localization of hydrogen peroxide accumulation during the ... · ing cross-linking reactions...
TRANSCRIPT

The Plant Cell, Vol. 9, 209-221, February 1997 O 1997 American Society of Plant Physiologists
Localization of Hydrogen Peroxide Accumulation during the Hypersensitive Reaction of Lettuce Cells to Pseudomonas syringae pv phaseolicola
Charles S. Bestwick,' Ian R. Brown, Mark H. R. Bennett, and John W. Mansfield Department of Biological Sciences, Wye College, University of London, Wye, Near Ashford, Kent TN25 5AH, United Kingdom
The active oxygen species hydrogen peroxide (H202) was detected cytochemically by its reaction with cerium chloride to produce electron-dense deposits of cerium perhydroxides. In uninoculated lettuce leaves, H202 was typically present within the secondary thickened walls of xylem vessels. Inoculation with wild-type cells of Pseudomonas syringae pv phaseolicola caused a rapid hypersensitive reaction (HR) during which highly localized accumulation of H202 was found in plant cell walls adjacent to attached bacteria. Quantitative analysis indicated a prolonged burst of H202 occurring be- tween 5 to 8 hr after inoculation in cells undergoing the HR during this example of non-host resistance. Cell wall alterations and papilla deposition, which occurred in response to both the wild-type strain and a nonpathogenic hrpD mutant, were not associated with intense staining for H202, unless the responding cell was undergoing the HR. Cata- lase treatment to decompose H,O, almost entirely eliminated staining, but 3-amino-l,2,4-triazole (catalase inhibitor) did not affect the pattern of distribution of H202 detected. H202 production was reduced more by the inhibition of plant per- oxidases (with potassium cyanide and sodium azide) than by inhibition of neutrophil-like NADPH oxidase (with diphenylene iodonium chloride). Results suggest that CeCI, reacts with excess H202 that is not rapidly metabolized dur- ing cross-linking reactions occurring in cell walls; such an excess of H202 in the early stages of the plant-bacterium interaction was only produced during the HR. The highly localized accumulation of H,Oz is consistent with its direct role as an antimicrobial agent and as the cause of localized membrane damage at sites of bacterial attachment.
INTRODUCTION
The active resistance of plants to colonization by bacteria and fungi is often expressed by the hypersensitive reaction (HR) of challenged plant cells (Ingram, 1978; Klement, 1982; Mansfield, 1990; Tenhaken et al., 1995). The HR can be rec- ognized as the rapid and localized death of cells in response to an avirulent pathogen; it has been observed during most interactions involving race-specific resistance and also in many examples of non-host resistance (Heath, 1989; Mansfield, 1990; Mansfield et al., 1997). A second form of resistance, more commonly found in non-host reactions, is the highly localized alteration of the cell wall at sites attacked by fungi or bacteria. Modification of the cell wall per se is often asso- ciated with the formation of a papilla or apposition at reac- tion sites (Ride, 1986; Nicholson and Hammerschmidt, 1992; Bestwick et al., 1995). In phytopathogenic bacteria, hyper- sensitive response and pathogenicity (hrp) genes determine the ability to multiply within susceptible plants and to cause
Current address: Division of Biochemical Sciences, Rowett Re- search Institute, Greenburn Road, Bucksburn, Aberdeen AB21 9SB, UK. To whom correspondence should be addressed.
a macroscopic HR in either resistant varieties of their host or in non-host plants (Bonas, 1994). Hrp mutants, although fail- ing to cause the HR, do cause defense responses in plants, including the alteration of adjacent cell walls and papilla for- mation (Jakobek and Lindgren, 1993; Bestwick et al., 1995; Brown et al., 1995; Fett and Jones, 1995).
The term active oxygen species (AOS; also referred to as reactive oxygen species or reactive oxygen intermediates) describes radicals and other nonradical but reactive species derived from oxygen. The rapid production and accumula- tion of AOS, primarily the superoxide anion (O,. -) and hy- drogen peroxide (H202), have been shown to occur in several plant-microbe interactions (for recent reviews, see Mehdy, 1994; Baker and Orlandi, 1995; Low and Merida, 1996). In aqueous solutions, O,' ~ exists in acid base equilib- rium with the protonated form, the hydroperoxyl radical HO,. (Sutherland, 1991). Both 0,'- and HO,. species un- dergo dismutation to yield H,O2. Increased AOS production may exert a toxic effect within cells via lipid peroxidation, protein degradationhodification, and DNA damage (re- viewed in Fridovich, 1986; Wolff et al., 1986; lmlay and Linn, 1988). In a number of studies, researchers have detected in- creased lipid peroxidation during the initial stages of the HR

21 O The Plant Cell
(Adám et al., 1989, 1995; Croft et al., 1990; May et al., 1996). Despite the temporal association of 0,. - production with lipid peroxidation, the superoxide anion, unlike Hop., is not capable by itself of performing H abstraction, and because it is charged, it is also unable to penetrate the hydrophobic in- terior of membranes (Halliwell and Gutteridge, 1989). H202 has a similarly low reactivity toward biological molecules. But in the presence of trace quantities of metal ions, H202 may participate in both the Fenton and Haber-Weiss reac- tions to form the highly reactive hydroxyl radical (HO’). For- mation of HO’ probably accounts for much of the in vivo toxicity associated with excessive H202 production. H,O, is relatively stable, can diffuse freely throughout the cell, and hence may be able to participate in HO’ formation at sites distant from its production (Baker and Orlandi, 1995).
Various enzyme systems have been proposed to generate AOS in plants. These include a neutrophil-analogous, mem- brane-bound NADPH oxidase (Doke, 1983; Levine et al., 1994; Auh and Murphy, 1995; Doke and Miura, 1995; Dwyer et al., 1996), lipoxygenase (Croft et al., 1990), and apoplastic peroxidases (Vera-Estrela et al., 1992; Bolwell et al., 1995). Adam et al. (1 995) have suggested that H202 accumulation may occur by a reduction in H202 scavenging and an in- crease in H,OZ production involving coordinated changes in peroxidase, superoxide dismutase, and catalase activities.
In many plants, treatment with inhibitors of protein syn- thesis has been found to prevent or delay the HR, implying a requirement for active metabolism within the plant cell lead- ing to cell collapse (Woods et al., 1989; He et al., 1993). The occurrence of lesion mimics (notably in Arabidopsis), which rapidly form HR-like lesions in the absence of an avirulent pathogen, suggests that the HR is under genetic control. The plant’s response may therefore represent a form of pro- grammed cell death (pcd; Dietrich et al., 1994; Greenberg et al., 1994). Levine et al. (1994) have suggested that H202 may initiate pcd events leading to the HR.
The most common form of pcd in animal cells is mani- fested in apoptosis. This descriptive term is used to define cells that demonstrate particular morphological and biochemical features associated with their death. Typical ultrastructural features of apoptosis include chromatin condensation, cell surface blebbing, chromatin migration to the nuclear mar- gins, nuclear fragmentation, and formation of several mem- brane-bound apoptotic bodies (Collins et al., 1992; Buja et al., 1993; Kerr et al., 1994). Biochemically, increased pro- tease and endonuclease activities and internucleosomal DNA cleavage have been reported (Arends and Wyllie, 1991; Martin et al., 1994). Some of these features have been asso- ciated with developmental and pathological cell death in plants (Mittler and Lam, 1995; Levine et al., 1996; Ryerson and Heath, 1996; Wang et al., 1996), but the HR often lacks the classic ultrastructural features associated with apopto- sis (Bestwick et al., 1995; Mittler and Lam, 1996). AOS have been shown to induce apoptosis in mammalian cells, but the involvement of AOS in the execution of apoptotic cell death is controversial (Buttke and Sandstrom, 1995; Jacobson, 1996).
In plants, H,Op production is also involved in the defen- sive modifications that occur within the walls of challenged cells (Bolwell, 1993; Wojtaszek et al., 1995). In some cases, alteration to cell wall structure may be an extremely rapid re- sponse. Bradley et al. (1992) identified a rapid H,O,-depen- dent insolubilization of a proline-rich protein and putative hydroxyproline-rich glycoprotein (HRGP) within 2 min of ap- plication of an elicitor to soybean cell suspensions. HRGPs have been shown to accumulate at sites of bacterial attach- ment to plant cell walls and also within the papillae that form adjacent to bacteria or sites of attempted fungal penetration (O’Connell et al., 1990; Bestwick et al., 1995; Brown et al., 1995). The accumulation of lignin or ligninlike materials has been observed in numerous incompatible interactions (Ride, 1986; Nicholson and Hammerschmidt, 1992). Polymeriza- tion of hydroxycinnamyl alcohols in the final stages of lignin biosynthesis and the intramolecular and intermolecular cross-linking of HRGPs are both catalyzed by isoperoxidase activities. In the presence of H202, peroxidase is also thought to mediate cross-linking between numerous other cell wall components (liyama et al., 1994). The detection of cross-linking within tissues is complicated by the highly lo- calized nature of reactions that occur at the subcellular level in plant cells (Bestwick et al., 1995; Brown et al., 1995). Al- terations to wall structure involving H20p may restrict diffu- sion of host-derived nutrients to the pathogen and pathogen-derived toxins to the host and alter the availability of wall linkages to cell wall-degrading enzymes produced by some potential pathogens (Friend, 1981; Brisson et al., 1994). The production of H202 may also lead to the develop- ment of an antimicrobial environment within the apoplasm (Peng and Kuc, 1992).
In addition to its proposed roles as (1) a direct cause of membrane damage, (2) a component of structural defense, (3) a signal molecule operating within the challenged cell, and (4) an antimicrobial compound, H202 has also been hy- pothesized to act as a signal molecule in the induction of systemic acquired resistance (Chen et al., 1993). There is, however, very little information on the precise localization of H202 production or accumulation during plant-pathogen in- teractions. Direct attempts to localize AOS by light micros- copy have included the use of nitro blue tetrazolium salt to detect 0,. ~ (Adám et al., 1989), potassium iodidektarch to detect H202 (Olson and Varner, 1993), and compounds that either fluoresce or have their fluorescence quenched when oxidized by H,Op (Yahraus et al., 1995). Detection of H,02 as a product of enzyme catalysis has formed the basis of the localization at the subcellular level of enzyme activities that generate the AOS in animals and plants, for example, NADH oxidase and glycolate oxidase (Briggs et al., 1975; Kausch et al., 1983; Kausch, 1987). The histochemical as- says used are based on the reaction of HP02 with CeCI, to produce electron-dense insoluble precipitates of cerium per- hydroxides, Ce[OH],OOH and Ce[OH],OOH. This ultrastruc- tural technique allows the precise localization of sites of H202 accumulation or production and has been used by

H,Op Localization and the HR 21 1
Czaninski et al. (1993) in a study of endogenous H,Op pro- duction in lignifying tissues.
Here, we report use of the CeCI, procedure to localize H,Op within lettuce cells challenged by wild-type and hrpD mutant strains of Pseudomonas syringae pv phaseolicola. Because both strains cause localized cell wall alterations, in- cluding HRGP and phenolic deposition, but only the wild type causes the HR (Bestwick et al., 1995), responses spe- cifically associated with the HR were determined. Our ex- periments reveal the highly localized accumulation of H202 occurring within regions of the cell wall adjacent to wild-type bacteria during the early stages of the HR. Results obtained support the proposals that H202 may have an antibacterial role and also may be a primary cause of membrane damage in plant cells undergoing the HR during non-host resistance.
RESULTS
Localization of H202 in Uninoculated Tissue
In noninoculated leaves or tissue away from bacterial colo- nies, precipitates of electron-dense cerium perhydroxides, indicating the presence H202, were located predominantly within the cell walls of xylem vessels with secondary thick-
ening and occasionally within the walls of surrounding cells (Figure 1A). Junctions between mesophyll cells, typically the sites of bacterial attachment to the plant cell wall, were usu- ally free of staining (Figure 1 B).
Localized Accumulation of H202 Is Detected at Reaction Sites
After injection, as inoculum droplets evaporated and were absorbed, bacterial cells were drawn into contact with plant cell walls. Wild-type and hrp mutant bacteria caused similar responses within the first 5 hr of inoculation. The plasma membrane adjacent to bacteria became convoluted, and amorphous material, initiating papilla formation, was depos- ited between the plant plasma membrane and cell wall. After 5 hr, deposition of HRGPs and phenolics was already ob- served in the altered cell walls adjacent to attached bacteria. Papillae continued to expand at sites inoculated with the hrp mutant, but vesiculation of the cytoplasm and cytoplasmic disorganization, indicative of the HR, occurred extensively only in plant cells next to wild-type bacteria, as reported in detail by Bestwick et al. (1995).
Preliminary observations, after CeCI, staining of samples 5 and 8 hr after inoculation, revealed H202 accumulation at reaction sites. Formation of the cerium perhydroxides was
Figure 1. Detection of H,Op in Uninoculated Tissue.
(A) Staining with CeCI, to form electron-dense deposits of cerium perhydroxides localized to spiral thickening of the xylem vessels and parts of the associated cell wall (arrows). Bar = 5 pm. (B) Absence of staining from a typical junction between two spongy mesophyll cells. Bar = 1 pm. c, chloroplast: CV, central vacuole; IS, intercellular space; XV, xylem vessel.

Figure 2. Localization of H2O2 Accumulation in Cells Adjacent to Wild-Type f . s. phaseokola.
(A) Electron-dense deposits of cerium perhydroxides extending from the site of attachment of wild-type bacteria (large arrow) into the surround- ing plant cell wall (small arrows). The small papillae formed were also intensely stained 8 hr after inoculation. Note that the cytoplasm of re- sponding cells is already vesiculated near the bacteria but that the nucleus (N), close to the reaction site, shows no signs of chromatin condensation or nuclear fragmentation (i.e., apoptotic features). Bar = 2 pm. (B) Restriction of H202 to one of a pair of adjacent mesophyll cells 5 hr after inoculation. Note that the one shared wall of two cells is intensely stained but that staining only extends into the individual wall of one cell (arrow). Bar = 1 pm. B, bacterium; c, chloroplast; CV, central vacuole; IS, intercellular space; m, mitochondrion; N, nucleus; V, vesiculation.

H,Op Localization and the HR 213
particularly striking in the plant cell walls adjacent to wild- type bacteria and was remarkably localized. The most dense deposits were found next to bacteria, and they extended with reducing intensity into the surrounding walls (Figure 2A). Staining was also frequently observed in material en- capsulating the bacteria. At some sites, although bacterial cells were lodged between two mesophyll cells, staining with CeCI, was detected predominantly in one cell, indicat- ing differential activation of the oxidative burst (Figure 2B).
Quantitative Analysis Reveals a Prolonged Burst of H202 during the HR
To assess H202 accumulation objectively, we developed a four-point scale to assess the appearance of deposits of ce- rium perhydroxides within the plant cell wall as follows: O, none; 1, faint and patchy; 2, dense but patchy; 3, dense and confluent deposits. The minimum degree of deposit as- signed to the categories is illustrated in Figures 3A to 3D. Figure 3 also provides additional data on the localization of deposits within responding cells.
The accumulation and persistence of H202 were then quantified at different times after inoculation, following as- signment of a total of at least 30 reaction sites to categories. The results presented in Table 1 clearly show that H202 ac- cumulation was an early response observed in the interac- tion with wild-type bacteria that caused the HR. A prolonged burst of H202 accumulation was apparent between 5 and 8 hr after inoculation with the wild-type strain. Analysis of data for the hrpD mutant and wild-type bacteria 5 and 8 hr after inoculation, using xz analysis of contingency tables pre- pared by grouping staining into low (categories O or 1) and high (categories 2 and 3) classes, revealed statistically sig- nificant differences between the strains at P < 0.001. The strong staining of a low percentage of cells challenged by the hrp mutant (after 8 and 16 hr) corresponded to the low numbers of cells found to undergo cytoplasmic collapse in this interaction (Bestwick et al., 1995).
The possibility that wild-type bacteria might produce H202 in the plant was discounted by experiments on the induction time for the HR. lnduction time, which was first demon- strated by Klement and Goodman (1967), defines the time during which protein synthesis must be performed by bacte- ria in the plant to induce a macroscopic HR. In the lettuce- P.s. phaseolicola interaction, the induction time is 2 to 2.5 hr, after which the prevention of bacterial protein synthesis does not stop the HR. Results presented in Table 2 show that when bacteria were inactivated by streptomycin injec- tion, H,Op continued to accumulate only if the induction time had passed, that is, if the HR continued to develop. The production of H202 was therefore shown to be a plant response.
In an attempt to determine the concentrations of H202 re- quired to caúse the staining observed in the cell wall, we in- jected noninoculated leaves with solutions of H202 ranging
from 1 FM to 3 M and immediately processed samples with CeCI,. Faint staining (equivalent to category 1) was ob- served after injection with 100 pM H,O,, and category 2 staining was frequently observed with concentrations in ex- cess of 100 mM. However, dense, category 3 staining was observed only with concentrations of or in excess of 1 M H202. It was necessary to process tissues within 5 min after the application of exogenous H202, because the ability to detect cerium perhydroxide formation in the cell wall rapidly declined after this time. Tissues injected with H202 were also examined by electron microscopy after 12 hr, and the number of collapsed cells was estimated. Treatment with 0.1 M H202 had caused cytoplasmic collapse in -0 to 10% of the mesophyll cells; 1 and 3 M H202 caused 25 to 30% and 80 to 90% cell collapse, respectively.
Papilla Formation 1s Not Associated with Major Accumulation of H20,
As previously reported (Bestwick et al., 1995), papilla devel- opment occurred in response to both wild-type and hrp mu- tant strains. Initially, papilla development was slightly more rapid in response to the wild-type bacterium; however, by 16 hr after inoculation, large papillae were more frequently observed in cells next to the hrp mutant. High levels of H2O2, indicated by either dense but patchy or confluent cerium per- hydroxide formation, were detected within small papillae produced in response to the wild-type strain during the early stages of the interaction (Figure 4A). In more developed pa- pillae (>0.3 pm thick), observed in the first 8 hr after inocu- lation, staining was often most intense in the outer layers of paramural deposits (Figure 4B). By 12 hr after inoculation, 50% of the lettuce cells next to wild-type bacteria had col- lapsed, and after 24 hr, the figure had increased to 85% (Bestwick et al., 1995). During the later stages of cell col- lapse (after 16 and 24 hr), staining was either absent from or located at the edge of papillae (Figure 4C). Although papillae adjacent to the hrpD mutant appeared morphologically identical to those formed after challenge with wild-type bac- teria throughout the time course, as shown in Figure 4D, they typically lacked dense deposits of cerium perhydrox- ides after staining with CeCI,.
Effects of lnhibitors on H202 Accumulation
Detecting deposits of cerium perhydroxides at reaction sites raised severa1 questions. For example, how specific was CeCI, staining for H,02, and was the accumulation of H,02 generated by peroxidase or NADPH oxidase activities? In attempts to answer these questions, CeCI, staining was ex- amined in the presence of catalase (to decompose H202), aminotriazole (ATZ; to inhibit catalase in the plant), potas- sium cyanide and sodium azide to inhibit peroxidases, and diphenylene iodonium chloride (DPI) to inhibit neutrophil-like

21 4 The Plant Cell
Figure 3. Minimum Intensity of Staining Used to Quantify H202 Accumulation in the Cell Wall as Detected by the Formation of Cerium Perhy- droxides at Reaction Sites.
(A) No staining. (B) Category 1 : faint and patchy staining. (C) Category 2: dense but patchy staining. . . (D) Category 3: dense and confluent staining. (A) and (B) show tissue 16 and 8 hr after inoculation with the hrpD mutant, respectively; (C) and (D) show tissue 8 and 5 hr after inoculation with wild-type P. s. phaseolicola, respectively. B, bacterium; CV, central vacuole; IS, intercellular space. Bars = 0.5 pm.

HzOp Localization and the HR 215
Table 1. Quantitative Assessment of Staining with CeCI, to Reveal H,Oz Accumulation within the Cell Wall Adjacent to Bacteria
Strain and Time after Inoculation
Percentage of Sites in Each Category of Staininga
O 1 2 3 0 1 2 3
(hr) (Without Aminotriazole) (With Aminotriazole)
Wild type 3 70 20 10 O 70 30 O O 5 10 10 2 78 7 12 12 69 8 40 10 5 45 30 3 14 53
1 6b 66 18 16 O 19 50 31 O
hrpD mutant 3 80 20 O O 83 14 3 O 5 52 24 8 8 63 19 3 16 8 58 14 14 14 64 6 19 11
16 77 3 17 3 80 9 O 11
=At least 30 sites were examined at each time from three replicate leaf samples. The minimum degrees of deposit assigned to each category of staining are as follows: O, none; 1, faint and patchy; 2, dense but patchy; and 3, confluent and dense staining. For illustra- tions, see Figure 3.
By 16 hr, most cells had collapsed.
NADPH oxidase (Levine et al., 1994; Bolwell et al., 1995; Murphy and Auh, 1996). Sites were assessed for CeCI, staining, and deposits were quantified. These data are sum- marized in Tables 1 and 2.
The specificity of CeCI, staining for H202 was demon- strated by the striking reduction observed after treatment with catalase. By contrast, ATZ did not cause major changes; however, at the later time points during the HR, in- creased staining in categories 1 and 2 was detected. Treat- ment with cyanide and azide effectively prevented intense staining, whereas DPI caused a less dramatic but also sta- tistically significant reduction (Table 2).
DISCUSSION
Increases in cerium perhydroxide production were clearly lo- calized to sites of bacterial attachment to lettuce cell walls. The inhibition of perhydroxide formation by exogenous cata- lase confirms that H202 was the species being detected. In plant cells, catalase is principally located in peroxisomes, al- though cytoplasmic forms have been reported. The catalase inhibitor ATZ rapidly diffuses into cells, and its failure to cause increased H202 accumulation in the cytoplasm or per- oxisomes demonstrates that reduced catalase activity does not necessarily lead to a corresponding increase in HzO, concentration. A similar conclusion has been reached in a number of studies (Ferguson and Dunning, 1986; Volk and
Feierabend, 1989; Amory et al., 1992). Amory et al. (1992), examining the failure of ATZ to induce H202 accumulation in pea seedlings, considered that any potential increase in de- tectable H202 must be prevented by its use in oxidative- reductive processes or scavenging by alternative antioxi- dant systems. The minor increase in H202 detection within lettuce tissues treated with ATZ 16 hr after inoculation, by which time cells had collapsed during the HR, suggests that H202 may be scavenged finally by catalase released from degrading peroxisomes.
H202 accumulation was localized to sites of wall alterations and papilla formation adjacent to attached bacteria. As such, it may be argued that H202 is produced in response to a requirement for peroxidase-mediated cross-linking reac- tions in the cell wall and papilla, both of which accumulate peroxidase substrates, such as HRGPs and phenolic com- pounds (Bestwick et al., 1995). However, by 16 hr after inoc- ulation, although papilla formation was far more prevalent in the interaction with the hrp mutant, H202 accumulation was much less than observed in response to the wild-type strain. Is it possible, therefore, that papillae formed in response to the hrp mutant are less cross-linked? Given the identical ap- pearance of papillae developing in response to Hrp- or Hrp+ bacteria and their similar impregnation with HRGPs and phenolic compounds (Bestwick et al., 1995), it appears that the same potential for cross-linking exists in both interactions.
The H,O,-dependent insolubilization of preexisting cell wall proteins has been reported in bean and soybean cells challenged with fungal elicitors and bacteria (Bradley et al., 1992; Brisson et al., 1994; Wojtaszek et al., 1995). In response to elicitors, such insolubilization is extremely rapid (Bradley et al., 1992). However, in response to avirulent bacteria, cross- linking is apparently much slower, occurring during the initial stages of visible hypersensitive cell collapse in soybean, some 24 hr after inoculation (Brisson et al., 1994). In soybean, as with other plant-pathogen interactions, the production of H202 in response to both elicitors and bacteria is a relatively rapid response (Levine et al., 1994; Baker and Orlandi, 1995). Increased AOS production is therefore not necessar- ily correlated with immediately detectable oxidative cross- linking. In the context of wall alterations, it will be interesting to localize sites of peroxidase activity relative to H,O, pro- duction and/or accumulation during these interactions.
We suggest that CeCI, is principally able to react with ex- cess H,O, that is not being rapidly metabolized during cross-linking reactions and that this excess H202 is a pri- mary factor in the development of membrane damage. Pre- viously, we were unable to follow the sequence of collapse observed in a few cells challenged with the hrp mutant, but now it appears that there is a correlation between the num- ber of cells collapsing (-10%; Bestwick et al., 1995) and the number of sites associated with high levels of H202.
Classically, two bursts of AOS production can be ob- served during incompatible plant-pathogen interactions. The first burst appears to be nonspecific, being produced in response to numerous elicitors, wounding, avirulent and
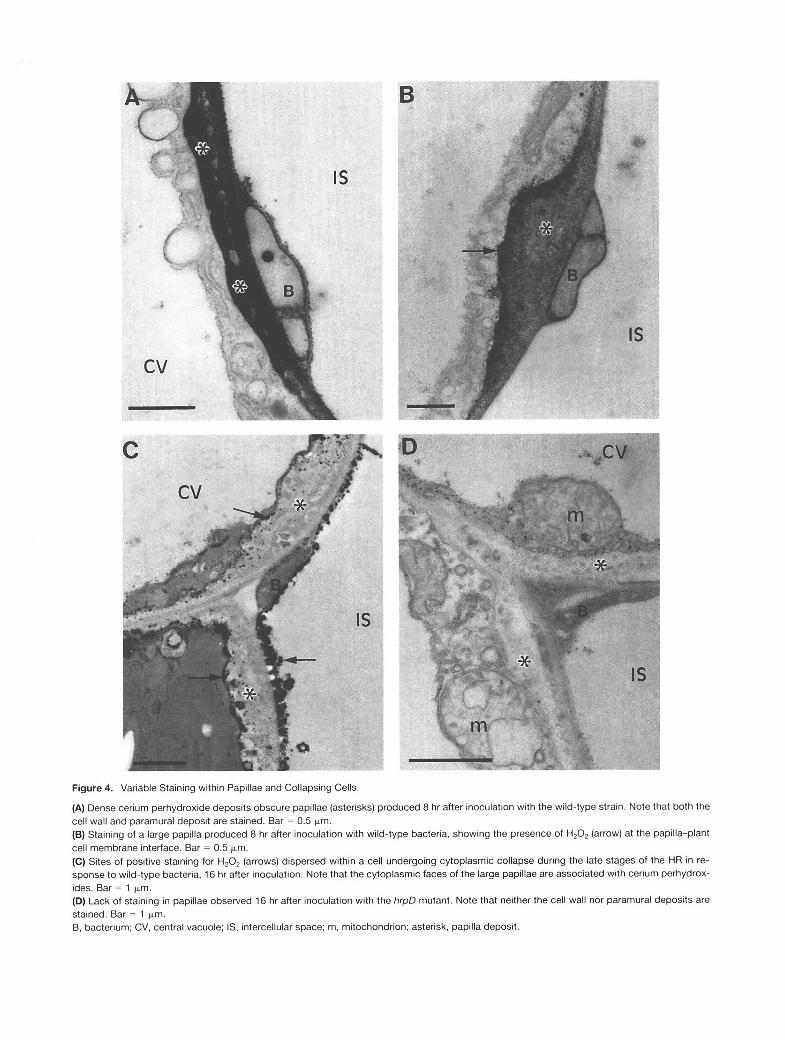
Figure 4. Variable Staining within Papillae and Collapsing Cells.
(A) Dense cerium perhydroxide deposits obscure papillae (asterisks) produced 8 hr after inoculation with the wild-type strain. Note that both the cell wall and paramural deposit are stained. Bar = 0.5 pm. (B) Staining of a large papilla produced 8 hr after inoculation with wild-type bacteria, showing the presence of H202 (arrow) at the papilla-plant cell membrane interface. Bar = 0.5 pm. (C) Sites of positive staining for H202 (arrows) dispersed within a cell undergoing cytoplasmic collapse during the late stages of the HR in re- sponse to wild-type bacteria, 16 hr after inoculation. Note that the cytoplasmic faces of the large papillae are associated with cerium perhydrox- ides. Bar = 1 pm. (D) Lack of staining in papillae observed 16 hr after inoculation with the hrpD mutant. Note that neither the cell wall nor paramural deposits are stained. Bar = 1 pm. 6, bacterium; CV, central vacuole; IS, intercellular space; m, mitochondrion; asterisk, papilla deposit.

H202 Localization and the HR 217
virulent pathogens, and hrp mutants (Doke, 1983; Aposto1 et al., 1989; Baker and Orlandi, 1995). The second burst was thought to be HR specific and a requirement for HR devel- opment; however, Glazener et al. (1996) have reported that hrmA mutants o f f . s. syringae, while failing to cause the HR in tobacco, do cause a biphasic oxidative burst. Further- more, Baker and Orlandi (1995) have pointed out that the levels of exogenous HzOz required to initiate plant cell death are far in excess of those observed in the HR. The conflict- ing evidence for a causal role for HzO2 in the HR may be partly reconciled in view of the highly localized accumulation of HzO2 detected in our work.
In noninoculated leaves, high concentrations of H202 are required to induce levels of staining similar to those ob- served during the HR. It is, however, difficult to define the in situ concentration of HZOz at sites of bacterial attachment. The detection of H202 will be influenced by various antioxi- dant mechanisms, and it is unlikely that the activity of such systems will remain unchanged throughout the development of the HR (Croft et al., 1990; Adam et al., 1995). Neverthe- less, the concentrations of H,Op required to induce cell col- lapse are comparable to those required to mimic the degree of cerium perhydroxide deposition observed during the HR. At reaction sites, concentrations of H2O2 may be sufficient to initiate toxic effects on the plasma membrane, leading to the distinctive vesiculation observed in ultrastructural studies (Bestwick et al., 1995). The availability and release of “cata- lytic” Fez+ from storage or transport proteins or iron-contain- ing enzymes may be important in regulating HzO2 toxicity to plant cells. A decrease in intracellular pH, providing condi- tions that are favorable for Fez+ release, has been reported during the HR (Baker and Orlandi, 1995) and may therefore enhance conversion of H202 to the active hydroxyl radical.
In lettuce cells, locally high levels of H,0, are maintained during the first 8 hr after inoculation in tissues undergoing the HR. During this period, cell viability was reduced to 50%, electrolyte leakage increased, and the first signs of cell decompartmentalization were detected (Bestwick et al., 1995). Oxidative stress is therefore probably maintained during the critical phases of membrane damage in lettuce. The maintenance of AOS production may be a more impor- tant factor in cell collapse than is a transient biphasic burst.
Variation in the level of H202 within individual cells pro- foundly affects the mechanism and type of cell death ob- served in mammalian cell types. Severe oxidative stress has been shown to induce necrosis directly, whereas lower lev- els of AOS induce apoptosis/pcd or even cell proliferation (Dypbukt et al., 1994). We suggest that during the HR, a specifically increased production of HzO2 may be viewed as part of a suicidal response, regardless of whether ultimate cell death is via a “toxic shock,” that is, caused directly by excessive production of H2O2 or indirectly through activation of pcd. It is also possible that HzO, released into the apo- plasm is directly toxic to the pathogen.
The source of H2O2 production is unclear, but its direc- tional accumulation in the cell wall and general absence
Table 2. Effects of Inhibitory Treatments on the Accumulation of H202 in the Cell Wall Adjacent to Wild-Type Bacteria 5 Hr after Inoculation
Percentage of Sites in Each Category of Staininga
Treatment o 1 2 3
None
Affecting bacteriab Streptomycin before induction
Streptomycin after induction
Heat-killed bacteria
timeC
timeC
Affecting the plantb Catalase KCN 1 mM KCN 3 mM Azide 1 mM Diphenylene iodonium chloride
8 PM
10 10 8 72
67 33 O O
8 O 8 8 4
82 18 O O
75 25 O O 41 16 8 5 67 33 O O 9 2 8 0 0 40 20 40 O
aCategories are as described in Table 1 and as illustrated in Figure 3. Statistical analysis using x2 contingency tests showed that all treatments, except for streptomycin after the induction time, signifi- cantly reduced the frequency of sites in categories 2 and 3 com- pared with O and 1 combined (P < 0.001). Major effects that were expected on either the bacteria or plant. Injection was with streptomycin before or after completion of the
induction time at 0.5 and 3 hr after inoculation, respectively.
from the cytoplasm suggest that the principal site is on the extracellular surface of the plasma membrane or within the apoplasm. The lack of staining in the cytoplasm does not re- flect an inability of CeCI, to enter plant cells. In both inocu- lated and noninoculated lettuce leaves, staining of the tonoplast was occasionally encountered (data not shown), demonstrating the potential penetration of CeCI, throughout the cytoplasm. Under similar incubation conditions, Kausch et al. (1 983) have reported deposition of cerium perhydrox- ides in peroxisomes of root parenchyma cells.
Studies of the HR in soybean and potato have suggested that the biphasic oxidative burst is the result of a mem- brane-bound neutrophil-like NADPH oxidase generating Oz. -, which rapidly dismutes to HzO2 (Doke, 1983; Levine et al., 1994; Doke and Miura, 1995; Dwyer et al., 1996). An elic- itor-induced oxidative burst in rose cells has also been cor- related with NADPH and NADH oxidase activities (Auh and Murphy, 1995; Murphy and Auh, 1996). In all of these stud- ies, the production of AOS was sensitive to inhibition by DPI, a potent inhibitor of neutrophil NADPH oxidase.
Although DPI inhibited H,02 production in lettuce, the peroxidase inhibitors cyanide and azide were much more ef- fective. Because neither azide nor cyanide inhibits neutro- phil-like NADPH oxidase activity (Bolwell et al., 1995;

218 The Plant Cell
Murphy and Auh, 1996), there appears to be a fundamen- tally different mechanism of H202 generation in lettuce from that suggested to operate in potato, rose, and soybean cells (Levine et al., 1994; Auh and Murphy, 1995; Doke and Miura, 1995; Murphy and Auh, 1996). It should be noted that DPI is also known to inhibit the activities of heme proteins, such as peroxidases (Deme et al., 1994; Dwyer et al., 1996). Peroxi- dases can generate H202 via H,O,-independent oxidation of a number of substrates, for example, NADH, NADPH, thiols, and certain phenols (Halliwell, 1978; Mader and Amberg- Fisher, 1982; Vianello and Macri, 1991; Pichorner et al., 1992; Bolwell et al., 1995). In bean cells, Bolwell et al. (1995) have presented evidence for the involvement of an apoplas- tic peroxidase activity in elicitor-induced H202 production that was insensitive to DPI but inhibited by cyanide. The in- volvement of peroxidase and components of the cell wall in the generation of AOS during the HR in lettuce merits further investigation. Perhaps significantly, protoplasts (from carrot) have been shown to be unable to generate an elicitor-induced oxidative burst (Bach et al., 1993).
The localized accumulation of H202 is consistent with the existence of a physically close link between the putative re- ceptor controlling recognition of the challenging bacterium and activation of AOS production. Confirmation of the mo- lecular organization of the signaling cascade leading to AOS production will require the identification of both the bacterial ligand and the plant’s receptor, which are both effective in generating the non-host HR in lettuce.
50 mM 3-(N-morpholino)propanesulfonic acid (Mops) at pH 7.2 for 1 hr. Tissues were then fixed in 1.25% (v/v) glutaraldehyde/l.25% (v/v) paraformaldehyde in 50 mM sodium cacodylate (CAB) buffer, pH 7.2, for 1 hr. After fixation, tissues were washed twice for 10 min in CAB buffer and postfixed for 45 min in 1 % (v/v) osmium tetroxide in CAB. Tissues were then washed again in CAB buffer (twice for 10 min) and dehydrated in a graded ethanol series (30, 50, 70, 80, and 90% [v/v] ethanol; 15 min each change and then three changes of 100% etha- nol of 20 min each in duration). Samples were transferred to two changes of propylene oxide of 20 min each in duration and progres- sively embedded in Eponaraldite (Agar Aids, Bishop’s Stortford, UK). Alternatively, tissues were dehydrated in a graded acetone series, transferred directly into acetone-resin mixtures, and progressively embedded. Tissues were then subjected to 12 hr in pure resin, fol- lowed by a change of fresh resin for 4 hr, and placed in blocks and polymerized at 60°C for 48 hr. Blocks were sectioned (70 to 90 nm) on a Reichert-Ultracut E microtome (Milton Keynes, UK), using a di- amond knife (Diatome, Bienne, Switzerland), and mounted on un- coated copper grids (300 mesh). Sections were examined using a transmission electron microscope (model 7000; Hitachi, Tokyo, Ja- pan) at an accelerating voltage of 75 kV.
Treatment with Inhibitors
Inoculated and noninoculated tissue segments were excised and in- cubated for 30 min in 50 mM Mops, pH 7.2, containing either 1 or 3 mM potassium cyanide, 1 mM sodium azide, 8 pM diphenylene io- donium chloride (DPI), 25 pg mL-l bovine liver catalase, or 20 pM 3-amino-I ,2,4-triazole (ATZ). Control tissues were incubated in Mops devoid of further additions. Tissues were then transferred to CeCI, solutions supplemented with inhibitors at the stated concen- trations, incubated for 1 hr, and processed for transmission electron microscopy as described above.
METHODS
Effect of Bacterial Viability on H202 Production Reagents
Unless stated otherwise, all reagents used were analytical or equiva- lent grade and supplied by BDH (Poole, Dorset, UK) or Sigma (Poole, Dorset, UK).
Maintenance of Plants and Bacteria
Lettuce plants (Lactuca sativa cv Diana) were grown from seed and inoculated by injection with suspensions of bacteria (1 O8 colony- forming units mL-l), as described by Bestwick et al. (1995). The strains of Pseudomonas syringae pv phaseolicola used were the wild-type race 6 strain S21-WT and the hrpD mutant S21-533 (Nollenburg et al., 1990).
Transmission Electron Microscopy
Samples were fixed, embedded, sectioned, and stained for conven- tional electron microscopy, as described by Bestwick et al. (1995). The histochemical method developed for localization of hydrogen peroxide (H202), based on the generation of cerium perhydroxides, was as follows. Tissue pieces (4 to 2 mm2) were excised from inoc- ulated leaf panels and incubated in freshly prepared 5 mM CeCI, in
The induction time for confluent expression of the hypersensitive re- action (HR) was determined by injecting aqueous solutions of the an- tibiotic streptomycin sulfate (200 pg mL-’) at 30-min intervals into leaf panels previously inoculated with bacteria, and the effect on confluent expression of the HR and H202 production was observed.
ACKNOWLEDGMENTS
This work was supported by a grant from the United Kingdom Agri- cultural and Food Research Council.
Received September 16, 1996; accepted December 13, 1996.
REFERENCES
Adám, A.L., Farkas, T., Somlyai, G., Hevesi, M., and Király, Z. (1 989). Consequence of 02- generation during bacterially induced

H202 Localization and the HR 21 9
hypersensitive reaction in tobacco: Deterioration of membrane lipids. Physiol. Mol. Plant Pathol. 34, 13-26.
Adám, A.L., Bestwick, C.S., Barna, B., and Mansfield, J.W. (1 995). Enzymes regulating the accumulation of active oxygen species during the hypersensitive reaction of bean to Pseudomo- nas syringae pv. phaseolicola. Planta 197, 240-249.
Amory, A.M., Ford, L., Pammenter, N.W., and Cresswell, C.F. (1992). The use of 3-amino-I ,2,4-triazole to investigate the short term effects of oxygen toxicity on carbon assimilation by Pisum sativum seedlings. Plant Cell Environ. 15, 655-663.
Apostol, I., Heinstein, P.F., and Low, P.S. (1989). Rapid stimulation of an oxidative burst during elicitation of cultured plant cells. Plant Physiol. 90, 109-1 16.
Arends, M.J., and Wyllie, A.H. (1991). Apoptosis: Mechanisms and roles in pathology. Int. Rev. Exp. Pathol. 32,223-254.
Auh, C.-K., and Murphy, T.M. (1995). Plasma membrane redox enzyme is involved in the synthesis of 02- and H202 by Phy- tophfhora elicitor-stimulated rose cells. Plant Physiol. 107,
Bach, M., Schnitzler, J.-P., and Seitz, U.H. (1 993). Elicitor-induced changes in Ca2+ influx, K+ efflux, and 4-hydroxybenzoic acid syn- thesis in protoplasts of Daucus carota L. Plant Physiol. 103,
Baker, C.J., and Orlandi, E.W. (1995). Active oxygen in plant/ pathogen interactions. Annu. Rev. Phytopathol. 33,299-321.
Bestwick, C.S., Bennett, M.H., and Mansfield, J.W. (1995). Hrp mutant of Pseudomonas syringae pv. phaseolicola induces cell wall alterations but not membrane damage leading to the HR in lettuce (Lactuca sativa). Plant Physiol. 108, 503-51 6.
Bolwell, G.P. (1993). Dynamic aspects of the plant extracellular matrix. Int. Rev. Cytol. 146, 261-324.
Bolwell, G.P., Butt, V.S., Davies, D.R., and Zimmerlin, A. (1995). The origin of the oxidative burst in plants. Free Rad. Res. 23,
Bonas, U. (1994). hrp genes of phytopathogenic bacteria. In Bacte- rial Pathogenesis of Plants and Animals: Molecular and Cellular Mechanisms, J.L. Dangl, ed (Heidelberg, Germany: Springer-Ver- lag), pp. 79-98.
Bradley, D.J., Kjellbom, P., and Lamb, C.J. (1992). Elicitor- and wound-induced oxidative cross-linking of a proline-rich plant cell wall protein: A novel, rapid defense response. Cell 70, 21-30.
Briggs, R.T., Drath, D.B., Darnovsky, M.L., and Karnovsky, M.J. (1975). Localization of NADH oxidase on the surface of human polymorphonuclear leukocytes by a new cytochemical method. J. Cell Rol. 67, 566-586.
Brisson, L.F., Tenhaken, R., and Lamb, C. (1994). Function of oxi- dative cross-linking of cell wall structural proteins in plant disease resistance. Plant Cell 6, 1703-1712.
Brown, I., Mansfield, J., and Bonas, U. (1995). hrp genes in Xanth- omonas campesfris pv. vesicatoria determine ability to suppress papilla deposition in pepper mesophyll cells. Mol. Plant-Microbe Interact. 8, 825-836.
Buja, M., Eigenbrodt, M.L., and Eigenbrodt, E.R. (1993). Apopto- sis and necrosis. Basic types and mechanisms of cell death. Arch. Pathol. Lab. Med. 117, 1208-1214.
1241-1 247.
407-412.
51 7-532.
Buttke, T.M., and Sandstrom, P.A. (1995). Redox regulation of pro- grammed cell death in lymphocytes. Free Rad. Res. 22, 389-397.
Chen, Z., Silva, R., and Klessig, D. (1993). Involvement of reactive oxygen species in the induction of systemic acquired resistance by salicylic acid in plants. Science 262, 1883-1 886.
Collins, R.J., Harmon, B.V., Gobe, G.C., and Kerr, J.F.R. (1992). lnternucleosomal DNA cleavage should not be the sole criterion for identifying apoptosis. Int. J. Radiat. Biol. 61, 451-453.
Croft, K.P.C., Voisey, C.R., and Slusarenko, A.J. (1990). Mecha- nism of hypersensitive cell collapse: Correlation of increased lipoxygenase activity with membrane damage in leaves of Phaseolus vulgaris (L.) inoculated with an avirulent race of Pseudomonas syringae pv. phaseolicola. Physiol. Mol. Plant Pathol. 36, 49-62.
Czaninski, Y., Sachot, R.M., and Catesson, A.M. (1993). Cytochemical localization of hydrogen peroxide in lignifying cell walls. Ann. Bot. 72, 547-550.
Deme, D., Doussiere, J., De Sandro, V., Dupuy, C., Pommier, J., and Virion, A. (1 994). The Ca2+/NADPH-dependent H202 genera- tor in thyroid plasma membrane: Inhibition by diphenyleneiodo- nium. Biochem. J. 301, 75-81.
Dietrich, R.A., Delaney, T.P., Uknes, S.J., Ward, E.R., Ryals, J.A., and Dangl, J.L. (1 994). Arabidopsis mutants simulating disease resistance response. Cell 77, 565-577.
Doke, N. (1 983). Involvement of superoxide anion generation in the hypersensitive response of potato tuber tissues to infection with an incompatible race of Phytophthora infestans and to the hyphal wall components. Physiol. Plant Pathol. 23, 345-358.
Doke, N., and Miura, Y. (1995). In vitro activation of NADPH-depen- dent 02- generating system in a plasma membrane-rich fraction of potato tuber tissues by treatment with an elicitor from Phytoph- thora infestam or with digitonin. Physiol. Mol. Plant Pathol. 46,
Dwyer, S.C., Legendre, L., Low, P.S., and Leto, T.L. (1996). Plant and human neutrophil oxidative burst complexes contain immu- nologically related proteins. Biochim. Biophys. Acta 1289,231-237.
Dypbukt, J.M., Ankarcrona, M., Burkitt, M., Sjoholm, A., Strom, K., Orrenius, S., and Nicotera, P. (1994). Different pro-oxidant levels stimulate growth, trigger apoptosis or produce necrosis of insulin secreting RINm5F cells. J. Biol. Chem. 269, 30553-30560.
Ferguson, I.B., and Dunning, S.J. (1986). Effect of 3-amino-I ,2,4- triazole, a catalase inhibitor, on peroxide content of suspension cultured pear fruit cells. Plant Sci. 43, 7-1 1.
Fett, W.F., and Jones, S.B. (1995). Microscopy of the interaction of hrp mutants of Pseudomonas syringae pv. phaseolicola with a non-host plant. Plant Sci. 107, 27-39.
Fridovich, 1. (1 986). Biological effects of the superoxide radical. Arch. Biochem. Biophys. 247, 1-1 1.
Friend, J. (1 981). Plant phenolics, lignification, and plant disease. Prog. Phytochem. 7, 197-261.
Glazener, J.A., Orlandi, E.W., and Baker, C.J. (1996). The active oxygen response of cell suspensions to incompatible bacteria is not sufficient to cause hypersensitive cell death. Plant Physiol.
Greenberg, J.T., Guo, A., Klessig, D.F., and Ausubel, F.M. (1994). Programmed cell death in plants: A pathogen-triggered response
17-28.
110,759-763.

220 The Plant Cell
activated coordinately with multiple defense functions. Cell 77,
Halliwell, B. (1 978). Lignin synthesis: The generation of hydrogen peroxide and superoxide by horseradish peroxidase and its stim- ulation by manganese (11) and phenols. Planta 140, 81-88.
Halliwell, B., and Gutteridge, J.M.C. (1989). Free Radicals in Biol- ogy and Medicine. (Oxford, UK: Oxford University Press).
He, S.Y., Huang, H.C., and Collmer, A. (1993). Pseudomonas syrin- gae pv. syringae Harpin Pss: A protein that is secreted via the Hrp pathway and elicits the hypersensitive response in plants. Cell 73, 1-20.
Heath, M.C. (1989). A comparison of fungal growth and plant responses in cowpea and bean cultivars inoculated with ure- diospores and basidiospores of the cowpea rust fungus. Physiol. Mol. Plant Pathol. 34, 415-426.
liyama, K., Lam, T.B.-T., and Stone, B.A. (1 994). Covalent cross- links in the cell wall. Plant Physiol. 104, 315-320.
Imlay, J.A., and Linn, S. (1988). DNA damage and oxygen radical toxicity. Science 240, 1302-1 304.
Ingram, D.S. (1978). Cell death and resistance to biotrophs. Ann. Appl. Biol. 89, 291-295.
Jacobson, M.D. (1 996). Reactive oxygen species and programmed cell death. Trends Biochem. Sci. 21, 83-86.
Jakobek, J.L., and Lindgren, P.B. (1 993). Generalized induction of defense responses in bean is not correlated with the induction of the hypersensitive reaction. Plant Cell 5, 49-56.
Kausch, A.P. (1987). Cerium precipitation. In Handbook of Plant Cytochemistry, Vol. 1, K.C. Vaughn, ed (Boca Raton, FL: CRC Press), pp. 25-36.
Kausch, A.P., Wagner, B.L., and Horner, H.T. (1983). Use of cerium chloride technique and energy dispersive x-ray microanal- ysis in plant peroxisome identification. Protoplasma 118, 1-9.
Kerr, J., Winterford, C., and Harmon, B. (1994). Morphological criteria for identifying apoptosis. In Cell Biology: A Laboratory Handbook, Vol. 2, J. Celis, ed (London: Academic Press), pp.
Klement, 2. (1 982). Hypersensitivity. In Phytopathogenic Procary- otes, M.S. Mount and G.H. Lacy, eds (New York: Academic Press), pp. 149-1 77.
Klement, Z., and Goodman, R.N. (1967). The role of the living cell and induction time in the hypersensitive reaction of the tobacco plant. Phytopathology 57, 322-323.
Levine, A., Tenhaken, R., Dixon, R., and Lamb, C. (1994). H2O2 from the oxidative burst orchestrates the plant hypersensitive dis- ease resistance response. Cell 79, 583-593.
Levine, A., Pennell, RI., Alvarez, M.E., Palmer, R., and Lamb, C. (1 996). Calcium-mediated apoptosis in a plant hypersensitive dis- ease resistance response Curr. Biol. 6, 427-437.
Low, P.S., and Merida, J.R. (1996). The oxidative burst in plant defense: Function and signal transduction. Physiol. Plant. 96,
Mader, M., and Amberg-Fisher, V. (1982). Role of peroxidase in the lignification of tobacco cells. I. Oxidation of nicotinamide ade- nine dinucleotide and formation of hydrogen peroxide by cell wall peroxidases. Plant Physiol. 70, 1128-1 131.
551 -563.
31 9-330.
533-542.
Mansfield, J.W. (1 990). Recognition and response in plant-fungus interactions. In Recognition and Response in Plant-Virus Interac- tions, R.S.S. Fraser, ed (Berlin: Springer-Verlag), pp. 31-52.
Mansfield, J.W., Bennett, M.H., Bestwick, C.S., and Woods-Tor, A.M. (1 997). Phenotypic expression of gene-for-gene interactions involving fungal and bacterial pathogens: Variation from recogni- tion to response. In The Gene-for-Gene Relationship in Host-Par- asite Interactions, I.R. Crute and E.B. Holub, eds (London: CAB International), pp. 265-291.
Martin, S.J., Green, D.R., and Cotter, T.G. (1994). Dicing with death: Dissecting the components of the apoptosis machinery. Trends Biochem. Sci. 19, 26-30.
May, M.A., Hammond-Kosack, K.E., and Jones, J.D.G. (1996). Involvement of reactive oxygen species, glutathione metabolism and lipid peroxidation in the Cf gene-dependent defense response of tomato cotyledons induced by race-specific elicitors from Cladosporium fulvum. Plant Physiol. 1 IO, 1367-1379.
Mehdy, M.C. (1994). Involvement of active oxygen species in plant defense against pathogens. Plant Physiol. 105, 467-472.
Mittler, R., and Lam, E. (1 995). Identification, characterization, and purification of a tobacco endonuclease activity induced upon hypersensitive response cell death. Plant Cell 7, 1951-1962.
Mittler, R., and Lam, E. (1 996). Sacrifice in the face of foes: Pathogen- induced programmed cell death in plants. Trends Microbiol.4,10-15.
Murphy, T.M., and Auh, C.-K. (1996). The superoxide synthases of plasma membrane preparations from cultured rose cells. Plant Physiol. 110, 621-629.
Nicholson, R.L., and Hammerschmidt, R. (1 992). Phenolic com- pounds and their role in disease resistance. Annu. Rev. Phyto- pathol. 30,369-380.
Nollenburg, M., Hevesi, M., Somylai, G., Rudolph, K., Klement, Z., and Kondorosi, A. (1990). Genetic and pathological charac- terization of path- and HR- mutant of Pseudomonas syringae pv. phaseolicola complemented by clones from a wild type genomic library. In Plant Pathogenic Bacteria: Proceedings of the Seventh International Conference on Plant Pathogenic Bacteria, Z. Klement, ed (Budapest, Hungary: Akademiai Kiado), pp. 363-369.
O’Connell, R.J., Brown, IR., Mansfield, J.W., Bailey, J.A., Mazau, D., Rumeau, D., and Esquerré-Tugayé, M.T. (1 990). Immunocy- tochemical localization of hydroxyproline-rich glycoproteins accu- mulating in melon and bean at sites of resistance to bacteria and fungi. Mol. Plant-Microbe Interact. 2, 33-40.
Olson, P.D., and Varner, J.E. (1993). Hydrogen peroxide and lignifi- cation. Plant J. 4, 887-892.
Peng, M., and Kuc, J. (1992). Peroxidase-generated hydrogen per- oxide as a source of antifungal activity in vitro and on tobacco leaf discs. Phytopathology 82, 696-699.
Pichorner, H., Couperus, A., Korori, S.A.A., and Ebermann, R. (1992). Plant peroxidase has thiol oxidase activity. Phytochemistry
Ride, J.P. (1986). Induced structural defenses in plants. In Natural Antimicrobial Systems in Plants and Animals, G.W. Gould, M.E. Rhodes-Roberts, A.K. Charnley, R.M. Cooper, and R.G. Board, eds (Bath, UK: Bath University Press), pp. 159-165.
Ryerson, D.E., and Heath, M.C. (1996). Cleavage of nuclear DNA into oligonucleosomal fragments during cell death induced by fungal infection or by abiotic treatments. Plant Cell 8, 393-402.
31,3371-3374.

H202 Localization and the HR 221
Sutherland, M.W. (1 991). The generation of oxygen radicals during host plant responses to infection. Physiol. Mol. Plant Pathol. 39, 79-93.
Tenhaken, R., Levine, A., Brisson, L.F., Dixon, R., and Lamb, C. (1 995). Function of the oxidative burst in hypersensitive disease resistance. Proc. Natl. Acad. Sci. USA 92,4158-41 63.
Vera-Estrella, R., Blumwald, E., and Higgins, V.J. (1992). Effect of specific elicitors of Cladosporium fulvum on tomato suspension cells. Plant Physiol. 99, 1208-1215.
Vianello, A., and Macri, F. (1991). Generation of superoxide anion and hydrogen peroxide at the surface of plant cells. J. Bioenerg. Biomembr. 23, 409-423.
Volk, S., and Feierabend, J. (1989). Photoinactivation of catalase at low temperature and its relevance to photosynthetic and peroxide metabolism in leaves. Plant Cell Environ. 12, 701-712.
Wang, H., Li, J., Bostock, R.M., and Gilchrist, D.G. (1996). Apop- tosis: A functional paradigm for programmed plant cell death
induced by a host-selective phytotoxin and invoked during devei- opment. Plant Cell 8, 375-391.
Wojtaszek, P., Trethowan, J., and Bolwell, G.P. (1995). Specificity in the immobilization of cell wall proteins in response to different elicitor molecules in suspension-cultured cells of French bean (Phaseolus vulgaris L.). Plant. Mol. Biol. 28, 1075-1087.
Wolff, S.P., Garner, A., and Dean, R.T. (1986). Free radicals, lipids and protein degradation. Trends Biochem. Sci. 11,27-31.
Woods, A.M., Fagg, J., and Mansfield, J.W. (1989). Effects of heat- shock and inhibitors of protein synthesis on irreversible mem- brane damage occurring during the hypersensitive reaction of Lactuca sativa L. to Bremia lactucae Regel. Physiol. Mol. Plant Pathol. 34, 531-544.
Yahraus, T., Chandra, S., Legendre, L., and Low, P.S. (1995). Evi- dence for a mechanically induced oxidative burst. Plant Physiol. 109,1259-1266.

DOI 10.1105/tpc.9.2.209 1997;9;209-221Plant Cell
C S Bestwick, I R Brown, M H Bennett and J W Mansfieldcells to Pseudomonas syringae pv phaseolicola.
Localization of hydrogen peroxide accumulation during the hypersensitive reaction of lettuce
This information is current as of April 29, 2020
Permissions X
https://www.copyright.com/ccc/openurl.do?sid=pd_hw1532298X&issn=1532298X&WT.mc_id=pd_hw1532298
eTOCs http://www.plantcell.org/cgi/alerts/ctmain
Sign up for eTOCs at:
CiteTrack Alerts http://www.plantcell.org/cgi/alerts/ctmain
Sign up for CiteTrack Alerts at:
Subscription Information http://www.aspb.org/publications/subscriptions.cfm
is available at:Plant Physiology and The Plant CellSubscription Information for
ADVANCING THE SCIENCE OF PLANT BIOLOGY © American Society of Plant Biologists