magna græcia salvatore venuta lna-i-mir-221 inhibitor...
TRANSCRIPT

1
A 13 mer LNA-i-miR-221 inhibitor restores drug-sensitivity in melphalan-refractory 1 multiple myeloma cells 2
Annamaria Gullà1, Maria Teresa Di Martino1, Maria Eugenia Gallo Cantafio1, Eugenio 3 Morelli1, Nicola Amodio1, Cirino Botta1, Maria Rita Pitari1, Santo Giovanni Lio2, Domenico 4 Britti1, Maria Angelica Stamato1, Teru Hideshima3, Nikhil C. Munshi3,4, Kenneth C. Anderson3, 5 Pierosandro Tagliaferri1 and Pierfrancesco Tassone1,5. 6 1Department of Experimental and Clinical Medicine, Magna Græcia University, Campus 7 Salvatore Venuta, Catanzaro, Italy; 2Pathology Unit, ‘‘Giovanni Paolo II’’ Hospital, Lamezia 8 Terme, Catanzaro, Italy, 3Jerome Lipper Multiple Myeloma Center, Department of Medical 9 Oncology, Dana-Farber Cancer Institute, Boston, MA, USA; 4VA Boston Healthcare System, 10 West Roxbury, Boston, MA, USA, 5Sbarro Institute for Cancer Research and Molecular 11 Medicine, Center for Biotechnology, College of Science and Technology, Temple University, 12 Philadelphia, PA, US. 13 Running title: LNA-i-miR-221 inhibitor overcome melphalan-resistance 14 Keywords: LNA-miR-221 inhibitors, melphalan, drug-resistance, microRNA, multiple 15 myeloma. 16 Financial support: This work has been supported by the Italian Association for Cancer 17 Research (AIRC), PI: PT. “Special Program Molecular Clinical Oncology - 5 per mille" n. 18 9980, 2010/15. This work has also been supported by a grant from NIH PO1-155258, RO1-19 124929, P50-100007, PO1-78378 and VA merit grant IO1-24467. Nicola Amodio was 20 supported by a "Fondazione Umberto Veronesi" Fellowship. 21 Corresponding author: Pierfrancesco Tassone, MD, Magna Graecia University, Viale 22 Europa, 88100 Catanzaro, Italy; E-mail: [email protected], Phone: +39-0961-3697029, Fax: 23 +39-0961-3697341. 24 Conflicts- of- interest disclosure: The authors declare no competing financial interests. 25 Text: 4955 words 26 Abstract: 210 words 27 Figures: 6 28
Research. on June 27, 2018. © 2015 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on November 2, 2015; DOI: 10.1158/1078-0432.CCR-15-0489

2
Statement of translational relevance 29 30
Disregulation of oncogenic microRNAs (miRNAs) is frequently involved in cancer progression 31 up to drug-resistant stage. We investigated the role of miR-221/222 in melphalan-resistance 32 in multiple myeloma (MM) cells. The translational relevance of our study relies in the 33 demonstration of in vivo efficacy of a novel 13 mer LNA-i-miR-221 inhibitor in restoring 34 melphalan-sensitivity of refractory MM cells by selective interference with relevant molecular 35 mechanisms of resistance. Importantly, LNA-i-miR-221 exerts anti-MM activity by itself and is 36 suitable for systemic delivery. Our findings provide therefore the rationale for clinical 37 investigation of LNA-i-miR-221 plus melphalan in drug-refractory stage of disease. 38
39 40 41 42 43 44 45 46 47 48 49 50 51 52
53 Research. on June 27, 2018. © 2015 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on November 2, 2015; DOI: 10.1158/1078-0432.CCR-15-0489

3
Abstract 54 55
Purpose: The onset of drug-resistance is a major cause of treatment failure in multiple 56 myeloma (MM). While increasing evidence is defining the role of microRNAs in mediating 57 drug-resistance, their potential activity as drug-sensitizing agents has not yet been 58 investigated in MM. 59 Experimental Design: Here we studied the potential utility of miR-221/222 inhibition in 60 sensitizing refractory MM cells to melphalan. 61 Results: MiR-221/222 expression inversely correlated with melphalan-sensitivity of MM cells. 62 Inhibition of miR-221/222 overcame melphalan-resistance and triggered apoptosis of MM 63 cells in vitro, in the presence or absence of human bone marrow stromal cells. Decreased 64 MM cell growth induced by inhibition of miR-221/222 plus melphalan was associated with a 65 marked upregulation of pro-apoptotic BBC3/PUMA protein, a miR-221/222 target, as well as 66 with modulation of drug influx-efflux transporters SLC7A5/LAT1 and the ATP-binding cassette 67 (ABC) transporter ABCC1/MRP1. Finally, in vivo treatment of SCID/NOD mice bearing human 68 melphalan-refractory MM xenografts with systemic LNA-i-miR-221 plus melphalan overcame 69 drug-resistance, evidenced by growth inhibition with significant antitumor effects together with 70 modulation of PUMA and ABCC1 in tumors retrieved from treated mice. 71 Conclusions: Taken together, our findings provide the proof of concept that LNA-i-miR-221 72 can reverse melphalan-resistance in preclinical models of MM, providing the framework for 73 clinical trials to overcome drug resistance and improve patient outcome in MM. 74 75 76 77
78 79 80 81 82 83 84
Research. on June 27, 2018. © 2015 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on November 2, 2015; DOI: 10.1158/1078-0432.CCR-15-0489

4
Introduction 85 86
Multiple Myeloma (MM) is characterized by the abnormal proliferation of malignant 87 plasma cells in the bone marrow (BM) (1, 2). Despite recent advances in MM biology (3), pre-88 clinical models (4-6), and translation of novel agents which have markedly improved the 89 outcome of MM patients, the development of drug-resistance remains an obstacle to long-90 term survival (7). MM commonly progresses to drug-refractory end-stage disease (8), and 91 novel therapeutic strategies are urgently needed. 92
For more than 30 years, melphalan has been the mainstay of MM treatment (9). 93 Presently its therapeutic value is in younger patients who undergo high-dose melphalan 94 (HDM) prior to autologous stem cell transplantation (ASCT), as well as in non-transplant 95 candidates or elderly patients as part of first-line combination regimens (10, 11). The use of 96 melphalan combination regimens with new agents, such as bortezomib or lenalidomide has 97 significantly prolonged progression-free (PFS) and overall survival (OS); however, 98 development of drug-resistance leads to relapse of disease (12). Recently a renewed 99 scientific interest on melphalan is emerging, and major efforts have been devoted to delineate 100 the mechanisms underlying primary or acquired melphalan-resistance (13). These efforts 101 have already led to the design of novel regimens to overcome melphalan-resistance or to 102 improve its anti-tumor activity (14). 103
Currently, there is a growing interest for the therapeutic potential of strategies aimed to 104 target microRNAs (miRNAs) network. MiRNAs are a class of short non-coding RNA that 105 function as post-transcriptional gene regulators. MiRNAs mainly act through complete or 106 partial binding to 3’ untranslated region (3’ UTR) of their mRNA targets, inducing either mRNA 107 degradation or translational repression (15). By targeting driver genes involved in critical 108 cellular pathways miRNAs can function as oncogenes or tumor suppressor genes, playing a 109 key role in tumorigenesis, as well as in cancer progression and aggressiveness (15). A variety 110 of studies have to date demonstrated the potential relevance of miRNA mimics/inhibitors as 111 therapeutic tools, and the promising results from the first Phase-2 trial in patients with HCV 112 infection treated with Locked Nucleic Acid (LNA)-miR-122 inhibitors have further stimulated 113 studies for the treatment of human cancer (16). In MM, miRNA-based strategies are presently 114 emerging as promising approaches (17-25). Moreover, recent findings have emphasized the 115 role of miRNAs in the development of drug-resistance in a variety of malignancies (26). In 116
Research. on June 27, 2018. © 2015 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on November 2, 2015; DOI: 10.1158/1078-0432.CCR-15-0489

5
particular, miRNAs have been shown to regulate drug efflux transporters, induction of 117 apoptosis, cell cycle progression, DNA repair mechanisms, and other alterations of drug 118 targets (27). Among miRNAs involved in development of drug-resistance, miR-221/222 plays 119 a key role: inhibition of miR-221/222 has been reported to overcome resistance to cisplatin 120 (28), tamoxifene (29), fulvestrant (30), temozolamide (31), tirosin Kinase Inhibitors (32), and 121 TRAIL (33) in a variety of cancers. We recently provided the first evidence that silencing of 122 miR-221/222 by specific inhibitors exerts anti-tumor activity in MM cells bearing t(4;14) 123 translocations in vitro and in vivo (34), and that naked LNA-inhibitors of miR-221 (LNA-i-miR-124 221) are suitable for systemic delivery in animals (35). Here we investigated the role of miR-125 221/222 in melphalan-refractory MM, and demonstrate restoration of melphalan-sensitivity in 126 refractory cells after exposure of MM cells to a novel 13 mer LNA-i-miR-221. Our findings 127 provide therefore the rationale for clinical trials investigating LNA-i-miR-221 plus melphalan in 128 drug-refractory MM. 129 130 131 132
Research. on June 27, 2018. © 2015 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on November 2, 2015; DOI: 10.1158/1078-0432.CCR-15-0489

6
Materials and Methods 133 134 Cell cultures, reagents and drugs. 135 Multiple Myeloma cell lines NCI-H929 t(4;14), RPMI-8226 t(14;16) and U266 t(11;14) were 136 purchased from DSMZ (Germany) which certified authentication performed by Short Tandem 137 Repeats DNA typing. These cells were immediately frozen and used from the original stock 138 within 6 months. Melphalan-resistant U266/LR7 t(11;14) cells were kindly provided by Dr. A. 139 Pandiella (University of Salamanca, Spain). AMO1 t(12;14) and bortezomib-resistant AMO1 140 Abzb t(12;14) cells were kindly provided by Dr. C Driessen (University of Tubingen, 141 Germany). U266/LR7, AMO1 and AMO1 Abzb were not further authenticated but confirmed 142 for the described drug-resistant phenotype. All cells were cultured in RPMI-1640 (Gibco®, 143 Life Technologies), as previously described (36, 37). Human stromal HS-5 cells were 144 purchased from ATCC, which certify authentication by Short Tandem Repeats profiling. Also 145 these cells were immediately frozen and used from the original stock within 6 months. HS-5 146 were cultured in Dulbecco’s modified Eagle’s medium (Gibco®, Life Technologies) 147 supplemented with 10% heath inactivated Fetal Bovine Serum (FBS) and 1% P/S 148 (Penicillin/Streptomycin). Following informed consent and Istitutional Ethical Comeettee 149 approval, peripheral blood mononuclear cells (PBMCs) and primary CD138+ MM cells from 150 BM aspirates of 3 MM patients, were isolated as previously described (38). LNA-i-miR-221 151 was designed and synthesized as previously described (35). Melphalan and Bortezomib were 152 purchased from Sigma Aldrich and Selleck Chemicals, respectively. 153 154 In vitro transfection of MM cells 155 Synthetic mirVana® miR-221 and miR-222 inhibitors or mimics were purchased (Life 156 Technologies); mirVana™ miRNA mimic and inhibitor Negative Control #1 (Life Technologies) 157 were used as experimental negative controls (NC). A total of 1 x 106 MM cells were 158 transfected at 100 nM miRNAs concentrations by the Neon® Transfection System (Life 159 Technologies) (1050 v, 2 pulse, 30 a); transfection efficiency, evaluated by flow-cytometry 160 analysis relative to a FAM dye–labeled anti-miR–negative control, reached 85% to 90%. 161 Similar conditions were applied for transfection of MM cells with Silencer® Select siRNA for 162 PUMA/BBC3 (siPUMA) or with Silencer® Select siRNA control (siCNT) (Life Technologies), 163
Research. on June 27, 2018. © 2015 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on November 2, 2015; DOI: 10.1158/1078-0432.CCR-15-0489

7
which was used at final concentration of 50 nM even in co-transfection experiments with 164 miRNAs inhibitors. 165 166 Virus generation and infection of Human Stromal HS-5 cells. 167 HS-5 cells cells stably expressing green fluorescent protein transgene were obtained as 168 previously described (39) (see Supplementary Methods for detailed information). 169 170 Reverse transcription and quantitative real-time PCR 171 Total RNA extraction from MM cells and quantitative real-time PCR were performed as 172 previously described (see Supplementary Methods for detailed information). (38) 173 174 Cell proliferation and survival assay 175 Cell growth inhibition was evaluated by Cell Counting Kit-8 (CCK-8) colorimetric assay 176 (Dojindo Molecular Technologies, Inc.), according to the manufacturer’s instructions. For 177 melphalan dose-response experiments, MM cells were seeded in 24-well plates at a density 178 of 2.5 x 105 cells per well in 1 ml of culture medium and incubated for 24 hours in the 179 presence of different µM melphalan concentrations; after incubation, MM cells were 180 inoculated in 96-well plates for CCK-8 assay. Final optical density (O.D.) was measured at 181 450 nm using GloMax (Promega). Wells without cells (culture medium alone) were used as 182 blank. For combination experiments with miRNAs, 1 x 106 electroporated cells with NC or 183 miR-221/222 inhibitors were incubated for 24 hours in 6-well plates; after harvesting, cells 184 were inoculated in 24-well plates at a density of 2.5 x 105 cells/ml and incubated in the 185 presence or absence of different µM melphalan concentrations. Twenty-four hours after 186 beginning drug exposure, cells were seeded in 96-well plates for CCK-8 assay and O.D. 187 measurement. For co-culture experiments, 2.5 x 105 U266/LR7 cells transfected with miR-188 221/222 inhibitors or NC were adhered to a monolayer of GFP+ HS-5 cells at 50% confluence 189 in 12-well plates for 24 h. After incubation, co-cultured cells were treated with 100 µM of 190 melphalan, and collected 24 hours later by gentle pipetting for 7-aminoactinomycin D (7-AAD) 191 staining and flow cytometry analysis. Each experiment was repeated at least three times. 192 Data represent the mean ± SD of at least 3 independent experiments. 193 194 195
Research. on June 27, 2018. © 2015 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on November 2, 2015; DOI: 10.1158/1078-0432.CCR-15-0489

8
Apoptosis detection by fluorescence microscopy 196 A total of 5 x 105 U266/LR7 cells transfected with NC or miR-221/222 inhibitors were 197 incubated for 24 hours in 12-well plates; after harvesting, each group was incubated in the 198 presence or absence of 100 µM melphalan. Twenty-four hours later, harvested cells were 199 plated in 96-well plates and visualized directly in the culture dishes after staining with Hoechst 200 33342 and Propidium Iodide (PI). Briefly, treated cells were washed and suspended in 201 phosphate-buffered saline (PBS) 1X at a density of 1 x 106 / ml with the addition of 5 µM 202 Hoechst 33342 (Sigma Aldrich). After incubation at 37° C for 15 minutes, cells were washed 203 and re-suspended in Binding Buffer 1X (BD Pharmigen) additions with 0.5 µM PI (BD 204 Pharmigen) and incubated at room temperature for 15 minutes avoiding light exposure. After 205 incubation, cells were washed and re-suspended in PBS for fluorescence microscopy 206 analysis using EVOS FLAuto Cell Imaging System (Life Technology). Representative fields of 207 cells were visualized (200X magnification). For apoptosis analysis by flow cytometry see 208 Supplementary Methods. 209 210 Luciferase reporter experiments. 211 Validation of miR-221/222 direct targeting of PUMA/BBC3 3’ UTR was performed as 212 previously described (see Supplementary Methods for detailed information). (38) 213 214 Protein extraction and Western blot analysis. 215 Total proteins were extracted with NP40 Cell Lysis Buffer (10 mmol/L Tris–HCl [pH 7.5], 150 216 mmol/L NaCl, 1% NP-40) (Life Technologies) with the addition of Halt Protease Inhibitor 217 Single-Use Cocktail 100 x (Thermo Scientific). For Western blot analysis, 50 µg per line of the 218 whole lysates were separated by electrophoresis on NuPAGE® precasted Gels (4%–12%) 219 (Invitrogen, Life Technologies) and electro-transferred to nitrocellulose membrane (Trans-Blot 220 Turbo Mini Nitrocellulose Transfer packs, Bio-Rad) using Trans-Blot Turbo Transfer System 221 (Bio-Rad). After protein transfer, the membranes were blotted with the primary antibodies 222 (see Supplementary Methods for detailed information). 223 224 Immune precipitation 225 Lysates were prepared by homogenizing retrieved tumor xenografts in NP-40 lysis buffer 226 containing Protease Inhibitor by using gentleMACS Dissociator (Miltenyi Biotec). Immune 227
Research. on June 27, 2018. © 2015 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on November 2, 2015; DOI: 10.1158/1078-0432.CCR-15-0489

9
precipitation was performed by incubating 1 mg lysates with 2 μg of antibody and 30 μl of 228 protein A/G-conjugated agarose beads (Santa Cruz Biotechnology) overnight at 4°C. The 229 precipitates were resuspended in loading buffer and resolved by NuPAGE electrophoresis, 230 followed by immune blotting afte electrotransfer. 231 232 Hoechst dye exclusion assay. 233 U266/LR7 cells were collected 48 hours after transfection with miR-221/222 inhibitors or NC 234 and resuspended in PBS (1X) at 1 x 106 / ml concentration. Cells were pre-incubated at 37°C 235 for 10 min in the presence or absence of verapamil 100 µM (Sigma Aldrich) to inhibit ABC 236 transporters and then were incubated for 90 min at 37°C with Hoechst 33342 5 µg/ml (Sigma 237 Aldrich), with intermittent shaking. Subsequently, cells were washed and re-suspended in ice-238 cold PBS (1X). 7-AAD was added (5 µl) to each tube prior to acquisition to exclude dead cells 239 from the analysis. The cells were filtered to obtain single cell suspension for flow cytometry 240 analysis performed by FACSAria III (Becton Dickinson). The Hoechst 33342 dye was excited 241 at 357 nm, and its fluorescence was dual-wavelength analyzed (402-446 nm for Hoechst 242 33342-Blue; 650-670 nm for Hoechst 33342-Red). 243 244 Animals and in vivo model of human MM. 245 Six to eight-week old female SCID/NOD mice (Harlan laboratories, Inc.) were housed and 246 monitored in Animal Research Facility at Magna Graecia University. All experimental 247 procedures and protocols had been approved by the Institutional Ethical Committee (Magna 248 Graecia University). In accordance with the institutional guidelines, mice were sacrificed when 249 their tumors reached 2 cm in diameter or in the event of paralysis or major compromise in 250 their quality of life. Specifically, we evaluated the in vivo ability of LNA-i-miR-221 (35), 251 specifically designed for systemic delivery, to enhance the anti-MM activity of melphalan. 252 Briefly, a cohort of 24 mice for each in vivo study were subcutaneously inoculated with 5 x 106 253 U266/LR7 cells in 100 µl of RPMI-1640 medium; treatments started when tumors became 254 measurable, approximately 3 weeks after cells were injected. In a first in vivo experiment, 255 mice were randomized to receive 4 different treatments: i) LNA-i-miR-221 intraperitoneally 256 (i.p.) injected at a dose of 25 mg/kg at day 1-4-8-15-22 from the beginning of the experiments; 257 ii) scrambled control with the same schedule; iii) 0.75 mg/kg of i.p. melphalan twice weekly for 258 3 weeks; iiii) melphalan plus LNA-i-miR-221 with the above described schedules. In a 259
Research. on June 27, 2018. © 2015 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on November 2, 2015; DOI: 10.1158/1078-0432.CCR-15-0489

10
subsequent in vivo study, treatment with LNA-i-miR-221 (25 mg/kg) plus melphalan (0.75 260 mg/kg) was given with for 4 consecutive days every 10 days. Tumor sizes were measured as 261 previously described (35). In vivo detection of the tumor mass in xenografted mice was 262 performed using IVIS LUMINA II Imaging System (Caliper Life sciences, Hopkinton, MA, 263 USA). At the end of observation, tumors were retrieved from animals and placed in 10 264 formalin for histology or stored at -80° C for protein analysis. 265 266 Histology and immunohistochemistry. 267 Retrieved tumors from animals were fixed in 4% buffered formaldehyde and 24 h later 268 washed, dehydrated, and embedded in paraffin. For light microscopy analysis by an optical 269 microscope Nikon i55 (Nikon Corporation, Tokyo, Japan), we performed staining with 270 Hematoxylin-Eosin on 4 µm tumors sections mounted on poly-lysine slides. For 271 immunohistochemistry staining, 2 µm thick tumor slices were de-paraffinized and pre-treated 272 with the Epitope Retrieval Solution 2 (EDTA-buffer pH 8.8) at 98°C for 20 min. After washing 273 steps, peroxidase blocking was carried out for 10 min using the Bond Polymer. All procedures 274 were performed using the Bond Max Automated Immunohistochemistry. Tissues were 275 washed and incubated with the primary antibody directed against Ki-67 (Dako, clone MIB-1; 276 1:150) or caspase-3 (Novocastra, clone JHM62; 1:500). Subsequently, tissues were 277 incubated with polymer for 10 min and developed with DAB-Chromogen for 10 min. Slides 278 were counterstained with hematoxylin. 279 280 Statistical analysis. 281 Each in vitro experiment was carried out at least 3 times, and all values are reported as 282 means ± SD. Comparisons between groups were made with Student t test, whereas statistical 283 significance of differences among multiple groups was determined by GraphPad software 284 (www.graphpad.com). Differences were considered significant with p < 0.05. The Synergistic 285 Index (SI) was determined as previously described (40) with the following formula: SI = (effect 286 induced by miR-221/222 inhibitors and melphalan in combination)/(effect of miR-221/222 287 inhibitors)+(effect of melphalan). Interactions were considered synergistic when SI was >1. 288 289 290
Research. on June 27, 2018. © 2015 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on November 2, 2015; DOI: 10.1158/1078-0432.CCR-15-0489

11
Results 291 292 1. Melphalan-induced modulation of miR-221/222 in MM cells. 293 We first evaluated the melphalan-sensitivity of our MM cell line panel including U266/S, NCI-294 H929, AMO1, RPMI-8226, AMO1 Abzb and U266/LR7. NCI-H929, AMO1 and U266/S were 295 sensitive at low melphalan concentrations; RPMI-8226 and the bortezomib-resistant AMO1 296 Abzb showed resistance at doses up to 20 µM, while the melphalan-resistant U266/LR7 cells 297 were resistant up to 200 µM drug concentration (IC50 not found) (Fig. 1a). In parallel, we 298 measured the miR-221/222 melphalan-induced expression in MM cells. As evaluated by q-299 RT-PCR, miR-221/222 expression after drug exposure inversely correlated with melphalan-300 sensitivity (Fig. 1b-c). In particular, melphalan-resistant U266/LR7 cells showed the highest 301 miR-221/222 expression after drug exposure. Significant miR-221 upregulation was also 302 observed in RPMI-8226 and AMO1 Abzb cells, which are moderately resistant to melphalan, 303 whereas those cell lines sensitive to melphalan including NCI-H929, U266/S, and AMO1 cells 304 did not show any change in miR-221/miR-222 expression following treatment. Moreover, q-305 RT-PCR analysis of miR-221 and miR-222 pre-melphalan treatment in U266/LR7 showed a 306 1.5-fold increase for miR-221 (Fig. S1a) and a 3.5-fold increase for miR-222 (Fig. S1b) 307 compared to the parental sensitive counterpart U266/S (Fig. S1). Altogether, these findings 308 demonstrate an inverse correlation of drug-induced miR-221/222 expression with melphalan 309 sensitivity of MM cells. 310 311 2. MiR-221/222 inhibition overcomes melphalan-resistance of MM cells in vitro. 312 We previous reported a strong in vitro and in vivo anti-tumor activity of miR-221/222 inhibitors 313 in MM cells bearing t(4;14) translocation (34). Since modulation of miR-221/222 seems to be 314 affected by melphalan treatment independently from chromosome translocation, we 315 investigated whether miR-221/222 plays a mechanistic role in MM cell melphalan-316 sensitivity/resistance regardless translocation status. To this aim, we first transfected 317 U266/LR7, RPMI-8226, and AMO1 Abzb melphalan-resistant cells with miR-221/222 318 inhibitors or scramble controls, and then exposed cells to increasing concentrations of 319 melphalan. As shown in Fig. 2a, miR-221/222 inhibitors synergistically enhanced the anti-MM 320 activity of melphalan in a dose-dependent manner. At a fixed concentration (100 nM) of miR-321 221/222 inhibitors with increasing doses of melphalan, the synergistic index was measured: 322
Research. on June 27, 2018. © 2015 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on November 2, 2015; DOI: 10.1158/1078-0432.CCR-15-0489

12
the highest synergistic effect was reached with 100 µM of melphalan (SI = 5.6) in U266/LR7 323 cells, and with 5 µM of melphalan (SI = 4.12) in RPMI-8226 cells, and in AMO1 Abzb cells (SI 324 = 1.36) (Table S1). Conversely, enforced expression of synthetic miR-221/222 mimics in 325 melphalan-sensitive U266/S cells, which previously we reported to exert a growth promoting 326 activity on MM cells (34), resulted in a reduced susceptibility to the drug (Fig. S2a). Moreover, 327 we evaluated whether miR-221/222 sensitizing effects to melphalan could be due to induction 328 of apoptosis. As shown in Fig. 2b using Annexin V/7-AAD assay, we found that the combined 329 treatment triggered greater apoptosis in all MM cell lines than each single agent. By western 330 blotting analysis, we showed activation/cleavage of caspase-8, caspase-9 and caspase-3, as 331 well as increased PARP cleavage, in U266/LR7 cells exposed to combination treatment (Fig. 332 2c, top panel). An increase of cleaved caspase-3 was also confirmed by western blotting in 333 RPMI-8226 cells (Fig. 2c, bottom panel). Conversely, enforced expression of synthetic miR-334 221/222 mimics in U266/S cells exposed to melphalan resulted in a decrease of cleaved 335 caspase-3 (Fig. S2b). Using Hoechst 33342 and PI staining, we showed induction of 336 apoptosis in cells after combination treatment, with a significant increase in late apoptosis 337 (Fig. 2d). Importantly, inhibition of miR-221/222 enhanced the cytotoxic effect of melphalan 338 even on primary CD138+ cells from 3 MM patients (Fig 3a). No effects on cell viability were 339 instead observed on PBMCs from 3 healthy donors (Fig 3b). We next studied whether 340 BMSCs protected against the synergistic combination therapy of miR-221/222 plus 341 melphalan. By q-RT-PCR, a slight increase of miR-221/222 expression (2 and 4-folds, 342 respectively) was detected in U266/LR7 cells adherent to the HS-5 BMSCs, as compared to 343 cells cultured alone (Fig. S3). Importantly, anti-MM activity of miR-221/222 inhibitors plus 344 melphalan against U266/LR7 cells adherent to HS5 BMSCs was similar to that against non 345 adherent U266/LR7 cells (Fig. 3c). Taken together, these findings demonstrate that miR-346 221/222 inhibitors enhance the therapeutic activity of melphalan in drug-resistant cells by 347 induction of apoptosis, even in the presence of BMSCs. 348 349 3. Genome-wide expression patterns triggered by miR-221/222 inhibitors plus 350 melphalan. 351 To analyze the transcriptome perturbations induced in MM cells by the combination of miR-352 221/222 inhibitors plus melphalan treatment, we performed a whole gene expression analysis 353
Research. on June 27, 2018. © 2015 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on November 2, 2015; DOI: 10.1158/1078-0432.CCR-15-0489

13
(GEP) by GeneChip® Affymetrix arrays. Data are available through GEO accession number 354 GSE66618. Since we demonstrated that inhibition of miR-221/222 restores sensitvity to 355 melphalan in resistant MM cells, we next examined whether this effect may be associated 356 with changes in the expression of genes involved in the most common mechanisms of drug-357 resistance. We performed a GEP on both untreated or drug-exposed melphalan-resistant 358 U266/LR7 cells and melphalan-sensitive parental U266/S MM cells. By hierarchical clustering 359 analysis, data points were grouped based on overall similarity in gene expression patterns. 360 Clustering analysis revealed a similar profile in melphalan-resistant U266/LR7 MM cells 361 treated with miR-221/222 inhibitors plus melphalan as was observed in the sensitive parental 362 cells U266/S cells treated with melphalan alone (data not shown). Accordingly, we next 363 performed fold change (FC) analysis, which yielded a list of 1487 genes modulated after 364 combination treatment in melphalan-resistant MM cells. Notably, differentially expressed 365 genes involved in apoptosis (PUMA/BBC3; BCL2L1), drug transport (ABCC1/MRP1, SLC7A8, 366 SLC7A5), and DNA repair mechanisms (XRCC4, PRKDC, CNOT6L, DDB2) were found (Fig. 367 S4a). By Ingenuity Pathway Analysis® (IPA) software, we evaluated the perturbation of 368 biological response pathways and regulatory networks. After IPA annotation, a list of 739 369 genes was used for “Core Analysis” and identification of different canonical pathways with 370 highly significant perturbation scores, primarily in the miR-221/222 inhibitors plus melphalan 371 treated U266/LR7 cells (p<0.05) (Fig. S4b). Specifically, IPA revealed modulation of 372 pathways: associated with alkylating agents-induced response (“NRF2-medited Oxidative 373 Stress Response”); induced by micro-environmental stimuli (“HGF Signaling”, “Chemokine 374 Signaling”, “IL-3 Signaling”, “Jak/Stat Signaling”, “STAT3 Pathway”); associated with tumor 375 proliferation (“PTEN Signaling”, “EIF2 Signaling” and “PI3K Signaling in B Lymphocytes”); 376 related to DNA repair mechanisms (“Cell Cycle:G2/M DNA Damage Checkpoint Regulation” 377 and “DNA Double-Strand Break Repair by Non-Homologous End Joining”); as well as 378 “Glucocorticoid Receptor Signaling”. Based on the IPA annotation analysis, we also 379 characterized genes involved in these signaling pathways (Fig. S4c, inserted table). 380 Interestingly, we detected downregulation of ABCC1, PIK3CB, XRCC4, PRKDC, AKT3 and 381 STAT6; as well as upregulation of CDKN1A and E2F7 after miR-221/222 inhibitor plus 382 melphalan treatment compared to either untreated- or either single agent treated- U266/LR7 383 cells. Altogether, these findings define a transcriptome profile that underlies the response to 384 the combination treatment in MM cells, suggesting further investigation of miR-221/222 385
Research. on June 27, 2018. © 2015 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on November 2, 2015; DOI: 10.1158/1078-0432.CCR-15-0489

14
involvement in resistance mechanisms. 386 387 4. PUMA/BBC3 is a target of miR-221/222 in MM cells. 388 We validated GEP data on XRCC4, CDKN1A, PRKDC, SLC7A8/LAT2, SLC7A5/LAT1, 389 ABCC1/MRP1, and PUMA/BBC3 by q-RT-PCR, (Fig. S5a-b). In silico search for target 390 prediction using mirDIP software indicates Bcl-2-binding component 3, PUMA/BBC3, as a 391 strongly predicted target of miR-221/222. On the basis of target prediction, we co-transfected 392 U266/LR7 cells with synthetic miR-221 or miR-222 mimics together with firefly luciferase 393 constructs, in which either the wild-type or mutant 3’ UTR of PUMA/BBC3 mRNA are cloned 394 downstream. A marked decrease in luciferase activity (42% for miR-221 and 38% for miR-395 222) indicated direct interactions between the miRNAs and PUMA/BBC3 3’UTR; moreover, 396 target gene repression was rescued by deletions in the mutant clone (Fig. 4a). These findings 397 provide validation of direct miR-221/222 targeting of 3’UTR of PUMA/BBC3 mRNA in MM 398 cells. 399 400 5. MiR-221/222 inhibitors upregulate PUMA/BBC3 protein levels triggering melphalan-401 sensitivity in drug-resistant MM cells. 402 We next analyzed PUMA/BBC3 expression both at the mRNA and protein levels in U266/LR7 403 cells transfected with miR-221/222 inhibitors and then exposed to melphalan. By western 404 blotting, a strong upregulation of PUMA/BBC3 protein was found in cells treated with miR-405 221/222 inhibitors plus melphalan; conversely, exposure to melphalan alone led to 406 upregulation of PUMA/BBC3 at mRNA, but not at protein, levels (Fig. 4b). Moreover, enforced 407 expression of synthetic miR-221/222 mimics in melphalan-treated U266/S cells abrogates 408 PUMA/BBC3 protein translation induced by melphalan alone (Fig. 4c). To evaluate the role of 409 PUMA overexpression in enhancing melphalan activity, we exposed the U266/LR7 cells 410 transfected with miR-221/222 inhibitors together with siRNA for PUMA/BBC3 (siPUMA) or 411 siRNA control (siCNT) to melphalan. Importantly, in resistant cell lines silenced for 412 PUMA/BBC3 (Fig. 4d, right panel), we found a significant antagonism (30%) of the sensitizing 413 activity of miR-221/222 inhibitors compared to co-transfection with siCNT (Fig. 4d, left panel). 414 As a further confirmation, PUMA silencing led to a 20% reduction of melphalan activity of in 415 drug-sensitive U266/S cells (Fig. 4e, right panel). Based on these findings, we conclude that 416
Research. on June 27, 2018. © 2015 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on November 2, 2015; DOI: 10.1158/1078-0432.CCR-15-0489

15
the sensitizing activity of miR-221/222 inhibitors to melphalan in MM cells is, at least in part, 417 dependent on PUMA/BBC3 protein upregulation. 418 419 420 6. MiR-221/222 inhibition modulates expression of drug influx-efflux transporters in MM 421 cells 422 Since GEP analysis showed modulation of drug transporter mRNA transcripts, we next 423 studied the effects of miR-221/222 inhibitors plus melphalan on their expression. L-Type 424 Amino Acid Transporter SLC7A5/LAT1 and ATP-binding cassette (ABC) transporter 425 ABCC1/MRP1 were evaluated at the protein level in miR221/222 inhibitor or scramble 426 transfected U266/LR7 cells. As shown in Fig 5a, miR-221/222 inhibition plus melphalan 427 resulted in a significant upregulation of SLC7A5/LAT1 protein, as well as a marked 428 downregulation of ABCC1/MRP1 protein (Fig. 5a). Moreover, based on the known ability of 429 ABCC1/MRP1 pump to exclude drugs as well as fluorescent dyes such as Hoechst 33342 430 (41), we next evaluated by Hoechst dye exclusion assay whether miR-221/222 inhibitor 431 treatment may affect dye cellular uptake. As expected, the cell fraction in U266/LR7 cells that 432 exhibits low level of Hoechst fluorescent intensity, identified as Side Population (SP), 433 decreased by 65% after treatment with miR-221/222 inhibitors, and by 74% after treatment 434 with ABC transporters inhibitor Verapamil (Fig. 5b,c). Taken together, these findings identified 435 a role of miR-221/222 in regulation of drug transporter expression. 436 437 438 7. LNA-i-miR-221 inhibitors overcome melphalan-induced drug resistance in MM cells 439 in vivo. 440 Finally, we evaluated the anti-tumor activity of the combination in SCID/NOD mice bearing 441 human melphalan-resistant MM xenografts in 2 independent in vivo studies. For each study, 442 animals were randomized to receive 4 different treatments. As shown in Fig. 6, i.p. treatment 443 with LNA-i-miR-221 (25 mg/kg) on days 1-4-8-15-22 plus melphalan (0.75 mg/kg) twice 444 weekly for 3 weeks overcame drug-resistance, as evidenced by a significant tumor growth 445 inhibition compared to control groups (p < 0.05) (Fig. 6a). In a subsequent in vivo study, we 446 optimized the treatment schedule to improve the sensitizing activity of LNA-i-miR-221 and to 447
Research. on June 27, 2018. © 2015 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on November 2, 2015; DOI: 10.1158/1078-0432.CCR-15-0489

16
adapt the LNA-i-miR-221 administration to a common melphalan schedule used in MM 448 patients. Specifically, the treatment with LNA-i-miR-221 plus melphalan was given for 4 449 consecutive days every 10 days. This approach strengthened the anti-tumor activity of the 450 combination (p < 0.001) (Fig.6b). Moreover, no changes in body weight or mice behaviors 451 were observed; H&E staining on retrieved organs from treated animals did not show any 452 organ toxicity (data not shown). Additionally and consistent with our in vitro findings, analysis 453 of retrieved tumors confirmed upregulation of PUMA/BBC3 protein (Fig. 6c bottom panel) and 454 inhibition of ABCC1/MRP1 protein (Fig. 6c top panel) in mice treated with LNA-i-miR-221 plus 455 mephalan. IHC analysis on excised tumors demonstrated extended necrosis following 456 combination treatment, with reduction of Ki-67 proliferation index (Fig. 6d). Altogether, these 457 in vivo findings further indicate the translational relevance of LNA-i-miR-221 as a sensitizing 458 agents in melphalan-resistant MM. 459 460 461 462
Research. on June 27, 2018. © 2015 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on November 2, 2015; DOI: 10.1158/1078-0432.CCR-15-0489

17
Discussion 463 464
Targeting the miRNA network is arising as a new promising strategy to overcome 465 drug-resistance of human cancer (42). We previously showed that inhibition of oncogenic 466 miR-221/222 in MM cell lines induces significant anti-MM activity via targeting key molecules 467 involved in cell proliferation and survival (34) and that LNA-inhibitors of miR-221 are suitable 468 for safe and effective systemic delivery in mice (35). We here report that in vitro inhibition of 469 miR-221/222 restores melphalan-sensitivity, inducing anti-proliferative and apoptotic cell 470 death in drug-refractory MM cells. More importantly, this anti-tumor activity has been also 471 shown in vivo following systemic treatment with LNA-i-miR-221 plus melphalan in SCID/NOD 472 mice bearing melphalan-resistant MM xenografts without any evidence of toxicity or side 473 effects in treated mice. Our results further support the role of miR-221/222 as crucial mediator 474 of tumor cell resistance to alkylating agents such as cisplatin and temozolamide (28, 31), and 475 we here provide proof-of principle that LNA-i-miR-221 is a potent sensitizing-agent in 476 melphalan-refractory MM. 477
To investigate the mechanistic role of miR-221/222 in melphalan-resistant MM, we first 478 assessed the correlation between miR-221/222 expression and the anti-proliferative activity of 479 melphalan. Decreased miR-221/222 expression after melphalan treatment in sensitive-cells 480 along with increased expression in resistant-cells suggested a correlation of miR-221/222 with 481 melphalan-resistance. This observation was also confirmed by the basal higher expression of 482 both miRNAs pre-melphalan treatment in U266/LR7 cells as compared to the parental 483 sensitive counterpart U266/S. Given that miR-221/222 expression has been shown to be 484 activated by transcription factor c-jun likely as DNA-damage response following radiation 485 therapy (43), it is tempting to hypothesize that a similar mechanism may occur in MM cells 486 after melphalan exposure. Follow-up studies will be carried out to clarify this point. 487
Indeed, inhibition of miR-221/222 markedly overcame melphalan-resistance in 488 U266/LR7, RPMI-8226 and AMO1 Abzb, evidenced by a strong synergism of these two 489 agents; conversely, overexpression of these miRNAs antagonizes drug-activity in melphalan-490 sensitive U266/S cells. Moreover, the synergistic effect of the combination affected the 491 survival of primary cells of MM patients without any cytotoxicity in PBMCs from healthy 492 donors, suggesting a low toxicity profile of our approach. In our in vitro experimental model, 493 miR-221/222 inhibitors enhance melphalan-induced apoptosis, as confirmed by activation of 494
Research. on June 27, 2018. © 2015 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on November 2, 2015; DOI: 10.1158/1078-0432.CCR-15-0489

18
Caspase cascade. Moreover, we found that upregulation of miR-221/222 induced by 495 adhesion of MM cells to BMSCs does not impair the effectiveness of this combination, 496 indicating that this treatment can overcome the role of the MM microenvironment in promoting 497 resistance to DNA-damaging agents (44). 498
Interestingly, functional analysis of genes and pathways significantly modulated in 499 resistant U266/LR7 cells demonstrated a remarkable similarity in gene expression signatures 500 of melphalan-resistant U266/LR7 MM cells treated with miR-221/222 inhibitors plus 501 melphalan compared with the sensitive parental U266/S cells treated with melphalan. 502 Notably, IPA analysis demonstrated significant perturbation of pathways of central relevance 503 in cellular response to alkylating agents, frequently correlated with drug-resistance such as 504 “NRF2-mediated Oxidative Stress Response” (45), “Cell Cycle:G2/M DNA Damage 505 Checkpoint Regulation”, “DNA Double-Strand Break Repair by Non-Homologous End Joining” 506 (46), as well as those pathways associated with tumor progression. Based on GEP data, we 507 focused on pro-apoptotic PUMA/BBC3, since its downregulation has been associated with 508 drug-resistance (47, 48) and miR-221/222-sensitizing activity to temozolamide (31). After 509 luciferase reporter assay validation of PUMA mRNA as a specific miR-221/222 target, we 510 demonstrated that miR-221/222 inhibitors induce mRNA and protein PUMA expression. 511 Conversely, PUMA silencing by siRNA transfection in melphalan-resistant U266/LR7 cells 512 antagonized the melphalan-sensitizing activity of miR-221/222 inhibitors. Of note, PUMA 513 silencing also led to a significant reduction of melphalan-activity in drug-sensitive U266/S 514 cells. Our results therefore highlight the role of PUMA as key player in miR-221/222-515 mediated induction of melphalan-resistance. 516
Several reports have emphasized the role of ATP-dependent ABC transporters which 517 rapidly efflux several drugs as crucial determinants of multidrug-resistance in cancer cells 518 (49). Of note, aberrant expression of influx and efflux transporters has been correlated with 519 altered sensitivity to melphalan (50). Interestingly, our GEP data suggested the activity of 520 miR-221/222 targeting ATP-dependent efflux MRP1/ABCC1 as well as influx SLC7A5/LAT1 521 transporters and we here confirmed that miR-221/222 inhibitors upregulate LAT1 and 522 downregulate ABCC1 at the protein level. Furthermore, a subset of cancer stem-like cells, 523 identified as “side population”, expresses these transporters which correlates with 524 chemoresistance and a high tumorigenic potential (41). In this light, we performed a functional 525 analysis to characterize cells that contribute to the SP phenotype, and we show a marked 526
Research. on June 27, 2018. © 2015 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on November 2, 2015; DOI: 10.1158/1078-0432.CCR-15-0489

19
decrease of SP in cells treated with miR-221/222 inhibitors. In silico search for target 527 prediction using mirDIP software did not indicate direct targeting of mRNAs and further 528 studies are necessary to solve the mechanism of miR-221/222 activity on LAT1 and ABCC1 529 transporters. Nonetheless our results are of translational relevance since they provide proof-530 of-concept that miR-221/222 inhibition can antagonize resistance to melphalan through 531 modulation of influx-efflux transporters. Finally, we validated our in vitro findings in murine 532 xenograft models with human melphalan-resistant MM cells in 2 independent experiments: a 533 stronger anti-tumor activity has been achieved with 4 consecutive days of treatment with LNA-534 i-miR-221 plus melphalan. This effect was associated with upregulation of PUMA/BBC3 and 535 downregulation of ABCC1 proteins in tumors harvested from treated animals. 536
In conclusion, the validation of LNA-i-miR-221 as a melphalan sensitizing-agent both 537 confirms the role of miR-221/222 in mediating drug resistance, and provides the framework 538 for combination clinical trials to sensitize or overcome resistance to melphalan in MM. 539
540 Disclaimer: 541 The Editor-in-Chief is an author on this article. In keeping with the AACR’s editorial policy, 542 the peer review of this submission was managed by a senior member of Cancer Research’s 543 editorial team; a member of the AACR Publications Committee rendered the final decision 544 concerning acceptability. 545 546
Research. on June 27, 2018. © 2015 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on November 2, 2015; DOI: 10.1158/1078-0432.CCR-15-0489

20
References 547 548 1. Palumbo A, Anderson K. Multiple myeloma. The New England journal of medicine. 2011 549 Mar 17;364(11):1046-60. 550 2. Munshi NC, Anderson KC. New strategies in the treatment of multiple myeloma. Clinical 551 cancer research : an official journal of the American Association for Cancer Research. 2013 Jul 552 1;19(13):3337-44. 553 3. Anderson KC, Carrasco RD. Pathogenesis of myeloma. Annual review of pathology. 554 2011;6:249-74. 555 4. Calimeri T, Battista E, Conforti F, Neri P, Di Martino MT, Rossi M, et al. A unique three-556 dimensional SCID-polymeric scaffold (SCID-synth-hu) model for in vivo expansion of human 557 primary multiple myeloma cells. Leukemia. 2011 Apr;25(4):707-11. 558 5. Tassone P, Neri P, Carrasco DR, Burger R, Goldmacher VS, Fram R, et al. A clinically 559 relevant SCID-hu in vivo model of human multiple myeloma. Blood. 2005 Jul 15;106(2):713-6. 560 6. Tassone P, Neri P, Burger R, Di Martino MT, Leone E, Amodio N, et al. Mouse models as a 561 translational platform for the development of new therapeutic agents in multiple myeloma. 562 Current cancer drug targets. 2012 Sep;12(7):814-22. 563 7. Rossi M, Di Martino MT, Morelli E, Leotta M, Rizzo A, Grimaldi A, et al. Molecular targets 564 for the treatment of multiple myeloma. Current cancer drug targets. 2012 Sep;12(7):757-67. 565 8. Anderson KC. Therapeutic advances in relapsed or refractory multiple myeloma. Journal 566 of the National Comprehensive Cancer Network : JNCCN. 2013 May;11(5 Suppl):676-9. 567 9. Bayraktar UD, Bashir Q, Qazilbash M, Champlin RE, Ciurea SO. Fifty years of melphalan use 568 in hematopoietic stem cell transplantation. Biology of blood and marrow transplantation : journal 569 of the American Society for Blood and Marrow Transplantation. 2013 Mar;19(3):344-56. 570 10. Child JA, Morgan GJ, Davies FE, Owen RG, Bell SE, Hawkins K, et al. High-dose 571 chemotherapy with hematopoietic stem-cell rescue for multiple myeloma. The New England 572 journal of medicine. 2003 May 8;348(19):1875-83. 573 11. Palumbo A, Attal M, Roussel M. Shifts in the therapeutic paradigm for patients newly 574 diagnosed with multiple myeloma: maintenance therapy and overall survival. Clinical cancer 575 research : an official journal of the American Association for Cancer Research. 2011 Mar 576 15;17(6):1253-63. 577 12. Palumbo A, Rajkumar SV, San Miguel JF, Larocca A, Niesvizky R, Morgan G, et al. 578 International Myeloma Working Group consensus statement for the management, treatment, and 579 supportive care of patients with myeloma not eligible for standard autologous stem-cell 580 transplantation. Journal of clinical oncology : official journal of the American Society of Clinical 581 Oncology. 2014 Feb 20;32(6):587-600. 582 13. Yarde DN, Oliveira V, Mathews L, Wang X, Villagra A, Boulware D, et al. Targeting the 583 Fanconi anemia/BRCA pathway circumvents drug resistance in multiple myeloma. Cancer 584 research. 2009 Dec 15;69(24):9367-75. 585 14. Chauhan D, Ray A, Viktorsson K, Spira J, Paba-Prada C, Munshi N, et al. In vitro and in vivo 586 antitumor activity of a novel alkylating agent, melphalan-flufenamide, against multiple myeloma 587 cells. Clinical cancer research : an official journal of the American Association for Cancer 588 Research. 2013 Jun 1;19(11):3019-31. 589 15. Croce CM. Causes and consequences of microRNA dysregulation in cancer. Nature reviews 590 Genetics. 2009 Oct;10(10):704-14. 591
Research. on June 27, 2018. © 2015 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on November 2, 2015; DOI: 10.1158/1078-0432.CCR-15-0489

21
16. Janssen HL, Reesink HW, Lawitz EJ, Zeuzem S, Rodriguez-Torres M, Patel K, et al. 592 Treatment of HCV infection by targeting microRNA. The New England journal of medicine. 2013 593 May 2;368(18):1685-94. 594 17. Di Martino MT, Leone E, Amodio N, Foresta U, Lionetti M, Pitari MR, et al. Synthetic miR-595 34a mimics as a novel therapeutic agent for multiple myeloma: in vitro and in vivo evidence. 596 Clinical cancer research : an official journal of the American Association for Cancer Research. 597 2012 Nov 15;18(22):6260-70. 598 18. Amodio N, Leotta M, Bellizzi D, Di Martino MT, D'Aquila P, Lionetti M, et al. DNA-599 demethylating and anti-tumor activity of synthetic miR-29b mimics in multiple myeloma. 600 Oncotarget. 2012 Oct;3(10):1246-58. 601 19. Amodio N, Di Martino MT, Foresta U, Leone E, Lionetti M, Leotta M, et al. miR-29b 602 sensitizes multiple myeloma cells to bortezomib-induced apoptosis through the activation of a 603 feedback loop with the transcription factor Sp1. Cell death & disease. 2012;3:e436. 604 20. Leone E, Morelli E, Di Martino MT, Amodio N, Foresta U, Gulla A, et al. Targeting miR-21 605 inhibits in vitro and in vivo multiple myeloma cell growth. Clinical cancer research : an official 606 journal of the American Association for Cancer Research. 2013 Apr 15;19(8):2096-106. 607 21. Amodio N, Bellizzi D, Leotta M, Raimondi L, Biamonte L, D'Aquila P, et al. miR-29b induces 608 SOCS-1 expression by promoter demethylation and negatively regulates migration of multiple 609 myeloma and endothelial cells. Cell Cycle. 2013 Dec 1;12(23):3650-62. 610 22. Di Martino MT, Campani V, Misso G, Gallo Cantafio ME, Gulla A, Foresta U, et al. In vivo 611 activity of miR-34a mimics delivered by stable nucleic acid lipid particles (SNALPs) against 612 multiple myeloma. PloS one. 2014;9(2):e90005. 613 23. Leotta M, Biamonte L, Raimondi L, Ronchetti D, Martino MT, Botta C, et al. A p53-614 dependent tumor suppressor network is induced by selective miR-125a-5p inhibition in multiple 615 myeloma cells in vitro. Journal of cellular physiology. 2014 May 13. 616 24. Raimondi L, Amodio N, Di Martino MT, Altomare E, Leotta M, Caracciolo D, et al. Targeting 617 of multiple myeloma-related angiogenesis by miR-199a-5p mimics: in vitro and in vivo anti-618 tumor activity. Oncotarget. 2014 May 30;5(10):3039-54. 619 25. Pitari MR, Rossi M, Amodio N, Botta C, Morelli E, Federico C, et al. Inhibition of miR-21 620 restores RANKL/OPG ratio in multiple myeloma-derived bone marrow stromal cells and impairs 621 the resorbing activity of mature osteoclasts. Oncotarget. 2015 Jun 24. 622 26. Garofalo M, Croce CM. MicroRNAs as therapeutic targets in chemoresistance. Drug 623 resistance updates : reviews and commentaries in antimicrobial and anticancer chemotherapy. 624 2013 Jul-Nov;16(3-5):47-59. 625 27. Zhu H, Wu H, Liu X, Evans BR, Medina DJ, Liu CG, et al. Role of MicroRNA miR-27a and miR-626 451 in the regulation of MDR1/P-glycoprotein expression in human cancer cells. Biochemical 627 pharmacology. 2008 Sep 1;76(5):582-8. 628 28. Zhao G, Cai C, Yang T, Qiu X, Liao B, Li W, et al. MicroRNA-221 induces cell survival and 629 cisplatin resistance through PI3K/Akt pathway in human osteosarcoma. PloS one. 630 2013;8(1):e53906. 631 29. Miller TE, Ghoshal K, Ramaswamy B, Roy S, Datta J, Shapiro CL, et al. MicroRNA-221/222 632 confers tamoxifen resistance in breast cancer by targeting p27Kip1. The Journal of biological 633 chemistry. 2008 Oct 31;283(44):29897-903. 634 30. Rao X, Di Leva G, Li M, Fang F, Devlin C, Hartman-Frey C, et al. MicroRNA-221/222 confers 635 breast cancer fulvestrant resistance by regulating multiple signaling pathways. Oncogene. 2011 636 Mar 3;30(9):1082-97. 637 Research. on June 27, 2018. © 2015 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on November 2, 2015; DOI: 10.1158/1078-0432.CCR-15-0489

22
31. Chen L, Zhang J, Han L, Zhang A, Zhang C, Zheng Y, et al. Downregulation of miR-221/222 638 sensitizes glioma cells to temozolomide by regulating apoptosis independently of p53 status. 639 Oncology reports. 2012 Mar;27(3):854-60. 640 32. Garofalo M, Romano G, Di Leva G, Nuovo G, Jeon YJ, Ngankeu A, et al. EGFR and MET 641 receptor tyrosine kinase-altered microRNA expression induces tumorigenesis and gefitinib 642 resistance in lung cancers. Nature medicine. 2012 Jan;18(1):74-82. 643 33. Garofalo M, Di Leva G, Romano G, Nuovo G, Suh SS, Ngankeu A, et al. miR-221&222 644 regulate TRAIL resistance and enhance tumorigenicity through PTEN and TIMP3 645 downregulation. Cancer cell. 2009 Dec 8;16(6):498-509. 646 34. Di Martino MT, Gulla A, Cantafio ME, Lionetti M, Leone E, Amodio N, et al. In vitro and in 647 vivo anti-tumor activity of miR-221/222 inhibitors in multiple myeloma. Oncotarget. 2013 648 Feb;4(2):242-55. 649 35. Di Martino MT, Gulla A, Gallo Cantafio ME, Altomare E, Amodio N, Leone E, et al. In vitro 650 and in vivo activity of a novel locked nucleic acid (LNA)-inhibitor-miR-221 against multiple 651 myeloma cells. PloS one. 2014;9(2):e89659. 652 36. Maiso P, Carvajal-Vergara X, Ocio EM, Lopez-Perez R, Mateo G, Gutierrez N, et al. The 653 histone deacetylase inhibitor LBH589 is a potent antimyeloma agent that overcomes drug 654 resistance. Cancer research. 2006 Jun 1;66(11):5781-9. 655 37. Ruckrich T, Kraus M, Gogel J, Beck A, Ovaa H, Verdoes M, et al. Characterization of the 656 ubiquitin-proteasome system in bortezomib-adapted cells. Leukemia. 2009 Jun;23(6):1098-105. 657 38. Morelli E, Leone E, Cantafio ME, Di Martino MT, Amodio N, Biamonte L, et al. Selective 658 targeting of IRF4 by synthetic microRNA-125b-5p mimics induces anti-multiple myeloma activity 659 in vitro and in vivo. Leukemia. 2015 May 19. 660 39. Amodio N, Scrima M, Palaia L, Salman AN, Quintiero A, Franco R, et al. Oncogenic role of 661 the E3 ubiquitin ligase NEDD4-1, a PTEN negative regulator, in non-small-cell lung carcinomas. 662 The American journal of pathology. 2010 Nov;177(5):2622-34. 663 40. Tassone P, Neri P, Burger R, Savino R, Shammas M, Catley L, et al. Combination therapy 664 with interleukin-6 receptor superantagonist Sant7 and dexamethasone induces antitumor effects 665 in a novel SCID-hu In vivo model of human multiple myeloma. Clinical cancer research : an official 666 journal of the American Association for Cancer Research. 2005 Jun 1;11(11):4251-8. 667 41. Hirschmann-Jax C, Foster AE, Wulf GG, Nuchtern JG, Jax TW, Gobel U, et al. A distinct "side 668 population" of cells with high drug efflux capacity in human tumor cells. Proceedings of the 669 National Academy of Sciences of the United States of America. 2004 Sep 28;101(39):14228-33. 670 42. Mishra PJ. The miRNA-drug resistance connection: a new era of personalized medicine 671 using noncoding RNA begins. Pharmacogenomics. 2012 Sep;13(12):1321-4. 672 43. Li W, Guo F, Wang P, Hong S, Zhang C. miR-221/222 confers radioresistance in 673 glioblastoma cells through activating Akt independent of PTEN status. Current molecular 674 medicine. 2014 Jan;14(1):185-95. 675 44. Hideshima T, Chauhan D, Podar K, Schlossman RL, Richardson P, Anderson KC. Novel 676 therapies targeting the myeloma cell and its bone marrow microenvironment. Seminars in 677 oncology. 2001 Dec;28(6):607-12. 678 45. Cho JM, Manandhar S, Lee HR, Park HM, Kwak MK. Role of the Nrf2-antioxidant system in 679 cytotoxicity mediated by anticancer cisplatin: implication to cancer cell resistance. Cancer letters. 680 2008 Feb 18;260(1-2):96-108. 681 46. Chen Q, Van der Sluis PC, Boulware D, Hazlehurst LA, Dalton WS. The FA/BRCA pathway is 682 involved in melphalan-induced DNA interstrand cross-link repair and accounts for melphalan 683 resistance in multiple myeloma cells. Blood. 2005 Jul 15;106(2):698-705. 684 Research. on June 27, 2018. © 2015 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on November 2, 2015; DOI: 10.1158/1078-0432.CCR-15-0489

23
47. Zhang LN, Li JY, Xu W. A review of the role of Puma, Noxa and Bim in the tumorigenesis, 685 therapy and drug resistance of chronic lymphocytic leukemia. Cancer gene therapy. 2013 686 Jan;20(1):1-7. 687 48. Yu J, Zhang L. No PUMA, no death: implications for p53-dependent apoptosis. Cancer cell. 688 2003 Oct;4(4):248-9. 689 49. Gottesman MM, Fojo T, Bates SE. Multidrug resistance in cancer: role of ATP-dependent 690 transporters. Nature reviews Cancer. 2002 Jan;2(1):48-58. 691 50. Kuhne A, Tzvetkov MV, Hagos Y, Lage H, Burckhardt G, Brockmoller J. Influx and efflux 692 transport as determinants of melphalan cytotoxicity: Resistance to melphalan in MDR1 693 overexpressing tumor cell lines. Biochemical pharmacology. 2009 Jul 1;78(1):45-53. 694 695 696 697 698 699 700 701 702 703 704 705 706 707 708 709 710 711 712 713 714 715 716 717 718 719 720 721 722 723 724 725 726 727 728 729 730 731 Research. on June 27, 2018. © 2015 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on November 2, 2015; DOI: 10.1158/1078-0432.CCR-15-0489

24
Figure legends 732 733
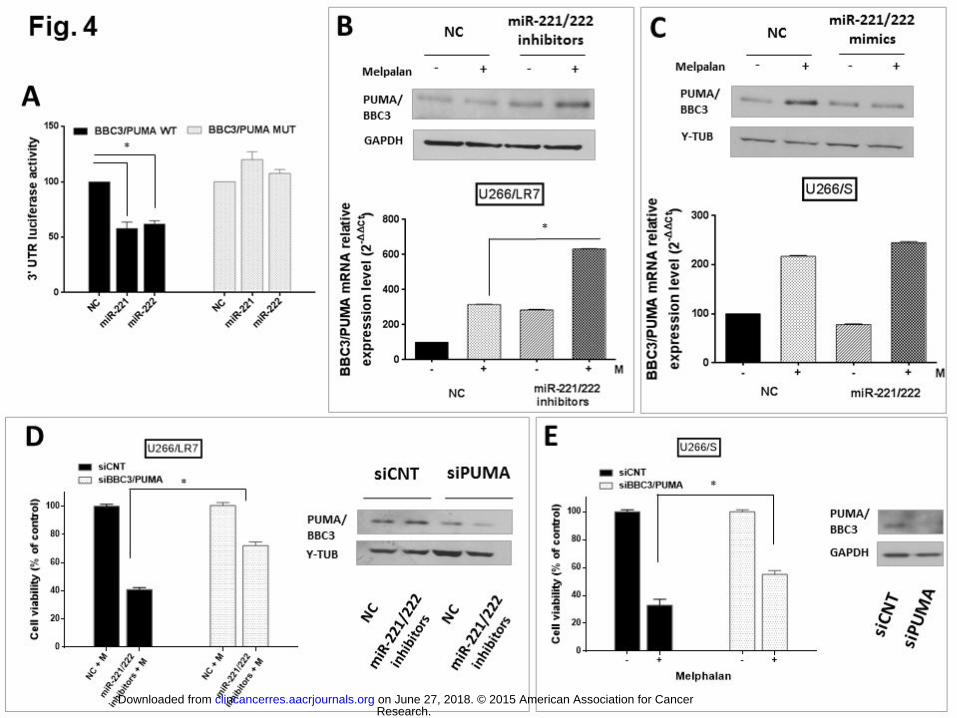
Fig. 1 Correlation of melphalan sensitivity with induction of miR-221 and miR-222 734 expression in MM cell lines. A) U266/LR7, U266/S, RPMI-8226, NCI-H929, AMO1 and 735 AMO1 Abzb cells were treated with increasing melphalan concentrations. Cell viability, 736 relative to the untreated controls, was measured after 24h by CCK-8 assay. Values represent 737 mean ± SD of 3 different experiments. B-C) qRT-PCR analysis of miR-221 (B) and miR-222 738 (C) expression was performed in all the six MM cell lines exposed to different melphalan 739 concentrations (0-10-20 µM). Raw Ct data were normalized to RNU44 houskeeping gene and 740 expressed as Ct values, calculated using the comparative cross threshold (Ct) method. 741 Values represent average ± SD of 3 independent experiments. P values (* p<0.05) were 742 obtained using two-tailed t test. 743 744 Fig. 2 Anti-proliferative and apoptotic effects in MM cells of miR-221/222 inhibitors 745 combined with melphalan. A) CCK-8 cell proliferation assay in U266/LR7, RPMI-8226 and 746 AMO1 Abzb tranfected with 100 nM of miR-221/222 inhibitors or scrambled Control (NC) and 747 then treated with increasing melphalan concentrations. Data shown are the average of three 748 independent experiments, and p values were obtained using two-tailed test. (* p<0.05). B) 749 Annexin V/7-AAD staining of U266/LR7, RPMI-8226 and AMO1 Abzb electroporated cells 750 (miR-221/222 inhibitors or NC) 24h after treatment with different melphalan concentrations. 751 The percentage of Annexin-V positive cells is plotted. Values represent the mean ± SD of 3 752 independent experiments. * p<0.05 C) Western blot analysis of total and cleaved Casp-8, 753 Casp-9, Casp-3 and Cleaved PARP in U266/LR7 cells (top panel) and Casp-3 levels in RPMI-754 8226 cells (bottom panel). Cells lines were transfected with 100 nM of miR-221/222 inhibitors 755 or NC and then exposed to melphalan (100µM for resistant U266/LR7 cells and 5µM for 756 RPMI-8226 cells). Loading control was performed using GAPD or γ-tubulin. D) Fluorescence 757 microscopy after Hoechst 33343 and PI staining of melphalan resistant U266/LR7 cells 758 transfected with miR-221/222 inhibitors or NC, and then exposed to 100 µM of melphalan. 759 DNA dye Hoechst stains the nucleus (blue color emission) of both viable and dead cells, 760 allowing identification of nuclear morphology. Apoptotic nuclei appear fragmented and 761 condensed, with greater signal intensity. Apoptotic and necrotic cells are identified as positive 762 PI staining cells (red color emission) with loss of membrane integrity. 763
Research. on June 27, 2018. © 2015 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on November 2, 2015; DOI: 10.1158/1078-0432.CCR-15-0489

25
764 Fig. 3 Effects of miR-221/222 inhibitors plus melphalan on primary cells and on MM cell 765 lines in the presence of BMSCs. A) CCK-8 cell proliferation assay in primary CD138+ MM 766 patients cells tranfected with 100 nM of miR-221/222 inhibitors or scrambled Control (NC) and 767 then treated with 100 uM of melphalan. Data shown are the average of two independent 768 experiments, and p values were obtained using two-tailed test. (* p<0.05). B) CCK-8 cell 769 proliferation assay in primary hPBMCs cells transfected with 100 nM of miR-221/222 770 inhibitors or scrambled Control (NC) and then treated with increased melphalan 771 concentrations. Data shown are the average of three independent experiments, and p values 772 were obtained using two-tailed test. (* p<0.05). C) 7-AAD staining of U266/LR7 cells 773 transfected with 100 nM of miR-221/222 inhibitors or NC, and then cultured in the presence or 774 absence of GFP+ hBMSCs. 24 hours after co-culture cells were treated with 100 µM of 775 melphalan; 24h later, they were collected and stained for 7-AAD flow cytometry analysis. The 776 percentage of 7-AAD positive cells is represented. Values represent the mean ± SD of 3 777 independent experiments. * p<0.05 778 779 780 Fig. 4 Inhibition of miR-221/222 enhances anti-MM activity of melphalan by modulation 781 of PUMA/BBC3 expression. A) Dual-luciferase assay of U266/LR7 cells co-transfected 782 either with NC or synthetic miR-221 or miR-222 mimics, together with firefly luciferase 783 constructs containing the wild-type or mutant 3’ UTR of PUMA/BBC3 mRNA. The firefly 784 luciferase activity was normalized to renilla luciferase activity. Data are represented as 785 relative luciferase activity of either miR-221 or miR-222 mimics electroporated cells as 786 compared to control. Values represent the mean ± SD of 3 independent experiments. * 787 p<0.05 B) q-RT-PCR (bottom panel) and immunoblot (top panel) of PUMA/BBC3 in resistant 788 U266/LR7 cells after transfection with NC or miR-221/222 inhibitors and treatment with 100 789 µM of melphalan. q-RT-PCR results are shown after normalization with GAPDH and Ct 790 calculation and represent an average ± SD of 3 independent experiments. Protein loading 791 control for immunoblot was performed using GAPDH. C) q-RT-PCR (bottom panel) and 792 immunoblot (top panel) of PUMA/BBC3 in sensitive U266/S cells after transfection with NC or 793 miR-221/222 mimics and treatment with 60 µM of melphalan. q-RT-PCR results are shown 794 after normalization with GAPDH and Ct calculation and represent an average ± SD of 3 795
Research. on June 27, 2018. © 2015 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on November 2, 2015; DOI: 10.1158/1078-0432.CCR-15-0489

26
independent experiments. Protein loading control for immunoblot was performed using γ -796 tubulin. D) CCK-8 cell proliferation assay on resistant U266/LR7 cells (left panel) treated with 797 either NC or miR-221/222 inhibitors co-transfected with siCNT or siBBC3/PUMA and 798 subsequently exposed to 100 µM of melphalan. Effective knockdown was confirmed by 799 immunoblot of PUMA/BBC3 protein and γ -tubulin normalization (right panel). Percentages of 800 growth inhibition are plotted compared to control. Values represent the mean ± SD of 3 801 independent experiments. * p<0.05 E) CCK-8 cell proliferation assay on sensitive U266/S (left 802 panel) transfected with siCNT or siBBC3/PUMA and treated with 60 µM of melphalan. 803 Effective knockdown was confirmed by immunoblot of PUMA/BBC3 protein and GAPDH 804 normalization (right panel). Cell viability is shown as percentage of control. Values represent 805 the mean ± SD of 3 independent experiments. * p<0.05 806 807 Fig. 5 Effects of miR-221/222 inhibitors on the expression of drug influx-efflux 808 transporters levels in resistant U266/LR7 cells. A) Western blot analysis of SLC7A5/LAT1 809 and ABCC1/MRP1 in U266/LR7 cells transfected with 100 nM of miR-221/222 inhibitors or 810 NC and then exposed to 100µM of melphalan. ABBC1/MRP1 was immunoprecipitated from 811 cell lysates (1mg) following incubation with 30 μl of protein A/G-conjugated agarose beads at 812 4°C overnight, and then detected by Western blot. Loading control was performed using γ-813 tubulin. B) Hoechst dye exclusion assay on U266/LR7 cells after treatment with either NC or 814 miR-221/222 inhibitors or NC with Verapamil. The blue circles show the Side population (SP) 815 with low levels of Hoechst fluorescence intensity. Dot plots represent one of three 816 independent experiments. C) Bar column representation of percentage of SP affected after 817 miR-221/222 inhibitors and Verapamil treatment as compared to NC. Values represent the 818 mean ± SD of 3 independent experiments. * p<0.05 819 820 Fig. 6 LNA-i-miR-221 enhances anti-MM activity of melphalan overcoming drug-821 resistance. For the in vivo study, mice xenografted with U266/LR7 cells were randomized in 822 4 groups of treatment. A) Treatments were: i) i.p. LNA-i-miR-221 (25 mg/kg) at day 1-4-8-15-823 22; ii) scrambled control with the same schedule; iii) i.p. melphalan (0.75 mg/kg) on days 2-5-824 9-12-16-19; iiii) melphalan plus LNA-i-miR-221 with the above described schedules. B) mice 825 were treated with: i) ip LNA-i-miR-221 (25 mg/kg) or ii) scramble control or iii) i.p melphalan 826 (0.75 mg/kg) or iiii) melphalan plus LNA-i-miR-221 for 4 consecutive days every 10 days (1-4, 827
Research. on June 27, 2018. © 2015 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on November 2, 2015; DOI: 10.1158/1078-0432.CCR-15-0489

27
15-18). Arrows indicate the days of treatment. Tumors were measured with an electronic 828 caliper every two days, and data are represented as averaged tumor volume ± SD of each 829 group. P values were obtained using two-tailed t test and calculated by comparing LNA-i-miR-830 221 plus melphalan group versus each of the others. The pictures inserted show the in vivo 831 detection of the tumor volume in a representative mouse of each group using IVIS LUMINA II 832 Imaging System. * p < 0.05 ** p < 0.001 B) Western blot analysis of PUMA/BBC3 and ABBC1 833 levels in lysates from a representative retrieved xenograft from each treatment group. 834 ABBC1/MRP1 was immunoprecipitated from cell lysates (1mg) following incubation with 30 μl 835 of protein A/G-conjugated agarose beads at 4°C overnight. Immunoprecipitated 836 ABCC1/MRP1 was detected by Western blot. Y-Tubulin was used as protein loading control. 837 C) H&E (400x) and Ki-67 (400x) immunohistochemical analysis of retrieved xenografted 838 tumors after different treatments. Representative images from each group are shown. 839 840 841 842 843 844 845 846 847 848 849 850 851 852 853 854 855 856 857 858
Research. on June 27, 2018. © 2015 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on November 2, 2015; DOI: 10.1158/1078-0432.CCR-15-0489

Research. on June 27, 2018. © 2015 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on November 2, 2015; DOI: 10.1158/1078-0432.CCR-15-0489

Research. on June 27, 2018. © 2015 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on November 2, 2015; DOI: 10.1158/1078-0432.CCR-15-0489

Research. on June 27, 2018. © 2015 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on November 2, 2015; DOI: 10.1158/1078-0432.CCR-15-0489
Research. on June 27, 2018. © 2015 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on November 2, 2015; DOI: 10.1158/1078-0432.CCR-15-0489

Research. on June 27, 2018. © 2015 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on November 2, 2015; DOI: 10.1158/1078-0432.CCR-15-0489

Research. on June 27, 2018. © 2015 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on November 2, 2015; DOI: 10.1158/1078-0432.CCR-15-0489

Published OnlineFirst November 2, 2015.Clin Cancer Res Annamaria Gulla', Maria T. Di Martino, Maria E. Gallo Cantafio, et al. melphalan-refractory multiple myeloma cellsA 13 mer LNA-i-miR-221 inhibitor restores drug-sensitivity in
Updated version
10.1158/1078-0432.CCR-15-0489doi:
Access the most recent version of this article at:
Material
Supplementary
http://clincancerres.aacrjournals.org/content/suppl/2015/10/31/1078-0432.CCR-15-0489.DC1
Access the most recent supplemental material at:
Manuscript
Authoredited. Author manuscripts have been peer reviewed and accepted for publication but have not yet been
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://clincancerres.aacrjournals.org/content/early/2015/10/31/1078-0432.CCR-15-0489To request permission to re-use all or part of this article, use this link
Research. on June 27, 2018. © 2015 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on November 2, 2015; DOI: 10.1158/1078-0432.CCR-15-0489