issn 0177-9214 - schloss-benrath.de · to 550 ka) and the soufriŁre hills (1011 m; upper...
TRANSCRIPT

ISSN 0177-9214
Ban
d 13
/200
6A
cta
Biol
ogic
a Be
nrod
is Acta Biologica Benrodis
Band 13 2006
und Mitteilungen aus demMuseum für Naturkunde in derStiftung Schloss und Park Benrath

Acta Biologica Benrodis 13 (2006)
Acta Biologica Benrodis
und Mitteilungen aus demMuseum für Naturkunde in der
Stiftung Schloss und Park Benrath
Band 13 � 2006
Gedruckt mitUnterstützung der StiftungSchloss und Park Benrath
sowiedem Freundeskreis
Museum für NaturkundeBenrath e.V.

Acta Biologica Benrodis+ Mitteilungen aus dem Museum für Naturkunde in der Stiftung Schloss und Park Benrath
Herausgeber
Prof Dr. H. GrevenInstitut für Zoomorphologie und Zellbiologie derHeinrich-Heine-Universität DüsseldorfUniversitätsstr.1D-40225 Düsseldorf
Dr. H. Heuwinkel Prof. Dr. R. WittigMuseum für Naturkunde Botanisches Institut derStiftung Schloss und Park Benrath Johann Wolfgang Goethe-UniversitätBenrather Schloßallee 102 Siesmayerstr. 70D-40597 Düsseldorf D-60323 Frankfurt am Main
Schriftleitung
Prof. Dr. H. GrevenInstitut für Zoomorphologie und Zellbiologie derHeinrich-Heine-Universität DüsseldorfUniversitätsstr. 1D-40252 Dü[email protected]
Produktion und Satz: Verlag Natur & Wissenschaft, Postfach 170209, D-42624 Solingen
ISBN 978-3-936616-51-4
Acta Biologica Benrodis erscheint einmal im Jahr mit ca. 120 Seiten Umfang pro Heft und kann vomMuseum für Naturkunde in der Stiftung Schloss und Park Benrath, Benrather Schlossallee 102,Schloss Benrath, D-40597 Düsseldorf, oder vom Verlag zum Abonnementspreis von 15,00 Euro(10,00 Euro für Studenten) pro Jahrgang (zuzüglich Versandkosten) oder im wissenschaftlichenZeitschriftenaustausch bezogen werden.
Titelfoto: Warnender Dreifarbglanzstar in einer Akazie. Aus dem Beitrag von H. Heuwinkel undU. Beigel-Heuwinkel (S. 249-252).Cover photo: Alarm calling Superb Starling in an Acacia. From the article of H. Heuwinkel und U.Beigel-Heuwinkel (S. 249-252).

Acta Biologica Benrodis 13 (2006)
Inhalt Band 13 – 2006
STEVENS, M., WALDMANN, G.: Endemics of the Lesser Antillean Islandof Montserrat, British West Indies/Endemiten der Insel Montserrat,Kleine Antillen (Britisch Westindien) ...............................................................................1
FUCHS, R., KUTZELNIGG, H., KEIL, P.: 20 Jahre Entwicklung der Flora undVegetation im NSG �Steinbruch Hofermühle-Süd� (GemeindeHeiligenhaus, Kreis Mettmann)/20 years development of flora andvegetation in the nature conservation area �Limestone-Quarry Hofermühle-Süd� (community Heiligenhaus, Mettmann district) ....................................................23
DIETRICH, W.: Pflanzliche Neubürger (Neophyten) im Raum Düsseldorf/Neuss(Deutschland, Nordrhein-Westfalen)/New plant species (neophytes) inthe Düsseldorf/Neuss area (Germany, North Rhine-Westphalia) ...............................51
FUCHS, R., KUTZELNIGG, H., FEIGE, G. B.: Seminatural ancient forest inurban agglomeration area �Ruhrgebiet�/Naturnahe Altwälder imBallungsraum Ruhrgebiet ................................................................................................91
WELTJE, L., OEHLMANN, J.: Effects of endocrine disrupting compoundsand temperature on the moulting frequency of the freshwater isopodAsellus aquaticus L. (Isopoda: Asellota)/Effekte von hormonähnlichwirksamen Substanzen und Temperatur auf die Häutungsfrequenzder Süßwasserassel Asellus aquaticus L. (Isopoda: Asellota) ........................................105
DIAS, A.N., HASSALL, M.: The abundance, life history and reproductiontraits of peracarid macro-decomposers in a Ria Formosa lagoon saltmarsh, Southern Portugal/Abundanz, �life history� und Charakteristikader Fortpflanzung von Makrozersetzern (Peracarida) in einer Salzwiese(Ria Formosa Lagune, Südportugal) ............................................................................117
ZIMMER, M.: The role of animal-microbe interactions in isopod ecologyand evolution/Die Bedeutung tierlich-mikrobieller Wechselwirkungenfür die Ökologie und Evolution von Asseln ...............................................................127
BEYL, F.E.: Morphometrische Untersuchungen am Gemüsespargel,Asparagus officinalis L./Morphometric studies in branches ofCommon Asparagus, Asparagus officinalis L. ...............................................................169
KREUELS, M.: Die Webspinnen (Arachnida: Araneae) aus Beifängen des NSGTeverener Heide (NRW, Kreis Heinsberg)/The spiders (Arachnidaa:Araneae) from bycatches of the nature reserve Teverener Heath (NorthRhine-Westphalia, district Heinsberg) ..........................................................................185

GREVEN, H., JÖMANN, N., CLEMEN, G.: Dentigerous bones, dentitionand dental laminae in the hynobiid salamander Ranodon sibiricusKessler, 1866 (Amphibia, Urodela)/Zahntragende Knochen,Bezahnung und Zahnleisten bei dem Hynobiiden Ranodon sibiricusKessler, 1866 (Amphibia, Urodela) ..............................................................................195
DASHDAMIROW, S.: A new genus and species of false scorpion fromVietnam showing remarkable chelal modifications (Archnida:Chelonetida) / Eine neue Gattung und Art von Pseudoskorpionenaus Vietnam mit bemerkenswerten Modifikationen der Chelae(Arachnida: Chelonethida) .............................................................................................219
TIEMANN, I., REHKÄMPER, G.: Rasseunterschiede bei Haushühnern (Gallusgallus f. d.) bei klassischer und operanter Konditionierung in derSkinner-Box/Breedspecific differences among domesticated chickens(Gallus gallus f. d.) revealed by classical and operant conditioning in theSkinner-Box ....................................................................................................................231
Kurze Mitteilungen/Short communications
SCHÜTT, H.: Wie kam die Nördliche Grundwasserschnecke Bythiospeumacicula moenanum (Hydrobiidae) an den Niederhein?/How came theNorthern Groundwater Snail Bythiospeum acicula moenanum(Hydrobiidae) to the Lower Rhine? ..............................................................................241
KRECHEL, R., BRAUN, TH., STEVENS, M.: Erstnachweis des Springfrosches(Rana dalmatina Bonaparte, 1840) im Regierungsbezirk Düsseldorf/First record of the Agile Frog (Rana dalmatina Bonaparte, 1840) in theadministrative district of Düsseldorf ...........................................................................245
HEUWINKEL, H., BEIGEL-HEUWINKEL, U.: Alarmrufe von Dreifarbglanzstaren(Lamprotornis superbus) bei Entdeckung eines Bodenfeindes/Alarm callsof Superb Starlings (Lamprotornis superbus) detecting a ground predator ...................249
Buchbesprechungen ............................................22, 50, 90, 116, 126, 194, 218, 230, 244, 253
Mitteilungen aus dem Museum für Naturkunde in der StiftungSchloss und Park Benrath ...........................................................................................M1

Endemics of the Lesser Antillean Island of Montserrat, British West Indies 1
Acta Biologica Benrodis 13 (2006)
Acta Biologica Benrodis 13 (2006): 1-21
Endemics of the Lesser Antillean Island of Montserrat,British West Indies
Endemiten der Insel Montserrat, Kleine Antillen (Britisch Westindien)
MICHAEL STEVENS1 & GEORG WALDMANN2
1 Institut für Zoomorphologie und Zellbiologie, Heinrich-Heine-Universität Düsseldorf,Universitätsstraße 1, D-40225 Düsseldorf, Germany; [email protected]
2 Am Trietenbroich 24, D-41352 Korschenbroich, Germany; [email protected]
Summary: Montserrat (British West Indies, Lesser Antilles), is part of the Caribbean biodiversityhotspot region with numerous endemic species including the following families: Lauraceae, Pipe-raceae, Theaceae, Marcgraviaceae, Clusiaceae, Elaeocarpaceae, Begoniaceae, Styracaceae, Melasto-mataceae, Rubiaceae, Cyclanthaceae, Bromeliacae, Agavaceae, Hesperiidae, Leptodactylidae, Gek-konidae, Polychrotidae, Teiidae, Psittacidae (fossil only), Trochilidae, Mimidae, Icteridae, Criceti-dae (fossil only) and Phyllostomidae. Natural hazards like hurricanes and recent volcanism probab-ly affect the survival of endemic species on Montserrat.
Lesser Antilles, Montserrat, single-island endemics, regional island endemics
Zusammenfassung: Die Karibikinsel Montserrat (Kleine Antillen; Britisch Westindien) liegtinmitten eines Zentrums globaler Biodiversität und beherbergt zahlreiche lokale und regionaleEndemiten aus folgenden Familien: Lauraceae, Piperaceae, Theaceae, Marcgraviaceae, Clusiaceae,Elaeocarpaceae, Begoniaceae, Styracaceae, Melastomataceae, Rubiaceae, Cyclanthaceae, Bromeli-acae, Agavaceae, Hesperiidae, Leptodactylidae, Gekkonidae, Polychrotidae, Teiidae, Psittacidae(nur fossil), Trochilidae, Mimidae, Icteridae, Cricetidae (nur fossil) und Phyllostomidae. Naturer-eignisse wie tropische Wirbelstürme oder rezenter Vulkanismus beeinflussen möglicherweise dasÜberleben von Endemiten auf Montserrat.
Kleine Antillen, Montserrat, Endemiten, regionale Endemiten
1. Introduction
The Caribbean Region is among the eightleading world biodiversity hotspots. Morethan 12,000 vascular plants and about 1,500vertebrates (excluding teleosts) have beendescribed from the Caribbean (DAVIS et al.1997; MYERS et al. 2000). Comprising only0.018 % of the Earth�s land surface, some2.3 % of all higher plants and 2.9 % verte-brates are endemic to the Caribbean (MYERS
et al. 2000). This paper attempts to list allendemic species of the island of Montserrat,both single-island endemics and regional is-
land endemics. Regional endemics currentlyknown, with a distribution on the LesserAntillean islands, in some cases reaching theVirgin Islands and Puerto Rico or Trinidadand Tobago are included.
2. The Lesser Antilles and the island ofMontserrat
The Caribbean island chain has a complexgeological and tectonical history. Recent islandshave been created by convergent plate boun-daries, tectonic uplifts, transform faults, ter-restrial and marine volcanic deposits (WEYL

2 MICHAEL STEVENS & GEORG WALDMANN
1966; MARTIN-KAYE 1969; ITURRALDE-VINENT
& MACPHEE 1999). The formation of moun-tains on the Greater Antilles during the earlyTertiary has been caused by uplift followingcompressional forces after the collision of theCaribbean Plate with the North AmericanPlate. The recent Greater Antillean islands dateback into the middle Eocene (ITURRALDE-VI-NENT & MACPHEE 1999). In contrast the for-mation of the Lesser Antillean islands was
initiated in the in mid- or late Cretaceous,due to subduction of the Atlantic Plate be-neath the Caribbean Plate. They are a chain ofoceanic islands of different age and origin(Fig. 1). According to MARTIN-KAYE (1969)the Lesser Antilles consist of two closely rela-ted arcs, both of volcanic construction. Theouter and discontinuous eastern arc (Lime-stone Caribees), extending from Grenada toSombrero, was formed in the early Tertiary.The islands of the western inner arc (VolcanicCaribees), ranging from Grenada to Saba, aredated into the later Miocene and Pliocene.Since the Eocene, marine sedimentary rocksare intercalated within subaerial volcanic se-quences. This indicates a complex history in-volving emergence, subsidence, and migrati-on of topographic elevations (ITURRALDE-VINENT & MACPHEE 1999). The present formof the western inner arc has been built upsince the Pliocene due to volcanic activity,which continues to the present day e.g. onMontserrat, St. Vincent, Martinique, Domi-nica and Guadeloupe. During the Andean(or Antillean) orogeny of the late Miocene toearly Pliocene the islands of Barbados andTobago were uplifted. Trinidad, in contrast,is part of South America.
The occurrence of mesozoic rocks of theAves Ridge and the Lesser Antilles suggest,that both structures were united during theCretaceous to the Eocene. Aves Ridge, now aremnant arc from the active Lesser Antilleanarc, was separated later. ITURRALDE-VINENT &MACPHEE (1999) have suggested a landbridgeduring the late Eocene and early Oligoceneconnecting South America and the GreaterAntilles, via the Aves Ridge and the adjacentLesser Antillean Islands. A general subsidencefollowed, ending the landspan phase, calledGAARlandia (Greater Antilles & Aves Ridge).
Montserrat is situated within the Lee-ward Islands of the Lesser Antilles in theCaribbean Sea. It is pear-shaped (Fig. 2), 18km long and 11 km wide with a terrestrialsurface of 104 km² (ECNAMP 1980). Se-veral volcanic peaks form today�s three
Fig. 1: The Lesser Antilles. The outer line repre-sents the coastline during the Pleistocene glacia-tion. It corresponds with 50 m below the actualsea level. The middle line shows the sea level oftoday. Terrestrial areas above 200 m NN are ex-pressed in black (modified from ROUGHGARDEN
1995).Abb. 1: Die Kleinen Antillen. Die äußere Liniezeigt den Küstenverlauf während der pleistozä-nen Vereisung. Sie entspricht heute einer Mee-restiefe von 50 m unter NN. Die mittlere Linieentspricht dem gegenwärtigen Küstenverlauf.Flächen oberhalb 200 m sind schwarz dargestellt(verändert nach ROUGHGARDEN 1995).

Endemics of the Lesser Antillean Island of Montserrat, British West Indies 3
Acta Biologica Benrodis 13 (2006)
Fig. 2: The topography of the island of Montserrat before 1995 (from POSSEKEL 1999).Abb. 2: Topographie der Insel Montserrat vor 1995 (aus POSSEKEL 1999).

4 MICHAEL STEVENS & GEORG WALDMANN
mountain ranges: the Silver Hills (403 m;upper Pliocene, 2,600 to 1,200 ka) in North,the Centre Hills (741 m; Pleistocene, 9,500to 550 ka) and the Soufrière Hills (1011 m;upper Pleistocene and late Quaternary, 170to present) in the South (HARFORD et al.2002). Montserrat�s climate is tropical witha relative low humidity. The mean annualtemperature is 26 °C with slight variationbetween 24 °C in January and 28 °C in Sep-tember. The mean annual precipitation va-ries between below 900 mm in the SilverHills, 1,638 mm in Plymouth, and 2,500mm in the Soufrière Hills (ECNAMP1980). Surviving primary rainforests andelfin forests are very limited due to the su-gar cane industry between the 17th and 19th
century. Secondary rainforests are now abun-dant in mountainous regions. At lower ele-vations a dry scrub woodland occurs (BEARD
1949).This small island is severely affected by
hurricanes. In September 1989 HurricaneHugo hit the Caribbean and defoliated 60 %of the forest of Montserrat and left slopesbare of vegetation (POSSEKEL 1999). In Ja-nuary 1992 elevated seismic activity begunand the Soufrière Hill Volcano became acti-ve again on July 18th 1995 (ROBERTSON et al.2000). Eruptive activity culminated onDecember 26th, 1997. During this event 55-60 million cubic meters of material wereremoved and a series of pyroclastic flowsdestroyed the landscape south-west of thevolcano, including the villages St. Patrick�sand the capital Plymouth. A chronology ofthe eruption is summarised by ROBERTSON
et al. (2000).At the present (December 2003), the vol-
cano remains active with minor explosions,rockfalls and ashfalls on a regular basis. Anew lava dome has been growing, consistingof 200 million cubic meters of lava. A cata-clysmical eruption towards the northwestcould lead to the complete evacuation ofhuman population of the entire island(STONE 2003) in case of its collapse.
3. Results
Our results summarize the published datafrom various authors before volcanic acti-vity has begun to limit biological fieldworksince 1995. Families listed follow the syste-matic order of biological textbooks. Withinfamilies the species are arranged alphabeti-cally. Every species is described with its dis-tribution. The distribution areal of everyspecies is given. Additionally, trivial namesand habitat description, distribution onMontserrat, habit (plants), flowering seasonare given if available.
3.1 Species and subspecies endemic toMontserrat
FlacourtiaceaeXylosma serratum (Sw.) Urb., Symb. Antill. 1:317 (1899)
Montserrat. Small tree to 6 m. The plantwas known to have a restricted distributionalong the track leading to the Great Alp Fallssouth of Chance�s Peak before 1995 (HOWARD
1974-1989; R. A. Howard in litt. in JOHNSON
1988). The vegetation cover in this localityhas been destroyed due to pyroclastic flows.Status of survival remains unclear.
RubiaceaeRondeletia buxifolia Vahl, Eclog. Amer. 2: 1 pl.12. (1798) � Privet
Montserrat. Shrub 1-2 m. (HOWARD 1974-1989; R. A. Howard in litt. in JOHNSON 1988).
OrchidaceaeEpidendrum montserratense Nir, Orchid. Antill.132 (2000)
Montserrat. Herb. Flowers in January-Feb-ruary. Inflorescence up to 0.6 m. Terrestrial,on volcanic bedrock or epiphytic on Loblolly(Pisonia fragrans; Mytaginaceae) (NIR 2000).
TheraphosidaeCyrtopholis femoralis Pocock, 1903 � Montser-rat Tarantula

Endemics of the Lesser Antillean Island of Montserrat, British West Indies 5
Acta Biologica Benrodis 13 (2006)
Montserrat (PERTRUNKEVITCH 1911).
StaphylinidaeOligolinus hubbardi Blackwelder, 1943
Montserrat (BLACKWELDER 1943).
Pseudolispinodes foveola Blackwelder, 1943Montserrat (BLACKWELDER 1943).
LycidaeThonalmus hubbardi Leng & Mutchler, 1922
Montserrat (LENG & MUTCHLER 1922).
Thonalmus sinuaticostis Leng & Mutchler, 1922Montserrat (LENG & MUTCHLER 1922).
CantharidaeTylocerus picipennis Leng & Mutchler, 1922
Montserrat (LENG & MUTCHLER 1922).
CucujidaeTelephanus nodicornis Nevermann, 1932
Montserrat (BLACKWELDER 1944-1947).
ColydiidaeMonoedus lecontei Fleutiaux, 1915Montserrat (LENG & MUTCHLER 1922).
ScarabaeidaePhyllophaga montserratense (Arrow, 1920)Montserrat (BLACKWELDER 1944-1947).
Phyllophaga cneda Saylor, 1940Montserrat (BLACKWELDER 1944-1947).
CerambycidaeCyrtinus hubbardi Fisher, 1926
Montserrat (CHEMSAK et al. 1992)
Strangalia benitoespinali Chalumeau, 1985Montserrat. St. George�s: Riley�s Estate, on
Chiny Bush (Philodendron giganteum; Araceae),400 m (CHALUMEAU 1985).
AnguidaeDiploglossus montisserati Underwood, 1964 �Montserrat Galliwasp
Montserrat. St. Peter: Woodlands Estate,Woodlands Springs (Runaway Ghaut). 180m (UNDERWOOD 1964) and Centre Hills (R.Crombie e-mail 16.5.2001). Mesophilic, nearstreams and springs, ground-dwelling, noc-turnal (SCHWARTZ & HENDERSON 1991).
GekkonidaeSphaerodactylus fantasticus ligniservulus King,1962 � Montserrat Dwarf Gekko
Montserrat. Mesophilic, in different ha-bitats from open settlements to forests(SCHWARTZ & HENDERSON 1991).
PolychrotidaeAnolis lividus Garman, 1887 � MontserratAnole
Montserrat. Mesophilic, in different habi-tats, diurnal (SCHWARTZ & HENDERSON 1991).
TeiidaeAmeiva pluvianotata pluvianotata Garman, 1888� Montserrat Ground Lizard
Montserrat. Mesophilic to xerophilic,mixed forest and pasture, on meadows andbeaches, diurnal (SCHWARTZ & HENDERSON
1991).
ColubridaeAlsophis antillensis manselli Parker, 1933 �Montserrat Racer
Montserrat. Xerophilic to mesophilic, rainforest, tropical deciduous forest, mountainpasture, gassy scrub, arid scrub, at the edge ofmangrove, lime grove, in dry stream and creekbeds, ground-dwelling, diurnal (SCHWARTZ &HENDERSON 1991).
TyphlopidaeTyphlops monastus monastus Thomas, 1966 �Montserrat Blindsnake
Montserrat. Xerophilic to mesophilic, oc-curs in open nor slightly forested areas, mesicand intermediate bush, open pastureland,cultivated clearings, open fern growths areaof human habitation (SCHWARTZ & HENDER-SON 1991).

6 MICHAEL STEVENS & GEORG WALDMANN
PsittacidaeAmazona spec. Undescribed species � FossilMontserrat Parrot
Montserrat. Trants archaeological site (WIL-LIAMS & STEADMAN 2001).
Ara spec. Undescribed species � Fossil Monts-errat Macaw
Montserrat. Trants archaeological site (WIL-LIAMS & STEADMAN 2001).
TurdidaeCichlherminia lherminieri lawrenci Cory, 1886 �Montserrat Forest Thrush
Montserrat. Moist mountain forests (RAF-FAELE et al. 1998).
IcteridaeIcterus oberi Lawrence, 1880 � MontserratOriole
Montserrat. Rare in Centre Hills and SouthSoufrière Hills. Mountain forests between230 - 800 m. Habitat loss due to volcanic erup-tions (BLANKENSHIP 1990; RAFFAELE et al. 1998;ARENDT et al. 1999; HILTON et al. 2003).
CricetidaeUndescribed species B � Fossil Rice RatMontserrat. Trants archaeological site (PRE-GILL et al. 1994).
PhyllostomidaeSturnira thomasi vulcanensis Genoways, 1998 �Thomas�s Epauletted Bat
Montserrat. St. George�s: Paradise Estate,1 km S Harris, 1,000 m. Presumably extinctdue to volcanic eruptions since 1997 (GENO-WAYS 1998; ADAMS & PEDERSEN 1998).
3.2 Species and subspecies on Montser-rat considered to be endemic to theLesser Antilles
PolypodiaceaeBlechnum binervatum (Poiret) Morton & Lel-linger, Am. Fern J. 57: 67 (1967)
St. Kitts, Montserrat, Guadeloupe, Domi-
nica, Martinique, St. Lucia (HOWARD 1974-1989; LAZAIRE et al. 1991).
Blechnum ryanii (Kaulf.) Hieron., Hedwigia 47:245 (1908)
Saba, St. Kitts, Nevis, Montserrat, Guade-loupe, Dominica, Martinique, St. Lucia, St.Vincent, Grenada (HOWARD 1974-1989; LA-ZAIRE et al. 1991).
Ctenidis meridionalis (Poiret) Ching, Sunyatse-nia 5: 250 (1940)
St. Eustatius, St. Kitts, Montserrat, Gua-deloupe, Martinique, (Grenada?) (HOWARD
1974-1989; LAZAIRE et al. 1991).
Elaphoglossum impressum (Fee) T. Moore, Ind.Fil. 10 (1857)Endemic to the Lesser Antilles? St. Kitts,Nevis, Montserrat, Guadeloupe, Dominica,Martinique, Grenada (HOWARD 1974-1989;LAZAIRE et al. 1991).
Grammitis eggersii (Baaker ex Hook.) Proctor,Brit. Fern Gaz. 9(3): 77 (1962)St. Kitts, Montserrat, Guadeloupe, Domini-ca, Martinique, St. Vincent, Grenada (HOWARD
1974-1989; LAZAIRE et al. 1991).
Thelypteris clypeolutata (Desvaux) Proctor,Rhodora 61: 306 (1959 [1960])St. Kitts, Nevis, Montserrat, Guadeloupe,Dominica, Martinique, St. Lucia, St. Vincent,Grenada (HOWARD 1974-1989; LAZAIRE et al.1991).
LauraceaeOcotea martinicensis Mez, Jahrb. Königl. Bot.Gart. Berlin 5: 270 (1889)
Montserrat, Guadeloupe, Dominica, Mar-tinique, St. Vincent, Grenada. Tree to 30 m.Rainforest, usually in clearings. Altitude (0-)100-600 m. Flowers from March-April (HO-WARD 1974-1989; FOURNET 2002).
Persea glaberrima Mez, Jahrb. Königl. Bot. Gart.Berlin 5: 144 (1889) � Sweetwood

Endemics of the Lesser Antillean Island of Montserrat, British West Indies 7
Acta Biologica Benrodis 13 (2006)
Montserrat, Guadeloupe, Dominica, Mar-tinique, St. Lucia. Tree to 7 m. (HOWARD 1974-1989; FOURNET 2002).
PiperaceaePeperomia myrtifolia (Vahl) A. Dietr. in L., Sp.Pl. ed. 6,1: 147 (1831)
St. Croix, St. Barthélémy, Saba, St. Eustati-us, St. Kitts, Antigua, Redonda, Montserrat,Guadeloupe, Les Saintes, Désirade, MarieGalante, Dominica, Martinique, St. Lucia, St.Vincent, Grenadines, Grenada, Barbados.Herb. Flowers throughout the year. Altitude0-500 (-1,000) m. Terrestrial, on rocks or epi-phytic, xerophilic (HOWARD 1974-1989; FOUR-NET 2002).
Piper dussii C. DC. in Urb., Symb. Antill. 3:190 (1902) � Doctor Bush
Nevis, Antigua, Montserrat, Guadeloupe,Marie Galante, Dominica, Martinique, St.Lucia, St. Vincent, Grenadines. Shrub or smalltree 0.8-3 m. Flowers all year round. Altitude60-800 m. In seasonal evergreen forests, be-low trees in the montane forest, on the mar-gins of ponds, rivers and waterfalls, forests,clearings and open spaces (HOWARD 1974-1989; FOURNET 2002).
Peperomia trifolia (L.) A. Dietr. in L., Sp. Pl. ed.6,1: 173 (1831)
Nevis, Montserrat, Dominica, St. Lucia, St.Vincent, Grenada. Small climbing herbaceousepiphyte. Flowers September-March (April).Altitude (100-) 200-1,500 m. Epiphytic orsaprophytic, mostly in degraded seasonal ever-green forest (HOWARD 1974-1989; FOURNET
2002).
TheaceaeFrezeria undulata (Sw.) Willd., Sp. Pl. 2: 1179(1799) � Mountain Parrot
Montserrat, Guadeloupe, Dominica, Mar-tinique, St. Lucia, St. Vincent, Grenada. Smalltree or stunted tree 1-8 m. Flowers March-July and October-December. Altitude 500-1,150 m. Upper horizon of the montane thi-
cket and elfin forest (HOWARD 1974-1989;FOURNET 2002).
Ternstroemia elliptica Sw., Prodr. 81 (1788)St. Kitts, Nevis, Antigua, Montserrat, Gu-
adeloupe, Dominica, Martinique, St. Vincent.Shrub or small tree 1.5-5 (-10) m. Altitude300-1200 m (HOWARD 1974-1989; FOURNET
2002).
MarcgraviaceaeMarcgravia umbellata L., Sp. Pl. 1: 503 (1753)
Saba, St. Kitts, Antigua, Guadeloupe, Do-minica, Martinique, St. Lucia, St. Vincent,Grenada. Liana, hemi-epiphytic. FlowersNovember-March. Altitude 150-950 m. Ra-inforest (HOWARD 1974-1989; FOURNET 2002).
ClusiaceaeClusia major L., Sp. Pl. 1: 509 (1753) � Coretor
St. Eustatius, St. Kitts, Nevis, Antigua,Montserrat, Guadeloupe, Désirade, MarieGalante, Dominica, Martinique, St. Lucia, St.Vincent. Tree, very often epiphytic to 20 m.Flowers April-September. Altitude (0-) 60-1,000 m. Seasonal evergreen forest and rain-forest, in both distributed below degradedtrees and in secondary forests (HOWARD 1974-1989; FOURNET 2002).
Marila racemosa Sw., Prodr. 84 (1788)St. Kitts, Antigua, Montserrat, Guadelou-
pe, Marie Galante, Dominica, Martinique, St.Lucia, St. Vincent. Large shrub to 2-4 m orsmall tree to 10 m. Flowers April-September.Altitude (50-) 200-1,050 m. Rainforest (HO-WARD 1974-1989; FOURNET 2002).
ElaeocarpaceaeSloanea dentata L., Sp. Pl. 1: 512 (1753)
St. Kitts, Montserrat, Guadeloupe, Domi-nica, Martinique. Very large tree to 30 m. Flo-wers from May-June. Altitude 200-700 m.Climax species of the montane forest andthe rainforest, often in the neighbourhoodof rivers (HOWARD 1974-1989; FOURNET
2002).

8 MICHAEL STEVENS & GEORG WALDMANN
Sloanea massoni Sw., Prodr. 82 (1788); FlowersInd. Occid. 2: 938 (1800)
Saba, St. Kitts, Montserrat, Guadeloupe,Dominica, Martinique St. Vincent, Grenada.Very large tree to 35 m. Flowers from June-August. Altitude 150-1,000 m. Climax spe-cies of the rainforest (HOWARD 1974-1989;FOURNET 2002).
SterculiaceaeSterculia caribaea R. Br. in Bennett, Pl. Jav.Rar. 228 (1852) � Wild Mahoe, Salt FishWood
Montserrat, Guadeloupe, Dominica, Mar-tinique, St. Lucia, St. Vincent. Tree to 15-35m. Flowers October-November and April-May. Altitude (0-) 200-1,000 m. In more orless degraded rainforest, often exposed towind. Sometimes occurring with Pterocarpus(HOWARD 1974-1989; FOURNET 2002).
MalvaceaeWercklea tulipiflora (Hook. f.) Fryx., J. ArnoldArbor. 62: 476 (1982)
Montserrat, Guadeloupe, Dominica, Mar-tinique, St. Lucia. Tree to 5-10 (-20) m. Flo-wers from November-January and March-May. Altitude 550-1,050 m. Degraded rainfo-rest or secondary rainforest (HOWARD 1974-1989; FOURNET 2002).
PassifloraceaePassiflora rotundifolia L., Sp. Pl. 2: 957 (1753)
Montserrat, Guadeloupe, Dominica,Martinique, St. Vincent, Grenada. Liana.Flowers March-April. Altitude 500-1,000m. Higher levels of the rainforest and dis-turbed rainforest (HOWARD 1974-1989;FOURNET 2002).
BegoniaceaeBegonia obliqua L., Sp. Pl. 2: 1056 (1753)
Montserrat, Guadeloupe, Dominica, Mar-tinique. Perennial herb to 2 m. Flowersthroughout the year. Altitude 450-900 m.Rainforest, below trees in moist places(HOWARD 1974-1989; FOURNET 2002).
Begonia retusa O.E. Schulz in Urb., Symb.Antill. 7: 20 (1911)
St. Barthélémy, Saba, St. Eustatius, St.Kitts, Nevis, Montserrat. Perennial herb to1.5 m (HOWARD 1974-1989; FOURNET 2002).
StyracaceaeStyrax glaber Sw., Prodr. 74 (1788) � Seguine
St. Kitts, Montserrat, Guadeloupe, MarieGalante, Dominica, Martinique, St. Lucia. Treeto 8 m. Flowers June-July and October-No-vember. Altitude (0?-) 250-650 m. Seasonalevergreen forest and rainforest (HOWARD
1974-1989; FOURNET 2002).
FabaceaeAndira sapindoides (DC.) Benth., J. Linn. Soc.,Bot. 4, suppl. 123 (1860) � Bastard Maho-gany
Antigua, Montserrat, Guadeloupe, Domi-nica, Martinique, St. Lucia, Grenada. Tree,smaller than 20 m (HOWARD 1974-1989; FOUR-NET 2002).
Galactia dubia DC., Prodr. 2: 238 (1825)Anguilla, St. Martin, St. Barthélémy, Bar-
buda, St. Eustatius, St. Kitts, Antigua,Montserrat, Guadeloupe, Désirade, MarieGalante, Martinique. Woody climber up to 4m. Altitude 0-200 m. Littoral woodland (of-ten calcareous), dry woodland and dry seaso-nal semi-evergreen forest (HOWARD 1974-1989; FOURNET 2002).
Galactia rubra (Jacq.) Urb., Symb. Antill. 2: 319(1900)
St. Martin, Saba, St. Eustatius, St. Kitts,Antigua, Montserrat, Guadeloupe, Domi-nica, Martinique. Vine up to 4 m, woody atbase. Flowers September-February (-April).Altitude 0-300 m. Shady thickets andhedges (HOWARD 1974-1989; FOURNET
2002).
MelastomataceaeBlakea pulverulenta Vahl, Symb. Bot. 3: 61(1794)

Endemics of the Lesser Antillean Island of Montserrat, British West Indies 9
Acta Biologica Benrodis 13 (2006)
Montserrat, Guadeloupe, Dominica, Mar-tinique, St. Vincent, Grenada. Shrub or smalltree or massive epiphyte, stems glabrous. Flo-wers June-October. Altitude 200-950 m. Inrainforest, often at margins of ravines andwaterfalls (HOWARD 1974-1989; FOURNET
2002).
Charianthus purpureus D. Don, Mem. Wern.Nat. Hist. Soc. 4: 329 (1823)
Saba, St. Kitts, Nevis, Montserrat, Guade-loupe, Dominica, Martinique, Grenada.Shrub 1-10 m. Altitude (150-) 600-1200 m(HOWARD 1974-1989; FOURNET 2002).
Henriettea lateriflora (Vahl) R. A. Howard &E. Kellog, J. Arnold Arbor. 67: 246 (1986)
Montserrat, Guadeloupe, Dominica, Mar-tinique, St. Lucia. Large shrub or small tree to4-5 (-8) m. Flowers June-August and Sep-tember-November. Altitude 500-1,100 m.Upper levels of the rainforest (HOWARD 1974-1989; FOURNET 2002).
Miconia coriacea (Sw.) DC., Prodr. 3: 189 (1828)Montserrat, Guadeloupe, Dominica. Small
dioceous shrub 0.8-1.8 m or small tree to 4m. Flowers throughout the year, mostly Feb-ruary-July. Altitude 1,000-1,467 m (HOWARD
1974-1989; FOURNET 2002).
Miconia cornifolia (Desr.) Naud., Ann. Sci. Nat.Bot. ser. 3, 16: 126 (1851)Montserrat, Guadeloupe, Les Saintes, Domi-nica, Martinique, St. Lucia, St. Vincent, Gre-nada, Barbados. Shrub to 3-8 m. Flowers May-July. Altitude (0-) 200-500 m. Rainforest. Ri-ver and pond margins (HOWARD 1974-1989;FOURNET 2002).
Miconia striata (Vahl) Cogn. in A. DC., Mo-nogr. Phan. 7: 765 (1891)
Antigua, Montserrat, Guadeloupe, Domi-nica, Martinique, St. Lucia. Shrub or small treeto 5 m. Flowers throughout the year, mostlyMay-November. Altitude 100-900 m. Savan-nas, woodland margins, river margins in
seasonal forest and rainforest (HOWARD 1974-1989; FOURNET 2002).
Tetrazygia discolor (L.) DC., Prodr. 3: 172 (1828)Saba, St. Kitts, Nevis, Antigua, Montser-
rat, Guadeloupe, Marie Galante, Dominica,Martinique, St. Lucia, St. Vincent, Grenada.Shrub to 3 m or small tree to 10 m. FlowersApril-June and September. Altitude 0-680 m.Littoral evergreen forest, lower levels and de-graded horizons of seasonal evergreen forestand rainforest (HOWARD 1974-1989; FOURNET
2002).
Tibouchina ornata (Sw.) Baillon, Adansonia 12:74 (1876
Saba, St. Kitts, Montserrat, Guadeloupe,St. Lucia. Dwarf shrub to 0.3 m. Flowersthroughout the year, mostly May-November.Altitude 1,000-1,467 m (HOWARD 1974-1989;FOURNET 2002).
AquifoliaceaeIlex dioica (Vahl) Griseb., Flowers Brit. W. In-dian Is. 147 (1859)
Nevis, Montserrat, Guadeloupe, Martini-que, St. Vincent, Grenada. Shrub or small treeto 8 m. Altitude 700-1,000 m (HOWARD 1974-1989; FOURNET 2002).
EuphorbiaceaePhyllanthus mimosoides Sw., Prodr. 27 (1788)
Montserrat, Guadeloupe, Dominica, Mar-tinique, Trinidad. Dwarf shrub or shrub to 5m. Flowers throughout the year, mostlyNovember-April. Altitude (10-) 150-950 m.Rainforest, often near rivers and in river beds(HOWARD 1974-1989; FOURNET 2002).
Sapium caribaeum Urb., Symb. Antill. 3: 308(1902) � Bird Lime
St. Kitts, Antigua, Montserrat, Guadelou-pe, Marie Galante, Dominica, Martinique, St.Lucia, St. Vincent, Grenada. Tree to 30 m.Flowers September-November. Altitude 0-1,000 m. Associated with Pterocarpus, in theupper horizon of seasonal evergreen forest,

10 MICHAEL STEVENS & GEORG WALDMANN
and secondary rainforest (HOWARD 1974-1989;FOURNET 2002).
SapindaceaePaullinia vespertilio Sw., Prodr. 64 (1788)
St. Kitts, Montserrat, Guadeloupe, Domi-nica, Martinique, St. Lucia, St. Vincent. Strongliana, reaches 10-15 m. Flowers (December)-January-March. Altitude 150-800 m. Rainfo-rest and seasonal evergreen forest (HOWARD
1974-1989; FOURNET 2002).
AraliaceaeSchefflera attenuata (Sw.) Frodin, Proc. Acad.Nat. Sci. Philadelphia 141: 315 (1989)
St. Kitts, Montserrat, Guadeloupe, Domi-nica, Martinique, St. Lucia, St. Vincent, Gre-nada. Small tree to 13 m. Flowers throug-hout the year. Altitude 600-1,467 m. Upperhorizon of the rainforest, montane forest(HOWARD 1974-1989; FOURNET 2002).
BoraginaceaeCordia nesophila I.M. Johnston, J. Arnold Ar-bor. 37: 289 (1956)
Barbuda, Saba, St. Eustatius, St. Kitts, An-tigua, Montserrat, Guadeloupe, Désirade, Ma-rie Galante, Martinique, Dominica, St. Lucia.Ramified shrub to 1-3 m. Flowers throug-hout the year. Altitude 0-30(-80) m. Dry slo-pes and rocky beaches. Very rare on peaty soil(HOWARD 1974-1989; FOURNET 2002).
Cordia reticulata Vahl, Eclog. Amer. 3: 5 (1807)Montserrat, Guadeloupe, Dominica, Mar-
tinique, St. Lucia. Shrub or small tree to 10m. Flowers throughout the year. Altitude 100-800 m. Seasonal evergreen forest, rainforestand upper horizon in degraded rainforest(HOWARD 1974-1989; FOURNET 2002).
VerbenaceaePetrea kohautiana C. Presl., Bot. Bemerk. 99
(1845) � Montserrat Wreath Plant, Sand Pa-per, Purple Wreath, St. John�s Vine, Petrea.
St. Kitts, Antigua, Montserrat, Guadelou-pe, Marie Galante, Dominica, Martinique, St.
Lucia, St. Vincent, Grenada, Trinidad. Strongliana, attached to the stems of large trees,dense leafage. Flowers January-October,occasionally throughout the year. Altitude0-200 (-600) m. Seasonal evergreen forest,lower levels of rainforest, rocky cliffs ofseasonal forest (HOWARD 1974-1989; FOUR-NET 2002).
AcanthaceaeJusticia eustachiana Jacq., Enum. Syst. Pl. 11(1760)
St. Barthélémy, Barbuda, Saba, St. Eusta-tius, Antigua, Redonda, Montserrat, Guade-loupe, Marie Galante. Herb to 1 m. Flowersmostly in August-January. Altitude 0-80 m.Generally below trees of the dry woodlandon sand or limestone (HOWARD 1974-1989;FOURNET 2002).
BignoniaceaeTabebuia pallida (Lindl.) Miers, Proc. Roy. Hort.Soc. 3: 199 (1863) � White Cedar, White Wood
Montserrat, Guadeloupe, Les Saintes, Do-minica, Martinique, St. Lucia, St. Vincent, Gre-nadines, Grenada, Barbados. Tree to 35 m.Flowers several times during the year. Alti-tude 0-200 (-500) m. Often in lower horizonof the seasonal evergreen forest (HOWARD
1974-1989; FOURNET 2002).
LobeliaceaeLobelia guadeloupensis Urb., Symb. Antill. 1: 454(1899)
(Montserrat?), Guadeloupe. Herb. FlowersJanuary-June. Altitude 800-1,200 m (HOWARD
1974-1989; FOURNET 2002).
RubiaceaePsychotria urbaniana Steyerm., Mem. New YorkBot. Gard. 23: 671 (1972)
Montserrat, Guadeloupe, Dominica, Mar-tinique, St. Vincent. Small shrub to 0.75-2 m.Flowers throughout the year. Altitude 150-900 m. Below trees of the rainforest. Veryrare in seasonal evergreen forest (HOWARD
1974-1989; FOURNET 2002).

Endemics of the Lesser Antillean Island of Montserrat, British West Indies 11
Acta Biologica Benrodis 13 (2006)
Spermacoce berteroana R.A. Howard, Phytolo-gia 65: 287 (1988)
Antigua, Montserrat, Guadeloupe, Domi-nica. Herb (HOWARD 1974-1989; FOURNET
2002).
AsteraceaeEupatorium dussii Urb., Symb. Antill. 5: 521(1908)
Montserrat, Guadeloupe. Shrub or smalltree to 1.5-4 m. Flowers March-May. Altitude600-1,000 m. Below trees of the higher levelsof rainforest up to montane forest (HOWARD
1974-1989; FOURNET 2002).
Eupatorium integrifolium Bert. ex Spreng., Syst.Veg. 3: 410 (1826)
St. Kitts, Montserrat, Guadeloupe, LesSaintes, Désirade, Marie Galante, Dominica.Ornamental shrub to 0.5-3 m. Flowers th-roughout the year. Altitude 0-80 m. Rockyand calcareous cliffs (HOWARD 1974-1989;FOURNET 2002).
Eupatorium trigonocarpum Griseb., Flowers Brit.W. Indian Is. 359 (1861)
St. Kitts, Montserrat, Guadeloupe, Domi-nica, Martinique, St. Lucia. Small shrub to 1-3 m. Flowers January-May. Altitude 300-1,100m. In higher levels of rainforest up to mon-tane forest (HOWARD 1974-1989; FOURNET
2002).
Mikania latifolia J. E. Smith in Rees, Cycl. 23Mikania 8 (1819)
Montserrat, Guadeloupe, Dominica, Mar-tinique, St. Lucia, St. Vincent, Barbados. Verystrong climbing liana. Flowers September-March. Altitude 150-900 m. In the rainforest(HOWARD 1974-1989; FOURNET 2002).
ArecaceaeSyagrus amara (Jacq.) Mart., Palmet. Orbign.132 (1847)
Montserrat, Guadeloupe, Dominica, Mar-tinique, St. Lucia, Trinidad. Tree to 15-20 m.Altitude 0-300 m. Seasonal evergreen forest
and lower elevations of rainforest (HOWARD
1974-1989; FOURNET 2002).
CyclanthaceaeAsplundia insignis (Duchass. ex Griseb.) Harl.,Acta Hort. Ber. 17(3): 42 (1954)
St. Kitts, Montserrat, Guadeloupe, Domi-nica, Martinique, St. Vincent, Grenada. Ter-restrial plant. Altitude 150-600 m. Rainforest.Slopes near rivers (HOWARD 1974-1989; FOUR-NET 2002).
Asplundia rigida (Aubl.) Harl., Acta Hort. Ber.17(3): 43 (1954)
St. Kitts, Montserrat, Guadeloupe, Domi-nica, Martinique, Grenada, St. Vincent, St. Lu-cia, Tobago, Trinidad. Strong liana, adheringby adventitious roots. In open ravines, rarelyterrestrial. Flowers February-May. Altitude150-900 m. Rainforest (HOWARD 1974-1989;FOURNET 2002).
PoaceaeIsachne disperma (Lam.) Doell in Mart., Flo-wers Bras. 2(2): 274 (1877)
St. Kitts, Nevis, Montserrat, Guadeloupe,Dominica, Martinique, Grenada, St. Lucia, St.Vincent, Tobago. Herb. Flowers throughoutthe year with interruptions. Altitude 400-1,100 m. Seasonal evergreen forest and rain-forest (HOWARD 1974-1989; FOURNET 2002).
BromeliaceaeAechmea smithiorum Mez in DC., Monogr.Phan. 9: 246 (1896)
Montserrat, Guadeloupe, Dominica, Mar-tinique, St. Lucia, St. Vincent, Grenada. Herb.Flowers throughout the year with interrupti-ons. Altitude 0-600 m. Seasonal evergreenforest and rainforest (HOWARD 1974-1989;FOURNET 2002).
Glomeropitcairnia penduliflora (Griseb.) Mez,Bull. Herb. Boiss. II 5: 232 (1905)
Montserrat, Guadeloupe, Dominica, Mar-tinique. Epiphytic or terrestrial herb, inflore-scence up to 1-2.5 m. Flowers December-

12 MICHAEL STEVENS & GEORG WALDMANN
June. Altitude 400-900 m. Upper elevationof the rainforest and montane forest (HO-WARD 1974-1989; FOURNET 2002).
Vriesea antillana L. B. Smith & Pittendr., J.Washington Acad. Sci. 43: 403 (1954)
Saba, St. Kitts, Montserrat, Guadeloupe,Dominica, Martinique. Acaulic epiphyte. Ra-inforest (HOWARD 1974-1989; FOURNET 2002).
Vriesea guadelupensis (E.G. Baker) Mez in DC,Monogr. Phan. 9: 595 (1896)
Antigua, Montserrat, Guadeloupe, Domi-nica. Epiphyte, tree-shaped to 0.85-1 m. Flo-wers June-September. Altitude 400-900 m(HOWARD 1974-1989; FOURNET 2002).
AgavaceaeAgave dussiana Trel., Mem. Nat. Acad. Sci.Washington 11: 26 (1913)
St. Barthélémy, Antigua, Montserrat, Gua-deloupe, Désirade, Martinique. Herb withacaul rosette. Draught resistant, in dry coastalelevations (HOWARD 1974-1989; FOURNET
2002).
Agave karatto Mill., Gard. Dict. Ed 8. Agaven° 6 (1768)
Barbuda, Saba, St. Kitts, Antigua, Monts-errat, Désirade. Herb with acaul rosette. Drau-ght resistant, in dry coastal elevations (HO-WARD 1974-1989; FOURNET 2002).
Furcraea tuberosa (Mill.) Ait. f. in Ait., Hort.Kew. ed. 2 2: 303 (1811)
St. Barthélémy, Saba, St. Eustatius, St.Kitts, Nevis, Antigua, Montserrat, Guade-loupe, Désirade, Marie Galante, Dominica,Martinique, St. Lucia, St. Vincent, Grenadi-nes, Grenada, Barbados. Herb. Flowers June-August. Altitude 0-250 m. Draught resistant,in dry coastal elevations (HOWARD 1974-1989;FOURNET 2002).
OrchidaceaeTolumnia urophylla (Lodd. ex Lindl.) Braem,Orchidee (Hamburg) 37(2): 59 (1986)
(Anguilla?), St. Martin, St. Barthélémy, Bar-buda, Saba, St. Eustatius, Nevis, Antigua,Montserrat, Guadeloupe, Les Saintes, Dési-rade, Marie Galante, Dominica. Epiphytic onlimestone or dry volcanic rock. FlowersMarch-June, but sometimes throughout theyear. Altitude 0-450 m.
Sometimes cultivated for ornamental re-asons, despite of its protection. Rarely esca-pes cultivation (HOWARD 1974-1989; FOUR-NET 2002).
PeripatidaePeripatus antiguensis Bouvier, 1899Antigua and Montserrat (READ 1988).
ButhidaeCentruroides pococki Sissom & Francke, 1983
St. Kitts, Nevis, Montserrat, Guadeloupe,Les Saintes, Désirade, Marie Galante (SISSOM
& FRANCKE 1983).
DiplocentridaeOieclus purvesii (Becker, 1880)
Saba, St. Eustatius, St. Kitts, Nevis, Bar-buda, Antigua, Montserrat, (Dominica?). St.Peter: Woodlands, under a stone in forest,210 m (LOURENÇO 1992; STEVENS & WALD-MANN 2001).
PhrynidaePhrynus goesii Thorell, 1889
Anguilla, St. Martin, St. Barthélémy, Saba,St. Eustatius, St. Kitts, Nevis, Antigua,Montserrat, Dominica, Martinique (QUINTE-RO 1981).
BlattidaePelmatosilpha purpurascens (Kirkby, 1903)
Antigua, Montserrat, Dominica. Plymouth(REHN & HEBARD 1927).
KalotermitidaeProcryptotermes leewardensis Scheffrahn &Krecek, 2001
Anguilla, St. Martin, St. Barthélémy, Saba,St. Eustatius, St. Kitts, Nevis, Barbuda, An-

Endemics of the Lesser Antillean Island of Montserrat, British West Indies 13
Acta Biologica Benrodis 13 (2006)
tigua, Montserrat, Guadeloupe, La Désirade,Marie-Galante. Spring Field (SCHEFFRAHN &KRECEK 2001).
StaphylinidaeErchomus sharpi (Cameron, 1922)
Antigua to St. Vincent (BLACKWELDER
1943).
Espeson crassulus Fauvel, 1863Montserrat to St. Vincent (BLACKWELDER
1943).
Espeson moratus Schaufuss, 1882St. Thomas, Montserrat, St. Lucia, St. Vin-
cent, Grenada (WOODRUFF et al. 1998).
Lithocharis sorocula Kraatz, 1859St. Croix, St. Kitts, Antigua, Montserrat,
Guadeloupe, Dominica, St. Lucia, St. Vin-cent, Grenada, Barbados (WOODRUFF et al.1998).
Neohypnus illucens (Erichson, 1829)Montserrat, Guadeloupe, Dominica, St.
Lucia, St. Vincent, Grenada (WOODRUFF et al.1998).
Thoracophorus simplex Wendeler, 1930Montserrat, Guadeloupe, Dominica, St.
Lucia, Grenada (WOODRUFF et al. 1998).
ScarabaeidaeAteuchus insulare Fleutiaux & Salle, 1889
Montserrat and Guadeloupe (BLACKWEL-DER 1944-1947).
NitidulidaeParalindria partia Olliff. 1883
Montserrat and Guadeloupe (LENG &MUTCHLER 1917, 1922).
TenebrionidaeCyrtosoma lherminieri (Chevrolat, 1844)
Montserrat, Guadeloupe, Dominica, St.Vincent, Grenada (LENG & MUTCHLER 1917;WOODRUFF et al. 1998).
ChrysomelidaeExora detritum (Fabricius, 1801)
Montserrat and Dominica (LENG & MUTCH-LER 1917).
CurculionidaeDiaprepes famelicus (Olivier, 1790) � LeewardIslands Sugarcane Root-borer
St. Barthélémy, Nevis, Montserrat, Gua-deloupe, Dominica, Martinique, Barbados.St. Anthony: Plymouth, St. George�s: Spa-nish Point, St. Anthony: Upper GalwaysEstate (BLACKWELDER 1944-1947, COOTER
1983).
Lachnopus villosipes (Boheman, 1834) � Gray-black Fiddler
St. Barthélémy, St. Eustatius, Montserrat(BLACKWELDER 1944-1947).
Litostylus pudens (Boheman, 1833) � Yellow-green Fiddler
St. Barthélémy, Antigua, Montserrat, Gua-deloupe (BLACKWELDER 1944-1947).
AnthophoridaeXylocopa (Neoxylocopa) caribaea Lepeltier, 1841
Anguilla, Montserrat, Guadeloupe, Domi-nica, Martinique, St. Lucia, St. Vincent (WOO-DRUFF et al. 1998).
NymphalidaeAnaea minor Hall, 1936 � Small Leafwing
Northern Lesser Antilles. Moister and dryforested habitats (SMITH et al. 1994).
HeliconiidaeDryas iulia warneri (Hall, 1936) � Flambeau
At least on St. Kitts and Montserrat. Inopen areas and the margins of woodland,less frequent in the highlands and generallyavoiding very dry regions (SMITH et al. 1994).
LycaenidaeElectrostrymon angerona (Godman & Salvin,1896)
Lesser Antilles (SMITH et al. 1994).

14 MICHAEL STEVENS & GEORG WALDMANN
Leptotes cassius chadwicki Comstock & Hun-tington, 1943 � Cassius Blue
Windward Islands to Barbuda. Open,moist or drier grassy or scrub land (SMITH etal. 1994).
Strymon acis acis (Drury, 1773) � Bartram�sScrub-Hairstreak
St. Martin to Dominica. Scattered and xericlocalities (SMITH et al. 1994).
PieridaeAppias drusilla comstocki Dillon, 1947 � Flori-da White
Montserrat to Dominica. Lowlands, mesicor dry woods (SMITH et al. 1994).
Eurema venusta emanona Dillon, 1947Montserrat to Grenada. Edges of forests
and woodlands (SMITH et al. 1994).
PapilionidaeBattus polydamas christopheranus (Hall, 1936) �Gold RimSt. Kitts and Montserrat, probably on Nevis.Gardens, low coastal country including be-achside scrub and dry woodland (SMITH et al.1994).
HesperiidaeAchlyodes mithridates minor Comstock, 1944 �Sickle-winged Skipper
Montserrat, Guadeloupe, Dominica.Wide choice of habitats, from mesic or xericwoodland and shady trails to open sunlitfields and other disturbed land (SMITH et al.1994).
Ephyriades brunnea domicinensis Bell & Com-stock, 1948 � Florida Duskywing
Montserrat to Dominica. Prefers open xe-ric scrub vegetation along roadsides, drystreambeds or disturbed fields and pinelands(SMITH et al. 1994).
Nyctelius nyctelius agari (Dillon, 1947) � Vio-let-banded Skipper
Antigua, Montserrat, Guadeloupe, Domi-nica, Martinique, St. Lucia, St. Vincent, Bar-bados, Grenada. Prefers lowland tropical ha-bitats, especially disturbed open fields oralong roadsides (SMITH et al. 1994).
Polites dictynna (Godman & Salvin, 1896)St. Kitts to Grenada. Open fields and mea-
dows, gardens and along roadsides and lowroadsides (SMITH et al. 1994).
Polygonus manueli punctus Bell & Comstock,1948 � Manuel�s Skipper
St. Martin to Grenada. Near freshwater andespecially in coastal habitats (SMITH et al. 1994).
Urbanus obscurus (Hewitson, 1867)Lesser Antilles. Forest paths, clearings,
roadsides, and disturbed land (SMITH et al.1994).
Wallengrenia ophites (Mabille, 1878) � SouthernBroken-Dash
St. Martin to St. Vincent, Trinidad. Alongthe edge of woods, on disturbed open landalong roadsides and in gardens and cultiva-ted groves (SMITH et al. 1994).
CulicidaeOchlerotatus (Howardina) busckii (Coquillett,1906)
St. Eustatius, Montserrat, Guadeloupe,Dominica, Martinique, Grenada (BELKIN &HEINEMANN 1976; WOODRUFF et al. 1998).Treeholes, flower bracts of Heliconia (Helico-nia caribaea; Heliconiaceae), cut Bamboo (Bam-busa vulgaris; Poaceae) and artificial container(tin can). Altitude 70-800 m (BELKIN & HEINE-MANN 1976).
Culex (Melanoconion) madininensis Senevet,1936
Nevis, St. Kitts, Montserrat, Guadeloupe,Dominica, Martinique (BELKIN & HEINEMANN
1976). Stream pools and mangrove swamp.Altitude 0-400 m (BELKIN & HEINEMANN
1976).

Endemics of the Lesser Antillean Island of Montserrat, British West Indies 15
Acta Biologica Benrodis 13 (2006)
Wyeomyia (Wyeomyia) grayii Theobald, 1901Montserrat, Guadeloupe, Dominica, Mar-
tinique, St. Lucia, Grenada (BELKIN & HEINE-MANN 1976). In epiphytic bromeliads (Vrieseaand Glomeropitcairnia; Bromeliaceae) in forests.Altitude 300-850 m (BELKIN & HEINEMANN
1976).
DrosophilidaeDrosophila pulchella Sturtevant, 1916
Montserrat and St. Vincent (GRIMALDI 1988)
Zygothrica insularis Grimaldi, 1987St. Kitts, Montserrat, Dominica, St. Vin-
cent, Trinidad (GRIMALDI 1988).
LeptodactylidaeEleutherodactylus johnstonei Barbour, 1914 �Lesser Antillean Eleuth
St. Martin, Saba, St. Eustatius, St. Kitts,Nevis, Barbuda, Antigua, Montserrat, Marti-nique, St. Lucia, St. Vincent, Grenada, Barba-dos; introduced on Jamaica, Anguilla, Gua-deloupe, Dominica, Grenadines. ThroughoutMontserrat. Mesic forests, rain forest, yards,gardens, nocturnal (BLANKENSHIP 1990; CENS-KY & KAISER 1999; SCHWARTZ & HENDERSON
1991).
Leptodactylus fallax Müller, 1926 � MountainChicken, on Dominica: Crapaud
Montserrat and Dominica. Extinct on St.Kitts, Guadeloupe, St. Lucia; presumablyextinct on Antigua. Montserrat leeward side:Centre Hills, St. George�s Hill, AmershamEstate. Mesic habitats, in woods, scrub, plan-tations on hillsides, wet palm grove, in rivervalley, nocturnal. Habitat loss due to volcaniceruptions (> 10 %) (BLANKENSHIP 1990;SCHWARTZ & HENDERSON 1991; DALTRY &GRAY 1999).
GekkonidaeSphaerodactylus fantasticus Dumérli & Bibron,1836 � Southern Leeward Dwarf Gekko
Montserrat, Guadeloupe and satellites, LesSaintes, Marie Galante, Désirade, Dominica.
Throughout Montserrat. Mesophilic, in dif-ferent habitats from open settlements to fo-rests (SCHWARTZ & HENDERSON 1991).
TeiidaeAmeiva pluvianotata Garman, 1888 � Monts-errat Ground Lizard
Redonda and Montserrat. ThroughoutMontserrat. Mesophilic to xerophilic, mixedforest and pasture, on meadows and beaches,diurnal (SCHWARTZ & HENDERSON 1991).
ColubridaeAlsophis antillensis Schlegel, 1937 � LeewardRacer
Montserrat, Guadeloupe and satellites, Ma-rie Galante, Dominica. Montserrat. Xerophi-lic to mesophilic, rain forest, tropical deci-duous forest, mountain pasture, gassy scrub,arid scrub, at the edge of mangrove, lime gro-ve, in dry stream and creek beds, ground-dwelling, diurnal (SCHWARTZ & HENDERSON
1991).
TyphlopidaeTyphlops monastus Thomas, 1966 � Montser-rat Blindsnake
St. Kitts, Nevis, Barbuda, Antigua,Montserrat. Xerophilic to mesophilic, occursin open nor slightly forested areas, mesic andintermediate bush, open pastureland, culti-vated clearings, open fern growths area ofhuman habitation (SCHWARTZ & HENDER-SON 1991).
ColumbidaeGeotrygon mystacea (Temminck, 1811) �Bridled Quail-Dove
Lesser Antilles to St. Lucia and Barbados;extremely rare on Puerto Rico. Uncommonresident. Dense mountain forests with thickundergrowth, though locally in coastal forests(RAFFAELE et al. 1998).
TrochilidaeEulampis jugularis (Linnaeus, 1766) � Purple-throated Carib

16 MICHAEL STEVENS & GEORG WALDMANN
St. Barthélémy to Grenada. Uncommonresident. Generally mountain forests and ba-nana plantations; occasionally and seasonallyat sea level (RAFFAELE et al. 1998).
Eulampis holosericeus (Linnaeus, 1758) � Green-throated Carib
Puerto Rico, Virgin Islands and the LesserAntilles. Gardens and forests at all elevations(RAFFAELE et al. 1998).
Orthorhyncus cristatus (Linnaeus, 1758) � An-tillean Crested Hummingbird
Puerto Rico, Virgin Islands and the LesserAntilles. Primarily lowland openings, gardens,forest edges and especially arid habitats, butalso mountain forests (RAFFAELE et al. 1998).
TurdidaeCichlherminia lherminieri (Lafresnaye, 1844) �Forest Thrush
Uncommon on Montserrat, Guadeloupe,Dominica; rare on St. Lucia. Moist mountainforests (RAFFAELE et al. 1998).
MimidaeMargarops fuscus (P. L. S. Müller, 1776) � Scaly-breasted Thrasher
Common on Saba to St. Vincent; rare onGrenada. Both moist and semi-arid forestsand woodlands (RAFFAELE et al. 1998).
Cinclocerthia ruficauda (Gould, 1766) � BrownTrembler
Common on Saba, Guadeloupe, Domi-nica; uncommon on St. Kitts, Nevis, Monts-errat, St. Lucia, St. Vincent; rare on Martini-que, Grenada. Wet forests on most islands,although occurs in secondary forests and dri-er woodlands (RAFFAELE et al. 1998).
EmberizidaeLoxigilla noctis (Linnaeus, 1766) � Lesser An-tillean Bullfinch
Saba to Grenada; expanded its range in the1970s to St. John and St. Croix. Shrubberygardens, thickets and forests understory at all
elevations. Sometimes mangroves, swampsand dry areas (RAFFAELE et al. 1998).
CricetidaeUndescribed species A � Fossil Rice Rat
Anguilla, St. Eustatius, St. Kitts, Monts-errat. Trants archaeological site (PREGILL et al.1994).
PhyllostomidaeArdops nichollsi montserratensis (Thomas, 1894)� Montserratian Tree Bat
St. Eustatius and Montserrat (BAKER &GENOWAYS 1978; KOOPMANN 1989). St. An-thony: Belham River Valley; St. George�s: Pa-radise Estate 1 km S Harris, Dyer�s RiverBridge, 0.25 km N Dyer�s; St. Peter: Water-works Estate, 0.5 km SE Happy Hill. Alti-tude 300-1,000 m (JONES & BAKER 1979;PIERSON et al. 1986; PEDERSEN et al. 1996;PEDERSEN 2001).
Chiroderma improvisum Baker & Genoways,1976 � Antillean White-lined BatMontserrat and Guadeloupe (BAKER & GE-NOWAYS 1978; KOOPMANN 1989).
St. George�s: Paradise Estate. Altitude 300-335 m (JONES & BAKER 1979; PIERSON et al.1986). Presumably extinct on Montserrat dueto Hurricane Hugo in 1989 (PEDERSEN et al.1996; ADAMS & PEDERSEN 1998).
Monophyllus plethodon luciae Millar, 1902 � Les-ser Antillean Long-tongued Bat
Anguilla to St. Vincent (BAKER & GENO-WAYS 1978; KOOPMANN 1989). St. George�s:Paradise Estate, 1 km S Harris; Paradise Ri-ver, 0.5 km E Harris; St. Peter: Town of St.Peter�s, Waterworks Estate, 0.5 km SE Hap-py Hill. Altitude 200-1,000 m (PIERSON et al.1986; PEDERSEN et al. 1996; PEDERSEN 2001).
Sturnira thomasi de la Torre & Schwartz, 1966� Thomas�s Epauletted Bat
Montserrat and Guadeloupe (BAKER & GE-NOWAYS 1978; KOOPMANN 1989; GENOWAYS
1998).

Endemics of the Lesser Antillean Island of Montserrat, British West Indies 17
Acta Biologica Benrodis 13 (2006)
4. Discussion
The wider Caribbean comprises of 263,500km², formerly covered with natural vegetati-on (MYERS et al. 2000). The remaining prima-ry vegetation is 11.3 % (29,840 km²). OnMontserrat almost no primary vegetationsurvived the deforestation due to Sugar CaneIndustry, the hurricane devastation in 1989and the ongoing volcanism since 1997 (BE-ARD 1949; PULSIPHER1986; STEVENS & WALD-MANN 2001). Humans have shaped signifi-cantly the island�s ecosystems and biogeogra-phy both in prehistoric and in recent times(MORGAN & WOODS 1986; WATTS 1987; KIM-BER 1988; WHITTAKER 1998; WILLIAMS &STEADMAN 2001).
The available data suggest that an averageof 6 endemic plants and 4 endemic verte-brates could be expected within 100 km² inthe Caribbean Region (MYERS et al. 2000).From Montserrat, 3 single-island endemic
phanerogames and 3 vertebrates are descri-bed (tab. 1). Further 63 vascular plants areconsidered to be local endemics of the Les-ser Antilles. 18 single-island endemic ani-mal species and additionally, 6 subspeciesare reported from Montserrat. 47 animalspecies and further 12 subspecies inhabitingMontserrat are restricted to the Lesser Antil-les. The knowledge of invertebrates exceptbutterflies and mosquitoes is far from beingcomplete (STEVENS & WALDMANN 2001),hence the actual number of endemic insectspecies is expected to be higher.
Our results reflect the published data fromvarious authors before volcanic activity hasbegun to limit biological fieldwork since 1995.The fate of two Montserratian endemics,Thomas�s Epauletted Bat (Sturnira thomasivulcanensis) and the plant Xylosma serratum re-mains unknown at present. The bat has notbeen spotted after 1997, while the plant wasknown to have a very limited distribution on
Tab. 1: Number and percentage of recent endemics (data from DAVIS et al. 1997; STEVENS &WALDMANN 2001)Tab. 1: Anzahl und Prozentsatz der rezenten Endemiten (Daten aus DAVIS et al. 1997; STEVENS &WALDMANN 2001)

18 MICHAEL STEVENS & GEORG WALDMANN
Chance�s Peak even before 1995. It cannot beruled out that volcanism has triggered theirextinction. Another regional endemic bat, theAntillean White-lined Bat (Chiroderma impro-visum) has not been observed since HurricaneHugo in 1989.
The following families comprise severalLesser Antillean endemics: Lauraceae, Piper-aceae, Theaceae, Marcgraviaceae, Clusiaceae,Elaeocarpaceae, Begoniaceae, Styracaceae, Me-lastomataceae, Rubiaceae, Cyclanthaceae, Bro-meliacae, Agavaceae , Hesperiidae, Leptodac-tylidae, Gekkonidae, Polychrotidae, Teiidae,fossil Psittacidae, Trochilidae, Mimidae, Ic-teridae, Cricetidae, and Phyllostomidae.
BEARD (1949) examined the endemism oftrees in the Lesser Antilles. He took all woo-dy plants into consideration that are capableof attaining a height of five meters at matu-rity. The proportions of endemism differfrom vegetation belt to vegetation belt. Man-grove flora does not contain endemics, dryzone flora comprises of 12 % endemics.About half of the trees of the montane andrainforest formations (52 % and 42 % respec-tively) are Lesser Antillean endemics. Alt-hough detailed information on the distribu-tion of most endemic animals is missing,most species occur in the montane elevation.
As expected, the flora and fauna consist ofpan-Caribbean, Greater Antillean, CentralAmerican, and Southern Continental elementsthat extend into the Lesser Antilles (KIMBER
1988). Pan-Caribbean genera are for example:Marcgravia, Miconia, Ocotea, Rondeletia (HOWARD
1973), Dryas, Leptotes, Strymon, Appias, Battus,Achlyodes, Nyctelius, Polites, Polygonus, Urbanus,Wallengrenia (MILLER & MILLER 2001). Examp-les of Central American origin are Anaea, Ephy-riades (MILLER & MILLER 2001); those of Cen-tral and South America are Tetrazygia (HOWARD
1973), Electrostrymon (MILLER & MILLER 2001),whereas Petrea is of South American origin(HOWARD 1973).
Some endemics are restricted to special ha-bitats, like rock pavement: Tabebuia pallida,volcanic and soufrière communities: Lobelia
guadeloupensis, Frezeria undulata, Crater Lakes:Miconia coriacea, Miconia cornifolia (HOWARD
1962, 1973).The unusual high level of biodiversity of
Montserrat requires increased conservation ef-forts (JOHNSON 1988; OLDFIELD & SHEPPARD
1997). Local authorities have launched a mo-nitoring program focussing on the Montser-rat Oriole (Icterus oberi) (ARENDT et al. 1999;HILTON et al. 2003) and the Mountain Chicken(Leptodactylus fallax) (DALTRY & GRAY 1999).Further monitoring is required to obtain ad-ditional data to prevent the impoverishmentregarding Montserrat�s biodiversity and to con-serve the island�s unique flora and fauna.
Acknowledgements
Fieldwork was funded by the Department ofInternational Development and the Foreignand Commonwealth Office/London. We areindebted to JACQUES FOURNET and JOHN S. P.BEARD, who generously have supported uswith unpublished data. RICHARD A. HOWARD
has been a huge moral support. We thankMICHAEL FIELD for his valuable support dur-ing our fieldwork. Former Governor ofMontserrat, ANTHONY ABBOTT, opened manydoors. We would like to thank CAROL andCEDRIC OSBORNE, INGRID and HARRY RAPUA-NO, LOU and SHIRLEY SPYCALLA for their kindhospitality. We thank the staff of the Mont-serrat Volcano Observatory for their kind co-operation allowing us to obtain data in theexclusion zone. BRIDGET BEATTIE has intro-duced us into local orchids. ALFRED �Roots-man� EDWARDS gave us insights into his uniqueethnobotanical knowledge. GERARD GRAY,Ministry of Agriculture, and the MontserratNational Trust have both supported our work.
References
ADAMS, R., PEDERSEN, S. C., 1998: The effects ofnatural disasters on bat populations on Mont-serrat, BWI: A 20 year history (Abstract). �American Zoologist 38, 52A

Endemics of the Lesser Antillean Island of Montserrat, British West Indies 19
Acta Biologica Benrodis 13 (2006)
ARENDT, W. J., GIBBONS, D. W., GRAY, G. A. L.,1999: Status of the volcanically threatenedMontserrat Oriole Icterus oberi and other fo-rest birds in Montserrat, West Indies. � BirdConservation International 9, 351-372
BAKER, R. J., GENOWAYS, H. H., 1978: Zoogeo-graphy of Antillean bats, pp. 53-97. In: Zo-ogeography in the Caribbean (F. B. GILL,ed.) � Special Publication of the Academyof Natural Sciences of Philadelphia 13, 1-128
BEARD, J. S., 1949: The natural vegetation of theWindward & Leeward Islands. � Oxford Fo-restry Memoir 21, 1-192
BELKIN, J. N., HEINEMANN, S. J., 1976: Collectionof the Project �Mosquitoes of Middle Ame-rica� 4. Leeward Islands: Anguilla (ANG),Antigua (ANT), Barbuda (BAR), Montserrat(MNT), Nevis (NVS), St. Kitts (KIT). �Mosquitoe Systematics 8, 123-162
BLACKWELDER, R. E., 1943: Monograph of theWest Indian beetles of the family Staphylini-dae. � Bulletin of the United States NationalMusem 182, 1-658
BLACKWELDER, R. E., 1944-1947: Checklist ofthe Coleopterous Insects of Mexico, CentralAmerica, the West Indies and South Ameri-ca. - Bulletin of the United States NationalMusem 185, 1-925
BLANKENSHIP, J. R., 1990: The Wildlife of Monts-errat. � Montserrat National Trust, Plymouth,Montserrat
CENSKY, E. J., KAISER, H., 1999: The Lesser An-tillean Fauna, pp. 180-222. In: CaribbeanAmphibians and Reptiles (B. I. CROTHER, ed.).� Academic Press, London
CHALUMEAU, F., 1985: Quelques Cerambycidae(Coleoptera) mimetiques des Petites Antilleset description d�une espece nouvelle. -L�Entomologiste 41, 147-152
CHEMSAK, J. A., LINDSLEY, E. G., NOGERA, F. A.,1992: Los Cerambycidae y Disteniidae deNorteamérica, Centroamérica y las Indias Oc-cidentales. � Listados Faunisticos de MéxicoII. Universidad National Autónoma de Mé-xico, México City
COOTER, J. (1983): A few insects from Montser-rat, West Indies. � Entomological Record andthe Journal of Variation 95, 185-186
DALTRY, J. C., GRAY, G., 1999: Effects of volca-nic activity on the endangered Mountain Chi-cken (Leptodactylus fallax). � Froglog 32, 1-2
DAVIS, S., HEYWOOD, V. H., HERRERA-MACBRIDE,O., VILLA-LOBOS, J., HAMILTON, A. C., 1997:Centres of Plant Diversity Vol. 3 The Ameri-cas. � IUCN Press, Cambridge
ECNAMP (EASTERN CARIBBEAN NATURAL AREAS
MANAGEMENT PROGRAMME) (ed.) 1980: Monts-errat preliminary data atlas. Survey of con-servation in the Lesser Antilles. � St. Croix,US Virgin Islands
FOURNET, J., 2002: Flore illustrée des phanéroga-mes de Guadeloupe et de Martinique. � Ciradet Gondwana editiones, Trinité et Mont-pellier
GENOWAYS, H. H., 1998: Two new subspecies ofbats of the genus Sturnira from the LesserAntilles, West Indies. � Occasional Papers ofthe Museum of the Texas Tech University176, 1-7
GRIMALDI, D. A., 1988: Relicts in the Drosophi-lidae (Diptera), pp. 183-213. In: Zoogeogra-phy of Caribbean Insects (J. K. LIEBHERR, ed.).� Cornell University Press, Ithaca
HARFORD, C. L., PINGLE, M. S., SPARKS, R. S. J.,YOUNG, S. R. 2002. The volcanic evolutionof Montserrat using 40Ar/39Ar geochronolo-gy, pp. 93-113. In: The Eruption of SoufriereHills Volcano, Montserrat, from 1995 to 1999(DRUITT, T.H., KOKELAAR, B.P. eds.). � Geolo-gical Society, London, Memoirs 21, 1-407
HILTON, G. M., ATKINSON, P.W., GRAY, G.A.L.,ARENDT, W. J., GIBBONS, D.W., 2003: Rapiddecline of the volcanically threatened Monts-errat oriole. � Biological Conservation 111,79-89
HOWARD, R. A., 1962: Volcanism and Vegetationin the Lesser Antilles. � Journal of the Ar-nold Arboretum of the Harvard University43, 279-313
HOWARD, R. A., 1973: The vegetation of theAntilles, pp. 1-19. In: Vegetation and vegeta-tional history of northern Latin America (A.GRAHAM, ed.). � Elsevier, Amsterdam
HOWARD, R. A., (ed.) 1974-1989: Flora of theLesser Antilles (Volumes 1-6). � Harvard Uni-versity Press, Jamaica Plain
ITURRALDE-VINENT, M. A., MACPHEE, R. D. E.,1999: Paleogeography of the Caribbean regi-on: Implications for cenozoic biogeography.� Bulletin of the American Museum of Na-tural History 238, 1-95
JOHNSON, T. H., 1988: Biodiversity and Conser-vation in the Caribbean. Profiles of selected

20 MICHAEL STEVENS & GEORG WALDMANN
Islands. � ICBP Monograph 1. ICBP PressCambridge
JONES, J. K. Jr., BAKER, R. J., 1979: Notes on acollection of bats from Montserrat, LesserAntilles. � Occasional Papers of the Museumof the Texas Tech University 60, 1-24
KIMBER, C. T., 1988: Martinique Revisited. TheChanging Plant Geographies of a West In-dian Island. � Texas A&M University Press,Austin
KOOPMAN, K. F., 1989: A review and analysis ofthe bats of the West Indies, pp. 635-643. In:Biogeography of the West Indies (C. A. WOODS,ed.). � Sandhill Crane Press, Gainesville
LAZAIRE, J.-J., VIVANT, J., SASTRE, C., 1991: LesPtérophytes de Guadeloupe: biodiversité,écologie, protection. � Bulletin de la SociétéBotnque de France 138, Actualités Botani-ques 2, 197-214
LENG, C. W., MUTCHLER, A. J., 1917: Supplementto preliminary list of the Coleoptera of theWest Indies. - Bulletin of the American Mu-seum of Natural History 37, 191-220.
LENG, C. W., MUTCHLER, A. J., 1922: The Lyci-dae, Lampyridae, and Cantharidae (Telepho-ridae) of the West Indies. � Bulletin of theAmerican Museum of Natural History 46,413-499
LOURENÇO, W. R., 1992: Les peuplements desScorpiones des Antilles; facteurs historiqueset écologiques en association avec les strate-giés biodémographiques. � Studies of the ne-otropical Fauna and Environment 27, 43-62
MARTIN-KAYE, P. H. A. (1969): A summary of thegeology of the Lesser Antilles. � OverseasGeology and Mineral Resources 10, 172-206
MILLER, J. Y., MILLER, L. D., 2001: New Perspec-tives on the biogeography of West Indianbutterflies: a vicariance model, pp. 127-156.In: Biogeography of the West Indies. Pat-terns and Perspectives (C. A.WOODS, F. E.SERGILE, eds.). � CRC Press, Boca Raton
MORGAN, G. S., WOODS, C. A., 1986: Extinctionand the zoogeography of West Indian landmammals. � Biological Journal of the Linne-an Society 28, 167-203
MYERS, N., MITTERMEIER, R. A., MITTERMEIER, C.G., DA FONSECA, G. A. B., KENT, J., 2000:Biodiversity hotspots for conservation prio-rities. � Nature 403, 853-858
NIR, M. A., 2000: Orchidaceae Antillanae. � DAGMedia Publishing, New York
OLDFIELD, S., SHEPPARD, C. 1997: Conservationof biodiversity and research needs in the UKDependent Territories. � Journal of AppliedEcology 34, 1111-1121
PEDERSEN, S. C., 2001: The impact of volcaniceruptions on the bat populations of Monts-errat (Abstract). � American Zoologist 40,1167A
PEDERSEN, S. C., GENOWAYS, H. H., FREEMAN, P.W., 1996: Notes on bats from Montserrat(Lesser Antilles) with comments concerningthe effects of Hurricane Hugo. � CaribbeanJounal of Science 32, 206-213
PIERSON, E. D., RAINEY, W. E., WARNER, R. M.,WHITE- WARNER, C. C., 1986: First record ofMonophyllus from Montserrat. � Mammalia 50,269-271
PERTRUNKEVITCH, A., 1911: A synonymic indexcatalogue of the spiders of North, Centraland South America with all adjacent Islands,Greenland, Bermuda, West Indies, Terra delFuego, Galapagos etc. � Bulletin of the Ame-rican Museum of Natural History 29, 1-892
POSSEKEL, A. K., 1999: Living with the unex-pected - Linking disaster recovery to sustai-nable development in Montserrat. � SpringerVerlag, Berlin, Heidelberg, New York, Tokyo
PREGILL, G. K., STEADMAN, D. W., WATTERS, D.R., 1994: Late Quaternary vertebrate fau-nas of the Lesser Antilles: historical compo-nents of Caribbean biogeography. � Bulletinof the Carnegie Museum of Natural Histo-ry 30, 1-51
PULSIPHER, L. M., 1986: Seventeenth CenturyMontserrat: an environmental impact state-ment. � Historical Geography Research Se-ries 17, 1-96
QUINTERO, D. Jr., 1981: Revision of amblypygidgenus Phrynus in the Americas (Amblypygi,Phrynidae). - Journal of Arachnology 9, 117-166
RAFFAELE, H., WILEY, J., GARRIDO, O., KEITH, A.,RAFFAELE, J., 1998: Birds of the West Indies.� C. Helm, London
READ, V. M. S., 1988: The Onychophora of Tri-nidad, Tobago and the Lesser Antilles. � Zoo-logical Journal of the Linnean Society 93,225-257
REHN, J. A. G., HEBARD, M., 1927: The Ortho-ptera of the West Indies. � Bulletin of theAmerican Museum of Natural History 54,1-320

Endemics of the Lesser Antillean Island of Montserrat, British West Indies 21
Acta Biologica Benrodis 13 (2006)
ROBERTSON, R. E. A., ASPINALL, W. P., HERD, R.A., NORTON, G. E., SPARKS, R. S. J., YOUNG, S.R., 2000: The 1995-1998 eruption of Souf-rière Hills Volcano, Montserrat, WI. � Philo-sophical Transactions of the Royal Societyof London A 358, 1619-1637
ROUGHGARDEN, J., 1995: Anolis Lizards of theCaribbean. � Oxford University Press, Ox-ford, New York, Tokyo
SCHEFFRAHN, R. H., KRECEK, J., 2001: New worldrevision of the termite genus Procryptotermes(Isoptera: Kalotermitidae). � Annals of theEntomological Societ of America 94, 503-539
SCHWARTZ, A., HENDERSON, R. W., 1991: Amphi-bians and Reptiles of the West Indies: De-scriptions, Distributions, and Natural Histo-ry. � University of Florida Press, Gainesville
SISSOM, W. D., FRANCKE, O. F., 1983: Redescrip-tion of Centruroides testaceus (DeGeer) anddescription of a new species from the LesserAntilles (Scorpiones: Buthidae). � Occasio-nal Papers of the Museum of the Texas TechUniversity 88, 1-13
SMITH, D. S., MILLER, L. D., MILLER, J. Y., 1994:The Butterflies of the West Indies and SouthFlorida. - Oxford University Press, Oxford,New York, Tokyo
STEVENS, M., WALDMANN, G., 2001: Animal Bio-diversity of the Lesser Antillean Island of
Montserrat (British West Indies): An Anno-tated Checklist of Terrestrial and FreshwaterAnimals. � Archiv Zoologischer Publikatio-nen, Nümbrecht 6, 1-149
STONE, R., 2003: Bracing for the big one onMontserrat. � Science 299, 2027-2030
UNDERWOOD, G. (1964): An anguid lizard fromthe Leeward islands. � Breviora 200, 1-10.
WATTS, D., 1987: The West Indies: Patterns ofDevelopment, Culture and EnvironmentalChange since 1492. � Cambridge UniversityPress, Cambridge
WEYL, R., 1966: Geologie der Antillen. � Ge-brüder Bornträger, Berlin
WHITTAKER, R. J., 1998: Island BiogeographyEcology, Evolution, and Conservation. �Oxford University Press, Oxford
WILLIAMS, M. I., STEADMAN, D. W., 2001: Thehistoric and prehistoric distribution of par-rots in the West Indies, pp. 355-368. In: Bio-geography of the West Indies. Patterns andPerspectives (C. A.WOODS, F. E. SERGILE,eds.). � CRC Press, Boca Raton
WOODRUFF, R. E., BECK, B. M., SKELLY, P. E.,SCHOTMAN, C. Y. L., THOMAS, M. C., 1998:Checklist and Bibliography of the Insects ofGrenada and the Grenadines. � Memoir No.2 Center for Systematic Entomolgy SandhillCrane Press, Gainesville
Accepted: 14.03.2004

22 MICHAEL STEVENS & GEORG WALDMANN
Book review
INDERJIT, MARC W. (ed. 2005): Invasive Plants: Ecological and Agricultural Aspects. �Birkhäuser, Basel, Boston, Berlin. 283 pages and numerous figures. Hardcover, 24.1 x 16.8cm. ISBN: 978-3-7643-7137-1. About 150.-- Euro
Biological invasion processes have a considerable impact on global biodiversity and natureconservation. The management of invasive species plays an important economic role. Thebook �Invasive Plants� is not a textbook but represents a collection of 17 contributions fromdifferent authors. The book is the result of a symposium held in December 2003 at theCentre for Environmental Management of Degraded Ecosystems (CEMDE), an interdisci-plinary research centre of the University of Delhi. The different chapters cover the followingthemes: Terminology of invasion processes (1), overview over different invasion ecologyhypotheses (2), theoretical approaches towards ecological concepts in terms of prediction ofinvaders, significance of residence time, replication in invasion and the relationship betweencommunity diversity and invasibility (3-7), invasion ecology of Centaurea diffusa, an invaderto the United States (8), regional plant management in the northwestern USA (9), allelo-pathy as a mechanism for resisting plant invasion with the case study of Polygonella myriophyl-la (10), agroecology and the management of weed invasion (11-14), economic, social andmanagement aspects (15-17). As almost all the authors are from USA and Canada, themajority of contributions emphasizes invasion processes from the North American pointof view. Only one contribution comes from the Czech Republic and two contributions arewith participation of Asian authors. A plus of this book is its register, which makes itpossible to look up words or themes of interest quickly.
Ulf Schmitz, Düsseldorf

20 Jahre Entwicklung der Flora und Vegetation im NSG �Steinbruch Hofermühle-Süd� 23
Acta Biologica Benrodis 13 (2006)
Acta Biologica Benrodis 13 (2006): 23-49
20 Jahre Entwicklung der Flora und Vegetation imNSG „Steinbruch Hofermühle-Süd“ (Gemeinde
Heiligenhaus, Kreis Mettmann)
20 years development of flora and vegetation in the natureconservation area “Limestone-Quarry Hofermühle-Süd”
(community Heiligenhaus, Mettmann district)
RENATE FUCHS1, HERFRIED KUTZELNIGG1 und PETER KEIL2
1Universität Duisburg-Essen, Campus Essen, Fachbereich Biologie undGeographie/Botanik, Universitätsstr. 5, D-45117 Essen, Germany
2Mühlenstr. 13, D-45473 Mülheim a.d.R., Germany
Zusammenfassung: 20 Jahre nach Unterschutzstellung des Kalksteinbruches �Hofermühle-Süd� erfolgt eine Bilanzierung der Bestandsentwicklung der Flora und Vegetation. Im Rahmendieser Erfolgskontrolle der Pflegemaßnahmen wurde das Steinbruchgelände im Jahr 2000erneut untersucht. Im Zeitraum von 1979 bis 2000 erhöhte sich stetig die erfasste Gesamtar-tenzahl auf nunmehr 325 Farn- und Blütenpflanzen. Unter den Neufunden sind zahlreichetypische, seltene und bemerkenswerte Taxa, die als Zielarten des Naturschutzgebietes aufge-fasst werden können. Die Vegetationsentwicklung zeigt in den Gehölzgesellschaften der Hang-bereiche und der Steinbruchsohle Tendenzen zum Waldmeister-Buchenwald (Galio-Fagetum);die Grünlandgesellschaften lassen sich zwanglos sehr artenreich ausgeprägten Glatthaferwie-sen-Gesellschaften (Arrhenatherion) zuordnen. Darüber hinaus wurden die wärmeliebendeHügelklee-Odermennig-Saumgesellschaft (Trifolio-Agrimonietum) und weitere bemerkens-werte Felsgrus- und Felsfugen-Gesellschaften nachgewiesen. Die Umsetzung der Pflege- undEntwicklungsmaßnahmen kann aus Sicht des Biotop- und Artenschutzes als erfolgreich ge-wertet werden.
Kalksteinbruch, Biotop- und Artenschutz, Flora, Vegetation
Summary: 20 years after constitution of the limestone quarry �Hofermühle-Süd� as a natureconservation area, the development of flora and vegetation was recorded. With respect to caresuccess controlling the area was again investigated in the year 2000. Within the period 1979-2000the total number of species continuously increase to 325 species of ferns and flowering plants.Many first recorded species are coenotic and ecological characteristic, rare, and remarkable, andcould be considered as target species of the nature conservation area. The wooden plant commu-nities in the range of inclinations and the stone pit bottom appear to develop into Galium odoratum-beach forest (Galio-Fagetum), the grassland vegetation seems to be related to species rich Arrhena-therum elatius-meadows (Arrhenatherion). Furthermore, the thermophilic Trifolium medium-Agrimo-nia-fringing community (Trifolio-Agrimonietum) and other remarkable fine gravel- and rock fis-sure-vegetations are recognized. From viewpoint of nature conservation it is obvious that the caremanagement of the area was successful.
Limestone quarry, natural conservation, flora, vegetation

24 RENATE FUCHS, HERFRIED KUTZELNIGG & PETER KEIL
1. Einleitung
Seit einigen Jahren betreut der Heiligenhau-ser Verein für wissenschaftliche Naturschutz-patenschaften e.V. in Kooperation mit eh-renamtlich arbeitenden Wissenschaftlern undder Unteren Landschaftsbehörde im KreisMettmann das Naturschutzgebiet �Stein-bruch Hofermühle-Süd�. Neben der fachwis-senschaftlichen Begleitung, z.B. von Pfleg-maßnahmen, besteht eine der wesentlichenAufgaben dieser Patenschaft und Kooperati-on darin, die Öffentlichkeit über die natur-schutzfachlichen Arbeiten in diesem Gebietund über dessen Entwicklung zu informie-ren. Ein erster, fachlich sehr breit angelegterÜberblick über naturschutzorientierte biolo-gische Arbeiten im Naturschutzgebiet �Stein-bruch Hofermühle-Süd� wurde bereits vonGERß (1995) in dieser Schriftenreihe gegeben.
Im Jahr 2000 wurde das Naturschutzge-biet erneut floristisch-vegetationskundlich be-arbeitet (FUCHS 2000). Ziel der vorliegendenPublikation ist es, auf der Grundlage dieserUntersuchungen unter Berücksichtigung al-ler bisher im Gebiet erhobenen floristisch-vegetationskundlichen Daten für den Ge-samtzeitraum von 20 Jahren die Entwicklungder Flora und Vegetation im NSG �Stein-bruch Hofermühle-Süd� aufzuzeigen, karto-graphisch darzustellen und unter natur-schutzfachlichen Gesichtspunkten hinsicht-lich der Effizienz der Pflege- und Entwick-lungsmaßnahmen zu diskutieren.
Nach der Eröffnung der Angerbachtalbahnvon Wülfrath nach Ratingen im Jahr 1903konnte der Steinbruch �Hofermühle-Süd� ab1906 bergbaulich erschlossen werden. Übereinen Zeitraum von etwa 30 Jahren wurdenhier Massenkalke aus dem Mitteldevon abge-baut. Das Ende des Abbaus wird für denZeitraum zwischen 1935-1940 angenommen.In den darauf folgenden Jahren diente derSteinbruch als Übungsfeld für die Artillerie,und als mit dem Ende des Zweiten Welt-kriegs keinerlei Bedarf einer Folgenutzungbestand, fiel die Fläche endgültig brach. 1979
untersuchte JACOB im Rahmen ihrer Staats-examensarbeit die Flora und Vegetation desGeländes und erkannte die hohe Funktiondes Steinbruches für Belange des Biotop- undArtenschutzes (JACOB 1979a, b). Auf ihr Be-treiben hin wurde das Gebiet �Hofermühle-Süd� 1984 unter Naturschutz gestellt (s.ADOLPHY 1995).
Mit dem 1988 von REINHOLD und ADOL-PHY (1988) vorgelegten Biotopmanagement-plan beginnt die Phase der Umsetzung vonPflege- und Entwicklungsmaßnahmen.Insbesondere die einsetzende Mahd und diegroßflächigen Entbuschungsmaßnahmen imNorden des Gebietes griffen in die seit derStilllegung des Steinbruchs weitestgehendfreie Sukzession ein und führten zu deutli-chen Veränderungen des floristischen Arten-inventars. 1992 erfolgte dann durch B. DÜLL
im Rahmen einer floristisch-vegetations-kundlichen Kartierung eine erste Erfolgskon-trolle (DÜLL 1992). Im Zusammenhang derGesamtdarstellung aller im Steinbruch durch-geführten biologischen Untersuchungen(GERß 1995) legte KUTZELNIGG (1995) einenÜberblick über Flora und Vegetation vor. Eshandelt sich um Auswertungen und Aktua-lisierungen bisheriger Erkenntnisse, die imRückblick auf die älteren BearbeitungenKommentare zu Neufunden und Verlustenenthält. Auf dieser Basis wurde im Jahre 2000die �Hofermühle-Süd� erneut floristisch-vegetationskundlich kartiert (FUCHS 2000).
2. Das Untersuchungsgebiet undMethoden
Der ehemalige Steinbruch �Hofermühle-Süd� befindet sich inmitten des Städtedrei-ecks Essen-Wuppertal-Düsseldorf (TK4607/43). In seiner näheren Umgebung wirder von den Städten Heiligenhaus, Wülfrath,Mettmann und Ratingen umschlossen (KreisMettmann). Er liegt in einer offenen, weitge-hend agrarisch genutzten Landschaft. Durchdichten Gehölzbewuchs, vor allem der Rand-bereiche, hebt sich der Steinbruch deutlich

20 Jahre Entwicklung der Flora und Vegetation im NSG �Steinbruch Hofermühle-Süd� 25
Acta Biologica Benrodis 13 (2006)
von der Umgebung ab. Hinter einem ausAbraum zusammengesetzten Wall schließtsich im Westen nahtlos der Auwald desZehnthof- und Angerbachs an, der zu einemkleinen Teil ebenfalls zum NSG gehört.
Das Gelände des mit einer Gesamtflächevon ca. 9 ha relativ kleinen Steinbruchs istüber einen Hohlweg erschlossen. Im Westenwird die Steinbruchsohle von zwei felsigenetwa 10-20 m hohen überwiegend bewalde-ten Abbruchkanten und einer dazugehörigenetwa 10-35 m breiten Berme umrahmt. Rela-tiv frei von Gehölzen sind diese felsigen Steil-wände sowie die Berme im Südwesten desGebietes. Die Steinbruchsohle dagegen wirdaufgrund regelmäßiger Pflegemaßnahmengroßflächig von Grünland eingenommen.Inmitten der Steinbruchsohle befindet sichder ehemalige Pumpensumpf, der in einerFelswanne von ca. 3 m Tiefe einen Wasser-körper aufweist.
Das NSG zählt naturräumlich zu den �Hei-ligenhausener Terrassen (3371 01)�, einem Teil-bereich der �Niederbergischen Höhenterrassen(3371 0)� im westlichen Bereich des regenrei-chen �Niederbergisch-Märkischen Hügellandes(3371)� (PAFFEN et al. 1963). Im SteinbruchHofermühle-Süd sind Massenkalke des Nord-randes des Velberter Sattels, des nördlichenGroßsattels des Rheinischen Schiefergebirgesaufgeschlossen, eine Schichtenfolge aus demUnteren Oberdevon, dem Dorpener Kalk. Vordem Abbau bedeckte, wie heute noch die an-grenzenden Hochflächen, quartärer Löß dieOberfläche (WUNSTORF 1929).
Der Niederbergische Raum, an dessenNord-Westgrenze sich das Untersuchungs-gebiet befindet, liegt großklimatisch im Über-gangsbereich des stark atlantisch geprägtenKlimas Nordwestdeutschlands zum stärkerkontinental beeinflussten Klima Mitteleuro-pas. Die Winter sind mild (Mitteltemperatu-ren Dez. und Jan. 1°-2°C), die Sommer rela-tiv kühl (Juli-August 17°-18°C). Die mittlereJahrestemperatur beträgt 9° bis 9,5°C, diemittleren Niederschläge 1000 bis 1100 mmim Jahr (MURL 1989). Das Lokalklima wird
durch die annähernd geschlossene Hohlformdes Steinbruches im Talbereich des Angerba-ches beeinflusst. Diese begünstigt bei aus-tauscharmen Wetterlagen die Bildung vonNebel, kleinräumigen Inversionswetterlagenund die Bildung eines lokalen Kalkluftsees.
Für die floristische Inventarisierung richtetsich die Nomenklatur der wissenschaftlichenPflanzennamen nach WISSKIRCHEN & HAEUP-LER (1998), die der Pflanzengesellschaftennach POTT (1995). Die Vegetationsaufnahmenwurden nach der Methode von BRAUN-BLAN-QUET (1964), modifiziert nach REICHELT &WILMANNS (1973), angefertigt.
3. Die Vegetation des NSG �SteinbruchHofermühle-Süd�
Vegetationskundlich stellt sich das NSG durchseine differenzierten Standortverhältnisse imSteinbruchbereich und durch die unterschied-lichen Pflegeregime als sehr vielfältig dar (Kar-te I). Das Angerbachtal und die Böschungs-bereiche im Steinbruch sind größtenteils be-waldet. Der naturnahe Abschnitt der Aue desAngerbaches wird dabei von einem artenrei-chen Hain-Sternmieren-Erlenauwald (Stella-rio-Alnetum) eingenommen (Tab. 1, Spalte 1).Bemerkenswert sind hier vor allem großeVorkommen von Corydalis cava. Innerhalbdes Steinbruches gliedern sich die Gehölzbe-stände zunächst in ältere, waldartige Bereichemit Baumarten wie Prunus avium, Fraxinusexcelsior, Acer pseudoplatanus und Carpinus be-tulus (vgl. Tab. 2, Spalte 1-11) sowie jüngereHochgebüsche mit hochwüchsigen Strauch-arten wie z. B. Crataegus monogyna, Crataegus×subsphaericea, Cornus sanguinea und Salix ca-prea (vgl. Tab. 2, Spalte 12-16). Unabhängigvon Alter und Struktur haben alle Gehölzge-sellschaften ihre typische, basenreiche Stand-orte anzeigende Artengarnitur mit beispiels-weise Adoxa moschatellina, Melica uniflora, Dac-tylis polygama, Lamium galeobdolon ssp. endtman-nii, Mercurialis perennis und Primula elatior ge-meinsam, welche auf den trockeneren bis fri-schen �rotbuchenwaldfähigen� Standorten �

26 RENATE FUCHS, HERFRIED KUTZELNIGG & PETER KEIL
plenium scolopendrium Anklänge zum Hirsch-zungen-Ahorn-Schluchtwald (Asplenio-Ace-retum) erkennen (Ein typischer Bestand istin Tab. 2, Spalte 1 dokumentiert). Darüberhinaus finden sich innerhalb des Steinbruch-geländes eine Vielzahl an Vorwald- und Ge-büschgesellschaften deren syntaxonomischeZugehörigkeit aufgrund des Fehlens vonCharakterarten nur schwer auszumachen ist.Tendenziell geht auch hier die Entwicklungin Richtung Galio-Fagetum; bei den sonnen-exponierten, auf trockenen Standorten sie-delnden Gebüschgesellschaften lässt sich eineTendenz zum Verband Berberidion(insbesondere mit Vorkommen von Rham-nus cathartica und Rosa arvensis) erkennen.
Ein Großteil der unteren Berme sowie Teil-bereiche der zweiten Berme im Südosten desNSG sind durch Pflegemaßnahmen offen-gehalten worden. Hier haben sich je nachStandortverhältnissen unterschiedliche, z. T.sehr artenreiche Grünlandgesellschaften ent-wickelt. Dominierend ist eine magere, arten-reiche Ausprägung der Glatthaferwiese, diein Teilbereichen auffällig viele Elemente derHalbtrockenrasen-Gesellschaften (Bromionerecti) sowie der Hügelklee-Odermennig-Saumgesellschaft (Trifolio-Agrimonietum)enthält (vgl. Tab. 3, Spalte 4-14). So findensich neben Linum catharticum und Euphrasiacf. nemorosa kleine Populationen von Dianthuscarthusianorum und Leontodon hispidus. Über-gänge zur feuchten Ausprägung der Wiesen-gesellschaft sind durch das Vorkommen vonCirsium palustre und Festuca arundinacea (Spal-te 1-10) gekennzeichnet, in der auch schwer-punktmäßig die Vorkommen von Ophioglos-sum vulgatum liegen (Spalte 4-7). Darüber hin-aus zeigt das Vorkommen von Cirsium arven-se, Heracleum sphondylium, Eupatorium cannabi-num, Tanacetum vulgare und Urtica dioica sowiedas teils dominierende Auftreten von Cala-magrostis epigejos die nährstoffreichere Ausprä-gung an (Spalte 1-2). Im Bereich der sonnen-exponierten Wald- und Gebüschränder cha-rakterisieren Vorkommen von Trifolium medi-um, Agrimonia eupatoria, Hypericum hirsutum
Spalte 1Flächen-Nr. W1Datum 04.00Größe der Aufnahmefläche [m²] 25Deckungsgrad Baumschicht [%] 40Deckungsgrad Strauchschicht [%] 5Deckungsgrad Krautschicht [%] 90
BaumschichtSalix fragilis 3Alnus glutinosa 3
StrauchschichtSambucus nigra 2bRubus idaeus +Rubus camptostachys +Ribes rubrum +
KrautschichtChrysosplenium oppositifolium 2aRanunculus ficaria 2aUrtica dioica 1Glechoma hederacea 1Brachypodium sylvaticum 1Circaea lutetiana 1Anemone nemorosa +Stachys sylvatica +Geum urbanum +Lamium endtmannii +Impatiens glandulifera +Athyrium filix-femina +Deschampsia cespitosa +Arum maculatum +Oxalis acetosella +Corydalis cava +
Tab. 1: Hain-Sternmieren-Erlenauenwald (Stel-lario nemorum-Alnetum glutinosae) im Bereichdes NSG �Steinbruch Hofermühle-Süd�.Tab. 1: Stellaria nemorum-Alnus glutinosa-flood-plain forest (Stellario nemorum-Alnetum gluti-nosae) in the nature conservation area �Lime-stone-Quarry Hofermühle-Süd� .
insbesondere durch das spontane Vorkom-men von Fagus sylvatica in der Krautschicht �die Entwicklungstendenz zum Waldmeister-Rotbuchenwald (Galio-Fagetum) anzeigt.
Im östlichen Abschnitt des Steinbruches �nordexponiert � lassen sich mit dem Vor-kommen von Polystichum aculeatum und As-

20 Jahre Entwicklung der Flora und Vegetation im NSG �Steinbruch Hofermühle-Süd� 27
Acta Biologica Benrodis 13 (2006)
Tab. 2: Waldgesellschaften und Hochgebüsche im NSG �Steinbruch Hofermühle-Süd�. Spalte 1:Waldgesellschaft mit Entwicklungstendenz zum Hirschzungen-Schluchtwald (Asplenio-Aceretum);Spalte 2-11: Waldgesellschaft mit Entwicklungstendenz zum Waldmeister-Buchenwald (Galio-odorati-Fagetum); Spalte 12-16: Hochgebüsche mit Entwicklungstendenz zum Waldmeister-Bu-chenwald (Galio odorati-Fagetum).Table 2: Forest and shrub communities in the nature conservation area �Limestone-Quarry Hof-ermühle-Süd�. Column 1: forest community with development tendencies to Asplenium scolopen-drium-Acer pseudoplatanus-gorge forest (Asplenio-Aceretum); Column 2-11: forest community withdevelopment tendencies to Galium odoratum-beech forest (Galio odorati-Fagetum); Column 12-16:shrub community with development tendencies to Galium odoratum-beech forest (Galio odorati-Fagetum).

28 RENATE FUCHS, HERFRIED KUTZELNIGG & PETER KEIL
Tab. 2: Fortsetzung.Tab. 2: Continued.
Tab. 3: Gesellschaften des Arrhenatherion im NSG �Steinbruch Hofermühle-Süd�. Spalte 4-14:Magere, artenreiche Ausprägung mit Elementen der Halbtrockenrasen-Gesellschaften (Bromionerecti) und der Hügelklee-Odermenning-Saumgesellschaft (Trifolio medii-Agrimonietum); Spalte1-10: Übergänge zur feuchten Ausprägung; Spalte 4-7: Schwerpunktmäßiges Vorkommen vonOphioglossum vulgatum; Spalte 1-2: Nährstoffreiche Ausprägung.Tab. 3: Plant communities of the Arrhenatherion in the nature conservation area �Limestone-Quarry Hofermühle-Süd�. Column 1-10: nutrient-poor species rich expression with floristic ele-ments of the mesoxerophytic meadows (Bromion erecti) and Trifolium medium-Agrimonia-fringingcommunity (Trifolio medii-Agrimonietum); column 4-7: transition to moist expression of Ophio-glossum vulgatum; column 1-2: nutrient-rich expression.

20 Jahre Entwicklung der Flora und Vegetation im NSG �Steinbruch Hofermühle-Süd� 29
Acta Biologica Benrodis 13 (2006)
Tab. 3: Fortsetzung.Tab. 3: Continued.

30 RENATE FUCHS, HERFRIED KUTZELNIGG & PETER KEIL

20 Jahre Entwicklung der Flora und Vegetation im NSG �Steinbruch Hofermühle-Süd� 31
Acta Biologica Benrodis 13 (2006)

32 RENATE FUCHS, HERFRIED KUTZELNIGG & PETER KEIL

20 Jahre Entwicklung der Flora und Vegetation im NSG �Steinbruch Hofermühle-Süd� 33
Acta Biologica Benrodis 13 (2006)

34 RENATE FUCHS, HERFRIED KUTZELNIGG & PETER KEIL

20 Jahre Entwicklung der Flora und Vegetation im NSG �Steinbruch Hofermühle-Süd� 35
Acta Biologica Benrodis 13 (2006)

36 RENATE FUCHS, HERFRIED KUTZELNIGG & PETER KEIL
Spalte 1 2Flächen-Nr. S1 S2Datum 06.00 07.00Größe der Aufnahmefläche [m²] 5 20Deckungsgrad [%] 100 100Median von �Stickstoff� 5,0 6,0Median von �Temperatur� 6,0 5,0
StrauchschichtCrataegus monogyna s.str. (3) .Crataegus ×subsphaericea (3) .Corylus avellana (3) .Rubus camptostachys 2a 2a
KrautschichtKennarten des Trifoliomedii-AgrimonietumTrifolium medium ssp. medium + .Hypericum hirsutum 2a .Agrimonia eupatoria 3 +Origanum vulgare 1 +Geranium robertianum + +Fragaria vesca + +Rosa arvensis . +
StickstoffzeigerEupatorium cannabinum . 2a
sowie Centaurium erythraea die Hügelklee-Odermennig-Saumgesellschaft (Trifolio-Agri-monietum) (s. Tab 4, Spalte 1-2). Hier findensich jedoch eng verzahnt mit stickstofflieben-den und feuchteanzeigenden Sippen alleÜbergänge zu den nitrophilen Hochstauden-gesellschaften (Aegopodion).
Kleinflächig abgeschobene Wuchsorte mitroher Gesteinsschuttbedeckung werden imFrühjahr von Erophila verna und Saxifragatridactylites besiedelt und bilden zusammenmit Arenaria serpyllifolia und Poa compressa dasArtenspektrum der Fingersteinbrech-Gesell-
Tab. 4: Saumgesellschaften im NSG �Steinbruch Hofermühle-Süd�. Spalte 1: Hügelklee-Oder-mennig-Saumgesellschaft (Trifolio medii-Agrimonietum); Spalte 2: Fragment der Hügelklee-Oder-mennig-Saumgesellschaft (Trifolio medii-Agrimonietum) mit Übergängen zur nitrophilen Hoch-staudengesellschaft.Table 4: Fringing communities in the nature conservation area �Limestone-Quarry Hofermühle-Süd�. Column 1: Trifolium medium-Agrimonia-fringing community (Trifolio medii-Agrimonietum);Column 2: fragment of the Trifolium medium-Agrimonia-fringing community (Trifolio medii-Agri-monietum) with transitions to nitrogen rich tall herb community.
Urtica dioica . +Galium aparine . +
BegleiterHolcus lanatus 1 +Rubus idaeus . 2aBrachypodium sylvaticum . 1Rubus caesius . 1Dryopteris filix-mas . +Geum urbanum . +Circaea lutetiana . +Epilobium angustifolium . +Ranunculus ficaria . +Veronica chamaedrys . +Cirsium palustre . +Pastinaca sativa . +Lathyrus pratensis . +Crataegus monogyna agg. juv. . +Galium album ssp. album . +Valeriana repens . +Epilobium parviflorum . +Centaurea jacea s. l. 2a .Glechoma hederacea 1 .Trisetum flavescens + .Quercus robur juv. + .Festuca arundinacea ssp. arundinacea + .Listera ovata + .
schaft (Saxifrago tridactylitis-Poetum com-pressae), die heute auf kaum einem Bahn-hof des Bergischen Landes und des Ruhrge-bietes fehlt (SCHÜTZ 1999; LESCHUS 1999).
Den tiefsten Punkt innerhalb des Steinbru-ches bildet ein kleines Grubengewässer miteiner beachtlichen Zonierung in der Verlan-dungsvegetation. Angefangen bei der sub-mersen Zone mit Vorkommen von Charavulgaris, Potamogeton berchtoldii und P. pusillusüber eine Schwimmblattzone mit Potamoge-ton natans und Lemna minor bis hin zu Röh-richtfragmenten und Initialstadien mit Iris

20 Jahre Entwicklung der Flora und Vegetation im NSG �Steinbruch Hofermühle-Süd� 37
Acta Biologica Benrodis 13 (2006)
pseudacorus, Alisma plantago-aquatica, Lycopuseuropaeus und Eleocharis palustris ssp. vulgariszeigt sich ein breites Spektrum der gewässer-begleitenden Flora.
In den wenigen noch offenen Felswändenfinden sich vereinzelt Vorkommen der xero-phytischen Farne Asplenium trichomanes (Abb.1) und A. ruta-muraria, deren gleichnamigeGesellschaft, das Asplenietum trichomano-ruta-murariae, als typische Kalk-Felsritzenge-sellschaft des Mittelgebirges hier zumindestlokal bestätigt werden kann (vgl. auch SCHOLZ
& LÖSCH 1994).Einen Überblick über die aktuellen Biotop-
typen und die Vegetation des NSG bietet Kar-te I. Dieser Karte folgen die kartographischenDarstellungen der Untersuchungen von B.DÜLL (1992) (Karte II) und REINHOLD & ADOL-PHY (1988) (Karte III). Aus Gründen der Über-sichtlichkeit werden in den Karten II und IIIdie Originalnummerierung der Aufnahmeflä-chen und die Bezeichnungen der Pflanzenge-sellschaften beibehalten, auch wenn von unsaus heutiger Sicht eine etwas andere syntaxo-nomische Auffassung, insbesondere der Ge-hölzgesellschaften, vertreten wird.
4. Ergebnisse der floristischen Untersu-chungen
Im Zeitraum zwischen (1971) 1979 und2000 wurde auf der lediglich etwa 9 ha gro-ßen Fläche die bemerkenswerte Anzahl von325 Pflanzensippen nachgewiesen, darunterzahlreiche in der Umgebung seltene undgefährdete Taxa (siehe Tab. 5). Diese Zahlist insofern besonders bemerkenswert, alsrelativ wenige Störzeiger die Artenzahl an-heben.
Neben einzelnen Angaben aus den frühen1970er Jahren, insbesondere zu einem ehe-maligen Vorkommen von Ophrys apifera (s.z.B. WOIKE & WOIKE 1988; ADOLPHY 1994)erfolgte im Jahre 1979 eine erste Zusammen-stellung der Flora des Steinbruches, die bereits109 Sippen enthält (JACOB 1979b).
Im Untersuchungsjahr 1985/86 belegenREINHOLD & ADOLPHY (1988) 184 Sippen,davon drei Pflanzenarten, die aufgrund ihrerSeltenheit sowie ihres Rückganges regional fürdas Süderbergland bzw. für den nordrhein-westfälischen Raum in der Roten-Liste NRWals gefährdet eingestuft werden, z.B. Euphra-sia cf. nemorosa und Myosotis ramosissima so-wie Dactylorhiza praetermissa, die für das Süd-erbergland sogar als �vom Aussterben be-droht� geführt wird (WOLFF-STRAUB et al.1999). Aus Sicht des Biotop- und Artenschut-zes sind darüber hinaus unter den typischenCharakterarten der Pflanzengemeinschaftender Hofermühle Linum catharticum, Cynosu-rus cristatus, Mercurialis perennis, Adoxa moscha-tellina, Chrysosplenium oppositifolium, Chrysosple-nium alternifolium, Corydalis cava und Potamo-genton natans als wertindizierende Taxa zunennen.
1992 nennt B. DÜLL 163 Sippen, daruntereine größere Zahl an Neufunden aus demBereich der Grünland- und Saumgesellschaf-ten, unter ihnen auch Sippen des magerenGrünlandes wie Leontodon autumnalis undLeontodon hispidus sowie das auf der Vorwarn-liste stehende Centaurium erythraea (DÜLL
1992). Unter den neu entdeckten Waldsip-
Abb. 1: Verbreitung von Asplenium trichomanes imNSG �Steinbruch Hofermühle-Süd�im Jahr 2000.Fig. 1: Distribution of Asplenium trichomanes inthe nature conservation area �Limestone-QuarryHofermühle-Süd� in 2000.

38 RENATE FUCHS, HERFRIED KUTZELNIGG & PETER KEIL
pen befinden sich Dactylis polygama und As-plenium scolopendrium, in der Roten Liste vonNRW in der Kategorie �3� � gefährdet ge-führt (WOLFF-STRAUB et al. 1999).
KUTZELNIGG zählt 1995 in seiner Artenlis-te 276 Sippen auf und kommentiert erstmalsneben weiteren Neufunden auch Verluste undfragliche Angaben. Für die Bewertung desSteinbruchs von besonderer Bedeutung sindhier z.B. die Neufunde von Potamogeton pusil-lus, Carlina vulgaris und Mespilus germanica(KUTZELNIGG 1995).
Während der hier vorgestellten Untersu-chung im Jahre 2000 konnten 272 Sippennachgewiesen werden, so dass die Gesamt-sippenzahl über einen Beobachtungszeitraumvon rund 20 Jahren nun 325 Taxa beträgt.Zu den interessanteren Neufunden im Jahr2000 zählen z.B. Polystichum aculeatum (Abb.2), Potamogeton berchtoldii sowie die bislangwohl übersehene Stellaria holostea.
Seit 1995 wird verstärkt auf das Vorkom-men von Kleinarten und stabilisierten Hyb-ridsippen geachtet. So zählt KUTZELNIGG (1995)u. a. Hypericum dubium, Betula ×aurata, Poa an-
gustifolia und Veronica sublobata auf. Währenddes Untersuchungsjahres 2000 wurden weite-re fünf Hybridsippen und zehn Kleinarten,darunter acht Rubus-Sippen erfasst.
Innerhalb der Gruppe der neophytischenSippen wurden 1985/86 zunächst Festuca bre-vipila, Impatiens glandulifera und der Garten-flüchter Vitis vinifera festgestellt. 1992 kamenBryonia dioica und Ribes alpinum neu hinzu.Hieracium aurantiacum, Impatiens parviflora, Sene-cio inaequidens, Narcissus pseudonarcissus (Gar-tenform), Aquilegia vulgaris (Gartenform) undein Exemplar von Heracleum mantegazzianumwerden im Jahr 1995 genannt. Im Jahr 2000wurden die Sippen Galinsoga ciliata und Impa-tiens parviflora neu beobachtet, während Hera-cleum mantegazzianum offensichtlich nach er-folgten Pflegemaßnahmen wieder ver-schwunden ist (s. auch Tab. 5).
Ein deutlicher Populationszuwachs konn-te für die im Steinbruch eingebrachte Listeraovata (Abb. 3) und insbesondere für Ophio-glossum vulgatum (Abb. 4) bemerkt werden.Listera ovata hat sich stark ausgebreitet und istmittlerweile auch auf der oberen Berme an-zutreffen, wo sie vermutlich nicht eingesetztwurde. Ophioglossum vulgatum trat im Jahr 2000massenhaft auf und konnte auf großer Flä-che auf der unteren Berme nachgewiesenwerden. Die Populationsgröße wird auf über10.000 Wedel geschätzt und zählt somit nachBENNERT (1999) zu den größeren im bun-desdeutschen Vergleich.
Aufgrund der Lage des Untersuchungsge-bietes am Nordwestrand des Mittelgebirgesim Übergang zum Norddeutschen Tieflandkommt dem Angerbachtal und dem Stein-bruch auch eine gewisse pflanzengeographi-sche Bedeutung zu. Bereits MÜLLER (1938) zeigtdies am Beispiel des Vorkommens von Bro-mus ramosus, die heute noch im Angerbachtalnachzuweisen ist und auch sehr selten im Stein-bruch-Gelände (leg. Jensen 1995) siedelt. Bro-mus ramosus erreicht hier ihren nordwestlichenmitteleuropäischen Arealrand (vgl. Karte beiHAEUPLER et al. 2003). Weitere aus chorologi-scher Sicht bemerkenswerte Taxa sind neben
Abb. 2: Verbreitung von Polystichum aculeatum imNSG �Steinbruch Hofermühle-Süd� im Jahr 2000.Fig. 2: Distribution of Polystichum aculeatum inthe nature conservation area �Limestone-QuarryHofermühle-Süd� in 2000.

20 Jahre Entwicklung der Flora und Vegetation im NSG �Steinbruch Hofermühle-Süd� 39
Acta Biologica Benrodis 13 (2006)
Asplenium trichomanes, die mit dem Vorkom-men auf Kalkfelsen in der Hofermühle Rand-vorkommen ihres geschlossenen Verbrei-tungsgebietes in Mitteleuropa bilden.
Wegen der unterschiedlichen Kartierungs-intensität der jeweiligen Bearbeiter lässt sichaus dem absoluten Sippenzuwachs nur be-dingt auf die Sukzessionsgeschwindigkeitschließen. Deutlicher wird dieses bei der Be-trachtung der absoluten Verluste an Taxa, dietrotz intensiver Nachsuche im Jahr 2000 nichtmehr bestätigt werden konnten. So zeigt sich,dass insbesondere in der Gruppe der jeweili-gen Neufunde viele Pflanzenarten enthaltensind, die sich offensichtlich nicht erfolgreichetablieren konnten und innerhalb eines kur-zen Zeitraumes wieder verschwunden sind(vgl. Tab. 5). Andere Taxa sind tatsächlich densich allmählich ändernden Standortbedingenbzw. der Pflege und der damit verbundenenVeränderung/Entwicklung der Pflanzenge-sellschaften �zum Opfer� gefallen und inner-halb des Steinbruches wieder verschwunden.Zu den Sippen, die seit 1995 nicht wiederbestätigt werden konnten, gehören z.B. diestenöken Taxa Dactylorhiza praetermissa, Soli-dago virgaurea sowie die standort- und gebiets-fremden Arten Alisma lanceolatum, Nymphoi-des peltata und Heracleum mantegazzianum.
5. Bestandsentwicklung und Pflegemaß-nahmen
Innerhalb der �Hofermühle-Süd� ist insbeson-dere die Entwicklung der nach Nordwest expo-nierten Wälder in Richtung einer �schlucht-waldähnlichen� Gesellschaft herauszustellen.Nach einem über 50-jährigen Entwicklungszeit-raum scheinen die Vorwaldgesellschaften einReifestadium erreicht zu haben, das neben dererfolgten Bodenentwicklung (s. HOFFROGGE &BURGHARDT 1997) ein günstiges Lokalklima ge-schaffen hat, welches die Ansiedlung solcherstandorttypischer Sippen wie Asplenium scolopend-rium (Abb. 5) und Polystichum aculeatum ermög-lichte. Ähnliche Entwicklungen sind in denräumlich benachbarten Kalksteinbrüchen Gru-
Abb. 3: Verbreitung von Listera ovata im NSG�Steinbruch Hofermühle-Süd� im Jahr 2000.Fig. 3: Distribution of Listera ovata in the natureconservation area �Limestone-Quarry Hofer-mühle-Süd� in 2000.
Abb. 4: Verbreitung von Ophioglossum vulgatum imNSG �Steinbruch Hofermühle-Süd� im Jahr 2000.Fig. 4: Distribution of Ophioglossum vulgatum inthe nature conservation area �Limestone-QuarryHofermühle-Süd� in 2000.
Mespilus germanica und der Moosart Rhytidia-delphus triquetrus (s. RISSE 1995, 2004 von denAutoren bestätigt), die Pteridophyta-Taxa Po-lystichum aculeatum, Asplenium scolopendrium und

40 RENATE FUCHS, HERFRIED KUTZELNIGG & PETER KEIL
be Voßbeck (Wuppertal-Dornap), Grube 7(Haan) und Fraunhofer Steinbruch (Neander-tal, Erkrath) zu beobachten (s. KORDGES 2001;RICHTER & LÖSCH 1999).
Eine positive Wirkung der Pflegemaßnah-men auf die Bestandsentwicklung der typi-schen Sippen des mageren Grünlandes kannbeispielsweise bei Centaurium erythraea, Carexpallescens, Dianthus carthusianorum (Abb. 6), Li-num catharticum, Leucanthemum vulgare s. str.,Campanula rapunculus, Euphrasia cf. nemorosaund Saxifraga tridactylites vermerkt werden.
Des Weiteren scheinen im Bereich der Teich-ufer und Makrophyten-Bestände positiveEntwicklungen statt zu finden. Nachdem diegebietsfremden Sippen Alisma lanceolata undNymphoides peltata, offensichtlich ursprünglicheingebracht, wieder verschwunden sind, zeigtsich eine artenreiche Zonierung, angefangenüber submerse Potamogeton berchtoldii-, P. pu-sillus-, Chara vulgaris-Beständen über eine Po-tamogeton natans-Schwimmblattdecke bis hinzu Röhrichtinitialstadien mit Alisma plantago-aquatica, Glyceria fluitans und Eleocharis palust-ris ssp. vulgaris.
Die Orchideenpopulationen von Epipactishelleborine und Listera ovata sind mit mehre-ren 100 Individuen bzw. mehreren DutzendIndividuen vertreten und bedürfen keinerbesonderen Pflegemaßnahmen. Epipactis hel-leborine ist in einer lockeren Bestandsdichte inallen Wäldern, Vorwäldern und Gebüschendes NSG anzutreffen.
Die Bestandsentwicklung der bundesweitals gefährdet eingestuften Ophioglossum vulga-tum ist ebenso positiv zu beurteilen. Auf-grund der nun erreichten Populationsgrößeübernimmt der Steinbruch �Hofermühle-Süd� eine besondere Bedeutung für denSchutz des seltenen Pteridophyten-Taxons (s.BENNERT 1999). In der Fachliteratur werdenkeine speziellen Pflegemaßnahmen empfoh-len. Unter den bisherigen Gegebenheiten hatsich diese Farnpflanze ausgesprochen gutentwickelt, demzufolge ist keine Änderungder Pflegemaßnahmen erforderlich oder wün-schenswert. Asplenium scolopendrium und Poly-stichum aculeatum bedürfen keiner besonde-ren Pflegemaßnahme, sie sollten jedoch nichtfreigestellt werden.
Abb. 6: Verbreitung von Dianthus carthusianorumim NSG �Steinbruch Hofermühle-Süd� im Jahr2000.Fig. 6: Distribution of Dianthus carthusianorumin the nature conservation area �Limestone-QuarryHofermühle-Süd� in 2000.
Abb. 5: Verbreitung von Asplenium scolopendriumim NSG �Steinbruch Hofermühle-Süd� im Jahr2000.Fig. 5: Distribution of Asplenium scolopendriumin the nature conservation area �Limestone-QuarryHofermühle-Süd� in 2000.

20 Jahre Entwicklung der Flora und Vegetation im NSG �Steinbruch Hofermühle-Süd� 41
Acta Biologica Benrodis 13 (2006)
Um weiterhin trocken-warme, nährstoff-arme Standorte für seltene, konkurrenzschwa-che Sippen (z. B. Saxifraga tridacylites) zu si-chern wird empfohlen, die randlichen Berei-che der auf der nördlichen Sohle befindlichenFelsmulden etwas großflächiger abzuschiebenund dies in regelmäßigen Abständen (alle 5-10 Jahre) zu wiederholen.
Insgesamt ist den bisher durchgeführtenPflege- und Entwicklungsmaßnahmen einepositive Wirkung zu attestieren.
6. Folgerungen für den Naturschutz
Die Veränderungen in der Flora des Stein-bruches �Hofermühle-Süd� markieren deut-lich den Sukzessionsverlauf und den Reife-prozess innerhalb der Grünland-, Hochstau-den- und Waldgesellschaften. Dies zeigenmarkant die positiven Populationsentwick-lungen von Ophioglossum vulgatum, Leontodonhispidus, Campanula rapunculus, Dianthus carthu-sianorum, Euphrasia cf. nemorosa und Listeraovata im Bereich der Magergrünlandgesellschaf-ten sowie von Asplenium scolopendrium, Poly-stichum aculeatum und Dactylis polygama imBereich der Waldgesellschaften. Nach einem50-jährigen Entwicklungszeitraum und fast15-jähriger Pflege hat sich vermutlich bereitsin den 1990er Jahren ein mehr oder wenigerkonstantes Artenspektrum von ca. 280 Sip-pen etabliert. Schwankungen in den angege-benen Artenzahlen begründen sich überwie-gend durch die Bearbeitungsintensität, z.B.in der Berücksichtigung von Kleinarten, Hy-bridsippen und schwer bestimmbarer Taxa,aber auch in dem intermittierenden Verhal-ten einer Reihe von Arten, die entsprechendnicht jedes Jahr beobachtet werden können.
Die große Anzahl und die z.T. bemer-kenswerten Populationsgrößen seltener undgefährdeter Taxa im Bereich des NSG �Stein-bruch Hofermühle-Süd� zeigt die hohe Be-deutung des Steinbruches für Belange desBiotop- und Artenschutzes, insbesonderefür solche konkurrenzschwachen Sippen ba-senreicher, nährstoffarmer und wärmebe-
günstigter Standorte, die in der Kulturland-schaft des Niederbergischen Raumes bereitsselten geworden sind. Dass Kalksteinbrü-che hier Refugialfunktionen für gefährdeteTaxa übernehmen, belegen eine Reihe vonUntersuchungen (z.B. KUPKA 1974; RISSE
1985; KEIL & KORDGES 1999; RICHTER &LÖSCH 1999; WOIKE & WOIKE 1999). Obimmer ein umfangreiches Pflegekonzept zurSicherung und Förderung bestimmter Taxabzw. Syntaxa notwendig ist, bleibt eine Fra-ge der Zielsetzung. Ohne Eingriffe und Pfle-ge greift innerhalb weniger Jahrzehnte dieSukzession in Richtung Waldentwicklung,die wie oben am Beispiel von Asplenium sco-lopendrium und Polystichum aculeatum gezeigtwurde, eine � auch unter Naturschutzaspek-ten � diskussionswürdige Entwicklungsva-riante darstellen kann. Zweifellos birgt eindurch Pflege bedingtes Mosaik unterschied-licher Biotopstrukturen ein höheres Besied-lungspotential, welches sich durch höhereArtenzahlen und potentiell durch den Nach-weis einer entsprechend größeren Anzahlgefährdeter Taxa niederschlägt. Im günsti-gen Fall sind beide Varianten � Sukzessionund Pflege � miteinander kombinierbar, sodass ein größtmögliches Entwicklungspo-tential besteht (s. z.B. TRANKLE 1997).Letztlich stellen die Pflegemaßnahmen mitder Förderung von Magergrünland, Wald-rändern und Hochstaudengesellschaften inder Hofermühle-Süd neben den Aspektendes Biotop- und Artenschutzes auch einenBeitrag zur Erhaltung der kleinbäuerlichenKulturlandschaft des Niederbergischen Rau-mes dar und sollten entsprechend fortge-führt werden.
Danksagung
Für die Überprüfung und Bestimmung kri-tischer Sippen danken wir den Herren GÖTZ
H. LOOS (Kamen) und Dr. KLAUS VAN DE
WEYER (Nettetal), für die kritische Durchsichtdes Manuskriptes Herrn Prof. Dr. R. WITTIG
(Frankfurt).

42 RENATE FUCHS, HERFRIED KUTZELNIGG & PETER KEIL
Literatur
ADOLPHY, K., 1994: Flora des Kreises Mettmann� unter besonderer Berücksichtigung vonSchutzgebieten. � Wupperdruck, Wuppertal.
ADOLPHY, K., 1995: Planerische Grundlagen undallgemeine Schutzziele des Naturschutzgebie-tes Hofermühle-Süd. � Acta Biologica Ben-rodis, Suppl. 2, 7-9.
BENNERT, H.W., 1999: Die seltenen und gefähr-deten Farnpflanzen Deutschlands. Biologie,Verbreitung, Schutz. � Landwirtschaftsver-lag, Bonn-Bad Godesberg.
BRAUN-BLANQUET, J., 1964: Pflanzensoziologie.3. Aufl. � Springer Verlag, Wien, New York.
GERß , W. (Hrsg.), 1995: Naturschutzorientiertebiologische Arbeiten im Naturschutzgebiet�Steinbruch Hofermühle-Süd� (GemeindeHeiligenhaus, Kreis Mettmann, Nordrhein-Westfalen). � Acta Biologica Benrodis,Suppl. 2, 1-129.
HAEUPLER, H., JAGEL, A., SCHUMACHER, W., 2003:Verbreitungsatlas der Farn- und Blütenpflan-zen in Nordrhein-Westfalen. � LÖBF NRW,Recklinghausen.
HOFFROGGE, E.H., BURGHARDT, W., 1997: Boden-bildung und chemische Eigenschaften desKalksteinbruches Hofermühle-Süd bei Hei-ligenhaus. � Deutsche Bodenkundliche Ge-sellschaft, Mitteilungen 84, 23-26.
KEIL, P., KORDGES, T., 1999: Verbreitung der Bie-nen-Ragwurz (Ophrys apifera Hudson) imNiederbergischen Raum (Wuppertal/Mett-mann). � Jahresberichte des Naturwissen-schaftlichen Vereins Wuppertal 52, 199-205.
KORDGES, T., 2001: Kalksteinbrüche in Wupper-tal-Dornap: Eingriffsflächen mit Refugial-funktionen für gefährdete Tier- und Pflan-zenarten, pp 33-52. In: Berichte zum Arten-und Biotopschutz Heft 1: Wildnis Wuppertal(Der Oberbürgermeister der Stadt Wupper-tal, ed.). � Wuppertal.
KUPKA, J., 1974: Die Kalkflora in der Umgebungvon Mettmann. � Jahresberichte des Naturwis-senschaftlichen Vereins Wuppertal 27, 104-118.
KUTZELNIGG, H., 1995: Flora und Vegetation imNaturschutzgebiet Hofermühle-Süd. � ActaBiologica Benrodis, Suppl. 2, 33-51.
LESCHUS, H., 1999: Flora der Bahnanlagen imnördlichen Bergischen Land. � Jahresberich-te des Naturwissenschaftlichen Vereins Wup-pertal 52, 121-198.
MÜLLER, J., 1938: Die botanische Erforschungdes bergischen Landes und das Bergische Her-barium. � Jahresberichte des Naturwissen-schaftlichen Vereins Wuppertal 17, 52-65.
MURL - Minister für Umwelt, Raumordnungund Landwirtschaft des Landes NRW, 1989:Klima-Atlas NRW � Düsseldorf.
PAFFEN, K., SCHÜTTLER, A., MÜLLER-MINY, H.,1963: Die Naturräumlichen Einheiten aufBlatt 108/109 Düsseldorf-Erkelenz. � Geo-graphische Landesaufnahme 1 : 200.000.Naturräumliche Gliederung Deutschlands.Bad Godesberg.
POTT, R., 1995: Die PflanzengesellschaftenDeutschlands. 2. Aufl. � Ulmer, Stuttgart.
REICHELT, G., WILMANNS, O., 1973: Vegetations-geographie. � Westermann, Braunschweig.
RICHTER, G., LÖSCH; R., 1999: Flora und Vegetati-on des Fraunhofer Steinbruchs, Erkrath, KreisMettmann. � Jahresberichte des Naturwissen-schaftlichen Vereins Wuppertal 52, 83-120.
RISSE, S., 1985: Schutzwürdige Biotope im RaumVelbert. Über die Moosflora der Feuchtge-biete, der ehemaligen Kalksteinbrüche undeiner alten Bergbauhalde im Gebiet der To-pographischen Karte 4608 (Velbert). � Ro-merike Berge, Zeitschrift für das BergischeLand 35, 41-47.
RISSE, S., 1995: Moose im NaturschutzgebietHofermühle-Süd. � Acta Biologica Benrodis,Suppl. 2, 53-57.
SCHOLZ, S.A., LÖSCH, R., 1994: Verbreitung undSoziologie der Mauerfugen-Vegetation im nie-derbergischen Teil des Kreises Mettmann. �Jahresberichte des NaturwissenschaftlichenVereins Wuppertal 47, 81-98.
SCHÜTZ, P., 1999: Kurzbeitrag zur Ausbreitungs-strategie des Dreifingersteinbrechs (Saxifraga tri-dactylites L.) und seiner künftiger Chancen im�Ruhrgebiet�. � Natur & Heimat 59, 11-14.
TRÄNKLE, U., 1997: Vergleichende Untersuchungzur Sukzession von Steinbrüchen in Südwest-deutschland und neue Ansätze für eine stand-orts- und naturschutzgerechte Renaturierung,pp 1-327. In: Steinbrüche und Naturschutz,Sukzession und Renaturierung (Poschlod, P.,Trankle, U., Böhmer, J. & Rahmann, H., eds).� Ecomed, Landsberg.
WISSKIRCHEN, R., HAEUPLER, H., 1998: Standard-liste der Farn- und Blütenpflanzen Deutsch-lands (Bundesamt für Naturschutz, Hrsg.). �Ulmer, Stuttgart.

20 Jahre Entwicklung der Flora und Vegetation im NSG �Steinbruch Hofermühle-Süd� 43
Acta Biologica Benrodis 13 (2006)
WOIKE, S., WOIKE, M., 1988: Das Neandertal. �Rheinische Landschaften 32, 1-43.
WOIKE, S., WOIKE, M., 1999: Steinbruch �Grube7� und der ehemalige Kalk-Schlammteich. �Der Kiebitz (Haan) 1/99, 4-14.
WOLFF-STRAUB, R., BÜSCHER, D., DIEKJOBST, H.,FASEL, P., FOERSTER, E., GÖDDE, R., JAGEL, A.,KAPLAN, K., KOSLOWSKI, I., KUTZELNIGG, H.,RAABE, U., SCHUMACHER, W., VANBERG, C., 1999:Rote Liste der gefährdeten Farn- und Blü-tenpflanzen (Pteridophyta et Spermatophy-ta) in Nordrhein-Westfalen. � LÖBF-Schrif-tenreihe 17, 75-171.
WUNSTORF, W., 1929: Geologische Karte vonPreußen und benachbarten Ländern Blatt2649 Kettwig (Preußische geologische Lan-desanstalt Berlin, Hrsg.). � Berlin.
Unveröffentlichte Arbeiten(Diese Arbeiten sind bei der Unteren Landschafts-behörde Kreis Mettmann bzw. bei den Autoreneinzusehen.)
DÜLL, B., 1992: Pflanzensoziologische Kartie-rung des NSG �Hofermühle-Süd�. � Gut-achten im Autrag des Heiligenhauser Vereinfür wissenschaftliche Naturschutzpatenschaf-ten e. V.
FUCHS, R., 2000: Flora und Vegetation im Na-turschutzgebiet Hofermühle-Süd. � Studien-arbeit im Fachbereich 9 der Gesamthoch-schule Essen.
JACOB, B., 1979a: Untersuchungen zur Vegetati-onsentwicklung in aufgelassenen Kalkstein-brüchen in der Umgebung von Mettmann. �Staatsexamensarbeit. Fachbereich Biologie,Gesamthochschule Wuppertal.
JACOB, B., 1979b: Biotopkataster Nordrhein-West-falen, Erhebungsbogen für den SteinbruchHofermühle + Artenliste.
REINHOLD, F.-O., ADOLPHY, K., 1988: Land-schaftsplan Kreis Mettmann. Biotopmanage-mentplan. Naturschutzgebiet �HofermühleSüd�.
Angenommen: 05.05.2004

44 RENATE FUCHS, HERFRIED KUTZELNIGG & PETER KEIL
Tab. 5: Gesamtartenliste NSG �Hofermühle-Süd� Nach: J = JACOB 1979a; A = REINHOLD undADOLPHY 1988; D = DÜLL 1992; K = KUTZELNIGG 1995, N = Nachträge unveröffentlicht aus denJahren 1995-1998, F = FUCHS 2000; * = unsicher oder offensichtliche Fehlbestimmung.Tab. 5: Total species of ferns and flowering plants in the nature conservation area �Hofermühle-Süd� according to: J = JACOB 1979a; A = REINHOLD und ADOLPHY 1988; D = DÜLL 1992; K =KUTZELNIGG 1995, N = unpublished postscripts from 1995-1998, F = FUCHS 2000; * = uncertainor obvious misidentification.

20 Jahre Entwicklung der Flora und Vegetation im NSG �Steinbruch Hofermühle-Süd� 45
Acta Biologica Benrodis 13 (2006)
Tab. 5: FortsetzungTab. 5: Continued.

46 RENATE FUCHS, HERFRIED KUTZELNIGG & PETER KEIL
Tab. 5: FortsetzungTab. 5: Continued.

20 Jahre Entwicklung der Flora und Vegetation im NSG �Steinbruch Hofermühle-Süd� 47
Acta Biologica Benrodis 13 (2006)
Tab. 5: FortsetzungTab. 5: Continued.

48 RENATE FUCHS, HERFRIED KUTZELNIGG & PETER KEIL
Tab. 5: FortsetzungTab. 5: Continued.
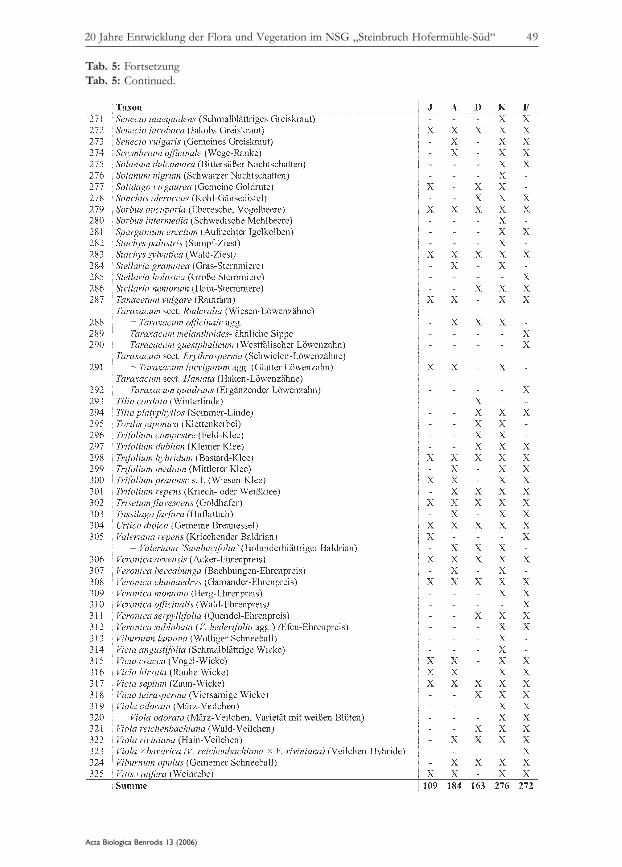
20 Jahre Entwicklung der Flora und Vegetation im NSG �Steinbruch Hofermühle-Süd� 49
Acta Biologica Benrodis 13 (2006)
Tab. 5: FortsetzungTab. 5: Continued.

50 RENATE FUCHS, HERFRIED KUTZELNIGG & PETER KEIL
Rezension
SEIFERT, BERNHARD (2007): Die Ameisen Mittel- und Nordeuropas. � lutra-Verlags- undVertriebsgesellschaft, Tauer. 368 Seiten mit zahlreichen Strichzeichnungen und Farbfotos.Gebunden, 15 x 21,5 cm. ISBN 978-3-936412-03-1. 39,-- Euro
Um erst keinen falschen Eindruck aufkommen zu lassen: Das ist ein wunderbares Buch, dasauch dem (interessierten) Laien einen Einstieg in die Biologie (heimischer) Ameisen ermög-licht, ihm aber nicht suggeriert, die Tiere ließen sich einfach bestimmen, möglichst noch nachfarbigen Fotos, wie sie dem Band in reichem Maße und in hervorragender Qualität beigege-ben sind.Das ein wenig wortreiche Vorwort schildert die Entstehungsgeschichte des vorliegendenBandes (das Buch hat einen erfolgreichen Vorläufer desselben Autors) und lamentiert überden heutigen Wissenschaftsbetrieb (s. auch die Einleitung S. 13) und die Probleme, die einemolekulare Systematik mit sich bringen könnte. Dies ist in der z.T. gebotenen Kürze undDiktion zumindest für den Laien unverständlich und für den Praktiker, für den das Buch jawohl in erster Linie geschrieben ist, nicht relevant und daher � entbehrlich.Der Rest kann sich sehen lassen: Auf 90 Seiten bekommt der Leser eine umfassende Einfüh-rung in Ameisen, ihre Organisation, Individualentwicklung, Lebensräume, Nester, Fort-pflanzungsbiologie (Schwarmflug, Begattung, Koloniegründung und Lebenszyklus), Er-nährungsstrategien und Feinde sowie in einige Sonderfälle wie Myrmecophilie und Bezie-hungen der Ameisen zu Bläulingen. Das Kernstück des Buches ist jedoch der spezielle Teilmit Ratschlägen für den ernsthaften Sammler, einer Liste der im Freiland lebenden Ameisen-arten Mittel-, Nord- und Nordwesteuropas sowie Deutschlands (114 Arten mit Angabenzur Gefährdung und ökologischer Kurzcharakterisierung), Angaben zur Präferenz vonAmeisenarten sowie mit einem akribisch verfassten dichotomen Bestimmungsschlüssel fürdie weiblichen Kasten mit zahlreichen Detailskizzen (S. 107-189) und mit nicht minderwertvollen Angaben zur �Biologie und Ökologie von Ameisenarten� (S. 191-333; ich dachteallerdings immer, die Ökologie sei ein Teil der Biologie?). Am Schluss folgen ein nützlichesGlossar, ein überaus umfangreiches Literaturverzeichnis (neben allgemeiner Literatur wer-den über 400 Arbeiten aufgelistet, die im Text zitiert sind), ein Register und Messvorschrif-ten für die Nutzer des Bestimmungsschlüssels. Druck und Einband sind ausgezeichnet.Eine Bereicherung nicht nur für Myrmecophile.
Hartmut Greven, Düsseldorf

Pflanzliche Neubürger (Neophyten) im Raum Düsseldorf/Neuss 51
Acta Biologica Benrodis 13 (2006)
Acta Biologica Benrodis 13 (2006): 51-89
Pflanzliche Neubürger (Neophyten) im RaumDüsseldorf/Neuss (Deutschland, Nordrhein-Westfalen)
New plant species (neophytes) in the Düsseldorf/Neuss area(Germany, North Rhine-Westphalia)
WERNER DIETRICH
Schloßallee 3a, D-50181 Bedburg, Germany
Zusammenfassung: Im dichtbesiedelten Raum der Rheinmetropolen Düsseldorf und Neusskommen zurzeit 252 Neophyten vor. Davon stammen 55 Arten aus Europa, 56 aus Eurasien, 25aus dem Mittelmeergebiet, 33 aus Asien, 58 aus Nordamerika und dem nördlichen Mittelamerika,21 aus Südamerika und dem südlichen Mittelamerika, 3 aus Afrika und eine Art aus Australien.Etwa 55 der 252 Arten sind als unbeständige Einwanderer anzusehen.
Neophyten im Raum Düsseldorf/Neuss, unbeständige Einwanderer
Summary: In the densely populated area of the capital cities Düsseldorf and Neuss 252 neophytesare currently present. 55 species came from Europe, 56 from Eurasia, 25 from the Mediterraneanregion, 33 from Asia, 58 from North America and the northern parts of Central America, 21 fromSouth America and the southern parts of Central America, three from Africa and a single speciesfrom Australia. Approximately 55 of the 252 species are occasional immigrants.
Neophytes in the Düsseldorf/Neuss area, occasional immigrants
1. Einleitung
Seit einigen Jahren wird der botanisch inter-essierte Wanderer häufig mit Pflanzen kon-frontiert, die das Bild altgewohnter Land-schaften drastisch verändern und überprägen.Lehrwanderungen im Heimatgebiet entfa-chen daher oft lebhafte Diskussionen um der-artige Neubürger aus aller Herren Länder. Soruft z.B. ein Exemplar des Riesenbärenklaus(Heracleum mantegazzianum), der mit demDrüsigen Springkraut (Impatiens glandulifera)verstellte Uferblick oder das Dickicht des Ja-panischen Staudenknöterichs (Reynoutria ja-ponica) beim Betrachter eine Mischung ausFaszination und Besorgnis hervor
Angesichts der sprunghaften Vermehrungvieler Neophyten tauchen Fragen auf, derenBeantwortung schwierig ist und zumeist län-gerfristiger Beobachtung bedarf. Was hat es
mit diesen Pflanzenarten auf sich? Müssenuns Neophyten angst machen? Wie sollenwir mit dieser Entwicklung umgehen?
Um den Begriff Neubürger (Neophyt)näher zu definieren, bedarf es einer zeitlichenRelation. Neu in Bezug auf die Arten derursprünglichen Vegetation (Apophyten) wa-ren auch schon die bei uns im Zusammen-hang mit der Neolithischen Revolution vor7000 Jahren, dem Beginn der Ackerbaukul-tur, eingeschleppten Arten (Archäophyten).Zwangsläufig wird somit der Bezugspunktfür den Beginn der Einbürgerung der in jün-gerer und jüngster Zeit neu hinzugekomme-nen Arten (Neophyten) künstlichen Charak-ter tragen müssen. Die großen überseeischenEntdeckungen ab Ende des 15. Jahrhundertsund der sich daraus ergebende globale Han-delsverkehr hatten zur Folge, dass Ausbrei-tungsschranken für biologische Arten zwi-

52 WERNER DIETRICH
schen den Kontinenten entfielen. Somit er-schien es zweckmäßig, das Datum der Ent-deckung der Neuen Welt (1492) zum Bezugs-punkt für die Einstufung als Neophyt zubestimmen (DÜLL & KÜTZELNIGG 1994).
Das Verständnis für die Eigenart unsererpflanzlichen Neubürger muß von der Tatsa-che ausgehen, dass jede Darstellung einerVegetationsgeschichte auf dem Prinzip derVeränderlichkeit beruht. So gesehen, wäreninnerhalb der Vegetation ständig neue For-men, �Neophyten� im weiteren Sinne, zubeobachten. Diese natürliche Entwicklungvollzieht sich jedoch, weitgehend unbemerktfür den Zeitgenossen, schrittweise und in lan-gen Zeiträumen. Was jedoch für die Flora imAllgemeinen gilt, kann nicht ohne weiteresauf die Neophyten im engeren Sinne über-tragen werden. Angesichts einer regelrechtenfloristischen Explosion � einer Entwicklung,die alles Bisherige in den Schatten stellt undderen Verursacher allein der Mensch ist � stelltsich nicht nur die Frage, was jetzt neu ist,sondern, vor allem, was anders ist. Die zubeobachtende Entwicklung besitzt eine völ-lig andere Dimension und ist nicht vergleich-bar mit der in Jahrhunderten vor sich gegan-genen Einwanderung der Archäophyten invorgeschichtlicher Zeit. Der heutige sprung-hafte Zuwachs fremder Florenelemente er-klärt sich aus der unbegrenzten Mobilität desMenschen, welche alle natürlichen Schrankenaufhebt. Die Dynamik dieser Entwicklunglässt schon heute keinen Zweifel darüber auf-kommen, dass dadurch irreversible Faktengeschaffen werden.
Wenn sich auch die Folgen solcher gravie-renden Umweltveränderungen vorerst nichtabschätzen lassen, so erfordern sie bereitsjetzt besondere Aufmerksamkeit. Je ernsterdiese Entwicklung genommen wird, destobesser wird man in der Lage sein, ihr sinn-voll zu begegnen. Es bleibt im Einzelfallabzuwägen, ob Gegenmaßnahmen zu einerausufernden Vermehrung bestimmter Neo-phyten notwendig oder überhaupt möglichsind.
Wie nicht anders zu erwarten, besiedeln alleNeophyten gestörte oder auf irgendeine Wei-se anthropogen beeinflußte Standorte, weilhier die einheimische Vegetation von vorn-herein beeinträchtigt ist. Zum Beispiel bietendie Uferbereiche und ehemaligen Auen ent-lang des Rheins und seiner Nebenflüsse imbehandelten Gebiet vielen Neuankömmlin-gen ideale Existenzmöglichkeiten. DrüsigesSpringkraut (Impatiens glandulifera) aus demHimalaya, Riesenbärenklau oder Herkules-staude (Heracleum mantegazzianum) aus demKaukasus, Topinambur (Helianthus tuberosus)aus Nordamerika, Goldrute (Solidago canaden-sis und S. gigantea), ebenfalls aus Nordameri-ka, und Japanischer Staudenknöterich (Reyn-outria japonica) aus Ostasien gehören inzwi-schen zum bleibenden und beherrschendenInventar. Auch Häfen, Industriebrachen undBahnanlagen sind wahre Fundgruben für denan exotischen Arten Interessierten. Überalldort sind Neophyten zum festen Bestandinnerhalb einheimischer Pflanzengesellschaf-ten geworden. Viele dieser Neubürger sindallgemein bekannt, so dass der Nichtfach-mann kaum auf die Idee käme, anzuneh-men, dass es das als lästiges Unkraut bekann-te Franzosenkraut (Galinsoga ciliata und G.parviflora) aus dem andinen Südamerika nichtschon seit jeher gegeben hätte. Ebenso sinddie Strahlenlose Kamille (Matricaria discoidea)aus dem nordöstlichen Asien, der PersischeEhrenpreis (Veronica persica) aus dem Vorde-ren Orient und das Drüsige Weidenröschen(Epilobium ciliatum) aus Nordamerika bei unsseit Jahrzehnten so allgegenwärtig wie ein-heimische Pflanzen.
In der vorliegenden Arbeit lege ich einekommentierte Artenliste der Neophyten imRaum Düsseldorf/Neuss vor, nehme aller-dings auch solche Arten als Neophyten in dieseListe auf, die in unserem Raum vorerst nurgelegentlich auftauchen. Die Artenliste beruhtauf eigenen Beobachtungen, bezieht aberauch sämtliche für den untersuchten Raumim Florenatlas von Nordrhein-Westfalen(HAEUPLER et al. 2003) als nicht indigen auf-

Pflanzliche Neubürger (Neophyten) im Raum Düsseldorf/Neuss 53
Acta Biologica Benrodis 13 (2006)
geführten Arten mit ein, soweit sie ab 1980kartiert wurden.
2. Hinweise zum Aufbau und Gebrauchder Pflanzenliste
Das behandelte Gebiet umfasst folgende to-pographischen Karten (TK 1:25 000): 4605(Krefeld, Quadranten 2 und 4), 4606 (Kai-serswerth), 4705 (Willich, Quadranten 2 und4), 4706 (Düsseldorf), 4707 (Mettmann),4806 (Neuss) und 4807 (Hilden).
Die Daten und Informationen zu den Ar-ten in der folgenden Liste wurden einer Viel-zahl von Publikationen entnommen, die mitwenigen Ausnahmen alle im Besitz des Ver-fassers sind. Die deutschen und wissenschaft-lichen Namen richten sich grundsätzlich nachdem SCHMEIL (SCHMEIL & FITSCHEN 2000). Wodies nicht möglich war, wurden ROTHMALER
(2002) und ZANDER (2002) zu Rate gezogen.Die Angaben zur Verbreitung und Herkunft,zur Einbürgerung und Einbürgerungsge-schichte sowie Inkulturnahme und Morpho-logie stammen aus folgenden Quellen: BÄR-TELS (2001), DÜLL & KUTZELNIGG (1994), FRAN-KE (1997), GARCKE (1972), HAEUPLER et al.(2003), HANF (1991), HECKER (1995), HEGI
(1908-1931), KÖRBER-GROHNE (1988),KRAUSCH (2003), KRÜSSMANN (1972, 1976,1977, 1978), LOHMEYER & SUKOPP (1992),OBERDORFER (2001), ROTHMALER (2002),SCHMEIL& FITSCHEN (2000), ZANDER (2002).Auch ökologische und pflanzensoziologischeDaten wurden den genannten Werken ent-nommen. Die Zuerkennung des Neophyten-status folgt HAEUPLER et al. (2003), HANF
(1991) und LOHMEYER & SUKOPP (1992).Die Hinweise auf Fundorte im Gebiet be-
ruhen auf eigenen Beobachtungen oder sindHAEUPLER et al. (2003) entnommen. Von deninsgesamt 1173 im Gebiet kartierten Artenwird dort 226 Arten der Status als Neophytzuerkannt, was etwa ein Fünftel der Gesamt-artenzahl ausmacht. Viele Zufallsfunde wür-den sich ergeben, wenn man die Hafen- undIndustrieanlagen systematisch absuchen wür-
de, wie es für den Neusser Hafen durch STIEG-LITZ (1980, 1981) geschehen ist. In HAEUPLER
& MUER (2000) sind die meisten Arten mithervorragenden Farbphotos abgebildet.
Über die etymologische Deutung und Er-klärung der wissenschaftlichen Pflanzenna-men gibt GENAUST (1996) Auskunft. Die eng-lischen Volksnamen der Pflanzen wurdenSTACE (1997) entnommen.
In der folgenden Liste sind die Arten nachihren Herkunfts- oder Verbreitungsgebietenin acht Gruppen geordnet: Europa, Eurasi-en, Mittelmeergebiet, Asien, Nordamerika/nördliches Mittelamerika, Südamerika/süd-liches Mittelamerika, Afrika und Australien.Die Reihenfolge der Arten innerhalb derGruppen ist alphabetisch nach den wissen-schaftlichen Namen. Wegen der klimatischenÄhnlichkeit und geographischen Nähe stam-men die meisten Neophyten aus Nordame-rika, dem Mittelmeergebiet und Asien, dage-gen nur wenige aus Südamerika, Afrika undAustralien.
3. Kommentierte Pflanzenliste
3.1. Europa
1. Aconitum napellus, Blauer Eisenhut (Ranun-culaceae, Hahnenfußgewächse).
Heimat: Mitteleuropa bis Südalpen; ausdau-erndes Kraut; hochmontane bis subalpine Ge-birgswälder, Hochstaudenfluren; in Nordrhein-Westfalen nicht einheimisch; aus Gartenabfäl-len gelegentlich verwildert; in Gärten auch an-dere Arten der Gattung und Hybriden, die eben-falls verwildern können; tödlich giftig in allenTeilen; hieß in früheren Zeiten aus naheliegen-den Gründen auch �Altsitzerkraut� (BECK-MANN & BECKMANN 1990), ebenso �Vegetabili-sches Arsenik� (HILLER & BICKERICH 1997); infrüheren Zeiten auch Pfeilgift (BECKMANN &BECKMANN 1990). Im Gebiet: z.B. HeltorferMark bei Angermund.2. Amaranthus bouchonii, Bouchons Fuchs-schwanz (Amaranthaceae, Fuchsschwanzge-wächse; Abb. 1).

54 WERNER DIETRICH
Heimat: vermutlich in Europa aus A. hyb-ridus agg. (S-Amerika) entstanden (SCHMITZ
2002); einjähriges Kraut in Maisäckern, aufRuderalstellen; Erstnachweis für Mitteleuro-pa 1925; am Niederrhein seit 1970er Jahre(SCHMITZ 2002). Im Gebiet: z. B. Himmel-geister Rheinbogen, auf dem Grind bei Dor-magen-Stürzelberg.3. Anchusa sempervirens, Ausdauernde Ochsen-zunge (Boraginaceae, Rauhblattgewächse).
Heimat: SW-Europa; ausdauerndes Kraut;Zierpflanze; im Gebiet: an der Erft bei HausEppinghoven (Neuss, TK 4806-1); fest einge-bürgert im Schlosspark Dyck bei Jüchen.4. Aquilegia vulgaris, Gewöhnliche Akelei (Ra-nunculaceae, Hahnenfußgewächse).
Heimat: Europa, Kanaren, Madeira, N-Afrika; ausdauerndes Kraut in sommerwar-men, lichten Laubmischwäldern, Gebüschen,Hecken, Wiesen, Halbtrockenrasen; Zier-pflanze und Gartenflüchtling; in Nordrhein-Westfalen nicht einheimisch; in der christli-chen Kunst oft auf Mariendarstellungen. ImGebiet: TK 4807-3.5. Armoracia rusticana, Meerrettich, Kren (Bras-sicaceae, Kreuzblütler).
Heimat: wahrscheinlich S-Rußland undöstliche Ukraine; ausdauerndes Kraut; keinSamenansatz, aber starke vegetative Vermeh-rung durch Wurzelsprosse; Neophyt seit 1594;seit alters Gewürz- und Heilpflanze (FURLEN-MEIER 1978). Der Dichter ADELBERT VON CHA-MISSO (1781-1838), Verfasser der Novelle �Pe-ter Schlemihls wundersame Geschichte� undKustos des Botanischen Gartens Berlin,schreibt 1827: �Das Kraut ist ein gutes milch-vermehrendes Futter für die Kühe.� Im Ge-biet: nicht selten.6. Aster x salignus, (= Aster lanceolatus x A.novi-belgii), Weidenblättrige Aster (Asteraceae,Korbblütler).
Heimat: Elternarten aus Nordamerika stam-mend, Bastard 18. Jahrhundert in Europa inKultur entstanden (ROTHMALER 2002); aus-dauerndes Kraut. Im Gebiet: TK 4705-4.7. Aubrieta deltoidea, Griechisches Blaukissen(Brassicaceae, Kreuzblütler).
Heimat: Sizilien, Balkan, Türkei; ausdau-erndes Kraut; Zierpflanze; entdeckt 1700 vondem französischen Botaniker Joseph Pittonde Tournefort auf Kreta; 1808 im Botani-schen Garten Berlin. Im Gebiet: in Fugen derRheinufermauer in Kaiserswerth.8. Barbarea intermedia, Mittleres Barbarakraut(Brassicaceae, Kreuzblütler).
Heimat: Europa (ROTHMALER 2002) oderW-Europa (GARCKE 1972); zweijährigesKraut auf Ruderalstellen, an Wegrändern undUfern, in Gärten und auf Äckern; Neophytseit 1865. Im Gebiet: TK 4706-3 .9. Cardaminopsis arenosa, Sand-Schaumkresse(Brassicaceae, Kreuzblütler).
Heimat: Mitteleuropa; ausdauerndes Krautauf sandigen und steinig-kiesigen Flächen;in Nordrhein-Westfalen nicht einheimisch;Neophyt im Nordwesten seit 1890. Im Ge-biet: TK 4606-2.10. Carex brizoides, Zittergras-Segge (Cypera-ceae, Sauergräser, Riedgräser).
Heimat: Europa mit Ausnahme der Briti-schen Inseln und Skandinavien; ausdauerndesKraut in staufeuchten Laubmischwäldern, anWaldwegen, auf waldnahen Feucht- und Naß-wiesen; in Nordrhein-Westfalen nicht einhei-misch; früher Streupflanze und als �Waldhaar�zum Polstern verwendet (ROTHMALER 2002).Im Gebiet: auf der Jücht im HimmelgeisterRheinbogen, Heltorfer Mark bei Angermund.11. Cochlearia danica, Dänisches Löffelkraut(Brassicaceae, Kreuzblütler).
Heimat: Küsten Nord- und Mitteleuro-pas; zweijähriges Kraut auf Salzwiesen derNord- und Ostseeküste; seit Anfang derNeunzigerjahre des vorigen Jahrhundertsauf den Mittelstreifen der Autobahnen desFlachlandes in Ausbreitung. Im Gebiet: TK4707-1, 4807-2.12. Conopodium majus, Französische Erdkas-tanie (Apiaceae, Doldenblütler).
Heimat: Atlantisches Europa bis Italien undKorsika; ausdauerndes Kraut in extensiv ge-nutzten Wiesen und Parkanlagen; die erbsen-bis haselnussgroßen Wurzelknollen sind ess-bar (HEGI 1926). Im Gebiet: TK 4606-4.

Pflanzliche Neubürger (Neophyten) im Raum Düsseldorf/Neuss 55
Acta Biologica Benrodis 13 (2006)
13. Corispermum leptopterum, SchmalflügeligerWanzensame (Chenopodiaceae, Gänsefußge-wächse).
Heimat: Mitteleuropa bis Ungarn; einjäh-riges Kraut auf Sandflächen; in Nordrhein-Westfalen nicht einheimisch; in Mitteleuropaaus dem Formenkreis der osteuropäischenC. hyssopifolium entstanden (SCHMITZ 2002);in Deutschland seit 1849; am Niederrhein seitden 1920er-Jahren (SCHMITZ 2002); wissen-schaftlicher Name: gr. koris = Bettwanze; dieflachen Früchte ähneln Wanzen. Im Gebiet:Himmelgeister Rheinbogen und auf demGrind bei Dormagen-Stürzelberg.14. Corydalis lutea, Gelber Lerchensporn (Fu-mariaceae, Erdrauchgewächse).
Heimat: Süd- und Zentralalpen; ausdau-erndes Kraut in Mauerspalten, Gärten undHecken; alte Zierpflanze seit dem Mittelalter;von GESSNER (1561) bereits als Gartenpflan-ze verzeichnet; erst seit dem 17. Jahrhundertallgemein in Gärten; wissenschaftlicher Name:gr. korys = Helm; die gespornte Blüte ähneltdem Kopf einer Haubenlerche. Im Gebiet:Kaiserpfalz in Kaiserswerth.15. Crepis taraxacifolia, Blasen-Pippau (Aster-aceae, Korbblütler).
Heimat: Europa, (auch NW-Afrika); zwei-jähriges Kraut in Wiesen und auf Ruderal-stellen; in Nordrhein-Westfalen nicht einhei-misch; Archäophyt in Baden-Württemberg,Rheinland-Pfalz, Südhessen. Im Gebiet: TK4606-3, 4807-3.16. Echinops exaltatus, Drüsenlose Kugeldis-tel (Asteraceae, Korbblütler).
Heimat: O- und SO-Europa; ausdauern-des Kraut; aus Gärten verwildert, auf Rude-ralstellen; Neophyt seit dem 19. Jahrhundert;Trachtpflanze für Bienen (ROTHMALER 2002).Im Gebiet: TK 4606-3.17. Erucastrum gallicum, Französische Hunds-rauke (Brassicaceae, Kreuzblütler).
Heimat: Europa; ein- bis zweijährigesKraut auf Ruderalstellen und nährstoffrei-chen Äckern; nicht einheimisch in Nordrhein-Westfalen, aber Ärchäophyt im Norden undOsten; wissenschaftlicher Name: lat. eruca =
Senfrauke (�Rucola�), die schon bei den Rö-mern ein beliebtes Küchengewürz war, aberauch Heilpflanze und potenzstärkendes Mit-tel, lat. �astrum� bedeutet ein vergröberndesSuffix. Über eine Nutzung von Erucastrumgallicum wird nirgends berichtet. Im Gebiet:TK 4606-1.18. Euphorbia x pseudovirgata (= E. esula x E.virgata), Scheinruten-Wolfsmilch (Euphorbi-aceae, Wolfsmilchgewächse).
Heimat: Europa; mehrjähriges Kraut auftrockenen Ruderalstellen. E. virgata kommtin Nordrhein-Westfalen nicht vor. E. x pseu-dovirgata ist im gemeinsamen Verbreitungs-gebiet der beiden Elternarten entstanden undvon dort verschleppt worden. Im Gebiet: TK4706-3.19. Festuca brevipila, Rauhblatt-Schafschwingel(Poaceae, Süßgräser).
Heimat: Frankreich und Mitteleuropa bisSkandinavien, O-Europa; ausdauerndes Grasin Trockenrasen; in Nordrhein-Westfalen nichteinheimisch; zur Saatgutgewinnung kultiviert;an Straßenböschungen und Dämmen ange-sät und aus den Ansaaten verwildert. Im Ge-biet: TK 4605-2, 4, 4606-1, 2, 3.20. Galanthus nivalis, Schneeglöckchen (Ama-ryllidaceae, Narzissengewächse).
Heimat: Europa; ausdauernde Zwiebel-pflanze in Wäldern und Gebüschen; Zierpflan-ze; Neophyt seit dem spätem 16. und frühen17. Jahrhundert. Alle Vorkommen nördlichder Alpen werden als nicht ursprünglich ange-sehen. Die in der ersten Hälfte des 16. Jahr-hunderts schreibenden Botaniker Brunfels,Bock und Fuchs, kannten das Schneeglöckchennoch nicht. Wissenschaftlicher Name: gr. gala= Milch, anthos = Blüte, lat. nivalis = schnee-weiß. Im Gebiet: nicht selten.21. Galeobdolon argentatum, Silberblatt-Gold-nessel (Lamiaceae, Lippenblütler).
Heimat: vermutlich in Kultur entstanden;ausdauerndes Kraut; Zierpflanze, oftmals inortsnahen Wäldern, Gebüschen und Anla-gen verwildert; bekannt seit 1873. Im Gebiet:z. B. Heltorfer Mark bei Angermund, EllerForst.

56 WERNER DIETRICH
22. Geranium pyrenaicum, Pyrenäen-Storchschna-bel (Geraniaceae, Storchschnabelgewächse).
Heimat: Sehr unterschiedliche Angaben:SO-Europa (SCHMEIL & FITSCHEN 2000), Mit-telmeergebiet bis N-Iran, W-Afrika (ZANDER
2002), Pyrenäen, Südalpen, Gebirge des Mit-telmeergebietes, Kaukasus (DÜLL & KUTZEL-NIGG 1994); einjähriges bis kurz ausdauern-des Kraut in Gebüschen und Gärten, aufWeiden und Äckern; Neophyt seit 1800. ImGebiet: häufig.23. Hieracium aurantiacum, Orangerotes Ha-bichtskraut (Asteraceae, Korbblütler).
Heimat: Europa (außer atlantisches Euro-pa); mehrjähriges Kraut in Bergwiesen, aufWeiden und in Silikat-Magerrasen; Zierpflan-ze, aus Gärten verwildert in Parkrasen undauf Ruderalstellen; in Kultur seit dem 16.Jahrhundert. Karl Förster, Altmeister der Stau-dengärtnerei, schreibt: �wirft überreichlich mitSämlingen um sich und wuchert unterirdischin schlimmster, nicht auszurottender Weise.�(KRAUSCH 2003). Im Gebiet: TK 4707-2,4807-1, 2, 4.24. Hyacinthoides hispanica, Spanisches Hasen-glöckchen (Liliaceae, Liliengewächse).
Heimat: SW-Europa (auch NW-Afrika);ausdauernde Zwiebelpflanze; Zierpflanze;1601 erstmalig abgebildet von Clusius; inDeutschland erstmals 1613 nachweislich inKultur. Im Gebiet: TK 4807-1, 2. (Wegenschwieriger Unterscheidbarkeit könnte es sichhierbei auch um H. non-scripta handeln.)25. Hyacinthoides non-scripta (= Scilla non-scrip-ta), Gemeines Hasenglöckchen (Liliaceae, Li-liengewächse).
Heimat: W-Europa; ausdauernde Zwiebel-pflanze in Laubwäldern und schattigen Hai-nen; in Kultur seit dem 16. Jahrhundert; inNordrhein-Westfalen ursprünglich nur imRaum Erkelenz, sonst verwildert; bastardiertmit der vorigen Art. Im Gebiet: selten.26. Iberis umbellata, Dolden-Schleifenblume(Brassicaceae, Kreuzblütler).
Heimat: S- und SO-Europa; einjährigesKraut; Gartenzierpflanze; um 1560 in Mittel-europa noch ziemlich selten; im 17. Jahrhun-
dert häufige Gartenblume; im Ruhrgebiet aufsonnenexponierten Hängen von Kohleab-raumhalden eingebürgert. Im Gebiet: aufdem Grind (TK 4806-2, 4807-1; 2.9.2004).27. Laburnum anagyroides, Gewöhnlicher Gold-regen (Fabaceae, Schmetterlingsblütler).
Heimat: S- und SO-Europa; bis 7 m ho-her Zierstrauch; auch in Eichen- und Kiefern-wäldern, an Waldrändern und in Gebüschen,auf Ruderalstellen; Neophyt seit 16. Jahrhun-dert; tödlich giftig in allen Pflanzenteilen; le-tale Dosis bei Kindern 3-4 Früchte oder 15-20 Samen (ALTMANN 1995); früher als Brech-und Abführmittel (GESSNER 1974). DerStrauch sollte keinesfalls in Kindereinrichtun-gen und auf Spielplätzen stehen. Holz schwer,hart und sehr gut polierfähig, verwendbar fürfeine Drechslerarbeiten, Messstäbe und Mu-sikinstrumente, früher Armbrustbogen (HE-CKER 1995). Nach HAEUPLER et al. (2003) han-delt es sich bei Verwilderungen überwiegendum Pflanzen, die als L. x watereri (= L. alpi-num x L. anagyroides, FITSCHEN 2002) in Gar-tencentern verkauft werden. Im Gebiet: TK4606-3, 4706-3, 4, 4806-3, 4807-3.28. Larix decidua, Europäische Lärche (Pin-aceae, Kieferngewächse).
Heimat: W- bis O-Europa; 35 � 40 (50) mhoher Baum; ursprünglich nur in den Alpen,sonst angepflanzt; Eiszeiteinwanderer aus Si-birien. CHAMISSO (1827): �Die Lärche liefertbesseres Harz als Kiefer, Fichte und Weißtan-ne. Der Venetianische Terpentin ist das Er-zeugnis dieses Baumes.� Im Gebiet: zerstreut.29. Lathyrus latifolius, Breitblättrige Platterb-se (Fabaceae, Schmetterlingsblütler).
Heimat: S-Europa, Mittelmeergebiet; aus-dauerndes Kraut, unbeständig verwildert inGebüsch- und Heckensäumen, auf Ruderal-stellen; in der zweiten Hälfte des 16. Jahr-hunderts als Zierpflanze nach Mitteleuropa;im Kräuterbuch von MATTHIOLUS & CAME-RARIUS (1586); im Hortus Eystettensis vonBESLER (1613). Im Gebiet: z. B. auf demCampus der Universität Düsseldorf.30. Lepidium graminifolium, Grasblättrige Kres-se (Brassicaceae, Kreuzblütler).

Pflanzliche Neubürger (Neophyten) im Raum Düsseldorf/Neuss 57
Acta Biologica Benrodis 13 (2006)
Heimat: Europa (außer Britische Inseln undSizilien), (auch N-Afrika und W-Türkei); zwei-jähriges Kraut an Weg- und Ackerrändern, aufBahngelände; in Nordrhein-Westfalen nichteinheimisch. Im Gebiet: TK 4605-2.31. Linaria repens, Gestreiftes Leinkraut (Scro-phulariaceae, Rachenblütler).
Heimat: W-Europa bis Oberitalien und N-Balkan; ausdauerndes Kraut; auf Bahngelän-de, an Straßen-, Wald- und Feldwegrändern,an Böschungen; in Nordrhein-Westfalen nichtursprünglich. Im Gebiet: TK 4807-2, 4.32. Lunaria annua, Judas-Silberblatt (Brassi-caceae, Kreuzblütler).
Heimat: SO-Europa; einjähriges bis kurzausdauerndes Kraut; Zierpflanze in Bauern-gärten; auch häufig verwildert auf Ruderal-stellen; in Kultur seit dem 16. Jahrhundert;Verwilderungen ab dem 19. Jahrhundert be-obachtet. Im Gebiet: nicht selten.33. Medicago x varia (= M. falcata x M. sativa),Bastard-Luzerne (Fabaceae, Schmetterlings-blütler).
Heimat: Europa; ausdauerndes Kraut;häufig kultiviert und verwildert; in Halbtro-ckenrasen, Wiesen, Gebüschsäumen und aufRuderalstellen; Futterpflanze. Im Gebiet:häufig.34. Mentha spicata, Grüne Minze (Lamiaceae,Lippenblütler).
Heimat: in Europa in Kultur entstanden;nicht einheimisch in Nordrhein-Westfalen; aus-dauerndes Kraut; verwildert an Gräben, Ufern,auf Äckern und Ruderalstellen. M. spicata isteine der Elternarten der Pfefferminze (M. x pipe-rita), die als Tripelbastard aus der Wasserminze(M. aquatica), der Roßminze (M. longifolia) undder genannten entstanden ist (FRANKE 1997).Die Krauseminze genannte Varietät crispata derGrünen Minze wird zuweilen in Gärten gezo-gen und war im 16. Jahrhundert sehr volks-tümlich als Gewürz- und Heilpflanze (HEGI
1927). Im Gebiet: häufig.35. Mentha x villosa, Zottige Minze (Lamiaceae,Lippenblütler).
Heimat: wahrscheinlich in Europa in Kul-tur entstanden; ausdauerndes Kraut; Eltern-
arten: Grüne Minze (M. spicata) und Rund-blättrige Minze (M. suaveolens); in Nordrhein-Westfalen nicht einheimisch; in der Varietätalopecuroides verwildert und eingebürgert aufnährstoffreichen Ruderalstellen; verbreitetsich nur vegetativ, da steril; wird heute alsTeepflanze häufiger gezogen als die Pfeffer-minze (M. x piperita; HAEUPLER et al. 2003).Im Gebiet: zerstreut.36. Narcissus pseudonarcissus, Gelbe Narzisse,Osterglocke (Amaryllidaceae, Narzissenge-wächse).
Heimat: W- und SW-Europa, westlichesMitteleuropa; ausdauernde Zwiebelpflanze;indigene Vorkommen nur aus der Rur-Eifelund dem deutsch-belgischen Grenzgebietbekannt; sonst Zierpflanze; viele Sorten undHybriden, welche auch verwildern; Zwiebelnfrüher als Brechmittel genutzt (GESSNER 1974).Bei Verzehr größerer Mengen, z. B. Verwechs-lung mit Speisezwiebeln (ALTMANN 1995),sind Kollaps und Lähmungserscheinungenmit tödlichem Ausgang möglich (HILLER &BICKERICH 1997). Im Gebiet: zerstreut.37. Oenothera x fallax, Täuschende Nachtkerze(Onagraceae, Nachtkerzengewächse; Abb. 2).
Heimat: in Europa als konstanter Bastardentstanden; Elternarten: die aus N-Amerikastammende Gewöhnliche Nachtkerze (O. bi-ennis) und die ebenso wie die TäuschendeNachtkerze in Europa entstandene Rotkel-chige Nachtkerze (O. glazioviana) (DIETRICH etal. 1997); zweijähriges Kraut. Im Gebiet: häu-fig entlang des Rheins.38. Oenothera glazioviana (= O. erythrosepala),Rotkelchige Nachtkerze (Onagraceae, Nacht-kerzengewächse; Abb. 3).
Heimat: in Europa (wahrscheinlich Eng-land) im 19. Jahrhundert durch Bastardierungvon aus N-Amerika stammenden Arten ent-standen; Elternarten: Gewöhnliche Nachtker-ze (O. biennis) und Großblütige Nachtkerze (O.grandiflora) (DIETRICH et al. 1997); zweijährigesKraut; wegen ihrer Schönheit sehr beliebteGartenpflanze. Im Gebiet: z. B. an der A 57zwischen Düsseldorf und Hildener Kreuz,auf dem Grind bei Dormagen-Stürzelberg.

58 WERNER DIETRICH
39. Onobrychis viciifolia, Futter-Esparsette (Fa-baceae, Schmetterlingsblütler).
Heimat: wahrscheinlich Europa; nach HA-EUPLER et al. (2003) in Nordrhein-Westfalennicht einheimisch; ausdauerndes Kraut; Kul-turpflanze; verwildert in halbtrockenen Ra-sen, trockenen Wiesen und auf Ruderalstel-len; an Straßenböschungen und Dämmen oftangesät. Im Gebiet: zerstreut.40. Picea abies, Fichte, Rottanne (Pinaceae, Kie-ferngewächse).
Heimat: in Europa ursprünglich nur in dersubalpinen Region der Gebirge; 30-50 mhoher Baum; durch Forstkultur weit verbrei-tet und sich durch Selbstaussaat verjüngend;CHAMISSO (1827) berichtet: �Die jungen Trie-be dieser und ähnlicher Arten werden vor-zugsweise von Seefahrern angewandt, umeine Art Bier zu brauen, welches zu den vor-züglichsten antiskorbutischen Mitteln gerech-net wird.� Im Gebiet: häufig.41. Populus x canadensis, Kanadische Pappel(Salicaceae, Weidengewächse).
Heimat: 1750 spontan in Frankreich ent-standen als Bastard aus der KanadischenSchwarzpappel (P. deltoides) und der einhei-mischen Schwarzpappel (P. nigra) (KRÜSS-MANN 1977); bis 30 m hoher Forst- und Zier-baum; verdrängt die Schwarzpappel wegenihrer Schnellwüchsigkeit; wird in der Zellulo-seindustrie genutzt (DÜLL & KUTZELNIGG
1994). Im Gebiet: häufig.42. Populus x canescens, Grau-Pappel (Salicaceae,Weidengewächse).
Heimat: Europa; 15-30 m hoher Baum inAuwäldern; auch Forst- und Parkbaum; na-türlicher Bastard aus der Silberpappel (P. alba)und der Zitterpappel, auch Espe genannt ( P.tremula); nach HAEUPLER et al. (2003) in Nord-rhein-Westfalen nicht einheimisch. Im Ge-biet: TK 4707-4, 4806-2, 4807-3.43. Potentilla intermedia, Mittleres Fingerkraut(Rosaceae, Rosengewächse).
Heimat: O-Europa; zweijähriges bis kurz aus-dauerndes Kraut auf trockenen und sandigenbis kiesigen Ruderalstellen; Neophyt seit 1825.Im Gebiet: TK 4605-4, 4606-3, 4705-2, 4807-4.
44. Reynoutria x bohemica (= Fallopia x bohemi-ca), Bastard-Staudenknöterich (Polygonaceae,Knöterichgewächse).
Heimat: in Europa entstandener sponta-ner Bastard; Elternarten: Japanischer Stauden-knöterich (R. japonica) und Sachalin-Stauden-knöterich (R. sachalinensis); ausdauerndesKraut; unabhängig von den Eltern an Fluß-ufern, Waldrändern und auf Ruderalstellenoder nur mit dem Japanischen Staudenknö-terich vergesellschaftet. Im Gebiet: auf demGrind (TK 4806-2; 2.9.2004).45. Rubus laciniatus, Schlitzblättrige Brombee-re (Rosaceae, Rosengewächse).
Heimat: wahrscheinlich nördliches gemä-ßigtes Europa; Strauch mit 3-5 m langenTrieben; Kulturpflanze, bekannt seit Mittedes 18. Jahrhunderts (meist als stachelloseChimäre); stachelige Form auf ärmeren san-digen Böden; häufig in Dünenanpflanzun-gen der Nordseeinseln; sehr ertragreichesBeerenobst; in Nordrhein-Westfalen vor al-lem in städtischen Bereichen angepflanzt,verwildert und zum Teil eingebürgert. ImGebiet: zerstreut.46. Rubus sciocharis, Schattenliebende Brom-beere (Rosaceae, Rosengewächse).
Heimat: W-Europa; Strauch mit 3-5 m lan-gen Trieben auf nährstoffreichen, kalkreichenBöden; aus holsteinischen Baumschulen ver-schleppt (ROTHMALER 2002). Im Gebiet: TK4707-4.47. Sempervivum tectorum, Echte Hauswurz(Crassulaceae, Dickblattgewächse).
Heimat: Gebirge SW-,W- und Mitteleuro-pas; sonst angepflanzt und verwildert; aus-dauerndes sukkulentes Kraut; früher wichti-ge Volksheil- und Zauberpflanze, z. B. gegenBlitz, deshalb oft auf Dächern angepflanzt;Karl d. Große (742-814) befahl im �Capitu-lare de villis� die Anpflanzung des �Jupiter-bart� auf den Dächern. CHAMISSO (1827):�Man ließ auch wider Ohrensausen und Taub-heit den Saft ins Ohr träufeln. Es mag man-chem um sein Gehör besorgten zum Trostgereicht haben, dass dort etwas zu seinerHeilung vorgenommen ward, und, mehr als

Pflanzliche Neubürger (Neophyten) im Raum Düsseldorf/Neuss 59
Acta Biologica Benrodis 13 (2006)
diesen Trost zu geben, vermag in vielem diewissenschaftliche Heilkunde nicht.� Im Ge-biet: TK 4807-3.48. Senecio vernalis, Frühlings-Greiskraut (As-teraceae, Korbblütler).
Heimat: aus O-Europa eingewandert undvielerorts zur �Landplage� geworden (RAUH
& HANF 1983); einjähriges Kraut auf mäßigtrockenen Sand- und Lehmböden sowie Ru-deralstellen (z. B. an Bahnanlagen); Neophytseit 1850; schädlich für das Vieh, für Pferdesogar als giftig geltend (CREMER et al. 1991).Im Gebiet: TK 4605-4, 4606-2.49. Sisymbrium volgense, Wolga-Rauke (Brassi-caceae, Kreuzblütler).
Heimat: mit Getreide aus SO-Rußland ein-geschleppt und eingebürgert; ausdauerndesKraut auf sandig-kiesigen Plätzen; Neophyt seit1880. Im Gebiet: TK 4605-4, 4606-3, 4705-2.50. Sorbus intermedia, Schwedische Eberesche(Rosaceae, Rosengewächse).
Heimat: Skandinavien, N-Deutschland,Polen, Baltikum; 5-15 m hoher, häufiger Stra-ßenbaum in Städten; seit langem in Kultur;aus Anpflanzungen verwildert. Im Gebiet:TK 4607-3, 4807-3.51. Spiraea x billardii, Billards Spierstrauch (Ro-saceae, Rosengewächse).
Heimat: in Frankreich entstanden. �Vor1854 von Billard in Fontenay-aux-Roses er-zielt. Wird leicht wüst und verwildert�(KRÜSSMANN 1978). Elternarten: Oregon-Spierstrauch (S. douglasii) aus dem westli-chen N-Amerika und Weiden-Spierstrauch(S. salicifolia) mit eurasischer Verbreitung;bis 1,5 (-2) m hoher Strauch. Im Gebiet:TK 4807-2.52. Symphoricarpos x chenaultii (= S. microphyl-lus x S. orbiculatus), Bastard-Korallenbeere(Caprifoliaceae, Geißblattgewächse).
Heimat: um 1910 von R. Chenault in Or-leans (Frankreich) erzielt. Die Heimat der ers-ten Elternart (S. microphyllus) ist Mexiko, dieder zweiten Art (S. orbiculatus) östliche USAund Mexiko. 1,5-2 m hoher und mindestensebenso breiter, stark wachsender, reich undlocker verzweigter Zierstrauch (BÄRTELS 2001,
KRÜSSMANN 1978). Im Gebiet: auf dem Grind(TK 4807-133114; 2.9.2004).53. Symphytum x uplandicum, Futter-Beinwell,Comfrey (Boraginaceae, Rauhblattgewächse).
Heimat: Bastard aus dem Gemeinen Bein-well (S. officinale) mit eurasischer Verbreitungund dem Rauhen Beinwell (S. asperum) ausdem Kaukasus (letzterer zu Beginn des 19.Jahrhunderts eingeführt); ausdauerndesKraut; Kulturpflanze; auf feuchten Ruderal-stellen und an Ufern verwildert. Sowohl derComfrey als auch der Rauhe Beinwell werdenals Futterpflanze (Schweinefutter) angebaut(DÜLL & KUTZELNIGG 1994). Im Gebiet: TK4806-1, 4807-4.54. Ulex europaeus, Stechginster (Fabaceae,Schmetterlingsblütler).
Heimat: W-Europa; sonst angpflanzt undverwildert; bis 2 m hoher Strauch auf Hal-den und Waldschlägen, an Waldrändern undBöschungen; bedeutende Pionierpflanze derBrachen und Heiden; in ozeanisch gemäßig-ten Zonen heute weltweit. Die erfolgreicheBekämpfung der Kaninchen in Englanddurch Myxomatose führte zu einem unge-hemmten Wachstum des Stechginsters, wo-durch weite Gebiete unbegehbar wurden(DÜLL & KUTZELNIGG 1994). Neophyt seitdem 18. Jahrhundert, jedoch nicht bestän-dig, da zu frostempfindlich. Im Gebiet: Mit-telstreifen der Münchner Straße in Düssel-dorf (ob noch?).55. Vicia villosa, Zottige Wicke (Fabaceae,Schmetterlingsblütler).
Heimat: O- und SO-Europa; einjährigesKraut auf Äckern, Wiesen und Ruderalstel-len; kultiviert als Futterpflanze und zur Stick-stoffanreicherung in Rekultivierungsgebieten;eingeführt und eingebürgert seit dem 19. Jahr-hundert, z. B. 1857 in Magdeburg. Im Ge-biet: TK 4606-3, 4707-4, 4806-1, 3, 4807-4.
3.2. Eurasien
56. Abutilon theophrasti, Samtpappel, GelbeSchönmalve (Malvaceae, Malvengewächse;Abb. 4).

60 WERNER DIETRICH
Heimat: östliches Mittelmeergebiet bis O-Asien; einjähriges Kraut; unbeständig beson-ders auf Müllkippen, in Häfen mit Öl- undGetreidemühlen sowie Vogelfutterstellen;erste Hälfte 16. Jh. in Deutschland eingeführt;1542 in Nürnberg; im 19. Jahrhundert häufi-ge Gartenpflanze. Im Gebiet: im Uferbereichdes Himmelgeister Rheinbogens.57. Aesculus hippocastanum, Roßkastanie (Hip-pocastanaceae, Roßkastaniengewächse).
Heimat: Balkan bis Himalaya; in Berg- undSchluchtwäldern; bis 25 m hoher Baum; ers-te Kunde in Mitteleuropa ein gemalter Frucht-zweig bei MATTHIOLUS (1565); hundert Jahrespäter in Leipzig angepflanzt; danach rascheAusbreitung; ab Ende 18. Jh. vielfältige Ver-wendung der öl- und stärkehaltigen Samen;später auch Verwendung in der Medizin. ImGebiet: häufig.58. Alchemilla mollis, Weicher Frauenmantel(Rosaceae, Rosengewächse).
Heimat: O-Europa bis Iran; ausdauerndesKraut; Zierpflanze; alte Bauerngartenpflan-ze; auch in Parks und auf Friedhöfen; gele-gentlich verwildert. Im Gebiet: TK 4706-3.59. Alnus incana, Grau-Erle (Betulaceae, Bir-kengewächse).Heimat: Europa, Kaukasus, W-Sibirien; 10(-25) m hoher Baum; in Nordrhein-Westfa-len nicht ursprünglich, nur angepflanzt alsVorholz zur Bodenbefestigung; verwildert inAuwäldern und an Flußufern. Im Gebiet:häufig.60. Amaranthus blitum, Aufsteigender Fuchs-schwanz (Amaranthaceae, Fuchsschwanzge-wächse; Abb. 5).
Heimat: Europa, Asien (auch Afrika); nachSCHMITZ (2002) S-Amerika, Madeira, Tenerif-fa, Ascension, Java; einjähriges Kraut aufRuderalstellen, in Gärten und auf Äckern;Erstnachweis für Mitteleuropa 1889; am Nie-derrhein seit den 1980er Jahren (SCHMITZ
2002); wird von HAEUPLER et al. (2003) fürNordrhein-Westfalen als Neophyt gewertet.Im Gebiet: TK 4806-2.61. Angelica archangelica, Echte Engelwurz,Garten-Engelwurz (Apiaceae, Doldenblütler).
Heimat: Europa, Kaukasus, W-Sibirien;ausdauerndes Kraut; in Nordrhein-Westfa-len nur die verwilderte Gartenform; alte Heil-und Gemüsepflanze. Im Gebiet: Himmel-geister Rheinbogen.62. Anthriscus caucalis, Hundskerbel (Apiaceae,Doldenblütler).
Heimat: Europa, W-Asien, Kaukasus (auchN-Afrika); einjähriges Kraut auf Ruderalstel-len, an Hecken; von LOHMEYER & SUKOPP
(1992), ROTHMALER (2002) und WILLERDING
(1986) für ganz Deutschland als Archäophytangesehen; von HAEUPLER et al. (2003) fürNordrhein-Westfalen als Neophyt gewertet.Im Gebiet: TK 4605-4, 4706-1, 3.63. Apera interrupta, Unterbrochener Wind-halm (Poaceae, Süßgräser).
Heimat: W-Europa bis Zentralasien (auchNW-Afrika); einjähriges Gras auf sandigenÄckern und Ruderalstellen; nach HAEUPLER
et al. (2003) auf Bahngelände und Industrie-brachen im Ruhrgebiet eingebürgert. Im Ge-biet: TK 4606-3, 4706-3.64. Asparagus officinalis, Gemüse-Spargel (Li-liaceae, Liliengewächse).
Heimat: Europa, Kaukasus, W-Sibirien; aus-dauerndes Kraut; Kulturpflanze; ab dem 16.Jahrhundert in Mitteleuropa angebaut; verwil-dert in Sandgebieten und der Uferregion desRheins. Im Gebiet: z. B. Himmelgeister Rhein-bogen, Grind bei Dormagen-Stürzelberg.65. Atriplex hortensis, Garten-Melde (Cheno-podiaceae, Gänsefußgewächse).
Heimat: Europa bis Zentralasien; einjähri-ges Kraut; alte Kulturpflanze. Die Garten-Melde wird in Wittgensteiner und Siegerlän-der Bauerngärten nach wie vor als Spinater-satz kultiviert (HAEUPLER et al. 2003). Auchverwildert auf dörflichen Ruderalstellen. ImGebiet: TK 4606-2, 4707-2, 4807-2.66. Atriplex oblongifolia, Langblättrige Melde(Chenopodiaceae, Gänsefußgewächse).
Heimat: östliches Mitteleuropa bis Zentral-asien; einjähriges Kraut; unbeständig verwildertauf Ruderalstellen. Im Gebiet: TK 4706-3.67. Berteroa incana, Graukresse (Brassicaceae,Kreuzblütler).

Pflanzliche Neubürger (Neophyten) im Raum Düsseldorf/Neuss 61
Acta Biologica Benrodis 13 (2006)
Abb. 1: Bouchons Fuchsschwanz (Amaranthusbouchonii).Fig. 1: Indehiscent Amaranth (Amaranthus bou-chonii).
Abb. 2: Täuschende Nachtkerze (Oenothera x fal-lax).Fig. 2: Intermediate Evening-primrose (Oenothe-ra x fallax).
Abb. 3: Rotkelchige Nachtkerze (Oenothera gla-zioviana).Fig. 3: Large-flowered Evening-primrose (Oeno-thera glazioviana).
Abb. 4: Samtpappel (Abutilon theophrasti).Fig. 4: Velvetleaf (Abutilon theophrasti).

62 WERNER DIETRICH
Heimat: O-Europa bis Zentralasien; ein-bis zweijähriges Kraut auf trockenen, sonni-gen Standorten und Sandfeldern; Erstfundein der Nähe von Exerzierplätzen; wahrschein-lich mit Pferdefutter aus dem Osten einge-schleppt; erster Nachweis 1769 aus Neuburgbei Wien. Im Gebiet: z. B. auf dem Mittel-streifen des Südrings in Düsseldorf, Him-melgeister Rheinbogen, auf dem Grind.68. Brassica nigra, Senf-Kohl, Schwarzer Senf(Brassicaceae, Kreuzblütler).
Heimat: Europa bis Kaukasus (auch nörd-liches Afrika); einjähriges Kraut; alte Kultur-pflanze; in Mitteleuropa heimisch seit dem16. Jahrhundert; wurde für den Innenstadt-bereich von Düsseldorf in einer archäologi-schen Ausgrabung für die Zeit um 1500 undspäter nachgewiesen (GELIUS-DIETRICH 2002/2003). Den Senfkörnern wie auch dem Senfwerden von alters her potenzanregende Wir-kungen zugesprochen; Klosterbrüdern wur-de deswegen später sogar der Senfgenuss ver-boten (RÄTSCH 1990). Medizinische Verwen-dung bis heute als Senfpackung aus gepul-verten Samen. Bis Ende des zweiten Welt-krieges waren die Körner Grundlage für diemeisten Senfsorten; heute nur angebaut, woHandernte sich noch lohnt, z. B. in Südita-lien und auf Sizilien (STOBART 1972); imRheintal verwildert und eingebürgert auf pe-riodisch überschwemmten Flußufern, Rude-ralstellen und Äckern. Im Gebiet: Himmel-geiser Rheinbogen und auf dem Grind beiDormagen-Stürzelberg.69. Bunias orientalis, Morgenländisches Za-ckenschötchen (Brassicaceae, Kreuzblütler).
Heimat: O-Europa bis W-Asien; einjähri-ges Kraut auf Getreideäckern, Schuttplätzenund an Wegrändern; Erstnachweis 1814 beiLüneburg; ähnlich wie die Graukresse(Berteroaincana) vorwiegend in der Nähe von Exer-zierplätzen beobachtet. Im Gebiet: z. B. Him-melgeister Rheinbogen, Neusser Hafen.70. Calystegia silvatica ssp. pulchra, Wald-Zaun-winde (Convolvulaceae, Windengewächse).
Heimat: NO-Asien oder als Gartenhybri-de in Europa entstanden (TUTIN et al. 1972);
ausdauerndes Kraut an Zäunen und Rude-ralstellen, auch Zierpflanze; Neophyt seit1871. Im Gebiet: TK 4606-2, 4, 4707-4.71. Cardamine hirsuta, Behaartes Schaumkraut(Brassicaceae, Kreuzblütler).
Heimat: Europa bis Himalaya (auch N-Af-rika); einjähriges allgegenwärtiges Unkraut mitexzessivem Samenaufkommen (kosmopo-litisch verwildert); nach HAEUPLER et al. (2003)in Nordrhein-Westfalen nicht einheimisch;wahrscheinlich über Baumschulen und Gärt-nereien eingeschleppt; zwischen 1975 und1985 explosionsartige Ausbreitung (DÜLL &KUTZELNIGG 1994). Im Gebiet: häufig.72. Cardaria draba, Pfeilkresse (Brassicaceae,Kreuzblütler).
Heimat: östliches Mitteleuropa bis Sibiri-en; ausdauerndes Kraut an Wegrändern,Bahndämmen und Ruderalstellen; einge-schleppt mit Saatgut und Futtergetreide(HANF 1991); erster Nachweis für Deutsch-land 1728 bei Ulm; lästiges Unkraut wegenvegetativer Vermehrung mit Wurzelsprossen.Im Gebiet: häufig.73. Carduus acanthoides, Weg-Distel (Astera-ceae, Korbblütler).
Heimat: Europa bis Kaukasus; zweijähri-ges Kraut auf Weiden und trockenen Rude-ralstellen; in Nordrhein-Westfalen nicht ein-heimisch. Im Gebiet: z. B. Rheinufer bei Dor-magen-Zons.74. Chaenorhinum minus, Orant (Scrophulari-aceae, Rachenblütler).
Heimat: Europa bis Zentralasien (auchNW-Afrika); einjähriges Kraut auf Äckernund Ruderalstellen, in Weinbergen; in Nord-rhein-Westfalen nicht einheimisch. Im Ge-biet: z. B. Kaiserswerth, HimmelgeisterRheinbogen, Düsseldorf-Hauptbahnhof,auf dem Grind.75. Chenopodium botrys, Klebriger Gänsefuß(Chenopodiaceae, Gänsefußgewächse).
Heimat: W-Europa bis Himalaya; einjähri-ges Kraut auf Ruderalstellen und wüstenPlätzen, so auch am Düsseldorfer Rheinufer;in Deutschland seit dem 19. Jahrhundert. ImGebiet: TK 4706-4, 4806-1,4807-3.

Pflanzliche Neubürger (Neophyten) im Raum Düsseldorf/Neuss 63
Acta Biologica Benrodis 13 (2006)
76. Chenopodium foliosum, Echter Erdbeerspi-nat (Chenopodiaceae, Gänsefußgewächse).
Heimat: Mittelmeergebiet bis Zentralasi-en; einjähriges Kraut; als Gemüsepflanze an-gebaut und selten auf Ruderalstellen verwil-dert; Neophyt seit 1910. Im Gebiet: NeusserHafen.77. Chenopodium opulifolium, Schneeballblätt-riger Gänsefuß (Chenopodiaceae, Gänsefuß-gewächse).
Heimat: Europa bis Zentralasien (auch N-Afrika); einjähriges Kraut auf Ruderalstellen;in Nordrhein-Westfalen nicht einheimisch.Im Gebiet: Neusser Hafen.78. Chenopodium strictum, Gestreifter Gänse-fuß (Chenopodiaceae, Gänsefußgewächse).
Heimat: Eurasien (auch Kanaren und Äthi-opien); einjähriges Kraut auf Ruderalstellen;in Nordrhein-Westfalen nicht einheimisch;Neophyt seit 1860. Im Gebiet: TK 4606-1,4705-2, 4706-1, 4806-1. 3.79. Cuscuta lupuliformis, Pappel-Seide (Con-volvulaceae, Windengewächse; Abb. 6).
Heimat: Europa bis O-Asien; einjährigesKraut im feuchten Ufergebüsch; Vollschma-rotzer auf Weide, Pappel, Holunder, Hop-fen, Beifuß und Kratzbeere. Im Gebiet: Him-melgeister Rheinbogen.80. Dipsacus laciniatus, Gelappte Karde (Dip-sacaceae, Kardengewächse).
Heimat: S-Europa bis Zentralasien; zwei-jähriges Kraut auf Wiesen, Ruderalflurenund an Waldrändern. Im Gebiet: TK 4606-2.81. Echinops sphaerocephalus, Bienen-Kugeldis-tel (Asteraceae, Korbblütler).
Heimat: O-Europa bis Zentralasien; aus-dauerndes Kraut auf Ruderalstellen; ab Mit-te des 16. Jahrhunderts in Deutschland; Zier-pflanze; Neophyt seit Ende des 19. Jahrhun-derts; Trachtpflanze für Bienen. Im Gebiet:auf dem Grind bei Dormagen-Stürzelberg.82. Euphorbia lathyris, Kreuzblütige Wolfs-milch (Euphorbiaceae, Wolfsmilchgewächse).
Heimat: Mittelmeergebiet und wärmeresAsien; zweijähriges Kraut; alte Heil- und Zier-pflanze; im �Capitulare� Karls d. Gr. unterdem Namen Lacteridas erwähnt. FUCHS (1543):
�Wechst fast in allen Gärten.� Verwildert aufRuderalstellen und in Gärten; nach ROTHMA-LER (2002) Archäophyt für ganz Deutschland;in Nordrhein-Westfalen aber nach HAEUPLER
et al. (2003) nicht einheimisch; vertreibt an-geblich Wühlmäuse. Der Milchsaft ist hochge-fährlich für die Augen (HILLER & BICKERICH
1997). Ableitung des wissenschaftlichen Gat-tungsnamens: zu Ehren des Euphorbos (ca.54 v. Chr.), dem Leibarzt des Königs Jubavon Numidien, der die Heilkräfte dieser Pflan-ze entdeckt haben soll. Im Gebiet: zerstreut.83. Galega officinalis, Geißraute (Fabaceae,Schmetterlingsblütler).
Heimat: östliches Mitteleuropa bis Pakistan;ausdauerndes Kraut; Zier- und Heilpflanze;verwildert auf Ruderalstellen und Brachen;Neophyt seit dem 19. Jahrhundert; als Vieh-futter zur Verbesserung der Milchleistung beiKühen geschätzt (LICHTENSTERN et al. 1994).Im Gebiet: TK 4606-2.84. Herniaria hirsuta, Behaartes Bruchkraut(Caryophyllaceae, Nelkengewächse).
Heimat: Europa, Türkei (auch N-Afrika);ausdauerndes Kraut auf sandigen Standor-ten; Heilpflanze bei Bruchleiden (Hernie =Leistenbruch) und als Blutreinigungsmittel(GESSNER 1974). Im Gebiet: TK 4807-1.85. Hesperis matronalis, Gewöhnliche Nachtvi-ole (Brassicaceae, Kreuzblütler).
Heimat: Europa bis Zentralasien; zweijäh-riges Kraut; alte Zierpflanze, besonders inBauerngärten; verwildert in Auwäldern, feuch-ten Gebüschen und auf Ruderalstellen; Ne-ophyt seit dem 16. Jahrhundert. Im Gebiet:TK 4807-4.86. Hippophae rhamnoides, Sanddorn (Elaea-gnaceae, Ölweidengewächse).
Heimat: Europa bis Himalaya und Mon-golei; Strauch oder bis 10 m hoher Baum aufKüstendünen und -steilufern und Kiefern-trockenwäldern; häufig angepflanzt und ver-wildert; wegen des hohen Gehalts an Vita-min A und C der Beeren vielfache Verwen-dung (5 g frische Beeren enthalten etwa denTagesbedarf eines Erwachsenen an VitaminC, VETVICKA 1985). Im Gebiet: zerstreut.
^ ^

64 WERNER DIETRICH
87. Kochia scoparia, Besen-Radmelde (Cheno-podiaceae, Gänsefußgewächse).
Heimat: SO-Europa bis Asien; einjährigesKraut; selten verwildert auf Müllkippen,Bahnanlagen und in Häfen; Neophyt seit1938. Im Gebiet: Neusser Hafen.88. Lactuca serriola, Kompass-Lattich, Stachel-Lattich (Asteraceae, Korbblütler).
Heimat: Europa bis Zentralasien (auch N-Afrika); ein- bis zweijähriges Kraut auf Ru-deralstellen und Brachen, in Weinbergen;Neophytenstatus unsicher; bereits von FUCHS
(1543) als �Wild Lattich� abgebildet und be-schrieben. Nach HEGI (1929) im Rheinlanderst im 19. Jahrhundert beobachtet. Im Ge-biet: häufig.89. Lepidium latifolium, Breitblättrige Kresse,Pfefferkraut (Brassicaceae, Kreuzblütler).
Heimat: Europa bis Himalaya (auch N-Af-rika); ausdauerndes Kraut; wild nur an salz-haltigen Stellen der Nord- und Ostseeküsteund des Binnenlandes; unbeständig verwil-dert auf Ruderalstellen und Brachen. CHA-MISSO (1827) erklärt: �Die Blätter, die bren-nend scharf schmecken, werden in einigenGegenden als Salatwürze gegessen.� Im Ge-biet: TK 4606-3, 4706-1.90. Lysimachia punctata, Punktierter Gilbwei-derich (Primulaceae, Primelgewächse).
Heimat: östliches Mitteleuropa bis Türkeiund Kaukasus; ausdauerndes Kraut; Bauern-gartenpflanze; verwildert auf Ruderalstellen,in Staudenfluren und Auewäldern; inDeutschland seit Ende des 16. Jahrhunderts;Neophyt seit dem 19. Jahrhundert; eingebür-gert durch sehr dominante vegetative Aus-breitung (HAEUPLER et al. 2003). Im Gebiet:z. B. Düsseldorf-Himmelgeist.91. Mercurialis annua, Einjähriges Bingelkraut(Euphorbiaceae, Wolfsmilchgewächse).
Heimat: Europa bis Kaukasus (auch Ka-naren, Azoren, N-Afrika); einjähriges Krautin Gärten, Weinbergen, auf Äckern undSchuttplätzen; wird für Nordrhein-Westfalenbei HAEUPLER & al. (2003) als Neophyt ge-wertet. Dagegen berichtet WILLERDING (1986)von subrezenten Funden aus der Römerzeit
(Neuss, Friesheim, Xanten) durch KNÖRZER
(1970, 1971); ebenso von CREMER et al. (1991)als alter Kulturbegleiter gewertet. Früher ge-bräuchliche Heilpflanze. CHAMISSO (1827): �Siewar ehedem officinell; sie ist nur schleimigund als unnützes Heilkraut aus den Apothe-ken verwiesen worden. Sie kann als ein ganzunschuldiges Gemüse gegessen werden.� ImGebiet: überall.92. Picris echioides, Wurmlattich (Asteraceae,Korbblütler).
Heimat: S-Europa bis Krim und Iran(auch Kanaren und N-Afrika); einjährigesKraut; unbeständig auf Äckern und Rude-ralstellen, in Gärten, an Ufern. Im Gebiet:TK 4606-3.93. Potentilla norvegica, Norwegisches Finger-kraut (Rosaceae, Rosengewächse).
Heimat: Europa bis O-Asien (auch N-Amerika); einjähriges bis kurz ausdauerndesKraut auf Ruderalstellen, trockengefallenenTeichböden, an Ufern; in Nordrhein-Westfa-len nicht einheimisch; ursprünglich nur vonThüringen bis Mecklenburg; sonst Neophytseit 1832. Im Gebiet: TK 4606-2, 4, 4706-4,4707-4, 4807-2, 4.94. Potentilla recta, Hohes Fingerkraut (Ro-saceae, Rosengewächse).
Heimat: O-Europa bis W-Asien; ausdau-erndes Kraut auf trockenen Ruderalstellen,sonnigen Hängen und Felsfluren, in Sand-trockenrasen; auch Zierpflanze. Im Gebiet:TK 4606-2, 3, 4, 4706-2, 3.95. Psyllium arenarium (= Plantago arenaria),Sand-Flohsame, Sand-Wegerich (Plantagi-naceae, Wegerichgewächse).
Heimat: Europa bis Zentralasien (auch N-Afrika); einjähriges Kraut; unbeständig aufsandigen Ruderalstellen; Neophyt für alle Bun-desländer; schleimhaltige Samen als Abführ-mittel (GESSNER 1974). Im Gebiet: z. B. aufdem Grind bei Dormagen-Stürzelberg.96. Rorippa austriaca, Östereichische Sumpf-kresse (Brassicaceae, Kreuzblütler).
Heimat: Mitteleuropa (ursprünglich nur anNeiße, Oder und Elbe) bis Zentralasien; aus-dauerndes Kraut auf periodisch überflute-

Pflanzliche Neubürger (Neophyten) im Raum Düsseldorf/Neuss 65
Acta Biologica Benrodis 13 (2006)
ten Flußufersäumen und Ruderalstellen;Neophyt seit Ende des 19. Jahrhunderts. ImGebiet: Rheinufer bei Kaiserswerth, auf demGrind.97. Rumex thyrsiflorus, Rispen-Ampfer (Poly-gonaceae, Knöterichgewächse).
Heimat: Skandinavien, östliches Mitteleu-ropa bis Balkan und Sibirien; ausdauerndesKraut auf Ruderalstellen, besonders inFlußtälern; im Rheinuferbereich vorherr-schend gegenüber dem Sauer-Ampfer (Ru-mex acetosa). Im Gebiet: Himmelgeist.98. Salsola kali ssp. tragus, Salzkraut (Cheno-podiaceae, Gänsefußgewächse).
Heimat: Eurasien (auch N-Afrika); einjähri-ges Kraut an Meeresstränden, in Küstendü-nen, auf Sandfeldern des Binnenlandes; salz-tolerant; Erstnachweis für Mitteleuropa 1812(SCHMITZ 2002); in Nordrhein-Westfalen nichteinheimisch. Im Gebiet: auf dem Grind, Neus-ser Hafen.99. Salvia verticillata, Quirlblütiger Salbei (La-miaceae, Lippenblütler).
Heimat: SO-Europa bis Kaukasus; ausdau-erndes Kraut in ruderal beeinflußten Halb-trockenrasen, auf mäßig trockenen Ruderal-stellen; Neophyt seit dem 16. Jahrhundert.Im Gebiet: z. B. an der Fleher Brücke aufDüsseldorfer Seite.100. Sanguisorba minor ssp. polygama, GrubigerWiesenknopf (Rosaceae, Rosengewächse).
Heimat: S-Europa bis Zentralasien (auchN-Afrika); ausdauerndes Kraut auf trocke-nen, sandigen Ruderalstellen, in lückigen Tro-ckenrasen; vor allem mit Rasenaussaaten ein-geschleppt; wie der einheimische Kleine Wie-senknopf (ssp. minor) wertvolle Futterpflan-ze. Im Gebiet: TK 4707-1, 2 ,4, 4806-2.101. Setaria italica, Kolbenhirse (Poaceae, Süß-gräser).
Heimat: unbekannt, weil alte Kulturpflan-ze; in Europa seit 3800 v. Chr. angebaut, heu-te nur als Vogelfutter; einjähriges Gras; vonHAEUPLER et al. (2003) als Neophyt für Nord-rhein-Westfalen angesehen, allerdings seit1900 nicht mehr nachgewiesen. Die Hirse derMärchen ist die Rispenhirse (Panicum miliace-
um), die früher bei uns als �Brot des armenMannes� angebaut wurde (FRANKE 1997). ImGebiet: ?102. Setaria verticillata, Wirtel-Borstenhirse(Poaceae, Süßgräser).
Heimat: Europa bis O-Asien, N-Amerika(var.verticillata); Europa, Kaukasus, W-Asien,N-Afrika (var. ambigua); einjähriges Gras inGärten, auf Äckern (Maisfelder) und Rude-ralstellen; nach HAEUPLER et al. (2003) in Nord-rhein-Westfalen Neophyt. Im Gebiet: TK4807-1, 3.103. Silybum marianum, Mariendistel (Astera-ceae, Korbblütler).
Heimat: S-Europa bis Zentralasien (auch N-Afrika, Kanaren, Madeira); zweijähriges Kraut;Gartenzierpflanze; verwildert auf Ruderalstel-len und Äckern; alte Heilpflanze: antagonisti-sche Wirkung gegen die Gifte Amanitin undPhalloidin des Knollenblätterpilzes (GESSNER
1974); auch Gemüsepflanze, z. B. Blütenköp-fe wie Artischocken genutzt (DÜLL & DÜLL
2003). Der deutsche Name bezieht sich aufdie weiße Zeichnung auf den Blättern, die nachder Legende von der Milch der Gottesmutterherrühren sollen (FURLENMEIER 1978). ImGebiet: z. B. auf Äckern bei Ratingen.104. Sisymbrium loeselii, Loesels Rauke (Bras-sicaceae, Kreuzblütler).
Heimat: SO-Europa bis O-Asien; ein- biszweijähriges Kraut auf Ruderalstellen; Neo-phyt seit dem 18. Jahrhundert; besiedelt erstin jüngerer Zeit Kulturland, wo sie die zu-nehmend bessere Nährstoffversorgung aus-nutzen kann (CREMER et al. 1991). Im Ge-biet: TK 4605-2, 4, 4606-3, 4706-3.105. Solanum luteum ssp. luteum (= S. villosum),Gelber Nachtschatten (Solanaceae, Nachtschat-tengewächse).
Heimat: Europa bis Zentralasien; einjähri-ges Kraut auf Ruderalstellen; in ganz Deutsch-land nicht einheimisch; giftig. Im Gebiet: TK4606-3, 4706-1, 4707-1, 4807-1, 2, 3.106. Syringa vulgaris, Gemeiner Flieder (Ole-aceae, Ölbaumgewächse).
Heimat: Balkan bis Kleinasien, nach HE-CKER (1995) nur Balkan; 2-6 (-10) m hoher

66 WERNER DIETRICH
Strauch oder Baum; Ziergehölz (ca. 900 Gar-tenformen); verwildert in trockenen, steini-gen Gebüschen, an Straßenrändern, auf Bahn-gelände und Mauern; eingeführt im 16. Jahr-hundert; erster Nachweis für Deutschland1588; Neophyt seit dem 18. Jahrhundert. Dieätherischen Öle der Blüten werden in der Par-fümindustrie gebraucht; das gut polierbarefeste Holz wird in der Drechslerei und Kunst-tischlerei verwendet (HECKER 1995). Im Ge-biet: z. B. TK 4807-3.107. Trifolium resupinatum, Persischer Klee (Fa-baceae, Schmetterlingsblütler).
Heimat: S-Europa bis Iran (auch N-Afri-ka); einjähriges Kraut; als Gründünger undFutterpflanze angebaut; unbeständig verwil-dert auf frischen Trittrasen, Ruderalstellen,an Ackerrändern; Neophyt seit dem 19. Jahr-hundert. Im Gebiet: zerstreut.108. Verbascum blattaria, Schaben-Königsker-ze (Scrophulariaceae, Rachenblütler).
Heimat: Europa bis W-Sibirien und Zen-tralasien; zweijähriges Kraut auf Ruderalstel-len und Brachen; in Nordrhein-Westfalennicht einheimisch; früher kultiviert gegenSchaben (ROTHMALER 2002). Im Gebiet: TK4605-4.109. Viola odorata, März-Veilchen (Violaceae,Veilchengewächse).
Heimat: Europa bis Iran und Kaukasus(auch NW-Afrika); ausdauerndes Kraut in lich-ten Laubgehölzen, an Bachufern und Wald-säumen; nach ROTHMALER (2002) Archäophytfür alle Bundesländer; gelangte spätestens imfrühen Mittelalter als Zier- und Arzneipflan-ze nach Mitteleuropa. Walahfried Strabonennt um 820 eine �viola nigella�; auch beiHildegard von Bingen (1098-1179) ist voneiner �viol� die Rede. Versteckt zwischen an-deren Pflanzen ist das Märzveilchen auf demum 1410 entstandenen �Paradiesgärtlein� ei-nes oberrheinischen Malers zu sehen(KRAUSCH 2003). Es gilt seit alters als Volks-heilpflanze (FURLENMEIER 1978, GESSNER
1974, LICHTENSTERN et al. 1994). CHAMISSO
(1827) berichtet: �Die Veilchen sind officinell,sie dienen aber nur, um andere Medicamente
zu färben.� Die Einstufung als Neophyt fürNordrhein-Westfalen durch HAEUPLER et al.(2003) bezieht sich wohl auf Verwilderun-gen und Einbürgerungen in neuerer Zeit. ImGebiet: häufig.110. Vulpia bromoides, Trespen-Fuchsschwin-gel (Poaceae, Süßgräser).
Heimat: Europa bis Kaukasus (auch N-Af-rika); heute fast weltweit eingebürgert; einjäh-riges Gras auf trockenen, sandigen bis lehmi-gen Ruderalstellen; in Nordrhein-Westfalennicht einheimisch. Im Gebiet: TK 4707-3.111. Vulpia myuros, Mäuseschwanz-Fuchs-schwingel (Poaceae, Süßgräser).
Heimat: Europa bis Himalaya (auch N-Af-rika); einjähriges Gras auf trockenen, sandi-gen bis kiesigen Ruderalstellen, z. B. aufBahngelände; in Nordrhein-Westfalen nichteinheimisch. Im Gebiet: häufig.
3.3. Mittelmeergebiet
112. Anthoxanthum aristatum (= A. puelii),Begranntes Ruchgras (Poaceae, Süßgräser).
Heimat: SW-Europa (auch Kanaren undMadeira); einjähriges Gras auf sandigen Äk-kern, Ruderalstellen und Brachen; in Nord-rhein-Westfalen nicht einheimisch; Neophytseit 1850; Ruchgrasarten duften wie auch derWaldmeister (Galium odoratum) nach Kuma-rin (HUBBARD 1985). Im Gebiet: TK 4606-3,4807-2.113. Antirrhinum majus, Großes Löwenmäul-chen (Scrophulariaceae, Rachenblütler).
Heimat: westliches Mittelmeergebiet; ein-jähriges Kraut; Zierpflanze; verwildert in Fels-spalten und an Mauern; in Deutschland seitMitte 16. Jh. Im Gebiet: z. B. Rheinufermau-er in Düsseldorf-Hamm.114. Brassica napus, Raps, Kohlrübe (Brassi-caceae, Kreuzblütler).
Heimat: alte Kulturpflanze, ursprünglichwahrscheinlich in S-Europa; ein- bis zweijäh-riges Kraut; verwildert auf Ruderalstellen; alteÖl- und Gemüsepflanze; für Rinder, Reheund Hasen in größerer Menge giftig (ROTH-MALER 2002). Durch spontane Kreuzung des

Pflanzliche Neubürger (Neophyten) im Raum Düsseldorf/Neuss 67
Acta Biologica Benrodis 13 (2006)
Wildrübsens (B. rapa) mit dem Wildkohl (B.oleracea) entstand der Raps (KÖRBER-GROH-NE 1988). Verwendung des Öls als Speiseölund für technische Zwecke, früher auch alsLampenöl (FRANKE et al. 1976). Im Gebiet:häufig.115. Calepina irregularis, Wendig (Brassicaceae,Kreuzblütler).
Heimat: östliches Mittelmeergebiet, S-Ruß-land; einjähriges Kraut; unbeständig aufBrachfeldern, Ruderalstellen und in Weinber-gen; Erstnachweis für Deutschland Ende des19. Jahrhunderts. Im Gebiet: Haltestelle Süd-park in Düsseldorf (LÖSCH mdl. Mitt.).116.Centranthus ruber, Spornblume (Valeria-naceae, Baldriangewächse).
Heimat: Mittelmeergebiet; ausdauerndesKraut; Zierpflanze; aus Gärten gelegentlichverwildert. Im Gebiet: z. B. Düsseldorf-Him-melgeist.117. Cerastium tomentosum, Filziges Hornkraut(Caryophyllaceae, Nelkengewächse).
Heimat: Mittel- und S-Italien; ausdauern-des Kraut; Zierpflanze in Steingärten; gele-gentlich verwildert; 1594 in den Niederlan-den als Gartenpflanze; in Deutschland erstseit dem 18. Jahrhundert (KRAUSCH 2003),nach ROTHMALER (2002) schon im 16. Jahr-hundert. Im Gebiet: z. B. Angermund.118. Colutea arborescens, Blasenstrauch, �Knall-schote� (Fabaceae, Schmetterlingsblütler).
Heimat: Mittelmeergebiet bis SW-Deutsch-land (KRAUSCH 2003); S-Europa bis Trans-kaukasus und N-Afrika (GARCKE 1972); 2-6m hoher Strauch; angepflanzt auf trockenenHügeln und in Gebüschen, auf Bahngelän-de;selten verwildert; um 1560 in deutschenGärten weit verbreitet (KRAUSCH 2003); Blät-ter wirken wie Sennesblätter als Abführmit-tel (CHAMISSO 1827, KRAUSCH 2003). Im Ge-biet: TK 4605-4.119. Coriandrum sativum, Koriander (Apiaceae,Doldenblütler).
Heimat: östliches Mittelmeergebiet; einjäh-riges Kraut; verwildert auf Brachen, lehmi-gen Äckern und Ruderalstellen; seit mindes-tens 3000 Jahren als Heil- und Gewürzpflan-
ze angebaut (QUATMANN 1996); schon in ägyp-tischen Papyri erwähnt (FRANKE 1997); seitdem 16. Jahrhundert in Deutschland ange-baut; wurde für den Zeitraum 1580-1620 imStadtgebiet Düsseldorf archäobotanisch nach-gewiesen (GELIUS-DIETRICH 2002/2003); giltseit alters als Aphrodisiacum. Schon Dios-corides (1. Jh. nach Chr.) empfahl den Kori-ander, in Wein getrunken, zur Vermehrungdes Samens (RÄTSCH 1990). Bestandteil desKarmelitergeistes (GESSNER 1974) und desCurrypulvers (FRANKE 1997). Im Gebiet: z.B. Düsseldorf-Himmelgeist.120. Cynodon dactylon, Hundszahngras, Ber-mudagras (Poaceae, Süßgräser).
Heimat: Mittelmeergebiet; heute kosmo-politisch verbreitet; ausdauerndes Gras aufsandigen Orten; wird von HAEUPLER et al.(2003) nicht als Neophyt gewertet; als Futter-gras in den Tropen angebaut (FRANKE 1997);ausgezeichnetes Rasengras; salztolerant, tritt-fest und trockenresistent (HUBBARD 1985,DÜLL & DÜLL 2003). Im Gebiet: Himmel-geister Rheinbogen, auf dem Grind.121. Digitaria sanguinalis ssp. pectinatiformis,Blut-Fingerhirse (Poaceae, Süßgräser).
Heimat: Mittelmeergebiet; die Art D. san-guinalis s. l. heute kosmopolitisch verbreitet;einjähriges Gras auf sandigen Ruderalstellen.Die ssp. sanguinalis gilt in ROTHMALER (2002)als Archäophyt für alle Bundesländer; dage-gen gilt die ssp. pectinatiformis zumindest inNordrhein-Westfalen als Neophyt. Im Ge-biet: auf dem Grind.122. Diplotaxis muralis, Mauer-Doppelsame(Brassicaceae, Kreuzblütler).
Heimat: Mittelmeergebiet; ein-bis zwei-jähriges Kraut auf Ruderalstellen und inHackkulturen (Weinberge, Äcker); Neophytseit dem 18. Jahrhundert. Im Gebiet: TK4606-3.123. Diplotaxis tenuifolia, SchmalblättrigerDoppelsame (Brassicaceae, Kreuzblütler).
Heimat: Mittelmeergebiet; ausdauerndesKraut auf Ruderalstellen, an Straßenrändern;Erstnachweise: 1740 bei Straßburg, 1808 beiKarlsruhe, 1818 am Niederrhein. Die Blätter

68 WERNER DIETRICH
werden in S-Frankreich als Salatwürze verwen-det (STOBART 1972). Im Gebiet: häufig.124. Eragrostis minor, Kleines Liebesgras (Poa-ceae, Süßgräser).
Heimat: Mittelmeergebiet; einjähriges Grasan Wegen, auf sandigen Stellen, Bahngelän-de und Schuttplätzen, im Straßenpflaster; Ne-ophyt seit 1782. Im Gebiet: z. B. Himmel-geister Rheinbogen, auf dem Grind, S-Bahn-höfe Bilk und Völklingerstraße in Düsseldorf.125. Foeniculum vulgare, Echter Fenchel (Api-aceae, Doldenblütler).
Heimat: Mittelmeergebiet (SCHMEIL & FIT-SCHEN 2000); Mittelmeergebiet bis Kauka-sus und Iran (ZANDER 2002); zweijährigesbis kurz ausdauerndes Kraut; Gewürz- undTeepflanze (var. dulce) und Gemüsepflanze(var. azoricum, Knollenfenchel); als Neophytvielerorts aus Ansaaten unbeständig verwil-dert in ruderal beeinflußten Rasengesell-schaften, auf Schutt und an Wegrändern, inWeinbergen; aber auch Archäophyt; von denBenediktinern im frühen Mittelalter nachMitteleuropa gebracht (TRONÍCKOVÁ 1985).Karl d. Gr. machte in seinem Capitulare deVillis den Anbau in den Bauerngärten zurPflicht (BECKMANN 1990). Wegen des Ge-halts an ätherischen Ölen seit alters vielseitigverwendete Heil- und Gewürzpflanze(FRANKE 1997, HLAVA & LANSKA 1977, LICH-TENSTERN et al. 1994, RAUH 1950, STOBART
1972). In Griechenland werden Fenchel undAnis (Pimpinella anisum) zur Herstellung vonOuzo verwendet (ALBERTS & MULLEN 2000).Im Gebiet: z. B. TK 4605-4, 4606-3, 4705-2,4706-1, 3, 4.126. Geranium purpureum, Purpur-Storchschna-bel (Geraniaceae, Storchschnabelgewächse).
Heimat: Mittelmeergebiet; in Deutschlandursprünglich in Baden-Württemberg, Hes-sen, Rheinland-Pfalz; einjähriges Kraut aufRuderalstellen (vornehmlich im Gleisschot-ter von Bahnanlagen); Neophyt seit 1890;Erstfund in Nordrhein-Westfalen 1996. ImGebiet: TK 4807-2.127. Hirschfeldia incana, Bastardsenf, Rempe(Brassicaceae, Kreuzblütler).
Heimat: Mittelmeergebiet; einjährigesKraut; unbeständig verwildert auf Industrie-und Bahngelände, auch sonstigen Ruderal-stellen sowie nährstoffreichen Äckern (beson-ders Kleefelder); Neophyt seit 1850. Im Ge-biet: TK 4706-4.128. Linaria cymbalaria (= Cymbalaria mura-lis), Zimbelkraut (Scrophulariaceae, Rachen-blütler).
Heimat: Mittelmeergebiet; einjähriges biskurz ausdauerndes Kraut in Mauer- und Fels-spalten; ursprünglich Gartenzierpflanze; Ne-ophyt seit dem 17. Jahrhundert. Im Gebiet:z. B. Kaiserpfalz in Düsseldorf-Kaiserswerth,Christophstraße in Düsseldorf.129. Lobularia maritima, Strand-Silberkraut,Strandkresse (Brassicaceae, Kreuzblütler).
Heimat: Mittelmeergebiet; einjähriges biskurz ausdauerndes Kraut; Zierpflanze inSteingärten; verwildert an Straßenrändern undMauern; in Mitteleuropa bekannt seit Endedes 16. Jahrhunderts; Neophyt seit der zwei-ten Hälfte des 19. Jahrhunderts, z. B. Bran-denburg 1884. Im Gebiet: z. B. auf demGrind (TK 4807-1; 2.9.2004).130. Lolium multiflorum, Italienisches Raygras(Poaceae, Süßgräser).
Heimat: Mittelmeergebiet; einjähriges biskurz ausdauerndes Gras; häufig als Grünland-aussaat; verwildert auf ruderal beeinflußtenÄckern und Wiesen; gutes Futtergras (DÜLL
& KUTZELNIGG 1994, FRANKE 1997). Im Ge-biet: häufig.131. Malus domestica, Kultur-Apfel (Rosaceae,Rosengewächse).
Heimat: nördliches Mittelmeergebiet; gro-ßer Strauch oder bis zu 10 m hoher Baum;Archäophyt seit der jüngeren Steinzeit (HE-CKER 1995); auch Neophyt bei rezenten Ver-wilderungen. Im Gebiet: z. B. Rheinufer beiDüsseldorf-Himmelgeist.132. Malva sylvestris ssp. mauritiana, Maureta-nische Malve (Malvaceae, Malvengewächse).
Heimat: Mittelmeergebiet; zweijähriges biskurz ausdauerndes Kraut auf Wildäckern. ImGebiet: z. B. spontan an Haus Christoph-straße 80 in Düsseldorf.
^

Pflanzliche Neubürger (Neophyten) im Raum Düsseldorf/Neuss 69
Acta Biologica Benrodis 13 (2006)
133. Oxalis corniculata, Horn-Sauerklee (Oxa-lidaceae, Sauerkleegewächse).
Heimat: Mittelmeergebiet (RAUH & HANF
1983); nach CREMER et al. (1991) N-Amerika;nach SEBALD et al. (1992) tropisches und sub-tropisches Asien, Afrika und Australien. ImGrabungshorizont 8.-13. Jahrhundert derAusgrabung Neusser Omnibusbahnhof (ab2001) wurde von GELIUS-DIETRICH (pers.Mitt.) im Jahre 2005 ein verkohlter Samenvon Oxalis corniculata gefunden. Somit ist derNeophytenstatus wohl nicht mehr aufrechtzu erhalten. Zumindest sind die Herkunfts-angaben N-Amerika und Australien damithinfällig. Einjähriges bis ausdauerndes Kraut;1576 als Bodendecker für Gräber und Beet-einfassungen nach Mitteleuropa eingeführt;1588 erstmalig wild bei Nürnberg beobach-tet; verwildert in Pflasterfugen, auf Kieswe-gen, in Parks und Gärten; lästiges Unkrautwegen hoher Samenproduktion. Im Gebiet:häufig.134 Phalaris canariensis, Kanariengras (Poaceae,Süßgräser).
Heimat: westliches und zentrales Mittel-meergebiet; einjähriges Gras; kultiviert als Vo-gelfutter; häufig verwildert auf Müllplätzen;in Italien seit 1532 in Kultur; in Deutschland1539 zum ersten Mal belegt (KRAUSCH 2003).Im Gebiet: TK 4706-3, 4.135. Reseda lutea, Gelber Wau (Resedaceae,Resedagewächse).
Heimat: Mittelmeergebiet; zweijährigesKraut auf trockenen Ruderalstellen und ex-tensiv genutzten Äckern; vermutlich Endedes 18. Jahrhunderts in Mitteleuropa aufge-taucht; verwildert entlang der Eisenbahnlini-en (AICHELE & SCHWEGLER 2000); nach HEGI
(1919) und HAEUPLER et al. (2003) teilweiseals Archäophyt zu betrachten. Im Gebiet:häufig.136. Tamarix parviflora, Kleinblütige Tamaris-ke (Tamaricaceae, Tamariskengewächse).
Heimat: östliches Mittelmeergebiet; dort inHecken und an Flußufern; bis 5 m hoher breit-buschiger Zierstrauch; seit 1853 in Kultur(KRÜSSMANN 1978). Im Gebiet: auf dem
Grind am rheinzugewandten Rand der Wei-denaue (TK 4807-133314; 15. 10. 2004); ca.1 m hohe Jungpflanze. Bisher in Nordrhein-Westfalen nicht als verwildert beobachtet.137. Trifolium incarnatum, Inkarnat-Klee (Fa-baceae, Schmetterlingsblütler).
Heimat: Mittelmeergebiet; ein- bis zwei-jähriges Kraut; als Gründünger angesät undunbeständig verwildert; Neophyt seit dem 19.Jahrhundert. Im Gebiet: z. B. TK 4707-4,4807-1.
3.4. Asien
138. Acorus calamus, Kalmus (Araceae, Aron-stabgewächse).
Heimat: Zentralasien bis O-Asien; ausdau-erndes Kraut an Gräben, Teichen, Ufern undAltwässern; als Arzneipflanze im 16. Jahr-hundert nach Mitteleuropa eingeführt; erst-mals 1557 in Prag; 1728 erstmals wild auseinem Graben bei Ulm gemeldet; gehört zuden ältesten Kulturpflanzen überhaupt; wur-de schon in vorindogermanischer Zeit ge-nutzt (BECKMANN 1990); aufgrund seinesGehalts an ätherischen Ölen seit alters vielsei-tig genutzte Heilpflanze (GESSNER 1974, LICH-TENSTERN et al. 1994, RÄTSCH 1990). HoheDosen von Kalmusöl steigern das Krebsrisi-ko (ALBERTS & MULLEN 2000). Auch Gewürz-pflanze, z. B. Bestandteil des Currypulvers(HLAVA & LANSKA 1977) sowie in der Likör-herstellung (FRANKE 1997). CHAMISSO (1827)berichtet: �Der Geruch des Kalmus soll dieHausinsekten vertreiben.� Im Gebiet: z. B.Rheinufer bei Düsseldorf-Kaiserswerth, aufdem Grind.139. Ailanthus altissima, Götterbaum (Sima-roubaceae, Bittereschengewächse).
Heimat: China, Korea; 20-25 m hoherBaum; in Europa Zierbaum seit 1750; Neo-phyt seit 1870; im Ruhrgebiet entlang der Ver-kehrswege mit zunehmender Einbürge-rungstendenz; Jungwuchs auffallend häufigim Windschatten von Brücken und ähnlichenBauwerken (HAEUPLER et al. 2003); unemp-findlich gegen Emissionen und Staubbelas-

70 WERNER DIETRICH
tung; eignet sich daher zur Anpflanzung inStädten und Industriegebieten (LUDWIG et al.2000); unterdrückt die übrige Vegetationdurch Abgabe chemischer Substanzen (Alle-lopathie, DÜLL & KUTZELNIGG 1994). DerName Ailanthus leitet sich von �Ailanto�,dem Volksnamen auf den Molukken, ab, wasso viel wie �Baum des Himmels� bedeutet(HECKER 1995). Die Schreibweise mit �th�ist falsch, da der Name nichts mit dem grie-chischen �anthos� (= Blüte) zu tun hat. ImGebiet: nicht selten.140. Buddleja davidii, Chinesischer Sommerflie-der, Schmetterlingsbusch (Buddlejaceae, Som-merfliedergewächse).
Heimat: W-China; 1-5 m hoher Strauch;1869 von David (1826-1900, französischerMissionar in China) entdeckt; erste Einfüh-rung nach Europa 1890; 1902 erstmals inBerlin; verwildert auf Schuttplätzen undBahngelände; Neophyt seit 1945; nach demzweiten Weltkrieg auffälligste Pflanze derTrümmergrundstücke (DÜLL & KUTZELNIGG
1994); wichtige Nektarquelle für Bienen undSchmetterlinge (LUDWIG et al. 2000). Im Ge-biet: häufig, z. B. Düsseldorfer Hauptbahn-hof, auf dem Grind.141. Cannabis sativa ssp. sativa, Hanf (Canna-baceae, Hanfgewächse).
Heimat: Asien; einjähriges Kraut; Archäo-phyt; wird für Nordrhein-Westfalen jedochals Neophyt gewertet; vielseitig genutzte alteKulturpflanze (Kleidung, Papier, Öl, Brenn-stoff, Nahrung, Baumaterial; Arzneimittel;auch Rauschdroge als Marihuana; HERER &BRÖCKERS 1995); in Deutschland umstritte-nes Anbauverbot; gelegentlich verwildert ausimportiertem Vogelfutter. Im Gebiet: TK4606-3.142. Cotoneaster horizontalis, Fächer-Zwergmis-pel (Rosaceae, Rosengewächse).
Heimat: W-China; niederliegender Zier-strauch, häufig als Bodendecker; 1880 nachEuropa eingeführt (VETVICKA 1985); ersterNachweis als Neophyt 1962. Im Gebiet: TK4606-3, 4706-3, 4707-1, 2. Nach HAEUPLER etal. (2003) existieren einige andere Arten mit
Einbürgerungstendenz, z. B. C. integrifolius,C. lucidus, C.bullatus, C. moupinensis.143. Cucumis sativus, Gurke (Cucurbitaceae,Kürbisgewächse).
Heimat: Indien; einjähriges Kraut; Kultur-pflanze. Für die Zeit um 1500 und später imInnenstadtbereich von Düsseldorf archäobo-tanisch nachgewiesen (GELIUS-DIETRICH
2002/2003). Mit 97% Wassergehalt ist dieGurke eine der wasserreichsten und kalorien-ärmsten Früchte. In manchen GegendenWestasiens führt die Landbevölkerung aufReisen oder bei der Feldarbeit stets einigeGurken als �Trinkwasserreservoir� mit sich(FRANKE et al. 1976). Im Gebiet: z. B. auf derJücht im Himmelgeister Rheinbogen.144. Datura ferox, Dorniger Stechapfel (Sola-naceae, Nachtschattengewächse).
Heimat: China; einjähriges Kraut. Im Ge-biet: Neusser Hafen (Erstfund 1979, STIEG-LITZ 1981).145. Fagopyrum esculentum, Echter Buchwei-zen, Heidekorn (Polygonaceae, Knöterichge-wächse).
Heimat: Turkestan, S-Sibirien, N-China;einjähriges Kraut; Kulturpflanze; gelegentlichverwildert auf Ruderalstellen; nach ROTHMA-LER (2002) Archäophyt, nach HAEUPLER et al.(2003) Neophyt. Gedeiht auch auf ärmstenSand- und Heideböden (RAUH 1950); auchals Trachtpflanze für Bienen. Im Gebiet: TK4705-2, 4706-3, 4807-2, 3.146. Fallopia baldschuanica (= F. aubertii), Sil-berregen (Polygonaceae, Knöterichgewächse).
Heimat: O-Asien; strauchige Kletterpflan-ze; Zierpflanze. 1883 von Albert Regel amFluße Voksh in Tatschikistan entdeckt. Nachdem ersten Weltkrieg allgemein in Garten-kultur; gelegentlich verwildert, aber selten ein-gebürgert. Im Gebiet: TK 4606-1, 2, 4706-3,4707-2.147. Glycine max, Sojabohne (Fabaceae,Schmetterlingsblütler).
Heimat: O-Sibirien, China, Mandschurei,Korea, Japan, Taiwan; einjähriges Kraut;Kulturpflanze; unbeständig eingeschleppt.Im Gebiet: z. B. im Neusser Hafen.
^^

Pflanzliche Neubürger (Neophyten) im Raum Düsseldorf/Neuss 71
Acta Biologica Benrodis 13 (2006)
148. Heracleum mantegazzianum, Riesen-Bären-klau, Herkulesstaude (Apiaceae, Doldenblüt-ler; Abb. 7).
Heimat: Kaukasus; ausdauerndes Kraut, bisüber 3 m hoch. 1890 von den Botanikern Som-mer und Levier aus dem Kaukasus als beson-dere Attraktion in den Alpengarten Correvonbei Genf gebracht; Erstfunde als Neophyt:1911 Dresden, 1912 Zürich, 1924 Schleswig-Holstein. Von Imkern bis 1990 als Bienenwei-de angesät; als Trachtpflanze jedoch uninteres-sant, da zur Blütezeit im Hochsommer genü-gend andere Bienenpflanzen verfügbar sind(LUDWIG et al. 2000). Mittlerweile größtenteilseingebürgert, sich entlang von Fließgewässern,Waldwegen und Verkehrswegen noch ständigausbreitend. Phototoxische Wirkung bei Haut-kontakt mit dem Saft der Pflanze (Hautaus-schlag, schwere Brandblasen, HILLER & BICKE-RICH 1997). Im Gebiet: z. B. HimmelgeisterRheinbogen, Urdenbacher Kämpe, S-Bahn-hof Völklingerstraße.149. Impatiens glandulifera, Drüsiges Spring-kraut (Balsaminaceae, Balsaminengewächse;Abb. 8).
Heimat: Himalaya; einjähriges bis über 2 mhohes Kraut; Gartenzierpflanze (�Bauernor-chidee�); wie die vorige beliebte Bienenfutter-pflanze (LUDWIG et al. 2000). 1837 in Dresdenangepflanzt; Erstnachweise als Neophyt: 1880auf der Pfaueninsel in Berlin-Wannsee, 1897Hamburg, 1912 in der Pfalz und in Tirol, 1917bei Garmisch; verwildert an Flußufern, in Au-wäldern; verdrängt die einheimische Vegetati-on. Im Gebiet: z. B. Kaiserswerth, Himmel-geister Rheinbogen, auf dem Grind.150. Impatiens parviflora, Kleinblütiges Spring-kraut (Balsaminaceae, Balsaminengewächse).
Heimat: O-Asien; einjähriges Kraut, dasseine Samen 3-4 m weit schleudert. 1837 imGroßen Garten in Dresden als Zierpflanzeausgesät. KLEIN (1973) berichtet: �Was einst-mals in den Parkanlagen von Dresden undGenf als Rarität bestaunt wurde, gilt heuteals unausrottbares Unkraut, das sich rück-sichtslos und schwungvoll freie Bahnschießt�. Dringt auch in natürliche Pflanzen-
gesellschaften feuchter Wälder ein und ver-drängt infolge seines massenhaften Auftre-tens die natürliche Vegetation (RAUH & HANF
1983). Im Gebiet: z. B. Urdenbacher Käm-pe, Himmelgeister Rheinbogen, auf demGrind.151. Larix kaempferi, Japanische Lärche (Pin-aceae, Kieferngewächse).
Heimat: Japan; bis 30 m hoher Forstbaum(in der Heimat bis 45 m); Ersteinführung1861; angepflanzt und sich natürlich verjün-gend; in Gebieten mit hoher Luftfeuchtig-keit der einheimischen Europäischen Lärche(Larix decidua) überlegen. Im Gebiet: TK4605-2, 4606-4.152. Lycium barbarum, Gemeiner Bocksdorn(Solanaceae, Nachtschattengewächse).
Heimat: China; bis 3,5 m hoher Zierstrauch;verwildert auf trockenen Ruderalstellen, inKüstendünen; in gärtnerischer Kultur seit1772; giftig. Im Gebiet: zerstreut.153. Matricaria discoidea, Strahllose Kamille(Asteraceae, Korbblütler.)
Heimat: NO-Asien; einjähriges Kraut; ein-gebürgert seit Mitte des 19. Jahrhunderts;zuerst in Botanischen Gärten, da sie wegender strahlenlosen Blütenköpfchen als Kurio-sum angesehen wurde; wird ähnlich verwen-det wie die Echte Kamille (Matricaria recuti-ta), hat aber keine entzündungshemmendeWirkung (LICHTENSTERN et al. 1994). Im Ge-biet: häufig.154. Panicum miliaceum, Echte Hirse (Poaceae,Süßgräser).
Heimat: Zentralasien; einjähriges Gras;Kulturpflanze seit der jüngeren Steinzeit; inMitteleuropa nur noch selten als Vogelfutterkultiviert; Archäophyt oder Neophyt aus re-zenten Ansaaten. Im Gebiet: TK 4606-3,4706-1, 3, 4807-2, 4.155. Phytolacca esculenta, Essbare Kermesbee-re (Phytolaccaceae, Kermesbeerengewächse).
Heimat: China, Korea, Japan; ausdauern-des Kraut; Kultur- und Zierpflanze; seltenverwildert; in der Heimat Gemüsepflanze;Neophyt seit dem 19. Jahrhundert. Im Ge-biet: TK 4606-3.

72 WERNER DIETRICH
Abb. 5: :Aufsteigender Fuchsschwanz (Amaran-thus blitum).Fig. 5: Guernsey Pigweed (Amaranthus blitum).
Abb. 6: Pappel-Seide (Cuscuta lupuliformis).Fig. 6: Hop Dodder (Cuscuta lupuliformis).
Abb. 7: Riesen-Bärenklau (Heracleum mantegaz-zianum).Fig. 7: Giant Hogweed (Heracleum mantegazzia-num).
Abb. 8: Drüsiges Springkraut (Impatiens glanduli-fera).Fig. 8: Indian Balsam (Impatiens glandulifera).

Pflanzliche Neubürger (Neophyten) im Raum Düsseldorf/Neuss 73
Acta Biologica Benrodis 13 (2006)
156. Polygonum polystachyum (= Aconogononpolystachyum), Himalaya-Knöterich (Polygo-naceae, Knöterichgewächse).
Heimat: Himalaya; ausdauerndes Kraut;Zierpflanze; aus Anpflanzungen auf vegeta-tivem Wege eingebürgert. Im Gebiet: TK4807-2.157. Prunus cerasifera, Kirschpflaume (Ro-saceae, Rosengewächse).
Heimat: W-Asien, Kaukasus; 5-8 m hoherStrauch; Park- und selten Obstgehölz; oft alsPfropfunterlage für andere Pflaumenartenverwendet; in aufgegebenem Gartenland häu-fig Austrieb aus der Pfropfunterlage; dringtauf diese Weise in benachbarte Hecken undGebüsche ein. Im Gebiet: z. B. am BrückerBach bei Wersten.158. Prunus domestica, Pflaume, Zwetschge(Rosaceae, Rosengewächse).
Heimat: Vorder- bis W-Asien; 6(-12) m ho-her Baum; alte euroasiatische Kulturpflanze;in zahlreichen Formen kultivierter Obst-baum; den Griechen seit 600-500 v. Chr. be-kannt; durch die Römer in die Gebiete nörd-lich der Alpen gelangt; zu allen Zeiten wohlschon verwildert, doch auch Neophyt (ROTH-MALER 2002), hervorgegangen aus Neuan-pflanzungen; von HAEUPLER et al. (2003) alsArchäophyt gewertet. Im Gebiet: zerstreut.159. Pyrus communis, Garten-Birne (Rosaceae,Rosengewächse).
Heimat: Vorderasien; bis 20 m hoher Obst-baum (mehr als 1500 Sorten); aus W-Asienzwischen 2000 und 1000 v. Chr. zu den Grie-chen und Römern gelangt; von diesen in derKaiserzeit über die Alpen gebracht (FRANKE etal. 1976); wertvolles, sehr hartes Nutzholz, viel-seitig verwendbar (z. B. für Fruchtpressen,Meßinstrumente und Druckstöcke, HECKER
1995). Bewertung des Neophytenstatus wiebei P. domestica. Im Gebiet: selten.160. Rapistrum rugosum, Runzliger Rapsdot-ter (Brassicaceae, Kreuzblütler).
Heimat: Vorderasien; einjähriges Kraut aufÄckern und Brachen; gilt für das Rheinlandals Neophyt (ROTHMALER 2002). Im Gebiet:Campus der Universität Düsseldorf.
161. Reynoutria japonica (= Fallopia japonica),Japanischer Staudenknöterich (Polygonaceae,Knöterichgewächse).
Heimat: O-Asien; ausdauerndes, bis über2 m hohes Kraut; Zierpflanze seit 1823;auch als Wildfutter angebaut (LUDWIG et al.2000); in feuchten Gebüschen und Bach-auen verwildert; unterdrückt die einheimi-sche Vegetation; Erstnachweise als Neo-phyt: 1844 in Westfalen, 1884 in Branden-burg (KRAUSCH 2003). Im Gebiet: z. B. Ur-denbacher Kämpe, Brücker Bach in Düs-seldorf-Wersten.162. Reynoutria sachalinensis (= Fallopia sachali-nensis), Sachalin-Staudenknöterich (Polygo-naceae, Knöterichgewächse).
Heimat: Sachalin, südliche Kurileninseln,N-Japan; ausdauerndes, bis 2 m hohes Kraut;Zierpflanze; 1853 entdeckt; 1855 im Botani-schen Garten St. Petersburg; 1863 in Eng-land; 1870 in Deutschland; verwildert an ähn-lichen Standorten wie vorige, aber wenigeragressiv als diese. Im Gebiet: häufig.163. Rosa rugosa, Kartoffel-Rose (Rosaceae,Rosengewächse).
Heimat: NO-Asien; 1-2 m hoher Zier-strauch; an Böschungen, Straßenrändernund auf Dünen zur Befestigung gepflanztund verwildert; entdeckt 1784; in Deutsch-land seit 1796; Erstnachweis als Neophyt1854; Hagebutten für Marmelade und Teeverwendbar (hoher Vitamin-C-Gehalt); inder Rosenzüchtung als Unterlage und Kreu-zungspartner verwendet (Kreuzungen ge-hören heute zu den modischsten Garten-rosensorten, VETVICKA 1985). Im Gebiet:im Rheinbereich selten verwildert, aber mitEinbürgerungstendenz. (HAEUPLER et al.2003).164. Rubus armeniacus, Armenische Brombee-re (Rosaceae, Rosengewächse).
Heimat: Kaukasus; Strauch; als Obstpflanzeverwildert in Siedlungsnähe, auf Bahn- undIndustriegelände; Neophyt seit vor 1900. ImGebiet: häufig.165. Rubus phoenicolasius, Japanische Weinbeere(Rosaceae, Rosengewächse).
^ ^

74 WERNER DIETRICH
Heimat: China, Korea, Japan; bis 3 m ho-her Strauch; Obstpflanze; seit 1876 in Europain Kultur; selten verwildert; wohlschmecken-de Früchte. Im Gebiet: Industriebrache beiDormagen-St. Peter in den 70er-Jahren.166. Sedum spurium, Zweifelhafte Fetthenne(Crassulaceae, Dickblattgewächse).
Heimat: Kaukasus, Türkei, N-Iran; ausdau-erndes Kraut; Zierpflanze; verwildert an Fel-sen und Mauern, auf sandigen und steinigenRuderalstellen; Neophyt seit dem 19. Jahrhun-dert. Im Gebiet: TK 4707-1, 4, 4806-4, 4807.167. Setaria faberi, Fabers Borstenhirse (Po-aceae, Süßgräser).
Heimat: China; einjähriges Gras; einge-schleppt mit Vogelfutter; unbeständig ver-wildert vor allem in Maisfeldern; Neophyt seit1960. Im Gebiet: TK 4706-3, 4807-1.168. Sisymbrium altissimum, Ungarische Rauke(Brassicaceae, Kreuzblütler).
Heimat: SW-Asien; einjähriges Kraut auftrockenen Ruderalstellen; Erstnachweise:1791 in Schlesien, seit etwa 1830 in Branden-burg. Im Gebiet: Neusser Hafen.169. Veronica filiformis, Faden-Ehrenpreis(Scrophulariaceae, Rachenblütler).
Heimat: Kaukasus bis Anatolien; ausdau-erndes Kraut; häufig flächendeckend in Wie-sen und Parkanlagen; 1780 nach Englandeingeführt; 1932 in dem Standardwerk Pa-reys Blumengärtnerei als �beste Staude fürRasenersatz in Steingärten und Alpenanla-gen� empfohlen (BONSTEDT 1932); Neophytseit ca 1930. Im Gebiet: z. B. SchloßparkBenrath, Botanischer Garten der UniversitätDüsseldorf.170. Veronica persica, Persischer Ehrenpreis(Scrophulariaceae, Rachenblütler).
Heimat: Gebirge SW-Asiens; einjährigesKraut in Gärten, auf Äckern und Ruderal-stellen; eines der häufigsten Unkräuter desWintergetreides; kann praktisch das ganzeJahr über keimen, blühen und fruchten; keimtbereits bei Temperaturen ab 2° C (CREMER etal. 1991); 1805 als Flüchtling aus dem Bota-nischen Garten Karlsruhe erstmals registriert.Im Gebiet: häufig.
3.5. Nordamerika/nördliches Mittel-amerika
171. Acer negundo, Eschen-Ahorn (Aceraceae,Ahorngewächse).
Heimat: N-Amerika; 6 (bis selten 25) mhoher Zierbaum; zweihäusig, windblütig,Bienentrachtpflanze; seit 1688 in Europa; abAnfang des 19. Jahrhunderts als Alleebaumwegen des schnellen Anfangswachstums;wurde in Amerika ähnlich wie der Zucker-ahorn (A. saccharum) zur Zuckergewinnunggenutzt (VETVICKA 1985). Im Gebiet: z. B.Himmelgeister Rheinbogen, auf dem Grind.172. Amaranthus albus, Weißer Fuchsschwanz(Amaranthaceae, Fuchsschwanzgewächse).
Heimat: südliches N-Amerika bis Mittel-amerika; einjähriges Kraut auf Bahngeländeund Ruderalstellen; Neophyt seit 1880; amNiederrhein seit den 1980er Jahren (SCHMITZ
2002). Im Gebiet: Himmelgeister Rheinbo-gen, auf dem Grind.173. Amaranthus blitoides, WestamerikanischerFuchsschwanz (Amaranthaceae, Fuchs-schwanzgewächse).
Heimat: westliches N-Amerika; einjährigesKraut auf Ruderalstellen; Neophyt seit 1893;am Niederrhein seit den 1990er Jahren (SCHMITZ
2002). Im Gebiet: Himmelgeister Rheinbogen,Rheinufer bei Kaiserswerth, auf dem Grind.174. Amaranthus retroflexus, RauhaarigerFuchsschwanz (Amaranthaceae, Fuchs-schwanzgewächse).
Heimat: wahrscheinlich N-Amerika; einjäh-riges Kraut auf Ruderalstellen, in Hackfrucht-äckern; Neophyt seit 1815; am Niederrhein seitden 1820er Jahren (SCHMITZ 2002). Im Gebiet:z. B. Himmelgeister Rheinbogen, Kaisers-werth, Urdenbacher Kämpe, auf dem Grind.175. Ambrosia artemisiifolia, Beifußblättrige Am-brosie, Traubenkraut (Asteraceae, Korbblütler).
Heimat: N-Amerika; einjähriges Kraut aufRuderalstellen, Verladeplätzen, in Hafenan-lagen; in Deutschland seit der zweiten Hälftedes 19. Jahrhunderts. Windblütig, gefährli-cher Allergieauslöser. Im Gebiet: Himmel-geister Rheinbogen, auf dem Grind.
^^

Pflanzliche Neubürger (Neophyten) im Raum Düsseldorf/Neuss 75
Acta Biologica Benrodis 13 (2006)
176. Ambrosia trifida, Dreilappige Ambrosie(Asteraceae, Korbblütler).
Heimat: USA, S-Kanada; einjähriges Krautauf Ruderalstellen, an Flußufern; Neophytseit 1900. Im Gebiet: Neusser Hafen.177. Amelanchier lamarckii (= A. canadensis),Kupfer-Felsenbirne (Rosaceae, Rosengewächse).
Heimat: nordöstliches N-Amerika; Zier-strauch; in der Heimat bis 10 m hoher Baum;Ersteinfuhr nach Europa 1623; Erstnachweisals Neophyt 1883. Im Gebiet: zerstreut.178. Aster lanceolatus, Lanzettblättrige Aster(Asteraceae, Korbblütler).
Heimat: östliches N-Amerika; ausdauerndesKraut; Zierpflanze; verwildert an Flußufern, inAuwäldern; eingeführt im 18. Jahrhundert;Neophyt seit dem 19. Jahrhundert. Im Gebiet:TK 4707-2, 3, 4806-2, 4807-2, 3.179. Aster novi-belgii, Glattblatt-Aster, (Aster-aceae, Korbblütler).
Heimat: küstennahes östliches N-Ameri-ka; mehrjähriges Kraut; Zierpflanze; Neo-phyt seit dem 18. Jahrhundert. Im Gebiet: z.B. Himmelgeister Rheinbogen, auf demGrind.180. Aster tradescantii (= A. parviflorus), Klein-blütige Aster (Asteraceae, Korbblütler).
Heimat: östliches N-Amerika; ausdauern-des Kraut; Zierpflanze; in Kultur seit 1640 inEngland; 1651 im herzoglich braunschweigi-schen Garten zu Hessen (KRAUSCH 2003);Neophyt seit 1736. Im Gebiet: auf demGrind.181. Bidens frondosa, Schwarzfrüchtiger Zwei-zahn (Asteraceae, Korbblütler).
Heimat: N-Amerika; einjähriges Kraut inUnkrautgesellschaften an Flußufern; Erstnach-weis für Deutschland 1894 (HANF 1991), nachROTHMALER (2002) 1891, nach LOHMEYER &SUKOPP (1992) 1736. Im Gebiet: z. B. Him-melgeister Rheinbogen, auf dem Grind.182. Claytonia perfoliata, Tellerkraut, Winter-portulak, Kubaspinat (Portulacaceae, Portu-lakgewächse).
Heimat: westliches N-Amerika; einjährigesKraut; aus Gärten verwildert in Grünanlagen,auf Friedhöfen und Ruderalstellen; als Salat
genutzt; Neophyt seit der zweiten Hälfte des19. Jahrhunderts. Im Gebiet: zerstreut.183. Cornus sericea (= C. stolonifera), Spros-sender Hartriegel (Cornaceae, Hartriegelge-wächse).
Heimat: N-Amerika; bis 2,5 m hoher Zier-strauch; Ersteinfuhr nach Europa 1656; Erst-nachweis als Neophyt 1861. Im Gebiet: TK4605-4, 4707-4, 4806-1, 3, 4807-4.184. Coronopus didymus, Zweiknotiger Krähen-fuß (Brassicaceae, Kreuzblütler).
Heimat: N-Amerika; einjähriges Kraut aufRuderalstellen, in Gärten; Erstnachweis als Ne-ophyt 1808. Im Gebiet: z. B. Neusser Hafen.185. Cuscuta campestris, NordamerikanischeSeide (Convolvulaceae, Windengewächse).
Heimat: südöstliches N-Amerika; einjährigesKraut in Klee- und Luzernefeldern; chlorophyll-loser Vollschmarotzer; Erstnachweise als Neo-phyt 1883, 1898. Im Gebiet: TK 4606-3.186. Cuscuta gronovii, Weiden-Seide (Convol-vulaceae, Windengewächse).
Heimat: östliches N-Amerika; einjährigesKraut; schmarotzt in Rheinufernähe aufWeiden, Brennesseln, Astern, Schafgarbe undanderen; Erstnachweis als Neophyt 1881. ImGebiet: TK 4606-2, 4, 4706-1, 3, 4807-3.187. Elodea canadensis, Kanadische Wasserpest(Hydrocharitaceae, Froschbissgewächse).
Heimat: N-Amerika; ausdauerndes zwei-häusiges Kraut; submers lebende Wasserpflan-ze in kühlen und klaren, stehenden und lang-sam fließenden Gewässern; ursprünglichAquarienpflanze; in Europa nur weiblichePflanzen, daher ausschließlich vegetative Ver-mehrung; in Europa seit Mitte des 19. Jahr-hundert; durch Gewässerverschmutzung starkzurückgegangen. Im gesamten Gebiet.188. Elodea nuttallii, Amerikanische Wasser-pest (Hydrocharitaceae, Froschbissgewächse).
Heimat: N-Amerika; ausdauerndes Kraut;submers lebende Wasserpflanze in stehen-den, auch verschmutzten Gewässern; zwei-häusig; in Europa nur männliche Pflanzen;ursprünglich Aquarienpflanze; hat gebietswei-se E. canadensis verdrängt (DÜLL & KUTZEL-NIGG 1994). Im gesamten Gebiet.

76 WERNER DIETRICH
Abb. 9: Zweijähriger Feinstrahl (Erigeron annu-us)Fig. 9: Tall Fleabane (Erigeron annuus).
Abb. 10: Riesen-Goldrute (Solidago gigantea).Fig. 10: Early Goldenrod (Solidago gigantea).
Abb. 11: Zucker-Spitzklette (Xanthium saccha-ratum).Fig. 11: Sugar Cocklebur (Xanthium saccharatum).
Abb. 12: Giftbeere (Nicandra physalodes).Fig. 12: Apple-of-Peru (Nicandra physalodes).

Pflanzliche Neubürger (Neophyten) im Raum Düsseldorf/Neuss 77
Acta Biologica Benrodis 13 (2006)
189. Epilobium ciliatum, Drüsiges Weidenrös-chen (Onagraceae, Nachtkerzengewächse).
Heimat: N-Amerika; ausdauerndes Kraut aufRuderalstellen, an Böschungen, in Gärten, aufBrachen und auf Kahlschlägen; Erstnachweisals Neophyt 1927. Im Gebiet: häufig.190. Erigeron annuus, Zweijähriger Feinstrahl(Asteraceae, Korbblütler; Abb. 9).
Heimat: Kanada, nördliche USA; zweijäh-riges, selten einjähriges oder ausdauerndesKraut in Auwäldern und Unkrautgesellschaf-ten; in Deutschland seit der zweiten Hälftedes 17. Jahrhunderts; verstärkte Ausbreitungseit Mitte des 19. Jahrhunderts. Im Gebiet:z. B. Himmelgeister Rheinbogen, auf demGrind, Heltorfer Mark bei Angermund.191. Erigeron canadensis (= Conyza canadensis),Kanadisches Berufkraut, Katzenschweif (As-teraceae, Korbblütler).
Heimat: N-Amerika; ein-bis zweijährigesKraut in Unkrautgesellschaften, Gärten undauf Kahlschlägen; Erstnachweis 1655 inFrankreich; schon im Laufe des 17. Jahrhun-derts über ganz Mitteleuropa sich ausbreitend;ungeheuere Vermehrung durch �Samen�(Achänen); bis zu 120 000 pro Pflanze (KLEIN
1973, RAUH & HANF 1983); Droge wird ge-gen starke Durchfälle verwendet (HOPPE 1975,LICHTENSTERN et al. 1994). Der Name Beruf-kraut bezieht sich auf das ähnliche Echte Be-rufkraut (Erigeron acris), das früher den Kin-dern in die Wiege gelegt wurde, um sie vor�Beschreien� oder �Berufen� zu schützen(CREMER et al. 1991). Im Gebiet: überall.192. Euphorbia maculata (= Chamaesyce macula-ta), Gefleckte Wolfsmilch (Euphorbiaceae,Wolfsmilchgewächse).
Heimat: N-Amerika; einjähriges Kraut aufBahn- und Industriegelände, auf Friedhö-fen, in Pflasterfugen; Neophyt seit 1877. ImGebiet: TK 4606-1.193. Helianthus annuus, Gemeine Sonnenblu-me (Asteraceae, Korbblütler).
Heimat: N-Amerika, auch Mexiko und Peru;einjähriges bis 3 m hohes Kraut auf Schuttplät-zen und ähnlichen Orten; im 16. Jahrhundertvon den Spaniern als Zierpflanze nach Europa
gebracht; Erstnachweis für Deutschland um1570; von den Ureinwohnern Amerikas bereitsvor 3000 Jahren kultiviert (QUATMANN 1996);wegen der ölreichen �Samen� (Achänen) heutein den gemäßigten Zonen in vielen Kulturfor-men angebaut; beliebte Zierpflanze; Vogelfut-ter. Im Gebiet: z. B. Rheinufer bei Kaiserswerth,Himmelgeister Rheinbogen, auf dem Grind.194. Helianthus tuberosus, Erdbirne, Topinam-bur, FRANKE (1997): Jerusalemartischocke,RAUH (1950): Pferdekartoffel (Asteraceae,Korbblütler).
Heimat: NO-Kanada, östliche und mittlereUSA; ausdauerndes Kraut auf Ruderalstellen,Brachen, an Flußufern; erst ab September blü-hend, da Kurztagpflanze; Früchte (Achänen)reifen bei uns nicht aus (DÜLL & KUTZELNIGG
1994); Nutzpflanze; Inulin anstelle von Stärkeals Speicherstoff in den Knollen; feldmäßigangebaut; Kraut als Viehfutter, grün oder si-liert; auch Knollen vorwiegend zur Tiefütte-rung;1607 nach Paris aus den französischenKolonien im östlichen N-Amerika; von dortrasche Verbreitung über Europa (KRAUSCH
2003); Erstnachweis als Neophyt für Deutsch-land um 1830. Die Bezeichnung Topinamburstammt vom Indianerstamm Tupinambus inKanada (TRONÍCKOVÁ 1985); nach MARZELL
(1972) soll dieser Indianerstamm jedoch inSüdamerika leben. Im Gebiet: HimmelgeisterRheinbogen, Kaiserswerth, auf dem Grind.195. Hordeum jubatum, Mähnen-Gerste (Po-aceae, Süßgräser).
Heimat: N-Amerika, Mexiko, (auch O-Asi-en, SCHMEIL & FITSCHEN 2000); einjährigesZiergras; verwildert auf Ruderalstellen, Tage-baukippen, salzbeeinflußte Küstenweiden;Neophyt seit dem 19. Jahrhundert. Im Ge-biet: TK 4706-3, 4707-4.196. Impatiens capensis, Orangefarbenes Spring-kraut (Balsaminaceae, Balsaminengewächse).
Heimat: N-Amerika; einjähriges Kraut anGewässerufern; Neophyt seit zweite Hälftedes 20. Jahrhunderts. Im Gebiet: Urdenba-cher Kämpe (HAEUPLER et al. 2003).197. Juncus tenuis, Zarte Binse (Juncaceae, Bin-sengewächse).
^

78 WERNER DIETRICH
Heimat: N-Amerika; ausdauerndes grasähn-liches Kraut, vor allem auf trittverdichtetenWegen; Neophyt seit 1834. Im Gebiet: häufig.198. Lemna minuta, Zierliche Wasserlinse (Lem-naceae, Wasserlinsengewächse).
Heimat: N-Amerika; ausdauernde Wasser-pflanze in stehenden und langsam fließen-den Gewässern; vermutlich Verschleppungdurch Wasservögel. Im Gebiet: TK 4806-1,3, 4807-3.199. Lepidium densiflorum, Dichtblütige Kres-se (Brassicaceae, Kreuzblütler).
Heimat: N-Amerika; einjähriges Kraut auftrockenen, sandigen Ruderalstellen; Neophytseit 1879; mit Baumwolle eingeschleppt(KLEIN 1973). Im Gebiet: z. B. Hauptbahn-hof Düsseldorf.200. Lepidium virginicum, Virginische Kresse(Brassicaceae, Kreuzblütler).
Heimat: N-Amerika; einjähriges Kraut aufRuderalstellen, in Gärten; Neophyt seit 1786.Im Gebiet: zerstreut.201. Lupinus polyphyllus, Vielblättrige Lupine(Fabaceae, Schmetterlingsblütler).
Heimat: westliches N-Amerika; mehrjäh-riges Kraut an Waldsäumen und Waldschlä-gen, an Böschungen; 1826 nach England ein-geführt; in Mitteleuropa seit 1837; Erstnach-weis als Neophyt 1890; als Wildfutter undGründünger sowie zur Böschungsbefesti-gung kultiviert; sät sich agressiv aus und ver-drängt heimische Vegetation; Samen giftig,vor allem für Pferde und Schafe in hohenDosen tödlich; auch Vergiftungen bei Kin-dern (ALTMANN 1995, STARY 1983). Im Ge-biet: häufig.202. Mahonia aquifolium, Mahonie (Berberi-daceae, Sauerdorngewächse).
Heimat: westliches N-Amerika; bis 1 m ho-her Zierstrauch; verwildert in stadtnahen, tro-ckenen Wäldern und Parks; 1838 in England;seit 1841 in Deutschland in Kultur; Erstnach-weis als Neophyt 1861; Beeren verwendbar fürMarmeladen und Süßmost (KRAUSCH 2003);Name Mahonia zu Ehren des amerikanischenGartenfachmanns Bernhard Mac Mahon(1775-1816). Im Gebiet: z. B. Eller Forst.
203. Mimulus guttatus, Gelbe Gauklerblume(Scrophulariaceae, Rachenblütler).
Heimat: westliches N-Amerika; ausdauern-de Zierpflanze; verwildert an Bach- und Fluß-ufern, an Gräben; Neophyt seit 1824. ImGebiet: TK 4605-4, 4606-3.204. Myriophyllum heterophyllum, Verschieden-blättriges Tausendblatt (Haloragaceae, Tau-sendblattgewächse).
Heimat: östliches N-Amerika; ausdau-ernde submerse Wasserpflanze in stehen-den Gewässern; Erstnachweis als Neophyt1930 (ROTHMALER 2002), 1940 (LOHMEYER
& SUKOPP 1992). Im Gebiet: z. B. spontanim Botanischen Garten der UniversitätDüsseldorf.205. Oenothera biennis, Gewöhnliche Nacht-kerze (Onagraceae, Nachtkerzengewächse).
Heimat: östliches N-Amerika; zweijährigesKraut an Bahndämmen, in Sandfeldern, aufRuderalstellen; eingeführt Anfang des 17.Jahrhunderts; Erstnachweise als Neophyt1720 in Hessen, 1769 in Schleswig-Holstein;Wurzeln früher wie Schwarzwurzeln verwen-det (Schinkenwurzel; DÜLL & KUTZELNIGG
1994); vielseitige medizinische Verwendungwegen des hohen Gehalts an Gamma-Lino-lensäure in den Samen (QUATMANN 1996). ImGebiet: z. B. Himmelgeister Rheinbogen, aufdem Grind.206. Oenothera parviflora, Kleinblütige Nacht-kerze (Onagraceae, Nachtkerzengewächse).
Heimat: östliches N-Amerika; zweijährigesKraut auf Ruderalstellen, an Flußufern; Neo-phyt seit Ende des 18. Jahrhunderts. ImGebiet: z. B. auf dem Grind.207. Oxalis fontana, Steifer Sauerklee (Oxali-daceae, Sauerkleegewächse).
Heimat: N-Amerika; ein- bis mehrjährigesKraut in Gärten, auf Äckern und Ruderal-stellen; eingeschleppt mit Tabak oder Kar-toffeln; Erstnachweise als Neophyt inDeutschland: 1807 in Schleswig-Holstein,1926 in Hannover, 1850 in Bayern; in Gemü-segärten lästiges Unkraut (RAUH & HANF
1983). Im Gebiet: häufig, z. B. Himmelgeis-ter Rheinbogen, auf dem Grind.

Pflanzliche Neubürger (Neophyten) im Raum Düsseldorf/Neuss 79
Acta Biologica Benrodis 13 (2006)
208. Panicum capillare, Haarstielige Hirse (Po-aceae, Süßgräser).
Heimat: N-Amerika; einjähriges Ziergras;verwildert in Gärten, auf Äckern, in Parkan-lagen, auf Ruderalstellen; Neophyt seit 1890.Im Gebiet: TK 4706-1, 3.209. Parthenocissus inserta (= P. vitacea), Ge-wöhnliche Jungfernrebe (Vitaceae, Weinre-bengewächse).
Heimat: N-Amerika; bis 8 m lang werden-de holzige Kletterpflanze; Zierpflanze zurMauerbegrünung; verwildert in Auwäldernund Gebüschen; in Deutschland vor 1800.Im Gebiet: zerstreut.210. Phacelia tanacetifolia, Büschelschön, Bie-nenfreund (Hydrophyllaceae, Wasserblattge-wächse).
Heimat: Kalifornien bis Mexiko; einjähri-ges Kraut, unbeständig verwildert auf Brach-flächen; ursprünglich als Zierpflanze einge-führt; 1832 in England, 1843 in Deutschland;heute oft kultiviert als Trachtpflanze für Bie-nen und zur Gründüngung. Im Gebiet: zer-streut, z. B. Himmelgeister Rheinbogen.211. Polygonum pensylvanicum, PensylvanischerKnöterich (Polygonaceae, Knöterichgewächse).
Heimat: N-Amerika; einjähriges Kraut anFlußufern und auf Ruderalstellen; einge-schleppt Ende 20. Jh. (WISSKIRCHEN &HAEUPLER 1998). Im Gebiet: HimmelgeisterRheinbogen, Neusser Hafen.212. Populus balsamifera, Balsam-Pappel (Sali-caceae, Weidengewächse).
Heimat: N-Amerika; bis 30 m hoher Zier-baum; Parkbaum; seit 1689 in gärtnerischerKultur; im Frühjahr auffällig duftend wegenstarker Harzabscheidung an den Knospen, die-se auch als Naturheilmittel verwendet (gemmaepopuli, VETVICKA 1985). Im Gebiet: zerstreut.213. Prunus serotina, Späte Traubenkirsche(Rosaceae, Rosengewächse).
Heimat: gemäßigtes nördliches und östli-ches N-Amerika; bis 30 m hoher Baum, beiuns oft nur strauchig wachsend; als Forst-baum angepflanzt und oft verwildert; inEuropa seit 1629; Neophyt seit der zweitenHälfte des 19. Jahrhunderts; starke vegetative
Vermehrung; bildet dichten Unterwuchs inWäldern, unterdrückt heimische Vegetation;ökologische Problempflanze (DÜLL & KUT-ZELNIGG 1994). Im Gebiet: häufig.214. Quercus rubra, Rot-Eiche (Fagaceae, Bu-chengewächse).
Heimat: N-Amerika; bis 25 (-40) m hoherBaum; seit 1724 in Kultur als Straßen- undParkbaum; häufig verwildert aus Naturverjün-gung; wichtigster fremdländischer Laubbaumder Forstwirtschaft (KRÜSSMANN 1978; DÜLL &KUTZELNIGG 1994). Im Gebiet: häufig.215. Rhus typhina, Essigbaum (Anacardiaceae,Sumachgewächse).
Heimat: östliches N-Amerika; bis 3 (-10)m hoher Strauch; Ziergehölz, in Kultur seit1629; verwildert auf Ruderalstellen. Im Ge-biet: zerstreut.216. Robinia pseudoacacia, Gewöhnliche Robi-nie, Scheinakazie (Fabaceae, Schmetterlings-blütler).
Heimat: N-Amerika; bis 25 m hoher Forst-,Park- und Straßenbaum; oft verwildert; 1601von dem Pariser Hofgärtner Jean Robin alsZierbaum eingeführt; seit 1635 in Deutsch-land, 1675 im Botanischen Garten Leipzig,1710 in Berlin; Erstnachweis als Neophyt1824; ökologische Problempflanze; in steterAusbreitung begriffen; durch Stickstoffanrei-cherung im Boden Förderung stickstofflie-bender Pflanzen; Verdrängung artenreicherVorvegetation. In Ostdeutschland wurdenwertvolle Trockenrasen durch Robinienauf-forstung zerstört (DÜLL & KUTZELNIGG 1994).Rinde und Laub giftig für Pferde (HILLER &BICKERICH 1997); Holz sehr fest und bieg-sam, kaum schwindend, vielseitig verwend-bar (HECKER 1995). Im Gebiet: häufig.217. Rudbeckia hirta, Rauher Sonnenhut (As-teraceae, Korbblütler).
Heimat: S-Kanada bis N-Mexiko; ein- bismehrjähriges Kraut; Zierpflanze; unbestän-dig verwildert auf Ruderalstandorten; Erst-nachweis als Neophyt 1860. Im Gebiet: TK4606-3, 4807-2.218. Rudbeckia laciniata, Schlitzblättriger Son-nenhut (Asteraceae, Korbblütler).
^^

80 WERNER DIETRICH
Heimat: NO-Kanada, östliche USA; aus-dauerndes Kraut; Zierpflanze; verwildertund eingebürgert in Staudenfluren, anFluß- und Bachufern; Ersteinfuhr um1650; Erstnachweis als Neophyt 1790. ImGebiet: 4706-4.219. Rumex triangulivalvis, Weidenblatt-Amp-fer (Polygonaceae, Knöterichgewächse).
Heimat: N-Amerika; ausdauerndes Krautan feuchten Ruderalstellen und Flußufern;Erstnachweis als Neophyt 1903. Im Gebiet:TK 4706-3.220. Shinnersia rivularis, Mexikanisches Eichen-blatt (Asteraceae, Korbblütler).
Heimat: Texas (nördöstliche Rio-Grande-Plains und südlicher Teil des Edwards Pla-teau), Coahuila in N-Mexiko (CORRELL &CORRELL 1975); ausdauernde Wasserpflanze;Warmwasserneophyt in der unteren Erft; nurblütenlose Bestände; in der Heimat am na-türlichen Standort auch terrestrisch wachsend.Im Gebiet: TK 4806-3. Vom Autor in derErft bei Grevenbroich-Wevelinghoven 1992beobachtet (TK 4905-2).221. Solanum carolinense, Carolina-Pferdenes-sel (Solanaceae, Nachtschattengewächse).
Heimat: östliches N-Amerika von Ontariobis Florida und Texas (RADFORD et al. 1968);ausdauerndes Kraut auf Ruderalstellen. ImGebiet: Neusser Hafen (14.09.1991).222. Solanum cornutum (= S. angustifolium, S.rostratum), Geschnäbelter Nachtschatten (So-lanaceae, Nachtschattengewächse).
Heimat: Kanada bis Mexiko; einjährigesKraut auf Ruderalstellen. Im Gebiet: NeusserHafen (Erstfund 1978, STIEGLITZ 1980).223. Solidago canadensis, Kanadische Goldrute(Asteraceae, Korbblütler).
Heimat: N-Amerika; ausdauerndes Kraut;Zierpflanze; verwildert in Auwäldern, anFlußufern, auf Ruderalstellen und Brachen;Ersteinfuhr nach Deutschland 1651; Erst-nachweis als Neophyt 1853. Im Gebiet: aufdem Grind.224. Solidago gigantea, Riesen-Goldrute (As-teraceae, Korbblütler; Abb. 10).
Heimat: N-Amerika; ausdauerndes Kraut;
Zierpflanze; verwildert an Standorten wie dievorige; Ersteinfuhr nach W-Europa 1758;Erstnachweis als Neophyt 1859; von Imkernals Bienentrachtpflanze ausgebracht, aber nurmäßiges Nektarangebot; gezieltes Ausbrin-gen daher abzulehnen (LUDWIG et al. 2000).Im Gebiet: Kaiserswerth, HimmelgeisterRheinbogen, auf dem Grind.225. Spiraea douglasii, Douglas� Spierstrauch(Rosaceae, Rosengewächse).
Heimat: westliches N-Amerika; bis 2,5 mhoher Zierstrauch, auch Heckenpflanze; ver-wildert vornehmlich auf Bahngelände; inMitteleuropa in Kultur seit 1827. Im Gebiet:TK 4606-3.226. Symphoricarpos albus, Schneebeere, �Knall-erbse� (Caprifoliaceae, Geißblattgewächse).
Heimat: westliches N-Amerika; bis 1,5 mhoher Zierstrauch; verwildert in siedlungsna-hen Gebüschen; Ersteinfuhr nach England1727, nach Deutschland in der zweiten Hälftedes 18. Jahrhunderts. Im Gebiet: zerstreut.227. Veronica peregrina, Amerikanischer Eh-renpreis (Scrophulariaceae, Rachenblütler).
Heimat: N- und S-Amerika; einjährigesKraut; Pionierpflanze auf feuchten Böden, inWegvertiefungen, an Ackerrändern, in Gärtne-reien und Baumschulen; in England seit 1680(SEBALD et al. 1996); Neophyt in Deutschlandseit 1838 (ROTHMALER 2002); nach LOHMEYER
& SUKOPP (1992) schon seit 1760. Im Gebiet:spontan im Botanischen Garten der Universi-tät Düsseldorf (Sommer 2004).228. Xanthium saccharatum, Zucker-Spitzklet-te (Asteraceae, Korbblütler; Abb. 11).
Heimat: N-Amerika; einjähriges Kraut aufRuderalstellen, an Flußufern. Neophyt seit1922. Im Gebiet: Himmelgeister Rheinbo-gen, auf dem Grind.
3.6. Südamerika/südliches Mittelamerika
229. Amaranthus patulus (= A. cruentus), Aus-gebreiteter Fuchsschwanz (Amaranthaceae,Fuchsschwanzgewächse).
Heimat: tropisches Amerika; einjährigesKraut auf Ruderalstellen; auch Zierpflanze. Im

Pflanzliche Neubürger (Neophyten) im Raum Düsseldorf/Neuss 81
Acta Biologica Benrodis 13 (2006)
Gebiet: Himmelgeister Rheinbogen (TK 4806-4; 1.9.2004); Neusser Hafen (STIEGLITZ 1981).230. Amaranthus powellii (= A. chlorostachys),Grünähriger Fuchsschwanz (Amaranthaceae,Fuchsschwanzgewächse).
Heimat: südliche USA, Mexiko (SCHMITZ
2002); einjähriges Kraut auf Ruderalstellenund Äckern; Erstnachweis als Neophyt inDeutschland 1889 im Hafen von Mannheimund Ludwigshafen; am Niederrhein seit denca. 1980er Jahren (SCHMITZ 2002). Im Gebiet:Kaiserswerth, Himmelgeister Rheinbogen,auf dem Grind.231. Amaranthus spinosus, Dorniger Amarant(Amaranthaceae, Fuchsschwanzgewächse).
Heimat: tropisches Amerika (?); heute inallen Tropen verbreitet; einjähriges Kraut aufRuderalstellen. Im Gebiet: Neusser Hafen(STIEGLITZ 1981).232. Anoda cristata, Gespornte Sternmalve(Malvaceae, Malvengewächse).
Heimat: tropisches S-Amerika bis südwest-liche USA; ein- bis mehrjähriges Kraut. ImGebiet: Neusser Hafen (STIEGLITZ 1981).233. Azolla filiculoides (= A. caroliniana), Al-genfarn (Azollaceae, Algenfarngewächse).
Heimat: subtropisches Amerika; ausdau-ernder Schwimmfarn auf warmen, windge-schützten, eutrophen, stehenden und strö-mungsarmen Fließgewässern; auch Aquari-enpflanze; Erstnachweis als Neophyt 1870;meist durch Aquarianer eingebracht; am Nie-derrhein eingebürgert, hier seit über zehn Jah-ren beobachtet und weiterhin in Ausbreitung(HAEUPLER et al. 2003). Im Gebiet: TK 4606,4806-1, 4807-1.234. Bromus catharticus (= B. unioloides, B. will-denowii), Pampas-Trespe (Poaceae, Süßgräser).
Heimat: S-Amerika; ein- bis zweijährigesGras auf Ruderalstellen (z. B. auf Bahnge-lände, in Häfen); Erstnachweis als Neophyt1889. Im Gebiet: z. B. TK 4706-3.235. Chenopodium ambrosioides, Wohlriechen-der Gänsefuß (Chenopodiaceae, Gänsefuß-gewächse).
Heimat: tropisches Amerika; einjährigesKraut auf Ruderalstellen; in Europa seit dem
17. Jahrhundert (LICHTENSTERN et al. 1994);vereinzelt auch angepflanzt; wichtige Arznei-und Gewürzpflanze der südamerikanischenIndianer; bei uns als Gewürz für Fleisch undGemüse verwendet (HLAVA & LANSKA 1977);in der Volksmedizin bei uns Wurmmittel,aber auch Magen- und Kräftigungsmittel(HOPPE 1975). Im Gebiet: auf dem Grind(TK 4806-2; 2.9.2004).236. Cucurbita maxima, Riesen-Kürbis (Cucur-bitaceae, Kürbisgewächse).
Heimat: tropisches Amerika; einjährigesKraut; Kulturpflanze; Futter- und Gemüse-pflanze (Früchte bis 100 kg schwer, FRANKE etal. 1976), auch Zierpflanze (Zierkürbis);im 16.Jahrhundert nach Europa gebracht (DE WIT
1964); wurde für die zweite Hälfte des 17. Jahr-hunderts archäobotanisch im Innenstadtbe-reich Düsseldorf nachgewiesen (GELIUS-DIET-RICH 2002/2003); verwildert auf Ruderalstel-len. Im Gebiet: auf dem Grind.237. Datura stramonium, Gewöhnlicher Stech-apfel (Solanaceae, Nachtschattengewächse).
Heimat: tropisches Amerika bis Mexiko;einjähriges Kraut auf sandigen Ruderalstel-len, Äckern und Brachen; in Europa seit 1584;wurde für den Zeitraum 1580-1620 archäo-botanisch für den Innenstadtbereich vonDüsseldorf nachgewiesen (GELIUS-DIETRICH
2002/2003). Erstnachweise als Neophyt im17. Jahrhundert; in allen Teilen der Pflanze,besonders in den Samen hoher Gehalt an dengiftigen Alkaloiden Hyoscyamin, Atropinund Scopolamin (ALTMANN 1995); vielfacherGebrauch und Mißbrauch; ca 20 Samen fürKinder tödlich (LICHTENSTERN et al. 1994,STARY 1983); wegen krampflösender Wirkungzur Herstellung sogenannter Asthmazigaret-ten verwendet (ALBERTS & MULLEN 2000);aber auch Bestandteil der Hexensalben(SCHERF 2002); früher nicht selten als Mord-und Selbstmordmittel (HILLER& BICKERICH
1997). Im Gebiet: z. B. Himmelgeister Rhein-bogen, auf dem Grind.238. Elodea densa (= Egeria densa), Dichtblätt-rige Wasserpest (Hydrocharitaceae, Froschbiss-gewächse).

82 WERNER DIETRICH
Heimat: tropisches Amerika bis Florida; aus-dauernde submers lebende Wasserpflanze;ursprünglich Aquarienpflanze; unbeständigverwildert in Warmwasserbereichen (Kanäle,Abwassergräben); Neophyt seit 1910. Im Ge-biet: TK 4806-1; zurzeit in der durch Sümp-fungswässer von Rhein-Braun erwärmten Erftfest eingebürgert (HAEUPLER et al. 2003).239. Galinsoga ciliata, Behaartes Franzosen-kraut (Asteraceae, Korbblütler).
Heimat: andines S-Amerika bis Mexiko;einjähriges Kraut auf Äckern und in Gärten;lästiges Unkraut; Neophyt seit Mitte des 19.Jahrhunderts. Im Gebiet: häufig.240. Galinsoga parviflora, Kleinblütiges Fran-zosenkraut (Asteraceae, Korbblütler).
Heimat: andines S-Amerika; einjährigesKraut; Vorkommen wie obige; bis zu 300000�Samen� (Achänen) pro Pflanze(KLEIN 1973);Neophyt seit Ende des 18. Jahrhunderts,Anfang des 19. Jahrhunderts aus BotanischenGärten verwildert (z. B. Berlin 1812, Erlan-gen 1821; MARZELL 1972). KLEIN (1973) be-richtet: �In zwei Fällen wurde die Ausbrei-tung des Franzosenkrauts der Nachwelt ganzpräzise aufgezeichnet. Pfarrer Hoffmann ausBudow in Pommern und Pfarrer Roth ausVegesack bei Bremen hatten es sich um 1800aus botanischen Gärten erbeten, um diesePflanze im eigenen Liebhabergarten zu ha-ben. Nicht lange, und es wurde auf den be-nachbarten Feldern zur Plage. Die Ausbrei-tung war nicht mehr aufzuhalten.� �Franzo-senkraut�, weil die Pflanze angeblich nach derBesetzung Osterodes durch die Franzosendort aufgetreten sein soll (DÜLL& KUTZEL-NIGG 1994). Im Gebiet: häufig.241. Myriophyllum aquaticum (= M. brasiliense),Brasilianisches Tausendblatt (Haloragaceae,Tausendblattgewächse).
Heimat: tropisches und subtropischesAmerika; ausdauernde Wasserpflanze. ImGebiet: Erftmündung (s. a. Nr. 233).242. Nicandra physalodes, Giftbeere (Solanaceae,Nachtschattengewächse; Abb. 12).
Heimat: Peru; einjähriges Kraut; Zierpflan-ze, verwildert auf Ruderalstellen; zwischen
1707 und 1712 von dem französischen Fran-ziskanerpater Louis Feuillée entdeckt; 1771 imBotanischen Garten Halle; Erstnachweis alsNeophyt um 1860; giftig. Im Gebiet: Him-melgeister Rheinbogen (TK 4806-4; 1.9.2004),auf dem Grind (TK 4806-2, 4807-1; 2.9.2004).243. Nicotiana rustica, Bauern-Tabak (Solana-ceae, Nachtschattengewächse).
Heimat: Peru (ROTHMALER 2002), Mittel-amerika (SCHMEIL & FITSCHEN 2000), Mittel-und nördliches S-Amerika (DÜLL & KUTZEL-NIGG 1994); einjähriges Kraut; Kulturpflanze;wurde für den Zeitraum 1580-1620 archäobo-tanisch für den Innenstadtbereich von Düs-seldorf nachgewiesen (GELIUS-DIETRICH 2002/2003); alte Bauerngartenpflanze; giftig; gele-gentlich verwildert. Im Gebiet: TK 4606-3.244. Panicum dichotomiflorum, SpätblühendeHirse (Poaceae, Süßgräser).
Heimat: S-Amerika; aus Argentinien mitWolle eingeschleppt; einjähriges Gras in Mais-äckern; Neophyt seit 1889. Im Gebiet: Neus-ser Hafen (STIEGLITZ 1981).245. Pharbitis hederacea (= Ipomoea hederacea),Efeu-Prunkwinde (Convolvulaceae, Winden-gewächse).
Heimat: tropisches Amerika; einjährigeswindendes Kraut; unbeständig einge-schleppt. Im Gebiet: Neusser Hafen (STIEG-LITZ 1981).246. Physalis peruviana, Kapstachelbeere (Sola-naceae, Nachtschattengewächse).
Heimat: Peru; einjähriges Kraut; Kultur-pflanze; selten verwildert; Verwendung:Frischobst, Marmelade, Gelee, in Getränken;unreife Früchte giftig (NOWACK & SCHULZ
1998); zu Beginn des 19. Jahrhunderts amKap der Guten Hoffnung eingeführt (daherder deutsche Name, FRANKE 1997, FRANKE etal. 1976). Im Gebiet: Himmelgeister Rhein-bogen, auf dem Grind.247. Solanum lycopersicum, Tomate (Solanaceae,Nachtschattengewächse).
Heimat: S-Amerika, besonders Peru; ein-jähriges Kraut, in der Natur ausdauernd; Kul-turpflanze; verwildert auf Ruderalstellen, ankiesigen Fluß- und Seeufern; den Indianern

Pflanzliche Neubürger (Neophyten) im Raum Düsseldorf/Neuss 83
Acta Biologica Benrodis 13 (2006)
Mexikos und Perus als Nutzpflanze bereitsbekannt; kam nach 1500 durch die Spaniermit der Kartoffel nach Europa; Erstnachweisals Neophyt 1584; in Italien, der �zweiten�Heimat der Tomate, schon um 1560 geges-sen; in Mitteleuropa zunächst Zierpflanze(�Liebesapfel�); Anbau als Gemüsepflanzehier noch um die Jahrhundertwende (18./19. Jahrhundert) verpönt, weil man sie fürgiftig hielt; erst seit etwa 100 Jahren hier alsNutzpflanze gebräuchlich; kam erst 1812 überEuropa nach N-Amerika; Züchtung vieler inGröße und Geschmack unterschiedlicher Sor-ten; Name leitet sich von dem mexikanischen�tomana� ab (STOBART 1972, FRANKE et al.1976, TRONÍCKOVÁ 1985, VOGEL 1991; DÜLL
et KUTZELNIGG 1994, FRANKE 1997); frühesteAbbildungen von Tomatenpflanzen baldnach ihrer Entdeckung Anfang 16. Jh. (KÖR-BER-GROHNE 1988). Im Gebiet: Himmelgeis-ter Rheinbogen, auf dem Grind.248. Solanum nitidibaccatum (= S. physalifoli-um), Glanzfrüchtiger Nachtschatten (Solana-ceae, Nachtschattengewächse).
Heimat: S-Amerika (Argentinien); einjäh-riges Kraut auf Ruderalstellen und Äckern;Neophyt seit 1880; giftig. Im Gebiet: z. B.auf dem Grind, Neusser Hafen.249. Solanum sarachoides, Saracho-Nachtschat-ten (Solanaceae, Nachtschattengewächse).
Heimat: S-Amerika (Brasilien); einjährigesKraut auf Ruderalstellen und Äckern; Neo-phyt seit 1903; giftig. Im Gebiet: TK 4606-3,4706-3, 4807-1.
3.7. Afrika
250. Ricinus communis, Rizinus, Wunderbaum(Euphorbiaceae, Wolfsmilchgewächse).
Heimat: tropisches Afrika; einjähriges Kraut;in der Heimat bis 12 m hoher Strauch oderBaum; bereits im Mittelalter in Europa; alsZierpflanze in Anlagen kultiviert; Neophyt ausrezenten Verwilderungen auf Ruderalstellen.Bereits im alten Ägypten und im Orient ver-wendete man das Öl der Samen zu kosmeti-schen, medizinischen und technischen Zwe-
cken (Brennöl); bis heute ähnlicher Gebrauch,sowohl als Kosmetikum, als Heilmittel (z. B.wirksames Abführmittel) und auch als hoch-wertiges Schmieröl (z. B. für Flugzeugtrieb-werke). Samen äußerst giftig wegen des Ge-halts an Ricin, das aber beim Kaltpressen nichtin das ungiftige Öl gelangt; tödliche Dosis 6-10 Samen (FRANKE et al. 1976; FURLENMEIER
1978; STARY 1983; LICHTENSTERN et al. 1994;ALTMANN 1995; FRANKE 1997; OBERDORFER
2001). Wissenschaftlicher Name wegen derÄhnlichkeit des Samens mit einer vollgesoge-nen Zecke (Ixodes ricinus, DÜLL& DÜLL 2003);deutscher Name Wunderbaum wegen der gro-ßen Blätter und des schnellen Wachstums(MARZELL 1977). Im Gebiet: Rheinaue (HAEU-PLER et al. 2003).251. Senecio inaequidens, Schmalblättriges Greis-kraut (Asteraceae, Korbblütler).
Heimat: S-Afrika; ausdauerndes Krautauf Ruderalstellen (Bahnanlagen, Straßen-ränder, Schuttplätze und ähnliche Orte);Neophyt seit 1889; seit den 60er Jahren ex-plosionsartige Ausbreitung über ganz Mit-teleuropa (KUHBIER 1996). Im Gebiet: all-gegenwärtig.252. Sorghum bicolor, Echte Mohrenhirse (Po-aceae, Süßgräser).
Heimat: wahrscheinlich Äquatorial-Afrika;einjähriges Gras; Kulturpflanze; unbeständigverwildert in Hafengelände oder auf Müllde-ponien; wichtigste Hirseart; Weltwirtschafts-pflanze; heute in allen wärmeren Ländernangebaut. Im Gebiet: Neusser Hafen (HAEU-PLER et al. 2003).
3.8. Australien
253. Chenopodium pumilio, Australischer Gän-sefuß (Chenopodiaceae, Gänsefußgewächse).
Heimat: Australien, Tasmanien; einjähri-ges Kraut auf sandigen Ruderalstellen; Erst-nachweis als Neophyt in Deutschland 1890;am Niederrhein Ende der 1980er/Anfangder 1990er Jahre (SCHMITZ 2002). Im Ge-biet: Himmelgeister Rheinbogen, auf demGrind.
^

84 WERNER DIETRICH
Danksagung
Mein Dank gilt Herrn Professor Dr. HARTMUT
GREVEN für die Möglichkeit der Publikation inActa Biologica Benrodis und meiner Frau, Dr.LISA GELIUS-DIETRICH, die mich auf vielen Ex-kursionen mit tatkräftiger Hilfe begleitet hat.
Literatur
AICHELE, D., SCHWEGLER, H.-W., 2000: Die Blü-tenpflanzen Mitteleuropas (2. Aufl.). � Fran-ckh-Kosmos, Stuttgart.
ALBERTS, A., MULLEN, P., 2000: PsychoaktivePflanzen, Pilze und Tiere. � Franckh-Kos-mos, Stuttgart.
ALTMANN, H., 1995: Giftpflanzen, Gifttiere. �BLV Verlagsgesellschaft, München.
BÄRTELS, A., 2001: Enzyklopädie der Gartenge-hölze. � Ulmer, Stuttgart.
BECKMANN, D., BECKMANN, B., 1990: Alraun, Bei-fuß und andere Hexenkräuter. � Campus-Verlag, Frankfurt, New York.
BESLER, B., 1613: Hortus Eystettensis. � Reprint,Schirmer Mosel, München 1988.
BONSTEDT, C. (ed.), 1932: Pareys Blumengärtne-rei. � Paul Parey, Berlin.
CHAMISSO, A. von, 1827: Übersicht der nutzbars-ten und der schädlichsten Gewächse, welchewild oder angebaut in Norddeutschland vor-kommen. � Dümmler, Berlin.
CORRELL, D. S., CORRELL, H.B., 1975: Aquaticand wetland plants of southeastern UnitedStates, Vol. 2. � University Press, Stanford.
CREMER, J., PARTZSCH, M., ZIMMERMANN, G.,SCHWÄR, CH., GOLTZ, H., 1991: Acker- undGartenwildkräuter. � Deutscher Landwirt-schaftsverlag, Berlin.
DE WIT, H.C.D., 1964: Knaurs Pflanzenreich inFarben, Band 1, Höhere Pflanzen 1. � Droe-mer, Zürich.
DIETRICH, W., WAGNER, W.L., RAVEN, P.H., 1997:Systematics of Oenothera Section OenotheraSubsection Oenothera. � Systematic BotanyMonographs 50, 1-234.
DÜLL, R., DÜLL, I., 2003: Geheimnisse der Mittel-meerflora. � IDH-Verlag, Bad Münstereifel.
DÜLL, R., KUTZELNIGG, H., 1994: Botanisch-ökologisches Exkursionstaschenbuch (5.Aufl.). � Quelle & Meyer, Heidelberg, Wies-baden.
FRANKE, W., 1997: Nutzpflanzenkunde (6. Aufl.).� Thieme, Stuttgart, New York.
FRANKE, G., HAMMER, H., HANELT, P., KETZ, H.-A.,NATHO, G., REINBOTHE, H., 1976: Früchte derErde. � Urania-Verlag, Leipzig, Jena, Berlin.
FUCHS, L., 1543: New Kreüterbuch. � M. Isen-grim, Basel.
FURLENMEIER, M., 1978: Wunderwelt der Heilpflan-zen. � Rheingauer Verlagsgesellschaft, Eltville.
GARCKE, A., 1972: Illustrierte Flora. Deutsch-land und angrenzende Gebiete (23. Aufl.). �Parey, Berlin, Hamburg.
GELIUS-DIETRICH, L., 2002/2003: BotanischeFunde und ihre Deutung aus der archäologi-schen Ausgrabung �Schlösser-Brauerei� in derDüsseldorfer Altstadt. � Acta Biologica Ben-rodis 12, 75-110.
GENAUST, H., 1996: Etymologisches Wörterbuchder botanischen Pflanzennamen (3. Aufl.). �Birkhäuser, Basel, Boston, Berlin.
GESSNER, K., 1561: Horti Germaniae. In: V. COR-DUS (ed.), Annotationes. � Argentorati, Strass-burg.
GESSNER, O., 1974: Gift- und Arzneipflanzen vonMitteleuropa (3. Aufl.). � C. Winter Univer-sitätsverlag, Heidelberg.
HAEUPLER, H., MUER, TH., 2000: Bildatlas derFarn- und Blütenpflanzen Deutschlands. �Ulmer, Stuttgart.
HAEUPLER, H., JAGEL, A., SCHUMACHER, W., 2003:Verbreitungsatlas der Farn- und Blütenpflan-zen in Nordrhein-Westfalen. � LÖBF Nord-rhein-Westfalen, Recklinghausen.
HANF, M., 1991: Neophyten � Neubürger in derPflanzenwelt Deutschlands. � BASF (Mittei-lungen für den Landbau 1), Limburgerhof.
HECKER, U., 1995: BLV Handbuch Bäume undSträucher. � BLV Verlagsgesellschaft, München.
HEGI, G., 1908-1931: Illustrierte Flora von Mit-teleuropa, 7 Bände (1. Aufl.). � J.F. Lehmanns,München.
HERER, J., BRÖCKERS, M., 1995: Die Wiederent-deckung der Nutzpflanze Hanf. � Heyne,München.
HILLER, K., BICKERICH, G., 1997: Das farbige Buchder Arznei- und Giftpflanzen. � Signa, Berlin.
HLAVA, B., LANSKA, D., 1977: Lexikon der Kü-chen- und Gewürzkräuter. � Nova Part,München.
HOPPE, H.A., 1975: Drogenkunde, Band 1, An-giospermen (8. Aufl.). � De Gruyter, Berlin,New York.

Pflanzliche Neubürger (Neophyten) im Raum Düsseldorf/Neuss 85
Acta Biologica Benrodis 13 (2006)
HUBBARD, C.E., 1985: Gräser. Beschreibung, Ver-breitung, Verwendung (2. Aufl.). � Ulmer,Stuttgart.
KLEIN, J.F., 1973: Unkraut verdirbt nicht. Ver-breitung der Art im Pflanzenreich. � Franckh-Kosmos, Stuttgart.
KNÖRZER, K.H., 1970: Römerzeitliche Pflanzen-funde aus Neuss. � Novaesium IV (= Limes-forschungen 10), Mann, Berlin.
KNÖRZER, K.H., 1971: Römerzeitliche Getrei-deunkräuter von kalkreichen Böden. � Rhei-nische Ausgrabungen 10, 467-481.
KÖRBER-GROHNE, U., 1988: Nutzpflanzen inDeutschland. Kulturgeschichte und Biologie(2. Aufl.). � Theiss, Stuttgart.
KRAUSCH, H.-D., 2003: �Kaiserkron und Paeoni-en rot...�. Entdeckung und Einführung unse-rer Gartenblumen. � Dölling und Galitz,Hamburg, München.
KRÜSSMANN, G., 1972: Handbuch der Nadelge-hölze. � Parey, Berlin, Hamburg.
KRÜSSMANN, G., 1976, 1977, 1978: Handbuchder Laubgehölze, 3 Bände (2. Aufl.). � Parey,Berlin, Hamburg.
KUHBIER, H., 1996: 100 Jahre Senecio inaequidens inBremen. � Abhandlungen des Naturwissen-schaftlichen Vereins zu Bremen 43/2, 531-536.
LICHTENSTERN, H. VOLAK, J., STODOLA, J., 1994:Das große Kräuterbuch der Gesundheit. �Gondrom, Bindlach.
LOHMEYER, W., SUKOPP, H., 1992: Agriophyten inder Vegetation Mitteleuropas. � Bundesfor-schungsanstalt für Naturschutz und Landschafts-ökologie, Heft 25, Bonn-Bad-Godesberg.
LUDWIG, M., GEBHARDT, H., LUDWIG, H.W.,SCHMIDT-FISCHER, S., 2000: Neue Tiere &Pflanzen in der heimischen Natur. � BLVVerlagsgesellschaft, München.
MARZELL, H., 1943-1979: Wörterbuch der deut-schen Pflanzennamen, 5 Bände. � Hirzel,Leipzig.
MATTHIOLUS, P.A., 1565: Commentarii in sex librosPedacii Dioscoridis. � V. Valgrisi, Venedig.
MATTHIOLUS, P.A., CAMERARIUS, J., 1586: De plan-tis epitome utilissima. � Feyerabend, Frank-furt/Main.
NOWACK, B., SCHULZ, B., 1998: Tropische Früch-te. Biologie, Verwendung, Anbau und Ernte.� BLV Verlagsgesellschaft, München.
OBERDORFER, E., 2001: PflanzensoziologischeExkursionsflora für Deutschland und angren-zende Gebiete (8. Aufl.). � Ulmer, Stuttgart.
QUATMANN, CH., 1996: Hausbuch der Kräuter. �Gondrom, Bindlach.
RADFORD, A.E., AHLES, H.E., BELL, C.R., 1968:Manual of the vascular flora of the Caroli-nas. � University of North Carolina Press,Chapel Hill.
RÄTSCH, CH., 1990: Pflanzen der Liebe. Aphro-disiaka in Mythos, Geschichte und Gegen-wart. � Hallwag, Bern, Stuttgart.
RAUH, W., 1950: Morphologie der Nutzpflanzen(2. Aufl.). � Quelle & Meyer, Heidelberg,Wiesbaden.
RAUH, W., HANF, M., 1983: Unkräuter (5.Aufl.). � Gebrüder Bornträger, Berlin,Stuttgart.
ROTHMALER, W. , JÄGER, E.J., WERNER, K. (eds.),2002: Exkursionsflora von Deutschland,Band 4 (9. Aufl.). � Spektrum AkademischerVerlag, Heidelberg, Berlin.
SCHERF, G., 2002: Zauberpflanzen, Hexenkräu-ter. Mythos und Magie heimischer Wild- undKulturpflanzen. � BLV Verlagsgesellschaft,München.
SCHMEIL, O., FITSCHEN, J. (SENGHAS, K., SEYBOLD,S., Hrsg.), 2000: Flora von Deutschland undangrenzender Länder (91. Aufl.). � Quelle &Meyer, Wiebelsheim.
SCHMITZ, U., 2002: Untersuchungen zum Vor-kommen und zur Ökologie neophytischerAmaranthaceae und Chenopodiaceae in derUfervegetation des Niederrheins. � Disser-tationes Botanicae 364. J. Cramer, Berlin,Stuttgart.
SEBALD, O., SEYBOLD, S., PHLIPPI, G., WÖRZ, A.,1992, 1996: Die Farn- und BlütenpflanzenBaden-Würrtembergs, Bände 4 und 5. � Ul-mer, Stuttgart.
STACE, C., 1997: New Flora of the British Isles(2. ed.).- University Press, Cambridge.
STARY, F., 1983: Giftpflanzen. � Dausien, Ha-nau/Main.
STIEGLITZ, W., 1980: Bemerkungen zur Adven-tivflora des Neusser Hafens. � Niederrheini-sches Jahrbuch 14, 121-128.
STIEGLITZ, W., 1981: Die Adventivflora desNeusser Hafens in den Jahren 1979 und1980. � Göttinger Floristische Rundbriefe15, 45-51.
STOBART, T., 1972: Lexikon der Gewürze. � Hör-nemann, Bonn.
TRONÍCKOVÁ, E., 1985: Gemüse. � Dausien, Ha-nau/Main.
^

86 WERNER DIETRICH
TUTIN, T.G., HEYWOOD, V.H., BURGES, N.A., MOO-RE, D.M., VALENTINE, S.M., WALTERS, S.M.,WEBB, D.A., 1972: Flora Europaea, Band 3. �University Press, Cambridge.
VETVICKA, V., 1985: Dausien�s großes Buch der Bäu-me und Sträucher. � Dausien, Hanau/Main.
VOGEL, S., 1991: Nutzpflanzen. � Mondadori,Verona.
WILLERDING, U., 1986: Zur Geschichte der Un-
5. Armoracia rusticana, Meerrettich64. Asparagus officinalis, Gemüse-Spargel178. Aster lanceolatus, Lanzettblättrige Aster179. Aster novi-belgii, Glattblatt-Aster6. Aster x salignus, Weidenblättrige Aster180. Aster tradescantii, Kleinblütige Aster65. Atriplex hortensis, Garten-Melde66. Atriplex oblongifolia, Langblättrige Melde7. Aubrieta deltoidea, Griechisches Blaukissen233. Azolla filicoloides, Algenfarn8. Barbarea intermedia, Mittleres Barbarakraut67. Berteroa incana, Graukresse181. Bidens frondosa, Schwarzfrüchtiger Zwei-
zahn114. Brassica napus, Raps68. Brassica nigra, Senf-Kohl234. Bromus catharticus, Pampas-Trespe140. Buddleja davidii, Chinesischer Sommerflie-
der69. Bunias orientalis, Morgenländisches Zacken-
schötchen115. Calepina irregularis, Wendig70. Calystegia silvatica ssp. pulchra141. Cannabis sativa ssp. sativa, Hanf71. Cardamine hirsuta, Behaartes Schaumkraut9. Cardaminopsis arenosa, Sand-Schaumkresse72. Cardaria draba, Pfeilkresse73. Carduus acanthoides, Weg-Distel10. Carex brizoides, Zittergras-Segge116. Centrantus ruber, Spornblume117. Cerastium tomentosum, Filziges Hornkraut74. Chaenorhinum minus, Orant235. Chenopodium ambrosioides, Wohlriechender
Gänsefuß75. Chenopodium botrys, Klebriger Gänsefuß76. Chenopodium foliosum, Echter Erdbeerspi-
nat
kräuter Mitteleuropas. � Wachholz, Neu-münster.
WISSKIRCHEN, R., & HAEUPLER, H., 1998: Stan-dardliste der Farn- und BlütenpflanzenDeutschlands. � Ulmer, Stuttgart.
ZANDER, R., ERHARDT, W., GÖTZ, E., BÖDEKER,N., & SEYBOLD, S., 2002: Handwörterbuchder Pflanzennamen (17. Aufl.). � Ulmer,Stuttgart.
Angenommen: 14.11.2004
56. Abutilon theophrasti, Samtpappel171. Acer negundo, Eschen-Ahorn1. Aconitum napellus, Blauer Eisenhut138. Acorus calamus, Kalmus57. Aesculus hippocastanum, Roßkastanie139. Ailanthus altissima, Götterbaum58. Alchemilla mollis, Weicher Frauenmantel59. Alnus incana, Grau-Erle172. Amaranthus albus, Weißer Fuchsschwanz173. Amaranthus blitoides, Westamerikanischer
Fuchsschwanz60. Amaranthus blitum, Aufsteigender Fuchs-
schwanz2. Amaranthus bouchonii, Bouchons Fuchs-
schwanz229. Amaranthus patulus, Ausgebreiteter Fuchs-
schwanz230. Amaranthus powellii, Grünähriger Fuchs-
schwanz174. Amaranthus retroflexus, Rauhaariger Fuchs-
schwanz231. Amaranthus spinosus, Dorniger Amarant175. Ambrosia artemisiifolia, Beifußblättrige
Ambrosie176. Ambrosia trifida, Dreilappige Ambrosie177. Amelanchier lamarckii, Kupfer-Felsenbirne3. Anchusa sempervirens, Ausdauernde Ochsen-
zunge61. Angelica archangelica, Echte Engelwurz232. Anoda cristata, Gepornte Sternmalve112. Anthoxanthum aristatum, Begranntes Ruch-
gras113 Antirrhinum majus, Großes Löwenmäulchen62. Anthriscus caucalis, Hundskerbel63. Apera interrupta, Unterbrochener Wind-
halm4. Aquilegia vulgaris, Gewöhnliche Akelei
Alphabetische Gesamtliste der Pflanzenarten
^^

Pflanzliche Neubürger (Neophyten) im Raum Düsseldorf/Neuss 87
Acta Biologica Benrodis 13 (2006)
77. Chenopodium opulifolium, Schneeballblättri-ger Gänsefuß
253. Chenopodium pumilio, Australischer Gänse-fuß
78. Chenopodium strictum, Gestreifter Gänsefuß182. Claytonia perfoliata, Tellerkraut11. Cochlearica danica, Dänisches Löffelkraut118. Colutea arborescens, Blasenstrauch12. Conopodium majus, Französische Erdkastanie119. Coriandrum sativum, Koriander13. Corispermum leptopterum, Schmalflügeliger
Wanzensame183. Cornus sericea, Sprossender Hartriegel184. Coronopus didymus, Zweiknotiger Krähenfuß14. Corydalis lutea, Gelber Lerchensporn142. Cotoneaster horizontalis, Fächer-Zwergmispel15. Crepis taraxacifolia, Blasen-Pippau143. Cucumis sativus, Gurke236. Cucurbita maxima, Riesen-Kürbis185. Cuscuta campestris, Nordamerikanische Seide186. Cuscuta gronovii, Weiden-Seide79. Cuscuta lupuliformis, Pappel-Seide120. Cynodon dactylon, Hundszahngras144. Datura ferox, Dorniger Stechapfel237. Datura stramonium, Gewöhnlicher Stech-
apfel121. Digitaria sanguinalis ssp. pectinatiformis, Blut-
Fingerhirse122. Diplotaxis muralis, Mauer-Doppelsame123. Diplotaxis tenuifolia, Schmalblättriger Dop-
pelsame80. Dipsacus laciniatus, Gelappte Karde16. Echinops exaltatus, Drüsenlose Kugeldistel81. Echinops sphaerocephalus, Bienen-Kugeldistel187. Elodea canadensis, Kanadische Wasserpest238. Elodea densa, Dichtblättrige Wasserpest188. Elodea nuttallii, Amerikanische Wasserpest189. Epilobium ciliatum, Drüsiges Weidenröschen124. Eragrostis minor, Kleines Liebesgras190. Erigeron annuus, Zweijähriger Feinstrahl191. Erigeron canadensis, Kanadisches Berufkraut17. Erucastrum gallicum, Französische Hunds-
rauke82. Euphorbia lathyris, Kreuzblütige Wolfsmilch192. Euphorbia maculata, Gefleckte Wolfsmilch18. Euphorbia x pseudovirgata, Scheinruten-
Wolfsmilch145. Fagopyrum esculentum, Echter Buchweizen146. Fallopia baldschuanica, Silberregen19. Festuca brevipila, Rauhblatt-Schafschwingel125. Foeniculum vulgare, Echter Fenchel20. Galanthus nivalis, Schneeglöckchen
83. Galega officinalis, Geisraute21. Galeobdolon argentatum, Silberblatt-Gold-
nessel239. Galinsoga ciliata, Behaartes Franzosenkraut240. Galinsoga parviflora, Kleinblütiges Franzo-
senkraut126. Geranium purpureum, Purpur-Storchschnabel22. Geranium pyrenaicum, Pyrenäen-Storch-
schnabel147. Glycine max, Sojabohne193. Helianthus annuus, Gemeine Sonnenblume194. Helianthus tuberosus, Erdbirne148. Heracleum mantegazzianum, Riesen-Bärenklau84. Herniaria hirsuta, Behaartes Bruchkraut85. Hesperis matronalis, Gewöhnliche Nacht-
viole23. Hieracium aurantiacum, Orangerotes Ha-
bichtskraut86. Hippophae rhamnoides, Sanddorn127. Hirschfeldia incana, Bastardsenf195. Hordeum jubatum, Mähnen-Gerste24. Hyacinthoides hispanica, Spanisches Hasen-
glöckchen25. Hyacinthoides non-scripta, Gemeines Hasen-
glöckchen26. Iberis umbellata, Dolden-Schleifenblume196. Impatiens capensis, Orangefarbenes Spring-
kraut149. Impatiens glandulifera, Drüsiges Spring-
kraut150. Impatiens parviflora, Kleinblütiges Spring-
kraut197. Juncus tenuis, Zarte Binse87. Kochia scoparia, Besen-Radmelde27. Laburnum anagyroides, Gewöhnlicher Gold-
regen88. Lactuca serriola, Kompass-Lattich28. Larix decidua, Europäische Lärche151. Larix kaempferi, Japanische Lärche29. Lathyrus latifolius, Breitblättrige Platterbse198. Lemna minuta, Zierliche Wasserlinse199. Lepidium densiflorum, Dichtblütige Kresse30. Lepidium graminifolium, Grasblättrige Kresse89. Lepidium latifolium, Breitblättrige Kresse200. Lepidium virginicum, Virginische Kresse128. Linaria cymbalaria, Zimbelkraut31. Linaria repens, Gestreiftes Leinkraut129. Lobularia maritima, Strand-Silberkraut130. Lolium multiflorum, Italienisches Raygras32. Lunaria annua, Judas-Silberblatt201. Lupinus polyphyllus, Vielblättrige Lupine152. Lycium barbarum, Gemeiner Bocksdorn

88 WERNER DIETRICH
90. Lysimachia punctata, Punktierter Gilbwei-derich
202. Mahonia aquifolium, Mahonie131. Malus domestica, Kultur-Apfel132. Malva sylvestris ssp. mauritiana, Mauretani-
sche Malve153. Matricaria discoidea, Strahllose Kamille33. Medicago x varia, Bastard-Luzerne34. Mentha spicata, Grüne Minze35. Mentha x villosa, Zottige Minze91. Mercurialis annua, Einjähriges Bingelkraut203. Mimulus guttatus, Gelbe Gauklerblume241. Myriophyllum aquaticum, Brasilianisches Tau-
sendblatt204. Myriophyllum heterophyllum, Verschieden-
blättriges Tausendblatt36. Narcissus pseudonarcissus, Gelbe Narzisse242. Nicandra physalodes, Giftbeere243. Nicotiana rustica, Bauern-Tabak205. Oenothera biennis, Gewöhnliche Nachtkerze37. Oenothera x fallax, Täuschende Nachtkerze38. Oenothera glazioviana, Rotkelchige Nacht-
kerze206. Oenothera parviflora, Kleinblütige Nachtkerze39. Onobrychis viciifolia, Futter-Esparsette133. Oxalis corniculata, Horn-Sauerklee207. Oxalis fontana, Steifer Sauerklee208. Panicum capillare, Haarstielige Hirse244. Panicum dichotomiflorum, Spätblühende Hirse154. Panicum miliaceum, Echte Hirse209. Parthenocissus inserta, Gewöhnliche Jung-
fernrebe210. Phacelia tanacetifolia, Büschelschön134. Phalaris canariensis, Kanariengras245. Pharbitis hederacea, Efeu-Prunkwinde246. Physalis peruviana, Kapstachelbeere155. Phytolacca esculenta, Essbare Kermesbeere40. Picea abies, Fichte92. Picris echioides, Wurmlattich211. Polygonum pensylvanicum, Pensylvanischer
Knöterich156. Polygonum polystachyum, Himalaya-Knöterich212. Populus balsamifera, Balsam-Pappel41. Populus x canadensis, Kanadische Pappel42. Populus x canescens, Grau-Pappel43. Potentilla intermedia, Mittleres Fingerkraut93. Potentilla norvegica, Norwegisches Fingerkraut94. Potentilla recta, Hohes Fingerkraut157. Prunus cerasifera, Kirschpflaume158. Prunus domestica, Pflaume213. Prunus serotina, Späte Traubenkirsche95. Psyllium arenarium, Sand-Flohsame
159. Pyrus communis, Garten-Birne214. Quercus rubra, Rot-Eiche160. Rapistrum rugosum, Runzliger Rapsdotter135. Reseda lutea, Gelber Wau44. Reynoutria x bohemica, Bastard-Staudenknö-
terich161. Reynoutria japonica, Japanischer Staudenknö-
terich162. Reynoutria sachalinensis, Sachalin-Stauden-
knöterich215. Rhus typhina, Essigbaum250. Ricinus communis, Rizinus216. Robinia pseudoacacia, Gewöhnliche Robinie96. Rorippa austriaca, Österreichische Sumpf-
kresse163. Rosa rugosa, Kartoffel-Rose164. Rubus armeniacus, Armenische Brombeere45. Rubus laciniatus, Schlitzblättrige Brombeere165. Rubus phoenicolasius, Japanische Weinbeere46. Rubus sciocharis, Schattenliebende Brom-
beere217. Rudbeckia hirta, Rauher Sonnenhut218. Rudbeckia laciniata, Schlitzblättriger Son-
nenhut97. Rumex thyrsiflorus, Rispen-Ampfer219. Rumex triangulivalvis, Weidenblatt-Ampfer98. Salsola kali ssp. tragus, Salzkraut99. Salvia verticillata, Quirlblütiger Salbei100. Sanguisorba minor ssp. polygama, Grubiger
Wiesenknopf166. Sedum spurium, Zweifelhafte Fetthenne47. Sempervivum tectorum, Echte Hauswurz251. Senecio inaequidens, Schmalblättriges Greis-
kraut48. Senecio vernalis, Frühlings-Greiskraut167. Setaria faberi, Fabers Borstenhirse101. Setaria italica, Kolbenhirse102. Setaria verticillata, Wirtel-Borstenhirse220. Shinnersia rivularis, Mexikanisches Eichen-
blatt103. Silybum marianum, Mariendistel168. Sisymbrium altissimum, Ungarische Rauke104. Sisymbrium loeselii, Loesels Rauke49. Sisymbrium volgense, Wolga-Rauke221. Solanum carolinense, Carolina-Pferdenessel222. Solanum cornutum, Geschnäbelter Nacht-
schatten105. Solanum luteum ssp. luteum, Gelber Nacht-
schatten247. Solanum lycopersicum, Tomate248. Solanum nitidibaccatum, Glanzfrüchtiger
Nachtschatten

Pflanzliche Neubürger (Neophyten) im Raum Düsseldorf/Neuss 89
Acta Biologica Benrodis 13 (2006)
249. Solanum sarachoides, Saracho-Nachtschatten223. Solidago canadensis, Kanadische Goldrute224. Solidago gigantea, Riesen-Goldrute50. Sorbus intermedia, Schwedische Eberesche252. Sorghum bicolor, Echte Moorenhirse51. Spiraea x billardii, Billards Spierstrauch225. Spiraea douglasii, Douglas� Spierstrauch226. Symphoricarpos albus, Schneebeere52. Symphoricarpos x chenaultii, Bastard-Koral-
lenbeere53. Symphytum x uplandicum, Futter-Beinwell106. Syringa vulgaris, Gemeiner Flieder136. Tamarix parviflora, Kleinblütige Tamariske
137. Trifolium incarnatum, Inkarnat-Klee107. Trifolium resupinatum, Persischer Klee54. Ulex europaeus, Stechginster108. Verbascum blattaria, Schaben-Königskerze169. Veronica filiformis, Faden-Ehrenpreis227. Veronica peregrina, Amerikanischer Ehrenpreis170. Veronica persica, Persischer Ehrenpreis55. Vicia villosa, Zottige Wicke109. Viola odorata, März-Veilchen110. Vulpia bromoides, Trespen-Fuchsschwingel111. Vulpia myuros, Mäuseschwanz-Fuchs-
schwingel228. Xanthium saccharatum, Zucker-Spitzklette

90 WERNER DIETRICH
Rezension
SAUERHOFF, FRIEDHELM (2003): Etymologisches Wörterbuch der Pflanzennamen. � Wis-senschaftliche Verlagsgesellschaft mbH, Stuttgart. 799 Seiten. Gebunden, 13,5 x 21,5 cm.ISBN 978-3-8047-1899-9, 98,-- Euro.
Der deutsche Name Brennnessel leitet sich von den bei Berührungen brennenden undnesselnden Eindrücken auf der Haut ab. Das scheint nahe liegend zu sein, ist aber falsch,denn �Nessel� kommt von Netz und bezieht sich darauf, dass früher aus den Bastfasern derBrennnessel ein leichtes, netzartiges Gewebe hergestellt wurde. So kann es dem Leser indiesem Buch häufiger gehen. Erstmals wird hier ausführlich die Herkunft von etwa 2300deutschen gebräuchlichen Pflanzennamen erläutert. Es werden sowohl ursprünglich hierheimische wie auch in den Gärten in den letzten Jahrzehnten vielfach angepflanzte Artenangeführt. Am besten schlägt man das Buch zuerst hinten auf. Dort findet sich nämlich dasVerzeichnis der deutschen Namen, wobei natürlich berücksichtigt wurde, dass zahlreichePflanzen unter mehreren Namen bekannt sind. So wird auch immer auf einen angenomme-nen Hauptnamen verwiesen. Warum aber bei manchen Verweisen der wissenschaftlicheName angegeben ist, bei anderen aber nicht, ist nicht einsichtig. Gelegentlich ist das Suchenetwas aufwändiger, wenn etwa der Bärlauch nicht hinter Bärlapp, sondern als �Bärenlauch,Bärlauch� hinter dem Bärenklau auftaucht. Bärlauch ist sicher der häufiger gebrauchte Name.Jedenfalls ist der Hauptteil des Buchs eben wegen dieser gelegentlichen Vieldeutigkeit derdeutschen Namen alphabetisch nach den wissenschaftlichen Namen geordnet (die ebenfallsein eigenes Register haben). Hier werden die Gattungsnamen, ihre deutschen Namen sowiederen Herkunft ausführlich erläutert. Immer angegeben sind auch die englischen und fran-zösischen Trivialnamen, gelegentlich auch die Namen in weiteren Sprachen, vor allem inSpanisch. Dort, wo der Volksmund eine Pflanzengattung mit mehreren Arten mit nureinem Namen belegt, wie beim Schilf, findet sich in der Regel nur die Erläuterung derGattung, sonst sind auch oft Arten angeführt. Die Erläuterungen sind nach kurzem Einle-sen gut verständlich und lassen viele vorher unverständliche Namen auf einmal verständlichwerden. Als Beispiel sei das Scharbockskraut Ranunculus ficaria genannt, denn Scharbock isteine andere Bezeichnung für Skorbut, eine Vitamin-C-Mangelkrankheit, und die Blätterenthalten viel Vitamin C. Aber schon danach fällt auch eine Lücke auf, so fehlt etwa (undsicher nicht als einziger) der Brennende Hahnenfuß Ranunculus flammula. Etwas unverständ-lich ist das Fehlen mancher Beschreibungen. Dann findet sich nur ein Hinweis auf Zander,das Handwörterbuch der Pflanzennamen (Zander, R., Encke, F., Buchheim, G. & Seybold,S. (1994): Handwörterbuch der Pflanzennamen, Verlag Eugen Ulmer, Stuttgart). Wenn mangerade bei einem Klassiker wie der Butterblume keine Antwort auf die Frage der Namens-entstehung findet, ist das etwas ärgerlich.Insgesamt ist das Buch für den, der an der Entstehung und Bedeutung deutscher Pflanzen-namen interessiert ist, und für den historisch arbeitenden Botaniker eine wertvolle Quelle.
Harro Hieronimus, Solingen

Seminatural ancient forest in urban agglomeration area "Ruhrgebiet" 91
Acta Biologica Benrodis 13 (2006)
Acta Biologica Benrodis 13 (2006): 91-104
Seminatural ancient forest in urban agglomerationarea “Ruhrgebiet”
Naturnahe Altwälder im Ballungsraum Ruhrgebiet
RENATE FUCHS, HERFRIED KUTZELNIGG & GUIDO BENNO FEIGE
Universität Duisburg-Essen, Campus Essen, Fachbereich Biologie undGeographie/Botanik, Universitätsstr. 5, D-45117 Essen, Germany
Summary: Through fieldwork, the influence of the neighbouring industrial area on flora andvegetation of the seminatural ancient forest district called �Duisburg-Mülheimer Wald� was inves-tigated. The urban as well as peri-urban woodland is characterized by red-oak, pine and spruceforests and a remarkable variety of different seminatural forest communities. In particular, thenumerous invasive alien plants (primarily plants escaped from cultivation) indicate the impairmentsand stress caused by the industrial area. Establishment of invasives is discussed considering thelocal problems in nature conservation and the floristic and vegetational importance of this semi-natural forest in urban agglomeration.
Urban woodland, seminatural forest, industrial area, invasive alien plants, ergasiophygophytes, Ruhrgebiet,Northrhine-Westphalia
Zusammenfassung: Im Rahmen einer floristisch-vegetationskundlichen Untersuchung des na-turnahen Altwaldgebiets �Duisburg-Mülheimer Wald� wurde der Frage der Auswirkungen des un-mittelbar benachbarten Industrieraumes nachgegangen. Der Wald im Kontakt bzw. in der Nähe dergroßstädtischen Bebauung zeichnet sich neben Rot-Eichen-, Kiefern- und Fichtenforste insbesonderedurch eine bemerkenswerten Vielfalt unterschiedlicher naturnaher Waldgesellschaften aus. Die hoheAnzahl gebietsfremder Pflanzenarten, hauptsächlich Garten- und Kulturflüchtlinge, zeigt jedochdeutlich die Beeinträchtigungen und Belastungen die vom Ballungsraum ausgehen. Im Hinblickauf die lokalen Probleme des Naturschutzes wird die floristische und vegetationskundliche Bedeu-tung der Etablierung von gebietsfremden Arten diskutiert.
Stadtwald, naturnahe Wälder, Industriegebiet, gebietsfremde Pflanzen, Ergasiophygophyten, Ruhrgebiet,Nordrhein-Westfalen
1. Introduction
In spite of developmental pressures on natu-ral environments since the beginning of in-dustrialization, a large seminatural forest dis-trict remains in the western part of the ur-ban agglomeration �Ruhrgebiet� between thecities Mülheim an der Ruhr and Duisburg. Avariety of different seminatural forest com-munities characterize the area which is called�Duisburg-Mülheimer Wald�, e.g. well de-
veloped remnants of lowland moors, birchswamps, alder swamp forests and oak-horn-beam woodlands. Considering the ecologi-cal conditions of the forest the occurencesof characteristic species is remarkably high.However, the influence of the neighbouringindustrial area is significant. In addition tothe well-known and much discussed impactsdue to air pollution (keyword: acid rain) (seeWITTIG 1999), there are also impacts relatedto the increasing number of people using

92 RENATE FUCHS, HERFRIED KUTZELNIGG & GUIDO BENNO FEIGE
the forest as a recreation area (keywords:trample damage and eutrophication) and theloss of forest habitat as a result of increas-ing building development. There are alsoindications of lowered ground water levels.Furthermore, a lot of alien plants, which haveescaped mainly from surrounding gardens,can be found in the flora within the wholeforest (KREFT 1993; WITTIG 2002). Soil aci-dification and mechanical stress have led toa decrease in species diversity within plantcommunities, eutrophication has occurreddue to the development of dominant popu-lations of nitrophytes, and plants escapedfrom cultivation are competing with nativespecies, thus affecting the ecological integ-rity of the forests.
The floristic and vegetational importanceof this natural forest district will be discussedconsidering the impairments and stressescaused by the area�s close proximity to anindustrial area. In particular, the phenomenonof establishment of invasive alien plants in-dependent of cultivation is assessed basedon the recently discovered taxa of Lysichitonamericanus and Onoclea sensibilis. Their origin,status and impact on aspects of environ-mental protection are under current dis-cussion (see FUCHS et al. 2003; FUCHS & KEIL
2004; ALBERTERNST & NAWRATH 2002).
2. Area under Investigation
The forest district �Duisburg-MülheimerWald� is located between Duisburg in theWest and Mülheim in the East. In the south,it ranges to downtown Ratingen (Mettmanndistrict). The large wooded area has been con-served as a result of belonging to the cen-turies-old territories of the aristocracy andclergy as well as its predominant historicaluse for hunting and horse breeding. Accord-ing to the geographical classification of natu-ral landscapes, the �Duisburg-MülheimerWald� is located on the border between �Nie-derrheinisches Tiefland� and �Süderberg-land�. The specific geographical position is
characterized by a distinct decrease of thetypical atlantic climate of the �Niederrhei-nisches Tiefland� with a mild, almost frost-free winter and a relatively cool and rainysummer (BURCKHARDT 1966; DINTER 1999).
The geological subsoil is composed ofslightly increasing terraces of sabulous andpebbly materials deposits of the Rhine inthe glacial epoch (BURCKHARDT 1966). Themain ingredients of the soils are sabulousand clayey sediments of the high tides, thepartly dune-like covers of wind-born sandas well as the floodplain sediments of thenumerous rivulets. These overlay the sabul-ous and pepply terraces, the materials ofthe till, tertiary clay as well as sandstone ofthe Upper Carboniferous. At only few lo-cations the tertiary clays appear at the sur-face. Therefore the morselled pattern ofsubsoil reflects the diversified geologicalmaterial.
3. Methods
The nomenclature of the scientific namesof the plants is based on WISSKIRCHEN &HAEUPLER (1998), DOSTÁL & KRAMER
(1984), WALTERS et al. (1984) and KOPERSKI
et al. (2000).In the following way the species spread
concerning the area of investigation is esti-mated and classified:ss � very rare: < 5%s � rare: < 10%sz � very dispersed: > 10%z � dispersed: > 25%zv � common: > 50%v � very common: > 75%
The concepts of status, time and rank ofnaturalization are based on SCHROEDER
(1969), DÜLL & KUTZELNIGG (1987), WISS-KIRCHEN & HAEUPLER (1998) and KEIL (1999):Native Species (I): indigenous speciesArchaeophytes (A): established before
1500 ADNeophytes (N): established after
1500 AD

Seminatural ancient forest in urban agglomeration area "Ruhrgebiet" 93
Acta Biologica Benrodis 13 (2006)
Industriophytes (NN): established after thebeginning of the in-dustrialization about1840
Agriophytes (AG): invasive plants estab-lished in natural/semi-natural vegeta-tion naturalized inde-pendently from culti-vation
Epecophytes (EÖ): invasive plants estab-lished in man-madehabitats, naturalizedbut dependent oncultivation
Ephemerophytes (U):casual invasive plantsCultivated plants (K):planted invasive spe-
cies, cultural relict inopen landscape
Synanthropic species (S): invasive plants withunknown rank of na-turalization
Ergasiophygophytes (EPG): invasive plants es-caped from cultivation
Akolutophytes (AK): man-made habitatscolonizing invasiveplants, immigratedwithout anthropo-genic influences
Xenophytes (XE): unintentionally intro-duced invasive
4. Results
Birch swamps, alder swamp forests, birch-oak and oak-hornbeam woodlands as wellas red beech forests belong to the relevantvegetation of the area under investigation(see FUCHS 2005). In addition to these semi-natural woodlands, the forest landscape iscomplemented by red-oak, pine and spruceforests.
The swamp forests can be split up intothe nutrient-poor habitat, which is colonisedby birch, and the comparatively rich habitatcolonised by alder. Whereas the birch swampsrepresent a highly endangered plant com-
munity, alder swamp forests are widely dis-tributed.
Following the graded index of humidityon drier and nutrient-poor habitats, it ispossible to differentiate the oak-birch-woodland. As a result of nutrient defi-ciency, the red beech is not able to growthere. Within the analysed wooded area,this plant community has merely been con-served as a relict.
The oak-hornbeam woodlands representthe most complex phytocoenosis of theinvestigated area. Based on the graded in-dex of alkalinity and nutrient balance aswell as the graded index of humidity, it ispossible to split up these plant commu-nities into six units.
The acidic beech forests can be classi-fied into a richer complex and the typicalform. The richer complex is characterisedby species requiring more nutrient and al-kaline rich soil like Milium effusum, Polygona-tum multiflorum, Anemone nemorosa and Dryo-pteris filix-mas the typical one, by the lack ofthese differentiating taxa.
In the context of the floristic-vegetationalinvestigation (FUCHS 2003), the number ofspecies was determined. The current list con-tains 538 species of ferns and floweringplants (including 13 cultivated plants) and 56mosses. The complete number of speciescorresponds to 49 % of the flora of theDuisburg area and its surroundings (DÜLL
& KUTZELNIGG 1987) and to 34 % of thecomplete flora of Northrhine-Westphalia(NRW) (RAABE et al. 1996). Thus the speciesdiversity indicates this woodland�s high flo-ristic quality. With 75 endangered taxa, theimportance of the �Duisburg-MülheimerWald� to the protection of habitats and spe-cies is very high, given the context of its loca-tion in an urban agglomeration.
Other unusual floristic features result fromits geographical location on the Northwest-ern edge of the �Rheinisch-WestfälischesMittelgebirge� (Süderbergland) in transitionto the �Niederrheinisches Tiefland�. Espe-

94 RENATE FUCHS, HERFRIED KUTZELNIGG & GUIDO BENNO FEIGE
cially noticeable is the accumulation of spe-cies which are usually spread in colline-sub-montane areas in Northrhine-Westfalia. Inthe investigated area, these species reach thelimit of their contiguous central Europeanarea of distribution, for example Blechnumspicant, Luzula luzuloides, Luzula sylvatica, Phe-gopteris connectilis, Hypericum pulchrum, Alche-milla xanthochlora and Oreopteris limbosperma.The occurrences of Poa chaixii, Calamagros-tis arundinacea and the moss Rhytidiadelphusloreus are remarkable. They are outside theircentral European area of distribution andtherefore constitute outposts. On the otherhand, there are some species that are usu-ally distributed in NRW lowlands such asCarex elata, Hydrocharis morsus-ranae, Mespilusgermanica and Hottonia palustris (HAEUPLER etal. 2003).
The preserved variety of endangered spe-cies (WOLF-STRAUB et al. 1999) may be dueto the fact that this large connected wood-land has never been used for modern agri-culture (BURCKHARDT 1966; TAPP 2000).Nevertheless, the human influences are sig-
nificant and include the impact of fossil fuelemissions and other pollutants, the presenceof alien woody plants and the establishmentof forest roads and paths and associatedtrampling damage. Last but not least, theproximity of the forest to urban areas makesthe forests vulnerable to invasion by alienplant species (through the deposition ofhorticultural wastes in these areas). The to-tal proportion of invasive alien plantsamounts to 33,2 % (179 taxa, neophytes,archaeophytes, including 13 cultivatedplants) (see tab. 1). Nevertheless the pro-portion of archaeophytes (6,0 %, 32 taxa)and neophytes (24,9 %, 134 taxa, neophytesincluding industriophytes, synanthrope taxa)is small compared to the city centers andindustrial fallows of the Ruhrgebiet (see forexample DETTMAR 1992: 11,7 % archaeo-phytes, 29 % neophytes; REIDL 1989: 9,5 %archaeophytes, 37,5 % neophytes). As ex-pected, many of these alien plants are erga-siophygophytes (22,2 %, 119 taxa), result-ing from the nearby settlements as explainedabove. The small amount of xenophytes
Tab. 1: Balance of vascular plants es-caped from cultivation (Ergasiophygo-phytes)Tab. 1: Sippenbilanz der Kulturflüch-ter (Ergasiophygophytes)
Tab. 2: Balance of vascularplants from the seminaturalforest �Duisburg-Mülhei-mer Wald�. Key of abbrevi-ation: EÖ: epecophytes; AG:agriophytes; A: archaeophy-tes; N: neophytes; NN: in-dustriophytes; EPG: ergasi-ophygophytes, XE: xeno-phytes; AK: akolutophytes
(status sensu SCHROEDER 1969, DÜLL & KUTZELNIGG 1987 WISSKIRCHEN & HAEUPLER (1998) andKEIL (1999)Tab. 2: Sippenbilanz der Gefäßpflanzen im Bereich des naturnahen Waldgebietes �Duisburg-Mül-heimer Wald� Erläuterung der Abkürzungen: EÖ: Epökophyten; AG: Agriophyten; A: Archaeo-phyten; N: Neophyten; NN: Industriophyen; EPG: Ergasiophygophyten, XE: Xenophyten; AK:Akolutophyten (status nach SCHROEDER 1969, DÜLL & KUTZELNIGG 1987, WISSKIRCHEN & HAEUP-LER (1998) und KEIL (1999)

Seminatural ancient forest in urban agglomeration area "Ruhrgebiet" 95
Acta Biologica Benrodis 13 (2006)
Tab. 3: Plants invaded from cultivation (Ergasiophygophytes) in the seminatural forest �Duisburg-Mülheimer Wald�. Abbreviations: Status: EÖ: epecophytes; AG: agriophytes; A: archaeophytes; N:neophytes; NN: industriophytes; EPG: ergasiophygophytes, XE: xenophytes; AK: akolutophytes(status sensu SCHROEDER 1969, DÜLL & KUTZELNIGG 1987, WISSKIRCHEN & HAEUPLER (1998) andKEIL (1999). Form of cultivation; F: silviculture (12 taxa); G: horticulture (70 taxa); (P): park (1taxon); (H): house garden (1 taxon); (T): pond plants (8 taxa); L: agriculture (5 taxa); S: other plan-ting (26 taxa); A: aquarium (3 taxa). Use: Z: ornamental plants (77 taxa); N: useful plants (38 taxa);H: medicinal plants (5 taxa); Aqu: aquarium plants (3 taxa).Tab. 3: Kulturflüchter (Ergasiophygophytes) im Bereich des naturnahen Waldgebietes �Duisburg-Mülheimer-Wald�. Erläuterung der Abkürzungen: Status: EÖ: Epökophyten; AG: Agriophyten; A:Archaeophyten; N: Neophyten; NN: Industriophyen; EPG: Ergasiophygophyten, XE: Xenophy-ten; AK: Akolutophyten (Status nach SCHROEDER 1969, DÜLL & KUTZELNIGG 1987, WISSKIRCHEN &HAEUPLER (1998) and KEIL (1999). Kulturform: F: Forstwirtschaft (12 Taxa); G: Gartenkultur (70Taxa); (P): Parks (1 Taxon); (H): Hausgarten (1 Taxon); (T): Gartenteich (8 Taxa); L: Landwirtschaft(5 Taxa); S: sonstige Anpflanzungen (26 Taxa); A: Aquarium (3 Taxa). Nutzungsart: Z: Zierpflanzen(77 Taxa); N: Nutzpflanzen (38 Taxa); H: Heilpflanzen (5=Taxa); Aqu: Aquarienpflanzen (3 Taxa).

96 RENATE FUCHS, HERFRIED KUTZELNIGG & GUIDO BENNO FEIGE
Tab. 3: Continued.Tab. 3: Fortsetzung.

Seminatural ancient forest in urban agglomeration area "Ruhrgebiet" 97
Acta Biologica Benrodis 13 (2006)
Tab. 3: Continued.Tab. 3: Fortsetzung.

98 RENATE FUCHS, HERFRIED KUTZELNIGG & GUIDO BENNO FEIGE
Tab. 3: Continued.Tab. 3: Fortsetzung.

Seminatural ancient forest in urban agglomeration area "Ruhrgebiet" 99
Acta Biologica Benrodis 13 (2006)
Fig. 1: Mountainbike course in a nature conservation area.Abb. 1: Mountainbike-Strecke innerhalb eines Naturschutzgebiets.
Fig. 2: Spring-aspect: Anemone nemorosa with subdominant Fallopia japonica (Mülheim-Saarn,Bühlsbach.Abb. 2: Frühlings-Aspekt: Anemone nemorosa mit subdominant Fallopia japonica (Mülheim-Saarn,Bühlsbach).

100 RENATE FUCHS, HERFRIED KUTZELNIGG & GUIDO BENNO FEIGE
Fig. 3: First record of an invasive alien fern-species, Onoclea sensibilis, in an semi natural alder swampforest (Duisburg, Weißbach) (see: FUCHS & KEIL 2004).Abb. 3: Erstnachweis einer neophytischen Farnpflanze, Onoclea sensiblis in einem naturnahen Erlen-bruchwald (Duisburg, Weißbach) (siehe: FUCHS & KEIL 2004).
Fig. 4: Lamium argentatum and Pachysandra terminalis naturalized in the �Duisburg-Mülheimer-Wald�.Abb. 4: Einbürgerungen von Lamium argentatum und Pachysandra terminalis im �Duisburg-Mülhei-mer-Wald.

Seminatural ancient forest in urban agglomeration area "Ruhrgebiet" 101
Acta Biologica Benrodis 13 (2006)
(5,6 %, 30 taxa) reflects the potential signi-ficance of traffic routes (highways, railwayline) by supplying diaspores of invasive ali-en plants (KOWARIK 2003).
The majority of alien plants escaped fromcultivation (ergasiophygophytes) after thebeginning of industrialization (see tab. 2).The percentages of these so-called indust-riophytes is more than three times greaterthan that of other neophyte types and even10 times higher than that of archaeophytes.The naturalization, in contrast, is inverselyproportional. Whereas 100% (8 taxa) of thearchaeophytes and 84,6 % (22 taxa) of theneophytes are already naturalized, the shareof the naturalized industriophytes is only38,8 % (33 taxa).
Regarding the life forms, 49,6 % (59 taxa)of the ergasiophygophytes are summergreen woody plants, 13 % (13 taxa) are ever-green woody plants and 4,2 % (5 taxa) aregrasses and sedges (see tab. 3). Most of theseergasiophygophytes are species used in hor-ticulture (70 taxa) and landscaping (26 taxa),whereas only very few (4 taxa) are derivedfrom agricultural uses. Interestingly, thereare 3 taxa originating from the aquariumtrade.
Most of the taxa escaped from cultivati-on (75) are ornamental plants. Only in a veryfew cases did medicinal plants (3 taxa) orplants of the aquarium trade run to seed.
5. Discussion
The surprising vascular species diversityof 538 vascular plants as well as 56mosses is quite unusual for a forest dis-trict in an urban agglomeration, but thishigh level is due, in part, to the high num-ber (134) of invasive species. When onecompares the number of species betweenthe current day and 200 years ago, thereis no species impoverishment. On thecontrary, it shows that the abundance ofspecies substantially increased. However,an increase in species diversity does notreflect the forest community�s health.Since the first floral recording in 1790(BÖNNINGER 1790) until today, 23 taxavanished and 61 vascular plants are highlyendangered or threatened by extinction(WOLFF-STRAUB et al. 1999).
Three reasons are crucial to understand-ing the significance of the increasing spe-cies diversity:
Tab. 3: Continued.Tab. 3: Fortsetzung.

102 RENATE FUCHS, HERFRIED KUTZELNIGG & GUIDO BENNO FEIGE
1. Forestry has created intermediate ha-bitat conditions favourable to the growthof many invasive alien plants. In particu-lar, the historical practice of drainingbirch an alder swamp forests has facili-tated the invasion of alien plants. Drain-age improves the soil�s air and nutrientbudgets in oligotrophic and wet habitatsand allowing establishment of mesophileplants. The changes within the plant com-munities may have been accelerated by theimpact of pollution. Which resulted fromindustrialization in the middle of the 19thcentury. More recently, the increase in at-mospheric nitrogen and acid, as well asindustrial dust deposition, have exten-sively changed the growth conditions (alsosee WITTIG 1999).
2. Since Second World War, the risingpopulation and the improving living con-ditions have equally lead to an increaseddemand for local recreation areas (see fig.1). The system of footpaths establishedin association with these demands tra-verses the forest district like a web. Simi-larly, a system of drainage channels criss-cross the area. Therefore, there is hardlyany undisturbed forest in the whole dis-trict. The system of footpaths and drai-nage channels have created linear struc-tures that serve as immigration corridorsfor invasive alien plants. Without thesestructures many alien plants would prob-ably not have invaded the centre of thisforest district.
3. Finally the proximity of the settle-ments accounts for a high influx of alienplant diaspores and short distribution dis-tances . Most of the noted invasive aliensplants have found their way into the for-est flora through garden garbage disposedinto the nearby woodland. In rare in-stances, these taxa are intentionally plantedinto the forest. Examples are species pro-moted by forestry like the woody plantsQuercus rubra, Prunus serotina and Acer pseu-doplatanus.
The most interesting taxa are those thatinvaded from cultivation and naturalizedindependently of human activit ies.These invasives have successfully becomean integral part of the flora. Prominentexamples are Fallopia sachalinensis, F. japo-nica (see Fig. 2), as well as Impatiens parvi-flora, which have been naturalized in theforest district «Duisburg-MülheimerWald» for decades. Extensive growth ofLamium argentatum and Impatiens glanduli-fera is found as well. Examples of somerecent invaders are Lysichiton americanus(see FUCHS et al. 2003), Onoclea sensibilis(see FUCHS & KEIL 2004, Fig. 3), and Pa-chysandra terminalis (see Fig. 4). While Fallo-pia sachalinensis, Impatiens parviflora, Lami-um argentatum and Pachysandra terminalis aremainly found in oak-hornbeam woo-dlands, Lysichiton americanus and Onocleasensibilis primarily settle in swamp forests.Whether any of the listed taxa threatennature conservation, especially of endan-gered species like Osmunda regalis or Carexelata, cannot clearly be concluded at pre-sent, but should be monitored.
Most woody species of the indigenousforest communities in Central Europe aredeciduous. Therefore, the large numberof invading taxa with evergreen leaves arepossibly benefiting from the global cli-mate change phenomenon. Commonevergreen invasives are Mahonia aquifoli-um, Prunus laurocerasus, Berberis julianae,Viburnum rhytidophyllum and Cotoneasterspecies and hybrids, especially Cotoneasterdivaricatus. In the last few years, however,many occurrences of other taxa have beendiscovered and the frequency of occur-rences is increasing (KEIL & LOOS 2004).In the forest district �Duisburg-Mülhei-mer Wald�, the first occurrences of Prunuslaurocerasus in relicts of birch swamps havebeen found. The expansion will be ofspecial interest for nature conservation,because it could impact the integrity ofthese forests.

Seminatural ancient forest in urban agglomeration area "Ruhrgebiet" 103
Acta Biologica Benrodis 13 (2006)
Acknowledgements
We would like to thank ANNA-MARIA GAIDA
(Queen Charlotte City, Canada), KATHRYN S.NICOLAI (Oberhausen) and IRIS BLAUM (Es-sen, Germany) for valuable suggestions andlinguistic corrections as well as GÖTZ HEIN-RICH LOOS (Kamen/Oberhausen, Germany)and RANDOLPH KRICKE (Essen, Germany) forcomments on the manuscript.
References
ALBERTERNST, B., NAWRATH S., 2002: Lysichitionamericanus neu in Kontinental-Europa � be-stehen Chancen für die Bekämpfung in derFrühphase der Einbürgerung? � Neobiota 1,91-99.
BÖNINGER, T.C., 1790: De Vegetabilibus Vene-natis et Speciatim de Plantis Venenatis AgriDuisburgensis. � Dissertatio Inauguralis Me-dica, Universität Duisburg (Typis Vid. Ben-thonianae), 1-133.
BURCKHARDT, H., 1966: Der Wald vor MülheimsToren. � Mülheimer Jahrbuch 1966: 38-68.
DETTMAR, J., 1992: Industrietypische Flora undVegetation im Ruhrgebiet. � Dissertationesbotanicae 191, 1-397.
DINTER, W., 1999: Naturräumliche Gliederung. In:Rote Liste der gefährdeten Farn- und Blüten-pflanzen (Pteridophyta et Spermatophyta) inNordrhein-Westfalen (Wolff-Straub, R, Büscher,D, Diekjobst, H, Fasel, P., Foerster, E., Götte,R., Jagel, A., Kaplan, K., Koslowski, I., Kutzel-nigg, H., Raabe, U., Schuhmacher, W., Vanberg,C.). � LÖBF-Schriftenreihe. 17, 29-36.
DOSTÁL, J., KRAMER, K.U., 1984: Athyriaceae. In:DOSTÁL, J., REICHSTEIN, T., FRASER-JENKINS, C.R.,KRAMER, K.U. (1984): Gustav Hegi. Bd. 1(1)/Illustrierte Flora von Mitteleuropa. Pteridophy-ta. 3. Aufl. � Parey, Berlin, Hamburg.
DÜLL, R., KUTZELNIGG, H., 1987: Punktkarten-flora von Duisburg und Umgebung, 2. Aufl.� IDH-Verlag, Rheurdt.
FUCHS, R., 2003: Floristisch-vegetationskundlicheUntersuchungen im Duisburg-MülheimerWald � unter besonderer Berücksichtigungder Moor-, Bruch- und Auwälder. � Dipl.-Arbeit, Univ. Duisburg-Essen.
FUCHS, R., 2005: Erlenbruchwald- (Alnion gluti-nosae) und Birkenbruchwald- (Betulion pu-
bescentis) Gesellschaften im Ruhrgebiet. �Tuexenia 25, in press.
FUCHS, R., KEIL, P., 2004: Onoclea sensibilis L. � derPerlfarn im Duisburg-Mülheimer Wald (West-liches Ruhrgebiet, Nordrhein-Westfalen). �Floristische Rundbriefe 37, 103-107.
FUCHS, R., KUTZELNIGG, H., FEIGE, G.B., KEIL, P.,2003: Verwilderte Vorkommen von Lysichitonamericanus Hultén & St. John (Araceae) in Du-isburg und Mülheim an der Ruhr. � Tuexenia22, 372-379.
HAEUPLER, H., JAGEL, A., SCHUHMACHER, W., 2003:Verbreitungsatlas der Farn- und Blütenpflan-zen in Nordrhein-Westfalen. � LÖBF NRW,Recklinghausen.
KEIL, P., 1999: Ökologie der gewässerbegleiten-den Agriophyten Angelica archangelica ssp. lito-ralis, Bidens frondosa und Rorippa austriaca imRuhrgebiet. � Dissertationes botanicae 321,1-186.
KEIL, P., LOOS, G. H., 2004: Ergasiophygophytictrees and shrubs in the Ruhrgebiet (WestGermany). In: Biological Invasions: Challen-ges for Science (Kühn, I., Klotz, S., eds.). �Neobiota 3, 90.
KOPERSKI, M, SAUER, M., BRAUN, W., GRADSTEIN,S. R., 2000: Referenzliste der Moose Deutsch-lands � Dokumentation unterschiedlicher ta-xonomischer Auffassungen. � Schriftenreihefür Vegetationskunde 34, 1-519.
KOWARIK, I., 2003: Biologische Invasionen. Ne-ophyten und Neozoen in Mitteleuropa. - Ul-mer, Stuttgart.
KREFT, H., 1993: Zur �Natur� urbaner Wälder,pp 103-114. In: Geoökologie � Beiträge zurForschung und Anwendung. Festschrift fürHans-Jürgen Klink (Hütter, M., Reinirkens,P., eds). � Brockmeyer, Bochum.
RAABE, U., FOERSTER, E., SCHUHMACHER, W.,WOLFF-STRAUB, R., 1996: Florenliste vonNordrhein-Westfalen. 3. verbesserte und er-weiterte Auflage. � LÖBF-Schriftenreihe 10.
REIDL, K., 1989: Floristische und vegetations-kundliche Untersuchungen als Grundlagenfür den Arten- und Biotopschutz in der Stadt- Dargestellt am Beispiel der Stadt Essen. �Diss. Univ.-Essen. Verlag G. Mainz, Aachen.
SCHROEDER, F.G. , 1969: Zur Klassifizierung derAnthropochoren. � Vegetatio 16, 225-238.
TAPP, I., 2000: Veränderungen in der natürlichenVegetation des Ruhrgebietes zwischen dem18. und 20. Jahrhundert an Hand von Land-

104 RENATE FUCHS, HERFRIED KUTZELNIGG & GUIDO BENNO FEIGE
kartenstudien. � Natur am Niederrhein (N.F.) 15(1), 3-32.
WALTERS, S.M., BRADY, A., BRICKELL, C.D., CUL-LEN, J., GREEN, P.S., LEWIS, J., MATTHEWS, V.A.,WEBB, D.A., YEO, P.F., ALEXANDER, J.C.M.(eds), 1984: The European Garden Flora.Vol. II. Monocotyledons (Part II). � Cam-bridge, London, New York, New Rochelle,Melbourne, Sydney.
WISSKIRCHEN, R., HAEUPLER, H., 1998: Standard-liste der Farn- und Blütenpflanzen Deutsch-lands. � Bundesamt für Naturschutz (Hrsg.).� Ulmer, Stuttgart.
WITTIG, R., 1999: Verbreitung und Standorte vonAnemone nemorosa und Polygonatum multiflorumin Buchenwäldern des Ruhrgebietes � Tue-xenia 19, 173-177.
WITTIG, R., 2002: Siedlungsvegetation. � Ulmer,Stuttgart.
WOLFF-STRAUB, R, BÜSCHER, D, DIEKJOBST, H, FA-SEL, P., FOERSTER, E., GÖTTE, R., JAGEL, A., KAP-LAN, K., KOSLOWSKI, I., KUTZELNIGG, H., RAABE,U., SCHUHMACHER, W., VANBERG, C., 1999: RoteListe der gefährdeten Farn- und Blütenpflan-zen (Pteridophyta et Spermatophyta) in Nord-rhein-Westfalen. � LÖBF-Schriftenreihe 17.
Accepted: 18.01.2005

Effects on the moulting frequency of Asellus aquaticus 105
Acta Biologica Benrodis 13 (2006)
Acta Biologica Benrodis 13 (2006): 105-115
Effects of endocrine disrupting compounds andtemperature on the moulting frequency of the
freshwater isopod Asellus aquaticus L. (Isopoda:Asellota)*
Effekte von hormonähnlich wirksamen Substanzen und Temperaturauf die Häutungsfrequenz der Süßwasserassel Asellus aquaticus L.
(Isopoda: Asellota)
LENNART WELTJE 1, 2 & JÖRG OEHLMANN 2
1 BASF AG, Agricultural Center – Ecotoxicology, APD/EE-Li425, Carl-Bosch-Straße 64,D-67117 Limburgerhof, Germany; [email protected] (corresponding author)
2 J.W. Goethe University, Department of Ecology and Evolution – Ecotoxicology,Siesmayerstraße 70, D-60323 Frankfurt am Main, Germany
Summary: The effects of the vertebrate endocrine disrupting compounds diuron, linuron, vinclo-zolin (pesticides), 17α-ethinylestradiol and tamoxifen (pharmaceuticals) on the moulting frequen-cy of the freshwater isopod Asellus aquaticus (L.) are described. In addition, the influence oftemperature on moulting frequency was studied. Since moulting is under the control of the steroidhormone 20-hydroxyecdysone, effects may be expected of substances known to influence thesteroid sex hormones (i.e. estradiol and testosterone) of vertebrates. Although the chemicalsinduced changes in moulting frequency (both stimulating and inhibiting effects were found),significant differences were not established. At 20 ºC, the moulting frequency was higher than at 15°C, but again the difference was not significant. Consequently, improvements in the experimentalset-up are discussed. It is concluded that experiments are best performed at a constant temperature,with young, isolated individuals of similar size and sex.
Pesticides, moulting frequency, Asellus aquaticus
Zusammenfassung: Effekte der wirbeltierhormonähnlich wirksamen Substanzen Diuron, Linu-ron, Vinclozolin (Pflanzenschutzmittel), 17α-Ethinylöstradiol und Tamoxifen (Arzneimittel) aufdie Häutungsfrequenz der Süßwasserassel Asellus aquaticus (L.) werden beschrieben. Zusätzlichwurde der Einfluss der Temperatur auf die Häutungsfrequenz untersucht. Da die Häutung über dasSteroidhormon 20-Hydroxyecdyson kontrolliert wird, können Effekte von Substanzen erwartetwerden, von denen bekannt ist, dass sie die steroiden Sexualhormone von Vertebraten (d.h. Östradiolund Testosteron) beeinflussen. Obwohl die getesteten Chemikalien einen Einfluss auf die Häutungs-frequenz hatten (sowohl fördernde als auch hemmende Effekte wurden beobachtet), wurden keinesignifikanten Unterschiede gefunden. Bei 20 °C war die Häutungsfrequenz höher als bei 15 °C, aberder Unterschied war ebenfalls nicht signifikant. Deshalb werden Verbesserungen des experimentellenAufbaus diskutiert. Daraus wird geschlossen, dass die Versuche am besten bei konstanter Temperaturmit jungen, isolierten Individuen von gleicher Größe und Geschlecht durchgeführt werden.
Pflanzenschutzmittel, Häutungsfrequenz, Asellus aquaticus* Contribution to the �First European Workshop on Isopods�, Düsseldorf 14.-16. November2003, Germany

106 LENNART WELTJE & JÖRG OEHLMANN
1. Introduction
1.1. Endocrine disruptions in invertebrates
At present, research on the disruption ofhormonal processes in animals is mainly fo-cussed on vertebrates and on in vitro systemsderived from them. The vertebrate endocrinesystem is quite well understood and the char-acterisation of its steroid hormones and as-sociated receptors is on a relatively high level.
A fair number of recent in vivo studies withchemicals known to interact with the verte-brate endocrine system (particularly the xeno-estrogens bisphenol A, 4-n-nonylphenol and17α-ethinylestradiol), have also demonstratedeffects in invertebrates, including molluscs,insects and crustaceans (BALDWIN et al. 1997;BROWN et al. 1999; OEHLMANN et al. 2000;HAHN et al. 2002; WATTS et al. 2002; VANDEN-BERGH et al. 2003; this list is not exhaustive).The difficulty with most of these invertebratestudies is to unequivocally prove that the ef-fect shown is due to the disturbance of anendocrine mechanism and not, for instance,by general toxicity (SEGNER et al. 2003). How-ever, such mechanistic proof is required,since it forms the very basis by which an en-docrine disrupting compound (EDC) is de-fined. To gather this proof, a reasonableamount of knowledge should be availableon the endocrinology of the organism understudy. In the case of invertebrates, this know-ledge is often insufficient.
There are three main issues that hamperthe interpretation of invertebrate EDC data:The first problem is the large diversity in endo-crine mechanisms, hormones and processes,existing within the invertebrate world, whichcomprises about thirty phyla, against onesingle phylum of vertebrates (DEFUR et al.1999). Many mechanisms of endocrine con-trol are simply not known or not fully inves-tigated yet. Secondly, vertebrate-type steroids(estradiol, testosterone, progesterone) havebeen detected in a number of invertebratespecies, e.g. in higher crustaceans (Malacostra-
ca), but it has not been established that theyfulfil a functional role (VANDENBERGH etal.2003; DEFUR et al. 1999). It may thereforebe unrealistic to expect vertebrate xeno-estro-gens to have estrogenic effects in crustaceans,but they could affect other endocrine pro-cesses, such as moulting (ZOU & FINGERMAN
1997). A possible working mechanism forsuch EDCs is an interaction with the ecdyste-roid receptor, and for bisphenol A and 17α-ethinylestradiol, ecdysteroid antagonistic ac-tivity has been shown in a Drosophila melano-gaster cell line (DINAN et al. 2001). Further, MU
& LEBLANC (2002) have shown that the devel-opmental toxicity of testosterone to thecrustacean Daphnia magna involves anti-ecdy-steroidal activity. DE LOOF & HUYBRECHTS
(1998) suggested in their review that ecdyste-roids, which are also involved in vitellogen-esis, may function as the counterparts of ver-tebrate sex steroids in insects. This impliesthat interactions of vertebrate EDCs with theecdysteroid receptor may elicit (anti-)estrogenictype of effects, not per se related to the pres-ence of an estrogen receptor. On the contrary,other invertebrate groups, notably molluscsand echinoderms, do seem to have a func-tional role for some vertebrate-type steroids,hence working mechanisms similar to thosein vertebrates may apply. Finally, the experi-mental design often does not allow for a de-finitive conclusion on the (endocrine) wor-king mechanism of a compound. Hence,many cases point towards a possibly endo-crine-mediated effect (e.g. increased reproduc-tive output, altered moulting behaviour,shifted sex-ratios), but evidence remains cir-cumstantial.
Although it is evident, from the meanwhile�classic� example of imposex in gastropods,caused by organotin compounds, such as tri-butyltin (TBT) and triphenyltin (TPhT) (foran overview see CSTEE, 1999), that endocri-ne disruption does occur in natural popula-tions of invertebrates, they have not receivedthe amount of attention they deserve withinthe framework of EDC research. Especially

Effects on the moulting frequency of Asellus aquaticus 107
Acta Biologica Benrodis 13 (2006)
if one considers that at least 95% of theknown animal species consists of inverte-brates, which comprise keystone species forthe functioning of ecosystems and food-webs, a greater effort should be undertakento investigate to what extent endocrine dis-ruption really affects invertebrates.
1.2. Isopods as model organisms toassess endocrine disruption
Isopods belong to the higher crustaceans(Malacostraca), the largest class of the sub-phylum Crustacea, which are part of the phy-lum Arthropoda. Insects are the other mainarthropod class, and in terms of speciesnumbers, arthropods are the most success-ful group of invertebrates, hence importantfor global biodiversity research. Arthropodsare a good starting point to investigate endo-crine disruption for a number of reasons:1) The endocrine system of insects is described
to a much greater extent than that of anyother invertebrate group, mainly because ofthe silkworm industry and pest control ofinsects (DEFUR et al. 1999). A substantialpart of the knowledge gathered on insectsis also applicable to crustaceans.
2) Insect pest control initiated the develop-ment of third generation pesticides, whichhave been designed to disrupt the actionof specific arthropod hormones, namelyecdysteroids and juvenile hormones. Thisimplies that there are specific positive con-trol substances available (agonists andantagonists), which are a necessary tool inexperiments, designed to evaluate theworking mechanism of suspected EDCs.
3) A unique androgenic gland and associatedandrogenic hormone (MARTIN et al. 1999)have been shown in isopods and otherMalacostraca. Androgenic hormone con-trols the development of primary andsecondary male characteristics and has anegative regulatory control on vitellogen-esis (DE LOOF & HUYBRECHTS 1998; DE-FUR et al. 1999).
4) Crustaceans have readily measurable end-points, such as moulting and regenerati-on, which are coupled processes underneuroendocrine control (WEIS et al. 1992;FINGERMAN et al. 1998).
5) A useful endpoint could be the forma-tion of oostegites in female isopods. Oo-stegites are ventral overlapping plates,forming the marsupium or brood pouch,which develop after a special moult, theso-called parturial moult, which preceedsfertilisation. The latter can only take placewhen the exoskeleton is still flexible inthe hours directly after moulting, and isthe main reason why male aquatic isopodsmonopolize females by carrying them ontheir ventral surface (a precopula behaviourcalled mate guarding) (ZIMMER 2001).
6) Field evidence exists for abnormal sexualdevelopment in amphipod (Malacostra-ca) populations (GROSS et al. 2001; NA-GEL et al. 2002). Various degrees of inter-sex have also been found in natural iso-pod populations (see CHARNIAUX-COTTON
1960), but a relation with environmentalchemicals has not been established.
Isopods occupy key positions as decom-posers in freshwater, terrestrial and marineecosystems and may thus provide sentinelspecies in endocrine disruptor research acrossall environments. This is further underlinedby the global occurrence of the species in thepresent paper, Asellus aquaticus, making it idealfor monitoring purposes (and such speciesare also available for terrestrial environments,for instance Armadillidium vulgare or Porcellioscaber). Finally, it should be added that litera-ture on isopods and endocrine disruptors isvirtually non-existent and presents thereforea completely open and challenging field.
1.3. Aim of the present work
The experiments described in this paperare the first in a series on the effects of endo-crine disruptors on freshwater isopods. Inthese experiments, the effects of two phar-

108 LENNART WELTJE & JÖRG OEHLMANN
maceuticals (tamoxifen and 17α-ethinylestra-diol) and three pesticides (the herbicides li-nuron and diuron and the fungicide vinclo-zolin) on the moulting frequency of Asellusaquaticus are investigated. The selected chemi-cals are known to interact with vertebrate re-ceptors for the steroids testosterone and estra-diol. The working mechanisms of thesechemicals are: estrogenic (17α-ethinylestradi-ol), anti-estrogenic (tamoxifen) and anti-an-drogenic (linuron, vinclozolin and diuron).Their endocrine effects come about by directi.e. receptor-mediated action, implying thatthese chemicals bind to the estrogen or to theandrogen receptor. In the case of vinclozo-lin, the effects are mainly due to two metabo-lites of vinclozolin which have greater affin-ity for the androgen receptor than vinclozo-lin itself. Since moulting in isopods is underthe control of the steroid hormone 20-hy-droxyecdysone (also known as β-ecdysone orecdysterone), effects may be expected fromexposure to these compounds.
Since it is known that temperature has aneffect on moulting, the moulting intervalgenerally decreases with temperature (BOU-CHON et al. 1992; MOMMERTZ 1993), we per-formed an additional experiment, whereinisopod moulting was studied at two diffe-rent temperatures, 15 ºC and 20 ºC. This ex-periment serves as a positive control, whichshould demonstrate that our lab-culturedisopods are sensitive to manipulations in en-vironmental parameters influencing themoulting frequency.
Because the aim of this work is to explorethe possibility of using freshwater isopodsfor the assessment of EDC effects, they canbe used to guide and improve future workon this topic.
2. Materials and methods
2.1. Chemicals
Vinclozolin, diuron and linuron werepurchased from Riedel-de Haën as Pestanal
standards (purity > 99%). Tamoxifen (pu-rity ≥ 99%) and 17α-ethinylestradiol (purity≥ 98%) were purchased from Sigma. Thestructures of these five substances are givenin figure 1. Ethanol, denatured with methyl-ethylketone (purity ≥ 99.5%, Merck, Darm-stadt) was used as a solvent. Chemicals thatwere used to prepare the artificial water (ac-cording to DUFT et al. 2003) were all of re-agent grade.
2.2. Organisms
Freshwater isopods (Asellus aquaticus) werecollected in the municipality of Zittau (Saxo-
Fig. 1: Chemical structures of the tested com-pounds. a) vinclozolin, b) diuron, c) linuron,d) tamoxifen, e) 17α-ethinylestradiol.Abb. 1: Strukturformeln der getesteten Substan-zen. a) Vinclozolin, b) Diuron, c) Linuron,d) Tamoxifen, e) 17α-Ethinylöstradiol.
a
b
c
d
e

Effects on the moulting frequency of Asellus aquaticus 109
Acta Biologica Benrodis 13 (2006)
ny, Germany) from a small drainage ditchflowing through agricultural land. Leaves oflocal alder trees (Alnus glutinosa) on which Asel-lus aquaticus feeds, were sampled to serve asfood and substrate in the laboratory massculture and in the experiments. The leaveswere analysed for heavy metals and polycyclicaromatic hydrocarbons (PAHs) (see below)to check the pollution state of the samplingsite and also the exposure history of the iso-pod population. The mass culture was estab-lished in glass aquaria (30 × 30 × 50 cm) con-taining gently aerated artificial water of pH8.2 ± 0.2 with a depth of 15 - 20 cm and wasmaintained at 15 °C in a climate room with aday-night rhythm of 16:8 h. The culture wasmainly fed with alder leaves, which alsoserved as a substrate. Additional food con-sisted of TetraPhyll (Melle, Germany), andoccasionally slices of cucumber and carrot.
2.3. Chemical analyses of the leaves
The field-collected alder (Alnus glutinosa)leaves with which the animals were fed in boththe cultures and the experiments, were ana-lysed to determine the concentration ofseventeen elements and also of the sixteenPAHs that are on the U.S. EPA�s list of prior-ity pollutants. The analyses were done to as-sure that the leaves had a normal (background)contamination and thus did not pose anextra chemical stress for the animals.
Analyses of the elements were as follows.The leaves were dried at 45°C overnight andmilled (Vibratory Disc Mill RS 1 of Specialtysteel, Retsch, Haan, Germany). About 0.3 gdry plant material was placed in a PTFE pres-sure vessel and 4 mL 65% HNO3 and 2 ml30% H2O2 (both Merck, Suprapur) wereadded. Digestion of the sample took place ina microwave (Ethos plus, MLS GmbH, Leut-kirch, Germany) which was heated in 3 minto 85°C, then in 5 min to 125°C, in 5 min to200°C and this temperature was held for 15min. After cooling down, the samples werebrought to a volume of 50 ml with Milli-Q
water in volumetric PMP flasks (VIT-LAB,Seeheim-Jugenheim, Germany). This wasdiluted two- and tenfold with Milli-Q wateror 3% HNO3 (Suprapur, Merck, Darmstadt,Germany) respectively, to achieve the requiredelemental intensities for the analyses. Micro-element concentrations in leave digests weremeasured with a quadrupole Perkin-ElmerElan 6000 ICP-MS equipped with a cross-flow nebulizer. Prior to the measurements,the ICP-MS was optimized for oxide forma-tion, by means of the ratio CeO+/Ce+, andfor formation of double-charged ions, bymeans of the Ba2+/Ba+ couple (for more de-tails on instrumental parameters see IVANOVA
et al. 2001). External calibration was per-formed with a multi-element standard solu-tion (Merck, Darmstadt, Germany). Eachsample was measured with three repetitionsto generate a measurement average with stan-dard deviation. Mathematical corrections forisotopic overlap were applied, using thebuilt-in Perkin-Elmer sofware routines.Macro-elements were measured with a Per-kin-Elmer Optima 3000 ICP-OES. The refe-rence material SRM 1515 Apple Leaves (Nati-onal Institute of Standards and Technology,Gaithersburg, MD, USA) was used for evalu-ation of the applied method.
Analyses of PAHs was done with HPLC,according to the procedure described for se-diment by DUFT et al. (2002) and followingDIN 38414 S 21 (DIN 1996). Prior to analy-sis, the leaves were freeze-dried (ALPHA 1-4,Christ, Osterode, Germany), milled (Vibra-tory Disc Mill RS 1 of Specialty steel, Retsch,Haan, Germany) and subjected to acceleratedsolvent extraction (ASE) using cyclohexane.
2.4. Experiments
The experiments with 17α-ethinylestradiol,vinclozolin, tamoxifen, diuron and linuronwere conducted simultaneously in a climateroom at 15 °C and shared three solvent (etha-nol) control treatments. Experiments wereconducted in glass beakers of 500 ml, contain-

110 LENNART WELTJE & JÖRG OEHLMANN
ing 300 ml of artificial water, 10 adult iso-pods (body length between 4 and 8 mm) anda 5 × 5 cm piece of alder leaf. For vinclozolin,tamoxifen, diuron and linuron, concentrationsof 10 ng/l and 10 µg/l were tested in dupli-cate (n = 2). For 17α-ethinylestradiol, concen-trations of 10 ng/l and 100 ng/l were tested(n = 1). Solutions were replaced at least onceevery week. Leaf pieces were replaced when inone of the treatments the leaf piece waseaten for about 75%. Over a period of 63days, daily observations were made on themoulting state of the animals. Exuviae anddead individuals were removed.
The temperature experiment was con-ducted at 15 °C and 20 °C. At each tempera-ture, one 12 wells plate (Nunc, Wiesbaden,Germany) was tested, with in each well 2 mlof artificial water, one juvenile isopod (bodylength < 3 mm) and a 1.5 × 1.5 cm piece ofalder leaf (n = 12). Solutions were replaced atleast once every week. Leaf pieces were replacedwhen in one of the treatments the leaf piecewas eaten for about 75%. Over a period of73 days, daily observations were made on themoulting state of each individual. Exuviaeand dead individuals were removed. In addi-tion, individuals were sexed and measured(pleotelson length and body length) at theend of this experiment or alternately on theday after they had died during the experiment.
2.5. Data handling and statistics
The moulting frequency was calculated as thenumber of moults per isopod per day. Datawere corrected for mortality in the vinclozo-lin, tamoxifen, diuron, linuron and17α-ethinylestradiol experiments, because tenindividuals were exposed simultaneously andthe exuviae could not be assigned to certainindividuals. In case of the temperature ex-periment, isopods could be studied indi-vidually. Individuals in this experiment thatdied within 14 days after the start of theexperiment were omitted from the calcula-tions.
An ANOVA with DUNNETT�s post testwas performed to test for substance effectsin comparison to the solvent control. Forthe temperature experiment a t-test wasused to test for significant differencesbetween the two temperatures. In this ex-periment, the pleotelson length and bodylength of all individuals was measured andthe correlation of these measures wasstudied. Since the data were normally dis-tributed (D�AGOSTINO and PEARSON omni-bus normality test), correlation was calcu-lated according to PEARSON. For all statisti-cal tests, P = 0.05. All calculations were per-formed with the software program Prism4.02 (GraphPad, San Diego, CA, USA).
Table 1: Concentrations (mg/kg dry wt) of 17elements with relative standard deviations in lea-ves of Alnus glutinosa, collected in 2002. Ele-ments in italics were measured with ICP-OES,others with ICP-MS.Tab. 1: Konzentrationen (mg/kg Trockengewicht)von 17 Elementen mit relativer Standardabwei-chung in Blättern von Alnus glutinosus (gesammelt2002). Elemente in Kursivschrift wurden mitICP-OES, die anderen mit ICP-MS bestimmt.

Effects on the moulting frequency of Asellus aquaticus 111
Acta Biologica Benrodis 13 (2006)
3. Results
The chemical analysis of the alder leaves (seetable 1 and 2) demonstrated that the sitewhere the isopods and the alder leaves werecollected has only a background pollution lev-el. Therefore, and as far as these analyses per-mit, we may conclude that the isopods haveno previous history of exposure to pollu-tion beyond a background level.
Exposure to the selected vertebrate endo-crine disruptors demonstrated effects onmoulting, albeit non significantly (P > 0.05,ANOVA, followed by Dunnett�s test). Theresults are presented in figs. 2 and 3. Expo-sure to the anti-androgenic pesticides diuron,linuron and vinclozolin induced highermoulting frequencies (linuron in a concentra-tion-dependent manner, see fig. 2), whereasthe anti-estrogenic drug tamoxifen loweredthe moulting frequency in a concentration-dependent manner (fig. 3). Effects of diuronand vinclozolin were slightly higher at
Table 2: Concentrations (mg/kg dry wt) of 16PAHs (listed in elution order) in leaves of Alnusglutinosa, collected in 2002; nd = not detected.Tab. 2: Konzentrationen (mg/kg Trockenge-wicht) von 16 PAHs (in der Reihenfolge ihrerElution aufgelistet) in Blättern von Alnus glutino-sus (gesammelt 2002); nd = nicht nachgewiesen.
Fig. 2: Effects of diuron, linuron and vinclozo-lin (10 ng/l and 10 µg/l) on the moulting fre-quency of freshwater isopods.Abb. 2: Effekte von Diuron, Linuron und Vin-clozolin (10 ng/l und 10 µg/l) auf die Häutungs-frequenz von Süsswasserasseln.
10 ng/l than at 10 µg/l. Exposure of the iso-pods to the estrogenic drug 17α-ethinylestra-diol at 10 ng/l lowered the moulting fre-quency, while at 100 ng/l moulting wasstimulated (fig. 3). Unfortunately, none ofthese results proved to be significant, due toseveral causes which will be dealt with in thenext section.

112 LENNART WELTJE & JÖRG OEHLMANN
As expected, the moulting frequency washigher at 20 ºC than at 15 ºC (fig. 4). However,also these results were not significant (P > 0.05,t-test). The pleotelson length and the bodylength of the animals in the temperature ex-periment were highly correlated (for pooleddata, Pearson r = 0.85, P < 0.0001), whichimplies that one of these measurements issufficient to characterise the size of the ani-mals. The relation between body length andpleotelson length is depicted in figure 5.
Fig. 3: Effects of tamoxifen (10 ng/l and 10µg/l) and of 17α-ethinylestradiol (10 ng/l and100 ng/L) on the moulting frequency of fresh-water isopods.Abb. 3: Effekte von Tamoxifen (10 ng/l und10 µg/l) und von 17α-Ethinylöstradiol (10 ng/lund 100 ng/l) auf die Häutungsfrequenz vonSüßwasserasseln.
Fig. 4: Effects of temperature (15 and 20 ºC) onthe moulting frequency of freshwater isopods.Abb. 4: Effekte der Temperatur (15 und 20 ºC)auf die Häutungsfrequenz von Süsswasserasseln.
Fig. 5: Relation between body length and pleo-telson length of juvenile freshwater isopods fromthe temperature experiment.Abb. 5: Zusammenhang zwischen Körperlängeund Länge des Pleotelsons von juvenilen Süß-wasserasseln aus dem Temperaturexperiment.
4. Discussion and conclusions
Because no significant differences for any ofthe treatments could be established, it is ob-vious that either the experimental set-up needsto be improved, or alternately, that moultingfrequency is not a sensitive endpoint regard-ing exposure to EDCs or to temperature dif-ferences.
The experimental set-up could certainly beimproved. The number of replicates, as wellas the homogeneity of the exposed isopodgroups, can be optimised. This implies totest a higher number of replicates; preferablyisolated individuals, of similar size and sex.Increasing the number of exposed individualswill lead to a higher statistical power of thetest, which is obviously needed for a variable

Effects on the moulting frequency of Asellus aquaticus 113
Acta Biologica Benrodis 13 (2006)
endpoint such as moulting. Isolating the iso-pods assures an exact recording of the inter-moult period of each individual and yieldsstatistically independent observations. Sincethe intermoult period increases with the ageof the individuals, the sensitivity of the testmay be increased by testing young, i.e. small,isopods. However, there was hardly any dif-ference between the moulting frequency ofthe control groups in the substance and tem-perature experiments (compare figs. 3 and 4;moulting frequency ~0.022 d-1), although theisopods in the temperature experiment weresmaller than the ones in the substance ex-periments. Because the sex of young isopodsis not easily determined, it is advisable to in-crease the number of exposed individuals,and evaluate the data for each sex separatelyafter the experiment has finished and the ani-mals have been sexed. The importance in dis-tinguishing the sexes is related to the diffe-rent moulting patterns of males and females(MOMMERTZ 1993). Whereas males have aregular moulting pattern, females have twotypes of moults which alternate; �normal�moults and so-called parturial moults in whichthe oostegites are formed. In addition, thetemperature should be controlled with someprecision, because this parameter will influ-ence the moulting behaviour of the exposedindividuals.
The effects of the chosen substances werestimulating in case of the anti-androgenicpesticides vinclozolin, diuron and linuron butalso in the highest concentration of the estro-genic drug 17α-ethinylestradiol. If interactionwith the ecdysteroid receptor is explainingthese observations, then the previously men-tioned compounds are acting as ecdysteroidagonists. Opposite effects were observed forthe anti-estrogenic drug tamoxifen, whichmay indicate ecdysteroid antagonism, butcould also be indicative for general toxicity,which has been reported for zinc effects onthe moulting frequency of the terrestrial iso-pod Porcellio scaber (DROBNE & �TRUS 1996).By co-exposure with an ecdysteroid agonist,
e.g. 20-hydroxyecdysone or tebufenozide, itmay be elucidated if there is an endocrineworking mechanism involved (see MU & LE-BLANC 2002).
Our observations agree with the anti-ec-dysteroid activity of testosterone in thecrustacean Daphnia magna (MU & LEBLANC
2002). The only exception is formed by theobserved lower moulting frequency in thelowest 17α-ethinylestradiol treatment,which does not fit into this series. How-ever, they may be explained by the slightecdysteroid antagonistic activity that17α-ethinylestradiol showed in an in vitrocell line (DINAN et al., 2001).
Overall, the work presented here did notproduce any conclusive evidence on the in-fluence of vertebrate EDCs on the moult-ing frequency of the freshwater isopod Asel-lus aquaticus. The results of the experimentswith 17α-ethinylestradiol and tamoxifen arehard to interpret, because of irregularitiesin the concentration-response relation andthe possible involvement of general toxi-city, respectively. On the other hand, theexperiments with the pesticides vinclozo-lin, diuron and linuron hint at ecdysteroidagonistic activity of these vertebrate anti-androgens. This should be investigated inmore detail in experiments with an im-proved lay-out.
Acknowledgements
The authors thank CONSTANZE STARK, SI-MONE ZIEBART, GERTRAUD WIRZINGER,CONNY SCHOLZ and CHRISTIAN VOGT for as-sistance in the laboratory and in the field.HEIKE HEIDENREICH and GERLINDE LIEPELT
are gratefully acknowledged for chemicalanalyses of the alder leaves. This researchhas been supported by a Marie-Curie Fel-lowship of the European Community pro-gramme Human Potential, under contractnumber HPMD-CT-2000-00039, fellow re-ference number HPMD-GH-00-00039-02to the first author.

114 LENNART WELTJE & JÖRG OEHLMANN
References
BALDWIN, W.S., GRAHAM, S.E., SHEA, D., LEBLANc,G.A., 1997: Metabolic androgenization of fe-male Daphnia magna by the xenoestrogen 4-nonylphenol. � Environmental Toxicology andChemistry 16: 1905-1911.
BOUCHON, D., SOUTY-GROSSET, C., MOCQUARD, J.-P.,CHENTOUFI, A., JUCHAULT. P., 1992: Photope-riodism and seasonal breeding in aquatic andterrestrial Eumalacostraca. � Invertebrate Re-production and Development 22: 203-212.
BROWN, R.J., CONRADI, M., DEPLEDGE, M.H., 1999:Long-term exposure to 4-nonylphenol affectssexual differentiation and growth of the am-phipod Corophium volutator (Pallas, 1766). �Science of the Total Environment 233, 77-88.
CHARNIAUX-COTTON, H., 1960: Sex determinati-on, pp 411-477. In: The physiology ofCrustacea, Vol. I, Metabolism and growth(Waterman, T.H., ed.). � Academic Press,New York.
CSTEE, 1999: Opinion on human and wildlifehealth effects of endocrine disrupting che-micals, with emphasis on wildlife and on eco-toxicology test methods. European Commis-sion, Scientific Committee on Toxicity, Eco-toxicity and the Environment. � (CSTEE),Brussels, Belgium.
DEFUR, P.L., CRANE, M., INGERSOLL, C.G., TATTERS-FIELD, L.J. (eds), 1999: Endocrine disruption ininvertebrates: endocrinology, testing, and assess-ment. � SETAC Press, Pensacola, FL, USA.
de Loof, A., Huybrechts, R., 1998: �Insects do nothave sex hormones�: a myth? - General andComparative Endocrinology 111: 245-260.
DIN, 1996: 38414. Bestimmung von 6 polyzycli-schen aromatischen Kohlenwasserstoffen(PAK) mittels Hochleistungs-Flüssigkeitschro-matographie (HPLC), p. 21. In: Deutsche Ver-fahren zur Wasser-, Abwasser- und Schlamm-untersuchung. Band 1. Loseblattsammlung.(Fachgruppe Wasserchemie der GDCh undNormenausschuss Wasserwesen (NAW) imDIN Deutsches Institut für Normung e.V., ed.).� VCH, Weinheim, Beuth, Berlin.
Dinan, L., Bourne, P., Whiting, P., Dhadialla, T.S.,Hutchinson, T.H., 2001: Screening of environ-mental contaminants for ecdysteroid agonist andantagonist activity using the Drosophila melano-gaster BII cell in vitro assay. � EnvironmentalToxicology and Chemistry 20, 2038-2046.
DROBNE, D., �TRUS, J., 1996: Moult frequency ofthe isopod Porcellio scaber, as a measure ofzinc-contaminated food. � EnvironmentalToxicology and Chemistry 15, 126-130.
DUFT, M., TILLMANN, M., OEHLMANN, J., 2002:Ökotoxikologische Sedimentkartierung dergrossen Flüsse Deutschlands. Report UBA-FB 000418. � UBA, Berlin, Germany.
DUFT, M., SCHULTE-OEHLMANN, U., WELTJE, L.,TILLMANN, M., & J. OEHLMANN, J., 2003: Sti-mulated embryo production as a parameterof estrogenic exposure via sediments in thefreshwater mudsnail Potamopyrgus antipodarum.- Aquatic Toxicology 64, 437-449.
FINGERMAN, M., JACKSON, N.C., NAGABHUSHANAM,R., 1998: Hormonally-regulated functions incrustaceans as biomarkers of environmentalpollution. � Comparative Biochemistry Phy-siology C 120, 343-350.
GROSS, M.Y., MAYCOCK, D.S., THORNDYKE, M.C.,MORRITT, D., CRANE, M., 2001: Abnormalitiesin sexual development of the amphipod Gam-marus pulex (L.) found below sewage treat-ment works. � Environmental Toxicology andChemistry 20, 1792-1797.
HAHN, T, SCHENK, K., SCHULZ, R., 2002: Envi-ronmental chemicals with known endocrinepotential affect yolk protein content in theaquatic insect Chironomus riparius. � Environ-mental Pollution 120, 525-528.
IVANOVA, J., KORHAMMER, S., DJINGOVA, R., HEI-DENREICH, H., MARKERT, B., 2001: Determina-tion of lanthanoids and some heavy andtoxic elements in plant certified reference ma-terials by inductively coupled plasma massspectrometry. � Spectrochimica Acta B, Ato-mic Spectroscopy 56, 3-12.
MARTIN, G., SOROKINE, O., MONIATTE, M., BU-LET, P., HETRU, C., VAN DORSSELAER, A.,1999: The structure of a glycosylated pro-tein hormone responsible for sex determi-nation in the isopod, Armadillidium vulgare.� European Journal of Biochemistry 262,727-736.
MOMMERTZ, S., 1993: Untersuchungen zum Se-xualverhalten von Asellus aquaticus L. � Sha-ker, Aachen, Germany.
MU, X., & LEBLANC, G.A., 2002: Developmentaltoxicity of testosterone in the crustaceanDaphnia magna involves anti-ecdysteroidalactivity. � General and Comparative Endo-crinology 129, 127-133.

Effects on the moulting frequency of Asellus aquaticus 115
Acta Biologica Benrodis 13 (2006)
NAGEL, R., KÖHLER, H.-R., TRIEBSKORN, R., JUNG-MANN, D., LADEWIG, V., LUDWICHOWSKI, K.U.,METAG, N., PREUSS, T., WOITSCHELLA, A., 2002:Effects of xeno-hormones in amphipods -from the molecular to the population level,pp 75-79. In: Second status seminar endocri-ne disruptors, Proceedings (Bundesministeri-um für Bildung und Forschung, ed.). � GSF,Munich, Germany.
OEHLMANN, J., SCHULTE-OEHLMANN, U., TILLMANN,M., MARKERT, B., 2000: Effects of endocrinedisruptors on prosobranch snails (Mollusca:Gastropoda) in the laboratory. Part I:Bisphenol A and octylphenol as xeno-estro-gens. � Ecotoxicology 9, 383-397.
SEGNER, H., CAROLL, K., FENSKE, M., JANSSEN, C.R.,MAACK, G., PASCOE, D., SCHÄFERS, C., VANDEN-BERGH, G.F., WATTS, M., WENZEL, A. 2003:Identification of endocrine-disrupting effectsin aquatic vertebrates and invertebrates: re-port from the European IDEA project. �Ecotoxicology and Environmental Safety 54,302-314.
VANDENBERGH, G.F., ADRIAENS, D., VERSLYKE, T.;JANSSEN, C.R., 2003: Effects of 17α-ethiny-lestradiol on sexual development of the am-phipod Hyalella azteca. � Ecotoxicology and.Environmental Safety 54, 216-222.
WATTS, M.M., PASCOE, D., CARROLL, K., 2002: Po-pulation responses of the freshwater amphipodGammarus pulex (L.) to an environmental estro-gen, 17α-ethinylestradiol. � EnvironmentalToxicology & Chemistry 21, 445-450.
WEIS, J.S., CRISTINI, A., K.R. RAO, K.R., 1992:Effects of pullutants on molting and regene-ration in Crustacea. � American Zoologist32, 495-500.
ZIMMER, M., 2001: Why do male terrestrial iso-pods (Isopoda: Oniscidea) not guard females?� Animal Behaviour 62, 815-821.
ZOU, E., FINGERMAN, M., 1997: Synthetic estro-genic agents do not interfere with sex diffe-rentiation but do inhibit molting of the cla-doceran Daphnia magna. � Bulletin of Envi-ronmental Contamination & Toxicology 58,596-602.
Accepted: 05.08.2005

116 LENNART WELTJE & JÖRG OEHLMANN
Book review
MAGURRAN, ANNE E. (2005): Evolutionary Ecology. The Trinidadian Guppy. OxfordSeries in Ecology and Evolution. � Oxford University Press, New York . 206 pages andnumerous figures. Paperback, 15.5 x 23.2 cm. ISBN 978-19-852786-2. 54.50 $
In 1997 A.E. Houde�s book �Sex, color, and mate choice in Guppies� was issued summa-rizing the work on sexual selection and mate choice in guppies. Now a further and highlywelcome synthesis of �what we know about the evolutionary ecology of the Trinidadianguppy� is available. Meanwhile populations of this small viviparous poeciliid have becomeone of the best studied �natural experiments� regarding the influence of predation pressureand the interrelations of ecology, behaviour and evolution. Hardly anyone seemed to havebeen made for this job as A.E. Magurran (University of St. Andrews, Scotland). Over theyears she and her students have decisively contributed and still contribute to this fascinatingtopic. The book is divided in eight chapters; some �key words� will be mentioned herein toindicate the wide range of subjects included: �1. Preview� contains an appreciation of earlyresearchers in this field, occasionally forgotten today, a definition of �evolutionary ecology inthe context of this book�, a brief introduction to guppy biology and a more precise descrip-tion of the objectives of the monograph. �2. Ecology of the guppy in Trinidad� describesthe �Trinidadian guppy system� and why it is attractive and suitable for evolutionary ecolo-gists, the various predators (fish, birds, other vertebrates, e.g. bats, invertebrates), relati-onships between productivity and phenotypic (and genotypic) variation, feeding behaviour,parasites and some dynamical aspects (density, sex-ratio). �3. Evading predators� deals withpredator-prey interactions, consequences of variation in predation risk, evidence for evoluti-on, significance of kinship and familiarity, learning of anti-predator skills, ontogenetic shifts,differences between the sexes. �4. Reproduction� summarizes the reproductive behaviour,female and male choice and mate choice in relation to risk, multiple mating, sperm compe-tition, cryptic choice and good genes. �5. Life history patterns� treats their evolution and therole of predators, intraspecific competition, seasonality, plasticity and predators and socialenvironment, temperature effects, aging and senescence. �6. Evolution of reproductiveisolation� contains a discussion of possible isolating barriers between river systems, poten-tial for allopatric speciation, pre-mating, post-mating, and post-zygotic isolation, sexualcoercion, learned mate recognition and reproductive isolation, sympatric speciation. �7. Con-serving a natural experiment� introduces guppies as a model species for conservation, paysthe attention to threatening by �homogenization� and the possible impact of scientists,population viability and consequences of artificial introductions. �8. Postscript and pro-spects� pleads for the guppy as experimental animal and outlines promising future direc-tions. Of course, chapters 2-7 end with a conclusion.This booklet is extremely informative, full of ideas and stimulating. At its end the reader willfind 37 pages with 796 references (not all are cited in the text): A must for all interested inbehavioural ecology and evolutionary biology and enthusiasts of the guppy.
Hartmut Greven, Düsseldorf

Abundance and life-histories of peracarid decomposers 117
Acta Biologica Benrodis 13 (2006)
Acta Biologica Benrodis 13 (2006): 117-125
The abundance, life history and reproduction traits ofperacarid macro-decomposers in a Ria Formosa
lagoon salt marsh, Southern Portugal*
Abundanz, „Life History“ und Charakteristika der Fortpflanzung vonMakrozersetzern (Peracarida) in einer Salzwiese (Ria Formosa
Lagune, Südportugal)
A. NATÁLIA DIAS1 & MARK HASSALL2
1Universidade do Algarve, Faculdade de Ciências do Mar e do Ambiente, Campusde Gambelas, P-8005-139, Faro, Portugal; present address: Centre of Ecology, Evo-
lution and Conservation, School of Environmental Sciences, University of East Anglia,Norwich NR4 7TY, United Kingdom; [email protected] (corresponding author)
2 Centre of Ecology, Evolution and Conservation, School of Environmental Sciences,University of East Anglia, Norwich, NR4 7TJ, United Kingdom; [email protected]
Summary : The abundances of four species of isopods and three of amphipods in a Ria Formosa saltmarsh in southern Portugal are compared. The three amphipods, Orchestia gammarellus, Orchestiamediterranea, and Talorchestia deshavesii were all more abundant, with densities of 358, 45 and 43 m-2
respectively, than three of the species of isopods, Porcellio lamellatus, Armadillidium album and Halophi-loscia couchii with densities of 36, 32, and 3 m-2 respectively, but the fourth species of isopod, Tylosponticus, was present in extremely high densities of 2950 m-2. The amphipods all had significantlyhigher growth rates than any of the isopods. Tylos ponticus grew significantly more slowly than any of theother species. The amphipods had shorter generation times and longer breeding periods, which werecontinuous for two of the species, than the isopods. They also have more broods than the isopods. Thiscombination of traits could help to explain the generally higher abundances of amphipods than isopodsin the highly productive but abiotically stressful conditions characteristic of salt marsh environments.
Density, relative growth rates, generation time, breeding phenology, trade-off, brood size, offspring size
Zusammenfassung: Die Abundanzen von vier Isopoden- und drei Amphipoden-Arten in einer Salz-wiese (Ria Formosa) in Südportugal werden miteinander verglichen. Die drei Amphipoden-Arten,Orchestia gammarellus, Orchestia mediterranea und Talorchestia deshavesii hatten höhere Abundanzen (358,45 und 43 m-2) als drei der Isopoden-Arten, Porcellio lamellatus, Armadillidium album und Halophilosciacouchii (36, 32 und 3 m-2 ). Die vierte Art, Tylos ponticus, erreichte sehr hohe Abundanzen (2950 m-2). AlleAmphipoden hatten signifikant höhere Wachstumsraten als die Isopoden. Tylos ponticus wuchs von allenArten am langsamsten. Die Amphipoden hatten eine kürzere Generationsdauer, eine längere Brutperi-ode und mehr Bruten als die Isopoden. Zwei Arten hatten kontinuierliche Brutperioden. Diese Eigen-schaften könnten die generell höheren Abundanzen der Amphipoden unter den hoch produktiven, aberim Hinblick auf die abiotischen Faktoren belastenden Bedingungen in Salzwiesen erklären.
Abundanzen, relative Wachstumsraten, Generationsdauer, Brutphänologie, trade off, Größe der Brut und desNachwuchses
*Contribution to the �First European Workshop on Isopods�, Düsseldorf, 14.-16. November2003, Germany

118 A. NATÁLIA DIAS & MARK HASSALL
1. Introduction
Salt marshes are highly productive but abio-tically stressful habitats (ODUM 1970; KNEIB
1984; CHAPMAN 1992; VERNBERG 1993). Thecombination of desiccation risk, osmoticvariability, and temperature fluctuations makethem harsher environments than either mar-ine or fully terrestrial habitats. However theyare predictably stressful. Such a combinationof high productivity and predictable stressimposes a distinctive suite of selection pres-sures on organisms living in salt marshes towhich they can potentially respond success-fully by developing life history strategies in-volving various different combinations ofresource acquisition and allocation strategies.
Under good growth conditions it can bepredicted that individuals with small off-spring, large broods and early reproductionwill be selected for and have a higher fitness,i.e., contribute more offspring to the nextgeneration than individuals with fewer largeoffspring (SOUTHWOOD 1988; MESSINA & FOX
2001). It is therefore logical to predict that aspecies of which the individuals have largerbroods of smaller offspring will becomemore abundant under good growth condi-tions, provided mortality is constant.
However, in environments in which mor-tality is not constant, further predictions basedon life history theory can be made. Where theratio of juvenile to adult mortality is high, itcan be predicted that individuals with smallerbroods of larger offspring and with an itero-parous breeding phenology, will be favouredby selection (SIBLY & CALOW 1985).
Where there is a combination of goodgrowth conditions but poor survivorship onespotentially a range of different combinationsof traits could be favoured by selection.
In this paper we compare the abundancesof seven species of peracarid macro-decom-posers in a salt marsh of the Ria Formosalagoon in Southern Portugal. Four of thespecies are isopods and three amphipods. Ona world wide scale isopods have been more
successful in colonising fully terrestrial habi-tats with over 5000 species reaching densitiesof up to 2150 m-2 in calcareous grasslands(SUTTON 1968). However, on a global scale,amphipods have been more successful in ex-ploiting intertidal habitats including the up-per reaches of salt marshes (DAHAL 1952;AVERILL 1976; ROBERTSON & LUCAS 1983;STENTON-DOZEY & GRIFFITHS 1983; INGLIS
1989). In this salt marsh both isopods andamphipods co-occur in the zone between theneap high tide level and spring high tide levelbut the different species do so at significantlydifferent densities. In this paper we hypoth-esise that the different success of the differ-ent species is related to differences in life his-tory traits and test this hypothesis by compar-ing their growth, development and reproduc-tive biology.
2. Material and Methods
2.1. Study area
The study area is a salt marsh system border-ing the Ria Formosa, a mesotidal lagoon onthe southern coast of Portugal separated fromthe Atlantic ocean by a 55 km long barrierisland system. The salt marshes in this la-goon are of the dry coast type (ADAM 1990)with the vegetation consisting predominantlyof Spartina maritima at lower levels, Sarcocor-nia spp. and Atriplex portulacoides L. in theintermediate region and Suaeda vera J. F. Gme-lin Suaeda maritima (L.) Dumort, Atriplex hali-mus L. and Limoniastrum monoptalum (L.) Bss.in the upper zone. Water salinity is approxi-mately 36 psu all year round (FALCÃO & VALE
1990).
2.2. Study species
Four species of terrestrial Oniscidea isopodsco-occur in the Ria Formosa salt marshes:Tylos ponticus Grebnitzky, 1874 (Tylidae),Porcellio lamellatus (Uljanin) Budde-Lund,1885 (Porcellionidae), Halophiloscia couchii

Abundance and life-histories of peracarid decomposers 119
Acta Biologica Benrodis 13 (2006)
(Kinahan, 1858) (Halophilosciidae) and Ar-madillidium album Dolfuss, 1887 (Armadillidi-idae). The four families to which these spe-cies belong show different morphological andphysiological adaptations to the terrestrialenvironment.
Porcellionidae and Armadillidiidae includespecies adapted to live in completely terres-trial habitats away from the coast and able tosurvive periodic desiccation of their environ-ment (SCHMALFUSS 1998). This is possible dueto a number of adaptations, e.g., changes inthe cuticle reducing transpiration, two to fivepairs of pleopodal lungs, together with otherqualities like shortening of the antennal fla-gellum (SCHMALFUSS 1989, 1998; HOPKIN 1991;ERHARD 1998). Tylidae, in contrast, still havean open water conducting system (SCHMAL-FUSS 1998), which only permits a limited cap-ture of water droplets from the environment.The Halophilosciidae is the only one ofthese families lacking pseudotrachea (SCHMAL-FUSS 1998). Consequently, species of these twofamilies are exclusively restricted to coastalmarine supra-littoral habitats, while Armadil-lidium album and Porcellio lamellatus have se-condarily returned to marine littoral habitats.
The amphipod assemblage that co-occurswith the isopods is composed of three spe-cies, Orchestia gammarellus (Pallas 1766), Orches-tia mediterranea mediterranea Costa 1853 and Tal-orchestia deshayesii (Audouin 1826). These spe-cies belong to the Talitridae family which do-minates the amphipod fauna in intertidal sys-tems, and alone contains truly terrestrial spe-cies (obligate land-dwellers) that are thoughtto have used salt marshes and/or estuaries asa route to colonization of the land (WILDISH
1988; RICHARDSON & SWAIN 2000).The adaptations of intertidal amphipods,
potentially convey an advantage for life onland, include strong pereiopods, strong de-velopment of the pleon, shortened uro-some, reduction of the third uropods andstiffening of uropods 1 and 2, which are suit-able for walking on land and supporting agreater weight than they would in water
(FRIEND & RICHARDSON 1986) and are associa-ted with an ability to jump (in air) using theposterior end of the body (BULYCHEVA 1957in FRIEND & RICHARDSON 1986). This abilityacts both as a means of locomotion and as ameans of escape from predators. The inter-tidal amphipods have also developed a ro-bust body form, presumably suited to life inthe high-energy littoral zone (FRIEND &RICHARDSON 1986). The enlargement of thegills may be due to an increase in the thick-ness of their cuticule to prevent desiccation(LINCOLN 1979) and it may provide a largersurface for ion uptake (BOUSFIELD person.commun. in MOORE & TAYLOR 1984).
The beachflea Orchestia gammarellus usuallyoccupies a fully saturated (> 95% R. H.) envi-ronment in the supralittoral, whether it be instrand-line wrack or under dense salt marshgrass swards (MORRITT 1998). It is distributedover a wide geographical area from Norwayand South-West Iceland in the North to theMediterranean Sea, the Canary Islands andthe Black Sea in the South. It also occurs onthe Atlantic coast of Canada and northernUSA (MOORE & FRANCIS 1986).
Orchestia mediterranea lives under stonesaround mean high water neap tide level, butalso occurs frequently in salt marshes (DEN
HARTOG 1963). Because O. mediterranea mi-grates on the shore so that its diurnal refugeis only covered by each recent tide, it is a trueeulittoral species. This species shows anupshore migration as tides change from neapto spring and a downshore migration astides change from spring to neap (WILDISH
1988). It is a Mediterranean-Atlantic speciesreaching its northern distribution limit in theIrish Sea and southern part of the North Sea,while its southern limit is in the Mediterrane-an Sea (DEN HARTOG 1963; LINCOLN 1979).
The sandhopper Talorchestia deshayesii in-habits sand beaches and occurs in non-tidalareas (Mediterranean and Baltic coasts) nearthe waterline as well as in areas with widetidal ranges (e.g., British Atlantic coasts) atthe base of the dune above high tide marks.

120 A. NATÁLIA DIAS & MARK HASSALL
It is distributed along the European coastsfrom southern Scandinavia to the Mediterra-nean Sea (NARDI et al. 2000).
2.3. Sampling methods
The animals were collected every sixth week atneap tide between November 1997 and July1999. Three different levels were sampled:(1) a lower stratum with compact mud at thesurface, corresponding to the neap high tidelevel (NHTL) dominated by Sarcocornia spp.and Atriplex portulacoides (3.15 m height and18.66 m distant from the high-water mark atneap tides); (2) an intermediate stratum withsandy soil and deposits of Zostera noltii Hor-nem, Spartina maritima and some algae (mainlyUlvales), which corresponds to the mean hightide level (MHTL) (3.31 m height and 22.63 mdistant from the high-water mark at neaptides); (3) an upper stratum with sandy soil,corresponding to the spring high tide level(SHTL) (3.90 m height and 28.23 m distantfrom the high-water mark at neap tides). Lit-ter and wrack deposits in the upper stratumwere rare and consisted mainly of dry leavesand stems of salt marsh plants.
Samples were taken by two methods: (a)pit-fall traps (plastic beakers 16 cm high x6 cm diameter) filled with very dilute forma-lin solution (0.01 %) left in place for 96 h (3replicates per level, randomly); (b) cores of113 cm2 to a depth of 20 cm sieved though a500 µm mesh sieve on site (15 replicates perlevel, randomly).
The following characteristics were recordedfor each specimen: body length (in 1 mm sizeclasses), sex, presence and number of eggs/young, ash free dry mass (AFDM). Ash freedry mass was obtained by pooling 1 to 20specimens of each size class for each core ortrap which were dried for 2 days at 60 ºC andashed for 3 h at 450 ºC.
Breeding season is defined as the time fromthe first appearance of ovigerous females tothe time of the last appearance of eggs/young in the marsupium. Reproductive activ-
ity is indicated by the percentage of femalescarrying eggs/young in the brood pouch.
2.4. Statistical analysis
Cohorts were identified by size frequencyanalysis using the Bhattacharya (1967)method in the FISAT software package(FAO-ICLARM Stock Assessment Tools).
Growth was calculated as relative growthrates, RGR (VAN EMDEN 1969), according tothe following equation:
RGR = (Log Lt1 - Log Lt0) / T
Where Lt0 is the length (mm) at the startof a given interval, Lt1 is the length at the endof that interval, and T is the number of yearsbetween the start (t0) and the end (t1).
3. Results
3.1. Abundance
The mean annual densities of all seven of thestudy species are shown in figure 1. Taking theaverage of mean high tide and spring high tidelevel densities, the isopod Tylos ponticus is themost abundant of all the species (2950 m-2).Judged on the criterion of its abundance, isclearly the most successful peracarid macro-de-composer in this salt marsh ecosystem. Thesecond most abundant peracarid was the am-phipod Orchestia gammarellus (358 m-2), follo-wed by the other two amphipods Orchestia me-diterranea (45 m-2) and Talorchestia deshayesii(43 m-2) both of which were more abundantthat any of the other three species of isopods(Porcellio lamellatus: 36 m-2, Armadillidium album:32 m-2, and Halophiloscia couchii: 3 m-2).
3.2. Relative Growth Rates, Adult BodySize, Development Rates, BreedingPhenology and Longevity
There is a consistent difference between mem-bers of the two orders in their relative growth

Abundance and life-histories of peracarid decomposers 121
Acta Biologica Benrodis 13 (2006)
rates (fig. 2). All species of amphipod grewfaster than any of the species of isopod. Ofall the species T. ponticus had conspicuouslythe lowest growth rate and the slowest deve-lopment rate taking two years to reach matu-
rity, compared with 12 months for the otherspecies of isopod and 3 to 7 months for theamphipods which all reached sexual maturitymore quickly than any of the isopods (tab. 1).The amphipods however, did not live as long
Fig. 1: Abundance of isopods and amphipods in the three sampling levels: NHTL � neap high tidelevel; MHTL � mean high tide level; SHTL � spring high tide level.Abb. 1: Abundanz der Isopoden auf den drei Merresniveaus, an denen gesammelt wurde: NHTL �Nipphochwasserr; MHTL � mittleres Hochwasser; SHTL - Springfluthochwasser.
Fig. 2: Relative growth rates (RGR) of isopods and amphipods. For most species +1. S.E. fallswithin the thickness of the lines; F6, 721 = 31959, P < 0.0001; all means significantly different to allother means at P < 0.05.Abb. 2: Relative Wachstumsraten (RGR) von Isopoden und Amphipoden. Für die meisten Artenfällt +1. S.E. in die Dicke der Linien. F6, 721 = 31959, P < 0,0001; alle Mittelwerte unterscheidensich mit P < 0.05 signifikant von allen anderen Mittelwerten.

122 A. NATÁLIA DIAS & MARK HASSALL
as the isopods which all had a lifespan ofover 12 months and for T. ponticus up tofour years.
T. ponticus had the highest mass at firstbreeding but was older at first sexual maturi-ty than O. mediterranea which was not so hea-vy when it first started breeding at six monthsold but reached a maximum mass of almostdouble that of T. ponticus.
The amphipods all had a longer breedingseason than any of the isopods (tab. 1). Or-chestia gammarellus and O. mediterranea bredcontinuously throughout the year with threeand six broods per year respectively, while T.deshayesii breeds from March through tillNovember (LOUIS 1980). The species with themost restricted breeding season was A. albumwhich only bred between May and July.
Tab. 1: Life history traits of isopods and amphipods in the Ria Formosa salt marsh. * source: VADER
& WOLF (1988), ** source: LOUIS (1980).Tab. 1: Life-history-Eigenschaften von Isopoden und Amphipoden in der Ria Formosa-Salzwiese.* Quelle: VADER & WOLF (1988), ** Quelle: LOUIS (1980)

Abundance and life-histories of peracarid decomposers 123
Acta Biologica Benrodis 13 (2006)
Only one of the seven species was poten-tially semelparous (tab. 1), this was Halophi-loscia couchii, all the amphipods had several,3-6, broods in their first year of life, Porcelliolamellatus bred twice in its first year and somesurvived long enough to also breed again intheir second season. Both T. ponticus and A.album bred in two seasons and some T. ponti-cus even also at the start of their fourth year.
3.3. Brood Size and Offspring Size
The two Orchestia species had the largestbroods of mean size 17 and 18 young brood-1
(tab. 1) while T. deshayesii has the smallestbrood of only 4 -7 young (LOUIS 1980). Tylosponticus had conspicuously the lowest size-specific fecundity of all the isopods but alsohad the largest individual offspring, whichwere ten times heavier than for the only otherspecies for which offspring mass data are avail-able, P. lamellatus.
4. Discussion
In the Ria Formosa salt marsh all three spe-cies of amphipods were more abundant thanthree out of the four isopod species withwhich they co-occurred. This is consistent withthe normal situation in salt marshes whereamphipods usually dominate the peracaridmacro-decomposer guild (DAHAL 1952; AVE-RILL 1976; ROBERTSON & LUCAS 1983; STEN-TON-DOZEY & GRIFFITHS 1983; INGLIS 1989)but raises the question of why are they betteradapted to exploit the exceptionally high pro-ductivity that is characteristic of salt marshesin general (ODUM 1970; CHAPMAN 1992; VERN-BERG 1993) than isopods are?
Examination of their life history and re-productive traits reveals some consistent dif-ferences between these members of the twoorders which may help to answer this questi-on. Most conspicuously the amphipods allhave higher relative growth rates than the iso-pods (fig. 1). This may reflect their consist-ently higher consumption rates as observed
by DIAS & HASSALL (2005) for these specieswith a range of foods that are available in thissalt marsh. These higher rates of resourceacquisition enable the amphipods to reach re-productive size at a younger age than the iso-pods and so to have a shorter generation time.When coupled with their longer, in the caseof two of the three species, continuous breed-ing season, extensive within season iteroparity,and larger broods, the reproductive potentialof the amphipods is considerably higher thanthat of the isopods.
It would thus appear that a combinationof higher resource acquisition rates and a re-source allocation strategy that leads to rapidgrowth, early maturation and high fecunditymay underpin the success of the amphipodsin exploiting the resources available in thissystem at higher levels of abundance. Theconspicuous exception to this generality is T.ponticus which, judged by the criterion ofabundance, is the most successful of all sevenof the peracarid species in this salt marsh.
Tylos ponticus has a completely contrastinglife history and reproductive strategy to boththe amphipods and to the other three iso-pods. It has the slowest growth rate, the lon-gest pre-reproductive period and generationtime of all seven species, it only has one broodper year and also has the lowest mass specificfecundity of any of the isopod species. Howthen can it achieve so much higher densitiesthan any of the other species? The answer tothis apparent enigma may lie in its reproduc-tive allocation strategy. Although its mass spe-cific fecundity is lower than that for the otherisopods, the relatively few offspring that ithas are bigger than those of any of the otherspecies studied, offspring ash free dry massbeing ten times higher than for the nextlargest isopod.
Thus although the number of new recruitsresulting from this resource allocation strategyis lower than the number released by thosespecies which have a short generation-high fe-cundity strategy, each individual offspring ismuch larger. This may be very significant in an

124 A. NATÁLIA DIAS & MARK HASSALL
environment that is subject to large fluctua-tions in temperature and moisture availability.In terrestrial isopods as little as 3% differencein offspring size can result in as much as 27%increase in survivorship of new recruits in theirfirst three weeks of life under fluctuating con-ditions or conditions of moisture stress, fur-thermore this survivorship advantage oflarger size increases with the magnitude of thedifference in offspring size (HASSALL et al. 2004).In the present interspecific comparison thereis over 1000% difference in size between theoffspring of T. ponticus and those of P. lamella-tus. It is thus reasonable to predict that theslow growth-low fecundity but large offspringsize strategy characteristic of T. ponticus will re-sult in a lower juvenile mortality which is cer-tainly the case in comparison to terrestrial spe-cies of isopod (DIAS et al. 2005) despite thehighly stressful environmental conditions thatcharacterise salt marsh habitats in general.
In conclusion this study has revealed a pos-sible explanation based on differences in lifehistory and reproductive strategies for the ge-neral phenomenon of amphipods oftenbeing more successful than isopods in ex-ploiting the very high productivity characteri-stic of salt marshes. It has however, alsoshown that exceptions to this broad genera-lisation exist and that by reducing juvenilemortality, as a result of changing the balancepoint in the resource allocation trade-offbetween brood size and offspring size, onespecies of isopod has achieved densities inthis abiotically stressful environment, thatexceed those of any population of fully ter-restrial isopod in the habitats to which theyare fully adapted.
Acknowledgements
We thank Dr. H. SCHMALFUSS who kindly gaveadvice on the phylogeny and evolution ofterrestrial isopods. The first author acknow-ledges financial support by the Fundação paraa Ciência e Tecnologia (grant PRAXIS XXI/BD/11039/97).
References
ADAM, P., 1990: Saltmarsh Ecology. � CambridgeUniversity Press, Cambridge.
AVERILL, P.H., 1976: The role of orchestid am-phipods in the breakdown of tidal marsh gras-ses. � M.Sc. thesis, University of Delaware,Delaware.
BHATTACHARYA, C.G., 1967: A simple method ofresolution of a distribution into Gaussiancomponents. � Biometrics 23, 115-135.
CHAPMAN, V.J., 1992: Ecosystems of the world �Wet coastal ecosystems (2nd ed.). � ElsevierScience Publishers B.V., Amsterdam.
DAHAL, E., 1952: Some aspects of the ecologyand zonation of the fauna on sandy beaches.� Oikos 4, 1-27.
DIAS, N, HASSALL, M., 2005: Food, feeding andgrowth rates of peralarid macrodecomposersin a Ria Formosa salt marsh, southern Portu-gal. � Journal of Experimental Marine Bio-logy and Ecology 325, 84-94.
DIAS, N., SPRUNG, M., HASSALL, M., 2005: Theabundance and life histories of terrestrial iso-pods in a salt marsh of the Ria Formosa la-goon system, southern Portugal. � MarineBiology 147, 1343-1352.
VAN EMDEN, H.F., 1969: Plant resistance to Myzuspersicae induced by a plant regulator and measu-red by aphid relative growth. � EntomologiaExperimentalis et Applicata 12, 125-131.
ERHARD, F., 1998: Phylogenetic relationshipswithin the Oniscidea (Crustacea, Isopoda). �Israel Journal of Zoology 44, 303-309.
FALCÃO, M., VALE, C., 1990: Study of the RioFormosa ecosystem; benthic nutrient re-mineralization and tidal variability of nu-trients in the water. � Hydrobiologia 207,137-146.
FRIEND, J.A., RICHARDSON, A.M.M., 1986: Biolo-gy of terrestrial amphipods. � Annual Re-view of Entomology 31, 25-48.
DEN HARTOG, C., 1963: The amphipods of thedeltaic region of the rivers Rhine, Meuse andScheldt in relation to the hydrography of thearea. Part II. The Talitridae. � NetherlandsJournal of Sea Research, 2, 40-67.
HASSALL, M., HELDON, A., GOLDSTON, A., CERANT,A., 2004: Ecotypic differentiation and phe-notypic plasticity in reproductive traits ofArmadillidium vulgare (Isopoda: Oniscidae). �Oecologia (submitted).

Abundance and life-histories of peracarid decomposers 125
Acta Biologica Benrodis 13 (2006)
HOPKIN, S., 1991: A key to the woodlice of Bri-tain and Ireland. � Field Studies 7, 599-650.
INGLIS, G., 1989: The colonization and degrada-tion of stranded Macrocystis pyrifera (L.) C.Ag. by the macrofauna of a New Zealandsandy beach. � Journal of Experimental Bio-logy and Ecology 125, 203-217.
KNEIB, R.T., 1984: Patterns of invertebrate dis-tribution and abundance in the intertidal saltmarsh: causes and questions. � Estuaries7(4A), 392-412.
LINCOLN, R.J., 1979: British Marine Amphipoda:Gammaridea. � British Museum (NaturalHistory), London.
LOUIS, M., 1980: Étude d�un peuplement mixted�Orchestia montagui Audouin et d�O. dehayeseiAudouin dans la baie de Bou Ismael (Algé-rie). � Bulletin d'Ecologie 11, 97-111.
MESSINA, F.J., FOX, C.W., 2001: Offspring sizeand number, pp. 113-127. In: EvolutionaryEcology: Concepts and Case Studies (Fox,C.W., Roff, D.A., & Fairbairn, D.T., eds) �Oxford University Press, Oxford.
MOORE, P.G., TAYLOR, A.C., 1984: Gill area rela-tionships in an ecological series of gammari-dean amphipods (Crustacea). � Journal ofExperimental Marine Biology and Ecology74, 179-186.
MOORE, P.G., FRANCIS, C.H., 1986: Notes on bree-ding periodicity and sex ratio of Orchestiagammarellus (Pallas) (Crustacea: Amphipoda)at Millport, Scotland. � Journal of Experi-mental Marine Biology and Ecology 95, 203-209.
MORRITT, D., 1998: Hygrokinetic responses oftalitrid amphipods. � Journal of CrustaceanBiology 18, 25-35.
NARDI, M., PERSSON, L.-E., & SCAPINI, F., 2000:Diel variation of visual response in Talitrussaltator and Talorchestia deshauesii (Crustacea:Amphipoda) from high latitude beaches onlow tidal amplitude. � Estuarine, Coastal andShelf Science 50, 333-340.
ODUM, W.E., 1970: Utilization of the direct gra-zing and plant detritus food chains by thestiped mullet, Mugil cephalus, pp. 222-240. In:Marine Food Chains (Steele, J.H., ed.). � Univ.California, Berkeley.
RICHARDSON, A.M.M., SWAIN, R., 2000: TerrestrialEvolution in Crustacea: The talitrid amphi-pod model, pp. 807-816. In: Crustacean Issues(12) � The Biodiversity Crisis and Crustacea.Proceedings of the Fourth InternationalCrustacean Congress (Klein, J.C.V.V., Schram,F.R., eds). � A.A. Balkema, Amsterdam.
ROBERTSON, A.I., LUCAS, J.S., 1983: Food choice,feeding rates, and the turnover of macrophy-te biomass by a surf-zone inhabiting amphi-pod. � Journal of Experimental Marine Bio-logy and Ecology 72, 99-124.
SCHMALFUSS, H., 1989: Phylogenetics in Onisci-dea. � Monitore Zoologico Italiano (N. S.)Monograph series 4, 3-27.
SCHMALFUSS, H., 1998: Evolutionary strategiesof the antennae in terrestrial isopods. � Jour-nal of Crustacean Biology 18, 10-24.
SIBLY, R.M., CALOW, P., 1985: Classification ofhabitats by selection pressures: a synthesis oflife-cycle and r/K theory: pp. 75-90. In: Be-havioural Ecology. Ecological Consequencesof Adaptive Behaviour. 25th Symposium ofthe Bristish Ecological Society, Reading 1984(Sibly, R. M., Smith, R. H., eds) � Blackwell,Oxford.
SOUTHWOOD, T.R.E., 1988: Tactics, strategies andtemplates. � Oikos 52, 3-18.
STENTON-DOZEY, J., & GRIFFITHS, C.L., 1983: Thefauna associated with kelp stranded on a sandybeach: pp. 557-568. In: Sandy beaches as eco-systems (McClachlan, A., Erasmus, T., eds). �Junk, The Hague.
SUTTON, S.L., 1968: The population dynamics ofTrichoniscus pusillus and Philoscia muscorum(Crustacea, Oniscoidea) in limestone grassland.� Journal of Animal Ecology 37, 425-444.
VADER, W., WOLF, L., 1988: Biotope and biologyof Armadillidium album Dollfuss, a terrestrialisopod of sandy beaches, in the SW Nether-lands. � Netherlands Journal of Sea Research22, 175-183.
VERNBERG, F.J., 1993: Salt-marsh processes: a re-view. � Environmental Toxicology and Che-mistry 12, 2167-2193.
WILDISH, D.J., 1988: Ecology and natural historyof aquatic Talitroidea. � Canadian Journal ofZoology 66, 2340-2359.
Accepted: 18.08.2005

126 A. NATÁLIA DIAS & MARK HASSALL
Rezension
WEHNER, RÜDIGER & GEHRING, WALTER (2007): Zoologie (24., vollständig überarbeiteteAuflage). � Georg Thieme Verlag, Stuttgart, New York. 954 Seiten, 1160 Abbildungen, 17Tabellen. Kartoniert, 19,7 x 23,8 cm. ISBN 978-3-13-367424-9. 49,95 Euro
Die �Zoologie� hat bis auf die Absicht, �jungen Biologen�(�)�Weite und Tiefe derProbleme unseres Faches vor Augen zu führen�, nahezu nichts mehr mit den ersten 17Auflagen des von A. Kühn verfassten �Grundriß der Allgemeinen Zoologie� gemein, derwegen seines relativ geringen Umfangs auch als �Kleiner Kühn� bezeichnet wurde. DieserGrundriss ist ab der 18. Auflage von R. Wehner und E. Hadorn und später von R. Wehnerund W. Gehring immer wieder aktualisiert worden und dabei notwendigerweise auch im-mer umfangreicher geworden. Nun liegt die 24. Auflage vor, d.h. das Buch hat seine Bewäh-rungsprobe längst bestanden und gehört zu den etablierten Lehrbüchern. Es ist jetzt groß-formatiger, umfangreicher, farbiger, bildreicher und noch übersichtlicher als die 23. Auflageund � sehr aktuell. Ob es tatsächlich spannend geschrieben ist (der Leser denkt bei diesemWort natürlich in erster Linie an einen Kriminalroman), wie der Klappentext anmerkt,möchte ich dahin gestellt sein lassen, allerdings ist auch der alte Text vielfach umgeschriebenworden und gewinnt dadurch zumindest an Präzision. Beim Vergleich mit der vorletztenAuflage zeigt sich, dass jedes Kapitel auch an Abbildungen hinzugewonnen hat. Was imEinzelnen anders oder neu ist, wird sich dem Leser (und dem Rezensenten) sicher erst nachund nach erschließen. Hervorzuheben ist allerdings auch die nun stringente Anwendungder phylogenetischen Systematik im Kapitel 12 �Vielfalt der Organismen�. Zwei kleineUngenauigkeiten, die mir beim ersten Durchblättern aufgefallen sind, könnten in der näch-sten Auflage leicht ausgemerzt werden könnten: Nicht nur Cichliden, sondern auch andereActinopterygii haben Schlundzähne (S. 650); die Darstellung der Chorda von Branchiostoma(S. 791) suggeriert einen Aufbau aus blasigen, turgeszenten Zellen anstatt aus Muskelplat-ten. Auffällige Neuerungen neben den bereits genannten sind: Rot markierte Abschnittebringen Kernsätze, die im Folgenden vertieft werden. Die Anzahl der Boxen (orangefarbigumrandet) mit Methodischem, Erklärendem und Ergänzendem ist beträchtlich erweitertworden. So genannte Plus-Boxen (grün eingefasst) bringen Historisches, Forschungsbei-spiele u.a.Insgesamt ist ein neues, modernes Lehrbuch entstanden, das sehr komprimiert und er-staunlich ausgewogen zoologisches Wissen in einer bewundernswerten Breite vermitteltund das sich ohne weiteres mit den umfangreichen amerikanischen Lehrbüchern, die zzt. inÜbersetzungen auf dem deutschen Buchmarkt zu haben sind, messen kann, das allerdingsweit über das im Grundstudium Gebotene hinausgeht, aber hier und auch später zumNachschlagen genutzt werden kann (nach wie vor ist das beigefügte Glossar hilfreich), undzwar ganz im Sinne A. Kühns, um �den jungen Biologen die über alle Spezialisierungen derheutigen biologischen Forschungsrichtungen hinausreichende Weite und Tiefe� zu zeigen.
Hartmut Greven, Düsseldorf

The role of animal-microbe interactions in isopod ecology and evolution 127
Acta Biologica Benrodis 13 (2006)
Acta Biologica Benrodis 13 (2006): 127-168
The role of animal-microbe interactions in isopodecology and evolution*
Die Bedeutung tierlich-mikrobieller Wechselwirkungen für dieÖkologie und Evolution von Asseln
MARTIN ZIMMER
Zoologisches Institut der Christian-Albrechts-Universität zu Kiel, Olshausenstr. 40,D-24098 Kiel, Germany; [email protected]
Summary: Due to their world-wide distribution in marine and terrestrial (as well as freshwater)habitats, the order Isopoda (Crustacea: Malacostraca: Peracarida) provides an excellent model forthe evolutionary ecology of terrestrialization.(1) Terrestrial isopods (Oniscidea) harbor endosymbiotic bacteria in their midgut glands (hepato-pancreas) that are lacking in marine isopods of the suborders Valvifera and Sphaeromatidea, con-sidered being (part of) a sister taxon of Oniscidea. Thus, these bacterial endosymbionts seem to besignificant in the context of living in terrestrial habitats and may have been important during thecourse of terrestrialization. In �truly terrestrial� species (Crinocheta), two different endosymbiontshave been characterized that are distantly related to known parasites and pathogens of the ordersRickettsiales and Mycoplasmatales, respectively. Both these endosymbionts form cytoplasmic ap-pendages that are in contact with the host epithelium and may serve in the exchange of nutrientsand information and or serve as holdfasts. In non-crinochete terrestrial isopods (Diplocheta, Tyli-da, Synocheta), hepatopancreatic bacteria belong to the genus Pseudomonas.Both marine and freshwater Asellota also harbor bacteria in their midgut glands. The lack of bac-teria in other marine suborders (as studied so far) may be due to antibiotic agents in these isopods.Based on the present findings, I propose a common (marine) ancestor of Asellota and Oniscideathat acquired the ability to harbor bacterial endosymbionts inside the hepatopancreas. While sym-biotic relationships remained unspecific in marine Asellota, they developed towards specific prima-ry symbioses with bacteria that aid in digesting cellulosic and phenolic compounds, and thus, faci-litate the utilization of terrestrial food sources in semi-terrestrial and terrestrial Oniscidea and infreshwater Asellota. I, further, hypothesize that later during early phylogeny of Crinocheta, primarysymbionts have been replaced by secondary endosymbionts that are still characteristic of recentCrinocheta.In contrast to previous studies, suggesting a role of hepatopancreatic bacteria in nutrition, our presentknowledge does not provide any evidence for crinochete symbionts to supply any digestive enzymesto their isopod host. However, Pseudomonas spp. are well-known to degrade both cellulosic and pheno-lic compounds. Thus, I hypothesize that, while primary symbionts of Oniscidea provide cellulasesand/or phenol oxidase, a transfer of cellulase and/or phenol oxidase genes from symbiont to hostoccurred in early Crinocheta, resulting in endogenous cellulase of evolutionarily bacterial origin. Be-sides (a) providing enzymes for the digestion of leaf litter, further possible contributions of hepato-pancreatic endosymbionts to their host�s physiological constitution and fitness include (b) increasingthe availability of nitrogen on a nitrogen-poor food source, (c) protecting their host from secondary(pathogenic) infection, (d) protecting their host from predatory attack, or (e) increasing fertility, mat-ing success and fecundity of their host � these hypotheses are briefly discussed.
*Contribution to the �First European Workshop on Isopods�, Düsseldorf, 14.-16. November 2003,Germany

128 MARTIN ZIMMER
(2) Terrestrial isopods interact with leaf litter-colonizing microbiota that they ingest along with theirmajor food source. While, however, it is well-documented that isopods gain from feeding on micro-bially inoculated leaf litter, reasons for this dependence are not well understood. Possibly, (a) micro-biota serve as supplementary high-quality food source and provide essential or otherwise limitingnutrients; (b) microbiota promote digestion of leaf litter itself, either prior to ingestion or during thegut passage; (c) microbiota simply act as indicators of easily digestible food sources of high quality.These explanations are not mutually exclusive, and the prevailing reason for preferentially consumingmicrobially inoculated leaf litter depends on both the species and developmental stage of the isopodand the nutritional context, i.e. the food source as such; recent results, however, indicate that cellu-lolytic capabilities of litter-colonizing microbiota [see (b)] may be less significant than previouslythought, while a role of litter-colonizing microbiota in indicating high-quality food [see (c)] is sup-ported. The ability to digestively utilize microbial cells as supplementary food [see (a)] depends on cellwall characteristics as indicated by gram-staining of the microbes, gram-positive bacteria beingdigested more effectively than gram-negative bacteria and fungi, and being preferred as food source.Despite numerous studies, the most recent ones using modern molecular techniques, it is still de-bated whether or not terrestrial isopods harbor resident gut microbes in their hindgut. Most hind-gut bacteria that may be candidates for hindgut residents appear to belong to gram-negative bac-terial taxa, and are taxonomically related to anaerobic species. Thus, we have to assume anoxicmicrohabitats in cuticular wrinkles. Further, the radial center of the hindgut is anoxic, too, allowingfor fermentative digestive processes, while the periphery of the hindgut lumen is largely oxic andoxidizing, thus, allowing for aerobic and oxidative digestive processes. These processes are pro-moted through cell compounds of ingested microbiota resulting in homeostatic maintenance of aslightly acidic pH that is optimal for the activity of involved enzymes. Potentially harmful effects ofphenolic food compounds that are likely under such conditions are counteracted through hydro-lytic enzymes and surfactants of microbial origin.In conclusion, our up-to-date knowledge as summarized and discussed herein strongly confirmsthe assumption that (terrestrial) isopods strongly depend on microbial activity and nutrients fortheir capability of digestively utilizing terrestrial leaf litter; on an evolutionary scale, this depend-ence may indicate the role that microbiota played during the course of terrestrialization, althoughthis aspect of isopod-microbe interactions is far from being understood.
Isopoda, terrestrialization, nutrition, digestion, endosymbiotic bacteria, symbiont-host interactions, leaf litter-coloniz-ing microorganisms, decomposition
Zusammenfassung: Aufgrund ihrer weltweiten Verbreitung in marinen und terrestrischen (sowielimnischen) Lebensräumen eignet sich die Ordnung Isopoda (Crustacea: Malacostraca: Peracarida)hervorragend als Modell für die Evolutionsökologie des Landgangs.(1) Landasseln (Oniscidea) beherbergen endosymbiotische Bakterien in ihren Mitteldarmdrüsen(Hepatopankreas), die ihren marinen Verwandten der Unterordnungen Valvifera und Sphaeromati-dea � beide als (Teil einer) Schwestergruppe der Oniscidea betrachtet � fehlen. Demnach scheinendiese bakteriellen Endosymbionten von Bedeutung für die Besiedlung terrestrischer Habitate zusein und könnten während der Terrestrialisierung eine wichtige Rolle übernommen haben. In �hö-heren Landasseln� (Crinocheta) konnten wir zwei verschiedene Endosymbionten charakterisie-ren, die entfernt mit bekannten Parasiten und Pathogenen der Ordnungen Rickettsiales und Myco-plasmatales verwandt sind. Beide bilden cytoplasmatische Zellfortsätze aus, die in Kontakt mit demWirtsepithel treten und entweder am Austausch von Nährstoffen und Information beteiligt sindoder als Verankerung dienen. In �niederen Landasseln� (Diplocheta, Tylida, Synocheta) gehörendie Endosymbionten in den Mitteldarmdrüsen zur Bakteriengattung Pseudomonas.Auch Asellota � sowohl marine als auch limnische � beherbergen Bakterien in ihren Mitteldarm-darmdrüsen. Das Fehlen dieser Bakterien in den bislang untersuchten anderen marinen Unterord-nungen ist möglicherweise auf antibiotische Agenzien in diesen Asseln zurückzuführen. Daherschlage ich einen gemeinsamen (marinen) Vorfahren der Asellota und Oniscidea mit der Fähigkeit,

The role of animal-microbe interactions in isopod ecology and evolution 129
Acta Biologica Benrodis 13 (2006)
bakterielle Symbionten im Hepatopankreas zu beherbergen vor. Während die daraus resultierendeEndosymbiose in marinen Asellota unspezifisch blieb, entwickelte sie sich in limnischen Asellotaund semi-terrestrischen � und später terrestrischen � Oniscidea zu einer primären spezifischenBeziehung mit Bakterien, die den Abbau cellulosischer und phenolischer Verbindungen ermögli-chen, und damit die Verdauung von Laubstreu erleichtern. Zudem vermute ich, dass später � wäh-rend der frühen Phylogenie der Crinocheta � primäre Endosymbionten durch sekundäre Endo-symbionten, die rezent in Crinocheta anzutreffen sind, ersetzt wurden.Im Gegensatz zu früheren Untersuchungen, die eine Rolle der Bakterien im Hepatopankreas bei Ver-dauungsvorgängen postulierten, gibt es nach heutigem Stand keinerlei Hinweise auf entsprechendeEnzyme in den Endosymbionten der Crinocheta. Pseudomonas spp. dagegen sind wohl bekannt für ihreFähigkeit, cellulosische und phenolische Verbindungen abzubauen. Ich stelle also die Hypothese auf,dass primäre Endosymbionten ihrem Wirt Cellulasen und/oder Phenoloxidasen zur Verfügung stellen,vor dem evolutiven Austausch der Symbionten aber ein Gentransfer zwischen primären Endosymbion-ten und Wirt stattfand, so dass �höhere Landasseln� endogen über diese Enzyme verfügen. Verschiede-ne Beiträge der Endosymbionten zur Konstitution ihres Wirtes sind vorstellbar: (a) die Lieferung vonVerdauungsenzymen, (b) eine erhöhte Stickstoffverfügbarkeit auf stickstoffarmer Nahrung, (c) Schutzvor (pathogenen) Sekundärinfektionen, (d) Schutz vor Fressfeinden, oder (e) Steigerung von Fertilität,Verpaarungserfolg und Fekundität. Diese Hypothesen werden kurz diskutiert.(2) Landasseln interagieren mit Streu besiedelnden Mikroorganismen, die sie gemeinsam mit ihrer Haupt-nahrungsquelle aufnehmen. Während aber wohl bekannt ist, dass Asseln davon profitieren, mikrobiellbesiedelte Streu zu fressen, sind die Gründe hierfür nur unzureichend verstanden. Möglicherweise (a)dienen Mikroorganismen als zusätzliche hochwertige Nahrungsquelle und liefern essenzielle oder an-derweitig limitierende Nährstoffe, (b) fördern Mikroorganismen die Verdauung der Laubstreu entwedervor der Konsumption oder während der Darmpassage oder (c) Mikroorganismen fungieren lediglich alsIndikatoren leicht verdaulicher Nahrung hoher Qualität. Diese Erklärungen schließen sich nicht gegen-seitig aus. Der Hauptgrund für eine Präferenz für mikrobiell besiedelte Laubstreu hängt sowohl von derArt und dem Stadium der Asseln, als auch von der Nahrungsquelle ab. Aktuelle Ergebnisse deutenjedoch an, dass cellulolytische Fähigkeiten Streu besiedelnder Mikroorganismen [siehe (b)] weniger be-deutend sind als bislang angenommen, während eine Rolle der Mikroorganismen als Indikatoren hoch-wertiger Nahrung [siehe (c)] wahrscheinlich erscheint. Die Fähigkeit der Assel, Streu besiedelnde Mikro-organismen als zusätzliche Nahrungsquelle zu nutzen [siehe (a)], hängt von deren Zellwandeigenschaf-ten, die durch Gramfärbung angezeigt werden, ab; gram-positive Bakterien werden effizienter verdautals gram-negative Bakterien und Pilze und zugleich als Nahrung bevorzugt.Trotz zahlreicher Untersuchungen mit klassischen und molekularbiologischen Methoden wird bisheute kontrovers diskutiert, ob Landasseln residente mikrobielle Enddarmbewohner beherbergen.Die meisten Kandidaten scheinen zu gram-negativen Taxa zu gehören und mit Anaerobiern ver-wandt zu sein. Demnach müssen wir anoxische Mikrohabitate in Kutikularfalten annehmen. Au-ßerdem ist auch das radiäre Zentrum des Enddarms anoxisch und ermöglicht fermentative Ver-dauungsprozesse, während die Bedingungen an der Peripherie oxisch und oxidierend sind, alsoaerobe und oxidative Verdauungsvorgänge ermöglichen. Derartige Prozesse werden durch pH-Homoiostasis infolge der Freisetzung mikrobieller Zellinhaltsstofe ermöglicht, die den pH-Wert ineinem optimalen Bereich für die entsprechenden Enzymaktivitäten konstant halten. Potenziell schäd-lichen Wirkungen phenolischer Nahrungsbestandteile, die unter solchen Bedingungen wahrscheinlichwären, wirken hydrolytische Enzyme und Detergenzien mikrobiellen Ursprungs entgegen.Zusammenfassend bestätigt unser heutiges Wissen, das in diesem Beitrag zusammengefasst unddiskutiert wird, die ausgeprägte Abhängigkeit der Landasseln von mikrobiellen Nährstoffen undEnzymen für ihre Fähigkeit, sich von Laubstreu zu ernähren. In evolutivem Maßstab mag dieseAbhängigkeit die bedeutende Rolle widerspiegeln, die Mikroorganismen während der Terrestriali-sierung spielten, obwohl dieser Aspekt tierlich-mikrobieller Interaktionen kaum verstanden ist.
Isopoda, Asseln, Landgang, Ernährung, Verdauung, endosymbiotische Bakterien, Wirt-Symbiont Wechselwirkun-gen, Laubstreu besiedelnde Mikroorganismen, Dekomposition

130 MARTIN ZIMMER
1. Introduction
Adaptive are those characteristics of an or-ganism that increase an individual�s fitnessrelative to that of its conspecifics. Thus, therespective characteristics will become moreand more frequent in the next generationsand will eventually become a common fea-ture of (almost) the entire population (e.g.FUTUYMA 1986).
Due to �formal constraints� (GOULD
1989; = �universal constraints� introducedby MAYNARD SMITH et al. 1985) and �histo-rical constraints� (GOULD 1989; = �localconstraints� of MAYNARD SMITH et al. 1985),however, adaptation is not necessarily com-plete or even optimal. On the other hand,other characteristics of organisms � havingbeen termed �pre-adaptation� (OSCHE
1962), �exaptation� GOULD & VRBA (1982),or �pre-disposition� (see SUDHAUS & REH-FELD 1992) � that are adaptive under recentconditions have been adopted from ances-tors to whom they may or may not havebeen adaptive when they developed. Someof these pre-dispositions may have beensignificant for the colonization of land viathe intertidal by animals with marine ance-stors. Due to the high number of recentmarine (ca 4,300), terrestrial (ca 3,500) andfreshwater (ca 650) species of isopods(Crustacea: Malacostraca: Peracarida), thisorder (Isopoda) provides a valuable modelin helping us understand the evolutionaryecology of terrestrialization.
The terrestrial environment exhibits nu-merous obstacles to hamper its coloniza-tion. Required adaptations comprise thestructural rigidity of the body, includingincreased strength of extremities, the struc-ture and function of respiratory organs,water budget and osmo- and thermoregu-lation, reproductive and developmental stra-tegies, and many more. In addition, terres-trial food sources (plants and parts of them,in case of herbivores and detritivores) dif-fer from aquatic food sources in terms of
physical and chemical characteristics. Sev-eral excellent reviews have been published(e.g., EDNEY 1954, 1968; HARTENSTEIN 1968;WIESER 1968, 1984; WARBURG 1968, 1987;SCHMALFUSS 1984, 1998; CAREFOOT 1993)that describe morphological, anatomical,physiological and ethological adaptationsand pre-dispositions of (terrestrial) isopods,but both reproductive (cf. ZIMMER 2001) andnutritive (cf. ZIMMER 2002a) biology of ter-restrial isopods have been regarded rarely.In particular, interactions of isopods withboth environmental and endosymbioticmicrobiota and their potential contributionsto digestive processes is far from being un-derstood. This review aims to summarizeour current knowledge extended by someunpublished data of the author and his co-workers on the significance of animal-mi-crobe interactions in ecology and evolutionof terrestrial isopods.
2. Terrestrial food sources
For most terrestrial detritivores, the carbonin plant tissue is not easily accessible, sincemost of it is part of recalcitrant cellulosemolecules (KOZLOVSKAJA & STRIGANOVA
1977; SZEGI 1988; BREZNAK & BRUNE 1994).The degradation of native (crystalline) cel-lulose is thought to usually be brought aboutby a complex of three enzyme classes(REESE & MANDELS 1971; ERIKSSON 1978;ERIKSSON & WOOD 1985) that have beendemonstrated in cellulolytic Protozoa (TRA-GER 1932; YAMIN & TRAGER 1979) and fun-gi (LJUNGDAHL & ERIKSSON 1985). Endo-β-1,4-glucanases (endocellulases, Cx-cellulases,EC 3.2.1.4) cleave inner β-1,4-glucosidicbounds and release oligosaccharides that,in turn, are cut into cellobiose by cellobio-hydrolases (exo-β-1,4-glucanases, exocellu-lases, C1-cellulases, EC 3.2.1.91). Cellobioseis the substrate of β-1,4-glucosidases (cello-biases, EC 3.2.1.21) that produce two mol-ecules of glucose. Since exocellulases couldbe demonstrated in only few bacteria (RAPP

The role of animal-microbe interactions in isopod ecology and evolution 131
Acta Biologica Benrodis 13 (2006)
& BEERMANN 1991), LJUNGDAHL & ERIKS-SON (1985) suggest the existence of onlytwo bacterial enzymes (corresponding tofungal endocellulases and cellobiases) thatappear to be cell wall-bound (WOOD &GARCIA-CAMPAYO 1990). Cellulolysis by an-aerobic bacteria � the radial center of theisopod hindgut is anoxic (ZIMMER & BRU-NE, 2005) � is brought about by cellulosomes(summarized in LESCHINE 1995), consistingof several enzymes with endo- and exocellu-lase activity and additional proteins that actas structural components or serve in bind-ing the substrate.
The degradation of cellulose is furtherconstrained through its close associationwith lignins (e.g. LJUNGDAHL & ERIKSSON
1985; BREZNAK & BRUNE 1994), high-mol-ecular phenolic polymers with a high de-gree of methoxylation (SWAIN 1979; HAGER-MAN & BUTLER 1991), that can only be de-graded through oxidation (BREZNAK &BRUNE 1994; TROJANOWSKI 2001). Numerousdifferent enzyme classes are known to beinvolved in oxidative degradation of phe-nolic compounds (ANDER & ERIKSSON 1976;WOOD 1980; MAYER 1987; CLAUS & FILIP
1990; SUMMERS & FELTON 1994; THURSTON
1994; FAURE et al. 1995), all of themcontaining a central metal ion (EVANS et al.1991).
ABE & HIGASHI (1991) summarized theirup-to-date knowledge by proposing thatmost (if not all) animals do not possess en-dogenous cellulases. However, HARTENSTEIN
(1964, 1982) had suggested earlier that sev-eral terrestrial isopods (Isopoda: Oniscidea)of the monophylum Crinocheta do producesuch enzymes [(HARTENSTEIN 1964: Oniscusasellus Linnaeus, 1758 (Oniscidae); HARTEN-STEIN 1982: Oniscus asellus, Trachelipus rathkii(Brandt, 1833) (Trachelipodidae), Armadil-lidium vulgare (Latreille, 1804) (Armadillidi-idae)]. On the other hand, subsequentstudies agreed with ABE & HIGASHI (1991)in that they suggested that woodlice dependupon cellulases produced by litter-coloniz-
ing microbiota (HASSALL & JENNINGS 1975;KOZLOVSKAJA & STRIGANOVA 1977), or atleast utilize fungal cellulases for cellulosedegradation (KUKOR & MARTIN 1986). Dur-ing their studies on the gut physiology ofPorcellio scaber Latreille, 1804 (Porcellionidae),ZIMMER & TOPP (1998a) discovered con-trasting data on cellulase activity and micro-bial counts in the hindgut: while the anter-ior hindgut � in accordance with findingsby HASSALL & JENNINGS (1975) in Philosciamuscorum (Scopoli, 1763) (�Philosciidae�) �exhibited the highest cellulolytic activity, italso contained the lowest density of bac-teria and fungi (ZIMMER & TOPP 1998a,b).Further, in contrast to HASSALL & JENNINGS
(1975), but in agreement with HARTENSTEIN
(1964a), cellulase activity was 3-30-timeshigher in the midgut glands (hepatopan-creas) than in the leaf litter, and thus, couldnot be attributed to enzymes ingested alongwith the food (ZIMMER & TOPP 1998a,b) ashad been suggested by HASSALL & JENNINGS
(1975). As had already been proposed ontheoretical grounds (HOPKIN & MARTIN
1982), ZIMMER & TOPP (1998b) concludedfrom results of feeding experiments withartificial diets and antibiotics that cellulasesare produced by bacterial endosymbiontsinhabiting the lumen of the hepatopancreas(fig. 1), or even by the isopod endogenously(cf. HARTENSTEIN 1964a). These enzymesshall, thus, be considered �functional en-dogenous� (cf. ZIMMER 2002a).
With respect to the origin of enzymes in-volved in the oxidative degradation of lig-nins and other phenolic compounds in theguts of invertebrates, there is little agree-ment either. Doubtlessly, phenol oxidasesare produced by arthropods, but these en-zymes are mostly required for molting (e.g.,ANDERSON 1985) and immune response (e.g.,GILLESPIE et al. 1997). Woodlice (Porcellioscaber and Oniscus asellus) produce endo-genous enzymes that are active in degrad-ing polycyclic aromatic hydrocarbons(PAHs) (DEKNECHT et al. 2001), and phe-

132 MARTIN ZIMMER
nolic leaf litter compounds are oxidizedduring the gut passage (NEUHAUSER & HAR-TENSTEIN 1976; ZIMMER & TOPP 1998c; ZIM-MER 1999). Questing for ecophysiological rea-sons for isopods storing vast amounts of cop-per in their midgut glands (discussed in ZIM-MER 2002a), ZIMMER & TOPP (1998c) per-formed feeding experiments with artificialdiets and antibiotics to observe correlationsbetween numbers of hepatopancreatic bac-teria and phenol oxidase activity in the hind-gut of Porcellio scaber; the copper store appearsto be significant for the production of cop-per-containing phenol oxidases (see fig. 1).
Leaf litter, the natural food source ofwoodlice, is usually densely colonized bymicrobiota, and woodlice were frequentlyproven to depend upon these microbiotawith respect to growth, reproduction andmortality; thus, woodlouse populations arestrongly influenced by the biomass and/or activity of leaf litter-colonizing micro-biota (UESBECK & TOPP 1995; ZIMMER &TOPP 1997a, 2000; KAUTZ et al. 2000).Hence, woodlice prefer feeding on denselycolonized leaf litter (GUNNARSSON 1987;SOMA & SAITO 1983; STÖCKLI 1990) that islocalized through olfaction (ZIMMER et al.
Fig. 1: Digestive processes in the hindgut of Porcellio scaber (Porcellionidae) (modified after ZIMMER
1998), including the ingestion of leaf litter, detoxification of ingestion phenolics (Detoxif.) in theforegut, digestion in the anterior hindgut through the activity of endogenous and bacterial en-zymes, absorption of nutrients and copper, microbial proliferation (Microb. prolif.) in the posteriorhindgut, and egestion of feces. See ZIMMER (2002a) for details.Abb. 1: Verdauungsprozesse im Enddarm von Porcellio scaber (Porcellionidae) (verändert nach ZIM-MER 1998), einschließlich der Aufnahme von Laubstreu, der Entgiftung phenolischer Inhaltsstoffe(Detoxif.) im Vorderdarm, der Verdauungsvorgänge im vorderen Enddarm durch die Aktivitätendogener und mikrobieller Enzyme, der Resorption von Nährstoffen und Kupfer, der mikrobiel-len Vermehrung (Microb. prolif.) im hinteren Enddarm, und der Ausscheidung von Faeces. Fürweitere Details, siehe ZIMMER (2002a).

The role of animal-microbe interactions in isopod ecology and evolution 133
Acta Biologica Benrodis 13 (2006)
1996). Possible reasons for the significanceof litter-colonizing microbiota can be sub-sumed as follows: (1) Microbial biomassserves as source of readily digestible (es-sential) nutrients (REYES & TIEDJE 1976;MARTIN 1984; SWIFT & BODDY 1984; GUN-NARSSON & TUNLID 1986; CAREFOOT
1984a,b; ULLRICH et al. 1991); (2) microbi-al extracellular enzymes that are ingestedalong with the leaf litter (partly) degradethe ingested lignocellulose (HASSALL & JEN-NINGS 1975; KUKOR & MARTIN 1986); (3)microbial procession of leaf litter removesrecalcitrant compounds (e.g., lignocellu-lose) prior to consumption by woodlice,thus facilitating digestion (HARTENSTEIN
1964a; RUSHTON & HASSALL 1983; HASSALL
et al. 1987).Without any doubt, microbial extracel-
lular enzymes are active in the hindgutof woodlice (fig. 1), but their role in di-gestive processes has been questioned(e.g., ZIMMER & TOPP 1998b,c). Recentresults suggest that litter-colonizingmicrobiota mainly serve as indicators ofhigh food quality to foraging woodlice(ZIMMER et al. 2003a), but obviously,digestive processes are also somehow pro-moted through ingested microbiota (fig.1). Possible reasons for this observationare (1) homeostasis of a gut pH (5.5-6.5)that is optimal for the activity of cellulasesand phenol oxidases through cellular com-pounds of digested microbiota (ZIMMER
& TOPP 1997b); (2) avoidance of toxiceffects of phenolic litter compounds,being likely under the prevailing pH con-ditions (BERENBAUM 1980; MARTIN & MAR-TIN 1983; SCHULTZ & LECHOWICZ 1986)through (a) hydrolysis of hydrolyzabletannins through enzymes of microbial ori-gin (ZIMMER 1999), or (b) precipitation ofphenolics through surfactants of possib-ly microbial origin (ZIMMER 1997).
The above-mentioned aspects of isopod-microbe interactions will be discussed in de-tail in turn.
3. Bacterial endosymbionts
3.1 General remarks
About 25 years after the first report of bac-teria in the midgut glands of terrestrial iso-pods (DONADEY & BESSE 1972), ZIMMER &TOPP (1998b,c) found correlations betweenbacterial numbers and enzymatic activitiesin the context of lignocellulose degradation(see fig. 1). Based on these results, they pro-posed that hepatopancreatic endosymbiontsof isopods facilitated or even made pos-sible the colonization of land through theircontribution to the digestive utilization ofterrestrial food sources (cf. ZIMMER et al.2001, 2002a). According to their originalhypothesis, these endosymbionts wouldhave been acquired once by a commonancestor of all oniscid isopods. In coinci-dence, bacterial endosymbionts have beenfound in midgut glands of numerous spe-cies of Oniscidea (tab. 1), but not in ma-rine Valvifera [ZIMMER et al. 2001: Idotea wos-nesenskii (Brandt, 1851) (Idoteidae); WANG,STINGL, BRUNE & ZIMMER, in review: Idoteabalthica (Pallas, 1772) (Idoteidae)] or Sphae-romatidea [SLEETER et al. 1978: Limnoria tri-punctata Menzies, 1951 (Limnoriidae); ZIM-MER et al. 2001: Gnorimosphaeroma oregonense(Dana, 1853) (Sphaeromatidae); ZIMMER &FRAUNE, unpubl.: Sphaeroma rugicauda)]. Incontrast, midgut glands of the freshwaterisopod, Asellus aquaticus (Linnaeus, 1758)(Asellota: Asellidae), harbor bacterial endo-symbionts that apparently provide cellulo-lytic and phenol-oxidizing enzymes (ZIM-MER & BARTHOLMÉ 2003). The authors con-clude from their results that Oniscidea andfreshwater Asellota � feeding on the samefood source, terrestrial leaf litter � acquiredsuch endosymbionts during convergent evo-lution. However, we recently found hepa-topancreatic bacteria in marine Asellota,Jaera albifrons Leach, 1814 (Janiridae) andEurycope cornuta Sars, 1864 (Eurycopidae)(ZIMMER & FRAUNE, unpubl.), suggesting a

134 MARTIN ZIMMER
Tab. 1: Selected characteristics of hepatopancreatic bacteria in different isopod species.Tab. 1: Ausgewählte Eigenschaften der Bakterien im Hepatopankreas verschiedenen Asselarten.
a: α-Proteobacteria; b: β-Proteobacteria; c: γ-Proteobacteria; d: Mollicutes; n.d.: not determined; ?:determination unsuccessful; 1: WANG, BRUNE & ZIMMER, in review; 2: ZIMMER & BARTHOLMÉ 2003;3: FRAUNE & ZIMMER, unpubl.; 4: WANG, STINGL, BRUNE & ZIMMER, in review; 5: ZIMMER et al. 2001;6: WANG & ZIMMER unpubl.; 7: HOPKIN & MARTIN, 1982; 8: WOOD & GRIFFITHS, 1988; 9: ULLRICH etal. 1991; 10: DONADEY & BESSE 1972; 11: HAMES & HOPKIN 1989; 12: ZIMMER 1998; 13: WANG et al.2004a; 14: WANG et al. 2004b.
common ancestor of Oniscidea and Asel-lota (see below).
With respect to cellulose hydrolysis andphenol oxidation in isopods, ZIMMER et al.(2002a) discuss two evolutionary scenarios:First, the ability to digest cellulose and oxi-dize phenolics may have been acquired si-multaneously with other physiological ad-
aptations (cf. CAREFOOT 1993) that allowedcolonization of land. In this case, these ad-aptations would be lacking from marine iso-pods. Second, the ability to digest celluloseand oxidize phenolics may have been a pre-disposition to the colonization of land. Inthis case, these capabilities would be pre-sent at least in some marine species. At least

The role of animal-microbe interactions in isopod ecology and evolution 135
Acta Biologica Benrodis 13 (2006)
those marine isopods that feed on brownalgae (Phaephycota) ingest consideraleamounts of phenolics (see ZIMMER et al.2001), while consumers of green algae(Chlorophycota) have to face relatively highcellulose contents of their food (VINOT etal. 1987; RUPEREZ & SAURA-CALIXTO 2001).Therefore, there would be a selective ad-vantage to the ability to digest cellulose andphenolics in many marine isopods (cf. ZIM-MER et al. 2001). If, however, the digestionof seaweed phenolics does not require theiroxidation � in contrast to lignins (seeabove) � the disadvantages of potentiallyharmful oxidation products (AHMAD 1992;APPEL 1993) would likely constrain theevolution of phenol-oxidizing capabilitiesin marine isopods. Comparing the inter-tidal Idotea wosnesenskii, Gnorimosphaeromaoregonense and the semi-terrestrial Ligia pal-lasii Brandt, 1833 (Oniscidea), we revealedevidence for the ability to hydrolyze cellu-lose and oxidize phenolics in intertidal andsemi-terrestrial isopods (ZIMMER et al.2002a), and ZIMMER et al. (2002b) concludefrom their comparison of semi-terrestrialand terrestrial isopods in a U.S. saltmarshthat phenolics in terrestrial leaf litter havenot been an obstacle to colonizing land andutilizing this food source. However, inter-tidal and semi-terrestrial isopods differ intheir abilitiy to both hydrolyze celluloseand oxidize phenolics, exhibiting a cleargradient of increasing digestive capabilitieswith terrestriality of the species� foodsources � Idotea wosnesenskii, preferentiallyfeeding on brown algae (Fucus spp.), do notoxidize phenolics, thus preventing poten-tially harmful effects of phenolic oxida-tion products.
On the other hand, as stated above, he-patopancreatic bacteria appear to be a syn-apomorphy of all Oniscidea (and possiblyAsellota: see below). Yet no Microcheta(Mesoniscidae) have been regarded so far,and data on Tylida (Tylidae) and Synochetaare still unpublished.
3.2 Characteristics and phylogeny ofbacterial endosymbionts
The up-to-date uncultured hepatopancreaticbacteria of terrestrial isopods have beenstudied most thoroughly in Porcellio scaber re-cently. Upon microscopic inspection, wefound three morphotypes of bacteria (tab.1); besides predominant species that couldbe detected frequently, there seem to be op-portunistic bacteria in Porcellio scaber (as wellas in other species). Molecular characteriz-ation (WANG et al. 2004a,b) revealed onlylittle similarity (< 81 %) of the 16S rRNAgenes of the two most frequent genotypeswith any other known bacterial species.According to phylogenetic analyses, endo-symbionts of Porcellio scaber cluster withRickettsiales (α-Proteobacteria) and Myco-plasmatales (Mollicutes), bacterial ordersmostly consisting of intracellular parasitesor pathogens in plants, animals and man.For rickettsial symbionts, we propose Can-didatus Hepatincola porcellionum (WANG etal. 2004a); for mycoplasmal symbionts, wepropose Candidatus Hepatoplasma crino-chetorum (WANG et al. 2004b).
Both Candidatus Hepatincola porcellio-num and Candidatus Hepatoplasma crino-chetorum live extracellularly in the hepa-topancreatic lumen, but are aggregated inthe periphery where they are attached tothe epithelium by means of cytoplasmicappendages (henceforth �stalks�) that in-sert into the spaces of the microvillousbrush border. (WANG et al. 2004a,b). Al-though the function of the stalks remainsunclear (see below), it is striking that twotaxonomically unrelated bacteria that sharethe same habitat independently develop afeature that is unknown from any of theirrelatives. According to STEINERT et al.(2000), the ability to attach to a host is oneof the prerequisites for establishing a sym-biotic relationship. Since the closest knownrelatives of hepatopancreatic bacteria areintracellular parasites or pathogens (see

136 MARTIN ZIMMER
above), we hypothesize that the stalks notonly serve in attaching to the host epithe-lium, but may represent an early evolu-tionary stage towards host cell invasion.Candidatus Hepatincola porcellionum andCandidatus Hepatoplasma crinochetorumwould then be representatives of an early-branching phylogenetic line of Rickett-siales and Mycoplasmatales, respectively,and the low sequence similarity corrobor-ates this assumption; Mycoplasma penetrans,a human pathogen that has first been iso-lated from AIDS patients, forms tip-likeappendages that insert deeply into infectedcells (e.g., LO et al. 1993; ROTTEM 2003).
Based on our limited knowledge of Can-didatus Hepatincola and Candidatus Hepato-plasma, we can only speculate on functionsof the stalk. In known prosthecate α-Pro-teobacteria (e.g., Caulobacter spp.; Caulobac-terales: Caulobacteraceae), stalks are in-volved in a complex life history with station-ary stages that are in close contact with thesubstrate in order to prevent drifting (HOLD
et al. 2001); bacterial endosymbionts ofPorcellio scaber could also utilize their stalksas holdfasts that insert in the microvillousbrush border of their hosts in order toavoid being washed off their habitat. Onthe other hand, well-studied symbiont-hostsystems (cf. SAVAGE 1972) suggest an ex-change of material and/or informationbetween closely associated symbionts andhost (WANG et al. 2004a,b). TOKUDA et al.(2000) concluded such an exchange from aclose association of endosymbiotic bacte-ria (Clostridium spp.) and microvilli of epi-thelial cells in the mesenteron of their ter-mite host (Nasutitermes takasagoensis; Isoptera:Termitidae).
Comparative studies of different speciesof Crinocheta provided evidence for Can-didatus Hepatincola being harbored by atleast three species: Porcellio scaber [individualsfrom Kiel and Cologne (Germany) andVancouver Island (B.C., Canada)], Oniscusasellus [Kiel, Cologne, Poitiers (France)], and
Trachelipus rathkii (Kiel), and CandidatusHepatoplasma also being present in Porcel-lio scaber, Oniscus asellus and Trachelipus rath-kii, but also in Philoscia muscorum (Kiel), Ar-madillidium vulgare (Kiel) and Alloniscus per-convexus Dana, 1856 (Alloniscidae) (Vancou-ver Island). Overall, however, CandidatusHepatoplasma was relatively rarely foundin Alloniscus perconvexus (n = 16; 25 %), Phi-loscia muscorum (n = 10; 60 %) and Armadil-lidium vulgare (n = 10; 10 %), but � if pre-sent � in high densities of 108-1011 per iso-pod (WANG & ZIMMER, unpubl.).
Considering the existence and identity ofhepatopancreatic endosymbionts a valuabletaxonomic feature (cf. HENNIG 1966; E1966;Brooks 1985), we propose a (preliminary)phylogenetic tree of the mentioned species(fig. 2a) that is in good agreement with treesbased on morphological (SCHMIDT & WÄ-GELE 2001: fig. 2b) and molecular data (MI-CHEL-SALZAT & BOUCHON 2000: fig. 2b;MATTERN & SCHLEGEL 2001: fig. 2c); it isonly the relation of Armadillidium (Armadil-lidiidae in SCHMIDT & WÄGELE 2001) to theother Crinocheta that distinguishes our treefrom that of SCHMIDT & WÄGELE (2001) andof MICHEL-SALZAT & BOUCHON (2000) inthat their tree would require a secondaryloss of Candidatus Hepatincola in Armadilli-dium if we considered the identity of hepa-topancreatic symbionts we found in thisspecies. In addition, our tree deviates fromthat by MATTERN & SCHLEGEL (2001) in thattheir tree would require a secondary lossof Candidatus Hepatincola in Philoscia, too.Thus, our phylogenetic tree of crinocheteisopods as based on microbiological cha-racteristics provides the simplest solutionto the interspecific distribution of endosym-biotic bacteria. However, in contrast tomorphology-based trees, the current pro-position does not consider numerous com-plex characteristics but, alike most molecu-lar trees, only a single feature, namely theidentity of hepatopancreatic endosym-bionts. Furthermore, we cannot exclude that

The role of animal-microbe interactions in isopod ecology and evolution 137
Acta Biologica Benrodis 13 (2006)
quent than the latter both in Porcellio scaber(20:80) and in Oniscus asellus (40:60). About30 % of all screened individuals of Oniscusasellus (n = 171) but only ca 10 % of allinvestigated Porcellio scaber (n = 265) did notharbor any bacteria in their hepatopancreas(tab. 2; WANG & ZIMMER, unpubl.; cf. WOOD
& GRIFFITHS 1988). Preliminary results of
Candidatus Hepatincola does exist in Ar-madillidium and/or Philoscia, but we were thusfar unable to detect this endosymbiont. De-tailed studies are needed and in progress inour laboratory.
Candidatus Hepatincola and CandidatusHepatoplasma never occurred together ina single specimen; the former was less fre-
Fig. 2: Phylogenetic tree ofsome families of Crinocheta; (a)as based on the identity of he-patopancreatic bacteria; (b) incomparison with a tree as pro-posed by MICHEL-SALZAT &BOUCHON (2000) and SCHMIDT &WÄGELE (2001); (c) in compari-son with a tree as proposed byMATTERN & SCHLEGEL (2001).Abb. 2: Stammbaum einigerFamilien der Crinocheta, (a)auf der Basis der Identität derBakterien im Hepatopankreas,(b) im Vergleich mit einemStammbaum nach MICHEL-SALZAT & BOUCHON (2000) undSCHMIDT & WÄGELE (2001),und (c) im Vergleich mit einemStammbaum nach MATTERN &SCHLEGEL (2001).

138 MARTIN ZIMMER
long-term studies in the laboratory (WANG
& ZIMMER, unpubl.) suggest that both lossand re-acquisition of hepatopancreatic bac-teria are possible and occur frequently inPorcellio scaber. However, seasonal changesin frequency or density of these endosym-bionts could be detected neither in Porcellioscaber nor in Oniscus asellus in the field (WANG
& ZIMMER, in prep.). In addition to our ob-servation of symbiont loss without any ob-vious harm, the relatively high number ofaposymbiotic isopod specimens (tab. 2) castsdoubt on the nutritive significance of he-patopancreatic bacteria (as will be discussedbelow) that had been proposed repeatedly(ZIMMER & TOPP 1998b,c; ZIMMER 1999;ZIMMER et al. 2001, 2002a; ZIMMER &BARTHOLMÉ 2003). On the other hand, theproportion of aposymbiotic Porcellio scaberin a given population appears at least in partto be controlled by food quality. When feed-ing on high-quality food (leaf litter of theraspberry, Rubus cf. fruticosus; Rosaceae), thenumber of symbiotic isopods decreases(WANG & ZIMMER, unpubl.). Possibly, thedegradation of lignocellulose or an addi-tional source of nitrogenous nutrients is notnecessary on this food source. Accordingto HASSALL (pers. comm.) three nutritionalstrategies of isopods are possible: (1) onhigh-quality food with high moisture, wood-lice simply squeeze out water-soluble nu-trients off the food and draw the nutritiousfluid into the hepatopancreas for resorp-tion of nutrients, while the remaining foodcompounds pass through the hindgut es-
sentially without any further nutritive utili-zation; (2) food of intermediate quality isenriched with digestive enzymes derivedfrom the hepatopancreas at the transitionof proventricle and hindgut and is digestedas efficiently as possible during a slowed-down gut passage; (3) since digestive util-ization of low-quality food is hardly pos-sible, this kind of food source is egestedvirtually undigested, and after microbial in-cubation and processing of feces they areutilized as food source with increased quali-ty (for a recent discussion of coprophagy,see KAUTZ et al. 2002).
Under favorable nutritional conditions(high-quality food; see above), the gain fromharboring bacterial endosymbionts might belower than costs (e.g., by contributing littleto digestion but utilizing food-derived nu-trients); according to a model by JOHNSTO-NE & BSHARY (2002), a host would be ex-pected to get rid of symbionts under suchconditions, particularly, if re-acquisition ofsymbionts is possible when conditionschange. On the opposite, one can deducethat digestively mutualistic symbionts arepromoted by their host when only low-qua-lity food is available, as had been assumedfor cockroaches (Blattopteroidea: Blattodea:Blattariae) and their intracellular endosym-bionts (BROOKS 1970) and could later beproven experimentally (GIJZEN et al. 1994)for Periplaneta americana and their symbioticciliates, Nyctotherus ovalis (Protista: Ciliopho-ra), that presumably provide cellulases. Thishypothesis would also explain why not all
Tab. 2: Percent of aposymbiotic individuals of different species of crinochete isopods (after WANG
& ZIMMER, in prep.).Tab. 2: Anteil aposymbiotischer Individuen verschiedener Crinocheta (nach WANG & ZIMMER, inVorb.).

The role of animal-microbe interactions in isopod ecology and evolution 139
Acta Biologica Benrodis 13 (2006)
woodlice harbor hepatopancreatic bacteria(see above). GOMULKIEWICZ et al. (2003)developed a model to demonstrate that in-teractions of two potential mutualists neednot necessarily be mutualistic at any giventime, at any given place; reciprocal adapta-tions (co-evolution) exhibit geographical va-riation (cf. THOMPSON & CUNNINGHAM 2002).Although our data show that both Candida-tus Hepatoplasma and Candidatus Hepatin-cola are abundant in Porcellio scaber and Oni-scus asellus from Germany, France and Ca-nada (WANG & ZIMMER, unpubl.: tab. 2), wedo not know anything about geographicalvariation in symbiont-host interactions orlocal adaptation of isopods and their sym-bionts (cf. LIVELY 1999).
To test the hypothesis of the commonancestor of all Oniscidea already harboringthe hepatopancreatic endosymbionts wefound in crinochete species as an early ad-aptation to a new habitat, we investigatedrepresentatives of the prototypal (SCHMAL-FUSS 1978, 1989; CAREFOOT & TAYLOR 1995)genus Ligia (Diplocheta: Ligiidae). Althoughwe found Pseudomonas n. sp. (γ-Proteobac-teria: Pseudomonadales) in midgut glandsof Ligia oceanica (Linnaeus, 1767) (WANG etal., in review), we were unable to demon-strate the existence of Candidatus Hepatin-cola or Candidatus Hepatoplasma. On theother hand, some individuals of both Porcel-lio scaber and Oniscus asellus (and Alloniscus per-convexus: ZIMMER & FRAUNE, unpubl.) alsocontain rod-shaped bacteria (HOPKIN &MARTIN 1982; WOOD & GRIFFITHS 1988;ULLRICH et al. 1991; ZIMMER 1998; tab. 1)that resemble Pseudomonas spp. and wereclassified as γ-Proteobacteria (WANG et al.,unpubl.). Rod-shaped bacteria in Ligidiumhypnorum (Cuvier, 1792) (Diplocheta: Ligii-dae), in Tylos ponticus Grebnitzsky, 1874 (Ty-lida: Tylidae) and in Trichoniscus pusillusBrandt, 1833 and Hyloniscus riparius (Koch,1838) (Synocheta: Trichoniscidae) were alsoidentified as representatives of the genusPseudomonas by hybridization of their 16S
rRNA genes with specific oligonucleotideprobes (ZIMMER & FRAUNE, unpubl.). Bycontrast, coccoid cells in Ligia pallasii (ZIM-MER et al. 2001; WANG & ZIMMER, unpubl.),in T. ponticus and in T. pusillus and H. riparius(ZIMMER & FRAUNE, unpubl.) as well as rod-shaped symbionts in the marine AsellotaJaera albifrons and Eurycope cornuta could thusfar not be determined (tab. 1). The coinci-dence between different phylogenetic lin-eages of Oniscidea (see fig. 3) with respectto the existence of Pseudomonas corrobora-tes our hypothesis concerning a single in-itial acquisition event of bacterial symbiontsin an early evolutionary stage of terrestria-lization. Possibly, the common ancestors ofthe Diplocheta (here: Ligia and Ligidium) andall other Oniscidea or even a common ma-rine ancestor of Oniscidea and Asellotasomehow developed the ability to harborbacteria in its midgut glands that getthere by chance, and implemented an un-specific endosymbiosis. In evolutionaryearly stages of Oniscidea, bacteria of thegenus Pseudomonas then established as spe-cific symbionts of terrestrial isopods thatare still present in Diplocheta, Tylida andSynocheta (and in some Crinocheta?) andmay be capable of degrading cellulose and/or phenolics. Based on a comparison of 16SrRNA gene sequences, we conclude that en-dosymbionts on Ligia oceanica derived frommarine pseudomonads. Thus, the acquisiti-on of the primary symbionts of Oniscideaprobably took place before the colonizationof terrestrial habitats, at least, however, nolater than in the intertidal (WANG et al., inreview). By contrast, marine Asellota appearto exhibit unspecific endosymbioses withdifferent bacterial species (WANG et al., inreview; cf. tab. 1). In Asellus aquaticus evolu-tion may � in convergence to Oniscidea �have led to specific symbioses that enablethe host to feed on leaf litter.
Hepatopancreatic bacteria have been de-monstrated in several aquatic crustaceans(MUSGROVE 1988; HARRIS et al. 1991; ZIM-

140 MARTIN ZIMMER
MER & BARTHOLMÉ 2003; WANG et al., inreview; ZIMMER & FRAUNE, unpubl.); thesymbionts of the freshwater crayfish, Para-nephrops zealandicus (Decapoda: Parastacidae),for instance, belong to the γ-proteobacteri-al genera Aeromonas (Aeromonadales), Cit-robacter and Hafnia (Enterobacteriales), andseem to provide cellulases for the digestionof leaf litter (MUSGROVE 1988). Freshwaterisopods of the species Asellus aquaticus alsoharbor endosymbiotic bacteria (see tab. 1)of the genera Rhodobacter (α-Proteobacteria:Rhodobacteriales), Burkholderia (β-Proteo-bacteria: Burkholderiales), Aeromonas, andRickettsiella (γ-Proetobacteria: Legionellales)in their midgut glands (WANG et al., in re-view). Both the high intraspecific diversityof bacterial endosymbionts and their tax-onomic identity apparently contradicts acommon evolutionary origin of endosym-
bioses in Asellota and Oniscidea and sug-gests convergent adaptations to terrestrialfood sources (leaf litter) in Asellus aquaticusand terrestrial isopods, unless we accept theidea of an intially unspecific symbiotic re-lationship in marine ancestors of Asellotaand Oniscidea that is still unspecific in ma-rine Asellota and evolved towards the func-tional endosymbiosis in freshwater Asello-ta. Feeding experiments actually providedevidence that (some of) the bacteria in A.aquaticus produce cellulases as well as phe-nol oxidases that are utilized for the diges-tion of leaf litter (ZIMMER & BARTHOLMÉ
2003). As discussed by ZIMMER (2002a) andZIMMER & BARTHOLMÉ (2003), we can, how-ever, not exclude the possibility of endo-genous cellulases in the tested isopods. Theconclusion by ZIMMER & TOPP (1998b,c) andZIMMER & BARTHOLMÉ (2003) are based on
Fig. 3: Phylogenetic tree of some suborders of Isopoda as based on the existence and identity ofendosymbiotic bacteria, with four evolutionary steps of symbiotic establishment; 1.: loss of anti-biotic agent and (passive) acquisition of hepatopancreatic bacteria in an unspecific endosymbiosis;2.: specific acquisition of Pseudomonas n. sp. in a common ancestor of all Oniscidea; 3.: acquisitionof Candidatus Hepatoplasma sp. in a common ancestor of all Crinocheta; 4.: acquisition of Candida-tus Hepatincola sp. (see also Figure 2).Abb. 3: Stammbaum einiger Unterordnungen der Isopoda, auf der Basis von Existenz und Identi-tät endosymbiotischer Bakterien, mit vier evolutiven Schritten: 1. Verlust antibiotischer Agentienund (passive) Akquirierung bakterieller Mitteldarmsymbionten, 2. spezifische Akquirierung vonPseudomonas n. sp. in einem gemeinsamen Vorfahren aller Oniscidea, 3. Akquirierung von CandidatusHepatoplasma sp. in einem gemeinsamen Vorfahren aller Crinocheta, 4. Akquirierung von Candida-tus Hepatincola sp. (s. Abbildung 2).

The role of animal-microbe interactions in isopod ecology and evolution 141
Acta Biologica Benrodis 13 (2006)
correlations between bacterial numbers andenymatic activities. According to HARTEN-STEIN (1964a), suggesting endogenous cel-lulases in crinochete isopods (see above),and to RAY & JULIAN (1952), providing evi-dence for endogenous cellulases in thewood-boring Limnoria lignorum (Rathke,1799) (Sphaeromatidea: Limnoriidae), it iswell possible that the experimental treat-ment simultaneously but independently re-duced both the number of hepatopancreaticbacteria and the release of endogenous cel-lulases into the gut lumen (for furtherdiscussion, see below).
The endosymbionts of Crinocheta makeus conclude that primary symbionts (Pseu-domonas) of Oniscidea were replaced (orsupplemented) by secondary symbionts(Candidatus Hepatoplasma and CandidatusHepatincola) soon after the branching ofCrinocheta (see fig. 3). Such an evolutionarytakeover has recently been demonstrated ex-perimentally in aphids (Hemiptera: Sternor-rhyncha: Aphidomorpha: Aphidina), replac-ing their primary symbionts, Buchnera aphi-dicola, by secondary symbionts (unknown γ-Proteobacteria), and has been discussed inthe context of establishing symbiotic rela-tionships (KOGA et al. 2003). Even in natu-ral systems, aphids harbor at least three dif-ferent secondary symbionts, belonging tothe genus Spiroplasma (Mollicutes: Mycoplas-matales) (FUKATSU et al. 2001), the γ-3 sub-division of the Proteobacteria and to thegenus Rickettsia (α-Proteobacteria: Rickett-siales) (MONTLLOR et al. 2002). Similarly, tse-tse flies (Glossina spp.; Diptera: Glossinidae)may lodge a commensal, Sodalis glossinidius(γ-Proteobacteria: Enterobacteriaceae) assecondary symbiont that belongs to thesame family as the primary symbiont, Wig-glesworthia glossinidia (DALE & MAUDLIN
1999). In dryophtherid beetles (Coleoptera:Curculionoidea) even three lineages of En-terobactericeae exist that colonized theirhost in independent evolutionary steps (cf.NARDON et al. 2003).
Applying a model by DOEBELI & DIECK-MANN (2000), I conclude that either the in-itial acquisiton of Pseudomonas sp. or the sub-sequent symbiotic takeover, possibly, led tothe branching and later speciation of Onis-cidea or Crinocheta, respectively, as has beendiscussed by BUCKLING & RAINEY (2002) ina broader context. LEONARDO & MUIRU
(2003) recently presented evidence for nu-tritional specialization being determined bythe identity of facultative bacterial sym-bionts in aphids.
In contrast to Asellota, marine Valviferaand Sphaeromatidea do not harbor hepato-pancreatic bacteria, although these animalslive in a microbe-rich environment and notbeing infected by bacterial endosymbiontsseems unlikely. SLEETER et al. (1978) pro-posed the production of antibiotics in thegut of the wood-boring Limnoria tripunctata(although ZACHARY & COLWELL (1979)found gut microbiota in this species). Thus,I extend the above hypothesis in that anti-biotics of endogenous or external origin inmarine isopods (except Asellota; see below)prevent the colonization of the midgutglands by bacteria (see fig. 3). HELLIO et al.(2000) found antibiotic agents in 9 out of16 species of marine macroalgae that mightstill be active in the guts of consumers (e.g.,isopods or amphipods) and may preventbacteria from conquering the hepatopan-creas. The initially unspecific(?) acquisitionof hepatopancreatic endosymbionts as a re-sult of the loss of antibiotic agents wouldthen simply be a consequence of an evolu-tionary change in feeding habits from con-suming fresh algae to preferring detritus.
In this context, PLANTE et al. (1990)discuss the significance of the habitat in de-termining an invertebrate�s role as host tomicrobial endosymbionts. In case of marineinvertebrates, the gut lumen and the envi-ronment exhibit virtually identical condi-tions in terms of ionic and osmotic levels,while this is not true for freshwater inver-tebrates. The gut of terrestrial invertebrates

142 MARTIN ZIMMER
provides a valuable aquatic habitat in a ter-restrial environment. Thus, the significanceof microbial gut symbionts can be explainedfrom different points of view: (1) Formarine microbiota, it is not advantageousto colonize the gut of an invertebrate in-stead of living in sea water, while the gutof a terrestrial invertebrate grants a morefavorable environment than soil or leaf lit-ter; (2) for terrestrial invertebrates, in turn,gut symbionts in a stable environment aremore reliable than soil or litter-colonizingmicrobes in a variable milieu, while this dif-ference is supposed to be insignificant formarine invertebrates (HARRIS 1993). Gutanatomy � in particular, the existence ofmidgut glands in shape of blind-endingtubes � can be interpreted as pre-disposi-tion (cf. HARRIS 1993).
Our current knowledge of endosymbioticbacteria in isopods makes the proposal of a(preliminary) phylogenetic tree of somesuborders of Isopoda possible (fig. 3); thistree contradicts most of the currentlydiscussed trees (WÄGELE 1989; BRUSCA &WILSON 1991; TABACARU & DANIELOPOL
1999) in that Asellota and Oniscidea areconsidered sister taxa here, forming a mo-nophylum that propose to be named Hepa-toconvivata (if the hosts provide nutrients)or Hepatohospita (if the hosts provide ahabitat rather than food). For the verifica-tion or rejection of this hypothesis and thusa decision upon the validity of this taxon,we urgently need additional data fromaquatic isopod suborders.
3.3 Symbiont-host interactions
Symbioses are generally accepted to act asevolutionary motor (MARGULIS & FESTER
1991; DOUGLAS 1992a; MAYNARD SMITH &SZATHMÁRY 1995; WATSON & POLLACK
2003). Further, it is frequently assumed thatmutualistic symbioses derive from parasiticones (BOUCHER et al. 1982; VANDERMEER
1983; PRICE 1991; YAMAMURA 1993; but see
NUISMER et al. 2003, for a different opin-ion), since the aim of a (vertically trans-mitted) parasite to be transferred to its host�soffspring conflicts the host�s interest ofkeeping its offspring free of parasites, andresults in an evolutionary arms race. Thesymbiont increases its transmission rate byreducing its virulence until both partnersgain from its transmission to the next gen-eration. If this is the case, the symbioticrelationship will be mutualistic (cf. THOMP-SON 1982). In addition, the exploitation ofthe host by the symbiont should not reducethe host�s longevity and fitness, because thiswould simultaneously reduce the fitness ofa vertically transmitted symbiont (ROUGH-GARDEN 1975). Even though � in spite ofinitial assumptions by ZIMMER & TOPP
(1998b,c) and ZIMMER et al. (2001, 2002a) �we could not yet unambiguously clarify whatkind of relation exists between isopods andtheir hepatopancreatic symbionts, their phy-logenetic affiliation with predominantly pa-rasitic and pathogenic bacteria is in agree-ment with this model.
Both aphids and their intracellular bac-terial endosymbionts, Buchnera aphidicola,evolved towards obligatory dependence oneach other (CHARLES & ISHIKAWA 1999;SHIGENOBU et al. 2000). Buchnera symbiontsare hosted in specialized cells, bacteriocytes,and provide essential amino acids to theirhost that feeds on nitrogen poor foodsources (see above) (WILKINSON & DOUGLAS
1996; WILKINSON & ISHIKAWA 2000; DOU-GLAS et al. 2001; BERNAYS & KLEIN 2002).As briefly discussed above, the transmissionof symbionts is critical in mutualistic sym-bioses, both for obligatory endosymbiontsand for their host. It is only through verti-cal transmission that a host can ensure thatnone of his progeny will stay aposymbiotic.Further, given that competition betweendifferent symbionts may exert negative ef-fects on the host (discussed in KOGA et al.2003), it seems advantageous to the host tokeep the diversity of symbionts low, and

The role of animal-microbe interactions in isopod ecology and evolution 143
Acta Biologica Benrodis 13 (2006)
the best way to achieve this aim is controlledvertical transmission (KORB & AANEN 2003).Symbiotic bacteria of the nudibranch, Den-drodoris nigra (Gastropoda: Ophistobranchia),are directly transferred from the vestibulargland, where they intrude between the epi-thelial microvilli, to the egg masses prior tospawning (KLUSSMANN-KOLB & BRODIE
1999). The transmission of yeast-like in-tracellular endosymbionts (Symbiotraphinaspp.) in the ceca of Stegobium paniceum andLasioderma serricorne (Coleoptera: Anobiidae)that provide vitamin B and sterols (PANT &FRAENKEL 1954) and detoxify harmful foodcompounds (DOWD 1989; SHEN & DOWD
1991) is brought about by smearing the eggsduring oviposition (NODA & KODAMA 1996).
Yet, many mutualists rely on horizontaltransmission (discussed in WILKINSON &SHERRATT 2001), and horizontal transfer ofsymbionts is the rule in sexually reproducinganimals with symbionts that are not harboredinside the reproductive tract (DOUGLAS 1995;but see YAMAMURA 1993). Larvae of ponyfish (Leiognathus nuchalis; Perciformes: Leio-gnathidae) hatch aposymbiotic and get in-fected by luminescent bacteria at an age ofat least 45 days (WADA et al. 1999). Similarly,aposymbiotic juveniles of lucinid mussels(Mollusca: Bivalvia) acquire sulfide-oxidizingintracellular gill symbionts (γ-Proteobacteria)horizontally. The flatworm, Convoluta roscof-fensis (Plathelminthes: Turbellaria), harborsendosymbiotic algae of the genus Tetraselmis(Chlorophyta), but host individuals differ inwhat species of Tetraselmis they accommo-date. Aposymbiotic juveniles take up algaewhile feeding and reject all but one spe-cies (DOUGLAS 1980). A similar trans-mission mechanism, with bacteria being in-gested along with the food, have been pro-posed for the sponge, Halichondria panicae(Porifera: Demospongiae), and its sym-bionts of the genus Rhodobacter (α-Proteo-bacteria) (ALTHOFF et al. 1998; discussedin STACKEBRANDT & PUKALL 1999; MÜLLER
1999).
Both Candidatus Hepatoplasma and Can-didatus Hepatincola, too, appear to be trans-mitted horizontally or even through abioticvectors in the environment: under sterileconditions in the laboratory, neither em-bryos nor 1d-old mancae I or 1wk-old man-cae II of Porcellio scaber harbored hepato-pancreatic bacteria (WANG et al., in review).If kept non-sterile, however, both embryosand mancae I are void of bacteria, too, whilebacteria can be found in the midgut glandsat the age of one week. Thus, either marsu-pial fluids are free of bacteria � contrastingpreliminary results by STEVENS & ZIMMER,unpubl. (in ZIMMER 2002a) and electron-mi-croscopic studies by STEVENS & GREVEN
(pers. comm.) � or embryos and marsupialmancae are unable to take up bacteria intotheir digestive tract. One possible obstacleto bacterial colonization of the midgutglands could be the filter system betweenproventricle and hepatopancreas that iscommon in Isopoda, Amphipoda and mostMalacostraca, preventing the influx of par-ticles into the glands (MARTIN 1964; SCHE-LOSKE 1976; OSHEL & STEELE 1988; WÄGE-LE 1989). According to WOOD & GRIFFITHS
(1988) particles larger than 1.17 µm in dia-meter are excluded, but HAMES & HOPKIN
(1989) observed a mesh size of just 40-50nm. However, it is questionable how thepressure should be built up that would beneeded to push fluids through a filter ofsuch small mesh size into the hepatopan-creas. STORCH (1987) discussed the possibil-ity of actively changing the mesh size ofthe proventricular filter. Yet, in this case,contamation of the midgut glands with lit-ter-colonizing microbiota would be likely.On the other hand, there must be some wayfor hepatopancreatic endosymbionts to passthe filter and enter the glands; this problemcould thus far not be solved (cf. ZIMMER
2002a).In the Spider Crab, Chionoectes opilio (Deca-
poda: Brachyura: Majidae), males transfer an-tibiotic agents with their sperm (BENHALIMA

144 MARTIN ZIMMER
& MORIYASU 2001), and comparable findingshave been reported for the fruitfly, Droso-phila melanogaster (LUNG et al. 2001). In iso-pods, too, such substances could be drawninto the marsupium to help prevent micro-bial contamination of marsupial embryos(but see above).
Horizontal transfer of symbionts via fecesor litter, as suggested by our findings inPorcellio scaber (see above), seems to be inef-ficient, since it cannot be ensured that alloffspring get access to (the correct) sym-bionts (see above). On the other hand, evenvertical transmission of (bacterial) sym-bionts is a bottleneck with respect to thenumber of transferred symbionts (MIRA &MORAN 2002), and the risk of enrichingharmful mutations in symbionts is higherthan in symbiont-host systems with hori-zontal transmission (RISPE & MORAN 2000).In this context, a comparison of CandidatusHepatincola with other Rickettsiales is in-teresting: while Wolbachia (see BOUCHON etal. 2005, in this issue) is usually transmittedvertically, horizontal transmission is typicalfor Rickettsia and Ehrlichia (ANDERSON &KARR 2001; SCHULENBURG et al. 2001); butWolbachia can be transferred horizontally,too (CORDEAUX et al. 2001).
Despite the risk inherent to horizontal trans-mitting, Candidatus Hepatincola or CandidatusHepatoplasma are present in 60-100 % (de-pending on the population and the season;see above) (WANG & ZIMMER, unpubl.).Thus, woodlice successfully detect and findbacteria in their environment (see also ZIM-MER et al. 1996), or the probability of en-countering and taking up these bacteria (ortheir propagules) is increased by the widedistribution in the isopods� environment (seeYAMAMURA 1993). Since bacteria inside thehepatopancreatic lumen actively proliferate(WANG et al., unpubl. TEM studies), we ex-pect surplus bacteria to be drawn to thehindgut and egested in feces (discussed inZIMMER 2002a). Extracellular luminescentendosymbionts of deep sea fish (Berycifor-
mes: flashlight fishes, Anomalopidae; pine-cone fishes, Monocentridae) (NEALSON etal. 1984) and of cuttlefish (Euprymna scolo-pes; Cephalopoda: Sepiidae) (LEE & RUBY
1994) are excreted at rates of 106-108 perday. Such a transfer mechanism could ex-plain the presence of Candidatus Hepatin-cola and Candidatus Hepatoplasma in mostcrinochete species tested thus far, if we as-sume that isopods but not their bacterialendosymbionts exhibit high specificity withrespect to their symbiotic partner (cf. FEIN-SINGER 1983). Up to now, however, we wereunable to detect either Candidatus Hepatin-cola or Candidatus Hepatoplasma using spe-cific fluorescence-labeled probes in thehindgut, or the feces of isopods or in leaflitter or soil (WANG & ZIMMER, unpubl.).Possibly, the symbionts are present as pro-pagules that cannot be detected with thistechnique. We currently try to detect themthrough PCR-amplification of their 16SrDNA using sequence-specific primers.
Neither in Porcellio scaber nor in Onicus asel-lus we ever found Candidatus Hepatincolaand Candidatus Hepatoplasma occurring to-gether (WANG & ZIMMER, unpubl.). Giventhat symbionts are transmitted horizontally(see above), an infection of an isopod withone of its potential endosymbionts seemsto preclude a secondary infection with theother one. Such interactions have been ex-plained in terms of competition betweensymbionts for their habitat (see KOGA et al.2003), but could also be due to induced de-fense of the host (cf. KURTZ & FRANZ 2003).
Based on our current knowledge we areto suppose that the initial inoculation ofjuveniles (as well as re-inoculation of apo-symbiotic adults; see above) depends on theuptake of potential endosymbionts with thefood, the most likely and reliable source ofsymbionts are probably feces of conspeci-fics. Beside its nutritional role (cf. KAUTZ etal. 2002), coprophagy, thus, seems to beadaptive in the context of acquiring hepa-topancreatic endosymbionts.

The role of animal-microbe interactions in isopod ecology and evolution 145
Acta Biologica Benrodis 13 (2006)
Symbiotic relationships between micro-biota and blood- or sapsuckers or detriti-vores (cf. BUCHNER 1965) have frequentlybeen described to include the transfer ofnutrients or a nutritive exploitation of sym-bionts by the host (e.g., BAUMANN & MO-RAN 1997; CAZEMIER et al. 1997; STEINERT
et al. 2000). Aphids (see above) and termi-tes (Blattopteroidea: Blattodea: Isoptera) areamong the best-studied symbiotic arthro-pods. Although both face the problem ofnitrogen-poor food source, termites � in con-trast to the sap-sucking aphids � are in ad-dition confronted with remarkable amountsof lignocellulose and other recalcitrant anddeterrent compounds of their natural foodsources (see BREZNAK & BRUNE 1994). Ho-wever, only some termite species rely ongut symbionts for the production of cellu-lases (cf. DOUGLAS 1992): lower termites(Mastotermitidae, Kalotermitidae, Hodoter-mitidae and Rhinotermitidae) harbor cellu-lolytic flagellates (Protista: Sarcomastigo-phora: Mastigophora) in their gut that pro-vide short-chained fatty acids to the nutri-tion of their host; cellulases of symbioticorigin are supplemented by endogenous cel-lulases (MARTIN 1984; TOKUDA et al. 2002;OHKUMA 2003). Higher termites (Termiti-dae) hydrolyze cellulose by endogenouscellulases that are produced in their midgut,but do not harbor cellulolytic protists (cf.OHKUMA 2003). Macrotermitinae, on theother hand, garden fungi (genus Termitomy-ces) in their colonies and feed on cellulase-rich nodules of the fruit bodies; cellulasesmaintain their activity during the gut passa-ge and contribute to cellulose digestion(MARTIN 1984). However, the nutritive sig-nificance of these enzymes for digestiveprocesses is unclear (cf. VEIVERS et al. 1991)as has also been stated with respect to cel-lulases in midguts of wood-feeding beetles(Coleoptera; e.g., Cerambycidae, Anobiidae,Bupestridae) or wood wasps (Hymenopte-ra: Symphyta: Siricoidea: Siricidae) (MARTIN
1991) and detritivorous stoneflies (Pteron-
arcys proteus; Plecoptera) or caddisflies (Py-cnopsyche luculenta; Trichoptera) (SINSABAUGH
et al. 1985).Although ABE & HIGASHI (1991) stated,
that �most animals [...] lack the ability toproduce the enzymes necessary for decom-posing cellulose� (p. 127), evidence for en-dogenous cellulases have been presented forthe marine isopod Limnoria lignorum (RAY &JULIAN 1952) and the freshwater amphipodGammarus pulex (MONK 1977; ZIMMER &BARTHOLMÉ 2003; but see BÄRLOCHER 1982,and BÄRLOCHER & PORTER 1986) as well asin the silverfish Thermobia domestica (Zygen-toma: Lepismatidae) (TREVES & MARTIN
1994) and the applesnail Ampullaria crossean(Mollusca: Gastropoda) (WANG et al. 2003).Further, parts of an active cellulase com-plex, namely β-glucosidases, have beendemonstrated in numerous invertebrates:Locusta migratoria (Orthopteroidea: Caelife-ra: Acrididae) (MORGAN 1975), Rhynchoscia-ra americana (Diptera: Nematocera: Sciaridae)(FERREIRA & TERRA 1983), Erinnyis ello (Le-pidoptera: Sphingidae) (SANTOS & TERRA
1985), Sitophilus oryzae (Coleoptera: Curcu-lionidae) (BAKER & WOO 1992), Abracris fla-volineata (Orthopteroidea: Caelifera: Acrid-idae) (MARANA et al. 1995), Spodoptera frugi-perda (Lepidoptera: Noctuidae) (MARANA etal. 2000). �Incomplete� (see above) cellu-lase complexes, consisting of endo-β-1,4-glucanases and β-glucosidases but lackingexocellulases and being capable of degrad-ing soluble but not native crystalline cellu-lose (see SKAMBRACKS & ZIMMER 1998), havealso been described: Trachelipus rathkii (KU-KOR & MARTIN 1986), Gammarus lacustris(Amphipoda: Gammaridea) (MCGRATH &MATTHEWS 2000), Macrotermes mülleri (Iso-ptera: Termitidae) (ROULAND et al. 1988),Panesthia cribrata (Blattopteroidea: Blattodea:Blattariae) (SCRIVENER & SLAYTOR 1994a).Although the degradative capacity of suchenzymes is lower than that of microbialcellulases (SLAYTOR 1992; SCRIVENER & SLAY-TOR 1994b), it is obviously sufficient to hy-

146 MARTIN ZIMMER
drolyze cellulose without microbial assis-tance (SLAYTOR 1992); the low efficacy iscompensated through high rates of produc-tion (SCRIVENER & SLAYTOR 1994b). Possibly,the supplementary uptake of fungal cellu-lases that provide critical components ofcellulase complexes is significant for theseanimals (MARTIN 1984). Neither ZIMMER &TOPP (1998b,c) nor ZIMMER & BARTHOLMÉ
(2003) distinguished between different com-ponents of the cellulase complex in theiranalyses of cellulase activity.
At least the fungal symbionts (genus Ter-mitomyces) of some termites (Macrotermesspp.) degrade lignins, thus facilitating cellu-lose digestion by the hosts (HYODO et al.2000, 2003); JOHJIMA et al. (2003) isolatedand characterized a peroxidase from Termi-tomyces albuminosus. Leaf-cutting ants (Hyme-noptera: Apocrita: Formicoidea: Myrmici-dae) and their gardened fungi show onlylittle overlap in their enzymes (D�ETTORE
et al. 2002), suggesting a stable dependenceof the hosts on their symbionts. No matterwhether or not cellulolytic or pectinolyticenyzmes are provided by the fungi (cf. AB-RIL & BUCHER 2002), the fungal activityseems to facilitate the ants� access to thenutritious cell contents of plant tissues. Job-sharing is also common in the marine aba-lone, Haliotis midae (Mollusca: Gastropoda:Prosobranchia), with resident gut bacteriadegrading cellulose (ERASMUS et al. 1997).In this same context, we are to interpret thesuggestion that cellulases and phenol oxi-dases in terrestrial isopods � and, as a re-sult of convergent evolution to feeding onterrestrial leaf litter, also in the freshwaterisopod, Asellus aquaticus (ZIMMER &BARTHOLMÉ 2003) � are provided by hepa-topancreatic bacteria (ZIMMER & TOPP
1998b,c; ZIMMER 1999; ZIMMER et al. 2001,2002a).
Whether or not bacterial endosymbiontsof isopods contribute cellulases and/or phe-nol oxidases can only be clarified by meansof genetic detection of the respective
genes. Relatives of Candidatus Hepatoplas-ma and Candidatus Hepatincola within theMollicutes and Rickettsiales, respectively, arenot known to produce any cellulolytic orphenol-oxidizing enyzmes which contradictsthe proposed function of these symbionts.However, even the closest relatives are sodistantly related that we cannot convincinglydeduce any physiological characteristics ofCandidatus Hepatoplasma or Candidatus He-patincola from what we know about myco-plasmas or rickettsias. ESHAM et al. (2000)were unable to predict the ability to degradelignins based on the phylogenetic affiliationof several α- and γ-Proteobacteria andgram-positive bacteria.
On the other hand, close relatives of en-dosymbionts in non-crinochete isopods (seeabove), e.g. Pseudomonas fluorescens, are capableof degrading both cellulose and hemicellu-lose and polycyclic aromatic hydrocarbons(see also Pseudomonas putida: PLAGGENBORG
et al. 2003). Similarly, relatives of hepato-pancreatic symbionts in Asellus aquaticus,Aeromonas spp. and Burkholderia spp. are well-known as xylanase and phenol oxidase pos-tive; these symbionts, thus, could well beinvolved in litter degradation in Asellus aqua-ticus (cf. ZIMMER & BARTHOLMÉ 2003). Basedon these, I hypothesize that the primarysymbiosis with Pseudomonas n. sp. provideddigestive enzymes for the degradation ofterrestrial food source to early semi-terres-trial isopods. Afterwards, prior to the evol-utionary loss of Pseudomonas and the take-over by Candidatus Hepatoplasma (and laterCandidatus Hepatincola), a gene transfertook place between symbiont and host, re-sulting in endogenous production of cellu-lase and/or phenol oxidase by the host (al-ternatively, the respective genes could havebeen transferred from the primary to thesecondary symbiont). Horizontal gene trans-fer beyond the species level is well-knownfrom both bacteria (e.g., JAENECKE et al.1996; KELLY & KADO 2002; HILL & WEIGHT-MAN 2003) and between prokaryotes and

The role of animal-microbe interactions in isopod ecology and evolution 147
Acta Biologica Benrodis 13 (2006)
eukaryotes (e.g., NESTER 2000; BIRD et al.2003; DOUGLAS & RAVEN 2003), and areconsidered significant for the evolution ofsymbiosis-specific capabilities of symbionts(FARRANT et al. 1997; TSUDA et al. 1999;NESTER 2000; DOUGLAS & RAVEN 2003; HILL
& WEIGHTMAN 2003; PARKER 2003; VAN
ELSAS et al. 2003). YAN et al. (1998) assumethat endogenous cellulases in plant-parasi-tic nematodes are evolutionarily of bacterialorigin. In some cases, close relatives of theproposed primary symbionts of Oniscidea,Pseudomonas putida (JAENECKE et al. 1996) andP. fluorescens (LILLEY et al. 2003), are involvedin such events; especially under nutrient-rich conditions, as prevail in the lumen ofthe midgut glands, the probability of inter-specific gene transfer is high (DROGE et al.1998) and could play an important role dur-ing the establishment of pathogens andparasites in their prospective host (WILSON
& SALYERS 2003).In contrast to termites, ingesting remark-
able amounts of lignocellulose and its deri-vates as well as other recalcitrant com-pounds (e.g., BREZNAK & BRUNE 1994),aphids, being sapsuckers, do not have to facemany recalcitrant or deterrent food com-pounds. Both, however, have in commonthat their food sources are extremely nitro-gen-limited so that they are in need of ad-ditional sources of nitrogen. In the gut ofTetraponera ants, VAN BORM et al. (2002) re-cently demonstrated nitrogen-fixing Pseudo-monas that share >97 % sequence similaritywith the endosymbionts of Ligia oceanica.The closely related Ligia pallasii mostly feedson stranded macroalgae (CAREFOOT 1973;PENNINGS et al. 2000) containing signifi-cantly less than 1 % N, and we assume asimilar nutritive behavior of Ligia oceanica.Thus, the nutritive significance of bacterialsymbionts could also be related to their ca-pability of making nitrogen available to theirN-limited host (discussed by NARDI et al.2002), as has been demonstrated repeatedlyfor the intracellular endosymbionts of
aphids (Buchnera aphidicola) and yeast-likesymbionts in the grasshopper Nilparvata lu-gens (Hemiptera: Auchenorrhyncha: Del-phacidae) (cf. DOUGLAS et al. 2001; WILKIN-SON & ISHIKAWA 2001; BERNAYS & KLEIN
2002). In addition, vitamins and essentialamino acids are produced and released byendosymbiotic microbiota and can be uti-lized by their hosts (DOUGLAS 1989, 1992).However, bacteria in the midgut glands ofLigia pallasii are apparently not closely re-lated to those in L. oceanica (WANG & ZIM-MER, unpubl.; tab. 1), questioning the evol-utionary significance of a potential nitro-gen source by means of endosymbiotic bac-teria.
Overall, our up-to-date knowledge assummarized herein (phylogenetic affiliationof symbionts, high proportion of aposym-biotic isopods without any obvious harm)does provide only weak arguments in favorof a nutritional significance of hepatopan-creatic bacteria. Further, the ability to hy-drolyze cellulose and oxidize phenolics ispresent (as a pre-disposition for the terres-trial habitat) in (some) Sphaeromatidea andValvifera (ZIMMER et al. 2002a). On the otherhand, despite their phylogenetic affiliation,nothing hints on a parasitic or pathogeniclife style of bacterial symbionts in Onisci-dea. Thus far, we can only speculate onthe nature of the symbiotic relationshipand the role symbionts play in the life oftheir host. I will discuss some hypothesesin turn.• Hepatopancreatic symbionts protect
their host from secondary infections by(microbial) pathogens or parasites.
Although woodlice are confronted with highdensities of (potentially pathogenic) micro-biota in their natural environment, themicrobial diversity in their midgut glands isremarkably low (see above). While the cuti-cular intima of the fore- and hindgut pro-tects from pathogens intruding the bodycavity, there is no such protection for themidgut epithelium; solely the proventricular

148 MARTIN ZIMMER
filters might act in this respect (see below).Most insects and crustaceans counteractpathogens through phenol oxidase activity(cf. GILLESPIE et al. 1997; see above). Al-though we proved phenol oxidation in thegut lumen of Porcellio scaber (ZIMMER & TOPP
1998c), alike Glyptonotus antarcticus Eights1852 (Valvifera) (SMITH & SÖDERHÄLL 1991)and presumably Asellus aquaticus (DEZFULI
2000), woodlice apparently do not possessphenol oxidases in their hemolymph (IRMAK
et al. 2005). Presumably, hemocyanine isinvolved here (for review, see DECKER &TUCZEK 2000) that also exerts antibacterialactivity (BOUCHON, pers. comm.; HERBINIÈRE,unpubl.). On the other hand, hepatopan-creatic bacteria may prevent the establish-ment of (microbial) parasites and pathogensin the hepatopancreatic lumen before theycan enter the body cavity, either as well-adapted, and thus superior, competitors, orthrough antibiotic agents; obviously, bothCandidatus Hepatoplasma and CandidatusHepatincola are capable of excluding theother potential symbiont from its habitat(see above). Comparable interactions betweenpotential symbionts have recently been de-scribed in the pea aphid, Acyrthosiphon pisum(KOGA et al. 2003). Besides producing aendo-β-(1-4)-glucanase (PASTOR et al. 2001),Paenibacillus sp. (Bacillales) exhibits a endo-β-(1-3)-glucanase that exerts fungicidal ef-fects through degrading the fungal cell wall(HONG & MENG 2003). Several unidentifiedPseudomonas strains produce both chitinasesand cellulases as well as additional enzymesthat act fungicidal (SINDHU & DADARWAL
2001), and some fluorescing soil pseudo-monads bring about a derivate of the phe-nolic phloroglucinol that also acts as fungi-cide (RAMETTE et al. 2003). Pseudonocardia-like (Actinomycetes) symbionts in leaf-cut-ting ants provide antibiotics that preventsparasitic fungi from intruding nutritionallysignificant fungal colonies (CURRIE et al.1999). However, thus far we were unableto detect any antibiotic activity in the hepa-
topancreas of Porcellio scaber (WANG & ZIM-MER, unpubl.).• Metabolites of hepatopancreatic sym-
bionts protect their hosts from preda-tors.
A close relative of Pseudomonas aeruginosa andthe symbionts in Ligia oceanica, Ligidium hyp-norum, Tylos ponticus, Hyloniscus riparius andTrichoniscus pusillus gives rise to the defen-sive compound pederine in the rove beetle,Paederus sabaeus (Coleoptera: Staphylinidae)(KELLNER 2001, 2002). Woodlice, too, exertchemical defense against predatory attack.Lobed glands in their uropods and the mar-gins of their epimera secret fluids uponstimulation that deter at least some inverte-brate predators (GORVETT 1952). Epimeralglands contain fluorescing compounds thatexhibit similar characteristics as those inZenker cells of the freshwater isopod, Asel-lus aquaticus (LORENZEN & ZIMMER, in prep.).Since lobed glands of terrestrial isopods areassumed homologous to the Zenker cellsof Asellidae (TER-POGHOSSIAN 1909), andbacterial symbionts in the gut of Asellusaquaticus may bring forth the fluorescenceof the Zenker cells (ZIMMER et al. 2002c),we hypothesize a correlation between bac-terial symbionts and the defensive functionof the lobed glands. However, experimen-tal evidence is lacking thus far.• Hepatopancreatic bacteria increase fer-
tility (production of oocytes or sperm)and reproductive success (mate choice)of their host.
If bacterial endosymbionts are (evolu-tionarily) advantageous to their host, wewill expect a positive effect on their host�sfitness. Aphids, for instance grow moreslowly if kept void of Buchnera (ADAMS etal. 1996). Experimental tests of selectiveadvantages to isopods from harboring he-patopancreatic bacteria in terms of highergrowth rates, higher fecundity, or highermating success thus far failed to demons-trate any gain to P. scaber (WANG & ZIM-MER, unpubl.).

The role of animal-microbe interactions in isopod ecology and evolution 149
Acta Biologica Benrodis 13 (2006)
4. Leaf litter-colonizing microbiota
Interactions of woodlice and litter-coloniz-ing microbiota can be classified either aspredator-prey interactions, i.e., ingestedmicrobiota are digested and utilized as food(e.g., REYES & TIEDJE 1976; KOZLOVSKAJA
& STRIGANOVA 1977; COUGHTREY et al. 1980;BECK & FRIEBE 1981; MÁRIALIGETI et al.1984; GUNNARSSON & TUNLID 1986; HAS-SALL et al. 1987; ZIMMER & TOPP 1998b), oras synergism or even mutualism with pro-motion of microbial biomass and/or acti-vity and distribution of microbial propa-gules (DUNGER 1958; STRIGANOVA 1967;HASSALL 1977; SOMA & SAITO 1983; HAS-SALL et al. 1987; TEUBEN & ROELOFSMA 1990;TEUBEN 1991; LUSSENHOP 1992; SZLÁVECZ
1993; VAN WEMSEN et al. 1993; KAUTZ &TOPP 2000; ZIMMER & TOPP 1999; ZIMMER
et al. 2002b) on one hand, and microbialprocessing of leaf litter prior to feeding bywoodlice (HARTENSTEIN 1964A; RUSHTON &HASSALL 1983; HASSALL et al. 1987) and/orduring the gut passage (HASSALL & JENNINGS
1975; KUKOR & MARTIN 1986) on the otherhand. With respect to the latter, it has beenshown repeatedly that consumption ofdensely colonized leaf litter is advantage-ous both on individual (UESBECK & TOPP
1995; ZIMMER & TOPP 1997a) and on popu-lation level (KAUTZ et al. 2000; ZIMMER &TOPP 1997a, 2000). Despite this generalagreement, the nutritive significance of lit-ter-colonizing microbiota is not well under-stood in detail (ZIMMER et al. 2003); the de-bated results and hypotheses can be sum-marized as follows:
(1) Digestion and utilization of micro-bial cells increase the nutritive value of leaflitter to consumers, since ingested micro-biota � upon digestion � serve as source ofeasily accessible (essential?) nutrients, suchas nitrogen (CAREFOOT 1984a,b; ULLRICH etal. 1991). However, other strategies thanutilizing microbiota as food source to copewith nitrogen limitation are known, too:
shrimp, Litopenaeus setiferus (Decapoda: Pe-naeidae), respond to protein deficiency withincreased production of proteolytic en-zymes (BRITO et al. 2000), and, similarly,aquatic detritivorous cranefly larvae (Di-ptera: Nematocera: Tipulidae) counteractthe low nitrogen availability in their foodwith extremely efficient protein degradationin their midgut with a correspondingly highpH level of 11.5 (MARTIN et al. 1980) foroptimal proteolytic activity and, at the sametime, minimal harm through phenolic com-pounds (cf. GRAÇA & BÄRLOCHER 1998); inaddition, β-1,4- and β-1,3-glucanases makethe digestion of ingested fungi, and accessto valuable, possibly even essential, nu-trients, possible. Woodlice (Trachelipus rath-kii) also exhibit these enzyme activities intheir hindgut, and thus, are capable of di-gesting fungal cells (see above; KUKOR &MARTIN 1986). Wood-boring isopods, Lim-noria spp., although presumably producingendogenous cellulases (RAY & JULIAN 1952),are in need of wood-colonizing fungi as asupplemental source of high-quality food(RAY 1959).
According to the literature (e.g., REYES &TIEDJE 1976; KOZLOVSKAJA & STRIGANOVA
1977; COUGHTREY et al. 1980; BECK & FRIEBE
1981; MÁRIALIGETI et al. 1984; GUNNARSSON
& TUNLID 1986; HASSALL et al. 1987), we knowthat ingested microbiota are digested in theanterior hindgut (ZIMMER & TOPP 1998b), andsurviving cells are egested in feces(COUGHTREY et al. 1980; HANLON & ANDER-SON 1980; INESON & ANDERSON 1985; HAS-SALL et al. 1987; ULLRICH et al. 1991) after ex-tensive proliferation (GUNNARSSON & TUNLID
1986; HASSALL et al. 1987) in the posteriorhindgut (ZIMMER & TOPP 1998b). Thus, micro-bial density of feces equals or surpasses thatof the ingested leaf litter (discussed in ZIM-MER 2002a). Due to the prevailing pH condi-tions in different gut sections (ZIMMER & TOPP
1997b; ZIMMER & BRUNE 2005), however, thispredominantly applies to bacteria but not tofungi (ZIMMER & TOPP 1998b).

150 MARTIN ZIMMER
Yeasts (Saccharomyces cerevisiae), accelerategrowth and increase fecundity of the milli-pede, Polydesmus angustus (Myriapoda: Diplo-poda), when experimentally supplementedto the diet (DAVID & CÉLÉRIER 1997); yeastcells are digested in the midgut of milli-pedes (BYZOV et al. 1993), but their ener-getic contribution to the consumers� nutri-tion is low, and DAVID & CÉLÉRIER (1997)assume them to be sources of nitrogen (seeabove). In earthworms (Lumbricus terrestris;Annelida: Oligochaeta), microbial cells arecleaved before they reach the foregut(SCHÖNHOLZER et al. 2002). During the sub-sequent gut passage, mostly Proteobacteria(gram-negative) of the α-, β- and γ-divi-sion are reduced in their number (SCHÖN-HOLZER et al. 2002), but overall there is anincrease in bacterial numbers from fore- tohindgut (SCHÖNHOLZER et al. 1999). By con-trast, it is chiefly δ-Proteobacteria that pro-liferate on feces (SCHÖNHOLZER et al. 2002).The mayfly, Ephemera danica (Ephemerop-tera), digests unselectively ingested bacterialspecies with different efficacies (AUSTIN &BAKER 1988): while gram-negative α-Pro-teobacteria � Aeromonas hydrophila (Aeromo-nadales) and Citrobacter freundii (Enterobac-teriales) � are digested effectively, Flavobac-terium spp. (Bacteroidetes = Flavobacterium-Flexibacter-Bacteroides-Cytophaga group:KIRCHMANN 2002; gram-negative) attach tothe hindgut wall. In Porcellio scaber, too, wefound specific digestibilities of differentmicrobial taxa (IHNEN & ZIMMER, in prep.).Thus, digestibility of actinomycetes [gram-positive: Streptomyces celluloflavus (Streptomy-cetaceae) and Pseudonocardia autotrophica(Pseudonocardiaceae)] is significantlyhigher that that of gram-negative bacteria[Pseudomonas fluorescens (α-Proteobacteria:Pseudomonaceae) and Myxococcus xanthus (δ-Proteobacteria: Myxococcaceae)] and fungi[Chaetomium globosum (Ascomycetes: Pyreno-mycetaceae) and Fusarium ventricosum (Deu-teromycetes)], possibly due to cell wall char-acteristics. Digestive specificity is, in addi-
tion, reflected by feeding preferences (IH-NEN & ZIMMER, in prep.). In preference tests,P. scaber consumed artificial diets in higherrates when inoculated with actinomycetesthan any other offered food, no matterwhether cellulolytic or non-cellulolyticmicrobiota were offered. Overall, microbialinoculation increased consumption ratesonly in case of low-quality food (cellulose),while consumption of diets of high quality(cf. ZIMMER & TOPP 1998b) was not alteredby microbial inoculation. These results arein agreement with findings by KAUTZ et al.(2002), indicating a decreasing nutritionalsignificance of feces-colonizing microbiotawith increasing quality of the initial leaf lit-ter material, but, on the other hand, contra-dict ZIMMER et al. (2003), concluding an in-creasing positive effect of litter-colonizingmicrobiota on digestion and utilization offood with increasing food quality. In anycase, our recent results (IHNEN & ZIMMER,in prep.; see above) indicate (i) that micro-biota obviously increase the attractivenessof low-quality food, but (ii) that microbialcellulases do not appear to be significant inthis context (see also ZIMMER et al. 2003),since the increased consumption rates dueto microbial inoculation did not depend onthe cellulolytic capability of actinomycetes.Actinomycetes also increased consumptionby Oniscus asellus, but the same was true fordifferent microbiota and high-quality food(IHNEN & ZIMMER, in prep.). As already sug-gested by ZIMMER & TOPP (2000), Porcellioscaber and Oniscus asellus obviously differ fromeach other with respect to nutritional re-quirements.
In addition to the above potential directnutritive gain, indirect ways of how ingestedmicrobiota positively influence digestiveprocesses have been described, too. Interrestrial isopods (Porcellio scaber), litter-co-lonizing microbiota appear to be involvedin hindgut pH homeostasis (ZIMMER & TOPP
1997b) and in detoxification of phenoliccompounds through providing surfactants

The role of animal-microbe interactions in isopod ecology and evolution 151
Acta Biologica Benrodis 13 (2006)
(ZIMMER 1997) and/or hydrolytic enzymes(ZIMMER 1999), releasing simple, harmlessphenolics and glucose. However, the releaseof glucose is not always advantageous, sin-ce most microbial β-glucosidases are inhi-bited by their product glucose (cf. DECKER
et al. 2001).Similarly, bacterial proliferation in the hind-
gut need not have positive effects on a con-sumer, since the gut lumen may simply actas an incubator, providing appropriate envi-ronmental conditions, without any gain frommicrobial activity to the consumer, particu-larly if microbial proliferation occurs in theposterior hindgut where no resorption ofnutrients takes place (as in terrestrial isopods:ZIMMER & TOPP 1998b). Woodlice appear togain from bacterial proliferation in the pos-terior hindgut through being coprophagous(discussed in ZIMMER 2002a). The advantagesof coprophagy to isopods have, however,been discussed controversially during the lastcouple of decades (summarized in ZIMMER
2002a; see also KAUTZ et al. 2002). WhileWIESER (1966) stressed the essential import-ance of coprophagy to survival of Porcellioscaber, others (WHITE 1968; COUGHTREY et al.1980; HASSALL & RUSHTON 1982, 1985; HOP-KIN & MARTIN 1984) maintained isopod po-pulations in the laboratory without access tofeces; SZLÁVECZ & MAIORANA (1998: Porcellioscaber) did not find any evidence for a nutri-tional advantage of being coprophagous, andthis behavior may be insignificant in the field(HASSALL & RUSHTON 1982; HOPKIN & MAR-TIN 1984; but see above). In the laboratory,the gain from feeding on feces is generallylow, but depends upon the quality of the in-itial leaf litter, increasing with decreasing lit-ter quality (KAUTZ et al. 2002). It is only incase of low-quality food (e.g., wide C:N ratio)that microbial activity or biomass adds to thenutritive value of the feces, while high-quali-ty litter is of higher value than feces. Thesefindings are in good agreement of HASSALL�s(pers. comm., see above) suggestion of differ-ent nutritive strategies in terrestrial isopods.
(2) Activity of microbial cells promotesdigestion of leaf litter by consumers, sinceextracellular microbial enzymes are utilizedfor digestive processes (HASSALL & JENNINGS
1975; KUKOR & MARTIN 1986), and micro-bial processing of the leaf litter prior toconsumption may be even more important.Woodlice prefer feeding on decomposingrather than fresh leaf litter (HARTENSTEIN
1964A; HASSALL et al. 1987; RUSHTON &HASSALL 1983), the former being reducedin toughness (RUSHTON & HASSALL 1983)and in phenolics and other repellents (KUI-TERS & SARINK 1986; POINSOT-BALAGUER etal. 1993; ZIMMER 2002b). BROOKS & BELL
(2001) state that fungal processing of thesubstrate plays a significant role in habitatchoice by the wood-boring Sphaeroma tere-brans Bate, 1866 (Sphaeromatidea: Sphae-romatidae).
Wood wasps, Sirex cyaneus (Hymenopte-ra: Symphyta: Siricoidea: Siricidae), digestwood with assistance of enzymes derivedfrom their fungal symbiont Amylostereumchailletii (Basidiomycetes) (KUKOR & MAR-TIN 1983). Unknown bacteria provide cel-lulases to desert millipedes (Myriapoda: Di-plopoda: Orthoporus ornatus und Comanchelussp.) (TAYLOR 1982). In marine blood-suckingisopods of the genus Gnathia (Gnathiidea:Gnathiidae) bacteria have been isolatedfrom the hindgut that belong to the generaPseudomonas (Pseudomonadales), Bacillus (Ba-cillales) und Streptococcus (Streptococcales)and presumably are involved in digestingfish blood (JUILF & WÄGELE 1987; DAVIES
1995). In natural microbial soil commu-nities, Pseudomonas fluorescens is one of themost effective degraders of humic acids(FILIP & BIELEK 2002), but complex bac-terial communities are much better in de-grading such substances than any testedisolate (ESHAM et al. 2000). By contrast, itis mostly fungi that are responsible for thedegradation of aromatic compounds inwood and leaf litter (cf. RABINOVICH et al.2004).

152 MARTIN ZIMMER
Enzymes of microbial origin may alsoplay a role in the digestion of ingestedmicrobiota. The degradation of chitin in thegut of fungus-feeding springtails, Folsomiacandida (Collembola: Isotomidae), is broughtabout by Xanthomonas maltophilia (γ-Proteo-bacteria: Pseudomonaceae) and Curtobac-terium sp. (Actinomycetales: Corynebacteri-aceae) (BORKOTT & INSAM 1990). In conse-quence, leaf litter that is inoculated with thesebacteria is preferred as food, but mechanismsof how springtails find and choose this foodsource remain unclear. In Porcellio scaber, vo-latile microbial metabolites are detected byolfaction and serve in finding food by for-aging isopods (ZIMMER et al. 1996).
(3) In both of the above cases, we wouldexpect increased digestibility by, as well ashigher growth rates of, consumers. Alterna-tively, litter-colonizing microbiota may serveas indicators of nutrient-rich, easily digestiblefood source that are used as cues while for-aging. Actually, Porcellio scaber responds posi-tively to volatile metabolites of litter-colo-nizing microbiota (ZIMMER et al. 1996). Ifso, we expect increased consumption ofdensely colonized litter, while digestion andgrowth may be affected only marginally. Re-cent results by ZIMMER et al. (2003) are infavor of the latter hypothesis: microbial in-oculation of experimental food increasedconsumption and slightly enhanced growthrates, but did not affect digestibility.
The millipede, Polydesmus angustus, by con-trast, did not show any effect of microbialcolonization (yeasts) of experimental foodon mortality, but both growth and fecun-dity were promoted significantly (DAVID &CÉLÉRIER 1997). These authors concludethat the microbial contribution to nutritionis qualitative rather than quantitative (seeabove). In contrast to our recent results(ZIMMER et al. 2003), previous studies re-vealed a positive effect of litter-colonizingmicrobiota on female fecundity and juve-nile mortality (ZIMMER & TOPP 1997a). Thus,the nutritional significance of litter-coloni-
zing microbiota is not only species-speci-fic, but also depends on the context. Bothextracellular microbial enzymes and essen-tial nutrients may promote some physiolo-gical processes that are related to growthand reproduction. In addition, the litter it-self appears to be of predominantly quan-titative importance. Such nutritional diffe-rences have been demonstrated in a com-parison of aquatic detritivores, Asellus aqua-ticus and Gammarus pulex; while isopodspreferentially graze fungal biomass off theleaf surface and thus directly depends onfungal biomass, amphipods consume fun-gally inoculated and processed leaf litter. Aswe demonstrated for terrestrial isopods (seeabove; IHNEN & ZIMMER, in prep.), the fun-gal species is of crucial significance for thenutritive value of the fungal biomass (BÄR-LOCHER & KENDRICK 1973a,b).
5. Gut microbiota and their environment
Until now is unclear, and still controver-sially debated, whether (species-specific?)resident gut microbes exist in terrestrialisopods, or whether the community of gutmicrobiota is a random assemblage thatdepends on the ingested food with mi-crobes simply being passive gut passengers.The abundance of Chryseobacterium sp.(Bacteroidetes) in the hindgut of the cock-roach, Periplaneta americana (Blatteroptoidea:Blattodea: Blattariae), alters with changingfood sources (DUGAS et al. 2001), and theabundance of Mycoplasma sp. in the gut ofthe Atlantic Salmon, Salmo salar (Salmon-idae), differs between populations fed ondifferent food sources (HOLBEN et al.2002). By contrast, the microbial commu-nity in the hindgut of Neotrypeaea (Callia-nassa) californiensis (Decapoda: Thalassin-idae) seems to be independent of nutri-tional conditions. Predominantly α-Proteo-bacteria, Bacteroidetes and gram-positivebacteria make up the resident gut micro-biota (LAU et al. 2002).

The role of animal-microbe interactions in isopod ecology and evolution 153
Acta Biologica Benrodis 13 (2006)
HASSALL & JENNINGS (1975) doubted theexistence of microbial residents in the iso-pod hindgut, while other authors disagreed(summarized in ZIMMER 2002a). In starvingcrinochete isopods (REYES & TIEDJE 1976;BECK & FRIEBE 1981; INESON & ANDERSON
1985), and during feeding on sterile arti-ficial diet (ZIMMER & TOPP 1998b; ZIMMER
1999), the number of gut microbes is dras-tically reduced. Both observations argueagainst resident gut microbes. CAREFOOT
(1984a), however, did not notice any changesin the microbial community of Ligia pallasiiafter five weeks of starving; two out of 21platable bacteria were only found in thehindgut but not on stranded macroalgae, thenatural food source of this species (SPENCE,unpubl.; cited in CAREFOOT 1984b).
GUNNARSSON & TUNLID (1986) assumedgram-positive bacteria to prevail in the hind-gut of O. asellus, and ULLRICH et al. (1991)characterized predominantly corynefor-mous Micrococcaceae (Firmicutes). Mostother investigators, however, mainly foundgram-negative bacteria. BEERSTECHER et al.(1954) isolated nitrogen-fixing Azotobactersp. (γ-Proteobacteria: Azotobacteriaceae) inthe hindgut of cf. Armadillidium vulgare.REYES & TIEDJE (1976) were unable to findsome of the (gram-negative) bacteria thatcolonized feces of Trachelipus rathkii in thelitter layer, while hindgut and feces weresimilar in species composition, and HASSALL
et al. (1987) isolated (cellulolytic?) Cytopha-ga spp. (Bacteroidetes) from feces of O. asel-lus that could not be detected in the leaflitter, either. As early as 1985, GRIFFITHS &WOOD (1985) found irregularly distributedrod-shaped bacteria in close association withthe (posterior) hindgut cuticle of Oniscusasellus that they affiliated to the genera Pseu-domonas (see also REYES & TIEDJE 1976: Tra-chelipus rathkii), Plesiomonas (Vibrionaceae)and Enterobacter (along with other Entero-bacteriaceae) of the gram-negative γ-Pro-teobacteria � at least some representativeof these genera are (facultatively) anaero-
bic (see below). GRIFFITHS & WOOD (1985)did not notice actinomycetes, and MÁRIA-LIGETI et al. (1984) proposed that thesegram-positive bacteria do not belong to anykind of gut community but are passive gutpassengers. DROBNE et al. (1996) describedby means of SEM filamentous and rod-shaped bacteria attached to the hindgut wallin Porcellio scaber, but did not provide anyinformation on whether or not these bac-teria were also detectable in feces and/orlitter. Later, KOSTANJI�EK et al. (2002) foundhigh bacterial diversity in the hindgut ofthis species and concluded from results ofmolecular studies that about half of thepresent species are not only unknown, butshare less than 80 % sequence similaritywith any known bacteria. About 70 % ofthese bacteria are, however, distantly relatedto typical (anaerobic) intestinal bacteria,namely Streptococci and Enterococci(Streptococcales; gram-positive) and Bac-teroides group (Bacteroidetes; gram-nega-tive), while the remaining species are relatedto soil bacteria such as Bacillus and Pseudo-monas. From these data, and from that somebacteria are specifically attached to cuticu-lar spines of the hindgut, KOSTANJI�EK etal. (2002) concluded that Porcellio scaber doharbor resident gut microbiota. Similarly,DROBNE (1995) described numerous bac-teria in association with the hindgut cuticleof Porcellio scaber � some of them havingbeen characterized by KOSTANJI�EK et al.(2002) � and Ligidium hypnorum (Diplocheta:Ligiidae), but not in Hyloniscus riparius (Syn-ocheta: Trichoniscidae). On the otherhand, freshly molted Porcellio scaber andLigidium hypnorum do not contain cuticle-attached bacteria in their hindgut (DROBNE
1995); she discusses this observation asactive protective mechanism by the iso-pod against microbial infection during asensitive phase with a unsklerotized gutcuticle, and thus, proposes re-acquisitionof specific bacteria from the environment(see above).

154 MARTIN ZIMMER
Despite the assumption of a relativelyshort gut passage of several hours (HARTEN-STEIN 1964a, in Oniscus asellus; ALIKHAN 1969,in Porcellio laevis; ZIMMER 1998, in Porcellio sca-ber; GRÜNWALD 1987, in different species), flu-orescent markers of the size of bacterial cellscould be detected as long as 16 days afteringestion by Porcellio scaber (CLEGG et al. 1996).Bacteria can be expected to be retained inand on cuticular structures (wrinkles andspines) and to remain in the gut for sometime as transient resident (KOSTANJ�EK et al.2002). Since some of the mentioned bacte-ria are taxonomically related to obligate ana-erobic species, we have to consider that theradial center of the hindgut, but not the pe-riphery is anoxic (ZIMMER & BRUNE 2005);thus, anoxic microhabitats in cuticularwrinkles are to be postulated.
As early as 1976, REYES & TIEDJE (1976)suggested hypoxic conditions to prevail inthe hindgut of Trachelipus rathkii due to oxy-gen consumption and limited gas diffusion,and MÁRIALIGETI et al. (1984) proposed fa-cultatively anaerobic microbiota (mostlyactinomycetes) to be the most stable gutpassengers. The assumption of aerobic ac-tivity and oxygen consumption is confirmedby in situ and in vivo measurements withmicroelectrodes, demonstrating an oxichindgut periphery but an anoxic hindgutcenter (ZIMMER & BRUNE 2005). Thus, theperiphery allows for aerobic and oxidativedigestive processes, while the radial centermay be a site of fermentative degradation.However, thus far, we could not detect anyfermentation products.
Optimal pH conditions for digestive pro-cesses (cellulose digestion and phenol ox-idation) in the anterior, and for bacterial pro-liferation in the posterior, hindgut as wellas coinciding redox potentials were also de-monstrated by utilizing microelectrodes insitu (ZIMMER & BRUNE 2005). Both cellu-lases and most known phenol oxidases ex-hibit maximal activity under these pH con-ditions (HARTENSTEIN 1964a; MAYER & HA-
REL 1979; WOOD 1980; ZIMMER & TOPP
1997b), and actually, pH gradients are re-flected by gradients of enzymatic activities(HARTENSTEIN 1964b; SALEEM & ALIKHAN
1974; HASSALL & JENNINGS 1975; ZIMMER
& TOPP 1998a,b; ZIMMER 1999). Excep-tionally high redoxpotentials of > +600 mVin the hindgut and the midgut glands facili-tate the aerobic and oxidative degradationof leaf litter compounds. Despite poten-tially harmful or toxic effects of phenoliclitter compounds at acidic pH conditions(BERENBAUM 1980; MARTIN & MARTIN 1983;SCHULTZ & LECHOWICZ 1986) and under ox-idizing conditions (APPEL 1993), a slightlyacidic gut pH of 5.0-6.0 in the anterior hind-gut (Crinocheta: ZIMMER & TOPP 1997b;ZIMMER & BRUNE 2005), thus, appears to beadaptive to the need of digesting the mostcommon leaf litter compounds. Further,surfactants (ZIMMER 1997) and tannin hy-drolysis (ZIMMER 1999) in the hindgut lu-men counteract potentially damage causedby phenolics. Proliferation of survivingbacteria in the posterior hindgut takes placeunder optimal pH conditions of 5.5-7.0(ZIMMER & TOPP 1998b). A rise in pH, thus,seems to be adaptive with respect to copro-phagy (see above), since favorable condi-tions for microbial processing of undigestedlitter compounds are provided in freshlydropped feces. Homeostasis of gut pH inthe hindgut lumen have been attributed tocell compounds of ingested and digestedmicrobiota (ZIMMER & TOPP 1997b). ZIM-MER & BRUNE (2005), by contrast, discusspH reduction as a consequence of fermen-tative digestive processes in the anoxic hind-gut center; the rise of pH in the posterior hind-gut would then be brought about by activeinflux of K+ ions as has been described forcaterpillars (DOW 1992).
While NICHOLLS (1931) in Ligia oceanicaalso found a pH gradient of 6.0 to 7.2, thepH level in the anterior hindgut of Tylosgranulatus Krauss, 1843 (Tylida: Tylidae)changes from 6.9 to 7.4 to drop to 6.8 again

The role of animal-microbe interactions in isopod ecology and evolution 155
Acta Biologica Benrodis 13 (2006)
in the posterior hindgut (KENSLEY 1974). Inthe hindgut of Trichoniscus pusillus it is solelythe most anterior section that exhibits ahigher pH (6.8) than the remaining regionswith pH 6.3-6.5 (ZIMMER & BRUNE 2005).Overall, the pH gradients are rather insig-nificant in the latter species as comparedwith Ligia oceanica and some studied Crino-cheta; furthermore, the gut pH is signifi-cantly higher in Ligia oceanica, Tylos granula-tus and Trichoniscus pusillus than in any ofthe studied Crinocheta. No pattern in he-patopancreatic pH can be brought in con-gruence with isopod phylogeny (ZIMMER &BRUNE 2005): values range from 6.1 in Oni-scus asellus and Porcellio scaber to 6.3 in Tylosgranulatus (KENSLEY 1974) and 6.5 in Tricho-niscus pusillus and Trachelipus rathkii. In ho-mogenates of the midgut glands WOOD &GRIFFITHS (1988) used colored indicators todetermine data of pH 4.7-5.4 and pH 5.4-6.1 in Oniscus asellus and Porcellio scaber. Thus,the nutritive and/or evolutionary significanceof these differences between phylogeneticlineages of Oniscidea remains unclear.
6. Conclusions
Although we do not yet fully understandisopod-microbe interactions in detail, it isobvious that microbiota are significant tothe ecology of terrestrial isopods, and mayhave been important evolutionarily in thecourse of terrestrialization. Possibly, theacquisition of hepatopancreatic symbiontsrendered woodlice less dependent on litter-colonizing microbiota. Although the nutri-tional utilization of terrestrial leaf litter ispromoted by litter-colonizing microbiotathrough their activity and their role as asupplementary nutrient source, and this mayhave facilitated the evolutionary coloniza-tion of land, woodlice appear to be able tofeed upon litter of low quality � low micro-bial density, wide C:N ratio, high structuraltoughness � during early stages of decom-position. This difference to other detri-
tivores, brought about by endosymbioticbacteria, may explain the evolutionary suc-cess of terrestrial isopods as detritivoresages after occupation of this niche byinsects, millipedes and eartworms, but thefunction(s) of hepatopancreatic bacteria aswell as how they interact with their host areonly beginning to be understood.
Acknowledgements
I am grateful to many people who contrib-uted to, and made it possible for me towrite, this review. My sincere thanks go tomy co-workers and students (as cited andreferred to in the text) having worked withme during the last couple of years; to WER-NER TOPP, my first supervisor in (isopod)ecology; to KLAUS LUNAU, my former in-structor to evolutionary ecology; to the headof my division, HEINZ BRENDELBERGER, forenabeling me to work on terrestrial (andaquatic) isopods; to HELMUT SCHMALFUSS, myteacher in isopod phylogeny; and to JOACHIM
KURTZ and JEAN PAUL DANKO for discus-sions on isopod ecology and evolution.
References
ABE, T., HIGASHI, M., 1991: Cellulose centeredperspective on terrestrial community struc-ture. � Oikos 60, 127-133.
ABRIL, B.A., BUCHER, E.H., 2002: Evidence thatthe fungus cultured by leaf-cutting ants doesnot metabolize cellulose. � Ecology Letters5, 325-328.
ADAMS, D., WILKINSON, T.L., DOUGLAS, A.E., 1996:The aphid-bacterial symbiosis: a comparisonbetween pea aphids and black bean aphids. �Entomologia Experimentalis at Applicata 80,275-278.
AHMAD, S., 1992: Biochemical defense of pro-oxidant plant allelochemicals by herbivorousinsects. � Biochemical Systematics and Eco-logy 20, 269-296.
ALIKHAN, M.A., 1969: The physiology of thewoodlouse, Porcellio laevis (Porcellionidae:Peracarida) � I: Studies on the gut epithe-lium cytology and its relation to maltase se-

156 MARTIN ZIMMER
cretion. � Canadian Journal of Zoology 47,65-75.
ALTHOFF, K., SCHÜTT, C., STEFFEN, R., BATEL, R.,MÜLLER, W.E.G., 1998: Evidence for a sym-biosis between bacteria of the genus Rhodo-bacter and the marine sponge Halichondria pa-nicea: harbor also for putatively toxic bacte-ria? � Marine Biology 130, 529-536.
ANDER, P., ERIKSSON, K.-E., 1976: The importanceof phenol oxidase activity in lignin degradationby the white-rot fungus Sporotrichum pulverulen-tum. � Archives of Microbiology 109, 1-8.
ANDERSON, S.O., 1985: Sclerotization and tanningof the cuticle, pp. 59-75. In: Comprehensiveinsect physiology, biochemistry and pharma-cology (Kerkut, G.A., Gilbert, L.I., eds). �Pergamon Press, Oxford.
ANDERSON, C.L., KARR, T.L., 2001: Wolbachia: evol-utionary novelty in a rickettsial bacteria. �BMC Evolutionary Biology 1, 10.
APPEL, H.M., 1993: Phenolics in ecological in-teractions: the importance of oxidation. �Journal of Chemical Ecology 19, 1521-1552.
AUSTIN, D.A., BAKER, J.H., 1988: Fate of bacteriaingested by larvae of the freshwater mayfly,Ephemera danica. � Microbial Ecology 15, 323-332.
BÄRLOCHER, F. 1982: The contribution of fungalenzymes to the digestion of leaves by Gam-marus fossarum Koch (Amphipoda). � Oecolo-gia 52, 1-4.
BÄRLOCHER, F., KENDRICK, B., 1973a: Fungi in thediet of Gammarus pseudolimnaeus (Amphipoda).� Oikos 24, 295-300.
BÄRLOCHER, F., KENDRICK, B., 1973b: Fungi andfood preferences of Gammarus pseudolimnaeus.� Archiv für Hydrobiologie 72, 501-516.
BÄRLOCHER, F., PORTER, C.W., 1986: Digestiveenzymes and feeding strategies of threestream invertebrates. � Journal of the NorthAmerican Benthological Society 5, 58-66.
BAKER, J.E., WOO, S.M., 1992: α-glucosidases inthe rice weevil, Sitophilus oryzae: purification,properties, and activity levels in wheat- andlegume-feeding strains. � Insect Biochemist-ry and Molecular Biology 22, 495-504.
BAUMANN, P., MORAN, N.A., 1997: Non-cultivab-le microorganisms from symbiotic associati-ons with insects and other hosts. � Antonievan Leeuwenhoek 72, 39-48.
BECK, L., FRIEBE, B., 1981: Verwertung von Koh-lenhydraten bei Onsicus asellus (Isopoda) und
Polydesmus angustus (Diplopoda). � Pedobiolo-gia 21, 19-29.
BEERSTECHER, E., CORNYN, J., VOLKMANN, C., CAR-DO, L., HARPER, R., 1954: Invertebrate nutriti-on I: a preliminary survey of the nutritionalrequirements of an isopod: Oniscus asellus. �Texas Reports on Biology and Medicine 12,207-210.
BENHALIMA, K., MORIYASU, M., 2001: Prevalenceof bacteria in the spermathecae of femalesnow crab, Chinoectes opilio (Brachyura: Maji-dae). � Hydrobiologia 449, 261-266.
BERENBAUM, M., 1980: Adaptive significance ofmidgut pH in larval Lepidoptera. � TheAmerican Naturalist 115, 138-146.
BERNAYS, E.A., KLEIN, B.A., 2002: Quantifying thesymbiont contribution to esssential aminoacids in aphids: the importance of tryptophanfor Uroleucon ambrosiae. � Physiological Ento-mology 27, 275-284.
BIRD, D.M., OPPERMAN, C.H., DAVIES, K.G., 2003:Interactions between bacteria and plant-pa-rasitic nematodes: now and then. � Internati-onal Journal of Parasitology 33, 1269-1276.
BORKOTT, H., INSAM, H., 1990: Symbiosis withbacteria enhances the use of chitin by thespringtail, Folsomia candida (Collembola). � Bio-logy and Fertility of Soils 9, 126-129.
BOUCHER, D.H., JAMES, S., KEELER, K.H., 1982:The ecology of mutualisms. � Annual Reviewof Ecology and Systematics 13, 315-347.
BREZNAK, J.A., BRUNE, A., 1994: Role of micro-organisms in the digestion of lignocelluloseby termites. � Annual Review of Entomology39, 453-487.
BRITO, R., CHIMAL, M.E., GAXIOLA, G., ROSAS, C.,2000: Growth, metabolic rate, and digestiveenzyme activity in the white shrimp Litope-naeus setiferus early postlarvae fed differentdiets. � Journal of Experimental Marine Bio-logy and Ecology 255, 21-36.
BROOKS, D.R., 1985: Historical ecology: a newapproach to studying the evolution of ecolo-gical associations. � Annals of the MissouriBotanical Garden 72, 660-680.
BROOKS, M.A., 1970: Comments on the classifi-cation of intracellular symbiotes of cock-roaches and a description of the species. � Jour-nal of Invertebrate Pathology 16, 249-258.
BROOKS, R.A., BELL, S.S., 2001: Colonization of adynamic substrate: factors influencing recruit-ment of the wood-boring isopod, Sphaeroma

The role of animal-microbe interactions in isopod ecology and evolution 157
Acta Biologica Benrodis 13 (2006)
terebrans, onto red mangrove (Rhizophora mang-le) prop roots. � Oecologia 127, 522-532.
BRUSCA, R.C., WILSON, G.D.F., 1991: A phyloge-netic analysis of the Isopoda with some clas-sificatory recommendations. � Memoirs ofthe Queensland Museum 31, 143-204.
BUCHNER, P., 1965: Endosymbiosis of animalswith plant microorganisms. � Interscience,New York.
BUCKLING, A., RAINEY, P.B., 2002: The role ofparasites in sympatric and allopatric host di-versification. � Nature 420, 496-499.
BYZOV, B.A., NGUYEN THANH, V., BABJEVA, I.P.,1993: Interrelationships between yeasts andsoil diplopods. � Soil Biology and Biochem-sitry 25, 1119-1126.
CAREFOOT, T.H., 1984: Studies on the nutrition ofthe supralitoral isopod Ligia pallasii using che-mically defined artificial diets: assessment ofvitamin, carbohydrate, fatty acid, cholesteroland mineral requirements. � Comparative Bi-ochemistry and Physiology A 79, 655-665.
CAREFOOT, T.H., 1984: Nutrition and growth ofLigia pallasii. � Symposia of the ZoologicalSociety of London 53, 455-467.
CAREFOOT, T.H., 1993: Physiology of terrestrialisopods. � Comparative Biochemistry andPhysiology A 106, 413-429.
CAREFOOT, T.H., TAYLOR, B.E., 1995: Ligia: a pro-totypal terrestrial isopod, pp. 47-60. In: Ter-restrial isopod biology (Alikhan, M.A., ed.). �Balkema, Rotterdam.
CAZEMIER, A.E., HACKSTEIN, J.H.P., OP DEN CAMP,H.J.M., ROSENBERG, J., VAN DER DRIFT C., 1997:Bacteria in the intestinal tract of differentspecies of arthropods. � Microbial Ecology33, 189-197.
CHARLES, H., ISHIKAWA, H., 1999: Physical andgenetic map of the genome of Buchnera, theprimary symbiont of the pea aphid Acyrthosi-phon pisum. � Journal of Molecular Evolution48, 142-150.
CLAUS, H., FILIP, Z., 1990: Effects of clays andother solids on the activity of phenoloxidasesproduced by some fungi and actinomycetes. �Soil Biology and Biochemistry 22, 483-488.
CLEGG, C.D., ANDERSON, J.M., LAPPIN-SCOTT,H.M., 1996: Biophysical processes affectingthe transit of a genetically-modied Pseudomo-nas fluorescens through the gut of the woodlousePorcellio scaber. � Soil Biology and Biochemist-ry 28, 997-1004
CORDAUX, R., MICHEL-SALZAT, A., BOUCHON, D.,2001: Wolbachia infection in crustaceans: no-vel hosts and potential routes for horizontaltransmission. � Journal of Evolutionary Bio-logy 14, 237-243.
COUGHTREY, P.J., MARTIN, M.H., CHARD, J., SHA-LES, S.W., 1980: Micro-organisms and metalretention in the woodlouse Oniscus asellus. �Soil Biology and Biochemistry 12, 23-27.
CURRIE, C.R., SCOTT, J.A., SUMMERBELL, R.C., MAL-LOCH, D., 1999: Fungus-growing ants use an-tibiotic-producing bacteria to control gardenparasites. � Nature 398, 701-704.
DALE, C., MAUDLIN, I., 1999: Sodalis gen. nov. andSodalis glossinidius sp. nov., a microaerophilicsecondary endosymbiont of the tsetse fly Glos-sina morsitans morsitans. � International Journalof Systematic Bacteriology 49,267-275.
DAVID, J.-F., CÉLÉRIER, M.-L., 1997: Effects ofyeast on the growth and the reproduction ofthe saprophagous millipede Polydesmus angus-tus (Diplopoda, Polydesmidae). � Biology andFertility of Soils 24, 66-69
DAVIES, A.J., 1995: Studies on the gut flora of thehaematophagous fish parasite Gnathia maxilla-ris Montagu. � Bulletin of the European Asso-ciation of Fish Pathology 15, 32-35.
DECKER, C.H., VISSER, J., SCHREIER, P., 2001: β-Glu-cosidase multiplicity from Aspergillus tubingen-sis CBS 643.92: purification and characteriza-tion of four β-glucosidases and their differen-tiation with respect to substrate specificity, glu-cose inhibition and acid tolerance. �Applied Microbiology and Biotechnology 55,157-163.
DECKER, H., TUCZEK, F., 2000: Tyrosinase/cate-choloxidase activity in hemocyanins: structu-ral basis and molecular mechanism. � Trendsin Biochemical Sciences 25, 392-397.
DE KNECHT, J.A., STROOMBERG, G.J., TUMP, C.,HELMS, M., VERWEIJ, R.A., COMMANDEUR, J.,VAN GESTEL, C.A., VAN STRAALEN, N.M., 2001:Characterization of enzymes involved in bi-otransformation of polycyclic hydrocarbonsin terrestrial isopods. � Environmental Toxi-cology and Chemistry 20, 1475-1464.
D�ETTORE, P., MORA, P., DIBANGOU, V., ROULAND,C., ERRARD, C., 2002: The role of the symbi-otic fungus in the digestive metabolismof two species of fungus-growing ants. �Journal of Comparative Physiology B 172,169-176.

158 MARTIN ZIMMER
DEZFULI, B.S., 2000: Host-parasite interfaces bet-ween Asellus aquaticus (Isopoda) and larvae ofAcanthocephalus anguillae (Acanthocephala). �Folia Parasitologica 47, 154-156.
DOBZHANSKY, T., 1973: Nothing in biology makessense except in the light of evolution. �American Biology Teacher 35, 125-129.
DOEBELI, M., DIECKMANN, U., 2000: Evolutionarybranching and sympatric speciation caused bydifferent types of ecological interactions. � TheAmerican Naturalist 156, S77-S101.
DONADEY, C., BESSE, G., 1972: Étude histologiqueultrastructurale et expérimentale des caecumsdigestifs de Porcellio dilatatus et Ligia oceanica(Crustacea, Isopoda). � Téthys 4, 145-162.
DOUGLAS, A.E., 1980: Establishment of the sym-biosis in Convoluta roscoffensis. � Journal of theMarine Biological Association UK 63, 419-434.
DOUGLAS, A.E., 1989: Mycetocyte symbiosis ininsects. � Biological Reviews 64, 409-434.
DOUGLAS, A.E., 1992a: Symbiosis in evolution. �Oxford Surveys in Environmental Biology 8,347-382.
DOUGLAS, A.E., 1992b: Microbial brokers ofinsect-plant interactions, pp. 329-336. In: Pro-ceedings of the 8th International Symposi-um on Insect-plant relationships (Menken,S.B.J., Visser, J.H., Harrewijn, P., eds). � Klu-wer Academic Publishers: Dordrecht.
DOUGLAS, A.E., 1995: The ecology of symbioticmicro-organisms. � Advances in EcologicalResearch 26, 69-103.
DOUGLAS, A.E., MINTO, L.B., WILKINSON, T.L.,2001: Quantifying nutrient production by themicrobial symbionts in an aphid. � Journal ofExperimental Biology 204, 349-358.
DOUGLAS, A.E., RAVEN, J.A., 2003: Genomes at theinterface between bacteria and organelles. �Philosophical Transactions of the Royal Socie-ty of London: Biological Sciences 358, 5-18.
DOW, J.A.T., 1992: pH gradients in lepidopteranmidgut. � Journal of Experimental Biology172, 355-375.
DOWD, P.F., 1989: In situ production of hydro-lytic detoxifying enzymes by symbiotic yeastsin the cigarette beetle (Coleoptera: Anobii-dae). � Journal of Economical Entomology82, 396-400.
DROBNE, D., 1995: Bacteria adherent to the hin-dgut of terrestrial isopods. � Acta Micro-biologica et Immunologica Hungarica 42,45-52.
DROBNE, D., PRIMOZ, Z., �TRUS, J., 1996: SEMAnalysis of midgut glands in isopod Porcellioscaber (Isopod, Crustacea) fed with metal con-taminated food. � Journal of Computer-As-sisted Microscopy 8, 249-250.
DROGE, M., PUHLER, A., SELBITSCHKA, W., 1998:Horizontal gene transfer as a biosafety issue:a natural phenomenon of public concern. �Journal of Biotechnology 64, 75-90.
DUGAS, J.E., ZUREK, L., PASTER, B.J., KEDDIE, B.A.,LEADBETTER, E.R., 2001: Isolation and charac-terization of a Chryseobacterium strain from thegut of the American cockroach, Periplanetaamericana. � Achives of Microbiology 175,259-262.
DUNGER, W., 1958: Über die Zersetzung derLaubstreu durch die Boden-Makrofauna imAuenwald. � Zoologisches Jahrbuch Systema-tik und Ökologie 86, 139-180.
EDNEY, E.B., 1954: Woodlice and the land habi-tat. � Biological Reviews 29, 185-220.
EDNEY, E.B., 1968: Transition from water to landin isopod crustaceans. � American Zoologist8, 309-326.
ERASMUS, J.H., COOK, P.A., COYNE, V.E., 1997: Therole of bacteria in the digestion of seaweedby the abalone Haliotis midae. � Aquaculture155, 377-386.
ERIKSSON, K.-E., 1978: Enzyme mechanisms in-volved in cellulose hydrolysis by the rot fun-gus Sporotrichum pulveruletum. � Biotechnologyand Bioengineering 20, 317-332.
ERIKSSON, K.-E., WOOD, T.M., 1985: Biogeradati-on of cellulose, pp. 469-504. In: Biosynthesisand biodegradation of wood components (Hi-guchi, T., ed.). � Academic Press: London.
ESHAM, E.C., YE, W., MORAN, M.A., 2000: Identi-fication and characterization of humic sub-stances-degrading bacterial isolates from anestuarine environment. � FEMS Microbiolo-gy Ecology 34, 103-111.
EVANS, C.S., GALLAGHER, I.M., ATKEY, P.T., WOOD,D.A., 1991: Localisation of degradative en-zymes in white-rot decay of lignocellulose. �Biodegradation 2, 93-106.
FARRANT, J.L., SANSONE, A., CANVIN, J.R., PALLEN,M.J., LANGFORD, P.R., WALLIS, T.S., DOUGAN,.G., KROLL, J.S., 1997: Bacterial copper- andzinc-cofactored superoxide dismutase cont-ributes to the pathogenesis of systemic sal-monellosis. � Molecular Microbiology 25,785-796.

The role of animal-microbe interactions in isopod ecology and evolution 159
Acta Biologica Benrodis 13 (2006)
FAURE, D., BOUILLANT, M.-L., BALLY, R., 1995:Comparative study of substrates and inhibi-tors of Azospirillum lipoferum and Pyriculariaoryzae laccases. � Applied and EnvironmentalMicrobiology 61, 1144-1146.
FEINSINGER, P., 1983: Coevolution and pollinati-on, pp. 282-310. In: Coevolution (Futuyma,D.J., Slatkin, M., eds). � Sinauer Associates,Sunderland (MA).
FERREIRA, C., TERRA, W.R., 1983: Physical and ki-netic properties of a plasma-membrane-boundα-D-glucosidase (cellobiase) from midgut cellsof an insect (Rhynchosciara americana larva). �Biochemistry Journal 213, 43-51.
FILIP, Z., BIELEK, P., 2002: Susceptibility of hu-mic acids from soils with various contents ofmetals to microbial utilization and transfor-mation. � Biology and Fertility of Soils 36,426-433.
FUKATSU, T., TSUCHIDA, T., NIKOH, N., KOGA, R.,2001: Spiroplasma symbiont of the pea aphid,Acyrthosiphon pisum (Insecta: Homoptera). �Applied and Environmental Microbiology 67,1284-1291.
FUTUYMA, D.J., 1986: Evolutionary Biology. � Si-nauer Associates, Sunderland (MA), USA.
GIJZEN, H.J., VAN DER DRIFT, C., BARUGAHARE, M.,OP DEN CAMP, H.J.M., 1994: Effect of hostdiet and hindgut microbial composition oncellulolytic activity in the hindgut of theAmerican Cockroach, Periplaneta americana. �Applied and Environmental Microbiology 60,1822-1826.
GILLESPIE, J.P., KANOST, M.R., TRENCZEK, T., 1997:Biological mediators of insect immunity. �Annual Review of Entomology 42, 611-643.
GOMULKIEWICZ, R., NUISMER, S.L., THOMPSON,J.N., 2003: Coevolution in variable mutua-lisms. � The American Naturalist 162, S80-S93.
GORVETT, H., 1952: The tegumental glands in theland Isopoda C � the lobed glands: the pro-perties of their secretion and their mode ofaction. � Quarterly Journal of MicroscopicSciences 93, 17-29.
GOULD, S.J., 1989: A developmental constraintin Cerion, with comments on the definition andinterpretation of constraint in evolution. �Evolution 43, 516-539.
GOULD, S.J., VRBA, E.S., 1982: Exaptation � a mis-sing term in the science of form. � Paleobio-logy 8, 4-15.
GRAÇA, M.A.S., BÄRLOCHER, F., 1998: Proteolyticgut enzymes in Tipula caloptera � interaction withphenolics. � Aquatic Insects 21, 11-18.
GRIFFITHS, B.S., WOOD, S., 1985: Microorganismsassociated with the hindgut of Oniscus asellus(Crustacea, Isopoda). � Pedobiologia 28, 377-381.
GRÜNWALD, M., 1987: Adaption und Dekompo-sitionsleistung von Landasseln (Isopoda,Oniscidea) an Standorten der Großen Bren-nessel (Urtica dioica L). � HochschulsammlungNaturwissenschaften � Biologie 20. Disser-tation, Universität Bayreuth.
GUNNARSSON, T., 1987: Selective feeding on amaple leaf by Oniscus asellus (Isopoda). � Pe-dobiologia 30, 161-165.
GUNNARSSON, T., TUNLID, A., 1986: Recycling offecal pellets in isopods: microorganisms andnitrogen compounds as potential food forOniscus asellus. � Soil Biology and Biochemist-ry 18, 595-600.
HAGERMAN, A.E., BUTLER, L.G., 1991: Tanninsand lignins, pp. 355-388. In: Herbivores: Theirinteractions with secondary plant metaboli-tes I: The chemical participants (Rosenthal,G.A., Berenbaum, M.R., eds). � AcademicPress, New York.
HANLON, R.D.G., ANDERSON, J.M., 1980: Influ-ence of macroarthropod feeding activities onmicroflora in the decomposing oak leaves. �Soil Biology and Biochemistry 12, 255-261.
HARRIS, J.M., 1993: The presence, nature, androle of gut microflora in aquatic inverte-brates: a synthesis. � Microbial Ecology 25,195-231.
HARRIS, J.M., SEIDERER, L.J., LUCAS, M.I., 1991:Gut microflora of two saltmarsh detritivo-re Thalassinid prawns, Ubogebia africana andCallianassa kraussi. � Microbial Ecology 21,63-82.
HARTENSTEIN, R., 1964a: Feeding, digestion, gly-cogen and the environmental conditions ofthe digestive system in Oniscus asellus. � Jour-nal of Insect Physiology 10, 611-621.
HARTENSTEIN, R., 1964b: Histochemical and spec-trophotometrical studies on several dehydro-genases of carbohydrate metabolism in Oni-scus asellus. � Journal of Insect Physiology 10,623-631.
HARTENSTEIN, R., 1968: Nitrogen metabolism inthe terrestrial isopod, Oniscus asellus. � Ameri-can Zoologist 8, 507-519.

160 MARTIN ZIMMER
HASSALL, M., 1977: Consumption of leaf litterby the terrestrial isopod Philoscia muscorumin relation to food availability in a dunegrassland ecosystem. � Ecological Bulletin45, 550-553.
HASSALL, M., JENNINGS, J.B., 1975: Adaptive featuresof gut structure and digestive physiology in theterrestrial isopod Philoscia muscorum (Scopoli)1763. � Biological Bulletin 149, 348-364.
HASSALL, M., RUSHTON, S.P., 1982: The role ofcoprophagy in the feeding strategies of ter-restrial isopods. � Oecologia 53, 374-381.
HASSALL, M., RUSHTON, S.P., 1985: The adaptivesignificance of coprophagous behaviour inthe terrestrial isopod Porcellio scaber. � Pedobi-ologia 28, 169-175.
HASSALL, M., TURNER, J.G., RANDS, M.R.W., 1987:Effects of terrestrial isopods on the decom-position of woodland leaf litter. � Oecologia72, 597-604.
HELLIO, C., BREMER, G., PONS, A.M., LE GAL, Y.,BOURGOUGNON, N., 2000: Inhibition of thedevelopment of microorganisms (bacteriaand fungi) by extracts of marine algae fromBrittany, France. � Applied Microbiology andBiotechnology 54, 543-549.
HENNIG, W., 1966: Phylogenetic systematics. �University of Illinois Press, Urbana.
HILL, K.E., WEIGHTMAN, A.J., 2003: Horizontaltrnasfer of dehalogenase genes on IncP1âplasmids during bacterial adaptation to degra-de α-halocarboxylic acids. � FEMS Microbi-ology Ecology 45, 273-282.
HOLBEN, W.E., WILLIAMS, P., SAARINEN, M., SÄR-KILAHTI, L.K., APAJALAHTI, J.H.A., 2001: Phy-logenetic analysis of intestinal microflora in-dicates a novel Mycoplasma phylotype in far-med and wild salmon. � Microbial Ecology44, 175-185.
HOLD, G.L., SMITH, E.A., RAPPÉ, M.S., MAAS, E.W.,MOORE, E.R.B., STROEMPL, C., STEPHEN, J.R.,PROSSER, J.I., BIRKBECK, H., GALLACHER, S.,2001: Characterisation of bacterial commu-nities associated with toxic and non-toxic di-noflagellates: Alexandrium spp. and Scrippsiellatrochoidea. � FEMS Microbiology Ecology 37,161-173.
HONG, T.-Y., MENG, M., 2003: Biochemical cha-racterization and antifungal activity of anendo-1,3-b-glucanase of Paenibacillus sp. iso-lated from garden soil. � Applied Microbio-logy and Biotechnology 61, 472�478
HOPKIN, S.P., MARTIN, M.H., 1982: The distributi-on of zinc, cadmium, lead and copper withinthe woodlouse Oniscus asellus (Crustacea, Iso-poda). � Oecologia 54, 227-232.
HOPKIN, S.P., MARTIN, M.H., 1984: Heavy metalsin woodlice. Symposia of the Zoological So-ciety of London 53, 143-166.
HYODO, F., INOUE, T., AZUMA, J.-I., TAYASU, I., ABE,T., 2000: Role of mutualistic fungus in lignindegradation in the fungus-growing termiteMacrotermes gilvus (Isoptera; Macrotermitinae).� Soil Biology and Biochemistry 32, 653-658
HYODO, F., TAYASU, I., INOUE, T., AZUMA, J.-I.,KUDO, T., ABE, T., 2003: Differential role ofsymbiotic fungi in lignin degradation and foodprovision for fungus-growing termites (Ma-crotermitinae: Isoptera). � Functional Eco-logy 17, 186-193.
INESON, P., ANDERSON, J.M., 1985: Aerobically iso-lated bacteria associated with the gut and fecesof the litter feeding macroarthropods Oniscusasellus and Glomeris marginata. � Soil Biologyand Biochemistry 17, 843-849.
JAENECKE, S., DE LORENZO, V., TIMMIS, K.N., DIAZ,E., 1996: A stringently controlled expressionsystem for analysing lateral gene transferbetween bacteria. � Molecular Microbiology21, 293-300.
JOHJIMA, T., OHKUMA, M., KUDO, T., 2003: Isola-tion and cDNA cloning of novel hydrogenperoxide-dependent phenol oxidase from thebasidiomycete Termitomyces albuminosus. � Ap-plied Microbiology and Biotechnology 61,220-225
JOHNSTONE, R.A., BSHARY, R., 2002: From parasi-tism to mutualism: partner control in asymme-tric interactions. � Ecology Letters 5, 634-639.
JUILFS, H.B., WÄGELE, J.W., 1987: Symbiotic bac-teria in the gut of the blood-sucking antarcticfish parasite Gnathia calva (Crustacea: Isopo-da). � Marine Biology 95, 493-499.
KAUTZ, G., ZIMMER, M., TOPP, W., 2000: Re-sponses of the parthenogenetic isopod, Tri-choniscus pusillus, to changes in food quality. �Pedobiologia 44, 75-85.
KAUTZ, G., ZIMMER, M., TOPP, W., 2002: DoesPorcellio scaber (Isopoda: Oniscidea) gain fromcoprophagy? � Soil Biology and Biochemist-ry 34, 1253�1259.
KAUTZ, G., TOPP, W., 2000: Acquisition of mic-robial communities and enhanced availabilityof soil nutrients by the isopod Porcellio scaber

The role of animal-microbe interactions in isopod ecology and evolution 161
Acta Biologica Benrodis 13 (2006)
(Latr.) (Isopoda: Oniscidea). � Biology andFertility of Soils 31, 102-107.
KELLNER, R.L.L., 2001: Suppression of pederinbiosynthesis through antibiotic elimination ofendosymbionts in Paederus sabaeus. � Journalof Insect Physiology 47, 475-483.
KELLNER, R.L.L., 2002: Molecular identificationof an endosymbiotic bacterium associatedwith pederin biosynthesis in Paederus sabaeus(Coleoptera: Staphylinidae). � Insect Bioche-mistry and Molecular Biology 32, 289-295.
KELLY, B.A., KADO, C.I., 2002: Agrobacterium-mediated T-DNA transfer and integrationinto the chromosome of Streptomyces lividans.� Molecular Plant Pathology 3, 125-134.
KENSLEY, B., 1974: Aspects of the biology andecology of the genus Tylos Latreille. � Annalsof the South African Museum 65, 401-471.
KIRCHMAN, D.L., 2002: The ecology of Cytopha-ga-Flavobacteria in aquatic environments. �FEMS Microbiology Ecology 39, 91-100.
KLUSSMANN-KOLB, A., BRODIE, G.D., 1999: Internalstorage and production of symbiotic bacteriain the reproductive system of a tropical marinegastropod. � Marine Biology 133, 443-447.
KOGA, R., TSUCHIDA, T., FUKATSU, T., 2003: Chan-ging partners in an obligate symbiosis: a fa-cultative endosymbiont can compensate forloss of the essential endosymbiont Buchnerain an aphid. � Proceedings of the Royal Soci-ety of London B 270, 2543-2550.
KORB, J., AANEN, D.K., 2003: The evolution ofuniparental transmission of fungal symbiontsin fungus-growing termites (Macrotermiti-nae). � Behavioral Ecology Sociobiology 53,65-71.
KOSTANJ�EK, R., �TRUS, J., AVGU�TIN, G., 2002:Genetic diversity of bacteria associated withthe hindgut of the terrestrial crustacean Porcel-lio scaber (Crustacea: Isopoda). � FEMS Mic-robiology Ecology 40, 171-179.
KOZLOVSKAJA, L.S., STRIGANOVA, B.R., 1977: Food,digestion and assimilation in desert woodliceand their relations to the soil microflora. �Ecological Bulletin 25, 240-245.
KUITERS, A.T., SARINK, H.M., 1986: Leaching ofphenolic compounds from leaf and needlelitter of several deciduous and coniferoustrees. � Soil Biology and Biochemistry 18,475-480.
KUKOR, J.J., MARTIN, M.M., 1983: Acquisition ofdigestive enzymes by siricid woodwasps from
their fungal symbionts. � Science 220, 1161-1163.
KUKOR, J.J., MARTIN, M.M., 1986: The effect ofacquired microbial enzymes on assimilationefficiency in the common woodlouse, Trache-oniscus rathkei. � Oecologia 69, 360-366.
KURTZ, J., FRANZ, K., 2003: Evidence for memo-ry in invertebrate immunity. � Nature 425, 37-38.
LAU, W.W.Y., JUMARS, P.A., ARMBRUST, E.V., 2002:Genetic diversity of attached bacteria in thehindgut of the deposit-feeding shrimp Neo-trypaea (formerly Callianassa) californiensis (De-capoda: Thalassinidae). � Microbial Ecology43, 455-466.
LEE, K.-H., RUBY, E.G., 1994: Effects of the squidhost on the abundance and distribution ofsymbiotic Vibrio fischeri in nature. �Applied and Environmental Microbiology 60,1565-1571..
LEONARDO, T.E., MUIRU, G.T., 2003: Facultativesymbionts are associated with host plant spe-cialization in pea aphid populations. � Pro-ceedings of the Royal Society of London B270, S209-S212
LESCHINE, S.B., 1995: Cellulose degradation inanaerobic environments. � Annual Review ofMicrobiology 49, 399-426.
LILLEY, A.K., BAILEY, M.J., BARR, M., KILSHAW, K.,TIMMS-WILSON, T.M., DAY, M.J., NORRIS, S.J.,JONES, T.H., GODFRAY, H.C.J., 2003: Populati-on dynamics and gene transfer in geneticallymodified bacteria in a model microcosm. �Molecular Ecology 12, 3097-3107.
LIVELY, C.M., 1999: Migration, virulence, andthe geographic mosaic of adaptation by pa-rasites. � The American Naturalist 153, S34-S47.
LJUNGDAHL, L.G., ERIKSSON, K.-E., 1985: Ecolo-gy of microbial cellulose degradation. � Ad-vances in Microbial Ecology 8, 237-299.
LO, S.C., HAYES, M.M., KOTANI, H., PIERCE, P.F.,WEAR, D.J., NEWTON, P.B., TULLY, J.G., SHIH,J.W., 1993: Adhesion to and invasion intomammalian cells by Mycoplasma penetrans: anewly isolated mycoplasma from patients withAIDS. � Modern Pathology 6, 276-280.
LUNG, O., KUO, L., WOLFNER, M.F., 2001: Droso-phila males transfer antibacterial proteins fromtheir accessory gland and ejaculatory duct totheir mates. � Journal of Insect Physiology47, 617-622.

162 MARTIN ZIMMER
LUSSENHOP, J., 1992: Mechanisms of microarth-ropod-microbial interactions in soil. � Advan-ces in Ecological Research 23, 1-33.
MANTER, H.W., 1966: Parasites of fishes as bio-logical indicators of recent and ancient con-ditions. In: Host-parasite relationships (Mc-Cauley, J.E., ed). � Oregon State UniversityPress, Corvallis.
MARANA, S.R., TERRA, W.R., FERREIRA, C., 1995:Midgut β-D-Glucosidases from Abracris fla-volineata (Orthoptera: Acrididae): physical pro-perties, substrate specificities and function. �Insect Biochemistry and Molecular Biology25, 835-843.
MARANA, S.R., TERRA, W.R., FERREIRA, C., 2000:Purification and properties of a β-glycosida-se purified from midgut cells of Spodoptera fru-giperda (Lepidoptera) larvae. � Insect Bioche-mistry and Molecular Biology 30, 1139-1146.
MARGULIS, L., FESTER, R. (eds), 1991: Symbiosesas a source of evolutionary innovation. � MITPress, Cambridge (MA, USA).
MÁRIALIGETI, K., JÁGER, K., SZABÓ, I.M., POBOZ-SNY, M., DZINGOV, A., 1984: The faecal acti-nomycete flora of Protracheoniscus amoenus(Woodlice; Isopoda). � Acta MicrobiologicaHungarica 31, 339-344.
MARTIN, A.L., 1964: The alimentary canal ofMarinogammarus obtusatus (Crustacea, Amphi-poda). � Proceedings of the Zoological Soci-ety of London 143, 525-544.
MARTIN, J.S., MARTIN, M.M., 1983: Tannin assaysin ecological studies: precipitation of Ribu-lose-1, 5-Bisphosphate Carboxylase/Oxyge-nase by tannic acid, quebracho, and oak folia-ge extracts. � Journal of Chemical Ecology9, 285-294.
MARTIN, M.M., 1984: The role of ingested enzy-mes in the digestive processes of insects, pp.155-172. In: Invertebrate-microbial interac-tions (Anderson, J.M., Rayner, A.D.M., Wal-ton, D.W.H., eds). � Cambridge UniversityPress, Cambridge.
MARTIN, M.M., 1991: The evolution of cellulosedigestion in insects. � Philosophical Transac-tions of the Royal Society of London B 333,281-288.
MARTIN, M.M., MARTIN, J.S., KUKOR, J.J., MERRITT,R.W., 1980: The digestion of protein and car-bohydrate by the stream detritivore, Tipulaabdominalis (Diptera, Tipulidae). � Oecologia46, 360-364.
MATTERN, D., SCHLEGEL, M., 2001: Molecular evo-lution of the small subunit ribosomal DNAin woodlice (Crustacea, Isopoda, Oniscidea)and implications for Oniscidean phylogeny.� Molecular Phylogenetics and Evolution, 18,54-65.
MAYER, A.M., 1987: Polyphenol oxidases in plants� recent progress. � Phytochemistry 26, 11-20.
MAYER, A.M., HAREL, E., 1979: Polyphenol oxi-dases in plants. � Phytochemistry 18, 193-215.
MAYNARD SMITH, J., SZATHMÁRY, E., 1995: Themajor transitions in evolution. � Freeman,San Francisco.
MAYNARD SMITH, J., BURIAN, R., KAUFFMAN, S.,ALBERCH, P., CAMPBELL, J., GOODWIN, B., LAN-DE, R., RAUP, D., WOLPERT, L., 1985: Develop-mental constraints and evolution. � Quarter-ly Review of Biology 60, 265-287.
MCGRATH, C.C., MATTHEWS, R.A., 2000: Cellula-se activity in the freshwater amphipod Gam-marus lacustris. � Journal of the North Ameri-can Benthological Society 19, 298-307.
MICHEL-SALZAT, A., BOUCHON, D., 2000: Phyloge-netic analysis of mitochondrial LSU rRNA inoniscids. � Comptes Rendus de l�Academie desSciences de Paris 323, 827-837
MIRA, A., MORAN, N.A., 2002: Estimating popu-lation size and transmission bottlenecks inmaternally transmitted endosymbiotic bacte-ria. � Microbial Ecology 44, 137-143.
MONK, D.C., 1977: The digestion of cellulose andother dietary components, and pH of the gutin the amphipod Gammarus pulex (L.). � Fresh-water Biololgy 7, 431-440.
MONTLLOR, C.B., MAXMEN, A., PURCELL, A.H., 2002:Facultative bacterial endosymbionts benefit peaaphids Acyrthosiphon pisum under heat stress. �Ecological Entomology 27, 189-195.
MORGAN, M.R.J., 1975: Relationship between gutcellobiase, lactase, ary-β-glucosidase, and aryl-β-galactosidase activities of Locusta migratoria.� Insect Biomchemistry 5, 609-617.
MÜLLER, W.E.G., 1999: 16S rRNA sequences asdiagnostic tools to elucidate potential symbi-otic relationships between bacteria and themarine sponge Halichondria panicea: reply toStackebrandt and Pukall (Mar Biol 133: 159-161). � Marine Biology 134, 213-215.
MUSGROVE, R.J., 1988: Digestive ability of thefreshwater crayfish Paranephrops zealandicus(Parastacidae) and the role of microbial en-zymes. � Freshwater Biology 19, 305-314.

The role of animal-microbe interactions in isopod ecology and evolution 163
Acta Biologica Benrodis 13 (2006)
NARDI, J.B., MACKIE, R.I., DAWSON, J.O., 2002:Could microbial symbionts of arthropod gutscontribute significantly to nitrogen fixationin terrestrial ecosystems? � Journal of InsectPhysiology 48, 751-763.
NARDON, P., CHARLES, H., DELOBEL, B., LEFEVRE,C., HEDDI, A., 2003: Symbiosis in the Dryo-phthoridae weevils (Coleoptera, Cuculionoi-dea): morphological variability of symbioticintracellular bacteria. � Symbiosis 34, 231-251.
NEALSON, K.H., HAYGOOD, M.G., TEBO, B.M.,ROMAN, M., MILLER, E., MCCOSKER, J.E., 1984:Contribution by symbiotically luminous fis-hes to the occurrence and bioluminescenceof luminous bacteria in sea water. � Micro-bial Ecology 10, 69-77.
NESTER, E.W., 2000: DNA and protein transfer frombacteria to eukaryotes � the Agrobacterium story.� Molecular Plant Pathology 1, 87-90.
NEUHAUSER, E., HARTENSTEIN, R., 1976: Degra-dation of phenol, cinnamic acid and quinicacid in the terrestrial crustacea, Oniscus asel-lus. � Soil Biology and Biochemistry 8, 95-98.
NICHOLLS, A.G., 1931: Studies on Ligia oceanica� II: The processes of feeding, digestionand absorption, with a description of thestructure of the foregut. � Journal of theMarine Biological Association of the UK17, 675-705.
NODA, H., KODAMA, K., 1996: Phylogenetic po-sition of yeastlike endosymbionts of anobiidbeetles. � Applied and Environmental Mic-robiology 62, 162-167.
NUISMER, S.L., GOMULKIEWICZ, R., MORGAN, M.T.,2003: Coevolution in Temporally VariableEnvironments. � The American Naturalist162, 195-204
OHKUMA, M., 2003: Termite symbiotic systems: ef-ficient bio-recycling of lignocellulose. � AppliedMicrobiology and Biotechnology 61, 1-9.
OSHEL, P.E., STEELE, D.H., 1988: SEM morpho-logy of the foreguts of gammaridean amphi-pods compared to Anaspides tasmaniae (Anas-pidacea: Anaspididae), Gnathophausia ingens(Mysidacea: Lophogastridae), and Idotea balthi-ca (Isopoda: Idoteidae). � Crustaceana Sup-plement 13, 209-219.
PANT, N.C., FRAENKEL, G., 1954: Studies on thesymbiotic yeasts of two insect species Lasio-derma serricorne F. and Stegobium paniceum L. �Biological Bulletin 107, 420-432.
PARKER, M.A., 2003: Genetic markers for analy-sing symbiotic relationships and lateral genetransfer in neotropical bradyrhizobia. � Mo-lecular Ecology 12, 2447-2455.
PASTOR, F.I.J., PUJOL, X., BLANCO, A., VIDAL, T.,TORRES, A.L., DÍAZ, P., 2001: Molecular clo-ning and characterization of a mulitdomainendoglucanase from Paenibacillus sp. BP-23:evaluation of its performance in pulp refining.� Applied Microbiology and Biotechnology55, 61-68.
PENNINGS, S.C., CAREFOOT, T.H., ZIMMER, M.,DANKO, J.P., ZIEGLER, A., 2000: Feeding pre-ferences of supralittoral isopods and amphi-pods. � Canadian Journal of Zoology 78,1918-1929.
PLANTE, C.J., JUMARS, P.A., BAROSS, J.A., 1990:Digestive associations between marine inver-tebrates and bacteria. � Annual Review inEcology and Systematics 21, 93-127.
POINSOT-BALAGUER, N., RACON, L., SADAKA, N.,LE PETIT, J., 1993: Effects of tannin com-pounds on two species of Collembola. � Eu-ropean Journal of Soil Biology 29, 13-16.
PRICE, P.W., 1991: The web of life: developmentover 3.8 billion years of trophic relationships,pp. 262-272. In: Symbiosis as a source of evo-lutionary innovation � speciation and mor-phogenesis (Margulis, L., Fester, R., eds). �Cambridge MIT Press, Cambridge.
RABINOVICH, M.L., BOLOBOVA, A.V., VASIL�CHENKO,L.G., 2004: Fungal decomposition of naturalaromatic structures and xenobiotics: A review.� Applied Biochemistry and Microbiology 40,1-17.
RAMETTE, A., MOËNNE-LOCCOZ, Y., DÉFAGO, G.,2003: Prevalence of fluorecent pseudomonadsproducing antifungal phloroglucinols and/orhydrogen cynide in soils naturally suppressiveor conducive to tobacco black root rot. �FEMS Microbiology Ecology 44, 35-43.
RAPP, P., BEERMANN, A., 1991: Bacterial cellula-ses, pp. 269-276. In: Biosynthesis and biode-gradation of cellulose (Haigler, C.H., Weimer,P.J., eds). � Marcel Dekker, New York.
RAY, D.L., 1959: Marine fungi and wood borerattack. � Proceedings of the American WoodPreservation Association 54, 1-7.
RAY, D.L., JULIAN, J.R., 1952: Occurrence of cel-lulase in Limnoria. � Nature 169, 32-33.
REESE, E.T., MANDELS, M., 1971: Degradation ofcellulose and its derivates � enzymic degra-

164 MARTIN ZIMMER
dation, pp. 1079-1094. In: High polymers 5(Bikales, N.M., Segal, L., eds). � Wiley-In-terscience: New York.
REYES, V.G., TIEDJE, J.M., 1976: Ecology of thegut microbiota of Tracheoniscus rathkei (Crus-tacea, Isopoda). � Pedobiologia 16, 67-74.
RISPE, C., MORAN, N.A., 2000: Accumulationof deleterious mutations in endosymbionts:Muller�s ratchet with two levels of selec-tion. � The American Naturalist 156, 425-441.
ROTTEM, S., 2003: Interaction of Mycoplasmas withhost cells. � Physiological Review 83, 417-432.
ROUGHGARDEN, J., 1975: Evolution of marinesymbioses � a simple cost-benefit model. �Ecology 56, 1201-1208.
RUPEREZ, P., SAURA-CALIXTO, F., 2001: Dietary andphysicochemical properties of edible Spanishseaweeds. � Zeitschrift für Lebensmittel Un-tersuchung und Forschung A 212, 349-354.
RUSHTON, S.P., HASSALL, M., 1983a: Food andfeeding rates of the terrestrial isopod Ar-madillidium vulgare (Latreille). � Oecologia 57,415-419.
SALEEM, M., ALIKHAN, M.A., 1974: The distribu-tion of phosphomonoesterases in the diges-tive gut of Porcellio laevis Latreille (Porcellio-nidae, Isopoda). � Comparative Biochemist-ry and Physiology 48A, 375-385.
SANTOS, C.D., TERRA, W.R., 1985: Physical pro-perties, substrate specificities and a probablemechanism for a α-D-glucosidase (cellobiase)from midgut cells of the cassava hornworm(Erinnyis ello). � Biochimica et Biophysica Acta831, 179-185.
SAVAGE, D.C., 1972: Survival on mucosal epithe-lia, epithelial penetration and growth in tissu-es of pathogenic bacteria, pp. 25-57. In: Mic-robiol pathogenicity of man and animals(Smith, H., Pearce, J.H., eds). � CambridgeUniversity Press, Cambridge
SCHELOSKE, H.W., 1976: Ist die Unterteilung desMagens der Malacostraca in �Cardia� und�Pylorus� berechtigt? � Zeitschrift für Zoo-logie: Systematik und Evolutionsforschung14, 253-280.
SCHMALFUSS, H., 1978: Ligia simoni: a model forthe evolution of terrestrial isopods. � Stutt-garter Beiträge zur Naturkunde 317, 1-5.
SCHMALFUSS, H., 1984: Eco-morphological stra-tegies in terrestrial isopods. � Symposia ofthe Zoological Society of London 53, 49-63.
SCHMALFUSS, H., 1998: Evolutionary strategies ofthe antennae in terrestrial isopods. � Journalof Crustacean Biology 18, 10-24.
SCHMIDT, C., WÄGELE, J.W., 2001: Morphology andevolution of respiratory structures in the ple-opod exopodites of terrestrial Isopoda(Crustacea, Isopoda, Oniscidea). � Acta Zo-ologica 82, 315-330.
SCHÖNHOLZER, F., HAHN, D., ZARDA, B., ZEYER, J.,2002: Automated image analysis and in situhybridization as tools to study bacterial po-pulations in food resources, gut and cast ofLumbricus terrestris L. � Journal of Microbio-logical Methods 48, 53-68.
SCHÖNHOLZER, F., HAHN, D., ZEYER, J., 1999: Ori-gins and fate of fungi and bacteria in the gut ofLumbricus terrestris L. studied by image analysis. �FEMS Microbiology Ecology 28, 235-248.
SCHULENBURG, GRAF VON DER J.H., HABIG, M.,SLOGGETT, J.J., WEBBERLEY, K.M., BERTRAND,D., HURST, G.D.D., MAJERUS, M.E.N., 2001:Incidence of male-killing Rickettsia spp. (α-Proteobacteria) in the Ten-Spot LadybirdBeetle Adalia decempunctata L (Coleoptera: Coc-cinellidae). � Applied and Environmental Mi-crobiology 67, 270-277.
SCHULTZ, J.C., LECHOWICZ, M.J., 1986: Host plant,larval age and feeding behavior influencemidgut pH in the Gypsy Moth (Lymantria dis-par L). � Oecologia 71, 133-137.
SCRIVENER, A.M., SLAYTOR, M., 1994a: Cellulosedigestion in Panesthia cribrata Saussure: doesfungal cellulase play a role? � ComparativeBiochemistry and Physiology 107B, 309-315.
SCRIVENER, A.M., SLAYTOR, M., 1994b: Propertiesof the endogenous cellulase from Panesthia crib-rata Saussure and purification of major endo-β-1,4-glucanase components. � Insect Biochemis-try and Molecular Biology 24, 223-231.
SHEN, S.K., DOWD, P.F., 1991: Detoxification spec-trum of the cigarette beetle symbiont Symbi-otaphrina kochii in culture. � Entomologia Ex-perimentalis et Applicata 60, 51-59.
SHIGENOBU, S., WATANABE, H., HATTORI, M., SA-KAKI, Y., ISHIKAWA, H., 2000: Genosequenceof the endocellular bacterial symbiont ofaphids Buchnera sp. APS. � Nature 407, 81-86.
SINDHU, S.S., DADARWAL, K.R., 2001: Chitinolyticand cellulolytic Pseudomonas sp. antagonistic tofungal pathogens enhances nodulation byMesorhizobium sp. cicer in chickpea. � Micro-biological Research 156, 353-358.

The role of animal-microbe interactions in isopod ecology and evolution 165
Acta Biologica Benrodis 13 (2006)
SINSABAUGH, R.L., LINKINS, A.E., BENFIELD, E.F.,1985: Cellulose digestion and assimilation bythree leaf-shredding aquatic insects. � Ecolo-gy 66, 1464-1471.
SKAMBRACKS, D., ZIMMER, M., 1998: Combinedmethods for the determination of microbialactivity in leaf litter. � European Journal ofSoil Biology 34, 105-110.
SLAYTOR, M., 1992: Cellulose digestion in termi-tes and cockroaches: what role do symbiontsplay? � Comparative Biochemistry and Phy-siology A 103, 775-784.
SLEETER, T.D., BOYLE, P.J., CUNDELL, A.M., MIT-CHELL, R., 1978: Relationships between mari-ne microorganisms and the wood-boring iso-pod Limnoria tripunctata. � Marine Biology 45,329-336.
SMITH, V.J., SÖDERHÄLL, K., 1991: A comparisonof phenoloxidase activity in the blood ofmarine invertebrates. � Developmental andComparative Immunology 15, 251-261.
SOMA, K., SAITO, T., 1983: Ecological studies ofsoil organisms with references to the decom-position of pine needles � II. Litter feedingand breakdown by the woodlouse, Porcellio sca-ber. � Plant and Soil 75, 139-151.
STACKEBRANDT, E., PUKALL, R., 1999: Derivingtaxonomic decisions from 16S rDNA: acase study: Response to Althoff et al. (MarBiol 130: 529-536). � Marine Biology 130,529-536.
STEINERT, M., HENTSCHEL, U., HACKER, J., 2000:Symbiosis and parthogenesis: evolution of themicrobe-host interaction. � Naturwissen-schaften 87, 1-11.
STÖCKLI, H., 1990: Das Unterscheidungsvermö-gen von Porcellio scaber (Crustacea, Isopoda)zwischen Blättern einer Baumart, unter Be-rücksichtigung der makroskopisch sichtbarenVerpilzung. � Pedobiologia 34, 191-205.
STORCH, V., 1987: Microscopic anatomy and ul-trastructure of the stomach of Porcellio scaber(Crustacea, Isopoda). � Zoomorphology 106,301-311.
SUDHAUS, W., REHFELD, K., 1992: Einführung indie Phylogenetik und Systematik. � GustavFischer, Stuttgart.
SUMMERS, C.B., FELTON, G.W., 1994: Prooxidanteffects of phenolic acids on the generalistherbivore Helicoverpa zea (Lepidoptera: Noc-tuidae): Potential mode of action for pheno-lic compounds in plant anti-herbivore che-
mistry. � Insect Biochemistry and MolecularBiology 24, 943-953.
STRIGANOVA, B.R., 1967: Über die Zersetzung vonüberwinterter Laubstreu durch Tausendfüßlerund Landasseln. � Pedobiologia 7, 125-134.
SWAIN, T., 1979: Tannins and lignins, pp. 657-718.In: Herbivores their interactions with se-condary plant metabolites. I: The chemicalparticipants (Rosenthal, G.A., Janzen, D.H.,eds). � Academic Press, San Diego.
SWIFT, M.J., BODDY, L., 1984: Animal-microbialinteractions in wood decomposition, pp. 89-131. In: Invertebrate-microbial interactions(Anderson, J.M., Rayner, A.D.M., WaltonD.W.H., eds). � Cambridge University Press,Cambridge.
SZEGI, J., 1988: Cellulose decomposition and soilfertility. � Akadémiai Kiadó, Budapest.
SZLÁVECZ, K., 1993: Needle litter consumption bytwo terrestrial isopods, Protracheoniscus amoenus(C.L. Koch), and Cylisticus convexus (de Geer) (Iso-poda, Oniscidea). � Pedobiologia 37, 57-64.
SZLÁVECZ, K., MAIORANA, V.C., 1998: Supplemen-tary food in the diet of the terrestrial isopodPorcellio scaber Latr (Isopoda: Oniscidea). � Is-rael Journal of Zoology 44, 413-422.
TABACARU, I., DANIELOPOL, D.L., 1999: Contributi-on a la connaissance de la phylogénie des Iso-poda (Crustacea). � Vie et Milieu 49, 163-176
TAYLOR, E.C., 1982: Role of aerobic microbialpopulations in cellulose digestion by desertmillipedes. � Applied and EnvironmentalMicrobiology 44, 281-293.
TER-POGHOSSIAN, A., 1909: Beiträge zur Kennt-nis der Exkretionsorgane der Isopoden. �Zeitschrift für Naturwissenschaften 81, 1-50.
TEUBEN, A., 1991: Nutrient availability and inter-actions between soil arthropods and micro-organisms during decomposition of conife-rous litter: a mesocosm study. � Biology andFertility of Soils 10, 256-266.
TEUBEN, A., ROELOFSMA, T.A.P.J., 1990: Dynamicinteractions between functional groups of soilarthropods and microorganisms during de-composition of coniferous litter in micro-cosm experiments. � Biology and Fertility ofSoils 9, 145-151.
THOMPSON, J.N., 1982: Interaction and Coevolu-tion. � Wiley, New York.
THOMPSON, J.N., CUNNINGHAM, B.M., 2002: Geo-graphic structure and dynamics of coevolu-tionary selection. � Nature 417, 735-738.

166 MARTIN ZIMMER
THURSTON, C.F., 1994: The structure and func-tion of fungal laccases. � Microbiology 140,19-26.
TOKUDA, G., SAITO, H., WATANABE, H., 2002: Adigestive β-glucosidase from the salivaryglands of the termite, Neotermes koshunensis(Shiraki): distributrion, characterization andisolation of its precursor cDNA by 5'- and3'-RACE amplifications with degenerate pri-mers. � Insect Biochemistry and MolecularBiology 32, 1681-1689.
TOKUDA, G., YAMAOKA, I., NODA, H., 2000: Lo-calization of symbiotic clostridia in the mixedsegment of the termite Nasutitermes takasago-ensis (Shiraki). � Applied and EnvirnmentalMicrobiology 66, 2199-2207.
TRAGER, W., 1932: A cellulase from the symbi-otic intestinal flagellates of termites and ofthe roach, Cryptocercus punctulatus. � Biochemi-cal Journal 26, 1762-1771.
TREVES, D.S., MARTIN, M.M., 1994: Cellulose di-gestion in primitive hexapods: effect of inge-sted antibiotics on gut microbial populationsand gut cellulase levels in the firebrat, Thermo-bia domestica (Zygentoma, Lepismatidae). � Jour-nal of Chemical Ecology 20, 2003-2020.
TROJANOWSKI, J., 2001: Biological degradation oflignin. � International Biodeterioration andBiodegradation 48, 213-218.
TSUDA, M., TAN, H.M., NISHI, A., FURUKAWA, K.,1999: Mobile catabolic genes in bacteria. �Journal of Bioscience and Bioengineering 87,401-410.
UESBECK, M., TOPP, W., 1995: The effect of leaflitter, microorganisms and Collembola on thefood allocation of Oniscus asellus (Isopoda), pp.121-132. In: Terrestrial Isopod Biology (Ali-khan, M.A., ed). � Balkema, Rotterdam.
ULLRICH, B., STORCH, V., SCHAIRER, H., 1991: Bacte-ria on the food, in the intestine and on the fa-eces of the woodlouse Oniscus asellus (Crustacea,Isopoda). � Pedobiologia 34, 41-51.
VAN BORM, S., BUSCHINGER, A., BOOMSMA, J.J.,BILLEN, J., 2002: Tetraponera ants have gut sym-bionts related to nitrogen-fixing root-nodulebacteria. � Proceedings of the Royal Societyof London B 269, 2023-2027.
VANDERMEER, J., 1983: The evolution of mutua-lism, pp. 221-232. In: Evoutionary ecology(23rd Symposium of the British EcologicalSociety Leeds 1982) (Shorrocks, B., ed). �Blackwell, Oxford.
VAN ELSAS, J.D., TURNER, S., BAILEY, M.J., 2003:Horizontal gene transfer in the phytosphere.� New Phytologist 157, 525-537.
VAN WEMSEN, J., VERHOEF, H.A., VAN STRAALEN,N.M., 1993: Litter degradation stage as a pri-me factor for isopod interaction with mine-ralization processes. � Soil Biology and Bio-chemistry 25, 1175-1183.
VEIVERS, P.C., MÜHLEMANN, R., SLAYTOR, M.,LEUTHOLD, R.H., BIGNEL, D.E., 1991: Diges-tion, diet and polytheism in two fungus-gro-wing termites: Macrotermes subhyalinus Ramburand M. michaelseni Sjøstedt. � Journal of InsectPhysiology 37, 675-682.
VINOT, C., DURAND, P., LECLERCQ, M., BOURGEAY
CAUSSE, M., 1987: Studies on the biochemicalcomposition of Undaria pinnatida with a viewto its utilization in human feeding. � Scienceof Alimentation 7, 589-601.
WADA, M., AZUMA, N., MIZUNO, N., KUROKURA, H.,1999: Transfer of symbiotic luminous bacte-ria from parental Leiognathus nuchalis to theiroffspring. � Marine Biology 135, 683-687.
WÄGELE, J.W., 1989: Evolution und phylogeneti-sches System der Isopoda � Stand der Forschungund neue Erkenntnisse. � Zoologica 140, 1-262
WANG, J., DING, M., LI, Y.-H., CHEN, Q.-X., XU,G.-J., ZHAO, F.-K., 2003: Isolation of a multi-functional endogenous cellulase gene frommollusc, Ampullaria crossean. � Acta Biochimi-ca et Biophysica Sinica 35, 941-946.
WANG, Y., BRUNE, A., ZIMMER, M., in review:Molecular identification of hepatopancreaticsymbionts in the freshwater isopod, Asellusaquaticus (Crustacea: Asellota).
WANG, Y., STINGL, U., ANTON-ERXLEBEN, F., GEIS-LER, S., BRUNE, A., ZIMMER, M., 2004b: �Candi-datus Hepatoplasma crinochetorum�, a new,stalk-forming lineage of Mollicutes coloni-zing the midgut glands of a terrestrial isopod.� Applied and Environmental Microbiology,70, 6166-6172.
WANG, Y., STINGL, U., ANTON-ERXLEBEN, F., ZIM-MER, M., BRUNE, A., 2004a: �Candidatus Hepa-tincola porcellionum� gen. nov., sp. nov., a new,stalk-forming lineage of Rickettsiales coloni-zing the midgut glands of a terrestrial isopod.� Archives of Microbiology, 181, 299-304.
WANG, Y., STINGL, U., BRUNE, A., ZIMMER, M., inreview: Symbiotic bacteria in the midgutglands of isopods (Crustacea: Isopoda): phy-logeny and evolution.

The role of animal-microbe interactions in isopod ecology and evolution 167
Acta Biologica Benrodis 13 (2006)
WARBURG, M.R., 1965: The evolutionary significanceof the ecological niche. � Oikos 16, 205-213.
WARBURG, M.R., 1987: Isopods and their terrest-rial environment. � Advances in EcologicalResearch 17, 187-242.
WATSON, R.A., POLLACK, J.B., 2003: A computati-onal model of symbiotic composition in evo-lutionary transitions. � BioSystems 69, 187-209.
WHITE, J.J., 1968: Bioenergetics of the woodlouseTracheoniscus rathkei Brandt in relation to litterdecomposition in a deciduous forest. � Eco-logy 49, 694-704.
WIESER, W., 1966: Copper and the role of isopo-ds in degradation of organic matter. � Sci-ence 153, 67-69.
WIESER, W., 1968: Aspects of nutrition and themetabolism of copper in isopods. � Ameri-can Zoologist 8, 495-506.
WILKINSON, D.M., SHERRATT, T.N., 2001: Horizon-tally acquired mutualisms, an unsolved pro-blem in ecology? � Oikos 92, 377-384.
WILKINSON, T.L., DOUGLAS, A.E., 1996: The im-pact of aposymbiosis on amino acid metabo-lism of pea aphids. � Entomologia Experi-mentalis at Applicata 80, 279-282.
WILKINSON, T.L., ISHIKAWA, H., 2000: Injection ofessential amino acids substitutes for bacterialsupply in aposymbiotic pea aphids (Acyrthosi-phon pisum). � Entomologia Experimentalis etApplicata 94, 85-91.
WILKINSON, T.L., ISHIKAWA, H., 2001: On the func-tional significance of symbiotic microorga-nisms in the Homoptera: a comparative stu-dy of Acyrthosiphon pisum and Nilaparvata lu-gens. � Physiological Entomology 26, 86-93.
WILSON, B.A., SALYERS, A.A., 2003: Is the evolutionof bacterial pathogens an out-of-body experi-ence? � Trends in Microbiology 11, 347-350.
WOOD, D.A., 1980: Production, purification andproperties of extracellular laccase of Agari-cus bisporus. � Journal of General Microbiolo-gy 117, 327-338.
WOOD, S., GRIFFITHS, B.S., 1988: Bacteria associa-ted with the hepatopancreas of the woodliceOniscus asellus and Porcellio scaber (Crustacea, Iso-poda). � Pedobiologia 31, 89-94.
WOOD, T.M., GARCIA-CAMPAYO, V., 1990: Enzy-mology of cellulose degradation. � Biodegra-dation 1, 147-161.
YAMAMURA, N., 1993: Diversity and evolution ofsymbiotic interactions, pp. 75-82. In: Biodi-
versity: an ecological perspective (ABE, T.,LEVIN, S.A., HIGASHI, M., eds). � Springer, NewYork.
YAMIN, M.A., TRAGER, W., 1979: Cellulolytic acti-vity of an axenically-cultivated termite flagel-late, Trichomitopsis termopsidis. � Journal of Ge-neral Microbiology 13, 417-420.
YAN, Y., SMANT, G., STOKKERMANS, J., QIN, L.,HELDER, J., BAUM, T., SCHOTS, A., DAVIS, E.,1998: Genomic organization of four β-1,4-endoglucanase genes in plant-parasitic cystnematodes and its evolutionary implications.� Gene 220, 61-70.
ZACHARY, A., COLWELL, R.R., 1979: Gut-associa-ted microflora of Limnoria tripunctata in mari-ne creosote-treated wood pilings. �Nature 282, 716-717.
ZIMMER, M., 1998: Wechselwirkungen zwischenVertretern der saprophagen Bodenmakrofau-na und saprotrophen Mikroorganismen. �Cuvillier, Göttingen.
ZIMMER, M., 1999: The fate and effects of inge-sted hydrolyzable tannins in Porcellio scaber. �Journal of Chemical Ecology 25, 611-628.
ZIMMER, M., 2001: Why do male terrestrial iso-pods (Isopoda: Oniscidea) not guard fema-les? Animal Behaviour 62, 815-821.
ZIMMER, M., 2002a: Nutrition in terrestrial iso-pods (Isopoda: Oniscidea): an evolutionary-ecological approach. � Biological Reviews 77,455-493.
ZIMMER, M., 2002b: Is decomposition of woodlandleaf litter influenced by its species richness? �Soil Biology and Biochemistry 34, 277-284.
ZIMMER, M., BARTHOLMÉ, S., 2003: Bacterial en-dosymbionts in Asellus aquaticus (Isopoda) andGammarus pulex (Amphipoda), and their con-tribution to digestion. � Limnology and Oce-anography 48, 2208-2213.
ZIMMER, M., BRUNE, A., 2005: Physiological pro-perties of the gut lumen of terrestrial isopo-ds (Isopoda: Oniscidea) � adaptive to diges-ting lignocellulose? � Journal of Comparati-ve Physiology B, 175, 275-283.
ZIMMER, M., TOPP, W., 1997a: Does leaf litterquality influence population parameters of thecommon woodlouse, Porcellio scaber Latr., 1804(Crustacea: Isopoda)? � Biology and FertilitySoils 24, 435-441.
ZIMMER M, TOPP W. 1997b: Homeostatic re-sponses in the gut of Porcellio scaber (Isopo-da: Oniscidea) optimize litter degradation.

168 MARTIN ZIMMER
� Journal of Comparative Physiology B 167,582-585.
ZIMMER, M., TOPP, W., 1998a: Do woodlice (Isopo-da: Oniscidea) produce endogenous cellulases?� Biology and Fertility of Soils 26, 155-156.
ZIMMER, M., TOPP, W., 1998b: Microorganismsand cellulose digestion in the gut of Porcellioscaber (Isopoda: Oniscidea). � Journal of Che-mical Ecology 24, 1397-1408.
ZIMMER, M., TOPP, W., 1998c: Nutritional biolo-gy of terrestrial isopods (Isopoda: Oniscidea):Copper revisited. � Israel Journal of Zoology44, 453-462.
ZIMMER M, TOPP W. 1999: Relations betweenwoodlice (Isopoda: Oniscidea), and microbialdensity and activity in the field. � Biology andFertility of Soils 30, 117-123
ZIMMER, M., TOPP, W., 2000: Species-specific uti-lization of food sources by sympatric wood-lice (Isopoda: Oniscidea). � Journal of Ani-mal Ecology 69, 1071-1082.
ZIMMER, M., KAUTZ, G., TOPP, W., 1996: Olfac-tion in terrestrial isopods (Isopoda: Onisci-dea): the responses of Porcellio scaber to theodour of litter. � European Journal of SoilBiology 32, 141-147.
ZIMMER, M., KAUTZ, G., TOPP, W., 2003: Leaf lit-ter-colonizing microbiota: supplementaryfood source or indicator of food quality forPorcellio scaber (Isopoda: Oniscidea)? � Euro-pean Journal of Soil Biology 39, 209-216.
ZIMMER, M., GEISLER, S., WALTER, S., BRENDEL-BERGER, H., 2002c: Fluorescence in Asellusaquaticus (Isopoda: Asellota): a first approach.� Evolutionary Ecology Research 4, 181-187.
ZIMMER, M., PENNINGS, S.C., BUCK, T.L., CARE-FOOT, T.H., 2002b: Species-specific patternsof litter processing by terrestrial isopods (Iso-poda: Oniscidea) in high intertidal salt mars-hes and coastal forests. � Functional Ecology16, 596-607.
ZIMMER, M., DANKO, J.P., PENNINGS, S.C., DAN-FORD, A.R., ZIEGLER, A., UGLOW, R.F., CARE-FOOT, T.H., 2001: Hepatopancreatic endosym-bionts in coastal isopods (Crustacea: Isopo-da), and their contribution to digestion. �Marine Biology 138, 955-963.
ZIMMER, M., DANKO, J.P., PENNINGS, S.C., DANFORD,A.R., CAREFOOT, T.H., ZIEGLER, A., UGLOW, R.F.,2002a: Cellulose digestion and phenol oxidati-on in coastal isopods (Crustacea: Isopoda). �Marine Biology 140, 1207-1213.
Accepted: 18.08.2005

Morphometrische Untersuchungen am Gemüsespargel, Asparagus officinalis L. 169
Acta Biologica Benrodis 13 (2006)
Acta Biologica Benrodis 13 (2006): 169-184
Morphometrische Untersuchungen amGemüsespargel, Asparagus officinalis L.*
Morphometric studies in branches of Common Asparagus,Asparagus officinalis L.
FRIEDRICH ERNST BEYHL
Pfingstweidstraße 9, D-60316 Frankfurt/Main, Germany
Zusammenfassung: Morphometrische Untersuchungen an oberirdischen Zweigen des Gemüse-spargels, Asparagus officinalis L., zeigen, dass das Zweigsystem dieser Spezies selbstähnlich ist, wobeidessen fraktale Dimensionen je nach der Untersuchungsmethode (Zollstockmethode, magere Git-termethode und fette Gittermethode) variieren (0,76360 bzw. 1,07968 bzw. 1,62382).
Asparagus officinalis L., Gemüsespargel, Selbstähnlichkeit, fraktaler Charakter, fraktale Dimensionen
Summary: Morphometric studies of the aerial branches of the Common Asparagus, Asparagusofficinalis L., demonstrate that its ramification system is self-similar and that its fractal dimensionsvary depending on the method of determination (yardstick method: 0.76360, lean box method:1.07968, or fat box method: 1.62382, resp.).
Asparagus officinalis L., Common Asparagus, selfsimilarity, fractal character, fractal dimensions
1. Einleitung
Die Luftsprosse des Gemüsespargels (Aspara-gus officinalis L.), die aus den unterirdischenTrieben entstehen, zeichnen sich durch einigeBesonderheiten aus. Auffällig ist die anmuti-ge, äußerst regelmäßige Verzweigung, die angewisse Farnwedel, Moose und Nadelhölzeroder an Bambussprosse und Blütenrispen vonGräsern erinnert. Der Hauptzweig, der aus derErde herauswächst, kann eine Höhe von über2 m erreichen, wird meistens aber nur halb sohoch. Im unteren Abschnitt ist er unverzweigt,besitzt aber chlorophyllfreie, brakteenartigeSchuppenblätter, die je einen zum Boden hingerichteten Stachel besitzen. Erst von einerbestimmten Höhe an trägt er spiralig ange-ordnete Seitenzweige, die jeweils der Achsel
eines derartigen Schuppenblattes entspringen.Von der Spitze des Hauptzweiges in Richtungzur Basis nehmen diese Seitenzweige an Län-ge zu. Auch sie sind meistens wiederum ver-zweigt und tragen in den Achseln von Schup-penblättern ebenfalls Seitenzweige zweiterOrdnung; auch deren Längen nehmen von derSpitze des jeweiligen Seitenzweiges erster Ord-nung, von dem sie abzweigen, in Richtung zudessen Basis zu. Die Seitenzweige zweiter Ord-nung tragen häufig ebenfalls Seitenzweige drit-ter Ordnung. Die Tendenz zur Verzweigungdes Hauptzweiges und der Nebenzweigenimmt von deren Spitze in Richtung zu ihrerBasis zu, so dass die jeweiligen Zweigspitzenunverzweigt sind. Auch der unterste, basaleTeil des Hauptzweiges ist unverzweigt, trägtaber ebenfalls Schuppenblätter.
*Diese Arbeit ist Herrn Dr. G. KUNKEL, Vélez Rubio, in Anerkennung seines umfang- und erfolgrei-chen botanischen Schaffens auf den Kanarischen Inseln gewidmet.

170 FRIEDRICH ERNST BEYHL
An Nadelhölzer, insbesondere an Lärchen,erinnern die nadelförmigen, blattartigen Or-gane des Spargels, die an den Zweigen inBüscheln angeordnet sind, und zwar an den-jenigen Abschnitten der Zweige, die keine Sei-tenzweige erster oder zweiter Ordnung tra-gen. Es handelt sich hierbei allerdings nichtum Blätter, sondern um Flachsprosse (Phyl-lokladien), die anstelle von Blättern die Pho-tosynthese durchführen (COONEY-SOVETTS &SATTLER 1986). Diese Büschel sind ihrerseitsals extrem verkürzte Kurztriebsysteme zu deu-ten, welche die Phyllokladien tragen (BEYHL
2004). Auch für die anderen Vertreter derGattung Asparagus sind derartige in Büschelnangeordnete Phyllokladien typisch. Bei meh-reren Asparagus-Arten (z. B. A. ferox) sind diePhyllokladien nicht weich und biegsam, son-dern steif, stachelspitzig und stechend undmachen zusammen mit den Brakteenstachelndie Pflanze äußerst wehrhaft!
Eine weitere Eigentümlichkeit dieser Gat-tung sind die so genannten Beitriebe, diepaarweise an den Verzweigungsstellen gebil-det werden; je ein derartiger Beitrieb stehtrechts oder links neben dem zugehörigenSeitenzweig, und zwar mit diesem gemein-sam in der Achsel des tragenden Schuppen-blattes. Bei einigen Vertretern, z. B. bei A.falcatus, gleichen die Beitriebe verkleinertenZweigen, indem sie ebenfalls Phyllokladien-Büschel tragen; bei einigen weiteren Arten, z.B. bei A. densiflorus, stellen diese Beitriebe da-gegen die Blütenstände dar. Beim Gemüse-spargel selbst besteht jeder Beitrieb aus je ei-ner einzelnen Blüte, wobei er in der Mitteeine Einschnürung trägt, so als ob er selbstwieder gestielt wäre. Die Natur dieser Ein-schnürung, die außer bei anderen Asparagus-Arten auch bei einigen (nicht allen!) Arten derGattung Dracaena an deren Blütenstielen auf-tritt (BEYHL 2004), ist bislang noch nicht be-friedigend geklärt worden. Möglicherweisehandelt es sich um einen weiteren Perianth-kreis, ähnlich wie er für den so genanntenCalyculus bei einigen Arten von Tofieldia dis-kutiert wird (REMIZOVA & SOKOLOFF 2003),
der aber bereits in einem frühen Entwick-lungsstadium �steckengeblieben� ist unddaher nicht mehr in Erscheinung tritt.
An solchen Verzweigungsstellen des Spar-geltriebes, welche zunächst zwar Seitenzwei-ge, aber keine blütentragende Beitriebe ent-wickelt haben, können sich übrigens nachAblauf der Blütezeit Beitriebe ausbilden, dieähnlich wie bei A. falcatus verkleinerte Zwei-ge darstellen und mit ihren Phyllokladien-büscheln zur Assimilation beitragen. Ins-besondere geschieht dies, wenn in der Zwi-schenzeit Teile des Vegetationskörpers in-folge von Beschädigungen (vor allem Ab-brechen) ausgefallen sind und nicht mehrzur Assimilation zur Verfügung stehen.Auch bei anderen Vertretern der Gattung be-obachtet man hin und wieder die nachträgli-che Ausbildung von Beitrieben an bereitsfertig ausgebildeten Schossen und deren in-tensives Wachstum, wodurch die ursprüng-liche, regelmäßige Zweigarchitektur starkverändert werden kann. Auch können dieuntersten, unverzweigten Blattknoten nochzu späterer Zeit, wenn der Aufbau des übri-gen Zweigsystems bereits abgeschlossen ist,aus ihren schuppenartigen Blättern Seiten-zweige und Beitriebe bilden. Es handelt sichhierbei also offensichtlich um Regenerations-vorgänge, als kompensatorische Reaktionauf Einflüsse der Umwelt.
Die feinen, nadelförmigen Phyllokladienverursachen ein filigranes Aussehen, die Zwei-ge liefern ein dekoratives, sich wiederholen-des Muster und das gesamte Zweigsystemerscheint als eine recht einfache geometrischeFigur, nämlich als ein Kegel. Da sich die Zwei-ge im Wind biegen und die Seitenzweigeschräg nach oben gebogen sind, wirkt einAsparagus-Spross äußerst anmutig und gra-ziös. Offensichtlich kann das menschliche Ge-hirn alle diese geometrischen Signale auf rela-tiv einfache und sichere Weise verarbeiten, umrasch zu dem Ergebnis �schön, ansprechend�zu kommen; und dies geschieht intuitiv, ohnebewußte Reflexion. Diese Fähigkeit zum ra-schen Erfassen geometrischer Formen scheint

Morphometrische Untersuchungen am Gemüsespargel, Asparagus officinalis L. 171
Acta Biologica Benrodis 13 (2006)
dem Menschen inhärent zu sein (DEHAENE
et al. 2006).Insgesamt wirkt also ein voll entwickelter
Spargeltrieb ästhetisch äußerst ansprechend,zumal auch die Berührung damit einen ähn-lichen taktilen Reiz ausübt wie die mit einembuschigen Säugerschwanz. Deshalb heißtauch auf Türkisch wild wachsender Spargel�ebenso wie auch Schachtelhalm (Equisetum sp.)und Tannenwedel (Hippuris vulgaris), tilkikuy-rugu, ,Fuchsschwanz� (HAUENSCHILD 1996),wörtlich �dem Fuchs sein Schwanz� (von til-ki ,Fuchs� und kuyruk ,Schwanz�). Und nichtumsonst sind verwandte Arten wie A. seta-ceus (syn. A. plumosus) und A. densiflorus (syn.A. sprengeri) wegen ihrer ornamentalen Ver-zweigung zu beliebten Zierpflanzen gewor-den (PERRY 1974), und zwar sowohl alsTopfpflanzen als auch als so genanntes Bin-degrün in der Floristik. Dies hat wiederumzu einer Verordnung der EuropäischenKommission geführt (ANONYMUS 1976),durch welche die Qualitätskriterien der ver-schiedenen Handelsklassen und derenKennzeichnung sowie die Inverkehrbrin-gung solchen Asparagus-Schnittgrüns ge-nauestens geregelt werden.
Die regelmäßige Art der Verzweigung desSpargels legt die Vermutung nahe, es könnesich hierbei um ein selbstähnliches Objekt(vgl. PEITGEN et al. 1992; DEUSSEN 2003) mitfraktalen Eigenschaften (MANDELBROT 1991)handeln, wobei Selbstähnlichkeit nicht un-bedingt zugleich auch fraktalen Charakter be-deutet (SCHMITTER 1989; KRAUSE 2000). ZurKlärung dieser Frage wurde die vorliegendeUntersuchung durchgeführt.
2. Material und Methoden
2.1. Untersuchungsmaterial
Als Untersuchungsmaterial dienten Zweigevon Spargelpflanzen, die an Wegrainen undGartenzäunen und auf aufgelassenen Gär-ten, Steuobstwiesen und Weiden in einemländlichen Gebiet im östlichen Teil von Frank-
furt am Main im Stadtteil Enkheim wild (bzw.verwildert) wachsen. Von diesen Pflanzenwurden nach dem Zufälligkeitsprinzip Zwei-ge ausgewählt und dem Standort zur Unter-suchung entnommen.
2.2. Messung von Zweiglängen
An einem der vorliegenden Zweige wurdendie Längen sämtlicher Seitenzweige LS ersterund zweiter Ordnung sowie die AbständeLH ihres Ursprungspunktes von der Spitzedes zugehörigen Zweiges niederer Ordnungvermessen. Bei der zu vermutenden Selbst-ähnlichkeit sollte der rechnerische Zusam-menhang zwischen den Längen der Seiten-zweige LS und deren jeweiligen Abständenvon der Spitze des zugehörigen, sie tragen-den Zweiges LH durch die Gleichung
LS = a + c � LHoder, nach Einführung einer Hilfsgröße ä =-a/c, durch die damit äquivalente Gleichung
LS = c � (LH - ä)wiedergegeben werden. Er wurde nach denkonventionellen Methoden der Statistik(THER 1965; WEBER 1972) durch lineare Re-gression ermittelt.
2.3. Untersuchung von Phyllokladien
An weiteren der zur Untersuchung vorliegen-den Spargelzweige wurden an 200 Phyllokla-dien-Büscheln (von mehreren verschiedenenSpargelpflanzen) die Zahlen der Phyllokladienpro Büschel abgezählt. Außerdem wurdenaus den abgemessenen Längen von 114 Phyl-lokladien (ebenfalls von mehreren verschie-denen Spargelpflanzen) deren durchschnitt-liche Längen ermittelt.
2.4. Untersuchung zum fraktalenCharakter von Spargelzweigen
Zur Messung der fraktalen Dimension(en)des Spargeltriebes wurden zwei unterschied-liche Methoden (siehe bei BAUMANN et al.1994) angewandt, nämlich die �Gittermetho-
^

172 FRIEDRICH ERNST BEYHL
de� (�box method�, �grid method�) und die�Zollstockmethode� (�yardstick method�).FALCONER (1993) und DE VRIES (1997) be-zeichnen die Gittermethode als �Kästchen-zählmethode� und diskutieren ihre Vor- undNachteile. Bei der Gittermethode wurde aufzwei verschiedene Weisen, nämlich nach derso genannten �fetten� Analyse und der sogenannten �mageren� Analyse (JESTCZEMSKI
1995) verfahren (s. u.). Dieser Autor bezeich-net allerdings nicht die Analyse selbst mit die-sen beiden Ausdrücken, sondern gebrauchtfür die auf die zwei Abzählweisen erhaltenenErgebnisse die Bezeichnungen �fette� und�magere Fraktale�. Bei CORBIT & GARBARY
(1995) sowie bei BERNTSON & SOLL (1995)heißt die �magere� Analyse �skeletonized�.Zum Prinzip aller dieser Untersuchungsme-thoden siehe Abb. 1.
Hierzu wurden ein möglichst großer ter-minaler Abschnitt eines Spargeltriebes sowieein von diesem abgetrennter Seitenzweig aufein Millimeterpapier des Formats DIN A3montiert, dieses Blatt wurde fotokopiert unddie Fotokopie ausgewertet.
Bei der Gittermethode wurde auf der Fo-tokopie die Anzahl N derjenigen Quadrategezählt, auf denen ein Teil des Spargeltriebeszu sehen war; diejenigen Quadrate, die kei-nen Spargel enthielten, blieben beim Zählenunberücksichtigt. Die Zählungen wurdenmanuell durchgeführt und zwar iterativ fürQuadrate mit den Seitenlängen s von je 2,00cm, 1,50 cm, 1,00 cm, 0,50 cm und 0,10 cm.Im ersten Durchgang, der �fetten� Analyse(Abb. 1 oben), wurden alle die jenigen Käst-chen berücksichtigt, welche sowohl Zweigeals auch Phyllokladien enthielten, wobei au-ßerdem auch noch die Dicke der Zweige mit-berücksichtigt wurde. Im zweitem Durch-gang, der �mageren� Analyse (Abb. 1 Mitte),wurden dagegen nur diejenigen Kästchen zurZählung verwendet, die Zweige enthielten;Kästchen, in denen nur Phyllokladien ent-halten waren, wurden dabei nicht mitgezähltund auch die Dicke der Zweige blieb unbe-rücksichtigt.
Abb. 1: Bestimmung der fraktalen DimensionDS. Vergleich dreier Methoden. Oben: Gitter-methode: fette Analyse; Mitte: Gittermetho-de: magere Analyse; unten: Zollstockmetho-de. � Jeweils links: Auszählung bei großerSchrittweite s; rechts: Auszählung bei kleinerSchrittweite s � bei der Zollstockmethode (un-ten) erkennt man neben dem Baumskelett die(etwas dicker gezeichneten) Streckenzüge der�Messlatte�.Fig. 1: Determination of fractal dimension DS.Comparison of three methods. Top: box me-thod: fat analysis; middle: box method: lean(or: skeletonized) analysis; bottom: yardstickanalysis. � Left: counting with large step lengths; right: counting with small step length s �with the yardstick method (below), one per-ceives, besides the tree skeleton, the lines ofthe �yardstick�, represented by thick lines.

Morphometrische Untersuchungen am Gemüsespargel, Asparagus officinalis L. 173
Acta Biologica Benrodis 13 (2006)
Bei der Zollstockmethode (Abb. 1 unten),die bei HASTINGS & SUGIHARA (1996) �Stech-zirkelmethode� heißt, wurden mit einemZirkel, der als Messlatte (�Zollstock�) dien-te, die Längen der Spargeltriebe abgegriffenund dabei die Anzahl N der Abgriffe ge-zählt, und zwar der Einfachheit halber eben-falls auf der Fotokopie. Auch diese Zählun-gen wurden manuell durchgeführt und zwarfür Längen s von je 3,00 cm, 2,50 cm, 2,00cm und 1,50 cm.
Die bei diesen iterativen Zählvorgängen ge-wonnenen Anzahlen N wurden zur Bestim-mung der fraktalen Dimension DS nach demSatz:
log N = b - DS � log s(vgl. MANDELBROT 1991; PEITGEN et al. 1992;HERRMANN 1994; DEUSSEN 2003) dazu ver-wendet und nach konventionellen Methodender Statistik (THER 1965; WEBER 1972) dieKorrelation zwischen den Logarithmen vonN und den Logarithmen von s berechnet;denn DS stellt in einer graphischen Darstel-lung (Abb. 1) von log N auf der Ordinategegen -log s auf der Abszisse die Steigungder hierbei entstehenden Geraden (bzw. desgeraden Abschnitts der Kurve) dar (BAUMANN
et al. 1994; CASERTA et al. 1995; vgl. JONES etal. 1993; EIBENSTEIN 1995). Hierbei wurden
die NEPERschen Logarithmen, also diejeni-gen zur Basis 10, verwendet, weil man mitihnen einfacher rechnen kann als mit den na-türlichen Logarithmen, also denen zur Basise (der EULERschen Zahl).
Derartige Untersuchungen wurden so-wohl für den Seitenzweig, den terminalenTeil des Haupttriebes und für beide zusam-mengenommen durchgeführt. Für jedeAnalysenmethode wurden aus den dabei ge-wonnenen drei Einzelwerten die zugehöri-gen Mittelwerte berechnet. Beim Vergleichdieser für die drei Methoden erhaltenen dreiMittelwerte wurden mittels des t-Testes(THER 1965) die statistischen Signifikanzenermittelt.
3. Ergebnisse
3.1. Untersuchung an Phyllokladien
Die Anzahlen der Phyllokladien je Büschelsind in Abbildung 2 oben zusammengestellt.Die durchschnittliche Phyllokladienzahl proBüschel beträgt 6,5 ± 2,4 (n = 200 Büschel).Am häufigsten kommen dabei Büschel von6 Phyllokladien vor; am zweithäufigsten sindBüschel mit 4 und 5 und am dritthäufigstensolche mit 7 Phyllokladien.
Tab. 1: Korrelation von Zweiglängen bei Asparagus officinalis. Die Parameter a, ä, c und z gehörenzu den Gleichungen: LS = a + c � LH = c � (LH - ä) und log LS = + z � log (LH - ä).Tab. 1: Correlation of branch lengths in Asparagus officinalis. Parameters a, ä, c, and z belong to theequations: LS = a + c � LH = c � (LH - ä) and log LS = + z � log (LH - ä).
Word explanations: Hauptzweig: main branch; Seitenzweig A�E: side branches A�E; SeitenzweigAa: side branch of sidebranch A

174 FRIEDRICH ERNST BEYHL
Abb. 2: Verzweigungsgeometrie beim Gemüsespargel, Asparagus officinalis. Oben: Anzahl vonPhyllokladien pro Phyllokladienbüschel (n = 200 Büschel); Mitte: Länge der Seitenzweige LS inAbhängigkeit von ihrem Abstand LH von der Spitze des Hauptzweiges (Kreis: Nebenzweige ersterOrdnung gegen Hauptzweig; Viereck und Dreieck: Nebenzweige zweiter Ordnung gegen je einenzugehörigen Nebenzweig erster Ordnung); unten: Schematischer Aufbau eines Phyllokladienbü-schels (von links nach rechts: 4, 5, 6, oder 7 Phyllokladien pro Büschel).Fig. 2: Branching geometry with Asparagus officinalis. Top: Numbers of phylloclades per phyllocla-de bunch (n = 200 bunches); middle: Length LS of subbranches in dependence of their distancesLH from the apex of the main branch (circle: Subbranch of first order against main branch;square and triangle: subbranches of second order against the corresponding subbranch of firstorder); bottom: Schematical drawing of the architecture of a phylloclade cluster (from left tworight:: clusters of 4, 5, 6, or 7 phylloclades).

Morphometrische Untersuchungen am Gemüsespargel, Asparagus officinalis L. 175
Acta Biologica Benrodis 13 (2006)
Die durchschnittliche Länge der Phyllokla-dien beträgt 1,76 ± 0,32 cm (n = 114).
3.2. Stellung der Seitenzweige entlangder Achse
Die Seitenzweige entspringen dem jeweili-gen Hauptzweig in einer fünfzähligen Spi-rale. Allerdings stehen manchmal je zweiSeitenzweige so eng beieinander, dass manfast von �Gegenständigkeit� sprechenkönnte.
3.3. Zweiglängen
Zwischen den Längen LS der Seitenzweigeund ihren Abständen LH von der Spitzedes sie tragenden Zweiges besteht eine li-neare Beziehung, und zwar für alle unter-suchten Zweige (Tab. 1, Abb. 2 Mitte). Überdie in der Tabelle ebenfalls angegebenenGrößen ä und z siehe oben (Abschnitt 2.2)und unten (Abschnitte 4.1 und 4.3.4).
Die in Tabelle 1 angegebenen SeitenzweigeA bis E sind nicht von der Spitze des Haupt-zweiges her, sondern von dessen Basis hergerechnet � nebenbei bemerkt: Der Haupt-zweig enthielt noch mehr Seitenzweige,insgesamt 49. Der Seitenzweig Aa ist einerder Seitenzweige zweiter Ordnung vom Sei-tenzweig A.
3.4. Fraktaler Charakter von Spargel-zweigen
Die Analysenergebnisse sind in Tabelle 2 undAbbildung 3 zusammengestellt. Von denWerten von DS wurden bei jeder Analysenme-thode für die beiden Teilzweige und den ge-samten Zweige die jeweiligen Mittelwerte samtden dazu gehörigen Standardabweichungenberechnet. Diese sind in in der Tabelle wieder-gegeben und betragen 1,62382 ± 0,06395 (Git-termethode: �fette Analyse�), 1,07968 ±0,04964 (Gittermethode: �magere Analyse�)und 0,76360 ± 0,03521 (Zollstockmethode).
4. Diskussion
4.1. Untersuchung der Phyllokladienund Phyllokladienbüschel
Die festgestellte durchschnittliche Länge derPhyllokladien bedarf keiner Diskussion.
Aus dem Mittelwert der Anzahl von Phyl-lokladien je Büscheln von 6,5 ± 2,4 und dar-aus, dass am häufigsten 4, 5, 6 oder 7 Phyllo-kladien zu einem Büschel angeordnet sind,lässt sich schließen, dass ein Verzweigungs-modus des zum Büschel zurückgebildetenKurztriebes hauptsächlich mit 4, 5, 6 oder 7Phyllokladien zugrunde liegt: Damit wärenz. B. vier Verzweigungsweisen denkbar (Abb.
Tab. 2: Fraktale Dimensionen Ds bei Asparagus officinalis. Weitere Erklärungen siehe im Text.Tab. 2: Fractal dimensionen Ds in Asparagus officinalis. Further specifications: see text.
Word explanations: Gittermethode: box method; Zollstockmethode: yardstick method; fette Ana-lyse: fat analysis; magere Analyse: lean analysis; kleiner Zweig: small branch; großer Zweig: bigbranch; Gesamtzweig: both branches taken together.

176 FRIEDRICH ERNST BEYHL
Abb. 3: Diagramme zur Bestim-mung der fraktalen DimensionenDS von Spargelzweigen. Oben:Gittermethode: fette Analyse;Mitte: Gittermethode: magereAnalyse; unten: Zollstock-Ana-lyse. � Dreieck: Seitenzweig;Raute: Hauptzweig ohne Seiten-zweig; Kreis: Hauptzweig mitSeitenzweig. � Man beachte, dassbei der Zollstock-Analyse die Ab-szisse einen anderen Maßstab be-sitzt als bei den beiden Gitter-Analysen!Fig. 3: Diagrams for the determi-nation of fractal dimensions DSof asparagus branches. Top: boxmethod: fat analysis; middle: boxmethod: lean analysis; bottom:yardstick analysis. � Triangle:subbranch; diamond: mainbranch without subbranch; cir-cle: main branch together withsubbranch. � N. b.: the abscissascale of the yardstick analysis dif-fers from that of the grid analysis!
2 unten), bei der die aufs Äußerste verkürzteHauptachse des die Phyllokladien tragendenKurztriebes entweder1.) zwei als Phyllokladien ausgebildete Bei-
triebe sowie einen beitrieblosen, als Phyl-lokladium ausgebildeten Seitentrieb be-sitzt und in ein endständiges Phyllokla-dium übergeht (Phyllokladienzahl 4) oder
2.) zwei als Phyllokladien ausgebildete Bei-triebe sowie zwei ebenfalls als Phyllokla-dien ausgebildete Seitentriebe besitzt undin ein endständiges Phyllokladium über-geht (Phyllokladienzahl 5) oder
3.) zwei als Phyllokladien ausgebildete Bei-triebe sowie einen als Phyllokladiumausgebildete Seitentrieb mit zwei Phyllo-kladien als Beitrieben besitzt und in ein
endständiges Phyllokladium übergeht(Phyllokladienzahl 6) oder
4.) zwei als Phyllokladien ausgebildete Bei-triebe sowie einen als Phyllokladium aus-gebildeten Seitentrieb mit zwei Phyllokla-dien als Beitrieben und zusätzlich nocheinen beitrieblosen, als Phyllokladiumausgebildeten Seitentrieb besitzt und inein endständiges Phyllokladium übergeht(Phyllokladienzahl 7).
Dabei sind alle diese Zweigorgane extremverkürzt; d. h., sie haben bereits im rudimen-tären Zustand ihr Wachstum eingestellt undbestehen praktisch nur noch aus den end-ständigen, allein sichtbaren Phyllokladien.Dies alles ist zur Zeit allerdings nur eine un-belegte Arbeitshypothese.

Morphometrische Untersuchungen am Gemüsespargel, Asparagus officinalis L. 177
Acta Biologica Benrodis 13 (2006)
4.2. Anordnung der Zweige entlang derAchse
Die fünfzählige Spirale, welche die Seitenzwei-ge am jeweiligen Hauptzweig bilden, ent-spricht einer normalen fünfzähligen Phyllo-taxis. Allerdings bedarf das Phänomen, dassmanchmal je zwei Seitenzweige sehr engbeieinander stehen, noch einer gesondertenUntersuchung.
4.3. Selbstähnlichkeit und fraktalerCharakter von Spargelzweigen
4.3.1. Verzweigungsgeometrie
Die Länge der Seitenzweige LS steht mit ih-rem Abstand LH von der Spitze des sie tra-genden Triebes in einem mathematisch gutcharakterisierten Zusammenhang: Die linea-re Regressionsrechnung ergibt die oben (Ab-schnitt 2.2.) angeführte Beziehung zwischendem Haupttrieb und den zugehörigen Ne-bentrieben, die auf eine einfache lineare Ab-hängigkeit zwischen den beiden Längen LSund LH nach der Gleichung schließen lässt.Dies gilt sowohl für den Haupttrieb mit sei-nen Seitentrieben erster Ordnung als auch fürdie untersuchten Seitentriebe mit ihren jewei-ligen Seitentrieben zweiter Ordnung (Tab. 1),so dass sich die entsprechenden Kurven über-lagern, wenn man sie in dasselbe Diagrammeinträgt (Abb. 2 Mitte). Die Konstante c liegtin der Nähe von 0,5, also von ½. Das heißt,die Seitenzweige sind etwa halb so lang wieihr Abstand von der Spitze des Hauptzwei-ges. Lediglich bei einigen spitzennahen (unddeshalb kürzeren) Seitenzweigen und bei denebenfalls viel kürzeren Seitenzweigen höhe-rer Ordnung liegt c unter 0,5.
Die in Abschnitt 2.2. eingeführte Hilfsgrö-ße ä ist also derjenige Abstand eines Seiten-zweiges von der Spitze des Hauptzweiges,für den die folgende Ungleichung
LS > 0 für LH > ägilt und stellt den theoretisch kleinsten Ab-stand von der Spitze dar, bei dem es über-
haupt zur Bildung eines Nebenzweiges kom-men kann; denn Seitenzweige mit negativenLängen sind natürlich realiter nicht möglich.
LS ist also proportional zu (LH - ä) mit derProportionalitätskonstante c; somit lässt sichder in der Einleitung verwendete Vergleichdes Spargeltriebes mit einem Kegel mathe-matisch belegen.
Diese Proportionalität zwischen der Längeder Seitenzweige und ihrem Abstand von derSpitze (oder einem Punkt unterhalb der Spit-ze) des Hauptzweiges ist auch bei einigenanderen Pflanzenarten anzutreffen (BEYHL,unveröff.).
Aus Tabelle 1, in welcher die Werte von äfür die untersuchten Zweige sowie der zuge-hörige Mittelwert angegeben sind, ergibt sich,dass ä mit im Durchschnitt ungefähr 2,5 cmwesentlich geringer ist als der tatsächliche Spit-zenabstand, bis zu dem die Hauptzweigekeine sichtbaren Seitenzweige ausgebildethaben und der ungefähr 16 cm beträgt (Abb.2 Mitte). Diese Diskrepanz ist zurzeit nichtzu erklären. Auch bei den Zweigenden meh-rerer Bambusarten, einiger Chenopodiaceen,von Pyracantha coccinea sowie von Solidago ca-nadensis findet man derartige Unstimmigkei-ten (BEYHL, unveröff.); sie sind also nicht aufden Spargel beschränkt.
Aus dieser Verzweigungsgeometrie geht dieSelbstähnlichkeit klar ersichtlich hervor: Je-der Seitenzweig der n-ten Ordung ist ein ver-kleinertes Abbild des Zweiges der (n-1)-tenOrdnung.
4.3.2. Fraktale Dimensionen DS desTriebsystems
Die bei den drei Analysemethoden ermittel-ten drei Mittelwerte von DS (Tab. 2) unter-scheiden sich statistisch signifikant vonein-ander mit p < 0,005 (t-Test).
4.3.2.1. Gittermethode
Die Gittermethode anhand der �magerenAnalyse� (Abb. 3 Mitte) ergab eine fraktale

178 FRIEDRICH ERNST BEYHL
Dimension DS von 1,08, so dass in diesemFall DS ziemlich nahe bei 1,0 liegt.
Anhand der �fetten Analyse� (Abb. 3oben), bei der die Stiele mitsamt den Phyl-lokladien berücksichtigt wurden, ermitteltsich für DS der Wert von 1,624 ± 0,064(n = 3). Nach derselben Methode sind vonCASTREJÓN PITA et al. (2002) ähnliche Unter-suchungen an Asparagus setaceus (syn. A. plu-mosus) durchgeführt worden, allerdings mit-tels eines Computerprogramms. Im Gegen-satz zum im dreidimensionalen Raum an-geordneten Verzweigungssystem des Gemü-sespargels ist das Verzweigungssystem vonA. setaceus in einer Ebene ausgebreitet unddaher leichter zu untersuchen. Diese Auto-ren fanden für DS einen Wert von 1,778 ±0,052 (Mittelwert ± Standardabweichungvon n = 9 berechneten Einzelwerten). Die-ser Wert liegt in ungefähr der gleichen Grö-ßenordnung (d. h. zwischen 1,5 und 2), un-terscheidet sich aber von unserem (1,624± 0,064) statistisch signifikant mit p < 0,005(t-Test: THER 1965).
Hierbei würde eine Dimension von 1 be-kanntlich der euklidische Dimension einerLinie und eine Dimension von 2 der einerFläche entsprechen (vgl. MANDELBROT 1991).Beim Abzählen würde der Wert 2 stets dannerreicht werden, wenn alle Kästchen gezähltworden wären, weil nämlich dann das unter-suchte Objekt vollständig flächenfüllend wäre.Dass die tatsächlich gefundene DimensionDS kleiner als 2 ist, hängt mit der Lakunarität(MANDELBROT 1994) fraktaler Objekte, d. h.mit deren mangelhafter Flächenfüllung, zu-sammen; denn es sind ja in Wirklichkeit Ob-jekte, welche aus Linien, d. h. eindimensio-nalen Strukturelementen, zusammengesetztsind. Dennoch ist ihre fraktale Dimensionhöher; sie liegt zwischen 1 (der euklidischenDimension für lineare Strukturen) und 2 (dereuklidischen Dimension für Flächenstruktu-ren) und deutet somit die unvollkommeneFlächenausfüllung an. Das Zweigsystem vonA. setaceus ist einer Fläche besser angenähertals das von A. officinalis; deshalb ist für die
erste Art DS auch näher am Wert 2 als bei derzweiten.
Andererseits übersteigt DS bei der �mage-ren Analyse� geringfügig den Wert 1, so dasses sich nicht um ein �lineares System� han-deln kann. Auf jeden Fall liegt DS hierbei nahean der Dimension eines linearen Systems, inengem Zusammenhang mit der Auffassungdes Untersuchungsobjektes als aus �mage-ren Linien� bestehend.
In der vorliegenden Untersuchung istnicht berücksichtigt worden, dass sich man-che Spargelzweige auf der Fotokopie über-decken. Wäre dieser Effekt berücksichtigtworden, dann hätten die Kästchen, auf de-nen sich mehrere Zweige überdecken, mehr-fach gezählt werden müssen und dann wä-ren insgesamt mehr Kästchen gezählt wor-den. Der Wert von DS wäre dann möglicher-weise höher geworden. Auf diese Weise lie-ße sich der Unterschied zwischen den Wer-ten von DS bei CASTREJÓN PITA et al. (2002)und unserer �fetten� Untersuchung wennauch nicht erklären, so aber doch plausibelmachen.
Sowohl in unserer �fetten� Analyse als auchbei der Untersuchung von CASTREJÓN PITA etal. (2002) sind die Phyllokladien mitberück-sichtigt worden, wodurch beim Abzählen derCharakter einer �unvollkommenen Fläche�resultierte. Bei der �mageren Analyse� wur-den nur die Stiele der Pflanze ohne Phyllokla-dien berücksichtigt. Diese Stiele stellen ja �wenn man von ihrer Dicke absieht, sie alsoeinfach als �Striche�, also Linien im geomet-rischen Sinn, betrachtet und beim Abzählenbehandelt � eindimensionale Strukturen dar.Deshalb bezeichnet DEUSSEN (2003) sie als�Baumskelette�.
Die Differenzen zwischen dem von unsgemessenen �fetten� Wert und dem �mage-ren� Wert von DS enthält möglicherweiseaußer der Dicke der Stiele auch noch den Bei-trag der Phyllokladien zum Gesamtsystem.
Ferner ist zu berücksichtigen, dass die An-zahl der Verzweigungsebenen bei natürlichenObjekten stets endlich ist, im Gegensatz zu

Morphometrische Untersuchungen am Gemüsespargel, Asparagus officinalis L. 179
Acta Biologica Benrodis 13 (2006)
den in der Literatur als Beispiele für Fraktaleangeführten Figuren (MANDELBROT 1991). Beidiesen ist die Anzahl der Verzweigungsebe-nen nämlich stets unendlich und das fraktaleObjekt dadurch unendlich oft ineinander ver-schachtelt. Das bedeutet, dass bei natürlichenObjekten die fraktale Iteration nach einer end-lichen Zahl von Schritten abbricht und dassdamit kein ideal-mathematisches Verhaltenerwartet werden kann (s. bei BERNTSON &STOLL 1995). Beim Spargeltrieb haben wirsogar nur 2-3 Verzweigungs-, d. h. auch nur2-3 Iterationsebenen vorliegen.
Auffälligerweise ist die fraktale Dimensiondes kleinen Seitenzweiges etwas kleiner als diedes großen und des Gesamtzweiges (Tab. 2).Hier deutet sich eine Multifraktalität des Spar-geltriebes an, wie sie auch in anderen frakta-len Naturobjekten vorkommen kann, dortaber viel stärker ausgeprägt ist (CRAWFORD &YOUNG 1990; JESTCZEMSKI 1995; SMITH & LAN-GE 1998).
4.3.2.2. Gittermethoden mit Flächenab-zählung
Zum Abzählen der �Kästchen� (d. h. derQuadrate) könnte man anstatt deren Seiten-längen s auch deren Flächeninhalte f mit
f = s²zugrundelegen und bekäme beim Abzählendie gleichen Werte N heraus. Wegen dieserBeziehung zwischen f und s gilt für DF � log fund DS � log s das Folgende:
DS � log s = DF � log f =DF � log (s ²) = 2 DF � log s
Demnach ist DF halb so groß wie DS, wassich in der Tat bei zusätzlichen Untersuchun-gen, über die hier nicht berichtet wird, erge-ben hat.
Anstatt mit quadratischen �Kästchen�könnte man das auf Vorhandensein fraktalerElemente zu untersuchende Terrain natür-lich auch mit anders gestalteten Flächenele-menten auslegen, z. B. mit Rechtecken, Tra-pezen, Rauten oder Dreiecken. Diese müs-sen nur die folgenden zwei Bedingungen er-
füllen: Sie müssen das zu untersuchende Ter-rain lückenlos und nicht-überlappend bede-cken und insgesamt ein wohldefiniertes, sichwiederholendes Muster aus endlich vielen undeindeutig abzählbaren Flächenelementen er-geben. Bei der Verwendung von Kreisen istdies beispielsweise nicht der Fall: Man kannzwar eine Fläche bekanntlich auf zweierleiWeise mit Kreisen des gleichen Radius in dich-ter Packung bedecken, nämlich so, dass jederKreis jeweils vier andere berührt, oder so, dasser jeweils sechs andere berührt; aber in beidenFällen überdecken die entstehenden Musterdie Gesamtfläche nur lückenhaft � �lakunär,porös� � (im ersten Fall nur zu ca. 78,5 %, imzweiten Fall nur zu ca. 90,7 % der Gesamtflä-che). Beim Abzählen fraktalhaltiger Flächen-elemente könnten hierbei u. U. einige unbe-rücksichtigt bleiben, weil sie in keinem derKreise liegen. Andernfalls müsste man dieKreise so anordnen, dass sie einander über-lappen; dann müßten über den Überlap-pungsgrad Absprachen getroffen werden undes bestünde außerdem die Gefahr, fraktalhal-tige Flächenelemente mehrfach zu zählen.Zwar lässt sich eine gegebene Fläche lücken-los mit unendlich vielen Kreisen von verschie-denen Radien, den so genannten Apolloni-schen Kreisen, überdecken (KASNER & SUP-NICK 1943), wobei man übrigens ebenfalls einSystem mit fraktalem Charakter erhält (HECHT
2000); aber bei einem solchen System ist eineAbzählung der Elemente nicht möglich.
Eine weitere Möglichkeit ist die, das auffraktale Elemente zu untersuchende Terrainmit konzentrischen Kreisen zu überdeckenund dann die ringförmigen Flächen zwischenden Kreisen zum Abzählen zu verwenden;aber auch bei diesem Verfahren gerät man inSchwierigkeiten: Damit alle Flächen zwischenje zwei Kreisen gleich groß sind und damitdas Auffinden darin enthaltener fraktaler Ele-mente gleich wahrscheinlich ist, müssen dieDifferenzen zwischen je zwei Kreisradien mitwachsendem Radius gesetzmäßig abnehmen,so dass jedes dieser Flächenelemente die glei-che Fläche umfasst. Außerdem hängt das Er-

180 FRIEDRICH ERNST BEYHL
gebnis einer solchen Auswertung stark da-von ab, an welcher Stelle man den gemeinsa-men Mittelpunkt aller Kreise annimmt. Denk-bar wären daher Auswertungen mit demComputer, bei denen solche Abzählprozes-se mit mehreren (je nach verlangter Genau-igkeit verschieden vielen), willkürlich ausge-suchten Kreiszentren durchgeführt werden� Verfahren, welche nicht mehr �von Hand�durchgeführt werden können. Eine derarti-ge Analyse fraktaler Systeme soll mit demNamen Kreis-Radius-Methode bezeichnetwerden (vgl. u.).
4.3.2.3. Zollstockmethode
Bei der lineare Abzählung mit der sog. �Zoll-stockmethode� (Abb. 3 unten) ergab sich fürDS ein Mittelwert unter 1, nämlich von 0, 764± 0, 036 (siehe Tab. 2).
Meines Wissens ist die vorliegende Zoll-stockanalyse die erste ihrer Art, die nicht aneiner geschlossenen Kurve, d. h. einer (rea-len oder geometrisch erzeugten) �Insel�,durchgeführt worden ist, sondern an einemverzweigten Kurvensystem (einem �Baum�im Sinne der Mathematik). Wie aus Abbil-dung 1 unten hervorgeht, kann man überdie Modalitäten der Auszählung durchausverschiedener Meinung sein. So kann mansich z. B. fragen, ob Zollstock-Abgriffe, dieüber das Ende des Baumes hinausgehen,oder solche, die bei Verzweigungen zweimalvom gleichen Ausgangspunkt angesetztwerden, überhaupt zulässig sind; denndadurch werden ja einige Strecken des zuuntersuchenden Baumes als zu lang ange-nommen oder mehrfach gezählt. Im vorlie-genden Fall wurden die �Doppelzählung�und das �Hinausgreifen� zugelassen � auchbei der Gitteranalyse werden ja diejenigenGitterelemente, in denen ein im Vergleichzur Gitterelementfläche nur winziges Teildes zu untersuchenden Objektes liegt, seitjeher bei der Abzählung berücksichtigt, ohnedass man diese vergleichsweise übermäßigeBewertung korrigiert!
4.3.2.4. Räumliche Fraktalanalyse
Aus diesen unterschiedlichen Ergebnissenunserer Untersuchung (DS < 1 bei der Zoll-stockmethode und zwischen 1 und 2 bei derGittermethode) stellt sich die weitergehendeFrage, ob dann, wenn man beim Abzählennicht ein Linienraster (�Zollstock-Abgriffe�)oder Flächenraster (�Kästchen�, �boxes�),sondern ein dreidimensionales Raster (�Wür-felchen�, �cubes�, �Kugeln�) verwendenwürde, DS zwischen 2 und 3 zu liegen käme,sich also mehr dem Charakter eines �unvoll-ständigen Volumens� annäherte. Dies mussallerdings nicht notwendigerweise der Fallsein, ist aufgrund der Zählergebnisse an drei-dimensionalen Objekten (s. nächsten Ab-schnitt) jedoch zu erwarten. Für derartigeAnalysen ist einfaches manuelles Abzählenunmöglich und die hierfür bereits bestehen-de Methode, nämlich die Masse-Radius-Ana-lyse, die hauptsächlich für verzweigte Adern-systeme angewandt wird (SERNETZ et al. 1995)und im Prinzip der oben erwähnten Kreis-Radius-Methode ähnelt, war mir nicht zu-gänglich.
4.3.3. Andere fraktale Naturobjekte
Eine Reihe anderer biologischer Objekte be-sitzt fraktale Dimensionen zwischen 1,5 und3 (Tab. 3). Hierbei handelt es sich um Nerven,Adern, Tracheen, Wurzeln und Pilzmycelien,also um fädige Strukturen bzw. Röhrensyste-me, die allesamt räumlich verzweigte Bäumedarstellen, sowie um Thalli (TATSUMI et al.1989; FITTER & STICKLAND 1992; CORBIT &GARBARY 1993; JONES et al. 1993; CASERTA et al.1995; DONNELLY et al. 1995; JESTCZEMSKI 1995;CANALS et al. 1998; CROSS 1998; SMITH & LANGE
1998; KNIFFKY et al. 2004). Die Präparations-methoden des Untersuchungsgutes und diebei den Untersuchungen angewendeten Ana-lyseverfahren, die zu den in der Tabelle zu-sammengestellten Ergebnissen geführt haben,sind voneinander verschieden, so dass sich dieUnterschiede bei DS leicht erklären lassen.

Morphometrische Untersuchungen am Gemüsespargel, Asparagus officinalis L. 181
Acta Biologica Benrodis 13 (2006)
Bei den von mir berechneten Mittelwertenund Standardabweichungen sind in Klam-mern die Anzahlen n der berücksichtigtenEinzelwerte angegeben. Bei den von den be-treffenden Autoren selbst angegebenen Mit-telwerten ist dagegen weder die Anzahl nder Einzelwerte angegeben noch ist es be-kannt, ob es sich um die Standardabweichun-gen oder um die Abweichungen der Mittel-werte handelt.
4.3.4. Beziehung zwischen LS und LH
Die aufgefundene direkte Proportionalität vonLS zu LH (siehe Abschnitt 4.3.1) legte zur Kon-trolle dieses Sachverhaltes auch eine nochmali-ge nichtlineare Korrelationsrechnung zwischenden beiden Größen gemäß der Gleichung
LS = c � (LH - ä)z
nahe. Hierbei erhält man für z einen Mittel-wert von 1,2740 ± 0,2912 (Tab. 1), der sich
Tab. 3: Andere biologische Objekte mit fraktalem Charakter. Weitere Angaben im Text.Tab. 3: Further biological objects with fractal character. Further specifications see text.
Word explanations: Arterien: arteries; Autoren: authors; Bronchien: bronchi; dendritisches Neu-ron: dendritic neuron; Fruchtkörper: fruiting body; Gehirn: brain; Luftsproß: aerial branch; Lunge:lung; Mycel: mycelium; Mycelstrang: mycelium string; Niere: kidney; Rückenmark: spinal chord;Wurzeln: roots. Autoren: [a]: CANALS et al. 1998; [b]: CASERTA et al. 1995; [c]: CASTREJÓN et al. 2002;[d]: CORBIT & GARBARY 1995; [e]: CROSS 1998; [f]: DONNELLY et al. 1995; [g]: FITTER & STICKLAND
1992; [h]: JESTCZEMSKI 1995; [i]: JONES et al. 1993; [j]: KNIFFKY et al. 2004; [k]: SMITH & LANGE 1998;[l]: TATSUMI et al. 1989.

182 FRIEDRICH ERNST BEYHL
von 1 (dem Wert von z bei direkter Proportio-nalität) nicht statistisch signifikant unterschei-det (t-Test), auch wenn einige Einzelwertestark davon abweichen. Insgesamt kann manalso durchaus von einer linearen Beziehungzwischen LS und LH ausgehen.
4.4. Einfluss der Methode auf dasErgebnis
Es bestehen also je nach Zählmethode Un-terschiede in der Dimension DS des Untersu-chungsobjektes: Verwendet man eine lineareUntersuchungsmethode, dann erhält man dieDimension eines �unvollkommenen� (d. h.lakunären) quasi-linearen Objektes, benutztman hingegen eine quadratische Untersu-chungsmethode, dann erscheint das Objektals �unvollkommen� (d. h. lakunär) quasi-planar, und zwar mehr an die planare Dimen-sion angenähert, wenn man außer den Stie-len des �Baumskelettes� zusätzlich derenDicke sowie die Flächenausfüllung durch diePhyllokladien mit berücksichtigt.
Hier liegt also ein eklatantes Beispiel dafürvor, wie man je nach Untersuchungsmetho-de verschiedene Ergebnisse erzielen kann undwie dabei ein ganz unterschiedliches Bild desuntersuchten Sachverhaltes entsteht. Im Be-reich der Biofraktale ist dies ein Novum:Bislang war es nur in der Atom- und Quan-tenphysik sowie in der Ethnologie bekannt,dass der Beobachter mit seiner Untersuchungin das von ihm zu untersuchende Gesche-hen eingreift und je nach angewandter Unter-suchungsmethode zu unterschiedlichen Er-gebnissen kommt. Kritik an unbedachterInterpretation von nicht durch Gegenkon-trollen zu Ende geführten Untersuchungenerscheint also immer angebracht.
4.5. Zusammenhang der Verzweigungs-struktur mit den Wachstumsprozessen
Es hat nicht an Versuchen gefehlt, Verzwei-gungs- und Wachstumsstrukturen vonPflanzen mit der Hilfe von Computerpro-
grammen zu simulieren; sie haben zu denso genannten LINDENMAYER-Systemen (L-Systemen) geführt (LINDENMAYER 1986 a,b; PRUSINKIEWICZ & HANAN 1989; PRUSIN-KIEWICZ & LINDENMAYER 1996) und damitletzten Endes zu Computerdarstellungennicht nur von einzelnen Pflanzen (z. B.NEUREITHER 1992; PRUSINKIEWICZ & LIN-DENMAYER 1996), sondern sogar von gan-zen mit Pflanzen besetzten Landschaften(PRUSINKIEWICZ & LINDENMAYER 1996;DEUSSEN 2003).
Die Steuerungsphänomene der Ontogene-se, also des Wachstums und der Differenzie-rung von pflanzlichen � und natürlich auchvon tierischen � Organismen, und zwar so-wohl der Gesamtorganismen als auch ihrerTeile, Organe und Gewebe, führen letzt-endlich zu den morphologischen Strukturen,welche man an voll entwickelten Organismenbeobachtet (BELL 1994). Sie werden von phy-sikochemischen und biochemischen Prozes-sen (Diffusion, Hemmung, Genexpression,Enzymreaktion) bewirkt, welche offensicht-lich letztlich in den mathematisch beschreib-baren Strukturen fertig entwickelter Lebewe-sen (TURING 1952; BUIS & BARTHOU 1985;WIRTZ 1997; BEYHL, in Vorber.) und derenGrößenverteilungen innerhalb einer Ge-meinschaft (KOHYAMA & HARA 1989; WIRTZ
1997) resultieren. Die fraktalen Gesetzmä-ßigkeiten, welche man bei morphometrischerUntersuchung der fertig entwickelten ma-kroskopischen Organe erkennt � oder zuerkennen glaubt � und welche deren Auf-bau beschreiben, sind somit das Resultatvon wesentlich komplizierteren Vorgängender Morphogenese.
Danksagung
Der Autor dankt Prau Prof. U. DUKOVA,Frankfurt/Main, Herrn Prof. A. KALIS, Frank-furt/Main, Frau Prof. M. PIEPENBRING, Frank-furt/Main, und Herrn Prof. R. WITTIG, Frank-furt/Main, für interessante und wichtige Dis-kussionshinweise.

Morphometrische Untersuchungen am Gemüsespargel, Asparagus officinalis L. 183
Acta Biologica Benrodis 13 (2006)
Literatur
ANONYMUS, 1976: Verordnung (EWG) Nr. 1155/76 der Kommission vom 18. Mai 1976 zurÄnderung von Anhang II der Verordnung(EWG) Nr. 316/68 betreffend die Quali-tätsnormen für Asparagus-Schnittgrün. �Amtsblatt Nt. L 130 vom 19/05/1976 derEuropäischen Gemeinschaften, S. 26-30;ohne Ortsangabe.
BAUMANN, G., BARTH, A., NONNENMACHER, T.F.,1994: Measuring fractal dimensions of cellcontours: Practical approaches and their li-mitations, pp.182-189. In: Fractals in Biolo-gy and Medicine, Bd. 1 (Nonnenmacher, T.F.,Losa, G.A., Weibel, E.R., eds). � BirkhäuserVerlag, Basel.
BELL, A.D., 1994: Illustrierte Morphologie der Blü-tenpflanzen. � Verlag Eugen Ulmer, Stuttgart.
BERNTSON, G.M., STOLL, P., 1995: Correcting forfinite spatial scales of self-similarity whencalculating the fractal dimensions of real-world-structures. � Proceedings of the Roy-al Society of London B 264, 1531-1537.
BEYHL, F.E., 2004: Der Kubanische Drachen-baum. � Mitteilungen der Deutschen Dend-rologischen Gesellschaft 89, 121-130.
BUIS, R., BARTHOU, H., 1985: Propositionsd´analyse des formes végétales: Fonctions decroissance organique et architecture del´organisme, pp. 295-304. In: IVe Séminairede l´École de Biologie Théorique. � Éditionsdu CNRS, Paris.
CANALS, M., OLIVARES, R., LABRA, F., CAPUTO, L.,RIVERA, A., NOVOA, F.F., 1998: Caracteriza-ción de la geometría fractal del árbol bron-quial en mamíferos. � Revista Chilena de Ana-tomía 16, 237-244.
CASERTA, F., ELDRED, W.D., FERNANDEZ, E., HAUS-MAN, R.E., STANFORD, L.R., BULDEREV, S.V.,SCHWARZER, S., STANLEY, H.E., 1995: Deter-mination of physiologically characterizedneurons in two and three dimensions. � Jour-nal of Neuroscience Methods 56, 133-144.
CASTREJÓN PITA, J.R., SARMIENTO GALÁN, A.,CASTREJÓN GARCÍA, R., 2002: Fractal dimensi-on and self-similarity in Asparagus plumosus. �Fractals 10, 429-434.
COONEY-SOVETTS, C., SATTLER, R., 1986: Phyllo-clade development in the Asparagaceae: anexample of homoeosis. � Botanical Journalof the Linnean Society 94, 327-371.
CORBIT, J.D., GARBARY, D.J., 1995: Fractal dimen-sion as a quantitative measure of complexityin plant development. � Proceedings of theRoyal Society of London B 262, 1-6.
CRAWFORD, J.W., YOUNG, I.M., 1990: A multiplescaled fractal tree. � Journal of theoreticalBiology 145, 199-206.
CROSS, S., 1998: Fractal geometry of the humanrenal arterial tree in development, health anddisease, pp. 294-313. In: Fractals in Biology andMedicine Bd. 1 (Nonnenmacher, T.F., Losa,G.A., Weibel, E.R., eds). � Birkhäuser, Basel.
DEHAENE, S., IZARD, V., PICA, P., SPELKE, E., 2006:Core knowledge of geometry in an Amazoni-an indigene group. � Science 311, 381-384.
DEUSSEN, O., 2003: Computergenerierte Pflanzen.Technik und Design digitaler Pflanzenwelten. �Springer-Verlag, Berlin, Heidelberg, New York.
DE VRIES, H., 1997: Skaleninvarianz in verzweig-ten Strukturen. Inaugural-Dissertation (Carlvon Ossietzky-Universität Oldenburg). �Shaker, Aachen.
DONNELLY, D. P., WILKINS, M. F., BODDY, L., 1995:An integrated image analysis approach for de-termining biomass, radial extent and box-count fractal dimension of macroscopic my-celial systems. � Binary, Computing in Micro-biology 7, 19-28.
EIBENSTEIN, M., 1995: Charakterisierung fetalerPlazentaarterien von Rind und Schaf alsFraktale. Inaugural-Dissertation (UniversitätGießen). � Gießen.
FALCONER, K.J., 1993: Fraktale Geometrie. Ma-thematische Grundlagen und Anwendungen.� Spektrum Akademischer Verlag, Heidelberg,Berlin, Oxford.
FITTER, A.H., STICKLAND, T.R., 1992: Fractal cha-racterization of root system architecture. �Functional Ecology 6, 632-635.
HASTINGS, H.M., SUGIHARA, G., 1996: Fraktale. einLeitfaden für Anwender. � Spektrum Akade-mischer Verlag, Heidelberg, Berlin, Oxford.
HAUENSCHILD, I., 1996: Tiermetaphorik in türk-sprachlichen Pflanzennamen. � HarrassowitzVerlag, Wiesbaden.
HECHT, C., 2000: Appolonian (sic!) packing andfractal shape of grains improving geomecha-nical properties in engeneering geology. � Pureand Applied Geophysics 157, 487-504.
HERRMANN, D., 1994: Algorithmen für Fraktale undChaostheorie. � Addison-Wesley, Bonn, Paris,Reading.

184 FRIEDRICH ERNST BEYHL
JESTCZEMSKI, F., 1995: Multifraktale und globaleCharakterisierung fetter inhomogener Frak-tale am Beispiel des arteriellen Blutgefäßsys-tems ausgewählter Organe. Inaugural-Disser-tation (Universität Gießen). � Gießen.
JONES, C.L., LONERGAN, G.T., MAINWARING, D.E.,1993: A rapid method for the fractal analysisof fungal colony growth using image proces-sing. � Binary, Computing in Microbiology 5,171-180.
KASNER, E., & SUPNICK, F., 1943: The Apolloni-an packing of circles. � Proceedings of theNational Academy of Sciences of the US 43,378-384.
KNIFFKY, K.-D., PAWLAK, M., VAHLE-HINZ, C.,2004: Fractal dimensions and dendritic bran-ching of neurons in the somatosensory tha-lamus, pp. 221-229 In: Fractals in Biologyand Medicine, Bd. 1 (Nonnenmacher, T.F.,Losa, G.A, Weibel, E.R., eds). � Birkhäuser,Basel.
KOHYHAMA, T., HARA, T., 1989: Frequency Distri-bution of Tree Growth Rate in Natural ForestStands. � Annals of Botany 64, 47�57.
KRAUSE, U., 2000: Fraktale Dimension: Messenvon Komplexität, Beitr. 2 (ohne Seitenanga-ben). In: Fraktale in Hydrologie und Biologie(Matthies, M., ed.). � Inst. f. Umweltsystem-forsch. Univ. Osnabrück, Osnabrück (= Bei-träge des Inst. f. Umweltsystemforschg. derUnivers. Osnabrück, Bd. 19).
LINDENMAYER, A., 1968 a: Mathematical modelsfor cellular interaction in development. � I:Filaments with one-sided inputs. � Journalof Theoretical Biology 18, 280-299.
LINDENMAYER, A., 1968 b: Mathematical modelsfor cellular interaction in development. � II:Simple and branching filaments with two-sided Inputs. � Journal of Theoretical Biolo-gy 18, 300-315.
MANDELBROT, B.B., 1991: Die fraktale Geomet-rie der Natur. � Birkhäuser Verlag, Basel.
MANDELBROT, B.B., 1994: A fractal´s lacunarity,and how it can be tuned and measured, pp. 9-21. In: Fractals in Biology and Medicine, Bd.1 (Nonnenmacher, T.F., Losa, G.A., Weibel,R., eds.). � Birkhäuser Verlag, Basel.
NEUREITHER, M., 1992: Rechnerische Generie-rung von Baumstrukturen, pp. 57-63. In: Ver-
zweigungen (F. Belger, ed.). � Sonderfor-schungsbereich 230, Stuttgart.
PEITGEN, H.-O., JÜRGENS, H., SAUPE, D., 1992:Bausteine des Chaos. Fraktale. � Klett-Cot-ta, Stuttgart; Springer-Verlag, Berlin, Heidel-berg, New York.
PERRY, F., Hrsg., 1974: Simon & Schuster´s Com-plete Guide to Plants & Flowers. � Simonand Schuster, New York.
PRUSINKIEWICZ, P., HANAN, J., 1989: Lindenmayersystems, fractals, and plants. � Lecture Notesin Biomathematics 79, 1-120.
PRUSINKIEWICZ, P., LINDENMAYER, A., 1996: TheAlgorithmic Beauty of Plants. � SpringerVerlag, New York, Berlin, Heidelberg.
REMIZOVA, M., SOKOLOFF, D., 2003: Inflorescenceand floral morphology in Tofieldia (Tofieldi-aceae) compared with Araceae, Acoraceaeand Alismatales s. str. � Botanische Jahrbü-cher für Systematik, Pflanzengeschichte undPflanzengeographie 124, 255-271.
SCHMITTER, E.D., 1989: Fraktale Geometrie. Einepraktische Einführung auf dem Computer. �Hofacker Verlag, Taufkirchen.
SERNETZ, M., JUSTEN, M., JESTCZEMSKI, F., 1995:Dispersive fractal characterisation of kidneyarteries by three-dimensional mass-radius ana-lysis. � Fractals 3, 879-891.
SMITH, T.G., LANGE, G.D., 1998: Biological cellu-lar morphology � fractal dimensions, lacuna-rity and multifractals, pp. 30-49. In: Fractalsin Biology and Medicine, Bd. 2 (Losa, G.A.,Merlini, D., Nonnenmacher,T.F., Weibel,E.R., eds.). � Birkhäuser Verlag, Basel.
TATSUMI, J., YAMAUCHI, A., KONO, Y., 1989: Frac-tal analysis of plant root systems. � Annalsof Botany 64, 499-503.
THER, L., 1965: Grundlagen der experimentellenArzneimittelforschung. � WissenschaftlicheVerlagsgesellschaft, Stuttgart.
TURING, A., 1952: The chemical basis of mor-phogenesis. � Philosophical Transactions B237, 37-72.
WIRTZ, K.-W., 1997: Self-regulation of plants:From individual to aggregated dynamics, pp.257-267. In: Self-Organization of ComplexStructures. From Individual to CollectiveDynamics (Schweitzer, F., ed.). � Gordonand Breach Science Publishers, Amsterdam.
Angenommen: 01.08.2006

Die Webspinnen (Arachnida: Araneae) aus Beifängen des NSG Teverener Heide 185
Acta Biologica Benrodis 13 (2006)
Acta Biologica Benrodis 13 (2006): 185-193
Die Webspinnen (Arachnida: Araneae) aus Beifängendes NSG Teverener Heide (NRW, Kreis Heinsberg)
The spiders (Arachnida: Araneae) from bycatches of the naturereserve Teverener Heath (North Rhine-Westphalia, district Heinsberg)
MARTIN KREUELS
AraDet, Alexander-Hammer-Weg 9, D-48161 Münster, Germany;[email protected]
Zusammenfassung: In der vorliegenden Arbeit werden die Webspinnen aus Beifängen des NSGTeverener Heide (Nordrhein-Westfalen, TK 5002) vorgestellt. Die Daten aus dem Beifangmaterial,dessen wissenschaftlicher Wert hervorgehoben wurde, liefern wichtige Indizien für die weitere Schutz-würdigkeit des Naturschutzgebiets. Es wird dafür plädiert, das Vorhandensein von Beifängen publikzu machen, um alte Aufsammlungen wissenschaftlich nutzen zu können. Neben zwei Neunachweisen(Eperigone trilobata, Xysticus ferrugineus) für Nordrhein-Westfalen konnten von insgesamt 87 Arten aus19 Familien 18 �Rote Listen�-Arten nachgewiesen werden. Die beiden Neunachweise und Tegenariapicta, Atypus affinis, Callilepis nocturna, Enoplognatha mordax, Haplodrassus dalmatensis und Pardosa proximawerden näher besprochen. Die aktuelle Diskussion des Klimawandels und seine möglichen Auswir-kungen auf die Fauna werden kurz anhand einiger Arten (E. trilobata, Haplodrassus dalmatensis, Pardosaproxima, Pirata tenuitarsis, T. picta, X. ferrugineus) skizziert und es wird dazu aufgerufen, regionaleArbeiten auch unter dem Gesichtspunkt der Arealverschiebung einzelner Arten zu betrachten.
Nordrhein-Westfalen, NSG Teverener Heide, Webspinnen, Beifänge, Klimawandel
Summary: In the present work, by-catch spiders from the Teverener Heath National Park (NorthRhine Westphalia, TK 5002) are presented. The data from the by-catch material, the value ofwhich has already been emphasized, provide important evidence regarding the further necessity topreserve the national park. We advocate for publishing the existence of by-catches, in order tomake old collections scientifically usable. Apart from two new finds (Eperigone trilobata, Xysticusferrugineus) for North Rhine-Westphalia, 18 Red List species were found among a total of 87species belonging to 19 families. The two new finds as well as Tegenaria picta, Atypus affinis, Callilepisnocturna, Enoplognatha mordax, Haplodrassus dalmatensis and Pardosa proxima are discussed in furtherdetail. The present debate concerning climate change and its possible consequences on fauna arebriefly outlined on the basis of some species (E. trilobata, Haplodrassus dalmatensis, Pardosa proxima,Pirata tenuitarsis, T. picta, X. ferrugineus). Furthermore, when considering regional works, we appealnot to forget the viewpoint of a possible area shift in individual species.
Northrhine-Westphalia, nature reserve Teverener Heath, spiders, bycatches, climate change
den bisher meist vernachlässigten Stellenwertvon zoologischen Beifängen nachzudenken,aus dem die vorliegende Arbeit erstellt wur-de. Neben einer ersten arachnologischen Da-tengrundlage für ein bisher nicht untersuch-tes Gebiet kann diese Arbeit auch erste Daten
1. Einführung
Eine vorläufige Artenliste der Webspinnen(Arachnida: Araneae) der Teverener Heidestellt nicht nur einen wissenschaftlichen Wertan sich da, sondern gibt auch Anlass, über

186 MARTIN KREUELS
zu den potenziellen Auswirkungen einer Kli-maänderung auf die Webspinnen und derenbiogeographische Verbreitung in Nordrhein-Westfalen liefern. Es werden drei Themenangesprochen: a) Artenbestand und Schutz-status der Spinnen des NSG Teverener Hei-de, b) potenzielle Klimaveränderung undmöglicher Einfluss auf die Biogeographie derSpinnenfauna und c) der wissenschaftlicheWert der Beifänge. Da alle drei Themen ge-nügend Material für eigene Arbeiten ergeben,kann der nachfolgende Text die genanntenAspekte nur anreißen. Vielleicht ergeben sichaber daraus zielgerichtete Untersuchungen, dienähere und weitergehende Erkenntnisse er-geben.
Das Naturschutzgebiet (NSG) und Fau-na-Flora-Habitat-Gebiet (FFH) TeverenerHeide liegt im südwestlichen Teil Nordrhein-Westfalens an der Grenze zu den Niederlan-den. Politisch gehört es zum Kreis Heins-berg, TK25 5002, MesstischblattbezeichnungGeilenkirchen. Nach mehreren Gebietsvergrö-ßerungen seit der unter Schutzstellung 1977weist das NSG seit 1995 eine Größe von ca.448 ha auf. Es liegt zwischen 82,5 m und 120m ü. NN und ist Teil der Schwalm-Nette-Platte, einer naturräumlichen Untereinheit desNiederrheinischen Tieflands (GRUBER 1999).
�Das NSG Teverener Heide bildet mitseinen im Kernbereich [�] natürlich erhal-tenen Binnendünenfeldern und den darineingebetteten Heidemoorkomplexen sowieden durch die Sand-, Kies- und Tongewin-nung abgegrabenen Flächen im Norden,Westen und Süden des Gebietes eine [�]vielfältig gestaltete Landschaft. Währendauf den Flugsandgebieten überwiegendKiefernbestände und die Heidemoore dasLandschaftsbild bestimmen, sind im Nor-den die Grünlandflächen mit den künst-lich entstandenen zahlreichen Seen undTeichen sowie im Westen und Süden diegroßen Offenlandgebiete mit den Sandtro-ckenrasen- und Heideflächen die das Land-schaftsbild vielfältig prägenden Elemente.�(GRUBER 1999, S. 7-9)
2. Material und Methoden
Die vorliegenden Beifänge der Untersuchungstammen aus dem Jahr 2004. Zur Erfassungder Arthropoden wurden Bodenfallen mit 5%iger Essigsäure als Fang- und Konservie-rungsflüssigkeit und Tetanal als Entspan-nungsmittel eingesetzt.
Nachfolgend werden die Fallenstandortekurz beschrieben, ohne deren Lokalität ge-nau zu nennen. Diese Maßnahme, die vomKreis Heinsberg gewünscht wurde, dient derSicherung seltener Pflanzen- und Tierbestän-de. Geschützte Arten können somit nichtdirekt aufgefunden werden. Die Kurzbe-schreibungen der Fallenstandorte stammenvon den Mitarbeitern des Hauses Wildenrath,die für die Betreuung der Untersuchungsflä-chen zuständig sind.
Fallenstandorte (KRÖNING in litt.)- F1 alte Besenheide: renaturierte (aufgefüll-
te), ehemalige Sandgrube, Boden schott-rig-sandig, zwischen Calluna-Pflanzen dichtmit Moosen und Flechten bedeckt;
- F2 Sandabgrabung: am Rande einer fastvegetationsfreien Sandfläche, einseitig be-grenzt durch aufkommende Kiefern;
- F3 Sanddüne, Waldrand: am Rande einerfast vegetationsfreien Stelle im Übergangs-bereich zu einer Sukzessionsfläche mit auf-kommenden Kiefern;
- F4 junge Besenheide: renaturierte (aufge-füllte), ehemalige Sandgrube; Boden fein-sandig, bei starkem Regen Staufeuchte; zwi-schen den Calluna-Pflanzen offener Boden;
- F5 Sandmagerrasen: renaturierte (aufgefüll-te), ehemalige Sandgrube; Boden feinsan-dig-lößig; geringe Vegetationsdichte;
- F6 Heidemoor (Wiggelewak): offener Be-reich am Rande eines Moorweihers auftorfigem Untergrund, durchgehend feuchtbis nass;
- F7 Heidemoor (Kuhhute): Verlandungs-zone am Rande eines Moorweihers, aufabgestorbenem Torfmooskörper, durchge-hend feucht bis nass, oberflächlich abge-trocknet;

Die Webspinnen (Arachnida: Araneae) aus Beifängen des NSG Teverener Heide 187
Acta Biologica Benrodis 13 (2006)
- F8 Grünland-Bestand: Fläche wird alsMähweide genutzt; Arrhenateretalia, stellen-weise mit Übergang zum Molinetalia, mitPoa trivialis, Agrostis tenuis und Holcus lana-tus.In die Ergebnistabelle (Tab. 1) wurden nur
die Arten in Schutz- und Bestandskategorieneingeordnet, die in der Roten Liste 2005(KREUELS & BUCHHOLZ 2006) Berücksichti-gung fanden. Die beiden Neunachweise fürNRW, Eperigone trilobata und Xysticus ferrugi-neus, werden zwar ebenfalls Kategorien zu-geordnet, waren aber zum Zeitpunkt derDrucklegung der Roten Liste 2005 für NRWnoch nicht bekannt. Die Artenliste enthält nurTiere, die adult waren oder im juvenilen Sta-dium einer Art eindeutig zugeordnet werdenkonnten. Die Taxonomie richtet sich nachNorman PLATNICK Version 5.5 (2000-2005)(http://research.amnh.org/entomology/spiders/catalog/index.html).
3. Ergebnisse
Die arachnologischen Beifänge enthielten 87Arten aus 19 Familien. 67 Arten (77 %) wur-den nach der Roten Liste als nicht gefährdeteingestuft. Sieben Arten (8,1 %) wurden derKategorie 3, sechs Arten (6,9 %) der Katego-rie R und sieben Arten (8,1 %) der KategorieV zugeordnet.
Die aktuelle Bestandssituation (H) dernachgewiesenen Arten spiegelt sich als hete-rogenes Bild wider: 28 (32,2 %) Arten sindsehr häufig und 27 (31 %) Arten häufig, 16Arten (18,4 %) sind mäßig häufig, 10 Arten(11,5 %) sind selten und sechs Arten (6,9 %)gelten als extrem selten.
Die langfristigen Bestandstrends (Tr_l-t)zeigen, dass 51 Arten (58,6 %) eher gleichbleibend in ihrer Entwicklung einzustufensind, wogegen 25 Arten (28,7 %) eine mäßi-ge Abnahme und 11 Arten (12,6 %) eine deut-liche Zunahme aufweisen.
Die kurzfristigen Bestandstrends (Tr_s-t)spiegeln ein ähnliches Bild wie die langfristi-gen Bestandstrends wider. 50 Arten (57,5 %)
sind eher gleich bleibend einzustufen, elf Ar-ten (12,6 %) zeigen eine deutliche Zunahmeund 25 Arten (28,7 %) eine Abnahme, derenAusmaß aber nicht abzuschätzen ist. Die Be-standsdiagnose für Atypus affinis wurde miteinem �?� versehen, da bei dieser Art Nach-weisprobleme (s.u., Diskussion) bestehen.
In Tabelle 1 werden alle nachgewiesen Ar-ten in alphabetischer Reihenfolge mit ihrerIndividuenzahl und ihrem momentanenSchutzstatus aufgeführt.
4. Diskussion
Die Aufsammlung der Webspinnen der Te-verener Heide spiegelt nur ein kleines zufälli-ges und punktuelles Spektrum wider, sie lie-fert aber eine Grundlage dafür, dass das Ge-biet arachnologisch als wertvoll zu bewertenist, da bereits der kleine Ausschnitt bemer-kenswerte Arten beinhaltet, darunter zweineue Nachweise für Nordrhein-Westfalen.Unterrepräsentiert sind die Arten der höhe-ren Straten und die nicht laufaktiven boden-gebundenen Arten, da mit den eingesetztenBodenfallen nur laufaktive Arten erfasst wer-den. Üblicherweise durch Handfänge nach-weisbare Arten waren nur in einem geringenMaße unter den Beifängen vorhanden. Zuerwarten ist die doppelte bis dreifache An-zahl an Arten durch den Einsatz gezielterHandfänge, durch eine längere Untersu-chungsdauer (2-3 Jahre) und Nachsuchenspezialisierter Arten.
Beifänge: Wissenschaftliche Untersuchun-gen in einem begrenzten Gebiet sind meistauf eine konkrete Fragestellung hin konzi-piert. Für die Bearbeitung von zoologischenFragestellungen werden Daten zu bestimm-ten Klassen benötigt. Dazu zählen z.B. An-gaben zum Vorkommen einer Art, ihre Akti-vitätszeiten im Tages- oder Jahresverlauf unddie Zahl der Individuen einer Art. Um aussa-gekräftige Daten zu erhalten, werden insbe-sondere für den Bereich der Arthropodenautomatisch fangende Falleneinrichtungenwie z.B. Bodenfallen eingesetzt. Dies ist ein

188 MARTIN KREUELS
Tab. 1: Arachnologische Nachweise aus dem NSG Teverener Heide incl. ihres Schutzstatus. RoteListe 2005 (aktueller Gefährdungsstatus): 3 = gefährdet; R = extrem seltene, nicht zurückgehendeArten; V = Gefährdung anzunehmen; H (aktuelle Bestandssituation): es = extrem selten; s = selten;mh = mäßig häufig; h = häufig; sh = sehr häufig; Tr_l-t (langfristiger Bestandstrend): < mäßigerRückgang; = gleichbleibend; > deutliche Zunahme; Tr_s-t (kurzfristiger Bestandstrend): = gleich-bleibend; ↑ deutliche Zunahme; (↓) Abnahme, Ausmaß unbekannt; ? Daten ungenügend; Zahlen:Männchen/Weibchen.Tab. 1: Arachnological records of the nature reserve Teverener Heath and the status of endanger-ment. Red Data Book 2005 (present status of endangerment): 3 = endangered; R = extremely rare,not declining species; V = endangerment is to be assumed; H (current numbers): es = extremelyrare; s = rare; mh = moderately frequent; h = frequent; sh = very frequent; Tr_l-t (long termpopulation trend): < moderate decline; = unchanged; > distinct increase; Tr_s-t (short term popu-lation trend): = unchanged; ↑ distinct increase; (↓) decrease, extent unkown; ? insufficient data;numbers: males/females.

Die Webspinnen (Arachnida: Araneae) aus Beifängen des NSG Teverener Heide 189
Acta Biologica Benrodis 13 (2006)
Tab. 1: Fortsetzung.Tab. 1: Continued.
etabliertes Verfahren, um Daten in Form vonArten zeitsparend und effektiv über größereZeiträume zu gewinnen. Neben der zu bear-beitenden Tiergruppe werden aber, da die an-gewandte Fangmethode nicht selektierendfängig ist, weitere Klassen erfasst, die nichtim Zentrum der geplanten Untersuchung ste-hen. Für diese mitgefangenen Arten (Beifän-ge) steht der Bearbeiter vor der Entscheidung,
diese aufzubewahren oder zu verwerfen. DieAufbewahrung der Beifänge ist gerade füreine geringe Anzahl Bearbeiter, wie dies fürviele Tier- und Pflanzengruppen der Fall ist,eine sinnvolle und effektive Nachweisergän-zung (vgl. auch HANNIG et al. 2005), da diezeitaufwändigen Geländearbeiten wie regelmä-ßige Fallenleerungen und Strukturaufnahmenentfallen. Die mittels Beifang gewonnenen

190 MARTIN KREUELS
Daten können in Zeiträumen bearbeitet wer-den, die nur einen Bruchteil der Arbeitszeitumfassen, die eine Materialbearbeitung ein-schließlich der Geländearbeiten in Anspruchnehmen würde.
Neben den Vorteilen gibt es allerdings auchNachteile: So werden zoologische Untersu-chungen meist auf konkrete Taxa hin konzi-piert. Die zusätzlich gewonnenen Beifängestimmen aber nicht gänzlich mit der Frage-stellung des primären Bearbeiters überein undstehen somit im Hintergrund, so dass dasMaterial aus den Beifängen unvollständig seinmuss. Denn nur der kompetente Bearbeiterweiß, wo er zusätzlich suchen muss, um bei-spielsweise stationäre oder versteckt lebendeArten zu finden. Dennoch liefern Beifängewichtige Anhaltspunkte zu einem Habitat odereiner Region, mit denen zukünftige zielgerich-tete Arbeiten fortgesetzt werden können.
Klimawandel: Durch die in der vorliegen-den Arbeit genannten Neunachweise ist derBrückenschlag zur aktuellen Diskussion zumThema Klimawandel möglich. Zukünftigsollten faunistische Arbeiten diesem AspektBerücksichtigung schenken. Gerade lokaleArbeiten liefern Daten, die Indizien liefernkönnen, die die Auswirkungen eines Klima-wandels auf die heimische Fauna belegen (SH-WARTZ 2002). Nach den aktuellen Hypothe-sen wird es zu einer Temperaturerhöhung miteiner gleichzeitigen Zunahme der Nieder-schlagsmenge kommen (LEUSCHNER & SCHIP-KA 2004). Dies kann bei der Fauna, nebenden spezifischen physiologischen Einflüssen,die hier nicht diskutiert werden, zu einer bio-geographischen Veränderung führen (KUHL-MANN in litt.). Hierbei liegt der Schwerpunktauf Arten, die in einem definierten Suchraum� hier NRW � eine Arealgrenze aufweisen.
Bei sechs nachgewiesenen Arten des NSGTeverener Heide liegt eine mehr oder wenigedeutliche Arealgrenze innerhalb NRWs vor.Dazu gehören Eperigone trilobata, Haplodras-sus dalmatensis, Pardosa proxima, Pirata tenuitar-sis, Tegenaria picta, Xysticus ferrugineus. Es darfdanach vermutet werden, dass diese Arten
zu einer potenziellen Ausbreitung fähig sind,wenn geeignete klimatische Bedingungen diesbegünstigen. Bei einer Art (Eperigone triloba-ta) kann bereits eine nordwärts gerichteteWanderung (DUMPERT & PLATEN 1985; KREU-ELS im Druck) belegt werden. Ob auch dieanderen genannten Arten dazu fähig sind,müssen zukünftige Untersuchungen ergeben.Vielleicht lassen sich aber schon auf derGrundlage vorhandener älterer Beifänge wei-tere Hinweise erkennen. Hierfür wäre es hilf-reich zu wissen, wo weiteres zur Auswertunggeeignetes Material lagert.
Nachfolgend werden nun einige interessan-te Arten hervorgehoben. Bei den Angabenzum Verbreitungsbild in NRW undDeutschland muss berücksichtigt werden,dass die biogeographischen Kenntnisse zuden Arten aufgrund der Bearbeiterdichte sehrlückenhaft sein können.
Tegenaria picta: Das Vorkommen von T.picta in Deutschland ist auf den Westen be-grenzt. Die Ostgrenze ist eine gedachte Linieentlang der Westküste Schleswig-Holsteinssüdlich zu den Alpen (ARACHNOLOGISCHE GE-SELLSCHAFT e.V. 2006). Nordgrenze ihrer Ver-breitung ist das Mittelgebirge in Nordrhein-Westfalen. T. picta wird an warmen Stellenunter Steinen und in niedriger Vegetationgefunden (KREUELS & BUCHHOLZ 2006).
Atypus affinis: Die nachgewiesene Vogels-pinne, die zu einer von drei Arten (A. affinis,A. piceus, A. muralis) in Deutschland gehört,stellt die häufigste der drei Arten dar. A. affi-nis ist über das gesamte Bundesgebiet ver-breitet (HIEBSCH & KAUSE 1976; ARACHNOLO-GISCHE GESELLSCHAFT e.V. 2006). Alle heimi-schen Vogelspinnen weisen eine sehr versteck-te Lebensweise auf (KREUELS & BUCHHOLZ
2006). Den Großteil ihres acht- bis neunjäh-rigen Lebens verbringen sie in einem finger-dicken Fangschlauch, der sich 30-50 cm in denBoden hinein erstreckt und mit einer Längevon 10-30 cm der Bodenoberfläche aufliegt.Durch äußere Einflüsse veralgt und ver-schmutzt der ursprünglich weiße Fang-schlauch und gleicht sich farblich der Umge-

Die Webspinnen (Arachnida: Araneae) aus Beifängen des NSG Teverener Heide 191
Acta Biologica Benrodis 13 (2006)
bung an. Hinzu kommt, dass die Vogelspin-ne meist unter überhängenden Kanten, dievon Gräsern überwachsen sind, ihren Fang-schlauch einrichtet, so dass der Grasvorhangmanuell an geeigneten Standorten durchsuchtwerden muss. Da nur die adulten Männchenwährend der Paarungszeit den Wohnschlauchverlassen, wird die Vogelspinne nur selten inBodenfallen nachgewiesen (Nachweispro-blem), so dass der vorliegende Fund einemglücklichen Zufall zu verdanken ist.
Callilepis nocturna: Die Plattbauchspinne C.nocturna ist eine seltene Art, die aber in deraktuellen Roten Liste (KREUELS & BUCHHOLZ
2006) keinen Schutzstatus zugesprochen be-kommen hat. Sie ist eine Zeigerart für offene,vegetationsfreie Standorte. DOER (2000) gehtdavon aus, dass die ameisenjagende Spinneursprünglich auf Binnendünenkomplexeund offene Standorte begrenzt war, bevor sieweitere Habitate wie z.B. Bergehalden imRuhrgebiet besiedelte.
In Deutschland lässt sich ein deutlichesNord-Südgefälle in ihrer Häufigkeit feststel-len. Es konnte nicht geklärt werden, ob die-ser Sachverhalt in ihrer tatsächlichen nördli-chen Seltenheit begründet liegt oder ob diesmit einem Nachweisproblem durch zu weni-ge Arachnologen im nördlichen Raum be-gründet werden muss (ARACHNOLOGISCHE
GESELLSCHAFT e.V. 2006). Neben ihrem auf-fälligen Erscheinungsbild weist sie eine inte-ressante Nahrungsbiologie (Nahrungsspezi-alist) auf, da sie Ameisen in die Fühlerbasisbeißt und diese damit unmittelbar lähmt(HELLER 1976; DOER 2000).
Eperigone trilobata: Diese Spinne ist ein Ein-wanderer aus den USA (KREUELS im Druck).Die Art wurde in der Literatur für NRW bishernicht genannt. Die Teverener Heide und einweiterer Fund am Rhein bei Düsseldorf (BLICK
in litt. 2003) sind die bisher einzigen Nachweisefür NRW. Es ist anzunehmen, dass diese Liny-phiidae weiter in Richtung Norden wandernwird. Bisherige Nachweise gelangen am Randesonnenexponierter Trockenstandorte im Über-gangsbereich zu Strauch- und Baumbeständen.
Enoplognatha mordax: E. mordax ist bis aufden südöstlichen Teil Deutschlands zwar imgesamten Bundesgebiet verbreitet, allerdingswurde sie bisher immer nur selten nachge-wiesen (ARACHNOLOGISCHE GESELLSCHAFT
e.V. 2006). Ihre Verbreitung in NRW scheintauf die westlichen Landesteile begrenzt zusein (KREUELS & BUCHHOLZ 2006). E. mor-dax wurde in der Roten Liste für NRW inKategorie 3 eingestuft mit einer sich verklei-nernden Population (KREUELS & BUCHHOLZ
2006). Ihre Habitatansprüche werden in derLiteratur unterschiedlich angegeben. Nachden vorliegenden Daten aus NRW ist E.mordax sowohl auf offenem Sandboden alsauch in Feldrainen von landwirtschaftlichenNutzflächen zu finden. Sie scheint aber wär-mere Standorte mit einer hohen Sonnen-einstrahlung zu bevorzugen (KREUELS &BUCHHOLZ 2006).
Haplodrassus dalmatensis: Das Verbreitungs-gebiet von H. dalmatensis in Deutschland istschwierig zu interpretieren. Einerseits er-streckt sich ein Streifen von zahlreichen Nach-weisen aus dem Gebiet Saarland/Luxem-burg/Oberrheingraben nach Nordosten inden Berliner Raum hinein, andererseits wur-de die Art auf den friesischen Inseln und ver-einzelt auch an der Ems nachgewiesen(ARACHNOLOGISCHE GESELLSCHAFT e.V. 2006).Weitere küstengebundene Nachweise ausBelgien stammen von BONTE et al. (2000).Zusätzliche Nachweise werden für Deutsch-land auf der Nachweiskarte der Arachnologi-schen Gesellschaft (ARACHNOLOGISCHE GE-SELLSCHAFT e.V. 2006) nicht wiedergegeben.Für Nordrhein-Westfalen sind erst zweiNachweise bekannt, aus dem hier bearbeite-ten Untersuchungsgebiet und ein Nachweisaus dem Raum Warstein im nördlichen Sau-erland in der Nähe von Bahngleisen. Betrach-tet man den breiten Nachweisstreifen, der sichvon Südwest nach Nordost durch Deutsch-land erstreckt, so liegen die nordrhein-west-fälischen Nachweise an der Nordgrenze desVerbreitungsschwerpunkts in Deutschland(KREUELS & BUCHHOLZ 2006). H. dalmatensis

192 MARTIN KREUELS
scheint wärmebegünstigte, offene Standortezu bevorzugen. In der Teverener Heide stam-men alle Nachweise von Sandflächen. STAUDT
(2000) weist darauf hin, dass für Deutsch-land keine primäre Bindung dieser Art anSandstandorte zu beobachten sei. Dies wi-derspricht den aktuellen Nachweisen für diefriesischen Inseln und den Erfahrungen derbelgischen Kollegen.Pardosa proxima: Bisher sind aus Deutschlandnur wenige gesicherte Nachweise dieser Wolfs-pinne bekannt (KREUELS 2001). In NRWscheint die Art auf die westlichen Landesteilebeschränkt zu sein (KREUELS & BUCHHOLZ
2006). Gesicherte Nachweise aus Nordrhein-Westfalen stammen aus dem westlichen undnordwestlichen Stadtgebiet von Münster undvon der Teverener Heide.Xysticus ferrugineus: Diese Art ist der zweiteNeunachweis dieser Untersuchung für NRW(vgl. KREUELS & BUCHHOLZ 2006). In Deutsch-land wurde diese Art bisher nur wenige Malegefunden (entlang der Saale in Sachsen-An-halt und Ost-Thüringen, Brandenburg undentlang des Mains in Rheinland-Pfalz undBayern (ARACHNOLOGISCHE GESELLSCHAFT e.V.2006)). Der Fund in der Teverener Heide stelltaktuell den westlichsten Nachweis in Deutsch-land dar. Nach BLICK et al. (2004) wurdenNachweise aus Mitteleuropa von den Kolle-gen der folgenden Ländern gemeldet: Belgi-en und Niederlande, Tschechische Republik,Slowakei und Polen. Nachweise aus derSchweiz und Österreich fehlen. Ein Vorkom-men für Frankreich und Luxemburg (FINCH
in litt.) kann vermutet werden, lag aber zurManuskripterstellung nicht vor. ROBERTS
(1998) beschreibt X. ferrugineus für trockeneHeideflächen und für Flächen mit kalklieben-der Vegetation. Angaben aus Belgien bestäti-gen diese Habitatansprüche (ALDERWEIRELDT
& MAES 1997).
Danksagung
Herzlich bedanke ich mich beim Kreis Heins-berg für die Erlaubnis, die Daten publizieren
zu dürfen. Den Mitarbeitern der Naturschutz-station Haus Wildenrath, insbesondere FrauPETRA KRÖNING, danke ich für die Bereitstel-lung des Fallenmaterials und weitergehendergebietsbetreffender Informationen. HerrnDR. MICHAEL KUHLMANN (Münster) danke ichfür die Diskussion zum Thema Klimawan-del und ihre möglichen Auswirkungen aufdie heimische Fauna. Herrn DR. OLIVER FINCH
(Oldenburg) und Herrn THEO BLICK (Hum-meltal) danke ich für ergänzende Daten zurBiogeographie einzelner Arten. Herrn SASCHA
BUCHHOLZ danke ich für die kritische Durch-sicht des Manuskripts.
Literatur
ALDERWEIRELDT, M., D. MAES, D., 1997: De spin-nenfauna van �De Vallei van de Grote Beek�en van de �Immertse Heide� in Heppen-Leo-poldsburg (Limburg, Belgie). � Nieuwsbriefvan de Belgische Arachnologische Vereniging12, 53-63.
ARACHNOLOGISCHE GESELLSCHAFT e.V., 2006: Nach-weiskarten der Spinnen(tiere) Deutschlands.� Internet: www.arages.de, Stand 07.06.2006.
BLICK, T., BOSMANS, R., BUCHAR, J., GAJDO�, P.,HÄNGGI, A., van HELSDINGEN, P., RU� ICKA, V.,STAREGA, W., THALER, K., 2004: Checklisteder Spinnen Mitteleuropas. (Arachnida: Ara-neae). Version 1. Dezember 2004. � Internet:http://www.arages.de/checklist.html#2004_Araneae
BONTE, D., MAELFAIT, J.-P., HOFFMANN, M., 2000:Seasonal and diurnal migration patterns ofthe spider (Araneae) fauna of coastal greydunes. � Ekologia (Bratislava) 19, 5-16.
DOER, D., 2000: Zum Vorkommen der Platt-bauchspinne Callilepis nocturna (Linnaeus,1758) (Araneae: Gnaphosidae) in NRW. �Natur und Heimat 60, 83-88.
DUMPERT, K., PLATEN, R., 1985: Zur Biologieeines Buchenwaldes, 4. Die Spinnenfauna. �carolinea 42, 75-106.
GRUBER, K., 1999: Das Naturschutzgebiet Teve-rener Heide. � Heinsberg, Kreis Heinsberg.
HANNIG, K., TERLUTTER, H., LÜCKMANN, J., 2005:Die Laufkäferfauna (Col., Carabidae) ausge-wählter Kalkmagerrasen des oberen Diemel-tals. � Natur und Heimat 65, 113-122.

Die Webspinnen (Arachnida: Araneae) aus Beifängen des NSG Teverener Heide 193
Acta Biologica Benrodis 13 (2006)
HELLER, G., 1976: Zum Beutefangverhalten derameisenfressenden Spinnen Callilepis nocturna(Arachnida: Araneae: Drassodidae). � Ento-mologica Germanica 3, 100-103.
HIEBSCH, H., KAUSE, R., 1976: Zur Verbreitungund Lebensweise von Atypus affinis EICH-WALD, 1830 in der Sächsischen Schweiz(Araneae, Atypidae). � Faunistische Abhand-lungen des Museums für Tierkunde Dresden6, 69-88.
KREUELS, M., 2001: Zweiter Nachweis der Wolfs-pinne Pardosa proxima (C.L.Koch, 1847) fürDeutschland in Münster (Arachnida: Arane-ae, Lycosidae). � Natur und Heimat 61, 1-4.
KREUELS, M., BUCHHOLZ, S., 2006: Ökologie,Verbreitung und Gefährdungsstatus der Web-spinnen Nordrhein-Westfalens: Erste über-arbeitete Fassung der Roten Liste der Web-spinnen (Arachnida: Araneae) mit ergänzen-
den ökologischen Angaben, ihrer Verbreitungin Nordrhein-Westfalen und den neuen Vor-gaben des BfN zum Gefährdungsstatus. � Ver-lag Wolf & Kreuels, Havixbeck
KREUELS, M. im Druck: Die amerikanische Bal-dachinspinne Eperigone trilobata (EMERTON,1882) breitet sich in Europa aus. � Naturund Heimat.
LEUSCHNER, C., SCHIPKA, F., 2004: Vorstudie: Kli-mawandel und Naturschutz in Deutschland.� Bundesamt für Naturschutz, Bonn.
ROBERTS, M. J., 1998: Spinnengids. - Tirion, Baarn.SHWARTZ, M., 2002: High carbon dioxide levels
can retard plant growth, study reveals. � Stan-ford Report 5, 1-3.
STAUDT, A., 2000: Neue und bemerkenswerteSpinnenfunde im Saarland und angrenzen-den Gebieten in den Jahren 1996-99. � Ab-handlungen der Delattinia 26, 5-22.
Angenommen: 15.09.2006

194 MARTIN KREUELS
Rezension
ULLRICH, HERBERT (2004): Schädel-Schicksale historischer Persönlichkeiten. � VerlagDr. Friedrich Pfeil, Stuttgart. 336 Seiten mit 266 SW-Abbildungen. Gebunden, 21 x 24,5 cm.ISBN 978-3-89937-055-3. 38,-- Euro.
Was fasziniert so an der Frage, was mit einem Schädel einer historischen Persönlichkeitpassiert ist? Diese Frage wird wohl nie so ganz geklärt werden können, vielleicht, weil derSchädel das Gehirn, das Zentrum unseres Denkens, beherbergt. Es gibt eine berühmtePlastik Goethes, auf der er sinnierend den Schädel Schillers in der Hand hält und im Juli2007 ist dieses Thema immer noch aktuell, denn es gibt, vom Autor ausführlich dargestellt,zwei Schiller zugeschriebene Schädel. Nun hat man das Grab von Schillers Sohn Ernstgeöffnet, um anhand von DNA-Untersuchungen endgültig festzustellen, welcher der bei-den Schädel wirklich Friedrich Schiller gehört.Dabei ist es noch keineswegs sicher, ob überhaupt einer der beiden Schädel von Schiller ist.Im Mozarteum in Salzburg ruht z.B. ein Schädel von Wolfgang Amadeus Mozart, dernachweislich nicht von Mozart stammt. Mozart wurde bekanntlich in einem Massengrab fürArme bestattet und derjenige, der das Grab später öffnete, hatte die Auswahl aus zahlreichenSchädeln, es war ein Massengrab. Diese und weitere Schädelschicksale sind nach einemausführlichen Prolog in folgenden Kapitel aufgeteilt: Komponisten und Maler; Dichter undDenker; Philosophen und Gelehrte; Heilige und Geistliche; Ritter und Hoffräulein; Feldher-ren und Admirale; Adlige und Bürger; Fürsten und Grafen; Kaiser und Könige; Großfür-sten und Zaren. Das Buch ist gut geschrieben, geradezu spannend, und bringt auch Persön-lichkeiten, die nicht jedermann bekannt sind (auch mir nicht, Rez.). Eine kurzweilige Lektürefür alle, die sich mit den Schädelknochen unserer Vorfahren einmal auf eine besonders Artbeschäftigen wollen.
Harro Hieronimus, Solingen

Dentition in Ranodon sibiricus Kessler 195
Acta Biologica Benrodis 13 (2006)
Acta Biologica Benrodis 13 (2006): 195-217
Dentigerous bones, dentition and dental laminae inthe hynobiid salamander Ranodon sibiricus Kessler,
1866 (Amphibia, Urodela)
Zahntragende Knochen, Bezahnung und Zahnleisten bei demHynobiiden Ranodon sibiricus Kessler, 1866 (Amphibia, Urodela)
HARTMUT GREVEN1, NORBERT JÖMANN2, GÜNTER CLEMEN2
1Institut für Zoomorphologie und Zellbiologie der Universität Düsseldorf,Universitätsstraße 1, D-40225 Düsseldorf, Germany; [email protected]
2Institut für Evolution und Ökologie der Tiere der Universität Münster, Hüfferstraße 1,D-48149 Münster, Germany
Summary: Among Urodela the hynobiids appear to be an ancient lineage, which seem to haveconserved several plesiomorphic characters. Such ancestral characters might also include dentalpattern and dentition. Using scanning electron microscopy and histological sections, the toothsystems of larvae, juveniles and adults of the hynobiid salamander Ranodon sibiricus were docu-mented and briefly compared with the tooth systems of other Urodela. Adults had three (upperjaw arcade, lower jaw arcade, and the paired vomers), larvae had five tooth systems, those retainedin the adults and the paired palatines and coronoids (splenials of some authors) including theirdental laminae. The palatine of our youngest larva bore two rows of teeth, all other dentigerousbones bore a single row of teeth. Post-metamorphosed specimens largely retained the larval pat-tern, i.e. single rows of teeth and replacements posterior to the tooth rows in labial direction.Attachment of teeth was horizontal and more or less pleurodont depending on the particular boneand numbers of previous dentitions. Succession of differently shaped tooth generations followedthe typical urodele pattern from larval �non-pedicellate� monocuspids via several, larval �subpedi-cellate� monocuspids to metamorphosed pedicellate bicuspids in a particular order. Even in theadult, bicuspidity of teeth and shaping of the zone of division separating the crown from thepedicle varied considerably. In larvae with a delayed metamorphosis the palatines were edentulous.In the upper jaw arcade no gap was seen in the tooth row between the two premaxillae and thepremaxillae and maxillae indicating a single continuous dental lamina; in the lower jaw the edentu-lous gap between the dentaries argues for two dental laminae. Histological sections reveal clearlyseparated vomerine and palatinal dental laminae in larvae. Already before regression of the palatineand the coronoid, dental laminae of the two bones became regressive, the coronoid lamina laterthan the palatinal one. Dental systems as realized in larval and metamorphosed R. sibiricus arewidespread among Urodela and appear to represent a highly conserved character.
Urodela, dentition, tooth systems, larval and transformed teeth, delayed metamorphosis, dental laminae, conser-ved character
Zusammenfassung: Innerhalb der Urodelen gelten Hynobiiden als altertümliche Gruppe, dieeine Reihe plesiomorpher Merkmale konserviert zu haben scheinen; dazu gehören wahrscheinlichauch das Bezahnungsmuster und die Bezahnung selbst. Anhand von rasterelektronenmikroskopi-schen Aufnahmen und histologischen Schnitten beschreiben wir noch einmal die Zahnsysteme voneinige Larven, Juvenilen und Adulti des Hynobiiden Ranodon sibiricus und vergleichen diese kurz mitden Zahnsystemen anderer Urodelen. Die Erwachsenen haben drei (Oberkieferbogen, Unterkie-

196 HARTMUT GREVEN, NORBERT JÖMANN & GÜNTER CLEMEN
ferbogen, paarige Vomeres), Larven jedoch fünf Zahnsysteme, und zwar die, welche vom Adultennach der Metamorphose zurückbehalten werden sowie die paarigen Palatina und Coronoide (dieSplenialia mancher Autoren) jeweils mit ihren Zahnleisten. Die Palatina der jüngsten uns zurVerfügung stehenden Larven trugen zwei Zahnzeilen, alle übrigen zahntragenden Knochen jedochnur eine Zahnzeile. Metamorphosierte Exemplare behalten weitgehend das larvale Muster bei, d.h.eine Zahnzeile sowie Zahnersatz in labialer Richtung hinter der Zahnzeile Die Verankerung derZähne ist in Abhängigkeit von Knochen und der Anzahl vorangegangener Dentitionen horizontalsowie mehr oder weniger pleurodont. Die Abfolge unterschiedlich gestalteter Zahngenerationenfolgt dem typischen bei Urodelen verwirklichtem Muster von larvalen einspitzigen Zähnen ohneabgesetzten Sockel über larvale einspitzige Zähne, bei denen die Grenze zwischen Sockel undKrone nur angedeutet ist, bis zu metamorphosierten zweispitzigen Zähnen mit einem durch eineRingnaht deutlich abgesetzten Sockel. Bei den Adulti sind Ausprägung der Zweispitzigkeit undErscheinungsbild der Ringnaht sehr variabel. Bei zwei Larven, deren Metamorphose offenbarverzögert war, waren die Palatina bereits zahnlos. Der Oberkieferbogen hat eine durchgehendeZahnzeile und daher wahrscheinlich auch eine Zahnleiste, welche die beiden Prämaxillaria sowiedie Grenze zwischen Prämaxillare und Maxillare lückenlos begleitet, während zwischen den beidenDentalia im Unterkiefer eine solche Lücke in der Zahnzeile vorhanden ist. Die Zahnleiste entlangdes larvalen Vomer und Palatinum ist deutlich getrennt. Bereits vor dem metamorphosebedingtenAbbau des Palatinum und des Coronoid werden deren Zahnleisten regressiv, und zwar die desCoronoids später als die des Palatinum. Die für larvale und metamorphosierte R. sibiricus hierbeschriebenen Zahnsysteme sind weitgehend identisch bei einer großen Zahl von Urodelen ver-wirklicht. Es scheint sich um hoch konservierte Merkmale zu handeln.
Urodela, Bezahnung, Zahnsysteme, larvale und metamorphosierte Zähne, verzögerte Metamorphose, Zahnlei-sten, konservierte Merkmale
1. Introduction
Previous studies have examined the patternof dentition of representative members ofextant paedomorphic and post-metamor-phosed Urodela to show the variation of thispattern, the diversity of tooth shapes duringontogeny (summarized in CLEMEN & GRE-VEN 1994) and most thoroughly studied byDAVIT-BEAL et al. (2006) focusing on ontoge-netical changes of a single tooth family (forreview see also DAVIT-BEAL et al. 2007), thecourse of dental laminae (not considered sys-tematically as yet; see also DAVIT-BEAL et al.2007) and the site of tooth replacement. Someof these features are considered to be valua-ble tools for phylogenetical considerations fora long time (for review see CLEMEN & GRE-VEN 1994). According to these studies three�tooth systems� can be distinguished in post-metamorphosed salamanders: (1) the upperjaw arcade consisting of the paired premaxillaand maxillae and the accompanying conti-
nuous dental lamina; (2) the lower jaw arcadeformed by the paired dentaries accompaniedby a dental lamina that is interrupted in thesymphyseal zone; and (3) the vomerine sy-stem consisting of the two vomers, each ac-companied by a dental lamina. In larvae andsome paedomorphic forms, two further sys-tems are present: the palatinal portions of thepaired palatopterygoids and the paired coro-noids each with their dental laminae. Teethwere typically monocuspid and �non pedicel-late� in early larvae and bicuspid and pedicel-late in transformed specimens. In this case thetooth consists of a crown that articulates witha basal pedicle; crown and pedicle develop inthe enamel organ. These patterns appear ratherconstant among Urodela; deviations primarilyconcern the vomerine system and the numberof tooth rows established on the bones.
Among Urodela, the Hynobiidae are cha-racterized by several plesiomorphic traits, e.g.,external fertilization and a separate angularbone in the lower jaw (e.g. SCHMALHAUSEN

Dentition in Ranodon sibiricus Kessler 197
Acta Biologica Benrodis 13 (2006)
1968; HECHT & EDWARDS 1977; LARSON &DIMMICK 1993; GAO & SHUBIN 2001; LARSON
et al. 2003) and are considered as an ancientlineage of extant urodeles. Within Hynobiidaemorphological analyses (ZHAO et al. 1988) andmolecular data (LARSON et al. 2003) are incon-sistent, but more recently complete sequencesof mitochondrial genomes indicate conver-gent development of several morphologicalcharacters in different hynobiid lineage. Fur-ther, Onychodactylus appears to be the sister-group of all other living hynobiids (ZHANG
et al. 2006).Skull ontogeny including the dentigerous
bones and dentitional pattern of the endan-gered (see IUCN 2006) hynobiid species, Ran-odon sibiricus, is amazingly well studied (see par-ticularly the summarizing work by LEBEDKINA
1979, now available in an English translation,LEBEDKINA 2005). VASSILIEVA & SMIRNOV (2001)re-examined LEBEDKINA's (l.c.) stained andcleared specimens that cover a nearly comple-te ontogenetic series with regard to the succes-sion of teeth from hatchlings to adults.Authors revealed the sequence of structurallydifferent tooth generations, i.e. �non-pedi-cellate�, monocuspid teeth precede �subpe-dicellate� monocuspids in larvae that startactive feeding, pedicellate teeth in and aftermetamorphosis, and pedicellate bicuspidteeth after metamorphosis, a pattern knownfrom a variety of transforming Urodela (seealso GREVEN 1989; CLEMEN & GREVEN 1994;DAVIT-BEAL et al. 2007). More recently JÖMANN
et al. (2005) published some supplementingdata on the skull, in particular of larvae witha delayed metamorphosis, and included a fewnotes on dentition.
In the present study a survey is given onsome aspects of dentitional ontogeny in R.sibiricus, which is based on the literature andexamination of the material used in our pre-vious study on cranial ontogeny (JÖMANN etal. 2005). We show dentition of all develop-mental stages available to us by means ofscanning electron micrographs despite the riskof reiteration. Unlike VASSILIEVA & SMIRNOV
(2001), we focus on the apex and the zone ofdivision that separates the crown from thepedicle and on the attachment of teeth to thedentigerous bones. In addition, we includesome histological sections to follow the cour-se of dental laminae in the mouth roof andbriefly touch on possible plesiomorphic traitsin the tooth systems.
2. Material and methods
The material examined corresponds largelyto that used in a previous article (JÖMANN etal. 2005, herein also a more detailed characteri-sation and staging of the specimens accord-ing to the tables of LEBEDKINA 1979, 2005;KUZMIN & THIESMEIER 2001). We studied (nr.1) larva (38 mm TL, stage 16); (nr. 2) larva atthe beginning of metamorphosis (41 mmTL, stage 16-17); (nr. 3) larva in metamor-phosis (60 mm TL, stage 19); (nr. 4) larva inadvanced metamorphosis (only the head wasavailable); (nr. 5, 6) larvae with delayed meta-morphosis (67 and 73 mm TL, stage 19); (nr.7) juvenile (75 mm, stage 20); (nr. 8) sub-adult (95 mm TL, stage 22); and (nr. 9) adult(approx. 112 mm TL).
Scanning electron microscopy (SEM): Den-tigerous bones were excised from the stainedand cleared specimens used in the previousarticle (JÖMANN et al. 2005) and the tissue wasremoved by 1% pancreatin in tetraborate buf-fer. The cleaning process was regularly control-led and the time needed for cleaning variedconsiderably. We did not use any other mace-rating agent to save the delicate bones andteeth especially in the younger stages. Never-theless, cleaning did not satisfyingly succeedin all preparations. During this procedure re-placement teeth and largely resorbed teethwere lost. Further, specimens may have suf-fered of some demineralisation.
A single head of a larva in an advancedstage of development (palatine teeth werealready missing) (larva nr. 4 a; nearly stage 18;no further data available and not listed above),fixed in 2.5% glutaraldehyde in 0.1 M cacody-

198 HARTMUT GREVEN, NORBERT JÖMANN & GÜNTER CLEMEN
late buffer and embedded in paraplast wasfreed from paraplast. All preparations weredehydrated in an ascending ethanol series, cri-tical-point dried, sputtered with gold andexamined in a Hitachi-Scanning ElectronMicroscope.
Heads of a larva in metamorphosis (larva3 a; nearly stage 19; no further data availableand not listed above) and a larva with de-layed (90 mm TL; larva 6 a; nearly stage 19)fixed in glutaraldehyde or neutral formalde-hyde, respectively, were embedded in para-plast, sectioned transversally at 7 µm andstained with Azan or Trichrom-Goldner(ROMEIS 1989).
3. Results
3.1. Larva (nr. 1) (Figs 1a-f)
Toothed bones were premaxillae, vomers,palatines (=anterior palatinal portion of thepalatopterygoid; homology with the palatineof other vertebrates is doubtful, e.g. SCHOCH,1998), dentals and coronoids.
Premaxillae (Fig. 1 a), vomers (Fig. 1 b),dentaries (Fig. 1 d), and coronoids (Fig. 1f)bore a single row of teeth (monostichousdentition), whereas the palatine has two rows(bistichous dentition) (Fig. 1 c).
Teeth of all bones were ankylosed with abroad base. The basis of dentary teeth wasthickened lingually (Fig. 1 d). Attachment va-ried from nearly acrodont, i.e teeth are attachedmore or less horizontal on the surface of thebone (vomer, coronoid, palatine) to slightlypleurodont, i.e. teeth are attached by one sideto the inner surface of the bone (dentary, pre-maxilla). Openings to the pulp varied in sizeand shape ranging from a large opening (pre-maxilla: Fig. 1a), and several small accesses(vomer: Fig. 1 b, palatine: Fig. 1 c; dentary:Fig. 1 d; coronoid: Fig. 1 f).
Teeth were monocuspid, undivided andwere curved more or less lingually with theexception of coronoid teeth, which werestraight (Fig. 1 f).
3.2. Larva at the beginning of metamor-phosis (nr. 2) (Figs 2a-i)
Dentigerous bones as in the previous stage.The disintegrating anterior and median por-tion of the palatine was toothless (Fig. 2 f).
All bones including the palatine bore asingle row of teeth. Size and shape of teethvary considerably even on the same bone (Fig.2). On the premaxilla and dentary the mostposterior teeth were smaller and sturdy, moreanteriorly, teeth were slender and curved moreor less inwards (Figs 2 a, c, g).
Ankylosis has changed to a pleurodont con-dition (premaxilla: Fig. 2 a; dentary: Fig. 1 g).In the latter the bases of teeth was wider thanthe shaft (Fig. 2 c). On the other bones anky-losis was horizontal and teeth were broad-based. Premaxillary teeth had a larger pulpalaccess (Fig. 2 b) than vomerine, dentary andcoronoid teeth, which showed severalsmaller openings.
Teeth were monocuspid and undivided.Teeth showed at best a globular ring be-tween the basis and the crown (e.g. dentary:Fig. 2 g).
3.3. Larvae in metamorphosis (nr. 3)(Figs 3a-h)
Premaxillae, maxillae, vomers, and dentariesbore a single row of teeth. Palatines and coro-noids were largely disintegrated. The palatineswere entirely toothless. The coronoid showedresorption pits (Fig. 3 g). Ankylosis of teethlargely as in the previous stage. Dentition ofmaxillaries was pleurodont (Fig. 3 a), that ofthe vomers slightly pleurodont due to rem-nants of previous dentitions (Fig. 3 e).
Pulpal accesses were large and few (premaxilla,maxilla, dentary: Figs 3 a, d, f) or smaller andmore numerous (vomer: Fig. 3 e).
Teeth of the premaxilla, maxilla and den-tary were monocuspid and incipiently divided(Fig. 3 a, f), teeth of the vomer, however,monocuspid and undivided (Fig. 3 e). Thezone of division was more distinct lingually

Dentition in Ranodon sibiricus Kessler 199
Acta Biologica Benrodis 13 (2006)
Figs 1 a-f: Larva nr. 1. a Premaxillary teeth, lingual view, with broad basis and slight pleurodontattachment. b Vomerine tooth, lingual view. c Palatine with two rows of teeth, bases touch oneanother, caudal view. d Dentary, lingual view; attachment of teeth is more or less pleurodont andteeth are widely separated. e Dentary, medio-lingual view; broad-based tooth. f Straight coronoidtooth with signs of basal resorption.Abb. 1 a-f: Larve nr. 1. a Prämaxillarzähne, lingual, mit breiter Basis und leicht pleurodonterVerankerung. b Vomerzahn, lingual. c Palatinum mit zwei Zahnzeilen. Die basalen Abschnitte derZähne berühren sich fast, caudal. d Dentale, lingual; mehr oder weniger pleurodonte Verankerungder weit voneinander getrennen Zähne. e Dentale, medio-lingual, mit breitbasigen Zähnen.f Gerader Zahn auf dem Coronoid mit Resorptionsnarben an der Basis.

200 HARTMUT GREVEN, NORBERT JÖMANN & GÜNTER CLEMEN
Figs 2 a-i: Larva at the onset of metamorphosis (nr. 2). a Premaxillary, lingual view; withpleurodont dentition. The more posterior teeth (left side) differ in size and shape from the medianones (right side). b Tooth of the median premaxilla, lingual view, with pulp opening (asterisk). cPosterior sturdy premaxillary tooth; the basis is thickened (arrow). d Anterior outgrowth of thevomerine plate (asterisk); the missing portion of the vomerine plate (arrowhead) is an artefact. eVomerine tooth, medio-lateral view, with enamel cap; prospective dividing zone (arrow). f Mono-stichous dentition of the palatine, top view, caudal; toothless anterior (arrowheads) and mediandisintegrating portion (arrow); bony bridge between the palatine and the pterygoid (asterisk). gDentary, lingual view; posterior teeth (arrow) are smaller as the anterior and more curved. hCoronoid, lingual view, with horizontally ankylosed teeth. i Coronoid tooth, lingual view, withboundary between the basis and the bone (arrow).Abb. 2 a-i: Larve am Beginn der Metamorphose (nr. 2). a Prämaxillare, lingual, mit pleurodontverankerten Zähnen. Die hintersten Zähne (links) unterscheiden sich von den mittleren (rechts). bZahn aus der Mitte des Prämaxillare, lingual; Öffnung zur Pulpa (Stern). c Hinterer untersetzter

Dentition in Ranodon sibiricus Kessler 201
Acta Biologica Benrodis 13 (2006)
than labially (Figs 3 a, b). The surface of theteeth became rougher in the direction of thedividing zone (dentary, Fig. 3 f). The singletooth established on the coronoid was mono-cuspid and undivided; the shaft above thebroad basis appeared waisted, perhaps a resultof a partial demineralisation (Figs 3 g, h).
Pedicles were bulged at the lingual side, butwere not broadened significantly (premaxilla,maxilla, dentary). They were distinctly separ-ated from the zone of ankylosis and did nottouch each other. Teeth were bent inwards,i.e. lingually (Figs 3 c and d), or were nearlystraight (dentary: Fig. 3 f).
3.4. Larva in an advanced stage of meta-morphosis (nr. 4 and 4 a) (Figs 4 a-i)
Number of rows, ankylosis of teeth andpulpal accesses as in the previous stage; coro-noids regressive and without teeth.
In the upper jaw dentition was continuouswithout a gap in the symphyseal zones be-tween the premaxillae and between prema-xilla/maxilla; vomers did not have contacteach other; their partes dentales were situatedin the midst of the bone and their laterocau-dal edges lied at the level of the choana (thatmark the lateral boundary of each vomer)(Figs 4 a, e-g).
Teeth were either bicuspid and divided (pre-maxilla: Figs 4 b, c), monocuspid and divided(maxilla: Fig. 4 d), monocuspid and inci-piently divided posteriorly or bicuspid anddivided anteriorly (dentary: Figs 4 h, i) ormonocuspid and undivided (vomer: Figs 4 e-g). On the dentary, the anterior teeth nearthe symphysis were larger and bicuspid. Pos-
teriorly teeth became smaller and were mono-cuspid and broad-based (Fig. 4 h, right side).
Each tooth had a large dominant lingualopening to the pulp (Fig. 4 d, h) or a largerand several small (vomer, Figs 4 e-f). Teethwere curved inwards; in the bicuspids, twocusps were of nearly equal size (Figs 4 c, i).
3.5. Larva with delayed metamorphosis(nr. 5 and 6) (Figs 5 a-l)
Premaxillae, maxillae, vomers, dentaries andcoronoids with a single row of teeth; pala-tine regressive without teeth. Dentition waspleurodont (premaxillae, maxillae, dentaries),slightly pleurodont (on the vomer; the parsdentalis was now situated in the posterior thirdof the vomer: Figs 5 e,d f) or broad-basedand nearly horizontal (coronoid: Fig. 5 k, l).Large pulpal accesses were seen in the teethattached in a pleurodont condition andsmaller and more numerous accesses in teethattached horizontally (Fig. 5 g). Teeth were allmonocuspid with an dividing zone indicatedby a globular surface (maxilla: figs 5 d). Trans-versal ridges (premaxilla: Fig. 5 a), incisions(dentary: Fig. 5 j) or a waist (Fig. 5 k, l) mightbe a result of partial demineralisation..
3.6. Juvenile specimen (nr. 7) (Figs 6 a-i)
Number of tooth rows, ankylosis of teethand pulpal accesses as in larva nr. 4 (seeabove); coronoids no longer present. Vomer-ine teeth shifted near the posterior edge ofthe bone (Fig. 6 f).
Teeth of all dentigerous bones were bi-cuspid and divided (premaxilla, maxilla: Figs
Prämaxillarzahn mit verdickter Basis (Pfeil). d Vorderer Bereich der Vomerplatte (Stern); ein Teilder Vomerplatte (Pfeilkopf) ist präparationsbedingt nicht mehr vorhanden. e Vomerzahn, medio-lateral, mit Schmelzkappe und prospektiver Ringnaht (Pfeil). f Einzeilige Bezahnung des Palati-num, Aufsicht; zahnloser vorderer (Pfeilköpfe) und mittlerer im Abbau begriffener Teil (Pfeil);Knochenbrücke zwischen Palatinum und Pterygoid (Stern). g Dentale, lingual; die hinteren Zähne(Pfeil) werden kleiner und sind gekrümmter. h Coronoid, lingual, mit horizontal verankertenZähnen. i Zahn auf dem Coronoid, lingual, mit deutlicher Grenze zwischen Basis und Knochen(Pfeil).

202 HARTMUT GREVEN, NORBERT JÖMANN & GÜNTER CLEMEN
Figs 3 a- h: Larva in metamorphosis (nr. 3). a Premaxillary, lingual view; attachment of the linguallycurved teeth is slightly pleurodont; teeth have a large pulpal access each (arrow). b Incipient dividingzone (arrowhead) of a premaxillary tooth, lingual view. c Apex of a maxillary tooth. d Maxillary teethwith large pulpal accesses (arrows); processus facialis maxillaris (asterisk). e Vomer, lingual view; labialremnants of previous dentitions (arrows); pars palatina (asterisk). f Dentary, lingual view; putativedivided (arrows) teeth are attached in a pleurodont condition; large pulpal openings (asterisks). g Singlecoronoid tooth, lingual view; resorption of the coronoid (sp) from posterior (below) to anterior (on thetop); prearticular (pa). h Detail of figure g. Straight �non-pedicellate� coronoid tooth.Abb. 3 a-h: Larve in der Metamorphose (nr. 3). a Prämaxillare, lingual; die Zähne sind lingualgekrümmt, pleurodont verankert und haben einen großen Zugang zur Pulpa (Pfeil). b Sich bildendeRingnaht (Pfeilkopf) eines Prämaxillarzahnes, lingual. c Spitze eines Maxillarzahns. d Maxillarzahnmit großen Pulpazugängen (Pfeile); processus facialis maxillaris (Stern). e Vomer, lingual: labiale Restefrüherer Dentitionen (Pfeile); pars palatina (Stern). f Dentale, lingual; die geteilten Zähne sind pleu-rodont verankert und haben große Pulpaöffnungen (Sterne). g Einzelner Zahn auf dem Coronoid,lingual; Resorption des Coronoids (co) von hinten (unten) nach vorn (oben); Präarticulare (pa). hAusschnitt aus Abbildung g. Gerader nicht zweigeteilter Zahn auf dem Coronoid.
Figs 4 a-i: Larvae in an advanced stage of metamorphosis (nr. 4, 4a); preparations with (a, c, g) andwithout (b, d-f, h, i) soft tissue. a Premaxillae and vomers, symphyseal region (arrow on the top) withcontinuous dentition, lingual view; vomers (arrowhead); gap between vomers (arrow); choana (aste-

Dentition in Ranodon sibiricus Kessler 203
Acta Biologica Benrodis 13 (2006)
risk). b Premaxillary teeth with incipient bicuspidity (arrowheads) and zone of division (arrow). c Abicuspid (arrow) and monocuspid tooth side by side in the upper jaw, frontal view. d Maxilla withlingually curved teeth attached in pleurodontous condition; processus facialis maxillaris (asterisk). eVomer; pars dentalis with remnants of previous dentitions (arrow) in the middle; vomerine plate (arrow-head) and pars palatina (asterisk). f Monocuspid vomerine teeth; pulpal access (asterisk); Howshiplacunae in the pedicle (arrow). g Monocuspid vomerine teeth penetrating the oral epithelium; taste buds(arrowheads), lingual. h Dentary, lingual view; the posterior teeth (arrows) are monocuspid, the anteriortooth (arrowhead) is bicuspid. i Apex of an anterior bicuspid dentary tooth with distinct enamel cap.Abb. 4 a-i: Larve in fortgeschrittenem Stadium der Metamorphose (nr. 4, 4a). Präparate mit (a, c, g) undohne (b, d-f, h, i) Weichgewebe. a Prämaxillaria und Vomeres; Symphysenregion (Pfeil oben) mit einerlückenlosen Bezahnung, lingual; Vomeres (Pfeilkopf); Lücke zwischen den Vomeres (Pfeil); Choane(Stern). b Prämaxillarzaähne mit Anzeichen von Zweispitzigkeit (Pfeilköpfe) und Ringnaht (Pfeil). cBenachbarter bicuspider (Pfeil) und monocuspider Zahn im Oberkiefer, von vorn. d Maxillare mitlingual gekrümmten, pleurodont verankerten Zähnen; processus facialis maxillaris (Stern). e Vomer; parsdentalis mit Resten früherer Dentitionen (Pfeil) inmitten der Vomerplatte (Pfeilkopf); pars palatina(Stern). f Monocuspide Vomerzähne; Pulpazugang (Stern); Howshipsche Lakunen im Sockel (Pfeil). gMonocuspider Vomerzahn durchdringt das Mundepithel; Geschmacksknospen (Pfeilköpfe). h Dentale,lingual; monocuspide, hintere Zähne (Pfeile), bicuspider vorderer Zahn (Pfeilkopf). i Dentale; Apexeines vorderen bicuspiden Zahnes mit deutlicher Schmelzkappe.

204 HARTMUT GREVEN, NORBERT JÖMANN & GÜNTER CLEMEN
Figs 5 a-l: Larvae with delayed metamorphosis (nr. 5 and 6). a Premaxillary teeth, lingual view. bMaxillary teeth, lingual view; a dividing zone can hardly be recognized; processus facialis maxillaris(asterisk). c Apex of a maxillary tooth. d Maxillary tooth with a lingual bulge (left side) andglobular surface (asterisk). e Vomer with pars dentalis (arrows) shifted more caudally; vomerineplate (arrowhead); pars palatina (asterisk) broken off (right side). f The anterior edge of thevomerine pars dentalis with remnants of previous dentitions, labial view. g Vomerine tooth with abroad basis and numerous small pulpal accesses. h Vomerine tooth with a worn-out apex. i Dentarywith pars dentalis; teeth attached in a pleurodont condition increase in height posteriorly. j Attach-ment of dentary teeth is pleurodontous, lingual view; zone of division (arrow). k Coronoid teeth,lingual view, are horizontally attached. l Waisted coronoid tooth, lingual view.Abb. 5 a-l: Larven mit verzögerter Metamorphose (nr. 5 und 6). a Prämaxillarzähne, lingual. bMaxilarzähne, lingual; eine Ringnaht ist kaum zu erkennen; processus facialis maxillaris (Stern). cApex eines Maxillarzahnes. d Maxillarzahn mit lingualer Ausbuchtung (links) und globulärer Ober-fläche (Stern). e Vomer mit pars dentalis (Pfeile) nach caudal versetzt; Vomerplatte (Pfeilkopf); parspalatina (Stern) gebrochen (rechts). f Vordere Kante der pars dentalis des Vomer mit Resten frühererDentitionen, labial. g Vomerzahn mit breiter Basis und zahlreichen Pulpazugängen. h Vomerzahnmit abgenutztem Apex. i Dentale mit pars dentalis; die pleurodont verankerten Zähne werden nachhinten hin größer. j Pleurodont verankerte Zähne, lingual. Ringnaht (Pfeil). k Die Zähne auf demCoronoid sind horizontal veankert. l Zahn auf dem Coronoid mit Taille, lingual.

Dentition in Ranodon sibiricus Kessler 205
Acta Biologica Benrodis 13 (2006)
Figs 6 a-i: Juvenile (nr. 7). a Premaxilla, lateral view; teeth have high, slender pedicles; dividingzone (arrowheads). b Premaxillary tooth with a small, labial (arrowhead) and a slightly largerlingual (arrow) sharp edged cusp. c Maxillary teeth with large pulpal accesses (arrowheads); proces-sus facialis maxillaris (asterisk). d Pediclelate (arrow) maxillary teeth, lingual view. e Linguallyrecurved maxillary teeth. f Vomer, lingual view; pars palatina (asterisk). g Slender vomerine teethwith a globular dividing zone (arrow), lingual view. h Dentary, lingual view; teeth have slender andlong pedicles. i Dentary teeth, labial view; note the varying aspect of the dividing zone (arrows).Abb. 6 a-i: Juvenis (nr. 7). a Prämaxillare, lateral; die Zähne besitzen hohe, schlanke Sockel;Ringnaht (Pfeilkpfe). b Prämaxillarzahn with kleiner, labialer (Pfeilkopf) und geringfügig größererlingualer (Pfeil) scharfkantigen Spitze. c Maxillarzahn mit großen Pulpazugängen (Pfeilköpfe);processus facialis maxillaris (Stern). d Maxillarzahn mit Sockel (Pfeil), lingual. e Lingual gekrümmterMaxiallarzahn. f Vomer, lingual; pars palatina (Stern). g Schlanker Vomerzahn mit globulärerRingnaht (Pfeil), lingual. h Dentale, lingual; die Zähne sind schlank und haben lange Sockel. iZähne auf dem Dentale, labial, mit unterschiedlich ausgeprägter Ringnaht (Pfeile).
6 a, b, c; dentary: Figs 6 h, i; vomer: Figs 6 f, g).The dividing zone in the upper jaw was lin-gually more distinct than labially (Figs 6 c-e).In dentary teeth the dividing zone showed adeep cleft at the labial side, whereas linguallythe cleft showed some material perhaps notor weakly mineralized (Figs 6 h, i). The divid-ing zone of vomerine teeth was characterized
by a globular ring (Figs 6 f, g). Apices of teethwere curved lingually and had two distinctcusps, the labial one was smaller and sharperthan the lingual one (Figs 6 b, i). In a labialview, slender pedicles towered above the an-terior edge of the bone (Figs 6 i). Blades ofcusps in vomerine teeth were not clearly con-spicuous (Fig. 6 g).

206 HARTMUT GREVEN, NORBERT JÖMANN & GÜNTER CLEMEN
Figs 7 a-k: Subadult specimen (nr. 8). a Premaxilla, lingual view; processus dorsalis praemaxillaris(asterisk). b Lingually curved premaxillary teeth with a small lingual dividing zone (arrowhead). cPremaxillary tooth, lateral view, with differently shaped cusps; labial blade (arrowhead). d Maxilla.e Maxillary teeth with a thin zone of division (arrowhead), lateral view. f Vomer; pars dentalis(black arrow) between the pars palatina (asterisk) and vomerine plate (arrowhead); gap for thechoana (white arrow). g Anterior portion of the vomerine pars dentalis with remnants of previousdentitions (asterisk), lateral view. h Vomerine tooth with a blunt lingual cusp (arrowhead). iDentary, lingual view, with ascending pars palatina (asterisk). j Dentary teeth are pleurodontouslyattached. k Dentary tooth with greatly reduced labial blade (arrow).Abb. 7 a-k: Subadultus (nr. 8). a Prämaxillare, lingual; processus dorsalis praemaxillaris (Stern). bLingual gekrümmte Prämaxillarzähne mit kleiner lingualen Ringnaht (Pfeilkopf). c Prämaxillarzahn,

Dentition in Ranodon sibiricus Kessler 207
Acta Biologica Benrodis 13 (2006)
3.7. Subadult (nr. 8) (Figs 7 a-k)
Number of tooth rows, ankylosis and cur-vature of teeth and pulpal accesses as in theprevious stage.
Teeth of all dentigerous bones were bi-cuspid and divided (Figs 7 a-k). The apex ofthe teeth had two cusps, the labial onesmaller than the lingual. However, propor-tion between the two cusps varied consider-ably (see Figs 7 c, h, j, k).
3.8. Adult (nr. 9) (Figs 8 a-k)
Number of tooth rows, anchorage and cur-vature of teeth and pulpal accesses as in theprevious stage. In the upper jaw only maxil-lary teeth were available.
All teeth were bicuspid and divided (Figs 8a-k). The dividing zone was deeper labiallythan lingually (Figs 8 d, e, h) showing a smallring like depression lingually (Fig. 8 k).
From the cusps, the labial one was signifi-cantly reduced (maxilla: Fig. 8 c; vomer: Fig. 8f); both cusps may have blades (maxilla: Fig.8 c). Teeth of the dentary showed a slightreduction of the labial blade; the lingual cuspwas rounded off (Fig. 8 i). Here teeth werestrongly recurved, so that the larger blade wasnearly horizontal (Fig. 8 j); the most posteri-or tooth differed from the other teeth by itsbroad pedicle, its sturdy shape and severalpulpal accesses (Fig. 8 l).
3.9. Dental laminae (Figs 9 a-h)
Dental laminae are two layered derivates ofthe stratum basale of the oropharyngeal epi-thelium. The face of the dental laminaeturned towards the oropharyngeal epithelium
was covered with dense connective tissue thatamalgamated with the tunica propria.
In the larvae in metamorphosis and withdelayed metamorphosis (both approximate-ly stage 19), the upper jaw arcade (premaxillaeand maxillae) had a single continuous dentallamina at the lingual side. Due to the lack ofan adequate histological series, this is con-cluded by the fact that the symphyseal zonesbetween the two premaxillae and the prema-xilla/maxilla were small and the tooth rowdid not show any gap.
Vomers possessed their own dental lami-na each. At the level of the choana (Fig. 9 a)and posterior to the choana the dental lami-na still produced tooth buds (Fig. 9b); beforedisappearing it became one-layered and toothbuds could not be seen (Fig. 9c). The pos-terior end of the vomerine dental lamina wasclearly separated from the palatinal dental la-mina by connective tissue (Fig. 9 d). The pala-tinal dental lamina began with a process en-tirely surrounded by connective tissue (Fig. 9e); more posteriorly its terminus fused withthe oral epithelium and was productive (Fig.9 f). Beyond the last tooth bud, the unpro-ductive dental lamina was shortened (Fig. 9g). Posteriorly, the dental lamina disappeared,although parts of the degenerating palatinewere still present (Fig. 9 h).
4. Discussion
4.1. Attachment and succession ofdifferently shaped tooth generations
Succession of differently shaped and dividedteeth described by VASSILIEVA & SMIRNOV
(2001) for Ranodon sibiricus and pictured here-in for some stages more thoroughly closely
lateral, mit unterschiedlichen Spitzen; labiale Schneide (Pfeilkopf). d Maxillare. e Maxillarzähne mitdünner Ringnahr (Pfeilkopf), lateral. f Vomer; pars dentalis (schwarzer Pfeil) zwischen pars palatina(Stern) und Vomerplatte (Pfeilkopf); Lücke für die Choane (weißer Pfeil). g Vorderer Teil der parsdentalis des Vomer mit Resten früherer Dentitionen (Stern), lateral. h Vomerzahn mit stumpferlingualer Spitze (Pfeilkopf). i Dentale, lingual mit aufsteigender pars palatina (Stern) .j Pleurodontverankerte Zähne des Dentale. k Zahn des Dentale mit stark reduzierter labialer Schneide (Pfeil).

208 HARTMUT GREVEN, NORBERT JÖMANN & GÜNTER CLEMEN
Figs 8 a-l: Adult specimen (nr. 9). a Maxillary, lingual view, and processus facialis maxillaris (aste-risk). b Slender pedicles of maxillary teeth, lingual view. c Maxillary tooth, lingual view, with twodifferent cusps. d Zone of division of a maxillary tooth, lingual view. e Zone of division of amaxillary tooth, labial view. f Apex of a vomerine tooth, lateral view. g Vomerine teeth withdistinct labial dividing zones (arrows); pars palatina (asterisk) and pars dentalis (arrowhead). hDividing zone of a vomerine tooth, lateral view. Note the incision labially (arrowhead). i Dentarytooth with a blunt lingual cusp (arrowhead). j Lingually curved dentary teeth; note the varyingbicuspidity. k Zone of division in a dentary tooth, lateral view. l The most posterior tooth of thedentary is sturdy, bicuspid and broad-based.Abb. 8 a-l: Adultus (nr. 9). a Maxillare, lingual, und processus facilais maxillaris (Stern). b SchlankeSockel der Maxillarzähne, lingual. c Maxillarzahn, lingual, mit zwei unterschiedlichen Spitzen. dRingnaht eines Maxillarzahns, lingual. e Ringnaht eines Maxillarzahns, labial. f Apex eines Vomer-zahns, lateral. g Vomerzähne mit deutlicher labialer Ringnaht (Pfeile); pars palatina (Stern) und parsdentalis (Pfeilkopf). h Ringnaht eines Vomerzahns, lateral. Man beachte den labialen Einschnitt(Pfeilkopf). i Zahn des Dentale mit stumpfer lingualer Spitze (Pfeilkopf). j Lingual gekrümmte

Dentition in Ranodon sibiricus Kessler 209
Acta Biologica Benrodis 13 (2006)
Figs 9 a-h: Dental laminae of the vomer (a-c), between vomer and palatine (d), and the palatine(e-h); cross sections. a Dental lamina (arrow) with associated replacement tooth (arrowhead);choana (asterisk). b Dental lamina at the level of the choana with tooth bud (arrow). c Beforedisappearing posteriorly the dental lamina becomes one-layered (arrow). d Between vomer (aste-risk) and palatine (arrow) a dental lamina is absent. e Free rostral end of the palatine dental laminain the subepithelial connective tissue (arrow). f Connection of the dental lamina with the oralepithelium (arrow); most anterior tooth bud (arrowhead). g Short dental lamina (arrow) near thepalatine (p) approx. 7 µm behind the last tooth bud. h In the section following g the dental laminais absent; palatine (p).Abb. 9 a-h: Zahnleisten des Vomer (a-c), zwischen Vomer und Palatinum (d) und Palatinim (e-h);Querschnitte. a Zahnleiste (Pfeil) mit Ersatzzahn (Pfeilkopf); Choane (Stern). b Zahnleiste inHöhe der Choane mit Zahnkeim (Pfeil). c Vor ihrem Verschwinden wird die Zahnleiste einschich-tig (Pfeil). d Zwischen Vomer (Stern) und Palatinum (Pfeile) fehlt die Zahnleiste. e Freies rostralesEnde der palatinalen Zahnleiste im subepithelialen Bindegewebe (Pfeil). f Verbindung der Zahn-leite mit dem Mundepithel (Pfeil); hinterster Zahnkeim (Pfeilkopf). g Kurze Zahnleiste (Pfeil)nahe des Palatinum (p) etwa 7 µm hinter dem letzten Zahnkeim. h Im Folgeschnitt ist die Zahn-leiste nicht mehr vorhanden; Palatinum (p.).
Zähne des Dentale mit unterschiedlicher Zweispitzigkeit. k Ringnaht eines Zahns auf dem Denta-le, lateral. l Der hinterste Zahn auf dem Dentale ist untersetzt, zweispitzig und breitbasig.

210 HARTMUT GREVEN, NORBERT JÖMANN & GÜNTER CLEMEN
corresponds to that seen in the majority oftransforming Urodela (CLEMEN & GREVEN
1994; see DAVIT-BEAL et al. 2006, 2007): Toothgerms appear at sites where the dentigerousbones will develop in most cases already dur-ing the embryonic period. Anchorage of the(functional) small, conical and undivided teethtakes place during the mineralization processof these bones (see also LEBEDKINA 2005)and teeth are attached to the bones horizon-tally (palatine, vomer, coronoid) or slightlypleurally (premaxilla, dentary). The mode ofankylosis appears to be dictated by the shapeof the dentigerous bone and by the numberof previous dentitions. In our material a pleu-rodont ankylosis was seen on the premaxil-lae, maxillae and dentaries, obviously rightfrom the start but with increasing pleurodontattachment in later stages. On the palatineand coronoid teeth were always horizontallyattached, on the flattened vomer, however,initially horizontally and later slightly pleuro-dont due to remnants of previous dentiti-ons (see also CLEMEN & GREVEN 1979). Ob-viously, there is the tendency to reduce andenlarge accesses to the pulp with an increa-sing pleurodont attachment. Broad-basedteeth seen on bones (e.g. dentaries, maxillae)bearing anteriorly a pleurodont dentition, al-ways were horizontally attached and showedseveral pulpal accesses. Generally, the conicalteeth of very young urodele larvae have a smallnon-vascularized pulp and a tubular dentine(BOLTE et al. 1996). Such teeth were sugge-sted to be conserved organs representing anancestral character for gnathostomes (SIRE etal. 2002). In the majority of Urodela these�early� larval teeth (undivided, conical, broad-based, monocuspid) are replaced by conicalmonocuspids that may gradually change froma �non-pedicellate� (a pedicle cannot be di-stinguished) to a pedicellate condition (thepedicle is separated from the crown by a moreor less complete distinct zone of division) viavarious intermediate stages named �subpe-dicellate� by some authors. Not until meta-morphosis teeth become replaced by fully
divided (=pedicellate), bicuspid teeth, andbicuspidity seems to develop also via inter-mediate stages. This was mentioned for in-stance in the articles of BENESKI & LARSEN
(1989 a, b) and (BOLTE & CLEMEN 1991) anddemonstrated unequivocally by DAVIT-BEAL
et al. (2006).Terminology of these three major ontoge-
netic stages is inconsistent in literature. BENE-SKI & LARSEN (1989 b) distinguished in theirSEM-pictures �for clarity and consistency� (p.166) 1) non-pedicellate teeth (stage I) with adistal monocuspid apex (covered entirely byenameloid) and a proximal shaft (not cover-ed by enameloid), 2) subpedicellate teeth (sta-ge II) with a distal monocuspid crown (onlyits apex is covered by enameloid) and a proxi-mal base separated from the crown by an ab-rupt increase in width and/or a lingual fi-brous zone of division, and 3) pedicellateteeth (stage III), with a distal crown consi-sting of the modified, i.e. bicuspid, apex, andshaft (the apex is entirely covered by enamel,the shaft only in its distal part), and a proxi-mal pedicle separated completely by an annu-lar zone of weakness (see Fig. 1 in BENESKI &LARSEN 1989 b; see also VASSILIEVA & SMIRO-NOV 2001). Apart from the fact that the apexof larval teeth probably possesses true ena-mel and enameloid instead of enameloid alo-ne (see BOLTE & CLEMEN 1992; BOLTE et al.1996; KOGAYA 1994, 1999; WISTUBA et al.2002), this terminology, although adoptedby various researchers, does not meet the realconditions and appears highly arbitrarily. Bydefinition non-pedicellate teeth do not showa dividing zone between the distal and proxi-mal part by LM and SEM. However, their�proximal shaft� or base definitely lacks den-tine tubules such as the pedicles of transfor-med teeth (BOLTE et al. 1996). That meansthat early larval teeth possess a true pedicle.Further, VASSILIEVA & SMIRNOV (2001) descri-be on the basis of their stained and clearedspecimens that mineralization of small non-pedicellate teeth begins from a single apicalmineralization centre and proceeds in apico-

Dentition in Ranodon sibiricus Kessler 211
Acta Biologica Benrodis 13 (2006)
basal direction, whereas mineralization of�subpedicellate� and pedicellate teeth startsfrom an apical and a basal centre moving inapico-basal and basal apical direction, respec-tively. There is evidence by TEM-micrographsthat also in early larvae two mineralizationfronts exist (Ambystoma mexicanum: WISTUBA
et al. 2002).Typically, in bicuspid teeth the lingual cusp
is elongated, but sharper than the labial one.Compared to the relative time the dividingzone develops, the change of the tooth crownfrom monocuspids to bicuspids seems totake place in a relatively shorter period aroundmetamorphosis.
Only in a few metamorphosed urodelesteeth deviate from the bicuspid type; e.g., insome Ambystoma species basic cusp shapesof pedicellate premaxillary teeth are disc orclub (see BENESKI & LARSEN, 1989a), which,however, all can be attributed to the bicuspidtype; male plethodontids have premaxillaryteeth that secondarily changed to cone-shapedmonocuspids under the influence of testo-sterone during the breeding season (e.g.,NOBLE & POPE 1929; EHMCKE et al. 2003;GREVEN et al. 2004).
In adult R. sibiricus the shape of the twocusps of teeth is rather variable, mainly regard-ing the lingual cusp, which probably cannotsolely be attributed to the fact that bicuspi-dity develops gradually on the different den-tigerous bones (see below). Also the zone ofdivision that creates the pedicellate conditionin metamorphosed urodele teeth and is con-sidered an apomorphic character of extantAmphibia (e.g., PARSONS & WILLIAMS 1962;DUELLMAN & TRUEB 1986; GREVEN 1989; AX
2001; the latter author also gives reasons whyto cancel the term �Lissamphibia� used forthe extant Amphibia) shows a considerablevariation. A transverse annular zone of divi-sion was lacking in our youngest larva (and islacking also in earlier stages; see VASSILIEVA &SMIRNOV 2001); in later stages this zone ap-pears to be indicated only by a more or lessglobular area around the tooth and curvature
of teeth might indicate the future region ofthe dividing zone (e.g., DAVIT-BEAT et al. 2006).Even in adults this zone was often remar-kably less distinct when compared to othermetamorphosed urodeles. We don�t know,whether this variability is a question of theage of the animals, i.e. fully metamorphosedand older adults may be expected to show amore conspicuous zone division as seen in avariety of Urodela, or caused by a heavier oreven secondary calcification of the collagenousfibres bridging the gap between crown andpedicle (MOURY et al. 1985; GREVEN et al. 1989;WISTUBA et al. 2002). Regarding the ontoge-netic sequence studied by VASSILIEVA & SMIR-NOV (2001), a secondary mineralization of thiszone is improbable. Generally, in R. sibiricusdevelopment of both characters (bicuspidityand zone of division) seems more variableand partly less conspicuous as in other Uro-dela.
4.2. The fate of dentigerous bonesduring ontogeny and direction of dentalreplacements
As in other Urodela, also in R. sibiricus de-velopment and replacement of the different-ly organized teeth takes place at different timesdepending on the particular dentigerous boneand its fate during further development.Tooth replacement in all dentigerous bones,typically is posterior to the tooth row in labialdirection. Development of dentigerousbones of R. sibiricus and their degradation orremodelling during metamorphosis wasthoroughly described by LEBEDKINA (sum-marized 1979, 2005, see also JÖMANN et al.2005); the various tooth generations and theirtemporally shifted development were illus-trated by VASSILIEVA & SMIRNOV (2001). Thelatter observations are deepened herein andbroadened with regard to the palatine andthe coronoid.
The typical sequence of differently shapedtooth generations has been used to classify�early� (see above), �late� larval teeth (mo-

212 HARTMUT GREVEN, NORBERT JÖMANN & GÜNTER CLEMEN
nocuspid, �subpedicellate�) and �trans-formed� or �metamorphosed� teeth (seeabove) in several Urodela including paedo-morphic species (e.g., CLEMEN & GREVEN 1977,1980, 1994; GREVEN & CLEMEN 1980). Initial-ly, larval dentigerous bones of R. sibiricus bear�non-pedicellate�, i.e., �early� larval teeth (seealso VASSILIEVA & SMIRNOV 2001). The laterthe bone develops in ontogeny, the moredeveloped teeth (in the direction of the �late�larval tooth) are established, i.e., very earlytooth stages are left out on bones developinglate in ontogenesis, e.g., on the maxilla. �Late�larval teeth are first established on the pre-maxilla, then on the dentary and later on thedeveloping maxilla. At this time vomerineand coronoid teeth (a single establishedtooth in larva 3) were from the �early� larvaltype. Palatinal teeth in larva 3 were, however,lacking. It was shown by VASSILIEVA & SMIR-NOV (2001) that the coronoid as well as thepalatinal dental laminae appear to be able toproduce short-lived �late� larval teeth(named by the authors �subpedicellate�). Ac-tually, we found �late� larval teeth on the co-ronoid of the specimens with delayed meta-morphosis. The discrepancies regarding thepalatine may be attributed to our limitedmaterial and the large variations suggestedfor the development of dentition in R. sibiri-cus (see the discussion in VASSILIEVA &SMIRNOW 2001).
Metamorphosed bicuspid teeth are firstproduced in the upper and lower jaw. All pre-maxillary and the most maxillary teeth of lar-va 4 in advanced metamorphosis had (inci-pient) bicuspids, whereas dentaries bore trans-formed teeth anteriorly and teeth of the �late�larval type posteriorly, i.e., transformation ofteeth takes place from the anterior to posteri-or. The vomers still had �early� larval teeth.After metamorphosis all systems still presentshow bicuspid divided teeth. In larvae withdelayed metamorphosis all systems presentat this time had �late� larval teeth.
Regression of the palatinal tooth systembegins relatively early. This is stressed by the
fact that in the larvae with delayed metamor-phosis palatinal teeth were totally absent (seeabove). After multiple hibernation as as-sumed for R. sibiricus larvae (KUZMIN & THIES-MEIER 2001; see the discussion in JÖMANN etal., 2005), the palatine is separated from thepterygoid, still fully developed, but free ofteeth. At this time the dental lamina of thecoronoid appeared still active.
Because of the relatively early degradationof these dental laminae a subsequent deliv-ery of new teeth ceases. This leads to a tooth-less palatine, either permanent when larvaewere hypopyhsectomized in time (Salaman-dra salamandra: CLEMEN 1978) or before com-plete resorption in course of metamorpho-sis when metamorphosis is delayed, e.g. bydeep temperatures (S. salamandra: CLEMEN
1978; R. sibiricus: JÖMANN et al. 2005; the pre-sent article). Degradation of the palatinalportion of the palatopterygoid is consideredas one of the key factors indicating the onsetof metamorphosis (REILLY 1986, 1987;REILLY & ALTIG 1996). Onset, however, maybe indicated earlier, e.g., by the less conspicu-ous degradation of dental laminae.
In brief, regarding chronology of tooth re-placements as well a disintegration of certaindentigerous bones, R. sibiricus follows thepattern realized in all urodeles hitherto exam-ined that undergo metamorphosis. The ty-pical spatial and temporal differences regard-ing development of fully transformed bi-cuspid teeth and their transitory stages (seeCLEMEN & GREVEN 1994 and the literaturecited above) in the various tooth systems isvery probably ascribed to different compe-tences of the tooth producing tissue and itsdifferent sensitivities to the metamorphosinghormone (CLEMEN 1988 a, b; GREVEN & CLE-MEN 1990).
Although briefly discussed in our previousstudy (JÖMANN et al. 2005), we again discussherein the changes during remodelling of thevomerine system around metamorphosis. Inmost Urodela, these changes are remarkableand have been long used for phylogenetic

Dentition in Ranodon sibiricus Kessler 213
Acta Biologica Benrodis 13 (2006)
considerations (e.g., LAURENT 1947; REGAL
1966; HECHT & EDWARDS 1977; ZHAO et al.1988; CLEMEN & GREVEN 1994).
Apart from the fact that the larval vomerof R. sibiricus bears several rows of teethrather than a single row (LEBEDKINA 1979,2005; VASSILIEVA & SMIRNOV 2001; see above)and that the vomerine tooth rows are lessparallel to the premaxillary arcade (see figuresin LEBEDKINA 2005 and JÖMANN et al. 2005),the changes of the vomerine system duringtransformation of R. sibiricus are less conspicu-ous as in many other Urodela (for review seeCLEMEN & GREVEN 1994). MetamorphosedR. sibiricus have a nearly transverse vomerinedentition more or less parallel to the upperjaw arcade with a tooth replacement posteri-or to the tooth row in labial direction, i.e. thelabial replacement pattern is retained in trans-formed specimens. In other hynobiids moreconspicuous changes occur (e.g., ZHAO et al.,1988; ADLER & ZHAO 1990; ZHANG et al. 2006).Here the partes dentales form a short anterior,transverse row and a longitudinal row direc-ted posteriorly resulting in replacements pos-terior (in labial direction) to the transversetooth row and lateral (in lingual direction) tothe longitudinal tooth rows (GREVEN & CLE-MEN 1985). The latter mode obviously hasevolved more than once in Urodela (see thediscussion in HECHT & EDWARDS 1977). R.sibiricus shares the transversely orientation ofthe vomerine dentition (and thus the pos-terior replacement of teeth) with the hyno-biid genera Liua, Batrachuperus, Pachyhynobius,Paradactylodon, and Onychodactylus (ZHAO etal. 1988; ZHANG et al. 2006), which belong tostream-type hynobiids and obviously do notform a natural group (LARSON et al. 2003;ZHANG et al. 2006). Transversely oriented vo-merine teeth were said to hinder escape ofprey in those species feeding by suction inwater (ZHANG et al. 2006). These functionalsuggestions do not contradict the assump-tion that a transverse tooth row with replace-ments in labial direction is a plesiomorphiccharacter of hynobiids and perhaps of all
Urodela (see the short discussion in JÖMANN
et al, 2005). Fossils from the Early Cretaceousrelated to hynobiids, such as Liaoxitriton havealso a transverse arrangement of the vomer-ine teeth (WANG 2004).
4.3. Multiple rows versus a single row ofteeth
In the youngest larva available larvae (stage16 according to LEBEDKINA�s table, l.c.; seeJÖMANN et al. 2005; late feeding larva accord-ing to VASSILIEVA & SMIRNOV 2001) we foundonly on the palatine clear signs of two rowsof teeth without resorption pits. Dentaries,premaxillae, and, when established, maxillaebear a single row of teeth throughout onto-geny (VASSILIEVA & SMIRNOV 2001; LEBEDKINA
1979, 2005; JÖMANN et al. 2005) and post-metamorphosed specimens have a single rowof teeth on all dentigerous bones. LEBED-KINA (1979, 2005) draws a single row of teethon all dentigerous bones up to stage 7 withthe exception of the coronoid (early feedinglarva; see VASSILIEVA & SMIRNOV 2001). In herstage 8 the palatine shows two rows of teethand two rows occur also on the vomer fromher stage 9 up to resorption in the premeta-morphic period (stage 17). Our youngestspecimen did not clearly show two rows ofteeth on the vomers. These findings showthat already in �late feeding larvae� resorp-tion of lateral margins of the vomer, butalso of the palatine and the coronoid has ledto the loss of teeth. In contrast to the find-ings of VASSILIEVA & SMIRNOW (2001) all teethof our specimen (stage 16) were from theearly larval type, i.e. monocuspid and entirelyundivided (see above). These discrepanciesmay be attributed to individual variation.
According to our results and the findingsof VASSILIEVA & SMIRNOV (2001), the sequenceof tooth row numbers in the tooth systemsof R. sibiricus is as follows (the first func-tional dentition of a dentigerous bones ne-cessarily comprises a single row): the upperjaw and the dentary bear a single row through-

214 HARTMUT GREVEN, NORBERT JÖMANN & GÜNTER CLEMEN
out ontogeny, the vomers bear up to threerows in larvae and a single row in metamor-phosed specimens. The coronoids bear upto three rows, and the palatines up to tworows that will be resorbed before metamor-phosis. In other Urodela deviations from thispattern occur mainly with respect to the pre-maxillae and palate (e.g. in Ambystomatidae,where the palate is partly retained after meta-morphosis) and vomer (e.g. in Plethodonti-dae) (see literature cited above).
Interestingly, the hynobiid, Onychodactylusfischeri, which is now considered as sistertaxon of all other extant hynobiids (LARSON
et al. 2003; ZHANG et al. 2006) and which lar-gely resembles R. sibiricus in the traits discus-sed herein, has a single row on all dentigerousbones already in early developmental stages(SMIRNOW & VASSILIEVA 2002).
4.4. Dental laminae
Knowledge of number and course of dentallaminae in larval and adult R. sibiricus is stillincomplete, but the available data and indi-rect evidence from the position and continu-ity of tooth rows allow for some conclusion.The upper jaw is accompanied by a conti-nuous dental lamina indicated by a conti-nuous tooth row (and tooth buds) in thesymphyseal region between the premaxillaeand at the transition of the premaxilla/ma-xilla. Between the dentaries here and in otherUrodela (e.g., GREVEN et al., 2002/2003) a gapin the tooth row may be indicative for a gapin the dental lamina. The paired coronoids,palatines and vomers are widely separated andhave their own continuous dental lamina each.This pattern exactly corresponds to that foundin the majority of extant Urodela. Exceptionshave been documented and discussed else-where (e.g., CLEMEN & GREVEN 1994).
The larvae with delayed metamorphosisshowed a fully developed palatinal portionof the palatopterygoid without teeth (see alsoJÖMANN et al. 2005). Thus, the dental laminamust have lost its capability to produce teeth
at the onset of the lengthened late larvalperiod. The dental laminae of the coronoidbetween vomer and palatine was, however,still active at this time (see the picture in JÖ-MANN et al. 2005).
4.5. Conclusions
As shown herein and in previous articles ofvarious authors, the pattern of dentition, thesuccession of differently shaped tooth gen-erations, the direction of replacements and thecourse and number of dental laminae (stillunknown in detail, but indirectly inferred fromgross morphology in part) of Ranodon. sibiri-cus, a representative of an ancient urodelelineage, fits very well in the pattern present inmany other Urodela. The remodelling mainlyof the vomerine system during metamorpho-sis appears comparatively inconspicuous. Wesuggest that many of the characters of thetooth systems summarized herein are highlyconserved plesiomorphic traits in Urodela.
Acknowledgements
The technical help of Mrs. MARTINA FASEL
(Münster) and Mr. MARCEL BRENNER (Düs-seldorf) is greatly appreciated.
References
ADLER, K., ZHAO, E., 1990: Studies on hynobiidsalamanders, with description of a newgenus. � Asiatic Herpetological Research 3,37-47.
AX, P., 2001: Das System der Metazoa. Ein Lehr-buch der phylogenetischen Systematik. Spek-trum Akademischer Verlag, Heidelberg.
BENESKI, J.H., LARSEN, J.H. Jr., 1989a: Interspe-cific, ontogenetic, and life history variationin the tooth morphology of mole salaman-ders (Amphibia, Urodela, and Ambystomati-dae). �Journal of Morphology 199, 53-69.
BENESKI, J.H., LARSEN, J.H. Jr., 1989b: Ontoge-netic alteration in the gross morphology ofDicamptodon and Rhyacotriton (Amphibia, Uro-dela, and Dicamptodontidae). � Journal ofMorphology 199, 165-174.

Dentition in Ranodon sibiricus Kessler 215
Acta Biologica Benrodis 13 (2006)
BOLTE, M., CLEMEN, G., 1991: Die Zahnformendes Unterkiefers beim adulten Ambystomamexicanum Shaw (Urodela: Ambystomatidae).� Acta Biologica Benrodis 3, 171-177.
BOLTE, M., CLEMEN, G., 1992: The enamel oflarval and adult teeth of Ambystoma mexica-num Shaw (Uodela: Ambystomatidae) � aSEM study. � Zoologischer Anzeiger 228,167-173.
BOLTE, M., KREFTING, E.-R., CLEMEN, G., 1996:Hard tissue of teeth and their calcium andphophate content in Ambystoma mexicanum(Urodela: Ambystomatidae). � Annals ofAnatomy 178, 71-80.
CLEMEN,G.,1978: Aufbau und Veränderungen derGaumenzahnleisten beim larvalen und meta-morphosierenden Salamandra salamandra (L.)(Salamandridae: Amphibia). � Zoomorpho-logie 90, 135-150.
CLEMEN, G., 1988a: Experimental analysis of thecapacity of dental laminae in Ambystoma me-xicanum Shaw. � Archives Biologiques (Bru-xelles) 99, 111-132.
CLEMEN, G., 1988 b: Competence and reactionsof early and late-larval dental laminae in ori-ginal and not-original dental systems of Am-bystoma mexicanum Shaw. � Archives Biolo-giques (Bruxelles) 99, 307-324.
CLEMEN, G., GREVEN, H., 1977: MorphologischeUntersuchungen an der Mundhöhle von Uro-delen. III. Die Munddachbezahnung vonAmbystoma mexicanum Cope (Ambystomati-dae: Amphibia). � Zoologische JahrbücherAbteilung für Anatomie und Ontogenie derTiere 8, 95-136.
CLEMEN, G., GREVEN, H., 1979: MorphologischeUntersuchungen an der Mundhöhle von Uro-delen. V. Die Munddachbezahnung von Tri-turus vulgaris (L.) (Salamandridae: Amphibia).� Zoologische Jahrbücher Abteilung für Ana-tomie und Ontogenie der Tiere 102, 170-186.
CLEMEN, G., GREVEN, H., 1980: MorphologischeUntersuchungen an der Mundhöhle von Uro-delen. VII. Die Munddachbezahnung vonAmphiuma (Amphiumidae: Amphibia). � Bon-ner zoologische Beiträge 31, 357-362.
CLEMEN, G., GREVEN, H., 1994: The buccal cavi-ty of larval and metamorphosed Salamandrasalamandra: Structural and developmental as-pects. � Mertensiella 4, 83-109.
DAVIT-BÉAL, T., ALLIZARD, F., SIRE, J.-Y., 2006:Morphological variations in a tooth family
through ontogeny in Pleurodeles waltl (Lissam-phibia, Caudata). � Journal of Morphology267, 1048-1065.
DAVIT-BÉAL, T., CHISAKA, H., DELGADO, S., SIRE,J.-Y., 2007: Amphibian teeth: current know-ledhe, unanswered questions, and some di-rections for future research. Biological Re-views 82: 49-81.
DUELLMAN, W.E., TRUEB, L., 1986: Biology ofAmphibians. � McGraw-Hill, New York.
EHMCKE, J., WISTUBA, J., CLEMEN, G., SCHLATT, S.,2003: Targeted expression of androgen re-ceptors in tooth-forming tissues of a neotro-pical salamander (Bolitoglossa schizodactyla)enables region-specific formation of dimor-phic types of teeth in the male. � Generaland Comparative Endocrinology 134, 26-35.
GAO, K.O., SHUBIN, N.H., 2001: Late Jurassicsalamanders from Northern China. � Nature410, 574-577.
GREVEN, H., 1989: Teeth of extant amphibian:morphology and some implications. � Fort-schritte der Zoologie 35, 451-455.
GREVEN, H., CLEMEN, G., 1980: Morphologicalstudies on the mouth cavity of urodeles. VI.The teeth of the upper jaw and the palate inAndrias davidianus (Blanchard) and A. japoni-cus (Temminck) (Cryotobranchidae: Amphi-bia). � Amphibia-Reptilia 1, 49-59.
GREVEN, H., HILLMANN, G., THEMANN, H., 1989:Ultrastructure of the dividing zone in teethof Salamandra salamandra (L.) (Amphibia,Urodela). � Fortschritte der Zoologie 35, 477-479.
GREVEN, H., CLEMEN, G., 1985: Morphologicalstudies on the mouth cavity of urodeles. VIII.The teeth of the upper jaw and the palate intwo Hynobius-species (Hynobiidae: Amphi-bia). �Zeitschrift für zoologisch-systemati-sche Evolutionsforschung 32, 136-147.
GREVEN, H., CLEMEN, G., 1990: Effects of hypo-physectomy on the structure of normal andectopically transplanted teeth in larval andadult urodeles. � Acta Embryologiae et Mor-phologiae Experimentalis, ns 1, 33-43.
GREVEN, H., SCHUBERT-JUNG, M., CLEMEN, G.,2002/2003: Notes on the skull of EuropeanSpeleomantes spp. (Plethodontidae, Urodela).� Acta Biologica Benrodis 12, 45-58.
GREVEN, H., SCHUBERT-JUNG, M., CLEMEN, G.,2004: The dentition of European Speleoman-tes spp. (Urodela, Plethodontidae) with spe-

216 HARTMUT GREVEN, NORBERT JÖMANN & GÜNTER CLEMEN
cial regard to sexual dimorphism. � Annalsof Anatomy 186, 33-43.
HECHT, M.K., EDWARDS, J.L., 1977: The metho-dology of phylogenetic inference above thespecies level, pp 3-51. In: Major patterns invertebrate evolution (Hecht, M.K., Goody,P.C., Hecht, B.M. , eds.). � New York, Ple-num Press.
IUCN Red List of threatened species., 2006:www.iucnredlist.org
JÖMANN, N., CLEMEN, G., GREVEN, H., 2005: No-tes on cranial ontogeny and delayed meta-morphosis in the hynobiid salamander Ran-odon sibiricus Kessler, 1866 (Urodela). � An-nals of Anatomy 187, 305-321.
KOGAYA, Y., 1994: Sulfated glycoconjugates inamelogenesis. � Progress in Histochemistryand Cytochemistry 29, 1-110.
KOGAYA, Y., 1999: Immunohistochemical locali-sation of amelogenin like proteins and type Icollagen and histochemical demonstration ofsulphated glycoconjugates in developing ena-meloid and enamel matrices of the larval uro-dele Triturus pyrrhogaster teeth. � Journal ofAnatomy 195, 455-464.
KUZMIN, S.L., THIESMEIER, B., 2001: Mountainsalamanders of the genus Ranodon . � Advan-ces of Amphibian Research of the FormerSoviet Union 6, 1-181.
LARSON, A., DIMMICK, W., 1993 : Phylogenetic rela-tionships of the salamander families: An analy-sis of congruence among morphological cha-racters. � Herpetological Monographs 7, 77-93
LARSON, A., WEISROCK, D.W., KOZAK, K.H., 2003:Phylogenetic systematics of salamanders (Am-phibia: Urodela), a review, pp 31-108. In:Reproductive Biology and phylogeny of Uro-dela (D.M. Sever, ed.). Vol 1 of the seriesReproductive Biology and Phylogeny (B.G.Jamieson, ed.). � Science Publishers, Inc.Enfield, USA.
LAURENT, R., 1947: La disposition des dents co-mediennes chez les row els superiors et sonimportance phylogenetique. �- Bulletin deMusée royal de Histoire naturelle Belgique23, 1-4.
LEBEDKINA, N.S., 1979: The evolution of amphi-bian skull. � Moscow, Nauk (in Russian)
LEBEDKINA, N.S., 2005: Evolution of the amphi-bian skull (translated by S.V. Smirnov). �Advances of Amphibian Research of theFormer Soviet Union 9, 1-265.
MOURY, J.D., CURTIS, S.K., PAV., D.I., 198:. Struc-ture of the radially asymmetrical uncalcifiedregion of the teeth of the red-backed sala-mander, Plethodon cinereus (Amphibia, Pletho-dontidae). � Journal of Morphology 185, 403-412.
NOBLE, G.K., POPE, S.H., 1929: The modificati-on of the cloaca and teeth of the adult sala-mander, Desmognathus, by testicular transplantsand by castration. � British Journal of Expe-rimental Biology 6, 399-411.
PARSONS, T.S., WILLIAMS, E.E., 1962: The teethof Amphibia and their relation to amphibianphylogeny. � Journal of Morphology 110, 375-389.
REGAL, P.J., 1966: Feeding specialisations and theclassification of terrestrial salamanders. �Evolution 20, 392-407.
REILLY, S.M., 1986: Ontogeny of cranial ossifi-cation in the eastern newt, Notophthalmus viri-descens (Caudata: Salamandridae), and its re-lationship to metamorphosis and neoteny. �Journal of Morphology 188, 215-326.
REILLY, S.M., 1987: Ontogeny of the hyobran-chial apparatus in the salamanders Ambysto-ma talpoideum (Ambystomatidae) and Noto-phthalmus viridescens (Salamandridae): the eco-logical morphology of two neotenic strate-gies. � Journal of Morphology 191, 201-214.
REILLY, S.M., ALTIG, R., 1996: Cranial ontogenyin Siren intermedia (Caudata: Sirenidae): pae-domorphic, metamorphic, and novel patternsof heterochrony. � Copeia 1996, 29-41.
ROMEIS, B., 1989: Mikroskopische Technik. � Ur-ban und Schwarzenberg, München, Wien,Baltimore.
SCHMALHAUSEN, I.I., 1968: The Origin of Terre-strial Vertebrates. � Academic Press, NewYork, London.
SCHOCH, R.R., 1998: Homology of cranial bonesin urodeles: the significance of developmen-tal data for fossil basal tetrapods. � NeuesJahrbuch für Geologie und Paläontologie,Monatshefte 11, 1-25.
SIRE, J.-Y., DAVIT-BEAL, T., DELGADO, S., VAN DER
HEYDEN, CH., HUYSSEUNE, A., 2002: First-ge-neration teeth in nonmammalian rowages:Evidence for a conserved ancestral charac-ter? � Microscopy Research and Technique59, 408-434.
SMIRNOV, S.V., VASSILIEVA, A.B., 2002: Skeletaland dental ontogeny in the long-tailed cla-

Dentition in Ranodon sibiricus Kessler 217
Acta Biologica Benrodis 13 (2006)
wed salamander, Onychodactylus fischeri (Uro-dela: Hynobiidae). � Russian Journal of Her-petology 9, 21-32.
VASSILIEVA, A.B., SMIRNOV, S.V., 2001: Develop-ment and morphology of the dentition in theAsian salamander, Ranodon sibiricus (Urodela:Hynobiidae). � Russian Journal of Herpeto-logy 8, 105-116.
WANG, Y., 2004: A new Mesozoic caudate (Liao-xitriton daohugouensis sp. nov.) from Inner Mon-golian China. � Chinese Science Bulletin 49,814-815.
WISTUBA, J., GREVEN, H., CLEMEN, G., 2002: De-velopment of larval and transformed teeth
in Ambystoma mexicanum (Urodela, Amphi-bia): an ultrastructural study. � Tissue & Cell34, 14-27.
ZHANG, P., CHEN, Y.-Q., ZHOU, H., LIU, Y.F.,WANG, X.-L., PAPENFUSS, T.J., WAKE, D.B.,QU, L.-H., 2006 : Phylogeny, evolution, andbiogeography of Asiatic salamanders (Hyn-obiidae). � Proceedings of the NationalAcademy of Sciences, USA 103, 7360-7365.
ZHAO, E., JIANG, Y., HU, Q., YANG, Y., 1988:Studies on Chinese Salamanders. � Societyfor the Study of Amphibians and Reptiles.Oxford, Ohio.
Accepted: 22.12. 2006

218 HARTMUT GREVEN, NORBERT JÖMANN & GÜNTER CLEMEN
Book review
LARINK, OTTO, WESTHEIDE, WILFRIED (2006): Coastal Plankton. Photo Guide for Euro-pean Seas. � Verlag Dr. Friedrich Pfeil, München, 143 pages, 60 colour plates with 649 micro-and macrophotograph. Paperback, 24 x 21 cm. ISBN 978-3-89937-062-1. 30.-- Euro
Photo guides are �in� and numerous are available for a variety of organisms living in the sea,in freshwater and on land. The present booklet is a joint effort written by two retiredscientists, who sifted through their micro- and micrographs taken on numerous excursionsand field courses in marine biology during their active time. They come out with a beautifullyillustrated guide on phytoplankton, protozoans, developmental stages and adults of zoo-plankton covering these organisms �mainly from the North Sea coastal area but also fromother European coastlines, including the western Mediterranean� (taken from the blurb). Itgoes without saying that the booklet is far from being �a comprehensive key� or a �textbookof planktology� or a determination key (see the foreword) � this was not the intention �,but it gives the reader a wonderful insight into a most diverse and beautiful world.The book begins with short introductory notes on �Phytoplankton diversity�, �Zooplank-ton diversity�, �Pelagic larvae � the meroplankton�, �Trophic relationships� and �Samplingand processing� and ends with �Literature� (96 citations including Haeckel�s �Kunstformender Natur� from 1904), some internet addresses of databases with information on marineplankton and a detailed �Index�. The main part (page 16-135) includes the colour plates and,on the opposite page, the legends, i.e. a short informative text on the biology of the taxonconsidered and its popular names in English, French, German, and Spanish. Authors treatso diverse organisms such as Bacillariophycea; various microalgae, Dinoflagellata includingthe famous Noctiluca, Choanaoflagellata, Foraminifera, �Radiolaria�, Ciliophora, Tintinni-na, Cnidaria, Ctenophora,, Platyhelminthes, Nemertini, Mollusca, Annelida, Crustacea,�Rotifera�, Phoronida, Bryozoa, Chaetognatha, Echinodermata with the early develop-ment of Psammechinus, Hemichordata, Tunicata, Acrania, and teleosts. Under �Miscellan-eous� even faecal pellets of copepods and Pinus sp. pollen are pictured. Micro- and macro-photographs are of high quality. The same applies to the printing.Leafing through the booklet I was enthusiastic about the various �Kunstformen der Natur�and I think that the reader and all, who love the fascinating microcosm of marine organismswill feel similarly.
Hartmut Greven, Düsseldorf

A new genus and species of false scorpion 219
Acta Biologica Benrodis 13 (2006)
Acta Biologica Benrodis 13 (2006): 219-229
A new genus and species of false scorpion fromVietnam showing remarkable chelal modifications
(Arachnida: Chelonethida)
Eine neue Gattung und Art von Pseudoskorpionen aus Vietnam mitbemerkenswerten Modifikationen der Chelae (Arachnida:
Chelonethida)
SELVIN DASHDAMIROV
Aquazoo – Löbbecke Museum, Kaiserswerther Straße 380, D-40474 Düsseldorf,Germany; [email protected]
Summary: A new pseudoscorpion genus and species (Caecogarypinus pectinodentatus gen. n., sp. n.) isdescribed from a lowland tropical forest in southern Vietnam. This new form is tentatively assignedto the family Garypinidae, being distinguished by a remarkable modification of the palpal chela, i.e.the presence of a pronounced basal comb on the dental ridge and an apparently reduced venomapparatus of the movable finger. Probably these animals capture small arthropods, which they holdand pierce with the modified chelae and/or sieve the substrate by means of the chelae to capturevery small prey. The phenotype of the specimens and their habitat (litter) suggest psammophily ofthe new species.
Pseudoscorpion, Garypinidae, taxonomy, new genus, new species, Vietnam
Zusammenfassung: Aus dem Regenwald Südvietnams wird eine neue Gattung und Art vonPseudoskorpionen (Caecogarypinus pectinodentatus gen. n., sp. n.) beschrieben, die möglicherweise zuden Garypinidae gehört und sich von anderen Pseudoskorpionen durch eine bemerkenswerteModifikation der Palpenscheren unterscheidet. Die Chelae zeigen einen auffälligen basalen Kammauf der bezahnten Leiste und einen offenbar reduzierten Giftapparat im beweglichen Finger.Wahrscheinlich jagen die Tiere kleine Arthropoden, die sie mit den modifizierten Chelae festhaltenund verletzen und/oder sieben das Substrat mit diesen aus, um sehr kleine Beute zu fangen. Dasäußere Erscheinungsbild sowie der Lebensraum der Tiere sprechen für Psammophilie.
Pseudoskorpione, Garypinidae, Taxonomie, neue Gattung, neue Art, Vietnam
1. Introduction
Through the kindness of Dr. SERGEI GOLO-VATCH and Mr. ALEXANDER ANICHKIN (bothMoscow), I have recently been privileged tostudy a rich collection of false scorpions fromVietnam made in 2003 to 2006. This collec-tion contains more than 25 species (ca. 1500specimens) and its detailed study is now inprogress.
The present paper treats only one of thespecies, which, by showing several remark-ably disjunct characters, definitely warrantsthe erection of a new genus. Furthermore,even placing it in a family has become prob-lematic, the more so as the available materialis very small and contains no males. Thisnew genus and species has tentatively beenattributed to the family Garypinidae Daday,1888. The status of this family is here accep-

220 SELVIN DASHDAMIROV
ted after JUDSON (2005) and STAHLAVSKÝ. Toconfirm or refute this assignment, additionalsamples, especially of males, are necessary.
2. Material and Methods
Type material is shared between the collectionsof the Zoological Museum of the MoscowState University (ZMMU) and Aquazoo �Löbbecke Museum, Düsseldorf (LB+No).Microscopic examinations were carried outusing a WILD M5 stereomicroscope, an Olym-pus BH2 with differential interference contrastattachment and an Olympus CX31, which wasalso used for taking measurements. Tempora-ry slide mounts were made in glycerol.
The following abbreviations are used inthe text. Tactile setae: B = basal; SB = sub-basal; ST = subterminal; T = terminal; IB =interior basal; ISB = interior sub-basal; IST= interior subterminal; IT = interior terminal;EB = exterior basal; ESB = exterior sub-basal;EST = exterior subterminal; ET = exteriorterminal. Cheliceral setae: GLS = galeal seta;ES = exterior seta; IS = interior seta; LS =lateral seta; SB = sub-basal seta; B = basalseta.
Both measurements and terminology hereaccepted are those after CHAMBERLIN (1931)and HARVEY (1992).
3. Systematics
Caecogarypinus gen. n.
Type species: Caecogarypinus pectinodentatussp. n.Diagnosis: Basically Olpioidea Banks, 1895,but totally blind and with a rudimentary ve-nom organ (both venom duct and venomtooth) on movable palpal finger. Carapace(sclerotised part) and pedipalps reddishbrown, latter more intensely so. Abdomenelongate, vermicular, being very similar to thatobserved, e.g., in Minniza Simon, 1881 (fam.Olpiidae Banks, 1895); distance between weak-ly pigmented, yellowish brown tergites
rather large; sternites weakly defined, smoothand whitish yellow. Both tergites and ster-nites weakly distinguished, with deep irregu-lar cuts; marginal setae simple, acuminate andalmost transparent; pleural membranes longi-tudinally rugulose. Caudal segments with 2-4 long and thin pseudotactile setae.
Carapace somewhat longer than posteriorbreadth; its shape usual for Garypinidae (i.e.a narrowed anterior part, or cucullus, poorly-developed transverse striae, a rounded caudalmargin and a modest number of slender,pointed, vestitural setae); margins of sclero-tised parts of sclerites indistinct; neither eyesnor even pigment patches; vestitural setaelong, very thin and transparent, prominentlyacuminate.
Chelicera with five setae on hand, setae Band SB shorter than other setae; movable fin-ger with one subterminal seta directly nearapex of finger. Movable finger without lo-bus, but with six (!) small but well visibleteeth in distal part. Lamina exterior present.Serrula, flagellum and galea more or less typi-cal for this superfamily.
Pedipalps moderately robust, mono-chrome; chela of palp with fine but well-vis-ible granulation in distal part of hand and atthe base of finger, this granulation extendingalso to base of movable finger. Fine granu-lation present also on median side of tro-chanter, of femur and of tibia. Chela denselysetose, with two different types of setation.Palpal fingers both laterally and medially withsemi-transparent, midway curved and distal-ly lanceolate setae located directly along dentalridge; distal ones shorter and less stronglybent; all strongly different from remaining,usual setae of palps. Dentition of chela ofusual garypiniid facies, except for teeth in ba-sal part of movable finger, 5�6 of which for-ming a comb-shaped structure. This groupof the teeth (= comb) combined with basalteeth of fixed finger overlap when both fin-gers are closed, so that of the movable fingerlies medial to the fixed finger. Several basalteeth of fixed finger forming a curved row,
^

A new genus and species of false scorpion 221
Acta Biologica Benrodis 13 (2006)
especially well-visible in ventral view. Tricho-bothriotaxy similar to that of Garypinus Da-day, 1888: IB-ISB-IST-IT-EB-ESB clusteredin basal half of fixed finger; however, IT,IST and ISB nearly forming a line in dorso-ventral direction. ET and EST located indistal half of fixed finger. On movable fin-ger, B-SB-ST assembled into one group, whileT located at about midway of finger. In fixedfinger, both venom duct and venom toothnormally developed, complete and distinct(transmission light microscopy); venom ductshort, nodus ramosus (nr) located betweenET and tip of finger. Venom duct of mov-able finger barely traceable even at strong ma-gnification, unusually slender, nr invisible,both possibly reduced.
Legs of typical facies, whitish yellow, aroli-um divided. Arolium distinctly longer thantarsal claws.
Male not known.Name: The complete absence of eyes or evenpigment spots, which is not characteristic ofmost of the genera composing the familyGarypinidae, as well as certain traits sharedwith Garypinus account for the name of thisnew genus (caecus, lat. = blind).Relations: Because of the lack of males thatare necessary to judge other taxonomicallyimportant traits, such as special fields of se-cretory setae on sternites VI-VIII (present inGarypinus and Serianus J.C, Chamberlin, 1930)and the genital structure, still remain unknownin Caecogarypinus gen. n, the assignment ofCaecogarypinus gen. n. to Garypinidae mustbe provisional. Even if this placement ap-pears to be correct, the new genus occupies ahighly isolated position within Garypinidae.Thus, the movable finger of the chelicerashows sufficiently clearly distinguishableteeth, whereas in most other genera of Ga-rypinidae this finger is terminally equippedwith only one subapical lobe, rarely two. Thecomplete absence of eyes or pigment spots,the unusual structure of the chela, namely,the presence of a group of tightly packed basalteeth forming a comb, the peculiar differen-
tiation of chelal setae, as well as the apparent-ly reduced venom apparatus of the movablefinger seem to be apomorphic characters ofthis taxon. Nevertheless, based on the tri-chobothriotaxy pattern, the shape and pro-portions of the pedipalps, the structure ofthe legs (especially the proportions of leg-I)and the presence of a divided arolium, thisnew genus rather closely resembles both Ga-rypinus and Serianus. However, in the absenceof male material, several important traits, suchas special sternal fields of secretory setae, aswell as the genital structure, still remainunknown in Caecogarypinus gen. n. This ham-pers any further speculations to be made con-cerning the status and affinities of this newgenus.
Caecogarypinus pectinodentatus sp.n.(Figs 1-3).
Holotype: & � ZMMU, Vietnam, Prov. BaRia-Vung Tau, Bihn Chau (Ap Bìhn Châu),10º31´N 107º28´E, sample 1, litter, 20.II.2005,leg. A. E. ANICHKIN.Paratypes: & � (LB No. 001), 1T (LB No 002),1T (ZMMU), Vietnam, Prov. Ba Ria-VungTau, Bihn Chau (Ap Bìhn Châu), 10º31´N107º28´E, sample 2, litter, 20.II.2005, leg.A.E. ANICHKIN.Description of the holotype (&) � (Figs 1A, C; 2 A, C, G, F, H, I; 3 A, E): Body long,vermicular, tegument almost transparent.Tergites and sternites weakly pigmented,light yellow, carapace light brown, palpsvividly reddish brown (chela more intenselyso), legs yellow-white. Carapace completelysmooth, without granulation, visible sclero-tised part 1.29 times longer than broad. Cu-cullus short and broad. Chaetotaxy of cara-pace: anterior margin with four setae, pos-terior margin with six setae (24 setae in total);formula of lyrifissures 2-2-2-2 (total eight),posterior pair located at the very edge of cara-pace. Tergites, except for two first ones, di-vided, but, tegument being almost transpar-ent, separating lines indistinct. Pleural mem-

222 SELVIN DASHDAMIROV
Figs 1 A-E: Caecogarypinus pectinodentatus sp.n. (A, C � &, holotype [ZMMU]; B, D, E � &, paratype[LB No 001]). A General aspect, dorsal view. B Left chelicera, dorsal view, showing dentition ofmovable finger. C Details of pleural membranes. D Tip of movable finger of chela, mesial view. EBase of movable finger of chela, lateral view.Abb. 1 A-E: Caecogarypinus pectinodentatus sp.n. (A, C � &, Holotyp [ZMMU]; B, D, E � &, Paratyp[LB No 001]). A Übersicht, dorsal. B Linke Chelicere, dorsal, mit der Bezahnung des beweglichenFingers. C Detail der Pleuralhaut. D Spitze des beweglichen Fingers der Chela, mesial. E Basis desbeweglichen Fingers der Chela, lateral.

A new genus and species of false scorpion 223
Acta Biologica Benrodis 13 (2006)
brane longitudinally rugulose. Tergal chaeto-taxy 7:6:8:8:8:8:8:8:10:14(10+4 long pseu-dotactile setae):12(8+4 long pseudotactilesetae):2. Chaetotaxy of sternites x:6(3+3):3mm-6-3mm:2mm-6-2mm:8:8:8:9:9:11(7+4 long pseudotactile setae):10 (8+2 longpseudotactile setae):2. These pseudotactilesetae, both on tergites and sternites, verystrongly narrowed at base and therefore ex-tremely flexible (Fig. 2G). Coxal chaetotaxy:P-11, manducatory process strongly nar-rowed, almost styliform, with one apical andone subapical, rather long seta and a very shortbut well-developed suboral seta near latter�sbase; I � 7; II � 6; III � 4-5; IV � 6. Chelicerawith five long acuminate setae in basal part;movable finger with one distal seta; serrulaexterior with 19 lamellae; flagellum with foursimple, blades without spinules. Galea withtwo long subterminal and one terminalbranch. Fixed finger with nine flat and smallteeth. Movable finger without subapical lobe,but with six tiny, but well-visible teeth. Palppattern and trichobothriotaxy as illustrated(Fig. 2C). Palp relatively robust, medial sur-face of trochanter, tibia and femur with veryfine and even granulation; hand with granu-lation in distal part; granulation also presentat base of fixed finger and a few grains pre-sent on ventral side at base of movable fin-ger, elsewhere quite smooth. Trichobothrio-taxy almost as in Garypinus: IB-ISB-IST-IT-EB-ESB clustered at base of fixed finger, EST-ET solitary in distal position of finger, SBalmost level to ST. Fingers rather thick indorsal view, especially so closer to base; pro-portions: trochanter 1.98 times as long asbroad; femur 2.46 times as long as broad;patella 2.15 times as long as broad; chela withpedicel 2.9 times, without pedicel 2.71 times,as long as broad, its chaetotaxy pattern, ve-nom apparatus and dentition as illustrated(Figs 3A, E). Hand with pedicel 1.85 times,without pedicel 1.62 times, as long as broad.Hand with pedicel 1.56 times, without pedi-cel 1.36 times, as long as movable finger.Movable finger of chela with 22 teeth, six
basal clustered into one large and tightlypacked group (comb) elevated well above thelevel of remaining dental ridge. Fixed fingerof chela with 25 marginal teeth, basal part ofdental ridge a little curved on lateral side. Thesetwo basal groups of teeth overlap when thefingers are closed, so that the comb-shapedbasal teeth of the movable finger enter the�scabbards� like the blade of a penknife. Bothfingers, in addition to a dense usual setation,with several strongly curved or bent, terminallylanceolate, almost transparent (especiallydistally) setae located directly along dentalridge, these setae only being visible using dif-ferential interference contrast. When chelal fin-gers are partly or fully closed, these stiff setae,which are located at an acute angle to the mainaxis of the fingers, overlap to create a kind of�cage�. Lamina defensor filamentary, presentin both fingers. A short venom duct wellvisible only in fixed finger, hardly traceableonly in distal part of movable finger, possib-ly rudimentary; diameter of channel aboutfour times less than that in fixed finger; nrinvisible in moveable finger, well visible infixed finger, situated there between T and tipof finger, attached to other 2-3 thick ducts(atrium), which pass into well visible venomglands. Leg-IV of typical garypiniid facies (Fig.2I), tibia and metatarsus with tactile setae, aswell as numerous, long, prominently acum-inate setae; proportions: femur+patella 2.86times as long as deep; tibia 3.47 times as longas deep, TS=0.50; metatarsus 1.81 times aslong as deep, TS=0.25; tarsus 2.78 times aslong as deep, tarsus 1.66 times as long asmetatarsus. Arolium bifurcate, with well-vi-sible denticulate edges. Claws shorter thanarolium.
Leg-I as illustrated (Fig. 2A). Proportions:femur+patella 3.1 times as long as deep; tibia3.23 times as long as deep; metatarsus 1.6times as long as deep; tarsus 2.94 times aslong as deep. Tarsus 1.66 times long as meta-tarsus.
Measurements (length/breadth or depth,in mm).

224 SELVIN DASHDAMIROV
Figs. 2 A-I: Caecogarypinus pectinodentatus sp.n. (A, C, G, F, H, I � &, holotype [ZMMU]; E � &,paratype [LB No 001]; B, D � tritonymph, paratype [LB No 002]). A Right leg-I, lateral view. BRight chela, lateral view, showing dentition, trichobothriotaxy and modified stiff setae on bothfingers (exclusive of vestitural setae). C Right palp, dorsal view. D Fingers of right chela, mesialview, showing dentition, trichobothriotaxy and modified stiff setae on both fingers (exclusive ofvestitural setae). E External chaetotaxy of the genital opercula (segments 2 and 3). F Base ofchelal fixed finger, ventral view. G Details of caudal segment, ventral view, showing pleuralmembranes and pseudotactile seta. H Arolium. I Right leg-IV, lateral view.Abb. 2 A-I: Caecogarypinus pectinodentatus sp.n. (A, C, G, F, H, I � &, Holotyp [ZMMU]; E � &,Paratyp [LB No 001]; B, D � Tritonymphe, Paratyp [LB No 002]). A Rechtes Bein-I, lateral. BRechte Chela, lateral, mit Bezahnung, Anordnung der Trichobothrien und modififierten, steifenSetae an beiden Fingern (mit Ausnahme der die Oberfläche bedeckenden Setae). C: RechterPalpus, dorsal. D: Finger der rechten Chela, mesial, mit Bezahnung, Anordnung der Trichobothrienund modifizierten, steifen Setae an beiden Fingern (mit Ausnahme die Oberfläche bedeckendenSetae). E: Anordnung der Chaetae auf den Opercula der Genitalöffnung (Segmente 2 und 3). FBasis der Chela des nicht beweglichen Fingers, ventral. G Detail des caudalen Segments, ventral,mit Pleuralhäuten und pseudotaktiler Seta. H Arolium. I Rechtes Bein-IV, lateral.

A new genus and species of false scorpion 225
Acta Biologica Benrodis 13 (2006)
Carapace 0.61/0.48. Palp: trochanter 0.35/0.18; femur 0.55/0.22; patella 0.55/0.25; che-la with pedicel 0.95/0.33; length of chela wi-thout pedicel 0.89; length of hand with pedi-cel 0.61, without pedicel 0.53; length of mo-vable finger 0.39. Leg IV: femur+patella 0.56/0.20; tibia 0.40/0.12; metatarsus 0.13/0.07;tarsus 0.18/0.06. Leg I: femur+patella 0.36/0.12; tibia 0.27/0.08; metatarsus 0.10/0.06;tarsus 0.16/0.05.
Name: Accounting for its comb-shaped teethin the basal portion of the movable chelalfinger (pecten, lat.=comb; dentatus, lat.= withteeth).
Description of the paratype (&&&&&) (Figs 1B,D, E; 2E; 3B-D, G-K):
Like holotype, except as follows. Palp: tro-chanter 1.89 times as long as broad; femur2.31 times as long as broad; patella 2.11 timesas long as broad; chela with pedicel 2.94 times,without pedicel 2.74 times, as long as broad,its chaetotaxy pattern, venom apparatus, den-tition and trichobothriotaxy as illustrated(Figs 1D, E, 3B, J). Hand with pedicel 1.84times, without pedicel 1.61 times, as long asbroad. Hand with pedicel 1.59 times, withoutpedicel 1.39 times, as long as movable finger.Movable finger of chela with 22 teeth, butonly five basal ones clustered into a comb.Fixed finger of chela with 25 marginal teeth.Fixed finger of this female lacking one tri-chobothrium (ET) on right chela. Both fin-gers also with strongly curved, terminallylanceolate setae. Lamina defensor filament-ary; present in both fingers.
Measurements (length/breadth, in mm).Palp: trochanter 0.32/0.17; femur 0.48/
0.21; patella 0.49/0.25; chela with pedicel 0.89/0.30; length of chela without pedicel 0.83;length of hand with pedicel 0.56, withoutpedicel 0.49; length of movable finger 0.35.
Description of the tritonymph (Figs 2B, D):Apart from a considerably smaller size, trito-nymph closely resembled the adults.
Chaetotaxy of carapace: anterior margin withfour setae, posterior margin with six setae(20 setae in total). Tergal chaetotaxy:6:6:6:6:8:7(?):8:8:8:12(8+4 long tactile se-tae):12(8+4 long tactile setae):2. Palp: trochan-ter 2.18 times as long as broad; femur 2.43times as long as broad; patella 2.31 times aslong as broad; chela with pedicel 3.10 times,without pedicel 2.91 times, as long as broad,its selected chaetotaxy pattern, venom appa-ratus, dentition and trichobothriotaxy as il-lustrated (figs. 2B, D). Hand with pedicel 1.86times, without pedicel 1.62 times, as long asbroad. Hand with pedicel 1.44 times, withoutpedicel 1.26 times, as long as movable finger.Movable finger of chela with 15 teeth, andonly four basal teeth, clustered in basal bulge.Fixed finger of chela with 22 marginal teeth.Both fingers with a smaller number of strong-ly curved, terminally lanceolate setae than inadult.
Measurements (length/breadth, in mm).Palp: trochanter 0.32/0.17; femur 0.48/
0.21; patella 0.49/0.25; chela with pedicel 0.89/0.30; length of chela without pedicel 0.83;length of hand with pedicel 0.56, withoutpedicel 0.49; length of movable finger 0.35.
3. Discussion
As noted above, one of the most striking fea-tures of the new genus and species describedherein is the structure of the palpal chela. Inparticular, in the basal portion of the movablefinger there is a comb-shaped dental groupwhich forms, together with the �scabbards�created by a curved dental ridge on the oppo-site finger, a kind of scissors when the fingersare closed. In addition, there are several pecu-liar stiff setae placed at an acute angle to thelongitudinal axis of the fingers that form akind of cage when the fingers are partly orcompletely closed. These strange structures canwell be presumed as being of utmost im-portance for searching and capturing a prey.
According to A.ANICHKIN (pers. comm.),the Bihn Chau Nature Reserve is a relict de-

226 SELVIN DASHDAMIROV
Figs 3 A-L: Caecogarypinus pectinodentatus sp.n. (A, E � &, holotype [ZMMU]; B-D, G-K � &,paratype [LB No 001]). A Tips of fingers of chela, lateral view, showing venom apparatus of fixedand hardly visible venom(?) duct of movable finger, respectively (nr � nodus ramosus). B Fingersof right chela, lateral view. C: Lamina exterior, lateral view. D Lamina interior. E Left chela,lateral view. F Tip of maxilla, showing chaetotaxy and apical process. G Suboral seta. H Carapaceand left chelicera, dorsal view. I Tips of fingers of chelicera, dorsal view, showing dentition. JFingers of right chela, mesal view, showing dentition, trichobothriotaxy, venom apparatus andmodified stiff setae on both fingers (exclusive of vestitural setae). K Basal group of teeth on

A new genus and species of false scorpion 227
Acta Biologica Benrodis 13 (2006)
Figs 4 A, B: Presumed ways of hunting. A Prey capture and retention. B �Sieving� the substrate.Sieved particles (small arrows), direction of movement (big arrow).Abb. 4 A, B: Mögliche Art und Weise des Beutefangs. A Ergreifen und Festhalten der Beuten. BAussieben des Substrats; ausgesiebte Partikel (kleine Pfeile), Bewegungsrichtung (großer Pfeil).
movable finger of chela, mesal view. L Modified stiff seta of movable finger of chela, showingcross section of seta.Abb. 3 A-L: Caecogarypinus pectinodentatus sp.n. (A, E � &, Holotyp [ZMMU]; B-D, G-K � &,Paratyp [LB No 001]). A Fingerspitzen der Chela, lateral, mit Giftapparat im unbeweglichen undkaum sichtbarem Giftgang (?) im beweglichen Finger (nr � nodus ramosus). B Finger der rechtenChela, lateral. C: Lamina exterior, lateral. D Lamina interior. E Linke Chela, lateral. F Spitze derMaxille mit Anordnung der Chaetae und apikalem Fortsatz. G Suborale Seta. H Carapax und linkeChelicere, dorsal. I Fingerspitzen der Chelicere, dorsal, mit Bezahnung. J Finger der rechten Chela,mesial, mit Bezahnung, Anordnung der Trichobothrien, Giftapparat und modifizierten, steifenSetae (mit Ausnahme der die Oberfläche bedeckenden Setae). K Basal gelegenen Zähne auf dembeweglichen Finger der Chela, mesial. L Modifizierte, steife Seta auf dem beweglichen Finger derChela mit den Querschnitt von Seta.

228 SELVIN DASHDAMIROV
ciduous dipterocarp forest located just at theshore of the South China Sea. Binh Chau isone of the few areas along the coastline ofVietnam to retain a significant cover of natu-ral forest. The principal reason for this maybe that the prevailing climate, which is charac-terised by low rainfall and an extended dryseason, makes the area unsuitable for perma-nent agriculture. The soil is loose large-grainquartz sandy to sandy clay, mainly of duneorigins. Due to heavy anthropogenic pres-sure the charcoal output is so heavy that inmany places the soil is still dark grey from theashes. At present, the forest is very light; theundergrowth is dense, a herb cover is mis-sing. At least in the dry season which lastsfrom December to April, when the animalswere collected, a rather thick leaf litter waspresent, in which Psocoptera (barklice), Ma-chiloidea (bristle-tails) and Blattodea (cock-roaches) dominated. Collembola (spring-tails)were few, oribatid mites, however, abundant.Since this false scorpion is obviously blind,soft-bodied and vermiform, living in the lit-ter and uppermost soil/sand, it seems veryreasonable to surmise its interstitial life-style.Mites and collembolans might well be amongits preys (Fig. 4A), in which case the �cage�formed by the modified stiff setae on bothfingers of the chela, as well as the �scissors�created by the dental ridges could well accountfor the way the animal hunts. The somewhatreduced venom apparatus in one of the fin-gers might be evidence of an evolutionarytrend, in which the killing or immobilisingfunction could have become shared betweenthe well-developed venom apparatus of theother finger and the �scissors�. Interestingly,a similar structure of the chelal fingers is ob-served in some other, quite distantly relatedfalse scorpions. Thus, Metatemnus superiorMuchmore, 1972 (fam. Atemnidae J.C. Cham-berlin, 1931), known as phoretic on somebeetles in Malaysia, shows a bulge at the baseof each of the chelal fingers (MUCHMORE
1972). However, in this case the �scissors�might well be advantageous not only for
hunting, but also for making a tighter clutchat the insect transport.
In Caecogarypinus pectinodentatus sp. n., a lifein the small crevices in the sand and in thelitter might well account for a frontal posi-tion of the shortened and stout palps usedfor probing. The �cage� could be helpful forsieving the substrate (Fig. 4B), as well as cap-turing and keeping a prey, while the �scis-sors� for cutting the prey, especially such hea-vily sclerotised organisms as oribatid mites.
Acknowledgements
I am deeply indebted to Mr. ALEXANDER
ANICHKIN (Vietnamese-Russian Tropical Cen-ter, Hanoi) and Dr. SERGEI GOLOVATCH (Mos-cow, Russia) for the loan of the material ex-amined here, to Prof. Dr. HARTMUT GREVEN
(Institut für Zoomorphologie und Zellbio-logie, Universität Düsseldorf) for helpful dis-cussions, to Dr. WOLFGANG GETTMANN
(Aquazoo � Löbbecke-Museum, Düsseldorf)for providing the necessary facilities to studythe material. I am again obliged to Dr. S.GOLOVATCH for checking the English of anearlier draft as well as for his critiques.
Literature
BANKS, N., 1895: Notes on the Pseudoscorpion-ida. � Journal of the New York Entomolog-ical Society 3, 1-13.
CHAMBERLIN, J.C., 1930: A synoptic classifica-tion of the false scorpions or chela-spin-ners, with a report on a cosmopolitan col-lection of the same. Part II. The Diplos-phyronida (Arachnida-Chelonethida). �Annals and Magazine of Natural History(10) 5, 1-48, 585-620.
CHAMBERLIN, J.C., 1931: The arachnid order Che-lonethida. � Stanford University Publications,Biological Sciences 7, 1-284.
DADAY, E., 1888a: A Magyar Nemzeti Muzeumálskorpióinak áttekintése. � TermészetrajziFüzetek 11, 111-136.
DADAY, E., 1888b: A Magyar Nemzeti Muzeumálskorpióinak áttekintése. � TermészetrajziFüzetek 11, 165-192.

A new genus and species of false scorpion 229
Acta Biologica Benrodis 13 (2006)
HARVEY, M.S., 1992: The phylogeny and syste-matics of the Pseudoscorpionida (Chelicera-ta: Arachnida), � Invertebrate Taxonomy 6,1373-1435.
JUDSON, M.L.I., 2005: Baltic amber fossil ofGarypinus electri Beier provides first videnceof phoresy in the pseudoscorpion family Ga-rypinidae (Arachnida, Chelonethi), � Arthro-poda selecta, Special issue 1 [2004], 127-131.
STAHLAVSKÝ, F., KRAL, J., HARVEY, M.S., HADDAD,C.R., 2006: A karyotype study on the pseu-
doscorpion families Geogarypidae, Garypini-dae and Olpiidae (Arachnida: Pseudoscorpio-nes). � European Journal of Entomology 103,277-289.
MUCHMORE, W.B., 1972: A phoretic Metatemnus(Pseudoscorpionida, Atemnidae) from Ma-laysia. � Entomological News 83, 11-14.
SIMON, E., 1881: Descriptions d�Arachnides nou-veaux d�Afrique (Chernetes de la BasseÉgypte rec. par M. Letourneux). � Bulletinde la Société zoologique de France 6, 1-15.
Accepted: 22.12.2006
^

230 SELVIN DASHDAMIROV
Rezension
WICHARD, WILFRIED & WEITSCHAT, WOLFGANG (2004): Im Bernsteinwald. � Gerstenberg-Verlag, Hildesheim. 168 Seiten mit zahlreichen Abbildungen. Gebunden, 25,5 x 25,3 cm.ISBN 978-3-8067-2551-39. 35,-- Euro
Poetisch ist der Titel, nicht minder poetisch ist der Inhalt. Schließlich kommen so namhafteDichter wie Martial und Ovid zu Wort und ich bin mir nicht sicher, ob spätere Autoren soschöne Worte über Bernstein gefunden haben wie diese beiden. Ein Zoologe und einGeologe haben ein Buch geschrieben, in dem ihre Liebe zu baltischem Bernstein deutlichwird und an der sie auch andere teilhaben lassen wollen, und dies tun sie mit einem ver-ständlichen Text und einer Fülle von eindrucksvollen Bildern, die zeigen, was alles an Tierenund Pflanzen in Bernstein eingeschlossen worden ist. Ein einführendes Kapitel (�Bernstein� Fenster in die Vergangenheit�) befasst sich kurz mit dem Ursprung des Bernsteins, mitFragen seiner �Wanderung�, mit dem baltischen Bernsteinwald und seiner möglichen Lageund Ausdehnung, mit Bernstein in eiszeitlichen Ablagerungen, mit der Frage, welche Bäu-me das Harz für den Bernstein produziert haben, mit der Entstehung von Inklusen etc.sowie mit früherer und heutiger Bernsteinforschung. Es folgen weitere Kapitel wie �Inklu-sensteine � goldene Gräber,�, �Szenen aus dem Bernsteinwald�, �Von Farnen, Moosen,Knospen, Blüten und Blättern�, �Tropische Exoten�, �Die Gewässerfauna des Bernstein-waldes� und �Wirbeltiere � eine Spurensuche�, in denen die Abbildungen (manchmal etwasübervergrößert und leider ohne Maßangaben) überwiegen, die aber stets mit einem infor-mativen Text versehen sind. Bereits die Kapitelüberschriften lassen das weite Spektrum vonOrganismen ahnen, die in Bernstein eingeschlossen sind. Im Anhang findet sich eine Auf-listung der Tiergruppen mit Angaben ihrer prozentualen Häufigkeit in baltischem Bern-stein. Darüber hinaus helfen ein kurzes Glossar, ein �Werkstattbericht� (über die Arbeits-weise von Bernsteinforschern), eine Auswahl weiterführender Literatur sowie eine Liste vonBernstein-Sammlungen und Bernstein-Museen in Europa weiter. Dies ist zunächst Rüst-zeug genug, um im Bernsteinwald auf Jagd zu gehen.
Hartmut Greven, Düsseldorf

Rassenunterschiede bei Haushühnern bei Konditionierung in der Skinner-Box 231
Acta Biologica Benrodis 13 (2006)
Acta Biologica Benrodis 13 (2006): 231-240
Rasseunterschiede bei Haushühnern (Gallus gallus f. d.)bei klassischer und operanter Konditionierung in der
Skinner-Box
Breedspecific differences among domesticated chickens (Gallusgallus f.d.) revealed by classical and operant conditioning in the
Skinner-Box
Inga TIEMANN 1,2 & Gerd REHKÄMPER 1
1 C. und O. Vogt Institut für Hirnforschung, Heinrich-Heine-Universität Düsseldorf,Universitätsstr. 1, D-40225 Düsseldorf, Germany
2 Wissenschaftlicher Geflügelhof des BDRG, Am Landwirtschaftsmuseum 10, D-41569 Rommerskirchen, Germany; [email protected]
Zusammenfassung: Vier Haushuhnrassen (Holländer Haubenhuhn, Bantam, Zwerg-Cochin undChabo) wurden während des Vortrainings (Shaping) und Autoshapings in der Skinner-Box beob-achtet. Dauer und Erfolgsquote beim Erreichen der Kriterien �Futteraufnahme� und �erste Pick-reaktion� wurden quantitativ ausgewertet. Ein Holländer Haubenhuhn schloss das Vortraining,sechs weitere Holländer Haubenhühner und ein Chabo das Autoshaping nicht erfolgreich ab.Signifikante Unterschiede zeigten sich im Vergleich des Zeitpunkts der ersten Pickreaktion und inder daraus resultierenden Anzahl notwendiger Wiederholungen des Autoshapings zwischen Hol-länder Haubenhühnern und Bantams auf der einen und Chabos und Zwerg-Cochin auf der anderenSeite. Die Holländer Haubenhühner erlernten die operante Konditionierung langsamer und damitschlechter als Tiere der Rassen Chabo und Zwerg-Cochin. Die Bantams nehmen etwa eine Mittel-stellung zwischen diesen beiden Gruppen ein. Diese verhaltensbiologischen Beobachtungen sindnicht mit der bekannten Vergrößerung lernrelevanter Hirnbereiche bei Holländer Haubenhühnernzu vereinbaren.
Haushühner, klassische und operante Konditionierung, Rasseunterschiede
Summary: Four breeds of the domestic chicken (White Crested Polands, Rosecomb Bantams,Peking Bantams and Japanese Bantams) were observed during shaping and autoshaping in anoperant conditioning chamber (Skinner-Box). Duration and rate of success were quantitativelyanalysed based on ethological criteria such as ingestion and first pecking. One White Crested Polishhen did not pass the pretraining and six White Crested Polands and one Japanese Bantam did notpass the autoshaping procedure. Comparing the point of first pecking and the resulting repetitionsof the autoshaping, significant differences appeared between White Crested Polands and Rose-comb Bantams on the one hand and Japanese Bantams and Peking Bantams on the other hand.White Crested Polands learned the operant conditioning slower and altogether worse than theJapanese Bantams and Peking Bantams. Rosecomb Bantams took a middle position between thetwo groups. These behavioural observations are not congruent with the known enlargement oflearning related brain areas in White Crested Polish chickens.
Domestic chickens, classical and operant conditioning, breedspecific differences

232 INGA TIEMANN & GERD REHKÄMPER
1. Einleitung
Domestikation wird als ein Prozess beschrie-ben, bei dem die Adaptation der Tiere an denMenschen und seine Umwelt im Vorder-grund steht. Diese Adaptation wird durcheine Kombination aus genetischen Verände-rungen über die Generationen hinweg unddurch eine umweltbedingte Individualent-wicklung gewährleistet, die sich in jeder Ge-neration wiederholt (PRICE 2002). Die Selek-tion durch den Menschen und individuelleErfahrungen beeinflussen das Tier. Die Sti-mulation durch die Umwelt, in Form vonHabituation an und Konditionierung durchStimuli, die mit dem Haltungsumfeld desTieres assoziiert sind, muss deshalb nebender selektionsfähigen Genetik berücksichtigtwerden. (OCHIENG-ODERO 1994).
Eines der ältesten Haustiere ist das Haus-huhn, das vom roten Dschungelhuhn Gallusgallus gallus abstammt und dessen Domesti-kation zwischen 5400 v. Chr. (CRAWFORD
1990) und 3200 v. Chr. (WOOD-GUSH 1959)einzuordnen ist. Der intensiven Nutzung desHaushuhns zur Lebensmittelgewinnungsteht die Haltung von Rassegeflügel als Frei-zeitgestaltung gegenüber. Zum jetzigen Zeit-punkt sind in Deutschland 189 Hühnerras-sen anerkannt, die nach Maßgabe eines Stan-dards (Deutscher Rassegeflügelstandard,BDRG E.V. 2006) gezüchtet werden. Wäh-rend für den Phänotyp definierte morpholo-gische Selektionskriterien vorliegen, sind Ver-haltensmerkmale nur in wenigen Rassen Teildes Zuchtziels (z.B. Bergische Kräher mit lan-gem Krähruf). Auffallend ist die rassespezi-fische Hirnzusammensetzung bei Haushüh-nern (REHKÄMPER et al. 2003), die als Anpas-sung an einen spezifischen Lebensraum zuverstehen ist (REHKÄMPER et al. 2000/2001).
Untersuchungen zur Rassespezifität desLernverhaltens stammen von SCHULZE-SCHOLZ (1961). Die Autorin stellte bei denvon ihr als unruhig beschriebenen Hauben-hühnern eine schlechte Lernleistung, für �ru-higere� Rassen (z.B. Chabo, Orpington) aber
eine bessere Lern- und Gedächtnisleistung festund begründete dies mit dem unterschiedli-chen Temperament der Rassen. REHKÄMPER
et al. (1995) beschrieben eine relative Vergrö-ßerung lernrelevanter Hirnareale (z.B. Nido-pallium) bei der Rasse �Chabo� und zeigten,dass Tiere dieser Rasse die operante Kondi-tionierung in einer Skinner-Box lernen kön-nen. Zur Konditionierung bedarf es einergrundlegenden ethologischen Plastizität, dieeine Adaptation an den Hausstand insoferndarstellt, als dass auch hier angepasste Ver-haltensweisen Erfolg versprechend sind.
Vergleichende Untersuchungen an Gehir-nen verschiedner Hühnerrasen haben be-trächtliche Unterschiede aufgezeigt (FRAHM
& REHKÄMPER 1998; REHKÄMPER et al. 2003).Namentlich bei Holländer Haubenhühnernwaren das Diencephalon und das Telence-phalon signifikant vergrößert; die Vergrö-ßerung des Telencephalons war vor allemauf die Vergrößerungen des Mesopalliumund des Hyperpallium densocellularum zu-rückzuführen. Diese Bereiche, die bei Hol-länder Haubenhühnern mit 405 mm3 fastdoppelt so groß sind wie bei Chabos (FRAHM
& REHKÄMPER 1998), sind nach MCCABE undHORN (1988) mit dem Lernverhalten asso-ziiert. Die Erwartungshaltung ist, dass mitder Vergrößerung dieser Areale eine höhereLernleistung einhergeht und Holländer Hau-benhühner schneller lernen als Vergleichs-rassen
Im Folgenden wird die Lernleistung vonHolländer Haubenhühnern mit der von an-deren Hühnerrassen (Bantam, Chabo, Zwerg-Cochin) verglichen. Als standardisierte Lern-situation ist die klassische und operante Kon-ditionierung in der Skinner-Box anzusehen.Dabei werden vorhandene Verhaltensweisenmit neuen Reizen durch Belohnung ver-knüpft. Die Verknüpfung selber ist als Ler-nen zu verstehen, so dass die Dauer bis zurVerknüpfung von Verhalten, Reiz und Be-lohnung als quantitatives Maß zur Lernfä-higkeit verwendet werden kann (LASHLEY
1929).

Rassenunterschiede bei Haushühnern bei Konditionierung in der Skinner-Box 233
Acta Biologica Benrodis 13 (2006)
2. Tiere, Material und Methode
2.1. Tiere und ihre Haltung
Die für den Rassevergleich verwendetenHennen gehörten den HaushuhnrassenBantam, Holländer Haubenhuhn, Chabound Zwerg-Cochin an (Abb. 1; zur Be-schreibung der rassespezifischen Merkma-le vgl. TEGETMEIER 1873; DÜRIGEN 1921).Die Hühner hatten, bevor das Experimentbegann, keine Erfahrung mit der Skinner-Box.
Die Hühner wurden nach Rassen getrenntund zu maximal je zehn Hennen in Volierenmit den Maßen 150x250x120 cm (HxBxT)untergebracht. Alle Tiere waren geschlechts-reif und individuell mit Ringen gekennzeich-net. Futter (handelsübliches Legemehl in Pel-let-Form) und Wasser standen ad libitum zurVerfügung. In der Trainingsphase war Futterallerdings nur für etwa 30 min im Anschlussan die Experimente verfügbar.
Um den Holländer Haubenhühnern eineuneingeschränkte Sichtfreiheit zu gewähren,wurden die Federn der Haube so beschnit-ten, dass nur der unterste Bereich des Fe-derschafts mit den Dunenstrahlen stehenblieb.
2.2. Skinner-Box
Alle Experimente wurden in der Skinner-Box durchgeführt (Abb. 2). Die Box bestandaus einem hölzernen Kasten mit den Ma-ß e n46 x 40 x 40 cm (H x B x T). Die Frontseiteder Box war mit drei Pickscheiben ausge-stattet, von denen nur die mittlere für dieExperimente genutzt wurde. Die Pickschei-ben waren aus Plastik, zweifach mit Folie(Letraset® opaque black) beklebt und wur-den von einem Diaprojektor angestrahlt.Ebenfalls an der Frontseite befand sich derFuttergeber. Die gegenüberliegende Seite waroffen; hier wurden die Tiere hineingesetztund nach dem Versuch entnommen. Die
Skinner-Box war während des Experimentsmit einem Netz verschlossen, um die Tieream Verlassen der Box zu hindern. Der Bo-den der Box konnte je nach verwendeterRasse in zwei verschiedene Höhen (7 cm oder15 cm) eingestellt werden, damit die TierePickscheibe (Höhe: 35 cm) und Futtergeber(Höhe: 21 cm) erreichen konnten. Die Tierestanden auf einer rutschfesten Gummimatte,die leicht zu reinigen war. Im Falle eines Pick-schlages wurde die Pickscheibe so ausgelenkt,dass sie eine Lichtschranke unterbrach. Dasentstehende elektrische Signal wurde als�Pick� vom Computer erfasst. Der Futterge-ber wurde durch einen Klappmechanismusreguliert, d.h. für das Tier (un-)zugänglichgemacht. Diese Apparatur wurde von einemComputer gesteuert, der auch die Daten spei-cherte.
Das Futter für den Futtergeber war Kü-kengrütze, bestehend aus Maisbruch, Weizenund Haferflocken (Firma Raiffeisen). In derBox konnte das Huhn mittels einer Kameramit angeschlossenem Monitor beobachtetwerden. Die in der Box angebrachten Lam-pen hatten unterschiedliche Funktionen. Eineder Lampen war das in Skinner-Boxen übli-che �Hauslicht�, welches während der gesam-ten Trainingszeit leuchtete, die andere das�Futterlicht�, das ausschließlich bei hochge-klapptem Futtergeber brannte.
2.3. Ablauf des Experiments
Alle Hennen durchliefen zuerst das Vortrai-ning (�Shaping�), in dem sie an die Skinner-Box gewöhnt wurden, bevor sich das Auto-shaping anschloss.
Unter �Shaping� versteht man eine Verhal-tensformung durch Verstärkung, d.h. einzel-ne Verhaltenskomponenten, die dem Lern-ziel entgegenkommen, werden belohnt undtreten daher in der Folge häufiger auf. DasAutoshaping folgt dem Vortraining und ent-spricht einer Selbstausformung des Verhal-tens durch das Individuum selbst; dabei kannes zu neuen Verhaltensweisen oder Verhal-

234 INGA TIEMANN & GERD REHKÄMPER
Abb. 1A-D: Kopfstudien von Hennen der untersuchten Rassen. A Schwarze Holländer Hauben-huhn-Henne mit weißer Vollhaube. B Schwarze Bantam-Henne. C Gelbe Chabo-Henne. D SchwarzeZwerg-Cochin-Henne.Figs. 1A-D: Heads of the hens of the observed breeds. A Black Polish hen with a white crest. BBlack Rosecomb Bantam. C Yellow Japanese Bantam. D Black Peking Bantam.
A B
C D
tenskombinationen kommen. In dieser Un-tersuchung ist das �geformte Verhalten� dasPicken auf die Pickscheibe.
Das Vortraining wurde in drei aufeinan-derfolgende Phasen (I-III) eingeteilt und täg-lich durchgeführt. In allen Phasen wurde diePickscheibe von dem Projektor angestrahlt;Haus- und Futterlicht brannten ebenfalls. InPhase I wurde das Huhn zum ersten Mal indie Box gesetzt. Auf dem fixierten Futterge-ber wurde � für das Huhn sichtbar � Futter(Kükengrütze) angehäuft und zudem Futterauf dem Boden verteilt. Konnte innerhalb
von 600 s auf dem Monitor beobachtet wer-den, dass das Tier Futter vom Boden auf-nahm, so galt die Phase I als erfolgreich durch-laufen. In Phase II lag kein Futter mehr aufdem Boden, doch war noch Kükengrütze aufdem fixierten Futtergeber aufgehäuft. Nahmdas Huhn innerhalb von 600 s dieses Futterauf, war auch Phase II erfolgreich abgeschlos-sen. In Phase III war das Futter nicht mehrdirekt sichtbar, sondern musste vom Tier ausdem fixierten Futtergeber aufgenommenwerden. War dies innerhalb von 600 s derFall, war auch diese Phase ebenfalls erfolg-

Rassenunterschiede bei Haushühnern bei Konditionierung in der Skinner-Box 235
Acta Biologica Benrodis 13 (2006)
reich beendet. Fraß das Huhn in einer derPhasen nicht, wurde diese am nächsten Tagwiederholt. War das Huhn nach drei Wieder-holungen einer Phase nicht erfolgreich, soschied es aus. Protokolliert wurden die An-zahl der täglich stattfindenden Sitzungen, diebenötigt wurden, bis alle drei Phasen erfolg-reich durchlaufen waren, und die Latenzzei-ten bis zur Futteraufnahme in den einzelnenPhasen.
Das Autoshaping, das am ersten Tag nacheinem erfolgreich abgeschlossenen Vortrai-ning begann, war in unterschiedliche Zeitin-tervalle gegliedert. Die aufeinanderfolgendeSequenz von Pause, Belohnungsverzöge-rung, Reizintervall und Futtergabe wird imWeiteren als �Durchgang� bezeichnet. Insge-samt 40 Durchgänge bildeten eine Sitzung,die einmal täglich stattfand. Ein Durchgangbegann mit einer Pause, in der die Pickschei-be nicht beleuchtet wurde. Die Länge der Pau-se betrug durchschnittlich 60 s. Nach Ablaufder Pause wurde die Pickscheibe durch denProjektor beleuchtet (Stimulus). Diese Be-leuchtungsphase wurde in zwei Intervalleuntergliedert, die Belohnungsverzögerung
(reinforcement-delay) und das Reizintervall.Die Belohnungsverzögerung war der Zeit-raum, in dem der Computer die Pickschlägeauf die Pickscheibe zählte, während im Reiz-intervall Pickschläge nicht registriert wurden.Das Picken des Huhns im Reizintervall undin der Pause hatte einen Abbruch des jeweili-gen Intervalls zur Folge, so dass schneller einerneuter Durchgang begann. Die Längen al-ler Intervalle konnten durch den Experimen-tator entsprechend der verschiedenen Phasendes Autoshapings verändert werden. Nachder Präsentation des Stimulus klappte derFuttergeber nach oben und das Huhn konn-te an das Futter gelangen. Nach Ablauf die-ser Futtergabe folgte erneut eine Pause. DasAutoshaping gliederte sich ebenfalls in dreiPhasen (I-III). Die Belohnungsverzögerungwurde von 0 s in Phase I und 2,5 s in Phase IIauf 5 s in Phase III erhöht. Das Reizintervallverringerte sich gleichzeitig so, dass beide In-tervalle (Stimuluspräsentation) zusammenimmer 8 s dauerten. Die Dauer der Fütte-rung, d.h. die Zeit, in der der Futtergeberhochgeklappt und das Futter zugänglich war,reduzierte sich von zunächst 5 s in Phase I
Abb. 2: Verwendete Apparatur (Skinner-Box, Computer, Projektor). Zeichnung: OPFERMANN-RÜNGELER.Fig. 2: Experimental setup (operant chamber, computer, projector). Drawing: OPFERMANN-RÜNGELER.

236 INGA TIEMANN & GERD REHKÄMPER
auf 3 s in den beiden folgenden Phasen. Pha-se I galt als beendet, wenn das Huhn daserste Mal auf die Pickscheibe pickte oder in-nerhalb von fünf Wiederholungen der Pha-se I nicht angefangen hatte auf die Pickscheibezu picken. In den folgenden beiden Phasenmusste ein Mindestwert von 20 Durchgängenerreicht werden, in denen gepickt wurde, be-vor sich die nächste Phase des Autoshapingsanschloss. Auch hierfür wurden die Sitzungenfünf Mal wiederholt, bevor das Autoshapingals nicht bestanden galt und das Tier ausge-schlossen wurde. Im Verlauf des Autosha-pings wurde der erste protokollierte Pick aufdie Pickscheibe zusammen mit der insgesamtbenötigten Anzahl an Sitzungen registriert.
2.4. Statistik
Die statistische Auswertung erfolgte mit SY-STAT (SPSS Inc.). Paarvergleiche wurden mitHilfe des Mann-Whitney U-Tests durchgeführt.Drei oder mehr Gruppen wurden mit Hilfe des
Kruskal-Wallis-Tests verglichen. Das Signifi-kanzniveau wurde bei α = 0,05 angesetzt.
Die Erfolgskriterien innerhalb der Teilex-perimente wurden nicht von allen Tieren er-reicht. Da die Sequenzen jedoch aufeinanderaufbauten, nahm daher zum Teil die Stich-probengröße, d.h. die Anzahl der Hennen,im Verlauf der Experimente ab. Insgesamtwurden acht Individuen der Rassen Chabo,Bantam und Zwerg-Cochin und 22 Hollän-der Haubenhühnern beobachtet.
3. Ergebnisse
Die durchschnittliche Anzahl der benötigtenSitzungen (eine Sitzung pro Tag, 24h) fürVortraining und Autoshaping der einzelnenRassen ist in Abbildung 3 dargestellt. Für diebenötigte Anzahl der Sitzungen bis zum er-folgreichen Abschluss des Vortrainings kön-nen keine signifikanten Unterschiede mitdem Kruskal-Wallis-Test (p = 0,916) zwi-schen den Rassen nachgewiesen werden.
Abb. 3: Anzahl der benötigten Tage bis zum Abschluss des Vortrainings (shapings) und des Autosha-pings pro Rasse (Mittelwerte mit Standardabweichungen); HH Holländer Haubenhühner, BT Ban-tam, CH Chabo, ZC Zwerg-Cochin. * p ≤ 0,05, ** p ≤ 0,01, *** p ≤ 0,001; für Details siehe Text.Fig. 3: Number of days needed to finish shaping and autoshaping of each breed (means andstandard deviations). HH White Crested Polands, BT Rosecomb Bantams, CH Japanese Bantams,ZC Peking Bantams. * p ≤ 0.05, ** p ≤ 0.01, *** p ≤ 0.001*; see text for details.

Rassenunterschiede bei Haushühnern bei Konditionierung in der Skinner-Box 237
Acta Biologica Benrodis 13 (2006)
Ein Holländer Haubenhuhn schloss dasVortraining nicht erfolgreich ab. Für die Be-rechnung des Autoshapings wurden deshalbdie folgenden Tierzahlen zugrunde gelegt:Holländer Weißhauben (n = 21), Chabo(n = 14), Bantam (n = 8), Zwerg-Cochin(n = 8). Im Autoshaping scheiterten ein Cha-bo und sechs Holländer Haubenhühner. Alleanderen Hennen waren erfolgreich.
Beim Autoshaping schwankten die Wertebis zum erfolgreichen Abschluss zwischendurchschnittlich 4,25±0,46 Sitzungen á 40Durchgängen bei den Zwerg-Cochin und6,71±2,05 Sitzungen bei den Holländer Hau-benhühnern. Diese Unterschiede sind hochsignifikant (Kruskal-Wallis-Test; p < 0,001).
Einzelvergleiche der Rassen, die mit demMann-Whitney U-Test berechnet wurden(vgl. Abb. 3), zeigen für das Autoshaping ei-nen hoch signifikanten Unterschied (p <0,001) zwischen den Rassen Holländer Hau-benhuhn und Chabo. Die Holländer Hau-benhühner benötigten mehr Sitzungen, bissie auf die Pickscheibe pickten und in Folgedessen das Autoshaping erfolgreich abschlos-
sen. Ein signifikanter Unterschied (p = 0,002)ist auch zwischen Holländer Haubenhühnernund Zwerg-Cochin vorhanden, wobei erneutdie Holländer Haubenhühner mehr Sitzun-gen benötigten. Auch die Bantam brauchtenmehr Sitzungen im Vergleich zu Chabos (p= 0,014) und Zwerg-Cochin (p = 0,03).
Abbildung 4 zeigt, dass der erste Pick jenach Rasse (Individuenanzahl s.o.) früher oderspäter im Verlauf des Autoshapings statt-fand. Dabei benötigten die Zwerg-Cochin mit64,4 (±18,57 Stabw.) Durchgängen wenigerDurchgänge als die Holländer Haubenhüh-ner mit 118 (±59,46 Stabw.) Durchgängen (p= 0,037). Auf die gleiche Weise unterschei-den sich Holländer Haubenhühner und Cha-bos (p = 0,012). Bantam ließen ebenfallsmehr Durchgänge vor ihrem ersten Pick aufdie Pickscheibe verstreichen als Chabos (p =0,012).
Auf der Basis von prozentualen Erwar-tungswerten (Gleichung 1) wurden die Hol-länder Haubenhühner gegen den generellenTrend der gesamten Gruppe (unabhängigvon Rassenzugehörigkeit) getestet.
Abb. 4: Anzahl benötigter Durchgänge pro Rasse bis zum 1. Pick (Mittelwertemit Standardabwei-chungen); HH Holländer Haubenhühner, BT Bantam, CH Chabo, ZC Zwerg-Cochin. * p ≤ 0,05,** p ≤ 0,01, *** p ≤ 0,001; für Details siehe Text.Fig. 4: Numbers of trails per breed before the 1st pick (means and standard deviations). HH WhiteCrested Polands, BT Rosecomb Bantams, CH Japanese Bantams, ZC Peking Bantams. * p ≤ 0.05,** p ≤ 0.01, *** p ≤ 0.001*; see text for details.

238 INGA TIEMANN & GERD REHKÄMPER
(1)
(2)
Holländer Haubenhühner weisen eine gro-ße Diskrepanz zwischen Erwartungswert undtatsächlicher Anzahl der Tiere auf, die dasAutoshaping erfolgreich beendet haben. Auf-grund der getesteten Individuenanzahl wärezu erwarten gewesen, dass 17 Holländer Hau-benhühner das Vortraining und Autosha-ping erfolgreich abgeschlossen, und vier Hen-nen dieser Rasse das Training nicht bestan-den hätten (Gleichung 2, gerundete Zahlen).Die Zahl von vier Hennen resultiert hierbeiaus der Subtraktion des Erwartungswertes(17) von der eingesetzten Grundgesamtheit(21 HH). Diesem Erwartungswert steht einempirischer Wert von sieben Hennen gegen-über, die Vortraining und Autoshaping nichterfolgreich abschlossen. Die Wahrscheinlich-keit dafür, dass bis zu sieben Hennen derHolländer Haubenhühner von 21 das Krite-rium des Autoshapings nicht erreichen, er-gibt P (x ≤ 7) = 0,0143. Damit ist die Abwei-chung des empirischen Wertes (sieben Hen-nen) vom Erwartungswert (3,706) signifikant(Binominaltest, p = 0,0143).
4. Diskussion
Ein Holländer Haubenhuhn schloss dasVortraining nicht erfolgreich ab, sechs weiterebeendeten das Autoshaping nicht mit demkonditionierten Picken auf die Pickscheibe.Eine Chabo-Henne wurde wegen mangeln-den Erfolges ebenfalls aus dem Autoshapingausgeschlossen, jedoch keine Henne der Ras-sen Bantam und Zwerg-Cochin. Alle Tiere,die das Autoshaping nicht bestanden, schei-terten an der Phase I, für deren erfolgreichenAbschluss das Picken auf die Pickscheibe ge-
fordert war. Das Ergebnis für die HolländerHaubenhühner unterschied sich im Test ge-gen einen Erwartungswert signifikant vomgenerellen Trend der Grundgesamtheit allerHennen. Die Anzahl der (täglich stattfinden-den) Sitzungen, die die Hennen einer Rassebenötigten, um das Autoshaping erfolgreichabzuschließen, war ebenfalls signifikant un-terschiedlich. Die Holländer Haubenhühnerbenötigten im Durchschnitt 118 Durchgän-ge bis zum ersten Pickschlag, während dieHennen der Rassen Chabo und Zwerg-Cochin durchschnittlich im 56. und 64.Durchgang pickten, d.h. Chabos und Zwerg-Cochin lernten die operante Konditionierungsignifikant schneller.
4.1. Vortraining (shaping)
Das Verfahren des Vortrainings ist eineVoraussetzung für die Konditionierung(BROWN & JENKINS 1968). Die Tiere werdenan die Versuchsapparatur gewöhnt; dabei wirdihnen freier Futterzugang gewährt. Die schritt-weise Gewöhnung an die Futteraufnahme ausdem Futtergeber war nicht problematisch.Ein Holländer Haubenhuhn nahm den Fut-tergeber nicht an und erfüllte daher die Krite-rien des Vortrainings nicht. Auffällig war, dassdiese Henne zwar in der ersten Sitzung (Vor-traingsphase 1) in der Box gefressen hat, da-nach jedoch nicht mehr. Möglicherweise han-delte es sich um eine besonders ängstlicheHenne.
4.2. Autoshaping
Beim Autoshaping wird das Picken als be-reits vorhandenes Verhalten mit der Pickschei-be verknüpft. Notwendig hierfür ist das zu-fällige Picken des Huhns auf die Scheibe undeine direkte Verstärkung dieses Verhaltensdurch eine Belohnung mit Futter. Anfäng-lich wurden die Hennen auf die Beleuchtungder Pickscheibe aufmerksam und beobachte-ten das Hervorklappen des Futtergebers. Imweiteren Verlauf fraßen die Hennen nach an-

Rassenunterschiede bei Haushühnern bei Konditionierung in der Skinner-Box 239
Acta Biologica Benrodis 13 (2006)
nähernd jeder Präsentation des Stimulus (Be-leuchtung der Pickscheibe) meist zuverlässigaus dem Futtergeber und hielten sich ver-mehrt an der Frontseite der Box auf. Letzt-lich folgte eine Pickbewegung in RichtungPickscheibe (klassische Konditionierung) undder erste Pick. Erst danach schließt sich dieoperante Konditionierung an, in der das er-worbene Verhalten durch Belohnung auf-rechterhalten wird.
4.3. Rasseunterschiede
Die hier dargestellten Unterschiede im Ver-halten der Rassen Holländer Haubenhuhn,Bantam, Chabo und Zwerg-Cochin ergän-zen morphologische und ethologische Be-funde, die ebenfalls Rasseunterschiede auf-gezeigt haben (ALTEVOGT 1951; SCHULZE-SC-HOLZ 1961; REHKÄMPER et al. 2003). ALTE-VOGT (1951) berichtet von einem �rassen-verschiedenen optischen Lernvermögen� beiHühnern (S. 103). Die Tatsache, dass sichdas Lernvermögen bei Bankiva, Zwerghüh-nern, Sperbern und Brahma parallel zurKörper- und damit zur absoluten Hirngrö-ße verbesserte, veranlasste den Autor einedirekte Korrelation zwischen Hirngröße undLernkapazität anzunehmen. Diese �Massen-regel� (LASHLEY 1931) ist anscheinend nichtauf die Holländer Haubenhühnern anzu-wenden. Entsprechend der vergrößerten lern-relevanten Hirnbereiche (s.o.) sollte mit die-ser Vergrößerung auch die Steigerung derLernleistung verbunden sein. Vergleicht manauf dieser Basis Holländer Haubenhühnermit Chabos oder Zwerg-Cochin, ist das of-fenbar nicht der Fall (vergl. REHKÄMPER et al.2003). Morphometrische Untersuchungendes Gehirns von Bantams liegen derzeitnoch nicht vor.
SCHULZE-SCHOLZ (1961) erklärte Lernunter-schiede bei verschiedenen Haushuhn-Rassenmit deren unterschiedlichem Temperament.Aktive Rassen, wie Englische Kämpfer undDeutsche Zwerge lernten Assoziationenschneller als ruhigere, wie z. B. Vertreter der
Rassen Orpington, Chabo und HolländerHaubenhühner. Zum Beispiel benötigten fürdas Erlernen des Deckelabwerfens vom Fut-ternapf Holländer Haubenhühner 17,7 undChabos 21,1 Tage (SCHULZE-SCHOLZ, 1953).Im Gegensatz dazu lernten Holländer Hau-benhühner im Vergleich zu Chabos undZwerg-Cochin die hier geforderte operanteKonditionierung langsamer und benötigtenmehr Zeit bis sie das gewünschte Verhaltenzeigten. Zudem schlossen mehr Individuendas Autoshaping nicht erfolgreich ab. ImHinblick auf das Lernen nehmen Bantamsnach unseren Ergebnissen eine Mittelstellungzwischen Holländer Haubenhühnern undChabos und Zwerg-Cochins ein.
SOSSINKA (1982) führt veränderte Tempe-ramente in Form von reduzierter lokomoto-rischer Aktivität und Vorsichtigkeit, wie auchdie schnelle Habituation domestizierter Vö-gel (Hühner, Tauben und Kanarienvögel) aufdie Domestikation zurück, da die Entstehungentsprechender Unterschiede im Besonderenauf Interaktionen des Tieres mit seiner spe-zifischen Umwelt zurückzuführen sind. Dierassespezifische Umwelt ist für Wahrneh-mungs- und Lernleistungen entscheidend(HEALY & JONES 2002).
Vor allem die Gesichtregion der Hühnermit Kamm, Schnabel und Kehllappen ist fürdas individuelle Erkennen ausschlaggebend(CANDLAND 1969). Holländer Haubenhühnerbesitzen keinen Kamm, dafür jedoch unüber-sehbare Veränderungen im Schädelbereich, diehauptsächlich auf die Ausbildung der Hau-be zurückgehen. Möglicherweise nehmen sichHolländer Haubenhühner untereinandernach anderen als den von CANDLAND (1969)postulierten Kriterien wahr und sind damitvielleicht anderen kognitiven Anforderungenausgesetzt. Untersuchungen hierzu fehlenjedoch bisher.
Die rassespezifische Vergrößerung vonArealen des Di- und Telencephalon bei Hol-länder Haubenhühnern sind (noch) nichtohne weiteres mit den hier mitgeteilten Er-gebnissen zum Lernvermögen zu vereinba-

240 INGA TIEMANN & GERD REHKÄMPER
ren. Festzuhalten ist jedoch, dass HolländerHaubenhühner im Vergleich zu anderenHühnerrassen viel schwerer zu konditionie-ren sind.
Literatur
ALTEVOGT, R., 1951: Vergleichend-psychologischeUntersuchungen an Hühnerrassen stark un-terschiedlicher Körpergröße. � Zeitschrift fürTierpsychologie 8, 75-109.
BROWN, P., JENKINS, H., 1968: Auto-Shaping ofthe pigeons key-peck. � Journal of the Expe-rimental Analysis of Behaviour 11, 1-8.
BUND DEUTSCHER RASSEGEFLÜGELZÜCHTER E.V.,2006: Rassegeflügel-Standard für Europa. �Howa Druck & Satz, Nürnberg.
CANDLAND, D., 1969: Discriminability of facialregions used by the domestic chicken in main-taining the social dominance order. � Journalof Comparative Physiology and Psychology69, 281-285.
CRAWFORD, R., 1990: Poultry Breeding and Ge-netics. � Elsevier, Amsterdam.
DÜRIGEN, B., 1921: Die Gefügelzucht. � 3. Aufl.,Paul Parey, Berlin.
FRAHM, H., REHKÄMPER, G., 1998: Allometriccomparison of the brain and brain structuresin the white crested polish chicken with un-crested domestic chicken breeds. � Brain, Be-haviour and Evolution 52, 292-307.
HEALY, S., JONES, C., 2002: Animal learning andmemory: an integration of cognition and ecol-ogy. � Zoology 105, 321-327.
LASHLEY, K.S., 1929: Brain Mechanisms and In-telligence. � University of Chicago Press,Chicago.
LASHLEY, K.S., 1931: Mass action in cerebralfunction. � Science, 73, 245-254.
MCCABE, B.J., HORN, G., 1988: Learning and me-mory: regional changes in N-methyl-D-aspar-tate receptors in the chick brain after im-printing. � Proceedings of the National Aca-demy of Science, USA 85, 2849-2853.
OCHIENG-ODERO, J., 1994: Does adaptation oc-cur in insect rearing systems, or is it a case ofselection, acclimatization and domestication?� Insect Science Applied 15, 1-7.
PRICE, E., 2002: Animal Domestication and Be-havior. � CAB International, Wallingford.
REHKÄMPER, G., FRAHM, H., STOLZE, C., 1995:Lernen bei erwachsenen Haushühnern derRasse Chabo. � Tierärztliche Umschau 50,826-832.
REHKÄMPER, G., WERNER, C., FRAHM, H., 2000/2001: Grundlagen, Stand und Perspektivender Haustierkunde. � Acta Biologica Ben-rodis 11, 95-120.
REHKÄMPER, G., KART, E., FRAHM, H., WERNER,C., 2003: Discontinuous variability of braincomposition among domestic chicken breeds.� Brain, Behaviour and Evolution 61, 59-69.
TEGETMEIER, W. B., 1873: The Poultry Book. �George Routledge and Sons, New York.
SCHULZE-SCHOLZ, I. 1953: Über den Einfluss ver-schiedener Verhaltensbesonderheiten auf dieLernfähigkeit von Haushühnern. � Unveröf-fentlichte Dissertation, Universität Kiel.
SCHULZE-SCHOLZ, I., 1961: Über den Einflussverschiedener Verhaltensbesonderheiten aufdie Lernfähigkeit von Haushühnern. � Zeit-schrift für wissenschaftliche Zoologie 168,209-280.
SOSSINKA, R., 1982: Domestication in birds, pp.373-403. In: Avian Biology (D. FARNER, ed.).� Academic Press, New York.
WOOD-GUSH, D., 1959: A history of the domes-tic chicken from antiquity to the 19th cen-tury. � Poultry Science 38, 321-326.
Angenommen: 29.12.2006

Wie kam die Nördliche Grundwasserschnecke an den Niederrhein? 241
Acta Biologica Benrodis 13 (2006)
Acta Biologica Benrodis 13 (2006): 241-243
Kurze Mitteilung/Short communication
Wie kam die Nördliche GrundwasserschneckeBythiospeum acicula moenanum (Hydrobiidae) an
den Niederrhein?
How came the Northern Groundwater Snail Bythiospeum aciculamoenanum (Hydrobiidae) to the Lower Rhine?
Hartwig SCHÜTT
Haydnstraße 50, D-40593 Düsseldorf, Germany; [email protected]
Summary: All species of the subterranean freshwater snails of the genus Bythiospeum are restrictedto S-Germany, only south of Frankfurt/M., in Bavaria, Baden-Württemberg, and Switzerland.Only two stygobiontic populations are known from the lower course of the River Rhine. Accor-ding to the historic geomorphologic development of the Rhine-valley, these populations may havebeen introduced via the so-called Lake Rhine-Hessonia of the early to middle Pleistocene.
Die kleine, subterran lebende Süßwasser-schnecke Bythiospeum acicula moenanum (FLACH,1886), (Mollusca Prosobranchia: Hydrobii-dae), die unter dem Synonym Paladilhiopsishusmanni von BOETTGER, 1963, aus demGrundwasserstrom der Ruhr bei Schwertebeschrieben wurde, ist die einzige Schneckedieser Gattung, die nördlich des RheinischenSchiefergebirges lebt. Alle anderen Arten derGattung Bythiospeum und die zahlreichen Po-pulationen leben, teilweise häufig, im Main-zer Becken oder in Süddeutschland sowie inder Schweiz (BOETERS 1984: 167, Karte 8); essind insgesamt vier Arten mit 13 Unterarten.Seit einer Revision durch ZILCH (1970) müs-sen die hier zu behandelnden Formen denNamen Bythiospeum acicula moenanum (FLACH,1886) tragen. Paladilhiopsis hat sich als eineandere, hauptsächlich in Frankreich vorkom-mende Gattung herausgestellt. Dieses wur-de schon durch BOLLING (1960, 1966) festge-stellt. Alle Arten der Gattung Bythiospeum le-ben als kleine kiemenatmende, heterosexuel-le Süßwassertiere in Sandlücken von Kiesbet-ten; sie sind farblos und blind. An stenother-me, interstitielle Biotope angepasst, werden
sie im Gebirge oft von Quellaustritten, Höh-len und Brunnen ans Licht befördert. ImGrundwasser des Flachlandes werden siebeim Kiesabbau in mäßiger Tiefe gefunden.
Der bis dahin nördlichste bekannte Fund-ort liegt am Südrand des Rheinischen Schie-fergebirges, der Speisbachquelle, 3 km nörd-lich von Assmannshausen am Rhein, undeinigen benachbarten Quellaustritten, wo diePopulationen unter dem Synonym Lartetiaclessini septentrionalis beschrieben wurden(SCHÜTT 1960). Es lag daher nahe anzuneh-men, dass diese sich aus den miozänen Hy-drobienschichten des Mainzer Beckens ent-wickelt hatten (SCHÜTT 1979), speziell den dortSchicht bildenden untermiozänen Hydrobiaelongata (FAUJAS, 1806), die im Mainzer Be-cken an zahlreichen Orten fossil gefundenwerden (WENZ 1926).
Umso überraschender war es, als sich imGrundwasser der Ruhr bei Schwerte in Westfa-len ähnliche Tiere wie in Assmannshausen fan-den. Die conchologische Ähnlichkeit mit denGrundwasser bewohnenden Arten der Bythio-speen im Mainzer Becken ist augenfällig (ZILCH
1970, T.16 F.10, 11). Später wurde eine ganz

242 HARTWIG SCHÜTT
aus dem Grundwasser des Mainzer Beckensbei Aschaffenburg bekannte Crustaceen derGattung Chappuisius auch im Grundwasserbei Bonn im Subterranstrom des Bröhlbachs,einem Nebenfluss der Sieg (HUSMANN 1964,Karte fig. 1), ohne diese Verbreitung erklärenzu können.
Es stellte sich also die Frage: Wie konnteeine Bythiospeum-Art als Grundwasserbewoh-ner von Süddeutschland über das RheinischeSchiefergebirge in die Norddeutsche Tiefebe-ne und an den Niederrhein kommen?Neuerdings scheint durch Rekonstruktion derEntwicklungsgeschichte des rheinischenFlusssystems (FRANZEN 2002) eine plausibleErklärung möglich: Es hat durch Anschlussdes Alpenrheins an den Oberrhein vor 0,8Millionen Jahren einen zusätzlichen Wasser-schub gegeben, der zum Entstehen einesStausees vor dem Rheinischen Schiefergebir-ge führte, dem sogenannten alt- bis mittel-pleistozänen Rheinhessensee (FRANZEN
2001). Der Mittelrhein-Canyon war bereits als
Abb. 1: Bythiospeum acicula moenanum (FLACH, 1886); lebendes Tier aus der Typlokalität von B.husmanni im Grundwasserstrom der Ruhr bei Schwerte. Foto: Dipl.-Ing. R. HANNEFORTH.Fig. 1: Bythiospeum acicula moenanum (FLACH, 1886); living specimen from the type locality of B. husmanniin the subterranean waterflow of river Ruhr near Schwerte. Foto: Dipl.-Ing. R. HANNEFORTH.
ähnliche Population im Grundwasser der Waal,eines Hauptarms des Rheindeltas in den Nie-derlanden, bei Kesteren, Gelderland, in 70 cmTiefe gefunden (KUIJPER & GITTENBERGER 1993,Karte fig.3). Die Tiere dieser holländischen Po-pulation wurden Bythiospeum husmanni (BOETT-GER, 1963) benannt, obwohl Bythiospeum aciculamoenanum (FLACH, 1886) valid ist. Sie leben inSiebbrunnen und Filtersandschichten (HUS-MANN, 1961) zusammen mit Ostracoden undGammariden und werden auch aus Peilrohrenund mit Rammpumpen erbeutet (HUSMANN
1971). Es sind Sandlückenbewohner eines grob-kiesigen Flussdeltas.
Gleichfalls aus dem Grundwasser der Ruhrwurde diese Art in Filtersanden des Wasser-werks Echthausen unterhalb der Möhne-Tal-sperre mit sehr individuenreicher Lebensge-meinschaft gefunden (RUMM 1999).
Diese Schnecken sind nicht die einzigen sty-gobionten Tiere im Niederrheingebiet, dieeine Beziehung zum Mainzer Becken haben.HAINE (1945) entdeckte zwei bis dahin nur

Wie kam die Nördliche Grundwasserschnecke an den Niederrhein? 243
Acta Biologica Benrodis 13 (2006)
antezedentes Tal vorhanden, konnte aber in-folge der tektonischen Hebung des Rheini-schen Schiefergebirges nur begrenzte Wasser-führung aufnehmen (SEMMEL 2001). Erst nachAuffüllen und zeitweisem Überlaufen desRheinhessensees wurde ein Abfluss und Ein-schneiden des Canyons bis in heutige Tiefeund Weite möglich (FRANZEN 2002). DiesenWeg könnte auch die am Nordrand des Main-zer Beckens subterran lebende Art Bythiospe-um acicula genommen haben, die sich dann indie interstitiellen Biotope von Ruhr und Waalausbreitete. Wahrscheinlich ist der aufgestau-te See auch für die Ausbreitung der Art amunteren Main verantwortlich und hat bei sei-ner Regression die bekannten Fundstellen der�Mainlartetien� bei Obernburg, Klingenbergund Miltenberg (BOLLING 1966) hinterlassen.
Aus diesen Überlegungen kann die Kon-sequenz gezogen werden, auch in den Kies-lagen des Rheinbettes im nördlichen Nord-rhein-Westfalen im Bereich der unteren Lip-pe und möglicherweise auch der Lahn aufdas Auftreten dieser Grundwasserschneckezu achten. Im Einzugsgebiet der Niers be-stehen jedoch infolge von Grundwasserab-senkungen wegen des Braunkohlenabbauskaum Erfolgsaussichten.
Literatur
BOETERS, H. D., 1984: Gedanken zu einer Revi-sion der Gattung Bythiospeum in Deutschland.� Mitteilungen der deutschen malakozoolo-gischen Gesellschaft 37, 142-171.
BOETTGER, C. R., 1963: Über das Auffinden ei-ner subterranen Schnecke aus der FamilieHydrobiidae im Grundwasser des Ruhrgebie-tes. � Archiv für Molluskenkunde 92, 45-48.
BOLLING, W., 1960: Kritische Bemerkungen zumGenus Lartetia. � Archiv für Molluskenkun-de 89, 153-156.
BOLLING, W., 1966: Beiträge zum Problem desGenus Bythiospeum BOURGUIGNAT. (Mollusca-Hydrobiidae. � Berichte der naturforschen-den Gesellschaft Bamberg 40, 21-102.
FRANZEN, J. L., 2001: Der Rheinhessensee � neueErkenntnisse, neue Fragen. � Natur undMuseum 131, 126-128.
FRANZEN, J. L., 2002: Versuch einer Rekonstruk-tion der Entwicklung des rheinischen Fluß-systems. � Natur und Museum 132: 408-423.
HAINE, E., 1946: Die Fauna des Grundwassersvon Bonn mit besonderer Berücksichtigungder Crustaceen, 1-144. � Inauguraldissertati-on, Bonn.
HUSMANN, S., 1961: Filtersandschichten inGrundwasseranreicherungs-Anlagen alskünstliche Biotope aquatiler Subterranorga-nismen. � Abhandlungen der Braunschweigi-schen Wissenschaftlichen Gesellschaft 18,163-181.
HUSMANN, S., 1964: Studien zur Ökologie undVerbreitung der Gattung Chappuisius KIEFER,1938. � Crustaceana 6, 179-194.
HUSMANN, S., 1971: Eine Rammpumpe zur Un-tersuchung des subaquatischen Stygorheals.�Gewässer und Abwässer 50/51, 115-119.
KUIJPER, W. J. & GITTENBERGER, E., 1993: Degrondwaterslag Bythiospeum husmanni (BOETT-GER, 1963) in Nederland (GastropodaProsobranchia: Hydrobiidae). � Basteria 57,89-94.
RUMM, P., 1999: Untersuchungen zum Abbaupartikulärer organischer Substanzen in einemLangsamsandfilter durch Metazoen am Bei-spiel von Niphargus fontanus. � Dissertation,Universität Oldenburg.
SCHÜTT, H., 1960: Das bisher nördlichste Larte-tienvorkommen. � Archiv für Molluskenkun-de 89, 77-78.
SCHÜTT, H., 1979: Die Bythiospeen-Fundorte amBinger Loch. � Club Conchylia Informatio-nen 9, 50-51.
SEMMEL, A., 2001: Warum ist es am Rhein soschön? Geowissenschaftliche Anmerkungenzum geplanten Weltkulturerbe Mittelrhein. �Natur und Museum 131, 115-125.
WENZ, W., 1926: Gastropoda extramarina terti-aria, 7 pars 32: 1863-2230. In: Fossilium Ca-talogus, 1, Animalia. � Junk, Berlin.
ZILCH, A., 1970: Die Typen und Typoide desNatur-Museums Senckenberg, 45: Mollusca,Hydrobiidae (1): Bythiospeum BOURGUIGNAT. �Archiv für Molluskenkunde 100, 319-346.
Angenommen: 01.02.2006

244 HARTWIG SCHÜTT
Rezension
WUKETITS, FRANZ M. (2003): ausgerottet � ausgestorben. über den Untergang von Ar-ten, Völkern und Sprachen. � S. Hirzel Verlag, Stuttgart, Leipzig. 2. Auflage. 240 Seiten, 14Abbildungen, 6 Tabellen. Gebunden, 13,5 x 22 cm. ISBN 978-3-7776-1259-1. 22,-- Euro.
Man hört häufig, Arten seien schon immer ausgestorben, das Artensterben sei aber noch nieso schnell und umfassend gewesen wie jetzt. Tatsache ist, dass es in der ErdgeschichteArtensterben in viel größerem Ausmaß gegeben hat, aber dass dafür noch nie ein einzigerOrganismus � in diesem Fall der Mensch � dafür verantwortlich war. Wuketits zählt inseinem neuen Buch nicht nur die Folgen von Erdkatastrophen auf, sondern betont auch,dass in unserer Zeit auch Völker, Kulturen und Sprachen untergehen. Dabei gehören Artenwie auch Kulturen nicht zwangsläufig zusammen. Sicher sind mit den hunderten ausgestor-benen (und ausgerotteten) Indianervölkern der USA, Brasiliens und Kolumbiens auchmindestens ebenso viele Sprachen ausgestorben; aber auch in unserer unmittelbaren Umge-bung erleben wir, dass Mundarten fast nur noch von älteren Leuten gesprochen werden.Wuketits will mit diesem Buch zeigen, dass das �große Sterben� nicht nur Tiere und Pflan-zen betrifft. Natürlich stellt er auch die Frage, ob uns das alles nicht gleichgültig sein könnteangesichts des ständigen Kommens und Gehens im Verlauf der Erdgeschichte. Aber seineethische Begründung, die Vielfalt zu bewahren, ist überzeugend. So ist dieses Buch einAppell an die Moral der Menschen und an eine Ethik des Bewahrens.
Harro Hieronimus, Solingen

Erstnachweis des Springfroschs (Rana dalmatica) im Regierungsbezirk Düsseldorf 245
Acta Biologica Benrodis 13 (2006)
Acta Biologica Benrodis 13 (2006): 245-248
Kurze Mitteilung/Short communication
Erstnachweis des Springfrosches (Rana dalmatinaBonaparte, 1840) im Regierungsbezirk Düsseldorf
First record of the Agile Frog (Rana dalmatina Bonaparte, 1840) inthe administrative district of Düsseldorf
RALF KRECHEL1, THOMAS BRAUN2 & MICHAEL STEVENS2,3
1 IVÖR Institut für Vegetationskunde, Ökologie und Raumplanung, VolmerswertherStraße 80-86, D-40225 Düsseldorf, Germany; [email protected]
2 Haus der Natur – Biologische Station im Rhein-Kreis Neuss e.V., KlosterKnechtsteden, D-41540 Dormagen, Germany; [email protected]
3 Institut für Zoomorphologie und Zellbiologie, Heinrich-Heine-Universität Düssel-dorf, Universitätsstraße 1, D-40225 Düsseldorf, Germany; stevensm@uni-
duesseldorf.de
Summary: Two spawning sites of the agile frog (Rana dalmatina) have been recorded in themunicipality of Rommerskirchen and one in the city of Dormagen (Rhine Country of Neuss,Administrative District of Düsseldorf, North Rhine-Westphalia). These spawning sites are consi-dered to be the most northern known in the Rhineland.
Der Springfrosch (Rana dalmatina BONAPAR-TE, 1840) ist eine europäische Art, derenVerbreitungsgebiet sich von den Pyrenäenund Frankreich über Südeuropa bis nachGriechenland und Rumänien erstreckt. DieVorkommen in Mittel- und Nordeuropasind mehr oder weniger deutlich separiert,wobei insbesondere die Vorkommen nörd-lich der Mainlinie stark voneinander iso-liert sind (STEINICKE et al. 2002). Dieses Ver-breitungsmuster wird als Relikt postglazi-aler Wärmeperioden betrachtet (GÜNTHER
et al. 1996). Aus dem nördlichen Rhein-land ist die Art nur aus der Niederrheini-schen Bucht bekannt (ARBEITSKREIS AMPHI-BIEN UND REPTILIEN NRW 2006). Dieses Ver-breitungsgebiet umfasst den GroßraumBonn einschließlich des Landkreises Ahr-weiler, die Voreifel, die Ruraue bis in denRaum Jülich sowie die Ville. Erstmaligkonnte die Reproduktion des Springfro-sches auch im Regierungsbezirk Düsseldorfdokumentiert werden.
In der Niederrheinischen Bucht zwischenDüsseldorf und Köln liegt das FFH-Gebiet�Knechtstedener Wald mit Chorbusch� (DE-4806-303). Es gehört sowohl zum Ballungs-raum Rhein-Ruhr als auch zur Köln-BonnerRheinebene. Dieser wertvolle Laubwald-Komplex hat eine Nord-Südausdehnungvon rund 12 km und eine maximale Breitevon 2 km. Er ist einer der größten unzer-schnittenen Lebensräume der Region. West-lich des Knechtstedener Waldes liegt in einerRandsenke entlang der Mittelterrasse eineehemals ausgedehnte Bruchlandschaft, diesich nach Meliorationsmaßnahmen heute alseine reich strukturierte, landwirtschaftlich ge-prägte Kulturlandschaft darstellt.
Am 17.03.2004 wurde an einem künstlichangelegten Tümpel auf dem Gelände desKlosters Knechtsteden ein Männchen desSpringfrosches gefangen, fotografiert und ver-messen (Länge: 5,4 cm, Gewicht: 17,9 g, FHL:0,30 mm, ZL: 0,74 mm, Quotient: 2,46).Daraufhin wurden in den folgenden Früh-

246 RALF KRECHEL, THOMAS BRAUN & MICHAEL STEVENS
jahren 2005 und 2006 im FFH-GebietKnechtstedener Wald mit Chorbusch undseinem Umfeld über 80 Kleingewässer aufmögliche Vorkommen des Springfroschesuntersucht. Insbesondere wurde eine inten-sive Laichballensuche durchgeführt. Dabeikonnte an drei ausdauernden Gewässern imFrixheimer Bruch, bei Gut Barbarastein undam ehemaligen Forsthaus in Dormagen-Stra-berg eine Reproduktion nachgewiesen wer-den (MTB 4906/1+2 Pulheim).
Frixheimer Bruch: Im Frixheimer Bruchwechseln kleinere Waldbestände, vorwiegendaus Hybrid-Pappeln, mit Grünland undAckerflächen ab. Innerhalb eines Wald-Ge-büsch-Komplexes liegt ein etwa Mitte der1980er Jahre angelegter kreisförmiger Weiher.Er besitzt einen Durchmesser von ca. 30 m beieiner Tiefe von über 80 cm im zentralen Be-reich. Die Ufer sind zum Teil dicht mit Gehöl-
zen, vorwiegend Weiden, bestanden. Entlangdes Ufers wächst kreisförmig ein Röhricht ausSchilf. Die zentrale Wasserfläche ist frei vonRöhrichtvegetation. Einige Uferabschnitte unddie zentralen Bereiche des Gewässers warenbei den Kartierungsarbeiten schlecht einseh-bar, dennoch konnten in den zugänglichenAbschnitten 13 Laichballen des Springfroschsregistriert werden (18.03.2005). Bei einemnächtlichen Verhör (19.03.2005) wurden 20-30 rufende Springfrösche und 3-5 rufendeGrasfrösche festgestellt. Darüber hinaus wur-den im Wasser zwei erwachsene Springfröscheund ein Totfund (Prädator: Iltis?) registriert.Der Springfrosch nutzt das Gewässer syntopmit fünf weiteren Amphibienarten: Von derErdkröte wurden im März 2005 > 30 Adulti,vom Grasfrosch > 30 Laichballen sowie vomTeichfrosch 3 Adulti festgestellt. Durch dasAusbringen von fünf Molchreusen wurden
Abb. 1: Erwachsener, männlicher Springfrosch, der am 17.03.2004 bei Kloster Knechtstedengefangen wurde (biometrische Daten im Text; Kantenlänge 1 cm).Fig. 1: Adult male agile frog captured near Knechtsteden monastery 17.03.2004 (biometrical dataare given in the text; length of edges 1 cm).

Erstnachweis des Springfroschs (Rana dalmatica) im Regierungsbezirk Düsseldorf 247
Acta Biologica Benrodis 13 (2006)
am 02.05.2005 darüber hinaus 33 Bergmol-che (18,15) und 58 Teichmolche (47,11) nach-gewiesen.
Gut Barbarastein: Die Landschaft um GutBarbarastein umfasst Ackerflächen, Grün-land, Gärten und Gehölzbestände. DasLaichgewässer ist ein mit rund 25 x 8 m Grö-ße und einer Tiefe von über 60 cm künstlichangelegter Folienweiher. Die Ufer sind be-sonnt und mit einem Röhricht bestanden,das vorwiegend aus Rohrkolben und Schilfbesteht. Hier konnten am 22.03.2005 neunLaichballen des Springfroschs festgestellt wer-den. Von beiden Probestellen wurden imAquarium Kaulquappen aufgezogen.
Ehemaliges Forsthaus Straberg: Die Um-gebung des ehemaligen Forsthauses bestehtaus dem Knechtstedener Wald, Ackerflächen,Gärten und der Ortschaft Straberg. Das Laich-gewässer ist ein mit rund 20 x 10 m Größeund einer Tiefe von über 150 cm ebenfallskünstlich angelegter Folienteich, der auch als
Badeteich genutzt wird. Die Ufer sind besonntund an wenigen Stellen mit einem Röhrichtbestanden, welches kurz vor dem Winter ober-halb der Wasserlinie gemäht wurde. Am27.03.2006 konnten hier ein Männchen undmindestens zwei Laichballen des Springfroschsfestgestellt werden. Darüber hinaus wurden350 Laichballen des Grasfrosches, ca. 400 Erd-kröten und ein Teichmolch nachgewiesen.
Die beiden erstgenannten Gewässer liegenrund 850 m voneinander entfernt auf demGebiet der Gemeinde Rommerskirchen, dasdritte im Stadtgebiet von Dormagen (alleRhein-Kreis Neuss). Diese drei Fundpunktesind die nördlichsten bekannten Laichgewäs-ser im Rheinland. Die nächsten Laichgewäs-ser liegen rund 11 km südlich im Rhein-Erft-Kreis im Bereich der nördlichsten Ausläuferder Ville (M. HACHTEL in lit.). Die Verbrei-tung und die Populationsgröße des Spring-froschs im Umfeld des Knechtstedener Wal-des werden weiter untersucht.
Abb. 2: Laichplatz im Frixheimer Broich (27.03.2006).Fig. 2: Spawning site in the Frixheimer Broich (27.03.2006).

248 RALF KRECHEL, THOMAS BRAUN & MICHAEL STEVENS
Literatur
ARBEITSKREIS AMPHIBIEN UND REPTILIEN NRW2006: http://www.herpetofauna-nrw.de/Arten/2.14Springfrosch.htm
GÜNTHER, R., PODLOUCKY, J., PODLOUCKY, R. 1996:Springfrosch � Rana dalmatina BONAPARTE,1840. S. 389-412. In: Die Amphibien und
Reptilien Deutschlands (R. Günther, ed.). �Gustav Fischer, Stuttgart.
STEINICKE, H., HENLE, K., GRUTTKE, H. 2002:Bewertung der Verantwortlichkeit Deutsch-lands für die Erhaltung von Amphibien- undReptilienarten. - Landwirtschaftsverlag, Mün-ster.
Angenommen: 19.04.2006

Alarmrufe von Dreifarbglanzstaren bei Entdeckung eines Bodenfeindes 249
Acta Biologica Benrodis 13 (2006)
Acta Biologica Benrodis 13 (2006): 249-252
Kurze Mitteilung/Short communication
Alarmrufe von Dreifarbglanzstaren (Lamprotornissuperbus) bei Entdeckung eines Bodenfeindes
Alarm calls of Superb Starlings (Lamprotornis superbus) detectinga ground predator
HUBERT HEUWINKEL & URSULA BEIGEL-HEUWINKEL !Museum für Naturkunde, Stiftung Schloss und Park Benrath, Benrather Schloßallee
102, D-40597 Düsseldorf
Summary: The behaviour and alarm calls of Superb Starlings in presence of a Puff adder aredescribed. When starlings had taken notice of the adder, a group of them assembled in a semicirclearound the adder calling continuously, whereas a single individual calling just as the birds in thegroup remained in the acacia, the former resting place of the group. These alarm calls were noisy,shrill and of considerable loudness. Frequencies with the highest sound intensity ranged from2,000 to 7,000 Hz.
Bei den Lautäußerungen der Singvögel handelt es sich zum einen um einfache oder hoch-komplexe Gesänge, die meistens der Revier-markierung, dem Anlocken von Weibchen,dem Paarzusammenhalt und weiteren sozia-len Funktionen dienen, zum anderen aberauch um Rufe, die nur in bestimmten Situa-tionen hervorgebracht werden. Neben Kon-takt-, Bettel-, Stimmfühlungs-, Stand-, Flug-rufen u.a. sind Alarm- oder Warnrufe fürVögel von besonderer Bedeutung, da sie ei-nerseits Artgenossen (intraspezifisch), aberauch Individuen anderer Arten (interspezi-fisch) vor Luft- oder Bodenfeinden warnen(Übersicht bei MARLER & SLABBEKOORN 2004). Alarmrufe auf Luftfeinde sind langgezogen,hochfrequent und schmalbandig; solche aku-stischen Signale sind schwer zu orten, so dassandere Vögel auf den Luftfeind aufmerksamwerden, der Rufer selbst aber nicht unbedingtvon ihm entdeckt wird. Alarmrufe auf Bo-denfeinde sind dagegen kürzer, tieferfrequent,breitbandig, laut und werden rasch wieder-holt. Da diese Rufe sich gut orten lassen, istder Bodenfeind für andere Vögel leicht zuentdecken (u.a. MARLER 1956; KLUMP & SHAL-TER 1984).
Während einer Safari durch Kenia im Fe-bruar/März 1980 wurden die Alarmrufe vonDreifarbglanzstaren auf eine Puffotter (Bitisarietans) aufgenommen sowie das Verhaltender warnenden Vögel mit Fotografien doku-mentiert. Für die Tonaufnahmen standen einNagra SN-Tonbandgerät, für Analysen einKay-Sonagraph 6061 B sowie ein B & K- Pe-gelschreiber Typ 2305 zur Verfügung.
Beim Abbau der Zelte am Navaisha-Seeam Morgen des 22.2.1980 gegen 7.30 Uhrentdeckten wir unter der Bodenplane einesder Zelte eine zusammengerollte Puffotter,die noch reglos war, da die Temperatur nuretwa 10 °C betrug. Die Puffotter wurde in-nerhalb kurzer Zeit von einer Gruppe vonDreifarbglanzstaren (Lamprotornis superbus)entdeckt, die sich in einer benachbarten Aka-zie aufhielten. Während ein warnender Vo-gel auf dem Baum sitzen blieb (Abb. 1),versammelten sich mehrere Glanzstare umdie Schlange und schrieen lauthals (Abb. 2).Der �Lärm� war so groß, dass einige We-bervögel in benachbarte Bäume einfielenund mit in das Geschrei einstimmten. Nach-

250 HUBERT HEUWINKEL & URSULA BEIGEL-HEUWINKEL
Abb. 1: Warnender Dreifarbglanzstar in einer Akazie.Fig. 1: Alarm calling Superb Starling in an Acacia.
Abb. 2: Ansammlung von Dreifarbglanzstaren, die Alarmrufe ausstoßen, um eine Puffotter.Fig. 2: A group of alarm calling Superb Starlings near the Puff adder.

Alarmrufe von Dreifarbglanzstaren bei Entdeckung eines Bodenfeindes 251
Acta Biologica Benrodis 13 (2006)
Abb. 3: Sonagramm der Alarmrufe, die Zeitmarken am unteren Rand des Sonagramms entsprechen0,05 s. Unten die dazugehörige relative Schalldruckpegelkurve.Fig. 3: Sonagram of alarm calls. Time marks at the lower edge 0.05 s. Below the relevant relativesound pressure curve.
Abb. 4: Sonagramm der letzten Alarmrufe aus Abbildung 3 im Frequenzbereich 160 -16000 Hz.Die Zeitmarken am oberen Sonagrammrand entsprechen 0,05 sec. Am Anfang des Sonagrammsüberdecken sich die Rufe zweier Individuen, am Ende ist ein Einzelruf dargestellt. Die Section-Analyse am Ende der Abbildung entspricht den Stellen im Sonagramm, die durch senkrechte Liniengekennzeichnet sind.Fig. 4: Sonagram of the last alarm calls in figure 3 in the frequency range of 160 - 16000 Hz. Timemarks at the upper edge 0.05 s. At the beginning of the sonagram the calls of two birds overlap. Atthe end a single call follows. The section analysis at the end of the figure corresponds to the verticallines in the sonagram.
dem die Schlange von uns unter äußersterVorsicht entfernt worden war, ebbten auchdie Alarmrufe ab und die warnenden Vögelverschwanden wieder in den umstehendenBäumen.
Eine während dieser Begebenheit entstan-dene Tonbandaufnahme enthält 262 Alarm-
rufe, die die Vögel vereinzelt oder zeitgleichund damit im Sonagramm einander über-deckend hervorbrachten (Abb. 3). Diese Bo-denfeind-Alarmrufe vermitteln demmenschlichen Ohr den Eindruck eines ge-räuschhaften, schrillen und recht lautenSchreis mit aggressiver Tönung. Abbildung

252 HUBERT HEUWINKEL & URSULA BEIGEL-HEUWINKEL
3 zeigt ein Sonagramm mit zwölf dieserAlarmlaute, die von mindestens zwei Indi-viduen stammen. Die beiden ersten Rufeüberlagern sich zum Teil, wobei der zweitevon einem Vogel stammt, der sich näher amMikrofon befand und daher entsprechendder Schalldruckpegelkurve um 13 dB lauterist als der erste; der zweite Ruf dauert mit0,5 s relativ lange.
Bei dem dritten Signal ist nicht eindeutigzu klären, ob es von einem oder zwei Rufernstammt; hier ist neben den beiden geräusch-haften frequenzspektralen Anteilen auch einklanghafter Anteil vorhanden, wie auch amEnde des siebten und achten Rufs. Der vierteRuf ist mit 0,05 s sehr kurz. Von den dreiletzten Rufen überlappen sich die beiden vor-letzten; sie stammen daher von zwei verschie-denen Individuen. Diese letzten Rufe ent-halten neben geräuschhaften auch klanghafteAnteile, wie die Obertonbänder zeigen.
Aus dem Sonagramm in Abbildung 4, daseine Frequenzanalyse von 160-16.000 Hzumfasst, geht hervor, dass die frequenzspek-tralen Anteile mit der höchsten Schallintensi-tät im Bereich von 2.000-7.000 Hz liegen, die
Alarmrufe aber auch im Bereich von 9.000-14.000 Hz deutliche frequenzspektrale An-teile enthalten. Ähnlich strukturierte Alarm-rufe sind auch von einer Reihe weiterer Vo-gelarten bekannt wie z. B. dem Teichrohrsän-ger (HEUWINKEL 1980).
Literatur
HEUWINKEL, H., 1980: Schalldruckpegel und Fre-quenzspektren der Gesänge von vier Rohr-sänger-Arten (Acrocephalus arundinanus, A. sci-rpaceus, A. schoenobaenus und A. palustris) inBeziehung zur Biotopakustik. � Inangural-Dissertation, Westfälische Wilhelms-Univer-sität zu Münster.
KLUMP, G., SHALTER, M.D., 1984: Acoustic behav-iour of birds and mammals in the predatorcontext. I. Factors affecting the structure ofalarm signals. II. The functional significanceand evolution of alarm signals. � Zeitschriftfür Tierpsychologie 66, 189-226.
MARLER, P., 1956: Über die Eigenschaften eini-ger tierlicher Rufe. � Journal für Ornitholo-gie 97: 220-227.
MARLER, P., SLABBEKOORN, H., 2004: Nature�sMusic � The Science of Birdsong. � ElsevierAcademic Press, London.
Angenommen: 22. 12. 2006

Totholz im collinen/montanen Laubwald 253
ACTA BIOLOGICA BENRODIS 13 (2006)
Rezensionen
ENGELMANN, WOLF-EBERHARD (Hrsg., 2005): Zootierhaltung. Tiere in menschlicher ObhutFische. � Verlag Harri Deutsch, Frankfurt am Main. 890 Seiten mit zahlreichen SW-Fotos.Kartoniert, 23 x 15,6 cm. ISBN 978-3-8171-1352-1. 49,80 Euro
ENGELMANN, WOLF-EBERHARD (Hrsg., 2006): Zootierhaltung. Tiere in menschlicher ObhutReptilien und Amphibien. � Verlag Harri Deutsch, Frankfurt am Main. 600 Seiten mitzahlreichen SW-Fotos und einer CD-ROM. Kartoniert, 22,8 x 16,2 cm. ISBN 978-3-8171-1743-7. 49,80 Euro
Das ist eine umfangreiche Gesamtschau, die ursprünglich wohl in erster Linie für die �Schau-aquaristik� und hier für die Ausbildung von Zootierpflegern gedacht war, dann aber lautVorwort auch �Nachschlagewerk zur Kurz- und Erstinformation für den Fachkollegen wie fürden interessierten Hobbyaquarianer� sein soll (Fische S. 16). Zahlreiche Helfer und Koautorenhaben an ihr mitgewirkt und eigene Erfahrungen und Beobachtungen mit einfließen lassen.Wo findet man schon z.B. so gebündelt Angaben zum Höchstalter von Fischen und Reptilienin menschlicher Obhut und anderes mehr? Darin liegt in erster Linie der Wert eines Buches, dassich mit keiner der angesprochenen Zielgruppen (Zootierpfleger, Fachkollegen) so recht iden-tifizieren kann und dessen Lektüre daher aus verschiedenen Gründen (noch) nicht ganz glück-lich macht. Darüber hinaus ist gerade auf diesem Gebiet � sieht man einmal von großen�Fischen� und �Reptilien� ab, die eben nur in �Schauaquarien� und �Schauterrarien� gehaltenwerden � die Konkurrenz riesig, denn an guten und vor allem auf die Belange einzelner Arteneingehenden Aquarien- und Terrarienbüchern ist kein Mangel.Im �Fischband� befasst sich ein erster Teil mit �Grundlagen der Schauaquaristik� (S. 21-137).Hier erfährt der Leser unter anderem etwas über die Geschichte und Entwicklung der Schauaqua-ristik, über Personen, die sich um diese verdient gemacht haben (hier findet man mit Recht nebenbekannten Größen wie A. E. Brehm, O. Heinroth u.a. auch M. Zahn, den ehemalige Leiter desLöbbecke Museums und Aquazoos in Düsseldorf, dessen Konzeption eine didaktische Glanz-leistung war), über (Groß-)Aquarienbau und -gestaltung, Aquarienbepflanzung, Themen, dieüber Aquarien vermittelt werden können, Biotopbecken, Aquarientechnik, Pflege und Sicher-heitsmaßnahmen, Fütterungskunde, Krankheiten und Quarantäne sowie Fang und Transport.Der weitaus umfangreichere zweite Teil �Systematik und Haltung� (S. 141-807) stellt Vertre-ter (in Auswahl und manchmal sehr pauschal; das ist der Preis, den man für ein eher enzyklo-pädisch angelegtes Werk zahlt) von 306 �Fischfamilien� vor. Die Behandlung der Fischefolgt stets einem weitgehend ähnlichen Muster. Vorgestellt werden von den Vertebratenausgehend die jeweils niedrigeren Taxa und aus den Familien jeweils typische Gattungenund Arten. Dazu gibt es Angaben zur Verbreitung, Lebensweise, Nahrung, Haltung undVermehrung und, falls bekannt, zum Höchstalter. Das ist auch so weit in Ordnung und dieLektüre vermittelt viel von der Erfahrung der jeweiligen Bearbeiter.Dennoch: Der Teufel liegt im Detail. Damit meine ich nicht so sehr die bei einem so umfangrei-chen Buch zwangsläufig sich einschleichenden Fehler oder Ungenauigkeiten, sondern auch dieTatsache, dass der Zootierpfleger bei der Lektüre mancher Passagen ohne weitere Erklärungenoder Kenntnisse aus der Zoologie einfach überfordert ist und der Fachkollege über mancheunpräzisen Formulierungen (hoffentlich) verwundert ist. Eine kleine Auswahl: Natürlich kanndas Buch �weder eine Bestimmungshilfe � noch ein Lehrbuch der Fischsystematik und Ich-

254 HEIKE KAPPES & WERNER TOPP
thyologie sein� (Einführung S. 18), doch das Taxon �Pisces� gibt es nun wirklich nicht mehr.Überhaupt wird m.E. ein ungeheurer Wert auf die Nennung der einzelnen Kategorien (Ord-nung, Unterstamm, Überklasse) gelegt. Hier sollte in zukünftigen Auflagen dem Leser (auch demZootierpfleger) klar gemacht werden, dass diese Kategorien nahezu ohne jeden Informations-wert sind. Und eigentlich ist auch das Schema von Branchiostoma (ohne weitere Erklärungen, z.B.Hirnbläschen, Gehirn etc.) in so einem Buch fehl am Platze, ebenso wie einige phylogenetischeSpekulationen, die zuwenig begründet sind. Die Termini �eilebendgebärend� (ovovivipar) und�echt lebendgebärend� (S. 154) sollten einer präziseren Definition weichen. Ob die �Neuralbö-gen� bei rezenten Neunaugen den Beginn (es handelt sich um rezente Organismen!) der Wirbel-bildung dokumentieren (S. 146), sei dahingestellt. Bei den �Osteichthyes� kann man nicht mehrQuastenflosser, Lungenfische und Störartige in einem Atemzug nennen (S. 219). Wenn schonGanoid- und Cosmoidschuppen erwähnt werden, sollte man auch deren Unterschiede nennen(S. 219). �Die Entwicklung der Eier erfolgt bereits im Mutterleib� (S. 222): Auch der angehendeZootierpfleger hätte eine präzisere Information verdient. Irreführend ist der Satz �An die Qua-stenflosser als direkte Vorfahren der Landwirbeltiere erinnern�� (S. 241, da denkt man sofort anLatimeria). Bei Halbschnabelhechten steht: �Die Embryonen entwickeln sich in einer sackartigenHöhlung des Eierstocks.� (S. 474) Was auch immer das ist, es ist zu verallgemeinernd. Dass beiAnableps anableps (der ist wohl gemeint) eine �dauerhafte Hälterung in reinem Süßwasser früheroder später zu Hauterkrankungen führt�, ist längst durch den Aquazoo in Düsseldorf widerlegtworden, der diese Tiere seit Generationen in reinem Süßwasser hält und züchtet (S. 485). Zu denGoodeiden liest man auf S. 493: �Besondere Nährstränge wachsen von den Embryonen zurEileiterwand, so dass sie von der Mutter ernährt werden (echtes Lebendgebären!).� Abgesehendavon, dass es nicht der Eileiter, sondern das Ovar ist, kann ich mir nicht allzu viel unter dieserAussage vorstellen. In einem Anhang gibt es einen Exkurs über giftige und elektrische Fischesowie über den Umgang mit diesen und ein Kapitel über Domestikation. Am Schluss werden dieAutoren vorgestellt. Das Literaturverzeichnis besteht überwiegend aus Sekundär- und Tertiärlite-ratur. Ihm folgt ein allgemeiner Index, ein Index der deutschen Fischnamen (gefallen haben mirder Amu-Darja-Pseudoschaufelstör, der Bastardgrunzer, der Wenigbestachelte Igelfisch und derUnbewaffnete Steinpicker) und ein Index der wissenschaftlichen Fischnamen. Leider finden sichim Text keine Literaturzitate, die es dem Leser ermöglichen würden, die eine oder andere Aussageeventuell etwas ausführlicher im Original nachzulesen, so z.B. über den Flug der Beilbäuche (S.335) oder die Zucht von Mormyriden und Messerfischen. Hier fehlen die sehr präzisen Angabenzur Zucht aus den Arbeiten von Kirschbaum und Mitarbeitern.Der Reptilien- und Amphibienband hat dieselbe(n) Zielgruppe(n). �Hier Basiswissen zu ver-mitteln, ist ein Hauptanliegen dieses Lehrbuchs. Daneben soll aber auch der Hauptkurator wieder Hobby-Terrarianer ein Nachschlagewerk in die Hand bekommen, das ihm zur Erstinfor-mation dient oder für den schnellen Überblick eine Hilfe sein kann.� (S. 12 Vorwort) Auch hierfinden sich zunächst Kapitel, die auf die Schauterraristik eingehen (Historie, Terrarium alsLebensraum, Pflege- und Sicherungsmaßnahmen, Fütterungskunde, Krankheiten, Fang, Trans-port) und wiederum der Hauptteil (s. 79-555) mit Systematik und Haltung. Insgesamt halte ichdiesen Band für weniger altmodisch und für viel präziser als den Fischband, vielleicht wegendes insgesamt etwas weniger zu bewältigenden Stoffes. Ähnlich wie Fische sollte man sich abermittlerweile doch langsam von liebgewordenen taxonomischen Einheiten trennen: Reptiliengibt es nun einmal nicht mehr. Die nähere Verwandtschaft der Krokodile mit Vögeln (S. 177)muss doch Konsequenzen haben. Irgendwann ist auch eine Tradition, und sei sie noch soliebenswürdig, obsolet. Für ein Lehrbuch, das die Bände ja nun erklärtermaßen auch seinsollen, ist die Abbildung des Amnioteneis unzureichend (S. 81). Wie ist eigentlich der Satz zu

Totholz im collinen/montanen Laubwald 255
ACTA BIOLOGICA BENRODIS 13 (2006)
verstehen: �Die Körperfarben werden im Wesentlichen durch die Chromatophoren (Farbträ-ger) in den Chromatoblasten (Farbzellen) der Unterhaut bestimmt.� (S. 82)? Für echte Vivipa-rie (S. 196) und Ovoviviparie (S. 84) gilt das oben Gesagte. Ob sich ein angehender Zootierpfle-ger etwas unter einem ZZ/ZW-Typ vorstellen kann? (S. 85) Auf Abb. 7.2 sind die Schleimdrü-sen der Amphibien als Körnerdrüsen und die Körnerdrüsen als Schleimdrüsen bezeichnet (S.427). Was sind �gestielte Zähne�? (S. 428) Schade, dass den Alpensalamandern nur �zusätzlichSubstanzen aus dem Körper zugeführt� (S. 429) werden. Diese zusätzlichen Substanzen sindEier und mütterliches Gewebe! Gymnophionen haben nicht schuppenähnliche Gebilde in derHaut, sondern tatsächlich Schuppen (S. 462). Im Text finden sich leider ebenfalls keine Litera-turangaben. Im Anhang erfährt der Leser unter anderem etwas über die Freiterrarienanlage imAlpenzoo Innsbruck und über die Autoren. Der Bildnachweis, ein Literaturverzeichnis, einallgemeiner Index, ein Index der deutschen Tiernamen (auch hier �erfreuen� erfundene deut-sche Namen wie Vierzehen-Handwühle, Talamanca-Raketenfrosch, Dunkelbäuchiger Herz-züngler usw.) und der wissenschaftlichen Tiernamen sowie eine schöne CD mit Farbbildernder im Text behandelten Reptilien und Amphibien beschließen diesen Band.Ich habe diese z.T. vielleicht kleinlichen Anmerkungen nicht gemacht, um diese Buchreihe zudesavouieren, sondern um aufzuzeigen, dass Nachschlagewerk und Lehrbuch wohl nur schwermiteinander zu vereinbaren sind. Ein basales Lehrbuch für angehende Zootierpfleger sollte vielmehr erklärend sein und biologische und durchaus auch phylogenetische Zusammenhängeetwas moderner, präziser und vor allen Dingen verständlich bieten � die einführenden Kapitel inbeiden Bänden setzen m.E. gehörige Kenntnisse in sehr spezieller vergleichender Anatomievoraus und sind teilweise ungenau �, dem Fachkollegen dürfte in dieser Beziehung wohl mehrzugemutet werden (können); er wird zudem Ungereimtheiten eher entdecken.Fazit: Lehrbücher sind diese beachtenswerten und verdienstvollen Bände (noch) nicht, brauch-bare Nachschlagewerke allemal. Man wünscht beiden weitere, geläuterte Auflagen
Hartmut Greven, Düsseldorf
EICHMANN, KLAUS (2005): Köhler�s Invention. � Birkhäuser Verlag, Basel, Berlin, Boston.223 Seiten. Gebunden, 23,2 x 16,2 cm. ISBN 978-3-7643-7173-9. 53,40 Euro
Zehn Jahre nach dem plötzlichen Tod des 38jährigen Georges Köhler hat Klaus Eichmannvorliegendes Buch zum Gedenken an seinen um sieben Jahre jüngeren Kollegen und Nobel-preisträger veröffentlicht. Es ist ein sehr persönliches Buch, gefüllt mit überraschenden Infor-mationen und Aussagen. Bereits der Titel des Buches �Köhlers Erfindung� lässt erkennen,dass es sich nicht nur um eine Biographie von Köhler handelt, sondern um die bahnbrechendeHybridoma-Technik, d.h. die Erzeugung von monoklonalen Antikörpern. Die Hybridoma-Geschichte wird in dem Buch in zwei große Abschnitte eingeteilt, die durch Köhlers kurzesLeben bestimmt sind: die Zeit vor dem Nobelpreis und die Zeit nach dem Nobelpreis.Eichmann beginnt mit einem kurzen geschichtlichen Abriss über das Antikörperproblem,d.h. die Aufgabe, die ungeheure Diversität der Antikörper schlüssig zu erklären, und endet mitdem Kapitel, dass auch heute das Antikörperproblem noch nicht gänzlich gelöst ist. Je nachNeigung und Vorbildung kann man das facettenreiche Buch unter drei verschiedenen, sichgegenseitig bedingenden Aspekten lesen. Der erste, nicht immer mühelose Weg führt durchdie Geschichte der wissenschaftlichen Fragestellungen zur Antikörperstruktur und -funktionund welche Ergebnisse im Laufe der Zeit gewonnen wurden. Dieser Stoff bildet den Hauptteil

256 HEIKE KAPPES & WERNER TOPP
des Buches. Es ist sicherlich, entgegen der ausgesprochenen Absicht des Autors, eine lehrreiche,subjektive Übersicht über den Fortschritt von Wissenschaft auf einem speziellen Gebiet derImmunologie. In dieser Zusammenschau bewertet Eichmann manchmal kurz oder auchausführlicher die Leistungen vieler seiner Kollegen im Umfeld von Georges Köhler. Manchmalkann man sich bei der Nennung von bekannten Namen des Eindrucks einer �mutual admiringsociety� nicht erwehren und sicherlich gibt es einige Immunologen, Biologen oder Chemiker,die sich gerne in dem Buch erwähnt fänden. Vielleicht wurde deshalb auf ein Personenregisterverzichtet und das 18 (!) Seiten lange Literaturverzeichnis für ausreichend empfunden. DieKapitel Post-Nobel science I und II sind etwas für Eingeweihte.Der zweite Gewinn beim Lesen des Buches ist ein Einblick in den Wissenschaftsbetrieb und dieArbeit nationaler wissenschaftlicher Institutionen, hier insbesondere die Entwicklung der Max-Planck-Gesellschaft. Was ist das Besondere an MPIs? Wie werden Institute gegründet und Wis-senschaftler für führende Posten gefunden? Welche Rolle spielen Gutachter dabei? Überraschenderfährt man, dass eine Gutachtermehrheit aus dem Ausland Georges Köhler für den Posten einesMPI-Direktors in Freiburg für ungeeignet hielt, was Klaus Eichmann letztlich abbiegen konnte.Man wird über das taktierende Spiel von Berufungsverhandlungen bei der Besetzung von akade-mischen Stellen in Europa aufgeklärt, über den Einfluss großer Namen bei Veröffentlichungen inSpitzenzeitschriften. Wie gehen Wissenschaftler miteinander um, wenn es um Anerkennungund Ehrungen geht? Wer war nun der eigentliche Erfinder der Hybridoma-Technik?Wissenschaft wird von Menschen gemacht, und diese Aussage führt zu dem dritten Genuss, denman aus Eichmanns Buch ziehen kann oder auch nicht. Der Autor frappiert den Leser mit sehrintimen und häufig erbarmungslosen Charakterisierungen prominenter Personen. Geniale Men-schen haben Stärken und Schwächen. Es gibt welche mit chaotischen Liebesaffären oder solche,die �das Trinken lieben�. Eichmann hält es für erwähnenswert, dass Köhler ein treuer Ehemannwar und sogar auf wissenschaftlichen Kongressen keine Affären hatte. Hat man je gehört, dass einVortrag während der Verleihung des Nobelpreises im betrunkenen Zustand gehalten wurde? Istes nicht zum Kopfschütteln, dass Köhler bei derselben Veranstaltung seinen Vortrag wie einAnfänger mit zu vielen Diapositiven befrachtet hatte? Nicht unerwartet lernt man sehr viel überden Menschen Georges Köhler; sogar ein ganzes Kapitel wird über �menschliche Beziehungen�geschrieben. Bemerkenswert ist die ablehnende Haltung von Köhlers Frau, etwas zur Erinnerungan ihren Mann beizutragen. Fasst man alles zusammen, was Eichmann in seinem Buch überKöhler zusammengetragen hat, so bleibt der zwiespältige Eindruck einer nicht sehr umgängli-chen Person zurück. Der Leser ist aufgefordert, sich eine eigene Meinung zu bilden. Sehr bewe-gend ist zum Schluss der Bericht über die Todesumstände von Köhler. Erwähnt sollte nochwerden, dass das Buch ein Faksimile der entscheidenden Experimente aus dem Jahr 1974 enthält,Photographien aus dem Leben Köhlers um und nach dem Nobelpreis, eine komplette Listeseiner Publikationen und seiner Ehrungen. Dem jungen Studenten oder Doktoranden mögenfolgende Eigenschaften Köhlers über eigene Unzulänglichkeiten hinwegtrösten. Köhler ist keinguter Schüler gewesen. Er hatte eine eigene Arbeitsmoral als Doktorand, d.h. nicht nachts arbeitenund nie an Wochenenden. Er war seelenverwandt mit seinem Mentor und MitnobelpreisträgerCesar Milstein, der sagte: Faulheit ist die Mutter guter Wissenschaft. Schöpferische Einfälle be-kommt man dann, wenn man nichts tun muss. (�Laziness is the mother of good science.Creation comes from moments when you don�t have anything to do.�) Was war das Geheimnisvon Georges Köhler? Man muss zur richtigen Zeit mit der richtigen Frage am richtigen Objekt amrichtigen Ort in der richtigen Arbeitsgruppe arbeiten und unerschütterlich an sich glauben.
Karl Heinz Glätzer, Düsseldorf

Mitteilungen aus dem Museum für Naturkunde in der Stiftung Schloss und Park Benrath M1
Acta Biologica Benrodis 13 (2006)
Mitteilungen aus demMuseum für Naturkunde, Stiftung Schloss und Park
Benrath
Freundeskreis Museum für Naturkunde Benrath e.V.
Im Berichtszeitraum unterstützte der Freundeskreis Sonderveranstaltungen, Ferien-programme für Kinder und die Herausgabe der Museumszeitschrift.
Im Jahre 2005 erschien der von RWE Power finanzierte 236 Seiten starke Supplement-band 10 von Acta Biologica Benrodis �Tiere und Pflanzen in der Rekultivierung � 40Jahre Freilandforschung im Rheinischen Braunkohlerevier� von Claus Albrecht, UlfDworschak, Thomas Esser, Horst Klein und Jochen Weglau (Ppbk. ISBN 978-3-936616-35-4, 8,- Euro; geb. ISBN 978-3-936616-36-1, 14,- Euro).
Für die museumspädagogische Arbeit beschaffte der Freundeskreis einen Laptop,einen Scanner sowie 20 Mikroskope.
Auf der Mitgliederversammlung am 09.12.2004 legte der Vorsitzende Herr HaraldMöhler sein Ehrenamt nieder, in das er auf der Mitgliederversammlung am 02.02.1999gewählt worden war, da er aus beruflichen Gründen Düsseldorf verlässt. Herr Möhlerhat in diesem Zeitraum, in dem das Naturkundliche Heimatmuseum Benrath in dieStiftung Schloss und Park Benrath unter dem Namen Museum für Naturkunde ein-gegliedert wurde, stets mit großem Interesse und Engagement bei der Weiterentwick-lung unseres Museums mitgewirkt. Ein besonders herausragendes Ereignis war dieEinwerbung von 400.000,00 DM bei der NRW-Stiftung für die Einrichtung einerDauerausstellung zum Thema �Flüsse und ihre Auen unter besonderer Berücksichti-gung des Rheins�. Für die Zusammenarbeit, die stets in harmonischer und freund-schaftlicher Atmosphäre verlief, wurde ihm ganz herzlich von den versammeltenMitgliedern gedankt. Als sein Nachfolger im Amt des Vorsitzenden wurde Herr RolfKlahold gewählt.
Der Freundeskreis dankt wie immer ganz herzlich allen Mitgliedern und Spendern,die das Museum ideell und finanziell bei der Wahrnehmung seiner vielfältigen Aufga-ben unterstützten, insbesondere der Klassik-Konzertgesellschaft e. V. und ihrem 1.Vorsitzenden Franz Lamprecht.
Für weitere Förderungen des Museums sind Spenden jeder Größenordnung stetssehr willkommen. Der Freundeskreis freut sich auch jederzeit über neue Mitglieder.
Freundeskreis Museum für Naturkunde Benrath e.V.,Benrather Schloßallee 102,40597 Düsseldorf-BenrathBankverbindung:Deutsche Bank AG Düsseldorf-Benrath, Konto 6523070 (BLZ 300 700 10)

M2
Verabschiedung von Herrn Harald Möhler (rechts) als Vorsitzender des Fördervereins �Freun-deskreis Museum für Naturkunde Benrath e.V.� durch Museumsleiter Dr. Hubert Heuwinkelauf der Mitgliederversammlung am 09.12.2004. Vorn rechts im Bild Herr Klaus Behrens,Schatzmeister des Fördervereins.
Teilnehmer des Internationalen Workshops �First European Workshop on Isopods� vor demEingang zum Vestibül des Schlosses Benrath am 15.11.2003.

Mitteilungen aus dem Museum für Naturkunde in der Stiftung Schloss und Park Benrath M3
Acta Biologica Benrodis 13 (2006)
Veranstaltungen und Ausstellungen
2003
12.01. - 13.03. Naturkundliche Begleitausstellung �Spinnen nur Spinnen? Biologievon Spinnentieren� zur Sonderausstellung �Verwoben� von Jean-Luc Bruniaux imMuseum für Europäische Gartenkunst. In Kooperation mit dem NABU Düsseldorf.
28.01. Abendvortrag �Halsbandsittiche in Düsseldorf � von Dipl. Biol. Tobias Krause.
05.02. Abendvortrag �Fallensteller, Wegelagerer und Meister der Pirsch� über die viel-fältigen Fangmethoden der Spinnen von Prof. Dr. Walter Kirchner.
12.02. Abendvortrag �Hochwasser-Katastrophe oder Segen für die Natur� von Dipl.Geogr. Holger Pieren
13.02. Abendvortrag �Zur Biologie der Spinnen� von Dr. Stephan Loksa
09.03./ 10.03. und 10.10. Abendvortrag über Fledermäuse und Fledermaus-Exkursionvon und mit Eberhard Menz
11.03. Filmvortrag �Aus dem Leben der Eisvögel� von Naturfilmer Otto Kieker
10.05./ 11.05. Nacht der Museen
Programm
Es spielt die New Dixie Band des Schloss-Gymnasiums Benrath, Leitung Dr. UdoZilkens�Eine naturkundliche Wanderung vom Rhein zum Bergischen Land im Museum�,Führung�Fliegende Edelsteine�, Filmvortrag von Otto Kieker�Blumen und Schmetterlinge in 3 D�, Diavortrag von Rolf Niggemeyer�Fledermäuse auf der Leinwand und live im Schlosspark� von und mit EberhardMenz�Die kleine Anfrage � Leonardo Quiz� von den Moderatoren Fatma Dasan und StefanBitterle des WDR 5
06.07. �Tag der offenen Tür�und Erlebnistag des Netzwerkes der naturkundlichen Museen, Infozentren und Bio-logischen Stationen der EUROGA-Region.Die Partner des net:natuur_cultuur präsentieren ihre Ausstellungen und Arbeitsge-biete im Innenhof des Museums für Naturkunde.
26.07. �Exkursion in die Welt der Spinnen� für Eltern mit Kindern mit Stefanie Egeling.

M4
14. - 16.11. �First European Workshop on Isopods�.
Vom 14. bis 16. November trafen sich im Museum für Naturkunde, Stiftung Schlossund Park Benrath zum ersten europäischen Workshop etwa 15 Wissenschaftler, diesich intensiv mit vermeintlichen Ekeltieren, den aus vielen Kellern bekannten Asseln,befassten.Die Themen reichten von Fortpflanzungs-Strategien bis hin zu Problemen der Nah-rungsaufnahme und der Nahrungsverarbeitung. Asseln haben Lebensräume im Meer,im Süßwasser und auf dem Lande besiedelt. Die landbewohnenden Asseln sind unteranderem am Abbau des Falllaubs (Streu) beteiligt.Dieser Workshop sollte dem Erfahrungsaustausch und einer intensiveren Kooperati-on dienen. Der Workshop wurde in Zusammenarbeit mit dem Museum für Natur-kunde, Stiftung Schloss und Park Benrath von Prof. Dr. M. Zimmer, Universität Kiel,sowie Prof. Dr. H. Greven und Dipl. Biol. M. Stevens, Universität Düsseldorf, ausge-richtet.
Programm
14.11.14.00 � H. Heuwinkel: Welcome to the �Museum für Naturkunde, Stiftung Schlossund Park Benrath�14.15 � H. Greven, M. Stevens, M. Zimmer: Welcome to the Workshop � Introduc-tionPart I: �Get to know your colleagues � who is doing what?�Reproduction in isopods � I: Evolutionary scaleChairperson: S. Taiti14.45 � E. Hornung: Review on the evolutionary aspects of oniscid (Isopoda) repro-ductive strategies15.45 � H. Greven: Are isopods brooding, viviparous, larviparous or pueriparous? �Reflections on isopod reproduction, and some insufficiently answered questions re-lated to it16.45 � M. Stevens: Behavioural and structural aspects of reproduction in Oniscusasellus: An account on current investigations17.45 � M. Zimmer, C. Henf, B. Leupolt: Reasons for and consequences of not guar-ding a mate15.11.Reproduction in isopods � II: Individual scaleChairperson: M. Stevens09.00 � M. Hassall, A. Helden, A. Goldson: Microevolution and phenotypic plasticityof reproductive traits in Armadillidium vulgare (Isopoda: Oniscidea)10.00 � D. Bouchon, Y. Caubet, M. Johnson: Host-parasite interactions: reproductivebehaviour of terrestrial crustaceans and Wolbachia11.00 L. Weltje, C. Vogt, J. Oehlmann: Endocrine disruption in terrestrial and fresh-water isopodsNutrition in isopodsChairperson: H. Schmalfuss

Mitteilungen aus dem Museum für Naturkunde in der Stiftung Schloss und Park Benrath M5
Acta Biologica Benrodis 13 (2006)
14.00 � Y. Wang, K. Ihnen, M. Zimmer: Evolutionary ecology of isopod nutrition:animal-microbe interactions15.00 � S. Farkas: Study on the nutrition biology of five woodlice species16.00 � N. Dias: Feeding rates and diet analysis of salt marsh isopodsPart II: �Cooperate with your colleagues�Proposals for joint projects and potential sources for financial supportChairperson: H. GrevenEverybody is invited to present project proposals and recent information on announce-ments17.00 � M. Hassall: Marie Curie Training courses17.30 � M. Zimmer: ESF � InterACT (and others)16.11.Part III: �Where does this workshop lead us to?�Final concluding roundtable discussionChairperson: M. Hassall
2004
07.02. Das Museum wurde 75 Jahre alt.
09.03. Filmvortrag �Der Eisvogel� von Tierfilmer Otto Kieker.
21.03. Sonderausstellung � Joseph Wolf (1820-1899) � Ein englischer Tiermaleraus dem Rheinland�. Die Eröffnung war die Auftaktveranstaltung zur Feier des 75-jährigen Bestehens des Museums.
Programm
Musikalische Begrüßung durch Schüler des Schloss-Gymnasiums BenrathGrußworte von Dr. Hubert Heuwinkel, Leiter des Museums für Naturkunde, undHarald Möhler, Vorsitzender des Freundeskreises des Museums für NaturkundeBenrath e.V.Einführende Worte von Dr. med. Karl Schulze-Hagen, Mönchengladbach:
�Joseph Wolf � ein englischer Tiermaler aus dem Rheinland�
Joseph Wolf, einer der bedeutendsten Tiermaler des 19. Jahrhunderts, revolutioniertedas Genre der Tiermalerei durch bis dahin unbekannte Malweisen. Von einfachenHolzkohle-Skizzen bis zu dramatischen Ölgemälden � nie verließ ihn sein einzigar-tiges Talent für das Festhalten des Wesentlichen.Nach Lehr- und Wanderjahren ließ sich Joseph Wolf 1848 in London nieder. Zuvorhatte er als ausgebildeter Lithograph und talentierter Künstler bedeutende Bildbändeillustriert. In London dann entwickelte er sich innerhalb weniger Jahre zu einem derbedeutendsten Tierillustratoren des 19. Jahrhunderts.Der Grund seiner Karriere lag in seinem nahezu photographischen Gedächtnis, mitdem er natürliche Verhaltensweisen einmal beobachteter Szenen virtuos zu Papierbrachte. Seine Technik nahm Elemente des Impressionismus vorweg, ohne die Exakt-

M6
Blick in einen der Sonderausstellungsräume. In der Vitrine sind neben dem Präparat eines Schuh-schnabels (Leihgabe des Zoologischen Forschungsinstituts und Museum Alexander König, Bonn),einer Schuhschnabel-Plastik von Joseph Pallenberg und einem Gerfalken-Präparat Lithogra-phien dieser Vogelarten von Joseph Wolf zu sehen sowie weitere gehängte Bilder an den Wänden.
Oben: Portraits von Joseph Wolfim jüngeren und fortgeschrittenenLebensalter. Unten: Plakat zurSonderausstellung �Joseph Wolf(1820-1899) � ein englischer Tier-maler aus dem Rheinland�, zu-gleich als Jubiläumsplakat zum 75-jährigen Bestehen des Museums.

Mitteilungen aus dem Museum für Naturkunde in der Stiftung Schloss und Park Benrath M7
Acta Biologica Benrodis 13 (2006)
heit und Akkuratesse naturwissenschaftlicher Dokumentation zu vernachlässigen.Zwar hatte Joseph Wolf bereits 1846 Abbildungen für populäre Bücher gefertigt, docherst in London machte sein Renommee ihn zum Illustrator der Wahl für Großwildjä-ger, Forscher und Dichter.Der Naturalismus Wolfs führte Bewegung und Landschaftseindruck ein, wie mandies vorher nicht kannte. So hingen Wolfs Bilder alsbald in der Royal Academy, inPrachtvillen und königlichen Palästen. Der Öffentlichkeit blieb sein Werk allerdingsfast unbekannt, da er es vermied, seine Bilder über Galerien zu verkaufen.Unter dem Titel �Joseph Wolf (1820-1899) Tiermaler � Animal Painter� liegt ein 361Seiten umfassender, opulenter Band in deutscher und englischer Sprache von denAutoren Dr. Karl Schulze-Hagen und Prof. Dr. Armin Geus vor (Basilisken Presse,Marburg, ISBN 3-925347-57-7).
25.03. Abendvortrag �Fledermäuse in 3 D� von Eberhard Menz
12.04. Sternenwanderung im abendlichen Schlosspark mit Werner Buschfeld
07.05. Abendvortrag über Fledermäuse mit anschließender Exkursion in den Schloss-park von und mit Eberhard Menz
08.05./ 09.05. Nacht der Museen
Programm
Musikalische Begrüßung durch die New Dixie Band des Schloss-Gymnasiums, Lei-tung Dr. Udo ZilkensFührung durch die Sonderausstellung �Joseph Wolf � ein englischer Tiermaler ausdem Rheinland��Ein bunter Zimmermann � Dendrocopus maior�, Filmvortrag von Otto Kieker�Falter der Nacht � Schmetterlinge des Tages�, Diavortrag in 3D von Rolf Niggemey-er�Fledermäuse auf der Leinwand und live im Schlosspark� von und mit EberhardMenz�Waldvöglein und Co. ganz groß � Wildorchideen�, Diavortrag in 3D von Rolf Nigge-meyer�Kleine Tierkunde zur Geisterstunde�, Heimliches und Unheimliches vorgetragenvon Markim Pause
04.07. �Tag der offenen Tür�Erlebnis- und Netzwerktag im Museum für Naturkunde zusammen mit NABU-Düsseldorf und net:natuur_cultuur ; 75 Jahre Museum für Naturkunde Benrath; 10Jahre Stadtverband NABU-Düsseldorf
Programm
Eröffnung der Sonderausstellung Natura 2000 der Landesanstalt für Ökologie, Bo-denordnung und Forsten, NRW

M8
11.00 Begrüßung im Innenhof durch Oberbürgermeister Joachim Erwin, Dr. HubertHeuwinkel, Museumsleiter, und Francisca Lienau, NABU-Vorsitzende11.30 Planwagenfahrt mit dem Pferdefuhrwerk um das Schloss12.00 �Schönheit der Natur in Düsseldorf und ihr Schutz�, Diavortrag von Dipl.Geogr. Holger Pieren, Biologische Station Urdenbacher Kämpe e.V.13.00 und 15.00 �Schmetterlinge in Düsseldorf�, Diavortrag von Ludwig Amen, NABU14.00 und16.00 �Zaunkönig � Vogel des Jahres 2004�, Diavortrag von Alfred Leisten,NABUDurchgehend: KinderaktivitätenStündlich: Museumsführungen
�Natura 2000 � Von der Vision zur Wirklichkeit�Die Ausstellung �Natura 2000� lässt ein spannendes Stück Naturschutzgeschichte �den wechselvollen Weg von der Vision zur Wirklichkeit eines europäischen Netz-werks für den Naturschutz � Revue passieren. Kernstück dieser Ausstellung der Lan-desanstalt für Ökologie, Bodenordnung und Forsten, NRW (LÖBF) ist die Präsenta-tion der Lebensräume, welche die Kriterien der so genannten Flora-Fauna-Habitat-Richtlinie (FFH) und EG- Vogelschutzrichtlinie erfüllen.Neun Lebensräume mit Gebietsbeispielen aus ganz NRW und den dort lebendenTieren und Pflanzen werden in Wort und Bild dargestellt. Die Ausstellung informiertunter anderem über die Stillgewässer des Vogelschutzgebietes �Unterer Niederrhein�mit ihren Libellen, Amphibien und Vogelarten und über die Wiesen der UrdenbacherKämpe, in denen zum Beispiel seltene Schmetterlinge zu Hause sind. Aber auchFließgewässer mit Auenwäldern, Heiden und Sandtrockenrasen, Moore und Moor-wälder, Magerrasen und Mischwälder werden präsentiert.Mit der FFH- und EG-Vogelschutzrichtlinie haben sich alle Mitgliedsstaaten verpflichtet,im zusammenwachsenden Europa einen gemeinsamen Beitrag zur Sicherung des Euro-päischen Naturerbes zu leisten. Bis heute wurden von NRW 513 FFH-Gebiete und 29EG-Vogelschutzgebiete an die Europäische Kommission gemeldet. Insgesamt wurdendamit rund 280 000 Hektar (8,2 % der Landesfläche) unter besonderen Schutz gestellt.
07.07. Abendvortrag über die FFH-Gebiete in der Stadt Düsseldorf von Dipl. Geogr.Holger Pieren.
19.09. Informationsstand des Museums für Naturkunde mit Exponaten aus dem Mu-seum mit dem Schwerpunkt Bienenkunde und Imkerei anlässlich der Festveranstal-tung zum 50-jährigen Bestehen der Bezirksvertretung 9 (Benrath) rund um das Rat-haus in Benrath.
11.10. Vorstellung �Zaunkönig � Vogel des Jahres 2004� durch UmweltdezernentinFrau Nieß-Mache im Museum für Naturkunde
10.11. Abendvortrag �Jahrhunderthochwasser � Jedes Jahr und immer höher?� vonDipl. Geogr. Holger Pieren
17.11. Abendvortrag �Dicke Luft in Düsseldorf? � Was Flechten und Moose uns überdie Luft sagen� von Dr. Norbert Staffer

Mitteilungen aus dem Museum für Naturkunde in der Stiftung Schloss und Park Benrath M9
Acta Biologica Benrodis 13 (2006)
Infostand des Museums für Naturkunde auf der Festveranstaltung zum 50-jährigen Bestehender Bezirksvertretung 9 rund um das Rathaus in Benrath am 19.09.2004 mit Museumspädago-gin Irmtrude Peters im Vordergrund.
Zum 75 jährigen Bestehendes Museums sowie dem 10jährigen Bestehen desNABU-StadtverbandesDüsseldorf wurde am04.07.2007 ein Tag der of-fenen Tür veranstaltet, indessen Verlauf sich etwa1500 Besucher einfanden.Von links MuseumsleiterDr. Hubert Heuwinkel,Oberbürgermeister JoachimErwin, NABU-VorsitzendeFrancisca Lienau, verdecktVorsitzender des Muse-umsfördervereins HaraldMöhler, StiftungsvorstandKlaus Krösche und Bezirks-verwaltungsstellenleiter derBV 9 Benrath WolfgangMehner.

M10
24.11. Abendvortrag �Faszination Niederrhein � mit allen Sinnen Natur erleben� undVorstellung des gleichnamigen Buches von den Autoren Dr. Georg Verbücheln undDr. Klaus van de Weyer (�Faszination Niederrhein, Mit allen Sinnen Natur erleben,Landschaften, Pflanzen und Tiere, Wanderungen, Kochen mit Kräutern�, Mercator-Verlag, ISBN 978-3-87463-353-1)
30.11. Abendvortragsreihe zum Thema: �Drei Jahre Deichrückverlegung im Mon-heimer Rheinbogen � Wie wirkt sich das Hochwasser auf Flora und Faunaaus?� In Kooperation mit der Biologischen Station Urdenbacher Kämpe e.V.
In den drei Jahren seit der Deichöffnung stand der Monheimer Rheinbogen schonmehrfach unter Wasser. Wie hat sich das auf die Flora und Fauna ausgewirkt? Vor undnach der Deichöffnung wurden, auf Anregung der Biologischen Station, verschiedeneTiergruppen und die Vegetation wissenschaftlich untersucht. An diesem Abend wer-den erste Ergebnisse von vier Monitoringuntersuchungen vorgestellt.Begrüßung und Einführung durch Dipl. Biol. Elke Löpke, Biologische Station Ur-denbacher Kämpe e.V.
Einführung in die Abendvortragsreihe �Drei Jahre Deichrückverlegung im Monheimer Rhein-bogen � Wie wirkt sich das Hochwasser auf Flora und Fauna aus?� von Elke Löpke, Leiterin derBiologischen Station Urdenbacher Kämpe e.V. am 30.11.2004.
�Vegetation des Monheimer Rheinbogens: Vergleich 1997 und 2003� von Dipl. Biol.Doris Riener�Bodentiere im Monheimer Rheinbogen vor und nach der Deichöffnung� von Dipl.Biol. Anke Struebig, Annika Dick, Prof. Dr. Werner Topp, Zoologisches Institut derUniversität Köln�Auswirkungen von winterlichen Hochwasser-Ereignissen auf Kleinsäuger (Insec-tivora, Rodentia) im Monheimer Rheinbogen� von Holger Meining�Entwicklung des Feldhasenbesatzes im Monheimer Rheinbogen� von Dr. HeinrichSpittler

Mitteilungen aus dem Museum für Naturkunde in der Stiftung Schloss und Park Benrath M11
Acta Biologica Benrodis 13 (2006)
Eröffnung der Sonderausstellung �Naslinge � Schöpfung der Phantasie? Phantasie der Schöp-fung?� am 30.01.2005 mit einer Einführung von Prof. Dr. Hartmut Greven.
Kleindiorama mit dem Nasling Eledonopsis suavis links und Nasobema lyricum rechts.

M12
2005
18.01. �Sternstunden � ein astronomischer Beobachtungsabend im Schlosspark� mitWerner Buschfeld
30.01. Eröffnung der Sonderausstellung �Naslinge � Schöpfung der Phantasie? �Phantasie der Schöpfung?�
Programm
Grußworte von Dr. Hubert Heuwinkel, Leiter des Museums für Naturkunde, undRolf Klahold, Vorsitzender des Freundeskreises des Museums für Naturkunde e.V.Einführende Worte von Prof. Dr. Hartmut Greven, Universität Düsseldorf
Umschlag von Harald Stümpkes grundlegendem Werk über die Naslinge (Rhinogradentia)

Mitteilungen aus dem Museum für Naturkunde in der Stiftung Schloss und Park Benrath M13
Acta Biologica Benrodis 13 (2006)
�Naslinge � Schöpfung der Phantasie? Phantasie der Schöpfung?�
Das Nasobem
Auf seinen Nasen schreiteteinher das Nasobem
von seinem Kind begleitet.Es steht noch nicht im Brehm.E steht noch nicht im Meyer.
Und auch im Brockhaus nicht.Es trat aus meiner Leyerzum erstenmal ans Licht.Auf seinen Nasen schreitet(wie schon gesagt) seitdem,von seinem Kind begleitet,
einher das Nasobem.
Christian Morgenstern (1871-1914)
Diese Verse sind der erste schriftliche Beleg dafür, dass Naslinge existieren. DerLyriker Christian Morgenstern, von keinem Geringeren stammt dieses Gedicht, warein gebildeter Mann und so nannte er diese besonderen Lebewesen �Nasobeme� ausnasus, lateinisch, die Nase, und bema, griechisch Schritt oder Gang.Was ist daran so erstaunlich? Eine überaus faszinierende Gruppe von Lebewesen wirdder �scientific community� (so pflegt sich die Gesamtheit derer zu nennen, die glau-ben, Wissenschaft zu betreiben) nicht über eine Originalarbeit vorgestellt, beispiels-weise in �Nature� und �Science� (um nur zwei der wissenschaftlichen Journale zunennen, die Aufsehen Erregendes bringen und in denen zu publizieren dem oder denAutoren (oft) höchstes Ansehen verschafft), und der interessierten Öffentlichkeit nichtüber allgemeinverständliche Artikel, beispielsweise in �Biologie in unserer Zeit� oderim �Kosmos�, sondern durch einen Dichter � einen (Sprach)künstler gewissermaßen�, und dazu noch in Reimen.Eine Zoologie in Reimen oder Versen zu schreiben, lag Morgenstern fern, obgleich vieleseiner Gedichte tiefes Interesse und Verständnis für biologische Sachverhalte zeigen. Soauch folgendes, in dem die umstrittene Frage, ob Tiere zählen können, behandelt wird:
Das Perlhuhn
Das Perlhuhn zählt: Eins, zwei, drei vier�Was zählt es wohl, das gute Tier,dort unter den dunklen Erlen?
Es zählt, vom Wissensdrang gejückt,(die es sowohl wie uns entzückt:)
die Anzahl seiner Perlen.
Christian Morgenstern (1871-1914)

M14
Aber das bringt uns vom Thema ab. Die erste (und leider auch letzte) zusammen-fassende wissenschaftliche Darstellung der Biologie der nur auf eine kleine, erst1941 entdeckte Südsee-Inselgruppe mit Namen Heieiei �die Inselgruppe wurdeversehentlich mit dem gesamten mittlerweile über Nasobeme zusammengetrage-nen Material bereits in den 50iger-Jahren bei einem atomaren Sprengversuch ver-senkt � beschränkten Naslinge, Nasobeme oder wissenschaftlich exakt Rhinogra-dentia, von rhinos griechisch Nase und gradi, lateinisch schreiten, erschien 1961 ausder Feder Dr. Harald Stümpkes alias Prof. Dr. G. Steiner, später Ordinarius fürZoologie an der Universität Karlsruhe und heute im wohlverdienten Ruhestand.Dieser ersten Darstellung folgten zahlreiche, allerdings unveränderte Auflagen inDeutsch, Englisch, Spanisch und sogar Japanisch. Die deutsche Ausgabe ist miteiner Gesamtauflage von nunmehr 55.000 ein Renner, der seinesgleichen in derwissenschaftlichen Literatur sucht, wenn man bedenkt, dass viele wissenschaftli-chen Zeitschriften gerade einmal eine Auflage von 300 Stück oder sogar wenigerhaben.Aufgrund ihrer besonderen Organisation und ihrer überaus bemerkenswerten ad-aptiven Radiation, die selbst die Darwinfinken alt aussehen lassen � ich kommenoch einmal auf dieses Problem zurück �, haben die Naslinge auch Eingang inmanche Lehrbücher der Zoologie gefunden (z.B. S. 543 in �Lehrbuch der Zoolo-gie, Band 2: Systematik�, herausgegeben von R. Siewing; Gustav Fischer Verlag,Stuttgart 1985). Dennoch sind sie heute nur einem verschwindend geringen Teilder jüngeren Biologen bekannt: Ein besonders eklatanter Fall wissenschaftlicherArroganz und Ignoranz den Leistungen anderer gegenüber, was ja dazu führt, das(auch in der Wissenschaft) das Rad jedes Jahr mehrmals von Neuem erfundenwird.Die wissenschaftliche Erforschung der Naslinge begann, folgt man den Ausfüh-rungen Dr. Stümpkes, erst mit der Ankunft des Biologen Dr. Einar Petterson-Skämtkvist auf dem Archipel im Jahre 1941.Von diesem Zeitpunkt an wurden zahl-reiche neue Arten beschrieben und die gesamte Gruppe sowie die einzelnen Artenerhielten wissenschaftliche Namen, die getreu einer jahrhundertealten Traditionnach bestimmten komplizierten, für den Philologen nicht immer nachvollzieh-baren Regeln dem Griechischen und dem Lateinischen entlehnt sind. Wer nunauch immer die gesamte Gruppe der Naslinge mit dem Namen Rhinogradentiabelegt hat, hat sich meiner Meinung nicht ganz fair Herrn Morgenstern gegenüberverhalten. Sein im einführenden Gedicht geschildertes Nasobem wurde späterNasobema lyricum, das Große Morgenstern-Nasobem, genannt, das zur Gruppe derVielnasen-Naslinge gehört (was ja auch unschwer auf dem Buchumschlag zu er-kennen ist).Die biologische Nomenklatur, also die Namensgebung, fordert seit dem großen Na-turforscher und übrigens auch bedeutenden Prosaschriftsteller Carl von Linné, dassjede Tier- und Pflanzenart mit zwei Begriffen gekennzeichnet wird, dem Gattungna-men � hier Nasobema � und dem Artepitheton, ein schmückendes Beiwort, das in derRegel den zu benennenden Organismus näher charakterisiert � hier lyricum, von latei-nisch lyricus, was eigentlich �lyrisch� bzw. �zur Leier oder zum Leierspiel gehörig�heißt, eine Anspielung auf das lyrische Gedicht Morgensterns.Es wäre sicher fairer gewesen, die ganze Gruppe nicht Rhinogradentia (sprachlich kor-rekt natürlich Rhinogradienta, doch ich erwähnte bereits, dass die Nomenklaturregeln

Mitteilungen aus dem Museum für Naturkunde in der Stiftung Schloss und Park Benrath M15
Acta Biologica Benrodis 13 (2006)
nicht immer philologisch nachvollziehbar sind) zu nennen, sondern Nasobemata, wasschließlich dasselbe bedeutet. Aber Wissenschaftler sind darauf angewiesen � das sindsie ihrem Ruf schuldig �, immer etwas Neues zu (er)finden und seien es nur Namen.Allerdings haben sich im Laufe der Zeit auch solche Wissenschaftler mit der Bio-logie der Rhinogradentia befasst, die über derlei profilneurotische Attitüden er-haben sind. Dazu gehörten so namhafte Forscher wie Bleedkopp, D�Epp, Bit-brain, Perischerzi und Schrimparsch � bitte haben Sie jetzt keine falschen Asso-ziationen, der Name leitet sich vom tschechischen sprymar = Spassmacher ab. Siealle sind, wie sich das auch gehört, in der Stümpkeschen Monographie gebührendberücksichtigt.In meiner kurzen Einführung in die heutige Ausstellung mit dem oben genanntenetwas sibyllinischen Titel will und kann ich, schon aus Zeitgründen, die wunderba-re Biologie der von Steiner geschaffenen Naslinge oder Rhinogradentia nicht aus-führlich behandeln. Dazu gibt es die bereits erwähnte Monographie von Stümpke,auch heute noch, nach über vierzig Jahren, lesenswert. Darüber hinaus gibt es füralle, die es noch genauer wissen wollen, eine Art Kommentar mit Rezensionen,Hintergründen und Erklärungen (vgl. K.D.S. Geeste: Stümpke�s Rhinogradentia.Versuch einer Analyse, Urban & Fischer, München, 2. Aufl. 1988), der, wie ich aller-dings vom Verlag erfahren habe, vergriffen ist.Wie sie schon aus dem bisherigen ahnen konnten, sind Naslinge Säugetiere, derennamengebende Nasen sich ausgehend von einer relativ einfachen in Einzahl vorhan-denen Urnase, die lediglich als Stützorgan diente, im Laufe der Evolution zu einemOrgan mit zum Teil vielfältigen Aufgaben, die z. T. weit über die Funktion als Atem-und Geruchsorgan hinausgehen, umgebildet hat. Die Nase wird zum Stütz- und Fort-bewegungsmittel, dient zur Lauterzeugung � ein Vertreter der schon erwähnten Viel-nasen-Naslinge, die Riesentatzelnase Rhinochilopus ingens (gewaltiger Nasentausend-füßler), soll sogar gelernt haben, Bachsche Orgelfugen mit Hilfe seiner modifiziertenNasen wiederzugeben.Man unterscheidet Einnasen-Naslinge (Monorrhina) mit den Urnaslingen, Weichna-sen und Nasenbeinlingen sowie die bereits erwähnten Vielnasen (Polyrrhina) mit denViernasen, Sechsnasen und Langschnauzennaslingen.Ich will einige der Vertreter kurz vorstellen; zwei davon auch anhand der Zeichnun-gen von H. Stümpke (für die Erlaubnis, diese zu verwenden, danke ich ganz herzlichdem Gustav Fischer Verlag), einen weiteren Vertreter anhand eines Fotos aus derAusstellung.Haeckels Urnasling (Archirrhinos haeckeli): Ursprünglichster Vertreter der Naslin-ge, der in schwer zugänglichen Bergwäldern lebt. Er ist etwa mausgroß, nachtaktiv,besitzt eine einzige Nase (Einnasen-Nasling) und läuft noch auf allen vier Beinen.Hat der Urnasling ein Insekt ergriffen, kippt er auf seine Nase, deren Ränder sichrasch ausbreiten � der austretende Nasenschleim sorgt für eine feste Haftung aufdem Boden � und führt die Beute mit allen vier Extremitäten zum Mund. Nach derMahlzeit wird der Kopfstand wieder aufgegeben.

M16
Schneuzender Schniefling (Emunctator sorbens): Die etwa rattengroßen Schnieflin-ge sind Einnasen-Naslinge, die in kleinen Gruppen am Ufer langsam fließender Bä-che leben. Sie lassen feine Fangfäden aus Nasenschleim ins Wasser hängen, an denenkleine Wassertiere kleben bleiben. Die Fangfäden mit der Beute werden in die Nasehochgezogen oder mit der langen Zunge abgeleckt. Der Schwanz besitzt eine Gift-drüsen und eine Klaue, mit der sich die Tiere verteidigen.Flugohr (Otopteryx volitans): Auf Bergwiesen. Einziger fliegender Nasenhopf mitenorm großen Ohren, verkümmertem Schwanz und besonders auffälliger Schil-lerfärbung. Zum Start springen die Tiere zunächst mit Hilfe ihrer Nase ähnlichwie der Nasenhopf Hopsorrhinus aureus (s.o.) in die Höhe, allerdings steiler, undschlagen dann die Ohren kräftig nach unten. Da der Flug aus dem Sprung derrückwärts hüpfenden Nasenhopfe abzuleiten ist, fliegt das Flugohr rückwärts underreicht dabei Höhen bis zu 20 m. Beute sind verschiedene Insekten, u.a. auchLibellen.Die Raubnase (Tyrannonasus imperator): Ein räuberischer Vielnasen-Nasling, der sichfast ausschließlich von dem erwähnten Nasobema lyricum ernährt. RaubtierähnlichesGebiss und Schwanz mit Giftklaue, mit der die Beute, wenn sie erst einmal eingeholtist, vergiftet wird.Wundernasenblümchen (Corbulonasus longicauda): Lebt in Kolonien auf Hahnenfuß-wiesen im Bergland. Die Jungen klettern an Blütenstängeln hoch. Ihr Schwanz strecktsich im Laufe der Entwicklung und bohrt sich in den Boden, so dass die Erwachsenen

Mitteilungen aus dem Museum für Naturkunde in der Stiftung Schloss und Park Benrath M17
Acta Biologica Benrodis 13 (2006)
unabhängig vom Pflanzenstängel auf ihrem Schwanz stehen. Die breiten, kurzen, blu-menblattartig den Mund umstehenden sechs Nasen und der buttermilchähnlicheMundgeruch der Tiere locken Insekten an, die dann durch schnelles Zusammenklap-pen der gespreizten Nasen gefangen werden.
Symbiose zwischen Saugmundnasenhopf (Hopsorrhinus mercator) und Säulennase(Columnifax lactans): Saugmundnasenhopfe können nicht wie andere Nasenhopfe (s.o.)geformte Nahrung zu sich nehmen � ihnen fehlen Zähne; zudem sind die Vorderbei-ne verkümmert � und sind daher auf die Säulennase angewiesen. Ein Saugmundnasen-hopf fängt z.B. einen Einsiedlerkrebs in der Uferzone, nähert sich damit einer Säulen-nase, übergibt dieser nach einem bestimmten Ritual, das die Abwehrreaktion (Stink-sekret) der Säulennase ausschaltet, die Beute und erhält im Tausch für diese Ware dieBrust, die sowohl bei Männchen als auch bei Weibchen von Columnifax lactans ständigMilch produziert.Flugohr (Otopteryx volitans): Auf Bergwiesen. Einziger fliegender Nasenhopf mit enormgroßen Ohren, verkümmertem Schwanz und besonders auffälliger Schillerfärbung.Zum Start springen die Tiere zunächst mit Hilfe ihrer Nase ähnlich wie der Nasen-hopf Hopsorrhinus aureus (s.o.) in die Höhe, allerdings steiler, und schlagen dann dieOhren kräftig nach unten. Da der Flug aus dem Sprung der rückwärts hüpfendenNasenhopfe abzuleiten ist, fliegt das Flugohr rückwärts und erreicht dabei Höhen biszu 20 m. Beute sind verschiedene Insekten, u.a. auch Libellen.

M18
Kopfstudie des Flugohrs (Otopteryx volitans). Foto: Katharina Greven

Mitteilungen aus dem Museum für Naturkunde in der Stiftung Schloss und Park Benrath M19
Acta Biologica Benrodis 13 (2006)
Kopfstudie des Orchideennaslings (Orchidiopsis rapax). Foto: Katharina Greven

M20
Die Raubnase (Tyrannonasus imperator): Ein räuberischer Vielnasen-Nasling, der sichfast ausschließlich von dem erwähnten Nasobema lyricum ernährt. RaubtierähnlichesGebiss und Schwanz mit Giftklaue, mit der die Beute, wenn sie erst einmal eingeholtist, vergiftet wird.Wundernasenblümchen (Corbulonasus longicauda): Lebt in Kolonien auf Hahnenfuß-wiesen im Bergland. Die Jungen klettern an Blütenstängeln hoch. Ihr Schwanz strecktsich im Laufe der Entwicklung und bohrt sich in den Boden, so dass die Erwachsenenunabhängig vom Pflanzenstängel auf ihrem Schwanz stehen. Die breiten, kurzen, blu-menblattartig den Mund umstehenden sechs Nasen und der buttermilchähnlicheMundgeruch der Tiere locken Insekten an, die dann durch schnelles Zusammenklap-pen der gespreizten Nasen gefangen werden.
Bereits diese Auswahl zeigt, dass sich die Naslinge im Laufe der Evolution auf ihrerkleinen Inselgruppe eine Fülle von ökologischen Nischen erobert haben, also eineadaptive Radiation durchlaufen. Dies bedarf für die Nichtbiologen unter ihnen einerkurzen Erklärung:Stellen sie sich vor, eine kleine Gruppe von Ihnen (natürlich im fortpflanzungsfähi-gen Alter) würde auf eine unbevölkerte Insel(gruppe) verschlagen. Sie könnten dortalle Ressourcen nutzen, wenn sie denn die Voraussetzungen mit sich brächten odernoch so flexibel wären, sich im Laufe der Zeit zu spezialisieren. Der eine würdeFischer, der andere Jäger, der dritte Gärtner usw. mit all den Anpassungen und Werk-zeugen, die für solche Berufe notwendig wären. Ihre Adresse wäre zwar dieselbe,nämlich besage Insel(gruppe), doch ihre Berufe wären verschieden, der Biologe wür-de sagen, sie haben verschiedene ökologische Nischen besetzt.Als Paradebeispiel für eine adaptive Radiation gelten die schon gerade erwähntenDarwinfinken auf den Galapagos-Inseln. Diese Inseln hatten aufgrund ihres vulkani-schen Ursprungs nie Kontakt mit irgendeiner anderen Landmasse. Folglich müssendie heute dort lebenden Pflanzen und Tiere im Laufe der Zeit eingewandert sein. Diedort heimischen 14 Finkenarten stammen offenbar von einer sehr wahrscheinlichKörner fressenden, weitgehend am Boden lebenden Art ab, die vor langer Zeit vomsüdamerikanischen Festland auf die Inseln verschlagen wurde. Man glaubt, dass eineheute noch in Ost-Ecuador lebende Finkenart (Catamblyrhynchus diadema) dieser Ur-form stark ähnelt.Langer Rede kurzer Sinn: Diese Finken fanden eine Umwelt mit zahlreichen Pflan-zen und einer Fülle von Insekten vor, so dass sie sich erfolgreich vermehren konntenund sich folglich zunehmend selbst Konkurrenz machten. Aus diesem Konkurrenz-verhältnis heraus entwickelten sich mehrere Arten von Körnerfressern, die sich je-weils auf verschiedene Samen spezialisiert haben, andere leben von Kakteen, anderevon Knospen und Früchten, wieder andere von Insekten. Das wird z.T. schon aus derGestalt der Schnäbel offenbar. Selbst der �Beruf� des Spechtes ist besetzt, da es eineArt gibt, die mit Hilfe von Kaktusstacheln Insekten aus den Bohrgängen im Holzherausholen. Jede Art hat eine besondere ökologische Nische besetzt.Wie viel vielfältiger und phantasievoller ist die Schöpfung doch mit den Naslingenumgegangen, die auf ihrer Inselgruppe meist mit Hilfe der Nase(n), aber auch auf-grund von Umbildungen anderer Organsysteme ganz unterschiedliche ökologischeNischen besiedeln konnten. Da gibt es � ich kann Ihnen das nicht alles vortragen �Parasiten, Schlickbewohner, die sich bis 30 cm eingraben und deren Nasen siphonar-

Mitteilungen aus dem Museum für Naturkunde in der Stiftung Schloss und Park Benrath M21
Acta Biologica Benrodis 13 (2006)
tig ausgezogen sind, und winzige nur millimeterlange Bewohner der kleinen Lückenzwischen Sandkörner, die praktisch nur noch aus Nasen bestehen (die so genanntenNur-Naslinge).Nun sind nur wenige Wissenschaftler wirklich in der Lage, wissenschaftliche Ergeb-nisse einem breiterem Publikum zugänglich zu machen. Oder können Sie etwas mitdem folgenden Satz aus der Stümpkeschen Monographie über die Rhinogradentiaanfangen. Es geht dabei um die Versteifung und gleichzeitige Elastizität der Nasendurch Schwellkörper, dem Nasenturgor also, der das Nasobema lyricum befähigt aufseinen Nasen zu schreiten (die kein knöchernes oder knorpeliges Skelett haben).
�Neben den Ampullae choanales spielen die die Ampullae pneumonales noch eine Rolle�.Sie sind inDreizahl jederseits vorhanden und sind Verteiler für die von den Ampullae choanales gelieferte Druckluft.Die Canales ramosi der Nasen selbst haben noch ein distales Orificium externum unterhalb der Nasen-spitze, das meist verschlossen ist, aber reflektorisch sehr schnell geöffnet werden kann��. (aus Stümpke,H.: Bau und Leben der Rhinogradentia, Gustav Fischer Verlag, Stuttgart 1989, S. 56)
Dies ist übrigens auch für einen gestandenen Wissenschaftler nahezu unverdaulich, essei denn, er ist hochgradiger Spezialist und arbeitet gerade auf diesem Gebiet.Es ist nun unser aller Anliegen, die Naslinge auch Ihnen nahe zu bringen und diesprimär mit Hilfe der heutigen Ausstellung, vielleicht später auch einmal mit einemSymposium oder, das klingt bescheidener, einem Workshop über diese wunderbarenTiere, zumal sich Naturwissenschaft, Philosophie und Dichtkunst wieder mit Rhi-nogradentiern zu befassen beginnen. Das ist nicht zuletzt auch dieser Ausstellung zuverdanken. Gerade die Gedichte über Naslinge � es gibt bereits einige: belehrendeVerse bis hin zu Kalauern �, erreichen bei weitem nicht die Qualität des Morgenstern-schen �Nasobem�. Hier zwei eher minderwertige Kostproben:
Grippe
Erst kürzlich in �nem Omnibussah ich ein Nasobem,
das litt stark unter Nasenfluss.Der Anblick war nicht schön.
Duranimus Strabo (geb. 1942)
Nasen
Die Nasen der Nasenaffensind manchmal so groß wie Karaffen.
Die Nasen der Nasenbärensind kaum länger, drauf möcht´ ich schwören.
Die Nasen des Nasobemsind jedoch unübertrefflich schön.
Duranimus Strabo (geb. 1942)

M22
Doch sollte unser ganzes Augenmerk auf die Ausstellung gerichtet sein: Sie ist auseiner Symbiose entstanden (dies ist bekanntlich eine Form des Zusammenlebens, beider alle beteiligten Partner profitieren), einer Symbiose zwischen der Stiftung SchlossBenrath und dem Naturkundemuseum Benrath einerseits und drei jungen Leutenandererseits. Von diesen hat sich jeder einzelne mit den ihm eigenen, vertrauten Mit-teln mit Naslingen auseinandergesetzt.Einer, Manfred Ruecker, ist bereits ein gestandener Komponist und bildender Künst-ler, der Musik und Psychologie an den Universitäten Siegen und Köln studiert hat, zurZeit gleichzeitig Student der Medienkunst bei Prof. David Larcher in Köln und Stu-dent der Bildhauerei bei Prof. Georg Herold hier an der Kunstakademie Düsseldorfist. Er hat bereits Preise bei Internationalen Musikwettbewerben eingeheimst. Ihnhaben die Rhinogradentier und deren verlorenes �Paradies� sowie das Morgenstern-sche Gedicht angeregt, ein 74 Minuten und 27 Sekunden dauerndes �akustisches Kunst-werk� zu schaffen, das nicht zwangsläufig zu dem eklatanten Fehlurteil Wilhelm Buschs(1832-1908) führen muss:
Musik wird oft nicht schön gefunden,weil sie stets mit Geräusch verbunden.
Die zweite im Bunde, Katharina Greven, hat diese Ausstellung initiiert und konzi-piert. Sie hat Freie Kunst/Fotografie an der Kunstakademie Düsseldorf studiert, istu.a. Meisterschülerin von Thomas Ruff. Sie hat schon einmal mit Manfred Rueckerzusammengearbeitet und im Rahmen von Ausstellungen der Ruff-Klasse, anderenSammelaustellungen und einer Einzelausstellung eigene Bilder zeigen können. Sieträgt Details von Naslingen bei, welche die Tiere realistischer erscheinen lassen alssie es in Wirklichkeit sind, wobei sich bei besonders schnellen Vertretern dieserTiere, manche Bewegungsunschärfen nicht vermeiden lassen.All dies wäre nicht möglich gewesen, gäbe es nicht Martina Fasel, die, mehr aus einervermeintlich exakten Wissenschaft kommend � sie hat in Münster Biologie studiertund ihre Diplomarbeit bei Prof. Dr. Pio Fioroni, Münster, geschrieben �, die Rhino-gradentia primär sich und ihrer Familie zur Freude liebevoll in ihrer natürlichenUmgebung nachgebildet hat. Das ist nicht leicht und erfordert subtile Kenntnisseausgefallener Anatomien, ein Gefühl für richtige Proportionen und viel Einfühlungs-vermögen in diese Schöpfungen der Phantasie � oder vielleicht doch Phantasien derSchöpfung?
Hartmut Greven, Düsseldorf

Mitteilungen aus dem Museum für Naturkunde in der Stiftung Schloss und Park Benrath M23
Acta Biologica Benrodis 13 (2006)
08.03. Abendvortrag �Aus dem Reich der geflügelten Jungfern � Lebensweise undLebensräume der Libellen� von Klaus Böhm.
10.03. Abendvortrag �Fledermäuse� mit anschließender Exkursion in den Schlossparkvon und mit Eberhard Menz
16.04./17.04. Nacht der Museen
Programm
Führung durch die Sonderausstellung �Naslinge � Schöpfung der Phantasie? Phanta-sie der Schöpfung?��Naslinge � Phantasie der Schöpfung? Schöpfung der Phantasie?� Vortrag von Prof.Dr. Hartmut Greven�Falter der Nacht�, Diavortrag von Rolf Niggemeyer�Fledermäuse auf der Leinwand und live im Schlosspark�, Diavortrag und Exkursionvon und mit Eberhard Menz�Nacht der Sterne� , Demonstrationen im Kleinplanetarium und Himmelsbeobach-tungen von und mit Werner Buschfeld
27.04. Vorstellung des Buches �Düsseldorf � das Naturerlebnis� von Michael Brock-erhoff und Thomas Buskamp (Fotos) im Museum für Naturkunde. (�Düsseldorf �ein Naturerlebnis�, Droste Verlag, ISBN 978-3-7700-1199-5)
21.05. Vogelkundliche Wanderung durch den frühmorgendlichen Schlosspark mit Dr.Hubert Heuwinkel
22.05. Naturgeschichtliche Radtour: �Auf den Spuren alter Rheinverläufe�, Exkursionmit Werner Buschfeld
28.05. Schlossparkführung von und mit Werner Buschfeld
03.07. �Tag der offenen Tür�, Tag der Begegnung und Netzwerktag net:natuur_cultuur im Museum für NaturkundeGrußworte des Bezirksvorstehers Heinz-Leo Schuth der Bezirksverwaltung 9 Ben-rath
25.08. Führung durch den Schlosspark unter natur- und kulturhistorischen Aspektenvon Dr. Hubert Heuwinkel
04.09. Informationsstand zum Tag des Denkmals auf Haus Bürgel, Urdenbacher Kämpe
13.09. Führung für Dr. Mühle, Dr. Kaden und Studentinnen und Studenten der Fach-richtung Geoökologie der Universität Potsdam im Museum und in den UrdenbacherKämpen von Dr. Hubert Heuwinkel
16.09. �Fledermäuse�, Vortrag und Exkursion von und mit Eberhard Menz

M24
07.11. Abendvortrag �Naturbeobachtungen am Neusiedler See� von Dipl. Geogr. Hol-ger Pieren
2006
25.01. Abendvortrag �Wilde Papageien in Düsseldorf � Zum Freileben des Halsband-sittichs im Rheinland� von Dipl. Biol. Tobias Krause
17.02. Vorstellung der museumspädagogischen Arbeit auf der Messe �Exponatec� inKöln
22.02. Abendvortrag �Jahrhunderthochwasser � Jedes Jahr und immer höher?� vonDipl. Geogr. Holger Pieren
22.03. Abendvortrag �Blumenküsser und Fliegenvögel � Aus dem Leben der Kolibris�von Dr. Christoph Hinkelmann
20.04. Abendvortrag �Geboren, um zu singen � Vogelstimmen, ihre akustische Struk-tur und ihre Funktion� von Dr. Hubert Heuwinkel im Wilhelm-Marx-Haus
26.04. Eröffnung der Sonderausstellung �Käfer � Könner und Strategen� (26.04.bis 30.07)mit dem gleichnamigen Abendvortrag von Edmund Wenzel
Das Wort Käfer leitet sich vom althochdeutschen Wort �kevar� ab � es ist die Bezeich-nung für eine einheitliche Tiergruppe (Ordnung) innerhalb der Klasse der Insekten.Käfer � die mit über fünf Millionen Arten individuenreichste Tiergruppe auf Erden �sind für manche Überraschung gut. Selbst viele Vertreter der nur rund 5000 Käferartenunserer Heimat erbringen Leistungen, die fast unvorstellbar sind.Selbst in dem vergleichsweise kleinen Schlosspark von Benrath, mit weniger als ei-nem Quadratkilometer Größe, umgeben von urbaner Struktur, konnten während ei-ner einjährigen Untersuchung mehr als 300 Käferarten nachgewiesen werden undzwar Arten, die hauptsächlich nur an Holz- und Holzpilzen leben, darunter für NRWäußerst seltene Käferarten. Es ist anzunehmen, dass auf dieser kleinen Enklave über600 Arten einen ihnen angemessenen Lebensraum gefunden haben.Die von Edmund Wenzel konzipierte und realisierte Ausstellung beleuchtete dieAnatomie sowie die Leistungen und Fähigkeiten der �Krabbeltiere�.
05.05. Abendvortrag �Fledermäuse� mit anschließender Exkursion im Schlossparkvon und mit Eberhard Menz.
06.05./ 07.05. Nacht der Museenund Eröffnung der Ausstellung �Wasserwerke� (bis 11.06.)

Mitteilungen aus dem Museum für Naturkunde in der Stiftung Schloss und Park Benrath M25
Acta Biologica Benrodis 13 (2006)
Besucher zur Eröffnung der Sonderausstellung �Käfer � Könner und Strategen� am 26.04.2006.
Eröffnung der Sonderausstellung �Wasserwerke� anlässlich der Nacht der Museen am 06.05.2006in Farben, Worten und Tönen mit Dr. Petra Urban, Gerd Keuenhof, Mareike Moeller undBärbel Hain.

M26
Programm
Im Museum für Naturkunde standen in diesem Jahr Wasser, Erde und Luft im Mittel-punkt. Wasser macht den Anfang, wenn �Spritziges� in Farben, Worten und Tönen vonDr. Petra Urban, Gerd Keuenhof, Mareike Moeller und Bärbel Hain �Wasserwerke�verheißt. Ausgestellt wurden bis zum 11.06. vierzig �Wasserbilder� (Pigmente auf Lein-wand, 40 x 40 cm) und Installationen zum Thema Wasser von Bärbel Hain.Wasser und Erde sind die Elemente der Amphibien, die Rolf Niggemeyer in einemspannenden Vortrag vorstellte.In der Luft bewegen sich die Fledermäuse, die man mit Eberhard Menz in einemDiavortrag mit anschließender Exkursion live erleben konnte.Um Mitternacht wurde der �Tanz der Galileischen Monde� bei einer Demonstrationim Kleinplanetarium und bei klarer Nacht als Himmelsbeobachtung ins Zentrum derAufmerksamkeit gerückt.
13.05. Ornithologische Frühexkursion durch den Schlosspark Benrath mit Dr. Hu-bert Heuwinkel
02.07. Tag der offenen Tür im Museum für Naturkunde, Tag der Begegnung desNABU-Stadtverbandes Düsseldorf und Netzwerktag der Museen und der Biologi-schen Stationen im net:natuur_cultuur, im Museum für Naturkunde.Grußworte der Umweltdezernentin der Stadt Düsseldorf, Frau Charlotte Nieß-Ma-che

Mitteilungen aus dem Museum für Naturkunde in der Stiftung Schloss und Park Benrath M27
Acta Biologica Benrodis 13 (2006)
Himmelsbeobachtungen in der Nacht der Museen am 06.05.2006 im Innenhof des Museumsmit Werner Buschfeld am Spiegelteleskop.
Ornithologische Frühexkursion durch den Schlosspark Benrath mit Dr. Hubert Heuwinkelam 13.05.2006.

M28
20.08. Eröffnung der Sonderausstellung �Faszinierende Welt der Spinnen� (20.08.bis 19.11)
Programm
Grußworte Dr. Hubert Heuwinkel, Leiter des Museums für Naturkunde
Einführung in die Ausstellung und Demonstration der lebenden Spinnen von Leihge-ber Jacek Pacyna (Polen)Diese Ausstellung gab Antworten darauf, ob die Geschöpfe mit acht Beinen, fünfAugenpaaren und Giftklauen wirklich unserem Schönheitssinn widersprechen. Beivielen Menschen erzeugt der Anblick von Spinnen Angst oder Ekel, sie haben eineArachnophobie. Mit den alten Vorurteilen gegenüber dem Leben und den Lebensge-wohnheiten der Spinnentiere konnte die Ausstellung �Faszinierende Welt der Spin-nen� aufräumen. Spinnen gehören zu den interessantesten und ältesten Tieren. Beein-druckend und faszinierend ist ihre unerschöpfliche Vielfalt an Formen und Farbenund die elegante Fortbewegung.Diese außergewöhnliche Ausstellung präsentierte über 30 lebende Arten von Spin-nentieren in 34 Glasterrarien. Darunter waren die aus Literatur und Film bekanntegrößte Spinne der Welt, die Riesenvogelspinne (Theraphosa leblondi), und eine der gif-tigsten Spinnen, die nur zwei Zentimeter große Schwarze Witwe (Latrodectus mactans).Auch Skorpione zählen zu den Spinnentieren. Von diesen war unter anderem derweltweit größte Skorpion überhaupt, der afrikanische Riesenskorpion (Pandinus impe-rator), zu sehen, der bis zu 20 cm lang werden kann..Die Ausstellung gab Besuchern die Gelegenheit, die Tiere hautnah zu beobachten undihnen direkt oder über Videoprojektion beim Fangen und Fressen der Beute zuzu-schauen. So verwunderte es nicht, dass diese Sonderausstellung mit über 7000 Besu-chern eine der am besten besuchten der letzten Jahre war.Auf einem Stich aus dem Werk �Metamorphosis Insectorum Surinamensum� (1705)der Naturforscherin Maria Sibylla Merian (1647-1717) ist unter anderem eine großeSpinne über einem toten Kolibri stehend abgebildet. Nach dieser Darstellung gabCarl von Linné (1707-1778), Begründer der modernen Systematik der Pflanzen undTiere, der Spinne über dem toten Vogel den Namen Vogelspinne. Nach heutigemWissensstand fressen jedoch nur sehr wenige Arten dieser Spinnengruppe tatsächlichVogelküken. Es sind zurzeit mehr als 800 Vogelspinnenarten bekannt.
03.09. Informationsstand zum Tag des Denkmals auf Haus Bürgel, Urdenbacher Käm-pe
09.09. Vortrag mit anschließender Exkursion in den Schlosspark �Die Bedeutung vonZecken als Überträger von Krankheitserregern� von und mit Prof. Dr. Heinz Mehl-horn, Universität Düsseldorf
11.06. Abendvortrag �Vogelspinnen zum Anfassen schön?!� von Dr. Stephan Loksa

Mitteilungen aus dem Museum für Naturkunde in der Stiftung Schloss und Park Benrath M29
Acta Biologica Benrodis 13 (2006)
�Besucherfülle� bei der Fütterung der Vogelspinnen durch Leihgeber Jacek Pacyna im Vor-tragssaal des Museums, die live auf eine Bildwand übertragen wurde.
Mehrere Male konn-ten während derAusstellungszeit Vo-gelspinnen bei derHäutung beobachtetwerden.

M30
Museumspädagogik
Wie in den Jahren zuvor wurden die museumspädagogischen Angebote wieder vonvielen Kindern, Schülern und Jugendlichen genutzt. Neben den zwei- bis dreistündi-gen Unterrichtseinheiten für Schüler aller Jahrgänge und Schultypen wurden auchFerienprogramme durchgeführt. Von 231 Schulklassen, die das Museum beispiels-weise im Jahre 2006 insgesamt besuchten, betreute der museumspädagogische Dienst181.Am Osterferienprogramm nahmen 431 und am Sommerferienprogramm 511 Kinderteil, außerdem wurden elf Kindergeburtstage im Museum gefeiert.Die Oster- und Sommerferienprogramme umfassten in den Jahren 2003 bis 2006 ne-ben der offenen Museumswerkstatt, in der Mal-, Rätsel-, Such- und Bastelbögen anjunge Besucher ab sechs Jahren ausgegeben wurden, folgende Sonderaktionen:
2003
�Eigenheim gesucht� � Kinder erfahren Interessantes über Tierbehausungen und bau-en Nistkästen unter der Anleitung des Jugend-Umwelt-Mobils der NAJU.
�Was macht die Amsel morgens um 8?� Ein Natur-Erlebnismorgen im Schlosspark.
�370 Mio. Jahre alte Elefantenzahnschnecke trifft modernen Homo sapiens im Jahr 2003�.Spurensuche in der Urzeit, selbstgemachte Fossilien zum Mitnehmen.
�Insektenkarriere � von der Putzfrau zur Kundschafterin�. Streifzug durch die Insek-tenwelt.
�Eine Art Umwelttheater�. Kinder lassen die Natur durch Wort und Ton erklingenund setzen sich in kleinen Szenen mit ihr auseinander.
�Fotojagd rund um das Naturkundemuseum�. Sehen lernen � fotografieren lernen.
�Naturerlebniswoche�
�Stadtökologie � was ist denn das?� Mit dem Jugend-Umwelt-Mobil der NAJU er-kunden und untersuchen Kinder die Natur im Stadtteil Benrath.
2004
�Sternstunden�. Ein Astro-Abend im Schlosspark
�Der Natur auf der Spur�. Lauschend, riechend, tastend und mit Adleraugen sehendgehen die Kinder mit dem Jugend-Umweltmobil der NAJU auf eine Entdeckungsrei-se durch den Schlosspark.
�Versteckspiel im Tierreich�. Einige Tricks der Tarnung werden u.a. anhand lebenderTiere aufgedeckt und in selbstgestalteten Bildern umgesetzt.

Mitteilungen aus dem Museum für Naturkunde in der Stiftung Schloss und Park Benrath M31
Acta Biologica Benrodis 13 (2006)
Teilnehmer des Osterferienprogramms 2005 für Kinder und Jugendliche im Vortragssaal.
�Ein Museum im Schuhkarton�, eine Sonderaktion im Sommerferienprogramm 2005 mitjungen Teilnehmern, Museumspädagogin Irmtrude Peters (hinten) und Museumsmitarbeite-rin Ina Vaassen (vorn).

M32
�Lebendige Unterwelt�. Bodenuntersuchungen mit dem Jugend-Umweltmobil derNAJU.
�Grüne Giganten � Rund um den Baum�. Kinder begegnen mit allen Sinnen denBäumen im Schlosspark und gestalten ein eigenes Baumbuch.
�Keine Angst vor Spinnen � Spinnen zum Anfassen�. Kinder erhalten Einblicke in dasLeben am seidenen Faden.
�Ein Museum im Schuhkarton�. Kinder lernen verschiedene Lebensräume heimi-scher Tiere kennen und gestalten ein eigenes Diorama.
�Mit den Augen sehen, mit der Kamera festhalten�. Ein Fotokurs rund um die Natur.
�Sechsfüßer zu Wasser, zu Land und in den Lüften�. Ein Streifzug durch die Insekten-welt.
�Fledermaushotels�. Kinder bauen Fledermauskästen und erfahren Wissenswertesüber das Leben der nicht �blutrünstigen� Tiere.
2005
�Insekten haltbar gemacht�. Kinder schauen dem Präparator über die Schulter undpräparieren eine echte Heuschrecke zum Mitnehmen.
�Tiere im Federkleid von A-Z�. Mit dem Jugend-Umweltmobil der NAJU beobach-ten Kinder heimische Vögel von der Amsel bis zum Zaunkönig, lernen Flugbildererkennen, Vogelstimmen zuordnen und erfahren Interessantes über das Vogelleben.
�Filz dir ein Tier � Marienkäfer, Schnecke und Co.� Im Jugend-Umweltmobil wirdFilz hergestellt und zum eigenen streichelweichen Tier für Zuhause geformt.
�Was wächst denn da � gelappt, gesägt, gefingert.� Kinder lernen während einer Exkur-sion in den Schlosspark einige Pflanzen und deren Standorte kennen.
�Hühnerlei � ein hautnahes Erlebnis mit dem Federvieh�. Der WissenschaftlicheGeflügelhof des Bundes Deutscher Rassegeflügelzüchter (BDRG) stellt verschiede-ne Hühnerrassen vor.
�Kleiner Fuchs und Admiral � wahre Verwandlungskünstler�. Kinder lernen heimi-sche Schmetterlingsarten kennen.
�Rheinfundstücke � Schnecken, Muscheln, Steine�. Während eines Streifzuges ent-lang des Rheinufers erfahren die Kinder Wissenswertes über Schnecken und Mu-scheln, Leben im Rhein und welche Geschichten Steine erzählen können.
�Streifzug durch die Insektenwelt�. Beobachtet wird alles, was sechs Beine hat.

Mitteilungen aus dem Museum für Naturkunde in der Stiftung Schloss und Park Benrath M33
Acta Biologica Benrodis 13 (2006)
2006
�Hühnerlei � Vielfalt beim Hühnervieh�
�Lebendiger und farbenreicher Waldboden�
�Der Urzeit auf der Spur. Kinder von heute begegnen der Tier- und Pflanzenwelt vongestern�
�Erleb dein grünes Wunder �Frühlingserwachen�
�Wildnisküche � Interessantes über wildes Gemüse und Kräuter�
�Eingewandert, ausgesetzt, geflüchtet � Neubürger in unserer Tier- und Pflanzen-welt�
�Kleine Wunder ganz groß�. Mikroskopierkurs für Kinder und Jugendliche ab 12Jahren.
�Unterkunft mit Vollpension� � Lebensraum Hecke
�Vor der Museumstür ist der Bär los�. Kinder begegnen einer Vielzahl von Tierartenund dem Braunen Bären.
�Pfui, Spinnen? � wahre Lebenskünstler!�
Hubert Heuwinkel, Düsseldorf

M34
Hinweise für Autoren
Acta biologica Benrodis veröffentlicht zoologische, botanische, geowissenschaftlicheund wissenschaftshistorische Originalarbeiten, kurze Mitteilungen und Übersichts-artikel in deutscher und englischer Sprache.
Die �Mitteilungen aus dem Museum für Naturkunde in der Stiftung Schloss undPark Benrath� informieren über Aktivitäten des Museums.
Manuskripte sind in doppelter Ausfertigung (Kopien) auf einseitig beschriebenen(zweifacher Zeilenabstand) DIN A 4-Seiten an einen der Herausgeber zu senden.
Nach Begutachtung � die Autoren können bei Einreichung des Manuskripts einenpotenziellen Gutachter benennen � und den eventuell notwendigen Änderungen soll-te das Manuskript auf Diskette (3 ½�) oder CD (PC-kompatibel, kein Mac) einge-reicht werden. Das Programm muss Word-kompatibel sein. Das Disketten- oderCD-Label sollte folgende Angaben enthalten: Autor, Titel der Arbeit, Dateiname undverwendetes Textprogramm. Text bitte nicht in Blocksatz und ohne Steuerzeichenschreiben.
Gliederung von Originalarbeiten: Titel in Deutsch und Englisch, Name und An-schrift des Autors/der Autoren, ausführliche Zusammenfassung sowie etwa fünf Schlüs-selwörter in Deutsch und Englisch, Einleitung, Material und Methoden, Ergebnisse,Diskussion, Danksagungen, Literatur. Fußnoten sind zu vermeiden.
Literatur ist folgendermaßen zu zitieren (Zeitschriftentitel bitte ausschreiben):
Im LiteraturverzeichnisGOETE, A., 1904: Über die Entwicklung der Hydromedusen. � Zoologischer Anzeiger27, 473-475HELB, H.W., DOWSETT-LEMAIRE, F., BERGMANN, H.H., CONRADS, K., 1985: Mixed sin-ging in European songbirds � a review. � Zeitschrift für Tierpsychologie 69, 27-41.LINDER, A., 1964: Statistische Methoden für Naturwissenschaftler, Mediziner und In-genieure. � Birkhäuser, Basel und Stuttgart.ANDREW, R.J., 1969: The effects of testosterone on avian vocalizations, pp 97-130. In:Bird Vocalizations (Hinde, R.A., ed.). � University Press, Cambridge.
Im laufenden TextGOETE (1904), WALGE & LUNAU (2003); (GOETE 1904; LINDER 1964; HELB et al. 1985;WALGE & LUNAU 2003)
Abbildungs- und Tabellenlegenden werden in Deutsch und Englisch auf gesondertenBlättern/in gesonderten Dateien erbeten.
Abbildungen sind als kontrastreiche Hochglanzfotos vorzugsweise zu Tafeln von 13cm Breite und höchstens 20 cm Höhe (Satzspiegel) zusammengestellt, als Strich-

Mitteilungen aus dem Museum für Naturkunde in der Stiftung Schloss und Park Benrath M35
Acta Biologica Benrodis 13 (2006)
zeichnungen, Farbfotos, Dias oder Dateien (TIFF-, JPEG- oder PDF-Dateien, Auf-lösung mindestens 300 dpi in Veröffentlichungsgröße), Tabellen und Grafiken alsdruckfertige Vorlagen in entsprechender Größe, besser als Word-, Excel-, TIFF- oderPDF-Datei einzureichen. Farbabbildungen sind gegen Kostenbeteiligung möglich.Mit Ausnahme von Dias werden die Vorlagen nur auf ausdrücklichen Wunsch zu-rückgeschickt.
Besonders umfangreiche Arbeiten können als Supplementbände gedruckt werden.Die Kosten hierfür sind bei der Schriftleitung zu erfragen.
Bei kurzen Mitteilungen unterbleibt eine Gliederung: Titel in Deutsch und Englisch,Name und Anschrift der Autoren, kurze Zusammenfassung (Englisch bei deutschemund Deutsch bei englischem Text mit Schlüsselwörtern), nicht weiter unterglieder-ter Text. Die Literatur wird wie in Originalarbeiten zitiert.
Die Autoren erhalten 30 Sonderdrucke ihrer Arbeit kostenlos, weitere gegen Berech-nung. Ebenfalls gegen Berechnung können Autoren eine druckfähige PDF-Datei ih-rer Arbeit bestellen, bei größeren Dateien erhalten Sie zusätzlich eine minimiertePDF-Datei. Die Dateien dürfen zum Download bereitgestellt werden.

M36