 1953
Grafner et al 1967
11
Coccidial species Host genus and
species
Life cycle
Location
Oocyst shape and
size
Oocyst
description
Sporocyst
description
Sporozoite
description
References
Eimeria ictidea Mustela
eversmanni
Mustela
nigripes
Mustela
putorius
Mustela
putorius furo
Homoxenous
Small intestine
Ovoid ndashellipsoid
L 13-27
W 13-21
OW 2 layers
M present
PG absent
OR absent
Ovoid
(irregular)
L 115
W65
SB present
SR present
- Hoare 1927 1935a
1935b
Jolley et al 1994
Litvenkova 1969
Svanbaev 1956
Tinar 1985
Williams et al 1988
1992
Eimeria irara Eira barbara
(tayra)
Homoxenous
Feces
Ovoid
L 21-25
W 18-20
OW outer
layer smooth
M absent
PG absent
OR absent
Ellipsoid
L 10-12
W 65
SB present
SR present
Elongate (one
end broader than
the other)
Carini amp da
Fonseca 1938
Eimeria melis Meles meles
(European
badger)
Homoxenous Ellipsoid
L 20plusmn018
W 157plusmn002
LW128plusmn0017
(112-15)
OW 2 layers
(outer
smooth)
M absent
PG present
OR present
Ovoid
L
119plusmn0018
W 65plusmn008
LW 183
(155-24)
SB present
L 90plusmn005
W 324plusmn0025
SRB present
Anwar et al 2000
Kotlan amp Pospesch
1933
Eimeria mustelae Mustela vison
Mustela nivalis
(snow weasel)
Homoxenous
Duodenumileum
Spherical or
Ellipsoid
L 18-26
W 14-24
OW 2 layers
M absent
PG present
OR absent
Ovoid
L 8
W 5
SB present
SR present
Broad at one
end and tapered
at other
L 7
W 3
Glebezdin 1978
Iwanoff-Gobzem
1934
Levine 1948
Musaev amp Veisov
1983
Tinar 1985
Eimeria sablii Martes zibellina
(sable)
Homoxenous
Gut
Spherical or
subspherical
L 112-126
W 112
OW 2 layers
M absent
OR absent
Ovoid
L 56
W 42
SR present
Elongate Nukerbaeva 1981
Eimeria sibirica Martes zibellina Homoxenous Ovoid
L avg 216
W avg 180
LW 1076
OW 2 layers
M absent
PG absent
OR absent
Ovoid
L 96-112
W 56-72
SR absent
Elongate Nukerbaeva 1981
Yakimoff amp
Gousseff 1934
Yakimoff amp
Terwinsky 1930
1931
12
Coccidial species Host genus and
species
Life cycle
Location
Oocyst shape and
size
Oocyst
description
Sporocyst
description
Sporozoite
description
References
Eimeria vison
(Eimeria
mustelae)
Mustela
putorius
Mustela
putorius furo
Mustela vison
Homoxenous
Small intestine
+- large intestine
Ovoid
L 17-22
W 9-18
OW 2 layers
M absent
OR
sometimes
present
Ovoid or
Piriform
L 10
W 55
SB absent
SR present
Curved or Club
shaped
L 9
W 25
Foreyt amp Todd 1976
Foreyt et al 1977
Kingscote 1934
1935
Levine 1948
McTaggart 1960
Nukerbaeva amp
Svanbaev
19731977
Tinar 1985
Umurzakov amp
Nukerbaeva 1985
Wolter 1961
Zimmermann 1959
Isospora africana Ictonyx libyca
(Libyan striped
weasel)
Homoxenous
Feces
Spherical
L 25-27
W 25-27
OW 2 layers
(outer
smooth)
M absent
PG absent
OR absent
Ovoid
L 15-17
W 10-12
SB absent
SR present
Elongate
L 135
W 3
Prasad 1961
Isospora altaica Mustela altaica
(mountain
weasel)
Homoxenous
Gut
Oval or spherical
L 280-336
W 252-280
LW 121 (111-
124)
OW 2 layers
M absent
PG absent
OR absent
Ovoid or
spherical
L 140-168
W 111-168
SR present
Svanbaev amp
Rachmatullina
1971
Isospora goussevi Mustela nivalis Homoxenous
Large intestine
Ovoid
L 224 (220-250)
W 174 (160-190)
LW 135 (133-
137)
OW 1 layer
PG present
OR present
Ovoid
L 120
(100-130)
W 70 (60-
80)
SB present
SR present
Elongate Musaev amp Veisov
1983
13
Coccidial species Host genus and
species
Life cycle
Location
Oocyst shape and
size
Oocyst
description
Sporocyst
description
Sporozoite
description
References
Isospora
hoogstraali
Ictonyx libyca Homoxenous
Feces
Ellipsoid
L 37-41
W 32-34
OW 2 layers
(outer
smooth)
M absent
PG some
OR absent
Ovoid
L 19-21
W 13-15
SB absent
SR present
Club-shaped
L 18-19
W 4-6
Prasad 1961
Isospora laidlawi Mustela
putorius
Mustela
putorius furo
Mustela vison
Homoxenous
Feces
Intestinal contents
Ovoid L
320-368
W 272-304
OW 2 layers
M absent
PG absent
OR absent
Ellipsoid
L 208
W 144
SB absent
SR present
Sausage shaped Foreyt et al 1977
Hoare 1927
Levine 1948
McTaggart 1960
Nukerbaeva amp
Svanbaev 1973
1974 1977
Tinar 1985
Isospora lutrae Lutra lutra
(European
otter)
Lutra
canadensis
(North
American river
otter)
Homoxenous Spherical
L 312 (275-32)
W 296 (28-31)
LW 104
(10-112)
OW 2 layers
(outer
smooth)
M absent
PG absent
OR absent
Ellipsoid
L 182 (17-
19)
W 144 (14-
16)
LW128
(12-14)
Sb absent
sSB absent
SR present
Spindle- shaped
L 124
W 25
SRB present
Torres et al 2000
Hoover et al 1985
Isospora
martessii
Martes zibellina Homoxenous
Gut
Ovoid short oval or
spherical
L 252 ndash 280 196
168
W 168 ndash 224 168
168
OW 2 layers
M absent
OR absent
Ovoid
L 112-168
W 84-112
SR present
Elongate Nukerbaeva 1981
Isospora melis Meles meles Homoxenous Ovoid
L 328plusmn034
W 269plusmn019
LW122 (110-
157)
OW 2 layers
(outer
smooth)
M absent
PG absent
OR absent
Ellipsoid
L
215plusmn0166
W 14plusmn012
LW 155
(133-185)
SR absent
Round at one
end other end
tapered
L 142plusmn116
W 40plusmn017
SRB absent
Anwar et al 2000
Glebezdin 1978
Kotlan amp Pospesch
1933
Pelleacuterdy 1955
14
Coccidial species Host genus and
species
Life cycle
Location
Oocyst shape and
size
Oocyst
description
Sporocyst
description
Sporozoite
description
References
Isospora
mustelae (nomen
nudum)
Martes martes Ovoid L
7 W
225
M present - - Galli-Valerio 1932
Isospora nivalis Mustela nivalis Homoxenous
Large intestine
Ovoid
L 206 (200-230)
W 184 (180-210)
LW 11 (109-111)
OW 1 layer
PG absent
OR absent
Ovoid
L 125
(120-130)
W 80 (70-
90)
SR present
Lemon or pear
shaped
Musaev amp Veisov
1983
Unnamed
ldquoCoccidiardquo^
Mustela
nigripes
Urinary bladder - - - - Jolley et al 1994
Unnamed
ldquoCoccidiardquo^
Mustela
nigripes
Trachea bronchus
bronchial glands
- - - - Jolley et al 1994
Unnamed
Eimeria sp^
Mustela
nigripes
Feces
intestinal contents
Ovoid
L 350-386
W 212-232
- - - Jolley et al 1994
Williams et al
1992
Unnamed
Eimeria sp^
Mustela
putorius furo
Small intestine - - - - Blankenship-Paris
et al 1993
Unnamed
Eimeria sp^
Mustela nivalis Homoxenous
Large intestine
Ovoid-ellipsoid L
2031 (1712-2162)
W 148 (1225-
1681)
LW 136 (121-16-
)
OW 1 layer
PG absent
OR absent
Ovoid or
pear-shaped
L 60-100
W 40-80
SR present
Elongate
L 50-90
W 30-70
Musaev amp Veisov
1983
Unnamed
Eimeria sp^
Martes martes
(marten)
Homoxenous Ovoid
L avg 216
W avg 180
LW 1076
OR absent 4 sporocysts
SR present
L 126
W 60
Yakimoff and
Gousseff 1934
Unnamed
Isospora sp^
Mustela
putorius furo
Feces - - - - Bell 1994
Unnamed
Isospora sp^
Mustela
putorius furo
Feces - - - - Bell 1994
Legend L = length W = width LW = length-width ratio avg = average OW = oocyst wall PG = polar granules M = micropyle SB = Stieda body sSB =
subStieda body OR = oocyst residuum SR = sporocyst residuum SRB = sporozoite refractile body ^ = species inquirendae - = no information provided by
author(s) = information obtained from secondary sources (primary reference could not be obtained) All measurements are in micrometers Bolded references
15
are those from which morphometric data were assembled Remaining references indicate other authors who have identified that parasite species in the same or
similar host
16
143 Eimeriid coccidia described from domestic ferrets
Three species of coccidia were originally described from 50 domestic ferrets (Mustela putorius
furo) Eimeria ictidea Eimeria furonis and Isospora (= Cystoisospora) laidlawi (Hoare 1927) All three
species were detected in feces from domestic ferrets at a research facility undergoing an outbreak of
canine distemper Sick ferrets appeared more frequently infected than healthy ones As per Hoare (1927)
none of the ferrets appeared to display clinical signs associated with protozoal infection For each
parasite the author described morphology of sporulated oocysts isolated from feces and sporulation time
(exogenous life stages) The pre-patent period (minimum duration of endogenous development) in an
inoculated naiumlve ferret was described only for E furonis and E ictidea due to insufficient sample size of
I (=C) laidlawi oocysts for an experimental infection trial Sporulation of oocysts occurred within 5-6
days for E furonis 3 days for E ictidea and 4 days for I (=C) laidlawi The sporulated oocysts of E
furonis were spherical with a double outer wall with a thin colourless outer layer and thick yellowish
inner layer no micropyle or residual body and measured on average 128 times 120 microm (length [L] 112-
144 width [W] 104-128 shape index [SI] 107) Unsporulated oocysts contained a zygote with a
diameter of 96 microm Sporocysts were spindle-shaped with one end constrictedblunted contained a
residual body and on average measured 8-88 times 4 microm Sporozoites were vermiform with one end
narrower than the other arranged head to tail and had a central nucleus a clear vacuole was identified in
some at the broad end The sporulated oocysts of E ictidea were oval or elliptical with a double outer
wall with a thin colourless outer layer and thick yellowish inner layer no micropyle or residual body
and measured on average 236 times 175 microm (L 184-272 W 128-208 shape index 135) The zygote in
unsporulated oocysts was elongate with a diameter of 15 times 12 microm when originally passed in feces but
became more spherical with time Sporocysts were irregularly oval with one end broad and the other
more constricted contained a residual body and on average measured 115 times 65 microm Sporozoites were
vermiform with one end narrower than the other arranged head to tail and had a central nucleus and a
clear vacuole at the broad end The sporulated oocysts of Isospora (=Cystoisospora) laidlawi were ovoid
with a double outer wall with a thin colourless outer layer and thick yellowish inner layer no micropyle
17
or residual body and measured on average 34 times 29 microm (L 320-368 W 272-304) Unsporulated
oocysts contained a spherical zygote with a diameter of 236 microm Two sporocysts were identified each
containing 4 sporozoites and no Stieda body sporocysts were elliptical contained a residual body and on
average measured on 208 times 144 microm Sporozoites were sausage shaped with one end slightly pointed
and had a central nucleus and a clear vacuole identified at the pointed end Sporozoites were arranged
with pointed ends all at the same pole of the sporocyst The pre-patent periods described for E furonis
and E ictidea were 6 days and 7 days respectively (Hoare 1927)
Since Hoarersquos initial description (Hoare 1927 Hoare 1935) multiple single case reports and
outbreaks of severe clinical disease associated with intestinal coccidiosis have been reported in domestic
ferrets Blankenship-Paris et al (1993) described a single case of a four-month-old domestic ferret that
presented depressed in thin body condition dehydrated and with pasty dark feces on the perineum This
ferret had been housed with its dam and another sibling neither dam nor sibling showed clinical signs of
enteric disease and both had negative fecal examination results on repeated evaluation Routine fecal
examination of the rest of the colony and necropsies on eight other ferrets in the colony revealed no
evidence of coccidial infection Enteric coccidiosis was determined to be the cause of disease in the four-
month-old ferret based on necropsy findings but the coccidia could not be speciated because diagnosis
was made on histologic findings only
Sledge et al ( 2011) described three separate outbreaks of severe enteric coccidiosis in domestic
ferrets from one ferret rescue centre (group 1) and two shelters (groups 2 and 3) all affected by the same
Eimeria sp The morphologic characteristics of sporulated oocysts were only described for group 1 no
coccidial oocysts were detected on direct smear or fecal flotation of diarrheic samples submitted from
groups 2 and 3 Oocysts were identified as spherical measuring 12-13 microm in diameter with four
sporocysts each containing two sporozoites Oocyst morphometrics histopathologic findings and nu 18S
rDNA partial sequences from all three groups were used collectively to confirm the coccidial species
identify in each outbreak as E furonis
18
Two cases of biliary coccidiosis with E furonis have been reported in domestic ferrets The first
was in a nine-week-old male ferret from a research facility (Williams Chimes amp Gardiner 1996) The
ferret presented with signs of hepatic disease and was negative for coccidia on fecal flotation and direct
smears Endogenous coccidial life stages were described from the gall bladder and liver on histologic
examination In tissue section the oocysts were oval to spherical and measured 125 times 120 microm Meronts
measured 108-130 times 89-93 microm and contained up to 16 merozoites The merozoites exhibited a double-
layered pellicle prominent conoid few rhoptries and many micronemes anterior to the nucleus Based on
the morphologic description of the life stages in this case the coccidia were identified by the authors as
an Eimeria species most likely E furonis Kaye et al (2015) described a second case of biliary
coccidiosis in an 18-month-old female pet domestic ferret with concurrent pure red cell aplasia In this
case all endogenous coccidial life stages were observed on histologic examination of the epithelium of
the extrahepatic biliary tree The oocysts were ovoid and measured 12 times 13 microm Meronts measured 12 times
15 microm and contained up to 16 merozoites each measuring 2 times 5 microm Based on the morphologic
description of the life stages in this case and nu 18S rDNA sequences the pathogen was also determined
to be E furonis Biliary coccidiosis has also been identified in mink (Mustela vison) with the etiologic
agent identified as Eimeria hiepei (Davis Chow amp Gorham 1953 Grafner Graubmann amp Dobbriner
1967)
Oocysts from Cystoisospora ohioensis have been reported from fecal samples collected from
healthy domestic ferret kits in a large American ferret breeding operation that were raised on the same
premise as juvenile domestic dogs (Patterson amp Fox 2007) The method of identification of this parasite
was not described by Patterson amp Fox A second similar institution reported the presence of a
Cystoisospora species also thought to be C ohioensis in routine fecal examination of their ferret colony
(Dr Bambi Jasmin personal communication) Coccidial identification in this case was performed by the
Animal Health Diagnostic Center at Cornell University The significance of these findings is unknown as
no clinical signs or histologic lesions have been described in domestic ferrets associated with shedding of
19
oocysts and the definitive host for C ohioensis is the domestic dog It is most likely that fecal
identification of C ohioensis represents a pseudoparasite in both of these cases or perhaps an
undescribed Cystoisospora sp that is morphologically indistinguishable from C ohioensis
It is difficult to estimate the prevalence of enteric coccidia within the North American domestic
ferret population Fecal samples submitted to university or large veterinary diagnostic laboratories from
domestic ferrets in Canada are uncommon and samples positive for coccidia appear infrequently (Dr
Donald Martin personal communication) Data from Idexx Vet Med Lab in Ludwigsburg Germany was
compiled to review the prevalence of coccidia and Giardia within fecal samples from domestic ferrets
(Pantchev et al 2011) The authors reported that of 284 fecal samples submitted from 2002-2004 18
(63) had detectable coccidial oocysts on fecal flotation Oocysts were identified based on morphologic
characteristics as E ictidea E furonis I (=C) laidlawi and another unidentified Isospora species
Comparative data from the same laboratory from 2009-2010 included sample submissions from 253
ferrets 21 (83) of which were positive for coccidial oocysts on fecal flotation Nine of the samples
were positive for E furonis three were positive with both E furonis and I(=C) laidlawi present eight
were positive only for I(=C) laidlawi and one sample contained both E furonis and E ictidea
identification in all cases was based on morphologic characteristics No statistically significant difference
in the occurrence of coccidial oocysts was detected when data from the two periods were compared
(Fisherrsquos exact test P=041) (Pantchev et al 2011)
144 Molecular characterization
Molecular characterization of Eimeria furonis was first performed by Abe et al (2008) using
oocysts purified from the feces of a single domestic ferret with clinical signs of coccidial enteritis Small
subunit ribosomal DNA (nu 18S rDNA) primers CYC1FE (5ʹ-TAC CCA ATG AAA ACA GTT T-3prime) and
CYC4RB (5prime-CGT CTT CAA ACC CCC TAC TG-3prime) were used to amplify a 347 base pair (bp) fragment
of nu 18S rDNA These primers were initially developed for molecular identification of Cyclospora
species but have since been shown to amplify nu 18S rDNA from several Eimeria species (Matsubayashi
20
et al 2005) The amplicon was sequenced (GenBank AB329724) and compared with previously
published partial nu 18S rDNA sequences from 40 Eimeria two Isospora and four Cyclospora species
The resulting phylogram grouped E furonis with E alabamensis (cattle) and E meleagrimitis (turkey) In
the same study the microscopic morphology of the oocysts was used to identify this coccidial species as
E furonis by comparison with published descriptions of E furonis E ictidea and E heipei by Hoare
(1927) Hoare (1935) and Grafner Graubmann amp Dobbriner (1967) respectively
Nuclear 18S rDNA was also used by Sledge et al (2011) for molecular identification of the
eimeriid coccidia implicated in the three distinct outbreaks of enteric disease in domestic ferrets As
described above initial identification and speciation of the coccidia was performed using morphologic
characteristics of the sporulated oocysts collected from feces in one of the three outbreaks being
investigated the oocysts were identified as E furonis Histologic sections of formalin fixed intestinal
segments from ferrets from each of the three outbreaks contained multiple coccidial life stages DNA was
then isolated from stored formalin-fixed tissues for further genetic analysis Using the partial nu 18S
rDNA gene sequence reported by Abe et al (2008) (GenBank AB329724) the following PCR primers
were created 5ʹ-ACA ATT GGA GGG CAA GTC TG-3ʹ and 5ʹ-GGCGAC AAG CCT GCT TGA AAC-
3ʹ PCR amplification produced a 247 bp amplicon from each of the three groups Analysis and
sequencing of amplicons from all three groups showed 100 homology to nucleic acid sequences
previously reported by Abe et al (2008) for the gene encoding E furonis nu 18S rDNA
Coccidia were identified within hepatobiliary lesions in a domestic ferret receiving
immunosuppressive therapy for red cell aplasia (Kaye et al 2015) DNA was extracted from frozen liver
and a 247 bp fragment of the nu 18S rDNA was amplified using the primers previously described by
Sledge et al (2011) and sequenced Kaye et al (2015) reported that the DNA sequence of the amplicon
was 100 homologous to the published nu 18S rDNA of E furonis and 95 homologous to the nu 18S
rDNA of E myoxi (rodent) E alabamensis (cattle) and I robini (avian)
21
145 Clinical signs of disease in domestic ferrets
Hoare (1927 1935b) in his initial descriptions of enteric coccidiosis in domestic ferrets
observed that clinical signs of intestinal disease were not evident The recent literature supports the
finding of subclinical disease but also describes signs ranging from mild transient diarrhea in young or
stressed animals to more severe disease with dehydration lethargy depression weight lossemaciation
inappetence and death (Blankenship-Paris et al 1993 Powers 2009 Sledge et al 2011 Hoefer et al
2012 Patterson et al 2014) Rectal prolapse has also been reported in ferrets with enteric coccidiosis
(Hillyer 1992 Hoefer et al 2012) In one study co-infection with coccidia and Lawsonia intracellularis
(Desulfovibrio sp) was diagnosed in 4 of 19 ferrets with proliferative bowel disease (Li et al 1996)
These ferrets presented with variable clinical signs including diarrhea lethargy anorexia weight loss
dehydration and emaciation
In the two reports of biliary coccidiosis clinical signs conformed to those expected with
hepatobiliary disease Williams et al (1996) described their case to have presented with emaciation poor
appetite abdominal distension and icterus Kaye et al (2015) described a one week history of lethargy
inappetence and icterus with serum biochemistry results consistent with cholestasis later clinical signs in
this case included melena anemia and cachexia
146 Gross necropsy and histologic findings
The pathology of enteric coccidiosis in domestic ferrets was described by Hoare (1927 1935b)
Two healthy domestic ferrets were experimentally inoculated one each with large numbers of mature
oocysts of either E furonis or E ictidea that were isolated during his initial work The inoculated ferrets
were killed humanely for histologic examination of intestinal sections at the time of first detection of fecal
oocyst shedding no clinical signs of coccidiosis were detected in these ferrets prior to death Infection
with E furonis resulted in invasion of the epithelium of the small intestine and rectum Within the small
intestine the parasites were concentrated in the tips of the villi but could be found to the level of the
22
opening of the crypts of Lieberkuumlhn In rectal sections life stages were limited to the epithelial ridges
between the openings of the glands of Lieberkuumlhn Organisms were located within the apical portion of
the epithelial cells and intensely infected regions exhibited multiple parasites within a single host cell
Both asexual and sexual life stages were present within the same sections Hoare (1927) described similar
histopathologic changes in naturally infected ferrets but the proportion of asexual versus sexual life
stages differed In natural infections sexual life stages were more numerous whereas in experimental
infections asexual life stages predominated these findings would be expected to correlate with the stage
of infection at which ferrets died or were humanely killed for tissue collection and would not be
reflective of differences between natural and experimental infection with this parasite Hoare also
described the morphology of the different endogenous stages including trophozoite (3-4 microm) merozoite
(stumpy sausage shaped L 3-4 microm W 2 microm) macrogamete (spherical 8 microm diameter with darkly
staining globular inclusions of reserve material) and microgamete (described as similar to those of other
Eimeria species) Two types of merogony are described from histologic sections the first with stumpy
merozoites as described above and the second with merozoites with elongated curved bodies and a
compact polar nucleus measuring 60 times 13 microm This second merogonic generation was observed almost
exclusively in the naturally infected ferrets and was associated with initiation of sexual differentiation and
reproduction
The pathology of experimental and non-experimental infection with E ictidea in domestic ferrets
was also described by Hoare (1927 1935b) Parasitic invasion of the epithelium was noted only in the
small intestine with patchy distribution of the parasite life stages throughout affected sections Within the
small intestinal villi the parasites were again concentrated in the tips of the villi with infected epithelial
cells never containing more than one parasite As each intracellular parasite grew it filled the entire host
cell displacing the nucleus to the base of the cell Predominantly sexual life stages were detected in tissue
sections with few asexual generations observed Interestingly the parasites were arranged into age
groups with forms of the same life stage grouped together within the affected epithelial sections this is in
23
contrast to E furonis where life stages of different maturities were found together in affected sections
Hoare described the morphology of the different endogenous stages of E ictidea including merozoites
(free within the lumen elongated vermiform with one pointed end and a nucleus located at the rounded
end 11 microm times 1 microm within the epithelium shortened and rounded 3-4 microm diameter) macrogametes
(elongated 20 times 7 microm occupying the entire host cell with darkly staining globular inclusions of reserve
material) and mature microgamonts (morphologically similar to those of other Eimeria species but larger
than those of E furonis) Of note a tissue reaction was observed specifically in association with more
developed life stages of E ictidea (eg mature meronts mature gamonts unsporulated oocysts) which
was not observed when cells contained earlier stages of development (eg trophozoites immature
gamonts) This tissue reaction was described by Hoare (1935a 1935b) as the development of an annular
constriction of the apical portion of the villus separating infected epithelial cells from unaffected cells
The constriction involved the epithelium but could also extend inwards into the core of the villus These
changes were associated with congestion of capillaries and extravasation of red blood cells within the
constricted segment and in some sections villar tip necrosis
In their case report of one domestic ferret Blankenship-Paris et al (1993) described the gross
pathologic lesions associated with intestinal coccidiosis in this case there was diffuse dilation and
reddening of the small intestine which was empty and the colon contained dark watery material
Histologic lesions were confined to the ileum and jejunum The jejunum exhibited thickening of the villi
with a crypt to villus ratio of 15 mild granulomatous inflammation in the lamina propria and large
numbers of coccidial meronts gamonts and oocysts within the enterocytes of the villar tips
The gross lesions described by Sledge et al (2011) from 20 domestic ferrets are as follows thin
body condition with moderate to marked dehydration perineal staining with diarrhea moderate dilation
of the small and large intestines and the presence of pasty tan to tarry black digesta within the distal small
intestine and colon Other findings in one to a small number of ferrets included enlarged pale tan livers
splenomegaly with dark red colouration and multiple superficial gastric or duodenal ulcers The
24
histologic lesions from 10 ferrets included moderate blunting and occasional fusion of jejunal and ileal
villi focal attenuation and erosion of the epithelium of the villar tips with exudation of fibrin neutrophils
and blood into the intestinal lumen in regions with severe erosion Intact epithelial cells at the villus tips
and rarely sloughed epithelial cells in the intestinal lumen contained numerous intracytoplasmic coccidia
representing a range of asexual and sexual life stages (meronts macrogamonts microgamonts and
oocysts) The subjacent lamina propria of the small intestine and of the large intestine exhibited moderate
lymphoplasmacytic infiltration with occasional neutrophils and congestion of blood vessels Marked
mucosal hemorrhage was identified in the most severely affected sections
Marked gross and histopathologic hepatobiliary lesions were described in a single ferret by
Williams et al (1996) On gross necropsy the liver was pale and enlarged with dilated firm bile ducts
and thickening of the gall bladder wall Similar gross necropsy findings were described by Kaye et al
(2015) marked dilation and mural thickening of the entire biliary tree (including gall bladder intrahepatic
and extrahepatic bile ducts) On histopathology Williams et al (1996) noted that the marked thickening
of the gallbladder wall was a result of cystic proliferation of mucosal glands which were separated by
tracts of fibrous connective tissue and marked granulomatous inflammation Liver sections exhibited
marked biliary hyperplasia marked periductular fibrosis and moderate periportal lymphoplasmacytic
cuffing There was multifocal papillary proliferation of bile duct epithelium and dilation of the bile ducts
and within the ductular lumens there were moderate numbers of lymphocytes and plasma cells small
numbers of degenerate neutrophils sloughed epithelial cells and debris All endogenous coccidial life
stages were present within the gall bladder and biliary epithelium with meronts visible in 20 of the
intact epithelial cells of the biliary tree and gallbladder and oocysts free within the lumen of the
intrahepatic bile ducts Similar lesions were present in the case described by Kaye et al (2015) and as
well as in juvenile and adult farmed mink (Mustela vison) with hepatobiliary coccidiosis (Davis Chow amp
Gorham 1953)
25
15 INTRODUCTION TO ENTERIC COCCIDIOSIS IN THE BLACK-FOOTED FERRET
151 Natural history and conservation of the black-footed ferret in North America
Black-footed ferrets are one of only three wild ferret species worldwide the other species are the
European polecat (Mustela putorius) and the Siberian polecat or steppe polecat (Mustela eversmanii)
They are the only native North American ferret species and the most endangered North American
carnivore They are nocturnal carnivores whose diet and lifestyle are highly dependent on local prairie
dog (Cynomys sp) populations Prairie dogs comprise almost exclusively the diet for the BFF who also
use the complex burrow systems made by prairie dogs to escape their predators and raise their young
(Santymire et al 2014 USFWS BFF Recovery Program 2017)
While formerly distributed throughout the North America prairie ecosystem BFF were
considered extinct by the late 1950s In 1964 a single population was discovered in Mellette County
South Dakota Progressive decline of this population in subsequent years resulted in the decision by
United States Fish and Wildlife Service (USFWS) to initiate a captive breeding program for the species
From 1971-1973 four females and five males were captured for this purpose Despite successful breeding
no kits survived and the last adult ferret in this captive colony died in 1979 at that time BFF were again
presumed extinct in the wild based on annual surveys of the initial capture site In 1981 a dead BFF was
discovered by a ranch dog outside of Meeteetse Wyoming allowing wildlife biologists to identify
another colony of BFF This colony flourished until 1985 when an outbreak of canine distemper in the
BFF population and an outbreak of sylvatic plague in the local prairie dog population resulted in sharp
population declines From 1985 through 1987 all 24 of the remaining BFF were trapped and brought into
captivity to re-initiate the captive breeding program Six ferrets in this initial group died of canine
distemper while in captivity and of the remaining 18 survivors 7 bred successfully to create the founding
population of the current captive breeding population Today this captive breeding population consists of
approximately 300 BFF distributed among multiple institutions (Santymire et al 2014)
26
Since 1986 this multi-institutional effort has been breeding BFF in captivity with reintroductions
back into the wild in 28 selected locations in Canada the USA and Mexico Currently six facilities
participate in the BFF Species Survival Plan (SSP) the Toronto Zoo USFWS National Black-footed
Ferret Conservation Center National Zoorsquos Smithsonian Conservation Biology Institute Louisville
Zoological Garden Cheyenne Mountain Zoo and the Phoenix Zoo (Black-footed Ferret Recovery
Implementation Team 2011) As of 2011 over 8000 BFF kits had been produced in captive breeding
facilities (Black-footed Ferret Recovery Implementation Team 2011)
Multiple infectious diseases pose a significant risk to the captive breeding and post-release
survival of BFF including canine distemper and sylvatic plague Coccidiosis is recognized as a cause of
significant juvenile morbidity and mortality in captive breeding programs and can result in significant
population losses (Bronson et al 2007 Santymire et al 2014 USFWS BFF Recovery Program 2017)
152 Coccidia identified from black-footed ferrets
Eimeria ictidea and Eimeria furonis have been identified in black-footed ferrets based on
morphologic criteria (Jolley et al 1994) Jolley et al examined fecal samples from six captive BFF during
a distemper outbreak as well as samples from wild BFF They described one medium-sized ovoid
tetrasporic dizoic oocyst with a double wall presence of a polar body and lacking both an oocyst residual
body and micropyle The oocysts measured 232 microm (range 182-274) by 155microm (range 130-162) with
a SI of 150 The sporocysts were elongate with the presence of both sporocyst residuum and a Stieda
body Sporozoites contained prominent refractile bodies at the posterior end and were aligned anterior to
posterior within sporocysts These oocysts were shed by all six captive ferrets On histopathology of
intestinal sections merogony and gametogony were observed within the villar epithelium throughout the
small intestine but were concentrated in the jejunum Two morphologically distinct meronts were
detected in these sections one at the villar tips which was larger and lacking in undifferentiated mass
and the other at the base of the villi or rarely in the intestinal crypts Gametogony was predominantly
27
observed at the villar tips and was noted throughout the small intestine These organisms were considered
consistent with Eimeria ictidea based on descriptions by Hoare (1927) from domestic ferrets
A second small spherical to subspherical tetrasporic dizoic oocyst was documented that had a
pink double wall a granular residual body and lacked both oocyst polar body and micropyle This
smaller oocyst measured 126plusmn12 microm (108-152) by 119plusmn09 microm (101-129) with a SI of 106 The
sporocysts were elongate with the presence of a Stieda body and sporozoites contained refractile bodies
Similar to the larger Eimeria species described above merogony and gametogony were observed within
the villar epithelium throughout the small intestine with endogenous developmental stages most
numerous in the jejunum The meronts were small with 16 or fewer merozoites Micro- and
macrogamonts were observed clustered within the apical third of the villar epithelium as were meronts
Jolley et al (1994) determined these small spherical oocysts to be consistent with Eimeria furonis as
described by Hoare (1927) from domestic ferrets
Jolley et al (1994) described a third type of coccidial oocyst occasionally detected in small
numbers within the BFF fecal samples the authors did not state whether this third type of oocyst was
recovered from wild or captive BFF The oocysts measured 370plusmn13 microm (350-386) by 223plusmn23 microm
(212-232) with a SI of 106 Attempts to sporulate collected oocysts were largely unsuccessful and
corresponding endogenous stages were not identified on histopathologic examination of necropsied
ferrets precluding further morphologic identification of the parasite It should be noted that coccidial
oocysts with similar measurements had not been detected in wild or captive prey species available for
ingestion by BFF (Jolley et al 1994)
Previous to this report coccidial oocysts had been isolated from the feces of BFF in two captive
populations (Carpenter amp Hillman 1979 Williams et al 1988) The abstract by Carpenter amp Hillman
(1979) did not describe the oocysts whereas Williams et al (1988) stated that two Eimeria species (one
with larger oocysts and one with smaller oocysts) were identified within the fecal samples but they were
28
not identified further Interestingly Williams et al reported both Eimeria species to be shed in the feces
of all ferrets concurrently affected by distemper and by approximately 30 of the clinically healthy
ferrets at the time of investigation
Non-enteric coccidia have been reported from captive BFF in one facility by two authors (Jolley
et al 1994 Williams et al 1988) Both reports presumably describing the same case(s) noted the
presence of endogenous coccidial life stages in histologic sections of respiratory tissue and merozoites of
an unidentified coccidium in impression smears of the urinary bladder from BFF diagnosed with canine
distemper Meronts were observed within the epithelium of the trachea a large bronchus and associated
bronchial glands Jolley et al (1994) described the lesions as occurring in the same ferret whereas in the
earlier report by Williams et al (1988) they are described as occurring in two different ferrets There have
been no subsequent published reports of systemic coccidiosis in black-footed ferrets and no cases have
been identified within the pathology database of the Toronto Zoo captive BFF population or by the
current SSP pathologist (Dr Michael M Garner personal communication)
There is a significant information gap regarding the pre-patent periods and pathogenicity of both
identified Eimeria species in BFF and studies to further characterize the eimeriid coccidia of the BFF are
lacking
153 Morbidity mortality and clinical signs associated with enteric coccidiosis in black-footed ferrets
The clinical signs of enteric coccidiosis in black-footed ferrets include mucoid to hemorrhagic
diarrhea abdominal discomfort lethargy appetite loss vomiting and dehydration In some cases sudden
death precedes the development of diarrhea Both adult and juvenile BFF are affected by the disease
which causes significant morbidity and mortality in captive populations (Bronson et al 2007) One
retrospective study of the captive BFF population at the Smithsonian National Zoological Park
determined that the most common cause of death in juvenile BFF (aged 30 days ndash 11 months) was
gastrointestinal pathology (524 of juvenile deaths) with 636 of these cases caused by enteric
29
coccidiosis (Bronson et al 2007) Despite the significance of this disease to the captive population its
effect on morbidity and mortality in wild BFF populations is unknown To the authorrsquos knowledge no
routine surveys of fecal parasites have been conducted on wild-born or captive released BFF during
yearly spotlighting events at ferret release sites However samples may be collected opportunistically if
fecal material is identified within the traps used to catch wild BFF during yearly surveys at release sites
Where fecal samples have been analyzed a 13 prevalence of coccidiosis has been identified in wild
born BFF (Dr Rachel Santymire personal communication) Fecal samples have been collected from BFF
at four release sites within the USA Wind Cave National Park (South Dakota) Badlands (South Dakota)
Conata Basin (South Dakota) and Aubrey Valley (Arizona) and positive samples were identified only at
the first site (Dr Rachel Santymire personal communication) Although radio-telemetry has been used at
some release sites to determine sources of mortality and factors involved in survival its use is not
widespread Furthermore the nocturnal and fossorial lifestyle of the BFF is a significant impediment to
the surveillance and monitoring of disease in this species
16 TREATMENT PREVENTION AND CONTROL OF INFECTION BY EIMERIA SPP
161 Current recommendations for treatment of eimeriid coccidia in carnivores
Described anticoccidial therapies for carnivores come from research in domestic cats and dogs
infected by Cystoisospora species these tissue coccidia (family Sarcocystidae) are only distantly related
to the Eimeria species infecting the BFF and other ferrets Current therapeutic recommendations by the
Companion Animal Parasite Council (CAPC 2013) for treatment of described Cystoisospora species
isolated from cats and dogs include the following amprolium (300-400 mg daily for 5 days in dogs 110-
200 mg daily for 7-12 days in dogs 60-100 mgkg daily for 7 days in cats) amproliumsulfadimethoxine
(150 mgkg amprolium and 25 mgkg sulfadimethoxine daily for 14 days in dogs) diclazuril (25 mgkg
for one dose in cats) furazolidone (8-20 mgkg 1-2 times daily for 5 days in dogs and cats) ponazuril (20
mgkg daily for 1-3 days in dogs and cats) quinacrine (10 mgkg daily for 5 days in cats)
30
sulfadimethoxine (50-60 mgkg daily for 5-20 days in dogs and cats) sulfadimethoxineormetoprim (55
mgkg sulfadimethoxine and 11 mgkg ormetoprim daily for 7-23 days in dogs) sulfaguanidine (150 or
200 mgkg daily for 6 days or 100-200 mgkg every 8 hours for 5 days in dogs and cats) toltrazuril (10-
30 mgkg daily for 1-3 days in dogs) trimethoprimsulfonamide (30-60 mgkg trimethoprim daily for 6
days if gt4kg 15-30 mgkg trimethoprim daily for 6 days if lt4kg) (CAPC 2013) Notably the use of all
drugs listed by the CAPC is considered off-label with the exception of sulfadimethoxine
162 Current recommendations for anticoccidial treatment and prophylaxis in domestic and black-
footed ferrets
1621 Domestic ferrets
Recommended daily oral treatment regimens for enteric coccidiosis in domestic ferrets include
amprolium (19 mgkg once daily 05 mgkg) decoquinate (05 mgkg) sulfadimethoxine (300 mgkg in
drinking water) or sulfadiazine-trimethoprim (30 mgkg once daily) all administered for a minimum of
two weeks (Bell 1994 Patterson amp Fox 2007 Patterson et al 2014) Both the aforementioned
coccidiostats amprolium and decoquinate are sold in large formats and are ideal for use in larger
operations such as breeding facilities research facilities or rescue centers Other anticoccidial therapies
used in domestic ferrets include toltrazuril (20 mgkg) and ponazuril (30-50 mgkg) once daily It should
be noted that all anticoccidial therapy used in domestic ferrets is considered off-label drug use
Multiple follow up fecal examinations should be performed after the treatment regimen is
complete and large groups may need to be treated multiple times Routine cage cleaning is also important
to decrease the environmental oocyst burden and prevent re-infection and in the case of coccidial
outbreaks ferrets should be transferred to clean cages multiple times during the course of anticoccidial
therapy Disinfectants such as bleach or quaternary ammonium compounds or dry heat should be used
for effective environmental decontamination (Patterson et al 2014)
31
1622 Species Survival Plan recommendations for black-footed ferrets
Treatment and prophylaxis of enteric coccidiosis with oral sulfadimethoxine was previously
recommended by the BFF Species Survival Plan (SSP) However due to a suspicion of decreasing
efficacy of treatment ponazuril has been recommended recently for treatment Due to the perceived
exquisite sensitivity of BFF to enteric coccidia the current SSP recommendation for treatment is oral
ponazuril at 30 mgkg once if ferrets are to be transported anesthetized stressed or are otherwise
suffering from another illness or injury (even in the absence of clinical signs or fecal shedding) The same
single oral dose of 30 mgkg is also recommended for kits at weaning (30-35 days of age) post weaning
(40-45 days of age) and prior to anesthesia for initial examination and vaccines (50-60 days of age)
Large crowded or otherwise stressed litters should be administered 30 mgkg orally once every 7-10 days
during the period of stress For treatment of coccidial diarrhea diagnosed by fecal examination 30 mgkg
orally once every 7 days for two doses or 50 mgkg orally once daily for 3 days in food (repeated in 7
days) is recommended In BFF with clinical signs of dehydration administration of subcutaneous or
intravenous fluid therapy has been performed Additional therapy with other antibiotics is sometimes
provided in cases with severe clinical signs or where secondary or primary bacterial enteritis is suspected
There is no pharmacokinetic or pharmacodynamic information available for the use of
anticoccidial drugs in BFF or other Mustelidae and thus it is unknown whether the current dose or
frequency of administration is truly appropriate for treatment of coccidiosis In 2 to 3-month-old piglets
administered a single dose of ponazuril orally at 5 mgkg peak serum concentration occurred at 42 hours
(36-48 hr) and elimination half-life was ~56 days (Zou et al 2014) In llamas administered ponazuril as
a single dose of 20 mgkg orally peak serum concentration occurred at 84 hours and elimination half-life
was ~56 days (Prado et al 2011) In domestic cows administered ponazuril as a single 5 mgkg dose
orally peak serum concentration occurred at 48 hours and elimination half-life was 58 hours (Dirikolu et
al 2009) The relevance of serum drug concentrations for treating an intestinal infection that lacks
extraintestinal life stages is likely minimal because the highest drug dose will reach the site of concern
(intestines) and systemic distribution is not required
32
Furthermore no safety or efficacy studies have been performed in any ferret species to validate
the current uses of either sulfadimethoxine or ponazuril for treatment nor have the current recommended
treatment lengths been validated However anecdotal information based on current usage would indicate
that they are safe at the current dosages and frequencies of administration as no adverse effects have been
reported A recent efficacy study in shelter dogs and cats showed that oral ponazuril (50 mgkg)
administered once daily for 3 days was effective for treatment of infection with Cystoisospora as
determined by a reduction in or cessation of fecal oocyst shedding at 4 and 8 days post treatment
Treatment efficacy in this study was inversely correlated to fecal oocyst counts at the initiation of
treatment (Litster et al 2014) Interestingly efficacy of this dose compared to the other two treatment
groups (single 50 mgkg or 20 mgkg oral dose) did not seem to differ but no statistical analysis was
performed Given the ubiquitous use of ponazuril in captive breeding facilities and concerns regarding
resistance of coccidia species to sulfadimethoxine therapy information on minimum effective doses and
dose regimes would be necessary to inform appropriate future SSP treatment and management plans and
to minimize development of drug resistance
17 VACCINES AGAINST COCCIDIA
171 Theory
The development of resistance of protozoal parasites to chemotherapeutic agents has resulted in a
shift towards the development of vaccines for the protection of domestic livestock Immunity to enteric
coccidiosis in avian and mammalian species involves both humoral and cell mediated responses Eimeria
spp infection in sheep rats poultry and other species generally results in a protective immune response
against subsequent re-infections (Catchpole et al 1993 Shi et al 2000) Interestingly this is not the case
for some host parasite interactions for example a recent report indicated that primary infection with E
ninakohlyakimovae in goat kids did not provide protective immunity against subsequent challenge with
the same parasite (Ruiz et al 2013)
33
Vaccines can be divided into four general categories live vaccines inactivatedkilled vaccines
subunit vaccines and recombinant vaccines Live vaccines are orally administered using small numbers of
infectious oocysts or oocysts from strains with low pathogenicity and result in patent but ideally sub-
clinical infections in the host that will elicit a protective immune response Such live vaccines can be
produced using attenuated forms of the pathogen of interest for example in chickens using ldquoprecociousrdquo
strains of Eimeria spp These precocious strains undergo a reduced number of merogonic replications
within the host cells and thus fewer oocysts are shed in the feces of vaccinated animals This reduction in
endogenous merogonic cycles reduces the amount of damage to the intestinal epithelium as well as
reducing the number of oocysts contaminating the environment
Another strategy has been to use live parasites with truncated life cycles An example of this is
the Toxoplasma gondii vaccine developed to prevent abortion in sheep This parasite was passaged
multiple times through a mouse host resulting in an inability to produce tissue cysts (Meeusen et al
2007) This is desirable as the cyst stage of this parasite normally inhibited by the immune system can be
reactivated during periods of stress or immunocompromise The potential drawbacks of live vaccines
include 1) the ability to produce and isolate adequate numbers of coccidial oocysts to meet vaccine
production requirements 2) the potential development of clinical disease in the host as a result of
inoculation 3) the need for all susceptible individuals to receive the vaccine simultaneously to prevent
fecal-oral inoculation of unvaccinated animals with high doses of the infective agent likely to be present
in a shared environment through fecal shedding
Inactivated vaccines are produced when the microbe of interest is killed via application of heat
radiation or chemical treatment prior to inoculation into the host species While safer because they cannot
induce disease in the inoculated patient inactivated vaccines stimulate a reduced immune response
compared with live vaccines and are consequently less effective Subunit vaccines contain single or
multiple antigens of importance in initiating the host immune response rather than the entire pathogen of
concern Subunit vaccines cannot induce disease in the immunized host but are more difficult to produce
34
because they require a detailed understanding of host immune response to infection Recombinant
vaccines involve the genetic modification of a vector (virus or bacteria) one capable of infecting the host
of interest to contain DNA of the pathogen of interest These vectors induce an immune response in the
vaccinated host but as with subunit vaccines cannot induce disease However recombinant vaccines are
again difficult to produce because they require an in depth understanding of the life cycle stages genes
and antigens targeted by the host immune response to infection There are currently no recombinant
vaccines marketed in Canada for use in veterinary medicine against protozoal disease
Creation of effective vaccines against protozoal parasites is complicated by parasite antigenic
diversity during the different life cycle stages and among protozoal species and strains of the same species
(Meeusen et al 2007) Although most parasites induce some level of immunity in their host species the
immunological response to different parasite life stages and species has been poorly characterized for
most coccidia Furthermore many parasites have developed mechanisms to evade host immune responses
or to continue survive and replicate in and transmission by previously infected hosts Our limited
understanding of the immune responses against coccidial antigens has restricted commercial vaccine
production to live or attenuated vaccines (Meeusen et al 2007)
A notable disadvantage of anticoccidial vaccines is that they need to be developed for each
coccidial species of interest because of the species-specific nature of the immune responses this is a
considerable limitation compared with anticoccidial drugs that can have a much wider spectrum of action
(Vermeulen 2005) While the requirement for mass production of vaccine is a limiting factor for vaccines
developed for the agricultural industry this drawback would be less important for production of a vaccine
to be used in an endangered species
172 Species successes in anticoccidial vaccination
The first successful immunization against coccidiosis was reported in 1918 in dogs (Hall amp
Wigdor) In this report a dog that had previously recovered from coccidial infection with Diplospora
35
bigemina was fed three increasing doses of live non-attenuated coccidial culture (at 14 32 and 48 days
post recovery from primary infection) which resulted in no development of clinical signs and no oocyst
shedding for 11mdash18 days after each challenge Subsequently immunization of dogs and cats against
coccidia with protection lasting up to seven months was reported by Andrews (1926) Immunization of
albino rats to eimeriid infection after administration of three or more sublethal doses of Eimeria
nieschulzi via gastric intubation was reported by Morehouse (1938) further experiments showed that
sporozoites did not enter the host intestinal epithelium in immunized rats given a challenge dose
(Morehouse 1938) Similar findings were reported in chickens immunized against Eimeria tenella that
had 50 fewer intra-epithelial sporozoites following challenge compared to naiumlve birds (Augustine and
Danforth 1986) Conversely chickens previously inoculated with Eimeria acervulina exhibited more
intracellular sporozoites after challenge than naiumlve birds but sporozoites were not observed to develop in
previously immunized birds (Augustine and Danforth 1986) These findings provide further evidence that
the immune response to Eimeria spp may differ among host species
Vaccination against Eimeria species has been most successful and is most widely used in the
poultry industry particularly in breeder and layer flocks Almost all vaccines marketed for poultry are
live vaccines (attenuated and non-attenuated) Vaccination against other apicomplexan parasites in
domestic mammals has also been achieved but has been generally less effective for disease prevention
and is less widely available Marketed killed and inactivated (attenuated) vaccines include those
containing killed tachyzoites of Neospora caninum for cattle (Neoguard Merck Animal Health) and
chemically inactivated merozoites of Sarcocystis neurona for horses (EPM Vaccine Fort Dodge ndash no
longer in production) A subunit vaccine for Babesia canis in dogs uses cultured antigen (Pirodog
Merial) Available live vaccines include a vaccine against Toxoplasma gondii in sheep (Ovilis Toxovax
Intervet) that uses an attenuated temperature sensitive strain (S48)
36
18 RESEARCH GOALS AND OBJECTIVES
181 Objectives
a) To determine and characterize (morphologically and molecularly) the enteric coccidial species
currently affecting the black-footed ferret population
b) To describe the natural history of enteric coccidiosis in captive black-footed ferrets including
pre-patent period shedding frequency and burdens and morbidity and mortality rates
c) To compare molecular morphologic and life history characteristics of enteric coccidial species
identified in domestic ferrets to those in black-footed ferrets
d) To validate domestic ferrets as an experimental model for intestinal coccidiosis in the black-
footed ferret
182 Hypotheses
a) Multiple Eimeria species will be isolated from the black-footed ferret population
b) The Eimeria species identified from black-footed ferrets will be the same as those previously
described in domestic ferrets
c) A single pathogenic Eimeria species will be implicated in recorded outbreaks of clinical
coccidiosis during the period of study
d) Domestic ferrets can act as an experimental model of intestinal coccidiosis for black-footed
ferrets
183 Applications
The goal of this project is to better characterize the enteric coccidia of the endangered black-
footed ferret in order to set the stage for improved disease prevention and treatment To the authorrsquos
knowledge this project is the first attempt to isolate and perform molecular characterization of the
coccidial species endemic in the black-footed ferret population This information will be used to compare
these species to known coccidia from domestic ferrets and other related mammals As experimental work
37
cannot be carried out on enteric coccidiosis in the BFF due to its endangered status if the domestic ferret
can be validated as an experimental model studies of the patterns of anticoccidial resistance and
development of immunity against Eimeria spp can be undertaken in vivo The ultimate goal would be the
development of an autogenous vaccine used to improve survival of ferret kits and reduce morbidity and
mortality associated with coccidiosis in BFF captive breeding programs Based on clinical experience
stressful life events such a breeding weaning and transfer between institutions appear to increase the risk
of coccidial outbreaks in adult BFF As such vaccination could assist in reducing disease outbreaks in
BFF associated with various management activities There is no data on the significance of coccidiosis in
wild populations and limited means of disease surveillance following release vaccination during captive-
rearing or pre-release conditioning of BFF would be an ideal method of reducing the potential effects of
this disease in released and free-living BFF Increasing the numbers of ferrets being released to the wild
and releasing ferrets immune to the subsequent threat of coccidiosis would support the goals of the
conservation initiative for the black-footed ferret
38
CHAPTER 2 MOLECULAR CHARACTERIZATION OF ENTERIC COCCIDIA FROM
DOMESTIC FERRETS (MUSTELA PUTORIUS FURO)
This chapter has been submitted for publication as
Adriana R Pastor Dale A Smith and John R Barta (2017) Molecular Characterization of Enteric
Coccidia from Domestic Ferrets (Mustela putorius furo) Vet Parasitol Regional Studies and Reports (In
review)
ABSTRACT
Combined morphometric and molecular characterization of coccidia that infect domestic ferrets
(Mustela putorius furo) was completed to improve the diagnostic specificity of enteric lsquococcidiosisrsquo in
this host Coccidia positive fecal samples (n=11) and formalin fixed paraffin embedded intestinal tissues
(n=3) from domestic ferrets were collected from diagnostic laboratories in Canada and Europe An
average of 35 and 13 domestic ferret fecal samples per year were coccidia-positive when tested by
Canadian and European diagnostic laboratories respectively during the period 2008-2015 Oocyst
morphometrics and sequence genotyping at two loci (nuclear 18S rDNA [nu 18S rDNA] and
mitochondrial cytochrome c oxidase subunit I [mt COI]) were conducted on all samples The first nu 18S
rDNA and mt COI sequences for Isospora (=Cystoisospora) laidlawi and the first mt COI sequence for
Eimeria furonis were generated during this study Phylogenetic analysis of the mitochondrial COI
sequences demonstrated that E furonis was most closely related to E cf ictidea isolated from a black-
footed ferret (Mustela nigripes) and that I (=C) laidlawi was closely related to C canis and C felis The
identifications provided by diagnostic laboratories of the specific parasite species present in a sample
showed poor agreement with their identifications based on genotyping obtained in this study Molecular
techniques appear to be essential for accurate determination of coccidial species responsible for individual
and group outbreaks of coccidiosis and for further understanding of eimeriid host-parasite relationships
Key words coccidia Cystoisospora laidlawi domestic ferret Eimeria furonis Eimeria ictidea Mustela
putorius furo
39
21 INTRODUCTION
Coccidia are host-specific parasites of the phylum Apicomplexa with greater than 2000 species
named to date (Duszynski et al 2000 Upton 2000) The eimeriorinid coccidia (suborder Eimeriorina)
include typical intestinal coccidia such as Eimeria Isospora and Cyclospora species belonging to the
family Eimeriidae as well as tissue (cyst-forming) coccidia such as Cystoisospora Besnoitia Toxoplasma
and Sarcocystis species that belong to the family Sarcocystidae (see Cox 1994)
Enteric coccidia affect both domestic ferrets (Mustela putorius furo) and their wild counterparts
In his initial descriptions of enteric coccidiosis in domestic ferrets Hoare (1927 1935b) did not observe
clinical signs of intestinal disease associated with infection More recently it has been recognized that
enteric coccidiosis can result in clinical signs ranging from mild transient diarrhea to more severe disease
with dehydration lethargy depression weight lossemaciation inappetence and death (Blankenship-Paris
et al 1993 Powers 2009 Sledge et al 2011 Hoefer et al 2012 Patterson et al 2014) Rectal prolapse
has also been reported in ferrets with enteric coccidiosis (Hillyer 1992 Hoefer et al 2012) Disease
appears to be most common in young or stressed animals In one study co-infection with coccidia and
Lawsonia intracellularis (Desulfovibrio sp) was diagnosed in 4 of 19 ferrets with proliferative bowel
disease (Li et al 1996) these ferrets presented with clinical signs including diarrhea lethargy anorexia
weight loss dehydration and emaciation Two cases of biliary coccidiosis have also been reported in
domestic ferrets infection was associated with biliary epithelial hyperplasia cholecystitis and
cholangiohepatitis (Williams et al 1996 Kaye et al 2015)
Three species of coccidia affecting domestic ferrets were originally described and named by
Hoare (1927) Eimeria ictidea Eimeria furonis and Isospora (=Cystoisospora) laidlawi The three
species were detected in feces from 50 domestic ferrets involved in an outbreak of canine distemper at a
research facility For each parasite the author described the morphology of sporulated oocysts isolated
from feces as well as sporulation time (exogenous life stages) All subsequent reports of morphologic
diagnoses of these coccidia have been based on Hoarersquos original descriptions The pre-patent period
40
(minimum duration of endogenous development) in inoculated naiumlve ferrets was described for E furonis
and E ictidea as 6 and 7 days respectively (Hoare 1935b) The pre-patent period for I (=C) laidlawi
was not determined because the number of oocysts available was insufficient for an experimental
infection trial
Hoare (1927) described the sporulated oocysts of E furonis as follows spherical double outer
wall with a thin colourless outer layer and a thick yellowish inner layer no micropyle or residual body
and measuring on average 128 times 120 microm (length [L] 112-144 width [W] 104-128 shape index [SI]
107) Sporocysts were spindle-shaped with one end constrictedblunted contained a residual body and
on average measured 8-88 times 4 microm Sporozoites were vermiform with one end narrower than the other
arranged head to tail and each had a central nucleus a clear vacuole was identified in some sporozoites at
their broad posterior end
The sporulated oocysts of E ictidea were described as follows oval or elliptical with a double
outer wall with a thin colourless outer layer and a thick yellowish inner layer no micropyle or residual
body and measuring on average 236 times 175 microm (L 184-272 W 128-208 SI 135) Sporocysts were
irregularly oval with one end broad and the other more constricted contained a residual body and
measured 115 times 65 microm on average Sporozoites were vermiform with one end narrower than the other
arranged head to tail in the sporocysts and had a central nucleus and a clear vacuole at their broad
posterior end
The sporulated oocysts of I (=C) laidlawi were ovoid with a double outer wall with a thin
colourless outer layer and a thick yellowish inner layer had no micropyle or residual body and measured
on average 34 times 29 microm (L 320-368 W 272-304) A SI of 117 can be calculated from the original
mean dimensions Two sporocysts were identified each containing four sporozoites and no Stieda body
sporocysts were elliptical contained a residual body and measured 208 times 144 microm on average
Sporozoites were sausage shaped with one end slightly pointed and had a central nucleus and a clear
41
vacuole identified at the pointed end Sporozoites were arranged with pointed ends all at the same pole of
the sporocyst
Oocysts identified as Cystoisospora ohioensis have been reported from fecal samples collected
from healthy domestic ferret kits in a large American ferret breeding operation that also housed juvenile
domestic dogs (Patterson and Fox 2007) The method of identification of this parasite was not described
A second similar institution reported the presence of Cystoisospora (=Isospora) species also thought to
be C ohioensis in routine fecal examination of their colony (Dr Bambi Jasmin personal
communication) Identification in this case was performed by the Animal Health Diagnostic Center at
Cornell University and was based on morphometrics using light microscopy The significance of these
findings is unknown but no clinical signs or histologic lesions were described in the ferrets shedding
these oocysts The definitive hosts for C ohioensis are canids including the domestic dog
More recently molecular techniques have been used for the more precise identification of
coccidia Nucleotide sequences like morphological features diverge over time under selective pressure
however recent evolutionary divergence among coccidia is more likely to be reflected in molecular as
compared to morphologic differences Thus nucleotide sequences that are more similar are inferred to be
more closely related and to have diverged more recently (Cox 1994)
Molecular characterization of ferret coccidia has only been performed for one species Eimeria
furonis Abe et al (2008) extracted DNA from oocysts from the feces of a single domestic ferret with
clinical signs resulting from coccidial enteritis Using primers initially developed for molecular
identification of Cyclospora species (see Matsubayashi et al 2005) small subunit ribosomal DNA (nu
18S rDNA) primers CYC1FE (5ʹ-TAC CCA ATG AAA ACA GTT T-3prime) and CYC4RB (5prime-CGT CTT
CAA ACC CCC TAC TG-3prime) were used to amplify a 347 base pair fragment of nu 18S rDNA The
amplicon was sequenced (GenBank AB329724) and compared with previously published partial nu 18S
rDNA sequences from 40 Eimeria two Isospora and four Cyclospora species The resulting phylogram
42
grouped E furonis with E alabamensis (cattle) and E meleagrimitis (turkey) In the same study the
microscopic morphology of the oocysts was used to identify this coccidial species as Eimeria furonis by
comparison with Hoarersquos (1927 1935b) published descriptions of Eimeria furonis and Eimeria ictidea
Sledge et al (2011) also used nu 18S rDNA to identify Eimeria furonis as the cause of three
distinct outbreaks of enteric disease in domestic ferrets Initial identification was performed using
morphometrics of sporulated oocysts collected from feces in one of the three outbreaks being
investigated Formalin fixed paraffin embedded intestinal segments from ferrets from each of the
outbreaks contained multiple coccidial life stages when examined by light microscopy PCR amplification
of a 247 base pair (bp) amplicon of the nu 18S rDNA was generated from DNA isolated from stored
formalin-fixed tissues for further genetic analysis Analysis and sequencing of amplicons from all three
groups showed 100 identity to sequences previously reported by Abe et al (2008) for the gene encoding
E furonis nu 18S
In 2015 Kaye et al identified coccidia within hepatobiliary lesions in a domestic ferret receiving
immunosuppressive therapy for red cell aplasia DNA was extracted from frozen liver and a fragment of
the nu 18S rDNA was amplified using the primers previously described by Sledge et al(2011) The
authors reported that the DNA sequence of the amplicon had 100 identity to the published nu 18S
rDNA sequence of E furonis and 95 identity to the nu 18S rDNA of E myoxi (rodent) E alabamensis
(cattle) and Isospora robini (avian) (Kaye et al 2015)
It is difficult to estimate the current prevalence of enteric coccidia within the North American
domestic ferret population and no studies have been conducted to do so Fecal samples submitted to
veterinary diagnostic laboratories from domestic ferrets in Canada are uncommon and samples positive
for coccidia appear infrequently (Dr Donald Martin personal communication) Conversely in Europe
the prevalence of coccidia within the domestic ferret population appears to be higher based on
submissions to a large veterinary diagnostic laboratory in Germany Data from Idexx Vet Med Lab in
Ludwigsburg Germany was compiled to review the prevalence of coccidia and Giardia within fecal
samples from domestic ferrets (Pantchev et al 2011) The authors reported that of 284 fecal samples
43
submitted from 2002-2004 18 (63) had detectable coccidial oocysts on fecal flotation Based on
morphologic characteristics oocysts were identified as E ictidea E furonis I laidlawi (herein referred
to as I (=C) laidlawi as noted above) and another unidentified Isospora species Comparative data from
the same laboratory from 2009-2010 included sample submissions from 253 ferrets 21 (83) of which
were positive for coccidial oocysts on fecal flotation Nine of the samples were identified as containing E
furonis three contained both E furonis and I (=C) laidlawi eight contained only I (=C) laidlawi and
one sample contained both E furonis and E ictidea identification in all cases was again based on oocyst
morphometrics No statistically significant difference in the occurrence of coccidial oocysts was detected
when data from the two periods were compared (Fisherrsquos exact test P=041) (Pantchev et al 2011)
The purpose of the present study was to perform a more detailed molecular characterization of the
coccidial species isolated from domestic ferrets to estimate prevalence of the different coccidial species
within the Canadian domestic ferret population and to associate morphologic and molecular
characteristics of a greater range of enteric coccidial species in order to improve diagnostic accuracy
22 MATERIALS amp METHODS
221 Fecal samples
Multiple diagnostic laboratories within Ontario Canada1 and a major European diagnostic
laboratory 2 were solicited for fecal samples from domestic ferrets shedding coccidial oocysts Fecal
samples were diagnosed positive for coccidia based on fecal flotation and light microscopic identification
of Eimeria or Cystoisospora species Eleven samples were collected during the study period (from 2014-
2017) and preserved in potassium dichromate (25 wv) eight from Europe and three from Canada
Centrifugal flotation with saturated salt solution (Ryley et al 1976) was used to isolate oocysts from fecal
samples for genomic DNA extraction Genomic DNA extraction and purification were performed using a
1 Animal Health Laboratory Guelph ON Antech Diagnostics Canada Ltd Mississauga ON IDEXX Canada
Markham ON 2 Vet Med Labor GmbH Division of IDEXX Laboratories Ludwigsburg Germany
44
QIAamp DNA Mini Kit (Qiagen Hilden Germany) according to manufacturerrsquos instructions After
addition of DNAzol to the samples (Qiagen Hilden Germany) samples were vortexed using 05 mm
glass beads (Biospec Products Inc Bartlesville OK USA) prior to extraction in order to fracture the
oocyst walls and release the sporocysts Concentrations of the resultant DNA were estimating using a
Nanodrop 2000 spectrophotometer (NanoDrop Products Wilmington DE USA) and stored at 4 degC for
immediate use or minus20 degC for later use
For each laboratory the number of domestic ferret fecal sample submissions numbers diagnosed
positive for coccidial oocysts and number of each coccidial species identified in positive samples were
tabulated for each of the years 2008-2015
222 Formalin fixed intestinal tissues
Major diagnostic pathology services across Canada3 were contacted to identify cases of enteric
coccidiosis identified on necropsy of domestic ferrets Cases were considered positive based on the
presence of asexual or sexual life stages of the parasites in intestinal sections The histologic sections on
each positive case were reviewed re-described and organisms measured (AP DAS) Gross necropsy
reports for all cases were also reviewed to identify any clinical correlates associated with enteric
coccidiosis DNA was extracted from ten 5-6 microm scrolls of formalin fixed paraffin embedded tissue
(FFPE) using the QIAamp DNA FFPE Tissue Kit (Qiagen) as per manufacturer instructions
223 Molecular characterization
Regions from the nu 18S rDNA and mitochondrial cytochrome c oxidase subunit I (mt COI)
DNA were amplified by polymerase chain reaction (PCR) from each sample using the primers listed in
Table 21 PCR amplification was performed for all samples in a volume of 25 microl containing ~100 ng of
3 Animal Health Centre Abbotsford BC Animal Health Laboratory Guelph ON Faculteacute de meacutedecine veacuteteacuterinaire
Universiteacute de Montreacuteal Saint-Hyacinthe QC Histovet Surgical Pathology Guelph ON IDEXX Canada
Markham ON Prairie Diagnostic Services Inc Saskatoon SK
45
genomic DNA 1times PCR buffer 15 mM MgCl2 02 mM deoxyribonucleotide triphosphates (dNTPs) 400
nM of each primer and 1 U of Invitrogen Platinum Taq DNA Polymerase (Thermo Fisher Scientific
Toronto ON Canada) Reactions were performed on a Bio-Rad T100 PCR thermal cycler (Bio-Rad
Laboratories Singapore) Samples were denatured and Taq polymerase activated at 95 degC for 3 min then
subjected to 35 cycles of 94 degC for 30s anneal at 50-62 degC (see Table 21 for specific anneal conditions
for the various primer pairs) for 30s and extension at 72 degC for 30-75s (see Table 21) followed by a
final extension at 72 degC for 7 min Suitable DNA (ie genomic DNA from an Eimeria or Sarcocystis sp)
was included in the PCR reactions to act as a positive control for the reaction chemistry All amplification
products were subjected to electrophoretic separation using 15 submarine agarose gel stained with
ethidium bromide and visualized on an ultraviolet transilluminator (Spectronics Corporation New York
NY USA) DNA band size was determined by comparison with a 1 kb DNA ladder (GeneRuler 1kb Plus
DNA ladder Thermo Fisher Scientific Waltham MA USA) Bands were excised with a new sterile
scalpel blade and PCR products were purified from the gel using a QIAquick Gel Extraction Kit (Qiagen)
PCR products were cycle sequenced using an ABI Prism 7000 Sequence Detection System (Applied
Biosystems Inc Foster City CA USA) by the Molecular Biology Unit of the Laboratory Services
Division University of Guelph (Guelph ON Canada) using the amplification primers to obtain
sequences in both directions The resulting chromatograms were aligned and analyzed with Geneious Ver
818 or later (Biomatters Limited Auckland New Zealand) and high quality consensus sequences
generated The resulting consensus sequences were searched from within Geneious against publically
available sequences on the BLAST server (blastncbinlmnihgovBlastcgi) using the blastn search
algorithm against the nrnt database (GenBank+EMBL+DDBJ+RefSeq ndash AA or DNA) Resultant new
nucleotide sequences were submitted to GenBank
46
224 Phylogenetic analysis
To determine the phylogenetic affinities of the newly obtained sequences with sequences from
related apicomplexan taxa representative nu 18S rDNA and mt COI sequences were downloaded from
GenBank with special reference to sequences from parasites that infect members of the order Carnivora
Nuclear 18S and mt COI sequences were aligned independently using MAFTT v7017 (Katoh et
al 2002) executed from within Geneious and then concatenated into a combined nu18S rDNA mt COI
dataset Multiple sequences from a single parasite were used to generate consensus sequences for each
locus as described by Ogedengbe et al (2017) Aligned sequences were trimmed to the length of the
largest newly generated nu 18S sequence Phylogenetic trees were generated using Bayesian Inference
(BI) using MrBayes Ver 326 (Huelsenbeck and Ronquist 2001) executed from within Geneious the
combined nu 18S and mt COI alignment was partitioned to permit locus-appropriate substitution models
to be applied to each partition For the nu 18S sequence partition the general time reversible (GTR)
substitution model (nst=6) with gamma rate variation (ie a GTR+G+I model) was applied For the mt
COI sequence partition the codon (M1) substitution model (using translation table 4 [ie lsquometmtrsquo]) was
used instead of the GTR with the remaining parameters remaining the same
The resulting tree was rooted using a pair of adeleid coccidia (Hepatozoon spp) as the taxonomic
outgroup All BI analyses were run for a chain length of 1000000 with tree sampling every 1000
following a burn-in of 100000 with default settings of 4 heated chains and heated chain temp of 02
47
23 RESULTS
231 Fresh fecal samples
From 2008-2015 inclusive the Canadian diagnostic parasitology laboratory4 received an average
of 1206 (range 81-160) domestic ferret fecal samples yearly the European parasitology laboratory5
received a yearly average of 230 samples (range 213-270) The number of fecal samples diagnosed as
positive for coccidial oocysts per year on fecal flotation during this time averaged 35 (range 0-8) and
130 (range 6-20) for the Canadian and European laboratories respectively The diagnosing laboratories
used oocyst morphometrics to identify the species of coccidia present Almost all coccidia-positive
submissions to the Canadian laboratory were identified as containing an I (=C) species based on light
microscopy Coccidia in only three samples from the Canadian laboratory were identified as E furonis
one in each of 2010 2012 and 2014 E ictidea was not identified in any samples submitted to the
Canadian laboratory Approximately equal numbers of coccidia-positive samples from the European
laboratory were identified as E furonis and I (=C) laidlawi each year Only two samples from the
European laboratory contained oocysts that were identified as Eimeria ictidea using morphometrics one
from each of 2011 and 2013 Laboratory submissions to both laboratories are summarized in Table 22
Twelve fecal samples preserved in potassium dichromate were received for analysis by the
authors Eleven samples had previously been identified as containing a single coccidial species five
containing E furonis two containing E ictidea and four containing I (=C) laidlawi A final sample had
been identified as containing a mix of E furonis and Cystoisospora canis Results of microscopic and
molecular characterization of these samples are summarized in Table 23
4 IDEXX Canada Markham ON
5 Vet Med Labor GmbH Division of IDEXX Laboratories Ludwigsburg Germany
48
232 Formalin fixed samples
Only three cases of coccidiosis were identified in domestic ferrets within the databases of the five
diagnostic laboratories that participated in the retrospective study Histologic sections of intestine were
received from these three cases which originated in Ontario6 and Quebec7 The Quebec sample (P2010-I)
was collected in 2010 and the Ontario samples (93-40404 and 17-008571) in 1993 and 2017 respectively
On gross necropsy the small intestinal contents of case P2010-I were described as pasty mucoid
yellow-brown feces with some blood For case 93-40404 the small intestines were described as empty
but melena was present within the terminal portion of the large intestine Scant intestinal contents and
dark brown fecal material in the colon were described in case 17-008571
In all cases endogenous developmental stages of coccidia were visible in histological sections
(Figure 21 is exemplary of the findings from one case) Hematoxylin and eosin stained sections from
P2010-I contained two affected regions of small intestine The intestinal mucosa of the first region
contained numerous asexual life stages and moderate numbers of sexual life stages as well as a small
number of oocysts free within the lumen The second section contained tissues that were poorly
preserved nonetheless 0-4 oocysts per 400times field were identifiable within the intestinal lumen Two
regions of affected small intestine were identified from 93-40404 after screening of all submitted sections
both contained low numbers of sexual and asexual endogenous stages Within one region there were small
numbers of meronts within the intestinal mucosa and lamina propria The second region had small
numbers of oocysts within cells of the epithelium and lamina propria as well as free within the intestinal
lumen In case 17-008571 multiple sections of jejunum contained numerous coccidian meronts gamonts
6 Animal Health Laboratory Guelph ON
7 Faculteacute de meacutedecine veacuteteacuterinaire Universiteacute de Montreacuteal Saint-Hyacinthe QC
49
and oocysts within intestinal villi within the ileum scattered epithelial cells also contained these various
life stages
Average length and width of oocysts were measured from slide sections for all cases For P2010-
I oocyst average length and width were determined from seven oocysts to be 94 microm (range 85-105) and
75 microm (range 69-84) respectively with a SI of 125 (range112-140) Average length and width of
oocysts measured from 93-40404 were determined from 5 oocysts to be 2814 microm (range 229-341) and
233 microm (range 180-308) respectively with a SI of 123 (range 110-146) For the third case 17-
008571 only 2 oocysts were identified and average length and width of oocysts measured 982 microm (range
973-992) and 845 microm (range 821-870) respectively with a SI of 116 (range 114-118)
233 Molecular characterization
DNA was successfully extracted from all twelve fecal samples and two cases with formalin fixed
tissue samples Attempts at amplification of DNA extracted from sample 93-40404 using the primer pairs
listed in Table 21 were unsuccessful Molecular identification results and GenBank accession numbers
for the remaining samples are summarized in Table 23 Both the nu 18S rDNA and mt COI sequences
from I (=C) laidlawi were unique when compared with available sequences from other Cystoisospora
species within the public databases However sequences from I (=C) laidlawi were most similar to
sequences from C canis and C felis and somewhat more divergent from sequences from members of the
C ohioensis species complex Two apparent genotypes of E furonis were identified based on nu 18S and
mt COI sequencing results Genotype 1 represented by EU sample 9014 had 100 identity to previously
published sequences of the nu 18S locus from two isolates from Japan (GenBank AB239130 and
AB329724) Genotype 2 represented by EU sample 907 and Canadian sample 17-008571 had 994
identity at the nu 18S locus (3 single nucleotide differences [SNDs] over 561 base pair region [bp]) to the
three sequences above belonging to E furonis genotype 1 Pairwise alignment of mt COI sequences from
both genotypes identified only 2 SNDs (996 pairwise identity over 513 bp region) Partial mt COI
50
sequences of E furonis from both genotypes were only distantly related (941 pairwise identity 30
SNDs over 513 bp and 905 pairwise identity 49 SNDs over 513 bp respectively) to publicly available
sequences from Eimeria ictidea from the black-footed ferret (Mustela nigripes) (GenBank KT203399)
and Eimeria mephitidis from the striped skunk (Mephitis mephitis) (GenBank KT203398) the only other
Eimeria species infecting members of the Carnivora for which sequence was available
234 Phylogenetic analysis
A phylogenetic reconstruction based on concatenated partial nu 18S rDNA and mt COI sequences
of E furonis I (=C) laidlawi and related coccidia is illustrated in Figure 22 The combined 18SCOI-
based tree demonstrates that the two Eimeria species from ferrets form a well-supported monophyletic
group that branches among a collection of other eimeriid coccidia that infect mammals The sarcocystid
parasite of the domestic ferret I (=C) laidlawi was found to group as the sister taxon to C canis that
together formed a monophyletic group with the closely related C felis all three of these closely related
Cystoisopora species possess comparatively large egg-shaped oocysts that are similar morphologically
24 DISCUSSION
The present work has generated the first nu 18S rDNA and mt COI sequences for Cystoisospora
laidlawi and the first mt COI sequence for Eimeria furonis both isolated from the domestic ferret In this
study histologic presence of organisms and microscopic identification of oocysts shed in feces have been
correlated with published and novel nu 18S and mt COI sequences
Eimeria ictidea was not identified in any Canadian sample and this coccidium was reported in
only 2 of 1840 fecal samples submitted from across the European Union (EU) to IDEXX Germany during
2008-2015 suggesting that E ictidea is not a frequent cause of enteric coccidiosis in domestic ferrets in
Canada or the EU
51
During the study period (2008-2015) almost twice as many domestic ferret fecal submissions
were made to the European as compared to the Canadian diagnostic laboratory however the prevalence
of coccidia-positive samples was similar The methodology used in this report cannot be used to
determine the actual prevalence of enteric coccidial infection (coccidiasis) or disease (coccidiosis) within
the domestic ferret population Fecal samples may be submitted to laboratories either as a result of
investigation into enteric disease or as part of a routine health examination Thus without historic
information accompanying each sample one can simply identify the proportion of positive samples and
compare the frequency of the finding of different coccidial species Prospective surveys of fecal samples
from healthy and sick domestic ferrets with greater sample size would be necessary to determine the true
prevalence of these parasites within the population and to infer their clinical significance
Comparatively few mitochondrial COI sequences have been generated for apicomplexan parasites
compared with other genetic loci the majority of published sequences obtained from Apicomplexa are
from nu 18S The disadvantage of using nu 18S rDNA sequences for parasite identification is that they
are poor at distinguishing among closely related eimeriid coccidia due to the highly conserved nature of
the nuclear ribosomal RNA locus In contrast mt COI sequences appear to be more useful for
distinguishing closely related coccidian species (Ogedengbe et al 2011) but are less useful than nu 18S
rDNA sequences for inferring more ancient relationships among more distantly related coccidia
Consequently the combined use of nu 18S rDNA and mt COI sequencing has been recommended for
improved species description and phylogenetic analysis (El-Sherry et al 2013) For these reasons both nu
18S and mt COI sequences were analysed in the present study
Despite adequate quantities of DNA extracted from the Ontario laboratory sample (93-40404)
successful amplification did not result with any primer pair (Table 21) Potential reasons for this include
degradation of formalin-fixed DNA into fragments too small for amplification with the desired primers
perhaps as a result of extended length of time in formalin prior to paraffin embedding or length of time
stored as FFPE tissue (23 years) or insufficient parasite DNA within the paraffin scrolls The primer pairs
52
used appear to be useful for most eimeriid coccidia (Ogedengbe 2015) and successfully amplified both
Eimeria species from DNA isolated from oocysts so it is unlikely that failure to amplify DNA from this
sample resulted from an inability of the primers used to recognize the parasite seen on section
Two genotypes of E furonis were identified in this study Genotype 1 was identified only from
samples originating from domestic ferrets in Europe but exhibited 100 identity based on nu 18S
sequencing with previously published sequences from both Japan and the USA Genotype 2 was
identified from samples originating from domestic ferrets in both Canada and Europe The small number
of single nucleotide differences between the two genotypes at two genetic loci in different genomes are
consistent with intraspecific variation (ie strain variation)
As might have been expected because of their morphological and host similarities nu 18S and mt
COI sequences of E furonis were determined to be most similar to an Eimeria species (E ictidea)
previously isolated from black-footed ferrets (Mustela nigripes) these eimeriid coccidia formed a
monophyletic group that was distinct from other eimeriid coccidia infecting mammals in the phylogenetic
analyses based on combined nu 18S rDNA and mt COI sequences Similarly the nu 18S rDNA and mt
COI sequences of I (=C) laidlawi are most similar to sequences from two other Cystoisospora species of
carnivores (C canis and C felis) that both have large egg-shaped oocysts comparable to those of I (=C)
laidlawi Both morphometrics and genotyping support the close relationships among these three
sarcocystid coccidia of carnivores These molecular data confirm that transfer of Isospora laidlawi to the
genus Cystoisospora by Barta et al (2005) is warranted
The previous light microscopic identifications of coccidial species in 3 of the 11 fecal samples
were not in agreement with the molecular findings These results were not surprising because light
microscopy has been shown to be an insensitive tool for distinguishing among apicomplexan parasites at
both the genus and species level Furthermore re-evaluation of these samples by the authors revealed that
many of the samples that were identified incorrectly based on morphometrics contained primarily
53
unsporulated oocysts making accurate identification based on microscopic appearance highly
challenging These findings further underscore the importance of molecular methods in accurate parasite
identification In the absence of molecular tools accurate measurement of oocyst size shape and
determination of SI can be useful for differentiating among species of Eimeria and Cystoisospora
however this can only be performed accurately on sporulated oocysts from feces Interestingly the size
and shape indices of oocysts of E furonis measured in histologic sections did not match those previously
described by Hoare (1927) for the same oocysts in feces despite molecular confirmation of identity
Thus measurements of oocysts in histologic sections are not recommended for use in coccidial
identification
Our observations highlight the utility of molecular methods for identifying enteric coccidia
infecting domestic ferrets and suggest that diagnoses based on morphological methods should perhaps be
limited to broad determinations of disease etiology (ie lsquococcidiosisrsquo or lsquococcidiasisrsquo) Using molecular
techniques we were able to differentiate morphologically similar coccidial species isolated from the feces
of domestic ferrets and specifically identify parasites seen in histological sections of ferret intestine
Molecular techniques thus appear to be essential for determining the coccidial species responsible for
individual and group outbreaks of coccidiosis and for further understanding of eimeriid host-parasite
relationships
ACKNOWLEDGEMENTS
Many thanks to Julia Whale and Alex Leveille for their assistance and encouragement during the course
of this project The authors would like to recognize the contributions of Dr Donald Martin (IDEXX
Canada) and Drs Nikola Pantchev and Majda Globokar (IDEXX Germany) for the contributions of data
and samples to this project The authors would also like to recognize the Laboratoire de Pathologie
(Service de diagnostic Faculteacute de meacutedecine veacuteteacuterinaire St Hyacinthe Quebec) and the Animal Health
Laboratory (Guelph Canada) for contributions of samples and data to this project Finally this project
was made possible through funding by the Toronto Zoo Residency Research Fund to DASAP and partial
54
funding from a Natural Sciences and Engineering Research Council of Canada (NSERC) Discovery Grant
(400566) to JRB
55
Table 21 Amplification primers for nuclear 18S rDNA and mitochondrial COI loci anneal temperatures (Ta) extension times and expected PCR
product sizes used in the identification of enteric coccidia from domestic ferrets (Mustela putorious furo)
Gene Target Primer Pairs Primer Sequence (5ʹ-3ʹ) Size (bp) Ta (degC) Anneal (sec) Reference
nu 18S rDNA CYC1FE TACCCAATGAAAACAGTTT 560 52 45 Matsubayashi et al (2005)
CYC4RB CGTCTTCAAACCCCCTACTG Matsubayashi et al (2005)
Cocci_18S_595F CCGCGGTAATTCCAGCTCCAAT 216 62 30 Present study
Cocci_18S_847R GCTGMAGTATTCAGGGCGACAA Present study
Lank_18S_224F TCATAGTAACCGAACGGATC 1080 54 60 Ogedengbe (2015)
Api_SSU_2733R CGGAATTAACCAGACAAATC Mathew et al (2000)
mt COI COI_10F GGWDSWGGWRYWGGWTGGAC 500 52 30 Ogedengbe et al (2011)
COI_500R CATRTGRTGDGCCCAWAC Ogedengbe et al (2011)
COI 272F CAATTCTAYGATGCCGCWTT 222 52 30 Present study
COI_500R CATRTGRTGDGCCCAWAC Ogedengbe et al (2011)
Sdae-COI_260F GATCTTTATGTTYTTRATGCC 890 50 75 Ogedengbe (2015)
Sdae-COI_1147R CATTACCCATAACYACACC Ogedengbe (2015)
56
Table 22 Summary of fecal samples from domestic ferrets (Mustela putorius furo) submitted to two diagnostic laboratories from 2008-2015
No fecal samples positive for coccidia
No samples submitted
(percentage positive)
No samples positive for
Cystoisospora sp
No samples positive for
Eimeria furonis
No samples positive for
Eimeria ictidea
Year Canada Europe Canada Europe Canada Europe Canada Europe
2008 3140 (21) 6214 (28) 3 2 0 4 0 0
2009 2160 (12) 14214 (65) 2 9 0 5 0 0
2010 8127 (63) 20213 (94) 7 10 1 10 0 0
2011 0114 (0) 17215 (79) 0 9 0 7 0 1
2012 3108 (28) 10231 (43) 2 4 1 6 0 0
2013 281 (25) 16270 (59) 2 13 0 2 0 1
2014 6127 (47) 12234 (51) 5 6 1 6 0 0
2015 4108 (37) 9249 (36) 4 3 0 6 0 0
Total 28 (29) 104 (56) 25 56 3 46 0 2
Average
year 35 130 31 70 04 58 00 03
Legend Numbers in brackets refer to the percent of the total number of fecal samples submitted
57
Table 23 Morphologic and molecular identification of coccidia from domestic ferrets (Mustela putorius furo)
Sample ID Source
External Lab Morphologic
Diagnosis Morphologic Diagnosis (ARP) Molecular Diagnosis
mt COI GenBank
Accession
nu 18S rDNA
GenBank Accession
93-40404 FFPE enteric coccidia Histologic sample - - -
P2010-I FFPE enteric coccidia Histologic sample E furonis Identical to MF774036 Identical to MF774678
17-008571 FFPE NP Histologic sample E furonis Same as MF774036 Same as MF774678
17-008571 feces NP E furonis E furonis MF774036 MF774678
907 feces E furonis E furonis E furonis MF774035 MF774679
938 feces I (=C) laidlawi no oocysts visualized I (=C) laidlawi MF774037 MF774677
952-A feces E ictidea Cystoisospora sp I (=C) laidlawi Identical to MF774037 Identical to MF774677
9958 feces E furonis no oocysts visualized E furonis Identical to MF774034 Identical to MF774680
9011 feces E furonis E furonis E furonis Identical to MF774035 Identical to MF774679
9014 feces E furonis E furonis E furonis MF774034 MF774680
9017 feces I (=C) laidlawi Cystoisospora sp I (=C) laidlawi Same as MF774037 Same as MF774677
9040 feces I (=C) laidlawi no oocysts visualized I (=C) laidlawi Same as MF774037 Same as MF774677
912-260 feces I (=C) laidlawi Cystoisospora sp I (=C) laidlawi Same as MF774037 Same as MF774677
CAN-2016-1 feces C canis + E furonis Cystoisospora sp I (=C) laidlawi MF774038 MF774676
Legend FFPE = formalin fixed paraffin embedded intestinal sections - = unsuccessful = morphologic diagnosis performed by JRB same as = 100 sequence
identity with listed GenBank entry over entire sequence length identical to = 100 sequence identity but shorter sequence than listed GenBank entry
58
Figure 21 Life stages of Eimeria furonis within the small intestinal epithelium of a domestic ferret
(Mustela putorius furo) Asexual life stages merozoites (black circle) Sexual life stages oocyst
(solid black arrow) macrogamonts (open arrows with labels) microgamont (dotted black arrow)
Hematoxylin and eosin staining scale bar = 25μm
25 microm
59
Figure 22 Phylogenetic relationships of coccidia (Eimeria ictidea Eimeria furonis and Isospora
(=Cystoisospora) laidlawi) from domestic (Mustela putorius furo) or black-footed (Mustela
nigripes) ferrets based on partial nuclear 18S rDNA and mitochondrial COI sequences of these
parasites and related apicomplexan parasites A summary of the sources of the molecular data for
the remaining taxa included in this phylogenetic analysis are found in Supplementary Table 1 of
Ogedengbe et al (2017) Bayesian support is indicated for each node horizontal distance is
proportional to hypothesized evolutionary change (scale indicates sequence divergence of 10)
60
CHAPTER 3 MORPHOLOGICAL AND MOLECULAR CHARACTERIZATION OF
ENTERIC COCCIDIA ISOLATED FROM BLACK-FOOTED FERRETS (MUSTELA
NIGRIPES)
ABSTRACT
Black-footed ferrets (BFF Mustela nigripes) are the only ferret species native to North America
and have been identified as endangered since 1967 Starting in 1986 a multi-institutional effort has been
breeding this species in captivity with successful reintroductions back into the wild Coccidiosis is
recognized as a cause of significant juvenile morbidity and mortality in captive breeding programs and
can result in significant population losses Little is known about the etiology of enteric coccidiosis in
BFF Coccidia positive fecal samples (n=12) and formalin fixed paraffin embedded intestinal tissues
(n=11) were obtained from BFF in the Toronto Zoo and Louisville Zoo Species Survival Plan (SSP)
populations Oocyst morphometrics and sequence genotyping at three loci (nuclear 18S rDNA
mitochondrial cytochrome c oxidase subunit I and mitochondrial cytochrome c oxidase subunit III) were
conducted Results suggest that the same Eimeria species E ictidea was the cause of enteric coccidiosis
in both SSP populations in both juvenile and adult age classes Wider research is indicated to determine
whether these findings are representative of the larger captive and wild BFF populations
31 INTRODUCTION
Black-footed ferrets (BFF) are one of only three wild ferret species worldwide the other two
being the European polecat (Mustela putorius) and the Siberian polecat or steppe polecat (Mustela
eversmanii) The BFF the only native North American ferret species was formerly distributed throughout
the North America prairie ecosystem but were considered extinct by the late 1950s In 1964 a single
population was discovered in Mellette County South Dakota Progressive decline of this population in
subsequent years resulted in the decision by United States Fish and Wildlife Service to initiate a captive
breeding program for the species From 1971-1973 four females and five males were captured for this
61
purpose Despite successful breeding no kits survived and the last adult ferret in this captive colony died
in 1979 BFF were again presumed extinct in the wild based on annual surveys of the initial capture site
In 1981 a dead BFF was discovered by a ranch dog outside of Meeteetse Wyoming allowing wildlife
biologists to identify another colony of BFF This colony flourished until 1985 when an outbreak of
canine distemper in this wild BFF population and an outbreak of sylvatic plague in the local prairie dog
population resulted in sharp population declines From 1985 through 1987 all 24 of the remaining BFF
were trapped and brought into captivity to re-initiate the captive breeding program Six ferrets in this
initial group died of canine distemper while in captivity and seven of the remaining eighteen survivors
are the founding population of the current captive breeding population Today this captive breeding
population consists of approximately 300 BFF distributed among multiple institutions (Santymire et al
2014)
Since 1986 a multi-institutional effort has been breeding BFF in captivity with reintroductions
back into the wild in selected locations in Canada the USA and Mexico Currently six facilities
participate in the BFF Species Survival Plan (SSP) the Toronto Zoo United States Fish and Wildlife
Services National Black-footed Ferret Conservation Center National Zoorsquos Smithsonian Conservation
Biology Institute Louisville Zoo Cheyenne Mountain Zoo and the Phoenix Zoo (Black-footed Ferret
Recovery Implementation Team 2011) In order to provide the best genetic matches BFF are transferred
among the six institutions for breeding Approximately 300-400 kits are produced annually between the
six SSP facilities with ~200 of these kits allocated for release to the wild yearly (Santymire et al 2014)
As of 2011 over 8000 BFF kits had been produced in captive breeding facilities (Black-footed Ferret
Recovery Implementation Team 2011)
Multiple infectious diseases pose a significant risk to the captive breeding and post-release
survival of BFF including canine distemper and sylvatic plague (Santymire et al 2014 USFWS BFF
Recovery Program 2017) Coccidiosis is recognized as a cause of significant juvenile morbidity and
mortality in captive breeding programs and can result in significant population losses (Bronson et al
62
2007 Santymire et al 2014 USFWS BFF Recovery Program 2017) While the effects of the disease on
the wild population are not clear a prevalence of approximately 13 has been reported based on fecal
samples collected from wild BFF born at release sites (Dr R Santymire personal commication)
Coccidia are eukaryotic host-specific parasites of the phylum Apicomplexa affecting numerous
hosts within a wide taxonomic range Two species of coccidia Eimeria ictidea Hoare 1927 and Eimeria
furonis Hoare 1927 have been identified in black-footed ferrets based on morphometrics (Jolley et al
1994) Jolley et al examined fecal samples from six captive BFF during a distemper outbreak as well as
samples from wild BFF They described one medium-sized ovoid eimeriid oocyst with a double wall
presence of a polar body and lacking both an oocyst residual body and micropyle Oocysts of this Eimeria
species (sp) measured 232 times 155 microm (range 182-274 times 130-162) with a shape index (SI) of 150 The
sporocysts were elongate with the presence of both sporocyst residuum and a Stieda body Sporozoites
contained prominent refractile bodies at the posterior end and were aligned anterior to posterior within
sporocysts These oocysts shed by all six captive ferrets were considered consistent with Eimeria ictidea
based on descriptions by Hoare (1927) On histopathologic examination of intestinal sections parasites
undergoing merogony and gamogony were observed within the villar epithelium throughout the small
intestine but were concentrated in the jejunum (Hoare 1935b) parasite life stages were not described
from other tissuesorgans
A second small spherical to subspherical eimeriid oocyst was also documented in the captive
ferrets by Jolley et al (1994) this second species had a pink double oocyst wall a granular residual body
and lacked both oocyst polar body and micropyle This smaller species measured 126 times 119 microm (range
108-152 times 101-129) with a SI of 106 The sporocysts were elongate and possessed a Stieda body and
sporozoites contained refractile bodies Similar to the larger Eimeria sp described above merogonic and
gamogonic stages were observed within the villar epithelium throughout the small intestine but were
most numerous in the jejunum Jolley et al (1994) concluded these small spherical oocysts were
consistent with E furonis described by Hoare (1927) from domestic ferrets
63
Jolley et al (1994) described a third type of coccidial oocyst occasionally detected in small
numbers within BFF fecal samples however the authors did not state whether this third oocyst
morphotype was recovered from wild or captive animals The oocysts measured 370 times 223 microm (range
350-386 times 212-232) with a SI of 106 Attempts to sporulate collected oocysts were unsuccessful and
corresponding endogenous stages were not identified on histopathologic examination of necropsied
ferrets precluding further morphologic identification of the parasite It should be noted that coccidial
oocysts with similar measurements had not been detected in wild or captive prey species available for
ingestion by BFF making it unlikely that this coccidial species would have been a pseudoparasite (Jolley
et al 1994)
Previous to this report by Jolley coccidial oocysts had been isolated from the feces of BFF in two
captive populations (Carpenter amp Hillman 1979 Williams et al 1988) The abstract by Carpenter amp
Hillman (1979) did not describe the oocysts whereas Williams et al (1988) stated that two Eimeria sp
(one with larger oocysts and one with smaller oocysts) were observed within the fecal samples but they
were not identified further Interestingly Williams et al reported both Eimeria sp to be shed in the feces
of all ferrets concurrently affected by distemper and by approximately 30 of the clinically healthy
ferrets at the time of investigation
Non-enteric coccidia have also been reported by two authors from captive BFF at one facility
(Jolley et al 1994 Williams et al 1988) Both reports which presumably described the same case(s)
noted the presence of endogenous coccidial life stages in histologic sections of respiratory tissue and
merozoites of an unidentified coccidium in impression smears of the urinary bladder from BFF diagnosed
with canine distemper Meronts were observed within the epithelium of the trachea a large bronchus and
associated bronchial glands In the later report Jolley et al (1994) described the lesions as occurring in the
same ferret whereas in the earlier report by Williams et al (1988) they are described as occurring in two
different ferrets Paraffin blocks containing formalin fixed tissues from these cases have since been
discarded precluding further attempts at parasite identification with molecular methods Subsequent to
64
these reports further cases of systemic coccidiosis in BFF have neither been published nor identified
within the pathology database of the Toronto Zoo captive BFF population nor by the current SSP
pathologist (Dr Michael M Garner personal communication)
Previous characterization of coccidia from black-footed ferrets has been based on host species
affected tissues in the host and morphometric characterization of life stages in histologic sections and
oocyst characteristics using light microscopy It is known that morphologically similar Eimeria species
are not necessarily conspecific and may vary in host specificity and pathogenicity Molecular
characterization is thus required to accurately identify coccidia to the species level No molecular
characterization of coccidian parasites from black-footed ferrets has been performed to date
There is a significant information gap regarding which parasite species are implicated in
morbidity and mortality events associated with enteric coccidiosis in BFF and whether different coccidia
are associated with this disease in adult versus juvenile age classes or in different SSP institutions
Studies to further characterize the eimeriid coccidia of the BFF are warranted to improve the management
of this disease in the captive population The objectives of this research were to morphologically and
molecularly characterize coccidia associated with enteric disease in BFF at the Toronto Zoo and in other
SSP facilities
32 MATERIALS AND METHODS
321 Fecal samples
Twelve fecal samples were collected during the study period (from 2014-2016) and preserved in
potassium dichromate (25 wv aqueous) seven from the Toronto Zoo and five from the Louisville Zoo
Centrifugal flotation with saturated salt solution (Ryley et al 1976) was used to isolate and concentrate
oocysts from fecal samples for light microscopic examination and genomic DNA extraction
One to two drops of the supernatant from the centrifugal flotation were placed directly on a slide
and beneath a coverslip The morphology and dimensions of sporulated oocysts were documented using a
65
Provis AX70 photomicroscope (Olympus Canada Richmond Hill ON Canada) fitted with a digital
imaging device (Infinity3-1C Lumenera Corporation Ottawa ON Canada) controlled using iSolution
Lite image analysis software (Hoskin Scientific Burlington ON Canada) operated at a total
magnification of 1000times Morphologic features noted for each oocyst included oocyst wall morphology
number of sporocysts presence or absence of a micropyle micropyle cap residual body and polar
granules For sporocysts size number of sporozoites per sporocyst and presence or absence of Stieda
body and sporocyst residuum were noted Alignment of sporozoites within the sporocyst and
presenceabsence of refractile bodies within sporozoites were also described The sporulated oocyst
length and width measurements (in microm) were then used to calculate the SI for each measured oocyst
Morphologic and morphometric features were compared to previously published values for E furonis and
E ictidea from domestic and black-footed ferrets
Genomic DNA extraction and purification were performed using a QIAamp DNA Mini Kit
(Qiagen Hilden Germany) according to manufacturerrsquos instructions as described in Chapter 2 (Materials
amp Methods)
322 Formalin fixed intestinal tissues
The pathology records of the Toronto Zoo were searched from 1993-2016 for cases of BFF
diagnosed with enteric coccidiosis on histopathology For each case slides of histologic sections from all
submitted intestinal segments were reviewed to confirm the presence of sexual andor asexual life stages
within the intestinal epithelium Scrolls (5-6 microm) were cut from the paraffin blocks containing affected
intestinal sections and DNA extracted from the formalin fixed paraffin-embedded tissue (FFPE) using
the QIAamp DNA FFPE Tissue Kit (Qiagen Toronto Ontario) as per manufacturerrsquos instructions
66
323 Molecular characterization
Molecular characterization of coccidial isolates was performed on oocysts purified from fresh
fecal samples (isolated as described above) that were collected from juvenile and adult ferrets from
2014-2016 and DNA extracted from FFPE samples of BFF intestine containing parasite life stages
Regions from the nuclear 18S (SSU) rDNA (nu 18S rDNA) mitochondrial cytochrome c oxidase
subunit I (mt COI) DNA and mitochondrial cytochrome c oxidase subunit III (mt COIII) DNA were
amplified by polymerase chain reaction (PCR) from each sample using the primers listed in Table 31 and
methodology described in the Materials amp Methods section of Chapter 2 Table 31 also contains the
specific anneal conditions used for the various primer pairs Genomic DNA from an Eimeria species of
poultry was included in the PCR reactions to act as a positive control for the reaction chemistry A
representative selection of the newly generated nucleotide sequences resulting from the above were
submitted to GenBank
DNA obtained from oocysts collected from fecal samples during the first year of the study (2014)
was used to generate a complete mitochondrial genome (see Chapter 6 for details) using primer pairs and
sequencing primers summarized in Table 31 All subsequent samples collected in 2015 and 2016 had
shorter mt COI and mt COIII sequences obtained to permit genotyping of all collected oocysts at these
two loci The location of each primer in the nu 18S mt COI and mt COIII genetic locus is illustrated in
Figure 31
33 RESULTS
From 2014-2016 coccidia-positive fecal samples were obtained from twelve BFF ferretsferret
groups from the Toronto Zoo and Louisville Zoo SSP populations (see Table 32) Nine samples were from
single housed adults between the ages of 1-5 years (63 MaleFemale) Two samples were from mixed
groups one pooled fecal sample from four adults (FERA-1 13 MF) and one fecal sample from a family
67
group consisting of a dam and five kits (23 MF) One fecal sample was collected from a juvenile male
ferret at the time of necropsy
Eleven BFF with enteric coccidiosis were identified in the Toronto Zoo necropsy reports from
1998-2016 and all were confirmed by histological re-evaluation (Table 32) Both juvenile (n=9 36 MF)
and adult ferrets (n=2 20 MF) were represented
331 Morphometric characterization
Twelve coccidia-positive fecal samples were identified from adult and juvenile BFF from 2014-
2016 by on site laboratories at either the Toronto Zoo or the Louisville Zoo Fecal flotation and light
microscopic re-examination of the samples identified coccidial oocysts in 10 of these 12 samples
Morphometric characterization was performed on six samples in which there was adequate
quantity and quality of sporulated oocysts for examination These included three samples from single-
housed adults one from a juvenile at the time of necropsy one of pooled feces from a group of adult
ferrets and one of pooled feces from a family group (dam and kits) Two of the three samples from
single-housed adults were from the same ferret on different dates in 2016 the dates of collection were
separated by a period in which shedding of oocysts was not identified on routine repeated fecal
examinations Oocysts were elliptical with a colourless double wall and contained four sporocysts each
with two sporozoites Sporocysts were ovoid and both Stieda body and residual body were present
Sporozoites exhibited an anterior to posterior alignment within the sporocysts and refractile bodies were
identified (Figure 32) Results for length width and shape index of sporulated oocysts including range
and average values are summarized in Table 33 and Figure 32 The average measurements based on the
results of all 148 oocysts measured were length 2398 microm (1859-3057) width 1855 microm (1373-2383)
and shape index 130 (101-160)
The same measurements were performed on 59 sporocysts from a single ferret (Noodle) and
results are as follows average length 1280 microm (898-1480) average width 738 microm (505-1028) and
average SI 176 (124-247) In one sample (Mohawk-2) sporozoites were visible free on the slide
68
Measurement of three of these provided an average length of 1068 microm (1044-1117) and an average
width of 341 microm (316-393)
332 Molecular characterization
Molecular characterization was successfully performed on oocysts from seven of 10 fecal
samples containing coccidial oocysts and FFPE tissue from nine of the 11 necropsy cases (see Table 32)
Attempts at amplification of DNA extracted from necropsy samples Z228-98 and Z137-14 using the
primer pairs listed in Table 31 were unsuccessful Similarly attempts at PCR and sequencing of DNA
extracted from fecal oocysts from two Toronto Zoo BFF Jenna and Ruckus were unsuccessful
Molecular identification results for the remaining samples are summarized in Table 32
Only one Eimeria species E ictidea was identified in all enteric coccidiosis cases diagnosed at
necropsy in both juvenile and adult BFF at the Toronto Zoo from 1998-2014 This same species was
identified in all Toronto and Louisville Zoo BFF fecal samples that were sequenced successfully (n=8)
with the exception of a single case from Louisville This Louisville ferret was identified as having a
rodent pseudoparasite (Eimeria species) in the submitted fecal sample the eimeriid pseudoparasite had
986 sequence identity at the mt COI locus to the murine coccidium Eimeria falciformis All sequences
generated for E ictidea exhibited 100 sequence identity at the mt COI and COIII loci
Novel nu 18S rDNA mt COI and mt COIII sequences were generated for E ictidea from both
geographic locations and deposited in GenBank (Accessions MF860826 MF860827 MF860823
MF860825 MF860822 MF860824) Sequences were compared to those previously published for related
eimeriid coccidia The nu 18S rDNA sequence from Eimeria ictidea isolated from the Toronto Zoo BFF
had 9736 identity (14 single nucleotide differences) to the previously published sequences from
isolates of E furonis from domestic ferrets (Mustela putorius furo) in Japan (GenBank AB239130 and
AB329724) and newly generated sequences from Canadian and European isolates (GenBank MF774678-
MF774680 see Chapter 2 and Figure 33) In contrast nu 18S rDNA sequence of E furonis from
domestic ferrets (see Chapter 2) showed 9953 to 100 identity (0 to 3 SND) to the Japanese
69
sequences Comparison of newly generated partial sequences of the mt COI region from E ictidea from
BFF to isolates of E furonis (GenBank MF774034-MF774036) from DF and E mephitidis (GenBank
KT203398) from the striped skunk (Mephitis mephitis) the only carnivore Eimeria sp for which a mt
COI sequence was previously available reveals only 9415 and 9084 sequence identity respectively
with these other Eimeria spp of carnivores (Figure 34)
34 DISCUSSION
This work presents the first nu 18S rDNA mt COI and mt COIII sequences (nu 18S rDNA -
MF860826 MF860827 mt COI - MF860823 MF860825 mt COIII - MF860822 MF860824) generated
from an intestinal eimeriid parasite of the BFF referred to here as E ictidea collected from multiple BFF
of different ages from two separate captive populations (Toronto Zoo Toronto Ontario Canada and
Louisville Zoo Louisville Kentucky USA)
The morphometric description of coccidial oocysts from BFF in this work are consistent with
previous descriptions of E ictidea from mustelids including BFF the Steppe polecat the European
polecat and domestic ferrets (Hoare 1927 Svanbaev 1956 Jolley et al 1994) Thus I propose the name
E ictidea for the enteric coccidium described from BFF reflecting the similarity in morphology host
species and location of infection in intestinal tissues yet recognizing the absence of species identification
by molecular techniques Molecular characterization of parasites that agree with the description of E
ictidea morphologically from various mustelid host species would allow not only for determination of
whether the parasites are conspecific but would also provide insight into the potential for cross-
transmission among related mustelid hosts
DNA extraction from FFPE samples allowed successful PCR and sequencing of small DNA
fragments (220 bp) in nine of the eleven cases in which the technique was attempted Age of the samples
did not appear to be the major factor associated with successful extraction of good quality DNA the two
samples for which it was unsuccessful were the most recent (2014) and oldest (1998) casesConsequently
it may be possible to use banked FFPE tissues from historic necropsy cases from other SSP institutions
70
and necropsies of wild-born or re-introduced ferrets to determine the identity of the coccidial species
underlying disease in these cases and to better characterize the disease in the greater BFF captive and wild
populations Williams et al (1988) were contacted regarding their historic FFPE samples but formalin
blocks were no longer available for these cases and thus comparisons could not be made Banked FFPE
samples were requested from other SSP institutions however the Convention on International Trade in
Endangered Species of Wild Fauna and Flora (CITES) restrictions on the international transport of DNA
from endangered species did not allow for sample acquisition during the period in which this research was
conducted
Evaluation of FFPE samples from the Toronto Zoo indicate that the same Eimeria species has
been implicated in deaths associated with enteric coccidiosis from 1999-2014 as well as episodes of
clinical disease in ferrets in the Toronto Zoo population from 2014-2016 Samples from coccidia-positive
BFF at the Louisville Zoo in 2016 also contained the same Eimeria species Finding the same parasite at
multiple SSP locations was expected because BFF are transferred among institutions on a yearly basis for
breeding and potential release Consequently these parasites have repeated opportunities to move
between institutions in infected hosts or on contaminated cage materials to become established at a new
location Furthermore the stress of transport and transfer to a new environment may precipitate shedding
of endemic coccidia and increase the risk of a coccidial outbreak this concern is reflected in the SSP
recommendations for prophylactic treatment of all BFF with anti-coccidial medication prior to shipment
(USFWS BFF Recovery Program 2017)
A single BFF from the Louisville SSP population not showing clinical signs consistent with
coccidiosis was identified as having a rodent Eimeria species in the submitted fecal sample
Morphometric characterization of oocysts in this sample was not performed due to the paucity of visible
oocysts however examination at 100times suggested that the oocysts in the sample were ovoid in shape and
of comparable size to oocysts identified in other BFF samples The finding of a rodent Eimeria in a BFF
fecal sample is not unexpected as whole rodents comprise a significant part of the captive BFF diet The
oocysts shed by the BFF were most likely acquired through ingestion of an infected prey item and thus
71
most likely represent pseudoparasitism Molecular characterization was however required to
differentiate this from a case of true enteric coccidiasis
Reports from the first captive BFF population derived from South Dakota indicate the presence
of an unidentified species of enteric coccidium (Carpenter and Hillman 1979) in this group before its
demise in 1979 No reports containing morphometric descriptions of the coccidia from this group were
found on literature review and all parasites of this group have been lost with their hosts All subsequent
reports on enteric coccidiosis in BFF are from ferrets derived from the second founder group from
Wyoming in the 1980s The frequent transfer of ferrets among SSP institutions within the captive
breeding program and to different release sites within North America would be expected to result in the
same Eimeria species being found in all populations The exception to this would be the potential for
cross-transfer of other eimeriid parasites to wild BFF from sympatric mustelid species such as the long-
tailed weasel (Mustela frenata) Jolley et al described two other species of enteric coccidia from this
second captive population in 1994 the first was similar to E furonis of domestic ferrets and the second a
large coccidian parasite of unknown genus Neither of these parasites was identified in the Toronto and
Louisville Zoo populations during the course of this study In order to determine whether these parasites
persist within the present-day BFF populations and their impact on this species more detailed
examination of coccidia-positive fecal samples from captive and wild BFF populations is recommended
Furthermore the molecular identification of enteric coccidia from historic and future necropsy samples of
wild and captive BFF could aid in determining the presence of and contribution to mortality events by
these additional coccidia species
ACKNOWLEDGEMENTS
The authors would like to recognize the Wildlife Health Centre staff at the Toronto Zoo for their
assistance with the collection of fecal samples from the BFF from 2014-2016 The authors would also like
to recognize the Louisville Zoo for their contribution of samples to this project Finally this project was
made possible through funding by the Toronto Zoo Residency Research Fund to DASAP and partial
72
funding from a Natural Sciences and Engineering Research Council of Canada (NSERC) Discovery
Grant (400566) to JRB
73
Table 31 Amplification primers used to sequence the nuclear 18S rDNA mitochondrial COI and COIII loci of Eimeria ictidea originating from
fecal and formalin-fixed paraffin embedded tissue samples from black-footed ferrets (Mustela nigripes) including anneal temeratures (Ta)
extension times and expected PCR product sizes
Figure 31 Nuclear and mitochondrial genetic loci targeted by primers listed in Table 31 and used to characterize Eimeria ictidea originating
from black-footed ferrets (Mustela nigripes)
Gene Target Primer Pairs Primer Sequence (5ʹ-3ʹ) Size (bp) Ta (degC) Anneal (sec) Reference
18SrDNA Sarco_18S_123F TATCAGCTTTCGACGGTAGTGTATT 1480 60 30 Ogedengbe et al (2016)
ERIB10_REV CTTCCGCAGGTTCACCTACGG
mt COI T_Eim_COI_272F CAATTCTAYGATGCCGCWTT 220 52 30 Chapter 2 (Table 21)
COX1-500R CATRTGRTGDGCCCAWAC Ogedengbe et al (2011)
COI-400F GGDTCAGGTRTTGGTTGGAC 800 52 60 El-Sherry et al (2013)
COI-1202R CAAKRAYHGCACCAAGAGATA El-Sherry et al (2013)
mt COIII WG-MT_4140F AGAAAACCTAAAATCATCATGT 1000 52 60 Ogedengbe et al (2015)
Eimeriid_CO3_799R AAGTGAGTTCGCATGTTTAC Ogedengbe et al (2015)
74
Figure 32 A+B) Features and cellular contents of Eimeria ictidea from a black-footed ferret
(Mustela nigripes) Legend Oocyst dotted thin white arrow = polar granule Sporocyst dotted thin
black arrow = Stieda body thick white arrow= sporozoite refractile body thick black arrow=
residuum scale bars as indicated C) Shape index length and width measurements of sporulated
oocysts of Eimeria ictidea from black-footed ferrets (Mustela nigripes) Legend times indicates the
mean Dotted oval indicates one standard deviation around the mean
10 microm 5 microm
75
Figure 33 Comparison of nuclear 18S rDNA sequence alignment of Eimeria ictidea from two black-footed ferrets (Mustela nigripes) to
newly generated (see Chapter 2) and published sequences of Eimeria furonis from domestic ferrets (Mustela putorius furo)
Figure 34 Comparison of mitochondrial cytochrome c oxidase subunit I sequence alignment of Eimeria ictidea from two black-footed
ferrets (Mustela nigripes) to sequences from other eimeriid parasites of carnivores
Identity
Eimeria mephitidis ndash KT2033981
Eimeria ictidea ndash MF860823 (Guanella ndash LZ)
Eimeria ictidea ndash MF860825 (Mystery ndash MTZ)
Eimeria furonis ndash MF774034 (Type 1)
Eimeria furonis ndash MF774035 (Type 2)
Identity
Eimeria ictidea ndash MF860827 (Guanella ndash LZ) Eimeria ictidea ndash MF860826 (Mystery ndash MTZ)
Eimeria furonis ndash MF774680 (Type 1) Eimeria furonis ndash AB3297241 (Type 1) Eimeria furonis ndash AB2391302 (Type 1)
Eimeria furonis ndash MF774678 (Type 2) Eimeria furonis ndash MF774679 (Type 2)
76
Table 32 Morphologic and molecular characterization of coccidia from fecal and FFPE necropsy samples from black-footed ferrets (Mustela
nigripes)
Sample ID Sample Source Year Age (years) Sex Source Microscopic Description^ Molecular Diagnosis
Z228-98 Toronto Zoo 1998 7 M FFPE rare asexual stages -
Z143-99 Toronto Zoo 1999 1 M FFPE sexual and asexual stages E ictidea
Z106-02 Toronto Zoo 2002 008 F FFPE rare asexual stages E ictidea
Z108-03 Toronto Zoo 2003 008 F FFPE sexual and asexual stages E ictidea
Z109-03 Toronto Zoo 2003 008 F FFPE sexual and asexual stages E ictidea
Z124-12 Toronto Zoo 2012 021 F FFPE sexual and asexual stages E ictidea
Z113-13 Toronto Zoo 2013 017 F FFPE sexual and asexual stages E ictidea
Z117-13 Toronto Zoo 2013 025 M FFPE sexual and asexual stages E ictidea
Z118-13 Toronto Zoo 2013 025 M FFPE sexual and asexual stages E ictidea
Z119-13 Toronto Zoo 2013 025 F FFPE sexual and asexual stages E ictidea
Z137-14 Toronto Zoo 2014 021 M FFPE
fresh feces
sexual and asexual stages
POS E cf ictidea
-
E ictidea
FERA_1 Toronto Zoo 2014 gt 1 MF fresh feces POS E cf ictidea E ictidea
Noodle Toronto Zoo 2015 1 M fresh feces POS E cf ictidea E ictidea
Ruckus Toronto Zoo 2016 3 F fresh feces POS -
Mystery Toronto Zoo 2016 1 M fresh feces POS E ictidea
Mohawk Toronto Zoo 2016 1 M fresh feces POS E cf ictidea E ictidea
Jenna Toronto Zoo 2016 1 F fresh feces POS -
Thrope Louisville Zoo 2016 3 M fresh feces NEG -
FloJean Louisville Zoo 2016 2 F fresh feces NEG -
Rigatoni Louisville Zoo 2016 5 M fresh feces POS rodent Eimeria
Guanella +kits Louisville Zoo 2016 2 015 F kits 2M3F fresh feces POS E cf ictidea E ictidea
Clive Louisville Zoo 2016 1 M fresh feces POS E ictidea
Legend FFPE= formalin fixed paraffin embedded Sex MF = samples from family groups containing both sexes - = PCR and sequencing unsuccessful ^ = life stages identified on
histologic section
= mitochondrial COI andor COIII sequencing results Presence (POS) or absence (NEG) of oocysts and morphological identification of oocysts based on measurements when
77
Table 33 Morphometric (length width shape index) characterization of Eimeria ictidea oocysts from fecal samples from black-footed ferrets
(Mustela nigripes)
Sample ID FERA - 1 ^ Z137-14 Noodle Mohawk -1 Mohawk - 2 Guanella Total
Number of oocysts 12 36 32 10 21 37 148
Length (microm) 2333 (2055-2583) 2456 (2111-2848) 2505 (2079-3008) 2779 (2590-3060) 2493 (2036-2822) 2139 (1859-2372) 2398 (1859-3057)
Width (microm) 1676 (1373-2180) 1835 (1643-2232) 1975 (1509-2360) 2253 (2092-2383) 1803 (1549-2017) 1751 (1610-1888) 1855 (1373-2383)
Shape index 135 (103-160) 134 (113-156) 127 (105-155) 124 (113-138) 139 (114-154) 122 (101-145) 130 (101-160)
Legend ^= mixed adult group = dam and kit group
78
CHAPTER 4 NATURAL HISTORY OF ENTERIC COCCIDIOSIS IN THE BLACK-
FOOTED FERRET (MUSTELA NIGRIPES)
ABSTRACT
Black-footed ferrets (BFF Mustela nigripes) the only native North American ferret species are
endangered throughout their former geographic range An intensive captive breeding program produces
animals to supplement re-established wild populations Coccidial enteritis is a major cause of morbidity in
young captive ferrets but the disease also affects adults Limited information is available on the
pathogenesis of intestinal coccidiosis in captive BFF and characterization of the natural history of the
disease for improved prevention and management is imperative The objectives of this research were to
determine morbidity and mortality rates in the Toronto Zoo captive BFF population as well as
characterizing the natural history of the disease in this species through evaluation of shedding patterns
body tissues affected pre-patent period and periods of enhanced host susceptibility to infection
Coccidia-associated mortality in BFF at the Toronto Zoo from 1997-2016 averaged 053 yearly in
adults (range 0-526) and 195 in juveniles (range 0-1667) Clinical signs and histologic lesions in
Toronto Zoo BFF were similar to those described in previous publications A seasonal influence on
oocyst shedding was identified in adult BFF and ferrets appeared to maintain persistent infection with E
ictidea shedding coccidia in multiple years A larger multi-institutional study is required to better
elucidate the natural history of enteric coccidiosis in this species
41 INTRODUCTION
Black-footed ferrets (BFF Mustela nigripes) are the only native North American ferret species
and are endangered throughout their former geographic range When the last remaining truly wild
population underwent serious decline as a result of disease the decision was made by the by United States
Fish and Wildlife Service to capture the remaining 24 animals and establish a captive breeding program
this occurred between 1985 and 1987 Only seven of the captured ferrets bred successfully and are the
79
founders of the current North American BFF population (USFWS BFF Recovery Program 2017) The
captive population which now numbers approximately 300 individuals is distributed among and managed
by six collaborating facilities these include the Toronto Zoo United States Fish and Wildlife Services
National Black-Footed Ferret Conservation Center National Zoorsquos Smithsonian Conservation Biology
Institute Louisville Zoo Cheyenne Mountain Zoo and the Phoenix Zoo (Black-footed Ferret Recovery
Implementation Team 2011 Santymire et al 2014) Since 1991 BFF have been released into
reintroduced into the wild at multiple sites within their former range and over 8000 BFF kits had been
produced in captive breeding facilities as of 2011(Black-footed Ferret Recovery Implementation Team
2011) Twenty-eight BFF reintroduction sites currently exist throughout North America however there
continues to be a need to support wild populations as only a four of the re-established groups are truly
self-sustaining
Enteric coccidiosis is recognized as a cause of significant morbidity and mortality in captive
breeding programs affecting both juvenile and adult animals (Bronson et al 2007 USFWS BFF
Recovery Program 2017) Two Eimeria species Eimeria ictidea and Eimeria furonis have been
identified from cases of entric coccidiosis in BFF (Jolley et al 1994) Jolley et al examined fecal samples
from both wild and captive BFF and provided detailed morphologic descriptions of the oocysts of both
Eimeria spp as well as descriptions of the intestinal pathology associated with infection Asexual and
sexual life stages of both of the aforementioned Eimeria spp were identified on histologic section within
the villar epithelium throughout the small intestine but were concentrated in the jejunum Intestinal
sections from BFF infected with E ictidea exhibited two morphologically distinct meronts one at the
villar tips which was larger and lacking in undifferentiated mass and the other at the base of the villi or
rarely in the intestinal crypts gamogony was predominantly observed at the villar tips and was noted
throughout the small intestine
80
Extraintestinal coccidia have also been reported from captive BFF at one facility (Jolley et al
1994 Williams et al 1988) The authors identified the presence of endogenous coccidial life stages in
histologic sections of respiratory tissue and in impression smears of the urinary bladder from BFF
diagnosed with canine distemper No subsequent reports of systemic coccidiosis in BFF have been
published or identified within the pathology database of the Toronto Zoo captive BFF population or by
the current SSP pathologist (Dr Michael M Garner personal communication)
Recent investigations into the etiologic agents of enteric coccidiosis in BFF at the Toronto Zoo
have identified a single Eimeria species associated with all cases of enteric coccidiosis and associated
mortality in juvenile and adult BFF from 1999-2016 Furthermore this pathogen was identified in fecal
samples based on morphologic and molecular characterization from adult and juvenile BFF in another
zoological collection (Louisville Zoo Kentucky USA) (see Chapter 3) This coccidium is
morphologically consistent with Hoarersquos original description of E ictidea (1927) and is referred to
henceforth as Eimeria ictidea
There is a significant information gap regarding the pathogenicity of E ictidea in BFF The
objectives of this research were to determine morbidity and mortality rates in the Toronto Zoo and
additional captive BFF SSP populations as well as characterizing the natural history of the disease in this
species through evaluation of shedding patterns body tissues affected pre-patent period and periods of
enhanced host susceptibility to infection
42 MATERIALS AND METHODS
421 Toronto Zoo BFF breeding program
At the Toronto Zoo black-footed ferret breeding program all adult ferrets are housed
individually with the exception of dams and kits After the birth of the kits dams are housed with their
offspring from whelp date until removal at approximately 4-6 months of age Routine monthly fecal
81
examinations (direct examination and flotation) are performed in house for all ferrets in the breeding
program based on SSP recommendations to evaluate for the presence of coccidia
422 Fecal oocyst evaluation
Family groups
From 2014-2016 daily fecal examination for coccidial oocysts was initiated for all group-housed
dams and kits In 2014 fecal samples were collected daily from all dams and kits from weaning (30 days
after whelping) to 72 days post whelping Based on 2014 data in 2015 this surveillance was extended
from weaning (35 days post whelping) to 135 days of age Furthermore fecal samples were collected
from the dam for an additional 14 days after removal of kits In 2016 no fecal samples were collected
from dam and kit groups at the Toronto Zoo but samples were submitted from one group of dam and kits
from another SSP population at the Louisville Zoo (Kentucky USA)
Adults
From 2015-2016 daily fecal samples were also collected from all adult ferrets identified as
shedding coccidial oocysts on their monthly routine fecal examination and from clinically ill BFF
Samples were collected for 10-14 days after initial positive sample identification In 2016 fecal samples
were also submitted from four coccidia-positive adult ferrets from the Louisville Zoo population samples
were collected for 7 days post initial identification of shedding
Individual fecal samples were analyzed via flotation using the McMaster method followed by
routine flotation in saturated salt solution (Dryden et al 2005) to determine the presence or absence of
oocysts and oocyst burden (oocysts per gram of feces OPG) Temporal trends in oocyst shedding were
monitored Coccidia-positive ferrets were evaluated visually on a daily basis for presence of clinical signs
consistent with infection Infected juvenile ferrets and adult ferrets were treated with oral ponazuril or
toltrazuril regardless of the presence of clinical signs as per the black-footed ferret SSP
recommendations Based on these recommendations ponazuril is typically administered orally at 30-50
82
mgkg once daily for 3-7 days until clinical signs have resolved or oocyst shedding has been significantly
reduced (USFWS BFF Recovery Program 2017)
423 Retrospective review of pathology records
The pathology records of the Toronto Zoo were searched from 1993-2016 for cases of BFF
diagnosed with enteric coccidiosis on histopathology For each case gross necropsy reports were
reviewed and slides of histologic sections from all submitted intestinal segments re-examined to confirm
the presence of sexual andor asexual life stages within the intestinal epithelium and describe the
histologic lesions associated with presence of the parasite life stages
424 Prospective modified necropsy protocol
During the study period 2014-2016 necropsy protocols for all BFF were modified to improve
detection of coccidial life stages and better to determine which portions of the intestinal tract were
affected The entire length of the intestine from duodenum to anus was measured and intestinal contents
were flushed with 12 mL of sterile saline into a sterile container Intestinal contents were preserved in
25 potassium dichromate solution (mixed 11 with intestinal contents vv) for molecular diagnostics
Paired 2-cm long intestinal samples were collected from all sections of small and large bowel duodenum
(1) jejunum (6) ileum (1) and colon (2) The eight small intestinal samples were collected at equal
distances from the pyloric sphincter to the beginning of the colon and the distance from the pylorus noted
for each Colon samples were taken at 25 and 75 of the length of the colon One sample from each
pair was preserved in Serra solution (100 ethanol (60 vv) 37 formaldehyde (30 vv) glacial
acetic acid (10 vv) and the second sample was frozen Representative tissues from all internal organs
as well as additional intestinal samples skin muscle and brain were also collected and preserved in 10
buffered formalin Histopathologic examination was performed on all tissues collected
83
425 Retrospective medical history review
Medical histories of all BFF held by the Toronto Zoo since the initiation of the SSP program were
reviewed for data on frequency of occurrence of shedding of coccidial oocysts in adults and juveniles as
well as any association of shedding with clinical signs and administration of anticoccidial treatment Data
was tabulated yearly for adult and juvenile ferrets to determine annual morbidity and mortality rates
associated with enteric coccidiosis Medical records and pathology reports were solicited from the other
SSP institutions to determine comparative morbidity and mortality rates associated with enteric
coccidiosis in BFF at other facilities Both morbidity and mortality rates were calculated as
incidenceattack rates with yearly adult population size or number of family groups (dam and kits) as the
denominator for morbidity rates and number of yearly deaths in each age class as the denominator for
mortality rates
43 RESULTS
431 Fecal oocyst evaluation and retrospective medical history review
Family groups
Fecal samples were collected from seven groups of dams and kits housed together at the Toronto
and the Louisville Zoos from 2014-2016 All data from first to last day of collection for all family groups
is listed in Appendix 1 selected pertinent data for each group is presented in Table 41 Five groups of
dams and kits were sampled in 2014 and one group in each of 2015 and 2016 Shedding occurred no
earlier than 55 days of kit-age in any of the groups and was identified from 55-81 days of age (Table 41
Table 42 Figure 41)
In 2014 fecal oocyst shedding was identified in three of the five surveyed groups In two of the
three groups (dams Poppy and Bumblefoot) changes to fecal colour and consistency were identified
concurrently with periods of oocyst shedding both groups shed higher numbers of oocysts than the other
dam and kit groups in 2014 and 2015 Both Poppy and Bumblefoot had had litters in the previous one and
84
two years prior to this study respectively based on medical record review these dams and their litters
were also diagnosed as shedding coccidial oocysts that were too numerous to count on direct exam and
fecal flotation Clinical signs in the previous years included dark tarry hemorrhagic or soft mucoid feces
and reduced appetite both groups received treatment with toltrazuril (Baycox Coccidiocide Solution
25 Bayer Inc Mississauga Canada) and trimethoprim sulfamethoxazole (Novo-Trimel Teva Canada
Ltd Scarborough Canada) (TMS) One of four kits from Poppyrsquos 2013 litter (Z113-13) died of enteric
coccidiosis three days after the group was diagnosed as shedding coccidial oocysts and the initiation of
treatment with TMS
In 2015 low grade fecal oocyst shedding (lt14 oocysts per gram of feces) without associated
clinical signs was noted in the Fiddlesticks group on three days during a seven day period from 63-69
days of kit-age and again for a single day at 128 days of kit-age The dam had been diagnosed and treated
for enteric coccidiosis in 2014 at which time she exhibited clinical signs of loose green feces to
hemorrhagic diarrhea lethargy and dehydration In 2016 she was diagnosed as shedding low numbers of
coccidia exhibited no clinical signs and did not receive treatment prior to resolution of shedding
In 2016 Guanella and kits shed oocysts over a nine day period and daily fecal oocyst shedding
ranged from 206 ndash 371714 OPG Combined treatment with ponazuril (first four days of shedding)
sulfadimethoxine injectable (first two days of shedding additional product information not available)
amoxicillin oral (first two days of shedding additional product information not available) penicillin
injectable (first two days of shedding additional product information not available) and subcutaneous
fluids (first two days of shedding additional product information not available) was administered to this
group Previous medical history was not available for this female for review
In 2014 and 2015 fecal oocyst shedding in all groups in the Toronto Zoo population started in the
three week period from the last week of July to mid-August In 2016 shedding was first identified in the
Louisville Zoo group in mid-July
85
Adults
Seven single-housed adult BFF (52 MF) were detected to have shed coccidia during the study
period (Table 43) Shedding periods lasted from 2-10 days and oocyst per gram counts ranged from 104
ndash 554274 (Table 44) Clinical signs were identified in four of the seven ferrets and consisted of
lossreduction of appetite (n=2) weight loss (n=1) lethargy (n=1) blood in feces (n=1) loose or runny
feces (n=3) soft mucoid feces (n=1) green colour of feces (n=2) Five of the seven adults received
treatment after detection of oocyst shedding two of which received treatment in the absence of clinical
signs Treatment consisted of oral toltrazuril in four cases toltrazuril in combination with trimethoprim
sulfamethoxazole in one case (Mohawk-A) and ponazuril and sulfadimethoxine (manufacturerrsquos
information not available) in one case (Clive) (Table 43)
Three of the adults in this study Mohawk Mystery and Jenna shed oocysts during multiple
different periods in 2016 Mohawk shed oocysts in May July and September of 2016 data from the first
two periods are reported in Tables 43 and 44 Mystery shed oocysts in June and July of 2016 Clinical
signs were observed only during the first shedding period and included poor appetite and hemorrhagic to
soft mucoid feces Jenna shed oocysts in July September and November of 2016 and again in February
and May of 2017 Although clinical signs were not detected in association with the initial period of
shedding in July 2016 (see Table 44) depressed mentation and hemorrhagic mucoid feces were identified
in the subsequent two shedding periods In both Mohawk and Jenna oocysts were not detected in feces on
multiple recheck and routine monthly fecal examination between shedding periods
Ruckus the fourth ferret shed low numbers of oocysts for two days in 2016 while housed alone
and had been reported to have shed oocysts during a 30 day period in 2014 while housed in a family
group with her kits Diarrhea to soft mucoid feces and loss of appetite were reported in 2014 Clinical
signs in 2016 consisted of small amounts of runny feces for a two day period two days after oocyst
shedding was no longer detected Treatment consisted of toltrazuril and TMS in 2014 and toltrazuril
again in 2016 Similarly Noodle shed low numbers of oocysts in July 2015 and had been identified as
shedding low numbers of oocysts during September of the previous year
86
The final ferret Rigatoni shed oocysts in feces in low numbers (54-732 OPG) over 5 days All
positive samples were pooled for molecular diagnostics and sequencing results showed 990 identity (7
single nucleotide differences SNDs) at the mt COI locus with a pair of rodent Eimeria species (ie
GenBank HM771682 JQ993704) (see Chapter 3) This ferret did not show clinical signs associated with
shedding and was not treated Previous medical history was not available for Clive the single adult ferret
from the Louisville Zoo
Based on sample collection dates and medical record review for the adult BFF for 2014-2016
shedding occurred during spring summer and fall with three ferrets shedding in May (Jenna Ruckus and
Mohawk) one ferret shedding in June (Mystery) three ferrets shedding in July (Mohawk Jenna Noodle)
one ferret shedding in August (Clive) two ferrets shedding in September (Noodle Jenna) and one ferret
shedding in November (Jenna)
432 Pathology
Eleven BFF with enteric coccidiosis were identified in the Toronto Zoo necropsy reports from
1993-2016 (Table 45) Cases were identified from 1998-2014 inclusive and all were confirmed by
histological re-evaluation (Chapter 3 Table 32) Both juvenile (n=9 36 MF) and adult ferrets (n=2 20
MF) were represented
Gross Pathology
Gross necropsy findings were similar across the 11 cases and included mucoid to fluid luminal
contents (n=7 636) beige to white pasty coating of the mucosal surface of the small intestine (n=6
545) and colon (n=4 364) gaseous dilation of intestinal segments (n=3 272) segmental enteritis
and hemorrhage (n=1 91) In one case Z228-98 no gross lesions were identified within the intestines
Impression smears of luminal contents or scrapings of intestinal mucosa were performed in four cases and
coccidia were identified in all four
Histopathology
87
Both sexual and asexual life stages were identified within the small intestinal segments in all
cases except Z228-98 and Z106-02 in which only rare asexual parasite life stages were identified (Table
45 Figure 42) Other histologic lesions seen in intestinal segments containing coccidia included
lymphoplasmacytic inflammation of the lamina propria (n=4) neutrophilic infiltration of the lamina
propria (n=2) villar necrosis (n=2) villar atrophy or blunting (n=3) and thrombi within the villar tips
(n=1)
Additional necropsy diagnoses included cholangiolar hyperplasia multiple hepatobiliary cysts
with suppuration renal adenocarcinoma apocrine gland adenocarcinoma (Z228-98) concurrent
clostridial enteritis (Z143-99) presumptive Salmonella sp septicemia (Z106-02) interstitial pneumonitis
(Z108-03) myocardial mineralization interstitial pneumonia and nephritis periportal hepatitis and
bacteremia (Z124-12) and suppurative esophagitis (Z137-14)
From 2014-2016 three black-footed ferrets were necropsied using the detailed protocol described
above Only one of the three cases Z137-14 was diagnosed with enteric coccidiosis based on
histopathology Two duodenal five jejunal two ileal and two colonic sections were collected at measured
lengths from the pylorus Parasite life stages were identified within the mucosal epithelium of all
intestinal segments extending from the distal duodenum (10-12 cm aboral to pylorus) through to the distal
colon (157-159 cm aboral from pylorus) The distal duodenal section contained asexual life stages only
with a single focus of epithelial cells containing meronts Sexual life stages (microgamonts
macrogamonts unsporulated oocysts) were identified within villar epithelial cells in all remaining
sections of the small intestine with numerous oocysts in the bowel lumen Mild lymphoplasmacytic
inflammation of the lamina propria was associated with the jejunal and ileal lesions and blunting of the
villi was identified within one jejunal segment The colonic sections contained small to moderate numbers
of sexual life stages identified within both superficial and deep crypt epithelium with occasional life
stages identified near the germinal cells Large numbers of oocysts and bacteria were identified within the
88
lumen of these colonic sections and both sections contained abscesses within the crypts The proximal
duodenum (0-2 cm) was the only section of the intestines not containing parasitic life stages
433 Morbidity and mortality
Annual morbidity rates for enteric coccidiosis at the Cheyenne Mountain Zoo and mortality rates
from enteric coccidiosis in Toronto Zoo BFF are summarized in Tables 46 and 47 During 2003-2016
yearly incidence of coccidiosis in adult BFF at the Cheyenne Mountain Zoo averaged 69 (range 0-
421) For family groups consisting of juvenile ferrets housed with their dams yearly incidence of
enteric coccidiosis averaged 115 (range 0-600)
From 1997-2016 coccidia-associated mortality in adult BFF at the Toronto Zoo averaged 053
yearly (range 0-526) with an average total mortality rate of 141 per year (range 0-526) For
juvenile ferrets (under 1 year of age) during the same period coccidia-associated mortality accounted for
an average of 133 of deaths yearly (range 0-100) with an overall average mortality rate of 170 per
year (range 0-3404) from all causes
Multiple additional SSP institutions provided partial medical and pathology data sets for use in
this study which were not sufficiently detailed to permit computation of morbidity and mortality rates for
those populations
44 DISCUSSION
The work described here supports previous clinical findings regarding the impact through both
morbidity and mortality associated with enteric coccidiosis in BFF No previous studies have been
undertaken to determine morbidity and mortality rates associated with enteric coccidiosis across BFF SSP
institutions
89
In a retrospective mortality study of captive BFF from 1984-2004 at the Smithsonianrsquos National
Zoological Park Bronson et al (2007) reported that gastrointestinal disease was the most common cause
of death in juvenile BFF (524) with 333 of juvenile mortality cases in the study caused by enteric
coccidiosis While the data is not directly comparable the findings reported here also reflect enteric
coccidiosis as a common cause of death in juveniles with increased mortality associated with the disease
compared with adult counterparts All Toronto Zoo mortalities in both juvenile and adult age classes for
which necropsy tissues were available have been attributed to infection with a single coccidia species
Eimeria ictidea (see Chapter 3)
Multiple SSP institutions provided partial medical and pathology data sets for use in this study
which were not sufficiently detailed to permit computation of morbidity and mortality rates for those
populations In future it would be useful to determine whether morbidity and mortality rates associated
with enteric coccidiosis vary among SSP institutions as this may allow for improved identification of
host parasite and environmental factors that increase risk
Clinical signs reported here are consistent with those described from both BFF and domestic
ferrets with enteric coccidiosis (Sledge et al 2011 Santymire et al 2014 USFWS BFF Recovery
Program 2017) Changes to fecal colour and consistency were the most common abnormalities identified
at the time of first detection of oocyst shedding While clinical signs in the cases described here do not
always correlate directly with the quantity of oocysts shed individuals shedding higher number of oocysts
showed clinical signs more frequently than those shedding lower numbers
In this study oocyst shedding from single-housed adult BFF ranged from 104 ndash 554274 oocysts
per gram Daily fecal samples produced from individual adult ferrets range in size from approximately 1-
16 grams In light of these findings during peak shedding from ~5times105 to 9times106 oocysts can be shed into
the environment in one day providing a massive infective dose The large numbers of oocysts shed
90
combined with confinement in a small enclosure space and hardiness of Eimeria oocysts in the
environment would be expected to markedly increase risk of infection in captive BFF
Oocyst shedding from family groups ranged from 0 ndash 371714 OPG The wide variability seen in
OPG counts between days as seen in Figure 41 and Table 41 may be accounted for by the staggered
initiation and resolution of shedding by different ferrets It is most likely that the source of infection in
these family groups is shedding by the dams some of which were identified to shed in multiple years
although environmental contamination cannot be excluded
Shedding in the adult ferrets was clustered during particular time periods specifically May July
and September If shedding is associated with stress and immunosuppression activities such as breeding
whelping electro-ejaculation of male ferrets and shipmenttransfer could act as stressors Whelping
which occurs primarily in May and June could also act as a stressor to other ferrets in the facility either
through social cues or as there would be associated changes in husbandry protocols The September
cluster could be associated with the transfer of ferrets between institutions kits are pulled during this time
and adult ferrets are moved among institutions thus changing the population dynamics of each SSP site A
large cluster of shedding was recorded in July With the exception of weaning of kits no other major
stressors are expected to occur during this time thus the increase in shedding by adults in this group
cannot be easily explained Interestingly shedding was not identified in ferrets from March through April
which is the typical breeding season and when ferrets are introduced for breeding a presumably stressful
time Shedding was not noted from December through April which could reflect reduced environmental
burdens due to low humidity levels as would be expected in a Nordic climate during the winter (which
would kill oocysts and thus block transmission) or may be consistent with reduced stress during this
period The results from single-housed adults are in contrast to the dam and juvenile ferret groups in
which oocyst shedding appeared to be correlated to a period of 55-81 days of kit age These results are
consistent with reports from other facilities of increased incidence of shedding by kits after 70 days of age
(USFWS BFF Recovery Program 2017)
91
Retrospective and prospective review of histologic sections of intestines from affected BFF at the
Toronto Zoo showed the presence parasitic life stages in epithelial cells of both the small and large
intestines Neither Hoare (1935b) nor Jolley et al (1994) mentioned the presence of parasites in the large
intestine of experimentally infected domestic ferrets or naturally infected BFF respectively In the study
described here asexual and sexual life stages were identified within the epithelial cells of the small
intestinal villi from base to tip and were most numerous in jejunum This matches the description by
Jolley et al (1994) however Hoare (1935a b) found E ictidea to be present primarily in the villar tips
Jolley et al (1994) also described two morphologically distinct meronts of E ictidea within the small
intestinal sections one at the villar tip that was larger and lacking in undifferentiated mass and the other
at the base of the villi or in the intestinal crypts these findings were not echoed in this study as
merogonic stages were identified throughout the intestinal epithelium from villus to the base of the crypt
and no visual differences between meronts in any location were identified Hoare (1935a b) also
described a resulting annular constriction of the villus separating the affected and non-affected segments
this constriction was neither seen in the cases described here nor mentioned by Jolley et al (1994)
Whether these differences result from E ictidea from BFF and E ictidea from domestic ferrets being
different parasites or from differences in tissue tropism of a single parasite in two different hosts cannot
be ascertained from the available information
Histologic lesions such as necrosis hemorrhage villar atrophy and inflammation associated with
the presence of parasitic life stages were rare These changes are normally elicited by the host immune
system (inflammation) and the parasite (cellular rupture to release life stages resulting in hemorrhage and
necrosis) in response to infection In light of the fact that acute death occurred in a number of these ferrets
(Z113-13 Z117-13 Z118-13 Z119-13 Z137-14) in the absence of secondary disease processes and
with the intestinal epithelium intact but containing myriad parasitic life stages an alternative mechanism
for mortality associated with the infection must be proposed It is possible that these parasites elaborate
exotoxins during their life cycle and when at high density result in sudden death of the host with minimal
92
tissue changes The presence of parasitic life stages occupying the majority of both small and large
intestinal epithelial cells could also potentially impair fluid and protein movement in and out of the
mucosa however clinical signs associated with malabsorptive diarrhea were not identified in any of these
cases The presence of bacteria within the blood or other organ tissues was not identified in any cases and
consequently sepsis is unlikely to be the cause of death
Black-footed ferrets appear to maintain persistent infection with E ictidea Adult BFF in the
Toronto Zoo population shed coccidia in multiple years and in two adult ferrets multiple times in the
same year While the coccidia seen in all cases were not confirmed as E ictidea using molecular
techniques morphologic similarities and a lack of additional Eimeria spp identified on molecular work
undertaken suggest that only one species of parasite is and has been present in the collection Two dams
that had been identified as infected based on routine fecal screening in previous years presumably acted as
the source of infection to their litters of kits in multiple years While continued environmental
contamination cannot be ruled out these findings imply a failure of the immune response of the BFF to
clear infection with E ictidea or even to protect against sufficient replication of organisms to result in
clinical disease
Based on clinical experience and review of the literature BFF appear to be much more sensitive
to infection with E ictidea compared with their domestic counterparts In domestic ferrets subclinical
shedding of oocysts appears to be the most common with rare reports in the literature of overt disease
and that only in juveniles (Blankenship-Paris et al 1993 Abe et al 2008) However a single report exists
of three separate clinical outbreaks of Eimeria furonis infection in domestic ferrets under intensive
management with increased morbidity and mortality affecting all ages classes (Sledge et al 2011) The
role of genetics in the apparent increased susceptibility of BFF to enteric coccidiosis is unknown but the
current captive BFF population is derived from seven founders and inbreeding depression or familial
genetic susceptibility may play a role in their increased susceptibility to disease caused by E ictidea
93
Black-footed ferrets diagnosed with enteric coccidiosis during the course of the study were
treated with either ponazuril or toltrazuril sulfonamide drugs or often a combination of the two groups
of therapeutic agents Toltrazuril and ponazuril are triazine coccidiocides with proven efficacy against
both asexual and sexual life stages of mammalian and avian Eimeria spp (Mehlhorn and Aspock 2008)
The sulfonamides are antimicrobial drugs that exhibit coccidiostatic or coccidiocidal effects depending on
dose they act by blocking folate synthesis and have effects on first and second generation meronts
(asexual life stages) as well as potentially acting on sexual life stages (Mehlhorn and Aspock 2008)
Based on the limited data available from this study and the fact that treatment was initiated in almost all
adult BFF and family groups at the time of oocyst detection regardless of the presence of clinical disease
the effects of treatment on duration of clinical signs cannot be effectively evaluated It appears
subjectively that adult ferrets treated with toltrazuril and in one case a combination of toltrazuril and
TMS showed reduction in oocyst shedding after 3-5 days of oral anti-coccidial therapy (see Table 43)
The effects of treatment with either sulfonamides or triazines would be expected to reduce oocyst
shedding consequently the duration and amount of oocyst shedding reported in this study may not
accurately characterize the natural course of disease
Perceived resistance to sulfa drugs has been reported from multiple SSP facilities In light of this
and their potential negative effects on ferret reproduction (eg prevention of embryo implantation in the
uterus and impairment of sperm development) sulfonamides are no longer recommended by the SSP for
treatment of coccidia in this species (USFWS BFF Recovery Program 2017) The frequent and
widespread use of triazines in the management of enteric coccidiosis in BFF presents a risk for
development of resistance to this drug class in the future Neither pharmacokinetic (PK) nor
pharmacodynamic (PD) studies have been published to validate the dose and frequency of dosage in
either class of drugs in BFF and consequently it is unclear whether this perceived failure of some ferrets
to respond to treatment is based on true resistance versus inappropriate dosing The only work evaluating
ponazuril in BFF evaluated serum levels of ponazuril after a single oral dose of 50 mgkg and reported
94
therapeutic levels for 10 days after administration (USFWS BFF Recovery Program 2017) No
information was provided on number or age of ferrets that participated in the study or on how the
determination of what were therapeutic levels was made Furthermore as life cycles of the coccidia
affecting BFF are limited to the gastrointestinal tract and do not exhibit tissue stages the validity and
usefulness of assessing blood levels of ponazuril in determining appropriate dosage and dose schedules is
questionable Further work to determine the PK and PD of triazines in ferrets is warranted to provide safe
and efficacious treatment and to reduce the risk of development of resistance Furthermore the creation
and validation of a model for enteric coccidiosis in a related species would allow for in vivo studies of
drug resistance
95
Figure 41 Oocyst per gram counts and shedding period of Eimeria ictidea from black-footed ferret
(Mustela nigripes) family groups from 2014-2016
0
50000
100000
150000
200000
250000
300000
350000
400000
29
33
37
41
45
49
53
57
61
65
69
73
77
81
85
89
93
97
10
1
10
5
10
9
11
3
11
7
12
1
12
5
12
9
13
3
13
7
14
1
14
5
14
9
OP
G
Age of Kits
Poppy Bumblefoot Calico Aubrey Ruckus Fiddlesticks Guanella
96
Figure 42 Small intestinal epithelium of a black-footed ferret (Mustela nigripes) containing sexual
life stages of Eimeria ictidea Legend Solid black arrow = oocyst Hatched arrow = macrogamont
Outlined arrow = microgamont Hematoxylin and eosin staining scale bar = 25 microm
25 microm
97
Table 41 Shedding of oocysts of Eimeria ictidea in black-footed ferret (Mustela nigripes) dam and kit
family groups from 2014-2016
Collection Year 2014 2014 2014 2014 2014 2015 2016
Age of kits (days) Poppy Bumblefoot Calico Aubrey Ruckus Fiddlesticks Guanella^
29 - - - 0 - - -
30 - 0 - 0 - - -
- - - - -
34 0 0 - 0 0 - -
35 0 0 - 0 0 0 -
- -
46 0 0 - 0 0 0 -
47 0 0 - 0 0 0 -
48 0 lt14 - 0 0 0 -
- -
53 0 0 - 0 0 0 +
54 0 0 - 0 0 0 +
55 0 0 - 0 0 0 3717146
56 0 0 - 0 - 0 1084436
57 0 0 - 0 - 0 64133
58 0 324 - 0 0 0 20654
59 0 0 - 0 0 0 0
60 0 446688 - 0 0 0 0
61 0 934828 - 0 0 0 41111
62 0 530469 - 0 0 0
63 0 1617131 - 0 0 lt14
64 0 8396357 - 114 0 0
65 0 234876 - 5368 0 lt14
66 0 374625 - 9455 0 0
67 lt14 2311575 - 39579 0 -
68 0 603563 - 125051 0 0
69 - 4103036 - 27067 0 lt14
70 2470327 7759974 - 0 0 0
71 1073085 - 1784 0 0
72 3430966 - 0 0 0
73 4459536 - 7326 0 0
74 4842212 - 2 0 0
75 3627307 - 0 0 0
76 5801885 - 0 0 0
77 5994 0 0 0 0
78 589835 0 0 0
79 24815 0 0 0
80 144016 0 0 0
81 97862 0 0 0
82 0 0 0
127 0 0
128 0 2843
136 0 0
150 0
Numbers of Oocysts Shed (oocysts per gram of feces)
Dam Identity
Legend lt 14 = oocyst positive samples with less than 14 oocyst per gram of feces - = no sample recorded for this date underline = last
sampling date + = coccidia present but OPG count not performed = Toronto Zoo ferret ^ = Louisville Zoo ferret thick outer border =
days treatment was received = range of sequential dates between previous and subsequent number during which OPG counts were
performed and samples contained 0 oocysts
98
Table 42 Summary of epidemiologic data for family groups of black-footed ferrets (Mustela nigripes)
shedding oocysts of Eimeria ictidea
Dam Identity
Poppy
2014
Bumblefoot
2014
Aubrey
2014
Fiddlesticks
2015
Guanella
^
2016
Number of kits 7 5 3 4 5
Kit age (days) at time of
shedding 70-81 58-70 64-73 63-69 54-61
Shedding period (days) 15 13 11 7 9
OPG min 0 0 0 0 206
OPG max 48442 83963 2707 lt14 371714
Clinical signs YES YES NO NO NO
Treated - YES YES NO YES
Legend = Toronto Zoo ferret ^ = Louisville Zoo ferret OPG = oocysts per gram of feces
- = missing data
Table 43 Shedding of oocysts of Eimeria ictidea in single-housed adult black-footed ferrets (Mustela nigripes) 2015-2016
Numbers of Oocysts Shed (oocysts per gram of feces)
Collection
Year 2015 2016 2016 2016 2016 2016 2016
Ferret Identity Noodle Ruckus Mohawk-A Mohawk-B Mystery Jenna Clive^
Age (years) 1 3 1 1 1 1 1
1084058 0 + + + + +
+ 0 42307650 6286676 + 183150 554274
857808 16650 12805238 7777929 + 215710 377920
1604894 16650 309690 139860 + 0 25808
377042 0 599400 119880 + - 37294
554445 0 34688 385579 117920 0 5363
26640 0 16650 0 0 0 7500
0 10406 0 0 0 1090
0 20813 0 0 0
- 0 1761 0
0 0 0 0
- 0
0
0
Legend lt 1 = oocyst positive samples with less than 1 oocyst per gram of feces underline = last sampling date + = coccidia present but OPG
count not performed = Toronto Zoo ferret ^ = Louisville Zoo ferret thick outer border = days treatment was received
100
Table 44 Summary of epidemiologic data for single housed adult black-footed ferrets (Mustela nigripes) shedding oocysts of Eimeria ictidea
Noodle Ruckus Mohawk-A Mohawk-B Mystery Jenna Clive^
Sex M F M M M F M
Age (years) 1 3 1 1 1 1 1
Shedding period (days) 7 2 9 6 10 4 8
OPG min 266 166 104 1199 1761 1831 1090
OPG max 10840 166 423076 77779 - 2157 554274
Clinical signs YES YES NO NO YES NO YES
Treated NO YES YES YES YES YES YES
Legend = Toronto Zoo ferret ^ = Louisville Zoo ferret M = male F= female OPG = oocysts per gram of feces - = missing data
NOTE Mohawk-A and Mohawk-B refer to two separate episodes of oocyst shedding by the same ferret
101
Table 45 Histologic findings from necropsies of black-footed ferrets (Mustela nigripes) with enteric coccidiosis
Number of Sections Affected
Ferret
ID Year
Age
(years) Sex Coccidia in Intestinal Sections Small Intestinea Large Intestinea
Z228-98 1998 7 M rare asexual stages S - 01 A - 11 S - 01 A - 01
Z143-99 1999 1 M sexual and asexual stages S - 24 A - 24 S - 03 A - 03
Z106-02 2002 008 F rare sexual stages S - 14 A - 04 S - 01 A - 01
Z108-03 2003 008 F sexual and asexual stages S - 12 A - 12 S - 01 A - 01
Z109-03 2003 008 F sexual and asexual stages S - 23 A - 23 S - 11 A - 11
Z124-12 2012 021 F sexual and asexual stages S - 24 A - 04 none
Z113-13 2013 017 F sexual and asexual stages S - 77 A - 07 none
Z117-13 2013 025 M sexual and asexual stages S - 56 A - 56 S - 11 A - 01
Z118-13 2013 025 M sexual and asexual stages S - 56 A - 46 S - 11 A - 01
Z119-13 2013 025 F sexual and asexual stages S - 45 A - 35 none
Z137-14 2014 021 M sexual and asexual stages S - 1011 A - 911 S - 22 A - 02
Legend a= xn where x is number of sections containing sexual or asexual lifestages n is the number of sections examined S = sexual life
stages A= asexual life stages
102
Table 46 Yearly incidence of coccidial infection in black-footed ferrets (Mustela nigripes) at the
Cheyenne Mountain Zoo
Cheyenne Mountain Zoo
Year Adult Family
2003 116 (625) -
2004 819 (4211) -
2005 021 (000) 14 (2500)
2006 021 (000) 07 (000)
2007 023 (000) 08 (000)
2008 224 (833) 14 (2500)
2009 025 (000) 06 (000)
2010 326 (1154) 07 (000)
2011 125 (400) 09 (000)
2012 125 (400) 08 (000)
2013 028 (000) 05 (000)
2014 430 (1333) 09 (000)
2015 035 (000) 35 (6000)
2016 - 27 (2857)
Mean annual () 689 1155
Legend - = missing data xn= where x is the number of ferrets shedding coccidial oocysts and n is the
total number of adult ferrets or family groups in a given year () = incidence expressed as a percentage
103
Table 47 Yearly mortality rate and incidence of mortality associated with coccidial infection in
black-footed ferrets (Mustela nigripes) at the Toronto Zoo
Total Mortality
Coccidia Other Causes
Year Kit Adult Kit Adult
1997 015 (000) 023 (000) 315 (2000) 423 (1739) 1998 038 (000) 119 (526) 838 (2105) 919 (4734) 1999 047 (000) 119 (526) 1647 (3404) 119 (526) 2000 034 (000) 015 (000) 434 (1176) 315 (2000) 2001 032 (000) 016 (000) 532 (1563) 116 (625) 2002 150 (200) 020 (000) 450 (800) 220 (1000) 2003 227 (741) 018 (000) 327 (1111) 118 (555) 2004 020 (000) 016 (000) 620 (3000) 216 (1250) 2005 016 (000) 015 (000) 416 (2500) 215 (1333) 2006 030 (000) 016 (000) 230 (667) 016 (000) 2007 019 (000) 015 (000) 419 (2105) 215 (1333) 2008 034 (000) 016 (000) 1134 (3235) 316 (1875) 2009 017 (000) 016 (000) 017 (000) 116 (625) 2010 017 (000) 016 (000) 317 (1765) 316 (1875) 2011 011 (000) 016 (000) 111 (909) 216 (1250) 2012 111 (909) 017 (000) 111 (909) 317 (1765) 2013 424 (1667) 017 (000) 424 (1667) 317 (1765) 2014 126 (384) 017 (000) 326 (1154) 217 (1176) 2015 04 (000) 017 (000) 04 (000) 217 (1176) 2016 011 (000) 017 (000) 211 (1818) 117 (588)
Mean annual () 195 053 1594 1359
Legend xn= where x is the number of ferrets that died with coccidial infection or of other
causes and n is the total number of adult ferrets or kits in a given year () = incidence expressed
as a percentage
104
CHAPTER 5 EVALUATING THE DOMESTIC FERRET (MUSTELA PUTORIUS FURO) AS
AN EXPERIMENTAL MODEL FOR ENTERIC COCCIDIOSIS IN THE BLACK-FOOTED
FERRET (MUSTELA NIGRIPES)
ABSTRACT
The purpose of this study was to determine whether the domestic ferret (Mustela putorius furo) is
susceptible to an isolate of Eimeria ictidea originating from black-footed ferrets (BFF Mustela nigripes)
and thus could act as a suitable experimental model in which to investigate the pathogenesis and
management of this disease A pilot study was performed with 10 male intact juvenile domestic ferrets
Ferrets were administered an oral inoculum containing either a high dose (1 times 106 oocysts) moderate
dose (5 times 104 oocysts) or saline control and observed for shedding of oocysts and development of
clinical signs Seven of ten ferrets developed patent infection all of which had received the high dose
inoculum The prepatent period was 7-9 days and duration of shedding varied from 1-7 days Clinical
signs were identified in six of the seven infected ferrets and were consistent with those previously
described for enteric coccidiosis in domestic and BFF Parasite life stages were identified within the
intestines of four of the seven ferrets with patent infection and were limited to the distal jejunum and
ileum The demonstrated ability to produce patent infections in domestic ferrets following oral inoculation
of a high dose of E ictidea (1 times 106 oocysts) isolated from BFF provides an avenue for future
experimental investigations into the control and treatment of enteric coccidiosis in this endangered
species
51 INTRODUCTION
Black-footed ferrets (BFF Mustela nigripes) are one of only three ferret species worldwide
While formerly distributed throughout the North American prairies black-footed ferrets were declared
extinct in the wild in the 1980s Since 1986 a multi-institutional consortium has been breeding this
species in captivity with reintroductions back into the wild within their historic range in selected locations
in Canada the USA and Mexico Introduced colonies of BFF are present in Arizona Colorado Kansas
105
Montana New Mexico South Dakota Utah Wyoming and Chihuahua (Mexico) Reintroduction
attempts in Saskatchewan Canada have been unsuccessful to date
Multiple infectious diseases pose a significant risk to the captive breeding and post-release
survival of BFF including distemper and sylvatic plague (Santymire et al 2014 USFWS BFF Recovery
Program 2017) Coccidiosis is a recognized cause of juvenile and adult morbidity and mortality in captive
breeding programs and can result in significant losses (Bronson et al 2007 Santymire et al 2014
USFWS BFF Recovery Program 2017) The effect of the disease on wild populations is unknown
Clinical signs of coccidiosis include mucoid to hemorrhagic diarrhea abdominal discomfort lethargy
appetite loss vomiting and dehydration Recent investigations into diseases affecting BFF at the Toronto
Zoo have identified a single Eimeria species E ictidea associated with all cases of enteric coccidiosis in
juvenile and adult BFF from 2014-2016 (see Chapter 3) This same Eimeria species was identified
retrospectively as the cause of juvenile and adult mortalities in previous years (1999 through 2014
inclusive) (Chapters 3 and 4) Furthermore this pathogen was identified in fecal samples based on
morphologic and molecular characterization from adult and juvenile BFF in another zoological collection
(Louisville Zoo Kentucky USA) (see Chapter 3)
Enteric coccidiosis also occurs in domestic ferrets (Mustela putorius furo) with three
morphologically distinct species of coccidia Eimeria ictidea Eimeria furonis and Isospora (=
Cystoisospora) laidlawi Both of the aforementioned Eimeria species have been identified in black-footed
ferrets based on morphologic criteria but molecular characterization was needed to confirm whether the
same species of parasite infects both ferret species (see Chapter 2 and 3) To this end nuclear and
mitochondrial sequences for E furonis and for I=(C) laidlawi were generated (Chapter 2) expanding the
existing limited sequence data from the nuclear 18S rRNA locus of Eimeria furonis Molecular
characterization of E ictidea from domestic ferrets was not possible because samples containing this
parasite were not available for study consequently it is unclear whether the same coccidium affects both
domestic and black-footed ferrets
106
There is no published information describing the pre-patent periods and pathogenicity of enteric
coccidia in BFF and given the conservation status of the BFF experimental work cannot be undertaken
in the natural host The purpose of this study was to determine whether the domestic ferret is susceptible
to E ictidea isolated from BFF if susceptible the domestic ferret could act as a suitable experimental
model in which to investigate the pathogenesis prevention and treatment of coccidiosis caused by E
ictidea
52 MATERIALS AND METHODS
521 Animal care
Ten juvenile male intact ferrets of 48 (n=6) or 50 (n=4) days of age were obtained from a
commercial source (Marshall BioResources North Rose New York USA) and were housed in the
University of Guelph Central Animal Facility Isolation Facility Ferret weights on arrival ranged from
334-475 g (average= 3928 g) All ferrets were housed individually in wire bottom cages of 813 times 1117
times 457 cm size and were divided in equal numbers between two non-adjoining rooms They received ad
libitum access to Envigo Teklad Certified Global Ferret Diet (Madison Wisconsin USA) and water
changed daily Room temperature was maintained at 18-21 degC and a 16 hour light 8 hour dark
photoperiod was provided All personnel working with the ferrets were required to wear personal
protective equipment including disposable facemasks gloves gowns and bouffant caps Shoes were
provided for use in each room This study was carried out in accordance with the recommendations in the
Canadian Council on Animal Care guidelines The protocol was approved by the Animal Care Committee
of the University of Guelph (Animal Use Protocol 3289) and by both the Animal Welfare Committee
and Animal Care and Research Committee of the Toronto Zoo
An initial physical examination and blood collection were performed on each ferret by the
principal investigator (ARP) one day after arrival to assess health status prior to enrollment in the study
Ferrets were mask induced with isoflurane (Isoflurane USP Fresenius Kabi Richmond Hill Ontario) in
107
oxygen placed on a heat disc (SnuggleSafe Lenric C21 Ltd Littlehampton United Kingdom) weighed
examined and blood was collected from the jugular vein for routine CBC and biochemical profiles
All ferrets subsequently underwent an acclimation period of two weeks During this time fecal
samples were collected daily from each ferret and examined for the presence of coccidial oocysts using a
standard salt flotation technique (Dryden et al 2005) to ensure that all individuals were free of coccidia
prior to initiation of experimental work Any ferret positive for coccidia was to be removed from the
study
522 Oocyst preparation
Oocysts used for inoculation originated from fecal samples from two naturally infected BFF These
samples were stored in potassium dichromate for four weeks prior to oocyst purification and use in this
infection trial Stored fecal samples were mixed with distilled water and passed through a small sieve to
remove debris The strained contents were transferred to a 50 mL conical vial and topped up to 50 mL
with additional distilled water Samples were centrifuged (Sorvall ST40R Centrifuge Thermo Scientific)
at 2800 rpm (1315 timesG) for 10 minutes at 12 degC A drop of supernatant was evaluated microscopically at
100times for the presence of oocysts If oocysts were observed the supernatant was poured off into a second
50 mL conical tube and again topped up to 50 mL with distilled water and re-centrifuged under the same
conditions Otherwise the supernatant was discarded The pellets from both the first and second tubes
were combined with saturated salt solution at a 14 ratio by volume Oocysts were floated in the salt
solution by centrifugation at 1500 rpm (377 timesG) for 10 minutes at 12 degC The top 5 mL of supernatant
were collected and transferred to a clean 50 mL conical tube topped up to 40 mL with distilled water and
washed via centrifugation at 2800 rpm (1315 timesG) for 10 minutes After the wash step the supernatant
was again checked for presence of oocysts and discarded if no oocysts were observed The pellet was
collected and the presence of oocysts confirmed by examination of a drop placed on a clean glass slide at
100times Once verified the contents of the pellet of concentrated oocysts was placed in a 250 mL storage
container and mixed with approximately 200 mL of sterile saline (09 sodium chloride Hospira
108
Montreal Quebec) prior to storage for two to four weeks at 4 degC until inoculation Prior to inoculation a
McMaster count was performed to determine the number of oocysts per mL in order to determine
appropriate volume of inoculum
523 Experimental infections
Part 1
Five ferrets were randomly assigned to each of the control and infection groups After the
acclimation period on day 0 four ferrets in the infection group were inoculated orally with a high dose
oocyst suspension (1 times 106 oocysts in 025 mL of saline) mixed into 1 mL of FerreTone Skin amp Coat
Supplement (United Pet Group Inc Blacksburg Virginia USA) a fifth ferret was inoculated with a
moderate dose oocyst suspension (5 times 104 oocysts in 025 mL of saline) in the same volume of FerreTone
Ferrets in the control group were inoculated with a placebo (025 mL of saline) in 1 mL of FerreTone
Inoculation was performed by offering the oocyst suspension or placebo to the ferrets in a plastic
container
Fecal samples were collected daily from each inoculated ferret for 14 days post-inoculation
Samples were analyzed via fecal flotation using the McMaster method followed by routine flotation in
saturated salt solution (Dryden et al 2005) to determine the presence or absence of oocysts and oocyst
burden (oocysts per gram of feces [OPG])Temporal trends in oocyst shedding were monitored Ferrets
were evaluated visually twice daily for the presence of clinical signs of coccidial disease The first of
every two ferrets identified to shed oocysts was to be humanely killed at the time of peak oocyst shedding
(ie the first day that fecal oocyst counts remained static or declined) and necropsied to confirm the
presence of and describe parasitic replication in the intestinal mucosa Any remaining animals that shed
oocysts were to be monitored throughout the 14 day period following inoculation in order to determine
the duration and intensity of oocyst shedding for these individuals the total number oocysts shed during
patency was determined
109
Part 2
All ferrets from the infection group that did not shed oocysts during Part 1 (n=4) and all but one
ferret from the previous control group (n=4) were orally inoculated with the high dose oocyst suspension
(1 times 106 in 1 ml of saline) mixed with an equal volume of FerreTone Consequently between phases 1
and 2 all but one ferret were inoculated at least once with the BFF coccidia in order to increase
experimental animal numbers and determine if ferret age played a role in susceptibility to infection
One ferret from the previous control group was inoculated with a lower dose (2 times 105 oocysts in
075 mL of 09 saline mixed with 1 mL FerreTone) of oocysts that had been collected from the single
domestic ferret that shed in Part 1 oocysts were purified as described above for the initial inocula Fecal
collection and analysis were performed as previously As in Part 1 one in every two ferrets sequentially
identified to be shedding oocysts in feces was killed humanely at the time of peak shedding and a
complete necropsy examination performed The remainder of the ferrets observed to be shedding were
monitored for the full 14 days of the trial after which they were killed humanely and necropsied and total
number of oocysts shed during patency was determined All ferrets that did not shed coccidial oocysts
during the infection trial were rehomed at the end of the trial
524 Animal welfare
Ferrets were evaluated twice daily for development of clinical signs of coccidial disease and any
animal showing clinical disease was to be treated as determined by a veterinarian with supportive care
including fluid therapy A grading system for clinical signs including intervention points and removal
criteria was created for use during daily evaluation (see Appendices 3 and 4) Animals whose clinical
signs could not be ameliorated without the use of specific anticoccidial therapy were to be euthanized
Should the inoculation in Part 1 have resulted in clinical disease that required extensive treatment andor
necessitated euthanasia a lower number of oocysts would be used for subsequent inoculation in Part 2
Ferrets to be euthanized were anesthetized by mask induction with isoflurane in oxygen a 1 mL blood
110
sample was collected from the cranial vena cava and then an intracardiac dose of potassium chloride (2
mEqkg) was administered
525 Hematology
Blood was collected from all ferrets under isoflurane anesthesia at the time of pre-trial health
examination and again at the time of humane killing Blood was collected via jugular venipuncture
initially due to the small size of the ferrets at arrival and then by cranial vena cava venipuncture or
cardiocentesis prior to euthanasia Complete blood count and serum biochemistries were performed by the
Animal Health Laboratory of the University of Guelph Guelph Ontario
526 Morphologic and molecular characterization
Morphologic and molecular characterization of oocysts shed by the domestic ferrets during the
course of the infection trial was performed to ensure that the ferrets were shedding the same species of
Eimeria with which they were inoculated Oocysts were concentrated from positive fecal samples as
described above A drop of concentrated oocyst solution was viewed photographed and measured at
400times and 600times for comparison with previously determined morphometrics of Eimeria ictidea oocysts
(Chapter 3)
Regions from the mitochondrial cytochrome c oxidase subunit I and III (mt COI and mt COIII)
DNA were amplified by polymerase chain reaction (PCR) from each sample using primer pairs
400F1202R and -172F799R respectively For all PCR reactions samples were denatured at 95 degC for 5
min then subjected to 35 cycles of 94 degC for 30s anneal at 52degC for 30s and extension at 72 degC for 60s
followed by a final extension at 72 degC for 7 min PCR gel electrophoresis and sequencing methods used
were as described in the Materials amp Methods section of Chapter 2
The resulting consensus sequences were searched from within Geneious against previous
sequences for E ictidea produced by the authors and against publically available sequences on the
111
BLAST server (blastncbinlmnihgovBlastcgi) using the blastn search algorithm against the nrnt
database (GenBank+EMBL+DDBJ+RefSeq ndash AA or DNA)
527 Necropsy protocol
All humanely killed ferrets underwent a complete necropsy (Appendix 6) using the modified
protocol described in Materials amp Methods section 424 of Chapter 4
53 RESULTS
Initial physical examination was unremarkable with the exception of mild to moderate bilateral
ceruminous discharge within the external ear canal of all ferrets Complete blood count and serum
biochemistry results for all ferrets were within normal reference intervals for juvenile domestic ferrets
(Appendices 2a and b) (Fox 2014) Six days after arrival a single ferret (103) in the control group
developed mild upper respiratory signs consisting of sneezing and clear nasal and ocular discharge these
clinical signs were associated with mild dehydration and decreased food and water consumption The
ferret was treated with subcutaneous fluid therapy (10 mL Plasmalyte-A subcutaneous Baxter Alliston
Ontario) heat and supportive care and all clinical signs resolved within three days This ferret was
deemed healthy to participate in the remainder of the clinical trial A second ferret (105) in the control
group developed unilateral purulent ocular discharge 14 days after arrival one day prior to placebo
inoculation The ferret was treated topically twice daily for five days with Isathal ophthalmic gel (fusidic
acid 10 mgg Dechra Veterinary Products Inc Point-Claire Quebec) and the discharge resolved but
reoccurred within 2 days of treatment cessation Ocular examination showed mild conjunctivitis but no
evidence of corneal lesions and fluorescein staining did not indicate the presence of corneal ulceration
The ferret was treated for an additional six days with tobramycin ophthalmic solution (3 mgmL Sandoz
Tobramycin 03 Boucherville Quebec) after which clinical signs resolved completely No coccidial
oocysts were shed in feces from any of the ferrets during the two week acclimation period
112
531 Oocyst shedding
All ferrets readily ingested the inoculum with either placebo or concentrated oocysts In Part 1
one ferret (203) in the infection group which had received the high dose (1 times 106 oocysts) shed oocysts
on day 8 and day 9 after inoculation (Tables 51 52) This ferret was 71 days of age at the time shedding
was initially identified The ferret was killed humanely 11 days post inoculation later than had been
outlined in the protocol as processing of fecal samples had been delayed by two days resulting in late
detection of oocyst shedding in this individual Oocysts were not identified in the feces of the three
remaining ferrets that received the high inoculation dose the single ferret that received the lower dose (5
times 104 oocysts) or in the ferrets within the control group
In Part 2 six of eight ferrets inoculated with the high dose (1 times 106 oocysts) shed oocysts during
the 14 day observation period (Tables 51 52) Four of these ferrets were from the previous control
group One of the ferrets previously inoculated with the high dose inoculum in Phase 1 that had not shed
oocysts did shed oocysts after being inoculated a second time with the same dose during Phase 2 The
ferret that had previously received the low dose (5 times 104 oocysts) of oocysts in Phase 1 also shed after
inoculation with the high dose in Phase 2 Three ferrets did not shed oocysts after high dose inoculation in
Phase 2 one of these had been part of the previous control group the other two had received the high
dose inoculation previously in Phase 1
The pre-patent period ranged from 7-9 days (Table 51) with equivalent numbers of ferrets
commencing shedding on each of days 7 through 9 All six ferrets were between 91 and 93 days of age at
the time shedding was initially identified Oocyst per gram counts and shedding trends for all individuals
are shown in Table 51 Total oocyst shedding during patency was lt14 oocysts 8904 oocysts and
172291 oocysts for ferrets 201 104 and 105 respectively The two ferrets for which the prepatent period
was 9 days only shed oocysts for one day and in low numbers Oocysts were not identified in the feces of
the three remaining ferrets two of which received the high inoculation dose (1 times 106 oocysts) and the
third that received the lower inoculation dose (2 times 105 oocysts) (Table 52)
113
532 Morphologic and molecular characterization
In all seven ferrets that shed oocysts the morphologic features and measurements (length width
shape index) of the shed oocysts were consistent with those of the E ictidea administered in the inoculum
(Figure 51) Molecular confirmation of the identity of the oocysts shed was successful in 3 out of the 7
ferrets (102 103 203) samples from the four remaining ferrets did not contain adequate quantity or
quality of DNA for confirmation
533 Clinical signs
Clinical signs associated with patent infection were identified in 6 of 7 ferrets (Table 52) These
signs included weight loss (n=5) diarrhea (n=1) mucoid soft feces (n=2) feces containing blood (n=2)
and malodorous feces (n=1) Appetite reduction was noted in two ferrets from the infection group in Part
1 between 6-8 days post infection however no oocyst shedding was detected from either ferret during
this time
534 Hematology
CBC and serum biochemistry values from ferrets collected during pre-trial health screening are
displayed in Appendices 1a and 1b Values obtained for ferrets euthanized during or after the
experimental trial are displayed in Appendices 3a and 3b Minor variances from reference range values
for CBC and serum biochemistry were identified in six of the seven ferrets with patent infection In all six
ferrets for which a complete serum biochemistry was obtained immediately prior to death creatinine
kinase (CK) values were elevated (see Appendix 3b) Ferret 103 exhibited a mild hypoalbuminemia (20
ref 24-40 gL) on ante-mortem serum biochemistry (see Appendix 3b)
114
535 Necropsy
No gross or histopathologic lesions were present and coccidia could not be identified in sections
of intestine from the single ferret (203) humanely killed in Part 1 For the ferrets humanely killed in Part
2 no evidence of diarrhea hematochezia or mucoid fecal material was identified grossly Ferret 105
killed at the termination of the experiment but still shedding low numbers of oocysts in its feces
exhibited a 7 cm region of congested mucosa within the distal jejunum
Coccidial life stages were identified in small intestinal sections from four of the seven ferrets that
were identified to shed oocysts at some point prior to necropsy (Figure 52 Tables 52 and 53) Affected
sections included jejunum in all four animals as well as ileum in one and were collected from 114 to 218
cm aboral from the pylorus (see Figure 53) Coccidia were not identified in sections of duodenum
proximal jejunum or large intestine however oocysts were identified within fecal material in the lumen
of the large intestine from one ferret (103) Of the 11-19 sections of intestine examined for each ferret the
number of sections containing parasites ranged from one to eight 1 section in ferret 102 2 sections in
ferret 201 4 sections in ferret 105 8 sections in ferret 103 A mix of sexual and asexual life stages was
observed within the enterocytes in small intestinal sections from ferret 103 the remainder of the ferrets
showed either asexual (102 201) or sexual (105) life stages in affected segments
Pathologic changes and additional histologic findings in small intestinal sections of ferrets with
enteric coccidia included rare regions of blunting of the villi and sloughing of the epithelium associated
with hemorrhage and inflammation The primary lesions identified were subjectively increased numbers
of eosinophils lymphocytes and plasma cells within the lamina propria of the small intestine and
similarly increased neutrophils lymphocytes and plasma cells within the lamina propria of the large
intestine Neutrophils were rarely present in intestinal crypts and glands
Other gross necropsy findings included two ferrets with renal cortico-medullary cysts and one
ferret with mild thickening of the esophageal mucosa midway along the esophageal length No histologic
changes were identified within a sample of esophagus taken from this region
115
54 DISCUSSION
The findings of this study show that domestic ferrets are susceptible to infection with the enteric
coccidium Eimeria ictidea isolated from black-footed ferrets Both morphometric and molecular
diagnostic methods were used to confirm that ferrets were shedding oocysts of the same species with
which they were inoculated Molecular characterization was successful in three of the seven ferrets that
developed patent infections and as no other coccidial species was identified during pre-trial observation
morphometry was considered to be confirmatory in the remaining four animals
We have referred to the eimeriid coccidium affecting BFF and used in this experimental trial as
E ictidea based on morphometric similarity of their oocysts with those of E ictidea as described from
domestic ferrets (see Chapter 3) There is limited published information on infection of domestic ferrets
with E ictidea outside of Hoarersquos original descriptions (1927 1935a b) which form the basis for all
subsequent identifications of E ictidea in domestic ferrets and in BFF Attempts to obtain exemplars of E
ictidea from domestic ferrets to characterize using molecular techniques were unsuccessful (Chapter 2)
Multiple diagnostic laboratories in Canada and Europe were solicited for coccidia-positive fecal samples
from domestic ferrets but no samples of E ictidea were received over a 4 year period (2014-2017)
Eimeria ictidea was identified based on microscopic examination in only two samples submitted to a
European diagnostic laboratory from 2008-2015 It is unproven whether the E ictidea described from
domestic ferrets and the E ictidea identified from black-footed ferrets and used in this experimental
work are the same or are simply morphologically indistinguishable Eimeria species However the
consistency in morphology host genus and location of infection within the intestinal tissues combined
with the successful cross-transmission of this parasite to domestic ferrets described in the present study
suggests they are likely conspecific
116
The pre-patent period (minimum duration of endogenous development) for infection with E
ictidea in the inoculated domestic ferrets ranged from 7-9 days (see Table 51) the pre-patent period for
this parasite in the BFF the natural host for this coccidium is unknown Hoare experimentally infected
naiumlve domestic ferrets with E ictidea derived from naturally occurring infection in this species (Hoare
1935b) The inoculated ferrets shed oocysts after a pre-patent period of 7 days consistent with the 7-9
days seen in the work described here with E ictidea
Shedding of oocysts was identified over a period of 1-7 days (see Table 51) and intensity ranged
from less than 14 up to 15624 OPG These results may be skewed with erroneously low duration of
shedding and number of oocysts shed as three of the seven ferrets were humanely killed at or prior to the
expected peak of oocyst shedding for tissue collection and histologic examination in order to increase the
probability of identifying parasite life stages within the intestinal sections Shedding periods were similar
to those identified in adult single-housed BFF which ranged from 2-9 days however oocyst per gram
counts from the domestic ferrets were consistently lower than OPG counts from BFF (104 - 554274
OPG) infected with the same parasite (see Chapter 4) Furthermore the total number of oocysts shed by
individual domestic ferrets (14 - 172291 oocysts) during patency was reduced compared to BFF despite
similar length of shedding period (see Chapter 4) The domestic ferrets in this study were naiumlve individual
juveniles whereas the BFF were adults 1-5 years of age some of which were showing clinical signs at
the time of oocyst shedding The relative influences of age species and previous exposure to the parasite
on our observations are unknown
Two different fecal flotation methods were used on all samples to increase the probability of
oocyst detection The McMaster method was used to provide accurate OPG counts for quantification of
oocyst shedding however this method had a minimum detection limit of ~13 oocysts per gram (1333
OPG calculated) because it is based on dilution of the initial sample with flotation media (saturated salt)
In samples with few oocysts oocysts may be missed or to be present in numbers below this detection
limit Routine salt flotation is in contrast performed using the entire sample allowing for detection of
117
small numbers of oocysts Consequently in cases where oocyst per gram counts were low shedding was
identified on routine salt flotation but not by the McMaster method and recorded as lsquopositiversquo but below
the detection limit of the enumeration method
Subclinical to clinical disease occurred in six of the seven ferrets that developed patent infection
with weight loss being the most frequent clinical sign Other clinical signs were typical of enteric
coccidiosis including diarrhea hematochezia and mucoid andor soft feces These clinical signs are
similar to those previously described for black-footed ferrets infected with this parasite (USFWS BFF
Recovery Program 2017 Chapter 4) and for domestic ferrets with enteric coccidiosis (Sledge et al
2011) Interestingly development of clinical disease was not described in the naiumlve domestic ferret
inoculated by Hoare (1935) with E ictidea derived from naturally occurring infection Based on review of
the literature severe clinical disease resulting from intestinal coccidiosis is rare in domestic ferrets
Black-footed ferrets however appear more susceptible to disease development and more frequently show
significant clinical signs No domestic ferret required treatment for clinical coccidiosis during the course
of this study It is possible that the more pronounced clinical signs associated with enteric coccidiosis in
BFF may result from the limited genetic diversity in a population derived from so few individuals and
increased susceptibility of BFF to other diseases such as sylvatic plague have been described in
comparison with their domestic counterparts (Williams et al 1994) Although unlikely recent acquisition
of E ictidea from a related host species such as the domestic ferret could have resulted in increased
pathogenicity and severity of clinical disease from infection with this parasite in BFF
Minor variances from reference range values for CBC and serum biochemistry were identified in
six of the seven ferrets with patent infection but only in one case (ferret 103) did this appear to be
correlated with infectiondisease In this animal a mild hypoalbuminemia (20 ref 24-40 gL) was noted
(Appendix 3b) On histopathology large regions of the small intestine contained parasite life stages
however inflammation lysis of epithelial cells and necrosis of affected areas that could be expected to
result in protein loss into the intestinal lumen were not identified
118
Creatine kinase values were elevated in all six ferrets for which a complete serum biochemistry
was obtained immediately prior to death these findings are consistent with release from CK rich tissues
such a skeletal muscle during venipuncture and manual restraint
The pathologic lesions identified within the intestinal sections of ferrets euthanized at the time of
oocyst shedding were similar to those identified in affected BFF but in most cases were less locally
extensive or widespread throughout the small intestine than those observed in necropsy cases of BFF (see
Chapter 4) Coccidia were seen in the histologic sections of four ferrets all three ferrets that were actively
shedding oocysts at the time of necropsy (102 103 105) and one of four in which necropsies were
performed after oocyst shedding had ceased (201)
Although the primary objective of the examination of histologic sections from affected ferrets
was to identify coccidial life stages attempts were made to describe the pathologic changes associated
with the presence of the parasite Despite necropsies being performed almost immediately after death and
the use of Serra solution fixative to improve parasite and tissue preservation the villi and villar
epithelium of the trimmed sections were frequently distorted or absent and consequently accurate
commentary on these areas was precluded and was made only on visible components of the lamina
propria and crypts or glands A deliberate decision was made not to kill and collect samples from the
saline inoculated control ferrets after Phase 1 and those not shedding oocysts during Phase 1 or 2 thus no
age-matched intestinal sections were available for comparison It is difficult to comment on the
significance of the inflammatory cells observed in the lamina propria of the small and large intestinal
segments or the proliferative rate of the crypt epithelium The only changes identified which may be
considered significant are the presence of neutrophils within the crypts and glands of small and large
intestinal sections respectively but these lesions were rare and not associated with the presence of
parasitic life stages
119
While no parasitic life stages were identified in intestinal sections from ferrets 104 203 and 205
one of the three ferrets ferret 104 exhibited lymphoplasmacytic inflammation and blunting of jejunal
villi in one section (36-38 cm aboral from pylorus) These findings might be expected as the most
extensive histologic lesions would occur associated with lysis of the intestinal epithelial cells as oocysts
were shed into the feces after which new intestinal epithelial cells would re-cover the denuded villar
surface Thus for those cases in which histologic examination was performed after shedding had ceased
presence of the protozoal life stages in the intestines would be expected to be significantly reduced or
absent
Hoare (1935a b) described a particular reaction to the presence of parasitic life stages of Eimeria
ictidea in the small intestine of domestic ferrets in which only the villar tips were affected and there was
resultant annular constriction of the villus separating the affected and non-affected segments These
particular changes were not identified in any of the ferrets in this study and had not been noted
retrospectively in naturally infected BFF (Chapter 4)
While patent infection and intestinal disease could be experimentally created in domestic ferrets
without equivalent experimental work in BFF it is difficult to fully compare the susceptibility to infection
and to the development of disease between the two species Eimeria species tend to be host specific thus
if E ictidea from BFF is not conspecific with E ictidea in domestic ferrets it might be expected that the
domestic ferret would be less susceptible to infection and the development of disease than is the BFF
Even if the two parasites are identical natural passaging through the BFF may alter affinity for the
domestic ferret
It appears that the infectious dose of oocysts of E ictidea derived from BFF required to initiate a
patent infection in domestic ferrets is high The ferrets that developed patent infections were administered
an inoculum containing 1 times 106 sporulated oocysts and even with this extremely high inoculating dose
only a proportion of inoculated ferrets became infected Neither the ferret that received the low dose
120
inoculum nor the ferret that received the passaged oocysts from Part 1 of the study shed oocysts during
the 14 day period post inoculation The latter finding was unexpected as fresh passaged oocysts would be
expected to contain larger proportions of viable oocysts and be comparatively more infective than oocysts
that had been stored for 2-4 weeks prior to inoculation
The only publication describing oral inoculation of Eimeria species in domestic ferrets (Hoare
1935b) did not quantify the number of oocysts administered A study performed in 16 farmed juvenile
mink (Mustela vison) administered 2000 sporulated oocysts of each of three coccidial species (I
laidlawi E vison and an unknown Eimeria species) resulting in patent infection with one of the three
species (Foreyt et al 1977) as determined by the presence of oocysts on fecal examination The authors
did not reveal which type of oocysts resulted in the infection
The number of oocysts required to result in infection in BFF is unknown Based on the
authorsrsquo observations of over 100000 oocysts per gram of feces being shed by black-footed ferrets into
cages of lt1m2 floor space (see Chapter 4) we estimated that under normal caged conditions animals
would likely ingest thousands of oocysts over a short period of time This was in part why a large number
of oocysts (up to 1000000 as available from our store of viable oocysts) was administered to each
domestic ferret in order to increase the probability that infection and shedding would result Furthermore
for the parasite to persist within the ferret population the total number of oocysts shed into the
environment would have to be several times higher than the infective dose required to generate a patent
infection otherwise the parasite would be expected to die out If E ictidea of domestic ferrets and E
ictidea of BFF are conspecific the difference in oocyst shedding between the host species during patent
infection could potentially explain the low prevalence of E ictidea reported from the domestic ferret
population (as seen in Chapter 2)
Both humoral and cell mediated immunity are involved in the immune response to coccidia The
role of maternal derived antibodies in combatting protozoal infection in mustelids has not been studied
121
but in carnivores maternal antibodies to viruses can last up to 16 weeks (Chappuis 1998) In poultry
maternal Eimeria-specific IgG is transferred via the egg yolk to offspring In one study breeding hens
were infected with a single species of Eimeria 28-39 days prior to lay Their hatched chicks were
challenged by inoculation with oocysts of the same and a related Eimeria species and showed reduced
oocyst shedding compared to age matched controls indicative of passive transfer of immunity (Smith et
al 1994) It is possible then that the presence or absence of maternal antibodies may be a factor in the
age at which ferrets are susceptible to coccidial infection and the development of disease The facility
from which the domestic ferrets were acquired has not previously detected Eimeria species on routine
fecal screening (Dr Bambi Jasmin personal communication) consequently it is unlikely that they would
have received maternal immunity to or been exposed to this parasite and thus can be considered to be
naive
Despite a theoretical lack of maternal immunity there did appear to be an effect of age on
susceptibility to infection In Part 1 when the ferrets were approximately 70 days of age only 1 of 4
ferrets inoculated with the high dose of oocysts developed patent infection In comparison in Part 2
when the ferrets were 91-93 days of age 3 of 4 ferrets that had been in the saline control group for Part 1
developed patent infections as did 2 of 4 ferrets that had been inoculated in Part 1 but had not shed
oocysts One of these previously inoculated ferrets was the individual that had received the lower dose of
oocysts Thus it appears that patent infection could be produced more easily in the older ferret kits
However the two ferrets that had previously been inoculated exhibited the shortest shedding periods (1
day) and lowest oocyst per gram counts and parasite life stages in these cases were rare (201) to absent
(205) on histologic examination of numerous sections of intestine It is thus possible that the primary
inoculation resulted in abbreviated infections or infections in which so few oocysts were shed that
infection was not detected Our observation of endogenous stages in the intestinal tissues of some of the
ferrets following cessation of oocyst shedding suggests that the pre-patent period and duration of patency
may vary considerably from animal to animal consequently it is possible that some of the kits would
122
have ultimately shed a few oocysts from the primary inoculum if followed beyond 14 days post-
inoculation Whether through an aborted or undetected infection previous exposure of these kits to the
parasite probably generated partial immunity against E ictidea and therefore the intensity of infection
upon challenge in previously exposed domestic ferret kits was reduced
The authors acknowledge the limitations of this initial pilot study however it was proven that
patent infection with E ictidea isolated from black-footed ferrets could be generated in a novel host the
domestic ferret Further studies will be required to investigate the effect of age on susceptibility to
infection as well as the possibility of development of immunity after exposure and its role in reducing
parasite replication and disease in subsequent infection With so few BFF in existence the use of BFF for
in vivo infection trials cannot be contemplated consequently refining the domestic ferret infection model
will be essential for carrying out research specifically intended to help manage coccidiosis in the
endangered black-footed ferret
Figure 51 Exogenous life stages of Eimeria ictidea shed from a domestic ferret (Mustela putorius
furo) experimentally inoculated with oocysts originating from black-footed ferrets (Mustela
nigripes) Unsporulated oocyst (solid black arrow) Sporulated oocyst (solid white arrow) Bright
field microscopy scale bar = 25 microm
25 microm
124
Figure 52 Life stages of Eimeria ictidea within the small intestinal epithelium of an experimentally
infected domestic ferret (Mustela putorius furo) A) Sexual life stages (micro- and macrogamonts
- white arrows) crowding the villar enterocytes all stages are found between the nucleus and
luminal surface of infected enterocytes Hematoxylin and eosin staining scale bar = 25microm B) At
higher magnification meronts (black arrows) and gamonts (open arrows) are crowded between the
enterocyte nuclei and brush border Hematoxylin and eosin staining scale bar = 25microm
25 microm
25 microm
125
Figure 53 Presence and location of sexual and asexual life stages of Eimeria ictidea within the
intestinal epithelium of domestic ferrets (Mustela putorius furo) (n=7) that developed patent
infection after experimental inoculation with oocysts originating from black-footed ferrets (Mustela
nigripes) Legend duod = duodenum jej= jejunum Sequential numbers for jejunal and colonic
sections represent the order aboral from the pylorus from which the samples were collected SI =
additional section(s) of small intestine whose aboral sequence was not recorded
0
1
2
3
duod jej 1 jej 2 jej 3 jej 4 jej 5 ileum colon 1 colon 2 SI
Nu
mb
er
of
Ferr
ets
Aff
ect
ed
Intestinal Section Containing Life Stages of Eimeria ictidea
Asexual life stages Sexual life stages
126
Table 51 Prepatent period and oocyst shedding patterns in domestic ferrets (Mustela putorius furo)
experimentally inoculated with 1 times 106 oocysts of Eimeria ictidea originating from black-footed ferret
(Mustela nigripes) that developed patent infections
Oocysts shed per gram of feces
Ferret Identity
Day post inoculation 102 103 104 105 201 203 205
1 0 0 0 0 0 0 0
2 0 0 0 0 0 0 0
3 0 0 0 0 0 0 0
4 0 0 0 0 0 0 0
5 0 0 0 0 0 0 0
6 0 0 0 0 0 0 0
7 lt 14 1807 0 0 0 0 0
8 11053 139 7091 0 156238 0
9 463 11733 lt 14 203 lt 14
10 578 7549 0 0 0
11 lt 14 0 0 0
12 0 lt 14 0
13 0 lt 14 0
14 0 lt 14 0
Legend ferret euthanized as of this date lt 14 = oocyst positive samples with less than 14 oocysts per
gram of feces
127
Table 52 Results of oral inoculation of domestic ferrets (Mustela putorius furo) with oocysts of Eimeria ictidea originating from black-footed
ferrets (Mustela nigripes)
Ferret
Identity
Results Part 1 Results Part 2
Inoculum Oocyst
Shedding
Presence
of clinical
disease
Coccidia
present in
sections
Inoculum Oocyst
Shedding
Presence
of clinical
disease
Coccidia
present in
sections
101 Saline N N - 2 times 105 oocystsa N N -
102 Saline N N - 1 times 106 oocysts Y N Y^
103 Saline N N - 1 times 106 oocysts Y Y Y^
104 Saline N N - 1 times 106 oocysts Y Y Nyen
105 Saline N N - 1 times 106 oocysts Y Y Yyen
201 1 times 106 oocysts N N - 1 times 106 oocysts Y Y Yyen
202 1 times 106 oocysts N N - 1 times 106 oocysts N N -
203 1 times 106 oocysts Y Y Nyen
204 1 times 106 oocysts N N - 1 times 106 oocysts N N -
205 5 times 104 oocysts N N - 1 times 106 oocysts Y Y N^
Legend includes any of the following weight loss diarrhea mucoid feces malodorous feces inappetence N = no Y= yes - = necropsy not
performed a = oocysts collected after passage through ferret 203 ^ = shedding oocysts at time of necropsy yen = not shedding oocysts at time of
necropsy
128
Table 53 Distribution of coccidial life stages in domestic ferrets (Mustela putorius furo) orally inoculated with oocysts of Eimeria ictidea
originating from black-footed ferrets (Mustela nigripes)
Ferret Identity
Intestinal level 102
103
104 105
201
203
205
Duodenum N N N N N N N
Jejunum 1 N N N N N N N
Jejunum 2 N N N N N N N
Jejunum 3 N S A N N N N N
Jejunum 4 N S A N N N N N
Jejunum 5 N S A N S A N N
Ileum N S A N N N N N
Colon 1 N Na N N N N N
Colon 2 N Na N N N N N
Unmeasured small intestinebc
S - 05
A - 15
S - 45
A - 25
S - 05
A - 05
S - 26
A - 06
S - 06
A - 06
S - 06
A - 06
S - 08
A - 08
Unmeasured large intestinebc
S - 01
A - 01
S - 01
A - 01
S - 01
A - 01
S - 02
A - 02
S - 01
A - 01 none
S - 01
A - 01
Legend N = no parasite life stages S = sexual life stages present A = asexual life stages present a= oocysts present in feces b=
additional sections of intestine for which the location measured from the pylorus was not obtained c= xn where x is number of
sections containing sexual or asexual lifestages n is the number of sections examined
129
CHAPTER 6 WHOLE MITOCHONDRIAL GENOME SEQUENCES OF TWO EIMERIA
SPECIES ISOLATED FROM DOMESTIC (MUSTELA PUTORIUS FURO) AND BLACK-
FOOTED FERRETS (MUSTELA NIGRIPES)
ABSTRACT
The complete mitochondrial (mt) genomes of Eimeria furonis and Eimeria ictidea (Eimeriidae
Coccidia Apicomplexa) originating from single fecal samples from a domestic (Mustela putorius furo)
and a black-footed ferret (Mustela nigripes) respectively were sequenced Both mt genomes were
circular-mapping with lengths of 6165 base pairs (Eimeria furonis - GenBank MF795598) and 6182
base pairs (Eimeria ictidea - GenBank KT203399) Genome organization and gene contents were
comparable with those of other publically available mt genomes from a variety of Eimeria species and
related coccidia there were three complete coding DNA sequence regions encoding cytochrome c
oxidase subunit I cytochrome c oxidase subunit III and cytochrome B and 33 regions encoding
fragmented rDNA Alignment of these mt genome sequences demonstrates a relatively high (945 340
single nucleotide differences [SNDs]) pairwise sequence identity between these Eimeria spp infecting
ferrets the majority of the SNDs resulted in synonymous codon changes with no changes to their protein
products Alignment of the newly sequenced mt genomes demonstrates and phylogenetic reconstructions
support the monophyly of these Eimeria spp of ferrets with another Eimeria sp of carnivores as the
sister taxon to this clade
61 INTRODUCTION
Coccidia are protozoal eukaryotic host-specific parasites of the phylum Apicomplexa and can
be divided into two major taxonomic suborders the eimerioirinid and adeleid coccidia The eimeriorinid
coccidia include both the typical intestinal coccidia (eg Eimeria Isospora Cyclospora) species
belonging to the family Eimeriidae as well as tissue or cyst-forming coccidia (eg Cystoisospora
130
Besnoitia Toxoplasma Sarcocystis) of the family Sarcocystidae (see Cox 1994) Ten species of
Eimeria and twelve species of Isospora (=Cystoisospora) have been described in the Mustelidae
(see Chapter 1 Table 11) Eimeria furonis has been reported in the European polecat (Mustela
putorius) domestic ferret (DF Mustela putorius furo) black-footed ferret (BFF Mustela nigripes) and
mink (Mustela vison) (Hoare 1927 Nukerbaeva and Svanbaev 1973 Jolley et al 1994) Eimeria ictidea
has been reported in the Steppe polecat (Mustela eversmanii) as well as the European polecat domestic
ferret and black-footed ferret (Hoare 1927 Svanbaev 1956 Jolley et al 1994) These reports are based on
the morphometric characteristics of oocysts identified in the feces of the aforementioned host species
without the use of molecular techniques to confirm specific parasite identities
Recently sequences of the mitochondrial cytochrome c oxidase subunit I gene (mt COI) and
nuclear small subunit ribosomal DNA (nu 18S rDNA) of E furonis originating from a domestic ferret (nu
18S rDNA GenBank MF774678-MF774680 mt COI GenBank MF774034-MF774036) and E ictidea
originating from a black-footed ferret (nu 18S rDNA GenBank MF860826-MF860827 mt COI
GenBank MF860823 MF860825) were generated (see Chapters 2 and 3) The parasite originating from
the black-footed ferret was identified as E ictidea based on morphologic similarity to the original
descriptions of E ictidea from domestic ferrets however sequence-based genotyping of E ictidea from
domestic ferrets has not been completed and consequently it has not been demonstrated unequivocally
that the two parasites are conspecific In the present work the complete mitochondrial genomes of E
furonis from the DF and E ictidea from the BFF are described and compared with the mitochondrial
genomes of related coccidia
62 MATERIALS amp METHODS
621 Parasites
Two isolates of morphologically distinct Eimeria species were used in this study Isolate one
identified morphologically and by nu 18S rDNA and mt COI sequences as Eimeria furonis was obtained
131
from a fecal sample from a DF that was submitted for routine ova and parasite examination to a European
diagnostic laboratory8 Isolate two identified morphologically and by nu 18S rDNA and mt COI
sequences as Eimeria ictidea was obtained from a fecal sample from a BFF and was collected during
routine cage cleaning in a captive breeding facility (see Chapters 3 and 4) Fecal collection techniques for
the BFF were reviewed and approved by both the Animal Welfare Committee and the Animal Care and
Research Committee of the Toronto Zoo
622 DNA isolation from coccidia in feces
Genomic DNA was isolated from fecal derived coccidial oocysts as described section 221 of the
Chapter 2 Materials amp Methods Parasite DNA concentration was estimated using a Nanodrop 2000
spectrophotometer (NanoDrop Products Wilmington DE USA) and DNA was stored at 4 degC for
immediate use or minus20 degC for later use
623 Whole genome sequencing
Mitochondrial whole genome amplification for both Eimeria species was initiated using sets of
mt-specific primers that generated overlapping polymerase chain reaction (PCR) fragments (Tables 61
and 62) PCR amplification was performed for all samples in a volume of 25 microl containing ~100 ng of
genomic DNA 1times PCR buffer 3 mM MgCl2 06 mM deoxyribonucleotide triphosphates (dNTPs) 500
nM of each primer and 4 U of Invitrogen Platinum Taq DNA Polymerase (Thermo Fisher Scientific
Toronto ON Canada) PCR reactions were performed on a Bio-Rad T100 PCR thermal cycler (Bio-Rad
Laboratories Singapore) using settings as described previously in the Materials amp Methods section of
Chapter 2 Table 61 details the specific anneal conditions used for the various primer pairs Genomic
DNA from either Eimeria maxima or Eimeria tenella acted as a positive control for the reaction
chemistry Gel electrophoresis purification and sequencing of the PCR amplification products were
8 Vet Med Labor GmbH Division of IDEXX Laboratories Ludwigsburg Germany
132
performed as described in Chapter 2 The resulting chromatograms were aligned and analyzed with
Geneious Ver 818 or later (Biomatters Limited Auckland New Zealand) and high quality consensus
sequences generated The completed mt genome sequences were annotated by comparison with
previously annotated mt genomes from other Eimeria species (eg Eimeria innocua - KR1082961) and
the annotated mt genomes deposited in GenBank
624 Phylogenetic analysis
To determine the phylogenetic affinities of the newly obtained sequences with sequences from
related apicomplexan taxa representative whole mt genome sequences from eimeriid coccidia were
downloaded from GenBank A complete mt genome sequence from an unnamed Choleoeimeria sp was
used to root the ingroup taxa several small genomic rearrangements in the Choleoeimeria sp sequence
required some rearrangement of the genome sequence to unambiguously align homologous regions across
the complete mt genomes
Whole genome sequences were aligned using MAFTT v7017 (Katoh et al 2002) executed from
within Geneious the resulting alignment was examined by eye to adjust start and stop codon positions in
aligned coding DNA sequence [CDS] regions (ie mt COI mitochondrial cytochrome c oxidase subunit
III gene [mtCOIII] mitochondrial cytochrome b gene [CytB]) Phylogenetic trees were generated using
Bayesian Inference (BI) using MrBayes Ver 326 (Huelsenbeck and Ronquist 2001) executed from
within Geneious The aligned complete mt genomes were partitioned into coding (ie CDS) and non-
coding regions so that region-specific models of nucleotide substitution could be applied Characters in
the non-coding region were analysed with the general time reversible (GTR) model (Tavareacute 1986) with
the following parameters nucmodel=4by4 nst=6 rates=invgamma (ie GTR+I+G) Characters in the
coding regions were analysed using the codon nucleotide model (ie lset nucmodel=codon rates=gamma
ngammacat=4) using metazoan mitochondrial translation (ie code=metmt)
133
All BI analyses were run for a chain length of 1000000 with tree sampling every 1000
following a burn-in of 100000 with default settings of 4 heated chains and heated chain temp of 02
63 RESULTS
The whole mt genome sequences of the single isolates of E furonis and E ictidea were
respectively 6165 base pairs (bp) (Figure 61 GenBank MF795598) and 6182 bp (Figure 62 GenBank
KT203399) Content and organization of both mt genomes consisted of three protein-coding genes (mt
COI mt COIII and CytB) interspersed with large and small subunit ribosomal DNA (rDNA) fragments
Details of the various CDS and rDNA fragments are summarized in Table 63 (for E furonis) and Table
64 (for E ictidea)
Pairwise alignment of the mt genome sequences from E furonis and E ictidea demonstrated a
relatively high pairwise sequence identity (946 333 single nucleotide differences [SNDs]) between
these two parasites The bulk of the SNDs (676 225333) were clustered within the three CDS regions
that encode CytB mt COI and mt COIII (see Figure 63 and Table 65) However the majority of these
SNDs (826 186225) were synonymous codon changes that resulted in no changes to the protein
products Only 41 SNDs were involved in 34 amino acid changes distributed among the three CDS
The 33 rDNA fragments comprised 2108 and 2109 bp respectively of the mt genomes of
E furonis and E ictidea Pairwise comparison of these rDNA fragments demonstrated high (986 30
SNDs) sequence identity between the two parasites The remaining 778 and 794 bp respectively of the
mt genomes of E furonis and E ictidea were intergenic stretches between the various rDNA and CDS
regions these intergenic regions were more variable that other regions of the genomes with 78 SNDs
(almost 10 sequence divergence) Additionally all indels were restricted to these variable intergenic
regions
The BI phylogeny generated from aligned complete mt genomes (Figure 64) supported the close
relationship between E furonis and E ictidea within a clade of Eimeria species that include the only three
sequences available for Eimeria spp of carnivores Eimeria mephitidis from the striped skunk (Mephitis
134
mephitis Family Mephitidae) was the sister taxon to the two Eimeria species of ferrets (Family
Mustelidae)
64 DISCUSSION
This work generated the first complete mt genomes from coccidia that infect domestic and black-
footed ferrets (Carnivora Mustelidae) Eimeria mephitidis from the striped skunk Mephitis mephitis
(Carnivora Mephitidae) is the only other Eimeria species from a carnivore for which a complete mt
genome has been reported
Comparatively few eimeriid coccidia only 26 Eimeria species have been described from
carnivores there are 14 named species from the mustelids four from the procyonids four from the ursids
three from the herpestids and one from the viverrids (Duszynski et al 2000) The majority of coccidia
that infect the digestive tract of carnivores belong to the family Sarcocystidae including monoxenous or
facultatively heteroxenous Cystoisospora species or heteroxenous parasites in the genera Sarcocystis
Hammondia and Neospora So far as is known none of the parasites in the Sarcocystidae possess typical
apicomplexan mt genomes with 3 complete CDS and many rDNA fragments (Ogedengbe 2015)
The mt genomes from the two Eimeria sp of mustelid origin demonstrate the same structural
organization (ie the order and number of CDS and rDNA fragments) and circular mapping as the mt
genomes from other Eimeria spp and other closely related eimeriid coccidia such as Isospora
Cyclospora and Lankesterella species Despite the ability of the eimeriid sequences to be mapped
circularly the physical form of Eimeria spp mt genomes may be a linear concatemer of multiple genome
copies as demonstrated for Eimeria tenella (Hikosaka et al 2011) As in the mt genomes of other
eimeriid coccidia (Ogedengbe et al 2013 2014) the CDS for mt COIII demonstrated the highest
sequence divergence between E furonis and E ictidea the mt COI CDS was somewhat more conserved
and CytB CDS demonstrated the fewest SNDs
As expected based on limited sequence divergence between E furonis and E ictidea a BI
phylogenetic analysis using aligned complete mt genome sequences generated a tree that placed these
135
two Eimeria species that infect mustelids within a well-supported monophyletic group The sister taxon
for these ferret parasites was the only other Eimeria species from carnivores for which a complete mt
genome is available E mephitidis which infects hosts belonging to a different family of carnivores
Eimeriid parasites that infect closely-related definitive hosts are commonly found in a single or limited
number of clades based on mitochondrial and nuclear genetic loci (Ogedengbe et al in press)
Sequencing of the mt genomes and at least one nuclear genetic locus (ie nu 18S rDNA) from additional
Eimeria species infecting carnivores will be required to determine if all carnivore-specific Eimeria
species share a common ancestor
136
Table 61 PCR primer pairs and resulting fragments used for sequencing the mitochondrial genome sequence of an isolate of Eimeria furonis
originating from a fecal sample from a domestic ferret (Mustela putorius furo)
Fragment Primer names Primer sequences (5ʹ-3ʹ) Size (bp) Anneal Temp References
1 WG-MT_4140F AGAAAACCTAAAATCATCATGT 1000 52 Ogedengbe et al (2015)
Eim_CO3_799R AAGTGAGTTCGCATGTTTAC Ogedengbe et al (2015)
2 Eim_COI_19F ACTGCYGCAAACCATAARGAA 1700 60 Present study
Api_LSUG_UNI_R AGATAGGGAACAAACTGYCTCAA Present study
3 WG_MT_5416F GGTCCAGATAAGCGATCTCATG 3400 53 Ogedengbe et al (2013)
Eim_COI_1436R CACATTGTGTTCARATAAGTTA Present study
4 WG-MT_6219F GCATCCATCTACAGCTGCGG 500 55 Ogedengbe et al (2013)
WG-MT_344R GTAGGAATCTRAATTCCCAACC Ogedengbe et al (2013)
5 Api_LSUE UNI_F AGGTGCTCAGGGTCTTACCG 500 55 Present study
WG_MT_63R CTGGTATGGATGGATAACACT Ogedengbe et al (2015)
6 Lank_COB-30F CCAGGCCAACTGAACTCGTT 1300 55 Present study
q_Eim_COI_221R GGCATAACTACAAAGAARATCATA Present study
7 Cocci_MT_WG_F TACACCTAGCCAACACGAT 1600 55 Ogedengbe et al (2014)
q_Eim_COI_221R GGCATAACTACAAAGAARATCATA Present study
137
Table 62 PCR primer pairs and resulting fragments used for sequencing the mitochondrial genome sequence of an isolate of Eimeria ictidea
originating from a fecal sample from a black-footed ferret (Mustela nigripes)
Fragment Primer names Primer sequences (5ʹ-3ʹ) Size (bp) Anneal Temp References
1 WG_MT_63R CTGGTATGGATGGATAACACT 2180 52 Ogedengbe et al (2015)
WG-MT_4140F AGAAAACCTAAAATCATCATGT Ogedengbe et al (2015)
2 Cocci_MT_WG_F TACACCTAGCCAACACGAT 1551 58 Ogedengbe et al (2014)
q_Eim_COI_221R GGCATAACTACAAAGAARATCATA Present study
3 WG-MT_3658F CTGGCGAGAAGGGAAGTGTG 1329 55 Ogedengbe et al (2013)
Eim_CO3_799R AAGTGAGTTCGCATGTTTAC Ogedengbe et al (2015)
4 Lank_COB-30F CCAGGCCAACTGAACTCGTT 3899 57 Present study
WG_MT_4072R GGTTGTTTCCATCTCGACTC Ogedengbe et al (2013)
138
Table 63 Coding regions within the mitochondrial genome of the eimeriid parasite Eimeria furonis from a domestic ferret (Mustela putorius faro)
139
Table 64 Coding regions of the mitochondrial genome of the eimeriid parasite Eimeria ictidea originating from a black-footed ferret (Mustela
nigripes)
Table 64 Features associated with the protein-coding regions of the mitochondrial genome of the eimeriid parasite Eimeria cf ictidea originating from a black-footed ferret (Mustela nigripes )
Protein coding regions (CDS) Sequence size (bp) Start position (bp) Stop position (bp) Direction Translation start codon Translation stop codon
Cytochrome c oxidase subunit I (COI) 1443 1343 2785 Forward ATG TAA
Cytochrome c oxidase subunit III (COIII) 756 4333 5088 Forward TTA TAA
Cytochrome b (CytB) 1080 226 1305 Forward ATG TAA
Ribosomal DNA fragments (rDNA) Product
SSUrRNA 46 17 62 forward RNA9 SSU8
SSUrRNA 77 104 180 forward SSUA SSU4 (partial)
SSUrRNA 35 181 215 forward RNA23t
LSUrRNA 20 2790 2809 forward RNA20 (partial) LSU
LSUrRNA 112 2835 2946 forward LSUF LSU11
LSUrRNA 106 2947 3052 forward LSUG LSU12
LSUrRNA 25 3087 3063 reverse LSU
SSUrRNA 39 3129 3167 forward RNA14 SSU1
LSUrRNA 16 3188 3173 reverse LSUC LSU4
SSUrRNA 33 3211 3243 forward SSU
SSUrRNA 61 3386 3326 reverse SSUF SSU12
LSUrRNA 74 3387 3460 forward RNA10 LSU13 (partial)
LSUrRNA 49 3476 3524 forward RNA11 LSU5
SSUrRNA 65 3533 3597 forward SSUD SSU10
SSUrRNA 40 3631 3670 forward RNA17 SSU3
SSUrRNA 30 3677 3706 forward RNA15 SSU
LSUrRNA 30 3711 3740 forward RNA13 LSU10
LSUrRNA 56 3749 3804 forward RNA6 LSU15
LSUrRNA 79 3899 3821 reverse LSUD LSU8
LSUrRNA 24 3931 3908 reverse RNA16 (partial)
SSUrRNA 92 4036 3945 reverse RNA8 SSU5
LSUrRNA 66 4054 4119 forward RNA2 LSU2
LSUrRNA 177 4316 4140 reverse LSUA LSU1
SSUrRNA 30 5125 5096 reverse RNA19 SSU7
LSUrRNA 88 5154 5241 forward RNA1 LSU6
LSUrRNA 26 5342 5317 reverse LSUB LSU3
LSUrRNA 73 5447 5375 reverse RNA3 LSU7
LSUrRNA 20 5479 5460 reverse RNA18 LSU14
SSUrRNA 119 5615 5497 reverse SSUB SSU6
LSUrRNA 80 5699 5620 reverse RNA7
LSUrRNA 188 5897 5710 reverse LSUE LSU9
SSUrRNA 33 6033 6001 reverse SSUE SSU11 (partial)
SSUrRNA 99 6156 6058 reverse RNA5SSU9
140
Table 65 Pairwise comparison of coding DNA and concatenated rDNA fragment sequences between the
mitochondrial genomes of Eimeria furonis originating from a domestic ferret (Mustela putorius furo) and
Eimeria ictidea originating from a black-footed ferret (Mustela nigripes)
Total length
(nucleotides)
Nucleotide
identity
Total amino
acids
Amino acid
identity
COI CDS 1443 934 (95) 480 975 (12)
COIII CDS 756 899 (76) 251 932 (17)
CytB CDS 1080 950 (54) 359 986 (5)
rDNA fragments 2109 985 (32) na na
Legend Numbers in brackets indicate the number of single nucleotide differences na = not
applicable
141
Figure 61 Circular mapping and organization of the mitochondrial genome content of Eimeria
furonis showing three protein-coding genes (COI COIII and CytB) interspersed with large and
small subunit rRNA fragments
142
Figure 62 Circular mapping and organization of the mitochondrial genome content of Eimeria
ictidea showing three protein-coding genes (COI COIII and CytB) interspersed with large and
small subunit rRNA fragments
143
Figure 63 Pairwise comparison of coding DNA and concatenated rDNA fragment sequences of
the mitochondrial genomes of Eimeria furonis and Eimeria ictidea Legend Yellow bands
correspond to coding DNA fragments Red arrows correspond to rDNA fragments Arrows of both
colours indicate the forward or reverse direction of these segments
144
Figure 64 Phylogenetic relationships of coccidia (Eimeria furonis and Eimeria ictidea) from
domestic (Mustela putorius furo) and black-footed ferrets (Mustela nigripes) based on the complete
mitochondrial genome sequences these Eimeria species and a selection of related apicomplexan
parasites
145
CHAPTER 7 CONCLUSIONS AND FUTURE DIRECTIONS
The primary objective of this project was to better characterize the enteric coccidia of the
endangered black-footed ferret (BFF) in order to set the stage for improved disease prevention
management and treatment To the authorrsquos knowledge this work was the first attempt to isolate and
perform molecular characterization of the coccidial species endemic in the black-footed ferret population
and to characterize the natural history of the disease in this host
Two Eimeria species Eimeria ictidea and Eimeria furonis have been described from BFF and
domestic ferrets A single Eimeria species morphologically resembling E ictidea of the domestic ferret
and referred to in this work as E ictidea was identified from all historic and active cases of enteric
coccidiosis in the Toronto Zoo BFF population The same species was identified in both BFF family
groups and single-housed adults that shed coccidia during the study period (2014-2016) as well as from
historic necropsy samples of juvenile and adult Toronto Zoo BFF from 1999-2014 Furthermore this
same species was identified from both family groups and single-housed adults shedding coccidia at an
additional BFF Species Survival Plan institution the Louisville Zoo in 2016 These findings are contrary
to previous published reports that indicated multiple coccidia species were affecting captive and wild
BFF In order to determine whether these additional parasites continue to exist within the present-day
BFF population and their impact on this species further examination of coccidia-positive fecal samples
from captive and wild BFF populations is recommended Moreover determination of which parasite
species impact BFF morbidity and mortality would allow for development of targeted therapies for
disease management
Coccidia-positive fecal samples and necropsy samples from domestic ferrets were collected from
multiple diagnostic laboratories in Canada and Europe for comparison with results from BFF Eimeria
furonis and Isospora (=Cystoisospora) laidlawi were identified from fecal parasitology reports from
2008-2015 and in fecal samples obtained prospectively from 2014-2016 from domestic ferrets submitted
to Canadian and European laboratories E furonis was also identified in necropsy samples from 2010 and
146
2017 from two Canadian diagnostic laboratories No samples containing E ictidea were submitted to the
Canadian laboratories however samples containing E ictidea were identified twice by the European
diagnostic laboratory with one sample in 2011 and a second in 2013 Consequently it appears that E
ictidea is rarely identified from domestic ferrets Since no samples of E ictidea were acquired from
domestic ferrets during the study period it remains undetermined whether E ictidea from domestic
ferrets and E ictidea of BFF are conspecific Future molecular characterization of E ictidea isolated from
various mustelid host species would allow not only for determination of whether the parasites are
conspecific but would also provide insight into the potential for cross-transmission of parasites between
related mustelid hosts
The identifications provided by diagnostic laboratories of the specific parasite species present in
the domestic ferret fecal samples showed poor agreement with their identifications based on genotyping
obtained in this study In the authorrsquos opinion molecular techniques are essential tools for determining
the specific coccidial species responsible for individual and group outbreaks of coccidiosis and for
further understanding of the eimeriid host-parasite relationships To this end the nu 18S rDNA region and
whole mitochondrial (mt) genomes of E ictidea and E furonis were sequenced these mt whole genome
sequences are the first for the Eimeria species of mustelids to be entered into GenBank These sequences
may provide suitable targets for the development of highly discriminatory PCR-based methods of
identification that could be applied to fecal tissue or even formalin-fixed paraffin-embedded samples
(see Chapters 2 and 3) Methodological improvements such as less expensive next generation sequencing
methods that could be applied to diagnostics will depend on the availability of high quality reference
sequences such as generated herein Furthermore evaluation of the evolution of the eimeriid coccidia
using phylogenetic analysis based on additional whole mitochondrial genome sequences may allow for
more accurate determinations of relationships between parasite species and timing of their divergence
from common ancestors
One of the goals of this research was to investigate the possibility of using the domestic ferret as
an experimental model to study enteric coccidiosis in the BFF Experimental work cannot be carried out
147
in the BFF due to its endangered status We showed that the domestic ferret is susceptible to infection
with Eimeria ictidea originating from BFF Infected domestic ferrets showed similar clinical signs and
pathologic lesions to BFF strengthening the possibility of their serving as a suitable model Future
investigations would include studies of the domestic ferretrsquos immune response to enteric coccidial
infection in vivo drug trials including pharmacokinetic pharmacodynamic and efficacy studies of
anticoccidial medications investigations into patterns of drug resistance in coccidial species of ferrets
and oral vaccine development
Ultimately the goal of research on coccidiosis in BFF may be the development of an autogenous
vaccine to improve the survival of ferret kits and reduce coccidiosis-related morbidity and mortality in
BFF captive breeding programs It may be possible to use the domestic ferret to select for E ictidea that
are more highly adapted to this host perhaps with a commensurate loss of virulence to its original host If
this were to be the case domestic ferret-derived coccidia could be used in BFF as vaccine organisms
Alternately methods such as use of a bioshuttle (live vaccination with coccidia followed by anticoccidial
treatment to limit pathogenicity) might be useful for generating long-lasting immunological protection in
BFF against E ictidea Improving immunity through vaccination could assist in reducing coccidial
shedding and disease in BFF associated with stressful life events such a breeding weaning and transfer
between institutions There is minimal available data on the significance of coccidiosis in wild
populations of BFF and limited means of disease surveillance post release however it is logical to
assume that the stresses associated with release to the wild might result in clinical disease as happens with
stressed BFF in captivity Reducing morbidity and mortality associated with coccidiosis in BFF could
result in increased numbers of ferrets being released to the wild and higher survival of released ferrets
increasing the number of BFF in the wild supports the goals of the conservation initiative for the black-
footed ferret and will support the longer-term survival and recovery of this species
148
REFERENCES
Abe N Tanoue T Ohta G Iseki M (2008) First record of Eimeria furonis infection in a ferret Japan with
notes on the usefulness of partial small subunit ribosomal RNA gene sequencing analysis for
discriminating among Eimeria species Parasitol Res 103967ndash70 doi 101007s00436-008-1037-x
Adl SM Simpson AGB Farmer MA et al (2005) The new higher level classification of eukaryotes with
emphasis on the taxonomy of protists J Eukaryot Microbiol 52399ndash451 doi 101111j1550-
7408200500053x
Andrews JM (1926) Coccidiosis in mammals Am J Hyg 6784ndash798
Augustine PC Danforth HD (1986) A study of the dynamics of the invasion of immunized birds by
Eimeria sporozoites Avian Dis 30347ndash351
Barta JR Schrenzel MD Carreno R Rideout BA (2005) The genus Atoxoplasma (Garnham 1950) as a
junior objective synonym of the genus Isospora (Schneider 1881) species infecting birds and
resurrection of Cystoisospora (Frenkel 1977) as the correct genus for Isospora species infecting
mammals J Parasitol 91726ndash727 doi 101645GE-33411
Bell JA (1994) Parasites of Domesticated Pet Ferrets Compend Contin Educ Pract Vet 16617ndash620
Black-footed Ferret Recovery Implementation Team (2011) Captive Breeding
httpwwwblackfootedferretorgcaptive-breeding Accessed 8 Jan 2014
Blankenship-Paris TL Chang J Bagnell CR (1993) Enteric coccidiosis in a ferret Lab Anim Sci 43361ndash
363
Bronson E Bush M Viner T et al (2007) Mortality of captive black-footed ferrets (Mustela nigripes) at
Smithsonianrsquos National Zoological Park 1989 ndash 2004 J Zoo Wildl Med 38169ndash176
149
CAPC (2013) Current Advice on Parasite Control Intestinal Parasites - Coccidia
httpwwwcapcvetorgcapc-recommendationscoccidia Accessed 6 Jun 2015
Carpenter JW Hillman CN (1979) Husbandry reproduction and veterinary care of captive ferrets In
1978 Proceedings of the Annual Meeting of the American Association of Zoo Veterinarians
Washington DC pp 36ndash47
Catchpole J Norton CC Gregory MW (1993) Immunisation of lambs against coccidosis Vet Rec
13256ndash59
Cavalier-Smith T (2014) Gregarine site-heterogeneous 18S rDNA trees revision of gregarine higher
classification and the evolutionary diversification of Sporozoa Eur J Protistol 50472ndash495 doi
101016jejop201407002
Chappuis G (1998) Neonatal immunity and immunisation in early age lessons from veterinary medicine
Vaccine 161468ndash1472
Cox FE (1994) The evolutionary expansion of the Sporozoa Int J Parasitol 241301ndash1316
Davis CL Chow TL Gorham JR (1953) Hepatic coccidiosis in mink Vet Med 48371ndash375
De Vos AJ (1970) Studies on the host range of ltigtEimeria chinchillaeltIgt de Vos and Van Der
Westhuizen 1968 Onderstepoort J Vet Res 3729ndash36
Dirikolu L Yohn R Garrett EF et al (2009) Detection quantifications and pharmacokinetics of
toltrazuril sulfone (Ponazuril) in cattle J Vet Pharmacol Ther 32280ndash288
Dryden MW Payne PA Ridley R Smith V (2005) Comparison of common fecal flotation techniques for
the recovery of parasite eggs and oocysts Vet Ther 615ndash28
Duszynski D Wilber PG (1997) A Guideline for the Preparation of Species Descriptions in the
Eimeriidae J Parasitol 83333ndash336
150
Duszynski DW Couch L Upton SJ (2000) The Coccidia of the World
httpbiologyunmeducoccidiacarniv2html Accessed 1 Sep 2017
El-Sherry S Ogedengbe ME Hafeez MA Barta JR (2013) Divergent nuclear 18S rDNA paralogs in a
turkey coccidium Eimeria meleagrimitis complicate molecular systematics and identification Int J
Parasitol 43679ndash685 doi 101016jijpara201303005
Evans HE An NQ (2014) Anatomy of the ferret In Fox JG Marini RP (eds) Biology and Diseases of the
Ferret 3rd edn Wiley Blackwell pp 23ndash67
Fayer R (1980) Epidemiology of Protozoan Infections The Coccidia Vet Parasitol 675ndash103
Foreyt WJ Todd AC Hartsough GR (1977) Anticoccidial Activity of Eight Compounds in Domestic
Mink AJVR 38391ndash394
Fox JG (2014) Normal Clinical and Biological Parameters In Fox JG Marini RP (eds) Biology and
Diseases of the Ferret 3rd edn John Wiley amp Sons Inc pp 157ndash185
Frenkel JK (1977) Besnoitia wallacei of Cats and Rodents With a Reclassification of Other Cyst-
Forming Isosporoid Coccidia J Parasitol 63611ndash628
Grafner G Graubmann HD Dobbriner W (1967) Leberkokzidiose beim Nerz (Lutreola vison Schreb)
hervorgerufen durch eine neue Kokzidienart Eimeria hiepei n sp Monatshefte fur
Veterinearmedizin 22696ndash700
Haberkorn A (1971) Zur Wirtsspezifitat yon Eimeria contorta nsp (Sporozoa Eimeriidae) Z
Parasitenkd 37303ndash314
Hall MC Wigdor M (1918) Canine coccidiosis with a note regarding other protozoan parasites from the
dog J Am Vet Med Assn 5364ndash76
Hikosaka K Nakai Y Watanabe YI et al (2011) Concatenated mitochondrial DNA of the coccidian
151
parasite Eimeria tenella Mitochondrion 11273ndash278
Hillyer E V (1992) Gastrointestinal diseases of ferrets (Mustela putorius furo) J Small Anim Med 244ndash
45
Hoare CA (1927) On the coccidia of the ferret Ann Trop Med Parasitol 21313ndash320
Hoare CA (1935a) A histopathological reaction of a special type on the part of the intestinal villi in ferret
coccidiosis Trans R Soc Trop Med Hyg 292
Hoare CA (1935b) The endogenous development of the coccidia of the ferret and the histopathological
reaction of the infected intestinal villi Ann Trop Med Parasitol 29111ndash122
Hoefer HL Fox JG Bell JA (2012) Gastrointestinal Diseases In Quesenberry KE Carpenter JW (eds)
Ferrets Rabbits and Rodents Clinical Medicine and Surgery 3rd edn Elsevier Saunders St Louis
Missouri pp 27ndash45
Huelsenbeck JP Ronquist F (2001) MrBayes Bayesian inference of phylogenetic trees Bioinformatics
17754ndash755 doi 101093bioinformatics178754
Jolley WR Kingston N Williams ES Lynn C (1994) Coccidia Giardia sp and a Physalopteran
Nematode Parasite from Black-footed Ferrets (Mustela nigripes) in Wyoming J Helminthol Soc
Washingt 6189ndash94
Katoh K Misawa K Kuma K Miyata T (2002) MAFFT a novel method for rapid multiple sequence
alignment based on fast Fourier transform Nucleic Acids Res 303059ndash3066 doi
101093nargkf436
Kaye SW Ossiboff RJ Noonan B et al (2015) Biliary coccidiosis associated with immunosuppressive
treatment of pure red cell aplasia in an adult ferret (Mustela putoris furo) J Exot Pet Med doi
101053jjepm201504012
152
Koepfli KP Deere K Slater GJ et al (2008) Multigene phylogeny of the Mustelidae resolving
relationships tempo and biogeographic history of a mammalian adaptive radiation BMC Biol 610
Lariviegravere S Jennings AP (2009) Family Mustelidae (weasels and relatives) In Wilson DE Mittermeier
RA (eds) Handbook of the Mammals of the World Volume 1 Carnivores Lynx Edicions
Barcelona pp 564ndash656
Levine ND Ivens V (1970) The coccidian parasites (Protozoa Sporozoa) of ruminants
Li X Pang J Fox JG (1996) Coinfection with intracellular Desulfovibrio species and coccidia in ferrets
with proliferative bowel disease Lab Anim Sci 46569ndash571
Litster AL Nichols J Hall K et al (2014) Use of ponazuril paste to treat coccidiosis in shelter-housed
cats and dogs Vet Parasitol 202319ndash325 doi 101016jvetpar201403003
Matsubayashi M Takami K Abe N et al (2005) Molecular characterization of crane coccidia Eimeria
gruis and E reichenowi found in feces of migratory cranes Parasitol Res 9780ndash83
Meeusen ENT Walker J Peters A et al (2007) Current status of veterinary vaccines Clin Microbiol Rev
20489ndash510 doi 101128CMR00005-07
Mehlhorn H Aspock H (eds) (2008) Coccidial Drugs In Encyclopedia of Parasitology 3rd edn Springer
Berlin Heidelberg New York pp 269ndash286
Morehouse NF (1938) The Reaction of the Immune Intestinal Epithelium of the Rat to Reingection wiht
Eimeria nieschulzi J Parasitol 24311ndash317
Nukerbaeva KK Svanbaev SK (1973) Coccidia of fur bearing mammals in Kazakhstan Vestn Selrsquoskokh
Nauk Kazakh 1250ndash54
Ogedengbe JD Hanner RH Barta JR (2011) DNA barcoding identifies Eimeria species and contributes to
the phylogenetics of coccidian parasites (Eimeriorina Apicomplexa Alveolata) Int J Parasitol
153
41843ndash850
Ogedengbe ME (2015) DNA Barcoding of Apicomplexa Mitochondrial Evolution across the Phylum
University of Guelph
Ogedengbe ME El-Sherry S Ogedengbe JD et al Whatrsquos in a name Phylogenies based on combined
mitochondrial and nuclear sequences conflict with morphologically defined genera in the eimeriid
coccidia (Apicomplexa)
Ogedengbe ME El-Sherry S Whale J Barta JR (2014) Complete mitochondrial genome sequences from
five Eimeria species (Apicomplexa Coccidia Eimeriidae) infecting domestic turkeys Parasit
Vectors 7335 doi 1011861756-3305-7-335
Ogedengbe ME Hafeez MA Barta JR (2013) Sequencing the complete mitochondrial genome of Eimeria
mitis strain USDA 50 (Apicomplexa Eimeriidae) suggests conserved start positions for mtCOI- and
mtCOIII-coding regions Parasitol Res 1124129ndash4136 doi 101007s00436-013-3604-z
Pantchev N Gassmann D Globokar-Vrhovec M (2011) Increasing numbers of Giardia (but not
coccidian) infections in ferrets 2002 to 2010 Vet Rec 168519 doi 101136vrd2962
Patterson M Fox JG (2007) Parasites of Ferrets In Baker DG (ed) Flynnrsquos Parasites of Laboratory
Animals 2nd edn Wiley Blackwell pp 501ndash508
Patterson MM Fox JG Eberhard ML (2014) Parasitic Diseases In Fox JG Marini RP (eds) Biology and
Diseases of the Ferret 3rd edn Wiley Blackwell pp 553ndash572
Powers L V (2009) Bacterial and parasitic diseases of ferrets Vet Clin North Am - Exot Anim Pract
12531ndash561 doi 101016jcvex200906001
Prado ME Ryman JT Boileau MJ et al (2011) Pharmacokinetics of ponazuril after oral administration to
healthy llamas (Llama glama) Am J Vet Res 721386ndash9
154
Ruiz A Muntildeoz MC Molina JM et al (2013) Primary infection of goats with Eimeria ninakohlyakimovae
does not provide protective immunity against high challenge infections Small Rumin Res 113258ndash
266 doi 101016jsmallrumres201301006
Ryley J Meade R Hazelhurst J Robinson T (1976) Methods in coccidiosis research separation of
oocysts from faeces Parasitology 73311ndash326
Santymire R Branvold-Faber H Marinari PE (2014) Recovery of the Black-Footed Ferret In Fox JG
Marini RP (eds) Biology and Diseases of the Ferret 3rd edn Wiley Blackwell pp 219ndash231
Shi MQ Huther S Burkhardt E Zahner H (2000) Immunity in rats against Eimeria separata oocyst
excretion effects on endogenous stages and local tissue response after primary and challenge
infections Parasitol Res 86891ndash898
Sledge DG Bolin SR Lim A et al (2011) Outbreaks of severe enteric disease associated with Eimeria
furonis infection in ferrets (Mustela putorius furo) of 3 densely populated groups J Am Vet Med
Assoc 2391584ndash1588 doi 102460javma239121584
Smith NC Wallach M Petracca M et al (1994) Maternal transfer of antibodies induced by infection with
Eimeria maxima partially protects chickens against challenge with Eimeria tenella Parasitology
109551ndash557
Svanbaev SK (1956) Materials on the fauna of coccidia of wild mammals in western Kazakhstan Tr
Instituta Zool Akad Nauk Kazachskoi SSR 5180ndash191
Tavareacute S (1986) Some probabilistic and statistical problems in the analysis of DNA sequences Am Math
Soc Lect Math Life Sci 1757ndash86 doi citeulike-article-id4801403
Tenter AM Barta JR Beveridge I et al (2002) The conceptual basis for a new classification of the
coccidia Int J Parasitol 32595ndash616
155
Upton SJ (2000) Suborder Eimeriorina Leacuteger 1911 In Lee JJ Leedale GF Bradbury P (eds) An
Illustrated Guide to the Protozoa vol 1 2nd edn Allen Press Lawrence Kansas pp 318ndash339
USFWS BFF Recovery Program (2017) Black-footed Ferret Managed Care Operations Manual
(BFFMCOM)
Vermeulen AN (2005) Vaccination against coccidial parasites The method of choice In Proceeding of
the 9th International Coccidiosis Conference
Williams BH Chimes MJ Gardiner CH (1996) Biliary coccidiosis in a ferret (Mustela putorius furo) Vet
Pathol 33437ndash439 doi 101177030098589603300412
Williams ES Mills K Kwiatkowski DR et al (1994) Plague in a Black-footed (Mustela nigripes)
presence J Wildl Dis 30581ndash585
Williams ES Thome ET Appel MJG Belitsky DW (1988) Canine Distemper in Black-Footed (Mustela
nigripes) from Wyoming J Wildl Dis 24385ndash398
Wozencraft WC (2005) Order Carnivora In Wilson DE Reeder DM (eds) Mammal Species of the
World A Taxonomic and Geographic Reference 3rd edn Johnrsquos Hopkins University Press
Baltimore Maryland p 2142
Yi-Fan C Le Y Yin D et al (2012) Emendation of 2 Isospora Species (Apicomplexa Eimeriidae)
Infecting the Steppe Polecat Mustela eversmanii Lesson 1827 in China to the Genus
Cystoisospora (Apicomplexa Sarcocystidae) Comp Parasitol 79147ndash152 doi 10165445311
Yu L Peng D Liu J et al (2011) On the phylogeny of Mustelidae subfamilies analysis of seventeen
nuclear non-coding loci and mitochondrial complete genomes BMC Evol Biol 1192 doi
1011861471-2148-11-92
Zou M Guo G Zhao Y Zhang Q (2014) Detection quantifications and pharmacokinetics of ponazuril in
156
healthy swine J Vet Pharmacol Ther 37598ndash602 doi 101111jvp12126
157
APPENDICES
158
Appendix 1 Shedding of oocysts of Eimeria ictidea in black-footed ferret (Mustela nigripes) dam and kit
family groups from 2014-2016
Collection Year 2014 2014 2014 2014 2014 2015 2016
Age of kits (days) Poppy Bumblefoot Calico Aubrey Ruckus Fiddlesticks Guanella^
29 - - - 0 - - -
30 - 0 - 0 - - -
31 - 0 - 0 - - -
32 - 0 - 0 - - -
33 - 0 - 0 - - -
34 0 0 - 0 0 - -
35 0 0 - 0 0 0 -
36 0 0 - 0 0 0 -
37 - 0 - 0 - 0 -
38 - 0 - 0 - 0 -
39 - 0 - 0 - 0 -
40 0 0 - 0 0 0 -
41 - 0 - 0 - 0 -
42 0 0 - 0 0 - -
43 0 0 - 0 0 0 -
44 0 0 - - 0 0 -
45 0 0 - 0 0 - -
46 0 0 - 0 0 0 -
47 0 0 - 0 0 0 -
48 0 lt14 - 0 0 0 -
49 0 0 - 0 0 0 -
50 0 0 - - 0 0 -
51 0 0 - 0 0 0 -
52 0 0 - 0 0 0 -
53 0 0 - 0 0 0 +
54 0 0 - 0 0 0 +
55 0 0 - 0 0 0 3717146
56 0 0 - 0 - 0 1084436
57 0 0 - 0 - 0 64133
58 0 324 - 0 0 0 20654
59 0 0 - 0 0 0 0
60 0 446688 - 0 0 0 0
61 0 934828 - 0 0 0 41111
62 0 530469 - 0 0 0
63 0 1617131 - 0 0 lt14
64 0 8396357 - 114 0 0
65 0 234876 - 5368 0 lt14
66 0 374625 - 9455 0 0
67 lt14 2311575 - 39579 0 -
68 0 603563 - 125051 0 0
69 - 4103036 - 27067 0 lt14
70 2470327 7759974 - 0 0 0
71 1073085 - 1784 0 0
72 3430966 - 0 0 0
73 4459536 - 7326 0 0
74 4842212 - 2 0 0
75 3627307 - 0 0 0
76 5801885 - 0 0 0
77 5994 0 0 0 0
78 589835 0 0 0
Numbers of Oocysts Shed (oocysts per gram of feces)
Dam Identity
159
Appendix 1 continued Shedding of oocysts of Eimeria ictidea in black-footed ferret (Mustela nigripes)
dam and kit family groups from 2014-2016
Collection Year 2014 2014 2014 2014 2014 2015 2016
Age of kits (days) Poppy Bumblefoot Calico Aubrey Ruckus Fiddlesticks Guanella^
79 24815 0 0 0
80 144016 0 0 0
81 97862 0 0 0
82 0 0 0
83 0 0 0
84 0 0 0
85 0 0 0
86 0 0 0
87 0 0 0
88 0 0 0
89 0 0 0
90 0 0 0
91 0 0 0
92 0 - -
93 0 - 0
94 0 - 0
95 0 0 -
96 0 - 0
97 0 0 -
98 0 0 0
99 0 0 0
100 0 0 0
101 0 0 0
102 0 0 0
103 0 0 0
104 0 0 0
105 0 0 -
106 0 0 0
107 0 0 0
108 - 0 0
109 0 0 -
110 0 0 0
111 0 0 -
112 0 0 0
113 0 0 -
114 0 0 -
115 0 0 -
116 0 0 -
117 0 -
118 0 -
119 0 0
120 0 0
121 0 0
122 0 -
Numbers of Oocysts Shed (oocysts per gram of feces)
Dam Identity
160
Appendix 1 continued Shedding of oocysts of Eimeria ictidea in black-footed ferret (Mustela nigripes)
dam and kit family groups from 2014-2016
Collection Year 2014 2014 2014 2014 2014 2015 2016
Age of kits (days) Poppy Bumblefoot Calico Aubrey Ruckus Fiddlesticks Guanella^
123 0 0
124 0 -
125 0 -
126 0 0
127 0 0
128 0 2843
129 0 0
130 0 -
131 0 0
132 0 0
133 0 0
134 0 0
135 0 0
136 0 0
137 0
138 0
139 0
140 0
141 0
142 0
143 0
144 0
145 0
146 -
147 0
148 0
149 0
150 0
Legend lt 14 = oocyst positive samples with less than 14 oocyst per gram of feces - = no sample recorded for this date
underline = last sampling date + = coccidia present but OPG count not performed = Toronto Zoo ferret ^ = Louisville Zoo
ferret thick outer border = days treatment was received
Numbers of Oocysts Shed (oocysts per gram of feces)
Dam Identity
161
Appendix 2a Hematology values for domestic ferrets (Mustela putorius furo) from 49-51 days of age
prior to experimental inoculation
Test Reference
Intervala
Reference
Intervalb
101 102 103 104 105 201 202 203 204 205
Age (days) adult 70 50 50 50 50 50 48 48 48 50 48
WBC (x 109L) 27-112 53-120 86 64 77 97 76 48 8 72 104 76
RBC (x 1012
L) 50-108 55ndash74 66 58 45 5 48 47 67 5 5 42
Hb (gL) 87-177 104ndash136 121 106 90 58 94 91 122 96 98 80
HCT (LL) 04 - 051 029ndash037 037 033 027 030 029 027 038 030 030 024
MCV (fL) 44-52 478ndash548 55 56 60 60 60 58 57 59 61 58
MCH (pg) 15-18 175ndash191 18 18 20 12 20 20 18 19 20 18
MCHC (gL) 325-362 347ndash370 331 327 328 196 325 337 321 327 324 331
RDW () 12-16 - 134 127 139 139 133 131 122 136 131 127
Platelets (x 109L) 54-695 629ndash775 777 329 529 480 640 402 534 336 527 363
MPV (fL) 5-10 - 78 78 96 75 66 74 76 81 74 82
TS Protein (gL) 49-76 - 54 51 - - - - - - - -
Seg Neuts (x 109L) 1-8 15ndash48 292 141 216 281 205 187 152 151 354 251
Lymphocytes (x 109L) 1-63 28ndash63 525 416 516 64 509 254 608 468 645 456
Monocytes (x 109L) 0-09 01ndash05 026 07 031 039 038 034 024 094 031 038
Eosinophils (x 109L) 0-13 01ndash06 009 013 008 01 008 005 016 007 010 015
Basophils (x 109L) 0-02 0 009 0 0 0 0 0 0 0 0 0
Polychromasia 2-5 - 5-10 2-5 10-15 10-15 10-15 10-15 2-5 10-15 10-15 5-10
Anisocytosis Occ 1+ 1+ 1+ 1+ 1+ 1+ 1+
HJ bodies rare rare rare rare rare
crenation Occ
poikilocytosis Occ Occ Occ
shift platelets Occ Occ
hemolysis Neg Neg Neg Neg Neg Neg Neg Neg Neg Neg Neg
lipemia mild mild mild mild mild Neg Neg Neg mild mild
Ferret Identity
Legend - = reference value unavailable bolded = outside reference range a = adult ferret reference ranges from Animal Health Laboratory
(University of Guelph Guelph Canada) b
= reference ranges for 10-week old ferrets (Fox JG 2014) Neg = negative Occ = occasional
162
Appendix 2b Serum biochemistry values for domestic ferrets (Mustela putorius furo) from 49-51 days of
age prior to experimental inoculation
Test Reference
Intervala
Reference
Intervalb
101 102 103 104 105 201 202 203 204 205
Age (days) adult 70 50 50 50 50 50 48 48 48 50 48
Calcium (mmolL) 185-242 253-302 239 233 241 244 24 221 242 234 253 242
Phosphorus (mmolL) 112-256 278-475 239 267 29 264 31 218 286 284 315 303
Magnesium (mmolL) 08-139 - 08 08 07 08 08 06 08 07 08 08
Sodium (mmolL) 147-159 146-154 149 149 149 149 148 144 153 148 152 152
Potassium (mmolL) 37-57 47-83 44 46 42 46 48 45 47 44 47 46
Chloride (mmolL) 111-129 115-121 110 112 115 113 113 110 119 117 117 120
Carbon dioxide (mmolL) 17-29 13-27 18 18 18 17 17 16 15 15 19 17
Anion gap (mmolL) 6 - 23 - 25 24 20 24 23 23 24 20 21 20
NaK ratio - - 34 32 35 32 31 32 33 34 32 33
Total protein (gL) 51-75 44-56 49 46 44 47 44 41 52 45 49 43
Albumin (gL) 24-40 26-32 29 26 28 28 28 25 28 27 29 25
Globulin (gL) 19-41 17-24 20 20 16 19 16 16 24 18 20 18
AG ratio 053-167 13ndash12 145 130 175 147 175 156 117 15 145 139
Blood urea nitrogen (mmolL) 45-153 71-139 83 89 107 11 136 73 119 105 118 135
Creatinine (umolL) 8-67 53-124 50 58 68 59 50 7 68 41 62 92
Glucose (mmolL) 32-91 64-138 47 54 54 42 55 53 57 56 52 59
Cholesterol (mmolL) 294-894 619-860 413 386 375 393 344 349 227 357 373 346
Total bilirubin (umolL) 2 to 7 - 1 L 0 L 0 0 0 1 0 0 0 0
Conjugated biilrubin (umolL) - 0-10 1 0 0 1 1 1 0 0 1 0
Free bilirubin (umolL) 0-2 0-15 0 0 0 0 0 0 0 0 0 0
ALKP (UL) 13-237 117ndash277 180 169 172 215 175 168 241 184 177 179
GGT (UL) 0-40 2ndash20 1 1 6 10 4 0 1 1 5 9
AST (UL) - 63ndash152 61 58 48 61 64 59 93 61 58 69
ALT (UL) 39-196 95ndash544 95 105 89 105 106 82 234 137 115 156
CK (UL) 74-294 - 513 330 496 560 530 492 793 539 479 724
Amylase (UL) - - 23 28 35 35 29 28 29 24 36 23
Lipase (UL) - - 65 63 60 64 62 56 67 69 60 68
Calculated osmo (mmolL) - - 298 300 301 301 303 289 311 300 308 311
Ferret Identity
Legend - = reference value unavailable bolded = outside reference range a = adult ferret reference ranges from Animal Health Laboratory
(University of Guelph Guelph Canada) b = reference ranges for 10-week old ferrets (Fox JG 2014)
163
Appendix 3a Hematology values for domestic ferrets (Mustela putorius furo) inoculated orally with
Eimeria ictidea
Test Reference
Intervala
Reference
Intervalb
102 103 104 105 201 203 205
Age (days) adult 98-112 92 91 99 99 97 74 92
Days post inocculation 8 7 15 15 15 11 10
WBC (x 109L) 27-112 52ndash150 94 124 81 85 88 85 142
RBC (x 1012
L) 50-108 62ndash92 71 62 73 60 65 63 65
Hb (gL) 87-177 127ndash159 122 110 122 98 103 114 102
HCT (LL) 04-051 030ndash043 037 033 037 031 032 035 032
MCV (fL) 44-52 50ndash54 53 53 52 51 50 55 49
MCH (pg) 15-18 16-21 17 18 17 16 16 18 16
MCHC (gL) 325-362 351ndash426 328 332 325 319 320 326 317
RDW () 12-16 - 122 126 129 129 145 127 145
Platelets (x 109L) 54-695 376ndash610 524 413 445 117 303 42 429
MPV (fL) 5-10 - 8 78 72 66 67 14 8
TS Protein (gL) 49-76 - 66 63 58 66 55 59 59
Seg Neuts (x 109L) 1-8 21ndash62 160 446 203 170 211 170 554
Lymphocytes (x 109L) 1-63 16ndash79 761 657 551 646 59 646 682
Monocytes (x 109L) 0-09 01ndash02 019 087 024 026 044 026 085
Eosinophils (x 109L) 0-13 03ndash09 0 037 032 009 026 009 099
Basophils (x 109L) 0-02 0 0 012 0 0 009 0 0
Polychromasia 2-5 - 1-3 0-2 0-2 1-3 1-3 2-5 5-10
Anisocytosis Occ Occ Occ
HJ bodies rare
crenation
rouleaux
poikilocytosis
shift platelets Occ Occ
hemolysis Neg
Ferret Identity
Legend - = reference value unavailable bolded = outside reference range a = adult ferret reference ranges from Animal Health
Laboratory (University of Guelph Guelph Canada) b
= reference ranges for 10-week old ferrets (Fox JG 2014) Neg = negative
Occ = occasional = many platelet clumps
164
Appendix 3b Serum biochemistry values for domestic ferrets (Mustela putorius furo) inoculated orally
with Eimeria ictidea
Test Reference
Intervala
Reference
Intervalb
102 103 104 105 201 203 205
Age (days) adult 98-112 92 91 99 99 97 74 92
Days post inocculation 8 7 15 15 15 11 10
Calcium (mmolL) 185-242 245-268 243 222 238 233 232 236 240
Phosphorus (mmolL) 112-256 200-323 292 223 248 298 268 288 306
Magnesium (mmolL) 08-139 - 06 08 08 08 09 08
Sodium (mmolL) 147-159 148-155 147 150 150 148 148 150
Potassium (mmolL) 37-57 45-55 42 41 51 41 50 44
Chloride (mmolL) 111-129 114-124 114 113 118 112 111 115
Carbon dioxide (mmolL) 17-29 16-24 16 21 19 20 12 18
Anion gap (mmolL) 6 - 23 - 21 20 18 20 30 21
NaK ratio - - 35 37 29 36 30 34
Total protein (gL) 51-75 49-64 52 55 55 49 56 57
Albumin (gL) 24-40 30-36 28 20 27 27 26 29 27
Globulin (gL) 19-41 19-30 32 28 28 23 27 30
AG ratio 053-167 11-17 063 096 096 113 107 090
Blood urea nitrogen (mmolL) 45-153 50-150 92 87 119 92 110 98 149
Creatinine (umolL) 8-67 706-1414 56 26 49 40 50 46 79
Glucose (mmolL) 32-91 688-943 52 59 64 71 58 17 66
Cholesterol (mmolL) 294-894 440-640 453 429 262 475 370 285
Total bilirubin (umolL) 2 to 7 - 0 0 0 0 1 1
Conjugated biilrubin (umolL)- - 1 0 0 0 0 0
Free bilirubin (umolL) 0-2 - 0 0 0 0 1 1
ALKP (UL) 13-237 41-181 124 120 213 120 146 170 196
GGT (UL) 0-40 1-2 2 1 3 0 0 2
AST (UL) - 47-128 48 95 104 66 100 100
ALT (UL) 39-196 78-279 133 110 140 203 158 183 281
CK (UL) 74-294 - 382 765 1190 578 680 930
Amylase (UL) - - 24 37 31 28 33 35
Lipase (UL) - - 60 65 72 72 86 79
Calculated osmo (mmolL) - - 296 305 305 300 296 309
Ferret Identity
Legend - = reference value unavailable bolded = outside reference range a = adult ferret reference ranges from Animal Health
Laboratory (University of Guelph Guelph Canada) b
= reference ranges for 10-week old ferrets (Fox JG 2014)
165
Appendix 4 Domestic ferret (Mustela putorius furo) weekly monitoring sheet
Mon
8 AM
Mon
4 PM
Tues
8 AM
Tues
4 PM
Wed
8 AM
Wed
4 PM
Thurs
8 AM
Thurs
8 PM
Fri
8 AM
Fri
4 PM
Sat
8 AM
Sat
4 PM
Sun
8AM
Sun
4 PM
Mentation
Weight (g)
Respiratory
Rate
Vomit
(+ ++ +++)
Diarrhea
(+ ++ +++)
Urination
(+ ++ +++)
Defecation
(+ ++ +++)
Food
offered
Food
remaining
Water remaining
(ml)
Treatments
Other
observations
Initials of
observer
166
Animal ID ________________________________________ Week ______________
Monitoring Criteria
Mentation BAR (bright alert responsive) QAR (quiet alert responsive) depressed lethargic quiet
Weight measured in grams to be performed once weekly (pre-inoculation) and once daily (post-inoculation)
Respiratory rate measured in breaths per minute
Vomit + (small amount) ++ (moderate amount) +++ (large amount)
Diarrhea + (small amount) ++ (moderate amount) +++ (large amount)
Urination + (small amount) ++ (normal amount) +++ (large amount)
Defecation + (one pile) ++ (two piles) +++ (3+ piles)
Food offered Y (yes) N (no)
Food remaining A (all) P (partial) N (none)
Please note any additional observations in the appropriate section
Monitoring Times
Pre-inoculation ndash ONCE daily at 8am during cage cleaningfeeding
Post-inoculation ndash TWICE daily at 8am and 4pm
Critical patients (as determined by veterinary examination) ndash 3-6 times daily (based on veterinarian recommendation) ndash switch to 24
hour care sheet
Intervention Points
If any ferret develops vomiting diarrhea goes off food or exhibits gt 3 weight loss both the PI and graduate student listed below are
to be notified and the animal will be assessed to determine further treatment plans
Removal Criteria
If an individual ferret develops severe gastrointestinal or systemic disease during the course of the study as characterized by abnormal
mentation marked dehydration 10 weight loss within 24-48 hours profuse diarrhea unresponsive to treatment anorexia of 2 or
more days or frequent vomiting unresponsive to treatment the individual will be removed from the study and humanely killed
167
Appendix 5 Domestic ferret (Mustela putorius furo) 24 hour intensive monitoring sheet
Animal ID ___________________________________________ Date___________________
0700 800 900 1000 1100 1200 1300 1400 1500 1600 1700 1800
Mentation
Weight (g)
Heart Rate
Respiratory Rate
Temperature (C)
Water offered
(mL)
Water consumed
(mL)
Food offered
(type and
amount in
grams)
Food consumed
(type and
amount in
grams)
Presence of urine
Presence of feces
168
Character of
feces
Presence of
vomit
Character of
vomit
Medications to
be administered
Other
observations
169
1900 2000 2100 2200 2300 2400 100 200 300 400 500 600
Mentation
Weight (g)
Heart Rate
Respiratory Rate
Temperature (C)
Water offered
(mL)
Water consumed
(mL)
Food offered
(type and amount
in grams)
Food consumed
(type and amount
in grams)
Presence of urine
Presence of feces
Character of
feces
Presence of vomit
Character of
vomit
170
Medications to be
administered
Other
observations
Monitoring Criteria
Mentation BAR (bright alert responsive) QAR (quiet alert responsive) depressed lethargic quiet
Weight measured in grams to be performed once weekly (pre-inocculation) and once daily (post-inocculation)
Respiratory rate measured in breaths per minute
Vomit + (small amount) ++ (moderate amount) +++ (large amount)
Diarrhea + (small amount) ++ (moderate amount) +++ (large amount)
Urination + (small amount) ++ (normal amount) +++ (large amount)
Defecation + (one pile) ++ (two piles) +++ (3+ piles)
171
Food offered Y (yes) N (no)
Food remaining A (all) P (partial) N (none)
Please note any additional observations in the appropriate section
Monitoring Times
Critical patients (as determined by veterinary examination) ndash 3-6 times daily (based on veterinarian recommendation) ndash switch to 24
hour care sheet
Intervention Points
If any ferret develops vomiting diarrhea goes off food or exhibits gt 3 weight loss both the PI and graduate student listed below are
to be notified and the animal will be assessed to determine further treatment plans
Removal Criteria
If an individual ferret develops severe gastrointestinal or systemic disease during the course of the study as characterized by abnormal
mentation marked dehydration 10 weight loss within 24-48 hours profuse diarrhea unresponsive to treatment anorexia of 2 or
more days or frequent vomiting unresponsive to treatment the individual will be removed from the study and humanely killed
172
Appendix 6 Domestic ferret (Mustela putorius furo) infection trial standard operating procedures
Daily Fecal Collection
1 Collection of all feces passed in a 24 hour period will be performed once daily for all ferrets from
July 13 to July 24 inclusive
2 CAF Isolation staff will collect all fecal material present at the time of daily cage cleaning and
place in individual pre-labelled plastic bags (one per cage)
3 The amount of non-fecal matter (bedding etc) collected should be as minimal as possible
4 CAF isolation staff will record fecal character observations daily for each sample by ticking the
appropriate box on the baggie label
5 Isolation staff will place samples in the necropsy cooler (4 degC) while awaiting collection by
summer student
6 Monday to Friday fecal samples will be collected by A Rodriguez and brought for processing to
the Barta Lab in Pathobiology Samples collected Saturday and Sunday will be put on hold in a
refrigerator (4degC) until pick up on Monday
Example Bag label
Ferret ID Date
Weight of Feces
Fecal Character Normal
Soft
Liquid
Bloody
Abnormal odour
Physical Examination and Health Assessments
1 Manual restraint by hand or in towel by CAF Isolation staff and project personnel If required
examination +- blood collection may be performed under general anesthesia with isoflurane
2 Physical examination body weight HR RR temperature to be performed by PIs (Adriana Pastor
and Dale Smith)
3 Blood collection (~1mL per ferret) is to be performed from the jugular vein if under manual
restraint or the cranial vena cava under anesthesia by PIs using a 25g needle and 1cc syringe
Blood will be collected into small heparinized tubes and submitted to AHL for
CBCBiochemistry
4 All physical examination findings will be recorded on the Exam Sheet
5 Any minor wounds will be treated as appropriate (to be determined by PIs)
Inoculation of Ferrets with Coccidia
1 All ferrets to be inoculated will have been confirmed negative on daily fecals for two weeks
173
2 Brief physical examination by PIs to confirm that the ferrets are healthy to continue in study
3 The concentrated oocyst solution in sterile saline (up to a volume of 1mL) will be combined with
up to 1 mL of FerreTone (or another highly palatable substance if more preferred by the ferrets)
and ferrets will be allowed to consume the mixture ad lib while being monitored
4 Control ferrets will be administered saline only (equal mL to oocyst solution) with 1 mL of
FerreTone (or other substance as used for experimental group)
5 If ferrets refuse to consume the mixture oral inoculation via syringe of concentrated oocyst
solution will be performed by PIs under manual restraint
6 If ferrets are resistant to manual restraint for oral inoculation then inoculation will be performed
by PIs under general anesthesia via gastric tube (8 Fr red rubber)
Euthanasia Protocol
1 Ferrets to be euthanized will be masked down with isoflurane under manual restraint (or in an
anesthetic chamber as judged most appropriate by the PI)
2 Once anesthetized the ferret will be weighed and any blood fecal or other samples will be
collected as required (as determined by PIs)
3 Once an adequate plane of anesthesia is obtained (as determined by PIs) potassium chloride at a
dose of 2 mEq K+kg will be administered either via vena caval puncture or cardiac puncture to
induce cardiac arrest
4 Presenceabsence of respirations heartbeat corneal reflex will be used to assess death
Necropsy Protocol
1 Measure the ferret from nose to tail base (body length)
2 Perform standard necropsy but start with gastrointestinal tract first then thoracic and abdominal
viscera
3 Gently flush the entire contents of the intestine with 12 mL saline into a sterile urine cup
Potassium dichromate should be added in a 11 volumevolume ratio to the same container and
mixed with the combined intestinal contents and saline Label the container with the ferretrsquos ID
ferret group ID date and place on appropriate shelf in the refrigerator (4 degC) in the Barta lab
4 Measure the length of the gut from duodenum to anus
5 Collect paired sections of intestine for histological sectioning and frozen These sections should
be collected along the entire length of the gut from duodenum to rectum (see below for GI length
calculations)
6 For histological sections cut a 2 cm long section of bowel open completely on one side place on
a pre-cut section of box-board and staple both ends to the board to create a flat section Place all
gastrointestinal sections (attached to the board) in Serra fixative solution (100 ethanol (60
VV) 37 formaldehyde (30 VV) glacial acetic acid (10 VV)) for fixation and trim in
cross (transverse) sections
7 Adjacent to each sample removed for histopathology remove another 2 cm section intact and
place in a labelled Whirl-pack for freezing
8 Box-board and pre-labelled Whirl-packs should be labelled in pencil and sharpie respectively
with the following information Animal ID Zoo pm number section of gut (information should
include the region of the gut - jejunum colon etc and the length from the pylorus to the section)
174
Ferret gastrointestinal length calculations (from Evans amp An 2014)
Adult ferret body length 36-41 cm
NB In domestic ferrets there is ~51 ratio of small intestine to body length
Adult domestic ferret GI lengths
Small intestine ndash 182-198 cm
Large intestine - ~10 cm long 06 cm diameter (colon ndash 7 cm rectum ndash 2 cm anus ndash 1 cm)
Based on the above information
Small intestine Six sections of the small intestine from duodenum to ileum will be collected each ~ 25-40
cm apart depending on the size of the ferret
Large intestine Collect two sections of colon (4 cm each) at 25 and 75 of the length of the colon from
the junction of the small and large intestine to the rectum Smaller ferrets may allow only on section
based on colon length
Laboratory SOPs
General notes on processing fecal samples
1 Samples will be collected from the isolation facility necropsy room cooler daily from Monday-
Friday for the entire 6 weeks of the project
2 Upon transfer to the lab each baggie will be weighed after filling to determine the amount of
feces (grams) in each bag and that number entered onto the label on the bag
3 All information regarding fecal processing will be entered onto the spreadsheet for the individual
ferret including ID weight of feces feces character date of sample collection date of sample
processing fecal flotation method presence or absence of oocysts oocyst quantification
Fecal Processing - Weeks 1+2 (acclimation period)
1 After weighing transfer the contents of one baggie to a small sieve on top of a paper cup
2 Fill a single glass container (for fecal flotation) with saturated salt solution
3 Slowly pour small amounts of the solution over the feces mix and strain liquid contents in the paper
cup using a tongue depressor
4 Dispose of the remaining fecal matter in the sieve
5 Pour the contents of the paper cup back into the glass container and cover slip for 5-7 minutes
6 Place coverslip on a clean glass slide and viewscan under microscope at 10x power for presence of
oocysts
7 Record all findings on the provided spreadsheet
8 Notify A Pastor if any oocysts are detected If detected contents of the slide should be flushed back
into the glass container with distilled water and contents of glass container should be transferred to a
50 mL plastic conical tube Clean to remove salt as per Barta lab SOP and mix with potassium
dichromate equal parts by volume to amount of oocysts in diH2O Place a checkmark on the lid of the
50 mL conical vial containing the remainder of the feces and store in the lab fridge at 4 degC
175
Fecal Processing - Weeks 3-7 (infection trial)
1 After weighing transfer the entire contents of one baggie to a small sieve on top of a paper cup
2 Slowly pour small amounts of distilled water over the feces (enough to wet) mix and strain liquid
contents in the paper cup using a tongue depressor until feces appear almost dry
3 Dispose of the remaining fecal matter in the sieve
4 Pour the contents of the paper cup into a 50 mL conical tube If samples are not going to be counted
on the same day then mix potassium dichromate (25 wv) 11 by volume with sieved fluid from
the paper cup and store at 4 degC (refrigerator) until ready to perform OPG counts
If proceeding with the count the same day
5 Pipette and place one drop of the sieved fecal fluid on a slide to determine approximate oocyst
concentration If oocysts rare to none ndash dilute 12 during step 6 if moderate numbers ndash dilute 19 if
too numerous to count ndash dilute 199 (or perform serial dilutions of 10x from initial 19 dilution)
6 Transfer x mL of the mixture from the 50 mL conical tube into a clean 15 mL conical tube and mix
with appropriate amount of saturated salt solution for desired dilution
7 Fill both sides of the McMaster counter chamber and count the number of oocysts per side (total for
one side= total number of oocysts from all 6 sections of the chamber )
8 Average the total counts from both sides
9 Use the following calculation to determine the oocyst per gram count
OocystsmL = oocysts counted times 666 times dilution (ie 3 if dilution 12)
OPG= oocystmL times total volume recorded at end of step 4
10 Record all findings on the provided spreadsheet
11 Notify A Pastor if any oocysts are detected If detected place a checkmark on the lid of the 50 mL
conical vial containing the remainder of the feces and store in the lab fridge at 4 degC
12 For all samples for which no oocysts are detected during steps 6-8 follow up with routine salt
flotation of the remainder of the fecal sample from step 6 (use instructions for fecal processing from
weeks 1-2)

coccidiosis in captive BFF Clinical signs and histopathologic changes associated with infection in BFF
were as described previously in the published literature Average yearly coccidia associated mortality
rates were 053 in adults and 195 in juveniles
Domestic ferrets were confirmed as experimental hosts of E ictidea isolated from BFF Seven of
10 juvenile DF inoculated with oocysts from a BFF developed patent infections and mild clinical disease
was observed in six of these seven Infection was confirmed via morphometric molecular and histologic
examination of samples from infected DF While much is still unknown about enteric coccidiosis in BFF
domestic ferrets provide a promising model for further investigation of this disease
iv
DEDICATION
For my mother Anna Pastorhellip
v
ACKNOWLEDGEMENTS
Itrsquos hard to believe that my residency and thesis have been completed and I have a lot of people
to thank for that
Dale and Graham the two people I wanted to be when I grew up thank you for your mentorship
for many years even before this program I know that you werenrsquot convinced that this project was DVSc
worthy when I first proposed it but Irsquom hoping that the results have changed your mind
I am sincerely grateful to all the members of my advisory committee Dale Smith John Barta and
Simon Hollamby for their insight support and interest in this project Dale you have been an exceptional
advisor I donrsquot know that I will ever get to your level but thank you for showing me that being a great
clinical zoo vet and pathologist are not mutually exclusive John thank you for spontaneously agreeing to
be my advisor when I came to you with this project proposal in my first semester for your energy and
enthusiasm and for supporting my widening interest in parasitology research
My heartfelt appreciation for the Toronto Zoo WHC veterinarians past and present Chris
Dutton Pauline Delnatte Simon Hollamby and Graham Crawshaw I have learned so much from all of
you that I will take forward into my future endeavours I appreciate the extra time you put in including
comps study sessions after-hours tecircte-agrave-tecirctes and the fact that your doors were open when I needed it
For the Toronto Zoo vet techs extraordinaire Michelle Lovering Cassia Devison Dawn
Mihailovic and Tasha Long ndash you have been indispensable during this program and there are not enough
words to express my gratitude
I would especially like to thank all the Wildlife Health Center staff (2013-2016) Mark Bongelli
Charles Guthrie Christine McKenzie Brian Telford Rick Vos Gerri Mintha Margaret Kolakowski
Andrew Lentini Rebecca Clark Lydia Attard Nigel Parr Paula Roberts Andrea Dada Mindy Waisglass
and Julie Digiandomenico for three very memorable years It is all of you that make the WHC such an
amazing place to be Irsquom not sure I have laughed so hard or so often as I did in that lunchroom and I hope
our paths will cross again
vi
I donrsquot think that I can truly express how thankful I am to Pathobiology laboratory technicians
Julie Cobean and Julia Whale Without your assistance patient teaching and friendship I would probably
still be screening fecal samples years from now and scratching my head as to how our lab protocols
actually work It is people like you who make sure graduate students become successful doctorates and I
canrsquot imagine Pathobio without you both in it
I would also like to thank my labmates in the Barta lab mdash Mian Hafeez Evelyn Rejman Rachel
Imai Perryn Kruth Ryan Snyder and Mosun Ogedengbe A special thank you goes to Alex Leveille
without whom my many adventures in parasitology research from coccidia to Babesia would not have
been as successful
To all the students who helped with ferret fecal sample processing data compilation and
necropsies Nathalie Ferriman Janessa Price Thisuri Eagalle Sarah Brisson thank you so much for your
hard work and excitement about my projecthellip even when it was very smelly
So many thanks to the amazing staff of Central Animal Facility - Linda Groocock Vicky Carson
Tony Cengija and Mary Fowler for the daily care and enrichment of my experimental ferrets Your
excitement about working with our ferrets and your assistance with all parts of the process helped made
this project a success
To Adriana Nielsen who was not only my better half but the other fifty percent of my brain for
several years It is your friendship fortitude and our endless phone conversations that got me through the
never-ending Toronto-Guelph commute and this program
To all the ldquoscope roomrdquo pathology co-residents past and present - thank you for being wonderful
friends and colleagues It is indeed rare to find so many amazing people in one place and I know this
program and my sanity would not have been the same without you
To the anatomic pathology faculty and senior graduate students - thank you for all the time
teaching and guidance you provided during my program While I canrsquot say that I have become an amazing
pathologist I can say that because of your mentorship I am a better diagnostician and the type of clinician
who asks better questions takes better samples and understands that you canrsquot ldquojust make a PCR for thatrdquo
vii
A special thank you to Tony van Dreumel who came out of retirement for a semester to try to teach the
Adrianas zoo pathology screening cases with you was always a pleasure
To all the lovely Histo Ladies PM room staff and the other AHL staff who helped me with
Toronto Zoo and HSC pathology cases along the way - I donrsquot think the anatomic path students could
survive without you Thank you for always smiling assisting and accommodating me even when I made
near-impossible processing requests during my weekly Guelph visits
I would also like to acknowledge and sincerely thank all the individuals who helped with resource
and sample acquisition for this project A special mention for those who went above and beyond because
of their interest in this project Don Duszynski who was instrumental in acquiring and then providing a
translator for many of the original mustelid Eimeria descriptions and Majda Globokar Nikola Pantchev
and Donald Martin who supplied my domestic ferret fecal samples and historical data
A shout-out to Julie Swenson Gary West and the Phoenix Zoo BFF team who fostered my love
of this endangered species and helped develop the idea for this project
As always I continue to go out into the world and pursue my dreams with the knowledge that I
have the support of my incredible family long-time friends and my partner Keith Morris I am so lucky
that my residency brought me home and that it afforded us all more time spent together For my aunt
Veronica Lacey who has never failed to believe in my potential and always pushed me to become an
academic ndash yoursquoll never get that PhD from me but I think this is pretty close Finally for my mother
Anna Pastor who never lived to see my greatest achievements but had absolute faith that I could reach
any goal I worked towardshellip this is for you
Finally none of this would have been possible without the generous support of the Toronto Zoo
Residency program and funding through the Barta Laboratory University of Guelph
Adriana Pastor
Toronto August 2017
viii
DECLARATION OF WORK PERFORMED
I declare that all the work reported in this thesis was performed by myself with the following
exceptions
Fecal samples were collected by personnel at the Toronto Zoo Louisville Zoo and participating
diagnostic laboratories
Fecal oocyst per gram counts (routine salt flotation and McMaster counts) were performed by
myself Julie Cobean Julia Whale Evelin Rejman Sarah Brisson Adriana Rodriguez and Perryn Kruth
Whole mitochondrial genome PCR and sequencing was performed by me in conjunction with
Julia Whale and Dr Mian Hafeez
Sequencing of PCR samples was performed at the University of Guelph Laboratory Services
(Guelph Ontario Canada) and results were obtained electronically
ix
TABLE OF CONTENTS
ABSTRACT ii
DEDICATION iv
ACKNOWLEDGEMENTS v
DECLARATION OF WORK PERFORMED viii
TABLE OF CONTENTS ix
LIST OF TABLES xiii
LIST OF FIGURES xiv
LIST OF APPENDICES xv
ABBREVIATIONS xvi
CHAPTER 1 LITERATURE REVIEW 1
11 INTRODUCTION 1
12 APICOMPLEXA 1
121 Brief introduction to apicomplexan pathogens 1
122 Life cycles of the Eimeria and Isospora species implicated in enteric coccidiosis 3
123 Methods of characterization 5
13 RECLASSIFICATION OF MAMMALIAN ISOSPORA 8
14 EIMERIID SPECIES CHARACTERIZED IN MUSTELIDS 8
141 The family Mustelidae 8
142 Eimeriid coccidia described from mustelids 9
143 Eimeriid coccidia described from domestic ferrets 16
144 Molecular characterization 19
145 Clinical signs of disease in domestic ferrets 21
146 Gross necropsy and histologic findings 21
15 INTRODUCTION TO ENTERIC COCCIDIOSIS IN THE BLACK-FOOTED FERRET
25
151 Natural history and conservation of the black-footed ferret in North America 25
152 Coccidia identified from black-footed ferrets 26
153 Morbidity mortality and clinical signs associated with enteric coccidiosis in black-footed
ferrets 28
16 TREATMENT PREVENTION AND CONTROL OF INFECTION BY EIMERIA SPP 29
161 Current recommendations for treatment of eimeriid coccidia in carnivores 29
x
162 Current recommendations for anticoccidial treatment and prophylaxis in domestic and
black-footed ferrets 30
17 VACCINES AGAINST COCCIDIA 32
171 Theory 32
172 Species successes in anticoccidial vaccination 34
18 RESEARCH GOALS AND OBJECTIVES 36
181 Objectives 36
182 Hypotheses 36
183 Applications 36
CHAPTER 2 MOLECULAR CHARACTERIZATION OF ENTERIC COCCIDIA FROM DOMESTIC
FERRETS (MUSTELA PUTORIUS FURO) 38
21 INTRODUCTION 39
22 MATERIALS amp METHODS 43
221 Fecal samples 43
222 Formalin fixed intestinal tissues 44
223 Molecular characterization 44
224 Phylogenetic analysis 46
23 RESULTS 47
231 Fresh fecal samples 47
232 Formalin fixed samples 48
233 Molecular characterization 49
234 Phylogenetic analysis 50
24 DISCUSSION 50
CHAPTER 3 MORPHOLOGICAL AND MOLECULAR CHARACTERIZATION OF ENTERIC
COCCIDIA ISOLATED FROM BLACK-FOOTED FERRETS (MUSTELA NIGRIPES) 60
31 INTRODUCTION 60
32 MATERIALS AND METHODS 64
321 Fecal samples 64
322 Formalin fixed intestinal tissues 65
323 Molecular characterization 66
33 RESULTS 66
331 Morphometric characterization 67
332 Molecular characterization 68
34 DISCUSSION 69
xi
CHAPTER 4 NATURAL HISTORY OF ENTERIC COCCIDIOSIS IN THE BLACK-FOOTED
FERRET (MUSTELA NIGRIPES) 78
41 INTRODUCTION 78
42 MATERIALS AND METHODS 80
421 Toronto Zoo BFF breeding program 80
422 Fecal oocyst evaluation 81
423 Retrospective review of pathology records 82
424 Prospective modified necropsy protocol 82
425 Retrospective medical history review 83
43 RESULTS 83
431 Fecal oocyst evaluation and retrospective medical history review 83
432 Pathology 86
433 Morbidity and mortality 88
44 DISCUSSION 88
CHAPTER 5 EVALUATING THE DOMESTIC FERRET (MUSTELA PUTORIUS FURO) AS AN
EXPERIMENTAL MODEL FOR ENTERIC COCCIDIOSIS IN THE BLACK-FOOTED FERRET
(MUSTELA NIGRIPES) 104
51 INTRODUCTION 104
52 MATERIALS AND METHODS 106
521 Animal care 106
522 Oocyst preparation 107
523 Experimental infections 108
524 Animal welfare 109
525 Hematology 110
526 Morphologic and molecular characterization 110
527 Necropsy protocol 111
53 RESULTS 111
531 Oocyst shedding 112
532 Morphologic and molecular characterization 113
533 Clinical signs 113
534 Hematology 113
535 Necropsy 114
54 DISCUSSION 115
xii
CHAPTER 6 WHOLE MITOCHONDRIAL GENOME SEQUENCES OF TWO EIMERIA SPECIES
ISOLATED FROM DOMESTIC (MUSTELA PUTORIUS FURO) AND BLACK- FOOTED FERRETS
(MUSTELA NIGRIPES) 129
61 INTRODUCTION 129
62 MATERIALS amp METHODS 130
621 Parasites 130
622 DNA isolation from coccidia in feces 131
623 Whole genome sequencing 131
624 Phylogenetic analysis 132
63 RESULTS 133
64 DISCUSSION 134
CHAPTER 7 CONCLUSIONS AND FUTURE DIRECTIONS 145
REFERENCES 148
APPENDICES 157
xiii
LIST OF TABLES
Table 11 Morphometrics of Eimeria and Isospora (=Cystoisospora) species affecting mustelids 10
Table 21 Amplification primers for nuclear 18S rDNA and mitochondrial COI loci used in the
identification of enteric coccidia from domestic ferrets 55
Table 22 Summary of fecal samples from domestic ferrets submitted to two diagnostic laboratories
from 2008-2015 56
Table 23 Morphologic and molecular identification of coccidia from domestic ferrets 57
Table 31 Amplification primers for nuclear 18S rDNA mitochondrial COI and COIII loci used in the
identification of coccidia from black-footed ferrets 73
Table 32 Morphologic and molecular characterization of coccidia from fecal and FFPE necropsy
samples from black-footed ferrets 76
Table 33 Morphometric characterization of Eimeria ictidea oocysts from black-footed ferrets 77
Table 41 Eimeria ictidea shedding in black-footed ferret dam and kit family groups - 2014-2016 97
Table 42 Epidemiologic data for family groups of black-footed ferrets shedding Eimeria ictidea 98
Table 43 Shedding of Eimeria ictidea in adult black-footed ferrets - 2015-2016 99
Table 44 Epidemiologic data for adult black-footed ferrets shedding Eimeria ictidea 100
Table 45 Histologic findings from black-footed ferrets with enteric coccidiosis 101
Table 46 Incidence of coccidial infections in black-footed ferrets at the Cheyenne Mountain Zoo 102
Table 47 Yearly mortality associated with coccidiosis in black-footed ferrets at the Toronto Zoo 103
Table 51 Prepatent period and oocyst shedding of Eimeria ictidea in experimentally infected
domestic ferrets 126
Table 52 Results of oral inoculation of domestic ferrets with oocysts of Eimeria ictidea 127
Table 53 Distribution of coccidial life stages in intestinal tract of domestic ferrets orally
inoculated with oocysts of Eimeria ictidea 128
Table 61 PCR primers used to sequence the mitochondrial genome of Eimeria furonis 136
Table 62 PCR primers used to sequence the mitochondrial genome of Eimeria ictidea 137
Table 63 Coding regions in the mitochondrial genome of Eimeria furonis from a domestic ferret 138
Table 64 Coding regions in the mitochondrial genome of Eimeria ictidea from a black-footed ferret 139
Table 65 Pairwise comparison of coding regions in the mitochondrial genomes of Eimeria furonis
and Eimeria ictidea 140
xiv
LIST OF FIGURES
Figure 11 Phylogeny of the Apicomplexa 2
Figure 12 Classical life cycle of coccidian parasites 4
Figure 13 Morphologic characteristics used for identification of eimeriid oocysts 6
Figure 21 Life stages of Eimeria furonis within the small intestine of a domestic ferret 58
Figure 22 Phylogenetic relationships of coccidia (Eimeria ictidea Eimeria furonis and Isospora
(=Cystoisospora) laidlawi) from domestic or black-footed ferrets 59
Figure 31 Nuclear and mitochondrial genetic loci targeted by primers listed in Table 31 73
Figure 32 Morphometrics of Eimeria ictidea from a black-footed ferret (Mustela nigripes) 74
Figure 33 Nuclear 18S rDNA sequences of Eimeria ictidea to newly generated (see Chapter 2) and
published sequences of Eimeria furonis 75
Figure 34 Mitochondrial cytochrome c oxidase subunit I sequences of Eimeria ictidea to sequences
from other eimeriid parasites of carnivores 75
Figure 41 Oocyst per gram counts and shedding period of Eimeria ictidea from black-footed ferret
family groups from 2014-2016 95
Figure 42 Sexual life stages of Eimeria ictidea in the small intestine of a black-footed ferret 96
Figure 51 Exogenous life stages of Eimeria ictidea 123
Figure 52 Endogenous life stages of Eimeria ictidea within the small intestine of an experimentally
infected domestic ferret 124
Figure 53 Distribution of sexual and asexual life stages of Eimeria ictidea along the intestinal tract
of experimentally infected domestic ferrets 125
Figure 61 Map of the mitochondrial genome of Eimeria furonis 141
Figure 62 Map of the mitochondrial genome of Eimeria ictidea 142
Figure 63 Comparison of the mitochondrial genomes of Eimeria furonis and Eimeria ictidea 143
Figure 64 Phylogenetic relationships of coccidia from domestic and black-footed ferrets based on
complete mitochondrial genome sequences 144
xv
LIST OF APPENDICES
Appendix 1 Shedding of oocysts of Eimeria ictidea in black-footed ferret (Mustela nigripes) dam and
kit family groups from 2014-2016 158
Appendix 2a Hematology values for domestic ferrets (Mustela putorius furo) from 49-51 days of
age prior to experimental inoculation 161
Appendix 2b Serum biochemistry values for domestic ferrets (Mustela putorius furo) from
49-51 days of age prior to experimental inoculation 162
Appendix 3a Hematology values for domestic ferrets (Mustela putorius furo) inoculated orally
with Eimeria ictidea 163
Appendix 3b Serum biochemistry values for domestic ferrets (Mustela putorius furo) inoculated
orally with Eimeria ictidea 164
Appendix 4 Domestic ferret (Mustela putorius furo) weekly monitoring sheet 165
Appendix 5 Domestic ferret (Mustela putorius furo) 24 hour intensive monitoring sheet 167
Appendix 6 Domestic ferret (Mustela putorius furo) infection trial standard operating procedures 172
xvi
ABBREVIATIONS
ATP Adenosine triphosphate
BFF Black-footed ferret(s)
BI Bayesian inference
bp Base pair
CAPC Companion Animal Parasitology Council
CDS Coding DNA sequence
CITES Convention on International Trade in Endangered Species of Wild Fauna and Flora
COI Cytochrome c oxidase subunit 1
COIII Cytochrome c oxidase subunit 3
CytB Cytochrome b
DF Domestic ferret(s)
DNA Deoxyribonucleic acid
FFPE Formalin-fixed paraffin embedded tissue
IUCN International Union on the Conservation of Nature
L Length
LSU Large subunit
mt Mitochondrial
NaOH Sodium hydroxide
nu Nuclear
OPG Oocyst per gram count
PCR Polymerase chain reaction
rDNA Ribosomal DNA
SI Shape index
SND Single nucleotide difference
SOP Standard operating procedure
sp spp Species (singular plural)
SSP Species Survival Plan
SSU Small subunit
TMS Trimethoprim sulfadimethoxine
USFWS United States Fish and Wildlife Service
W Width
1
CHAPTER 1 LITERATURE REVIEW
11 INTRODUCTION
Black-footed ferrets (Mustela nigripes) are one of three wild ferret species worldwide Although
formerly distributed throughout the North American prairies black-footed ferrets (BFF) had been
extirpated from the majority of their range by the 1970s and were declared extinct in the wild in 1987
Since 1986 a multi-institutional effort has been breeding this species in captivity with reintroduction back
into the wild at select sites within Canada the USA and Mexico
Coccidial enteritis is a major cause of death in young captive black-footed ferrets (Bronson et al
2007) but coccidiosis can affect all age classes (personal observation) As a result fewer captive-bred
ferrets may be reared successfully for release to the wild The significance of coccidiosis in wild ferrets is
unknown Consequently the prevention and control of coccidial outbreaks is an important part of black-
footed ferret captive breeding programs and management This research is intended to improve the in situ
and ex situ health of the black-footed ferret through the provision of a better understanding of the
pathogenesis of enteric coccidiosis in this species and to pave the way for the investigation of novel
methods for disease treatment and control
12 APICOMPLEXA
121 Brief introduction to apicomplexan pathogens
The phylum Apicomplexa comprises a large number of eukaryotic intracellular parasitic
organisms many of which are of importance to human and veterinary medicine As indicated by their
name these parasites are characterized by the presence of an apical complex at the anterior aspect of the
infective stage of the life-cycle (Tenter et al 2002) The taxonomic classifications of members of the
Apicomplexa continue to be in a state of flux (reviewed by Adl et al 2005 Cavalier-Smith 2014 Tenter
et al 2002) For this reason a more simplified taxonomic structure has been used in this review (see
2
Figure 11) The subclass Coccidia is a speciose group within the Apicomplexa with most genera falling
into one of two major coccidian suborders within the Eucoccidiorida To date greater than 2000 species
of coccidia have been named (Duszynski Upton amp Couch nd Upton 2000) The adeleid coccidia
(suborder Adeleorina) include monoxenous (single host) and heteroxenous (multiple hosts) parasites in
genera such as Adelea Haemogregarina Hepatozoon and Karyolysus The eimeriorinid coccidia
(suborder Eimeriorina) include the typical intestinal coccidia such as Eimeria Isospora and Cyclospora
species belonging to the family Eimeriidae as well as tissue (cyst forming) coccidia such as
Cystoisospora Besnoitia Toxoplasma and Sarcocystis species that belong to the family Sarcocystidae
(Cox 1994)
Figure 11 Phylogeny of the Apicomplexa Numbers on branches and thickness indicate diversity
(ie named species) Taxonomic groupings demonstrated by the phylogenetic tree (1) subclass
Coccidia (2) suborder Adeleorina (3) suborder Eimeriorina (4) family Eimeriidae and (5) family
Sarcocystidae Adapted from Šlapeta J Morin-Adeline V (2011) Apicomplexa Levine 1970
Sporozoa Leucart 1879 httptolweborgApicomplexa2446 in The Tree of Life Web Project
httptolweborg
2
1
3
4
5
3
122 Life cycles of the Eimeria and Isospora species implicated in enteric coccidiosis
The life cycle of Eimeria species is considered the classical coccidian life cycle which is
typically completed in one host (monoxenous) with many Eimeria species parasitizing only a single host
species (stenoxenous) (Figure 12) The life cycle has two main phases of development one that takes
place within the host (endogenous) and the other that takes places outside of the host (exogenous)
Classically the endogenous stages of the Eimeria life cycle take place within the intestinal epithelium
however some Eimeria species undergo extraintestinal endogenous development such as Eimeria stiedae
in rabbits which replicates within the epithelium of the biliary tree During the exogenous phase of the
life cycle unsporulated oocysts that are shed in the feces of the host sporulate within the environment
resulting in the formation of four sporocysts within each oocyst (tetrasporocystic) Each sporocyst
contains two sporozoites (dizoic) Sporulation is affected by three main factors temperature moisture and
aerobic conditions (Fayer 1980)
Once ingested by the host the wall of the sporulated oocyst is broken to release sporocysts from
which the sporozoites (infective stage) excyst The freed sporozoites penetrate the intestinal epithelial
cells and undergo multiple mitotic divisions to form a single multinucleate meront The meront then
undergoes simultaneous cytokinesis to form first generation merozoites which leave the host cell to infect
new cells and undergo further asexual replications The undifferentiated uninucleate tissue stage of the
parasite within the intestinal epithelial cell is called a trophozoite The number of cycles of asexual
replication (merogony) is predetermined after which the last generation of merozoites penetrate host cells
and undergo sexual differentiation into male and female gamonts (gametogony) Each microgamont
(male) undergoes simultaneous fission to produce numerous motile microgametes each macrogamont
(female) develops into a single mature macrogamete Fertilization of a macrogamete by a motile
microgamete results in formation of a zygote that is rapidly enclosed in a thick wall to form an
unsporulated oocyst Oocysts are shed with the hostrsquos feces into the environment where they are
protected from desiccation and chemical disinfection by the oocyst wall Traditionally Eimeria species
4
have been differentiated based on the host species or host genus affected the site of endogenous life cycle
development and the microscopic cellular characteristics of the different life stages Interestingly
experimental cross infection of Eimeria species from their natural host to a novel host of a taxonomically
similar species has been successful in some cases (De Vos 1970 Levine and Ivens 1970 Haberkorn
1971) challenging the notion that Eimeria are truly stenoxenous parasites
Figure 12 Classical life cycle of coccidian parasites This apicomplexan life cycle includes both
sexual and asexual development The three processes in the life cycle are merogony (asexual
replication A-D) followed by gametogony (formation of gametes E-H) within the digestive tract
of the host with release of unsporulated oocysts (I) Exogenous sporogony (I-L) results in the
production of infective sporulated oocysts (L) Adapted from Barta 2001 with permission of the
author
The life cycle of Isospora spp is similar to that of species in the genus Eimeria (see Figure 12)
but the number of sporocysts and sporozoites differ sporulated oocysts contain two sporocysts (disporic)
5
each of which contains four sporozoites (tetrazoic) These characteristics are not unique to Isospora spp
because diasporic tetrazoic sporulated oocysts are also found in the genera Besnoitia Frenkelia
Hammondia Sarcocystis and Toxoplasma However the sporocysts in the latter parasites are
morphologically distinct in that they lack Stieda bodies
123 Methods of characterization
1231 Morphological features
Historically eimeriid coccidia have been classified based on the cellular morphology of the
different life stages (particularly the morphometrics of sporulated oocysts) where these stages occur in
the host and apparent host specificity (frequently assumed and not tested experimentally) The
morphological features and dimensions of oocysts and their components are important diagnostic features
because of the availability of these stages in clinical specimens these characteristics can include size
(length [L] width [W] shape index [SI=LW]) number of sporocysts wall morphology
presenceabsence of a micropyle micropyle cap residual body or polar granules for oocysts size number
of sporozoites wall morphology presenceabsence of Stieda body subStieda body paraStieda body or
residual body for sporocysts and presenceabsence of refractile bodies for sporozoites (see Figure 13)
Pertinent life cycle information includes type of life cycle (monoxenous versus heteroxenous) tissue
sites of merogony and gametogony (intestinal versus extraintestinal) and the presence or absence of
extraintestinal hypobiotic stages (eg dormozoites or hypnozoites) Further information used to
characterize coccidia that form tissue cysts generally includes details on life stages in the definitive and
intermediate hosts location and morphology of tissue cysts route(s) of transmission among host species
and morphologic descriptions of merozoites (eg tachyzoites or bradyzoites) in tissue culture
6
Figure 13 Morphologic characteristics used for identification of eimeriid oocysts 1) Oocyst in cross
section ol - oocyst length or - oocyst residual body ow - oocyst width pg - polar granule row -rough
outer wall 2) The top of a hypothetical oocyst mcd - depth of the micropyle cap mcw - width of the
micropyle cap mw - width of the micropyle sow - smooth outer wall 3) Sporocyst in cross section
psb - paraStieda body sb - Stieda body sl - sporocyst length sp - sporozoite sr - sporocyst residual
body srb - sporozoite refractile body ssb - subStieda body sw - sporocyst width From Duszynski D
Wilber PG (1997) A guideline for the preparation of species descriptions in the Eimeriidae Journal of
Parasitology 83(2)333-336 reproduced with permission of Allen Press Publishing Services
1232 Molecular characterization (genetic loci and methods)
More recently molecular techniques have been used to infer phylogenetic or evolutionary
relationships among coccidia and to reclassify taxonomic assignments to better reflect the evolutionary
history of these parasites Molecular data can be more informative than phenotypic data because recent
evolutionary divergence among coccidia is unlikely to be reflected in morphologic differences but may
be detectable using molecular data The principle behind the use of molecular sequencing to describe
evolutionary relationships is that nucleotide sequences like morphological features diverge over time
under selective pressure however nucleotide sequences evolve at a more regular rate than do
morphologic characteristics Phenotypic data is thus less likely to detect recent evolutionary divergence
Sequences that are more similar are inferred to be more closely related and to have diverged more
recently (Cox 1994) Molecular characterization can be performed using DNA RNA or protein
sequences Most of the early molecular phylogenetic analyses of coccidia performed used ribosomal RNA
sequences usually by PCR amplification of ribosomal DNA (rDNA) in the nuclear genome of the
7
parasites Ribosomes contain both small and large RNA subunits in eukaryotes the large ribosomal RNA
consists of two forms 5S and 28S while the small ribosomal RNA exists only as 18S Sequences from
several genetic loci have been used for characterization of parasites most commonly 18S rDNA 28S
rDNA and ribosomal internal transcribed spacer regions (ITS) from the nuclear genome and more
recently mitochondrial cytochrome c oxidase subunits I (COI) and III (COIII) however sequencing of
nuclear 18S rDNA (nu 18S rDNA) has been the most prevalent in the literature by far Early attempts to
use 5S RNA sequences formed unlikely phylogenies and too few 28S ribosomal DNA sequences have
been obtained to make this locus useful (Cox 1994 Tenter et al 2002) The disadvantage of nu 18S
rDNA is that it is comparatively poor at distinguishing among closely related eimeriid coccidial species
because of its conserved nature but for that reason the nu18S rDNA locus is useful for inferring
relationships among species with greater evolutionary divergence Although only exploited recently
because of the paucity of suitable PCR primers the mitochondrial COI locus appears to be more useful
for distinguishing closely related eimeriid coccidia (Ogedengbe Hanner amp Barta 2011) but COI
sequences are less useful for inferring more ancient relationships between highly divergent coccidial
species Consequently the combined use of nu 18S rDNA and mitochondrial COI sequencing has been
recommended for improved species description and phylogenetic analysis (El-Sherry et al 2013)
Molecular characterization has also been used for diagnostic purposes and is well-suited to the
identification of coccidia when information on host specificity parasite life cycle and life stages is not
available as the molecular (genetic) data is the same for a given parasite during each of its life cycle
stages This information can be particularly useful in identifying the relationship between different life
stages of heteroxenous parasites collected from different hosts (intermediate definitive) Furthermore for
previously unidentified coccidia or those for which limited information is available molecular
characterization could be used to predict likely definitive hosts or parasite life cycle traits based on
phylogenetic relationships to other known species
8
13 RECLASSIFICATION OF MAMMALIAN ISOSPORA
Recommendations have been made to reclassify the avian and mammalian Isospora into two
separate genera based on life cycle molecular phylogenetic studies and morphologic description of
sporulated oocysts (Frenkel 1977 Barta et al 2005) Due to their classical coccidian life cycle presence
of Stieda bodies within sporocysts and close phylogenetic association with Eimeria species the avian
Atoxoplasma and Isospora have been retained in the genus Isospora (see Barta et al 2005) Conversely
the presence of tissue life cycle stages lack of Stieda bodies within sporocysts and close phylogenetic
association with other genera within the family Sarcocystidae have required many mammalian Isospora
to be reclassified as members of the genus Cystoisospora Frenkel 1977 (Frenkel 1977 Barta et al 2005)
Consequently for the remainder of this thesis Isospora species from mustelids will be referred to as
Isospora (=Cystoisospora) to reflect their probable generic association
14 EIMERIID SPECIES CHARACTERIZED IN MUSTELIDS
141 The family Mustelidae
The family Mustelidae within the order Carnivora comprises a group of approximately 59
carnivorous mammalian species within 22 genera Native mustelids are found in terrestrial and aquatic
environments on almost every continent with the exception of Australia and Antarctica The Mustelidae
are classically divided into two subfamilies as defined by Wozencraft (2005) 1) Mustelinae (weasels
mink ferrets marten wolverine) the larger subfamily including the following genera Arctonyx Eira
Galictis Gulo Ictonyx Lyncodon Martes Meles Mellivora Melogale Mustela Neovison Poecilogale
Taxidea and Vormela and 2) Lutrinae (otters) including seven genera Aonyx Enhydra Hydrictis
Lontra Lutra Lutrogale and Pteronura More recently molecular data suggest the Mustelidae should be
separated into eight subfamilies although this is not universally accepted (Koepfli et al 2008 Lariviegravere
and Jennings 2009 Yu et al 2011)
9
142 Eimeriid coccidia described from mustelids
Ten named Eimeria species and twelve named Isospora (=Cystoisospora) species have been
described in the Mustelidae and are summarized in Table 11 This table includes information on host
range life cycle and detailed morphologic data used to identify and classify the individual parasites Two
coccidial parasites isolated from the Libyan striped weasel (Ictonyx libyca) and the European polecat
(Mustela putorius) initially ascribed to the genus Isospora Isospora zorillae and Isospora putori
respectively have since been reclassified as Sarcocystis spp (see footnote to Table 2 of Yi-Fan et al
2012)
10
Table 11 Morphologic characteristics of Eimeria and Isospora (=Cystoisospora) species affecting mustelids
Coccidial species Host genus and
species
Life cycle
Location
Oocyst shape and
size
Oocyst
description
Sporocyst
description
Sporozoite
description
References
Cytoisospora
eversmanni
Mustela
eversmanii
(Steppe polecat)
Mustela
putorius
(European
polecat)
Homoxenous L185 (16ndash20)
W 148 (16ndash12)
LW 13 (11ndash16)
M absent
PG absent
OR absent
L 115
(10ndash135)
W 98
(9ndash11)
LW 12
(11ndash15)
SB absent
SR present
SRB present Yi-Fan et al 2012
Svanbaev 1956
Nukerbaeva amp
Svanbaev 1973
1977
Cystoisospora
pavlovskyi
Mustela
eversmanii
Mustela
putorius
Homoxenous L 322 (29ndash36)
W 273 (265ndash285)
LW 12 (11ndash14)
M absent
PG absent
OR absent
L 195
(18ndash21)
W 144
(12ndash15)
LW 14
(12ndash15)
SB absent
SR present
SRB present Yi-Fan et al 2012
Svanbaev 1956
Nukerbaeva amp
Svanbaev 1973
1977
Eimeria
baskanica^
Mustela
erminae
(ermine)
Homoxenous Oval with tapered
ends
L 112-126
W 84-98
M absent
PG absent
OR present
SR absent Bean shaped Nukerbaeva amp
Svanbaev 1977
Eimeria furonis Mustela
putorius
Mustela
putorius furo
(dom ferret)
Mustela
nigripes (BFF)
Mustela vison
(mink)
Homoxenous
Small intestine
rectum (H 1927)
Jejunumileum (BP
1993)
Spherical ndash
subspherical
L 11-14
W 10-13
OW 2 layers
M absent
PG absent
OR absent
Spindloid
L 8-9
W 4
SB present
SR present
Vermiform Blankenship-Paris
et al 1993
Hoare 1927 1935b
Jolley et al 1994
Nukerbaeva amp
Svanbaev
19731977
Williams et al 1988
1992 1996
Eimeria hiepei Mustela vison Homoxenous
Bile duct
Spherical
L 13-17
W 13-17
OW 2 layers
(outer
smooth)
M absent
PG absent
OR absent
L 6
W 4
SB absent
SR absent
Banana shaped Davis et al 1953
Grafner et al 1967
11
Coccidial species Host genus and
species
Life cycle
Location
Oocyst shape and
size
Oocyst
description
Sporocyst
description
Sporozoite
description
References
Eimeria ictidea Mustela
eversmanni
Mustela
nigripes
Mustela
putorius
Mustela
putorius furo
Homoxenous
Small intestine
Ovoid ndashellipsoid
L 13-27
W 13-21
OW 2 layers
M present
PG absent
OR absent
Ovoid
(irregular)
L 115
W65
SB present
SR present
- Hoare 1927 1935a
1935b
Jolley et al 1994
Litvenkova 1969
Svanbaev 1956
Tinar 1985
Williams et al 1988
1992
Eimeria irara Eira barbara
(tayra)
Homoxenous
Feces
Ovoid
L 21-25
W 18-20
OW outer
layer smooth
M absent
PG absent
OR absent
Ellipsoid
L 10-12
W 65
SB present
SR present
Elongate (one
end broader than
the other)
Carini amp da
Fonseca 1938
Eimeria melis Meles meles
(European
badger)
Homoxenous Ellipsoid
L 20plusmn018
W 157plusmn002
LW128plusmn0017
(112-15)
OW 2 layers
(outer
smooth)
M absent
PG present
OR present
Ovoid
L
119plusmn0018
W 65plusmn008
LW 183
(155-24)
SB present
L 90plusmn005
W 324plusmn0025
SRB present
Anwar et al 2000
Kotlan amp Pospesch
1933
Eimeria mustelae Mustela vison
Mustela nivalis
(snow weasel)
Homoxenous
Duodenumileum
Spherical or
Ellipsoid
L 18-26
W 14-24
OW 2 layers
M absent
PG present
OR absent
Ovoid
L 8
W 5
SB present
SR present
Broad at one
end and tapered
at other
L 7
W 3
Glebezdin 1978
Iwanoff-Gobzem
1934
Levine 1948
Musaev amp Veisov
1983
Tinar 1985
Eimeria sablii Martes zibellina
(sable)
Homoxenous
Gut
Spherical or
subspherical
L 112-126
W 112
OW 2 layers
M absent
OR absent
Ovoid
L 56
W 42
SR present
Elongate Nukerbaeva 1981
Eimeria sibirica Martes zibellina Homoxenous Ovoid
L avg 216
W avg 180
LW 1076
OW 2 layers
M absent
PG absent
OR absent
Ovoid
L 96-112
W 56-72
SR absent
Elongate Nukerbaeva 1981
Yakimoff amp
Gousseff 1934
Yakimoff amp
Terwinsky 1930
1931
12
Coccidial species Host genus and
species
Life cycle
Location
Oocyst shape and
size
Oocyst
description
Sporocyst
description
Sporozoite
description
References
Eimeria vison
(Eimeria
mustelae)
Mustela
putorius
Mustela
putorius furo
Mustela vison
Homoxenous
Small intestine
+- large intestine
Ovoid
L 17-22
W 9-18
OW 2 layers
M absent
OR
sometimes
present
Ovoid or
Piriform
L 10
W 55
SB absent
SR present
Curved or Club
shaped
L 9
W 25
Foreyt amp Todd 1976
Foreyt et al 1977
Kingscote 1934
1935
Levine 1948
McTaggart 1960
Nukerbaeva amp
Svanbaev
19731977
Tinar 1985
Umurzakov amp
Nukerbaeva 1985
Wolter 1961
Zimmermann 1959
Isospora africana Ictonyx libyca
(Libyan striped
weasel)
Homoxenous
Feces
Spherical
L 25-27
W 25-27
OW 2 layers
(outer
smooth)
M absent
PG absent
OR absent
Ovoid
L 15-17
W 10-12
SB absent
SR present
Elongate
L 135
W 3
Prasad 1961
Isospora altaica Mustela altaica
(mountain
weasel)
Homoxenous
Gut
Oval or spherical
L 280-336
W 252-280
LW 121 (111-
124)
OW 2 layers
M absent
PG absent
OR absent
Ovoid or
spherical
L 140-168
W 111-168
SR present
Svanbaev amp
Rachmatullina
1971
Isospora goussevi Mustela nivalis Homoxenous
Large intestine
Ovoid
L 224 (220-250)
W 174 (160-190)
LW 135 (133-
137)
OW 1 layer
PG present
OR present
Ovoid
L 120
(100-130)
W 70 (60-
80)
SB present
SR present
Elongate Musaev amp Veisov
1983
13
Coccidial species Host genus and
species
Life cycle
Location
Oocyst shape and
size
Oocyst
description
Sporocyst
description
Sporozoite
description
References
Isospora
hoogstraali
Ictonyx libyca Homoxenous
Feces
Ellipsoid
L 37-41
W 32-34
OW 2 layers
(outer
smooth)
M absent
PG some
OR absent
Ovoid
L 19-21
W 13-15
SB absent
SR present
Club-shaped
L 18-19
W 4-6
Prasad 1961
Isospora laidlawi Mustela
putorius
Mustela
putorius furo
Mustela vison
Homoxenous
Feces
Intestinal contents
Ovoid L
320-368
W 272-304
OW 2 layers
M absent
PG absent
OR absent
Ellipsoid
L 208
W 144
SB absent
SR present
Sausage shaped Foreyt et al 1977
Hoare 1927
Levine 1948
McTaggart 1960
Nukerbaeva amp
Svanbaev 1973
1974 1977
Tinar 1985
Isospora lutrae Lutra lutra
(European
otter)
Lutra
canadensis
(North
American river
otter)
Homoxenous Spherical
L 312 (275-32)
W 296 (28-31)
LW 104
(10-112)
OW 2 layers
(outer
smooth)
M absent
PG absent
OR absent
Ellipsoid
L 182 (17-
19)
W 144 (14-
16)
LW128
(12-14)
Sb absent
sSB absent
SR present
Spindle- shaped
L 124
W 25
SRB present
Torres et al 2000
Hoover et al 1985
Isospora
martessii
Martes zibellina Homoxenous
Gut
Ovoid short oval or
spherical
L 252 ndash 280 196
168
W 168 ndash 224 168
168
OW 2 layers
M absent
OR absent
Ovoid
L 112-168
W 84-112
SR present
Elongate Nukerbaeva 1981
Isospora melis Meles meles Homoxenous Ovoid
L 328plusmn034
W 269plusmn019
LW122 (110-
157)
OW 2 layers
(outer
smooth)
M absent
PG absent
OR absent
Ellipsoid
L
215plusmn0166
W 14plusmn012
LW 155
(133-185)
SR absent
Round at one
end other end
tapered
L 142plusmn116
W 40plusmn017
SRB absent
Anwar et al 2000
Glebezdin 1978
Kotlan amp Pospesch
1933
Pelleacuterdy 1955
14
Coccidial species Host genus and
species
Life cycle
Location
Oocyst shape and
size
Oocyst
description
Sporocyst
description
Sporozoite
description
References
Isospora
mustelae (nomen
nudum)
Martes martes Ovoid L
7 W
225
M present - - Galli-Valerio 1932
Isospora nivalis Mustela nivalis Homoxenous
Large intestine
Ovoid
L 206 (200-230)
W 184 (180-210)
LW 11 (109-111)
OW 1 layer
PG absent
OR absent
Ovoid
L 125
(120-130)
W 80 (70-
90)
SR present
Lemon or pear
shaped
Musaev amp Veisov
1983
Unnamed
ldquoCoccidiardquo^
Mustela
nigripes
Urinary bladder - - - - Jolley et al 1994
Unnamed
ldquoCoccidiardquo^
Mustela
nigripes
Trachea bronchus
bronchial glands
- - - - Jolley et al 1994
Unnamed
Eimeria sp^
Mustela
nigripes
Feces
intestinal contents
Ovoid
L 350-386
W 212-232
- - - Jolley et al 1994
Williams et al
1992
Unnamed
Eimeria sp^
Mustela
putorius furo
Small intestine - - - - Blankenship-Paris
et al 1993
Unnamed
Eimeria sp^
Mustela nivalis Homoxenous
Large intestine
Ovoid-ellipsoid L
2031 (1712-2162)
W 148 (1225-
1681)
LW 136 (121-16-
)
OW 1 layer
PG absent
OR absent
Ovoid or
pear-shaped
L 60-100
W 40-80
SR present
Elongate
L 50-90
W 30-70
Musaev amp Veisov
1983
Unnamed
Eimeria sp^
Martes martes
(marten)
Homoxenous Ovoid
L avg 216
W avg 180
LW 1076
OR absent 4 sporocysts
SR present
L 126
W 60
Yakimoff and
Gousseff 1934
Unnamed
Isospora sp^
Mustela
putorius furo
Feces - - - - Bell 1994
Unnamed
Isospora sp^
Mustela
putorius furo
Feces - - - - Bell 1994
Legend L = length W = width LW = length-width ratio avg = average OW = oocyst wall PG = polar granules M = micropyle SB = Stieda body sSB =
subStieda body OR = oocyst residuum SR = sporocyst residuum SRB = sporozoite refractile body ^ = species inquirendae - = no information provided by
author(s) = information obtained from secondary sources (primary reference could not be obtained) All measurements are in micrometers Bolded references
15
are those from which morphometric data were assembled Remaining references indicate other authors who have identified that parasite species in the same or
similar host
16
143 Eimeriid coccidia described from domestic ferrets
Three species of coccidia were originally described from 50 domestic ferrets (Mustela putorius
furo) Eimeria ictidea Eimeria furonis and Isospora (= Cystoisospora) laidlawi (Hoare 1927) All three
species were detected in feces from domestic ferrets at a research facility undergoing an outbreak of
canine distemper Sick ferrets appeared more frequently infected than healthy ones As per Hoare (1927)
none of the ferrets appeared to display clinical signs associated with protozoal infection For each
parasite the author described morphology of sporulated oocysts isolated from feces and sporulation time
(exogenous life stages) The pre-patent period (minimum duration of endogenous development) in an
inoculated naiumlve ferret was described only for E furonis and E ictidea due to insufficient sample size of
I (=C) laidlawi oocysts for an experimental infection trial Sporulation of oocysts occurred within 5-6
days for E furonis 3 days for E ictidea and 4 days for I (=C) laidlawi The sporulated oocysts of E
furonis were spherical with a double outer wall with a thin colourless outer layer and thick yellowish
inner layer no micropyle or residual body and measured on average 128 times 120 microm (length [L] 112-
144 width [W] 104-128 shape index [SI] 107) Unsporulated oocysts contained a zygote with a
diameter of 96 microm Sporocysts were spindle-shaped with one end constrictedblunted contained a
residual body and on average measured 8-88 times 4 microm Sporozoites were vermiform with one end
narrower than the other arranged head to tail and had a central nucleus a clear vacuole was identified in
some at the broad end The sporulated oocysts of E ictidea were oval or elliptical with a double outer
wall with a thin colourless outer layer and thick yellowish inner layer no micropyle or residual body
and measured on average 236 times 175 microm (L 184-272 W 128-208 shape index 135) The zygote in
unsporulated oocysts was elongate with a diameter of 15 times 12 microm when originally passed in feces but
became more spherical with time Sporocysts were irregularly oval with one end broad and the other
more constricted contained a residual body and on average measured 115 times 65 microm Sporozoites were
vermiform with one end narrower than the other arranged head to tail and had a central nucleus and a
clear vacuole at the broad end The sporulated oocysts of Isospora (=Cystoisospora) laidlawi were ovoid
with a double outer wall with a thin colourless outer layer and thick yellowish inner layer no micropyle
17
or residual body and measured on average 34 times 29 microm (L 320-368 W 272-304) Unsporulated
oocysts contained a spherical zygote with a diameter of 236 microm Two sporocysts were identified each
containing 4 sporozoites and no Stieda body sporocysts were elliptical contained a residual body and on
average measured on 208 times 144 microm Sporozoites were sausage shaped with one end slightly pointed
and had a central nucleus and a clear vacuole identified at the pointed end Sporozoites were arranged
with pointed ends all at the same pole of the sporocyst The pre-patent periods described for E furonis
and E ictidea were 6 days and 7 days respectively (Hoare 1927)
Since Hoarersquos initial description (Hoare 1927 Hoare 1935) multiple single case reports and
outbreaks of severe clinical disease associated with intestinal coccidiosis have been reported in domestic
ferrets Blankenship-Paris et al (1993) described a single case of a four-month-old domestic ferret that
presented depressed in thin body condition dehydrated and with pasty dark feces on the perineum This
ferret had been housed with its dam and another sibling neither dam nor sibling showed clinical signs of
enteric disease and both had negative fecal examination results on repeated evaluation Routine fecal
examination of the rest of the colony and necropsies on eight other ferrets in the colony revealed no
evidence of coccidial infection Enteric coccidiosis was determined to be the cause of disease in the four-
month-old ferret based on necropsy findings but the coccidia could not be speciated because diagnosis
was made on histologic findings only
Sledge et al ( 2011) described three separate outbreaks of severe enteric coccidiosis in domestic
ferrets from one ferret rescue centre (group 1) and two shelters (groups 2 and 3) all affected by the same
Eimeria sp The morphologic characteristics of sporulated oocysts were only described for group 1 no
coccidial oocysts were detected on direct smear or fecal flotation of diarrheic samples submitted from
groups 2 and 3 Oocysts were identified as spherical measuring 12-13 microm in diameter with four
sporocysts each containing two sporozoites Oocyst morphometrics histopathologic findings and nu 18S
rDNA partial sequences from all three groups were used collectively to confirm the coccidial species
identify in each outbreak as E furonis
18
Two cases of biliary coccidiosis with E furonis have been reported in domestic ferrets The first
was in a nine-week-old male ferret from a research facility (Williams Chimes amp Gardiner 1996) The
ferret presented with signs of hepatic disease and was negative for coccidia on fecal flotation and direct
smears Endogenous coccidial life stages were described from the gall bladder and liver on histologic
examination In tissue section the oocysts were oval to spherical and measured 125 times 120 microm Meronts
measured 108-130 times 89-93 microm and contained up to 16 merozoites The merozoites exhibited a double-
layered pellicle prominent conoid few rhoptries and many micronemes anterior to the nucleus Based on
the morphologic description of the life stages in this case the coccidia were identified by the authors as
an Eimeria species most likely E furonis Kaye et al (2015) described a second case of biliary
coccidiosis in an 18-month-old female pet domestic ferret with concurrent pure red cell aplasia In this
case all endogenous coccidial life stages were observed on histologic examination of the epithelium of
the extrahepatic biliary tree The oocysts were ovoid and measured 12 times 13 microm Meronts measured 12 times
15 microm and contained up to 16 merozoites each measuring 2 times 5 microm Based on the morphologic
description of the life stages in this case and nu 18S rDNA sequences the pathogen was also determined
to be E furonis Biliary coccidiosis has also been identified in mink (Mustela vison) with the etiologic
agent identified as Eimeria hiepei (Davis Chow amp Gorham 1953 Grafner Graubmann amp Dobbriner
1967)
Oocysts from Cystoisospora ohioensis have been reported from fecal samples collected from
healthy domestic ferret kits in a large American ferret breeding operation that were raised on the same
premise as juvenile domestic dogs (Patterson amp Fox 2007) The method of identification of this parasite
was not described by Patterson amp Fox A second similar institution reported the presence of a
Cystoisospora species also thought to be C ohioensis in routine fecal examination of their ferret colony
(Dr Bambi Jasmin personal communication) Coccidial identification in this case was performed by the
Animal Health Diagnostic Center at Cornell University The significance of these findings is unknown as
no clinical signs or histologic lesions have been described in domestic ferrets associated with shedding of
19
oocysts and the definitive host for C ohioensis is the domestic dog It is most likely that fecal
identification of C ohioensis represents a pseudoparasite in both of these cases or perhaps an
undescribed Cystoisospora sp that is morphologically indistinguishable from C ohioensis
It is difficult to estimate the prevalence of enteric coccidia within the North American domestic
ferret population Fecal samples submitted to university or large veterinary diagnostic laboratories from
domestic ferrets in Canada are uncommon and samples positive for coccidia appear infrequently (Dr
Donald Martin personal communication) Data from Idexx Vet Med Lab in Ludwigsburg Germany was
compiled to review the prevalence of coccidia and Giardia within fecal samples from domestic ferrets
(Pantchev et al 2011) The authors reported that of 284 fecal samples submitted from 2002-2004 18
(63) had detectable coccidial oocysts on fecal flotation Oocysts were identified based on morphologic
characteristics as E ictidea E furonis I (=C) laidlawi and another unidentified Isospora species
Comparative data from the same laboratory from 2009-2010 included sample submissions from 253
ferrets 21 (83) of which were positive for coccidial oocysts on fecal flotation Nine of the samples
were positive for E furonis three were positive with both E furonis and I(=C) laidlawi present eight
were positive only for I(=C) laidlawi and one sample contained both E furonis and E ictidea
identification in all cases was based on morphologic characteristics No statistically significant difference
in the occurrence of coccidial oocysts was detected when data from the two periods were compared
(Fisherrsquos exact test P=041) (Pantchev et al 2011)
144 Molecular characterization
Molecular characterization of Eimeria furonis was first performed by Abe et al (2008) using
oocysts purified from the feces of a single domestic ferret with clinical signs of coccidial enteritis Small
subunit ribosomal DNA (nu 18S rDNA) primers CYC1FE (5ʹ-TAC CCA ATG AAA ACA GTT T-3prime) and
CYC4RB (5prime-CGT CTT CAA ACC CCC TAC TG-3prime) were used to amplify a 347 base pair (bp) fragment
of nu 18S rDNA These primers were initially developed for molecular identification of Cyclospora
species but have since been shown to amplify nu 18S rDNA from several Eimeria species (Matsubayashi
20
et al 2005) The amplicon was sequenced (GenBank AB329724) and compared with previously
published partial nu 18S rDNA sequences from 40 Eimeria two Isospora and four Cyclospora species
The resulting phylogram grouped E furonis with E alabamensis (cattle) and E meleagrimitis (turkey) In
the same study the microscopic morphology of the oocysts was used to identify this coccidial species as
E furonis by comparison with published descriptions of E furonis E ictidea and E heipei by Hoare
(1927) Hoare (1935) and Grafner Graubmann amp Dobbriner (1967) respectively
Nuclear 18S rDNA was also used by Sledge et al (2011) for molecular identification of the
eimeriid coccidia implicated in the three distinct outbreaks of enteric disease in domestic ferrets As
described above initial identification and speciation of the coccidia was performed using morphologic
characteristics of the sporulated oocysts collected from feces in one of the three outbreaks being
investigated the oocysts were identified as E furonis Histologic sections of formalin fixed intestinal
segments from ferrets from each of the three outbreaks contained multiple coccidial life stages DNA was
then isolated from stored formalin-fixed tissues for further genetic analysis Using the partial nu 18S
rDNA gene sequence reported by Abe et al (2008) (GenBank AB329724) the following PCR primers
were created 5ʹ-ACA ATT GGA GGG CAA GTC TG-3ʹ and 5ʹ-GGCGAC AAG CCT GCT TGA AAC-
3ʹ PCR amplification produced a 247 bp amplicon from each of the three groups Analysis and
sequencing of amplicons from all three groups showed 100 homology to nucleic acid sequences
previously reported by Abe et al (2008) for the gene encoding E furonis nu 18S rDNA
Coccidia were identified within hepatobiliary lesions in a domestic ferret receiving
immunosuppressive therapy for red cell aplasia (Kaye et al 2015) DNA was extracted from frozen liver
and a 247 bp fragment of the nu 18S rDNA was amplified using the primers previously described by
Sledge et al (2011) and sequenced Kaye et al (2015) reported that the DNA sequence of the amplicon
was 100 homologous to the published nu 18S rDNA of E furonis and 95 homologous to the nu 18S
rDNA of E myoxi (rodent) E alabamensis (cattle) and I robini (avian)
21
145 Clinical signs of disease in domestic ferrets
Hoare (1927 1935b) in his initial descriptions of enteric coccidiosis in domestic ferrets
observed that clinical signs of intestinal disease were not evident The recent literature supports the
finding of subclinical disease but also describes signs ranging from mild transient diarrhea in young or
stressed animals to more severe disease with dehydration lethargy depression weight lossemaciation
inappetence and death (Blankenship-Paris et al 1993 Powers 2009 Sledge et al 2011 Hoefer et al
2012 Patterson et al 2014) Rectal prolapse has also been reported in ferrets with enteric coccidiosis
(Hillyer 1992 Hoefer et al 2012) In one study co-infection with coccidia and Lawsonia intracellularis
(Desulfovibrio sp) was diagnosed in 4 of 19 ferrets with proliferative bowel disease (Li et al 1996)
These ferrets presented with variable clinical signs including diarrhea lethargy anorexia weight loss
dehydration and emaciation
In the two reports of biliary coccidiosis clinical signs conformed to those expected with
hepatobiliary disease Williams et al (1996) described their case to have presented with emaciation poor
appetite abdominal distension and icterus Kaye et al (2015) described a one week history of lethargy
inappetence and icterus with serum biochemistry results consistent with cholestasis later clinical signs in
this case included melena anemia and cachexia
146 Gross necropsy and histologic findings
The pathology of enteric coccidiosis in domestic ferrets was described by Hoare (1927 1935b)
Two healthy domestic ferrets were experimentally inoculated one each with large numbers of mature
oocysts of either E furonis or E ictidea that were isolated during his initial work The inoculated ferrets
were killed humanely for histologic examination of intestinal sections at the time of first detection of fecal
oocyst shedding no clinical signs of coccidiosis were detected in these ferrets prior to death Infection
with E furonis resulted in invasion of the epithelium of the small intestine and rectum Within the small
intestine the parasites were concentrated in the tips of the villi but could be found to the level of the
22
opening of the crypts of Lieberkuumlhn In rectal sections life stages were limited to the epithelial ridges
between the openings of the glands of Lieberkuumlhn Organisms were located within the apical portion of
the epithelial cells and intensely infected regions exhibited multiple parasites within a single host cell
Both asexual and sexual life stages were present within the same sections Hoare (1927) described similar
histopathologic changes in naturally infected ferrets but the proportion of asexual versus sexual life
stages differed In natural infections sexual life stages were more numerous whereas in experimental
infections asexual life stages predominated these findings would be expected to correlate with the stage
of infection at which ferrets died or were humanely killed for tissue collection and would not be
reflective of differences between natural and experimental infection with this parasite Hoare also
described the morphology of the different endogenous stages including trophozoite (3-4 microm) merozoite
(stumpy sausage shaped L 3-4 microm W 2 microm) macrogamete (spherical 8 microm diameter with darkly
staining globular inclusions of reserve material) and microgamete (described as similar to those of other
Eimeria species) Two types of merogony are described from histologic sections the first with stumpy
merozoites as described above and the second with merozoites with elongated curved bodies and a
compact polar nucleus measuring 60 times 13 microm This second merogonic generation was observed almost
exclusively in the naturally infected ferrets and was associated with initiation of sexual differentiation and
reproduction
The pathology of experimental and non-experimental infection with E ictidea in domestic ferrets
was also described by Hoare (1927 1935b) Parasitic invasion of the epithelium was noted only in the
small intestine with patchy distribution of the parasite life stages throughout affected sections Within the
small intestinal villi the parasites were again concentrated in the tips of the villi with infected epithelial
cells never containing more than one parasite As each intracellular parasite grew it filled the entire host
cell displacing the nucleus to the base of the cell Predominantly sexual life stages were detected in tissue
sections with few asexual generations observed Interestingly the parasites were arranged into age
groups with forms of the same life stage grouped together within the affected epithelial sections this is in
23
contrast to E furonis where life stages of different maturities were found together in affected sections
Hoare described the morphology of the different endogenous stages of E ictidea including merozoites
(free within the lumen elongated vermiform with one pointed end and a nucleus located at the rounded
end 11 microm times 1 microm within the epithelium shortened and rounded 3-4 microm diameter) macrogametes
(elongated 20 times 7 microm occupying the entire host cell with darkly staining globular inclusions of reserve
material) and mature microgamonts (morphologically similar to those of other Eimeria species but larger
than those of E furonis) Of note a tissue reaction was observed specifically in association with more
developed life stages of E ictidea (eg mature meronts mature gamonts unsporulated oocysts) which
was not observed when cells contained earlier stages of development (eg trophozoites immature
gamonts) This tissue reaction was described by Hoare (1935a 1935b) as the development of an annular
constriction of the apical portion of the villus separating infected epithelial cells from unaffected cells
The constriction involved the epithelium but could also extend inwards into the core of the villus These
changes were associated with congestion of capillaries and extravasation of red blood cells within the
constricted segment and in some sections villar tip necrosis
In their case report of one domestic ferret Blankenship-Paris et al (1993) described the gross
pathologic lesions associated with intestinal coccidiosis in this case there was diffuse dilation and
reddening of the small intestine which was empty and the colon contained dark watery material
Histologic lesions were confined to the ileum and jejunum The jejunum exhibited thickening of the villi
with a crypt to villus ratio of 15 mild granulomatous inflammation in the lamina propria and large
numbers of coccidial meronts gamonts and oocysts within the enterocytes of the villar tips
The gross lesions described by Sledge et al (2011) from 20 domestic ferrets are as follows thin
body condition with moderate to marked dehydration perineal staining with diarrhea moderate dilation
of the small and large intestines and the presence of pasty tan to tarry black digesta within the distal small
intestine and colon Other findings in one to a small number of ferrets included enlarged pale tan livers
splenomegaly with dark red colouration and multiple superficial gastric or duodenal ulcers The
24
histologic lesions from 10 ferrets included moderate blunting and occasional fusion of jejunal and ileal
villi focal attenuation and erosion of the epithelium of the villar tips with exudation of fibrin neutrophils
and blood into the intestinal lumen in regions with severe erosion Intact epithelial cells at the villus tips
and rarely sloughed epithelial cells in the intestinal lumen contained numerous intracytoplasmic coccidia
representing a range of asexual and sexual life stages (meronts macrogamonts microgamonts and
oocysts) The subjacent lamina propria of the small intestine and of the large intestine exhibited moderate
lymphoplasmacytic infiltration with occasional neutrophils and congestion of blood vessels Marked
mucosal hemorrhage was identified in the most severely affected sections
Marked gross and histopathologic hepatobiliary lesions were described in a single ferret by
Williams et al (1996) On gross necropsy the liver was pale and enlarged with dilated firm bile ducts
and thickening of the gall bladder wall Similar gross necropsy findings were described by Kaye et al
(2015) marked dilation and mural thickening of the entire biliary tree (including gall bladder intrahepatic
and extrahepatic bile ducts) On histopathology Williams et al (1996) noted that the marked thickening
of the gallbladder wall was a result of cystic proliferation of mucosal glands which were separated by
tracts of fibrous connective tissue and marked granulomatous inflammation Liver sections exhibited
marked biliary hyperplasia marked periductular fibrosis and moderate periportal lymphoplasmacytic
cuffing There was multifocal papillary proliferation of bile duct epithelium and dilation of the bile ducts
and within the ductular lumens there were moderate numbers of lymphocytes and plasma cells small
numbers of degenerate neutrophils sloughed epithelial cells and debris All endogenous coccidial life
stages were present within the gall bladder and biliary epithelium with meronts visible in 20 of the
intact epithelial cells of the biliary tree and gallbladder and oocysts free within the lumen of the
intrahepatic bile ducts Similar lesions were present in the case described by Kaye et al (2015) and as
well as in juvenile and adult farmed mink (Mustela vison) with hepatobiliary coccidiosis (Davis Chow amp
Gorham 1953)
25
15 INTRODUCTION TO ENTERIC COCCIDIOSIS IN THE BLACK-FOOTED FERRET
151 Natural history and conservation of the black-footed ferret in North America
Black-footed ferrets are one of only three wild ferret species worldwide the other species are the
European polecat (Mustela putorius) and the Siberian polecat or steppe polecat (Mustela eversmanii)
They are the only native North American ferret species and the most endangered North American
carnivore They are nocturnal carnivores whose diet and lifestyle are highly dependent on local prairie
dog (Cynomys sp) populations Prairie dogs comprise almost exclusively the diet for the BFF who also
use the complex burrow systems made by prairie dogs to escape their predators and raise their young
(Santymire et al 2014 USFWS BFF Recovery Program 2017)
While formerly distributed throughout the North America prairie ecosystem BFF were
considered extinct by the late 1950s In 1964 a single population was discovered in Mellette County
South Dakota Progressive decline of this population in subsequent years resulted in the decision by
United States Fish and Wildlife Service (USFWS) to initiate a captive breeding program for the species
From 1971-1973 four females and five males were captured for this purpose Despite successful breeding
no kits survived and the last adult ferret in this captive colony died in 1979 at that time BFF were again
presumed extinct in the wild based on annual surveys of the initial capture site In 1981 a dead BFF was
discovered by a ranch dog outside of Meeteetse Wyoming allowing wildlife biologists to identify
another colony of BFF This colony flourished until 1985 when an outbreak of canine distemper in the
BFF population and an outbreak of sylvatic plague in the local prairie dog population resulted in sharp
population declines From 1985 through 1987 all 24 of the remaining BFF were trapped and brought into
captivity to re-initiate the captive breeding program Six ferrets in this initial group died of canine
distemper while in captivity and of the remaining 18 survivors 7 bred successfully to create the founding
population of the current captive breeding population Today this captive breeding population consists of
approximately 300 BFF distributed among multiple institutions (Santymire et al 2014)
26
Since 1986 this multi-institutional effort has been breeding BFF in captivity with reintroductions
back into the wild in 28 selected locations in Canada the USA and Mexico Currently six facilities
participate in the BFF Species Survival Plan (SSP) the Toronto Zoo USFWS National Black-footed
Ferret Conservation Center National Zoorsquos Smithsonian Conservation Biology Institute Louisville
Zoological Garden Cheyenne Mountain Zoo and the Phoenix Zoo (Black-footed Ferret Recovery
Implementation Team 2011) As of 2011 over 8000 BFF kits had been produced in captive breeding
facilities (Black-footed Ferret Recovery Implementation Team 2011)
Multiple infectious diseases pose a significant risk to the captive breeding and post-release
survival of BFF including canine distemper and sylvatic plague Coccidiosis is recognized as a cause of
significant juvenile morbidity and mortality in captive breeding programs and can result in significant
population losses (Bronson et al 2007 Santymire et al 2014 USFWS BFF Recovery Program 2017)
152 Coccidia identified from black-footed ferrets
Eimeria ictidea and Eimeria furonis have been identified in black-footed ferrets based on
morphologic criteria (Jolley et al 1994) Jolley et al examined fecal samples from six captive BFF during
a distemper outbreak as well as samples from wild BFF They described one medium-sized ovoid
tetrasporic dizoic oocyst with a double wall presence of a polar body and lacking both an oocyst residual
body and micropyle The oocysts measured 232 microm (range 182-274) by 155microm (range 130-162) with
a SI of 150 The sporocysts were elongate with the presence of both sporocyst residuum and a Stieda
body Sporozoites contained prominent refractile bodies at the posterior end and were aligned anterior to
posterior within sporocysts These oocysts were shed by all six captive ferrets On histopathology of
intestinal sections merogony and gametogony were observed within the villar epithelium throughout the
small intestine but were concentrated in the jejunum Two morphologically distinct meronts were
detected in these sections one at the villar tips which was larger and lacking in undifferentiated mass
and the other at the base of the villi or rarely in the intestinal crypts Gametogony was predominantly
27
observed at the villar tips and was noted throughout the small intestine These organisms were considered
consistent with Eimeria ictidea based on descriptions by Hoare (1927) from domestic ferrets
A second small spherical to subspherical tetrasporic dizoic oocyst was documented that had a
pink double wall a granular residual body and lacked both oocyst polar body and micropyle This
smaller oocyst measured 126plusmn12 microm (108-152) by 119plusmn09 microm (101-129) with a SI of 106 The
sporocysts were elongate with the presence of a Stieda body and sporozoites contained refractile bodies
Similar to the larger Eimeria species described above merogony and gametogony were observed within
the villar epithelium throughout the small intestine with endogenous developmental stages most
numerous in the jejunum The meronts were small with 16 or fewer merozoites Micro- and
macrogamonts were observed clustered within the apical third of the villar epithelium as were meronts
Jolley et al (1994) determined these small spherical oocysts to be consistent with Eimeria furonis as
described by Hoare (1927) from domestic ferrets
Jolley et al (1994) described a third type of coccidial oocyst occasionally detected in small
numbers within the BFF fecal samples the authors did not state whether this third type of oocyst was
recovered from wild or captive BFF The oocysts measured 370plusmn13 microm (350-386) by 223plusmn23 microm
(212-232) with a SI of 106 Attempts to sporulate collected oocysts were largely unsuccessful and
corresponding endogenous stages were not identified on histopathologic examination of necropsied
ferrets precluding further morphologic identification of the parasite It should be noted that coccidial
oocysts with similar measurements had not been detected in wild or captive prey species available for
ingestion by BFF (Jolley et al 1994)
Previous to this report coccidial oocysts had been isolated from the feces of BFF in two captive
populations (Carpenter amp Hillman 1979 Williams et al 1988) The abstract by Carpenter amp Hillman
(1979) did not describe the oocysts whereas Williams et al (1988) stated that two Eimeria species (one
with larger oocysts and one with smaller oocysts) were identified within the fecal samples but they were
28
not identified further Interestingly Williams et al reported both Eimeria species to be shed in the feces
of all ferrets concurrently affected by distemper and by approximately 30 of the clinically healthy
ferrets at the time of investigation
Non-enteric coccidia have been reported from captive BFF in one facility by two authors (Jolley
et al 1994 Williams et al 1988) Both reports presumably describing the same case(s) noted the
presence of endogenous coccidial life stages in histologic sections of respiratory tissue and merozoites of
an unidentified coccidium in impression smears of the urinary bladder from BFF diagnosed with canine
distemper Meronts were observed within the epithelium of the trachea a large bronchus and associated
bronchial glands Jolley et al (1994) described the lesions as occurring in the same ferret whereas in the
earlier report by Williams et al (1988) they are described as occurring in two different ferrets There have
been no subsequent published reports of systemic coccidiosis in black-footed ferrets and no cases have
been identified within the pathology database of the Toronto Zoo captive BFF population or by the
current SSP pathologist (Dr Michael M Garner personal communication)
There is a significant information gap regarding the pre-patent periods and pathogenicity of both
identified Eimeria species in BFF and studies to further characterize the eimeriid coccidia of the BFF are
lacking
153 Morbidity mortality and clinical signs associated with enteric coccidiosis in black-footed ferrets
The clinical signs of enteric coccidiosis in black-footed ferrets include mucoid to hemorrhagic
diarrhea abdominal discomfort lethargy appetite loss vomiting and dehydration In some cases sudden
death precedes the development of diarrhea Both adult and juvenile BFF are affected by the disease
which causes significant morbidity and mortality in captive populations (Bronson et al 2007) One
retrospective study of the captive BFF population at the Smithsonian National Zoological Park
determined that the most common cause of death in juvenile BFF (aged 30 days ndash 11 months) was
gastrointestinal pathology (524 of juvenile deaths) with 636 of these cases caused by enteric
29
coccidiosis (Bronson et al 2007) Despite the significance of this disease to the captive population its
effect on morbidity and mortality in wild BFF populations is unknown To the authorrsquos knowledge no
routine surveys of fecal parasites have been conducted on wild-born or captive released BFF during
yearly spotlighting events at ferret release sites However samples may be collected opportunistically if
fecal material is identified within the traps used to catch wild BFF during yearly surveys at release sites
Where fecal samples have been analyzed a 13 prevalence of coccidiosis has been identified in wild
born BFF (Dr Rachel Santymire personal communication) Fecal samples have been collected from BFF
at four release sites within the USA Wind Cave National Park (South Dakota) Badlands (South Dakota)
Conata Basin (South Dakota) and Aubrey Valley (Arizona) and positive samples were identified only at
the first site (Dr Rachel Santymire personal communication) Although radio-telemetry has been used at
some release sites to determine sources of mortality and factors involved in survival its use is not
widespread Furthermore the nocturnal and fossorial lifestyle of the BFF is a significant impediment to
the surveillance and monitoring of disease in this species
16 TREATMENT PREVENTION AND CONTROL OF INFECTION BY EIMERIA SPP
161 Current recommendations for treatment of eimeriid coccidia in carnivores
Described anticoccidial therapies for carnivores come from research in domestic cats and dogs
infected by Cystoisospora species these tissue coccidia (family Sarcocystidae) are only distantly related
to the Eimeria species infecting the BFF and other ferrets Current therapeutic recommendations by the
Companion Animal Parasite Council (CAPC 2013) for treatment of described Cystoisospora species
isolated from cats and dogs include the following amprolium (300-400 mg daily for 5 days in dogs 110-
200 mg daily for 7-12 days in dogs 60-100 mgkg daily for 7 days in cats) amproliumsulfadimethoxine
(150 mgkg amprolium and 25 mgkg sulfadimethoxine daily for 14 days in dogs) diclazuril (25 mgkg
for one dose in cats) furazolidone (8-20 mgkg 1-2 times daily for 5 days in dogs and cats) ponazuril (20
mgkg daily for 1-3 days in dogs and cats) quinacrine (10 mgkg daily for 5 days in cats)
30
sulfadimethoxine (50-60 mgkg daily for 5-20 days in dogs and cats) sulfadimethoxineormetoprim (55
mgkg sulfadimethoxine and 11 mgkg ormetoprim daily for 7-23 days in dogs) sulfaguanidine (150 or
200 mgkg daily for 6 days or 100-200 mgkg every 8 hours for 5 days in dogs and cats) toltrazuril (10-
30 mgkg daily for 1-3 days in dogs) trimethoprimsulfonamide (30-60 mgkg trimethoprim daily for 6
days if gt4kg 15-30 mgkg trimethoprim daily for 6 days if lt4kg) (CAPC 2013) Notably the use of all
drugs listed by the CAPC is considered off-label with the exception of sulfadimethoxine
162 Current recommendations for anticoccidial treatment and prophylaxis in domestic and black-
footed ferrets
1621 Domestic ferrets
Recommended daily oral treatment regimens for enteric coccidiosis in domestic ferrets include
amprolium (19 mgkg once daily 05 mgkg) decoquinate (05 mgkg) sulfadimethoxine (300 mgkg in
drinking water) or sulfadiazine-trimethoprim (30 mgkg once daily) all administered for a minimum of
two weeks (Bell 1994 Patterson amp Fox 2007 Patterson et al 2014) Both the aforementioned
coccidiostats amprolium and decoquinate are sold in large formats and are ideal for use in larger
operations such as breeding facilities research facilities or rescue centers Other anticoccidial therapies
used in domestic ferrets include toltrazuril (20 mgkg) and ponazuril (30-50 mgkg) once daily It should
be noted that all anticoccidial therapy used in domestic ferrets is considered off-label drug use
Multiple follow up fecal examinations should be performed after the treatment regimen is
complete and large groups may need to be treated multiple times Routine cage cleaning is also important
to decrease the environmental oocyst burden and prevent re-infection and in the case of coccidial
outbreaks ferrets should be transferred to clean cages multiple times during the course of anticoccidial
therapy Disinfectants such as bleach or quaternary ammonium compounds or dry heat should be used
for effective environmental decontamination (Patterson et al 2014)
31
1622 Species Survival Plan recommendations for black-footed ferrets
Treatment and prophylaxis of enteric coccidiosis with oral sulfadimethoxine was previously
recommended by the BFF Species Survival Plan (SSP) However due to a suspicion of decreasing
efficacy of treatment ponazuril has been recommended recently for treatment Due to the perceived
exquisite sensitivity of BFF to enteric coccidia the current SSP recommendation for treatment is oral
ponazuril at 30 mgkg once if ferrets are to be transported anesthetized stressed or are otherwise
suffering from another illness or injury (even in the absence of clinical signs or fecal shedding) The same
single oral dose of 30 mgkg is also recommended for kits at weaning (30-35 days of age) post weaning
(40-45 days of age) and prior to anesthesia for initial examination and vaccines (50-60 days of age)
Large crowded or otherwise stressed litters should be administered 30 mgkg orally once every 7-10 days
during the period of stress For treatment of coccidial diarrhea diagnosed by fecal examination 30 mgkg
orally once every 7 days for two doses or 50 mgkg orally once daily for 3 days in food (repeated in 7
days) is recommended In BFF with clinical signs of dehydration administration of subcutaneous or
intravenous fluid therapy has been performed Additional therapy with other antibiotics is sometimes
provided in cases with severe clinical signs or where secondary or primary bacterial enteritis is suspected
There is no pharmacokinetic or pharmacodynamic information available for the use of
anticoccidial drugs in BFF or other Mustelidae and thus it is unknown whether the current dose or
frequency of administration is truly appropriate for treatment of coccidiosis In 2 to 3-month-old piglets
administered a single dose of ponazuril orally at 5 mgkg peak serum concentration occurred at 42 hours
(36-48 hr) and elimination half-life was ~56 days (Zou et al 2014) In llamas administered ponazuril as
a single dose of 20 mgkg orally peak serum concentration occurred at 84 hours and elimination half-life
was ~56 days (Prado et al 2011) In domestic cows administered ponazuril as a single 5 mgkg dose
orally peak serum concentration occurred at 48 hours and elimination half-life was 58 hours (Dirikolu et
al 2009) The relevance of serum drug concentrations for treating an intestinal infection that lacks
extraintestinal life stages is likely minimal because the highest drug dose will reach the site of concern
(intestines) and systemic distribution is not required
32
Furthermore no safety or efficacy studies have been performed in any ferret species to validate
the current uses of either sulfadimethoxine or ponazuril for treatment nor have the current recommended
treatment lengths been validated However anecdotal information based on current usage would indicate
that they are safe at the current dosages and frequencies of administration as no adverse effects have been
reported A recent efficacy study in shelter dogs and cats showed that oral ponazuril (50 mgkg)
administered once daily for 3 days was effective for treatment of infection with Cystoisospora as
determined by a reduction in or cessation of fecal oocyst shedding at 4 and 8 days post treatment
Treatment efficacy in this study was inversely correlated to fecal oocyst counts at the initiation of
treatment (Litster et al 2014) Interestingly efficacy of this dose compared to the other two treatment
groups (single 50 mgkg or 20 mgkg oral dose) did not seem to differ but no statistical analysis was
performed Given the ubiquitous use of ponazuril in captive breeding facilities and concerns regarding
resistance of coccidia species to sulfadimethoxine therapy information on minimum effective doses and
dose regimes would be necessary to inform appropriate future SSP treatment and management plans and
to minimize development of drug resistance
17 VACCINES AGAINST COCCIDIA
171 Theory
The development of resistance of protozoal parasites to chemotherapeutic agents has resulted in a
shift towards the development of vaccines for the protection of domestic livestock Immunity to enteric
coccidiosis in avian and mammalian species involves both humoral and cell mediated responses Eimeria
spp infection in sheep rats poultry and other species generally results in a protective immune response
against subsequent re-infections (Catchpole et al 1993 Shi et al 2000) Interestingly this is not the case
for some host parasite interactions for example a recent report indicated that primary infection with E
ninakohlyakimovae in goat kids did not provide protective immunity against subsequent challenge with
the same parasite (Ruiz et al 2013)
33
Vaccines can be divided into four general categories live vaccines inactivatedkilled vaccines
subunit vaccines and recombinant vaccines Live vaccines are orally administered using small numbers of
infectious oocysts or oocysts from strains with low pathogenicity and result in patent but ideally sub-
clinical infections in the host that will elicit a protective immune response Such live vaccines can be
produced using attenuated forms of the pathogen of interest for example in chickens using ldquoprecociousrdquo
strains of Eimeria spp These precocious strains undergo a reduced number of merogonic replications
within the host cells and thus fewer oocysts are shed in the feces of vaccinated animals This reduction in
endogenous merogonic cycles reduces the amount of damage to the intestinal epithelium as well as
reducing the number of oocysts contaminating the environment
Another strategy has been to use live parasites with truncated life cycles An example of this is
the Toxoplasma gondii vaccine developed to prevent abortion in sheep This parasite was passaged
multiple times through a mouse host resulting in an inability to produce tissue cysts (Meeusen et al
2007) This is desirable as the cyst stage of this parasite normally inhibited by the immune system can be
reactivated during periods of stress or immunocompromise The potential drawbacks of live vaccines
include 1) the ability to produce and isolate adequate numbers of coccidial oocysts to meet vaccine
production requirements 2) the potential development of clinical disease in the host as a result of
inoculation 3) the need for all susceptible individuals to receive the vaccine simultaneously to prevent
fecal-oral inoculation of unvaccinated animals with high doses of the infective agent likely to be present
in a shared environment through fecal shedding
Inactivated vaccines are produced when the microbe of interest is killed via application of heat
radiation or chemical treatment prior to inoculation into the host species While safer because they cannot
induce disease in the inoculated patient inactivated vaccines stimulate a reduced immune response
compared with live vaccines and are consequently less effective Subunit vaccines contain single or
multiple antigens of importance in initiating the host immune response rather than the entire pathogen of
concern Subunit vaccines cannot induce disease in the immunized host but are more difficult to produce
34
because they require a detailed understanding of host immune response to infection Recombinant
vaccines involve the genetic modification of a vector (virus or bacteria) one capable of infecting the host
of interest to contain DNA of the pathogen of interest These vectors induce an immune response in the
vaccinated host but as with subunit vaccines cannot induce disease However recombinant vaccines are
again difficult to produce because they require an in depth understanding of the life cycle stages genes
and antigens targeted by the host immune response to infection There are currently no recombinant
vaccines marketed in Canada for use in veterinary medicine against protozoal disease
Creation of effective vaccines against protozoal parasites is complicated by parasite antigenic
diversity during the different life cycle stages and among protozoal species and strains of the same species
(Meeusen et al 2007) Although most parasites induce some level of immunity in their host species the
immunological response to different parasite life stages and species has been poorly characterized for
most coccidia Furthermore many parasites have developed mechanisms to evade host immune responses
or to continue survive and replicate in and transmission by previously infected hosts Our limited
understanding of the immune responses against coccidial antigens has restricted commercial vaccine
production to live or attenuated vaccines (Meeusen et al 2007)
A notable disadvantage of anticoccidial vaccines is that they need to be developed for each
coccidial species of interest because of the species-specific nature of the immune responses this is a
considerable limitation compared with anticoccidial drugs that can have a much wider spectrum of action
(Vermeulen 2005) While the requirement for mass production of vaccine is a limiting factor for vaccines
developed for the agricultural industry this drawback would be less important for production of a vaccine
to be used in an endangered species
172 Species successes in anticoccidial vaccination
The first successful immunization against coccidiosis was reported in 1918 in dogs (Hall amp
Wigdor) In this report a dog that had previously recovered from coccidial infection with Diplospora
35
bigemina was fed three increasing doses of live non-attenuated coccidial culture (at 14 32 and 48 days
post recovery from primary infection) which resulted in no development of clinical signs and no oocyst
shedding for 11mdash18 days after each challenge Subsequently immunization of dogs and cats against
coccidia with protection lasting up to seven months was reported by Andrews (1926) Immunization of
albino rats to eimeriid infection after administration of three or more sublethal doses of Eimeria
nieschulzi via gastric intubation was reported by Morehouse (1938) further experiments showed that
sporozoites did not enter the host intestinal epithelium in immunized rats given a challenge dose
(Morehouse 1938) Similar findings were reported in chickens immunized against Eimeria tenella that
had 50 fewer intra-epithelial sporozoites following challenge compared to naiumlve birds (Augustine and
Danforth 1986) Conversely chickens previously inoculated with Eimeria acervulina exhibited more
intracellular sporozoites after challenge than naiumlve birds but sporozoites were not observed to develop in
previously immunized birds (Augustine and Danforth 1986) These findings provide further evidence that
the immune response to Eimeria spp may differ among host species
Vaccination against Eimeria species has been most successful and is most widely used in the
poultry industry particularly in breeder and layer flocks Almost all vaccines marketed for poultry are
live vaccines (attenuated and non-attenuated) Vaccination against other apicomplexan parasites in
domestic mammals has also been achieved but has been generally less effective for disease prevention
and is less widely available Marketed killed and inactivated (attenuated) vaccines include those
containing killed tachyzoites of Neospora caninum for cattle (Neoguard Merck Animal Health) and
chemically inactivated merozoites of Sarcocystis neurona for horses (EPM Vaccine Fort Dodge ndash no
longer in production) A subunit vaccine for Babesia canis in dogs uses cultured antigen (Pirodog
Merial) Available live vaccines include a vaccine against Toxoplasma gondii in sheep (Ovilis Toxovax
Intervet) that uses an attenuated temperature sensitive strain (S48)
36
18 RESEARCH GOALS AND OBJECTIVES
181 Objectives
a) To determine and characterize (morphologically and molecularly) the enteric coccidial species
currently affecting the black-footed ferret population
b) To describe the natural history of enteric coccidiosis in captive black-footed ferrets including
pre-patent period shedding frequency and burdens and morbidity and mortality rates
c) To compare molecular morphologic and life history characteristics of enteric coccidial species
identified in domestic ferrets to those in black-footed ferrets
d) To validate domestic ferrets as an experimental model for intestinal coccidiosis in the black-
footed ferret
182 Hypotheses
a) Multiple Eimeria species will be isolated from the black-footed ferret population
b) The Eimeria species identified from black-footed ferrets will be the same as those previously
described in domestic ferrets
c) A single pathogenic Eimeria species will be implicated in recorded outbreaks of clinical
coccidiosis during the period of study
d) Domestic ferrets can act as an experimental model of intestinal coccidiosis for black-footed
ferrets
183 Applications
The goal of this project is to better characterize the enteric coccidia of the endangered black-
footed ferret in order to set the stage for improved disease prevention and treatment To the authorrsquos
knowledge this project is the first attempt to isolate and perform molecular characterization of the
coccidial species endemic in the black-footed ferret population This information will be used to compare
these species to known coccidia from domestic ferrets and other related mammals As experimental work
37
cannot be carried out on enteric coccidiosis in the BFF due to its endangered status if the domestic ferret
can be validated as an experimental model studies of the patterns of anticoccidial resistance and
development of immunity against Eimeria spp can be undertaken in vivo The ultimate goal would be the
development of an autogenous vaccine used to improve survival of ferret kits and reduce morbidity and
mortality associated with coccidiosis in BFF captive breeding programs Based on clinical experience
stressful life events such a breeding weaning and transfer between institutions appear to increase the risk
of coccidial outbreaks in adult BFF As such vaccination could assist in reducing disease outbreaks in
BFF associated with various management activities There is no data on the significance of coccidiosis in
wild populations and limited means of disease surveillance following release vaccination during captive-
rearing or pre-release conditioning of BFF would be an ideal method of reducing the potential effects of
this disease in released and free-living BFF Increasing the numbers of ferrets being released to the wild
and releasing ferrets immune to the subsequent threat of coccidiosis would support the goals of the
conservation initiative for the black-footed ferret
38
CHAPTER 2 MOLECULAR CHARACTERIZATION OF ENTERIC COCCIDIA FROM
DOMESTIC FERRETS (MUSTELA PUTORIUS FURO)
This chapter has been submitted for publication as
Adriana R Pastor Dale A Smith and John R Barta (2017) Molecular Characterization of Enteric
Coccidia from Domestic Ferrets (Mustela putorius furo) Vet Parasitol Regional Studies and Reports (In
review)
ABSTRACT
Combined morphometric and molecular characterization of coccidia that infect domestic ferrets
(Mustela putorius furo) was completed to improve the diagnostic specificity of enteric lsquococcidiosisrsquo in
this host Coccidia positive fecal samples (n=11) and formalin fixed paraffin embedded intestinal tissues
(n=3) from domestic ferrets were collected from diagnostic laboratories in Canada and Europe An
average of 35 and 13 domestic ferret fecal samples per year were coccidia-positive when tested by
Canadian and European diagnostic laboratories respectively during the period 2008-2015 Oocyst
morphometrics and sequence genotyping at two loci (nuclear 18S rDNA [nu 18S rDNA] and
mitochondrial cytochrome c oxidase subunit I [mt COI]) were conducted on all samples The first nu 18S
rDNA and mt COI sequences for Isospora (=Cystoisospora) laidlawi and the first mt COI sequence for
Eimeria furonis were generated during this study Phylogenetic analysis of the mitochondrial COI
sequences demonstrated that E furonis was most closely related to E cf ictidea isolated from a black-
footed ferret (Mustela nigripes) and that I (=C) laidlawi was closely related to C canis and C felis The
identifications provided by diagnostic laboratories of the specific parasite species present in a sample
showed poor agreement with their identifications based on genotyping obtained in this study Molecular
techniques appear to be essential for accurate determination of coccidial species responsible for individual
and group outbreaks of coccidiosis and for further understanding of eimeriid host-parasite relationships
Key words coccidia Cystoisospora laidlawi domestic ferret Eimeria furonis Eimeria ictidea Mustela
putorius furo
39
21 INTRODUCTION
Coccidia are host-specific parasites of the phylum Apicomplexa with greater than 2000 species
named to date (Duszynski et al 2000 Upton 2000) The eimeriorinid coccidia (suborder Eimeriorina)
include typical intestinal coccidia such as Eimeria Isospora and Cyclospora species belonging to the
family Eimeriidae as well as tissue (cyst-forming) coccidia such as Cystoisospora Besnoitia Toxoplasma
and Sarcocystis species that belong to the family Sarcocystidae (see Cox 1994)
Enteric coccidia affect both domestic ferrets (Mustela putorius furo) and their wild counterparts
In his initial descriptions of enteric coccidiosis in domestic ferrets Hoare (1927 1935b) did not observe
clinical signs of intestinal disease associated with infection More recently it has been recognized that
enteric coccidiosis can result in clinical signs ranging from mild transient diarrhea to more severe disease
with dehydration lethargy depression weight lossemaciation inappetence and death (Blankenship-Paris
et al 1993 Powers 2009 Sledge et al 2011 Hoefer et al 2012 Patterson et al 2014) Rectal prolapse
has also been reported in ferrets with enteric coccidiosis (Hillyer 1992 Hoefer et al 2012) Disease
appears to be most common in young or stressed animals In one study co-infection with coccidia and
Lawsonia intracellularis (Desulfovibrio sp) was diagnosed in 4 of 19 ferrets with proliferative bowel
disease (Li et al 1996) these ferrets presented with clinical signs including diarrhea lethargy anorexia
weight loss dehydration and emaciation Two cases of biliary coccidiosis have also been reported in
domestic ferrets infection was associated with biliary epithelial hyperplasia cholecystitis and
cholangiohepatitis (Williams et al 1996 Kaye et al 2015)
Three species of coccidia affecting domestic ferrets were originally described and named by
Hoare (1927) Eimeria ictidea Eimeria furonis and Isospora (=Cystoisospora) laidlawi The three
species were detected in feces from 50 domestic ferrets involved in an outbreak of canine distemper at a
research facility For each parasite the author described the morphology of sporulated oocysts isolated
from feces as well as sporulation time (exogenous life stages) All subsequent reports of morphologic
diagnoses of these coccidia have been based on Hoarersquos original descriptions The pre-patent period
40
(minimum duration of endogenous development) in inoculated naiumlve ferrets was described for E furonis
and E ictidea as 6 and 7 days respectively (Hoare 1935b) The pre-patent period for I (=C) laidlawi
was not determined because the number of oocysts available was insufficient for an experimental
infection trial
Hoare (1927) described the sporulated oocysts of E furonis as follows spherical double outer
wall with a thin colourless outer layer and a thick yellowish inner layer no micropyle or residual body
and measuring on average 128 times 120 microm (length [L] 112-144 width [W] 104-128 shape index [SI]
107) Sporocysts were spindle-shaped with one end constrictedblunted contained a residual body and
on average measured 8-88 times 4 microm Sporozoites were vermiform with one end narrower than the other
arranged head to tail and each had a central nucleus a clear vacuole was identified in some sporozoites at
their broad posterior end
The sporulated oocysts of E ictidea were described as follows oval or elliptical with a double
outer wall with a thin colourless outer layer and a thick yellowish inner layer no micropyle or residual
body and measuring on average 236 times 175 microm (L 184-272 W 128-208 SI 135) Sporocysts were
irregularly oval with one end broad and the other more constricted contained a residual body and
measured 115 times 65 microm on average Sporozoites were vermiform with one end narrower than the other
arranged head to tail in the sporocysts and had a central nucleus and a clear vacuole at their broad
posterior end
The sporulated oocysts of I (=C) laidlawi were ovoid with a double outer wall with a thin
colourless outer layer and a thick yellowish inner layer had no micropyle or residual body and measured
on average 34 times 29 microm (L 320-368 W 272-304) A SI of 117 can be calculated from the original
mean dimensions Two sporocysts were identified each containing four sporozoites and no Stieda body
sporocysts were elliptical contained a residual body and measured 208 times 144 microm on average
Sporozoites were sausage shaped with one end slightly pointed and had a central nucleus and a clear
41
vacuole identified at the pointed end Sporozoites were arranged with pointed ends all at the same pole of
the sporocyst
Oocysts identified as Cystoisospora ohioensis have been reported from fecal samples collected
from healthy domestic ferret kits in a large American ferret breeding operation that also housed juvenile
domestic dogs (Patterson and Fox 2007) The method of identification of this parasite was not described
A second similar institution reported the presence of Cystoisospora (=Isospora) species also thought to
be C ohioensis in routine fecal examination of their colony (Dr Bambi Jasmin personal
communication) Identification in this case was performed by the Animal Health Diagnostic Center at
Cornell University and was based on morphometrics using light microscopy The significance of these
findings is unknown but no clinical signs or histologic lesions were described in the ferrets shedding
these oocysts The definitive hosts for C ohioensis are canids including the domestic dog
More recently molecular techniques have been used for the more precise identification of
coccidia Nucleotide sequences like morphological features diverge over time under selective pressure
however recent evolutionary divergence among coccidia is more likely to be reflected in molecular as
compared to morphologic differences Thus nucleotide sequences that are more similar are inferred to be
more closely related and to have diverged more recently (Cox 1994)
Molecular characterization of ferret coccidia has only been performed for one species Eimeria
furonis Abe et al (2008) extracted DNA from oocysts from the feces of a single domestic ferret with
clinical signs resulting from coccidial enteritis Using primers initially developed for molecular
identification of Cyclospora species (see Matsubayashi et al 2005) small subunit ribosomal DNA (nu
18S rDNA) primers CYC1FE (5ʹ-TAC CCA ATG AAA ACA GTT T-3prime) and CYC4RB (5prime-CGT CTT
CAA ACC CCC TAC TG-3prime) were used to amplify a 347 base pair fragment of nu 18S rDNA The
amplicon was sequenced (GenBank AB329724) and compared with previously published partial nu 18S
rDNA sequences from 40 Eimeria two Isospora and four Cyclospora species The resulting phylogram
42
grouped E furonis with E alabamensis (cattle) and E meleagrimitis (turkey) In the same study the
microscopic morphology of the oocysts was used to identify this coccidial species as Eimeria furonis by
comparison with Hoarersquos (1927 1935b) published descriptions of Eimeria furonis and Eimeria ictidea
Sledge et al (2011) also used nu 18S rDNA to identify Eimeria furonis as the cause of three
distinct outbreaks of enteric disease in domestic ferrets Initial identification was performed using
morphometrics of sporulated oocysts collected from feces in one of the three outbreaks being
investigated Formalin fixed paraffin embedded intestinal segments from ferrets from each of the
outbreaks contained multiple coccidial life stages when examined by light microscopy PCR amplification
of a 247 base pair (bp) amplicon of the nu 18S rDNA was generated from DNA isolated from stored
formalin-fixed tissues for further genetic analysis Analysis and sequencing of amplicons from all three
groups showed 100 identity to sequences previously reported by Abe et al (2008) for the gene encoding
E furonis nu 18S
In 2015 Kaye et al identified coccidia within hepatobiliary lesions in a domestic ferret receiving
immunosuppressive therapy for red cell aplasia DNA was extracted from frozen liver and a fragment of
the nu 18S rDNA was amplified using the primers previously described by Sledge et al(2011) The
authors reported that the DNA sequence of the amplicon had 100 identity to the published nu 18S
rDNA sequence of E furonis and 95 identity to the nu 18S rDNA of E myoxi (rodent) E alabamensis
(cattle) and Isospora robini (avian) (Kaye et al 2015)
It is difficult to estimate the current prevalence of enteric coccidia within the North American
domestic ferret population and no studies have been conducted to do so Fecal samples submitted to
veterinary diagnostic laboratories from domestic ferrets in Canada are uncommon and samples positive
for coccidia appear infrequently (Dr Donald Martin personal communication) Conversely in Europe
the prevalence of coccidia within the domestic ferret population appears to be higher based on
submissions to a large veterinary diagnostic laboratory in Germany Data from Idexx Vet Med Lab in
Ludwigsburg Germany was compiled to review the prevalence of coccidia and Giardia within fecal
samples from domestic ferrets (Pantchev et al 2011) The authors reported that of 284 fecal samples
43
submitted from 2002-2004 18 (63) had detectable coccidial oocysts on fecal flotation Based on
morphologic characteristics oocysts were identified as E ictidea E furonis I laidlawi (herein referred
to as I (=C) laidlawi as noted above) and another unidentified Isospora species Comparative data from
the same laboratory from 2009-2010 included sample submissions from 253 ferrets 21 (83) of which
were positive for coccidial oocysts on fecal flotation Nine of the samples were identified as containing E
furonis three contained both E furonis and I (=C) laidlawi eight contained only I (=C) laidlawi and
one sample contained both E furonis and E ictidea identification in all cases was again based on oocyst
morphometrics No statistically significant difference in the occurrence of coccidial oocysts was detected
when data from the two periods were compared (Fisherrsquos exact test P=041) (Pantchev et al 2011)
The purpose of the present study was to perform a more detailed molecular characterization of the
coccidial species isolated from domestic ferrets to estimate prevalence of the different coccidial species
within the Canadian domestic ferret population and to associate morphologic and molecular
characteristics of a greater range of enteric coccidial species in order to improve diagnostic accuracy
22 MATERIALS amp METHODS
221 Fecal samples
Multiple diagnostic laboratories within Ontario Canada1 and a major European diagnostic
laboratory 2 were solicited for fecal samples from domestic ferrets shedding coccidial oocysts Fecal
samples were diagnosed positive for coccidia based on fecal flotation and light microscopic identification
of Eimeria or Cystoisospora species Eleven samples were collected during the study period (from 2014-
2017) and preserved in potassium dichromate (25 wv) eight from Europe and three from Canada
Centrifugal flotation with saturated salt solution (Ryley et al 1976) was used to isolate oocysts from fecal
samples for genomic DNA extraction Genomic DNA extraction and purification were performed using a
1 Animal Health Laboratory Guelph ON Antech Diagnostics Canada Ltd Mississauga ON IDEXX Canada
Markham ON 2 Vet Med Labor GmbH Division of IDEXX Laboratories Ludwigsburg Germany
44
QIAamp DNA Mini Kit (Qiagen Hilden Germany) according to manufacturerrsquos instructions After
addition of DNAzol to the samples (Qiagen Hilden Germany) samples were vortexed using 05 mm
glass beads (Biospec Products Inc Bartlesville OK USA) prior to extraction in order to fracture the
oocyst walls and release the sporocysts Concentrations of the resultant DNA were estimating using a
Nanodrop 2000 spectrophotometer (NanoDrop Products Wilmington DE USA) and stored at 4 degC for
immediate use or minus20 degC for later use
For each laboratory the number of domestic ferret fecal sample submissions numbers diagnosed
positive for coccidial oocysts and number of each coccidial species identified in positive samples were
tabulated for each of the years 2008-2015
222 Formalin fixed intestinal tissues
Major diagnostic pathology services across Canada3 were contacted to identify cases of enteric
coccidiosis identified on necropsy of domestic ferrets Cases were considered positive based on the
presence of asexual or sexual life stages of the parasites in intestinal sections The histologic sections on
each positive case were reviewed re-described and organisms measured (AP DAS) Gross necropsy
reports for all cases were also reviewed to identify any clinical correlates associated with enteric
coccidiosis DNA was extracted from ten 5-6 microm scrolls of formalin fixed paraffin embedded tissue
(FFPE) using the QIAamp DNA FFPE Tissue Kit (Qiagen) as per manufacturer instructions
223 Molecular characterization
Regions from the nu 18S rDNA and mitochondrial cytochrome c oxidase subunit I (mt COI)
DNA were amplified by polymerase chain reaction (PCR) from each sample using the primers listed in
Table 21 PCR amplification was performed for all samples in a volume of 25 microl containing ~100 ng of
3 Animal Health Centre Abbotsford BC Animal Health Laboratory Guelph ON Faculteacute de meacutedecine veacuteteacuterinaire
Universiteacute de Montreacuteal Saint-Hyacinthe QC Histovet Surgical Pathology Guelph ON IDEXX Canada
Markham ON Prairie Diagnostic Services Inc Saskatoon SK
45
genomic DNA 1times PCR buffer 15 mM MgCl2 02 mM deoxyribonucleotide triphosphates (dNTPs) 400
nM of each primer and 1 U of Invitrogen Platinum Taq DNA Polymerase (Thermo Fisher Scientific
Toronto ON Canada) Reactions were performed on a Bio-Rad T100 PCR thermal cycler (Bio-Rad
Laboratories Singapore) Samples were denatured and Taq polymerase activated at 95 degC for 3 min then
subjected to 35 cycles of 94 degC for 30s anneal at 50-62 degC (see Table 21 for specific anneal conditions
for the various primer pairs) for 30s and extension at 72 degC for 30-75s (see Table 21) followed by a
final extension at 72 degC for 7 min Suitable DNA (ie genomic DNA from an Eimeria or Sarcocystis sp)
was included in the PCR reactions to act as a positive control for the reaction chemistry All amplification
products were subjected to electrophoretic separation using 15 submarine agarose gel stained with
ethidium bromide and visualized on an ultraviolet transilluminator (Spectronics Corporation New York
NY USA) DNA band size was determined by comparison with a 1 kb DNA ladder (GeneRuler 1kb Plus
DNA ladder Thermo Fisher Scientific Waltham MA USA) Bands were excised with a new sterile
scalpel blade and PCR products were purified from the gel using a QIAquick Gel Extraction Kit (Qiagen)
PCR products were cycle sequenced using an ABI Prism 7000 Sequence Detection System (Applied
Biosystems Inc Foster City CA USA) by the Molecular Biology Unit of the Laboratory Services
Division University of Guelph (Guelph ON Canada) using the amplification primers to obtain
sequences in both directions The resulting chromatograms were aligned and analyzed with Geneious Ver
818 or later (Biomatters Limited Auckland New Zealand) and high quality consensus sequences
generated The resulting consensus sequences were searched from within Geneious against publically
available sequences on the BLAST server (blastncbinlmnihgovBlastcgi) using the blastn search
algorithm against the nrnt database (GenBank+EMBL+DDBJ+RefSeq ndash AA or DNA) Resultant new
nucleotide sequences were submitted to GenBank
46
224 Phylogenetic analysis
To determine the phylogenetic affinities of the newly obtained sequences with sequences from
related apicomplexan taxa representative nu 18S rDNA and mt COI sequences were downloaded from
GenBank with special reference to sequences from parasites that infect members of the order Carnivora
Nuclear 18S and mt COI sequences were aligned independently using MAFTT v7017 (Katoh et
al 2002) executed from within Geneious and then concatenated into a combined nu18S rDNA mt COI
dataset Multiple sequences from a single parasite were used to generate consensus sequences for each
locus as described by Ogedengbe et al (2017) Aligned sequences were trimmed to the length of the
largest newly generated nu 18S sequence Phylogenetic trees were generated using Bayesian Inference
(BI) using MrBayes Ver 326 (Huelsenbeck and Ronquist 2001) executed from within Geneious the
combined nu 18S and mt COI alignment was partitioned to permit locus-appropriate substitution models
to be applied to each partition For the nu 18S sequence partition the general time reversible (GTR)
substitution model (nst=6) with gamma rate variation (ie a GTR+G+I model) was applied For the mt
COI sequence partition the codon (M1) substitution model (using translation table 4 [ie lsquometmtrsquo]) was
used instead of the GTR with the remaining parameters remaining the same
The resulting tree was rooted using a pair of adeleid coccidia (Hepatozoon spp) as the taxonomic
outgroup All BI analyses were run for a chain length of 1000000 with tree sampling every 1000
following a burn-in of 100000 with default settings of 4 heated chains and heated chain temp of 02
47
23 RESULTS
231 Fresh fecal samples
From 2008-2015 inclusive the Canadian diagnostic parasitology laboratory4 received an average
of 1206 (range 81-160) domestic ferret fecal samples yearly the European parasitology laboratory5
received a yearly average of 230 samples (range 213-270) The number of fecal samples diagnosed as
positive for coccidial oocysts per year on fecal flotation during this time averaged 35 (range 0-8) and
130 (range 6-20) for the Canadian and European laboratories respectively The diagnosing laboratories
used oocyst morphometrics to identify the species of coccidia present Almost all coccidia-positive
submissions to the Canadian laboratory were identified as containing an I (=C) species based on light
microscopy Coccidia in only three samples from the Canadian laboratory were identified as E furonis
one in each of 2010 2012 and 2014 E ictidea was not identified in any samples submitted to the
Canadian laboratory Approximately equal numbers of coccidia-positive samples from the European
laboratory were identified as E furonis and I (=C) laidlawi each year Only two samples from the
European laboratory contained oocysts that were identified as Eimeria ictidea using morphometrics one
from each of 2011 and 2013 Laboratory submissions to both laboratories are summarized in Table 22
Twelve fecal samples preserved in potassium dichromate were received for analysis by the
authors Eleven samples had previously been identified as containing a single coccidial species five
containing E furonis two containing E ictidea and four containing I (=C) laidlawi A final sample had
been identified as containing a mix of E furonis and Cystoisospora canis Results of microscopic and
molecular characterization of these samples are summarized in Table 23
4 IDEXX Canada Markham ON
5 Vet Med Labor GmbH Division of IDEXX Laboratories Ludwigsburg Germany
48
232 Formalin fixed samples
Only three cases of coccidiosis were identified in domestic ferrets within the databases of the five
diagnostic laboratories that participated in the retrospective study Histologic sections of intestine were
received from these three cases which originated in Ontario6 and Quebec7 The Quebec sample (P2010-I)
was collected in 2010 and the Ontario samples (93-40404 and 17-008571) in 1993 and 2017 respectively
On gross necropsy the small intestinal contents of case P2010-I were described as pasty mucoid
yellow-brown feces with some blood For case 93-40404 the small intestines were described as empty
but melena was present within the terminal portion of the large intestine Scant intestinal contents and
dark brown fecal material in the colon were described in case 17-008571
In all cases endogenous developmental stages of coccidia were visible in histological sections
(Figure 21 is exemplary of the findings from one case) Hematoxylin and eosin stained sections from
P2010-I contained two affected regions of small intestine The intestinal mucosa of the first region
contained numerous asexual life stages and moderate numbers of sexual life stages as well as a small
number of oocysts free within the lumen The second section contained tissues that were poorly
preserved nonetheless 0-4 oocysts per 400times field were identifiable within the intestinal lumen Two
regions of affected small intestine were identified from 93-40404 after screening of all submitted sections
both contained low numbers of sexual and asexual endogenous stages Within one region there were small
numbers of meronts within the intestinal mucosa and lamina propria The second region had small
numbers of oocysts within cells of the epithelium and lamina propria as well as free within the intestinal
lumen In case 17-008571 multiple sections of jejunum contained numerous coccidian meronts gamonts
6 Animal Health Laboratory Guelph ON
7 Faculteacute de meacutedecine veacuteteacuterinaire Universiteacute de Montreacuteal Saint-Hyacinthe QC
49
and oocysts within intestinal villi within the ileum scattered epithelial cells also contained these various
life stages
Average length and width of oocysts were measured from slide sections for all cases For P2010-
I oocyst average length and width were determined from seven oocysts to be 94 microm (range 85-105) and
75 microm (range 69-84) respectively with a SI of 125 (range112-140) Average length and width of
oocysts measured from 93-40404 were determined from 5 oocysts to be 2814 microm (range 229-341) and
233 microm (range 180-308) respectively with a SI of 123 (range 110-146) For the third case 17-
008571 only 2 oocysts were identified and average length and width of oocysts measured 982 microm (range
973-992) and 845 microm (range 821-870) respectively with a SI of 116 (range 114-118)
233 Molecular characterization
DNA was successfully extracted from all twelve fecal samples and two cases with formalin fixed
tissue samples Attempts at amplification of DNA extracted from sample 93-40404 using the primer pairs
listed in Table 21 were unsuccessful Molecular identification results and GenBank accession numbers
for the remaining samples are summarized in Table 23 Both the nu 18S rDNA and mt COI sequences
from I (=C) laidlawi were unique when compared with available sequences from other Cystoisospora
species within the public databases However sequences from I (=C) laidlawi were most similar to
sequences from C canis and C felis and somewhat more divergent from sequences from members of the
C ohioensis species complex Two apparent genotypes of E furonis were identified based on nu 18S and
mt COI sequencing results Genotype 1 represented by EU sample 9014 had 100 identity to previously
published sequences of the nu 18S locus from two isolates from Japan (GenBank AB239130 and
AB329724) Genotype 2 represented by EU sample 907 and Canadian sample 17-008571 had 994
identity at the nu 18S locus (3 single nucleotide differences [SNDs] over 561 base pair region [bp]) to the
three sequences above belonging to E furonis genotype 1 Pairwise alignment of mt COI sequences from
both genotypes identified only 2 SNDs (996 pairwise identity over 513 bp region) Partial mt COI
50
sequences of E furonis from both genotypes were only distantly related (941 pairwise identity 30
SNDs over 513 bp and 905 pairwise identity 49 SNDs over 513 bp respectively) to publicly available
sequences from Eimeria ictidea from the black-footed ferret (Mustela nigripes) (GenBank KT203399)
and Eimeria mephitidis from the striped skunk (Mephitis mephitis) (GenBank KT203398) the only other
Eimeria species infecting members of the Carnivora for which sequence was available
234 Phylogenetic analysis
A phylogenetic reconstruction based on concatenated partial nu 18S rDNA and mt COI sequences
of E furonis I (=C) laidlawi and related coccidia is illustrated in Figure 22 The combined 18SCOI-
based tree demonstrates that the two Eimeria species from ferrets form a well-supported monophyletic
group that branches among a collection of other eimeriid coccidia that infect mammals The sarcocystid
parasite of the domestic ferret I (=C) laidlawi was found to group as the sister taxon to C canis that
together formed a monophyletic group with the closely related C felis all three of these closely related
Cystoisopora species possess comparatively large egg-shaped oocysts that are similar morphologically
24 DISCUSSION
The present work has generated the first nu 18S rDNA and mt COI sequences for Cystoisospora
laidlawi and the first mt COI sequence for Eimeria furonis both isolated from the domestic ferret In this
study histologic presence of organisms and microscopic identification of oocysts shed in feces have been
correlated with published and novel nu 18S and mt COI sequences
Eimeria ictidea was not identified in any Canadian sample and this coccidium was reported in
only 2 of 1840 fecal samples submitted from across the European Union (EU) to IDEXX Germany during
2008-2015 suggesting that E ictidea is not a frequent cause of enteric coccidiosis in domestic ferrets in
Canada or the EU
51
During the study period (2008-2015) almost twice as many domestic ferret fecal submissions
were made to the European as compared to the Canadian diagnostic laboratory however the prevalence
of coccidia-positive samples was similar The methodology used in this report cannot be used to
determine the actual prevalence of enteric coccidial infection (coccidiasis) or disease (coccidiosis) within
the domestic ferret population Fecal samples may be submitted to laboratories either as a result of
investigation into enteric disease or as part of a routine health examination Thus without historic
information accompanying each sample one can simply identify the proportion of positive samples and
compare the frequency of the finding of different coccidial species Prospective surveys of fecal samples
from healthy and sick domestic ferrets with greater sample size would be necessary to determine the true
prevalence of these parasites within the population and to infer their clinical significance
Comparatively few mitochondrial COI sequences have been generated for apicomplexan parasites
compared with other genetic loci the majority of published sequences obtained from Apicomplexa are
from nu 18S The disadvantage of using nu 18S rDNA sequences for parasite identification is that they
are poor at distinguishing among closely related eimeriid coccidia due to the highly conserved nature of
the nuclear ribosomal RNA locus In contrast mt COI sequences appear to be more useful for
distinguishing closely related coccidian species (Ogedengbe et al 2011) but are less useful than nu 18S
rDNA sequences for inferring more ancient relationships among more distantly related coccidia
Consequently the combined use of nu 18S rDNA and mt COI sequencing has been recommended for
improved species description and phylogenetic analysis (El-Sherry et al 2013) For these reasons both nu
18S and mt COI sequences were analysed in the present study
Despite adequate quantities of DNA extracted from the Ontario laboratory sample (93-40404)
successful amplification did not result with any primer pair (Table 21) Potential reasons for this include
degradation of formalin-fixed DNA into fragments too small for amplification with the desired primers
perhaps as a result of extended length of time in formalin prior to paraffin embedding or length of time
stored as FFPE tissue (23 years) or insufficient parasite DNA within the paraffin scrolls The primer pairs
52
used appear to be useful for most eimeriid coccidia (Ogedengbe 2015) and successfully amplified both
Eimeria species from DNA isolated from oocysts so it is unlikely that failure to amplify DNA from this
sample resulted from an inability of the primers used to recognize the parasite seen on section
Two genotypes of E furonis were identified in this study Genotype 1 was identified only from
samples originating from domestic ferrets in Europe but exhibited 100 identity based on nu 18S
sequencing with previously published sequences from both Japan and the USA Genotype 2 was
identified from samples originating from domestic ferrets in both Canada and Europe The small number
of single nucleotide differences between the two genotypes at two genetic loci in different genomes are
consistent with intraspecific variation (ie strain variation)
As might have been expected because of their morphological and host similarities nu 18S and mt
COI sequences of E furonis were determined to be most similar to an Eimeria species (E ictidea)
previously isolated from black-footed ferrets (Mustela nigripes) these eimeriid coccidia formed a
monophyletic group that was distinct from other eimeriid coccidia infecting mammals in the phylogenetic
analyses based on combined nu 18S rDNA and mt COI sequences Similarly the nu 18S rDNA and mt
COI sequences of I (=C) laidlawi are most similar to sequences from two other Cystoisospora species of
carnivores (C canis and C felis) that both have large egg-shaped oocysts comparable to those of I (=C)
laidlawi Both morphometrics and genotyping support the close relationships among these three
sarcocystid coccidia of carnivores These molecular data confirm that transfer of Isospora laidlawi to the
genus Cystoisospora by Barta et al (2005) is warranted
The previous light microscopic identifications of coccidial species in 3 of the 11 fecal samples
were not in agreement with the molecular findings These results were not surprising because light
microscopy has been shown to be an insensitive tool for distinguishing among apicomplexan parasites at
both the genus and species level Furthermore re-evaluation of these samples by the authors revealed that
many of the samples that were identified incorrectly based on morphometrics contained primarily
53
unsporulated oocysts making accurate identification based on microscopic appearance highly
challenging These findings further underscore the importance of molecular methods in accurate parasite
identification In the absence of molecular tools accurate measurement of oocyst size shape and
determination of SI can be useful for differentiating among species of Eimeria and Cystoisospora
however this can only be performed accurately on sporulated oocysts from feces Interestingly the size
and shape indices of oocysts of E furonis measured in histologic sections did not match those previously
described by Hoare (1927) for the same oocysts in feces despite molecular confirmation of identity
Thus measurements of oocysts in histologic sections are not recommended for use in coccidial
identification
Our observations highlight the utility of molecular methods for identifying enteric coccidia
infecting domestic ferrets and suggest that diagnoses based on morphological methods should perhaps be
limited to broad determinations of disease etiology (ie lsquococcidiosisrsquo or lsquococcidiasisrsquo) Using molecular
techniques we were able to differentiate morphologically similar coccidial species isolated from the feces
of domestic ferrets and specifically identify parasites seen in histological sections of ferret intestine
Molecular techniques thus appear to be essential for determining the coccidial species responsible for
individual and group outbreaks of coccidiosis and for further understanding of eimeriid host-parasite
relationships
ACKNOWLEDGEMENTS
Many thanks to Julia Whale and Alex Leveille for their assistance and encouragement during the course
of this project The authors would like to recognize the contributions of Dr Donald Martin (IDEXX
Canada) and Drs Nikola Pantchev and Majda Globokar (IDEXX Germany) for the contributions of data
and samples to this project The authors would also like to recognize the Laboratoire de Pathologie
(Service de diagnostic Faculteacute de meacutedecine veacuteteacuterinaire St Hyacinthe Quebec) and the Animal Health
Laboratory (Guelph Canada) for contributions of samples and data to this project Finally this project
was made possible through funding by the Toronto Zoo Residency Research Fund to DASAP and partial
54
funding from a Natural Sciences and Engineering Research Council of Canada (NSERC) Discovery Grant
(400566) to JRB
55
Table 21 Amplification primers for nuclear 18S rDNA and mitochondrial COI loci anneal temperatures (Ta) extension times and expected PCR
product sizes used in the identification of enteric coccidia from domestic ferrets (Mustela putorious furo)
Gene Target Primer Pairs Primer Sequence (5ʹ-3ʹ) Size (bp) Ta (degC) Anneal (sec) Reference
nu 18S rDNA CYC1FE TACCCAATGAAAACAGTTT 560 52 45 Matsubayashi et al (2005)
CYC4RB CGTCTTCAAACCCCCTACTG Matsubayashi et al (2005)
Cocci_18S_595F CCGCGGTAATTCCAGCTCCAAT 216 62 30 Present study
Cocci_18S_847R GCTGMAGTATTCAGGGCGACAA Present study
Lank_18S_224F TCATAGTAACCGAACGGATC 1080 54 60 Ogedengbe (2015)
Api_SSU_2733R CGGAATTAACCAGACAAATC Mathew et al (2000)
mt COI COI_10F GGWDSWGGWRYWGGWTGGAC 500 52 30 Ogedengbe et al (2011)
COI_500R CATRTGRTGDGCCCAWAC Ogedengbe et al (2011)
COI 272F CAATTCTAYGATGCCGCWTT 222 52 30 Present study
COI_500R CATRTGRTGDGCCCAWAC Ogedengbe et al (2011)
Sdae-COI_260F GATCTTTATGTTYTTRATGCC 890 50 75 Ogedengbe (2015)
Sdae-COI_1147R CATTACCCATAACYACACC Ogedengbe (2015)
56
Table 22 Summary of fecal samples from domestic ferrets (Mustela putorius furo) submitted to two diagnostic laboratories from 2008-2015
No fecal samples positive for coccidia
No samples submitted
(percentage positive)
No samples positive for
Cystoisospora sp
No samples positive for
Eimeria furonis
No samples positive for
Eimeria ictidea
Year Canada Europe Canada Europe Canada Europe Canada Europe
2008 3140 (21) 6214 (28) 3 2 0 4 0 0
2009 2160 (12) 14214 (65) 2 9 0 5 0 0
2010 8127 (63) 20213 (94) 7 10 1 10 0 0
2011 0114 (0) 17215 (79) 0 9 0 7 0 1
2012 3108 (28) 10231 (43) 2 4 1 6 0 0
2013 281 (25) 16270 (59) 2 13 0 2 0 1
2014 6127 (47) 12234 (51) 5 6 1 6 0 0
2015 4108 (37) 9249 (36) 4 3 0 6 0 0
Total 28 (29) 104 (56) 25 56 3 46 0 2
Average
year 35 130 31 70 04 58 00 03
Legend Numbers in brackets refer to the percent of the total number of fecal samples submitted
57
Table 23 Morphologic and molecular identification of coccidia from domestic ferrets (Mustela putorius furo)
Sample ID Source
External Lab Morphologic
Diagnosis Morphologic Diagnosis (ARP) Molecular Diagnosis
mt COI GenBank
Accession
nu 18S rDNA
GenBank Accession
93-40404 FFPE enteric coccidia Histologic sample - - -
P2010-I FFPE enteric coccidia Histologic sample E furonis Identical to MF774036 Identical to MF774678
17-008571 FFPE NP Histologic sample E furonis Same as MF774036 Same as MF774678
17-008571 feces NP E furonis E furonis MF774036 MF774678
907 feces E furonis E furonis E furonis MF774035 MF774679
938 feces I (=C) laidlawi no oocysts visualized I (=C) laidlawi MF774037 MF774677
952-A feces E ictidea Cystoisospora sp I (=C) laidlawi Identical to MF774037 Identical to MF774677
9958 feces E furonis no oocysts visualized E furonis Identical to MF774034 Identical to MF774680
9011 feces E furonis E furonis E furonis Identical to MF774035 Identical to MF774679
9014 feces E furonis E furonis E furonis MF774034 MF774680
9017 feces I (=C) laidlawi Cystoisospora sp I (=C) laidlawi Same as MF774037 Same as MF774677
9040 feces I (=C) laidlawi no oocysts visualized I (=C) laidlawi Same as MF774037 Same as MF774677
912-260 feces I (=C) laidlawi Cystoisospora sp I (=C) laidlawi Same as MF774037 Same as MF774677
CAN-2016-1 feces C canis + E furonis Cystoisospora sp I (=C) laidlawi MF774038 MF774676
Legend FFPE = formalin fixed paraffin embedded intestinal sections - = unsuccessful = morphologic diagnosis performed by JRB same as = 100 sequence
identity with listed GenBank entry over entire sequence length identical to = 100 sequence identity but shorter sequence than listed GenBank entry
58
Figure 21 Life stages of Eimeria furonis within the small intestinal epithelium of a domestic ferret
(Mustela putorius furo) Asexual life stages merozoites (black circle) Sexual life stages oocyst
(solid black arrow) macrogamonts (open arrows with labels) microgamont (dotted black arrow)
Hematoxylin and eosin staining scale bar = 25μm
25 microm
59
Figure 22 Phylogenetic relationships of coccidia (Eimeria ictidea Eimeria furonis and Isospora
(=Cystoisospora) laidlawi) from domestic (Mustela putorius furo) or black-footed (Mustela
nigripes) ferrets based on partial nuclear 18S rDNA and mitochondrial COI sequences of these
parasites and related apicomplexan parasites A summary of the sources of the molecular data for
the remaining taxa included in this phylogenetic analysis are found in Supplementary Table 1 of
Ogedengbe et al (2017) Bayesian support is indicated for each node horizontal distance is
proportional to hypothesized evolutionary change (scale indicates sequence divergence of 10)
60
CHAPTER 3 MORPHOLOGICAL AND MOLECULAR CHARACTERIZATION OF
ENTERIC COCCIDIA ISOLATED FROM BLACK-FOOTED FERRETS (MUSTELA
NIGRIPES)
ABSTRACT
Black-footed ferrets (BFF Mustela nigripes) are the only ferret species native to North America
and have been identified as endangered since 1967 Starting in 1986 a multi-institutional effort has been
breeding this species in captivity with successful reintroductions back into the wild Coccidiosis is
recognized as a cause of significant juvenile morbidity and mortality in captive breeding programs and
can result in significant population losses Little is known about the etiology of enteric coccidiosis in
BFF Coccidia positive fecal samples (n=12) and formalin fixed paraffin embedded intestinal tissues
(n=11) were obtained from BFF in the Toronto Zoo and Louisville Zoo Species Survival Plan (SSP)
populations Oocyst morphometrics and sequence genotyping at three loci (nuclear 18S rDNA
mitochondrial cytochrome c oxidase subunit I and mitochondrial cytochrome c oxidase subunit III) were
conducted Results suggest that the same Eimeria species E ictidea was the cause of enteric coccidiosis
in both SSP populations in both juvenile and adult age classes Wider research is indicated to determine
whether these findings are representative of the larger captive and wild BFF populations
31 INTRODUCTION
Black-footed ferrets (BFF) are one of only three wild ferret species worldwide the other two
being the European polecat (Mustela putorius) and the Siberian polecat or steppe polecat (Mustela
eversmanii) The BFF the only native North American ferret species was formerly distributed throughout
the North America prairie ecosystem but were considered extinct by the late 1950s In 1964 a single
population was discovered in Mellette County South Dakota Progressive decline of this population in
subsequent years resulted in the decision by United States Fish and Wildlife Service to initiate a captive
breeding program for the species From 1971-1973 four females and five males were captured for this
61
purpose Despite successful breeding no kits survived and the last adult ferret in this captive colony died
in 1979 BFF were again presumed extinct in the wild based on annual surveys of the initial capture site
In 1981 a dead BFF was discovered by a ranch dog outside of Meeteetse Wyoming allowing wildlife
biologists to identify another colony of BFF This colony flourished until 1985 when an outbreak of
canine distemper in this wild BFF population and an outbreak of sylvatic plague in the local prairie dog
population resulted in sharp population declines From 1985 through 1987 all 24 of the remaining BFF
were trapped and brought into captivity to re-initiate the captive breeding program Six ferrets in this
initial group died of canine distemper while in captivity and seven of the remaining eighteen survivors
are the founding population of the current captive breeding population Today this captive breeding
population consists of approximately 300 BFF distributed among multiple institutions (Santymire et al
2014)
Since 1986 a multi-institutional effort has been breeding BFF in captivity with reintroductions
back into the wild in selected locations in Canada the USA and Mexico Currently six facilities
participate in the BFF Species Survival Plan (SSP) the Toronto Zoo United States Fish and Wildlife
Services National Black-footed Ferret Conservation Center National Zoorsquos Smithsonian Conservation
Biology Institute Louisville Zoo Cheyenne Mountain Zoo and the Phoenix Zoo (Black-footed Ferret
Recovery Implementation Team 2011) In order to provide the best genetic matches BFF are transferred
among the six institutions for breeding Approximately 300-400 kits are produced annually between the
six SSP facilities with ~200 of these kits allocated for release to the wild yearly (Santymire et al 2014)
As of 2011 over 8000 BFF kits had been produced in captive breeding facilities (Black-footed Ferret
Recovery Implementation Team 2011)
Multiple infectious diseases pose a significant risk to the captive breeding and post-release
survival of BFF including canine distemper and sylvatic plague (Santymire et al 2014 USFWS BFF
Recovery Program 2017) Coccidiosis is recognized as a cause of significant juvenile morbidity and
mortality in captive breeding programs and can result in significant population losses (Bronson et al
62
2007 Santymire et al 2014 USFWS BFF Recovery Program 2017) While the effects of the disease on
the wild population are not clear a prevalence of approximately 13 has been reported based on fecal
samples collected from wild BFF born at release sites (Dr R Santymire personal commication)
Coccidia are eukaryotic host-specific parasites of the phylum Apicomplexa affecting numerous
hosts within a wide taxonomic range Two species of coccidia Eimeria ictidea Hoare 1927 and Eimeria
furonis Hoare 1927 have been identified in black-footed ferrets based on morphometrics (Jolley et al
1994) Jolley et al examined fecal samples from six captive BFF during a distemper outbreak as well as
samples from wild BFF They described one medium-sized ovoid eimeriid oocyst with a double wall
presence of a polar body and lacking both an oocyst residual body and micropyle Oocysts of this Eimeria
species (sp) measured 232 times 155 microm (range 182-274 times 130-162) with a shape index (SI) of 150 The
sporocysts were elongate with the presence of both sporocyst residuum and a Stieda body Sporozoites
contained prominent refractile bodies at the posterior end and were aligned anterior to posterior within
sporocysts These oocysts shed by all six captive ferrets were considered consistent with Eimeria ictidea
based on descriptions by Hoare (1927) On histopathologic examination of intestinal sections parasites
undergoing merogony and gamogony were observed within the villar epithelium throughout the small
intestine but were concentrated in the jejunum (Hoare 1935b) parasite life stages were not described
from other tissuesorgans
A second small spherical to subspherical eimeriid oocyst was also documented in the captive
ferrets by Jolley et al (1994) this second species had a pink double oocyst wall a granular residual body
and lacked both oocyst polar body and micropyle This smaller species measured 126 times 119 microm (range
108-152 times 101-129) with a SI of 106 The sporocysts were elongate and possessed a Stieda body and
sporozoites contained refractile bodies Similar to the larger Eimeria sp described above merogonic and
gamogonic stages were observed within the villar epithelium throughout the small intestine but were
most numerous in the jejunum Jolley et al (1994) concluded these small spherical oocysts were
consistent with E furonis described by Hoare (1927) from domestic ferrets
63
Jolley et al (1994) described a third type of coccidial oocyst occasionally detected in small
numbers within BFF fecal samples however the authors did not state whether this third oocyst
morphotype was recovered from wild or captive animals The oocysts measured 370 times 223 microm (range
350-386 times 212-232) with a SI of 106 Attempts to sporulate collected oocysts were unsuccessful and
corresponding endogenous stages were not identified on histopathologic examination of necropsied
ferrets precluding further morphologic identification of the parasite It should be noted that coccidial
oocysts with similar measurements had not been detected in wild or captive prey species available for
ingestion by BFF making it unlikely that this coccidial species would have been a pseudoparasite (Jolley
et al 1994)
Previous to this report by Jolley coccidial oocysts had been isolated from the feces of BFF in two
captive populations (Carpenter amp Hillman 1979 Williams et al 1988) The abstract by Carpenter amp
Hillman (1979) did not describe the oocysts whereas Williams et al (1988) stated that two Eimeria sp
(one with larger oocysts and one with smaller oocysts) were observed within the fecal samples but they
were not identified further Interestingly Williams et al reported both Eimeria sp to be shed in the feces
of all ferrets concurrently affected by distemper and by approximately 30 of the clinically healthy
ferrets at the time of investigation
Non-enteric coccidia have also been reported by two authors from captive BFF at one facility
(Jolley et al 1994 Williams et al 1988) Both reports which presumably described the same case(s)
noted the presence of endogenous coccidial life stages in histologic sections of respiratory tissue and
merozoites of an unidentified coccidium in impression smears of the urinary bladder from BFF diagnosed
with canine distemper Meronts were observed within the epithelium of the trachea a large bronchus and
associated bronchial glands In the later report Jolley et al (1994) described the lesions as occurring in the
same ferret whereas in the earlier report by Williams et al (1988) they are described as occurring in two
different ferrets Paraffin blocks containing formalin fixed tissues from these cases have since been
discarded precluding further attempts at parasite identification with molecular methods Subsequent to
64
these reports further cases of systemic coccidiosis in BFF have neither been published nor identified
within the pathology database of the Toronto Zoo captive BFF population nor by the current SSP
pathologist (Dr Michael M Garner personal communication)
Previous characterization of coccidia from black-footed ferrets has been based on host species
affected tissues in the host and morphometric characterization of life stages in histologic sections and
oocyst characteristics using light microscopy It is known that morphologically similar Eimeria species
are not necessarily conspecific and may vary in host specificity and pathogenicity Molecular
characterization is thus required to accurately identify coccidia to the species level No molecular
characterization of coccidian parasites from black-footed ferrets has been performed to date
There is a significant information gap regarding which parasite species are implicated in
morbidity and mortality events associated with enteric coccidiosis in BFF and whether different coccidia
are associated with this disease in adult versus juvenile age classes or in different SSP institutions
Studies to further characterize the eimeriid coccidia of the BFF are warranted to improve the management
of this disease in the captive population The objectives of this research were to morphologically and
molecularly characterize coccidia associated with enteric disease in BFF at the Toronto Zoo and in other
SSP facilities
32 MATERIALS AND METHODS
321 Fecal samples
Twelve fecal samples were collected during the study period (from 2014-2016) and preserved in
potassium dichromate (25 wv aqueous) seven from the Toronto Zoo and five from the Louisville Zoo
Centrifugal flotation with saturated salt solution (Ryley et al 1976) was used to isolate and concentrate
oocysts from fecal samples for light microscopic examination and genomic DNA extraction
One to two drops of the supernatant from the centrifugal flotation were placed directly on a slide
and beneath a coverslip The morphology and dimensions of sporulated oocysts were documented using a
65
Provis AX70 photomicroscope (Olympus Canada Richmond Hill ON Canada) fitted with a digital
imaging device (Infinity3-1C Lumenera Corporation Ottawa ON Canada) controlled using iSolution
Lite image analysis software (Hoskin Scientific Burlington ON Canada) operated at a total
magnification of 1000times Morphologic features noted for each oocyst included oocyst wall morphology
number of sporocysts presence or absence of a micropyle micropyle cap residual body and polar
granules For sporocysts size number of sporozoites per sporocyst and presence or absence of Stieda
body and sporocyst residuum were noted Alignment of sporozoites within the sporocyst and
presenceabsence of refractile bodies within sporozoites were also described The sporulated oocyst
length and width measurements (in microm) were then used to calculate the SI for each measured oocyst
Morphologic and morphometric features were compared to previously published values for E furonis and
E ictidea from domestic and black-footed ferrets
Genomic DNA extraction and purification were performed using a QIAamp DNA Mini Kit
(Qiagen Hilden Germany) according to manufacturerrsquos instructions as described in Chapter 2 (Materials
amp Methods)
322 Formalin fixed intestinal tissues
The pathology records of the Toronto Zoo were searched from 1993-2016 for cases of BFF
diagnosed with enteric coccidiosis on histopathology For each case slides of histologic sections from all
submitted intestinal segments were reviewed to confirm the presence of sexual andor asexual life stages
within the intestinal epithelium Scrolls (5-6 microm) were cut from the paraffin blocks containing affected
intestinal sections and DNA extracted from the formalin fixed paraffin-embedded tissue (FFPE) using
the QIAamp DNA FFPE Tissue Kit (Qiagen Toronto Ontario) as per manufacturerrsquos instructions
66
323 Molecular characterization
Molecular characterization of coccidial isolates was performed on oocysts purified from fresh
fecal samples (isolated as described above) that were collected from juvenile and adult ferrets from
2014-2016 and DNA extracted from FFPE samples of BFF intestine containing parasite life stages
Regions from the nuclear 18S (SSU) rDNA (nu 18S rDNA) mitochondrial cytochrome c oxidase
subunit I (mt COI) DNA and mitochondrial cytochrome c oxidase subunit III (mt COIII) DNA were
amplified by polymerase chain reaction (PCR) from each sample using the primers listed in Table 31 and
methodology described in the Materials amp Methods section of Chapter 2 Table 31 also contains the
specific anneal conditions used for the various primer pairs Genomic DNA from an Eimeria species of
poultry was included in the PCR reactions to act as a positive control for the reaction chemistry A
representative selection of the newly generated nucleotide sequences resulting from the above were
submitted to GenBank
DNA obtained from oocysts collected from fecal samples during the first year of the study (2014)
was used to generate a complete mitochondrial genome (see Chapter 6 for details) using primer pairs and
sequencing primers summarized in Table 31 All subsequent samples collected in 2015 and 2016 had
shorter mt COI and mt COIII sequences obtained to permit genotyping of all collected oocysts at these
two loci The location of each primer in the nu 18S mt COI and mt COIII genetic locus is illustrated in
Figure 31
33 RESULTS
From 2014-2016 coccidia-positive fecal samples were obtained from twelve BFF ferretsferret
groups from the Toronto Zoo and Louisville Zoo SSP populations (see Table 32) Nine samples were from
single housed adults between the ages of 1-5 years (63 MaleFemale) Two samples were from mixed
groups one pooled fecal sample from four adults (FERA-1 13 MF) and one fecal sample from a family
67
group consisting of a dam and five kits (23 MF) One fecal sample was collected from a juvenile male
ferret at the time of necropsy
Eleven BFF with enteric coccidiosis were identified in the Toronto Zoo necropsy reports from
1998-2016 and all were confirmed by histological re-evaluation (Table 32) Both juvenile (n=9 36 MF)
and adult ferrets (n=2 20 MF) were represented
331 Morphometric characterization
Twelve coccidia-positive fecal samples were identified from adult and juvenile BFF from 2014-
2016 by on site laboratories at either the Toronto Zoo or the Louisville Zoo Fecal flotation and light
microscopic re-examination of the samples identified coccidial oocysts in 10 of these 12 samples
Morphometric characterization was performed on six samples in which there was adequate
quantity and quality of sporulated oocysts for examination These included three samples from single-
housed adults one from a juvenile at the time of necropsy one of pooled feces from a group of adult
ferrets and one of pooled feces from a family group (dam and kits) Two of the three samples from
single-housed adults were from the same ferret on different dates in 2016 the dates of collection were
separated by a period in which shedding of oocysts was not identified on routine repeated fecal
examinations Oocysts were elliptical with a colourless double wall and contained four sporocysts each
with two sporozoites Sporocysts were ovoid and both Stieda body and residual body were present
Sporozoites exhibited an anterior to posterior alignment within the sporocysts and refractile bodies were
identified (Figure 32) Results for length width and shape index of sporulated oocysts including range
and average values are summarized in Table 33 and Figure 32 The average measurements based on the
results of all 148 oocysts measured were length 2398 microm (1859-3057) width 1855 microm (1373-2383)
and shape index 130 (101-160)
The same measurements were performed on 59 sporocysts from a single ferret (Noodle) and
results are as follows average length 1280 microm (898-1480) average width 738 microm (505-1028) and
average SI 176 (124-247) In one sample (Mohawk-2) sporozoites were visible free on the slide
68
Measurement of three of these provided an average length of 1068 microm (1044-1117) and an average
width of 341 microm (316-393)
332 Molecular characterization
Molecular characterization was successfully performed on oocysts from seven of 10 fecal
samples containing coccidial oocysts and FFPE tissue from nine of the 11 necropsy cases (see Table 32)
Attempts at amplification of DNA extracted from necropsy samples Z228-98 and Z137-14 using the
primer pairs listed in Table 31 were unsuccessful Similarly attempts at PCR and sequencing of DNA
extracted from fecal oocysts from two Toronto Zoo BFF Jenna and Ruckus were unsuccessful
Molecular identification results for the remaining samples are summarized in Table 32
Only one Eimeria species E ictidea was identified in all enteric coccidiosis cases diagnosed at
necropsy in both juvenile and adult BFF at the Toronto Zoo from 1998-2014 This same species was
identified in all Toronto and Louisville Zoo BFF fecal samples that were sequenced successfully (n=8)
with the exception of a single case from Louisville This Louisville ferret was identified as having a
rodent pseudoparasite (Eimeria species) in the submitted fecal sample the eimeriid pseudoparasite had
986 sequence identity at the mt COI locus to the murine coccidium Eimeria falciformis All sequences
generated for E ictidea exhibited 100 sequence identity at the mt COI and COIII loci
Novel nu 18S rDNA mt COI and mt COIII sequences were generated for E ictidea from both
geographic locations and deposited in GenBank (Accessions MF860826 MF860827 MF860823
MF860825 MF860822 MF860824) Sequences were compared to those previously published for related
eimeriid coccidia The nu 18S rDNA sequence from Eimeria ictidea isolated from the Toronto Zoo BFF
had 9736 identity (14 single nucleotide differences) to the previously published sequences from
isolates of E furonis from domestic ferrets (Mustela putorius furo) in Japan (GenBank AB239130 and
AB329724) and newly generated sequences from Canadian and European isolates (GenBank MF774678-
MF774680 see Chapter 2 and Figure 33) In contrast nu 18S rDNA sequence of E furonis from
domestic ferrets (see Chapter 2) showed 9953 to 100 identity (0 to 3 SND) to the Japanese
69
sequences Comparison of newly generated partial sequences of the mt COI region from E ictidea from
BFF to isolates of E furonis (GenBank MF774034-MF774036) from DF and E mephitidis (GenBank
KT203398) from the striped skunk (Mephitis mephitis) the only carnivore Eimeria sp for which a mt
COI sequence was previously available reveals only 9415 and 9084 sequence identity respectively
with these other Eimeria spp of carnivores (Figure 34)
34 DISCUSSION
This work presents the first nu 18S rDNA mt COI and mt COIII sequences (nu 18S rDNA -
MF860826 MF860827 mt COI - MF860823 MF860825 mt COIII - MF860822 MF860824) generated
from an intestinal eimeriid parasite of the BFF referred to here as E ictidea collected from multiple BFF
of different ages from two separate captive populations (Toronto Zoo Toronto Ontario Canada and
Louisville Zoo Louisville Kentucky USA)
The morphometric description of coccidial oocysts from BFF in this work are consistent with
previous descriptions of E ictidea from mustelids including BFF the Steppe polecat the European
polecat and domestic ferrets (Hoare 1927 Svanbaev 1956 Jolley et al 1994) Thus I propose the name
E ictidea for the enteric coccidium described from BFF reflecting the similarity in morphology host
species and location of infection in intestinal tissues yet recognizing the absence of species identification
by molecular techniques Molecular characterization of parasites that agree with the description of E
ictidea morphologically from various mustelid host species would allow not only for determination of
whether the parasites are conspecific but would also provide insight into the potential for cross-
transmission among related mustelid hosts
DNA extraction from FFPE samples allowed successful PCR and sequencing of small DNA
fragments (220 bp) in nine of the eleven cases in which the technique was attempted Age of the samples
did not appear to be the major factor associated with successful extraction of good quality DNA the two
samples for which it was unsuccessful were the most recent (2014) and oldest (1998) casesConsequently
it may be possible to use banked FFPE tissues from historic necropsy cases from other SSP institutions
70
and necropsies of wild-born or re-introduced ferrets to determine the identity of the coccidial species
underlying disease in these cases and to better characterize the disease in the greater BFF captive and wild
populations Williams et al (1988) were contacted regarding their historic FFPE samples but formalin
blocks were no longer available for these cases and thus comparisons could not be made Banked FFPE
samples were requested from other SSP institutions however the Convention on International Trade in
Endangered Species of Wild Fauna and Flora (CITES) restrictions on the international transport of DNA
from endangered species did not allow for sample acquisition during the period in which this research was
conducted
Evaluation of FFPE samples from the Toronto Zoo indicate that the same Eimeria species has
been implicated in deaths associated with enteric coccidiosis from 1999-2014 as well as episodes of
clinical disease in ferrets in the Toronto Zoo population from 2014-2016 Samples from coccidia-positive
BFF at the Louisville Zoo in 2016 also contained the same Eimeria species Finding the same parasite at
multiple SSP locations was expected because BFF are transferred among institutions on a yearly basis for
breeding and potential release Consequently these parasites have repeated opportunities to move
between institutions in infected hosts or on contaminated cage materials to become established at a new
location Furthermore the stress of transport and transfer to a new environment may precipitate shedding
of endemic coccidia and increase the risk of a coccidial outbreak this concern is reflected in the SSP
recommendations for prophylactic treatment of all BFF with anti-coccidial medication prior to shipment
(USFWS BFF Recovery Program 2017)
A single BFF from the Louisville SSP population not showing clinical signs consistent with
coccidiosis was identified as having a rodent Eimeria species in the submitted fecal sample
Morphometric characterization of oocysts in this sample was not performed due to the paucity of visible
oocysts however examination at 100times suggested that the oocysts in the sample were ovoid in shape and
of comparable size to oocysts identified in other BFF samples The finding of a rodent Eimeria in a BFF
fecal sample is not unexpected as whole rodents comprise a significant part of the captive BFF diet The
oocysts shed by the BFF were most likely acquired through ingestion of an infected prey item and thus
71
most likely represent pseudoparasitism Molecular characterization was however required to
differentiate this from a case of true enteric coccidiasis
Reports from the first captive BFF population derived from South Dakota indicate the presence
of an unidentified species of enteric coccidium (Carpenter and Hillman 1979) in this group before its
demise in 1979 No reports containing morphometric descriptions of the coccidia from this group were
found on literature review and all parasites of this group have been lost with their hosts All subsequent
reports on enteric coccidiosis in BFF are from ferrets derived from the second founder group from
Wyoming in the 1980s The frequent transfer of ferrets among SSP institutions within the captive
breeding program and to different release sites within North America would be expected to result in the
same Eimeria species being found in all populations The exception to this would be the potential for
cross-transfer of other eimeriid parasites to wild BFF from sympatric mustelid species such as the long-
tailed weasel (Mustela frenata) Jolley et al described two other species of enteric coccidia from this
second captive population in 1994 the first was similar to E furonis of domestic ferrets and the second a
large coccidian parasite of unknown genus Neither of these parasites was identified in the Toronto and
Louisville Zoo populations during the course of this study In order to determine whether these parasites
persist within the present-day BFF populations and their impact on this species more detailed
examination of coccidia-positive fecal samples from captive and wild BFF populations is recommended
Furthermore the molecular identification of enteric coccidia from historic and future necropsy samples of
wild and captive BFF could aid in determining the presence of and contribution to mortality events by
these additional coccidia species
ACKNOWLEDGEMENTS
The authors would like to recognize the Wildlife Health Centre staff at the Toronto Zoo for their
assistance with the collection of fecal samples from the BFF from 2014-2016 The authors would also like
to recognize the Louisville Zoo for their contribution of samples to this project Finally this project was
made possible through funding by the Toronto Zoo Residency Research Fund to DASAP and partial
72
funding from a Natural Sciences and Engineering Research Council of Canada (NSERC) Discovery
Grant (400566) to JRB
73
Table 31 Amplification primers used to sequence the nuclear 18S rDNA mitochondrial COI and COIII loci of Eimeria ictidea originating from
fecal and formalin-fixed paraffin embedded tissue samples from black-footed ferrets (Mustela nigripes) including anneal temeratures (Ta)
extension times and expected PCR product sizes
Figure 31 Nuclear and mitochondrial genetic loci targeted by primers listed in Table 31 and used to characterize Eimeria ictidea originating
from black-footed ferrets (Mustela nigripes)
Gene Target Primer Pairs Primer Sequence (5ʹ-3ʹ) Size (bp) Ta (degC) Anneal (sec) Reference
18SrDNA Sarco_18S_123F TATCAGCTTTCGACGGTAGTGTATT 1480 60 30 Ogedengbe et al (2016)
ERIB10_REV CTTCCGCAGGTTCACCTACGG
mt COI T_Eim_COI_272F CAATTCTAYGATGCCGCWTT 220 52 30 Chapter 2 (Table 21)
COX1-500R CATRTGRTGDGCCCAWAC Ogedengbe et al (2011)
COI-400F GGDTCAGGTRTTGGTTGGAC 800 52 60 El-Sherry et al (2013)
COI-1202R CAAKRAYHGCACCAAGAGATA El-Sherry et al (2013)
mt COIII WG-MT_4140F AGAAAACCTAAAATCATCATGT 1000 52 60 Ogedengbe et al (2015)
Eimeriid_CO3_799R AAGTGAGTTCGCATGTTTAC Ogedengbe et al (2015)
74
Figure 32 A+B) Features and cellular contents of Eimeria ictidea from a black-footed ferret
(Mustela nigripes) Legend Oocyst dotted thin white arrow = polar granule Sporocyst dotted thin
black arrow = Stieda body thick white arrow= sporozoite refractile body thick black arrow=
residuum scale bars as indicated C) Shape index length and width measurements of sporulated
oocysts of Eimeria ictidea from black-footed ferrets (Mustela nigripes) Legend times indicates the
mean Dotted oval indicates one standard deviation around the mean
10 microm 5 microm
75
Figure 33 Comparison of nuclear 18S rDNA sequence alignment of Eimeria ictidea from two black-footed ferrets (Mustela nigripes) to
newly generated (see Chapter 2) and published sequences of Eimeria furonis from domestic ferrets (Mustela putorius furo)
Figure 34 Comparison of mitochondrial cytochrome c oxidase subunit I sequence alignment of Eimeria ictidea from two black-footed
ferrets (Mustela nigripes) to sequences from other eimeriid parasites of carnivores
Identity
Eimeria mephitidis ndash KT2033981
Eimeria ictidea ndash MF860823 (Guanella ndash LZ)
Eimeria ictidea ndash MF860825 (Mystery ndash MTZ)
Eimeria furonis ndash MF774034 (Type 1)
Eimeria furonis ndash MF774035 (Type 2)
Identity
Eimeria ictidea ndash MF860827 (Guanella ndash LZ) Eimeria ictidea ndash MF860826 (Mystery ndash MTZ)
Eimeria furonis ndash MF774680 (Type 1) Eimeria furonis ndash AB3297241 (Type 1) Eimeria furonis ndash AB2391302 (Type 1)
Eimeria furonis ndash MF774678 (Type 2) Eimeria furonis ndash MF774679 (Type 2)
76
Table 32 Morphologic and molecular characterization of coccidia from fecal and FFPE necropsy samples from black-footed ferrets (Mustela
nigripes)
Sample ID Sample Source Year Age (years) Sex Source Microscopic Description^ Molecular Diagnosis
Z228-98 Toronto Zoo 1998 7 M FFPE rare asexual stages -
Z143-99 Toronto Zoo 1999 1 M FFPE sexual and asexual stages E ictidea
Z106-02 Toronto Zoo 2002 008 F FFPE rare asexual stages E ictidea
Z108-03 Toronto Zoo 2003 008 F FFPE sexual and asexual stages E ictidea
Z109-03 Toronto Zoo 2003 008 F FFPE sexual and asexual stages E ictidea
Z124-12 Toronto Zoo 2012 021 F FFPE sexual and asexual stages E ictidea
Z113-13 Toronto Zoo 2013 017 F FFPE sexual and asexual stages E ictidea
Z117-13 Toronto Zoo 2013 025 M FFPE sexual and asexual stages E ictidea
Z118-13 Toronto Zoo 2013 025 M FFPE sexual and asexual stages E ictidea
Z119-13 Toronto Zoo 2013 025 F FFPE sexual and asexual stages E ictidea
Z137-14 Toronto Zoo 2014 021 M FFPE
fresh feces
sexual and asexual stages
POS E cf ictidea
-
E ictidea
FERA_1 Toronto Zoo 2014 gt 1 MF fresh feces POS E cf ictidea E ictidea
Noodle Toronto Zoo 2015 1 M fresh feces POS E cf ictidea E ictidea
Ruckus Toronto Zoo 2016 3 F fresh feces POS -
Mystery Toronto Zoo 2016 1 M fresh feces POS E ictidea
Mohawk Toronto Zoo 2016 1 M fresh feces POS E cf ictidea E ictidea
Jenna Toronto Zoo 2016 1 F fresh feces POS -
Thrope Louisville Zoo 2016 3 M fresh feces NEG -
FloJean Louisville Zoo 2016 2 F fresh feces NEG -
Rigatoni Louisville Zoo 2016 5 M fresh feces POS rodent Eimeria
Guanella +kits Louisville Zoo 2016 2 015 F kits 2M3F fresh feces POS E cf ictidea E ictidea
Clive Louisville Zoo 2016 1 M fresh feces POS E ictidea
Legend FFPE= formalin fixed paraffin embedded Sex MF = samples from family groups containing both sexes - = PCR and sequencing unsuccessful ^ = life stages identified on
histologic section
= mitochondrial COI andor COIII sequencing results Presence (POS) or absence (NEG) of oocysts and morphological identification of oocysts based on measurements when
77
Table 33 Morphometric (length width shape index) characterization of Eimeria ictidea oocysts from fecal samples from black-footed ferrets
(Mustela nigripes)
Sample ID FERA - 1 ^ Z137-14 Noodle Mohawk -1 Mohawk - 2 Guanella Total
Number of oocysts 12 36 32 10 21 37 148
Length (microm) 2333 (2055-2583) 2456 (2111-2848) 2505 (2079-3008) 2779 (2590-3060) 2493 (2036-2822) 2139 (1859-2372) 2398 (1859-3057)
Width (microm) 1676 (1373-2180) 1835 (1643-2232) 1975 (1509-2360) 2253 (2092-2383) 1803 (1549-2017) 1751 (1610-1888) 1855 (1373-2383)
Shape index 135 (103-160) 134 (113-156) 127 (105-155) 124 (113-138) 139 (114-154) 122 (101-145) 130 (101-160)
Legend ^= mixed adult group = dam and kit group
78
CHAPTER 4 NATURAL HISTORY OF ENTERIC COCCIDIOSIS IN THE BLACK-
FOOTED FERRET (MUSTELA NIGRIPES)
ABSTRACT
Black-footed ferrets (BFF Mustela nigripes) the only native North American ferret species are
endangered throughout their former geographic range An intensive captive breeding program produces
animals to supplement re-established wild populations Coccidial enteritis is a major cause of morbidity in
young captive ferrets but the disease also affects adults Limited information is available on the
pathogenesis of intestinal coccidiosis in captive BFF and characterization of the natural history of the
disease for improved prevention and management is imperative The objectives of this research were to
determine morbidity and mortality rates in the Toronto Zoo captive BFF population as well as
characterizing the natural history of the disease in this species through evaluation of shedding patterns
body tissues affected pre-patent period and periods of enhanced host susceptibility to infection
Coccidia-associated mortality in BFF at the Toronto Zoo from 1997-2016 averaged 053 yearly in
adults (range 0-526) and 195 in juveniles (range 0-1667) Clinical signs and histologic lesions in
Toronto Zoo BFF were similar to those described in previous publications A seasonal influence on
oocyst shedding was identified in adult BFF and ferrets appeared to maintain persistent infection with E
ictidea shedding coccidia in multiple years A larger multi-institutional study is required to better
elucidate the natural history of enteric coccidiosis in this species
41 INTRODUCTION
Black-footed ferrets (BFF Mustela nigripes) are the only native North American ferret species
and are endangered throughout their former geographic range When the last remaining truly wild
population underwent serious decline as a result of disease the decision was made by the by United States
Fish and Wildlife Service to capture the remaining 24 animals and establish a captive breeding program
this occurred between 1985 and 1987 Only seven of the captured ferrets bred successfully and are the
79
founders of the current North American BFF population (USFWS BFF Recovery Program 2017) The
captive population which now numbers approximately 300 individuals is distributed among and managed
by six collaborating facilities these include the Toronto Zoo United States Fish and Wildlife Services
National Black-Footed Ferret Conservation Center National Zoorsquos Smithsonian Conservation Biology
Institute Louisville Zoo Cheyenne Mountain Zoo and the Phoenix Zoo (Black-footed Ferret Recovery
Implementation Team 2011 Santymire et al 2014) Since 1991 BFF have been released into
reintroduced into the wild at multiple sites within their former range and over 8000 BFF kits had been
produced in captive breeding facilities as of 2011(Black-footed Ferret Recovery Implementation Team
2011) Twenty-eight BFF reintroduction sites currently exist throughout North America however there
continues to be a need to support wild populations as only a four of the re-established groups are truly
self-sustaining
Enteric coccidiosis is recognized as a cause of significant morbidity and mortality in captive
breeding programs affecting both juvenile and adult animals (Bronson et al 2007 USFWS BFF
Recovery Program 2017) Two Eimeria species Eimeria ictidea and Eimeria furonis have been
identified from cases of entric coccidiosis in BFF (Jolley et al 1994) Jolley et al examined fecal samples
from both wild and captive BFF and provided detailed morphologic descriptions of the oocysts of both
Eimeria spp as well as descriptions of the intestinal pathology associated with infection Asexual and
sexual life stages of both of the aforementioned Eimeria spp were identified on histologic section within
the villar epithelium throughout the small intestine but were concentrated in the jejunum Intestinal
sections from BFF infected with E ictidea exhibited two morphologically distinct meronts one at the
villar tips which was larger and lacking in undifferentiated mass and the other at the base of the villi or
rarely in the intestinal crypts gamogony was predominantly observed at the villar tips and was noted
throughout the small intestine
80
Extraintestinal coccidia have also been reported from captive BFF at one facility (Jolley et al
1994 Williams et al 1988) The authors identified the presence of endogenous coccidial life stages in
histologic sections of respiratory tissue and in impression smears of the urinary bladder from BFF
diagnosed with canine distemper No subsequent reports of systemic coccidiosis in BFF have been
published or identified within the pathology database of the Toronto Zoo captive BFF population or by
the current SSP pathologist (Dr Michael M Garner personal communication)
Recent investigations into the etiologic agents of enteric coccidiosis in BFF at the Toronto Zoo
have identified a single Eimeria species associated with all cases of enteric coccidiosis and associated
mortality in juvenile and adult BFF from 1999-2016 Furthermore this pathogen was identified in fecal
samples based on morphologic and molecular characterization from adult and juvenile BFF in another
zoological collection (Louisville Zoo Kentucky USA) (see Chapter 3) This coccidium is
morphologically consistent with Hoarersquos original description of E ictidea (1927) and is referred to
henceforth as Eimeria ictidea
There is a significant information gap regarding the pathogenicity of E ictidea in BFF The
objectives of this research were to determine morbidity and mortality rates in the Toronto Zoo and
additional captive BFF SSP populations as well as characterizing the natural history of the disease in this
species through evaluation of shedding patterns body tissues affected pre-patent period and periods of
enhanced host susceptibility to infection
42 MATERIALS AND METHODS
421 Toronto Zoo BFF breeding program
At the Toronto Zoo black-footed ferret breeding program all adult ferrets are housed
individually with the exception of dams and kits After the birth of the kits dams are housed with their
offspring from whelp date until removal at approximately 4-6 months of age Routine monthly fecal
81
examinations (direct examination and flotation) are performed in house for all ferrets in the breeding
program based on SSP recommendations to evaluate for the presence of coccidia
422 Fecal oocyst evaluation
Family groups
From 2014-2016 daily fecal examination for coccidial oocysts was initiated for all group-housed
dams and kits In 2014 fecal samples were collected daily from all dams and kits from weaning (30 days
after whelping) to 72 days post whelping Based on 2014 data in 2015 this surveillance was extended
from weaning (35 days post whelping) to 135 days of age Furthermore fecal samples were collected
from the dam for an additional 14 days after removal of kits In 2016 no fecal samples were collected
from dam and kit groups at the Toronto Zoo but samples were submitted from one group of dam and kits
from another SSP population at the Louisville Zoo (Kentucky USA)
Adults
From 2015-2016 daily fecal samples were also collected from all adult ferrets identified as
shedding coccidial oocysts on their monthly routine fecal examination and from clinically ill BFF
Samples were collected for 10-14 days after initial positive sample identification In 2016 fecal samples
were also submitted from four coccidia-positive adult ferrets from the Louisville Zoo population samples
were collected for 7 days post initial identification of shedding
Individual fecal samples were analyzed via flotation using the McMaster method followed by
routine flotation in saturated salt solution (Dryden et al 2005) to determine the presence or absence of
oocysts and oocyst burden (oocysts per gram of feces OPG) Temporal trends in oocyst shedding were
monitored Coccidia-positive ferrets were evaluated visually on a daily basis for presence of clinical signs
consistent with infection Infected juvenile ferrets and adult ferrets were treated with oral ponazuril or
toltrazuril regardless of the presence of clinical signs as per the black-footed ferret SSP
recommendations Based on these recommendations ponazuril is typically administered orally at 30-50
82
mgkg once daily for 3-7 days until clinical signs have resolved or oocyst shedding has been significantly
reduced (USFWS BFF Recovery Program 2017)
423 Retrospective review of pathology records
The pathology records of the Toronto Zoo were searched from 1993-2016 for cases of BFF
diagnosed with enteric coccidiosis on histopathology For each case gross necropsy reports were
reviewed and slides of histologic sections from all submitted intestinal segments re-examined to confirm
the presence of sexual andor asexual life stages within the intestinal epithelium and describe the
histologic lesions associated with presence of the parasite life stages
424 Prospective modified necropsy protocol
During the study period 2014-2016 necropsy protocols for all BFF were modified to improve
detection of coccidial life stages and better to determine which portions of the intestinal tract were
affected The entire length of the intestine from duodenum to anus was measured and intestinal contents
were flushed with 12 mL of sterile saline into a sterile container Intestinal contents were preserved in
25 potassium dichromate solution (mixed 11 with intestinal contents vv) for molecular diagnostics
Paired 2-cm long intestinal samples were collected from all sections of small and large bowel duodenum
(1) jejunum (6) ileum (1) and colon (2) The eight small intestinal samples were collected at equal
distances from the pyloric sphincter to the beginning of the colon and the distance from the pylorus noted
for each Colon samples were taken at 25 and 75 of the length of the colon One sample from each
pair was preserved in Serra solution (100 ethanol (60 vv) 37 formaldehyde (30 vv) glacial
acetic acid (10 vv) and the second sample was frozen Representative tissues from all internal organs
as well as additional intestinal samples skin muscle and brain were also collected and preserved in 10
buffered formalin Histopathologic examination was performed on all tissues collected
83
425 Retrospective medical history review
Medical histories of all BFF held by the Toronto Zoo since the initiation of the SSP program were
reviewed for data on frequency of occurrence of shedding of coccidial oocysts in adults and juveniles as
well as any association of shedding with clinical signs and administration of anticoccidial treatment Data
was tabulated yearly for adult and juvenile ferrets to determine annual morbidity and mortality rates
associated with enteric coccidiosis Medical records and pathology reports were solicited from the other
SSP institutions to determine comparative morbidity and mortality rates associated with enteric
coccidiosis in BFF at other facilities Both morbidity and mortality rates were calculated as
incidenceattack rates with yearly adult population size or number of family groups (dam and kits) as the
denominator for morbidity rates and number of yearly deaths in each age class as the denominator for
mortality rates
43 RESULTS
431 Fecal oocyst evaluation and retrospective medical history review
Family groups
Fecal samples were collected from seven groups of dams and kits housed together at the Toronto
and the Louisville Zoos from 2014-2016 All data from first to last day of collection for all family groups
is listed in Appendix 1 selected pertinent data for each group is presented in Table 41 Five groups of
dams and kits were sampled in 2014 and one group in each of 2015 and 2016 Shedding occurred no
earlier than 55 days of kit-age in any of the groups and was identified from 55-81 days of age (Table 41
Table 42 Figure 41)
In 2014 fecal oocyst shedding was identified in three of the five surveyed groups In two of the
three groups (dams Poppy and Bumblefoot) changes to fecal colour and consistency were identified
concurrently with periods of oocyst shedding both groups shed higher numbers of oocysts than the other
dam and kit groups in 2014 and 2015 Both Poppy and Bumblefoot had had litters in the previous one and
84
two years prior to this study respectively based on medical record review these dams and their litters
were also diagnosed as shedding coccidial oocysts that were too numerous to count on direct exam and
fecal flotation Clinical signs in the previous years included dark tarry hemorrhagic or soft mucoid feces
and reduced appetite both groups received treatment with toltrazuril (Baycox Coccidiocide Solution
25 Bayer Inc Mississauga Canada) and trimethoprim sulfamethoxazole (Novo-Trimel Teva Canada
Ltd Scarborough Canada) (TMS) One of four kits from Poppyrsquos 2013 litter (Z113-13) died of enteric
coccidiosis three days after the group was diagnosed as shedding coccidial oocysts and the initiation of
treatment with TMS
In 2015 low grade fecal oocyst shedding (lt14 oocysts per gram of feces) without associated
clinical signs was noted in the Fiddlesticks group on three days during a seven day period from 63-69
days of kit-age and again for a single day at 128 days of kit-age The dam had been diagnosed and treated
for enteric coccidiosis in 2014 at which time she exhibited clinical signs of loose green feces to
hemorrhagic diarrhea lethargy and dehydration In 2016 she was diagnosed as shedding low numbers of
coccidia exhibited no clinical signs and did not receive treatment prior to resolution of shedding
In 2016 Guanella and kits shed oocysts over a nine day period and daily fecal oocyst shedding
ranged from 206 ndash 371714 OPG Combined treatment with ponazuril (first four days of shedding)
sulfadimethoxine injectable (first two days of shedding additional product information not available)
amoxicillin oral (first two days of shedding additional product information not available) penicillin
injectable (first two days of shedding additional product information not available) and subcutaneous
fluids (first two days of shedding additional product information not available) was administered to this
group Previous medical history was not available for this female for review
In 2014 and 2015 fecal oocyst shedding in all groups in the Toronto Zoo population started in the
three week period from the last week of July to mid-August In 2016 shedding was first identified in the
Louisville Zoo group in mid-July
85
Adults
Seven single-housed adult BFF (52 MF) were detected to have shed coccidia during the study
period (Table 43) Shedding periods lasted from 2-10 days and oocyst per gram counts ranged from 104
ndash 554274 (Table 44) Clinical signs were identified in four of the seven ferrets and consisted of
lossreduction of appetite (n=2) weight loss (n=1) lethargy (n=1) blood in feces (n=1) loose or runny
feces (n=3) soft mucoid feces (n=1) green colour of feces (n=2) Five of the seven adults received
treatment after detection of oocyst shedding two of which received treatment in the absence of clinical
signs Treatment consisted of oral toltrazuril in four cases toltrazuril in combination with trimethoprim
sulfamethoxazole in one case (Mohawk-A) and ponazuril and sulfadimethoxine (manufacturerrsquos
information not available) in one case (Clive) (Table 43)
Three of the adults in this study Mohawk Mystery and Jenna shed oocysts during multiple
different periods in 2016 Mohawk shed oocysts in May July and September of 2016 data from the first
two periods are reported in Tables 43 and 44 Mystery shed oocysts in June and July of 2016 Clinical
signs were observed only during the first shedding period and included poor appetite and hemorrhagic to
soft mucoid feces Jenna shed oocysts in July September and November of 2016 and again in February
and May of 2017 Although clinical signs were not detected in association with the initial period of
shedding in July 2016 (see Table 44) depressed mentation and hemorrhagic mucoid feces were identified
in the subsequent two shedding periods In both Mohawk and Jenna oocysts were not detected in feces on
multiple recheck and routine monthly fecal examination between shedding periods
Ruckus the fourth ferret shed low numbers of oocysts for two days in 2016 while housed alone
and had been reported to have shed oocysts during a 30 day period in 2014 while housed in a family
group with her kits Diarrhea to soft mucoid feces and loss of appetite were reported in 2014 Clinical
signs in 2016 consisted of small amounts of runny feces for a two day period two days after oocyst
shedding was no longer detected Treatment consisted of toltrazuril and TMS in 2014 and toltrazuril
again in 2016 Similarly Noodle shed low numbers of oocysts in July 2015 and had been identified as
shedding low numbers of oocysts during September of the previous year
86
The final ferret Rigatoni shed oocysts in feces in low numbers (54-732 OPG) over 5 days All
positive samples were pooled for molecular diagnostics and sequencing results showed 990 identity (7
single nucleotide differences SNDs) at the mt COI locus with a pair of rodent Eimeria species (ie
GenBank HM771682 JQ993704) (see Chapter 3) This ferret did not show clinical signs associated with
shedding and was not treated Previous medical history was not available for Clive the single adult ferret
from the Louisville Zoo
Based on sample collection dates and medical record review for the adult BFF for 2014-2016
shedding occurred during spring summer and fall with three ferrets shedding in May (Jenna Ruckus and
Mohawk) one ferret shedding in June (Mystery) three ferrets shedding in July (Mohawk Jenna Noodle)
one ferret shedding in August (Clive) two ferrets shedding in September (Noodle Jenna) and one ferret
shedding in November (Jenna)
432 Pathology
Eleven BFF with enteric coccidiosis were identified in the Toronto Zoo necropsy reports from
1993-2016 (Table 45) Cases were identified from 1998-2014 inclusive and all were confirmed by
histological re-evaluation (Chapter 3 Table 32) Both juvenile (n=9 36 MF) and adult ferrets (n=2 20
MF) were represented
Gross Pathology
Gross necropsy findings were similar across the 11 cases and included mucoid to fluid luminal
contents (n=7 636) beige to white pasty coating of the mucosal surface of the small intestine (n=6
545) and colon (n=4 364) gaseous dilation of intestinal segments (n=3 272) segmental enteritis
and hemorrhage (n=1 91) In one case Z228-98 no gross lesions were identified within the intestines
Impression smears of luminal contents or scrapings of intestinal mucosa were performed in four cases and
coccidia were identified in all four
Histopathology
87
Both sexual and asexual life stages were identified within the small intestinal segments in all
cases except Z228-98 and Z106-02 in which only rare asexual parasite life stages were identified (Table
45 Figure 42) Other histologic lesions seen in intestinal segments containing coccidia included
lymphoplasmacytic inflammation of the lamina propria (n=4) neutrophilic infiltration of the lamina
propria (n=2) villar necrosis (n=2) villar atrophy or blunting (n=3) and thrombi within the villar tips
(n=1)
Additional necropsy diagnoses included cholangiolar hyperplasia multiple hepatobiliary cysts
with suppuration renal adenocarcinoma apocrine gland adenocarcinoma (Z228-98) concurrent
clostridial enteritis (Z143-99) presumptive Salmonella sp septicemia (Z106-02) interstitial pneumonitis
(Z108-03) myocardial mineralization interstitial pneumonia and nephritis periportal hepatitis and
bacteremia (Z124-12) and suppurative esophagitis (Z137-14)
From 2014-2016 three black-footed ferrets were necropsied using the detailed protocol described
above Only one of the three cases Z137-14 was diagnosed with enteric coccidiosis based on
histopathology Two duodenal five jejunal two ileal and two colonic sections were collected at measured
lengths from the pylorus Parasite life stages were identified within the mucosal epithelium of all
intestinal segments extending from the distal duodenum (10-12 cm aboral to pylorus) through to the distal
colon (157-159 cm aboral from pylorus) The distal duodenal section contained asexual life stages only
with a single focus of epithelial cells containing meronts Sexual life stages (microgamonts
macrogamonts unsporulated oocysts) were identified within villar epithelial cells in all remaining
sections of the small intestine with numerous oocysts in the bowel lumen Mild lymphoplasmacytic
inflammation of the lamina propria was associated with the jejunal and ileal lesions and blunting of the
villi was identified within one jejunal segment The colonic sections contained small to moderate numbers
of sexual life stages identified within both superficial and deep crypt epithelium with occasional life
stages identified near the germinal cells Large numbers of oocysts and bacteria were identified within the
88
lumen of these colonic sections and both sections contained abscesses within the crypts The proximal
duodenum (0-2 cm) was the only section of the intestines not containing parasitic life stages
433 Morbidity and mortality
Annual morbidity rates for enteric coccidiosis at the Cheyenne Mountain Zoo and mortality rates
from enteric coccidiosis in Toronto Zoo BFF are summarized in Tables 46 and 47 During 2003-2016
yearly incidence of coccidiosis in adult BFF at the Cheyenne Mountain Zoo averaged 69 (range 0-
421) For family groups consisting of juvenile ferrets housed with their dams yearly incidence of
enteric coccidiosis averaged 115 (range 0-600)
From 1997-2016 coccidia-associated mortality in adult BFF at the Toronto Zoo averaged 053
yearly (range 0-526) with an average total mortality rate of 141 per year (range 0-526) For
juvenile ferrets (under 1 year of age) during the same period coccidia-associated mortality accounted for
an average of 133 of deaths yearly (range 0-100) with an overall average mortality rate of 170 per
year (range 0-3404) from all causes
Multiple additional SSP institutions provided partial medical and pathology data sets for use in
this study which were not sufficiently detailed to permit computation of morbidity and mortality rates for
those populations
44 DISCUSSION
The work described here supports previous clinical findings regarding the impact through both
morbidity and mortality associated with enteric coccidiosis in BFF No previous studies have been
undertaken to determine morbidity and mortality rates associated with enteric coccidiosis across BFF SSP
institutions
89
In a retrospective mortality study of captive BFF from 1984-2004 at the Smithsonianrsquos National
Zoological Park Bronson et al (2007) reported that gastrointestinal disease was the most common cause
of death in juvenile BFF (524) with 333 of juvenile mortality cases in the study caused by enteric
coccidiosis While the data is not directly comparable the findings reported here also reflect enteric
coccidiosis as a common cause of death in juveniles with increased mortality associated with the disease
compared with adult counterparts All Toronto Zoo mortalities in both juvenile and adult age classes for
which necropsy tissues were available have been attributed to infection with a single coccidia species
Eimeria ictidea (see Chapter 3)
Multiple SSP institutions provided partial medical and pathology data sets for use in this study
which were not sufficiently detailed to permit computation of morbidity and mortality rates for those
populations In future it would be useful to determine whether morbidity and mortality rates associated
with enteric coccidiosis vary among SSP institutions as this may allow for improved identification of
host parasite and environmental factors that increase risk
Clinical signs reported here are consistent with those described from both BFF and domestic
ferrets with enteric coccidiosis (Sledge et al 2011 Santymire et al 2014 USFWS BFF Recovery
Program 2017) Changes to fecal colour and consistency were the most common abnormalities identified
at the time of first detection of oocyst shedding While clinical signs in the cases described here do not
always correlate directly with the quantity of oocysts shed individuals shedding higher number of oocysts
showed clinical signs more frequently than those shedding lower numbers
In this study oocyst shedding from single-housed adult BFF ranged from 104 ndash 554274 oocysts
per gram Daily fecal samples produced from individual adult ferrets range in size from approximately 1-
16 grams In light of these findings during peak shedding from ~5times105 to 9times106 oocysts can be shed into
the environment in one day providing a massive infective dose The large numbers of oocysts shed
90
combined with confinement in a small enclosure space and hardiness of Eimeria oocysts in the
environment would be expected to markedly increase risk of infection in captive BFF
Oocyst shedding from family groups ranged from 0 ndash 371714 OPG The wide variability seen in
OPG counts between days as seen in Figure 41 and Table 41 may be accounted for by the staggered
initiation and resolution of shedding by different ferrets It is most likely that the source of infection in
these family groups is shedding by the dams some of which were identified to shed in multiple years
although environmental contamination cannot be excluded
Shedding in the adult ferrets was clustered during particular time periods specifically May July
and September If shedding is associated with stress and immunosuppression activities such as breeding
whelping electro-ejaculation of male ferrets and shipmenttransfer could act as stressors Whelping
which occurs primarily in May and June could also act as a stressor to other ferrets in the facility either
through social cues or as there would be associated changes in husbandry protocols The September
cluster could be associated with the transfer of ferrets between institutions kits are pulled during this time
and adult ferrets are moved among institutions thus changing the population dynamics of each SSP site A
large cluster of shedding was recorded in July With the exception of weaning of kits no other major
stressors are expected to occur during this time thus the increase in shedding by adults in this group
cannot be easily explained Interestingly shedding was not identified in ferrets from March through April
which is the typical breeding season and when ferrets are introduced for breeding a presumably stressful
time Shedding was not noted from December through April which could reflect reduced environmental
burdens due to low humidity levels as would be expected in a Nordic climate during the winter (which
would kill oocysts and thus block transmission) or may be consistent with reduced stress during this
period The results from single-housed adults are in contrast to the dam and juvenile ferret groups in
which oocyst shedding appeared to be correlated to a period of 55-81 days of kit age These results are
consistent with reports from other facilities of increased incidence of shedding by kits after 70 days of age
(USFWS BFF Recovery Program 2017)
91
Retrospective and prospective review of histologic sections of intestines from affected BFF at the
Toronto Zoo showed the presence parasitic life stages in epithelial cells of both the small and large
intestines Neither Hoare (1935b) nor Jolley et al (1994) mentioned the presence of parasites in the large
intestine of experimentally infected domestic ferrets or naturally infected BFF respectively In the study
described here asexual and sexual life stages were identified within the epithelial cells of the small
intestinal villi from base to tip and were most numerous in jejunum This matches the description by
Jolley et al (1994) however Hoare (1935a b) found E ictidea to be present primarily in the villar tips
Jolley et al (1994) also described two morphologically distinct meronts of E ictidea within the small
intestinal sections one at the villar tip that was larger and lacking in undifferentiated mass and the other
at the base of the villi or in the intestinal crypts these findings were not echoed in this study as
merogonic stages were identified throughout the intestinal epithelium from villus to the base of the crypt
and no visual differences between meronts in any location were identified Hoare (1935a b) also
described a resulting annular constriction of the villus separating the affected and non-affected segments
this constriction was neither seen in the cases described here nor mentioned by Jolley et al (1994)
Whether these differences result from E ictidea from BFF and E ictidea from domestic ferrets being
different parasites or from differences in tissue tropism of a single parasite in two different hosts cannot
be ascertained from the available information
Histologic lesions such as necrosis hemorrhage villar atrophy and inflammation associated with
the presence of parasitic life stages were rare These changes are normally elicited by the host immune
system (inflammation) and the parasite (cellular rupture to release life stages resulting in hemorrhage and
necrosis) in response to infection In light of the fact that acute death occurred in a number of these ferrets
(Z113-13 Z117-13 Z118-13 Z119-13 Z137-14) in the absence of secondary disease processes and
with the intestinal epithelium intact but containing myriad parasitic life stages an alternative mechanism
for mortality associated with the infection must be proposed It is possible that these parasites elaborate
exotoxins during their life cycle and when at high density result in sudden death of the host with minimal
92
tissue changes The presence of parasitic life stages occupying the majority of both small and large
intestinal epithelial cells could also potentially impair fluid and protein movement in and out of the
mucosa however clinical signs associated with malabsorptive diarrhea were not identified in any of these
cases The presence of bacteria within the blood or other organ tissues was not identified in any cases and
consequently sepsis is unlikely to be the cause of death
Black-footed ferrets appear to maintain persistent infection with E ictidea Adult BFF in the
Toronto Zoo population shed coccidia in multiple years and in two adult ferrets multiple times in the
same year While the coccidia seen in all cases were not confirmed as E ictidea using molecular
techniques morphologic similarities and a lack of additional Eimeria spp identified on molecular work
undertaken suggest that only one species of parasite is and has been present in the collection Two dams
that had been identified as infected based on routine fecal screening in previous years presumably acted as
the source of infection to their litters of kits in multiple years While continued environmental
contamination cannot be ruled out these findings imply a failure of the immune response of the BFF to
clear infection with E ictidea or even to protect against sufficient replication of organisms to result in
clinical disease
Based on clinical experience and review of the literature BFF appear to be much more sensitive
to infection with E ictidea compared with their domestic counterparts In domestic ferrets subclinical
shedding of oocysts appears to be the most common with rare reports in the literature of overt disease
and that only in juveniles (Blankenship-Paris et al 1993 Abe et al 2008) However a single report exists
of three separate clinical outbreaks of Eimeria furonis infection in domestic ferrets under intensive
management with increased morbidity and mortality affecting all ages classes (Sledge et al 2011) The
role of genetics in the apparent increased susceptibility of BFF to enteric coccidiosis is unknown but the
current captive BFF population is derived from seven founders and inbreeding depression or familial
genetic susceptibility may play a role in their increased susceptibility to disease caused by E ictidea
93
Black-footed ferrets diagnosed with enteric coccidiosis during the course of the study were
treated with either ponazuril or toltrazuril sulfonamide drugs or often a combination of the two groups
of therapeutic agents Toltrazuril and ponazuril are triazine coccidiocides with proven efficacy against
both asexual and sexual life stages of mammalian and avian Eimeria spp (Mehlhorn and Aspock 2008)
The sulfonamides are antimicrobial drugs that exhibit coccidiostatic or coccidiocidal effects depending on
dose they act by blocking folate synthesis and have effects on first and second generation meronts
(asexual life stages) as well as potentially acting on sexual life stages (Mehlhorn and Aspock 2008)
Based on the limited data available from this study and the fact that treatment was initiated in almost all
adult BFF and family groups at the time of oocyst detection regardless of the presence of clinical disease
the effects of treatment on duration of clinical signs cannot be effectively evaluated It appears
subjectively that adult ferrets treated with toltrazuril and in one case a combination of toltrazuril and
TMS showed reduction in oocyst shedding after 3-5 days of oral anti-coccidial therapy (see Table 43)
The effects of treatment with either sulfonamides or triazines would be expected to reduce oocyst
shedding consequently the duration and amount of oocyst shedding reported in this study may not
accurately characterize the natural course of disease
Perceived resistance to sulfa drugs has been reported from multiple SSP facilities In light of this
and their potential negative effects on ferret reproduction (eg prevention of embryo implantation in the
uterus and impairment of sperm development) sulfonamides are no longer recommended by the SSP for
treatment of coccidia in this species (USFWS BFF Recovery Program 2017) The frequent and
widespread use of triazines in the management of enteric coccidiosis in BFF presents a risk for
development of resistance to this drug class in the future Neither pharmacokinetic (PK) nor
pharmacodynamic (PD) studies have been published to validate the dose and frequency of dosage in
either class of drugs in BFF and consequently it is unclear whether this perceived failure of some ferrets
to respond to treatment is based on true resistance versus inappropriate dosing The only work evaluating
ponazuril in BFF evaluated serum levels of ponazuril after a single oral dose of 50 mgkg and reported
94
therapeutic levels for 10 days after administration (USFWS BFF Recovery Program 2017) No
information was provided on number or age of ferrets that participated in the study or on how the
determination of what were therapeutic levels was made Furthermore as life cycles of the coccidia
affecting BFF are limited to the gastrointestinal tract and do not exhibit tissue stages the validity and
usefulness of assessing blood levels of ponazuril in determining appropriate dosage and dose schedules is
questionable Further work to determine the PK and PD of triazines in ferrets is warranted to provide safe
and efficacious treatment and to reduce the risk of development of resistance Furthermore the creation
and validation of a model for enteric coccidiosis in a related species would allow for in vivo studies of
drug resistance
95
Figure 41 Oocyst per gram counts and shedding period of Eimeria ictidea from black-footed ferret
(Mustela nigripes) family groups from 2014-2016
0
50000
100000
150000
200000
250000
300000
350000
400000
29
33
37
41
45
49
53
57
61
65
69
73
77
81
85
89
93
97
10
1
10
5
10
9
11
3
11
7
12
1
12
5
12
9
13
3
13
7
14
1
14
5
14
9
OP
G
Age of Kits
Poppy Bumblefoot Calico Aubrey Ruckus Fiddlesticks Guanella
96
Figure 42 Small intestinal epithelium of a black-footed ferret (Mustela nigripes) containing sexual
life stages of Eimeria ictidea Legend Solid black arrow = oocyst Hatched arrow = macrogamont
Outlined arrow = microgamont Hematoxylin and eosin staining scale bar = 25 microm
25 microm
97
Table 41 Shedding of oocysts of Eimeria ictidea in black-footed ferret (Mustela nigripes) dam and kit
family groups from 2014-2016
Collection Year 2014 2014 2014 2014 2014 2015 2016
Age of kits (days) Poppy Bumblefoot Calico Aubrey Ruckus Fiddlesticks Guanella^
29 - - - 0 - - -
30 - 0 - 0 - - -
- - - - -
34 0 0 - 0 0 - -
35 0 0 - 0 0 0 -
- -
46 0 0 - 0 0 0 -
47 0 0 - 0 0 0 -
48 0 lt14 - 0 0 0 -
- -
53 0 0 - 0 0 0 +
54 0 0 - 0 0 0 +
55 0 0 - 0 0 0 3717146
56 0 0 - 0 - 0 1084436
57 0 0 - 0 - 0 64133
58 0 324 - 0 0 0 20654
59 0 0 - 0 0 0 0
60 0 446688 - 0 0 0 0
61 0 934828 - 0 0 0 41111
62 0 530469 - 0 0 0
63 0 1617131 - 0 0 lt14
64 0 8396357 - 114 0 0
65 0 234876 - 5368 0 lt14
66 0 374625 - 9455 0 0
67 lt14 2311575 - 39579 0 -
68 0 603563 - 125051 0 0
69 - 4103036 - 27067 0 lt14
70 2470327 7759974 - 0 0 0
71 1073085 - 1784 0 0
72 3430966 - 0 0 0
73 4459536 - 7326 0 0
74 4842212 - 2 0 0
75 3627307 - 0 0 0
76 5801885 - 0 0 0
77 5994 0 0 0 0
78 589835 0 0 0
79 24815 0 0 0
80 144016 0 0 0
81 97862 0 0 0
82 0 0 0
127 0 0
128 0 2843
136 0 0
150 0
Numbers of Oocysts Shed (oocysts per gram of feces)
Dam Identity
Legend lt 14 = oocyst positive samples with less than 14 oocyst per gram of feces - = no sample recorded for this date underline = last
sampling date + = coccidia present but OPG count not performed = Toronto Zoo ferret ^ = Louisville Zoo ferret thick outer border =
days treatment was received = range of sequential dates between previous and subsequent number during which OPG counts were
performed and samples contained 0 oocysts
98
Table 42 Summary of epidemiologic data for family groups of black-footed ferrets (Mustela nigripes)
shedding oocysts of Eimeria ictidea
Dam Identity
Poppy
2014
Bumblefoot
2014
Aubrey
2014
Fiddlesticks
2015
Guanella
^
2016
Number of kits 7 5 3 4 5
Kit age (days) at time of
shedding 70-81 58-70 64-73 63-69 54-61
Shedding period (days) 15 13 11 7 9
OPG min 0 0 0 0 206
OPG max 48442 83963 2707 lt14 371714
Clinical signs YES YES NO NO NO
Treated - YES YES NO YES
Legend = Toronto Zoo ferret ^ = Louisville Zoo ferret OPG = oocysts per gram of feces
- = missing data
Table 43 Shedding of oocysts of Eimeria ictidea in single-housed adult black-footed ferrets (Mustela nigripes) 2015-2016
Numbers of Oocysts Shed (oocysts per gram of feces)
Collection
Year 2015 2016 2016 2016 2016 2016 2016
Ferret Identity Noodle Ruckus Mohawk-A Mohawk-B Mystery Jenna Clive^
Age (years) 1 3 1 1 1 1 1
1084058 0 + + + + +
+ 0 42307650 6286676 + 183150 554274
857808 16650 12805238 7777929 + 215710 377920
1604894 16650 309690 139860 + 0 25808
377042 0 599400 119880 + - 37294
554445 0 34688 385579 117920 0 5363
26640 0 16650 0 0 0 7500
0 10406 0 0 0 1090
0 20813 0 0 0
- 0 1761 0
0 0 0 0
- 0
0
0
Legend lt 1 = oocyst positive samples with less than 1 oocyst per gram of feces underline = last sampling date + = coccidia present but OPG
count not performed = Toronto Zoo ferret ^ = Louisville Zoo ferret thick outer border = days treatment was received
100
Table 44 Summary of epidemiologic data for single housed adult black-footed ferrets (Mustela nigripes) shedding oocysts of Eimeria ictidea
Noodle Ruckus Mohawk-A Mohawk-B Mystery Jenna Clive^
Sex M F M M M F M
Age (years) 1 3 1 1 1 1 1
Shedding period (days) 7 2 9 6 10 4 8
OPG min 266 166 104 1199 1761 1831 1090
OPG max 10840 166 423076 77779 - 2157 554274
Clinical signs YES YES NO NO YES NO YES
Treated NO YES YES YES YES YES YES
Legend = Toronto Zoo ferret ^ = Louisville Zoo ferret M = male F= female OPG = oocysts per gram of feces - = missing data
NOTE Mohawk-A and Mohawk-B refer to two separate episodes of oocyst shedding by the same ferret
101
Table 45 Histologic findings from necropsies of black-footed ferrets (Mustela nigripes) with enteric coccidiosis
Number of Sections Affected
Ferret
ID Year
Age
(years) Sex Coccidia in Intestinal Sections Small Intestinea Large Intestinea
Z228-98 1998 7 M rare asexual stages S - 01 A - 11 S - 01 A - 01
Z143-99 1999 1 M sexual and asexual stages S - 24 A - 24 S - 03 A - 03
Z106-02 2002 008 F rare sexual stages S - 14 A - 04 S - 01 A - 01
Z108-03 2003 008 F sexual and asexual stages S - 12 A - 12 S - 01 A - 01
Z109-03 2003 008 F sexual and asexual stages S - 23 A - 23 S - 11 A - 11
Z124-12 2012 021 F sexual and asexual stages S - 24 A - 04 none
Z113-13 2013 017 F sexual and asexual stages S - 77 A - 07 none
Z117-13 2013 025 M sexual and asexual stages S - 56 A - 56 S - 11 A - 01
Z118-13 2013 025 M sexual and asexual stages S - 56 A - 46 S - 11 A - 01
Z119-13 2013 025 F sexual and asexual stages S - 45 A - 35 none
Z137-14 2014 021 M sexual and asexual stages S - 1011 A - 911 S - 22 A - 02
Legend a= xn where x is number of sections containing sexual or asexual lifestages n is the number of sections examined S = sexual life
stages A= asexual life stages
102
Table 46 Yearly incidence of coccidial infection in black-footed ferrets (Mustela nigripes) at the
Cheyenne Mountain Zoo
Cheyenne Mountain Zoo
Year Adult Family
2003 116 (625) -
2004 819 (4211) -
2005 021 (000) 14 (2500)
2006 021 (000) 07 (000)
2007 023 (000) 08 (000)
2008 224 (833) 14 (2500)
2009 025 (000) 06 (000)
2010 326 (1154) 07 (000)
2011 125 (400) 09 (000)
2012 125 (400) 08 (000)
2013 028 (000) 05 (000)
2014 430 (1333) 09 (000)
2015 035 (000) 35 (6000)
2016 - 27 (2857)
Mean annual () 689 1155
Legend - = missing data xn= where x is the number of ferrets shedding coccidial oocysts and n is the
total number of adult ferrets or family groups in a given year () = incidence expressed as a percentage
103
Table 47 Yearly mortality rate and incidence of mortality associated with coccidial infection in
black-footed ferrets (Mustela nigripes) at the Toronto Zoo
Total Mortality
Coccidia Other Causes
Year Kit Adult Kit Adult
1997 015 (000) 023 (000) 315 (2000) 423 (1739) 1998 038 (000) 119 (526) 838 (2105) 919 (4734) 1999 047 (000) 119 (526) 1647 (3404) 119 (526) 2000 034 (000) 015 (000) 434 (1176) 315 (2000) 2001 032 (000) 016 (000) 532 (1563) 116 (625) 2002 150 (200) 020 (000) 450 (800) 220 (1000) 2003 227 (741) 018 (000) 327 (1111) 118 (555) 2004 020 (000) 016 (000) 620 (3000) 216 (1250) 2005 016 (000) 015 (000) 416 (2500) 215 (1333) 2006 030 (000) 016 (000) 230 (667) 016 (000) 2007 019 (000) 015 (000) 419 (2105) 215 (1333) 2008 034 (000) 016 (000) 1134 (3235) 316 (1875) 2009 017 (000) 016 (000) 017 (000) 116 (625) 2010 017 (000) 016 (000) 317 (1765) 316 (1875) 2011 011 (000) 016 (000) 111 (909) 216 (1250) 2012 111 (909) 017 (000) 111 (909) 317 (1765) 2013 424 (1667) 017 (000) 424 (1667) 317 (1765) 2014 126 (384) 017 (000) 326 (1154) 217 (1176) 2015 04 (000) 017 (000) 04 (000) 217 (1176) 2016 011 (000) 017 (000) 211 (1818) 117 (588)
Mean annual () 195 053 1594 1359
Legend xn= where x is the number of ferrets that died with coccidial infection or of other
causes and n is the total number of adult ferrets or kits in a given year () = incidence expressed
as a percentage
104
CHAPTER 5 EVALUATING THE DOMESTIC FERRET (MUSTELA PUTORIUS FURO) AS
AN EXPERIMENTAL MODEL FOR ENTERIC COCCIDIOSIS IN THE BLACK-FOOTED
FERRET (MUSTELA NIGRIPES)
ABSTRACT
The purpose of this study was to determine whether the domestic ferret (Mustela putorius furo) is
susceptible to an isolate of Eimeria ictidea originating from black-footed ferrets (BFF Mustela nigripes)
and thus could act as a suitable experimental model in which to investigate the pathogenesis and
management of this disease A pilot study was performed with 10 male intact juvenile domestic ferrets
Ferrets were administered an oral inoculum containing either a high dose (1 times 106 oocysts) moderate
dose (5 times 104 oocysts) or saline control and observed for shedding of oocysts and development of
clinical signs Seven of ten ferrets developed patent infection all of which had received the high dose
inoculum The prepatent period was 7-9 days and duration of shedding varied from 1-7 days Clinical
signs were identified in six of the seven infected ferrets and were consistent with those previously
described for enteric coccidiosis in domestic and BFF Parasite life stages were identified within the
intestines of four of the seven ferrets with patent infection and were limited to the distal jejunum and
ileum The demonstrated ability to produce patent infections in domestic ferrets following oral inoculation
of a high dose of E ictidea (1 times 106 oocysts) isolated from BFF provides an avenue for future
experimental investigations into the control and treatment of enteric coccidiosis in this endangered
species
51 INTRODUCTION
Black-footed ferrets (BFF Mustela nigripes) are one of only three ferret species worldwide
While formerly distributed throughout the North American prairies black-footed ferrets were declared
extinct in the wild in the 1980s Since 1986 a multi-institutional consortium has been breeding this
species in captivity with reintroductions back into the wild within their historic range in selected locations
in Canada the USA and Mexico Introduced colonies of BFF are present in Arizona Colorado Kansas
105
Montana New Mexico South Dakota Utah Wyoming and Chihuahua (Mexico) Reintroduction
attempts in Saskatchewan Canada have been unsuccessful to date
Multiple infectious diseases pose a significant risk to the captive breeding and post-release
survival of BFF including distemper and sylvatic plague (Santymire et al 2014 USFWS BFF Recovery
Program 2017) Coccidiosis is a recognized cause of juvenile and adult morbidity and mortality in captive
breeding programs and can result in significant losses (Bronson et al 2007 Santymire et al 2014
USFWS BFF Recovery Program 2017) The effect of the disease on wild populations is unknown
Clinical signs of coccidiosis include mucoid to hemorrhagic diarrhea abdominal discomfort lethargy
appetite loss vomiting and dehydration Recent investigations into diseases affecting BFF at the Toronto
Zoo have identified a single Eimeria species E ictidea associated with all cases of enteric coccidiosis in
juvenile and adult BFF from 2014-2016 (see Chapter 3) This same Eimeria species was identified
retrospectively as the cause of juvenile and adult mortalities in previous years (1999 through 2014
inclusive) (Chapters 3 and 4) Furthermore this pathogen was identified in fecal samples based on
morphologic and molecular characterization from adult and juvenile BFF in another zoological collection
(Louisville Zoo Kentucky USA) (see Chapter 3)
Enteric coccidiosis also occurs in domestic ferrets (Mustela putorius furo) with three
morphologically distinct species of coccidia Eimeria ictidea Eimeria furonis and Isospora (=
Cystoisospora) laidlawi Both of the aforementioned Eimeria species have been identified in black-footed
ferrets based on morphologic criteria but molecular characterization was needed to confirm whether the
same species of parasite infects both ferret species (see Chapter 2 and 3) To this end nuclear and
mitochondrial sequences for E furonis and for I=(C) laidlawi were generated (Chapter 2) expanding the
existing limited sequence data from the nuclear 18S rRNA locus of Eimeria furonis Molecular
characterization of E ictidea from domestic ferrets was not possible because samples containing this
parasite were not available for study consequently it is unclear whether the same coccidium affects both
domestic and black-footed ferrets
106
There is no published information describing the pre-patent periods and pathogenicity of enteric
coccidia in BFF and given the conservation status of the BFF experimental work cannot be undertaken
in the natural host The purpose of this study was to determine whether the domestic ferret is susceptible
to E ictidea isolated from BFF if susceptible the domestic ferret could act as a suitable experimental
model in which to investigate the pathogenesis prevention and treatment of coccidiosis caused by E
ictidea
52 MATERIALS AND METHODS
521 Animal care
Ten juvenile male intact ferrets of 48 (n=6) or 50 (n=4) days of age were obtained from a
commercial source (Marshall BioResources North Rose New York USA) and were housed in the
University of Guelph Central Animal Facility Isolation Facility Ferret weights on arrival ranged from
334-475 g (average= 3928 g) All ferrets were housed individually in wire bottom cages of 813 times 1117
times 457 cm size and were divided in equal numbers between two non-adjoining rooms They received ad
libitum access to Envigo Teklad Certified Global Ferret Diet (Madison Wisconsin USA) and water
changed daily Room temperature was maintained at 18-21 degC and a 16 hour light 8 hour dark
photoperiod was provided All personnel working with the ferrets were required to wear personal
protective equipment including disposable facemasks gloves gowns and bouffant caps Shoes were
provided for use in each room This study was carried out in accordance with the recommendations in the
Canadian Council on Animal Care guidelines The protocol was approved by the Animal Care Committee
of the University of Guelph (Animal Use Protocol 3289) and by both the Animal Welfare Committee
and Animal Care and Research Committee of the Toronto Zoo
An initial physical examination and blood collection were performed on each ferret by the
principal investigator (ARP) one day after arrival to assess health status prior to enrollment in the study
Ferrets were mask induced with isoflurane (Isoflurane USP Fresenius Kabi Richmond Hill Ontario) in
107
oxygen placed on a heat disc (SnuggleSafe Lenric C21 Ltd Littlehampton United Kingdom) weighed
examined and blood was collected from the jugular vein for routine CBC and biochemical profiles
All ferrets subsequently underwent an acclimation period of two weeks During this time fecal
samples were collected daily from each ferret and examined for the presence of coccidial oocysts using a
standard salt flotation technique (Dryden et al 2005) to ensure that all individuals were free of coccidia
prior to initiation of experimental work Any ferret positive for coccidia was to be removed from the
study
522 Oocyst preparation
Oocysts used for inoculation originated from fecal samples from two naturally infected BFF These
samples were stored in potassium dichromate for four weeks prior to oocyst purification and use in this
infection trial Stored fecal samples were mixed with distilled water and passed through a small sieve to
remove debris The strained contents were transferred to a 50 mL conical vial and topped up to 50 mL
with additional distilled water Samples were centrifuged (Sorvall ST40R Centrifuge Thermo Scientific)
at 2800 rpm (1315 timesG) for 10 minutes at 12 degC A drop of supernatant was evaluated microscopically at
100times for the presence of oocysts If oocysts were observed the supernatant was poured off into a second
50 mL conical tube and again topped up to 50 mL with distilled water and re-centrifuged under the same
conditions Otherwise the supernatant was discarded The pellets from both the first and second tubes
were combined with saturated salt solution at a 14 ratio by volume Oocysts were floated in the salt
solution by centrifugation at 1500 rpm (377 timesG) for 10 minutes at 12 degC The top 5 mL of supernatant
were collected and transferred to a clean 50 mL conical tube topped up to 40 mL with distilled water and
washed via centrifugation at 2800 rpm (1315 timesG) for 10 minutes After the wash step the supernatant
was again checked for presence of oocysts and discarded if no oocysts were observed The pellet was
collected and the presence of oocysts confirmed by examination of a drop placed on a clean glass slide at
100times Once verified the contents of the pellet of concentrated oocysts was placed in a 250 mL storage
container and mixed with approximately 200 mL of sterile saline (09 sodium chloride Hospira
108
Montreal Quebec) prior to storage for two to four weeks at 4 degC until inoculation Prior to inoculation a
McMaster count was performed to determine the number of oocysts per mL in order to determine
appropriate volume of inoculum
523 Experimental infections
Part 1
Five ferrets were randomly assigned to each of the control and infection groups After the
acclimation period on day 0 four ferrets in the infection group were inoculated orally with a high dose
oocyst suspension (1 times 106 oocysts in 025 mL of saline) mixed into 1 mL of FerreTone Skin amp Coat
Supplement (United Pet Group Inc Blacksburg Virginia USA) a fifth ferret was inoculated with a
moderate dose oocyst suspension (5 times 104 oocysts in 025 mL of saline) in the same volume of FerreTone
Ferrets in the control group were inoculated with a placebo (025 mL of saline) in 1 mL of FerreTone
Inoculation was performed by offering the oocyst suspension or placebo to the ferrets in a plastic
container
Fecal samples were collected daily from each inoculated ferret for 14 days post-inoculation
Samples were analyzed via fecal flotation using the McMaster method followed by routine flotation in
saturated salt solution (Dryden et al 2005) to determine the presence or absence of oocysts and oocyst
burden (oocysts per gram of feces [OPG])Temporal trends in oocyst shedding were monitored Ferrets
were evaluated visually twice daily for the presence of clinical signs of coccidial disease The first of
every two ferrets identified to shed oocysts was to be humanely killed at the time of peak oocyst shedding
(ie the first day that fecal oocyst counts remained static or declined) and necropsied to confirm the
presence of and describe parasitic replication in the intestinal mucosa Any remaining animals that shed
oocysts were to be monitored throughout the 14 day period following inoculation in order to determine
the duration and intensity of oocyst shedding for these individuals the total number oocysts shed during
patency was determined
109
Part 2
All ferrets from the infection group that did not shed oocysts during Part 1 (n=4) and all but one
ferret from the previous control group (n=4) were orally inoculated with the high dose oocyst suspension
(1 times 106 in 1 ml of saline) mixed with an equal volume of FerreTone Consequently between phases 1
and 2 all but one ferret were inoculated at least once with the BFF coccidia in order to increase
experimental animal numbers and determine if ferret age played a role in susceptibility to infection
One ferret from the previous control group was inoculated with a lower dose (2 times 105 oocysts in
075 mL of 09 saline mixed with 1 mL FerreTone) of oocysts that had been collected from the single
domestic ferret that shed in Part 1 oocysts were purified as described above for the initial inocula Fecal
collection and analysis were performed as previously As in Part 1 one in every two ferrets sequentially
identified to be shedding oocysts in feces was killed humanely at the time of peak shedding and a
complete necropsy examination performed The remainder of the ferrets observed to be shedding were
monitored for the full 14 days of the trial after which they were killed humanely and necropsied and total
number of oocysts shed during patency was determined All ferrets that did not shed coccidial oocysts
during the infection trial were rehomed at the end of the trial
524 Animal welfare
Ferrets were evaluated twice daily for development of clinical signs of coccidial disease and any
animal showing clinical disease was to be treated as determined by a veterinarian with supportive care
including fluid therapy A grading system for clinical signs including intervention points and removal
criteria was created for use during daily evaluation (see Appendices 3 and 4) Animals whose clinical
signs could not be ameliorated without the use of specific anticoccidial therapy were to be euthanized
Should the inoculation in Part 1 have resulted in clinical disease that required extensive treatment andor
necessitated euthanasia a lower number of oocysts would be used for subsequent inoculation in Part 2
Ferrets to be euthanized were anesthetized by mask induction with isoflurane in oxygen a 1 mL blood
110
sample was collected from the cranial vena cava and then an intracardiac dose of potassium chloride (2
mEqkg) was administered
525 Hematology
Blood was collected from all ferrets under isoflurane anesthesia at the time of pre-trial health
examination and again at the time of humane killing Blood was collected via jugular venipuncture
initially due to the small size of the ferrets at arrival and then by cranial vena cava venipuncture or
cardiocentesis prior to euthanasia Complete blood count and serum biochemistries were performed by the
Animal Health Laboratory of the University of Guelph Guelph Ontario
526 Morphologic and molecular characterization
Morphologic and molecular characterization of oocysts shed by the domestic ferrets during the
course of the infection trial was performed to ensure that the ferrets were shedding the same species of
Eimeria with which they were inoculated Oocysts were concentrated from positive fecal samples as
described above A drop of concentrated oocyst solution was viewed photographed and measured at
400times and 600times for comparison with previously determined morphometrics of Eimeria ictidea oocysts
(Chapter 3)
Regions from the mitochondrial cytochrome c oxidase subunit I and III (mt COI and mt COIII)
DNA were amplified by polymerase chain reaction (PCR) from each sample using primer pairs
400F1202R and -172F799R respectively For all PCR reactions samples were denatured at 95 degC for 5
min then subjected to 35 cycles of 94 degC for 30s anneal at 52degC for 30s and extension at 72 degC for 60s
followed by a final extension at 72 degC for 7 min PCR gel electrophoresis and sequencing methods used
were as described in the Materials amp Methods section of Chapter 2
The resulting consensus sequences were searched from within Geneious against previous
sequences for E ictidea produced by the authors and against publically available sequences on the
111
BLAST server (blastncbinlmnihgovBlastcgi) using the blastn search algorithm against the nrnt
database (GenBank+EMBL+DDBJ+RefSeq ndash AA or DNA)
527 Necropsy protocol
All humanely killed ferrets underwent a complete necropsy (Appendix 6) using the modified
protocol described in Materials amp Methods section 424 of Chapter 4
53 RESULTS
Initial physical examination was unremarkable with the exception of mild to moderate bilateral
ceruminous discharge within the external ear canal of all ferrets Complete blood count and serum
biochemistry results for all ferrets were within normal reference intervals for juvenile domestic ferrets
(Appendices 2a and b) (Fox 2014) Six days after arrival a single ferret (103) in the control group
developed mild upper respiratory signs consisting of sneezing and clear nasal and ocular discharge these
clinical signs were associated with mild dehydration and decreased food and water consumption The
ferret was treated with subcutaneous fluid therapy (10 mL Plasmalyte-A subcutaneous Baxter Alliston
Ontario) heat and supportive care and all clinical signs resolved within three days This ferret was
deemed healthy to participate in the remainder of the clinical trial A second ferret (105) in the control
group developed unilateral purulent ocular discharge 14 days after arrival one day prior to placebo
inoculation The ferret was treated topically twice daily for five days with Isathal ophthalmic gel (fusidic
acid 10 mgg Dechra Veterinary Products Inc Point-Claire Quebec) and the discharge resolved but
reoccurred within 2 days of treatment cessation Ocular examination showed mild conjunctivitis but no
evidence of corneal lesions and fluorescein staining did not indicate the presence of corneal ulceration
The ferret was treated for an additional six days with tobramycin ophthalmic solution (3 mgmL Sandoz
Tobramycin 03 Boucherville Quebec) after which clinical signs resolved completely No coccidial
oocysts were shed in feces from any of the ferrets during the two week acclimation period
112
531 Oocyst shedding
All ferrets readily ingested the inoculum with either placebo or concentrated oocysts In Part 1
one ferret (203) in the infection group which had received the high dose (1 times 106 oocysts) shed oocysts
on day 8 and day 9 after inoculation (Tables 51 52) This ferret was 71 days of age at the time shedding
was initially identified The ferret was killed humanely 11 days post inoculation later than had been
outlined in the protocol as processing of fecal samples had been delayed by two days resulting in late
detection of oocyst shedding in this individual Oocysts were not identified in the feces of the three
remaining ferrets that received the high inoculation dose the single ferret that received the lower dose (5
times 104 oocysts) or in the ferrets within the control group
In Part 2 six of eight ferrets inoculated with the high dose (1 times 106 oocysts) shed oocysts during
the 14 day observation period (Tables 51 52) Four of these ferrets were from the previous control
group One of the ferrets previously inoculated with the high dose inoculum in Phase 1 that had not shed
oocysts did shed oocysts after being inoculated a second time with the same dose during Phase 2 The
ferret that had previously received the low dose (5 times 104 oocysts) of oocysts in Phase 1 also shed after
inoculation with the high dose in Phase 2 Three ferrets did not shed oocysts after high dose inoculation in
Phase 2 one of these had been part of the previous control group the other two had received the high
dose inoculation previously in Phase 1
The pre-patent period ranged from 7-9 days (Table 51) with equivalent numbers of ferrets
commencing shedding on each of days 7 through 9 All six ferrets were between 91 and 93 days of age at
the time shedding was initially identified Oocyst per gram counts and shedding trends for all individuals
are shown in Table 51 Total oocyst shedding during patency was lt14 oocysts 8904 oocysts and
172291 oocysts for ferrets 201 104 and 105 respectively The two ferrets for which the prepatent period
was 9 days only shed oocysts for one day and in low numbers Oocysts were not identified in the feces of
the three remaining ferrets two of which received the high inoculation dose (1 times 106 oocysts) and the
third that received the lower inoculation dose (2 times 105 oocysts) (Table 52)
113
532 Morphologic and molecular characterization
In all seven ferrets that shed oocysts the morphologic features and measurements (length width
shape index) of the shed oocysts were consistent with those of the E ictidea administered in the inoculum
(Figure 51) Molecular confirmation of the identity of the oocysts shed was successful in 3 out of the 7
ferrets (102 103 203) samples from the four remaining ferrets did not contain adequate quantity or
quality of DNA for confirmation
533 Clinical signs
Clinical signs associated with patent infection were identified in 6 of 7 ferrets (Table 52) These
signs included weight loss (n=5) diarrhea (n=1) mucoid soft feces (n=2) feces containing blood (n=2)
and malodorous feces (n=1) Appetite reduction was noted in two ferrets from the infection group in Part
1 between 6-8 days post infection however no oocyst shedding was detected from either ferret during
this time
534 Hematology
CBC and serum biochemistry values from ferrets collected during pre-trial health screening are
displayed in Appendices 1a and 1b Values obtained for ferrets euthanized during or after the
experimental trial are displayed in Appendices 3a and 3b Minor variances from reference range values
for CBC and serum biochemistry were identified in six of the seven ferrets with patent infection In all six
ferrets for which a complete serum biochemistry was obtained immediately prior to death creatinine
kinase (CK) values were elevated (see Appendix 3b) Ferret 103 exhibited a mild hypoalbuminemia (20
ref 24-40 gL) on ante-mortem serum biochemistry (see Appendix 3b)
114
535 Necropsy
No gross or histopathologic lesions were present and coccidia could not be identified in sections
of intestine from the single ferret (203) humanely killed in Part 1 For the ferrets humanely killed in Part
2 no evidence of diarrhea hematochezia or mucoid fecal material was identified grossly Ferret 105
killed at the termination of the experiment but still shedding low numbers of oocysts in its feces
exhibited a 7 cm region of congested mucosa within the distal jejunum
Coccidial life stages were identified in small intestinal sections from four of the seven ferrets that
were identified to shed oocysts at some point prior to necropsy (Figure 52 Tables 52 and 53) Affected
sections included jejunum in all four animals as well as ileum in one and were collected from 114 to 218
cm aboral from the pylorus (see Figure 53) Coccidia were not identified in sections of duodenum
proximal jejunum or large intestine however oocysts were identified within fecal material in the lumen
of the large intestine from one ferret (103) Of the 11-19 sections of intestine examined for each ferret the
number of sections containing parasites ranged from one to eight 1 section in ferret 102 2 sections in
ferret 201 4 sections in ferret 105 8 sections in ferret 103 A mix of sexual and asexual life stages was
observed within the enterocytes in small intestinal sections from ferret 103 the remainder of the ferrets
showed either asexual (102 201) or sexual (105) life stages in affected segments
Pathologic changes and additional histologic findings in small intestinal sections of ferrets with
enteric coccidia included rare regions of blunting of the villi and sloughing of the epithelium associated
with hemorrhage and inflammation The primary lesions identified were subjectively increased numbers
of eosinophils lymphocytes and plasma cells within the lamina propria of the small intestine and
similarly increased neutrophils lymphocytes and plasma cells within the lamina propria of the large
intestine Neutrophils were rarely present in intestinal crypts and glands
Other gross necropsy findings included two ferrets with renal cortico-medullary cysts and one
ferret with mild thickening of the esophageal mucosa midway along the esophageal length No histologic
changes were identified within a sample of esophagus taken from this region
115
54 DISCUSSION
The findings of this study show that domestic ferrets are susceptible to infection with the enteric
coccidium Eimeria ictidea isolated from black-footed ferrets Both morphometric and molecular
diagnostic methods were used to confirm that ferrets were shedding oocysts of the same species with
which they were inoculated Molecular characterization was successful in three of the seven ferrets that
developed patent infections and as no other coccidial species was identified during pre-trial observation
morphometry was considered to be confirmatory in the remaining four animals
We have referred to the eimeriid coccidium affecting BFF and used in this experimental trial as
E ictidea based on morphometric similarity of their oocysts with those of E ictidea as described from
domestic ferrets (see Chapter 3) There is limited published information on infection of domestic ferrets
with E ictidea outside of Hoarersquos original descriptions (1927 1935a b) which form the basis for all
subsequent identifications of E ictidea in domestic ferrets and in BFF Attempts to obtain exemplars of E
ictidea from domestic ferrets to characterize using molecular techniques were unsuccessful (Chapter 2)
Multiple diagnostic laboratories in Canada and Europe were solicited for coccidia-positive fecal samples
from domestic ferrets but no samples of E ictidea were received over a 4 year period (2014-2017)
Eimeria ictidea was identified based on microscopic examination in only two samples submitted to a
European diagnostic laboratory from 2008-2015 It is unproven whether the E ictidea described from
domestic ferrets and the E ictidea identified from black-footed ferrets and used in this experimental
work are the same or are simply morphologically indistinguishable Eimeria species However the
consistency in morphology host genus and location of infection within the intestinal tissues combined
with the successful cross-transmission of this parasite to domestic ferrets described in the present study
suggests they are likely conspecific
116
The pre-patent period (minimum duration of endogenous development) for infection with E
ictidea in the inoculated domestic ferrets ranged from 7-9 days (see Table 51) the pre-patent period for
this parasite in the BFF the natural host for this coccidium is unknown Hoare experimentally infected
naiumlve domestic ferrets with E ictidea derived from naturally occurring infection in this species (Hoare
1935b) The inoculated ferrets shed oocysts after a pre-patent period of 7 days consistent with the 7-9
days seen in the work described here with E ictidea
Shedding of oocysts was identified over a period of 1-7 days (see Table 51) and intensity ranged
from less than 14 up to 15624 OPG These results may be skewed with erroneously low duration of
shedding and number of oocysts shed as three of the seven ferrets were humanely killed at or prior to the
expected peak of oocyst shedding for tissue collection and histologic examination in order to increase the
probability of identifying parasite life stages within the intestinal sections Shedding periods were similar
to those identified in adult single-housed BFF which ranged from 2-9 days however oocyst per gram
counts from the domestic ferrets were consistently lower than OPG counts from BFF (104 - 554274
OPG) infected with the same parasite (see Chapter 4) Furthermore the total number of oocysts shed by
individual domestic ferrets (14 - 172291 oocysts) during patency was reduced compared to BFF despite
similar length of shedding period (see Chapter 4) The domestic ferrets in this study were naiumlve individual
juveniles whereas the BFF were adults 1-5 years of age some of which were showing clinical signs at
the time of oocyst shedding The relative influences of age species and previous exposure to the parasite
on our observations are unknown
Two different fecal flotation methods were used on all samples to increase the probability of
oocyst detection The McMaster method was used to provide accurate OPG counts for quantification of
oocyst shedding however this method had a minimum detection limit of ~13 oocysts per gram (1333
OPG calculated) because it is based on dilution of the initial sample with flotation media (saturated salt)
In samples with few oocysts oocysts may be missed or to be present in numbers below this detection
limit Routine salt flotation is in contrast performed using the entire sample allowing for detection of
117
small numbers of oocysts Consequently in cases where oocyst per gram counts were low shedding was
identified on routine salt flotation but not by the McMaster method and recorded as lsquopositiversquo but below
the detection limit of the enumeration method
Subclinical to clinical disease occurred in six of the seven ferrets that developed patent infection
with weight loss being the most frequent clinical sign Other clinical signs were typical of enteric
coccidiosis including diarrhea hematochezia and mucoid andor soft feces These clinical signs are
similar to those previously described for black-footed ferrets infected with this parasite (USFWS BFF
Recovery Program 2017 Chapter 4) and for domestic ferrets with enteric coccidiosis (Sledge et al
2011) Interestingly development of clinical disease was not described in the naiumlve domestic ferret
inoculated by Hoare (1935) with E ictidea derived from naturally occurring infection Based on review of
the literature severe clinical disease resulting from intestinal coccidiosis is rare in domestic ferrets
Black-footed ferrets however appear more susceptible to disease development and more frequently show
significant clinical signs No domestic ferret required treatment for clinical coccidiosis during the course
of this study It is possible that the more pronounced clinical signs associated with enteric coccidiosis in
BFF may result from the limited genetic diversity in a population derived from so few individuals and
increased susceptibility of BFF to other diseases such as sylvatic plague have been described in
comparison with their domestic counterparts (Williams et al 1994) Although unlikely recent acquisition
of E ictidea from a related host species such as the domestic ferret could have resulted in increased
pathogenicity and severity of clinical disease from infection with this parasite in BFF
Minor variances from reference range values for CBC and serum biochemistry were identified in
six of the seven ferrets with patent infection but only in one case (ferret 103) did this appear to be
correlated with infectiondisease In this animal a mild hypoalbuminemia (20 ref 24-40 gL) was noted
(Appendix 3b) On histopathology large regions of the small intestine contained parasite life stages
however inflammation lysis of epithelial cells and necrosis of affected areas that could be expected to
result in protein loss into the intestinal lumen were not identified
118
Creatine kinase values were elevated in all six ferrets for which a complete serum biochemistry
was obtained immediately prior to death these findings are consistent with release from CK rich tissues
such a skeletal muscle during venipuncture and manual restraint
The pathologic lesions identified within the intestinal sections of ferrets euthanized at the time of
oocyst shedding were similar to those identified in affected BFF but in most cases were less locally
extensive or widespread throughout the small intestine than those observed in necropsy cases of BFF (see
Chapter 4) Coccidia were seen in the histologic sections of four ferrets all three ferrets that were actively
shedding oocysts at the time of necropsy (102 103 105) and one of four in which necropsies were
performed after oocyst shedding had ceased (201)
Although the primary objective of the examination of histologic sections from affected ferrets
was to identify coccidial life stages attempts were made to describe the pathologic changes associated
with the presence of the parasite Despite necropsies being performed almost immediately after death and
the use of Serra solution fixative to improve parasite and tissue preservation the villi and villar
epithelium of the trimmed sections were frequently distorted or absent and consequently accurate
commentary on these areas was precluded and was made only on visible components of the lamina
propria and crypts or glands A deliberate decision was made not to kill and collect samples from the
saline inoculated control ferrets after Phase 1 and those not shedding oocysts during Phase 1 or 2 thus no
age-matched intestinal sections were available for comparison It is difficult to comment on the
significance of the inflammatory cells observed in the lamina propria of the small and large intestinal
segments or the proliferative rate of the crypt epithelium The only changes identified which may be
considered significant are the presence of neutrophils within the crypts and glands of small and large
intestinal sections respectively but these lesions were rare and not associated with the presence of
parasitic life stages
119
While no parasitic life stages were identified in intestinal sections from ferrets 104 203 and 205
one of the three ferrets ferret 104 exhibited lymphoplasmacytic inflammation and blunting of jejunal
villi in one section (36-38 cm aboral from pylorus) These findings might be expected as the most
extensive histologic lesions would occur associated with lysis of the intestinal epithelial cells as oocysts
were shed into the feces after which new intestinal epithelial cells would re-cover the denuded villar
surface Thus for those cases in which histologic examination was performed after shedding had ceased
presence of the protozoal life stages in the intestines would be expected to be significantly reduced or
absent
Hoare (1935a b) described a particular reaction to the presence of parasitic life stages of Eimeria
ictidea in the small intestine of domestic ferrets in which only the villar tips were affected and there was
resultant annular constriction of the villus separating the affected and non-affected segments These
particular changes were not identified in any of the ferrets in this study and had not been noted
retrospectively in naturally infected BFF (Chapter 4)
While patent infection and intestinal disease could be experimentally created in domestic ferrets
without equivalent experimental work in BFF it is difficult to fully compare the susceptibility to infection
and to the development of disease between the two species Eimeria species tend to be host specific thus
if E ictidea from BFF is not conspecific with E ictidea in domestic ferrets it might be expected that the
domestic ferret would be less susceptible to infection and the development of disease than is the BFF
Even if the two parasites are identical natural passaging through the BFF may alter affinity for the
domestic ferret
It appears that the infectious dose of oocysts of E ictidea derived from BFF required to initiate a
patent infection in domestic ferrets is high The ferrets that developed patent infections were administered
an inoculum containing 1 times 106 sporulated oocysts and even with this extremely high inoculating dose
only a proportion of inoculated ferrets became infected Neither the ferret that received the low dose
120
inoculum nor the ferret that received the passaged oocysts from Part 1 of the study shed oocysts during
the 14 day period post inoculation The latter finding was unexpected as fresh passaged oocysts would be
expected to contain larger proportions of viable oocysts and be comparatively more infective than oocysts
that had been stored for 2-4 weeks prior to inoculation
The only publication describing oral inoculation of Eimeria species in domestic ferrets (Hoare
1935b) did not quantify the number of oocysts administered A study performed in 16 farmed juvenile
mink (Mustela vison) administered 2000 sporulated oocysts of each of three coccidial species (I
laidlawi E vison and an unknown Eimeria species) resulting in patent infection with one of the three
species (Foreyt et al 1977) as determined by the presence of oocysts on fecal examination The authors
did not reveal which type of oocysts resulted in the infection
The number of oocysts required to result in infection in BFF is unknown Based on the
authorsrsquo observations of over 100000 oocysts per gram of feces being shed by black-footed ferrets into
cages of lt1m2 floor space (see Chapter 4) we estimated that under normal caged conditions animals
would likely ingest thousands of oocysts over a short period of time This was in part why a large number
of oocysts (up to 1000000 as available from our store of viable oocysts) was administered to each
domestic ferret in order to increase the probability that infection and shedding would result Furthermore
for the parasite to persist within the ferret population the total number of oocysts shed into the
environment would have to be several times higher than the infective dose required to generate a patent
infection otherwise the parasite would be expected to die out If E ictidea of domestic ferrets and E
ictidea of BFF are conspecific the difference in oocyst shedding between the host species during patent
infection could potentially explain the low prevalence of E ictidea reported from the domestic ferret
population (as seen in Chapter 2)
Both humoral and cell mediated immunity are involved in the immune response to coccidia The
role of maternal derived antibodies in combatting protozoal infection in mustelids has not been studied
121
but in carnivores maternal antibodies to viruses can last up to 16 weeks (Chappuis 1998) In poultry
maternal Eimeria-specific IgG is transferred via the egg yolk to offspring In one study breeding hens
were infected with a single species of Eimeria 28-39 days prior to lay Their hatched chicks were
challenged by inoculation with oocysts of the same and a related Eimeria species and showed reduced
oocyst shedding compared to age matched controls indicative of passive transfer of immunity (Smith et
al 1994) It is possible then that the presence or absence of maternal antibodies may be a factor in the
age at which ferrets are susceptible to coccidial infection and the development of disease The facility
from which the domestic ferrets were acquired has not previously detected Eimeria species on routine
fecal screening (Dr Bambi Jasmin personal communication) consequently it is unlikely that they would
have received maternal immunity to or been exposed to this parasite and thus can be considered to be
naive
Despite a theoretical lack of maternal immunity there did appear to be an effect of age on
susceptibility to infection In Part 1 when the ferrets were approximately 70 days of age only 1 of 4
ferrets inoculated with the high dose of oocysts developed patent infection In comparison in Part 2
when the ferrets were 91-93 days of age 3 of 4 ferrets that had been in the saline control group for Part 1
developed patent infections as did 2 of 4 ferrets that had been inoculated in Part 1 but had not shed
oocysts One of these previously inoculated ferrets was the individual that had received the lower dose of
oocysts Thus it appears that patent infection could be produced more easily in the older ferret kits
However the two ferrets that had previously been inoculated exhibited the shortest shedding periods (1
day) and lowest oocyst per gram counts and parasite life stages in these cases were rare (201) to absent
(205) on histologic examination of numerous sections of intestine It is thus possible that the primary
inoculation resulted in abbreviated infections or infections in which so few oocysts were shed that
infection was not detected Our observation of endogenous stages in the intestinal tissues of some of the
ferrets following cessation of oocyst shedding suggests that the pre-patent period and duration of patency
may vary considerably from animal to animal consequently it is possible that some of the kits would
122
have ultimately shed a few oocysts from the primary inoculum if followed beyond 14 days post-
inoculation Whether through an aborted or undetected infection previous exposure of these kits to the
parasite probably generated partial immunity against E ictidea and therefore the intensity of infection
upon challenge in previously exposed domestic ferret kits was reduced
The authors acknowledge the limitations of this initial pilot study however it was proven that
patent infection with E ictidea isolated from black-footed ferrets could be generated in a novel host the
domestic ferret Further studies will be required to investigate the effect of age on susceptibility to
infection as well as the possibility of development of immunity after exposure and its role in reducing
parasite replication and disease in subsequent infection With so few BFF in existence the use of BFF for
in vivo infection trials cannot be contemplated consequently refining the domestic ferret infection model
will be essential for carrying out research specifically intended to help manage coccidiosis in the
endangered black-footed ferret
Figure 51 Exogenous life stages of Eimeria ictidea shed from a domestic ferret (Mustela putorius
furo) experimentally inoculated with oocysts originating from black-footed ferrets (Mustela
nigripes) Unsporulated oocyst (solid black arrow) Sporulated oocyst (solid white arrow) Bright
field microscopy scale bar = 25 microm
25 microm
124
Figure 52 Life stages of Eimeria ictidea within the small intestinal epithelium of an experimentally
infected domestic ferret (Mustela putorius furo) A) Sexual life stages (micro- and macrogamonts
- white arrows) crowding the villar enterocytes all stages are found between the nucleus and
luminal surface of infected enterocytes Hematoxylin and eosin staining scale bar = 25microm B) At
higher magnification meronts (black arrows) and gamonts (open arrows) are crowded between the
enterocyte nuclei and brush border Hematoxylin and eosin staining scale bar = 25microm
25 microm
25 microm
125
Figure 53 Presence and location of sexual and asexual life stages of Eimeria ictidea within the
intestinal epithelium of domestic ferrets (Mustela putorius furo) (n=7) that developed patent
infection after experimental inoculation with oocysts originating from black-footed ferrets (Mustela
nigripes) Legend duod = duodenum jej= jejunum Sequential numbers for jejunal and colonic
sections represent the order aboral from the pylorus from which the samples were collected SI =
additional section(s) of small intestine whose aboral sequence was not recorded
0
1
2
3
duod jej 1 jej 2 jej 3 jej 4 jej 5 ileum colon 1 colon 2 SI
Nu
mb
er
of
Ferr
ets
Aff
ect
ed
Intestinal Section Containing Life Stages of Eimeria ictidea
Asexual life stages Sexual life stages
126
Table 51 Prepatent period and oocyst shedding patterns in domestic ferrets (Mustela putorius furo)
experimentally inoculated with 1 times 106 oocysts of Eimeria ictidea originating from black-footed ferret
(Mustela nigripes) that developed patent infections
Oocysts shed per gram of feces
Ferret Identity
Day post inoculation 102 103 104 105 201 203 205
1 0 0 0 0 0 0 0
2 0 0 0 0 0 0 0
3 0 0 0 0 0 0 0
4 0 0 0 0 0 0 0
5 0 0 0 0 0 0 0
6 0 0 0 0 0 0 0
7 lt 14 1807 0 0 0 0 0
8 11053 139 7091 0 156238 0
9 463 11733 lt 14 203 lt 14
10 578 7549 0 0 0
11 lt 14 0 0 0
12 0 lt 14 0
13 0 lt 14 0
14 0 lt 14 0
Legend ferret euthanized as of this date lt 14 = oocyst positive samples with less than 14 oocysts per
gram of feces
127
Table 52 Results of oral inoculation of domestic ferrets (Mustela putorius furo) with oocysts of Eimeria ictidea originating from black-footed
ferrets (Mustela nigripes)
Ferret
Identity
Results Part 1 Results Part 2
Inoculum Oocyst
Shedding
Presence
of clinical
disease
Coccidia
present in
sections
Inoculum Oocyst
Shedding
Presence
of clinical
disease
Coccidia
present in
sections
101 Saline N N - 2 times 105 oocystsa N N -
102 Saline N N - 1 times 106 oocysts Y N Y^
103 Saline N N - 1 times 106 oocysts Y Y Y^
104 Saline N N - 1 times 106 oocysts Y Y Nyen
105 Saline N N - 1 times 106 oocysts Y Y Yyen
201 1 times 106 oocysts N N - 1 times 106 oocysts Y Y Yyen
202 1 times 106 oocysts N N - 1 times 106 oocysts N N -
203 1 times 106 oocysts Y Y Nyen
204 1 times 106 oocysts N N - 1 times 106 oocysts N N -
205 5 times 104 oocysts N N - 1 times 106 oocysts Y Y N^
Legend includes any of the following weight loss diarrhea mucoid feces malodorous feces inappetence N = no Y= yes - = necropsy not
performed a = oocysts collected after passage through ferret 203 ^ = shedding oocysts at time of necropsy yen = not shedding oocysts at time of
necropsy
128
Table 53 Distribution of coccidial life stages in domestic ferrets (Mustela putorius furo) orally inoculated with oocysts of Eimeria ictidea
originating from black-footed ferrets (Mustela nigripes)
Ferret Identity
Intestinal level 102
103
104 105
201
203
205
Duodenum N N N N N N N
Jejunum 1 N N N N N N N
Jejunum 2 N N N N N N N
Jejunum 3 N S A N N N N N
Jejunum 4 N S A N N N N N
Jejunum 5 N S A N S A N N
Ileum N S A N N N N N
Colon 1 N Na N N N N N
Colon 2 N Na N N N N N
Unmeasured small intestinebc
S - 05
A - 15
S - 45
A - 25
S - 05
A - 05
S - 26
A - 06
S - 06
A - 06
S - 06
A - 06
S - 08
A - 08
Unmeasured large intestinebc
S - 01
A - 01
S - 01
A - 01
S - 01
A - 01
S - 02
A - 02
S - 01
A - 01 none
S - 01
A - 01
Legend N = no parasite life stages S = sexual life stages present A = asexual life stages present a= oocysts present in feces b=
additional sections of intestine for which the location measured from the pylorus was not obtained c= xn where x is number of
sections containing sexual or asexual lifestages n is the number of sections examined
129
CHAPTER 6 WHOLE MITOCHONDRIAL GENOME SEQUENCES OF TWO EIMERIA
SPECIES ISOLATED FROM DOMESTIC (MUSTELA PUTORIUS FURO) AND BLACK-
FOOTED FERRETS (MUSTELA NIGRIPES)
ABSTRACT
The complete mitochondrial (mt) genomes of Eimeria furonis and Eimeria ictidea (Eimeriidae
Coccidia Apicomplexa) originating from single fecal samples from a domestic (Mustela putorius furo)
and a black-footed ferret (Mustela nigripes) respectively were sequenced Both mt genomes were
circular-mapping with lengths of 6165 base pairs (Eimeria furonis - GenBank MF795598) and 6182
base pairs (Eimeria ictidea - GenBank KT203399) Genome organization and gene contents were
comparable with those of other publically available mt genomes from a variety of Eimeria species and
related coccidia there were three complete coding DNA sequence regions encoding cytochrome c
oxidase subunit I cytochrome c oxidase subunit III and cytochrome B and 33 regions encoding
fragmented rDNA Alignment of these mt genome sequences demonstrates a relatively high (945 340
single nucleotide differences [SNDs]) pairwise sequence identity between these Eimeria spp infecting
ferrets the majority of the SNDs resulted in synonymous codon changes with no changes to their protein
products Alignment of the newly sequenced mt genomes demonstrates and phylogenetic reconstructions
support the monophyly of these Eimeria spp of ferrets with another Eimeria sp of carnivores as the
sister taxon to this clade
61 INTRODUCTION
Coccidia are protozoal eukaryotic host-specific parasites of the phylum Apicomplexa and can
be divided into two major taxonomic suborders the eimerioirinid and adeleid coccidia The eimeriorinid
coccidia include both the typical intestinal coccidia (eg Eimeria Isospora Cyclospora) species
belonging to the family Eimeriidae as well as tissue or cyst-forming coccidia (eg Cystoisospora
130
Besnoitia Toxoplasma Sarcocystis) of the family Sarcocystidae (see Cox 1994) Ten species of
Eimeria and twelve species of Isospora (=Cystoisospora) have been described in the Mustelidae
(see Chapter 1 Table 11) Eimeria furonis has been reported in the European polecat (Mustela
putorius) domestic ferret (DF Mustela putorius furo) black-footed ferret (BFF Mustela nigripes) and
mink (Mustela vison) (Hoare 1927 Nukerbaeva and Svanbaev 1973 Jolley et al 1994) Eimeria ictidea
has been reported in the Steppe polecat (Mustela eversmanii) as well as the European polecat domestic
ferret and black-footed ferret (Hoare 1927 Svanbaev 1956 Jolley et al 1994) These reports are based on
the morphometric characteristics of oocysts identified in the feces of the aforementioned host species
without the use of molecular techniques to confirm specific parasite identities
Recently sequences of the mitochondrial cytochrome c oxidase subunit I gene (mt COI) and
nuclear small subunit ribosomal DNA (nu 18S rDNA) of E furonis originating from a domestic ferret (nu
18S rDNA GenBank MF774678-MF774680 mt COI GenBank MF774034-MF774036) and E ictidea
originating from a black-footed ferret (nu 18S rDNA GenBank MF860826-MF860827 mt COI
GenBank MF860823 MF860825) were generated (see Chapters 2 and 3) The parasite originating from
the black-footed ferret was identified as E ictidea based on morphologic similarity to the original
descriptions of E ictidea from domestic ferrets however sequence-based genotyping of E ictidea from
domestic ferrets has not been completed and consequently it has not been demonstrated unequivocally
that the two parasites are conspecific In the present work the complete mitochondrial genomes of E
furonis from the DF and E ictidea from the BFF are described and compared with the mitochondrial
genomes of related coccidia
62 MATERIALS amp METHODS
621 Parasites
Two isolates of morphologically distinct Eimeria species were used in this study Isolate one
identified morphologically and by nu 18S rDNA and mt COI sequences as Eimeria furonis was obtained
131
from a fecal sample from a DF that was submitted for routine ova and parasite examination to a European
diagnostic laboratory8 Isolate two identified morphologically and by nu 18S rDNA and mt COI
sequences as Eimeria ictidea was obtained from a fecal sample from a BFF and was collected during
routine cage cleaning in a captive breeding facility (see Chapters 3 and 4) Fecal collection techniques for
the BFF were reviewed and approved by both the Animal Welfare Committee and the Animal Care and
Research Committee of the Toronto Zoo
622 DNA isolation from coccidia in feces
Genomic DNA was isolated from fecal derived coccidial oocysts as described section 221 of the
Chapter 2 Materials amp Methods Parasite DNA concentration was estimated using a Nanodrop 2000
spectrophotometer (NanoDrop Products Wilmington DE USA) and DNA was stored at 4 degC for
immediate use or minus20 degC for later use
623 Whole genome sequencing
Mitochondrial whole genome amplification for both Eimeria species was initiated using sets of
mt-specific primers that generated overlapping polymerase chain reaction (PCR) fragments (Tables 61
and 62) PCR amplification was performed for all samples in a volume of 25 microl containing ~100 ng of
genomic DNA 1times PCR buffer 3 mM MgCl2 06 mM deoxyribonucleotide triphosphates (dNTPs) 500
nM of each primer and 4 U of Invitrogen Platinum Taq DNA Polymerase (Thermo Fisher Scientific
Toronto ON Canada) PCR reactions were performed on a Bio-Rad T100 PCR thermal cycler (Bio-Rad
Laboratories Singapore) using settings as described previously in the Materials amp Methods section of
Chapter 2 Table 61 details the specific anneal conditions used for the various primer pairs Genomic
DNA from either Eimeria maxima or Eimeria tenella acted as a positive control for the reaction
chemistry Gel electrophoresis purification and sequencing of the PCR amplification products were
8 Vet Med Labor GmbH Division of IDEXX Laboratories Ludwigsburg Germany
132
performed as described in Chapter 2 The resulting chromatograms were aligned and analyzed with
Geneious Ver 818 or later (Biomatters Limited Auckland New Zealand) and high quality consensus
sequences generated The completed mt genome sequences were annotated by comparison with
previously annotated mt genomes from other Eimeria species (eg Eimeria innocua - KR1082961) and
the annotated mt genomes deposited in GenBank
624 Phylogenetic analysis
To determine the phylogenetic affinities of the newly obtained sequences with sequences from
related apicomplexan taxa representative whole mt genome sequences from eimeriid coccidia were
downloaded from GenBank A complete mt genome sequence from an unnamed Choleoeimeria sp was
used to root the ingroup taxa several small genomic rearrangements in the Choleoeimeria sp sequence
required some rearrangement of the genome sequence to unambiguously align homologous regions across
the complete mt genomes
Whole genome sequences were aligned using MAFTT v7017 (Katoh et al 2002) executed from
within Geneious the resulting alignment was examined by eye to adjust start and stop codon positions in
aligned coding DNA sequence [CDS] regions (ie mt COI mitochondrial cytochrome c oxidase subunit
III gene [mtCOIII] mitochondrial cytochrome b gene [CytB]) Phylogenetic trees were generated using
Bayesian Inference (BI) using MrBayes Ver 326 (Huelsenbeck and Ronquist 2001) executed from
within Geneious The aligned complete mt genomes were partitioned into coding (ie CDS) and non-
coding regions so that region-specific models of nucleotide substitution could be applied Characters in
the non-coding region were analysed with the general time reversible (GTR) model (Tavareacute 1986) with
the following parameters nucmodel=4by4 nst=6 rates=invgamma (ie GTR+I+G) Characters in the
coding regions were analysed using the codon nucleotide model (ie lset nucmodel=codon rates=gamma
ngammacat=4) using metazoan mitochondrial translation (ie code=metmt)
133
All BI analyses were run for a chain length of 1000000 with tree sampling every 1000
following a burn-in of 100000 with default settings of 4 heated chains and heated chain temp of 02
63 RESULTS
The whole mt genome sequences of the single isolates of E furonis and E ictidea were
respectively 6165 base pairs (bp) (Figure 61 GenBank MF795598) and 6182 bp (Figure 62 GenBank
KT203399) Content and organization of both mt genomes consisted of three protein-coding genes (mt
COI mt COIII and CytB) interspersed with large and small subunit ribosomal DNA (rDNA) fragments
Details of the various CDS and rDNA fragments are summarized in Table 63 (for E furonis) and Table
64 (for E ictidea)
Pairwise alignment of the mt genome sequences from E furonis and E ictidea demonstrated a
relatively high pairwise sequence identity (946 333 single nucleotide differences [SNDs]) between
these two parasites The bulk of the SNDs (676 225333) were clustered within the three CDS regions
that encode CytB mt COI and mt COIII (see Figure 63 and Table 65) However the majority of these
SNDs (826 186225) were synonymous codon changes that resulted in no changes to the protein
products Only 41 SNDs were involved in 34 amino acid changes distributed among the three CDS
The 33 rDNA fragments comprised 2108 and 2109 bp respectively of the mt genomes of
E furonis and E ictidea Pairwise comparison of these rDNA fragments demonstrated high (986 30
SNDs) sequence identity between the two parasites The remaining 778 and 794 bp respectively of the
mt genomes of E furonis and E ictidea were intergenic stretches between the various rDNA and CDS
regions these intergenic regions were more variable that other regions of the genomes with 78 SNDs
(almost 10 sequence divergence) Additionally all indels were restricted to these variable intergenic
regions
The BI phylogeny generated from aligned complete mt genomes (Figure 64) supported the close
relationship between E furonis and E ictidea within a clade of Eimeria species that include the only three
sequences available for Eimeria spp of carnivores Eimeria mephitidis from the striped skunk (Mephitis
134
mephitis Family Mephitidae) was the sister taxon to the two Eimeria species of ferrets (Family
Mustelidae)
64 DISCUSSION
This work generated the first complete mt genomes from coccidia that infect domestic and black-
footed ferrets (Carnivora Mustelidae) Eimeria mephitidis from the striped skunk Mephitis mephitis
(Carnivora Mephitidae) is the only other Eimeria species from a carnivore for which a complete mt
genome has been reported
Comparatively few eimeriid coccidia only 26 Eimeria species have been described from
carnivores there are 14 named species from the mustelids four from the procyonids four from the ursids
three from the herpestids and one from the viverrids (Duszynski et al 2000) The majority of coccidia
that infect the digestive tract of carnivores belong to the family Sarcocystidae including monoxenous or
facultatively heteroxenous Cystoisospora species or heteroxenous parasites in the genera Sarcocystis
Hammondia and Neospora So far as is known none of the parasites in the Sarcocystidae possess typical
apicomplexan mt genomes with 3 complete CDS and many rDNA fragments (Ogedengbe 2015)
The mt genomes from the two Eimeria sp of mustelid origin demonstrate the same structural
organization (ie the order and number of CDS and rDNA fragments) and circular mapping as the mt
genomes from other Eimeria spp and other closely related eimeriid coccidia such as Isospora
Cyclospora and Lankesterella species Despite the ability of the eimeriid sequences to be mapped
circularly the physical form of Eimeria spp mt genomes may be a linear concatemer of multiple genome
copies as demonstrated for Eimeria tenella (Hikosaka et al 2011) As in the mt genomes of other
eimeriid coccidia (Ogedengbe et al 2013 2014) the CDS for mt COIII demonstrated the highest
sequence divergence between E furonis and E ictidea the mt COI CDS was somewhat more conserved
and CytB CDS demonstrated the fewest SNDs
As expected based on limited sequence divergence between E furonis and E ictidea a BI
phylogenetic analysis using aligned complete mt genome sequences generated a tree that placed these
135
two Eimeria species that infect mustelids within a well-supported monophyletic group The sister taxon
for these ferret parasites was the only other Eimeria species from carnivores for which a complete mt
genome is available E mephitidis which infects hosts belonging to a different family of carnivores
Eimeriid parasites that infect closely-related definitive hosts are commonly found in a single or limited
number of clades based on mitochondrial and nuclear genetic loci (Ogedengbe et al in press)
Sequencing of the mt genomes and at least one nuclear genetic locus (ie nu 18S rDNA) from additional
Eimeria species infecting carnivores will be required to determine if all carnivore-specific Eimeria
species share a common ancestor
136
Table 61 PCR primer pairs and resulting fragments used for sequencing the mitochondrial genome sequence of an isolate of Eimeria furonis
originating from a fecal sample from a domestic ferret (Mustela putorius furo)
Fragment Primer names Primer sequences (5ʹ-3ʹ) Size (bp) Anneal Temp References
1 WG-MT_4140F AGAAAACCTAAAATCATCATGT 1000 52 Ogedengbe et al (2015)
Eim_CO3_799R AAGTGAGTTCGCATGTTTAC Ogedengbe et al (2015)
2 Eim_COI_19F ACTGCYGCAAACCATAARGAA 1700 60 Present study
Api_LSUG_UNI_R AGATAGGGAACAAACTGYCTCAA Present study
3 WG_MT_5416F GGTCCAGATAAGCGATCTCATG 3400 53 Ogedengbe et al (2013)
Eim_COI_1436R CACATTGTGTTCARATAAGTTA Present study
4 WG-MT_6219F GCATCCATCTACAGCTGCGG 500 55 Ogedengbe et al (2013)
WG-MT_344R GTAGGAATCTRAATTCCCAACC Ogedengbe et al (2013)
5 Api_LSUE UNI_F AGGTGCTCAGGGTCTTACCG 500 55 Present study
WG_MT_63R CTGGTATGGATGGATAACACT Ogedengbe et al (2015)
6 Lank_COB-30F CCAGGCCAACTGAACTCGTT 1300 55 Present study
q_Eim_COI_221R GGCATAACTACAAAGAARATCATA Present study
7 Cocci_MT_WG_F TACACCTAGCCAACACGAT 1600 55 Ogedengbe et al (2014)
q_Eim_COI_221R GGCATAACTACAAAGAARATCATA Present study
137
Table 62 PCR primer pairs and resulting fragments used for sequencing the mitochondrial genome sequence of an isolate of Eimeria ictidea
originating from a fecal sample from a black-footed ferret (Mustela nigripes)
Fragment Primer names Primer sequences (5ʹ-3ʹ) Size (bp) Anneal Temp References
1 WG_MT_63R CTGGTATGGATGGATAACACT 2180 52 Ogedengbe et al (2015)
WG-MT_4140F AGAAAACCTAAAATCATCATGT Ogedengbe et al (2015)
2 Cocci_MT_WG_F TACACCTAGCCAACACGAT 1551 58 Ogedengbe et al (2014)
q_Eim_COI_221R GGCATAACTACAAAGAARATCATA Present study
3 WG-MT_3658F CTGGCGAGAAGGGAAGTGTG 1329 55 Ogedengbe et al (2013)
Eim_CO3_799R AAGTGAGTTCGCATGTTTAC Ogedengbe et al (2015)
4 Lank_COB-30F CCAGGCCAACTGAACTCGTT 3899 57 Present study
WG_MT_4072R GGTTGTTTCCATCTCGACTC Ogedengbe et al (2013)
138
Table 63 Coding regions within the mitochondrial genome of the eimeriid parasite Eimeria furonis from a domestic ferret (Mustela putorius faro)
139
Table 64 Coding regions of the mitochondrial genome of the eimeriid parasite Eimeria ictidea originating from a black-footed ferret (Mustela
nigripes)
Table 64 Features associated with the protein-coding regions of the mitochondrial genome of the eimeriid parasite Eimeria cf ictidea originating from a black-footed ferret (Mustela nigripes )
Protein coding regions (CDS) Sequence size (bp) Start position (bp) Stop position (bp) Direction Translation start codon Translation stop codon
Cytochrome c oxidase subunit I (COI) 1443 1343 2785 Forward ATG TAA
Cytochrome c oxidase subunit III (COIII) 756 4333 5088 Forward TTA TAA
Cytochrome b (CytB) 1080 226 1305 Forward ATG TAA
Ribosomal DNA fragments (rDNA) Product
SSUrRNA 46 17 62 forward RNA9 SSU8
SSUrRNA 77 104 180 forward SSUA SSU4 (partial)
SSUrRNA 35 181 215 forward RNA23t
LSUrRNA 20 2790 2809 forward RNA20 (partial) LSU
LSUrRNA 112 2835 2946 forward LSUF LSU11
LSUrRNA 106 2947 3052 forward LSUG LSU12
LSUrRNA 25 3087 3063 reverse LSU
SSUrRNA 39 3129 3167 forward RNA14 SSU1
LSUrRNA 16 3188 3173 reverse LSUC LSU4
SSUrRNA 33 3211 3243 forward SSU
SSUrRNA 61 3386 3326 reverse SSUF SSU12
LSUrRNA 74 3387 3460 forward RNA10 LSU13 (partial)
LSUrRNA 49 3476 3524 forward RNA11 LSU5
SSUrRNA 65 3533 3597 forward SSUD SSU10
SSUrRNA 40 3631 3670 forward RNA17 SSU3
SSUrRNA 30 3677 3706 forward RNA15 SSU
LSUrRNA 30 3711 3740 forward RNA13 LSU10
LSUrRNA 56 3749 3804 forward RNA6 LSU15
LSUrRNA 79 3899 3821 reverse LSUD LSU8
LSUrRNA 24 3931 3908 reverse RNA16 (partial)
SSUrRNA 92 4036 3945 reverse RNA8 SSU5
LSUrRNA 66 4054 4119 forward RNA2 LSU2
LSUrRNA 177 4316 4140 reverse LSUA LSU1
SSUrRNA 30 5125 5096 reverse RNA19 SSU7
LSUrRNA 88 5154 5241 forward RNA1 LSU6
LSUrRNA 26 5342 5317 reverse LSUB LSU3
LSUrRNA 73 5447 5375 reverse RNA3 LSU7
LSUrRNA 20 5479 5460 reverse RNA18 LSU14
SSUrRNA 119 5615 5497 reverse SSUB SSU6
LSUrRNA 80 5699 5620 reverse RNA7
LSUrRNA 188 5897 5710 reverse LSUE LSU9
SSUrRNA 33 6033 6001 reverse SSUE SSU11 (partial)
SSUrRNA 99 6156 6058 reverse RNA5SSU9
140
Table 65 Pairwise comparison of coding DNA and concatenated rDNA fragment sequences between the
mitochondrial genomes of Eimeria furonis originating from a domestic ferret (Mustela putorius furo) and
Eimeria ictidea originating from a black-footed ferret (Mustela nigripes)
Total length
(nucleotides)
Nucleotide
identity
Total amino
acids
Amino acid
identity
COI CDS 1443 934 (95) 480 975 (12)
COIII CDS 756 899 (76) 251 932 (17)
CytB CDS 1080 950 (54) 359 986 (5)
rDNA fragments 2109 985 (32) na na
Legend Numbers in brackets indicate the number of single nucleotide differences na = not
applicable
141
Figure 61 Circular mapping and organization of the mitochondrial genome content of Eimeria
furonis showing three protein-coding genes (COI COIII and CytB) interspersed with large and
small subunit rRNA fragments
142
Figure 62 Circular mapping and organization of the mitochondrial genome content of Eimeria
ictidea showing three protein-coding genes (COI COIII and CytB) interspersed with large and
small subunit rRNA fragments
143
Figure 63 Pairwise comparison of coding DNA and concatenated rDNA fragment sequences of
the mitochondrial genomes of Eimeria furonis and Eimeria ictidea Legend Yellow bands
correspond to coding DNA fragments Red arrows correspond to rDNA fragments Arrows of both
colours indicate the forward or reverse direction of these segments
144
Figure 64 Phylogenetic relationships of coccidia (Eimeria furonis and Eimeria ictidea) from
domestic (Mustela putorius furo) and black-footed ferrets (Mustela nigripes) based on the complete
mitochondrial genome sequences these Eimeria species and a selection of related apicomplexan
parasites
145
CHAPTER 7 CONCLUSIONS AND FUTURE DIRECTIONS
The primary objective of this project was to better characterize the enteric coccidia of the
endangered black-footed ferret (BFF) in order to set the stage for improved disease prevention
management and treatment To the authorrsquos knowledge this work was the first attempt to isolate and
perform molecular characterization of the coccidial species endemic in the black-footed ferret population
and to characterize the natural history of the disease in this host
Two Eimeria species Eimeria ictidea and Eimeria furonis have been described from BFF and
domestic ferrets A single Eimeria species morphologically resembling E ictidea of the domestic ferret
and referred to in this work as E ictidea was identified from all historic and active cases of enteric
coccidiosis in the Toronto Zoo BFF population The same species was identified in both BFF family
groups and single-housed adults that shed coccidia during the study period (2014-2016) as well as from
historic necropsy samples of juvenile and adult Toronto Zoo BFF from 1999-2014 Furthermore this
same species was identified from both family groups and single-housed adults shedding coccidia at an
additional BFF Species Survival Plan institution the Louisville Zoo in 2016 These findings are contrary
to previous published reports that indicated multiple coccidia species were affecting captive and wild
BFF In order to determine whether these additional parasites continue to exist within the present-day
BFF population and their impact on this species further examination of coccidia-positive fecal samples
from captive and wild BFF populations is recommended Moreover determination of which parasite
species impact BFF morbidity and mortality would allow for development of targeted therapies for
disease management
Coccidia-positive fecal samples and necropsy samples from domestic ferrets were collected from
multiple diagnostic laboratories in Canada and Europe for comparison with results from BFF Eimeria
furonis and Isospora (=Cystoisospora) laidlawi were identified from fecal parasitology reports from
2008-2015 and in fecal samples obtained prospectively from 2014-2016 from domestic ferrets submitted
to Canadian and European laboratories E furonis was also identified in necropsy samples from 2010 and
146
2017 from two Canadian diagnostic laboratories No samples containing E ictidea were submitted to the
Canadian laboratories however samples containing E ictidea were identified twice by the European
diagnostic laboratory with one sample in 2011 and a second in 2013 Consequently it appears that E
ictidea is rarely identified from domestic ferrets Since no samples of E ictidea were acquired from
domestic ferrets during the study period it remains undetermined whether E ictidea from domestic
ferrets and E ictidea of BFF are conspecific Future molecular characterization of E ictidea isolated from
various mustelid host species would allow not only for determination of whether the parasites are
conspecific but would also provide insight into the potential for cross-transmission of parasites between
related mustelid hosts
The identifications provided by diagnostic laboratories of the specific parasite species present in
the domestic ferret fecal samples showed poor agreement with their identifications based on genotyping
obtained in this study In the authorrsquos opinion molecular techniques are essential tools for determining
the specific coccidial species responsible for individual and group outbreaks of coccidiosis and for
further understanding of the eimeriid host-parasite relationships To this end the nu 18S rDNA region and
whole mitochondrial (mt) genomes of E ictidea and E furonis were sequenced these mt whole genome
sequences are the first for the Eimeria species of mustelids to be entered into GenBank These sequences
may provide suitable targets for the development of highly discriminatory PCR-based methods of
identification that could be applied to fecal tissue or even formalin-fixed paraffin-embedded samples
(see Chapters 2 and 3) Methodological improvements such as less expensive next generation sequencing
methods that could be applied to diagnostics will depend on the availability of high quality reference
sequences such as generated herein Furthermore evaluation of the evolution of the eimeriid coccidia
using phylogenetic analysis based on additional whole mitochondrial genome sequences may allow for
more accurate determinations of relationships between parasite species and timing of their divergence
from common ancestors
One of the goals of this research was to investigate the possibility of using the domestic ferret as
an experimental model to study enteric coccidiosis in the BFF Experimental work cannot be carried out
147
in the BFF due to its endangered status We showed that the domestic ferret is susceptible to infection
with Eimeria ictidea originating from BFF Infected domestic ferrets showed similar clinical signs and
pathologic lesions to BFF strengthening the possibility of their serving as a suitable model Future
investigations would include studies of the domestic ferretrsquos immune response to enteric coccidial
infection in vivo drug trials including pharmacokinetic pharmacodynamic and efficacy studies of
anticoccidial medications investigations into patterns of drug resistance in coccidial species of ferrets
and oral vaccine development
Ultimately the goal of research on coccidiosis in BFF may be the development of an autogenous
vaccine to improve the survival of ferret kits and reduce coccidiosis-related morbidity and mortality in
BFF captive breeding programs It may be possible to use the domestic ferret to select for E ictidea that
are more highly adapted to this host perhaps with a commensurate loss of virulence to its original host If
this were to be the case domestic ferret-derived coccidia could be used in BFF as vaccine organisms
Alternately methods such as use of a bioshuttle (live vaccination with coccidia followed by anticoccidial
treatment to limit pathogenicity) might be useful for generating long-lasting immunological protection in
BFF against E ictidea Improving immunity through vaccination could assist in reducing coccidial
shedding and disease in BFF associated with stressful life events such a breeding weaning and transfer
between institutions There is minimal available data on the significance of coccidiosis in wild
populations of BFF and limited means of disease surveillance post release however it is logical to
assume that the stresses associated with release to the wild might result in clinical disease as happens with
stressed BFF in captivity Reducing morbidity and mortality associated with coccidiosis in BFF could
result in increased numbers of ferrets being released to the wild and higher survival of released ferrets
increasing the number of BFF in the wild supports the goals of the conservation initiative for the black-
footed ferret and will support the longer-term survival and recovery of this species
148
REFERENCES
Abe N Tanoue T Ohta G Iseki M (2008) First record of Eimeria furonis infection in a ferret Japan with
notes on the usefulness of partial small subunit ribosomal RNA gene sequencing analysis for
discriminating among Eimeria species Parasitol Res 103967ndash70 doi 101007s00436-008-1037-x
Adl SM Simpson AGB Farmer MA et al (2005) The new higher level classification of eukaryotes with
emphasis on the taxonomy of protists J Eukaryot Microbiol 52399ndash451 doi 101111j1550-
7408200500053x
Andrews JM (1926) Coccidiosis in mammals Am J Hyg 6784ndash798
Augustine PC Danforth HD (1986) A study of the dynamics of the invasion of immunized birds by
Eimeria sporozoites Avian Dis 30347ndash351
Barta JR Schrenzel MD Carreno R Rideout BA (2005) The genus Atoxoplasma (Garnham 1950) as a
junior objective synonym of the genus Isospora (Schneider 1881) species infecting birds and
resurrection of Cystoisospora (Frenkel 1977) as the correct genus for Isospora species infecting
mammals J Parasitol 91726ndash727 doi 101645GE-33411
Bell JA (1994) Parasites of Domesticated Pet Ferrets Compend Contin Educ Pract Vet 16617ndash620
Black-footed Ferret Recovery Implementation Team (2011) Captive Breeding
httpwwwblackfootedferretorgcaptive-breeding Accessed 8 Jan 2014
Blankenship-Paris TL Chang J Bagnell CR (1993) Enteric coccidiosis in a ferret Lab Anim Sci 43361ndash
363
Bronson E Bush M Viner T et al (2007) Mortality of captive black-footed ferrets (Mustela nigripes) at
Smithsonianrsquos National Zoological Park 1989 ndash 2004 J Zoo Wildl Med 38169ndash176
149
CAPC (2013) Current Advice on Parasite Control Intestinal Parasites - Coccidia
httpwwwcapcvetorgcapc-recommendationscoccidia Accessed 6 Jun 2015
Carpenter JW Hillman CN (1979) Husbandry reproduction and veterinary care of captive ferrets In
1978 Proceedings of the Annual Meeting of the American Association of Zoo Veterinarians
Washington DC pp 36ndash47
Catchpole J Norton CC Gregory MW (1993) Immunisation of lambs against coccidosis Vet Rec
13256ndash59
Cavalier-Smith T (2014) Gregarine site-heterogeneous 18S rDNA trees revision of gregarine higher
classification and the evolutionary diversification of Sporozoa Eur J Protistol 50472ndash495 doi
101016jejop201407002
Chappuis G (1998) Neonatal immunity and immunisation in early age lessons from veterinary medicine
Vaccine 161468ndash1472
Cox FE (1994) The evolutionary expansion of the Sporozoa Int J Parasitol 241301ndash1316
Davis CL Chow TL Gorham JR (1953) Hepatic coccidiosis in mink Vet Med 48371ndash375
De Vos AJ (1970) Studies on the host range of ltigtEimeria chinchillaeltIgt de Vos and Van Der
Westhuizen 1968 Onderstepoort J Vet Res 3729ndash36
Dirikolu L Yohn R Garrett EF et al (2009) Detection quantifications and pharmacokinetics of
toltrazuril sulfone (Ponazuril) in cattle J Vet Pharmacol Ther 32280ndash288
Dryden MW Payne PA Ridley R Smith V (2005) Comparison of common fecal flotation techniques for
the recovery of parasite eggs and oocysts Vet Ther 615ndash28
Duszynski D Wilber PG (1997) A Guideline for the Preparation of Species Descriptions in the
Eimeriidae J Parasitol 83333ndash336
150
Duszynski DW Couch L Upton SJ (2000) The Coccidia of the World
httpbiologyunmeducoccidiacarniv2html Accessed 1 Sep 2017
El-Sherry S Ogedengbe ME Hafeez MA Barta JR (2013) Divergent nuclear 18S rDNA paralogs in a
turkey coccidium Eimeria meleagrimitis complicate molecular systematics and identification Int J
Parasitol 43679ndash685 doi 101016jijpara201303005
Evans HE An NQ (2014) Anatomy of the ferret In Fox JG Marini RP (eds) Biology and Diseases of the
Ferret 3rd edn Wiley Blackwell pp 23ndash67
Fayer R (1980) Epidemiology of Protozoan Infections The Coccidia Vet Parasitol 675ndash103
Foreyt WJ Todd AC Hartsough GR (1977) Anticoccidial Activity of Eight Compounds in Domestic
Mink AJVR 38391ndash394
Fox JG (2014) Normal Clinical and Biological Parameters In Fox JG Marini RP (eds) Biology and
Diseases of the Ferret 3rd edn John Wiley amp Sons Inc pp 157ndash185
Frenkel JK (1977) Besnoitia wallacei of Cats and Rodents With a Reclassification of Other Cyst-
Forming Isosporoid Coccidia J Parasitol 63611ndash628
Grafner G Graubmann HD Dobbriner W (1967) Leberkokzidiose beim Nerz (Lutreola vison Schreb)
hervorgerufen durch eine neue Kokzidienart Eimeria hiepei n sp Monatshefte fur
Veterinearmedizin 22696ndash700
Haberkorn A (1971) Zur Wirtsspezifitat yon Eimeria contorta nsp (Sporozoa Eimeriidae) Z
Parasitenkd 37303ndash314
Hall MC Wigdor M (1918) Canine coccidiosis with a note regarding other protozoan parasites from the
dog J Am Vet Med Assn 5364ndash76
Hikosaka K Nakai Y Watanabe YI et al (2011) Concatenated mitochondrial DNA of the coccidian
151
parasite Eimeria tenella Mitochondrion 11273ndash278
Hillyer E V (1992) Gastrointestinal diseases of ferrets (Mustela putorius furo) J Small Anim Med 244ndash
45
Hoare CA (1927) On the coccidia of the ferret Ann Trop Med Parasitol 21313ndash320
Hoare CA (1935a) A histopathological reaction of a special type on the part of the intestinal villi in ferret
coccidiosis Trans R Soc Trop Med Hyg 292
Hoare CA (1935b) The endogenous development of the coccidia of the ferret and the histopathological
reaction of the infected intestinal villi Ann Trop Med Parasitol 29111ndash122
Hoefer HL Fox JG Bell JA (2012) Gastrointestinal Diseases In Quesenberry KE Carpenter JW (eds)
Ferrets Rabbits and Rodents Clinical Medicine and Surgery 3rd edn Elsevier Saunders St Louis
Missouri pp 27ndash45
Huelsenbeck JP Ronquist F (2001) MrBayes Bayesian inference of phylogenetic trees Bioinformatics
17754ndash755 doi 101093bioinformatics178754
Jolley WR Kingston N Williams ES Lynn C (1994) Coccidia Giardia sp and a Physalopteran
Nematode Parasite from Black-footed Ferrets (Mustela nigripes) in Wyoming J Helminthol Soc
Washingt 6189ndash94
Katoh K Misawa K Kuma K Miyata T (2002) MAFFT a novel method for rapid multiple sequence
alignment based on fast Fourier transform Nucleic Acids Res 303059ndash3066 doi
101093nargkf436
Kaye SW Ossiboff RJ Noonan B et al (2015) Biliary coccidiosis associated with immunosuppressive
treatment of pure red cell aplasia in an adult ferret (Mustela putoris furo) J Exot Pet Med doi
101053jjepm201504012
152
Koepfli KP Deere K Slater GJ et al (2008) Multigene phylogeny of the Mustelidae resolving
relationships tempo and biogeographic history of a mammalian adaptive radiation BMC Biol 610
Lariviegravere S Jennings AP (2009) Family Mustelidae (weasels and relatives) In Wilson DE Mittermeier
RA (eds) Handbook of the Mammals of the World Volume 1 Carnivores Lynx Edicions
Barcelona pp 564ndash656
Levine ND Ivens V (1970) The coccidian parasites (Protozoa Sporozoa) of ruminants
Li X Pang J Fox JG (1996) Coinfection with intracellular Desulfovibrio species and coccidia in ferrets
with proliferative bowel disease Lab Anim Sci 46569ndash571
Litster AL Nichols J Hall K et al (2014) Use of ponazuril paste to treat coccidiosis in shelter-housed
cats and dogs Vet Parasitol 202319ndash325 doi 101016jvetpar201403003
Matsubayashi M Takami K Abe N et al (2005) Molecular characterization of crane coccidia Eimeria
gruis and E reichenowi found in feces of migratory cranes Parasitol Res 9780ndash83
Meeusen ENT Walker J Peters A et al (2007) Current status of veterinary vaccines Clin Microbiol Rev
20489ndash510 doi 101128CMR00005-07
Mehlhorn H Aspock H (eds) (2008) Coccidial Drugs In Encyclopedia of Parasitology 3rd edn Springer
Berlin Heidelberg New York pp 269ndash286
Morehouse NF (1938) The Reaction of the Immune Intestinal Epithelium of the Rat to Reingection wiht
Eimeria nieschulzi J Parasitol 24311ndash317
Nukerbaeva KK Svanbaev SK (1973) Coccidia of fur bearing mammals in Kazakhstan Vestn Selrsquoskokh
Nauk Kazakh 1250ndash54
Ogedengbe JD Hanner RH Barta JR (2011) DNA barcoding identifies Eimeria species and contributes to
the phylogenetics of coccidian parasites (Eimeriorina Apicomplexa Alveolata) Int J Parasitol
153
41843ndash850
Ogedengbe ME (2015) DNA Barcoding of Apicomplexa Mitochondrial Evolution across the Phylum
University of Guelph
Ogedengbe ME El-Sherry S Ogedengbe JD et al Whatrsquos in a name Phylogenies based on combined
mitochondrial and nuclear sequences conflict with morphologically defined genera in the eimeriid
coccidia (Apicomplexa)
Ogedengbe ME El-Sherry S Whale J Barta JR (2014) Complete mitochondrial genome sequences from
five Eimeria species (Apicomplexa Coccidia Eimeriidae) infecting domestic turkeys Parasit
Vectors 7335 doi 1011861756-3305-7-335
Ogedengbe ME Hafeez MA Barta JR (2013) Sequencing the complete mitochondrial genome of Eimeria
mitis strain USDA 50 (Apicomplexa Eimeriidae) suggests conserved start positions for mtCOI- and
mtCOIII-coding regions Parasitol Res 1124129ndash4136 doi 101007s00436-013-3604-z
Pantchev N Gassmann D Globokar-Vrhovec M (2011) Increasing numbers of Giardia (but not
coccidian) infections in ferrets 2002 to 2010 Vet Rec 168519 doi 101136vrd2962
Patterson M Fox JG (2007) Parasites of Ferrets In Baker DG (ed) Flynnrsquos Parasites of Laboratory
Animals 2nd edn Wiley Blackwell pp 501ndash508
Patterson MM Fox JG Eberhard ML (2014) Parasitic Diseases In Fox JG Marini RP (eds) Biology and
Diseases of the Ferret 3rd edn Wiley Blackwell pp 553ndash572
Powers L V (2009) Bacterial and parasitic diseases of ferrets Vet Clin North Am - Exot Anim Pract
12531ndash561 doi 101016jcvex200906001
Prado ME Ryman JT Boileau MJ et al (2011) Pharmacokinetics of ponazuril after oral administration to
healthy llamas (Llama glama) Am J Vet Res 721386ndash9
154
Ruiz A Muntildeoz MC Molina JM et al (2013) Primary infection of goats with Eimeria ninakohlyakimovae
does not provide protective immunity against high challenge infections Small Rumin Res 113258ndash
266 doi 101016jsmallrumres201301006
Ryley J Meade R Hazelhurst J Robinson T (1976) Methods in coccidiosis research separation of
oocysts from faeces Parasitology 73311ndash326
Santymire R Branvold-Faber H Marinari PE (2014) Recovery of the Black-Footed Ferret In Fox JG
Marini RP (eds) Biology and Diseases of the Ferret 3rd edn Wiley Blackwell pp 219ndash231
Shi MQ Huther S Burkhardt E Zahner H (2000) Immunity in rats against Eimeria separata oocyst
excretion effects on endogenous stages and local tissue response after primary and challenge
infections Parasitol Res 86891ndash898
Sledge DG Bolin SR Lim A et al (2011) Outbreaks of severe enteric disease associated with Eimeria
furonis infection in ferrets (Mustela putorius furo) of 3 densely populated groups J Am Vet Med
Assoc 2391584ndash1588 doi 102460javma239121584
Smith NC Wallach M Petracca M et al (1994) Maternal transfer of antibodies induced by infection with
Eimeria maxima partially protects chickens against challenge with Eimeria tenella Parasitology
109551ndash557
Svanbaev SK (1956) Materials on the fauna of coccidia of wild mammals in western Kazakhstan Tr
Instituta Zool Akad Nauk Kazachskoi SSR 5180ndash191
Tavareacute S (1986) Some probabilistic and statistical problems in the analysis of DNA sequences Am Math
Soc Lect Math Life Sci 1757ndash86 doi citeulike-article-id4801403
Tenter AM Barta JR Beveridge I et al (2002) The conceptual basis for a new classification of the
coccidia Int J Parasitol 32595ndash616
155
Upton SJ (2000) Suborder Eimeriorina Leacuteger 1911 In Lee JJ Leedale GF Bradbury P (eds) An
Illustrated Guide to the Protozoa vol 1 2nd edn Allen Press Lawrence Kansas pp 318ndash339
USFWS BFF Recovery Program (2017) Black-footed Ferret Managed Care Operations Manual
(BFFMCOM)
Vermeulen AN (2005) Vaccination against coccidial parasites The method of choice In Proceeding of
the 9th International Coccidiosis Conference
Williams BH Chimes MJ Gardiner CH (1996) Biliary coccidiosis in a ferret (Mustela putorius furo) Vet
Pathol 33437ndash439 doi 101177030098589603300412
Williams ES Mills K Kwiatkowski DR et al (1994) Plague in a Black-footed (Mustela nigripes)
presence J Wildl Dis 30581ndash585
Williams ES Thome ET Appel MJG Belitsky DW (1988) Canine Distemper in Black-Footed (Mustela
nigripes) from Wyoming J Wildl Dis 24385ndash398
Wozencraft WC (2005) Order Carnivora In Wilson DE Reeder DM (eds) Mammal Species of the
World A Taxonomic and Geographic Reference 3rd edn Johnrsquos Hopkins University Press
Baltimore Maryland p 2142
Yi-Fan C Le Y Yin D et al (2012) Emendation of 2 Isospora Species (Apicomplexa Eimeriidae)
Infecting the Steppe Polecat Mustela eversmanii Lesson 1827 in China to the Genus
Cystoisospora (Apicomplexa Sarcocystidae) Comp Parasitol 79147ndash152 doi 10165445311
Yu L Peng D Liu J et al (2011) On the phylogeny of Mustelidae subfamilies analysis of seventeen
nuclear non-coding loci and mitochondrial complete genomes BMC Evol Biol 1192 doi
1011861471-2148-11-92
Zou M Guo G Zhao Y Zhang Q (2014) Detection quantifications and pharmacokinetics of ponazuril in
156
healthy swine J Vet Pharmacol Ther 37598ndash602 doi 101111jvp12126
157
APPENDICES
158
Appendix 1 Shedding of oocysts of Eimeria ictidea in black-footed ferret (Mustela nigripes) dam and kit
family groups from 2014-2016
Collection Year 2014 2014 2014 2014 2014 2015 2016
Age of kits (days) Poppy Bumblefoot Calico Aubrey Ruckus Fiddlesticks Guanella^
29 - - - 0 - - -
30 - 0 - 0 - - -
31 - 0 - 0 - - -
32 - 0 - 0 - - -
33 - 0 - 0 - - -
34 0 0 - 0 0 - -
35 0 0 - 0 0 0 -
36 0 0 - 0 0 0 -
37 - 0 - 0 - 0 -
38 - 0 - 0 - 0 -
39 - 0 - 0 - 0 -
40 0 0 - 0 0 0 -
41 - 0 - 0 - 0 -
42 0 0 - 0 0 - -
43 0 0 - 0 0 0 -
44 0 0 - - 0 0 -
45 0 0 - 0 0 - -
46 0 0 - 0 0 0 -
47 0 0 - 0 0 0 -
48 0 lt14 - 0 0 0 -
49 0 0 - 0 0 0 -
50 0 0 - - 0 0 -
51 0 0 - 0 0 0 -
52 0 0 - 0 0 0 -
53 0 0 - 0 0 0 +
54 0 0 - 0 0 0 +
55 0 0 - 0 0 0 3717146
56 0 0 - 0 - 0 1084436
57 0 0 - 0 - 0 64133
58 0 324 - 0 0 0 20654
59 0 0 - 0 0 0 0
60 0 446688 - 0 0 0 0
61 0 934828 - 0 0 0 41111
62 0 530469 - 0 0 0
63 0 1617131 - 0 0 lt14
64 0 8396357 - 114 0 0
65 0 234876 - 5368 0 lt14
66 0 374625 - 9455 0 0
67 lt14 2311575 - 39579 0 -
68 0 603563 - 125051 0 0
69 - 4103036 - 27067 0 lt14
70 2470327 7759974 - 0 0 0
71 1073085 - 1784 0 0
72 3430966 - 0 0 0
73 4459536 - 7326 0 0
74 4842212 - 2 0 0
75 3627307 - 0 0 0
76 5801885 - 0 0 0
77 5994 0 0 0 0
78 589835 0 0 0
Numbers of Oocysts Shed (oocysts per gram of feces)
Dam Identity
159
Appendix 1 continued Shedding of oocysts of Eimeria ictidea in black-footed ferret (Mustela nigripes)
dam and kit family groups from 2014-2016
Collection Year 2014 2014 2014 2014 2014 2015 2016
Age of kits (days) Poppy Bumblefoot Calico Aubrey Ruckus Fiddlesticks Guanella^
79 24815 0 0 0
80 144016 0 0 0
81 97862 0 0 0
82 0 0 0
83 0 0 0
84 0 0 0
85 0 0 0
86 0 0 0
87 0 0 0
88 0 0 0
89 0 0 0
90 0 0 0
91 0 0 0
92 0 - -
93 0 - 0
94 0 - 0
95 0 0 -
96 0 - 0
97 0 0 -
98 0 0 0
99 0 0 0
100 0 0 0
101 0 0 0
102 0 0 0
103 0 0 0
104 0 0 0
105 0 0 -
106 0 0 0
107 0 0 0
108 - 0 0
109 0 0 -
110 0 0 0
111 0 0 -
112 0 0 0
113 0 0 -
114 0 0 -
115 0 0 -
116 0 0 -
117 0 -
118 0 -
119 0 0
120 0 0
121 0 0
122 0 -
Numbers of Oocysts Shed (oocysts per gram of feces)
Dam Identity
160
Appendix 1 continued Shedding of oocysts of Eimeria ictidea in black-footed ferret (Mustela nigripes)
dam and kit family groups from 2014-2016
Collection Year 2014 2014 2014 2014 2014 2015 2016
Age of kits (days) Poppy Bumblefoot Calico Aubrey Ruckus Fiddlesticks Guanella^
123 0 0
124 0 -
125 0 -
126 0 0
127 0 0
128 0 2843
129 0 0
130 0 -
131 0 0
132 0 0
133 0 0
134 0 0
135 0 0
136 0 0
137 0
138 0
139 0
140 0
141 0
142 0
143 0
144 0
145 0
146 -
147 0
148 0
149 0
150 0
Legend lt 14 = oocyst positive samples with less than 14 oocyst per gram of feces - = no sample recorded for this date
underline = last sampling date + = coccidia present but OPG count not performed = Toronto Zoo ferret ^ = Louisville Zoo
ferret thick outer border = days treatment was received
Numbers of Oocysts Shed (oocysts per gram of feces)
Dam Identity
161
Appendix 2a Hematology values for domestic ferrets (Mustela putorius furo) from 49-51 days of age
prior to experimental inoculation
Test Reference
Intervala
Reference
Intervalb
101 102 103 104 105 201 202 203 204 205
Age (days) adult 70 50 50 50 50 50 48 48 48 50 48
WBC (x 109L) 27-112 53-120 86 64 77 97 76 48 8 72 104 76
RBC (x 1012
L) 50-108 55ndash74 66 58 45 5 48 47 67 5 5 42
Hb (gL) 87-177 104ndash136 121 106 90 58 94 91 122 96 98 80
HCT (LL) 04 - 051 029ndash037 037 033 027 030 029 027 038 030 030 024
MCV (fL) 44-52 478ndash548 55 56 60 60 60 58 57 59 61 58
MCH (pg) 15-18 175ndash191 18 18 20 12 20 20 18 19 20 18
MCHC (gL) 325-362 347ndash370 331 327 328 196 325 337 321 327 324 331
RDW () 12-16 - 134 127 139 139 133 131 122 136 131 127
Platelets (x 109L) 54-695 629ndash775 777 329 529 480 640 402 534 336 527 363
MPV (fL) 5-10 - 78 78 96 75 66 74 76 81 74 82
TS Protein (gL) 49-76 - 54 51 - - - - - - - -
Seg Neuts (x 109L) 1-8 15ndash48 292 141 216 281 205 187 152 151 354 251
Lymphocytes (x 109L) 1-63 28ndash63 525 416 516 64 509 254 608 468 645 456
Monocytes (x 109L) 0-09 01ndash05 026 07 031 039 038 034 024 094 031 038
Eosinophils (x 109L) 0-13 01ndash06 009 013 008 01 008 005 016 007 010 015
Basophils (x 109L) 0-02 0 009 0 0 0 0 0 0 0 0 0
Polychromasia 2-5 - 5-10 2-5 10-15 10-15 10-15 10-15 2-5 10-15 10-15 5-10
Anisocytosis Occ 1+ 1+ 1+ 1+ 1+ 1+ 1+
HJ bodies rare rare rare rare rare
crenation Occ
poikilocytosis Occ Occ Occ
shift platelets Occ Occ
hemolysis Neg Neg Neg Neg Neg Neg Neg Neg Neg Neg Neg
lipemia mild mild mild mild mild Neg Neg Neg mild mild
Ferret Identity
Legend - = reference value unavailable bolded = outside reference range a = adult ferret reference ranges from Animal Health Laboratory
(University of Guelph Guelph Canada) b
= reference ranges for 10-week old ferrets (Fox JG 2014) Neg = negative Occ = occasional
162
Appendix 2b Serum biochemistry values for domestic ferrets (Mustela putorius furo) from 49-51 days of
age prior to experimental inoculation
Test Reference
Intervala
Reference
Intervalb
101 102 103 104 105 201 202 203 204 205
Age (days) adult 70 50 50 50 50 50 48 48 48 50 48
Calcium (mmolL) 185-242 253-302 239 233 241 244 24 221 242 234 253 242
Phosphorus (mmolL) 112-256 278-475 239 267 29 264 31 218 286 284 315 303
Magnesium (mmolL) 08-139 - 08 08 07 08 08 06 08 07 08 08
Sodium (mmolL) 147-159 146-154 149 149 149 149 148 144 153 148 152 152
Potassium (mmolL) 37-57 47-83 44 46 42 46 48 45 47 44 47 46
Chloride (mmolL) 111-129 115-121 110 112 115 113 113 110 119 117 117 120
Carbon dioxide (mmolL) 17-29 13-27 18 18 18 17 17 16 15 15 19 17
Anion gap (mmolL) 6 - 23 - 25 24 20 24 23 23 24 20 21 20
NaK ratio - - 34 32 35 32 31 32 33 34 32 33
Total protein (gL) 51-75 44-56 49 46 44 47 44 41 52 45 49 43
Albumin (gL) 24-40 26-32 29 26 28 28 28 25 28 27 29 25
Globulin (gL) 19-41 17-24 20 20 16 19 16 16 24 18 20 18
AG ratio 053-167 13ndash12 145 130 175 147 175 156 117 15 145 139
Blood urea nitrogen (mmolL) 45-153 71-139 83 89 107 11 136 73 119 105 118 135
Creatinine (umolL) 8-67 53-124 50 58 68 59 50 7 68 41 62 92
Glucose (mmolL) 32-91 64-138 47 54 54 42 55 53 57 56 52 59
Cholesterol (mmolL) 294-894 619-860 413 386 375 393 344 349 227 357 373 346
Total bilirubin (umolL) 2 to 7 - 1 L 0 L 0 0 0 1 0 0 0 0
Conjugated biilrubin (umolL) - 0-10 1 0 0 1 1 1 0 0 1 0
Free bilirubin (umolL) 0-2 0-15 0 0 0 0 0 0 0 0 0 0
ALKP (UL) 13-237 117ndash277 180 169 172 215 175 168 241 184 177 179
GGT (UL) 0-40 2ndash20 1 1 6 10 4 0 1 1 5 9
AST (UL) - 63ndash152 61 58 48 61 64 59 93 61 58 69
ALT (UL) 39-196 95ndash544 95 105 89 105 106 82 234 137 115 156
CK (UL) 74-294 - 513 330 496 560 530 492 793 539 479 724
Amylase (UL) - - 23 28 35 35 29 28 29 24 36 23
Lipase (UL) - - 65 63 60 64 62 56 67 69 60 68
Calculated osmo (mmolL) - - 298 300 301 301 303 289 311 300 308 311
Ferret Identity
Legend - = reference value unavailable bolded = outside reference range a = adult ferret reference ranges from Animal Health Laboratory
(University of Guelph Guelph Canada) b = reference ranges for 10-week old ferrets (Fox JG 2014)
163
Appendix 3a Hematology values for domestic ferrets (Mustela putorius furo) inoculated orally with
Eimeria ictidea
Test Reference
Intervala
Reference
Intervalb
102 103 104 105 201 203 205
Age (days) adult 98-112 92 91 99 99 97 74 92
Days post inocculation 8 7 15 15 15 11 10
WBC (x 109L) 27-112 52ndash150 94 124 81 85 88 85 142
RBC (x 1012
L) 50-108 62ndash92 71 62 73 60 65 63 65
Hb (gL) 87-177 127ndash159 122 110 122 98 103 114 102
HCT (LL) 04-051 030ndash043 037 033 037 031 032 035 032
MCV (fL) 44-52 50ndash54 53 53 52 51 50 55 49
MCH (pg) 15-18 16-21 17 18 17 16 16 18 16
MCHC (gL) 325-362 351ndash426 328 332 325 319 320 326 317
RDW () 12-16 - 122 126 129 129 145 127 145
Platelets (x 109L) 54-695 376ndash610 524 413 445 117 303 42 429
MPV (fL) 5-10 - 8 78 72 66 67 14 8
TS Protein (gL) 49-76 - 66 63 58 66 55 59 59
Seg Neuts (x 109L) 1-8 21ndash62 160 446 203 170 211 170 554
Lymphocytes (x 109L) 1-63 16ndash79 761 657 551 646 59 646 682
Monocytes (x 109L) 0-09 01ndash02 019 087 024 026 044 026 085
Eosinophils (x 109L) 0-13 03ndash09 0 037 032 009 026 009 099
Basophils (x 109L) 0-02 0 0 012 0 0 009 0 0
Polychromasia 2-5 - 1-3 0-2 0-2 1-3 1-3 2-5 5-10
Anisocytosis Occ Occ Occ
HJ bodies rare
crenation
rouleaux
poikilocytosis
shift platelets Occ Occ
hemolysis Neg
Ferret Identity
Legend - = reference value unavailable bolded = outside reference range a = adult ferret reference ranges from Animal Health
Laboratory (University of Guelph Guelph Canada) b
= reference ranges for 10-week old ferrets (Fox JG 2014) Neg = negative
Occ = occasional = many platelet clumps
164
Appendix 3b Serum biochemistry values for domestic ferrets (Mustela putorius furo) inoculated orally
with Eimeria ictidea
Test Reference
Intervala
Reference
Intervalb
102 103 104 105 201 203 205
Age (days) adult 98-112 92 91 99 99 97 74 92
Days post inocculation 8 7 15 15 15 11 10
Calcium (mmolL) 185-242 245-268 243 222 238 233 232 236 240
Phosphorus (mmolL) 112-256 200-323 292 223 248 298 268 288 306
Magnesium (mmolL) 08-139 - 06 08 08 08 09 08
Sodium (mmolL) 147-159 148-155 147 150 150 148 148 150
Potassium (mmolL) 37-57 45-55 42 41 51 41 50 44
Chloride (mmolL) 111-129 114-124 114 113 118 112 111 115
Carbon dioxide (mmolL) 17-29 16-24 16 21 19 20 12 18
Anion gap (mmolL) 6 - 23 - 21 20 18 20 30 21
NaK ratio - - 35 37 29 36 30 34
Total protein (gL) 51-75 49-64 52 55 55 49 56 57
Albumin (gL) 24-40 30-36 28 20 27 27 26 29 27
Globulin (gL) 19-41 19-30 32 28 28 23 27 30
AG ratio 053-167 11-17 063 096 096 113 107 090
Blood urea nitrogen (mmolL) 45-153 50-150 92 87 119 92 110 98 149
Creatinine (umolL) 8-67 706-1414 56 26 49 40 50 46 79
Glucose (mmolL) 32-91 688-943 52 59 64 71 58 17 66
Cholesterol (mmolL) 294-894 440-640 453 429 262 475 370 285
Total bilirubin (umolL) 2 to 7 - 0 0 0 0 1 1
Conjugated biilrubin (umolL)- - 1 0 0 0 0 0
Free bilirubin (umolL) 0-2 - 0 0 0 0 1 1
ALKP (UL) 13-237 41-181 124 120 213 120 146 170 196
GGT (UL) 0-40 1-2 2 1 3 0 0 2
AST (UL) - 47-128 48 95 104 66 100 100
ALT (UL) 39-196 78-279 133 110 140 203 158 183 281
CK (UL) 74-294 - 382 765 1190 578 680 930
Amylase (UL) - - 24 37 31 28 33 35
Lipase (UL) - - 60 65 72 72 86 79
Calculated osmo (mmolL) - - 296 305 305 300 296 309
Ferret Identity
Legend - = reference value unavailable bolded = outside reference range a = adult ferret reference ranges from Animal Health
Laboratory (University of Guelph Guelph Canada) b
= reference ranges for 10-week old ferrets (Fox JG 2014)
165
Appendix 4 Domestic ferret (Mustela putorius furo) weekly monitoring sheet
Mon
8 AM
Mon
4 PM
Tues
8 AM
Tues
4 PM
Wed
8 AM
Wed
4 PM
Thurs
8 AM
Thurs
8 PM
Fri
8 AM
Fri
4 PM
Sat
8 AM
Sat
4 PM
Sun
8AM
Sun
4 PM
Mentation
Weight (g)
Respiratory
Rate
Vomit
(+ ++ +++)
Diarrhea
(+ ++ +++)
Urination
(+ ++ +++)
Defecation
(+ ++ +++)
Food
offered
Food
remaining
Water remaining
(ml)
Treatments
Other
observations
Initials of
observer
166
Animal ID ________________________________________ Week ______________
Monitoring Criteria
Mentation BAR (bright alert responsive) QAR (quiet alert responsive) depressed lethargic quiet
Weight measured in grams to be performed once weekly (pre-inoculation) and once daily (post-inoculation)
Respiratory rate measured in breaths per minute
Vomit + (small amount) ++ (moderate amount) +++ (large amount)
Diarrhea + (small amount) ++ (moderate amount) +++ (large amount)
Urination + (small amount) ++ (normal amount) +++ (large amount)
Defecation + (one pile) ++ (two piles) +++ (3+ piles)
Food offered Y (yes) N (no)
Food remaining A (all) P (partial) N (none)
Please note any additional observations in the appropriate section
Monitoring Times
Pre-inoculation ndash ONCE daily at 8am during cage cleaningfeeding
Post-inoculation ndash TWICE daily at 8am and 4pm
Critical patients (as determined by veterinary examination) ndash 3-6 times daily (based on veterinarian recommendation) ndash switch to 24
hour care sheet
Intervention Points
If any ferret develops vomiting diarrhea goes off food or exhibits gt 3 weight loss both the PI and graduate student listed below are
to be notified and the animal will be assessed to determine further treatment plans
Removal Criteria
If an individual ferret develops severe gastrointestinal or systemic disease during the course of the study as characterized by abnormal
mentation marked dehydration 10 weight loss within 24-48 hours profuse diarrhea unresponsive to treatment anorexia of 2 or
more days or frequent vomiting unresponsive to treatment the individual will be removed from the study and humanely killed
167
Appendix 5 Domestic ferret (Mustela putorius furo) 24 hour intensive monitoring sheet
Animal ID ___________________________________________ Date___________________
0700 800 900 1000 1100 1200 1300 1400 1500 1600 1700 1800
Mentation
Weight (g)
Heart Rate
Respiratory Rate
Temperature (C)
Water offered
(mL)
Water consumed
(mL)
Food offered
(type and
amount in
grams)
Food consumed
(type and
amount in
grams)
Presence of urine
Presence of feces
168
Character of
feces
Presence of
vomit
Character of
vomit
Medications to
be administered
Other
observations
169
1900 2000 2100 2200 2300 2400 100 200 300 400 500 600
Mentation
Weight (g)
Heart Rate
Respiratory Rate
Temperature (C)
Water offered
(mL)
Water consumed
(mL)
Food offered
(type and amount
in grams)
Food consumed
(type and amount
in grams)
Presence of urine
Presence of feces
Character of
feces
Presence of vomit
Character of
vomit
170
Medications to be
administered
Other
observations
Monitoring Criteria
Mentation BAR (bright alert responsive) QAR (quiet alert responsive) depressed lethargic quiet
Weight measured in grams to be performed once weekly (pre-inocculation) and once daily (post-inocculation)
Respiratory rate measured in breaths per minute
Vomit + (small amount) ++ (moderate amount) +++ (large amount)
Diarrhea + (small amount) ++ (moderate amount) +++ (large amount)
Urination + (small amount) ++ (normal amount) +++ (large amount)
Defecation + (one pile) ++ (two piles) +++ (3+ piles)
171
Food offered Y (yes) N (no)
Food remaining A (all) P (partial) N (none)
Please note any additional observations in the appropriate section
Monitoring Times
Critical patients (as determined by veterinary examination) ndash 3-6 times daily (based on veterinarian recommendation) ndash switch to 24
hour care sheet
Intervention Points
If any ferret develops vomiting diarrhea goes off food or exhibits gt 3 weight loss both the PI and graduate student listed below are
to be notified and the animal will be assessed to determine further treatment plans
Removal Criteria
If an individual ferret develops severe gastrointestinal or systemic disease during the course of the study as characterized by abnormal
mentation marked dehydration 10 weight loss within 24-48 hours profuse diarrhea unresponsive to treatment anorexia of 2 or
more days or frequent vomiting unresponsive to treatment the individual will be removed from the study and humanely killed
172
Appendix 6 Domestic ferret (Mustela putorius furo) infection trial standard operating procedures
Daily Fecal Collection
1 Collection of all feces passed in a 24 hour period will be performed once daily for all ferrets from
July 13 to July 24 inclusive
2 CAF Isolation staff will collect all fecal material present at the time of daily cage cleaning and
place in individual pre-labelled plastic bags (one per cage)
3 The amount of non-fecal matter (bedding etc) collected should be as minimal as possible
4 CAF isolation staff will record fecal character observations daily for each sample by ticking the
appropriate box on the baggie label
5 Isolation staff will place samples in the necropsy cooler (4 degC) while awaiting collection by
summer student
6 Monday to Friday fecal samples will be collected by A Rodriguez and brought for processing to
the Barta Lab in Pathobiology Samples collected Saturday and Sunday will be put on hold in a
refrigerator (4degC) until pick up on Monday
Example Bag label
Ferret ID Date
Weight of Feces
Fecal Character Normal
Soft
Liquid
Bloody
Abnormal odour
Physical Examination and Health Assessments
1 Manual restraint by hand or in towel by CAF Isolation staff and project personnel If required
examination +- blood collection may be performed under general anesthesia with isoflurane
2 Physical examination body weight HR RR temperature to be performed by PIs (Adriana Pastor
and Dale Smith)
3 Blood collection (~1mL per ferret) is to be performed from the jugular vein if under manual
restraint or the cranial vena cava under anesthesia by PIs using a 25g needle and 1cc syringe
Blood will be collected into small heparinized tubes and submitted to AHL for
CBCBiochemistry
4 All physical examination findings will be recorded on the Exam Sheet
5 Any minor wounds will be treated as appropriate (to be determined by PIs)
Inoculation of Ferrets with Coccidia
1 All ferrets to be inoculated will have been confirmed negative on daily fecals for two weeks
173
2 Brief physical examination by PIs to confirm that the ferrets are healthy to continue in study
3 The concentrated oocyst solution in sterile saline (up to a volume of 1mL) will be combined with
up to 1 mL of FerreTone (or another highly palatable substance if more preferred by the ferrets)
and ferrets will be allowed to consume the mixture ad lib while being monitored
4 Control ferrets will be administered saline only (equal mL to oocyst solution) with 1 mL of
FerreTone (or other substance as used for experimental group)
5 If ferrets refuse to consume the mixture oral inoculation via syringe of concentrated oocyst
solution will be performed by PIs under manual restraint
6 If ferrets are resistant to manual restraint for oral inoculation then inoculation will be performed
by PIs under general anesthesia via gastric tube (8 Fr red rubber)
Euthanasia Protocol
1 Ferrets to be euthanized will be masked down with isoflurane under manual restraint (or in an
anesthetic chamber as judged most appropriate by the PI)
2 Once anesthetized the ferret will be weighed and any blood fecal or other samples will be
collected as required (as determined by PIs)
3 Once an adequate plane of anesthesia is obtained (as determined by PIs) potassium chloride at a
dose of 2 mEq K+kg will be administered either via vena caval puncture or cardiac puncture to
induce cardiac arrest
4 Presenceabsence of respirations heartbeat corneal reflex will be used to assess death
Necropsy Protocol
1 Measure the ferret from nose to tail base (body length)
2 Perform standard necropsy but start with gastrointestinal tract first then thoracic and abdominal
viscera
3 Gently flush the entire contents of the intestine with 12 mL saline into a sterile urine cup
Potassium dichromate should be added in a 11 volumevolume ratio to the same container and
mixed with the combined intestinal contents and saline Label the container with the ferretrsquos ID
ferret group ID date and place on appropriate shelf in the refrigerator (4 degC) in the Barta lab
4 Measure the length of the gut from duodenum to anus
5 Collect paired sections of intestine for histological sectioning and frozen These sections should
be collected along the entire length of the gut from duodenum to rectum (see below for GI length
calculations)
6 For histological sections cut a 2 cm long section of bowel open completely on one side place on
a pre-cut section of box-board and staple both ends to the board to create a flat section Place all
gastrointestinal sections (attached to the board) in Serra fixative solution (100 ethanol (60
VV) 37 formaldehyde (30 VV) glacial acetic acid (10 VV)) for fixation and trim in
cross (transverse) sections
7 Adjacent to each sample removed for histopathology remove another 2 cm section intact and
place in a labelled Whirl-pack for freezing
8 Box-board and pre-labelled Whirl-packs should be labelled in pencil and sharpie respectively
with the following information Animal ID Zoo pm number section of gut (information should
include the region of the gut - jejunum colon etc and the length from the pylorus to the section)
174
Ferret gastrointestinal length calculations (from Evans amp An 2014)
Adult ferret body length 36-41 cm
NB In domestic ferrets there is ~51 ratio of small intestine to body length
Adult domestic ferret GI lengths
Small intestine ndash 182-198 cm
Large intestine - ~10 cm long 06 cm diameter (colon ndash 7 cm rectum ndash 2 cm anus ndash 1 cm)
Based on the above information
Small intestine Six sections of the small intestine from duodenum to ileum will be collected each ~ 25-40
cm apart depending on the size of the ferret
Large intestine Collect two sections of colon (4 cm each) at 25 and 75 of the length of the colon from
the junction of the small and large intestine to the rectum Smaller ferrets may allow only on section
based on colon length
Laboratory SOPs
General notes on processing fecal samples
1 Samples will be collected from the isolation facility necropsy room cooler daily from Monday-
Friday for the entire 6 weeks of the project
2 Upon transfer to the lab each baggie will be weighed after filling to determine the amount of
feces (grams) in each bag and that number entered onto the label on the bag
3 All information regarding fecal processing will be entered onto the spreadsheet for the individual
ferret including ID weight of feces feces character date of sample collection date of sample
processing fecal flotation method presence or absence of oocysts oocyst quantification
Fecal Processing - Weeks 1+2 (acclimation period)
1 After weighing transfer the contents of one baggie to a small sieve on top of a paper cup
2 Fill a single glass container (for fecal flotation) with saturated salt solution
3 Slowly pour small amounts of the solution over the feces mix and strain liquid contents in the paper
cup using a tongue depressor
4 Dispose of the remaining fecal matter in the sieve
5 Pour the contents of the paper cup back into the glass container and cover slip for 5-7 minutes
6 Place coverslip on a clean glass slide and viewscan under microscope at 10x power for presence of
oocysts
7 Record all findings on the provided spreadsheet
8 Notify A Pastor if any oocysts are detected If detected contents of the slide should be flushed back
into the glass container with distilled water and contents of glass container should be transferred to a
50 mL plastic conical tube Clean to remove salt as per Barta lab SOP and mix with potassium
dichromate equal parts by volume to amount of oocysts in diH2O Place a checkmark on the lid of the
50 mL conical vial containing the remainder of the feces and store in the lab fridge at 4 degC
175
Fecal Processing - Weeks 3-7 (infection trial)
1 After weighing transfer the entire contents of one baggie to a small sieve on top of a paper cup
2 Slowly pour small amounts of distilled water over the feces (enough to wet) mix and strain liquid
contents in the paper cup using a tongue depressor until feces appear almost dry
3 Dispose of the remaining fecal matter in the sieve
4 Pour the contents of the paper cup into a 50 mL conical tube If samples are not going to be counted
on the same day then mix potassium dichromate (25 wv) 11 by volume with sieved fluid from
the paper cup and store at 4 degC (refrigerator) until ready to perform OPG counts
If proceeding with the count the same day
5 Pipette and place one drop of the sieved fecal fluid on a slide to determine approximate oocyst
concentration If oocysts rare to none ndash dilute 12 during step 6 if moderate numbers ndash dilute 19 if
too numerous to count ndash dilute 199 (or perform serial dilutions of 10x from initial 19 dilution)
6 Transfer x mL of the mixture from the 50 mL conical tube into a clean 15 mL conical tube and mix
with appropriate amount of saturated salt solution for desired dilution
7 Fill both sides of the McMaster counter chamber and count the number of oocysts per side (total for
one side= total number of oocysts from all 6 sections of the chamber )
8 Average the total counts from both sides
9 Use the following calculation to determine the oocyst per gram count
OocystsmL = oocysts counted times 666 times dilution (ie 3 if dilution 12)
OPG= oocystmL times total volume recorded at end of step 4
10 Record all findings on the provided spreadsheet
11 Notify A Pastor if any oocysts are detected If detected place a checkmark on the lid of the 50 mL
conical vial containing the remainder of the feces and store in the lab fridge at 4 degC
12 For all samples for which no oocysts are detected during steps 6-8 follow up with routine salt
flotation of the remainder of the fecal sample from step 6 (use instructions for fecal processing from
weeks 1-2)

iv
DEDICATION
For my mother Anna Pastorhellip
v
ACKNOWLEDGEMENTS
Itrsquos hard to believe that my residency and thesis have been completed and I have a lot of people
to thank for that
Dale and Graham the two people I wanted to be when I grew up thank you for your mentorship
for many years even before this program I know that you werenrsquot convinced that this project was DVSc
worthy when I first proposed it but Irsquom hoping that the results have changed your mind
I am sincerely grateful to all the members of my advisory committee Dale Smith John Barta and
Simon Hollamby for their insight support and interest in this project Dale you have been an exceptional
advisor I donrsquot know that I will ever get to your level but thank you for showing me that being a great
clinical zoo vet and pathologist are not mutually exclusive John thank you for spontaneously agreeing to
be my advisor when I came to you with this project proposal in my first semester for your energy and
enthusiasm and for supporting my widening interest in parasitology research
My heartfelt appreciation for the Toronto Zoo WHC veterinarians past and present Chris
Dutton Pauline Delnatte Simon Hollamby and Graham Crawshaw I have learned so much from all of
you that I will take forward into my future endeavours I appreciate the extra time you put in including
comps study sessions after-hours tecircte-agrave-tecirctes and the fact that your doors were open when I needed it
For the Toronto Zoo vet techs extraordinaire Michelle Lovering Cassia Devison Dawn
Mihailovic and Tasha Long ndash you have been indispensable during this program and there are not enough
words to express my gratitude
I would especially like to thank all the Wildlife Health Center staff (2013-2016) Mark Bongelli
Charles Guthrie Christine McKenzie Brian Telford Rick Vos Gerri Mintha Margaret Kolakowski
Andrew Lentini Rebecca Clark Lydia Attard Nigel Parr Paula Roberts Andrea Dada Mindy Waisglass
and Julie Digiandomenico for three very memorable years It is all of you that make the WHC such an
amazing place to be Irsquom not sure I have laughed so hard or so often as I did in that lunchroom and I hope
our paths will cross again
vi
I donrsquot think that I can truly express how thankful I am to Pathobiology laboratory technicians
Julie Cobean and Julia Whale Without your assistance patient teaching and friendship I would probably
still be screening fecal samples years from now and scratching my head as to how our lab protocols
actually work It is people like you who make sure graduate students become successful doctorates and I
canrsquot imagine Pathobio without you both in it
I would also like to thank my labmates in the Barta lab mdash Mian Hafeez Evelyn Rejman Rachel
Imai Perryn Kruth Ryan Snyder and Mosun Ogedengbe A special thank you goes to Alex Leveille
without whom my many adventures in parasitology research from coccidia to Babesia would not have
been as successful
To all the students who helped with ferret fecal sample processing data compilation and
necropsies Nathalie Ferriman Janessa Price Thisuri Eagalle Sarah Brisson thank you so much for your
hard work and excitement about my projecthellip even when it was very smelly
So many thanks to the amazing staff of Central Animal Facility - Linda Groocock Vicky Carson
Tony Cengija and Mary Fowler for the daily care and enrichment of my experimental ferrets Your
excitement about working with our ferrets and your assistance with all parts of the process helped made
this project a success
To Adriana Nielsen who was not only my better half but the other fifty percent of my brain for
several years It is your friendship fortitude and our endless phone conversations that got me through the
never-ending Toronto-Guelph commute and this program
To all the ldquoscope roomrdquo pathology co-residents past and present - thank you for being wonderful
friends and colleagues It is indeed rare to find so many amazing people in one place and I know this
program and my sanity would not have been the same without you
To the anatomic pathology faculty and senior graduate students - thank you for all the time
teaching and guidance you provided during my program While I canrsquot say that I have become an amazing
pathologist I can say that because of your mentorship I am a better diagnostician and the type of clinician
who asks better questions takes better samples and understands that you canrsquot ldquojust make a PCR for thatrdquo
vii
A special thank you to Tony van Dreumel who came out of retirement for a semester to try to teach the
Adrianas zoo pathology screening cases with you was always a pleasure
To all the lovely Histo Ladies PM room staff and the other AHL staff who helped me with
Toronto Zoo and HSC pathology cases along the way - I donrsquot think the anatomic path students could
survive without you Thank you for always smiling assisting and accommodating me even when I made
near-impossible processing requests during my weekly Guelph visits
I would also like to acknowledge and sincerely thank all the individuals who helped with resource
and sample acquisition for this project A special mention for those who went above and beyond because
of their interest in this project Don Duszynski who was instrumental in acquiring and then providing a
translator for many of the original mustelid Eimeria descriptions and Majda Globokar Nikola Pantchev
and Donald Martin who supplied my domestic ferret fecal samples and historical data
A shout-out to Julie Swenson Gary West and the Phoenix Zoo BFF team who fostered my love
of this endangered species and helped develop the idea for this project
As always I continue to go out into the world and pursue my dreams with the knowledge that I
have the support of my incredible family long-time friends and my partner Keith Morris I am so lucky
that my residency brought me home and that it afforded us all more time spent together For my aunt
Veronica Lacey who has never failed to believe in my potential and always pushed me to become an
academic ndash yoursquoll never get that PhD from me but I think this is pretty close Finally for my mother
Anna Pastor who never lived to see my greatest achievements but had absolute faith that I could reach
any goal I worked towardshellip this is for you
Finally none of this would have been possible without the generous support of the Toronto Zoo
Residency program and funding through the Barta Laboratory University of Guelph
Adriana Pastor
Toronto August 2017
viii
DECLARATION OF WORK PERFORMED
I declare that all the work reported in this thesis was performed by myself with the following
exceptions
Fecal samples were collected by personnel at the Toronto Zoo Louisville Zoo and participating
diagnostic laboratories
Fecal oocyst per gram counts (routine salt flotation and McMaster counts) were performed by
myself Julie Cobean Julia Whale Evelin Rejman Sarah Brisson Adriana Rodriguez and Perryn Kruth
Whole mitochondrial genome PCR and sequencing was performed by me in conjunction with
Julia Whale and Dr Mian Hafeez
Sequencing of PCR samples was performed at the University of Guelph Laboratory Services
(Guelph Ontario Canada) and results were obtained electronically
ix
TABLE OF CONTENTS
ABSTRACT ii
DEDICATION iv
ACKNOWLEDGEMENTS v
DECLARATION OF WORK PERFORMED viii
TABLE OF CONTENTS ix
LIST OF TABLES xiii
LIST OF FIGURES xiv
LIST OF APPENDICES xv
ABBREVIATIONS xvi
CHAPTER 1 LITERATURE REVIEW 1
11 INTRODUCTION 1
12 APICOMPLEXA 1
121 Brief introduction to apicomplexan pathogens 1
122 Life cycles of the Eimeria and Isospora species implicated in enteric coccidiosis 3
123 Methods of characterization 5
13 RECLASSIFICATION OF MAMMALIAN ISOSPORA 8
14 EIMERIID SPECIES CHARACTERIZED IN MUSTELIDS 8
141 The family Mustelidae 8
142 Eimeriid coccidia described from mustelids 9
143 Eimeriid coccidia described from domestic ferrets 16
144 Molecular characterization 19
145 Clinical signs of disease in domestic ferrets 21
146 Gross necropsy and histologic findings 21
15 INTRODUCTION TO ENTERIC COCCIDIOSIS IN THE BLACK-FOOTED FERRET
25
151 Natural history and conservation of the black-footed ferret in North America 25
152 Coccidia identified from black-footed ferrets 26
153 Morbidity mortality and clinical signs associated with enteric coccidiosis in black-footed
ferrets 28
16 TREATMENT PREVENTION AND CONTROL OF INFECTION BY EIMERIA SPP 29
161 Current recommendations for treatment of eimeriid coccidia in carnivores 29
x
162 Current recommendations for anticoccidial treatment and prophylaxis in domestic and
black-footed ferrets 30
17 VACCINES AGAINST COCCIDIA 32
171 Theory 32
172 Species successes in anticoccidial vaccination 34
18 RESEARCH GOALS AND OBJECTIVES 36
181 Objectives 36
182 Hypotheses 36
183 Applications 36
CHAPTER 2 MOLECULAR CHARACTERIZATION OF ENTERIC COCCIDIA FROM DOMESTIC
FERRETS (MUSTELA PUTORIUS FURO) 38
21 INTRODUCTION 39
22 MATERIALS amp METHODS 43
221 Fecal samples 43
222 Formalin fixed intestinal tissues 44
223 Molecular characterization 44
224 Phylogenetic analysis 46
23 RESULTS 47
231 Fresh fecal samples 47
232 Formalin fixed samples 48
233 Molecular characterization 49
234 Phylogenetic analysis 50
24 DISCUSSION 50
CHAPTER 3 MORPHOLOGICAL AND MOLECULAR CHARACTERIZATION OF ENTERIC
COCCIDIA ISOLATED FROM BLACK-FOOTED FERRETS (MUSTELA NIGRIPES) 60
31 INTRODUCTION 60
32 MATERIALS AND METHODS 64
321 Fecal samples 64
322 Formalin fixed intestinal tissues 65
323 Molecular characterization 66
33 RESULTS 66
331 Morphometric characterization 67
332 Molecular characterization 68
34 DISCUSSION 69
xi
CHAPTER 4 NATURAL HISTORY OF ENTERIC COCCIDIOSIS IN THE BLACK-FOOTED
FERRET (MUSTELA NIGRIPES) 78
41 INTRODUCTION 78
42 MATERIALS AND METHODS 80
421 Toronto Zoo BFF breeding program 80
422 Fecal oocyst evaluation 81
423 Retrospective review of pathology records 82
424 Prospective modified necropsy protocol 82
425 Retrospective medical history review 83
43 RESULTS 83
431 Fecal oocyst evaluation and retrospective medical history review 83
432 Pathology 86
433 Morbidity and mortality 88
44 DISCUSSION 88
CHAPTER 5 EVALUATING THE DOMESTIC FERRET (MUSTELA PUTORIUS FURO) AS AN
EXPERIMENTAL MODEL FOR ENTERIC COCCIDIOSIS IN THE BLACK-FOOTED FERRET
(MUSTELA NIGRIPES) 104
51 INTRODUCTION 104
52 MATERIALS AND METHODS 106
521 Animal care 106
522 Oocyst preparation 107
523 Experimental infections 108
524 Animal welfare 109
525 Hematology 110
526 Morphologic and molecular characterization 110
527 Necropsy protocol 111
53 RESULTS 111
531 Oocyst shedding 112
532 Morphologic and molecular characterization 113
533 Clinical signs 113
534 Hematology 113
535 Necropsy 114
54 DISCUSSION 115
xii
CHAPTER 6 WHOLE MITOCHONDRIAL GENOME SEQUENCES OF TWO EIMERIA SPECIES
ISOLATED FROM DOMESTIC (MUSTELA PUTORIUS FURO) AND BLACK- FOOTED FERRETS
(MUSTELA NIGRIPES) 129
61 INTRODUCTION 129
62 MATERIALS amp METHODS 130
621 Parasites 130
622 DNA isolation from coccidia in feces 131
623 Whole genome sequencing 131
624 Phylogenetic analysis 132
63 RESULTS 133
64 DISCUSSION 134
CHAPTER 7 CONCLUSIONS AND FUTURE DIRECTIONS 145
REFERENCES 148
APPENDICES 157
xiii
LIST OF TABLES
Table 11 Morphometrics of Eimeria and Isospora (=Cystoisospora) species affecting mustelids 10
Table 21 Amplification primers for nuclear 18S rDNA and mitochondrial COI loci used in the
identification of enteric coccidia from domestic ferrets 55
Table 22 Summary of fecal samples from domestic ferrets submitted to two diagnostic laboratories
from 2008-2015 56
Table 23 Morphologic and molecular identification of coccidia from domestic ferrets 57
Table 31 Amplification primers for nuclear 18S rDNA mitochondrial COI and COIII loci used in the
identification of coccidia from black-footed ferrets 73
Table 32 Morphologic and molecular characterization of coccidia from fecal and FFPE necropsy
samples from black-footed ferrets 76
Table 33 Morphometric characterization of Eimeria ictidea oocysts from black-footed ferrets 77
Table 41 Eimeria ictidea shedding in black-footed ferret dam and kit family groups - 2014-2016 97
Table 42 Epidemiologic data for family groups of black-footed ferrets shedding Eimeria ictidea 98
Table 43 Shedding of Eimeria ictidea in adult black-footed ferrets - 2015-2016 99
Table 44 Epidemiologic data for adult black-footed ferrets shedding Eimeria ictidea 100
Table 45 Histologic findings from black-footed ferrets with enteric coccidiosis 101
Table 46 Incidence of coccidial infections in black-footed ferrets at the Cheyenne Mountain Zoo 102
Table 47 Yearly mortality associated with coccidiosis in black-footed ferrets at the Toronto Zoo 103
Table 51 Prepatent period and oocyst shedding of Eimeria ictidea in experimentally infected
domestic ferrets 126
Table 52 Results of oral inoculation of domestic ferrets with oocysts of Eimeria ictidea 127
Table 53 Distribution of coccidial life stages in intestinal tract of domestic ferrets orally
inoculated with oocysts of Eimeria ictidea 128
Table 61 PCR primers used to sequence the mitochondrial genome of Eimeria furonis 136
Table 62 PCR primers used to sequence the mitochondrial genome of Eimeria ictidea 137
Table 63 Coding regions in the mitochondrial genome of Eimeria furonis from a domestic ferret 138
Table 64 Coding regions in the mitochondrial genome of Eimeria ictidea from a black-footed ferret 139
Table 65 Pairwise comparison of coding regions in the mitochondrial genomes of Eimeria furonis
and Eimeria ictidea 140
xiv
LIST OF FIGURES
Figure 11 Phylogeny of the Apicomplexa 2
Figure 12 Classical life cycle of coccidian parasites 4
Figure 13 Morphologic characteristics used for identification of eimeriid oocysts 6
Figure 21 Life stages of Eimeria furonis within the small intestine of a domestic ferret 58
Figure 22 Phylogenetic relationships of coccidia (Eimeria ictidea Eimeria furonis and Isospora
(=Cystoisospora) laidlawi) from domestic or black-footed ferrets 59
Figure 31 Nuclear and mitochondrial genetic loci targeted by primers listed in Table 31 73
Figure 32 Morphometrics of Eimeria ictidea from a black-footed ferret (Mustela nigripes) 74
Figure 33 Nuclear 18S rDNA sequences of Eimeria ictidea to newly generated (see Chapter 2) and
published sequences of Eimeria furonis 75
Figure 34 Mitochondrial cytochrome c oxidase subunit I sequences of Eimeria ictidea to sequences
from other eimeriid parasites of carnivores 75
Figure 41 Oocyst per gram counts and shedding period of Eimeria ictidea from black-footed ferret
family groups from 2014-2016 95
Figure 42 Sexual life stages of Eimeria ictidea in the small intestine of a black-footed ferret 96
Figure 51 Exogenous life stages of Eimeria ictidea 123
Figure 52 Endogenous life stages of Eimeria ictidea within the small intestine of an experimentally
infected domestic ferret 124
Figure 53 Distribution of sexual and asexual life stages of Eimeria ictidea along the intestinal tract
of experimentally infected domestic ferrets 125
Figure 61 Map of the mitochondrial genome of Eimeria furonis 141
Figure 62 Map of the mitochondrial genome of Eimeria ictidea 142
Figure 63 Comparison of the mitochondrial genomes of Eimeria furonis and Eimeria ictidea 143
Figure 64 Phylogenetic relationships of coccidia from domestic and black-footed ferrets based on
complete mitochondrial genome sequences 144
xv
LIST OF APPENDICES
Appendix 1 Shedding of oocysts of Eimeria ictidea in black-footed ferret (Mustela nigripes) dam and
kit family groups from 2014-2016 158
Appendix 2a Hematology values for domestic ferrets (Mustela putorius furo) from 49-51 days of
age prior to experimental inoculation 161
Appendix 2b Serum biochemistry values for domestic ferrets (Mustela putorius furo) from
49-51 days of age prior to experimental inoculation 162
Appendix 3a Hematology values for domestic ferrets (Mustela putorius furo) inoculated orally
with Eimeria ictidea 163
Appendix 3b Serum biochemistry values for domestic ferrets (Mustela putorius furo) inoculated
orally with Eimeria ictidea 164
Appendix 4 Domestic ferret (Mustela putorius furo) weekly monitoring sheet 165
Appendix 5 Domestic ferret (Mustela putorius furo) 24 hour intensive monitoring sheet 167
Appendix 6 Domestic ferret (Mustela putorius furo) infection trial standard operating procedures 172
xvi
ABBREVIATIONS
ATP Adenosine triphosphate
BFF Black-footed ferret(s)
BI Bayesian inference
bp Base pair
CAPC Companion Animal Parasitology Council
CDS Coding DNA sequence
CITES Convention on International Trade in Endangered Species of Wild Fauna and Flora
COI Cytochrome c oxidase subunit 1
COIII Cytochrome c oxidase subunit 3
CytB Cytochrome b
DF Domestic ferret(s)
DNA Deoxyribonucleic acid
FFPE Formalin-fixed paraffin embedded tissue
IUCN International Union on the Conservation of Nature
L Length
LSU Large subunit
mt Mitochondrial
NaOH Sodium hydroxide
nu Nuclear
OPG Oocyst per gram count
PCR Polymerase chain reaction
rDNA Ribosomal DNA
SI Shape index
SND Single nucleotide difference
SOP Standard operating procedure
sp spp Species (singular plural)
SSP Species Survival Plan
SSU Small subunit
TMS Trimethoprim sulfadimethoxine
USFWS United States Fish and Wildlife Service
W Width
1
CHAPTER 1 LITERATURE REVIEW
11 INTRODUCTION
Black-footed ferrets (Mustela nigripes) are one of three wild ferret species worldwide Although
formerly distributed throughout the North American prairies black-footed ferrets (BFF) had been
extirpated from the majority of their range by the 1970s and were declared extinct in the wild in 1987
Since 1986 a multi-institutional effort has been breeding this species in captivity with reintroduction back
into the wild at select sites within Canada the USA and Mexico
Coccidial enteritis is a major cause of death in young captive black-footed ferrets (Bronson et al
2007) but coccidiosis can affect all age classes (personal observation) As a result fewer captive-bred
ferrets may be reared successfully for release to the wild The significance of coccidiosis in wild ferrets is
unknown Consequently the prevention and control of coccidial outbreaks is an important part of black-
footed ferret captive breeding programs and management This research is intended to improve the in situ
and ex situ health of the black-footed ferret through the provision of a better understanding of the
pathogenesis of enteric coccidiosis in this species and to pave the way for the investigation of novel
methods for disease treatment and control
12 APICOMPLEXA
121 Brief introduction to apicomplexan pathogens
The phylum Apicomplexa comprises a large number of eukaryotic intracellular parasitic
organisms many of which are of importance to human and veterinary medicine As indicated by their
name these parasites are characterized by the presence of an apical complex at the anterior aspect of the
infective stage of the life-cycle (Tenter et al 2002) The taxonomic classifications of members of the
Apicomplexa continue to be in a state of flux (reviewed by Adl et al 2005 Cavalier-Smith 2014 Tenter
et al 2002) For this reason a more simplified taxonomic structure has been used in this review (see
2
Figure 11) The subclass Coccidia is a speciose group within the Apicomplexa with most genera falling
into one of two major coccidian suborders within the Eucoccidiorida To date greater than 2000 species
of coccidia have been named (Duszynski Upton amp Couch nd Upton 2000) The adeleid coccidia
(suborder Adeleorina) include monoxenous (single host) and heteroxenous (multiple hosts) parasites in
genera such as Adelea Haemogregarina Hepatozoon and Karyolysus The eimeriorinid coccidia
(suborder Eimeriorina) include the typical intestinal coccidia such as Eimeria Isospora and Cyclospora
species belonging to the family Eimeriidae as well as tissue (cyst forming) coccidia such as
Cystoisospora Besnoitia Toxoplasma and Sarcocystis species that belong to the family Sarcocystidae
(Cox 1994)
Figure 11 Phylogeny of the Apicomplexa Numbers on branches and thickness indicate diversity
(ie named species) Taxonomic groupings demonstrated by the phylogenetic tree (1) subclass
Coccidia (2) suborder Adeleorina (3) suborder Eimeriorina (4) family Eimeriidae and (5) family
Sarcocystidae Adapted from Šlapeta J Morin-Adeline V (2011) Apicomplexa Levine 1970
Sporozoa Leucart 1879 httptolweborgApicomplexa2446 in The Tree of Life Web Project
httptolweborg
2
1
3
4
5
3
122 Life cycles of the Eimeria and Isospora species implicated in enteric coccidiosis
The life cycle of Eimeria species is considered the classical coccidian life cycle which is
typically completed in one host (monoxenous) with many Eimeria species parasitizing only a single host
species (stenoxenous) (Figure 12) The life cycle has two main phases of development one that takes
place within the host (endogenous) and the other that takes places outside of the host (exogenous)
Classically the endogenous stages of the Eimeria life cycle take place within the intestinal epithelium
however some Eimeria species undergo extraintestinal endogenous development such as Eimeria stiedae
in rabbits which replicates within the epithelium of the biliary tree During the exogenous phase of the
life cycle unsporulated oocysts that are shed in the feces of the host sporulate within the environment
resulting in the formation of four sporocysts within each oocyst (tetrasporocystic) Each sporocyst
contains two sporozoites (dizoic) Sporulation is affected by three main factors temperature moisture and
aerobic conditions (Fayer 1980)
Once ingested by the host the wall of the sporulated oocyst is broken to release sporocysts from
which the sporozoites (infective stage) excyst The freed sporozoites penetrate the intestinal epithelial
cells and undergo multiple mitotic divisions to form a single multinucleate meront The meront then
undergoes simultaneous cytokinesis to form first generation merozoites which leave the host cell to infect
new cells and undergo further asexual replications The undifferentiated uninucleate tissue stage of the
parasite within the intestinal epithelial cell is called a trophozoite The number of cycles of asexual
replication (merogony) is predetermined after which the last generation of merozoites penetrate host cells
and undergo sexual differentiation into male and female gamonts (gametogony) Each microgamont
(male) undergoes simultaneous fission to produce numerous motile microgametes each macrogamont
(female) develops into a single mature macrogamete Fertilization of a macrogamete by a motile
microgamete results in formation of a zygote that is rapidly enclosed in a thick wall to form an
unsporulated oocyst Oocysts are shed with the hostrsquos feces into the environment where they are
protected from desiccation and chemical disinfection by the oocyst wall Traditionally Eimeria species
4
have been differentiated based on the host species or host genus affected the site of endogenous life cycle
development and the microscopic cellular characteristics of the different life stages Interestingly
experimental cross infection of Eimeria species from their natural host to a novel host of a taxonomically
similar species has been successful in some cases (De Vos 1970 Levine and Ivens 1970 Haberkorn
1971) challenging the notion that Eimeria are truly stenoxenous parasites
Figure 12 Classical life cycle of coccidian parasites This apicomplexan life cycle includes both
sexual and asexual development The three processes in the life cycle are merogony (asexual
replication A-D) followed by gametogony (formation of gametes E-H) within the digestive tract
of the host with release of unsporulated oocysts (I) Exogenous sporogony (I-L) results in the
production of infective sporulated oocysts (L) Adapted from Barta 2001 with permission of the
author
The life cycle of Isospora spp is similar to that of species in the genus Eimeria (see Figure 12)
but the number of sporocysts and sporozoites differ sporulated oocysts contain two sporocysts (disporic)
5
each of which contains four sporozoites (tetrazoic) These characteristics are not unique to Isospora spp
because diasporic tetrazoic sporulated oocysts are also found in the genera Besnoitia Frenkelia
Hammondia Sarcocystis and Toxoplasma However the sporocysts in the latter parasites are
morphologically distinct in that they lack Stieda bodies
123 Methods of characterization
1231 Morphological features
Historically eimeriid coccidia have been classified based on the cellular morphology of the
different life stages (particularly the morphometrics of sporulated oocysts) where these stages occur in
the host and apparent host specificity (frequently assumed and not tested experimentally) The
morphological features and dimensions of oocysts and their components are important diagnostic features
because of the availability of these stages in clinical specimens these characteristics can include size
(length [L] width [W] shape index [SI=LW]) number of sporocysts wall morphology
presenceabsence of a micropyle micropyle cap residual body or polar granules for oocysts size number
of sporozoites wall morphology presenceabsence of Stieda body subStieda body paraStieda body or
residual body for sporocysts and presenceabsence of refractile bodies for sporozoites (see Figure 13)
Pertinent life cycle information includes type of life cycle (monoxenous versus heteroxenous) tissue
sites of merogony and gametogony (intestinal versus extraintestinal) and the presence or absence of
extraintestinal hypobiotic stages (eg dormozoites or hypnozoites) Further information used to
characterize coccidia that form tissue cysts generally includes details on life stages in the definitive and
intermediate hosts location and morphology of tissue cysts route(s) of transmission among host species
and morphologic descriptions of merozoites (eg tachyzoites or bradyzoites) in tissue culture
6
Figure 13 Morphologic characteristics used for identification of eimeriid oocysts 1) Oocyst in cross
section ol - oocyst length or - oocyst residual body ow - oocyst width pg - polar granule row -rough
outer wall 2) The top of a hypothetical oocyst mcd - depth of the micropyle cap mcw - width of the
micropyle cap mw - width of the micropyle sow - smooth outer wall 3) Sporocyst in cross section
psb - paraStieda body sb - Stieda body sl - sporocyst length sp - sporozoite sr - sporocyst residual
body srb - sporozoite refractile body ssb - subStieda body sw - sporocyst width From Duszynski D
Wilber PG (1997) A guideline for the preparation of species descriptions in the Eimeriidae Journal of
Parasitology 83(2)333-336 reproduced with permission of Allen Press Publishing Services
1232 Molecular characterization (genetic loci and methods)
More recently molecular techniques have been used to infer phylogenetic or evolutionary
relationships among coccidia and to reclassify taxonomic assignments to better reflect the evolutionary
history of these parasites Molecular data can be more informative than phenotypic data because recent
evolutionary divergence among coccidia is unlikely to be reflected in morphologic differences but may
be detectable using molecular data The principle behind the use of molecular sequencing to describe
evolutionary relationships is that nucleotide sequences like morphological features diverge over time
under selective pressure however nucleotide sequences evolve at a more regular rate than do
morphologic characteristics Phenotypic data is thus less likely to detect recent evolutionary divergence
Sequences that are more similar are inferred to be more closely related and to have diverged more
recently (Cox 1994) Molecular characterization can be performed using DNA RNA or protein
sequences Most of the early molecular phylogenetic analyses of coccidia performed used ribosomal RNA
sequences usually by PCR amplification of ribosomal DNA (rDNA) in the nuclear genome of the
7
parasites Ribosomes contain both small and large RNA subunits in eukaryotes the large ribosomal RNA
consists of two forms 5S and 28S while the small ribosomal RNA exists only as 18S Sequences from
several genetic loci have been used for characterization of parasites most commonly 18S rDNA 28S
rDNA and ribosomal internal transcribed spacer regions (ITS) from the nuclear genome and more
recently mitochondrial cytochrome c oxidase subunits I (COI) and III (COIII) however sequencing of
nuclear 18S rDNA (nu 18S rDNA) has been the most prevalent in the literature by far Early attempts to
use 5S RNA sequences formed unlikely phylogenies and too few 28S ribosomal DNA sequences have
been obtained to make this locus useful (Cox 1994 Tenter et al 2002) The disadvantage of nu 18S
rDNA is that it is comparatively poor at distinguishing among closely related eimeriid coccidial species
because of its conserved nature but for that reason the nu18S rDNA locus is useful for inferring
relationships among species with greater evolutionary divergence Although only exploited recently
because of the paucity of suitable PCR primers the mitochondrial COI locus appears to be more useful
for distinguishing closely related eimeriid coccidia (Ogedengbe Hanner amp Barta 2011) but COI
sequences are less useful for inferring more ancient relationships between highly divergent coccidial
species Consequently the combined use of nu 18S rDNA and mitochondrial COI sequencing has been
recommended for improved species description and phylogenetic analysis (El-Sherry et al 2013)
Molecular characterization has also been used for diagnostic purposes and is well-suited to the
identification of coccidia when information on host specificity parasite life cycle and life stages is not
available as the molecular (genetic) data is the same for a given parasite during each of its life cycle
stages This information can be particularly useful in identifying the relationship between different life
stages of heteroxenous parasites collected from different hosts (intermediate definitive) Furthermore for
previously unidentified coccidia or those for which limited information is available molecular
characterization could be used to predict likely definitive hosts or parasite life cycle traits based on
phylogenetic relationships to other known species
8
13 RECLASSIFICATION OF MAMMALIAN ISOSPORA
Recommendations have been made to reclassify the avian and mammalian Isospora into two
separate genera based on life cycle molecular phylogenetic studies and morphologic description of
sporulated oocysts (Frenkel 1977 Barta et al 2005) Due to their classical coccidian life cycle presence
of Stieda bodies within sporocysts and close phylogenetic association with Eimeria species the avian
Atoxoplasma and Isospora have been retained in the genus Isospora (see Barta et al 2005) Conversely
the presence of tissue life cycle stages lack of Stieda bodies within sporocysts and close phylogenetic
association with other genera within the family Sarcocystidae have required many mammalian Isospora
to be reclassified as members of the genus Cystoisospora Frenkel 1977 (Frenkel 1977 Barta et al 2005)
Consequently for the remainder of this thesis Isospora species from mustelids will be referred to as
Isospora (=Cystoisospora) to reflect their probable generic association
14 EIMERIID SPECIES CHARACTERIZED IN MUSTELIDS
141 The family Mustelidae
The family Mustelidae within the order Carnivora comprises a group of approximately 59
carnivorous mammalian species within 22 genera Native mustelids are found in terrestrial and aquatic
environments on almost every continent with the exception of Australia and Antarctica The Mustelidae
are classically divided into two subfamilies as defined by Wozencraft (2005) 1) Mustelinae (weasels
mink ferrets marten wolverine) the larger subfamily including the following genera Arctonyx Eira
Galictis Gulo Ictonyx Lyncodon Martes Meles Mellivora Melogale Mustela Neovison Poecilogale
Taxidea and Vormela and 2) Lutrinae (otters) including seven genera Aonyx Enhydra Hydrictis
Lontra Lutra Lutrogale and Pteronura More recently molecular data suggest the Mustelidae should be
separated into eight subfamilies although this is not universally accepted (Koepfli et al 2008 Lariviegravere
and Jennings 2009 Yu et al 2011)
9
142 Eimeriid coccidia described from mustelids
Ten named Eimeria species and twelve named Isospora (=Cystoisospora) species have been
described in the Mustelidae and are summarized in Table 11 This table includes information on host
range life cycle and detailed morphologic data used to identify and classify the individual parasites Two
coccidial parasites isolated from the Libyan striped weasel (Ictonyx libyca) and the European polecat
(Mustela putorius) initially ascribed to the genus Isospora Isospora zorillae and Isospora putori
respectively have since been reclassified as Sarcocystis spp (see footnote to Table 2 of Yi-Fan et al
2012)
10
Table 11 Morphologic characteristics of Eimeria and Isospora (=Cystoisospora) species affecting mustelids
Coccidial species Host genus and
species
Life cycle
Location
Oocyst shape and
size
Oocyst
description
Sporocyst
description
Sporozoite
description
References
Cytoisospora
eversmanni
Mustela
eversmanii
(Steppe polecat)
Mustela
putorius
(European
polecat)
Homoxenous L185 (16ndash20)
W 148 (16ndash12)
LW 13 (11ndash16)
M absent
PG absent
OR absent
L 115
(10ndash135)
W 98
(9ndash11)
LW 12
(11ndash15)
SB absent
SR present
SRB present Yi-Fan et al 2012
Svanbaev 1956
Nukerbaeva amp
Svanbaev 1973
1977
Cystoisospora
pavlovskyi
Mustela
eversmanii
Mustela
putorius
Homoxenous L 322 (29ndash36)
W 273 (265ndash285)
LW 12 (11ndash14)
M absent
PG absent
OR absent
L 195
(18ndash21)
W 144
(12ndash15)
LW 14
(12ndash15)
SB absent
SR present
SRB present Yi-Fan et al 2012
Svanbaev 1956
Nukerbaeva amp
Svanbaev 1973
1977
Eimeria
baskanica^
Mustela
erminae
(ermine)
Homoxenous Oval with tapered
ends
L 112-126
W 84-98
M absent
PG absent
OR present
SR absent Bean shaped Nukerbaeva amp
Svanbaev 1977
Eimeria furonis Mustela
putorius
Mustela
putorius furo
(dom ferret)
Mustela
nigripes (BFF)
Mustela vison
(mink)
Homoxenous
Small intestine
rectum (H 1927)
Jejunumileum (BP
1993)
Spherical ndash
subspherical
L 11-14
W 10-13
OW 2 layers
M absent
PG absent
OR absent
Spindloid
L 8-9
W 4
SB present
SR present
Vermiform Blankenship-Paris
et al 1993
Hoare 1927 1935b
Jolley et al 1994
Nukerbaeva amp
Svanbaev
19731977
Williams et al 1988
1992 1996
Eimeria hiepei Mustela vison Homoxenous
Bile duct
Spherical
L 13-17
W 13-17
OW 2 layers
(outer
smooth)
M absent
PG absent
OR absent
L 6
W 4
SB absent
SR absent
Banana shaped Davis et al 1953
Grafner et al 1967
11
Coccidial species Host genus and
species
Life cycle
Location
Oocyst shape and
size
Oocyst
description
Sporocyst
description
Sporozoite
description
References
Eimeria ictidea Mustela
eversmanni
Mustela
nigripes
Mustela
putorius
Mustela
putorius furo
Homoxenous
Small intestine
Ovoid ndashellipsoid
L 13-27
W 13-21
OW 2 layers
M present
PG absent
OR absent
Ovoid
(irregular)
L 115
W65
SB present
SR present
- Hoare 1927 1935a
1935b
Jolley et al 1994
Litvenkova 1969
Svanbaev 1956
Tinar 1985
Williams et al 1988
1992
Eimeria irara Eira barbara
(tayra)
Homoxenous
Feces
Ovoid
L 21-25
W 18-20
OW outer
layer smooth
M absent
PG absent
OR absent
Ellipsoid
L 10-12
W 65
SB present
SR present
Elongate (one
end broader than
the other)
Carini amp da
Fonseca 1938
Eimeria melis Meles meles
(European
badger)
Homoxenous Ellipsoid
L 20plusmn018
W 157plusmn002
LW128plusmn0017
(112-15)
OW 2 layers
(outer
smooth)
M absent
PG present
OR present
Ovoid
L
119plusmn0018
W 65plusmn008
LW 183
(155-24)
SB present
L 90plusmn005
W 324plusmn0025
SRB present
Anwar et al 2000
Kotlan amp Pospesch
1933
Eimeria mustelae Mustela vison
Mustela nivalis
(snow weasel)
Homoxenous
Duodenumileum
Spherical or
Ellipsoid
L 18-26
W 14-24
OW 2 layers
M absent
PG present
OR absent
Ovoid
L 8
W 5
SB present
SR present
Broad at one
end and tapered
at other
L 7
W 3
Glebezdin 1978
Iwanoff-Gobzem
1934
Levine 1948
Musaev amp Veisov
1983
Tinar 1985
Eimeria sablii Martes zibellina
(sable)
Homoxenous
Gut
Spherical or
subspherical
L 112-126
W 112
OW 2 layers
M absent
OR absent
Ovoid
L 56
W 42
SR present
Elongate Nukerbaeva 1981
Eimeria sibirica Martes zibellina Homoxenous Ovoid
L avg 216
W avg 180
LW 1076
OW 2 layers
M absent
PG absent
OR absent
Ovoid
L 96-112
W 56-72
SR absent
Elongate Nukerbaeva 1981
Yakimoff amp
Gousseff 1934
Yakimoff amp
Terwinsky 1930
1931
12
Coccidial species Host genus and
species
Life cycle
Location
Oocyst shape and
size
Oocyst
description
Sporocyst
description
Sporozoite
description
References
Eimeria vison
(Eimeria
mustelae)
Mustela
putorius
Mustela
putorius furo
Mustela vison
Homoxenous
Small intestine
+- large intestine
Ovoid
L 17-22
W 9-18
OW 2 layers
M absent
OR
sometimes
present
Ovoid or
Piriform
L 10
W 55
SB absent
SR present
Curved or Club
shaped
L 9
W 25
Foreyt amp Todd 1976
Foreyt et al 1977
Kingscote 1934
1935
Levine 1948
McTaggart 1960
Nukerbaeva amp
Svanbaev
19731977
Tinar 1985
Umurzakov amp
Nukerbaeva 1985
Wolter 1961
Zimmermann 1959
Isospora africana Ictonyx libyca
(Libyan striped
weasel)
Homoxenous
Feces
Spherical
L 25-27
W 25-27
OW 2 layers
(outer
smooth)
M absent
PG absent
OR absent
Ovoid
L 15-17
W 10-12
SB absent
SR present
Elongate
L 135
W 3
Prasad 1961
Isospora altaica Mustela altaica
(mountain
weasel)
Homoxenous
Gut
Oval or spherical
L 280-336
W 252-280
LW 121 (111-
124)
OW 2 layers
M absent
PG absent
OR absent
Ovoid or
spherical
L 140-168
W 111-168
SR present
Svanbaev amp
Rachmatullina
1971
Isospora goussevi Mustela nivalis Homoxenous
Large intestine
Ovoid
L 224 (220-250)
W 174 (160-190)
LW 135 (133-
137)
OW 1 layer
PG present
OR present
Ovoid
L 120
(100-130)
W 70 (60-
80)
SB present
SR present
Elongate Musaev amp Veisov
1983
13
Coccidial species Host genus and
species
Life cycle
Location
Oocyst shape and
size
Oocyst
description
Sporocyst
description
Sporozoite
description
References
Isospora
hoogstraali
Ictonyx libyca Homoxenous
Feces
Ellipsoid
L 37-41
W 32-34
OW 2 layers
(outer
smooth)
M absent
PG some
OR absent
Ovoid
L 19-21
W 13-15
SB absent
SR present
Club-shaped
L 18-19
W 4-6
Prasad 1961
Isospora laidlawi Mustela
putorius
Mustela
putorius furo
Mustela vison
Homoxenous
Feces
Intestinal contents
Ovoid L
320-368
W 272-304
OW 2 layers
M absent
PG absent
OR absent
Ellipsoid
L 208
W 144
SB absent
SR present
Sausage shaped Foreyt et al 1977
Hoare 1927
Levine 1948
McTaggart 1960
Nukerbaeva amp
Svanbaev 1973
1974 1977
Tinar 1985
Isospora lutrae Lutra lutra
(European
otter)
Lutra
canadensis
(North
American river
otter)
Homoxenous Spherical
L 312 (275-32)
W 296 (28-31)
LW 104
(10-112)
OW 2 layers
(outer
smooth)
M absent
PG absent
OR absent
Ellipsoid
L 182 (17-
19)
W 144 (14-
16)
LW128
(12-14)
Sb absent
sSB absent
SR present
Spindle- shaped
L 124
W 25
SRB present
Torres et al 2000
Hoover et al 1985
Isospora
martessii
Martes zibellina Homoxenous
Gut
Ovoid short oval or
spherical
L 252 ndash 280 196
168
W 168 ndash 224 168
168
OW 2 layers
M absent
OR absent
Ovoid
L 112-168
W 84-112
SR present
Elongate Nukerbaeva 1981
Isospora melis Meles meles Homoxenous Ovoid
L 328plusmn034
W 269plusmn019
LW122 (110-
157)
OW 2 layers
(outer
smooth)
M absent
PG absent
OR absent
Ellipsoid
L
215plusmn0166
W 14plusmn012
LW 155
(133-185)
SR absent
Round at one
end other end
tapered
L 142plusmn116
W 40plusmn017
SRB absent
Anwar et al 2000
Glebezdin 1978
Kotlan amp Pospesch
1933
Pelleacuterdy 1955
14
Coccidial species Host genus and
species
Life cycle
Location
Oocyst shape and
size
Oocyst
description
Sporocyst
description
Sporozoite
description
References
Isospora
mustelae (nomen
nudum)
Martes martes Ovoid L
7 W
225
M present - - Galli-Valerio 1932
Isospora nivalis Mustela nivalis Homoxenous
Large intestine
Ovoid
L 206 (200-230)
W 184 (180-210)
LW 11 (109-111)
OW 1 layer
PG absent
OR absent
Ovoid
L 125
(120-130)
W 80 (70-
90)
SR present
Lemon or pear
shaped
Musaev amp Veisov
1983
Unnamed
ldquoCoccidiardquo^
Mustela
nigripes
Urinary bladder - - - - Jolley et al 1994
Unnamed
ldquoCoccidiardquo^
Mustela
nigripes
Trachea bronchus
bronchial glands
- - - - Jolley et al 1994
Unnamed
Eimeria sp^
Mustela
nigripes
Feces
intestinal contents
Ovoid
L 350-386
W 212-232
- - - Jolley et al 1994
Williams et al
1992
Unnamed
Eimeria sp^
Mustela
putorius furo
Small intestine - - - - Blankenship-Paris
et al 1993
Unnamed
Eimeria sp^
Mustela nivalis Homoxenous
Large intestine
Ovoid-ellipsoid L
2031 (1712-2162)
W 148 (1225-
1681)
LW 136 (121-16-
)
OW 1 layer
PG absent
OR absent
Ovoid or
pear-shaped
L 60-100
W 40-80
SR present
Elongate
L 50-90
W 30-70
Musaev amp Veisov
1983
Unnamed
Eimeria sp^
Martes martes
(marten)
Homoxenous Ovoid
L avg 216
W avg 180
LW 1076
OR absent 4 sporocysts
SR present
L 126
W 60
Yakimoff and
Gousseff 1934
Unnamed
Isospora sp^
Mustela
putorius furo
Feces - - - - Bell 1994
Unnamed
Isospora sp^
Mustela
putorius furo
Feces - - - - Bell 1994
Legend L = length W = width LW = length-width ratio avg = average OW = oocyst wall PG = polar granules M = micropyle SB = Stieda body sSB =
subStieda body OR = oocyst residuum SR = sporocyst residuum SRB = sporozoite refractile body ^ = species inquirendae - = no information provided by
author(s) = information obtained from secondary sources (primary reference could not be obtained) All measurements are in micrometers Bolded references
15
are those from which morphometric data were assembled Remaining references indicate other authors who have identified that parasite species in the same or
similar host
16
143 Eimeriid coccidia described from domestic ferrets
Three species of coccidia were originally described from 50 domestic ferrets (Mustela putorius
furo) Eimeria ictidea Eimeria furonis and Isospora (= Cystoisospora) laidlawi (Hoare 1927) All three
species were detected in feces from domestic ferrets at a research facility undergoing an outbreak of
canine distemper Sick ferrets appeared more frequently infected than healthy ones As per Hoare (1927)
none of the ferrets appeared to display clinical signs associated with protozoal infection For each
parasite the author described morphology of sporulated oocysts isolated from feces and sporulation time
(exogenous life stages) The pre-patent period (minimum duration of endogenous development) in an
inoculated naiumlve ferret was described only for E furonis and E ictidea due to insufficient sample size of
I (=C) laidlawi oocysts for an experimental infection trial Sporulation of oocysts occurred within 5-6
days for E furonis 3 days for E ictidea and 4 days for I (=C) laidlawi The sporulated oocysts of E
furonis were spherical with a double outer wall with a thin colourless outer layer and thick yellowish
inner layer no micropyle or residual body and measured on average 128 times 120 microm (length [L] 112-
144 width [W] 104-128 shape index [SI] 107) Unsporulated oocysts contained a zygote with a
diameter of 96 microm Sporocysts were spindle-shaped with one end constrictedblunted contained a
residual body and on average measured 8-88 times 4 microm Sporozoites were vermiform with one end
narrower than the other arranged head to tail and had a central nucleus a clear vacuole was identified in
some at the broad end The sporulated oocysts of E ictidea were oval or elliptical with a double outer
wall with a thin colourless outer layer and thick yellowish inner layer no micropyle or residual body
and measured on average 236 times 175 microm (L 184-272 W 128-208 shape index 135) The zygote in
unsporulated oocysts was elongate with a diameter of 15 times 12 microm when originally passed in feces but
became more spherical with time Sporocysts were irregularly oval with one end broad and the other
more constricted contained a residual body and on average measured 115 times 65 microm Sporozoites were
vermiform with one end narrower than the other arranged head to tail and had a central nucleus and a
clear vacuole at the broad end The sporulated oocysts of Isospora (=Cystoisospora) laidlawi were ovoid
with a double outer wall with a thin colourless outer layer and thick yellowish inner layer no micropyle
17
or residual body and measured on average 34 times 29 microm (L 320-368 W 272-304) Unsporulated
oocysts contained a spherical zygote with a diameter of 236 microm Two sporocysts were identified each
containing 4 sporozoites and no Stieda body sporocysts were elliptical contained a residual body and on
average measured on 208 times 144 microm Sporozoites were sausage shaped with one end slightly pointed
and had a central nucleus and a clear vacuole identified at the pointed end Sporozoites were arranged
with pointed ends all at the same pole of the sporocyst The pre-patent periods described for E furonis
and E ictidea were 6 days and 7 days respectively (Hoare 1927)
Since Hoarersquos initial description (Hoare 1927 Hoare 1935) multiple single case reports and
outbreaks of severe clinical disease associated with intestinal coccidiosis have been reported in domestic
ferrets Blankenship-Paris et al (1993) described a single case of a four-month-old domestic ferret that
presented depressed in thin body condition dehydrated and with pasty dark feces on the perineum This
ferret had been housed with its dam and another sibling neither dam nor sibling showed clinical signs of
enteric disease and both had negative fecal examination results on repeated evaluation Routine fecal
examination of the rest of the colony and necropsies on eight other ferrets in the colony revealed no
evidence of coccidial infection Enteric coccidiosis was determined to be the cause of disease in the four-
month-old ferret based on necropsy findings but the coccidia could not be speciated because diagnosis
was made on histologic findings only
Sledge et al ( 2011) described three separate outbreaks of severe enteric coccidiosis in domestic
ferrets from one ferret rescue centre (group 1) and two shelters (groups 2 and 3) all affected by the same
Eimeria sp The morphologic characteristics of sporulated oocysts were only described for group 1 no
coccidial oocysts were detected on direct smear or fecal flotation of diarrheic samples submitted from
groups 2 and 3 Oocysts were identified as spherical measuring 12-13 microm in diameter with four
sporocysts each containing two sporozoites Oocyst morphometrics histopathologic findings and nu 18S
rDNA partial sequences from all three groups were used collectively to confirm the coccidial species
identify in each outbreak as E furonis
18
Two cases of biliary coccidiosis with E furonis have been reported in domestic ferrets The first
was in a nine-week-old male ferret from a research facility (Williams Chimes amp Gardiner 1996) The
ferret presented with signs of hepatic disease and was negative for coccidia on fecal flotation and direct
smears Endogenous coccidial life stages were described from the gall bladder and liver on histologic
examination In tissue section the oocysts were oval to spherical and measured 125 times 120 microm Meronts
measured 108-130 times 89-93 microm and contained up to 16 merozoites The merozoites exhibited a double-
layered pellicle prominent conoid few rhoptries and many micronemes anterior to the nucleus Based on
the morphologic description of the life stages in this case the coccidia were identified by the authors as
an Eimeria species most likely E furonis Kaye et al (2015) described a second case of biliary
coccidiosis in an 18-month-old female pet domestic ferret with concurrent pure red cell aplasia In this
case all endogenous coccidial life stages were observed on histologic examination of the epithelium of
the extrahepatic biliary tree The oocysts were ovoid and measured 12 times 13 microm Meronts measured 12 times
15 microm and contained up to 16 merozoites each measuring 2 times 5 microm Based on the morphologic
description of the life stages in this case and nu 18S rDNA sequences the pathogen was also determined
to be E furonis Biliary coccidiosis has also been identified in mink (Mustela vison) with the etiologic
agent identified as Eimeria hiepei (Davis Chow amp Gorham 1953 Grafner Graubmann amp Dobbriner
1967)
Oocysts from Cystoisospora ohioensis have been reported from fecal samples collected from
healthy domestic ferret kits in a large American ferret breeding operation that were raised on the same
premise as juvenile domestic dogs (Patterson amp Fox 2007) The method of identification of this parasite
was not described by Patterson amp Fox A second similar institution reported the presence of a
Cystoisospora species also thought to be C ohioensis in routine fecal examination of their ferret colony
(Dr Bambi Jasmin personal communication) Coccidial identification in this case was performed by the
Animal Health Diagnostic Center at Cornell University The significance of these findings is unknown as
no clinical signs or histologic lesions have been described in domestic ferrets associated with shedding of
19
oocysts and the definitive host for C ohioensis is the domestic dog It is most likely that fecal
identification of C ohioensis represents a pseudoparasite in both of these cases or perhaps an
undescribed Cystoisospora sp that is morphologically indistinguishable from C ohioensis
It is difficult to estimate the prevalence of enteric coccidia within the North American domestic
ferret population Fecal samples submitted to university or large veterinary diagnostic laboratories from
domestic ferrets in Canada are uncommon and samples positive for coccidia appear infrequently (Dr
Donald Martin personal communication) Data from Idexx Vet Med Lab in Ludwigsburg Germany was
compiled to review the prevalence of coccidia and Giardia within fecal samples from domestic ferrets
(Pantchev et al 2011) The authors reported that of 284 fecal samples submitted from 2002-2004 18
(63) had detectable coccidial oocysts on fecal flotation Oocysts were identified based on morphologic
characteristics as E ictidea E furonis I (=C) laidlawi and another unidentified Isospora species
Comparative data from the same laboratory from 2009-2010 included sample submissions from 253
ferrets 21 (83) of which were positive for coccidial oocysts on fecal flotation Nine of the samples
were positive for E furonis three were positive with both E furonis and I(=C) laidlawi present eight
were positive only for I(=C) laidlawi and one sample contained both E furonis and E ictidea
identification in all cases was based on morphologic characteristics No statistically significant difference
in the occurrence of coccidial oocysts was detected when data from the two periods were compared
(Fisherrsquos exact test P=041) (Pantchev et al 2011)
144 Molecular characterization
Molecular characterization of Eimeria furonis was first performed by Abe et al (2008) using
oocysts purified from the feces of a single domestic ferret with clinical signs of coccidial enteritis Small
subunit ribosomal DNA (nu 18S rDNA) primers CYC1FE (5ʹ-TAC CCA ATG AAA ACA GTT T-3prime) and
CYC4RB (5prime-CGT CTT CAA ACC CCC TAC TG-3prime) were used to amplify a 347 base pair (bp) fragment
of nu 18S rDNA These primers were initially developed for molecular identification of Cyclospora
species but have since been shown to amplify nu 18S rDNA from several Eimeria species (Matsubayashi
20
et al 2005) The amplicon was sequenced (GenBank AB329724) and compared with previously
published partial nu 18S rDNA sequences from 40 Eimeria two Isospora and four Cyclospora species
The resulting phylogram grouped E furonis with E alabamensis (cattle) and E meleagrimitis (turkey) In
the same study the microscopic morphology of the oocysts was used to identify this coccidial species as
E furonis by comparison with published descriptions of E furonis E ictidea and E heipei by Hoare
(1927) Hoare (1935) and Grafner Graubmann amp Dobbriner (1967) respectively
Nuclear 18S rDNA was also used by Sledge et al (2011) for molecular identification of the
eimeriid coccidia implicated in the three distinct outbreaks of enteric disease in domestic ferrets As
described above initial identification and speciation of the coccidia was performed using morphologic
characteristics of the sporulated oocysts collected from feces in one of the three outbreaks being
investigated the oocysts were identified as E furonis Histologic sections of formalin fixed intestinal
segments from ferrets from each of the three outbreaks contained multiple coccidial life stages DNA was
then isolated from stored formalin-fixed tissues for further genetic analysis Using the partial nu 18S
rDNA gene sequence reported by Abe et al (2008) (GenBank AB329724) the following PCR primers
were created 5ʹ-ACA ATT GGA GGG CAA GTC TG-3ʹ and 5ʹ-GGCGAC AAG CCT GCT TGA AAC-
3ʹ PCR amplification produced a 247 bp amplicon from each of the three groups Analysis and
sequencing of amplicons from all three groups showed 100 homology to nucleic acid sequences
previously reported by Abe et al (2008) for the gene encoding E furonis nu 18S rDNA
Coccidia were identified within hepatobiliary lesions in a domestic ferret receiving
immunosuppressive therapy for red cell aplasia (Kaye et al 2015) DNA was extracted from frozen liver
and a 247 bp fragment of the nu 18S rDNA was amplified using the primers previously described by
Sledge et al (2011) and sequenced Kaye et al (2015) reported that the DNA sequence of the amplicon
was 100 homologous to the published nu 18S rDNA of E furonis and 95 homologous to the nu 18S
rDNA of E myoxi (rodent) E alabamensis (cattle) and I robini (avian)
21
145 Clinical signs of disease in domestic ferrets
Hoare (1927 1935b) in his initial descriptions of enteric coccidiosis in domestic ferrets
observed that clinical signs of intestinal disease were not evident The recent literature supports the
finding of subclinical disease but also describes signs ranging from mild transient diarrhea in young or
stressed animals to more severe disease with dehydration lethargy depression weight lossemaciation
inappetence and death (Blankenship-Paris et al 1993 Powers 2009 Sledge et al 2011 Hoefer et al
2012 Patterson et al 2014) Rectal prolapse has also been reported in ferrets with enteric coccidiosis
(Hillyer 1992 Hoefer et al 2012) In one study co-infection with coccidia and Lawsonia intracellularis
(Desulfovibrio sp) was diagnosed in 4 of 19 ferrets with proliferative bowel disease (Li et al 1996)
These ferrets presented with variable clinical signs including diarrhea lethargy anorexia weight loss
dehydration and emaciation
In the two reports of biliary coccidiosis clinical signs conformed to those expected with
hepatobiliary disease Williams et al (1996) described their case to have presented with emaciation poor
appetite abdominal distension and icterus Kaye et al (2015) described a one week history of lethargy
inappetence and icterus with serum biochemistry results consistent with cholestasis later clinical signs in
this case included melena anemia and cachexia
146 Gross necropsy and histologic findings
The pathology of enteric coccidiosis in domestic ferrets was described by Hoare (1927 1935b)
Two healthy domestic ferrets were experimentally inoculated one each with large numbers of mature
oocysts of either E furonis or E ictidea that were isolated during his initial work The inoculated ferrets
were killed humanely for histologic examination of intestinal sections at the time of first detection of fecal
oocyst shedding no clinical signs of coccidiosis were detected in these ferrets prior to death Infection
with E furonis resulted in invasion of the epithelium of the small intestine and rectum Within the small
intestine the parasites were concentrated in the tips of the villi but could be found to the level of the
22
opening of the crypts of Lieberkuumlhn In rectal sections life stages were limited to the epithelial ridges
between the openings of the glands of Lieberkuumlhn Organisms were located within the apical portion of
the epithelial cells and intensely infected regions exhibited multiple parasites within a single host cell
Both asexual and sexual life stages were present within the same sections Hoare (1927) described similar
histopathologic changes in naturally infected ferrets but the proportion of asexual versus sexual life
stages differed In natural infections sexual life stages were more numerous whereas in experimental
infections asexual life stages predominated these findings would be expected to correlate with the stage
of infection at which ferrets died or were humanely killed for tissue collection and would not be
reflective of differences between natural and experimental infection with this parasite Hoare also
described the morphology of the different endogenous stages including trophozoite (3-4 microm) merozoite
(stumpy sausage shaped L 3-4 microm W 2 microm) macrogamete (spherical 8 microm diameter with darkly
staining globular inclusions of reserve material) and microgamete (described as similar to those of other
Eimeria species) Two types of merogony are described from histologic sections the first with stumpy
merozoites as described above and the second with merozoites with elongated curved bodies and a
compact polar nucleus measuring 60 times 13 microm This second merogonic generation was observed almost
exclusively in the naturally infected ferrets and was associated with initiation of sexual differentiation and
reproduction
The pathology of experimental and non-experimental infection with E ictidea in domestic ferrets
was also described by Hoare (1927 1935b) Parasitic invasion of the epithelium was noted only in the
small intestine with patchy distribution of the parasite life stages throughout affected sections Within the
small intestinal villi the parasites were again concentrated in the tips of the villi with infected epithelial
cells never containing more than one parasite As each intracellular parasite grew it filled the entire host
cell displacing the nucleus to the base of the cell Predominantly sexual life stages were detected in tissue
sections with few asexual generations observed Interestingly the parasites were arranged into age
groups with forms of the same life stage grouped together within the affected epithelial sections this is in
23
contrast to E furonis where life stages of different maturities were found together in affected sections
Hoare described the morphology of the different endogenous stages of E ictidea including merozoites
(free within the lumen elongated vermiform with one pointed end and a nucleus located at the rounded
end 11 microm times 1 microm within the epithelium shortened and rounded 3-4 microm diameter) macrogametes
(elongated 20 times 7 microm occupying the entire host cell with darkly staining globular inclusions of reserve
material) and mature microgamonts (morphologically similar to those of other Eimeria species but larger
than those of E furonis) Of note a tissue reaction was observed specifically in association with more
developed life stages of E ictidea (eg mature meronts mature gamonts unsporulated oocysts) which
was not observed when cells contained earlier stages of development (eg trophozoites immature
gamonts) This tissue reaction was described by Hoare (1935a 1935b) as the development of an annular
constriction of the apical portion of the villus separating infected epithelial cells from unaffected cells
The constriction involved the epithelium but could also extend inwards into the core of the villus These
changes were associated with congestion of capillaries and extravasation of red blood cells within the
constricted segment and in some sections villar tip necrosis
In their case report of one domestic ferret Blankenship-Paris et al (1993) described the gross
pathologic lesions associated with intestinal coccidiosis in this case there was diffuse dilation and
reddening of the small intestine which was empty and the colon contained dark watery material
Histologic lesions were confined to the ileum and jejunum The jejunum exhibited thickening of the villi
with a crypt to villus ratio of 15 mild granulomatous inflammation in the lamina propria and large
numbers of coccidial meronts gamonts and oocysts within the enterocytes of the villar tips
The gross lesions described by Sledge et al (2011) from 20 domestic ferrets are as follows thin
body condition with moderate to marked dehydration perineal staining with diarrhea moderate dilation
of the small and large intestines and the presence of pasty tan to tarry black digesta within the distal small
intestine and colon Other findings in one to a small number of ferrets included enlarged pale tan livers
splenomegaly with dark red colouration and multiple superficial gastric or duodenal ulcers The
24
histologic lesions from 10 ferrets included moderate blunting and occasional fusion of jejunal and ileal
villi focal attenuation and erosion of the epithelium of the villar tips with exudation of fibrin neutrophils
and blood into the intestinal lumen in regions with severe erosion Intact epithelial cells at the villus tips
and rarely sloughed epithelial cells in the intestinal lumen contained numerous intracytoplasmic coccidia
representing a range of asexual and sexual life stages (meronts macrogamonts microgamonts and
oocysts) The subjacent lamina propria of the small intestine and of the large intestine exhibited moderate
lymphoplasmacytic infiltration with occasional neutrophils and congestion of blood vessels Marked
mucosal hemorrhage was identified in the most severely affected sections
Marked gross and histopathologic hepatobiliary lesions were described in a single ferret by
Williams et al (1996) On gross necropsy the liver was pale and enlarged with dilated firm bile ducts
and thickening of the gall bladder wall Similar gross necropsy findings were described by Kaye et al
(2015) marked dilation and mural thickening of the entire biliary tree (including gall bladder intrahepatic
and extrahepatic bile ducts) On histopathology Williams et al (1996) noted that the marked thickening
of the gallbladder wall was a result of cystic proliferation of mucosal glands which were separated by
tracts of fibrous connective tissue and marked granulomatous inflammation Liver sections exhibited
marked biliary hyperplasia marked periductular fibrosis and moderate periportal lymphoplasmacytic
cuffing There was multifocal papillary proliferation of bile duct epithelium and dilation of the bile ducts
and within the ductular lumens there were moderate numbers of lymphocytes and plasma cells small
numbers of degenerate neutrophils sloughed epithelial cells and debris All endogenous coccidial life
stages were present within the gall bladder and biliary epithelium with meronts visible in 20 of the
intact epithelial cells of the biliary tree and gallbladder and oocysts free within the lumen of the
intrahepatic bile ducts Similar lesions were present in the case described by Kaye et al (2015) and as
well as in juvenile and adult farmed mink (Mustela vison) with hepatobiliary coccidiosis (Davis Chow amp
Gorham 1953)
25
15 INTRODUCTION TO ENTERIC COCCIDIOSIS IN THE BLACK-FOOTED FERRET
151 Natural history and conservation of the black-footed ferret in North America
Black-footed ferrets are one of only three wild ferret species worldwide the other species are the
European polecat (Mustela putorius) and the Siberian polecat or steppe polecat (Mustela eversmanii)
They are the only native North American ferret species and the most endangered North American
carnivore They are nocturnal carnivores whose diet and lifestyle are highly dependent on local prairie
dog (Cynomys sp) populations Prairie dogs comprise almost exclusively the diet for the BFF who also
use the complex burrow systems made by prairie dogs to escape their predators and raise their young
(Santymire et al 2014 USFWS BFF Recovery Program 2017)
While formerly distributed throughout the North America prairie ecosystem BFF were
considered extinct by the late 1950s In 1964 a single population was discovered in Mellette County
South Dakota Progressive decline of this population in subsequent years resulted in the decision by
United States Fish and Wildlife Service (USFWS) to initiate a captive breeding program for the species
From 1971-1973 four females and five males were captured for this purpose Despite successful breeding
no kits survived and the last adult ferret in this captive colony died in 1979 at that time BFF were again
presumed extinct in the wild based on annual surveys of the initial capture site In 1981 a dead BFF was
discovered by a ranch dog outside of Meeteetse Wyoming allowing wildlife biologists to identify
another colony of BFF This colony flourished until 1985 when an outbreak of canine distemper in the
BFF population and an outbreak of sylvatic plague in the local prairie dog population resulted in sharp
population declines From 1985 through 1987 all 24 of the remaining BFF were trapped and brought into
captivity to re-initiate the captive breeding program Six ferrets in this initial group died of canine
distemper while in captivity and of the remaining 18 survivors 7 bred successfully to create the founding
population of the current captive breeding population Today this captive breeding population consists of
approximately 300 BFF distributed among multiple institutions (Santymire et al 2014)
26
Since 1986 this multi-institutional effort has been breeding BFF in captivity with reintroductions
back into the wild in 28 selected locations in Canada the USA and Mexico Currently six facilities
participate in the BFF Species Survival Plan (SSP) the Toronto Zoo USFWS National Black-footed
Ferret Conservation Center National Zoorsquos Smithsonian Conservation Biology Institute Louisville
Zoological Garden Cheyenne Mountain Zoo and the Phoenix Zoo (Black-footed Ferret Recovery
Implementation Team 2011) As of 2011 over 8000 BFF kits had been produced in captive breeding
facilities (Black-footed Ferret Recovery Implementation Team 2011)
Multiple infectious diseases pose a significant risk to the captive breeding and post-release
survival of BFF including canine distemper and sylvatic plague Coccidiosis is recognized as a cause of
significant juvenile morbidity and mortality in captive breeding programs and can result in significant
population losses (Bronson et al 2007 Santymire et al 2014 USFWS BFF Recovery Program 2017)
152 Coccidia identified from black-footed ferrets
Eimeria ictidea and Eimeria furonis have been identified in black-footed ferrets based on
morphologic criteria (Jolley et al 1994) Jolley et al examined fecal samples from six captive BFF during
a distemper outbreak as well as samples from wild BFF They described one medium-sized ovoid
tetrasporic dizoic oocyst with a double wall presence of a polar body and lacking both an oocyst residual
body and micropyle The oocysts measured 232 microm (range 182-274) by 155microm (range 130-162) with
a SI of 150 The sporocysts were elongate with the presence of both sporocyst residuum and a Stieda
body Sporozoites contained prominent refractile bodies at the posterior end and were aligned anterior to
posterior within sporocysts These oocysts were shed by all six captive ferrets On histopathology of
intestinal sections merogony and gametogony were observed within the villar epithelium throughout the
small intestine but were concentrated in the jejunum Two morphologically distinct meronts were
detected in these sections one at the villar tips which was larger and lacking in undifferentiated mass
and the other at the base of the villi or rarely in the intestinal crypts Gametogony was predominantly
27
observed at the villar tips and was noted throughout the small intestine These organisms were considered
consistent with Eimeria ictidea based on descriptions by Hoare (1927) from domestic ferrets
A second small spherical to subspherical tetrasporic dizoic oocyst was documented that had a
pink double wall a granular residual body and lacked both oocyst polar body and micropyle This
smaller oocyst measured 126plusmn12 microm (108-152) by 119plusmn09 microm (101-129) with a SI of 106 The
sporocysts were elongate with the presence of a Stieda body and sporozoites contained refractile bodies
Similar to the larger Eimeria species described above merogony and gametogony were observed within
the villar epithelium throughout the small intestine with endogenous developmental stages most
numerous in the jejunum The meronts were small with 16 or fewer merozoites Micro- and
macrogamonts were observed clustered within the apical third of the villar epithelium as were meronts
Jolley et al (1994) determined these small spherical oocysts to be consistent with Eimeria furonis as
described by Hoare (1927) from domestic ferrets
Jolley et al (1994) described a third type of coccidial oocyst occasionally detected in small
numbers within the BFF fecal samples the authors did not state whether this third type of oocyst was
recovered from wild or captive BFF The oocysts measured 370plusmn13 microm (350-386) by 223plusmn23 microm
(212-232) with a SI of 106 Attempts to sporulate collected oocysts were largely unsuccessful and
corresponding endogenous stages were not identified on histopathologic examination of necropsied
ferrets precluding further morphologic identification of the parasite It should be noted that coccidial
oocysts with similar measurements had not been detected in wild or captive prey species available for
ingestion by BFF (Jolley et al 1994)
Previous to this report coccidial oocysts had been isolated from the feces of BFF in two captive
populations (Carpenter amp Hillman 1979 Williams et al 1988) The abstract by Carpenter amp Hillman
(1979) did not describe the oocysts whereas Williams et al (1988) stated that two Eimeria species (one
with larger oocysts and one with smaller oocysts) were identified within the fecal samples but they were
28
not identified further Interestingly Williams et al reported both Eimeria species to be shed in the feces
of all ferrets concurrently affected by distemper and by approximately 30 of the clinically healthy
ferrets at the time of investigation
Non-enteric coccidia have been reported from captive BFF in one facility by two authors (Jolley
et al 1994 Williams et al 1988) Both reports presumably describing the same case(s) noted the
presence of endogenous coccidial life stages in histologic sections of respiratory tissue and merozoites of
an unidentified coccidium in impression smears of the urinary bladder from BFF diagnosed with canine
distemper Meronts were observed within the epithelium of the trachea a large bronchus and associated
bronchial glands Jolley et al (1994) described the lesions as occurring in the same ferret whereas in the
earlier report by Williams et al (1988) they are described as occurring in two different ferrets There have
been no subsequent published reports of systemic coccidiosis in black-footed ferrets and no cases have
been identified within the pathology database of the Toronto Zoo captive BFF population or by the
current SSP pathologist (Dr Michael M Garner personal communication)
There is a significant information gap regarding the pre-patent periods and pathogenicity of both
identified Eimeria species in BFF and studies to further characterize the eimeriid coccidia of the BFF are
lacking
153 Morbidity mortality and clinical signs associated with enteric coccidiosis in black-footed ferrets
The clinical signs of enteric coccidiosis in black-footed ferrets include mucoid to hemorrhagic
diarrhea abdominal discomfort lethargy appetite loss vomiting and dehydration In some cases sudden
death precedes the development of diarrhea Both adult and juvenile BFF are affected by the disease
which causes significant morbidity and mortality in captive populations (Bronson et al 2007) One
retrospective study of the captive BFF population at the Smithsonian National Zoological Park
determined that the most common cause of death in juvenile BFF (aged 30 days ndash 11 months) was
gastrointestinal pathology (524 of juvenile deaths) with 636 of these cases caused by enteric
29
coccidiosis (Bronson et al 2007) Despite the significance of this disease to the captive population its
effect on morbidity and mortality in wild BFF populations is unknown To the authorrsquos knowledge no
routine surveys of fecal parasites have been conducted on wild-born or captive released BFF during
yearly spotlighting events at ferret release sites However samples may be collected opportunistically if
fecal material is identified within the traps used to catch wild BFF during yearly surveys at release sites
Where fecal samples have been analyzed a 13 prevalence of coccidiosis has been identified in wild
born BFF (Dr Rachel Santymire personal communication) Fecal samples have been collected from BFF
at four release sites within the USA Wind Cave National Park (South Dakota) Badlands (South Dakota)
Conata Basin (South Dakota) and Aubrey Valley (Arizona) and positive samples were identified only at
the first site (Dr Rachel Santymire personal communication) Although radio-telemetry has been used at
some release sites to determine sources of mortality and factors involved in survival its use is not
widespread Furthermore the nocturnal and fossorial lifestyle of the BFF is a significant impediment to
the surveillance and monitoring of disease in this species
16 TREATMENT PREVENTION AND CONTROL OF INFECTION BY EIMERIA SPP
161 Current recommendations for treatment of eimeriid coccidia in carnivores
Described anticoccidial therapies for carnivores come from research in domestic cats and dogs
infected by Cystoisospora species these tissue coccidia (family Sarcocystidae) are only distantly related
to the Eimeria species infecting the BFF and other ferrets Current therapeutic recommendations by the
Companion Animal Parasite Council (CAPC 2013) for treatment of described Cystoisospora species
isolated from cats and dogs include the following amprolium (300-400 mg daily for 5 days in dogs 110-
200 mg daily for 7-12 days in dogs 60-100 mgkg daily for 7 days in cats) amproliumsulfadimethoxine
(150 mgkg amprolium and 25 mgkg sulfadimethoxine daily for 14 days in dogs) diclazuril (25 mgkg
for one dose in cats) furazolidone (8-20 mgkg 1-2 times daily for 5 days in dogs and cats) ponazuril (20
mgkg daily for 1-3 days in dogs and cats) quinacrine (10 mgkg daily for 5 days in cats)
30
sulfadimethoxine (50-60 mgkg daily for 5-20 days in dogs and cats) sulfadimethoxineormetoprim (55
mgkg sulfadimethoxine and 11 mgkg ormetoprim daily for 7-23 days in dogs) sulfaguanidine (150 or
200 mgkg daily for 6 days or 100-200 mgkg every 8 hours for 5 days in dogs and cats) toltrazuril (10-
30 mgkg daily for 1-3 days in dogs) trimethoprimsulfonamide (30-60 mgkg trimethoprim daily for 6
days if gt4kg 15-30 mgkg trimethoprim daily for 6 days if lt4kg) (CAPC 2013) Notably the use of all
drugs listed by the CAPC is considered off-label with the exception of sulfadimethoxine
162 Current recommendations for anticoccidial treatment and prophylaxis in domestic and black-
footed ferrets
1621 Domestic ferrets
Recommended daily oral treatment regimens for enteric coccidiosis in domestic ferrets include
amprolium (19 mgkg once daily 05 mgkg) decoquinate (05 mgkg) sulfadimethoxine (300 mgkg in
drinking water) or sulfadiazine-trimethoprim (30 mgkg once daily) all administered for a minimum of
two weeks (Bell 1994 Patterson amp Fox 2007 Patterson et al 2014) Both the aforementioned
coccidiostats amprolium and decoquinate are sold in large formats and are ideal for use in larger
operations such as breeding facilities research facilities or rescue centers Other anticoccidial therapies
used in domestic ferrets include toltrazuril (20 mgkg) and ponazuril (30-50 mgkg) once daily It should
be noted that all anticoccidial therapy used in domestic ferrets is considered off-label drug use
Multiple follow up fecal examinations should be performed after the treatment regimen is
complete and large groups may need to be treated multiple times Routine cage cleaning is also important
to decrease the environmental oocyst burden and prevent re-infection and in the case of coccidial
outbreaks ferrets should be transferred to clean cages multiple times during the course of anticoccidial
therapy Disinfectants such as bleach or quaternary ammonium compounds or dry heat should be used
for effective environmental decontamination (Patterson et al 2014)
31
1622 Species Survival Plan recommendations for black-footed ferrets
Treatment and prophylaxis of enteric coccidiosis with oral sulfadimethoxine was previously
recommended by the BFF Species Survival Plan (SSP) However due to a suspicion of decreasing
efficacy of treatment ponazuril has been recommended recently for treatment Due to the perceived
exquisite sensitivity of BFF to enteric coccidia the current SSP recommendation for treatment is oral
ponazuril at 30 mgkg once if ferrets are to be transported anesthetized stressed or are otherwise
suffering from another illness or injury (even in the absence of clinical signs or fecal shedding) The same
single oral dose of 30 mgkg is also recommended for kits at weaning (30-35 days of age) post weaning
(40-45 days of age) and prior to anesthesia for initial examination and vaccines (50-60 days of age)
Large crowded or otherwise stressed litters should be administered 30 mgkg orally once every 7-10 days
during the period of stress For treatment of coccidial diarrhea diagnosed by fecal examination 30 mgkg
orally once every 7 days for two doses or 50 mgkg orally once daily for 3 days in food (repeated in 7
days) is recommended In BFF with clinical signs of dehydration administration of subcutaneous or
intravenous fluid therapy has been performed Additional therapy with other antibiotics is sometimes
provided in cases with severe clinical signs or where secondary or primary bacterial enteritis is suspected
There is no pharmacokinetic or pharmacodynamic information available for the use of
anticoccidial drugs in BFF or other Mustelidae and thus it is unknown whether the current dose or
frequency of administration is truly appropriate for treatment of coccidiosis In 2 to 3-month-old piglets
administered a single dose of ponazuril orally at 5 mgkg peak serum concentration occurred at 42 hours
(36-48 hr) and elimination half-life was ~56 days (Zou et al 2014) In llamas administered ponazuril as
a single dose of 20 mgkg orally peak serum concentration occurred at 84 hours and elimination half-life
was ~56 days (Prado et al 2011) In domestic cows administered ponazuril as a single 5 mgkg dose
orally peak serum concentration occurred at 48 hours and elimination half-life was 58 hours (Dirikolu et
al 2009) The relevance of serum drug concentrations for treating an intestinal infection that lacks
extraintestinal life stages is likely minimal because the highest drug dose will reach the site of concern
(intestines) and systemic distribution is not required
32
Furthermore no safety or efficacy studies have been performed in any ferret species to validate
the current uses of either sulfadimethoxine or ponazuril for treatment nor have the current recommended
treatment lengths been validated However anecdotal information based on current usage would indicate
that they are safe at the current dosages and frequencies of administration as no adverse effects have been
reported A recent efficacy study in shelter dogs and cats showed that oral ponazuril (50 mgkg)
administered once daily for 3 days was effective for treatment of infection with Cystoisospora as
determined by a reduction in or cessation of fecal oocyst shedding at 4 and 8 days post treatment
Treatment efficacy in this study was inversely correlated to fecal oocyst counts at the initiation of
treatment (Litster et al 2014) Interestingly efficacy of this dose compared to the other two treatment
groups (single 50 mgkg or 20 mgkg oral dose) did not seem to differ but no statistical analysis was
performed Given the ubiquitous use of ponazuril in captive breeding facilities and concerns regarding
resistance of coccidia species to sulfadimethoxine therapy information on minimum effective doses and
dose regimes would be necessary to inform appropriate future SSP treatment and management plans and
to minimize development of drug resistance
17 VACCINES AGAINST COCCIDIA
171 Theory
The development of resistance of protozoal parasites to chemotherapeutic agents has resulted in a
shift towards the development of vaccines for the protection of domestic livestock Immunity to enteric
coccidiosis in avian and mammalian species involves both humoral and cell mediated responses Eimeria
spp infection in sheep rats poultry and other species generally results in a protective immune response
against subsequent re-infections (Catchpole et al 1993 Shi et al 2000) Interestingly this is not the case
for some host parasite interactions for example a recent report indicated that primary infection with E
ninakohlyakimovae in goat kids did not provide protective immunity against subsequent challenge with
the same parasite (Ruiz et al 2013)
33
Vaccines can be divided into four general categories live vaccines inactivatedkilled vaccines
subunit vaccines and recombinant vaccines Live vaccines are orally administered using small numbers of
infectious oocysts or oocysts from strains with low pathogenicity and result in patent but ideally sub-
clinical infections in the host that will elicit a protective immune response Such live vaccines can be
produced using attenuated forms of the pathogen of interest for example in chickens using ldquoprecociousrdquo
strains of Eimeria spp These precocious strains undergo a reduced number of merogonic replications
within the host cells and thus fewer oocysts are shed in the feces of vaccinated animals This reduction in
endogenous merogonic cycles reduces the amount of damage to the intestinal epithelium as well as
reducing the number of oocysts contaminating the environment
Another strategy has been to use live parasites with truncated life cycles An example of this is
the Toxoplasma gondii vaccine developed to prevent abortion in sheep This parasite was passaged
multiple times through a mouse host resulting in an inability to produce tissue cysts (Meeusen et al
2007) This is desirable as the cyst stage of this parasite normally inhibited by the immune system can be
reactivated during periods of stress or immunocompromise The potential drawbacks of live vaccines
include 1) the ability to produce and isolate adequate numbers of coccidial oocysts to meet vaccine
production requirements 2) the potential development of clinical disease in the host as a result of
inoculation 3) the need for all susceptible individuals to receive the vaccine simultaneously to prevent
fecal-oral inoculation of unvaccinated animals with high doses of the infective agent likely to be present
in a shared environment through fecal shedding
Inactivated vaccines are produced when the microbe of interest is killed via application of heat
radiation or chemical treatment prior to inoculation into the host species While safer because they cannot
induce disease in the inoculated patient inactivated vaccines stimulate a reduced immune response
compared with live vaccines and are consequently less effective Subunit vaccines contain single or
multiple antigens of importance in initiating the host immune response rather than the entire pathogen of
concern Subunit vaccines cannot induce disease in the immunized host but are more difficult to produce
34
because they require a detailed understanding of host immune response to infection Recombinant
vaccines involve the genetic modification of a vector (virus or bacteria) one capable of infecting the host
of interest to contain DNA of the pathogen of interest These vectors induce an immune response in the
vaccinated host but as with subunit vaccines cannot induce disease However recombinant vaccines are
again difficult to produce because they require an in depth understanding of the life cycle stages genes
and antigens targeted by the host immune response to infection There are currently no recombinant
vaccines marketed in Canada for use in veterinary medicine against protozoal disease
Creation of effective vaccines against protozoal parasites is complicated by parasite antigenic
diversity during the different life cycle stages and among protozoal species and strains of the same species
(Meeusen et al 2007) Although most parasites induce some level of immunity in their host species the
immunological response to different parasite life stages and species has been poorly characterized for
most coccidia Furthermore many parasites have developed mechanisms to evade host immune responses
or to continue survive and replicate in and transmission by previously infected hosts Our limited
understanding of the immune responses against coccidial antigens has restricted commercial vaccine
production to live or attenuated vaccines (Meeusen et al 2007)
A notable disadvantage of anticoccidial vaccines is that they need to be developed for each
coccidial species of interest because of the species-specific nature of the immune responses this is a
considerable limitation compared with anticoccidial drugs that can have a much wider spectrum of action
(Vermeulen 2005) While the requirement for mass production of vaccine is a limiting factor for vaccines
developed for the agricultural industry this drawback would be less important for production of a vaccine
to be used in an endangered species
172 Species successes in anticoccidial vaccination
The first successful immunization against coccidiosis was reported in 1918 in dogs (Hall amp
Wigdor) In this report a dog that had previously recovered from coccidial infection with Diplospora
35
bigemina was fed three increasing doses of live non-attenuated coccidial culture (at 14 32 and 48 days
post recovery from primary infection) which resulted in no development of clinical signs and no oocyst
shedding for 11mdash18 days after each challenge Subsequently immunization of dogs and cats against
coccidia with protection lasting up to seven months was reported by Andrews (1926) Immunization of
albino rats to eimeriid infection after administration of three or more sublethal doses of Eimeria
nieschulzi via gastric intubation was reported by Morehouse (1938) further experiments showed that
sporozoites did not enter the host intestinal epithelium in immunized rats given a challenge dose
(Morehouse 1938) Similar findings were reported in chickens immunized against Eimeria tenella that
had 50 fewer intra-epithelial sporozoites following challenge compared to naiumlve birds (Augustine and
Danforth 1986) Conversely chickens previously inoculated with Eimeria acervulina exhibited more
intracellular sporozoites after challenge than naiumlve birds but sporozoites were not observed to develop in
previously immunized birds (Augustine and Danforth 1986) These findings provide further evidence that
the immune response to Eimeria spp may differ among host species
Vaccination against Eimeria species has been most successful and is most widely used in the
poultry industry particularly in breeder and layer flocks Almost all vaccines marketed for poultry are
live vaccines (attenuated and non-attenuated) Vaccination against other apicomplexan parasites in
domestic mammals has also been achieved but has been generally less effective for disease prevention
and is less widely available Marketed killed and inactivated (attenuated) vaccines include those
containing killed tachyzoites of Neospora caninum for cattle (Neoguard Merck Animal Health) and
chemically inactivated merozoites of Sarcocystis neurona for horses (EPM Vaccine Fort Dodge ndash no
longer in production) A subunit vaccine for Babesia canis in dogs uses cultured antigen (Pirodog
Merial) Available live vaccines include a vaccine against Toxoplasma gondii in sheep (Ovilis Toxovax
Intervet) that uses an attenuated temperature sensitive strain (S48)
36
18 RESEARCH GOALS AND OBJECTIVES
181 Objectives
a) To determine and characterize (morphologically and molecularly) the enteric coccidial species
currently affecting the black-footed ferret population
b) To describe the natural history of enteric coccidiosis in captive black-footed ferrets including
pre-patent period shedding frequency and burdens and morbidity and mortality rates
c) To compare molecular morphologic and life history characteristics of enteric coccidial species
identified in domestic ferrets to those in black-footed ferrets
d) To validate domestic ferrets as an experimental model for intestinal coccidiosis in the black-
footed ferret
182 Hypotheses
a) Multiple Eimeria species will be isolated from the black-footed ferret population
b) The Eimeria species identified from black-footed ferrets will be the same as those previously
described in domestic ferrets
c) A single pathogenic Eimeria species will be implicated in recorded outbreaks of clinical
coccidiosis during the period of study
d) Domestic ferrets can act as an experimental model of intestinal coccidiosis for black-footed
ferrets
183 Applications
The goal of this project is to better characterize the enteric coccidia of the endangered black-
footed ferret in order to set the stage for improved disease prevention and treatment To the authorrsquos
knowledge this project is the first attempt to isolate and perform molecular characterization of the
coccidial species endemic in the black-footed ferret population This information will be used to compare
these species to known coccidia from domestic ferrets and other related mammals As experimental work
37
cannot be carried out on enteric coccidiosis in the BFF due to its endangered status if the domestic ferret
can be validated as an experimental model studies of the patterns of anticoccidial resistance and
development of immunity against Eimeria spp can be undertaken in vivo The ultimate goal would be the
development of an autogenous vaccine used to improve survival of ferret kits and reduce morbidity and
mortality associated with coccidiosis in BFF captive breeding programs Based on clinical experience
stressful life events such a breeding weaning and transfer between institutions appear to increase the risk
of coccidial outbreaks in adult BFF As such vaccination could assist in reducing disease outbreaks in
BFF associated with various management activities There is no data on the significance of coccidiosis in
wild populations and limited means of disease surveillance following release vaccination during captive-
rearing or pre-release conditioning of BFF would be an ideal method of reducing the potential effects of
this disease in released and free-living BFF Increasing the numbers of ferrets being released to the wild
and releasing ferrets immune to the subsequent threat of coccidiosis would support the goals of the
conservation initiative for the black-footed ferret
38
CHAPTER 2 MOLECULAR CHARACTERIZATION OF ENTERIC COCCIDIA FROM
DOMESTIC FERRETS (MUSTELA PUTORIUS FURO)
This chapter has been submitted for publication as
Adriana R Pastor Dale A Smith and John R Barta (2017) Molecular Characterization of Enteric
Coccidia from Domestic Ferrets (Mustela putorius furo) Vet Parasitol Regional Studies and Reports (In
review)
ABSTRACT
Combined morphometric and molecular characterization of coccidia that infect domestic ferrets
(Mustela putorius furo) was completed to improve the diagnostic specificity of enteric lsquococcidiosisrsquo in
this host Coccidia positive fecal samples (n=11) and formalin fixed paraffin embedded intestinal tissues
(n=3) from domestic ferrets were collected from diagnostic laboratories in Canada and Europe An
average of 35 and 13 domestic ferret fecal samples per year were coccidia-positive when tested by
Canadian and European diagnostic laboratories respectively during the period 2008-2015 Oocyst
morphometrics and sequence genotyping at two loci (nuclear 18S rDNA [nu 18S rDNA] and
mitochondrial cytochrome c oxidase subunit I [mt COI]) were conducted on all samples The first nu 18S
rDNA and mt COI sequences for Isospora (=Cystoisospora) laidlawi and the first mt COI sequence for
Eimeria furonis were generated during this study Phylogenetic analysis of the mitochondrial COI
sequences demonstrated that E furonis was most closely related to E cf ictidea isolated from a black-
footed ferret (Mustela nigripes) and that I (=C) laidlawi was closely related to C canis and C felis The
identifications provided by diagnostic laboratories of the specific parasite species present in a sample
showed poor agreement with their identifications based on genotyping obtained in this study Molecular
techniques appear to be essential for accurate determination of coccidial species responsible for individual
and group outbreaks of coccidiosis and for further understanding of eimeriid host-parasite relationships
Key words coccidia Cystoisospora laidlawi domestic ferret Eimeria furonis Eimeria ictidea Mustela
putorius furo
39
21 INTRODUCTION
Coccidia are host-specific parasites of the phylum Apicomplexa with greater than 2000 species
named to date (Duszynski et al 2000 Upton 2000) The eimeriorinid coccidia (suborder Eimeriorina)
include typical intestinal coccidia such as Eimeria Isospora and Cyclospora species belonging to the
family Eimeriidae as well as tissue (cyst-forming) coccidia such as Cystoisospora Besnoitia Toxoplasma
and Sarcocystis species that belong to the family Sarcocystidae (see Cox 1994)
Enteric coccidia affect both domestic ferrets (Mustela putorius furo) and their wild counterparts
In his initial descriptions of enteric coccidiosis in domestic ferrets Hoare (1927 1935b) did not observe
clinical signs of intestinal disease associated with infection More recently it has been recognized that
enteric coccidiosis can result in clinical signs ranging from mild transient diarrhea to more severe disease
with dehydration lethargy depression weight lossemaciation inappetence and death (Blankenship-Paris
et al 1993 Powers 2009 Sledge et al 2011 Hoefer et al 2012 Patterson et al 2014) Rectal prolapse
has also been reported in ferrets with enteric coccidiosis (Hillyer 1992 Hoefer et al 2012) Disease
appears to be most common in young or stressed animals In one study co-infection with coccidia and
Lawsonia intracellularis (Desulfovibrio sp) was diagnosed in 4 of 19 ferrets with proliferative bowel
disease (Li et al 1996) these ferrets presented with clinical signs including diarrhea lethargy anorexia
weight loss dehydration and emaciation Two cases of biliary coccidiosis have also been reported in
domestic ferrets infection was associated with biliary epithelial hyperplasia cholecystitis and
cholangiohepatitis (Williams et al 1996 Kaye et al 2015)
Three species of coccidia affecting domestic ferrets were originally described and named by
Hoare (1927) Eimeria ictidea Eimeria furonis and Isospora (=Cystoisospora) laidlawi The three
species were detected in feces from 50 domestic ferrets involved in an outbreak of canine distemper at a
research facility For each parasite the author described the morphology of sporulated oocysts isolated
from feces as well as sporulation time (exogenous life stages) All subsequent reports of morphologic
diagnoses of these coccidia have been based on Hoarersquos original descriptions The pre-patent period
40
(minimum duration of endogenous development) in inoculated naiumlve ferrets was described for E furonis
and E ictidea as 6 and 7 days respectively (Hoare 1935b) The pre-patent period for I (=C) laidlawi
was not determined because the number of oocysts available was insufficient for an experimental
infection trial
Hoare (1927) described the sporulated oocysts of E furonis as follows spherical double outer
wall with a thin colourless outer layer and a thick yellowish inner layer no micropyle or residual body
and measuring on average 128 times 120 microm (length [L] 112-144 width [W] 104-128 shape index [SI]
107) Sporocysts were spindle-shaped with one end constrictedblunted contained a residual body and
on average measured 8-88 times 4 microm Sporozoites were vermiform with one end narrower than the other
arranged head to tail and each had a central nucleus a clear vacuole was identified in some sporozoites at
their broad posterior end
The sporulated oocysts of E ictidea were described as follows oval or elliptical with a double
outer wall with a thin colourless outer layer and a thick yellowish inner layer no micropyle or residual
body and measuring on average 236 times 175 microm (L 184-272 W 128-208 SI 135) Sporocysts were
irregularly oval with one end broad and the other more constricted contained a residual body and
measured 115 times 65 microm on average Sporozoites were vermiform with one end narrower than the other
arranged head to tail in the sporocysts and had a central nucleus and a clear vacuole at their broad
posterior end
The sporulated oocysts of I (=C) laidlawi were ovoid with a double outer wall with a thin
colourless outer layer and a thick yellowish inner layer had no micropyle or residual body and measured
on average 34 times 29 microm (L 320-368 W 272-304) A SI of 117 can be calculated from the original
mean dimensions Two sporocysts were identified each containing four sporozoites and no Stieda body
sporocysts were elliptical contained a residual body and measured 208 times 144 microm on average
Sporozoites were sausage shaped with one end slightly pointed and had a central nucleus and a clear
41
vacuole identified at the pointed end Sporozoites were arranged with pointed ends all at the same pole of
the sporocyst
Oocysts identified as Cystoisospora ohioensis have been reported from fecal samples collected
from healthy domestic ferret kits in a large American ferret breeding operation that also housed juvenile
domestic dogs (Patterson and Fox 2007) The method of identification of this parasite was not described
A second similar institution reported the presence of Cystoisospora (=Isospora) species also thought to
be C ohioensis in routine fecal examination of their colony (Dr Bambi Jasmin personal
communication) Identification in this case was performed by the Animal Health Diagnostic Center at
Cornell University and was based on morphometrics using light microscopy The significance of these
findings is unknown but no clinical signs or histologic lesions were described in the ferrets shedding
these oocysts The definitive hosts for C ohioensis are canids including the domestic dog
More recently molecular techniques have been used for the more precise identification of
coccidia Nucleotide sequences like morphological features diverge over time under selective pressure
however recent evolutionary divergence among coccidia is more likely to be reflected in molecular as
compared to morphologic differences Thus nucleotide sequences that are more similar are inferred to be
more closely related and to have diverged more recently (Cox 1994)
Molecular characterization of ferret coccidia has only been performed for one species Eimeria
furonis Abe et al (2008) extracted DNA from oocysts from the feces of a single domestic ferret with
clinical signs resulting from coccidial enteritis Using primers initially developed for molecular
identification of Cyclospora species (see Matsubayashi et al 2005) small subunit ribosomal DNA (nu
18S rDNA) primers CYC1FE (5ʹ-TAC CCA ATG AAA ACA GTT T-3prime) and CYC4RB (5prime-CGT CTT
CAA ACC CCC TAC TG-3prime) were used to amplify a 347 base pair fragment of nu 18S rDNA The
amplicon was sequenced (GenBank AB329724) and compared with previously published partial nu 18S
rDNA sequences from 40 Eimeria two Isospora and four Cyclospora species The resulting phylogram
42
grouped E furonis with E alabamensis (cattle) and E meleagrimitis (turkey) In the same study the
microscopic morphology of the oocysts was used to identify this coccidial species as Eimeria furonis by
comparison with Hoarersquos (1927 1935b) published descriptions of Eimeria furonis and Eimeria ictidea
Sledge et al (2011) also used nu 18S rDNA to identify Eimeria furonis as the cause of three
distinct outbreaks of enteric disease in domestic ferrets Initial identification was performed using
morphometrics of sporulated oocysts collected from feces in one of the three outbreaks being
investigated Formalin fixed paraffin embedded intestinal segments from ferrets from each of the
outbreaks contained multiple coccidial life stages when examined by light microscopy PCR amplification
of a 247 base pair (bp) amplicon of the nu 18S rDNA was generated from DNA isolated from stored
formalin-fixed tissues for further genetic analysis Analysis and sequencing of amplicons from all three
groups showed 100 identity to sequences previously reported by Abe et al (2008) for the gene encoding
E furonis nu 18S
In 2015 Kaye et al identified coccidia within hepatobiliary lesions in a domestic ferret receiving
immunosuppressive therapy for red cell aplasia DNA was extracted from frozen liver and a fragment of
the nu 18S rDNA was amplified using the primers previously described by Sledge et al(2011) The
authors reported that the DNA sequence of the amplicon had 100 identity to the published nu 18S
rDNA sequence of E furonis and 95 identity to the nu 18S rDNA of E myoxi (rodent) E alabamensis
(cattle) and Isospora robini (avian) (Kaye et al 2015)
It is difficult to estimate the current prevalence of enteric coccidia within the North American
domestic ferret population and no studies have been conducted to do so Fecal samples submitted to
veterinary diagnostic laboratories from domestic ferrets in Canada are uncommon and samples positive
for coccidia appear infrequently (Dr Donald Martin personal communication) Conversely in Europe
the prevalence of coccidia within the domestic ferret population appears to be higher based on
submissions to a large veterinary diagnostic laboratory in Germany Data from Idexx Vet Med Lab in
Ludwigsburg Germany was compiled to review the prevalence of coccidia and Giardia within fecal
samples from domestic ferrets (Pantchev et al 2011) The authors reported that of 284 fecal samples
43
submitted from 2002-2004 18 (63) had detectable coccidial oocysts on fecal flotation Based on
morphologic characteristics oocysts were identified as E ictidea E furonis I laidlawi (herein referred
to as I (=C) laidlawi as noted above) and another unidentified Isospora species Comparative data from
the same laboratory from 2009-2010 included sample submissions from 253 ferrets 21 (83) of which
were positive for coccidial oocysts on fecal flotation Nine of the samples were identified as containing E
furonis three contained both E furonis and I (=C) laidlawi eight contained only I (=C) laidlawi and
one sample contained both E furonis and E ictidea identification in all cases was again based on oocyst
morphometrics No statistically significant difference in the occurrence of coccidial oocysts was detected
when data from the two periods were compared (Fisherrsquos exact test P=041) (Pantchev et al 2011)
The purpose of the present study was to perform a more detailed molecular characterization of the
coccidial species isolated from domestic ferrets to estimate prevalence of the different coccidial species
within the Canadian domestic ferret population and to associate morphologic and molecular
characteristics of a greater range of enteric coccidial species in order to improve diagnostic accuracy
22 MATERIALS amp METHODS
221 Fecal samples
Multiple diagnostic laboratories within Ontario Canada1 and a major European diagnostic
laboratory 2 were solicited for fecal samples from domestic ferrets shedding coccidial oocysts Fecal
samples were diagnosed positive for coccidia based on fecal flotation and light microscopic identification
of Eimeria or Cystoisospora species Eleven samples were collected during the study period (from 2014-
2017) and preserved in potassium dichromate (25 wv) eight from Europe and three from Canada
Centrifugal flotation with saturated salt solution (Ryley et al 1976) was used to isolate oocysts from fecal
samples for genomic DNA extraction Genomic DNA extraction and purification were performed using a
1 Animal Health Laboratory Guelph ON Antech Diagnostics Canada Ltd Mississauga ON IDEXX Canada
Markham ON 2 Vet Med Labor GmbH Division of IDEXX Laboratories Ludwigsburg Germany
44
QIAamp DNA Mini Kit (Qiagen Hilden Germany) according to manufacturerrsquos instructions After
addition of DNAzol to the samples (Qiagen Hilden Germany) samples were vortexed using 05 mm
glass beads (Biospec Products Inc Bartlesville OK USA) prior to extraction in order to fracture the
oocyst walls and release the sporocysts Concentrations of the resultant DNA were estimating using a
Nanodrop 2000 spectrophotometer (NanoDrop Products Wilmington DE USA) and stored at 4 degC for
immediate use or minus20 degC for later use
For each laboratory the number of domestic ferret fecal sample submissions numbers diagnosed
positive for coccidial oocysts and number of each coccidial species identified in positive samples were
tabulated for each of the years 2008-2015
222 Formalin fixed intestinal tissues
Major diagnostic pathology services across Canada3 were contacted to identify cases of enteric
coccidiosis identified on necropsy of domestic ferrets Cases were considered positive based on the
presence of asexual or sexual life stages of the parasites in intestinal sections The histologic sections on
each positive case were reviewed re-described and organisms measured (AP DAS) Gross necropsy
reports for all cases were also reviewed to identify any clinical correlates associated with enteric
coccidiosis DNA was extracted from ten 5-6 microm scrolls of formalin fixed paraffin embedded tissue
(FFPE) using the QIAamp DNA FFPE Tissue Kit (Qiagen) as per manufacturer instructions
223 Molecular characterization
Regions from the nu 18S rDNA and mitochondrial cytochrome c oxidase subunit I (mt COI)
DNA were amplified by polymerase chain reaction (PCR) from each sample using the primers listed in
Table 21 PCR amplification was performed for all samples in a volume of 25 microl containing ~100 ng of
3 Animal Health Centre Abbotsford BC Animal Health Laboratory Guelph ON Faculteacute de meacutedecine veacuteteacuterinaire
Universiteacute de Montreacuteal Saint-Hyacinthe QC Histovet Surgical Pathology Guelph ON IDEXX Canada
Markham ON Prairie Diagnostic Services Inc Saskatoon SK
45
genomic DNA 1times PCR buffer 15 mM MgCl2 02 mM deoxyribonucleotide triphosphates (dNTPs) 400
nM of each primer and 1 U of Invitrogen Platinum Taq DNA Polymerase (Thermo Fisher Scientific
Toronto ON Canada) Reactions were performed on a Bio-Rad T100 PCR thermal cycler (Bio-Rad
Laboratories Singapore) Samples were denatured and Taq polymerase activated at 95 degC for 3 min then
subjected to 35 cycles of 94 degC for 30s anneal at 50-62 degC (see Table 21 for specific anneal conditions
for the various primer pairs) for 30s and extension at 72 degC for 30-75s (see Table 21) followed by a
final extension at 72 degC for 7 min Suitable DNA (ie genomic DNA from an Eimeria or Sarcocystis sp)
was included in the PCR reactions to act as a positive control for the reaction chemistry All amplification
products were subjected to electrophoretic separation using 15 submarine agarose gel stained with
ethidium bromide and visualized on an ultraviolet transilluminator (Spectronics Corporation New York
NY USA) DNA band size was determined by comparison with a 1 kb DNA ladder (GeneRuler 1kb Plus
DNA ladder Thermo Fisher Scientific Waltham MA USA) Bands were excised with a new sterile
scalpel blade and PCR products were purified from the gel using a QIAquick Gel Extraction Kit (Qiagen)
PCR products were cycle sequenced using an ABI Prism 7000 Sequence Detection System (Applied
Biosystems Inc Foster City CA USA) by the Molecular Biology Unit of the Laboratory Services
Division University of Guelph (Guelph ON Canada) using the amplification primers to obtain
sequences in both directions The resulting chromatograms were aligned and analyzed with Geneious Ver
818 or later (Biomatters Limited Auckland New Zealand) and high quality consensus sequences
generated The resulting consensus sequences were searched from within Geneious against publically
available sequences on the BLAST server (blastncbinlmnihgovBlastcgi) using the blastn search
algorithm against the nrnt database (GenBank+EMBL+DDBJ+RefSeq ndash AA or DNA) Resultant new
nucleotide sequences were submitted to GenBank
46
224 Phylogenetic analysis
To determine the phylogenetic affinities of the newly obtained sequences with sequences from
related apicomplexan taxa representative nu 18S rDNA and mt COI sequences were downloaded from
GenBank with special reference to sequences from parasites that infect members of the order Carnivora
Nuclear 18S and mt COI sequences were aligned independently using MAFTT v7017 (Katoh et
al 2002) executed from within Geneious and then concatenated into a combined nu18S rDNA mt COI
dataset Multiple sequences from a single parasite were used to generate consensus sequences for each
locus as described by Ogedengbe et al (2017) Aligned sequences were trimmed to the length of the
largest newly generated nu 18S sequence Phylogenetic trees were generated using Bayesian Inference
(BI) using MrBayes Ver 326 (Huelsenbeck and Ronquist 2001) executed from within Geneious the
combined nu 18S and mt COI alignment was partitioned to permit locus-appropriate substitution models
to be applied to each partition For the nu 18S sequence partition the general time reversible (GTR)
substitution model (nst=6) with gamma rate variation (ie a GTR+G+I model) was applied For the mt
COI sequence partition the codon (M1) substitution model (using translation table 4 [ie lsquometmtrsquo]) was
used instead of the GTR with the remaining parameters remaining the same
The resulting tree was rooted using a pair of adeleid coccidia (Hepatozoon spp) as the taxonomic
outgroup All BI analyses were run for a chain length of 1000000 with tree sampling every 1000
following a burn-in of 100000 with default settings of 4 heated chains and heated chain temp of 02
47
23 RESULTS
231 Fresh fecal samples
From 2008-2015 inclusive the Canadian diagnostic parasitology laboratory4 received an average
of 1206 (range 81-160) domestic ferret fecal samples yearly the European parasitology laboratory5
received a yearly average of 230 samples (range 213-270) The number of fecal samples diagnosed as
positive for coccidial oocysts per year on fecal flotation during this time averaged 35 (range 0-8) and
130 (range 6-20) for the Canadian and European laboratories respectively The diagnosing laboratories
used oocyst morphometrics to identify the species of coccidia present Almost all coccidia-positive
submissions to the Canadian laboratory were identified as containing an I (=C) species based on light
microscopy Coccidia in only three samples from the Canadian laboratory were identified as E furonis
one in each of 2010 2012 and 2014 E ictidea was not identified in any samples submitted to the
Canadian laboratory Approximately equal numbers of coccidia-positive samples from the European
laboratory were identified as E furonis and I (=C) laidlawi each year Only two samples from the
European laboratory contained oocysts that were identified as Eimeria ictidea using morphometrics one
from each of 2011 and 2013 Laboratory submissions to both laboratories are summarized in Table 22
Twelve fecal samples preserved in potassium dichromate were received for analysis by the
authors Eleven samples had previously been identified as containing a single coccidial species five
containing E furonis two containing E ictidea and four containing I (=C) laidlawi A final sample had
been identified as containing a mix of E furonis and Cystoisospora canis Results of microscopic and
molecular characterization of these samples are summarized in Table 23
4 IDEXX Canada Markham ON
5 Vet Med Labor GmbH Division of IDEXX Laboratories Ludwigsburg Germany
48
232 Formalin fixed samples
Only three cases of coccidiosis were identified in domestic ferrets within the databases of the five
diagnostic laboratories that participated in the retrospective study Histologic sections of intestine were
received from these three cases which originated in Ontario6 and Quebec7 The Quebec sample (P2010-I)
was collected in 2010 and the Ontario samples (93-40404 and 17-008571) in 1993 and 2017 respectively
On gross necropsy the small intestinal contents of case P2010-I were described as pasty mucoid
yellow-brown feces with some blood For case 93-40404 the small intestines were described as empty
but melena was present within the terminal portion of the large intestine Scant intestinal contents and
dark brown fecal material in the colon were described in case 17-008571
In all cases endogenous developmental stages of coccidia were visible in histological sections
(Figure 21 is exemplary of the findings from one case) Hematoxylin and eosin stained sections from
P2010-I contained two affected regions of small intestine The intestinal mucosa of the first region
contained numerous asexual life stages and moderate numbers of sexual life stages as well as a small
number of oocysts free within the lumen The second section contained tissues that were poorly
preserved nonetheless 0-4 oocysts per 400times field were identifiable within the intestinal lumen Two
regions of affected small intestine were identified from 93-40404 after screening of all submitted sections
both contained low numbers of sexual and asexual endogenous stages Within one region there were small
numbers of meronts within the intestinal mucosa and lamina propria The second region had small
numbers of oocysts within cells of the epithelium and lamina propria as well as free within the intestinal
lumen In case 17-008571 multiple sections of jejunum contained numerous coccidian meronts gamonts
6 Animal Health Laboratory Guelph ON
7 Faculteacute de meacutedecine veacuteteacuterinaire Universiteacute de Montreacuteal Saint-Hyacinthe QC
49
and oocysts within intestinal villi within the ileum scattered epithelial cells also contained these various
life stages
Average length and width of oocysts were measured from slide sections for all cases For P2010-
I oocyst average length and width were determined from seven oocysts to be 94 microm (range 85-105) and
75 microm (range 69-84) respectively with a SI of 125 (range112-140) Average length and width of
oocysts measured from 93-40404 were determined from 5 oocysts to be 2814 microm (range 229-341) and
233 microm (range 180-308) respectively with a SI of 123 (range 110-146) For the third case 17-
008571 only 2 oocysts were identified and average length and width of oocysts measured 982 microm (range
973-992) and 845 microm (range 821-870) respectively with a SI of 116 (range 114-118)
233 Molecular characterization
DNA was successfully extracted from all twelve fecal samples and two cases with formalin fixed
tissue samples Attempts at amplification of DNA extracted from sample 93-40404 using the primer pairs
listed in Table 21 were unsuccessful Molecular identification results and GenBank accession numbers
for the remaining samples are summarized in Table 23 Both the nu 18S rDNA and mt COI sequences
from I (=C) laidlawi were unique when compared with available sequences from other Cystoisospora
species within the public databases However sequences from I (=C) laidlawi were most similar to
sequences from C canis and C felis and somewhat more divergent from sequences from members of the
C ohioensis species complex Two apparent genotypes of E furonis were identified based on nu 18S and
mt COI sequencing results Genotype 1 represented by EU sample 9014 had 100 identity to previously
published sequences of the nu 18S locus from two isolates from Japan (GenBank AB239130 and
AB329724) Genotype 2 represented by EU sample 907 and Canadian sample 17-008571 had 994
identity at the nu 18S locus (3 single nucleotide differences [SNDs] over 561 base pair region [bp]) to the
three sequences above belonging to E furonis genotype 1 Pairwise alignment of mt COI sequences from
both genotypes identified only 2 SNDs (996 pairwise identity over 513 bp region) Partial mt COI
50
sequences of E furonis from both genotypes were only distantly related (941 pairwise identity 30
SNDs over 513 bp and 905 pairwise identity 49 SNDs over 513 bp respectively) to publicly available
sequences from Eimeria ictidea from the black-footed ferret (Mustela nigripes) (GenBank KT203399)
and Eimeria mephitidis from the striped skunk (Mephitis mephitis) (GenBank KT203398) the only other
Eimeria species infecting members of the Carnivora for which sequence was available
234 Phylogenetic analysis
A phylogenetic reconstruction based on concatenated partial nu 18S rDNA and mt COI sequences
of E furonis I (=C) laidlawi and related coccidia is illustrated in Figure 22 The combined 18SCOI-
based tree demonstrates that the two Eimeria species from ferrets form a well-supported monophyletic
group that branches among a collection of other eimeriid coccidia that infect mammals The sarcocystid
parasite of the domestic ferret I (=C) laidlawi was found to group as the sister taxon to C canis that
together formed a monophyletic group with the closely related C felis all three of these closely related
Cystoisopora species possess comparatively large egg-shaped oocysts that are similar morphologically
24 DISCUSSION
The present work has generated the first nu 18S rDNA and mt COI sequences for Cystoisospora
laidlawi and the first mt COI sequence for Eimeria furonis both isolated from the domestic ferret In this
study histologic presence of organisms and microscopic identification of oocysts shed in feces have been
correlated with published and novel nu 18S and mt COI sequences
Eimeria ictidea was not identified in any Canadian sample and this coccidium was reported in
only 2 of 1840 fecal samples submitted from across the European Union (EU) to IDEXX Germany during
2008-2015 suggesting that E ictidea is not a frequent cause of enteric coccidiosis in domestic ferrets in
Canada or the EU
51
During the study period (2008-2015) almost twice as many domestic ferret fecal submissions
were made to the European as compared to the Canadian diagnostic laboratory however the prevalence
of coccidia-positive samples was similar The methodology used in this report cannot be used to
determine the actual prevalence of enteric coccidial infection (coccidiasis) or disease (coccidiosis) within
the domestic ferret population Fecal samples may be submitted to laboratories either as a result of
investigation into enteric disease or as part of a routine health examination Thus without historic
information accompanying each sample one can simply identify the proportion of positive samples and
compare the frequency of the finding of different coccidial species Prospective surveys of fecal samples
from healthy and sick domestic ferrets with greater sample size would be necessary to determine the true
prevalence of these parasites within the population and to infer their clinical significance
Comparatively few mitochondrial COI sequences have been generated for apicomplexan parasites
compared with other genetic loci the majority of published sequences obtained from Apicomplexa are
from nu 18S The disadvantage of using nu 18S rDNA sequences for parasite identification is that they
are poor at distinguishing among closely related eimeriid coccidia due to the highly conserved nature of
the nuclear ribosomal RNA locus In contrast mt COI sequences appear to be more useful for
distinguishing closely related coccidian species (Ogedengbe et al 2011) but are less useful than nu 18S
rDNA sequences for inferring more ancient relationships among more distantly related coccidia
Consequently the combined use of nu 18S rDNA and mt COI sequencing has been recommended for
improved species description and phylogenetic analysis (El-Sherry et al 2013) For these reasons both nu
18S and mt COI sequences were analysed in the present study
Despite adequate quantities of DNA extracted from the Ontario laboratory sample (93-40404)
successful amplification did not result with any primer pair (Table 21) Potential reasons for this include
degradation of formalin-fixed DNA into fragments too small for amplification with the desired primers
perhaps as a result of extended length of time in formalin prior to paraffin embedding or length of time
stored as FFPE tissue (23 years) or insufficient parasite DNA within the paraffin scrolls The primer pairs
52
used appear to be useful for most eimeriid coccidia (Ogedengbe 2015) and successfully amplified both
Eimeria species from DNA isolated from oocysts so it is unlikely that failure to amplify DNA from this
sample resulted from an inability of the primers used to recognize the parasite seen on section
Two genotypes of E furonis were identified in this study Genotype 1 was identified only from
samples originating from domestic ferrets in Europe but exhibited 100 identity based on nu 18S
sequencing with previously published sequences from both Japan and the USA Genotype 2 was
identified from samples originating from domestic ferrets in both Canada and Europe The small number
of single nucleotide differences between the two genotypes at two genetic loci in different genomes are
consistent with intraspecific variation (ie strain variation)
As might have been expected because of their morphological and host similarities nu 18S and mt
COI sequences of E furonis were determined to be most similar to an Eimeria species (E ictidea)
previously isolated from black-footed ferrets (Mustela nigripes) these eimeriid coccidia formed a
monophyletic group that was distinct from other eimeriid coccidia infecting mammals in the phylogenetic
analyses based on combined nu 18S rDNA and mt COI sequences Similarly the nu 18S rDNA and mt
COI sequences of I (=C) laidlawi are most similar to sequences from two other Cystoisospora species of
carnivores (C canis and C felis) that both have large egg-shaped oocysts comparable to those of I (=C)
laidlawi Both morphometrics and genotyping support the close relationships among these three
sarcocystid coccidia of carnivores These molecular data confirm that transfer of Isospora laidlawi to the
genus Cystoisospora by Barta et al (2005) is warranted
The previous light microscopic identifications of coccidial species in 3 of the 11 fecal samples
were not in agreement with the molecular findings These results were not surprising because light
microscopy has been shown to be an insensitive tool for distinguishing among apicomplexan parasites at
both the genus and species level Furthermore re-evaluation of these samples by the authors revealed that
many of the samples that were identified incorrectly based on morphometrics contained primarily
53
unsporulated oocysts making accurate identification based on microscopic appearance highly
challenging These findings further underscore the importance of molecular methods in accurate parasite
identification In the absence of molecular tools accurate measurement of oocyst size shape and
determination of SI can be useful for differentiating among species of Eimeria and Cystoisospora
however this can only be performed accurately on sporulated oocysts from feces Interestingly the size
and shape indices of oocysts of E furonis measured in histologic sections did not match those previously
described by Hoare (1927) for the same oocysts in feces despite molecular confirmation of identity
Thus measurements of oocysts in histologic sections are not recommended for use in coccidial
identification
Our observations highlight the utility of molecular methods for identifying enteric coccidia
infecting domestic ferrets and suggest that diagnoses based on morphological methods should perhaps be
limited to broad determinations of disease etiology (ie lsquococcidiosisrsquo or lsquococcidiasisrsquo) Using molecular
techniques we were able to differentiate morphologically similar coccidial species isolated from the feces
of domestic ferrets and specifically identify parasites seen in histological sections of ferret intestine
Molecular techniques thus appear to be essential for determining the coccidial species responsible for
individual and group outbreaks of coccidiosis and for further understanding of eimeriid host-parasite
relationships
ACKNOWLEDGEMENTS
Many thanks to Julia Whale and Alex Leveille for their assistance and encouragement during the course
of this project The authors would like to recognize the contributions of Dr Donald Martin (IDEXX
Canada) and Drs Nikola Pantchev and Majda Globokar (IDEXX Germany) for the contributions of data
and samples to this project The authors would also like to recognize the Laboratoire de Pathologie
(Service de diagnostic Faculteacute de meacutedecine veacuteteacuterinaire St Hyacinthe Quebec) and the Animal Health
Laboratory (Guelph Canada) for contributions of samples and data to this project Finally this project
was made possible through funding by the Toronto Zoo Residency Research Fund to DASAP and partial
54
funding from a Natural Sciences and Engineering Research Council of Canada (NSERC) Discovery Grant
(400566) to JRB
55
Table 21 Amplification primers for nuclear 18S rDNA and mitochondrial COI loci anneal temperatures (Ta) extension times and expected PCR
product sizes used in the identification of enteric coccidia from domestic ferrets (Mustela putorious furo)
Gene Target Primer Pairs Primer Sequence (5ʹ-3ʹ) Size (bp) Ta (degC) Anneal (sec) Reference
nu 18S rDNA CYC1FE TACCCAATGAAAACAGTTT 560 52 45 Matsubayashi et al (2005)
CYC4RB CGTCTTCAAACCCCCTACTG Matsubayashi et al (2005)
Cocci_18S_595F CCGCGGTAATTCCAGCTCCAAT 216 62 30 Present study
Cocci_18S_847R GCTGMAGTATTCAGGGCGACAA Present study
Lank_18S_224F TCATAGTAACCGAACGGATC 1080 54 60 Ogedengbe (2015)
Api_SSU_2733R CGGAATTAACCAGACAAATC Mathew et al (2000)
mt COI COI_10F GGWDSWGGWRYWGGWTGGAC 500 52 30 Ogedengbe et al (2011)
COI_500R CATRTGRTGDGCCCAWAC Ogedengbe et al (2011)
COI 272F CAATTCTAYGATGCCGCWTT 222 52 30 Present study
COI_500R CATRTGRTGDGCCCAWAC Ogedengbe et al (2011)
Sdae-COI_260F GATCTTTATGTTYTTRATGCC 890 50 75 Ogedengbe (2015)
Sdae-COI_1147R CATTACCCATAACYACACC Ogedengbe (2015)
56
Table 22 Summary of fecal samples from domestic ferrets (Mustela putorius furo) submitted to two diagnostic laboratories from 2008-2015
No fecal samples positive for coccidia
No samples submitted
(percentage positive)
No samples positive for
Cystoisospora sp
No samples positive for
Eimeria furonis
No samples positive for
Eimeria ictidea
Year Canada Europe Canada Europe Canada Europe Canada Europe
2008 3140 (21) 6214 (28) 3 2 0 4 0 0
2009 2160 (12) 14214 (65) 2 9 0 5 0 0
2010 8127 (63) 20213 (94) 7 10 1 10 0 0
2011 0114 (0) 17215 (79) 0 9 0 7 0 1
2012 3108 (28) 10231 (43) 2 4 1 6 0 0
2013 281 (25) 16270 (59) 2 13 0 2 0 1
2014 6127 (47) 12234 (51) 5 6 1 6 0 0
2015 4108 (37) 9249 (36) 4 3 0 6 0 0
Total 28 (29) 104 (56) 25 56 3 46 0 2
Average
year 35 130 31 70 04 58 00 03
Legend Numbers in brackets refer to the percent of the total number of fecal samples submitted
57
Table 23 Morphologic and molecular identification of coccidia from domestic ferrets (Mustela putorius furo)
Sample ID Source
External Lab Morphologic
Diagnosis Morphologic Diagnosis (ARP) Molecular Diagnosis
mt COI GenBank
Accession
nu 18S rDNA
GenBank Accession
93-40404 FFPE enteric coccidia Histologic sample - - -
P2010-I FFPE enteric coccidia Histologic sample E furonis Identical to MF774036 Identical to MF774678
17-008571 FFPE NP Histologic sample E furonis Same as MF774036 Same as MF774678
17-008571 feces NP E furonis E furonis MF774036 MF774678
907 feces E furonis E furonis E furonis MF774035 MF774679
938 feces I (=C) laidlawi no oocysts visualized I (=C) laidlawi MF774037 MF774677
952-A feces E ictidea Cystoisospora sp I (=C) laidlawi Identical to MF774037 Identical to MF774677
9958 feces E furonis no oocysts visualized E furonis Identical to MF774034 Identical to MF774680
9011 feces E furonis E furonis E furonis Identical to MF774035 Identical to MF774679
9014 feces E furonis E furonis E furonis MF774034 MF774680
9017 feces I (=C) laidlawi Cystoisospora sp I (=C) laidlawi Same as MF774037 Same as MF774677
9040 feces I (=C) laidlawi no oocysts visualized I (=C) laidlawi Same as MF774037 Same as MF774677
912-260 feces I (=C) laidlawi Cystoisospora sp I (=C) laidlawi Same as MF774037 Same as MF774677
CAN-2016-1 feces C canis + E furonis Cystoisospora sp I (=C) laidlawi MF774038 MF774676
Legend FFPE = formalin fixed paraffin embedded intestinal sections - = unsuccessful = morphologic diagnosis performed by JRB same as = 100 sequence
identity with listed GenBank entry over entire sequence length identical to = 100 sequence identity but shorter sequence than listed GenBank entry
58
Figure 21 Life stages of Eimeria furonis within the small intestinal epithelium of a domestic ferret
(Mustela putorius furo) Asexual life stages merozoites (black circle) Sexual life stages oocyst
(solid black arrow) macrogamonts (open arrows with labels) microgamont (dotted black arrow)
Hematoxylin and eosin staining scale bar = 25μm
25 microm
59
Figure 22 Phylogenetic relationships of coccidia (Eimeria ictidea Eimeria furonis and Isospora
(=Cystoisospora) laidlawi) from domestic (Mustela putorius furo) or black-footed (Mustela
nigripes) ferrets based on partial nuclear 18S rDNA and mitochondrial COI sequences of these
parasites and related apicomplexan parasites A summary of the sources of the molecular data for
the remaining taxa included in this phylogenetic analysis are found in Supplementary Table 1 of
Ogedengbe et al (2017) Bayesian support is indicated for each node horizontal distance is
proportional to hypothesized evolutionary change (scale indicates sequence divergence of 10)
60
CHAPTER 3 MORPHOLOGICAL AND MOLECULAR CHARACTERIZATION OF
ENTERIC COCCIDIA ISOLATED FROM BLACK-FOOTED FERRETS (MUSTELA
NIGRIPES)
ABSTRACT
Black-footed ferrets (BFF Mustela nigripes) are the only ferret species native to North America
and have been identified as endangered since 1967 Starting in 1986 a multi-institutional effort has been
breeding this species in captivity with successful reintroductions back into the wild Coccidiosis is
recognized as a cause of significant juvenile morbidity and mortality in captive breeding programs and
can result in significant population losses Little is known about the etiology of enteric coccidiosis in
BFF Coccidia positive fecal samples (n=12) and formalin fixed paraffin embedded intestinal tissues
(n=11) were obtained from BFF in the Toronto Zoo and Louisville Zoo Species Survival Plan (SSP)
populations Oocyst morphometrics and sequence genotyping at three loci (nuclear 18S rDNA
mitochondrial cytochrome c oxidase subunit I and mitochondrial cytochrome c oxidase subunit III) were
conducted Results suggest that the same Eimeria species E ictidea was the cause of enteric coccidiosis
in both SSP populations in both juvenile and adult age classes Wider research is indicated to determine
whether these findings are representative of the larger captive and wild BFF populations
31 INTRODUCTION
Black-footed ferrets (BFF) are one of only three wild ferret species worldwide the other two
being the European polecat (Mustela putorius) and the Siberian polecat or steppe polecat (Mustela
eversmanii) The BFF the only native North American ferret species was formerly distributed throughout
the North America prairie ecosystem but were considered extinct by the late 1950s In 1964 a single
population was discovered in Mellette County South Dakota Progressive decline of this population in
subsequent years resulted in the decision by United States Fish and Wildlife Service to initiate a captive
breeding program for the species From 1971-1973 four females and five males were captured for this
61
purpose Despite successful breeding no kits survived and the last adult ferret in this captive colony died
in 1979 BFF were again presumed extinct in the wild based on annual surveys of the initial capture site
In 1981 a dead BFF was discovered by a ranch dog outside of Meeteetse Wyoming allowing wildlife
biologists to identify another colony of BFF This colony flourished until 1985 when an outbreak of
canine distemper in this wild BFF population and an outbreak of sylvatic plague in the local prairie dog
population resulted in sharp population declines From 1985 through 1987 all 24 of the remaining BFF
were trapped and brought into captivity to re-initiate the captive breeding program Six ferrets in this
initial group died of canine distemper while in captivity and seven of the remaining eighteen survivors
are the founding population of the current captive breeding population Today this captive breeding
population consists of approximately 300 BFF distributed among multiple institutions (Santymire et al
2014)
Since 1986 a multi-institutional effort has been breeding BFF in captivity with reintroductions
back into the wild in selected locations in Canada the USA and Mexico Currently six facilities
participate in the BFF Species Survival Plan (SSP) the Toronto Zoo United States Fish and Wildlife
Services National Black-footed Ferret Conservation Center National Zoorsquos Smithsonian Conservation
Biology Institute Louisville Zoo Cheyenne Mountain Zoo and the Phoenix Zoo (Black-footed Ferret
Recovery Implementation Team 2011) In order to provide the best genetic matches BFF are transferred
among the six institutions for breeding Approximately 300-400 kits are produced annually between the
six SSP facilities with ~200 of these kits allocated for release to the wild yearly (Santymire et al 2014)
As of 2011 over 8000 BFF kits had been produced in captive breeding facilities (Black-footed Ferret
Recovery Implementation Team 2011)
Multiple infectious diseases pose a significant risk to the captive breeding and post-release
survival of BFF including canine distemper and sylvatic plague (Santymire et al 2014 USFWS BFF
Recovery Program 2017) Coccidiosis is recognized as a cause of significant juvenile morbidity and
mortality in captive breeding programs and can result in significant population losses (Bronson et al
62
2007 Santymire et al 2014 USFWS BFF Recovery Program 2017) While the effects of the disease on
the wild population are not clear a prevalence of approximately 13 has been reported based on fecal
samples collected from wild BFF born at release sites (Dr R Santymire personal commication)
Coccidia are eukaryotic host-specific parasites of the phylum Apicomplexa affecting numerous
hosts within a wide taxonomic range Two species of coccidia Eimeria ictidea Hoare 1927 and Eimeria
furonis Hoare 1927 have been identified in black-footed ferrets based on morphometrics (Jolley et al
1994) Jolley et al examined fecal samples from six captive BFF during a distemper outbreak as well as
samples from wild BFF They described one medium-sized ovoid eimeriid oocyst with a double wall
presence of a polar body and lacking both an oocyst residual body and micropyle Oocysts of this Eimeria
species (sp) measured 232 times 155 microm (range 182-274 times 130-162) with a shape index (SI) of 150 The
sporocysts were elongate with the presence of both sporocyst residuum and a Stieda body Sporozoites
contained prominent refractile bodies at the posterior end and were aligned anterior to posterior within
sporocysts These oocysts shed by all six captive ferrets were considered consistent with Eimeria ictidea
based on descriptions by Hoare (1927) On histopathologic examination of intestinal sections parasites
undergoing merogony and gamogony were observed within the villar epithelium throughout the small
intestine but were concentrated in the jejunum (Hoare 1935b) parasite life stages were not described
from other tissuesorgans
A second small spherical to subspherical eimeriid oocyst was also documented in the captive
ferrets by Jolley et al (1994) this second species had a pink double oocyst wall a granular residual body
and lacked both oocyst polar body and micropyle This smaller species measured 126 times 119 microm (range
108-152 times 101-129) with a SI of 106 The sporocysts were elongate and possessed a Stieda body and
sporozoites contained refractile bodies Similar to the larger Eimeria sp described above merogonic and
gamogonic stages were observed within the villar epithelium throughout the small intestine but were
most numerous in the jejunum Jolley et al (1994) concluded these small spherical oocysts were
consistent with E furonis described by Hoare (1927) from domestic ferrets
63
Jolley et al (1994) described a third type of coccidial oocyst occasionally detected in small
numbers within BFF fecal samples however the authors did not state whether this third oocyst
morphotype was recovered from wild or captive animals The oocysts measured 370 times 223 microm (range
350-386 times 212-232) with a SI of 106 Attempts to sporulate collected oocysts were unsuccessful and
corresponding endogenous stages were not identified on histopathologic examination of necropsied
ferrets precluding further morphologic identification of the parasite It should be noted that coccidial
oocysts with similar measurements had not been detected in wild or captive prey species available for
ingestion by BFF making it unlikely that this coccidial species would have been a pseudoparasite (Jolley
et al 1994)
Previous to this report by Jolley coccidial oocysts had been isolated from the feces of BFF in two
captive populations (Carpenter amp Hillman 1979 Williams et al 1988) The abstract by Carpenter amp
Hillman (1979) did not describe the oocysts whereas Williams et al (1988) stated that two Eimeria sp
(one with larger oocysts and one with smaller oocysts) were observed within the fecal samples but they
were not identified further Interestingly Williams et al reported both Eimeria sp to be shed in the feces
of all ferrets concurrently affected by distemper and by approximately 30 of the clinically healthy
ferrets at the time of investigation
Non-enteric coccidia have also been reported by two authors from captive BFF at one facility
(Jolley et al 1994 Williams et al 1988) Both reports which presumably described the same case(s)
noted the presence of endogenous coccidial life stages in histologic sections of respiratory tissue and
merozoites of an unidentified coccidium in impression smears of the urinary bladder from BFF diagnosed
with canine distemper Meronts were observed within the epithelium of the trachea a large bronchus and
associated bronchial glands In the later report Jolley et al (1994) described the lesions as occurring in the
same ferret whereas in the earlier report by Williams et al (1988) they are described as occurring in two
different ferrets Paraffin blocks containing formalin fixed tissues from these cases have since been
discarded precluding further attempts at parasite identification with molecular methods Subsequent to
64
these reports further cases of systemic coccidiosis in BFF have neither been published nor identified
within the pathology database of the Toronto Zoo captive BFF population nor by the current SSP
pathologist (Dr Michael M Garner personal communication)
Previous characterization of coccidia from black-footed ferrets has been based on host species
affected tissues in the host and morphometric characterization of life stages in histologic sections and
oocyst characteristics using light microscopy It is known that morphologically similar Eimeria species
are not necessarily conspecific and may vary in host specificity and pathogenicity Molecular
characterization is thus required to accurately identify coccidia to the species level No molecular
characterization of coccidian parasites from black-footed ferrets has been performed to date
There is a significant information gap regarding which parasite species are implicated in
morbidity and mortality events associated with enteric coccidiosis in BFF and whether different coccidia
are associated with this disease in adult versus juvenile age classes or in different SSP institutions
Studies to further characterize the eimeriid coccidia of the BFF are warranted to improve the management
of this disease in the captive population The objectives of this research were to morphologically and
molecularly characterize coccidia associated with enteric disease in BFF at the Toronto Zoo and in other
SSP facilities
32 MATERIALS AND METHODS
321 Fecal samples
Twelve fecal samples were collected during the study period (from 2014-2016) and preserved in
potassium dichromate (25 wv aqueous) seven from the Toronto Zoo and five from the Louisville Zoo
Centrifugal flotation with saturated salt solution (Ryley et al 1976) was used to isolate and concentrate
oocysts from fecal samples for light microscopic examination and genomic DNA extraction
One to two drops of the supernatant from the centrifugal flotation were placed directly on a slide
and beneath a coverslip The morphology and dimensions of sporulated oocysts were documented using a
65
Provis AX70 photomicroscope (Olympus Canada Richmond Hill ON Canada) fitted with a digital
imaging device (Infinity3-1C Lumenera Corporation Ottawa ON Canada) controlled using iSolution
Lite image analysis software (Hoskin Scientific Burlington ON Canada) operated at a total
magnification of 1000times Morphologic features noted for each oocyst included oocyst wall morphology
number of sporocysts presence or absence of a micropyle micropyle cap residual body and polar
granules For sporocysts size number of sporozoites per sporocyst and presence or absence of Stieda
body and sporocyst residuum were noted Alignment of sporozoites within the sporocyst and
presenceabsence of refractile bodies within sporozoites were also described The sporulated oocyst
length and width measurements (in microm) were then used to calculate the SI for each measured oocyst
Morphologic and morphometric features were compared to previously published values for E furonis and
E ictidea from domestic and black-footed ferrets
Genomic DNA extraction and purification were performed using a QIAamp DNA Mini Kit
(Qiagen Hilden Germany) according to manufacturerrsquos instructions as described in Chapter 2 (Materials
amp Methods)
322 Formalin fixed intestinal tissues
The pathology records of the Toronto Zoo were searched from 1993-2016 for cases of BFF
diagnosed with enteric coccidiosis on histopathology For each case slides of histologic sections from all
submitted intestinal segments were reviewed to confirm the presence of sexual andor asexual life stages
within the intestinal epithelium Scrolls (5-6 microm) were cut from the paraffin blocks containing affected
intestinal sections and DNA extracted from the formalin fixed paraffin-embedded tissue (FFPE) using
the QIAamp DNA FFPE Tissue Kit (Qiagen Toronto Ontario) as per manufacturerrsquos instructions
66
323 Molecular characterization
Molecular characterization of coccidial isolates was performed on oocysts purified from fresh
fecal samples (isolated as described above) that were collected from juvenile and adult ferrets from
2014-2016 and DNA extracted from FFPE samples of BFF intestine containing parasite life stages
Regions from the nuclear 18S (SSU) rDNA (nu 18S rDNA) mitochondrial cytochrome c oxidase
subunit I (mt COI) DNA and mitochondrial cytochrome c oxidase subunit III (mt COIII) DNA were
amplified by polymerase chain reaction (PCR) from each sample using the primers listed in Table 31 and
methodology described in the Materials amp Methods section of Chapter 2 Table 31 also contains the
specific anneal conditions used for the various primer pairs Genomic DNA from an Eimeria species of
poultry was included in the PCR reactions to act as a positive control for the reaction chemistry A
representative selection of the newly generated nucleotide sequences resulting from the above were
submitted to GenBank
DNA obtained from oocysts collected from fecal samples during the first year of the study (2014)
was used to generate a complete mitochondrial genome (see Chapter 6 for details) using primer pairs and
sequencing primers summarized in Table 31 All subsequent samples collected in 2015 and 2016 had
shorter mt COI and mt COIII sequences obtained to permit genotyping of all collected oocysts at these
two loci The location of each primer in the nu 18S mt COI and mt COIII genetic locus is illustrated in
Figure 31
33 RESULTS
From 2014-2016 coccidia-positive fecal samples were obtained from twelve BFF ferretsferret
groups from the Toronto Zoo and Louisville Zoo SSP populations (see Table 32) Nine samples were from
single housed adults between the ages of 1-5 years (63 MaleFemale) Two samples were from mixed
groups one pooled fecal sample from four adults (FERA-1 13 MF) and one fecal sample from a family
67
group consisting of a dam and five kits (23 MF) One fecal sample was collected from a juvenile male
ferret at the time of necropsy
Eleven BFF with enteric coccidiosis were identified in the Toronto Zoo necropsy reports from
1998-2016 and all were confirmed by histological re-evaluation (Table 32) Both juvenile (n=9 36 MF)
and adult ferrets (n=2 20 MF) were represented
331 Morphometric characterization
Twelve coccidia-positive fecal samples were identified from adult and juvenile BFF from 2014-
2016 by on site laboratories at either the Toronto Zoo or the Louisville Zoo Fecal flotation and light
microscopic re-examination of the samples identified coccidial oocysts in 10 of these 12 samples
Morphometric characterization was performed on six samples in which there was adequate
quantity and quality of sporulated oocysts for examination These included three samples from single-
housed adults one from a juvenile at the time of necropsy one of pooled feces from a group of adult
ferrets and one of pooled feces from a family group (dam and kits) Two of the three samples from
single-housed adults were from the same ferret on different dates in 2016 the dates of collection were
separated by a period in which shedding of oocysts was not identified on routine repeated fecal
examinations Oocysts were elliptical with a colourless double wall and contained four sporocysts each
with two sporozoites Sporocysts were ovoid and both Stieda body and residual body were present
Sporozoites exhibited an anterior to posterior alignment within the sporocysts and refractile bodies were
identified (Figure 32) Results for length width and shape index of sporulated oocysts including range
and average values are summarized in Table 33 and Figure 32 The average measurements based on the
results of all 148 oocysts measured were length 2398 microm (1859-3057) width 1855 microm (1373-2383)
and shape index 130 (101-160)
The same measurements were performed on 59 sporocysts from a single ferret (Noodle) and
results are as follows average length 1280 microm (898-1480) average width 738 microm (505-1028) and
average SI 176 (124-247) In one sample (Mohawk-2) sporozoites were visible free on the slide
68
Measurement of three of these provided an average length of 1068 microm (1044-1117) and an average
width of 341 microm (316-393)
332 Molecular characterization
Molecular characterization was successfully performed on oocysts from seven of 10 fecal
samples containing coccidial oocysts and FFPE tissue from nine of the 11 necropsy cases (see Table 32)
Attempts at amplification of DNA extracted from necropsy samples Z228-98 and Z137-14 using the
primer pairs listed in Table 31 were unsuccessful Similarly attempts at PCR and sequencing of DNA
extracted from fecal oocysts from two Toronto Zoo BFF Jenna and Ruckus were unsuccessful
Molecular identification results for the remaining samples are summarized in Table 32
Only one Eimeria species E ictidea was identified in all enteric coccidiosis cases diagnosed at
necropsy in both juvenile and adult BFF at the Toronto Zoo from 1998-2014 This same species was
identified in all Toronto and Louisville Zoo BFF fecal samples that were sequenced successfully (n=8)
with the exception of a single case from Louisville This Louisville ferret was identified as having a
rodent pseudoparasite (Eimeria species) in the submitted fecal sample the eimeriid pseudoparasite had
986 sequence identity at the mt COI locus to the murine coccidium Eimeria falciformis All sequences
generated for E ictidea exhibited 100 sequence identity at the mt COI and COIII loci
Novel nu 18S rDNA mt COI and mt COIII sequences were generated for E ictidea from both
geographic locations and deposited in GenBank (Accessions MF860826 MF860827 MF860823
MF860825 MF860822 MF860824) Sequences were compared to those previously published for related
eimeriid coccidia The nu 18S rDNA sequence from Eimeria ictidea isolated from the Toronto Zoo BFF
had 9736 identity (14 single nucleotide differences) to the previously published sequences from
isolates of E furonis from domestic ferrets (Mustela putorius furo) in Japan (GenBank AB239130 and
AB329724) and newly generated sequences from Canadian and European isolates (GenBank MF774678-
MF774680 see Chapter 2 and Figure 33) In contrast nu 18S rDNA sequence of E furonis from
domestic ferrets (see Chapter 2) showed 9953 to 100 identity (0 to 3 SND) to the Japanese
69
sequences Comparison of newly generated partial sequences of the mt COI region from E ictidea from
BFF to isolates of E furonis (GenBank MF774034-MF774036) from DF and E mephitidis (GenBank
KT203398) from the striped skunk (Mephitis mephitis) the only carnivore Eimeria sp for which a mt
COI sequence was previously available reveals only 9415 and 9084 sequence identity respectively
with these other Eimeria spp of carnivores (Figure 34)
34 DISCUSSION
This work presents the first nu 18S rDNA mt COI and mt COIII sequences (nu 18S rDNA -
MF860826 MF860827 mt COI - MF860823 MF860825 mt COIII - MF860822 MF860824) generated
from an intestinal eimeriid parasite of the BFF referred to here as E ictidea collected from multiple BFF
of different ages from two separate captive populations (Toronto Zoo Toronto Ontario Canada and
Louisville Zoo Louisville Kentucky USA)
The morphometric description of coccidial oocysts from BFF in this work are consistent with
previous descriptions of E ictidea from mustelids including BFF the Steppe polecat the European
polecat and domestic ferrets (Hoare 1927 Svanbaev 1956 Jolley et al 1994) Thus I propose the name
E ictidea for the enteric coccidium described from BFF reflecting the similarity in morphology host
species and location of infection in intestinal tissues yet recognizing the absence of species identification
by molecular techniques Molecular characterization of parasites that agree with the description of E
ictidea morphologically from various mustelid host species would allow not only for determination of
whether the parasites are conspecific but would also provide insight into the potential for cross-
transmission among related mustelid hosts
DNA extraction from FFPE samples allowed successful PCR and sequencing of small DNA
fragments (220 bp) in nine of the eleven cases in which the technique was attempted Age of the samples
did not appear to be the major factor associated with successful extraction of good quality DNA the two
samples for which it was unsuccessful were the most recent (2014) and oldest (1998) casesConsequently
it may be possible to use banked FFPE tissues from historic necropsy cases from other SSP institutions
70
and necropsies of wild-born or re-introduced ferrets to determine the identity of the coccidial species
underlying disease in these cases and to better characterize the disease in the greater BFF captive and wild
populations Williams et al (1988) were contacted regarding their historic FFPE samples but formalin
blocks were no longer available for these cases and thus comparisons could not be made Banked FFPE
samples were requested from other SSP institutions however the Convention on International Trade in
Endangered Species of Wild Fauna and Flora (CITES) restrictions on the international transport of DNA
from endangered species did not allow for sample acquisition during the period in which this research was
conducted
Evaluation of FFPE samples from the Toronto Zoo indicate that the same Eimeria species has
been implicated in deaths associated with enteric coccidiosis from 1999-2014 as well as episodes of
clinical disease in ferrets in the Toronto Zoo population from 2014-2016 Samples from coccidia-positive
BFF at the Louisville Zoo in 2016 also contained the same Eimeria species Finding the same parasite at
multiple SSP locations was expected because BFF are transferred among institutions on a yearly basis for
breeding and potential release Consequently these parasites have repeated opportunities to move
between institutions in infected hosts or on contaminated cage materials to become established at a new
location Furthermore the stress of transport and transfer to a new environment may precipitate shedding
of endemic coccidia and increase the risk of a coccidial outbreak this concern is reflected in the SSP
recommendations for prophylactic treatment of all BFF with anti-coccidial medication prior to shipment
(USFWS BFF Recovery Program 2017)
A single BFF from the Louisville SSP population not showing clinical signs consistent with
coccidiosis was identified as having a rodent Eimeria species in the submitted fecal sample
Morphometric characterization of oocysts in this sample was not performed due to the paucity of visible
oocysts however examination at 100times suggested that the oocysts in the sample were ovoid in shape and
of comparable size to oocysts identified in other BFF samples The finding of a rodent Eimeria in a BFF
fecal sample is not unexpected as whole rodents comprise a significant part of the captive BFF diet The
oocysts shed by the BFF were most likely acquired through ingestion of an infected prey item and thus
71
most likely represent pseudoparasitism Molecular characterization was however required to
differentiate this from a case of true enteric coccidiasis
Reports from the first captive BFF population derived from South Dakota indicate the presence
of an unidentified species of enteric coccidium (Carpenter and Hillman 1979) in this group before its
demise in 1979 No reports containing morphometric descriptions of the coccidia from this group were
found on literature review and all parasites of this group have been lost with their hosts All subsequent
reports on enteric coccidiosis in BFF are from ferrets derived from the second founder group from
Wyoming in the 1980s The frequent transfer of ferrets among SSP institutions within the captive
breeding program and to different release sites within North America would be expected to result in the
same Eimeria species being found in all populations The exception to this would be the potential for
cross-transfer of other eimeriid parasites to wild BFF from sympatric mustelid species such as the long-
tailed weasel (Mustela frenata) Jolley et al described two other species of enteric coccidia from this
second captive population in 1994 the first was similar to E furonis of domestic ferrets and the second a
large coccidian parasite of unknown genus Neither of these parasites was identified in the Toronto and
Louisville Zoo populations during the course of this study In order to determine whether these parasites
persist within the present-day BFF populations and their impact on this species more detailed
examination of coccidia-positive fecal samples from captive and wild BFF populations is recommended
Furthermore the molecular identification of enteric coccidia from historic and future necropsy samples of
wild and captive BFF could aid in determining the presence of and contribution to mortality events by
these additional coccidia species
ACKNOWLEDGEMENTS
The authors would like to recognize the Wildlife Health Centre staff at the Toronto Zoo for their
assistance with the collection of fecal samples from the BFF from 2014-2016 The authors would also like
to recognize the Louisville Zoo for their contribution of samples to this project Finally this project was
made possible through funding by the Toronto Zoo Residency Research Fund to DASAP and partial
72
funding from a Natural Sciences and Engineering Research Council of Canada (NSERC) Discovery
Grant (400566) to JRB
73
Table 31 Amplification primers used to sequence the nuclear 18S rDNA mitochondrial COI and COIII loci of Eimeria ictidea originating from
fecal and formalin-fixed paraffin embedded tissue samples from black-footed ferrets (Mustela nigripes) including anneal temeratures (Ta)
extension times and expected PCR product sizes
Figure 31 Nuclear and mitochondrial genetic loci targeted by primers listed in Table 31 and used to characterize Eimeria ictidea originating
from black-footed ferrets (Mustela nigripes)
Gene Target Primer Pairs Primer Sequence (5ʹ-3ʹ) Size (bp) Ta (degC) Anneal (sec) Reference
18SrDNA Sarco_18S_123F TATCAGCTTTCGACGGTAGTGTATT 1480 60 30 Ogedengbe et al (2016)
ERIB10_REV CTTCCGCAGGTTCACCTACGG
mt COI T_Eim_COI_272F CAATTCTAYGATGCCGCWTT 220 52 30 Chapter 2 (Table 21)
COX1-500R CATRTGRTGDGCCCAWAC Ogedengbe et al (2011)
COI-400F GGDTCAGGTRTTGGTTGGAC 800 52 60 El-Sherry et al (2013)
COI-1202R CAAKRAYHGCACCAAGAGATA El-Sherry et al (2013)
mt COIII WG-MT_4140F AGAAAACCTAAAATCATCATGT 1000 52 60 Ogedengbe et al (2015)
Eimeriid_CO3_799R AAGTGAGTTCGCATGTTTAC Ogedengbe et al (2015)
74
Figure 32 A+B) Features and cellular contents of Eimeria ictidea from a black-footed ferret
(Mustela nigripes) Legend Oocyst dotted thin white arrow = polar granule Sporocyst dotted thin
black arrow = Stieda body thick white arrow= sporozoite refractile body thick black arrow=
residuum scale bars as indicated C) Shape index length and width measurements of sporulated
oocysts of Eimeria ictidea from black-footed ferrets (Mustela nigripes) Legend times indicates the
mean Dotted oval indicates one standard deviation around the mean
10 microm 5 microm
75
Figure 33 Comparison of nuclear 18S rDNA sequence alignment of Eimeria ictidea from two black-footed ferrets (Mustela nigripes) to
newly generated (see Chapter 2) and published sequences of Eimeria furonis from domestic ferrets (Mustela putorius furo)
Figure 34 Comparison of mitochondrial cytochrome c oxidase subunit I sequence alignment of Eimeria ictidea from two black-footed
ferrets (Mustela nigripes) to sequences from other eimeriid parasites of carnivores
Identity
Eimeria mephitidis ndash KT2033981
Eimeria ictidea ndash MF860823 (Guanella ndash LZ)
Eimeria ictidea ndash MF860825 (Mystery ndash MTZ)
Eimeria furonis ndash MF774034 (Type 1)
Eimeria furonis ndash MF774035 (Type 2)
Identity
Eimeria ictidea ndash MF860827 (Guanella ndash LZ) Eimeria ictidea ndash MF860826 (Mystery ndash MTZ)
Eimeria furonis ndash MF774680 (Type 1) Eimeria furonis ndash AB3297241 (Type 1) Eimeria furonis ndash AB2391302 (Type 1)
Eimeria furonis ndash MF774678 (Type 2) Eimeria furonis ndash MF774679 (Type 2)
76
Table 32 Morphologic and molecular characterization of coccidia from fecal and FFPE necropsy samples from black-footed ferrets (Mustela
nigripes)
Sample ID Sample Source Year Age (years) Sex Source Microscopic Description^ Molecular Diagnosis
Z228-98 Toronto Zoo 1998 7 M FFPE rare asexual stages -
Z143-99 Toronto Zoo 1999 1 M FFPE sexual and asexual stages E ictidea
Z106-02 Toronto Zoo 2002 008 F FFPE rare asexual stages E ictidea
Z108-03 Toronto Zoo 2003 008 F FFPE sexual and asexual stages E ictidea
Z109-03 Toronto Zoo 2003 008 F FFPE sexual and asexual stages E ictidea
Z124-12 Toronto Zoo 2012 021 F FFPE sexual and asexual stages E ictidea
Z113-13 Toronto Zoo 2013 017 F FFPE sexual and asexual stages E ictidea
Z117-13 Toronto Zoo 2013 025 M FFPE sexual and asexual stages E ictidea
Z118-13 Toronto Zoo 2013 025 M FFPE sexual and asexual stages E ictidea
Z119-13 Toronto Zoo 2013 025 F FFPE sexual and asexual stages E ictidea
Z137-14 Toronto Zoo 2014 021 M FFPE
fresh feces
sexual and asexual stages
POS E cf ictidea
-
E ictidea
FERA_1 Toronto Zoo 2014 gt 1 MF fresh feces POS E cf ictidea E ictidea
Noodle Toronto Zoo 2015 1 M fresh feces POS E cf ictidea E ictidea
Ruckus Toronto Zoo 2016 3 F fresh feces POS -
Mystery Toronto Zoo 2016 1 M fresh feces POS E ictidea
Mohawk Toronto Zoo 2016 1 M fresh feces POS E cf ictidea E ictidea
Jenna Toronto Zoo 2016 1 F fresh feces POS -
Thrope Louisville Zoo 2016 3 M fresh feces NEG -
FloJean Louisville Zoo 2016 2 F fresh feces NEG -
Rigatoni Louisville Zoo 2016 5 M fresh feces POS rodent Eimeria
Guanella +kits Louisville Zoo 2016 2 015 F kits 2M3F fresh feces POS E cf ictidea E ictidea
Clive Louisville Zoo 2016 1 M fresh feces POS E ictidea
Legend FFPE= formalin fixed paraffin embedded Sex MF = samples from family groups containing both sexes - = PCR and sequencing unsuccessful ^ = life stages identified on
histologic section
= mitochondrial COI andor COIII sequencing results Presence (POS) or absence (NEG) of oocysts and morphological identification of oocysts based on measurements when
77
Table 33 Morphometric (length width shape index) characterization of Eimeria ictidea oocysts from fecal samples from black-footed ferrets
(Mustela nigripes)
Sample ID FERA - 1 ^ Z137-14 Noodle Mohawk -1 Mohawk - 2 Guanella Total
Number of oocysts 12 36 32 10 21 37 148
Length (microm) 2333 (2055-2583) 2456 (2111-2848) 2505 (2079-3008) 2779 (2590-3060) 2493 (2036-2822) 2139 (1859-2372) 2398 (1859-3057)
Width (microm) 1676 (1373-2180) 1835 (1643-2232) 1975 (1509-2360) 2253 (2092-2383) 1803 (1549-2017) 1751 (1610-1888) 1855 (1373-2383)
Shape index 135 (103-160) 134 (113-156) 127 (105-155) 124 (113-138) 139 (114-154) 122 (101-145) 130 (101-160)
Legend ^= mixed adult group = dam and kit group
78
CHAPTER 4 NATURAL HISTORY OF ENTERIC COCCIDIOSIS IN THE BLACK-
FOOTED FERRET (MUSTELA NIGRIPES)
ABSTRACT
Black-footed ferrets (BFF Mustela nigripes) the only native North American ferret species are
endangered throughout their former geographic range An intensive captive breeding program produces
animals to supplement re-established wild populations Coccidial enteritis is a major cause of morbidity in
young captive ferrets but the disease also affects adults Limited information is available on the
pathogenesis of intestinal coccidiosis in captive BFF and characterization of the natural history of the
disease for improved prevention and management is imperative The objectives of this research were to
determine morbidity and mortality rates in the Toronto Zoo captive BFF population as well as
characterizing the natural history of the disease in this species through evaluation of shedding patterns
body tissues affected pre-patent period and periods of enhanced host susceptibility to infection
Coccidia-associated mortality in BFF at the Toronto Zoo from 1997-2016 averaged 053 yearly in
adults (range 0-526) and 195 in juveniles (range 0-1667) Clinical signs and histologic lesions in
Toronto Zoo BFF were similar to those described in previous publications A seasonal influence on
oocyst shedding was identified in adult BFF and ferrets appeared to maintain persistent infection with E
ictidea shedding coccidia in multiple years A larger multi-institutional study is required to better
elucidate the natural history of enteric coccidiosis in this species
41 INTRODUCTION
Black-footed ferrets (BFF Mustela nigripes) are the only native North American ferret species
and are endangered throughout their former geographic range When the last remaining truly wild
population underwent serious decline as a result of disease the decision was made by the by United States
Fish and Wildlife Service to capture the remaining 24 animals and establish a captive breeding program
this occurred between 1985 and 1987 Only seven of the captured ferrets bred successfully and are the
79
founders of the current North American BFF population (USFWS BFF Recovery Program 2017) The
captive population which now numbers approximately 300 individuals is distributed among and managed
by six collaborating facilities these include the Toronto Zoo United States Fish and Wildlife Services
National Black-Footed Ferret Conservation Center National Zoorsquos Smithsonian Conservation Biology
Institute Louisville Zoo Cheyenne Mountain Zoo and the Phoenix Zoo (Black-footed Ferret Recovery
Implementation Team 2011 Santymire et al 2014) Since 1991 BFF have been released into
reintroduced into the wild at multiple sites within their former range and over 8000 BFF kits had been
produced in captive breeding facilities as of 2011(Black-footed Ferret Recovery Implementation Team
2011) Twenty-eight BFF reintroduction sites currently exist throughout North America however there
continues to be a need to support wild populations as only a four of the re-established groups are truly
self-sustaining
Enteric coccidiosis is recognized as a cause of significant morbidity and mortality in captive
breeding programs affecting both juvenile and adult animals (Bronson et al 2007 USFWS BFF
Recovery Program 2017) Two Eimeria species Eimeria ictidea and Eimeria furonis have been
identified from cases of entric coccidiosis in BFF (Jolley et al 1994) Jolley et al examined fecal samples
from both wild and captive BFF and provided detailed morphologic descriptions of the oocysts of both
Eimeria spp as well as descriptions of the intestinal pathology associated with infection Asexual and
sexual life stages of both of the aforementioned Eimeria spp were identified on histologic section within
the villar epithelium throughout the small intestine but were concentrated in the jejunum Intestinal
sections from BFF infected with E ictidea exhibited two morphologically distinct meronts one at the
villar tips which was larger and lacking in undifferentiated mass and the other at the base of the villi or
rarely in the intestinal crypts gamogony was predominantly observed at the villar tips and was noted
throughout the small intestine
80
Extraintestinal coccidia have also been reported from captive BFF at one facility (Jolley et al
1994 Williams et al 1988) The authors identified the presence of endogenous coccidial life stages in
histologic sections of respiratory tissue and in impression smears of the urinary bladder from BFF
diagnosed with canine distemper No subsequent reports of systemic coccidiosis in BFF have been
published or identified within the pathology database of the Toronto Zoo captive BFF population or by
the current SSP pathologist (Dr Michael M Garner personal communication)
Recent investigations into the etiologic agents of enteric coccidiosis in BFF at the Toronto Zoo
have identified a single Eimeria species associated with all cases of enteric coccidiosis and associated
mortality in juvenile and adult BFF from 1999-2016 Furthermore this pathogen was identified in fecal
samples based on morphologic and molecular characterization from adult and juvenile BFF in another
zoological collection (Louisville Zoo Kentucky USA) (see Chapter 3) This coccidium is
morphologically consistent with Hoarersquos original description of E ictidea (1927) and is referred to
henceforth as Eimeria ictidea
There is a significant information gap regarding the pathogenicity of E ictidea in BFF The
objectives of this research were to determine morbidity and mortality rates in the Toronto Zoo and
additional captive BFF SSP populations as well as characterizing the natural history of the disease in this
species through evaluation of shedding patterns body tissues affected pre-patent period and periods of
enhanced host susceptibility to infection
42 MATERIALS AND METHODS
421 Toronto Zoo BFF breeding program
At the Toronto Zoo black-footed ferret breeding program all adult ferrets are housed
individually with the exception of dams and kits After the birth of the kits dams are housed with their
offspring from whelp date until removal at approximately 4-6 months of age Routine monthly fecal
81
examinations (direct examination and flotation) are performed in house for all ferrets in the breeding
program based on SSP recommendations to evaluate for the presence of coccidia
422 Fecal oocyst evaluation
Family groups
From 2014-2016 daily fecal examination for coccidial oocysts was initiated for all group-housed
dams and kits In 2014 fecal samples were collected daily from all dams and kits from weaning (30 days
after whelping) to 72 days post whelping Based on 2014 data in 2015 this surveillance was extended
from weaning (35 days post whelping) to 135 days of age Furthermore fecal samples were collected
from the dam for an additional 14 days after removal of kits In 2016 no fecal samples were collected
from dam and kit groups at the Toronto Zoo but samples were submitted from one group of dam and kits
from another SSP population at the Louisville Zoo (Kentucky USA)
Adults
From 2015-2016 daily fecal samples were also collected from all adult ferrets identified as
shedding coccidial oocysts on their monthly routine fecal examination and from clinically ill BFF
Samples were collected for 10-14 days after initial positive sample identification In 2016 fecal samples
were also submitted from four coccidia-positive adult ferrets from the Louisville Zoo population samples
were collected for 7 days post initial identification of shedding
Individual fecal samples were analyzed via flotation using the McMaster method followed by
routine flotation in saturated salt solution (Dryden et al 2005) to determine the presence or absence of
oocysts and oocyst burden (oocysts per gram of feces OPG) Temporal trends in oocyst shedding were
monitored Coccidia-positive ferrets were evaluated visually on a daily basis for presence of clinical signs
consistent with infection Infected juvenile ferrets and adult ferrets were treated with oral ponazuril or
toltrazuril regardless of the presence of clinical signs as per the black-footed ferret SSP
recommendations Based on these recommendations ponazuril is typically administered orally at 30-50
82
mgkg once daily for 3-7 days until clinical signs have resolved or oocyst shedding has been significantly
reduced (USFWS BFF Recovery Program 2017)
423 Retrospective review of pathology records
The pathology records of the Toronto Zoo were searched from 1993-2016 for cases of BFF
diagnosed with enteric coccidiosis on histopathology For each case gross necropsy reports were
reviewed and slides of histologic sections from all submitted intestinal segments re-examined to confirm
the presence of sexual andor asexual life stages within the intestinal epithelium and describe the
histologic lesions associated with presence of the parasite life stages
424 Prospective modified necropsy protocol
During the study period 2014-2016 necropsy protocols for all BFF were modified to improve
detection of coccidial life stages and better to determine which portions of the intestinal tract were
affected The entire length of the intestine from duodenum to anus was measured and intestinal contents
were flushed with 12 mL of sterile saline into a sterile container Intestinal contents were preserved in
25 potassium dichromate solution (mixed 11 with intestinal contents vv) for molecular diagnostics
Paired 2-cm long intestinal samples were collected from all sections of small and large bowel duodenum
(1) jejunum (6) ileum (1) and colon (2) The eight small intestinal samples were collected at equal
distances from the pyloric sphincter to the beginning of the colon and the distance from the pylorus noted
for each Colon samples were taken at 25 and 75 of the length of the colon One sample from each
pair was preserved in Serra solution (100 ethanol (60 vv) 37 formaldehyde (30 vv) glacial
acetic acid (10 vv) and the second sample was frozen Representative tissues from all internal organs
as well as additional intestinal samples skin muscle and brain were also collected and preserved in 10
buffered formalin Histopathologic examination was performed on all tissues collected
83
425 Retrospective medical history review
Medical histories of all BFF held by the Toronto Zoo since the initiation of the SSP program were
reviewed for data on frequency of occurrence of shedding of coccidial oocysts in adults and juveniles as
well as any association of shedding with clinical signs and administration of anticoccidial treatment Data
was tabulated yearly for adult and juvenile ferrets to determine annual morbidity and mortality rates
associated with enteric coccidiosis Medical records and pathology reports were solicited from the other
SSP institutions to determine comparative morbidity and mortality rates associated with enteric
coccidiosis in BFF at other facilities Both morbidity and mortality rates were calculated as
incidenceattack rates with yearly adult population size or number of family groups (dam and kits) as the
denominator for morbidity rates and number of yearly deaths in each age class as the denominator for
mortality rates
43 RESULTS
431 Fecal oocyst evaluation and retrospective medical history review
Family groups
Fecal samples were collected from seven groups of dams and kits housed together at the Toronto
and the Louisville Zoos from 2014-2016 All data from first to last day of collection for all family groups
is listed in Appendix 1 selected pertinent data for each group is presented in Table 41 Five groups of
dams and kits were sampled in 2014 and one group in each of 2015 and 2016 Shedding occurred no
earlier than 55 days of kit-age in any of the groups and was identified from 55-81 days of age (Table 41
Table 42 Figure 41)
In 2014 fecal oocyst shedding was identified in three of the five surveyed groups In two of the
three groups (dams Poppy and Bumblefoot) changes to fecal colour and consistency were identified
concurrently with periods of oocyst shedding both groups shed higher numbers of oocysts than the other
dam and kit groups in 2014 and 2015 Both Poppy and Bumblefoot had had litters in the previous one and
84
two years prior to this study respectively based on medical record review these dams and their litters
were also diagnosed as shedding coccidial oocysts that were too numerous to count on direct exam and
fecal flotation Clinical signs in the previous years included dark tarry hemorrhagic or soft mucoid feces
and reduced appetite both groups received treatment with toltrazuril (Baycox Coccidiocide Solution
25 Bayer Inc Mississauga Canada) and trimethoprim sulfamethoxazole (Novo-Trimel Teva Canada
Ltd Scarborough Canada) (TMS) One of four kits from Poppyrsquos 2013 litter (Z113-13) died of enteric
coccidiosis three days after the group was diagnosed as shedding coccidial oocysts and the initiation of
treatment with TMS
In 2015 low grade fecal oocyst shedding (lt14 oocysts per gram of feces) without associated
clinical signs was noted in the Fiddlesticks group on three days during a seven day period from 63-69
days of kit-age and again for a single day at 128 days of kit-age The dam had been diagnosed and treated
for enteric coccidiosis in 2014 at which time she exhibited clinical signs of loose green feces to
hemorrhagic diarrhea lethargy and dehydration In 2016 she was diagnosed as shedding low numbers of
coccidia exhibited no clinical signs and did not receive treatment prior to resolution of shedding
In 2016 Guanella and kits shed oocysts over a nine day period and daily fecal oocyst shedding
ranged from 206 ndash 371714 OPG Combined treatment with ponazuril (first four days of shedding)
sulfadimethoxine injectable (first two days of shedding additional product information not available)
amoxicillin oral (first two days of shedding additional product information not available) penicillin
injectable (first two days of shedding additional product information not available) and subcutaneous
fluids (first two days of shedding additional product information not available) was administered to this
group Previous medical history was not available for this female for review
In 2014 and 2015 fecal oocyst shedding in all groups in the Toronto Zoo population started in the
three week period from the last week of July to mid-August In 2016 shedding was first identified in the
Louisville Zoo group in mid-July
85
Adults
Seven single-housed adult BFF (52 MF) were detected to have shed coccidia during the study
period (Table 43) Shedding periods lasted from 2-10 days and oocyst per gram counts ranged from 104
ndash 554274 (Table 44) Clinical signs were identified in four of the seven ferrets and consisted of
lossreduction of appetite (n=2) weight loss (n=1) lethargy (n=1) blood in feces (n=1) loose or runny
feces (n=3) soft mucoid feces (n=1) green colour of feces (n=2) Five of the seven adults received
treatment after detection of oocyst shedding two of which received treatment in the absence of clinical
signs Treatment consisted of oral toltrazuril in four cases toltrazuril in combination with trimethoprim
sulfamethoxazole in one case (Mohawk-A) and ponazuril and sulfadimethoxine (manufacturerrsquos
information not available) in one case (Clive) (Table 43)
Three of the adults in this study Mohawk Mystery and Jenna shed oocysts during multiple
different periods in 2016 Mohawk shed oocysts in May July and September of 2016 data from the first
two periods are reported in Tables 43 and 44 Mystery shed oocysts in June and July of 2016 Clinical
signs were observed only during the first shedding period and included poor appetite and hemorrhagic to
soft mucoid feces Jenna shed oocysts in July September and November of 2016 and again in February
and May of 2017 Although clinical signs were not detected in association with the initial period of
shedding in July 2016 (see Table 44) depressed mentation and hemorrhagic mucoid feces were identified
in the subsequent two shedding periods In both Mohawk and Jenna oocysts were not detected in feces on
multiple recheck and routine monthly fecal examination between shedding periods
Ruckus the fourth ferret shed low numbers of oocysts for two days in 2016 while housed alone
and had been reported to have shed oocysts during a 30 day period in 2014 while housed in a family
group with her kits Diarrhea to soft mucoid feces and loss of appetite were reported in 2014 Clinical
signs in 2016 consisted of small amounts of runny feces for a two day period two days after oocyst
shedding was no longer detected Treatment consisted of toltrazuril and TMS in 2014 and toltrazuril
again in 2016 Similarly Noodle shed low numbers of oocysts in July 2015 and had been identified as
shedding low numbers of oocysts during September of the previous year
86
The final ferret Rigatoni shed oocysts in feces in low numbers (54-732 OPG) over 5 days All
positive samples were pooled for molecular diagnostics and sequencing results showed 990 identity (7
single nucleotide differences SNDs) at the mt COI locus with a pair of rodent Eimeria species (ie
GenBank HM771682 JQ993704) (see Chapter 3) This ferret did not show clinical signs associated with
shedding and was not treated Previous medical history was not available for Clive the single adult ferret
from the Louisville Zoo
Based on sample collection dates and medical record review for the adult BFF for 2014-2016
shedding occurred during spring summer and fall with three ferrets shedding in May (Jenna Ruckus and
Mohawk) one ferret shedding in June (Mystery) three ferrets shedding in July (Mohawk Jenna Noodle)
one ferret shedding in August (Clive) two ferrets shedding in September (Noodle Jenna) and one ferret
shedding in November (Jenna)
432 Pathology
Eleven BFF with enteric coccidiosis were identified in the Toronto Zoo necropsy reports from
1993-2016 (Table 45) Cases were identified from 1998-2014 inclusive and all were confirmed by
histological re-evaluation (Chapter 3 Table 32) Both juvenile (n=9 36 MF) and adult ferrets (n=2 20
MF) were represented
Gross Pathology
Gross necropsy findings were similar across the 11 cases and included mucoid to fluid luminal
contents (n=7 636) beige to white pasty coating of the mucosal surface of the small intestine (n=6
545) and colon (n=4 364) gaseous dilation of intestinal segments (n=3 272) segmental enteritis
and hemorrhage (n=1 91) In one case Z228-98 no gross lesions were identified within the intestines
Impression smears of luminal contents or scrapings of intestinal mucosa were performed in four cases and
coccidia were identified in all four
Histopathology
87
Both sexual and asexual life stages were identified within the small intestinal segments in all
cases except Z228-98 and Z106-02 in which only rare asexual parasite life stages were identified (Table
45 Figure 42) Other histologic lesions seen in intestinal segments containing coccidia included
lymphoplasmacytic inflammation of the lamina propria (n=4) neutrophilic infiltration of the lamina
propria (n=2) villar necrosis (n=2) villar atrophy or blunting (n=3) and thrombi within the villar tips
(n=1)
Additional necropsy diagnoses included cholangiolar hyperplasia multiple hepatobiliary cysts
with suppuration renal adenocarcinoma apocrine gland adenocarcinoma (Z228-98) concurrent
clostridial enteritis (Z143-99) presumptive Salmonella sp septicemia (Z106-02) interstitial pneumonitis
(Z108-03) myocardial mineralization interstitial pneumonia and nephritis periportal hepatitis and
bacteremia (Z124-12) and suppurative esophagitis (Z137-14)
From 2014-2016 three black-footed ferrets were necropsied using the detailed protocol described
above Only one of the three cases Z137-14 was diagnosed with enteric coccidiosis based on
histopathology Two duodenal five jejunal two ileal and two colonic sections were collected at measured
lengths from the pylorus Parasite life stages were identified within the mucosal epithelium of all
intestinal segments extending from the distal duodenum (10-12 cm aboral to pylorus) through to the distal
colon (157-159 cm aboral from pylorus) The distal duodenal section contained asexual life stages only
with a single focus of epithelial cells containing meronts Sexual life stages (microgamonts
macrogamonts unsporulated oocysts) were identified within villar epithelial cells in all remaining
sections of the small intestine with numerous oocysts in the bowel lumen Mild lymphoplasmacytic
inflammation of the lamina propria was associated with the jejunal and ileal lesions and blunting of the
villi was identified within one jejunal segment The colonic sections contained small to moderate numbers
of sexual life stages identified within both superficial and deep crypt epithelium with occasional life
stages identified near the germinal cells Large numbers of oocysts and bacteria were identified within the
88
lumen of these colonic sections and both sections contained abscesses within the crypts The proximal
duodenum (0-2 cm) was the only section of the intestines not containing parasitic life stages
433 Morbidity and mortality
Annual morbidity rates for enteric coccidiosis at the Cheyenne Mountain Zoo and mortality rates
from enteric coccidiosis in Toronto Zoo BFF are summarized in Tables 46 and 47 During 2003-2016
yearly incidence of coccidiosis in adult BFF at the Cheyenne Mountain Zoo averaged 69 (range 0-
421) For family groups consisting of juvenile ferrets housed with their dams yearly incidence of
enteric coccidiosis averaged 115 (range 0-600)
From 1997-2016 coccidia-associated mortality in adult BFF at the Toronto Zoo averaged 053
yearly (range 0-526) with an average total mortality rate of 141 per year (range 0-526) For
juvenile ferrets (under 1 year of age) during the same period coccidia-associated mortality accounted for
an average of 133 of deaths yearly (range 0-100) with an overall average mortality rate of 170 per
year (range 0-3404) from all causes
Multiple additional SSP institutions provided partial medical and pathology data sets for use in
this study which were not sufficiently detailed to permit computation of morbidity and mortality rates for
those populations
44 DISCUSSION
The work described here supports previous clinical findings regarding the impact through both
morbidity and mortality associated with enteric coccidiosis in BFF No previous studies have been
undertaken to determine morbidity and mortality rates associated with enteric coccidiosis across BFF SSP
institutions
89
In a retrospective mortality study of captive BFF from 1984-2004 at the Smithsonianrsquos National
Zoological Park Bronson et al (2007) reported that gastrointestinal disease was the most common cause
of death in juvenile BFF (524) with 333 of juvenile mortality cases in the study caused by enteric
coccidiosis While the data is not directly comparable the findings reported here also reflect enteric
coccidiosis as a common cause of death in juveniles with increased mortality associated with the disease
compared with adult counterparts All Toronto Zoo mortalities in both juvenile and adult age classes for
which necropsy tissues were available have been attributed to infection with a single coccidia species
Eimeria ictidea (see Chapter 3)
Multiple SSP institutions provided partial medical and pathology data sets for use in this study
which were not sufficiently detailed to permit computation of morbidity and mortality rates for those
populations In future it would be useful to determine whether morbidity and mortality rates associated
with enteric coccidiosis vary among SSP institutions as this may allow for improved identification of
host parasite and environmental factors that increase risk
Clinical signs reported here are consistent with those described from both BFF and domestic
ferrets with enteric coccidiosis (Sledge et al 2011 Santymire et al 2014 USFWS BFF Recovery
Program 2017) Changes to fecal colour and consistency were the most common abnormalities identified
at the time of first detection of oocyst shedding While clinical signs in the cases described here do not
always correlate directly with the quantity of oocysts shed individuals shedding higher number of oocysts
showed clinical signs more frequently than those shedding lower numbers
In this study oocyst shedding from single-housed adult BFF ranged from 104 ndash 554274 oocysts
per gram Daily fecal samples produced from individual adult ferrets range in size from approximately 1-
16 grams In light of these findings during peak shedding from ~5times105 to 9times106 oocysts can be shed into
the environment in one day providing a massive infective dose The large numbers of oocysts shed
90
combined with confinement in a small enclosure space and hardiness of Eimeria oocysts in the
environment would be expected to markedly increase risk of infection in captive BFF
Oocyst shedding from family groups ranged from 0 ndash 371714 OPG The wide variability seen in
OPG counts between days as seen in Figure 41 and Table 41 may be accounted for by the staggered
initiation and resolution of shedding by different ferrets It is most likely that the source of infection in
these family groups is shedding by the dams some of which were identified to shed in multiple years
although environmental contamination cannot be excluded
Shedding in the adult ferrets was clustered during particular time periods specifically May July
and September If shedding is associated with stress and immunosuppression activities such as breeding
whelping electro-ejaculation of male ferrets and shipmenttransfer could act as stressors Whelping
which occurs primarily in May and June could also act as a stressor to other ferrets in the facility either
through social cues or as there would be associated changes in husbandry protocols The September
cluster could be associated with the transfer of ferrets between institutions kits are pulled during this time
and adult ferrets are moved among institutions thus changing the population dynamics of each SSP site A
large cluster of shedding was recorded in July With the exception of weaning of kits no other major
stressors are expected to occur during this time thus the increase in shedding by adults in this group
cannot be easily explained Interestingly shedding was not identified in ferrets from March through April
which is the typical breeding season and when ferrets are introduced for breeding a presumably stressful
time Shedding was not noted from December through April which could reflect reduced environmental
burdens due to low humidity levels as would be expected in a Nordic climate during the winter (which
would kill oocysts and thus block transmission) or may be consistent with reduced stress during this
period The results from single-housed adults are in contrast to the dam and juvenile ferret groups in
which oocyst shedding appeared to be correlated to a period of 55-81 days of kit age These results are
consistent with reports from other facilities of increased incidence of shedding by kits after 70 days of age
(USFWS BFF Recovery Program 2017)
91
Retrospective and prospective review of histologic sections of intestines from affected BFF at the
Toronto Zoo showed the presence parasitic life stages in epithelial cells of both the small and large
intestines Neither Hoare (1935b) nor Jolley et al (1994) mentioned the presence of parasites in the large
intestine of experimentally infected domestic ferrets or naturally infected BFF respectively In the study
described here asexual and sexual life stages were identified within the epithelial cells of the small
intestinal villi from base to tip and were most numerous in jejunum This matches the description by
Jolley et al (1994) however Hoare (1935a b) found E ictidea to be present primarily in the villar tips
Jolley et al (1994) also described two morphologically distinct meronts of E ictidea within the small
intestinal sections one at the villar tip that was larger and lacking in undifferentiated mass and the other
at the base of the villi or in the intestinal crypts these findings were not echoed in this study as
merogonic stages were identified throughout the intestinal epithelium from villus to the base of the crypt
and no visual differences between meronts in any location were identified Hoare (1935a b) also
described a resulting annular constriction of the villus separating the affected and non-affected segments
this constriction was neither seen in the cases described here nor mentioned by Jolley et al (1994)
Whether these differences result from E ictidea from BFF and E ictidea from domestic ferrets being
different parasites or from differences in tissue tropism of a single parasite in two different hosts cannot
be ascertained from the available information
Histologic lesions such as necrosis hemorrhage villar atrophy and inflammation associated with
the presence of parasitic life stages were rare These changes are normally elicited by the host immune
system (inflammation) and the parasite (cellular rupture to release life stages resulting in hemorrhage and
necrosis) in response to infection In light of the fact that acute death occurred in a number of these ferrets
(Z113-13 Z117-13 Z118-13 Z119-13 Z137-14) in the absence of secondary disease processes and
with the intestinal epithelium intact but containing myriad parasitic life stages an alternative mechanism
for mortality associated with the infection must be proposed It is possible that these parasites elaborate
exotoxins during their life cycle and when at high density result in sudden death of the host with minimal
92
tissue changes The presence of parasitic life stages occupying the majority of both small and large
intestinal epithelial cells could also potentially impair fluid and protein movement in and out of the
mucosa however clinical signs associated with malabsorptive diarrhea were not identified in any of these
cases The presence of bacteria within the blood or other organ tissues was not identified in any cases and
consequently sepsis is unlikely to be the cause of death
Black-footed ferrets appear to maintain persistent infection with E ictidea Adult BFF in the
Toronto Zoo population shed coccidia in multiple years and in two adult ferrets multiple times in the
same year While the coccidia seen in all cases were not confirmed as E ictidea using molecular
techniques morphologic similarities and a lack of additional Eimeria spp identified on molecular work
undertaken suggest that only one species of parasite is and has been present in the collection Two dams
that had been identified as infected based on routine fecal screening in previous years presumably acted as
the source of infection to their litters of kits in multiple years While continued environmental
contamination cannot be ruled out these findings imply a failure of the immune response of the BFF to
clear infection with E ictidea or even to protect against sufficient replication of organisms to result in
clinical disease
Based on clinical experience and review of the literature BFF appear to be much more sensitive
to infection with E ictidea compared with their domestic counterparts In domestic ferrets subclinical
shedding of oocysts appears to be the most common with rare reports in the literature of overt disease
and that only in juveniles (Blankenship-Paris et al 1993 Abe et al 2008) However a single report exists
of three separate clinical outbreaks of Eimeria furonis infection in domestic ferrets under intensive
management with increased morbidity and mortality affecting all ages classes (Sledge et al 2011) The
role of genetics in the apparent increased susceptibility of BFF to enteric coccidiosis is unknown but the
current captive BFF population is derived from seven founders and inbreeding depression or familial
genetic susceptibility may play a role in their increased susceptibility to disease caused by E ictidea
93
Black-footed ferrets diagnosed with enteric coccidiosis during the course of the study were
treated with either ponazuril or toltrazuril sulfonamide drugs or often a combination of the two groups
of therapeutic agents Toltrazuril and ponazuril are triazine coccidiocides with proven efficacy against
both asexual and sexual life stages of mammalian and avian Eimeria spp (Mehlhorn and Aspock 2008)
The sulfonamides are antimicrobial drugs that exhibit coccidiostatic or coccidiocidal effects depending on
dose they act by blocking folate synthesis and have effects on first and second generation meronts
(asexual life stages) as well as potentially acting on sexual life stages (Mehlhorn and Aspock 2008)
Based on the limited data available from this study and the fact that treatment was initiated in almost all
adult BFF and family groups at the time of oocyst detection regardless of the presence of clinical disease
the effects of treatment on duration of clinical signs cannot be effectively evaluated It appears
subjectively that adult ferrets treated with toltrazuril and in one case a combination of toltrazuril and
TMS showed reduction in oocyst shedding after 3-5 days of oral anti-coccidial therapy (see Table 43)
The effects of treatment with either sulfonamides or triazines would be expected to reduce oocyst
shedding consequently the duration and amount of oocyst shedding reported in this study may not
accurately characterize the natural course of disease
Perceived resistance to sulfa drugs has been reported from multiple SSP facilities In light of this
and their potential negative effects on ferret reproduction (eg prevention of embryo implantation in the
uterus and impairment of sperm development) sulfonamides are no longer recommended by the SSP for
treatment of coccidia in this species (USFWS BFF Recovery Program 2017) The frequent and
widespread use of triazines in the management of enteric coccidiosis in BFF presents a risk for
development of resistance to this drug class in the future Neither pharmacokinetic (PK) nor
pharmacodynamic (PD) studies have been published to validate the dose and frequency of dosage in
either class of drugs in BFF and consequently it is unclear whether this perceived failure of some ferrets
to respond to treatment is based on true resistance versus inappropriate dosing The only work evaluating
ponazuril in BFF evaluated serum levels of ponazuril after a single oral dose of 50 mgkg and reported
94
therapeutic levels for 10 days after administration (USFWS BFF Recovery Program 2017) No
information was provided on number or age of ferrets that participated in the study or on how the
determination of what were therapeutic levels was made Furthermore as life cycles of the coccidia
affecting BFF are limited to the gastrointestinal tract and do not exhibit tissue stages the validity and
usefulness of assessing blood levels of ponazuril in determining appropriate dosage and dose schedules is
questionable Further work to determine the PK and PD of triazines in ferrets is warranted to provide safe
and efficacious treatment and to reduce the risk of development of resistance Furthermore the creation
and validation of a model for enteric coccidiosis in a related species would allow for in vivo studies of
drug resistance
95
Figure 41 Oocyst per gram counts and shedding period of Eimeria ictidea from black-footed ferret
(Mustela nigripes) family groups from 2014-2016
0
50000
100000
150000
200000
250000
300000
350000
400000
29
33
37
41
45
49
53
57
61
65
69
73
77
81
85
89
93
97
10
1
10
5
10
9
11
3
11
7
12
1
12
5
12
9
13
3
13
7
14
1
14
5
14
9
OP
G
Age of Kits
Poppy Bumblefoot Calico Aubrey Ruckus Fiddlesticks Guanella
96
Figure 42 Small intestinal epithelium of a black-footed ferret (Mustela nigripes) containing sexual
life stages of Eimeria ictidea Legend Solid black arrow = oocyst Hatched arrow = macrogamont
Outlined arrow = microgamont Hematoxylin and eosin staining scale bar = 25 microm
25 microm
97
Table 41 Shedding of oocysts of Eimeria ictidea in black-footed ferret (Mustela nigripes) dam and kit
family groups from 2014-2016
Collection Year 2014 2014 2014 2014 2014 2015 2016
Age of kits (days) Poppy Bumblefoot Calico Aubrey Ruckus Fiddlesticks Guanella^
29 - - - 0 - - -
30 - 0 - 0 - - -
- - - - -
34 0 0 - 0 0 - -
35 0 0 - 0 0 0 -
- -
46 0 0 - 0 0 0 -
47 0 0 - 0 0 0 -
48 0 lt14 - 0 0 0 -
- -
53 0 0 - 0 0 0 +
54 0 0 - 0 0 0 +
55 0 0 - 0 0 0 3717146
56 0 0 - 0 - 0 1084436
57 0 0 - 0 - 0 64133
58 0 324 - 0 0 0 20654
59 0 0 - 0 0 0 0
60 0 446688 - 0 0 0 0
61 0 934828 - 0 0 0 41111
62 0 530469 - 0 0 0
63 0 1617131 - 0 0 lt14
64 0 8396357 - 114 0 0
65 0 234876 - 5368 0 lt14
66 0 374625 - 9455 0 0
67 lt14 2311575 - 39579 0 -
68 0 603563 - 125051 0 0
69 - 4103036 - 27067 0 lt14
70 2470327 7759974 - 0 0 0
71 1073085 - 1784 0 0
72 3430966 - 0 0 0
73 4459536 - 7326 0 0
74 4842212 - 2 0 0
75 3627307 - 0 0 0
76 5801885 - 0 0 0
77 5994 0 0 0 0
78 589835 0 0 0
79 24815 0 0 0
80 144016 0 0 0
81 97862 0 0 0
82 0 0 0
127 0 0
128 0 2843
136 0 0
150 0
Numbers of Oocysts Shed (oocysts per gram of feces)
Dam Identity
Legend lt 14 = oocyst positive samples with less than 14 oocyst per gram of feces - = no sample recorded for this date underline = last
sampling date + = coccidia present but OPG count not performed = Toronto Zoo ferret ^ = Louisville Zoo ferret thick outer border =
days treatment was received = range of sequential dates between previous and subsequent number during which OPG counts were
performed and samples contained 0 oocysts
98
Table 42 Summary of epidemiologic data for family groups of black-footed ferrets (Mustela nigripes)
shedding oocysts of Eimeria ictidea
Dam Identity
Poppy
2014
Bumblefoot
2014
Aubrey
2014
Fiddlesticks
2015
Guanella
^
2016
Number of kits 7 5 3 4 5
Kit age (days) at time of
shedding 70-81 58-70 64-73 63-69 54-61
Shedding period (days) 15 13 11 7 9
OPG min 0 0 0 0 206
OPG max 48442 83963 2707 lt14 371714
Clinical signs YES YES NO NO NO
Treated - YES YES NO YES
Legend = Toronto Zoo ferret ^ = Louisville Zoo ferret OPG = oocysts per gram of feces
- = missing data
Table 43 Shedding of oocysts of Eimeria ictidea in single-housed adult black-footed ferrets (Mustela nigripes) 2015-2016
Numbers of Oocysts Shed (oocysts per gram of feces)
Collection
Year 2015 2016 2016 2016 2016 2016 2016
Ferret Identity Noodle Ruckus Mohawk-A Mohawk-B Mystery Jenna Clive^
Age (years) 1 3 1 1 1 1 1
1084058 0 + + + + +
+ 0 42307650 6286676 + 183150 554274
857808 16650 12805238 7777929 + 215710 377920
1604894 16650 309690 139860 + 0 25808
377042 0 599400 119880 + - 37294
554445 0 34688 385579 117920 0 5363
26640 0 16650 0 0 0 7500
0 10406 0 0 0 1090
0 20813 0 0 0
- 0 1761 0
0 0 0 0
- 0
0
0
Legend lt 1 = oocyst positive samples with less than 1 oocyst per gram of feces underline = last sampling date + = coccidia present but OPG
count not performed = Toronto Zoo ferret ^ = Louisville Zoo ferret thick outer border = days treatment was received
100
Table 44 Summary of epidemiologic data for single housed adult black-footed ferrets (Mustela nigripes) shedding oocysts of Eimeria ictidea
Noodle Ruckus Mohawk-A Mohawk-B Mystery Jenna Clive^
Sex M F M M M F M
Age (years) 1 3 1 1 1 1 1
Shedding period (days) 7 2 9 6 10 4 8
OPG min 266 166 104 1199 1761 1831 1090
OPG max 10840 166 423076 77779 - 2157 554274
Clinical signs YES YES NO NO YES NO YES
Treated NO YES YES YES YES YES YES
Legend = Toronto Zoo ferret ^ = Louisville Zoo ferret M = male F= female OPG = oocysts per gram of feces - = missing data
NOTE Mohawk-A and Mohawk-B refer to two separate episodes of oocyst shedding by the same ferret
101
Table 45 Histologic findings from necropsies of black-footed ferrets (Mustela nigripes) with enteric coccidiosis
Number of Sections Affected
Ferret
ID Year
Age
(years) Sex Coccidia in Intestinal Sections Small Intestinea Large Intestinea
Z228-98 1998 7 M rare asexual stages S - 01 A - 11 S - 01 A - 01
Z143-99 1999 1 M sexual and asexual stages S - 24 A - 24 S - 03 A - 03
Z106-02 2002 008 F rare sexual stages S - 14 A - 04 S - 01 A - 01
Z108-03 2003 008 F sexual and asexual stages S - 12 A - 12 S - 01 A - 01
Z109-03 2003 008 F sexual and asexual stages S - 23 A - 23 S - 11 A - 11
Z124-12 2012 021 F sexual and asexual stages S - 24 A - 04 none
Z113-13 2013 017 F sexual and asexual stages S - 77 A - 07 none
Z117-13 2013 025 M sexual and asexual stages S - 56 A - 56 S - 11 A - 01
Z118-13 2013 025 M sexual and asexual stages S - 56 A - 46 S - 11 A - 01
Z119-13 2013 025 F sexual and asexual stages S - 45 A - 35 none
Z137-14 2014 021 M sexual and asexual stages S - 1011 A - 911 S - 22 A - 02
Legend a= xn where x is number of sections containing sexual or asexual lifestages n is the number of sections examined S = sexual life
stages A= asexual life stages
102
Table 46 Yearly incidence of coccidial infection in black-footed ferrets (Mustela nigripes) at the
Cheyenne Mountain Zoo
Cheyenne Mountain Zoo
Year Adult Family
2003 116 (625) -
2004 819 (4211) -
2005 021 (000) 14 (2500)
2006 021 (000) 07 (000)
2007 023 (000) 08 (000)
2008 224 (833) 14 (2500)
2009 025 (000) 06 (000)
2010 326 (1154) 07 (000)
2011 125 (400) 09 (000)
2012 125 (400) 08 (000)
2013 028 (000) 05 (000)
2014 430 (1333) 09 (000)
2015 035 (000) 35 (6000)
2016 - 27 (2857)
Mean annual () 689 1155
Legend - = missing data xn= where x is the number of ferrets shedding coccidial oocysts and n is the
total number of adult ferrets or family groups in a given year () = incidence expressed as a percentage
103
Table 47 Yearly mortality rate and incidence of mortality associated with coccidial infection in
black-footed ferrets (Mustela nigripes) at the Toronto Zoo
Total Mortality
Coccidia Other Causes
Year Kit Adult Kit Adult
1997 015 (000) 023 (000) 315 (2000) 423 (1739) 1998 038 (000) 119 (526) 838 (2105) 919 (4734) 1999 047 (000) 119 (526) 1647 (3404) 119 (526) 2000 034 (000) 015 (000) 434 (1176) 315 (2000) 2001 032 (000) 016 (000) 532 (1563) 116 (625) 2002 150 (200) 020 (000) 450 (800) 220 (1000) 2003 227 (741) 018 (000) 327 (1111) 118 (555) 2004 020 (000) 016 (000) 620 (3000) 216 (1250) 2005 016 (000) 015 (000) 416 (2500) 215 (1333) 2006 030 (000) 016 (000) 230 (667) 016 (000) 2007 019 (000) 015 (000) 419 (2105) 215 (1333) 2008 034 (000) 016 (000) 1134 (3235) 316 (1875) 2009 017 (000) 016 (000) 017 (000) 116 (625) 2010 017 (000) 016 (000) 317 (1765) 316 (1875) 2011 011 (000) 016 (000) 111 (909) 216 (1250) 2012 111 (909) 017 (000) 111 (909) 317 (1765) 2013 424 (1667) 017 (000) 424 (1667) 317 (1765) 2014 126 (384) 017 (000) 326 (1154) 217 (1176) 2015 04 (000) 017 (000) 04 (000) 217 (1176) 2016 011 (000) 017 (000) 211 (1818) 117 (588)
Mean annual () 195 053 1594 1359
Legend xn= where x is the number of ferrets that died with coccidial infection or of other
causes and n is the total number of adult ferrets or kits in a given year () = incidence expressed
as a percentage
104
CHAPTER 5 EVALUATING THE DOMESTIC FERRET (MUSTELA PUTORIUS FURO) AS
AN EXPERIMENTAL MODEL FOR ENTERIC COCCIDIOSIS IN THE BLACK-FOOTED
FERRET (MUSTELA NIGRIPES)
ABSTRACT
The purpose of this study was to determine whether the domestic ferret (Mustela putorius furo) is
susceptible to an isolate of Eimeria ictidea originating from black-footed ferrets (BFF Mustela nigripes)
and thus could act as a suitable experimental model in which to investigate the pathogenesis and
management of this disease A pilot study was performed with 10 male intact juvenile domestic ferrets
Ferrets were administered an oral inoculum containing either a high dose (1 times 106 oocysts) moderate
dose (5 times 104 oocysts) or saline control and observed for shedding of oocysts and development of
clinical signs Seven of ten ferrets developed patent infection all of which had received the high dose
inoculum The prepatent period was 7-9 days and duration of shedding varied from 1-7 days Clinical
signs were identified in six of the seven infected ferrets and were consistent with those previously
described for enteric coccidiosis in domestic and BFF Parasite life stages were identified within the
intestines of four of the seven ferrets with patent infection and were limited to the distal jejunum and
ileum The demonstrated ability to produce patent infections in domestic ferrets following oral inoculation
of a high dose of E ictidea (1 times 106 oocysts) isolated from BFF provides an avenue for future
experimental investigations into the control and treatment of enteric coccidiosis in this endangered
species
51 INTRODUCTION
Black-footed ferrets (BFF Mustela nigripes) are one of only three ferret species worldwide
While formerly distributed throughout the North American prairies black-footed ferrets were declared
extinct in the wild in the 1980s Since 1986 a multi-institutional consortium has been breeding this
species in captivity with reintroductions back into the wild within their historic range in selected locations
in Canada the USA and Mexico Introduced colonies of BFF are present in Arizona Colorado Kansas
105
Montana New Mexico South Dakota Utah Wyoming and Chihuahua (Mexico) Reintroduction
attempts in Saskatchewan Canada have been unsuccessful to date
Multiple infectious diseases pose a significant risk to the captive breeding and post-release
survival of BFF including distemper and sylvatic plague (Santymire et al 2014 USFWS BFF Recovery
Program 2017) Coccidiosis is a recognized cause of juvenile and adult morbidity and mortality in captive
breeding programs and can result in significant losses (Bronson et al 2007 Santymire et al 2014
USFWS BFF Recovery Program 2017) The effect of the disease on wild populations is unknown
Clinical signs of coccidiosis include mucoid to hemorrhagic diarrhea abdominal discomfort lethargy
appetite loss vomiting and dehydration Recent investigations into diseases affecting BFF at the Toronto
Zoo have identified a single Eimeria species E ictidea associated with all cases of enteric coccidiosis in
juvenile and adult BFF from 2014-2016 (see Chapter 3) This same Eimeria species was identified
retrospectively as the cause of juvenile and adult mortalities in previous years (1999 through 2014
inclusive) (Chapters 3 and 4) Furthermore this pathogen was identified in fecal samples based on
morphologic and molecular characterization from adult and juvenile BFF in another zoological collection
(Louisville Zoo Kentucky USA) (see Chapter 3)
Enteric coccidiosis also occurs in domestic ferrets (Mustela putorius furo) with three
morphologically distinct species of coccidia Eimeria ictidea Eimeria furonis and Isospora (=
Cystoisospora) laidlawi Both of the aforementioned Eimeria species have been identified in black-footed
ferrets based on morphologic criteria but molecular characterization was needed to confirm whether the
same species of parasite infects both ferret species (see Chapter 2 and 3) To this end nuclear and
mitochondrial sequences for E furonis and for I=(C) laidlawi were generated (Chapter 2) expanding the
existing limited sequence data from the nuclear 18S rRNA locus of Eimeria furonis Molecular
characterization of E ictidea from domestic ferrets was not possible because samples containing this
parasite were not available for study consequently it is unclear whether the same coccidium affects both
domestic and black-footed ferrets
106
There is no published information describing the pre-patent periods and pathogenicity of enteric
coccidia in BFF and given the conservation status of the BFF experimental work cannot be undertaken
in the natural host The purpose of this study was to determine whether the domestic ferret is susceptible
to E ictidea isolated from BFF if susceptible the domestic ferret could act as a suitable experimental
model in which to investigate the pathogenesis prevention and treatment of coccidiosis caused by E
ictidea
52 MATERIALS AND METHODS
521 Animal care
Ten juvenile male intact ferrets of 48 (n=6) or 50 (n=4) days of age were obtained from a
commercial source (Marshall BioResources North Rose New York USA) and were housed in the
University of Guelph Central Animal Facility Isolation Facility Ferret weights on arrival ranged from
334-475 g (average= 3928 g) All ferrets were housed individually in wire bottom cages of 813 times 1117
times 457 cm size and were divided in equal numbers between two non-adjoining rooms They received ad
libitum access to Envigo Teklad Certified Global Ferret Diet (Madison Wisconsin USA) and water
changed daily Room temperature was maintained at 18-21 degC and a 16 hour light 8 hour dark
photoperiod was provided All personnel working with the ferrets were required to wear personal
protective equipment including disposable facemasks gloves gowns and bouffant caps Shoes were
provided for use in each room This study was carried out in accordance with the recommendations in the
Canadian Council on Animal Care guidelines The protocol was approved by the Animal Care Committee
of the University of Guelph (Animal Use Protocol 3289) and by both the Animal Welfare Committee
and Animal Care and Research Committee of the Toronto Zoo
An initial physical examination and blood collection were performed on each ferret by the
principal investigator (ARP) one day after arrival to assess health status prior to enrollment in the study
Ferrets were mask induced with isoflurane (Isoflurane USP Fresenius Kabi Richmond Hill Ontario) in
107
oxygen placed on a heat disc (SnuggleSafe Lenric C21 Ltd Littlehampton United Kingdom) weighed
examined and blood was collected from the jugular vein for routine CBC and biochemical profiles
All ferrets subsequently underwent an acclimation period of two weeks During this time fecal
samples were collected daily from each ferret and examined for the presence of coccidial oocysts using a
standard salt flotation technique (Dryden et al 2005) to ensure that all individuals were free of coccidia
prior to initiation of experimental work Any ferret positive for coccidia was to be removed from the
study
522 Oocyst preparation
Oocysts used for inoculation originated from fecal samples from two naturally infected BFF These
samples were stored in potassium dichromate for four weeks prior to oocyst purification and use in this
infection trial Stored fecal samples were mixed with distilled water and passed through a small sieve to
remove debris The strained contents were transferred to a 50 mL conical vial and topped up to 50 mL
with additional distilled water Samples were centrifuged (Sorvall ST40R Centrifuge Thermo Scientific)
at 2800 rpm (1315 timesG) for 10 minutes at 12 degC A drop of supernatant was evaluated microscopically at
100times for the presence of oocysts If oocysts were observed the supernatant was poured off into a second
50 mL conical tube and again topped up to 50 mL with distilled water and re-centrifuged under the same
conditions Otherwise the supernatant was discarded The pellets from both the first and second tubes
were combined with saturated salt solution at a 14 ratio by volume Oocysts were floated in the salt
solution by centrifugation at 1500 rpm (377 timesG) for 10 minutes at 12 degC The top 5 mL of supernatant
were collected and transferred to a clean 50 mL conical tube topped up to 40 mL with distilled water and
washed via centrifugation at 2800 rpm (1315 timesG) for 10 minutes After the wash step the supernatant
was again checked for presence of oocysts and discarded if no oocysts were observed The pellet was
collected and the presence of oocysts confirmed by examination of a drop placed on a clean glass slide at
100times Once verified the contents of the pellet of concentrated oocysts was placed in a 250 mL storage
container and mixed with approximately 200 mL of sterile saline (09 sodium chloride Hospira
108
Montreal Quebec) prior to storage for two to four weeks at 4 degC until inoculation Prior to inoculation a
McMaster count was performed to determine the number of oocysts per mL in order to determine
appropriate volume of inoculum
523 Experimental infections
Part 1
Five ferrets were randomly assigned to each of the control and infection groups After the
acclimation period on day 0 four ferrets in the infection group were inoculated orally with a high dose
oocyst suspension (1 times 106 oocysts in 025 mL of saline) mixed into 1 mL of FerreTone Skin amp Coat
Supplement (United Pet Group Inc Blacksburg Virginia USA) a fifth ferret was inoculated with a
moderate dose oocyst suspension (5 times 104 oocysts in 025 mL of saline) in the same volume of FerreTone
Ferrets in the control group were inoculated with a placebo (025 mL of saline) in 1 mL of FerreTone
Inoculation was performed by offering the oocyst suspension or placebo to the ferrets in a plastic
container
Fecal samples were collected daily from each inoculated ferret for 14 days post-inoculation
Samples were analyzed via fecal flotation using the McMaster method followed by routine flotation in
saturated salt solution (Dryden et al 2005) to determine the presence or absence of oocysts and oocyst
burden (oocysts per gram of feces [OPG])Temporal trends in oocyst shedding were monitored Ferrets
were evaluated visually twice daily for the presence of clinical signs of coccidial disease The first of
every two ferrets identified to shed oocysts was to be humanely killed at the time of peak oocyst shedding
(ie the first day that fecal oocyst counts remained static or declined) and necropsied to confirm the
presence of and describe parasitic replication in the intestinal mucosa Any remaining animals that shed
oocysts were to be monitored throughout the 14 day period following inoculation in order to determine
the duration and intensity of oocyst shedding for these individuals the total number oocysts shed during
patency was determined
109
Part 2
All ferrets from the infection group that did not shed oocysts during Part 1 (n=4) and all but one
ferret from the previous control group (n=4) were orally inoculated with the high dose oocyst suspension
(1 times 106 in 1 ml of saline) mixed with an equal volume of FerreTone Consequently between phases 1
and 2 all but one ferret were inoculated at least once with the BFF coccidia in order to increase
experimental animal numbers and determine if ferret age played a role in susceptibility to infection
One ferret from the previous control group was inoculated with a lower dose (2 times 105 oocysts in
075 mL of 09 saline mixed with 1 mL FerreTone) of oocysts that had been collected from the single
domestic ferret that shed in Part 1 oocysts were purified as described above for the initial inocula Fecal
collection and analysis were performed as previously As in Part 1 one in every two ferrets sequentially
identified to be shedding oocysts in feces was killed humanely at the time of peak shedding and a
complete necropsy examination performed The remainder of the ferrets observed to be shedding were
monitored for the full 14 days of the trial after which they were killed humanely and necropsied and total
number of oocysts shed during patency was determined All ferrets that did not shed coccidial oocysts
during the infection trial were rehomed at the end of the trial
524 Animal welfare
Ferrets were evaluated twice daily for development of clinical signs of coccidial disease and any
animal showing clinical disease was to be treated as determined by a veterinarian with supportive care
including fluid therapy A grading system for clinical signs including intervention points and removal
criteria was created for use during daily evaluation (see Appendices 3 and 4) Animals whose clinical
signs could not be ameliorated without the use of specific anticoccidial therapy were to be euthanized
Should the inoculation in Part 1 have resulted in clinical disease that required extensive treatment andor
necessitated euthanasia a lower number of oocysts would be used for subsequent inoculation in Part 2
Ferrets to be euthanized were anesthetized by mask induction with isoflurane in oxygen a 1 mL blood
110
sample was collected from the cranial vena cava and then an intracardiac dose of potassium chloride (2
mEqkg) was administered
525 Hematology
Blood was collected from all ferrets under isoflurane anesthesia at the time of pre-trial health
examination and again at the time of humane killing Blood was collected via jugular venipuncture
initially due to the small size of the ferrets at arrival and then by cranial vena cava venipuncture or
cardiocentesis prior to euthanasia Complete blood count and serum biochemistries were performed by the
Animal Health Laboratory of the University of Guelph Guelph Ontario
526 Morphologic and molecular characterization
Morphologic and molecular characterization of oocysts shed by the domestic ferrets during the
course of the infection trial was performed to ensure that the ferrets were shedding the same species of
Eimeria with which they were inoculated Oocysts were concentrated from positive fecal samples as
described above A drop of concentrated oocyst solution was viewed photographed and measured at
400times and 600times for comparison with previously determined morphometrics of Eimeria ictidea oocysts
(Chapter 3)
Regions from the mitochondrial cytochrome c oxidase subunit I and III (mt COI and mt COIII)
DNA were amplified by polymerase chain reaction (PCR) from each sample using primer pairs
400F1202R and -172F799R respectively For all PCR reactions samples were denatured at 95 degC for 5
min then subjected to 35 cycles of 94 degC for 30s anneal at 52degC for 30s and extension at 72 degC for 60s
followed by a final extension at 72 degC for 7 min PCR gel electrophoresis and sequencing methods used
were as described in the Materials amp Methods section of Chapter 2
The resulting consensus sequences were searched from within Geneious against previous
sequences for E ictidea produced by the authors and against publically available sequences on the
111
BLAST server (blastncbinlmnihgovBlastcgi) using the blastn search algorithm against the nrnt
database (GenBank+EMBL+DDBJ+RefSeq ndash AA or DNA)
527 Necropsy protocol
All humanely killed ferrets underwent a complete necropsy (Appendix 6) using the modified
protocol described in Materials amp Methods section 424 of Chapter 4
53 RESULTS
Initial physical examination was unremarkable with the exception of mild to moderate bilateral
ceruminous discharge within the external ear canal of all ferrets Complete blood count and serum
biochemistry results for all ferrets were within normal reference intervals for juvenile domestic ferrets
(Appendices 2a and b) (Fox 2014) Six days after arrival a single ferret (103) in the control group
developed mild upper respiratory signs consisting of sneezing and clear nasal and ocular discharge these
clinical signs were associated with mild dehydration and decreased food and water consumption The
ferret was treated with subcutaneous fluid therapy (10 mL Plasmalyte-A subcutaneous Baxter Alliston
Ontario) heat and supportive care and all clinical signs resolved within three days This ferret was
deemed healthy to participate in the remainder of the clinical trial A second ferret (105) in the control
group developed unilateral purulent ocular discharge 14 days after arrival one day prior to placebo
inoculation The ferret was treated topically twice daily for five days with Isathal ophthalmic gel (fusidic
acid 10 mgg Dechra Veterinary Products Inc Point-Claire Quebec) and the discharge resolved but
reoccurred within 2 days of treatment cessation Ocular examination showed mild conjunctivitis but no
evidence of corneal lesions and fluorescein staining did not indicate the presence of corneal ulceration
The ferret was treated for an additional six days with tobramycin ophthalmic solution (3 mgmL Sandoz
Tobramycin 03 Boucherville Quebec) after which clinical signs resolved completely No coccidial
oocysts were shed in feces from any of the ferrets during the two week acclimation period
112
531 Oocyst shedding
All ferrets readily ingested the inoculum with either placebo or concentrated oocysts In Part 1
one ferret (203) in the infection group which had received the high dose (1 times 106 oocysts) shed oocysts
on day 8 and day 9 after inoculation (Tables 51 52) This ferret was 71 days of age at the time shedding
was initially identified The ferret was killed humanely 11 days post inoculation later than had been
outlined in the protocol as processing of fecal samples had been delayed by two days resulting in late
detection of oocyst shedding in this individual Oocysts were not identified in the feces of the three
remaining ferrets that received the high inoculation dose the single ferret that received the lower dose (5
times 104 oocysts) or in the ferrets within the control group
In Part 2 six of eight ferrets inoculated with the high dose (1 times 106 oocysts) shed oocysts during
the 14 day observation period (Tables 51 52) Four of these ferrets were from the previous control
group One of the ferrets previously inoculated with the high dose inoculum in Phase 1 that had not shed
oocysts did shed oocysts after being inoculated a second time with the same dose during Phase 2 The
ferret that had previously received the low dose (5 times 104 oocysts) of oocysts in Phase 1 also shed after
inoculation with the high dose in Phase 2 Three ferrets did not shed oocysts after high dose inoculation in
Phase 2 one of these had been part of the previous control group the other two had received the high
dose inoculation previously in Phase 1
The pre-patent period ranged from 7-9 days (Table 51) with equivalent numbers of ferrets
commencing shedding on each of days 7 through 9 All six ferrets were between 91 and 93 days of age at
the time shedding was initially identified Oocyst per gram counts and shedding trends for all individuals
are shown in Table 51 Total oocyst shedding during patency was lt14 oocysts 8904 oocysts and
172291 oocysts for ferrets 201 104 and 105 respectively The two ferrets for which the prepatent period
was 9 days only shed oocysts for one day and in low numbers Oocysts were not identified in the feces of
the three remaining ferrets two of which received the high inoculation dose (1 times 106 oocysts) and the
third that received the lower inoculation dose (2 times 105 oocysts) (Table 52)
113
532 Morphologic and molecular characterization
In all seven ferrets that shed oocysts the morphologic features and measurements (length width
shape index) of the shed oocysts were consistent with those of the E ictidea administered in the inoculum
(Figure 51) Molecular confirmation of the identity of the oocysts shed was successful in 3 out of the 7
ferrets (102 103 203) samples from the four remaining ferrets did not contain adequate quantity or
quality of DNA for confirmation
533 Clinical signs
Clinical signs associated with patent infection were identified in 6 of 7 ferrets (Table 52) These
signs included weight loss (n=5) diarrhea (n=1) mucoid soft feces (n=2) feces containing blood (n=2)
and malodorous feces (n=1) Appetite reduction was noted in two ferrets from the infection group in Part
1 between 6-8 days post infection however no oocyst shedding was detected from either ferret during
this time
534 Hematology
CBC and serum biochemistry values from ferrets collected during pre-trial health screening are
displayed in Appendices 1a and 1b Values obtained for ferrets euthanized during or after the
experimental trial are displayed in Appendices 3a and 3b Minor variances from reference range values
for CBC and serum biochemistry were identified in six of the seven ferrets with patent infection In all six
ferrets for which a complete serum biochemistry was obtained immediately prior to death creatinine
kinase (CK) values were elevated (see Appendix 3b) Ferret 103 exhibited a mild hypoalbuminemia (20
ref 24-40 gL) on ante-mortem serum biochemistry (see Appendix 3b)
114
535 Necropsy
No gross or histopathologic lesions were present and coccidia could not be identified in sections
of intestine from the single ferret (203) humanely killed in Part 1 For the ferrets humanely killed in Part
2 no evidence of diarrhea hematochezia or mucoid fecal material was identified grossly Ferret 105
killed at the termination of the experiment but still shedding low numbers of oocysts in its feces
exhibited a 7 cm region of congested mucosa within the distal jejunum
Coccidial life stages were identified in small intestinal sections from four of the seven ferrets that
were identified to shed oocysts at some point prior to necropsy (Figure 52 Tables 52 and 53) Affected
sections included jejunum in all four animals as well as ileum in one and were collected from 114 to 218
cm aboral from the pylorus (see Figure 53) Coccidia were not identified in sections of duodenum
proximal jejunum or large intestine however oocysts were identified within fecal material in the lumen
of the large intestine from one ferret (103) Of the 11-19 sections of intestine examined for each ferret the
number of sections containing parasites ranged from one to eight 1 section in ferret 102 2 sections in
ferret 201 4 sections in ferret 105 8 sections in ferret 103 A mix of sexual and asexual life stages was
observed within the enterocytes in small intestinal sections from ferret 103 the remainder of the ferrets
showed either asexual (102 201) or sexual (105) life stages in affected segments
Pathologic changes and additional histologic findings in small intestinal sections of ferrets with
enteric coccidia included rare regions of blunting of the villi and sloughing of the epithelium associated
with hemorrhage and inflammation The primary lesions identified were subjectively increased numbers
of eosinophils lymphocytes and plasma cells within the lamina propria of the small intestine and
similarly increased neutrophils lymphocytes and plasma cells within the lamina propria of the large
intestine Neutrophils were rarely present in intestinal crypts and glands
Other gross necropsy findings included two ferrets with renal cortico-medullary cysts and one
ferret with mild thickening of the esophageal mucosa midway along the esophageal length No histologic
changes were identified within a sample of esophagus taken from this region
115
54 DISCUSSION
The findings of this study show that domestic ferrets are susceptible to infection with the enteric
coccidium Eimeria ictidea isolated from black-footed ferrets Both morphometric and molecular
diagnostic methods were used to confirm that ferrets were shedding oocysts of the same species with
which they were inoculated Molecular characterization was successful in three of the seven ferrets that
developed patent infections and as no other coccidial species was identified during pre-trial observation
morphometry was considered to be confirmatory in the remaining four animals
We have referred to the eimeriid coccidium affecting BFF and used in this experimental trial as
E ictidea based on morphometric similarity of their oocysts with those of E ictidea as described from
domestic ferrets (see Chapter 3) There is limited published information on infection of domestic ferrets
with E ictidea outside of Hoarersquos original descriptions (1927 1935a b) which form the basis for all
subsequent identifications of E ictidea in domestic ferrets and in BFF Attempts to obtain exemplars of E
ictidea from domestic ferrets to characterize using molecular techniques were unsuccessful (Chapter 2)
Multiple diagnostic laboratories in Canada and Europe were solicited for coccidia-positive fecal samples
from domestic ferrets but no samples of E ictidea were received over a 4 year period (2014-2017)
Eimeria ictidea was identified based on microscopic examination in only two samples submitted to a
European diagnostic laboratory from 2008-2015 It is unproven whether the E ictidea described from
domestic ferrets and the E ictidea identified from black-footed ferrets and used in this experimental
work are the same or are simply morphologically indistinguishable Eimeria species However the
consistency in morphology host genus and location of infection within the intestinal tissues combined
with the successful cross-transmission of this parasite to domestic ferrets described in the present study
suggests they are likely conspecific
116
The pre-patent period (minimum duration of endogenous development) for infection with E
ictidea in the inoculated domestic ferrets ranged from 7-9 days (see Table 51) the pre-patent period for
this parasite in the BFF the natural host for this coccidium is unknown Hoare experimentally infected
naiumlve domestic ferrets with E ictidea derived from naturally occurring infection in this species (Hoare
1935b) The inoculated ferrets shed oocysts after a pre-patent period of 7 days consistent with the 7-9
days seen in the work described here with E ictidea
Shedding of oocysts was identified over a period of 1-7 days (see Table 51) and intensity ranged
from less than 14 up to 15624 OPG These results may be skewed with erroneously low duration of
shedding and number of oocysts shed as three of the seven ferrets were humanely killed at or prior to the
expected peak of oocyst shedding for tissue collection and histologic examination in order to increase the
probability of identifying parasite life stages within the intestinal sections Shedding periods were similar
to those identified in adult single-housed BFF which ranged from 2-9 days however oocyst per gram
counts from the domestic ferrets were consistently lower than OPG counts from BFF (104 - 554274
OPG) infected with the same parasite (see Chapter 4) Furthermore the total number of oocysts shed by
individual domestic ferrets (14 - 172291 oocysts) during patency was reduced compared to BFF despite
similar length of shedding period (see Chapter 4) The domestic ferrets in this study were naiumlve individual
juveniles whereas the BFF were adults 1-5 years of age some of which were showing clinical signs at
the time of oocyst shedding The relative influences of age species and previous exposure to the parasite
on our observations are unknown
Two different fecal flotation methods were used on all samples to increase the probability of
oocyst detection The McMaster method was used to provide accurate OPG counts for quantification of
oocyst shedding however this method had a minimum detection limit of ~13 oocysts per gram (1333
OPG calculated) because it is based on dilution of the initial sample with flotation media (saturated salt)
In samples with few oocysts oocysts may be missed or to be present in numbers below this detection
limit Routine salt flotation is in contrast performed using the entire sample allowing for detection of
117
small numbers of oocysts Consequently in cases where oocyst per gram counts were low shedding was
identified on routine salt flotation but not by the McMaster method and recorded as lsquopositiversquo but below
the detection limit of the enumeration method
Subclinical to clinical disease occurred in six of the seven ferrets that developed patent infection
with weight loss being the most frequent clinical sign Other clinical signs were typical of enteric
coccidiosis including diarrhea hematochezia and mucoid andor soft feces These clinical signs are
similar to those previously described for black-footed ferrets infected with this parasite (USFWS BFF
Recovery Program 2017 Chapter 4) and for domestic ferrets with enteric coccidiosis (Sledge et al
2011) Interestingly development of clinical disease was not described in the naiumlve domestic ferret
inoculated by Hoare (1935) with E ictidea derived from naturally occurring infection Based on review of
the literature severe clinical disease resulting from intestinal coccidiosis is rare in domestic ferrets
Black-footed ferrets however appear more susceptible to disease development and more frequently show
significant clinical signs No domestic ferret required treatment for clinical coccidiosis during the course
of this study It is possible that the more pronounced clinical signs associated with enteric coccidiosis in
BFF may result from the limited genetic diversity in a population derived from so few individuals and
increased susceptibility of BFF to other diseases such as sylvatic plague have been described in
comparison with their domestic counterparts (Williams et al 1994) Although unlikely recent acquisition
of E ictidea from a related host species such as the domestic ferret could have resulted in increased
pathogenicity and severity of clinical disease from infection with this parasite in BFF
Minor variances from reference range values for CBC and serum biochemistry were identified in
six of the seven ferrets with patent infection but only in one case (ferret 103) did this appear to be
correlated with infectiondisease In this animal a mild hypoalbuminemia (20 ref 24-40 gL) was noted
(Appendix 3b) On histopathology large regions of the small intestine contained parasite life stages
however inflammation lysis of epithelial cells and necrosis of affected areas that could be expected to
result in protein loss into the intestinal lumen were not identified
118
Creatine kinase values were elevated in all six ferrets for which a complete serum biochemistry
was obtained immediately prior to death these findings are consistent with release from CK rich tissues
such a skeletal muscle during venipuncture and manual restraint
The pathologic lesions identified within the intestinal sections of ferrets euthanized at the time of
oocyst shedding were similar to those identified in affected BFF but in most cases were less locally
extensive or widespread throughout the small intestine than those observed in necropsy cases of BFF (see
Chapter 4) Coccidia were seen in the histologic sections of four ferrets all three ferrets that were actively
shedding oocysts at the time of necropsy (102 103 105) and one of four in which necropsies were
performed after oocyst shedding had ceased (201)
Although the primary objective of the examination of histologic sections from affected ferrets
was to identify coccidial life stages attempts were made to describe the pathologic changes associated
with the presence of the parasite Despite necropsies being performed almost immediately after death and
the use of Serra solution fixative to improve parasite and tissue preservation the villi and villar
epithelium of the trimmed sections were frequently distorted or absent and consequently accurate
commentary on these areas was precluded and was made only on visible components of the lamina
propria and crypts or glands A deliberate decision was made not to kill and collect samples from the
saline inoculated control ferrets after Phase 1 and those not shedding oocysts during Phase 1 or 2 thus no
age-matched intestinal sections were available for comparison It is difficult to comment on the
significance of the inflammatory cells observed in the lamina propria of the small and large intestinal
segments or the proliferative rate of the crypt epithelium The only changes identified which may be
considered significant are the presence of neutrophils within the crypts and glands of small and large
intestinal sections respectively but these lesions were rare and not associated with the presence of
parasitic life stages
119
While no parasitic life stages were identified in intestinal sections from ferrets 104 203 and 205
one of the three ferrets ferret 104 exhibited lymphoplasmacytic inflammation and blunting of jejunal
villi in one section (36-38 cm aboral from pylorus) These findings might be expected as the most
extensive histologic lesions would occur associated with lysis of the intestinal epithelial cells as oocysts
were shed into the feces after which new intestinal epithelial cells would re-cover the denuded villar
surface Thus for those cases in which histologic examination was performed after shedding had ceased
presence of the protozoal life stages in the intestines would be expected to be significantly reduced or
absent
Hoare (1935a b) described a particular reaction to the presence of parasitic life stages of Eimeria
ictidea in the small intestine of domestic ferrets in which only the villar tips were affected and there was
resultant annular constriction of the villus separating the affected and non-affected segments These
particular changes were not identified in any of the ferrets in this study and had not been noted
retrospectively in naturally infected BFF (Chapter 4)
While patent infection and intestinal disease could be experimentally created in domestic ferrets
without equivalent experimental work in BFF it is difficult to fully compare the susceptibility to infection
and to the development of disease between the two species Eimeria species tend to be host specific thus
if E ictidea from BFF is not conspecific with E ictidea in domestic ferrets it might be expected that the
domestic ferret would be less susceptible to infection and the development of disease than is the BFF
Even if the two parasites are identical natural passaging through the BFF may alter affinity for the
domestic ferret
It appears that the infectious dose of oocysts of E ictidea derived from BFF required to initiate a
patent infection in domestic ferrets is high The ferrets that developed patent infections were administered
an inoculum containing 1 times 106 sporulated oocysts and even with this extremely high inoculating dose
only a proportion of inoculated ferrets became infected Neither the ferret that received the low dose
120
inoculum nor the ferret that received the passaged oocysts from Part 1 of the study shed oocysts during
the 14 day period post inoculation The latter finding was unexpected as fresh passaged oocysts would be
expected to contain larger proportions of viable oocysts and be comparatively more infective than oocysts
that had been stored for 2-4 weeks prior to inoculation
The only publication describing oral inoculation of Eimeria species in domestic ferrets (Hoare
1935b) did not quantify the number of oocysts administered A study performed in 16 farmed juvenile
mink (Mustela vison) administered 2000 sporulated oocysts of each of three coccidial species (I
laidlawi E vison and an unknown Eimeria species) resulting in patent infection with one of the three
species (Foreyt et al 1977) as determined by the presence of oocysts on fecal examination The authors
did not reveal which type of oocysts resulted in the infection
The number of oocysts required to result in infection in BFF is unknown Based on the
authorsrsquo observations of over 100000 oocysts per gram of feces being shed by black-footed ferrets into
cages of lt1m2 floor space (see Chapter 4) we estimated that under normal caged conditions animals
would likely ingest thousands of oocysts over a short period of time This was in part why a large number
of oocysts (up to 1000000 as available from our store of viable oocysts) was administered to each
domestic ferret in order to increase the probability that infection and shedding would result Furthermore
for the parasite to persist within the ferret population the total number of oocysts shed into the
environment would have to be several times higher than the infective dose required to generate a patent
infection otherwise the parasite would be expected to die out If E ictidea of domestic ferrets and E
ictidea of BFF are conspecific the difference in oocyst shedding between the host species during patent
infection could potentially explain the low prevalence of E ictidea reported from the domestic ferret
population (as seen in Chapter 2)
Both humoral and cell mediated immunity are involved in the immune response to coccidia The
role of maternal derived antibodies in combatting protozoal infection in mustelids has not been studied
121
but in carnivores maternal antibodies to viruses can last up to 16 weeks (Chappuis 1998) In poultry
maternal Eimeria-specific IgG is transferred via the egg yolk to offspring In one study breeding hens
were infected with a single species of Eimeria 28-39 days prior to lay Their hatched chicks were
challenged by inoculation with oocysts of the same and a related Eimeria species and showed reduced
oocyst shedding compared to age matched controls indicative of passive transfer of immunity (Smith et
al 1994) It is possible then that the presence or absence of maternal antibodies may be a factor in the
age at which ferrets are susceptible to coccidial infection and the development of disease The facility
from which the domestic ferrets were acquired has not previously detected Eimeria species on routine
fecal screening (Dr Bambi Jasmin personal communication) consequently it is unlikely that they would
have received maternal immunity to or been exposed to this parasite and thus can be considered to be
naive
Despite a theoretical lack of maternal immunity there did appear to be an effect of age on
susceptibility to infection In Part 1 when the ferrets were approximately 70 days of age only 1 of 4
ferrets inoculated with the high dose of oocysts developed patent infection In comparison in Part 2
when the ferrets were 91-93 days of age 3 of 4 ferrets that had been in the saline control group for Part 1
developed patent infections as did 2 of 4 ferrets that had been inoculated in Part 1 but had not shed
oocysts One of these previously inoculated ferrets was the individual that had received the lower dose of
oocysts Thus it appears that patent infection could be produced more easily in the older ferret kits
However the two ferrets that had previously been inoculated exhibited the shortest shedding periods (1
day) and lowest oocyst per gram counts and parasite life stages in these cases were rare (201) to absent
(205) on histologic examination of numerous sections of intestine It is thus possible that the primary
inoculation resulted in abbreviated infections or infections in which so few oocysts were shed that
infection was not detected Our observation of endogenous stages in the intestinal tissues of some of the
ferrets following cessation of oocyst shedding suggests that the pre-patent period and duration of patency
may vary considerably from animal to animal consequently it is possible that some of the kits would
122
have ultimately shed a few oocysts from the primary inoculum if followed beyond 14 days post-
inoculation Whether through an aborted or undetected infection previous exposure of these kits to the
parasite probably generated partial immunity against E ictidea and therefore the intensity of infection
upon challenge in previously exposed domestic ferret kits was reduced
The authors acknowledge the limitations of this initial pilot study however it was proven that
patent infection with E ictidea isolated from black-footed ferrets could be generated in a novel host the
domestic ferret Further studies will be required to investigate the effect of age on susceptibility to
infection as well as the possibility of development of immunity after exposure and its role in reducing
parasite replication and disease in subsequent infection With so few BFF in existence the use of BFF for
in vivo infection trials cannot be contemplated consequently refining the domestic ferret infection model
will be essential for carrying out research specifically intended to help manage coccidiosis in the
endangered black-footed ferret
Figure 51 Exogenous life stages of Eimeria ictidea shed from a domestic ferret (Mustela putorius
furo) experimentally inoculated with oocysts originating from black-footed ferrets (Mustela
nigripes) Unsporulated oocyst (solid black arrow) Sporulated oocyst (solid white arrow) Bright
field microscopy scale bar = 25 microm
25 microm
124
Figure 52 Life stages of Eimeria ictidea within the small intestinal epithelium of an experimentally
infected domestic ferret (Mustela putorius furo) A) Sexual life stages (micro- and macrogamonts
- white arrows) crowding the villar enterocytes all stages are found between the nucleus and
luminal surface of infected enterocytes Hematoxylin and eosin staining scale bar = 25microm B) At
higher magnification meronts (black arrows) and gamonts (open arrows) are crowded between the
enterocyte nuclei and brush border Hematoxylin and eosin staining scale bar = 25microm
25 microm
25 microm
125
Figure 53 Presence and location of sexual and asexual life stages of Eimeria ictidea within the
intestinal epithelium of domestic ferrets (Mustela putorius furo) (n=7) that developed patent
infection after experimental inoculation with oocysts originating from black-footed ferrets (Mustela
nigripes) Legend duod = duodenum jej= jejunum Sequential numbers for jejunal and colonic
sections represent the order aboral from the pylorus from which the samples were collected SI =
additional section(s) of small intestine whose aboral sequence was not recorded
0
1
2
3
duod jej 1 jej 2 jej 3 jej 4 jej 5 ileum colon 1 colon 2 SI
Nu
mb
er
of
Ferr
ets
Aff
ect
ed
Intestinal Section Containing Life Stages of Eimeria ictidea
Asexual life stages Sexual life stages
126
Table 51 Prepatent period and oocyst shedding patterns in domestic ferrets (Mustela putorius furo)
experimentally inoculated with 1 times 106 oocysts of Eimeria ictidea originating from black-footed ferret
(Mustela nigripes) that developed patent infections
Oocysts shed per gram of feces
Ferret Identity
Day post inoculation 102 103 104 105 201 203 205
1 0 0 0 0 0 0 0
2 0 0 0 0 0 0 0
3 0 0 0 0 0 0 0
4 0 0 0 0 0 0 0
5 0 0 0 0 0 0 0
6 0 0 0 0 0 0 0
7 lt 14 1807 0 0 0 0 0
8 11053 139 7091 0 156238 0
9 463 11733 lt 14 203 lt 14
10 578 7549 0 0 0
11 lt 14 0 0 0
12 0 lt 14 0
13 0 lt 14 0
14 0 lt 14 0
Legend ferret euthanized as of this date lt 14 = oocyst positive samples with less than 14 oocysts per
gram of feces
127
Table 52 Results of oral inoculation of domestic ferrets (Mustela putorius furo) with oocysts of Eimeria ictidea originating from black-footed
ferrets (Mustela nigripes)
Ferret
Identity
Results Part 1 Results Part 2
Inoculum Oocyst
Shedding
Presence
of clinical
disease
Coccidia
present in
sections
Inoculum Oocyst
Shedding
Presence
of clinical
disease
Coccidia
present in
sections
101 Saline N N - 2 times 105 oocystsa N N -
102 Saline N N - 1 times 106 oocysts Y N Y^
103 Saline N N - 1 times 106 oocysts Y Y Y^
104 Saline N N - 1 times 106 oocysts Y Y Nyen
105 Saline N N - 1 times 106 oocysts Y Y Yyen
201 1 times 106 oocysts N N - 1 times 106 oocysts Y Y Yyen
202 1 times 106 oocysts N N - 1 times 106 oocysts N N -
203 1 times 106 oocysts Y Y Nyen
204 1 times 106 oocysts N N - 1 times 106 oocysts N N -
205 5 times 104 oocysts N N - 1 times 106 oocysts Y Y N^
Legend includes any of the following weight loss diarrhea mucoid feces malodorous feces inappetence N = no Y= yes - = necropsy not
performed a = oocysts collected after passage through ferret 203 ^ = shedding oocysts at time of necropsy yen = not shedding oocysts at time of
necropsy
128
Table 53 Distribution of coccidial life stages in domestic ferrets (Mustela putorius furo) orally inoculated with oocysts of Eimeria ictidea
originating from black-footed ferrets (Mustela nigripes)
Ferret Identity
Intestinal level 102
103
104 105
201
203
205
Duodenum N N N N N N N
Jejunum 1 N N N N N N N
Jejunum 2 N N N N N N N
Jejunum 3 N S A N N N N N
Jejunum 4 N S A N N N N N
Jejunum 5 N S A N S A N N
Ileum N S A N N N N N
Colon 1 N Na N N N N N
Colon 2 N Na N N N N N
Unmeasured small intestinebc
S - 05
A - 15
S - 45
A - 25
S - 05
A - 05
S - 26
A - 06
S - 06
A - 06
S - 06
A - 06
S - 08
A - 08
Unmeasured large intestinebc
S - 01
A - 01
S - 01
A - 01
S - 01
A - 01
S - 02
A - 02
S - 01
A - 01 none
S - 01
A - 01
Legend N = no parasite life stages S = sexual life stages present A = asexual life stages present a= oocysts present in feces b=
additional sections of intestine for which the location measured from the pylorus was not obtained c= xn where x is number of
sections containing sexual or asexual lifestages n is the number of sections examined
129
CHAPTER 6 WHOLE MITOCHONDRIAL GENOME SEQUENCES OF TWO EIMERIA
SPECIES ISOLATED FROM DOMESTIC (MUSTELA PUTORIUS FURO) AND BLACK-
FOOTED FERRETS (MUSTELA NIGRIPES)
ABSTRACT
The complete mitochondrial (mt) genomes of Eimeria furonis and Eimeria ictidea (Eimeriidae
Coccidia Apicomplexa) originating from single fecal samples from a domestic (Mustela putorius furo)
and a black-footed ferret (Mustela nigripes) respectively were sequenced Both mt genomes were
circular-mapping with lengths of 6165 base pairs (Eimeria furonis - GenBank MF795598) and 6182
base pairs (Eimeria ictidea - GenBank KT203399) Genome organization and gene contents were
comparable with those of other publically available mt genomes from a variety of Eimeria species and
related coccidia there were three complete coding DNA sequence regions encoding cytochrome c
oxidase subunit I cytochrome c oxidase subunit III and cytochrome B and 33 regions encoding
fragmented rDNA Alignment of these mt genome sequences demonstrates a relatively high (945 340
single nucleotide differences [SNDs]) pairwise sequence identity between these Eimeria spp infecting
ferrets the majority of the SNDs resulted in synonymous codon changes with no changes to their protein
products Alignment of the newly sequenced mt genomes demonstrates and phylogenetic reconstructions
support the monophyly of these Eimeria spp of ferrets with another Eimeria sp of carnivores as the
sister taxon to this clade
61 INTRODUCTION
Coccidia are protozoal eukaryotic host-specific parasites of the phylum Apicomplexa and can
be divided into two major taxonomic suborders the eimerioirinid and adeleid coccidia The eimeriorinid
coccidia include both the typical intestinal coccidia (eg Eimeria Isospora Cyclospora) species
belonging to the family Eimeriidae as well as tissue or cyst-forming coccidia (eg Cystoisospora
130
Besnoitia Toxoplasma Sarcocystis) of the family Sarcocystidae (see Cox 1994) Ten species of
Eimeria and twelve species of Isospora (=Cystoisospora) have been described in the Mustelidae
(see Chapter 1 Table 11) Eimeria furonis has been reported in the European polecat (Mustela
putorius) domestic ferret (DF Mustela putorius furo) black-footed ferret (BFF Mustela nigripes) and
mink (Mustela vison) (Hoare 1927 Nukerbaeva and Svanbaev 1973 Jolley et al 1994) Eimeria ictidea
has been reported in the Steppe polecat (Mustela eversmanii) as well as the European polecat domestic
ferret and black-footed ferret (Hoare 1927 Svanbaev 1956 Jolley et al 1994) These reports are based on
the morphometric characteristics of oocysts identified in the feces of the aforementioned host species
without the use of molecular techniques to confirm specific parasite identities
Recently sequences of the mitochondrial cytochrome c oxidase subunit I gene (mt COI) and
nuclear small subunit ribosomal DNA (nu 18S rDNA) of E furonis originating from a domestic ferret (nu
18S rDNA GenBank MF774678-MF774680 mt COI GenBank MF774034-MF774036) and E ictidea
originating from a black-footed ferret (nu 18S rDNA GenBank MF860826-MF860827 mt COI
GenBank MF860823 MF860825) were generated (see Chapters 2 and 3) The parasite originating from
the black-footed ferret was identified as E ictidea based on morphologic similarity to the original
descriptions of E ictidea from domestic ferrets however sequence-based genotyping of E ictidea from
domestic ferrets has not been completed and consequently it has not been demonstrated unequivocally
that the two parasites are conspecific In the present work the complete mitochondrial genomes of E
furonis from the DF and E ictidea from the BFF are described and compared with the mitochondrial
genomes of related coccidia
62 MATERIALS amp METHODS
621 Parasites
Two isolates of morphologically distinct Eimeria species were used in this study Isolate one
identified morphologically and by nu 18S rDNA and mt COI sequences as Eimeria furonis was obtained
131
from a fecal sample from a DF that was submitted for routine ova and parasite examination to a European
diagnostic laboratory8 Isolate two identified morphologically and by nu 18S rDNA and mt COI
sequences as Eimeria ictidea was obtained from a fecal sample from a BFF and was collected during
routine cage cleaning in a captive breeding facility (see Chapters 3 and 4) Fecal collection techniques for
the BFF were reviewed and approved by both the Animal Welfare Committee and the Animal Care and
Research Committee of the Toronto Zoo
622 DNA isolation from coccidia in feces
Genomic DNA was isolated from fecal derived coccidial oocysts as described section 221 of the
Chapter 2 Materials amp Methods Parasite DNA concentration was estimated using a Nanodrop 2000
spectrophotometer (NanoDrop Products Wilmington DE USA) and DNA was stored at 4 degC for
immediate use or minus20 degC for later use
623 Whole genome sequencing
Mitochondrial whole genome amplification for both Eimeria species was initiated using sets of
mt-specific primers that generated overlapping polymerase chain reaction (PCR) fragments (Tables 61
and 62) PCR amplification was performed for all samples in a volume of 25 microl containing ~100 ng of
genomic DNA 1times PCR buffer 3 mM MgCl2 06 mM deoxyribonucleotide triphosphates (dNTPs) 500
nM of each primer and 4 U of Invitrogen Platinum Taq DNA Polymerase (Thermo Fisher Scientific
Toronto ON Canada) PCR reactions were performed on a Bio-Rad T100 PCR thermal cycler (Bio-Rad
Laboratories Singapore) using settings as described previously in the Materials amp Methods section of
Chapter 2 Table 61 details the specific anneal conditions used for the various primer pairs Genomic
DNA from either Eimeria maxima or Eimeria tenella acted as a positive control for the reaction
chemistry Gel electrophoresis purification and sequencing of the PCR amplification products were
8 Vet Med Labor GmbH Division of IDEXX Laboratories Ludwigsburg Germany
132
performed as described in Chapter 2 The resulting chromatograms were aligned and analyzed with
Geneious Ver 818 or later (Biomatters Limited Auckland New Zealand) and high quality consensus
sequences generated The completed mt genome sequences were annotated by comparison with
previously annotated mt genomes from other Eimeria species (eg Eimeria innocua - KR1082961) and
the annotated mt genomes deposited in GenBank
624 Phylogenetic analysis
To determine the phylogenetic affinities of the newly obtained sequences with sequences from
related apicomplexan taxa representative whole mt genome sequences from eimeriid coccidia were
downloaded from GenBank A complete mt genome sequence from an unnamed Choleoeimeria sp was
used to root the ingroup taxa several small genomic rearrangements in the Choleoeimeria sp sequence
required some rearrangement of the genome sequence to unambiguously align homologous regions across
the complete mt genomes
Whole genome sequences were aligned using MAFTT v7017 (Katoh et al 2002) executed from
within Geneious the resulting alignment was examined by eye to adjust start and stop codon positions in
aligned coding DNA sequence [CDS] regions (ie mt COI mitochondrial cytochrome c oxidase subunit
III gene [mtCOIII] mitochondrial cytochrome b gene [CytB]) Phylogenetic trees were generated using
Bayesian Inference (BI) using MrBayes Ver 326 (Huelsenbeck and Ronquist 2001) executed from
within Geneious The aligned complete mt genomes were partitioned into coding (ie CDS) and non-
coding regions so that region-specific models of nucleotide substitution could be applied Characters in
the non-coding region were analysed with the general time reversible (GTR) model (Tavareacute 1986) with
the following parameters nucmodel=4by4 nst=6 rates=invgamma (ie GTR+I+G) Characters in the
coding regions were analysed using the codon nucleotide model (ie lset nucmodel=codon rates=gamma
ngammacat=4) using metazoan mitochondrial translation (ie code=metmt)
133
All BI analyses were run for a chain length of 1000000 with tree sampling every 1000
following a burn-in of 100000 with default settings of 4 heated chains and heated chain temp of 02
63 RESULTS
The whole mt genome sequences of the single isolates of E furonis and E ictidea were
respectively 6165 base pairs (bp) (Figure 61 GenBank MF795598) and 6182 bp (Figure 62 GenBank
KT203399) Content and organization of both mt genomes consisted of three protein-coding genes (mt
COI mt COIII and CytB) interspersed with large and small subunit ribosomal DNA (rDNA) fragments
Details of the various CDS and rDNA fragments are summarized in Table 63 (for E furonis) and Table
64 (for E ictidea)
Pairwise alignment of the mt genome sequences from E furonis and E ictidea demonstrated a
relatively high pairwise sequence identity (946 333 single nucleotide differences [SNDs]) between
these two parasites The bulk of the SNDs (676 225333) were clustered within the three CDS regions
that encode CytB mt COI and mt COIII (see Figure 63 and Table 65) However the majority of these
SNDs (826 186225) were synonymous codon changes that resulted in no changes to the protein
products Only 41 SNDs were involved in 34 amino acid changes distributed among the three CDS
The 33 rDNA fragments comprised 2108 and 2109 bp respectively of the mt genomes of
E furonis and E ictidea Pairwise comparison of these rDNA fragments demonstrated high (986 30
SNDs) sequence identity between the two parasites The remaining 778 and 794 bp respectively of the
mt genomes of E furonis and E ictidea were intergenic stretches between the various rDNA and CDS
regions these intergenic regions were more variable that other regions of the genomes with 78 SNDs
(almost 10 sequence divergence) Additionally all indels were restricted to these variable intergenic
regions
The BI phylogeny generated from aligned complete mt genomes (Figure 64) supported the close
relationship between E furonis and E ictidea within a clade of Eimeria species that include the only three
sequences available for Eimeria spp of carnivores Eimeria mephitidis from the striped skunk (Mephitis
134
mephitis Family Mephitidae) was the sister taxon to the two Eimeria species of ferrets (Family
Mustelidae)
64 DISCUSSION
This work generated the first complete mt genomes from coccidia that infect domestic and black-
footed ferrets (Carnivora Mustelidae) Eimeria mephitidis from the striped skunk Mephitis mephitis
(Carnivora Mephitidae) is the only other Eimeria species from a carnivore for which a complete mt
genome has been reported
Comparatively few eimeriid coccidia only 26 Eimeria species have been described from
carnivores there are 14 named species from the mustelids four from the procyonids four from the ursids
three from the herpestids and one from the viverrids (Duszynski et al 2000) The majority of coccidia
that infect the digestive tract of carnivores belong to the family Sarcocystidae including monoxenous or
facultatively heteroxenous Cystoisospora species or heteroxenous parasites in the genera Sarcocystis
Hammondia and Neospora So far as is known none of the parasites in the Sarcocystidae possess typical
apicomplexan mt genomes with 3 complete CDS and many rDNA fragments (Ogedengbe 2015)
The mt genomes from the two Eimeria sp of mustelid origin demonstrate the same structural
organization (ie the order and number of CDS and rDNA fragments) and circular mapping as the mt
genomes from other Eimeria spp and other closely related eimeriid coccidia such as Isospora
Cyclospora and Lankesterella species Despite the ability of the eimeriid sequences to be mapped
circularly the physical form of Eimeria spp mt genomes may be a linear concatemer of multiple genome
copies as demonstrated for Eimeria tenella (Hikosaka et al 2011) As in the mt genomes of other
eimeriid coccidia (Ogedengbe et al 2013 2014) the CDS for mt COIII demonstrated the highest
sequence divergence between E furonis and E ictidea the mt COI CDS was somewhat more conserved
and CytB CDS demonstrated the fewest SNDs
As expected based on limited sequence divergence between E furonis and E ictidea a BI
phylogenetic analysis using aligned complete mt genome sequences generated a tree that placed these
135
two Eimeria species that infect mustelids within a well-supported monophyletic group The sister taxon
for these ferret parasites was the only other Eimeria species from carnivores for which a complete mt
genome is available E mephitidis which infects hosts belonging to a different family of carnivores
Eimeriid parasites that infect closely-related definitive hosts are commonly found in a single or limited
number of clades based on mitochondrial and nuclear genetic loci (Ogedengbe et al in press)
Sequencing of the mt genomes and at least one nuclear genetic locus (ie nu 18S rDNA) from additional
Eimeria species infecting carnivores will be required to determine if all carnivore-specific Eimeria
species share a common ancestor
136
Table 61 PCR primer pairs and resulting fragments used for sequencing the mitochondrial genome sequence of an isolate of Eimeria furonis
originating from a fecal sample from a domestic ferret (Mustela putorius furo)
Fragment Primer names Primer sequences (5ʹ-3ʹ) Size (bp) Anneal Temp References
1 WG-MT_4140F AGAAAACCTAAAATCATCATGT 1000 52 Ogedengbe et al (2015)
Eim_CO3_799R AAGTGAGTTCGCATGTTTAC Ogedengbe et al (2015)
2 Eim_COI_19F ACTGCYGCAAACCATAARGAA 1700 60 Present study
Api_LSUG_UNI_R AGATAGGGAACAAACTGYCTCAA Present study
3 WG_MT_5416F GGTCCAGATAAGCGATCTCATG 3400 53 Ogedengbe et al (2013)
Eim_COI_1436R CACATTGTGTTCARATAAGTTA Present study
4 WG-MT_6219F GCATCCATCTACAGCTGCGG 500 55 Ogedengbe et al (2013)
WG-MT_344R GTAGGAATCTRAATTCCCAACC Ogedengbe et al (2013)
5 Api_LSUE UNI_F AGGTGCTCAGGGTCTTACCG 500 55 Present study
WG_MT_63R CTGGTATGGATGGATAACACT Ogedengbe et al (2015)
6 Lank_COB-30F CCAGGCCAACTGAACTCGTT 1300 55 Present study
q_Eim_COI_221R GGCATAACTACAAAGAARATCATA Present study
7 Cocci_MT_WG_F TACACCTAGCCAACACGAT 1600 55 Ogedengbe et al (2014)
q_Eim_COI_221R GGCATAACTACAAAGAARATCATA Present study
137
Table 62 PCR primer pairs and resulting fragments used for sequencing the mitochondrial genome sequence of an isolate of Eimeria ictidea
originating from a fecal sample from a black-footed ferret (Mustela nigripes)
Fragment Primer names Primer sequences (5ʹ-3ʹ) Size (bp) Anneal Temp References
1 WG_MT_63R CTGGTATGGATGGATAACACT 2180 52 Ogedengbe et al (2015)
WG-MT_4140F AGAAAACCTAAAATCATCATGT Ogedengbe et al (2015)
2 Cocci_MT_WG_F TACACCTAGCCAACACGAT 1551 58 Ogedengbe et al (2014)
q_Eim_COI_221R GGCATAACTACAAAGAARATCATA Present study
3 WG-MT_3658F CTGGCGAGAAGGGAAGTGTG 1329 55 Ogedengbe et al (2013)
Eim_CO3_799R AAGTGAGTTCGCATGTTTAC Ogedengbe et al (2015)
4 Lank_COB-30F CCAGGCCAACTGAACTCGTT 3899 57 Present study
WG_MT_4072R GGTTGTTTCCATCTCGACTC Ogedengbe et al (2013)
138
Table 63 Coding regions within the mitochondrial genome of the eimeriid parasite Eimeria furonis from a domestic ferret (Mustela putorius faro)
139
Table 64 Coding regions of the mitochondrial genome of the eimeriid parasite Eimeria ictidea originating from a black-footed ferret (Mustela
nigripes)
Table 64 Features associated with the protein-coding regions of the mitochondrial genome of the eimeriid parasite Eimeria cf ictidea originating from a black-footed ferret (Mustela nigripes )
Protein coding regions (CDS) Sequence size (bp) Start position (bp) Stop position (bp) Direction Translation start codon Translation stop codon
Cytochrome c oxidase subunit I (COI) 1443 1343 2785 Forward ATG TAA
Cytochrome c oxidase subunit III (COIII) 756 4333 5088 Forward TTA TAA
Cytochrome b (CytB) 1080 226 1305 Forward ATG TAA
Ribosomal DNA fragments (rDNA) Product
SSUrRNA 46 17 62 forward RNA9 SSU8
SSUrRNA 77 104 180 forward SSUA SSU4 (partial)
SSUrRNA 35 181 215 forward RNA23t
LSUrRNA 20 2790 2809 forward RNA20 (partial) LSU
LSUrRNA 112 2835 2946 forward LSUF LSU11
LSUrRNA 106 2947 3052 forward LSUG LSU12
LSUrRNA 25 3087 3063 reverse LSU
SSUrRNA 39 3129 3167 forward RNA14 SSU1
LSUrRNA 16 3188 3173 reverse LSUC LSU4
SSUrRNA 33 3211 3243 forward SSU
SSUrRNA 61 3386 3326 reverse SSUF SSU12
LSUrRNA 74 3387 3460 forward RNA10 LSU13 (partial)
LSUrRNA 49 3476 3524 forward RNA11 LSU5
SSUrRNA 65 3533 3597 forward SSUD SSU10
SSUrRNA 40 3631 3670 forward RNA17 SSU3
SSUrRNA 30 3677 3706 forward RNA15 SSU
LSUrRNA 30 3711 3740 forward RNA13 LSU10
LSUrRNA 56 3749 3804 forward RNA6 LSU15
LSUrRNA 79 3899 3821 reverse LSUD LSU8
LSUrRNA 24 3931 3908 reverse RNA16 (partial)
SSUrRNA 92 4036 3945 reverse RNA8 SSU5
LSUrRNA 66 4054 4119 forward RNA2 LSU2
LSUrRNA 177 4316 4140 reverse LSUA LSU1
SSUrRNA 30 5125 5096 reverse RNA19 SSU7
LSUrRNA 88 5154 5241 forward RNA1 LSU6
LSUrRNA 26 5342 5317 reverse LSUB LSU3
LSUrRNA 73 5447 5375 reverse RNA3 LSU7
LSUrRNA 20 5479 5460 reverse RNA18 LSU14
SSUrRNA 119 5615 5497 reverse SSUB SSU6
LSUrRNA 80 5699 5620 reverse RNA7
LSUrRNA 188 5897 5710 reverse LSUE LSU9
SSUrRNA 33 6033 6001 reverse SSUE SSU11 (partial)
SSUrRNA 99 6156 6058 reverse RNA5SSU9
140
Table 65 Pairwise comparison of coding DNA and concatenated rDNA fragment sequences between the
mitochondrial genomes of Eimeria furonis originating from a domestic ferret (Mustela putorius furo) and
Eimeria ictidea originating from a black-footed ferret (Mustela nigripes)
Total length
(nucleotides)
Nucleotide
identity
Total amino
acids
Amino acid
identity
COI CDS 1443 934 (95) 480 975 (12)
COIII CDS 756 899 (76) 251 932 (17)
CytB CDS 1080 950 (54) 359 986 (5)
rDNA fragments 2109 985 (32) na na
Legend Numbers in brackets indicate the number of single nucleotide differences na = not
applicable
141
Figure 61 Circular mapping and organization of the mitochondrial genome content of Eimeria
furonis showing three protein-coding genes (COI COIII and CytB) interspersed with large and
small subunit rRNA fragments
142
Figure 62 Circular mapping and organization of the mitochondrial genome content of Eimeria
ictidea showing three protein-coding genes (COI COIII and CytB) interspersed with large and
small subunit rRNA fragments
143
Figure 63 Pairwise comparison of coding DNA and concatenated rDNA fragment sequences of
the mitochondrial genomes of Eimeria furonis and Eimeria ictidea Legend Yellow bands
correspond to coding DNA fragments Red arrows correspond to rDNA fragments Arrows of both
colours indicate the forward or reverse direction of these segments
144
Figure 64 Phylogenetic relationships of coccidia (Eimeria furonis and Eimeria ictidea) from
domestic (Mustela putorius furo) and black-footed ferrets (Mustela nigripes) based on the complete
mitochondrial genome sequences these Eimeria species and a selection of related apicomplexan
parasites
145
CHAPTER 7 CONCLUSIONS AND FUTURE DIRECTIONS
The primary objective of this project was to better characterize the enteric coccidia of the
endangered black-footed ferret (BFF) in order to set the stage for improved disease prevention
management and treatment To the authorrsquos knowledge this work was the first attempt to isolate and
perform molecular characterization of the coccidial species endemic in the black-footed ferret population
and to characterize the natural history of the disease in this host
Two Eimeria species Eimeria ictidea and Eimeria furonis have been described from BFF and
domestic ferrets A single Eimeria species morphologically resembling E ictidea of the domestic ferret
and referred to in this work as E ictidea was identified from all historic and active cases of enteric
coccidiosis in the Toronto Zoo BFF population The same species was identified in both BFF family
groups and single-housed adults that shed coccidia during the study period (2014-2016) as well as from
historic necropsy samples of juvenile and adult Toronto Zoo BFF from 1999-2014 Furthermore this
same species was identified from both family groups and single-housed adults shedding coccidia at an
additional BFF Species Survival Plan institution the Louisville Zoo in 2016 These findings are contrary
to previous published reports that indicated multiple coccidia species were affecting captive and wild
BFF In order to determine whether these additional parasites continue to exist within the present-day
BFF population and their impact on this species further examination of coccidia-positive fecal samples
from captive and wild BFF populations is recommended Moreover determination of which parasite
species impact BFF morbidity and mortality would allow for development of targeted therapies for
disease management
Coccidia-positive fecal samples and necropsy samples from domestic ferrets were collected from
multiple diagnostic laboratories in Canada and Europe for comparison with results from BFF Eimeria
furonis and Isospora (=Cystoisospora) laidlawi were identified from fecal parasitology reports from
2008-2015 and in fecal samples obtained prospectively from 2014-2016 from domestic ferrets submitted
to Canadian and European laboratories E furonis was also identified in necropsy samples from 2010 and
146
2017 from two Canadian diagnostic laboratories No samples containing E ictidea were submitted to the
Canadian laboratories however samples containing E ictidea were identified twice by the European
diagnostic laboratory with one sample in 2011 and a second in 2013 Consequently it appears that E
ictidea is rarely identified from domestic ferrets Since no samples of E ictidea were acquired from
domestic ferrets during the study period it remains undetermined whether E ictidea from domestic
ferrets and E ictidea of BFF are conspecific Future molecular characterization of E ictidea isolated from
various mustelid host species would allow not only for determination of whether the parasites are
conspecific but would also provide insight into the potential for cross-transmission of parasites between
related mustelid hosts
The identifications provided by diagnostic laboratories of the specific parasite species present in
the domestic ferret fecal samples showed poor agreement with their identifications based on genotyping
obtained in this study In the authorrsquos opinion molecular techniques are essential tools for determining
the specific coccidial species responsible for individual and group outbreaks of coccidiosis and for
further understanding of the eimeriid host-parasite relationships To this end the nu 18S rDNA region and
whole mitochondrial (mt) genomes of E ictidea and E furonis were sequenced these mt whole genome
sequences are the first for the Eimeria species of mustelids to be entered into GenBank These sequences
may provide suitable targets for the development of highly discriminatory PCR-based methods of
identification that could be applied to fecal tissue or even formalin-fixed paraffin-embedded samples
(see Chapters 2 and 3) Methodological improvements such as less expensive next generation sequencing
methods that could be applied to diagnostics will depend on the availability of high quality reference
sequences such as generated herein Furthermore evaluation of the evolution of the eimeriid coccidia
using phylogenetic analysis based on additional whole mitochondrial genome sequences may allow for
more accurate determinations of relationships between parasite species and timing of their divergence
from common ancestors
One of the goals of this research was to investigate the possibility of using the domestic ferret as
an experimental model to study enteric coccidiosis in the BFF Experimental work cannot be carried out
147
in the BFF due to its endangered status We showed that the domestic ferret is susceptible to infection
with Eimeria ictidea originating from BFF Infected domestic ferrets showed similar clinical signs and
pathologic lesions to BFF strengthening the possibility of their serving as a suitable model Future
investigations would include studies of the domestic ferretrsquos immune response to enteric coccidial
infection in vivo drug trials including pharmacokinetic pharmacodynamic and efficacy studies of
anticoccidial medications investigations into patterns of drug resistance in coccidial species of ferrets
and oral vaccine development
Ultimately the goal of research on coccidiosis in BFF may be the development of an autogenous
vaccine to improve the survival of ferret kits and reduce coccidiosis-related morbidity and mortality in
BFF captive breeding programs It may be possible to use the domestic ferret to select for E ictidea that
are more highly adapted to this host perhaps with a commensurate loss of virulence to its original host If
this were to be the case domestic ferret-derived coccidia could be used in BFF as vaccine organisms
Alternately methods such as use of a bioshuttle (live vaccination with coccidia followed by anticoccidial
treatment to limit pathogenicity) might be useful for generating long-lasting immunological protection in
BFF against E ictidea Improving immunity through vaccination could assist in reducing coccidial
shedding and disease in BFF associated with stressful life events such a breeding weaning and transfer
between institutions There is minimal available data on the significance of coccidiosis in wild
populations of BFF and limited means of disease surveillance post release however it is logical to
assume that the stresses associated with release to the wild might result in clinical disease as happens with
stressed BFF in captivity Reducing morbidity and mortality associated with coccidiosis in BFF could
result in increased numbers of ferrets being released to the wild and higher survival of released ferrets
increasing the number of BFF in the wild supports the goals of the conservation initiative for the black-
footed ferret and will support the longer-term survival and recovery of this species
148
REFERENCES
Abe N Tanoue T Ohta G Iseki M (2008) First record of Eimeria furonis infection in a ferret Japan with
notes on the usefulness of partial small subunit ribosomal RNA gene sequencing analysis for
discriminating among Eimeria species Parasitol Res 103967ndash70 doi 101007s00436-008-1037-x
Adl SM Simpson AGB Farmer MA et al (2005) The new higher level classification of eukaryotes with
emphasis on the taxonomy of protists J Eukaryot Microbiol 52399ndash451 doi 101111j1550-
7408200500053x
Andrews JM (1926) Coccidiosis in mammals Am J Hyg 6784ndash798
Augustine PC Danforth HD (1986) A study of the dynamics of the invasion of immunized birds by
Eimeria sporozoites Avian Dis 30347ndash351
Barta JR Schrenzel MD Carreno R Rideout BA (2005) The genus Atoxoplasma (Garnham 1950) as a
junior objective synonym of the genus Isospora (Schneider 1881) species infecting birds and
resurrection of Cystoisospora (Frenkel 1977) as the correct genus for Isospora species infecting
mammals J Parasitol 91726ndash727 doi 101645GE-33411
Bell JA (1994) Parasites of Domesticated Pet Ferrets Compend Contin Educ Pract Vet 16617ndash620
Black-footed Ferret Recovery Implementation Team (2011) Captive Breeding
httpwwwblackfootedferretorgcaptive-breeding Accessed 8 Jan 2014
Blankenship-Paris TL Chang J Bagnell CR (1993) Enteric coccidiosis in a ferret Lab Anim Sci 43361ndash
363
Bronson E Bush M Viner T et al (2007) Mortality of captive black-footed ferrets (Mustela nigripes) at
Smithsonianrsquos National Zoological Park 1989 ndash 2004 J Zoo Wildl Med 38169ndash176
149
CAPC (2013) Current Advice on Parasite Control Intestinal Parasites - Coccidia
httpwwwcapcvetorgcapc-recommendationscoccidia Accessed 6 Jun 2015
Carpenter JW Hillman CN (1979) Husbandry reproduction and veterinary care of captive ferrets In
1978 Proceedings of the Annual Meeting of the American Association of Zoo Veterinarians
Washington DC pp 36ndash47
Catchpole J Norton CC Gregory MW (1993) Immunisation of lambs against coccidosis Vet Rec
13256ndash59
Cavalier-Smith T (2014) Gregarine site-heterogeneous 18S rDNA trees revision of gregarine higher
classification and the evolutionary diversification of Sporozoa Eur J Protistol 50472ndash495 doi
101016jejop201407002
Chappuis G (1998) Neonatal immunity and immunisation in early age lessons from veterinary medicine
Vaccine 161468ndash1472
Cox FE (1994) The evolutionary expansion of the Sporozoa Int J Parasitol 241301ndash1316
Davis CL Chow TL Gorham JR (1953) Hepatic coccidiosis in mink Vet Med 48371ndash375
De Vos AJ (1970) Studies on the host range of ltigtEimeria chinchillaeltIgt de Vos and Van Der
Westhuizen 1968 Onderstepoort J Vet Res 3729ndash36
Dirikolu L Yohn R Garrett EF et al (2009) Detection quantifications and pharmacokinetics of
toltrazuril sulfone (Ponazuril) in cattle J Vet Pharmacol Ther 32280ndash288
Dryden MW Payne PA Ridley R Smith V (2005) Comparison of common fecal flotation techniques for
the recovery of parasite eggs and oocysts Vet Ther 615ndash28
Duszynski D Wilber PG (1997) A Guideline for the Preparation of Species Descriptions in the
Eimeriidae J Parasitol 83333ndash336
150
Duszynski DW Couch L Upton SJ (2000) The Coccidia of the World
httpbiologyunmeducoccidiacarniv2html Accessed 1 Sep 2017
El-Sherry S Ogedengbe ME Hafeez MA Barta JR (2013) Divergent nuclear 18S rDNA paralogs in a
turkey coccidium Eimeria meleagrimitis complicate molecular systematics and identification Int J
Parasitol 43679ndash685 doi 101016jijpara201303005
Evans HE An NQ (2014) Anatomy of the ferret In Fox JG Marini RP (eds) Biology and Diseases of the
Ferret 3rd edn Wiley Blackwell pp 23ndash67
Fayer R (1980) Epidemiology of Protozoan Infections The Coccidia Vet Parasitol 675ndash103
Foreyt WJ Todd AC Hartsough GR (1977) Anticoccidial Activity of Eight Compounds in Domestic
Mink AJVR 38391ndash394
Fox JG (2014) Normal Clinical and Biological Parameters In Fox JG Marini RP (eds) Biology and
Diseases of the Ferret 3rd edn John Wiley amp Sons Inc pp 157ndash185
Frenkel JK (1977) Besnoitia wallacei of Cats and Rodents With a Reclassification of Other Cyst-
Forming Isosporoid Coccidia J Parasitol 63611ndash628
Grafner G Graubmann HD Dobbriner W (1967) Leberkokzidiose beim Nerz (Lutreola vison Schreb)
hervorgerufen durch eine neue Kokzidienart Eimeria hiepei n sp Monatshefte fur
Veterinearmedizin 22696ndash700
Haberkorn A (1971) Zur Wirtsspezifitat yon Eimeria contorta nsp (Sporozoa Eimeriidae) Z
Parasitenkd 37303ndash314
Hall MC Wigdor M (1918) Canine coccidiosis with a note regarding other protozoan parasites from the
dog J Am Vet Med Assn 5364ndash76
Hikosaka K Nakai Y Watanabe YI et al (2011) Concatenated mitochondrial DNA of the coccidian
151
parasite Eimeria tenella Mitochondrion 11273ndash278
Hillyer E V (1992) Gastrointestinal diseases of ferrets (Mustela putorius furo) J Small Anim Med 244ndash
45
Hoare CA (1927) On the coccidia of the ferret Ann Trop Med Parasitol 21313ndash320
Hoare CA (1935a) A histopathological reaction of a special type on the part of the intestinal villi in ferret
coccidiosis Trans R Soc Trop Med Hyg 292
Hoare CA (1935b) The endogenous development of the coccidia of the ferret and the histopathological
reaction of the infected intestinal villi Ann Trop Med Parasitol 29111ndash122
Hoefer HL Fox JG Bell JA (2012) Gastrointestinal Diseases In Quesenberry KE Carpenter JW (eds)
Ferrets Rabbits and Rodents Clinical Medicine and Surgery 3rd edn Elsevier Saunders St Louis
Missouri pp 27ndash45
Huelsenbeck JP Ronquist F (2001) MrBayes Bayesian inference of phylogenetic trees Bioinformatics
17754ndash755 doi 101093bioinformatics178754
Jolley WR Kingston N Williams ES Lynn C (1994) Coccidia Giardia sp and a Physalopteran
Nematode Parasite from Black-footed Ferrets (Mustela nigripes) in Wyoming J Helminthol Soc
Washingt 6189ndash94
Katoh K Misawa K Kuma K Miyata T (2002) MAFFT a novel method for rapid multiple sequence
alignment based on fast Fourier transform Nucleic Acids Res 303059ndash3066 doi
101093nargkf436
Kaye SW Ossiboff RJ Noonan B et al (2015) Biliary coccidiosis associated with immunosuppressive
treatment of pure red cell aplasia in an adult ferret (Mustela putoris furo) J Exot Pet Med doi
101053jjepm201504012
152
Koepfli KP Deere K Slater GJ et al (2008) Multigene phylogeny of the Mustelidae resolving
relationships tempo and biogeographic history of a mammalian adaptive radiation BMC Biol 610
Lariviegravere S Jennings AP (2009) Family Mustelidae (weasels and relatives) In Wilson DE Mittermeier
RA (eds) Handbook of the Mammals of the World Volume 1 Carnivores Lynx Edicions
Barcelona pp 564ndash656
Levine ND Ivens V (1970) The coccidian parasites (Protozoa Sporozoa) of ruminants
Li X Pang J Fox JG (1996) Coinfection with intracellular Desulfovibrio species and coccidia in ferrets
with proliferative bowel disease Lab Anim Sci 46569ndash571
Litster AL Nichols J Hall K et al (2014) Use of ponazuril paste to treat coccidiosis in shelter-housed
cats and dogs Vet Parasitol 202319ndash325 doi 101016jvetpar201403003
Matsubayashi M Takami K Abe N et al (2005) Molecular characterization of crane coccidia Eimeria
gruis and E reichenowi found in feces of migratory cranes Parasitol Res 9780ndash83
Meeusen ENT Walker J Peters A et al (2007) Current status of veterinary vaccines Clin Microbiol Rev
20489ndash510 doi 101128CMR00005-07
Mehlhorn H Aspock H (eds) (2008) Coccidial Drugs In Encyclopedia of Parasitology 3rd edn Springer
Berlin Heidelberg New York pp 269ndash286
Morehouse NF (1938) The Reaction of the Immune Intestinal Epithelium of the Rat to Reingection wiht
Eimeria nieschulzi J Parasitol 24311ndash317
Nukerbaeva KK Svanbaev SK (1973) Coccidia of fur bearing mammals in Kazakhstan Vestn Selrsquoskokh
Nauk Kazakh 1250ndash54
Ogedengbe JD Hanner RH Barta JR (2011) DNA barcoding identifies Eimeria species and contributes to
the phylogenetics of coccidian parasites (Eimeriorina Apicomplexa Alveolata) Int J Parasitol
153
41843ndash850
Ogedengbe ME (2015) DNA Barcoding of Apicomplexa Mitochondrial Evolution across the Phylum
University of Guelph
Ogedengbe ME El-Sherry S Ogedengbe JD et al Whatrsquos in a name Phylogenies based on combined
mitochondrial and nuclear sequences conflict with morphologically defined genera in the eimeriid
coccidia (Apicomplexa)
Ogedengbe ME El-Sherry S Whale J Barta JR (2014) Complete mitochondrial genome sequences from
five Eimeria species (Apicomplexa Coccidia Eimeriidae) infecting domestic turkeys Parasit
Vectors 7335 doi 1011861756-3305-7-335
Ogedengbe ME Hafeez MA Barta JR (2013) Sequencing the complete mitochondrial genome of Eimeria
mitis strain USDA 50 (Apicomplexa Eimeriidae) suggests conserved start positions for mtCOI- and
mtCOIII-coding regions Parasitol Res 1124129ndash4136 doi 101007s00436-013-3604-z
Pantchev N Gassmann D Globokar-Vrhovec M (2011) Increasing numbers of Giardia (but not
coccidian) infections in ferrets 2002 to 2010 Vet Rec 168519 doi 101136vrd2962
Patterson M Fox JG (2007) Parasites of Ferrets In Baker DG (ed) Flynnrsquos Parasites of Laboratory
Animals 2nd edn Wiley Blackwell pp 501ndash508
Patterson MM Fox JG Eberhard ML (2014) Parasitic Diseases In Fox JG Marini RP (eds) Biology and
Diseases of the Ferret 3rd edn Wiley Blackwell pp 553ndash572
Powers L V (2009) Bacterial and parasitic diseases of ferrets Vet Clin North Am - Exot Anim Pract
12531ndash561 doi 101016jcvex200906001
Prado ME Ryman JT Boileau MJ et al (2011) Pharmacokinetics of ponazuril after oral administration to
healthy llamas (Llama glama) Am J Vet Res 721386ndash9
154
Ruiz A Muntildeoz MC Molina JM et al (2013) Primary infection of goats with Eimeria ninakohlyakimovae
does not provide protective immunity against high challenge infections Small Rumin Res 113258ndash
266 doi 101016jsmallrumres201301006
Ryley J Meade R Hazelhurst J Robinson T (1976) Methods in coccidiosis research separation of
oocysts from faeces Parasitology 73311ndash326
Santymire R Branvold-Faber H Marinari PE (2014) Recovery of the Black-Footed Ferret In Fox JG
Marini RP (eds) Biology and Diseases of the Ferret 3rd edn Wiley Blackwell pp 219ndash231
Shi MQ Huther S Burkhardt E Zahner H (2000) Immunity in rats against Eimeria separata oocyst
excretion effects on endogenous stages and local tissue response after primary and challenge
infections Parasitol Res 86891ndash898
Sledge DG Bolin SR Lim A et al (2011) Outbreaks of severe enteric disease associated with Eimeria
furonis infection in ferrets (Mustela putorius furo) of 3 densely populated groups J Am Vet Med
Assoc 2391584ndash1588 doi 102460javma239121584
Smith NC Wallach M Petracca M et al (1994) Maternal transfer of antibodies induced by infection with
Eimeria maxima partially protects chickens against challenge with Eimeria tenella Parasitology
109551ndash557
Svanbaev SK (1956) Materials on the fauna of coccidia of wild mammals in western Kazakhstan Tr
Instituta Zool Akad Nauk Kazachskoi SSR 5180ndash191
Tavareacute S (1986) Some probabilistic and statistical problems in the analysis of DNA sequences Am Math
Soc Lect Math Life Sci 1757ndash86 doi citeulike-article-id4801403
Tenter AM Barta JR Beveridge I et al (2002) The conceptual basis for a new classification of the
coccidia Int J Parasitol 32595ndash616
155
Upton SJ (2000) Suborder Eimeriorina Leacuteger 1911 In Lee JJ Leedale GF Bradbury P (eds) An
Illustrated Guide to the Protozoa vol 1 2nd edn Allen Press Lawrence Kansas pp 318ndash339
USFWS BFF Recovery Program (2017) Black-footed Ferret Managed Care Operations Manual
(BFFMCOM)
Vermeulen AN (2005) Vaccination against coccidial parasites The method of choice In Proceeding of
the 9th International Coccidiosis Conference
Williams BH Chimes MJ Gardiner CH (1996) Biliary coccidiosis in a ferret (Mustela putorius furo) Vet
Pathol 33437ndash439 doi 101177030098589603300412
Williams ES Mills K Kwiatkowski DR et al (1994) Plague in a Black-footed (Mustela nigripes)
presence J Wildl Dis 30581ndash585
Williams ES Thome ET Appel MJG Belitsky DW (1988) Canine Distemper in Black-Footed (Mustela
nigripes) from Wyoming J Wildl Dis 24385ndash398
Wozencraft WC (2005) Order Carnivora In Wilson DE Reeder DM (eds) Mammal Species of the
World A Taxonomic and Geographic Reference 3rd edn Johnrsquos Hopkins University Press
Baltimore Maryland p 2142
Yi-Fan C Le Y Yin D et al (2012) Emendation of 2 Isospora Species (Apicomplexa Eimeriidae)
Infecting the Steppe Polecat Mustela eversmanii Lesson 1827 in China to the Genus
Cystoisospora (Apicomplexa Sarcocystidae) Comp Parasitol 79147ndash152 doi 10165445311
Yu L Peng D Liu J et al (2011) On the phylogeny of Mustelidae subfamilies analysis of seventeen
nuclear non-coding loci and mitochondrial complete genomes BMC Evol Biol 1192 doi
1011861471-2148-11-92
Zou M Guo G Zhao Y Zhang Q (2014) Detection quantifications and pharmacokinetics of ponazuril in
156
healthy swine J Vet Pharmacol Ther 37598ndash602 doi 101111jvp12126
157
APPENDICES
158
Appendix 1 Shedding of oocysts of Eimeria ictidea in black-footed ferret (Mustela nigripes) dam and kit
family groups from 2014-2016
Collection Year 2014 2014 2014 2014 2014 2015 2016
Age of kits (days) Poppy Bumblefoot Calico Aubrey Ruckus Fiddlesticks Guanella^
29 - - - 0 - - -
30 - 0 - 0 - - -
31 - 0 - 0 - - -
32 - 0 - 0 - - -
33 - 0 - 0 - - -
34 0 0 - 0 0 - -
35 0 0 - 0 0 0 -
36 0 0 - 0 0 0 -
37 - 0 - 0 - 0 -
38 - 0 - 0 - 0 -
39 - 0 - 0 - 0 -
40 0 0 - 0 0 0 -
41 - 0 - 0 - 0 -
42 0 0 - 0 0 - -
43 0 0 - 0 0 0 -
44 0 0 - - 0 0 -
45 0 0 - 0 0 - -
46 0 0 - 0 0 0 -
47 0 0 - 0 0 0 -
48 0 lt14 - 0 0 0 -
49 0 0 - 0 0 0 -
50 0 0 - - 0 0 -
51 0 0 - 0 0 0 -
52 0 0 - 0 0 0 -
53 0 0 - 0 0 0 +
54 0 0 - 0 0 0 +
55 0 0 - 0 0 0 3717146
56 0 0 - 0 - 0 1084436
57 0 0 - 0 - 0 64133
58 0 324 - 0 0 0 20654
59 0 0 - 0 0 0 0
60 0 446688 - 0 0 0 0
61 0 934828 - 0 0 0 41111
62 0 530469 - 0 0 0
63 0 1617131 - 0 0 lt14
64 0 8396357 - 114 0 0
65 0 234876 - 5368 0 lt14
66 0 374625 - 9455 0 0
67 lt14 2311575 - 39579 0 -
68 0 603563 - 125051 0 0
69 - 4103036 - 27067 0 lt14
70 2470327 7759974 - 0 0 0
71 1073085 - 1784 0 0
72 3430966 - 0 0 0
73 4459536 - 7326 0 0
74 4842212 - 2 0 0
75 3627307 - 0 0 0
76 5801885 - 0 0 0
77 5994 0 0 0 0
78 589835 0 0 0
Numbers of Oocysts Shed (oocysts per gram of feces)
Dam Identity
159
Appendix 1 continued Shedding of oocysts of Eimeria ictidea in black-footed ferret (Mustela nigripes)
dam and kit family groups from 2014-2016
Collection Year 2014 2014 2014 2014 2014 2015 2016
Age of kits (days) Poppy Bumblefoot Calico Aubrey Ruckus Fiddlesticks Guanella^
79 24815 0 0 0
80 144016 0 0 0
81 97862 0 0 0
82 0 0 0
83 0 0 0
84 0 0 0
85 0 0 0
86 0 0 0
87 0 0 0
88 0 0 0
89 0 0 0
90 0 0 0
91 0 0 0
92 0 - -
93 0 - 0
94 0 - 0
95 0 0 -
96 0 - 0
97 0 0 -
98 0 0 0
99 0 0 0
100 0 0 0
101 0 0 0
102 0 0 0
103 0 0 0
104 0 0 0
105 0 0 -
106 0 0 0
107 0 0 0
108 - 0 0
109 0 0 -
110 0 0 0
111 0 0 -
112 0 0 0
113 0 0 -
114 0 0 -
115 0 0 -
116 0 0 -
117 0 -
118 0 -
119 0 0
120 0 0
121 0 0
122 0 -
Numbers of Oocysts Shed (oocysts per gram of feces)
Dam Identity
160
Appendix 1 continued Shedding of oocysts of Eimeria ictidea in black-footed ferret (Mustela nigripes)
dam and kit family groups from 2014-2016
Collection Year 2014 2014 2014 2014 2014 2015 2016
Age of kits (days) Poppy Bumblefoot Calico Aubrey Ruckus Fiddlesticks Guanella^
123 0 0
124 0 -
125 0 -
126 0 0
127 0 0
128 0 2843
129 0 0
130 0 -
131 0 0
132 0 0
133 0 0
134 0 0
135 0 0
136 0 0
137 0
138 0
139 0
140 0
141 0
142 0
143 0
144 0
145 0
146 -
147 0
148 0
149 0
150 0
Legend lt 14 = oocyst positive samples with less than 14 oocyst per gram of feces - = no sample recorded for this date
underline = last sampling date + = coccidia present but OPG count not performed = Toronto Zoo ferret ^ = Louisville Zoo
ferret thick outer border = days treatment was received
Numbers of Oocysts Shed (oocysts per gram of feces)
Dam Identity
161
Appendix 2a Hematology values for domestic ferrets (Mustela putorius furo) from 49-51 days of age
prior to experimental inoculation
Test Reference
Intervala
Reference
Intervalb
101 102 103 104 105 201 202 203 204 205
Age (days) adult 70 50 50 50 50 50 48 48 48 50 48
WBC (x 109L) 27-112 53-120 86 64 77 97 76 48 8 72 104 76
RBC (x 1012
L) 50-108 55ndash74 66 58 45 5 48 47 67 5 5 42
Hb (gL) 87-177 104ndash136 121 106 90 58 94 91 122 96 98 80
HCT (LL) 04 - 051 029ndash037 037 033 027 030 029 027 038 030 030 024
MCV (fL) 44-52 478ndash548 55 56 60 60 60 58 57 59 61 58
MCH (pg) 15-18 175ndash191 18 18 20 12 20 20 18 19 20 18
MCHC (gL) 325-362 347ndash370 331 327 328 196 325 337 321 327 324 331
RDW () 12-16 - 134 127 139 139 133 131 122 136 131 127
Platelets (x 109L) 54-695 629ndash775 777 329 529 480 640 402 534 336 527 363
MPV (fL) 5-10 - 78 78 96 75 66 74 76 81 74 82
TS Protein (gL) 49-76 - 54 51 - - - - - - - -
Seg Neuts (x 109L) 1-8 15ndash48 292 141 216 281 205 187 152 151 354 251
Lymphocytes (x 109L) 1-63 28ndash63 525 416 516 64 509 254 608 468 645 456
Monocytes (x 109L) 0-09 01ndash05 026 07 031 039 038 034 024 094 031 038
Eosinophils (x 109L) 0-13 01ndash06 009 013 008 01 008 005 016 007 010 015
Basophils (x 109L) 0-02 0 009 0 0 0 0 0 0 0 0 0
Polychromasia 2-5 - 5-10 2-5 10-15 10-15 10-15 10-15 2-5 10-15 10-15 5-10
Anisocytosis Occ 1+ 1+ 1+ 1+ 1+ 1+ 1+
HJ bodies rare rare rare rare rare
crenation Occ
poikilocytosis Occ Occ Occ
shift platelets Occ Occ
hemolysis Neg Neg Neg Neg Neg Neg Neg Neg Neg Neg Neg
lipemia mild mild mild mild mild Neg Neg Neg mild mild
Ferret Identity
Legend - = reference value unavailable bolded = outside reference range a = adult ferret reference ranges from Animal Health Laboratory
(University of Guelph Guelph Canada) b
= reference ranges for 10-week old ferrets (Fox JG 2014) Neg = negative Occ = occasional
162
Appendix 2b Serum biochemistry values for domestic ferrets (Mustela putorius furo) from 49-51 days of
age prior to experimental inoculation
Test Reference
Intervala
Reference
Intervalb
101 102 103 104 105 201 202 203 204 205
Age (days) adult 70 50 50 50 50 50 48 48 48 50 48
Calcium (mmolL) 185-242 253-302 239 233 241 244 24 221 242 234 253 242
Phosphorus (mmolL) 112-256 278-475 239 267 29 264 31 218 286 284 315 303
Magnesium (mmolL) 08-139 - 08 08 07 08 08 06 08 07 08 08
Sodium (mmolL) 147-159 146-154 149 149 149 149 148 144 153 148 152 152
Potassium (mmolL) 37-57 47-83 44 46 42 46 48 45 47 44 47 46
Chloride (mmolL) 111-129 115-121 110 112 115 113 113 110 119 117 117 120
Carbon dioxide (mmolL) 17-29 13-27 18 18 18 17 17 16 15 15 19 17
Anion gap (mmolL) 6 - 23 - 25 24 20 24 23 23 24 20 21 20
NaK ratio - - 34 32 35 32 31 32 33 34 32 33
Total protein (gL) 51-75 44-56 49 46 44 47 44 41 52 45 49 43
Albumin (gL) 24-40 26-32 29 26 28 28 28 25 28 27 29 25
Globulin (gL) 19-41 17-24 20 20 16 19 16 16 24 18 20 18
AG ratio 053-167 13ndash12 145 130 175 147 175 156 117 15 145 139
Blood urea nitrogen (mmolL) 45-153 71-139 83 89 107 11 136 73 119 105 118 135
Creatinine (umolL) 8-67 53-124 50 58 68 59 50 7 68 41 62 92
Glucose (mmolL) 32-91 64-138 47 54 54 42 55 53 57 56 52 59
Cholesterol (mmolL) 294-894 619-860 413 386 375 393 344 349 227 357 373 346
Total bilirubin (umolL) 2 to 7 - 1 L 0 L 0 0 0 1 0 0 0 0
Conjugated biilrubin (umolL) - 0-10 1 0 0 1 1 1 0 0 1 0
Free bilirubin (umolL) 0-2 0-15 0 0 0 0 0 0 0 0 0 0
ALKP (UL) 13-237 117ndash277 180 169 172 215 175 168 241 184 177 179
GGT (UL) 0-40 2ndash20 1 1 6 10 4 0 1 1 5 9
AST (UL) - 63ndash152 61 58 48 61 64 59 93 61 58 69
ALT (UL) 39-196 95ndash544 95 105 89 105 106 82 234 137 115 156
CK (UL) 74-294 - 513 330 496 560 530 492 793 539 479 724
Amylase (UL) - - 23 28 35 35 29 28 29 24 36 23
Lipase (UL) - - 65 63 60 64 62 56 67 69 60 68
Calculated osmo (mmolL) - - 298 300 301 301 303 289 311 300 308 311
Ferret Identity
Legend - = reference value unavailable bolded = outside reference range a = adult ferret reference ranges from Animal Health Laboratory
(University of Guelph Guelph Canada) b = reference ranges for 10-week old ferrets (Fox JG 2014)
163
Appendix 3a Hematology values for domestic ferrets (Mustela putorius furo) inoculated orally with
Eimeria ictidea
Test Reference
Intervala
Reference
Intervalb
102 103 104 105 201 203 205
Age (days) adult 98-112 92 91 99 99 97 74 92
Days post inocculation 8 7 15 15 15 11 10
WBC (x 109L) 27-112 52ndash150 94 124 81 85 88 85 142
RBC (x 1012
L) 50-108 62ndash92 71 62 73 60 65 63 65
Hb (gL) 87-177 127ndash159 122 110 122 98 103 114 102
HCT (LL) 04-051 030ndash043 037 033 037 031 032 035 032
MCV (fL) 44-52 50ndash54 53 53 52 51 50 55 49
MCH (pg) 15-18 16-21 17 18 17 16 16 18 16
MCHC (gL) 325-362 351ndash426 328 332 325 319 320 326 317
RDW () 12-16 - 122 126 129 129 145 127 145
Platelets (x 109L) 54-695 376ndash610 524 413 445 117 303 42 429
MPV (fL) 5-10 - 8 78 72 66 67 14 8
TS Protein (gL) 49-76 - 66 63 58 66 55 59 59
Seg Neuts (x 109L) 1-8 21ndash62 160 446 203 170 211 170 554
Lymphocytes (x 109L) 1-63 16ndash79 761 657 551 646 59 646 682
Monocytes (x 109L) 0-09 01ndash02 019 087 024 026 044 026 085
Eosinophils (x 109L) 0-13 03ndash09 0 037 032 009 026 009 099
Basophils (x 109L) 0-02 0 0 012 0 0 009 0 0
Polychromasia 2-5 - 1-3 0-2 0-2 1-3 1-3 2-5 5-10
Anisocytosis Occ Occ Occ
HJ bodies rare
crenation
rouleaux
poikilocytosis
shift platelets Occ Occ
hemolysis Neg
Ferret Identity
Legend - = reference value unavailable bolded = outside reference range a = adult ferret reference ranges from Animal Health
Laboratory (University of Guelph Guelph Canada) b
= reference ranges for 10-week old ferrets (Fox JG 2014) Neg = negative
Occ = occasional = many platelet clumps
164
Appendix 3b Serum biochemistry values for domestic ferrets (Mustela putorius furo) inoculated orally
with Eimeria ictidea
Test Reference
Intervala
Reference
Intervalb
102 103 104 105 201 203 205
Age (days) adult 98-112 92 91 99 99 97 74 92
Days post inocculation 8 7 15 15 15 11 10
Calcium (mmolL) 185-242 245-268 243 222 238 233 232 236 240
Phosphorus (mmolL) 112-256 200-323 292 223 248 298 268 288 306
Magnesium (mmolL) 08-139 - 06 08 08 08 09 08
Sodium (mmolL) 147-159 148-155 147 150 150 148 148 150
Potassium (mmolL) 37-57 45-55 42 41 51 41 50 44
Chloride (mmolL) 111-129 114-124 114 113 118 112 111 115
Carbon dioxide (mmolL) 17-29 16-24 16 21 19 20 12 18
Anion gap (mmolL) 6 - 23 - 21 20 18 20 30 21
NaK ratio - - 35 37 29 36 30 34
Total protein (gL) 51-75 49-64 52 55 55 49 56 57
Albumin (gL) 24-40 30-36 28 20 27 27 26 29 27
Globulin (gL) 19-41 19-30 32 28 28 23 27 30
AG ratio 053-167 11-17 063 096 096 113 107 090
Blood urea nitrogen (mmolL) 45-153 50-150 92 87 119 92 110 98 149
Creatinine (umolL) 8-67 706-1414 56 26 49 40 50 46 79
Glucose (mmolL) 32-91 688-943 52 59 64 71 58 17 66
Cholesterol (mmolL) 294-894 440-640 453 429 262 475 370 285
Total bilirubin (umolL) 2 to 7 - 0 0 0 0 1 1
Conjugated biilrubin (umolL)- - 1 0 0 0 0 0
Free bilirubin (umolL) 0-2 - 0 0 0 0 1 1
ALKP (UL) 13-237 41-181 124 120 213 120 146 170 196
GGT (UL) 0-40 1-2 2 1 3 0 0 2
AST (UL) - 47-128 48 95 104 66 100 100
ALT (UL) 39-196 78-279 133 110 140 203 158 183 281
CK (UL) 74-294 - 382 765 1190 578 680 930
Amylase (UL) - - 24 37 31 28 33 35
Lipase (UL) - - 60 65 72 72 86 79
Calculated osmo (mmolL) - - 296 305 305 300 296 309
Ferret Identity
Legend - = reference value unavailable bolded = outside reference range a = adult ferret reference ranges from Animal Health
Laboratory (University of Guelph Guelph Canada) b
= reference ranges for 10-week old ferrets (Fox JG 2014)
165
Appendix 4 Domestic ferret (Mustela putorius furo) weekly monitoring sheet
Mon
8 AM
Mon
4 PM
Tues
8 AM
Tues
4 PM
Wed
8 AM
Wed
4 PM
Thurs
8 AM
Thurs
8 PM
Fri
8 AM
Fri
4 PM
Sat
8 AM
Sat
4 PM
Sun
8AM
Sun
4 PM
Mentation
Weight (g)
Respiratory
Rate
Vomit
(+ ++ +++)
Diarrhea
(+ ++ +++)
Urination
(+ ++ +++)
Defecation
(+ ++ +++)
Food
offered
Food
remaining
Water remaining
(ml)
Treatments
Other
observations
Initials of
observer
166
Animal ID ________________________________________ Week ______________
Monitoring Criteria
Mentation BAR (bright alert responsive) QAR (quiet alert responsive) depressed lethargic quiet
Weight measured in grams to be performed once weekly (pre-inoculation) and once daily (post-inoculation)
Respiratory rate measured in breaths per minute
Vomit + (small amount) ++ (moderate amount) +++ (large amount)
Diarrhea + (small amount) ++ (moderate amount) +++ (large amount)
Urination + (small amount) ++ (normal amount) +++ (large amount)
Defecation + (one pile) ++ (two piles) +++ (3+ piles)
Food offered Y (yes) N (no)
Food remaining A (all) P (partial) N (none)
Please note any additional observations in the appropriate section
Monitoring Times
Pre-inoculation ndash ONCE daily at 8am during cage cleaningfeeding
Post-inoculation ndash TWICE daily at 8am and 4pm
Critical patients (as determined by veterinary examination) ndash 3-6 times daily (based on veterinarian recommendation) ndash switch to 24
hour care sheet
Intervention Points
If any ferret develops vomiting diarrhea goes off food or exhibits gt 3 weight loss both the PI and graduate student listed below are
to be notified and the animal will be assessed to determine further treatment plans
Removal Criteria
If an individual ferret develops severe gastrointestinal or systemic disease during the course of the study as characterized by abnormal
mentation marked dehydration 10 weight loss within 24-48 hours profuse diarrhea unresponsive to treatment anorexia of 2 or
more days or frequent vomiting unresponsive to treatment the individual will be removed from the study and humanely killed
167
Appendix 5 Domestic ferret (Mustela putorius furo) 24 hour intensive monitoring sheet
Animal ID ___________________________________________ Date___________________
0700 800 900 1000 1100 1200 1300 1400 1500 1600 1700 1800
Mentation
Weight (g)
Heart Rate
Respiratory Rate
Temperature (C)
Water offered
(mL)
Water consumed
(mL)
Food offered
(type and
amount in
grams)
Food consumed
(type and
amount in
grams)
Presence of urine
Presence of feces
168
Character of
feces
Presence of
vomit
Character of
vomit
Medications to
be administered
Other
observations
169
1900 2000 2100 2200 2300 2400 100 200 300 400 500 600
Mentation
Weight (g)
Heart Rate
Respiratory Rate
Temperature (C)
Water offered
(mL)
Water consumed
(mL)
Food offered
(type and amount
in grams)
Food consumed
(type and amount
in grams)
Presence of urine
Presence of feces
Character of
feces
Presence of vomit
Character of
vomit
170
Medications to be
administered
Other
observations
Monitoring Criteria
Mentation BAR (bright alert responsive) QAR (quiet alert responsive) depressed lethargic quiet
Weight measured in grams to be performed once weekly (pre-inocculation) and once daily (post-inocculation)
Respiratory rate measured in breaths per minute
Vomit + (small amount) ++ (moderate amount) +++ (large amount)
Diarrhea + (small amount) ++ (moderate amount) +++ (large amount)
Urination + (small amount) ++ (normal amount) +++ (large amount)
Defecation + (one pile) ++ (two piles) +++ (3+ piles)
171
Food offered Y (yes) N (no)
Food remaining A (all) P (partial) N (none)
Please note any additional observations in the appropriate section
Monitoring Times
Critical patients (as determined by veterinary examination) ndash 3-6 times daily (based on veterinarian recommendation) ndash switch to 24
hour care sheet
Intervention Points
If any ferret develops vomiting diarrhea goes off food or exhibits gt 3 weight loss both the PI and graduate student listed below are
to be notified and the animal will be assessed to determine further treatment plans
Removal Criteria
If an individual ferret develops severe gastrointestinal or systemic disease during the course of the study as characterized by abnormal
mentation marked dehydration 10 weight loss within 24-48 hours profuse diarrhea unresponsive to treatment anorexia of 2 or
more days or frequent vomiting unresponsive to treatment the individual will be removed from the study and humanely killed
172
Appendix 6 Domestic ferret (Mustela putorius furo) infection trial standard operating procedures
Daily Fecal Collection
1 Collection of all feces passed in a 24 hour period will be performed once daily for all ferrets from
July 13 to July 24 inclusive
2 CAF Isolation staff will collect all fecal material present at the time of daily cage cleaning and
place in individual pre-labelled plastic bags (one per cage)
3 The amount of non-fecal matter (bedding etc) collected should be as minimal as possible
4 CAF isolation staff will record fecal character observations daily for each sample by ticking the
appropriate box on the baggie label
5 Isolation staff will place samples in the necropsy cooler (4 degC) while awaiting collection by
summer student
6 Monday to Friday fecal samples will be collected by A Rodriguez and brought for processing to
the Barta Lab in Pathobiology Samples collected Saturday and Sunday will be put on hold in a
refrigerator (4degC) until pick up on Monday
Example Bag label
Ferret ID Date
Weight of Feces
Fecal Character Normal
Soft
Liquid
Bloody
Abnormal odour
Physical Examination and Health Assessments
1 Manual restraint by hand or in towel by CAF Isolation staff and project personnel If required
examination +- blood collection may be performed under general anesthesia with isoflurane
2 Physical examination body weight HR RR temperature to be performed by PIs (Adriana Pastor
and Dale Smith)
3 Blood collection (~1mL per ferret) is to be performed from the jugular vein if under manual
restraint or the cranial vena cava under anesthesia by PIs using a 25g needle and 1cc syringe
Blood will be collected into small heparinized tubes and submitted to AHL for
CBCBiochemistry
4 All physical examination findings will be recorded on the Exam Sheet
5 Any minor wounds will be treated as appropriate (to be determined by PIs)
Inoculation of Ferrets with Coccidia
1 All ferrets to be inoculated will have been confirmed negative on daily fecals for two weeks
173
2 Brief physical examination by PIs to confirm that the ferrets are healthy to continue in study
3 The concentrated oocyst solution in sterile saline (up to a volume of 1mL) will be combined with
up to 1 mL of FerreTone (or another highly palatable substance if more preferred by the ferrets)
and ferrets will be allowed to consume the mixture ad lib while being monitored
4 Control ferrets will be administered saline only (equal mL to oocyst solution) with 1 mL of
FerreTone (or other substance as used for experimental group)
5 If ferrets refuse to consume the mixture oral inoculation via syringe of concentrated oocyst
solution will be performed by PIs under manual restraint
6 If ferrets are resistant to manual restraint for oral inoculation then inoculation will be performed
by PIs under general anesthesia via gastric tube (8 Fr red rubber)
Euthanasia Protocol
1 Ferrets to be euthanized will be masked down with isoflurane under manual restraint (or in an
anesthetic chamber as judged most appropriate by the PI)
2 Once anesthetized the ferret will be weighed and any blood fecal or other samples will be
collected as required (as determined by PIs)
3 Once an adequate plane of anesthesia is obtained (as determined by PIs) potassium chloride at a
dose of 2 mEq K+kg will be administered either via vena caval puncture or cardiac puncture to
induce cardiac arrest
4 Presenceabsence of respirations heartbeat corneal reflex will be used to assess death
Necropsy Protocol
1 Measure the ferret from nose to tail base (body length)
2 Perform standard necropsy but start with gastrointestinal tract first then thoracic and abdominal
viscera
3 Gently flush the entire contents of the intestine with 12 mL saline into a sterile urine cup
Potassium dichromate should be added in a 11 volumevolume ratio to the same container and
mixed with the combined intestinal contents and saline Label the container with the ferretrsquos ID
ferret group ID date and place on appropriate shelf in the refrigerator (4 degC) in the Barta lab
4 Measure the length of the gut from duodenum to anus
5 Collect paired sections of intestine for histological sectioning and frozen These sections should
be collected along the entire length of the gut from duodenum to rectum (see below for GI length
calculations)
6 For histological sections cut a 2 cm long section of bowel open completely on one side place on
a pre-cut section of box-board and staple both ends to the board to create a flat section Place all
gastrointestinal sections (attached to the board) in Serra fixative solution (100 ethanol (60
VV) 37 formaldehyde (30 VV) glacial acetic acid (10 VV)) for fixation and trim in
cross (transverse) sections
7 Adjacent to each sample removed for histopathology remove another 2 cm section intact and
place in a labelled Whirl-pack for freezing
8 Box-board and pre-labelled Whirl-packs should be labelled in pencil and sharpie respectively
with the following information Animal ID Zoo pm number section of gut (information should
include the region of the gut - jejunum colon etc and the length from the pylorus to the section)
174
Ferret gastrointestinal length calculations (from Evans amp An 2014)
Adult ferret body length 36-41 cm
NB In domestic ferrets there is ~51 ratio of small intestine to body length
Adult domestic ferret GI lengths
Small intestine ndash 182-198 cm
Large intestine - ~10 cm long 06 cm diameter (colon ndash 7 cm rectum ndash 2 cm anus ndash 1 cm)
Based on the above information
Small intestine Six sections of the small intestine from duodenum to ileum will be collected each ~ 25-40
cm apart depending on the size of the ferret
Large intestine Collect two sections of colon (4 cm each) at 25 and 75 of the length of the colon from
the junction of the small and large intestine to the rectum Smaller ferrets may allow only on section
based on colon length
Laboratory SOPs
General notes on processing fecal samples
1 Samples will be collected from the isolation facility necropsy room cooler daily from Monday-
Friday for the entire 6 weeks of the project
2 Upon transfer to the lab each baggie will be weighed after filling to determine the amount of
feces (grams) in each bag and that number entered onto the label on the bag
3 All information regarding fecal processing will be entered onto the spreadsheet for the individual
ferret including ID weight of feces feces character date of sample collection date of sample
processing fecal flotation method presence or absence of oocysts oocyst quantification
Fecal Processing - Weeks 1+2 (acclimation period)
1 After weighing transfer the contents of one baggie to a small sieve on top of a paper cup
2 Fill a single glass container (for fecal flotation) with saturated salt solution
3 Slowly pour small amounts of the solution over the feces mix and strain liquid contents in the paper
cup using a tongue depressor
4 Dispose of the remaining fecal matter in the sieve
5 Pour the contents of the paper cup back into the glass container and cover slip for 5-7 minutes
6 Place coverslip on a clean glass slide and viewscan under microscope at 10x power for presence of
oocysts
7 Record all findings on the provided spreadsheet
8 Notify A Pastor if any oocysts are detected If detected contents of the slide should be flushed back
into the glass container with distilled water and contents of glass container should be transferred to a
50 mL plastic conical tube Clean to remove salt as per Barta lab SOP and mix with potassium
dichromate equal parts by volume to amount of oocysts in diH2O Place a checkmark on the lid of the
50 mL conical vial containing the remainder of the feces and store in the lab fridge at 4 degC
175
Fecal Processing - Weeks 3-7 (infection trial)
1 After weighing transfer the entire contents of one baggie to a small sieve on top of a paper cup
2 Slowly pour small amounts of distilled water over the feces (enough to wet) mix and strain liquid
contents in the paper cup using a tongue depressor until feces appear almost dry
3 Dispose of the remaining fecal matter in the sieve
4 Pour the contents of the paper cup into a 50 mL conical tube If samples are not going to be counted
on the same day then mix potassium dichromate (25 wv) 11 by volume with sieved fluid from
the paper cup and store at 4 degC (refrigerator) until ready to perform OPG counts
If proceeding with the count the same day
5 Pipette and place one drop of the sieved fecal fluid on a slide to determine approximate oocyst
concentration If oocysts rare to none ndash dilute 12 during step 6 if moderate numbers ndash dilute 19 if
too numerous to count ndash dilute 199 (or perform serial dilutions of 10x from initial 19 dilution)
6 Transfer x mL of the mixture from the 50 mL conical tube into a clean 15 mL conical tube and mix
with appropriate amount of saturated salt solution for desired dilution
7 Fill both sides of the McMaster counter chamber and count the number of oocysts per side (total for
one side= total number of oocysts from all 6 sections of the chamber )
8 Average the total counts from both sides
9 Use the following calculation to determine the oocyst per gram count
OocystsmL = oocysts counted times 666 times dilution (ie 3 if dilution 12)
OPG= oocystmL times total volume recorded at end of step 4
10 Record all findings on the provided spreadsheet
11 Notify A Pastor if any oocysts are detected If detected place a checkmark on the lid of the 50 mL
conical vial containing the remainder of the feces and store in the lab fridge at 4 degC
12 For all samples for which no oocysts are detected during steps 6-8 follow up with routine salt
flotation of the remainder of the fecal sample from step 6 (use instructions for fecal processing from
weeks 1-2)

v
ACKNOWLEDGEMENTS
Itrsquos hard to believe that my residency and thesis have been completed and I have a lot of people
to thank for that
Dale and Graham the two people I wanted to be when I grew up thank you for your mentorship
for many years even before this program I know that you werenrsquot convinced that this project was DVSc
worthy when I first proposed it but Irsquom hoping that the results have changed your mind
I am sincerely grateful to all the members of my advisory committee Dale Smith John Barta and
Simon Hollamby for their insight support and interest in this project Dale you have been an exceptional
advisor I donrsquot know that I will ever get to your level but thank you for showing me that being a great
clinical zoo vet and pathologist are not mutually exclusive John thank you for spontaneously agreeing to
be my advisor when I came to you with this project proposal in my first semester for your energy and
enthusiasm and for supporting my widening interest in parasitology research
My heartfelt appreciation for the Toronto Zoo WHC veterinarians past and present Chris
Dutton Pauline Delnatte Simon Hollamby and Graham Crawshaw I have learned so much from all of
you that I will take forward into my future endeavours I appreciate the extra time you put in including
comps study sessions after-hours tecircte-agrave-tecirctes and the fact that your doors were open when I needed it
For the Toronto Zoo vet techs extraordinaire Michelle Lovering Cassia Devison Dawn
Mihailovic and Tasha Long ndash you have been indispensable during this program and there are not enough
words to express my gratitude
I would especially like to thank all the Wildlife Health Center staff (2013-2016) Mark Bongelli
Charles Guthrie Christine McKenzie Brian Telford Rick Vos Gerri Mintha Margaret Kolakowski
Andrew Lentini Rebecca Clark Lydia Attard Nigel Parr Paula Roberts Andrea Dada Mindy Waisglass
and Julie Digiandomenico for three very memorable years It is all of you that make the WHC such an
amazing place to be Irsquom not sure I have laughed so hard or so often as I did in that lunchroom and I hope
our paths will cross again
vi
I donrsquot think that I can truly express how thankful I am to Pathobiology laboratory technicians
Julie Cobean and Julia Whale Without your assistance patient teaching and friendship I would probably
still be screening fecal samples years from now and scratching my head as to how our lab protocols
actually work It is people like you who make sure graduate students become successful doctorates and I
canrsquot imagine Pathobio without you both in it
I would also like to thank my labmates in the Barta lab mdash Mian Hafeez Evelyn Rejman Rachel
Imai Perryn Kruth Ryan Snyder and Mosun Ogedengbe A special thank you goes to Alex Leveille
without whom my many adventures in parasitology research from coccidia to Babesia would not have
been as successful
To all the students who helped with ferret fecal sample processing data compilation and
necropsies Nathalie Ferriman Janessa Price Thisuri Eagalle Sarah Brisson thank you so much for your
hard work and excitement about my projecthellip even when it was very smelly
So many thanks to the amazing staff of Central Animal Facility - Linda Groocock Vicky Carson
Tony Cengija and Mary Fowler for the daily care and enrichment of my experimental ferrets Your
excitement about working with our ferrets and your assistance with all parts of the process helped made
this project a success
To Adriana Nielsen who was not only my better half but the other fifty percent of my brain for
several years It is your friendship fortitude and our endless phone conversations that got me through the
never-ending Toronto-Guelph commute and this program
To all the ldquoscope roomrdquo pathology co-residents past and present - thank you for being wonderful
friends and colleagues It is indeed rare to find so many amazing people in one place and I know this
program and my sanity would not have been the same without you
To the anatomic pathology faculty and senior graduate students - thank you for all the time
teaching and guidance you provided during my program While I canrsquot say that I have become an amazing
pathologist I can say that because of your mentorship I am a better diagnostician and the type of clinician
who asks better questions takes better samples and understands that you canrsquot ldquojust make a PCR for thatrdquo
vii
A special thank you to Tony van Dreumel who came out of retirement for a semester to try to teach the
Adrianas zoo pathology screening cases with you was always a pleasure
To all the lovely Histo Ladies PM room staff and the other AHL staff who helped me with
Toronto Zoo and HSC pathology cases along the way - I donrsquot think the anatomic path students could
survive without you Thank you for always smiling assisting and accommodating me even when I made
near-impossible processing requests during my weekly Guelph visits
I would also like to acknowledge and sincerely thank all the individuals who helped with resource
and sample acquisition for this project A special mention for those who went above and beyond because
of their interest in this project Don Duszynski who was instrumental in acquiring and then providing a
translator for many of the original mustelid Eimeria descriptions and Majda Globokar Nikola Pantchev
and Donald Martin who supplied my domestic ferret fecal samples and historical data
A shout-out to Julie Swenson Gary West and the Phoenix Zoo BFF team who fostered my love
of this endangered species and helped develop the idea for this project
As always I continue to go out into the world and pursue my dreams with the knowledge that I
have the support of my incredible family long-time friends and my partner Keith Morris I am so lucky
that my residency brought me home and that it afforded us all more time spent together For my aunt
Veronica Lacey who has never failed to believe in my potential and always pushed me to become an
academic ndash yoursquoll never get that PhD from me but I think this is pretty close Finally for my mother
Anna Pastor who never lived to see my greatest achievements but had absolute faith that I could reach
any goal I worked towardshellip this is for you
Finally none of this would have been possible without the generous support of the Toronto Zoo
Residency program and funding through the Barta Laboratory University of Guelph
Adriana Pastor
Toronto August 2017
viii
DECLARATION OF WORK PERFORMED
I declare that all the work reported in this thesis was performed by myself with the following
exceptions
Fecal samples were collected by personnel at the Toronto Zoo Louisville Zoo and participating
diagnostic laboratories
Fecal oocyst per gram counts (routine salt flotation and McMaster counts) were performed by
myself Julie Cobean Julia Whale Evelin Rejman Sarah Brisson Adriana Rodriguez and Perryn Kruth
Whole mitochondrial genome PCR and sequencing was performed by me in conjunction with
Julia Whale and Dr Mian Hafeez
Sequencing of PCR samples was performed at the University of Guelph Laboratory Services
(Guelph Ontario Canada) and results were obtained electronically
ix
TABLE OF CONTENTS
ABSTRACT ii
DEDICATION iv
ACKNOWLEDGEMENTS v
DECLARATION OF WORK PERFORMED viii
TABLE OF CONTENTS ix
LIST OF TABLES xiii
LIST OF FIGURES xiv
LIST OF APPENDICES xv
ABBREVIATIONS xvi
CHAPTER 1 LITERATURE REVIEW 1
11 INTRODUCTION 1
12 APICOMPLEXA 1
121 Brief introduction to apicomplexan pathogens 1
122 Life cycles of the Eimeria and Isospora species implicated in enteric coccidiosis 3
123 Methods of characterization 5
13 RECLASSIFICATION OF MAMMALIAN ISOSPORA 8
14 EIMERIID SPECIES CHARACTERIZED IN MUSTELIDS 8
141 The family Mustelidae 8
142 Eimeriid coccidia described from mustelids 9
143 Eimeriid coccidia described from domestic ferrets 16
144 Molecular characterization 19
145 Clinical signs of disease in domestic ferrets 21
146 Gross necropsy and histologic findings 21
15 INTRODUCTION TO ENTERIC COCCIDIOSIS IN THE BLACK-FOOTED FERRET
25
151 Natural history and conservation of the black-footed ferret in North America 25
152 Coccidia identified from black-footed ferrets 26
153 Morbidity mortality and clinical signs associated with enteric coccidiosis in black-footed
ferrets 28
16 TREATMENT PREVENTION AND CONTROL OF INFECTION BY EIMERIA SPP 29
161 Current recommendations for treatment of eimeriid coccidia in carnivores 29
x
162 Current recommendations for anticoccidial treatment and prophylaxis in domestic and
black-footed ferrets 30
17 VACCINES AGAINST COCCIDIA 32
171 Theory 32
172 Species successes in anticoccidial vaccination 34
18 RESEARCH GOALS AND OBJECTIVES 36
181 Objectives 36
182 Hypotheses 36
183 Applications 36
CHAPTER 2 MOLECULAR CHARACTERIZATION OF ENTERIC COCCIDIA FROM DOMESTIC
FERRETS (MUSTELA PUTORIUS FURO) 38
21 INTRODUCTION 39
22 MATERIALS amp METHODS 43
221 Fecal samples 43
222 Formalin fixed intestinal tissues 44
223 Molecular characterization 44
224 Phylogenetic analysis 46
23 RESULTS 47
231 Fresh fecal samples 47
232 Formalin fixed samples 48
233 Molecular characterization 49
234 Phylogenetic analysis 50
24 DISCUSSION 50
CHAPTER 3 MORPHOLOGICAL AND MOLECULAR CHARACTERIZATION OF ENTERIC
COCCIDIA ISOLATED FROM BLACK-FOOTED FERRETS (MUSTELA NIGRIPES) 60
31 INTRODUCTION 60
32 MATERIALS AND METHODS 64
321 Fecal samples 64
322 Formalin fixed intestinal tissues 65
323 Molecular characterization 66
33 RESULTS 66
331 Morphometric characterization 67
332 Molecular characterization 68
34 DISCUSSION 69
xi
CHAPTER 4 NATURAL HISTORY OF ENTERIC COCCIDIOSIS IN THE BLACK-FOOTED
FERRET (MUSTELA NIGRIPES) 78
41 INTRODUCTION 78
42 MATERIALS AND METHODS 80
421 Toronto Zoo BFF breeding program 80
422 Fecal oocyst evaluation 81
423 Retrospective review of pathology records 82
424 Prospective modified necropsy protocol 82
425 Retrospective medical history review 83
43 RESULTS 83
431 Fecal oocyst evaluation and retrospective medical history review 83
432 Pathology 86
433 Morbidity and mortality 88
44 DISCUSSION 88
CHAPTER 5 EVALUATING THE DOMESTIC FERRET (MUSTELA PUTORIUS FURO) AS AN
EXPERIMENTAL MODEL FOR ENTERIC COCCIDIOSIS IN THE BLACK-FOOTED FERRET
(MUSTELA NIGRIPES) 104
51 INTRODUCTION 104
52 MATERIALS AND METHODS 106
521 Animal care 106
522 Oocyst preparation 107
523 Experimental infections 108
524 Animal welfare 109
525 Hematology 110
526 Morphologic and molecular characterization 110
527 Necropsy protocol 111
53 RESULTS 111
531 Oocyst shedding 112
532 Morphologic and molecular characterization 113
533 Clinical signs 113
534 Hematology 113
535 Necropsy 114
54 DISCUSSION 115
xii
CHAPTER 6 WHOLE MITOCHONDRIAL GENOME SEQUENCES OF TWO EIMERIA SPECIES
ISOLATED FROM DOMESTIC (MUSTELA PUTORIUS FURO) AND BLACK- FOOTED FERRETS
(MUSTELA NIGRIPES) 129
61 INTRODUCTION 129
62 MATERIALS amp METHODS 130
621 Parasites 130
622 DNA isolation from coccidia in feces 131
623 Whole genome sequencing 131
624 Phylogenetic analysis 132
63 RESULTS 133
64 DISCUSSION 134
CHAPTER 7 CONCLUSIONS AND FUTURE DIRECTIONS 145
REFERENCES 148
APPENDICES 157
xiii
LIST OF TABLES
Table 11 Morphometrics of Eimeria and Isospora (=Cystoisospora) species affecting mustelids 10
Table 21 Amplification primers for nuclear 18S rDNA and mitochondrial COI loci used in the
identification of enteric coccidia from domestic ferrets 55
Table 22 Summary of fecal samples from domestic ferrets submitted to two diagnostic laboratories
from 2008-2015 56
Table 23 Morphologic and molecular identification of coccidia from domestic ferrets 57
Table 31 Amplification primers for nuclear 18S rDNA mitochondrial COI and COIII loci used in the
identification of coccidia from black-footed ferrets 73
Table 32 Morphologic and molecular characterization of coccidia from fecal and FFPE necropsy
samples from black-footed ferrets 76
Table 33 Morphometric characterization of Eimeria ictidea oocysts from black-footed ferrets 77
Table 41 Eimeria ictidea shedding in black-footed ferret dam and kit family groups - 2014-2016 97
Table 42 Epidemiologic data for family groups of black-footed ferrets shedding Eimeria ictidea 98
Table 43 Shedding of Eimeria ictidea in adult black-footed ferrets - 2015-2016 99
Table 44 Epidemiologic data for adult black-footed ferrets shedding Eimeria ictidea 100
Table 45 Histologic findings from black-footed ferrets with enteric coccidiosis 101
Table 46 Incidence of coccidial infections in black-footed ferrets at the Cheyenne Mountain Zoo 102
Table 47 Yearly mortality associated with coccidiosis in black-footed ferrets at the Toronto Zoo 103
Table 51 Prepatent period and oocyst shedding of Eimeria ictidea in experimentally infected
domestic ferrets 126
Table 52 Results of oral inoculation of domestic ferrets with oocysts of Eimeria ictidea 127
Table 53 Distribution of coccidial life stages in intestinal tract of domestic ferrets orally
inoculated with oocysts of Eimeria ictidea 128
Table 61 PCR primers used to sequence the mitochondrial genome of Eimeria furonis 136
Table 62 PCR primers used to sequence the mitochondrial genome of Eimeria ictidea 137
Table 63 Coding regions in the mitochondrial genome of Eimeria furonis from a domestic ferret 138
Table 64 Coding regions in the mitochondrial genome of Eimeria ictidea from a black-footed ferret 139
Table 65 Pairwise comparison of coding regions in the mitochondrial genomes of Eimeria furonis
and Eimeria ictidea 140
xiv
LIST OF FIGURES
Figure 11 Phylogeny of the Apicomplexa 2
Figure 12 Classical life cycle of coccidian parasites 4
Figure 13 Morphologic characteristics used for identification of eimeriid oocysts 6
Figure 21 Life stages of Eimeria furonis within the small intestine of a domestic ferret 58
Figure 22 Phylogenetic relationships of coccidia (Eimeria ictidea Eimeria furonis and Isospora
(=Cystoisospora) laidlawi) from domestic or black-footed ferrets 59
Figure 31 Nuclear and mitochondrial genetic loci targeted by primers listed in Table 31 73
Figure 32 Morphometrics of Eimeria ictidea from a black-footed ferret (Mustela nigripes) 74
Figure 33 Nuclear 18S rDNA sequences of Eimeria ictidea to newly generated (see Chapter 2) and
published sequences of Eimeria furonis 75
Figure 34 Mitochondrial cytochrome c oxidase subunit I sequences of Eimeria ictidea to sequences
from other eimeriid parasites of carnivores 75
Figure 41 Oocyst per gram counts and shedding period of Eimeria ictidea from black-footed ferret
family groups from 2014-2016 95
Figure 42 Sexual life stages of Eimeria ictidea in the small intestine of a black-footed ferret 96
Figure 51 Exogenous life stages of Eimeria ictidea 123
Figure 52 Endogenous life stages of Eimeria ictidea within the small intestine of an experimentally
infected domestic ferret 124
Figure 53 Distribution of sexual and asexual life stages of Eimeria ictidea along the intestinal tract
of experimentally infected domestic ferrets 125
Figure 61 Map of the mitochondrial genome of Eimeria furonis 141
Figure 62 Map of the mitochondrial genome of Eimeria ictidea 142
Figure 63 Comparison of the mitochondrial genomes of Eimeria furonis and Eimeria ictidea 143
Figure 64 Phylogenetic relationships of coccidia from domestic and black-footed ferrets based on
complete mitochondrial genome sequences 144
xv
LIST OF APPENDICES
Appendix 1 Shedding of oocysts of Eimeria ictidea in black-footed ferret (Mustela nigripes) dam and
kit family groups from 2014-2016 158
Appendix 2a Hematology values for domestic ferrets (Mustela putorius furo) from 49-51 days of
age prior to experimental inoculation 161
Appendix 2b Serum biochemistry values for domestic ferrets (Mustela putorius furo) from
49-51 days of age prior to experimental inoculation 162
Appendix 3a Hematology values for domestic ferrets (Mustela putorius furo) inoculated orally
with Eimeria ictidea 163
Appendix 3b Serum biochemistry values for domestic ferrets (Mustela putorius furo) inoculated
orally with Eimeria ictidea 164
Appendix 4 Domestic ferret (Mustela putorius furo) weekly monitoring sheet 165
Appendix 5 Domestic ferret (Mustela putorius furo) 24 hour intensive monitoring sheet 167
Appendix 6 Domestic ferret (Mustela putorius furo) infection trial standard operating procedures 172
xvi
ABBREVIATIONS
ATP Adenosine triphosphate
BFF Black-footed ferret(s)
BI Bayesian inference
bp Base pair
CAPC Companion Animal Parasitology Council
CDS Coding DNA sequence
CITES Convention on International Trade in Endangered Species of Wild Fauna and Flora
COI Cytochrome c oxidase subunit 1
COIII Cytochrome c oxidase subunit 3
CytB Cytochrome b
DF Domestic ferret(s)
DNA Deoxyribonucleic acid
FFPE Formalin-fixed paraffin embedded tissue
IUCN International Union on the Conservation of Nature
L Length
LSU Large subunit
mt Mitochondrial
NaOH Sodium hydroxide
nu Nuclear
OPG Oocyst per gram count
PCR Polymerase chain reaction
rDNA Ribosomal DNA
SI Shape index
SND Single nucleotide difference
SOP Standard operating procedure
sp spp Species (singular plural)
SSP Species Survival Plan
SSU Small subunit
TMS Trimethoprim sulfadimethoxine
USFWS United States Fish and Wildlife Service
W Width
1
CHAPTER 1 LITERATURE REVIEW
11 INTRODUCTION
Black-footed ferrets (Mustela nigripes) are one of three wild ferret species worldwide Although
formerly distributed throughout the North American prairies black-footed ferrets (BFF) had been
extirpated from the majority of their range by the 1970s and were declared extinct in the wild in 1987
Since 1986 a multi-institutional effort has been breeding this species in captivity with reintroduction back
into the wild at select sites within Canada the USA and Mexico
Coccidial enteritis is a major cause of death in young captive black-footed ferrets (Bronson et al
2007) but coccidiosis can affect all age classes (personal observation) As a result fewer captive-bred
ferrets may be reared successfully for release to the wild The significance of coccidiosis in wild ferrets is
unknown Consequently the prevention and control of coccidial outbreaks is an important part of black-
footed ferret captive breeding programs and management This research is intended to improve the in situ
and ex situ health of the black-footed ferret through the provision of a better understanding of the
pathogenesis of enteric coccidiosis in this species and to pave the way for the investigation of novel
methods for disease treatment and control
12 APICOMPLEXA
121 Brief introduction to apicomplexan pathogens
The phylum Apicomplexa comprises a large number of eukaryotic intracellular parasitic
organisms many of which are of importance to human and veterinary medicine As indicated by their
name these parasites are characterized by the presence of an apical complex at the anterior aspect of the
infective stage of the life-cycle (Tenter et al 2002) The taxonomic classifications of members of the
Apicomplexa continue to be in a state of flux (reviewed by Adl et al 2005 Cavalier-Smith 2014 Tenter
et al 2002) For this reason a more simplified taxonomic structure has been used in this review (see
2
Figure 11) The subclass Coccidia is a speciose group within the Apicomplexa with most genera falling
into one of two major coccidian suborders within the Eucoccidiorida To date greater than 2000 species
of coccidia have been named (Duszynski Upton amp Couch nd Upton 2000) The adeleid coccidia
(suborder Adeleorina) include monoxenous (single host) and heteroxenous (multiple hosts) parasites in
genera such as Adelea Haemogregarina Hepatozoon and Karyolysus The eimeriorinid coccidia
(suborder Eimeriorina) include the typical intestinal coccidia such as Eimeria Isospora and Cyclospora
species belonging to the family Eimeriidae as well as tissue (cyst forming) coccidia such as
Cystoisospora Besnoitia Toxoplasma and Sarcocystis species that belong to the family Sarcocystidae
(Cox 1994)
Figure 11 Phylogeny of the Apicomplexa Numbers on branches and thickness indicate diversity
(ie named species) Taxonomic groupings demonstrated by the phylogenetic tree (1) subclass
Coccidia (2) suborder Adeleorina (3) suborder Eimeriorina (4) family Eimeriidae and (5) family
Sarcocystidae Adapted from Šlapeta J Morin-Adeline V (2011) Apicomplexa Levine 1970
Sporozoa Leucart 1879 httptolweborgApicomplexa2446 in The Tree of Life Web Project
httptolweborg
2
1
3
4
5
3
122 Life cycles of the Eimeria and Isospora species implicated in enteric coccidiosis
The life cycle of Eimeria species is considered the classical coccidian life cycle which is
typically completed in one host (monoxenous) with many Eimeria species parasitizing only a single host
species (stenoxenous) (Figure 12) The life cycle has two main phases of development one that takes
place within the host (endogenous) and the other that takes places outside of the host (exogenous)
Classically the endogenous stages of the Eimeria life cycle take place within the intestinal epithelium
however some Eimeria species undergo extraintestinal endogenous development such as Eimeria stiedae
in rabbits which replicates within the epithelium of the biliary tree During the exogenous phase of the
life cycle unsporulated oocysts that are shed in the feces of the host sporulate within the environment
resulting in the formation of four sporocysts within each oocyst (tetrasporocystic) Each sporocyst
contains two sporozoites (dizoic) Sporulation is affected by three main factors temperature moisture and
aerobic conditions (Fayer 1980)
Once ingested by the host the wall of the sporulated oocyst is broken to release sporocysts from
which the sporozoites (infective stage) excyst The freed sporozoites penetrate the intestinal epithelial
cells and undergo multiple mitotic divisions to form a single multinucleate meront The meront then
undergoes simultaneous cytokinesis to form first generation merozoites which leave the host cell to infect
new cells and undergo further asexual replications The undifferentiated uninucleate tissue stage of the
parasite within the intestinal epithelial cell is called a trophozoite The number of cycles of asexual
replication (merogony) is predetermined after which the last generation of merozoites penetrate host cells
and undergo sexual differentiation into male and female gamonts (gametogony) Each microgamont
(male) undergoes simultaneous fission to produce numerous motile microgametes each macrogamont
(female) develops into a single mature macrogamete Fertilization of a macrogamete by a motile
microgamete results in formation of a zygote that is rapidly enclosed in a thick wall to form an
unsporulated oocyst Oocysts are shed with the hostrsquos feces into the environment where they are
protected from desiccation and chemical disinfection by the oocyst wall Traditionally Eimeria species
4
have been differentiated based on the host species or host genus affected the site of endogenous life cycle
development and the microscopic cellular characteristics of the different life stages Interestingly
experimental cross infection of Eimeria species from their natural host to a novel host of a taxonomically
similar species has been successful in some cases (De Vos 1970 Levine and Ivens 1970 Haberkorn
1971) challenging the notion that Eimeria are truly stenoxenous parasites
Figure 12 Classical life cycle of coccidian parasites This apicomplexan life cycle includes both
sexual and asexual development The three processes in the life cycle are merogony (asexual
replication A-D) followed by gametogony (formation of gametes E-H) within the digestive tract
of the host with release of unsporulated oocysts (I) Exogenous sporogony (I-L) results in the
production of infective sporulated oocysts (L) Adapted from Barta 2001 with permission of the
author
The life cycle of Isospora spp is similar to that of species in the genus Eimeria (see Figure 12)
but the number of sporocysts and sporozoites differ sporulated oocysts contain two sporocysts (disporic)
5
each of which contains four sporozoites (tetrazoic) These characteristics are not unique to Isospora spp
because diasporic tetrazoic sporulated oocysts are also found in the genera Besnoitia Frenkelia
Hammondia Sarcocystis and Toxoplasma However the sporocysts in the latter parasites are
morphologically distinct in that they lack Stieda bodies
123 Methods of characterization
1231 Morphological features
Historically eimeriid coccidia have been classified based on the cellular morphology of the
different life stages (particularly the morphometrics of sporulated oocysts) where these stages occur in
the host and apparent host specificity (frequently assumed and not tested experimentally) The
morphological features and dimensions of oocysts and their components are important diagnostic features
because of the availability of these stages in clinical specimens these characteristics can include size
(length [L] width [W] shape index [SI=LW]) number of sporocysts wall morphology
presenceabsence of a micropyle micropyle cap residual body or polar granules for oocysts size number
of sporozoites wall morphology presenceabsence of Stieda body subStieda body paraStieda body or
residual body for sporocysts and presenceabsence of refractile bodies for sporozoites (see Figure 13)
Pertinent life cycle information includes type of life cycle (monoxenous versus heteroxenous) tissue
sites of merogony and gametogony (intestinal versus extraintestinal) and the presence or absence of
extraintestinal hypobiotic stages (eg dormozoites or hypnozoites) Further information used to
characterize coccidia that form tissue cysts generally includes details on life stages in the definitive and
intermediate hosts location and morphology of tissue cysts route(s) of transmission among host species
and morphologic descriptions of merozoites (eg tachyzoites or bradyzoites) in tissue culture
6
Figure 13 Morphologic characteristics used for identification of eimeriid oocysts 1) Oocyst in cross
section ol - oocyst length or - oocyst residual body ow - oocyst width pg - polar granule row -rough
outer wall 2) The top of a hypothetical oocyst mcd - depth of the micropyle cap mcw - width of the
micropyle cap mw - width of the micropyle sow - smooth outer wall 3) Sporocyst in cross section
psb - paraStieda body sb - Stieda body sl - sporocyst length sp - sporozoite sr - sporocyst residual
body srb - sporozoite refractile body ssb - subStieda body sw - sporocyst width From Duszynski D
Wilber PG (1997) A guideline for the preparation of species descriptions in the Eimeriidae Journal of
Parasitology 83(2)333-336 reproduced with permission of Allen Press Publishing Services
1232 Molecular characterization (genetic loci and methods)
More recently molecular techniques have been used to infer phylogenetic or evolutionary
relationships among coccidia and to reclassify taxonomic assignments to better reflect the evolutionary
history of these parasites Molecular data can be more informative than phenotypic data because recent
evolutionary divergence among coccidia is unlikely to be reflected in morphologic differences but may
be detectable using molecular data The principle behind the use of molecular sequencing to describe
evolutionary relationships is that nucleotide sequences like morphological features diverge over time
under selective pressure however nucleotide sequences evolve at a more regular rate than do
morphologic characteristics Phenotypic data is thus less likely to detect recent evolutionary divergence
Sequences that are more similar are inferred to be more closely related and to have diverged more
recently (Cox 1994) Molecular characterization can be performed using DNA RNA or protein
sequences Most of the early molecular phylogenetic analyses of coccidia performed used ribosomal RNA
sequences usually by PCR amplification of ribosomal DNA (rDNA) in the nuclear genome of the
7
parasites Ribosomes contain both small and large RNA subunits in eukaryotes the large ribosomal RNA
consists of two forms 5S and 28S while the small ribosomal RNA exists only as 18S Sequences from
several genetic loci have been used for characterization of parasites most commonly 18S rDNA 28S
rDNA and ribosomal internal transcribed spacer regions (ITS) from the nuclear genome and more
recently mitochondrial cytochrome c oxidase subunits I (COI) and III (COIII) however sequencing of
nuclear 18S rDNA (nu 18S rDNA) has been the most prevalent in the literature by far Early attempts to
use 5S RNA sequences formed unlikely phylogenies and too few 28S ribosomal DNA sequences have
been obtained to make this locus useful (Cox 1994 Tenter et al 2002) The disadvantage of nu 18S
rDNA is that it is comparatively poor at distinguishing among closely related eimeriid coccidial species
because of its conserved nature but for that reason the nu18S rDNA locus is useful for inferring
relationships among species with greater evolutionary divergence Although only exploited recently
because of the paucity of suitable PCR primers the mitochondrial COI locus appears to be more useful
for distinguishing closely related eimeriid coccidia (Ogedengbe Hanner amp Barta 2011) but COI
sequences are less useful for inferring more ancient relationships between highly divergent coccidial
species Consequently the combined use of nu 18S rDNA and mitochondrial COI sequencing has been
recommended for improved species description and phylogenetic analysis (El-Sherry et al 2013)
Molecular characterization has also been used for diagnostic purposes and is well-suited to the
identification of coccidia when information on host specificity parasite life cycle and life stages is not
available as the molecular (genetic) data is the same for a given parasite during each of its life cycle
stages This information can be particularly useful in identifying the relationship between different life
stages of heteroxenous parasites collected from different hosts (intermediate definitive) Furthermore for
previously unidentified coccidia or those for which limited information is available molecular
characterization could be used to predict likely definitive hosts or parasite life cycle traits based on
phylogenetic relationships to other known species
8
13 RECLASSIFICATION OF MAMMALIAN ISOSPORA
Recommendations have been made to reclassify the avian and mammalian Isospora into two
separate genera based on life cycle molecular phylogenetic studies and morphologic description of
sporulated oocysts (Frenkel 1977 Barta et al 2005) Due to their classical coccidian life cycle presence
of Stieda bodies within sporocysts and close phylogenetic association with Eimeria species the avian
Atoxoplasma and Isospora have been retained in the genus Isospora (see Barta et al 2005) Conversely
the presence of tissue life cycle stages lack of Stieda bodies within sporocysts and close phylogenetic
association with other genera within the family Sarcocystidae have required many mammalian Isospora
to be reclassified as members of the genus Cystoisospora Frenkel 1977 (Frenkel 1977 Barta et al 2005)
Consequently for the remainder of this thesis Isospora species from mustelids will be referred to as
Isospora (=Cystoisospora) to reflect their probable generic association
14 EIMERIID SPECIES CHARACTERIZED IN MUSTELIDS
141 The family Mustelidae
The family Mustelidae within the order Carnivora comprises a group of approximately 59
carnivorous mammalian species within 22 genera Native mustelids are found in terrestrial and aquatic
environments on almost every continent with the exception of Australia and Antarctica The Mustelidae
are classically divided into two subfamilies as defined by Wozencraft (2005) 1) Mustelinae (weasels
mink ferrets marten wolverine) the larger subfamily including the following genera Arctonyx Eira
Galictis Gulo Ictonyx Lyncodon Martes Meles Mellivora Melogale Mustela Neovison Poecilogale
Taxidea and Vormela and 2) Lutrinae (otters) including seven genera Aonyx Enhydra Hydrictis
Lontra Lutra Lutrogale and Pteronura More recently molecular data suggest the Mustelidae should be
separated into eight subfamilies although this is not universally accepted (Koepfli et al 2008 Lariviegravere
and Jennings 2009 Yu et al 2011)
9
142 Eimeriid coccidia described from mustelids
Ten named Eimeria species and twelve named Isospora (=Cystoisospora) species have been
described in the Mustelidae and are summarized in Table 11 This table includes information on host
range life cycle and detailed morphologic data used to identify and classify the individual parasites Two
coccidial parasites isolated from the Libyan striped weasel (Ictonyx libyca) and the European polecat
(Mustela putorius) initially ascribed to the genus Isospora Isospora zorillae and Isospora putori
respectively have since been reclassified as Sarcocystis spp (see footnote to Table 2 of Yi-Fan et al
2012)
10
Table 11 Morphologic characteristics of Eimeria and Isospora (=Cystoisospora) species affecting mustelids
Coccidial species Host genus and
species
Life cycle
Location
Oocyst shape and
size
Oocyst
description
Sporocyst
description
Sporozoite
description
References
Cytoisospora
eversmanni
Mustela
eversmanii
(Steppe polecat)
Mustela
putorius
(European
polecat)
Homoxenous L185 (16ndash20)
W 148 (16ndash12)
LW 13 (11ndash16)
M absent
PG absent
OR absent
L 115
(10ndash135)
W 98
(9ndash11)
LW 12
(11ndash15)
SB absent
SR present
SRB present Yi-Fan et al 2012
Svanbaev 1956
Nukerbaeva amp
Svanbaev 1973
1977
Cystoisospora
pavlovskyi
Mustela
eversmanii
Mustela
putorius
Homoxenous L 322 (29ndash36)
W 273 (265ndash285)
LW 12 (11ndash14)
M absent
PG absent
OR absent
L 195
(18ndash21)
W 144
(12ndash15)
LW 14
(12ndash15)
SB absent
SR present
SRB present Yi-Fan et al 2012
Svanbaev 1956
Nukerbaeva amp
Svanbaev 1973
1977
Eimeria
baskanica^
Mustela
erminae
(ermine)
Homoxenous Oval with tapered
ends
L 112-126
W 84-98
M absent
PG absent
OR present
SR absent Bean shaped Nukerbaeva amp
Svanbaev 1977
Eimeria furonis Mustela
putorius
Mustela
putorius furo
(dom ferret)
Mustela
nigripes (BFF)
Mustela vison
(mink)
Homoxenous
Small intestine
rectum (H 1927)
Jejunumileum (BP
1993)
Spherical ndash
subspherical
L 11-14
W 10-13
OW 2 layers
M absent
PG absent
OR absent
Spindloid
L 8-9
W 4
SB present
SR present
Vermiform Blankenship-Paris
et al 1993
Hoare 1927 1935b
Jolley et al 1994
Nukerbaeva amp
Svanbaev
19731977
Williams et al 1988
1992 1996
Eimeria hiepei Mustela vison Homoxenous
Bile duct
Spherical
L 13-17
W 13-17
OW 2 layers
(outer
smooth)
M absent
PG absent
OR absent
L 6
W 4
SB absent
SR absent
Banana shaped Davis et al 1953
Grafner et al 1967
11
Coccidial species Host genus and
species
Life cycle
Location
Oocyst shape and
size
Oocyst
description
Sporocyst
description
Sporozoite
description
References
Eimeria ictidea Mustela
eversmanni
Mustela
nigripes
Mustela
putorius
Mustela
putorius furo
Homoxenous
Small intestine
Ovoid ndashellipsoid
L 13-27
W 13-21
OW 2 layers
M present
PG absent
OR absent
Ovoid
(irregular)
L 115
W65
SB present
SR present
- Hoare 1927 1935a
1935b
Jolley et al 1994
Litvenkova 1969
Svanbaev 1956
Tinar 1985
Williams et al 1988
1992
Eimeria irara Eira barbara
(tayra)
Homoxenous
Feces
Ovoid
L 21-25
W 18-20
OW outer
layer smooth
M absent
PG absent
OR absent
Ellipsoid
L 10-12
W 65
SB present
SR present
Elongate (one
end broader than
the other)
Carini amp da
Fonseca 1938
Eimeria melis Meles meles
(European
badger)
Homoxenous Ellipsoid
L 20plusmn018
W 157plusmn002
LW128plusmn0017
(112-15)
OW 2 layers
(outer
smooth)
M absent
PG present
OR present
Ovoid
L
119plusmn0018
W 65plusmn008
LW 183
(155-24)
SB present
L 90plusmn005
W 324plusmn0025
SRB present
Anwar et al 2000
Kotlan amp Pospesch
1933
Eimeria mustelae Mustela vison
Mustela nivalis
(snow weasel)
Homoxenous
Duodenumileum
Spherical or
Ellipsoid
L 18-26
W 14-24
OW 2 layers
M absent
PG present
OR absent
Ovoid
L 8
W 5
SB present
SR present
Broad at one
end and tapered
at other
L 7
W 3
Glebezdin 1978
Iwanoff-Gobzem
1934
Levine 1948
Musaev amp Veisov
1983
Tinar 1985
Eimeria sablii Martes zibellina
(sable)
Homoxenous
Gut
Spherical or
subspherical
L 112-126
W 112
OW 2 layers
M absent
OR absent
Ovoid
L 56
W 42
SR present
Elongate Nukerbaeva 1981
Eimeria sibirica Martes zibellina Homoxenous Ovoid
L avg 216
W avg 180
LW 1076
OW 2 layers
M absent
PG absent
OR absent
Ovoid
L 96-112
W 56-72
SR absent
Elongate Nukerbaeva 1981
Yakimoff amp
Gousseff 1934
Yakimoff amp
Terwinsky 1930
1931
12
Coccidial species Host genus and
species
Life cycle
Location
Oocyst shape and
size
Oocyst
description
Sporocyst
description
Sporozoite
description
References
Eimeria vison
(Eimeria
mustelae)
Mustela
putorius
Mustela
putorius furo
Mustela vison
Homoxenous
Small intestine
+- large intestine
Ovoid
L 17-22
W 9-18
OW 2 layers
M absent
OR
sometimes
present
Ovoid or
Piriform
L 10
W 55
SB absent
SR present
Curved or Club
shaped
L 9
W 25
Foreyt amp Todd 1976
Foreyt et al 1977
Kingscote 1934
1935
Levine 1948
McTaggart 1960
Nukerbaeva amp
Svanbaev
19731977
Tinar 1985
Umurzakov amp
Nukerbaeva 1985
Wolter 1961
Zimmermann 1959
Isospora africana Ictonyx libyca
(Libyan striped
weasel)
Homoxenous
Feces
Spherical
L 25-27
W 25-27
OW 2 layers
(outer
smooth)
M absent
PG absent
OR absent
Ovoid
L 15-17
W 10-12
SB absent
SR present
Elongate
L 135
W 3
Prasad 1961
Isospora altaica Mustela altaica
(mountain
weasel)
Homoxenous
Gut
Oval or spherical
L 280-336
W 252-280
LW 121 (111-
124)
OW 2 layers
M absent
PG absent
OR absent
Ovoid or
spherical
L 140-168
W 111-168
SR present
Svanbaev amp
Rachmatullina
1971
Isospora goussevi Mustela nivalis Homoxenous
Large intestine
Ovoid
L 224 (220-250)
W 174 (160-190)
LW 135 (133-
137)
OW 1 layer
PG present
OR present
Ovoid
L 120
(100-130)
W 70 (60-
80)
SB present
SR present
Elongate Musaev amp Veisov
1983
13
Coccidial species Host genus and
species
Life cycle
Location
Oocyst shape and
size
Oocyst
description
Sporocyst
description
Sporozoite
description
References
Isospora
hoogstraali
Ictonyx libyca Homoxenous
Feces
Ellipsoid
L 37-41
W 32-34
OW 2 layers
(outer
smooth)
M absent
PG some
OR absent
Ovoid
L 19-21
W 13-15
SB absent
SR present
Club-shaped
L 18-19
W 4-6
Prasad 1961
Isospora laidlawi Mustela
putorius
Mustela
putorius furo
Mustela vison
Homoxenous
Feces
Intestinal contents
Ovoid L
320-368
W 272-304
OW 2 layers
M absent
PG absent
OR absent
Ellipsoid
L 208
W 144
SB absent
SR present
Sausage shaped Foreyt et al 1977
Hoare 1927
Levine 1948
McTaggart 1960
Nukerbaeva amp
Svanbaev 1973
1974 1977
Tinar 1985
Isospora lutrae Lutra lutra
(European
otter)
Lutra
canadensis
(North
American river
otter)
Homoxenous Spherical
L 312 (275-32)
W 296 (28-31)
LW 104
(10-112)
OW 2 layers
(outer
smooth)
M absent
PG absent
OR absent
Ellipsoid
L 182 (17-
19)
W 144 (14-
16)
LW128
(12-14)
Sb absent
sSB absent
SR present
Spindle- shaped
L 124
W 25
SRB present
Torres et al 2000
Hoover et al 1985
Isospora
martessii
Martes zibellina Homoxenous
Gut
Ovoid short oval or
spherical
L 252 ndash 280 196
168
W 168 ndash 224 168
168
OW 2 layers
M absent
OR absent
Ovoid
L 112-168
W 84-112
SR present
Elongate Nukerbaeva 1981
Isospora melis Meles meles Homoxenous Ovoid
L 328plusmn034
W 269plusmn019
LW122 (110-
157)
OW 2 layers
(outer
smooth)
M absent
PG absent
OR absent
Ellipsoid
L
215plusmn0166
W 14plusmn012
LW 155
(133-185)
SR absent
Round at one
end other end
tapered
L 142plusmn116
W 40plusmn017
SRB absent
Anwar et al 2000
Glebezdin 1978
Kotlan amp Pospesch
1933
Pelleacuterdy 1955
14
Coccidial species Host genus and
species
Life cycle
Location
Oocyst shape and
size
Oocyst
description
Sporocyst
description
Sporozoite
description
References
Isospora
mustelae (nomen
nudum)
Martes martes Ovoid L
7 W
225
M present - - Galli-Valerio 1932
Isospora nivalis Mustela nivalis Homoxenous
Large intestine
Ovoid
L 206 (200-230)
W 184 (180-210)
LW 11 (109-111)
OW 1 layer
PG absent
OR absent
Ovoid
L 125
(120-130)
W 80 (70-
90)
SR present
Lemon or pear
shaped
Musaev amp Veisov
1983
Unnamed
ldquoCoccidiardquo^
Mustela
nigripes
Urinary bladder - - - - Jolley et al 1994
Unnamed
ldquoCoccidiardquo^
Mustela
nigripes
Trachea bronchus
bronchial glands
- - - - Jolley et al 1994
Unnamed
Eimeria sp^
Mustela
nigripes
Feces
intestinal contents
Ovoid
L 350-386
W 212-232
- - - Jolley et al 1994
Williams et al
1992
Unnamed
Eimeria sp^
Mustela
putorius furo
Small intestine - - - - Blankenship-Paris
et al 1993
Unnamed
Eimeria sp^
Mustela nivalis Homoxenous
Large intestine
Ovoid-ellipsoid L
2031 (1712-2162)
W 148 (1225-
1681)
LW 136 (121-16-
)
OW 1 layer
PG absent
OR absent
Ovoid or
pear-shaped
L 60-100
W 40-80
SR present
Elongate
L 50-90
W 30-70
Musaev amp Veisov
1983
Unnamed
Eimeria sp^
Martes martes
(marten)
Homoxenous Ovoid
L avg 216
W avg 180
LW 1076
OR absent 4 sporocysts
SR present
L 126
W 60
Yakimoff and
Gousseff 1934
Unnamed
Isospora sp^
Mustela
putorius furo
Feces - - - - Bell 1994
Unnamed
Isospora sp^
Mustela
putorius furo
Feces - - - - Bell 1994
Legend L = length W = width LW = length-width ratio avg = average OW = oocyst wall PG = polar granules M = micropyle SB = Stieda body sSB =
subStieda body OR = oocyst residuum SR = sporocyst residuum SRB = sporozoite refractile body ^ = species inquirendae - = no information provided by
author(s) = information obtained from secondary sources (primary reference could not be obtained) All measurements are in micrometers Bolded references
15
are those from which morphometric data were assembled Remaining references indicate other authors who have identified that parasite species in the same or
similar host
16
143 Eimeriid coccidia described from domestic ferrets
Three species of coccidia were originally described from 50 domestic ferrets (Mustela putorius
furo) Eimeria ictidea Eimeria furonis and Isospora (= Cystoisospora) laidlawi (Hoare 1927) All three
species were detected in feces from domestic ferrets at a research facility undergoing an outbreak of
canine distemper Sick ferrets appeared more frequently infected than healthy ones As per Hoare (1927)
none of the ferrets appeared to display clinical signs associated with protozoal infection For each
parasite the author described morphology of sporulated oocysts isolated from feces and sporulation time
(exogenous life stages) The pre-patent period (minimum duration of endogenous development) in an
inoculated naiumlve ferret was described only for E furonis and E ictidea due to insufficient sample size of
I (=C) laidlawi oocysts for an experimental infection trial Sporulation of oocysts occurred within 5-6
days for E furonis 3 days for E ictidea and 4 days for I (=C) laidlawi The sporulated oocysts of E
furonis were spherical with a double outer wall with a thin colourless outer layer and thick yellowish
inner layer no micropyle or residual body and measured on average 128 times 120 microm (length [L] 112-
144 width [W] 104-128 shape index [SI] 107) Unsporulated oocysts contained a zygote with a
diameter of 96 microm Sporocysts were spindle-shaped with one end constrictedblunted contained a
residual body and on average measured 8-88 times 4 microm Sporozoites were vermiform with one end
narrower than the other arranged head to tail and had a central nucleus a clear vacuole was identified in
some at the broad end The sporulated oocysts of E ictidea were oval or elliptical with a double outer
wall with a thin colourless outer layer and thick yellowish inner layer no micropyle or residual body
and measured on average 236 times 175 microm (L 184-272 W 128-208 shape index 135) The zygote in
unsporulated oocysts was elongate with a diameter of 15 times 12 microm when originally passed in feces but
became more spherical with time Sporocysts were irregularly oval with one end broad and the other
more constricted contained a residual body and on average measured 115 times 65 microm Sporozoites were
vermiform with one end narrower than the other arranged head to tail and had a central nucleus and a
clear vacuole at the broad end The sporulated oocysts of Isospora (=Cystoisospora) laidlawi were ovoid
with a double outer wall with a thin colourless outer layer and thick yellowish inner layer no micropyle
17
or residual body and measured on average 34 times 29 microm (L 320-368 W 272-304) Unsporulated
oocysts contained a spherical zygote with a diameter of 236 microm Two sporocysts were identified each
containing 4 sporozoites and no Stieda body sporocysts were elliptical contained a residual body and on
average measured on 208 times 144 microm Sporozoites were sausage shaped with one end slightly pointed
and had a central nucleus and a clear vacuole identified at the pointed end Sporozoites were arranged
with pointed ends all at the same pole of the sporocyst The pre-patent periods described for E furonis
and E ictidea were 6 days and 7 days respectively (Hoare 1927)
Since Hoarersquos initial description (Hoare 1927 Hoare 1935) multiple single case reports and
outbreaks of severe clinical disease associated with intestinal coccidiosis have been reported in domestic
ferrets Blankenship-Paris et al (1993) described a single case of a four-month-old domestic ferret that
presented depressed in thin body condition dehydrated and with pasty dark feces on the perineum This
ferret had been housed with its dam and another sibling neither dam nor sibling showed clinical signs of
enteric disease and both had negative fecal examination results on repeated evaluation Routine fecal
examination of the rest of the colony and necropsies on eight other ferrets in the colony revealed no
evidence of coccidial infection Enteric coccidiosis was determined to be the cause of disease in the four-
month-old ferret based on necropsy findings but the coccidia could not be speciated because diagnosis
was made on histologic findings only
Sledge et al ( 2011) described three separate outbreaks of severe enteric coccidiosis in domestic
ferrets from one ferret rescue centre (group 1) and two shelters (groups 2 and 3) all affected by the same
Eimeria sp The morphologic characteristics of sporulated oocysts were only described for group 1 no
coccidial oocysts were detected on direct smear or fecal flotation of diarrheic samples submitted from
groups 2 and 3 Oocysts were identified as spherical measuring 12-13 microm in diameter with four
sporocysts each containing two sporozoites Oocyst morphometrics histopathologic findings and nu 18S
rDNA partial sequences from all three groups were used collectively to confirm the coccidial species
identify in each outbreak as E furonis
18
Two cases of biliary coccidiosis with E furonis have been reported in domestic ferrets The first
was in a nine-week-old male ferret from a research facility (Williams Chimes amp Gardiner 1996) The
ferret presented with signs of hepatic disease and was negative for coccidia on fecal flotation and direct
smears Endogenous coccidial life stages were described from the gall bladder and liver on histologic
examination In tissue section the oocysts were oval to spherical and measured 125 times 120 microm Meronts
measured 108-130 times 89-93 microm and contained up to 16 merozoites The merozoites exhibited a double-
layered pellicle prominent conoid few rhoptries and many micronemes anterior to the nucleus Based on
the morphologic description of the life stages in this case the coccidia were identified by the authors as
an Eimeria species most likely E furonis Kaye et al (2015) described a second case of biliary
coccidiosis in an 18-month-old female pet domestic ferret with concurrent pure red cell aplasia In this
case all endogenous coccidial life stages were observed on histologic examination of the epithelium of
the extrahepatic biliary tree The oocysts were ovoid and measured 12 times 13 microm Meronts measured 12 times
15 microm and contained up to 16 merozoites each measuring 2 times 5 microm Based on the morphologic
description of the life stages in this case and nu 18S rDNA sequences the pathogen was also determined
to be E furonis Biliary coccidiosis has also been identified in mink (Mustela vison) with the etiologic
agent identified as Eimeria hiepei (Davis Chow amp Gorham 1953 Grafner Graubmann amp Dobbriner
1967)
Oocysts from Cystoisospora ohioensis have been reported from fecal samples collected from
healthy domestic ferret kits in a large American ferret breeding operation that were raised on the same
premise as juvenile domestic dogs (Patterson amp Fox 2007) The method of identification of this parasite
was not described by Patterson amp Fox A second similar institution reported the presence of a
Cystoisospora species also thought to be C ohioensis in routine fecal examination of their ferret colony
(Dr Bambi Jasmin personal communication) Coccidial identification in this case was performed by the
Animal Health Diagnostic Center at Cornell University The significance of these findings is unknown as
no clinical signs or histologic lesions have been described in domestic ferrets associated with shedding of
19
oocysts and the definitive host for C ohioensis is the domestic dog It is most likely that fecal
identification of C ohioensis represents a pseudoparasite in both of these cases or perhaps an
undescribed Cystoisospora sp that is morphologically indistinguishable from C ohioensis
It is difficult to estimate the prevalence of enteric coccidia within the North American domestic
ferret population Fecal samples submitted to university or large veterinary diagnostic laboratories from
domestic ferrets in Canada are uncommon and samples positive for coccidia appear infrequently (Dr
Donald Martin personal communication) Data from Idexx Vet Med Lab in Ludwigsburg Germany was
compiled to review the prevalence of coccidia and Giardia within fecal samples from domestic ferrets
(Pantchev et al 2011) The authors reported that of 284 fecal samples submitted from 2002-2004 18
(63) had detectable coccidial oocysts on fecal flotation Oocysts were identified based on morphologic
characteristics as E ictidea E furonis I (=C) laidlawi and another unidentified Isospora species
Comparative data from the same laboratory from 2009-2010 included sample submissions from 253
ferrets 21 (83) of which were positive for coccidial oocysts on fecal flotation Nine of the samples
were positive for E furonis three were positive with both E furonis and I(=C) laidlawi present eight
were positive only for I(=C) laidlawi and one sample contained both E furonis and E ictidea
identification in all cases was based on morphologic characteristics No statistically significant difference
in the occurrence of coccidial oocysts was detected when data from the two periods were compared
(Fisherrsquos exact test P=041) (Pantchev et al 2011)
144 Molecular characterization
Molecular characterization of Eimeria furonis was first performed by Abe et al (2008) using
oocysts purified from the feces of a single domestic ferret with clinical signs of coccidial enteritis Small
subunit ribosomal DNA (nu 18S rDNA) primers CYC1FE (5ʹ-TAC CCA ATG AAA ACA GTT T-3prime) and
CYC4RB (5prime-CGT CTT CAA ACC CCC TAC TG-3prime) were used to amplify a 347 base pair (bp) fragment
of nu 18S rDNA These primers were initially developed for molecular identification of Cyclospora
species but have since been shown to amplify nu 18S rDNA from several Eimeria species (Matsubayashi
20
et al 2005) The amplicon was sequenced (GenBank AB329724) and compared with previously
published partial nu 18S rDNA sequences from 40 Eimeria two Isospora and four Cyclospora species
The resulting phylogram grouped E furonis with E alabamensis (cattle) and E meleagrimitis (turkey) In
the same study the microscopic morphology of the oocysts was used to identify this coccidial species as
E furonis by comparison with published descriptions of E furonis E ictidea and E heipei by Hoare
(1927) Hoare (1935) and Grafner Graubmann amp Dobbriner (1967) respectively
Nuclear 18S rDNA was also used by Sledge et al (2011) for molecular identification of the
eimeriid coccidia implicated in the three distinct outbreaks of enteric disease in domestic ferrets As
described above initial identification and speciation of the coccidia was performed using morphologic
characteristics of the sporulated oocysts collected from feces in one of the three outbreaks being
investigated the oocysts were identified as E furonis Histologic sections of formalin fixed intestinal
segments from ferrets from each of the three outbreaks contained multiple coccidial life stages DNA was
then isolated from stored formalin-fixed tissues for further genetic analysis Using the partial nu 18S
rDNA gene sequence reported by Abe et al (2008) (GenBank AB329724) the following PCR primers
were created 5ʹ-ACA ATT GGA GGG CAA GTC TG-3ʹ and 5ʹ-GGCGAC AAG CCT GCT TGA AAC-
3ʹ PCR amplification produced a 247 bp amplicon from each of the three groups Analysis and
sequencing of amplicons from all three groups showed 100 homology to nucleic acid sequences
previously reported by Abe et al (2008) for the gene encoding E furonis nu 18S rDNA
Coccidia were identified within hepatobiliary lesions in a domestic ferret receiving
immunosuppressive therapy for red cell aplasia (Kaye et al 2015) DNA was extracted from frozen liver
and a 247 bp fragment of the nu 18S rDNA was amplified using the primers previously described by
Sledge et al (2011) and sequenced Kaye et al (2015) reported that the DNA sequence of the amplicon
was 100 homologous to the published nu 18S rDNA of E furonis and 95 homologous to the nu 18S
rDNA of E myoxi (rodent) E alabamensis (cattle) and I robini (avian)
21
145 Clinical signs of disease in domestic ferrets
Hoare (1927 1935b) in his initial descriptions of enteric coccidiosis in domestic ferrets
observed that clinical signs of intestinal disease were not evident The recent literature supports the
finding of subclinical disease but also describes signs ranging from mild transient diarrhea in young or
stressed animals to more severe disease with dehydration lethargy depression weight lossemaciation
inappetence and death (Blankenship-Paris et al 1993 Powers 2009 Sledge et al 2011 Hoefer et al
2012 Patterson et al 2014) Rectal prolapse has also been reported in ferrets with enteric coccidiosis
(Hillyer 1992 Hoefer et al 2012) In one study co-infection with coccidia and Lawsonia intracellularis
(Desulfovibrio sp) was diagnosed in 4 of 19 ferrets with proliferative bowel disease (Li et al 1996)
These ferrets presented with variable clinical signs including diarrhea lethargy anorexia weight loss
dehydration and emaciation
In the two reports of biliary coccidiosis clinical signs conformed to those expected with
hepatobiliary disease Williams et al (1996) described their case to have presented with emaciation poor
appetite abdominal distension and icterus Kaye et al (2015) described a one week history of lethargy
inappetence and icterus with serum biochemistry results consistent with cholestasis later clinical signs in
this case included melena anemia and cachexia
146 Gross necropsy and histologic findings
The pathology of enteric coccidiosis in domestic ferrets was described by Hoare (1927 1935b)
Two healthy domestic ferrets were experimentally inoculated one each with large numbers of mature
oocysts of either E furonis or E ictidea that were isolated during his initial work The inoculated ferrets
were killed humanely for histologic examination of intestinal sections at the time of first detection of fecal
oocyst shedding no clinical signs of coccidiosis were detected in these ferrets prior to death Infection
with E furonis resulted in invasion of the epithelium of the small intestine and rectum Within the small
intestine the parasites were concentrated in the tips of the villi but could be found to the level of the
22
opening of the crypts of Lieberkuumlhn In rectal sections life stages were limited to the epithelial ridges
between the openings of the glands of Lieberkuumlhn Organisms were located within the apical portion of
the epithelial cells and intensely infected regions exhibited multiple parasites within a single host cell
Both asexual and sexual life stages were present within the same sections Hoare (1927) described similar
histopathologic changes in naturally infected ferrets but the proportion of asexual versus sexual life
stages differed In natural infections sexual life stages were more numerous whereas in experimental
infections asexual life stages predominated these findings would be expected to correlate with the stage
of infection at which ferrets died or were humanely killed for tissue collection and would not be
reflective of differences between natural and experimental infection with this parasite Hoare also
described the morphology of the different endogenous stages including trophozoite (3-4 microm) merozoite
(stumpy sausage shaped L 3-4 microm W 2 microm) macrogamete (spherical 8 microm diameter with darkly
staining globular inclusions of reserve material) and microgamete (described as similar to those of other
Eimeria species) Two types of merogony are described from histologic sections the first with stumpy
merozoites as described above and the second with merozoites with elongated curved bodies and a
compact polar nucleus measuring 60 times 13 microm This second merogonic generation was observed almost
exclusively in the naturally infected ferrets and was associated with initiation of sexual differentiation and
reproduction
The pathology of experimental and non-experimental infection with E ictidea in domestic ferrets
was also described by Hoare (1927 1935b) Parasitic invasion of the epithelium was noted only in the
small intestine with patchy distribution of the parasite life stages throughout affected sections Within the
small intestinal villi the parasites were again concentrated in the tips of the villi with infected epithelial
cells never containing more than one parasite As each intracellular parasite grew it filled the entire host
cell displacing the nucleus to the base of the cell Predominantly sexual life stages were detected in tissue
sections with few asexual generations observed Interestingly the parasites were arranged into age
groups with forms of the same life stage grouped together within the affected epithelial sections this is in
23
contrast to E furonis where life stages of different maturities were found together in affected sections
Hoare described the morphology of the different endogenous stages of E ictidea including merozoites
(free within the lumen elongated vermiform with one pointed end and a nucleus located at the rounded
end 11 microm times 1 microm within the epithelium shortened and rounded 3-4 microm diameter) macrogametes
(elongated 20 times 7 microm occupying the entire host cell with darkly staining globular inclusions of reserve
material) and mature microgamonts (morphologically similar to those of other Eimeria species but larger
than those of E furonis) Of note a tissue reaction was observed specifically in association with more
developed life stages of E ictidea (eg mature meronts mature gamonts unsporulated oocysts) which
was not observed when cells contained earlier stages of development (eg trophozoites immature
gamonts) This tissue reaction was described by Hoare (1935a 1935b) as the development of an annular
constriction of the apical portion of the villus separating infected epithelial cells from unaffected cells
The constriction involved the epithelium but could also extend inwards into the core of the villus These
changes were associated with congestion of capillaries and extravasation of red blood cells within the
constricted segment and in some sections villar tip necrosis
In their case report of one domestic ferret Blankenship-Paris et al (1993) described the gross
pathologic lesions associated with intestinal coccidiosis in this case there was diffuse dilation and
reddening of the small intestine which was empty and the colon contained dark watery material
Histologic lesions were confined to the ileum and jejunum The jejunum exhibited thickening of the villi
with a crypt to villus ratio of 15 mild granulomatous inflammation in the lamina propria and large
numbers of coccidial meronts gamonts and oocysts within the enterocytes of the villar tips
The gross lesions described by Sledge et al (2011) from 20 domestic ferrets are as follows thin
body condition with moderate to marked dehydration perineal staining with diarrhea moderate dilation
of the small and large intestines and the presence of pasty tan to tarry black digesta within the distal small
intestine and colon Other findings in one to a small number of ferrets included enlarged pale tan livers
splenomegaly with dark red colouration and multiple superficial gastric or duodenal ulcers The
24
histologic lesions from 10 ferrets included moderate blunting and occasional fusion of jejunal and ileal
villi focal attenuation and erosion of the epithelium of the villar tips with exudation of fibrin neutrophils
and blood into the intestinal lumen in regions with severe erosion Intact epithelial cells at the villus tips
and rarely sloughed epithelial cells in the intestinal lumen contained numerous intracytoplasmic coccidia
representing a range of asexual and sexual life stages (meronts macrogamonts microgamonts and
oocysts) The subjacent lamina propria of the small intestine and of the large intestine exhibited moderate
lymphoplasmacytic infiltration with occasional neutrophils and congestion of blood vessels Marked
mucosal hemorrhage was identified in the most severely affected sections
Marked gross and histopathologic hepatobiliary lesions were described in a single ferret by
Williams et al (1996) On gross necropsy the liver was pale and enlarged with dilated firm bile ducts
and thickening of the gall bladder wall Similar gross necropsy findings were described by Kaye et al
(2015) marked dilation and mural thickening of the entire biliary tree (including gall bladder intrahepatic
and extrahepatic bile ducts) On histopathology Williams et al (1996) noted that the marked thickening
of the gallbladder wall was a result of cystic proliferation of mucosal glands which were separated by
tracts of fibrous connective tissue and marked granulomatous inflammation Liver sections exhibited
marked biliary hyperplasia marked periductular fibrosis and moderate periportal lymphoplasmacytic
cuffing There was multifocal papillary proliferation of bile duct epithelium and dilation of the bile ducts
and within the ductular lumens there were moderate numbers of lymphocytes and plasma cells small
numbers of degenerate neutrophils sloughed epithelial cells and debris All endogenous coccidial life
stages were present within the gall bladder and biliary epithelium with meronts visible in 20 of the
intact epithelial cells of the biliary tree and gallbladder and oocysts free within the lumen of the
intrahepatic bile ducts Similar lesions were present in the case described by Kaye et al (2015) and as
well as in juvenile and adult farmed mink (Mustela vison) with hepatobiliary coccidiosis (Davis Chow amp
Gorham 1953)
25
15 INTRODUCTION TO ENTERIC COCCIDIOSIS IN THE BLACK-FOOTED FERRET
151 Natural history and conservation of the black-footed ferret in North America
Black-footed ferrets are one of only three wild ferret species worldwide the other species are the
European polecat (Mustela putorius) and the Siberian polecat or steppe polecat (Mustela eversmanii)
They are the only native North American ferret species and the most endangered North American
carnivore They are nocturnal carnivores whose diet and lifestyle are highly dependent on local prairie
dog (Cynomys sp) populations Prairie dogs comprise almost exclusively the diet for the BFF who also
use the complex burrow systems made by prairie dogs to escape their predators and raise their young
(Santymire et al 2014 USFWS BFF Recovery Program 2017)
While formerly distributed throughout the North America prairie ecosystem BFF were
considered extinct by the late 1950s In 1964 a single population was discovered in Mellette County
South Dakota Progressive decline of this population in subsequent years resulted in the decision by
United States Fish and Wildlife Service (USFWS) to initiate a captive breeding program for the species
From 1971-1973 four females and five males were captured for this purpose Despite successful breeding
no kits survived and the last adult ferret in this captive colony died in 1979 at that time BFF were again
presumed extinct in the wild based on annual surveys of the initial capture site In 1981 a dead BFF was
discovered by a ranch dog outside of Meeteetse Wyoming allowing wildlife biologists to identify
another colony of BFF This colony flourished until 1985 when an outbreak of canine distemper in the
BFF population and an outbreak of sylvatic plague in the local prairie dog population resulted in sharp
population declines From 1985 through 1987 all 24 of the remaining BFF were trapped and brought into
captivity to re-initiate the captive breeding program Six ferrets in this initial group died of canine
distemper while in captivity and of the remaining 18 survivors 7 bred successfully to create the founding
population of the current captive breeding population Today this captive breeding population consists of
approximately 300 BFF distributed among multiple institutions (Santymire et al 2014)
26
Since 1986 this multi-institutional effort has been breeding BFF in captivity with reintroductions
back into the wild in 28 selected locations in Canada the USA and Mexico Currently six facilities
participate in the BFF Species Survival Plan (SSP) the Toronto Zoo USFWS National Black-footed
Ferret Conservation Center National Zoorsquos Smithsonian Conservation Biology Institute Louisville
Zoological Garden Cheyenne Mountain Zoo and the Phoenix Zoo (Black-footed Ferret Recovery
Implementation Team 2011) As of 2011 over 8000 BFF kits had been produced in captive breeding
facilities (Black-footed Ferret Recovery Implementation Team 2011)
Multiple infectious diseases pose a significant risk to the captive breeding and post-release
survival of BFF including canine distemper and sylvatic plague Coccidiosis is recognized as a cause of
significant juvenile morbidity and mortality in captive breeding programs and can result in significant
population losses (Bronson et al 2007 Santymire et al 2014 USFWS BFF Recovery Program 2017)
152 Coccidia identified from black-footed ferrets
Eimeria ictidea and Eimeria furonis have been identified in black-footed ferrets based on
morphologic criteria (Jolley et al 1994) Jolley et al examined fecal samples from six captive BFF during
a distemper outbreak as well as samples from wild BFF They described one medium-sized ovoid
tetrasporic dizoic oocyst with a double wall presence of a polar body and lacking both an oocyst residual
body and micropyle The oocysts measured 232 microm (range 182-274) by 155microm (range 130-162) with
a SI of 150 The sporocysts were elongate with the presence of both sporocyst residuum and a Stieda
body Sporozoites contained prominent refractile bodies at the posterior end and were aligned anterior to
posterior within sporocysts These oocysts were shed by all six captive ferrets On histopathology of
intestinal sections merogony and gametogony were observed within the villar epithelium throughout the
small intestine but were concentrated in the jejunum Two morphologically distinct meronts were
detected in these sections one at the villar tips which was larger and lacking in undifferentiated mass
and the other at the base of the villi or rarely in the intestinal crypts Gametogony was predominantly
27
observed at the villar tips and was noted throughout the small intestine These organisms were considered
consistent with Eimeria ictidea based on descriptions by Hoare (1927) from domestic ferrets
A second small spherical to subspherical tetrasporic dizoic oocyst was documented that had a
pink double wall a granular residual body and lacked both oocyst polar body and micropyle This
smaller oocyst measured 126plusmn12 microm (108-152) by 119plusmn09 microm (101-129) with a SI of 106 The
sporocysts were elongate with the presence of a Stieda body and sporozoites contained refractile bodies
Similar to the larger Eimeria species described above merogony and gametogony were observed within
the villar epithelium throughout the small intestine with endogenous developmental stages most
numerous in the jejunum The meronts were small with 16 or fewer merozoites Micro- and
macrogamonts were observed clustered within the apical third of the villar epithelium as were meronts
Jolley et al (1994) determined these small spherical oocysts to be consistent with Eimeria furonis as
described by Hoare (1927) from domestic ferrets
Jolley et al (1994) described a third type of coccidial oocyst occasionally detected in small
numbers within the BFF fecal samples the authors did not state whether this third type of oocyst was
recovered from wild or captive BFF The oocysts measured 370plusmn13 microm (350-386) by 223plusmn23 microm
(212-232) with a SI of 106 Attempts to sporulate collected oocysts were largely unsuccessful and
corresponding endogenous stages were not identified on histopathologic examination of necropsied
ferrets precluding further morphologic identification of the parasite It should be noted that coccidial
oocysts with similar measurements had not been detected in wild or captive prey species available for
ingestion by BFF (Jolley et al 1994)
Previous to this report coccidial oocysts had been isolated from the feces of BFF in two captive
populations (Carpenter amp Hillman 1979 Williams et al 1988) The abstract by Carpenter amp Hillman
(1979) did not describe the oocysts whereas Williams et al (1988) stated that two Eimeria species (one
with larger oocysts and one with smaller oocysts) were identified within the fecal samples but they were
28
not identified further Interestingly Williams et al reported both Eimeria species to be shed in the feces
of all ferrets concurrently affected by distemper and by approximately 30 of the clinically healthy
ferrets at the time of investigation
Non-enteric coccidia have been reported from captive BFF in one facility by two authors (Jolley
et al 1994 Williams et al 1988) Both reports presumably describing the same case(s) noted the
presence of endogenous coccidial life stages in histologic sections of respiratory tissue and merozoites of
an unidentified coccidium in impression smears of the urinary bladder from BFF diagnosed with canine
distemper Meronts were observed within the epithelium of the trachea a large bronchus and associated
bronchial glands Jolley et al (1994) described the lesions as occurring in the same ferret whereas in the
earlier report by Williams et al (1988) they are described as occurring in two different ferrets There have
been no subsequent published reports of systemic coccidiosis in black-footed ferrets and no cases have
been identified within the pathology database of the Toronto Zoo captive BFF population or by the
current SSP pathologist (Dr Michael M Garner personal communication)
There is a significant information gap regarding the pre-patent periods and pathogenicity of both
identified Eimeria species in BFF and studies to further characterize the eimeriid coccidia of the BFF are
lacking
153 Morbidity mortality and clinical signs associated with enteric coccidiosis in black-footed ferrets
The clinical signs of enteric coccidiosis in black-footed ferrets include mucoid to hemorrhagic
diarrhea abdominal discomfort lethargy appetite loss vomiting and dehydration In some cases sudden
death precedes the development of diarrhea Both adult and juvenile BFF are affected by the disease
which causes significant morbidity and mortality in captive populations (Bronson et al 2007) One
retrospective study of the captive BFF population at the Smithsonian National Zoological Park
determined that the most common cause of death in juvenile BFF (aged 30 days ndash 11 months) was
gastrointestinal pathology (524 of juvenile deaths) with 636 of these cases caused by enteric
29
coccidiosis (Bronson et al 2007) Despite the significance of this disease to the captive population its
effect on morbidity and mortality in wild BFF populations is unknown To the authorrsquos knowledge no
routine surveys of fecal parasites have been conducted on wild-born or captive released BFF during
yearly spotlighting events at ferret release sites However samples may be collected opportunistically if
fecal material is identified within the traps used to catch wild BFF during yearly surveys at release sites
Where fecal samples have been analyzed a 13 prevalence of coccidiosis has been identified in wild
born BFF (Dr Rachel Santymire personal communication) Fecal samples have been collected from BFF
at four release sites within the USA Wind Cave National Park (South Dakota) Badlands (South Dakota)
Conata Basin (South Dakota) and Aubrey Valley (Arizona) and positive samples were identified only at
the first site (Dr Rachel Santymire personal communication) Although radio-telemetry has been used at
some release sites to determine sources of mortality and factors involved in survival its use is not
widespread Furthermore the nocturnal and fossorial lifestyle of the BFF is a significant impediment to
the surveillance and monitoring of disease in this species
16 TREATMENT PREVENTION AND CONTROL OF INFECTION BY EIMERIA SPP
161 Current recommendations for treatment of eimeriid coccidia in carnivores
Described anticoccidial therapies for carnivores come from research in domestic cats and dogs
infected by Cystoisospora species these tissue coccidia (family Sarcocystidae) are only distantly related
to the Eimeria species infecting the BFF and other ferrets Current therapeutic recommendations by the
Companion Animal Parasite Council (CAPC 2013) for treatment of described Cystoisospora species
isolated from cats and dogs include the following amprolium (300-400 mg daily for 5 days in dogs 110-
200 mg daily for 7-12 days in dogs 60-100 mgkg daily for 7 days in cats) amproliumsulfadimethoxine
(150 mgkg amprolium and 25 mgkg sulfadimethoxine daily for 14 days in dogs) diclazuril (25 mgkg
for one dose in cats) furazolidone (8-20 mgkg 1-2 times daily for 5 days in dogs and cats) ponazuril (20
mgkg daily for 1-3 days in dogs and cats) quinacrine (10 mgkg daily for 5 days in cats)
30
sulfadimethoxine (50-60 mgkg daily for 5-20 days in dogs and cats) sulfadimethoxineormetoprim (55
mgkg sulfadimethoxine and 11 mgkg ormetoprim daily for 7-23 days in dogs) sulfaguanidine (150 or
200 mgkg daily for 6 days or 100-200 mgkg every 8 hours for 5 days in dogs and cats) toltrazuril (10-
30 mgkg daily for 1-3 days in dogs) trimethoprimsulfonamide (30-60 mgkg trimethoprim daily for 6
days if gt4kg 15-30 mgkg trimethoprim daily for 6 days if lt4kg) (CAPC 2013) Notably the use of all
drugs listed by the CAPC is considered off-label with the exception of sulfadimethoxine
162 Current recommendations for anticoccidial treatment and prophylaxis in domestic and black-
footed ferrets
1621 Domestic ferrets
Recommended daily oral treatment regimens for enteric coccidiosis in domestic ferrets include
amprolium (19 mgkg once daily 05 mgkg) decoquinate (05 mgkg) sulfadimethoxine (300 mgkg in
drinking water) or sulfadiazine-trimethoprim (30 mgkg once daily) all administered for a minimum of
two weeks (Bell 1994 Patterson amp Fox 2007 Patterson et al 2014) Both the aforementioned
coccidiostats amprolium and decoquinate are sold in large formats and are ideal for use in larger
operations such as breeding facilities research facilities or rescue centers Other anticoccidial therapies
used in domestic ferrets include toltrazuril (20 mgkg) and ponazuril (30-50 mgkg) once daily It should
be noted that all anticoccidial therapy used in domestic ferrets is considered off-label drug use
Multiple follow up fecal examinations should be performed after the treatment regimen is
complete and large groups may need to be treated multiple times Routine cage cleaning is also important
to decrease the environmental oocyst burden and prevent re-infection and in the case of coccidial
outbreaks ferrets should be transferred to clean cages multiple times during the course of anticoccidial
therapy Disinfectants such as bleach or quaternary ammonium compounds or dry heat should be used
for effective environmental decontamination (Patterson et al 2014)
31
1622 Species Survival Plan recommendations for black-footed ferrets
Treatment and prophylaxis of enteric coccidiosis with oral sulfadimethoxine was previously
recommended by the BFF Species Survival Plan (SSP) However due to a suspicion of decreasing
efficacy of treatment ponazuril has been recommended recently for treatment Due to the perceived
exquisite sensitivity of BFF to enteric coccidia the current SSP recommendation for treatment is oral
ponazuril at 30 mgkg once if ferrets are to be transported anesthetized stressed or are otherwise
suffering from another illness or injury (even in the absence of clinical signs or fecal shedding) The same
single oral dose of 30 mgkg is also recommended for kits at weaning (30-35 days of age) post weaning
(40-45 days of age) and prior to anesthesia for initial examination and vaccines (50-60 days of age)
Large crowded or otherwise stressed litters should be administered 30 mgkg orally once every 7-10 days
during the period of stress For treatment of coccidial diarrhea diagnosed by fecal examination 30 mgkg
orally once every 7 days for two doses or 50 mgkg orally once daily for 3 days in food (repeated in 7
days) is recommended In BFF with clinical signs of dehydration administration of subcutaneous or
intravenous fluid therapy has been performed Additional therapy with other antibiotics is sometimes
provided in cases with severe clinical signs or where secondary or primary bacterial enteritis is suspected
There is no pharmacokinetic or pharmacodynamic information available for the use of
anticoccidial drugs in BFF or other Mustelidae and thus it is unknown whether the current dose or
frequency of administration is truly appropriate for treatment of coccidiosis In 2 to 3-month-old piglets
administered a single dose of ponazuril orally at 5 mgkg peak serum concentration occurred at 42 hours
(36-48 hr) and elimination half-life was ~56 days (Zou et al 2014) In llamas administered ponazuril as
a single dose of 20 mgkg orally peak serum concentration occurred at 84 hours and elimination half-life
was ~56 days (Prado et al 2011) In domestic cows administered ponazuril as a single 5 mgkg dose
orally peak serum concentration occurred at 48 hours and elimination half-life was 58 hours (Dirikolu et
al 2009) The relevance of serum drug concentrations for treating an intestinal infection that lacks
extraintestinal life stages is likely minimal because the highest drug dose will reach the site of concern
(intestines) and systemic distribution is not required
32
Furthermore no safety or efficacy studies have been performed in any ferret species to validate
the current uses of either sulfadimethoxine or ponazuril for treatment nor have the current recommended
treatment lengths been validated However anecdotal information based on current usage would indicate
that they are safe at the current dosages and frequencies of administration as no adverse effects have been
reported A recent efficacy study in shelter dogs and cats showed that oral ponazuril (50 mgkg)
administered once daily for 3 days was effective for treatment of infection with Cystoisospora as
determined by a reduction in or cessation of fecal oocyst shedding at 4 and 8 days post treatment
Treatment efficacy in this study was inversely correlated to fecal oocyst counts at the initiation of
treatment (Litster et al 2014) Interestingly efficacy of this dose compared to the other two treatment
groups (single 50 mgkg or 20 mgkg oral dose) did not seem to differ but no statistical analysis was
performed Given the ubiquitous use of ponazuril in captive breeding facilities and concerns regarding
resistance of coccidia species to sulfadimethoxine therapy information on minimum effective doses and
dose regimes would be necessary to inform appropriate future SSP treatment and management plans and
to minimize development of drug resistance
17 VACCINES AGAINST COCCIDIA
171 Theory
The development of resistance of protozoal parasites to chemotherapeutic agents has resulted in a
shift towards the development of vaccines for the protection of domestic livestock Immunity to enteric
coccidiosis in avian and mammalian species involves both humoral and cell mediated responses Eimeria
spp infection in sheep rats poultry and other species generally results in a protective immune response
against subsequent re-infections (Catchpole et al 1993 Shi et al 2000) Interestingly this is not the case
for some host parasite interactions for example a recent report indicated that primary infection with E
ninakohlyakimovae in goat kids did not provide protective immunity against subsequent challenge with
the same parasite (Ruiz et al 2013)
33
Vaccines can be divided into four general categories live vaccines inactivatedkilled vaccines
subunit vaccines and recombinant vaccines Live vaccines are orally administered using small numbers of
infectious oocysts or oocysts from strains with low pathogenicity and result in patent but ideally sub-
clinical infections in the host that will elicit a protective immune response Such live vaccines can be
produced using attenuated forms of the pathogen of interest for example in chickens using ldquoprecociousrdquo
strains of Eimeria spp These precocious strains undergo a reduced number of merogonic replications
within the host cells and thus fewer oocysts are shed in the feces of vaccinated animals This reduction in
endogenous merogonic cycles reduces the amount of damage to the intestinal epithelium as well as
reducing the number of oocysts contaminating the environment
Another strategy has been to use live parasites with truncated life cycles An example of this is
the Toxoplasma gondii vaccine developed to prevent abortion in sheep This parasite was passaged
multiple times through a mouse host resulting in an inability to produce tissue cysts (Meeusen et al
2007) This is desirable as the cyst stage of this parasite normally inhibited by the immune system can be
reactivated during periods of stress or immunocompromise The potential drawbacks of live vaccines
include 1) the ability to produce and isolate adequate numbers of coccidial oocysts to meet vaccine
production requirements 2) the potential development of clinical disease in the host as a result of
inoculation 3) the need for all susceptible individuals to receive the vaccine simultaneously to prevent
fecal-oral inoculation of unvaccinated animals with high doses of the infective agent likely to be present
in a shared environment through fecal shedding
Inactivated vaccines are produced when the microbe of interest is killed via application of heat
radiation or chemical treatment prior to inoculation into the host species While safer because they cannot
induce disease in the inoculated patient inactivated vaccines stimulate a reduced immune response
compared with live vaccines and are consequently less effective Subunit vaccines contain single or
multiple antigens of importance in initiating the host immune response rather than the entire pathogen of
concern Subunit vaccines cannot induce disease in the immunized host but are more difficult to produce
34
because they require a detailed understanding of host immune response to infection Recombinant
vaccines involve the genetic modification of a vector (virus or bacteria) one capable of infecting the host
of interest to contain DNA of the pathogen of interest These vectors induce an immune response in the
vaccinated host but as with subunit vaccines cannot induce disease However recombinant vaccines are
again difficult to produce because they require an in depth understanding of the life cycle stages genes
and antigens targeted by the host immune response to infection There are currently no recombinant
vaccines marketed in Canada for use in veterinary medicine against protozoal disease
Creation of effective vaccines against protozoal parasites is complicated by parasite antigenic
diversity during the different life cycle stages and among protozoal species and strains of the same species
(Meeusen et al 2007) Although most parasites induce some level of immunity in their host species the
immunological response to different parasite life stages and species has been poorly characterized for
most coccidia Furthermore many parasites have developed mechanisms to evade host immune responses
or to continue survive and replicate in and transmission by previously infected hosts Our limited
understanding of the immune responses against coccidial antigens has restricted commercial vaccine
production to live or attenuated vaccines (Meeusen et al 2007)
A notable disadvantage of anticoccidial vaccines is that they need to be developed for each
coccidial species of interest because of the species-specific nature of the immune responses this is a
considerable limitation compared with anticoccidial drugs that can have a much wider spectrum of action
(Vermeulen 2005) While the requirement for mass production of vaccine is a limiting factor for vaccines
developed for the agricultural industry this drawback would be less important for production of a vaccine
to be used in an endangered species
172 Species successes in anticoccidial vaccination
The first successful immunization against coccidiosis was reported in 1918 in dogs (Hall amp
Wigdor) In this report a dog that had previously recovered from coccidial infection with Diplospora
35
bigemina was fed three increasing doses of live non-attenuated coccidial culture (at 14 32 and 48 days
post recovery from primary infection) which resulted in no development of clinical signs and no oocyst
shedding for 11mdash18 days after each challenge Subsequently immunization of dogs and cats against
coccidia with protection lasting up to seven months was reported by Andrews (1926) Immunization of
albino rats to eimeriid infection after administration of three or more sublethal doses of Eimeria
nieschulzi via gastric intubation was reported by Morehouse (1938) further experiments showed that
sporozoites did not enter the host intestinal epithelium in immunized rats given a challenge dose
(Morehouse 1938) Similar findings were reported in chickens immunized against Eimeria tenella that
had 50 fewer intra-epithelial sporozoites following challenge compared to naiumlve birds (Augustine and
Danforth 1986) Conversely chickens previously inoculated with Eimeria acervulina exhibited more
intracellular sporozoites after challenge than naiumlve birds but sporozoites were not observed to develop in
previously immunized birds (Augustine and Danforth 1986) These findings provide further evidence that
the immune response to Eimeria spp may differ among host species
Vaccination against Eimeria species has been most successful and is most widely used in the
poultry industry particularly in breeder and layer flocks Almost all vaccines marketed for poultry are
live vaccines (attenuated and non-attenuated) Vaccination against other apicomplexan parasites in
domestic mammals has also been achieved but has been generally less effective for disease prevention
and is less widely available Marketed killed and inactivated (attenuated) vaccines include those
containing killed tachyzoites of Neospora caninum for cattle (Neoguard Merck Animal Health) and
chemically inactivated merozoites of Sarcocystis neurona for horses (EPM Vaccine Fort Dodge ndash no
longer in production) A subunit vaccine for Babesia canis in dogs uses cultured antigen (Pirodog
Merial) Available live vaccines include a vaccine against Toxoplasma gondii in sheep (Ovilis Toxovax
Intervet) that uses an attenuated temperature sensitive strain (S48)
36
18 RESEARCH GOALS AND OBJECTIVES
181 Objectives
a) To determine and characterize (morphologically and molecularly) the enteric coccidial species
currently affecting the black-footed ferret population
b) To describe the natural history of enteric coccidiosis in captive black-footed ferrets including
pre-patent period shedding frequency and burdens and morbidity and mortality rates
c) To compare molecular morphologic and life history characteristics of enteric coccidial species
identified in domestic ferrets to those in black-footed ferrets
d) To validate domestic ferrets as an experimental model for intestinal coccidiosis in the black-
footed ferret
182 Hypotheses
a) Multiple Eimeria species will be isolated from the black-footed ferret population
b) The Eimeria species identified from black-footed ferrets will be the same as those previously
described in domestic ferrets
c) A single pathogenic Eimeria species will be implicated in recorded outbreaks of clinical
coccidiosis during the period of study
d) Domestic ferrets can act as an experimental model of intestinal coccidiosis for black-footed
ferrets
183 Applications
The goal of this project is to better characterize the enteric coccidia of the endangered black-
footed ferret in order to set the stage for improved disease prevention and treatment To the authorrsquos
knowledge this project is the first attempt to isolate and perform molecular characterization of the
coccidial species endemic in the black-footed ferret population This information will be used to compare
these species to known coccidia from domestic ferrets and other related mammals As experimental work
37
cannot be carried out on enteric coccidiosis in the BFF due to its endangered status if the domestic ferret
can be validated as an experimental model studies of the patterns of anticoccidial resistance and
development of immunity against Eimeria spp can be undertaken in vivo The ultimate goal would be the
development of an autogenous vaccine used to improve survival of ferret kits and reduce morbidity and
mortality associated with coccidiosis in BFF captive breeding programs Based on clinical experience
stressful life events such a breeding weaning and transfer between institutions appear to increase the risk
of coccidial outbreaks in adult BFF As such vaccination could assist in reducing disease outbreaks in
BFF associated with various management activities There is no data on the significance of coccidiosis in
wild populations and limited means of disease surveillance following release vaccination during captive-
rearing or pre-release conditioning of BFF would be an ideal method of reducing the potential effects of
this disease in released and free-living BFF Increasing the numbers of ferrets being released to the wild
and releasing ferrets immune to the subsequent threat of coccidiosis would support the goals of the
conservation initiative for the black-footed ferret
38
CHAPTER 2 MOLECULAR CHARACTERIZATION OF ENTERIC COCCIDIA FROM
DOMESTIC FERRETS (MUSTELA PUTORIUS FURO)
This chapter has been submitted for publication as
Adriana R Pastor Dale A Smith and John R Barta (2017) Molecular Characterization of Enteric
Coccidia from Domestic Ferrets (Mustela putorius furo) Vet Parasitol Regional Studies and Reports (In
review)
ABSTRACT
Combined morphometric and molecular characterization of coccidia that infect domestic ferrets
(Mustela putorius furo) was completed to improve the diagnostic specificity of enteric lsquococcidiosisrsquo in
this host Coccidia positive fecal samples (n=11) and formalin fixed paraffin embedded intestinal tissues
(n=3) from domestic ferrets were collected from diagnostic laboratories in Canada and Europe An
average of 35 and 13 domestic ferret fecal samples per year were coccidia-positive when tested by
Canadian and European diagnostic laboratories respectively during the period 2008-2015 Oocyst
morphometrics and sequence genotyping at two loci (nuclear 18S rDNA [nu 18S rDNA] and
mitochondrial cytochrome c oxidase subunit I [mt COI]) were conducted on all samples The first nu 18S
rDNA and mt COI sequences for Isospora (=Cystoisospora) laidlawi and the first mt COI sequence for
Eimeria furonis were generated during this study Phylogenetic analysis of the mitochondrial COI
sequences demonstrated that E furonis was most closely related to E cf ictidea isolated from a black-
footed ferret (Mustela nigripes) and that I (=C) laidlawi was closely related to C canis and C felis The
identifications provided by diagnostic laboratories of the specific parasite species present in a sample
showed poor agreement with their identifications based on genotyping obtained in this study Molecular
techniques appear to be essential for accurate determination of coccidial species responsible for individual
and group outbreaks of coccidiosis and for further understanding of eimeriid host-parasite relationships
Key words coccidia Cystoisospora laidlawi domestic ferret Eimeria furonis Eimeria ictidea Mustela
putorius furo
39
21 INTRODUCTION
Coccidia are host-specific parasites of the phylum Apicomplexa with greater than 2000 species
named to date (Duszynski et al 2000 Upton 2000) The eimeriorinid coccidia (suborder Eimeriorina)
include typical intestinal coccidia such as Eimeria Isospora and Cyclospora species belonging to the
family Eimeriidae as well as tissue (cyst-forming) coccidia such as Cystoisospora Besnoitia Toxoplasma
and Sarcocystis species that belong to the family Sarcocystidae (see Cox 1994)
Enteric coccidia affect both domestic ferrets (Mustela putorius furo) and their wild counterparts
In his initial descriptions of enteric coccidiosis in domestic ferrets Hoare (1927 1935b) did not observe
clinical signs of intestinal disease associated with infection More recently it has been recognized that
enteric coccidiosis can result in clinical signs ranging from mild transient diarrhea to more severe disease
with dehydration lethargy depression weight lossemaciation inappetence and death (Blankenship-Paris
et al 1993 Powers 2009 Sledge et al 2011 Hoefer et al 2012 Patterson et al 2014) Rectal prolapse
has also been reported in ferrets with enteric coccidiosis (Hillyer 1992 Hoefer et al 2012) Disease
appears to be most common in young or stressed animals In one study co-infection with coccidia and
Lawsonia intracellularis (Desulfovibrio sp) was diagnosed in 4 of 19 ferrets with proliferative bowel
disease (Li et al 1996) these ferrets presented with clinical signs including diarrhea lethargy anorexia
weight loss dehydration and emaciation Two cases of biliary coccidiosis have also been reported in
domestic ferrets infection was associated with biliary epithelial hyperplasia cholecystitis and
cholangiohepatitis (Williams et al 1996 Kaye et al 2015)
Three species of coccidia affecting domestic ferrets were originally described and named by
Hoare (1927) Eimeria ictidea Eimeria furonis and Isospora (=Cystoisospora) laidlawi The three
species were detected in feces from 50 domestic ferrets involved in an outbreak of canine distemper at a
research facility For each parasite the author described the morphology of sporulated oocysts isolated
from feces as well as sporulation time (exogenous life stages) All subsequent reports of morphologic
diagnoses of these coccidia have been based on Hoarersquos original descriptions The pre-patent period
40
(minimum duration of endogenous development) in inoculated naiumlve ferrets was described for E furonis
and E ictidea as 6 and 7 days respectively (Hoare 1935b) The pre-patent period for I (=C) laidlawi
was not determined because the number of oocysts available was insufficient for an experimental
infection trial
Hoare (1927) described the sporulated oocysts of E furonis as follows spherical double outer
wall with a thin colourless outer layer and a thick yellowish inner layer no micropyle or residual body
and measuring on average 128 times 120 microm (length [L] 112-144 width [W] 104-128 shape index [SI]
107) Sporocysts were spindle-shaped with one end constrictedblunted contained a residual body and
on average measured 8-88 times 4 microm Sporozoites were vermiform with one end narrower than the other
arranged head to tail and each had a central nucleus a clear vacuole was identified in some sporozoites at
their broad posterior end
The sporulated oocysts of E ictidea were described as follows oval or elliptical with a double
outer wall with a thin colourless outer layer and a thick yellowish inner layer no micropyle or residual
body and measuring on average 236 times 175 microm (L 184-272 W 128-208 SI 135) Sporocysts were
irregularly oval with one end broad and the other more constricted contained a residual body and
measured 115 times 65 microm on average Sporozoites were vermiform with one end narrower than the other
arranged head to tail in the sporocysts and had a central nucleus and a clear vacuole at their broad
posterior end
The sporulated oocysts of I (=C) laidlawi were ovoid with a double outer wall with a thin
colourless outer layer and a thick yellowish inner layer had no micropyle or residual body and measured
on average 34 times 29 microm (L 320-368 W 272-304) A SI of 117 can be calculated from the original
mean dimensions Two sporocysts were identified each containing four sporozoites and no Stieda body
sporocysts were elliptical contained a residual body and measured 208 times 144 microm on average
Sporozoites were sausage shaped with one end slightly pointed and had a central nucleus and a clear
41
vacuole identified at the pointed end Sporozoites were arranged with pointed ends all at the same pole of
the sporocyst
Oocysts identified as Cystoisospora ohioensis have been reported from fecal samples collected
from healthy domestic ferret kits in a large American ferret breeding operation that also housed juvenile
domestic dogs (Patterson and Fox 2007) The method of identification of this parasite was not described
A second similar institution reported the presence of Cystoisospora (=Isospora) species also thought to
be C ohioensis in routine fecal examination of their colony (Dr Bambi Jasmin personal
communication) Identification in this case was performed by the Animal Health Diagnostic Center at
Cornell University and was based on morphometrics using light microscopy The significance of these
findings is unknown but no clinical signs or histologic lesions were described in the ferrets shedding
these oocysts The definitive hosts for C ohioensis are canids including the domestic dog
More recently molecular techniques have been used for the more precise identification of
coccidia Nucleotide sequences like morphological features diverge over time under selective pressure
however recent evolutionary divergence among coccidia is more likely to be reflected in molecular as
compared to morphologic differences Thus nucleotide sequences that are more similar are inferred to be
more closely related and to have diverged more recently (Cox 1994)
Molecular characterization of ferret coccidia has only been performed for one species Eimeria
furonis Abe et al (2008) extracted DNA from oocysts from the feces of a single domestic ferret with
clinical signs resulting from coccidial enteritis Using primers initially developed for molecular
identification of Cyclospora species (see Matsubayashi et al 2005) small subunit ribosomal DNA (nu
18S rDNA) primers CYC1FE (5ʹ-TAC CCA ATG AAA ACA GTT T-3prime) and CYC4RB (5prime-CGT CTT
CAA ACC CCC TAC TG-3prime) were used to amplify a 347 base pair fragment of nu 18S rDNA The
amplicon was sequenced (GenBank AB329724) and compared with previously published partial nu 18S
rDNA sequences from 40 Eimeria two Isospora and four Cyclospora species The resulting phylogram
42
grouped E furonis with E alabamensis (cattle) and E meleagrimitis (turkey) In the same study the
microscopic morphology of the oocysts was used to identify this coccidial species as Eimeria furonis by
comparison with Hoarersquos (1927 1935b) published descriptions of Eimeria furonis and Eimeria ictidea
Sledge et al (2011) also used nu 18S rDNA to identify Eimeria furonis as the cause of three
distinct outbreaks of enteric disease in domestic ferrets Initial identification was performed using
morphometrics of sporulated oocysts collected from feces in one of the three outbreaks being
investigated Formalin fixed paraffin embedded intestinal segments from ferrets from each of the
outbreaks contained multiple coccidial life stages when examined by light microscopy PCR amplification
of a 247 base pair (bp) amplicon of the nu 18S rDNA was generated from DNA isolated from stored
formalin-fixed tissues for further genetic analysis Analysis and sequencing of amplicons from all three
groups showed 100 identity to sequences previously reported by Abe et al (2008) for the gene encoding
E furonis nu 18S
In 2015 Kaye et al identified coccidia within hepatobiliary lesions in a domestic ferret receiving
immunosuppressive therapy for red cell aplasia DNA was extracted from frozen liver and a fragment of
the nu 18S rDNA was amplified using the primers previously described by Sledge et al(2011) The
authors reported that the DNA sequence of the amplicon had 100 identity to the published nu 18S
rDNA sequence of E furonis and 95 identity to the nu 18S rDNA of E myoxi (rodent) E alabamensis
(cattle) and Isospora robini (avian) (Kaye et al 2015)
It is difficult to estimate the current prevalence of enteric coccidia within the North American
domestic ferret population and no studies have been conducted to do so Fecal samples submitted to
veterinary diagnostic laboratories from domestic ferrets in Canada are uncommon and samples positive
for coccidia appear infrequently (Dr Donald Martin personal communication) Conversely in Europe
the prevalence of coccidia within the domestic ferret population appears to be higher based on
submissions to a large veterinary diagnostic laboratory in Germany Data from Idexx Vet Med Lab in
Ludwigsburg Germany was compiled to review the prevalence of coccidia and Giardia within fecal
samples from domestic ferrets (Pantchev et al 2011) The authors reported that of 284 fecal samples
43
submitted from 2002-2004 18 (63) had detectable coccidial oocysts on fecal flotation Based on
morphologic characteristics oocysts were identified as E ictidea E furonis I laidlawi (herein referred
to as I (=C) laidlawi as noted above) and another unidentified Isospora species Comparative data from
the same laboratory from 2009-2010 included sample submissions from 253 ferrets 21 (83) of which
were positive for coccidial oocysts on fecal flotation Nine of the samples were identified as containing E
furonis three contained both E furonis and I (=C) laidlawi eight contained only I (=C) laidlawi and
one sample contained both E furonis and E ictidea identification in all cases was again based on oocyst
morphometrics No statistically significant difference in the occurrence of coccidial oocysts was detected
when data from the two periods were compared (Fisherrsquos exact test P=041) (Pantchev et al 2011)
The purpose of the present study was to perform a more detailed molecular characterization of the
coccidial species isolated from domestic ferrets to estimate prevalence of the different coccidial species
within the Canadian domestic ferret population and to associate morphologic and molecular
characteristics of a greater range of enteric coccidial species in order to improve diagnostic accuracy
22 MATERIALS amp METHODS
221 Fecal samples
Multiple diagnostic laboratories within Ontario Canada1 and a major European diagnostic
laboratory 2 were solicited for fecal samples from domestic ferrets shedding coccidial oocysts Fecal
samples were diagnosed positive for coccidia based on fecal flotation and light microscopic identification
of Eimeria or Cystoisospora species Eleven samples were collected during the study period (from 2014-
2017) and preserved in potassium dichromate (25 wv) eight from Europe and three from Canada
Centrifugal flotation with saturated salt solution (Ryley et al 1976) was used to isolate oocysts from fecal
samples for genomic DNA extraction Genomic DNA extraction and purification were performed using a
1 Animal Health Laboratory Guelph ON Antech Diagnostics Canada Ltd Mississauga ON IDEXX Canada
Markham ON 2 Vet Med Labor GmbH Division of IDEXX Laboratories Ludwigsburg Germany
44
QIAamp DNA Mini Kit (Qiagen Hilden Germany) according to manufacturerrsquos instructions After
addition of DNAzol to the samples (Qiagen Hilden Germany) samples were vortexed using 05 mm
glass beads (Biospec Products Inc Bartlesville OK USA) prior to extraction in order to fracture the
oocyst walls and release the sporocysts Concentrations of the resultant DNA were estimating using a
Nanodrop 2000 spectrophotometer (NanoDrop Products Wilmington DE USA) and stored at 4 degC for
immediate use or minus20 degC for later use
For each laboratory the number of domestic ferret fecal sample submissions numbers diagnosed
positive for coccidial oocysts and number of each coccidial species identified in positive samples were
tabulated for each of the years 2008-2015
222 Formalin fixed intestinal tissues
Major diagnostic pathology services across Canada3 were contacted to identify cases of enteric
coccidiosis identified on necropsy of domestic ferrets Cases were considered positive based on the
presence of asexual or sexual life stages of the parasites in intestinal sections The histologic sections on
each positive case were reviewed re-described and organisms measured (AP DAS) Gross necropsy
reports for all cases were also reviewed to identify any clinical correlates associated with enteric
coccidiosis DNA was extracted from ten 5-6 microm scrolls of formalin fixed paraffin embedded tissue
(FFPE) using the QIAamp DNA FFPE Tissue Kit (Qiagen) as per manufacturer instructions
223 Molecular characterization
Regions from the nu 18S rDNA and mitochondrial cytochrome c oxidase subunit I (mt COI)
DNA were amplified by polymerase chain reaction (PCR) from each sample using the primers listed in
Table 21 PCR amplification was performed for all samples in a volume of 25 microl containing ~100 ng of
3 Animal Health Centre Abbotsford BC Animal Health Laboratory Guelph ON Faculteacute de meacutedecine veacuteteacuterinaire
Universiteacute de Montreacuteal Saint-Hyacinthe QC Histovet Surgical Pathology Guelph ON IDEXX Canada
Markham ON Prairie Diagnostic Services Inc Saskatoon SK
45
genomic DNA 1times PCR buffer 15 mM MgCl2 02 mM deoxyribonucleotide triphosphates (dNTPs) 400
nM of each primer and 1 U of Invitrogen Platinum Taq DNA Polymerase (Thermo Fisher Scientific
Toronto ON Canada) Reactions were performed on a Bio-Rad T100 PCR thermal cycler (Bio-Rad
Laboratories Singapore) Samples were denatured and Taq polymerase activated at 95 degC for 3 min then
subjected to 35 cycles of 94 degC for 30s anneal at 50-62 degC (see Table 21 for specific anneal conditions
for the various primer pairs) for 30s and extension at 72 degC for 30-75s (see Table 21) followed by a
final extension at 72 degC for 7 min Suitable DNA (ie genomic DNA from an Eimeria or Sarcocystis sp)
was included in the PCR reactions to act as a positive control for the reaction chemistry All amplification
products were subjected to electrophoretic separation using 15 submarine agarose gel stained with
ethidium bromide and visualized on an ultraviolet transilluminator (Spectronics Corporation New York
NY USA) DNA band size was determined by comparison with a 1 kb DNA ladder (GeneRuler 1kb Plus
DNA ladder Thermo Fisher Scientific Waltham MA USA) Bands were excised with a new sterile
scalpel blade and PCR products were purified from the gel using a QIAquick Gel Extraction Kit (Qiagen)
PCR products were cycle sequenced using an ABI Prism 7000 Sequence Detection System (Applied
Biosystems Inc Foster City CA USA) by the Molecular Biology Unit of the Laboratory Services
Division University of Guelph (Guelph ON Canada) using the amplification primers to obtain
sequences in both directions The resulting chromatograms were aligned and analyzed with Geneious Ver
818 or later (Biomatters Limited Auckland New Zealand) and high quality consensus sequences
generated The resulting consensus sequences were searched from within Geneious against publically
available sequences on the BLAST server (blastncbinlmnihgovBlastcgi) using the blastn search
algorithm against the nrnt database (GenBank+EMBL+DDBJ+RefSeq ndash AA or DNA) Resultant new
nucleotide sequences were submitted to GenBank
46
224 Phylogenetic analysis
To determine the phylogenetic affinities of the newly obtained sequences with sequences from
related apicomplexan taxa representative nu 18S rDNA and mt COI sequences were downloaded from
GenBank with special reference to sequences from parasites that infect members of the order Carnivora
Nuclear 18S and mt COI sequences were aligned independently using MAFTT v7017 (Katoh et
al 2002) executed from within Geneious and then concatenated into a combined nu18S rDNA mt COI
dataset Multiple sequences from a single parasite were used to generate consensus sequences for each
locus as described by Ogedengbe et al (2017) Aligned sequences were trimmed to the length of the
largest newly generated nu 18S sequence Phylogenetic trees were generated using Bayesian Inference
(BI) using MrBayes Ver 326 (Huelsenbeck and Ronquist 2001) executed from within Geneious the
combined nu 18S and mt COI alignment was partitioned to permit locus-appropriate substitution models
to be applied to each partition For the nu 18S sequence partition the general time reversible (GTR)
substitution model (nst=6) with gamma rate variation (ie a GTR+G+I model) was applied For the mt
COI sequence partition the codon (M1) substitution model (using translation table 4 [ie lsquometmtrsquo]) was
used instead of the GTR with the remaining parameters remaining the same
The resulting tree was rooted using a pair of adeleid coccidia (Hepatozoon spp) as the taxonomic
outgroup All BI analyses were run for a chain length of 1000000 with tree sampling every 1000
following a burn-in of 100000 with default settings of 4 heated chains and heated chain temp of 02
47
23 RESULTS
231 Fresh fecal samples
From 2008-2015 inclusive the Canadian diagnostic parasitology laboratory4 received an average
of 1206 (range 81-160) domestic ferret fecal samples yearly the European parasitology laboratory5
received a yearly average of 230 samples (range 213-270) The number of fecal samples diagnosed as
positive for coccidial oocysts per year on fecal flotation during this time averaged 35 (range 0-8) and
130 (range 6-20) for the Canadian and European laboratories respectively The diagnosing laboratories
used oocyst morphometrics to identify the species of coccidia present Almost all coccidia-positive
submissions to the Canadian laboratory were identified as containing an I (=C) species based on light
microscopy Coccidia in only three samples from the Canadian laboratory were identified as E furonis
one in each of 2010 2012 and 2014 E ictidea was not identified in any samples submitted to the
Canadian laboratory Approximately equal numbers of coccidia-positive samples from the European
laboratory were identified as E furonis and I (=C) laidlawi each year Only two samples from the
European laboratory contained oocysts that were identified as Eimeria ictidea using morphometrics one
from each of 2011 and 2013 Laboratory submissions to both laboratories are summarized in Table 22
Twelve fecal samples preserved in potassium dichromate were received for analysis by the
authors Eleven samples had previously been identified as containing a single coccidial species five
containing E furonis two containing E ictidea and four containing I (=C) laidlawi A final sample had
been identified as containing a mix of E furonis and Cystoisospora canis Results of microscopic and
molecular characterization of these samples are summarized in Table 23
4 IDEXX Canada Markham ON
5 Vet Med Labor GmbH Division of IDEXX Laboratories Ludwigsburg Germany
48
232 Formalin fixed samples
Only three cases of coccidiosis were identified in domestic ferrets within the databases of the five
diagnostic laboratories that participated in the retrospective study Histologic sections of intestine were
received from these three cases which originated in Ontario6 and Quebec7 The Quebec sample (P2010-I)
was collected in 2010 and the Ontario samples (93-40404 and 17-008571) in 1993 and 2017 respectively
On gross necropsy the small intestinal contents of case P2010-I were described as pasty mucoid
yellow-brown feces with some blood For case 93-40404 the small intestines were described as empty
but melena was present within the terminal portion of the large intestine Scant intestinal contents and
dark brown fecal material in the colon were described in case 17-008571
In all cases endogenous developmental stages of coccidia were visible in histological sections
(Figure 21 is exemplary of the findings from one case) Hematoxylin and eosin stained sections from
P2010-I contained two affected regions of small intestine The intestinal mucosa of the first region
contained numerous asexual life stages and moderate numbers of sexual life stages as well as a small
number of oocysts free within the lumen The second section contained tissues that were poorly
preserved nonetheless 0-4 oocysts per 400times field were identifiable within the intestinal lumen Two
regions of affected small intestine were identified from 93-40404 after screening of all submitted sections
both contained low numbers of sexual and asexual endogenous stages Within one region there were small
numbers of meronts within the intestinal mucosa and lamina propria The second region had small
numbers of oocysts within cells of the epithelium and lamina propria as well as free within the intestinal
lumen In case 17-008571 multiple sections of jejunum contained numerous coccidian meronts gamonts
6 Animal Health Laboratory Guelph ON
7 Faculteacute de meacutedecine veacuteteacuterinaire Universiteacute de Montreacuteal Saint-Hyacinthe QC
49
and oocysts within intestinal villi within the ileum scattered epithelial cells also contained these various
life stages
Average length and width of oocysts were measured from slide sections for all cases For P2010-
I oocyst average length and width were determined from seven oocysts to be 94 microm (range 85-105) and
75 microm (range 69-84) respectively with a SI of 125 (range112-140) Average length and width of
oocysts measured from 93-40404 were determined from 5 oocysts to be 2814 microm (range 229-341) and
233 microm (range 180-308) respectively with a SI of 123 (range 110-146) For the third case 17-
008571 only 2 oocysts were identified and average length and width of oocysts measured 982 microm (range
973-992) and 845 microm (range 821-870) respectively with a SI of 116 (range 114-118)
233 Molecular characterization
DNA was successfully extracted from all twelve fecal samples and two cases with formalin fixed
tissue samples Attempts at amplification of DNA extracted from sample 93-40404 using the primer pairs
listed in Table 21 were unsuccessful Molecular identification results and GenBank accession numbers
for the remaining samples are summarized in Table 23 Both the nu 18S rDNA and mt COI sequences
from I (=C) laidlawi were unique when compared with available sequences from other Cystoisospora
species within the public databases However sequences from I (=C) laidlawi were most similar to
sequences from C canis and C felis and somewhat more divergent from sequences from members of the
C ohioensis species complex Two apparent genotypes of E furonis were identified based on nu 18S and
mt COI sequencing results Genotype 1 represented by EU sample 9014 had 100 identity to previously
published sequences of the nu 18S locus from two isolates from Japan (GenBank AB239130 and
AB329724) Genotype 2 represented by EU sample 907 and Canadian sample 17-008571 had 994
identity at the nu 18S locus (3 single nucleotide differences [SNDs] over 561 base pair region [bp]) to the
three sequences above belonging to E furonis genotype 1 Pairwise alignment of mt COI sequences from
both genotypes identified only 2 SNDs (996 pairwise identity over 513 bp region) Partial mt COI
50
sequences of E furonis from both genotypes were only distantly related (941 pairwise identity 30
SNDs over 513 bp and 905 pairwise identity 49 SNDs over 513 bp respectively) to publicly available
sequences from Eimeria ictidea from the black-footed ferret (Mustela nigripes) (GenBank KT203399)
and Eimeria mephitidis from the striped skunk (Mephitis mephitis) (GenBank KT203398) the only other
Eimeria species infecting members of the Carnivora for which sequence was available
234 Phylogenetic analysis
A phylogenetic reconstruction based on concatenated partial nu 18S rDNA and mt COI sequences
of E furonis I (=C) laidlawi and related coccidia is illustrated in Figure 22 The combined 18SCOI-
based tree demonstrates that the two Eimeria species from ferrets form a well-supported monophyletic
group that branches among a collection of other eimeriid coccidia that infect mammals The sarcocystid
parasite of the domestic ferret I (=C) laidlawi was found to group as the sister taxon to C canis that
together formed a monophyletic group with the closely related C felis all three of these closely related
Cystoisopora species possess comparatively large egg-shaped oocysts that are similar morphologically
24 DISCUSSION
The present work has generated the first nu 18S rDNA and mt COI sequences for Cystoisospora
laidlawi and the first mt COI sequence for Eimeria furonis both isolated from the domestic ferret In this
study histologic presence of organisms and microscopic identification of oocysts shed in feces have been
correlated with published and novel nu 18S and mt COI sequences
Eimeria ictidea was not identified in any Canadian sample and this coccidium was reported in
only 2 of 1840 fecal samples submitted from across the European Union (EU) to IDEXX Germany during
2008-2015 suggesting that E ictidea is not a frequent cause of enteric coccidiosis in domestic ferrets in
Canada or the EU
51
During the study period (2008-2015) almost twice as many domestic ferret fecal submissions
were made to the European as compared to the Canadian diagnostic laboratory however the prevalence
of coccidia-positive samples was similar The methodology used in this report cannot be used to
determine the actual prevalence of enteric coccidial infection (coccidiasis) or disease (coccidiosis) within
the domestic ferret population Fecal samples may be submitted to laboratories either as a result of
investigation into enteric disease or as part of a routine health examination Thus without historic
information accompanying each sample one can simply identify the proportion of positive samples and
compare the frequency of the finding of different coccidial species Prospective surveys of fecal samples
from healthy and sick domestic ferrets with greater sample size would be necessary to determine the true
prevalence of these parasites within the population and to infer their clinical significance
Comparatively few mitochondrial COI sequences have been generated for apicomplexan parasites
compared with other genetic loci the majority of published sequences obtained from Apicomplexa are
from nu 18S The disadvantage of using nu 18S rDNA sequences for parasite identification is that they
are poor at distinguishing among closely related eimeriid coccidia due to the highly conserved nature of
the nuclear ribosomal RNA locus In contrast mt COI sequences appear to be more useful for
distinguishing closely related coccidian species (Ogedengbe et al 2011) but are less useful than nu 18S
rDNA sequences for inferring more ancient relationships among more distantly related coccidia
Consequently the combined use of nu 18S rDNA and mt COI sequencing has been recommended for
improved species description and phylogenetic analysis (El-Sherry et al 2013) For these reasons both nu
18S and mt COI sequences were analysed in the present study
Despite adequate quantities of DNA extracted from the Ontario laboratory sample (93-40404)
successful amplification did not result with any primer pair (Table 21) Potential reasons for this include
degradation of formalin-fixed DNA into fragments too small for amplification with the desired primers
perhaps as a result of extended length of time in formalin prior to paraffin embedding or length of time
stored as FFPE tissue (23 years) or insufficient parasite DNA within the paraffin scrolls The primer pairs
52
used appear to be useful for most eimeriid coccidia (Ogedengbe 2015) and successfully amplified both
Eimeria species from DNA isolated from oocysts so it is unlikely that failure to amplify DNA from this
sample resulted from an inability of the primers used to recognize the parasite seen on section
Two genotypes of E furonis were identified in this study Genotype 1 was identified only from
samples originating from domestic ferrets in Europe but exhibited 100 identity based on nu 18S
sequencing with previously published sequences from both Japan and the USA Genotype 2 was
identified from samples originating from domestic ferrets in both Canada and Europe The small number
of single nucleotide differences between the two genotypes at two genetic loci in different genomes are
consistent with intraspecific variation (ie strain variation)
As might have been expected because of their morphological and host similarities nu 18S and mt
COI sequences of E furonis were determined to be most similar to an Eimeria species (E ictidea)
previously isolated from black-footed ferrets (Mustela nigripes) these eimeriid coccidia formed a
monophyletic group that was distinct from other eimeriid coccidia infecting mammals in the phylogenetic
analyses based on combined nu 18S rDNA and mt COI sequences Similarly the nu 18S rDNA and mt
COI sequences of I (=C) laidlawi are most similar to sequences from two other Cystoisospora species of
carnivores (C canis and C felis) that both have large egg-shaped oocysts comparable to those of I (=C)
laidlawi Both morphometrics and genotyping support the close relationships among these three
sarcocystid coccidia of carnivores These molecular data confirm that transfer of Isospora laidlawi to the
genus Cystoisospora by Barta et al (2005) is warranted
The previous light microscopic identifications of coccidial species in 3 of the 11 fecal samples
were not in agreement with the molecular findings These results were not surprising because light
microscopy has been shown to be an insensitive tool for distinguishing among apicomplexan parasites at
both the genus and species level Furthermore re-evaluation of these samples by the authors revealed that
many of the samples that were identified incorrectly based on morphometrics contained primarily
53
unsporulated oocysts making accurate identification based on microscopic appearance highly
challenging These findings further underscore the importance of molecular methods in accurate parasite
identification In the absence of molecular tools accurate measurement of oocyst size shape and
determination of SI can be useful for differentiating among species of Eimeria and Cystoisospora
however this can only be performed accurately on sporulated oocysts from feces Interestingly the size
and shape indices of oocysts of E furonis measured in histologic sections did not match those previously
described by Hoare (1927) for the same oocysts in feces despite molecular confirmation of identity
Thus measurements of oocysts in histologic sections are not recommended for use in coccidial
identification
Our observations highlight the utility of molecular methods for identifying enteric coccidia
infecting domestic ferrets and suggest that diagnoses based on morphological methods should perhaps be
limited to broad determinations of disease etiology (ie lsquococcidiosisrsquo or lsquococcidiasisrsquo) Using molecular
techniques we were able to differentiate morphologically similar coccidial species isolated from the feces
of domestic ferrets and specifically identify parasites seen in histological sections of ferret intestine
Molecular techniques thus appear to be essential for determining the coccidial species responsible for
individual and group outbreaks of coccidiosis and for further understanding of eimeriid host-parasite
relationships
ACKNOWLEDGEMENTS
Many thanks to Julia Whale and Alex Leveille for their assistance and encouragement during the course
of this project The authors would like to recognize the contributions of Dr Donald Martin (IDEXX
Canada) and Drs Nikola Pantchev and Majda Globokar (IDEXX Germany) for the contributions of data
and samples to this project The authors would also like to recognize the Laboratoire de Pathologie
(Service de diagnostic Faculteacute de meacutedecine veacuteteacuterinaire St Hyacinthe Quebec) and the Animal Health
Laboratory (Guelph Canada) for contributions of samples and data to this project Finally this project
was made possible through funding by the Toronto Zoo Residency Research Fund to DASAP and partial
54
funding from a Natural Sciences and Engineering Research Council of Canada (NSERC) Discovery Grant
(400566) to JRB
55
Table 21 Amplification primers for nuclear 18S rDNA and mitochondrial COI loci anneal temperatures (Ta) extension times and expected PCR
product sizes used in the identification of enteric coccidia from domestic ferrets (Mustela putorious furo)
Gene Target Primer Pairs Primer Sequence (5ʹ-3ʹ) Size (bp) Ta (degC) Anneal (sec) Reference
nu 18S rDNA CYC1FE TACCCAATGAAAACAGTTT 560 52 45 Matsubayashi et al (2005)
CYC4RB CGTCTTCAAACCCCCTACTG Matsubayashi et al (2005)
Cocci_18S_595F CCGCGGTAATTCCAGCTCCAAT 216 62 30 Present study
Cocci_18S_847R GCTGMAGTATTCAGGGCGACAA Present study
Lank_18S_224F TCATAGTAACCGAACGGATC 1080 54 60 Ogedengbe (2015)
Api_SSU_2733R CGGAATTAACCAGACAAATC Mathew et al (2000)
mt COI COI_10F GGWDSWGGWRYWGGWTGGAC 500 52 30 Ogedengbe et al (2011)
COI_500R CATRTGRTGDGCCCAWAC Ogedengbe et al (2011)
COI 272F CAATTCTAYGATGCCGCWTT 222 52 30 Present study
COI_500R CATRTGRTGDGCCCAWAC Ogedengbe et al (2011)
Sdae-COI_260F GATCTTTATGTTYTTRATGCC 890 50 75 Ogedengbe (2015)
Sdae-COI_1147R CATTACCCATAACYACACC Ogedengbe (2015)
56
Table 22 Summary of fecal samples from domestic ferrets (Mustela putorius furo) submitted to two diagnostic laboratories from 2008-2015
No fecal samples positive for coccidia
No samples submitted
(percentage positive)
No samples positive for
Cystoisospora sp
No samples positive for
Eimeria furonis
No samples positive for
Eimeria ictidea
Year Canada Europe Canada Europe Canada Europe Canada Europe
2008 3140 (21) 6214 (28) 3 2 0 4 0 0
2009 2160 (12) 14214 (65) 2 9 0 5 0 0
2010 8127 (63) 20213 (94) 7 10 1 10 0 0
2011 0114 (0) 17215 (79) 0 9 0 7 0 1
2012 3108 (28) 10231 (43) 2 4 1 6 0 0
2013 281 (25) 16270 (59) 2 13 0 2 0 1
2014 6127 (47) 12234 (51) 5 6 1 6 0 0
2015 4108 (37) 9249 (36) 4 3 0 6 0 0
Total 28 (29) 104 (56) 25 56 3 46 0 2
Average
year 35 130 31 70 04 58 00 03
Legend Numbers in brackets refer to the percent of the total number of fecal samples submitted
57
Table 23 Morphologic and molecular identification of coccidia from domestic ferrets (Mustela putorius furo)
Sample ID Source
External Lab Morphologic
Diagnosis Morphologic Diagnosis (ARP) Molecular Diagnosis
mt COI GenBank
Accession
nu 18S rDNA
GenBank Accession
93-40404 FFPE enteric coccidia Histologic sample - - -
P2010-I FFPE enteric coccidia Histologic sample E furonis Identical to MF774036 Identical to MF774678
17-008571 FFPE NP Histologic sample E furonis Same as MF774036 Same as MF774678
17-008571 feces NP E furonis E furonis MF774036 MF774678
907 feces E furonis E furonis E furonis MF774035 MF774679
938 feces I (=C) laidlawi no oocysts visualized I (=C) laidlawi MF774037 MF774677
952-A feces E ictidea Cystoisospora sp I (=C) laidlawi Identical to MF774037 Identical to MF774677
9958 feces E furonis no oocysts visualized E furonis Identical to MF774034 Identical to MF774680
9011 feces E furonis E furonis E furonis Identical to MF774035 Identical to MF774679
9014 feces E furonis E furonis E furonis MF774034 MF774680
9017 feces I (=C) laidlawi Cystoisospora sp I (=C) laidlawi Same as MF774037 Same as MF774677
9040 feces I (=C) laidlawi no oocysts visualized I (=C) laidlawi Same as MF774037 Same as MF774677
912-260 feces I (=C) laidlawi Cystoisospora sp I (=C) laidlawi Same as MF774037 Same as MF774677
CAN-2016-1 feces C canis + E furonis Cystoisospora sp I (=C) laidlawi MF774038 MF774676
Legend FFPE = formalin fixed paraffin embedded intestinal sections - = unsuccessful = morphologic diagnosis performed by JRB same as = 100 sequence
identity with listed GenBank entry over entire sequence length identical to = 100 sequence identity but shorter sequence than listed GenBank entry
58
Figure 21 Life stages of Eimeria furonis within the small intestinal epithelium of a domestic ferret
(Mustela putorius furo) Asexual life stages merozoites (black circle) Sexual life stages oocyst
(solid black arrow) macrogamonts (open arrows with labels) microgamont (dotted black arrow)
Hematoxylin and eosin staining scale bar = 25μm
25 microm
59
Figure 22 Phylogenetic relationships of coccidia (Eimeria ictidea Eimeria furonis and Isospora
(=Cystoisospora) laidlawi) from domestic (Mustela putorius furo) or black-footed (Mustela
nigripes) ferrets based on partial nuclear 18S rDNA and mitochondrial COI sequences of these
parasites and related apicomplexan parasites A summary of the sources of the molecular data for
the remaining taxa included in this phylogenetic analysis are found in Supplementary Table 1 of
Ogedengbe et al (2017) Bayesian support is indicated for each node horizontal distance is
proportional to hypothesized evolutionary change (scale indicates sequence divergence of 10)
60
CHAPTER 3 MORPHOLOGICAL AND MOLECULAR CHARACTERIZATION OF
ENTERIC COCCIDIA ISOLATED FROM BLACK-FOOTED FERRETS (MUSTELA
NIGRIPES)
ABSTRACT
Black-footed ferrets (BFF Mustela nigripes) are the only ferret species native to North America
and have been identified as endangered since 1967 Starting in 1986 a multi-institutional effort has been
breeding this species in captivity with successful reintroductions back into the wild Coccidiosis is
recognized as a cause of significant juvenile morbidity and mortality in captive breeding programs and
can result in significant population losses Little is known about the etiology of enteric coccidiosis in
BFF Coccidia positive fecal samples (n=12) and formalin fixed paraffin embedded intestinal tissues
(n=11) were obtained from BFF in the Toronto Zoo and Louisville Zoo Species Survival Plan (SSP)
populations Oocyst morphometrics and sequence genotyping at three loci (nuclear 18S rDNA
mitochondrial cytochrome c oxidase subunit I and mitochondrial cytochrome c oxidase subunit III) were
conducted Results suggest that the same Eimeria species E ictidea was the cause of enteric coccidiosis
in both SSP populations in both juvenile and adult age classes Wider research is indicated to determine
whether these findings are representative of the larger captive and wild BFF populations
31 INTRODUCTION
Black-footed ferrets (BFF) are one of only three wild ferret species worldwide the other two
being the European polecat (Mustela putorius) and the Siberian polecat or steppe polecat (Mustela
eversmanii) The BFF the only native North American ferret species was formerly distributed throughout
the North America prairie ecosystem but were considered extinct by the late 1950s In 1964 a single
population was discovered in Mellette County South Dakota Progressive decline of this population in
subsequent years resulted in the decision by United States Fish and Wildlife Service to initiate a captive
breeding program for the species From 1971-1973 four females and five males were captured for this
61
purpose Despite successful breeding no kits survived and the last adult ferret in this captive colony died
in 1979 BFF were again presumed extinct in the wild based on annual surveys of the initial capture site
In 1981 a dead BFF was discovered by a ranch dog outside of Meeteetse Wyoming allowing wildlife
biologists to identify another colony of BFF This colony flourished until 1985 when an outbreak of
canine distemper in this wild BFF population and an outbreak of sylvatic plague in the local prairie dog
population resulted in sharp population declines From 1985 through 1987 all 24 of the remaining BFF
were trapped and brought into captivity to re-initiate the captive breeding program Six ferrets in this
initial group died of canine distemper while in captivity and seven of the remaining eighteen survivors
are the founding population of the current captive breeding population Today this captive breeding
population consists of approximately 300 BFF distributed among multiple institutions (Santymire et al
2014)
Since 1986 a multi-institutional effort has been breeding BFF in captivity with reintroductions
back into the wild in selected locations in Canada the USA and Mexico Currently six facilities
participate in the BFF Species Survival Plan (SSP) the Toronto Zoo United States Fish and Wildlife
Services National Black-footed Ferret Conservation Center National Zoorsquos Smithsonian Conservation
Biology Institute Louisville Zoo Cheyenne Mountain Zoo and the Phoenix Zoo (Black-footed Ferret
Recovery Implementation Team 2011) In order to provide the best genetic matches BFF are transferred
among the six institutions for breeding Approximately 300-400 kits are produced annually between the
six SSP facilities with ~200 of these kits allocated for release to the wild yearly (Santymire et al 2014)
As of 2011 over 8000 BFF kits had been produced in captive breeding facilities (Black-footed Ferret
Recovery Implementation Team 2011)
Multiple infectious diseases pose a significant risk to the captive breeding and post-release
survival of BFF including canine distemper and sylvatic plague (Santymire et al 2014 USFWS BFF
Recovery Program 2017) Coccidiosis is recognized as a cause of significant juvenile morbidity and
mortality in captive breeding programs and can result in significant population losses (Bronson et al
62
2007 Santymire et al 2014 USFWS BFF Recovery Program 2017) While the effects of the disease on
the wild population are not clear a prevalence of approximately 13 has been reported based on fecal
samples collected from wild BFF born at release sites (Dr R Santymire personal commication)
Coccidia are eukaryotic host-specific parasites of the phylum Apicomplexa affecting numerous
hosts within a wide taxonomic range Two species of coccidia Eimeria ictidea Hoare 1927 and Eimeria
furonis Hoare 1927 have been identified in black-footed ferrets based on morphometrics (Jolley et al
1994) Jolley et al examined fecal samples from six captive BFF during a distemper outbreak as well as
samples from wild BFF They described one medium-sized ovoid eimeriid oocyst with a double wall
presence of a polar body and lacking both an oocyst residual body and micropyle Oocysts of this Eimeria
species (sp) measured 232 times 155 microm (range 182-274 times 130-162) with a shape index (SI) of 150 The
sporocysts were elongate with the presence of both sporocyst residuum and a Stieda body Sporozoites
contained prominent refractile bodies at the posterior end and were aligned anterior to posterior within
sporocysts These oocysts shed by all six captive ferrets were considered consistent with Eimeria ictidea
based on descriptions by Hoare (1927) On histopathologic examination of intestinal sections parasites
undergoing merogony and gamogony were observed within the villar epithelium throughout the small
intestine but were concentrated in the jejunum (Hoare 1935b) parasite life stages were not described
from other tissuesorgans
A second small spherical to subspherical eimeriid oocyst was also documented in the captive
ferrets by Jolley et al (1994) this second species had a pink double oocyst wall a granular residual body
and lacked both oocyst polar body and micropyle This smaller species measured 126 times 119 microm (range
108-152 times 101-129) with a SI of 106 The sporocysts were elongate and possessed a Stieda body and
sporozoites contained refractile bodies Similar to the larger Eimeria sp described above merogonic and
gamogonic stages were observed within the villar epithelium throughout the small intestine but were
most numerous in the jejunum Jolley et al (1994) concluded these small spherical oocysts were
consistent with E furonis described by Hoare (1927) from domestic ferrets
63
Jolley et al (1994) described a third type of coccidial oocyst occasionally detected in small
numbers within BFF fecal samples however the authors did not state whether this third oocyst
morphotype was recovered from wild or captive animals The oocysts measured 370 times 223 microm (range
350-386 times 212-232) with a SI of 106 Attempts to sporulate collected oocysts were unsuccessful and
corresponding endogenous stages were not identified on histopathologic examination of necropsied
ferrets precluding further morphologic identification of the parasite It should be noted that coccidial
oocysts with similar measurements had not been detected in wild or captive prey species available for
ingestion by BFF making it unlikely that this coccidial species would have been a pseudoparasite (Jolley
et al 1994)
Previous to this report by Jolley coccidial oocysts had been isolated from the feces of BFF in two
captive populations (Carpenter amp Hillman 1979 Williams et al 1988) The abstract by Carpenter amp
Hillman (1979) did not describe the oocysts whereas Williams et al (1988) stated that two Eimeria sp
(one with larger oocysts and one with smaller oocysts) were observed within the fecal samples but they
were not identified further Interestingly Williams et al reported both Eimeria sp to be shed in the feces
of all ferrets concurrently affected by distemper and by approximately 30 of the clinically healthy
ferrets at the time of investigation
Non-enteric coccidia have also been reported by two authors from captive BFF at one facility
(Jolley et al 1994 Williams et al 1988) Both reports which presumably described the same case(s)
noted the presence of endogenous coccidial life stages in histologic sections of respiratory tissue and
merozoites of an unidentified coccidium in impression smears of the urinary bladder from BFF diagnosed
with canine distemper Meronts were observed within the epithelium of the trachea a large bronchus and
associated bronchial glands In the later report Jolley et al (1994) described the lesions as occurring in the
same ferret whereas in the earlier report by Williams et al (1988) they are described as occurring in two
different ferrets Paraffin blocks containing formalin fixed tissues from these cases have since been
discarded precluding further attempts at parasite identification with molecular methods Subsequent to
64
these reports further cases of systemic coccidiosis in BFF have neither been published nor identified
within the pathology database of the Toronto Zoo captive BFF population nor by the current SSP
pathologist (Dr Michael M Garner personal communication)
Previous characterization of coccidia from black-footed ferrets has been based on host species
affected tissues in the host and morphometric characterization of life stages in histologic sections and
oocyst characteristics using light microscopy It is known that morphologically similar Eimeria species
are not necessarily conspecific and may vary in host specificity and pathogenicity Molecular
characterization is thus required to accurately identify coccidia to the species level No molecular
characterization of coccidian parasites from black-footed ferrets has been performed to date
There is a significant information gap regarding which parasite species are implicated in
morbidity and mortality events associated with enteric coccidiosis in BFF and whether different coccidia
are associated with this disease in adult versus juvenile age classes or in different SSP institutions
Studies to further characterize the eimeriid coccidia of the BFF are warranted to improve the management
of this disease in the captive population The objectives of this research were to morphologically and
molecularly characterize coccidia associated with enteric disease in BFF at the Toronto Zoo and in other
SSP facilities
32 MATERIALS AND METHODS
321 Fecal samples
Twelve fecal samples were collected during the study period (from 2014-2016) and preserved in
potassium dichromate (25 wv aqueous) seven from the Toronto Zoo and five from the Louisville Zoo
Centrifugal flotation with saturated salt solution (Ryley et al 1976) was used to isolate and concentrate
oocysts from fecal samples for light microscopic examination and genomic DNA extraction
One to two drops of the supernatant from the centrifugal flotation were placed directly on a slide
and beneath a coverslip The morphology and dimensions of sporulated oocysts were documented using a
65
Provis AX70 photomicroscope (Olympus Canada Richmond Hill ON Canada) fitted with a digital
imaging device (Infinity3-1C Lumenera Corporation Ottawa ON Canada) controlled using iSolution
Lite image analysis software (Hoskin Scientific Burlington ON Canada) operated at a total
magnification of 1000times Morphologic features noted for each oocyst included oocyst wall morphology
number of sporocysts presence or absence of a micropyle micropyle cap residual body and polar
granules For sporocysts size number of sporozoites per sporocyst and presence or absence of Stieda
body and sporocyst residuum were noted Alignment of sporozoites within the sporocyst and
presenceabsence of refractile bodies within sporozoites were also described The sporulated oocyst
length and width measurements (in microm) were then used to calculate the SI for each measured oocyst
Morphologic and morphometric features were compared to previously published values for E furonis and
E ictidea from domestic and black-footed ferrets
Genomic DNA extraction and purification were performed using a QIAamp DNA Mini Kit
(Qiagen Hilden Germany) according to manufacturerrsquos instructions as described in Chapter 2 (Materials
amp Methods)
322 Formalin fixed intestinal tissues
The pathology records of the Toronto Zoo were searched from 1993-2016 for cases of BFF
diagnosed with enteric coccidiosis on histopathology For each case slides of histologic sections from all
submitted intestinal segments were reviewed to confirm the presence of sexual andor asexual life stages
within the intestinal epithelium Scrolls (5-6 microm) were cut from the paraffin blocks containing affected
intestinal sections and DNA extracted from the formalin fixed paraffin-embedded tissue (FFPE) using
the QIAamp DNA FFPE Tissue Kit (Qiagen Toronto Ontario) as per manufacturerrsquos instructions
66
323 Molecular characterization
Molecular characterization of coccidial isolates was performed on oocysts purified from fresh
fecal samples (isolated as described above) that were collected from juvenile and adult ferrets from
2014-2016 and DNA extracted from FFPE samples of BFF intestine containing parasite life stages
Regions from the nuclear 18S (SSU) rDNA (nu 18S rDNA) mitochondrial cytochrome c oxidase
subunit I (mt COI) DNA and mitochondrial cytochrome c oxidase subunit III (mt COIII) DNA were
amplified by polymerase chain reaction (PCR) from each sample using the primers listed in Table 31 and
methodology described in the Materials amp Methods section of Chapter 2 Table 31 also contains the
specific anneal conditions used for the various primer pairs Genomic DNA from an Eimeria species of
poultry was included in the PCR reactions to act as a positive control for the reaction chemistry A
representative selection of the newly generated nucleotide sequences resulting from the above were
submitted to GenBank
DNA obtained from oocysts collected from fecal samples during the first year of the study (2014)
was used to generate a complete mitochondrial genome (see Chapter 6 for details) using primer pairs and
sequencing primers summarized in Table 31 All subsequent samples collected in 2015 and 2016 had
shorter mt COI and mt COIII sequences obtained to permit genotyping of all collected oocysts at these
two loci The location of each primer in the nu 18S mt COI and mt COIII genetic locus is illustrated in
Figure 31
33 RESULTS
From 2014-2016 coccidia-positive fecal samples were obtained from twelve BFF ferretsferret
groups from the Toronto Zoo and Louisville Zoo SSP populations (see Table 32) Nine samples were from
single housed adults between the ages of 1-5 years (63 MaleFemale) Two samples were from mixed
groups one pooled fecal sample from four adults (FERA-1 13 MF) and one fecal sample from a family
67
group consisting of a dam and five kits (23 MF) One fecal sample was collected from a juvenile male
ferret at the time of necropsy
Eleven BFF with enteric coccidiosis were identified in the Toronto Zoo necropsy reports from
1998-2016 and all were confirmed by histological re-evaluation (Table 32) Both juvenile (n=9 36 MF)
and adult ferrets (n=2 20 MF) were represented
331 Morphometric characterization
Twelve coccidia-positive fecal samples were identified from adult and juvenile BFF from 2014-
2016 by on site laboratories at either the Toronto Zoo or the Louisville Zoo Fecal flotation and light
microscopic re-examination of the samples identified coccidial oocysts in 10 of these 12 samples
Morphometric characterization was performed on six samples in which there was adequate
quantity and quality of sporulated oocysts for examination These included three samples from single-
housed adults one from a juvenile at the time of necropsy one of pooled feces from a group of adult
ferrets and one of pooled feces from a family group (dam and kits) Two of the three samples from
single-housed adults were from the same ferret on different dates in 2016 the dates of collection were
separated by a period in which shedding of oocysts was not identified on routine repeated fecal
examinations Oocysts were elliptical with a colourless double wall and contained four sporocysts each
with two sporozoites Sporocysts were ovoid and both Stieda body and residual body were present
Sporozoites exhibited an anterior to posterior alignment within the sporocysts and refractile bodies were
identified (Figure 32) Results for length width and shape index of sporulated oocysts including range
and average values are summarized in Table 33 and Figure 32 The average measurements based on the
results of all 148 oocysts measured were length 2398 microm (1859-3057) width 1855 microm (1373-2383)
and shape index 130 (101-160)
The same measurements were performed on 59 sporocysts from a single ferret (Noodle) and
results are as follows average length 1280 microm (898-1480) average width 738 microm (505-1028) and
average SI 176 (124-247) In one sample (Mohawk-2) sporozoites were visible free on the slide
68
Measurement of three of these provided an average length of 1068 microm (1044-1117) and an average
width of 341 microm (316-393)
332 Molecular characterization
Molecular characterization was successfully performed on oocysts from seven of 10 fecal
samples containing coccidial oocysts and FFPE tissue from nine of the 11 necropsy cases (see Table 32)
Attempts at amplification of DNA extracted from necropsy samples Z228-98 and Z137-14 using the
primer pairs listed in Table 31 were unsuccessful Similarly attempts at PCR and sequencing of DNA
extracted from fecal oocysts from two Toronto Zoo BFF Jenna and Ruckus were unsuccessful
Molecular identification results for the remaining samples are summarized in Table 32
Only one Eimeria species E ictidea was identified in all enteric coccidiosis cases diagnosed at
necropsy in both juvenile and adult BFF at the Toronto Zoo from 1998-2014 This same species was
identified in all Toronto and Louisville Zoo BFF fecal samples that were sequenced successfully (n=8)
with the exception of a single case from Louisville This Louisville ferret was identified as having a
rodent pseudoparasite (Eimeria species) in the submitted fecal sample the eimeriid pseudoparasite had
986 sequence identity at the mt COI locus to the murine coccidium Eimeria falciformis All sequences
generated for E ictidea exhibited 100 sequence identity at the mt COI and COIII loci
Novel nu 18S rDNA mt COI and mt COIII sequences were generated for E ictidea from both
geographic locations and deposited in GenBank (Accessions MF860826 MF860827 MF860823
MF860825 MF860822 MF860824) Sequences were compared to those previously published for related
eimeriid coccidia The nu 18S rDNA sequence from Eimeria ictidea isolated from the Toronto Zoo BFF
had 9736 identity (14 single nucleotide differences) to the previously published sequences from
isolates of E furonis from domestic ferrets (Mustela putorius furo) in Japan (GenBank AB239130 and
AB329724) and newly generated sequences from Canadian and European isolates (GenBank MF774678-
MF774680 see Chapter 2 and Figure 33) In contrast nu 18S rDNA sequence of E furonis from
domestic ferrets (see Chapter 2) showed 9953 to 100 identity (0 to 3 SND) to the Japanese
69
sequences Comparison of newly generated partial sequences of the mt COI region from E ictidea from
BFF to isolates of E furonis (GenBank MF774034-MF774036) from DF and E mephitidis (GenBank
KT203398) from the striped skunk (Mephitis mephitis) the only carnivore Eimeria sp for which a mt
COI sequence was previously available reveals only 9415 and 9084 sequence identity respectively
with these other Eimeria spp of carnivores (Figure 34)
34 DISCUSSION
This work presents the first nu 18S rDNA mt COI and mt COIII sequences (nu 18S rDNA -
MF860826 MF860827 mt COI - MF860823 MF860825 mt COIII - MF860822 MF860824) generated
from an intestinal eimeriid parasite of the BFF referred to here as E ictidea collected from multiple BFF
of different ages from two separate captive populations (Toronto Zoo Toronto Ontario Canada and
Louisville Zoo Louisville Kentucky USA)
The morphometric description of coccidial oocysts from BFF in this work are consistent with
previous descriptions of E ictidea from mustelids including BFF the Steppe polecat the European
polecat and domestic ferrets (Hoare 1927 Svanbaev 1956 Jolley et al 1994) Thus I propose the name
E ictidea for the enteric coccidium described from BFF reflecting the similarity in morphology host
species and location of infection in intestinal tissues yet recognizing the absence of species identification
by molecular techniques Molecular characterization of parasites that agree with the description of E
ictidea morphologically from various mustelid host species would allow not only for determination of
whether the parasites are conspecific but would also provide insight into the potential for cross-
transmission among related mustelid hosts
DNA extraction from FFPE samples allowed successful PCR and sequencing of small DNA
fragments (220 bp) in nine of the eleven cases in which the technique was attempted Age of the samples
did not appear to be the major factor associated with successful extraction of good quality DNA the two
samples for which it was unsuccessful were the most recent (2014) and oldest (1998) casesConsequently
it may be possible to use banked FFPE tissues from historic necropsy cases from other SSP institutions
70
and necropsies of wild-born or re-introduced ferrets to determine the identity of the coccidial species
underlying disease in these cases and to better characterize the disease in the greater BFF captive and wild
populations Williams et al (1988) were contacted regarding their historic FFPE samples but formalin
blocks were no longer available for these cases and thus comparisons could not be made Banked FFPE
samples were requested from other SSP institutions however the Convention on International Trade in
Endangered Species of Wild Fauna and Flora (CITES) restrictions on the international transport of DNA
from endangered species did not allow for sample acquisition during the period in which this research was
conducted
Evaluation of FFPE samples from the Toronto Zoo indicate that the same Eimeria species has
been implicated in deaths associated with enteric coccidiosis from 1999-2014 as well as episodes of
clinical disease in ferrets in the Toronto Zoo population from 2014-2016 Samples from coccidia-positive
BFF at the Louisville Zoo in 2016 also contained the same Eimeria species Finding the same parasite at
multiple SSP locations was expected because BFF are transferred among institutions on a yearly basis for
breeding and potential release Consequently these parasites have repeated opportunities to move
between institutions in infected hosts or on contaminated cage materials to become established at a new
location Furthermore the stress of transport and transfer to a new environment may precipitate shedding
of endemic coccidia and increase the risk of a coccidial outbreak this concern is reflected in the SSP
recommendations for prophylactic treatment of all BFF with anti-coccidial medication prior to shipment
(USFWS BFF Recovery Program 2017)
A single BFF from the Louisville SSP population not showing clinical signs consistent with
coccidiosis was identified as having a rodent Eimeria species in the submitted fecal sample
Morphometric characterization of oocysts in this sample was not performed due to the paucity of visible
oocysts however examination at 100times suggested that the oocysts in the sample were ovoid in shape and
of comparable size to oocysts identified in other BFF samples The finding of a rodent Eimeria in a BFF
fecal sample is not unexpected as whole rodents comprise a significant part of the captive BFF diet The
oocysts shed by the BFF were most likely acquired through ingestion of an infected prey item and thus
71
most likely represent pseudoparasitism Molecular characterization was however required to
differentiate this from a case of true enteric coccidiasis
Reports from the first captive BFF population derived from South Dakota indicate the presence
of an unidentified species of enteric coccidium (Carpenter and Hillman 1979) in this group before its
demise in 1979 No reports containing morphometric descriptions of the coccidia from this group were
found on literature review and all parasites of this group have been lost with their hosts All subsequent
reports on enteric coccidiosis in BFF are from ferrets derived from the second founder group from
Wyoming in the 1980s The frequent transfer of ferrets among SSP institutions within the captive
breeding program and to different release sites within North America would be expected to result in the
same Eimeria species being found in all populations The exception to this would be the potential for
cross-transfer of other eimeriid parasites to wild BFF from sympatric mustelid species such as the long-
tailed weasel (Mustela frenata) Jolley et al described two other species of enteric coccidia from this
second captive population in 1994 the first was similar to E furonis of domestic ferrets and the second a
large coccidian parasite of unknown genus Neither of these parasites was identified in the Toronto and
Louisville Zoo populations during the course of this study In order to determine whether these parasites
persist within the present-day BFF populations and their impact on this species more detailed
examination of coccidia-positive fecal samples from captive and wild BFF populations is recommended
Furthermore the molecular identification of enteric coccidia from historic and future necropsy samples of
wild and captive BFF could aid in determining the presence of and contribution to mortality events by
these additional coccidia species
ACKNOWLEDGEMENTS
The authors would like to recognize the Wildlife Health Centre staff at the Toronto Zoo for their
assistance with the collection of fecal samples from the BFF from 2014-2016 The authors would also like
to recognize the Louisville Zoo for their contribution of samples to this project Finally this project was
made possible through funding by the Toronto Zoo Residency Research Fund to DASAP and partial
72
funding from a Natural Sciences and Engineering Research Council of Canada (NSERC) Discovery
Grant (400566) to JRB
73
Table 31 Amplification primers used to sequence the nuclear 18S rDNA mitochondrial COI and COIII loci of Eimeria ictidea originating from
fecal and formalin-fixed paraffin embedded tissue samples from black-footed ferrets (Mustela nigripes) including anneal temeratures (Ta)
extension times and expected PCR product sizes
Figure 31 Nuclear and mitochondrial genetic loci targeted by primers listed in Table 31 and used to characterize Eimeria ictidea originating
from black-footed ferrets (Mustela nigripes)
Gene Target Primer Pairs Primer Sequence (5ʹ-3ʹ) Size (bp) Ta (degC) Anneal (sec) Reference
18SrDNA Sarco_18S_123F TATCAGCTTTCGACGGTAGTGTATT 1480 60 30 Ogedengbe et al (2016)
ERIB10_REV CTTCCGCAGGTTCACCTACGG
mt COI T_Eim_COI_272F CAATTCTAYGATGCCGCWTT 220 52 30 Chapter 2 (Table 21)
COX1-500R CATRTGRTGDGCCCAWAC Ogedengbe et al (2011)
COI-400F GGDTCAGGTRTTGGTTGGAC 800 52 60 El-Sherry et al (2013)
COI-1202R CAAKRAYHGCACCAAGAGATA El-Sherry et al (2013)
mt COIII WG-MT_4140F AGAAAACCTAAAATCATCATGT 1000 52 60 Ogedengbe et al (2015)
Eimeriid_CO3_799R AAGTGAGTTCGCATGTTTAC Ogedengbe et al (2015)
74
Figure 32 A+B) Features and cellular contents of Eimeria ictidea from a black-footed ferret
(Mustela nigripes) Legend Oocyst dotted thin white arrow = polar granule Sporocyst dotted thin
black arrow = Stieda body thick white arrow= sporozoite refractile body thick black arrow=
residuum scale bars as indicated C) Shape index length and width measurements of sporulated
oocysts of Eimeria ictidea from black-footed ferrets (Mustela nigripes) Legend times indicates the
mean Dotted oval indicates one standard deviation around the mean
10 microm 5 microm
75
Figure 33 Comparison of nuclear 18S rDNA sequence alignment of Eimeria ictidea from two black-footed ferrets (Mustela nigripes) to
newly generated (see Chapter 2) and published sequences of Eimeria furonis from domestic ferrets (Mustela putorius furo)
Figure 34 Comparison of mitochondrial cytochrome c oxidase subunit I sequence alignment of Eimeria ictidea from two black-footed
ferrets (Mustela nigripes) to sequences from other eimeriid parasites of carnivores
Identity
Eimeria mephitidis ndash KT2033981
Eimeria ictidea ndash MF860823 (Guanella ndash LZ)
Eimeria ictidea ndash MF860825 (Mystery ndash MTZ)
Eimeria furonis ndash MF774034 (Type 1)
Eimeria furonis ndash MF774035 (Type 2)
Identity
Eimeria ictidea ndash MF860827 (Guanella ndash LZ) Eimeria ictidea ndash MF860826 (Mystery ndash MTZ)
Eimeria furonis ndash MF774680 (Type 1) Eimeria furonis ndash AB3297241 (Type 1) Eimeria furonis ndash AB2391302 (Type 1)
Eimeria furonis ndash MF774678 (Type 2) Eimeria furonis ndash MF774679 (Type 2)
76
Table 32 Morphologic and molecular characterization of coccidia from fecal and FFPE necropsy samples from black-footed ferrets (Mustela
nigripes)
Sample ID Sample Source Year Age (years) Sex Source Microscopic Description^ Molecular Diagnosis
Z228-98 Toronto Zoo 1998 7 M FFPE rare asexual stages -
Z143-99 Toronto Zoo 1999 1 M FFPE sexual and asexual stages E ictidea
Z106-02 Toronto Zoo 2002 008 F FFPE rare asexual stages E ictidea
Z108-03 Toronto Zoo 2003 008 F FFPE sexual and asexual stages E ictidea
Z109-03 Toronto Zoo 2003 008 F FFPE sexual and asexual stages E ictidea
Z124-12 Toronto Zoo 2012 021 F FFPE sexual and asexual stages E ictidea
Z113-13 Toronto Zoo 2013 017 F FFPE sexual and asexual stages E ictidea
Z117-13 Toronto Zoo 2013 025 M FFPE sexual and asexual stages E ictidea
Z118-13 Toronto Zoo 2013 025 M FFPE sexual and asexual stages E ictidea
Z119-13 Toronto Zoo 2013 025 F FFPE sexual and asexual stages E ictidea
Z137-14 Toronto Zoo 2014 021 M FFPE
fresh feces
sexual and asexual stages
POS E cf ictidea
-
E ictidea
FERA_1 Toronto Zoo 2014 gt 1 MF fresh feces POS E cf ictidea E ictidea
Noodle Toronto Zoo 2015 1 M fresh feces POS E cf ictidea E ictidea
Ruckus Toronto Zoo 2016 3 F fresh feces POS -
Mystery Toronto Zoo 2016 1 M fresh feces POS E ictidea
Mohawk Toronto Zoo 2016 1 M fresh feces POS E cf ictidea E ictidea
Jenna Toronto Zoo 2016 1 F fresh feces POS -
Thrope Louisville Zoo 2016 3 M fresh feces NEG -
FloJean Louisville Zoo 2016 2 F fresh feces NEG -
Rigatoni Louisville Zoo 2016 5 M fresh feces POS rodent Eimeria
Guanella +kits Louisville Zoo 2016 2 015 F kits 2M3F fresh feces POS E cf ictidea E ictidea
Clive Louisville Zoo 2016 1 M fresh feces POS E ictidea
Legend FFPE= formalin fixed paraffin embedded Sex MF = samples from family groups containing both sexes - = PCR and sequencing unsuccessful ^ = life stages identified on
histologic section
= mitochondrial COI andor COIII sequencing results Presence (POS) or absence (NEG) of oocysts and morphological identification of oocysts based on measurements when
77
Table 33 Morphometric (length width shape index) characterization of Eimeria ictidea oocysts from fecal samples from black-footed ferrets
(Mustela nigripes)
Sample ID FERA - 1 ^ Z137-14 Noodle Mohawk -1 Mohawk - 2 Guanella Total
Number of oocysts 12 36 32 10 21 37 148
Length (microm) 2333 (2055-2583) 2456 (2111-2848) 2505 (2079-3008) 2779 (2590-3060) 2493 (2036-2822) 2139 (1859-2372) 2398 (1859-3057)
Width (microm) 1676 (1373-2180) 1835 (1643-2232) 1975 (1509-2360) 2253 (2092-2383) 1803 (1549-2017) 1751 (1610-1888) 1855 (1373-2383)
Shape index 135 (103-160) 134 (113-156) 127 (105-155) 124 (113-138) 139 (114-154) 122 (101-145) 130 (101-160)
Legend ^= mixed adult group = dam and kit group
78
CHAPTER 4 NATURAL HISTORY OF ENTERIC COCCIDIOSIS IN THE BLACK-
FOOTED FERRET (MUSTELA NIGRIPES)
ABSTRACT
Black-footed ferrets (BFF Mustela nigripes) the only native North American ferret species are
endangered throughout their former geographic range An intensive captive breeding program produces
animals to supplement re-established wild populations Coccidial enteritis is a major cause of morbidity in
young captive ferrets but the disease also affects adults Limited information is available on the
pathogenesis of intestinal coccidiosis in captive BFF and characterization of the natural history of the
disease for improved prevention and management is imperative The objectives of this research were to
determine morbidity and mortality rates in the Toronto Zoo captive BFF population as well as
characterizing the natural history of the disease in this species through evaluation of shedding patterns
body tissues affected pre-patent period and periods of enhanced host susceptibility to infection
Coccidia-associated mortality in BFF at the Toronto Zoo from 1997-2016 averaged 053 yearly in
adults (range 0-526) and 195 in juveniles (range 0-1667) Clinical signs and histologic lesions in
Toronto Zoo BFF were similar to those described in previous publications A seasonal influence on
oocyst shedding was identified in adult BFF and ferrets appeared to maintain persistent infection with E
ictidea shedding coccidia in multiple years A larger multi-institutional study is required to better
elucidate the natural history of enteric coccidiosis in this species
41 INTRODUCTION
Black-footed ferrets (BFF Mustela nigripes) are the only native North American ferret species
and are endangered throughout their former geographic range When the last remaining truly wild
population underwent serious decline as a result of disease the decision was made by the by United States
Fish and Wildlife Service to capture the remaining 24 animals and establish a captive breeding program
this occurred between 1985 and 1987 Only seven of the captured ferrets bred successfully and are the
79
founders of the current North American BFF population (USFWS BFF Recovery Program 2017) The
captive population which now numbers approximately 300 individuals is distributed among and managed
by six collaborating facilities these include the Toronto Zoo United States Fish and Wildlife Services
National Black-Footed Ferret Conservation Center National Zoorsquos Smithsonian Conservation Biology
Institute Louisville Zoo Cheyenne Mountain Zoo and the Phoenix Zoo (Black-footed Ferret Recovery
Implementation Team 2011 Santymire et al 2014) Since 1991 BFF have been released into
reintroduced into the wild at multiple sites within their former range and over 8000 BFF kits had been
produced in captive breeding facilities as of 2011(Black-footed Ferret Recovery Implementation Team
2011) Twenty-eight BFF reintroduction sites currently exist throughout North America however there
continues to be a need to support wild populations as only a four of the re-established groups are truly
self-sustaining
Enteric coccidiosis is recognized as a cause of significant morbidity and mortality in captive
breeding programs affecting both juvenile and adult animals (Bronson et al 2007 USFWS BFF
Recovery Program 2017) Two Eimeria species Eimeria ictidea and Eimeria furonis have been
identified from cases of entric coccidiosis in BFF (Jolley et al 1994) Jolley et al examined fecal samples
from both wild and captive BFF and provided detailed morphologic descriptions of the oocysts of both
Eimeria spp as well as descriptions of the intestinal pathology associated with infection Asexual and
sexual life stages of both of the aforementioned Eimeria spp were identified on histologic section within
the villar epithelium throughout the small intestine but were concentrated in the jejunum Intestinal
sections from BFF infected with E ictidea exhibited two morphologically distinct meronts one at the
villar tips which was larger and lacking in undifferentiated mass and the other at the base of the villi or
rarely in the intestinal crypts gamogony was predominantly observed at the villar tips and was noted
throughout the small intestine
80
Extraintestinal coccidia have also been reported from captive BFF at one facility (Jolley et al
1994 Williams et al 1988) The authors identified the presence of endogenous coccidial life stages in
histologic sections of respiratory tissue and in impression smears of the urinary bladder from BFF
diagnosed with canine distemper No subsequent reports of systemic coccidiosis in BFF have been
published or identified within the pathology database of the Toronto Zoo captive BFF population or by
the current SSP pathologist (Dr Michael M Garner personal communication)
Recent investigations into the etiologic agents of enteric coccidiosis in BFF at the Toronto Zoo
have identified a single Eimeria species associated with all cases of enteric coccidiosis and associated
mortality in juvenile and adult BFF from 1999-2016 Furthermore this pathogen was identified in fecal
samples based on morphologic and molecular characterization from adult and juvenile BFF in another
zoological collection (Louisville Zoo Kentucky USA) (see Chapter 3) This coccidium is
morphologically consistent with Hoarersquos original description of E ictidea (1927) and is referred to
henceforth as Eimeria ictidea
There is a significant information gap regarding the pathogenicity of E ictidea in BFF The
objectives of this research were to determine morbidity and mortality rates in the Toronto Zoo and
additional captive BFF SSP populations as well as characterizing the natural history of the disease in this
species through evaluation of shedding patterns body tissues affected pre-patent period and periods of
enhanced host susceptibility to infection
42 MATERIALS AND METHODS
421 Toronto Zoo BFF breeding program
At the Toronto Zoo black-footed ferret breeding program all adult ferrets are housed
individually with the exception of dams and kits After the birth of the kits dams are housed with their
offspring from whelp date until removal at approximately 4-6 months of age Routine monthly fecal
81
examinations (direct examination and flotation) are performed in house for all ferrets in the breeding
program based on SSP recommendations to evaluate for the presence of coccidia
422 Fecal oocyst evaluation
Family groups
From 2014-2016 daily fecal examination for coccidial oocysts was initiated for all group-housed
dams and kits In 2014 fecal samples were collected daily from all dams and kits from weaning (30 days
after whelping) to 72 days post whelping Based on 2014 data in 2015 this surveillance was extended
from weaning (35 days post whelping) to 135 days of age Furthermore fecal samples were collected
from the dam for an additional 14 days after removal of kits In 2016 no fecal samples were collected
from dam and kit groups at the Toronto Zoo but samples were submitted from one group of dam and kits
from another SSP population at the Louisville Zoo (Kentucky USA)
Adults
From 2015-2016 daily fecal samples were also collected from all adult ferrets identified as
shedding coccidial oocysts on their monthly routine fecal examination and from clinically ill BFF
Samples were collected for 10-14 days after initial positive sample identification In 2016 fecal samples
were also submitted from four coccidia-positive adult ferrets from the Louisville Zoo population samples
were collected for 7 days post initial identification of shedding
Individual fecal samples were analyzed via flotation using the McMaster method followed by
routine flotation in saturated salt solution (Dryden et al 2005) to determine the presence or absence of
oocysts and oocyst burden (oocysts per gram of feces OPG) Temporal trends in oocyst shedding were
monitored Coccidia-positive ferrets were evaluated visually on a daily basis for presence of clinical signs
consistent with infection Infected juvenile ferrets and adult ferrets were treated with oral ponazuril or
toltrazuril regardless of the presence of clinical signs as per the black-footed ferret SSP
recommendations Based on these recommendations ponazuril is typically administered orally at 30-50
82
mgkg once daily for 3-7 days until clinical signs have resolved or oocyst shedding has been significantly
reduced (USFWS BFF Recovery Program 2017)
423 Retrospective review of pathology records
The pathology records of the Toronto Zoo were searched from 1993-2016 for cases of BFF
diagnosed with enteric coccidiosis on histopathology For each case gross necropsy reports were
reviewed and slides of histologic sections from all submitted intestinal segments re-examined to confirm
the presence of sexual andor asexual life stages within the intestinal epithelium and describe the
histologic lesions associated with presence of the parasite life stages
424 Prospective modified necropsy protocol
During the study period 2014-2016 necropsy protocols for all BFF were modified to improve
detection of coccidial life stages and better to determine which portions of the intestinal tract were
affected The entire length of the intestine from duodenum to anus was measured and intestinal contents
were flushed with 12 mL of sterile saline into a sterile container Intestinal contents were preserved in
25 potassium dichromate solution (mixed 11 with intestinal contents vv) for molecular diagnostics
Paired 2-cm long intestinal samples were collected from all sections of small and large bowel duodenum
(1) jejunum (6) ileum (1) and colon (2) The eight small intestinal samples were collected at equal
distances from the pyloric sphincter to the beginning of the colon and the distance from the pylorus noted
for each Colon samples were taken at 25 and 75 of the length of the colon One sample from each
pair was preserved in Serra solution (100 ethanol (60 vv) 37 formaldehyde (30 vv) glacial
acetic acid (10 vv) and the second sample was frozen Representative tissues from all internal organs
as well as additional intestinal samples skin muscle and brain were also collected and preserved in 10
buffered formalin Histopathologic examination was performed on all tissues collected
83
425 Retrospective medical history review
Medical histories of all BFF held by the Toronto Zoo since the initiation of the SSP program were
reviewed for data on frequency of occurrence of shedding of coccidial oocysts in adults and juveniles as
well as any association of shedding with clinical signs and administration of anticoccidial treatment Data
was tabulated yearly for adult and juvenile ferrets to determine annual morbidity and mortality rates
associated with enteric coccidiosis Medical records and pathology reports were solicited from the other
SSP institutions to determine comparative morbidity and mortality rates associated with enteric
coccidiosis in BFF at other facilities Both morbidity and mortality rates were calculated as
incidenceattack rates with yearly adult population size or number of family groups (dam and kits) as the
denominator for morbidity rates and number of yearly deaths in each age class as the denominator for
mortality rates
43 RESULTS
431 Fecal oocyst evaluation and retrospective medical history review
Family groups
Fecal samples were collected from seven groups of dams and kits housed together at the Toronto
and the Louisville Zoos from 2014-2016 All data from first to last day of collection for all family groups
is listed in Appendix 1 selected pertinent data for each group is presented in Table 41 Five groups of
dams and kits were sampled in 2014 and one group in each of 2015 and 2016 Shedding occurred no
earlier than 55 days of kit-age in any of the groups and was identified from 55-81 days of age (Table 41
Table 42 Figure 41)
In 2014 fecal oocyst shedding was identified in three of the five surveyed groups In two of the
three groups (dams Poppy and Bumblefoot) changes to fecal colour and consistency were identified
concurrently with periods of oocyst shedding both groups shed higher numbers of oocysts than the other
dam and kit groups in 2014 and 2015 Both Poppy and Bumblefoot had had litters in the previous one and
84
two years prior to this study respectively based on medical record review these dams and their litters
were also diagnosed as shedding coccidial oocysts that were too numerous to count on direct exam and
fecal flotation Clinical signs in the previous years included dark tarry hemorrhagic or soft mucoid feces
and reduced appetite both groups received treatment with toltrazuril (Baycox Coccidiocide Solution
25 Bayer Inc Mississauga Canada) and trimethoprim sulfamethoxazole (Novo-Trimel Teva Canada
Ltd Scarborough Canada) (TMS) One of four kits from Poppyrsquos 2013 litter (Z113-13) died of enteric
coccidiosis three days after the group was diagnosed as shedding coccidial oocysts and the initiation of
treatment with TMS
In 2015 low grade fecal oocyst shedding (lt14 oocysts per gram of feces) without associated
clinical signs was noted in the Fiddlesticks group on three days during a seven day period from 63-69
days of kit-age and again for a single day at 128 days of kit-age The dam had been diagnosed and treated
for enteric coccidiosis in 2014 at which time she exhibited clinical signs of loose green feces to
hemorrhagic diarrhea lethargy and dehydration In 2016 she was diagnosed as shedding low numbers of
coccidia exhibited no clinical signs and did not receive treatment prior to resolution of shedding
In 2016 Guanella and kits shed oocysts over a nine day period and daily fecal oocyst shedding
ranged from 206 ndash 371714 OPG Combined treatment with ponazuril (first four days of shedding)
sulfadimethoxine injectable (first two days of shedding additional product information not available)
amoxicillin oral (first two days of shedding additional product information not available) penicillin
injectable (first two days of shedding additional product information not available) and subcutaneous
fluids (first two days of shedding additional product information not available) was administered to this
group Previous medical history was not available for this female for review
In 2014 and 2015 fecal oocyst shedding in all groups in the Toronto Zoo population started in the
three week period from the last week of July to mid-August In 2016 shedding was first identified in the
Louisville Zoo group in mid-July
85
Adults
Seven single-housed adult BFF (52 MF) were detected to have shed coccidia during the study
period (Table 43) Shedding periods lasted from 2-10 days and oocyst per gram counts ranged from 104
ndash 554274 (Table 44) Clinical signs were identified in four of the seven ferrets and consisted of
lossreduction of appetite (n=2) weight loss (n=1) lethargy (n=1) blood in feces (n=1) loose or runny
feces (n=3) soft mucoid feces (n=1) green colour of feces (n=2) Five of the seven adults received
treatment after detection of oocyst shedding two of which received treatment in the absence of clinical
signs Treatment consisted of oral toltrazuril in four cases toltrazuril in combination with trimethoprim
sulfamethoxazole in one case (Mohawk-A) and ponazuril and sulfadimethoxine (manufacturerrsquos
information not available) in one case (Clive) (Table 43)
Three of the adults in this study Mohawk Mystery and Jenna shed oocysts during multiple
different periods in 2016 Mohawk shed oocysts in May July and September of 2016 data from the first
two periods are reported in Tables 43 and 44 Mystery shed oocysts in June and July of 2016 Clinical
signs were observed only during the first shedding period and included poor appetite and hemorrhagic to
soft mucoid feces Jenna shed oocysts in July September and November of 2016 and again in February
and May of 2017 Although clinical signs were not detected in association with the initial period of
shedding in July 2016 (see Table 44) depressed mentation and hemorrhagic mucoid feces were identified
in the subsequent two shedding periods In both Mohawk and Jenna oocysts were not detected in feces on
multiple recheck and routine monthly fecal examination between shedding periods
Ruckus the fourth ferret shed low numbers of oocysts for two days in 2016 while housed alone
and had been reported to have shed oocysts during a 30 day period in 2014 while housed in a family
group with her kits Diarrhea to soft mucoid feces and loss of appetite were reported in 2014 Clinical
signs in 2016 consisted of small amounts of runny feces for a two day period two days after oocyst
shedding was no longer detected Treatment consisted of toltrazuril and TMS in 2014 and toltrazuril
again in 2016 Similarly Noodle shed low numbers of oocysts in July 2015 and had been identified as
shedding low numbers of oocysts during September of the previous year
86
The final ferret Rigatoni shed oocysts in feces in low numbers (54-732 OPG) over 5 days All
positive samples were pooled for molecular diagnostics and sequencing results showed 990 identity (7
single nucleotide differences SNDs) at the mt COI locus with a pair of rodent Eimeria species (ie
GenBank HM771682 JQ993704) (see Chapter 3) This ferret did not show clinical signs associated with
shedding and was not treated Previous medical history was not available for Clive the single adult ferret
from the Louisville Zoo
Based on sample collection dates and medical record review for the adult BFF for 2014-2016
shedding occurred during spring summer and fall with three ferrets shedding in May (Jenna Ruckus and
Mohawk) one ferret shedding in June (Mystery) three ferrets shedding in July (Mohawk Jenna Noodle)
one ferret shedding in August (Clive) two ferrets shedding in September (Noodle Jenna) and one ferret
shedding in November (Jenna)
432 Pathology
Eleven BFF with enteric coccidiosis were identified in the Toronto Zoo necropsy reports from
1993-2016 (Table 45) Cases were identified from 1998-2014 inclusive and all were confirmed by
histological re-evaluation (Chapter 3 Table 32) Both juvenile (n=9 36 MF) and adult ferrets (n=2 20
MF) were represented
Gross Pathology
Gross necropsy findings were similar across the 11 cases and included mucoid to fluid luminal
contents (n=7 636) beige to white pasty coating of the mucosal surface of the small intestine (n=6
545) and colon (n=4 364) gaseous dilation of intestinal segments (n=3 272) segmental enteritis
and hemorrhage (n=1 91) In one case Z228-98 no gross lesions were identified within the intestines
Impression smears of luminal contents or scrapings of intestinal mucosa were performed in four cases and
coccidia were identified in all four
Histopathology
87
Both sexual and asexual life stages were identified within the small intestinal segments in all
cases except Z228-98 and Z106-02 in which only rare asexual parasite life stages were identified (Table
45 Figure 42) Other histologic lesions seen in intestinal segments containing coccidia included
lymphoplasmacytic inflammation of the lamina propria (n=4) neutrophilic infiltration of the lamina
propria (n=2) villar necrosis (n=2) villar atrophy or blunting (n=3) and thrombi within the villar tips
(n=1)
Additional necropsy diagnoses included cholangiolar hyperplasia multiple hepatobiliary cysts
with suppuration renal adenocarcinoma apocrine gland adenocarcinoma (Z228-98) concurrent
clostridial enteritis (Z143-99) presumptive Salmonella sp septicemia (Z106-02) interstitial pneumonitis
(Z108-03) myocardial mineralization interstitial pneumonia and nephritis periportal hepatitis and
bacteremia (Z124-12) and suppurative esophagitis (Z137-14)
From 2014-2016 three black-footed ferrets were necropsied using the detailed protocol described
above Only one of the three cases Z137-14 was diagnosed with enteric coccidiosis based on
histopathology Two duodenal five jejunal two ileal and two colonic sections were collected at measured
lengths from the pylorus Parasite life stages were identified within the mucosal epithelium of all
intestinal segments extending from the distal duodenum (10-12 cm aboral to pylorus) through to the distal
colon (157-159 cm aboral from pylorus) The distal duodenal section contained asexual life stages only
with a single focus of epithelial cells containing meronts Sexual life stages (microgamonts
macrogamonts unsporulated oocysts) were identified within villar epithelial cells in all remaining
sections of the small intestine with numerous oocysts in the bowel lumen Mild lymphoplasmacytic
inflammation of the lamina propria was associated with the jejunal and ileal lesions and blunting of the
villi was identified within one jejunal segment The colonic sections contained small to moderate numbers
of sexual life stages identified within both superficial and deep crypt epithelium with occasional life
stages identified near the germinal cells Large numbers of oocysts and bacteria were identified within the
88
lumen of these colonic sections and both sections contained abscesses within the crypts The proximal
duodenum (0-2 cm) was the only section of the intestines not containing parasitic life stages
433 Morbidity and mortality
Annual morbidity rates for enteric coccidiosis at the Cheyenne Mountain Zoo and mortality rates
from enteric coccidiosis in Toronto Zoo BFF are summarized in Tables 46 and 47 During 2003-2016
yearly incidence of coccidiosis in adult BFF at the Cheyenne Mountain Zoo averaged 69 (range 0-
421) For family groups consisting of juvenile ferrets housed with their dams yearly incidence of
enteric coccidiosis averaged 115 (range 0-600)
From 1997-2016 coccidia-associated mortality in adult BFF at the Toronto Zoo averaged 053
yearly (range 0-526) with an average total mortality rate of 141 per year (range 0-526) For
juvenile ferrets (under 1 year of age) during the same period coccidia-associated mortality accounted for
an average of 133 of deaths yearly (range 0-100) with an overall average mortality rate of 170 per
year (range 0-3404) from all causes
Multiple additional SSP institutions provided partial medical and pathology data sets for use in
this study which were not sufficiently detailed to permit computation of morbidity and mortality rates for
those populations
44 DISCUSSION
The work described here supports previous clinical findings regarding the impact through both
morbidity and mortality associated with enteric coccidiosis in BFF No previous studies have been
undertaken to determine morbidity and mortality rates associated with enteric coccidiosis across BFF SSP
institutions
89
In a retrospective mortality study of captive BFF from 1984-2004 at the Smithsonianrsquos National
Zoological Park Bronson et al (2007) reported that gastrointestinal disease was the most common cause
of death in juvenile BFF (524) with 333 of juvenile mortality cases in the study caused by enteric
coccidiosis While the data is not directly comparable the findings reported here also reflect enteric
coccidiosis as a common cause of death in juveniles with increased mortality associated with the disease
compared with adult counterparts All Toronto Zoo mortalities in both juvenile and adult age classes for
which necropsy tissues were available have been attributed to infection with a single coccidia species
Eimeria ictidea (see Chapter 3)
Multiple SSP institutions provided partial medical and pathology data sets for use in this study
which were not sufficiently detailed to permit computation of morbidity and mortality rates for those
populations In future it would be useful to determine whether morbidity and mortality rates associated
with enteric coccidiosis vary among SSP institutions as this may allow for improved identification of
host parasite and environmental factors that increase risk
Clinical signs reported here are consistent with those described from both BFF and domestic
ferrets with enteric coccidiosis (Sledge et al 2011 Santymire et al 2014 USFWS BFF Recovery
Program 2017) Changes to fecal colour and consistency were the most common abnormalities identified
at the time of first detection of oocyst shedding While clinical signs in the cases described here do not
always correlate directly with the quantity of oocysts shed individuals shedding higher number of oocysts
showed clinical signs more frequently than those shedding lower numbers
In this study oocyst shedding from single-housed adult BFF ranged from 104 ndash 554274 oocysts
per gram Daily fecal samples produced from individual adult ferrets range in size from approximately 1-
16 grams In light of these findings during peak shedding from ~5times105 to 9times106 oocysts can be shed into
the environment in one day providing a massive infective dose The large numbers of oocysts shed
90
combined with confinement in a small enclosure space and hardiness of Eimeria oocysts in the
environment would be expected to markedly increase risk of infection in captive BFF
Oocyst shedding from family groups ranged from 0 ndash 371714 OPG The wide variability seen in
OPG counts between days as seen in Figure 41 and Table 41 may be accounted for by the staggered
initiation and resolution of shedding by different ferrets It is most likely that the source of infection in
these family groups is shedding by the dams some of which were identified to shed in multiple years
although environmental contamination cannot be excluded
Shedding in the adult ferrets was clustered during particular time periods specifically May July
and September If shedding is associated with stress and immunosuppression activities such as breeding
whelping electro-ejaculation of male ferrets and shipmenttransfer could act as stressors Whelping
which occurs primarily in May and June could also act as a stressor to other ferrets in the facility either
through social cues or as there would be associated changes in husbandry protocols The September
cluster could be associated with the transfer of ferrets between institutions kits are pulled during this time
and adult ferrets are moved among institutions thus changing the population dynamics of each SSP site A
large cluster of shedding was recorded in July With the exception of weaning of kits no other major
stressors are expected to occur during this time thus the increase in shedding by adults in this group
cannot be easily explained Interestingly shedding was not identified in ferrets from March through April
which is the typical breeding season and when ferrets are introduced for breeding a presumably stressful
time Shedding was not noted from December through April which could reflect reduced environmental
burdens due to low humidity levels as would be expected in a Nordic climate during the winter (which
would kill oocysts and thus block transmission) or may be consistent with reduced stress during this
period The results from single-housed adults are in contrast to the dam and juvenile ferret groups in
which oocyst shedding appeared to be correlated to a period of 55-81 days of kit age These results are
consistent with reports from other facilities of increased incidence of shedding by kits after 70 days of age
(USFWS BFF Recovery Program 2017)
91
Retrospective and prospective review of histologic sections of intestines from affected BFF at the
Toronto Zoo showed the presence parasitic life stages in epithelial cells of both the small and large
intestines Neither Hoare (1935b) nor Jolley et al (1994) mentioned the presence of parasites in the large
intestine of experimentally infected domestic ferrets or naturally infected BFF respectively In the study
described here asexual and sexual life stages were identified within the epithelial cells of the small
intestinal villi from base to tip and were most numerous in jejunum This matches the description by
Jolley et al (1994) however Hoare (1935a b) found E ictidea to be present primarily in the villar tips
Jolley et al (1994) also described two morphologically distinct meronts of E ictidea within the small
intestinal sections one at the villar tip that was larger and lacking in undifferentiated mass and the other
at the base of the villi or in the intestinal crypts these findings were not echoed in this study as
merogonic stages were identified throughout the intestinal epithelium from villus to the base of the crypt
and no visual differences between meronts in any location were identified Hoare (1935a b) also
described a resulting annular constriction of the villus separating the affected and non-affected segments
this constriction was neither seen in the cases described here nor mentioned by Jolley et al (1994)
Whether these differences result from E ictidea from BFF and E ictidea from domestic ferrets being
different parasites or from differences in tissue tropism of a single parasite in two different hosts cannot
be ascertained from the available information
Histologic lesions such as necrosis hemorrhage villar atrophy and inflammation associated with
the presence of parasitic life stages were rare These changes are normally elicited by the host immune
system (inflammation) and the parasite (cellular rupture to release life stages resulting in hemorrhage and
necrosis) in response to infection In light of the fact that acute death occurred in a number of these ferrets
(Z113-13 Z117-13 Z118-13 Z119-13 Z137-14) in the absence of secondary disease processes and
with the intestinal epithelium intact but containing myriad parasitic life stages an alternative mechanism
for mortality associated with the infection must be proposed It is possible that these parasites elaborate
exotoxins during their life cycle and when at high density result in sudden death of the host with minimal
92
tissue changes The presence of parasitic life stages occupying the majority of both small and large
intestinal epithelial cells could also potentially impair fluid and protein movement in and out of the
mucosa however clinical signs associated with malabsorptive diarrhea were not identified in any of these
cases The presence of bacteria within the blood or other organ tissues was not identified in any cases and
consequently sepsis is unlikely to be the cause of death
Black-footed ferrets appear to maintain persistent infection with E ictidea Adult BFF in the
Toronto Zoo population shed coccidia in multiple years and in two adult ferrets multiple times in the
same year While the coccidia seen in all cases were not confirmed as E ictidea using molecular
techniques morphologic similarities and a lack of additional Eimeria spp identified on molecular work
undertaken suggest that only one species of parasite is and has been present in the collection Two dams
that had been identified as infected based on routine fecal screening in previous years presumably acted as
the source of infection to their litters of kits in multiple years While continued environmental
contamination cannot be ruled out these findings imply a failure of the immune response of the BFF to
clear infection with E ictidea or even to protect against sufficient replication of organisms to result in
clinical disease
Based on clinical experience and review of the literature BFF appear to be much more sensitive
to infection with E ictidea compared with their domestic counterparts In domestic ferrets subclinical
shedding of oocysts appears to be the most common with rare reports in the literature of overt disease
and that only in juveniles (Blankenship-Paris et al 1993 Abe et al 2008) However a single report exists
of three separate clinical outbreaks of Eimeria furonis infection in domestic ferrets under intensive
management with increased morbidity and mortality affecting all ages classes (Sledge et al 2011) The
role of genetics in the apparent increased susceptibility of BFF to enteric coccidiosis is unknown but the
current captive BFF population is derived from seven founders and inbreeding depression or familial
genetic susceptibility may play a role in their increased susceptibility to disease caused by E ictidea
93
Black-footed ferrets diagnosed with enteric coccidiosis during the course of the study were
treated with either ponazuril or toltrazuril sulfonamide drugs or often a combination of the two groups
of therapeutic agents Toltrazuril and ponazuril are triazine coccidiocides with proven efficacy against
both asexual and sexual life stages of mammalian and avian Eimeria spp (Mehlhorn and Aspock 2008)
The sulfonamides are antimicrobial drugs that exhibit coccidiostatic or coccidiocidal effects depending on
dose they act by blocking folate synthesis and have effects on first and second generation meronts
(asexual life stages) as well as potentially acting on sexual life stages (Mehlhorn and Aspock 2008)
Based on the limited data available from this study and the fact that treatment was initiated in almost all
adult BFF and family groups at the time of oocyst detection regardless of the presence of clinical disease
the effects of treatment on duration of clinical signs cannot be effectively evaluated It appears
subjectively that adult ferrets treated with toltrazuril and in one case a combination of toltrazuril and
TMS showed reduction in oocyst shedding after 3-5 days of oral anti-coccidial therapy (see Table 43)
The effects of treatment with either sulfonamides or triazines would be expected to reduce oocyst
shedding consequently the duration and amount of oocyst shedding reported in this study may not
accurately characterize the natural course of disease
Perceived resistance to sulfa drugs has been reported from multiple SSP facilities In light of this
and their potential negative effects on ferret reproduction (eg prevention of embryo implantation in the
uterus and impairment of sperm development) sulfonamides are no longer recommended by the SSP for
treatment of coccidia in this species (USFWS BFF Recovery Program 2017) The frequent and
widespread use of triazines in the management of enteric coccidiosis in BFF presents a risk for
development of resistance to this drug class in the future Neither pharmacokinetic (PK) nor
pharmacodynamic (PD) studies have been published to validate the dose and frequency of dosage in
either class of drugs in BFF and consequently it is unclear whether this perceived failure of some ferrets
to respond to treatment is based on true resistance versus inappropriate dosing The only work evaluating
ponazuril in BFF evaluated serum levels of ponazuril after a single oral dose of 50 mgkg and reported
94
therapeutic levels for 10 days after administration (USFWS BFF Recovery Program 2017) No
information was provided on number or age of ferrets that participated in the study or on how the
determination of what were therapeutic levels was made Furthermore as life cycles of the coccidia
affecting BFF are limited to the gastrointestinal tract and do not exhibit tissue stages the validity and
usefulness of assessing blood levels of ponazuril in determining appropriate dosage and dose schedules is
questionable Further work to determine the PK and PD of triazines in ferrets is warranted to provide safe
and efficacious treatment and to reduce the risk of development of resistance Furthermore the creation
and validation of a model for enteric coccidiosis in a related species would allow for in vivo studies of
drug resistance
95
Figure 41 Oocyst per gram counts and shedding period of Eimeria ictidea from black-footed ferret
(Mustela nigripes) family groups from 2014-2016
0
50000
100000
150000
200000
250000
300000
350000
400000
29
33
37
41
45
49
53
57
61
65
69
73
77
81
85
89
93
97
10
1
10
5
10
9
11
3
11
7
12
1
12
5
12
9
13
3
13
7
14
1
14
5
14
9
OP
G
Age of Kits
Poppy Bumblefoot Calico Aubrey Ruckus Fiddlesticks Guanella
96
Figure 42 Small intestinal epithelium of a black-footed ferret (Mustela nigripes) containing sexual
life stages of Eimeria ictidea Legend Solid black arrow = oocyst Hatched arrow = macrogamont
Outlined arrow = microgamont Hematoxylin and eosin staining scale bar = 25 microm
25 microm
97
Table 41 Shedding of oocysts of Eimeria ictidea in black-footed ferret (Mustela nigripes) dam and kit
family groups from 2014-2016
Collection Year 2014 2014 2014 2014 2014 2015 2016
Age of kits (days) Poppy Bumblefoot Calico Aubrey Ruckus Fiddlesticks Guanella^
29 - - - 0 - - -
30 - 0 - 0 - - -
- - - - -
34 0 0 - 0 0 - -
35 0 0 - 0 0 0 -
- -
46 0 0 - 0 0 0 -
47 0 0 - 0 0 0 -
48 0 lt14 - 0 0 0 -
- -
53 0 0 - 0 0 0 +
54 0 0 - 0 0 0 +
55 0 0 - 0 0 0 3717146
56 0 0 - 0 - 0 1084436
57 0 0 - 0 - 0 64133
58 0 324 - 0 0 0 20654
59 0 0 - 0 0 0 0
60 0 446688 - 0 0 0 0
61 0 934828 - 0 0 0 41111
62 0 530469 - 0 0 0
63 0 1617131 - 0 0 lt14
64 0 8396357 - 114 0 0
65 0 234876 - 5368 0 lt14
66 0 374625 - 9455 0 0
67 lt14 2311575 - 39579 0 -
68 0 603563 - 125051 0 0
69 - 4103036 - 27067 0 lt14
70 2470327 7759974 - 0 0 0
71 1073085 - 1784 0 0
72 3430966 - 0 0 0
73 4459536 - 7326 0 0
74 4842212 - 2 0 0
75 3627307 - 0 0 0
76 5801885 - 0 0 0
77 5994 0 0 0 0
78 589835 0 0 0
79 24815 0 0 0
80 144016 0 0 0
81 97862 0 0 0
82 0 0 0
127 0 0
128 0 2843
136 0 0
150 0
Numbers of Oocysts Shed (oocysts per gram of feces)
Dam Identity
Legend lt 14 = oocyst positive samples with less than 14 oocyst per gram of feces - = no sample recorded for this date underline = last
sampling date + = coccidia present but OPG count not performed = Toronto Zoo ferret ^ = Louisville Zoo ferret thick outer border =
days treatment was received = range of sequential dates between previous and subsequent number during which OPG counts were
performed and samples contained 0 oocysts
98
Table 42 Summary of epidemiologic data for family groups of black-footed ferrets (Mustela nigripes)
shedding oocysts of Eimeria ictidea
Dam Identity
Poppy
2014
Bumblefoot
2014
Aubrey
2014
Fiddlesticks
2015
Guanella
^
2016
Number of kits 7 5 3 4 5
Kit age (days) at time of
shedding 70-81 58-70 64-73 63-69 54-61
Shedding period (days) 15 13 11 7 9
OPG min 0 0 0 0 206
OPG max 48442 83963 2707 lt14 371714
Clinical signs YES YES NO NO NO
Treated - YES YES NO YES
Legend = Toronto Zoo ferret ^ = Louisville Zoo ferret OPG = oocysts per gram of feces
- = missing data
Table 43 Shedding of oocysts of Eimeria ictidea in single-housed adult black-footed ferrets (Mustela nigripes) 2015-2016
Numbers of Oocysts Shed (oocysts per gram of feces)
Collection
Year 2015 2016 2016 2016 2016 2016 2016
Ferret Identity Noodle Ruckus Mohawk-A Mohawk-B Mystery Jenna Clive^
Age (years) 1 3 1 1 1 1 1
1084058 0 + + + + +
+ 0 42307650 6286676 + 183150 554274
857808 16650 12805238 7777929 + 215710 377920
1604894 16650 309690 139860 + 0 25808
377042 0 599400 119880 + - 37294
554445 0 34688 385579 117920 0 5363
26640 0 16650 0 0 0 7500
0 10406 0 0 0 1090
0 20813 0 0 0
- 0 1761 0
0 0 0 0
- 0
0
0
Legend lt 1 = oocyst positive samples with less than 1 oocyst per gram of feces underline = last sampling date + = coccidia present but OPG
count not performed = Toronto Zoo ferret ^ = Louisville Zoo ferret thick outer border = days treatment was received
100
Table 44 Summary of epidemiologic data for single housed adult black-footed ferrets (Mustela nigripes) shedding oocysts of Eimeria ictidea
Noodle Ruckus Mohawk-A Mohawk-B Mystery Jenna Clive^
Sex M F M M M F M
Age (years) 1 3 1 1 1 1 1
Shedding period (days) 7 2 9 6 10 4 8
OPG min 266 166 104 1199 1761 1831 1090
OPG max 10840 166 423076 77779 - 2157 554274
Clinical signs YES YES NO NO YES NO YES
Treated NO YES YES YES YES YES YES
Legend = Toronto Zoo ferret ^ = Louisville Zoo ferret M = male F= female OPG = oocysts per gram of feces - = missing data
NOTE Mohawk-A and Mohawk-B refer to two separate episodes of oocyst shedding by the same ferret
101
Table 45 Histologic findings from necropsies of black-footed ferrets (Mustela nigripes) with enteric coccidiosis
Number of Sections Affected
Ferret
ID Year
Age
(years) Sex Coccidia in Intestinal Sections Small Intestinea Large Intestinea
Z228-98 1998 7 M rare asexual stages S - 01 A - 11 S - 01 A - 01
Z143-99 1999 1 M sexual and asexual stages S - 24 A - 24 S - 03 A - 03
Z106-02 2002 008 F rare sexual stages S - 14 A - 04 S - 01 A - 01
Z108-03 2003 008 F sexual and asexual stages S - 12 A - 12 S - 01 A - 01
Z109-03 2003 008 F sexual and asexual stages S - 23 A - 23 S - 11 A - 11
Z124-12 2012 021 F sexual and asexual stages S - 24 A - 04 none
Z113-13 2013 017 F sexual and asexual stages S - 77 A - 07 none
Z117-13 2013 025 M sexual and asexual stages S - 56 A - 56 S - 11 A - 01
Z118-13 2013 025 M sexual and asexual stages S - 56 A - 46 S - 11 A - 01
Z119-13 2013 025 F sexual and asexual stages S - 45 A - 35 none
Z137-14 2014 021 M sexual and asexual stages S - 1011 A - 911 S - 22 A - 02
Legend a= xn where x is number of sections containing sexual or asexual lifestages n is the number of sections examined S = sexual life
stages A= asexual life stages
102
Table 46 Yearly incidence of coccidial infection in black-footed ferrets (Mustela nigripes) at the
Cheyenne Mountain Zoo
Cheyenne Mountain Zoo
Year Adult Family
2003 116 (625) -
2004 819 (4211) -
2005 021 (000) 14 (2500)
2006 021 (000) 07 (000)
2007 023 (000) 08 (000)
2008 224 (833) 14 (2500)
2009 025 (000) 06 (000)
2010 326 (1154) 07 (000)
2011 125 (400) 09 (000)
2012 125 (400) 08 (000)
2013 028 (000) 05 (000)
2014 430 (1333) 09 (000)
2015 035 (000) 35 (6000)
2016 - 27 (2857)
Mean annual () 689 1155
Legend - = missing data xn= where x is the number of ferrets shedding coccidial oocysts and n is the
total number of adult ferrets or family groups in a given year () = incidence expressed as a percentage
103
Table 47 Yearly mortality rate and incidence of mortality associated with coccidial infection in
black-footed ferrets (Mustela nigripes) at the Toronto Zoo
Total Mortality
Coccidia Other Causes
Year Kit Adult Kit Adult
1997 015 (000) 023 (000) 315 (2000) 423 (1739) 1998 038 (000) 119 (526) 838 (2105) 919 (4734) 1999 047 (000) 119 (526) 1647 (3404) 119 (526) 2000 034 (000) 015 (000) 434 (1176) 315 (2000) 2001 032 (000) 016 (000) 532 (1563) 116 (625) 2002 150 (200) 020 (000) 450 (800) 220 (1000) 2003 227 (741) 018 (000) 327 (1111) 118 (555) 2004 020 (000) 016 (000) 620 (3000) 216 (1250) 2005 016 (000) 015 (000) 416 (2500) 215 (1333) 2006 030 (000) 016 (000) 230 (667) 016 (000) 2007 019 (000) 015 (000) 419 (2105) 215 (1333) 2008 034 (000) 016 (000) 1134 (3235) 316 (1875) 2009 017 (000) 016 (000) 017 (000) 116 (625) 2010 017 (000) 016 (000) 317 (1765) 316 (1875) 2011 011 (000) 016 (000) 111 (909) 216 (1250) 2012 111 (909) 017 (000) 111 (909) 317 (1765) 2013 424 (1667) 017 (000) 424 (1667) 317 (1765) 2014 126 (384) 017 (000) 326 (1154) 217 (1176) 2015 04 (000) 017 (000) 04 (000) 217 (1176) 2016 011 (000) 017 (000) 211 (1818) 117 (588)
Mean annual () 195 053 1594 1359
Legend xn= where x is the number of ferrets that died with coccidial infection or of other
causes and n is the total number of adult ferrets or kits in a given year () = incidence expressed
as a percentage
104
CHAPTER 5 EVALUATING THE DOMESTIC FERRET (MUSTELA PUTORIUS FURO) AS
AN EXPERIMENTAL MODEL FOR ENTERIC COCCIDIOSIS IN THE BLACK-FOOTED
FERRET (MUSTELA NIGRIPES)
ABSTRACT
The purpose of this study was to determine whether the domestic ferret (Mustela putorius furo) is
susceptible to an isolate of Eimeria ictidea originating from black-footed ferrets (BFF Mustela nigripes)
and thus could act as a suitable experimental model in which to investigate the pathogenesis and
management of this disease A pilot study was performed with 10 male intact juvenile domestic ferrets
Ferrets were administered an oral inoculum containing either a high dose (1 times 106 oocysts) moderate
dose (5 times 104 oocysts) or saline control and observed for shedding of oocysts and development of
clinical signs Seven of ten ferrets developed patent infection all of which had received the high dose
inoculum The prepatent period was 7-9 days and duration of shedding varied from 1-7 days Clinical
signs were identified in six of the seven infected ferrets and were consistent with those previously
described for enteric coccidiosis in domestic and BFF Parasite life stages were identified within the
intestines of four of the seven ferrets with patent infection and were limited to the distal jejunum and
ileum The demonstrated ability to produce patent infections in domestic ferrets following oral inoculation
of a high dose of E ictidea (1 times 106 oocysts) isolated from BFF provides an avenue for future
experimental investigations into the control and treatment of enteric coccidiosis in this endangered
species
51 INTRODUCTION
Black-footed ferrets (BFF Mustela nigripes) are one of only three ferret species worldwide
While formerly distributed throughout the North American prairies black-footed ferrets were declared
extinct in the wild in the 1980s Since 1986 a multi-institutional consortium has been breeding this
species in captivity with reintroductions back into the wild within their historic range in selected locations
in Canada the USA and Mexico Introduced colonies of BFF are present in Arizona Colorado Kansas
105
Montana New Mexico South Dakota Utah Wyoming and Chihuahua (Mexico) Reintroduction
attempts in Saskatchewan Canada have been unsuccessful to date
Multiple infectious diseases pose a significant risk to the captive breeding and post-release
survival of BFF including distemper and sylvatic plague (Santymire et al 2014 USFWS BFF Recovery
Program 2017) Coccidiosis is a recognized cause of juvenile and adult morbidity and mortality in captive
breeding programs and can result in significant losses (Bronson et al 2007 Santymire et al 2014
USFWS BFF Recovery Program 2017) The effect of the disease on wild populations is unknown
Clinical signs of coccidiosis include mucoid to hemorrhagic diarrhea abdominal discomfort lethargy
appetite loss vomiting and dehydration Recent investigations into diseases affecting BFF at the Toronto
Zoo have identified a single Eimeria species E ictidea associated with all cases of enteric coccidiosis in
juvenile and adult BFF from 2014-2016 (see Chapter 3) This same Eimeria species was identified
retrospectively as the cause of juvenile and adult mortalities in previous years (1999 through 2014
inclusive) (Chapters 3 and 4) Furthermore this pathogen was identified in fecal samples based on
morphologic and molecular characterization from adult and juvenile BFF in another zoological collection
(Louisville Zoo Kentucky USA) (see Chapter 3)
Enteric coccidiosis also occurs in domestic ferrets (Mustela putorius furo) with three
morphologically distinct species of coccidia Eimeria ictidea Eimeria furonis and Isospora (=
Cystoisospora) laidlawi Both of the aforementioned Eimeria species have been identified in black-footed
ferrets based on morphologic criteria but molecular characterization was needed to confirm whether the
same species of parasite infects both ferret species (see Chapter 2 and 3) To this end nuclear and
mitochondrial sequences for E furonis and for I=(C) laidlawi were generated (Chapter 2) expanding the
existing limited sequence data from the nuclear 18S rRNA locus of Eimeria furonis Molecular
characterization of E ictidea from domestic ferrets was not possible because samples containing this
parasite were not available for study consequently it is unclear whether the same coccidium affects both
domestic and black-footed ferrets
106
There is no published information describing the pre-patent periods and pathogenicity of enteric
coccidia in BFF and given the conservation status of the BFF experimental work cannot be undertaken
in the natural host The purpose of this study was to determine whether the domestic ferret is susceptible
to E ictidea isolated from BFF if susceptible the domestic ferret could act as a suitable experimental
model in which to investigate the pathogenesis prevention and treatment of coccidiosis caused by E
ictidea
52 MATERIALS AND METHODS
521 Animal care
Ten juvenile male intact ferrets of 48 (n=6) or 50 (n=4) days of age were obtained from a
commercial source (Marshall BioResources North Rose New York USA) and were housed in the
University of Guelph Central Animal Facility Isolation Facility Ferret weights on arrival ranged from
334-475 g (average= 3928 g) All ferrets were housed individually in wire bottom cages of 813 times 1117
times 457 cm size and were divided in equal numbers between two non-adjoining rooms They received ad
libitum access to Envigo Teklad Certified Global Ferret Diet (Madison Wisconsin USA) and water
changed daily Room temperature was maintained at 18-21 degC and a 16 hour light 8 hour dark
photoperiod was provided All personnel working with the ferrets were required to wear personal
protective equipment including disposable facemasks gloves gowns and bouffant caps Shoes were
provided for use in each room This study was carried out in accordance with the recommendations in the
Canadian Council on Animal Care guidelines The protocol was approved by the Animal Care Committee
of the University of Guelph (Animal Use Protocol 3289) and by both the Animal Welfare Committee
and Animal Care and Research Committee of the Toronto Zoo
An initial physical examination and blood collection were performed on each ferret by the
principal investigator (ARP) one day after arrival to assess health status prior to enrollment in the study
Ferrets were mask induced with isoflurane (Isoflurane USP Fresenius Kabi Richmond Hill Ontario) in
107
oxygen placed on a heat disc (SnuggleSafe Lenric C21 Ltd Littlehampton United Kingdom) weighed
examined and blood was collected from the jugular vein for routine CBC and biochemical profiles
All ferrets subsequently underwent an acclimation period of two weeks During this time fecal
samples were collected daily from each ferret and examined for the presence of coccidial oocysts using a
standard salt flotation technique (Dryden et al 2005) to ensure that all individuals were free of coccidia
prior to initiation of experimental work Any ferret positive for coccidia was to be removed from the
study
522 Oocyst preparation
Oocysts used for inoculation originated from fecal samples from two naturally infected BFF These
samples were stored in potassium dichromate for four weeks prior to oocyst purification and use in this
infection trial Stored fecal samples were mixed with distilled water and passed through a small sieve to
remove debris The strained contents were transferred to a 50 mL conical vial and topped up to 50 mL
with additional distilled water Samples were centrifuged (Sorvall ST40R Centrifuge Thermo Scientific)
at 2800 rpm (1315 timesG) for 10 minutes at 12 degC A drop of supernatant was evaluated microscopically at
100times for the presence of oocysts If oocysts were observed the supernatant was poured off into a second
50 mL conical tube and again topped up to 50 mL with distilled water and re-centrifuged under the same
conditions Otherwise the supernatant was discarded The pellets from both the first and second tubes
were combined with saturated salt solution at a 14 ratio by volume Oocysts were floated in the salt
solution by centrifugation at 1500 rpm (377 timesG) for 10 minutes at 12 degC The top 5 mL of supernatant
were collected and transferred to a clean 50 mL conical tube topped up to 40 mL with distilled water and
washed via centrifugation at 2800 rpm (1315 timesG) for 10 minutes After the wash step the supernatant
was again checked for presence of oocysts and discarded if no oocysts were observed The pellet was
collected and the presence of oocysts confirmed by examination of a drop placed on a clean glass slide at
100times Once verified the contents of the pellet of concentrated oocysts was placed in a 250 mL storage
container and mixed with approximately 200 mL of sterile saline (09 sodium chloride Hospira
108
Montreal Quebec) prior to storage for two to four weeks at 4 degC until inoculation Prior to inoculation a
McMaster count was performed to determine the number of oocysts per mL in order to determine
appropriate volume of inoculum
523 Experimental infections
Part 1
Five ferrets were randomly assigned to each of the control and infection groups After the
acclimation period on day 0 four ferrets in the infection group were inoculated orally with a high dose
oocyst suspension (1 times 106 oocysts in 025 mL of saline) mixed into 1 mL of FerreTone Skin amp Coat
Supplement (United Pet Group Inc Blacksburg Virginia USA) a fifth ferret was inoculated with a
moderate dose oocyst suspension (5 times 104 oocysts in 025 mL of saline) in the same volume of FerreTone
Ferrets in the control group were inoculated with a placebo (025 mL of saline) in 1 mL of FerreTone
Inoculation was performed by offering the oocyst suspension or placebo to the ferrets in a plastic
container
Fecal samples were collected daily from each inoculated ferret for 14 days post-inoculation
Samples were analyzed via fecal flotation using the McMaster method followed by routine flotation in
saturated salt solution (Dryden et al 2005) to determine the presence or absence of oocysts and oocyst
burden (oocysts per gram of feces [OPG])Temporal trends in oocyst shedding were monitored Ferrets
were evaluated visually twice daily for the presence of clinical signs of coccidial disease The first of
every two ferrets identified to shed oocysts was to be humanely killed at the time of peak oocyst shedding
(ie the first day that fecal oocyst counts remained static or declined) and necropsied to confirm the
presence of and describe parasitic replication in the intestinal mucosa Any remaining animals that shed
oocysts were to be monitored throughout the 14 day period following inoculation in order to determine
the duration and intensity of oocyst shedding for these individuals the total number oocysts shed during
patency was determined
109
Part 2
All ferrets from the infection group that did not shed oocysts during Part 1 (n=4) and all but one
ferret from the previous control group (n=4) were orally inoculated with the high dose oocyst suspension
(1 times 106 in 1 ml of saline) mixed with an equal volume of FerreTone Consequently between phases 1
and 2 all but one ferret were inoculated at least once with the BFF coccidia in order to increase
experimental animal numbers and determine if ferret age played a role in susceptibility to infection
One ferret from the previous control group was inoculated with a lower dose (2 times 105 oocysts in
075 mL of 09 saline mixed with 1 mL FerreTone) of oocysts that had been collected from the single
domestic ferret that shed in Part 1 oocysts were purified as described above for the initial inocula Fecal
collection and analysis were performed as previously As in Part 1 one in every two ferrets sequentially
identified to be shedding oocysts in feces was killed humanely at the time of peak shedding and a
complete necropsy examination performed The remainder of the ferrets observed to be shedding were
monitored for the full 14 days of the trial after which they were killed humanely and necropsied and total
number of oocysts shed during patency was determined All ferrets that did not shed coccidial oocysts
during the infection trial were rehomed at the end of the trial
524 Animal welfare
Ferrets were evaluated twice daily for development of clinical signs of coccidial disease and any
animal showing clinical disease was to be treated as determined by a veterinarian with supportive care
including fluid therapy A grading system for clinical signs including intervention points and removal
criteria was created for use during daily evaluation (see Appendices 3 and 4) Animals whose clinical
signs could not be ameliorated without the use of specific anticoccidial therapy were to be euthanized
Should the inoculation in Part 1 have resulted in clinical disease that required extensive treatment andor
necessitated euthanasia a lower number of oocysts would be used for subsequent inoculation in Part 2
Ferrets to be euthanized were anesthetized by mask induction with isoflurane in oxygen a 1 mL blood
110
sample was collected from the cranial vena cava and then an intracardiac dose of potassium chloride (2
mEqkg) was administered
525 Hematology
Blood was collected from all ferrets under isoflurane anesthesia at the time of pre-trial health
examination and again at the time of humane killing Blood was collected via jugular venipuncture
initially due to the small size of the ferrets at arrival and then by cranial vena cava venipuncture or
cardiocentesis prior to euthanasia Complete blood count and serum biochemistries were performed by the
Animal Health Laboratory of the University of Guelph Guelph Ontario
526 Morphologic and molecular characterization
Morphologic and molecular characterization of oocysts shed by the domestic ferrets during the
course of the infection trial was performed to ensure that the ferrets were shedding the same species of
Eimeria with which they were inoculated Oocysts were concentrated from positive fecal samples as
described above A drop of concentrated oocyst solution was viewed photographed and measured at
400times and 600times for comparison with previously determined morphometrics of Eimeria ictidea oocysts
(Chapter 3)
Regions from the mitochondrial cytochrome c oxidase subunit I and III (mt COI and mt COIII)
DNA were amplified by polymerase chain reaction (PCR) from each sample using primer pairs
400F1202R and -172F799R respectively For all PCR reactions samples were denatured at 95 degC for 5
min then subjected to 35 cycles of 94 degC for 30s anneal at 52degC for 30s and extension at 72 degC for 60s
followed by a final extension at 72 degC for 7 min PCR gel electrophoresis and sequencing methods used
were as described in the Materials amp Methods section of Chapter 2
The resulting consensus sequences were searched from within Geneious against previous
sequences for E ictidea produced by the authors and against publically available sequences on the
111
BLAST server (blastncbinlmnihgovBlastcgi) using the blastn search algorithm against the nrnt
database (GenBank+EMBL+DDBJ+RefSeq ndash AA or DNA)
527 Necropsy protocol
All humanely killed ferrets underwent a complete necropsy (Appendix 6) using the modified
protocol described in Materials amp Methods section 424 of Chapter 4
53 RESULTS
Initial physical examination was unremarkable with the exception of mild to moderate bilateral
ceruminous discharge within the external ear canal of all ferrets Complete blood count and serum
biochemistry results for all ferrets were within normal reference intervals for juvenile domestic ferrets
(Appendices 2a and b) (Fox 2014) Six days after arrival a single ferret (103) in the control group
developed mild upper respiratory signs consisting of sneezing and clear nasal and ocular discharge these
clinical signs were associated with mild dehydration and decreased food and water consumption The
ferret was treated with subcutaneous fluid therapy (10 mL Plasmalyte-A subcutaneous Baxter Alliston
Ontario) heat and supportive care and all clinical signs resolved within three days This ferret was
deemed healthy to participate in the remainder of the clinical trial A second ferret (105) in the control
group developed unilateral purulent ocular discharge 14 days after arrival one day prior to placebo
inoculation The ferret was treated topically twice daily for five days with Isathal ophthalmic gel (fusidic
acid 10 mgg Dechra Veterinary Products Inc Point-Claire Quebec) and the discharge resolved but
reoccurred within 2 days of treatment cessation Ocular examination showed mild conjunctivitis but no
evidence of corneal lesions and fluorescein staining did not indicate the presence of corneal ulceration
The ferret was treated for an additional six days with tobramycin ophthalmic solution (3 mgmL Sandoz
Tobramycin 03 Boucherville Quebec) after which clinical signs resolved completely No coccidial
oocysts were shed in feces from any of the ferrets during the two week acclimation period
112
531 Oocyst shedding
All ferrets readily ingested the inoculum with either placebo or concentrated oocysts In Part 1
one ferret (203) in the infection group which had received the high dose (1 times 106 oocysts) shed oocysts
on day 8 and day 9 after inoculation (Tables 51 52) This ferret was 71 days of age at the time shedding
was initially identified The ferret was killed humanely 11 days post inoculation later than had been
outlined in the protocol as processing of fecal samples had been delayed by two days resulting in late
detection of oocyst shedding in this individual Oocysts were not identified in the feces of the three
remaining ferrets that received the high inoculation dose the single ferret that received the lower dose (5
times 104 oocysts) or in the ferrets within the control group
In Part 2 six of eight ferrets inoculated with the high dose (1 times 106 oocysts) shed oocysts during
the 14 day observation period (Tables 51 52) Four of these ferrets were from the previous control
group One of the ferrets previously inoculated with the high dose inoculum in Phase 1 that had not shed
oocysts did shed oocysts after being inoculated a second time with the same dose during Phase 2 The
ferret that had previously received the low dose (5 times 104 oocysts) of oocysts in Phase 1 also shed after
inoculation with the high dose in Phase 2 Three ferrets did not shed oocysts after high dose inoculation in
Phase 2 one of these had been part of the previous control group the other two had received the high
dose inoculation previously in Phase 1
The pre-patent period ranged from 7-9 days (Table 51) with equivalent numbers of ferrets
commencing shedding on each of days 7 through 9 All six ferrets were between 91 and 93 days of age at
the time shedding was initially identified Oocyst per gram counts and shedding trends for all individuals
are shown in Table 51 Total oocyst shedding during patency was lt14 oocysts 8904 oocysts and
172291 oocysts for ferrets 201 104 and 105 respectively The two ferrets for which the prepatent period
was 9 days only shed oocysts for one day and in low numbers Oocysts were not identified in the feces of
the three remaining ferrets two of which received the high inoculation dose (1 times 106 oocysts) and the
third that received the lower inoculation dose (2 times 105 oocysts) (Table 52)
113
532 Morphologic and molecular characterization
In all seven ferrets that shed oocysts the morphologic features and measurements (length width
shape index) of the shed oocysts were consistent with those of the E ictidea administered in the inoculum
(Figure 51) Molecular confirmation of the identity of the oocysts shed was successful in 3 out of the 7
ferrets (102 103 203) samples from the four remaining ferrets did not contain adequate quantity or
quality of DNA for confirmation
533 Clinical signs
Clinical signs associated with patent infection were identified in 6 of 7 ferrets (Table 52) These
signs included weight loss (n=5) diarrhea (n=1) mucoid soft feces (n=2) feces containing blood (n=2)
and malodorous feces (n=1) Appetite reduction was noted in two ferrets from the infection group in Part
1 between 6-8 days post infection however no oocyst shedding was detected from either ferret during
this time
534 Hematology
CBC and serum biochemistry values from ferrets collected during pre-trial health screening are
displayed in Appendices 1a and 1b Values obtained for ferrets euthanized during or after the
experimental trial are displayed in Appendices 3a and 3b Minor variances from reference range values
for CBC and serum biochemistry were identified in six of the seven ferrets with patent infection In all six
ferrets for which a complete serum biochemistry was obtained immediately prior to death creatinine
kinase (CK) values were elevated (see Appendix 3b) Ferret 103 exhibited a mild hypoalbuminemia (20
ref 24-40 gL) on ante-mortem serum biochemistry (see Appendix 3b)
114
535 Necropsy
No gross or histopathologic lesions were present and coccidia could not be identified in sections
of intestine from the single ferret (203) humanely killed in Part 1 For the ferrets humanely killed in Part
2 no evidence of diarrhea hematochezia or mucoid fecal material was identified grossly Ferret 105
killed at the termination of the experiment but still shedding low numbers of oocysts in its feces
exhibited a 7 cm region of congested mucosa within the distal jejunum
Coccidial life stages were identified in small intestinal sections from four of the seven ferrets that
were identified to shed oocysts at some point prior to necropsy (Figure 52 Tables 52 and 53) Affected
sections included jejunum in all four animals as well as ileum in one and were collected from 114 to 218
cm aboral from the pylorus (see Figure 53) Coccidia were not identified in sections of duodenum
proximal jejunum or large intestine however oocysts were identified within fecal material in the lumen
of the large intestine from one ferret (103) Of the 11-19 sections of intestine examined for each ferret the
number of sections containing parasites ranged from one to eight 1 section in ferret 102 2 sections in
ferret 201 4 sections in ferret 105 8 sections in ferret 103 A mix of sexual and asexual life stages was
observed within the enterocytes in small intestinal sections from ferret 103 the remainder of the ferrets
showed either asexual (102 201) or sexual (105) life stages in affected segments
Pathologic changes and additional histologic findings in small intestinal sections of ferrets with
enteric coccidia included rare regions of blunting of the villi and sloughing of the epithelium associated
with hemorrhage and inflammation The primary lesions identified were subjectively increased numbers
of eosinophils lymphocytes and plasma cells within the lamina propria of the small intestine and
similarly increased neutrophils lymphocytes and plasma cells within the lamina propria of the large
intestine Neutrophils were rarely present in intestinal crypts and glands
Other gross necropsy findings included two ferrets with renal cortico-medullary cysts and one
ferret with mild thickening of the esophageal mucosa midway along the esophageal length No histologic
changes were identified within a sample of esophagus taken from this region
115
54 DISCUSSION
The findings of this study show that domestic ferrets are susceptible to infection with the enteric
coccidium Eimeria ictidea isolated from black-footed ferrets Both morphometric and molecular
diagnostic methods were used to confirm that ferrets were shedding oocysts of the same species with
which they were inoculated Molecular characterization was successful in three of the seven ferrets that
developed patent infections and as no other coccidial species was identified during pre-trial observation
morphometry was considered to be confirmatory in the remaining four animals
We have referred to the eimeriid coccidium affecting BFF and used in this experimental trial as
E ictidea based on morphometric similarity of their oocysts with those of E ictidea as described from
domestic ferrets (see Chapter 3) There is limited published information on infection of domestic ferrets
with E ictidea outside of Hoarersquos original descriptions (1927 1935a b) which form the basis for all
subsequent identifications of E ictidea in domestic ferrets and in BFF Attempts to obtain exemplars of E
ictidea from domestic ferrets to characterize using molecular techniques were unsuccessful (Chapter 2)
Multiple diagnostic laboratories in Canada and Europe were solicited for coccidia-positive fecal samples
from domestic ferrets but no samples of E ictidea were received over a 4 year period (2014-2017)
Eimeria ictidea was identified based on microscopic examination in only two samples submitted to a
European diagnostic laboratory from 2008-2015 It is unproven whether the E ictidea described from
domestic ferrets and the E ictidea identified from black-footed ferrets and used in this experimental
work are the same or are simply morphologically indistinguishable Eimeria species However the
consistency in morphology host genus and location of infection within the intestinal tissues combined
with the successful cross-transmission of this parasite to domestic ferrets described in the present study
suggests they are likely conspecific
116
The pre-patent period (minimum duration of endogenous development) for infection with E
ictidea in the inoculated domestic ferrets ranged from 7-9 days (see Table 51) the pre-patent period for
this parasite in the BFF the natural host for this coccidium is unknown Hoare experimentally infected
naiumlve domestic ferrets with E ictidea derived from naturally occurring infection in this species (Hoare
1935b) The inoculated ferrets shed oocysts after a pre-patent period of 7 days consistent with the 7-9
days seen in the work described here with E ictidea
Shedding of oocysts was identified over a period of 1-7 days (see Table 51) and intensity ranged
from less than 14 up to 15624 OPG These results may be skewed with erroneously low duration of
shedding and number of oocysts shed as three of the seven ferrets were humanely killed at or prior to the
expected peak of oocyst shedding for tissue collection and histologic examination in order to increase the
probability of identifying parasite life stages within the intestinal sections Shedding periods were similar
to those identified in adult single-housed BFF which ranged from 2-9 days however oocyst per gram
counts from the domestic ferrets were consistently lower than OPG counts from BFF (104 - 554274
OPG) infected with the same parasite (see Chapter 4) Furthermore the total number of oocysts shed by
individual domestic ferrets (14 - 172291 oocysts) during patency was reduced compared to BFF despite
similar length of shedding period (see Chapter 4) The domestic ferrets in this study were naiumlve individual
juveniles whereas the BFF were adults 1-5 years of age some of which were showing clinical signs at
the time of oocyst shedding The relative influences of age species and previous exposure to the parasite
on our observations are unknown
Two different fecal flotation methods were used on all samples to increase the probability of
oocyst detection The McMaster method was used to provide accurate OPG counts for quantification of
oocyst shedding however this method had a minimum detection limit of ~13 oocysts per gram (1333
OPG calculated) because it is based on dilution of the initial sample with flotation media (saturated salt)
In samples with few oocysts oocysts may be missed or to be present in numbers below this detection
limit Routine salt flotation is in contrast performed using the entire sample allowing for detection of
117
small numbers of oocysts Consequently in cases where oocyst per gram counts were low shedding was
identified on routine salt flotation but not by the McMaster method and recorded as lsquopositiversquo but below
the detection limit of the enumeration method
Subclinical to clinical disease occurred in six of the seven ferrets that developed patent infection
with weight loss being the most frequent clinical sign Other clinical signs were typical of enteric
coccidiosis including diarrhea hematochezia and mucoid andor soft feces These clinical signs are
similar to those previously described for black-footed ferrets infected with this parasite (USFWS BFF
Recovery Program 2017 Chapter 4) and for domestic ferrets with enteric coccidiosis (Sledge et al
2011) Interestingly development of clinical disease was not described in the naiumlve domestic ferret
inoculated by Hoare (1935) with E ictidea derived from naturally occurring infection Based on review of
the literature severe clinical disease resulting from intestinal coccidiosis is rare in domestic ferrets
Black-footed ferrets however appear more susceptible to disease development and more frequently show
significant clinical signs No domestic ferret required treatment for clinical coccidiosis during the course
of this study It is possible that the more pronounced clinical signs associated with enteric coccidiosis in
BFF may result from the limited genetic diversity in a population derived from so few individuals and
increased susceptibility of BFF to other diseases such as sylvatic plague have been described in
comparison with their domestic counterparts (Williams et al 1994) Although unlikely recent acquisition
of E ictidea from a related host species such as the domestic ferret could have resulted in increased
pathogenicity and severity of clinical disease from infection with this parasite in BFF
Minor variances from reference range values for CBC and serum biochemistry were identified in
six of the seven ferrets with patent infection but only in one case (ferret 103) did this appear to be
correlated with infectiondisease In this animal a mild hypoalbuminemia (20 ref 24-40 gL) was noted
(Appendix 3b) On histopathology large regions of the small intestine contained parasite life stages
however inflammation lysis of epithelial cells and necrosis of affected areas that could be expected to
result in protein loss into the intestinal lumen were not identified
118
Creatine kinase values were elevated in all six ferrets for which a complete serum biochemistry
was obtained immediately prior to death these findings are consistent with release from CK rich tissues
such a skeletal muscle during venipuncture and manual restraint
The pathologic lesions identified within the intestinal sections of ferrets euthanized at the time of
oocyst shedding were similar to those identified in affected BFF but in most cases were less locally
extensive or widespread throughout the small intestine than those observed in necropsy cases of BFF (see
Chapter 4) Coccidia were seen in the histologic sections of four ferrets all three ferrets that were actively
shedding oocysts at the time of necropsy (102 103 105) and one of four in which necropsies were
performed after oocyst shedding had ceased (201)
Although the primary objective of the examination of histologic sections from affected ferrets
was to identify coccidial life stages attempts were made to describe the pathologic changes associated
with the presence of the parasite Despite necropsies being performed almost immediately after death and
the use of Serra solution fixative to improve parasite and tissue preservation the villi and villar
epithelium of the trimmed sections were frequently distorted or absent and consequently accurate
commentary on these areas was precluded and was made only on visible components of the lamina
propria and crypts or glands A deliberate decision was made not to kill and collect samples from the
saline inoculated control ferrets after Phase 1 and those not shedding oocysts during Phase 1 or 2 thus no
age-matched intestinal sections were available for comparison It is difficult to comment on the
significance of the inflammatory cells observed in the lamina propria of the small and large intestinal
segments or the proliferative rate of the crypt epithelium The only changes identified which may be
considered significant are the presence of neutrophils within the crypts and glands of small and large
intestinal sections respectively but these lesions were rare and not associated with the presence of
parasitic life stages
119
While no parasitic life stages were identified in intestinal sections from ferrets 104 203 and 205
one of the three ferrets ferret 104 exhibited lymphoplasmacytic inflammation and blunting of jejunal
villi in one section (36-38 cm aboral from pylorus) These findings might be expected as the most
extensive histologic lesions would occur associated with lysis of the intestinal epithelial cells as oocysts
were shed into the feces after which new intestinal epithelial cells would re-cover the denuded villar
surface Thus for those cases in which histologic examination was performed after shedding had ceased
presence of the protozoal life stages in the intestines would be expected to be significantly reduced or
absent
Hoare (1935a b) described a particular reaction to the presence of parasitic life stages of Eimeria
ictidea in the small intestine of domestic ferrets in which only the villar tips were affected and there was
resultant annular constriction of the villus separating the affected and non-affected segments These
particular changes were not identified in any of the ferrets in this study and had not been noted
retrospectively in naturally infected BFF (Chapter 4)
While patent infection and intestinal disease could be experimentally created in domestic ferrets
without equivalent experimental work in BFF it is difficult to fully compare the susceptibility to infection
and to the development of disease between the two species Eimeria species tend to be host specific thus
if E ictidea from BFF is not conspecific with E ictidea in domestic ferrets it might be expected that the
domestic ferret would be less susceptible to infection and the development of disease than is the BFF
Even if the two parasites are identical natural passaging through the BFF may alter affinity for the
domestic ferret
It appears that the infectious dose of oocysts of E ictidea derived from BFF required to initiate a
patent infection in domestic ferrets is high The ferrets that developed patent infections were administered
an inoculum containing 1 times 106 sporulated oocysts and even with this extremely high inoculating dose
only a proportion of inoculated ferrets became infected Neither the ferret that received the low dose
120
inoculum nor the ferret that received the passaged oocysts from Part 1 of the study shed oocysts during
the 14 day period post inoculation The latter finding was unexpected as fresh passaged oocysts would be
expected to contain larger proportions of viable oocysts and be comparatively more infective than oocysts
that had been stored for 2-4 weeks prior to inoculation
The only publication describing oral inoculation of Eimeria species in domestic ferrets (Hoare
1935b) did not quantify the number of oocysts administered A study performed in 16 farmed juvenile
mink (Mustela vison) administered 2000 sporulated oocysts of each of three coccidial species (I
laidlawi E vison and an unknown Eimeria species) resulting in patent infection with one of the three
species (Foreyt et al 1977) as determined by the presence of oocysts on fecal examination The authors
did not reveal which type of oocysts resulted in the infection
The number of oocysts required to result in infection in BFF is unknown Based on the
authorsrsquo observations of over 100000 oocysts per gram of feces being shed by black-footed ferrets into
cages of lt1m2 floor space (see Chapter 4) we estimated that under normal caged conditions animals
would likely ingest thousands of oocysts over a short period of time This was in part why a large number
of oocysts (up to 1000000 as available from our store of viable oocysts) was administered to each
domestic ferret in order to increase the probability that infection and shedding would result Furthermore
for the parasite to persist within the ferret population the total number of oocysts shed into the
environment would have to be several times higher than the infective dose required to generate a patent
infection otherwise the parasite would be expected to die out If E ictidea of domestic ferrets and E
ictidea of BFF are conspecific the difference in oocyst shedding between the host species during patent
infection could potentially explain the low prevalence of E ictidea reported from the domestic ferret
population (as seen in Chapter 2)
Both humoral and cell mediated immunity are involved in the immune response to coccidia The
role of maternal derived antibodies in combatting protozoal infection in mustelids has not been studied
121
but in carnivores maternal antibodies to viruses can last up to 16 weeks (Chappuis 1998) In poultry
maternal Eimeria-specific IgG is transferred via the egg yolk to offspring In one study breeding hens
were infected with a single species of Eimeria 28-39 days prior to lay Their hatched chicks were
challenged by inoculation with oocysts of the same and a related Eimeria species and showed reduced
oocyst shedding compared to age matched controls indicative of passive transfer of immunity (Smith et
al 1994) It is possible then that the presence or absence of maternal antibodies may be a factor in the
age at which ferrets are susceptible to coccidial infection and the development of disease The facility
from which the domestic ferrets were acquired has not previously detected Eimeria species on routine
fecal screening (Dr Bambi Jasmin personal communication) consequently it is unlikely that they would
have received maternal immunity to or been exposed to this parasite and thus can be considered to be
naive
Despite a theoretical lack of maternal immunity there did appear to be an effect of age on
susceptibility to infection In Part 1 when the ferrets were approximately 70 days of age only 1 of 4
ferrets inoculated with the high dose of oocysts developed patent infection In comparison in Part 2
when the ferrets were 91-93 days of age 3 of 4 ferrets that had been in the saline control group for Part 1
developed patent infections as did 2 of 4 ferrets that had been inoculated in Part 1 but had not shed
oocysts One of these previously inoculated ferrets was the individual that had received the lower dose of
oocysts Thus it appears that patent infection could be produced more easily in the older ferret kits
However the two ferrets that had previously been inoculated exhibited the shortest shedding periods (1
day) and lowest oocyst per gram counts and parasite life stages in these cases were rare (201) to absent
(205) on histologic examination of numerous sections of intestine It is thus possible that the primary
inoculation resulted in abbreviated infections or infections in which so few oocysts were shed that
infection was not detected Our observation of endogenous stages in the intestinal tissues of some of the
ferrets following cessation of oocyst shedding suggests that the pre-patent period and duration of patency
may vary considerably from animal to animal consequently it is possible that some of the kits would
122
have ultimately shed a few oocysts from the primary inoculum if followed beyond 14 days post-
inoculation Whether through an aborted or undetected infection previous exposure of these kits to the
parasite probably generated partial immunity against E ictidea and therefore the intensity of infection
upon challenge in previously exposed domestic ferret kits was reduced
The authors acknowledge the limitations of this initial pilot study however it was proven that
patent infection with E ictidea isolated from black-footed ferrets could be generated in a novel host the
domestic ferret Further studies will be required to investigate the effect of age on susceptibility to
infection as well as the possibility of development of immunity after exposure and its role in reducing
parasite replication and disease in subsequent infection With so few BFF in existence the use of BFF for
in vivo infection trials cannot be contemplated consequently refining the domestic ferret infection model
will be essential for carrying out research specifically intended to help manage coccidiosis in the
endangered black-footed ferret
Figure 51 Exogenous life stages of Eimeria ictidea shed from a domestic ferret (Mustela putorius
furo) experimentally inoculated with oocysts originating from black-footed ferrets (Mustela
nigripes) Unsporulated oocyst (solid black arrow) Sporulated oocyst (solid white arrow) Bright
field microscopy scale bar = 25 microm
25 microm
124
Figure 52 Life stages of Eimeria ictidea within the small intestinal epithelium of an experimentally
infected domestic ferret (Mustela putorius furo) A) Sexual life stages (micro- and macrogamonts
- white arrows) crowding the villar enterocytes all stages are found between the nucleus and
luminal surface of infected enterocytes Hematoxylin and eosin staining scale bar = 25microm B) At
higher magnification meronts (black arrows) and gamonts (open arrows) are crowded between the
enterocyte nuclei and brush border Hematoxylin and eosin staining scale bar = 25microm
25 microm
25 microm
125
Figure 53 Presence and location of sexual and asexual life stages of Eimeria ictidea within the
intestinal epithelium of domestic ferrets (Mustela putorius furo) (n=7) that developed patent
infection after experimental inoculation with oocysts originating from black-footed ferrets (Mustela
nigripes) Legend duod = duodenum jej= jejunum Sequential numbers for jejunal and colonic
sections represent the order aboral from the pylorus from which the samples were collected SI =
additional section(s) of small intestine whose aboral sequence was not recorded
0
1
2
3
duod jej 1 jej 2 jej 3 jej 4 jej 5 ileum colon 1 colon 2 SI
Nu
mb
er
of
Ferr
ets
Aff
ect
ed
Intestinal Section Containing Life Stages of Eimeria ictidea
Asexual life stages Sexual life stages
126
Table 51 Prepatent period and oocyst shedding patterns in domestic ferrets (Mustela putorius furo)
experimentally inoculated with 1 times 106 oocysts of Eimeria ictidea originating from black-footed ferret
(Mustela nigripes) that developed patent infections
Oocysts shed per gram of feces
Ferret Identity
Day post inoculation 102 103 104 105 201 203 205
1 0 0 0 0 0 0 0
2 0 0 0 0 0 0 0
3 0 0 0 0 0 0 0
4 0 0 0 0 0 0 0
5 0 0 0 0 0 0 0
6 0 0 0 0 0 0 0
7 lt 14 1807 0 0 0 0 0
8 11053 139 7091 0 156238 0
9 463 11733 lt 14 203 lt 14
10 578 7549 0 0 0
11 lt 14 0 0 0
12 0 lt 14 0
13 0 lt 14 0
14 0 lt 14 0
Legend ferret euthanized as of this date lt 14 = oocyst positive samples with less than 14 oocysts per
gram of feces
127
Table 52 Results of oral inoculation of domestic ferrets (Mustela putorius furo) with oocysts of Eimeria ictidea originating from black-footed
ferrets (Mustela nigripes)
Ferret
Identity
Results Part 1 Results Part 2
Inoculum Oocyst
Shedding
Presence
of clinical
disease
Coccidia
present in
sections
Inoculum Oocyst
Shedding
Presence
of clinical
disease
Coccidia
present in
sections
101 Saline N N - 2 times 105 oocystsa N N -
102 Saline N N - 1 times 106 oocysts Y N Y^
103 Saline N N - 1 times 106 oocysts Y Y Y^
104 Saline N N - 1 times 106 oocysts Y Y Nyen
105 Saline N N - 1 times 106 oocysts Y Y Yyen
201 1 times 106 oocysts N N - 1 times 106 oocysts Y Y Yyen
202 1 times 106 oocysts N N - 1 times 106 oocysts N N -
203 1 times 106 oocysts Y Y Nyen
204 1 times 106 oocysts N N - 1 times 106 oocysts N N -
205 5 times 104 oocysts N N - 1 times 106 oocysts Y Y N^
Legend includes any of the following weight loss diarrhea mucoid feces malodorous feces inappetence N = no Y= yes - = necropsy not
performed a = oocysts collected after passage through ferret 203 ^ = shedding oocysts at time of necropsy yen = not shedding oocysts at time of
necropsy
128
Table 53 Distribution of coccidial life stages in domestic ferrets (Mustela putorius furo) orally inoculated with oocysts of Eimeria ictidea
originating from black-footed ferrets (Mustela nigripes)
Ferret Identity
Intestinal level 102
103
104 105
201
203
205
Duodenum N N N N N N N
Jejunum 1 N N N N N N N
Jejunum 2 N N N N N N N
Jejunum 3 N S A N N N N N
Jejunum 4 N S A N N N N N
Jejunum 5 N S A N S A N N
Ileum N S A N N N N N
Colon 1 N Na N N N N N
Colon 2 N Na N N N N N
Unmeasured small intestinebc
S - 05
A - 15
S - 45
A - 25
S - 05
A - 05
S - 26
A - 06
S - 06
A - 06
S - 06
A - 06
S - 08
A - 08
Unmeasured large intestinebc
S - 01
A - 01
S - 01
A - 01
S - 01
A - 01
S - 02
A - 02
S - 01
A - 01 none
S - 01
A - 01
Legend N = no parasite life stages S = sexual life stages present A = asexual life stages present a= oocysts present in feces b=
additional sections of intestine for which the location measured from the pylorus was not obtained c= xn where x is number of
sections containing sexual or asexual lifestages n is the number of sections examined
129
CHAPTER 6 WHOLE MITOCHONDRIAL GENOME SEQUENCES OF TWO EIMERIA
SPECIES ISOLATED FROM DOMESTIC (MUSTELA PUTORIUS FURO) AND BLACK-
FOOTED FERRETS (MUSTELA NIGRIPES)
ABSTRACT
The complete mitochondrial (mt) genomes of Eimeria furonis and Eimeria ictidea (Eimeriidae
Coccidia Apicomplexa) originating from single fecal samples from a domestic (Mustela putorius furo)
and a black-footed ferret (Mustela nigripes) respectively were sequenced Both mt genomes were
circular-mapping with lengths of 6165 base pairs (Eimeria furonis - GenBank MF795598) and 6182
base pairs (Eimeria ictidea - GenBank KT203399) Genome organization and gene contents were
comparable with those of other publically available mt genomes from a variety of Eimeria species and
related coccidia there were three complete coding DNA sequence regions encoding cytochrome c
oxidase subunit I cytochrome c oxidase subunit III and cytochrome B and 33 regions encoding
fragmented rDNA Alignment of these mt genome sequences demonstrates a relatively high (945 340
single nucleotide differences [SNDs]) pairwise sequence identity between these Eimeria spp infecting
ferrets the majority of the SNDs resulted in synonymous codon changes with no changes to their protein
products Alignment of the newly sequenced mt genomes demonstrates and phylogenetic reconstructions
support the monophyly of these Eimeria spp of ferrets with another Eimeria sp of carnivores as the
sister taxon to this clade
61 INTRODUCTION
Coccidia are protozoal eukaryotic host-specific parasites of the phylum Apicomplexa and can
be divided into two major taxonomic suborders the eimerioirinid and adeleid coccidia The eimeriorinid
coccidia include both the typical intestinal coccidia (eg Eimeria Isospora Cyclospora) species
belonging to the family Eimeriidae as well as tissue or cyst-forming coccidia (eg Cystoisospora
130
Besnoitia Toxoplasma Sarcocystis) of the family Sarcocystidae (see Cox 1994) Ten species of
Eimeria and twelve species of Isospora (=Cystoisospora) have been described in the Mustelidae
(see Chapter 1 Table 11) Eimeria furonis has been reported in the European polecat (Mustela
putorius) domestic ferret (DF Mustela putorius furo) black-footed ferret (BFF Mustela nigripes) and
mink (Mustela vison) (Hoare 1927 Nukerbaeva and Svanbaev 1973 Jolley et al 1994) Eimeria ictidea
has been reported in the Steppe polecat (Mustela eversmanii) as well as the European polecat domestic
ferret and black-footed ferret (Hoare 1927 Svanbaev 1956 Jolley et al 1994) These reports are based on
the morphometric characteristics of oocysts identified in the feces of the aforementioned host species
without the use of molecular techniques to confirm specific parasite identities
Recently sequences of the mitochondrial cytochrome c oxidase subunit I gene (mt COI) and
nuclear small subunit ribosomal DNA (nu 18S rDNA) of E furonis originating from a domestic ferret (nu
18S rDNA GenBank MF774678-MF774680 mt COI GenBank MF774034-MF774036) and E ictidea
originating from a black-footed ferret (nu 18S rDNA GenBank MF860826-MF860827 mt COI
GenBank MF860823 MF860825) were generated (see Chapters 2 and 3) The parasite originating from
the black-footed ferret was identified as E ictidea based on morphologic similarity to the original
descriptions of E ictidea from domestic ferrets however sequence-based genotyping of E ictidea from
domestic ferrets has not been completed and consequently it has not been demonstrated unequivocally
that the two parasites are conspecific In the present work the complete mitochondrial genomes of E
furonis from the DF and E ictidea from the BFF are described and compared with the mitochondrial
genomes of related coccidia
62 MATERIALS amp METHODS
621 Parasites
Two isolates of morphologically distinct Eimeria species were used in this study Isolate one
identified morphologically and by nu 18S rDNA and mt COI sequences as Eimeria furonis was obtained
131
from a fecal sample from a DF that was submitted for routine ova and parasite examination to a European
diagnostic laboratory8 Isolate two identified morphologically and by nu 18S rDNA and mt COI
sequences as Eimeria ictidea was obtained from a fecal sample from a BFF and was collected during
routine cage cleaning in a captive breeding facility (see Chapters 3 and 4) Fecal collection techniques for
the BFF were reviewed and approved by both the Animal Welfare Committee and the Animal Care and
Research Committee of the Toronto Zoo
622 DNA isolation from coccidia in feces
Genomic DNA was isolated from fecal derived coccidial oocysts as described section 221 of the
Chapter 2 Materials amp Methods Parasite DNA concentration was estimated using a Nanodrop 2000
spectrophotometer (NanoDrop Products Wilmington DE USA) and DNA was stored at 4 degC for
immediate use or minus20 degC for later use
623 Whole genome sequencing
Mitochondrial whole genome amplification for both Eimeria species was initiated using sets of
mt-specific primers that generated overlapping polymerase chain reaction (PCR) fragments (Tables 61
and 62) PCR amplification was performed for all samples in a volume of 25 microl containing ~100 ng of
genomic DNA 1times PCR buffer 3 mM MgCl2 06 mM deoxyribonucleotide triphosphates (dNTPs) 500
nM of each primer and 4 U of Invitrogen Platinum Taq DNA Polymerase (Thermo Fisher Scientific
Toronto ON Canada) PCR reactions were performed on a Bio-Rad T100 PCR thermal cycler (Bio-Rad
Laboratories Singapore) using settings as described previously in the Materials amp Methods section of
Chapter 2 Table 61 details the specific anneal conditions used for the various primer pairs Genomic
DNA from either Eimeria maxima or Eimeria tenella acted as a positive control for the reaction
chemistry Gel electrophoresis purification and sequencing of the PCR amplification products were
8 Vet Med Labor GmbH Division of IDEXX Laboratories Ludwigsburg Germany
132
performed as described in Chapter 2 The resulting chromatograms were aligned and analyzed with
Geneious Ver 818 or later (Biomatters Limited Auckland New Zealand) and high quality consensus
sequences generated The completed mt genome sequences were annotated by comparison with
previously annotated mt genomes from other Eimeria species (eg Eimeria innocua - KR1082961) and
the annotated mt genomes deposited in GenBank
624 Phylogenetic analysis
To determine the phylogenetic affinities of the newly obtained sequences with sequences from
related apicomplexan taxa representative whole mt genome sequences from eimeriid coccidia were
downloaded from GenBank A complete mt genome sequence from an unnamed Choleoeimeria sp was
used to root the ingroup taxa several small genomic rearrangements in the Choleoeimeria sp sequence
required some rearrangement of the genome sequence to unambiguously align homologous regions across
the complete mt genomes
Whole genome sequences were aligned using MAFTT v7017 (Katoh et al 2002) executed from
within Geneious the resulting alignment was examined by eye to adjust start and stop codon positions in
aligned coding DNA sequence [CDS] regions (ie mt COI mitochondrial cytochrome c oxidase subunit
III gene [mtCOIII] mitochondrial cytochrome b gene [CytB]) Phylogenetic trees were generated using
Bayesian Inference (BI) using MrBayes Ver 326 (Huelsenbeck and Ronquist 2001) executed from
within Geneious The aligned complete mt genomes were partitioned into coding (ie CDS) and non-
coding regions so that region-specific models of nucleotide substitution could be applied Characters in
the non-coding region were analysed with the general time reversible (GTR) model (Tavareacute 1986) with
the following parameters nucmodel=4by4 nst=6 rates=invgamma (ie GTR+I+G) Characters in the
coding regions were analysed using the codon nucleotide model (ie lset nucmodel=codon rates=gamma
ngammacat=4) using metazoan mitochondrial translation (ie code=metmt)
133
All BI analyses were run for a chain length of 1000000 with tree sampling every 1000
following a burn-in of 100000 with default settings of 4 heated chains and heated chain temp of 02
63 RESULTS
The whole mt genome sequences of the single isolates of E furonis and E ictidea were
respectively 6165 base pairs (bp) (Figure 61 GenBank MF795598) and 6182 bp (Figure 62 GenBank
KT203399) Content and organization of both mt genomes consisted of three protein-coding genes (mt
COI mt COIII and CytB) interspersed with large and small subunit ribosomal DNA (rDNA) fragments
Details of the various CDS and rDNA fragments are summarized in Table 63 (for E furonis) and Table
64 (for E ictidea)
Pairwise alignment of the mt genome sequences from E furonis and E ictidea demonstrated a
relatively high pairwise sequence identity (946 333 single nucleotide differences [SNDs]) between
these two parasites The bulk of the SNDs (676 225333) were clustered within the three CDS regions
that encode CytB mt COI and mt COIII (see Figure 63 and Table 65) However the majority of these
SNDs (826 186225) were synonymous codon changes that resulted in no changes to the protein
products Only 41 SNDs were involved in 34 amino acid changes distributed among the three CDS
The 33 rDNA fragments comprised 2108 and 2109 bp respectively of the mt genomes of
E furonis and E ictidea Pairwise comparison of these rDNA fragments demonstrated high (986 30
SNDs) sequence identity between the two parasites The remaining 778 and 794 bp respectively of the
mt genomes of E furonis and E ictidea were intergenic stretches between the various rDNA and CDS
regions these intergenic regions were more variable that other regions of the genomes with 78 SNDs
(almost 10 sequence divergence) Additionally all indels were restricted to these variable intergenic
regions
The BI phylogeny generated from aligned complete mt genomes (Figure 64) supported the close
relationship between E furonis and E ictidea within a clade of Eimeria species that include the only three
sequences available for Eimeria spp of carnivores Eimeria mephitidis from the striped skunk (Mephitis
134
mephitis Family Mephitidae) was the sister taxon to the two Eimeria species of ferrets (Family
Mustelidae)
64 DISCUSSION
This work generated the first complete mt genomes from coccidia that infect domestic and black-
footed ferrets (Carnivora Mustelidae) Eimeria mephitidis from the striped skunk Mephitis mephitis
(Carnivora Mephitidae) is the only other Eimeria species from a carnivore for which a complete mt
genome has been reported
Comparatively few eimeriid coccidia only 26 Eimeria species have been described from
carnivores there are 14 named species from the mustelids four from the procyonids four from the ursids
three from the herpestids and one from the viverrids (Duszynski et al 2000) The majority of coccidia
that infect the digestive tract of carnivores belong to the family Sarcocystidae including monoxenous or
facultatively heteroxenous Cystoisospora species or heteroxenous parasites in the genera Sarcocystis
Hammondia and Neospora So far as is known none of the parasites in the Sarcocystidae possess typical
apicomplexan mt genomes with 3 complete CDS and many rDNA fragments (Ogedengbe 2015)
The mt genomes from the two Eimeria sp of mustelid origin demonstrate the same structural
organization (ie the order and number of CDS and rDNA fragments) and circular mapping as the mt
genomes from other Eimeria spp and other closely related eimeriid coccidia such as Isospora
Cyclospora and Lankesterella species Despite the ability of the eimeriid sequences to be mapped
circularly the physical form of Eimeria spp mt genomes may be a linear concatemer of multiple genome
copies as demonstrated for Eimeria tenella (Hikosaka et al 2011) As in the mt genomes of other
eimeriid coccidia (Ogedengbe et al 2013 2014) the CDS for mt COIII demonstrated the highest
sequence divergence between E furonis and E ictidea the mt COI CDS was somewhat more conserved
and CytB CDS demonstrated the fewest SNDs
As expected based on limited sequence divergence between E furonis and E ictidea a BI
phylogenetic analysis using aligned complete mt genome sequences generated a tree that placed these
135
two Eimeria species that infect mustelids within a well-supported monophyletic group The sister taxon
for these ferret parasites was the only other Eimeria species from carnivores for which a complete mt
genome is available E mephitidis which infects hosts belonging to a different family of carnivores
Eimeriid parasites that infect closely-related definitive hosts are commonly found in a single or limited
number of clades based on mitochondrial and nuclear genetic loci (Ogedengbe et al in press)
Sequencing of the mt genomes and at least one nuclear genetic locus (ie nu 18S rDNA) from additional
Eimeria species infecting carnivores will be required to determine if all carnivore-specific Eimeria
species share a common ancestor
136
Table 61 PCR primer pairs and resulting fragments used for sequencing the mitochondrial genome sequence of an isolate of Eimeria furonis
originating from a fecal sample from a domestic ferret (Mustela putorius furo)
Fragment Primer names Primer sequences (5ʹ-3ʹ) Size (bp) Anneal Temp References
1 WG-MT_4140F AGAAAACCTAAAATCATCATGT 1000 52 Ogedengbe et al (2015)
Eim_CO3_799R AAGTGAGTTCGCATGTTTAC Ogedengbe et al (2015)
2 Eim_COI_19F ACTGCYGCAAACCATAARGAA 1700 60 Present study
Api_LSUG_UNI_R AGATAGGGAACAAACTGYCTCAA Present study
3 WG_MT_5416F GGTCCAGATAAGCGATCTCATG 3400 53 Ogedengbe et al (2013)
Eim_COI_1436R CACATTGTGTTCARATAAGTTA Present study
4 WG-MT_6219F GCATCCATCTACAGCTGCGG 500 55 Ogedengbe et al (2013)
WG-MT_344R GTAGGAATCTRAATTCCCAACC Ogedengbe et al (2013)
5 Api_LSUE UNI_F AGGTGCTCAGGGTCTTACCG 500 55 Present study
WG_MT_63R CTGGTATGGATGGATAACACT Ogedengbe et al (2015)
6 Lank_COB-30F CCAGGCCAACTGAACTCGTT 1300 55 Present study
q_Eim_COI_221R GGCATAACTACAAAGAARATCATA Present study
7 Cocci_MT_WG_F TACACCTAGCCAACACGAT 1600 55 Ogedengbe et al (2014)
q_Eim_COI_221R GGCATAACTACAAAGAARATCATA Present study
137
Table 62 PCR primer pairs and resulting fragments used for sequencing the mitochondrial genome sequence of an isolate of Eimeria ictidea
originating from a fecal sample from a black-footed ferret (Mustela nigripes)
Fragment Primer names Primer sequences (5ʹ-3ʹ) Size (bp) Anneal Temp References
1 WG_MT_63R CTGGTATGGATGGATAACACT 2180 52 Ogedengbe et al (2015)
WG-MT_4140F AGAAAACCTAAAATCATCATGT Ogedengbe et al (2015)
2 Cocci_MT_WG_F TACACCTAGCCAACACGAT 1551 58 Ogedengbe et al (2014)
q_Eim_COI_221R GGCATAACTACAAAGAARATCATA Present study
3 WG-MT_3658F CTGGCGAGAAGGGAAGTGTG 1329 55 Ogedengbe et al (2013)
Eim_CO3_799R AAGTGAGTTCGCATGTTTAC Ogedengbe et al (2015)
4 Lank_COB-30F CCAGGCCAACTGAACTCGTT 3899 57 Present study
WG_MT_4072R GGTTGTTTCCATCTCGACTC Ogedengbe et al (2013)
138
Table 63 Coding regions within the mitochondrial genome of the eimeriid parasite Eimeria furonis from a domestic ferret (Mustela putorius faro)
139
Table 64 Coding regions of the mitochondrial genome of the eimeriid parasite Eimeria ictidea originating from a black-footed ferret (Mustela
nigripes)
Table 64 Features associated with the protein-coding regions of the mitochondrial genome of the eimeriid parasite Eimeria cf ictidea originating from a black-footed ferret (Mustela nigripes )
Protein coding regions (CDS) Sequence size (bp) Start position (bp) Stop position (bp) Direction Translation start codon Translation stop codon
Cytochrome c oxidase subunit I (COI) 1443 1343 2785 Forward ATG TAA
Cytochrome c oxidase subunit III (COIII) 756 4333 5088 Forward TTA TAA
Cytochrome b (CytB) 1080 226 1305 Forward ATG TAA
Ribosomal DNA fragments (rDNA) Product
SSUrRNA 46 17 62 forward RNA9 SSU8
SSUrRNA 77 104 180 forward SSUA SSU4 (partial)
SSUrRNA 35 181 215 forward RNA23t
LSUrRNA 20 2790 2809 forward RNA20 (partial) LSU
LSUrRNA 112 2835 2946 forward LSUF LSU11
LSUrRNA 106 2947 3052 forward LSUG LSU12
LSUrRNA 25 3087 3063 reverse LSU
SSUrRNA 39 3129 3167 forward RNA14 SSU1
LSUrRNA 16 3188 3173 reverse LSUC LSU4
SSUrRNA 33 3211 3243 forward SSU
SSUrRNA 61 3386 3326 reverse SSUF SSU12
LSUrRNA 74 3387 3460 forward RNA10 LSU13 (partial)
LSUrRNA 49 3476 3524 forward RNA11 LSU5
SSUrRNA 65 3533 3597 forward SSUD SSU10
SSUrRNA 40 3631 3670 forward RNA17 SSU3
SSUrRNA 30 3677 3706 forward RNA15 SSU
LSUrRNA 30 3711 3740 forward RNA13 LSU10
LSUrRNA 56 3749 3804 forward RNA6 LSU15
LSUrRNA 79 3899 3821 reverse LSUD LSU8
LSUrRNA 24 3931 3908 reverse RNA16 (partial)
SSUrRNA 92 4036 3945 reverse RNA8 SSU5
LSUrRNA 66 4054 4119 forward RNA2 LSU2
LSUrRNA 177 4316 4140 reverse LSUA LSU1
SSUrRNA 30 5125 5096 reverse RNA19 SSU7
LSUrRNA 88 5154 5241 forward RNA1 LSU6
LSUrRNA 26 5342 5317 reverse LSUB LSU3
LSUrRNA 73 5447 5375 reverse RNA3 LSU7
LSUrRNA 20 5479 5460 reverse RNA18 LSU14
SSUrRNA 119 5615 5497 reverse SSUB SSU6
LSUrRNA 80 5699 5620 reverse RNA7
LSUrRNA 188 5897 5710 reverse LSUE LSU9
SSUrRNA 33 6033 6001 reverse SSUE SSU11 (partial)
SSUrRNA 99 6156 6058 reverse RNA5SSU9
140
Table 65 Pairwise comparison of coding DNA and concatenated rDNA fragment sequences between the
mitochondrial genomes of Eimeria furonis originating from a domestic ferret (Mustela putorius furo) and
Eimeria ictidea originating from a black-footed ferret (Mustela nigripes)
Total length
(nucleotides)
Nucleotide
identity
Total amino
acids
Amino acid
identity
COI CDS 1443 934 (95) 480 975 (12)
COIII CDS 756 899 (76) 251 932 (17)
CytB CDS 1080 950 (54) 359 986 (5)
rDNA fragments 2109 985 (32) na na
Legend Numbers in brackets indicate the number of single nucleotide differences na = not
applicable
141
Figure 61 Circular mapping and organization of the mitochondrial genome content of Eimeria
furonis showing three protein-coding genes (COI COIII and CytB) interspersed with large and
small subunit rRNA fragments
142
Figure 62 Circular mapping and organization of the mitochondrial genome content of Eimeria
ictidea showing three protein-coding genes (COI COIII and CytB) interspersed with large and
small subunit rRNA fragments
143
Figure 63 Pairwise comparison of coding DNA and concatenated rDNA fragment sequences of
the mitochondrial genomes of Eimeria furonis and Eimeria ictidea Legend Yellow bands
correspond to coding DNA fragments Red arrows correspond to rDNA fragments Arrows of both
colours indicate the forward or reverse direction of these segments
144
Figure 64 Phylogenetic relationships of coccidia (Eimeria furonis and Eimeria ictidea) from
domestic (Mustela putorius furo) and black-footed ferrets (Mustela nigripes) based on the complete
mitochondrial genome sequences these Eimeria species and a selection of related apicomplexan
parasites
145
CHAPTER 7 CONCLUSIONS AND FUTURE DIRECTIONS
The primary objective of this project was to better characterize the enteric coccidia of the
endangered black-footed ferret (BFF) in order to set the stage for improved disease prevention
management and treatment To the authorrsquos knowledge this work was the first attempt to isolate and
perform molecular characterization of the coccidial species endemic in the black-footed ferret population
and to characterize the natural history of the disease in this host
Two Eimeria species Eimeria ictidea and Eimeria furonis have been described from BFF and
domestic ferrets A single Eimeria species morphologically resembling E ictidea of the domestic ferret
and referred to in this work as E ictidea was identified from all historic and active cases of enteric
coccidiosis in the Toronto Zoo BFF population The same species was identified in both BFF family
groups and single-housed adults that shed coccidia during the study period (2014-2016) as well as from
historic necropsy samples of juvenile and adult Toronto Zoo BFF from 1999-2014 Furthermore this
same species was identified from both family groups and single-housed adults shedding coccidia at an
additional BFF Species Survival Plan institution the Louisville Zoo in 2016 These findings are contrary
to previous published reports that indicated multiple coccidia species were affecting captive and wild
BFF In order to determine whether these additional parasites continue to exist within the present-day
BFF population and their impact on this species further examination of coccidia-positive fecal samples
from captive and wild BFF populations is recommended Moreover determination of which parasite
species impact BFF morbidity and mortality would allow for development of targeted therapies for
disease management
Coccidia-positive fecal samples and necropsy samples from domestic ferrets were collected from
multiple diagnostic laboratories in Canada and Europe for comparison with results from BFF Eimeria
furonis and Isospora (=Cystoisospora) laidlawi were identified from fecal parasitology reports from
2008-2015 and in fecal samples obtained prospectively from 2014-2016 from domestic ferrets submitted
to Canadian and European laboratories E furonis was also identified in necropsy samples from 2010 and
146
2017 from two Canadian diagnostic laboratories No samples containing E ictidea were submitted to the
Canadian laboratories however samples containing E ictidea were identified twice by the European
diagnostic laboratory with one sample in 2011 and a second in 2013 Consequently it appears that E
ictidea is rarely identified from domestic ferrets Since no samples of E ictidea were acquired from
domestic ferrets during the study period it remains undetermined whether E ictidea from domestic
ferrets and E ictidea of BFF are conspecific Future molecular characterization of E ictidea isolated from
various mustelid host species would allow not only for determination of whether the parasites are
conspecific but would also provide insight into the potential for cross-transmission of parasites between
related mustelid hosts
The identifications provided by diagnostic laboratories of the specific parasite species present in
the domestic ferret fecal samples showed poor agreement with their identifications based on genotyping
obtained in this study In the authorrsquos opinion molecular techniques are essential tools for determining
the specific coccidial species responsible for individual and group outbreaks of coccidiosis and for
further understanding of the eimeriid host-parasite relationships To this end the nu 18S rDNA region and
whole mitochondrial (mt) genomes of E ictidea and E furonis were sequenced these mt whole genome
sequences are the first for the Eimeria species of mustelids to be entered into GenBank These sequences
may provide suitable targets for the development of highly discriminatory PCR-based methods of
identification that could be applied to fecal tissue or even formalin-fixed paraffin-embedded samples
(see Chapters 2 and 3) Methodological improvements such as less expensive next generation sequencing
methods that could be applied to diagnostics will depend on the availability of high quality reference
sequences such as generated herein Furthermore evaluation of the evolution of the eimeriid coccidia
using phylogenetic analysis based on additional whole mitochondrial genome sequences may allow for
more accurate determinations of relationships between parasite species and timing of their divergence
from common ancestors
One of the goals of this research was to investigate the possibility of using the domestic ferret as
an experimental model to study enteric coccidiosis in the BFF Experimental work cannot be carried out
147
in the BFF due to its endangered status We showed that the domestic ferret is susceptible to infection
with Eimeria ictidea originating from BFF Infected domestic ferrets showed similar clinical signs and
pathologic lesions to BFF strengthening the possibility of their serving as a suitable model Future
investigations would include studies of the domestic ferretrsquos immune response to enteric coccidial
infection in vivo drug trials including pharmacokinetic pharmacodynamic and efficacy studies of
anticoccidial medications investigations into patterns of drug resistance in coccidial species of ferrets
and oral vaccine development
Ultimately the goal of research on coccidiosis in BFF may be the development of an autogenous
vaccine to improve the survival of ferret kits and reduce coccidiosis-related morbidity and mortality in
BFF captive breeding programs It may be possible to use the domestic ferret to select for E ictidea that
are more highly adapted to this host perhaps with a commensurate loss of virulence to its original host If
this were to be the case domestic ferret-derived coccidia could be used in BFF as vaccine organisms
Alternately methods such as use of a bioshuttle (live vaccination with coccidia followed by anticoccidial
treatment to limit pathogenicity) might be useful for generating long-lasting immunological protection in
BFF against E ictidea Improving immunity through vaccination could assist in reducing coccidial
shedding and disease in BFF associated with stressful life events such a breeding weaning and transfer
between institutions There is minimal available data on the significance of coccidiosis in wild
populations of BFF and limited means of disease surveillance post release however it is logical to
assume that the stresses associated with release to the wild might result in clinical disease as happens with
stressed BFF in captivity Reducing morbidity and mortality associated with coccidiosis in BFF could
result in increased numbers of ferrets being released to the wild and higher survival of released ferrets
increasing the number of BFF in the wild supports the goals of the conservation initiative for the black-
footed ferret and will support the longer-term survival and recovery of this species
148
REFERENCES
Abe N Tanoue T Ohta G Iseki M (2008) First record of Eimeria furonis infection in a ferret Japan with
notes on the usefulness of partial small subunit ribosomal RNA gene sequencing analysis for
discriminating among Eimeria species Parasitol Res 103967ndash70 doi 101007s00436-008-1037-x
Adl SM Simpson AGB Farmer MA et al (2005) The new higher level classification of eukaryotes with
emphasis on the taxonomy of protists J Eukaryot Microbiol 52399ndash451 doi 101111j1550-
7408200500053x
Andrews JM (1926) Coccidiosis in mammals Am J Hyg 6784ndash798
Augustine PC Danforth HD (1986) A study of the dynamics of the invasion of immunized birds by
Eimeria sporozoites Avian Dis 30347ndash351
Barta JR Schrenzel MD Carreno R Rideout BA (2005) The genus Atoxoplasma (Garnham 1950) as a
junior objective synonym of the genus Isospora (Schneider 1881) species infecting birds and
resurrection of Cystoisospora (Frenkel 1977) as the correct genus for Isospora species infecting
mammals J Parasitol 91726ndash727 doi 101645GE-33411
Bell JA (1994) Parasites of Domesticated Pet Ferrets Compend Contin Educ Pract Vet 16617ndash620
Black-footed Ferret Recovery Implementation Team (2011) Captive Breeding
httpwwwblackfootedferretorgcaptive-breeding Accessed 8 Jan 2014
Blankenship-Paris TL Chang J Bagnell CR (1993) Enteric coccidiosis in a ferret Lab Anim Sci 43361ndash
363
Bronson E Bush M Viner T et al (2007) Mortality of captive black-footed ferrets (Mustela nigripes) at
Smithsonianrsquos National Zoological Park 1989 ndash 2004 J Zoo Wildl Med 38169ndash176
149
CAPC (2013) Current Advice on Parasite Control Intestinal Parasites - Coccidia
httpwwwcapcvetorgcapc-recommendationscoccidia Accessed 6 Jun 2015
Carpenter JW Hillman CN (1979) Husbandry reproduction and veterinary care of captive ferrets In
1978 Proceedings of the Annual Meeting of the American Association of Zoo Veterinarians
Washington DC pp 36ndash47
Catchpole J Norton CC Gregory MW (1993) Immunisation of lambs against coccidosis Vet Rec
13256ndash59
Cavalier-Smith T (2014) Gregarine site-heterogeneous 18S rDNA trees revision of gregarine higher
classification and the evolutionary diversification of Sporozoa Eur J Protistol 50472ndash495 doi
101016jejop201407002
Chappuis G (1998) Neonatal immunity and immunisation in early age lessons from veterinary medicine
Vaccine 161468ndash1472
Cox FE (1994) The evolutionary expansion of the Sporozoa Int J Parasitol 241301ndash1316
Davis CL Chow TL Gorham JR (1953) Hepatic coccidiosis in mink Vet Med 48371ndash375
De Vos AJ (1970) Studies on the host range of ltigtEimeria chinchillaeltIgt de Vos and Van Der
Westhuizen 1968 Onderstepoort J Vet Res 3729ndash36
Dirikolu L Yohn R Garrett EF et al (2009) Detection quantifications and pharmacokinetics of
toltrazuril sulfone (Ponazuril) in cattle J Vet Pharmacol Ther 32280ndash288
Dryden MW Payne PA Ridley R Smith V (2005) Comparison of common fecal flotation techniques for
the recovery of parasite eggs and oocysts Vet Ther 615ndash28
Duszynski D Wilber PG (1997) A Guideline for the Preparation of Species Descriptions in the
Eimeriidae J Parasitol 83333ndash336
150
Duszynski DW Couch L Upton SJ (2000) The Coccidia of the World
httpbiologyunmeducoccidiacarniv2html Accessed 1 Sep 2017
El-Sherry S Ogedengbe ME Hafeez MA Barta JR (2013) Divergent nuclear 18S rDNA paralogs in a
turkey coccidium Eimeria meleagrimitis complicate molecular systematics and identification Int J
Parasitol 43679ndash685 doi 101016jijpara201303005
Evans HE An NQ (2014) Anatomy of the ferret In Fox JG Marini RP (eds) Biology and Diseases of the
Ferret 3rd edn Wiley Blackwell pp 23ndash67
Fayer R (1980) Epidemiology of Protozoan Infections The Coccidia Vet Parasitol 675ndash103
Foreyt WJ Todd AC Hartsough GR (1977) Anticoccidial Activity of Eight Compounds in Domestic
Mink AJVR 38391ndash394
Fox JG (2014) Normal Clinical and Biological Parameters In Fox JG Marini RP (eds) Biology and
Diseases of the Ferret 3rd edn John Wiley amp Sons Inc pp 157ndash185
Frenkel JK (1977) Besnoitia wallacei of Cats and Rodents With a Reclassification of Other Cyst-
Forming Isosporoid Coccidia J Parasitol 63611ndash628
Grafner G Graubmann HD Dobbriner W (1967) Leberkokzidiose beim Nerz (Lutreola vison Schreb)
hervorgerufen durch eine neue Kokzidienart Eimeria hiepei n sp Monatshefte fur
Veterinearmedizin 22696ndash700
Haberkorn A (1971) Zur Wirtsspezifitat yon Eimeria contorta nsp (Sporozoa Eimeriidae) Z
Parasitenkd 37303ndash314
Hall MC Wigdor M (1918) Canine coccidiosis with a note regarding other protozoan parasites from the
dog J Am Vet Med Assn 5364ndash76
Hikosaka K Nakai Y Watanabe YI et al (2011) Concatenated mitochondrial DNA of the coccidian
151
parasite Eimeria tenella Mitochondrion 11273ndash278
Hillyer E V (1992) Gastrointestinal diseases of ferrets (Mustela putorius furo) J Small Anim Med 244ndash
45
Hoare CA (1927) On the coccidia of the ferret Ann Trop Med Parasitol 21313ndash320
Hoare CA (1935a) A histopathological reaction of a special type on the part of the intestinal villi in ferret
coccidiosis Trans R Soc Trop Med Hyg 292
Hoare CA (1935b) The endogenous development of the coccidia of the ferret and the histopathological
reaction of the infected intestinal villi Ann Trop Med Parasitol 29111ndash122
Hoefer HL Fox JG Bell JA (2012) Gastrointestinal Diseases In Quesenberry KE Carpenter JW (eds)
Ferrets Rabbits and Rodents Clinical Medicine and Surgery 3rd edn Elsevier Saunders St Louis
Missouri pp 27ndash45
Huelsenbeck JP Ronquist F (2001) MrBayes Bayesian inference of phylogenetic trees Bioinformatics
17754ndash755 doi 101093bioinformatics178754
Jolley WR Kingston N Williams ES Lynn C (1994) Coccidia Giardia sp and a Physalopteran
Nematode Parasite from Black-footed Ferrets (Mustela nigripes) in Wyoming J Helminthol Soc
Washingt 6189ndash94
Katoh K Misawa K Kuma K Miyata T (2002) MAFFT a novel method for rapid multiple sequence
alignment based on fast Fourier transform Nucleic Acids Res 303059ndash3066 doi
101093nargkf436
Kaye SW Ossiboff RJ Noonan B et al (2015) Biliary coccidiosis associated with immunosuppressive
treatment of pure red cell aplasia in an adult ferret (Mustela putoris furo) J Exot Pet Med doi
101053jjepm201504012
152
Koepfli KP Deere K Slater GJ et al (2008) Multigene phylogeny of the Mustelidae resolving
relationships tempo and biogeographic history of a mammalian adaptive radiation BMC Biol 610
Lariviegravere S Jennings AP (2009) Family Mustelidae (weasels and relatives) In Wilson DE Mittermeier
RA (eds) Handbook of the Mammals of the World Volume 1 Carnivores Lynx Edicions
Barcelona pp 564ndash656
Levine ND Ivens V (1970) The coccidian parasites (Protozoa Sporozoa) of ruminants
Li X Pang J Fox JG (1996) Coinfection with intracellular Desulfovibrio species and coccidia in ferrets
with proliferative bowel disease Lab Anim Sci 46569ndash571
Litster AL Nichols J Hall K et al (2014) Use of ponazuril paste to treat coccidiosis in shelter-housed
cats and dogs Vet Parasitol 202319ndash325 doi 101016jvetpar201403003
Matsubayashi M Takami K Abe N et al (2005) Molecular characterization of crane coccidia Eimeria
gruis and E reichenowi found in feces of migratory cranes Parasitol Res 9780ndash83
Meeusen ENT Walker J Peters A et al (2007) Current status of veterinary vaccines Clin Microbiol Rev
20489ndash510 doi 101128CMR00005-07
Mehlhorn H Aspock H (eds) (2008) Coccidial Drugs In Encyclopedia of Parasitology 3rd edn Springer
Berlin Heidelberg New York pp 269ndash286
Morehouse NF (1938) The Reaction of the Immune Intestinal Epithelium of the Rat to Reingection wiht
Eimeria nieschulzi J Parasitol 24311ndash317
Nukerbaeva KK Svanbaev SK (1973) Coccidia of fur bearing mammals in Kazakhstan Vestn Selrsquoskokh
Nauk Kazakh 1250ndash54
Ogedengbe JD Hanner RH Barta JR (2011) DNA barcoding identifies Eimeria species and contributes to
the phylogenetics of coccidian parasites (Eimeriorina Apicomplexa Alveolata) Int J Parasitol
153
41843ndash850
Ogedengbe ME (2015) DNA Barcoding of Apicomplexa Mitochondrial Evolution across the Phylum
University of Guelph
Ogedengbe ME El-Sherry S Ogedengbe JD et al Whatrsquos in a name Phylogenies based on combined
mitochondrial and nuclear sequences conflict with morphologically defined genera in the eimeriid
coccidia (Apicomplexa)
Ogedengbe ME El-Sherry S Whale J Barta JR (2014) Complete mitochondrial genome sequences from
five Eimeria species (Apicomplexa Coccidia Eimeriidae) infecting domestic turkeys Parasit
Vectors 7335 doi 1011861756-3305-7-335
Ogedengbe ME Hafeez MA Barta JR (2013) Sequencing the complete mitochondrial genome of Eimeria
mitis strain USDA 50 (Apicomplexa Eimeriidae) suggests conserved start positions for mtCOI- and
mtCOIII-coding regions Parasitol Res 1124129ndash4136 doi 101007s00436-013-3604-z
Pantchev N Gassmann D Globokar-Vrhovec M (2011) Increasing numbers of Giardia (but not
coccidian) infections in ferrets 2002 to 2010 Vet Rec 168519 doi 101136vrd2962
Patterson M Fox JG (2007) Parasites of Ferrets In Baker DG (ed) Flynnrsquos Parasites of Laboratory
Animals 2nd edn Wiley Blackwell pp 501ndash508
Patterson MM Fox JG Eberhard ML (2014) Parasitic Diseases In Fox JG Marini RP (eds) Biology and
Diseases of the Ferret 3rd edn Wiley Blackwell pp 553ndash572
Powers L V (2009) Bacterial and parasitic diseases of ferrets Vet Clin North Am - Exot Anim Pract
12531ndash561 doi 101016jcvex200906001
Prado ME Ryman JT Boileau MJ et al (2011) Pharmacokinetics of ponazuril after oral administration to
healthy llamas (Llama glama) Am J Vet Res 721386ndash9
154
Ruiz A Muntildeoz MC Molina JM et al (2013) Primary infection of goats with Eimeria ninakohlyakimovae
does not provide protective immunity against high challenge infections Small Rumin Res 113258ndash
266 doi 101016jsmallrumres201301006
Ryley J Meade R Hazelhurst J Robinson T (1976) Methods in coccidiosis research separation of
oocysts from faeces Parasitology 73311ndash326
Santymire R Branvold-Faber H Marinari PE (2014) Recovery of the Black-Footed Ferret In Fox JG
Marini RP (eds) Biology and Diseases of the Ferret 3rd edn Wiley Blackwell pp 219ndash231
Shi MQ Huther S Burkhardt E Zahner H (2000) Immunity in rats against Eimeria separata oocyst
excretion effects on endogenous stages and local tissue response after primary and challenge
infections Parasitol Res 86891ndash898
Sledge DG Bolin SR Lim A et al (2011) Outbreaks of severe enteric disease associated with Eimeria
furonis infection in ferrets (Mustela putorius furo) of 3 densely populated groups J Am Vet Med
Assoc 2391584ndash1588 doi 102460javma239121584
Smith NC Wallach M Petracca M et al (1994) Maternal transfer of antibodies induced by infection with
Eimeria maxima partially protects chickens against challenge with Eimeria tenella Parasitology
109551ndash557
Svanbaev SK (1956) Materials on the fauna of coccidia of wild mammals in western Kazakhstan Tr
Instituta Zool Akad Nauk Kazachskoi SSR 5180ndash191
Tavareacute S (1986) Some probabilistic and statistical problems in the analysis of DNA sequences Am Math
Soc Lect Math Life Sci 1757ndash86 doi citeulike-article-id4801403
Tenter AM Barta JR Beveridge I et al (2002) The conceptual basis for a new classification of the
coccidia Int J Parasitol 32595ndash616
155
Upton SJ (2000) Suborder Eimeriorina Leacuteger 1911 In Lee JJ Leedale GF Bradbury P (eds) An
Illustrated Guide to the Protozoa vol 1 2nd edn Allen Press Lawrence Kansas pp 318ndash339
USFWS BFF Recovery Program (2017) Black-footed Ferret Managed Care Operations Manual
(BFFMCOM)
Vermeulen AN (2005) Vaccination against coccidial parasites The method of choice In Proceeding of
the 9th International Coccidiosis Conference
Williams BH Chimes MJ Gardiner CH (1996) Biliary coccidiosis in a ferret (Mustela putorius furo) Vet
Pathol 33437ndash439 doi 101177030098589603300412
Williams ES Mills K Kwiatkowski DR et al (1994) Plague in a Black-footed (Mustela nigripes)
presence J Wildl Dis 30581ndash585
Williams ES Thome ET Appel MJG Belitsky DW (1988) Canine Distemper in Black-Footed (Mustela
nigripes) from Wyoming J Wildl Dis 24385ndash398
Wozencraft WC (2005) Order Carnivora In Wilson DE Reeder DM (eds) Mammal Species of the
World A Taxonomic and Geographic Reference 3rd edn Johnrsquos Hopkins University Press
Baltimore Maryland p 2142
Yi-Fan C Le Y Yin D et al (2012) Emendation of 2 Isospora Species (Apicomplexa Eimeriidae)
Infecting the Steppe Polecat Mustela eversmanii Lesson 1827 in China to the Genus
Cystoisospora (Apicomplexa Sarcocystidae) Comp Parasitol 79147ndash152 doi 10165445311
Yu L Peng D Liu J et al (2011) On the phylogeny of Mustelidae subfamilies analysis of seventeen
nuclear non-coding loci and mitochondrial complete genomes BMC Evol Biol 1192 doi
1011861471-2148-11-92
Zou M Guo G Zhao Y Zhang Q (2014) Detection quantifications and pharmacokinetics of ponazuril in
156
healthy swine J Vet Pharmacol Ther 37598ndash602 doi 101111jvp12126
157
APPENDICES
158
Appendix 1 Shedding of oocysts of Eimeria ictidea in black-footed ferret (Mustela nigripes) dam and kit
family groups from 2014-2016
Collection Year 2014 2014 2014 2014 2014 2015 2016
Age of kits (days) Poppy Bumblefoot Calico Aubrey Ruckus Fiddlesticks Guanella^
29 - - - 0 - - -
30 - 0 - 0 - - -
31 - 0 - 0 - - -
32 - 0 - 0 - - -
33 - 0 - 0 - - -
34 0 0 - 0 0 - -
35 0 0 - 0 0 0 -
36 0 0 - 0 0 0 -
37 - 0 - 0 - 0 -
38 - 0 - 0 - 0 -
39 - 0 - 0 - 0 -
40 0 0 - 0 0 0 -
41 - 0 - 0 - 0 -
42 0 0 - 0 0 - -
43 0 0 - 0 0 0 -
44 0 0 - - 0 0 -
45 0 0 - 0 0 - -
46 0 0 - 0 0 0 -
47 0 0 - 0 0 0 -
48 0 lt14 - 0 0 0 -
49 0 0 - 0 0 0 -
50 0 0 - - 0 0 -
51 0 0 - 0 0 0 -
52 0 0 - 0 0 0 -
53 0 0 - 0 0 0 +
54 0 0 - 0 0 0 +
55 0 0 - 0 0 0 3717146
56 0 0 - 0 - 0 1084436
57 0 0 - 0 - 0 64133
58 0 324 - 0 0 0 20654
59 0 0 - 0 0 0 0
60 0 446688 - 0 0 0 0
61 0 934828 - 0 0 0 41111
62 0 530469 - 0 0 0
63 0 1617131 - 0 0 lt14
64 0 8396357 - 114 0 0
65 0 234876 - 5368 0 lt14
66 0 374625 - 9455 0 0
67 lt14 2311575 - 39579 0 -
68 0 603563 - 125051 0 0
69 - 4103036 - 27067 0 lt14
70 2470327 7759974 - 0 0 0
71 1073085 - 1784 0 0
72 3430966 - 0 0 0
73 4459536 - 7326 0 0
74 4842212 - 2 0 0
75 3627307 - 0 0 0
76 5801885 - 0 0 0
77 5994 0 0 0 0
78 589835 0 0 0
Numbers of Oocysts Shed (oocysts per gram of feces)
Dam Identity
159
Appendix 1 continued Shedding of oocysts of Eimeria ictidea in black-footed ferret (Mustela nigripes)
dam and kit family groups from 2014-2016
Collection Year 2014 2014 2014 2014 2014 2015 2016
Age of kits (days) Poppy Bumblefoot Calico Aubrey Ruckus Fiddlesticks Guanella^
79 24815 0 0 0
80 144016 0 0 0
81 97862 0 0 0
82 0 0 0
83 0 0 0
84 0 0 0
85 0 0 0
86 0 0 0
87 0 0 0
88 0 0 0
89 0 0 0
90 0 0 0
91 0 0 0
92 0 - -
93 0 - 0
94 0 - 0
95 0 0 -
96 0 - 0
97 0 0 -
98 0 0 0
99 0 0 0
100 0 0 0
101 0 0 0
102 0 0 0
103 0 0 0
104 0 0 0
105 0 0 -
106 0 0 0
107 0 0 0
108 - 0 0
109 0 0 -
110 0 0 0
111 0 0 -
112 0 0 0
113 0 0 -
114 0 0 -
115 0 0 -
116 0 0 -
117 0 -
118 0 -
119 0 0
120 0 0
121 0 0
122 0 -
Numbers of Oocysts Shed (oocysts per gram of feces)
Dam Identity
160
Appendix 1 continued Shedding of oocysts of Eimeria ictidea in black-footed ferret (Mustela nigripes)
dam and kit family groups from 2014-2016
Collection Year 2014 2014 2014 2014 2014 2015 2016
Age of kits (days) Poppy Bumblefoot Calico Aubrey Ruckus Fiddlesticks Guanella^
123 0 0
124 0 -
125 0 -
126 0 0
127 0 0
128 0 2843
129 0 0
130 0 -
131 0 0
132 0 0
133 0 0
134 0 0
135 0 0
136 0 0
137 0
138 0
139 0
140 0
141 0
142 0
143 0
144 0
145 0
146 -
147 0
148 0
149 0
150 0
Legend lt 14 = oocyst positive samples with less than 14 oocyst per gram of feces - = no sample recorded for this date
underline = last sampling date + = coccidia present but OPG count not performed = Toronto Zoo ferret ^ = Louisville Zoo
ferret thick outer border = days treatment was received
Numbers of Oocysts Shed (oocysts per gram of feces)
Dam Identity
161
Appendix 2a Hematology values for domestic ferrets (Mustela putorius furo) from 49-51 days of age
prior to experimental inoculation
Test Reference
Intervala
Reference
Intervalb
101 102 103 104 105 201 202 203 204 205
Age (days) adult 70 50 50 50 50 50 48 48 48 50 48
WBC (x 109L) 27-112 53-120 86 64 77 97 76 48 8 72 104 76
RBC (x 1012
L) 50-108 55ndash74 66 58 45 5 48 47 67 5 5 42
Hb (gL) 87-177 104ndash136 121 106 90 58 94 91 122 96 98 80
HCT (LL) 04 - 051 029ndash037 037 033 027 030 029 027 038 030 030 024
MCV (fL) 44-52 478ndash548 55 56 60 60 60 58 57 59 61 58
MCH (pg) 15-18 175ndash191 18 18 20 12 20 20 18 19 20 18
MCHC (gL) 325-362 347ndash370 331 327 328 196 325 337 321 327 324 331
RDW () 12-16 - 134 127 139 139 133 131 122 136 131 127
Platelets (x 109L) 54-695 629ndash775 777 329 529 480 640 402 534 336 527 363
MPV (fL) 5-10 - 78 78 96 75 66 74 76 81 74 82
TS Protein (gL) 49-76 - 54 51 - - - - - - - -
Seg Neuts (x 109L) 1-8 15ndash48 292 141 216 281 205 187 152 151 354 251
Lymphocytes (x 109L) 1-63 28ndash63 525 416 516 64 509 254 608 468 645 456
Monocytes (x 109L) 0-09 01ndash05 026 07 031 039 038 034 024 094 031 038
Eosinophils (x 109L) 0-13 01ndash06 009 013 008 01 008 005 016 007 010 015
Basophils (x 109L) 0-02 0 009 0 0 0 0 0 0 0 0 0
Polychromasia 2-5 - 5-10 2-5 10-15 10-15 10-15 10-15 2-5 10-15 10-15 5-10
Anisocytosis Occ 1+ 1+ 1+ 1+ 1+ 1+ 1+
HJ bodies rare rare rare rare rare
crenation Occ
poikilocytosis Occ Occ Occ
shift platelets Occ Occ
hemolysis Neg Neg Neg Neg Neg Neg Neg Neg Neg Neg Neg
lipemia mild mild mild mild mild Neg Neg Neg mild mild
Ferret Identity
Legend - = reference value unavailable bolded = outside reference range a = adult ferret reference ranges from Animal Health Laboratory
(University of Guelph Guelph Canada) b
= reference ranges for 10-week old ferrets (Fox JG 2014) Neg = negative Occ = occasional
162
Appendix 2b Serum biochemistry values for domestic ferrets (Mustela putorius furo) from 49-51 days of
age prior to experimental inoculation
Test Reference
Intervala
Reference
Intervalb
101 102 103 104 105 201 202 203 204 205
Age (days) adult 70 50 50 50 50 50 48 48 48 50 48
Calcium (mmolL) 185-242 253-302 239 233 241 244 24 221 242 234 253 242
Phosphorus (mmolL) 112-256 278-475 239 267 29 264 31 218 286 284 315 303
Magnesium (mmolL) 08-139 - 08 08 07 08 08 06 08 07 08 08
Sodium (mmolL) 147-159 146-154 149 149 149 149 148 144 153 148 152 152
Potassium (mmolL) 37-57 47-83 44 46 42 46 48 45 47 44 47 46
Chloride (mmolL) 111-129 115-121 110 112 115 113 113 110 119 117 117 120
Carbon dioxide (mmolL) 17-29 13-27 18 18 18 17 17 16 15 15 19 17
Anion gap (mmolL) 6 - 23 - 25 24 20 24 23 23 24 20 21 20
NaK ratio - - 34 32 35 32 31 32 33 34 32 33
Total protein (gL) 51-75 44-56 49 46 44 47 44 41 52 45 49 43
Albumin (gL) 24-40 26-32 29 26 28 28 28 25 28 27 29 25
Globulin (gL) 19-41 17-24 20 20 16 19 16 16 24 18 20 18
AG ratio 053-167 13ndash12 145 130 175 147 175 156 117 15 145 139
Blood urea nitrogen (mmolL) 45-153 71-139 83 89 107 11 136 73 119 105 118 135
Creatinine (umolL) 8-67 53-124 50 58 68 59 50 7 68 41 62 92
Glucose (mmolL) 32-91 64-138 47 54 54 42 55 53 57 56 52 59
Cholesterol (mmolL) 294-894 619-860 413 386 375 393 344 349 227 357 373 346
Total bilirubin (umolL) 2 to 7 - 1 L 0 L 0 0 0 1 0 0 0 0
Conjugated biilrubin (umolL) - 0-10 1 0 0 1 1 1 0 0 1 0
Free bilirubin (umolL) 0-2 0-15 0 0 0 0 0 0 0 0 0 0
ALKP (UL) 13-237 117ndash277 180 169 172 215 175 168 241 184 177 179
GGT (UL) 0-40 2ndash20 1 1 6 10 4 0 1 1 5 9
AST (UL) - 63ndash152 61 58 48 61 64 59 93 61 58 69
ALT (UL) 39-196 95ndash544 95 105 89 105 106 82 234 137 115 156
CK (UL) 74-294 - 513 330 496 560 530 492 793 539 479 724
Amylase (UL) - - 23 28 35 35 29 28 29 24 36 23
Lipase (UL) - - 65 63 60 64 62 56 67 69 60 68
Calculated osmo (mmolL) - - 298 300 301 301 303 289 311 300 308 311
Ferret Identity
Legend - = reference value unavailable bolded = outside reference range a = adult ferret reference ranges from Animal Health Laboratory
(University of Guelph Guelph Canada) b = reference ranges for 10-week old ferrets (Fox JG 2014)
163
Appendix 3a Hematology values for domestic ferrets (Mustela putorius furo) inoculated orally with
Eimeria ictidea
Test Reference
Intervala
Reference
Intervalb
102 103 104 105 201 203 205
Age (days) adult 98-112 92 91 99 99 97 74 92
Days post inocculation 8 7 15 15 15 11 10
WBC (x 109L) 27-112 52ndash150 94 124 81 85 88 85 142
RBC (x 1012
L) 50-108 62ndash92 71 62 73 60 65 63 65
Hb (gL) 87-177 127ndash159 122 110 122 98 103 114 102
HCT (LL) 04-051 030ndash043 037 033 037 031 032 035 032
MCV (fL) 44-52 50ndash54 53 53 52 51 50 55 49
MCH (pg) 15-18 16-21 17 18 17 16 16 18 16
MCHC (gL) 325-362 351ndash426 328 332 325 319 320 326 317
RDW () 12-16 - 122 126 129 129 145 127 145
Platelets (x 109L) 54-695 376ndash610 524 413 445 117 303 42 429
MPV (fL) 5-10 - 8 78 72 66 67 14 8
TS Protein (gL) 49-76 - 66 63 58 66 55 59 59
Seg Neuts (x 109L) 1-8 21ndash62 160 446 203 170 211 170 554
Lymphocytes (x 109L) 1-63 16ndash79 761 657 551 646 59 646 682
Monocytes (x 109L) 0-09 01ndash02 019 087 024 026 044 026 085
Eosinophils (x 109L) 0-13 03ndash09 0 037 032 009 026 009 099
Basophils (x 109L) 0-02 0 0 012 0 0 009 0 0
Polychromasia 2-5 - 1-3 0-2 0-2 1-3 1-3 2-5 5-10
Anisocytosis Occ Occ Occ
HJ bodies rare
crenation
rouleaux
poikilocytosis
shift platelets Occ Occ
hemolysis Neg
Ferret Identity
Legend - = reference value unavailable bolded = outside reference range a = adult ferret reference ranges from Animal Health
Laboratory (University of Guelph Guelph Canada) b
= reference ranges for 10-week old ferrets (Fox JG 2014) Neg = negative
Occ = occasional = many platelet clumps
164
Appendix 3b Serum biochemistry values for domestic ferrets (Mustela putorius furo) inoculated orally
with Eimeria ictidea
Test Reference
Intervala
Reference
Intervalb
102 103 104 105 201 203 205
Age (days) adult 98-112 92 91 99 99 97 74 92
Days post inocculation 8 7 15 15 15 11 10
Calcium (mmolL) 185-242 245-268 243 222 238 233 232 236 240
Phosphorus (mmolL) 112-256 200-323 292 223 248 298 268 288 306
Magnesium (mmolL) 08-139 - 06 08 08 08 09 08
Sodium (mmolL) 147-159 148-155 147 150 150 148 148 150
Potassium (mmolL) 37-57 45-55 42 41 51 41 50 44
Chloride (mmolL) 111-129 114-124 114 113 118 112 111 115
Carbon dioxide (mmolL) 17-29 16-24 16 21 19 20 12 18
Anion gap (mmolL) 6 - 23 - 21 20 18 20 30 21
NaK ratio - - 35 37 29 36 30 34
Total protein (gL) 51-75 49-64 52 55 55 49 56 57
Albumin (gL) 24-40 30-36 28 20 27 27 26 29 27
Globulin (gL) 19-41 19-30 32 28 28 23 27 30
AG ratio 053-167 11-17 063 096 096 113 107 090
Blood urea nitrogen (mmolL) 45-153 50-150 92 87 119 92 110 98 149
Creatinine (umolL) 8-67 706-1414 56 26 49 40 50 46 79
Glucose (mmolL) 32-91 688-943 52 59 64 71 58 17 66
Cholesterol (mmolL) 294-894 440-640 453 429 262 475 370 285
Total bilirubin (umolL) 2 to 7 - 0 0 0 0 1 1
Conjugated biilrubin (umolL)- - 1 0 0 0 0 0
Free bilirubin (umolL) 0-2 - 0 0 0 0 1 1
ALKP (UL) 13-237 41-181 124 120 213 120 146 170 196
GGT (UL) 0-40 1-2 2 1 3 0 0 2
AST (UL) - 47-128 48 95 104 66 100 100
ALT (UL) 39-196 78-279 133 110 140 203 158 183 281
CK (UL) 74-294 - 382 765 1190 578 680 930
Amylase (UL) - - 24 37 31 28 33 35
Lipase (UL) - - 60 65 72 72 86 79
Calculated osmo (mmolL) - - 296 305 305 300 296 309
Ferret Identity
Legend - = reference value unavailable bolded = outside reference range a = adult ferret reference ranges from Animal Health
Laboratory (University of Guelph Guelph Canada) b
= reference ranges for 10-week old ferrets (Fox JG 2014)
165
Appendix 4 Domestic ferret (Mustela putorius furo) weekly monitoring sheet
Mon
8 AM
Mon
4 PM
Tues
8 AM
Tues
4 PM
Wed
8 AM
Wed
4 PM
Thurs
8 AM
Thurs
8 PM
Fri
8 AM
Fri
4 PM
Sat
8 AM
Sat
4 PM
Sun
8AM
Sun
4 PM
Mentation
Weight (g)
Respiratory
Rate
Vomit
(+ ++ +++)
Diarrhea
(+ ++ +++)
Urination
(+ ++ +++)
Defecation
(+ ++ +++)
Food
offered
Food
remaining
Water remaining
(ml)
Treatments
Other
observations
Initials of
observer
166
Animal ID ________________________________________ Week ______________
Monitoring Criteria
Mentation BAR (bright alert responsive) QAR (quiet alert responsive) depressed lethargic quiet
Weight measured in grams to be performed once weekly (pre-inoculation) and once daily (post-inoculation)
Respiratory rate measured in breaths per minute
Vomit + (small amount) ++ (moderate amount) +++ (large amount)
Diarrhea + (small amount) ++ (moderate amount) +++ (large amount)
Urination + (small amount) ++ (normal amount) +++ (large amount)
Defecation + (one pile) ++ (two piles) +++ (3+ piles)
Food offered Y (yes) N (no)
Food remaining A (all) P (partial) N (none)
Please note any additional observations in the appropriate section
Monitoring Times
Pre-inoculation ndash ONCE daily at 8am during cage cleaningfeeding
Post-inoculation ndash TWICE daily at 8am and 4pm
Critical patients (as determined by veterinary examination) ndash 3-6 times daily (based on veterinarian recommendation) ndash switch to 24
hour care sheet
Intervention Points
If any ferret develops vomiting diarrhea goes off food or exhibits gt 3 weight loss both the PI and graduate student listed below are
to be notified and the animal will be assessed to determine further treatment plans
Removal Criteria
If an individual ferret develops severe gastrointestinal or systemic disease during the course of the study as characterized by abnormal
mentation marked dehydration 10 weight loss within 24-48 hours profuse diarrhea unresponsive to treatment anorexia of 2 or
more days or frequent vomiting unresponsive to treatment the individual will be removed from the study and humanely killed
167
Appendix 5 Domestic ferret (Mustela putorius furo) 24 hour intensive monitoring sheet
Animal ID ___________________________________________ Date___________________
0700 800 900 1000 1100 1200 1300 1400 1500 1600 1700 1800
Mentation
Weight (g)
Heart Rate
Respiratory Rate
Temperature (C)
Water offered
(mL)
Water consumed
(mL)
Food offered
(type and
amount in
grams)
Food consumed
(type and
amount in
grams)
Presence of urine
Presence of feces
168
Character of
feces
Presence of
vomit
Character of
vomit
Medications to
be administered
Other
observations
169
1900 2000 2100 2200 2300 2400 100 200 300 400 500 600
Mentation
Weight (g)
Heart Rate
Respiratory Rate
Temperature (C)
Water offered
(mL)
Water consumed
(mL)
Food offered
(type and amount
in grams)
Food consumed
(type and amount
in grams)
Presence of urine
Presence of feces
Character of
feces
Presence of vomit
Character of
vomit
170
Medications to be
administered
Other
observations
Monitoring Criteria
Mentation BAR (bright alert responsive) QAR (quiet alert responsive) depressed lethargic quiet
Weight measured in grams to be performed once weekly (pre-inocculation) and once daily (post-inocculation)
Respiratory rate measured in breaths per minute
Vomit + (small amount) ++ (moderate amount) +++ (large amount)
Diarrhea + (small amount) ++ (moderate amount) +++ (large amount)
Urination + (small amount) ++ (normal amount) +++ (large amount)
Defecation + (one pile) ++ (two piles) +++ (3+ piles)
171
Food offered Y (yes) N (no)
Food remaining A (all) P (partial) N (none)
Please note any additional observations in the appropriate section
Monitoring Times
Critical patients (as determined by veterinary examination) ndash 3-6 times daily (based on veterinarian recommendation) ndash switch to 24
hour care sheet
Intervention Points
If any ferret develops vomiting diarrhea goes off food or exhibits gt 3 weight loss both the PI and graduate student listed below are
to be notified and the animal will be assessed to determine further treatment plans
Removal Criteria
If an individual ferret develops severe gastrointestinal or systemic disease during the course of the study as characterized by abnormal
mentation marked dehydration 10 weight loss within 24-48 hours profuse diarrhea unresponsive to treatment anorexia of 2 or
more days or frequent vomiting unresponsive to treatment the individual will be removed from the study and humanely killed
172
Appendix 6 Domestic ferret (Mustela putorius furo) infection trial standard operating procedures
Daily Fecal Collection
1 Collection of all feces passed in a 24 hour period will be performed once daily for all ferrets from
July 13 to July 24 inclusive
2 CAF Isolation staff will collect all fecal material present at the time of daily cage cleaning and
place in individual pre-labelled plastic bags (one per cage)
3 The amount of non-fecal matter (bedding etc) collected should be as minimal as possible
4 CAF isolation staff will record fecal character observations daily for each sample by ticking the
appropriate box on the baggie label
5 Isolation staff will place samples in the necropsy cooler (4 degC) while awaiting collection by
summer student
6 Monday to Friday fecal samples will be collected by A Rodriguez and brought for processing to
the Barta Lab in Pathobiology Samples collected Saturday and Sunday will be put on hold in a
refrigerator (4degC) until pick up on Monday
Example Bag label
Ferret ID Date
Weight of Feces
Fecal Character Normal
Soft
Liquid
Bloody
Abnormal odour
Physical Examination and Health Assessments
1 Manual restraint by hand or in towel by CAF Isolation staff and project personnel If required
examination +- blood collection may be performed under general anesthesia with isoflurane
2 Physical examination body weight HR RR temperature to be performed by PIs (Adriana Pastor
and Dale Smith)
3 Blood collection (~1mL per ferret) is to be performed from the jugular vein if under manual
restraint or the cranial vena cava under anesthesia by PIs using a 25g needle and 1cc syringe
Blood will be collected into small heparinized tubes and submitted to AHL for
CBCBiochemistry
4 All physical examination findings will be recorded on the Exam Sheet
5 Any minor wounds will be treated as appropriate (to be determined by PIs)
Inoculation of Ferrets with Coccidia
1 All ferrets to be inoculated will have been confirmed negative on daily fecals for two weeks
173
2 Brief physical examination by PIs to confirm that the ferrets are healthy to continue in study
3 The concentrated oocyst solution in sterile saline (up to a volume of 1mL) will be combined with
up to 1 mL of FerreTone (or another highly palatable substance if more preferred by the ferrets)
and ferrets will be allowed to consume the mixture ad lib while being monitored
4 Control ferrets will be administered saline only (equal mL to oocyst solution) with 1 mL of
FerreTone (or other substance as used for experimental group)
5 If ferrets refuse to consume the mixture oral inoculation via syringe of concentrated oocyst
solution will be performed by PIs under manual restraint
6 If ferrets are resistant to manual restraint for oral inoculation then inoculation will be performed
by PIs under general anesthesia via gastric tube (8 Fr red rubber)
Euthanasia Protocol
1 Ferrets to be euthanized will be masked down with isoflurane under manual restraint (or in an
anesthetic chamber as judged most appropriate by the PI)
2 Once anesthetized the ferret will be weighed and any blood fecal or other samples will be
collected as required (as determined by PIs)
3 Once an adequate plane of anesthesia is obtained (as determined by PIs) potassium chloride at a
dose of 2 mEq K+kg will be administered either via vena caval puncture or cardiac puncture to
induce cardiac arrest
4 Presenceabsence of respirations heartbeat corneal reflex will be used to assess death
Necropsy Protocol
1 Measure the ferret from nose to tail base (body length)
2 Perform standard necropsy but start with gastrointestinal tract first then thoracic and abdominal
viscera
3 Gently flush the entire contents of the intestine with 12 mL saline into a sterile urine cup
Potassium dichromate should be added in a 11 volumevolume ratio to the same container and
mixed with the combined intestinal contents and saline Label the container with the ferretrsquos ID
ferret group ID date and place on appropriate shelf in the refrigerator (4 degC) in the Barta lab
4 Measure the length of the gut from duodenum to anus
5 Collect paired sections of intestine for histological sectioning and frozen These sections should
be collected along the entire length of the gut from duodenum to rectum (see below for GI length
calculations)
6 For histological sections cut a 2 cm long section of bowel open completely on one side place on
a pre-cut section of box-board and staple both ends to the board to create a flat section Place all
gastrointestinal sections (attached to the board) in Serra fixative solution (100 ethanol (60
VV) 37 formaldehyde (30 VV) glacial acetic acid (10 VV)) for fixation and trim in
cross (transverse) sections
7 Adjacent to each sample removed for histopathology remove another 2 cm section intact and
place in a labelled Whirl-pack for freezing
8 Box-board and pre-labelled Whirl-packs should be labelled in pencil and sharpie respectively
with the following information Animal ID Zoo pm number section of gut (information should
include the region of the gut - jejunum colon etc and the length from the pylorus to the section)
174
Ferret gastrointestinal length calculations (from Evans amp An 2014)
Adult ferret body length 36-41 cm
NB In domestic ferrets there is ~51 ratio of small intestine to body length
Adult domestic ferret GI lengths
Small intestine ndash 182-198 cm
Large intestine - ~10 cm long 06 cm diameter (colon ndash 7 cm rectum ndash 2 cm anus ndash 1 cm)
Based on the above information
Small intestine Six sections of the small intestine from duodenum to ileum will be collected each ~ 25-40
cm apart depending on the size of the ferret
Large intestine Collect two sections of colon (4 cm each) at 25 and 75 of the length of the colon from
the junction of the small and large intestine to the rectum Smaller ferrets may allow only on section
based on colon length
Laboratory SOPs
General notes on processing fecal samples
1 Samples will be collected from the isolation facility necropsy room cooler daily from Monday-
Friday for the entire 6 weeks of the project
2 Upon transfer to the lab each baggie will be weighed after filling to determine the amount of
feces (grams) in each bag and that number entered onto the label on the bag
3 All information regarding fecal processing will be entered onto the spreadsheet for the individual
ferret including ID weight of feces feces character date of sample collection date of sample
processing fecal flotation method presence or absence of oocysts oocyst quantification
Fecal Processing - Weeks 1+2 (acclimation period)
1 After weighing transfer the contents of one baggie to a small sieve on top of a paper cup
2 Fill a single glass container (for fecal flotation) with saturated salt solution
3 Slowly pour small amounts of the solution over the feces mix and strain liquid contents in the paper
cup using a tongue depressor
4 Dispose of the remaining fecal matter in the sieve
5 Pour the contents of the paper cup back into the glass container and cover slip for 5-7 minutes
6 Place coverslip on a clean glass slide and viewscan under microscope at 10x power for presence of
oocysts
7 Record all findings on the provided spreadsheet
8 Notify A Pastor if any oocysts are detected If detected contents of the slide should be flushed back
into the glass container with distilled water and contents of glass container should be transferred to a
50 mL plastic conical tube Clean to remove salt as per Barta lab SOP and mix with potassium
dichromate equal parts by volume to amount of oocysts in diH2O Place a checkmark on the lid of the
50 mL conical vial containing the remainder of the feces and store in the lab fridge at 4 degC
175
Fecal Processing - Weeks 3-7 (infection trial)
1 After weighing transfer the entire contents of one baggie to a small sieve on top of a paper cup
2 Slowly pour small amounts of distilled water over the feces (enough to wet) mix and strain liquid
contents in the paper cup using a tongue depressor until feces appear almost dry
3 Dispose of the remaining fecal matter in the sieve
4 Pour the contents of the paper cup into a 50 mL conical tube If samples are not going to be counted
on the same day then mix potassium dichromate (25 wv) 11 by volume with sieved fluid from
the paper cup and store at 4 degC (refrigerator) until ready to perform OPG counts
If proceeding with the count the same day
5 Pipette and place one drop of the sieved fecal fluid on a slide to determine approximate oocyst
concentration If oocysts rare to none ndash dilute 12 during step 6 if moderate numbers ndash dilute 19 if
too numerous to count ndash dilute 199 (or perform serial dilutions of 10x from initial 19 dilution)
6 Transfer x mL of the mixture from the 50 mL conical tube into a clean 15 mL conical tube and mix
with appropriate amount of saturated salt solution for desired dilution
7 Fill both sides of the McMaster counter chamber and count the number of oocysts per side (total for
one side= total number of oocysts from all 6 sections of the chamber )
8 Average the total counts from both sides
9 Use the following calculation to determine the oocyst per gram count
OocystsmL = oocysts counted times 666 times dilution (ie 3 if dilution 12)
OPG= oocystmL times total volume recorded at end of step 4
10 Record all findings on the provided spreadsheet
11 Notify A Pastor if any oocysts are detected If detected place a checkmark on the lid of the 50 mL
conical vial containing the remainder of the feces and store in the lab fridge at 4 degC
12 For all samples for which no oocysts are detected during steps 6-8 follow up with routine salt
flotation of the remainder of the fecal sample from step 6 (use instructions for fecal processing from
weeks 1-2)

vi
I donrsquot think that I can truly express how thankful I am to Pathobiology laboratory technicians
Julie Cobean and Julia Whale Without your assistance patient teaching and friendship I would probably
still be screening fecal samples years from now and scratching my head as to how our lab protocols
actually work It is people like you who make sure graduate students become successful doctorates and I
canrsquot imagine Pathobio without you both in it
I would also like to thank my labmates in the Barta lab mdash Mian Hafeez Evelyn Rejman Rachel
Imai Perryn Kruth Ryan Snyder and Mosun Ogedengbe A special thank you goes to Alex Leveille
without whom my many adventures in parasitology research from coccidia to Babesia would not have
been as successful
To all the students who helped with ferret fecal sample processing data compilation and
necropsies Nathalie Ferriman Janessa Price Thisuri Eagalle Sarah Brisson thank you so much for your
hard work and excitement about my projecthellip even when it was very smelly
So many thanks to the amazing staff of Central Animal Facility - Linda Groocock Vicky Carson
Tony Cengija and Mary Fowler for the daily care and enrichment of my experimental ferrets Your
excitement about working with our ferrets and your assistance with all parts of the process helped made
this project a success
To Adriana Nielsen who was not only my better half but the other fifty percent of my brain for
several years It is your friendship fortitude and our endless phone conversations that got me through the
never-ending Toronto-Guelph commute and this program
To all the ldquoscope roomrdquo pathology co-residents past and present - thank you for being wonderful
friends and colleagues It is indeed rare to find so many amazing people in one place and I know this
program and my sanity would not have been the same without you
To the anatomic pathology faculty and senior graduate students - thank you for all the time
teaching and guidance you provided during my program While I canrsquot say that I have become an amazing
pathologist I can say that because of your mentorship I am a better diagnostician and the type of clinician
who asks better questions takes better samples and understands that you canrsquot ldquojust make a PCR for thatrdquo
vii
A special thank you to Tony van Dreumel who came out of retirement for a semester to try to teach the
Adrianas zoo pathology screening cases with you was always a pleasure
To all the lovely Histo Ladies PM room staff and the other AHL staff who helped me with
Toronto Zoo and HSC pathology cases along the way - I donrsquot think the anatomic path students could
survive without you Thank you for always smiling assisting and accommodating me even when I made
near-impossible processing requests during my weekly Guelph visits
I would also like to acknowledge and sincerely thank all the individuals who helped with resource
and sample acquisition for this project A special mention for those who went above and beyond because
of their interest in this project Don Duszynski who was instrumental in acquiring and then providing a
translator for many of the original mustelid Eimeria descriptions and Majda Globokar Nikola Pantchev
and Donald Martin who supplied my domestic ferret fecal samples and historical data
A shout-out to Julie Swenson Gary West and the Phoenix Zoo BFF team who fostered my love
of this endangered species and helped develop the idea for this project
As always I continue to go out into the world and pursue my dreams with the knowledge that I
have the support of my incredible family long-time friends and my partner Keith Morris I am so lucky
that my residency brought me home and that it afforded us all more time spent together For my aunt
Veronica Lacey who has never failed to believe in my potential and always pushed me to become an
academic ndash yoursquoll never get that PhD from me but I think this is pretty close Finally for my mother
Anna Pastor who never lived to see my greatest achievements but had absolute faith that I could reach
any goal I worked towardshellip this is for you
Finally none of this would have been possible without the generous support of the Toronto Zoo
Residency program and funding through the Barta Laboratory University of Guelph
Adriana Pastor
Toronto August 2017
viii
DECLARATION OF WORK PERFORMED
I declare that all the work reported in this thesis was performed by myself with the following
exceptions
Fecal samples were collected by personnel at the Toronto Zoo Louisville Zoo and participating
diagnostic laboratories
Fecal oocyst per gram counts (routine salt flotation and McMaster counts) were performed by
myself Julie Cobean Julia Whale Evelin Rejman Sarah Brisson Adriana Rodriguez and Perryn Kruth
Whole mitochondrial genome PCR and sequencing was performed by me in conjunction with
Julia Whale and Dr Mian Hafeez
Sequencing of PCR samples was performed at the University of Guelph Laboratory Services
(Guelph Ontario Canada) and results were obtained electronically
ix
TABLE OF CONTENTS
ABSTRACT ii
DEDICATION iv
ACKNOWLEDGEMENTS v
DECLARATION OF WORK PERFORMED viii
TABLE OF CONTENTS ix
LIST OF TABLES xiii
LIST OF FIGURES xiv
LIST OF APPENDICES xv
ABBREVIATIONS xvi
CHAPTER 1 LITERATURE REVIEW 1
11 INTRODUCTION 1
12 APICOMPLEXA 1
121 Brief introduction to apicomplexan pathogens 1
122 Life cycles of the Eimeria and Isospora species implicated in enteric coccidiosis 3
123 Methods of characterization 5
13 RECLASSIFICATION OF MAMMALIAN ISOSPORA 8
14 EIMERIID SPECIES CHARACTERIZED IN MUSTELIDS 8
141 The family Mustelidae 8
142 Eimeriid coccidia described from mustelids 9
143 Eimeriid coccidia described from domestic ferrets 16
144 Molecular characterization 19
145 Clinical signs of disease in domestic ferrets 21
146 Gross necropsy and histologic findings 21
15 INTRODUCTION TO ENTERIC COCCIDIOSIS IN THE BLACK-FOOTED FERRET
25
151 Natural history and conservation of the black-footed ferret in North America 25
152 Coccidia identified from black-footed ferrets 26
153 Morbidity mortality and clinical signs associated with enteric coccidiosis in black-footed
ferrets 28
16 TREATMENT PREVENTION AND CONTROL OF INFECTION BY EIMERIA SPP 29
161 Current recommendations for treatment of eimeriid coccidia in carnivores 29
x
162 Current recommendations for anticoccidial treatment and prophylaxis in domestic and
black-footed ferrets 30
17 VACCINES AGAINST COCCIDIA 32
171 Theory 32
172 Species successes in anticoccidial vaccination 34
18 RESEARCH GOALS AND OBJECTIVES 36
181 Objectives 36
182 Hypotheses 36
183 Applications 36
CHAPTER 2 MOLECULAR CHARACTERIZATION OF ENTERIC COCCIDIA FROM DOMESTIC
FERRETS (MUSTELA PUTORIUS FURO) 38
21 INTRODUCTION 39
22 MATERIALS amp METHODS 43
221 Fecal samples 43
222 Formalin fixed intestinal tissues 44
223 Molecular characterization 44
224 Phylogenetic analysis 46
23 RESULTS 47
231 Fresh fecal samples 47
232 Formalin fixed samples 48
233 Molecular characterization 49
234 Phylogenetic analysis 50
24 DISCUSSION 50
CHAPTER 3 MORPHOLOGICAL AND MOLECULAR CHARACTERIZATION OF ENTERIC
COCCIDIA ISOLATED FROM BLACK-FOOTED FERRETS (MUSTELA NIGRIPES) 60
31 INTRODUCTION 60
32 MATERIALS AND METHODS 64
321 Fecal samples 64
322 Formalin fixed intestinal tissues 65
323 Molecular characterization 66
33 RESULTS 66
331 Morphometric characterization 67
332 Molecular characterization 68
34 DISCUSSION 69
xi
CHAPTER 4 NATURAL HISTORY OF ENTERIC COCCIDIOSIS IN THE BLACK-FOOTED
FERRET (MUSTELA NIGRIPES) 78
41 INTRODUCTION 78
42 MATERIALS AND METHODS 80
421 Toronto Zoo BFF breeding program 80
422 Fecal oocyst evaluation 81
423 Retrospective review of pathology records 82
424 Prospective modified necropsy protocol 82
425 Retrospective medical history review 83
43 RESULTS 83
431 Fecal oocyst evaluation and retrospective medical history review 83
432 Pathology 86
433 Morbidity and mortality 88
44 DISCUSSION 88
CHAPTER 5 EVALUATING THE DOMESTIC FERRET (MUSTELA PUTORIUS FURO) AS AN
EXPERIMENTAL MODEL FOR ENTERIC COCCIDIOSIS IN THE BLACK-FOOTED FERRET
(MUSTELA NIGRIPES) 104
51 INTRODUCTION 104
52 MATERIALS AND METHODS 106
521 Animal care 106
522 Oocyst preparation 107
523 Experimental infections 108
524 Animal welfare 109
525 Hematology 110
526 Morphologic and molecular characterization 110
527 Necropsy protocol 111
53 RESULTS 111
531 Oocyst shedding 112
532 Morphologic and molecular characterization 113
533 Clinical signs 113
534 Hematology 113
535 Necropsy 114
54 DISCUSSION 115
xii
CHAPTER 6 WHOLE MITOCHONDRIAL GENOME SEQUENCES OF TWO EIMERIA SPECIES
ISOLATED FROM DOMESTIC (MUSTELA PUTORIUS FURO) AND BLACK- FOOTED FERRETS
(MUSTELA NIGRIPES) 129
61 INTRODUCTION 129
62 MATERIALS amp METHODS 130
621 Parasites 130
622 DNA isolation from coccidia in feces 131
623 Whole genome sequencing 131
624 Phylogenetic analysis 132
63 RESULTS 133
64 DISCUSSION 134
CHAPTER 7 CONCLUSIONS AND FUTURE DIRECTIONS 145
REFERENCES 148
APPENDICES 157
xiii
LIST OF TABLES
Table 11 Morphometrics of Eimeria and Isospora (=Cystoisospora) species affecting mustelids 10
Table 21 Amplification primers for nuclear 18S rDNA and mitochondrial COI loci used in the
identification of enteric coccidia from domestic ferrets 55
Table 22 Summary of fecal samples from domestic ferrets submitted to two diagnostic laboratories
from 2008-2015 56
Table 23 Morphologic and molecular identification of coccidia from domestic ferrets 57
Table 31 Amplification primers for nuclear 18S rDNA mitochondrial COI and COIII loci used in the
identification of coccidia from black-footed ferrets 73
Table 32 Morphologic and molecular characterization of coccidia from fecal and FFPE necropsy
samples from black-footed ferrets 76
Table 33 Morphometric characterization of Eimeria ictidea oocysts from black-footed ferrets 77
Table 41 Eimeria ictidea shedding in black-footed ferret dam and kit family groups - 2014-2016 97
Table 42 Epidemiologic data for family groups of black-footed ferrets shedding Eimeria ictidea 98
Table 43 Shedding of Eimeria ictidea in adult black-footed ferrets - 2015-2016 99
Table 44 Epidemiologic data for adult black-footed ferrets shedding Eimeria ictidea 100
Table 45 Histologic findings from black-footed ferrets with enteric coccidiosis 101
Table 46 Incidence of coccidial infections in black-footed ferrets at the Cheyenne Mountain Zoo 102
Table 47 Yearly mortality associated with coccidiosis in black-footed ferrets at the Toronto Zoo 103
Table 51 Prepatent period and oocyst shedding of Eimeria ictidea in experimentally infected
domestic ferrets 126
Table 52 Results of oral inoculation of domestic ferrets with oocysts of Eimeria ictidea 127
Table 53 Distribution of coccidial life stages in intestinal tract of domestic ferrets orally
inoculated with oocysts of Eimeria ictidea 128
Table 61 PCR primers used to sequence the mitochondrial genome of Eimeria furonis 136
Table 62 PCR primers used to sequence the mitochondrial genome of Eimeria ictidea 137
Table 63 Coding regions in the mitochondrial genome of Eimeria furonis from a domestic ferret 138
Table 64 Coding regions in the mitochondrial genome of Eimeria ictidea from a black-footed ferret 139
Table 65 Pairwise comparison of coding regions in the mitochondrial genomes of Eimeria furonis
and Eimeria ictidea 140
xiv
LIST OF FIGURES
Figure 11 Phylogeny of the Apicomplexa 2
Figure 12 Classical life cycle of coccidian parasites 4
Figure 13 Morphologic characteristics used for identification of eimeriid oocysts 6
Figure 21 Life stages of Eimeria furonis within the small intestine of a domestic ferret 58
Figure 22 Phylogenetic relationships of coccidia (Eimeria ictidea Eimeria furonis and Isospora
(=Cystoisospora) laidlawi) from domestic or black-footed ferrets 59
Figure 31 Nuclear and mitochondrial genetic loci targeted by primers listed in Table 31 73
Figure 32 Morphometrics of Eimeria ictidea from a black-footed ferret (Mustela nigripes) 74
Figure 33 Nuclear 18S rDNA sequences of Eimeria ictidea to newly generated (see Chapter 2) and
published sequences of Eimeria furonis 75
Figure 34 Mitochondrial cytochrome c oxidase subunit I sequences of Eimeria ictidea to sequences
from other eimeriid parasites of carnivores 75
Figure 41 Oocyst per gram counts and shedding period of Eimeria ictidea from black-footed ferret
family groups from 2014-2016 95
Figure 42 Sexual life stages of Eimeria ictidea in the small intestine of a black-footed ferret 96
Figure 51 Exogenous life stages of Eimeria ictidea 123
Figure 52 Endogenous life stages of Eimeria ictidea within the small intestine of an experimentally
infected domestic ferret 124
Figure 53 Distribution of sexual and asexual life stages of Eimeria ictidea along the intestinal tract
of experimentally infected domestic ferrets 125
Figure 61 Map of the mitochondrial genome of Eimeria furonis 141
Figure 62 Map of the mitochondrial genome of Eimeria ictidea 142
Figure 63 Comparison of the mitochondrial genomes of Eimeria furonis and Eimeria ictidea 143
Figure 64 Phylogenetic relationships of coccidia from domestic and black-footed ferrets based on
complete mitochondrial genome sequences 144
xv
LIST OF APPENDICES
Appendix 1 Shedding of oocysts of Eimeria ictidea in black-footed ferret (Mustela nigripes) dam and
kit family groups from 2014-2016 158
Appendix 2a Hematology values for domestic ferrets (Mustela putorius furo) from 49-51 days of
age prior to experimental inoculation 161
Appendix 2b Serum biochemistry values for domestic ferrets (Mustela putorius furo) from
49-51 days of age prior to experimental inoculation 162
Appendix 3a Hematology values for domestic ferrets (Mustela putorius furo) inoculated orally
with Eimeria ictidea 163
Appendix 3b Serum biochemistry values for domestic ferrets (Mustela putorius furo) inoculated
orally with Eimeria ictidea 164
Appendix 4 Domestic ferret (Mustela putorius furo) weekly monitoring sheet 165
Appendix 5 Domestic ferret (Mustela putorius furo) 24 hour intensive monitoring sheet 167
Appendix 6 Domestic ferret (Mustela putorius furo) infection trial standard operating procedures 172
xvi
ABBREVIATIONS
ATP Adenosine triphosphate
BFF Black-footed ferret(s)
BI Bayesian inference
bp Base pair
CAPC Companion Animal Parasitology Council
CDS Coding DNA sequence
CITES Convention on International Trade in Endangered Species of Wild Fauna and Flora
COI Cytochrome c oxidase subunit 1
COIII Cytochrome c oxidase subunit 3
CytB Cytochrome b
DF Domestic ferret(s)
DNA Deoxyribonucleic acid
FFPE Formalin-fixed paraffin embedded tissue
IUCN International Union on the Conservation of Nature
L Length
LSU Large subunit
mt Mitochondrial
NaOH Sodium hydroxide
nu Nuclear
OPG Oocyst per gram count
PCR Polymerase chain reaction
rDNA Ribosomal DNA
SI Shape index
SND Single nucleotide difference
SOP Standard operating procedure
sp spp Species (singular plural)
SSP Species Survival Plan
SSU Small subunit
TMS Trimethoprim sulfadimethoxine
USFWS United States Fish and Wildlife Service
W Width
1
CHAPTER 1 LITERATURE REVIEW
11 INTRODUCTION
Black-footed ferrets (Mustela nigripes) are one of three wild ferret species worldwide Although
formerly distributed throughout the North American prairies black-footed ferrets (BFF) had been
extirpated from the majority of their range by the 1970s and were declared extinct in the wild in 1987
Since 1986 a multi-institutional effort has been breeding this species in captivity with reintroduction back
into the wild at select sites within Canada the USA and Mexico
Coccidial enteritis is a major cause of death in young captive black-footed ferrets (Bronson et al
2007) but coccidiosis can affect all age classes (personal observation) As a result fewer captive-bred
ferrets may be reared successfully for release to the wild The significance of coccidiosis in wild ferrets is
unknown Consequently the prevention and control of coccidial outbreaks is an important part of black-
footed ferret captive breeding programs and management This research is intended to improve the in situ
and ex situ health of the black-footed ferret through the provision of a better understanding of the
pathogenesis of enteric coccidiosis in this species and to pave the way for the investigation of novel
methods for disease treatment and control
12 APICOMPLEXA
121 Brief introduction to apicomplexan pathogens
The phylum Apicomplexa comprises a large number of eukaryotic intracellular parasitic
organisms many of which are of importance to human and veterinary medicine As indicated by their
name these parasites are characterized by the presence of an apical complex at the anterior aspect of the
infective stage of the life-cycle (Tenter et al 2002) The taxonomic classifications of members of the
Apicomplexa continue to be in a state of flux (reviewed by Adl et al 2005 Cavalier-Smith 2014 Tenter
et al 2002) For this reason a more simplified taxonomic structure has been used in this review (see
2
Figure 11) The subclass Coccidia is a speciose group within the Apicomplexa with most genera falling
into one of two major coccidian suborders within the Eucoccidiorida To date greater than 2000 species
of coccidia have been named (Duszynski Upton amp Couch nd Upton 2000) The adeleid coccidia
(suborder Adeleorina) include monoxenous (single host) and heteroxenous (multiple hosts) parasites in
genera such as Adelea Haemogregarina Hepatozoon and Karyolysus The eimeriorinid coccidia
(suborder Eimeriorina) include the typical intestinal coccidia such as Eimeria Isospora and Cyclospora
species belonging to the family Eimeriidae as well as tissue (cyst forming) coccidia such as
Cystoisospora Besnoitia Toxoplasma and Sarcocystis species that belong to the family Sarcocystidae
(Cox 1994)
Figure 11 Phylogeny of the Apicomplexa Numbers on branches and thickness indicate diversity
(ie named species) Taxonomic groupings demonstrated by the phylogenetic tree (1) subclass
Coccidia (2) suborder Adeleorina (3) suborder Eimeriorina (4) family Eimeriidae and (5) family
Sarcocystidae Adapted from Šlapeta J Morin-Adeline V (2011) Apicomplexa Levine 1970
Sporozoa Leucart 1879 httptolweborgApicomplexa2446 in The Tree of Life Web Project
httptolweborg
2
1
3
4
5
3
122 Life cycles of the Eimeria and Isospora species implicated in enteric coccidiosis
The life cycle of Eimeria species is considered the classical coccidian life cycle which is
typically completed in one host (monoxenous) with many Eimeria species parasitizing only a single host
species (stenoxenous) (Figure 12) The life cycle has two main phases of development one that takes
place within the host (endogenous) and the other that takes places outside of the host (exogenous)
Classically the endogenous stages of the Eimeria life cycle take place within the intestinal epithelium
however some Eimeria species undergo extraintestinal endogenous development such as Eimeria stiedae
in rabbits which replicates within the epithelium of the biliary tree During the exogenous phase of the
life cycle unsporulated oocysts that are shed in the feces of the host sporulate within the environment
resulting in the formation of four sporocysts within each oocyst (tetrasporocystic) Each sporocyst
contains two sporozoites (dizoic) Sporulation is affected by three main factors temperature moisture and
aerobic conditions (Fayer 1980)
Once ingested by the host the wall of the sporulated oocyst is broken to release sporocysts from
which the sporozoites (infective stage) excyst The freed sporozoites penetrate the intestinal epithelial
cells and undergo multiple mitotic divisions to form a single multinucleate meront The meront then
undergoes simultaneous cytokinesis to form first generation merozoites which leave the host cell to infect
new cells and undergo further asexual replications The undifferentiated uninucleate tissue stage of the
parasite within the intestinal epithelial cell is called a trophozoite The number of cycles of asexual
replication (merogony) is predetermined after which the last generation of merozoites penetrate host cells
and undergo sexual differentiation into male and female gamonts (gametogony) Each microgamont
(male) undergoes simultaneous fission to produce numerous motile microgametes each macrogamont
(female) develops into a single mature macrogamete Fertilization of a macrogamete by a motile
microgamete results in formation of a zygote that is rapidly enclosed in a thick wall to form an
unsporulated oocyst Oocysts are shed with the hostrsquos feces into the environment where they are
protected from desiccation and chemical disinfection by the oocyst wall Traditionally Eimeria species
4
have been differentiated based on the host species or host genus affected the site of endogenous life cycle
development and the microscopic cellular characteristics of the different life stages Interestingly
experimental cross infection of Eimeria species from their natural host to a novel host of a taxonomically
similar species has been successful in some cases (De Vos 1970 Levine and Ivens 1970 Haberkorn
1971) challenging the notion that Eimeria are truly stenoxenous parasites
Figure 12 Classical life cycle of coccidian parasites This apicomplexan life cycle includes both
sexual and asexual development The three processes in the life cycle are merogony (asexual
replication A-D) followed by gametogony (formation of gametes E-H) within the digestive tract
of the host with release of unsporulated oocysts (I) Exogenous sporogony (I-L) results in the
production of infective sporulated oocysts (L) Adapted from Barta 2001 with permission of the
author
The life cycle of Isospora spp is similar to that of species in the genus Eimeria (see Figure 12)
but the number of sporocysts and sporozoites differ sporulated oocysts contain two sporocysts (disporic)
5
each of which contains four sporozoites (tetrazoic) These characteristics are not unique to Isospora spp
because diasporic tetrazoic sporulated oocysts are also found in the genera Besnoitia Frenkelia
Hammondia Sarcocystis and Toxoplasma However the sporocysts in the latter parasites are
morphologically distinct in that they lack Stieda bodies
123 Methods of characterization
1231 Morphological features
Historically eimeriid coccidia have been classified based on the cellular morphology of the
different life stages (particularly the morphometrics of sporulated oocysts) where these stages occur in
the host and apparent host specificity (frequently assumed and not tested experimentally) The
morphological features and dimensions of oocysts and their components are important diagnostic features
because of the availability of these stages in clinical specimens these characteristics can include size
(length [L] width [W] shape index [SI=LW]) number of sporocysts wall morphology
presenceabsence of a micropyle micropyle cap residual body or polar granules for oocysts size number
of sporozoites wall morphology presenceabsence of Stieda body subStieda body paraStieda body or
residual body for sporocysts and presenceabsence of refractile bodies for sporozoites (see Figure 13)
Pertinent life cycle information includes type of life cycle (monoxenous versus heteroxenous) tissue
sites of merogony and gametogony (intestinal versus extraintestinal) and the presence or absence of
extraintestinal hypobiotic stages (eg dormozoites or hypnozoites) Further information used to
characterize coccidia that form tissue cysts generally includes details on life stages in the definitive and
intermediate hosts location and morphology of tissue cysts route(s) of transmission among host species
and morphologic descriptions of merozoites (eg tachyzoites or bradyzoites) in tissue culture
6
Figure 13 Morphologic characteristics used for identification of eimeriid oocysts 1) Oocyst in cross
section ol - oocyst length or - oocyst residual body ow - oocyst width pg - polar granule row -rough
outer wall 2) The top of a hypothetical oocyst mcd - depth of the micropyle cap mcw - width of the
micropyle cap mw - width of the micropyle sow - smooth outer wall 3) Sporocyst in cross section
psb - paraStieda body sb - Stieda body sl - sporocyst length sp - sporozoite sr - sporocyst residual
body srb - sporozoite refractile body ssb - subStieda body sw - sporocyst width From Duszynski D
Wilber PG (1997) A guideline for the preparation of species descriptions in the Eimeriidae Journal of
Parasitology 83(2)333-336 reproduced with permission of Allen Press Publishing Services
1232 Molecular characterization (genetic loci and methods)
More recently molecular techniques have been used to infer phylogenetic or evolutionary
relationships among coccidia and to reclassify taxonomic assignments to better reflect the evolutionary
history of these parasites Molecular data can be more informative than phenotypic data because recent
evolutionary divergence among coccidia is unlikely to be reflected in morphologic differences but may
be detectable using molecular data The principle behind the use of molecular sequencing to describe
evolutionary relationships is that nucleotide sequences like morphological features diverge over time
under selective pressure however nucleotide sequences evolve at a more regular rate than do
morphologic characteristics Phenotypic data is thus less likely to detect recent evolutionary divergence
Sequences that are more similar are inferred to be more closely related and to have diverged more
recently (Cox 1994) Molecular characterization can be performed using DNA RNA or protein
sequences Most of the early molecular phylogenetic analyses of coccidia performed used ribosomal RNA
sequences usually by PCR amplification of ribosomal DNA (rDNA) in the nuclear genome of the
7
parasites Ribosomes contain both small and large RNA subunits in eukaryotes the large ribosomal RNA
consists of two forms 5S and 28S while the small ribosomal RNA exists only as 18S Sequences from
several genetic loci have been used for characterization of parasites most commonly 18S rDNA 28S
rDNA and ribosomal internal transcribed spacer regions (ITS) from the nuclear genome and more
recently mitochondrial cytochrome c oxidase subunits I (COI) and III (COIII) however sequencing of
nuclear 18S rDNA (nu 18S rDNA) has been the most prevalent in the literature by far Early attempts to
use 5S RNA sequences formed unlikely phylogenies and too few 28S ribosomal DNA sequences have
been obtained to make this locus useful (Cox 1994 Tenter et al 2002) The disadvantage of nu 18S
rDNA is that it is comparatively poor at distinguishing among closely related eimeriid coccidial species
because of its conserved nature but for that reason the nu18S rDNA locus is useful for inferring
relationships among species with greater evolutionary divergence Although only exploited recently
because of the paucity of suitable PCR primers the mitochondrial COI locus appears to be more useful
for distinguishing closely related eimeriid coccidia (Ogedengbe Hanner amp Barta 2011) but COI
sequences are less useful for inferring more ancient relationships between highly divergent coccidial
species Consequently the combined use of nu 18S rDNA and mitochondrial COI sequencing has been
recommended for improved species description and phylogenetic analysis (El-Sherry et al 2013)
Molecular characterization has also been used for diagnostic purposes and is well-suited to the
identification of coccidia when information on host specificity parasite life cycle and life stages is not
available as the molecular (genetic) data is the same for a given parasite during each of its life cycle
stages This information can be particularly useful in identifying the relationship between different life
stages of heteroxenous parasites collected from different hosts (intermediate definitive) Furthermore for
previously unidentified coccidia or those for which limited information is available molecular
characterization could be used to predict likely definitive hosts or parasite life cycle traits based on
phylogenetic relationships to other known species
8
13 RECLASSIFICATION OF MAMMALIAN ISOSPORA
Recommendations have been made to reclassify the avian and mammalian Isospora into two
separate genera based on life cycle molecular phylogenetic studies and morphologic description of
sporulated oocysts (Frenkel 1977 Barta et al 2005) Due to their classical coccidian life cycle presence
of Stieda bodies within sporocysts and close phylogenetic association with Eimeria species the avian
Atoxoplasma and Isospora have been retained in the genus Isospora (see Barta et al 2005) Conversely
the presence of tissue life cycle stages lack of Stieda bodies within sporocysts and close phylogenetic
association with other genera within the family Sarcocystidae have required many mammalian Isospora
to be reclassified as members of the genus Cystoisospora Frenkel 1977 (Frenkel 1977 Barta et al 2005)
Consequently for the remainder of this thesis Isospora species from mustelids will be referred to as
Isospora (=Cystoisospora) to reflect their probable generic association
14 EIMERIID SPECIES CHARACTERIZED IN MUSTELIDS
141 The family Mustelidae
The family Mustelidae within the order Carnivora comprises a group of approximately 59
carnivorous mammalian species within 22 genera Native mustelids are found in terrestrial and aquatic
environments on almost every continent with the exception of Australia and Antarctica The Mustelidae
are classically divided into two subfamilies as defined by Wozencraft (2005) 1) Mustelinae (weasels
mink ferrets marten wolverine) the larger subfamily including the following genera Arctonyx Eira
Galictis Gulo Ictonyx Lyncodon Martes Meles Mellivora Melogale Mustela Neovison Poecilogale
Taxidea and Vormela and 2) Lutrinae (otters) including seven genera Aonyx Enhydra Hydrictis
Lontra Lutra Lutrogale and Pteronura More recently molecular data suggest the Mustelidae should be
separated into eight subfamilies although this is not universally accepted (Koepfli et al 2008 Lariviegravere
and Jennings 2009 Yu et al 2011)
9
142 Eimeriid coccidia described from mustelids
Ten named Eimeria species and twelve named Isospora (=Cystoisospora) species have been
described in the Mustelidae and are summarized in Table 11 This table includes information on host
range life cycle and detailed morphologic data used to identify and classify the individual parasites Two
coccidial parasites isolated from the Libyan striped weasel (Ictonyx libyca) and the European polecat
(Mustela putorius) initially ascribed to the genus Isospora Isospora zorillae and Isospora putori
respectively have since been reclassified as Sarcocystis spp (see footnote to Table 2 of Yi-Fan et al
2012)
10
Table 11 Morphologic characteristics of Eimeria and Isospora (=Cystoisospora) species affecting mustelids
Coccidial species Host genus and
species
Life cycle
Location
Oocyst shape and
size
Oocyst
description
Sporocyst
description
Sporozoite
description
References
Cytoisospora
eversmanni
Mustela
eversmanii
(Steppe polecat)
Mustela
putorius
(European
polecat)
Homoxenous L185 (16ndash20)
W 148 (16ndash12)
LW 13 (11ndash16)
M absent
PG absent
OR absent
L 115
(10ndash135)
W 98
(9ndash11)
LW 12
(11ndash15)
SB absent
SR present
SRB present Yi-Fan et al 2012
Svanbaev 1956
Nukerbaeva amp
Svanbaev 1973
1977
Cystoisospora
pavlovskyi
Mustela
eversmanii
Mustela
putorius
Homoxenous L 322 (29ndash36)
W 273 (265ndash285)
LW 12 (11ndash14)
M absent
PG absent
OR absent
L 195
(18ndash21)
W 144
(12ndash15)
LW 14
(12ndash15)
SB absent
SR present
SRB present Yi-Fan et al 2012
Svanbaev 1956
Nukerbaeva amp
Svanbaev 1973
1977
Eimeria
baskanica^
Mustela
erminae
(ermine)
Homoxenous Oval with tapered
ends
L 112-126
W 84-98
M absent
PG absent
OR present
SR absent Bean shaped Nukerbaeva amp
Svanbaev 1977
Eimeria furonis Mustela
putorius
Mustela
putorius furo
(dom ferret)
Mustela
nigripes (BFF)
Mustela vison
(mink)
Homoxenous
Small intestine
rectum (H 1927)
Jejunumileum (BP
1993)
Spherical ndash
subspherical
L 11-14
W 10-13
OW 2 layers
M absent
PG absent
OR absent
Spindloid
L 8-9
W 4
SB present
SR present
Vermiform Blankenship-Paris
et al 1993
Hoare 1927 1935b
Jolley et al 1994
Nukerbaeva amp
Svanbaev
19731977
Williams et al 1988
1992 1996
Eimeria hiepei Mustela vison Homoxenous
Bile duct
Spherical
L 13-17
W 13-17
OW 2 layers
(outer
smooth)
M absent
PG absent
OR absent
L 6
W 4
SB absent
SR absent
Banana shaped Davis et al 1953
Grafner et al 1967
11
Coccidial species Host genus and
species
Life cycle
Location
Oocyst shape and
size
Oocyst
description
Sporocyst
description
Sporozoite
description
References
Eimeria ictidea Mustela
eversmanni
Mustela
nigripes
Mustela
putorius
Mustela
putorius furo
Homoxenous
Small intestine
Ovoid ndashellipsoid
L 13-27
W 13-21
OW 2 layers
M present
PG absent
OR absent
Ovoid
(irregular)
L 115
W65
SB present
SR present
- Hoare 1927 1935a
1935b
Jolley et al 1994
Litvenkova 1969
Svanbaev 1956
Tinar 1985
Williams et al 1988
1992
Eimeria irara Eira barbara
(tayra)
Homoxenous
Feces
Ovoid
L 21-25
W 18-20
OW outer
layer smooth
M absent
PG absent
OR absent
Ellipsoid
L 10-12
W 65
SB present
SR present
Elongate (one
end broader than
the other)
Carini amp da
Fonseca 1938
Eimeria melis Meles meles
(European
badger)
Homoxenous Ellipsoid
L 20plusmn018
W 157plusmn002
LW128plusmn0017
(112-15)
OW 2 layers
(outer
smooth)
M absent
PG present
OR present
Ovoid
L
119plusmn0018
W 65plusmn008
LW 183
(155-24)
SB present
L 90plusmn005
W 324plusmn0025
SRB present
Anwar et al 2000
Kotlan amp Pospesch
1933
Eimeria mustelae Mustela vison
Mustela nivalis
(snow weasel)
Homoxenous
Duodenumileum
Spherical or
Ellipsoid
L 18-26
W 14-24
OW 2 layers
M absent
PG present
OR absent
Ovoid
L 8
W 5
SB present
SR present
Broad at one
end and tapered
at other
L 7
W 3
Glebezdin 1978
Iwanoff-Gobzem
1934
Levine 1948
Musaev amp Veisov
1983
Tinar 1985
Eimeria sablii Martes zibellina
(sable)
Homoxenous
Gut
Spherical or
subspherical
L 112-126
W 112
OW 2 layers
M absent
OR absent
Ovoid
L 56
W 42
SR present
Elongate Nukerbaeva 1981
Eimeria sibirica Martes zibellina Homoxenous Ovoid
L avg 216
W avg 180
LW 1076
OW 2 layers
M absent
PG absent
OR absent
Ovoid
L 96-112
W 56-72
SR absent
Elongate Nukerbaeva 1981
Yakimoff amp
Gousseff 1934
Yakimoff amp
Terwinsky 1930
1931
12
Coccidial species Host genus and
species
Life cycle
Location
Oocyst shape and
size
Oocyst
description
Sporocyst
description
Sporozoite
description
References
Eimeria vison
(Eimeria
mustelae)
Mustela
putorius
Mustela
putorius furo
Mustela vison
Homoxenous
Small intestine
+- large intestine
Ovoid
L 17-22
W 9-18
OW 2 layers
M absent
OR
sometimes
present
Ovoid or
Piriform
L 10
W 55
SB absent
SR present
Curved or Club
shaped
L 9
W 25
Foreyt amp Todd 1976
Foreyt et al 1977
Kingscote 1934
1935
Levine 1948
McTaggart 1960
Nukerbaeva amp
Svanbaev
19731977
Tinar 1985
Umurzakov amp
Nukerbaeva 1985
Wolter 1961
Zimmermann 1959
Isospora africana Ictonyx libyca
(Libyan striped
weasel)
Homoxenous
Feces
Spherical
L 25-27
W 25-27
OW 2 layers
(outer
smooth)
M absent
PG absent
OR absent
Ovoid
L 15-17
W 10-12
SB absent
SR present
Elongate
L 135
W 3
Prasad 1961
Isospora altaica Mustela altaica
(mountain
weasel)
Homoxenous
Gut
Oval or spherical
L 280-336
W 252-280
LW 121 (111-
124)
OW 2 layers
M absent
PG absent
OR absent
Ovoid or
spherical
L 140-168
W 111-168
SR present
Svanbaev amp
Rachmatullina
1971
Isospora goussevi Mustela nivalis Homoxenous
Large intestine
Ovoid
L 224 (220-250)
W 174 (160-190)
LW 135 (133-
137)
OW 1 layer
PG present
OR present
Ovoid
L 120
(100-130)
W 70 (60-
80)
SB present
SR present
Elongate Musaev amp Veisov
1983
13
Coccidial species Host genus and
species
Life cycle
Location
Oocyst shape and
size
Oocyst
description
Sporocyst
description
Sporozoite
description
References
Isospora
hoogstraali
Ictonyx libyca Homoxenous
Feces
Ellipsoid
L 37-41
W 32-34
OW 2 layers
(outer
smooth)
M absent
PG some
OR absent
Ovoid
L 19-21
W 13-15
SB absent
SR present
Club-shaped
L 18-19
W 4-6
Prasad 1961
Isospora laidlawi Mustela
putorius
Mustela
putorius furo
Mustela vison
Homoxenous
Feces
Intestinal contents
Ovoid L
320-368
W 272-304
OW 2 layers
M absent
PG absent
OR absent
Ellipsoid
L 208
W 144
SB absent
SR present
Sausage shaped Foreyt et al 1977
Hoare 1927
Levine 1948
McTaggart 1960
Nukerbaeva amp
Svanbaev 1973
1974 1977
Tinar 1985
Isospora lutrae Lutra lutra
(European
otter)
Lutra
canadensis
(North
American river
otter)
Homoxenous Spherical
L 312 (275-32)
W 296 (28-31)
LW 104
(10-112)
OW 2 layers
(outer
smooth)
M absent
PG absent
OR absent
Ellipsoid
L 182 (17-
19)
W 144 (14-
16)
LW128
(12-14)
Sb absent
sSB absent
SR present
Spindle- shaped
L 124
W 25
SRB present
Torres et al 2000
Hoover et al 1985
Isospora
martessii
Martes zibellina Homoxenous
Gut
Ovoid short oval or
spherical
L 252 ndash 280 196
168
W 168 ndash 224 168
168
OW 2 layers
M absent
OR absent
Ovoid
L 112-168
W 84-112
SR present
Elongate Nukerbaeva 1981
Isospora melis Meles meles Homoxenous Ovoid
L 328plusmn034
W 269plusmn019
LW122 (110-
157)
OW 2 layers
(outer
smooth)
M absent
PG absent
OR absent
Ellipsoid
L
215plusmn0166
W 14plusmn012
LW 155
(133-185)
SR absent
Round at one
end other end
tapered
L 142plusmn116
W 40plusmn017
SRB absent
Anwar et al 2000
Glebezdin 1978
Kotlan amp Pospesch
1933
Pelleacuterdy 1955
14
Coccidial species Host genus and
species
Life cycle
Location
Oocyst shape and
size
Oocyst
description
Sporocyst
description
Sporozoite
description
References
Isospora
mustelae (nomen
nudum)
Martes martes Ovoid L
7 W
225
M present - - Galli-Valerio 1932
Isospora nivalis Mustela nivalis Homoxenous
Large intestine
Ovoid
L 206 (200-230)
W 184 (180-210)
LW 11 (109-111)
OW 1 layer
PG absent
OR absent
Ovoid
L 125
(120-130)
W 80 (70-
90)
SR present
Lemon or pear
shaped
Musaev amp Veisov
1983
Unnamed
ldquoCoccidiardquo^
Mustela
nigripes
Urinary bladder - - - - Jolley et al 1994
Unnamed
ldquoCoccidiardquo^
Mustela
nigripes
Trachea bronchus
bronchial glands
- - - - Jolley et al 1994
Unnamed
Eimeria sp^
Mustela
nigripes
Feces
intestinal contents
Ovoid
L 350-386
W 212-232
- - - Jolley et al 1994
Williams et al
1992
Unnamed
Eimeria sp^
Mustela
putorius furo
Small intestine - - - - Blankenship-Paris
et al 1993
Unnamed
Eimeria sp^
Mustela nivalis Homoxenous
Large intestine
Ovoid-ellipsoid L
2031 (1712-2162)
W 148 (1225-
1681)
LW 136 (121-16-
)
OW 1 layer
PG absent
OR absent
Ovoid or
pear-shaped
L 60-100
W 40-80
SR present
Elongate
L 50-90
W 30-70
Musaev amp Veisov
1983
Unnamed
Eimeria sp^
Martes martes
(marten)
Homoxenous Ovoid
L avg 216
W avg 180
LW 1076
OR absent 4 sporocysts
SR present
L 126
W 60
Yakimoff and
Gousseff 1934
Unnamed
Isospora sp^
Mustela
putorius furo
Feces - - - - Bell 1994
Unnamed
Isospora sp^
Mustela
putorius furo
Feces - - - - Bell 1994
Legend L = length W = width LW = length-width ratio avg = average OW = oocyst wall PG = polar granules M = micropyle SB = Stieda body sSB =
subStieda body OR = oocyst residuum SR = sporocyst residuum SRB = sporozoite refractile body ^ = species inquirendae - = no information provided by
author(s) = information obtained from secondary sources (primary reference could not be obtained) All measurements are in micrometers Bolded references
15
are those from which morphometric data were assembled Remaining references indicate other authors who have identified that parasite species in the same or
similar host
16
143 Eimeriid coccidia described from domestic ferrets
Three species of coccidia were originally described from 50 domestic ferrets (Mustela putorius
furo) Eimeria ictidea Eimeria furonis and Isospora (= Cystoisospora) laidlawi (Hoare 1927) All three
species were detected in feces from domestic ferrets at a research facility undergoing an outbreak of
canine distemper Sick ferrets appeared more frequently infected than healthy ones As per Hoare (1927)
none of the ferrets appeared to display clinical signs associated with protozoal infection For each
parasite the author described morphology of sporulated oocysts isolated from feces and sporulation time
(exogenous life stages) The pre-patent period (minimum duration of endogenous development) in an
inoculated naiumlve ferret was described only for E furonis and E ictidea due to insufficient sample size of
I (=C) laidlawi oocysts for an experimental infection trial Sporulation of oocysts occurred within 5-6
days for E furonis 3 days for E ictidea and 4 days for I (=C) laidlawi The sporulated oocysts of E
furonis were spherical with a double outer wall with a thin colourless outer layer and thick yellowish
inner layer no micropyle or residual body and measured on average 128 times 120 microm (length [L] 112-
144 width [W] 104-128 shape index [SI] 107) Unsporulated oocysts contained a zygote with a
diameter of 96 microm Sporocysts were spindle-shaped with one end constrictedblunted contained a
residual body and on average measured 8-88 times 4 microm Sporozoites were vermiform with one end
narrower than the other arranged head to tail and had a central nucleus a clear vacuole was identified in
some at the broad end The sporulated oocysts of E ictidea were oval or elliptical with a double outer
wall with a thin colourless outer layer and thick yellowish inner layer no micropyle or residual body
and measured on average 236 times 175 microm (L 184-272 W 128-208 shape index 135) The zygote in
unsporulated oocysts was elongate with a diameter of 15 times 12 microm when originally passed in feces but
became more spherical with time Sporocysts were irregularly oval with one end broad and the other
more constricted contained a residual body and on average measured 115 times 65 microm Sporozoites were
vermiform with one end narrower than the other arranged head to tail and had a central nucleus and a
clear vacuole at the broad end The sporulated oocysts of Isospora (=Cystoisospora) laidlawi were ovoid
with a double outer wall with a thin colourless outer layer and thick yellowish inner layer no micropyle
17
or residual body and measured on average 34 times 29 microm (L 320-368 W 272-304) Unsporulated
oocysts contained a spherical zygote with a diameter of 236 microm Two sporocysts were identified each
containing 4 sporozoites and no Stieda body sporocysts were elliptical contained a residual body and on
average measured on 208 times 144 microm Sporozoites were sausage shaped with one end slightly pointed
and had a central nucleus and a clear vacuole identified at the pointed end Sporozoites were arranged
with pointed ends all at the same pole of the sporocyst The pre-patent periods described for E furonis
and E ictidea were 6 days and 7 days respectively (Hoare 1927)
Since Hoarersquos initial description (Hoare 1927 Hoare 1935) multiple single case reports and
outbreaks of severe clinical disease associated with intestinal coccidiosis have been reported in domestic
ferrets Blankenship-Paris et al (1993) described a single case of a four-month-old domestic ferret that
presented depressed in thin body condition dehydrated and with pasty dark feces on the perineum This
ferret had been housed with its dam and another sibling neither dam nor sibling showed clinical signs of
enteric disease and both had negative fecal examination results on repeated evaluation Routine fecal
examination of the rest of the colony and necropsies on eight other ferrets in the colony revealed no
evidence of coccidial infection Enteric coccidiosis was determined to be the cause of disease in the four-
month-old ferret based on necropsy findings but the coccidia could not be speciated because diagnosis
was made on histologic findings only
Sledge et al ( 2011) described three separate outbreaks of severe enteric coccidiosis in domestic
ferrets from one ferret rescue centre (group 1) and two shelters (groups 2 and 3) all affected by the same
Eimeria sp The morphologic characteristics of sporulated oocysts were only described for group 1 no
coccidial oocysts were detected on direct smear or fecal flotation of diarrheic samples submitted from
groups 2 and 3 Oocysts were identified as spherical measuring 12-13 microm in diameter with four
sporocysts each containing two sporozoites Oocyst morphometrics histopathologic findings and nu 18S
rDNA partial sequences from all three groups were used collectively to confirm the coccidial species
identify in each outbreak as E furonis
18
Two cases of biliary coccidiosis with E furonis have been reported in domestic ferrets The first
was in a nine-week-old male ferret from a research facility (Williams Chimes amp Gardiner 1996) The
ferret presented with signs of hepatic disease and was negative for coccidia on fecal flotation and direct
smears Endogenous coccidial life stages were described from the gall bladder and liver on histologic
examination In tissue section the oocysts were oval to spherical and measured 125 times 120 microm Meronts
measured 108-130 times 89-93 microm and contained up to 16 merozoites The merozoites exhibited a double-
layered pellicle prominent conoid few rhoptries and many micronemes anterior to the nucleus Based on
the morphologic description of the life stages in this case the coccidia were identified by the authors as
an Eimeria species most likely E furonis Kaye et al (2015) described a second case of biliary
coccidiosis in an 18-month-old female pet domestic ferret with concurrent pure red cell aplasia In this
case all endogenous coccidial life stages were observed on histologic examination of the epithelium of
the extrahepatic biliary tree The oocysts were ovoid and measured 12 times 13 microm Meronts measured 12 times
15 microm and contained up to 16 merozoites each measuring 2 times 5 microm Based on the morphologic
description of the life stages in this case and nu 18S rDNA sequences the pathogen was also determined
to be E furonis Biliary coccidiosis has also been identified in mink (Mustela vison) with the etiologic
agent identified as Eimeria hiepei (Davis Chow amp Gorham 1953 Grafner Graubmann amp Dobbriner
1967)
Oocysts from Cystoisospora ohioensis have been reported from fecal samples collected from
healthy domestic ferret kits in a large American ferret breeding operation that were raised on the same
premise as juvenile domestic dogs (Patterson amp Fox 2007) The method of identification of this parasite
was not described by Patterson amp Fox A second similar institution reported the presence of a
Cystoisospora species also thought to be C ohioensis in routine fecal examination of their ferret colony
(Dr Bambi Jasmin personal communication) Coccidial identification in this case was performed by the
Animal Health Diagnostic Center at Cornell University The significance of these findings is unknown as
no clinical signs or histologic lesions have been described in domestic ferrets associated with shedding of
19
oocysts and the definitive host for C ohioensis is the domestic dog It is most likely that fecal
identification of C ohioensis represents a pseudoparasite in both of these cases or perhaps an
undescribed Cystoisospora sp that is morphologically indistinguishable from C ohioensis
It is difficult to estimate the prevalence of enteric coccidia within the North American domestic
ferret population Fecal samples submitted to university or large veterinary diagnostic laboratories from
domestic ferrets in Canada are uncommon and samples positive for coccidia appear infrequently (Dr
Donald Martin personal communication) Data from Idexx Vet Med Lab in Ludwigsburg Germany was
compiled to review the prevalence of coccidia and Giardia within fecal samples from domestic ferrets
(Pantchev et al 2011) The authors reported that of 284 fecal samples submitted from 2002-2004 18
(63) had detectable coccidial oocysts on fecal flotation Oocysts were identified based on morphologic
characteristics as E ictidea E furonis I (=C) laidlawi and another unidentified Isospora species
Comparative data from the same laboratory from 2009-2010 included sample submissions from 253
ferrets 21 (83) of which were positive for coccidial oocysts on fecal flotation Nine of the samples
were positive for E furonis three were positive with both E furonis and I(=C) laidlawi present eight
were positive only for I(=C) laidlawi and one sample contained both E furonis and E ictidea
identification in all cases was based on morphologic characteristics No statistically significant difference
in the occurrence of coccidial oocysts was detected when data from the two periods were compared
(Fisherrsquos exact test P=041) (Pantchev et al 2011)
144 Molecular characterization
Molecular characterization of Eimeria furonis was first performed by Abe et al (2008) using
oocysts purified from the feces of a single domestic ferret with clinical signs of coccidial enteritis Small
subunit ribosomal DNA (nu 18S rDNA) primers CYC1FE (5ʹ-TAC CCA ATG AAA ACA GTT T-3prime) and
CYC4RB (5prime-CGT CTT CAA ACC CCC TAC TG-3prime) were used to amplify a 347 base pair (bp) fragment
of nu 18S rDNA These primers were initially developed for molecular identification of Cyclospora
species but have since been shown to amplify nu 18S rDNA from several Eimeria species (Matsubayashi
20
et al 2005) The amplicon was sequenced (GenBank AB329724) and compared with previously
published partial nu 18S rDNA sequences from 40 Eimeria two Isospora and four Cyclospora species
The resulting phylogram grouped E furonis with E alabamensis (cattle) and E meleagrimitis (turkey) In
the same study the microscopic morphology of the oocysts was used to identify this coccidial species as
E furonis by comparison with published descriptions of E furonis E ictidea and E heipei by Hoare
(1927) Hoare (1935) and Grafner Graubmann amp Dobbriner (1967) respectively
Nuclear 18S rDNA was also used by Sledge et al (2011) for molecular identification of the
eimeriid coccidia implicated in the three distinct outbreaks of enteric disease in domestic ferrets As
described above initial identification and speciation of the coccidia was performed using morphologic
characteristics of the sporulated oocysts collected from feces in one of the three outbreaks being
investigated the oocysts were identified as E furonis Histologic sections of formalin fixed intestinal
segments from ferrets from each of the three outbreaks contained multiple coccidial life stages DNA was
then isolated from stored formalin-fixed tissues for further genetic analysis Using the partial nu 18S
rDNA gene sequence reported by Abe et al (2008) (GenBank AB329724) the following PCR primers
were created 5ʹ-ACA ATT GGA GGG CAA GTC TG-3ʹ and 5ʹ-GGCGAC AAG CCT GCT TGA AAC-
3ʹ PCR amplification produced a 247 bp amplicon from each of the three groups Analysis and
sequencing of amplicons from all three groups showed 100 homology to nucleic acid sequences
previously reported by Abe et al (2008) for the gene encoding E furonis nu 18S rDNA
Coccidia were identified within hepatobiliary lesions in a domestic ferret receiving
immunosuppressive therapy for red cell aplasia (Kaye et al 2015) DNA was extracted from frozen liver
and a 247 bp fragment of the nu 18S rDNA was amplified using the primers previously described by
Sledge et al (2011) and sequenced Kaye et al (2015) reported that the DNA sequence of the amplicon
was 100 homologous to the published nu 18S rDNA of E furonis and 95 homologous to the nu 18S
rDNA of E myoxi (rodent) E alabamensis (cattle) and I robini (avian)
21
145 Clinical signs of disease in domestic ferrets
Hoare (1927 1935b) in his initial descriptions of enteric coccidiosis in domestic ferrets
observed that clinical signs of intestinal disease were not evident The recent literature supports the
finding of subclinical disease but also describes signs ranging from mild transient diarrhea in young or
stressed animals to more severe disease with dehydration lethargy depression weight lossemaciation
inappetence and death (Blankenship-Paris et al 1993 Powers 2009 Sledge et al 2011 Hoefer et al
2012 Patterson et al 2014) Rectal prolapse has also been reported in ferrets with enteric coccidiosis
(Hillyer 1992 Hoefer et al 2012) In one study co-infection with coccidia and Lawsonia intracellularis
(Desulfovibrio sp) was diagnosed in 4 of 19 ferrets with proliferative bowel disease (Li et al 1996)
These ferrets presented with variable clinical signs including diarrhea lethargy anorexia weight loss
dehydration and emaciation
In the two reports of biliary coccidiosis clinical signs conformed to those expected with
hepatobiliary disease Williams et al (1996) described their case to have presented with emaciation poor
appetite abdominal distension and icterus Kaye et al (2015) described a one week history of lethargy
inappetence and icterus with serum biochemistry results consistent with cholestasis later clinical signs in
this case included melena anemia and cachexia
146 Gross necropsy and histologic findings
The pathology of enteric coccidiosis in domestic ferrets was described by Hoare (1927 1935b)
Two healthy domestic ferrets were experimentally inoculated one each with large numbers of mature
oocysts of either E furonis or E ictidea that were isolated during his initial work The inoculated ferrets
were killed humanely for histologic examination of intestinal sections at the time of first detection of fecal
oocyst shedding no clinical signs of coccidiosis were detected in these ferrets prior to death Infection
with E furonis resulted in invasion of the epithelium of the small intestine and rectum Within the small
intestine the parasites were concentrated in the tips of the villi but could be found to the level of the
22
opening of the crypts of Lieberkuumlhn In rectal sections life stages were limited to the epithelial ridges
between the openings of the glands of Lieberkuumlhn Organisms were located within the apical portion of
the epithelial cells and intensely infected regions exhibited multiple parasites within a single host cell
Both asexual and sexual life stages were present within the same sections Hoare (1927) described similar
histopathologic changes in naturally infected ferrets but the proportion of asexual versus sexual life
stages differed In natural infections sexual life stages were more numerous whereas in experimental
infections asexual life stages predominated these findings would be expected to correlate with the stage
of infection at which ferrets died or were humanely killed for tissue collection and would not be
reflective of differences between natural and experimental infection with this parasite Hoare also
described the morphology of the different endogenous stages including trophozoite (3-4 microm) merozoite
(stumpy sausage shaped L 3-4 microm W 2 microm) macrogamete (spherical 8 microm diameter with darkly
staining globular inclusions of reserve material) and microgamete (described as similar to those of other
Eimeria species) Two types of merogony are described from histologic sections the first with stumpy
merozoites as described above and the second with merozoites with elongated curved bodies and a
compact polar nucleus measuring 60 times 13 microm This second merogonic generation was observed almost
exclusively in the naturally infected ferrets and was associated with initiation of sexual differentiation and
reproduction
The pathology of experimental and non-experimental infection with E ictidea in domestic ferrets
was also described by Hoare (1927 1935b) Parasitic invasion of the epithelium was noted only in the
small intestine with patchy distribution of the parasite life stages throughout affected sections Within the
small intestinal villi the parasites were again concentrated in the tips of the villi with infected epithelial
cells never containing more than one parasite As each intracellular parasite grew it filled the entire host
cell displacing the nucleus to the base of the cell Predominantly sexual life stages were detected in tissue
sections with few asexual generations observed Interestingly the parasites were arranged into age
groups with forms of the same life stage grouped together within the affected epithelial sections this is in
23
contrast to E furonis where life stages of different maturities were found together in affected sections
Hoare described the morphology of the different endogenous stages of E ictidea including merozoites
(free within the lumen elongated vermiform with one pointed end and a nucleus located at the rounded
end 11 microm times 1 microm within the epithelium shortened and rounded 3-4 microm diameter) macrogametes
(elongated 20 times 7 microm occupying the entire host cell with darkly staining globular inclusions of reserve
material) and mature microgamonts (morphologically similar to those of other Eimeria species but larger
than those of E furonis) Of note a tissue reaction was observed specifically in association with more
developed life stages of E ictidea (eg mature meronts mature gamonts unsporulated oocysts) which
was not observed when cells contained earlier stages of development (eg trophozoites immature
gamonts) This tissue reaction was described by Hoare (1935a 1935b) as the development of an annular
constriction of the apical portion of the villus separating infected epithelial cells from unaffected cells
The constriction involved the epithelium but could also extend inwards into the core of the villus These
changes were associated with congestion of capillaries and extravasation of red blood cells within the
constricted segment and in some sections villar tip necrosis
In their case report of one domestic ferret Blankenship-Paris et al (1993) described the gross
pathologic lesions associated with intestinal coccidiosis in this case there was diffuse dilation and
reddening of the small intestine which was empty and the colon contained dark watery material
Histologic lesions were confined to the ileum and jejunum The jejunum exhibited thickening of the villi
with a crypt to villus ratio of 15 mild granulomatous inflammation in the lamina propria and large
numbers of coccidial meronts gamonts and oocysts within the enterocytes of the villar tips
The gross lesions described by Sledge et al (2011) from 20 domestic ferrets are as follows thin
body condition with moderate to marked dehydration perineal staining with diarrhea moderate dilation
of the small and large intestines and the presence of pasty tan to tarry black digesta within the distal small
intestine and colon Other findings in one to a small number of ferrets included enlarged pale tan livers
splenomegaly with dark red colouration and multiple superficial gastric or duodenal ulcers The
24
histologic lesions from 10 ferrets included moderate blunting and occasional fusion of jejunal and ileal
villi focal attenuation and erosion of the epithelium of the villar tips with exudation of fibrin neutrophils
and blood into the intestinal lumen in regions with severe erosion Intact epithelial cells at the villus tips
and rarely sloughed epithelial cells in the intestinal lumen contained numerous intracytoplasmic coccidia
representing a range of asexual and sexual life stages (meronts macrogamonts microgamonts and
oocysts) The subjacent lamina propria of the small intestine and of the large intestine exhibited moderate
lymphoplasmacytic infiltration with occasional neutrophils and congestion of blood vessels Marked
mucosal hemorrhage was identified in the most severely affected sections
Marked gross and histopathologic hepatobiliary lesions were described in a single ferret by
Williams et al (1996) On gross necropsy the liver was pale and enlarged with dilated firm bile ducts
and thickening of the gall bladder wall Similar gross necropsy findings were described by Kaye et al
(2015) marked dilation and mural thickening of the entire biliary tree (including gall bladder intrahepatic
and extrahepatic bile ducts) On histopathology Williams et al (1996) noted that the marked thickening
of the gallbladder wall was a result of cystic proliferation of mucosal glands which were separated by
tracts of fibrous connective tissue and marked granulomatous inflammation Liver sections exhibited
marked biliary hyperplasia marked periductular fibrosis and moderate periportal lymphoplasmacytic
cuffing There was multifocal papillary proliferation of bile duct epithelium and dilation of the bile ducts
and within the ductular lumens there were moderate numbers of lymphocytes and plasma cells small
numbers of degenerate neutrophils sloughed epithelial cells and debris All endogenous coccidial life
stages were present within the gall bladder and biliary epithelium with meronts visible in 20 of the
intact epithelial cells of the biliary tree and gallbladder and oocysts free within the lumen of the
intrahepatic bile ducts Similar lesions were present in the case described by Kaye et al (2015) and as
well as in juvenile and adult farmed mink (Mustela vison) with hepatobiliary coccidiosis (Davis Chow amp
Gorham 1953)
25
15 INTRODUCTION TO ENTERIC COCCIDIOSIS IN THE BLACK-FOOTED FERRET
151 Natural history and conservation of the black-footed ferret in North America
Black-footed ferrets are one of only three wild ferret species worldwide the other species are the
European polecat (Mustela putorius) and the Siberian polecat or steppe polecat (Mustela eversmanii)
They are the only native North American ferret species and the most endangered North American
carnivore They are nocturnal carnivores whose diet and lifestyle are highly dependent on local prairie
dog (Cynomys sp) populations Prairie dogs comprise almost exclusively the diet for the BFF who also
use the complex burrow systems made by prairie dogs to escape their predators and raise their young
(Santymire et al 2014 USFWS BFF Recovery Program 2017)
While formerly distributed throughout the North America prairie ecosystem BFF were
considered extinct by the late 1950s In 1964 a single population was discovered in Mellette County
South Dakota Progressive decline of this population in subsequent years resulted in the decision by
United States Fish and Wildlife Service (USFWS) to initiate a captive breeding program for the species
From 1971-1973 four females and five males were captured for this purpose Despite successful breeding
no kits survived and the last adult ferret in this captive colony died in 1979 at that time BFF were again
presumed extinct in the wild based on annual surveys of the initial capture site In 1981 a dead BFF was
discovered by a ranch dog outside of Meeteetse Wyoming allowing wildlife biologists to identify
another colony of BFF This colony flourished until 1985 when an outbreak of canine distemper in the
BFF population and an outbreak of sylvatic plague in the local prairie dog population resulted in sharp
population declines From 1985 through 1987 all 24 of the remaining BFF were trapped and brought into
captivity to re-initiate the captive breeding program Six ferrets in this initial group died of canine
distemper while in captivity and of the remaining 18 survivors 7 bred successfully to create the founding
population of the current captive breeding population Today this captive breeding population consists of
approximately 300 BFF distributed among multiple institutions (Santymire et al 2014)
26
Since 1986 this multi-institutional effort has been breeding BFF in captivity with reintroductions
back into the wild in 28 selected locations in Canada the USA and Mexico Currently six facilities
participate in the BFF Species Survival Plan (SSP) the Toronto Zoo USFWS National Black-footed
Ferret Conservation Center National Zoorsquos Smithsonian Conservation Biology Institute Louisville
Zoological Garden Cheyenne Mountain Zoo and the Phoenix Zoo (Black-footed Ferret Recovery
Implementation Team 2011) As of 2011 over 8000 BFF kits had been produced in captive breeding
facilities (Black-footed Ferret Recovery Implementation Team 2011)
Multiple infectious diseases pose a significant risk to the captive breeding and post-release
survival of BFF including canine distemper and sylvatic plague Coccidiosis is recognized as a cause of
significant juvenile morbidity and mortality in captive breeding programs and can result in significant
population losses (Bronson et al 2007 Santymire et al 2014 USFWS BFF Recovery Program 2017)
152 Coccidia identified from black-footed ferrets
Eimeria ictidea and Eimeria furonis have been identified in black-footed ferrets based on
morphologic criteria (Jolley et al 1994) Jolley et al examined fecal samples from six captive BFF during
a distemper outbreak as well as samples from wild BFF They described one medium-sized ovoid
tetrasporic dizoic oocyst with a double wall presence of a polar body and lacking both an oocyst residual
body and micropyle The oocysts measured 232 microm (range 182-274) by 155microm (range 130-162) with
a SI of 150 The sporocysts were elongate with the presence of both sporocyst residuum and a Stieda
body Sporozoites contained prominent refractile bodies at the posterior end and were aligned anterior to
posterior within sporocysts These oocysts were shed by all six captive ferrets On histopathology of
intestinal sections merogony and gametogony were observed within the villar epithelium throughout the
small intestine but were concentrated in the jejunum Two morphologically distinct meronts were
detected in these sections one at the villar tips which was larger and lacking in undifferentiated mass
and the other at the base of the villi or rarely in the intestinal crypts Gametogony was predominantly
27
observed at the villar tips and was noted throughout the small intestine These organisms were considered
consistent with Eimeria ictidea based on descriptions by Hoare (1927) from domestic ferrets
A second small spherical to subspherical tetrasporic dizoic oocyst was documented that had a
pink double wall a granular residual body and lacked both oocyst polar body and micropyle This
smaller oocyst measured 126plusmn12 microm (108-152) by 119plusmn09 microm (101-129) with a SI of 106 The
sporocysts were elongate with the presence of a Stieda body and sporozoites contained refractile bodies
Similar to the larger Eimeria species described above merogony and gametogony were observed within
the villar epithelium throughout the small intestine with endogenous developmental stages most
numerous in the jejunum The meronts were small with 16 or fewer merozoites Micro- and
macrogamonts were observed clustered within the apical third of the villar epithelium as were meronts
Jolley et al (1994) determined these small spherical oocysts to be consistent with Eimeria furonis as
described by Hoare (1927) from domestic ferrets
Jolley et al (1994) described a third type of coccidial oocyst occasionally detected in small
numbers within the BFF fecal samples the authors did not state whether this third type of oocyst was
recovered from wild or captive BFF The oocysts measured 370plusmn13 microm (350-386) by 223plusmn23 microm
(212-232) with a SI of 106 Attempts to sporulate collected oocysts were largely unsuccessful and
corresponding endogenous stages were not identified on histopathologic examination of necropsied
ferrets precluding further morphologic identification of the parasite It should be noted that coccidial
oocysts with similar measurements had not been detected in wild or captive prey species available for
ingestion by BFF (Jolley et al 1994)
Previous to this report coccidial oocysts had been isolated from the feces of BFF in two captive
populations (Carpenter amp Hillman 1979 Williams et al 1988) The abstract by Carpenter amp Hillman
(1979) did not describe the oocysts whereas Williams et al (1988) stated that two Eimeria species (one
with larger oocysts and one with smaller oocysts) were identified within the fecal samples but they were
28
not identified further Interestingly Williams et al reported both Eimeria species to be shed in the feces
of all ferrets concurrently affected by distemper and by approximately 30 of the clinically healthy
ferrets at the time of investigation
Non-enteric coccidia have been reported from captive BFF in one facility by two authors (Jolley
et al 1994 Williams et al 1988) Both reports presumably describing the same case(s) noted the
presence of endogenous coccidial life stages in histologic sections of respiratory tissue and merozoites of
an unidentified coccidium in impression smears of the urinary bladder from BFF diagnosed with canine
distemper Meronts were observed within the epithelium of the trachea a large bronchus and associated
bronchial glands Jolley et al (1994) described the lesions as occurring in the same ferret whereas in the
earlier report by Williams et al (1988) they are described as occurring in two different ferrets There have
been no subsequent published reports of systemic coccidiosis in black-footed ferrets and no cases have
been identified within the pathology database of the Toronto Zoo captive BFF population or by the
current SSP pathologist (Dr Michael M Garner personal communication)
There is a significant information gap regarding the pre-patent periods and pathogenicity of both
identified Eimeria species in BFF and studies to further characterize the eimeriid coccidia of the BFF are
lacking
153 Morbidity mortality and clinical signs associated with enteric coccidiosis in black-footed ferrets
The clinical signs of enteric coccidiosis in black-footed ferrets include mucoid to hemorrhagic
diarrhea abdominal discomfort lethargy appetite loss vomiting and dehydration In some cases sudden
death precedes the development of diarrhea Both adult and juvenile BFF are affected by the disease
which causes significant morbidity and mortality in captive populations (Bronson et al 2007) One
retrospective study of the captive BFF population at the Smithsonian National Zoological Park
determined that the most common cause of death in juvenile BFF (aged 30 days ndash 11 months) was
gastrointestinal pathology (524 of juvenile deaths) with 636 of these cases caused by enteric
29
coccidiosis (Bronson et al 2007) Despite the significance of this disease to the captive population its
effect on morbidity and mortality in wild BFF populations is unknown To the authorrsquos knowledge no
routine surveys of fecal parasites have been conducted on wild-born or captive released BFF during
yearly spotlighting events at ferret release sites However samples may be collected opportunistically if
fecal material is identified within the traps used to catch wild BFF during yearly surveys at release sites
Where fecal samples have been analyzed a 13 prevalence of coccidiosis has been identified in wild
born BFF (Dr Rachel Santymire personal communication) Fecal samples have been collected from BFF
at four release sites within the USA Wind Cave National Park (South Dakota) Badlands (South Dakota)
Conata Basin (South Dakota) and Aubrey Valley (Arizona) and positive samples were identified only at
the first site (Dr Rachel Santymire personal communication) Although radio-telemetry has been used at
some release sites to determine sources of mortality and factors involved in survival its use is not
widespread Furthermore the nocturnal and fossorial lifestyle of the BFF is a significant impediment to
the surveillance and monitoring of disease in this species
16 TREATMENT PREVENTION AND CONTROL OF INFECTION BY EIMERIA SPP
161 Current recommendations for treatment of eimeriid coccidia in carnivores
Described anticoccidial therapies for carnivores come from research in domestic cats and dogs
infected by Cystoisospora species these tissue coccidia (family Sarcocystidae) are only distantly related
to the Eimeria species infecting the BFF and other ferrets Current therapeutic recommendations by the
Companion Animal Parasite Council (CAPC 2013) for treatment of described Cystoisospora species
isolated from cats and dogs include the following amprolium (300-400 mg daily for 5 days in dogs 110-
200 mg daily for 7-12 days in dogs 60-100 mgkg daily for 7 days in cats) amproliumsulfadimethoxine
(150 mgkg amprolium and 25 mgkg sulfadimethoxine daily for 14 days in dogs) diclazuril (25 mgkg
for one dose in cats) furazolidone (8-20 mgkg 1-2 times daily for 5 days in dogs and cats) ponazuril (20
mgkg daily for 1-3 days in dogs and cats) quinacrine (10 mgkg daily for 5 days in cats)
30
sulfadimethoxine (50-60 mgkg daily for 5-20 days in dogs and cats) sulfadimethoxineormetoprim (55
mgkg sulfadimethoxine and 11 mgkg ormetoprim daily for 7-23 days in dogs) sulfaguanidine (150 or
200 mgkg daily for 6 days or 100-200 mgkg every 8 hours for 5 days in dogs and cats) toltrazuril (10-
30 mgkg daily for 1-3 days in dogs) trimethoprimsulfonamide (30-60 mgkg trimethoprim daily for 6
days if gt4kg 15-30 mgkg trimethoprim daily for 6 days if lt4kg) (CAPC 2013) Notably the use of all
drugs listed by the CAPC is considered off-label with the exception of sulfadimethoxine
162 Current recommendations for anticoccidial treatment and prophylaxis in domestic and black-
footed ferrets
1621 Domestic ferrets
Recommended daily oral treatment regimens for enteric coccidiosis in domestic ferrets include
amprolium (19 mgkg once daily 05 mgkg) decoquinate (05 mgkg) sulfadimethoxine (300 mgkg in
drinking water) or sulfadiazine-trimethoprim (30 mgkg once daily) all administered for a minimum of
two weeks (Bell 1994 Patterson amp Fox 2007 Patterson et al 2014) Both the aforementioned
coccidiostats amprolium and decoquinate are sold in large formats and are ideal for use in larger
operations such as breeding facilities research facilities or rescue centers Other anticoccidial therapies
used in domestic ferrets include toltrazuril (20 mgkg) and ponazuril (30-50 mgkg) once daily It should
be noted that all anticoccidial therapy used in domestic ferrets is considered off-label drug use
Multiple follow up fecal examinations should be performed after the treatment regimen is
complete and large groups may need to be treated multiple times Routine cage cleaning is also important
to decrease the environmental oocyst burden and prevent re-infection and in the case of coccidial
outbreaks ferrets should be transferred to clean cages multiple times during the course of anticoccidial
therapy Disinfectants such as bleach or quaternary ammonium compounds or dry heat should be used
for effective environmental decontamination (Patterson et al 2014)
31
1622 Species Survival Plan recommendations for black-footed ferrets
Treatment and prophylaxis of enteric coccidiosis with oral sulfadimethoxine was previously
recommended by the BFF Species Survival Plan (SSP) However due to a suspicion of decreasing
efficacy of treatment ponazuril has been recommended recently for treatment Due to the perceived
exquisite sensitivity of BFF to enteric coccidia the current SSP recommendation for treatment is oral
ponazuril at 30 mgkg once if ferrets are to be transported anesthetized stressed or are otherwise
suffering from another illness or injury (even in the absence of clinical signs or fecal shedding) The same
single oral dose of 30 mgkg is also recommended for kits at weaning (30-35 days of age) post weaning
(40-45 days of age) and prior to anesthesia for initial examination and vaccines (50-60 days of age)
Large crowded or otherwise stressed litters should be administered 30 mgkg orally once every 7-10 days
during the period of stress For treatment of coccidial diarrhea diagnosed by fecal examination 30 mgkg
orally once every 7 days for two doses or 50 mgkg orally once daily for 3 days in food (repeated in 7
days) is recommended In BFF with clinical signs of dehydration administration of subcutaneous or
intravenous fluid therapy has been performed Additional therapy with other antibiotics is sometimes
provided in cases with severe clinical signs or where secondary or primary bacterial enteritis is suspected
There is no pharmacokinetic or pharmacodynamic information available for the use of
anticoccidial drugs in BFF or other Mustelidae and thus it is unknown whether the current dose or
frequency of administration is truly appropriate for treatment of coccidiosis In 2 to 3-month-old piglets
administered a single dose of ponazuril orally at 5 mgkg peak serum concentration occurred at 42 hours
(36-48 hr) and elimination half-life was ~56 days (Zou et al 2014) In llamas administered ponazuril as
a single dose of 20 mgkg orally peak serum concentration occurred at 84 hours and elimination half-life
was ~56 days (Prado et al 2011) In domestic cows administered ponazuril as a single 5 mgkg dose
orally peak serum concentration occurred at 48 hours and elimination half-life was 58 hours (Dirikolu et
al 2009) The relevance of serum drug concentrations for treating an intestinal infection that lacks
extraintestinal life stages is likely minimal because the highest drug dose will reach the site of concern
(intestines) and systemic distribution is not required
32
Furthermore no safety or efficacy studies have been performed in any ferret species to validate
the current uses of either sulfadimethoxine or ponazuril for treatment nor have the current recommended
treatment lengths been validated However anecdotal information based on current usage would indicate
that they are safe at the current dosages and frequencies of administration as no adverse effects have been
reported A recent efficacy study in shelter dogs and cats showed that oral ponazuril (50 mgkg)
administered once daily for 3 days was effective for treatment of infection with Cystoisospora as
determined by a reduction in or cessation of fecal oocyst shedding at 4 and 8 days post treatment
Treatment efficacy in this study was inversely correlated to fecal oocyst counts at the initiation of
treatment (Litster et al 2014) Interestingly efficacy of this dose compared to the other two treatment
groups (single 50 mgkg or 20 mgkg oral dose) did not seem to differ but no statistical analysis was
performed Given the ubiquitous use of ponazuril in captive breeding facilities and concerns regarding
resistance of coccidia species to sulfadimethoxine therapy information on minimum effective doses and
dose regimes would be necessary to inform appropriate future SSP treatment and management plans and
to minimize development of drug resistance
17 VACCINES AGAINST COCCIDIA
171 Theory
The development of resistance of protozoal parasites to chemotherapeutic agents has resulted in a
shift towards the development of vaccines for the protection of domestic livestock Immunity to enteric
coccidiosis in avian and mammalian species involves both humoral and cell mediated responses Eimeria
spp infection in sheep rats poultry and other species generally results in a protective immune response
against subsequent re-infections (Catchpole et al 1993 Shi et al 2000) Interestingly this is not the case
for some host parasite interactions for example a recent report indicated that primary infection with E
ninakohlyakimovae in goat kids did not provide protective immunity against subsequent challenge with
the same parasite (Ruiz et al 2013)
33
Vaccines can be divided into four general categories live vaccines inactivatedkilled vaccines
subunit vaccines and recombinant vaccines Live vaccines are orally administered using small numbers of
infectious oocysts or oocysts from strains with low pathogenicity and result in patent but ideally sub-
clinical infections in the host that will elicit a protective immune response Such live vaccines can be
produced using attenuated forms of the pathogen of interest for example in chickens using ldquoprecociousrdquo
strains of Eimeria spp These precocious strains undergo a reduced number of merogonic replications
within the host cells and thus fewer oocysts are shed in the feces of vaccinated animals This reduction in
endogenous merogonic cycles reduces the amount of damage to the intestinal epithelium as well as
reducing the number of oocysts contaminating the environment
Another strategy has been to use live parasites with truncated life cycles An example of this is
the Toxoplasma gondii vaccine developed to prevent abortion in sheep This parasite was passaged
multiple times through a mouse host resulting in an inability to produce tissue cysts (Meeusen et al
2007) This is desirable as the cyst stage of this parasite normally inhibited by the immune system can be
reactivated during periods of stress or immunocompromise The potential drawbacks of live vaccines
include 1) the ability to produce and isolate adequate numbers of coccidial oocysts to meet vaccine
production requirements 2) the potential development of clinical disease in the host as a result of
inoculation 3) the need for all susceptible individuals to receive the vaccine simultaneously to prevent
fecal-oral inoculation of unvaccinated animals with high doses of the infective agent likely to be present
in a shared environment through fecal shedding
Inactivated vaccines are produced when the microbe of interest is killed via application of heat
radiation or chemical treatment prior to inoculation into the host species While safer because they cannot
induce disease in the inoculated patient inactivated vaccines stimulate a reduced immune response
compared with live vaccines and are consequently less effective Subunit vaccines contain single or
multiple antigens of importance in initiating the host immune response rather than the entire pathogen of
concern Subunit vaccines cannot induce disease in the immunized host but are more difficult to produce
34
because they require a detailed understanding of host immune response to infection Recombinant
vaccines involve the genetic modification of a vector (virus or bacteria) one capable of infecting the host
of interest to contain DNA of the pathogen of interest These vectors induce an immune response in the
vaccinated host but as with subunit vaccines cannot induce disease However recombinant vaccines are
again difficult to produce because they require an in depth understanding of the life cycle stages genes
and antigens targeted by the host immune response to infection There are currently no recombinant
vaccines marketed in Canada for use in veterinary medicine against protozoal disease
Creation of effective vaccines against protozoal parasites is complicated by parasite antigenic
diversity during the different life cycle stages and among protozoal species and strains of the same species
(Meeusen et al 2007) Although most parasites induce some level of immunity in their host species the
immunological response to different parasite life stages and species has been poorly characterized for
most coccidia Furthermore many parasites have developed mechanisms to evade host immune responses
or to continue survive and replicate in and transmission by previously infected hosts Our limited
understanding of the immune responses against coccidial antigens has restricted commercial vaccine
production to live or attenuated vaccines (Meeusen et al 2007)
A notable disadvantage of anticoccidial vaccines is that they need to be developed for each
coccidial species of interest because of the species-specific nature of the immune responses this is a
considerable limitation compared with anticoccidial drugs that can have a much wider spectrum of action
(Vermeulen 2005) While the requirement for mass production of vaccine is a limiting factor for vaccines
developed for the agricultural industry this drawback would be less important for production of a vaccine
to be used in an endangered species
172 Species successes in anticoccidial vaccination
The first successful immunization against coccidiosis was reported in 1918 in dogs (Hall amp
Wigdor) In this report a dog that had previously recovered from coccidial infection with Diplospora
35
bigemina was fed three increasing doses of live non-attenuated coccidial culture (at 14 32 and 48 days
post recovery from primary infection) which resulted in no development of clinical signs and no oocyst
shedding for 11mdash18 days after each challenge Subsequently immunization of dogs and cats against
coccidia with protection lasting up to seven months was reported by Andrews (1926) Immunization of
albino rats to eimeriid infection after administration of three or more sublethal doses of Eimeria
nieschulzi via gastric intubation was reported by Morehouse (1938) further experiments showed that
sporozoites did not enter the host intestinal epithelium in immunized rats given a challenge dose
(Morehouse 1938) Similar findings were reported in chickens immunized against Eimeria tenella that
had 50 fewer intra-epithelial sporozoites following challenge compared to naiumlve birds (Augustine and
Danforth 1986) Conversely chickens previously inoculated with Eimeria acervulina exhibited more
intracellular sporozoites after challenge than naiumlve birds but sporozoites were not observed to develop in
previously immunized birds (Augustine and Danforth 1986) These findings provide further evidence that
the immune response to Eimeria spp may differ among host species
Vaccination against Eimeria species has been most successful and is most widely used in the
poultry industry particularly in breeder and layer flocks Almost all vaccines marketed for poultry are
live vaccines (attenuated and non-attenuated) Vaccination against other apicomplexan parasites in
domestic mammals has also been achieved but has been generally less effective for disease prevention
and is less widely available Marketed killed and inactivated (attenuated) vaccines include those
containing killed tachyzoites of Neospora caninum for cattle (Neoguard Merck Animal Health) and
chemically inactivated merozoites of Sarcocystis neurona for horses (EPM Vaccine Fort Dodge ndash no
longer in production) A subunit vaccine for Babesia canis in dogs uses cultured antigen (Pirodog
Merial) Available live vaccines include a vaccine against Toxoplasma gondii in sheep (Ovilis Toxovax
Intervet) that uses an attenuated temperature sensitive strain (S48)
36
18 RESEARCH GOALS AND OBJECTIVES
181 Objectives
a) To determine and characterize (morphologically and molecularly) the enteric coccidial species
currently affecting the black-footed ferret population
b) To describe the natural history of enteric coccidiosis in captive black-footed ferrets including
pre-patent period shedding frequency and burdens and morbidity and mortality rates
c) To compare molecular morphologic and life history characteristics of enteric coccidial species
identified in domestic ferrets to those in black-footed ferrets
d) To validate domestic ferrets as an experimental model for intestinal coccidiosis in the black-
footed ferret
182 Hypotheses
a) Multiple Eimeria species will be isolated from the black-footed ferret population
b) The Eimeria species identified from black-footed ferrets will be the same as those previously
described in domestic ferrets
c) A single pathogenic Eimeria species will be implicated in recorded outbreaks of clinical
coccidiosis during the period of study
d) Domestic ferrets can act as an experimental model of intestinal coccidiosis for black-footed
ferrets
183 Applications
The goal of this project is to better characterize the enteric coccidia of the endangered black-
footed ferret in order to set the stage for improved disease prevention and treatment To the authorrsquos
knowledge this project is the first attempt to isolate and perform molecular characterization of the
coccidial species endemic in the black-footed ferret population This information will be used to compare
these species to known coccidia from domestic ferrets and other related mammals As experimental work
37
cannot be carried out on enteric coccidiosis in the BFF due to its endangered status if the domestic ferret
can be validated as an experimental model studies of the patterns of anticoccidial resistance and
development of immunity against Eimeria spp can be undertaken in vivo The ultimate goal would be the
development of an autogenous vaccine used to improve survival of ferret kits and reduce morbidity and
mortality associated with coccidiosis in BFF captive breeding programs Based on clinical experience
stressful life events such a breeding weaning and transfer between institutions appear to increase the risk
of coccidial outbreaks in adult BFF As such vaccination could assist in reducing disease outbreaks in
BFF associated with various management activities There is no data on the significance of coccidiosis in
wild populations and limited means of disease surveillance following release vaccination during captive-
rearing or pre-release conditioning of BFF would be an ideal method of reducing the potential effects of
this disease in released and free-living BFF Increasing the numbers of ferrets being released to the wild
and releasing ferrets immune to the subsequent threat of coccidiosis would support the goals of the
conservation initiative for the black-footed ferret
38
CHAPTER 2 MOLECULAR CHARACTERIZATION OF ENTERIC COCCIDIA FROM
DOMESTIC FERRETS (MUSTELA PUTORIUS FURO)
This chapter has been submitted for publication as
Adriana R Pastor Dale A Smith and John R Barta (2017) Molecular Characterization of Enteric
Coccidia from Domestic Ferrets (Mustela putorius furo) Vet Parasitol Regional Studies and Reports (In
review)
ABSTRACT
Combined morphometric and molecular characterization of coccidia that infect domestic ferrets
(Mustela putorius furo) was completed to improve the diagnostic specificity of enteric lsquococcidiosisrsquo in
this host Coccidia positive fecal samples (n=11) and formalin fixed paraffin embedded intestinal tissues
(n=3) from domestic ferrets were collected from diagnostic laboratories in Canada and Europe An
average of 35 and 13 domestic ferret fecal samples per year were coccidia-positive when tested by
Canadian and European diagnostic laboratories respectively during the period 2008-2015 Oocyst
morphometrics and sequence genotyping at two loci (nuclear 18S rDNA [nu 18S rDNA] and
mitochondrial cytochrome c oxidase subunit I [mt COI]) were conducted on all samples The first nu 18S
rDNA and mt COI sequences for Isospora (=Cystoisospora) laidlawi and the first mt COI sequence for
Eimeria furonis were generated during this study Phylogenetic analysis of the mitochondrial COI
sequences demonstrated that E furonis was most closely related to E cf ictidea isolated from a black-
footed ferret (Mustela nigripes) and that I (=C) laidlawi was closely related to C canis and C felis The
identifications provided by diagnostic laboratories of the specific parasite species present in a sample
showed poor agreement with their identifications based on genotyping obtained in this study Molecular
techniques appear to be essential for accurate determination of coccidial species responsible for individual
and group outbreaks of coccidiosis and for further understanding of eimeriid host-parasite relationships
Key words coccidia Cystoisospora laidlawi domestic ferret Eimeria furonis Eimeria ictidea Mustela
putorius furo
39
21 INTRODUCTION
Coccidia are host-specific parasites of the phylum Apicomplexa with greater than 2000 species
named to date (Duszynski et al 2000 Upton 2000) The eimeriorinid coccidia (suborder Eimeriorina)
include typical intestinal coccidia such as Eimeria Isospora and Cyclospora species belonging to the
family Eimeriidae as well as tissue (cyst-forming) coccidia such as Cystoisospora Besnoitia Toxoplasma
and Sarcocystis species that belong to the family Sarcocystidae (see Cox 1994)
Enteric coccidia affect both domestic ferrets (Mustela putorius furo) and their wild counterparts
In his initial descriptions of enteric coccidiosis in domestic ferrets Hoare (1927 1935b) did not observe
clinical signs of intestinal disease associated with infection More recently it has been recognized that
enteric coccidiosis can result in clinical signs ranging from mild transient diarrhea to more severe disease
with dehydration lethargy depression weight lossemaciation inappetence and death (Blankenship-Paris
et al 1993 Powers 2009 Sledge et al 2011 Hoefer et al 2012 Patterson et al 2014) Rectal prolapse
has also been reported in ferrets with enteric coccidiosis (Hillyer 1992 Hoefer et al 2012) Disease
appears to be most common in young or stressed animals In one study co-infection with coccidia and
Lawsonia intracellularis (Desulfovibrio sp) was diagnosed in 4 of 19 ferrets with proliferative bowel
disease (Li et al 1996) these ferrets presented with clinical signs including diarrhea lethargy anorexia
weight loss dehydration and emaciation Two cases of biliary coccidiosis have also been reported in
domestic ferrets infection was associated with biliary epithelial hyperplasia cholecystitis and
cholangiohepatitis (Williams et al 1996 Kaye et al 2015)
Three species of coccidia affecting domestic ferrets were originally described and named by
Hoare (1927) Eimeria ictidea Eimeria furonis and Isospora (=Cystoisospora) laidlawi The three
species were detected in feces from 50 domestic ferrets involved in an outbreak of canine distemper at a
research facility For each parasite the author described the morphology of sporulated oocysts isolated
from feces as well as sporulation time (exogenous life stages) All subsequent reports of morphologic
diagnoses of these coccidia have been based on Hoarersquos original descriptions The pre-patent period
40
(minimum duration of endogenous development) in inoculated naiumlve ferrets was described for E furonis
and E ictidea as 6 and 7 days respectively (Hoare 1935b) The pre-patent period for I (=C) laidlawi
was not determined because the number of oocysts available was insufficient for an experimental
infection trial
Hoare (1927) described the sporulated oocysts of E furonis as follows spherical double outer
wall with a thin colourless outer layer and a thick yellowish inner layer no micropyle or residual body
and measuring on average 128 times 120 microm (length [L] 112-144 width [W] 104-128 shape index [SI]
107) Sporocysts were spindle-shaped with one end constrictedblunted contained a residual body and
on average measured 8-88 times 4 microm Sporozoites were vermiform with one end narrower than the other
arranged head to tail and each had a central nucleus a clear vacuole was identified in some sporozoites at
their broad posterior end
The sporulated oocysts of E ictidea were described as follows oval or elliptical with a double
outer wall with a thin colourless outer layer and a thick yellowish inner layer no micropyle or residual
body and measuring on average 236 times 175 microm (L 184-272 W 128-208 SI 135) Sporocysts were
irregularly oval with one end broad and the other more constricted contained a residual body and
measured 115 times 65 microm on average Sporozoites were vermiform with one end narrower than the other
arranged head to tail in the sporocysts and had a central nucleus and a clear vacuole at their broad
posterior end
The sporulated oocysts of I (=C) laidlawi were ovoid with a double outer wall with a thin
colourless outer layer and a thick yellowish inner layer had no micropyle or residual body and measured
on average 34 times 29 microm (L 320-368 W 272-304) A SI of 117 can be calculated from the original
mean dimensions Two sporocysts were identified each containing four sporozoites and no Stieda body
sporocysts were elliptical contained a residual body and measured 208 times 144 microm on average
Sporozoites were sausage shaped with one end slightly pointed and had a central nucleus and a clear
41
vacuole identified at the pointed end Sporozoites were arranged with pointed ends all at the same pole of
the sporocyst
Oocysts identified as Cystoisospora ohioensis have been reported from fecal samples collected
from healthy domestic ferret kits in a large American ferret breeding operation that also housed juvenile
domestic dogs (Patterson and Fox 2007) The method of identification of this parasite was not described
A second similar institution reported the presence of Cystoisospora (=Isospora) species also thought to
be C ohioensis in routine fecal examination of their colony (Dr Bambi Jasmin personal
communication) Identification in this case was performed by the Animal Health Diagnostic Center at
Cornell University and was based on morphometrics using light microscopy The significance of these
findings is unknown but no clinical signs or histologic lesions were described in the ferrets shedding
these oocysts The definitive hosts for C ohioensis are canids including the domestic dog
More recently molecular techniques have been used for the more precise identification of
coccidia Nucleotide sequences like morphological features diverge over time under selective pressure
however recent evolutionary divergence among coccidia is more likely to be reflected in molecular as
compared to morphologic differences Thus nucleotide sequences that are more similar are inferred to be
more closely related and to have diverged more recently (Cox 1994)
Molecular characterization of ferret coccidia has only been performed for one species Eimeria
furonis Abe et al (2008) extracted DNA from oocysts from the feces of a single domestic ferret with
clinical signs resulting from coccidial enteritis Using primers initially developed for molecular
identification of Cyclospora species (see Matsubayashi et al 2005) small subunit ribosomal DNA (nu
18S rDNA) primers CYC1FE (5ʹ-TAC CCA ATG AAA ACA GTT T-3prime) and CYC4RB (5prime-CGT CTT
CAA ACC CCC TAC TG-3prime) were used to amplify a 347 base pair fragment of nu 18S rDNA The
amplicon was sequenced (GenBank AB329724) and compared with previously published partial nu 18S
rDNA sequences from 40 Eimeria two Isospora and four Cyclospora species The resulting phylogram
42
grouped E furonis with E alabamensis (cattle) and E meleagrimitis (turkey) In the same study the
microscopic morphology of the oocysts was used to identify this coccidial species as Eimeria furonis by
comparison with Hoarersquos (1927 1935b) published descriptions of Eimeria furonis and Eimeria ictidea
Sledge et al (2011) also used nu 18S rDNA to identify Eimeria furonis as the cause of three
distinct outbreaks of enteric disease in domestic ferrets Initial identification was performed using
morphometrics of sporulated oocysts collected from feces in one of the three outbreaks being
investigated Formalin fixed paraffin embedded intestinal segments from ferrets from each of the
outbreaks contained multiple coccidial life stages when examined by light microscopy PCR amplification
of a 247 base pair (bp) amplicon of the nu 18S rDNA was generated from DNA isolated from stored
formalin-fixed tissues for further genetic analysis Analysis and sequencing of amplicons from all three
groups showed 100 identity to sequences previously reported by Abe et al (2008) for the gene encoding
E furonis nu 18S
In 2015 Kaye et al identified coccidia within hepatobiliary lesions in a domestic ferret receiving
immunosuppressive therapy for red cell aplasia DNA was extracted from frozen liver and a fragment of
the nu 18S rDNA was amplified using the primers previously described by Sledge et al(2011) The
authors reported that the DNA sequence of the amplicon had 100 identity to the published nu 18S
rDNA sequence of E furonis and 95 identity to the nu 18S rDNA of E myoxi (rodent) E alabamensis
(cattle) and Isospora robini (avian) (Kaye et al 2015)
It is difficult to estimate the current prevalence of enteric coccidia within the North American
domestic ferret population and no studies have been conducted to do so Fecal samples submitted to
veterinary diagnostic laboratories from domestic ferrets in Canada are uncommon and samples positive
for coccidia appear infrequently (Dr Donald Martin personal communication) Conversely in Europe
the prevalence of coccidia within the domestic ferret population appears to be higher based on
submissions to a large veterinary diagnostic laboratory in Germany Data from Idexx Vet Med Lab in
Ludwigsburg Germany was compiled to review the prevalence of coccidia and Giardia within fecal
samples from domestic ferrets (Pantchev et al 2011) The authors reported that of 284 fecal samples
43
submitted from 2002-2004 18 (63) had detectable coccidial oocysts on fecal flotation Based on
morphologic characteristics oocysts were identified as E ictidea E furonis I laidlawi (herein referred
to as I (=C) laidlawi as noted above) and another unidentified Isospora species Comparative data from
the same laboratory from 2009-2010 included sample submissions from 253 ferrets 21 (83) of which
were positive for coccidial oocysts on fecal flotation Nine of the samples were identified as containing E
furonis three contained both E furonis and I (=C) laidlawi eight contained only I (=C) laidlawi and
one sample contained both E furonis and E ictidea identification in all cases was again based on oocyst
morphometrics No statistically significant difference in the occurrence of coccidial oocysts was detected
when data from the two periods were compared (Fisherrsquos exact test P=041) (Pantchev et al 2011)
The purpose of the present study was to perform a more detailed molecular characterization of the
coccidial species isolated from domestic ferrets to estimate prevalence of the different coccidial species
within the Canadian domestic ferret population and to associate morphologic and molecular
characteristics of a greater range of enteric coccidial species in order to improve diagnostic accuracy
22 MATERIALS amp METHODS
221 Fecal samples
Multiple diagnostic laboratories within Ontario Canada1 and a major European diagnostic
laboratory 2 were solicited for fecal samples from domestic ferrets shedding coccidial oocysts Fecal
samples were diagnosed positive for coccidia based on fecal flotation and light microscopic identification
of Eimeria or Cystoisospora species Eleven samples were collected during the study period (from 2014-
2017) and preserved in potassium dichromate (25 wv) eight from Europe and three from Canada
Centrifugal flotation with saturated salt solution (Ryley et al 1976) was used to isolate oocysts from fecal
samples for genomic DNA extraction Genomic DNA extraction and purification were performed using a
1 Animal Health Laboratory Guelph ON Antech Diagnostics Canada Ltd Mississauga ON IDEXX Canada
Markham ON 2 Vet Med Labor GmbH Division of IDEXX Laboratories Ludwigsburg Germany
44
QIAamp DNA Mini Kit (Qiagen Hilden Germany) according to manufacturerrsquos instructions After
addition of DNAzol to the samples (Qiagen Hilden Germany) samples were vortexed using 05 mm
glass beads (Biospec Products Inc Bartlesville OK USA) prior to extraction in order to fracture the
oocyst walls and release the sporocysts Concentrations of the resultant DNA were estimating using a
Nanodrop 2000 spectrophotometer (NanoDrop Products Wilmington DE USA) and stored at 4 degC for
immediate use or minus20 degC for later use
For each laboratory the number of domestic ferret fecal sample submissions numbers diagnosed
positive for coccidial oocysts and number of each coccidial species identified in positive samples were
tabulated for each of the years 2008-2015
222 Formalin fixed intestinal tissues
Major diagnostic pathology services across Canada3 were contacted to identify cases of enteric
coccidiosis identified on necropsy of domestic ferrets Cases were considered positive based on the
presence of asexual or sexual life stages of the parasites in intestinal sections The histologic sections on
each positive case were reviewed re-described and organisms measured (AP DAS) Gross necropsy
reports for all cases were also reviewed to identify any clinical correlates associated with enteric
coccidiosis DNA was extracted from ten 5-6 microm scrolls of formalin fixed paraffin embedded tissue
(FFPE) using the QIAamp DNA FFPE Tissue Kit (Qiagen) as per manufacturer instructions
223 Molecular characterization
Regions from the nu 18S rDNA and mitochondrial cytochrome c oxidase subunit I (mt COI)
DNA were amplified by polymerase chain reaction (PCR) from each sample using the primers listed in
Table 21 PCR amplification was performed for all samples in a volume of 25 microl containing ~100 ng of
3 Animal Health Centre Abbotsford BC Animal Health Laboratory Guelph ON Faculteacute de meacutedecine veacuteteacuterinaire
Universiteacute de Montreacuteal Saint-Hyacinthe QC Histovet Surgical Pathology Guelph ON IDEXX Canada
Markham ON Prairie Diagnostic Services Inc Saskatoon SK
45
genomic DNA 1times PCR buffer 15 mM MgCl2 02 mM deoxyribonucleotide triphosphates (dNTPs) 400
nM of each primer and 1 U of Invitrogen Platinum Taq DNA Polymerase (Thermo Fisher Scientific
Toronto ON Canada) Reactions were performed on a Bio-Rad T100 PCR thermal cycler (Bio-Rad
Laboratories Singapore) Samples were denatured and Taq polymerase activated at 95 degC for 3 min then
subjected to 35 cycles of 94 degC for 30s anneal at 50-62 degC (see Table 21 for specific anneal conditions
for the various primer pairs) for 30s and extension at 72 degC for 30-75s (see Table 21) followed by a
final extension at 72 degC for 7 min Suitable DNA (ie genomic DNA from an Eimeria or Sarcocystis sp)
was included in the PCR reactions to act as a positive control for the reaction chemistry All amplification
products were subjected to electrophoretic separation using 15 submarine agarose gel stained with
ethidium bromide and visualized on an ultraviolet transilluminator (Spectronics Corporation New York
NY USA) DNA band size was determined by comparison with a 1 kb DNA ladder (GeneRuler 1kb Plus
DNA ladder Thermo Fisher Scientific Waltham MA USA) Bands were excised with a new sterile
scalpel blade and PCR products were purified from the gel using a QIAquick Gel Extraction Kit (Qiagen)
PCR products were cycle sequenced using an ABI Prism 7000 Sequence Detection System (Applied
Biosystems Inc Foster City CA USA) by the Molecular Biology Unit of the Laboratory Services
Division University of Guelph (Guelph ON Canada) using the amplification primers to obtain
sequences in both directions The resulting chromatograms were aligned and analyzed with Geneious Ver
818 or later (Biomatters Limited Auckland New Zealand) and high quality consensus sequences
generated The resulting consensus sequences were searched from within Geneious against publically
available sequences on the BLAST server (blastncbinlmnihgovBlastcgi) using the blastn search
algorithm against the nrnt database (GenBank+EMBL+DDBJ+RefSeq ndash AA or DNA) Resultant new
nucleotide sequences were submitted to GenBank
46
224 Phylogenetic analysis
To determine the phylogenetic affinities of the newly obtained sequences with sequences from
related apicomplexan taxa representative nu 18S rDNA and mt COI sequences were downloaded from
GenBank with special reference to sequences from parasites that infect members of the order Carnivora
Nuclear 18S and mt COI sequences were aligned independently using MAFTT v7017 (Katoh et
al 2002) executed from within Geneious and then concatenated into a combined nu18S rDNA mt COI
dataset Multiple sequences from a single parasite were used to generate consensus sequences for each
locus as described by Ogedengbe et al (2017) Aligned sequences were trimmed to the length of the
largest newly generated nu 18S sequence Phylogenetic trees were generated using Bayesian Inference
(BI) using MrBayes Ver 326 (Huelsenbeck and Ronquist 2001) executed from within Geneious the
combined nu 18S and mt COI alignment was partitioned to permit locus-appropriate substitution models
to be applied to each partition For the nu 18S sequence partition the general time reversible (GTR)
substitution model (nst=6) with gamma rate variation (ie a GTR+G+I model) was applied For the mt
COI sequence partition the codon (M1) substitution model (using translation table 4 [ie lsquometmtrsquo]) was
used instead of the GTR with the remaining parameters remaining the same
The resulting tree was rooted using a pair of adeleid coccidia (Hepatozoon spp) as the taxonomic
outgroup All BI analyses were run for a chain length of 1000000 with tree sampling every 1000
following a burn-in of 100000 with default settings of 4 heated chains and heated chain temp of 02
47
23 RESULTS
231 Fresh fecal samples
From 2008-2015 inclusive the Canadian diagnostic parasitology laboratory4 received an average
of 1206 (range 81-160) domestic ferret fecal samples yearly the European parasitology laboratory5
received a yearly average of 230 samples (range 213-270) The number of fecal samples diagnosed as
positive for coccidial oocysts per year on fecal flotation during this time averaged 35 (range 0-8) and
130 (range 6-20) for the Canadian and European laboratories respectively The diagnosing laboratories
used oocyst morphometrics to identify the species of coccidia present Almost all coccidia-positive
submissions to the Canadian laboratory were identified as containing an I (=C) species based on light
microscopy Coccidia in only three samples from the Canadian laboratory were identified as E furonis
one in each of 2010 2012 and 2014 E ictidea was not identified in any samples submitted to the
Canadian laboratory Approximately equal numbers of coccidia-positive samples from the European
laboratory were identified as E furonis and I (=C) laidlawi each year Only two samples from the
European laboratory contained oocysts that were identified as Eimeria ictidea using morphometrics one
from each of 2011 and 2013 Laboratory submissions to both laboratories are summarized in Table 22
Twelve fecal samples preserved in potassium dichromate were received for analysis by the
authors Eleven samples had previously been identified as containing a single coccidial species five
containing E furonis two containing E ictidea and four containing I (=C) laidlawi A final sample had
been identified as containing a mix of E furonis and Cystoisospora canis Results of microscopic and
molecular characterization of these samples are summarized in Table 23
4 IDEXX Canada Markham ON
5 Vet Med Labor GmbH Division of IDEXX Laboratories Ludwigsburg Germany
48
232 Formalin fixed samples
Only three cases of coccidiosis were identified in domestic ferrets within the databases of the five
diagnostic laboratories that participated in the retrospective study Histologic sections of intestine were
received from these three cases which originated in Ontario6 and Quebec7 The Quebec sample (P2010-I)
was collected in 2010 and the Ontario samples (93-40404 and 17-008571) in 1993 and 2017 respectively
On gross necropsy the small intestinal contents of case P2010-I were described as pasty mucoid
yellow-brown feces with some blood For case 93-40404 the small intestines were described as empty
but melena was present within the terminal portion of the large intestine Scant intestinal contents and
dark brown fecal material in the colon were described in case 17-008571
In all cases endogenous developmental stages of coccidia were visible in histological sections
(Figure 21 is exemplary of the findings from one case) Hematoxylin and eosin stained sections from
P2010-I contained two affected regions of small intestine The intestinal mucosa of the first region
contained numerous asexual life stages and moderate numbers of sexual life stages as well as a small
number of oocysts free within the lumen The second section contained tissues that were poorly
preserved nonetheless 0-4 oocysts per 400times field were identifiable within the intestinal lumen Two
regions of affected small intestine were identified from 93-40404 after screening of all submitted sections
both contained low numbers of sexual and asexual endogenous stages Within one region there were small
numbers of meronts within the intestinal mucosa and lamina propria The second region had small
numbers of oocysts within cells of the epithelium and lamina propria as well as free within the intestinal
lumen In case 17-008571 multiple sections of jejunum contained numerous coccidian meronts gamonts
6 Animal Health Laboratory Guelph ON
7 Faculteacute de meacutedecine veacuteteacuterinaire Universiteacute de Montreacuteal Saint-Hyacinthe QC
49
and oocysts within intestinal villi within the ileum scattered epithelial cells also contained these various
life stages
Average length and width of oocysts were measured from slide sections for all cases For P2010-
I oocyst average length and width were determined from seven oocysts to be 94 microm (range 85-105) and
75 microm (range 69-84) respectively with a SI of 125 (range112-140) Average length and width of
oocysts measured from 93-40404 were determined from 5 oocysts to be 2814 microm (range 229-341) and
233 microm (range 180-308) respectively with a SI of 123 (range 110-146) For the third case 17-
008571 only 2 oocysts were identified and average length and width of oocysts measured 982 microm (range
973-992) and 845 microm (range 821-870) respectively with a SI of 116 (range 114-118)
233 Molecular characterization
DNA was successfully extracted from all twelve fecal samples and two cases with formalin fixed
tissue samples Attempts at amplification of DNA extracted from sample 93-40404 using the primer pairs
listed in Table 21 were unsuccessful Molecular identification results and GenBank accession numbers
for the remaining samples are summarized in Table 23 Both the nu 18S rDNA and mt COI sequences
from I (=C) laidlawi were unique when compared with available sequences from other Cystoisospora
species within the public databases However sequences from I (=C) laidlawi were most similar to
sequences from C canis and C felis and somewhat more divergent from sequences from members of the
C ohioensis species complex Two apparent genotypes of E furonis were identified based on nu 18S and
mt COI sequencing results Genotype 1 represented by EU sample 9014 had 100 identity to previously
published sequences of the nu 18S locus from two isolates from Japan (GenBank AB239130 and
AB329724) Genotype 2 represented by EU sample 907 and Canadian sample 17-008571 had 994
identity at the nu 18S locus (3 single nucleotide differences [SNDs] over 561 base pair region [bp]) to the
three sequences above belonging to E furonis genotype 1 Pairwise alignment of mt COI sequences from
both genotypes identified only 2 SNDs (996 pairwise identity over 513 bp region) Partial mt COI
50
sequences of E furonis from both genotypes were only distantly related (941 pairwise identity 30
SNDs over 513 bp and 905 pairwise identity 49 SNDs over 513 bp respectively) to publicly available
sequences from Eimeria ictidea from the black-footed ferret (Mustela nigripes) (GenBank KT203399)
and Eimeria mephitidis from the striped skunk (Mephitis mephitis) (GenBank KT203398) the only other
Eimeria species infecting members of the Carnivora for which sequence was available
234 Phylogenetic analysis
A phylogenetic reconstruction based on concatenated partial nu 18S rDNA and mt COI sequences
of E furonis I (=C) laidlawi and related coccidia is illustrated in Figure 22 The combined 18SCOI-
based tree demonstrates that the two Eimeria species from ferrets form a well-supported monophyletic
group that branches among a collection of other eimeriid coccidia that infect mammals The sarcocystid
parasite of the domestic ferret I (=C) laidlawi was found to group as the sister taxon to C canis that
together formed a monophyletic group with the closely related C felis all three of these closely related
Cystoisopora species possess comparatively large egg-shaped oocysts that are similar morphologically
24 DISCUSSION
The present work has generated the first nu 18S rDNA and mt COI sequences for Cystoisospora
laidlawi and the first mt COI sequence for Eimeria furonis both isolated from the domestic ferret In this
study histologic presence of organisms and microscopic identification of oocysts shed in feces have been
correlated with published and novel nu 18S and mt COI sequences
Eimeria ictidea was not identified in any Canadian sample and this coccidium was reported in
only 2 of 1840 fecal samples submitted from across the European Union (EU) to IDEXX Germany during
2008-2015 suggesting that E ictidea is not a frequent cause of enteric coccidiosis in domestic ferrets in
Canada or the EU
51
During the study period (2008-2015) almost twice as many domestic ferret fecal submissions
were made to the European as compared to the Canadian diagnostic laboratory however the prevalence
of coccidia-positive samples was similar The methodology used in this report cannot be used to
determine the actual prevalence of enteric coccidial infection (coccidiasis) or disease (coccidiosis) within
the domestic ferret population Fecal samples may be submitted to laboratories either as a result of
investigation into enteric disease or as part of a routine health examination Thus without historic
information accompanying each sample one can simply identify the proportion of positive samples and
compare the frequency of the finding of different coccidial species Prospective surveys of fecal samples
from healthy and sick domestic ferrets with greater sample size would be necessary to determine the true
prevalence of these parasites within the population and to infer their clinical significance
Comparatively few mitochondrial COI sequences have been generated for apicomplexan parasites
compared with other genetic loci the majority of published sequences obtained from Apicomplexa are
from nu 18S The disadvantage of using nu 18S rDNA sequences for parasite identification is that they
are poor at distinguishing among closely related eimeriid coccidia due to the highly conserved nature of
the nuclear ribosomal RNA locus In contrast mt COI sequences appear to be more useful for
distinguishing closely related coccidian species (Ogedengbe et al 2011) but are less useful than nu 18S
rDNA sequences for inferring more ancient relationships among more distantly related coccidia
Consequently the combined use of nu 18S rDNA and mt COI sequencing has been recommended for
improved species description and phylogenetic analysis (El-Sherry et al 2013) For these reasons both nu
18S and mt COI sequences were analysed in the present study
Despite adequate quantities of DNA extracted from the Ontario laboratory sample (93-40404)
successful amplification did not result with any primer pair (Table 21) Potential reasons for this include
degradation of formalin-fixed DNA into fragments too small for amplification with the desired primers
perhaps as a result of extended length of time in formalin prior to paraffin embedding or length of time
stored as FFPE tissue (23 years) or insufficient parasite DNA within the paraffin scrolls The primer pairs
52
used appear to be useful for most eimeriid coccidia (Ogedengbe 2015) and successfully amplified both
Eimeria species from DNA isolated from oocysts so it is unlikely that failure to amplify DNA from this
sample resulted from an inability of the primers used to recognize the parasite seen on section
Two genotypes of E furonis were identified in this study Genotype 1 was identified only from
samples originating from domestic ferrets in Europe but exhibited 100 identity based on nu 18S
sequencing with previously published sequences from both Japan and the USA Genotype 2 was
identified from samples originating from domestic ferrets in both Canada and Europe The small number
of single nucleotide differences between the two genotypes at two genetic loci in different genomes are
consistent with intraspecific variation (ie strain variation)
As might have been expected because of their morphological and host similarities nu 18S and mt
COI sequences of E furonis were determined to be most similar to an Eimeria species (E ictidea)
previously isolated from black-footed ferrets (Mustela nigripes) these eimeriid coccidia formed a
monophyletic group that was distinct from other eimeriid coccidia infecting mammals in the phylogenetic
analyses based on combined nu 18S rDNA and mt COI sequences Similarly the nu 18S rDNA and mt
COI sequences of I (=C) laidlawi are most similar to sequences from two other Cystoisospora species of
carnivores (C canis and C felis) that both have large egg-shaped oocysts comparable to those of I (=C)
laidlawi Both morphometrics and genotyping support the close relationships among these three
sarcocystid coccidia of carnivores These molecular data confirm that transfer of Isospora laidlawi to the
genus Cystoisospora by Barta et al (2005) is warranted
The previous light microscopic identifications of coccidial species in 3 of the 11 fecal samples
were not in agreement with the molecular findings These results were not surprising because light
microscopy has been shown to be an insensitive tool for distinguishing among apicomplexan parasites at
both the genus and species level Furthermore re-evaluation of these samples by the authors revealed that
many of the samples that were identified incorrectly based on morphometrics contained primarily
53
unsporulated oocysts making accurate identification based on microscopic appearance highly
challenging These findings further underscore the importance of molecular methods in accurate parasite
identification In the absence of molecular tools accurate measurement of oocyst size shape and
determination of SI can be useful for differentiating among species of Eimeria and Cystoisospora
however this can only be performed accurately on sporulated oocysts from feces Interestingly the size
and shape indices of oocysts of E furonis measured in histologic sections did not match those previously
described by Hoare (1927) for the same oocysts in feces despite molecular confirmation of identity
Thus measurements of oocysts in histologic sections are not recommended for use in coccidial
identification
Our observations highlight the utility of molecular methods for identifying enteric coccidia
infecting domestic ferrets and suggest that diagnoses based on morphological methods should perhaps be
limited to broad determinations of disease etiology (ie lsquococcidiosisrsquo or lsquococcidiasisrsquo) Using molecular
techniques we were able to differentiate morphologically similar coccidial species isolated from the feces
of domestic ferrets and specifically identify parasites seen in histological sections of ferret intestine
Molecular techniques thus appear to be essential for determining the coccidial species responsible for
individual and group outbreaks of coccidiosis and for further understanding of eimeriid host-parasite
relationships
ACKNOWLEDGEMENTS
Many thanks to Julia Whale and Alex Leveille for their assistance and encouragement during the course
of this project The authors would like to recognize the contributions of Dr Donald Martin (IDEXX
Canada) and Drs Nikola Pantchev and Majda Globokar (IDEXX Germany) for the contributions of data
and samples to this project The authors would also like to recognize the Laboratoire de Pathologie
(Service de diagnostic Faculteacute de meacutedecine veacuteteacuterinaire St Hyacinthe Quebec) and the Animal Health
Laboratory (Guelph Canada) for contributions of samples and data to this project Finally this project
was made possible through funding by the Toronto Zoo Residency Research Fund to DASAP and partial
54
funding from a Natural Sciences and Engineering Research Council of Canada (NSERC) Discovery Grant
(400566) to JRB
55
Table 21 Amplification primers for nuclear 18S rDNA and mitochondrial COI loci anneal temperatures (Ta) extension times and expected PCR
product sizes used in the identification of enteric coccidia from domestic ferrets (Mustela putorious furo)
Gene Target Primer Pairs Primer Sequence (5ʹ-3ʹ) Size (bp) Ta (degC) Anneal (sec) Reference
nu 18S rDNA CYC1FE TACCCAATGAAAACAGTTT 560 52 45 Matsubayashi et al (2005)
CYC4RB CGTCTTCAAACCCCCTACTG Matsubayashi et al (2005)
Cocci_18S_595F CCGCGGTAATTCCAGCTCCAAT 216 62 30 Present study
Cocci_18S_847R GCTGMAGTATTCAGGGCGACAA Present study
Lank_18S_224F TCATAGTAACCGAACGGATC 1080 54 60 Ogedengbe (2015)
Api_SSU_2733R CGGAATTAACCAGACAAATC Mathew et al (2000)
mt COI COI_10F GGWDSWGGWRYWGGWTGGAC 500 52 30 Ogedengbe et al (2011)
COI_500R CATRTGRTGDGCCCAWAC Ogedengbe et al (2011)
COI 272F CAATTCTAYGATGCCGCWTT 222 52 30 Present study
COI_500R CATRTGRTGDGCCCAWAC Ogedengbe et al (2011)
Sdae-COI_260F GATCTTTATGTTYTTRATGCC 890 50 75 Ogedengbe (2015)
Sdae-COI_1147R CATTACCCATAACYACACC Ogedengbe (2015)
56
Table 22 Summary of fecal samples from domestic ferrets (Mustela putorius furo) submitted to two diagnostic laboratories from 2008-2015
No fecal samples positive for coccidia
No samples submitted
(percentage positive)
No samples positive for
Cystoisospora sp
No samples positive for
Eimeria furonis
No samples positive for
Eimeria ictidea
Year Canada Europe Canada Europe Canada Europe Canada Europe
2008 3140 (21) 6214 (28) 3 2 0 4 0 0
2009 2160 (12) 14214 (65) 2 9 0 5 0 0
2010 8127 (63) 20213 (94) 7 10 1 10 0 0
2011 0114 (0) 17215 (79) 0 9 0 7 0 1
2012 3108 (28) 10231 (43) 2 4 1 6 0 0
2013 281 (25) 16270 (59) 2 13 0 2 0 1
2014 6127 (47) 12234 (51) 5 6 1 6 0 0
2015 4108 (37) 9249 (36) 4 3 0 6 0 0
Total 28 (29) 104 (56) 25 56 3 46 0 2
Average
year 35 130 31 70 04 58 00 03
Legend Numbers in brackets refer to the percent of the total number of fecal samples submitted
57
Table 23 Morphologic and molecular identification of coccidia from domestic ferrets (Mustela putorius furo)
Sample ID Source
External Lab Morphologic
Diagnosis Morphologic Diagnosis (ARP) Molecular Diagnosis
mt COI GenBank
Accession
nu 18S rDNA
GenBank Accession
93-40404 FFPE enteric coccidia Histologic sample - - -
P2010-I FFPE enteric coccidia Histologic sample E furonis Identical to MF774036 Identical to MF774678
17-008571 FFPE NP Histologic sample E furonis Same as MF774036 Same as MF774678
17-008571 feces NP E furonis E furonis MF774036 MF774678
907 feces E furonis E furonis E furonis MF774035 MF774679
938 feces I (=C) laidlawi no oocysts visualized I (=C) laidlawi MF774037 MF774677
952-A feces E ictidea Cystoisospora sp I (=C) laidlawi Identical to MF774037 Identical to MF774677
9958 feces E furonis no oocysts visualized E furonis Identical to MF774034 Identical to MF774680
9011 feces E furonis E furonis E furonis Identical to MF774035 Identical to MF774679
9014 feces E furonis E furonis E furonis MF774034 MF774680
9017 feces I (=C) laidlawi Cystoisospora sp I (=C) laidlawi Same as MF774037 Same as MF774677
9040 feces I (=C) laidlawi no oocysts visualized I (=C) laidlawi Same as MF774037 Same as MF774677
912-260 feces I (=C) laidlawi Cystoisospora sp I (=C) laidlawi Same as MF774037 Same as MF774677
CAN-2016-1 feces C canis + E furonis Cystoisospora sp I (=C) laidlawi MF774038 MF774676
Legend FFPE = formalin fixed paraffin embedded intestinal sections - = unsuccessful = morphologic diagnosis performed by JRB same as = 100 sequence
identity with listed GenBank entry over entire sequence length identical to = 100 sequence identity but shorter sequence than listed GenBank entry
58
Figure 21 Life stages of Eimeria furonis within the small intestinal epithelium of a domestic ferret
(Mustela putorius furo) Asexual life stages merozoites (black circle) Sexual life stages oocyst
(solid black arrow) macrogamonts (open arrows with labels) microgamont (dotted black arrow)
Hematoxylin and eosin staining scale bar = 25μm
25 microm
59
Figure 22 Phylogenetic relationships of coccidia (Eimeria ictidea Eimeria furonis and Isospora
(=Cystoisospora) laidlawi) from domestic (Mustela putorius furo) or black-footed (Mustela
nigripes) ferrets based on partial nuclear 18S rDNA and mitochondrial COI sequences of these
parasites and related apicomplexan parasites A summary of the sources of the molecular data for
the remaining taxa included in this phylogenetic analysis are found in Supplementary Table 1 of
Ogedengbe et al (2017) Bayesian support is indicated for each node horizontal distance is
proportional to hypothesized evolutionary change (scale indicates sequence divergence of 10)
60
CHAPTER 3 MORPHOLOGICAL AND MOLECULAR CHARACTERIZATION OF
ENTERIC COCCIDIA ISOLATED FROM BLACK-FOOTED FERRETS (MUSTELA
NIGRIPES)
ABSTRACT
Black-footed ferrets (BFF Mustela nigripes) are the only ferret species native to North America
and have been identified as endangered since 1967 Starting in 1986 a multi-institutional effort has been
breeding this species in captivity with successful reintroductions back into the wild Coccidiosis is
recognized as a cause of significant juvenile morbidity and mortality in captive breeding programs and
can result in significant population losses Little is known about the etiology of enteric coccidiosis in
BFF Coccidia positive fecal samples (n=12) and formalin fixed paraffin embedded intestinal tissues
(n=11) were obtained from BFF in the Toronto Zoo and Louisville Zoo Species Survival Plan (SSP)
populations Oocyst morphometrics and sequence genotyping at three loci (nuclear 18S rDNA
mitochondrial cytochrome c oxidase subunit I and mitochondrial cytochrome c oxidase subunit III) were
conducted Results suggest that the same Eimeria species E ictidea was the cause of enteric coccidiosis
in both SSP populations in both juvenile and adult age classes Wider research is indicated to determine
whether these findings are representative of the larger captive and wild BFF populations
31 INTRODUCTION
Black-footed ferrets (BFF) are one of only three wild ferret species worldwide the other two
being the European polecat (Mustela putorius) and the Siberian polecat or steppe polecat (Mustela
eversmanii) The BFF the only native North American ferret species was formerly distributed throughout
the North America prairie ecosystem but were considered extinct by the late 1950s In 1964 a single
population was discovered in Mellette County South Dakota Progressive decline of this population in
subsequent years resulted in the decision by United States Fish and Wildlife Service to initiate a captive
breeding program for the species From 1971-1973 four females and five males were captured for this
61
purpose Despite successful breeding no kits survived and the last adult ferret in this captive colony died
in 1979 BFF were again presumed extinct in the wild based on annual surveys of the initial capture site
In 1981 a dead BFF was discovered by a ranch dog outside of Meeteetse Wyoming allowing wildlife
biologists to identify another colony of BFF This colony flourished until 1985 when an outbreak of
canine distemper in this wild BFF population and an outbreak of sylvatic plague in the local prairie dog
population resulted in sharp population declines From 1985 through 1987 all 24 of the remaining BFF
were trapped and brought into captivity to re-initiate the captive breeding program Six ferrets in this
initial group died of canine distemper while in captivity and seven of the remaining eighteen survivors
are the founding population of the current captive breeding population Today this captive breeding
population consists of approximately 300 BFF distributed among multiple institutions (Santymire et al
2014)
Since 1986 a multi-institutional effort has been breeding BFF in captivity with reintroductions
back into the wild in selected locations in Canada the USA and Mexico Currently six facilities
participate in the BFF Species Survival Plan (SSP) the Toronto Zoo United States Fish and Wildlife
Services National Black-footed Ferret Conservation Center National Zoorsquos Smithsonian Conservation
Biology Institute Louisville Zoo Cheyenne Mountain Zoo and the Phoenix Zoo (Black-footed Ferret
Recovery Implementation Team 2011) In order to provide the best genetic matches BFF are transferred
among the six institutions for breeding Approximately 300-400 kits are produced annually between the
six SSP facilities with ~200 of these kits allocated for release to the wild yearly (Santymire et al 2014)
As of 2011 over 8000 BFF kits had been produced in captive breeding facilities (Black-footed Ferret
Recovery Implementation Team 2011)
Multiple infectious diseases pose a significant risk to the captive breeding and post-release
survival of BFF including canine distemper and sylvatic plague (Santymire et al 2014 USFWS BFF
Recovery Program 2017) Coccidiosis is recognized as a cause of significant juvenile morbidity and
mortality in captive breeding programs and can result in significant population losses (Bronson et al
62
2007 Santymire et al 2014 USFWS BFF Recovery Program 2017) While the effects of the disease on
the wild population are not clear a prevalence of approximately 13 has been reported based on fecal
samples collected from wild BFF born at release sites (Dr R Santymire personal commication)
Coccidia are eukaryotic host-specific parasites of the phylum Apicomplexa affecting numerous
hosts within a wide taxonomic range Two species of coccidia Eimeria ictidea Hoare 1927 and Eimeria
furonis Hoare 1927 have been identified in black-footed ferrets based on morphometrics (Jolley et al
1994) Jolley et al examined fecal samples from six captive BFF during a distemper outbreak as well as
samples from wild BFF They described one medium-sized ovoid eimeriid oocyst with a double wall
presence of a polar body and lacking both an oocyst residual body and micropyle Oocysts of this Eimeria
species (sp) measured 232 times 155 microm (range 182-274 times 130-162) with a shape index (SI) of 150 The
sporocysts were elongate with the presence of both sporocyst residuum and a Stieda body Sporozoites
contained prominent refractile bodies at the posterior end and were aligned anterior to posterior within
sporocysts These oocysts shed by all six captive ferrets were considered consistent with Eimeria ictidea
based on descriptions by Hoare (1927) On histopathologic examination of intestinal sections parasites
undergoing merogony and gamogony were observed within the villar epithelium throughout the small
intestine but were concentrated in the jejunum (Hoare 1935b) parasite life stages were not described
from other tissuesorgans
A second small spherical to subspherical eimeriid oocyst was also documented in the captive
ferrets by Jolley et al (1994) this second species had a pink double oocyst wall a granular residual body
and lacked both oocyst polar body and micropyle This smaller species measured 126 times 119 microm (range
108-152 times 101-129) with a SI of 106 The sporocysts were elongate and possessed a Stieda body and
sporozoites contained refractile bodies Similar to the larger Eimeria sp described above merogonic and
gamogonic stages were observed within the villar epithelium throughout the small intestine but were
most numerous in the jejunum Jolley et al (1994) concluded these small spherical oocysts were
consistent with E furonis described by Hoare (1927) from domestic ferrets
63
Jolley et al (1994) described a third type of coccidial oocyst occasionally detected in small
numbers within BFF fecal samples however the authors did not state whether this third oocyst
morphotype was recovered from wild or captive animals The oocysts measured 370 times 223 microm (range
350-386 times 212-232) with a SI of 106 Attempts to sporulate collected oocysts were unsuccessful and
corresponding endogenous stages were not identified on histopathologic examination of necropsied
ferrets precluding further morphologic identification of the parasite It should be noted that coccidial
oocysts with similar measurements had not been detected in wild or captive prey species available for
ingestion by BFF making it unlikely that this coccidial species would have been a pseudoparasite (Jolley
et al 1994)
Previous to this report by Jolley coccidial oocysts had been isolated from the feces of BFF in two
captive populations (Carpenter amp Hillman 1979 Williams et al 1988) The abstract by Carpenter amp
Hillman (1979) did not describe the oocysts whereas Williams et al (1988) stated that two Eimeria sp
(one with larger oocysts and one with smaller oocysts) were observed within the fecal samples but they
were not identified further Interestingly Williams et al reported both Eimeria sp to be shed in the feces
of all ferrets concurrently affected by distemper and by approximately 30 of the clinically healthy
ferrets at the time of investigation
Non-enteric coccidia have also been reported by two authors from captive BFF at one facility
(Jolley et al 1994 Williams et al 1988) Both reports which presumably described the same case(s)
noted the presence of endogenous coccidial life stages in histologic sections of respiratory tissue and
merozoites of an unidentified coccidium in impression smears of the urinary bladder from BFF diagnosed
with canine distemper Meronts were observed within the epithelium of the trachea a large bronchus and
associated bronchial glands In the later report Jolley et al (1994) described the lesions as occurring in the
same ferret whereas in the earlier report by Williams et al (1988) they are described as occurring in two
different ferrets Paraffin blocks containing formalin fixed tissues from these cases have since been
discarded precluding further attempts at parasite identification with molecular methods Subsequent to
64
these reports further cases of systemic coccidiosis in BFF have neither been published nor identified
within the pathology database of the Toronto Zoo captive BFF population nor by the current SSP
pathologist (Dr Michael M Garner personal communication)
Previous characterization of coccidia from black-footed ferrets has been based on host species
affected tissues in the host and morphometric characterization of life stages in histologic sections and
oocyst characteristics using light microscopy It is known that morphologically similar Eimeria species
are not necessarily conspecific and may vary in host specificity and pathogenicity Molecular
characterization is thus required to accurately identify coccidia to the species level No molecular
characterization of coccidian parasites from black-footed ferrets has been performed to date
There is a significant information gap regarding which parasite species are implicated in
morbidity and mortality events associated with enteric coccidiosis in BFF and whether different coccidia
are associated with this disease in adult versus juvenile age classes or in different SSP institutions
Studies to further characterize the eimeriid coccidia of the BFF are warranted to improve the management
of this disease in the captive population The objectives of this research were to morphologically and
molecularly characterize coccidia associated with enteric disease in BFF at the Toronto Zoo and in other
SSP facilities
32 MATERIALS AND METHODS
321 Fecal samples
Twelve fecal samples were collected during the study period (from 2014-2016) and preserved in
potassium dichromate (25 wv aqueous) seven from the Toronto Zoo and five from the Louisville Zoo
Centrifugal flotation with saturated salt solution (Ryley et al 1976) was used to isolate and concentrate
oocysts from fecal samples for light microscopic examination and genomic DNA extraction
One to two drops of the supernatant from the centrifugal flotation were placed directly on a slide
and beneath a coverslip The morphology and dimensions of sporulated oocysts were documented using a
65
Provis AX70 photomicroscope (Olympus Canada Richmond Hill ON Canada) fitted with a digital
imaging device (Infinity3-1C Lumenera Corporation Ottawa ON Canada) controlled using iSolution
Lite image analysis software (Hoskin Scientific Burlington ON Canada) operated at a total
magnification of 1000times Morphologic features noted for each oocyst included oocyst wall morphology
number of sporocysts presence or absence of a micropyle micropyle cap residual body and polar
granules For sporocysts size number of sporozoites per sporocyst and presence or absence of Stieda
body and sporocyst residuum were noted Alignment of sporozoites within the sporocyst and
presenceabsence of refractile bodies within sporozoites were also described The sporulated oocyst
length and width measurements (in microm) were then used to calculate the SI for each measured oocyst
Morphologic and morphometric features were compared to previously published values for E furonis and
E ictidea from domestic and black-footed ferrets
Genomic DNA extraction and purification were performed using a QIAamp DNA Mini Kit
(Qiagen Hilden Germany) according to manufacturerrsquos instructions as described in Chapter 2 (Materials
amp Methods)
322 Formalin fixed intestinal tissues
The pathology records of the Toronto Zoo were searched from 1993-2016 for cases of BFF
diagnosed with enteric coccidiosis on histopathology For each case slides of histologic sections from all
submitted intestinal segments were reviewed to confirm the presence of sexual andor asexual life stages
within the intestinal epithelium Scrolls (5-6 microm) were cut from the paraffin blocks containing affected
intestinal sections and DNA extracted from the formalin fixed paraffin-embedded tissue (FFPE) using
the QIAamp DNA FFPE Tissue Kit (Qiagen Toronto Ontario) as per manufacturerrsquos instructions
66
323 Molecular characterization
Molecular characterization of coccidial isolates was performed on oocysts purified from fresh
fecal samples (isolated as described above) that were collected from juvenile and adult ferrets from
2014-2016 and DNA extracted from FFPE samples of BFF intestine containing parasite life stages
Regions from the nuclear 18S (SSU) rDNA (nu 18S rDNA) mitochondrial cytochrome c oxidase
subunit I (mt COI) DNA and mitochondrial cytochrome c oxidase subunit III (mt COIII) DNA were
amplified by polymerase chain reaction (PCR) from each sample using the primers listed in Table 31 and
methodology described in the Materials amp Methods section of Chapter 2 Table 31 also contains the
specific anneal conditions used for the various primer pairs Genomic DNA from an Eimeria species of
poultry was included in the PCR reactions to act as a positive control for the reaction chemistry A
representative selection of the newly generated nucleotide sequences resulting from the above were
submitted to GenBank
DNA obtained from oocysts collected from fecal samples during the first year of the study (2014)
was used to generate a complete mitochondrial genome (see Chapter 6 for details) using primer pairs and
sequencing primers summarized in Table 31 All subsequent samples collected in 2015 and 2016 had
shorter mt COI and mt COIII sequences obtained to permit genotyping of all collected oocysts at these
two loci The location of each primer in the nu 18S mt COI and mt COIII genetic locus is illustrated in
Figure 31
33 RESULTS
From 2014-2016 coccidia-positive fecal samples were obtained from twelve BFF ferretsferret
groups from the Toronto Zoo and Louisville Zoo SSP populations (see Table 32) Nine samples were from
single housed adults between the ages of 1-5 years (63 MaleFemale) Two samples were from mixed
groups one pooled fecal sample from four adults (FERA-1 13 MF) and one fecal sample from a family
67
group consisting of a dam and five kits (23 MF) One fecal sample was collected from a juvenile male
ferret at the time of necropsy
Eleven BFF with enteric coccidiosis were identified in the Toronto Zoo necropsy reports from
1998-2016 and all were confirmed by histological re-evaluation (Table 32) Both juvenile (n=9 36 MF)
and adult ferrets (n=2 20 MF) were represented
331 Morphometric characterization
Twelve coccidia-positive fecal samples were identified from adult and juvenile BFF from 2014-
2016 by on site laboratories at either the Toronto Zoo or the Louisville Zoo Fecal flotation and light
microscopic re-examination of the samples identified coccidial oocysts in 10 of these 12 samples
Morphometric characterization was performed on six samples in which there was adequate
quantity and quality of sporulated oocysts for examination These included three samples from single-
housed adults one from a juvenile at the time of necropsy one of pooled feces from a group of adult
ferrets and one of pooled feces from a family group (dam and kits) Two of the three samples from
single-housed adults were from the same ferret on different dates in 2016 the dates of collection were
separated by a period in which shedding of oocysts was not identified on routine repeated fecal
examinations Oocysts were elliptical with a colourless double wall and contained four sporocysts each
with two sporozoites Sporocysts were ovoid and both Stieda body and residual body were present
Sporozoites exhibited an anterior to posterior alignment within the sporocysts and refractile bodies were
identified (Figure 32) Results for length width and shape index of sporulated oocysts including range
and average values are summarized in Table 33 and Figure 32 The average measurements based on the
results of all 148 oocysts measured were length 2398 microm (1859-3057) width 1855 microm (1373-2383)
and shape index 130 (101-160)
The same measurements were performed on 59 sporocysts from a single ferret (Noodle) and
results are as follows average length 1280 microm (898-1480) average width 738 microm (505-1028) and
average SI 176 (124-247) In one sample (Mohawk-2) sporozoites were visible free on the slide
68
Measurement of three of these provided an average length of 1068 microm (1044-1117) and an average
width of 341 microm (316-393)
332 Molecular characterization
Molecular characterization was successfully performed on oocysts from seven of 10 fecal
samples containing coccidial oocysts and FFPE tissue from nine of the 11 necropsy cases (see Table 32)
Attempts at amplification of DNA extracted from necropsy samples Z228-98 and Z137-14 using the
primer pairs listed in Table 31 were unsuccessful Similarly attempts at PCR and sequencing of DNA
extracted from fecal oocysts from two Toronto Zoo BFF Jenna and Ruckus were unsuccessful
Molecular identification results for the remaining samples are summarized in Table 32
Only one Eimeria species E ictidea was identified in all enteric coccidiosis cases diagnosed at
necropsy in both juvenile and adult BFF at the Toronto Zoo from 1998-2014 This same species was
identified in all Toronto and Louisville Zoo BFF fecal samples that were sequenced successfully (n=8)
with the exception of a single case from Louisville This Louisville ferret was identified as having a
rodent pseudoparasite (Eimeria species) in the submitted fecal sample the eimeriid pseudoparasite had
986 sequence identity at the mt COI locus to the murine coccidium Eimeria falciformis All sequences
generated for E ictidea exhibited 100 sequence identity at the mt COI and COIII loci
Novel nu 18S rDNA mt COI and mt COIII sequences were generated for E ictidea from both
geographic locations and deposited in GenBank (Accessions MF860826 MF860827 MF860823
MF860825 MF860822 MF860824) Sequences were compared to those previously published for related
eimeriid coccidia The nu 18S rDNA sequence from Eimeria ictidea isolated from the Toronto Zoo BFF
had 9736 identity (14 single nucleotide differences) to the previously published sequences from
isolates of E furonis from domestic ferrets (Mustela putorius furo) in Japan (GenBank AB239130 and
AB329724) and newly generated sequences from Canadian and European isolates (GenBank MF774678-
MF774680 see Chapter 2 and Figure 33) In contrast nu 18S rDNA sequence of E furonis from
domestic ferrets (see Chapter 2) showed 9953 to 100 identity (0 to 3 SND) to the Japanese
69
sequences Comparison of newly generated partial sequences of the mt COI region from E ictidea from
BFF to isolates of E furonis (GenBank MF774034-MF774036) from DF and E mephitidis (GenBank
KT203398) from the striped skunk (Mephitis mephitis) the only carnivore Eimeria sp for which a mt
COI sequence was previously available reveals only 9415 and 9084 sequence identity respectively
with these other Eimeria spp of carnivores (Figure 34)
34 DISCUSSION
This work presents the first nu 18S rDNA mt COI and mt COIII sequences (nu 18S rDNA -
MF860826 MF860827 mt COI - MF860823 MF860825 mt COIII - MF860822 MF860824) generated
from an intestinal eimeriid parasite of the BFF referred to here as E ictidea collected from multiple BFF
of different ages from two separate captive populations (Toronto Zoo Toronto Ontario Canada and
Louisville Zoo Louisville Kentucky USA)
The morphometric description of coccidial oocysts from BFF in this work are consistent with
previous descriptions of E ictidea from mustelids including BFF the Steppe polecat the European
polecat and domestic ferrets (Hoare 1927 Svanbaev 1956 Jolley et al 1994) Thus I propose the name
E ictidea for the enteric coccidium described from BFF reflecting the similarity in morphology host
species and location of infection in intestinal tissues yet recognizing the absence of species identification
by molecular techniques Molecular characterization of parasites that agree with the description of E
ictidea morphologically from various mustelid host species would allow not only for determination of
whether the parasites are conspecific but would also provide insight into the potential for cross-
transmission among related mustelid hosts
DNA extraction from FFPE samples allowed successful PCR and sequencing of small DNA
fragments (220 bp) in nine of the eleven cases in which the technique was attempted Age of the samples
did not appear to be the major factor associated with successful extraction of good quality DNA the two
samples for which it was unsuccessful were the most recent (2014) and oldest (1998) casesConsequently
it may be possible to use banked FFPE tissues from historic necropsy cases from other SSP institutions
70
and necropsies of wild-born or re-introduced ferrets to determine the identity of the coccidial species
underlying disease in these cases and to better characterize the disease in the greater BFF captive and wild
populations Williams et al (1988) were contacted regarding their historic FFPE samples but formalin
blocks were no longer available for these cases and thus comparisons could not be made Banked FFPE
samples were requested from other SSP institutions however the Convention on International Trade in
Endangered Species of Wild Fauna and Flora (CITES) restrictions on the international transport of DNA
from endangered species did not allow for sample acquisition during the period in which this research was
conducted
Evaluation of FFPE samples from the Toronto Zoo indicate that the same Eimeria species has
been implicated in deaths associated with enteric coccidiosis from 1999-2014 as well as episodes of
clinical disease in ferrets in the Toronto Zoo population from 2014-2016 Samples from coccidia-positive
BFF at the Louisville Zoo in 2016 also contained the same Eimeria species Finding the same parasite at
multiple SSP locations was expected because BFF are transferred among institutions on a yearly basis for
breeding and potential release Consequently these parasites have repeated opportunities to move
between institutions in infected hosts or on contaminated cage materials to become established at a new
location Furthermore the stress of transport and transfer to a new environment may precipitate shedding
of endemic coccidia and increase the risk of a coccidial outbreak this concern is reflected in the SSP
recommendations for prophylactic treatment of all BFF with anti-coccidial medication prior to shipment
(USFWS BFF Recovery Program 2017)
A single BFF from the Louisville SSP population not showing clinical signs consistent with
coccidiosis was identified as having a rodent Eimeria species in the submitted fecal sample
Morphometric characterization of oocysts in this sample was not performed due to the paucity of visible
oocysts however examination at 100times suggested that the oocysts in the sample were ovoid in shape and
of comparable size to oocysts identified in other BFF samples The finding of a rodent Eimeria in a BFF
fecal sample is not unexpected as whole rodents comprise a significant part of the captive BFF diet The
oocysts shed by the BFF were most likely acquired through ingestion of an infected prey item and thus
71
most likely represent pseudoparasitism Molecular characterization was however required to
differentiate this from a case of true enteric coccidiasis
Reports from the first captive BFF population derived from South Dakota indicate the presence
of an unidentified species of enteric coccidium (Carpenter and Hillman 1979) in this group before its
demise in 1979 No reports containing morphometric descriptions of the coccidia from this group were
found on literature review and all parasites of this group have been lost with their hosts All subsequent
reports on enteric coccidiosis in BFF are from ferrets derived from the second founder group from
Wyoming in the 1980s The frequent transfer of ferrets among SSP institutions within the captive
breeding program and to different release sites within North America would be expected to result in the
same Eimeria species being found in all populations The exception to this would be the potential for
cross-transfer of other eimeriid parasites to wild BFF from sympatric mustelid species such as the long-
tailed weasel (Mustela frenata) Jolley et al described two other species of enteric coccidia from this
second captive population in 1994 the first was similar to E furonis of domestic ferrets and the second a
large coccidian parasite of unknown genus Neither of these parasites was identified in the Toronto and
Louisville Zoo populations during the course of this study In order to determine whether these parasites
persist within the present-day BFF populations and their impact on this species more detailed
examination of coccidia-positive fecal samples from captive and wild BFF populations is recommended
Furthermore the molecular identification of enteric coccidia from historic and future necropsy samples of
wild and captive BFF could aid in determining the presence of and contribution to mortality events by
these additional coccidia species
ACKNOWLEDGEMENTS
The authors would like to recognize the Wildlife Health Centre staff at the Toronto Zoo for their
assistance with the collection of fecal samples from the BFF from 2014-2016 The authors would also like
to recognize the Louisville Zoo for their contribution of samples to this project Finally this project was
made possible through funding by the Toronto Zoo Residency Research Fund to DASAP and partial
72
funding from a Natural Sciences and Engineering Research Council of Canada (NSERC) Discovery
Grant (400566) to JRB
73
Table 31 Amplification primers used to sequence the nuclear 18S rDNA mitochondrial COI and COIII loci of Eimeria ictidea originating from
fecal and formalin-fixed paraffin embedded tissue samples from black-footed ferrets (Mustela nigripes) including anneal temeratures (Ta)
extension times and expected PCR product sizes
Figure 31 Nuclear and mitochondrial genetic loci targeted by primers listed in Table 31 and used to characterize Eimeria ictidea originating
from black-footed ferrets (Mustela nigripes)
Gene Target Primer Pairs Primer Sequence (5ʹ-3ʹ) Size (bp) Ta (degC) Anneal (sec) Reference
18SrDNA Sarco_18S_123F TATCAGCTTTCGACGGTAGTGTATT 1480 60 30 Ogedengbe et al (2016)
ERIB10_REV CTTCCGCAGGTTCACCTACGG
mt COI T_Eim_COI_272F CAATTCTAYGATGCCGCWTT 220 52 30 Chapter 2 (Table 21)
COX1-500R CATRTGRTGDGCCCAWAC Ogedengbe et al (2011)
COI-400F GGDTCAGGTRTTGGTTGGAC 800 52 60 El-Sherry et al (2013)
COI-1202R CAAKRAYHGCACCAAGAGATA El-Sherry et al (2013)
mt COIII WG-MT_4140F AGAAAACCTAAAATCATCATGT 1000 52 60 Ogedengbe et al (2015)
Eimeriid_CO3_799R AAGTGAGTTCGCATGTTTAC Ogedengbe et al (2015)
74
Figure 32 A+B) Features and cellular contents of Eimeria ictidea from a black-footed ferret
(Mustela nigripes) Legend Oocyst dotted thin white arrow = polar granule Sporocyst dotted thin
black arrow = Stieda body thick white arrow= sporozoite refractile body thick black arrow=
residuum scale bars as indicated C) Shape index length and width measurements of sporulated
oocysts of Eimeria ictidea from black-footed ferrets (Mustela nigripes) Legend times indicates the
mean Dotted oval indicates one standard deviation around the mean
10 microm 5 microm
75
Figure 33 Comparison of nuclear 18S rDNA sequence alignment of Eimeria ictidea from two black-footed ferrets (Mustela nigripes) to
newly generated (see Chapter 2) and published sequences of Eimeria furonis from domestic ferrets (Mustela putorius furo)
Figure 34 Comparison of mitochondrial cytochrome c oxidase subunit I sequence alignment of Eimeria ictidea from two black-footed
ferrets (Mustela nigripes) to sequences from other eimeriid parasites of carnivores
Identity
Eimeria mephitidis ndash KT2033981
Eimeria ictidea ndash MF860823 (Guanella ndash LZ)
Eimeria ictidea ndash MF860825 (Mystery ndash MTZ)
Eimeria furonis ndash MF774034 (Type 1)
Eimeria furonis ndash MF774035 (Type 2)
Identity
Eimeria ictidea ndash MF860827 (Guanella ndash LZ) Eimeria ictidea ndash MF860826 (Mystery ndash MTZ)
Eimeria furonis ndash MF774680 (Type 1) Eimeria furonis ndash AB3297241 (Type 1) Eimeria furonis ndash AB2391302 (Type 1)
Eimeria furonis ndash MF774678 (Type 2) Eimeria furonis ndash MF774679 (Type 2)
76
Table 32 Morphologic and molecular characterization of coccidia from fecal and FFPE necropsy samples from black-footed ferrets (Mustela
nigripes)
Sample ID Sample Source Year Age (years) Sex Source Microscopic Description^ Molecular Diagnosis
Z228-98 Toronto Zoo 1998 7 M FFPE rare asexual stages -
Z143-99 Toronto Zoo 1999 1 M FFPE sexual and asexual stages E ictidea
Z106-02 Toronto Zoo 2002 008 F FFPE rare asexual stages E ictidea
Z108-03 Toronto Zoo 2003 008 F FFPE sexual and asexual stages E ictidea
Z109-03 Toronto Zoo 2003 008 F FFPE sexual and asexual stages E ictidea
Z124-12 Toronto Zoo 2012 021 F FFPE sexual and asexual stages E ictidea
Z113-13 Toronto Zoo 2013 017 F FFPE sexual and asexual stages E ictidea
Z117-13 Toronto Zoo 2013 025 M FFPE sexual and asexual stages E ictidea
Z118-13 Toronto Zoo 2013 025 M FFPE sexual and asexual stages E ictidea
Z119-13 Toronto Zoo 2013 025 F FFPE sexual and asexual stages E ictidea
Z137-14 Toronto Zoo 2014 021 M FFPE
fresh feces
sexual and asexual stages
POS E cf ictidea
-
E ictidea
FERA_1 Toronto Zoo 2014 gt 1 MF fresh feces POS E cf ictidea E ictidea
Noodle Toronto Zoo 2015 1 M fresh feces POS E cf ictidea E ictidea
Ruckus Toronto Zoo 2016 3 F fresh feces POS -
Mystery Toronto Zoo 2016 1 M fresh feces POS E ictidea
Mohawk Toronto Zoo 2016 1 M fresh feces POS E cf ictidea E ictidea
Jenna Toronto Zoo 2016 1 F fresh feces POS -
Thrope Louisville Zoo 2016 3 M fresh feces NEG -
FloJean Louisville Zoo 2016 2 F fresh feces NEG -
Rigatoni Louisville Zoo 2016 5 M fresh feces POS rodent Eimeria
Guanella +kits Louisville Zoo 2016 2 015 F kits 2M3F fresh feces POS E cf ictidea E ictidea
Clive Louisville Zoo 2016 1 M fresh feces POS E ictidea
Legend FFPE= formalin fixed paraffin embedded Sex MF = samples from family groups containing both sexes - = PCR and sequencing unsuccessful ^ = life stages identified on
histologic section
= mitochondrial COI andor COIII sequencing results Presence (POS) or absence (NEG) of oocysts and morphological identification of oocysts based on measurements when
77
Table 33 Morphometric (length width shape index) characterization of Eimeria ictidea oocysts from fecal samples from black-footed ferrets
(Mustela nigripes)
Sample ID FERA - 1 ^ Z137-14 Noodle Mohawk -1 Mohawk - 2 Guanella Total
Number of oocysts 12 36 32 10 21 37 148
Length (microm) 2333 (2055-2583) 2456 (2111-2848) 2505 (2079-3008) 2779 (2590-3060) 2493 (2036-2822) 2139 (1859-2372) 2398 (1859-3057)
Width (microm) 1676 (1373-2180) 1835 (1643-2232) 1975 (1509-2360) 2253 (2092-2383) 1803 (1549-2017) 1751 (1610-1888) 1855 (1373-2383)
Shape index 135 (103-160) 134 (113-156) 127 (105-155) 124 (113-138) 139 (114-154) 122 (101-145) 130 (101-160)
Legend ^= mixed adult group = dam and kit group
78
CHAPTER 4 NATURAL HISTORY OF ENTERIC COCCIDIOSIS IN THE BLACK-
FOOTED FERRET (MUSTELA NIGRIPES)
ABSTRACT
Black-footed ferrets (BFF Mustela nigripes) the only native North American ferret species are
endangered throughout their former geographic range An intensive captive breeding program produces
animals to supplement re-established wild populations Coccidial enteritis is a major cause of morbidity in
young captive ferrets but the disease also affects adults Limited information is available on the
pathogenesis of intestinal coccidiosis in captive BFF and characterization of the natural history of the
disease for improved prevention and management is imperative The objectives of this research were to
determine morbidity and mortality rates in the Toronto Zoo captive BFF population as well as
characterizing the natural history of the disease in this species through evaluation of shedding patterns
body tissues affected pre-patent period and periods of enhanced host susceptibility to infection
Coccidia-associated mortality in BFF at the Toronto Zoo from 1997-2016 averaged 053 yearly in
adults (range 0-526) and 195 in juveniles (range 0-1667) Clinical signs and histologic lesions in
Toronto Zoo BFF were similar to those described in previous publications A seasonal influence on
oocyst shedding was identified in adult BFF and ferrets appeared to maintain persistent infection with E
ictidea shedding coccidia in multiple years A larger multi-institutional study is required to better
elucidate the natural history of enteric coccidiosis in this species
41 INTRODUCTION
Black-footed ferrets (BFF Mustela nigripes) are the only native North American ferret species
and are endangered throughout their former geographic range When the last remaining truly wild
population underwent serious decline as a result of disease the decision was made by the by United States
Fish and Wildlife Service to capture the remaining 24 animals and establish a captive breeding program
this occurred between 1985 and 1987 Only seven of the captured ferrets bred successfully and are the
79
founders of the current North American BFF population (USFWS BFF Recovery Program 2017) The
captive population which now numbers approximately 300 individuals is distributed among and managed
by six collaborating facilities these include the Toronto Zoo United States Fish and Wildlife Services
National Black-Footed Ferret Conservation Center National Zoorsquos Smithsonian Conservation Biology
Institute Louisville Zoo Cheyenne Mountain Zoo and the Phoenix Zoo (Black-footed Ferret Recovery
Implementation Team 2011 Santymire et al 2014) Since 1991 BFF have been released into
reintroduced into the wild at multiple sites within their former range and over 8000 BFF kits had been
produced in captive breeding facilities as of 2011(Black-footed Ferret Recovery Implementation Team
2011) Twenty-eight BFF reintroduction sites currently exist throughout North America however there
continues to be a need to support wild populations as only a four of the re-established groups are truly
self-sustaining
Enteric coccidiosis is recognized as a cause of significant morbidity and mortality in captive
breeding programs affecting both juvenile and adult animals (Bronson et al 2007 USFWS BFF
Recovery Program 2017) Two Eimeria species Eimeria ictidea and Eimeria furonis have been
identified from cases of entric coccidiosis in BFF (Jolley et al 1994) Jolley et al examined fecal samples
from both wild and captive BFF and provided detailed morphologic descriptions of the oocysts of both
Eimeria spp as well as descriptions of the intestinal pathology associated with infection Asexual and
sexual life stages of both of the aforementioned Eimeria spp were identified on histologic section within
the villar epithelium throughout the small intestine but were concentrated in the jejunum Intestinal
sections from BFF infected with E ictidea exhibited two morphologically distinct meronts one at the
villar tips which was larger and lacking in undifferentiated mass and the other at the base of the villi or
rarely in the intestinal crypts gamogony was predominantly observed at the villar tips and was noted
throughout the small intestine
80
Extraintestinal coccidia have also been reported from captive BFF at one facility (Jolley et al
1994 Williams et al 1988) The authors identified the presence of endogenous coccidial life stages in
histologic sections of respiratory tissue and in impression smears of the urinary bladder from BFF
diagnosed with canine distemper No subsequent reports of systemic coccidiosis in BFF have been
published or identified within the pathology database of the Toronto Zoo captive BFF population or by
the current SSP pathologist (Dr Michael M Garner personal communication)
Recent investigations into the etiologic agents of enteric coccidiosis in BFF at the Toronto Zoo
have identified a single Eimeria species associated with all cases of enteric coccidiosis and associated
mortality in juvenile and adult BFF from 1999-2016 Furthermore this pathogen was identified in fecal
samples based on morphologic and molecular characterization from adult and juvenile BFF in another
zoological collection (Louisville Zoo Kentucky USA) (see Chapter 3) This coccidium is
morphologically consistent with Hoarersquos original description of E ictidea (1927) and is referred to
henceforth as Eimeria ictidea
There is a significant information gap regarding the pathogenicity of E ictidea in BFF The
objectives of this research were to determine morbidity and mortality rates in the Toronto Zoo and
additional captive BFF SSP populations as well as characterizing the natural history of the disease in this
species through evaluation of shedding patterns body tissues affected pre-patent period and periods of
enhanced host susceptibility to infection
42 MATERIALS AND METHODS
421 Toronto Zoo BFF breeding program
At the Toronto Zoo black-footed ferret breeding program all adult ferrets are housed
individually with the exception of dams and kits After the birth of the kits dams are housed with their
offspring from whelp date until removal at approximately 4-6 months of age Routine monthly fecal
81
examinations (direct examination and flotation) are performed in house for all ferrets in the breeding
program based on SSP recommendations to evaluate for the presence of coccidia
422 Fecal oocyst evaluation
Family groups
From 2014-2016 daily fecal examination for coccidial oocysts was initiated for all group-housed
dams and kits In 2014 fecal samples were collected daily from all dams and kits from weaning (30 days
after whelping) to 72 days post whelping Based on 2014 data in 2015 this surveillance was extended
from weaning (35 days post whelping) to 135 days of age Furthermore fecal samples were collected
from the dam for an additional 14 days after removal of kits In 2016 no fecal samples were collected
from dam and kit groups at the Toronto Zoo but samples were submitted from one group of dam and kits
from another SSP population at the Louisville Zoo (Kentucky USA)
Adults
From 2015-2016 daily fecal samples were also collected from all adult ferrets identified as
shedding coccidial oocysts on their monthly routine fecal examination and from clinically ill BFF
Samples were collected for 10-14 days after initial positive sample identification In 2016 fecal samples
were also submitted from four coccidia-positive adult ferrets from the Louisville Zoo population samples
were collected for 7 days post initial identification of shedding
Individual fecal samples were analyzed via flotation using the McMaster method followed by
routine flotation in saturated salt solution (Dryden et al 2005) to determine the presence or absence of
oocysts and oocyst burden (oocysts per gram of feces OPG) Temporal trends in oocyst shedding were
monitored Coccidia-positive ferrets were evaluated visually on a daily basis for presence of clinical signs
consistent with infection Infected juvenile ferrets and adult ferrets were treated with oral ponazuril or
toltrazuril regardless of the presence of clinical signs as per the black-footed ferret SSP
recommendations Based on these recommendations ponazuril is typically administered orally at 30-50
82
mgkg once daily for 3-7 days until clinical signs have resolved or oocyst shedding has been significantly
reduced (USFWS BFF Recovery Program 2017)
423 Retrospective review of pathology records
The pathology records of the Toronto Zoo were searched from 1993-2016 for cases of BFF
diagnosed with enteric coccidiosis on histopathology For each case gross necropsy reports were
reviewed and slides of histologic sections from all submitted intestinal segments re-examined to confirm
the presence of sexual andor asexual life stages within the intestinal epithelium and describe the
histologic lesions associated with presence of the parasite life stages
424 Prospective modified necropsy protocol
During the study period 2014-2016 necropsy protocols for all BFF were modified to improve
detection of coccidial life stages and better to determine which portions of the intestinal tract were
affected The entire length of the intestine from duodenum to anus was measured and intestinal contents
were flushed with 12 mL of sterile saline into a sterile container Intestinal contents were preserved in
25 potassium dichromate solution (mixed 11 with intestinal contents vv) for molecular diagnostics
Paired 2-cm long intestinal samples were collected from all sections of small and large bowel duodenum
(1) jejunum (6) ileum (1) and colon (2) The eight small intestinal samples were collected at equal
distances from the pyloric sphincter to the beginning of the colon and the distance from the pylorus noted
for each Colon samples were taken at 25 and 75 of the length of the colon One sample from each
pair was preserved in Serra solution (100 ethanol (60 vv) 37 formaldehyde (30 vv) glacial
acetic acid (10 vv) and the second sample was frozen Representative tissues from all internal organs
as well as additional intestinal samples skin muscle and brain were also collected and preserved in 10
buffered formalin Histopathologic examination was performed on all tissues collected
83
425 Retrospective medical history review
Medical histories of all BFF held by the Toronto Zoo since the initiation of the SSP program were
reviewed for data on frequency of occurrence of shedding of coccidial oocysts in adults and juveniles as
well as any association of shedding with clinical signs and administration of anticoccidial treatment Data
was tabulated yearly for adult and juvenile ferrets to determine annual morbidity and mortality rates
associated with enteric coccidiosis Medical records and pathology reports were solicited from the other
SSP institutions to determine comparative morbidity and mortality rates associated with enteric
coccidiosis in BFF at other facilities Both morbidity and mortality rates were calculated as
incidenceattack rates with yearly adult population size or number of family groups (dam and kits) as the
denominator for morbidity rates and number of yearly deaths in each age class as the denominator for
mortality rates
43 RESULTS
431 Fecal oocyst evaluation and retrospective medical history review
Family groups
Fecal samples were collected from seven groups of dams and kits housed together at the Toronto
and the Louisville Zoos from 2014-2016 All data from first to last day of collection for all family groups
is listed in Appendix 1 selected pertinent data for each group is presented in Table 41 Five groups of
dams and kits were sampled in 2014 and one group in each of 2015 and 2016 Shedding occurred no
earlier than 55 days of kit-age in any of the groups and was identified from 55-81 days of age (Table 41
Table 42 Figure 41)
In 2014 fecal oocyst shedding was identified in three of the five surveyed groups In two of the
three groups (dams Poppy and Bumblefoot) changes to fecal colour and consistency were identified
concurrently with periods of oocyst shedding both groups shed higher numbers of oocysts than the other
dam and kit groups in 2014 and 2015 Both Poppy and Bumblefoot had had litters in the previous one and
84
two years prior to this study respectively based on medical record review these dams and their litters
were also diagnosed as shedding coccidial oocysts that were too numerous to count on direct exam and
fecal flotation Clinical signs in the previous years included dark tarry hemorrhagic or soft mucoid feces
and reduced appetite both groups received treatment with toltrazuril (Baycox Coccidiocide Solution
25 Bayer Inc Mississauga Canada) and trimethoprim sulfamethoxazole (Novo-Trimel Teva Canada
Ltd Scarborough Canada) (TMS) One of four kits from Poppyrsquos 2013 litter (Z113-13) died of enteric
coccidiosis three days after the group was diagnosed as shedding coccidial oocysts and the initiation of
treatment with TMS
In 2015 low grade fecal oocyst shedding (lt14 oocysts per gram of feces) without associated
clinical signs was noted in the Fiddlesticks group on three days during a seven day period from 63-69
days of kit-age and again for a single day at 128 days of kit-age The dam had been diagnosed and treated
for enteric coccidiosis in 2014 at which time she exhibited clinical signs of loose green feces to
hemorrhagic diarrhea lethargy and dehydration In 2016 she was diagnosed as shedding low numbers of
coccidia exhibited no clinical signs and did not receive treatment prior to resolution of shedding
In 2016 Guanella and kits shed oocysts over a nine day period and daily fecal oocyst shedding
ranged from 206 ndash 371714 OPG Combined treatment with ponazuril (first four days of shedding)
sulfadimethoxine injectable (first two days of shedding additional product information not available)
amoxicillin oral (first two days of shedding additional product information not available) penicillin
injectable (first two days of shedding additional product information not available) and subcutaneous
fluids (first two days of shedding additional product information not available) was administered to this
group Previous medical history was not available for this female for review
In 2014 and 2015 fecal oocyst shedding in all groups in the Toronto Zoo population started in the
three week period from the last week of July to mid-August In 2016 shedding was first identified in the
Louisville Zoo group in mid-July
85
Adults
Seven single-housed adult BFF (52 MF) were detected to have shed coccidia during the study
period (Table 43) Shedding periods lasted from 2-10 days and oocyst per gram counts ranged from 104
ndash 554274 (Table 44) Clinical signs were identified in four of the seven ferrets and consisted of
lossreduction of appetite (n=2) weight loss (n=1) lethargy (n=1) blood in feces (n=1) loose or runny
feces (n=3) soft mucoid feces (n=1) green colour of feces (n=2) Five of the seven adults received
treatment after detection of oocyst shedding two of which received treatment in the absence of clinical
signs Treatment consisted of oral toltrazuril in four cases toltrazuril in combination with trimethoprim
sulfamethoxazole in one case (Mohawk-A) and ponazuril and sulfadimethoxine (manufacturerrsquos
information not available) in one case (Clive) (Table 43)
Three of the adults in this study Mohawk Mystery and Jenna shed oocysts during multiple
different periods in 2016 Mohawk shed oocysts in May July and September of 2016 data from the first
two periods are reported in Tables 43 and 44 Mystery shed oocysts in June and July of 2016 Clinical
signs were observed only during the first shedding period and included poor appetite and hemorrhagic to
soft mucoid feces Jenna shed oocysts in July September and November of 2016 and again in February
and May of 2017 Although clinical signs were not detected in association with the initial period of
shedding in July 2016 (see Table 44) depressed mentation and hemorrhagic mucoid feces were identified
in the subsequent two shedding periods In both Mohawk and Jenna oocysts were not detected in feces on
multiple recheck and routine monthly fecal examination between shedding periods
Ruckus the fourth ferret shed low numbers of oocysts for two days in 2016 while housed alone
and had been reported to have shed oocysts during a 30 day period in 2014 while housed in a family
group with her kits Diarrhea to soft mucoid feces and loss of appetite were reported in 2014 Clinical
signs in 2016 consisted of small amounts of runny feces for a two day period two days after oocyst
shedding was no longer detected Treatment consisted of toltrazuril and TMS in 2014 and toltrazuril
again in 2016 Similarly Noodle shed low numbers of oocysts in July 2015 and had been identified as
shedding low numbers of oocysts during September of the previous year
86
The final ferret Rigatoni shed oocysts in feces in low numbers (54-732 OPG) over 5 days All
positive samples were pooled for molecular diagnostics and sequencing results showed 990 identity (7
single nucleotide differences SNDs) at the mt COI locus with a pair of rodent Eimeria species (ie
GenBank HM771682 JQ993704) (see Chapter 3) This ferret did not show clinical signs associated with
shedding and was not treated Previous medical history was not available for Clive the single adult ferret
from the Louisville Zoo
Based on sample collection dates and medical record review for the adult BFF for 2014-2016
shedding occurred during spring summer and fall with three ferrets shedding in May (Jenna Ruckus and
Mohawk) one ferret shedding in June (Mystery) three ferrets shedding in July (Mohawk Jenna Noodle)
one ferret shedding in August (Clive) two ferrets shedding in September (Noodle Jenna) and one ferret
shedding in November (Jenna)
432 Pathology
Eleven BFF with enteric coccidiosis were identified in the Toronto Zoo necropsy reports from
1993-2016 (Table 45) Cases were identified from 1998-2014 inclusive and all were confirmed by
histological re-evaluation (Chapter 3 Table 32) Both juvenile (n=9 36 MF) and adult ferrets (n=2 20
MF) were represented
Gross Pathology
Gross necropsy findings were similar across the 11 cases and included mucoid to fluid luminal
contents (n=7 636) beige to white pasty coating of the mucosal surface of the small intestine (n=6
545) and colon (n=4 364) gaseous dilation of intestinal segments (n=3 272) segmental enteritis
and hemorrhage (n=1 91) In one case Z228-98 no gross lesions were identified within the intestines
Impression smears of luminal contents or scrapings of intestinal mucosa were performed in four cases and
coccidia were identified in all four
Histopathology
87
Both sexual and asexual life stages were identified within the small intestinal segments in all
cases except Z228-98 and Z106-02 in which only rare asexual parasite life stages were identified (Table
45 Figure 42) Other histologic lesions seen in intestinal segments containing coccidia included
lymphoplasmacytic inflammation of the lamina propria (n=4) neutrophilic infiltration of the lamina
propria (n=2) villar necrosis (n=2) villar atrophy or blunting (n=3) and thrombi within the villar tips
(n=1)
Additional necropsy diagnoses included cholangiolar hyperplasia multiple hepatobiliary cysts
with suppuration renal adenocarcinoma apocrine gland adenocarcinoma (Z228-98) concurrent
clostridial enteritis (Z143-99) presumptive Salmonella sp septicemia (Z106-02) interstitial pneumonitis
(Z108-03) myocardial mineralization interstitial pneumonia and nephritis periportal hepatitis and
bacteremia (Z124-12) and suppurative esophagitis (Z137-14)
From 2014-2016 three black-footed ferrets were necropsied using the detailed protocol described
above Only one of the three cases Z137-14 was diagnosed with enteric coccidiosis based on
histopathology Two duodenal five jejunal two ileal and two colonic sections were collected at measured
lengths from the pylorus Parasite life stages were identified within the mucosal epithelium of all
intestinal segments extending from the distal duodenum (10-12 cm aboral to pylorus) through to the distal
colon (157-159 cm aboral from pylorus) The distal duodenal section contained asexual life stages only
with a single focus of epithelial cells containing meronts Sexual life stages (microgamonts
macrogamonts unsporulated oocysts) were identified within villar epithelial cells in all remaining
sections of the small intestine with numerous oocysts in the bowel lumen Mild lymphoplasmacytic
inflammation of the lamina propria was associated with the jejunal and ileal lesions and blunting of the
villi was identified within one jejunal segment The colonic sections contained small to moderate numbers
of sexual life stages identified within both superficial and deep crypt epithelium with occasional life
stages identified near the germinal cells Large numbers of oocysts and bacteria were identified within the
88
lumen of these colonic sections and both sections contained abscesses within the crypts The proximal
duodenum (0-2 cm) was the only section of the intestines not containing parasitic life stages
433 Morbidity and mortality
Annual morbidity rates for enteric coccidiosis at the Cheyenne Mountain Zoo and mortality rates
from enteric coccidiosis in Toronto Zoo BFF are summarized in Tables 46 and 47 During 2003-2016
yearly incidence of coccidiosis in adult BFF at the Cheyenne Mountain Zoo averaged 69 (range 0-
421) For family groups consisting of juvenile ferrets housed with their dams yearly incidence of
enteric coccidiosis averaged 115 (range 0-600)
From 1997-2016 coccidia-associated mortality in adult BFF at the Toronto Zoo averaged 053
yearly (range 0-526) with an average total mortality rate of 141 per year (range 0-526) For
juvenile ferrets (under 1 year of age) during the same period coccidia-associated mortality accounted for
an average of 133 of deaths yearly (range 0-100) with an overall average mortality rate of 170 per
year (range 0-3404) from all causes
Multiple additional SSP institutions provided partial medical and pathology data sets for use in
this study which were not sufficiently detailed to permit computation of morbidity and mortality rates for
those populations
44 DISCUSSION
The work described here supports previous clinical findings regarding the impact through both
morbidity and mortality associated with enteric coccidiosis in BFF No previous studies have been
undertaken to determine morbidity and mortality rates associated with enteric coccidiosis across BFF SSP
institutions
89
In a retrospective mortality study of captive BFF from 1984-2004 at the Smithsonianrsquos National
Zoological Park Bronson et al (2007) reported that gastrointestinal disease was the most common cause
of death in juvenile BFF (524) with 333 of juvenile mortality cases in the study caused by enteric
coccidiosis While the data is not directly comparable the findings reported here also reflect enteric
coccidiosis as a common cause of death in juveniles with increased mortality associated with the disease
compared with adult counterparts All Toronto Zoo mortalities in both juvenile and adult age classes for
which necropsy tissues were available have been attributed to infection with a single coccidia species
Eimeria ictidea (see Chapter 3)
Multiple SSP institutions provided partial medical and pathology data sets for use in this study
which were not sufficiently detailed to permit computation of morbidity and mortality rates for those
populations In future it would be useful to determine whether morbidity and mortality rates associated
with enteric coccidiosis vary among SSP institutions as this may allow for improved identification of
host parasite and environmental factors that increase risk
Clinical signs reported here are consistent with those described from both BFF and domestic
ferrets with enteric coccidiosis (Sledge et al 2011 Santymire et al 2014 USFWS BFF Recovery
Program 2017) Changes to fecal colour and consistency were the most common abnormalities identified
at the time of first detection of oocyst shedding While clinical signs in the cases described here do not
always correlate directly with the quantity of oocysts shed individuals shedding higher number of oocysts
showed clinical signs more frequently than those shedding lower numbers
In this study oocyst shedding from single-housed adult BFF ranged from 104 ndash 554274 oocysts
per gram Daily fecal samples produced from individual adult ferrets range in size from approximately 1-
16 grams In light of these findings during peak shedding from ~5times105 to 9times106 oocysts can be shed into
the environment in one day providing a massive infective dose The large numbers of oocysts shed
90
combined with confinement in a small enclosure space and hardiness of Eimeria oocysts in the
environment would be expected to markedly increase risk of infection in captive BFF
Oocyst shedding from family groups ranged from 0 ndash 371714 OPG The wide variability seen in
OPG counts between days as seen in Figure 41 and Table 41 may be accounted for by the staggered
initiation and resolution of shedding by different ferrets It is most likely that the source of infection in
these family groups is shedding by the dams some of which were identified to shed in multiple years
although environmental contamination cannot be excluded
Shedding in the adult ferrets was clustered during particular time periods specifically May July
and September If shedding is associated with stress and immunosuppression activities such as breeding
whelping electro-ejaculation of male ferrets and shipmenttransfer could act as stressors Whelping
which occurs primarily in May and June could also act as a stressor to other ferrets in the facility either
through social cues or as there would be associated changes in husbandry protocols The September
cluster could be associated with the transfer of ferrets between institutions kits are pulled during this time
and adult ferrets are moved among institutions thus changing the population dynamics of each SSP site A
large cluster of shedding was recorded in July With the exception of weaning of kits no other major
stressors are expected to occur during this time thus the increase in shedding by adults in this group
cannot be easily explained Interestingly shedding was not identified in ferrets from March through April
which is the typical breeding season and when ferrets are introduced for breeding a presumably stressful
time Shedding was not noted from December through April which could reflect reduced environmental
burdens due to low humidity levels as would be expected in a Nordic climate during the winter (which
would kill oocysts and thus block transmission) or may be consistent with reduced stress during this
period The results from single-housed adults are in contrast to the dam and juvenile ferret groups in
which oocyst shedding appeared to be correlated to a period of 55-81 days of kit age These results are
consistent with reports from other facilities of increased incidence of shedding by kits after 70 days of age
(USFWS BFF Recovery Program 2017)
91
Retrospective and prospective review of histologic sections of intestines from affected BFF at the
Toronto Zoo showed the presence parasitic life stages in epithelial cells of both the small and large
intestines Neither Hoare (1935b) nor Jolley et al (1994) mentioned the presence of parasites in the large
intestine of experimentally infected domestic ferrets or naturally infected BFF respectively In the study
described here asexual and sexual life stages were identified within the epithelial cells of the small
intestinal villi from base to tip and were most numerous in jejunum This matches the description by
Jolley et al (1994) however Hoare (1935a b) found E ictidea to be present primarily in the villar tips
Jolley et al (1994) also described two morphologically distinct meronts of E ictidea within the small
intestinal sections one at the villar tip that was larger and lacking in undifferentiated mass and the other
at the base of the villi or in the intestinal crypts these findings were not echoed in this study as
merogonic stages were identified throughout the intestinal epithelium from villus to the base of the crypt
and no visual differences between meronts in any location were identified Hoare (1935a b) also
described a resulting annular constriction of the villus separating the affected and non-affected segments
this constriction was neither seen in the cases described here nor mentioned by Jolley et al (1994)
Whether these differences result from E ictidea from BFF and E ictidea from domestic ferrets being
different parasites or from differences in tissue tropism of a single parasite in two different hosts cannot
be ascertained from the available information
Histologic lesions such as necrosis hemorrhage villar atrophy and inflammation associated with
the presence of parasitic life stages were rare These changes are normally elicited by the host immune
system (inflammation) and the parasite (cellular rupture to release life stages resulting in hemorrhage and
necrosis) in response to infection In light of the fact that acute death occurred in a number of these ferrets
(Z113-13 Z117-13 Z118-13 Z119-13 Z137-14) in the absence of secondary disease processes and
with the intestinal epithelium intact but containing myriad parasitic life stages an alternative mechanism
for mortality associated with the infection must be proposed It is possible that these parasites elaborate
exotoxins during their life cycle and when at high density result in sudden death of the host with minimal
92
tissue changes The presence of parasitic life stages occupying the majority of both small and large
intestinal epithelial cells could also potentially impair fluid and protein movement in and out of the
mucosa however clinical signs associated with malabsorptive diarrhea were not identified in any of these
cases The presence of bacteria within the blood or other organ tissues was not identified in any cases and
consequently sepsis is unlikely to be the cause of death
Black-footed ferrets appear to maintain persistent infection with E ictidea Adult BFF in the
Toronto Zoo population shed coccidia in multiple years and in two adult ferrets multiple times in the
same year While the coccidia seen in all cases were not confirmed as E ictidea using molecular
techniques morphologic similarities and a lack of additional Eimeria spp identified on molecular work
undertaken suggest that only one species of parasite is and has been present in the collection Two dams
that had been identified as infected based on routine fecal screening in previous years presumably acted as
the source of infection to their litters of kits in multiple years While continued environmental
contamination cannot be ruled out these findings imply a failure of the immune response of the BFF to
clear infection with E ictidea or even to protect against sufficient replication of organisms to result in
clinical disease
Based on clinical experience and review of the literature BFF appear to be much more sensitive
to infection with E ictidea compared with their domestic counterparts In domestic ferrets subclinical
shedding of oocysts appears to be the most common with rare reports in the literature of overt disease
and that only in juveniles (Blankenship-Paris et al 1993 Abe et al 2008) However a single report exists
of three separate clinical outbreaks of Eimeria furonis infection in domestic ferrets under intensive
management with increased morbidity and mortality affecting all ages classes (Sledge et al 2011) The
role of genetics in the apparent increased susceptibility of BFF to enteric coccidiosis is unknown but the
current captive BFF population is derived from seven founders and inbreeding depression or familial
genetic susceptibility may play a role in their increased susceptibility to disease caused by E ictidea
93
Black-footed ferrets diagnosed with enteric coccidiosis during the course of the study were
treated with either ponazuril or toltrazuril sulfonamide drugs or often a combination of the two groups
of therapeutic agents Toltrazuril and ponazuril are triazine coccidiocides with proven efficacy against
both asexual and sexual life stages of mammalian and avian Eimeria spp (Mehlhorn and Aspock 2008)
The sulfonamides are antimicrobial drugs that exhibit coccidiostatic or coccidiocidal effects depending on
dose they act by blocking folate synthesis and have effects on first and second generation meronts
(asexual life stages) as well as potentially acting on sexual life stages (Mehlhorn and Aspock 2008)
Based on the limited data available from this study and the fact that treatment was initiated in almost all
adult BFF and family groups at the time of oocyst detection regardless of the presence of clinical disease
the effects of treatment on duration of clinical signs cannot be effectively evaluated It appears
subjectively that adult ferrets treated with toltrazuril and in one case a combination of toltrazuril and
TMS showed reduction in oocyst shedding after 3-5 days of oral anti-coccidial therapy (see Table 43)
The effects of treatment with either sulfonamides or triazines would be expected to reduce oocyst
shedding consequently the duration and amount of oocyst shedding reported in this study may not
accurately characterize the natural course of disease
Perceived resistance to sulfa drugs has been reported from multiple SSP facilities In light of this
and their potential negative effects on ferret reproduction (eg prevention of embryo implantation in the
uterus and impairment of sperm development) sulfonamides are no longer recommended by the SSP for
treatment of coccidia in this species (USFWS BFF Recovery Program 2017) The frequent and
widespread use of triazines in the management of enteric coccidiosis in BFF presents a risk for
development of resistance to this drug class in the future Neither pharmacokinetic (PK) nor
pharmacodynamic (PD) studies have been published to validate the dose and frequency of dosage in
either class of drugs in BFF and consequently it is unclear whether this perceived failure of some ferrets
to respond to treatment is based on true resistance versus inappropriate dosing The only work evaluating
ponazuril in BFF evaluated serum levels of ponazuril after a single oral dose of 50 mgkg and reported
94
therapeutic levels for 10 days after administration (USFWS BFF Recovery Program 2017) No
information was provided on number or age of ferrets that participated in the study or on how the
determination of what were therapeutic levels was made Furthermore as life cycles of the coccidia
affecting BFF are limited to the gastrointestinal tract and do not exhibit tissue stages the validity and
usefulness of assessing blood levels of ponazuril in determining appropriate dosage and dose schedules is
questionable Further work to determine the PK and PD of triazines in ferrets is warranted to provide safe
and efficacious treatment and to reduce the risk of development of resistance Furthermore the creation
and validation of a model for enteric coccidiosis in a related species would allow for in vivo studies of
drug resistance
95
Figure 41 Oocyst per gram counts and shedding period of Eimeria ictidea from black-footed ferret
(Mustela nigripes) family groups from 2014-2016
0
50000
100000
150000
200000
250000
300000
350000
400000
29
33
37
41
45
49
53
57
61
65
69
73
77
81
85
89
93
97
10
1
10
5
10
9
11
3
11
7
12
1
12
5
12
9
13
3
13
7
14
1
14
5
14
9
OP
G
Age of Kits
Poppy Bumblefoot Calico Aubrey Ruckus Fiddlesticks Guanella
96
Figure 42 Small intestinal epithelium of a black-footed ferret (Mustela nigripes) containing sexual
life stages of Eimeria ictidea Legend Solid black arrow = oocyst Hatched arrow = macrogamont
Outlined arrow = microgamont Hematoxylin and eosin staining scale bar = 25 microm
25 microm
97
Table 41 Shedding of oocysts of Eimeria ictidea in black-footed ferret (Mustela nigripes) dam and kit
family groups from 2014-2016
Collection Year 2014 2014 2014 2014 2014 2015 2016
Age of kits (days) Poppy Bumblefoot Calico Aubrey Ruckus Fiddlesticks Guanella^
29 - - - 0 - - -
30 - 0 - 0 - - -
- - - - -
34 0 0 - 0 0 - -
35 0 0 - 0 0 0 -
- -
46 0 0 - 0 0 0 -
47 0 0 - 0 0 0 -
48 0 lt14 - 0 0 0 -
- -
53 0 0 - 0 0 0 +
54 0 0 - 0 0 0 +
55 0 0 - 0 0 0 3717146
56 0 0 - 0 - 0 1084436
57 0 0 - 0 - 0 64133
58 0 324 - 0 0 0 20654
59 0 0 - 0 0 0 0
60 0 446688 - 0 0 0 0
61 0 934828 - 0 0 0 41111
62 0 530469 - 0 0 0
63 0 1617131 - 0 0 lt14
64 0 8396357 - 114 0 0
65 0 234876 - 5368 0 lt14
66 0 374625 - 9455 0 0
67 lt14 2311575 - 39579 0 -
68 0 603563 - 125051 0 0
69 - 4103036 - 27067 0 lt14
70 2470327 7759974 - 0 0 0
71 1073085 - 1784 0 0
72 3430966 - 0 0 0
73 4459536 - 7326 0 0
74 4842212 - 2 0 0
75 3627307 - 0 0 0
76 5801885 - 0 0 0
77 5994 0 0 0 0
78 589835 0 0 0
79 24815 0 0 0
80 144016 0 0 0
81 97862 0 0 0
82 0 0 0
127 0 0
128 0 2843
136 0 0
150 0
Numbers of Oocysts Shed (oocysts per gram of feces)
Dam Identity
Legend lt 14 = oocyst positive samples with less than 14 oocyst per gram of feces - = no sample recorded for this date underline = last
sampling date + = coccidia present but OPG count not performed = Toronto Zoo ferret ^ = Louisville Zoo ferret thick outer border =
days treatment was received = range of sequential dates between previous and subsequent number during which OPG counts were
performed and samples contained 0 oocysts
98
Table 42 Summary of epidemiologic data for family groups of black-footed ferrets (Mustela nigripes)
shedding oocysts of Eimeria ictidea
Dam Identity
Poppy
2014
Bumblefoot
2014
Aubrey
2014
Fiddlesticks
2015
Guanella
^
2016
Number of kits 7 5 3 4 5
Kit age (days) at time of
shedding 70-81 58-70 64-73 63-69 54-61
Shedding period (days) 15 13 11 7 9
OPG min 0 0 0 0 206
OPG max 48442 83963 2707 lt14 371714
Clinical signs YES YES NO NO NO
Treated - YES YES NO YES
Legend = Toronto Zoo ferret ^ = Louisville Zoo ferret OPG = oocysts per gram of feces
- = missing data
Table 43 Shedding of oocysts of Eimeria ictidea in single-housed adult black-footed ferrets (Mustela nigripes) 2015-2016
Numbers of Oocysts Shed (oocysts per gram of feces)
Collection
Year 2015 2016 2016 2016 2016 2016 2016
Ferret Identity Noodle Ruckus Mohawk-A Mohawk-B Mystery Jenna Clive^
Age (years) 1 3 1 1 1 1 1
1084058 0 + + + + +
+ 0 42307650 6286676 + 183150 554274
857808 16650 12805238 7777929 + 215710 377920
1604894 16650 309690 139860 + 0 25808
377042 0 599400 119880 + - 37294
554445 0 34688 385579 117920 0 5363
26640 0 16650 0 0 0 7500
0 10406 0 0 0 1090
0 20813 0 0 0
- 0 1761 0
0 0 0 0
- 0
0
0
Legend lt 1 = oocyst positive samples with less than 1 oocyst per gram of feces underline = last sampling date + = coccidia present but OPG
count not performed = Toronto Zoo ferret ^ = Louisville Zoo ferret thick outer border = days treatment was received
100
Table 44 Summary of epidemiologic data for single housed adult black-footed ferrets (Mustela nigripes) shedding oocysts of Eimeria ictidea
Noodle Ruckus Mohawk-A Mohawk-B Mystery Jenna Clive^
Sex M F M M M F M
Age (years) 1 3 1 1 1 1 1
Shedding period (days) 7 2 9 6 10 4 8
OPG min 266 166 104 1199 1761 1831 1090
OPG max 10840 166 423076 77779 - 2157 554274
Clinical signs YES YES NO NO YES NO YES
Treated NO YES YES YES YES YES YES
Legend = Toronto Zoo ferret ^ = Louisville Zoo ferret M = male F= female OPG = oocysts per gram of feces - = missing data
NOTE Mohawk-A and Mohawk-B refer to two separate episodes of oocyst shedding by the same ferret
101
Table 45 Histologic findings from necropsies of black-footed ferrets (Mustela nigripes) with enteric coccidiosis
Number of Sections Affected
Ferret
ID Year
Age
(years) Sex Coccidia in Intestinal Sections Small Intestinea Large Intestinea
Z228-98 1998 7 M rare asexual stages S - 01 A - 11 S - 01 A - 01
Z143-99 1999 1 M sexual and asexual stages S - 24 A - 24 S - 03 A - 03
Z106-02 2002 008 F rare sexual stages S - 14 A - 04 S - 01 A - 01
Z108-03 2003 008 F sexual and asexual stages S - 12 A - 12 S - 01 A - 01
Z109-03 2003 008 F sexual and asexual stages S - 23 A - 23 S - 11 A - 11
Z124-12 2012 021 F sexual and asexual stages S - 24 A - 04 none
Z113-13 2013 017 F sexual and asexual stages S - 77 A - 07 none
Z117-13 2013 025 M sexual and asexual stages S - 56 A - 56 S - 11 A - 01
Z118-13 2013 025 M sexual and asexual stages S - 56 A - 46 S - 11 A - 01
Z119-13 2013 025 F sexual and asexual stages S - 45 A - 35 none
Z137-14 2014 021 M sexual and asexual stages S - 1011 A - 911 S - 22 A - 02
Legend a= xn where x is number of sections containing sexual or asexual lifestages n is the number of sections examined S = sexual life
stages A= asexual life stages
102
Table 46 Yearly incidence of coccidial infection in black-footed ferrets (Mustela nigripes) at the
Cheyenne Mountain Zoo
Cheyenne Mountain Zoo
Year Adult Family
2003 116 (625) -
2004 819 (4211) -
2005 021 (000) 14 (2500)
2006 021 (000) 07 (000)
2007 023 (000) 08 (000)
2008 224 (833) 14 (2500)
2009 025 (000) 06 (000)
2010 326 (1154) 07 (000)
2011 125 (400) 09 (000)
2012 125 (400) 08 (000)
2013 028 (000) 05 (000)
2014 430 (1333) 09 (000)
2015 035 (000) 35 (6000)
2016 - 27 (2857)
Mean annual () 689 1155
Legend - = missing data xn= where x is the number of ferrets shedding coccidial oocysts and n is the
total number of adult ferrets or family groups in a given year () = incidence expressed as a percentage
103
Table 47 Yearly mortality rate and incidence of mortality associated with coccidial infection in
black-footed ferrets (Mustela nigripes) at the Toronto Zoo
Total Mortality
Coccidia Other Causes
Year Kit Adult Kit Adult
1997 015 (000) 023 (000) 315 (2000) 423 (1739) 1998 038 (000) 119 (526) 838 (2105) 919 (4734) 1999 047 (000) 119 (526) 1647 (3404) 119 (526) 2000 034 (000) 015 (000) 434 (1176) 315 (2000) 2001 032 (000) 016 (000) 532 (1563) 116 (625) 2002 150 (200) 020 (000) 450 (800) 220 (1000) 2003 227 (741) 018 (000) 327 (1111) 118 (555) 2004 020 (000) 016 (000) 620 (3000) 216 (1250) 2005 016 (000) 015 (000) 416 (2500) 215 (1333) 2006 030 (000) 016 (000) 230 (667) 016 (000) 2007 019 (000) 015 (000) 419 (2105) 215 (1333) 2008 034 (000) 016 (000) 1134 (3235) 316 (1875) 2009 017 (000) 016 (000) 017 (000) 116 (625) 2010 017 (000) 016 (000) 317 (1765) 316 (1875) 2011 011 (000) 016 (000) 111 (909) 216 (1250) 2012 111 (909) 017 (000) 111 (909) 317 (1765) 2013 424 (1667) 017 (000) 424 (1667) 317 (1765) 2014 126 (384) 017 (000) 326 (1154) 217 (1176) 2015 04 (000) 017 (000) 04 (000) 217 (1176) 2016 011 (000) 017 (000) 211 (1818) 117 (588)
Mean annual () 195 053 1594 1359
Legend xn= where x is the number of ferrets that died with coccidial infection or of other
causes and n is the total number of adult ferrets or kits in a given year () = incidence expressed
as a percentage
104
CHAPTER 5 EVALUATING THE DOMESTIC FERRET (MUSTELA PUTORIUS FURO) AS
AN EXPERIMENTAL MODEL FOR ENTERIC COCCIDIOSIS IN THE BLACK-FOOTED
FERRET (MUSTELA NIGRIPES)
ABSTRACT
The purpose of this study was to determine whether the domestic ferret (Mustela putorius furo) is
susceptible to an isolate of Eimeria ictidea originating from black-footed ferrets (BFF Mustela nigripes)
and thus could act as a suitable experimental model in which to investigate the pathogenesis and
management of this disease A pilot study was performed with 10 male intact juvenile domestic ferrets
Ferrets were administered an oral inoculum containing either a high dose (1 times 106 oocysts) moderate
dose (5 times 104 oocysts) or saline control and observed for shedding of oocysts and development of
clinical signs Seven of ten ferrets developed patent infection all of which had received the high dose
inoculum The prepatent period was 7-9 days and duration of shedding varied from 1-7 days Clinical
signs were identified in six of the seven infected ferrets and were consistent with those previously
described for enteric coccidiosis in domestic and BFF Parasite life stages were identified within the
intestines of four of the seven ferrets with patent infection and were limited to the distal jejunum and
ileum The demonstrated ability to produce patent infections in domestic ferrets following oral inoculation
of a high dose of E ictidea (1 times 106 oocysts) isolated from BFF provides an avenue for future
experimental investigations into the control and treatment of enteric coccidiosis in this endangered
species
51 INTRODUCTION
Black-footed ferrets (BFF Mustela nigripes) are one of only three ferret species worldwide
While formerly distributed throughout the North American prairies black-footed ferrets were declared
extinct in the wild in the 1980s Since 1986 a multi-institutional consortium has been breeding this
species in captivity with reintroductions back into the wild within their historic range in selected locations
in Canada the USA and Mexico Introduced colonies of BFF are present in Arizona Colorado Kansas
105
Montana New Mexico South Dakota Utah Wyoming and Chihuahua (Mexico) Reintroduction
attempts in Saskatchewan Canada have been unsuccessful to date
Multiple infectious diseases pose a significant risk to the captive breeding and post-release
survival of BFF including distemper and sylvatic plague (Santymire et al 2014 USFWS BFF Recovery
Program 2017) Coccidiosis is a recognized cause of juvenile and adult morbidity and mortality in captive
breeding programs and can result in significant losses (Bronson et al 2007 Santymire et al 2014
USFWS BFF Recovery Program 2017) The effect of the disease on wild populations is unknown
Clinical signs of coccidiosis include mucoid to hemorrhagic diarrhea abdominal discomfort lethargy
appetite loss vomiting and dehydration Recent investigations into diseases affecting BFF at the Toronto
Zoo have identified a single Eimeria species E ictidea associated with all cases of enteric coccidiosis in
juvenile and adult BFF from 2014-2016 (see Chapter 3) This same Eimeria species was identified
retrospectively as the cause of juvenile and adult mortalities in previous years (1999 through 2014
inclusive) (Chapters 3 and 4) Furthermore this pathogen was identified in fecal samples based on
morphologic and molecular characterization from adult and juvenile BFF in another zoological collection
(Louisville Zoo Kentucky USA) (see Chapter 3)
Enteric coccidiosis also occurs in domestic ferrets (Mustela putorius furo) with three
morphologically distinct species of coccidia Eimeria ictidea Eimeria furonis and Isospora (=
Cystoisospora) laidlawi Both of the aforementioned Eimeria species have been identified in black-footed
ferrets based on morphologic criteria but molecular characterization was needed to confirm whether the
same species of parasite infects both ferret species (see Chapter 2 and 3) To this end nuclear and
mitochondrial sequences for E furonis and for I=(C) laidlawi were generated (Chapter 2) expanding the
existing limited sequence data from the nuclear 18S rRNA locus of Eimeria furonis Molecular
characterization of E ictidea from domestic ferrets was not possible because samples containing this
parasite were not available for study consequently it is unclear whether the same coccidium affects both
domestic and black-footed ferrets
106
There is no published information describing the pre-patent periods and pathogenicity of enteric
coccidia in BFF and given the conservation status of the BFF experimental work cannot be undertaken
in the natural host The purpose of this study was to determine whether the domestic ferret is susceptible
to E ictidea isolated from BFF if susceptible the domestic ferret could act as a suitable experimental
model in which to investigate the pathogenesis prevention and treatment of coccidiosis caused by E
ictidea
52 MATERIALS AND METHODS
521 Animal care
Ten juvenile male intact ferrets of 48 (n=6) or 50 (n=4) days of age were obtained from a
commercial source (Marshall BioResources North Rose New York USA) and were housed in the
University of Guelph Central Animal Facility Isolation Facility Ferret weights on arrival ranged from
334-475 g (average= 3928 g) All ferrets were housed individually in wire bottom cages of 813 times 1117
times 457 cm size and were divided in equal numbers between two non-adjoining rooms They received ad
libitum access to Envigo Teklad Certified Global Ferret Diet (Madison Wisconsin USA) and water
changed daily Room temperature was maintained at 18-21 degC and a 16 hour light 8 hour dark
photoperiod was provided All personnel working with the ferrets were required to wear personal
protective equipment including disposable facemasks gloves gowns and bouffant caps Shoes were
provided for use in each room This study was carried out in accordance with the recommendations in the
Canadian Council on Animal Care guidelines The protocol was approved by the Animal Care Committee
of the University of Guelph (Animal Use Protocol 3289) and by both the Animal Welfare Committee
and Animal Care and Research Committee of the Toronto Zoo
An initial physical examination and blood collection were performed on each ferret by the
principal investigator (ARP) one day after arrival to assess health status prior to enrollment in the study
Ferrets were mask induced with isoflurane (Isoflurane USP Fresenius Kabi Richmond Hill Ontario) in
107
oxygen placed on a heat disc (SnuggleSafe Lenric C21 Ltd Littlehampton United Kingdom) weighed
examined and blood was collected from the jugular vein for routine CBC and biochemical profiles
All ferrets subsequently underwent an acclimation period of two weeks During this time fecal
samples were collected daily from each ferret and examined for the presence of coccidial oocysts using a
standard salt flotation technique (Dryden et al 2005) to ensure that all individuals were free of coccidia
prior to initiation of experimental work Any ferret positive for coccidia was to be removed from the
study
522 Oocyst preparation
Oocysts used for inoculation originated from fecal samples from two naturally infected BFF These
samples were stored in potassium dichromate for four weeks prior to oocyst purification and use in this
infection trial Stored fecal samples were mixed with distilled water and passed through a small sieve to
remove debris The strained contents were transferred to a 50 mL conical vial and topped up to 50 mL
with additional distilled water Samples were centrifuged (Sorvall ST40R Centrifuge Thermo Scientific)
at 2800 rpm (1315 timesG) for 10 minutes at 12 degC A drop of supernatant was evaluated microscopically at
100times for the presence of oocysts If oocysts were observed the supernatant was poured off into a second
50 mL conical tube and again topped up to 50 mL with distilled water and re-centrifuged under the same
conditions Otherwise the supernatant was discarded The pellets from both the first and second tubes
were combined with saturated salt solution at a 14 ratio by volume Oocysts were floated in the salt
solution by centrifugation at 1500 rpm (377 timesG) for 10 minutes at 12 degC The top 5 mL of supernatant
were collected and transferred to a clean 50 mL conical tube topped up to 40 mL with distilled water and
washed via centrifugation at 2800 rpm (1315 timesG) for 10 minutes After the wash step the supernatant
was again checked for presence of oocysts and discarded if no oocysts were observed The pellet was
collected and the presence of oocysts confirmed by examination of a drop placed on a clean glass slide at
100times Once verified the contents of the pellet of concentrated oocysts was placed in a 250 mL storage
container and mixed with approximately 200 mL of sterile saline (09 sodium chloride Hospira
108
Montreal Quebec) prior to storage for two to four weeks at 4 degC until inoculation Prior to inoculation a
McMaster count was performed to determine the number of oocysts per mL in order to determine
appropriate volume of inoculum
523 Experimental infections
Part 1
Five ferrets were randomly assigned to each of the control and infection groups After the
acclimation period on day 0 four ferrets in the infection group were inoculated orally with a high dose
oocyst suspension (1 times 106 oocysts in 025 mL of saline) mixed into 1 mL of FerreTone Skin amp Coat
Supplement (United Pet Group Inc Blacksburg Virginia USA) a fifth ferret was inoculated with a
moderate dose oocyst suspension (5 times 104 oocysts in 025 mL of saline) in the same volume of FerreTone
Ferrets in the control group were inoculated with a placebo (025 mL of saline) in 1 mL of FerreTone
Inoculation was performed by offering the oocyst suspension or placebo to the ferrets in a plastic
container
Fecal samples were collected daily from each inoculated ferret for 14 days post-inoculation
Samples were analyzed via fecal flotation using the McMaster method followed by routine flotation in
saturated salt solution (Dryden et al 2005) to determine the presence or absence of oocysts and oocyst
burden (oocysts per gram of feces [OPG])Temporal trends in oocyst shedding were monitored Ferrets
were evaluated visually twice daily for the presence of clinical signs of coccidial disease The first of
every two ferrets identified to shed oocysts was to be humanely killed at the time of peak oocyst shedding
(ie the first day that fecal oocyst counts remained static or declined) and necropsied to confirm the
presence of and describe parasitic replication in the intestinal mucosa Any remaining animals that shed
oocysts were to be monitored throughout the 14 day period following inoculation in order to determine
the duration and intensity of oocyst shedding for these individuals the total number oocysts shed during
patency was determined
109
Part 2
All ferrets from the infection group that did not shed oocysts during Part 1 (n=4) and all but one
ferret from the previous control group (n=4) were orally inoculated with the high dose oocyst suspension
(1 times 106 in 1 ml of saline) mixed with an equal volume of FerreTone Consequently between phases 1
and 2 all but one ferret were inoculated at least once with the BFF coccidia in order to increase
experimental animal numbers and determine if ferret age played a role in susceptibility to infection
One ferret from the previous control group was inoculated with a lower dose (2 times 105 oocysts in
075 mL of 09 saline mixed with 1 mL FerreTone) of oocysts that had been collected from the single
domestic ferret that shed in Part 1 oocysts were purified as described above for the initial inocula Fecal
collection and analysis were performed as previously As in Part 1 one in every two ferrets sequentially
identified to be shedding oocysts in feces was killed humanely at the time of peak shedding and a
complete necropsy examination performed The remainder of the ferrets observed to be shedding were
monitored for the full 14 days of the trial after which they were killed humanely and necropsied and total
number of oocysts shed during patency was determined All ferrets that did not shed coccidial oocysts
during the infection trial were rehomed at the end of the trial
524 Animal welfare
Ferrets were evaluated twice daily for development of clinical signs of coccidial disease and any
animal showing clinical disease was to be treated as determined by a veterinarian with supportive care
including fluid therapy A grading system for clinical signs including intervention points and removal
criteria was created for use during daily evaluation (see Appendices 3 and 4) Animals whose clinical
signs could not be ameliorated without the use of specific anticoccidial therapy were to be euthanized
Should the inoculation in Part 1 have resulted in clinical disease that required extensive treatment andor
necessitated euthanasia a lower number of oocysts would be used for subsequent inoculation in Part 2
Ferrets to be euthanized were anesthetized by mask induction with isoflurane in oxygen a 1 mL blood
110
sample was collected from the cranial vena cava and then an intracardiac dose of potassium chloride (2
mEqkg) was administered
525 Hematology
Blood was collected from all ferrets under isoflurane anesthesia at the time of pre-trial health
examination and again at the time of humane killing Blood was collected via jugular venipuncture
initially due to the small size of the ferrets at arrival and then by cranial vena cava venipuncture or
cardiocentesis prior to euthanasia Complete blood count and serum biochemistries were performed by the
Animal Health Laboratory of the University of Guelph Guelph Ontario
526 Morphologic and molecular characterization
Morphologic and molecular characterization of oocysts shed by the domestic ferrets during the
course of the infection trial was performed to ensure that the ferrets were shedding the same species of
Eimeria with which they were inoculated Oocysts were concentrated from positive fecal samples as
described above A drop of concentrated oocyst solution was viewed photographed and measured at
400times and 600times for comparison with previously determined morphometrics of Eimeria ictidea oocysts
(Chapter 3)
Regions from the mitochondrial cytochrome c oxidase subunit I and III (mt COI and mt COIII)
DNA were amplified by polymerase chain reaction (PCR) from each sample using primer pairs
400F1202R and -172F799R respectively For all PCR reactions samples were denatured at 95 degC for 5
min then subjected to 35 cycles of 94 degC for 30s anneal at 52degC for 30s and extension at 72 degC for 60s
followed by a final extension at 72 degC for 7 min PCR gel electrophoresis and sequencing methods used
were as described in the Materials amp Methods section of Chapter 2
The resulting consensus sequences were searched from within Geneious against previous
sequences for E ictidea produced by the authors and against publically available sequences on the
111
BLAST server (blastncbinlmnihgovBlastcgi) using the blastn search algorithm against the nrnt
database (GenBank+EMBL+DDBJ+RefSeq ndash AA or DNA)
527 Necropsy protocol
All humanely killed ferrets underwent a complete necropsy (Appendix 6) using the modified
protocol described in Materials amp Methods section 424 of Chapter 4
53 RESULTS
Initial physical examination was unremarkable with the exception of mild to moderate bilateral
ceruminous discharge within the external ear canal of all ferrets Complete blood count and serum
biochemistry results for all ferrets were within normal reference intervals for juvenile domestic ferrets
(Appendices 2a and b) (Fox 2014) Six days after arrival a single ferret (103) in the control group
developed mild upper respiratory signs consisting of sneezing and clear nasal and ocular discharge these
clinical signs were associated with mild dehydration and decreased food and water consumption The
ferret was treated with subcutaneous fluid therapy (10 mL Plasmalyte-A subcutaneous Baxter Alliston
Ontario) heat and supportive care and all clinical signs resolved within three days This ferret was
deemed healthy to participate in the remainder of the clinical trial A second ferret (105) in the control
group developed unilateral purulent ocular discharge 14 days after arrival one day prior to placebo
inoculation The ferret was treated topically twice daily for five days with Isathal ophthalmic gel (fusidic
acid 10 mgg Dechra Veterinary Products Inc Point-Claire Quebec) and the discharge resolved but
reoccurred within 2 days of treatment cessation Ocular examination showed mild conjunctivitis but no
evidence of corneal lesions and fluorescein staining did not indicate the presence of corneal ulceration
The ferret was treated for an additional six days with tobramycin ophthalmic solution (3 mgmL Sandoz
Tobramycin 03 Boucherville Quebec) after which clinical signs resolved completely No coccidial
oocysts were shed in feces from any of the ferrets during the two week acclimation period
112
531 Oocyst shedding
All ferrets readily ingested the inoculum with either placebo or concentrated oocysts In Part 1
one ferret (203) in the infection group which had received the high dose (1 times 106 oocysts) shed oocysts
on day 8 and day 9 after inoculation (Tables 51 52) This ferret was 71 days of age at the time shedding
was initially identified The ferret was killed humanely 11 days post inoculation later than had been
outlined in the protocol as processing of fecal samples had been delayed by two days resulting in late
detection of oocyst shedding in this individual Oocysts were not identified in the feces of the three
remaining ferrets that received the high inoculation dose the single ferret that received the lower dose (5
times 104 oocysts) or in the ferrets within the control group
In Part 2 six of eight ferrets inoculated with the high dose (1 times 106 oocysts) shed oocysts during
the 14 day observation period (Tables 51 52) Four of these ferrets were from the previous control
group One of the ferrets previously inoculated with the high dose inoculum in Phase 1 that had not shed
oocysts did shed oocysts after being inoculated a second time with the same dose during Phase 2 The
ferret that had previously received the low dose (5 times 104 oocysts) of oocysts in Phase 1 also shed after
inoculation with the high dose in Phase 2 Three ferrets did not shed oocysts after high dose inoculation in
Phase 2 one of these had been part of the previous control group the other two had received the high
dose inoculation previously in Phase 1
The pre-patent period ranged from 7-9 days (Table 51) with equivalent numbers of ferrets
commencing shedding on each of days 7 through 9 All six ferrets were between 91 and 93 days of age at
the time shedding was initially identified Oocyst per gram counts and shedding trends for all individuals
are shown in Table 51 Total oocyst shedding during patency was lt14 oocysts 8904 oocysts and
172291 oocysts for ferrets 201 104 and 105 respectively The two ferrets for which the prepatent period
was 9 days only shed oocysts for one day and in low numbers Oocysts were not identified in the feces of
the three remaining ferrets two of which received the high inoculation dose (1 times 106 oocysts) and the
third that received the lower inoculation dose (2 times 105 oocysts) (Table 52)
113
532 Morphologic and molecular characterization
In all seven ferrets that shed oocysts the morphologic features and measurements (length width
shape index) of the shed oocysts were consistent with those of the E ictidea administered in the inoculum
(Figure 51) Molecular confirmation of the identity of the oocysts shed was successful in 3 out of the 7
ferrets (102 103 203) samples from the four remaining ferrets did not contain adequate quantity or
quality of DNA for confirmation
533 Clinical signs
Clinical signs associated with patent infection were identified in 6 of 7 ferrets (Table 52) These
signs included weight loss (n=5) diarrhea (n=1) mucoid soft feces (n=2) feces containing blood (n=2)
and malodorous feces (n=1) Appetite reduction was noted in two ferrets from the infection group in Part
1 between 6-8 days post infection however no oocyst shedding was detected from either ferret during
this time
534 Hematology
CBC and serum biochemistry values from ferrets collected during pre-trial health screening are
displayed in Appendices 1a and 1b Values obtained for ferrets euthanized during or after the
experimental trial are displayed in Appendices 3a and 3b Minor variances from reference range values
for CBC and serum biochemistry were identified in six of the seven ferrets with patent infection In all six
ferrets for which a complete serum biochemistry was obtained immediately prior to death creatinine
kinase (CK) values were elevated (see Appendix 3b) Ferret 103 exhibited a mild hypoalbuminemia (20
ref 24-40 gL) on ante-mortem serum biochemistry (see Appendix 3b)
114
535 Necropsy
No gross or histopathologic lesions were present and coccidia could not be identified in sections
of intestine from the single ferret (203) humanely killed in Part 1 For the ferrets humanely killed in Part
2 no evidence of diarrhea hematochezia or mucoid fecal material was identified grossly Ferret 105
killed at the termination of the experiment but still shedding low numbers of oocysts in its feces
exhibited a 7 cm region of congested mucosa within the distal jejunum
Coccidial life stages were identified in small intestinal sections from four of the seven ferrets that
were identified to shed oocysts at some point prior to necropsy (Figure 52 Tables 52 and 53) Affected
sections included jejunum in all four animals as well as ileum in one and were collected from 114 to 218
cm aboral from the pylorus (see Figure 53) Coccidia were not identified in sections of duodenum
proximal jejunum or large intestine however oocysts were identified within fecal material in the lumen
of the large intestine from one ferret (103) Of the 11-19 sections of intestine examined for each ferret the
number of sections containing parasites ranged from one to eight 1 section in ferret 102 2 sections in
ferret 201 4 sections in ferret 105 8 sections in ferret 103 A mix of sexual and asexual life stages was
observed within the enterocytes in small intestinal sections from ferret 103 the remainder of the ferrets
showed either asexual (102 201) or sexual (105) life stages in affected segments
Pathologic changes and additional histologic findings in small intestinal sections of ferrets with
enteric coccidia included rare regions of blunting of the villi and sloughing of the epithelium associated
with hemorrhage and inflammation The primary lesions identified were subjectively increased numbers
of eosinophils lymphocytes and plasma cells within the lamina propria of the small intestine and
similarly increased neutrophils lymphocytes and plasma cells within the lamina propria of the large
intestine Neutrophils were rarely present in intestinal crypts and glands
Other gross necropsy findings included two ferrets with renal cortico-medullary cysts and one
ferret with mild thickening of the esophageal mucosa midway along the esophageal length No histologic
changes were identified within a sample of esophagus taken from this region
115
54 DISCUSSION
The findings of this study show that domestic ferrets are susceptible to infection with the enteric
coccidium Eimeria ictidea isolated from black-footed ferrets Both morphometric and molecular
diagnostic methods were used to confirm that ferrets were shedding oocysts of the same species with
which they were inoculated Molecular characterization was successful in three of the seven ferrets that
developed patent infections and as no other coccidial species was identified during pre-trial observation
morphometry was considered to be confirmatory in the remaining four animals
We have referred to the eimeriid coccidium affecting BFF and used in this experimental trial as
E ictidea based on morphometric similarity of their oocysts with those of E ictidea as described from
domestic ferrets (see Chapter 3) There is limited published information on infection of domestic ferrets
with E ictidea outside of Hoarersquos original descriptions (1927 1935a b) which form the basis for all
subsequent identifications of E ictidea in domestic ferrets and in BFF Attempts to obtain exemplars of E
ictidea from domestic ferrets to characterize using molecular techniques were unsuccessful (Chapter 2)
Multiple diagnostic laboratories in Canada and Europe were solicited for coccidia-positive fecal samples
from domestic ferrets but no samples of E ictidea were received over a 4 year period (2014-2017)
Eimeria ictidea was identified based on microscopic examination in only two samples submitted to a
European diagnostic laboratory from 2008-2015 It is unproven whether the E ictidea described from
domestic ferrets and the E ictidea identified from black-footed ferrets and used in this experimental
work are the same or are simply morphologically indistinguishable Eimeria species However the
consistency in morphology host genus and location of infection within the intestinal tissues combined
with the successful cross-transmission of this parasite to domestic ferrets described in the present study
suggests they are likely conspecific
116
The pre-patent period (minimum duration of endogenous development) for infection with E
ictidea in the inoculated domestic ferrets ranged from 7-9 days (see Table 51) the pre-patent period for
this parasite in the BFF the natural host for this coccidium is unknown Hoare experimentally infected
naiumlve domestic ferrets with E ictidea derived from naturally occurring infection in this species (Hoare
1935b) The inoculated ferrets shed oocysts after a pre-patent period of 7 days consistent with the 7-9
days seen in the work described here with E ictidea
Shedding of oocysts was identified over a period of 1-7 days (see Table 51) and intensity ranged
from less than 14 up to 15624 OPG These results may be skewed with erroneously low duration of
shedding and number of oocysts shed as three of the seven ferrets were humanely killed at or prior to the
expected peak of oocyst shedding for tissue collection and histologic examination in order to increase the
probability of identifying parasite life stages within the intestinal sections Shedding periods were similar
to those identified in adult single-housed BFF which ranged from 2-9 days however oocyst per gram
counts from the domestic ferrets were consistently lower than OPG counts from BFF (104 - 554274
OPG) infected with the same parasite (see Chapter 4) Furthermore the total number of oocysts shed by
individual domestic ferrets (14 - 172291 oocysts) during patency was reduced compared to BFF despite
similar length of shedding period (see Chapter 4) The domestic ferrets in this study were naiumlve individual
juveniles whereas the BFF were adults 1-5 years of age some of which were showing clinical signs at
the time of oocyst shedding The relative influences of age species and previous exposure to the parasite
on our observations are unknown
Two different fecal flotation methods were used on all samples to increase the probability of
oocyst detection The McMaster method was used to provide accurate OPG counts for quantification of
oocyst shedding however this method had a minimum detection limit of ~13 oocysts per gram (1333
OPG calculated) because it is based on dilution of the initial sample with flotation media (saturated salt)
In samples with few oocysts oocysts may be missed or to be present in numbers below this detection
limit Routine salt flotation is in contrast performed using the entire sample allowing for detection of
117
small numbers of oocysts Consequently in cases where oocyst per gram counts were low shedding was
identified on routine salt flotation but not by the McMaster method and recorded as lsquopositiversquo but below
the detection limit of the enumeration method
Subclinical to clinical disease occurred in six of the seven ferrets that developed patent infection
with weight loss being the most frequent clinical sign Other clinical signs were typical of enteric
coccidiosis including diarrhea hematochezia and mucoid andor soft feces These clinical signs are
similar to those previously described for black-footed ferrets infected with this parasite (USFWS BFF
Recovery Program 2017 Chapter 4) and for domestic ferrets with enteric coccidiosis (Sledge et al
2011) Interestingly development of clinical disease was not described in the naiumlve domestic ferret
inoculated by Hoare (1935) with E ictidea derived from naturally occurring infection Based on review of
the literature severe clinical disease resulting from intestinal coccidiosis is rare in domestic ferrets
Black-footed ferrets however appear more susceptible to disease development and more frequently show
significant clinical signs No domestic ferret required treatment for clinical coccidiosis during the course
of this study It is possible that the more pronounced clinical signs associated with enteric coccidiosis in
BFF may result from the limited genetic diversity in a population derived from so few individuals and
increased susceptibility of BFF to other diseases such as sylvatic plague have been described in
comparison with their domestic counterparts (Williams et al 1994) Although unlikely recent acquisition
of E ictidea from a related host species such as the domestic ferret could have resulted in increased
pathogenicity and severity of clinical disease from infection with this parasite in BFF
Minor variances from reference range values for CBC and serum biochemistry were identified in
six of the seven ferrets with patent infection but only in one case (ferret 103) did this appear to be
correlated with infectiondisease In this animal a mild hypoalbuminemia (20 ref 24-40 gL) was noted
(Appendix 3b) On histopathology large regions of the small intestine contained parasite life stages
however inflammation lysis of epithelial cells and necrosis of affected areas that could be expected to
result in protein loss into the intestinal lumen were not identified
118
Creatine kinase values were elevated in all six ferrets for which a complete serum biochemistry
was obtained immediately prior to death these findings are consistent with release from CK rich tissues
such a skeletal muscle during venipuncture and manual restraint
The pathologic lesions identified within the intestinal sections of ferrets euthanized at the time of
oocyst shedding were similar to those identified in affected BFF but in most cases were less locally
extensive or widespread throughout the small intestine than those observed in necropsy cases of BFF (see
Chapter 4) Coccidia were seen in the histologic sections of four ferrets all three ferrets that were actively
shedding oocysts at the time of necropsy (102 103 105) and one of four in which necropsies were
performed after oocyst shedding had ceased (201)
Although the primary objective of the examination of histologic sections from affected ferrets
was to identify coccidial life stages attempts were made to describe the pathologic changes associated
with the presence of the parasite Despite necropsies being performed almost immediately after death and
the use of Serra solution fixative to improve parasite and tissue preservation the villi and villar
epithelium of the trimmed sections were frequently distorted or absent and consequently accurate
commentary on these areas was precluded and was made only on visible components of the lamina
propria and crypts or glands A deliberate decision was made not to kill and collect samples from the
saline inoculated control ferrets after Phase 1 and those not shedding oocysts during Phase 1 or 2 thus no
age-matched intestinal sections were available for comparison It is difficult to comment on the
significance of the inflammatory cells observed in the lamina propria of the small and large intestinal
segments or the proliferative rate of the crypt epithelium The only changes identified which may be
considered significant are the presence of neutrophils within the crypts and glands of small and large
intestinal sections respectively but these lesions were rare and not associated with the presence of
parasitic life stages
119
While no parasitic life stages were identified in intestinal sections from ferrets 104 203 and 205
one of the three ferrets ferret 104 exhibited lymphoplasmacytic inflammation and blunting of jejunal
villi in one section (36-38 cm aboral from pylorus) These findings might be expected as the most
extensive histologic lesions would occur associated with lysis of the intestinal epithelial cells as oocysts
were shed into the feces after which new intestinal epithelial cells would re-cover the denuded villar
surface Thus for those cases in which histologic examination was performed after shedding had ceased
presence of the protozoal life stages in the intestines would be expected to be significantly reduced or
absent
Hoare (1935a b) described a particular reaction to the presence of parasitic life stages of Eimeria
ictidea in the small intestine of domestic ferrets in which only the villar tips were affected and there was
resultant annular constriction of the villus separating the affected and non-affected segments These
particular changes were not identified in any of the ferrets in this study and had not been noted
retrospectively in naturally infected BFF (Chapter 4)
While patent infection and intestinal disease could be experimentally created in domestic ferrets
without equivalent experimental work in BFF it is difficult to fully compare the susceptibility to infection
and to the development of disease between the two species Eimeria species tend to be host specific thus
if E ictidea from BFF is not conspecific with E ictidea in domestic ferrets it might be expected that the
domestic ferret would be less susceptible to infection and the development of disease than is the BFF
Even if the two parasites are identical natural passaging through the BFF may alter affinity for the
domestic ferret
It appears that the infectious dose of oocysts of E ictidea derived from BFF required to initiate a
patent infection in domestic ferrets is high The ferrets that developed patent infections were administered
an inoculum containing 1 times 106 sporulated oocysts and even with this extremely high inoculating dose
only a proportion of inoculated ferrets became infected Neither the ferret that received the low dose
120
inoculum nor the ferret that received the passaged oocysts from Part 1 of the study shed oocysts during
the 14 day period post inoculation The latter finding was unexpected as fresh passaged oocysts would be
expected to contain larger proportions of viable oocysts and be comparatively more infective than oocysts
that had been stored for 2-4 weeks prior to inoculation
The only publication describing oral inoculation of Eimeria species in domestic ferrets (Hoare
1935b) did not quantify the number of oocysts administered A study performed in 16 farmed juvenile
mink (Mustela vison) administered 2000 sporulated oocysts of each of three coccidial species (I
laidlawi E vison and an unknown Eimeria species) resulting in patent infection with one of the three
species (Foreyt et al 1977) as determined by the presence of oocysts on fecal examination The authors
did not reveal which type of oocysts resulted in the infection
The number of oocysts required to result in infection in BFF is unknown Based on the
authorsrsquo observations of over 100000 oocysts per gram of feces being shed by black-footed ferrets into
cages of lt1m2 floor space (see Chapter 4) we estimated that under normal caged conditions animals
would likely ingest thousands of oocysts over a short period of time This was in part why a large number
of oocysts (up to 1000000 as available from our store of viable oocysts) was administered to each
domestic ferret in order to increase the probability that infection and shedding would result Furthermore
for the parasite to persist within the ferret population the total number of oocysts shed into the
environment would have to be several times higher than the infective dose required to generate a patent
infection otherwise the parasite would be expected to die out If E ictidea of domestic ferrets and E
ictidea of BFF are conspecific the difference in oocyst shedding between the host species during patent
infection could potentially explain the low prevalence of E ictidea reported from the domestic ferret
population (as seen in Chapter 2)
Both humoral and cell mediated immunity are involved in the immune response to coccidia The
role of maternal derived antibodies in combatting protozoal infection in mustelids has not been studied
121
but in carnivores maternal antibodies to viruses can last up to 16 weeks (Chappuis 1998) In poultry
maternal Eimeria-specific IgG is transferred via the egg yolk to offspring In one study breeding hens
were infected with a single species of Eimeria 28-39 days prior to lay Their hatched chicks were
challenged by inoculation with oocysts of the same and a related Eimeria species and showed reduced
oocyst shedding compared to age matched controls indicative of passive transfer of immunity (Smith et
al 1994) It is possible then that the presence or absence of maternal antibodies may be a factor in the
age at which ferrets are susceptible to coccidial infection and the development of disease The facility
from which the domestic ferrets were acquired has not previously detected Eimeria species on routine
fecal screening (Dr Bambi Jasmin personal communication) consequently it is unlikely that they would
have received maternal immunity to or been exposed to this parasite and thus can be considered to be
naive
Despite a theoretical lack of maternal immunity there did appear to be an effect of age on
susceptibility to infection In Part 1 when the ferrets were approximately 70 days of age only 1 of 4
ferrets inoculated with the high dose of oocysts developed patent infection In comparison in Part 2
when the ferrets were 91-93 days of age 3 of 4 ferrets that had been in the saline control group for Part 1
developed patent infections as did 2 of 4 ferrets that had been inoculated in Part 1 but had not shed
oocysts One of these previously inoculated ferrets was the individual that had received the lower dose of
oocysts Thus it appears that patent infection could be produced more easily in the older ferret kits
However the two ferrets that had previously been inoculated exhibited the shortest shedding periods (1
day) and lowest oocyst per gram counts and parasite life stages in these cases were rare (201) to absent
(205) on histologic examination of numerous sections of intestine It is thus possible that the primary
inoculation resulted in abbreviated infections or infections in which so few oocysts were shed that
infection was not detected Our observation of endogenous stages in the intestinal tissues of some of the
ferrets following cessation of oocyst shedding suggests that the pre-patent period and duration of patency
may vary considerably from animal to animal consequently it is possible that some of the kits would
122
have ultimately shed a few oocysts from the primary inoculum if followed beyond 14 days post-
inoculation Whether through an aborted or undetected infection previous exposure of these kits to the
parasite probably generated partial immunity against E ictidea and therefore the intensity of infection
upon challenge in previously exposed domestic ferret kits was reduced
The authors acknowledge the limitations of this initial pilot study however it was proven that
patent infection with E ictidea isolated from black-footed ferrets could be generated in a novel host the
domestic ferret Further studies will be required to investigate the effect of age on susceptibility to
infection as well as the possibility of development of immunity after exposure and its role in reducing
parasite replication and disease in subsequent infection With so few BFF in existence the use of BFF for
in vivo infection trials cannot be contemplated consequently refining the domestic ferret infection model
will be essential for carrying out research specifically intended to help manage coccidiosis in the
endangered black-footed ferret
Figure 51 Exogenous life stages of Eimeria ictidea shed from a domestic ferret (Mustela putorius
furo) experimentally inoculated with oocysts originating from black-footed ferrets (Mustela
nigripes) Unsporulated oocyst (solid black arrow) Sporulated oocyst (solid white arrow) Bright
field microscopy scale bar = 25 microm
25 microm
124
Figure 52 Life stages of Eimeria ictidea within the small intestinal epithelium of an experimentally
infected domestic ferret (Mustela putorius furo) A) Sexual life stages (micro- and macrogamonts
- white arrows) crowding the villar enterocytes all stages are found between the nucleus and
luminal surface of infected enterocytes Hematoxylin and eosin staining scale bar = 25microm B) At
higher magnification meronts (black arrows) and gamonts (open arrows) are crowded between the
enterocyte nuclei and brush border Hematoxylin and eosin staining scale bar = 25microm
25 microm
25 microm
125
Figure 53 Presence and location of sexual and asexual life stages of Eimeria ictidea within the
intestinal epithelium of domestic ferrets (Mustela putorius furo) (n=7) that developed patent
infection after experimental inoculation with oocysts originating from black-footed ferrets (Mustela
nigripes) Legend duod = duodenum jej= jejunum Sequential numbers for jejunal and colonic
sections represent the order aboral from the pylorus from which the samples were collected SI =
additional section(s) of small intestine whose aboral sequence was not recorded
0
1
2
3
duod jej 1 jej 2 jej 3 jej 4 jej 5 ileum colon 1 colon 2 SI
Nu
mb
er
of
Ferr
ets
Aff
ect
ed
Intestinal Section Containing Life Stages of Eimeria ictidea
Asexual life stages Sexual life stages
126
Table 51 Prepatent period and oocyst shedding patterns in domestic ferrets (Mustela putorius furo)
experimentally inoculated with 1 times 106 oocysts of Eimeria ictidea originating from black-footed ferret
(Mustela nigripes) that developed patent infections
Oocysts shed per gram of feces
Ferret Identity
Day post inoculation 102 103 104 105 201 203 205
1 0 0 0 0 0 0 0
2 0 0 0 0 0 0 0
3 0 0 0 0 0 0 0
4 0 0 0 0 0 0 0
5 0 0 0 0 0 0 0
6 0 0 0 0 0 0 0
7 lt 14 1807 0 0 0 0 0
8 11053 139 7091 0 156238 0
9 463 11733 lt 14 203 lt 14
10 578 7549 0 0 0
11 lt 14 0 0 0
12 0 lt 14 0
13 0 lt 14 0
14 0 lt 14 0
Legend ferret euthanized as of this date lt 14 = oocyst positive samples with less than 14 oocysts per
gram of feces
127
Table 52 Results of oral inoculation of domestic ferrets (Mustela putorius furo) with oocysts of Eimeria ictidea originating from black-footed
ferrets (Mustela nigripes)
Ferret
Identity
Results Part 1 Results Part 2
Inoculum Oocyst
Shedding
Presence
of clinical
disease
Coccidia
present in
sections
Inoculum Oocyst
Shedding
Presence
of clinical
disease
Coccidia
present in
sections
101 Saline N N - 2 times 105 oocystsa N N -
102 Saline N N - 1 times 106 oocysts Y N Y^
103 Saline N N - 1 times 106 oocysts Y Y Y^
104 Saline N N - 1 times 106 oocysts Y Y Nyen
105 Saline N N - 1 times 106 oocysts Y Y Yyen
201 1 times 106 oocysts N N - 1 times 106 oocysts Y Y Yyen
202 1 times 106 oocysts N N - 1 times 106 oocysts N N -
203 1 times 106 oocysts Y Y Nyen
204 1 times 106 oocysts N N - 1 times 106 oocysts N N -
205 5 times 104 oocysts N N - 1 times 106 oocysts Y Y N^
Legend includes any of the following weight loss diarrhea mucoid feces malodorous feces inappetence N = no Y= yes - = necropsy not
performed a = oocysts collected after passage through ferret 203 ^ = shedding oocysts at time of necropsy yen = not shedding oocysts at time of
necropsy
128
Table 53 Distribution of coccidial life stages in domestic ferrets (Mustela putorius furo) orally inoculated with oocysts of Eimeria ictidea
originating from black-footed ferrets (Mustela nigripes)
Ferret Identity
Intestinal level 102
103
104 105
201
203
205
Duodenum N N N N N N N
Jejunum 1 N N N N N N N
Jejunum 2 N N N N N N N
Jejunum 3 N S A N N N N N
Jejunum 4 N S A N N N N N
Jejunum 5 N S A N S A N N
Ileum N S A N N N N N
Colon 1 N Na N N N N N
Colon 2 N Na N N N N N
Unmeasured small intestinebc
S - 05
A - 15
S - 45
A - 25
S - 05
A - 05
S - 26
A - 06
S - 06
A - 06
S - 06
A - 06
S - 08
A - 08
Unmeasured large intestinebc
S - 01
A - 01
S - 01
A - 01
S - 01
A - 01
S - 02
A - 02
S - 01
A - 01 none
S - 01
A - 01
Legend N = no parasite life stages S = sexual life stages present A = asexual life stages present a= oocysts present in feces b=
additional sections of intestine for which the location measured from the pylorus was not obtained c= xn where x is number of
sections containing sexual or asexual lifestages n is the number of sections examined
129
CHAPTER 6 WHOLE MITOCHONDRIAL GENOME SEQUENCES OF TWO EIMERIA
SPECIES ISOLATED FROM DOMESTIC (MUSTELA PUTORIUS FURO) AND BLACK-
FOOTED FERRETS (MUSTELA NIGRIPES)
ABSTRACT
The complete mitochondrial (mt) genomes of Eimeria furonis and Eimeria ictidea (Eimeriidae
Coccidia Apicomplexa) originating from single fecal samples from a domestic (Mustela putorius furo)
and a black-footed ferret (Mustela nigripes) respectively were sequenced Both mt genomes were
circular-mapping with lengths of 6165 base pairs (Eimeria furonis - GenBank MF795598) and 6182
base pairs (Eimeria ictidea - GenBank KT203399) Genome organization and gene contents were
comparable with those of other publically available mt genomes from a variety of Eimeria species and
related coccidia there were three complete coding DNA sequence regions encoding cytochrome c
oxidase subunit I cytochrome c oxidase subunit III and cytochrome B and 33 regions encoding
fragmented rDNA Alignment of these mt genome sequences demonstrates a relatively high (945 340
single nucleotide differences [SNDs]) pairwise sequence identity between these Eimeria spp infecting
ferrets the majority of the SNDs resulted in synonymous codon changes with no changes to their protein
products Alignment of the newly sequenced mt genomes demonstrates and phylogenetic reconstructions
support the monophyly of these Eimeria spp of ferrets with another Eimeria sp of carnivores as the
sister taxon to this clade
61 INTRODUCTION
Coccidia are protozoal eukaryotic host-specific parasites of the phylum Apicomplexa and can
be divided into two major taxonomic suborders the eimerioirinid and adeleid coccidia The eimeriorinid
coccidia include both the typical intestinal coccidia (eg Eimeria Isospora Cyclospora) species
belonging to the family Eimeriidae as well as tissue or cyst-forming coccidia (eg Cystoisospora
130
Besnoitia Toxoplasma Sarcocystis) of the family Sarcocystidae (see Cox 1994) Ten species of
Eimeria and twelve species of Isospora (=Cystoisospora) have been described in the Mustelidae
(see Chapter 1 Table 11) Eimeria furonis has been reported in the European polecat (Mustela
putorius) domestic ferret (DF Mustela putorius furo) black-footed ferret (BFF Mustela nigripes) and
mink (Mustela vison) (Hoare 1927 Nukerbaeva and Svanbaev 1973 Jolley et al 1994) Eimeria ictidea
has been reported in the Steppe polecat (Mustela eversmanii) as well as the European polecat domestic
ferret and black-footed ferret (Hoare 1927 Svanbaev 1956 Jolley et al 1994) These reports are based on
the morphometric characteristics of oocysts identified in the feces of the aforementioned host species
without the use of molecular techniques to confirm specific parasite identities
Recently sequences of the mitochondrial cytochrome c oxidase subunit I gene (mt COI) and
nuclear small subunit ribosomal DNA (nu 18S rDNA) of E furonis originating from a domestic ferret (nu
18S rDNA GenBank MF774678-MF774680 mt COI GenBank MF774034-MF774036) and E ictidea
originating from a black-footed ferret (nu 18S rDNA GenBank MF860826-MF860827 mt COI
GenBank MF860823 MF860825) were generated (see Chapters 2 and 3) The parasite originating from
the black-footed ferret was identified as E ictidea based on morphologic similarity to the original
descriptions of E ictidea from domestic ferrets however sequence-based genotyping of E ictidea from
domestic ferrets has not been completed and consequently it has not been demonstrated unequivocally
that the two parasites are conspecific In the present work the complete mitochondrial genomes of E
furonis from the DF and E ictidea from the BFF are described and compared with the mitochondrial
genomes of related coccidia
62 MATERIALS amp METHODS
621 Parasites
Two isolates of morphologically distinct Eimeria species were used in this study Isolate one
identified morphologically and by nu 18S rDNA and mt COI sequences as Eimeria furonis was obtained
131
from a fecal sample from a DF that was submitted for routine ova and parasite examination to a European
diagnostic laboratory8 Isolate two identified morphologically and by nu 18S rDNA and mt COI
sequences as Eimeria ictidea was obtained from a fecal sample from a BFF and was collected during
routine cage cleaning in a captive breeding facility (see Chapters 3 and 4) Fecal collection techniques for
the BFF were reviewed and approved by both the Animal Welfare Committee and the Animal Care and
Research Committee of the Toronto Zoo
622 DNA isolation from coccidia in feces
Genomic DNA was isolated from fecal derived coccidial oocysts as described section 221 of the
Chapter 2 Materials amp Methods Parasite DNA concentration was estimated using a Nanodrop 2000
spectrophotometer (NanoDrop Products Wilmington DE USA) and DNA was stored at 4 degC for
immediate use or minus20 degC for later use
623 Whole genome sequencing
Mitochondrial whole genome amplification for both Eimeria species was initiated using sets of
mt-specific primers that generated overlapping polymerase chain reaction (PCR) fragments (Tables 61
and 62) PCR amplification was performed for all samples in a volume of 25 microl containing ~100 ng of
genomic DNA 1times PCR buffer 3 mM MgCl2 06 mM deoxyribonucleotide triphosphates (dNTPs) 500
nM of each primer and 4 U of Invitrogen Platinum Taq DNA Polymerase (Thermo Fisher Scientific
Toronto ON Canada) PCR reactions were performed on a Bio-Rad T100 PCR thermal cycler (Bio-Rad
Laboratories Singapore) using settings as described previously in the Materials amp Methods section of
Chapter 2 Table 61 details the specific anneal conditions used for the various primer pairs Genomic
DNA from either Eimeria maxima or Eimeria tenella acted as a positive control for the reaction
chemistry Gel electrophoresis purification and sequencing of the PCR amplification products were
8 Vet Med Labor GmbH Division of IDEXX Laboratories Ludwigsburg Germany
132
performed as described in Chapter 2 The resulting chromatograms were aligned and analyzed with
Geneious Ver 818 or later (Biomatters Limited Auckland New Zealand) and high quality consensus
sequences generated The completed mt genome sequences were annotated by comparison with
previously annotated mt genomes from other Eimeria species (eg Eimeria innocua - KR1082961) and
the annotated mt genomes deposited in GenBank
624 Phylogenetic analysis
To determine the phylogenetic affinities of the newly obtained sequences with sequences from
related apicomplexan taxa representative whole mt genome sequences from eimeriid coccidia were
downloaded from GenBank A complete mt genome sequence from an unnamed Choleoeimeria sp was
used to root the ingroup taxa several small genomic rearrangements in the Choleoeimeria sp sequence
required some rearrangement of the genome sequence to unambiguously align homologous regions across
the complete mt genomes
Whole genome sequences were aligned using MAFTT v7017 (Katoh et al 2002) executed from
within Geneious the resulting alignment was examined by eye to adjust start and stop codon positions in
aligned coding DNA sequence [CDS] regions (ie mt COI mitochondrial cytochrome c oxidase subunit
III gene [mtCOIII] mitochondrial cytochrome b gene [CytB]) Phylogenetic trees were generated using
Bayesian Inference (BI) using MrBayes Ver 326 (Huelsenbeck and Ronquist 2001) executed from
within Geneious The aligned complete mt genomes were partitioned into coding (ie CDS) and non-
coding regions so that region-specific models of nucleotide substitution could be applied Characters in
the non-coding region were analysed with the general time reversible (GTR) model (Tavareacute 1986) with
the following parameters nucmodel=4by4 nst=6 rates=invgamma (ie GTR+I+G) Characters in the
coding regions were analysed using the codon nucleotide model (ie lset nucmodel=codon rates=gamma
ngammacat=4) using metazoan mitochondrial translation (ie code=metmt)
133
All BI analyses were run for a chain length of 1000000 with tree sampling every 1000
following a burn-in of 100000 with default settings of 4 heated chains and heated chain temp of 02
63 RESULTS
The whole mt genome sequences of the single isolates of E furonis and E ictidea were
respectively 6165 base pairs (bp) (Figure 61 GenBank MF795598) and 6182 bp (Figure 62 GenBank
KT203399) Content and organization of both mt genomes consisted of three protein-coding genes (mt
COI mt COIII and CytB) interspersed with large and small subunit ribosomal DNA (rDNA) fragments
Details of the various CDS and rDNA fragments are summarized in Table 63 (for E furonis) and Table
64 (for E ictidea)
Pairwise alignment of the mt genome sequences from E furonis and E ictidea demonstrated a
relatively high pairwise sequence identity (946 333 single nucleotide differences [SNDs]) between
these two parasites The bulk of the SNDs (676 225333) were clustered within the three CDS regions
that encode CytB mt COI and mt COIII (see Figure 63 and Table 65) However the majority of these
SNDs (826 186225) were synonymous codon changes that resulted in no changes to the protein
products Only 41 SNDs were involved in 34 amino acid changes distributed among the three CDS
The 33 rDNA fragments comprised 2108 and 2109 bp respectively of the mt genomes of
E furonis and E ictidea Pairwise comparison of these rDNA fragments demonstrated high (986 30
SNDs) sequence identity between the two parasites The remaining 778 and 794 bp respectively of the
mt genomes of E furonis and E ictidea were intergenic stretches between the various rDNA and CDS
regions these intergenic regions were more variable that other regions of the genomes with 78 SNDs
(almost 10 sequence divergence) Additionally all indels were restricted to these variable intergenic
regions
The BI phylogeny generated from aligned complete mt genomes (Figure 64) supported the close
relationship between E furonis and E ictidea within a clade of Eimeria species that include the only three
sequences available for Eimeria spp of carnivores Eimeria mephitidis from the striped skunk (Mephitis
134
mephitis Family Mephitidae) was the sister taxon to the two Eimeria species of ferrets (Family
Mustelidae)
64 DISCUSSION
This work generated the first complete mt genomes from coccidia that infect domestic and black-
footed ferrets (Carnivora Mustelidae) Eimeria mephitidis from the striped skunk Mephitis mephitis
(Carnivora Mephitidae) is the only other Eimeria species from a carnivore for which a complete mt
genome has been reported
Comparatively few eimeriid coccidia only 26 Eimeria species have been described from
carnivores there are 14 named species from the mustelids four from the procyonids four from the ursids
three from the herpestids and one from the viverrids (Duszynski et al 2000) The majority of coccidia
that infect the digestive tract of carnivores belong to the family Sarcocystidae including monoxenous or
facultatively heteroxenous Cystoisospora species or heteroxenous parasites in the genera Sarcocystis
Hammondia and Neospora So far as is known none of the parasites in the Sarcocystidae possess typical
apicomplexan mt genomes with 3 complete CDS and many rDNA fragments (Ogedengbe 2015)
The mt genomes from the two Eimeria sp of mustelid origin demonstrate the same structural
organization (ie the order and number of CDS and rDNA fragments) and circular mapping as the mt
genomes from other Eimeria spp and other closely related eimeriid coccidia such as Isospora
Cyclospora and Lankesterella species Despite the ability of the eimeriid sequences to be mapped
circularly the physical form of Eimeria spp mt genomes may be a linear concatemer of multiple genome
copies as demonstrated for Eimeria tenella (Hikosaka et al 2011) As in the mt genomes of other
eimeriid coccidia (Ogedengbe et al 2013 2014) the CDS for mt COIII demonstrated the highest
sequence divergence between E furonis and E ictidea the mt COI CDS was somewhat more conserved
and CytB CDS demonstrated the fewest SNDs
As expected based on limited sequence divergence between E furonis and E ictidea a BI
phylogenetic analysis using aligned complete mt genome sequences generated a tree that placed these
135
two Eimeria species that infect mustelids within a well-supported monophyletic group The sister taxon
for these ferret parasites was the only other Eimeria species from carnivores for which a complete mt
genome is available E mephitidis which infects hosts belonging to a different family of carnivores
Eimeriid parasites that infect closely-related definitive hosts are commonly found in a single or limited
number of clades based on mitochondrial and nuclear genetic loci (Ogedengbe et al in press)
Sequencing of the mt genomes and at least one nuclear genetic locus (ie nu 18S rDNA) from additional
Eimeria species infecting carnivores will be required to determine if all carnivore-specific Eimeria
species share a common ancestor
136
Table 61 PCR primer pairs and resulting fragments used for sequencing the mitochondrial genome sequence of an isolate of Eimeria furonis
originating from a fecal sample from a domestic ferret (Mustela putorius furo)
Fragment Primer names Primer sequences (5ʹ-3ʹ) Size (bp) Anneal Temp References
1 WG-MT_4140F AGAAAACCTAAAATCATCATGT 1000 52 Ogedengbe et al (2015)
Eim_CO3_799R AAGTGAGTTCGCATGTTTAC Ogedengbe et al (2015)
2 Eim_COI_19F ACTGCYGCAAACCATAARGAA 1700 60 Present study
Api_LSUG_UNI_R AGATAGGGAACAAACTGYCTCAA Present study
3 WG_MT_5416F GGTCCAGATAAGCGATCTCATG 3400 53 Ogedengbe et al (2013)
Eim_COI_1436R CACATTGTGTTCARATAAGTTA Present study
4 WG-MT_6219F GCATCCATCTACAGCTGCGG 500 55 Ogedengbe et al (2013)
WG-MT_344R GTAGGAATCTRAATTCCCAACC Ogedengbe et al (2013)
5 Api_LSUE UNI_F AGGTGCTCAGGGTCTTACCG 500 55 Present study
WG_MT_63R CTGGTATGGATGGATAACACT Ogedengbe et al (2015)
6 Lank_COB-30F CCAGGCCAACTGAACTCGTT 1300 55 Present study
q_Eim_COI_221R GGCATAACTACAAAGAARATCATA Present study
7 Cocci_MT_WG_F TACACCTAGCCAACACGAT 1600 55 Ogedengbe et al (2014)
q_Eim_COI_221R GGCATAACTACAAAGAARATCATA Present study
137
Table 62 PCR primer pairs and resulting fragments used for sequencing the mitochondrial genome sequence of an isolate of Eimeria ictidea
originating from a fecal sample from a black-footed ferret (Mustela nigripes)
Fragment Primer names Primer sequences (5ʹ-3ʹ) Size (bp) Anneal Temp References
1 WG_MT_63R CTGGTATGGATGGATAACACT 2180 52 Ogedengbe et al (2015)
WG-MT_4140F AGAAAACCTAAAATCATCATGT Ogedengbe et al (2015)
2 Cocci_MT_WG_F TACACCTAGCCAACACGAT 1551 58 Ogedengbe et al (2014)
q_Eim_COI_221R GGCATAACTACAAAGAARATCATA Present study
3 WG-MT_3658F CTGGCGAGAAGGGAAGTGTG 1329 55 Ogedengbe et al (2013)
Eim_CO3_799R AAGTGAGTTCGCATGTTTAC Ogedengbe et al (2015)
4 Lank_COB-30F CCAGGCCAACTGAACTCGTT 3899 57 Present study
WG_MT_4072R GGTTGTTTCCATCTCGACTC Ogedengbe et al (2013)
138
Table 63 Coding regions within the mitochondrial genome of the eimeriid parasite Eimeria furonis from a domestic ferret (Mustela putorius faro)
139
Table 64 Coding regions of the mitochondrial genome of the eimeriid parasite Eimeria ictidea originating from a black-footed ferret (Mustela
nigripes)
Table 64 Features associated with the protein-coding regions of the mitochondrial genome of the eimeriid parasite Eimeria cf ictidea originating from a black-footed ferret (Mustela nigripes )
Protein coding regions (CDS) Sequence size (bp) Start position (bp) Stop position (bp) Direction Translation start codon Translation stop codon
Cytochrome c oxidase subunit I (COI) 1443 1343 2785 Forward ATG TAA
Cytochrome c oxidase subunit III (COIII) 756 4333 5088 Forward TTA TAA
Cytochrome b (CytB) 1080 226 1305 Forward ATG TAA
Ribosomal DNA fragments (rDNA) Product
SSUrRNA 46 17 62 forward RNA9 SSU8
SSUrRNA 77 104 180 forward SSUA SSU4 (partial)
SSUrRNA 35 181 215 forward RNA23t
LSUrRNA 20 2790 2809 forward RNA20 (partial) LSU
LSUrRNA 112 2835 2946 forward LSUF LSU11
LSUrRNA 106 2947 3052 forward LSUG LSU12
LSUrRNA 25 3087 3063 reverse LSU
SSUrRNA 39 3129 3167 forward RNA14 SSU1
LSUrRNA 16 3188 3173 reverse LSUC LSU4
SSUrRNA 33 3211 3243 forward SSU
SSUrRNA 61 3386 3326 reverse SSUF SSU12
LSUrRNA 74 3387 3460 forward RNA10 LSU13 (partial)
LSUrRNA 49 3476 3524 forward RNA11 LSU5
SSUrRNA 65 3533 3597 forward SSUD SSU10
SSUrRNA 40 3631 3670 forward RNA17 SSU3
SSUrRNA 30 3677 3706 forward RNA15 SSU
LSUrRNA 30 3711 3740 forward RNA13 LSU10
LSUrRNA 56 3749 3804 forward RNA6 LSU15
LSUrRNA 79 3899 3821 reverse LSUD LSU8
LSUrRNA 24 3931 3908 reverse RNA16 (partial)
SSUrRNA 92 4036 3945 reverse RNA8 SSU5
LSUrRNA 66 4054 4119 forward RNA2 LSU2
LSUrRNA 177 4316 4140 reverse LSUA LSU1
SSUrRNA 30 5125 5096 reverse RNA19 SSU7
LSUrRNA 88 5154 5241 forward RNA1 LSU6
LSUrRNA 26 5342 5317 reverse LSUB LSU3
LSUrRNA 73 5447 5375 reverse RNA3 LSU7
LSUrRNA 20 5479 5460 reverse RNA18 LSU14
SSUrRNA 119 5615 5497 reverse SSUB SSU6
LSUrRNA 80 5699 5620 reverse RNA7
LSUrRNA 188 5897 5710 reverse LSUE LSU9
SSUrRNA 33 6033 6001 reverse SSUE SSU11 (partial)
SSUrRNA 99 6156 6058 reverse RNA5SSU9
140
Table 65 Pairwise comparison of coding DNA and concatenated rDNA fragment sequences between the
mitochondrial genomes of Eimeria furonis originating from a domestic ferret (Mustela putorius furo) and
Eimeria ictidea originating from a black-footed ferret (Mustela nigripes)
Total length
(nucleotides)
Nucleotide
identity
Total amino
acids
Amino acid
identity
COI CDS 1443 934 (95) 480 975 (12)
COIII CDS 756 899 (76) 251 932 (17)
CytB CDS 1080 950 (54) 359 986 (5)
rDNA fragments 2109 985 (32) na na
Legend Numbers in brackets indicate the number of single nucleotide differences na = not
applicable
141
Figure 61 Circular mapping and organization of the mitochondrial genome content of Eimeria
furonis showing three protein-coding genes (COI COIII and CytB) interspersed with large and
small subunit rRNA fragments
142
Figure 62 Circular mapping and organization of the mitochondrial genome content of Eimeria
ictidea showing three protein-coding genes (COI COIII and CytB) interspersed with large and
small subunit rRNA fragments
143
Figure 63 Pairwise comparison of coding DNA and concatenated rDNA fragment sequences of
the mitochondrial genomes of Eimeria furonis and Eimeria ictidea Legend Yellow bands
correspond to coding DNA fragments Red arrows correspond to rDNA fragments Arrows of both
colours indicate the forward or reverse direction of these segments
144
Figure 64 Phylogenetic relationships of coccidia (Eimeria furonis and Eimeria ictidea) from
domestic (Mustela putorius furo) and black-footed ferrets (Mustela nigripes) based on the complete
mitochondrial genome sequences these Eimeria species and a selection of related apicomplexan
parasites
145
CHAPTER 7 CONCLUSIONS AND FUTURE DIRECTIONS
The primary objective of this project was to better characterize the enteric coccidia of the
endangered black-footed ferret (BFF) in order to set the stage for improved disease prevention
management and treatment To the authorrsquos knowledge this work was the first attempt to isolate and
perform molecular characterization of the coccidial species endemic in the black-footed ferret population
and to characterize the natural history of the disease in this host
Two Eimeria species Eimeria ictidea and Eimeria furonis have been described from BFF and
domestic ferrets A single Eimeria species morphologically resembling E ictidea of the domestic ferret
and referred to in this work as E ictidea was identified from all historic and active cases of enteric
coccidiosis in the Toronto Zoo BFF population The same species was identified in both BFF family
groups and single-housed adults that shed coccidia during the study period (2014-2016) as well as from
historic necropsy samples of juvenile and adult Toronto Zoo BFF from 1999-2014 Furthermore this
same species was identified from both family groups and single-housed adults shedding coccidia at an
additional BFF Species Survival Plan institution the Louisville Zoo in 2016 These findings are contrary
to previous published reports that indicated multiple coccidia species were affecting captive and wild
BFF In order to determine whether these additional parasites continue to exist within the present-day
BFF population and their impact on this species further examination of coccidia-positive fecal samples
from captive and wild BFF populations is recommended Moreover determination of which parasite
species impact BFF morbidity and mortality would allow for development of targeted therapies for
disease management
Coccidia-positive fecal samples and necropsy samples from domestic ferrets were collected from
multiple diagnostic laboratories in Canada and Europe for comparison with results from BFF Eimeria
furonis and Isospora (=Cystoisospora) laidlawi were identified from fecal parasitology reports from
2008-2015 and in fecal samples obtained prospectively from 2014-2016 from domestic ferrets submitted
to Canadian and European laboratories E furonis was also identified in necropsy samples from 2010 and
146
2017 from two Canadian diagnostic laboratories No samples containing E ictidea were submitted to the
Canadian laboratories however samples containing E ictidea were identified twice by the European
diagnostic laboratory with one sample in 2011 and a second in 2013 Consequently it appears that E
ictidea is rarely identified from domestic ferrets Since no samples of E ictidea were acquired from
domestic ferrets during the study period it remains undetermined whether E ictidea from domestic
ferrets and E ictidea of BFF are conspecific Future molecular characterization of E ictidea isolated from
various mustelid host species would allow not only for determination of whether the parasites are
conspecific but would also provide insight into the potential for cross-transmission of parasites between
related mustelid hosts
The identifications provided by diagnostic laboratories of the specific parasite species present in
the domestic ferret fecal samples showed poor agreement with their identifications based on genotyping
obtained in this study In the authorrsquos opinion molecular techniques are essential tools for determining
the specific coccidial species responsible for individual and group outbreaks of coccidiosis and for
further understanding of the eimeriid host-parasite relationships To this end the nu 18S rDNA region and
whole mitochondrial (mt) genomes of E ictidea and E furonis were sequenced these mt whole genome
sequences are the first for the Eimeria species of mustelids to be entered into GenBank These sequences
may provide suitable targets for the development of highly discriminatory PCR-based methods of
identification that could be applied to fecal tissue or even formalin-fixed paraffin-embedded samples
(see Chapters 2 and 3) Methodological improvements such as less expensive next generation sequencing
methods that could be applied to diagnostics will depend on the availability of high quality reference
sequences such as generated herein Furthermore evaluation of the evolution of the eimeriid coccidia
using phylogenetic analysis based on additional whole mitochondrial genome sequences may allow for
more accurate determinations of relationships between parasite species and timing of their divergence
from common ancestors
One of the goals of this research was to investigate the possibility of using the domestic ferret as
an experimental model to study enteric coccidiosis in the BFF Experimental work cannot be carried out
147
in the BFF due to its endangered status We showed that the domestic ferret is susceptible to infection
with Eimeria ictidea originating from BFF Infected domestic ferrets showed similar clinical signs and
pathologic lesions to BFF strengthening the possibility of their serving as a suitable model Future
investigations would include studies of the domestic ferretrsquos immune response to enteric coccidial
infection in vivo drug trials including pharmacokinetic pharmacodynamic and efficacy studies of
anticoccidial medications investigations into patterns of drug resistance in coccidial species of ferrets
and oral vaccine development
Ultimately the goal of research on coccidiosis in BFF may be the development of an autogenous
vaccine to improve the survival of ferret kits and reduce coccidiosis-related morbidity and mortality in
BFF captive breeding programs It may be possible to use the domestic ferret to select for E ictidea that
are more highly adapted to this host perhaps with a commensurate loss of virulence to its original host If
this were to be the case domestic ferret-derived coccidia could be used in BFF as vaccine organisms
Alternately methods such as use of a bioshuttle (live vaccination with coccidia followed by anticoccidial
treatment to limit pathogenicity) might be useful for generating long-lasting immunological protection in
BFF against E ictidea Improving immunity through vaccination could assist in reducing coccidial
shedding and disease in BFF associated with stressful life events such a breeding weaning and transfer
between institutions There is minimal available data on the significance of coccidiosis in wild
populations of BFF and limited means of disease surveillance post release however it is logical to
assume that the stresses associated with release to the wild might result in clinical disease as happens with
stressed BFF in captivity Reducing morbidity and mortality associated with coccidiosis in BFF could
result in increased numbers of ferrets being released to the wild and higher survival of released ferrets
increasing the number of BFF in the wild supports the goals of the conservation initiative for the black-
footed ferret and will support the longer-term survival and recovery of this species
148
REFERENCES
Abe N Tanoue T Ohta G Iseki M (2008) First record of Eimeria furonis infection in a ferret Japan with
notes on the usefulness of partial small subunit ribosomal RNA gene sequencing analysis for
discriminating among Eimeria species Parasitol Res 103967ndash70 doi 101007s00436-008-1037-x
Adl SM Simpson AGB Farmer MA et al (2005) The new higher level classification of eukaryotes with
emphasis on the taxonomy of protists J Eukaryot Microbiol 52399ndash451 doi 101111j1550-
7408200500053x
Andrews JM (1926) Coccidiosis in mammals Am J Hyg 6784ndash798
Augustine PC Danforth HD (1986) A study of the dynamics of the invasion of immunized birds by
Eimeria sporozoites Avian Dis 30347ndash351
Barta JR Schrenzel MD Carreno R Rideout BA (2005) The genus Atoxoplasma (Garnham 1950) as a
junior objective synonym of the genus Isospora (Schneider 1881) species infecting birds and
resurrection of Cystoisospora (Frenkel 1977) as the correct genus for Isospora species infecting
mammals J Parasitol 91726ndash727 doi 101645GE-33411
Bell JA (1994) Parasites of Domesticated Pet Ferrets Compend Contin Educ Pract Vet 16617ndash620
Black-footed Ferret Recovery Implementation Team (2011) Captive Breeding
httpwwwblackfootedferretorgcaptive-breeding Accessed 8 Jan 2014
Blankenship-Paris TL Chang J Bagnell CR (1993) Enteric coccidiosis in a ferret Lab Anim Sci 43361ndash
363
Bronson E Bush M Viner T et al (2007) Mortality of captive black-footed ferrets (Mustela nigripes) at
Smithsonianrsquos National Zoological Park 1989 ndash 2004 J Zoo Wildl Med 38169ndash176
149
CAPC (2013) Current Advice on Parasite Control Intestinal Parasites - Coccidia
httpwwwcapcvetorgcapc-recommendationscoccidia Accessed 6 Jun 2015
Carpenter JW Hillman CN (1979) Husbandry reproduction and veterinary care of captive ferrets In
1978 Proceedings of the Annual Meeting of the American Association of Zoo Veterinarians
Washington DC pp 36ndash47
Catchpole J Norton CC Gregory MW (1993) Immunisation of lambs against coccidosis Vet Rec
13256ndash59
Cavalier-Smith T (2014) Gregarine site-heterogeneous 18S rDNA trees revision of gregarine higher
classification and the evolutionary diversification of Sporozoa Eur J Protistol 50472ndash495 doi
101016jejop201407002
Chappuis G (1998) Neonatal immunity and immunisation in early age lessons from veterinary medicine
Vaccine 161468ndash1472
Cox FE (1994) The evolutionary expansion of the Sporozoa Int J Parasitol 241301ndash1316
Davis CL Chow TL Gorham JR (1953) Hepatic coccidiosis in mink Vet Med 48371ndash375
De Vos AJ (1970) Studies on the host range of ltigtEimeria chinchillaeltIgt de Vos and Van Der
Westhuizen 1968 Onderstepoort J Vet Res 3729ndash36
Dirikolu L Yohn R Garrett EF et al (2009) Detection quantifications and pharmacokinetics of
toltrazuril sulfone (Ponazuril) in cattle J Vet Pharmacol Ther 32280ndash288
Dryden MW Payne PA Ridley R Smith V (2005) Comparison of common fecal flotation techniques for
the recovery of parasite eggs and oocysts Vet Ther 615ndash28
Duszynski D Wilber PG (1997) A Guideline for the Preparation of Species Descriptions in the
Eimeriidae J Parasitol 83333ndash336
150
Duszynski DW Couch L Upton SJ (2000) The Coccidia of the World
httpbiologyunmeducoccidiacarniv2html Accessed 1 Sep 2017
El-Sherry S Ogedengbe ME Hafeez MA Barta JR (2013) Divergent nuclear 18S rDNA paralogs in a
turkey coccidium Eimeria meleagrimitis complicate molecular systematics and identification Int J
Parasitol 43679ndash685 doi 101016jijpara201303005
Evans HE An NQ (2014) Anatomy of the ferret In Fox JG Marini RP (eds) Biology and Diseases of the
Ferret 3rd edn Wiley Blackwell pp 23ndash67
Fayer R (1980) Epidemiology of Protozoan Infections The Coccidia Vet Parasitol 675ndash103
Foreyt WJ Todd AC Hartsough GR (1977) Anticoccidial Activity of Eight Compounds in Domestic
Mink AJVR 38391ndash394
Fox JG (2014) Normal Clinical and Biological Parameters In Fox JG Marini RP (eds) Biology and
Diseases of the Ferret 3rd edn John Wiley amp Sons Inc pp 157ndash185
Frenkel JK (1977) Besnoitia wallacei of Cats and Rodents With a Reclassification of Other Cyst-
Forming Isosporoid Coccidia J Parasitol 63611ndash628
Grafner G Graubmann HD Dobbriner W (1967) Leberkokzidiose beim Nerz (Lutreola vison Schreb)
hervorgerufen durch eine neue Kokzidienart Eimeria hiepei n sp Monatshefte fur
Veterinearmedizin 22696ndash700
Haberkorn A (1971) Zur Wirtsspezifitat yon Eimeria contorta nsp (Sporozoa Eimeriidae) Z
Parasitenkd 37303ndash314
Hall MC Wigdor M (1918) Canine coccidiosis with a note regarding other protozoan parasites from the
dog J Am Vet Med Assn 5364ndash76
Hikosaka K Nakai Y Watanabe YI et al (2011) Concatenated mitochondrial DNA of the coccidian
151
parasite Eimeria tenella Mitochondrion 11273ndash278
Hillyer E V (1992) Gastrointestinal diseases of ferrets (Mustela putorius furo) J Small Anim Med 244ndash
45
Hoare CA (1927) On the coccidia of the ferret Ann Trop Med Parasitol 21313ndash320
Hoare CA (1935a) A histopathological reaction of a special type on the part of the intestinal villi in ferret
coccidiosis Trans R Soc Trop Med Hyg 292
Hoare CA (1935b) The endogenous development of the coccidia of the ferret and the histopathological
reaction of the infected intestinal villi Ann Trop Med Parasitol 29111ndash122
Hoefer HL Fox JG Bell JA (2012) Gastrointestinal Diseases In Quesenberry KE Carpenter JW (eds)
Ferrets Rabbits and Rodents Clinical Medicine and Surgery 3rd edn Elsevier Saunders St Louis
Missouri pp 27ndash45
Huelsenbeck JP Ronquist F (2001) MrBayes Bayesian inference of phylogenetic trees Bioinformatics
17754ndash755 doi 101093bioinformatics178754
Jolley WR Kingston N Williams ES Lynn C (1994) Coccidia Giardia sp and a Physalopteran
Nematode Parasite from Black-footed Ferrets (Mustela nigripes) in Wyoming J Helminthol Soc
Washingt 6189ndash94
Katoh K Misawa K Kuma K Miyata T (2002) MAFFT a novel method for rapid multiple sequence
alignment based on fast Fourier transform Nucleic Acids Res 303059ndash3066 doi
101093nargkf436
Kaye SW Ossiboff RJ Noonan B et al (2015) Biliary coccidiosis associated with immunosuppressive
treatment of pure red cell aplasia in an adult ferret (Mustela putoris furo) J Exot Pet Med doi
101053jjepm201504012
152
Koepfli KP Deere K Slater GJ et al (2008) Multigene phylogeny of the Mustelidae resolving
relationships tempo and biogeographic history of a mammalian adaptive radiation BMC Biol 610
Lariviegravere S Jennings AP (2009) Family Mustelidae (weasels and relatives) In Wilson DE Mittermeier
RA (eds) Handbook of the Mammals of the World Volume 1 Carnivores Lynx Edicions
Barcelona pp 564ndash656
Levine ND Ivens V (1970) The coccidian parasites (Protozoa Sporozoa) of ruminants
Li X Pang J Fox JG (1996) Coinfection with intracellular Desulfovibrio species and coccidia in ferrets
with proliferative bowel disease Lab Anim Sci 46569ndash571
Litster AL Nichols J Hall K et al (2014) Use of ponazuril paste to treat coccidiosis in shelter-housed
cats and dogs Vet Parasitol 202319ndash325 doi 101016jvetpar201403003
Matsubayashi M Takami K Abe N et al (2005) Molecular characterization of crane coccidia Eimeria
gruis and E reichenowi found in feces of migratory cranes Parasitol Res 9780ndash83
Meeusen ENT Walker J Peters A et al (2007) Current status of veterinary vaccines Clin Microbiol Rev
20489ndash510 doi 101128CMR00005-07
Mehlhorn H Aspock H (eds) (2008) Coccidial Drugs In Encyclopedia of Parasitology 3rd edn Springer
Berlin Heidelberg New York pp 269ndash286
Morehouse NF (1938) The Reaction of the Immune Intestinal Epithelium of the Rat to Reingection wiht
Eimeria nieschulzi J Parasitol 24311ndash317
Nukerbaeva KK Svanbaev SK (1973) Coccidia of fur bearing mammals in Kazakhstan Vestn Selrsquoskokh
Nauk Kazakh 1250ndash54
Ogedengbe JD Hanner RH Barta JR (2011) DNA barcoding identifies Eimeria species and contributes to
the phylogenetics of coccidian parasites (Eimeriorina Apicomplexa Alveolata) Int J Parasitol
153
41843ndash850
Ogedengbe ME (2015) DNA Barcoding of Apicomplexa Mitochondrial Evolution across the Phylum
University of Guelph
Ogedengbe ME El-Sherry S Ogedengbe JD et al Whatrsquos in a name Phylogenies based on combined
mitochondrial and nuclear sequences conflict with morphologically defined genera in the eimeriid
coccidia (Apicomplexa)
Ogedengbe ME El-Sherry S Whale J Barta JR (2014) Complete mitochondrial genome sequences from
five Eimeria species (Apicomplexa Coccidia Eimeriidae) infecting domestic turkeys Parasit
Vectors 7335 doi 1011861756-3305-7-335
Ogedengbe ME Hafeez MA Barta JR (2013) Sequencing the complete mitochondrial genome of Eimeria
mitis strain USDA 50 (Apicomplexa Eimeriidae) suggests conserved start positions for mtCOI- and
mtCOIII-coding regions Parasitol Res 1124129ndash4136 doi 101007s00436-013-3604-z
Pantchev N Gassmann D Globokar-Vrhovec M (2011) Increasing numbers of Giardia (but not
coccidian) infections in ferrets 2002 to 2010 Vet Rec 168519 doi 101136vrd2962
Patterson M Fox JG (2007) Parasites of Ferrets In Baker DG (ed) Flynnrsquos Parasites of Laboratory
Animals 2nd edn Wiley Blackwell pp 501ndash508
Patterson MM Fox JG Eberhard ML (2014) Parasitic Diseases In Fox JG Marini RP (eds) Biology and
Diseases of the Ferret 3rd edn Wiley Blackwell pp 553ndash572
Powers L V (2009) Bacterial and parasitic diseases of ferrets Vet Clin North Am - Exot Anim Pract
12531ndash561 doi 101016jcvex200906001
Prado ME Ryman JT Boileau MJ et al (2011) Pharmacokinetics of ponazuril after oral administration to
healthy llamas (Llama glama) Am J Vet Res 721386ndash9
154
Ruiz A Muntildeoz MC Molina JM et al (2013) Primary infection of goats with Eimeria ninakohlyakimovae
does not provide protective immunity against high challenge infections Small Rumin Res 113258ndash
266 doi 101016jsmallrumres201301006
Ryley J Meade R Hazelhurst J Robinson T (1976) Methods in coccidiosis research separation of
oocysts from faeces Parasitology 73311ndash326
Santymire R Branvold-Faber H Marinari PE (2014) Recovery of the Black-Footed Ferret In Fox JG
Marini RP (eds) Biology and Diseases of the Ferret 3rd edn Wiley Blackwell pp 219ndash231
Shi MQ Huther S Burkhardt E Zahner H (2000) Immunity in rats against Eimeria separata oocyst
excretion effects on endogenous stages and local tissue response after primary and challenge
infections Parasitol Res 86891ndash898
Sledge DG Bolin SR Lim A et al (2011) Outbreaks of severe enteric disease associated with Eimeria
furonis infection in ferrets (Mustela putorius furo) of 3 densely populated groups J Am Vet Med
Assoc 2391584ndash1588 doi 102460javma239121584
Smith NC Wallach M Petracca M et al (1994) Maternal transfer of antibodies induced by infection with
Eimeria maxima partially protects chickens against challenge with Eimeria tenella Parasitology
109551ndash557
Svanbaev SK (1956) Materials on the fauna of coccidia of wild mammals in western Kazakhstan Tr
Instituta Zool Akad Nauk Kazachskoi SSR 5180ndash191
Tavareacute S (1986) Some probabilistic and statistical problems in the analysis of DNA sequences Am Math
Soc Lect Math Life Sci 1757ndash86 doi citeulike-article-id4801403
Tenter AM Barta JR Beveridge I et al (2002) The conceptual basis for a new classification of the
coccidia Int J Parasitol 32595ndash616
155
Upton SJ (2000) Suborder Eimeriorina Leacuteger 1911 In Lee JJ Leedale GF Bradbury P (eds) An
Illustrated Guide to the Protozoa vol 1 2nd edn Allen Press Lawrence Kansas pp 318ndash339
USFWS BFF Recovery Program (2017) Black-footed Ferret Managed Care Operations Manual
(BFFMCOM)
Vermeulen AN (2005) Vaccination against coccidial parasites The method of choice In Proceeding of
the 9th International Coccidiosis Conference
Williams BH Chimes MJ Gardiner CH (1996) Biliary coccidiosis in a ferret (Mustela putorius furo) Vet
Pathol 33437ndash439 doi 101177030098589603300412
Williams ES Mills K Kwiatkowski DR et al (1994) Plague in a Black-footed (Mustela nigripes)
presence J Wildl Dis 30581ndash585
Williams ES Thome ET Appel MJG Belitsky DW (1988) Canine Distemper in Black-Footed (Mustela
nigripes) from Wyoming J Wildl Dis 24385ndash398
Wozencraft WC (2005) Order Carnivora In Wilson DE Reeder DM (eds) Mammal Species of the
World A Taxonomic and Geographic Reference 3rd edn Johnrsquos Hopkins University Press
Baltimore Maryland p 2142
Yi-Fan C Le Y Yin D et al (2012) Emendation of 2 Isospora Species (Apicomplexa Eimeriidae)
Infecting the Steppe Polecat Mustela eversmanii Lesson 1827 in China to the Genus
Cystoisospora (Apicomplexa Sarcocystidae) Comp Parasitol 79147ndash152 doi 10165445311
Yu L Peng D Liu J et al (2011) On the phylogeny of Mustelidae subfamilies analysis of seventeen
nuclear non-coding loci and mitochondrial complete genomes BMC Evol Biol 1192 doi
1011861471-2148-11-92
Zou M Guo G Zhao Y Zhang Q (2014) Detection quantifications and pharmacokinetics of ponazuril in
156
healthy swine J Vet Pharmacol Ther 37598ndash602 doi 101111jvp12126
157
APPENDICES
158
Appendix 1 Shedding of oocysts of Eimeria ictidea in black-footed ferret (Mustela nigripes) dam and kit
family groups from 2014-2016
Collection Year 2014 2014 2014 2014 2014 2015 2016
Age of kits (days) Poppy Bumblefoot Calico Aubrey Ruckus Fiddlesticks Guanella^
29 - - - 0 - - -
30 - 0 - 0 - - -
31 - 0 - 0 - - -
32 - 0 - 0 - - -
33 - 0 - 0 - - -
34 0 0 - 0 0 - -
35 0 0 - 0 0 0 -
36 0 0 - 0 0 0 -
37 - 0 - 0 - 0 -
38 - 0 - 0 - 0 -
39 - 0 - 0 - 0 -
40 0 0 - 0 0 0 -
41 - 0 - 0 - 0 -
42 0 0 - 0 0 - -
43 0 0 - 0 0 0 -
44 0 0 - - 0 0 -
45 0 0 - 0 0 - -
46 0 0 - 0 0 0 -
47 0 0 - 0 0 0 -
48 0 lt14 - 0 0 0 -
49 0 0 - 0 0 0 -
50 0 0 - - 0 0 -
51 0 0 - 0 0 0 -
52 0 0 - 0 0 0 -
53 0 0 - 0 0 0 +
54 0 0 - 0 0 0 +
55 0 0 - 0 0 0 3717146
56 0 0 - 0 - 0 1084436
57 0 0 - 0 - 0 64133
58 0 324 - 0 0 0 20654
59 0 0 - 0 0 0 0
60 0 446688 - 0 0 0 0
61 0 934828 - 0 0 0 41111
62 0 530469 - 0 0 0
63 0 1617131 - 0 0 lt14
64 0 8396357 - 114 0 0
65 0 234876 - 5368 0 lt14
66 0 374625 - 9455 0 0
67 lt14 2311575 - 39579 0 -
68 0 603563 - 125051 0 0
69 - 4103036 - 27067 0 lt14
70 2470327 7759974 - 0 0 0
71 1073085 - 1784 0 0
72 3430966 - 0 0 0
73 4459536 - 7326 0 0
74 4842212 - 2 0 0
75 3627307 - 0 0 0
76 5801885 - 0 0 0
77 5994 0 0 0 0
78 589835 0 0 0
Numbers of Oocysts Shed (oocysts per gram of feces)
Dam Identity
159
Appendix 1 continued Shedding of oocysts of Eimeria ictidea in black-footed ferret (Mustela nigripes)
dam and kit family groups from 2014-2016
Collection Year 2014 2014 2014 2014 2014 2015 2016
Age of kits (days) Poppy Bumblefoot Calico Aubrey Ruckus Fiddlesticks Guanella^
79 24815 0 0 0
80 144016 0 0 0
81 97862 0 0 0
82 0 0 0
83 0 0 0
84 0 0 0
85 0 0 0
86 0 0 0
87 0 0 0
88 0 0 0
89 0 0 0
90 0 0 0
91 0 0 0
92 0 - -
93 0 - 0
94 0 - 0
95 0 0 -
96 0 - 0
97 0 0 -
98 0 0 0
99 0 0 0
100 0 0 0
101 0 0 0
102 0 0 0
103 0 0 0
104 0 0 0
105 0 0 -
106 0 0 0
107 0 0 0
108 - 0 0
109 0 0 -
110 0 0 0
111 0 0 -
112 0 0 0
113 0 0 -
114 0 0 -
115 0 0 -
116 0 0 -
117 0 -
118 0 -
119 0 0
120 0 0
121 0 0
122 0 -
Numbers of Oocysts Shed (oocysts per gram of feces)
Dam Identity
160
Appendix 1 continued Shedding of oocysts of Eimeria ictidea in black-footed ferret (Mustela nigripes)
dam and kit family groups from 2014-2016
Collection Year 2014 2014 2014 2014 2014 2015 2016
Age of kits (days) Poppy Bumblefoot Calico Aubrey Ruckus Fiddlesticks Guanella^
123 0 0
124 0 -
125 0 -
126 0 0
127 0 0
128 0 2843
129 0 0
130 0 -
131 0 0
132 0 0
133 0 0
134 0 0
135 0 0
136 0 0
137 0
138 0
139 0
140 0
141 0
142 0
143 0
144 0
145 0
146 -
147 0
148 0
149 0
150 0
Legend lt 14 = oocyst positive samples with less than 14 oocyst per gram of feces - = no sample recorded for this date
underline = last sampling date + = coccidia present but OPG count not performed = Toronto Zoo ferret ^ = Louisville Zoo
ferret thick outer border = days treatment was received
Numbers of Oocysts Shed (oocysts per gram of feces)
Dam Identity
161
Appendix 2a Hematology values for domestic ferrets (Mustela putorius furo) from 49-51 days of age
prior to experimental inoculation
Test Reference
Intervala
Reference
Intervalb
101 102 103 104 105 201 202 203 204 205
Age (days) adult 70 50 50 50 50 50 48 48 48 50 48
WBC (x 109L) 27-112 53-120 86 64 77 97 76 48 8 72 104 76
RBC (x 1012
L) 50-108 55ndash74 66 58 45 5 48 47 67 5 5 42
Hb (gL) 87-177 104ndash136 121 106 90 58 94 91 122 96 98 80
HCT (LL) 04 - 051 029ndash037 037 033 027 030 029 027 038 030 030 024
MCV (fL) 44-52 478ndash548 55 56 60 60 60 58 57 59 61 58
MCH (pg) 15-18 175ndash191 18 18 20 12 20 20 18 19 20 18
MCHC (gL) 325-362 347ndash370 331 327 328 196 325 337 321 327 324 331
RDW () 12-16 - 134 127 139 139 133 131 122 136 131 127
Platelets (x 109L) 54-695 629ndash775 777 329 529 480 640 402 534 336 527 363
MPV (fL) 5-10 - 78 78 96 75 66 74 76 81 74 82
TS Protein (gL) 49-76 - 54 51 - - - - - - - -
Seg Neuts (x 109L) 1-8 15ndash48 292 141 216 281 205 187 152 151 354 251
Lymphocytes (x 109L) 1-63 28ndash63 525 416 516 64 509 254 608 468 645 456
Monocytes (x 109L) 0-09 01ndash05 026 07 031 039 038 034 024 094 031 038
Eosinophils (x 109L) 0-13 01ndash06 009 013 008 01 008 005 016 007 010 015
Basophils (x 109L) 0-02 0 009 0 0 0 0 0 0 0 0 0
Polychromasia 2-5 - 5-10 2-5 10-15 10-15 10-15 10-15 2-5 10-15 10-15 5-10
Anisocytosis Occ 1+ 1+ 1+ 1+ 1+ 1+ 1+
HJ bodies rare rare rare rare rare
crenation Occ
poikilocytosis Occ Occ Occ
shift platelets Occ Occ
hemolysis Neg Neg Neg Neg Neg Neg Neg Neg Neg Neg Neg
lipemia mild mild mild mild mild Neg Neg Neg mild mild
Ferret Identity
Legend - = reference value unavailable bolded = outside reference range a = adult ferret reference ranges from Animal Health Laboratory
(University of Guelph Guelph Canada) b
= reference ranges for 10-week old ferrets (Fox JG 2014) Neg = negative Occ = occasional
162
Appendix 2b Serum biochemistry values for domestic ferrets (Mustela putorius furo) from 49-51 days of
age prior to experimental inoculation
Test Reference
Intervala
Reference
Intervalb
101 102 103 104 105 201 202 203 204 205
Age (days) adult 70 50 50 50 50 50 48 48 48 50 48
Calcium (mmolL) 185-242 253-302 239 233 241 244 24 221 242 234 253 242
Phosphorus (mmolL) 112-256 278-475 239 267 29 264 31 218 286 284 315 303
Magnesium (mmolL) 08-139 - 08 08 07 08 08 06 08 07 08 08
Sodium (mmolL) 147-159 146-154 149 149 149 149 148 144 153 148 152 152
Potassium (mmolL) 37-57 47-83 44 46 42 46 48 45 47 44 47 46
Chloride (mmolL) 111-129 115-121 110 112 115 113 113 110 119 117 117 120
Carbon dioxide (mmolL) 17-29 13-27 18 18 18 17 17 16 15 15 19 17
Anion gap (mmolL) 6 - 23 - 25 24 20 24 23 23 24 20 21 20
NaK ratio - - 34 32 35 32 31 32 33 34 32 33
Total protein (gL) 51-75 44-56 49 46 44 47 44 41 52 45 49 43
Albumin (gL) 24-40 26-32 29 26 28 28 28 25 28 27 29 25
Globulin (gL) 19-41 17-24 20 20 16 19 16 16 24 18 20 18
AG ratio 053-167 13ndash12 145 130 175 147 175 156 117 15 145 139
Blood urea nitrogen (mmolL) 45-153 71-139 83 89 107 11 136 73 119 105 118 135
Creatinine (umolL) 8-67 53-124 50 58 68 59 50 7 68 41 62 92
Glucose (mmolL) 32-91 64-138 47 54 54 42 55 53 57 56 52 59
Cholesterol (mmolL) 294-894 619-860 413 386 375 393 344 349 227 357 373 346
Total bilirubin (umolL) 2 to 7 - 1 L 0 L 0 0 0 1 0 0 0 0
Conjugated biilrubin (umolL) - 0-10 1 0 0 1 1 1 0 0 1 0
Free bilirubin (umolL) 0-2 0-15 0 0 0 0 0 0 0 0 0 0
ALKP (UL) 13-237 117ndash277 180 169 172 215 175 168 241 184 177 179
GGT (UL) 0-40 2ndash20 1 1 6 10 4 0 1 1 5 9
AST (UL) - 63ndash152 61 58 48 61 64 59 93 61 58 69
ALT (UL) 39-196 95ndash544 95 105 89 105 106 82 234 137 115 156
CK (UL) 74-294 - 513 330 496 560 530 492 793 539 479 724
Amylase (UL) - - 23 28 35 35 29 28 29 24 36 23
Lipase (UL) - - 65 63 60 64 62 56 67 69 60 68
Calculated osmo (mmolL) - - 298 300 301 301 303 289 311 300 308 311
Ferret Identity
Legend - = reference value unavailable bolded = outside reference range a = adult ferret reference ranges from Animal Health Laboratory
(University of Guelph Guelph Canada) b = reference ranges for 10-week old ferrets (Fox JG 2014)
163
Appendix 3a Hematology values for domestic ferrets (Mustela putorius furo) inoculated orally with
Eimeria ictidea
Test Reference
Intervala
Reference
Intervalb
102 103 104 105 201 203 205
Age (days) adult 98-112 92 91 99 99 97 74 92
Days post inocculation 8 7 15 15 15 11 10
WBC (x 109L) 27-112 52ndash150 94 124 81 85 88 85 142
RBC (x 1012
L) 50-108 62ndash92 71 62 73 60 65 63 65
Hb (gL) 87-177 127ndash159 122 110 122 98 103 114 102
HCT (LL) 04-051 030ndash043 037 033 037 031 032 035 032
MCV (fL) 44-52 50ndash54 53 53 52 51 50 55 49
MCH (pg) 15-18 16-21 17 18 17 16 16 18 16
MCHC (gL) 325-362 351ndash426 328 332 325 319 320 326 317
RDW () 12-16 - 122 126 129 129 145 127 145
Platelets (x 109L) 54-695 376ndash610 524 413 445 117 303 42 429
MPV (fL) 5-10 - 8 78 72 66 67 14 8
TS Protein (gL) 49-76 - 66 63 58 66 55 59 59
Seg Neuts (x 109L) 1-8 21ndash62 160 446 203 170 211 170 554
Lymphocytes (x 109L) 1-63 16ndash79 761 657 551 646 59 646 682
Monocytes (x 109L) 0-09 01ndash02 019 087 024 026 044 026 085
Eosinophils (x 109L) 0-13 03ndash09 0 037 032 009 026 009 099
Basophils (x 109L) 0-02 0 0 012 0 0 009 0 0
Polychromasia 2-5 - 1-3 0-2 0-2 1-3 1-3 2-5 5-10
Anisocytosis Occ Occ Occ
HJ bodies rare
crenation
rouleaux
poikilocytosis
shift platelets Occ Occ
hemolysis Neg
Ferret Identity
Legend - = reference value unavailable bolded = outside reference range a = adult ferret reference ranges from Animal Health
Laboratory (University of Guelph Guelph Canada) b
= reference ranges for 10-week old ferrets (Fox JG 2014) Neg = negative
Occ = occasional = many platelet clumps
164
Appendix 3b Serum biochemistry values for domestic ferrets (Mustela putorius furo) inoculated orally
with Eimeria ictidea
Test Reference
Intervala
Reference
Intervalb
102 103 104 105 201 203 205
Age (days) adult 98-112 92 91 99 99 97 74 92
Days post inocculation 8 7 15 15 15 11 10
Calcium (mmolL) 185-242 245-268 243 222 238 233 232 236 240
Phosphorus (mmolL) 112-256 200-323 292 223 248 298 268 288 306
Magnesium (mmolL) 08-139 - 06 08 08 08 09 08
Sodium (mmolL) 147-159 148-155 147 150 150 148 148 150
Potassium (mmolL) 37-57 45-55 42 41 51 41 50 44
Chloride (mmolL) 111-129 114-124 114 113 118 112 111 115
Carbon dioxide (mmolL) 17-29 16-24 16 21 19 20 12 18
Anion gap (mmolL) 6 - 23 - 21 20 18 20 30 21
NaK ratio - - 35 37 29 36 30 34
Total protein (gL) 51-75 49-64 52 55 55 49 56 57
Albumin (gL) 24-40 30-36 28 20 27 27 26 29 27
Globulin (gL) 19-41 19-30 32 28 28 23 27 30
AG ratio 053-167 11-17 063 096 096 113 107 090
Blood urea nitrogen (mmolL) 45-153 50-150 92 87 119 92 110 98 149
Creatinine (umolL) 8-67 706-1414 56 26 49 40 50 46 79
Glucose (mmolL) 32-91 688-943 52 59 64 71 58 17 66
Cholesterol (mmolL) 294-894 440-640 453 429 262 475 370 285
Total bilirubin (umolL) 2 to 7 - 0 0 0 0 1 1
Conjugated biilrubin (umolL)- - 1 0 0 0 0 0
Free bilirubin (umolL) 0-2 - 0 0 0 0 1 1
ALKP (UL) 13-237 41-181 124 120 213 120 146 170 196
GGT (UL) 0-40 1-2 2 1 3 0 0 2
AST (UL) - 47-128 48 95 104 66 100 100
ALT (UL) 39-196 78-279 133 110 140 203 158 183 281
CK (UL) 74-294 - 382 765 1190 578 680 930
Amylase (UL) - - 24 37 31 28 33 35
Lipase (UL) - - 60 65 72 72 86 79
Calculated osmo (mmolL) - - 296 305 305 300 296 309
Ferret Identity
Legend - = reference value unavailable bolded = outside reference range a = adult ferret reference ranges from Animal Health
Laboratory (University of Guelph Guelph Canada) b
= reference ranges for 10-week old ferrets (Fox JG 2014)
165
Appendix 4 Domestic ferret (Mustela putorius furo) weekly monitoring sheet
Mon
8 AM
Mon
4 PM
Tues
8 AM
Tues
4 PM
Wed
8 AM
Wed
4 PM
Thurs
8 AM
Thurs
8 PM
Fri
8 AM
Fri
4 PM
Sat
8 AM
Sat
4 PM
Sun
8AM
Sun
4 PM
Mentation
Weight (g)
Respiratory
Rate
Vomit
(+ ++ +++)
Diarrhea
(+ ++ +++)
Urination
(+ ++ +++)
Defecation
(+ ++ +++)
Food
offered
Food
remaining
Water remaining
(ml)
Treatments
Other
observations
Initials of
observer
166
Animal ID ________________________________________ Week ______________
Monitoring Criteria
Mentation BAR (bright alert responsive) QAR (quiet alert responsive) depressed lethargic quiet
Weight measured in grams to be performed once weekly (pre-inoculation) and once daily (post-inoculation)
Respiratory rate measured in breaths per minute
Vomit + (small amount) ++ (moderate amount) +++ (large amount)
Diarrhea + (small amount) ++ (moderate amount) +++ (large amount)
Urination + (small amount) ++ (normal amount) +++ (large amount)
Defecation + (one pile) ++ (two piles) +++ (3+ piles)
Food offered Y (yes) N (no)
Food remaining A (all) P (partial) N (none)
Please note any additional observations in the appropriate section
Monitoring Times
Pre-inoculation ndash ONCE daily at 8am during cage cleaningfeeding
Post-inoculation ndash TWICE daily at 8am and 4pm
Critical patients (as determined by veterinary examination) ndash 3-6 times daily (based on veterinarian recommendation) ndash switch to 24
hour care sheet
Intervention Points
If any ferret develops vomiting diarrhea goes off food or exhibits gt 3 weight loss both the PI and graduate student listed below are
to be notified and the animal will be assessed to determine further treatment plans
Removal Criteria
If an individual ferret develops severe gastrointestinal or systemic disease during the course of the study as characterized by abnormal
mentation marked dehydration 10 weight loss within 24-48 hours profuse diarrhea unresponsive to treatment anorexia of 2 or
more days or frequent vomiting unresponsive to treatment the individual will be removed from the study and humanely killed
167
Appendix 5 Domestic ferret (Mustela putorius furo) 24 hour intensive monitoring sheet
Animal ID ___________________________________________ Date___________________
0700 800 900 1000 1100 1200 1300 1400 1500 1600 1700 1800
Mentation
Weight (g)
Heart Rate
Respiratory Rate
Temperature (C)
Water offered
(mL)
Water consumed
(mL)
Food offered
(type and
amount in
grams)
Food consumed
(type and
amount in
grams)
Presence of urine
Presence of feces
168
Character of
feces
Presence of
vomit
Character of
vomit
Medications to
be administered
Other
observations
169
1900 2000 2100 2200 2300 2400 100 200 300 400 500 600
Mentation
Weight (g)
Heart Rate
Respiratory Rate
Temperature (C)
Water offered
(mL)
Water consumed
(mL)
Food offered
(type and amount
in grams)
Food consumed
(type and amount
in grams)
Presence of urine
Presence of feces
Character of
feces
Presence of vomit
Character of
vomit
170
Medications to be
administered
Other
observations
Monitoring Criteria
Mentation BAR (bright alert responsive) QAR (quiet alert responsive) depressed lethargic quiet
Weight measured in grams to be performed once weekly (pre-inocculation) and once daily (post-inocculation)
Respiratory rate measured in breaths per minute
Vomit + (small amount) ++ (moderate amount) +++ (large amount)
Diarrhea + (small amount) ++ (moderate amount) +++ (large amount)
Urination + (small amount) ++ (normal amount) +++ (large amount)
Defecation + (one pile) ++ (two piles) +++ (3+ piles)
171
Food offered Y (yes) N (no)
Food remaining A (all) P (partial) N (none)
Please note any additional observations in the appropriate section
Monitoring Times
Critical patients (as determined by veterinary examination) ndash 3-6 times daily (based on veterinarian recommendation) ndash switch to 24
hour care sheet
Intervention Points
If any ferret develops vomiting diarrhea goes off food or exhibits gt 3 weight loss both the PI and graduate student listed below are
to be notified and the animal will be assessed to determine further treatment plans
Removal Criteria
If an individual ferret develops severe gastrointestinal or systemic disease during the course of the study as characterized by abnormal
mentation marked dehydration 10 weight loss within 24-48 hours profuse diarrhea unresponsive to treatment anorexia of 2 or
more days or frequent vomiting unresponsive to treatment the individual will be removed from the study and humanely killed
172
Appendix 6 Domestic ferret (Mustela putorius furo) infection trial standard operating procedures
Daily Fecal Collection
1 Collection of all feces passed in a 24 hour period will be performed once daily for all ferrets from
July 13 to July 24 inclusive
2 CAF Isolation staff will collect all fecal material present at the time of daily cage cleaning and
place in individual pre-labelled plastic bags (one per cage)
3 The amount of non-fecal matter (bedding etc) collected should be as minimal as possible
4 CAF isolation staff will record fecal character observations daily for each sample by ticking the
appropriate box on the baggie label
5 Isolation staff will place samples in the necropsy cooler (4 degC) while awaiting collection by
summer student
6 Monday to Friday fecal samples will be collected by A Rodriguez and brought for processing to
the Barta Lab in Pathobiology Samples collected Saturday and Sunday will be put on hold in a
refrigerator (4degC) until pick up on Monday
Example Bag label
Ferret ID Date
Weight of Feces
Fecal Character Normal
Soft
Liquid
Bloody
Abnormal odour
Physical Examination and Health Assessments
1 Manual restraint by hand or in towel by CAF Isolation staff and project personnel If required
examination +- blood collection may be performed under general anesthesia with isoflurane
2 Physical examination body weight HR RR temperature to be performed by PIs (Adriana Pastor
and Dale Smith)
3 Blood collection (~1mL per ferret) is to be performed from the jugular vein if under manual
restraint or the cranial vena cava under anesthesia by PIs using a 25g needle and 1cc syringe
Blood will be collected into small heparinized tubes and submitted to AHL for
CBCBiochemistry
4 All physical examination findings will be recorded on the Exam Sheet
5 Any minor wounds will be treated as appropriate (to be determined by PIs)
Inoculation of Ferrets with Coccidia
1 All ferrets to be inoculated will have been confirmed negative on daily fecals for two weeks
173
2 Brief physical examination by PIs to confirm that the ferrets are healthy to continue in study
3 The concentrated oocyst solution in sterile saline (up to a volume of 1mL) will be combined with
up to 1 mL of FerreTone (or another highly palatable substance if more preferred by the ferrets)
and ferrets will be allowed to consume the mixture ad lib while being monitored
4 Control ferrets will be administered saline only (equal mL to oocyst solution) with 1 mL of
FerreTone (or other substance as used for experimental group)
5 If ferrets refuse to consume the mixture oral inoculation via syringe of concentrated oocyst
solution will be performed by PIs under manual restraint
6 If ferrets are resistant to manual restraint for oral inoculation then inoculation will be performed
by PIs under general anesthesia via gastric tube (8 Fr red rubber)
Euthanasia Protocol
1 Ferrets to be euthanized will be masked down with isoflurane under manual restraint (or in an
anesthetic chamber as judged most appropriate by the PI)
2 Once anesthetized the ferret will be weighed and any blood fecal or other samples will be
collected as required (as determined by PIs)
3 Once an adequate plane of anesthesia is obtained (as determined by PIs) potassium chloride at a
dose of 2 mEq K+kg will be administered either via vena caval puncture or cardiac puncture to
induce cardiac arrest
4 Presenceabsence of respirations heartbeat corneal reflex will be used to assess death
Necropsy Protocol
1 Measure the ferret from nose to tail base (body length)
2 Perform standard necropsy but start with gastrointestinal tract first then thoracic and abdominal
viscera
3 Gently flush the entire contents of the intestine with 12 mL saline into a sterile urine cup
Potassium dichromate should be added in a 11 volumevolume ratio to the same container and
mixed with the combined intestinal contents and saline Label the container with the ferretrsquos ID
ferret group ID date and place on appropriate shelf in the refrigerator (4 degC) in the Barta lab
4 Measure the length of the gut from duodenum to anus
5 Collect paired sections of intestine for histological sectioning and frozen These sections should
be collected along the entire length of the gut from duodenum to rectum (see below for GI length
calculations)
6 For histological sections cut a 2 cm long section of bowel open completely on one side place on
a pre-cut section of box-board and staple both ends to the board to create a flat section Place all
gastrointestinal sections (attached to the board) in Serra fixative solution (100 ethanol (60
VV) 37 formaldehyde (30 VV) glacial acetic acid (10 VV)) for fixation and trim in
cross (transverse) sections
7 Adjacent to each sample removed for histopathology remove another 2 cm section intact and
place in a labelled Whirl-pack for freezing
8 Box-board and pre-labelled Whirl-packs should be labelled in pencil and sharpie respectively
with the following information Animal ID Zoo pm number section of gut (information should
include the region of the gut - jejunum colon etc and the length from the pylorus to the section)
174
Ferret gastrointestinal length calculations (from Evans amp An 2014)
Adult ferret body length 36-41 cm
NB In domestic ferrets there is ~51 ratio of small intestine to body length
Adult domestic ferret GI lengths
Small intestine ndash 182-198 cm
Large intestine - ~10 cm long 06 cm diameter (colon ndash 7 cm rectum ndash 2 cm anus ndash 1 cm)
Based on the above information
Small intestine Six sections of the small intestine from duodenum to ileum will be collected each ~ 25-40
cm apart depending on the size of the ferret
Large intestine Collect two sections of colon (4 cm each) at 25 and 75 of the length of the colon from
the junction of the small and large intestine to the rectum Smaller ferrets may allow only on section
based on colon length
Laboratory SOPs
General notes on processing fecal samples
1 Samples will be collected from the isolation facility necropsy room cooler daily from Monday-
Friday for the entire 6 weeks of the project
2 Upon transfer to the lab each baggie will be weighed after filling to determine the amount of
feces (grams) in each bag and that number entered onto the label on the bag
3 All information regarding fecal processing will be entered onto the spreadsheet for the individual
ferret including ID weight of feces feces character date of sample collection date of sample
processing fecal flotation method presence or absence of oocysts oocyst quantification
Fecal Processing - Weeks 1+2 (acclimation period)
1 After weighing transfer the contents of one baggie to a small sieve on top of a paper cup
2 Fill a single glass container (for fecal flotation) with saturated salt solution
3 Slowly pour small amounts of the solution over the feces mix and strain liquid contents in the paper
cup using a tongue depressor
4 Dispose of the remaining fecal matter in the sieve
5 Pour the contents of the paper cup back into the glass container and cover slip for 5-7 minutes
6 Place coverslip on a clean glass slide and viewscan under microscope at 10x power for presence of
oocysts
7 Record all findings on the provided spreadsheet
8 Notify A Pastor if any oocysts are detected If detected contents of the slide should be flushed back
into the glass container with distilled water and contents of glass container should be transferred to a
50 mL plastic conical tube Clean to remove salt as per Barta lab SOP and mix with potassium
dichromate equal parts by volume to amount of oocysts in diH2O Place a checkmark on the lid of the
50 mL conical vial containing the remainder of the feces and store in the lab fridge at 4 degC
175
Fecal Processing - Weeks 3-7 (infection trial)
1 After weighing transfer the entire contents of one baggie to a small sieve on top of a paper cup
2 Slowly pour small amounts of distilled water over the feces (enough to wet) mix and strain liquid
contents in the paper cup using a tongue depressor until feces appear almost dry
3 Dispose of the remaining fecal matter in the sieve
4 Pour the contents of the paper cup into a 50 mL conical tube If samples are not going to be counted
on the same day then mix potassium dichromate (25 wv) 11 by volume with sieved fluid from
the paper cup and store at 4 degC (refrigerator) until ready to perform OPG counts
If proceeding with the count the same day
5 Pipette and place one drop of the sieved fecal fluid on a slide to determine approximate oocyst
concentration If oocysts rare to none ndash dilute 12 during step 6 if moderate numbers ndash dilute 19 if
too numerous to count ndash dilute 199 (or perform serial dilutions of 10x from initial 19 dilution)
6 Transfer x mL of the mixture from the 50 mL conical tube into a clean 15 mL conical tube and mix
with appropriate amount of saturated salt solution for desired dilution
7 Fill both sides of the McMaster counter chamber and count the number of oocysts per side (total for
one side= total number of oocysts from all 6 sections of the chamber )
8 Average the total counts from both sides
9 Use the following calculation to determine the oocyst per gram count
OocystsmL = oocysts counted times 666 times dilution (ie 3 if dilution 12)
OPG= oocystmL times total volume recorded at end of step 4
10 Record all findings on the provided spreadsheet
11 Notify A Pastor if any oocysts are detected If detected place a checkmark on the lid of the 50 mL
conical vial containing the remainder of the feces and store in the lab fridge at 4 degC
12 For all samples for which no oocysts are detected during steps 6-8 follow up with routine salt
flotation of the remainder of the fecal sample from step 6 (use instructions for fecal processing from
weeks 1-2)