measurements of the concentration of active sites in preparations of yeast alcohol dehydrogenase
TRANSCRIPT

Eur. J. Biocheni. 42,31-36 (1974)
Measurements of the Concentration of Active Sites in Preparations of Yeast Alcohol Dehydrogenase
Mark DICEINSON Department of Biochemistry, University of Hull
(Received August 14, 1973)
Studies on the inactivation of yeast alcohol dehydrogenase by iodoacetate indicate that under certain conditions reaction of two equivalents of reagent produces a 90--95O/, loss of activity. Such observations suggest two essential thiol groups per molecule. Measurements of NADH binding using three independent methods indicate a similar number of coenzyme binding sites.
Studies on the inactivation of yeast alcohol dehydrogenase with iodoacetamide have indicated that reaction of 4 groups/molecule of enzyme abol- ished all enzymic activity [1,-31. Since the enzyme is comprised of four very similar, if not identical, subunits [4,5], the experiments with iodoacetamide lead to the expectation that the enzyme contains four active sites and a t least four coenzyme binding sites. Early studies of NADH binding by the enzyme [6] using the ultracentrifugal separation technique yielded estimates of around 3.6 NADH binding sites/molecule. However, more recent studies [7] using methods which can be carried out much more rapidly and which are thus less subject to errors arising from the blank reaction of NADH with the enzyme, have given estimates in the range 2.1-2.8 coenzyme binding sites/molecule of molecular weight 141 000.
A study of the inactivation of yeast alcohol dehydrogenase by iodoacetate showed that reaction of 0.95 equivalent of reagent produced a loss of some 40-450/, of the initial enzymic activity. This result indicated only about 2 essential thiol groups/ molecule and implied that only 2 of the 4 thiol groups which react readily with iodoacetamide control activity. The experiment with iodoacetate indicated a close correlation between the number of essential thiol residues and the lowest estimates [7] of coenzyme binding capacity. The implications of this observa- tion are very important in any attempt to understand the relationship between structure and activity of the enzyme. Accordingly attempts have been made to study the reaction of alcohol dehydrogenase with iodoacetate in detail. In addition the coenzyme binding characteristics of the enzyme have been re- examined.
Enzyme. Yeast alcohol dehydrogenase (EC 1.1.1.1).
MATERIALS AND MXTHODS Reagents
NADH (fluorimetric grade) was purchased from Boehringer Corp (London) Ltd. (London, W.5, U.K.). Radioactive NADH was prepared by the method of Dalziel [8] from 50 mg NADf containing 0.25 mCi [4-3H]nicotinamide adenine dinucleotide (Radio- chemical Centre, Amersham, Bucks, U.K.). Analysis of the product showed that the material was pure by the normal criteria. However, chromatography on DEAE-cellulose a t pH 7.0 indicated that 7O/, of the radioactivity resided in material other than NADH. This presumably came from some radioactive impur- ity in the radioactive NAD+ which was carried through the preparation procedure. Iodoacetamide, iodo- acetic acid and acetamide were obtained from Bri- tish Drug Houses Ltd (Poole, Dorset, U.K.). Iodo- acetamide was recrystallised from 50 (v/v) ethanol and iodoacetic acid was recrystallised from carbon tetrachloride. Iod~[l-~~C]acetsmide and iodo[2-14C]- acetic acid were obtained from The Radiochemical Centre (Amersham, Bucks, U.K.). Alcohol dehydro- genase was prepared from baker's yeast as previously described [3,7].
All solutions were prepared with glass distilled water. For experiments involving zinc analyses the water was further treated by passage through a mixed-bed ion-exchange resin. All buffer solutions except those used in zinc analyses contained 0.3 mM EDTA.
Enzyme Activity and Protein concentration
Both of these quantities were determined an previously described. A specific activity of 360 units/ mg in the standard assay corresponds to a molecular- activity of 34000 a t 20 "C in the assay of Wallenfels
Eur. J. Biochem. 41 (1974)

32 Active Sites of Alcohol Dehydrogenase
and Sund [9] based on a molecular weight of 141 000. Enzyme of this activity contained 33 thiol groups/ molecule when assayed with p-chloromercuribenzoate a t pH 7.0 in M/15 phosphate buffer using the condi- tions given by Wallenfels and Sund [9]. The enzyme used here compares very favourably with the best preparations of Buhner and Sund [lo] and Wallen- fels and Sund 191 which had molecular activities of 32 000 and 27 000, respectively.
Protein molar concentrations are calculated throughout using a molecular weight of 141 000 [lo]. In previous work from this laboratory [3,7] a molec- ular weight of 149000 was used derived from sedi- mentation equilibrium experiments a t 10 "C but using a value of B = 0.743 ml/g determined at 25 "C. If a corrected value of 8 is used assuming a variation of 0.0008 ml x g-l x deg-l which seems a reasonable figure for a globular protein [I l l , an estimate for the molecular weight of 142000 is obtained.
Inactivation Experiments with I~do[Z-~*C]acetate and lodo[I - 14 Clacetamide
Enzyme at a concentration of 10 mg/ml in phos- phate buffer I = 0.1, pH 7.0 was incubated a t 0 "C with a 16-fold molar excess of iodo[2-14C]acetate or i~do[l-~~C]acetamide. One experiment with iodo- acetate also contained 460 pM NADH. 0.8-ml sam- ples were withdrawn a t appropriate times after the incubation was started and the reaction stopped by filtration through a column (21 x 1.5 cm) of G-25 Sephadex. The column was thermostatted a t 0 "C and was equilibrated with phosphate buffer, pH 7.0 I = 0.1. On elution from the column, samples were assayed for enzymic activity and radioactivity. Radioactivity measurements were made with liquid scintillation counting. Filtration through the columns took approximately 10 min, the reaction probably being stopped in half of that time. There was thus only a small uncertainty in the total reaction times.
NADH Binding Studies The coenzyme binding capacity of the enzyme
was studied by the gel filtration method of Hummel and Dreyer 1121 and the steady-state dialysis techni- que of Colowick and Womack [13]. Gel filtration was carried out much as before [3,7] using 90 nmol enzyme with NADH concentrations in the range 9-360 pM. In the present experiments, however, NADH concentrations in the enzyme- bearing frac- tions were measured following the change in absorb- ance a t 340nm following the addition of an excess of acetaldehyde. The steady-state dialysis experi- ments were carried out in an apparatus of similar dimensions to that described by Colowick and Wo- mack [13]. Enzyme a t a concentration of 8.8 mg/ml in phosphate buffer pH 7.0, I = 0.1 a t 22 "C was
placed in the upper chamber and aliquots of first radioactive and then unlabelled NADH were sub- sequently added. Buffer was pumped through the lower chamber a t a constant rate of 1.7 ml/min. The radioactivity of samples was determined by liquid scintillation counting. The enzyme was com- pletely stable throughout the experiment. As has been noted earlier some 7 O / , of the radioactivity of the radioactive NADH preparation was in material other than NADH. Further, control experiments indicated that on mixing enzyme with the radioactive NADH, some 7O/, of the coenzyme was lost in the first 10 min in a blank reaction [7], presumably by oxidation to NAD+. Further loss was negligible since the reaction was much slower after the initial period and the NADH concentration was thereafter progressively increased in the course of the experi- ment. Under conditions used here the dissociation constant of NAD+ from the enzyme is some 30 times greater than that for NADH+ [3]. In the calculation of results it has been assumed that the binding of NAD+ to the enzyme was negligible and that the other 70/, radiochemical impurity did not bind either. Thus 150/, of the total radioactivity was assumed to be freely diffusible a t all times. The correction led to an increase of estimated bound coenzyme concentra- tions, which resulted in an increase in the total estimated NADH binding capacity of the enzyme by about 10°/,.
RESULTS AND DISCUSSION The fall in specific activity of solutions of yeast
alcohol dehydrogenase (67 pM) after incubation for 4 h a t 25 "C and pH 7, with various concentrations of excess of iodoacetate is shown in Fig. 1. The reac- tions were complete in 4 h. Enzyme incubated under similar conditions, but in the absence of iodoacetate, was stable for the whole of the incubation period. It is apparent from Fig.1 that 0.95 equivalent of iodoacetate produced a 40°/, loss of enzymic activity whilst 1.9 equivalents produced a 75O/, loss. It is known that reaction of yeast alcohol dehydrogenase with iodoacetate results in the carboxymethylation of certain specific cysteine residues [5 ] . The initial slope of Fig.1 suggests a maximum of 2.3-2.4 cysteine residues per 141 000 g protein which control enzymic activity.
The conclusion arising from the results in Fig.1 is surprising. Incubation of similar preparations of enzyme with iodoacetamide under otherwise identical conditions to those obtained in the experiments of Fig.1 were consistent with a situation where in- activation of the enzyme paralleled the alkylation of 4 equivalent cysteine residues [3]. This finding was in good agreement with earlier work [1,2] using iodo- acetamide and agreed with studies [5] showing that the enzyme contains 4 cysteine residues which react
Eur. J. Biochem. 41 (1974)
M. Dickinson
400
- c aJ .- - 2 300 a
F . 3 - 5 200 ._ >
" m u c
.- c
.-
._ y 100 a ul
0
1 0
2 4 6 8 Iodoacetate /enzyme (rnol/ rnol)
Fig. 1. Inactivation of yeast alcohol dehydrogenase (67 p M ) after incubation for 4 h at 25 " C in phosphate buffer p H 7.0, I = 0.1 with stated concentrations of excess of iodoacetate
readily with iodoacetate. The results of Fig.1 can only be accommodated within the framework of existing data if of the four reactive cysteine residues, the two controlling activity are slightly more reac- tive towards iodoacetate than the alternate pair which do not affect activity. All must be equally reactive towards iodoacetamide.
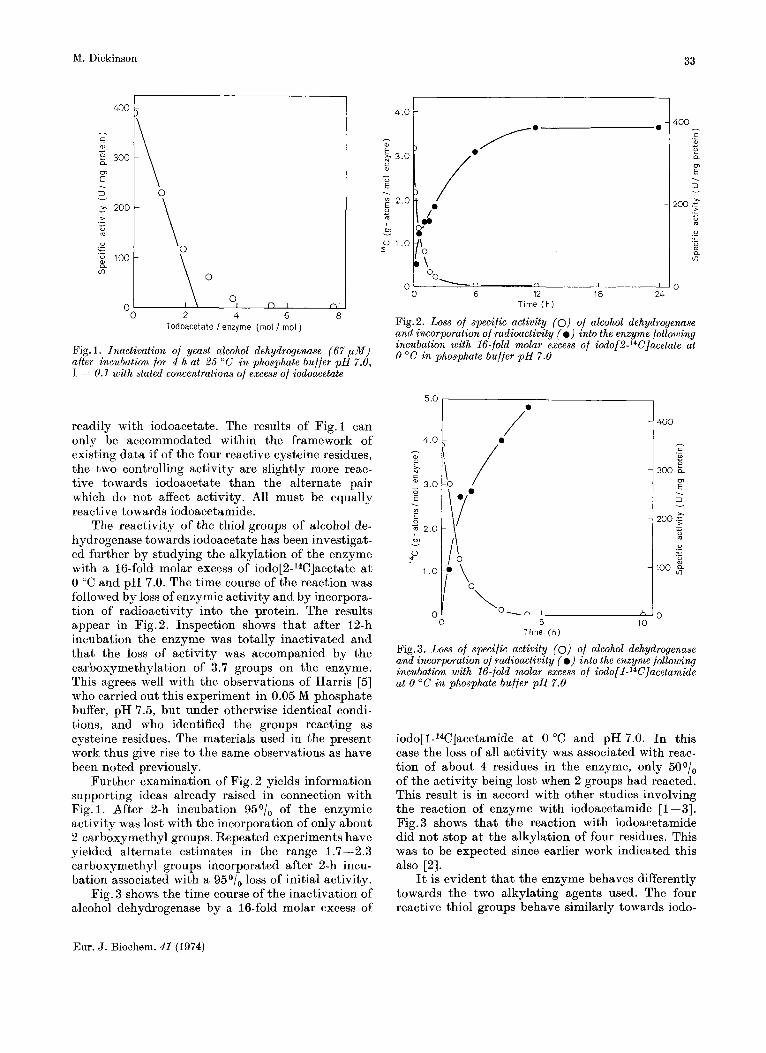
The reactivity of the thiol groups of alcohol de- hydrogenase towards iodoacetate has been investigat- ed further by studying the alkylation of the enzyme with a 16-fold molar excess of iodo[Z-W]acetate a t 0 "C and pH 7.0. The time course of the reaction was followed by loss of enzymic activity and by incorpora- tion of radioactivity into the protein. The results appear in Fig.2. Inspection shows that after 12-h incubation the enzyme was totally inactivated and that the loss of activity was accompanied by the carboxymethylation of 3.7 groups on the enzyme. This agrees well with the observations of Harris [5] who carried out this experiment in 0.05 M phosphate buffer, pH 7.5 , but under otherwise identical condi- tions, and who identified the groups reacting as cysteine residues. The materials used in the present work thus give rise to the same observations as have been noted previously.
Further examination of Fig. 2 yields information supporting ideas already raised in connection with Fig.1. After 2-h incubation 95O/, of the enzymic activity was lost with the incorporation of only about 2 carboxymethyl groups. Repeated experiments have yielded alternate estimates in the range 1.7-2.3 carboxymethyl groups incorporated after 2-h incu- bation associated with a 950/, loss of initial activity.
Fig. 3 shows the time course of the inactivation of alcohol dehydrogenase by a 16-fold molar excess of
4.0 3-
"0 - I I
0 6 12 18 26 Time ( h )
33
- 400 c a, ._ I e E
3
a m . -
200 p ._ - " m U
U
a
._ + .-
u)
3
Fig.2. Loss of specific activity (0) of alcohol dehydrogenase and incorporation of radioactivity (e) into the enzyme following incubation with 16-fold molar excess of iod0(2-~~C]acetate at 0 "C in phosphate buffer p H 7.0
Time ( h )
Fig.3. Loss of specific activity (0) of alcohol dehydrogenase and incorporation of radioactivity ( 0 ) into the enzyme following incubation with 16-fold molar excess of i~do[l- '~C]acetamide at 0 "C in phosphate buffer pH 7.0
i~do[l-~~C]acetamide a t 0 "C and pH 7.0. In this case the loss of all activity was associated with reac- tion of about 4 residues in the enzyme, only 50°/, of the activity being lost when 2 groups had reacted. This result is in accord with other studies involving the reaction of enzyme with iodoacetamide [l-31. Fig. 3 shows that the reaction with iodoacetamide did not stop a t the alkylation of four residues. This was to be expected since earlier work indicated this also [2].
It is evident that the enzyme behaves differently towards the two alkylating agents used. The four reactive thiol groups behave similarly towards iodo-
Eur. J. Biochem. 41 (1974)

34 Active Sites of Alcohol Dehydrogenase
3 .o
2 .o
r
1 .o
0 0 0.05 0.10
F l [ N A D H ] (pM- ' )
Fig.4. Binding of N A D H by alcohol dehydrogenase at pH 7.0 studied by gel filtration at 25 "C ( 0 ) and by the steady-state dialysis method at 22 "C (0). p = molecules NADH bound/ molecule enzyme
acetamide while iodoacetate distinguishes them into two classes, the more reactive ones being those affecting enzymic activity. Whitehead and Rabin [l], showed that with both iodoacetate and iodo- acetamide the loss of enzymic activity in the pres- ence of excess reagent is a pseudo first-order process. This has been confirmed here over periods covering 5-95O/, of the total reaction. These findings indicate that the two essential cysteine residues are indepen- dent and identical with respect to both alkylating agents.
The finding of about 2 cysteine residues per molecule controlling enzymic activity suggests that there should be a similar number of coenzyme binding sites and catalytic sites. Detailed coenzyme binding studies have already been made with these enzyme preparations and estimates in the range 2.05-2.8 binding sites/molecule of molecular weight 141 000 were obtained [7] depending upon the method used for the determination. There was thus quite a spread of values but emphasis was placed on the higher estimates suggesting an integral value of 3 binding sites/molecule. Comparison with the results of iodo- acetate inactivation now imply that the lowest estimates were the more reliable.
The NADH binding studies using the gel filtra- tion method have now been repeated using an im- proved technique in which bound NADH concentra- tions were estimated in the peak fractions by oxida- tion with acetaldehyde and alcohol dehydrogenase. The results of these experiments appear in Fig.4 in the form of a plot of molecules of NADH bound/ molecule of enzyme (7) versus this quantity divided by the free NADH concentration (?/"ADHI). The plot indicates a maximum of 2.05 NADH binding sites/molecule and a dissociation constant of 11.5 pM for the E * NADH complex.
600
- 5 E
2 - 400 r 3 0
1
I 0
200
C
I i r
I
d I I I I 20 40 60 80
Fraction n u m b e r
Fig.5. Time course of the dialysis of N A D H at 22 ' C in phosphate buffer pH 7.0 in the presence ( 0 ) or absence (0) of 8.8 mglml alcohol dehydrogenase. At the first arrow radioactive NADH at a specific activity of approx. 3 mCi/mmol was added to raise the NADH concentration to 105 pM. At sub- sequent arrows the NADH concentration was raised to 162, 218, 332,446, 674 and 2724 pM by the addition of unlabelled NADH
A separate study of NADH binding has been carried out using the steady-state dialysis method of Colowick and Womack [13]. Fig.5 shows how the radioactivity of the buffer passing the dialysis membrane increased on addition of an initial aliquot of radioactive NADH to alcohol dehydrogenase and subsequently on addition of aliquots of unlabelled NADH. Also shown is the radioactivity measure- ment relating to the steady-state dialysis rate of radioactive NADH a t the same initial concentration as before but in the absence of enzyme. The data of Fig. 5 on suitable manipulation give rise to the second titration curve appearing in Fig. 4.
NADH binding studies using fluorescence titra- tion [7], gel filtration and steady-state dialysis methods now agree. The higher values found in previous gel filtration experiments [3,7] were due to a failure to allow for the effects of high enzyme concentration on determinations of bound NADH using direct absorbance measurements a t 340 nm. The estimate of 2.4-2.8 sites/molecules capable of forming the enzyme * pyridine-3-aldehyde-adenine- dinucleotide * hydroxylamine complex [7] is now the only estimate which does not agree. This estimate was obtained using an absorption coefficient for the complex a t 315nm of 6.0 mM-l cm-l [14]. The accuracy of that value is unconfirmed. The present work suggests a somewhat larger figure.
Whitehead and Rabin [l] showed that the pres- ence of coenzyme protects alcohol dehydrogenase
Eur. J. Biochem. 41 (1974)

M. Dickinson 35
from inactivation by iodoacetamide. The finding here of four cysteine residues/molecule reacting with iodoacetate, only two of which are essential, leads to the expectation that in the presence of NADH only the inessential thiols will be alkylated. This expectation was tested in an experiment in which enzyme (60 mg in a final volume of 6.4 ml) was in- cubated with a 16-fold molar excess of iodoacetate in the presence of 460 pM NADH a t 0 "C. As anti- cipated the presence of coenzyme markedly protected the enzyme. After 10.5 h the activity of the prepara- tion had only fallen by about 15O/,. At the same time there was an incorporation of 0.96 equivalents of carboxymethyl group. In the absence of coenzyme (see Fig.2) reaction of 1 equivalent of iodoacetate produced about a 50°/, loss of activity. Obviously the more reactive cysteine residues were protected by coenzyme and the nonessential ones were tending to react first. The later stages of the reaction were less clear cut, there being a progressive loss of enzyme activity as the level of carboxymethyl group in- corporation rose to 4.0 groups/molecule after 2.4 days with the loss of 95O/, of the activity. The results of the experiment provide qualified support for expecta- tions based on the earlier experiments.
It is well established that yeast alcohol dehydro- genase is a tetramer composed of subunits of approx- imate molecular weight 35000 [5]. Sequence work on a 21-residue peptide around the iodoacetate-reacting thiol groups indicated that the four subunits are very similar if not identical. The suggestion of only two catalytic sites arising from this work raises certain interesting possibilities.
Inactivation experiments with iodoacetate show that two pairs of cysteine residues do not reside in the same environment. The difference may arise because the enzyme is built up from two types of similar but different subunits or because of partial degradation of the enzyme in preparation. Alter- natively i t may result from a quaternary structure which requires chemically identical subunits to main- tain different conformations. It is probable that the observation of only two NADH binding sites/ molecule does not arise from extreme negative co- operativity in which two of the active sites become inoperative when the other two bind coenzyme. It seems perhaps unlikely that iodoacetate should induce a similar structural change in the enzyme to that induced by coenzyme. Thus the experiments with iodoacetate imply that only two active sites pre-exist on the enzyme.
The possibility that the present observations arise as an artefact in the enzyme isolation procedure cannot be dismissed although efforts have been made to eliminate some obvious possibilities. Preparations of enzyme appear perfectly homogenous in sedimenta- tion and sedimentation equilibrium experiments in the ultracentrifuge and contain no significant amount
of inactive protein as judged by electrophoresis on polyacrylamide gels [7]. Isolation of the enzyme by a different procedure yielded enzyme of about the same specific activity and with the same coenzyme binding capacity 171. The specific activities of the enzyme preparations compare very favourably (see Materials and Methods) with those of other workers [9,10] who have shown that loss of sulphydryl groups from these preparations by oxidation results in the loss of enzy- mic activity. The enzyme is reported to contain 4-5 atoms zinc/molecule protein and zinc is essential for enzymic activity [4,15,16]. Our preparations when analysed using a very slight modification of the dithizone method of Cremona and Singer [17] after dialysis against zinc-free buffers and dry ashing a t 500 "C for 24 h gave estimates of 4.3-5.0 g-atoms zinc/l41000 g protein. There was thus no evidence that the enzyme preparations used here were partly denatured but this must remain a possibility. If the enzyme is partly denatured than it seems probable that the preparations used by other workers hitherto were similarly affected. One may note here that yeast aldehyde dehydrogenase, prepared by a method designed to eliminate the possibility of partial degradation in preparation, exhibits only two NADH binding sites/molecule despite its tetrameric structure [18]. Perhaps yeast alcohol and aldehyde dehydrogenases provide two examples of an unusual quaternary structure.
It is interesting to note a t this point some obser- vations made in a study of the binding of l-anilino-8- naphthalene sulphonate by the enzyme [19]. There appears to be about 2 binding sites/molecule for the dye which are not identical with active sites as jud- ged by failure of coenzyme to completely displace the dye and by the fact that the inhibitor constant Ki is significantly greater than the dissociation constant of the enzyme - dye complex. It seems possible that these observations are a reflection of the same struc- tural features in the enzyme which gives rise to only two active sites.
I wish to thank Mr M. Girling for technical assistance and the Science Research Council for financial support.
REFERENCES 1.
2.
3. 4.
5. 6.
7. 8. 9.
Whitehead. E. P. & Rabin. B. R. (1964) Biochem. J . > ,
90, 532-539. Rabin. B. R.. Ruiz Cruz, J., Watts, D. C. & Whitehead,
E. P. (1964) Biochem. J. 90, 539-542. Dickinson, F. M. (1972) Biochem. J. 126, 133-138. Kani. J . H. R. & Vallee. B. L. (1960) J . Biol. Chem. 235, ~ 588-3192. Harris, J. I. (1964) Nature (Lond.) 203, 30-34. Hayes, J . E. & Velick, S. F. (1954) J . Biol. Chem. 207,
225-244. Dickinson, F. M. (1970) Biochem. J. 120, 821-830. Dalziel, K. (1961) Biochem. J. 84, 240-244. Wallenfels, K. & Sund, H. (1957) Biochem. 2. 329, 17-
30.
Eiir. J. Biochem. 41 (1974)

36 M. Dickinson: Active Sites of Alcohol Dehydrogenase
10. Buhner, M. & Sund, H. (1969) Eur. J . Biochem. 11, 73-
11. Reithel, F. J. & Sakura, J. D. (1963) J . Phys. Chem. 67,
12. Hummel, J. P. & Dreyer, W. J. (1962) Biochim. Biophys.
13. Colowick, S. P. 8: Womack, F. C. (1968) J . Bid. Chem.
14. Van Eys, J., Ciotti, M. &I. & Kaplan, N. 0. (1957) Bio-
15. Vallee, B. L. & Hoch, F. L. (1955) Proc. Natl. Acad. Sci.
16. Wallenfels, K., Sund, H., Faessler, A. & Burchard, W.
17. Cremona, T. & Singer, T. P. (1964) J . Biol. Chem. 239,
18. Bradbury, S. L. & Jakoby, W. B. (1971) J . Biol. Chem.
19. Dickinson, F. M. (1971) FEBS. Lett. 15, 17-20.
79. U . S. A . 41, 327-338.
2497- 2498.
A&, 63, 530-532. 1466-1473.
(1957) Biochem. 2. 329, 31-40.
244, 774-777. 246, 6929-6932.
chim. Biophys. Actu, 23, 581-58 .
M. Dickinson, Department of Biochemistry, University of Hull, Hull, Great Britain, HU6 7RX
Eur. J. Biochem. 41 (1974)