macroalgae on shallow tropical reefs reduce the availability of
TRANSCRIPT

BULLETIN OF MARINE SCIENCE. 88(4):1019–1033. 2012http://dx.doi.org/10.5343/bms.2011.1084
1019Bulletin of Marine Science© 2012 Rosenstiel School of Marine & Atmospheric Science of the University of Miami
Macroalgae on shallow tropical reefs reduce the availability of reflected light
for use in coral photosynthesis
Nicholas B Colvard and Peter J Edmunds
abstract
we tested the hypothesis that light reflected from dense growths of macroalgae [i.e., a low reflectance (lr) surface] vs heavily grazed surfaces [i.e., a high reflectance (hr) surface] has no effect on the photophysiology of scleractinian corals on the shallow (3-m depth) back reef of Moorea, french polynesia. underwater light was measured using a cosine-corrected sensor that detected photosynthetically active radiation (par, between 400 and 700 nm), and an underwater spectrophotometer that measured spectral reflectance, r(λ), with nanometer wavelength resolution. The effect of reflected light on corals was assessed by incubating small colonies of massive Porites spp. and Pocillopora verrucosa (ellis and solander, 1786) for 27 d on artificial hr and lr surfaces, and measuring the effective photochemical efficiency (Δf⁄fm') of Symbiodinium on the sides of colonies adjacent to the benthos. at noon on a cloudless day, upwelling photon irradiance (eu) from the benthos was reduced 20% over lr vs hr surfaces, and lr surfaces reduced r(λ) 50%–60% for par. for massive Porites spp., Δf⁄fm' did not differ between hr and lr surfaces, but for P. verrucosa, Δf⁄fm' increased 60% on lr compared to hr surfaces. The increase in Δf⁄fm' suggests that P. verrucosa modifies its photophysiological performance in tissue adjacent to lr surfaces to increase the efficiency with which light can be utilized in photochemical pathways.
one striking change affecting coral reefs over the last half century has been the widespread decline in cover of scleractinian corals (gardner et al. 2003, bellwood et al. 2004, hughes et al. 2011), and a shift to dominance by alternate taxa (bruno et al. 2009, norström et al. 2009), typically macroalgae (McManus and polsenberg 2004, aronson and precht 2006). while documentation of these trends has featured prom-inently in coral reef literature (pandolfi et al. 2005, Kuffner et al. 2006, norström et al. 2009), these efforts have overlooked the significance of alternate taxa in altering the color (i.e., the reflective properties) of the benthos relative to reefs that are heavily grazed (e.g., fig. 5 in hoegh-guldberg et al. 2007).
on reefs dominated by scleractinians, space on hard substrata that is not occupied by coral is typically exploited by algal turf (carpenter 1986, tanner 1995), crustose coralline algae (cca; Klumpp and McKinnon 1992), or sessile invertebrates (chadwick and Morrow 2011, colvard and edmunds 2011). The benthos on such reefs is generally distinguished by pastel-colored corals, pale branch apices and coral colony margins (fabricius 2006), and pink cca. in contrast, a reef dominated by macroalgae has dense growths of a variety of algae that can include Amansia, Dictyota, Halimeda, and Sargassum (connor and adey 1977, adjeroud and salvat 1996, adjeroud 1997, Mcclanahan et al. 1999) that are dark as a result of brown (phaeophycea) and green algae (chlorophycea) pigments.
coRAl Reef pApeR

BULLETIN OF MARINE SCIENCE. VOL 88, NO 4. 20121020
remote sensing of coral reefs has enabled the description of regional-scale differ-ences in the benthic community structure based on the reflective characteristics of the benthos (dickey et al. 2006, lesser and Mobley 2007, hochberg 2011). in con-trast, in situ analysis of underwater light using spectrometry equipment has allowed underwater light microenvironments to be quantified on a submeter scale (lesser 2000, Joyce and phinn 2003). at this scale, light microenvironments are created by the attenuation and reflectance of sunlight at the air-water interface (wilkinson 2004), the optical properties of seawater (smith and Mobley 2008), the topographic complexity of the reef surface (dove et al. 2008), and differences in the reflectance of benthic organisms (adjeroud and salvat 1996, hochberg et al. 2003, werdell and roesler 2003, hochberg 2011, Kaniewska et al. 2011). although reflectance of ben-thic surfaces is likely to have a strong effect on the ambient light regime of shallow marine environments, these effects pose challenges for measurement because un-derwater reflectance is subject to high variability attributed to a diversity of bio-logical agents and physical principles (Maritorena et al. 1994, Joyce and phinn 2003). Thus, empirical measurements of underwater reflectance are valuable in describing relative differences among treatments, but they do not provide absolute measures that can be extrapolated directly to other systems.
brakel (1979) was among the first to describe depth-dependent light attenuation on a coral reef, and his analysis revealed the potential for variation in the angular dis-tribution of irradiance to affect coral photosynthesis. Though the acclimatization of corals to different irradiances has been studied for decades (redalje 1976, falkowski and dubinsky 1981), the response of corals to varying light regimes continues to be a rich field for investigation (ulstrup et al. 2006, Mass et al. 2010, dubinsky and falkowski 2011). however, recent changes in the community structure of coral reefs may provide a new context for such studies because the declines in coral cover that have taken place over the last few decades (Mccook 1999, bruno et al. 2009) lead to changes that affect the way light interacts with the benthos. for instance, the loss of coral favors a reduction in three-dimensional structure (McManus and polsenberg 2004, alvarez-filip et al. 2009) that reduces the availability of light microenviron-ments, and often leads to an increased abundance of macroalgae that alters the light available to corals beneath their canopy (birrell et al. 2008). changes in the com-munity structure of coral reefs also are likely to affect the reflectance of the benthos, yet this topic has not been investigated in detail (but see Maritorena et al. 1994, Joyce and phinn 2003). given the importance of light to tropical corals (dubinsky and falkowski 2011), it is likely that changes in the underwater light regime attributed to benthic reflectance will influence coral performance.
The goals of the present study were to test the effects of benthic macroalgae in altering the light regime on a coral reef, and to determine whether these effects influ-ence the photophysiology of reef corals. first, light reflected from benthic surfaces on a shallow reef was characterized through a contrast of areas at a single depth that were dominated either by macroalgae [hereafter, defined as a low reflectance (lr) surface] or coral, cca, and carbonate pavement [hereafter, defined as a high reflectance (hr) surface]. second, the effect of lr and hr surfaces on the effective photochemical efficiency of psii (Δf⁄fm') of Symbiodinium within corals was quanti-fied with the objective of evaluating how differences in irradiance (the radiant flux density emanating from a surface per unit solid angle, after campbell and norman 1998) and spectral reflectance (the ratio of reflected radiant flux to incident radiant

colvard and edmunds: change in spectral composition of reef from macroalgae 1021
flux; hochberg et al. 2003, Kaniewska et al. 2011) affect coral photophysiology. we define light microenvironments on a scale of centimeters-to-meters for both down-welling and upwelling photon irradiance (i.e., reflected) from the substratum, and do not address large-scale (i.e., over kilometers) processes that are typically described by remote sensing techniques (lesser and Mobley 2007, hochberg 2011).
Methods
study sites and environment.—downwelling photon irradiance, ed, upwelling photon irradiance, eu, and relative spectral reflectance, r(λ) (Mobley 1994, Mobley and sundman 2003), from benthic surfaces were measured on a shallow (3-m depth) back reef on the north shore of Moorea, french polynesia (17°30´s, 149°50´w). between 2005 and 2007—just before the present analysis was conducted—this habitat was characterized by low-profile carbonate pavement where 6%–68% of the benthos was sand, 14%–39% a combination of cca and bare space (that often cannot be distinguished reliably in digital images), <17% turf, and <2% macroalgae (edmunds et al. 2010). coral bommies are spread throughout this habitat, and on these surfaces coral cover reaches 8%–43% (edmunds et al. 2010) and is composed mostly of Porites spp. (6%–30% cover), Montipora spp. (0%–27% cover), and Pocillopora spp. (0%–6% cover; adjeroud 1997, edmunds et al. 2010). dense aggregates of the macroalgae Amansia rhodantha (J. agardh, 1841), Sargassum spp., and Turbinaria ornata (J. agardh, 1848) are scattered among the coral colonies (adjeroud and salvat 1996, adjeroud 1997).
downwelling and upwelling photon irradiance.—we defined ed as the rate of in-cident radiant energy received in quanta m−2 s−1, and eu as the energy reflected per unit per time (quanta m−2 s−1) from a given surface (e.g., light reflected from the substratum; Mobley 1994). The underwater light regime was recorded at 3-m depth using a cosine-corrected pho-tosynthetically active radiation (par) sensor fitted to a pulse amplitude modulation fluo-rometer (diving paM, waltz, gmbh) and calibrated against an underwater quantum sensor (li-192, li-cor, lincoln, nebraska, usa). The light sensor on the diving paM was used to measure the ed (μmol quanta m−2 s−1) in the par range. par was recorded from 12:00 to 13:00 hrs on nine cloudless days from 23 april to 20 May, 2009, with the cosine-corrected sensor oriented first, at 0° from vertical to record ed, and second, at 135° from vertical to quantify eu. to complete these measurements, the par sensor was held approximately 10 cm above the benthos, and recordings were made from locations selected at random on benthic surfaces. twenty such recordings were made for both the 0° and 135° orientations and were made above a contiguous area of reef extending over approximately 20 m2. using this procedure, for each day that measurements were made (n = 9 d), there were approximately 20 recordings of ed and eu collected for each orientation of the sensor. These measurements were used to characterize ed as well as the light reflected from each substratum type (e.g., hr and lr surfaces) and were used to characterize the underwater regime of par levels within the back reef. Multiple re-cordings every day were necessary to reduce the variability in the measured underwater irra-diance due to light scattering and attenuation from the water column (Maritorena et al. 1994).
underwater relative spectral reflectance.—to evaluate r(λ) (radiance integrated over unit wavelength; campbell and norman 1998, Kaniewska et al. 2011) of benthic surfaces at 3-m depth, a spectrophotometer (usb2000, ocean optics, dunedin, fl, usa) was operat-ed from a laptop computer (asus eee, taipei, taiwan), running ooibase32 software (ocean optics, dunedin, fl, usa). This instrument was contained in a custom underwater housing and connected using a vacuum feed-through (ocean optics, dunedin, fl, usa) to a 2-m long, 400-µm diameter fiber optic cable. This cable was used to sample light reflected from hr and lr surfaces with 0.33 nm wavelength resolution between 400 and 700 nm (i.e., par). The spectrophotometer used a 2048-element linear silicon ccd array detector to quantify photons of light separated by wavelength, and was configured to integrate light over 4 ms. The

BULLETIN OF MARINE SCIENCE. VOL 88, NO 4. 20121022
manufacturers settings were used to generate a single spectrum from the average of five con-secutive discrete spectral acquisitions, with each spectrum smoothed using a moving average derived from the intensities at six wavelengths. r(λ) was measured using the “scope mode” of the instrument, which displays the spectral irradiance in raw arbitrary counts for each pixel in the array, scaled from 0 to 4000. to account for changes in sunlight intensity, the spectro-photometer signal was adjusted to the current light condition using both a dark reference (a spectrum taken with the light path blocked) and a white reference (a diffuse reflectance stan-dard of matte white plastic) prior to all r(λ) measurements.
to quantify r(λ), the fiber optic cable was positioned in a holder that supported the tip of the sensor 1.0 cm above the substratum and at 135° from vertical (fig. 1), so that measure-ments were not affected by the shadow of the operator (Mazel 1997). The collection angle of the sensor was 26.5°, and thus light intercepted by the downward-pointing sensor was re-flected entirely off the benthos. The light sensitive tip of this sensor was not fitted with a co-sine-corrected filter, as is customary for measurements of reflectance in bio-optical literature (dickey et al. 2006, smith and Mobley 2008) because we sought to use the narrow collection angle of the instrument to sample small areas (0.80 cm2) of benthos. This was advantageous because it ensured that the sensor measured light emitted from the surface at which it was pointed, rather than harvesting light over a larger collection angle and surfaces other than that of interest.
one limitation of measuring light using a sensor with a narrow collection angle is that it samples light from a small area, particularly when it is close to the surface of interest. on surfaces with a high degree of reflectance heterogeneity, a small sampling area could result in measurements of r(λ) having a high variance. however, in our application, preliminary experiments revealed that the reflectance of reef surface were relatively homogeneous on the scale investigated, with measurements of r(λ) differing by <15% for multiple placement of the sensor over substrata that were selected to represent a single benthic category.
r(λ) of hr and lr surfaces was quantified on 13 and 15 May, 2009, with measurements taken between 11:00 and 14:00 hrs to reduce the solar zenith angle and lessen the possibility of shading from adjacent objects. six replicate scans of reflected light from each substratum were generated daily, with each replicate composed of five sequential smoothed scans (each lasting 5–7 s) that were averaged by wavelength. averaging at this stage reduced the variabil-ity in underwater irradiance caused by light flecking, surface ripples, and probe placement (barron et al. 2000, devlin et al. 2008, smith and Mobley 2008). r(λ) (dimensionless) was calculated following Kaniewska et al. (2011) using:
r(λ) = (i − id) ⁄ (i0 − id) (eq. 1)
where i = sample measurement, and id = dark reference standard, and i0 = white reference standard. This calculation of r(λ) accounted for variability in the ambient light environment, and relies on measurements of in arbitrary units. r(λ) is presented as percent reflectance, calculated as r(λ) × 100% (chittka et al. 1994). r(λ) was measured with sequential determina-tions of (i − id) and (i0 − id), which was dictated by instrument limitations; all spectral mea-surements were recorded <5 s apart.
Manipulative experiment.—Juvenile colonies (≤40-mm diameter) of massive Porites spp. (as described in edmunds 2008) and small branches (50–80 mm long) of Pocillopora verrucosa (ellis and solander, 1786) were used to test the photophysiological response of corals to hr and lr surfaces. Massive Porites spp. and P. verrucosa were chosen because they are common in the back reef of Moorea (n colvard, pers obs).
Juvenile massive Porites spp. and branch tips of P. verrucosa were collected from 3-m depth in the back reef on 20 april, 2009, and prepared as nubbins (birkeland 1976). colonies of massive Porites spp. were assumed to be genetically unique as they originated from the settle-ment of planula larvae of sexual origin; genetic independence of P. verrucosa branches was sought by collecting one branch per colony. twenty massive Porites spp. and 20 branches of

colvard and edmunds: change in spectral composition of reef from macroalgae 1023
P. verrucosa were prepared as nubbins and assigned haphazardly to one of two treatments contrasting the effects of lr and hr surfaces. immediately following collection, corals were placed in a shaded aquarium at the uc berkeley richard b gump south pacific research station to prevent photophysiological stress attributed to exposure to surface light intensi-ties. corals were fastened to 100-cm2 acrylic plates (one per plate) painted with matte paint to mimic the light reflected from natural lr and hr surfaces in the back reef. lr plates were painted brown to mimic Turbinaria ornata (J. agardh, 1848), one of the dominant macroal-gae in the back reef of Moorea (adjeroud and salvat 1996, adjeroud 1997; rc carpenter, csu northridge, pers comm), and hr plates were painted light beige to mimic heavily grazed reef surfaces and carbonate pavement. to evaluate the effectiveness of these mimics, r(λ) of painted plates was compared to the corresponding natural surfaces by a randomized block anova, with sampling days treated as a block, and surfaces a fixed factor.
on 23 april, 2009, the acrylic plates and attached corals were nailed in positions selected haphazardly in a 20-m2 area of carbonate pavement at 3-m depth in the back reef of Moorea. on 20 May, 2009 (after 27 d of treatment), Δf⁄fm' of the coral nubbins on each surface was evaluated. Δf⁄fm' is calculated from (fm' − f') ⁄ fm', where fm' is the maximal fluorescence yield in actinic light, and f' is the fluorescence yield in actinic light (cosgrove and borowitzka 2010). Δf⁄fm' measures the efficiency of light energy utilization by psii in a light-adapted state, and it was measured using a diving paM (walz, gmbh), fitted with a fiber optic probe (5.5-mm diameter) supported by an aluminum clip that held the sensor tip 1.0 cm above the coral tissue and at 60° to the sample plane. The settings of the diving paM were held constant
Figure 1. Diagram showing the sources of light affecting small corals, and the position of the sen-sors used in the analysis of light and photophysiological performance. In the shallow back reef of Moorea, small corals receive light from (A) downwelling and (B) reflective sources, both of which were quantified with a cosine-corrected quantum sensor. To analyze the spectral composition of light, the tip of the sampling cable of the spectrophotometer was positioned at 135° from the verti-cal axis to sample light reflected off the benthos (C). To measure photophysiological performance, the sensor of the Diving PAM was positioned approximately 2 cm above the benthos on the side of corals [i.e., on marginal tissue (D)] to measure effective photochemical efficiency of PSII of Symbiodinium (ΔF⁄F
m’). The high reflective surface is representative of a sandy or carbonate reef
rock substratum, whereas the low reflective surface is representative of a substratum dominated by macroalgae.

BULLETIN OF MARINE SCIENCE. VOL 88, NO 4. 20121024
at a gain of 6 and a measuring intensity of 6 for all recordings, and all measurements were completed on a single day between 12:00 and 13:00 hrs to reduce the effect of variation in short-term light history prior to sampling (brown et al. 1999, Kaniewska et al. 2008). to stan-dardize the measurements of Δf⁄fm' with regard to treatments, measurements were taken on the sides of the nubbins, approximately 2 cm above the acrylic plate (hereafter described as “marginal” tissue). This position was selected to detect a photophysiological response of the Symbiodinium to the light reflected from the adjacent substrata. downwelling irradiance, as well as Symbiodinium genotypes, can vary across the surface of coral colonies (rowan and Knowlton 1995, iglesias-prieto et al. 2004, dove et al. 2008), and therefore the results of the present analysis are likely to apply only to coral tissue on the margins of the colony.
statistical analysis.—to describe the light reflected from natural surfaces, r(λ) for hr and lr surfaces was integrated within five wavelength bins selected to characterize the ab-sorptivity of the photosynthetically active pigment chlorophyll a (420–440 and 650–670 nm), as well as the accessory pigments of Symbiodinium (Jeffrey and haxo 1968): xanthophylls (440–460 nm), carotenoids (520–540 nm), and phycobilins (570–590 nm). This analytical approach is similar to that used to quantify the color of flowers through r(λ) (chittka et al. 1994, Mcewen and vamosi 2010). for the purposes of the present study we used three wave-length bins to detect variation in r(λ) that was biologically meaningful to Symbiodinium. r(λ) was compared between hr and lr surfaces using a randomized block anova, in which sampling day was treated as a block, and surfaces a fixed factor. The same statistical model was used in the manipulative experiment to compare the r(λ) of the hr and lr acrylic tiles. a one-way anova was used to compare Δf⁄fm' of corals on hr vs lr surfaces. all statistical analyses were completed with systat versions 11 and 12, and the statistical assumptions of normality and homoscedasticity were evaluated through graphical analyses of residuals.
results
downwelling and upwelling photon irradiance.—The cosine-corrected par sensor demonstrated that the underwater ed did not differ in intensity over hr and lr surfaces (anova: P > 0.05). however, underwater eu differed between sur-faces (anova: P < 0.001), with a mean intensity of 687 (se 20) μmol quanta m−2 s−1 reflected from hr, and 609 (se 19) μmol quanta m−2 s−1 reflected from lr surfaces (n = 9 d, fig. 2). overall, when r(λ) was quantified with a cosine corrected sensor, lr surfaces reflected 11% less light than hr surfaces.
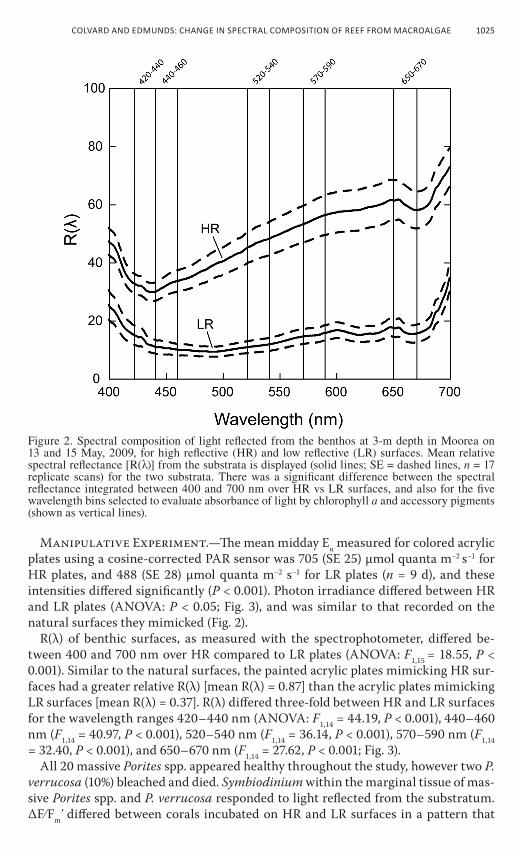
underwater spectral reflectance.—r(λ) of hr surfaces [mean r(λ) = 0.76] differed significantly from that of lr surfaces [mean r(λ) = 0.36; anova: F1,13 = 11.17, P = 0.005]. when light was measured with the underwater spectrophotom-eter, mean r(λ) was about 52% lower for lr compared to hr surfaces. There were also significant differences between hr and lr surfaces for the select wavelength ranges (i.e., bins). for wavelengths corresponding to the two absorption peaks for chlorophyll a, the integrated intensity of light between 420 nm and 440 nm differed between hr and lr surfaces (anova: F1,13 = 21.46, P < 0.001) as it did between 650 nm and 670 nm (F1,13 = 32.46, P < 0.001); in both cases, >60% more light was reflected from hr compared to lr surfaces. for the wavelength bins corresponding to the absorption of chlorophyll accessory pigments, integrated light intensity differed be-tween hr and lr surfaces for the wavelengths 440–460 nm (anova: F1,13 = 28.72, P < 0.001), 520–540 nm (F1,13 = 29.71, P < 0.001), and 570–590 nm (F1,13 = 26.39, P < 0.001; fig. 2).
colvard and edmunds: change in spectral composition of reef from macroalgae 1025
Manipulative experiment.—The mean midday eu measured for colored acrylic plates using a cosine-corrected par sensor was 705 (se 25) µmol quanta m−2 s−1 for hr plates, and 488 (se 28) µmol quanta m−2 s−1 for lr plates (n = 9 d), and these intensities differed significantly (P < 0.001). photon irradiance differed between hr and lr plates (anova: P < 0.05; fig. 3), and was similar to that recorded on the natural surfaces they mimicked (fig. 2).
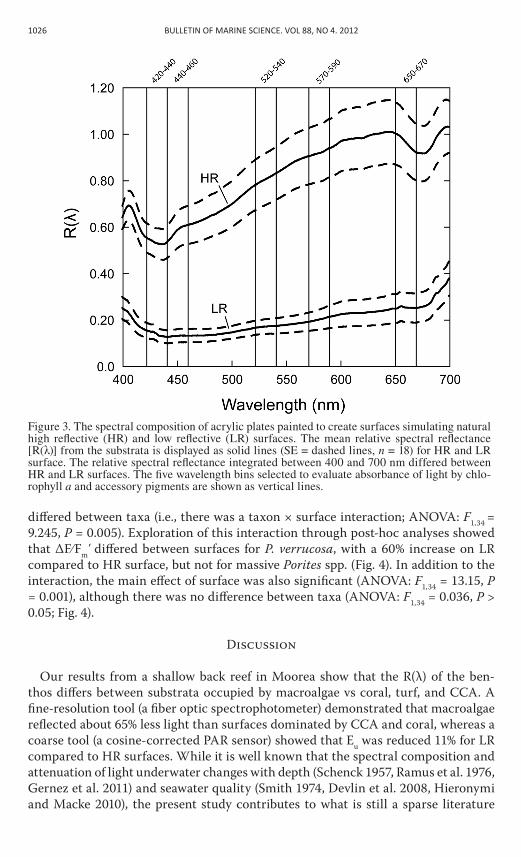
r(λ) of benthic surfaces, as measured with the spectrophotometer, differed be-tween 400 and 700 nm over hr compared to lr plates (anova: F1,15 = 18.55, P < 0.001). similar to the natural surfaces, the painted acrylic plates mimicking hr sur-faces had a greater relative r(λ) [mean r(λ) = 0.87] than the acrylic plates mimicking lr surfaces [mean r(λ) = 0.37]. r(λ) differed three-fold between hr and lr surfaces for the wavelength ranges 420–440 nm (anova: F1,14 = 44.19, P < 0.001), 440–460 nm (F1,14 = 40.97, P < 0.001), 520–540 nm (F1,14 = 36.14, P < 0.001), 570–590 nm (F1,14 = 32.40, P < 0.001), and 650–670 nm (F1,14 = 27.62, P < 0.001; fig. 3).
all 20 massive Porites spp. appeared healthy throughout the study, however two P. verrucosa (10%) bleached and died. Symbiodinium within the marginal tissue of mas-sive Porites spp. and P. verrucosa responded to light reflected from the substratum. Δf⁄fm' differed between corals incubated on hr and lr surfaces in a pattern that
Figure 2. Spectral composition of light reflected from the benthos at 3-m depth in Moorea on 13 and 15 May, 2009, for high reflective (HR) and low reflective (LR) surfaces. Mean relative spectral reflectance [R(λ)] from the substrata is displayed (solid lines; SE = dashed lines, n = 17 replicate scans) for the two substrata. There was a significant difference between the spectral reflectance integrated between 400 and 700 nm over HR vs LR surfaces, and also for the five wavelength bins selected to evaluate absorbance of light by chlorophyll a and accessory pigments (shown as vertical lines).
BULLETIN OF MARINE SCIENCE. VOL 88, NO 4. 20121026
differed between taxa (i.e., there was a taxon × surface interaction; anova: F1,34 = 9.245, P = 0.005). exploration of this interaction through post-hoc analyses showed that Δf⁄fm' differed between surfaces for P. verrucosa, with a 60% increase on lr compared to hr surface, but not for massive Porites spp. (fig. 4). in addition to the interaction, the main effect of surface was also significant (anova: F1,34 = 13.15, P = 0.001), although there was no difference between taxa (anova: F1,34 = 0.036, P > 0.05; fig. 4).
discussion
our results from a shallow back reef in Moorea show that the r(λ) of the ben-thos differs between substrata occupied by macroalgae vs coral, turf, and cca. a fine-resolution tool (a fiber optic spectrophotometer) demonstrated that macroalgae reflected about 65% less light than surfaces dominated by cca and coral, whereas a coarse tool (a cosine-corrected par sensor) showed that eu was reduced 11% for lr compared to hr surfaces. while it is well known that the spectral composition and attenuation of light underwater changes with depth (schenck 1957, ramus et al. 1976, gernez et al. 2011) and seawater quality (smith 1974, devlin et al. 2008, hieronymi and Macke 2010), the present study contributes to what is still a sparse literature
Figure 3. The spectral composition of acrylic plates painted to create surfaces simulating natural high reflective (HR) and low reflective (LR) surfaces. The mean relative spectral reflectance [R(λ)] from the substrata is displayed as solid lines (SE = dashed lines, n = 18) for HR and LR surface. The relative spectral reflectance integrated between 400 and 700 nm differed between HR and LR surfaces. The five wavelength bins selected to evaluate absorbance of light by chlo-rophyll a and accessory pigments are shown as vertical lines.

colvard and edmunds: change in spectral composition of reef from macroalgae 1027
describing how the underwater light regime on shallow coral reefs is affected by the reflective properties of the benthos (but see Maritorena et al. 1994, Joyce and phinn 2003, Mobley and sundman 2003, hedley et al. 2004).
additionally, we show the potential for benthos-mediated variation in eu and r(λ) to affect the performance of reef corals. as r(λ) is reduced by macroalgae compared to coral, turf, and cca, the temporal trend for rising macroalgal cover on coral reefs (McManus and polsenberg 2004, Kuffner et al. 2006, norström et al. 2009) is likely to be associated with reduced near-bottom light intensities. while the light-related implications of this trend are unclear, the reductions in reflected light motivate at least two predictions regarding the effects on corals. first, corals might have a selec-tive advantage on surfaces dominated by macroalgae if they can maintain high rates of photosynthesis at low irradiances. second, it is possible that macroalgal dominat-ed areas of reef could serve as a spatial refuge for corals that are sensitive to damage at high light intensities.
based on the aforementioned predictions and the present results for Δf⁄fm', it is unlikely that small colonies of massive Porites spp. and P. verrucosa on shallow reefs will be negatively affected by the impacts of macroalgae on reflected light. not only was Δf⁄fm' in the marginal tissues of both taxa resistant to the negative effects of reduced reflected light intensities over lr surfaces, but for P. verrucosa, Δf⁄fm' in-creased on lr compared to hr surfaces. Therefore Symbiodinium in P. verrucosa may benefit from lr surfaces where they lose less light energy to non-photosynthetic pathways (i.e., non-photosynthetic quenching, npQ; cosgrove and borowitzka 2010) compared to conspecifics on hr surfaces. while this pattern cannot be interpreted as evidence that photosynthetic carbon fixation is higher in marginal tissues of small P. verrucosa on lr compared to hr surfaces (enriquez and borowitzka 2010), nev-ertheless, it demonstrates that the reductions in light reflected from macroalgae can have functional implications for coral photophysiology.
Figure 4. Effective photochemical efficiency of PSII (ΔF⁄Fm') of Porites spp. (n = 10 for each treat-ment) and Pocillopora verrucosa (n = 9 for each treatment) after 27 d on acrylic plates painted to mimic high reflective (HR) and low reflective (LR) surfaces. Incubations were completed at 3-m depth in the back reef, and all measurements were obtained between 12:00 and 13:00 hrs. ΔF⁄F
m’
was affected significantly by a taxon × surface interaction, as well as surface type.

BULLETIN OF MARINE SCIENCE. VOL 88, NO 4. 20121028
in the context of the well-known capacity for acclimatization to shade in corals that involves changes in pigment concentration, thylakoid structures, and symbiont densities (falkowski and dubinsky 1981, iglesias-prieto and trench 1994, dubinsky and falkowski 2011), it is intriguing that Symbiodinium in the marginal tissue of massive Porites spp. were unresponsive to reduced intensities of eu. however, this finding may not be without precedent because at least one other poritid [Porites cy-lindrica (dana, 1846)], has a limited capacity to acclimate to shade (anthony and fabricius 2000). Moreover, because there are indications that massive Porites spp. is functionally unequal to other corals (loya et al. 2001, fabricius 2011), it may not respond to shade in similar ways to other corals. The Symbiodinium offer one reason why massive Porites spp. and P. verrucosa might differ in their response to reflected light, because massive Porites spp. in Moorea contain c15 Symbiodinium (edmunds et al. in press), whereas P. verrucosa contain c1c Symbiodinium throughout the pacific (laJeunesse et al. 2004). The response of Symbiodinium to varying light re-gimes differs among genetic variants of this taxon (hennige et al. 2009), although to our knowledge, the phenotypes of c1c and c15 have not been compared.
by analyzing the light regime by wavelength bins corresponding to the peak ab-sorptivity of chlorophyll a and its accessory pigments, we sought to explore the mechanisms through which variation in r(λ) could affect coral photophysiology. The contrast of light reflected from hr and lr surfaces revealed a two-fold difference in the light intensity for the wavelength bins corresponding to the absorptivity of chlorophyll a, with hr surfaces reflecting more light than lr surfaces. There were also differences between hr and lr surfaces for the wavelength bins corresponding to the chlorophyll accessory pigments, with hr surfaces reflecting three- to four-fold more light within these bins than lr surfaces. These differences between hr and lr surfaces are likely to be a function of the absorption of light for photosynthesis by macroalgae (Mccook et al. 2001, birrell et al. 2008). by analyzing reflected light in discrete wavelength bins, our results suggest that light reflected from hr substrata may have the potential to affect the photophysiology of the Symbiodinium in two ways: (1) by providing a greater amount of light, and (2) providing a greater propor-tion of wavelengths that excite chlorophyll a as well as the accessory pigments within these algae.
as many coral reefs are experiencing a decline in scleractinian cover (gardner et al. 2003, bellwood et al. 2004, hughes et al. 2011) and an increase in abundance of macroalgae (tanner 1995, aronson and precht 2006, norström et al. 2009), evaluat-ing the consequences of these changes is an important priority to understand the functional implications of algal dominance on tropical reefs (Mccook et al. 2001). The present analysis shows that macroalgae can affect the near-bottom light regime on coral reefs in ways that have not previously been emphasized (Mccook et al. 2001, birrell et al. 2008), and demonstrate that these effects can influence the photophysi-ology of coral tissue located close to these benthic surfaces. it is important to note, however, that our study utilized small corals that were either juvenile colonies or branch fragments, and therefore it is unknown to what extent our findings can be extrapolated to other corals or to other reefs. to elucidate the ecological relevance of the aforementioned patterns, it will be necessary to conduct similar studies with larger colonies, to evaluate the impacts of adjacent surfaces to whole colonies (not just the marginal tissue), and to evaluate the extent to which the reflectance properties of meter-scale areas of reef can impact the reflectance properties of centimeter-scale

colvard and edmunds: change in spectral composition of reef from macroalgae 1029
areas of reef (as investigated in the present study), and the light regime within the tissue where the Symbiodinium are located (Kaniewska et al. 2011).
acknowledgments
This material is based upon work supported by the us national science foundation through the Moorea coral reef, long-term ecological research program (grants oce 04-17412 and 10-26851), and gifts from the gordon and betty Moore foundation; it was completed under a research permit issued by the french polynesian Ministry of research. we are grateful to n davies and the staff of the uc berkeley, richard b gump south pacific research station for making our visits to Moorea productive and enjoyable, and c cameron, w goldenheim, v Moriarty, M Johnson, and s swanson for field assistance. we would also like to thank three anonymous reviewers as well as b helmuth for providing comments that improved an earlier draft of this paper. This manuscript was submitted in partial fulfillment for the Ms degree at california state university, northridge. This is contribution number 183 of the csun Marine biology program.
literature cited
adjeroud M. 1997. long-term changes of epibenthic macrofauna communities in a closed la-goon (taiaro atoll, french polynesia): 1972–1994. hydrobiologia. 356:11–19. http://dx.doi.org/10.1023/a:1003052515305
adjeroud M, salvat b. 1996. spatial patterns in biodiversity of a fringing reef community along opunohu bay, Moorea, french polynesia. bull Mar sci. 59:175–187.
alvarez-filip l, dulvy nK, gill Ja, côté iM, watkinson ar. 2009. flattening of caribbean coral reefs: region-wide declines in architectural complexity. proc roy soc b-biol sci. 276(1669):3019–3025.
anthony Krn, fabricius Ke. 2000. shifting roles of heterotrophy and autotrophy in coral ener-getics under varying turbidity. J exp Mar biol ecol. 252:221–253. http://dx.doi.org/10.1016/s0022-0981(00)00237-9
aronson r, precht w. 2006. conservation, precaution, and caribbean reefs. coral reefs. 25:441–450. http://dx.doi.org/10.1007/s00338-006-0122-9
barron Mg, little ee, calfee r, diamond s. 2000. Quantifying solar spectral irradiance in aquatic habitats for the assessment of photoenhanced toxicity. environ toxicol chem. 19:920–925. http://dx.doi.org/10.1002/etc.5620190419
bellwood dr, hughes tp, folke c, nyster M. 2004. confronting the coral reef crisis. nature. 429:827–833. pMid:15215854. http://dx.doi.org/10.1038/nature02691
birkeland c. 1976. an experimental method of studying corals during early stages of growth. Micronesica-series. 12:319–322.
birrell c, Mccook l, willis bl, harrington l. 2008. chemical effects of macroalgae on lar-val settlement of the broadcast spawning coral Acropora millepora. Mar ecol prog ser. 362:129–137. http://dx.doi.org/10.3354/meps07524
brakel wh. 1979. small-scale spatial variation in light available to coral reef benthos: quantum irradiance measurements from a Jamaican reef. bull Mar sci. 29:406–413.
brown be, ambarsari i, warner Me, fitt wK, dunne rp, gibb sw, cummings dg. 1999. diurnal changes in photochemical efficiency and xanthophyll concentrations in shallow water reef corals: evidence for photoinhibition and photoprotection. coral reefs. 18:99–105. http://dx.doi.org/10.1007/s003380050163
bruno Jf, sweatman h, precht wf, selig er, schutte vgw. 2009. assessing evidence of phase shifts from coral to macroalgal dominance on coral reefs. ecology. 90:1478–1484. pMid:19569362. http://dx.doi.org/10.1890/08-1781.1

BULLETIN OF MARINE SCIENCE. VOL 88, NO 4. 20121030
campbell gs, norman JM. 1998. an introduction to environmental biophysics. new york: springer-verlag new york, inc. http://dx.doi.org/10.1007/978-1-4612-1626-1
carpenter rc. 1986. partitioning herbivory and its effects on coral reef algal communities. ecol Monogr. 56(4):345–363. http://dx.doi.org/10.2307/1942551
chadwick ne, Morrow KM. 2011. competition among sessile organisms on coral reefs. In: dubinsky Z, stambler n, editors. coral reefs: an ecosystem in transition. springer netherlands. p. 347–371. http://dx.doi.org/10.1007/978-94-007-0114-4_20
chittka l, shmida a, troje n, Menzel r. 1994. ultraviolet as a component of flower reflections, and the color-perception of hymenoptera. vision res. 34(11):1489–1508. http://dx.doi.org/10.1016/0042-6989(94)90151-1
colvard nb, edmunds pJ. 2011. decadal-scale changes in abundance of non-scleractinian in-vertebrates on a caribbean coral reef. J exp Mar biol ecol. 397(2):153–160. http://dx.doi.org/10.1016/j.jembe.2010.11.015
connor Jl, adey wh. 1977. The benthic algal composition, standing crop and productivity of a caribbean algal ridge. atoll res bull. 211:1–40. http://dx.doi.org/10.5479/si.00775630.211.1
cosgrove J, borowitzka Ma. 2010. chlorophyll fluorescence terminology: an introduction. In: suggett dJ, prášil o, borowitzka Ma, editors. chlorophyll a fluorescence in aquatic sci-ences: methods and applications. springer netherlands. p. 1–17.
devlin MJ, barry J, Mills dK, gowen rJ, foden J, sivyer d, tett p. 2008. relationships between suspended particulate material, light attenuation and secchi depth in uK marine waters. estuar coast shelf sci. 79(3):429–439. http://dx.doi.org/10.1016/j.ecss.2008.04.024
dickey t, lewis M, chang g. 2006. optical oceanography: recent advances and future di-rections using global remote sensing and in situ observations. rev geophys. 44(1). http://dx.doi.org/10.1029/2003rg000148
dove sg, lovell c, fine M, deckenback J, hoegh-guldberg ove, iglesias-prieto r, anthony Krn. 2008. host pigments: potential facilitators of photosynthesis in coral symbioses. plant cell environ. 31(11):1523–1533. http://dx.doi.org/10.1111/j.1365-3040.2008.01852.x
dubinsky Z, falkowski p. 2011. light as a source of information and energy in zooxanthel-late corals. In: dubinsky Z, stambler n, editors. coral reefs: an ecosystem in transition. springer netherlands. p. 107–118. http://dx.doi.org/10.1007/978-94-007-0114-4_8
edmunds p. 2008. The effects of temperature on the growth of juvenile scleractinian corals. Mar biol. 154:153–162. http://dx.doi.org/10.1007/s00227-008-0910-6
edmunds pJ, leichter JJ, adjeroud M. 2010. landscape-scale variation in coral recruitment in Moorea, french polynesia. Mar ecol prog ser. 414:75–89. http://dx.doi.org/10.3354/meps08728
edmunds pJ, putnam hM, gates rd. in press. photophysiological consequences of vertical stratification of Symbiodinium in tissue of the coral Porites lutea. biol bull.
enríquez s, borowitzka Ma. 2010. The use of the fluorescence signal in studies of seagrasses and macroalgae. In: suggett dJ, prášil o, borowitzka Ma, editors. chlorophyll a fluores-cence in aquatic sciences: methods and applications. springer netherlands. p. 187–208.
fabricius Ke. 2006. effects of irradiance, flow, and colony pigmentation on the temperature microenvironment around corals: implications for coral bleaching? limnol oceanogr. 51:30–37. http://dx.doi.org/10.4319/lo.2006.51.1.0030
fabricius Ke. 2011. factors determining the resilience of coral reefs to eutrophication: a review and conceptual model. In: dubinsky Z, stambler n, editors. coral reefs: an ecosystem in tran-sition. springer netherlands. p. 493–505. http://dx.doi.org/10.1007/978-94-007-0114-4_28
falkowski pg, dubinsky Z. 1981. light-shade adaptation of Stylophora pistillata, a hermatypic coral from the gulf of eilat. nature. 289:172–174. http://dx.doi.org/10.1038/289172a0
gardner t, côté i, gill J, grant a, watkinson a. 2003. long-term, region-wide declines in caribbean corals. science. 301:958–960. pMid:12869698. http://dx.doi.org/10.1126/science.1086050

colvard and edmunds: change in spectral composition of reef from macroalgae 1031
gernez p, stramski d, darecki M. 2011. vertical changes in the probability distribution of downward irradiance within the near-surface ocean under sunny conditions. J geophys res. 116:c00h07. http://dx.doi.org/10.1029/2011Jc007156
hedley J, Mumby p, Joyce K, phinn s. 2004. spectral unmixing of coral reef benthos under ideal conditions. coral reefs. 23:60–73. http://dx.doi.org/10.1007/s00338-003-0354-x
hennige s, suggett d, warner M, Mcdougall K, smith d. 2009. photobiology of Symbiodinium revisited: bio-physical and bio-optical signatures. coral reefs. 28:179–195. http://dx.doi.org/10.1007/s00338-008-0444-x
hieronymi M, Macke a. 2010. spatiotemporal underwater light field fluctuations in the open ocean. J eur opt soc-rapid. 5:1–8.
hochberg eJ. 2011. remote sensing of coral reef processes. In: dubinsky Z, stambler n, edi-tors. coral reefs: an ecosystem in transition. springer netherlands. p. 25–35. http://dx.doi.org/10.1007/978-94-007-0114-4_3
hochberg eJ, atkinson MJ, andréfouët s. 2003. spectral reflectance of coral reef bottom-types worldwide and implications for coral reef remote sensing. remote sens environ. 85:159–173. http://dx.doi.org/10.1016/s0034-4257(02)00201-8
hoegh-guldberg o, Mumby pJ, hooten aJ, steneck rs, greenfield p, gomez e, harvell cd, sale pf, edwards aJ, caldeira K, et al. 2007. coral reefs under rapid climate change and ocean acidification. science. 318:1737–1742. pMid:18079392. http://dx.doi.org/10.1126/science.1152509
hughes t, bellwood d, baird a, brodie J, bruno J, pandolfi J. 2011. shifting base-lines, declin-ing coral cover, and the erosion of reef resilience: comment on sweatman et al. (2011). coral reefs. 30:653–660. http://dx.doi.org/10.1007/s00338-011-0787-6
iglesias-prieto r, beltran vh, laJeunesse tc, reyes-bonilla h, Thome pe. 2004. different al-gal symbionts explain the vertical distribution of dominant reef corals in the eastern pacific. proc r soc london. 271:1757–1763. pMid:15306298. pMcid:1691786. http://dx.doi.org/10.1098/rspb.2004.2757
iglesias-prieto r, trench rK. 1994. acclimation and adaptation to irradiance in symbiotic di-noflagellates. i. responses of the photosynthetic unit to changes in photon flux density. Mar ecol prog ser. 113:163–175. http://dx.doi.org/10.3354/meps113163
Jeffrey sw, haxo ft. 1968. photosynthetic pigments of symbiotic dinoflagellates (zooxanthel-lae) from corals and clams. biol bull. 135:149–165. http://dx.doi.org/10.2307/1539622
Joyce Ke, phinn sr. 2003. hyperspectral analysis of chlorophyll content and photosynthetic capacity of coral reef substrates. limnol oceanogr. 48:489–496. http://dx.doi.org/10.4319/lo.2003.48.1_part_2.0489
Kaniewska p, anthony K, hoegh-guldberg o. 2008. variation in colony geometry modulates internal light levels in branching corals Acropora humilis and Stylophora pistillata. Mar biol. 155:649–660. http://dx.doi.org/10.1007/s00227-008-1061-5
Kaniewska p, Magnusson sh, anthony K, reef r, Kühl M, hoegh-guldberg o. 2011. importance of macro- versus microstructure in modulating light levels inside coral colo-nies. J phycol. 47:846–860. http://dx.doi.org/10.1111/j.1529-8817.2011.01021.x
Klumpp dw, McKnnon ad. 1992. community structure, biomass and productivity of epilithic algal communities on the great barrier reef: dynamics at different spatial scales. Mar ecol prog ser. 86:77–89. http://dx.doi.org/10.3354/meps086077
Kuffner ib, walters lJ, becerro Ma, paul vJ, ritson-williams r, beach Ks. 2006. inhibition of coral recruitment by macroalgae and cyanobacteria. Mar ecol prog ser. 323:107–117. http://dx.doi.org/10.3354/meps323107
laJeunesse tc, Thornhill dJ, cox ef, stanton fg, fitt wK, schmidt gw. 2004. high diversity and host specificity observed among symbiotic dinoflagellates in reef coral communities from hawaii. coral reefs. 23:596–603.
lesser Mp. 2000. depth-dependent photoacclimatization to solar ultraviolet radiation in the caribbean coral Montastraea faveolata. Mar ecol prog ser. 192:137–151. http://dx.doi.org/10.3354/meps192137

BULLETIN OF MARINE SCIENCE. VOL 88, NO 4. 20121032
lesser Mp, Mobley c. 2007. bathymetry, water optical properties, and benthic classification of coral reefs using hyperspectral remote sensing imagery. coral reefs. 26:819–829. http://dx.doi.org/10.1007/s00338-007-0271-5
loya y, sakai K, yamazato K, nakano y, sambali h, van woesik r. 2001. coral bleaching: the win-ners and the losers. ecol lett. 4:122–131. http://dx.doi.org/10.1046/j.1461-0248.2001.00203.x
Maritorena s, Morel a, gentili b. 1994. diffuse reflectance of oceanic shallow waters: influ-ence of water depth and bottom albedo. limnol oceanogr. 39:1689–1703. http://dx.doi.org/10.4319/lo.1994.39.7.1689
Mass t, Kline di, roopin M, veal cJ, cohen s, iluz d, levy o. 2010. The spectral quality of light is a key driver of photosynthesis and photoadaptation in Stylophora pistillata colo-nies from different depths in the red sea. J exp biol. 213(23):4084–4091. pMid:21075950. http://dx.doi.org/10.1242/jeb.039891
Mazel ch. 1997. diver-operated instrument for in situ measurement of spectral fluorescence and reflectance of benthic marine organisms and substrates. opt eng. 36(9):2612–2617. http://dx.doi.org/10.1117/1.601486
Mcclanahan tr, hendrick v, rodrigues MJ, polunin nvc. 1999. varying responses of herbiv-orous and invertebrate-feeding fishes to macroalgal reduction on a coral reef. coral reefs. 18:195–203. http://dx.doi.org/10.1007/s003380050181
Mccook lJ. 1999. Macroalgae, nutrients and phase shifts on coral reefs: scientific issues and management consequences for the great barrier reef. coral reefs. 18:357–367. http://dx.doi.org/10.1007/s003380050213
Mccook lJ, Jompa J, diaz-pulido g. 2001. competition between corals and algae on coral reefs: a review of evidence and mechanisms. coral reefs. 19:400–417. http://dx.doi.org/10.1007/s003380000129
Mcewen Jr, vamosi Jc. 2010. floral colour versus phylogeny in structuring subalpine flower-ing communities. proc r soc b. 277(1696):2957–2965. pMid:20484236. pMcid:2982023. http://dx.doi.org/10.1098/rspb.2010.0501
McManus Jw, polsenberg Jf. 2004. coral-algal phase shifts on coral reefs: ecological and environmental aspects. prog oceanogr. 60(2–4):263–279. http://dx.doi.org/10.1016/j.pocean.2004.02.014
Mobley cd. 1994. light and water: radiative transfer in natural waters. academic press san diego, ca.
Mobley cd, sundman lK. 2003. effects of optically shallow bottoms on upwelling radianc-es: inhomogeneous and sloping bottoms. limnol oceanogr. 48:329–336. http://dx.doi.org/10.4319/lo.2003.48.1_part_2.0329
norström av, nyström M, lokrantz J, folke c. 2009. alternative states on coral reefs: beyond coral macroalgal phase shifts. Mar ecol prog ser. 376:295–306. http://dx.doi.org/10.3354/meps07815
pandolfi JM, Jackson Jbc, baron n, bradbury rh, guzman hM, hughes tp, Kappel cv, Micheli f, ogden Jc, possingham hp, et al. 2005. are us coral reefs on the slippery slope to slime? science. 307:1725–1726. pMid:15774744. http://dx.doi.org/10.1126/science.1104258
ramus J, beale si, Mauzerall d, howard Kl. 1976. changes in photosynthetic pigment con-centration in seaweeds as a function of water depth. Mar biol. 37:223–229. http://dx.doi.org/10.1007/bf00387607
redalje r. 1976. light adaptation strategies of hermatypic corals. pac sci. 30(3):212–212.rowan r, Knowlton n. 1995. intraspecific diversity and ecological zonation in coral-algal sym-
biosis. proc natl acad sci-biol. 92(7):2850–2853.schenck Jh. 1957. on the focusing of sunlight by ocean waves. J opt soc am. 47:653–657.
http://dx.doi.org/10.1364/Josa.47.000653smith rc. 1974. structure of solar radiation in the upper layer of the sea. new york: academic.
p. 95–119.smith rc, Mobley cd. 2008. underwater light. In: bjorn lo, editor. photobiology: the science
of life and light. springer new york. p. 131–138.

colVARD AND eDMUNDS: cHANGe IN SpecTRAl coMpoSITIoN of Reef fRoM MAcRoAlGAe 1033
tanner Je. 1995. competition between scleractinian corals and macroalgae: an experimental investigation of coral growth, survival and reproduction. J exp Mar biol ecol. 190(2):151–168. http://dx.doi.org/10.1016/0022-0981(95)00027-o
ulstrup K, ralph p, larkum a, Kühl M. 2006. intra-colonial variability in light acclimation of zooxanthellae in coral tissues of Pocillopora damicornis. Mar biol. 149:1325–1335. http://dx.doi.org/10.1007/s00227-006-0286-4
werdell pJ, roesler cs. 2003. remote assessment of benthic substrate composition in shal-low waters using multispectral refl ectance. limnol oceanogr. 48:557–567. http://dx.doi.org/10.4319/lo.2003.48.1_part_2.0557
wilkinson cr. 2004. status of coral reefs of the world: 2004. townsville, australia: australian institute of Marine science.
date suBmitted: 1 august, 2011.date accepted: 30 July, 2012.availaBle online: 23 august, 2012.
addresses: (nbc) Department of Biological Sciences, University of South Carolina, 715 Sumter St. Columbia, South Carolina 29208. (pJe) Department of Biology, California State University, Northridge, 18111 Nordhoff Street, Northridge, California 91330-8303. corresponding author: (nbc) Email: <[email protected]>.