late holocene environmental changes inferred from diatom, chironomid, and pollen assemblages in an...
TRANSCRIPT

MOUNTAIN LAKES
Late Holocene environmental changes inferred from diatom,chironomid, and pollen assemblages in an Andean lakein Central Chile, Lake Laja (36�S)
Roberto Urrutia • Alberto Araneda • Laura Torres • Fabiola Cruces •
Caterina Vivero • Fernando Torrejon • Ricardo Barra • Nathalie Fagel •
Burkhard Scharf
Published online: 28 April 2010
� Springer Science+Business Media B.V. 2010
Abstract A sediment core encompassing the last
2000 years was extracted from Lake Laja, Chile,
(36�540S, 71�050W) using an Uwitec drilling plat-
form. The sediment was subsampled for loss on
ignition, nutrients, biogenic silica, and biological
proxies (diatoms, chironomids, pollen). The sedimen-
tary profile was characterized by several coarse
volcanic layers. Loss on ignition, nutrients, and
biogenic silica showed an increasing trend that
suggests a recent shift to a higher trophic status.
Diatom assemblages also suggested higher nutrient
content with increased abundances of Aulacoseira
granulata, A. distans, and Asterionella formosa. At
the same time, a marked change in the benthic and
facultative planktonic taxa may be associated with
cooling. This period of change coincides with the
European Little Ice Age (LIA). The chironomid
profile showed four key zones distinguished largely
by changes in the abundance of Tanytarsini, Para-
chironomus, and Macropelopia. Like diatoms, chir-
onomids also seemed to reflect a shift to higher
trophic conditions in the upper part reflected by
increasing abundance of taxa such as Tribelos/
Phaenopsectra, Cricotopus/Orthocladius, and Abla-
besmyia. The most striking feature in the chironomid
assemblage is the abundance of Podonominae,
Parapsectrocladius, and Limnophyes/Compterosmit-
tia, which could be associated with a cold-dry period
between 1500 and 1900 AD in Lake Laja (the period
of the European LIA). Pollen assemblages indicated
fluctuations in humidity through changes in Nothof-
agus dombeyi-type, Poaceae, and Ephedra, and we
inferred a strong human impact over the last
100 years from the appearance of Plantago and
increased levels of Poaceae and Asteraceae subf.
Cichorioidae. Finally, the three proxies showed the
occurrence of a cold-dry event in Lake Laja (*1550–
1900 AD), which roughly coincides with the
Guest editors: Hilde Eggermont, Martin Kernan & Koen
Martens / Global change impacts on mountain lakes
R. Urrutia (&) � A. Araneda � L. Torres �F. Cruces � C. Vivero � F. Torrejon � R. Barra
Group of Paleolimnological Studies (GEP), Aquatic
Systems Research Unit, Environmental Sciences Center
EULA-Chile, University of Concepcion, Barrio
Universitario s/n, Concepcion, Chile
e-mail: [email protected]
L. Torres
Department of Basic Sciences, Education School,
Campus Los Angeles, University of Concepcion,
Juan Antonio Coloma 0201, Los Angeles, Chile
F. Cruces
Faculty of Natural and Oceanographic Sciences,
Department of Botany, University of Concepcion,
Concepcion, Chile
N. Fagel
Clays and Paleoclimate Research Unit, Department
of Geology, University of Liege, Liege, Belgium
B. Scharf
Centre for Environmental Research, UFZ-Centrum,
Mardeburg, Germany
123
Hydrobiologia (2010) 648:207–225
DOI 10.1007/s10750-010-0264-1

European LIA. However the data from this research,
does not prove neither rejects the existence of the
occurrence of the MWP in the central Andes.
Keywords Late Holocene � MWP �LIA � Chilean Andes � Lake sediments �Multiproxy approach
Introduction
High resolution paleoclimate reconstructions of the
last 1000 years have become very important in recent
decades. According to Esper et al. (2005), under-
standing the correct amplitude of major climatic
episodes in the past millennium is ‘‘critical for
predicting future trends.’’ This is because evidence
showing that the amplitudes of major historical
climate episodes were as great as or even greater
than twentieth century global warming would indi-
cate the continued importance of the role played by
natural drivers in forcing temperature changes.
Among the climate events of the last millennium,
the Medieval Warm Period (MWP), or Medieval
Climate Anomaly (Bradley et al., 2003) which
occurred from *900 to 1300 AD and the Little Ice
Age (LIA; *1350–1850 AD) have gained much
attention. However, despite the considerable research
effort, the magnitude and timing of these events are
still debated, particularly the timing and synchronic-
ity (Lamb, 1977; Bradley & Jones, 1993; Mann et al.,
1998; Grove, 2001; Ogilvie & Jonsson, 2001; Mann,
2002; Soon et al., 2003; Goosse et al., 2005), and
some researchers even doubt the global expression of
such events (Winkler, 2004; McKinzey et al., 2004).
Hughes & Diaz (1994) indicated that, in some
areas of the world, temperatures were relatively high
during the MWP. However, other studies have shown
that temperatures during this period were slightly
above current levels (Huang & Pollack, 1997; Loehle,
2007), and temperatures at other sites such as
southeastern US and southern Europe did not differ
from those of previous or subsequent centuries
(Goosse et al., 2005). Crowley & Lowery (2000)
pointed out an asynchrony in the worldwide expres-
sion of this warm event. Moreover, Bradley et al.
(2003) rejected the idea of a warm event during
medieval times, indicating the need to disentangle
natural climate variability from anthropogenic
forcing and, thus, the necessity for more reconstruc-
tions of the last millennium.
High resolution climatic studies of Late Holocene
climate events such as the LIA and MWP are
common for the Northern Hemisphere (Jones et al.,
2001; Soon et al., 2003; Goosse et al., 2005; Torres
et al., 2008). Comparatively little research has been
published about these events in South America (e.g.,
Villalba, 1994; Lamy et al., 2001; Koch & Kilian,
2005; Bertrand et al., 2005; Araneda et al., 2007;
Moy et al., 2008), and this limits our ability to
understand and predict future global climate changes
(Villalba, 1990; Bradley & Jones, 1993; Nesje &
Dahl, 2003).
Several different approaches have been used to try
to identify the LIA in the Southern Hemisphere.
Thompson et al. (1986) used the Quelccaya ice core
(Southern Peru) to place the LIA between ca. AD
1530 and 1900, with a peak around AD 1800–1820.
Villalba (1994) relied on tree-ring records from
Northern Patagonia to identify two cold periods
(AD 900–1070 and 1270–1660) either side of a warm
period (AD 1080–1250) that was coincident with the
MWP.
Cioccale (1999) used documentary data to deter-
mine that the LIA in central Argentina comprised two
cold pulses interrupted by an intermediate period.
The first pulse lasted from the beginning of the
fifteenth to the end of the sixteenth century, and the
second and most important pulse from the early
eighteenth until the start of the nineteenth century.
Kreutz et al. (1997) found evidence of a synchronous
onset of the LIA when comparing results from studies
of West Antarctic ice cores with the Greenland Ice
Sheet Project Two (GISP2).
In Chile, Lamy et al. (2001) used humidity proxies
in marine sediments to show the existence of the
MWP and LIA in southern Chile (41�S). Bertrand
et al. (2005) studied sedimentological proxies in Lake
Puyehue (40�S) to reveal wet climate conditions
between AD 1490 and 1700; they associated these
with the onset of the European LIA. Araneda et al.
(2007) analyzed historical data and found evidence of
a cold period lasting from AD 1766 to 1898 that
peaked between 1857 and 1871; this would have been
the last pulse of the LIA in Northern Patagonia.
However, these authors did not record a warm event
corresponding to the MWP. Recently, Moy et al.
(2008) used stable isotopes to identify two periods of
208 Hydrobiologia (2010) 648:207–225
123

enhanced evaporation (900–550 and 400–50 cal yr
BP), the first one coincident with the MWP and the
second broadly agreeing with the timing of the LIA.
As indicated above, most information about the
occurrence of LIA and MWP in Chile stems from the
southern part of the country, with relatively few
studies undertaken in central Chile. Furthermore,
most of these studies relied on a single proxy.
Accordingly, the aim of this research was to assess
the occurrence, timing, and expression of the LIA and
MWP in the Andes of central Chile using a multipr-
oxy approach including diatoms, pollen, chironomids,
and chemical parameters in the sediment record of
Lake Laja.
Materials and methods
Study area
Lake Laja (36�540S; 71�050W; Fig. 1) is located at
1360 m a.s.l. in the Chilean Andes. It has a maximum
depth of 135 m, a surface area of 87 km2, an approx-
imate length of 35 km from north (Punta Chillan) to
south (El Pino Bay), and a maximum width of 7 km in
the area of Antuco Volcano. The lake was created by a
volcanic avalanche that dammed the natural valley of
the Laja River around 9700 ± 600 yr BP (Thiele et al.,
1998). The average annual precipitation is 2172 mm,
occurring mostly between May and August (fall-
winter) and falling mainly as snow. The driest months
are January, February, and March (summer). The
lowest temperatures occur in June (on average,
-0.3�C), and the highest in January (on average,
13.6�C, Torres et al., 2008).
Current vegetation in the Lake Laja basin is
composed of Austrocedrus chilensis (D. Don) Pic.
Ser. et Bizz, Lomatia hirsuta (Lam.) Diels ex Macbr,
Maytenus boaria Mol., Schinus polygamus (Cav.)
Cabrera, Colletia ulicina Gill. et Hook., Aristotelia
chilensis (Mol.) Stuntz, Orites myrtoidea (Poepp. and
Endl.) Benth. and Hooker, and Ephedra chilensis K.
Presl (Donoso, 1982; Hoffmann, 1982; Veblen &
Schlegel, 1982; Torres et al., 2008).
Sampling and sedimentological analyses
Before selecting Lake Laja as the sampling site,
bathymetric and seismological studies were
performed using a 300 J sparker equipment from
the Renard Center of Marine Geology, University of
Ghent, Belgium. This allowed us to select an
appropriate site for sediment coring. A 522-cm core
was taken from the deepest part of the lake (Fig. 1)
using an Uwitec platform, but only the section
covering the last 2000 years was analyzed (402 cm).
Grain size measurements, performed on bulk
sediment using a laser diffraction particle analyzer
Malvern Mastersizer 2000, detected a range between
0.02 and 2000 lm. Samples were introduced into a
100-ml deionized water tank free of additive disper-
sants, mixed with a 2000 rpm stirrer, and crumbled
with ultrasonic waves. The grain size was classified
according to Wentworth (1922). Organic matter and
carbonates were determined by loss on ignition. For
this, approximately 1 g of sediment was dried for
24 h at 105�C and then was weighed after heating to
550�C to determine carbon content (Boyle, 2002).
After this, the samples were heated to 1000�C for 2 h
and then weighed again in order to obtain the
carbonates present in the sediment.
Biogenic silica was determined in two stages.
First, the organic matter was eliminated and the
compound was extracted from the sediment matrix
Fig. 1 Study area showing the core sampling site and the
bathymetric map of Lake Laja
Hydrobiologia (2010) 648:207–225 209
123

with chlorhydric acid 1 N, hydrogen peroxide 10%,
and sodium carbonate 2 M. Then, the dissolved silica
was quantified according to the modified Mortlock &
Froelich (1989) molybdenum blue method, determin-
ing absorbance with molecular spectrophotometry at
810 nm wavelength.
Samples for determination of total phosphorous
were prepared with sulfuric acid 30% and potassium
peroxydisulfate solution. In this determination, PO4
was quantified with the colorimetric molybdenum
blue method (Jackson, 1964) using a molecular
absorption spectrophotometer at 890 nm wavelength.
Geochronology
The first 26 cm of the sediment core was analyzed for210Pb and 137Cs activity; ages were determined using
the Constant Rate of Supply model (CRS; Appleby &
Oldfield, 1978). In order to date the entire sediment
sequence from Lake Laja, four samples were ana-
lyzed by AMS radiocarbon dating and the calibrated
ages were obtained using the program CALIB 5.0.2
(McCormac et al., 2004). A detailed description of
the dating models applied to 210Pb and AMS dates
can be found in Torres et al. (2008).
Biological proxies
Diatoms
Diatoms were analyzed according to Battarbee
(1986), 0.1 g of dry sediment was oxidised with
H2O2 and permanent slides were mounted with Hyrax
resin (I.R. = 1.7). The diatom concentration was
estimated by adding a microsphere solution to the
samples (Battarbee & Kneen, 1982), and about 500–
600 diatom valves were counted and identified for
each sample using standard taxonomies (Rivera,
1970, 1974; Rivera et al., 1973, 1982; Krammer &
Lange-Bertalot, 1991–2000).
Chironomids
The chironomid analysis consisted of deflocculating
4 ml of wet sediment in KOH 10% for 15 min at
70�C and passing it through a 90-lm sieve. The
remains were then transferred to a Bogorov counting
tray where head capsules were picked out with
entomological forceps. Each head capsule was
dehydrated in 80 and 100% ethanol and then mounted
in Euparal, ventral side up. Chironomids were
identified using a Zeiss microscope at magnifications
of 25, 40, or 1009, and the keys of Hofmann (1971),
Rieradevall & Brooks (2001), Wiederholm (1983),
Epler (2001), and Paggi (2001).
Pollen
The pollen analysis was performed following Erdt-
man (1960). The sediment samples were processed
using concentrated HF and acetolysis. Permanent
samples were mounted in Gelatin–Glycerin, and 350
pollen grains were counted for each centimeter.
Pollen was identified following Marticorena (1968)
and Heusser (1971). The results are shown in
percentage diagrams; the terrestrial pollen estimate
includes arboreal and non-arboreal taxa, excluding
fern spores.
Statistical methods
To distinguish different associations along the profile,
a stratigraphically constrained sum-of-squares cluster
analysis (CONISS) was applied to the percentage
values of the different proxies using Tilia and Tilia
Graph (Grimm, 1987) programs. Diagrams were also
constructed using Tilia, Tilia Graph (Grimm, 1991).
A detrended correspondence analysis (DCA) was also
applied to the chironomid, pollen, and diatom
assemblages in order to assess the compositional
structure and taxa turnover throughout the profile
(Birks, 1998; Langdon et al., 2004) with the program
R and the vegan and rioja packages (Juggins, 2009;
Oksanen et al., 2005).
Results
Geochronology
The activity of 210Pb in the upper section of Lake
Laja was discussed thoroughly in Quiroz et al.
(2005), but, in general terms, the activity of unsup-
ported 210Pb shows important fluctuations along the
sediment profile. It was low in the upper part, had an
important peak around 5 cm, and then generally
tended to decline toward the deepest layers of the
sediment. Despite these important fluctuations, it was
210 Hydrobiologia (2010) 648:207–225
123
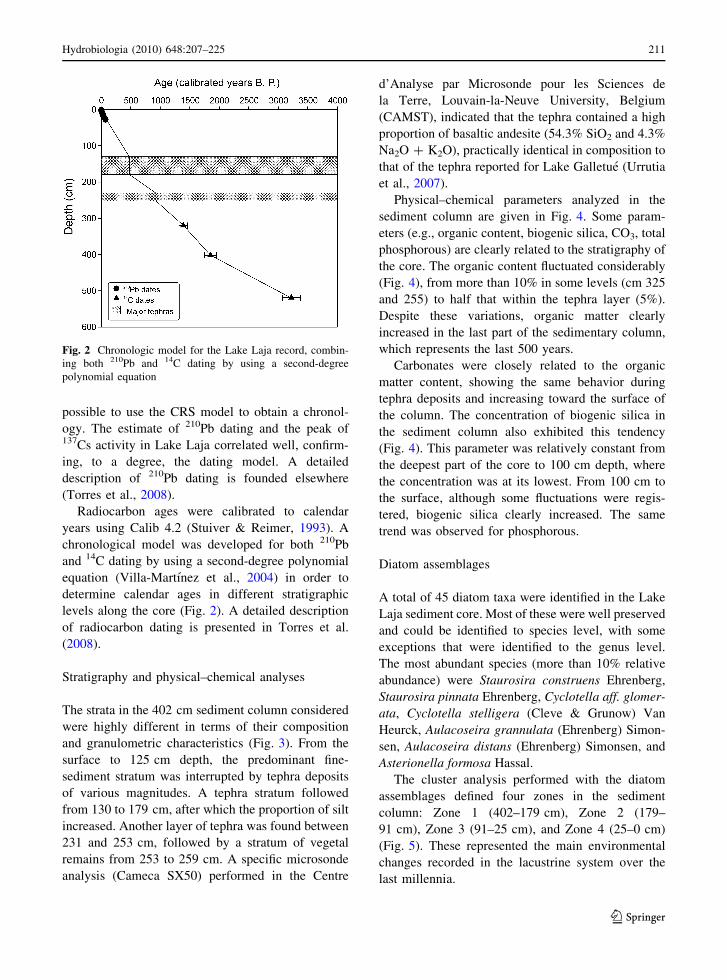
possible to use the CRS model to obtain a chronol-
ogy. The estimate of 210Pb dating and the peak of137Cs activity in Lake Laja correlated well, confirm-
ing, to a degree, the dating model. A detailed
description of 210Pb dating is founded elsewhere
(Torres et al., 2008).
Radiocarbon ages were calibrated to calendar
years using Calib 4.2 (Stuiver & Reimer, 1993). A
chronological model was developed for both 210Pb
and 14C dating by using a second-degree polynomial
equation (Villa-Martınez et al., 2004) in order to
determine calendar ages in different stratigraphic
levels along the core (Fig. 2). A detailed description
of radiocarbon dating is presented in Torres et al.
(2008).
Stratigraphy and physical–chemical analyses
The strata in the 402 cm sediment column considered
were highly different in terms of their composition
and granulometric characteristics (Fig. 3). From the
surface to 125 cm depth, the predominant fine-
sediment stratum was interrupted by tephra deposits
of various magnitudes. A tephra stratum followed
from 130 to 179 cm, after which the proportion of silt
increased. Another layer of tephra was found between
231 and 253 cm, followed by a stratum of vegetal
remains from 253 to 259 cm. A specific microsonde
analysis (Cameca SX50) performed in the Centre
d’Analyse par Microsonde pour les Sciences de
la Terre, Louvain-la-Neuve University, Belgium
(CAMST), indicated that the tephra contained a high
proportion of basaltic andesite (54.3% SiO2 and 4.3%
Na2O ? K2O), practically identical in composition to
that of the tephra reported for Lake Galletue (Urrutia
et al., 2007).
Physical–chemical parameters analyzed in the
sediment column are given in Fig. 4. Some param-
eters (e.g., organic content, biogenic silica, CO3, total
phosphorous) are clearly related to the stratigraphy of
the core. The organic content fluctuated considerably
(Fig. 4), from more than 10% in some levels (cm 325
and 255) to half that within the tephra layer (5%).
Despite these variations, organic matter clearly
increased in the last part of the sedimentary column,
which represents the last 500 years.
Carbonates were closely related to the organic
matter content, showing the same behavior during
tephra deposits and increasing toward the surface of
the column. The concentration of biogenic silica in
the sediment column also exhibited this tendency
(Fig. 4). This parameter was relatively constant from
the deepest part of the core to 100 cm depth, where
the concentration was at its lowest. From 100 cm to
the surface, although some fluctuations were regis-
tered, biogenic silica clearly increased. The same
trend was observed for phosphorous.
Diatom assemblages
A total of 45 diatom taxa were identified in the Lake
Laja sediment core. Most of these were well preserved
and could be identified to species level, with some
exceptions that were identified to the genus level.
The most abundant species (more than 10% relative
abundance) were Staurosira construens Ehrenberg,
Staurosira pinnata Ehrenberg, Cyclotella aff. glomer-
ata, Cyclotella stelligera (Cleve & Grunow) Van
Heurck, Aulacoseira grannulata (Ehrenberg) Simon-
sen, Aulacoseira distans (Ehrenberg) Simonsen, and
Asterionella formosa Hassal.
The cluster analysis performed with the diatom
assemblages defined four zones in the sediment
column: Zone 1 (402–179 cm), Zone 2 (179–
91 cm), Zone 3 (91–25 cm), and Zone 4 (25–0 cm)
(Fig. 5). These represented the main environmental
changes recorded in the lacustrine system over the
last millennia.
Fig. 2 Chronologic model for the Lake Laja record, combin-
ing both 210Pb and 14C dating by using a second-degree
polynomial equation
Hydrobiologia (2010) 648:207–225 211
123

Zone D-I (402–179 cm)
The diatom assemblages are dominated by Staurosira
pinnata, S. construens, and C. stelligera (Fig. 5), with
maximum abundances of 39, 33, and 30%, respec-
tively. Diatom valves were absent in the sediment
between 253 and 231 and 299–296 cm; these strata
correspond to tephra deposits. Diatom concentrations
fluctuate between 3.13 9 105 and 1.72 9 106 valves
per gram of dry sediment.
Zone D-II (175–91 cm)
This zone is dominated by the same species as D-I
and a thick layer of tephra lies between 179 and
130 cm. The total diatom concentration increases
immediately after the tephra layer and peaked at the
end of this zone. The diatom concentration in this
section ranges from 3.16 9 105 to 4.42 9 106 valves
per gram of dry sediment.
Zone D-III (91–25 cm)
The diatom composition changes significantly within
this zone, which is dominated by the centric species
C. stelligera and C. aff. glomerata, with maximum
abundances of 60% and 50%, respectively (Fig. 5).
The abundance of the Staurosira genera declines
drastically, and Aulacoseira distans appears in the
sediment record for the first time, reaching a max-
imum abundance of 11.6%. The total diatom con-
centration continues to increase, fluctuating between
1.32 9 106 and 5.83 9 106 valves per gram of dry
sediment.
Fig. 3 Stratigraphy and grain size (Phi) of the sedimentary column
212 Hydrobiologia (2010) 648:207–225
123

Zone D-IV (25–0 cm)
In this zone diatom assemblages change significantly.
First, Aulacoseira distans (80.6%) and then Asterio-
nella formosa (85.1%) displace the dominant species
from Zone D-III. Abundances of A. granulata
(23.9%) remain high (Fig. 5). C. aff. glomerata and
C. stelligera abundances decrease sharply, and the
former returns to the abundance levels found in zones
D-I and D-II. Synedra species (S. rumpens (Kutzing)
Carlson, S. ulna (Nitzsch) Lange-Bertalot y S. ulna
var. acus (Kutzing) Lange-Bertalot), initially very
scarce, increases during this period. Diatom concen-
trations which peaked at 8.36 9 106 valves per gram
of dry sediment at the beginning of this zone start to
decrease from 15 cm upwards and reached
9.06 9 105 valves per gram of dry sediment at the
surface.
Fig. 4 Physical–chemical
characterization of the
sediment column. The
major sand layers are also
shown (light-grey color)
Fig. 5 Diatom stratigraphy of Lake Laja showing the dominant taxa, abundances (%), total concentration, and biogenic silica in the
different zones identified by the cluster analysis
Hydrobiologia (2010) 648:207–225 213
123

Chironomid assemblages
A total of 5599 chironomid head capsules from Lake
Laja were sorted into 30 different taxa. The most
abundant subfamily was Chironominae (54.2%),
composed of Tanytarsini (30%) and Chironomini
(24%), followed by Orthocladiinae (23.4%), Tany-
podinae (19.7%), and finally Podonominae and
Diamesinae (both at 1.4%); unidentified remains
made up 4% of the total. Two morphotypes of the
Tanytarsini tribe were identified according to the
shape of their antennal pedestal (types A and B).
Type A was defined by a pedestal with a short,
rounded spur, whereas type B presented a more
pronounced spur, generally at an obtuse angle with
respect to the antennal pedestal. Figure 6 shows the
chironomid assemblages of Lake Laja. The CONISS
analysis identified four zones in the assemblage: Zone
C-I (402–248 cm), Zone C-II (247–180 cm), Zone C-
III (180–98 cm), and Zone C-IV (98–0 cm).
Zone C-I (402–175 cm)
Ablabesmyia abundances were high during this
period, especially toward the beginning of the zone,
exceeding 20% total abundance before declining
toward the end of the zone. Cricotopus/Orthocladius
and Dicrotendipes taxa were very abundant exceed-
ing 18% of the total in some layers but the
Tanytarsini as a group were the most abundant in
this zone. Similarly, Podonominae abundances are
also high in this zone, and abundances of Limno-
phyes/Compterosmittia, Macropelopia, Parapsectro-
cladius, and Tanytarsina type B were important. The
number of head capsules per gram of wet sediment
remained around five.
Zone C-II (175–98 cm)
At the beginning of this zone Ablablesmyia is absent
but it increases toward the upper part of this zone.
Other taxa abundant in the previous zone such as
Cricotopus/Orthocladius and Diamesinae, decrease
to zero abundance. Toward the end of this zone other
taxa also recover (e.g., Parapsectrocladius, Tanytar-
sini group). This zone is characterized by the thickest
tephra layer of the all profile.
Zone C-III (98–20 cm)
In this zone there is a marked change in comparison
with to the previous zone (C-II), with an increase in
taxa that previously had very low abundances such as
Macropelopia and Parachironomus, which exhibit
their highest abundances throughout the profile within
this zone. Other taxa that also increase in abundance
(but with a lower magnitude) are Tribelos/Phaenop-
sectra, Podonominae, Polypedilum, Limnophyes,
Djalmabista. Clearly the increase in the abundance
of these taxa could be related to the re-establishment
of previous lake conditions after the deposition of the
tephra layer together with a possible increase in
nutrient inputs. Other taxa record a decrease in
abundance compared to the previous zone (Tanytar-
sina type B, Parapsectrocladius, Ablabesmyia).
Zone C-IV (20–0 cm)
In this zone there are important changes in taxa that
previously exhibited very high abundances. Macro-
pelopia and Parachironomus which had abundances
of almost 60% in the zone C-III both decreased until
absent within this zone. Other taxa decreasing to zero
abundances include Monodiamesa, Limnophyes/
Compterosmittia, Parapsectrocladius, and Djalmaba-
tista. The top of the core shows important increases in
Ablabesmyia, Apedilum, Cricotopus/Orthocladius,
and Tribelos/Phaenopsectra, the latter reaching its
highest abundance.
Pollen
The pollen assemblages were divided into four zones
(Fig. 7). Figure 7 shows the total lack of arboreal
(AP) and non-arboreal pollen (NAP) in some strata.
Those sections probably correspond to sediments that
arrived suddenly at the lake bottom (landslides/tephra
falls) triggered by seismic or volcanic events, which
would explain the absence of pollen.
Zone P-I (402–120 cm)
In this zone, the assemblages were characterized by
significant proportions of AP (mean = 60.7%), with
the most important taxa being N. dombeyi-
type (mean = 36.3%), N. obliqua-type (mean =
17.6%), and E. chilensis (mean = 21.7%). Poaceae
214 Hydrobiologia (2010) 648:207–225
123

and Austrocedrus chilensis are also present but at low
abundances. Within this zone Caryophyllaceae
reached its highest abundance throughout the profile.
In the upper part of the zone the tephra layer is
clearly distinguishable and is characterized by the
complete absence of pollen. The total pollen concen-
tration fluctuated between 93,285 pollen grains g-1 at
340 cm and 22,875 pollen grains g-1 at 234 cm.
Zone P-II (120–63 cm)
This zone was dominated by N. dombeyi-type
(mean = 37.3%), E. chilensis (mean = 23.1%), and
N. obliqua-type (mean = 12.1%). The assemblage
was dominated by AP (mean = 55.3%). This period
was characterized by a substantial rise in Poaceae
(12.9%) and a decline in E. chilensis in the upper part
of the zone. The total pollen concentration ranged
between 137,402 pollen grains g-1 (101 cm) and
17,560 pollen grains g-1 (77 cm).
Zone P-III (63–41 cm)
This zone was characterized by significant proportions
of AP (mean = 50.8%). N. dombeyi-type decreased
(mean = 32.8%) and N. obliqua-type averaged 5%.
The Poaceae increased to 20%, whereas E. chilensis
decreased to 18.2%. The arboreal taxa Austrocedrus
chilensis exhibited an increase toward the upper part of
the zone. The highest total pollen concentration value
was 39,314 pollen grains g-1 (53 cm) and the lowest
was 7,554 pollen grains g-1 (43 cm).
Zone P-IV (41–0 cm)
This zone was dominated by AP (mean = 57.3%),
specifically N. dombeyi-type (mean = 35%), N. ob-
liqua-type (mean = 12.5%), and A. chilensis
(mean = 9.4%). Poacaea and E. chilensis declined
subsequently in the upper part of the zone. Total
pollen concentration values increased with respect to
the previous zone, reaching a maximum of 184,770
pollen grains g-1 at 20 cm and a minimum of 8,347
pollen grains g-1 at 2 cm.
Discussion
Diatoms
Both diatom assemblages and the chemical parame-
ters of Lake Laja have shown important variations
Fig. 6 Chironomid stratigraphy from Lake Laja showing the abundances (%) of dominant taxa across zones identified by cluster
analysis. Te timing of the LIA is shown between dashed lines and major sand layers in light-grey
Hydrobiologia (2010) 648:207–225 215
123

over the last 2000 years. These changes may be
related to different anthropogenic and natural
forcings.
The elevated abundances of Aulacoseira distans
and Aulacoseira granulata in the upper part of the
sediment record reflect increased lake productivity in
recent decades as evidenced by the increase in S. ulna
and S. rumpens. Bracco et al. (2005) suggest that A.
distans and A. granulata appear when the trophic
condition of a lake increases (e.g., increased organic
matter or phosphorous). Furthermore, the species
belonging to this genus are considered to be oppor-
tunistic (Willen, 1991), which accounts for their
higher abundances in recent times, where important
reductions in the lake level have occurred (Mardones
& Vargas, 2005; Cruces et al., 2006), which in turn
increases the nutrient concentration. The occurrence
and increase of A. formosa and Stephanodiscus sp. in
Zone IV also indicate elevated nutrients in the lake
(e.g., phosphorous, silica), as both species have been
recorded in environments with medium to high
nutrient levels (Anderson, 1990; Alefs & Muller,
1999; Clerk et al., 2000; Merilainen et al., 2000;
Bradbury et al., 2004).
Together, the mineralogical characteristics of the
sediment and the diatom assemblages indicate that
three strata (130–179, 231–253, 296–299) with pre-
dominantly minerogenic environments were depos-
ited in a short time. These sediments would have
originated from the volcanic activity of the Antuco-
Sierra Velluda complex located to the south west of
the lake. Even thought Cruces et al. (2006) indicate
that at least 11 volcanic events have been historically
recognized for this complex over the last 400 years,
not all of these were recorded in the sediment,
probably due to variations in the wind direction at the
time of each eruption. Hence in the sediment of Lake
Laja we can identify seven volcanic events with the
thickest below the 130 cm.
The low concentrations of organic matter, carbon-
ates, and biogenic silica in the tephra layers of the
profile result from the dilution generated by the
sudden input of volcanic material. The peak in organic
content (253–259 cm) was generated by the vegeta-
tion remains in this stratum from the watershed.
The volcanic events that affected Lake Laja are
likely to have affected the diatom assemblages.
According to different authors (Hickman & Schweger,
1991; Hickman & Reasoner, 1998) both Staurosira
construens and Staurosira pinnata have been cited as
the first colonizers following strong perturbations to
lakes but are subject to changes in abundances after
volcanic events. However, unlike the findings of
Cruces et al. (2006) at Lake Galletue, the occurrence
of these both species in Lake Laja seems to be
unaffected by the volcanic eruption. Rather, the high
abundances of these benthic species in zones I and II
suggests lower nutrient levels in the water column, as
Fig. 7 Pollen percentage diagram. AP arboreal pollen, NAP non-arboreal pollen. LIA temporal window is shown by the area of
dashed lines, and major sand layers in light-grey
216 Hydrobiologia (2010) 648:207–225
123

indicated by Brugam et al. (1998) for Lake Michigan.
This idea is backed by the low phosphorous concen-
trations found in these two zones.
Climate fluctuations may also have been associ-
ated with more abundant Staurosira species. Previous
studies have indicated that such species could reflect
more humid, colder conditions (Smol, 1988; Laing
et al., 1999). However, the strong volcanic influence
on Lake Laja hampered our ability to disentangle the
response of the diatoms to climate fluctuations from
their response to other forcings (e.g., volcanic and
anthropogenic activity). Furthermore, the efficacy of
diatoms for reconstructing past temperatures is con-
founded by changes in pH or nutrients (Wolin &
Duthie, 1999). Even though volcanic eruptions can
cause pH changes in the lake due to sulfur deposition
(Birks & Lotter, 1994), in the case of Lake Laja,
diatom assemblages did not show any effect. Unfor-
tunately, no calibration data set has been developed
for pH reconstruction in Chile, and it is not possible
to infer any change in this variable. Rather, diatoms
seem to be more sensitive to nutrient fluctuations.
Changes in assemblages within Zone III resulted
mainly from changes in the water body itself,
probably due to heightened nutrient concentrations
triggered by a reduction in the lake level. This
generated a shift from the previously dominant diatom
taxa to Cyclotella, which reached its maximum
abundances in this zone. According to some studies
(Gasse et al., 1989; Marciniak, 1990; Owen et al.,
1982), changes in the planktonic/non-planktonic ratio
can be used to indicate water level fluctuations.
Benthic and epiphytic diatoms increase when lake
levels are low whereas planktonic species are more
numerous at high water levels. However, depending
on the lake bathymetry, it is possible to find an inverse
relationship between depth and planktonic species
abundance (Wolin & Duthie, 1999). In lakes with
steeply shelving shores such as Lake Laja (Melnick
et al., 2006), a drop in lake level reduces the coastal
zones and, thereby, the representation of taxa from
this type of environment (e.g., Staurosira), at the same
time augmenting planktonic species.
Finally, in the upper part of the sediment record
(Zone IV), the increase in benthic-littoral species
coincided with a decline in the abundance of
planktonic taxa. These changes are the result of
recent hydrological changes in the lake, which is now
used to generate hydroelectric power (Mardones &
Vargas, 2005). In this same period diatom species
compositions suggest a rise in lake productivity (e.g.,
A. formosa, A. granulata, and Stephanodiscus sp.), as
reflected by the increase of organic matter, phospho-
rous and biogenic silica.
Chironomids
The Lake Laja sediment record revealed a 2000-year
sequence of environmental changes that are reflected
in the chironomid assemblages. CONISS analyses
produced four zones determined largely by Tanytar-
sina A and B, Cricotopus/Orthocladius, Parachiron-
omus, Parapsectrocladius, Macropelopia, and
Ablabesmyia.
In Chile, little information is available on the
ecology of these groups (e.g., Brundin, 1966; Arenas,
1995; Andersen, 1996), reducing the utility of these
organisms for reconstructing past environmental
changes. Nevertheless, some researchers have made
paleoenvironmental inferences by transferring the
ecological typology of holarctic chironomids to local
representatives of the same genera (Verschuren &
Eggermont, 2006).
The predominance of Parachironomus in the most
recent part of the record suggests a higher trophic
status of the lake for that period. According to Lotter
et al. (1998), Parachironomus is very abundant in
eutrophic and hypereutrophic lakes in the Alps.
Toward the end of the record here, levels of
Ablabesmyia, Cricotopus/Ortocladius, and Tribelos/
Phaenopsectra are also greater. Rae (1989) found
higher proportions of Ablabesmyia and Tribelos in
fluvial environments with elevated organic loads in
Ohio (USA), whereas Brooks et al. (2001) deter-
mined a slight increase of Cricotopus toward the end
of the Betton Pool (England) record, which was
congruent with an increase of total phosphorous in
the same record. This information, combined with the
chemical parameters of the sediment (e.g., organic
matter, carbonates, and total phosphorous all increas-
ing toward the end of the core), suggest that Lake
Laja has been more productive during the last
*450 years, and particularly over the last
*80 years.
All previous climate reconstruction studies under-
taken in Chile using chironomids as a paleoclimate
proxy have focused on the Late-glacial period
(Massaferro & Brooks, 2002; Massaferro et al.,
Hydrobiologia (2010) 648:207–225 217
123

2005, 2009). Although some records extend to the
present, they lack the temporal resolution required
to analyze changes in the last millennium such as
the LIA. During the LIA, lower temperatures and
glacier advances were registered across the world
(McDermott et al., 2001; Esper et al., 2002; Soon
et al., 2003; Rabatel et al., 2005; Polissar et al., 2006)
and, despite some differences in timing, it is gener-
ally accepted that colder conditions predominated
between 1550 and 1850 in the North Atlantic region
(Jones & Bradley, 1992; Bradley, 2000).
Parachironomus and Macropelopia, which were
previously related to an increase in the lake’s trophic
status, were very abundant in Lake Laja during the
LIA. In the same period, Parapsectrocladius, Podo-
nominae, Limnophyes/Compterosmittia and to a
lesser degree, Parakiefferiella and Monodiamesa also
increased slightly. Brundin (1966), Massaferro &
Brooks (2002), and Massaferro et al. (2005) indicated
that the presence of Podonominae is associated with
cold, glacier-fed waters whereas Massaferro & Corley
(1998) and Porinchu & Cwynar (2002) noted that
Parakiefferiella is related to cold, well-oxygenated
waters. High Limnophyes abundances, associated with
a higher proportion of littoral zone, were indicative of
water level fluctuations (Hofmann, 1998). Hence, the
occurrence of this taxon within the LIA allows us to
infer colder and probably drier conditions. This
conclusion was further reinforced by the occurrence
of taxa typical of more productive conditions, since
nutrient concentrations can increase in a lake with a
lower volume, thereby making it more productive
(Massaferro & Brooks, 2002).
Although chironomids are a useful tool for recon-
structing past climate changes in the Late-glacial,
their utility as a proxy for climate changes during the
Holocene is less clear as the magnitude of compo-
sitional changes in the Holocene are not as great.
However, some studies have shown that chironomids
can also record low-amplitude climate changes.
Hence, Brooks & Birks (2001) used a chironomid-
based temperature reconstruction to indicate a cool-
ing period of about 1.5�C corresponding with to the
LIA in Scotland. Brooks & Birks (2004) also reported
evidence of environmental changes in Norway that
could be associated with the LIA. Likewise, Velle
et al. (2005) found increased abundances of typical
cold-environment taxa such as Heterotrissocladius
and Sergentia in a Holocene sediment sequence from
south central Norway, providing some evidence of
the LIA in that area.
Pollen
The palynological records from Lake Laja show
important changes during the Late Holocene. Accord-
ing to the pollen record, N. dombeyi-type, N. obliqua-
type, E. chilensis, and Poaceae were the most
important taxa over the last 2000 cal yr BP, suggest-
ing alternating drier and wetter conditions during this
period. The presence of Plantago and increasing
levels of Poaceae indicate an important human
influence during the first half of the twentieth century
(Fig. 7).
A drier period is suggested by increases of E.
chilensis, an arid-adapted species (Marticorena &
Rodrıguez, 1995; Ickert-Bond et al., 2003), around
660 years AD (Zone I). This dry period was followed
by more humid conditions between 660 AD and 1561
AD, as indicated by the rise of N. dombeyi-type and
decline of Poaceae and E. chilensis.
After 933 AD, when the MWP was evident in
Europe, the Lake Laja pollen assemblage implies the
prevalence of conditions more humid than seen at
present. However, the low pollen resolution for that
period impedes verifying the occurrence of such
humid conditions. The stratigraphy in this zone is
very diverse, possibly reflecting a greater influence of
runoff from the watershed, suggesting higher precip-
itation during this period. Unfortunately, little
research has been undertaken for this period in
central Chile, limiting potential comparisons. None-
theless, Lamy et al. (2001) reported declining rainfall
during the MWP in southern Chile and Haberzettl
et al. (2005) suggesting the same for southeastern
Patagonia.
The higher relative abundance of Poaceae and E.
chilensis (the latter at the beginning of Zone II)
between 1450 and 1766 AD and the pollen concen-
trations of Chenopodiaceae and Asteraceae subf.
asteroidae suggest the beginning of a drier period that
peaked from 1766 to 1894 AD (Zone III and the
beginning of zone IV), with a concomitant decrease
in N. dombeyi-type and N. obliqua-type.
The pollen of N. obliqua-type is associated with
humid conditions (Rodrıguez et al., 1983). When
taken together with the tree growth of N. pumilio in
the Lake Laja watershed (a species that forms part of
218 Hydrobiologia (2010) 648:207–225
123

N. dombeyi-type), this record shows a positive
correlation with annual precipitation under a Medi-
terranean-type climate where water availability is a
major limiting factor (Lara et al., 2001, 2005).
The occurrence of Poaceae is normally associated
with human activity but could also be related to lower
precipitation (Bush, 2000). Given the very low
human population in the area at that time, we believe
that Poaceae responded more to changes in precip-
itation than to anthropogenic factors. Therefore, and
according to previous authors, we think that lower
precipitation levels drove an increase in Poaceae
between 1766 and 1894 AD, while the peak between
1900 and 1950 could be associated with anthropo-
genic influence in Lake Laja. A detailed description
of the human influence in the study area is found
elsewhere (Torres et al., 2008).
This dry period may be associated with the
manifestation of the LIA in central Chile. This was
not a single event but rather was characterised by
humid periods. Evidence of this comes from the high
prevalence of Ephedra chilensis, an arid-adapted
species (Marticorena & Rodrıguez, 1995; Ickert-
Bond et al., 2003), interspersed with a few peaks of
N. dombeyi-type (which according the current vege-
tation, would correspond to N. pumilio), an indicator
species of higher precipitation (Lara et al., 2001,
2005).
According to Jenny et al. (2002), the LIA in
central Chile (32�–38�S) began with a humid period
(1300–1700 AD) and ended with a drier period
(1700–1850 AD). Bertrand et al. (2005), working at
Lake Puyehue (40�S), highlighted a humid period
(1490–1700 AD) followed by a drier period (1700–
1900 AD). Only Lamy et al. (2001) found a different
trend, with heightened rainfall during the LIA around
41�S. These findings are in rough agreement with the
drier conditions reported in our results for the LIA in
Lake Laja, which occurred between ca. 1550 and
1900 AD. As stated earlier, the LIA was probably not
a single homogenous event, but comprised different
cold pulses that were not necessarily synchronous
(Ciocale, 1999; Araneda et al., 2009).
The most important changes in Zone IV could be
related to human impacts. NAP, specifically from
Poaceae and E. chilensis, increased at the beginning
of this period and then declined in the upper part
of this zone (1968–2001 AD). N. dombeyi-type and
N. obliqua-type increased simultaneously, whereas
Plantago appeared in the core for the first time
(Fig. 7).
Between 1938 and 1968, the human impacts on the
Lake Laja basin expanded as a result of hydropower
plant construction, tourism, and recreational activities
(Nardini & Montoya, 1993). We think that the
increase in Poaceae from 1938 to 1968 resulted
mainly from intensified human activity. During these
years, E. chilensis was rare, and it is possible that the
species cannot survive in disturbed ecosystems
(Torres et al., 2008). Finally, the palynological
changes detected in the upper part of the core may
be a response to reduced anthropogenic intervention
related to the end of hydropower plant construction
(Nardini & Montoya, 1993) and the founding of
Parque Nacional Laguna del Laja in 1957.
Combined analysis
One advantage of multiproxy reconstructions is they
offer the possibility of determining inherent climate
variation (Battarbee, 2000; Larocque & Bigler,
2004). Moreover, a multiproxy approach decreases
the introduction of errors associated with the ‘‘over-
simplification’’ of relationships between environmen-
tal variables and organisms, which occurs with
reconstructions based on just one proxy (Ammann
et al., 2000; Birks et al., 2000; Bigler et al., 2002).
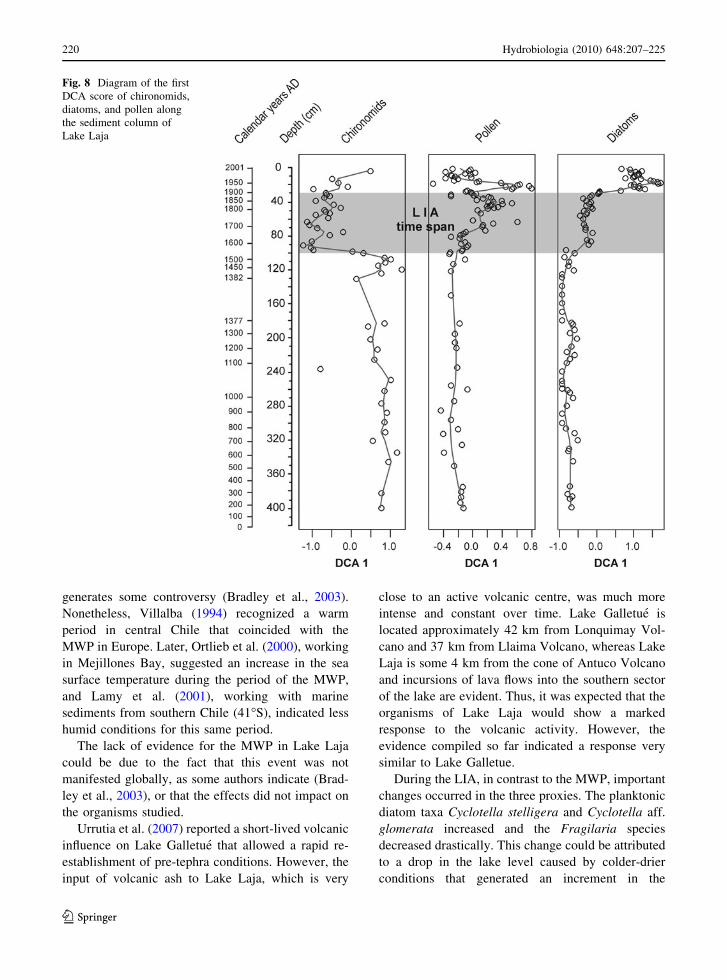
All three proxies underwent important changes, as
seen in the record of the last 2000 years from Lake
Laja. During the first thousand years of the record
where the volcanic sediments are more evident, the
three proxies do not show any major change as
reflected in the DCA scores; however, this should be
tested in a quantitative way in a further study (e.g.,
Lotter & Birks, 1993). Conversely, in the last
millennium significant changes occurred in the three
proxies (DCA scores) that coincide with the mani-
festation of LIA in Europe (Fig. 8), while in the last
century, important changes are recognized mostly in
diatoms, which reflects heightened anthropogenic
activity in the basin.
The first part of the record coincides with the
timing of the MWP (*800–1300 AD; Trouet et al.,
2009). However, no responses from the three proxies
were observed that might indicate climatic conditions
associated with the MWP. In Chile, few studies have
described the occurrence of the MWP, and the
existence of this period as a global phenomenon
Hydrobiologia (2010) 648:207–225 219
123
generates some controversy (Bradley et al., 2003).
Nonetheless, Villalba (1994) recognized a warm
period in central Chile that coincided with the
MWP in Europe. Later, Ortlieb et al. (2000), working
in Mejillones Bay, suggested an increase in the sea
surface temperature during the period of the MWP,
and Lamy et al. (2001), working with marine
sediments from southern Chile (41�S), indicated less
humid conditions for this same period.
The lack of evidence for the MWP in Lake Laja
could be due to the fact that this event was not
manifested globally, as some authors indicate (Brad-
ley et al., 2003), or that the effects did not impact on
the organisms studied.
Urrutia et al. (2007) reported a short-lived volcanic
influence on Lake Galletue that allowed a rapid re-
establishment of pre-tephra conditions. However, the
input of volcanic ash to Lake Laja, which is very
close to an active volcanic centre, was much more
intense and constant over time. Lake Galletue is
located approximately 42 km from Lonquimay Vol-
cano and 37 km from Llaima Volcano, whereas Lake
Laja is some 4 km from the cone of Antuco Volcano
and incursions of lava flows into the southern sector
of the lake are evident. Thus, it was expected that the
organisms of Lake Laja would show a marked
response to the volcanic activity. However, the
evidence compiled so far indicated a response very
similar to Lake Galletue.
During the LIA, in contrast to the MWP, important
changes occurred in the three proxies. The planktonic
diatom taxa Cyclotella stelligera and Cyclotella aff.
glomerata increased and the Fragilaria species
decreased drastically. This change could be attributed
to a drop in the lake level caused by colder-drier
conditions that generated an increment in the
Fig. 8 Diagram of the first
DCA score of chironomids,
diatoms, and pollen along
the sediment column of
Lake Laja
220 Hydrobiologia (2010) 648:207–225
123

planktonic species. Normally, a drop in the lake level
would cause these types of diatoms to decline in
abundance and benthic species to increase (Fritz,
1990; Moser et al. 1996; Caballero & Ortega, 1998;
Fritz et al., 1999; Telford et al., 1999). Such changes
have been reported at millennial-decadal scales,
inferring changes in lake levels when planktonic
communities are replaced by benthic and epiphytic
diatoms (Barker et al., 2002; Cumming et al., 2002;
Xue et al., 2003). But as was stated above, the
opposite response seems to had occurred lake Laja as
a result of the morphometry of the lake basin.
Chironomids also showed important changes dur-
ing the LIA, including increased abundances of
Macropelopia and Parachironomus taxa, the latter
being an indicator of greater trophic conditions.
Increased levels of Podonominae and Limnophyes
were also recorded, related to cold temperatures and
lower lake levels, respectively. These variations
denote a prevalence of colder, drier conditions over
the period of the LIA in Lake Laja. This is reflected
by the abundance of taxa which favor higher nutrient
levels, since lower lake levels tend to concentrate
nutrients.
The occurrence of cold, dry conditions was also
corroborated by the pollen record. More pollen was
found from Poaceae, E. chilensis, Chenopodiaceae,
and Asteraceae subf. Asteroidae, all taxa that indicate
dry conditions. At the same time, N. dombeyi-type
and N. obliqua-type decreased; according to Lara
et al. (2001, 2005), these taxa flourish under humid
conditions, again supporting the likelihood that the
lake was characterised by cold-dry conditions during
this period.
Finally, the effect of anthropogenic activity on the
lacustrine ecosystem and its watershed was observed
in the upper layers of the sediment column. Changes
recorded in the aquatic communities (diatoms and
chironomids) after the cold-dry condition (LIA
manifestation) were indicative of an increase in the
productivity of the lake associated with the hydro-
logical changes caused by the use of the lake for the
generation of hydroelectric energy (Cruces et al.,
2006). Human activity was also reflected in the pollen
profile in the form of increased NAP, specifically
Poaceae, and the appearance of Plantago in the first
strata along with a decline in the pollen of N.
dombeyi-type and E. chilensis. The latter species is
sensitive to anthropogenic intervention and has low
survival rates in heavily perturbed ecosystems (Tor-
res et al., 2008).
Conclusions
– The sedimentary record shows a clear volcanic
influence, most evident in the first 1000 years;
however, the organisms studied did not show a
marked response. Thus, the volcanic events are
likely to have had a more instantaneous rather
than long-term effects.
– No evidence of the occurrence of the MWP was
found, either because the event was not global, or
its magnitude was not sufficient regionally to
cause a change. Therefore, it is not possible to
prove or reject reliably the existence of this event
in the central Andes.
– The three proxies indicate the occurrence of a
cold, dry event in Lake Laja centered around
1550–1900 AD, coinciding roughly with the
occurrence of the LIA in Europe. Pollen and
chironomids were the most sensitive proxies to
this change; the diatom response was less direct
and associated more with changes in the lake
level and/or its trophic state.
– In the most recent part of the record, both diatoms
and chironomids indicate an increased trophic state
of the lake due to greater anthropogenic interven-
tion in the watershed. This is further reflected,
although to a lesser degree, in the pollen profile.
– Finally, we note the importance of using a
multiproxy approach to study past environmental
conditions (climate and others), since this dimin-
ished the bias associated with ‘‘oversimplifying’’
the relationships between the environmental
variables and the organisms.
Acknowledgments This research was funded by Fondecyt
projects No 1070508, No 1080294, and 11080158, and WBI
Wallonie-Chile cooperation project, FNRS, and ULg fundings.
References
Alefs, J. & J. Muller, 1999. Differences in the eutrophication
dynamics of Ammersee and Starnberger See (Southern
Germany), reflected by the diatom succession in varve-
dated sediments. Journal of Paleolimnology 21: 395–407.
Hydrobiologia (2010) 648:207–225 221
123

Ammann, B., H. Birks, S. Brooks, U. Eicher, U. von Grafen-
stein, W. Hofmann, G. Lemdahl, J. Schwander, K. To-
bolski & L. Wick, 2000. Quantification of biotic responses
to rapid climatic changes around the Younger Dryas – a
synthesis. Palaeogeography Palaeoclimatology Palaeoe-
cology 159: 313–347.
Andersen, T., 1996. A new species of Monodiamesa Kieffer,
1922 from Southern Chile (Diptera: Chironomidae: Pro-
diamesinae). Revista Chilena de Entomologıa 23: 43–49.
Anderson, N., 1990. The biostratigraphy and taxonomy of
small Stephanodiscus and Cyclostephanos species (Bac-
illariophyceae) in a eutrophic lake and their ecological
implications. British Phycological Journal 25: 217–235.
Appleby, P. G. & F. Oldfield, 1978. The calculation of lead-
210 dates assuming a constant rate of supply of unsup-
ported 210Pb to the sediment. Catena 5: 1–8.
Araneda, A., F. Torrejon, M. Aguayo, L. Torres, F. Cruces, M.
Cisternas & R. Urrutia, 2007. Historical record of San
Rafael glacier advances (North Patagonian Icefield):
another clue for Little Ice Age timing in Souther Chile.
The Holocene 17: 987–998.
Araneda, A., F. Torrejon, M. Aguayo, I. Alvial, C. Mendoza &
R. Urrutia, 2009. Historical records of Cipreses glacier
(34�S): combining documentary-inferred ‘Little Ice Age’
evidence from Southern and Central Chile. The Holocene
19: 1173–1183.
Arenas, J., 1995. Composicion y distribucion del macrozoo-
bentos del curso principal del rıo Biobıo, Chile. Medio
Ambiente 12: 39–50.
Barker, P., R. Telford, F. Gasse & R. Thevenon, 2002. Late
Pleistocene and Holocene palaeohydrology of Lake Ru-
kwa, Tanzania, inferred from diatom analysis. Palaeoge-
ography, Palaeoclimatology, Palaeoecology 187: 295–
305.
Battarbee, R. W., 1986. Diatom analysis. In Berglund, B. E.
(ed.), Handbook of Holocene Palaeoecology and Pala-
eohydrology. Wiley, Chichester: 527–570.
Battarbee, R. W., 2000. Palaeolimnological approaches to
climate change, with special regard to the biological
record. Quaternary Science Reviews 19: 107–124.
Battarbee, R. & M. Kneen, 1982. The use of electronically
counted microspheres in absolute diatom analysis. Lim-
nology and Oceanography 27: 184–188.
Bertrand, S., X. Boes, J. Castiaux, F. Charlet, R. Urrutia, C.
Espinoza, G. Lepoint, B. Charlier & N. Fagel, 2005.
Temporal evolution of sediment supply in Lago Puyehue
(Southern Chile) during the last 600 yr and its climatic
significance. Quaternary Research 64: 163–175.
Bigler, C., I. Larocque, S. Peglar, H. Birks & R. Hall, 2002.
Quantitative multiproxy assessment of long-term patterns
of Holocene environmental change from a small lake
near Abisko, northern Sweden. The Holocene 12: 481–
496.
Birks, H. J. B., 1998. Numerical tools in palaeolimnology –
progress, potentialities, and problems. Journal of Paleo-
limnology 20: 307–332.
Birks, H. J. B. & A. F. Lotter, 1994. The impact of the Laacher
See Volcano (11, 000 yr B.P.) on terrestrial vegetation
and diatoms. Journal of Paleolimnology 11: 313–322.
Birks, H. J. B., R. Battarbee & H. Birks, 2000. The develop-
ment of the aquatic ecosystem at Krakenes Lake, western
Norway, during the late glacial and early Holocene – a
synthesis. Journal of Paleolimnology 23: 91–114.
Boyle, J. F., 2002. Mineralogical and geochemical indicator
techniques. In Last, W. & J. P. Smol (eds), Tracking
Environmental Change Using Lake Sediments. Springer-
Verlag, New York, USA: 83–141.
Bracco, R., H. Inda, L. del Puerto, C. Castineira, P. Sprech-
mann & F. Garcıa-Rodrıguez, 2005. Relationships
between Holocene sea-level variations, trophic develop-
ment, and climatic change in Negra Lagoon, Southern
Uruguay. Journal of Paleolimnology 33: 253–263.
Bradbury, J., S. Colman & R. Reynolds, 2004. The history of
recent limnological changes and human impact on Upper
Klamath Lake, Oregon. Journal of Paleolimnology 31:
151–165.
Bradley, R. S., 2000. Climate paradigms for the last millen-
nium. PAGES Newsletters 8: 2–3.
Bradley, R. & P. D. Jones, 1993. Little Ice Age summer
temperature variations: their nature and relevance to
recent global warming. The Holocene 3: 367–376.
Bradley, R., M. Hughes & H. Diaz, 2003. Climate in medieval
time. Science 302: 404–405.
Brooks, S. J. & H. J. B. Birks, 2001. Chironomid-inferred
temperatures from Lateglacial and Holocene sites in
north-west Europe: progress and problems. Quaternary
Science Reviews 20: 1723–1741.
Brooks, S. J. & H. J. B. Birks, 2004. The dynamics of Chiro-
nomidae (Insecta: Diptera) assemblages in response to
environmental change during the past 700 years on
Svalbard. Journal of Paleolimnology 31: 483–498.
Brooks, S. J., H. Bennion & H. J. B. Birks, 2001. Tracking lake
trophic history with a chironomid-total phosphorus
inference model. Freshwater Biology 46: 513–533.
Brugam, R. B., K. McKeever & L. Kolesa, 1998. A diatom-
inferred water depth reconstruction for an Upper Peninsula,
Michigan, lake. Journal of Paleolimnology 20: 267–276.
Brundin, L., 1966. Transarctic relationships and their signifi-
cance, as evidenced by chironomid midges, with a
monograph of the subfamilies Podonominae and Aph-
roteniinae and the austral Heptagyiae. Kung Svenska
Vetenskapsakademiens Handinglar 11: 1–472.
Bush, M., 2000. Deriving response matrices from Central
American modern pollen rain. Quaternary Research 54:
132–143.
Caballero, M. & B. Ortega, 1998. Lake levels since about
40000 years ago at Lake Chalco, near Mexico City.
Quaternary Research 50: 69–79.
Cioccale, M. A., 1999. Climatic fluctuations in the central
region of Argentina in the last 1000 years. QuaternaryInternational 62: 35–47.
Clerk, S., R. Hall, R. Quinlan & J. Smol, 2000. Quantitative
inferences of past hypolimnetic anoxia and nutrient levels
from a Canadian Precambrian Shield lake. Journal of
Paleolimnology 23: 319–336.
Crowley, T. J. & T. S. Lowery, 2000. How warm was the
medieval warm period? Ambio 29: 51–54.
Cruces, F., R. Urrutia, O. Parra, A. Araneda, H. Treutler, S.
Bertrand, N. Fagel, L. Torres, R. Barra & L. Chirinos,
2006. Changes in diatom assemblages in an Andean lake
in response to a recent volcanic event. Archiv fur Hy-
drobiologie 165: 23–35.
222 Hydrobiologia (2010) 648:207–225
123

Cumming, B., K. Laird, J. Bennett, J. Smol & A. Salomon,
2002. Persistent millennial-scale shifts in moisture
regimes in western Canada during the past six millennia.
Proceedings of the National Academy of Sciences of the
United States of America 25: 16117–16121.
Donoso, C., 1982. Resena ecologica de los bosques mediter-
raneos de Chile. Bosque 4: 117–146.
Epler, J. H., 2001. Identification Manual for the larval Chiro-
nomidae (Diptera) of North and South Carolina. A Guide
to the Taxonomy of the Midges of the Southeastern
United States, Including Florida. North Carolina Depart-
ment of Environment and Natural Resources, Florida: 526
pp.
Erdtman, G., 1960. The acetolysis method. A revised
description. Svensk Botanisk Tidskrift 54: 561–564.
Esper, J., E. Cook & F. H. Schweingruber, 2002. Low-fre-
quency signals in long tree-ring chronologies for recon-
structing past temperature variability. Science 295: 2250–
2253.
Esper, J., R. J. S. Wilson, D. C. Frank, A. Moberg, H. Wanner
& J. Luterbacher, 2005. Climate: past ranges and future
changes. Quaternary Science Reviews 24: 2164–2166.
Fritz, S., 1990. Twentieth-century salinity and water-level
fluctuations in Devil’s Lake, North Dakota, test of a dia-
tom-based transfer function. Limnology and Oceanogra-
phy 35: 1771–1781.
Fritz, S., B. Cumming, F. Gasse & K. Laird, 1999. Diatoms as
indicators of hydrologic and climatic change in saline
lakes. In Stoermer, E. F. & J. P. Smol (eds), The Diatoms:
Applications to the Environmental and Earth Sciences.
Cambridge University Press, Cambridge, MA: 41–72.
Gasse, F., V. Ledee, M. Massault & J.-C. Fontes, 1989. Water
level fluctuations of Lake Tanganika in phase with oce-
anic changes during the last glaciation and deglaciation.
Nature 342: 57–59.
Goosse, H., H. Renssen, A. Timmermann & R. Bradley, 2005.
Internal and forced climate variability during the last
millennium: a model-data comparison using ensemble
simulations. Quaternary Science Reviews 24: 1345–1360.
Grimm, E., 1987. CONISS: a Fortran 77 program for strati-
graphically constrained cluster analysis by the method of
incremental sum of squares. Computational Geosciences
13: 13–35.
Grimm, E., 1991. Tilia and Tilia Graph. Illinois State Museum,
Springfield.
Grove, J. M., 2001. The initiation of the ‘Little Ice Age’ in
regions around the North Atlantic. Climatic Change 48:
53–82.
Haberzettl, T., M. Fey, A. Lucke, N. Maidana, C. Mayr, C.
Ohlendorf, F. Schabitz, G. Schleser, M. Wille & B. Zo-
litschka, 2005. Climatically induced lake level changes
during the last two millennia as reflected in sediments of
Laguna Potrok Aike, southern Patagonia (Santa Cruz,
Argentina). Journal of Paleolimnology 33: 283–302.
Heusser, C., 1971. Pollen and Spores of Chile. Modern types of
the Pteridophytas, Gimnospermae and Angiospermae.
University of Arizona, Arizona, AZ.
Hickman, M. & M. Reasoner, 1998. Late quaternary diatom
responses to vegetation and climate change in a subalpine
lake in Banff National Park, Alberta. Journal of Paleo-
limnology 20: 253–265.
Hickman, M. & C. E. Schweger, 1991. A palaeoenvironmental
study of Fairfax Lake, a small lake situated in the Rocky
Mountain Foothills of west-central Alberta. Journal of
Paleolimnology 6: 1–15.
Hoffmann, A., 1982. Flora silvestre de Chile. Fundacion
Claudio Gay, Zona Araucana (Santiago de Chile).
Hofmann, W., 1971. Zur taxonomie und palokologie subfoss-
iler Chironomiden (Dipt.) in seesedimenten. Ergebnisse
der Limnologie 6: 1–50.
Hofmann, W., 1998. Cladocerans and chironomids as indica-
tors of lake level changes in north temperate lakes. Journal
of Paleolimnology 19: 55–62.
Huang, S. & H. N. Pollack, 1997. Late quaternary temperature
changes seen in world-wide continental heat flow mea-
surements. Geophysical Research Letters 24: 1947–1950.
Hughes, M. K. & H. F. Diaz, 1994. Was there a ‘‘Medieval
warm period’’ and if so where and when? Climatic
Change 26: 109–142.
Ickert-Bond, S., J. Skvarla & W. Chissoe, 2003. Pollen
dimorphism in Ephedra L. (Ephedraceae). Review of
Palaeobotany and Palynology 124: 325–334.
Jackson, M., 1964. Analisis quımico de suelos, Vol. 1. Edici-
ones Omega S.A., Barcelona, Espana: 367 pp.
Jenny, B., B. Valero-Garces, R. Villa-Martinez, R. Urrutia, M.
Geyh & H. Veit, 2002. Early to mid-Holocene aridity in
central Chile and the southern westerlies: the Laguna
Aculeo record (34�S). Quaternary Research 58: 160–170.
Jones, P. D. & R. S. Bradley, 1992. Climatic variations over the
last 500 years. In Bradley, R. S. & P. D. Jones (eds),
Climate since AD 1500. Routledge, London: 649–665.
Jones, P. D., T. J. Osborn & K. R. Briffa, 2001. The evolution of
climate over the last millennium. Science 292: 662–667.
Juggins, S., 2009. Rioja: an R package for the analysis of
quaternary science data, Version 0.5-3. (http://cran.r-project.org/package=rioja).
Koch, J. & R. Kilian, 2005. ‘‘Little Ice Age’’ glacier fluctua-
tions, Gran Campo Nevado, southernmost Chile. The
Holocene 15: 20–28.
Krammer, K. & H. Lange-Bertalot, 1991–2000. Bacillario-
phyceae. In Ettl, H., J. Gerloff, H. Heyning & D. Molle-
nhauser (eds) Subwasserflora von Mitteleuropa, Vol. 2.
Fischer, Stuttgart: 1–4.
Kreutz, K. J., P. A. Mayewski, L. D. Meeker, M. S. Twickler,
S. I. Whitlow & I. I. Pittalwala, 1997. Bipolar changes in
atmospheric circulation during the Little Ice Age. Science
277: 1294–1296.
Laing, T., K. Ruhland & J. Smol, 1999. Past environmental and
climate changes related to tree-line shifts inferred from
fossil diatoms from a lake near the Lena River Delta,
Siberia. The Holocene 9: 547–557.
Lamb, H. H., 1977. Climate: present, past and future, Vol. 2:
Climatic history and the future. Methuen, London.
Lamy, F., D. Hebbeln, U. Rohl & G. Wefer, 2001. Holocene
rainfall variability in southern Chile: a marine record of
latitudinal shifts of the southern westerlies. Earth and
Planetary Science Letters 185: 369–382.
Langdon, P. G., K. E. Barber & S. H. Lomas-Clarke, 2004.
Reconstructing climate and environmental change in
northern England through chironomid and pollen analy-
ses: evidence from Talkin Tarn, Cumbria. Journal of Pa-
leolimnology 32: 197–213.
Hydrobiologia (2010) 648:207–225 223
123

Lara, A., J. Aravena, R. Villalba, A. Wolodarsky-Franke, B.
Luckman & R. Wilson, 2001. Dendroclimatology of high-
elevation Nothofagus pumilio forests at their northern
distribution limit in the central Andes of Chile. Canadian
Journal of Forest Research 31: 925–936.
Lara, A., R. Villalba, A. Wolodarsky-Franke, J. Aravena, B.
Luckman & E. Cuq, 2005. Spatial and temporal variation
in Nothofagus pumilio growth at tree line along its lati-
tudinal range (35�400–55�S) in the Chilean Andes. Journal
of Biogeography 32: 879–893.
Larocque, I. C. & C. Bigler, 2004. Similarities and discrep-
ancies between chironomid- and diatom-inferred temper-
ature reconstructions through the Holocene at Lake 850,
northern Sweden. Quaternary International 122: 109–121.
Loehle, C., 2007. A 2000-year global temperature reconstruc-
tion based on non-tree-ring proxies. Energy and Envi-
ronment 18: 1049–1058.
Lotter, A. F. & H. J. B. Birks, 1993. The impact of the Laacher
See tephra on terrestrial and aquatic ecosystems in the
Black Forest, southern Germany. Journal of Quaternary
Science 8: 263–276.
Lotter, A. F., H. J. B. Birks, W. Hofmann & A. Marchetto,
1998. Modern diatom, cladocera, chironomid, and
chrysophyte cyst assemblages as quantitative indicators
for the reconstruction of past environmental conditions in
the Alps. II. Nutrients. Journal of Paleolimnology 19:
443–463.
Mann, M. E., 2002. Little ice age. In MacCracken, M. C. & J.
S. Perry (eds), The Earth System: Physical and Chemical
Dimensions of Global Environmental Change. Wiley,
New York: 504–509.
Mann, M. E., R. S. Bradley & M. K. Hughes, 1998. Global-
scale temperature patterns and climate forcing over the
past six centuries. Nature 392: 779–787.
Marciniak, B., 1990. Late glacial and Holocene diatoms in
sediments of the Bledowo Lake (Central Poland). In Si-
mola H (ed.) Proceedings of the 10th Diatom Symposium.
Koeltz Scientific Books, Koenigstein: 379–390.
Mardones, M. & J. Vargas, 2005. Efectos hidrologicos de los
usos electrico y agrıcola en la cuenca del rıo Laja (Chile
centro-sur). Revista de Geografıa Norte Grande 33: 89–
102.
Marticorena, C., 1968. Granos de polen de plantas chilenas,
Vol. 17. Gayana Botanica, Chile: 34 pp.
Marticorena, C. & R. Rodrıguez, 1995. Flora de Chile, Vol 1.
Pteridophyta-Gimnospermae. Universidad de Concepcion,
Concepcion, Chile: 351 pp.
Massaferro, J. & S. J. Brooks, 2002. Response of chironomids
to late quaternary environmental change in the Taitao
peninsula, southern Chile. Journal of Quaternary Science
17: 101–111.
Massaferro, J. & J. Corley, 1998. Environmental disturbance
and chironomid paleodiversity: 15 kyr BP of history at
Lake Mascardi, Patagonia, Argentina. Aquatic Conserva-
tion: Marine and Freshwater Ecosystems 8: 315–323.
Massaferro, J., S. J. Brooks & S. G. Haberle, 2005. The
dynamics of chironomid assemblages and vegetation
during the Late quaternary at Laguna Facil, Chonos
Archipelago, southern Chile. Quaternary Science Reviews
24: 2510–2522.
Massaferro, J., P. Moreno, G. Denton, M. Vandergoes & A.
Dieffenbacher-Krall, 2009. Chironomid and pollen evi-
dence for climate fluctuations during the Last Glacial
Termination in NW Patagonia. Quaternary Science
Reviews 28: 517–525.
McCormac, F., A. Hogg, P. Blackwell, C. Buck, T. Higham &
P. Reimer, 2004. SHCal04 southern hemisphere calibra-
tion 0–11.0 cal Kyr BP. Radiocarbon 46: 1087–1092.
McDermott, F., D. Mattey & C. Hawkesworth, 2001. Centen-
nial-scale Holocene climate variability revealed by high-
resolution speleotherm 18O record from SW Ireland.
Science 294: 1328–1331.
McKinzey, K. M., W. Lawson, D. Kelly & A. Hubbard, 2004.
A revised Little Ice Age chronology of the Franz Josef
Glacier, Westland, New Zealand. Journal of the Royal
Society of New Zealand 34: 381–394.
Melnick, D., F. Charlet, H. P. Echtler & M. De Batist, 2006.
Incipient axial collapse of the Main Cordillera and strain
partitioning gradient between the central and Patagonian
Andes, Lago Laja, Chile. Tectonics 25: 1–22.
Merilainen, J., J. Hynynen, A. Teppo, A. Palomaki, K. Gran-
berg & P. Reinikainen, 2000. Importance of diffuse
nutrient loading and lake level changes to the eutrophi-
cation of an originally oligotrophic boreal lake: a palae-
olimnological diatom and chironomid analysis. Journal of
Paleolimnology 24: 251–270.
Mortlock, R. & P. Froelich, 1989. A simple method for the
rapid determination of biogenic opal in pelagic marine
sediments. Deep Sea Research 36: 1415–1426.
Moser, K., G. MacDonald & J. Smol, 1996. Applications of
freshwater diatoms to geographical research. Progress in
Physical Geography 20: 21–52.
Moy, C., R. Dunbar, P. Moreno, J.-P. Francois, R. Villa-
Martınez, D. Mucciarone, T. Guilderson & R. Garreaud,
2008. Isotopic evidence for hydrologic change related to
the westerlies in SW Patagonia, Chile, during the last
millennium. Quternary Science Reviews 27: 1335–1349.
Nardini, A. & D. Montoya, 1993. Planteamiento de un modelo
decisional para la gestion integrada del sistema lago Laja-
rıo Laja (con respecto al proyecto ‘‘Canal Laja-Dig-
uillın’’), Vol. 3. Monografıa cientıficas, Centro Eula-
Chile.
Nesje, A. & S. O. Dahl, 2003. The ‘‘Little ice Age’’ – only
temperature? The Holocene 13: 139–145.
Ogilvie, A. E. J. & T. Jonsson, 2001. ‘Little Ice Age’ research:
a perspective from Iceland. Climate Change 48: 9–52.
Oksanen, J., R. Kindt, P. Legendre & R. B. O’Hara, 2005.
Vegan: community ecology package version 1.15-3.
http://cc.oulu.fi/*jarioksa/.
Ortlieb, L., R. Escribano, R. Follegati, O. Zuniga, I. Kong, L.
Rodriguez, J. Valdes, N. Guzman & P. Iratchet, 2000.
Recording of ocean-climate changes during the last 2,
000 years in a hypoxic marine environment off northern
Chile (23�S). Revista Chilena de Historia Natural 73(2):
221–242.
Owen, R., J. Barthelme, R. Renaut & A. Vincens, 1982. Pa-
leolimnology and archaeology of Holocene deposits
north-east of Lake Turkana, Nenya. Nature 298: 523–529.
Paggi, A., 2001. Diptera: chironomidae. In Fernandez, H. R. &
E. Domınguez (eds), Guıa para la determinacion de los
224 Hydrobiologia (2010) 648:207–225
123

artropodos bentonicos Sudamericanos. Editorial Univers-
itaria de Tucuman, Tucuman, Argentina: 167–194.
Polissar, P. J., M. B. Abbott, A. P. Wolfe, M. Bezada, V. Rull
& R. S. Bradley, 2006. Solar modulation of Little Ice Age
climate in the tropical Andes. Proceedings of the National
Academy of Sciences of the United States of America
103: 8937–8942.
Porinchu, D. F. & L. C. Cwynar, 2002. Late-Quaternary history
of midge communities and climate from a tundra site near
the lower Lena River, Northeast Siberia. Journal of Pa-
leolimnology 27: 59–69.
Quiroz, R., P. Popp, R. Urrutia, C. Bauer, A. Araneda, H.-C.
Treutler & R. Barra, 2005. PAH fluxes in the Laja Lake of
south central-Chile Andes over the last 50 years: evidence
from a dated sediment core. Science of the Total Envi-
ronment 349: 150–160.
Rabatel, A., V. Jomelli, P. Naveau, B. Francou & D. Grancher,
2005. Dating of Little Ice Age glacier fluctuations in the
tropical Andes: Charquini glaciers, Bolivia, 16�S.
Comptes Rendur Geoscience 337: 1311–1322.
Rae, J. G., 1989. Chironomid midges as indicators of organic
pollution in the Scioto River basin, Ohio. Ohio Journal of
Science 1: 5–9.
Rieradevall, M. & S. J. Brooks, 2001. An identification guide
to subfossil Tanypodinae larvae (Insecta: Diptera: Chiro-
nomidae) based on cephalic setation. Journal of Paleo-
limnology 25: 81–99.
Rivera, P., 1970. Diatomeas de los lagos Ranco, Laja y Laguna
Chica de San Pedro Vol. 20. Gayana Botanica, Chile: 23
pp.
Rivera, P., O. Parra & M. Gonzalez, 1973. Fitoplancton del
Estero Lenga, Vol. 23. Gayana Botanica, Chile: 93 pp.
Rivera, P., 1974. Diatomeas de agua dulce de Concepcion y
alrededores Vol. 28. Gayana Botanica, Chile: 134 pp.
Rivera, P., O. Parra, M. Gonzalez, V. Dellarossa & M. Orell-
ana, 1982. Manual taxonomico del fitoplancton de aguas
continentales. Editorial Universidad de Concepcion,
Concepcion: 97.
Rodrıguez, R., O. Matthei & M. Quezada, 1983. Flora Arborea
de Chile. Editorial de la Universidad de Concepcion,
Concepcion, Chile.
Smol, J., 1988. Paleoclimate proxy from freshwater arctic
diatoms. Verhandlungen der Internationalen Vereinigung
fur Limnologie 23: 837–844.
Soon, W., S. Baliunas, C. Idso, S. Idso & D. Legates, 2003.
Reconstructing climatic and environmental changes of the
past 1000 years: a reappraisal. Energy and Environment
14(2–3): 233–299.
Stuiver, M. & P. J. Reimer, 1993. Extended 14C database and
revised CALIB radiocarbon calibration program. Radio-
carbon 35: 215–230.
Telford, R. J., H. F. Lamb & U. Mohammed, 1999. Diatom-
derived paleoconductivity estimates from Lake Awassa,
Ethiopia: evidence for pulsed inflows of saline ground-
water? Journal of Paleolimnology 21: 409–421.
Thiele, R., H. Moreno, S. Elgueta, A. Lahsen, S. Rebolledo &
M. Petit-Breuilh, 1998. Evolucion geologico-geomor-
fologica cuaternaria del tramo superior del valle del rıo
Laja. Revista Geologica de Chile 25(2): 229–253.
Thompson, L. G., E. Mosley-Thompson, W. Dansgaard & P.
M. Grootes, 1986. The Little Ice Age as recorded in the
stratigraphy of the tropical Quelccaya ice cap. Science
234: 361–364.
Torres, L., O. Parra, A. Araneda, R. Urrutia, F. Cruces & L.
Chirinos, 2008. Vegetational and climatic history during
the late Holocene in Lake Laja basin (central Chile)
inferred from sedimentary pollen record. Reviews of
Palaeobotany and Palynology 149: 18–28.
Trouet, V., J. Esper, N. Graham, A. Baker, J. Scourse & D.
Frank, 2009. Persistent positive North Atlantic oscillation
mode dominated the medieval climate anomaly. Science
324: 78–80.
Urrutia, R., A. Araneda, F. Cruces, L. Torres, L. Chirinos, H.-
C. Treutler, N. Fagel, S. Bertrand, I. Alvial, R. Barra & E.
Chapron, 2007. Changes in diatom, pollen, and chirono-
mid assemblages in response to a recent volcanic event in
Lake Galletue0 (Chilean Andes). Limnologica 37: 49–62.
Veblen, T. & F. Schlegel, 1982. Resena ecologica de los
bosques del sur de Chile. Bosque 4(2): 73–115.
Velle, G., J. Larsen, W. Eide, S. M. Peglar & H. J. Birks, 2005.
Holocene environmental history and climate of Ratasjøen,
a low-alpine lake in south-central Norway. Journal of
Paleolimnology 33: 129–153.
Verschuren, D. & H. Eggermont, 2006. Quaternary paleo-
ecology of aquatic Diptera in tropical and Southern
Hemisphere regions, with special reference to the Chiro-
nomidae. Quaternary Science Reviews 25: 1926–1947.
Villalba, R., 1990. Climatic fluctuations in Northern Patagonia
during the last 1000 years as inferred from tree-ring
records. Quaternary Research 34: 346–360.
Villalba, R., 1994. Tree-ring and glacial evidence for the
medieval warm epoch and the little ice age in Southern
South America. Climatic Change 26: 183–197.
Villa-Martınez, R., C. Villagran & B. Jenny, 2004. Pollen
evidence for late-Holocene climatic variability at Laguna
de Aculeo, Central Chile (lat. 34�S). The Holocene 14:
361–367.
Wentworth, C. K., 1922. A scale of grade and class terms for
clastic sediments. Journal of Geology 30: 377–392.
Wiederholm, T., 1983. Chironomidae of the holartic region.
Keys and diagnoses. Part I – Larvae. Entomologica
Scandinavica Supplement 19: 1–457.
Willen, E., 1991. Planktonic diatoms-an ecological review.
Algological Studies 62: 69–106.
Winkler, S., 2004. Lichenometric dating of the ‘Little Ice Age’
maximum in Mt Cook National Park, Southern Alps, New
Zealand. The Holocene 14: 911–920.
Wolin, J. & H. Duthie, 1999. Diatoms as indicators of water
level change in freshwater lakes. In Stoermer, E. F. & J. P.
Smol (eds), The Diatoms: Applications to the Environ-
mental and Earth Sciences. Cambridge University Press,
Cambridge: 183–204.
Xue, B., W. Qu, S. Wang, Y. Ma & M. D. Dickman, 2003.
Lake level changes documented by sediment properties
and diatom of Hulun Lake, China since the late Glacial.
Hydrobiologia 498: 133–141.
Hydrobiologia (2010) 648:207–225 225
123