low sfrp1 expression correlates with poor prognosis...
TRANSCRIPT

Research Article
Low SFRP1 Expression Correlates with PoorPrognosis and Promotes Cell Invasion byActivating the Wnt/b-Catenin Signaling Pathwayin NPCXian-Yue Ren1, Guan-Qun Zhou1,Wei Jiang2, Ying Sun1, Ya-Fei Xu1, Ying-Qin Li1,Xin-Ran Tang1, Xin Wen1, Qing-Mei He1, Xiao-Jing Yang1, Na Liu1, and Jun Ma1
Abstract
Distant metastasis remains the predominant mode of treat-ment failure in nasopharyngeal carcinoma (NPC). Unfortu-nately, the molecular events underlying NPC metastasisremain poorly understood. Secreted frizzled-related protein1 (SFRP1) plays an important role in tumorigenesis andprogression. However, little is known about the function andmechanism of SFRP1 in NPC. Immunohistochemistry wasused to determine SFRP1 expression levels in patients withNPC. SFRP1 function was evaluated using MTT, colony for-mation, wound-healing, Transwell assays, and in vivo models.The methylation level of SFRP1 in NPC cells was examinedusing bisulfate pyrosequencing; the Wnt/b-catenin signalingpathway genes were studied using Western blotting. Com-pared with patients with high SFRP1 expression, patients withlow SFRP1 expression had worse overall survival [HR, 2.32;95% confidence interval (CI), 1.36–3.94; P ¼ 0.002], disease-
free survival (HR, 1.98; 95% CI, 1.23–3.18; P ¼ 0.005), anddistant metastasis-free survival (HR, 2.07; 95% CI, 1.19–3.59;P ¼ 0.009). Multivariate Cox regression analysis indicatedthat SFRP1 was an independent prognostic factor. Further-more, SFRP1 was significantly downregulated in NPC celllines. SFRP1 overexpression suppressed NPC cell proliferation,migration, and invasion in vitro and lung colonization in vivo.SFRP1 expression was restored after treatment with a demeth-ylation agent, and the SFRP1 promoter region was hyper-methylated in NPC cells. b-Catenin, c-Myc, and cyclin D1were downregulated after SFRP1 restoration, which suggestedthat SFRP1 suppressed growth and metastasis by inhibitingthe Wnt/b-catenin signaling pathway in NPC. SFRP1 providesfurther insight into NPC progression and may provide noveltherapeutic targets for NPC treatment. Cancer Prev Res; 8(10);968–77. �2015 AACR.
IntroductionNasopharyngeal carcinoma (NPC) is one of the leading
malignancies of the head and neck; prevalence is low (0.5cases per 100,000 individuals) in Europe and the United States,but the incidence rate is high (20–50 cases per 100,000 people)in southern China (1). Improvements in intensity-modulatedradiotherapy and chemotherapy have improved local control.However, the outcome remains poor for approximately 30%of patients with NPC. Distant metastasis remains the main
reason for treatment failure in NPC (2). Despite great effortsover the past decades to better understand the molecularmechanisms underlying migration and invasion in NPC, bio-markers that can accurately identify patients with high risk ofdistant metastasis remain absent. Therefore, it is urgent toidentify new molecular prognostic markers that can predictdistant metastasis and guide individualized treatment ofpatients with NPC.
Metastasis is an important event in cancer development andprogression that contributes to the majority of cancer deaths (3).Deregulation of the signaling pathways involved in cell adhesionand migration is likely to lead to tumor metastasis. Currently,several signaling pathways have been identified as being asso-ciated with tumor metastasis, one of which is the Wnt/b-cateninsignaling pathway (4, 5). The Wnt/b-catenin signaling pathwayis associated with many cell biologic processes such as differ-entiation, proliferation, and migration (6–8). Overactivation ofthe Wnt/b-catenin signaling pathway contributes to tumorigen-esis, proliferation, and migration in several human cancers, suchas hepatocellular carcinoma (9), colon cancer (10), and breastcancer (11). Therefore, therapeutic agents that alter abnormallyactivated Wnt signaling pathway genes provide potential ther-apeutic targets for cancer therapies (12–14). Secreted frizzled-related protein 1 (SFRP1), a Wnt antagonist on 8p11, plays animportant regulatory role in cellular biologic processes (15, 16).
1SunYat-senUniversityCancerCenter; StateKeyLaboratoryofOncol-ogy, SouthChina;Collaborative InnovationCenterofCancerMedicine,Guangzhou, PR China. 2Department of Radiation Oncology, GuilinMedical University Affiliated Hospital, Guilin, PR China.
Note: Supplementary data for this article are available at Cancer PreventionResearch Online (http://cancerprevres.aacrjournals.org/).
X.-Y. Ren, G.-Q. Zhou, and W. Jiang contributed equally to this article.
Corresponding Authors: Jun Ma, State Key Laboratory of Oncology in SouthChina, Sun Yat-sen University Cancer Center, 651 Dongfeng Road East, Guangz-hou 510060, PR China. Phone: 86-20-87343469; Fax: 86-20-87343295; E-mail:[email protected]; and Na Liu. Phone: 86-20-87342370; Fax: 86-20-87343295; E-mail: [email protected]
doi: 10.1158/1940-6207.CAPR-14-0369
�2015 American Association for Cancer Research.
CancerPreventionResearch
Cancer Prev Res; 8(10) October 2015968
Research. on May 20, 2018. © 2015 American Association for Cancercancerpreventionresearch.aacrjournals.org Downloaded from
Published OnlineFirst August 14, 2015; DOI: 10.1158/1940-6207.CAPR-14-0369
It blocks Wnt signaling by hindering Wnt–receptor interactionsvia the N-terminal cysteine-rich domain homologous to Frizzledproteins. SFRP1 suppresses tumor proliferation and metastasisin several cancers, such as breast (17) and colorectal carcinomas(18). More importantly, SFRP1 has been reported to be associ-ated with cancer therapy. Recently, it has been reported thatSFRP1 could suppress NPC cells proliferation in vitro (19).However, it remains unclear whether SFRP1 plays a direct rolein NPC metastasis.
Our research was designed to investigate SFRP1 expressionlevels in NPC tissues and its correlation with the clinicalcharacteristics of a cohort of patients with NPC. Furthermore,we explored the potential roles of SFRP1 in NPC cell migrationand invasion to better understand the mechanism of NPCmetastasis, which may provide a novel therapeutic target fortreating NPC.
Materials and MethodsClinical specimens
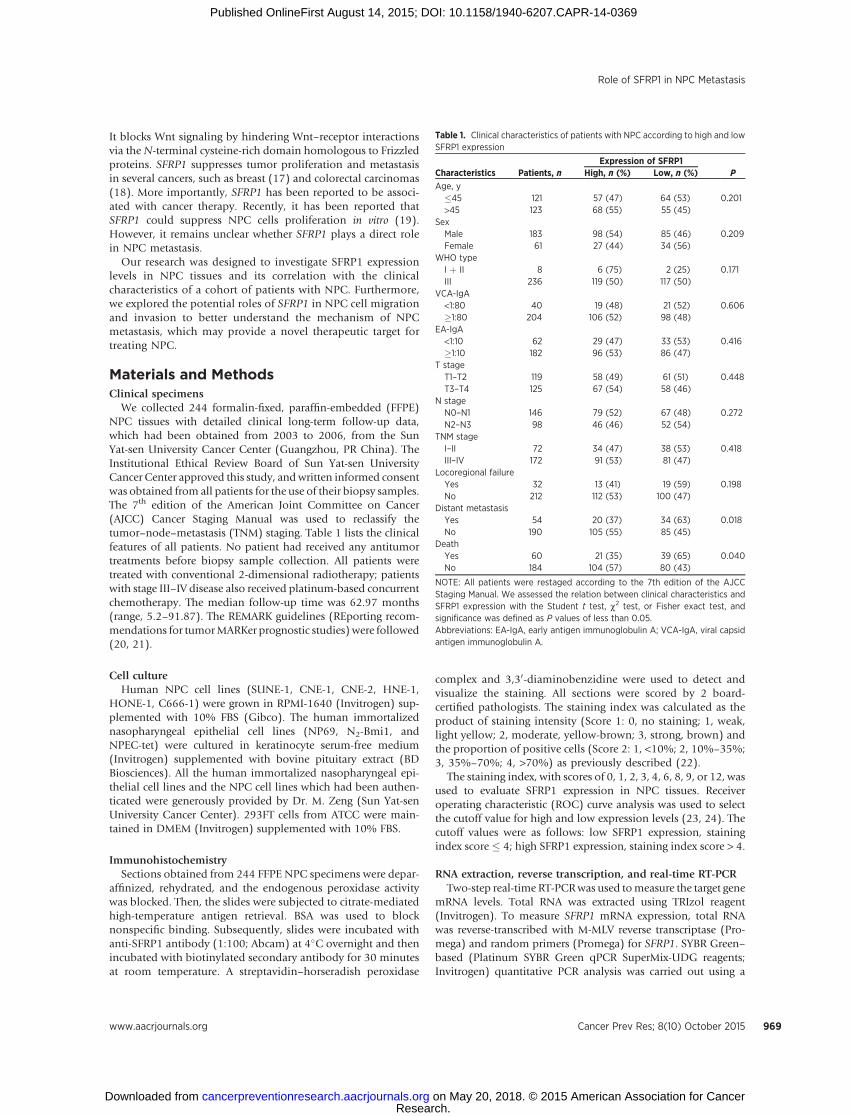
We collected 244 formalin-fixed, paraffin-embedded (FFPE)NPC tissues with detailed clinical long-term follow-up data,which had been obtained from 2003 to 2006, from the SunYat-sen University Cancer Center (Guangzhou, PR China). TheInstitutional Ethical Review Board of Sun Yat-sen UniversityCancer Center approved this study, and written informed consentwas obtained from all patients for the use of their biopsy samples.The 7th edition of the American Joint Committee on Cancer(AJCC) Cancer Staging Manual was used to reclassify thetumor–node–metastasis (TNM) staging. Table 1 lists the clinicalfeatures of all patients. No patient had received any antitumortreatments before biopsy sample collection. All patients weretreated with conventional 2-dimensional radiotherapy; patientswith stage III–IV disease also received platinum-based concurrentchemotherapy. The median follow-up time was 62.97 months(range, 5.2–91.87). The REMARK guidelines (REporting recom-mendations for tumorMARKer prognostic studies) were followed(20, 21).
Cell cultureHuman NPC cell lines (SUNE-1, CNE-1, CNE-2, HNE-1,
HONE-1, C666-1) were grown in RPMI-1640 (Invitrogen) sup-plemented with 10% FBS (Gibco). The human immortalizednasopharyngeal epithelial cell lines (NP69, N2-Bmi1, andNPEC-tet) were cultured in keratinocyte serum-free medium(Invitrogen) supplemented with bovine pituitary extract (BDBiosciences). All the human immortalized nasopharyngeal epi-thelial cell lines and the NPC cell lines which had been authen-ticated were generously provided by Dr. M. Zeng (Sun Yat-senUniversity Cancer Center). 293FT cells from ATCC were main-tained in DMEM (Invitrogen) supplemented with 10% FBS.
ImmunohistochemistrySections obtained from 244 FFPE NPC specimens were depar-
affinized, rehydrated, and the endogenous peroxidase activitywas blocked. Then, the slides were subjected to citrate-mediatedhigh-temperature antigen retrieval. BSA was used to blocknonspecific binding. Subsequently, slides were incubated withanti-SFRP1 antibody (1:100; Abcam) at 4�C overnight and thenincubated with biotinylated secondary antibody for 30 minutesat room temperature. A streptavidin–horseradish peroxidase
complex and 3,30-diaminobenzidine were used to detect andvisualize the staining. All sections were scored by 2 board-certified pathologists. The staining index was calculated as theproduct of staining intensity (Score 1: 0, no staining; 1, weak,light yellow; 2, moderate, yellow-brown; 3, strong, brown) andthe proportion of positive cells (Score 2: 1, <10%; 2, 10%–35%;3, 35%–70%; 4, >70%) as previously described (22).
The staining index, with scores of 0, 1, 2, 3, 4, 6, 8, 9, or 12, wasused to evaluate SFRP1 expression in NPC tissues. Receiveroperating characteristic (ROC) curve analysis was used to selectthe cutoff value for high and low expression levels (23, 24). Thecutoff values were as follows: low SFRP1 expression, stainingindex score � 4; high SFRP1 expression, staining index score > 4.
RNA extraction, reverse transcription, and real-time RT-PCRTwo-step real-time RT-PCRwas used tomeasure the target gene
mRNA levels. Total RNA was extracted using TRIzol reagent(Invitrogen). To measure SFRP1 mRNA expression, total RNAwas reverse-transcribed with M-MLV reverse transcriptase (Pro-mega) and random primers (Promega) for SFRP1. SYBR Green–based (Platinum SYBR Green qPCR SuperMix-UDG reagents;Invitrogen) quantitative PCR analysis was carried out using a
Table 1. Clinical characteristics of patients with NPC according to high and lowSFRP1 expression
Expression of SFRP1Characteristics Patients, n High, n (%) Low, n (%) P
Age, y�45 121 57 (47) 64 (53) 0.201>45 123 68 (55) 55 (45)
SexMale 183 98 (54) 85 (46) 0.209Female 61 27 (44) 34 (56)
WHO typeI þ II 8 6 (75) 2 (25) 0.171III 236 119 (50) 117 (50)
VCA-IgA<1:80 40 19 (48) 21 (52) 0.606�1:80 204 106 (52) 98 (48)
EA-IgA<1:10 62 29 (47) 33 (53) 0.416�1:10 182 96 (53) 86 (47)
T stageT1–T2 119 58 (49) 61 (51) 0.448T3–T4 125 67 (54) 58 (46)
N stageN0–N1 146 79 (52) 67 (48) 0.272N2–N3 98 46 (46) 52 (54)
TNM stageI–II 72 34 (47) 38 (53) 0.418III–IV 172 91 (53) 81 (47)
Locoregional failureYes 32 13 (41) 19 (59) 0.198No 212 112 (53) 100 (47)
Distant metastasisYes 54 20 (37) 34 (63) 0.018No 190 105 (55) 85 (45)
DeathYes 60 21 (35) 39 (65) 0.040No 184 104 (57) 80 (43)
NOTE: All patients were restaged according to the 7th edition of the AJCCStaging Manual. We assessed the relation between clinical characteristics andSFRP1 expression with the Student t test, c2 test, or Fisher exact test, andsignificance was defined as P values of less than 0.05.Abbreviations: EA-IgA, early antigen immunoglobulin A; VCA-IgA, viral capsidantigen immunoglobulin A.
Role of SFRP1 in NPC Metastasis
www.aacrjournals.org Cancer Prev Res; 8(10) October 2015 969
Research. on May 20, 2018. © 2015 American Association for Cancercancerpreventionresearch.aacrjournals.org Downloaded from
Published OnlineFirst August 14, 2015; DOI: 10.1158/1940-6207.CAPR-14-0369

CFX96 Touch sequence detection system (Bio-Rad). Real-timeRT-PCR primers for SFRP1 were 50-ATCTCTGTGCCAGCGAGTTT-30
(forward, F) and 50-GGCTTCTTCTTCTTGGGGAC-30 (reverse, R).GAPDH was used as the endogenous control for SFRP1, and thecomparative threshold cycle (2�DDCT) equation was used tocalculate the relative expression level.
Plasmid construction and transfectionThe pcDNA3.1-vector, pcDNA3.1-SFRP1, pSin-EF2-puro-vec-
tor, and pSin-EF2-puro-SFRP1 plasmids were obtained fromLand.Hua Gene Biosciences. All plasmids were verified by DNAsequencing before use. The pcDNA3.1-vector and pSin-EF2-puro-vector plasmids were used as controls.
The pSin-EF2-puro-SFRP1 or pSin-EF2-puro-vector plasmidswere transfected into 293FT cells using Lipofectamine 2000reagent (Invitrogen) to generate stably transfected cell lines.Lentiviral particles were harvested and infected into SUNE-1 cells48hours later. After selectionwithpuromycin (Sigma-Aldrich) for2 weeks, real-time RT-PCR and Western blotting were used tovalidate the stably transfected cells.
MTT assay and colony formation assayFor MTT assay, CNE-2, SUNE-1, and NP69 cells after transfect-
ing with plasmids or siRNAs were seeded at 1,000 cells per well in96-well plates. The cell viability was measured at 490 nm with aspectrophotometric plate reader at 1, 2, 3, 4, and 5 days. Forcolony formation assay, cells were plated at 400 cells per well in6-well plates and cultured for 7 or 12 days. Colonies were countedunder the inverted microscope after fixing in 4% paraformalde-hyde and staining with 0.5% crystal violet.
Wound-healing assayCNE-2, SUNE-1, and NP69 cells were transfected with plas-
mids or siRNAs and grown to near confluence in 6-well plates.After 24-hour serum starving, the monolayers were scratchedusing a sterile 200-mL tip, followed by washing with serum-freemedium to remove the detached cells. Then, the cells werecultured without serum over the next 24 hours. Images of cellsmigrating at the corresponding wound sites were captured at0 and 24 hours using an inverted microscope (40�).
Transwell migration and invasion assaysTranswell chambers (8-mmpores; Corning) in which the upper
surface of the membrane were coated with or without Matrigel(BD Biosciences) before cell seeding were used to test the cellmigration or invasive ability. The dishes were placed in a cellculture incubator for 1 hour at 37�C. CNE-2, SUNE-1, and NP69cells (5� 104 or 1� 105) that had been transfected with plasmidsor siRNAs and suspended in serum-free medium were plated intheupper chambers. The lower chamberswerefilledwithmediumsupplemented with 10% FBS. After 12- or 24-hour incubation,cells that had migrated or invaded were fixed, stained, andcounted under an inverted microscope (100�).
In vivo xenografted tumor modelForty BALB/c nudemice (4–6weeks old, male) were purchased
from the Medical Experimental Animal Center of GuangdongProvince (Guangzhou, China), 5mice in each group. In total, 5�105 CNE-2 cells or 1 � 106 SUNE-1 cells stably overexpressingvector or SFRP1 that had been resuspended in 200 mL PBS were
injected into the dorsal flank and the tail veins of the mice. Fortumor growth model, tumor size was measured for 2 or 4 weeks.Then, the tumors were dissected and weighted after the micesacrificing. For lung colonization model, after 6 or 8 weeks, themicewere sacrificed and the lung tissueswere harvested,fixed, andparaffin-embedded before 5-mmtissue sectionswere obtained. Allsections were stained with hematoxylin and eosin (H&E) forexamination. All animal research was conducted in accordancewith the detailed rules approved by the Animal Care and UseEthnic Committee. All efforts were made to minimize animalsuffering.
DNA isolation and bisulfite pyrosequencing analysisCNE-2 and SUNE-1 cellswere treatedwithorwithout 10mmol/L
DAC (Sigma-Aldrich) for 72 hours, with the drug being replacedevery 24hours. AnEZ1DNATissueKit (Qiagen)wasused to isolatecellular gDNA. The gDNA (1–2 mg) was treated with sodiumbisulfite using an EpiTect Bisulfite kit (Qiagen) according to themanufacturer's instructions. The bisulfite pyrosequencingprimers were designed using PyroMark Assay Design Software2.0 (Qiagen). The primer sequences were as follows: PCR primers:50-GTTAAAATTAAGGGTTTTTATTAGGGTAGA-30 (F); 50-TCACT-CCCAACTCTCCAAAACT-30 (R); sequencing primer: 50-TAACAA-AAAAACTTCTATTCC-30. The PyroMark Q96 System (Qiagen) wasused for the sequencing reaction and for quantifying methylationlevels.
Western blottingRIPA buffer containing protease inhibitor cocktail (FDbio
Science) was used for cell lysis. Total proteins were separatedusing SDS-PAGE and then transferred to polyvinylidene fluoridemembranes (Millipore). The membranes were hybridized withvarious primary antibodies overnight at 4�C, followed by incu-bationwith species-matched secondary antibodies. Detectionwasachieved using enhanced chemiluminescence.
The following antibodies were used: SFRP1 (1:2,000; Abcam);b-catenin (1:500; Proteintech); c-Myc (1:2,000; Proteintech);cyclin D1 (1:500; Proteintech); GAPDH (1:5,000; Epitomics);and anti-rabbit IgG antibody (1:5,000; Epitomics). GAPDH wasused as the endogenous control.
Statistical analysisAll statistical analysis was performed using SPSS 16.0 software
(SPSS Inc.); differences with P < 0.05 were considered statisticallysignificant. Data were representative of 3 independent experi-ments and presented as the mean � SD. The Student t test, Fisherexact test, or the c2 test were used to compare groups. Survivalcurves were estimated using the Kaplan–Meier method and uni-variate analysis. Multivariate Cox regression analysis with back-ward stepwise approach was used to test for independent prog-nostic factors.
ResultsRelationship between SFRP1 expression and clinicalcharacteristics of NPC patients
To examine whether the SFRP1 protein expression level isrelated to the clinical features of patients with NPC, we appliedimmunohistochemistry to 244 paraffin-embedded NPC tissuesamples. Figure 1A–D depicts representative SFRP1 stainingin NPC tissue. SFRP1 protein levels were detected in 221 NPC
Ren et al.
Cancer Prev Res; 8(10) October 2015 Cancer Prevention Research970
Research. on May 20, 2018. © 2015 American Association for Cancercancerpreventionresearch.aacrjournals.org Downloaded from
Published OnlineFirst August 14, 2015; DOI: 10.1158/1940-6207.CAPR-14-0369

specimens; 119 of 244 (48.8%) patients with NPC had repre-sentatively low SFRP1 expression. More patients with lowSFRP1 expression died (P ¼ 0.04) and developed distantmetastasis (P ¼ 0.018) than those with high SFRP1 expression(Table 1). No statistically significant correlations betweenSFRP1 expression and patient age, sex, World Health Organi-zation type, viral capsid antigen immunoglobulin A (VCA-IgA),early antigen immunoglobulin A (EA-IgA), T stage, N stage,TNM stage, or locoregional failure were observed.
Low SFRP1 expression was associated with worse survival inNPC
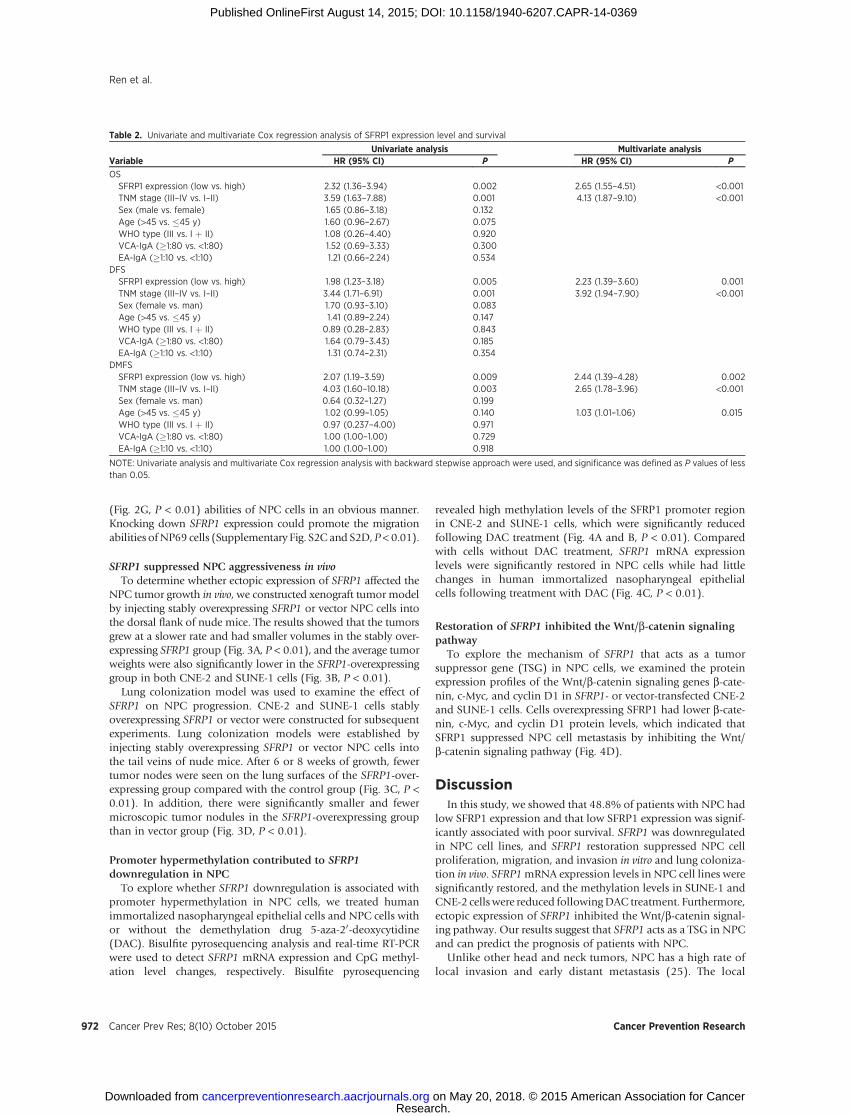
Kaplan–Meier analysis was used to evaluate overall survival(OS), disease-free survival (DFS), and distant metastasis–freesurvival (DMFS) in patients with NPC based on the SFRP1protein expression level. Patients with NPC with low SFRP1expression had significantly poorer OS [HR, 2.32; 95% confi-dence interval (CI), 1.36–3.94; P ¼ 0.002], DFS (HR, 1.98; 95%CI, 1.23–3.18; P ¼ 0.005), and DMFS (HR, 2.07; 95% CI, 1.19–3.59; P ¼ 0.009) than those with high SFRP1 expression (Fig.1E–G). Multivariate Cox regression analyses indicated thatpatients with low SFRP1 expression and advanced disease hadworse OS, DFS, and DMFS than those with high SFRP1 expres-sion, suggesting that both SFRP1 expression and TNM stage are
independent prognostic indicators for survival in patients withNPC (Table 2).
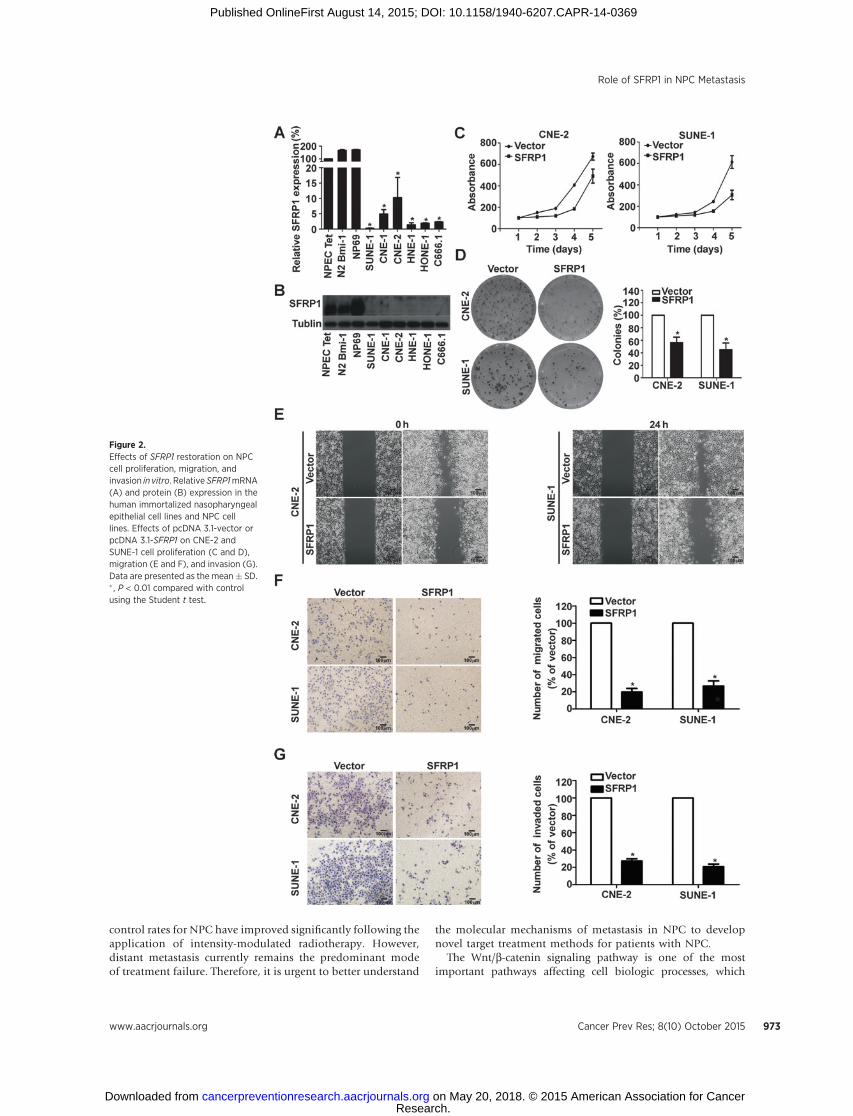
SFRP1 suppressed NPC cell proliferation in vitroWe examined SFRP1 mRNA and protein levels in 3 normal
nasopharyngeal epithelial cell lines and 6 NPC cell lines usingreal-time RT-PCR and Western blotting and found that both theSFRP1 mRNA and protein expression levels were significantlydownregulated in NPC cell lines (Fig. 2A and B, P < 0.01). MTTand colony formation assays demonstrated that ectopic expres-sion of SFRP1 (Supplementary Fig. S1, P < 0.01) could suppressthe cell viability and proliferation in CNE-2 and SUNE-1 cells(Fig. 2C andD, P < 0.05). SFRP1 silence (Supplementary Fig. S2A,P < 0.01) could promote the cell proliferation in NP69 cells(Supplementary Fig. S2B, P < 0.05).
SFRP1 suppressed NPC cell migration and invasion in vitroThe wound-healing and Transwell assays were used to study the
roles of SFRP1 in NPC cell migration and invasion. The wound-healing assaydemonstrated that themigration ability ofCNE-2 andSUNE-1 cells transfected with SFRP1 was much lower than that incells transfected with plasmid vector (Fig. 2E). Restoring SFRP1expression suppressed themigration (Fig. 2F,P<0.01) and invasive
Figure 1.SFRP1 expression levels and survival of patients with NPC. A–D, immunohistochemical detection of SFRP1 expression in 244 patients diagnosed with NPC.A, negative staining (400�). B, weak staining: light yellow (400�). C, moderate staining: yellow brown (400�). D, strong staining: brown (400�). E–G,SFRP1 downregulation is associated with (E) poor OS, (F) DFS, and (G) DMFS. HR values were calculated using unadjusted Cox regression analysis.P values were calculated using the log-rank test.
Role of SFRP1 in NPC Metastasis
www.aacrjournals.org Cancer Prev Res; 8(10) October 2015 971
Research. on May 20, 2018. © 2015 American Association for Cancercancerpreventionresearch.aacrjournals.org Downloaded from
Published OnlineFirst August 14, 2015; DOI: 10.1158/1940-6207.CAPR-14-0369
(Fig. 2G, P < 0.01) abilities of NPC cells in an obvious manner.Knocking down SFRP1 expression could promote the migrationabilities ofNP69 cells (Supplementary Fig. S2C and S2D, P < 0.01).
SFRP1 suppressed NPC aggressiveness in vivoTo determine whether ectopic expression of SFRP1 affected the
NPC tumor growth in vivo, we constructed xenograft tumor modelby injecting stably overexpressing SFRP1 or vector NPC cells intothe dorsal flank of nude mice. The results showed that the tumorsgrew at a slower rate and had smaller volumes in the stably over-expressing SFRP1 group (Fig. 3A, P < 0.01), and the average tumorweights were also significantly lower in the SFRP1-overexpressinggroup in both CNE-2 and SUNE-1 cells (Fig. 3B, P < 0.01).
Lung colonization model was used to examine the effect ofSFRP1 on NPC progression. CNE-2 and SUNE-1 cells stablyoverexpressing SFRP1 or vector were constructed for subsequentexperiments. Lung colonization models were established byinjecting stably overexpressing SFRP1 or vector NPC cells intothe tail veins of nude mice. After 6 or 8 weeks of growth, fewertumor nodes were seen on the lung surfaces of the SFRP1-over-expressing group compared with the control group (Fig. 3C, P <0.01). In addition, there were significantly smaller and fewermicroscopic tumor nodules in the SFRP1-overexpressing groupthan in vector group (Fig. 3D, P < 0.01).
Promoter hypermethylation contributed to SFRP1downregulation in NPC
To explore whether SFRP1 downregulation is associated withpromoter hypermethylation in NPC cells, we treated humanimmortalized nasopharyngeal epithelial cells and NPC cells withor without the demethylation drug 5-aza-20-deoxycytidine(DAC). Bisulfite pyrosequencing analysis and real-time RT-PCRwere used to detect SFRP1 mRNA expression and CpG methyl-ation level changes, respectively. Bisulfite pyrosequencing
revealed high methylation levels of the SFRP1 promoter regionin CNE-2 and SUNE-1 cells, which were significantly reducedfollowing DAC treatment (Fig. 4A and B, P < 0.01). Comparedwith cells without DAC treatment, SFRP1 mRNA expressionlevels were significantly restored in NPC cells while had littlechanges in human immortalized nasopharyngeal epithelialcells following treatment with DAC (Fig. 4C, P < 0.01).
Restoration of SFRP1 inhibited the Wnt/b-catenin signalingpathway
To explore the mechanism of SFRP1 that acts as a tumorsuppressor gene (TSG) in NPC cells, we examined the proteinexpression profiles of the Wnt/b-catenin signaling genes b-cate-nin, c-Myc, and cyclin D1 in SFRP1- or vector-transfected CNE-2and SUNE-1 cells. Cells overexpressing SFRP1 had lower b-cate-nin, c-Myc, and cyclin D1 protein levels, which indicated thatSFRP1 suppressed NPC cell metastasis by inhibiting the Wnt/b-catenin signaling pathway (Fig. 4D).
DiscussionIn this study, we showed that 48.8% of patients with NPC had
low SFRP1 expression and that low SFRP1 expression was signif-icantly associated with poor survival. SFRP1 was downregulatedin NPC cell lines, and SFRP1 restoration suppressed NPC cellproliferation, migration, and invasion in vitro and lung coloniza-tion in vivo. SFRP1mRNA expression levels in NPC cell lines weresignificantly restored, and the methylation levels in SUNE-1 andCNE-2 cells were reduced followingDAC treatment. Furthermore,ectopic expression of SFRP1 inhibited the Wnt/b-catenin signal-ing pathway. Our results suggest that SFRP1 acts as a TSG in NPCand can predict the prognosis of patients with NPC.
Unlike other head and neck tumors, NPC has a high rate oflocal invasion and early distant metastasis (25). The local
Table 2. Univariate and multivariate Cox regression analysis of SFRP1 expression level and survival
Univariate analysis Multivariate analysisVariable HR (95% CI) P HR (95% CI) P
OSSFRP1 expression (low vs. high) 2.32 (1.36–3.94) 0.002 2.65 (1.55–4.51) <0.001TNM stage (III–IV vs. I–II) 3.59 (1.63–7.88) 0.001 4.13 (1.87–9.10) <0.001Sex (male vs. female) 1.65 (0.86–3.18) 0.132Age (>45 vs. �45 y) 1.60 (0.96–2.67) 0.075WHO type (III vs. I þ II) 1.08 (0.26–4.40) 0.920VCA-IgA (�1:80 vs. <1:80) 1.52 (0.69–3.33) 0.300EA-IgA (�1:10 vs. <1:10) 1.21 (0.66–2.24) 0.534
DFSSFRP1 expression (low vs. high) 1.98 (1.23–3.18) 0.005 2.23 (1.39–3.60) 0.001TNM stage (III–IV vs. I–II) 3.44 (1.71–6.91) 0.001 3.92 (1.94–7.90) <0.001Sex (female vs. man) 1.70 (0.93–3.10) 0.083Age (>45 vs. �45 y) 1.41 (0.89–2.24) 0.147WHO type (III vs. I þ II) 0.89 (0.28–2.83) 0.843VCA-IgA (�1:80 vs. <1:80) 1.64 (0.79–3.43) 0.185EA-IgA (�1:10 vs. <1:10) 1.31 (0.74–2.31) 0.354
DMFSSFRP1 expression (low vs. high) 2.07 (1.19–3.59) 0.009 2.44 (1.39–4.28) 0.002TNM stage (III–IV vs. I–II) 4.03 (1.60–10.18) 0.003 2.65 (1.78–3.96) <0.001Sex (female vs. man) 0.64 (0.32–1.27) 0.199Age (>45 vs. �45 y) 1.02 (0.99–1.05) 0.140 1.03 (1.01–1.06) 0.015WHO type (III vs. I þ II) 0.97 (0.237–4.00) 0.971VCA-IgA (�1:80 vs. <1:80) 1.00 (1.00–1.00) 0.729EA-IgA (�1:10 vs. <1:10) 1.00 (1.00–1.00) 0.918
NOTE: Univariate analysis and multivariate Cox regression analysis with backward stepwise approach were used, and significance was defined as P values of lessthan 0.05.
Ren et al.
Cancer Prev Res; 8(10) October 2015 Cancer Prevention Research972
Research. on May 20, 2018. © 2015 American Association for Cancercancerpreventionresearch.aacrjournals.org Downloaded from
Published OnlineFirst August 14, 2015; DOI: 10.1158/1940-6207.CAPR-14-0369
control rates for NPC have improved significantly following theapplication of intensity-modulated radiotherapy. However,distant metastasis currently remains the predominant modeof treatment failure. Therefore, it is urgent to better understand
the molecular mechanisms of metastasis in NPC to developnovel target treatment methods for patients with NPC.
The Wnt/b-catenin signaling pathway is one of the mostimportant pathways affecting cell biologic processes, which
Figure 2.Effects of SFRP1 restoration on NPCcell proliferation, migration, andinvasion in vitro. Relative SFRP1mRNA(A) and protein (B) expression in thehuman immortalized nasopharyngealepithelial cell lines and NPC celllines. Effects of pcDNA 3.1-vector orpcDNA 3.1-SFRP1 on CNE-2 andSUNE-1 cell proliferation (C and D),migration (E and F), and invasion (G).Data are presented as the mean� SD.� , P < 0.01 compared with controlusing the Student t test.
Role of SFRP1 in NPC Metastasis
www.aacrjournals.org Cancer Prev Res; 8(10) October 2015 973
Research. on May 20, 2018. © 2015 American Association for Cancercancerpreventionresearch.aacrjournals.org Downloaded from
Published OnlineFirst August 14, 2015; DOI: 10.1158/1940-6207.CAPR-14-0369

include differentiation, proliferation, and adhesion (4). In nor-mal conditions, signaling begins with the Wnt ligand binding toFrizzled proteins and lipoprotein receptor–related proteins 5 and6 receptors. Then, b-catenin accumulates and functions as atranscription cofactor with T-cell factor/lymphoid enhancer fac-
tor, which regulates the expression of target genes such as c-mycand cyclin D1. Abnormal activation of the Wnt/b-catenin signal-ing pathway is a common event in several human cancers,including NPC (26–28), and the abnormal methylation statusof Wnt antagonists such as Dickkopf proteins, Wnt inhibitory
Figure 3.SFRP1 suppressed NPC cellaggressiveness in vivo. A,representative picture of tumorsformed and the growth curves oftumor volumes. B, tumor weight. C,representative images ofmacroscopictumor colonization and growth in thelung tissues and quantification of theaverage number of tumor nodes onthe lung surface; arrows indicatetumor nodes. D, representativeimages of H&E-stained lung sections(100�) and quantification of theaverage number ofmicroscopic tumornodes in the lungs based onpathologic analysis of the H&E-stained sections. Data are presentedas the mean � SD. �P < 0.01 (Studentt test).
Ren et al.
Cancer Prev Res; 8(10) October 2015 Cancer Prevention Research974
Research. on May 20, 2018. © 2015 American Association for Cancercancerpreventionresearch.aacrjournals.org Downloaded from
Published OnlineFirst August 14, 2015; DOI: 10.1158/1940-6207.CAPR-14-0369

factor 1, and SFRPs may contribute to it (29–32). An SFRP familymember, SFRP1, can block Wnt/b-catenin signaling by hinderingWnt–receptor interactions via an N-terminal cysteine-richdomain homologous to Frizzled proteins (33). SFRP1 is hyper-methylated and downregulated in various cancers, includingbreast (34), colorectal (35), and ovarian cancer (36). SFRP1hypermethylation and downregulation are also associated withpoor prognosis in several tumors (34, 37, 38).Moreover, SFRP1 isassociated with tumor chemotherapy, and some antitumor drugsinhibit cell growth through the re-expression of SFRP1 (39–41).However, emerging evidence has indicated that SFRP1 may alsobe highly expressed in carcinomas and promote tumor prolifer-ation or migration, such as in basal-like breast cancer (42), gastriccancer (43), and metastatic renal carcinoma (44). Recently, it hasbeen reported that SFRP1 could suppress NPC cell proliferation invitro (19). Notably, there is no research on SFRP1 in NPC metas-tasis. It remains unclear whether SFRP1 plays a direct role in NPC
development and progression and whether it can predict distantmetastasis and be a potential biomarker to guide individualizedtreatment of patients with NPC.
Our study demonstrates that SFRP1 is downregulated in NPCcell lines, and 48.8% of clinical specimens showed representativelow SFRP1 protein expression levels. Kaplan–Meier analysisshowed that patients with NPC with low SFRP1 expression hadsignificantly poorerOS,DFS, andDMFS rates than thosewithhighSFRP1 expression. Multivariate Cox regression analysis indicatedthat high SFRP1 expression was an independent prognostic indi-cator for patients with NPC. These results suggest that the SFRP1expression level can predict the prognosis of patients with NPCand may provide a new therapy target for individualized treat-ment of patients with NPC.
SFRP1 acts as a suppressor gene in several tumor types byregulating cell proliferation, migration, and invasion. In thisresearch, we used CNE-2 and SUNE-1 cells overexpressing SFRP1
Figure 4.SFRP1 downregulation was associated with promoter methylation. SFRP1 restoration downregulated Wnt/b-catenin target genes. A and B, bisulfitepyrosequencing analysis of the SFRP1 promoter region (A) and average methylation level (B) for 3 CpG sites in DAC-treated (DACþ) or -untreated (DAC�)CNE-2 and SUNE-1 cells. Data are presented as the mean � SD. � , P < 0.01 compared with control (Student t test). C, relative SFRP1 mRNA expressionin DAC-treated (DACþ) or -untreated (DAC�) human immortalized nasopharyngeal epithelial cell lines and NPC cell lines. D, Western blot analysis ofWnt/b-catenin target gene expression in CNE-2 and SUNE-1 cells. GAPDH was used as the loading control.
Role of SFRP1 in NPC Metastasis
www.aacrjournals.org Cancer Prev Res; 8(10) October 2015 975
Research. on May 20, 2018. © 2015 American Association for Cancercancerpreventionresearch.aacrjournals.org Downloaded from
Published OnlineFirst August 14, 2015; DOI: 10.1158/1940-6207.CAPR-14-0369

to investigate the function of SFRP1 in NPC. First, we examinedthe effects of SFRP1 on NPC cells using MTT, colony formation,wound-healing, and Transwell assays, which showed that cellproliferation, invasion, and migration abilities were significantlysuppressed in cells overexpressing SFRP1. The role of SFRP1 inNPC proliferation in vitro was consistent with Li and colleagues(19). Nude mice xenografts tumor model and lung colonizationmodel were used to investigate the effects of SFRP1 in vivo andrevealed that SFRP1 overexpression inhibited tumor aggres-siveness of NPC. Taken together, these results demonstrate thatSFRP1 suppresses NPC cell proliferation, migration, and invasionin vitro and lung colonization in vivo.
Epigenetic alterations of TSGs have been reported in thedevelopment of several cancers. Hypermethylation of theSFRP1 promoter has been recognized as a common reason forSFRP1 downregulation in tumors. Therefore, we treated NPCcells with the demethylation agent DAC to explore the mech-anism of SFRP1 downregulation in NPC. DAC treatment sig-nificantly restored SFRP1 mRNA expression levels; bisulfatepyrosequencing analysis demonstrated that the methylationstatus of SFRP1 was reduced in CNE-2 and SUNE-1 cells afterDAC treatment. In summary, SFRP1was downregulated in NPCdue to hypermethylation of the SFRP1 promoter region.
As SFRP1 is a Wnt/b-catenin signaling pathway antagonist, weexamined changes to the downstream target genes using Westernblotting. b-Catenin, c-Myc, and cyclin D1 were all downregulatedafter SFRP1 restoration in NPC cells, which suggests that SFRP1might suppress NPC proliferation and metastasis via the Wnt/b-catenin signaling pathway.
In conclusion, SFRP1 downregulation may result from epi-genetic silencing and is associated with worse survival in NPC.Ectopic SFRP1 expression suppresses cell proliferation, migra-tion, and invasion by the blocking Wnt/b-catenin signalingpathway. Our findings also highlight the possibility that SFRP1
is a potential molecular biomarker for predicting the prognosisof patients with NPC and may provide novel therapeuticstrategies in NPC.
Disclosure of Potential Conflicts of InterestNo potential conflicts of interest were disclosed.
Authors' ContributionsConception and design: X.-Y. Ren, G.-Q. Zhou, W. Jiang, Y. Sun, N. Liu, J. MaDevelopment of methodology: X.-Y. Ren, G.-Q. Zhou, W. Jiang, Q.-M. HeAcquisition of data (provided animals, acquired and managed patients,provided facilities, etc.): X.-Y. Ren, G.-Q. Zhou, Y.-F. Xu, Y.-Q. Li, X.-R. Tang,Q.-M. He, N. Liu, J. MaAnalysis and interpretation of data (e.g., statistical analysis, biostatistics,computational analysis):X.-Y. Ren,W. Jiang, Y.-Q. Li, X.Wen, X.-J. Yang,N. Liu,J. MaWriting, review, and/or revision of the manuscript: X.-Y. Ren, N. Liu, J. MaAdministrative, technical, or material support (i.e., reporting or organizingdata, constructing databases): X.-Y. Ren, X.-R. Tang, J. MaStudy supervision: X.-Y. Ren, N. Liu, J. Ma
Grant SupportThis work was supported by grants from the National Natural Science
Foundation of China (Nos. 81402532, G.-Q. Zhou; 81372409, Y. Sun); theNatural Science Foundation of Guangdong Province (No. S2013010012220,Y. Sun); the Science and Technology Project of Guangzhou City, China(No. 132000507, Y. Sun); the Science and Technology Project of GuangzhouCity, China (No. 14570006, J. Ma); the Health & Medical CollaborativeInnovation Project of Guangzhou City, China (No. 201400000001, J. Ma);and the National Science & Technology Pillar Program during the TwelfthFive-year Plan Period (No. 2014BAI09B10, J. Ma).
The costs of publication of this article were defrayed in part by the paymentof page charges. This article must therefore be hereby marked advertisementin accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
Received October 22, 2014; revised July 24, 2015; accepted July 24, 2015;published OnlineFirst August 14, 2015.
References1. Liu N, Chen NY, Cui RX, Li WF, Li Y, Wei RR, et al. Pronostic value of a
microRNAsignature innasopharyngeal carcinoma: amicroRNAexpressionanalysis. Lancet Oncol 2012;13:633–41.
2. Lai SZ, Li WF, Chen L, Luo W, Chen YY, Liu LZ, et al. How does intensity-modulated radiotherapy versus conventional two-dimensional radiother-apy influence the treatment results in nasopharyngeal carcinoma patients?Int J Radiat Oncol Biol Phys 2011;80:661–8.
3. Steeg PS. Tumor metastasis: mechanistic insights and clinical challenges.Nat Med 2006;12:895–904.
4. Nelson WJ, Nusse R. Convergence of Wnt, beta-catenin, and cadherinpathways. Science 2004;303:1483–7.
5. Chou J, Lin YC, Kim J, You L, Xu Z, He B, et al. Nasopharyngeal carcinoma–review of the molecular mechanisms of tumorigenesis. Head Neck2008;30:946–63.
6. Kirstetter P, Anderson K, Porse BT, Jacobsen SE, Nerlov C. Activation ofthe canonical Wnt pathway leads to loss of hematopoietic stem cellrepopulation and multilineage differentiation block. Na Immunol2006;7:1048–56.
7. Kamino M, Kishida M, Kibe T, Ikoma K, Iijima M, Hirano H, et al. Wnt-5asignaling is correlated with infiltrative activity in human glioma by induc-ing cellular migration and MMP-2. Cancer Sci 2011;102:540–8.
8. Prunier C, Hocevar BA, Howe PH. Wnt signaling: physiology and pathol-ogy. Growth Factors 2004;22:141–50.
9. Wands JR, Kim M. WNT/beta-catenin signaling and hepatocellular carci-noma. Hepatology 2014;60:452–4.
10. Stewart DJ. Wnt signaling pathway in non-small cell lung cancer. J NatlCancer Inst 2014;106:djt356.
11. DeyN, Barwick BG,MorenoCS,Ordanic-KodaniM, Chen Z,Oprea-Ilies G,et al. Wnt signaling in triple negative breast cancer is associated withmetastasis. BMC Cancer 2013;13:537.
12. Anastas JN, Moon RT. WNT signalling pathways as therapeutic targets incancer. Nat Rev Cancer 2013;13:11–26.
13. Kahn M. Can we safely target the WNT pathway? Nat Rev Drug Discov2014;13:513–32.
14. Herbst A, Kolligs FT. Wnt signaling as a therapeutic target for cancer.Methods Mol Biol 2007;361:63–91.
15. MacDonald BT, Tamai K, He X. Wnt/beta-catenin signaling: components,mechanisms, and diseases. Dev Cell 2009;17:9–26.
16. Caldwell GM, Jones C, Gensberg K, Jan S, Hardy RG, Byrd P, et al. The Wntantagonist sFRP1 in colorectal tumorigenesis. Cancer Res 2004;64:883–8.
17. Gauger KJ, Chenausky KL, Murray ME, Schneider SS. SFRP1 reductionresults in an increased sensitivity to TGF-beta signaling. BMC Cancer2011;11:59.
18. Nojima M, Suzuki H, Toyota M, Watanabe Y, Maruyama R, Sasaki S,et al. Frequent epigenetic inactivation of SFRP genes and constitutiveactivation of Wnt signaling in gastric cancer. Oncogene 2007;26:4699–713.
19. Li L, Zhang Y, Fan Y, Sun K, Su X, Du Z, et al. Characterization of thenasopharyngeal carcinoma methylome identifies aberrant disruption ofkey signaling pathways and methylated tumor suppressor genes. Epige-nomics 2015:7:155–73.
20. McShane LM, AltmanDG, Sauerbrei W, Taube SE, GionM, Clark GM, et al.Reporting recommendations for tumor marker prognostic studies. J Clin-oncol 2005;23:9067–72.
Cancer Prev Res; 8(10) October 2015 Cancer Prevention Research976
Ren et al.
Research. on May 20, 2018. © 2015 American Association for Cancercancerpreventionresearch.aacrjournals.org Downloaded from
Published OnlineFirst August 14, 2015; DOI: 10.1158/1940-6207.CAPR-14-0369

21. McShane LM, AltmanDG, SauerbreiW, Taube SE, GionM, Clark GM, et al.Reporting recommendations for tumor marker prognostic studies(REMARK). J Natl Cancer Inst 2005;97:1180–4.
22. Liu N, Tang LL, Sun Y, Cui RX, Wang HY, Huang BJ, et al. MiR-29csuppresses invasion and metastasis by targeting TIAM1 in nasopharyngealcarcinoma. Cancer Lett 2013;329:181–8.
23. Hanley JA, McNeil BJ. The meaning and use of the area under a receiveroperating characteristic (ROC) curve. Radiology 1982;143:29–36.
24. Zweig MH, Campbell G. Receiver-operating characteristic (ROC) plots: afundamental evaluation tool in clinical medicine. Clin Chem 1993;39:561–77.
25. Lee AW, Poon YF, FooW, LawSC, Cheung FK, ChanDK, et al. Retrospectiveanalysis of 5037 patients with nasopharyngeal carcinoma treated during1976–1985: overall survival and patterns of failure. Int J Radiat Oncol BiolPhys 1992;23:261–70.
26. QingLingZ, LiNaY, Li L, ShuangW, YuFang Y, YiD, et al. LMP1 antagonizesWNT/beta-catenin signalling through inhibition of WTX and promotesnasopharyngeal dysplasia but not tumourigenesis in LMP1(B95-8) trans-genic mice. J Pathol 2011;223:574–83.
27. Shu XS, Geng H, Li L, Ying J, Ma C, Wang Y, et al. The epigenetic modifierPRDM5 functions as a tumor suppressor through modulating WNT/beta-catenin signaling and is frequently silenced in multiple tumors. PLoS One2011;6:e27346.
28. Xu L, Jiang Y, Zheng J, Xie G, Li J, Shi L, et al. Aberrant expression of beta-catenin and E-cadherin is correlated with poor prognosis of nasopharyn-geal cancer. Hum Pathol 2013;44:1357–64.
29. Guo Y, Guo W, Chen Z, Kuang G, Yang Z, Dong Z. Hypermethylation andaberrant expression of Wnt-antagonist family genes in gastric cardiaadenocarcinoma. Neoplasma 2011;58:110–7.
30. Kim JT, Li J, Jang ER, Gulhati P, Rychahou PG, Napier DL, et al.Deregulation of Wnt/beta-catenin signaling through genetic or epige-netic alterations in human neuroendocrine tumors. Carcinogenesis2013;34:953–61.
31. Aguilera O, Fraga MF, Ballestar E, Paz MF, Herranz M, Espada J, et al.Epigenetic inactivation of the Wnt antagonist DICKKOPF-1 (DKK-1) genein human colorectal cancer. Oncogene 2006;25:4116–21.
32. Mitsui Y, Yasumoto H, Nagami T, Hiraki M, Arichi N, Ishikawa N, et al.Extracellular activation of Wnt signaling through epigenetic dysregulationof Wnt inhibitory factor-1 (Wif-1) is associated with pathogenesis ofadrenocortical tumor. Oncotarget 2014;5:2198–207.
33. Suzuki H, Watkins DN, Jair KW, Schuebel KE, Markowitz SD, Chen WD,et al. Epigenetic inactivation of SFRP genes allows constitutive WNTsignaling in colorectal cancer. Nat Genet 2004;36:417–22.
34. Yang ZQ, Liu G, Bollig-Fischer A, Haddad R, Tarca AL, Ethier SP. Meth-ylation-associated silencing of SFRP1 with an 8p11-12 amplificationinhibits canonical and non-canonical WNT pathways in breast cancers.Int J Cancer 2009;125:1613–21.
35. Rawson JB,MannoM,MrkonjicM,DaftaryD, Dicks E, BuchananDD, et al.Promoter methylation of Wnt antagonists DKK1 and SFRP1 is associatedwith opposing tumor subtypes in two large populations of colorectalcancer patients. Carcinogenesis 2011;32:741–7.
36. Flanagan JM, Wilhelm-Benartzi CS, Metcalf M, Kaye SB, Brown R.Association of somatic DNA methylation variability with progres-sion-free survival and toxicity in ovarian cancer patients. Ann Oncol2013;24:2813–8.
37. Cheng CK, Li L, Cheng SH, Ng K, Chan NP, Ip RK, et al. Secreted-frizzledrelated protein 1 is a transcriptional repression target of the t(8;21) fusionprotein in acute myeloid leukemia. Blood 2011;118:6638–48.
38. Kandimalla R, van Tilborg AA, Zwarthoff EC. DNA methylation-basedbiomarkers in bladder cancer. Nat Rev Urol 2013;10:327–35.
39. Cooper SJ, von Roemeling CA, Kang KH, Marlow LA, Grebe SK, MenefeeME, et al. Reexpression of tumor suppressor, sFRP1, leads to antitumorsynergy of combined HDAC andmethyltransferase inhibitors in chemore-sistant cancers. Mol Cancer Ther 2012;11:2105–15.
40. Bernemann C, Hulsewig C, Ruckert C, Schafer S, Blumel L, Hempel G, et al.Influence of secreted frizzled receptor protein 1 (SFRP1) on neoadjuvantchemotherapy in triple negative breast cancer does not rely on WNTsignaling. Mol Cancer 2014;13:174.
41. Dave K, Panchal H. Review on chemogenomics approach: interpretingantagonist activity of secreted frizzled-related protein 1 in glaucomadisease with in-silico docking. Curr Topics Med Chem 2012;12:1834–42.
42. Jeong YJ, Jeong HY, Bong JG, Park SH, Oh HK. Low methylation levels ofthe SFRP1 gene are associated with the basal-like subtype of breast cancer.Oncol Rep 2013;29:1946–54.
43. Qu Y, Ray PS, Li J, Cai Q, Bagaria SP, Moran C, et al. High levels of secretedfrizzled-related protein 1 correlate with poor prognosis and promotetumourigenesis in gastric cancer. Eur J Cancer 2013;49:3718–28.
44. GumzML, ZouH, Kreinest PA, Childs AC, Belmonte LS, LeGrand SN, et al.Secreted frizzled-related protein 1 loss contributes to tumor phenotype ofclear cell renal cell carcinoma. Clin Cancer Res 2007;13:4740–9.
www.aacrjournals.org Cancer Prev Res; 8(10) October 2015 977
Role of SFRP1 in NPC Metastasis
Research. on May 20, 2018. © 2015 American Association for Cancercancerpreventionresearch.aacrjournals.org Downloaded from
Published OnlineFirst August 14, 2015; DOI: 10.1158/1940-6207.CAPR-14-0369

2015;8:968-977. Published OnlineFirst August 14, 2015.Cancer Prev Res Xian-Yue Ren, Guan-Qun Zhou, Wei Jiang, et al. Pathway in NPC
-Catenin SignalingβPromotes Cell Invasion by Activating the Wnt/Low SFRP1 Expression Correlates with Poor Prognosis and
Updated version
10.1158/1940-6207.CAPR-14-0369doi:
Access the most recent version of this article at:
Material
Supplementary
1
http://cancerpreventionresearch.aacrjournals.org/content/suppl/2015/08/08/1940-6207.CAPR-14-0369.DCAccess the most recent supplemental material at:
Cited articles
http://cancerpreventionresearch.aacrjournals.org/content/8/10/968.full#ref-list-1
This article cites 44 articles, 7 of which you can access for free at:
Citing articles
http://cancerpreventionresearch.aacrjournals.org/content/8/10/968.full#related-urls
This article has been cited by 2 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications Department at
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cancerpreventionresearch.aacrjournals.org/content/8/10/968To request permission to re-use all or part of this article, use this link
Research. on May 20, 2018. © 2015 American Association for Cancercancerpreventionresearch.aacrjournals.org Downloaded from
Published OnlineFirst August 14, 2015; DOI: 10.1158/1940-6207.CAPR-14-0369