mandibular growth retardation as a cause of cleft palat ien mice...
TRANSCRIPT

J. Embryol. exp. Morph. Vol. 38, pp. 227-238, 1977 2 2 7
Printed in Great Britain
Mandibular growthretardation as a cause of cleft palate in micehomozygous for the chondrodysplasia gene
By ROBERT E. SEEGMILLER1 AND F.CLARKE FRASERFrom the Department of Zoology, Brigham Young University, Provo, Utah,
and the Department of Biology, McGill University, Montreal
SUMMARYDefective chondrogenesis in C57BL mice homozygous for the chondrodysplasia gene leads
to deformity of limbs, ribs, trachea, mandible and palate. Since formation of the secondarypalate depends upon coordinated development of several craniofacial structures, the presenceof micrognathia and cleft palate in cho/cho newborn mice suggested a cause-and-effectrelation between these two deformities. To determine whether or not lower jaw shorteningcoincided with the time of palate closure, heads from mutant and control littermates previouslyrated morphologically were examined in median sagittal section. Of six parameters analyzed,growth rates for mutant mandible and anterior vertical dimension were significantly less thanthose of controls from the beginning of control palate closure. Since there is evidence that in-trinsic shelf force is normal, these observations suggested that, during palatogenesis, growthretardation of Meckel's cartilage did not allow forward displacement of the tongue, and thatthe consequent failure to straighten the tongue impaired shelf movement. The data supportthe concept that growth of Meckel's cartilage is necessary for normal palate formation.
INTRODUCTION
Although the precise role played by the lower jaw in development of the sec-ondary palate is not known, evidence is increasing that mandibular growthretardation during embryogenesis can be a contributing factor to cleft palate(Cohlan, 1953; Thalhammer & Heller-Szollosy, 1955; Asling et al. 1960;Fitch, 1961; Latham, 1965; Schwarz & Chaudry, 1968; Fraser, 1969; Burdiet al. 1972; Ohyama, Pruzansky, Heinze & Parris (1974). A causal relation,however, cannot be determined by the mere association at birth of micrognathiaand cleft palate but only by demonstrating that mandibular growth retarda-tion coincides with palate development. Indeed, Shih, Trasler & Fraser (1974)have shown that a teratogen (vitamin A) that causes micrognathia and cleftpalate, causes the micrognathia after palate closure in the control is complete, so
1 Author's address: Department of Zoology, Brigham Young University, Provo, Utah84602, U.S.A.

228 R. E. S E E G M I L L E R A N D F . C . F R A S E R
that the micrognathia does not appear to cause the cleft palate in this case. However, this may not apply to all cases of cleft palate with micrognathia.
Mice with completely penetrant, recessive chondrodysplasia {chojcho) (Seegmiller, Fraser & Sheldon, 1971; Seegmiller, Ferguson & Sheldon, 1972) are born with micrognathia and cleft palate and are therefore a suitable model for study of the role of the lower jaw in palate formation. From sagittal measurements of craniofacial structures associated with palate closure, this study suggests that mandibular growth retardation resulting from defective chondrogenesis is the principal cause of cleft palate in cho mice.
M A T E R I A L S A N D M E T H O D S
Mature heterozygous female mice {choj ±) of the C57/BL/Fr strain were placed overnight with heterozygous males. The presence of a copulation plug the following morning indicated that mating had occurred and that the embryos were approximately 6 h old (day 0/6, Snell, Fekete, Hummel & Law, 1940). Between days 13/20 and 15/4, pregnant uteri were excised and preserved in Bouin's fixative. After fixation for 48-72 h the fetuses were removed from the uterus and rated morphologically (MR) according to Walker & Crain's method (1960). The heads were removed and further processed for examination by light microscopy. Sagittal sections were cut at 10 //m and stained with toluidine blue. Mutants were easily identified by reduced staining intensity of the cartilage matrix (Seegmiller et al. 1971). Midsagittal sections showing thyroglossal duct and nasal septum were photographed at low magnification using a Zeiss photo-microscope with a plan 1 x objective. The overall enlargement from tissue to print was approximately 35 x . Reference points were marked on acetate matte overlays (Fig. 1) and measurements of the head were made in millimeters or degrees by Hart, Smiley & Dixon's method (1972). These measurements permitted correlation of palate closure stage (PS 0-7, Trasler, 1965) with the following parameters: maxillary and mandibular growth, changing angulation of the cranial base and lower jaw, and changing vertical dimension of anterior and posterior regions of the mouth. Palate closure stage was determined by examining serial sagittal sections for the position of the shelves.
Most fetuses were collected over a period of 2 years. Twenty-five litters were examined, from which most morphological stages from MR 0-15 were represented. The number of controls recovered was 90; the number of mutants was 19, which approached the 25 % expected frequency for a recessive gene. Because the MR of the 19 mutants was random, there were stages which consisted of few or no mutants. For statistical purpose, morphological stages were therefore grouped, such that each group was represented by at least two mutant fetuses (Table 1). Mutant and control littermates generally were of the same group. A comparison of the curves before and after grouping suggested that this procedure did not affect the shape of the curves. Differences were tested by the Mann-

Chondrodysplasia, micrognathia and cleft palate in mouse 229
Anteriorvertical
Fig. 1. Median sagittal section of a day-14 (MR 4) control fetal head showingposition of tongue and mandible prior to palate closure. Labels indicate the para-meters measured in millimeters or degrees. Enlarged approximately 35 x .
Whitney U test (Siegel, 1956); a probability of less than 5 % was consideredsignificant.
RESULTS
Palate closure stage vs. morphological age
Palate shelves of control fetuses at MR 0-3 were vertically orientated (PS 0). AtMR 4 and 5 the shelves were either vertical (PS 0) or in the early stages of shelfmovement (PS 1-3). Between MR 6 and 11 the shelves were generally horizontal(PS 4) and many were fusing (PS 5-6). At MR 13-15 closure of the secondarypalate was complete (stage 7). Timing of palate closure was also plotted againstchronological age. Fetuses recovered late on day 13 or early on day 14 generallyhad open palates; those recovered between days 14/6 and 14/12 were at inter-mediate stages of palate closure; and those recovered late on day 14 or early onday 15 had closed palates. Palate closure time for control C57BL mice was similarto that reported by Trasler (1965) and Shih et al. (1974), although the shape of thecurve deviated slightly, perhaps due to our use of tissue sections rather thanwhole heads to determine palate stage.
Mutant fetuses were randomly distributed from MR 0-15. A comparison ofmean MR for control and cho mice within litters suggested that the cho genedid not affect the MR. Mutant heads examined in sagittal section consistentlyshowed palate shelves in the vertical position. Although there was no evidenceof movement to the horizontal position, the shelves were capable of moving

Tab
le 1
. Cra
niof
acia
l di
men
sion
s {t
aken
from
x
35
prin
ts)
for
norm
al a
nd c
ho fe
tuse
s du
ring
pal
atog
enes
is
MR
f
0-2
3-4
5-7
8-11
13-1
5
* C
on
trol
Gro
up
Co
ntr
ol
cho
cho,
% c
ontr
ol
Co
ntr
ol
cho
cho,
%
co
ntr
ol
Co
ntr
ol
cho
cho,
% c
on
tro
l
Co
ntr
ol
cho
cho,
% c
on
tro
l
Co
ntr
ol
cho
cho,
% c
on
tro
l
an
d rh
n m
ean
s diffe
r*
N 19 4 33 2 17 4 7 5 14 4
;d si
enif
n
(mm
)
94-1
±2
-291
-8 +
5-3
97-6
112-
6± 1
-210
7-5
±6
-695
-5
114-
1 +
1-6
113-
5 +
5-6
99-5
123-
0 ±
2-9
120-
6 ±
4-7
98
0
142-
8 ±
2-2
141-
2 ±
6-5
98-9
=an
tlv a
t th
e. 0
05
Cra
nia
l ba
se(d
egre
es)
3C
±S
.E.
151-
0± 1
114
9-0
±2
-798
-7
159-
5±0-
715
6-0
±2
-097
-8
15
9-8
±l-
015
4-5
±5
-396
-6
163-
7 ±
0-8
16
3-6
±l-
899
-9
169-
8 ±
1-7
16
8-5
+1
-299
-2
nro
hah
ilit
v l
evel
Man
dibu
lar
(mm
)
52
-6±
ll51
-8±
2-4
98-5
67
-0±
l-0
58
0±
41
86-6
*
69-5
±1
-261
-2 +
3-3
88-1
*
79
-9±
l-7
68-0
+ 2
-08
51
94-8
±2
-478
-8 +
1-1
83-1
J
det
erm
ined
hv
ti
Ant
erio
r ve
rt.
(mm
)
5-5±
O-8
4-5
±l-
381
-8
16-0
±0-
78-
5±2-
553
-1*
19
-7±
l-0
12-2
±2-
061
-9*
23-6
±0-
614
-0±
2-3
59-3
*
31-1
± 1
-319
-5 ±
0-4
62-7
J
ie
Man
n—W
hitn
e/
Pos
teri
or
vert
.(m
m)
21-8
±0-
42
20
±0-
710
0-9
22-9
±0
-521
-8±
1-5
95-2
24-3
±0
-522
-2 ±
0-6
91-4
22-4
±0
-62
1-l
±0
-994
-2
26-6
±1
-529
-8 ±
0-8
11
20
v TT
tes
t
Pos
teri
or a
ngle
(deg
rees
)
38-1
±0
-637
-5±
2-4
98-4
38-0
±0-
93
60
±2
094
-7
40-3
±0
-841
-5±
7-0
1030
37-6
±0
-732
-6 ±
1-7
86-7
*
38
-6±
l-5
38-0
+ 3
-298
-4
w in W O >—i r m & p m 1*
gy
py
,
y
yt
Tw
o t
o f
our
morp
holo
gic
al g
roups
wer
e co
mbi
ned i
n o
rder
to
inc
reas
e th
e s
ampl
e si
ze.
Tot
al n
umbe
r of
contr
ol
fetu
ses
equa
ls 9
0, m
utan
tseq
ual
s 19.
% C
on
tro
l an
d c
ho
mea
ns
diff
ered
sig
nifi
cant
ly a
t th
e 0
01 p
robab
ilit
y l
evel
.
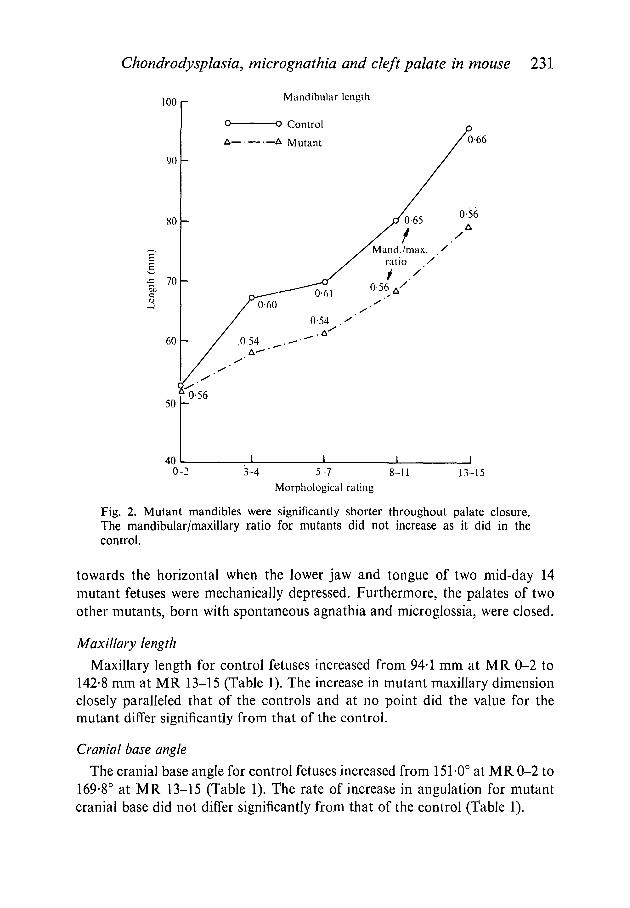
Chondrodysplasia, micrognathia and cleft palate in mouse 231
100
90
80
- 70
Mandibular length
Control
A A Mutant
0-56A
Mand./max. .ratio /t /
o-56 y
0-66
0-2 3-4 5-7
Morphological rating
8-11 13-15
Fig. 2. Mutant mandibles were significantly shorter throughout palate closure.The mandibular/maxillary ratio for mutants did not increase as it did in thecontrol.
towards the horizontal when the lower jaw and tongue of two mid-day 14mutant fetuses were mechanically depressed. Furthermore, the palates of twoother mutants, born with spontaneous agnathia and microglossia, were closed.
Maxillary length
Maxillary length for control fetuses increased from 94-1 mm at MR 0-2 to142-8 mm at MR 13-15 (Table 1). The increase in mutant maxillary dimensionclosely paralleled that of the controls and at no point did the value for themutant differ significantly from that of the control.
Cranial base angle
The cranial base angle for control fetuses increased from 151-0° at MR 0-2 to169-8° at MR 13-15 (Table 1). The rate of increase in angulation for mutantcranial base did not differ significantly from that of the control (Table 1).

232 R. E. SEEGMILLER AND F. C. FRASER
Fig. 3. (A) Day-15 (MR 15) control midsagittal section showing horizontal shelf,relatively straight tongue, and a line extending through hyoid cartilage, Meckel'scartilage and apex of lower jaw. Force from primary palate is directed through theanterior lower jaw (arrow). Distance between hyoid and Meckel's cartilages isindicated (50 mm is representative of MR 15 fetuses). (B) Day-15 (MR 15) mutantmidsagittal section of cleft palate fetus showing curvature of tongue, and shorteningof mandible as measured from hyoid to Meckel's cartilage (35 mm, representative). Aline extended through hyoid and Meckel's cartilages traverses above the apex of thelower jaw, through the tongue and maxilla. The line of force (arrow) from the primarypalate is directed through the tip of the tongue but anterior to the lower jaw.
Mandible length
Mandibular dimension for control fetuses increased from 52-6 mm at MR 0-2to 94-8 mm at MR 13-15 (Table 1, Fig. 2), an increase of approximately 1-8times. Mutant mandibular dimension increased from 51-8 mm at MR 0-2 to

Chondrodysplasia, micrognathia and cleft palate in mouse 233
78-8 mm at MR 13-15 (Table 1), an increase of approximately 1-5 times. Man-dibular shortening was significant and present as early as MR 3-4 (86-6 % ofcontrol, Table 1), during the commencement of palate closure. At MR 13-15,i.e. after palate closure, mutant mandibular dimension was 83-1 % of control.Thus throughout palate closure, mutant lower jaws were consistently and signifi-cantly shorter than those of the controls (Fig. 2). However, the mandible anteriorto Meckel's cartilage, which is comprised of soft tissue, was not reduced inlength, and the distance between Meckel's cartilage and the hyoid cartilage wasonly 70 % of the control measurement (Fig. 3 A, B). This suggests that the reduc-tion in mandibular length occurs primarily through retardation in growth ofMeckel's cartilage.
Mandibular growth retardation was further demonstrated by comparingmutant and control for mandibular/maxillary ratio (Fig. 2). Whereas the ratiofor control increased from 0-56 at MR 0-2 to 0-66 at MR 13-15, the ratio forcho fetuses was approximately 0-56 throughout palate closure.
Anterior vertical dimensionThis parameter represents the degree to which the mandible rotates as the
mouth opens. For control fetuses this measurement increased from 5-5 mm atMR 0-2 to 31-1 mm at MR 13-15 (Table 1). Mutant anterior vertical dimen-sion increased from 4-5 mm at MR 0-2 to 19-5 mm at MR 13-15. The differencebetween mutant and control at MR 0-2 was not statistically significant whereasthe difference thereafter was (Table 1). The greatest difference was observed atMR 3-4 when mutant vertical dimension was 53-1 % of control.
Posterior vertical dimensionThis parameter for control fetuses ranged from 21 -8 mm at MR 0-2 to 26-6 mm
at MR 13-15 (Table 1). Mutant posterior vertical dimension increased similarlyfrom 22-0 mm at MR 0 to 29-8 mm at MR 15. At all stages the mutant did notdiffer significantly from control for posterior vertical dimension.
Posterior angle
This dimension for control fetuses was 38-1 at MR 0-2 and 38-6 at MR 13-15(Table 1) and for cho fetuses it was 37-5 at MR 0-2 and 38-0 at MR 13-15. Thedeviation from control was significant only at MR 8-11.
DISCUSSION
During the time of active chondrogenesis in normal C57BL fetuses the palatineshelves reorientate themselves from vertical to horizontal, and fuse above thetongue to form the secondary palate. The mechanism for shelf movement is notcompletely understood. It may involve, among other things, extension of thecartilaginous cranial base (Harris, 1964; Verrusio, 1970; Long, Larsson &Lohmander, 1973) and contraction of actin-like proteins within the shelves

234 R. E. SEEGMILLER AND F. C. FRASER
(Lessard, Wee & Zimmerman, 1974; Babiarz, Allenspach & Zimmerman, 1975;Wee, Wolfson & Zimmerman, 1976).
Evidence from the present study suggested that shelf force in cho fetuses isnormal. The mutant's shelves before closure appeared histologically andmorphologically normal, and upon surgical removal of the tongue and lowerjaw they rotated horizontally. They were also fused in cho fetuses bornwith spontaneous agnathia and microglossia. These observations suggestedthat the underlying cause of cleft palate in cho fetuses was not related to intrin-sic shelf force,
Before palate closure the tongue lies between the shelves and behind theprimary palate. Afterwards the tongue lies below the shelves, which have becomehorizontal, and below the primary palate. Although the evidence is controver-sial (see Larsson (1974) for review), tongue displacement seems necessary forshelf rotation and probably involves tongue-primary-palate contact (Fraser,1968) and possibly mouth opening reflexes (Walker, 1969; Walker & Ross, 1972).
The lower jaw has been associated with tongue displacement because of itsattachment to the tongue and because both undergo rapid growth during thetime of shelf rotation (Sicher, 1915; Zeiler, Weinstein & Gibson, 1964; Hart,Smiley & Dixon, 1969; Burdi & Silvey, 1969; Wragg, Klein, Steinvorth &Warpeha, 1970; Diewert, 1974). In the present study the lower jaw of controlfetuses nearly doubled its length between MR 0 and 15 (Fig. 2), stages whichimmediately precede and follow palate closure.
Further support for the developmental relation between jaw growth, tonguedisplacement and palate closure comes from several examples of cleft palate thatare associated with mandibular shortening or ankylosis (Sisken, Gluecksohn-Waelsch, 1959; Deuschel & Kalter, 1962; Latham, 1965; Nanda, 1970; Ohyamaet al. 1974). Burdi et al. (1972) observed that ' . . . a majority of syndromes inwhich micrognathia is a feature also include cleft palate as a feature, whereasmany syndromes involving cleft palate do not include micrognathia. Thisprovides indirect evidence that micrognathia of Pierre Robin [and of cho]syndrome is a contributory cause to the cleft palate'.
The cho mandibular dimension was 98-5 % of control prior to shelf rotation(MR 0-2), 86-6 % during the initial stages of palate closure (MR 3-4) and 83-1 %after closure (MR 13-15, Table 1). Since the average distance between hyoidand Meckel's cartilage for cho fetuses at MR 13-15 was 70 % of control (Fig. 3),and since the mandibular skeleton during palate closure is comprised predom-inantly of well-developed cartilage (extensive intramembranous bone formationhas yet to occur), defective chondrogenesis alone apparently accounted formandibular growth retardation.
Growth retardation was further demonstrated by comparing mutant andcontrol fetuses for mandibular maxillary ratio. The ratio for control increasedfrom 0-56 at 0-2 to 0-66 at MR 13-15 (Fig. 2). A similar increase, to the pointof transient prognathism, was reported during palate closure in lower animals

Chondrodysplasia, micrognathia and cleft palate in mouse 235
and human beings (Polzl, 1905; Sicher, 1915; Zeiler, Weinstein & Gibson,1964; Wragg et al 1970; and Die wart, 1974), and was interpreted as acceleratedgrowth of the lower jaw. The ratio for cho fetuses at MR 0-2 was also 0-56 butno increase during palate closure was observed. Although the mutant's man-dibular dimension showed an absolute increase from 51-8 mm at MR 0-2 to78-8 mm at MR 13-15 (Table 1), the absence of an increase relative to maxillarydimension can be interpreted as no acceleration of growth during the criticaltime for palate closure.
Electron microscopy provided evidence that the cho gene affects only chondro-genic tissues (Seegmiller et al. 1971, 1972). The presence of large collagen fibrilsin the matrix of Meckel's cartilage (unpublished) confirmed that cartilage of thelower jaw was also defective. This supports the hypothesis that defective chon-drogenesis of Meckel's cartilage causes mandibular growth retardation. Theabsence of statistically significant deviations from control for cranial base angleand maxillary dimension (Table 1) suggested that mandibular growth retardationalone, acting through the tongue, causes cleft palate in cho fetuses.
The diminished anterior vertical dimension of mutant fetuses (Table 1) presentat MR 3-4, and the (underlying) distortion of the tongue and mandible duringpalate closure suggest a mechanism by which the mandibular growth deficiencycauses cleft palate. A line drawn through the control lower jaw at the beginningof palate closure extends from the posterior cranial base through hyoid andMeckel's cartilage and the apex of the lower jaw (Fig. 1). The tongue is onlyslightly curved and positioned posterior to the primary palate. After palateclosure the tongue is relatively straight and lies below the primary and secon-dary palate. Despite the morphological changes in the tongue and secondarypalate, a line traversing the lower jaw passed through both cartilages andapex (Fig. 3 A), as it did before closure. In the mutant, prior to palate closure,this line extended similarly through all structures of the lower jaw. However,after the normal closure time it did not extend through the apex of the lower jawbut above it, traversing the tongue and primary palate (Fig. 3B). The tongueremained curved and between the vertical palate shelves. It is suggested that, inthe mutant when the tongue is not carried forward by the extension of Meckel'scartilage, and does not come to lie under the primary palate, it is pressed upagainst the anterior part of the cranial base and adopts its curvature. Its tip there-fore points downward instead of anteriorly, and depresses the anterior part ofthe lower jaw which, anterior to Meckel's cartilage, is still only soft tissue. Thiswould result in the body of the tongue being arched upwards into the spacebetween the shelves, which would increase the obstruction of the tongue to shelfmovement so much that they would not reach the horizontal position in time tofuse.
The above suggests that accelerated growth of Meckel's cartilage plays acritical role in palate formation; the anterior tip of the cartilage, moving anter-iorly, may bring the tongue to lie under the primary palate. This would permit

236 R. E. SEEGMILLER AND F. C. FRASER
the tongue to be straightened and cause its tip to lie between the primary palateand tip and the lower jaw. Thus, the entire lower jaw would be depressed ina hinge-like fashion, thereby increasing the anterior vertical dimension, andcreating space for the shelves to move into (Eraser, 1969).
The concept that Meeker s cartilage enhances growth of the lower jaw andthereby plays a special role in palate development is further supported by theuniqueness of mandibular ossification. Meckel's cartilage forms approximately1 day before palate closure (Bhaskar, 1953). Not until after palate closure andformation of Meckel's cartilage does significant ossification occur, and then itoccurs lateral to the cartilage. The mandible is not an endochondrial bone andtherefore does not require Meckel's cartilage for its formation, although Meckel'scartilage may contribute to mandibular shaping (Hamilton, Boyd & Mossman,1964). This suggests that Meckel's cartilage functions during the critical stagesof palate closure to provide a point of attachment to the tongue, and to lengthenand provide rigidity to the lower jaw.
Our current understanding of the cho gene is that through defective collagen-proteoglycan interaction (Stephens & Seegmiller, 1976) it expresses itself only inhyaline cartilage causing interstitial growth retardation and loss of matrixrigidity. Growth of the lower jaw is retarded primarily because of the presence ofMeckel's cartilage. This study supports the concept that mandibular growthretardation during palate closure prevents tongue displacement necessary forshelf movement and is therefore the primary cause of cleft palate in the chomutant.
This research was supported by a Basil O'Connor Starter Research Grant from the NationalFoundation - March of Dimes and by the Medical Research Council of Canada. Technicalassistance of Vaunda Barrus and Devon Hiatt is gratefully acknowledged.
REFERENCES
ASLING, C. W., NELSON, M. M., DOUGHERTY, H. L., WRIGHT, H. V. & EVANS, H. M. (1960).The development of cleft palate resulting from maternal pteroylglutamic (folic) aciddeficiency during the latter half of gestation in rats. Surgery ofGynec. Obstet. I l l , 19-28.
BABIARZ, B., ALLENSPACH, A. & ZIMMERMAN, E. (1975). Ultrastructural evidence of contrac-tile systems in mouse palates prior to rotation. Devi Biol. 47, 32-44.
BHASKAR, S. N. (1953). Growth pattern of the rat mandible from 13 days insemination age to30 days after birth. Am. J. Anat. 92, 1-53.
BURDI, A. R. & SILVEY, R. G. (1969). Sexual differences in closure of the human palatalshelves. Cleft Pal. 6, 1-7.
BURDI, A., FEINGOLD, M., LARSSON, K. S., LECK, I., ZIMMERMAN, E. F. & FRASER, F. C.(1972). Etiology and pathogenesis of congenital cleft lip and cleft palate, an NJDR state ofthe art report. Teratology 6, 255-268.
COHLAN, S. (1953). Excessive intake of vitamin A as a cause of congenital anomalies in the rat.Science, N. Y. 117, 535-536.
DEUSCHLE, F. M. & KALTER, H. (1962). Observations on the mandible in association withdefects of the lip and palate. /. dent. Res. 41, 1085-1095.
DIEWERT, V. M. (1974). A cephalometric study of orofacial structures during secondarypalate closure in the rat. Archs Oral Biol. 19, 303-315.

Chondrodysplasia, micrognathia and cleft palate in mouse 237FITCH, N. (1961). Development of cleft palate in mice homozygous for the shorthead muta-
tion. /. Morph. 109, 151-157.FRASER, F. C. (1968). Workshop on embryology of cleft lip and cleft palate. Teratology 1,
353-358.FRASER, F. C. (1969). Gene-environment interactions in the production of cleft palate. In
Methods for Teratological Studies in Experimental Animals and Man. (ed. H. Nishimura &J. R. Miller), pp. 34-49. Tokyo: Shoin.
HAMILTON, W. J., BOYD, J. D. & MOSSMAN, H. W. (1964). Human Embryology, p. 389.Cambridge: W. Heffer and Sons Ltd.
HARRIS, J. W. S. (1964). Oligohydramnios and cortisone-induced cleft palate. Nature, Lond.203, 533-534.
HART, J. S., SMILEY, G. R. & DIXON, A. D. (1969). Sagittal growth of the craniofacial com-plex in normal embryonic mice. Archs Oral Biol. 14, 995-997.
HART, J., SMILEY, G. & DIXON, A. (1972). Sagittal growth trends of the craniofacial complexduring formation of the secondary palate in mice. Teratology 6, 43-50.
LARSSON, K. S. (1974). Mechanism of cleft palate formation. In Congenital Defects, (ed. D. T.Janerich, R. G. Skalko & I. H. Porter), pp. 255-273. New York: Academic Press.
LATHAM, R. A. (1966). The pathogenesis of cleft palate associated with the Pierre Robinsyndrome. An analysis of a seventeen-week human fetus. Br. J. Plastic Surg. Vol. 19:205-214.
LESSARD, J. L., WEE, E. L. & ZIMMERMAN, E. F. (1974). Presence of contractile proteins inmouse fetal palate prior to shelf elevation. Teratology 9, 113-126.
LONG, S. Y., LARSSON, K. S. & LOHMANDER, S. (1973). Cell proliferation in the cranial base ofA/J mice with 6-AN-induced cleft palate. Teratology 8, 127-138.
NANDA, R. (1970). The role of sulfated mucopolysaccharides in cleft palate production.Teratology 3, 237-244.
OHYAMA, K., PRUZANSKY, S., HEINZE, W. &PARRIS, P. (1974). Discriminate function analysisin the diagnosis of Pierre Robin syndrome. Teratology 9, A-32.
POLZL, A. (1905). Zur Entwicklungsgechichte des menchlichen Gaumens. Anat. Heftell, 245-281.
SCHWARTZ, D. & CHAUDRY, A. (1968). Planimetric studies of mandibles in A/Jax mice bornwith cortisone-induced cleft palates. /. dent. Res. 47, 725-732.
SEEGMILLER, R., FRASER, F. C. & SHELDON, H. (1971). A new chondrodystrophic mutant inmice: electron microscopy of normal and abnormal chondrogenesis. / . Cell Biol. 48, 580-593.
SEEGMILLER, R., FERGUSON, C. C. & SHELDON, H. (1972). Studies on cartilage. VI. A genet-ically determined defect in tracheal cartilage. /. ultrastruct. Res. 98, 288-301.
SHIH, L., TRASLER, D. & FRASER, F. (1974). Relation of mandible growth to palate closure inmice. Teratology 9, 191-202.
SICHER, H. (1915). Die Entwickelung des sekundaren Gaumens beim Menschen. Anat. Am.47, 513-523, 545-562.
SIEGEL, S. (1956). Non-parametric statistics for the Behavioral Sciences, pp. 117-277. NewYork: McGraw Hill.
SISKEN, B. & GLUECKSOHN-WAELSCH, S. (1959). A developmental study of the mutationphocomelia in the mouse. /. exp Zool. 142, 623-641.
SNELL, G., FEKETE, E., HUMMEL, K. & LAW, L. (1940). The relation of mating, ovulation andthe estrus smear in the house mouse to time of day. Anat Rec. 76, 39-54.
STEPHENS, T. D. & SEEGMILLER, R. E. (1976). Normal production of cartilage glycosamino-glycon in mice homozygous for the chondrodysplasia gene. Teratology 19, 317-326.
THALHAMMER, O. & HELLER-SZOLLOSY, E. (1955). Ekogene Bildungsfehler (Miszbildungen)durch lostinjektion bei der graviden Maus. Z. Kinderhailk 16, 351-365.
TRASLER, D. G. (1965). Strain differences in susceptibility to teratogenesis. Survey of spon-taneously occurring malformations in mice. In Teratology: Principles and Techniques (ed.J. G. Wilson & J. Warkany), pp. 38-55. University of Chicago. .
VERRUSIO, A. (1970). A mechanism for closure of the secondary palate. Teratology 3, 17-20.WALKER, B. E. (1969). Correlation of embryonic movement with palate closure in mice.
Teratology 2, 191-198.l6 EMB 38

238 R. E. SEEGMILLER AND F. C. FRASER
WALKER, B. E. & CRAIN, B. (1960). Effects of hypervitamosis A on palate development in twostrains of mice. Am. J. Anat. 107, 49-8.
WALKER, B. E. & Ross, L. M. (1972). Observation of palatine shelves in living rabbit embryos.Teratology 5, 97-102.
WEE, E. L., WOLFSON, L. G. & ZIMMERMAN, E. F. (1976). Palate shelf movement in mouseembryo culture: evidence for skeletal and smooth muscle contractility. Devi Biol. 48, 91-103.
WRAGG, L. E., KLEIN, M., STEINVORTH, G. & WARPEHA, R. (1970). Facial growth accommo-dating secondary palate closure in rat and man. Archs Oral Biol. 15, 705-719.
2EILER, K. B., WEINSTEIN, S. & GIBSON, R. D. (1964). A study of the morphology and thetime of closure of the palate in the albino rat. Archs Oral Biol. 9, 545-665.
(Received 14 September 1976)