ion-binding properties of the dehydrin erd14 are dependent upon
TRANSCRIPT

Phosphorylation-dependent ion-binding by ERD14
1
Ion-binding properties of the dehydrin ERD14 are dependent
upon phosphorylation
Muath K. Alsheikh 1, Bruce J. Heyen2, and Stephen K. Randall 1
1 Department of Biology, Indiana University - Purdue University Indianapolis, 723 West
Michigan Street, Indianapolis, Indiana 46202-5132
2 Departments of Biology and Chemistry, Tabor College, 400 South Jefferson, Hillsboro,
Kansas 67063
Corresponding author:
Dr. Stephen K. Randall, Department of Biology
Indiana University - Purdue University Indianapolis
723 West Michigan Street
Indianapolis, IN 46202-5132
Telephone # (317) 274-0592
Fax # (317) 274-2846
E-Mail [email protected]
Copyright 2003 by The American Society for Biochemistry and Molecular Biology, Inc.
JBC Papers in Press. Published on August 13, 2003 as Manuscript M307151200 by guest on A
pril 14, 2018http://w
ww
.jbc.org/D
ownloaded from

Phosphorylation-dependent ion-binding by ERD14
2
Running title: Phosphorylation-dependent ion-binding by ERD14
by guest on April 14, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Phosphorylation-dependent ion-binding by ERD14
3
Summary
The ERD14 protein (Early Response to Dehydration) is a member of the dehydrin
family of proteins which accumulate in response to dehydration-related environmental
stresses. Here we show the Arabidopsis dehydrin, ERD14, possesses ion-binding
properties. ERD14 is an in vitro substrate of casein kinase II; the phosphorylation
resulting both in a shift in apparent molecular mass on SDS-PAGE gels, and increased
calcium binding activity. The phosphorylated protein bound significantly more calcium
than the non-phosphorylated protein, with a dissociation constant of 120 uM, and 2.86
mole of calcium bound per mole of protein. ERD14 is phosphorylated by extracts of
cold-treated tissues, suggesting that the phosphorylation status of this protein might be
modulated by cold-regulated kinases or phosphatases. Calcium-binding properties of
ERD14 purified from Arabidopsis extracts were comparable to phosphorylated E. coli
expressed ERD14. Approximately two moles of phosphate were incorporated per mole
of ERD14, indicating a minimum of two phosphorylation sites. Western blot analyses
confirmed that threonine and serine are possible phosphorylation sites on ERD14.
Utilizing MALDI-TOF/MS, we identified five phosphorylated peptides that were present
in both in vivo and in vitro phosphorylated ERD14. Our results suggest that the poly-
serine (“S”) domain is most likely the site of phosphorylation in ERD14 responsible for
the activation of calcium binding.
Keywords: dehydrin, calcium, cold-stress, dehydration stress, plant
by guest on April 14, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Phosphorylation-dependent ion-binding by ERD14
4
Introduction.
Environmental stresses such as cold and drought have significant impact on plant
growth and development; hence agricultural productivity. Plants have evolved a wide
variety of molecular responses to enable them to survive severe abiotic stresses (1,2).
Among these responses is the alteration of expression of a family of genes which encode
dehydrins, a subfamily of the group II Late Embryogenesis Abundant mRNAs (LEAs).
Levels of dehydrins are increased in response to low temperature, drought, osmotic
stress, or are ABA-induced (3-8). In Arabidopsis, dehydrins can be characterized as the
acidic dehydrins, pI 4.6-6.4 (COR47, ERD10, ERD14, NP_195554, NP_195624, and
X91920), that are generally highly enriched in glutamic acid, and the neutral/basic
dehydrins, pI 7.6-9.8 (RAB18, XERO1, and XERO2) that are enriched in glycine.
Dehydrin proteins remain soluble following boiling, are extremely hydrophilic, and can
be defined by the presence of at least one lysine-rich consensus sequence, the “K”
domain (EKKGIMDKIKEKLPG), which is similar to class A2 amphipathic -helical
domains found in lipid-binding proteins (7,9). Some dehydrins meet the criteria for
hydrophillins (10). Additionally, many dehydrins contain the “S” domain, a Ser-rich
sequence (3,11-13). Although their biochemical and physiological roles are still unclear,
it has been suggested that dehydrins may play a role in stabilizing proteins or membrane
structures under environmental stresses through interactions with an amphipathic -helix
(9).
Calcium levels control a variety of plant developmental and signal transduction
processes (14,15), and in particular, calcium signaling is requisite for plant responses to
by guest on April 14, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Phosphorylation-dependent ion-binding by ERD14
5
environmental signals (16-19). A transient change in cytosolic calcium levels is one of
the initial responses of some plants to adverse environmental conditions such as low
temperature, drought and salinity (16,19-23). Several studies have shown that the ability
of plants to withstand calcium level changes is essential for them to survive different
abiotic stresses (16,24,25). Typically, cytosolic free calcium is maintained at
submicromolecular levels (~200 nM) by homeostatic mechanisms involving a variety of
calcium channels, pumps and secondary transporters in a variety of cell organelles, such
as vacuole, endoplasmic reticulum, chloroplasts and mitochondria, and cell wall
(14,15,26). Intracellular calcium homeostasis is also maintained in plant cells by a variety
of calcium-binding proteins, such as calsequestrin (27,28), calnexin (29), and calreticulin
(30). Calcium levels in the cytosol increases by several orders of magnitude upon
signaling, however, sustained elevation of calcium can be toxic (31). It has been
suggested that cold-induced [Ca2+]cyt increases are caused by calcium leakage across
membranes; initiated at the plasma membrane and perpetuated by calcium release mainly
from vacuole and endoplasmic reticulum (16,17,32).
The connection between stress-induced increases in cytosolic calcium and the
accumulation of dehydrins, has suggested to us a potential physiological role for
dehydrins in cold stress management could be ion binding. It was recently determined by
our laboratory that a celery protein, similar in sequence to dehydrins (VCaB45), binds
calcium (33), and it has been reported that another dehydrin-like protein found in castor
bean has metal binding capability (34). The present study is a characterization of a known
by guest on April 14, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Phosphorylation-dependent ion-binding by ERD14
6
Arabidopsis dehydrin, ERD14, its ion binding activity, and its regulation by
phosphorylation.
Experimental Procedures
Plant materials and extractions
Arabidopsis thaliana (Columbia ecotype) seedlings were grown with 16/8 hours
light/dark period at 20°C. Low temperature treatment was performed on 3-4 week old
plants for 2 days 4.7°C. Total protein extracts of Arabidopsis plants were obtained from
cold-treated and control tissues (33). In some cases low density membranes were isolated
from cold-treated Arabidopsis thaliana in the presence of protease and phosphatase
inhibitors as described in Heyen et al. (2002). Triton X-100 (0.2% w/w) was used to
release the contents of the membrane vesicles (33). Supernatants derived from the
permeabilized membranes were heat-treated in an 85°C water bath for 20 minutes and
quickly cooled in an ethanol bath (–50°C) for 5 minutes. The soluble fraction was then
obtained following ultracentrifugation for 1 hour at 100,000 g at 4°C. Anion exchange
chromatography was performed on a MonoQ column (Pharmacia) in 20 mM Tris-HCl
(pH 8.2 at 4°C).
ERD14 cloning and expression
A lambda-PRL2 expression library (obtained from Ohio State Plant Resource
Center) was subcloned into the pHEP2 vector (50) using the Pst I and Xba I sites.
Bacterial colonies were screened on replicate lifts following induction with IPTG as
reported previously (50), with the antibody raised against the celery dehydrin-like
protein, VCaB45 (33), using a secondary antibody of anti-mouse/alkaline phosphatase.
by guest on April 14, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Phosphorylation-dependent ion-binding by ERD14
7
Blots were developed with the colorimetric substrate, NBT/BCIP. The fusion protein
was expected to be approximately 5 kDa greater than ERD14 due to the inclusion of v-ras
sequences (50). However, the induced ERD14 was present as two bands. A minor band
(of the expected size for the fusion protein) and an approximately 5 kDa smaller band
(major band) were obtained. The major band was purified and was sequenced by Edman
degradation from the amino terminus. This revealed that the v-ras portion of the fusion
was missing, and additionally the two N-terminal amino acids (methionine-alanine) of
ERD14 had been removed by proteolysis. The presence of the predicted amino-terminal
tryptic peptide (lacking met-ala) was also confirmed by MALDI-TOF/MS. This product,
ERD14 lacking the amino terminal two residues, was used for all the studies described
here.
Purification of ERD14
Induced cells (1 mM isopropyl B-D-thiogalactopyranoside) were harvested and
resuspended in 2X homogenizing buffer in the presence of protease inhibitors (33).
Samples were sonicated for 5 minutes, and heat-treated as described before. Lysates were
then centrifuged at 4,700 g for 15 minutes at 4°C. Supernatants were treated with 0.2%
protamine sulfate to precipitate DNA, and ultracentrifuged for 1 hour at 100,000 g at 4°C.
The high speed supernatant was fractionated by anion exchange chromatography on a 40
mL packed bed of diethyl aminoethyl (DEAE) Sepharose (Pharmacia) (1 mL/min flow
rate). ERD14 fractions were further concentrated on a MonoQ column (Pharmacia).
by guest on April 14, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Phosphorylation-dependent ion-binding by ERD14
8
Western blotting
Antibodies raised against phosphoserine and phosphothreonine were obtained
from Zymed Laboratories Inc. and Cell Signaling Technology, respectively. Anti-
dehydrin antiserum was kindly supplied by Dr. Timothy J. Close (3). Samples were
separated by 10% SDS-PAGE, and electroblotted onto nitrocellulose membrane (Protran,
Mid-West Scientific). For anti-dehydrin antibody, membranes were blocked with 5%
non-fat dry milk in phosphate-buffered saline. When anti-phosphoserine and anti-
phosphothreonine were used, membranes were blocked with 5% Tween-20 and 5% non-
fat dry milk / 0.1% Tween-20, respectively, in Tris-buffered saline. Anti-rabbit and anti-
mouse immunoglobulin G (goat) conjugated to horseradish peroxidase (Sigma) was used
as secondary antibodies. Antibody detection procedures were with SuperSignal West
Pico reagent (Pierce).
ERD14 in vitro phosphorylation and dephosphorylation
Purified E. coli expressed ERD14 (approximately 7.5 µg) was incubated with 10
units of casein kinase II, in 1X kinase buffer (BIOMOL Research Laboratories, Inc.), and
1 mM ATP at 30°C in a total of 10 µL. In some cases, 1 mM Staurosporine (BIOMOL
Research Laboratories, Inc.) was added after 30 min to inhibit protein kinase activity.
Dephosphorylation was performed with shrimp alkaline phosphatase (Roche) (33).
Reactions were stopped by the addition of SDS-PAGE sample buffer and were boiled for
5 min. For the 32P incorporation experiments, purified ERD14 was phosphorylated in the
presence of 0.8 mM ATP (0.17 µCi of [g-32P] ATP). 32P incorporation was visualized
with a PhosphoImager (STORM 840, Molecular Dynamics). In some cases, reactions
by guest on April 14, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Phosphorylation-dependent ion-binding by ERD14
9
were terminated with 25% trichloroacetic acid (TCA) and precipitates were collected on
GFA glass fiber filters (Mid-West Scientific), washed with 5% TCA and then 100%
acetone. In such cases, the incorporated 32P was determined by liquid scintillation
counting.
Phosphorylation of ERD14 by A. thaliana protein extract
Extracts from whole A. thaliana plants treated for 2 days at 4°C or 20°C were
incubated with or without the bacterial purified ERD14 (final concentration of 95 ng/µL),
0.1 mM ATP containing [g-32P] ATP (1 µCi) in 50 mM Hepes, pH 7.2, 0.5 mM CaCl2, 10
mM MgCl2, and 10 mM NaF at 30°C. Reactions were stopped by adding SDS-PAGE
loading buffer and boiling for 5 minutes.
Calcium-binding activity assays
Calcium-binding activity was determined by ligand blotting and equilibrium
dialysis (33). Equilibrium dialysis assays were performed with Teflon Micro Dialyzer
cells (QuizSep, Mid-West Scientific) and Spectra/Por MWCO 6-8,000 membranes
(Spectrum Laboratories, Inc.).
MALDI-TOF mass spectrometry
Protein samples (puried from E. coli) were mixed with an equal volume of 8 M
urea and incubated for 30 minutes at 55 degrees. To reduce the urea concentration to 1 M,
10 mM ammonium bicarbonate buffer, pH 8.5, was added. Protein was then digested
overnight with shaking at 37°C with 1 mg of freshly prepared sequencing grade trypsin
(Promega, V511A) dissolved in 50 mM acetic acid. A C18 Zip-Tip (Millipore, USA) was
by guest on April 14, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Phosphorylation-dependent ion-binding by ERD14
10
used to remove urea and desalt the digestion solution. ERD14 purified from Arabidopsis
was separated on 10% SDS-PAGE gels, and coomassie-stained. Protein bands
corresponding to ERD14 were excised and manually processed. Samples were destained
with 50% acetonitrile in water for 15 minutes at 20°C, 50% acetonitrile in 50 mM
ammonium bicarbonate for 15 minutes at 20°C, and 100% acetonitrile for 10 minutes at
20°C. After removing the destain solution, 10 mM DTT in 100 mM ammonium
bicarbonate was added to cover the gel pieces and incubated for 45 minuutes at 50°C. For
alkylation, DTT was replaced with 55 mM iodoacetamide in 50 mM ammonium
bicarbonate and the samples incubated for 30 minutes in the dark at 20°C. Gel pieces
were rinsed twice with 50 mM ammonium bicarbonate for 15 minutes at 20°C. Protein
bands were digested overnight at 37°C with 150 ng of freshly prepared sequencing grade
trypsin. Equal volumes of water and 100 mM ammonium bicarbonate were added to the
digestion solution to give a final trypsin solution of 1 ng/uL. Following a C18 Zip-Tip
(Millipore, USA) purification, the protein solution was mixed with equal volume of a -
cyano-4-hydroxycinnamic acid matrix (Sigma Chemical Co.) and analyzed by MALDI-
TOF/MS (MALDI LR‰, Micromass, UK, located at the IUPUI Proteomic Core Facility).
The mass of peptides obtained from each band were compared with the computer-
generated list of deduced tryptic fragments to identify the 80 Da shift corresponding to
phosphorylation; using the FindMod site (http://ca.expasy.org/tools/findmod) for single
phosphorylation sites and a Command Prompt program written to find multiple
phosphorylations on single peptide fragments. Genomic Solutions‰
(http://165.219.84.5/index.html) web site was used to search for protein identification
by guest on April 14, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Phosphorylation-dependent ion-binding by ERD14
11
(ProFound‰) and was used to identify the theoretical phosphorylation sites on ERD14
(Protein Association Work Station, PAWS‰).
Results
In vitro phosphorylation and calcium binding of ERD14
ERD14 was cloned from an Arabidopsis expression library, using antibody raised
against celery VCaB45, a dehydrin-like protein (33). The apparent molecular mass of the
purified E. coli expressed ERD14, as estimated by 10% SDS-PAGE was 37 kDa (See
Figures 1, 2, and 3B). The molecular mass of ERD14, deduced from the amino acid
sequence is 20.9 kDa. The mass of the protein used in these studies is predicted to be
20.6 kDa (lacks the amino terminal two amino acids). The anomalous migration
resulting in the overestimation of mass by SDS-PAGE is commonly found in other
dehydrins (5,33,35-38).
Our previous work with a dehydrin-like protein (33) and the presence of potential
casein kinase II phosphorylation sites on ERD14 and other dehydrins suggested that
ERD14 might be a substrate for CKII. Indeed, treatment of the purified E. coli expressed
ERD14 with CKII resulted in a shift in the apparent molecular mass on SDS-PAGE gels
(Figures 1A, and 2); with an apparent increase of approximately 5 kDa. The shifted
by guest on April 14, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Phosphorylation-dependent ion-binding by ERD14
12
polypeptides were confirmed to be ERD14 with a dehydrin specific antibody, raised
against the conserved K-domain of dehydrins (Figure 1A). The ERD14 gel shift was
nearly completed after a 30 minutes phosphorylation (Figure 1A). Dephosphorylation of
CKII phosphorylated ERD14 with SAP (shrimp alkaline phosphatase), resulted in a
quantitative return to the smaller molecular mass (Figure 2). We have utilized this gel
shifting phenomena to routinely assess the phosphorylation status of ERD14, as
demonstrated for other phosphorylated proteins (33,39,40). To quantitate ERD14
phosphorylation, ERD14 was phosphorylated with CKII in the presence of [g-32P]-ATP.
A rapid increase in phosphorylation of ERD14 was observed in the first 30 minutes of
CKII treatment, and approached a plateau after 1 hour. Phosphorylation of ERD14
correlated temporally with the gel shifting of ERD14 on SDS-PAGE gels (Figure 1B).
To determine whether the E. coli expressed ERD14 bound calcium and whether
this activity was regulated by phosphorylation, ERD14 was in vitro phosphorylated with
CKII and separated on SDS-PAGE. Calcium-binding activity was then assessed by
calcium (45CaCl2) ligand blots and visualized by PhosphoImaging. Figure 1C
demonstrates that E. coli expressed ERD14 binds calcium only when phosphorylated.
Quantitative analysis of the calcium ligand blot showed an increase in ERD14 calcium
binding with increased phosphorylation and that ERD14 calcium-binding activity
approached saturation after 1 hour (Figure 1C). Calcium binding, as measured by
calcium-ligand blot (Figure 1C), correlated well with the gel shifting of ERD14 on SDS-
PAGE gels (Figure 1A) and with the extent of phosphorylation (Figure 1B).
To analyze the extent of phosphorylation of the E. coli expressed ERD14, CKII-
phosphorylated ERD14 (with [g-32P]-ATP) was precipitated with TCA and collected on
by guest on April 14, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Phosphorylation-dependent ion-binding by ERD14
13
glass fiber filters. Approximately two moles of phosphate were incorporated per mole of
ERD14 after 3 hours phosphorylation (Figure 3A), indicating at least two possible
phosphorylation sites on ERD14. No further 32P was incorporated into ERD14 up to 5
hours of phosphorylation. In some experiments, fresh CKII was added after 3 hours
(Alsheikh and Randall, unpublished) with no further incorporation observed. Western
blots probed with antibodies raised against phosphoserine and phosphothreonine,
indicated that both threonine and serine were possible phosphorylation sites on ERD14
(Figure 3B). Since phosphothreonine was detected only after three hours
phosphorylation; it is unlikely the phosphorylation requisite for the gel shifting and
calcium-binding activation was due to phosphorylation on a threonine. Western blots
probed with anti-phosphoserine showed a subtle increase in serine phosphorylation over
time (15-120 minutes). However, the anti-phosphoserine antibodies used were not
specific for serine-phosphorylation (Sigma, Cat. No. P3430, and Zymed, Cat. No. 61-
8100), as they reacted with non-phosphorylated ERD14.
Because dehydrins seem to lack significant secondary structure and have been
considered to be intrinsically unstructured proteins (IUP’s) (41); we thought it was
possible that activation of calcium-binding could be due to a rapid phosphorylation but
require a slower induced structural change. To test this hypothesis ERD14 was treated for
30 minutes with CKII, further phosphorylation was prevented by adding staurosporine;
then the incubation was continued. No further increase in calcium-binding occurred with
further incubation in the presence of staurosporine. These results suggest that gel shifting
requires an enzymatically active kinase and that calcium-binding activity of the purified
ERD14 does not require slow phosphorylation-dependent conformational changes.
by guest on April 14, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Phosphorylation-dependent ion-binding by ERD14
14
The quantitative effect of the phosphorylation on ERD14 calcium binding activity
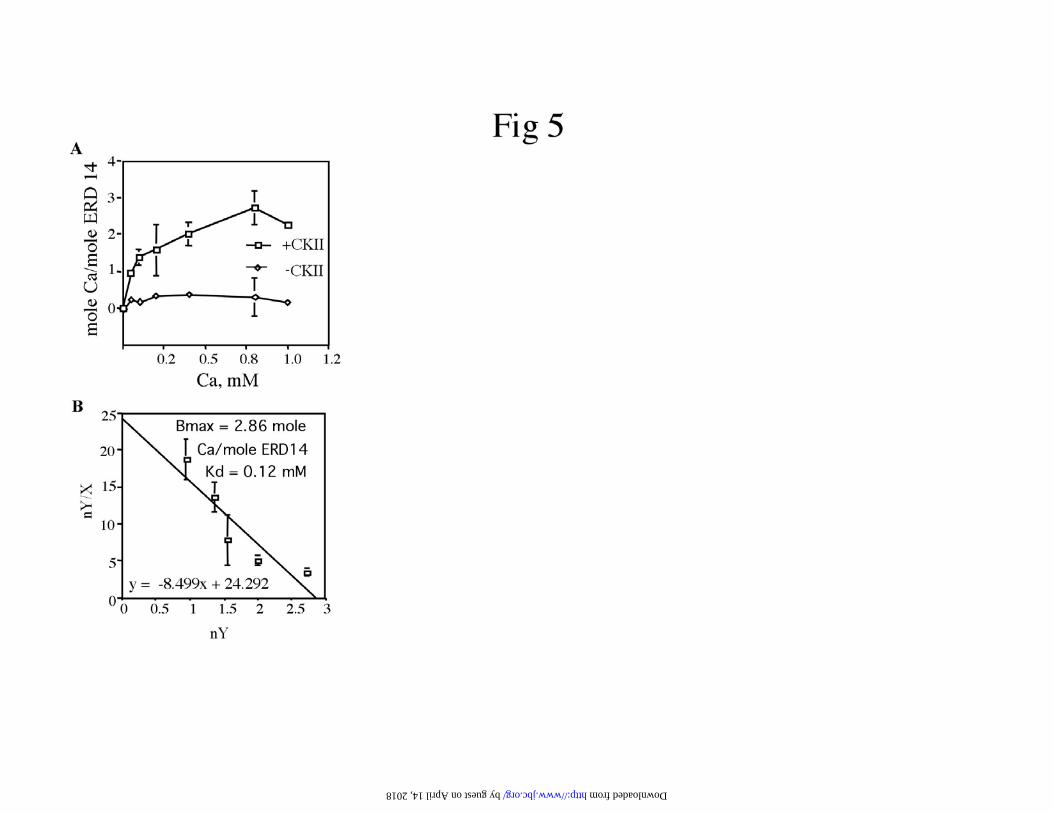
is illustrated by equilibrium dialysis experiments, which show saturable binding of
calcium to native ERD14 (Figure 5). Scatchard plot analysis (Figure 5B) indicated that
CKII-phosphorylated ERD14 binds approximately 3 moles of calcium per mole of
ERD14 with relatively low affinity, having an apparent Kd for calcium of 120 uM.
We examined inhibition of calcium binding by various ions (K+, Mg2+, Zn2+,
Mn2+, Fe2+, Fe3+, and La3+) at 0.25 mM final concentration (Figure 6). K+ and the
trivalent Fe3+, had little affect on calcium binding to ERD14. In contrast, the divalent
cations, Mg2+, Zn2+, Mn2+, or Fe2+ did significantly reduce ERD14 calcium-binding
activity. In the presence of the trivalent cation, lanthanum, calcium binding to ERD14
was strongly inhibited.
In vivo phosphorylation and calcium binding of ERD14
The accumulation of dehydrins in response to low temperature has been
previously reported (5,8,33,35). Here we wanted to determine whether the dehydrin
ERD14 phosphorylation could be modulated by a cold-regulated kinase. E. coli
expressed ERD14 was phosphorylated in the presence of [ g-32P]-ATP with extracts from
4°C or 20°C-treated Arabidopsis plants. In the absence of added ERD14, no
incorporation of [32P] into extract proteins was observed. In the presence of ERD14,
phosphorylation was detected at the expected mass of ERD14. A subtle increase of
kinase activity toward ERD14 in cold-treated Arabidopsis extracts was observed
(Figure7A and B). To determine whether the calcium-binding activity of ERD14 derived
from Arabidopsis extracts is comparable to E. coli expressed ERD14, we isolated ERD14
by guest on April 14, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Phosphorylation-dependent ion-binding by ERD14
15
from 4 week-old cold-treated Arabidopsis plants. ERD14 was purified from low density
membranes which were subsequently permeabilized with 0.2% w/w Triton X-100,
followed by heat treatment and anion exchange chromatography. Peak fractions were
analyzed by SDS-PAGE and western blotting (Figure 8A). A band corresponding to
ERD14 was excised and analyzed by MALDI-TOF/MS. ERD14 identification was
confirmed by comparison of the observed peptides to the SWISS-PROT nr database
(ProFound‰ web site in Genomic Solution‰, http://65.219.84.5/index.html). The best
match was found to be the Arabidopsis dehydrin ERD14, with a Z-value of 1.86, (with
44% coverage) and a probability of 1.0e+000. Similar to E. coli expressed ERD14,
treatment of Arabidopsis ERD14 with shrimp alkaline phosphatase (SAP) resulted in a
shift to a smaller molecular mass on SDS-PAGE gels and greatly reduced the ability of
this protein to bind calcium (Figure 8B). These results indicated that the Arabidopsis
ERD14 is in vivo phosphorylated in cold-treated plants, and its calcium-binding activity
is phosphorylation dependent.
Analysis of phosphorylation sites in ERD14
E. coli expressed ERD14 is phosphorylated by CKII at a minimum of two
phosphorylation sites (~1.6 moles phosphate incorporated per mole of ERD14) (Figure
3A). Further, based on the kinetics of phosphorylation (Figure 1A, B and Figure 3A), it
appeared that one site was phosphorylated rapidly while a second was phosphorylated
more slowly. Purified E. coli expressed ERD14 was phosphorylated with casein kinase II
for 0, 30, and 180 minutes and was then analysed by MALDI-TOF/MS. Mass
spectrometric analysis consistently identified one phosphorylated peptide (peptide # IV)
by guest on April 14, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Phosphorylation-dependent ion-binding by ERD14
16
from the in vitro phosphorylated ERD14 (Figure 9), though several others were observed
in more than one experiment. We compared phospho-peptides from the in vitro
phosphorylated ERD14 to that of the in vivo phosphorylated ERD14 derived from cold-
treated Arabidopsis plants. Since both the in vivo and in vitro phosphorylated ERD14
proteins showed alterations in both gel shifting and calcium-binding, we reasoned that the
common phosphorylated peptides would be likely candidates for containing the phospho-
peptide responsible for calcium-binding. Four phosphorylated peptides (#’s II, IV, V,
and VII) were consistently identified (Figure 9) on the in vivo phosphorylated
Arabidopsis ERD14 and were found consistently in common to both in vitro and in vivo
phosphorylated ERD14. Phosphorylation of peptide VII was deduced not to be important
for calcium-binding activity as the phosphate was not removed from that site by SAP
treatment even though this treatment abrogated calcium binding (Figure 8). The
remaining peptides were dephosphorylated after treatment of Arabidopsis ERD14 with
SAP. Of these only three phospho-peptides (#’s II, IV, and V), appeared to temporally
correlate with the kinetics of the gel shifts and calcium-binding; that is, their presence
was consistently observed after only 30 minutes phosphorylation. Because of the lack of
a significant amount of phosphothreonine detectable at short times (see Figure 3B), we
concluded that the peptide V was not a likely candidate responsible for the shifting and
calcium-binding activation. Of the two remaining phospho-peptides only peptide IV,
containing the “S” domain, was identified in every experiment, and whose presence
consistently correlated with calcium-binding activity. The ERD14 “S”-domain (peptide
IV) has four potential phosphorylation sites for CKII. Analyses at 30 minutes
phosphorylation of E. coli expressed ERD14 with CKII indicated two phosphorylated
by guest on April 14, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Phosphorylation-dependent ion-binding by ERD14
17
serines in the “S”-domain, and three phosphorylation sites after three hours of
phosphorylation. MALDI-TOF analyses of purified in vivo phosphorylated Arabidopsis
ERD14 also revealed up to three phosphorylated serines in the “S”-domain.
by guest on April 14, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Phosphorylation-dependent ion-binding by ERD14
18
Discussion
ERD14 was previously identified as a cDNA encoding a member of the dehydrin
family of proteins (42,43). Although the expression patterns and transcriptional
regulation of dehydrins have been well characterized (5,6,8,42-44), the biochemical
function of these proteins and their physiological role in plants during stress conditions
are not fully understood. Here, we have shown that ERD14 possesses ion-binding activity
which is dependent on its phosphorylation status. Interaction of dehydrins on
immobilized metal-ion affinity chromatography columns (38) was consistent with the
postulate that dehydrins might interact with ions (45). Recently a phloem-localized
dehydrin (ITP) from castor bean was shown to bind Fe3+
,and was postulated to play a
role in iron transport (34). In that study, it was shown that the iron transport protein
(ITP), preferentially binds Fe3+
over Fe2+
and complexes with Cu2+
, Zn2+
, and Mn2+
in
vitro. In another case, a vacuole-localized celery dehydrin (VCaB45) was shown to bind
Ca2+
(33) and was postulated to function as a calcium buffer or as a calcium-dependent
chaperone. In both cases, proteins were isolated from plant tissues in an active ion-
binding form (likely phosphorylated). In this study we advance this understanding by
showing that the Arabidopsis dehydrin ERD14 is capable of binding a variety of ions and
that ion-binding (calcium in particular) is dependent upon ERD14’s phosphorylation
state. Arabidopsis ERD14 is phosphorylated in vivo. Further we have localized the
activating phosphorylation site to be contained within the “S” domain, a common feature
of most dehydrins.
by guest on April 14, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Phosphorylation-dependent ion-binding by ERD14
19
Of all the ions tested for interaction with ERD14, it seems most likely that the
binding of calcium and magnesium to ERD14 are the most physiologically relevant.
Intraorganellar levels of calcium and magnesium may exceed 1 mM and cytoplasmic
levels of these ions reach uM concentrations under a variety of conditions. Thus, the
observed Kd of 120 uM for calcium suggests the possibility for reversible binding at near
physiological conditions. ERD14 binds about 3 moles of calcium per mole, significantly
less than the approximately 25 moles bound to VCaB45 (33). No information is available
for the number of Fe3+
binding sites for the dehydrin-like ITP. Unlike ITP, the inhibition
of calcium binding suggests that ERD14 may bind Fe2+
preferentially over Fe3+
. The Kd
for the highest affinity calcium-binding sites for ERD14 (120 uM) and VCaB45 (200
uM) are similar. Known calcium-binding proteins such as calreticulin, calsequestrin,
calnexin, and calmegin use the negatively charged acidic amino acid rich-region at the
carboxyl terminal (C-domain) quarter of the protein to bind calcium at high capacity and
low affinity (0.3-2 mM) (46). Calreticulin also binds calcium with high affinity in a
proline-rich domain (~1 uM) (46). Generally, calcium-binding domains include the
presence of glycine or proline and the presence of acidic or hydroxylated residues.
ERD14, an acidic protein with a pI of 5.28, which is highly enriched in glutamic acid
(constitutes 21% of the protein); has several regions which meet these general calcium-
binding characteristics.
We demonstrated that the dehydrin ERD14 is phosphorylated in planta and can be
phosphorylated in vitro with casein kinase II. The phosphorylation state of the “S”
domain likely regulates ion binding. Like other dehydrins, ERD14 has many potential
by guest on April 14, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Phosphorylation-dependent ion-binding by ERD14
20
phosphorylation sites. Both within and outside the “S” domain there are multiple
consensus phosphorylation sites for the protein kinase CKII. When E. coli expressed
ERD14 is phosphorylated with CKII, phosphorylation approaches saturation at 2 moles
of phosphate per mole of ERD14. Mass spectrometry revealed several phosphorylated
peptides in common to the purified Arabidopsis ERD14 and CKII-phosphorylated E. coli
expressed ERD14. Only one phospho-peptide, SDSSSSSSSEEEGSDGEK, was found
consistently by MALDI, and was always correlated with the characteristics of the gel
shifts and calcium binding. It is intriguing that this site and the regions immediately
surrounding it are highly conserved in a number of Arabidopsis dehydrins. Since the
phosphothreonine specific antibodies revealed no phosphothreonine that correlated with
the ability of ERD14 to bind calcium; we have concluded that the “S” domain contains
the activating phosphorylation site. A similar site has been shown to be phosphorylated in
maize dehydrin, Rab17 (11-13). In this case, it was concluded to be important for nuclear
targeting or membrane association. The commonality between in vivo and in vitro
phosphorylated peptides on ERD14 suggests that CKII may be responsible for in vivo
phosphorylation of ERD14. We observed that the activity of an endogenous kinase
capable of phosphorylating ERD14 is increased following cold stress (Figure 7);
suggesting that this might be a cold modulated kinase, perhaps CKII. It is unlikely that
this increase reflects an alteration in phosphatase activity, as phosphatase inhibitors were
present in the assay. Accompanying phosphorylation was a shift in apparent mass.
Whether under normal growth conditions or under cold stress conditions, Arabidopsis
ERD14 was always present in the phosphorylated state and always had calcium-binding
by guest on April 14, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Phosphorylation-dependent ion-binding by ERD14
21
activity (data not shown). This suggests that the phosphorylation is constitutive and might
not be used as a means to modulate ion-binding activity under these conditions.
The physiological function of the cold-induced dehydrins remains unknown. It
was recently shown (9) that an acidic dehydrin, DHN1 binds preferentially to acidic
phospholipids. In addition, phosphorylation of this same protein appears to be requisite
for targeting to nucleus (13). These latter results are consistent with a possible role for
dehydrins in stabilizing membranes in stress conditions (7,47). It was of interest to
determine whether calcium or phosphorylation of the dehydrins had an impact on lipid
interactions. A logical extension from our work was to determine whether membrane
association of dehydrins might be dependent on calcium. While low concentrations of
Triton X-100 readily remove ERD14 from membranes (a routine purification step used in
this paper) we have been unable to remove ERD14 from the membranes with EGTA
(data not shown). This is consistent with the conclusion drawn by Koag et al, (2002), that
dehydrins may interact with membranes through a lipid-binding class A2 a-helix
structure. Ion binding is likely not requisite for membrane interaction and we speculate
that the calcium (or ion-) binding has an independent function. We have proposed that an
alternative function of the dehydrins might be as calcium-dependent protein or
carbohydrate chaperones, a function similar to that performed by calreticulin and
calnexin (48,49). The similarity in structure of the chaperones and ERD14, number of
calcium-binding sites, and affinity make this postulate attractive. This hypothesis has not
yet been tested.
This study provides evidence that ion binding, most likely to be calcium, is one of
the major biochemical functions of the Arabidopsis ERD14 and perhaps of the other
by guest on April 14, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Phosphorylation-dependent ion-binding by ERD14
22
acidic dehydrins. Further, the ability of the acidic dehydrins, in particular ERD14, to bind
calcium is regulated by their phosphorylation state. On the basis of these results, we
hypothesize that ERD14 and perhaps other dehydrins could act as either calcium buffers,
conferred by their high calcium binding capacity, or have calcium-dependent chaperone-
like activity similar to the function of calreticulin and calnexin. However, further studies
are under way to evaluate which of the models is the likely biochemical function of
ERD14.
Acknowledgements
We extend our thanks to Dr. Mu Wang and Mr. Ross Cocklin (Proteomics Core Facility,
Indiana University), for consultations on the mass spectrometric data. We are also
grateful to Dr. Osama Alshaykh (Packet Video Corporation, CA) for developing the
software used to identify phosphorylated peptides in the mass spectra data. by guest on April 14, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Phosphorylation-dependent ion-binding by ERD14
23
References
1. Palva, E. T., Tahtiharju, S. T., Tamminen, I., Puhakainen, T., Laitinen, R.,
Svensson, J., Helenius, E., and Heino, P. (2002) JIRCAS Working Report, 9-15
2. Kim, K. N., Cheong, Y. H., Grant, J. J., Pandey, G. K., and Luan, S. (2003) Plant
Cell 15, 411-423
3. Close, T. J., Fenton, R. D., and Moonan, F. (1993) Plant Mol. Biol. 23, 279-286
4. Close, T. J., Fenton, R. D., Yang, A., Asghar, R., DeMason, D. A., Crone, D. E.,
Meyer, N. C., and Moonan, F. (1993) Curr. Top. Plant Physiol. 10, 104-118
5. Welin, B. V., Olson, A., Nylander, M., and Palva, E. T. (1994) Plant Mol. Biol.
26, 131-144
6. Shinozaki, K., and Yamaguchi-Shinozaki, K. (1996) Curr. Opin. Biotech. 7, 161-
167
7. Close, T. J. (1997) Physiol. Plant. 100, 291-296
8. Nylander, M., Svensson, J., Palva, E. T., and Welin, B. V. (2001) Plant Mol. Biol.
45, 263-279
9. Koag, M. C., Fenton, R. D., Wilkens, S., and Close, T. J. (2003) Plant Physiol.
131, 309-316
10. Garay-Arroyo, A., Colmenero-Flores, J. M., Garciarrubio, A., and Covarrubias,
A. A. (2000) J. Biol. Chem. 275, 5668-5674
11. Vilardell, J., Goday, A., Freire, M. A., Torrent, M., Martinez, M. C., Torne, J. M.,
and Pages, M. (1990) Plant Mol. Biol. 14, 423-432
12. Plana, M., Itarte, E., Eritja, R., Goday, A., Pages, M., and Martinez, M. C. (1991)
J. Biol. Chem. 266, 22510-22514
by guest on April 14, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Phosphorylation-dependent ion-binding by ERD14
24
13. Jensen, A. B., Goday, A., Figueras, M., Jessop, A. C., and Pages, M. (1998) Plant
J. 13, 691-697
14. Bush, D. S. (1995) Annu. Rev. Plant Physiol. Plant Mol. Biol. 46, 95-122
15. Sanders, D., Brownlee, C., and Harper, J. F. (1999) Plant Cell 11, 691-706
16. Knight, H., Trewavas, A. J., and Knight, M. R. (1996) Plant Cell 8, 489-503
17. Polisensky, D. H., and Braam, J. (1996) Plant Physiol. 111, 1271-1279
18. Sedbrook, J. C., Kronebusch, P. J., Borisy, G. G., Trewavas, A. J., and Masson, P.
H. (1996) Plant Physiol. 111, 243-257
19. Knight, H., Brandt, S., and Knight, M. R. (1998) Plant J. 16, 681-687
20. Bush, D. S. (1993) Plant Physiol. 103, 7-13
21. Ding, J. P., and Pickard, B. G. (1993) Plant J. 3, 713-720
22. Knight, H., Trewavas, A. J., and Knight, M. R. (1997) Plant J. 12, 1067-1078
23. Lewis, B. D., Karlin-Neumann, G., Davis, R. W., and Spalding, E. P. (1997)
Plant Physiol. 114, 1327-1334
24. Minorsky, P. V. (1989) Plant Cell Environ. 12, 119-135
25. Minorsky, P. V., and Spanswick, R. M. (1989) Plant Cell Environ. 12, 137-143
26. Sze, H., Liang, F., Hwang, I., Curran, A. C., and Harper, J. F. (2000) Annu. Rev.
Plant Physiol. Plant Mol. Biol. 51, 433-462
27. Xing, T., Williams, L. E., Nelson, S. J., East, J. M., and Hall, J. L. (1994)
Protoplasma 179, 158-165
28. Persson, S., Wyatt, S. E., Love, J., Thompson, W. F., Robertson, D., and Boss, W.
F. (2001) Plant Physiol. 126, 1092-1104
29. Li, X., Su, R. T., Hsu, H. T., and Sze, H. (1998) Plant Cell 10, 119-130
by guest on April 14, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Phosphorylation-dependent ion-binding by ERD14
25
30. Nelson, D. E., Glaunsinger, B., and Bohnert, H. J. (1997) Plant Physiol. 114, 29-
37
31. Hepler, P. K., and Wayne, R. O. (1985) Annu. Rev. Plant Physiol. Plant Mol.
Biol. 36, 397-439
32. Clapham, D. E. (1995) Cell 80, 259-268
33. Heyen, B. J., Alsheikh, M. K., Smith, E. A., Torvik, C. F., Seals, D. F., and
Randall, S. K. (2002) Plant Physiol. 130, 675-687
34. Kruger, C., Berkowitz, O., Stephan, U. W., and Hell, R. (2002) J. Biol. Chem.
277, 25062-25069
35. Gilmour, S. J., Artus, N. N., and Thomashow, M. F. (1992) Plant Mol. Biol. 18,
13-21
36. Ceccardi, T. L., Meyer, N. C., and Close, T. J. (1994) Protein Expr. Purif. 5, 266-
269
37. Welin, B. V., Olson, A., and Palva, E. T. (1995) Plant Mol. Biol. 29, 391-395
38. Svensson, J., Palva, E. T., and Welin, B. (2000) Protein Expr. Purif. 20, 169-178
39. Kitta, K., Clement, S. A., Remeika, J., Blumberg, J. B., and Suzuki, Y. J. (2001)
Biochem. J. 359, 375-380
40. Rivedal, E., and Opsahl, H. (2001) Carcinogenesis 22, 1543-1550
41. Tompa, P. (2002) Trends Biochem. Sci. 27, 527-533
42. Kiyosue, T., Yamaguchi-Shinozaki, K., and Shinozaki, K. (1994) Plant Mol. Biol.
25, 791-798
43. Kiyosue, T., Yamaguchi-Shinozaki, K., and Shinozaki, K. (1994) Plant Cell
Physiol. 35, 225-231
by guest on April 14, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Phosphorylation-dependent ion-binding by ERD14
26
44. Ingram, J., and Bartels, D. (1996) Annu. Rev. Plant Physiol. Plant Mol. Biol. 47,
377-403
45. Dure, L. (1993) Curr. Top. Plant Physiol. 10, 91-103
46. Corbett, E. F., and Michalak, M. (2000) Trends Biochem. Sci. 25, 307-311
47. Kaye, C., and Guy, C. L. (1995) Sci. Prog. 78 ( Pt 4), 271-299
48. Nigam, S. K., Goldberg, A. L., Ho, S., Rohde, M. F., Bush, K. T., and Sherman,
M. (1994) J. Biol. Chem. 269, 1744-1749
49. Peterson, J. R., Ora, A., Van, P. N., and Helenius, A. (1995) Mol. Biol. Cell 6,
1173-1184
50. Biermann, B. J., Morehead, T. A., Tate, S. E., Price, J. R., Randall, S. K., and
Crowell, D. N. (1994) J. Biol. Chem. 269, 25251-25254
by guest on April 14, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Phosphorylation-dependent ion-binding by ERD14
27
Footnotes
1 The abbreviations used are: ERD14, early response to dehydration; CKII, casein kinase
II; MALDI-TOF/MS, matrix assisted laser desorption ionization-time of flight/mass
spectrometry.
by guest on April 14, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Phosphorylation-dependent ion-binding by ERD14
28
Figure Legends
Figure 1. Treatment of ERD14 with casein kinase II (CKII) results in a shift in apparent
molecular mass and the shift correlates with phosphorylation and calcium-binding
activities. Purified E. coli expressed ERD14 was treated with CKII for the indicated
times. Reactions terminated by the addition of hot SDS-PAGE sample buffer were
analyzed on SDS-PAGE gel. Panel A. Apparent mass shifting of the phosphorylated
ERD14. SDS-PAGE gels were coomassie-stained (CO) or transferred onto nitrocellulose
membranes and probed with anti-dehydrin (WB). Panel B. [32P] Phosphorus/ERD14
incorporation. ERD14 was in vitro phosphorylated with CKII in the presence of [g-32P]
ATP. [32P] Phosphorus was visualized with a STORM PhosphoImager. Panel C.
Calcium-binding activity of ERD14 as measured by ligand blot. SDS-PAGE gels were
transferred onto nitrocellulose membranes and incubated in 200-300 nM 45calcium
chloride as described in Heyen et al. (2002). 32P and 45Ca incorporation were visualized
and quantitated using a STORM PhosphoImager. Gels are from one typical experiment,
quantitative plots are average of three experiments. Standard deviations are shown as
error bars.
Figure 2. Phosphorylation and apparent mass shift of ERD14 by CKII is reversible.
Purified E. coli expressed ERD14 was treated for 3 hours at 30°C with or without casein
kinase II (CKII). One portion of the phosphorylated ERD14 was then incubated for 20
minutes at 37°C in the presence or absence of 5 units of SAP. Following SDS-PAGE
analyses, gels were stained with Coomassie Brilliant Blue.
by guest on April 14, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Phosphorylation-dependent ion-binding by ERD14
29
Figure 3. Panel A. Purified E. coli expressed ERD14 is phosphorylated on at least two
sites. ERD14 was in vitro phosphorylated at 30°C with casein kinase II in the presence of
[g-32
P] ATP. At the indicated times, reactions were terminated with 25% trichloroacetic
acid. Precipitates were collected on GFA glass fiber filters and washed with 5% TCA and
100% acetone. 32
P incorporation was determined by liquid scintillation counting. Data are
average of three experiments. Standard deviations are shown as error bars. Panel B.
Purified E. coli expressed ERD14 is phosphorylated by casein kinase II (CKII) in
threonine and serine sites. Following SDS-PAGE, blots were probed with anti-
phosphothreonine (Top Panel) and anti-phosphoserine (Bottom Panel).
Figure 4. Phosphorylation activates calcium-binding activity of ERD14. Purified E. coli
expressed ERD14 was in vitro phosphorylated for 3 hours with casein kinase II (CKII).
In one case (+ST), 1 mM staurosporine was added at 30 minutes to inhibit protein kinase
activity. Calcium-binding activity (at 0.25 mM) was estimated by equilibrium dialysis
(33). Standard deviations are indicated as error bars. Data are average of three
experiments. Inset: The corresponding gel shift as visualized by Coomassie staining.
Figure 5. Calcium-binding properties of ERD14. Panel A. Calcium binding of the
purified E. coli expressed ERD14, as estimated by equilibrium dialysis when non-
phosphorylated (-CKII) and phosphorylated (+CKII). Panel B. Scatchard plot indicating
maximal binding sites (Bmax) (2.86 mole Ca++/ mole phosphorylated ERD14) and Kd
(0.12 mM). The data was plotted as nY/X vs nY; where nY is the moles of calcium
by guest on April 14, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Phosphorylation-dependent ion-binding by ERD14
30
bound per mole of ERD14 and X is the concentration of free calcium. Standard
deviations are shown as error bars. Data are average of three experiments.
Figure 6. Inhibition of ERD14 calcium-binding activity by cations. Purified E. coli
expressed ERD14 was in vitro phosphorylated for 3 hours with casein kinase II. Calcium
binding was estimated by equilibrium dialysis. Calcium binding was performed in the
presence of 0.25 mM calcium chloride. Competition was examined by adding additional
0.25 mM of the indicated metal ion. Standard deviations are indicated as error bars. Data
are average of three experiments.
Figure 7. ERD14 is phosphorylated by Arabidopsis protein extracts. Panel A. Extracts
from plants treated for 2 days at 4°C or 20°C were incubated with or without purified E.
coli expressed ERD14, 0.8 mM ATP, and [g-32P] ATP (1 µCi). At the indicated times
(min), aliquots were analyzed by SDS-PAGE and STORM PhosphoImaging. Panel B.
Quantitation of the gel. Standard deviations are shown as error bar. Data are average of
three experiments.
Figure 8. ERD14 from cold-treated Arabidopsis plants is phosphorylated. Panel A. Peak
fractions obtained from the MonoQ column (fractions 26, 27, 28) are compared to
ERD14 and Phosphorylated ERD14 (P-ERD14). The band seen in fraction # 27 was
excised and confirmed to be ERD14 by MALDI-TOF mass spectroscopy. Panel B.
Treatment of the Arabidopsis ERD14 (fraction # 27 in A) with SAP results in a shift to a
smaller apparent molecular mass and a loss in calcium-binding activity. ERD14 was
by guest on April 14, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Phosphorylation-dependent ion-binding by ERD14
31
incubated for 20 minutes at 37°C in the presence or absence of SAP. To obtain sufficient
sample, proteins were separated on a 2 mm thick 12% SDS-PAGE gel, and analyzed by
coomassie staining of gels (CO), western blotting (WB) using anti-dehydrin antibody, or
by calcium-ligand blot (CB). 45Calcium was visualized with the STORM PhosphoImager.
Figure 9. Matrix assisted laser desorption ionization-time of flight/mass spectrometry
(MALDI-TOF/MS) identification of the phospho-peptides in ERD14. Purified E. coli
expressed ERD14 was phosphorylated with casein kinase II for the indicated times and
tryptic digested. ERD14, isolated from 4 week-old Arabidopsis seedlings treated for 2
days at 4°C, was incubated for 20 minutes at 37°C in the presence or absence of SAP.
Following SDS-PAGE, ERD14 bands were stained, excised, destained, reduced,
alkylated, trypsin-digested, and extracted as described in Experimental Procedures.
ERD14 digested solutions were mixed with equal volume of a -cyano-4-
hydroxycinnamic acid matrix and analyzed by MALDI-TOF/MS. ProFound‰ and
PAWS‰ in the Genomic Solutions‰ (http://165.219.84.5/index.html) web site were used
for protein identification and to identify the theoretical phosphorylation sites on ERD14
with four tryptic digestion missed cleavages, respectively. Panel A. ERD14 amino acid
sequence. Bold letters indicate the predicted phosphorylation sites for protein casein
kinase II determined by PhosphoBase v2.0 (www.cbs.dtu.dk/databases/PhosphoBase/).
Lines and Roman numerals show phospho-peptide positions and designated peptide
numbers. K-domains are indicated by dotted underlines. Panel B. Phosphorylation sites
in E. coli expressed and Arabidopsis ERD14. Results are from three experiments; a, b,
and c. Fragments obtained from each reaction were compared with the computer-
by guest on April 14, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Phosphorylation-dependent ion-binding by ERD14
32
generated list to identify the 80, 160, 240, and 320 Da shift corresponding to
phosphorylation sites 1, 2, 3, and 4, respectively. The sequences shown are the minimum
sized tryptic fragments (except peptide IV), however, alternative peptides (up to four
missed tryptic cleavages) were examined for possible phosphorylation modifications and
included in the analysis. Panel C. The ion spectra map depicting the monoisotopic profile
of the phospho-peptides (II, IV, V, and VII) found in the ERD14 isolated from
Arabidopsis plants (not treated with SAP) acquired by MALDI-TOF/MS. Peptide II
(mass/z was 1584.72) has no missed cleavages, and one phosphorylation; peptide IV
(mass/z was 2186.77) has one missed cleavage (indicated by (R)) and three
phosphorylations; peptide V (mass/z was 982.48) has no missed cleavage and one
phosphorylation; and peptide VII (mass/z was 995.4) has no missed cleavage and two
phosphorylations. In parentheses are the peptide numbers on ERD14 amino acid
sequence (from Panel A). x-axis is the mass-to-charge value of the ion (m/z), and y-axis
represents peak relative intensity.
by guest on April 14, 2018
http://ww
w.jbc.org/
Dow
nloaded from









Muath K. Alsheikh, Bruce J. Heyen and Stephen K. RandallIon-binding properties of the dehydrin ERD14 are dependent upon phosphorylation
published online August 13, 2003J. Biol. Chem.
10.1074/jbc.M307151200Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
by guest on April 14, 2018
http://ww
w.jbc.org/
Dow
nloaded from