involvement of hsp70, a stress protein, in the resistance of long-term culture of pc12 cells against...
TRANSCRIPT

Arch Toxicol (2010) 84:699–708
DOI 10.1007/s00204-010-0565-1MOLECULAR TOXICOLOGY
Involvement of Hsp70, a stress protein, in the resistance of long-term culture of PC12 cells against sodium nitroprusside (SNP)-induced cell death
Carmen Romero · Juana Benedí · Angel Villar · Sagrario Martín-Aragón
Received: 29 March 2010 / Accepted: 31 May 2010 / Published online: 18 June 2010© Springer-Verlag 2010
Abstract Sodium nitroprusside (SNP)-treated PC12 cellline is being used in our laboratory as a cell model of nitricoxide (NO)-mediated damage for in vitro evaluation ofpotential neuroprotective compounds, thus cell response toSNP must be standardized to gain reproducible data. TheNO-donor SNP has been shown to induce cell death at highconcentrations in undiVerentiated PC12 cells. DiVerenceswere found in sensitivity to SNP between cells from short-and long-term cultured cells. After 24-h exposure to 100–500 �M SNP, a decrease of cell viability was observed inboth short- (17, 21 and 23rd passages) and long-term cul-tures (46, 49 and 50th passages), with IC50 values of 312.72and 462.90 �M, respectively. In cells from early passages,SNP-induced cell death was accompanied by signiWcantincreases of LDH leakage, nitrite production, malondialde-hyde (MDA) levels, catalase (CAT) activity, cleavage ofpoly(ADP-ribose)polymerase (PARP) and caspase-3 acti-vation in comparison with those from late passages. Fur-thermore, untreated and SNP-treated cells from long-termcultures displayed an increase of the stress protein Hsp70levels when compared with those from short-term cultures.Up-regulated levels of Hsp70 may be associated with cellsurvival. Therefore, cells may acquire a certain resistanceto SNP-induced toxicity associated with an increase in cellpassage-dependent Hsp70. The protein Hsp70 might modu-late the cellular response to the toxic insult by increasingCAT and GSH-Px activities and decreasing caspase-3 acti-
vation. Finally, it is crucial for the standardization of thiscell model of neurotoxicity, at least in part, the use of PC12cells in an optimum and reliable range of passages.
Keywords Resistance · Short-term culture · Long-term culture · Hsp70 · Sodium nitroprusside (SNP) · PC12 cell line
Introduction
PC12 cell line is a rat adrenal gland pheochromocytomacell line which has been widely used as a model for study-ing the survival of neuronal cells or antioxidant protection(Heo et al. 2002; Sasaki et al. 2007). In the present study,PC12 cell line was chosen since as it has low viability bynitric oxide (NO) and gradually develops a characteristicpattern associated with apoptosis events. The interest in NOlies in its involvement in many pathophysiologies of thecentral nervous system (CNS) including brain ischemia,inXammation and neurodegeneration (Jung et al. 2007).Therefore, models of NO-mediated cytotoxicity could bepartially useful tools to evaluate strategies for prevention ortreating those pathologies.
Sodium nitroprusside (SNP), a NO donor, is a water-sol-uble iron nitrosyl complex which in basic research has beenused to mainly investigate the cytotoxic eVects of nitrosa-tive stress (Cardaci et al. 2008). Altogether, SNP-treatedPC12 cell line is presented here as a cell model of NO-med-iated damage for further in vitro evaluation of potentialneuroprotective compounds with reported antioxidant prop-erties (unpublished results). In fact, it is our aim to developand use cell line models of free-radical-mediated toxicitywhich mimics or resembles, with clear limitations, oxida-tive damage-related neuropathologies (Molina-Jimenez
C. Romero · J. Benedí · A. Villar · S. Martín-Aragón (&)Pharmacology Department, School of Pharmacy, Complutense University of Madrid, Plaza Ramón y Cajal s/n, 28040 Madrid, Spaine-mail: [email protected]
123

700 Arch Toxicol (2010) 84:699–708
et al. 2003; Benedi et al. 2004; Molina-Jimenez et al. 2004,2005; Sanchez-Reus et al. 2005) as an alternative to the useof experimental animals for testing pharmacologicalsubstances (Martin-Aragon et al. 1997a, b, 1998, 2001;Sanchez-Reus et al. 2007).
It is worth it to consider that the metabolic and geneticcharacteristics of permanently growing cell lines diVerremarkably depending on the culture passage (Saminathanet al. 2009), and it is especially well known that the proper-ties of the PC12 cells vary after long-term culture (Yuyamaet al. 2001). Thus, it could be suspected that cell responseto toxins may vary through serial passages.
A number of mechanisms are involved in cell resistanceor loss of cell sensitivity to toxin-, stressor-, or drug-induced apoptosis, and we previously reported some ofthem with clinical implications (Martin-Aragon et al. 2000;Fu et al. 2001; Majlessipour et al. 2001).
Among the responsible mechanisms of PC12 cell resis-tance against NO, the expression of the neuronal isoform ofNOS, but not eNOS and iNOS, has been associated with theprotection against NO-induced cytotoxicity in long-termcultures (Yuyama et al. 2001).
On the other hand, heat shock proteins (Hsps) are themajor components of the intracellular defense system(Papp et al. 2003) and are reported to reduce inXammationand apoptosis in a variety of brain insults (Sinn et al.2007). Increased expression of some Hsps, e.g., Hsp70,occurs after exposure of cells to environmental and physi-ological stressors such as non-lethal elevations in tempera-ture or oxidants (Gorman et al. 1999). Hsps has beenshown to protect cells against apoptosis (Yang et al. 2008)by interfering with caspase activation at several diVerentlevels (Samali and Orrenius 1998; Lee et al. 2008). More-over, the mechanisms by which Hsp70 protects cells fromnecrosis may involve maintenance of cellular redox bal-ance and/or prevention of cellular damage as a result ofATP depletion (Quigney et al. 2003). In addition, Hsp70has been identiWed as a defense molecule against NO sinceoverexpression of Hsp70 confers resistance against SNP-induced NO cytotoxicity in rat insulinoma cells (Bellmannet al. 1996).
The need for standardizing experimental conditions inthe continuously growing cell line PC12 in order to gainrelevant and reproducible results led us to study the diVer-ences in cell response and sensitivity to SNP between short-and long-term cultures and therefore, Wnd the optimum andreliable range of cell passages.
On the assumption that the stress protein Hsp70 protectscells from NO-mediated cell death and its induction isrelated to an increase in cell survival, we hypothesized thatloss of sensitivity in PC12 cells to SNP in late passages isdue to an up-regulation of Hsp70 levels which might modu-late the cellular response to the toxic insult.
Materials and methods
Materials and reagents
All materials for cell line culture were obtained from Gibco(Invitrogen; CA, USA). Monoclonal anti-Hsp70, monoclo-nal anti-�-actin and horseradish peroxidase (HRP)-conju-gated antibodies were from Sigma–Aldrich (St. Louis, MO,USA). Monoclonal anti-PARP antibody and Xuorescentprobe for caspase-3 activity were purchased from BD Bio-sciences (San Jose, CA, USA). All other chemicals wereobtained from Sigma–Aldrich (St. Louis, MO, USA).
Cell culture and drug treatment
PC12 cells were kindly donated by Dr. Fando from theBiochemistry and Biology Department at the University ofAlcalá (Alcalá de Henares, Spain). Cells were culturedboth short-term (16–30th passages) and long-term (over45th passage) and maintained and grown routinely in Dul-becco’s modiWed Eagle’s medium (DMEM) supplementedwith 10% heat-inactivated horse serum (HS) and 5% heat-inactivated fetal bovine serum (FBS), 50 �g/ml gentamy-cin and 0.2 mM piruvate at 37°C in a humidiWedatmosphere of 5% CO2. During the logarithmic growthphase, cells were digested with trypsin at a concentrationof 0.25% and then were resuspended in the medium andseeded at appropriate densities on plates according to eachexperimental scale. Experiments were always carried out24 h after cells were seeded. For treatments, cells wereincubated in DMEM containing 1.0% HS and 0.5% FBS.Stock solutions of SNP were made in phosphate buVersaline (PBS). Subsequent dilutions of SNP were made incell medium (1.0% HS and 0.5% FBS) for cell treatments.In a set of pilot experiments, cells were treated with SNP atconcentrations ranging from 100 to 500 �M for variousintervals, 24, 48 and 72 h, respectively. Concentrations of200 and 400 �M SNP, respectively, as controls were usedin an extensive study of the markers of cell injury up to24-h exposure. For inhibition of Hsp70 synthesis, cellswere pre-treated with 5 �M quercetin for 3 h and thenincubated with SNP for 24 h.
MTT assay for cell viability
Methylthiazolyldiphenyl-tetrazolium (MTT) bromidereduction was used to examine the metabolic activity of thecells. The MTT assay is based on the reduction of the yel-lowish MTT to the insoluble and dark blue formazan byviable and metabolically active cells (Mosmann 1983).PC12 cells were subcultured in 96-well plates at a seedingdensity of 2 £ 104 cells/well. For assessing the cytotoxiceVect of SNP, PC12 cells were incubated with 200 and
123

Arch Toxicol (2010) 84:699–708 701
400 �M SNP for 24 h. Then, MTT dissolved in PBS, wasadded into each well to get the Wnal concentration of MTTat 0.5 mg/ml and incubation continued at 37°C for 1 h. Themedium with MTT solution was removed, and the forma-zan crystals were dissolved with DMSO. The optical den-sity (OD) which is proportional to metabolic activity ofcells was measured at 550 nm using a Digiscan MicroplateReader (Assys Hitech Kornenburg, Austria). Cell viabilitywas expressed as a percentage of cells untreated whichserved as the control group (untreated) and was designatedas 100%. MTT assay was also performed in cells underquercetin treatment.
Lactate dehydrogenase (LDH) assay
LDH (EC 1.1.1.27) is of a great value as an in vitro markerfor cellular toxicity. LDH released into the culture mediumwas measured by monitoring Xuorometrically the produc-tion of NAD+ from NADH during the conversion of pyru-vate to lactate (Imahashi 1968). For assessing the plasmamembrane integration upon the exposure to SNP, PC12cells were subcultured in 96-well plates at a seeding densityof 2 £ 104 cells/well. Twenty-four hours later cells wereincubated with 200 and 400 �M SNP for 24 h. The cellsupernatant (100 �l) was incubated with 0.18 mM NADPHand 0.60 mM pyruvate in 50 mM phosphate buVer (pH7.4). The rate of NAD+ formation was monitored for 1 minby reading the Xuorescence in a FLx800 microplate Xuores-cence reader (Bio-tek Instruments, Inc.) with the excitationWlter set at 360 nm and the emission Wlter set at 460 nm.LDH leakage was expressed as a percentage of untreated orcontrol cells which was designated as 100%.
Measurement of reactive oxygen species (ROS)
Intracellular accumulation of ROS was measured Xuoro-metrically. The non-Xuorescent dye 2�,7�-dichloroXuores-cin diacetate (DCFA-DA), used as a molecular probe, isfreely permeable to cells. The DCFA-DA is hydrolyzed to2�,7�-dichloroXuorescin (DCFH) inside the cells, where itconverts upon interaction with ROS to dichloroXuorescin(DCF), a green Xuorescent dye (LeBel et al. 1992). Cellswere plated at a density of 4 £ 104 cells/well in non-Xuo-rescent 96-well plates. Forty-eight hours later, each wellwas loaded with 20 �M DCFA-DA. Half an hour later, cellswere washed with PBS and 200 and 400 �M SNP wasadded to the cells, respectively. The Xuorescence causedafter 120-min exposure of the cells to SNP was measured.DCF Xuorescence reading of the wells was conducted usinga FLx800 microplate Xuorescence reader (Bio-tek, Instru-ments, Inc.) with the excitation Wlter set at 485 nm and theemission Wlter set at 530 nm. Data points were taken every15 min.
QuantiWcation of nitrites
The presence of nitrite, a stable oxidized product of NO, wasdetermined in phenol red-free cell medium by the Griessreagent (1% sulfanamide and 0.1% N-(1-naphtyl) ethylenedi-amine dihydrochloride in 2.5% H3PO4) (Miranda et al.2001). Griess’s reaction entails formation of a chromophorefrom the diazotization of sulfanilamide by acidic nitrite fol-lowed by coupling with bicyclic amines such as N-1-(naph-tyl) ethyendiamine. PC12 cells were plated at a seedingdensity of 2 £ 104 cells/well in 96-well plates. Twenty-fourhours later cells were incubated with 200 and 400 �M SNPfor 24 h. Afterward, 100 �l of cell culture supernatant wasremoved and combined with 100 �l of Griess reagent in a96-well plate followed by spectrophotometric measurementat 550 nm using a Digiscan Microplate Reader (Assys HitechKornenburg, Austria). Nitrite concentration in the superna-tants was determined by interpolation from a sodium nitritestandard curve. Nitrite content was expressed as a percentageof untreated cells which was designated as 100%.
Superoxide dismutase (SOD) activity
SOD activity was determined in cell lysates by means ofinhibition of pyrogallol autoxidation (Marklund and Marklund1974). The reaction mixture consisted of 0.2 mM pyro-gallol in air-equilibrated 50 mM Tris-cacodylic acid buVerpH 8.2, containing 1 mM diethylenetriaminepentaaceticacid. The rate of autoxidation was taken from the increasein absorbance at 420 nm and 25°C. One unit of the enzymeis generally deWned as the amount of the enzyme whichinhibits the reaction by 50%. SOD activity was expressedas International Units (IU) per milligram of protein.Enzyme activity was measured in an Uvikon-930 spectro-photometer (Kontron).
Catalase (CAT) activity
CAT was assayed in Triton X-100 (l%)-treated superna-tants from cell lysates following the disappearance of15 mM H2O2 in 50 mM phosphate buVer, pH 7.4, at240 nm for 1 min (Aebi 1984). One unit of enzyme decom-poses one �mol of H2O2 per minute at 25°C and pH 7.4.The activity was calculated using a molar extinction coeY-cient of 7.1 £ 10¡3 M¡1 cm¡1. CAT activity was expressedas International Units (IU) per milligram of protein (�molH2O2/min mg prot). Enzyme activity was measured in anUvikon-930 spectrophotometer (Kontron).
Glutathione Peroxidase (GSH-Px) activity
GSH-Px activity was measured in cell lysates according tothe method of Paglia and Valentine (Paglia and Valentine
123

702 Arch Toxicol (2010) 84:699–708
1967). GSH-Px catalyzes the oxidation of the 4 mM GSHby cumene hydroperoxide as substrate, in 50 mM phos-phate buVer, pH 7.4, including 4 mM sodium azide (inorder to inhibit catalase activity). The oxidized glutathioneproduced is converted to the reduced form in the presenceof 0.22 mM NADPH, 1 mM EDTA and 0.702 IU GR. Theconcomitant oxidation of NADPH was followed by thedecrease in absorbance at 340 nm and 25°C. One enzymeunit is deWned as the oxidation of 1 �mol NADPH per min-ute using an extinction coeYcient of 6.22 mM¡1 cm¡1. Theenzyme activity was expressed as mU per milligram of pro-tein (nmol NADPH/min mg prot). Enzyme activity wasmeasured in an Uvikon-930 spectrophotometer (Kontron).
Cell lysates for determining lipid peroxidation, PARP cleavage, Hsp70 levels and caspase-3 activation
PC12 cells in logarithmic growth phase were digested withtrypsin at a concentration of 0.25% and then resuspended inthe medium and subcultured in Petri dishes (100 £ 15 mm)at a seeding density of 1.5 £ 105 cells/ml. After 24 h, thecells were treated with 200 and 400 �M SNP, respectively,for 24 h. Then the cells were harvested, washed with PBS(pH 7.4) twice and recovered by centrifugation at 1000£gfor 5 min. The cell pellet was resuspended in PBS and cen-trifuged at 1000£g for 10 min at 4°C to remove the super-natant. After centrifugation, cell pellet samples were lysedin a lysis buVer consisting of 10 mM Tris–HCl (pH 7.5)with 0.5% CHAPS, 1 mM Cl2Mg, 1 mM EGTA, 1 mMEDTA, 10% glycerol, 5 mM �-mercaptoethanol, 1 mMDTT, 1 mM phenylmethylsulfonyl Xuoride (PMSF),100 �M leupeptin and 1 �M pepstatin. After incubation for30 min on ice, the cell lysate was cleared by centrifugationat 13000g for 5 min. Protein determinations were per-formed in the supernatant according to the bicinchonic acidmethod (Hill and Straka 1988).
Estimation of lipid peroxidation (MDA)
Malondialdehyde (MDA) was determined in cell lysatesfrom untreated or SNP-treated PC12 cells by HPLC asdescribed previously by our group (Bermejo et al. 1997).HPLC (Varian model 2510) was equipped with a SPD-M10AVP model diode array detector (Shimadzu). Thestationary phase was a Hypersil C18 column (ShandonScientiWc, UK; 25 cm 9 4.6 mm, 5 lm) that was thermostatedat 40°C. The mobile phase was PO4HNa2 and myristilam-monium bromide buVer (pH 7.4)/acetonitrile. The Xow ratewas maintained isocratically at 0.5 ml/min. The absorbanceof the eluent was monitored at 268 nm. Before injection,cell lysate samples were cleaned up by precipitation withacetonitrile in order to remove most of the protein content.Finally, the supernatant solutions were Wltered (0.2-�m
pore diameter membrane) and directly injected into the col-umn. Injection volumes were 10 �l. The MDA peak in thechromatogram was identiWed by comparison with a refer-ence chromatogram of freshly prepared free MDA. Theconcentration of MDA was expressed as nmol MDA/mgprotein.
Western blot analysis
The degree of poly(ADP-ribose)polymerase (PARP)cleavage and levels of Hsp70 were measured by Westernblot. Cells were incubated with 200 and 400 �M SNP,respectively, for 24 h. Afterward, cells were collected forcell lysis. When needed cytosolic lysates were alsoobtained. An equal amount of protein from each cell lysatesample was boiled and electrophoresed under reducingconditions in a sodium dodecyl sulphate (SDS)/polyacryl-amide gel. Transfer of proteins was carried out to a PVDFmembrane for 1 h. Non-speciWc binding was inhibited byincubation the membranes in blocking buVer containing10% non-fat dried milk for 1 h. Afterward, membraneswere exposed to a monoclonal anti-PARP (1:20000), and amonoclonal anti-Hsp70 (1:200000) antibodies, respec-tively, overnight at 4°C. Membranes were incubated withhorseradish peroxidase (HRP)-conjugated secondaryantibodies followed by detection with enhanced chemilu-minescence (ECL) (Amersham Bioscience Corp., Piscata-way, NJ). The intensity of the bands was analyzed andcalculated by the Multigenius gel documentation and anal-ysis system (Syngene, Synoptics Limited, Cambridge) andwas normalized to �-actin. Membranes were stripped andreprobed with monoclonal mouse anti-�-actin (1:50000) asprimary antibody and goat anti-mouse-HRP (1:40000) assecondary antibody.
Caspase-3 activity assay
To determine the eVects of SNP on apoptosis, a Xuoro-metric assay of caspase-3 activity was conducted asdescribed elsewhere (Fiorucci et al. 1999). Cell lysates forthis assay were obtained from diVerent periods of incuba-tion with 200 and 400 �M SNP (3, 6, 9, 12 and 24 h,respectively) in order to Wnd the highest peak of caspaseactivation. BrieXy, 50 �g of cellular protein (from whole-cell lysates or cytosolic lysates) was incubated with20 �M caspase-3 substrate (Ac-DEVD-AMC, CPP32) for2 h at 37°C. Substrate cleavage was measured with aFLx800 microplate Xuorescence reader (Bio-tek Instru-ments, Inc.) with the excitation Wlter set at 360 nm and theemission Wlter set at 460 nm. The activity of caspase wascalculated from the cleavage of the speciWc Xuorogenicsubstrate (AC-DEVD-AMC). The data were presented asarbitrary units of Xuorescence.
123
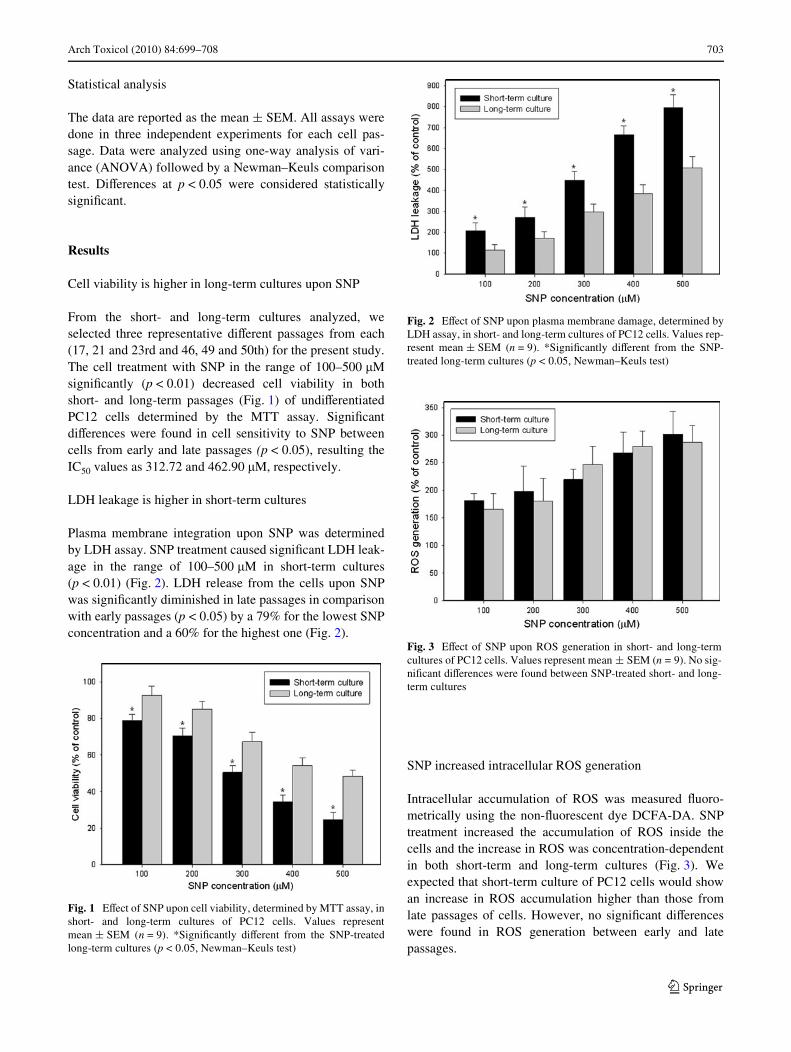
Arch Toxicol (2010) 84:699–708 703
Statistical analysis
The data are reported as the mean § SEM. All assays weredone in three independent experiments for each cell pas-sage. Data were analyzed using one-way analysis of vari-ance (ANOVA) followed by a Newman–Keuls comparisontest. DiVerences at p < 0.05 were considered statisticallysigniWcant.
Results
Cell viability is higher in long-term cultures upon SNP
From the short- and long-term cultures analyzed, weselected three representative diVerent passages from each(17, 21 and 23rd and 46, 49 and 50th) for the present study.The cell treatment with SNP in the range of 100–500 �MsigniWcantly (p < 0.01) decreased cell viability in bothshort- and long-term passages (Fig. 1) of undiVerentiatedPC12 cells determined by the MTT assay. SigniWcantdiVerences were found in cell sensitivity to SNP betweencells from early and late passages (p < 0.05), resulting theIC50 values as 312.72 and 462.90 �M, respectively.
LDH leakage is higher in short-term cultures
Plasma membrane integration upon SNP was determinedby LDH assay. SNP treatment caused signiWcant LDH leak-age in the range of 100–500 �M in short-term cultures(p < 0.01) (Fig. 2). LDH release from the cells upon SNPwas signiWcantly diminished in late passages in comparisonwith early passages (p < 0.05) by a 79% for the lowest SNPconcentration and a 60% for the highest one (Fig. 2).
SNP increased intracellular ROS generation
Intracellular accumulation of ROS was measured Xuoro-metrically using the non-Xuorescent dye DCFA-DA. SNPtreatment increased the accumulation of ROS inside thecells and the increase in ROS was concentration-dependentin both short-term and long-term cultures (Fig. 3). Weexpected that short-term culture of PC12 cells would showan increase in ROS accumulation higher than those fromlate passages of cells. However, no signiWcant diVerenceswere found in ROS generation between early and latepassages.
Fig. 1 EVect of SNP upon cell viability, determined by MTT assay, inshort- and long-term cultures of PC12 cells. Values representmean § SEM (n = 9). *SigniWcantly diVerent from the SNP-treatedlong-term cultures (p < 0.05, Newman–Keuls test)
Fig. 2 EVect of SNP upon plasma membrane damage, determined byLDH assay, in short- and long-term cultures of PC12 cells. Values rep-resent mean § SEM (n = 9). *SigniWcantly diVerent from the SNP-treated long-term cultures (p < 0.05, Newman–Keuls test)
Fig. 3 EVect of SNP upon ROS generation in short- and long-termcultures of PC12 cells. Values represent mean § SEM (n = 9). No sig-niWcant diVerences were found between SNP-treated short- and long-term cultures
123

704 Arch Toxicol (2010) 84:699–708
Nitrite accumulation is lower in long-term cultures
Stimulation of PC12 cells with 200 and 400 �M SNP,respectively, for 24 h caused a signiWcant increase of nitriteproduction (the stable metabolite of NO) compared toun-stimulated cells (p < 0.01) in early passages (Fig. 4).Nitrite accumulation in medium upon SNP treatment oflong-term cultures was decreased when compared withthose from short-term cultures by a 34% for 200 �M SNPtreatment and a 50% for the 400 �M SNP.
MDA levels are lower in long-term cultures
Intracellular MDA was used as a suitable marker for lipidperoxidation induced by oxidative damage. SNP treatmentat 200 or 400 �M signiWcantly increased MDA formationby 4.28 or 6.02-folds in comparison with untreated cells(p < 0.05) in early passages (Fig. 5). SNP-induced MDAformation upon treatment with 200 and 400 �M SNP wasdecreased in late passages when comparing with early pas-sages. MDA levels upon SNP treatment at 200 or 400 �Mincreased signiWcantly by 2.02 or 2.90-folds in comparisonwith untreated cells (p < 0.05) in late passages (Fig. 5).
CAT and GSH-Px activities are increased in long-term cultures
The activities of SOD, CAT and GSH-Px in PC12 cellsuntreated and cells upon 200 and 400 �M SNP treatmentsare presented in Table 1. No signiWcant diVerences werefound in SOD activity between short- and long-term cul-tures. SOD activity was unaVected by incubation with SNP,in both early and late passages. However, an increase in
CAT and GSH-Px activities were observed in untreated latepassages in comparison with early passages (p < 0.05).SNP treatment increased signiWcantly CAT activity com-pared to untreated cells in early passages (p < 0.05). More-over, an increase in CAT and GSH-Px activities wereobserved in SNP-treated cells from late passages. Theincrease in CAT activity upon 200 and 400 �M SNP treat-ments was much higher in late passages than early passageswhen compared with untreated cells.
PARP cleavage is higher in short-term cultures
PARP cleavage has been used as a marker of cell deathsince PARP is hydrolyzed by active caspase-3 during apop-tosis, yielding an active 85-kD fragment (Hanzel andVerstraeten 2009). SNP-treated PC12 cells from early pas-sages have shown an important and higher decrease in thePARP proform determined by western blot in comparisonwith late passages (Fig. 6). The bands of PARP proformcorresponding to 200 and 400 �M were almost undetect-able by chemiluminescence for the short-term culture.However, although there was cleavage of PARP upon SNPtreatment in long-term cultures, the extent of hydrolysiswas lower being the bands of the PARP proform detectableand approximately one-third the intensity of the untreatedcontrol band.
SNP increased caspase-3 activation in short-term cultures
Caspases are cysteine proteases that cleave a critical set ofcellular proteins to initiate the apoptotic signal (Thornberryand Lazebnik 1998). Because caspase-3 is an executioner
Fig. 4 EVect of SNP upon nitrite production in short- and long-termcultures of PC12 cells. Values represent mean § SEM (n = 9). *Sig-niWcantly diVerent from the SNP-treated long-term cultures (p < 0.05,Newman–Keuls test)
Fig. 5 EVect of SNP upon MDA formation in short- and long-termcultures of PC12 cells. Values represent mean § SEM (n = 9). *Sig-niWcantly diVerent from the untreated short-term cultures (p < 0.05,Newman-Keuls test). #SigniWcantly diVerent from the untreated long-term cultures (p < 0.05, Newman–Keuls test). For both SNP concen-trations, no signiWcant diVerences were found between short- andlong-term cultures
123

Arch Toxicol (2010) 84:699–708 705
of apoptosis induced by a variety of stimuli, we examinedwhether NO generated by SNP activated caspase-3 in PC12cell. Since no diVerences among treatments in caspase-3activity from whole-cell lysates were found, cytosoliclysates were obtained for further assays. SNP at 200 or 400�M for 6 h increased the caspase-3 activity approximatelyby a 1100 or 1675% in comparison with the basal levels ofcontrol cells (p < 0.05) (Fig. 7). The activation of caspase-3after 200 or 400 �M SNP treatment for 6 h was signiW-cantly attenuated in late passages in comparison with earlypassages by 7.41 and 9.81-folds, respectively (p < 0.05).No signiWcant diVerences were found between both SNPconcentrations and between both passages in caspase-3activity after 24-h SNP treatment (data not shown).Besides, we examined the eVect of quercetin, an Hsp70inhibitor (Khomenko et al. 2007; Sasaki et al. 2007), oncaspase activation of SNP-treated PC12 cells in order totest whether Hsp70 inhibition could account for an increasein SNP-induced caspase activation. For this purpose, PC12cells were pre-incubated with 5 �M quercetin for 3 h andthen SNP was added to the cells for 24 h. Caspase activitywas increased in the short- and long-term cultures, respec-tively, upon the single 3-h incubation with quercetin incomparison with untreated cells, by 2.5 and 3.5-folds(p < 0.05). Caspase activity was increased in long-term cul-tures upon quercetin plus SNP treatment when comparedwith single 200 �M SNP and 400 �M treatments by 5.05and 3.24-folds, respectively (p < 0.05) (Fig. 7).
Hsp70 levels are increased in long-term cultures
Although PC12 cells from short-term cultures were notstress-stimulated in this study, they weakly expressedHsp70 protein (Fig. 8). The basal levels of Hsp70 in long-term cultures from untreated PC12 were 2.8-folds higherthan those from early passages. SNP treatment of cells fromearly passages slightly increased Hsp70 levels at 200 and400 �M. However, high levels were observed in Hsp70after SNP treatment of long-term cultures, being theincrease higher at 200 �M by 2.4-folds in comparison withuntreated cells (Fig. 8).
SNP-induced cell death in early and late passages upon quercetin pre-treatment
We examined the eVect of quercetin on cell viability ofSNP-treated PC12 cells in order to test whether Hsp70 inhi-bition could account for an increase in SNP-induced cell
Fig. 6 EVect of SNP upon PARP cleavage in short- and long-term cul-tures of PC12 cells. Equal amounts of cell lysates (10 �g) obtainedfrom cells grown under the conditions described in “Cell culture anddrug treatment” were resolved by SDS–PAGE, transferred to a PVDFmembrane and then probed with an anti-PARP antibody (proform).The proteins were then visualized using an ECL detection system.Actin was used as an internal control
Table 1 EVect of SNP upon SOD, CAT and GSH-Px activities in short- and long-term cultures of PC12 cells
Values represent mean § SEM (n = 9). *SigniWcantly diVerent from the untreated short-term cultures (p < 0.05, Newman–Keuls test). #SigniW-cantly diVerent from the untreated long-term cultures (p < 0.05, Newman–Keuls test)
Experimental condition Untreated cells 200 �M SNP 400 �M SNP
Short-term Long-term Short-term Long-term Short-term Long-term
SOD (IU/mg prot) 8.73 § 0.65 6.75 § 0.81 7.22 § 0.95 8.49 § 0.95 8.92 § 1.07 9.35 § 1.23
CAT (IU/mg prot) 4.72 § 0.63 7.79 § 1.47 8.37 § 0.71* 18.85 § 2.02# 12.23 § 1.83* 21.92 § 2.64#
GSH-Px (mU/min mg prot) 49.31 § 1.27 53.41 § 2.64 52.26 § 2.06* 62.09 § 3.11# 50.57 § 1.92* 68.45 § 3.25#
Fig. 7 EVect of SNP and/or pre-incubation with 5 �M quercetin (priorSNP treatment) upon caspase-3 activation in short- and long-term cul-tures of PC12 cells. Caspase-3 activation was determined in cytosoliccell lysates from PC12 cells. Values represent mean § SEM (n = 9).*SigniWcantly diVerent from the SNP-treated long-term cultures(p < 0.05, Newman-Keuls test). #SigniWcantly diVerent from the onlySNP-treated long-term cultures (p < 0.05, Newman–Keuls test)
123

706 Arch Toxicol (2010) 84:699–708
death. For this purpose, PC12 cells were preincubated with5 �M quercetin for 3 h and then SNP was added to the cellsfor 24 h. Cell viability diminished in the short-term culturesupon the single 3-h incubation with quercetin in compari-son with untreated cells; however, no signiWcant variationsin cell viability were observed upon quercetin plus 24-hSNP treatment when compared with the single SNP treat-ments. Cell viability diminished in long-term cultures uponquercetin plus SNP treatment when compared with singleSNP treatments (Fig. 9).
Discussion
Sodium nitroprusside (SNP)-treated PC12 cell line is beingused in our laboratory as a cell model of nitric oxide (NO)-mediated damage for in vitro evaluation of potential neuro-
protective compounds. NO is involved in many pathologiesof the central nervous system (CNS) (Jung et al. 2007);therefore, models of NO-mediated cytotoxicity may par-tially serve as tools to evaluate potential active compoundsfor further treatments.
In our concern about the integrity of this cell model, wehave considered that PC12 cell lines derived from diVerentlaboratories have shown distinct diVerences in size, shape,adherence and clumping behavior, which varied in responseto diVerent culture media, growth substrates and nervegrowth factor (Dixon et al. 2005).
Interestingly, in our laboratory work through the time,we have found diVerences in sensitivity to SNP betweencells from short- and long-term cultured of PC12 cells. Thenumber of cell passages discriminates between young andaged cells. Our data from the markers of cell death haverevealed that aged cells evade from the apoptosis inducedby SNP. However, the capability for apoptosis is main-tained in aged PC12 cells as the sensitivity to SNP-trig-gered apoptosis remained in a certain extent and was SNPconcentration-dependent for most of the parameters evalu-ated.
After a number of serial passages, the basal levels ofHsp70 start increasing in untreated PC12 cells which mightbe indicative of an adaptive response to cumulative intra-cellular stress during aging and an important mechanism ofcytoprotection as described elsewhere in cultured HT22mouse hippocampal neurons (Khomenko et al. 2007). Thisalteration in Hsp70 levels has been accompanied by animproved survival ability of the cells against SNP toxicityas well as a lesser damage of cell membrane as observed byLDH leakage.
In performing our experiments, we have always beencautious to carry out cell treatments with SNP 24 h aftercells were seeded since cellular glutathione contentincreases rapidly during the Wrst 24 h of subculture, includ-ing the lag and early log phases of growth, and decreasescontinuously thereafter, thus cells exposed to toxins atdiVerent times during the logarithmic growth phase mightshow diVerential sensitivity (Kang and Enger 1990).
In the absence of heat shock treatment, our PC12 cellsfrom short-term cultures weakly expressed the Hsp70 pro-tein. However, higher levels were found in long-term cul-tures, both in untreated and SNP-treated cells. The highestlevels of Hsp70 were found at 200 �M SNP indicating thatHsp70 induction was not SNP concentration-dependent inPC12 cells which already possessed signiWcant high basallevels of Hsp70. No SNP concentration-dependent changeswere found either in Hsp70 levels in short-term cultures.The treatment of cells with an Hsp70 inhibitor, quercetin,may have demonstrated, on one hand, that late passages ofuntreated cells possesses enough levels of Hsp70 to copewith the Hsp inhibition and remain viable in comparison
Fig. 8 EVect of SNP upon Hsp70 levels in short- and long-term cul-tures of PC12 cells. Equal amounts of cell lysates (10 �g) obtainedfrom cells grown under the conditions described in “Cell culture anddrug treatment” were resolved by SDS–PAGE, transferred to a PVDFmembrane and then probed with an anti-Hsp70 antibody. The proteinswere then visualized using an ECL detection system. Actin was usedas an internal control
Fig. 9 EVect of pre-incubation with 5 �M quercetin prior SNP treat-ment upon cell viability, determined by MTT assay, in short- and long-term cultures of PC12 cells. Values represent mean § SEM (n = 9).*SigniWcantly diVerent from the untreated short-term cultures(p < 0.05, Newman–Keuls test). #SigniWcantly diVerent from the200 �M SNP-treated long-term cultures (p < 0.05, Newman–Keulstest). ##SigniWcantly diVerent from the 400 �M SNP-treated long-termcultures (p < 0.05, Newman–Keuls test)
123

Arch Toxicol (2010) 84:699–708 707
with early passage of untreated cells which undergo a cer-tain extent of cell death, and on the other hand, that cellsfrom late passages could probably become more sensitiveto SNP, at least partially, as far as Hsp70 levels were dimin-ished and caspase was activated. Thus, up-regulated levelsof Hsp70 may be associated with cell survival in our exper-imental conditions, spite that activation of the heat shockresponse is not necessarily neuroprotective (Fonager et al.2002). Cytoprotection against SNP-induced NO cytotoxic-ity as a result of Hsp70 overexpression has been describedelsewhere in rat insulinoma cells (Bellmann et al. 1996)which could be in agreement with the cyprotection we haveseen here.
In our present study, the most important eVect of thehigh levels of Hsp70 in late passages of SNP-treated PC12cells could be an increase in CAT and GSH-Px activitiesand a decrease of caspase-3 activation in comparison withearly passages. It is reasonable that Hsp70 could interactwith antioxidant enzymes and caspase proteins and regulateits activity. On the other hand, Hsp70 does not seem toexert its protective action by scavenging ROS since no sig-niWcant diVerences were found in ROS generation betweenearly and late passages upon SNP treatment. It is, of course,possible that a combination of these alterations explains thestate of lower sensitivity in long-term cultures of the PC12cell line against SNP.
As a result of the increase antioxidant capacity of long-term cultures, nitrite production and MDA content werelower than those from early passages. Therefore, elevatedvalues of activity of these endogenous antioxidant enzymescould be involved in prevention of free-radical production,lipid peroxidation and cellular toxicity as a result of treat-ment with SNP as described elsewhere with other neurotox-ins for the nervous system (Hom et al. 1997).
Moreover, the degree of PARP cleavage, a substrate ofcaspase-3, observed in early and late passages was relatedto the extent of caspase-3 activation which was muchhigher in the short-term cultures. Therefore, the apoptosismachinery could be less functional in long-term culturesdue to the interference of the Hsp70 protein (Samali andOrrenius 1998; Lee et al. 2008). Our Wndings are in accor-dance with the previous reports describing that Hsp70inhibits activation of caspase-3 (Mosser et al. 1997;Eftekharzadeh et al. 2010).
The need for standardizing experimental conditions inthe permanently growing PC12 cell line which serve us as amodel for drug testing demands for selecting early passagesto gain relevant and reproducible results. Through serialpassages cells start aging and adapt to oxidative stress andthis phenomenon may lead to misestimate subsequent cellresponses to toxins, which is dependent on cellular antioxi-dant capacity or redox state, determined partially by theheat shock proteins. Taken together, our data suggest that
increased expression of Hsps may lead researchers to underestimate drug toxicity in long-term cultures of PC12 cells.
In conclusion, we observed that long-term cultures ofPC12 cells have a certain extent of resistance to SNP-induced cell death, suggesting that the stress protein Hsp70plays a role in the cytoprotective eVect, being involved atleast in part in the cellular resistance to SNP, a cytotoxicagent, which causes cell death in vitro. Other mechanisms,of course, might have contributed to the neuroprotection ofPC12 long-term cultures against SNP which need furtherinvestigation as stabilization of intracellular energy levelsand maintenance of correct protein folding due to theknown chaperone function of Hsps as well as the reorgani-zation of cytoskeletal structures.
Acknowledgments Funds for this study were granted by the NationalResearch Foundation of the Spanish Ministry of Health (FIS;PI021745).
ConXict of Interest statement The authors declare that there are noconXicts of interest.
References
Aebi H (1984) Catalase in vitro. Methods Enzymol 105:121–126Bellmann K, Jaattela M, Wissing D, Burkart V, Kolb H (1996) Heat
shock protein hsp70 overexpression confers resistance againstnitric oxide. FEBS Lett 391:185–188
Benedi J, Arroyo R, Romero C, Martin-Aragon S, Villar AM (2004)Antioxidant properties and protective eVects of a standardizedextract of Hypericum perforatum on hydrogen peroxide-inducedoxidative damage in PC12 cells. Life Sci 75:1263–1276
Bermejo P, Gomez-Serranillos P, Santos J, Pastor E, Gil P, Martin-Aragon S (1997) Determination of malonaldehyde inAlzheimer’s disease: a comparative study of high-performanceliquid chromatography and thiobarbituric acid test. Gerontology43:218–222
Cardaci S, Filomeni G, Rotilio G, Ciriolo MR (2008) Reactive oxygenspecies mediate p53 activation and apoptosis induced by sodiumnitroprusside in SH-SY5Y cells. Mol Pharmacol 74:1234–1245
Dixon DN, Loxley RA, Barron A, Cleary S, Phillips JK (2005) Com-parative studies of PC12 and mouse pheochromocytoma-derivedrodent cell lines as models for the study of neuroendocrine sys-tems. In Vitro Cell Dev Biol Anim 41:197–206
Eftekharzadeh B, Khodagholi F, Abdia A, Maghsoudia N (2010) Algi-nate protects NT2 neurons against H2O2-induced neurotoxicity.Carbohydr Polym 79:1063–1072
Fiorucci S, Santucci L, Federici B, Antonelli E, Distrutti E, Morelli O(1999) Nitric oxide-releasing NSAIDs inhibit interleukin-1betaconverting enzyme-like cysteine proteases and protect endothelialcells from apoptosis induced by TNFalpha. Aliment PharmacolTher 13:421–435
Fonager J, Beedholm R, Clark BF, Rattan SI (2002) Mild stress-in-duced stimulation of heat-shock protein synthesis and improvedfunctional ability of human Wbroblasts undergoing aging in vitro.Exp Gerontol 37:1223–1228
Fu CH, Martin-Aragon S, Weinberg KI, Ardi VC, Danenberg PV,Avramis VI (2001) Reversal of cytosine arabinoside (ara-C)resistance by the synergistic combination of 6-thioguanine plusara-C plus PEG-asparaginase (TGAP) in human leukemia lines
123

708 Arch Toxicol (2010) 84:699–708
lacking or expressing p53 protein. Cancer Chemother Pharmacol48:123–133
Gorman AM, Heavey B, Creagh E, Cotter TG, Samali A (1999) Anti-oxidant-mediated inhibition of the heat shock response leads toapoptosis. FEBS Lett 445:98–102
Hanzel CE, Verstraeten SV (2009) Tl(I) and Tl(III) activate both mito-chondrial and extrinsic pathways of apoptosis in rat pheochromo-cytoma (PC12) cells. Toxicol Appl Pharmacol 236:59–70
Heo HJ, Cho HY, Hong B, Kim HK, Heo TR, Kim EK (2002) Ursolicacid of Origanum majorana L. reduces Abeta-induced oxidativeinjury. Mol Cells 13:5–11
Hill HD, Straka JG (1988) Protein determination using bicinchoninicacid in the presence of sulfhydryl reagents. Anal Biochem170(1):203–208
Hom DG, Jiang D, Hong EJ, Mo JQ, Andersen JK (1997) Elevatedexpression of glutathione peroxidase in PC12 cells results in pro-tection against methamphetamine but not MPTP toxicity. BrainRes Mol 46:154–160
Imahashi D (1968) Quantitation of LDH isoenzymes by the Xuorimet-ric and colorimetric methods. Can J Med Technol 30:235–248
Jung JY, Han CR, Jeong YJ, Kim HJ, Lim HS, Lee KH (2007) Epigal-locatechin gallate inhibits nitric oxide-induced apoptosis in ratPC12 cells. Neurosci Lett 411:222–227
Kang YJ, Enger MD (1990) Cadmium cytotoxicity correlates with thechanges in glutathione content that occur during the logarithmicgrowth phase of A549–T27 cells. Toxicol Lett 51:23–28
Khomenko IP, Bakhtina LY, Zelenina OM, Kruglov SV, ManukhinaEB, Bayda LA (2007) Role of heat shock proteins HSP70 andHSP32 in the protective eVect of adaptation of cultured HT22 hip-pocampal cells to oxidative stress. Bull Exp Biol Med 144:174–177
LeBel CP, Ischiropoulos H, Bondy SC (1992) Evaluation of the probe2’, 7’-dichloroXuorescin as an indicator of reactive oxygen spe-cies formation and oxidative stress. Chem Res Toxicol 5:227–231
Lee JY, Kim YJ, Kim TY, Koh JY, Kim YH (2008) Essential role forzinc-triggered p75NTR activation in preconditioning neuropro-tection. J Neurosci 28:10919–10927
Majlessipour F, Kwock R, Martin-Aragon S, Weinberg KI, AvramisVI (2001) Development of a double-drug-resistant human leuke-mia model to cytosine arabinoside and L-asparaginase: evaluationof cross-resistance to other treatment modalities. Anticancer Res21:11–22
Marklund S, Marklund G (1974) Involvement of the superoxide anionradical in the autoxidation of pyrogallol and a convenient assayfor superoxide dismutase. Eur J Biochem 47:469–474
Martin-Aragon S, Benedi JM, Villar AM (1997a) EVects of fraxetin onglutathione redox status. Z Naturforsch C 52:55–59
Martin-Aragon S, Benedi JM, Villar AM (1997b) ModiWcations onantioxidant capacity and lipid peroxidation in mice under fraxetintreatment. J Pharm Pharmacol 49:49–52
Martin-Aragon S, Benedi JM, Villar AM (1998) EVects of the antiox-idant (6, 7-dihydroxycoumarin) esculetin on the glutathione sys-tem and lipid peroxidation in mice. Gerontology 44:21–25
Martin-Aragon S, Mukherjee SK, Taylor BJ, Ivy SP, Fu CH, Ardi VC(2000) Cytosine arabinoside (ara-C) resistance confers cross-resistance or collateral sensitivity to other classes of anti-leuke-mic drugs. Anticancer Res 20:139–150
Martin-Aragon S, de las Heras B, Sanchez-Reus MI, Benedi J (2001)Pharmacological modiWcation of endogenous antioxidantenzymes by ursolic acid on tetrachloride-induced liver damage inrats and primary cultures of rat hepatocytes. Exp Toxicol Pathol53:199–206
Miranda KM, Espey MG, Wink DA (2001) A rapid, simple spectro-photometric method for simultaneous detection of nitrate andnitrite. Nitric Oxide 5:62–71
Molina-Jimenez MF, Sanchez-Reus MI, Benedi J (2003) EVect offraxetin and myricetin on rotenone-induced cytotoxicity inSH-SY5Y cells: comparison with N-acetylcysteine. Eur JPharmacol 472:81–87
Molina-Jimenez MF, Sanchez-Reus MI, Andres D, Cascales M,Benedi J (2004) Neuroprotective eVect of fraxetin and myricetinagainst rotenone-induced apoptosis in neuroblastoma cells. BrainRes 1009:9–16
Molina-Jimenez MF, Sanchez-Reus MI, Cascales M, Andres D,Benedi J (2005) EVect of fraxetin on antioxidant defense andstress proteins in human neuroblastoma cell model of rotenone neuro-toxicity. Comparative study with myricetin and N-acetylcysteine.Toxicol Appl Pharmacol 209:214–225
Mosmann T (1983) Rapid colorimetric assay for cellular growth andsurvival: application to proliferation and cytotoxicity assays. JImmunol Methods 65:55–63
Mosser DD, Caron AW, Bourget L, Denis-Larose C, Massie B (1997)Role of the human heat shock protein hsp70 in protection againststress-induced apoptosis. Mol Cell Biol 17:5317–5327
Paglia DE, Valentine WN (1967) Studies on the quantitative and qual-itative characterization of erythrocyte glutathione peroxidase. JLab Clin Med 70:158–169
Papp E, Nardai G, Soti C, Csermely P (2003) Molecular chaperones,stress proteins and redox homeostasis. Biofactors 17:249–257
Quigney DJ, Gorman AM, Samali A (2003) Heat shock protects PC12cells against MPP + toxicity. Brain Res 993:133–139
Samali A, Orrenius S (1998) Heat shock proteins: regulators of stressresponse and apoptosis. Cell Stress Chaperones 3:228–236
Saminathan R, Pachiappan A, Feng L, Rowan EG, GopalakrishnakoneP (2009) Transcriptome proWling of neuronal model cell PC12from rat pheochromocytoma. Cell Mol Neurobiol 29:533–548
Sanchez-Reus MI, Peinado II, Molina-Jimenez MF, Benedi J (2005)Fraxetin prevents rotenone-induced apoptosis by induction ofendogenous glutathione in human neuroblastoma cells. NeurosciRes 53:48–56
Sanchez-Reus MI, Gomez del Rio MA, Iglesias I, Elorza M, SlowingK, Benedi J (2007) Standardized Hypericum perforatum reducesoxidative stress and increases gene expression of antioxidantenzymes on rotenone-exposed rats. Neuropharmacology 52:606–616
Sasaki M, Nakamura H, Tsuchiya S, Horie S, Kashiwayanagi M, SaitoT (2007) Quercetin-induced PC12 cell death accompanied bycaspase-mediated DNA fragmentation. Biol Pharm Bull 30:682–686
Sinn DI, Chu K, Lee ST, Song EC, Jung KH, Kim EH (2007) Pharma-cological induction of heat shock protein exerts neuroprotectiveeVects in experimental intracerebral hemorrhage. Brain Res1135:167–176
Thornberry NA, Lazebnik Y (1998) Caspases: enemies within. Science281:1312–1316
Yang L, Liu X, Hao J, Yang Y, Zhao M, Zuo J (2008) Glucose-regu-lated protein 75 suppresses apoptosis induced by glucose depriva-tion in PC12 cells through inhibition of Bax conformationalchange. Acta Biochim Biophys Sin (Shanghai) 40:339–348
Yuyama K, Yamamoto H, Nakamura K, Kato T, Sora I, Yamamoto T(2001) Resistance of PC12 cells against nitric oxide (NO)-in-duced toxicity in long-term culture: implication of neuronal NOsynthase expression. Neurosci Lett 309:169–172
123