lipid rafts signaling detected by fret-based molecular
TRANSCRIPT

REVIEW
Lipid rafts signaling detected by FRET-based molecularbiosensors
Jihye Seong
Received: 24 April 2015 / Accepted: 30 April 2015 / Published online: 12 May 2015
� The Korean Society for Applied Biological Chemistry 2015
Abstract Cells recognize and process various extracel-
lular signals via transmembrane receptors and membrane-
associated signaling molecules. Therefore, the precise
regulation of these signaling events at the plasma mem-
brane, which occur spatiotemporally, is necessary for
proper cellular functions such as cell proliferation, migra-
tion, and survival. The plasma membrane may contain
dynamic microdomains called lipid rafts, which are sug-
gested to be crucial for the regulation of efficient and
specific signaling pathways. However, because of the
limitation in methodologies, the specific molecular
mechanisms underlying dynamic signaling events at lipid
rafts are largely unknown. This review discusses the tra-
ditional biochemical methods for the visualization of lipid
rafts and the related signaling events at these mi-
crodomains. In addition, the review reports on fluorescence
resonance energy transfer (FRET)-based molecular
biosensors with lipid rafts targeting sequences as a pow-
erful tool for live-cell imaging of spatiotemporal signaling
events at lipid rafts. In particular, examples of dynamic
lipid rafts signaling mechanisms visualized by FRET-based
biosensors in live cells are covered in the last section.
Keywords Acylation � Biosensor � Fluorescenceresonance energy transfer � Lipid rafts � Live-cell imaging �Prenylation � Signaling
Introduction
Lipid rafts at plasma membrane
According to the fluid mosaic model proposed by Singer
and Nicolson in 1972, the plasma membrane was thought
to be a 2D phospholipid bilayer containing peripheral and
integral proteins floating freely in this bilayer (Singer and
Nicolson 1972). Since then, however, studies in a model
lipid bilayer system and in living cells have reported that
the lipid bilayer is not a static homogenous fluid. These
studies indicate that the plasma membrane contains deter-
gent-resistant microdomains of more ordered states
(Thompson and Tillack 1985; Brown and Rose 1992; Sil-
vius et al. 1996; Schroeder et al. 1998). After two decades,
Simons and colleagues postulated the ‘‘lipid rafts hy-
pothesis’’, defining that these membrane microdomains
may float around the surrounding lipid fluid like a raft
(Simons and Ikonen 1997). Lipid rafts are enriched with
sphingolipids and cholesterol, and they may be more
tightly packed because of these lipid compositions. These
distinct physicochemical properties may allow the segre-
gated distribution of membrane-associated proteins in and
outside the lipid rafts. For example, glycosylphos-
phatidylinositol (GPI)-anchored proteins and saturated long
fatty acids are shown to be strongly attached to lipid rafts
microdomains (Brown and London 1998; Simons and
Toomre 2000). The selective segregation of specific sig-
naling proteins is involved in the tight regulation of signal
transduction on the plasma membrane. For example, in T
cell antigen receptor signaling (Horejsi 2003), neurotrans-
mitter signaling (Allen et al. 2007), and membrane traf-
ficking (Hanzal-Bayer and Hancock 2007), extracellular
signals have been suggested to be sorted and transferred
inside the cells more efficiently through the clustering of
J. Seong (&)
Center for Neuro-Medicine, Brain Science Institute, Korea
Institute of Science and Technology (KIST), Hwarangno 14-gil
5, Seongbuk-gu, Seoul 136-791, Republic of Korea
e-mail: [email protected]
123
J Korean Soc Appl Biol Chem (2015) 58(5):629–636 Online ISSN 2234-344X
DOI 10.1007/s13765-015-0082-2 Print ISSN 1738-2203

lipid rafts. Moreover, lipid rafts are believed to be of var-
ious sizes (from 10 nm to submicrometer range) and highly
dynamic in their assembly/disassembly in response to ex-
tracellular signals. Considering that the resolution of con-
ventional microscopy is several hundred micrometers, it
has been a major challenge to clearly understand the
mechanisms underlying the involvement of these small,
heterogeneous, and highly dynamic microdomains in the
regulation of signal transduction pathways in live cells.
Visualization of lipid rafts
To understand the highly dynamic features of membrane
microdomains, numerous studies have been tried to visu-
alize lipid rafts using fluorescent lipid/protein analogs or
lipid-binding toxins/antibodies (Ishitsuka et al. 2005).
For example, fluorescein–polyethylene glycol–cholesterol,
Cy5-dimyristoyl-sn-glycero-phosphatidylethanolamine, and
GPI linked to green fluorescent protein (GFP) have been
used as fluorescent analogs of lipid rafts components:
cholesterol, saturated long fatty acids, and GPI-associated
proteins, respectively (Schutz et al. 2000; Sato et al. 2004;
Sharma et al. 2004). In addition, non-toxic recombinant
derivatives of pore-forming toxins, such as lysenin, cholera
toxin subunit B, and aerolysin, have been used for the lipid
raft markers because they specifically bind to sphin-
gomyelin, glycosphingolipid GM1, and GPI-associated
proteins, respectively (Kenworthy et al. 2000; Shakor et al.
2003; Skocaj et al. 2013).
These fluorescent analogs and probes are useful tools to
visualize lipid rafts microdomains, although the size of the
individual raft seems to be beyond the resolution of the
current conventional microscopy. Super-resolution imaging
techniques have recently begun to be applied to imaging lipid
rafts, and they include stimulated emission depletion
(STED), photoactivated localization microscopy (PALM),
and stochastic optical reconstruction microscopy (STORM)
(Simons and Gerl 2010; Owen et al. 2012; Owen and Gaus
2013). Various cutting-edge optical methods have also been
applied such as fluorescence resonance energy transfer
(FRET), fluorescence lifetime imaging (FLIM), fluorescence
recovery after photobleaching (FRAP), and fluorescence
correlation spectroscopy (FCS) (Lagerholm et al. 2005; Rao
and Mayor 2005; de Almeida et al. 2009). These optical
techniques are expected to provide more accurate informa-
tion of lipid rafts dynamics and the spatiotemporal signaling
events at these microdomains.
Biochemical methods to study lipid rafts signaling
The functions of lipid rafts in signal transduction have been
studied using several biochemical methods. First, using the
detergent-resistant property of lipid rafts, these membrane
fractions can be extracted with cold non-ionic detergent
(e.g., Triton X-100 at 4 �C), and then separated by sucrose
gradient ultracentrifugation (Simons and Ikonen 1997;
Hooper 1999). In fact, lipid rafts are also called detergent-
resistant membranes (DRMs). The distribution of signaling
proteins in separated DRMs can be measured by im-
munoblotting. However, this method has been controver-
sial because the results are largely affected by conditions of
the extraction (e.g., the types and concentrations of deter-
gents) (Hooper 1999). The proteins weakly attached to
lipid rafts may not be successfully isolated by the detergent
extraction method, and it is also difficult to capture the
dynamic distribution of signaling proteins, which occurs
during signal transduction in live cells.
Another typical method to study lipid rafts signaling is
to deplete cholesterol with methyl-b-cyclodextrin (MbCD)to measure the effect of rafts disruption (Simons and
Toomre 2000). Other approaches to disrupt the integrity of
lipid rafts have also been applied such as the inhibition of
cholesterol biosynthesis by Lovastatin, the addition of ex-
ogenous cholesterol, and the sequestration of cholesterol by
filipin or saponin (Simons and Toomre 2000). However,
these methods may promote some nonspecific artifacts, as
cholesterol is not simply the major component of lipid rafts
but is also involved in many cellular functions. In addition,
MbCD may work nonspecifically on other hydrophobic
molecules (Zidovetzki and Levitan 2007). These methods
are also limited to detecting dynamic signaling events
spatiotemporally at lipid rafts in live cells.
FRET-based molecular biosensor in lipid rafts signaling
FRET-based molecular biosensor
Fluorescent lipid/protein analogs or probes are useful tools
to investigate the distribution of proteins of interest on
membrane microdomains. However, simple fluorescent
tagging is not enough to visualize real activity of target
proteins on lipid rafts. Traditional biochemical assays also
have limitations for the detection of dynamic subcellular
protein activities in live cells. To visualize real-time activity
of signaling proteins in live cells, genetically encoded
molecular biosensors have been developed based on FRET
(Zhang et al. 2002; Aoki et al. 2013). FRET is a physical
phenomenon observed between donor and acceptor FPs at a
proximal distance within 10 nm. In addition to the proximal
distance, the emission spectrum of the donor FP should
overlap with the excitation spectrum of the effector FP to
achieve successful FRET. For example, the emission spec-
trum of cyan FP (CFP) largely overlap with the excitation
spectrum of yellow FP (YFP), thus they are widely used as a
FRET pair (Zhang et al. 2002; Aoki et al. 2013).
630 J Korean Soc Appl Biol Chem (2015) 58(5):629–636
123

Various FRET-based biosensors can be designed to vi-
sualize different signaling events (Zhang et al. 2002; Gaits
and Hahn 2003; Aoki et al. 2013). For example, molecular
biosensors detecting posttranslational modifications, such
as phosphorylation and methylation, can be designed to
include a specific substrate and its sensory domain between
a FRET pair. Some ligand-sensing biosensors contain only
a sensory domain between a FRET pair, and upon ligand
binding to the sensory domain, its conformation is altered
to cause FRET changes. Protease biosensors have a specific
substrate site for cleavage between a FRET pair. All of
these FRET biosensors are designed to induce huge con-
formational changes after specifically sensing the target
signaling events, resulting in significant changes in the
FRET level. Thus, by detecting FRET level changes, we
can monitor the dynamic signaling events in live cells. In
addition to these intra-molecular FRET biosensors, mole-
cular biosensors based on inter-molecular FRET have also
been developed to visualize the interactions/proximity be-
tween two target proteins, with each fused to either the
donor or acceptor FPs. Therefore, we can now monitor
real-time molecular interactions and dynamic signaling
events in live cells by measuring the changes in the FRET
level of these genetically encoded molecular biosensors.
Lipid rafts targeting signals
Genetically encoded FRET biosensors can be further tar-
geted to the subcellular compartments of the cell to detect
local activities of signaling molecules in live cells. Thus,
the integration of the lipid rafts targeting motif with FRET
biosensors allows the visualization of real-time signaling
events at lipid rafts. Zacharias et al. have shown that acy-
lated, but not prenylated, monomeric FPs can be parti-
tioned into membrane microdomains (Melkonian et al.
1999; Zacharias et al. 2002). When monomeric CFP or
YFP are genetically fused to the acylation sequences for
myristoylation and palmitoylation, which are from the Src
family kinase member Lyn kinase, efficient clustering be-
tween CFP and YFP is detected by FRET and this can be
completely inhibited by MbCD. In contrast, clustering of
monomeric CFP and YFP with the prenylation sequence
(i.e., the geranylgeranylation motif derived from Rho
GTPase) does not respond to MbCD treatment. In addition,
the prenylated FP is not clustered with the acylated FP,
suggesting that different lipid modifications can be used as
targeting signals to different membrane microdomains.
These results also indicate that the acylation, but not the
prenylation, motif is sufficient to target a protein at lipid
raft microdomains. Therefore, the addition of lipid
modification sequences to FRET biosensors can be applied
to the visualization of dynamic signaling events at mem-
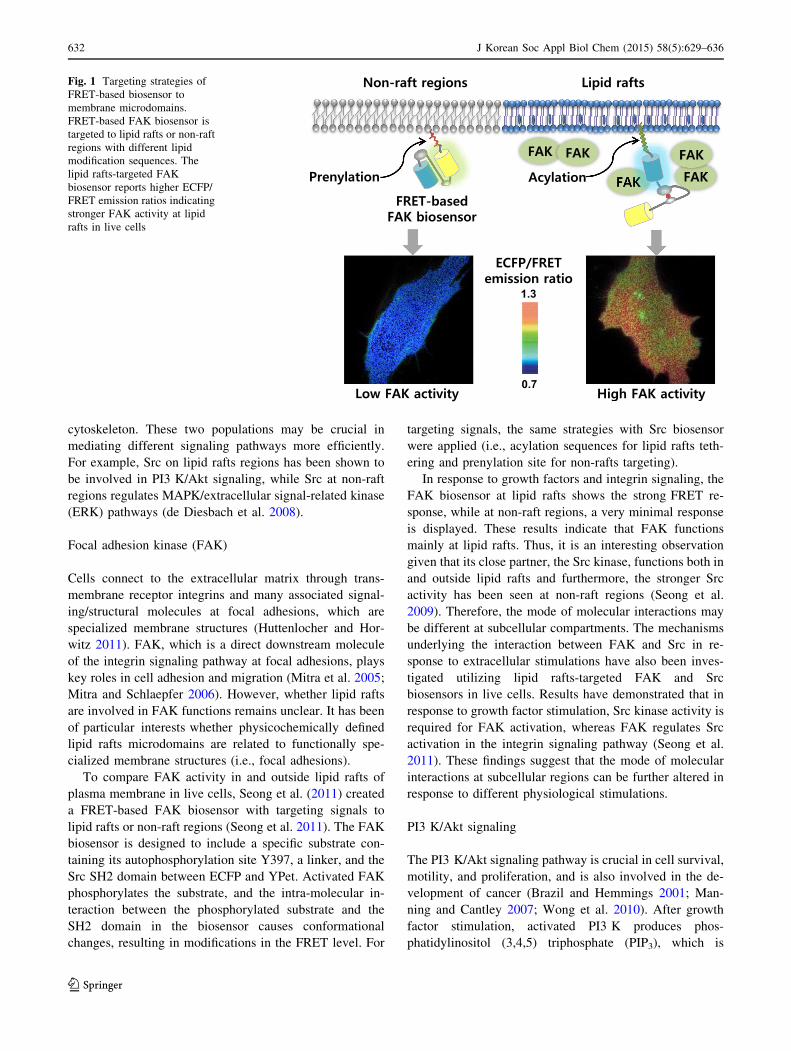
brane microdomains in living cells (Fig. 1).
FRET-based visualization of signaling events at lipid
rafts
Src kinase
Src kinase plays important roles in many cellular functions
such as cell migration, proliferation, and survival, and is
often found to be hyper-activated in many cancers (Thomas
and Brugge 1997; Martin 2001; Mitra and Schlaepfer
2006). Src kinases interact with various downstream sig-
naling molecules, including p130CAS, paxillin, focal ad-
hesion kinase, Ras/Mitogen-activated protein kinase
(MAPK), and phosphatidylinositol 3-kinase (PI3 K)/Akt.
Therefore, the tight regulation of spatiotemporal Src ac-
tivity is required to control these multiple signaling events
accurately and more efficiently. In response to various
extracellular signals such as growth factors, Src has been
suggested to be activated near the plasma membrane
(Sandilands et al. 2004). However, whether Src resides in
lipid rafts remains controversial, most likely because of the
limitation of previous tools to investigate signaling events
in these dynamic subcellular regions.
To visualize Src activity in and outside lipid rafts, Seong
et al. (2009) constructed FRET-based Src biosensors that
target these regions (Wang et al. 2005; Seong et al. 2009).
The Src biosensor is composed of a Src substrate peptide
derived from p130CAS, a flexible linker, and the SH2
domain between enhanced CFP (ECFP) and a YFP variant
(YPet). When active Src kinase phosphorylates the sub-
strate, the phosphorylated tyrosine in the substrate binds to
the nearby SH2 domain in the biosensor. This intra-mole-
cular interaction would cause a huge conformational
change of the biosensor, and the strong FRET between
ECFP and YPet would be significantly decreased. There-
fore, by measuring changes in the FRET level, real-time
Src activation in live cells can be visualized. This biosensor
is further targeted to lipid rafts or non-raft regions with
different lipid modifications (i.e., acylation or prenylation
sequences) as discussed in the previous section (Zacharias
et al. 2002).
Surprisingly, the results utilizing these subcellular-tar-
geted FRET-based Src biosensors showed the faster and
stronger Src activation outside lipid rafts in response to
growth factors and chemical stimulations (Seong et al.
2009). The quantitative analysis of Src activation by cal-
culating kinetic factors confirmed different kinetics of Src
activations in and outside lipid rafts (Seong et al. 2009).
Further experiments have revealed that relatively slower
Src activation on lipid rafts is dependent on actin-mediated
transportation, while fast Src activation in non-raft regions
is independent of the actin cytoskeleton (Seong et al.
2009). These results suggest the existence of two distinct
Src populations that are differentially regulated by the
J Korean Soc Appl Biol Chem (2015) 58(5):629–636 631
123
cytoskeleton. These two populations may be crucial in
mediating different signaling pathways more efficiently.
For example, Src on lipid rafts regions has been shown to
be involved in PI3 K/Akt signaling, while Src at non-raft
regions regulates MAPK/extracellular signal-related kinase
(ERK) pathways (de Diesbach et al. 2008).
Focal adhesion kinase (FAK)
Cells connect to the extracellular matrix through trans-
membrane receptor integrins and many associated signal-
ing/structural molecules at focal adhesions, which are
specialized membrane structures (Huttenlocher and Hor-
witz 2011). FAK, which is a direct downstream molecule
of the integrin signaling pathway at focal adhesions, plays
key roles in cell adhesion and migration (Mitra et al. 2005;
Mitra and Schlaepfer 2006). However, whether lipid rafts
are involved in FAK functions remains unclear. It has been
of particular interests whether physicochemically defined
lipid rafts microdomains are related to functionally spe-
cialized membrane structures (i.e., focal adhesions).
To compare FAK activity in and outside lipid rafts of
plasma membrane in live cells, Seong et al. (2011) created
a FRET-based FAK biosensor with targeting signals to
lipid rafts or non-raft regions (Seong et al. 2011). The FAK
biosensor is designed to include a specific substrate con-
taining its autophosphorylation site Y397, a linker, and the
Src SH2 domain between ECFP and YPet. Activated FAK
phosphorylates the substrate, and the intra-molecular in-
teraction between the phosphorylated substrate and the
SH2 domain in the biosensor causes conformational
changes, resulting in modifications in the FRET level. For
targeting signals, the same strategies with Src biosensor
were applied (i.e., acylation sequences for lipid rafts teth-
ering and prenylation site for non-rafts targeting).
In response to growth factors and integrin signaling, the
FAK biosensor at lipid rafts shows the strong FRET re-
sponse, while at non-raft regions, a very minimal response
is displayed. These results indicate that FAK functions
mainly at lipid rafts. Thus, it is an interesting observation
given that its close partner, the Src kinase, functions both in
and outside lipid rafts and furthermore, the stronger Src
activity has been seen at non-raft regions (Seong et al.
2009). Therefore, the mode of molecular interactions may
be different at subcellular compartments. The mechanisms
underlying the interaction between FAK and Src in re-
sponse to extracellular stimulations have also been inves-
tigated utilizing lipid rafts-targeted FAK and Src
biosensors in live cells. Results have demonstrated that in
response to growth factor stimulation, Src kinase activity is
required for FAK activation, whereas FAK regulates Src
activation in the integrin signaling pathway (Seong et al.
2011). These findings suggest that the mode of molecular
interactions at subcellular regions can be further altered in
response to different physiological stimulations.
PI3 K/Akt signaling
The PI3 K/Akt signaling pathway is crucial in cell survival,
motility, and proliferation, and is also involved in the de-
velopment of cancer (Brazil and Hemmings 2001; Man-
ning and Cantley 2007; Wong et al. 2010). After growth
factor stimulation, activated PI3 K produces phos-
phatidylinositol (3,4,5) triphosphate (PIP3), which is
Fig. 1 Targeting strategies of
FRET-based biosensor to
membrane microdomains.
FRET-based FAK biosensor is
targeted to lipid rafts or non-raft
regions with different lipid
modification sequences. The
lipid rafts-targeted FAK
biosensor reports higher ECFP/
FRET emission ratios indicating
stronger FAK activity at lipid
rafts in live cells
632 J Korean Soc Appl Biol Chem (2015) 58(5):629–636
123

negatively regulated by phosphatase and tensin homolog
deleted on chromosome 10 (PTEN). PIP3 then recruits Akt
to the plasma membrane, which is then phosphorylated by
phosphoinositide-dependent kinase 1 (PDK1), and is sub-
sequently fully activated by multiple proteins including
mammalian target of rapamycin complex (mTORC). Lipid
rafts have been suggested to be involved in PI3 K/Akt
signaling, but the regulatory mechanisms of dynamic Akt
activation at membrane microdomains are unclear.
Gao and Zhang (2008) constructed a FRET-based Akt
biosensor consisting of a substrate from FOXO1 and its
binding domain forkhead-associated domain (FHA1) be-
tween the FRET pair Cerulean and cpV E172 (Gao and
Zhang 2008). This Akt biosensor has been further targeted
in or outside lipid rafts by different lipid modifications. In
response to platelet-derived growth factors (PDGF) or in-
sulin-like growth factor-1 (IGF-1), these Akt biosensors
have reported the activation of Akt, both in and outside
lipid rafts. However, Akt activation in lipid rafts has been
shown to be faster and stronger than that of outside lipid
rafts.
In addition, when MbCD, which disrupts lipid rafts in-
tegrity, was treated, the PDGF-stimulated Akt activity at
lipid rafts, but not at non-raft regions, was decreased,
suggesting two independent Akt populations at membrane
microdomains. For IGF-1 stimulation, however, MbCDinhibits Akt activation both in and outside lipid rafts, and
interestingly the Akt activation at non-raft regions is
completely blocked. These findings suggest that IGF-1-
induced Akt activation at non-raft regions may be depen-
dent on the one at lipid rafts. Thus, stimulation by different
growth factors may result in different mechanisms of Akt
activation at membrane microdomains.
Gao et al. (2011) further investigated the regulatory
mechanisms of the compartmentalized activities of PI3 K/
Akt at membrane microdomains (Gao et al. 2011). This
study investigated at which membrane microdomains the
Akt-related signaling molecules are localized to regulate
the spatiotemporal activation of Akt. Therefore, they con-
structed a FRET-based biosensor to detect the activity of
PDK1, an upstream signaling molecule for Akt activation.
The PDK1 biosensor is composed of the full-length PDK1
molecule flanked by a FRET pair, ECFP and Citrine, and is
designed to directly detect the conformational change of
PDK1 by FRET measurement. The biosensor is targeted in
or outside lipid rafts with lipid modification sequences.
Results from these FRET-based PDK1 biosensors have
indicated that the activation of PDK1 is exclusively ob-
served in lipid rafts, which may explain the higher Akt
activity at this region. In contrast, PTEN, a negative
regulator for Akt activity, has been shown to be mainly
located at non-raft regions, and the altered distribution of
PTEN inhibits proper Akt activation. These also support
the regulatory mechanism of the higher Akt activity at lipid
rafts. Therefore, the precise spatiotemporal activities of
related signaling molecules are crucial for the proper sig-
naling of PI3 K/Akt.
Cyclic adenosine monophosphate (cAMP)/protein
kinase A (PKA)
Cyclic adenosine monophosphate is a second messenger
converted from adenosine triphosphate by adenylyl cyclase
(AC), which is activated by the G-protein-coupled receptor
(GPCR) on the plasma membrane (Chin et al. 2002).
Protein kinase A is a traditional target of the second mes-
senger cAMP and is involved in many cellular functions
such as gene expression and proliferation (Tasken and
Aandahl 2004). A-kinase anchoring protein (AKAP) (e.g.,
Ezrin) is known to localize PKA to appropriate subcellular
regions, which is crucial for PKA functions. However, how
PKA activity is regulated in and outside lipid rafts of the
plasma membrane is unclear.
To visualize PKA activity, Depry et al. (2011) devel-
oped the FRET-based PKA biosensor, which is composed
of the PKA substrate and forkhead-associated domain
(FHA1) between Cerulean and cpVenus (Depry et al.
2011). This FRET biosensor has also been further targeted
to membrane microdomains by lipid modifications. Results
from this study show that the basal PKA activity at resting
state is higher at lipid rafts than non-raft regions, and this
activity is dependent on PKA localization and lipid rafts
integrity. Interestingly, the disruption of lipid rafts by
MbCD induces a stronger and prolonged PKA response,
and this finding suggests that lipid rafts may be involved in
the proper desensitization of the b-adrenergic receptor
(bAR)-induced cAMP response (Depry et al. 2011). In fact,
bAR is shown to be localized at lipid rafts (Depry et al.
2011). It is possible that the disruption of lipid rafts may
cause the slower desensitization of this GPCR, because
lipid rafts are also suggested to be involved in endocytosis
and membrane trafficking (Hanzal-Bayer and Hancock
2007).
cAMP/exchange proteins directly activated by cAMP
(Epac)
The production of cAMP also regulates other signaling
molecules such as Epac (Bos 2003). Epac is a recently
found guanine exchange factor (GEF) that works on the
small GTPase, Rap1. It plays important roles in insulin
secretion, vascular permeability, and cardiac contraction.
The activation of Epac results from its conformational
changes upon cAMP binding. Therefore, spatiotemporal
distribution of cAMP is likely to control the cAMP-induced
Epac signaling pathway at subcellular levels.
J Korean Soc Appl Biol Chem (2015) 58(5):629–636 633
123
To visualize the distribution of cAMP at membrane
microdomains in live cells, DiPilato et al. (2004) designed
a FRET-based cAMP biosensor, which is composed of full-
length Epac1 between ECFP and citrine (DiPilato et al.
2004). Findings from this biosensor system have shown
that increased production of cAMP directly binds to Epac1
in the biosensor, causing the conformational change of the
biosensor and the subsequent changes in the FRET level.
This biosensor has been further targeted to lipid rafts by
acylation sequences (DiPilato and Zhang 2009). Upon ac-
tivation of the GPCR, bAR, this biosensor shows local andtransient increases in cAMP production in live cells. In-
terestingly, this cAMP activity is enhanced by the treat-
ment of MbCD, suggesting that lipid rafts may be involved
in the desensitization of bAR-induced cAMP signaling.
The prostaglandin E receptor is another GPCR located in
non-raft regions, and with the treatment of MbCD has been
shown not to enhance the prostaglandin E receptor-induced
cAMP response (DiPilato and Zhang 2009). Therefore, the
negative role of lipid rafts in cAMP signaling may be de-
pendent on the type and/or distribution of GPCR.
Agarwal et al. (2014) also constructed a FRET-based
cAMP biosensor. This biosensor is composed of Epac2
between EYFP and ECFP, and is targeted to lipid rafts or
non-raft regions of the plasma membrane by lipid modifi-
cation sequences (Agarwal et al. 2014). Results from this
group have revealed that basal cAMP activity is sig-
nificantly higher in non-raft regions at plasma membrane,
because the biosensors tethered at non-raft regions show
more sensitive FRET response to the inhibition of AC, a
positive regulator of cAMP. The FRET response of the
biosensors at non-raft regions also show a less sensitive
response to the inhibition of phosphodiesterase (PDE),
which is involved in the cAMP metabolism. Therefore,
different levels of cAMP production in and outside lipid
rafts may be due to the compartmentalized regulation of
the positive/negative regulatory molecules such as AC and
PDE.
Rac1
Rac1, a member of Rho GTPases, regulates cell adhesion
and motility through cytoskeletal reorganization (Jaffe and
Hall 2005). GDP-bound Rac1 is inactive and associated
with GDP-dissociation inhibitor (GDI) in the cytosol. Rac1
gets activated when GTP exchange factors (GEFs) replaces
GDP to GTP, and the dissociation of GDI exposes its
membrane targeting signals, leading Rac1 to plasma
membrane. Rac1-bound GTP can be hydrolyzed to GDP by
GTPase-activating protein (GAP), inactivating Rac1. The
precise location of Rac1 is required for the correct inter-
action with downstream signaling molecules. A previous
study has suggested that Rac1 may function in lipid raft
microdomains (del Pozo et al. 2004).
To investigate the distribution of Rac1 at membrane
microdomains of live cells, Moissoglu et al. (2014) de-
signed a FRET-based system consisting of GFP-tagged
Rac1 and mCherry fluorescence proteins fused to acyla-
tion/prenylation sequences (Moissoglu et al. 2014). In this
system, the acylated or prenylated mCherry FP is readily
located in or outside lipid rafts. When the activated Rac1
translocates to each microdomain of the plasma mem-
brane, FRET would be observed between Rac1-attached
GFP and mCherry that is tethered in or outside lipid rafts.
The FRET results have shown that Rac1 is located both in
lipid rafts and non-raft regions on the plasma membrane.
Further studies with a Rac GEF (Tiam) and a Rac GAP
(b2-chimaerin) have indicated that GEF activates both
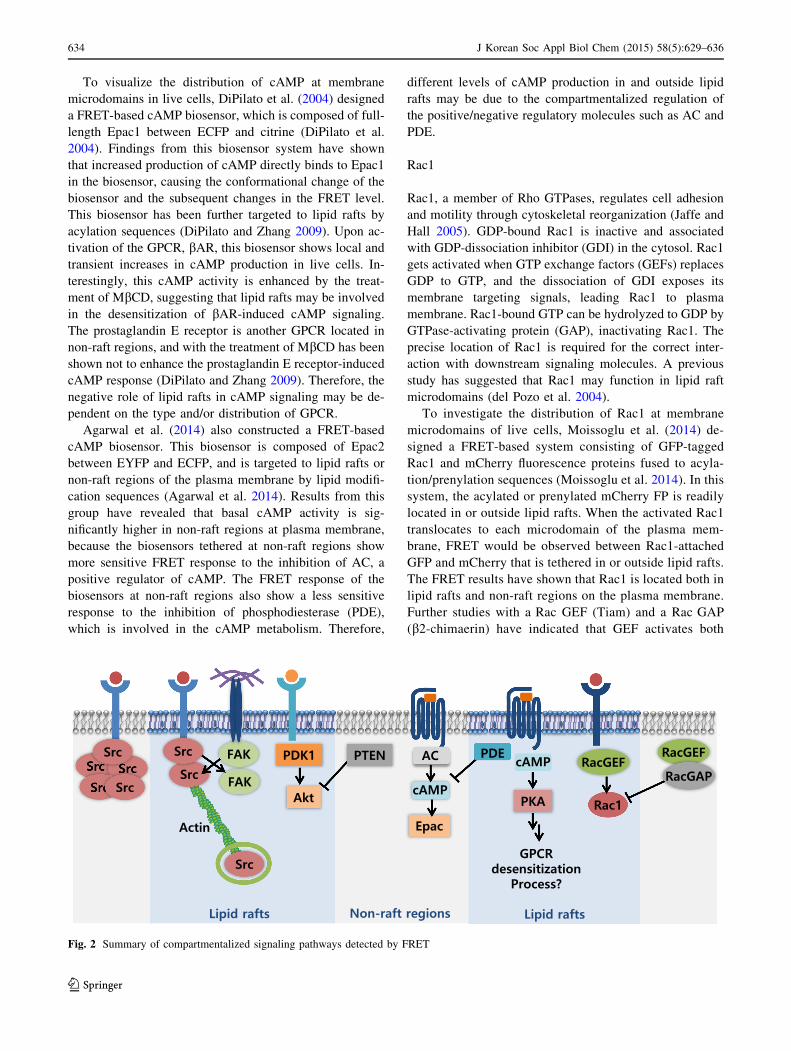
Fig. 2 Summary of compartmentalized signaling pathways detected by FRET
634 J Korean Soc Appl Biol Chem (2015) 58(5):629–636
123

Rac1 populations in and outside lipid rafts, while GAP
preferentially inactivates the Rac1 population in non-raft
regions. Thus, the selective function of the negative
regulator in the membrane compartments may contribute
to the preferential Rac1 activity that occurs at lipid rafts.
These FRET imaging system findings provide an insight
into the molecular mechanism of the differential Rac1
activity at membrane microdomains in live cells.
Conclusion and perspectives
In this review, several lipid rafts signaling pathways visu-
alized by FRET biosensors were discussed (Summarized in
Fig. 2). FRET-based molecular biosensors have been used
as powerful tools to visualize real-time signaling events in
live cells with high spatiotemporal resolutions. Therefore,
the integration of these biosensors with subcellular target-
ing signals (i.e., acylation/prenylation sequences) success-
fully provides a unique strategy to precisely monitor
dynamic signaling events and to study the molecular
regulatory mechanisms at these membrane microdomains.
FRET biosensors with lipid modification sequences pro-
vide a much deeper understanding of the distribution,
functional activities, and regulatory mechanisms of crucial
signaling molecules (e.g., Src, FAK, PI3 K/Akt, cAMP/
PKA, and Rac1) at membrane microdomains. These sig-
naling pathways are also related to diseases such as can-
cers. Therefore, subcellular-targeted FRET biosensors can
contribute to the development of new diagnostic tools, drug
screening methods, and therapeutic strategies. Recent ad-
vances in super-resolution optical techniques such as STED
microscopy, PALM, and STORM are promising to visu-
alize nanometer-sized individual lipid raft and the local
signaling pathways more accurately. The combination of
these new optical techniques with FRET-based biosensors
will provide a more in-depth understanding of lipid rafts
signaling pathways with super spatiotemporal resolutions.
Acknowledgments This work is supported by the Korea Institute of
Science and Technology (KIST-2E25240, 2E25473).
Conflict Interests The author declares no competing interests.
References
Agarwal SR, Yang PC, Rice M, Singer CA, Nikolaev VO, Lohse MJ,
Harvey RD (2014) Role of membrane microdomains in
compartmentation of cAMP signaling. PLoS ONE 9:e95835
Allen JA, Halverson-Tamboli RA, Rasenick MM (2007) Lipid raft
microdomains and neurotransmitter signalling. Nat Rev Neu-
rosci 8:128–140
Aoki K, Kamioka Y, Matsuda M (2013) Fluorescence resonance
energy transfer imaging of cell signaling from in vitro to in vivo:
basis of biosensor construction, live imaging, and image
processing. Dev Growth Differ 55:515–522
Bos JL (2003) Epac: a new cAMP target and new avenues in cAMP
research. Nat Rev Mol Cell Bio 4:733–738
Brazil DP, Hemmings BA (2001) Ten years of protein kinase B
signalling: a hard Akt to follow. Trends Biochem Sci
26:657–664
Brown DA, London E (1998) Functions of lipid rafts in biological
membranes. Annu Rev Cell Dev Biol 14:111–136
Brown DA, Rose JK (1992) Sorting of GPI-anchored proteins to
glycolipid-enriched membrane subdomains during transport to
the apical cell surface. Cell 68:533–544
Chin KV, Yang WL, Ravatn R, Kita T, Reitman E, Vettori D, Cvijic
ME, Shin KV, Iacono L (2002) Reinventing the wheel of cyclic
AMP—novel mechanisms of cAMP signaling. Ann Ny Acad Sci
968:49–64
de Almeida RF, Loura LM, Prieto M (2009) Membrane lipid domains
and rafts: current applications of fluorescence lifetime spec-
troscopy and imaging. Chem Phys Lipids 157:61–77
de Diesbach P, Medts T, Carpentier S, D’Auria L, Van Der Smissen
P, Platek A,Mettlen M, Caplanusi A, van den Hove MF, Tyteca
D, Courtoy PJ (2008) Differential subcellular membrane recruit-
ment of Src may specify its downstream signalling. Exp Cell Res
314:1465–1479
del Pozo MA, Alderson NB, Kiosses WB, Chiang HH, Anderson RG,
Schwartz MA (2004) Integrins regulate Rac targeting by
internalization of membrane domains. Science 303:839–842
Depry C, Allen MD, Zhang J (2011) Visualization of PKA activity in
plasma membrane microdomains. Mol BioSyst 7:52–58
DiPilato LM, Zhang J (2009) The role of membrane microdomains in
shaping beta2-adrenergic receptor-mediated cAMP dynamics.
Mol BioSyst 5:832–837
DiPilato LM, Cheng X, Zhang J (2004) Fluorescent indicators of
cAMP and Epac activation reveal differential dynamics of
cAMP signaling within discrete subcellular compartments. Proc
Natl Acad Sci USA 101:16513–16518
Gaits F, Hahn K (2003) Shedding light on cell signaling: interpre-
tation of FRET biosensors Science’s STKE : signal transduction
knowledge environment 2003:PE3
Gao X, Zhang J (2008) Spatiotemporal analysis of differential Akt
regulation in plasma membrane microdomains. Mol Biol Cell
19:4366–4373
Gao X, Lowry PR, Zhou X, Depry C, Wei Z, Wong GW, Zhang J
(2011) PI3 K/Akt signaling requires spatial compartmentaliza-
tion in plasma membrane microdomains. Proc Natl Acad Sci
USA 108:14509–14514
Hanzal-Bayer MF, Hancock JF (2007) Lipid rafts and membrane
traffic. FEBS Lett 581:2098–2104
Hooper NM (1999) Detergent-insoluble glycosphingolipid/choles-
terol-rich membrane domains, lipid rafts and caveolae (review).
Mol Membr Biol 16:145–156
Horejsi V (2003) The roles of membrane microdomains (rafts) in T
cell activation. Immunol Rev 191:148–164
Huttenlocher A, Horwitz AR (2011) Integrins in cell migration. Csh
Perspect Biol 3. doi: 10.1101/cshperspect.a005074
Ishitsuka R, Sato SB, Kobayashi T (2005) Imaging lipid rafts.
J Biochem 137:249–254
Jaffe AB, Hall A (2005) Rho GTPases: biochemistry and biology.
Ann Rev cell Dev Biol 21:247–269
Kenworthy AK, Petranova N, Edidin M (2000) High-resolution FRET
microscopy of cholera toxin B-subunit and GPI-anchored
proteins in cell plasma membranes. Mol Biol Cell 11:1645–1655
Lagerholm BC, Weinreb GE, Jacobson K, Thompson NL (2005)
Detecting microdomains in intact cell membranes. Ann Rev
Phys Chem 56:309–336
J Korean Soc Appl Biol Chem (2015) 58(5):629–636 635
123

Manning BD, Cantley LC (2007) AKT/PKB signaling: navigating
downstream. Cell 129:1261–1274
Martin GS (2001) The hunting of the Src. Nat Rev Mol Cell Biol
2:467–475
Melkonian KA, Ostermeyer AG, Chen JZ, Roth MG, Brown DA
(1999) Role of lipid modifications in targeting proteins to
detergent-resistant membrane rafts Many raft proteins are
acylated, while few are prenylated. J Biol Chem 274:3910–3917
Mitra SK, Schlaepfer DD (2006) Integrin-regulated FAK-Src signal-
ing in normal and cancer cells. Curr Opin Cell Biol 18:516–523
Mitra SK, Hanson DA, Schlaepfer DD (2005) Focal adhesion kinase:
in command and control of cell motility. Nat Rev Mol Cell Bio
6:56–68
Moissoglu K, Kiessling V, Wan C, Hoffman BD, Norambuena A,
Tamm LK, Schwartz MA (2014) Regulation of Rac1 transloca-
tion and activation by membrane domains and their boundaries.
J Cell Sci 127:2565–2576
Owen DM, Gaus K (2013) Imaging lipid domains in cell membranes:
the advent of super-resolution fluorescence microscopy. Front
Plant Sci 4:503. doi:10.3389/fpls.2013.00503
Owen DM, Magenau A, Williamson D, Gaus K (2012) The lipid raft
hypothesis revisited–new insights on raft composition and
function from super-resolution fluorescence microscopy. BioEs-
says 34:739–747
Rao M, Mayor S (2005) Use of forster’s resonance energy transfer
microscopy to study lipid rafts. Biochim et Biophys Acta
1746:221–233
Sandilands E, Cans C, Fincham VJ, Brunton VG, Mellor H,
Prendergast GC, Norman JC, Superti-Furga G, Frame MC
(2004) RhoB and actin polymerization coordinate Src activation
with endosome-mediated delivery to the membrane. Dev Cell
7:855–869
Sato SB, Ishii K, Makino A, Iwabuchi K, Yamaji-Hasegawa A,
Senoh Y, Nagaoka I, Sakuraba H, Kobayashi T (2004) Distri-
bution and transport of cholesterol-rich membrane domains
monitored by a membrane-impermeant fluorescent polyethylene
glycol-derivatized cholesterol. J Biol Chem 279:23790–23796
Schroeder RJ, Ahmed SN, Zhu Y, London E, Brown DA (1998)
Cholesterol and sphingolipid enhance the Triton X-100 in-
solubility of glycosylphosphatidylinositol-anchored proteins by
promoting the formation of detergent-insoluble ordered mem-
brane domains. J Biol Chem 273:1150–1157
Schutz GJ, Kada G, Pastushenko VP, Schindler H (2000) Properties
of lipid microdomains in a muscle cell membrane visualized by
single molecule microscopy. EMBO J 19:892–901
Seong J, Lu S, Ouyang M, Huang H, Zhang J, Frame MC, Wang Y
(2009) Visualization of Src activity at different compartments of
the plasma membrane by FRET imaging. Chem Biol 16:48–57
Seong J, Ouyang M, Kim T, Sun J, Wen PC, Lu S, Zhuo Y,Llewel-
lyn NM, Schlaepfer DD, Guan JL, Chien S, Wang Y (2011)
Detection of focal adhesion kinase activation at membrane
microdomains by fluorescence resonance energy transfer. Nat
Commun 2:406. doi:10.1038/ncomms1414
Shakor AB, Czurylo EA, Sobota A (2003) Lysenin, a unique
sphingomyelin-binding protein. FEBS Lett 542:1–6
Sharma P, Varma R, Sarasij RC, Ira, Gousset K, Krishnamoorthy
G, Rao M, Mayor S (2004) Nanoscale organization of multiple
GPI-anchored proteins in living cell membranes. Cell
116:577–589
Silvius JR, del Giudice D, Lafleur M (1996) Cholesterol at different
bilayer concentrations can promote or antagonize lateral segre-
gation of phospholipids of differing acyl chain length. Biochem-
istry 35:15198–15208
Simons K, Gerl MJ (2010) Revitalizing membrane rafts: new tools
and insights. Nat Rev Mol Cell Biol 11:688–699
Simons K, Ikonen E (1997) Functional rafts in cell membranes.
Nature 387:569–572
Simons K, Toomre D (2000) Lipid rafts and signal transduction. Nat
Rev Mol Cell Biol 1:31–39
Singer SJ, Nicolson GL (1972) The fluid mosaic model of the
structure of cell membranes. Science 175:720–731
Skocaj M, Bakrac B, Krizaj I, Macek P, Anderluh G, Sepcic K (2013)
The sensing of membrane microdomains based on pore-forming
toxins. Curr Med Chem 20:491–501
Tasken K, Aandahl EM (2004) Localized effects of cAMP mediated
by distinct routes of protein kinase A. Physiol Rev 84:137–167
Thomas SM, Brugge JS (1997) Cellular functions regulated by Src
family kinases. Annu Rev Cell Dev Biol 13:513–609
Thompson TE, Tillack TW (1985) Organization of glycosphin-
golipids in bilayers and plasma membranes of mammalian cells.
Annu Rev Biophys Biophys Chem 14:361–386
Wang Y, Botvinick EL, Zhao Y, Berns MW, Usami S, Tsien RY,
Chien S (2005) Visualizing the mechanical activation of Src.
Nature 434:1040–1045
Wong KK, Engelman JA, Cantley LC (2010) Targeting the PI3 K
signaling pathway in cancer. Curr Opin Genet Dev 20:87–90
Zacharias DA, Violin JD, Newton AC, Tsien RY (2002) Partitioning
of lipid-modified monomeric GFPs into membrane mi-
crodomains of live cells. Science 296:913–916
Zhang J, Campbell RE, Ting AY, Tsien RY (2002) Creating new
fluorescent probes for cell biology. Nat Rev Mol Cell Biol
3:906–918
Zidovetzki R, Levitan I (2007) Use of cyclodextrins to manipulate
plasma membrane cholesterol content: evidence, misconceptions
and control strategies. Biochim Biophys Acta 1768:1311–1324
636 J Korean Soc Appl Biol Chem (2015) 58(5):629–636
123