isolated rat hepatocyte couplets in short-term culture: structural characteristics and plasma...
TRANSCRIPT

0270-9139/87/0702-0216$02.00/0 HEPATOLOCY Copyright 0 1987 by the Amencan Association for the Study of Liver Diseases
Vol. 7, No. 2, pp. 216223, 1987 Printed in U.S. A.
Original Articles
Isolated Rat Hepatocyte Couplets in Short-Term Culture: Structural Characteristics and Plasma Membrane
Reorganization
ANIL GAUTAM, 01-CHENG NG AND JAMES L. BOYER Department of Medicine and Liver Research Center, Yak University School of Medicine, New Haven, Connecticut 06510
Studies of canalicular bile secretion have been limited due to lack of direct access to the canalicular lumen. Isolated rat hepatocyte couplets, consisting of two hep- atocytes enclosing a canalicular space, have been pro- posed as a primary secretory unit that may be useful for direct studies of unmodified canalicular bile secretion. The present study examines their structural character- istics and plasma membrane reorganization. The cana- licular space of freshly isolated hepatocyte couplets is freely permeable to ruthenium red, but within 4 hr the junctional borders reseal in most couplets which then exclude ruthenium red from the luminal area. These resealed spaces expand in 61.8 f 10% of couplets as secretion is elaborated and after 4 hr in monolayer culture, 12.7 f 4.7% of the canalicular spaces are dila- ted to diameters greater than 3 pm. Normal-appearing microvilli line these canalicular membranes in the ma- jority of dilated spaces as assessed by electron micros- copy. Immediately after isolation, Mg++-ATPase, a his- tochemical marker for canalicular membranes, is lo- cated as a discrete band corresponding to the normal in uiuo circumferential distribution of the canalicular membrane domain. However, this pattern of Mg++-ATP- ase staining rapidly diminishes and reorganizes at the remaining canalicular pole within several hours. This membrane reorganization is a microfilament-dependent process, since it is inhibited by cytochalasin D but not by colchicine, an inhibitor of microtubular function or cycloheximide, an inhibitor of new protein synthesis.
Thus, 3 to 4 hr after isolation, the isolated hepatocyte couplet has reorganized its canalicular membrane, re- taining polarity of its excretory domain only at the remaining junctional contacts between adjacent hepa- tocytes. Secretion is elaborated into the ruthenium red impermeant canalicular lumen permitting direct studies of canalicular bile secretion by micropuncture, electro- physiologic and quantitative microscopic techniques.
Received April 18,1986, accepted November 20,1986. This work was supported in part by USPHS Grants AM25636,
AM01180 and Liver Center Grant 1 P30 AM 34989. Parts of this work were presented at the Annual Meeting of the
American Association of Physicians (TM~s . Assoc. Am. Phys. 1985; 9821-29) and the American Gastroenterology Association and have appeared in abstract form (Clin. Res. 1985; 33602A and Gastroenter- ology 1985; 88A660).
Address reprint requests to. Anil Gautam, M.D., Liver Research Center, lOs0 LMP, Yale University School of Medicine, 333 Cedar Street, New Haven, Connecticut 06510.
The primary secretion of bile is elaborated into small canalicular spaces that are formed between adjacent hepatocytes and encircle these parenchymal cells in a belt-like fashion, interconnecting throughout the hepatic lobule in a three-dimensional chicken-wire like mesh. The tiny (1 to 2 pm in diameter) size and inaccessibility of these canalicular spaces have prevented direct sam- pling of this secretion as well as the application of more classical electrophysiological methods to study mecha- nisms of canalicular bile formation. Instead, the study of bile secretory function has relied on a number of indirect methods that sample bile from biliary cannulas in intact animals or isolated perfused liver preparations at sites distal to the canaliculus and after contact with and probable modification by cells lining the bile ductules. A more direct approach involves the use of canalicular membrane vesicles to study the driving forces for the transport of solutes across this excretory domain, but the intact cell is disrupted in this method, limiting its use- fulness.
Recently, a new model has been described for the study of bile secretory function that consists of isolated rat hepatocyte couplets (IRHC) in short-term monolayer culture (1,2). This technique permits direct study of the primary canalicular secretion in isolated intact hepato- cytes without the modifying influence of bile ductular cells, blood flow or hormonal effects. As originally ob- served by Oshio and co-workers (l), during isolation of rat hepatocytes by collagenase perfusion, a number of hepatocytes fail to dissociate completely and remain coupled together. Some of these isolated hepatocyte pairs enclose a canalicular space which reseals when cells are placed into short-term monolayer cultures. Continued secretion into the space results in periodic expansions followed by spontaneous collapse or contraction (1, 3). Recent work from our laboratory (2) has demonstrated that microelectrodes can be inserted into the expanded canalicular space through one cell, enabling the recording of transepithelial potential profiles. In addition, IRHC transport fluorescein from the medium and concentrate this organic anion within the canalicular vacuoles (2). Furthermore, addition of the choleretic bile salt, tauro- cholate, to the external media stimulates secretion into the canalicular space (2, 3). Measurement of electrical resistances are consistent with tight junction conduct-
216

Vol. 7, No. 2, 1987 CANALICULAR BILE SECRETION IN ISOLATED HEPATOCYTE COUPLETS 217
ances in leaky epithelia (4), whereas canalicular reten- tion of fluorescein and reductions in the canalicular space volume in response to externally imposed hyperosmotic sucrose gradients suggest that the canalicular vacuole is bounded by membranes that are selectively permeable to small ions but not to larger osmotically active substances (2). Finally, the IRHC is capable of regulation of intra- cellular pH following intracellular acidification with NKCl(5) .
These studies indicate that the IRHC model functions as a primary unit of canalicular bile secretion and should be a useful model for elucidating mechanisms of canalic- ular bile formation with electrophysiological and other quantitative techniques.
Despite these advantages, the IRHC differs in several important respeda from the normal intact liver. First, the canalicular space is a closed vacuole rather than an open tube. Second, the canalicular membranes are in a dynamic state, where this membrane surface is undergo- ing a reorganization to the new excretory pole of the cell during the 4 to 8 hr after being placed in cultures. In this report, we examine the structural reorganization of the canalicular membrane of this unique hepatocyte couplet- secretory unit in greater detail.
MATERIALS AND METHODS IRHCs are prepared in the Liver Center Core Laboratory for
Hepatmyte Isolation and Culture from fed, male Sprague- Dawley rats (200 to 250 gm; Camm Laboratories, Wayne, NJ) by the nonrecirculating collagenase perfusion method of Seglen (6) as modified in our laboratory (2, 7). The concentration of collegenase (Type 1, Sigma Chemical Co., St. Louis, Mo.) is decreased to 0.05% (w/v) in a total volume of 150 ml of Hank's buffer containing 4 mM calcium, 0.9 mM magnesium, 26 mM eodium bicarbonate and 0.8 unit of trypsin inhibitor (Sigma)/ unit of tryptic activity in the collagenam. The final cell suspen- sion is filtered through a 30 pm nylon mesh (Tetko, Elmaford, NY) to obtain a cell population relatively enriched in hepato- cyte couplets (31.6 f 8.6% of cells were IRHC in 138 consecu- tive isolations). Intermediate steps to purify hepatmytea from nonparenchymal cells are performed in hbovitz-15 (L-15) medium using unit gravity sedimentations for three successive 15-min periods to selectively sediment hepatocytes. The result- ing cell preparations are >95% hepatocytes and 91.7 * 2.9% exclude trypan blue consistently (n = 231 isolations). Prepa- ratio- in which 4 6 % cells exclude trypan blue are diecarded. For short-term monolayer culture, the final cell suspension is plated onto glass coverslip fragments in 35 x 10 mm plastic tissue culture dishes at a cell concentration of 0.75 X l@ per ml of L-15 medium (GIBCO, Grand Island, NY) containing 50 units penicillin and 50 ccg streptomycin per ml. Calf serum and hormones were omitted from the culture medium to prevent flattening of cells onto the glass cover slips and to assess cell function in a more defined medium.
Isolated couplets were examined by both light and electron microscopy. Cells were fued with 1.5% glutaraldehyde in 0.1 M sodium cacodylate buffer (pH 7.4), immediately aRer isola- tion and then at 1,2,4,6, and 8 hr after incubation at 37°C.
Fixed cells are washed overnight at 4'C in 0.1 M sucrose in 0.1 M cacodylate buffer and, for electron microscopy studies, postsxea for 45 min with 1% osmium tetraoxide in the same bder containing 1% potassium ferrocyanide at 4°C. After en bloc staining with 0.5% uranyl acetate for 1 hr, specimens are dehydrated in ethanol and embedded in epon. The epon capsule is inverted and placed flat on the cover slip. After polymeriza-
tion, the cover slip is cracked, sections are cut in a plane parallel to the culture layer and stained with uranyl acetate and lead citrate. Specimens are examined with a Zeiss 10B electron microscope at an accelerating voltage of 80 kV.
Permeability of the tight junctions is assessed by exposing the isolated couplets at various times after isolation to the electron-dense dye, ruthenium red, which coats cell membranes (8).The ruthenium red concentration is adjusted to 1,OOO ppm in 2.5% glutaraldehyde fixative buffered in 0.1 M cacodylate buffer (pH 7.4) (8). After washing overnight with 0.1 M sodium cacodylate buffer (pH 7.4) containing 1,OOO ppm of ruthenium red, the specimen is postfixed with 2% osmium tetraoxide for 3 hr in the dark. Specimens are further processed with the transmission electron microscopic technique as above.
Preliminary evidence indicates that canalicular membrane elements tend to reorganize from their in uitro circumferential belt-like configuration on the cell surface toward the remaining canalidar pole between the two adjoining hepatmytea in IRHC (2). To study this membrane reorganization in isolated couplets, the marker enzyme, Mg++-ATPase, was used to local- ize the canalicular membrane domains by the Wachstein-Mei- sel histochemical method with 1 mM tetramisole to inhibit background alkaline phosphatase activity (9). Cells were ex- amined at various time points after initial plating and photo- graphed in a Zeiss IM35 microscope fitted with double differ- ential contrast (Nomarski) optics. The determinants of this process of membrane reorganization were studied by using 1 to 5 pg per ml cytochalasin D (Sigma), lo-' M colchicine (Sigma) and cycloheximide (5 x M) to inhibit mic ro fhen t (lo), microtubular (11, 12) and protein synthetic (13) functions, respectively. In the colchicine group, rats were pretreated with i.v. colchicine (0.2 mg per 100 gm body weight) 2 hr prior to cell isolation (11), and lo-' M colchicine was included through- out the purification and culture steps. Cytochalasin D and cycloheximide were added to the culture media immediately after the cells were isolated.
Thiamine pyrophosphatase was assessed histochemically by the lead capture method in separate experiments (14) as an additional marker for the trans side of the Golgi membranes and the canalicular membranes. Viability of hepatmytea in culture was assessed at 4 and 8 hr by lactate dehydrogenase release into the medium (la, by trypan blue exclusion and also corroborated by measurements of electrical potentials of hep- atocytes and their ability to regulate intracellular pH (5).
To quantitate canalidar size, a micrometer was fitted into the eyepiece, and the number of canaliculi greater or less than a maximum diameter of 3 pm were counted. A Neofluar 63/ 1.26-oil objective, x10 eyepieces and an Achromatic-aplanatic condenser with a 1.4 Pol condenser (oil-immersion) were used in the optical system, which yields a lateral resolution of 0.25 to 0.3 pm and a depth of focus of 0.3 pm with Nomarski optics.
Mg++-ATPase activity was assayed biochemcially by the method of Scharschmidt et al. (16) in cell samples harvested immediately after isolation as well as in cells obtained from monolayer cultures at 1, 2, 4 and 8 hr after initial plating. At defined times the monolayers were washed twice and then scraped into Krebs-Henseleit buffer and sonicated for assays of Mg++-ATPase activity and protein concentrations by the Lowry technique (17).
To determine if Mg++-ATPam is newly synthesized and inserted into the remaining canalicular pole membranes, the effect of cycloheximide (5 X lO-'M) and cytochalasin D on biochemically assayable Mg++-ATPase activity was compared to control, untreated hepatocyte couplets.
Startieticd Methale. All values represent arithmetic m e m with standard deviations. Data from each treatment group WBS evaluated in a "blinded" manner and compared to controls using the Student's t test.

218 GAUTAM, NG AND BOYER HEPATOLOGY
RESULTS CeU Viability. Isolated hepatocytes maintained ex-
cellent viability for 4 to 8 hr after isolation and attach- ment to glass cover slips (90.6 f 3.1% of attached cells excluded trypan blue 4 hr after plating n = 46 experi- ments). Lactate dehydrogenase was released into the supernatant medium during the first hour and amounted to 14.9 f 6.1% (n = 10) of total cellular activity at the time of plating. However, subsequent release of lactate dehydrogenase over the next 4 hr was minimal, and total supernatant activity was still 17.3 f 5.6% of total cellular activity at plating (n = 10).
The light microscopic appearance of several IRHC with variably dilated canalicular spaces between the ad- jacent hepatocytes after 4 hr in culture is illustrated in Figure 1. The isolated liver cells remain spherical over the 8-hr culture period and, in the absence of added serum to the media, usually do not flatten onto the coverslips.
Canalicular Spcrces. Couplets (232 k 58) were counted in 11 separate isolations after 4 hr in culture and separated into three groups according to canalicular size: Group I = canalicular lumen greater than 3 pm; Group I1 = canalicular lumen less than 3 pm, and Group I11 = couplets without expanded canalicular lumen. All canalicular spaces were collapsed and less than 1 pm in maximum diameter immediately after isolation. How- ever, after 4 hr in culture, a majority of couplets dem- onstrated canalicular dilatation. Forty-eight f 17% of IRHC demonstrated canaliculi that were moderately di- lated (less than 3 pm), whereas 12.7 k 4.7% were widely dilated (>3 pm). Notably, after 4 hr, 39.2 f 12% of the examined couplets either remained collapsed or were in a collapsed phase following a cycle of expansion.
The electron microscopic appearance of a couplet is illustrated in Figure 2 and shows the ultrastructure of the hepatocytes to be essentially normal. Vacuoles are usually increased in freshly isolated hepatocytes but are
FIG. 1. Light microscopic appearance of IRHC 4 hr after isolation and plating onto glass cover slips in L-15 medium (original magnifica- tion, X 740). The canalicular spaces are variably expanded. Nomarski Optics were used.
8
FIG. 2. Electron microscopic appearance of an isolated couplet after 4 hr in culture. Sections were cut parallel to [(A) original magnification, x 4,4001 or perpendicular to the planes of the cover slip [(B) original magnification, X 5,0001. Arrows = original position of cover slip.
largely absent in cells maintained for 4 hr in culture, suggesting that vacuoles may reflect cell damage during the isolation process.
Thin sections, cut parallel to the plane of the cover slip (Figure 2A) and perpendicular to the cover slip (Figure 2B), further demonstrate the essentially spheri- cal configuration of the coupled hepatocytes afer 4 hr in culture and provide a three-dimensional perspective of the couplet structure and the enclosed canalicular space. Canalicular microvilli are normal in appearance, and the lumen is moderately dilated. A similar appearance was noted in 40 of 55 dilated canalicular spaces examined in detail by electron microscopy in five separate experi- ments (Figure 3A). In contrast, 15 of 55 dilated spaces appeared cholestatic (Figure 3B), as characterized by canalicular membranes that were devoid of microvilli over most of their surface with thickening of the perican- alicular ectoplasm.
Permeability of the Canalicular Space to Ruthe- nium Red. Couplets were exposed to ruthenium red immediately after isolation and at various times in cul- ture. Examination of the fixed samples (n = 4) by elec- tron microscopy revealed that ruthenium red freely pen- etrated 100% of 30 canalicular spaces in freshly isolated

Vol. 7, No. 2, 1987 CANALICULAR BILE SECRETION IN ISOLATED HEPATOCYTE COUPLETS 219
FIG. 3. (A) Normal c~alicular spece encloeed between adjacent hepatocytes in M isolated couplet at 4 hr. High power electron micros- copy ( 0 x i g i ~ I maenifieation, X 8.800). Note microvilli liniDg tbe can- alicular membrane, pen- gob and tight junctions ( a r m s ) , l h t i n g the canalicular epace. (B) Choleatatic canalicular apace with flattsned microvilli and increaee in pericanalicular ectophm (0- magnificntion, X 8,400).
hepatocyte couplets (Figure 4A). In contrast, the dye penetrated expanded canalicular spaces in only 8% of 26 couplets with dilated canaliculi exposed to ruthenium red after 4 to 8 hr in culture (Table 1). The electron- dense material coated the intercellular spaces only up to the level of the tight junction and was excluded from the canalicular luminal area. These findings suggest that the canalicular lumen is initially open and freely commun- ciates with the bathing medium in couplets freshly dis- sociated fiwm neighboring hepatocytes as would be ex- pected. However, with time in culture, the junctional complex apparently reseals in some couplets and remains relatively impermeant to large solutes such as ruthenium red as the lumen expands with secretion. In contrast, 33% of 24 collapwd canalicular spaces were permeable to ruthenium red after 4 to 8 hr in culture (Figure 5B). Canaliculi that are freely permeable to ruthenium red at 4 to 8 hr occur either because their junctional contacts that border the canalicular space initially failed to reseal or because they are in a collapsed phase between secre- tory cycles.
Canalicular Membrane Rearrangement. Hieto- chemical staining of Mg++-ATPase was used a~ a marker for canalicular membrane domains. Immediately follow- ing isolation, the hepatocyte couplets display band-like circumferential staining on the cell surface, similar to the in uiuo configuration of bile canaliculi. By electron microscopy, freshly isolated couplets display dieCrete membrane areas with the appearance of c a n a l i h membrane domains (Figure 6). These "hemicanaliculi" or canalicular remnants represent attachment aitea to neighboring hepatocytes in uioo from which the couplet is now dissociated. A small amount of Mg++-ATPase
FIG. 4. Permeability of the canalicular limiting membrauea to ruthe- nium red. (A) Fmhly isolated couplet: NtheniUm red is freely perme- able into the canalicular apace (arrow) (or ighl m&h, X l0,ooO). (B) Ieolated couplet 4 hr &r culture. Ruthenium red in exchded from the expanded canaliculnr space but c r ~ b tbe lateral intercellular spaces up to the tight junction8 (WrOW8) (0- tication, x 10,290).
TABLE 1. Permeability of the d c u l a r lumen to ruthemium red
Freshly iaolated couplets 100 0
Couplets cultured 4-8 hr (n = 30)
Dilated canalicular lu- 8 92
Collapsed lumen 33 67 men (n = 26)
(n = 24)
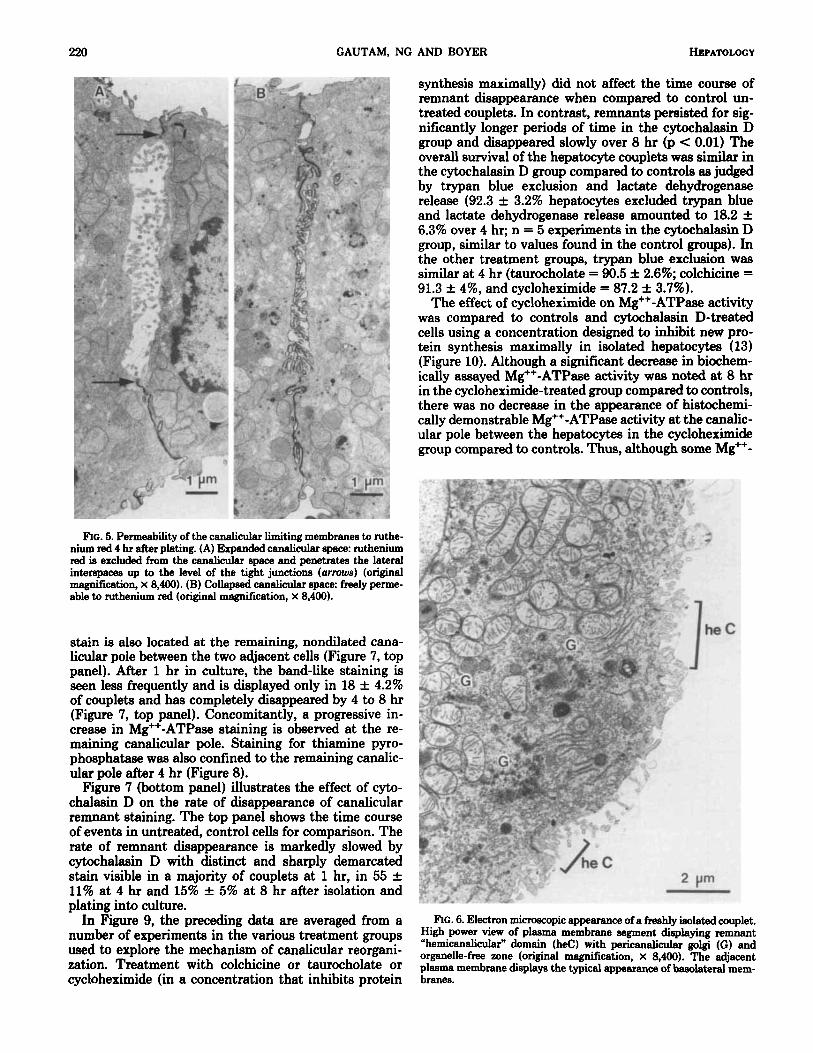
220 GAUTAM, NG AND BOYER HEPATOLOGY
synthesis maximally) did not affect the time course of remnant disappearance when compared to control un- treated couplets. In contrast, remnants persisted for sig- nificantly longer periods of time in the cytochalasin D group and disappeared slowly over 8 hr (p < 0.01) The overall survival of the hepatocyte couplets was similar in the cytochalasin D group compared to controls as judged by trypan blue exclusion and lactate dehydrogenase release (92.3 f 3.2% hepatocytes excluded trypan blue and lactate dehydrogenase release amounted to 18.2 f 6.3% over 4 hr; n = 5 experiments in the cytochalasin D group, similar to values found in the control groups). In the other treatment groups, trypan blue exclusion was similar at 4 hr (taurocholate = 90.5 f 2.6%; colchicine = 91.3 f 4%, and cycloheximide = 87.2 f 3.7%).
The effect of cycloheximide on Mg++-ATPase activity was compared to controls and cytochalasin D-treated cells using a concentration designed to inhibit new pro- tein synthesis maximally in isolated hepatocytes (13) (Figure 10). Although a significant decrease in biochem- ically assayed Mg++-ATPase activity was noted at 8 hr in the cycloheximide-treated group compared to controls, there was no decrease in the appearance of histochemi- cally demonstrable Mg++-ATPase activity at the canalic- ular pole between the hepatocytes in the cycloheximide group compared to controls. Thus, although some Mg++-
FIG. 5. Permeability of the canalicular limiting membranes to Nthe- nium red 4 hr after plating. (A) Expanded canalicular apace: ruthenium red is excluded from the cnnalicular space and penetrates the lateral intempaces up to the level of the tight junctions (arrows) (original magnification, x 8,400). (B) Collapsed canalicular space: freely perme- able to NtheniUm red (original magnification, X 8,400).
stain is also located at the remaining, nondilated cana- licular pole between the two adjacent cells (Figure 7, top panel). After 1 hr in culture, the band-like staining is seen less frequently and is displayed only in 18 f 4.2% of couplets and has completely disappeared by 4 to 8 hr (Figure 7, top panel). Concomitantly, a progressive in- crease in Mg++-ATPase staining is observed at the re- maining canalicular pole. Staining for thiamine pyro- phosphatase was also confined to the remaining canalic- ular pole after 4 hr (Figure 8).
Figure 7 (bottom panel) illustrates the effect of cyto- chalasin D on the rate of disappearance of canalicular remnant staining. The top panel shows the time course of events in untreated, control cells for comparison. The rate of remnant disappearance is markedly slowed by cytochalasin D with distinct and sharply demarcated stain visible in a majority of couplets at 1 hr, in 55 f 11% at 4 hr and 15% f 5% at 8 hr after isolation and plating into culture.
In Figure 9, the preceding data are averaged from a number of experiments in the various treatment groups used to explore the mechanism of canalicular reorgani- zation. Treatment with colchicine or taurocholate or cycloheximide (in a concentration that inhibits protein
FIG. 6. Electron microscopic appearance of a freshly isolated couplet. High power view of plasma membrane segment displaying remnant “hemicanalicular” domain ( h e ) with pericanalicular golgi (G) and organelle-free zone (original magnification, x 8,400). The adjacent plasma membrane displays the typical appearance of basolateral mem- branes.

Vol. 7, No. 2,1987 CANALICULAR BILE SECRETION IN ISOLATED HEPATOCYTE COUPLETS 221
Ftc. 7. Hietochemical localization of the canalicular marker enzyme, Mg"-ATPase, in hlated hepatmyte couplets. The number in each panel indicates the hours in culture prior to fixation for hietochemistry. (Toppanel) Untreated, control couplate in L-15 medium. (Bottom panel) Cells plated in medium containing 5 pg per ml cytochalasin D. Original megnircation. X 586.
- . I 1 , , >-
F ~ G . 8. Thiamine pyrophoephatase staining of isolated coupleta 4 hr after plating. (A) Low power view (original magnification, X 162) and (B) higher power light microecopic view (original mannificetinn, X 426) displaying staining predominantly a t or near the remainii canalicular pole.
ATPase is synthesized during the 4 to 8 hr in monolayer culture, the bulk of activity located at the canalicular pole must represent a microfilament-dependent reorga- nization of Mg++-ATPase and presumably other canalic- ular membrane elements from the canalicular remnants to the surviving canalicular pole.
0 CONTROL (n.9) = CYTOCHALASIN D (5pqlrnl; n.6) A COLCHICINE ( 1 0 - 5 ~ ; n.5) A CYCLOHEXIMIDE ( 5 ~ 1 0 ' ~ M ; n.6) o TAUROCHOLATE ( I O ~ M ; n.4) -
W z
F c
IY) Lkl
1 8.5
0' ' 0.5 2.5 4.5
TIME IN CULTURE (hrs) RG. 9. Rate of di68ppe-e of can&cular remnant &*-AW-
staining with time in culture in control coupleta (0) and in the presence of 5 pg per ml cytochalaaii D 0.10 pA4 taurocholate (01, lod M colchicii (A) and 5 x lo4 Mcycloheximide (A). Each point rspresente the percentage of coupleta displaying remnant atainln9. bors indicate mean f S.D. Cytochalaei D significently aelayed the d i q - pearance of canalieular remnants compared to controls at 1,2,4H and 8% hr after culture (p < 0.01 at each point).
DISCUSSION Isolated hepatocyte couplets consist of two nondiseo-
ciated isolated hepatocytes in a monolayer culture sys- tem, which enclose a canalicular space. Secretion occurs with time to distend the canalicular lumen in a milieu entirely free of hormonal or blood flow effecte or the

222 GAUTAM, NG AND BOYER HEPATOLOGY
r
t 0 CONTROL
I .o 0 CYCLOHEXIMIDE (5xIO"M) Y
I A CYTOCHALASIN D ( 5pg / m l )
0 ' ' 1 1 I
.5 4.5 8.5 T I M E AFTER ISOLATION (hrs)
lation of Mg++-ATPase at the remaining canalicular pole. Whether this process of rearrangement represents lateral diffusion of canalicular membrane elements into baso- lateral domains with subsequent relocalization at the remaining canalicular pole or transcellular migration via a membrane vesicle-mediated transport process or a com- bination of both mechanisms is presently unknown. However, by analogy to other cell systems, notably toad bladder epithelial cells which also display extensive re- organization of differentiated membrane domains after isolation, the dependency of canalicular remnant disap- pearance on microfilament function might favor lateral intramembranous dislocation as the initial event (18).
Thus, isolated hepatocyte couplets in short-term cul- ture undergo within l hr a rapid reorganization of cana- licular remnants to the remaining canalicular pole (Fig- ure 11). This process results in a highly polarized config- uration with bile canalicular membranes of the cell now organized entirely at the pole of the cell that retained junctional contacts between the two adjacent hepato- cytes. With time in culture, the canalicular space expands in size as its membrane area appears to increase and as secretion accumulates in the lumen. Figure 11 is a three- dimensional schematic of the cell couplet immediately
FIG. 10. Biochemically assayable Mg++-ATPase activity (cmoles per - after isolation and following reorganization of the a p i d mg protein per hr) in ieolated couplets at the designated times after culture in control, untreated couplets (0) and in the presence of 5 pg per ml cytochalasin D (A) and 5 X lo4 M cycloheximide (0). Range &bars indikte mean f S.D.
-
modifying influence of nonparenchymal liver cells or bile dudular cells. This cell preparation thus represents a unique experimental model to study hepatic bile secre- tory processes. Organic anions are transported into the lumen, and secretion is modified by osmotic gradients (2). An intracanalicular potential of -5 to -6 mV is maintained as determined by direct micropuncture stud- ies. Thus, the hepatocyte couplet system appears to possess electrophysiological and functional characteris- tics of a primary bile secretory unit.
Immediately after isolation, the freshly dissociated couplets retain structural evidence that they were pre- viously attached to neighboring liver cells, since canalic- ular elements can be found on the cell surface distributed in a normal belt-like arrangement. These elements, iden- tified by Mg++-ATPase staining, correspond to the cir- cumferential configuration of canaliculi in uiuo and thus represent the specialized membrane domains that are the excretory poles of the liver cell. However, in culture, these canalicular remnants disappear relatively rapidly, presumably as a consequence of disruption of tight junc- tions that normally would separate the canalicular do- mains from adjacent lateral surfaces of the hepatocyte plasma membrane. This process appears to depend on microfilament but not on microtubular function, since cytochalasin D inhibits the loss of the canalicular rem- nants but colchicine does not. In contrast, at the same time, there is progressive accumulation of Mg++-ATPase and thiamine pyrophosphatase (as assessed histochemi- cally) at the remaining canalicular pole between the cells. This process,appears to represent a rearrangement of
membrane and expansion of the canalicular space. Immediately after the cell couplet is isolated, the re-
maining canalicular space is completely open to the culture medium as judged by penetration of ruthenium red freely into the luminal area as well as by the relatively
I I
FIG. 11. Schematic three-dimensional representation of a freshly isolated couplet (top right) displaying "hemicanaliculi" or canalicular membrane remnants on the cell surface and a fully reorganized couplet displaying an expanded canalicular space encircled by tight junctions. The insert (bottom left) show a cutaway of the bottom half of the canalicular mace to demonstrate orientation of the membranes limiting preexisting membrane elements rather than denouo syn-
thesis, since cycloheximide failed to inhibit the accumu- the canalicuiar space.

Vol. 7, No. 2, 1987 CANALICULAR BILE SECRETION IN ISOLATED HEPATOCYTE COUPLETS 223
small (<1 pm), nondilated canaliculi observed at this early time point. However, the junctional complexes of most couplets apparently reseal with time, and the lumen then expands as secretion is elaborated. That these can- alicular spaces are impermeant to solutes of higher mo- lecular weight is also noted when fluorescein (molecular weight = 334 daltons) is injected by iontophoresis through a microelectrode and remains confined within the canalicular lumen (2). However, the junctional com- plexes are freely permeable to small ions, since addition of sucrose to the media rapidly decreases the size of the canalicular lumen, and electrical measurements demon- strate that the junctional pathway has a low electrical resistance and is relativley nonselective to Na+, K+ and C1- (4). Junctional domains can be visualized by electron microscopy, and preliminary freeze fracture studies in- dicate that there are normal-appearing tight junctional elements as well as gap junctions in 4-hr-old hepatmyte couplets. Electrical measurements indicate that the he- patocyte couplets are also tightly coupled, suggesting a functional role for the gap junctions (2,4).
After 4 to 8 hr in culture, approximately 30 to 40% of couplets fail to display distended canalicular spaces and 33% of collapsed canaliculi are permeable to ruthenium red. Some of these nondistended canaliculi may result because the couplets failed to adequately reseal their canalicular lumen but others probably represent canalic- uli that periodically contract or collapse and lose their sealing properties. Oshio and Phillips and Graf et al. (1, 2) originally reported periodic (both partial and com- plete) collapses of the canalicular lumen in hepatocyte couplets that represented either a primary contractile process or a periodic leak/rupture of the limiting mem- branes and collapse of the lumen under pressure of accumulating secretion. The contraction/collapse cycles are followed by periods of expansion, presumably as the transiently disrupted membranes reseal, similar to the rapid assembly and reformation of tight junctions follow- ing transient disruption noted in tissue/organ culture conditions in other epithelia (19, 20).
A few of the widely dilated canaliculi examined by electron microscopy display a cholestatic appearance of the canalicular membrane. Whether these canaliculi are a normal occurrence towards the end of an expansion cycle and occur as a result of steady pressure build-up from accumulating secretions prior to a spontaneous collapae/contraction or represent injured canaliculi is not known. The majority of couplets, however, display nor- mal canalicular membrane morphology by electron mi- crogcopy and contain microvilli lining the surface mem- brane.
Thus, in short-term culture, freshly isolated, nondis- aociated hepatocyte pairs reorganize structurally within 1 to 4 hr into hepatocyte couplets with a polar orientation towards the remaining canaliculus. The canalicular space reseals and becomes impermeant to solutes in the 300 to 500 dalton molecular weight range. This space then
distends with secretion in this unique model which rep- resents the primary unit for canalicular bile secretion. Isolated hepatocyte couplets, therefore, represent a novel opportunity to study primary mechanisms of canalicular bile formation by a variety of techniques, including c b - ical electrophysiologic and quantitative microscopy methods that are impossible to perform in the intact organ.
1.
2.
3.
4.
5.
6.
7.
8.
9.
10.
11.
12.
13.
14.
15.
16.
17.
18.
19.
20.
REFERENCES &hi0 C, Phillips MJ. Contractility of bile canalid implications for liver function. Science 1981; 2121041-1042. Graf J , Gautam A, Boyer JL. Isolated rat hepatocyb couplets: a primary unit for electrophysiological studies of bile secretory func- tion. Pmc Natl Acad Sci USA 1984,81:6516-6520. Miyairi M. Ohio C, Watanabe S, et al. Taurocholate accelerates bile canalicular contractions in isolated rat hepatocytes. Castm- enterobgy 1984; 87788-792. Craf J, Henderson RM, Krumpholz B, et al. Cell membrane and transepithelial voltages and resistances in isolated rat hepa- couplets. J Membrane Biol1987; 92 (in press). Henderson RJ, Graf J , Boyer JL. Na-H exchange regulates intra- cellular pH in isolated rat hepatocyte couplets. Am J Physiol1987; 252GlWG113. w e n PO. Preparation of isolated rat liver cells. Meth Cell Biol 1976; 1329-83. Blitzer BL, Ratooeh SL, Donovan CB, et al. Effecta of inhibitors of Na+-coupled ion transport on bile acid uptake by isolated rat hepatocytea. Am J Physiol1982; 24kG48453. Waneon J-C, Drochomas P, Moeselmane R, et al. Adult rat hepa- tocytes in primary monolayer culture. J Cell Biol 1977; 74864- 877. Wachstein G, Meisel E. Histochemistry of hepatic phoephatases at physiologic pH. Am J Clin Pathol 1957; 2713-23. Phillips MJ, &hi0 C, Miyairi M, et al. Intrahepatic choleetaeie as a canalicuh motility disorder: evidence using cytochalaein. Lab Invest 1983,48:205-211. Kacich RL, Renston RH, Jones AL. Effecte of cytochalaain D and colchicine on the uptake, translocation and biliary secretion of horeeradiah peroxidase and ( “ c ) - d u m taurocholate in the rat. Gastroenterology 1983,86:386-394. Gebhardt R. Participation of microtubules and microfilaments in the transcellular biliary secretion of immunoglobulin A in primary cultures of rat hepatocytes. Experientia 1% 402269-271. Kovacs AL, w e n PO. Inhibition of hepatmytic protein dsgnda- tion by methylaminopurines and inhibitors of protein eynthesi. Biochem Biophys Acta 1981; 676213-220. Sandetrom B. A comparison between histochemically demonstrable Golgi-associated and bile canalicular TDP-ase activity in liver, Histochemie 1970; 23289-294. Wacker WEC, Ulmer DD, Vallee BL. Metalloenzymea and myo- cardial infarction. 11. Malic and lactic dehydro(leneee activih and zinc concentration in wrum. N End J Med 1966,256:449-460. Scharschmidt BF, Keeffe EB, Blankenahip NM. et al. Validation of a recording spectrophotometric method for measurement of membrane associated Mg++ and Na+,K+-ATPaee activity. J Lab Clin Med 1979; 93:790-799. Lowry OH, Roeebrough HJ, Farr AL, et al. Protein mesnuement with the Folin phenol reagent. J Biol Chem 1951; 193:266-275. Fujimoto T, Ogawa K. Cell membrane polarity in diwociated frog urinary bladder epithelial cells. J Histochem Cytochem 1983;
Meldolesi J, Castiglioni G, Panna R, et al. Ca++-dependent diaas- sembly and reassembly of occluding junctions in guinea pig pan- creatic acinar cells. Effecta of drugs. J Cell Biol197& 7%156-172. Kachar B. DaSilva P. Rapid massive assembly of tight junction strands. Science 1981; 213541-544.
31~131-138.