lect. 6-1 bch 443 biochemistry of specialized tissues 2. fibrous proteins
TRANSCRIPT

Lect. 6-1
BCH 443BCH 443Biochemistry ofBiochemistry of
Specialized TissuesSpecialized Tissues
2. Fibrous Proteins2. Fibrous Proteins

Lect. 6-2
Fibrous vs. Globular Proteins
1. Compact protein structure Extended protein structure
2. Soluble in water (or in lipid Insoluble in water (or in lipid bilayers) bilayers)
3. Secondary structure is complex Secondary structure is simplewith a mixture of -helix, -sheet based on one type onlyand loop structures 4. Quaternary structure is held Quaternary structure is usually together by noncovalent forces held together by covalent bridges
5. Functions in all aspects of Functions in structure of the body metabolism (enzymes, transport, or cell (tendons, bones, muscle, immune protection, hormones, etc). ligaments, hair, skin)
Globular Fibrous

Lect. 6-3
Fibrous Proteins
Fibrous proteins have high -helix or -sheet content. Most are structural proteins.
Examples include:
• Collagen
• Elastin
• Keratin
• Fibroin

Lect. 6-4
Fibrous Proteins
• Much or most of the polypeptide chain is parallel to a single axis
• Fibrous proteins are often mechanically strong & highly cross-linked
• Fibrous proteins are usually insoluble
• Usually play a structural role

Lect. 6-5
COLLAGENWhat makes collagen a strong
tensile protein?

Lect. 6-6
Questions?
1. How would you define the structure of a collagen molecule?
2. What are the dimensions of a collagen molecule?
3. What are the dimensions of a collagen fibril?
4. State the most important amino acids in collagen and explain their importance.
5. What is the periodicity of collagen? Why does it happen?

Lect. 6-7
Collagen Background
The collagens are the most abundant proteins in the body.• They occur in connective tissues where tensile strength is needed.• Examples: skin, tendons, cartilage, bones.
Tensile strength results from the use of:
(a) The triple helix secondary structure(b) The assembly of tropocollagen subunits into a fibre(c) Chemical cross linking to strengthen the fibre

Lect. 6-8
Secondary structure - the triple helix
Collagen is formed from tropocollagen subunits. The triple helix in tropocollagen is highly extended and strong.
Features: (1) Three separate polypeptide chains arranged as a left-handed helix (note that an -helix is right-handed).(2) 3.3 residues per turn (3) Each chain forms hydrogen bonds with the other two: STRENGTH!

Lect. 6-9
Collagen A Triple Helix
Principal component of connective tissue (tendons, cartilage, bones, teeth)
Basic unit is tropocollagen:
• Three intertwined polypeptide chains (1000 amino acid residues each)
• MW = 285,000
• 300 nm long, 1.4 nm diameter
• Unique amino acid composition

Lect. 6-10
Collagen Amino Acid Composition
• Nearly one residue out of three is Gly
• Proline content is unusually high
• Many modified amino acids present:
– 4-hydroxyproline
– 3-hydroxyproline
– 5-hydroxylysine
• Pro and HyPro together make 30% of res.

Lect. 6-11
Collagen Amino Acid Sequence
AA sequence of C-terminal region of bovine type-I collagen

Lect. 6-12
Hydroxylated residues found in collagen

Lect. 6-13
Biosynthesis of hydroxyPro and hydroxyLys requires O2 and ascorbic acid (vitamin C). Vit. C deficiency leads to disorders in bone, skin and teeth.

Lect. 6-14
The Collagen Triple Helix
• The unusual amino acid composition of collagen is not favorable for helices OR -sheets
• But it is ideally suited for the collagen triple helix: three intertwined helical strands
• Much more extended than helix, with a rise per residue of 2.9 Angstroms
• 3.3 residues per turn
• Long stretches of Gly-Pro-Pro-HyPro

Lect. 6-15
In collagen triple helix H-bonds form between separate chains. In -helix H-bonds formed between residues of the same chain.

Lect. 6-16
Collagen Fibers
• Fibers are formed by staggered arrays of tropocollagens
• Banding pattern in EMs with 68 nm repeat
• Since tropocollagens are 300 nm long, there must be 40 nm gaps between adjacent tropocollagens (5 x 68 = 340 Angstroms)
• 40 nm gaps are called "hole regions" - they contain carbohydrate and are thought to be nucleation sites for bone formation
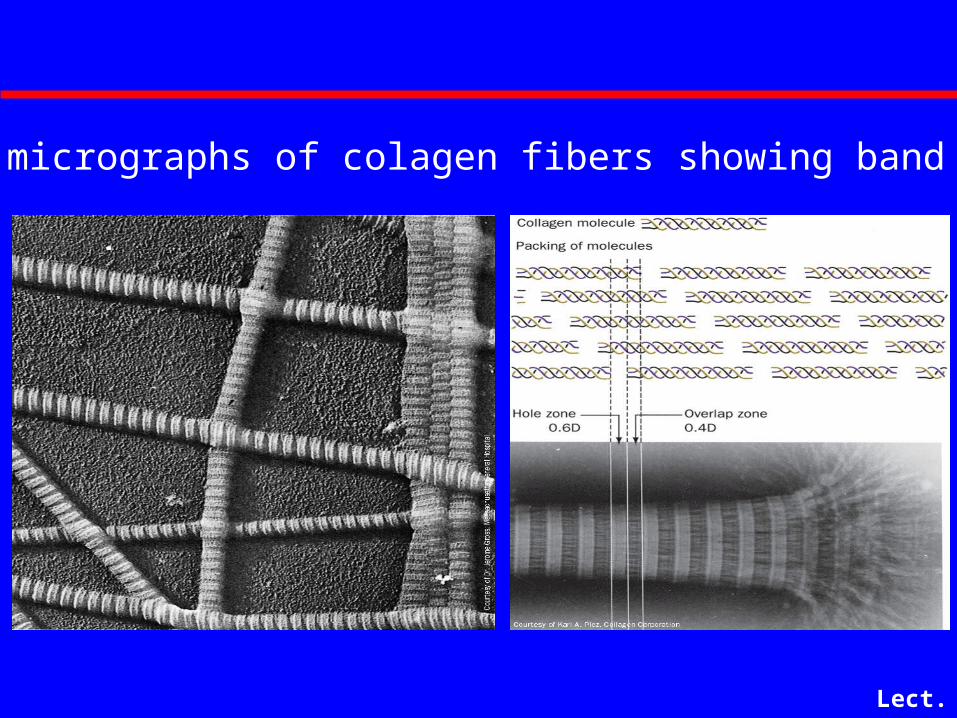
Lect. 6-17
Electron micrographs of colagen fibers showing band pattern

Lect. 6-18
Structure of collagen fibers
•(a) and (b) the primary and secondary structure
•(c) lower magnification emphasizes the triple-helix
• (d) tropocollagen molecules align side by side to form collagen fiber
19
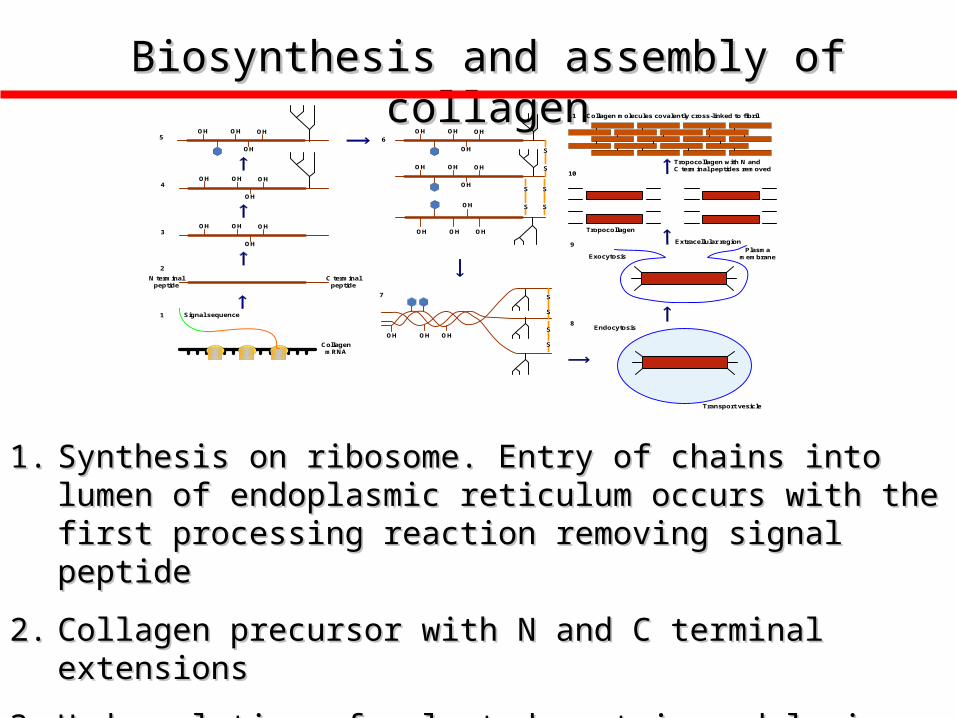
Biosynthesis and assembly of collagenBiosynthesis and assembly of collagen
1.1. Synthesis on ribosome. Entry of chains into lumen of endoplasmic Synthesis on ribosome. Entry of chains into lumen of endoplasmic reticulum occurs with the first processing reaction removing signal reticulum occurs with the first processing reaction removing signal peptidepeptide
2.2. Collagen precursor with N and C terminal extensionsCollagen precursor with N and C terminal extensions
3.3. Hydroxylation of selected protein and lysinesHydroxylation of selected protein and lysines
OH OH
OH
OH
1 Signal sequence
OH OH
OH
OH
OH OH
OH
OH
Collagen mRNA
2
3
4
5OH OH
OH
OH
OH OH
OH
OH
OH OH
OH
OH
S
S
S
S
S
S
C terminalpeptide
N terminalpeptide
S
S
S
S
OH OH OH
6
7
Endocytosis
Transport vesicle
Plasmamembrane
Extracellular region
Exocytosis
Tropocollagen
Tropocollagen with N andC terminal peptides removed
Collagen molecules covalently cross-linked to fibril11
10
9
8

Lect. 6-20
Biosynthesis and assembly of collagen (Con’t)
4. Addition of Asn-linked oligosaccharides to collagen
5. Initial glycosylation of hydroxylyine residues
6. Alignment of three polypeptide chains and formation of inter-chain disulfide bridges
7. Formation of triple helical procollagen
8. Transfer by endocytosis to transport vesicle
9. Exocytosis transfers triple helix to extracellular phase
10. Removal of N and C terminal propeptides by specific peptidase
11. Lateral association of collagen molecules coupled to covalent cross linking creates fibril

Lect. 6-21
Structural Basis of Collagen Triple Helix
• Every third residue faces the crowded center of the helix only Gly fits
• Pro and HyPro suit the constraints of phi and psi
• Interchain H-bonds involving HyPro stabilize helix
• Fibrils are strengthened by intrachain lysine-lysine and interchain hydroxypyridinium cross links

Lect. 6-22
Biosynthesis of Aldol Cross-links in Collagen
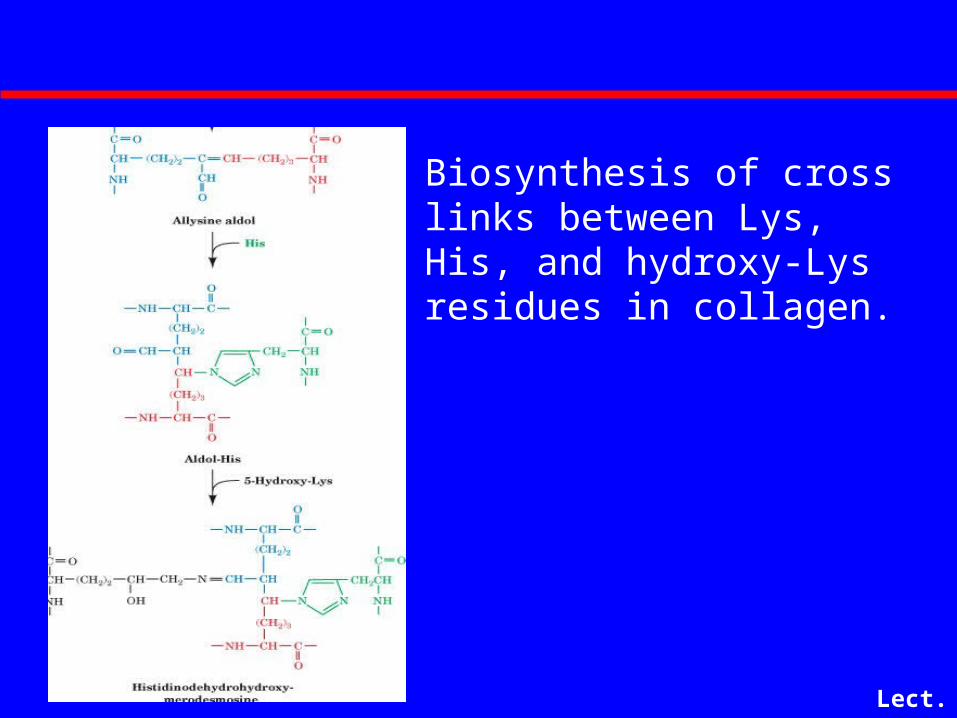
Lect. 6-23
Biosynthesis of cross links between Lys, His, and hydroxy-Lys residues in collagen.

Lect. 6-24
The Major Collagen Groups
In humans at least there are 19 different collagens. Within these 19 structural types four major classes are generally identified.

Lect. 6-25
Classification of Collagens
Type Chains Tissue Found Characteristics
I a1(I)2, a2(I) Bone, skin, tendons
Low carbohydrate; <10%Hydroxylysines per chain
II a1(II)3 Cartilage, vitreous
10% carbohydrate; >20 hydroxylysines per chain
III a1(III)3 Blood vessels, scar tissue, uterine wall

Lect. 6-26
Classification Continued
IV [a1(IV)3
a2(IV)3]
Basement membrane
lens capsule
High carbohydrate,
>40 hydroxylysines per chain
V [a1(V)2a2(V)]
[a1(V)3]
[a1(V)a2(V)a3(V)]
Cell surface and exoskeleton
High carbohydrate, high glycine and hydroxylysine
VI Aortic intima, placenta, kidney
Low mol.weight, equal amounts of hydroxylysine and hydroxyproline

Lect. 6-27
Thermal Denaturation Curve
In normal collagens the transition midpoint temperature or Tm is related to the normal body temperature of the organism and for animal is above 40 oC as shown in blue line in the graph..
Tm
Triple helix stabilization is through HyPro and formation of H bonds with neighboring chains.

Lect. 6-28
DISORDERS OF COLLAGEN DEPOSITION

Lect. 6-29
Disorders of Collagen Deposition
• Disorders of collagen deposition– insufficient collagen content– presence of chemically and/or morphologically
abnormal collagen– excessive collagen content– insufficient collagen resorption– excessive collagen resorption

Lect. 6-30
Disorders of Collagen Deposition
• Genetic abnormalities of collagen– mutations that lead to aminoacid deletions or
additions– deficient synthesis of a portion– disorders in post-translational modification
(hydroxylation of lysine, hydroxylation of proline)– defects in enzymes essential for post-translational
modification

Lect. 6-31
Disorders of Collagen Deposition
• Collagen is the building block; thus, its disorders lead to significant deterioration in the mechanical integrity of tissues
• Several disorders– Ehlers-Danlos syndrome
– Osteogenesis Imperfecta
– Marfan syndrome

Lect. 6-32
ELASTIN Three factors make it
stretchy and elastic

Lect. 6-33
Elastin
Elastin can stretch several times - then return to the original starting size
Elastin is found in large arteries (the aorta), ligaments, and the lung wall. (1) The subunits of elastin are called tropoelastin –
molecules 1, 2, 3 and 4.
The crosslinking of tropoelastin via lysine residues results in a stable starting network of elastin (i.e. when not stretched).
Either desmosine (4 Lys) links 4 molecules of tropoelastin, or lysinonorleucine (2 Lys) links 2 tropoelastin molecules.
It is clinically relevant in cardiovascular disease and lung emphysema

Lect. 6-34
Desmosine is formed from 4 lysines, 3 of which are oxidised.
Desmosine
CH
NH
CO
CH2 CH2 CHOCH2 CH
NH
CO
CH2CH2CHO CH2
Lysine
Allysine
CH
NH CO
CH2
CH2
CH2
CH2
NH3+
CH
NHCO
CH2
CH2
CH2
CHOAllysine
Allysine
CH
NH
CO
CH2 CH2 CH
NH
CO
CH2CH2
CH
NH CO
CH2
CH2
CH2
CH2
N+
CH
NHCO
CH2
CH2
CH2
CC
C
C
C
Desmosine

Lect. 6-35
(2) Amino acid composition of elastin33% Gly 10% Pro and Hyp 23% Ala 13% ValHence 79% of the residues come from 4 amino acids.There are large hydrophobic peptides rich in Ala, Val, Ile and Leu. As these sidechains do not interact with each other by hydrogen bonds, they enable the core of elastin to separate and stretch easily.
Elastin
(3) Secondary structure of elastinA different type of helix structure from those in the a-helix is present. This is able to stretch and relax like a coiled spring. So elastin is elastic!This is constructed from a helix of repeated b-turns based on the sequence Val.Pro.Gly.Val, and is called the-spiral.

Lect. 6-36
Elastin• Abundant in ligaments, lungs, artery walls, skin.
• Provides tissues with ability to stretch in all directions without tearing.
• Contains predominantly small hydrophobic residues: 1/3 Gly, 1/3 Ala + Val, many Pro but no hydroxyPro or hydroxyLys.
• Lacks regular secondary structure.
• Has unordered coil structure that is highly cross-linked into 3-dimensional network of fibers to provide rubber-like elasticity.

Lect. 6-37
Elastin• Cross-links formed from allyysine (aldehyde derivative
of Lys)
• Extracellular Lys oxidase specific for Lys-Ala-Ala-Lys and Lys-(Ala)3-Lys sequences
• Lys + 3 allysine combine to from desmosine or isodesmosine cross-links responsible for yellow color of elastin
• Also forms lysinorleucine cross-links from 2 allysine, as in collagen.
• Cross-links responsible for elasticity & insolubility

Lect. 6-38
CH2 (CH2)3
(CH2)2 CH2
H2CCH2
H2C CH2
N+
(CH2)3 (CH2)3
(CH2)3 (CH2)3
CC
C CH2
NH2
H
H H
O
OO
(CH2)3 (CH2)3
(CH2)3 (CH2)3
CH2 CH2
NH2 NH2
CH2 CH2
NH2 NH2Lysine amino
oxidase
Aldol condensations
Desmosinecross-link
Biosynthesis of desmosine and isodesmosine cross-links unique to elastin

Lect. 6-39
KERATIN
-Keratins are found in mammals -Keratins are found as a left-handed super
helix -Keratins are found in birds and reptiles -Keratins are analogs to the silk fibroin
structures produced by spiders and silkworms

Lect. 6-40
-KERATINTwo reasons why this is a tough protective fibrous
protein

Lect. 6-41
-Keratin is found in hair, nails, outer layer of skin. It forms almost the entire dry weight of these materials.
(1) The entire secondary structure is a dimer of two -helices.
It is rich in amino acids that favours -helix formation (Phe, Ile, Val, Met, Ala)These hydrophobic side chains are on the -helix surface-explaining its insolubility.
It is also rich in Cys residues.

Lect. 6-42
Structure of dimer of two -helices.

Lect. 6-43
Proposed structure for -keratin intermediate filaments
• Two monomers (a) pair via a parallel coiled-coil to form 50- nm-long dimer (b)
• These then associate to form 1st protofilament (c)
• These then associate to form protofibril (d)
• Regular spacing of 25 nm along the fibers is accounted for by overlap
Lect. 6-44
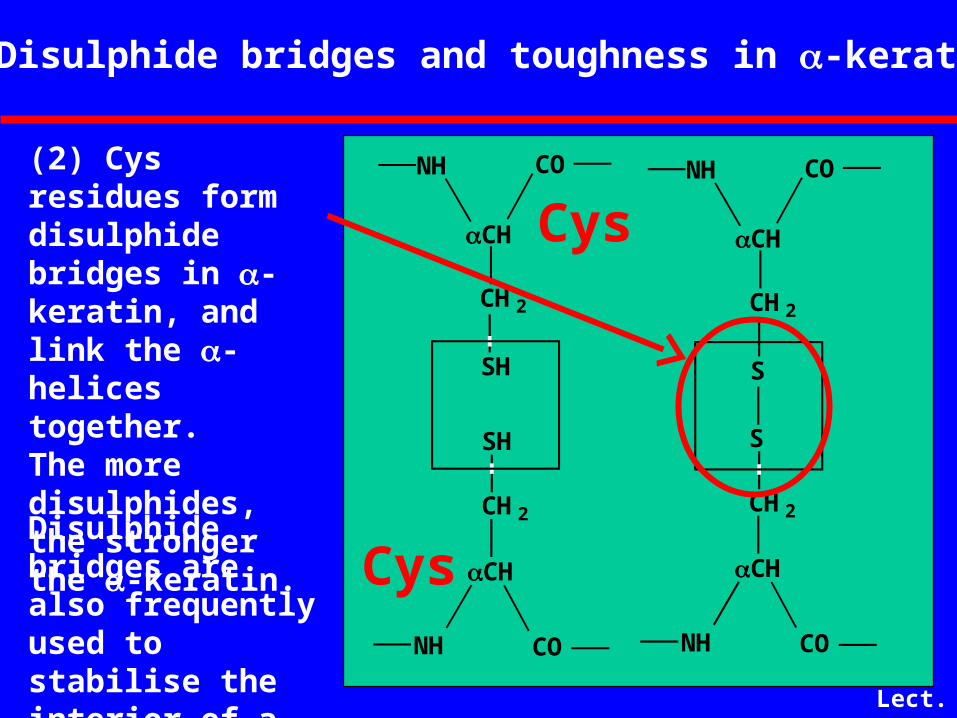
Disulphide bridges and toughness in -keratin
Disulphide bridges are also frequently used to stabilise the interior of a globular protein.
CH
NH CO
CH2
SH
CH
NH CO
CH2
SH
CH
NH CO
CH2
S
CH
NH CO
CH2
S
(2) Cys residues form disulphide bridges in -keratin, and link the -helices together. The more disulphides, the stronger the -keratin.
Cys
Cys

Lect. 6-45
Quaternary structure -keratin of
•The association of long parallel a-helices also gives toughness to -keratin.
UP-TO-DATE MODEL
Proto-filament of anti-parallel dimers
The up-to-date view is that TWO parallel -helices supercoil around each other to form a dimer. Then each dimer associates antiparallel with two other dimers to form the protofibril.
The association of four protofibrils forms a four-stranded rope. These successive overlaps explain why -keratin is such a tough protein.
Clinical relevance in skin diseases: psioriasis – the overproduction of -keratin
•The incorrect explanation of -keratin structure states that THREE -helices supercoil around each other to form a protofibril, and that the association of 2 and 9 protofibrils forms a hair microfibril. •Lippincott’s Fig 3.31 on page 45 is wrong!

Lect. 6-46
-Keratin Structure

Lect. 6-47
Fibroin
• Fibroins are the silk proteins. They also form the spider webs
• Made with a -sheet structures (M6.12) with Gly on one face and Ala/Ser on the other
• Fibroins contain repeats of [Gly-Ala-Gly-Ala-Gly-Ser-Gly-Ala-Ala-Gly-(Ser-Gly-Ala-Gly-Ala-Gly)8]
• The -sheet structures stack on top of each other (M6.12b)
• Bulky regions with valine and tyrosine interrupt the -sheet and allow the stretchiness

Lect. 6-48
Structure of silk fibroin
• (a)Three dimension view of the stacked -sheets
• (b) Interdigitation of Al or Ser and Gly side chain
• The plane of the section is perpendicular to the folded sheets