isolation electron microscopic observations ... · reticulate bodies (rb) of chlamydia psittaci...
TRANSCRIPT

Vol. 145, No. 1JOURNAL OF BACTERIOLOGY, Jan. 1981, p. 605-6120021-9193/81/010605-08$02.00/0
Isolation and Electron Microscopic Observations ofIntracytoplasmic Inclusions Containing Chlamydia psittaci
AKIRA MATSUMOTO
Department ofMicrobiology, Kawasaki Medical School, Kurashiki, Okayama 701-01, Japan
Intracytoplasmic inclusions containing Chlamydia psittaci were isolated by anewly established method. Infected L-cells at 20 h after infection were suspendedin 0.25 M sucrose-tris(hydroxymethyl)aminomethane buffer containing ethylene-diaminetetraacetic acid, homogenized in a Dounce tissue grinder, and filteredthrough a 2,000-mesh screen. Isolated inclusions were stabilized in 5% bovineserum albumin in 10 mM tris(hydroxymethyl)aminomethane buffer. Electronmicroscopic observations revealed the presence of surface projections on thevegetative, reticulate bodies and a direct connection between the reticulate bodiesand the inclusion membrane by means of projections.
Cell envelopes isolated from the infectious,elementary bodies (EB) and the developmental,reticulate bodies (RB) of Chlamydia psittacihave been previously studied by electron mi-croscopy. These observations revealed surfaceprojections with roughly hexagonal arrange-ments on a limited surface area on EB (8-10).When isolated cell walls were stained negatively,the sites of these projections coincided with therosettes and B structures seen on the concavesurface exposed by cleavage of frozen EB (7, 8).No projections were found on RB surfaces. Ex-amination by the freeze-replica technique dem-onstrated some groups of fine particles on thesmooth convex surface of the intracytoplasmicinclusions which contained only RB in samplesharvested at 18 h after infection (7). The ar-rangement of the fine particles was roughly hex-agonal and was similar to that of the projectionson EB, suggesting the presence of the projec-tions on the RB surface and structural interre-lation between RB and the inclusion membraneby means of the projections. The projectionswere, however, never seen in inclusions in situin thin sections. Experiments were then designedto examine inclusions isolated in native form,since the projections on EB apparently possessvery low electron opacity and are hard to stainspecifically.The isolation of intact inclusions and a de-
scription of their morphology, together with ob-servations on the interrelation between RB andinclusion membrane, are reported in this paper.
MATERIALS AND METHODSChlamydial strain and its propagation. The
meningopneumonitis strain of Chlamydiapsittaci andL-cells in suspension culture were used throughoutthis experiment. The propagation of the organisms
was carried out by the method of Tamura and Higashi(16).
Isolation of inclusions. L-ceUs inoculated with 5to 10 inclusion-forming units per cell were cultivatedin a Spinner culture bottle at 37°C for 20 h andcollected with low-speed centrifugation (350 x g).After washing twice in the phosphate-buffered salineof Dulbecco and Vogt (4), but lacking Ca and Mg, thecells, approximately 108 in number, were suspended in10 ml of 0.25 M sucrose-ET buffer solution (1 mMEDTA in 10 mM Tris buffer, pH 7.4) and kept in icefor 60 min. The ceU suspension was then transferredto a Dounce tissue grinder SS, 15 ml in size (KontesGlass Co., Vineland, N.J.) and homogenized gently for20 strokes. The homogenate was taken in a 10-mlsyringe and filtered through a stainless-steel, 2,000-mesh screen (Twilled Dutch Weave, purchased fromTaiyo Wire Cloth Co. Ltd., Osaka, Japan) set in astainless-steel Millipore membrane filter holder 2.5 cmin diameter. The filtrate was then spun down at 1,500x g for 5 min in a Kubota K-80 table type centrifuge.The peUet obtained was washed twice with 0.25 Msucrose in 10 mM Tris buffer. This procedure wasmonitored with a phase-contrast light microscope.
Electron microscopy. For thin-sectioning tech-nique, the samples in the peUets were fixed in 2.5%glutaraldehyde in phosphate-buffered saline lackingCa and Mg or in Karnovsky formaldehyde-glutaral-dehyde mixture (M. J. Karnovsky, J. Cel Biol. 27:137A, 1965) for 60 min at room temperature. The fixedsamples were treated with tannic acid as previouslyreported to enhance the electron opacity of the spec-kiaens, especially the fine surface projections in thinsections (9). After several washings in distilled water,the samples were fixed again in 1% osmium tetroxidein 0.1 M cacodylate buffer, dehydrated in an ethanolseries, and embedded in the low-viscosity embeddingmedium described by Spurr (14). Thin sections werecut with a glass knife on a Porter-Blum microtomeMT2-B and doubly stained with 2% uranyl acetate in70% ethanol and lead citrate solution (13).
For freeze-replica preparations, the samples of in-fected cells and isolated inclusions, in pellets prefixed
605
on April 30, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

606 MATSUMOTO
in glutaraldehyde or Karnovsky fixative and impreg-nated with 10% glycerol, were frozen in Freon 22 atliquid nitrogen temperature and then transferred intoliquid nitrogen. The replica was prepared in a BalzersBAF301 freeze-replica apparatus without sublimationof ice. The replica membrane was treated with com-mercial bleaching solution containing sodium hypo-chlorite, rinsed several times in distilled water, andthen collected on a copper grid.
All specimens were examined with a Hitachi H-500
J. BACTERIOL.
electron microscope at an accelerating voltage of 75kV.
RESULTSFigure 1 presents phase-contrast light micro-
graphs showing the process of inclusion isola-tion. Figure la represents the cells kept in 0.25M sucrose-ET buffer for 60 min. The cells ap-pear somewhat swollen, and the inclusions can
FIG. 1. Phase-contrast micrographs of the process of inclusion isolation. The process is shown alphabeti-cally (a to f). (a) L-cells kept in 0.25Msucrose-ET buffer for 60 min in ice. Inclusions are clearly seen as thebright bodies (arrows). (b) Cells after a five-stroke homogenization in a Dounce tissuegrinder. Some inclusionsare released, but the majority of the cells retain their shape. (c) Fraction after 20 strokes of homogenization.The cells are disintegrated and make large aggregates. Many inclusions and organelles are released. (d)Filtrate of the 23-stroke homogenate through the screen. The large aggregates are removed. Many inclusionsand much small cell debris are obtained. (e) Suspension ofpellet in 0.25M sucrose-Tris buffer, obtained bycentrifugation at 1,500 x g for 5 min. Small debris still remains. (f) Suspension in 0.25M sucrose-Tris bufferafter two washings by centrifugation at 1,500 x g for 5 min. Many inclusions in high purity are obtained. Allmicrographs are shown at the same magnification. Bar, 10 um.
on April 30, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

INCLUSIONS CONTAINING C. PSITTACI 607
be clearly seen as the bright bodies in the centralregion of cytoplasm (arrows). After a five-strokehomogenization, some inclusions were released,but the majority of the cells retained their shapeand the inclusions were still contained in eachcell (Fig. lb). When the homogenization wascontinued up to 20 strokes, the cells were almostcompletely disintegrated (c). Many inclusionsand cell organelles were mixed in amid the largecell debris, which tended to make large aggre-gates. These large aggregates were efficientlyremoved by the filtration through the screen,although many small organelles and much mem-branous debris remained in this fraction (d). Thesuspension of the pellet obtained from the frac-tion after centrifugation contained mainly inclu-sions (e). The purity of this fraction did not,however, seem sufficient for morphological ex-amination, especially for the freeze-replica tech-nique, since small particulate contaminants werestill present. After two washings in 0.25 M su-crose-Tris buffer, the small contaminants weremostly removed (f). When the inclusions werecollected into a pellet, cut into thin sections, andfollowed by electron microscopic examination,they showed severe damage in inclusion mem-brane and RB (Fig. 2). The inclusion membranewas broken at many places and lacked continu-ity. RB appeared to be remarkably shrunk andlacked their native morphology, although thefraction was rich in inclusions.To determine the conditions under which the
inclusion membrane and RB can retain theirnative morphology, the isolated inclusions in asuspension were divided equivalently into sevenparts, then spun down at 1,500 x g for 5 min,simultaneously. Each pellet was suspended, in0.25 M sucrose, 0.5, 1, 2.5, and 5% bovine serum,and 5 and 10% fetal calf serum, respectively. Allsolutions tested were prepared with 10 mM Trisbuffer. The suspensions were incubated at roomtemperature. The turbidity of the supernatantsof the 1,500 x g, 5-min centrifugation was as-sayed at 0, 30, 60, and 120 min after incubation,with the expectation that the turbidity of thesupernatant would indicate the degree of RBliberation due to the disintegration of the inclu-sion membrane in the suspension. The results(Fig. 3) indicated that bovine serum albuminwas capable, in proportion to its concentrationin the suspension, of stabilizing the inclusionmembrane, and that 5% bovine serum albumineffectively maintained the integrity of inclusions.It is noteworthy that the turbidity in 0.25%sucrose showed rapid increase, indicating thatthe inclusion membrane was unstable in thissolution, which has been widely used for theisolation of organelles from various types of cellsand tissues. The inclusions in 5% bovine serum
FIG. 2. Thin-sectioned inclusions in 0.25 M su-crose- Tris buffer. Inclusion membranes are broken atmanyplaces (arrows). RB are shrunk and form irreg-ular shapes. Bar, 1 fum.
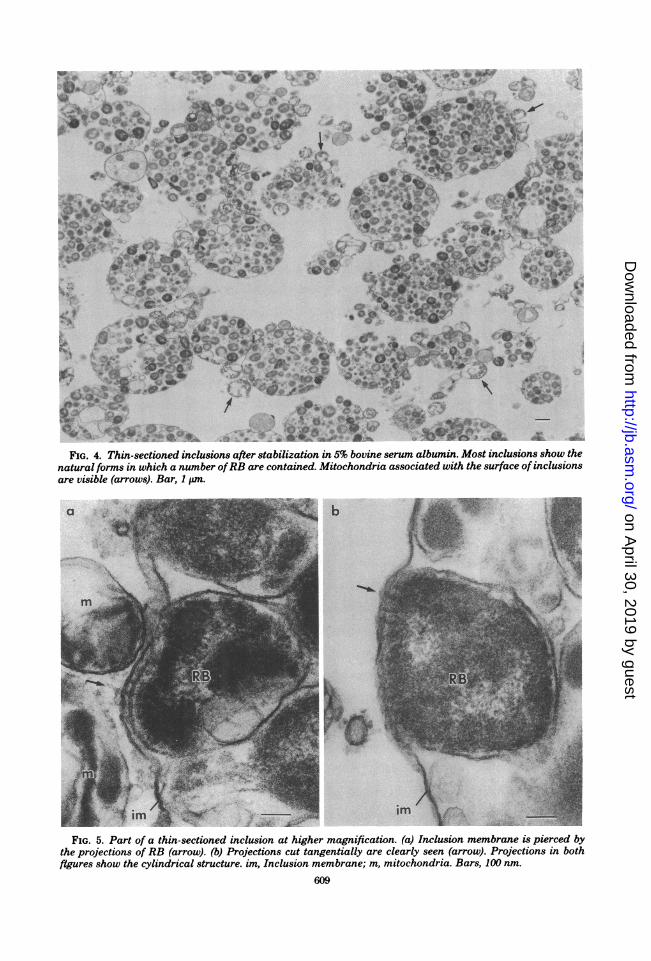
were examined in thin sections by electron mi-croscopy.As shown in Fig. 4, each inclusion was round
in shape, and the RB that were surrounded bythe inclusion membrane appeared to retain theirnatural shapes. At higher magnification, it ap-peared that some RB were closely connected tothe inside surface of the inclusion membrane bymeans of fine projections, cylindrical in shapeand 10 to 13 nm in diameter, which appeared topierce the inclusion membrane (Fig. 5a). Figure5b illustrates the projections sectioned tangen-tially, showing RB adhering closely to the innersurface of inclusion membrane with the projec-tions.
Figures 6 and 7 illustrate the freeze-replicaimages of the inclusions in situ. In Fig. 6, showinga convex surface of partially fractured inclusionmembrane, some particles are seen on the top ofthe RB in the inclusion (arrows). Large convexand concave surfaces in Fig. 7 show two differentfaces exposed by the cleavage of inclusion mem-brane of neighboring inclusions. No RB profileis seen in each inclusion, but both surfaces showclear ruggedness along RB outlines. On the con-
VOL. 145, 1981
on April 30, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

608 MATSUMOTO
likely that the fine particles on the convex sur-face of the inclusion membrane are RB surface
0. 25M Sucrose projections, which pierce the inclusion mem-brane.
It was noticed that many mitochondria wereassociated with the outer surface of inclusions,with minimum spacing of 5 nm (Fig. 9). How-ever, the relation between the location of mito-chondria and the projections is not clear.
.2 *°6 _ 0. 5% BSA
Z~.06-I%BSA
~~~.04~~~~~~5%FCS
.02
2. 5% BSA
, 5% BSA30 60 120
Time( minFIG. 3. Effects of bovine serum albumin (BSA),
fetal calf serum (FCS), and 0.25M sucrose solutionsin 10 mM Tris buffer on the stability of the isolatedinclusions. Bovine serum albumin was capable ofstabilizing the inclusion membrane in proportion toits concentration, whereas no difference between 5and 10% fetal calfserum was observed. The turbidityin 0.25M sucrose increased rapidly, showing break-age of the inclusion membrane and release ofRB.
vex surface (conv), several groups of particles,which seem to be the same particles as thoseshown in Fig. 6, are seen (arrows). These parti-cles are also the same as those I reported previ-ously (7). On the concave surface (conc), fineparticle-like depressions can be seen (arrows),which correspond to the pattern of the particleson the convex surface. These results indicatethat the particles on the convex surface and thedepressions on the concave surface are identicalto the structures seen in the inclusion mem-
brane. The convex surface, which may corre-
spond to the "E face" (2) of the host plasmamembrane, completely lacks membrane parti-cles, whereas many membrane particles 8 to 10nm in diameter are seen on the concave surface,which may be the "P face" (2) ofthe host plasmamembrane. The freeze-replica of the isolatedinclusions also showed a convex or concave sur-face, on which several of these structures, as
shown in Fig. 6 and 7, were seen in a regularlyarrayed pattem. Figure 8 illustrates the convex
surface of an isolated inclusion on which thestructures in groups are clearly seen. Each struc-ture is surrounded by six structures lying 40 to50 nm apart. From these results, it appears very
DISCUSSIONThe key points for the isolation of inclusions
in their native form seem to be as follows: (i) useof0.25M sucrose-ET buffer, in which the plasmamembrane swells and becomes unstable ingentle homogenization; (ii) use of the Douncetissue grinder, with which swollen plasma mem-brane is broken; (iii) use of the 2,000-meshscreen, with which the larger aggregates were
readily removed from the homogenate. The ac-
tual pore size of the screen is not known. Whena normal L-cell suspension (108/10 ml) was fil-tered in the same way, only about 2 ml of thesuspension, with 103 to 104 cells per ml, wasobtained and the majority of L cells remained,showing that the pores in the screen were
stopped up with the cells. The pore size of thescreen seems, therefore, to be a little bit lessthan 10,um in diameter. Direct measurement ofthe pore size by electron microscopy failed be-cause of the Twilled Dutch Weave.Many attempts to isolate the inclusions from
the homogenate by various techniques such as
differential, sucrose-cushioning, and density gra-dient centrifugations failed.The inclusion membrane is very unstable even
in 0.25M sucrose solution, although this solutionhas been widely used for the isolation of variousorganelles of animal cells. Addition of bovineserum albumin effectively stabilized the integ-rity of inclusions. The purity of the inclusions inthe final fraction was, at least as seen by phase-contrast microscopy, very high. However, thefraction still contained some cell debris. Addi-tional treatment may be necessary for use of theinclusion-rich fraction in further experiments.We previously reported that the projections
on EB, treated with tannic acid and thinly sec-
tioned, are cylindrical in structure (9), and thatone edge of each projection is connected withthe cytoplasmic membrane while the other edgeprojects beyond the cell wall through fine holes,which are seen as rosettes in negatively stainedcell wall preparations and as B structures on theinner surface of the EB cell wall by either freeze-fracture or freeze-etching technique (8). The Bstructures are identical to the craters of EB andRB recently reported by Louis et al. (6), but the
Ec
0
0
C
to
C
J. BACTERIOL.
on April 30, 2019 by guest
http://jb.asm.org/
Dow
nloaded from
-8 * aw-t iF -*w i .*> &wej
's,'t;b0''gCXot,,< Jk 4Uk l'
.,4t,4 1 * * * , &.
V U
t.§ 4 ( T n tsv * ^^+ B + C)
FIG. 4. Thin-sectioned inclusions after stabilization in 5% bovine serum albumin. Most inclusions show thenatural forms in which a number ofRB are contained. Mitochondria associated with the surface of inclusionsare visible (arrows). Bar, 1 Lm.
FIG. 5. Part of a thin-sectioned inclusion at higher magnification. (a) Inclusion membrane is pierced bythe projections of RB (arrow). (b) Projections cut tangentially are clearly seen (arrow). Projections in bothfigures show the cylindrical structure im Inclusion membrane; m, mitochondria. Bars, 100 nm.
609
on April 30, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

610 MATSUMOTO
FIG. 6. Freeze-replica image of an inclusion in a host cell. The inclusion membrane is partially cleavedand shows a convex surface, on which several particles are seen on the top of the RB within the inclusion(arrows). RB are also cleaved and show convex and concave surfaces. The thick arrow at bottom indicatesthe shadowing direction. Bar, 100 nm.
FIG. 7. Freeze-replica image of two neighboring inclusions in a host cell. Particles (arrows) in groups areseen on the convex surface (conv). Fine depressions (arrows) in a similar pattern are seen on the concavesurface (conc). Both surfaces show ruggedness along RB outlines. The thick arrow at bottom indicates theshadowing direction. Bar, 1 ,um.
location of the crater is, according to their expla-nation, on the fractured face of the cytoplasmicmembrane of the organism. This may be due toincorrect application of technique. With theiridea, it may be difficult to make clear the factthat the B structures are located on a surfacepossessing the hexagonally arrayed subunitswhich compose the inner surface of the EB cell
wall and are visualized only after optimum etch-ing (7).The results obtained in the present experi-
ment strongly suggest that RB also have surfaceprojections and that RB are connected directlywith the host cytoplasm through the canals ofthe projections. In this manner, RB within aninclusion may regularly connect with the host
J. BACTERIOL.
on April 30, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

INCLUSIONS CONTAINING C. PSITTACI 611
cytoplasm during their multiplication and inter-change information with the host cell. A .The relation between loci of mitochondria and Im
those of the projections was not clear. Further- /more, it was not clear whether mitochondriatouched the inclusion membrane. Considering mthe obligate parasitism of Chlamydia organisms,the significance of the attachment of mitochon-dria remains an important problem.From many studies of the plasma membrane
of many types of cells by means of freeze-replicatechnique, it may be generalized that both P and R tE faces, according to the nomenclature by Bran-ton et al. (2), are covered by the membraneparticles, but the E face is studded with fewermembrane particles than the P face (1, 3, 12, 17). , yThe morphology of the inclusion membrane re-vealed by the freeze-replica technique appar-ently differs from that of the plasma membrane.The inclusion membrane is derived from theplasma membrane of the host cell as EB isphagocytized (5). Therefore, the convex surfaceof the inclusion membrane corresponds to the E
FIG. 9. Thin-sectioned inclusion. Many mitochon-dria are closely associated with the surface of theinclusion. Bar, 1 ,um. Inset shows the association ofmitochondria and inclusion membrane at higher
i(_;,> magnification; m, mitochondria; im, inclusion mem-j9-'t, brane. Bar, 100 nm.
face, and the P face is exposed as the concavesurface. No membrane particles are encounteredon the convex surface, whereas many membraneparticles are seen on the concave surface. The
___<, inclusion membrane seems to be modified in itsnature during the multiplication of these orga-nisms. Biochemical study by Stokes also mdi-
" g ;gCcates this modification by Chlamydia-directedglycosylation (15). The modified nature of theseglyclmembranes is also suggested by the instability
_ai.of the inclusions in 0.25 M sucrose solution.
ACKNOWLEDGMENTSI thank N. Higashi, Kawasaki Medical School, and G. P.s_- ar/!<>--'.1,-*.Manire, University of North Carolina, for helpful and stimu-
_=asl ^ * ,, - ^ *42 ,; ^ < lating discussion.This work was supported by Grant-in-Aid for Scientific
Research 357149 from the Ministry of Education, Science and-
i~
J .' Culture, Japan.FIG. 8. Freeze-replica image of the convex surface LITERATURE CITED
ofan isolated inclusion. The particles in groups show 1. Benedetti, E. L., I. Dunia, and A. Diawara. 1973. Thea pattern similar to that on the convex surface of the organization of the plasma membrane in mammalianinclusion shown in Fig. 7. The thick arrow indicates cells. Eur. J. Cancer 9:263-272.the shadowing direction. Bar, 1 Jim. 2. Branton, D., N. B. Gilula, M. J. Karnovsky, H. Moor,
VOL. 145, 1981
on April 30, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

612 MATSUMOTO
K. Miihlethaler, D. H. Northcote, L. Packer, B.Satir, P. Satir, V. Speth, L. A. Staehlin, R. L.Steere, and R. S. Weinstein. 1975. Freeze-etchingnomenclature. Science 190:54-56.
3. Chalcroft, J. P., and S. Bullivant. 1970. An interpreta-tion of liver cell membrane and junction structure basedon observation of freeze-fracture replicas of both sidesof the fracture. J. Cell Biol. 47:49-60.
4. Dulbecco, A., and M. Vogt. 1954. One-step growth curveof Western encephalomyelitis virus grown in vitro andanalysis of the virus yield from single cell. J. Exp. Med.99:183-199.
5. Higashi, N. 1965. Electron microscopic studies on themode of reproduction of trachoma virus and psittacosisvirus in cell cultures. Exp. Mol. Pathol. 4:24-39.
6. Louis, C., G. Nicolas, F. Eb, J.-F. Lefebvre, and J.Orfila. 1980. Modifications of envelope of Chiamydiapsittaci during its developmental cycle: freeze-fracturestudy of complementary replica. J. Bacteriol. 141:868-875.
7. Matsumoto, A. 1973. Fine structures of cell envelopes ofChiamydia organisms as revealed by freeze-etching andnegative staining techniques. J. Bacteriol. 116:1355-1363.
8. Matsumoto, A. 1979. Recent progress of electron micros-copy in microbiology and its development in future:from a study of the obligate intracellular parasites,Chiamydia organisms. J. Electron Microsc. 28(Suppl.):s57-s64.
9. Matsumoto, A., E. Fujiwara, and N. Higashi. 1976.Observations of the surface projections of infectioussmall cell of Chiamydia psittaci in thin sections. J.Electron Microsc. 25:169-170.
10. Matsumoto, A., and N. Higashi. 1975. Morphology ofthe envelopes of Chlamydia organisms as revealed byfreeze-etching technique and scanning electron micros-copy. Ann. Rep. Inst. Virus Res. Kyoto Univ. 18:51-61.
11. Matsumoto, A., and G. P. Manire. 1970. Electron mi-croscopic observations on the fine structure of cell wallsof Chlamydia psittaci. J. Bacteriol. 104:1332-1337.
12. Miihlethaler, K. 1971. Studies on freeze-etching of cellmembrane. Intern. Rev. Cytol. 31:1-19.
13. Reynolds, E. S. 1963. The use of lead citrate at high pHas an electron-opaque stain in electron microscopy. J.Cell Biol. 17:208-212.
14. Spurr, A. R. 1969. A low-viscosity epoxy resin embeddingmedium for electron microscopy. J. Ultrastruct. Res.26:31-43.
15. Stokes, G. V. 1974. Cycloheximide-resistant glycosylationin L cells infected with Chiamydia psittaci. Infect.Immun. 9:497-499.
16. Tamura, A., and N. Higashi. 1963. Purification andchemical composition of meningopneumonitis virus. Vi-rology 20:596-604.
17. Wehrli, E. K., K. Muihlethaler, and H. Moor. 1970.Membrane structure as seen with a double replicamethod for freeze fracturing. Exp. Cell Res. 59:336-339.
J. BACTERIOL.
on April 30, 2019 by guest
http://jb.asm.org/
Dow
nloaded from