king s research portal ofset-shiftinginefficienciesthathavepreviouslybeenlinkedwithdistinctneural...
TRANSCRIPT

King’s Research Portal
DOI:10.1371/journal.pone.0124027
Document VersionPublisher's PDF, also known as Version of record
Link to publication record in King's Research Portal
Citation for published version (APA):Lao-Kaim, N. P., Fonville, L., Giampietro, V. P., Williams, S. C. R., Simmons, A., & Tchanturia, K. (2015).Aberrant function of learning and cognitive control networks underlie inefficient cognitive flexibility in AnorexiaNervosa: A cross-sectional fMRI study. PloS one, 10(5), [e0124027]. DOI: 10.1371/journal.pone.0124027
Citing this paperPlease note that where the full-text provided on King's Research Portal is the Author Accepted Manuscript or Post-Print version this maydiffer from the final Published version. If citing, it is advised that you check and use the publisher's definitive version for pagination,volume/issue, and date of publication details. And where the final published version is provided on the Research Portal, if citing you areagain advised to check the publisher's website for any subsequent corrections.
General rightsCopyright and moral rights for the publications made accessible in the Research Portal are retained by the authors and/or other copyrightowners and it is a condition of accessing publications that users recognize and abide by the legal requirements associated with these rights.
•Users may download and print one copy of any publication from the Research Portal for the purpose of private study or research.•You may not further distribute the material or use it for any profit-making activity or commercial gain•You may freely distribute the URL identifying the publication in the Research Portal
Take down policyIf you believe that this document breaches copyright please contact [email protected] providing details, and we will remove access tothe work immediately and investigate your claim.
Download date: 15. Jun. 2018

RESEARCH ARTICLE
Aberrant Function of Learning and CognitiveControl Networks Underlie InefficientCognitive Flexibility in Anorexia Nervosa: ACross-Sectional fMRI StudyNick P. Lao-Kaim1, Leon Fonville1, Vincent P. Giampietro2, Steven C. R. Williams2,3,Andrew Simmons2,3, Kate Tchanturia1*
1 King’s College London, Institute of Psychiatry, Department of Psychological Medicine, London, UnitedKingdom, 2 King’s College London, Institute of Psychiatry, Department of Neuroimaging, London UnitedKingdom, 3 NIHR Biomedical Research Centre for Mental Health at South London and Maudsley NHSFoundation Trust and Institute of Psychiatry, King’s College London, London, United Kingdom
Abstract
Objectives
People with Anorexia Nervosa exhibit difficulties flexibly adjusting behaviour in response to
environmental changes. This has previously been attributed to problematic behavioural
shifting, characterised by a decrease in fronto-striatal activity. Additionally, alterations of
instrumental learning, which relies on fronto-striatal networks, may contribute to the obser-
vation of inflexible behaviour. The authors sought to investigate the neural correlates of cog-
nitive flexibility and learning in Anorexia Nervosa.
Method
Thirty-two adult females with Anorexia Nervosa and thirty-two age-matched female control
participants completed the Wisconsin Card Sorting Task whilst undergoing functional mag-
netic resonance imaging. Event-related analysis permitted the comparison of cognitive shift
trials against those requiring maintenance of rule-sets and allowed assessment of trials
representing learning.
Results
Although both groups performed similarly, we found significant interactions in the left middle
frontal gyrus, precuneus and superior parietal lobule whereby blood-oxygenated-level de-
pendent response was higher in Anorexia Nervosa patients during shifting but lower when
maintaining rule-sets, as compared to healthy controls. During learning, posterior cingulate
cortex activity in healthy controls decreased whilst increasing in the Anorexia Nervosa
group, whereas the right precuneus exhibited the opposite pattern. Furthermore, learning
was associated with lower blood-oxygenated-level dependent response in the caudate
body, as compared to healthy controls.
PLOS ONE | DOI:10.1371/journal.pone.0124027 May 13, 2015 1 / 16
OPEN ACCESS
Citation: Lao-Kaim NP, Fonville L, Giampietro VP,Williams SCR, Simmons A, Tchanturia K (2015)Aberrant Function of Learning and Cognitive ControlNetworks Underlie Inefficient Cognitive Flexibility inAnorexia Nervosa: A Cross-Sectional fMRI Study.PLoS ONE 10(5): e0124027. doi:10.1371/journal.pone.0124027
Academic Editor: J Bruce Morton, University ofWestern Ontario, CANADA
Received: October 22, 2014
Accepted: March 9, 2015
Published: May 13, 2015
Copyright: © 2015 Lao-Kaim et al. This is an openaccess article distributed under the terms of theCreative Commons Attribution License, which permitsunrestricted use, distribution, and reproduction in anymedium, provided the original author and source arecredited.
Data Availability Statement: All relevant data arewithin the paper and its Supporting Information files
Funding: This work was supported by the SwissAnorexia Foundation, the Psychiatry Research Trust,the NIHR Biomedical Research Centre for MentalHealth at South London and Maudsley NHSFoundation Trust and the Institute of Psychiatry,King's College London. The funders had no role instudy design, data collection and analysis, decision topublish, or preparation of the manuscript.

Conclusions
People with Anorexia Nervosa display widespread changes in executive function. Whilst
cognitive flexibility appears to be associated with aberrant functioning of the fronto-parietal
control network that mediates between internally and externally directed cognition, fronto-
striatal alterations, particularly within the caudate body, were associated with instrumental
learning. Together, this shows how perseverative tendencies could be a substrate of multi-
ple high-order processes that may contribute to the maintenance of Anorexia Nervosa.
IntroductionAnorexia nervosa (AN) is a severe mental illness associated with a lifetime prevalence of 0.9%in females and 0.3% in males [1], and carries the highest mortality rate among psychiatric con-ditions [2]. It has been suggested that persistent dietary restriction, intense fear of gainingweight and disturbances in bodily perception may be caused and maintained by a combinationof socio-emotional and cognitive abnormalities [3]. In particular, people with AN demonstratean inability to advantageously alter their current behaviour in response to changes in theenvironment [4,5]. This lack of cognitive flexibility remains present after recovery [4,6], is in-dependent of body mass index (BMI: weight/height2) and illness duration [5,7] and has beenobserved in unaffected sisters [5,6,8] as well as in children whose mothers suffer from AN [9].As such, inefficient cognitive flexibility has been marked as a possible endophenotype of ANthat facilitates persistent appetitive control [3] and could contribute to its development [10].
Using a functional magnetic resonance imaging (fMRI) coupled with a target-detectiontask, Zastrow et al. [11] found decreased activation in AN in the fronto-striatal network whenperforming behavioural shifts independently of cognitive shift requirement. These findingsare corroborated by an activation likelihood estimation meta-analysis of nine voxel-basedmorphometry studies showing decreased grey matter volume in the right caudate and right len-tiform nucleus in currently ill AN [12], and decreased ACC grey matter in recovered partici-pants [13]. In addition, Zastrow et al. demonstrated dominant activation of the right middlefrontal gyrus and bilateral temporoparietal junction in AN, regions that form the fronto-parie-tal control network (FPCN; [14,15]). These observations suggest that inefficient cognitive set-shifting in ANmay in fact be attributed to problematic behavioural shifting, characterised bygreater cognitive supervisory control and aberrant function of motivation-related circuitry.However, neuropsychological studies report that set-shifting inefficiencies in AN are character-ised by an increase in perseverative errors [4,6], whilst no change is observed in the number ofloss-of-set/failure-to-maintain-set scores [4]. Since loss-of-sets occur when participants errone-ously shift category following positive feedback, avoiding such errors requires efficient beha-vioural shifting regardless of the ability to shift between abstract rules. The absence of cognitiveset-shifting differences by Zastrow et al. could therefore be methodological; the majority of tri-als categorised as requiring a cognitive shift did not occur immediately following a rule change.Indeed, a recent fMRI study using the Wisconsin Card Sorting Test (WCST) found differentialactivation during shifting as opposed to maintaining set in the inferior frontal gyrus and bilat-eral parahippocampal gyrus [16], demonstrating functional alterations associated with cogni-tive flexibility above that of behavioural flexibility.
The current study aimed to assess cortical function in AN during cognitive flexibility usingevent-related fMRI. We chose to use the WCST because it permits investigation of separableforms of set-shifting inefficiencies that have previously been linked with distinct neural
Cognitive Flexibility in Anorexia Nervosa: An fMRI study
PLOS ONE | DOI:10.1371/journal.pone.0124027 May 13, 2015 2 / 16
Competing Interests: This work was supported bythe Swiss Anorexia Foundation, the PsychiatryResearch Trust, the NIHR Biomedical ResearchCentre for Mental Health at South London andMaudsley NHS Foundation Trust and the Institute ofPsychiatry, King's College London. The funders hadno role in study design, data collection and analysis,decision to publish, or preparation of the manuscriptand their support does not alter the authors'adherence to all PLOS ONE policies on sharing dataand materials.

patterns. For instance, failure to respond appropriately to negative feedback has been associat-ed with decreased regional cerebral bloodflow in the middle frontal gyrus whereas the inabilityto keep in mind previously unsuccessful strategies was related to decreased regional cerebralbloodflow in the inferior parietal lobule [17]. Such characterisation may therefore help to im-prove understanding of the pathophysiology associated with inefficient cognitive flexibilityin AN.
The WCST also allows measurement of cortical function during instrumental learning andreinforcement, arguably vital processes for efficient flexible behaviour. Indeed, abnormal activi-ty within the dorsal striatum, which plays a major role in forming stimulus-response and ac-tion-outcome associations [18], has been observed during cognitive-behavioural flexibility[11,16] as well as during a reward-based guessing game that in healthy individuals relies onventral striatal function [19]. Furthermore, people with AN exhibit deficits of implicit categorylearning, which may be the result of disrupted dorsal striatal dopaminergic pathways [20].
We hypothesised that patients with anorexia would 1) perform poorly on the WCST, partic-ularly with regards to perseverative errors, 2) exhibit abnormal fronto-striatal activity duringlearning and reinforcement and 3) utilise the fronto-parietal control network to a greater de-gree than healthy controls (HC) during cognitive set-shifting.
Methods and Materials
ParticipantsA total of 32 women with AN (Age range = 18–41; years of illness = 1–25) were recruited fromthe South London and Maudsley (SLaM) eating disorder service and from a B-EAT communitysample (http://www.b-eat.co.uk/) (Inpatient = 10; Outpatient = 9; Day-care = 6; Communi-ty = 7). All AN participants had been diagnosed by eating disorder clinicians as fulfillingDSM-IV criteria and included 24 restrictive and 8 binge/purge subtypes. Fifteen were taking�1 psychoactive medications during the study (S1 Table). 32 age-matched HC women (Agerange = 22–46) with a BMI of>18.5, EDE-Q score of<3 and no personal history of or first de-gree relative with a psychiatric illness were recruited from the local community. Exclusion cri-teria for all participants included a history of brain trauma and/or neurological problems (e.g.epilepsy), pregnancy, claustrophobia, inadequate use of the English language, colour blindness,non-corrected visual impairment and metallic implants.
Prior to scanning, all participants were measured to calculate their current BMI. The re-search version of the structured clinical interview for DSM-IV disorders (SCID-I;[21])screened for current Axis I psychopathology. IQ was estimated using the Revised NationalAdult Reading Test (NART-R;[22]). Self-report questionnaires included the 36-item EatingDisorder Examination Questionnaire (EDE-Q;[23]), 14-item Hospital Anxiety and DepressionScale (HADS;[24]) and 12-item Cognitive Flexibility Scale (CFS;[25]).
Ethics StatementThis study was carried out in accordance with the Declaration of Helsinki under approval ofthe National Research Ethics Committee, London (Ref 11/LO/0952). Written informed con-sent was obtained following complete description of the study.
Wisconsin Card Sorting TestParticipants performed a modified version of the WCST during the scan. At the start of eachtrial, a single ‘stimulus’ card was presented in the centre of a grey screen, surrounded by four‘reference’ cards (Fig 1). ‘References’ were chosen at random at the beginning of each
Cognitive Flexibility in Anorexia Nervosa: An fMRI study
PLOS ONE | DOI:10.1371/journal.pone.0124027 May 13, 2015 3 / 16

experimental run from a deck of 64 cards that differed along three dimensions; shape (dia-mond, square, triangle, cross), colour (blue, red, green, yellow) and quantity (one, two, three,four), such that each ‘reference’ was unique on every dimension. ‘Reference’ cards remainedunchanged throughout a participants’ experimental run. The remaining 60 cards were used todraw a ‘stimulus’ at random for each trial.
Participants had 3000ms from the start of each trial to match the ‘stimulus’ card to one ofthe four ‘references’ using a 4-way joystick, according to either shape, colour or quantity. Thechosen ‘reference’ was then immediately outlined for 2000ms; in green if correct, in red if in-correct, and the ‘stimulus’ card was replaced with corresponding written feedback. This wasfollowed by a rest period lasting until the trial had elapsed 6000ms, at which point a new trialstarted (Fig 1). Participants were never directly informed of the correct sorting rule and hadto respond based on feedback given at the point of response. The sorting rule at time zero ofthe experimental run was randomly selected and changed without notice when the participantattained 8 consecutive correct trials (set), in which case the other two dimensions had a50% chance of becoming the new sorting rule. The task consisted of 100 trials, each lasting
Fig 1. Time course of aWCST trial and events of interest for behavioural and fMRI analysis. Eachexample presents the current rule chosen by the program (‘correct rule’) with concurrent trial-by-trialparticipant responses (‘chosen rule’), illustrating different WCST ‘event types’. Note that for event-relatedfMRI analysis, each event is modelled as the period between two consecutive responses and the ‘event type’is defined by the combination of the two responses. EXAMPLE A: I) Efficient Shifts (Eff-Sh) were whenparticipants changed sorting rules following negative feedback to one that had not been previously tested. II)Stuck-in-set perseverations (SiS-P) occurred if the same sorting rule that was incorrect in the previous trialwas applied in the subsequent trial. III) First correct sort (F-Corr) of a new set. IV) Second correct sort(S-Corr) follows first correct sort. V) fMRI baseline (Baseline) was designated as trials 3–8 of a string of 8consecutive correct sorts. EXAMPLE B: VI) Recurrent perseverations (Rec-P) constituted shifts to anothersorting rule following incorrect feedback, but to one that had been tested two trials previously and already fedback as incorrect. EXAMPLE C: VII) Error Set trials (Error Set) were correct sorts not included as baselinetrials due to occurrence of ‘loss of set’ errors within that particular set. VIII) Loss of set (Loss of set) trials areshifts to a different rule following positive feedback.
doi:10.1371/journal.pone.0124027.g001
Cognitive Flexibility in Anorexia Nervosa: An fMRI study
PLOS ONE | DOI:10.1371/journal.pone.0124027 May 13, 2015 4 / 16

6 seconds, making the total experimental time 10 minutes. Participants were trained prior toscanning to ensure they understood task requirements.
Reaction time and responses were recorded during scanning and classified as discrete eventsaccording to previous literature (Fig 1). Notably, we included two forms of perseveration:stuck-in-set (SiS-P) and recurrent (Rec-P) [17], and adopted the suggestion of distinguishingloss-of-set from efficient shifts (Eff-Sh), both traditionally classified as non-perseverative errors[26]. The former is indicative of a disruption in task maintenance. The latter represents adap-tive cognitive shifting. First and second correct trials (F-Corr and S-Corr respectively) were alsoconsidered separately for fMRI analysis as they represent learning and reinforcement process-es. A high-level baseline of consecutive correct trials was used in order to subtract behaviouralflexibility processes from events of interest. Error set, omission, anticipatory and ambiguoustrials not conforming to selected event criteria were coded as nuisance covariates.
fMRI AcquisitionfMRI data were acquired on a 1.5-Tesla GE Signa HDx system running 14m5 software (GeneralElectric Medical Systems, Wisconsin) at the Centre for Neuroimaging Sciences of the Institute ofPsychiatry, King’s College London. A body coil was used for RF transmission and an 8-channelhead coil for RF reception. T2�-weighted gradient echo echo-planar images (GE-EPI) depictingblood-oxygen-level-dependent (BOLD) contrast were acquired in the axial plane, parallel to theanterior commissure—posterior commissure (AC-PC) line, with the following parameters: repe-tition time (TR) = 2000ms, echo time (TE) = 40ms, flip angle = 70°, slice thickness = 5mm, slicegap = 0.5mm, field of view = 24x24cm, matrix size = 64x64. Whole-brain coverage was acquiredwith 25 slices and 300 T2�-weighted whole-brain volumes were acquired for each subject.
To facilitate co-registration of fMRI data in standard space, a whole-brain high-resolutionGE-EPI volume consisting of 43 slices parallel to the AC-PC line was acquired for each partici-pant (TR = 3000ms, TE = 40ms, flip angle = 90°, slice thickness = 3mm, slice gap = 0.3mm,FoV = 24x24cm, matrix size = 128x128). Data quality was assured using an automated qualitycontrol procedure [27].
Demographic, Clinical and Performance AnalysisDemographic, clinical and performance data were analysed using Statistical Package for the So-cial Sciences (SPSS 20.0) [28]. Parametric Student t-tests were used to assess between-groupdifferences unless Shapiro-Wilk test indicated non-normality of data, in which case non-parametric Mann-Whitney U comparisons were conducted. Performance measures includedtotal number of correct trials, completed sets (8 consecutive correct trials), stuck-in-set persev-erations, recurrent perseverations and loss-of-set errors. Reaction times for these events werealso analysed, as well as for first correct, second correct and efficient shift trials.
fMRI AnalysisfMRI data were analysed with XBAM v4.1, developed at King’s College London’s Institute ofPsychiatry, which utilises a non-parametric permutation-based approach to minimize assump-tions and reduce the effect of outliers (c.f. http://brainmap.co.uk).
Individual brain activation mapsData were first processed to minimize motion related artefacts [29]. Following realignment,images were smoothed using an 8.8mm full-width half-maximum Gaussian filter, chosen toimprove signal-to-noise ratio over the spatial neighbourhood of each voxel. Responses to each
Cognitive Flexibility in Anorexia Nervosa: An fMRI study
PLOS ONE | DOI:10.1371/journal.pone.0124027 May 13, 2015 5 / 16

event (‘Baseline’, ‘Eff-Sh’, ‘F-Corr’, ‘S-Corr’, ‘Sis-P’ and ‘Rec-P’) were then detected by time-se-ries analysis using a linear model in which each component of the experimental design wasconvolved separately with a pair of Poisson kernels (λ = 4 and 8 seconds) to allow variability inthe haemodynamic delay. Events were modelled as continuous periods between two sequentialparticipant responses, where the type of event was defined by the combination of these two re-sponses (see Fig 1). The best fit between the weighted sum of these convolutions and the time-series at each voxel was computed using the constrained BOLD effect model [30]. A goodnessof fit statistic was then computed as the ratio of the sum of squares of deviations from themean image intensity resulting from the model (over the whole time-series) to the sum ofsquares of deviations resulting from the residuals (SSQ ratio).
Following computation of the observed SSQ ratio at each voxel, the data were permuted bythe wavelet-based method [31]. Repeated application of this method at each voxel followed byre-computation of the SSQ ratio from the permuted data allows (by combination of resultsover all intracerebral voxels) the data-driven calculation of the null distribution of SSQ ratiosunder the assumption of no experimentally determined response. Using this distributionmakes it possible to calculate the critical value of SSQ ratio needed to threshold the maps atany desired type I error rate.
Group activation maps. The observed and permuted SSQ ratio maps for each individualwere transformed into the standard space of Talairach and Tournoux [32] using a two-stagewarping procedure [33]. First, the average image intensity map was computed for each individ-ual over the course of the experiment. The transformations required to map this image to thestructural scan for each individual and then from structural space to the Talairach template arethen calculated by maximising the correlation between the images at each stage. These trans-formations are then applied to the SSQ ratio maps. Group activation maps were then computedby determining the median SSQ ratio at each voxel (over all individuals) in the observed andpermuted data map. Computing intra and inter participant variations in effect separately con-stitutes a mixed effect approach, which is desirable in fMRI. Detection of activated voxels wasextended from voxel to 3D cluster-level using the method described by Bullmore et al. [34]. Re-sulting cluster-level maps were then thresholded to ensure<1 expected type I error cluster permap. Unlike Bonferroni-based multiple comparison correction procedures, use of the false dis-covery rate (FDR) ensures adequate control over the type I error rate whilst preventing over-in-flation of type II error [35].
Group and event comparisons. Between and within-group comparisons were performedby fitting the data at each intracerebral voxel at which all subjects have non-zero data using alinear model of the type:
Y ¼ aþ bX þ e
where ‘Y’ is the vector of SSQ ratios for each individual, ‘X’ is the contrast matrix for the partic-ular inter-group/inter-event contrasts required, ‘a’ is the mean effect across all individuals inthe various groups/events, ‘b’ is the computed group/event difference and ‘e’ is a vector of re-sidual errors. The model is fitted by minimising the sum of absolute deviations rather than thesums of squares to reduce outlier effects. The null distribution of ‘b’ is computed by permutingdata between groups or events (assuming the null hypothesis of no effect of group membershipor WCST event) and refitting the above model 50 times at each voxel and combining the dataover all intracerebral voxels. F-Corr vs. S-Corr (learning) and F-Corr vs. Eff-Sh (cognitive flexi-bility) by group interactions were performed using split-plot analyses of variance (ANOVA) inorder to ascertain whether any brain regions exhibit group dependency in differential event-re-lated activation patterns.
Cognitive Flexibility in Anorexia Nervosa: An fMRI study
PLOS ONE | DOI:10.1371/journal.pone.0124027 May 13, 2015 6 / 16
Results
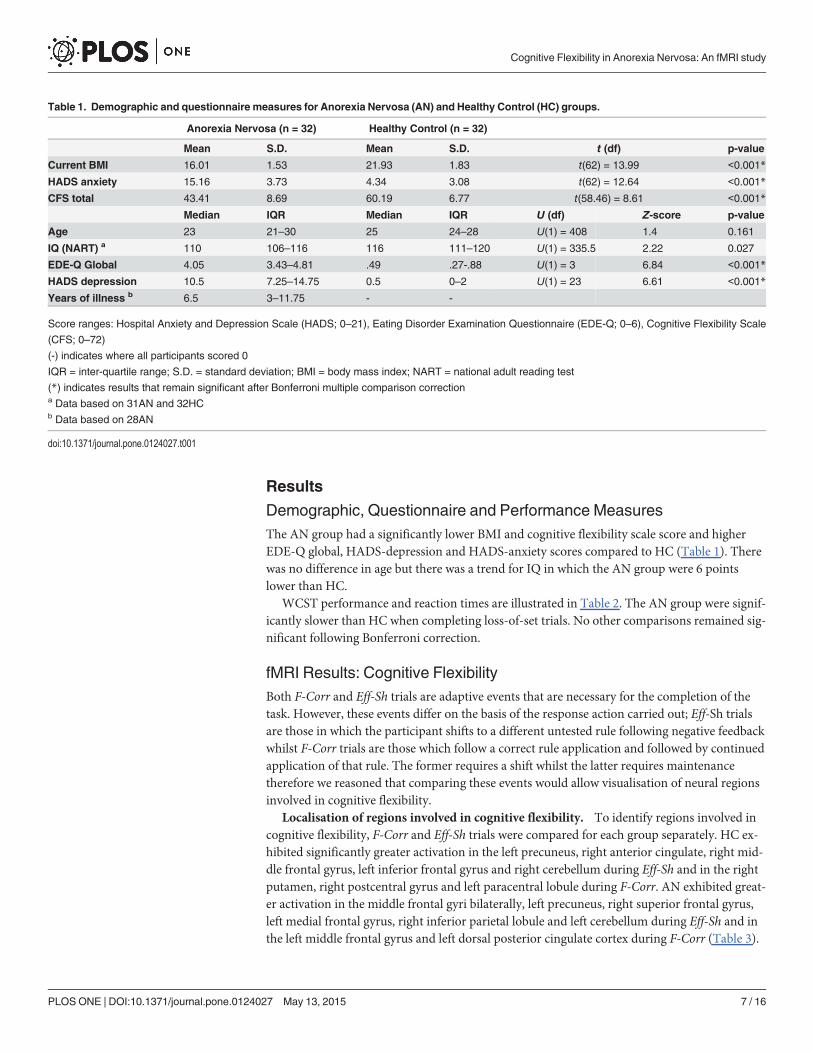
Demographic, Questionnaire and Performance MeasuresThe AN group had a significantly lower BMI and cognitive flexibility scale score and higherEDE-Q global, HADS-depression and HADS-anxiety scores compared to HC (Table 1). Therewas no difference in age but there was a trend for IQ in which the AN group were 6 pointslower than HC.
WCST performance and reaction times are illustrated in Table 2. The AN group were signif-icantly slower than HC when completing loss-of-set trials. No other comparisons remained sig-nificant following Bonferroni correction.
fMRI Results: Cognitive FlexibilityBoth F-Corr and Eff-Sh trials are adaptive events that are necessary for the completion of thetask. However, these events differ on the basis of the response action carried out; Eff-Sh trialsare those in which the participant shifts to a different untested rule following negative feedbackwhilst F-Corr trials are those which follow a correct rule application and followed by continuedapplication of that rule. The former requires a shift whilst the latter requires maintenancetherefore we reasoned that comparing these events would allow visualisation of neural regionsinvolved in cognitive flexibility.
Localisation of regions involved in cognitive flexibility. To identify regions involved incognitive flexibility, F-Corr and Eff-Sh trials were compared for each group separately. HC ex-hibited significantly greater activation in the left precuneus, right anterior cingulate, right mid-dle frontal gyrus, left inferior frontal gyrus and right cerebellum during Eff-Sh and in the rightputamen, right postcentral gyrus and left paracentral lobule during F-Corr. AN exhibited great-er activation in the middle frontal gyri bilaterally, left precuneus, right superior frontal gyrus,left medial frontal gyrus, right inferior parietal lobule and left cerebellum during Eff-Sh and inthe left middle frontal gyrus and left dorsal posterior cingulate cortex during F-Corr (Table 3).
Table 1. Demographic and questionnaire measures for Anorexia Nervosa (AN) and Healthy Control (HC) groups.
Anorexia Nervosa (n = 32) Healthy Control (n = 32)
Mean S.D. Mean S.D. t (df) p-value
Current BMI 16.01 1.53 21.93 1.83 t(62) = 13.99 <0.001*
HADS anxiety 15.16 3.73 4.34 3.08 t(62) = 12.64 <0.001*
CFS total 43.41 8.69 60.19 6.77 t(58.46) = 8.61 <0.001*
Median IQR Median IQR U (df) Z-score p-value
Age 23 21–30 25 24–28 U(1) = 408 1.4 0.161
IQ (NART) a 110 106–116 116 111–120 U(1) = 335.5 2.22 0.027
EDE-Q Global 4.05 3.43–4.81 .49 .27-.88 U(1) = 3 6.84 <0.001*
HADS depression 10.5 7.25–14.75 0.5 0–2 U(1) = 23 6.61 <0.001*
Years of illness b 6.5 3–11.75 - -
Score ranges: Hospital Anxiety and Depression Scale (HADS; 0–21), Eating Disorder Examination Questionnaire (EDE-Q; 0–6), Cognitive Flexibility Scale
(CFS; 0–72)
(-) indicates where all participants scored 0
IQR = inter-quartile range; S.D. = standard deviation; BMI = body mass index; NART = national adult reading test
(*) indicates results that remain significant after Bonferroni multiple comparison correctiona Data based on 31AN and 32HCb Data based on 28AN
doi:10.1371/journal.pone.0124027.t001
Cognitive Flexibility in Anorexia Nervosa: An fMRI study
PLOS ONE | DOI:10.1371/journal.pone.0124027 May 13, 2015 7 / 16

Group differences in first correct and efficient shift trials. No differences were foundwhen comparing AN and HC on F-Corr trials. There were however differences when compar-ing groups on Eff-Sh trials; AN participants exhibited greater activation in the left angulargyrus and left precuneus (Table 3). There were no areas of greater activation in the HC group.
Cognitive flexibility interaction. A group (AN, HC) by event (F-Corr, Eff-Sh) interactionrevealed that the AN group exhibited greater activation during Eff-Sh trials but lesser activationduring F-Corr trials in the left middle frontal gyrus, left inferior precuneus and left superior pa-rietal lobule, as compared to the HC group (Fig 2A).
Only a percentage of participants committed SiS-P (50%) and Rec-P (37.5%) errors over thecourse of the task with these participants also showing low event frequency (S2 Table). Thismeant that there was insufficient power to conduct whole-brain analyses on these event types.Non-parametric voxel statistics (SSQ ratios) for SiS-P and Rec-P were therefore extracted fromsignificant clusters resulting from the cognitive flexibility interaction analysis and plotted inFig 2B.
fMRI Results: LearningFirst correct (F-Corr) and second correct (S-Corr) trials both represent events in which the par-ticipant has applied the correct sorting rule. Although a requisite of F-Corr is maintenance andapplication of the rule in the next trial, the period modelled may be associated with uncertainty;first after because the correct rule is found through trial and error and second because the par-ticipant may not be sure of the rule even a correct response. As such, the event following
Table 2. WCST performance and reaction timemeasures for Anorexia Nervosa (AN) and Healthy Control (HC) groups.
Anorexia Nervosa(n = 32)
Healthy Control(n = 32)
Performance Score Median IQR Median IQR U (df) Z-Score p-value
First Correct 11 10.25–11 11 11–11 U(1) = 495 0.26 .796
Second Correct 11 10–11 11 10–11 U(1) = 475.5 0.54 .586
Completed Sets 8 7–9 9.5 8–10 U(1) = 334.5 2.45 .014
Efficient Shift 14 13–15.75 14 13–15 U(1) = 421 1.25 .213
Total Correct 80 75–83 81 79–84 U(1) = 399.5 1.52 .129
SiS-Perseveration .5 0–2.75 1 0–1.75 U(1) = 508.5 0.05 .960
Rec-Perseveration 0 0–1.75 0 0–1 U(1) = 493.5 0.29 .774
Loss-of-Set 2 1–3 1 1–2.75 U(1) = 430 1.12 .261
Mean Reaction Time (ms) Mean S.D. Mean S.D. t (df) p-valueFirst Correct 1473.9 252.9 1374.3 234.2 t(62) = 1.64 .107
Second Correct 1355.1 191.1 1328.6 163.8 t(62) = 0.59 .553
Completed Set 1417.3 278.9 1260.4 195.3 t(62) = 2.61 .011
Efficient Shift 1426.8 221.2 1288.7 200.0 t(62) = 2.62 .011
SiS-Perseveration a 1409.9 562.4 1257.4 314.3 t(32) = 0.99 .329
Rec-Perseveration b 1283.4 443.4 1394.4 385.6 t(21) = 0.64 .527
Loss-of-Set c 1531.9 401.7 1212.7 269.3 t(43.85) = 3.35 .002 *
IQR = inter-quartile range; S.D. = standard deviation
(*) indicates results that remain significant after Bonferroni multiple comparison correctiona Data based on 16AN and 16HC; the remaining participants committed 0 errorsb Data based on 11AN and 13HC; the remaining participants committed 0 errorsc Data based on 26AN and 25HC; the remaining participants committed 0 errors
doi:10.1371/journal.pone.0124027.t002
Cognitive Flexibility in Anorexia Nervosa: An fMRI study
PLOS ONE | DOI:10.1371/journal.pone.0124027 May 13, 2015 8 / 16

repeated application (S-Corr) represents verification of what participants believe to be the cor-rect rule. We reasoned that comparing these events would allow assessment of neural mecha-nisms involved in learning.
Group differences in learning trials. To assess BOLD response during learning, AN andHC groups were compared on F-Corr and S-Corr trials. As previously stated, there were no dif-ferences when comparing groups on F-Corr trials. For S-Corr however, the HC group exhibitedsignificantly greater activation in the right precuneus, left caudate body and right cerebellum ascompared to AN. The AN group exhibited significantly greater activation in the right posteriorcingulate cortex as compared to HC (Fig 3B).
Learning interaction. A group (AN, HC) by event (F-Corr, S-Corr) interaction revealedthat the AN group had increasing activation in the right posterior cingulate cortex from F-Corrto S-Corr whereas the HC group exhibited a decreasing trend. Oppositely, AN show decreasingactivation in the right precuneus from F-Corr to S-Corr whilst HC exhibited an increasingtrend (Fig 3A).
Table 3. Regions showing a significant difference between Efficient Shift (Eff-Sh) and First Correct (F-Corr) trials.
Peak TalairachCoordinates
Region BA Side x y z Cluster Size (voxels) p-value
Healthy Control: Eff-Sh > F-CorrPrecuneus 7 L 0 -74 42 152 0.000178
Cerebellum - R 47 -59 -29 108 0.000178
Anterior Cingulate Gyrus 32 R 4 19 42 53 0.003018
Inferior Frontal Gyrus 47 L -47 22 -2 62 0.005859
Middle Frontal Gyrus 9 R 51 15 31 35 0.00799
Healthy Control: F-Corr > Eff-ShParacentral Lobule 31 L 0 -30 42 127 0.001584
Putamen - R 14 7 4 42 0.005631
Postcentral Gyrus 3 R 36 -30 53 45 0.006335
Anorexia Nervosa: Eff-Sh > F-CorrPrecuneus 7 L -4 -63 42 302 0.000179
Middle Frontal Gyrus 10 L -43 44 9 119 0.000359
9 R 43 30 26 103 0.000359
6 L -29 7 53 39 0.003406
Cerebellum - L -33 -56 -24 118 0.000359
Medial Frontal Gyrus 8 L 0 22 42 40 0.003048
Inferior Parietal Lobule 40 R 36 -48 37 55 0.003227
Superior Frontal Gyrus 8 R 36 15 48 31 0.00502
Anorexia Nervosa: F-Corr > Eff-ShPosterior Cingulate Gyrus 31 L -11 -30 42 781 0.000183
Medial Frontal Gyrus 9 L -4 37 26 52 0.007156
Eff-Sh: AN > HC
Precuneus 7 L -7 -63 48 60 0.000765
Angular Gyrus 39 L -29 -48 26 27 0.003825
Efficient Shift (Eff-Sh); First Correct (F-Corr); Anorexia Nervosa (AN); Healthy Controls (HC)
All listed regions survived correction for multiple comparisons.
doi:10.1371/journal.pone.0124027.t003
Cognitive Flexibility in Anorexia Nervosa: An fMRI study
PLOS ONE | DOI:10.1371/journal.pone.0124027 May 13, 2015 9 / 16

DiscussionThe current study used the Wisconsin Card Sorting Test (WCST) in conjunction with whole-brain event-related fMRI to elucidate the neural underpinnings of instrumental learning andcognitive flexibility in AN. With regards to behavioural measures, whilst AN attained lowerscores on the cognitive flexibility scale and a measure of general WCST performance (complet-ed sets), we did not detect differences with regards to perseverations. Previous neuropsycholog-ical studies using the WCST consistently identify heightened perseverative tendency in AN
Fig 2. fMRI results of cognitive flexibility comparisons. (A): Axial slices and line graphs showingsignificant group (Anorexia Nervosa, Healthy Control) x event (Efficient Shift [Eff-Sh], First Correct [F-Corr])interactions of BOLD response (SSQ value) in the left middle frontal gyrus (BA9, Peak activation Talairachcoordinates = -40, 22, 26, cluster size = 14, p = 0.0069), left inferior precuneus (BA7, Peak activationTalairach coordinates = -25, -56, 26, cluster size = 34, p = 0.0029) and left superior parietal lobule extendinginto the precuneus (BA7, Peak activation Talairach coordinates = -18, -67, 48, cluster size = 17, p = 0.0078).(B): Sagittal slices and bar charts illustrating the BOLD response during SiS-P and Rec-P and correspondingactivity during Eff-Sh in the left middle frontal gyrus, left inferior precuneus and left superior parietal lobule.Functional data were thresholded to yield less than 1 false positive cluster per map and overlaid onto a highresolution single subject T1 structural image in Talairach space.
doi:10.1371/journal.pone.0124027.g002
Cognitive Flexibility in Anorexia Nervosa: An fMRI study
PLOS ONE | DOI:10.1371/journal.pone.0124027 May 13, 2015 10 / 16

[4,6]. However, neuropsychological WCST protocol refrains from providing informationabout the rules embedded within the task whilst training sessions are needed to ensure ade-quate performance in fMRI settings. Indeed, a similar fMRI experiment using the WCST alsofailed to find perseverative error differences [16]; hence prior strategic knowledge could be re-sponsible for the difference in results.
Despite this, lower activation was found in the left caudate body in AN as compared to HCafter consolidating a new rule (S-Corr), although no difference was found after first correct tri-als (F-Corr). This was somewhat surprising, given that previous fMRI investigations havefound hyperactivity during various executive processes, including reward and cognitiveflexibility [16,19] as well as when presented with aversive food stimuli [36]. In addition to
Fig 3. fMRI results of learning comparisons. (A): Coronal, sagittal and axial views of regions showingsignificant group (Anorexia Nervosa, Healthy Control) by event (First Correct [F-Corr], Second Correct[S-Corr]) interactions of BOLD response (SSQ value) in the right posterior cingulate cortex (BA31, Peakactivation Talairach coordinates = 14, -56, 20, cluster size = 26, p = 0.0071) and right precuneus (BA4, Peakactivation Talairach coordinates = 29, -33, 42, cluster size = 33, p = 0.0026). (B): Significant difference inactivity between Anorexia Nervosa and Healthy Control groups during Second Correct trials (S-Corr) in theright cerebellum (Peak activation Talairach coordinates = 7, -30, -24, cluster size = 36, p = 0.0031), leftcaudate body (Peak activation Talairach coordinates = -22, -26, 26, cluster size = 30, p = 0.003), rightprecuneus (BA31, Peak activation Talairach coordinates = 22, -30, 42, cluster size = 21, p = 0.0017) and rightposterior cingulate cortex (BA31, Peak activation Talairach coordinates = 11, -56, 20, cluster size = 37,p = 0.0019). Functional data were thresholded to yield less than 1 false positive cluster and overlaid onto ahigh resolution single subject T1 structural image in Talairach space.
doi:10.1371/journal.pone.0124027.g003
Cognitive Flexibility in Anorexia Nervosa: An fMRI study
PLOS ONE | DOI:10.1371/journal.pone.0124027 May 13, 2015 11 / 16

facilitating planning and decision-making processes, the caudate nucleus is important forinstrumental learning, specifically supporting goal-directed action through formation of ac-tion-outcome contingencies [18]. To our knowledge, no studies have directly assessed corticalfunction during learning processes in AN, although one neuropsychological study provided anindication of disrupted striatal dopamine pathways by showing poorer performance on an im-plicit category learning task [20]. Since both groups committed an equal number of loss-of-seterrors, it is unlikely that decreased caudate activation affected adequate formation of action-outcome contingencies. However, one study of the marmoset medial striatum, an area analo-gous to the caudate nucleus, showed that focal lesions to this structure cause impairments inreversal learning when goal values and/or task requirements change, whilst the ability to estab-lish new action-outcome contingencies remains intact [37]. Indeed, our AN group exhibited atrend for slower reaction times for shift trials following negative feedback which cannot be ex-plained by general motor deficits as reaction time differences were not widespread, indicating adifficulty in shifting set when prompted. Thus, it is possible that aberrant caudate activationmay affect the ability of AN patients to reverse learned associations, which may subsequentlyproduce perseverative tendencies.
We found that activity in the anterior portion of the right precuneus decreased from thefirst to second correct trials in AN whereas the opposite pattern occurred in HC. This regionexhibits high functional connectivity with the superior parietal cortex, paracentral lobule andmotor cortex, constituting a network related to sensorimotor processing [38]. Although theexact function of the right precuneus in sensorimotor functions remains elusive, the superiorparietal cortex has been found to respond when detection of stimulus features is necessary toupdate stimulus-response associations [39,40]. Decreasing activation could therefore indicatethat people with AN are quicker to assume stimulus-response contingencies than HC. In addi-tion, a recent fMRI study revealed that activity in the caudate, which receives projections fromthe superior parietal cortex [41], decreases steadily for successive trials following initial acquisi-tion [42]. Although we cannot comment on the activation trajectory of the caudate from firstto second correct trials, the fact that it is lower in the second correct trial for AN lends supportto this theory of quicker acquisition. Further work is needed to clarify the relative contributionsand mutual dependency of aberrant caudate and precuneus activation to learning in AN.
We found a greater increase in activity in AN in the left precuneus and middle frontal gyruswhen shifting but greater decrease when maintaining set, as compared to controls. This is inline with Zastrow et al. [11] who demonstrated dominant activation of the fronto-parietal con-trol network (FPCN) in AN during behavioural shifts.We add here that differential FPCN acti-vation is also evident during cognitive set-shifting and further, that perseverative errors werecharacterised by a failure to activate these regions. Interestingly, this lack of activation occurredfor both stuck-in-set and recurrent perseverations, suggesting that it is independent of cogni-tive set-shifting as well as feedback, although due to lack of power this finding should be treatedas preliminary. We also found a similar activation difference in the superior parietal lobule ex-tending into the precuneus. This area is part of the dorsal attention network (DAN; [43]),which as aforementioned is linked with perceptual shifting [39,40]. The FPCN shows a high de-gree of between-network interconnectivity with the DAN and default mode network (DMN),acting as an intermediary that delegates cortical resources towards either externally or internal-ly focussed goal-directed cognition respectively [15,43]. Taken together, these findings indicatethat cognitive set-shifting places greater functional demands on people with AN and supportcurrent opinion of greater supervisory cognitive control when carrying out external goals[11,19].
Cognitive Flexibility in Anorexia Nervosa: An fMRI study
PLOS ONE | DOI:10.1371/journal.pone.0124027 May 13, 2015 12 / 16

LimitationsOur design used a high-level baseline in an attempt to isolate processes occurring in additionto behavioural flexibility. However, Zastrow et al. [11] showed that the BOLD response associ-ated with behavioural flexibility is different between AN and HC, hence our results may beconfounded by our baseline. Although we took this into account by restricting the selection ofbaseline trials to those that occurred within sets completed with 100% accuracy, and primarilyusing interaction analyses to visualise the baseline, care must be taken when interpreting the re-sults presented here. We also note that the use of F-Corr and S-Corr trials to represent learningin the context of cognitive flexibility may be overly simplistic and therefore our results may notbe wholly generalizable. However, we maintain that our findings indicate the involvement oflearning circuitry in the pathophysiology of AN and thus outline the importance of furtherneuroimaging investigation.
Perseverative errors were infrequent which due to the noise inherent in fMRI data, causedour analysis to lack power. The WCST is often used in behavioural studies to characterise cog-nitive flexibility in AN with particular regard to perseveration, therefore it is important to elab-orate on its neural bases. The WCST was not designed for use in fMRI settings and it ispossible that pre-scan practice sessions together with short task duration resulted in low eventcount and general lack of behavioural differences. Decreasing the number of trials required tocomplete a set could increase perseverative error scores whilst keeping the risk of fatigue lowand using the practice sessions to measure performance may allow a more valid quantificationof cognitive flexibility inefficiencies without confounding practice effects. Alternatively, ablock-design method such as that employed by Lie et al. [44] could be used to isolate compo-nents of the task via subtraction logic, although this may not allow direct assessment of allWCST event types. Lastly, we found that depression and anxiety scores were higher in AN,however, research suggests that up to 94% of patients with an eating disorder also suffer fromat least one comorbid mood disorder [45], therefore we opted to analyse a cohort that reflectsclinical presentation.
ConclusionsPoor cognitive flexibility in AN is characterised by aberrant functioning of the fronto-parietalcontrol network which may contribute to a dysregulation in the way resources are directed be-tween external and internal goal-directed cognition. We corroborate previous work showingno fronto-striatal differences during cognitive flexibility over and above that of behaviouralflexibility. However, the finding of altered caudate activation during learning could indicatethat people with AN are unable to maintain and/or reverse acquired action-outcome associa-tions. Aberrant learning circuitry may therefore contribute to the observed tendency for peoplewith AN to perseverate and as such, merits further investigation.
Supporting InformationS1 Table. Number of Anorexia Nervosa participants taking psychoactive medications at thetime of study.(DOCX)
S2 Table. Descriptive statistics of SiS-P, Rec-P and Eff-Sh events occurring in Anorexia Ner-vosa and Healthy Control groups for perseveration analysis.(DOCX)
Cognitive Flexibility in Anorexia Nervosa: An fMRI study
PLOS ONE | DOI:10.1371/journal.pone.0124027 May 13, 2015 13 / 16
AcknowledgmentsWe would like to thank Dr. Helen Davies and Naima Lounes for data collection and Dr. OwenO’Daly for statistical support.
Author ContributionsConceived and designed the experiments: KT. Performed the experiments: KT LF. Analyzedthe data: NPLK LF. Contributed reagents/materials/analysis tools: VPG. Wrote the paper:NPLK AS SCRW.
References1. Hudson JI, Hiripi E, Pope HG, Kessler RC (2007) The prevalence and correlates of eating disorders in
the National Comorbidity Survey Replication. Biol Psychiatry 61: 348–358. PMID: 16815322
2. Harris EC, Barraclough B (1998) Excess mortality of mental disorder. Br J Psychiatry 173: 11–53.PMID: 9850203
3. Schmidt U, Treasure J (2006) Anorexia nervosa: valued and visible. A cognitive-interpersonal mainte-nance model and its implications for research and practice. Br J Clin Psychol 45: 343–366. PMID:17147101
4. Tchanturia K, Davies H, Roberts M, Harrison A, Nakazato M, Schmidt U, et al. (2012) Poor CognitiveFlexibility in Eating Disorders: Examining the Evidence using the Wisconsin Card Sorting Task. PlosOne 7: e28331. doi: 10.1371/journal.pone.0028331 PMID: 22253689
5. Roberts ME, Tchanturia K, Treasure JL (2010) Exploring the neurocognitive signature of poor set-shift-ing in anorexia and bulimia nervosa. Journal of Psychiatric Research 44: 964–970. doi: 10.1016/j.jpsychires.2010.03.001 PMID: 20398910
6. Tenconi E, Santonastaso P, Degortes D, Bosello R, Titton F, Mapelli D, et al. (2010) Set-shifting abili-ties, central coherence, and handedness in anorexia nervosa patients, their unaffected siblings andhealthy controls: Exploring putative endophenotypes. World Journal of Biological Psychiatry 11: 813–823. doi: 10.3109/15622975.2010.483250 PMID: 20486870
7. Tchanturia K, Harrison A, Davies H, Roberts M, Oldershaw A, Nakazato M, et al. (2011) Cognitive flexi-bility and clinical severity in eating disorders. Plos One 6: e20462. doi: 10.1371/journal.pone.0020462PMID: 21698277
8. Holliday J, Tchanturia K, Landau S, Collier D, Treasure J (2005) Is impaired set-shifting an endopheno-type of anorexia nervosa? Am J Psychiatry 162: 2269–2275. PMID: 16330590
9. Kothari R, Solmi F, Treasure J, Micali N (2013) The neuropsychological profile of children at high risk ofdeveloping an eating disorder. Psychological Medicine 43: 1543–1554. doi: 10.1017/S0033291712002188 PMID: 23021014
10. Anderluh MB, Tchanturia K, Rabe-Hesketh S, Treasure J (2003) Childhood obsessive-compulsive per-sonality traits in adult women with eating disorders: defining a broader eating disorder phenotype. Am JPsychiatry 160: 242–247. PMID: 12562569
11. Zastrow A, Kaiser S, Stippich C, Walther S, HerzogW, Tchanturia K, et al. (2009) Neural correlates ofimpaired cognitive-behavioral flexibility in anorexia nervosa. Am J Psychiatry 166: 608–616. doi: 10.1176/appi.ajp.2008.08050775 PMID: 19223435
12. Titova OE, Hjorth OC, Schioth HB, Brooks SJ (2013) Anorexia nervosa is linked to reduced brain struc-ture in reward and somatosensory regions: a meta-analysis of VBM studies. Bmc Psychiatry 13: 110.doi: 10.1186/1471-244X-13-110 PMID: 23570420
13. Muhlau M, Gaser C, Ilg R, Conrad B, Leibl C, Cebulla MH, et al. (2007) Gray matter decrease of the an-terior cingulate cortex in anorexia nervosa. Am J Psychiatry 164: 1850–1857. PMID: 18056240
14. Dang LC, O'Neil JP, Jagust WJ (2012) Dopamine supports coupling of attention-related networks. Jour-nal of Neuroscience 32: 9582–9587. doi: 10.1523/JNEUROSCI.0909-12.2012 PMID: 22787044
15. Spreng RN, StevensWD, Chamberlain JP, Gilmore AW, Schacter DL (2010) Default network activity,coupled with the frontoparietal control network, supports goal-directed cognition. Neuroimage 53: 303–317. doi: 10.1016/j.neuroimage.2010.06.016 PMID: 20600998
16. Sato Y, Saito N, Utsumi A, Aizawa E, Shoji T, IzumiyamaM, et al. (2013): Neural Basis of ImpairedCognitive Flexibility in Patients with Anorexia Nervosa. Plos One 8: e61108. doi: 10.1371/journal.pone.0061108 PMID: 23675408
Cognitive Flexibility in Anorexia Nervosa: An fMRI study
PLOS ONE | DOI:10.1371/journal.pone.0124027 May 13, 2015 14 / 16

17. Nagahama Y, Okina T, Suzuki N, Nabatame H, Matsuda M (2005): The cerebral correlates of differenttypes of perseveration in theWisconsin Card Sorting Test. Journal of Neurology, Neurosurgery andPsychiatry 76: 169–175. PMID: 15654026
18. Grahn JA, Parkinson JA, Owen AM (2008) The cognitive functions of the caudate nucleus. Prog Neuro-biol 86: 141–155. doi: 10.1016/j.pneurobio.2008.09.004 PMID: 18824075
19. Wagner A, Aizenstein H, Venkatraman VK, Fudge J, May JC, Mazurkewicz L, et al. (2007) Altered re-ward processing in women recovered from anorexia nervosa. Am J Psychiatry 164: 1842–1849. PMID:18056239
20. Shott ME, Filoteo JV, Jappe LM, Pryor T, MaddoxWT, Rollin MD, et al. (2012) Altered implicit categorylearning in anorexia nervosa. Neuropsychology 26: 191–201. doi: 10.1037/a0026771 PMID:22201300
21. First M, Spitzer R, Gibbon M, Williams J (1997) Structured clinical interview for DSM-IV axis I disorders.Washington, DC: American Psychiatric Press.
22. Nelson HE, Willison JR (1991) The revised National Adult Reading Test (NART): Test Manual. Wind-sor, UK: NFER Nelson.
23. Fairburn CG, Beglin SJ (1994) Assessment of Eating Disorders—Interview or Self-Report Question-naire. International Journal of Eating Disorders 16: 363–370. PMID: 7866415
24. Zigmond AS, Snaith RP (1983): The hospital anxiety and depression scale. Acta Psychiatr Scand 67:361–370. PMID: 6880820
25. Martin MM, Rubin RB (1995) A NewMeasure of Cognitive Flexibility. Psychological Reports 76: 623–626.
26. Barcelo F, Knight RT (2002) Both random and perseverative errors underlie WCST deficits in prefrontalpatients. Neuropsychologia 40: 349–356.
27. Simmons A, Moore E, Williams SCR (1999) Quality control for functional magnetic resonance imagingusing automated data analysis and Shewhart charting. Magnetic Resonance in Medicine 41: 1274–1278. PMID: 10371463
28. IBM-Corp (2011) SPSS Statistics for Windows, Version 20.0. Armonk, NY: IBM Corp.
29. Bullmore ET, Brammer MJ, Rabe-Hesketh S, Curtis VA, Morris RG, Williams SCR, et al. (1999) Meth-ods for diagnosis and treatment of stimulus-correlated motion in generic brain activation studies usingfMRI. Hum Brain Mapp 7: 38–48. PMID: 9882089
30. Friman O, Borga M, Lundberg P, Knutsson H (2003) Adaptive analysis of fMRI data. Neuroimage 19:837–845. PMID: 12880812
31. Bullmore E, Long C, Suckling J, Fadili J, Calvert G, Zelaya F, et al. (2001) Colored noise and computa-tional inference in neurophysiological (fMRI) time series analysis: Resampling methods in time andwavelet domains. Hum Brain Mapp 12: 61–78. PMID: 11169871
32. Talairach J, Tournoux P (1988) Co-planar stereotactic atlas of the human brain. Stuttgart: Thieme.
33. Brammer MJ, Bullmore ET, Simmons A, Williams SC, Grasby PM, Howard RJ, et al. (1997) Genericbrain activation mapping in functional magnetic resonance imaging: a nonparametric approach. MagnReson Imaging 15: 763–770. PMID: 9309607
34. Bullmore ET, Suckling J, Overmeyer S, Rabe-Hesketh S, Taylor E, Brammer MJ (1999) Global, voxel,and cluster tests, by theory and permutation, for a difference between two groups of structural MR im-ages of the brain. Ieee Transactions on Medical Imaging 18: 32–42. PMID: 10193695
35. Genovese CR, Lazar NA, Nichols T (2002) Thresholding of statistical maps in functional neuroimagingusing the false discovery rate. Neuroimage 15: 870–878. PMID: 11906227
36. Cowdrey FA, Park RJ, Harmer CJ, McCabe C (2011) Increased neural processing of rewarding andaversive food stimuli in recovered anorexia nervosa. Biol Psychiatry 70: 736–743. doi: 10.1016/j.biopsych.2011.05.028 PMID: 21714958
37. Clarke HF, Robbins TW, Roberts AC (2008) Lesions of the Medial Striatum in Monkeys Produce Per-severative Impairments during Reversal Learning Similar to Those Produced by Lesions of the Orbito-frontal Cortex. Journal of Neuroscience 28: 10972–10982. doi: 10.1523/JNEUROSCI.1521-08.2008PMID: 18945905
38. Margulies DS, Vincent JL, Kelly C, Lohmann G, Uddin LQ, Biswal BB, et al. (2009) Precuneus sharesintrinsic functional architecture in humans and monkeys. Proc Natl Acad Sci U S A 106: 20069–20074.doi: 10.1073/pnas.0905314106 PMID: 19903877
39. Barber AD, Carter CS (2005) Cognitive control involved in overcoming prepotent response tendenciesand switching between tasks. Cerebral Cortex 15: 899–912. PMID: 15459082
Cognitive Flexibility in Anorexia Nervosa: An fMRI study
PLOS ONE | DOI:10.1371/journal.pone.0124027 May 13, 2015 15 / 16

40. Ravizza SM, Carter CS (2008) Shifting set about task switching: behavioral and neural evidence for dis-tinct forms of cognitive flexibility. Neuropsychologia 46: 2924–2935. doi: 10.1016/j.neuropsychologia.2008.06.006 PMID: 18601941
41. Yeterian EH, Pandya DN (1993) Striatal connections of the parietal association cortices in rhesus mon-keys. J Comp Neurol 332: 175–197.
42. Eliassen JC, LamyM, Allendorfer JB, Boespflug E, Bullard DP, Smith MS, et al. (2012) Selective rolefor striatal and prefrontal regions in processing first trial feedback during single-trial associative learn-ing. Brain Res 1458: 56–66. doi: 10.1016/j.brainres.2012.04.010 PMID: 22560503
43. Spreng RN, Sepulcre J, Turner GR, StevensWD, Schacter DL (2013) Intrinsic Architecture Underlyingthe Relations among the Default, Dorsal Attention, and Frontoparietal Control Networks of the HumanBrain. Journal of Cognitive Neuroscience 25: 74–86. doi: 10.1162/jocn_a_00281 PMID: 22905821
44. Lie CH, Specht K, Marshall JC, Fink GR (2006) Using fMRI to decompose the neural processes under-lying the Wisconsin Card Sorting Test. Neuroimage 30: 1038–1049. PMID: 16414280
45. Blinder BJ, Cumella EJ, Sanathara VA (2006) Psychiatric comorbidities of female inpatients with eatingdisorders. PsychosomMed 68: 454–462. PMID: 16738079
Cognitive Flexibility in Anorexia Nervosa: An fMRI study
PLOS ONE | DOI:10.1371/journal.pone.0124027 May 13, 2015 16 / 16