mechanism of electron transfera - oregon state university
TRANSCRIPT

AN ABSTRACT OF THE DISSERTATION OF
Shoutao Xu for the degree of Doctor of Philosophy in Biological and Ecological
Engineering presented on June 15 2012
Title Bacterial Community Analysis New Exoelectrogen Isolation and Enhanced
Performance of Microbial Electrochemical Systems Using Nano-Decorated Anodes
Abstract approved
Hong Liu Frank WR Chaplen
Microbial electrochemical systems (MESs) have attracted much research attention in
recent years due to their promising applications in renewable energy generation
bioremediation and wastewater treatment In a MES microorganisms interact with
electrodes via electrons catalyzing oxidation and reduction reactions at the anode and the
cathode
The bacterial community of a high power mixed consortium MESs (maximum power
density is 65Wm2) was analyzed by using denature gradient gel electrophoresis (DGGE)
and 16S DNA clone library methods The bacterial DGGE profiles were relatively
complex (more than 10 bands) but only three brightly dominant bands in DGGE results
These results indicated there are three dominant bacterial species in mixed consortium
MFCs The 16S DNA clone library method results revealed that the predominant
bacterial species in mixed culture is Geobacter sp (66) Arcobacter sp and Citrobacter
sp These three bacterial species reached to 88 of total bacterial species This result is
consistent with the DGGE result which showed that three bright bands represented three
dominant bacterial species
Exoelectrogenic bacterial strain SX-1 was isolated from a mediator-less microbial
fuel cell by conventional plating techniques with ferric citrate as electron acceptor under
anaerobic conditions Phylogenetic analysis of the 16S rDNA sequence revealed that it
was related to the members of Citrobacter genus with Citrobacter sp sdy-48 being the
most closely related species The bacterial strain SX-1 produced electricity from citrate
acetate glucose sucrose glycerol and lactose in MFCs with the highest current density
of 205 mAm2 generated from citrate Cyclic voltammetry analysis indicated that
membrane associated proteins may play an important role in facilitating electron transfer
from the bacteria to the electrode This is the first study that demonstrates that
Citrobacter species can transfer electrons to extracellular electron acceptors Citrobacter
strain SX-1 is capable of generating electricity from a wide range of substrates in MFCs
This finding increases the known diversity of power generating exoelectrogens and
provids a new strain to explore the mechanisms of extracellular electron transfer from
bacteria to electrode The wide range of substrate utilization by SX-1 increases the
application potential of MFCs in renewable energy generation and waste treatment
Anode properties are critical for the performance of microbial electrolysis cells
(MECs) Inexpensive Fe nanoparticle modified graphite disks were used as anodes to
preliminarily investigate the effects of nanoparticles on the performance of Shewanella
oneidensis MR-1 in MECs Results demonstrated that average current densities
produced with Fe nanoparticle decorated anodes were up to 59-fold higher than plain
graphite anodes Whole genome microarray analysis of the gene expression showed that
genes encoding biofilm formation were significantly up-regulated as a response to
nanoparticle decorated anodes Increased expression of genes related to nanowires
flavins and c-type cytochromes indicate that enhanced mechanisms of electron transfer
to the anode may also have contributed to the observed increases in current density The
majority of the remaining differentially expressed genes were associated with electron
transport and anaerobic metabolism demonstrating a systemic response to increased
power loads
The carbon nanotube (CNT) is another form of nano materials Carbon nanotube
(CNT) modified graphite disks were used as anodes to investigate the effects of
nanostructures on the performance S oneidensis MR-1 in microbial electrolysis cells
(MECs) The current densities produced with CNT decorated anodes were up to 56-fold
higher than plain graphite anodes Global transcriptome analysis showed that cytochrome
c genes associated with extracellular electron transfer are up-expressed by CNT
decorated anodes which is the leading factor to contribute current increase in CNT
decorated anode MECs The up regulated genes encoded to flavin also contribute to
current enhancement in CNT decorated anode MECs
copyCopyright by Shoutao Xu
June 15 2012
All Rights Reserved
Bacterial Community Analysis New Exoelectrogen Isolation and Enhanced Performance
of Microbial Electrochemical Systems Using Nano-Decorated Anodes
by
Shoutao Xu
A DISSERTATION
Submitted to
Oregon State University
in partial fulfillment of
the requirements for the
degree of
Doctor of Philosophy
Presented June 15 2012
Commencement June 2013
Doctor of Philosophy dissertation of Shoutao Xu presented on June 15 2012
APPROVED
Co-Major Professor representing Biological and Ecological Engineering
Co-Major Professor representing Biological and Ecological Engineering
Head of the Department of Biological and Ecological Engineering
Dean of the Graduate School
I understand that my dissertation will become part of the permanent collection of
Oregon State University libraries My signature below authorizes release of my
dissertation to any reader upon request
Shoutao Xu Author
ACKNOWLEDGEMENTS
I would like to thank all people who have helped and inspired me during my
doctoral study Foremost I would like to express my sincere gratitude to my advisors Dr
Hong Liu and Dr Frank Chaplen for their continuous support of my PhD study and the
research of Microbial Fuel Cells at Oregon State University Hong inspired me to devote
myself to the field of Bio-energy with her great patience and enthusiasm towards
scientific educations Frank was always available and willing to help me with my study
especially during the period of Hongrsquos sabbatical leave Thanks to his kindness and
assistance my study at OSU became smooth and rewarding
Besides my advisors I would like to thank everyone in my dissertation committee Dr
Martin Schuster Dr Clare Reimers and Dr Mark Dolan Due to their encouragement
and insightful comments on my research I could always have the courage and knowledge
to overcome difficulties in my research I benefited greatly from Martinrsquos valuable
suggestions on my writing skills and his generous help regarding microarray data analysis
Clarersquos advice helped me build a solid foundation of Electrochemistry Also it was a
great honor to have Mark as my committee member His teaching gave me an insight into
the world of environmental engineering and enhanced the depth and width of my research
I thanked Dr Yanzhen Fan for always giving me guidance about the designs of the
reactors in my experiments
It was also important for me to say thanks to my current and previous lab-mates
Keaton Lesnik Kuhuan Chien Cheng Li Corale Abourached Anthony Janicek
Hongqiang Hu Jeremy Chignell Yudith Nieto and Wengguo Wu I had a wonderful
time enjoying doing research with them Their friendship and help made me confident of
my ability to do research as a scientist In addition I cherished and appreciated the
friendly environment in our BEE department Faculty members especially Dr John Bolte
Dr John Selker Dr Roger Ely Dr Ganti Murthy and Desiree Tullos made BEE an
excellent department for our students
My deepest gratitude went to my father Fuqin Wang and my mother Jinzhi Wang
for their endless care love and support throughout my life Also I would like to say
thanks to my best-loved wife Songhua Zhu She always supported me and never
complained that I could spend so little time accompanying with her This dissertation was
also for my lovely son Gabriel Hong-Yi Xu
TABLE OF CONTENTS
Page
1 General Introductionhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 1
2 Bacterial Community Analysis of Mixed Consortium in Microbial
Electrochemical Systemshelliphelliphelliphellip 13
3 New Exoelectrogen Citrobacter sp SX-1 Isolation and
Characterizationhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 27
4
Enhanced Performance and Mechanism Study of Microbial Electrolysis
Cells using Fe Nanoparticles Decorated Anodeshelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 45
5 Global Transcriptome Analysis of Response of Shewanella oneidensis
MR-1 to CNT Nanostructure Decorated Anodes in Microbial
Electrochemical System
66
6 Summaryhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 80
7 Bibliography helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 82
8 Appendices helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 93
LIST OF FIGURES
Figure Page
1-1 Generalized schematics (a) Microbial fuel cell (MFCs)(b) Microbial
electrolysis cell (MECs) helliphelliphelliphelliphelliphelliphelliphelliphelliphellip helliphelliphelliphellip
2
1-2 Mechanisms for extracellular electron transport in a MFCs anode 7
2-1 Polarization curves of high power mixed culture MFCshelliphelliphelliphelliphelliphellip 20
2-2 DGGE result of high power generation mixed culture MFCshelliphelliphellip 21
2-3 Bacterial species and percentage of bacterial community of mixed
culture MFCs helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip
22
2-4 Phylogenetic trees of bacterial species from mixed culture MFCs 22
2-5 Dominant bacterial species identification in DGGEhelliphelliphelliphelliphelliphelliphelliphellip 23
2-6 Current density generated by different dominant isolateshelliphelliphelliphelliphelliphellip 24
3-1 Phylogenetic tree of strain SX-1 and closely related species based on
16S rDNA sequenceshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip
35
3-2 SEM image of planktonic cells of Citrobacter sp SX-1helliphelliphelliphelliphelliphellip 35
3-3 Electricity generation by Citrobacter sp SX-1 in a single chamber
MFCs with sodium citrate helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip
36
3-4 Electricity generation of Citrobacter sp SX-1 using different
substrates in single chamber air-cathode MFCs at 1 KΩhelliphelliphellip
37
3-5 Power and voltage generation by Citrobacter sp SX-1 as a function
of current density helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip
38
3-6 Four current generating stages for CV analysis helliphelliphelliphelliphelliphelliphelliphellip 39
3-7 Cyclic Voltammetry analysis of of Citrobacter sp SX-1 biofilms helliphellip 41
4-1 SEM images of Fe NPs decorated anodeshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 50
4-2 Effects of Fe-NP decorated anodes on the current density in MECs helliphellip 51
4-3 Differentially expressed genes grouped by functional classification in
Fe NP decorated anodeshelliphelliphelliphelliphelliphellip helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip
53
5-1 SEM images of CNT decorated anodeshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 70
5-2 Effects of CNT decorated anodes on the current density in MECshelliphellip 73
5-3 Differentially expressed genes grouped by functional classification in
CNT decorated anodes helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip
74
LIST OF TABLES
Table
1-1
4-1
4-2
4-3
Taxa of bacteria current density generated and reactor configuration
used in microbial electrochemical cell experimentshelliphelliphelliphellip
Expression levels of genes related to biofilm formationhelliphelliphelliphelliphellip
Genes related to anaerobic growth and electron transfer with
significantly change expression levelhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip
Expression level of flavin and cytochrome geneshelliphelliphelliphelliphelliphelliphelliphellip
Page
4
55
56
58
4-4
5-1
5-2
Other genes with significantly changed expression level
Expression level of cytochrome c as response to CNT decorated anodes
Expression level of genes related to flavin synthesis as response to CNT
decorated anode in CNT decorated anodeshelliphellip
59
76
78
1
1
Bacterial Community Analysis New Exoelectrogen Isolation and Enhanced Performance
of Microbial Electrochemical Systems Using Nano-Decorated Anodes
Chapter 1
General Introduction
11 Microbial Fuel Cells and Microbial Electrolysis Cells
Microbial electrochemical systems (MESs) have drawn the attention of researchers
in recent years due to their promising applications in a variety of scientific fields such as
renewable energy generation bioremediation and wastewater treatment In a MES
microorganisms interact with electrodes catalyzing oxidation and reduction reactions at
the anode and the cathode
The most-described type of MESs is the microbial fuel cells (MFCs) in which
useful power is generated directly using the catalytic action of active microorganisms
(Kim 2002 Cheng and Logan 2007 Cheng et al 2006) In a typical two-chamber
MFCs (Figure 1a) organic matter is oxidized by electrochemically active
microorganisms in the anode chamber to release protons and electrons Protons diffuse
into the cathode chamber through a proton exchange membrane Meanwhile electrons
are transferred to the anode through various mechanisms (Logan et al 2006 Lovley
2
2006 Rabaey et al 2003) and then travel to the cathode where normally they combine
with oxygen and protons to form water
Another common used type of MES is the microbial electrolysis cell (MECs) in
which hydrogen is produced instead of electricity by applying a circuit voltage to the
MFCs system and maintaining anaerobic conditions in the cathode chamber (Liu et al
2005 Logan et al 2008 Rozendal et al 2006 Schaller et al 2009) (Figure 1b) A
cathode potential of at least - 410 mV (vs standard hydrogen electrode at pH 70) is
required to produce hydrogen at the cathode Therefore a theoretical potential of 110 mV
(ie 410-300 mV) is needed but voltages of gt02 V are normally applied due to various
overpotentials
Fig 1-1 Generalized schematics (a) Microbial fuel cells (MFCs) where electricity is
captured through the resistance R (b) Microbial electrolysis cells (MECs) where
hydrogen gas is produced in the cathode chamber R for resistor PS for power supply and
PEM for proton exchange membrane
The fundamental feature shared by microbial electrochemical systems (MESs)
(MECs and MFCs) is the microbe-catalyzed electron transfer from organic matter to
electrodes (anodes) (Rebaey et al 2004) Therefore different types of MESs can be
PEM
R
H+
e- e-
Anode
Bacte
ria
a
Cath
ode
H2O
PEM
PS
H+
e- e-
Anode
Bacte
ria
b
Cath
ode
H2
3
utilized to investigate the phenomena and mechanisms of interactions between microbes
and electrodes
12 Bacteria and Electron Transfer Mechanisms in MESs
The electrochemically active bacteria in MFCs are thought to be iron-reducing
bacteria such as Shewanella and Geobacter species (Gorby et al 2006) They have great
importance in the natural environment principally in metal oxidation and reduction
However recent studies have shown that the diversity of bacterial communities is much
greater than these model iron reducers on MFCs anodes (Reimers et al 2001 Liu et al
2004b Kim et al 2005) Over 20 electrochemically active bacterial species those can
transfer electrons exocellularly to electrodes have been reported in the past 10 years
(Logan 2009 Liu et al 2010) These bacteria have been categorized into diverse genetic
groups so far including alphaproteobacteria (Rhodopseudomonas Ochrobactrum and
Acidiphilium) (Malki et al 2008 Xing et al 2008 Zuo et al 2008) betaproteobacteria
(Rhodoferax) (Chaudhuri et al 2003) gammaproteobacteria (Shewanella Pseudomonas
Klebsiella Enterobacter and Aeronomas) (Kim et al 2002 Pham et al 2003 Rabaey et
al 2005 Bretschger et al 2007 Zhang et al 2008 Chung et al 2009 Rezaei et al 2009)
deltaproteobacteria (Geobacter Geopsychrobacter Desulfuromonas and Desulfobulbus)
(Bond et al 2002 Bond and Lovley 2003 Holmes et al 2004ab 2006)
Epsilonproteobacteria (Arcobacter) (Fedorovich et al 2009) Firmicutes (Clostridium and
Thermincola) (Park et al 2001 Wrighton et al 2008) Acidobacteria (Geothrix) (Bond
and Lovley 2005) and actinobacteria (Propionibacterium) (Wang et al 2008) A wider
range of electrochemically active bacteria are expected to be discovered
4
Table 1 Taxa of bacteria current density generated and reactor configuration used in
microbial electrochemical cell experiments (Liu et al 2010)
The electrochemically active bacterial species that possess the ability to transfer
electrons outside of the cell are called exoelectrogens in the MESs research field The
different exoelectrogens have demonstrated a wide-ranging power generation ability in
MFCs For example Shewanella oneidensis MR-1 has demonstrated the ability to
generate 0018 Am2 current density in single chamber MFCs while the Geobacter
Taxon Microorganisms Current Density (Am2
)
α-proteobacteria
Rhodopseudomonas palustris DX-1 003
Ochrobactrum anthropi YZ-1 071
Acidiphilium sp 32sup5 300
β-proteobacteria Rhodoferax ferrireducens 0031
γ-proteobacteria
Shewanella putrefaciens IR-1 0016
Shewanella oneidensis DSP10 0013
Shewanella oneidensis MR-1 0018
Pseudomonas aeruginosa KRA3 0017
Escherichia coli K12 HB101 100
Klebsiella pneumoniae L17 120
Enterobacter cloacae 013
Aeromonas hydrophila PA3 030
δ-proteobacteria
Geobacter metallireducens 065
Geobacter sulfurreducens 800
Desulfuromonas acetoxidans 0005
Geopsychrobacter
Electrodiphilus strain A2
0066
Desulfobulbus propionicus 003
Firmicutes
Lactococcus lactis 003
Thermincola sp strain Jr 020
Clostridium butyricum EG3 022
Thermincola ferriacetica Z-0001 040
Brevibacillus spp PTH1 0009
Desulfitobacterium hafniense DCB2 110
Acidobacteria Geothrix fermentans 0097
Actinobacteria Propionibacterium freudenreichiiET-3 120
5
sulfurreducens species has generated current densities as high as 80 Am2 which is 400
times higher than the one generated by Shewanella oneidensis MR-1 However mixed-
culture communities have generated much higher power densities than their pure-culture
counterparts in MFCs perhaps due to synergistic interactions within the anode bacterial
communities and the participation of currently unknown bacteria species and mechanisms
(Jung and Regan 2007) The dominant bacteria species vary in mixed cultures bacteria
communities in MFCs due to the enrichment of different substrates
Traditional methods of extroelectrogen study depend on cultivation hampered novel
exoelectrogens discovery because the inadequacy of defined media underestimates the
actual microbial diversity in MFCs Slow growth rate and unknown growth requirements
of anaerobic microorganisms make some anaerobic extroelectrogens cultivation difficult
However these limitations have been overcome by using molecular biological methods
based on DNARNA analysis Molecular biological techniques are now widely applied to
assess the diversity of microbial communities by analyzing the 16S rDNA sequence The
most commonly used molecular biological techniques for bacterial community analysis
include denaturing gradient gel electrophoresis (DGGE) restriction fragment length
polymorphism (RFLP) and 16S rDNA sequencing These methods are much less time
consuming than traditional isolation and cultivation methods
Isolated exoelectrogens were utilized to explore the mechanism of electron transfer
to the anode However the mechanisms of electron transfer to extracellular electron
acceptors are not well understood Three mechanisms have been proposed for exocellular
transport of electrons by exoelectrogens to anodes in MFCs so far Some exoelectrogenic
bacteria can transfer electrons to electrodes through soluble redox compounds (Bond and
6
Lovley 2005) These compounds include artificial mediators and mediators secreted by
exoelectrogenic bacteria A variety of chemicals have been used to facilitate the shuttling
of electrons from inside of cell to electrodes outside the cell These exogenous mediators
include neutral red (Park et al 1999) anthraquinone-2-6 disulfonate (AQDS) thionin
potassium ferricyanide (Bond et al 2002) methyl viologen and others (Logan 2004
Rabaey and Verstraete 2005) Some bacteria can secrete their own chemical mediator
for example Pseudomonas aeruginosa secretes pyocyanin and phenazine-1-carboxamide
to transfer electrons toward the MFCs anode (Rabaey et al 2005) Another evidence for
mediator production by bacteria is Geothrix fermentans When the medium was replaced
in a MFCs that had stable power generation with this bacteria power dropped by 50
and required 10 days to resume the original level
Some bacteria can directly transfer electrons to anodes via outer cell membrane
proteins (Myers and Myers 1992) The outer cell membrane protein cytochrome c is
thought to play a critical role in to transferring electrons to anodes Ly et al (2011)
isolated the haem protein cytochrome c and demonstrated that electric field effects may
be functional for the natural redox processes of cytochrome c in the respiratory chain
Shewanella oneidensis is the example of a bacterium that is capable of transfer electron to
anode via outer cell membrane proteins When Shewanella oneidensis adhered to an iron
surface the greater force has showed benefits to grow cells because closer contact
required for electron transfer from cell bound cytochromes (Lower et al 2001)
7
Fig1-2 Mechanisms for extracellular electron transport in an MFCs anode (1) direct
contact (top in green) (2) by nanowires (middle in purple) and (3) self-produced
mediators (bottom in blue) (Logan 2009)
More and more evidence supports the involvement of bacterial nanowires in
extracellular electron transport (Reguera et al 2005 2006 Gorby et al 2006) Nanowires
are conductive appendages produced by both Geobacter and Shewanella species (Gorby
and Beveridge 2005) The conductivity of the appendages was examined and confirmed
by using conductive scanning tunneling microscopy (STM) (Gorby et al 2006)
Nanowires can carry electrons from the cell to the anode surface of MFCs
The solid component of the extracellular biofilm matrix has high efficiency on
extracellular electron transfer compared with other extracellular electron transfer
8
mechanisms and recently Torres et al (2010) hypothesized that the solid component of
the extracellular biofilm matrix formed by exoelectogens is another way bacteria transfer
electrons to electrodes This hypothesis was based on kinetic analysis of each EET
mechanism reported in available literature (Torres et al 2010)
13 Anode electrodes
In MFCsMECs anode electrodes are a critical component because exoelectrogens
adhere to the surface of anodes to transfer electrons to the electrode The characteristics
of anodes have significant effects on electron transfer rate from bacteria to anode
electrodes in MFCs The requirements of an anode material are it should be highly
conductive non-corrosive have a high specific surface area (area per volume) high
porosity be non-fouling inexpensive and easily scaled to larger sizes Of these
properties the most important one that is different from other biofilm reactors is that the
material must be electrically conductive Normally they are made of various carbon
materials including carbon fiber carbon clothe and carbon paper due to their stability
high conductivity and high specific surface-area Nevertheless they have little
electrocatalytic activity for the anode microbial reactions and thus a modification of the
carbon materials is the main approach for improving their performance Consequently
there is a great need to develop a new type of anode material for MFCsMECs
It is a great challenge to develop a new anode material to further increase the power
density of a MFCs The nature of the catalytic mechanism of a MFCrsquos anode involves not
only a biological but also an electrocatalytic process An optimal nanostructure with a
high specific surface area favorable for both catalytic processes could play a critical role
in improving the power density of the MFCs such a structure needs to host the bacteria
9
with high bioactivity while enhancing the electron-transfer rate Schroumlder et al (2008)
employed PANI to modify a platinum anode for MFCs and achieved a current density 1
order of magnitude higher than the previously reported value PANIinorganic
composites are also reported to have better conductivity Qiao et al also applied a new
mesoporous TiO2 electrode material with uniform nanopore distribution and a high
specific surface area to anode in comparison to previously reported work with E coli
MFCs the composite anode delivers 2-fold higher power density (Qiao et al 2008)
Thus it has great potential for use as the anode in a high-power MFCs and may be a new
approach for improving performance of MFCs
14 Other parts of MESs
141 Membranes and ion transport
The ion exchange membrane is another one of the critical components in two-
chamber MESs systems It separates anode and cathode chambers and at the same time
maintains the electron neutrality of the system ie transport of electrons to the cathode
needs to be compensated by transport of an equal amount of positive charge to the
cathode chamber A proton exchange membrane (PEM) such as Nafion a per-fluorinated
sulphonic acid membrane (PFSAs) consisting of a hydrophobic fluorocarbon backbone to
which hydrophilic sulfonate groups (-SO3-) are attached is commonly used in chemical
fuel cell systems For MFCs systems however mainly cation species like Na+ and K
+
other than proton are often responsible for the dominant transport of positive charge
through the cation exchange membrane (CEM) to maintain electroneutrality due to the
low proton concentration in any aqueous medium with near neutral pH (Rozendal et al
2006) Consequently the pH increases in the cathode chamber due to the consumption of
10
protons and decreases in the anode chamber because of the accumulation of protons
(Rozendal et al 2007) This pH change in two-chamber systems results in a decrease of
the cathode potential and performance The application of anion exchange membrane
(AEM) has also been explored in MFCs systems (Cheng and Logan 2007 Kim and
Logan 2007) where it has been proposed that protons are transferred via pH buffers like
phosphate anions
142 Cathodes and Catalysts
The cathode is another challenge for making MFCs commercially available
technology because the chemical reaction that occurs at the cathode is difficult to
engineer as the electrons protons and oxygen must all meet at a catalyst in a tri-phase
reaction (solid catalyst air and water) The catalyst must be on a conductive surface and
must be exposed to both water and air so that protons and electrons in these different
phases can reach the same point The most commonly used material for a cathode is
commercially available carbon paper pre-loaded with a Pt catalyst on one side When it is
used in a MFC the side that contains the catalyst faces the water and the uncoated side
faces air To reduce the high cathode cost associated with platinum catalyst other precious-
metal-free cathodes such as (NiMo and NiW) were developed by electrodepositing on a
carbon fiber They have achieved comparable performance with Pt catalyst with same
loading at a much lower cathode fabrication cost (Hu 2010)
The requirements of cathode for MECs are quite similar to the requirements of
cathode for MFCs but easier than cathode for MFCs for the manufacturing process
because the cathode in MECs is not necessarily to exposed to air Therefore it can be
made of the exact same materials of cathode in MFCs except a waterproof layer
11
Recently some researchers have attempted to apply microorganisms as a biocatalyst to
precede the combination of electron with oxygen in the cathodes
15 Dissertation overview
Low power densities in MESs limit practical applications The improvement of
MESs performance requires a detailed understanding of the physiology and ecology of
microorganisms in MESs including the mechanism of electron transfer to the anode from
the microorganism
This dissertation focuses on the problem of the low power density of MESs The
bacterial community structure of a high power generated mixed culture communities in
MFCs will be identified firstly and then one of major exoelectrogens will be isolated and
characterized This information will be helpful to understand the physiology and ecology
of exoelectrogens in MESs Consequently they will be beneficial to improve power
density of MFCs Lastly the nanostructure decorated anodes in MECs will be utilized to
improve the power density The power enhancement mechanism will be explored by
using a whole genome microarray They are presented here as four papers
In the first paper the cultivation independent molecular biological techniques
denaturing gradient gel electrophoresis (DGGE) and 16S rDNA clone library are utilized
to analyze the bacterial community structure of a higher power mixed culture MFCs The
analyzed results provide fundamental information for isolating the dominant bacteria in
mixed culture MFCs Two of dominant bacterial species has been isolated and used aone
to test power generation in MFCs The possible interaction among different bacterial
species in mixed culture is discussed
12
In the second paper one isolated exoelectrogenic bacterial strain SX-1 is
characterized It is identified as a member of the Citrobacter genus and power generation
is tested ultilizing a wide range of different substrates The electron transfer mechanism is
explored using Cyclic Voltammetry (CV) This study increases the known diversity of
power generating exoelectrogens and provides a new strain to explore the mechanisms of
extracellular electron transfer from bacteria to electrodes
The third paper shows effects on MESs performance by Fe nanoparticle decorated
anodes in the MESs The average current density produced with Fe nanoparticle
decorated anodes increased up to 59-fold higher than plain graphite anodes A whole
genome microarray is utilized to analyze the possible mechanism of enhanced current
density as responded to nanoparticle decorated anodes
The fourth paper describes the effects of carbon nanotube (CNT) modified anode on
the performance S oneidensis MR-1 in MESs Results demonstrate that current densities
produced with CNT decorated anodes are up to 56-fold higher than plain graphite anodes
The possible mechanisms of enhanced current density by CNT decorated are explored
13
Chapter 2
Bacterial community analysis of mixed consortium in higher
power density MESs
Shoutao Xu and Hong Liu
1 Introduction
The improvement in the performance of mixed culture MFCs requires an
understanding of the ecology in microbial communities of MFCs Many researchers have
attempted to characterize microbial populations and activities to elucidate the behaviors
and ecology of microorganisms in MFCs (Wrighton et al 2008 Fedorovich et al 2009)
In order to study the microbial ecology of the mixed culture in a MFC and select the
appropriate isolation medium for dominant bacterial species in the mixed culture the
fundamental steps are to analyze the bacterial diversity in the mixed culture of MFCs and
identity the dominant bacterial species in bacteria communities in MFCs
14
As for the identification of bacterial communities typically there are two general
methods The first method for identification of bacterial community is the traditional
cultivation processes using selective nutrients to promote the growth of different types of
bacteria within the samples (Amman et al 2000) The community structure can then be
assessed by identifying the isolates from the dominant colonies that were cultured This
can often be costly and laborious as each isolate has to be further studied by examining
its physiology taxonomy and reactivity to stains (Adwards et al 1989)
The second method relies on utilizing molecular techniques to analyze bacterial
community DNA Several molecular methods involving the extraction and analysis of
DNA from entire bacterial communities are used to identify genetic fingerprints of
bacteria These methods including the cloning and sequencing of 16S rDNA automated
ribosomal intergenic spacer analysis (ARISA) terminal restriction fragment length
polymorphism (T-RFLP) and denaturing gradient gel electrophoresis (DGGE) generate
profiles of bacterial community structures They can rapidly assess complex communities
from various environments (Amman et al 2000)
Due to the conservative characteristic of 16S rDNA in bacteria during the process
of evolution 16S rDNA sequencing can be used to identify different species of bacteria
Therefore the molecular techniques of denaturing gradient gel electrophoresis (DGGE)
with PCR and 16S rDNA clone library are used for analysis of the microbial diversity
These methods are more convenient and save time compared to traditional
isolationcultivation methods for microorganism analysis
In this work a biofilm bacterial community from an anode of a MFCs wase studied
by using denaturing gradient gel electrophoresis (DGGE) of PCR-amplified partial 16S
15
rRNA genes followed by cloning and sequencing of 16S rDNA The results provided
essential information for dominant bacterial isolation in mixed culture MFCs
2 Materials and methods
21 High power generation of mixed culture MFCs
Mixed cultures were originally inoculated from domestic wastewater (Corvallis
Wastewater Treatment Plant Corvallis OR) to MFCs after operating for 2 years using a
defined medium solution (Lovely 2002) with sodium acetate as the carbon source A
new MFCs was inoculated from the operating MFCs The polarization curves were
performed to measure the power generation when maximal stable power were established
(normally 3-5 batches post-inoculation) and the genomic DNA from the microbial
biofilm on the anode was extracted under the sterile conditions for further bacterial
community analysis
22 Denaturing gradient gel electrophoresis (DGGE)
DGGE with PCR is a method of analysis of bacterial community composition
based on different GC percentage of 16s rDNA in different bacterial species PCR-DGGE
comprises three steps (i) extraction of total community DNA from the sample (ii) PCR-
controlled amplification using specific oligonucleotide primers and (iii) separation of the
amplicons using DGGE For this purpose a reproducible and efficient method for total
DNA extraction is indispensable and needs to be evaluated and optimized depending on
the nature of the sample In the subsequent PCR step multiple PCR primer sets with
different resolution can be used In most PCR-DGGE applications on bacteria universal
or specific primers are targeting the 16S rRNA gene Upon electrophoresis of the PCR
16
amplicons and gel staining (using ethidium bromide silver staining or SYBR green)
DGGE gels are digitally captured and further analyzed using computer software packages
The richness of DNA fragment bands in DGGE indicates the bacterial diversity of the
sample
The detailed steps are as followed Biofilms were scratched from the anodes of high
power generated MFCs fed with sodium acetate Bacterial genomic DNA was extracted
from the biofilm samples using the DNeasy blood and tissue kit (Qiagen) according to the
manufacturerrsquos instructions The universal primer set was used to amplify the 16S rDNA
from the extracted genomic DNA then the V3 region of bacterial 16S ribosomal DNA
(rDNA) was amplified with GC-clamp primer (Muyzer et al 1993) for DGGE PCR
amplification was performed in a thermocycler DGGE of the PCR products was carried
out in a DcodeTM
Universal Mutation Detection System) The 8 (wv) polyacrylamide
gels contains from 30 to 55 denaturing gradients Electrophoresis was conducted
using a 1timesTAE (Tris-Acetate-EDTA) buffer at 130V and 60degC for 5 hours After
electrophoresis the gel was stained with ethidium bromide in 1timesTAE buffer for 15
minutes and washed in 1timesTAE buffer for 10 minutes The fragments were visualized
under a UV transilluminator The richness of single band in DGGE gel picture
preliminarily showed that the bacterial diversity in mixed culture since the single band in
gel represents one bacterial species
23 16S rDNA clone library construction method
16S rDNA-DGGE fingerprinting has proven to be particularly useful as an initial
investigation into bacterial communities and is suitable for identifying the predominant
bacteria that comprise 1 or more of cells within a given sample In the 16s rDNA clone
17
library construction process the first step is the extratction of the total genomic DNA
then the genomic DNA of the mixed culture biofilm was used as template for PCR
amplification of approximately 1500 bp of 16S rDNA with universal primers The PCR
products of 16S rDNA were purified and inverted into pGEM-T Easy vector system
before they were transformed into competent E coli The transformed cells were spread
on Luria-Bertani (LB) plates containing the selective antibiotic ampicillin and X-Gal and
incubated overnight at 37 OC after the more than 80 inverted 16s DNA fragment E coli
colonies were picked up and inoculated in the 1-3 ml liquid LB medium and left to grow
for 16 hours The plasmid DNA were extracted and sequenced The sequences were
compared directly to all known sequences deposited in GenBank databases using the
basic local alignment search tool (BLAST)
24 Bioinformatics Analysis
16S rDNA sequencing results of more than 80 colones were queried against the
GenBank and Ribosomal Database Project (RDP) databases using BLAST and
SEQUATCH algorithms (Cole et al 2007 Wang et al 2007 Cole et al 2009) The
neighbor-joining trees were constructed with the Molecular Evolutionary Genetics
Analysis package (MEGA Version 4) based on 1000 bootstrap analysis (Tamura et al
2007)
25 Dominant bacterial species band in DGGE identification
The DGGE results gave the information of dominant bacterial richness of bacterial
species in mixed culture based on the theory that single band represents one bacterial
species which showed the diversity of bacterial community in the mixed culture of a
MFC 16S rDNA clone library results provided the whole picture of bacterial community
18
including the bacterial species name and bacterial species percentage in the mixed culture
The known pure bacterial species which have been sequenced can be used as markers to
identify dominant bacterial species in DGGE The whole experimental procedure is
similar to the procedure of the mixed culture DGGE steps The first step is to extract the
collect the mixed culture biofilm genomic DNA and pure bacterial species marker
genomic DNA Then the universal primer set was used to amplify the 16S rDNA from
the extracted genomic DNA After that the V3 region of bacterial 16S ribosomal DNA
(rDNA) was amplified with GC-clamp primer (Muyzer et al 1993) for DGGE The
subsequent steps were similar to the mixed culture DGGE experimental procedure The
dominant bacterial bands in DGGE were determined by comparisons of the single band
position in the mixed culture DGGE with single pure bacterial species location in DGGE
26 Dominant bacterial species isolation and power generation
The bacterial community of mixed culture in MFCs were predominantly composed
of Geobacter sp Arcobacter sp and Citrobacter sp Two dominant bacterial species
have been isolated
Citrobacter sp SX-1 was isolated from the anode biofilm by using a solid agar
medium containing NH4Cl 6mM KCl 2mM CH3COONa (electron donor) 30 mM
FeC6H5O7 (electron acceptor) 20mM minerals and vitamins (Lovley and Phillips 1988)
Single colonies were picked up after culturing on the plate for 36 hours at 30 degC and
transferred two times on the agar plate for purification Then the isolates grew in a liquid
medium solution in anaerobic tubes containing the same constituents as the solid
medium All isolation process was operated in a glove box anaerobic chamber (Coy
Laboratory Products Grass Lake MI)
19
Arcobacter sp SX-2 was isolated from the anode biofilm of a MFC fed with sodium
acetate by using the same method of isolation of pure bacterial species Citrobacter XS-1
except using Trypticase Soy Agar with 5 Sheep Blood solid (from American type
culture collection (ATCC)) agar medium instead of Citrobacter sp medium Single
colonies were picked up after culturing on the plate for 36 hours at 30 degC and transferred
twice on the agar plate for purification Then the isolates grew in a liquid medium
solution microaerobically
Geobacter sp have been trying to isolate by using three different methods They are
the colony pickup after different condition enrichment Goebacter medium isolation and
dilution to extinction by multiple channel mini MFCs isolation however the Geobacter
sp has not been isolated yet due to some limits so far But they provide valuable
information for further isolation of Geobacter sp
Single chamber MFCs were used to evaluate power generation by different isolates
The MFCs were constructed as described previously (Liu and Logan 2004) and modified
with 3 cm2 carbon cloth anodes and 7 cm
2 carbon clothPt cathodes The total liquid
volume of each MFCs was about 15 ml with electrode spacing about 17 cm All MFCs
were operated in an autoclaved closed plastic box and sterile cotton was attached to the
outer surface of the air cathodes to prevent contamination A MFCs without bacterial
culture was used as control MFCs were inoculated with 3 ml late exponential phase
cultures of SX-1 in the medium solution reported previously (Liu and Logan 2004)
3 Results and discussion
31 Power production by mixed culture MFCs
20
A polarization curve was used to characterize current as a function of voltage in the
MFCs The polarization curves are performed by varying a series of external resistances
The results are shown in Figure 21 With MFCs acclimated to 25 Ω external resistance
the maximum power density was achieved at 65Wm2 based on the polarization data At
this point the current density is 21mAcm2 The power density was three times higher
than previous report (20 Wm2) using phosphate buffer (Fan et al 2007) Power density
was also 2 times higher than (28 m Wm2) using a bicarbonate buffer ( Fan et al 2007)
32 Bacterial community structure assessed by PCR-DGGE
Bacterial DGGE profiles for higher power generating MFCs mixed culture setups
were relatively complex (more than 10 bands) However there are three brightly stained
bands and other less intense bands (Fig 2-2) These results indicated there were probably
three dominant bacterial species in mixed culture MFCs
010
020
030
040
050
060
070
2
3
4
5
6
7
04 05 05 06 07 08 10 11 13 15 21 22 23 24
Volta
ge (
V)
Pow
er
densi
ty (
Wm
2)
Current density (mAcm2)
Power
Voltage
power density 65wm2 at current
density208 mAcm2
Figure 2-1 Polarization curves of high power mixed culture MFCs
21
33 16S rDNA clone library results
A total of 83 colonies were sequenced for 16S rDNA clone library construction They
represented the bacterial community structure in mixed culture in MFCs The results
revealed that the predominant bacterial species in mixed culture are Geobacter sp (66
of the mixed culture) Arcobacter sp (12 of the mixed culture) and Citrobacter sp (11
of the mixed culture) Those three genuses comprised 88 of the total bacterial species
(Fig 23) This result is consistent with the DGGE result which showed that three bright
bands in the DGGE gel represented three dominant bacterial species in the mixed culture
The phylogenetic analysis of the colonies (83 colony sequences) is shown in Figure 24
Figure 22 DGGE result of high power generation mixed culture MFC (1 mixed
culture 2 pure culture control)
1 2
22
34 Dominant bacterial band in DGGE identification
Geobacter
66
Arcobacter
12
others
6
Pseudomonas
1
Citrobacter
11
Clostridium
2
Anaerovorax
2 Geobacter
Arcobacter
Citrobacter
Clostridium
Anaerovorax
Pseudomonas
others
Figure 23 Bacterial species and percentage of bacterial community of mixed
culture MFC lt 1 Alcaligensgt
lt1 Commamonasgt
lt1 Pseudomonas aeruginosagt
lt1 Klebsiellagt
lt11 Citrobactergt
lt1 Wolinellagt
lt12 Arobactergt
lt2 Anaeroboraxgt
lt2 Clostridium gt
lt1 unclear bacteriagt
lt66Geobactergt
100
100
85
100
100
82
99
87
100
002
Figure 24 Phylogenetic trees of bacterial species from mixed culture MFC
23
The dominant bacterial species including Goebacter sp Citrobacter sp and Arcobacter
sp in the mixed culture are shown in Figure 25 The upper band is Arcobacter sp and
Citrobacter sp is located in the bottom band of DGGE Geobacter sp is the middle one
The analysis of the bacterial community using denaturing gradient gel electrophoresis
(DGGE) of PCR-amplified 16S rRNA gene fragments and 16S rDNA clone library
construction method showed great phylogenetic diversity of mixed culture in MFCs with
the identification of sequences derived from bacteria of the taxa deltaproteobacteria
(Geobacter sp) gammaproteobacteria (Citrobacter sp) and epsilonproteobacteria
(Arcobacter sp) The result is consistent with the conclusion that among the isolated
exoelectrogens gammaproteobacteria and deltaprotebactia appear to be dominant among
others (Parot et al 2009 Liu et al 2010)
35 The dominant bacterial species isolation and power generation
Figure 25 Dominant bacterial species identification in DGGE
A B C D
A Citrobacter sp
B Geobacter sp
C Arobacter butzmeri
D mixed culture control
24
Two dominant bacterial species Citrobacter sp XS-1 and Arcobacter sp XS-2 have been
isolated and tested for power generation The power densities generated by the two
isolates are shown in the Fig 26 Current densities generated by isolate Citrobacter sp
and Arcobacter sp were 98 mAm2 and 20 mAm
2 respectively which were much less
than that (21Am2) generated by the mixed culture from which they were isolated
Figure 26 Current density generated by different dominant isolates
The maximum current density generated by mixed culture is much higher than the
one produced by pure bacterial species Citrobacter sp XS-1 and Arcobacter sp XS-2 in
the same structure MFCs Although the Geobacter sp in our mixed culture has not been
isolated a previous study showed that maximum current density generated by Geobacter
sp was 080 Am2
(Bond and Lovley 2003) The current density results seem partially
supporting the conclusion that mixed-culture generated higher power densities than their
pure-culture counterparts in MFCs (Jung and Regan 2007) This result suggests there
may be synergistic interactions within the anode bacterial communities
0
500
1000
1500
2000
2500
Mixed culture Citrobacter sp Arcobacter Sp
Cu
rre
nt
de
nsi
ty (
mA
m2)
25
Geobacter sp are strictly anaerobic bacteria which were enriched in most of the anode
biofilms (George 2005) Citrobacter species are facultative anaerobic bacteria (George
2005) which can be found in a wide variety of habitats including in soil water and
wastewater Arcobacter sp are micro-aerobic bacteria including both environmental
nonpathogens and opportunistic human pathogens They are able to grow in aerobic
conditions but in the optimal growth under micro-aerobic conditions (George 2005)
Arcobacter sp has been discovered in the MFCs by Fedorovich (Fedorovich et al 2009)
It is interesting that the mixed culture bacterial community composed of primarily these
three bacterial species has demonstrated to generate high power at non-strictly anaerobic
condition while leading dominant bacterial species Geobacter sp (66) is strict
anaerobic bacteria and generated powder in the MFCs only under strict anaerobic
condition (George 2005) Therefore it is possible that microaerobic bacteria such as
Arcobacter sp and facultative anaerobic bacteria Citrobacter sp can facilitate to create
anaerobic condition for Geobacter sp when they utilized oxygen for growing in the
mixed culture Syntrophic communities study have showed that anaerobic bacteria and
methanogenic archaea form compact microbial structures that operate like an organ rather
than a set of microorganisms functioning independently (Stams and Plugge 2009) Some
substrates have been degradeted within these communities while they are not able to be
fermented by individual species alone interspecies electron transfer also have been
presented in these communities (Stams and Plugge 2009) Summers et al (2010) also
discovered that direct exchange of electrons happened within coculture of Geobacter
metallireducens and Geobacter sulfurreducens Therefore we believe that there are
26
more complex synergistic interactions between different bacteria species in mixed culture
in the MFCs
Acknowledgements
The authors acknowledge support from the US National Science Foundation
(CBET 0955124) We also thank Keaton Lesnik for the review of this manuscript
27
Chapter 3
New Exoelectrogen Citrobacter sp SX-1 Isolated from a Microbial Fuel Cell
Shoutao Xu and Hong Liu
Published in
Journal of Applied Microbiology 111(5)1108-1115 (2011)
ABSTRACT
Exoelectrogenic bacterial strain SX-1 was isolated from a mediator-less microbial fuel
cell by conventional plating techniques with ferric citrate as electron acceptor under
anaerobic condition Phylogenetic analysis of the 16S rDNA sequence revealed that it
28
was related to the members of Citrobacter genus with Citrobacter sp sdy-48 being the
most closely related species The bacterial strain SX-1 produced electricity from citrate
acetate glucose sucrose glycerol and lactose in MFCs with the highest current density
of 205 mA m-2
generated from citrate Cyclic voltammetry analysis indicated that
membrane associated proteins may play an important role in facilitating the electrons
transferring from bacteria to electrode This is the first study that demonstrates that
Citrobacter species can transfer electrons to extracellular electron acceptors Citrobacter
strain SX-1 is capable of generating electricity from a wide range of substrates in MFCs
This finding increases the known diversity of power generating exoelectrogens and
provided a new strain to explore the mechanisms of extracellular electron transfer from
bacteria to electrode The wide range of substrate utilization by SX-1 increases the
application potential of MFCs in renewable energy generation and waste treatment
Keywords Microbial fuel cell Exoelectrogen Citrobacter sp SX-1 Extracellular
electron transfer
29
Introduction
Microbial Fuel Cells (MFCs) technology has become an active research area
recently as a promising approach for renewable energy generation wastewater treatment
and bioremediation (Rabaey and Verstraete 2005 Lovley 2006 Fan et al 2008 Logan
2009 Liu et al 2010) The key feature of MFCs system is the microbe-catalyzed electron
transfer from organic matter to anodes Many studies have shown the presence of diverse
bacterial communities on MFCs anodes (Reimers et al 2001 Liu et al 2004b Kim et al
2005) Over 20 exoelectrogens ie microbes that can transfer electrons exocellularly to
electrodes have been reported in the past 10 years (Logan 2009 Liu et al 2010) The
exoelectrogens so far belong to diverse genetic groups including alphaproteobacteria
(Rhodopseudomonas Ochrobactrum and Acidiphilium) (Malki et al 2008 Xing et al
2008 Zuo et al 2008) betaproteobacteria (Rhodoferax) (Chaudhuri et al 2003)
gammaproteobacteria (Shewanella Pseudomonas Klebsiella Enterobacter and
Aeronomas) (Kim et al 2002 Pham et al 2003 Rabaey et al 2005 Bretschger et al
2007 Zhang et al 2008 Chung et al 2009 Rezaei et al 2009) deltaproteobacteria
(Geobacter Geopsychrobacter Desulfuromonas and Desulfobulbus) (Bond et al 2002
Bond and Lovley 2003 Holmes et al 2004ab 2006) Epsilonproteobacteria (Arcobacter)
(Fedorovich et al 2009) Firmicutes (Clostridium and Thermincola) (Park et al 2001
Wrighton et al 2008) Acidobacteria (Geothrix) (Bond and Lovley 2005) and
actinobacteria (Propionibacterium) (Wang et al 2008) A wider range of
exoelectrogenic species are expected to be discovered
Three mechanisms have been proposed for exocellular transport of electrons by
exoelectrogens without artificial mediators Some exoelectrogenic bacteria can transfer
30
electrons to electrodes through soluble redox compounds excreted by microorganisms
(Bond and Lovley 2005 Rabaey et al 2005) and others can directly transfer electrons to
anodes via outer cell membrane proteins (Myers and Myers 1992) Recently more and
more evidence supports the involvement of bacterial nanowires in extracellular electron
transport (Reguera et al 2005 2006 Gorby et al 2006) In spite of the discovery of
many bacterial species that can transfer the electrons to electrode without the need of
artificial mediators the investigation of extracellular electron transfer mechanisms was
mainly focused on a few species such as those from Geobacter and Shewanella genera
(Kim et al 1999 2002 Reguera et al 2005 2006 Gorby et al 2006) The electron
transfer mechanisms for many of the isolated exoelectrogens species are still not well
studied
In the present study we isolated a new exoelectrogen strain SX-1 from a MFC a
strain phylogenetically related to Citrobacter sp Power generation from various carbon
sources by this strain was evaluated using single chamber MFCs Plausible extracellular
electron transfer mechanisms were also discussed based on the characterization of anodic
biofilms by cyclic voltammetry (CV)
Materials and methods
Bacterial strain SX-1 isolation
Bacterial strain SX-1 was isolated from the anode biofilm of a MFC fed with sodium
acetate operated in fed-batch mode over a period of six months The original source of
the inoculum is wastewater from a local waste water treatment plant Bacterial cells were
released from the carbon cloth anode by shaking the cloth in a sterile flask with 15 mL
sterile 1 NaCl solution and glass beads (2 mm diameter) The suspension was then
31
serially diluted from 10 times to 105
times and plated on a petri dish with a solid agar
medium containing NH4Cl 6mM KCl 2mM CH3COONa (electron donor) 30 mM
FeC6H5O7 (electron acceptor) 20mM minerals and vitamins (Lovley and Phillips 1988)
Single colony was randomly selected after culturing on the plate for 36 hours at 30 degC
and purified on a new agar plate following a procedure reported previously (Chung and
Okabe 2009) Then the isolate was picked up and grew in a liquid medium solution in
anaerobic tubes containing the same constituents as the solid medium for further analysis
All isolation process was operated in a glove box anaerobic chamber (Coy Laboratory
Products Grass Lake MI)
16S rDNA sequencing and phylogenetic analysis
Bacterial genomic DNA was extracted from the isolated strain SX-1 using the DNeasy
tissue Kits (Qiagen Valencia CA) according to the manufacturerrsquos instructions The 16S
rDNA was amplified by PCR using a pair of universal primers 27F (5rsquo-
AGAGTTTGATCMTGGCTCAG-3rsquo) and 1492R (5rsquo-GGTTACCTTTGTTACGACTT-
3rsquo) (Park et al 2001) The conditions used for PCR were described previously
(Ferna ndez et al 1999) PCR products were purified ligated and cloned according to a
procedure described in a previous report (Xing et al 2008) Plasmids were isolated from
randomly selected clone colonies with a QIAprep Spin Miniprep kit (Qiagen Valencia
CA) and three plasmid inserts were then sequenced in both directions using an ABI 3730
DNA sequencer (Applied Biosystems Carlsbad CA)United States) Due to the identical
16S rDNA sequence of the three colones only one was queried against the GenBank and
Ribosomal Database Project (RDP) databases using BLAST and SEQUATCH algorithms
(Cole et al 2007 Wang et al 2007 Cole et al 2009) The 16S rDNA sequence of
32
strain SX-1 and closely related type strains were aligned using CLUSTALX software
(Thompson et al 1997) The neighbor-joining trees were constructed with the Molecular
Evolutionary Genetics Analysis package (MEGA Version 4) based on 1000 bootstrap
analysis (Tamura et al 2007)
SEM
The morphologies of SX-1 cells growing overnight in anaerobic tubes were examined
with a scanning electron microscope (FEI Quanta 600 FEG FEI Company Hillsboro
OR) The suspended cells were fixed in a 25 glutaraldehyde and 01M phosphate
buffer solution and dehydrated with a graded ethanol series from 30 to 100 After
dehydration the samples were dried in a critical point dryer and then sputter-coated with
AuPd for SEM examination (Liu and Logan 2004)
Evaluation of power generation by SX-1 in MFCs
Single chamber MFCs were used to evaluate power generation by SX-1 using various
substrates The MFCs were constructed as described previously (Liu and Logan 2004)
and modified with 3 cm2 carbon cloth anodes and 7cm
2 carbon clothPt cathodes The
total liquid volume of each MFCs was about 15 ml with electrode spacing about 17 cm
All MFCs were operated in an autoclaved closed plastic box and sterile cotton was
attached to the outer surface of the air cathodes to prevent contamination A MFC
without bacterial culture was used as control MFCs were inoculated with 3 ml late
exponential phase cultures of SX-1 in the medium solution reported previously (Liu and
Logan 2004) Seven substrates including citrate acetate glucose sucrose glycerol and
lactose were evaluated individually for power generation in a fed-batch mode in a
temperature control chamber (30 plusmn2degC) For each substrate at least 8 batches were run to
33
investigate the effect of biofilm formation on current output at fixed external resistance of
1kΩ and the same initial substrate concentration of 30 mM Phosphate buffer (100mM)
was used to maintain the solution pH and conductivity The MFCs with sodium citrate as
electron donors were also examined for maximum power output by varying the external
resistance from 20 kΩ to 4 kΩ A data acquisition system was used to record voltage data
during the MFCs operation (Keithley 2700 Keithley Instruments Inc Cleveland OH) It
took about 15 to 30 minutes for the MFCs to stabilize depending on the external
resistance At each resistance we collected at least five data at the steady condition to
make the polarization curves Averaged voltages were used to calculate the power density
(mW m-2
) according to P=IVA where I was the current V was voltage and A was cross-
sectional area of the anode
CV analysis
CV was used to characterize the oxidation and reduction reactions on the anodic surface
of the MFCs fed with citrate and operated at 7 KΩ This external resistance was selected
because the maximum power density was obtained at this resistance based on the
polarization experiment We assumed the biofilm on the anode of MFCs were well-
developed when stable power output was obtained after 3 batches of operation The
MFCs were then used directly for CV analysis at four current generating stages of the
fourth batch (1) initial exponential current increasing stage (middle point of stage about
4 h after media change) (2) current plateau stage (middle point of stage about 8 h after
media change) and (3) current decreasing stage (middle point of stage about 16h after
media change) (4) right after the complete replacement of medium solution (Figure 6)
The anode was used as working electrode the cathode as counter electrode and an
34
AgAgCl electrode was selected as the reference The CV curves were scanned from 200
to -600 mV at a rate of 5 mV s-1
using a potentiostat (G300 Gamry Instrument
IncWarminster PA) Control experiment was also conducted using new anode (without
biofilm) and new medium solution
Nucleotide sequence accession number
The 16S r DNA sequence determined in this study has been deposited in the GenBank
database under accession number HQ845373
Results
Identification of the strain SX-1
An almost-complete 16S rDNA sequence (1475 bp) of strain SX-1 was obtained and
subjected to comparative analysis with the 16S rDNA of closely related reference strains
A phylogenetic analysis revealed that strain SX-1 should be assigned to the genus
Citrobacter and was most closely related to Citrobacter sp sdy-48 EJ463782 (990
sequence similarity) and Citrobacter sp yy-21 FJ463779 (975 sequence similarity)
These three strains formed a distinct sub cluster in the neighbor-joining in which the new
isolate and Citrobacter sp sdy-48 EJ463782 formed a distinct subline (Figure 31)
The SEM image illustrated that the strain SX-1 is a rod-shaped bacterium 03-06 microm
wide and 12-17 microm long (Figure 32) Strain SX-1 colonies on nutrient agar appeared 2-
3 mm in diameter smooth moist opaque with a shiny surface after 5 days incubation
under anaerobic condition The fact that strain SX-1 grew both aerobically and
anaerobically using FeC6H5O7 as electron acceptor suggests that the strain is facultatively
anaerobic Strain SX-1 showed robust growth on citrate as a carbon source These D
35
properties were similar to those of genus Citrobacter described in Bergeyrsquos Manual of
Systematic Bacteriology (George 2005)
Figure 31 phylogenetic tree of strain SX-1 and closely related species based on 16S
rDNA sequences The tree was constructed using the neighbor-joining method
Figure 32 SEM image of planktonic cells of Citrobacter sp SX-1
Strain SX-1
Citrobacter sp sdy-48 (FJ463782)
Citrobacter sp yy-21 (FJ463779)
Citrobacter koseri E639 (ATCC 25408)
Citrobater koseri CDC 3613-63(AF025372)
Citrobacter koseri CDC 8132-86(AF025366)
Salmonella enterica SL483 (CP001138)
Salmonella enterica AKU12601(AY696668)
Shigella sonnei Ss046(CP000038)
E coli C2 (AF403733)
E coli E24377A(CP000800)88
100
100
76
44
92
53
36
0001
1 microm
36
Electricity production by strain SX-1 in MFCs
The isolated strain SX-1 was first tested for its ability to generate current from sodium
citrate at 1kΩ Current gradually increased to a maximum 50 mA m-2
after cell
inoculation and then decreased (Figure 33) After the MFCs was refilled with new
substrate solution the current recovered rapidly and reached a higher level than the first
batch After 4 batches operation the maximum current output of each batch became
stable The highest current density achieved in the MFCs with Citrobacter sp SX-1 at 1
KΩ was 98 mA m-2
(Figure 33)
Figure 33 Electricity generation by Citrobacter sp SX-1 in a single chamber MFCs
with sodium citrate (30 mM) as substrate at 1 KΩ
0
20
40
60
80
100
120
0 50 100 150 200
Curr
ent
den
sity
(m
A m
-2)
Time (h)
37
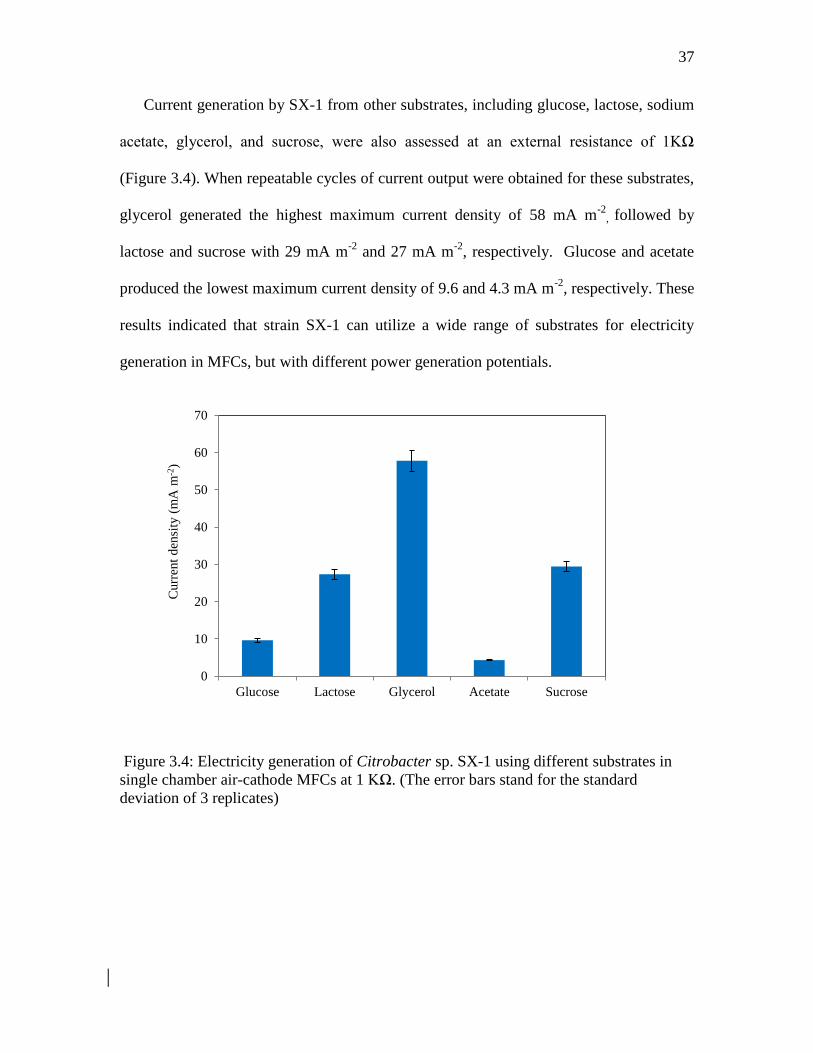
Current generation by SX-1 from other substrates including glucose lactose sodium
acetate glycerol and sucrose were also assessed at an external resistance of 1KΩ
(Figure 34) When repeatable cycles of current output were obtained for these substrates
glycerol generated the highest maximum current density of 58 mA m-2
followed by
lactose and sucrose with 29 mA m-2
and 27 mA m-2
respectively Glucose and acetate
produced the lowest maximum current density of 96 and 43 mA m-2
respectively These
results indicated that strain SX-1 can utilize a wide range of substrates for electricity
generation in MFCs but with different power generation potentials
Figure 34 Electricity generation of Citrobacter sp SX-1 using different substrates in
single chamber air-cathode MFCs at 1 KΩ (The error bars stand for the standard
deviation of 3 replicates)
0
10
20
30
40
50
60
70
Glucose Lactose Glycerol Acetate Sucrose
Curr
ent
den
sity
(m
A m
-2)
38
Polarization experiment was further conducted to determine the maximum power
density generated by strain SX-1 in the single chamber MFCs with citrate as substrate A
maximum power density of 881 mW m-2
was obtained at current density of 205 mA m-2
at an external resistance of 7 KΩ (Figure 35)
Figure 35 Power and voltage generation by Citrobacter sp SX-1 as a function of
current density using sodium citrate (30 mM) as substrate The error bars stand for the
standard deviation of the 3 voltagepower outputs obtained in three MFCs
20
30
40
50
60
70
80
90
100
01
02
03
04
05
06
07
80 110 140 170 200 230
Po
wer
den
sity
(m
W m
-2)
Vo
ltag
e (V
)
Current density (mA m-2)
Voltage
Power
39
Figure 36 Four current generating stages for CV analysis current increasing stage (stage
1) current plateau stage (stage 2) current decreasing stage (stage 3) and right after the
replacement of medium solution (stage 4) The MFCs was operated at 7 KΩ with sodium
citrate (30 mM) as substrate
Cyclic Voltammetry
To determine the presence of redox active compounds produced by SX-1 and the
location of these compounds CV scan of the anodic biofilms at four current output stages
and the supernatant of MFCs medium solution at the end of the batch experiment were
performed Reduction peaks at range of -310 ndash -350 mV and oxidation peaks at range -
100 ndash -150 mV were observed at all current generating stages (Figure 36 Figure 37A)
suggesting the presence of redox active compounds may involve in extracellular electron
transfer by Citrobacter sp SX-1 The amplitude of the peaks increased according to the
growth stage of the batch and the highest peaks were present after the current plateau
stage which indicated the redox active compounds mainly were secreted in the current
plateau stage While the current density at stage 3 (deceasing stage) was lower than that
at stage 2 (current plateau stage) the redox peak at stage 3 was higher than that in stage 2
0
50
100
150
200
250
50 55 60 65 70 75 80 85
Curr
ent
den
sity
(m
A m
-2)
Time (h)
A
A
Stage 1
Stage 2 Stage 3
Stage 4
40
A plausible explanation for this pattern was that redox active compounds were
continuously being secreted and accumulated after the current peak was reached
resulting in more redox compounds present after the plateau stage But since most of the
carbon source had already been being used up many of the redox compounds at this
stage had not been as active as those at current plateau stage due to much less electrons
were available to be transferred The bigger peak amplitude in stage 4 than in stage 1
indicates that the redox active compounds were continuously being secreted and
accumulated in the biofilm even after stable power was generated at 7 kΩ The slightly
change of the peak locations at different stages was possible due to the solution chemistry
change during the current generation process or the slightly location change of reference
electrode during the measurement (Figure 37B) When the medium solution in the MFCs
was replaced by fresh medium at the end of the batch the oxidization and reduction
peaks were still presented in the CV (Figure 37B) but no peaks were observed when the
supernatant and fresh mediums were tested in a MFCs with no biofilm (Figure 37C)
These results suggest the compounds involved in the electron transfer were located not in
the supernatant (solution) but in the biofilm
41
Figure 37 Cyclic Voltammetry analysis of Citrobacter sp SX-1 biofilms at (A) current
increasing stage current plateau stage and decreasing stage (end of the batch) (B) end of
the batch and replaced with fresh medium and (C) controls
-40E-04
-30E-04
-20E-04
-10E-04
00E+00
10E-04
20E-04
30E-04
-060 -050 -040 -030 -020 -010 000 010 020
Curr
ent
(A)
Potential (V) vs AgAgCl
Current plateau stage
Current increasing stage
Current decreasing stage
-40E-04
-30E-04
-20E-04
-10E-04
00E+00
10E-04
20E-04
30E-04
-060 -050 -040 -030 -020 -010 000 010 020
Curr
ent
(A)
Potential (V) vs AgAgCl
Current decreasing stage
Biofilm with fresh medium
-40E-04
-35E-04
-30E-04
-25E-04
-20E-04
-15E-04
-10E-04
-50E-05
00E+00
-060 -050 -040 -030 -020 -010 000 010 020
Curr
ent(
A)
Potential (V) vs AgAgCl
Supernatant
Fresh medium
A
B
C
B C
42
Discussion
Citrobacter species belonging to Gammaproteobacteria Enterobacteriales
Enterobacteriaceae are facultative anaerobic bacteria (George 2005) They can grow with
oxygen as an electron acceptor but also with a variety of electron accepters such as Fe (III)
in the absence of oxygen (George 2005) Citrobacter species can be found in a wide
variety of habitats including in soil water and wastewater Several Citrobacter species
have been studied as important bioremediation bacterium for heavy metal removals
sulfate reduction phenol degradation and chlorophenol degradation (Macaskie et al 1995
Narde et al 2004 Qiu et al 2009) Citrobacter species have also been found on the
anodic biofilm of a diesel-degrading MFCs (Morris et al 2009) However there was no
study reported that Citrobacter species can transfer electrons to extracellular electron
acceptors This finding that Citrobacter strain SX-1 can generate electricity in MFCs
increased the diversity of power generating exoelectrogens and provided a new strain to
explore the mechanisms of extracellular electron transfer from bacteria to electrode
Among the isolated exoelectrogens gammaproteobacteria and deltaprotebactia appear to
be dominant among others (Parot et al 2009 Liu et al 2010) The isolated strain
Citrobacter sp SX-1 phylogenetically belongs to gammaproteobacteria which provides
another evidence to support this observation
Most exoelectrogens utilize a limited range of substrates for power generation For
example Shewanella oniedensis MR-1 can oxidize lactate under anaerobic condition but
cannot utilize acetate for electricity generation (Kim et al 2002) Geobacter
sulfurreducens can completely oxidize acetate for power generation but it cannot utilize
simple sugars (Bond and Lovley 2003 Caccavo et al 1994)) In comparison strain SX-1
43
not only can oxidize citrate for power generation but also utilize a wide range of
substrates for power generation including glycerol glucose lactose sucrose and acetate
Interestingly the higher current density generated by strain SX-1 from glycerol a main
by-product of biodiesel production process suggests that strain SX-1 may be potentially
used for harvesting energy from biodiesel wastes using MFCs However the current
density generated by strain SX-1 (205 mA m-2
) is lower than that (805 mA m-2
at 1 kΩ)
generated by the mixed culture from which the SX-1 was isolated indicating the
existence of other higher power generating bacteria andor complex ecology in the mixed
culture community
Understanding mechanisms of microbial extracellular electron transfer is critical for
enhancing the electron transfer rate from bacteria to electrode through metabolic or
genetic engineering of exoelectrogens Since the spent supernatant solution of MFCs run
with SX-1 showed no redox properties the redox compounds produced by SX-1 may
have been retained in the biofilm Alternatively SX-1 may utilize one or both of the
other two known extracellular electron transfer mechanisms For example it is possible
that strain SX-1 transfers electrons to the anode via cell-bound outer membrane proteins
The two distinct oxidation and reduction peaks observed at -100mv- -150 mV and -310--
350 mV of the SX-1 biofilm have a midpoint potential range from -205mV to -250mV
which is close to the redox potential of some c-type cytochromes (Field et al 2000 Meitl
et al 2009) which are well-known to play an important role in extracellular electron
trasnfer (Eggleston et al 2008 Field et al 2000 Meitl et al 2009) Field et al (2000)
reported that the c-type cytochromes CymA had the midpoint potentials of -229 mV
Meitl et al (2009) and Eggleston et al (2008) reported that the c- type cytochromes
44
OmcA in Shewanella oneidensis MR-1 had the midpoint potentials of -201 mV and -208
mV
While itrsquos possible that SX-1 may also have conductive appendages to enhance the
extracellular transfer electron similar to the nanowires discovered in some Geobacter
and Shewanella species (Reguera et al 2005 2006 Gorby et al 2006) further
investigation on the presence of these appendages on the cell surface of SX-1 is needed
Acknowledgements
The authors acknowledge support from the US National Science Foundation
(CBET 0955124) We also thank Teresa Sawyer for her helps with the SEM analysis and
Jeremy Chignell and Yanzhen Fan for their review of this manuscript
45
Chapter 4
Enhanced Performance and Mechanism Study of Microbial Electrolysis Cells
Using Fe Nanoparticles Decorated Anodes
Shoutao Xu Hong LiuYanzhen Fan Rebecca Schaller Jun Jiao Frank Chaplen
Published in
Applied Microbiology and biotechnology 93(2)871-880 (2012)
46
ABSTRACT
Anode properties are critical for performance of microbial electrolysis cells (MECs) In
the present study Fe nanoparticle modified graphite disks were used as anodes to
investigate the effects of nanoparticles on the performance of Shewanella oneidensis MR-
1 in MECs Results demonstrated that average current densities produced with Fe
nanoparticle decorated anodes were up to 56-fold higher than plain graphite anodes
Whole genome microarray analysis of the gene expression showed that genes encoding
biofilm formation were significantly up-regulated as response to nanoparticle decorated
anodes Increased expression of genes related to nanowires flavins and c-type
cytochromes indicate that enhanced mechanisms of electron transfer to the anode may
also have contributed to the observed increases in current density The majority of the
remaining differentially expressed genes were associated with electron transport and
anaerobic metabolism demonstrating a systemic response to increased power loads
Keywords Microbial electrochemical system microbial fuel cell microbial electrolysis
cell nanotechnology differential gene expression DNA microarray
47
Introduction
Microbial electrochemical systems (MESs) have been intensively studied since Lewis
achieved practical advances in this field (Logan 2007) however they attracted much
research attention in recent years due to their promising applications in renewable energy
generation bioremediation and wastewater treatment In a MES microorganisms
interact with electrodes via electrons catalyzing oxidation and reduction reactions at the
anode and the cathode The most-described type of MESs is microbial fuel cells (MFCs)
in which useful power is generated from electron donors typically biodegradable organic
materials (Logan et al 2006) Various novel MESs have recently been developed to
produce hydrogen (microbial electrolysis cells (MECs)) (Liu et al 2005 Logan et al
2008 Rozendal et al 2006 Schaller et al 2009) to power an autonomous sensor or
sensor network (Ringeisen et al 2006) to reduce bioremediation targets (Lovley 2006
Shea et al 2008) Sukkasem et al 2008) and to desalinate water (Cao et al 2009) The
key feature shared by these systems is the microbe-catalyzed electron transfer from
organic matter to electrodes (anodes) (Rabaey et al 2004) Enhancing the anodic current
output which highly depends on the performance of the electrodes is critical for the
successful application of all these processes (Logan et al 2007 Park and Zeilus 2002
2003)
Nanomaterials have received much attention from researchers in the context of
microbiology due to their unique physical electrical and chemical properties which
facilitate the study of interactions between bacteria and surfaces (Liu 2006) Previous
studies have demonstrated that electrodes decorated with different nanostructures such
as carbon nanotubes nanostructured polyanilinetitanium dioxide composite and titania
48
nanotubes have the potential to enhance power generation in MFCs (Morozan et al 2007
Qiao et al 2008 Quan et al 2005) Furthermore our recent study shows nanoparticle
(NP) decorated anodes greatly increased the electrochemical electron transfer rate in
MFCs (Fan et al 2011) However Fe NPs as an ideal candidate for decorating electrodes
because of respectively low price and high conductivity compared to other materials has
not been focused to study in MECs
S oneidensis MR-1 an important electrochemically active bacterial strain has been
exploited for the electricity generation in MFCs (Logan et al 2007 Logan 2009 Park
and Zeikus 2002) The availability of genome sequence for this strain makes it possible to
use transcriptome assays to globally measure the responses to different growth conditions
and environmental stresses (Murray et al 2001) S oneidensis MR-1 gene expression
response to heat shock cold shock Cr (VI) and U (VI) reduction chromate stress and
iron and acid tolerance have been studied in the past several years (Bencheikh-Latmani et
al 2005 Brown et al 2006 Gao et al2006 Gao et al 2004 Yang et al 2008) However
no studies have been focused on the Shewanella gene expression response to NPs in
MESs
The mechanism of increased electron transfer rate exhibited by nanoparticle decorated
electrodes is not well understood yet In the present study graphite disks decorated with
Fe NPs were used as anodes to explore the effects of nanostructures on current generation
in a multi-anode MECs DNA microarrays were utilized to investigate differences in the
global gene expression profile of S oneidensis MR-1 grown on plain versus Fe NPs
decorated anodes
Materials and methods
49
Bacterial cultures
S oneidensis MR-1 was purchased from American Type Culture collection (ATCC
700550) and stored as freezer stocks at -80degC Stock cells of S oneidensis MR-1 was
grown aerobically using a medium containing 30 gL Trypticase Soy Broth (BD 211825
Becton Dickinson Company Franklin lakes NJ) at 30degC Cells in early stationary
growth phase (optical density of 15-16 at 600nm (Spectrophotometer UV-1700
Shimadzu Columbia MD) were washed two times then injected into the chamber of the
MECs for current production Trypticase Soy Broth medium with 30 mM sodium lactate
as electron donor was used in MECs Phosphate buffer (100mM) was used to maintain
the solution pH 7 and solution conductivity at 15 mScm
Characterization of nanostructured anodes
Superfine isomolded graphite disks (16 cm x 05 cm GraphiteStorecom Inc) were
polished with ultrafine sand paper (2000 grit 3M Company) as the base for the NP
decorated and control anodes Fe NPs decorated anodes were fabricated by thermal
annealing of metal thin film coated graphite disks Thin films of Fe were firstly deposited
on the polished graphite disk by using sputter coating for 95 min and the samples then
were annealed in the Chemical vapor deposition (CVD) at 750degC for 20 min to form Fe
NPs Fe-NPs were produced with a diameter of 200plusmn100 nm and a density range
(distance between NPs) of 200plusmn100 nm with a coating time of 95 min SEM images of
Fe NPs decorated anode and control surfaces were shown in the Figure 41
50
Figure 41 SEM images of Fe NPs decorated anodes (A) Graphite disk control (B) Fe
Nanoparticle decorated
Multiple channel MECs construction and operation
A MECs with removable multiple anodes with each effective anode area 07 cm2
was
constructed and used to evaluate the effects of nanostructure on current density of
according to a previous report (Fan et al 2011) The cathode was made of wet-proof
(30) carbon cloth (type B E-TEK Division Inc USA) coated with PtC (20 E-TEK
Division Inc USA) following a previously reported procedure (Liu et al 2005) The
final platinum loading was 05 mgcm2 per projected cathode area The size of cathode
(150 cm2) was 25 times larger than the total effective area of the 8 graphite disk anodes
(56 cm2) to prevent cathode limitations on the performance of the MECs system All
MECs with Fe-NP decorated anodes and control anodes were tested for current
generation for 24 h with medium (without bacteria) before injecting the bacterial cells in
order to determine whether the Fe-NP decorated anodes demonstrate chemical current
generation compared to control anodes All the testing anodes were set up in the MECs
B
500 nm
A
500 nm
51
at once so that the S oneidensis MR-1 culture was applied to all of them simultaneously
Short electrode spacing (17 cm) was used in the MECs design to reduce the internal
resistance A voltage of 06 V was applied the MECs for current generation after the cells
of S oneidensis MR-1 were inoculated into MECs at early stationary phase A multimeter
with a data acquisition system (2700 Keithly USA) was used to monitor the current
change by measuring the voltage (every 300 seconds) drop through a resistor at 300 ohm
Fig 4 2 Effects of Fe-NP decorated anodes on the current density in MECs Control is
plain graphite disk anode Fe-NP is the anode with Fe-NP decoration
Microarray analysis
Biofilms for whole gene microarray analysis were aseptically removed from the plain
and Fe-NP decorated anodes of MECs after 110 hours further incubation at 30oC when
current density obviously started to decease Total RNA was extracted using Trizol
(Ambion inc AustinTX) following the manufacturerrsquos instructions Integrity of the
52
RNA samples was confirmed using the Agilent 2100 Bioanalyzer (Agilent Technology
Inc Palo Alto CA) The same amount of RNA (200 ng) from each sample was used for
further analysis after amplification by using the MessageAmpTM
II-Bacteria Prokaryotic
RNA Amplification kit (Ambion AustinTX) according to the manufacturerrsquos
instructions All of the 4295 genes (resource from NCBI) represented on the S oneidensis
MR-1 whole-gene microarrays were ordered from Roche NimbleGen Inc (Madison WI)
Biological triplicates of biofilms on the Fe-NP decorated anodes and control were
analyzed respectively cDNA synthesis labeling and hybridization were carried out by
the CGRB Core Laboratories at Oregon State University DNASTAR ArrayStarTM
3
software was used to identify genes that were up- or down-regulated more than 2-fold
when grown on the nanoparticle modified anodes using the unpaired two sample t-test
with a cutoff p-value of 005 The complete microarray data set generated in this study is
deposited for public access in the Gene Expression Omnibus
(httpwwwncbinlmnihgovgeo) under accession number GSE31535
Results
Enhancement of current generation by using NP decorated anodes in MECs
The current density generated by Fe-NP decorated anode MECs with S onidensis MR-1
increased to approximately 43 microAcm2 20 h following inoculation and then slowly
decreased Figure 42 shows that the maximum current density achieved by NP
decorated anodes was 83 times higher than that (51 microAcm2) generated by the control
(plain graphite disk) The average current density improvement of 110 hours was 59
times of that generated by the control The current densities generated in MECs in the
53
absence of bacteria were negligible (03 microAcm2) which demonstrated that the current
enhancement observed with the Fe-NP decorated anodes was biologically-derived
Global transcriptome analysis
Whole-genome DNA microarrays were used to attain a comprehensive general
overview
of the transcriptional response to Fe-NP decorated anodes by S oneidensis MR-1 in MEC
Of a total 4295 genes assayed 392 (188 induced and 204 repressed) exhibited significant
(P lt 005) differential expression at a 2-fold (log (expression ratio 1) level in 3
replicates in response to Fe-NPs These
total gene numbers present 9 of the 4295 open
reading frames (ORFs) presented on the array
Fig 43 Differentially expressed genes grouped by functional classification according to
the TIGR S oneidensis genome database (wwwtigrorg) Columns 1 amino acid
biosynthesis 2 biosynthesis of cofactors prosthetic groups and carriers 3 cell
envelope 4 cellular processes 5 central intermediary metabolism 6 DNA metabolism
7 energy metabolism 8 fatty acid and phospholipid metabolism 9 other categories 10
protein fate 11 protein synthesis 12 purines pyrimidines nucleosides and nucleotides
13 regulatory functions 14 signal transduction 15 transcription 16 transport and
binding proteins 17 unknown function 18 hypothetical proteins
Up-Regulated
-75
-50
-25
0
25
50
75
100
125
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18
Down-Regulated
Num
ber
of
Gen
es
54
Figure 43 summarizes the overall genomic response of S oneidensis MR-1 to Fe-NP
decorated anodes by grouping the differentially expressed genes into their functional role
categories based on The Institute for Genomic Researchrsquos annotation (Version 8) of the
MR-1 genome sequence The wide distribution of putative functional roles attributed to
the differentially expressed genes indicated the extent of the molecular response of
Soniedenis MR-1 to the NP decorated anodes Among genes with defined function a
large number of up-regulated genes were associated with cellular processes (group 4)
energy metabolism (group 7) other categories (group 9) and hypothetical proteins (group
18) While those genes encoding proteins involved in amino acid biosynthesis (group 1)
cellular processes (group 4) substrate transport (group 16) and hypothetical proteins
(group 18) were among the most down-regulated genes Two groups of genes that were
the most noticeable among the functional gene groups genes related to energy
metabolism (group 7) and genes related to amino acid biosynthesis (group 1) The ratios
of up-regulated genes to down-regulated genes in these functional groupings were much
higher than other functional gene groupings there were 21 up-regulated genes and 7
down-regulated genes in energy metabolism gene group and 2 up-regulated and 22 down
regulated genes in the amino acid biosynthesis group
Genes related to biofilm formation
In order to determine whether there was a correlation between the observed biofilm
enhancements by NP decorated anodes and the genes related to biofilm formation the
significant modified genes related to biofilm formation were examined and summarized
in the Table 41 These included SO_3223 and SO_3228 genes encoding the flagellum
proteins (Thormann et al 2004) which has critical impacts on initial attachment to the
55
surface as swimming motility functions were up-regulated over 10 and 09 fold
respectively The gene clusters SO_4178-SO_4180 which are thought to be essential for
formation of a three-dimensional biofilm were up-regulated from 12 to 13 fold
(Thormann et al 2004) In addition the genes SO_4105 encoding IV pilus which is
known to represent a critical factor for biofilm formation on a biotic surfaces (Thormann
et al 2004) were also induced 08 fold (Table 41) The genes related to flagellum
motility (SO_4286 SO_1529 and SO_1530) were also induced slightly (Thormann et al
2004)
Table 41 Expression levels of genes related to biofilm formation
Locus Tag Gene product descriptions Fold change
SO_3228 flagellar basal-body MS-ring and collar protein FliF 10
SO_3223 flagellar hook-length control protein FliK 09
SO_4103 MshA minor pilin protein MshD 05
SO_4105 MSHA major pilin protein MshA 08
SO_4178 expressed protein of unknown function MxdC 12
SO_4179 inner membrane family 2 glycosyltransferase MxdB 13
SO_4180 diguanylate cyclase-like protein MxdA 10
a fold changes were presented after log2
b positive number represented up-regulated and negative represented down-regulated
Genes related to energy metabolism
A closer consideration of energy metabolism genes was undertaken as anaerobic
metabolism for electron generation and electron transport functions may play critical
roles in enhanced current density generation in MECs Genes associated with energy
56
metabolism and with significantly modified gene expression levels are summarized in the
Table 42 Several of the up-regulated genes were related to formate dehydrogenase
proteins Most notably four genes related to formate dehydrogenase were up-regulated
more than 2-fold SO_0102 SO_0103 SO_0104 and SO_0107 The other genes
expression related to formate dehydrogenase proteins SO_4513 SO_4514 SO_4515 also
significantly increased Another interesting gene is that encoding the cytochrome c
oxidase protein gene SO_4609 which was up-regulated 11 fold while the gene
SO_0479 related periplasmic octaheme cytochrome c MccA was down-regulated
However SO_1519 the gene encoding lactate dehydrogenase was down-regulated 12
fold despite lactate being the primary carbon source in the media
Table 4 2 Genes related to anaerobic growth and electron transfer with significantly change expression level
Locus Tag Gene product descriptions Fold
change
SO_0101 nitrate-inducible formate dehydrogenase molybdopterin-binding
subunit FdnG
18
SO_0102 formate dehydrogenase nitrate-inducible iron-sulfur subunit 21
SO_0103 formate dehydrogenase nitrate-inducible cytochrome b556 subunit 24
SO_0104 formate dehydrogenase accessory protein FdhE 22
SO_0107 formate dehydrogenase accessory protein fdhD 24
SO_0397 quinolfumarate reductase menaquinol-oxidizing subunit FrdC_2 12
SO_0452 thioredoxin 2 15
SO_0903 Na-translocating NADH-quinone reductase subunit B NqrB_1 12
SO_1012 NADH-ubiquinone oxidoreductase subunit K NuoK 12
SO_1013 NADH-ubiquinone oxidoreductase subunit J NuoJ 12
SO_1363 hydroxylamine reductase 13
57
SO_2417 ferredoxin cofactor maintenance protein YfaE 10
SO_3922 formate dehydrogenase cytochrome b Fdh -12
SO_4513 formate dehydrogenase molybdopterin-binding subunit _2 15
SO_4514 formate dehydrogenase FeS subunit FdhB_2 12
SO_4515 formate dehydrogenase cytochrome b subunit FdhC_2 14
SO_4609 aa3 type cytochrome c oxidase subunit III CoxC 11
SO_0479 periplasmic octaheme cytochrome c MccA -12
SO_0847 periplasmic nitrate reductase ferredoxin component NapG -15
SO_1519 L-lactate dehydrogenase iron-sulfur cluster-binding protein LldF -10
SO_1251 ferredoxin 4Fe-4S -11
SO_37411 hypothetical inner membrane protein -11
a fold changes were presented after log2
b positive number represented up-regulated and negative represented down-regulated
Flavin and cytochrome related genes
Flavins can be secreted by Shewanella species as electron shuttle to facilitate
extracellular electron transfer (Cansterin et al 2008) C-type cytochromes play the
important roles on the process of extracellular electron transfer (Shi et al 2007) The
genes related to flavins synthesis and the genes encoding the cytochromes electron
transport proteins were shown in the table 43 the genes SO_1414 and SO_3058 related
to flavins synthesis was up-regulated to 03 and 01 folds respectively The genes related
to s C-type cytochromes SO_0135 SO_4105 SO_0169 SO_1778 are increased to 10
08 05 04 01 folds respectively However most of genes have shown no significant
changes (less one-fold change) as response to nano particle decorated anode in MECs
58
Table 43 Expression level of flavin and cytochrome genes
Locus Tag Gene product descriptions Fold
change
SO_1414 flavocytochrome c flavin subunit putative 03
SO_3468 riboflavin synthase subunit alpha -02
SO_3058 flavocytochrome c flavin subunit 01
SO_4105 MSHA pilin protein MshA 08
SO_0169 general secretion pathway protein GspG 05
SO_1778 outer membrane decaheme cytochrome c lipoprotein MtrC 04
SO_1779 outer membrane decaheme cytochrome c OmcA 01
SO_0135 lipoprotein of unknown function DUF333 10
SO_0136 conserved hypothetical inner membrane protein 01
a fold changes were presented after log2
b positive number represented up-regulated and negative represented down-regulated
Other genes with significantly modified expression levels
These genes over 15 fold change and possibly related to current enhancements were
shown in Table 44 SO_3104 (expressed inner membrane protein) was over expressed
and the gene SO_2194 which encodes outer membrane protein (OmpA family protein)
was repressed as response to nanoparticle decorated anode however it is unclear the
relationships of this modified membrane protein genes with the current enhancement
Another interesting phenomenon was possible co-regulation of several gene clusters
indicating possible operon associations This includes three sets of genes that were
induced more than 15 fold SO_0108SO_0109 and SO_1042SO_1043SO_1044 The
59
consistency of expression of these genes under the NP conditions provides basic evidence
to support operon structure However the correlation of these significant changed gene
with current density enhance are unclear which showed there are unknown multiple and
complex responses of S oneidensis MR-1 to nanoparticle anode of MECs
Table 44 Other genes with significantly changed expression levels
Locus Tag Gene product descriptions Fold change
SO_0101 nitrate-inducible formate dehydrogenase molybdopterin-binding
subunit FdnG
18
SO_0108 integral membrane protein of unknown function DUF39 YedE 17
SO_0109 SirA family protein YedF 17
SO_0275 N-acetyl-gamma-glutamyl-phosphate reductase -31
SO_0277 ornithine carbamoyltransferase -21
SO_0279 argininosuccinate lyase -24
SO_0404 zinc dependent metalloprotease domain lipoprotein 21
SO_0956 alkyl hydroperoxide reductase F subunit -20
SO_1042 amino acid ABC transporter ATP-binding protein -18
SO_1043 amino acid ABC transporter permease protein -27
SO_1044 amino acid ABC transporter periplasmic amino acid-binding protein -27
SO_1072 chitin-binding protein putative 21
SO_1405 transglutaminase family protein 20
SO_1822 TonB-dependent receptor putative -22
SO_2069
1-(5-phosphoribosyl)-5-[(5-phosphoribosylamino)
methylideneamino] imidazole-4-carboxamide isomerase
-19
SO_2070 amidotransferase HisH -26
SO_2071 imidazole glycerol-phosphate dehydratasehistidinol phosphatase -27
60
SO_2072 histidinol-phosphate aminotransferase -28
SO_2073 histidinol dehydrogenase -30
SO_2194 OmpA family protein -36
SO_2195 inter-alpha-trypsin inhibitor domain protein -27
SO_2767 asparagine synthetase B -26
SO_2945 prophage LambdaSo tail fiber protein 20
SO_2963 prophage LambdaSo major capsid protein HK97 family 20
SO_2987 prophage LambdaSo hypothetical protein 19
SO_2988 prophage LambdaSo expressed protein of unknown function 18
SO_3104 expressed inner membrane protein 131
SO_3408 conserved hypothetical inner membrane protein -18
SO_3585 azoreductase putative -18
SO_3586 glyoxalase family protein -17
SO_3687 curli production assemblytransport component CsgE putative -18
SO_38191 hypothetical ammonia permease -19
SO_4014 AcrBAcrDAcrF family protein 18
SO_40151 type I secretion system membrane fusion protein RND family 18
SO_4054 510-methylenetetrahydrofolate reductase 18
SO_4245 N-acetylglutamate synthase -22
SO_45251 hypothetical transcriptional regulator LysR family -17
SO_4527 integral membrane domain protein -19
SO_4705 transcriptional regulator putative -19
a fold changes were presented after log2
b positive number represented up-regulated and negative represented down-regulated
Discussion
61
The addition of NP to anodes significantly impacts current densities with the elemental
composition being a critical factor The chemical composition of NPs significantly affects
the current enhancement in MECs (Fan et al 2011) because the chemical composition of
NP properties is a critical factor in determining the conductivity which significantly
affects the efficiency of electron transfer from bacteria to electrode Fe is an ideal
candidate for NP formation for current enhancement in MESs because of high
conductivity and low price compared to gold and other conductive materials Gold NPs
has also been studies for enhancement of electricity conductivity (Bao et al 2008)
However the disadvantages of the high price of gold limit their practical application in
MECs (Fan et al 2011) The different size and density of the same composition of NP
also has essential effects on the current density enhancement (Fan et al 2011) In this
study the results showed the current density enhancement with Fe-NP size range of
200plusmn100 nm and density range of 200plusmn100 nm is considerable enhancement consequence
There are a number of reports of studies of current density enhancement in MFCs
by nanostructured anodes (Fan et al 2011 Qiao et al 2008 Sharma et al 2008 Tsai et al
2009) Based on bacterial behavior on nanostructured anode surface Qiao et al (2008)
believes that Escherichia coli cells on the nanostructured electrode surface produce hair-
like structures similar to pili that could facilitate the electron transfer between the cells
and electrode The production of hair-like structures is believed to be stimulated by
nanostructures and could play the key role on current density enhancement in pili-
producing bacteria in MFCs (Qiao et al 2008) S oneidensis can produce electrically
conductive bacterial nanowires which have similar structure to the hair-like structures in
Ecoli (Gorby et al 2006 Reguera et al 2005) A number of possible genes related
62
nanowires of S oneidensis have been identified including mshA (SO_4105) and gspG
(SO_0169) (Konstantinidis et al 2009) They are up-regulated 08 and 05 fold in the
current study respectively even though they are not significant induced This result
supported the increased nanowire genes had contributions to enhance current density in
MFCs by NPs decorated anodes
Based on the previous research on electron transfer mechanisms for S oneidensis it is
known that self-produced mediators (flavins) and multihaem c-type cytochromes (c-Cyts)
may play critical roles in the electron transfer of S oneidensis to extracellular insoluble
electrode (Canstein et al 2008 Myers and Myers 1992 Scott and Nealson 1994 Shi et al
2007) The flavins secreted by S oneidensis can facilitate to transfer electrons from
bacteria to solid anode electrodes by acting as extracellular electron shuttles (Canstein et
al 2008) In this study the genes encoding flavins (SO_1414 SO_3468 and SO_3058)
had no significant increase (less than 1 fold change) as a response to nanoparticle
decorated anode in MECs which suggests that increased flavin gene expression does not
significantly contribute to the current density enhancements in nano-particle decorated
anode The genes encoding cytochromes electron transport proteins (SO_4104 SO_0417
SO_177879 SO_013536) had no significant increase (less than 1-fold change)
However those genes were up-regulated slightly These results suggest the increased
expression flavin and c type cytochromes genes had partial contributions even not
significantly to enhance current density in MFCs by NPs decorated anodes
The thickness of bacterial biofilm on the anode can affect the power generation in
MFCs (Biffinger et al 2007 Heydorn et al 2000) since increased biofilm thickness on
the anode represents more active bacteria involving in the process of electron generation
63
and transfer in MFCs Biofilm formation related genes included flagellar genes IV pilus
genes and other genes The flagellar related genes SO_3228 SO_3229 (fliB and fliK) and
IV pilus genes SO_4105 (mshA) were induced from 19 to 09 fold the genes (SO_4178
-4180) related to formation of a three-dimensional biofilm were up-regulated from 10 to
13 folds even though the genes related bacterial motility protein (SO_ 4286 SO_4287
SO_1529 and SO_1530) are induced slightly (less than 0 5 fold) However the motility
genes most probably play a role for initial biofilm attachment on the anode surfaces
(Thormann et al 2004 Thormann et al 2006) with the rest of biofilm genes being more
essential for biofilm maintenance in mature cultures as were presented at harvest in this
study This enhancement of biofilm density under conditions of increase current density
has been observed for gold NP decorated anodes using confocal light microscopy (data
not shown) but has not yet been confirmed for Fe NP decorated anodes In any event an
increase in the expression level of genes related biofilm formation therefore also
supported the possibility of enhanced biofilm formation on NPs decorated anodes thus
facilitating electron transfer from bacteria to anodes
Beliaev et al described a model of the Fe reduction pathway in S oneidensis MR-1 using
solid substrates as the electron acceptor (Beliaev et al 2001) This model proposes that
electrons are generated and released in cytoplasm then transferred to the quinone pool
eg menaquinone in the cytoplasmic membrane The reduced menaquinone in turn
reduces CymA and from there electrons are then transferred to a periplasmic shuttle (eg
CctA) and then to MtrA a periplasmic soluble decaheme cytochrome c located in
associated with the outer membrane via interaction with MtrB Finally surface displayed
outer membrane proteins decaheme cytochromes (OmcA MtrC or MtrF) can transfer
64
electron to Fe or other solid substrates (Beliaev et al 2001)) Bencheikh-Latmani et al
demonstrated that the same electron transport pathway may be used for more than one
electron acceptor through studies of the response of S oneidensis MR-1 to U(VI) and
Cr(VI) under anaerobic conditions particularly focused on the critical genes
(SO_1777mtrA SO_1776mtrB SO_1778mtrC) (Bencheikh-Latmani et al 2005)
However our studies showed that these genes had no significant expression changes in
response to nanostructured anodes Contrastingly one of expressed inner membrane
protein genes SO_3104 and one of cytochromes genes SO_4609 encoding cytochrome c
oxidase subunit III demonstrated a 13-fold and 11-fold up-regulation respectively in this
study while the gene SO_2194 which encodes outer membrane protein (OmpA family
protein) was down regulated 36 fold Our results therefore suggest that S oneidensis
MR-1 does not utilize the same set of core critical genes (SO_1777 mtrA SO_1776
mtrB SO_1778 mtrC) products to transfer electrons in all instances This is the similar
result with the study of Bretschger et al (2007) which indicated that the electron
transport system in S oneidensis MR-1 is complex with several different proteins able to
participate in electron transfer to the anode of MECs
It should also be mentioned that the gene (SO_1519) encoding lactate
dehydrogenase repressed (-12 fold) here suggesting that lactate was depleted at the time
of cell harvest from the MECs and that other pathways of anaerobiosis had been activated
including those associated with amino acid uptake and consumption the growth media
was a complex mixture including yeast extract In particular the catabolic pathway for
histidine of S oneidensis MR-1 a pathway that may formate as an intermediate is up-
regulated with the genes for histidine ammonia lyase SO_0098 SO_3057 and SO_4374
65
up-regulated 13 11 and 11-fold respectively The possible production of formate
during histidine degradation may in turn explain the high levels of formate
dehydrogenates as evidenced by upregulation of SO_0102 SO_0103 SO_0104 and
SO_0107 annotated as formate dehydrogenase proteins These genes had the highest
levels of up-regulated expression as response to Fe NP decorated anodes (greater than 2-
fold)
Acknowledgements
This research was partially supported by the US National Science Foundation CBET
0828544 and the funds from ONAMIDOD (ARL-DOD Cooperative Agreement
W911NF-07-2-0083) We thank Barbara Gvakharia and Caprice Rosato for helpful
suggestions We also thank Margaret Romine of Pacific Northwest Laboratories for
valuable comments on manuscripts We also thank anonymous reviewers for significant
suggested improvements imparted as part of prior review of this manuscript
66
Chapter 5
Global Transcriptome Analysis of the Response of Shewanella oneidensis MR-1 to
Carbon Nanotube Decorated Anodes in Microbial Electrochemical Systems
Shoutao Xu Yanzhen Fan Rebecca Schaller Frank Chaplen Jun Jiao Hong Liu
67
Abstract
Shewanella oneidensis MR-1 is an important model microorganism for metabolic studies
on the effects of different environmental factors because of its diverse respiratory
capabilities Carbon nanotube (CNT) modified graphite disks were used as anodes to
investigate the effects of nanomatrials on the performance of S oneidensis MR-1 in
microbial electrolysis cells (MECs) The current densities produced with CNT decorated
anodes were on average 56-fold higher than plain graphite anodes Whole genome
microarray analysis of gene expression showed that up-regulation of cytochromes c genes
associated with extracellular electron transfer are strongly correlated to current increases
in CNT decorated anodes Genes involved in flavin biosynthesis also contribute to
current increase in CNT decorated anode MECs
Keywords Microbial electrochemical system microbial fuel cell microbial
electrolysis cell carbon nanotube gene expression DNA microarray
68
Introduction
Shewanella oneidensis MR-1 is an important model microorganism for metabolic
studies of the effects of different environmental factors because of its diverse respiratory
capabilities It has been used for transcriptome analysis to investigate the responses to
different growth conditions and environmental stresses (Murray et al 2001) Gene
expression patterns under different conditions such as heat shock cold shock Cr (VI) U
(VI) reduction chromate stress iron and acid tolerance have been studied previously
(Bencheikh-Latmani et al 2005 Brown et al 2006 Gao et al2006 Gao et al 2004
Yang et al 2008) More recently it has been exploited as a model species for power
generation in microbial electrochemical systems (MESs)which have potential
applications in renewable energy generation bioremediation and wastewater treatment
(Logan et al 2007 Logan 2009 Park and Zeikus 2002 Fan et al 2011 Xu et al 2012)
In a MES electrochemically active microorganisms oxidize organic matter in the an
ode chamber to release electrons Electrons are then transferred to the anode electrode thr
ough various mechanisms (Logan et al 2006 Lovley 2006 Rabaey et al 2003) and fina
lly travel to the cathode electrode and combine with the terminal electron acceptor The
key feature of MESs is the microbe-catalyzed electron transfer from the organic matter to
the anode (Rabaey et al 2004) Enhancing the current output which highly depends on
the performance of the anode electrode is critical for the successful application of MESs
(Logan et al 2007 Park and Zeilus 2002 2003) Several nanomaterials including TiO2
gold (Au) and palladium (Pd) nanoparticles have been utilized to decorate electrodes to
enhance power density in MESs (Qiao et al 2008 Quan et al 2005 Fan et al 2011) The
possible mechanisms for increased current densities have been studied by using Fe
69
nanoparticle-decorated anodes (Xu et al 2012)
Carbon nanotubes (CNT) are cylindrically shaped nanomaterials with extremely
high surface area excellent electrical conductivity and chemical inertness (He et al
2005a Serp et al 2003) These unique properties make CNT a promising electrode
material (Liang et al 2008) The biocompatibility of microorganisms and carbon
nanostructures has been evaluated (Morozan et al 2007) The effects of CNT on anodic
biofilms have been tested in MFCs (Liang et al 2011) However no studies have been re
ported on global transcriptome analysis for the response of S oneidensis MR-1 to CNT
decorated anode in MECs
In the present study the graphite disks decorated with CNT were used as anodes to
investigate the effects of nanomaterials on current generation in multi-anode MECs
DNA microarrays were used to analyze differences in the global gene expression profile
of S oneidensis MR-1 grown on plain versus CNT decorated anodes
Materials and methods
Bacterial cultures S oneidensis MR-1 was purchased from American Type Culture
collection (ATCC 700550) and stored as freezer stocks at -80degC Stock S oneidensis
MR-1 cells were grown aerobically using M1 medium at 30degC Cells in early stationary
growth phase (optical density of 15-16 at 600nm (Spectrophotometer UV-1700
Shimadzu Columbia MD) were injected into the chamber of the MECs for current
production Sodium lactate (final concentration 30mM) was added as the additional
electron donor
Fabrication and characterization of nanostructured anodes Superfine isomolded
graphite disks (16 cm x 05 cm GraphiteStorecom Inc) were polished with ultrafine
70
sand paper (2000 grit 3M Company) as the base for the CNT decorated and control
anodes Aligned multi-walled carbon nanotube samples were synthesized via plasma
enhanced chemical vapor deposition (PECVD) Samples superfine isomolded graphite
pillars with a 16 cm diameter and 05 cm thickness were sputtered with a thin film of Ni
in a CRC 300 magnetron sputter coater for 1 to 2 min They were then placed in the
PECVD reactor where the Ni was annealed into small catalyst islands C2H2 was bled
into the chamber for growth and an NH3 plasma was used to vertically align the growth
of the multi-walled carbon nanotubes (MWCNTs) The temperature and time were varied
between 725degC and 775oC and 15 to 60 min to control the length of the tubes and density
of samples Samples were then analyzed in a FEI Sirion field emission scanning electron
microscope (FESEM) SEM images of CNT decorated anode and control surfaces were
shown in the Figure 51
Figure 51 SEM images of CNT decorated anodes (A) Graphite disk control (B) CNT
decorated anode
Multiple channel MECs construction and operation A MECs with multiple
removable anodes each with an effective anode area of 07 cm2
was constructed and used
to evaluate the effects of nanostructures on the current density of according to a previous
B A
71
study (Fan et al 2011) The cathode was made of wet-proof (30) carbon cloth (type B
E-TEK Division Inc USA) coated with PtC (20 E-TEK Division Inc USA)
following a previously reported procedure (Liu et al 2005) The final platinum loading
was 05 mgcm2 per projected cathode area The size of the cathode (150 cm
2) was 25
times larger than the total effective area of the 8 graphite disk anodes (56 cm2) to prevent
cathode limitations on the performance of the MECs system All MECs with CNT
decorated anodes and control anodes were tested for current generation for 24 h with
sterile a medium (without bacteria) before injecting the bacterial cells in order to
determine whether the CNT decorated anodes demonstrate chemical current generation
compared to control anodes All the testing anodes were set up in the MECs
simultaneously so that the S oneidensis MR-1 culture was applied to all of them
simultaneously Short electrode spacing (17 cm) was used in the MECs design to reduce
the internal resistance A voltage of 06 V was applied the MECs for current generation
after the cells of S oneidensis MR-1 were inoculated into the MECs in the early
stationary phase A multimeter with a data acquisition system (2700 Keithly USA) was
used to monitor the current change by measuring the voltage drop through a resistor
Microarray analysis Biofilms for whole gene microarray analysis were aseptically
removed from the plain and CNT decorated anodes of MECs after 80 hours further
incubation at 30oC when current density obviously reached to a stable phase Total RNA
was extracted using Trizol (Ambion inc AustinTX) following the manufacturerrsquos
instructions Integrity of the RNA samples was confirmed using the Agilent 2100
Bioanalyzer (Agilent Technology Inc Palo Alto CA) Total RNA (200 ng) from each
sample was amplified using the MessageAmpTM
II-Bacteria Prokaryotic RNA
72
Amplification kit (Ambion AustinTX) according to the manufacturerrsquos instructions The
S oneidensis MR-1 whole-gene microarrays representing all of the 4295 genes (resource
from NCBI) were ordered from Roche NimbleGen Inc (Madison WI) Biological
triplicates of biofilms on the CNT decorated anodes and control were analyzed cDNA
synthesis labeling and hybridization were carried out by the CGRB Core Laboratories at
Oregon State University DNASTAR ArrayStarTM
3 software was used to identify genes
that were up- or down-regulated more than 2-fold when grown on the nanoparticle
modified anodes using the unpaired two sample t-test with a cutoff p-value of 005 with
bonferroni correction
Results and discussion
Enhancement of current generation using CNT decorated anodes in MECs The
current density generated by CNT decorated anode MECs with S onidensis MR-1
increased to 36 microAcm2 10 hours post-inoculation and reached a maximum of 40 microAcm
2
at 29 hours inoculation then gradually decreased The current density stabilized at
30microAcm2 after 78 h operation Figure 52 shows that the current density curve generated
by CNT decorated anode in MECs The average current density generated by CNT
decorated anodes was 56 times that of the control The current density generated in the
MECs without bacteria was negligible (data not shown) This result demonstrates that the
current enhancement observed with the CNT decorated anodes was biologically-derived
A
73
Figure 52 Effects of CNT decorated anodes on the current density in MECs (3 replicates)
Control is plain graphite disk anode CNT is the anode with CNT decoration (Error bars
represent standard deviation)
It has been reported that nanostructured decorated anodes have significant impacts on
current densities (Fan et al 2011 Qiao et al 2006 Sharma 2008 Tsai et al 2009) The
CNT decorated anodes had significantly enhanced current densities in MECs in this
study This result is consistent with previous studies Interestingly the current density
curve generated by CNT decorated anode MECs is quite different from the current
density curve generated by Au NP and Pd NP decorated anodes using same bacterial
species (Fan et al 2011) In the Au and Pd NP decorated anode MECs the current
density curve was not significantly increased in the beginning compared with the control
but gradually increased after the inoculation the current density reached the maximum
current density more than 50 hours after the inoculation However the current density
generated by CNT decorated anode MECs started to increase significantly immediately
after the inoculation It reached a maximum current density 29 hours post-inoculation
-1
6
13
20
27
34
41
48
0 10 20 30 40 50 60 70 80
Curr
ent
den
sity
(micro
Ac
m2)
Time (Hours)
Control
CNT
74
These results indicate that there might be different current density enhancement
mechanisms between CNT with other metal NP decorated anodes in MECs
Global transcriptome analysis Whole-genome DNA microarrays were used to attain a
comprehensive general overview of the transcriptional response of S oneidensis
MR-1 of
S oneidensis MR-1 to CNT decorated anodes in MECs In a total of 4295 genes analyzed
457 genes (250 genes up-regulated and 207 genes down-regulated) exhibited significantly
(P lt 005) changed expression at a 2-fold level in at least 3 replicates in the response to
CNT The total number of the regulated genes present 11 of the 4295 open reading
frames (ORFs) presented on the microarray The total number of genes at significant
expression levels in response to CNT decorated anodes is close to that to Fe NP decorated
anodes in MECs (392 9) ( Xu et al 2012)
Fig 53 Differentially expressed genes grouped by functional classification according to
the TIGR S oneidensis genome database (wwwtigrorg) Columns 1 amino acid
biosynthesis 2 biosynthesis of cofactors prosthetic groups and carriers 3 cell
envelope 4 cellular processes 5 central intermediary metabolism 6 DNA metabolism
7 energy metabolism 8 fatty acid and phospholipid metabolism 9 other categories 10
protein fate 11 protein synthesis 12 purines pyrimidines nucleosides and nucleotides
-80
-60
-40
-20
0
20
40
60
80
100
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18
Nu
mb
er
of
Ge
ne
s
Up-regulated
Down-regulated
75
13 regulatory functions 14 signal transduction 15 transcription 16 transport and
binding proteins 17 unknown function 18 hypothetical proteins
The overall transcript genomic response of S oneidensis MR-1 to CNT decorated
anodes was summarized in Figure 53 by grouping the differentially expressed genes into
their functional role categories based on The Institute for Genomic Researchrsquos annotation
(Version 8) of the MR-1 genome sequence The wide distribution of putative functional
roles attributed to the differentially expressed genes indicated the extent of the molecular
response of Soniedenis MR-1 to the CNT decorated anodes A large number of up-
regulated genes were presented in genes associated with biosynthesis of cofactors
prosthetic groups and carriers (group 2) cell envelope (group 3) protein fate (group 10)
protein synthesis (group 11) and purines pyrimidines nucleosides and nucleotides
(group 12) in this study The number of up-regulated genes was two times more than that
of down-regulated genes in each functional group Contrastingly in the genomic response
S oneidensis MR-1 to Fe NP decorated anodes (chapter 4) most of up-regulated genes
were associated with cellular processes (group 4) energy metabolism (group 7) other
categories (group 9) and hypothetical proteins (group 18) The down-regulated genes
presented in the amino acid biosynthesis (group 1) cellular processes (group 4) substrate
transport (group 16) and hypothetical proteins (group 18) While those genes encoding
proteins involved in amino acid biosynthesis (group 1) central intermediary metabolism
( group 5) DNA metabolism( group 6) fatty acid and phospholipid metabolism (group
8) and other categories (group 9) were among the most down-regulated genes in this
study However the two groups of genes that were the most noticeable between the
genomic response S oneidensis MR-1 to CNT and Fe NP decorated anodes which they
are the genes related to energy metabolism (group 7) and hypothetical proteins (group
76
18) These results indicated there are different and common genomic responses to S
oneidensis MR-1 on CNT and Fe NP decorated anodes in MECs
Electron transfer related genes Based on previous electron transfer mechanism
studies on Shewanella a serial of group proteins collectively described as the Mtr
pathway play the role to transfer electron rom the inner bacterial body to the outer
membrane (Beliaev et al 2001 Shi et al 2007 Brutinel and Gralnick 2012) In the Mtr
pathway firstly electrons generated by bacteria received by CymA a tetraheme c-type
cytochrome anchored in the inner membrane then electrons were transferred to MtrA a
periplasmic decaheme c-type cytochrome After that electrons were transferred from
MtrA to MtrC an outer-membrane decaheme c-type cytochrome is facilitated by MtrB a
non-heme containing integral outer-membrane proteinOuter-membrane decaheme c-type
cytochrome MtrC and OmcA are in change of electron transfer to exreacellular electron
acceptor (Shi et al 2007 Brutinel and Gralnick 2012 ) These genes encoding electron
transport proteins are shown in the table 51 The critical genes SO_1777 mtrA SO_1776
mtrB SO_1778 mtrC are unregulated 41 to 80 fold in response to CNT decorated
anodes other cytochrome c genes related to electron transfer in MtrA pathway also are
significantly increased in response to CNT decorated anodes These genes encoding outer
membrane proteins have different level increases supporting that the increase of
cytochrome c gene expression contributes to the current enhancements of MECs as
response to CNT decorated anodes
77
Table 51 Expression level of cytochrome c as response to CNT decorated anodes
Locus Tag Gene product descriptions Fold change CNT
SO_0165 general secretion pathway protein GspC 31
SO_0167 general secretion pathway protein GspE 45
SO_1776 outer membrane protein precursor MtrB 41
SO_1777 outer membrane decaheme cytochrome c
lipoprotein MtrA
80
SO_1778 outer membrane decaheme cytochrome c
lipoprotein MtrC
43
SO_1779 outer membrane decaheme cytochrome c
OmcA
63
SO_0135 lipoprotein of unknown function DUF333 22
SO_0136 conserved hypothetical inner membrane protein 24
Flavin related genes It is known that self-produced mediators play critical roles in the
electron transfer of S oneidensis to extracellular insoluble electrode (Canstein von et al 2
008 Myers et al 1992 Scott et al 1994 Shi et al 2007) It has been proved that the
flavins secreted by S oneidensis can facilitate electron transfer from bacteria to solid
anode electrodes by acting as extracellular electron shuttles (Cansterin von et al 2008) T
wo critical genes (SO_1414 and SO_3468) related to flavin synthesis were shown increa
sed significantly (more than 2 folds) increase in this study (Table 52) This result support
s the hypothesis that flavin production increases as the response to CNT decorated anodes
in MECs have significant contribution to the current density enhancements
78
Table 52 Expression levels of genes related to flavin synthesis as response to CNT
decorated anode
Locus Tag Gene product descriptions Fold change CNT
SO_1414 flavocytochrome c flavin subunit putative 22
SO_3468 riboflavin synthase subunit alpha 20
The amount of bacterial biofilm biomass on the anode can affect the power
generation in MFCs (Biffinger et al 2007 Heydorn et al 2000) Biofilm formation
related genes include flagellar gene IV pilus genes and other genes the flagellar related
genes (fliB and fliK) and IV pilus genes (mshA) were induced only from 105 to 134
fold as response of S oneidensis MR-1 to CNT decorated anodes in this study No
biofilm-associated genes presented significant changes in expression levels in this study
indicating there is no direct connections between biofilm enhancements to CNT
decorated anodes in this study This result is consistent with the Liang et al (2011)
conclusion that using CNT enhanced the electrochemical activity of anodic biofilms but
did not result in a significant increase of biomass in the anodic biofilms
Our microarray results showed two significantly up-regulated gene groups the
genes encoding proteins localized on the outer membrane and the genes involved in
flavin biosynthesis contributed to current density enhancement by CNT decorated anodes
Among 457 significantly changed genes to CNT in this study there are also a relatively
large number of genes encoding proteins with unknown functions which are either up- or
down-regulated in the response to CNT decorated anodes which indicated more
79
complicated responses of S oneidensis MR-1 to CNT decorated anodes in MECs and
further study is needed
80
Chapter 6
Summary
Microbial electrochemical systems (MESs) with a mixed culture initially inoculated
from Corvallis wastewater treatment plant have been studied for more than 6 years for
varying purposes including power generation hydrogen production heavy metal
removal and wastewater treatment Experiment results have shown that the mixed culture
is quite stable with excellent performance in MESs Our community analysis using
denaturing gradient gel electrophoresis (DGGE) and 16S rDNA clone library construction
suggests that the mixed culture is composed predominantly of Geobacter sp (66)
Arcobacter sp (12) and Citrobacter sp (11) These results not only improved our
understanding of the mixed culture community but also guided our studies on the
cultivation and isolation of the different bacterial species
Exoelectrogenic Citrobacter sp SX-1 was first isolated and characterized from a
MFC operated with the mixed culture Citrobacter sp SX-1 has been demonstrated to
produce electricity from wide range of different substrates including citrate acetate
glucose sucrose glycerol and lactosein MFCs Cyclic voltammetry analysis indicated
that membrane associated proteins may play an important role in facilitating the electrons
transferring from bacteria to electrode The strain SX-1 increased the known diversity of
power generating exoelectrogens and provided a unique bacterial species for study in
renewable energy generation and waste treatment
81
To enhance the electron transfer from bacteria to anode nano decorated anodes
including Fe NP and CNT were developed and characterized and evaluated in MECs
using Shewanella oneidensis MR-1 as a model species Both nanostructures have
significantly increased current density compared with the control Whole genome
microarray analysis elucidated the possible mechanisms of power enhancement in
response to these nano-decorated anodes
These results benefit to understanding of physiology and ecology of mixed
cultures in MFCs and improve the efficiency of current generation in MESs which will
facilitate the viability of niche applications for MESs in near future
82
Bibliography
1 Amman R Ludwig W (2000) Ribosomal RNA-targeted nucleic acid probes for
studies in microbial ecology FEMS Microbiol Ecol 24 555ndash565
1 Bao SJ Li CM Zang JF Cui XQ Qiao Y Guo J (2008) Novel Nanoporous TiO2
Electrode for Directly Electrochemistry of glucose oxidase Adv Funct Mater
18(4) 591-599
2 Beliaev AS Saffarini DA McLaughlin JL Hunnicutt D (2001) MtrC an outer
membrane decahaem c cytochrome required for metal reduction in Shewanella
putrefaciens MR-1 Mol Microbiol 39722-30
3 Bencheikh-Latmani R Williams SM Haucke L Criddle CS Wu L Criddle CS
Zhou J Tebo BM (2005) Global transcriptional profiling of Shewanella oneidensis
MR-1 during Cr(VI) and U(VI) reduction Appl Environ Microbiol 71 7453ndash7460
4 Biffinger JC Pietron J Ray R Little B and Ringeisen BR (2007) A biofilm
enhanced miniature microbial fuel cell using Shewanella oneidensis DSP10 and
oxygen reduction cathodes Biosens Bioelectron 221672ndash1679
5 Bodelier PLE Roslev P Henckel T FenzelP (2000) Stimulation by ammonium
based fertilizers of methane oxidation in soil around rice root Nature 40 421ndash424
6 Bond DR Holmes DE Tender LM Lovley DR (2002) Electrode-reducing
microorganisms that harvest energy from marine sediments Science 295 483-485
7 Bond DR Lovley DR (2003) Electricity production by Geobacter sulfurreducens
attached to electrodes Appl Environ Microbiol 69 1548-1555
8 Bond DR Lovley DR (2005) Evidence for involvement of an electron shuttle in
electricity generation by Geothrix fermentans Appl Environ Microbiol 71 2186-
2189
9 Bretschger O Obraztsova A Sturm CA Chang IS Gorby YA Reed SB Culley
DE Reardon CL Barua S Romine MF Zhou J Beliaev AS Bouhenni R
Saffarini D Mansfeld F Kim BH Fredrickson JK Nealson KH (2007) Current
production and metal oxide reduction by Shewanella oneidensis MR-1 wildtype and
mutants Appl Environ Microbiol 73 7003ndash7012
10 Brown SD Thompson MR Verberkmoes NC Chourey K Shah M Zhou J
Hettich RL and Thompson DK (2006) Molecular dynamics of the Shewanella
oneidensis response to chromate stress Mol Cell Proteomics 51054-1071
11 Brutinel ED Gralnick JA (2012) Shuttling happens soluble flavin mediators of
extracellular electron transfer in Shewanella Appl Microbiol Biotechnol 93(1) 41-8
12 Butler JE Kaufmann F Coppi MV Nuacutentildeez C Lovley DR (2004) MacA a diheme
c-type cytochrome involved in Fe(III) reduction by Geobacter sulfurreducens J
Bacteriol 186(12) 4042ndash4045
83
13 Caccavo F Lonergan DJ Lovley DR Davis M Stolz JF McInerney MJ (1994)
Geobacter sulfurreducens sp nov a hydrogen- and acetate oxidizing dissimilatory
metal-reducing microorganism Appl Environ Microbiol 60 3752ndash3759
14 Canstein von H Ogawa J Shimizu S Lloyd JR (2008) Secretion of flavins by
Shewanella species and their role in extracellular electron transfer Appl Environ
Microbiol 74 615ndash623
15 Cao X Huang X Liang P Xiao K Zhou Y Zhang X Logan BE (2009) A New
Method for Water Desalination Using Microbial Desalination Cells Environ Sci
Technol 43(18)7148-7152
16 Chaudhuri SK Lovley DR (2003) Electricity generation by direct oxidation of
glucose in mediatorless microbial fuel cells Nat Biotechnol 21 1229-1232
17 Cheng S Liu H Logan B E (2006) Increased power generation in a continuous
flow MFC with advective flow through the porous anode and reduced electrode
spacing Environ Sci Technol 402426-2432
18 Cheng S Logan B E (2007) Sustainable and efficient biohydrogen production via
electrohydrogenesis PNAS 104 18871-18873
19 Chignell J (2010) Energy Recovery from Biodiesel Waste Performance of
Microbial Electrochemical Systems on Glycerol Thesis
20 Chung K Okabe S (2009) Characterization of electrochemical activity of a strain
ISO2-3 phylogenetically related to Aeromonas sp isolated from a glucose-fed
microbial fuel cell Biotechnol Bioeng 104(5) 901-910
21 Cole JR Chai B Farris RJ Wang Q Kulam-Syed-Mohideen AS McGarrell
DM Bandela AM Cardenas E Garrity GM Tiedje JM (2007) The ribosomal
database project (RDP-II) introducing myRDP space and quality controlled public
data Nucleic Acids Res 35 (Database issue) D169-D172 doi 101093nargkl889
22 Cole JR Wang Q Cardena E Fish J Chai B Farris RJ Kulam-Syed-Mohideen
AS McGarrell DM Marsh T Garrity GM Tiedje JM (2009) The Ribosomal
Database Project improved alignments and new tools for rRNA analysis Nucleic
Acids Res 37 (Database issue) D141-D145 doi 101093nargkn879
23 Dey A De S De A De SK (2004) Characterization and dielectric properties of
polyaniline-TiO2 nanocomposites Nanotechnol 15 1277-1283
24 Edwards U Rogall T Bloumlcker H Emde M Boumlttger EC (1989) Isolation and
direct complete nucleotide determination of entire genesmdashcharacterization of a gene
coding for 16S-ribosomal RNA NucleicAcids Res 17 7843ndash7853
25 Eggleston CM Voumlroumls J Shi L Lower BH Droubay TC Colberg PJS (2008)
Binding and direct electrochemistry of OmcA an outer-membrane cytochrome from
an iron reducing bacterium with oxide electrodes a candidate biofuel cell system
Inorg Chim Acta 361769ndash777
26 Fan Y Sharbrough E Liu H (2008) Quantification of the Internal Resistance
Distribution of Microbial Fuel Cells Environ Sci Technol 42 8101ndash8107
84
27 Fan Y Xu S Schaller R Jiao J Chaplen F Liu H (2011) Nanoparticle decorated
anodes for enhanced current generation in microbial electrochemical
cells Biosensors and Bioelectronics 26 (5)1908ndash1912
28 Fan Y Hu H Liu H (2007)Sustainable power generation in microbial fuel cells
using bicarbonate buffer and proton transfer mechanisms Environ Sci
Technol 41(23) 8154-8
29 Fedorovich V Knighton MC Pagaling E Ward FB Free A Goryanin I (2009)
Novel electrochemically active bacterium phylogenetically related to Arcobacter
butzleri isolated from a microbial fuel cell Appl Environ Microbiol 75(23) 7326-
7334
30 Ferna ndez A Huang S Seston S Xing J Hickey R Criddle C Tiedje J (1999)
How stable is stable Function versus community composition Appl Environ
Microbiol 65 3697ndash3704
31 Field SJ Dobbin PS Cheesman MR Watmough NJ Thomson AJ Richardson
DJ (2000) Purification and magneto-optical spectroscopic characterization of
cytoplasmic membrane and outer membrane multiheme c-type cytochromes from
Shewanella frigidimarina NCIMB400 J Biol Chem 275 8515ndash8522
32 Freguia S Masuda M Tsujimura S Kano K (2009) Lactococcus lactis catalyses
electricity generation at microbial fuel cell anodes via excretion of a soluble quinone
Bioelectrochemistry 7614ndash18
33 Fricke K Harnisch F and Schroumlder U (2008) On the use of cyclic voltammetry for
the study of the anodic electron transfer in microbial fuel cells Energy Environ
Sci 1 144ndash147
34 Gao H Wang Y Liu X Yan T Wu L Alm E Arkin A Thompson DK and
Zhou J (2004) Global transcriptome analysis of the heat shock response of
Shewanella oneidensis J Bacteriol 1867796-7803
35 Gao H Yang ZK Wu L Thompson DK Zhou J (2006) Global transcriptome
analysis of the cold shock response of Shewanella oneidensis MR-1 and mutational
analysis of its classical cold shock proteins J Bacteriol 1884560-4569
36 George MG (2005) Bergeyrsquos Manual of Systematic Bacteriology Volume Two The
Proteobacteria Part B The Gammaproteobacteria Second Edition New York
Springer 651p
37 Gonzaacutelez-Toril E Llobet-Brossa E Casamayor EO Amann R Amils R (2003)
Microbial ecology of an extreme acidic environment the Tinto river Appl Environ
Microbiol 69(8) 4853ndash4865
38 Gorby YA Yanina S Mclean JS Rosso KM Moyles D Dohnalkova A
Beveridge TJ Chang IS Kim BH Kim KS Culley DE Reed SB Romine M
F Saffarini DA Hill EA Shi L Elias DA Kennedy DW Pinchuk G
Watanabe K Ishii S Logan B Nealson KH Fredrickson JK (2006) Electrically
conductive bacterial nanowires produced by Shewanella oneidensis strain MR-1 and
other microorganisms Proc Natl Acad Sci USA 10311358-11363
85
39 He Z Wagner N Minteer SD Angenent LT (2006) An upflow microbial fuel cell
with an interior cathodes assessment of the internal resistance by impedance
spectroscopy Environ Sci Technol 40 5212-5217
40 Heydorn A Nielsen AT Hentzer M Sternberg C Givskov M Ersboslashll BK Molin
S (2000) Quantification of biofilm structures by the novel computer program
COMSTAT Microbiol 1462395-2407
41 Holmes DE Bond DR Lovley DR (2004a) Electron transfer by Desulfobulbus
propionicus to Fe(III) and graphite electrodes Appl Environ Microbiol 701234ndash
1237
42 Holmes DE Chaudhuri SK Nevin KP Mehta T Methe BA Liu A Ward JE
Woodard TL Webster J Lovley DR (2006) Microarray and genetic analysis of
electron transfer to electrodes in Geobacter sulfurreducens Environ Microbiol
81805-1815
43 Holmes DE Nicoll JS Bond DR Lovley DR (2004b) Potential role of a novel
psychrotolerant member of the family Geobacteraceae Geopsychrobacter
electrodiphilus gen nov sp nov in electricity production by a marine sediment
fuel cell Appl Environ Microbiol 70 6023ndash6030
44 Holmes DE Chaudhuri SK Nevin KP Mehta T Metheacute BA Liu A Ward JE
Woodard TL Webster J Lovley DR (2006) microarray and genetic analysis of
electron transfer to electrodes in Geobacter sulfurreducens Environ Microbial
8(10)1805-1815
45 Hu H (2009) Enhancing Hydrogen Production in Microbial Electrolysis Cells
through Development of Platinum-free Cathode and Improvement of Reactor
Dissertation
46 Ince BK Ayman O N Turker G Ccedilelikkol S Ince O (2010) Microbial ecology of
anaerobic reactors for treatment of alcohol industry wastewaters a review Current
research technology and education topics in applied microbiology and microbial
ecology 988-999
47 Jung S Regan JR (2007) Comparison of anode bacterial communities and
performance in microbial fuel cells with different electron donors Appl Microbiol
Biotechnol 77393ndash 402
48 Kim BH Kim HJ Hyun MS Park DH (1999) Direct electrode reaction of Fe(III)
reducing bacterium Shwwanella putrefaciens J Microbial Biotechnol 9127-131
49 Kim GT Hyun MS Chang IS Kim HJ Park HS Kim BH Kim SD Wimpenny
JW Weightman AJ (2005) Dissimilatory Fe(III) reduction by an electrochemically
active lactic acid bacterium phylogenetically related to Enterococcus gallinarum
isolated from submerged soil J Appl Microbiol 99(4) 978ndash987
50 Kim GT Webster G Wimpenny JW Kim BH Kim HJ Weightman AJ (2006)
Bacterial community structure compartmentalization and activity in a microbial fuel
cell J Appl Microbiol 101(3) 698ndash710
86
51 Kim HJ Park HS Hyun MS Chang IS Kim M Kim BH (2002) A mediator-
less microbial fuel cell using a metal reducing bacterium Shewanella putrefaciens
Enzyme Microb Technol 30 145-152
52 Kim JR Min B Logan BE (2005) Evaluation of procedures to acclimate a
microbial fuel cell for electricity production Appl Microbiol Biotechnol 68(1)23-30
53 Kim JR Cheng S Oh SE Logan BE (2007) Power generation using different
cation anion and ultrafiltration membranes in microbial fuel cells Environ Sci
Technol 41 1004-1009
54 Konstantinidis KT Serres MH Romine MF Rodrigues JL Auchtung J McCue
LA Lipton MS Obraztsova A Giometti CS Nealson KH Fredrickson JK
Tiedje JM (2009) Comparative systems biology across an evolutionary gradient
within the Shewanella genus Proc Natl Acad Sci U S A 106(37)15909-15914
55 Lee J Phung NT Chang IS Kim BH Sung HC (2003) Use of acetate for
enrichment of electrochemically active microorganisms and their 16S rDNA
analyses FEMS Microbiol Lett 223185ndash191
56 Liang P Wang H Xia X Huang X Mo Y Cao X Fan M(2011)Carbon
nanotube powders as electrode modifier to enhance the activity of anodic biofilm
in microbial fuel cells Biosens Bioelectron 26(6)3000-4
57 Lies DP Hernandez ME Kappler A Mielke RE Gralnick JA Newman DK
(2005) Shewanaella oneidensis MR-1 uses overlapping pathways for iron reduction
at a distance and by direct contact under conditions relevant for biofilms Appl
Environ Microbiol 71 4414-4426
58 Liu H and Logan B E (2004) Electricity generation using an air-cathode single
chamber microbial fuel cell in the presence and absence of a proton exchange
membrane Environ Sci Technol 38 4040-4046
59 Liu H Grot S Logan BE (2005) Electrochemically assisted microbial production
of hydrogen from acetate Environ Sci Technol 39 4317 ndash 320
60 Liu H Hu H Chignell J Fan Y (2010) Microbial Electrolysis Novel Technology
for Hydrogen Production from Biomass Biofuels 1(1) 129ndash142
61 Liu H Ramnarayanan R Logan BE (2004) Production of electricity during
wastewater treatment using a single chamber microbial fuel cell Environ Sci Technol
382281-2285
62 Liu WT Marsh TL Cheng H Forney LJ (1997) Characterization of microbial
diversity by determining terminal restriction fragment length polymorphisms of gene
encoding 16S rRNA Appl Environ Microbiol 63 4516ndash4522
63 Logan B E Hamelers B Rozendal R Schroder U Keller J Freguia S Aelterman
P Verstraete W Rabaey K (2006) Microbial fuel cells methodology and
technology Environ Sci Technol 40 (17) 5181-5192
64 Logan B E and Regan JM (2006) Electricity-producing bacterial communities in
microbial fuel cells Trends Microbiol 14512ndash518
87
65 Logan B Cheng S Watson V Estadt G (2007) Graphite fiber brush anodes for
increased power production in air-cathode microbial fuel cells Environ Sci Technol
41(9)3341-3346
66 Logan BE (2009) Exoelectrogenic bacteria that power microbial fuel cells Nat Rev
Microbiol 5 375-381
67 Logan BE Call D Cheng S Hamelers HV Sleutels TH Jeremiasse AW
Rozendal RA (2008) Microbial electrolysis cells for high yield hydrogen gas
production from organic matter Environ Sci Technol 42 8630-8640
68 Logan BE Hamelers B Rozendal R Schroder U Keller J Freguia S Aelterman
P Verstraete W Rabaey K (2006) Microbial fuel cells methodology and
technology Environ SciTechnol 40 5181-5192
69 Lovely DR (2006) Bug juice harvesting electricity with microorganisms Nature
Review| Microbiology 4 497-508
70 Lovley DR Phillips EJ (1988) Novel Mode of Microbial Energy Metabolism
Organic Carbon Oxidation Coupled to Dissimilatory Reduction of Iron or
Manganese Appl Environ Microbiol 54(6) 1472-1480
71 Ly HK Sezer M Wisitruangsakul N Feng JJ Kranich A Millo D Weidinger
IM Zebger I Murgida DH Hildebrandt P (2011) Surface-enhanced vibrational
spectroscopy for probing transient interactions of proteins with biomimetic interfaces
electric field effects on structure dynamics and function of cytochrome c FEBS J
278(9)1382-90
72 Macaskie LE Hewitt CJ Shearer JA Kent CA (1995) Biomass production for
the removal of heavy metals from aqueous solutions at low pH using growth-
decoupled cells of a Citrobacter sp Int Biodeter Biodegr 3573ndash92
73 Malki M De Lacey AL Rodriguez N Amils R Fernandez VM (2008)
Preferential use of an anode as an electron acceptor by an acidophilic bacterium in
the presence of oxygen Appl Environ Microbiol 74 4472ndash4476
74 Meitl LA Eggleston CM Colberg PJS Khare N Reardon CL Shi L (2009)
Electrochemical interaction of Shewanella oneidensis MR-1 and its outer membrane
cytochromes OmcA and MtrC with hematite electrodes Geochim Cosmochim
Ac 73(18) 5292-5307
75 Morozan A Stamatin L and Nastase
F (2007) The biocompatibility
microorganisms-carbon nanostructures for applications in microbial fuel cells Phys
Stat Sol 6 1797-1803
76 Morris JM Jin S Crimi B Pruden A (2009) Microbial fuel cell in enhancing
anaerobic biodegradation of diesel Chem Eng J 146 161ndash167
77 Murray AE Lies D Li G Nealson K Zhou J Tiedje JM (2001) DNADNA
hybridization to microarrays reveals gene-specific differences between closely
related microbial genomes Proc Natl Acad Sci U S A 98 9853-9858
78 Muyzer G de Waal EC Uitterlinden AG (1993) Profiling of complex microbial
communities by denaturing gradient gel electrophoresis analysis of polymerase chain
88
reaction amplified genes coding for 16S rRNA Appl Environ Microbiol 59 695ndash
700
79 Myers CR Myers JM (1992) Localization of cytochromes to the outer membrane
of anaerobically grown Shewanella putrefacians MR-1 J Bacteriol 174 3429ndash3438
80 Narde GK Kapley A Purohit HJ (2004) Isolation and characterization of
Citrobacter strain HPC255 for broad-range substrate specificity for chlorophenols
Curr Microbiol 48(6) 419-423
81 Okabe S Ito T Sugita K Satoh H (2005) Succession of internal sulfur cycles and
sulfur-oxidizing bacterial communities in microaerophilic wastewater biofilms Appl
Environ Microbiol 71(5) 2520-2529
82 Park DH and Zeikus JG (2000) Electricity generation in microbial fuel cells using
neutral red as an electronophore Appl Environ Microbial 66 292-1297
83 Park DH and Zeikus JG (2002) Impact of electrode composition on electricity
generation in a single-compartment fuel cell using Shewanella putrefaciens Appl
Microbiol Biotechnol 5958ndash56
84 Park DH and Zeikus JG (2003) Improved fuel cell and electrode designs for
producing electricity from microbial degradation Biotechnol Bioeng 81 348ndash355
85 Park HS Kim BH Kim HS Kim HJ Kim GT Kim M Chang IS Park Y
K Chang HI (2001) A novel electrochemically active and Fe(III)-reducing
bacterium phylogenetically related to Clostridium butyricum isolated from a
microbial fuel cell Anaerobe 7 297-306
86 Parot S Nercessian O Delia ML Achouak W Bergel A (2009) Electrochemical
checking of aerobic isolates from electrochemically active biofilms formed in
compost J Appl Microbiol 106(4) 1350-1359
87 Pham CA Jung SJ Phung NT Lee J Chang IS Kim BH Yi H Chun J
(2003) A novel electrochemically active and Fe(III)-reducing bacterium
phylogenetically related to Aeromonas hydrophila isolated from a microbial fuel cell
FEMS Microbiol Lett 223 129-134
88 Qiao Y Bao SJ Li CM Cui XQ Lu ZS Guo J (2008) Nanostructured
polyanilinetitanium dioxide composite anode for microbial fuel cells ACS Nano 2
113-119
89 Qiu R Zhao B Liu J Huang X Li Q Brewer E Wang S Shi N (2009) Sulfate
reduction and copper precipitation by a Citrobacter sp isolated from a mining area J
Hazard Mater 1641310-1315
90 Quan X Yang SG Ruan XL and Zhao HM (2005) Preparation of titania
nanotubes and their environmental applications as electrode Environ Sci Technol 39
3770ndash3775
91 Rabaey K Boon N Hofte M Verstraete W (2005) Microbial phenazine
production enhances electron transfer in biofuel cells Environ Sci Technol 393401ndash
3408
89
92 Rabaey K Boon N Siciliano S D Verhaege M Verstraete W (2004) Biofuel cells
select for microbial consortia that self-mediate electron transfer Appl Environ
Microbiol 70(9) 5373ndash5382
93 Rabaey K Lissens G Siciliano S D Verstraete W (2003) Amicrobial fuel cell
capable of converting glucose to electricity at high rate and efficiency Biotechnol
Lett 25 1531-15
94 Rabaey K Verstraete W (2005) Microbial fuel cells novel biotechnology for
energy generation Trends Biotechnol 23(6) 291-298
95 Reguera G McCarthy KD Mehta T Nicoll JS Tuominen MT Lovley DR
(2005) Extracellular electron transfer via microbial nanowires Nature 435 1098-
1101
96 Reguera G Nevin KP Nicoll JS Covalla SF Woodard TL Lovley DR (2006)
Biofilm and nanowire production leads to increased current in Geobacter
sulfurreducens fuel cells Appl Environ Microbiol 72 7345-7348
97 Reimers CE Tender LM Fertig S Wang W (2001) Harvesting energy from the
marine sediment-water interface Environ Sci Technol 35192-195
98 Rezaei F Xing D Wagner R Regan JM Richard TM Logan BE (2009)
Simultaneous cellulose degradation and electricity production by Enterobacter
cloacae in a microbial fuel cell Appl Microbiol Biotechnol 75 3673ndash3678
99 Richter H Nevin KP Jia H Lowy DA Lovley DR Tender LM (2009) Cyclic
voltammetry of biofilms of wild type and mutant Geobacter sulfurreducens on fuel
cell anodes indicates possible roles of OmcB OmcZ type IV pili and protons in
extracellular electron transfer Energy Environ Sci 2 506ndash516
100 Riesenfeld SC Schloss DP Handelsman J(2004) Metagenomics Genomic
Analysis of Microbial Communities Annual Review Genetics 38525-552
101 Rozendal RA Hamelers HV Buisman CJ (2006) Effects of membrane cation
transport on pH and microbial fuel cell performance Environ Sci Technol 40
5206-5211
102 Rozendal RA Hamelers HVM Euverink GJW Metz SJ Buisman CJN (2006)
Principle and perspectives of hydrogen production through biocatalyzed electrolysis
Int J Hydrogen Energ 311632-1640
103 Rozendal RA Hamelers HV Molenkamp RJ Buisman CJ (2007) Performance of
single chamber biocatalyzed electrolysis with different types of ion exchange
membrances Water Res 41 1984-1994
104 Schluumlter A Bekel T Diaz NN Dondrup M Eichenlaub R Gartemann
KH Krahn I Krause L Kroumlmeke H Kruse O Mussgnug JH Neuweger
H Niehaus K Puumlhler A Runte KJ Szczepanowski R Tauch A Tilker
A Viehoumlver P Goesmann A (2008) The metagenome of a biogas-producing
microbial community of a production-scale biogas plant fermenter analyzed by the
454-pyrosequencing technology J Biotechnol136 77-90
90
105 Scott J H and Nealson KH (1994) A biochemical study of the intermediary carbon
metabolism of Shewanella putrefaciens J Bacteriol 176(11) 3408ndash3411
106 Sekiguchi H Tomioka H Nakahara T and Uchiyama H (2001) A single band
does not always represent single bacterial strains in denaturing gradient gel
electrophoresis analysis Biotechnology Letters23 1205ndash1208
107 Sharma T Leela M Reddy A Chandra TS Ramaprabhu S (2008) Development
of carbon nanotubes and nanofluids based microbial fuel cell Int J Hydrogen Energ
336749ndash6754
108 Shea C Clauwaert P Verstraete W Nerenberg R (2008) Adapting a denitrifying
biocathode for perchlorate reduction Water Sci Technol 58 (10) 1941-1946
109 Shi L Squier TC Zachara JM Fredrickson JK (2007) Respiration of metal (hydr)
oxides by Shewanella and Geobacter a key role for multihaem c-type cytochromes
Mol Microbiol 6512-20
110 Stams AJ and Plugge CM(2009)Electron transfer in syntrophic communities
of anaerobic bacteria and archaea Nat Rev Microbiol7(8)568-77
111 Sukkasem C Xu S Park S Boonsawang P Liu H (2008) Effect of Nitrate on the
Performance of Single Chamber Air Cathode Microbial Fuel Cells Water Res 42(19)
4743-4750
112 Summers ZM Fogarty HE Leang C Franks AE Malvankar NS Lovley DR
(2010)Direct exchange of electrons within aggregates of an evolved syntrophic
coculture of anaerobic bacteria Science 330 (6009)1413-5
113 Szczepanowski R Bekel T Goesmann A Krause L Kroumlmeke H Kaiser
O Eichler W Puumlhler A Schluumlter A (2008) Insight into the plasmid metagenome
of wastewater treatment plant bacteria showing reduced susceptibility to
antimicrobial drugs analysed by the 454-pyrosequencing technology J
Biotechnol 136 54-64
114 Tamura K Dudley J Nei M Kumar S (2007) MEGA4 Molecular Evolutionary
Genetics Analysis (MEGA) software version 40 MolBiol Evol 24(8) 1596-1599
115 Thompson JD Gibson TJ Plewniak F Jeanmougin F Higgins DG (1997) The
CLUSTAL_X windows interface flexible strategies for multiple sequence alignment
aided by quality analysis tools Nucleic Acids Res 25 4876-4882
116 Thormann KM Duttler S Saville RM Hyodo M Shukla S Hayakawa Y
Spormann AM (2006) Control of formation and cellular detachment from
Shewanella oneidensis MR-1 biofilms by cyclic di-GMP J Bacteriol 1882681-2691
117 Thormann KM Saville RM Shukla S Pelletier DA Spormann AM (2004)
Initial Phases of biofilm formation in Shewanella oneidensis MR-1 J Bacteriol
186(23) 8096-104
118 Torres CI Kato Marcus A Rittmann BE (2008b) Proton transport inside the
biofilm limits electrical current generation by anode-respiring bacteria Biotechnol
Bioeng 100 872ndash881
91
119 Torres CI Marcus AK Lee HS Parameswaran P Krajmalnik-Brown R Rittmann
BE (2010) A kinetic perspective on extracellular electron transfer by anode-
respiring bacteria FEMS Microbiol Rev 234(1)3-17
120 Torres CI Marcus AK Parameswaran P Rittmann BE (2008a) Kinetic
experiments for evaluating the NernstndashMonod model for anode-respiring bacteria
(ARB) in a biofilm anode Environ Sci Technol 42 6593ndash6597
121 Tsai HY Wu Ch-Ch Lee Ch-Y Shih EP (2009) Microbial fuel cell performance
of multiwall carbon nanotubes on carbon cloth as electrodes J Power Sources
194199-205
122 Tyson GW Chapman J Hugenholtz P Allen EE Ram RJ Richardson
PM Solovyev VV Rubin EM Rokhsar DS Banfield JF(2004) Community
structure and metabolism through reconstruction of microbial genomes from the
environment Nature42837-43
123 Venkateswaran K Moser DP Dollhopf ME Lies DP Saffarini D A MacGregor
BJ Ringelberg DB White DC Nishijima M Sano H Burghardt J Stackebrandt
E Nealson KH (1999) Polyphasic taxonomy of the genus Shewanella and
description of Shewanella oneidensis sp Int J Syst Bacteriol 49705-724
124 von Canstein H Ogawa J Shimizu S Lloyd JR (2008) secretion of Flavins
by Shewanella Species and Their Role in Extracellular Electron Transfer Appl
Environ Microbiol 74 3615-623
125 Wang A Liu W Cheng Sh Xing D Zhou J Logan BE (2009) Source of methane
and methods to control its formation in single chamber microbial electrolysis cells
Int J Hydrogen Energ 34 3653-3658
126 Wang J Ma T Zhao L Lv J Li G Liang F Liu R (2008) PCRndashDGGE method
for analyzing the bacterial community in a high temperature petroleum reservoir
World J Microbiol Biotechnol 2 9694-9702
127 Wang Q Garrity GM Tiedje JM Cole JR (2007) Naiumlve Bayesian Classifier for
Rapid Assignment of rRNA Sequences into the New Bacterial Taxonomy Appl
Environ Microbiol 73(16) 5261-5267
128 Wang YF Masuda M Tsulimura S Kano K (2008) Electrochemical regulation of
the end-product profile in Propionibacterium freudenreichii ET-3 with an
endogenous mediator Biotechnol Bioeng 101(3) 579ndash586
129 Wrighton KC Agbo P Warnecke F Weber KA Brodie EL DeSantis TZ
Hugenholtz P Andersen GL Coates JD (2008) A novel ecological role of the
Firmicutes identified in thermophilic microbial fuel cells ISME J 2 1146ndash1156
130 Xing D Zuo Y Cheng S Regan JM Logan BE (2008) Electricity generation by
Rhodopseudomonas palustris DX-1 Environ Sci Technol 42 4146-4151
131 Xu S Liu H Fan Y Schaller R Jiao J Chaplen F (2012) Enhanced performance
and mechanism study of microbial electrolysis cells using Fenanoparticle-decorated
anodes Appl Microbiol Biotechnol 93(2)871-880
92
132 Yang Y Harris DP Luo F Wu L Parsons AB Palumbo AV Zhou J (2008)
Characterization of the Shewanella oneidensis Fur gene roles in iron and acid
tolerance response BMC Genomics 9 Suppl 1S11
133 Zhang L Zhou S Zhuang L Li W Zhang J Lu N Deng L (2008) Microbial
fuel cell based on Klebsiella pneumoniae biofilm Electrochem Commun10 1641ndash
1643
134 Zuo Y Xing DF Regan JM Logan BE (2008) Isolation of the exoelectrogenic
bacterium Ochrobactrum anthropi YZ-1 by using a U-tube microbial fuel cell Appl
Environ Microbiol 74 3130-3137
93
APPENDICES
List of published papers during PhD study
1 Shoutao Xu Hong Liu Yanzhen Fan Rebecca Schaller Jun Jiao and Frank
Chaplen (2012) Enhanced performance and mechanism study
of microbial electrolysis cells using Fe nanoparticle-decorated anodes Applied
Microbiology and Biotechnology 93(2)871-880
2 Shoutao Xu Hong Liu (2011) New exoelectrogen Citrobacter sp SX-1 isolated
from a microbial fuel cell Journal of Applied Microbiology111(5)1108-1115
3 Yanzhen Fan Shoutao Xu Rebecca Schaller Jun Jiao Frank Chaplen Hong Liu
(2011) Nanoparticle decorated anodes for enhanced current generation in microbial
electrochemical cells Biosensors and Bioelectronics 26(5) 1908-1912
4 Tunc Catal Shoutao Xu Kaichang Li Hakan Bermek Hong Liu
(2008)
Electricity generation from polyalcohols in single-chamber microbial fuel
cells Biosensors and Bioelectronics 24(4)849-854
5 Chontisa Sukkasema Shoutao Xu Sunhwa Park Piyarat Boonsawang Hong Liu
(2008) Effect of nitrate on the performance of single chamber air cathode
microbial fuel cells Water research 424743-4750
6 Rebecca Schaller Yanzhen Fan Shoutao Xu Alan Fern Frank Chaplen Hong
Liu and Jun Jiao (2009) Vertically Aligned Multi-walled Carbon Nanotube
Decorated Anodes for Microbial Fuel Cells Proceedings of Materials Research
Society 2009 1170 R05-13

bacterial species in mixed culture is Geobacter sp (66) Arcobacter sp and Citrobacter
sp These three bacterial species reached to 88 of total bacterial species This result is
consistent with the DGGE result which showed that three bright bands represented three
dominant bacterial species
Exoelectrogenic bacterial strain SX-1 was isolated from a mediator-less microbial
fuel cell by conventional plating techniques with ferric citrate as electron acceptor under
anaerobic conditions Phylogenetic analysis of the 16S rDNA sequence revealed that it
was related to the members of Citrobacter genus with Citrobacter sp sdy-48 being the
most closely related species The bacterial strain SX-1 produced electricity from citrate
acetate glucose sucrose glycerol and lactose in MFCs with the highest current density
of 205 mAm2 generated from citrate Cyclic voltammetry analysis indicated that
membrane associated proteins may play an important role in facilitating electron transfer
from the bacteria to the electrode This is the first study that demonstrates that
Citrobacter species can transfer electrons to extracellular electron acceptors Citrobacter
strain SX-1 is capable of generating electricity from a wide range of substrates in MFCs
This finding increases the known diversity of power generating exoelectrogens and
provids a new strain to explore the mechanisms of extracellular electron transfer from
bacteria to electrode The wide range of substrate utilization by SX-1 increases the
application potential of MFCs in renewable energy generation and waste treatment
Anode properties are critical for the performance of microbial electrolysis cells
(MECs) Inexpensive Fe nanoparticle modified graphite disks were used as anodes to
preliminarily investigate the effects of nanoparticles on the performance of Shewanella
oneidensis MR-1 in MECs Results demonstrated that average current densities
produced with Fe nanoparticle decorated anodes were up to 59-fold higher than plain
graphite anodes Whole genome microarray analysis of the gene expression showed that
genes encoding biofilm formation were significantly up-regulated as a response to
nanoparticle decorated anodes Increased expression of genes related to nanowires
flavins and c-type cytochromes indicate that enhanced mechanisms of electron transfer
to the anode may also have contributed to the observed increases in current density The
majority of the remaining differentially expressed genes were associated with electron
transport and anaerobic metabolism demonstrating a systemic response to increased
power loads
The carbon nanotube (CNT) is another form of nano materials Carbon nanotube
(CNT) modified graphite disks were used as anodes to investigate the effects of
nanostructures on the performance S oneidensis MR-1 in microbial electrolysis cells
(MECs) The current densities produced with CNT decorated anodes were up to 56-fold
higher than plain graphite anodes Global transcriptome analysis showed that cytochrome
c genes associated with extracellular electron transfer are up-expressed by CNT
decorated anodes which is the leading factor to contribute current increase in CNT
decorated anode MECs The up regulated genes encoded to flavin also contribute to
current enhancement in CNT decorated anode MECs
copyCopyright by Shoutao Xu
June 15 2012
All Rights Reserved
Bacterial Community Analysis New Exoelectrogen Isolation and Enhanced Performance
of Microbial Electrochemical Systems Using Nano-Decorated Anodes
by
Shoutao Xu
A DISSERTATION
Submitted to
Oregon State University
in partial fulfillment of
the requirements for the
degree of
Doctor of Philosophy
Presented June 15 2012
Commencement June 2013
Doctor of Philosophy dissertation of Shoutao Xu presented on June 15 2012
APPROVED
Co-Major Professor representing Biological and Ecological Engineering
Co-Major Professor representing Biological and Ecological Engineering
Head of the Department of Biological and Ecological Engineering
Dean of the Graduate School
I understand that my dissertation will become part of the permanent collection of
Oregon State University libraries My signature below authorizes release of my
dissertation to any reader upon request
Shoutao Xu Author
ACKNOWLEDGEMENTS
I would like to thank all people who have helped and inspired me during my
doctoral study Foremost I would like to express my sincere gratitude to my advisors Dr
Hong Liu and Dr Frank Chaplen for their continuous support of my PhD study and the
research of Microbial Fuel Cells at Oregon State University Hong inspired me to devote
myself to the field of Bio-energy with her great patience and enthusiasm towards
scientific educations Frank was always available and willing to help me with my study
especially during the period of Hongrsquos sabbatical leave Thanks to his kindness and
assistance my study at OSU became smooth and rewarding
Besides my advisors I would like to thank everyone in my dissertation committee Dr
Martin Schuster Dr Clare Reimers and Dr Mark Dolan Due to their encouragement
and insightful comments on my research I could always have the courage and knowledge
to overcome difficulties in my research I benefited greatly from Martinrsquos valuable
suggestions on my writing skills and his generous help regarding microarray data analysis
Clarersquos advice helped me build a solid foundation of Electrochemistry Also it was a
great honor to have Mark as my committee member His teaching gave me an insight into
the world of environmental engineering and enhanced the depth and width of my research
I thanked Dr Yanzhen Fan for always giving me guidance about the designs of the
reactors in my experiments
It was also important for me to say thanks to my current and previous lab-mates
Keaton Lesnik Kuhuan Chien Cheng Li Corale Abourached Anthony Janicek
Hongqiang Hu Jeremy Chignell Yudith Nieto and Wengguo Wu I had a wonderful
time enjoying doing research with them Their friendship and help made me confident of
my ability to do research as a scientist In addition I cherished and appreciated the
friendly environment in our BEE department Faculty members especially Dr John Bolte
Dr John Selker Dr Roger Ely Dr Ganti Murthy and Desiree Tullos made BEE an
excellent department for our students
My deepest gratitude went to my father Fuqin Wang and my mother Jinzhi Wang
for their endless care love and support throughout my life Also I would like to say
thanks to my best-loved wife Songhua Zhu She always supported me and never
complained that I could spend so little time accompanying with her This dissertation was
also for my lovely son Gabriel Hong-Yi Xu
TABLE OF CONTENTS
Page
1 General Introductionhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 1
2 Bacterial Community Analysis of Mixed Consortium in Microbial
Electrochemical Systemshelliphelliphelliphellip 13
3 New Exoelectrogen Citrobacter sp SX-1 Isolation and
Characterizationhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 27
4
Enhanced Performance and Mechanism Study of Microbial Electrolysis
Cells using Fe Nanoparticles Decorated Anodeshelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 45
5 Global Transcriptome Analysis of Response of Shewanella oneidensis
MR-1 to CNT Nanostructure Decorated Anodes in Microbial
Electrochemical System
66
6 Summaryhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 80
7 Bibliography helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 82
8 Appendices helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 93
LIST OF FIGURES
Figure Page
1-1 Generalized schematics (a) Microbial fuel cell (MFCs)(b) Microbial
electrolysis cell (MECs) helliphelliphelliphelliphelliphelliphelliphelliphelliphellip helliphelliphelliphellip
2
1-2 Mechanisms for extracellular electron transport in a MFCs anode 7
2-1 Polarization curves of high power mixed culture MFCshelliphelliphelliphelliphelliphellip 20
2-2 DGGE result of high power generation mixed culture MFCshelliphelliphellip 21
2-3 Bacterial species and percentage of bacterial community of mixed
culture MFCs helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip
22
2-4 Phylogenetic trees of bacterial species from mixed culture MFCs 22
2-5 Dominant bacterial species identification in DGGEhelliphelliphelliphelliphelliphelliphelliphellip 23
2-6 Current density generated by different dominant isolateshelliphelliphelliphelliphelliphellip 24
3-1 Phylogenetic tree of strain SX-1 and closely related species based on
16S rDNA sequenceshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip
35
3-2 SEM image of planktonic cells of Citrobacter sp SX-1helliphelliphelliphelliphelliphellip 35
3-3 Electricity generation by Citrobacter sp SX-1 in a single chamber
MFCs with sodium citrate helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip
36
3-4 Electricity generation of Citrobacter sp SX-1 using different
substrates in single chamber air-cathode MFCs at 1 KΩhelliphelliphellip
37
3-5 Power and voltage generation by Citrobacter sp SX-1 as a function
of current density helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip
38
3-6 Four current generating stages for CV analysis helliphelliphelliphelliphelliphelliphelliphellip 39
3-7 Cyclic Voltammetry analysis of of Citrobacter sp SX-1 biofilms helliphellip 41
4-1 SEM images of Fe NPs decorated anodeshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 50
4-2 Effects of Fe-NP decorated anodes on the current density in MECs helliphellip 51
4-3 Differentially expressed genes grouped by functional classification in
Fe NP decorated anodeshelliphelliphelliphelliphelliphellip helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip
53
5-1 SEM images of CNT decorated anodeshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 70
5-2 Effects of CNT decorated anodes on the current density in MECshelliphellip 73
5-3 Differentially expressed genes grouped by functional classification in
CNT decorated anodes helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip
74
LIST OF TABLES
Table
1-1
4-1
4-2
4-3
Taxa of bacteria current density generated and reactor configuration
used in microbial electrochemical cell experimentshelliphelliphelliphellip
Expression levels of genes related to biofilm formationhelliphelliphelliphelliphellip
Genes related to anaerobic growth and electron transfer with
significantly change expression levelhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip
Expression level of flavin and cytochrome geneshelliphelliphelliphelliphelliphelliphelliphellip
Page
4
55
56
58
4-4
5-1
5-2
Other genes with significantly changed expression level
Expression level of cytochrome c as response to CNT decorated anodes
Expression level of genes related to flavin synthesis as response to CNT
decorated anode in CNT decorated anodeshelliphellip
59
76
78
1
1
Bacterial Community Analysis New Exoelectrogen Isolation and Enhanced Performance
of Microbial Electrochemical Systems Using Nano-Decorated Anodes
Chapter 1
General Introduction
11 Microbial Fuel Cells and Microbial Electrolysis Cells
Microbial electrochemical systems (MESs) have drawn the attention of researchers
in recent years due to their promising applications in a variety of scientific fields such as
renewable energy generation bioremediation and wastewater treatment In a MES
microorganisms interact with electrodes catalyzing oxidation and reduction reactions at
the anode and the cathode
The most-described type of MESs is the microbial fuel cells (MFCs) in which
useful power is generated directly using the catalytic action of active microorganisms
(Kim 2002 Cheng and Logan 2007 Cheng et al 2006) In a typical two-chamber
MFCs (Figure 1a) organic matter is oxidized by electrochemically active
microorganisms in the anode chamber to release protons and electrons Protons diffuse
into the cathode chamber through a proton exchange membrane Meanwhile electrons
are transferred to the anode through various mechanisms (Logan et al 2006 Lovley
2
2006 Rabaey et al 2003) and then travel to the cathode where normally they combine
with oxygen and protons to form water
Another common used type of MES is the microbial electrolysis cell (MECs) in
which hydrogen is produced instead of electricity by applying a circuit voltage to the
MFCs system and maintaining anaerobic conditions in the cathode chamber (Liu et al
2005 Logan et al 2008 Rozendal et al 2006 Schaller et al 2009) (Figure 1b) A
cathode potential of at least - 410 mV (vs standard hydrogen electrode at pH 70) is
required to produce hydrogen at the cathode Therefore a theoretical potential of 110 mV
(ie 410-300 mV) is needed but voltages of gt02 V are normally applied due to various
overpotentials
Fig 1-1 Generalized schematics (a) Microbial fuel cells (MFCs) where electricity is
captured through the resistance R (b) Microbial electrolysis cells (MECs) where
hydrogen gas is produced in the cathode chamber R for resistor PS for power supply and
PEM for proton exchange membrane
The fundamental feature shared by microbial electrochemical systems (MESs)
(MECs and MFCs) is the microbe-catalyzed electron transfer from organic matter to
electrodes (anodes) (Rebaey et al 2004) Therefore different types of MESs can be
PEM
R
H+
e- e-
Anode
Bacte
ria
a
Cath
ode
H2O
PEM
PS
H+
e- e-
Anode
Bacte
ria
b
Cath
ode
H2
3
utilized to investigate the phenomena and mechanisms of interactions between microbes
and electrodes
12 Bacteria and Electron Transfer Mechanisms in MESs
The electrochemically active bacteria in MFCs are thought to be iron-reducing
bacteria such as Shewanella and Geobacter species (Gorby et al 2006) They have great
importance in the natural environment principally in metal oxidation and reduction
However recent studies have shown that the diversity of bacterial communities is much
greater than these model iron reducers on MFCs anodes (Reimers et al 2001 Liu et al
2004b Kim et al 2005) Over 20 electrochemically active bacterial species those can
transfer electrons exocellularly to electrodes have been reported in the past 10 years
(Logan 2009 Liu et al 2010) These bacteria have been categorized into diverse genetic
groups so far including alphaproteobacteria (Rhodopseudomonas Ochrobactrum and
Acidiphilium) (Malki et al 2008 Xing et al 2008 Zuo et al 2008) betaproteobacteria
(Rhodoferax) (Chaudhuri et al 2003) gammaproteobacteria (Shewanella Pseudomonas
Klebsiella Enterobacter and Aeronomas) (Kim et al 2002 Pham et al 2003 Rabaey et
al 2005 Bretschger et al 2007 Zhang et al 2008 Chung et al 2009 Rezaei et al 2009)
deltaproteobacteria (Geobacter Geopsychrobacter Desulfuromonas and Desulfobulbus)
(Bond et al 2002 Bond and Lovley 2003 Holmes et al 2004ab 2006)
Epsilonproteobacteria (Arcobacter) (Fedorovich et al 2009) Firmicutes (Clostridium and
Thermincola) (Park et al 2001 Wrighton et al 2008) Acidobacteria (Geothrix) (Bond
and Lovley 2005) and actinobacteria (Propionibacterium) (Wang et al 2008) A wider
range of electrochemically active bacteria are expected to be discovered
4
Table 1 Taxa of bacteria current density generated and reactor configuration used in
microbial electrochemical cell experiments (Liu et al 2010)
The electrochemically active bacterial species that possess the ability to transfer
electrons outside of the cell are called exoelectrogens in the MESs research field The
different exoelectrogens have demonstrated a wide-ranging power generation ability in
MFCs For example Shewanella oneidensis MR-1 has demonstrated the ability to
generate 0018 Am2 current density in single chamber MFCs while the Geobacter
Taxon Microorganisms Current Density (Am2
)
α-proteobacteria
Rhodopseudomonas palustris DX-1 003
Ochrobactrum anthropi YZ-1 071
Acidiphilium sp 32sup5 300
β-proteobacteria Rhodoferax ferrireducens 0031
γ-proteobacteria
Shewanella putrefaciens IR-1 0016
Shewanella oneidensis DSP10 0013
Shewanella oneidensis MR-1 0018
Pseudomonas aeruginosa KRA3 0017
Escherichia coli K12 HB101 100
Klebsiella pneumoniae L17 120
Enterobacter cloacae 013
Aeromonas hydrophila PA3 030
δ-proteobacteria
Geobacter metallireducens 065
Geobacter sulfurreducens 800
Desulfuromonas acetoxidans 0005
Geopsychrobacter
Electrodiphilus strain A2
0066
Desulfobulbus propionicus 003
Firmicutes
Lactococcus lactis 003
Thermincola sp strain Jr 020
Clostridium butyricum EG3 022
Thermincola ferriacetica Z-0001 040
Brevibacillus spp PTH1 0009
Desulfitobacterium hafniense DCB2 110
Acidobacteria Geothrix fermentans 0097
Actinobacteria Propionibacterium freudenreichiiET-3 120
5
sulfurreducens species has generated current densities as high as 80 Am2 which is 400
times higher than the one generated by Shewanella oneidensis MR-1 However mixed-
culture communities have generated much higher power densities than their pure-culture
counterparts in MFCs perhaps due to synergistic interactions within the anode bacterial
communities and the participation of currently unknown bacteria species and mechanisms
(Jung and Regan 2007) The dominant bacteria species vary in mixed cultures bacteria
communities in MFCs due to the enrichment of different substrates
Traditional methods of extroelectrogen study depend on cultivation hampered novel
exoelectrogens discovery because the inadequacy of defined media underestimates the
actual microbial diversity in MFCs Slow growth rate and unknown growth requirements
of anaerobic microorganisms make some anaerobic extroelectrogens cultivation difficult
However these limitations have been overcome by using molecular biological methods
based on DNARNA analysis Molecular biological techniques are now widely applied to
assess the diversity of microbial communities by analyzing the 16S rDNA sequence The
most commonly used molecular biological techniques for bacterial community analysis
include denaturing gradient gel electrophoresis (DGGE) restriction fragment length
polymorphism (RFLP) and 16S rDNA sequencing These methods are much less time
consuming than traditional isolation and cultivation methods
Isolated exoelectrogens were utilized to explore the mechanism of electron transfer
to the anode However the mechanisms of electron transfer to extracellular electron
acceptors are not well understood Three mechanisms have been proposed for exocellular
transport of electrons by exoelectrogens to anodes in MFCs so far Some exoelectrogenic
bacteria can transfer electrons to electrodes through soluble redox compounds (Bond and
6
Lovley 2005) These compounds include artificial mediators and mediators secreted by
exoelectrogenic bacteria A variety of chemicals have been used to facilitate the shuttling
of electrons from inside of cell to electrodes outside the cell These exogenous mediators
include neutral red (Park et al 1999) anthraquinone-2-6 disulfonate (AQDS) thionin
potassium ferricyanide (Bond et al 2002) methyl viologen and others (Logan 2004
Rabaey and Verstraete 2005) Some bacteria can secrete their own chemical mediator
for example Pseudomonas aeruginosa secretes pyocyanin and phenazine-1-carboxamide
to transfer electrons toward the MFCs anode (Rabaey et al 2005) Another evidence for
mediator production by bacteria is Geothrix fermentans When the medium was replaced
in a MFCs that had stable power generation with this bacteria power dropped by 50
and required 10 days to resume the original level
Some bacteria can directly transfer electrons to anodes via outer cell membrane
proteins (Myers and Myers 1992) The outer cell membrane protein cytochrome c is
thought to play a critical role in to transferring electrons to anodes Ly et al (2011)
isolated the haem protein cytochrome c and demonstrated that electric field effects may
be functional for the natural redox processes of cytochrome c in the respiratory chain
Shewanella oneidensis is the example of a bacterium that is capable of transfer electron to
anode via outer cell membrane proteins When Shewanella oneidensis adhered to an iron
surface the greater force has showed benefits to grow cells because closer contact
required for electron transfer from cell bound cytochromes (Lower et al 2001)
7
Fig1-2 Mechanisms for extracellular electron transport in an MFCs anode (1) direct
contact (top in green) (2) by nanowires (middle in purple) and (3) self-produced
mediators (bottom in blue) (Logan 2009)
More and more evidence supports the involvement of bacterial nanowires in
extracellular electron transport (Reguera et al 2005 2006 Gorby et al 2006) Nanowires
are conductive appendages produced by both Geobacter and Shewanella species (Gorby
and Beveridge 2005) The conductivity of the appendages was examined and confirmed
by using conductive scanning tunneling microscopy (STM) (Gorby et al 2006)
Nanowires can carry electrons from the cell to the anode surface of MFCs
The solid component of the extracellular biofilm matrix has high efficiency on
extracellular electron transfer compared with other extracellular electron transfer
8
mechanisms and recently Torres et al (2010) hypothesized that the solid component of
the extracellular biofilm matrix formed by exoelectogens is another way bacteria transfer
electrons to electrodes This hypothesis was based on kinetic analysis of each EET
mechanism reported in available literature (Torres et al 2010)
13 Anode electrodes
In MFCsMECs anode electrodes are a critical component because exoelectrogens
adhere to the surface of anodes to transfer electrons to the electrode The characteristics
of anodes have significant effects on electron transfer rate from bacteria to anode
electrodes in MFCs The requirements of an anode material are it should be highly
conductive non-corrosive have a high specific surface area (area per volume) high
porosity be non-fouling inexpensive and easily scaled to larger sizes Of these
properties the most important one that is different from other biofilm reactors is that the
material must be electrically conductive Normally they are made of various carbon
materials including carbon fiber carbon clothe and carbon paper due to their stability
high conductivity and high specific surface-area Nevertheless they have little
electrocatalytic activity for the anode microbial reactions and thus a modification of the
carbon materials is the main approach for improving their performance Consequently
there is a great need to develop a new type of anode material for MFCsMECs
It is a great challenge to develop a new anode material to further increase the power
density of a MFCs The nature of the catalytic mechanism of a MFCrsquos anode involves not
only a biological but also an electrocatalytic process An optimal nanostructure with a
high specific surface area favorable for both catalytic processes could play a critical role
in improving the power density of the MFCs such a structure needs to host the bacteria
9
with high bioactivity while enhancing the electron-transfer rate Schroumlder et al (2008)
employed PANI to modify a platinum anode for MFCs and achieved a current density 1
order of magnitude higher than the previously reported value PANIinorganic
composites are also reported to have better conductivity Qiao et al also applied a new
mesoporous TiO2 electrode material with uniform nanopore distribution and a high
specific surface area to anode in comparison to previously reported work with E coli
MFCs the composite anode delivers 2-fold higher power density (Qiao et al 2008)
Thus it has great potential for use as the anode in a high-power MFCs and may be a new
approach for improving performance of MFCs
14 Other parts of MESs
141 Membranes and ion transport
The ion exchange membrane is another one of the critical components in two-
chamber MESs systems It separates anode and cathode chambers and at the same time
maintains the electron neutrality of the system ie transport of electrons to the cathode
needs to be compensated by transport of an equal amount of positive charge to the
cathode chamber A proton exchange membrane (PEM) such as Nafion a per-fluorinated
sulphonic acid membrane (PFSAs) consisting of a hydrophobic fluorocarbon backbone to
which hydrophilic sulfonate groups (-SO3-) are attached is commonly used in chemical
fuel cell systems For MFCs systems however mainly cation species like Na+ and K
+
other than proton are often responsible for the dominant transport of positive charge
through the cation exchange membrane (CEM) to maintain electroneutrality due to the
low proton concentration in any aqueous medium with near neutral pH (Rozendal et al
2006) Consequently the pH increases in the cathode chamber due to the consumption of
10
protons and decreases in the anode chamber because of the accumulation of protons
(Rozendal et al 2007) This pH change in two-chamber systems results in a decrease of
the cathode potential and performance The application of anion exchange membrane
(AEM) has also been explored in MFCs systems (Cheng and Logan 2007 Kim and
Logan 2007) where it has been proposed that protons are transferred via pH buffers like
phosphate anions
142 Cathodes and Catalysts
The cathode is another challenge for making MFCs commercially available
technology because the chemical reaction that occurs at the cathode is difficult to
engineer as the electrons protons and oxygen must all meet at a catalyst in a tri-phase
reaction (solid catalyst air and water) The catalyst must be on a conductive surface and
must be exposed to both water and air so that protons and electrons in these different
phases can reach the same point The most commonly used material for a cathode is
commercially available carbon paper pre-loaded with a Pt catalyst on one side When it is
used in a MFC the side that contains the catalyst faces the water and the uncoated side
faces air To reduce the high cathode cost associated with platinum catalyst other precious-
metal-free cathodes such as (NiMo and NiW) were developed by electrodepositing on a
carbon fiber They have achieved comparable performance with Pt catalyst with same
loading at a much lower cathode fabrication cost (Hu 2010)
The requirements of cathode for MECs are quite similar to the requirements of
cathode for MFCs but easier than cathode for MFCs for the manufacturing process
because the cathode in MECs is not necessarily to exposed to air Therefore it can be
made of the exact same materials of cathode in MFCs except a waterproof layer
11
Recently some researchers have attempted to apply microorganisms as a biocatalyst to
precede the combination of electron with oxygen in the cathodes
15 Dissertation overview
Low power densities in MESs limit practical applications The improvement of
MESs performance requires a detailed understanding of the physiology and ecology of
microorganisms in MESs including the mechanism of electron transfer to the anode from
the microorganism
This dissertation focuses on the problem of the low power density of MESs The
bacterial community structure of a high power generated mixed culture communities in
MFCs will be identified firstly and then one of major exoelectrogens will be isolated and
characterized This information will be helpful to understand the physiology and ecology
of exoelectrogens in MESs Consequently they will be beneficial to improve power
density of MFCs Lastly the nanostructure decorated anodes in MECs will be utilized to
improve the power density The power enhancement mechanism will be explored by
using a whole genome microarray They are presented here as four papers
In the first paper the cultivation independent molecular biological techniques
denaturing gradient gel electrophoresis (DGGE) and 16S rDNA clone library are utilized
to analyze the bacterial community structure of a higher power mixed culture MFCs The
analyzed results provide fundamental information for isolating the dominant bacteria in
mixed culture MFCs Two of dominant bacterial species has been isolated and used aone
to test power generation in MFCs The possible interaction among different bacterial
species in mixed culture is discussed
12
In the second paper one isolated exoelectrogenic bacterial strain SX-1 is
characterized It is identified as a member of the Citrobacter genus and power generation
is tested ultilizing a wide range of different substrates The electron transfer mechanism is
explored using Cyclic Voltammetry (CV) This study increases the known diversity of
power generating exoelectrogens and provides a new strain to explore the mechanisms of
extracellular electron transfer from bacteria to electrodes
The third paper shows effects on MESs performance by Fe nanoparticle decorated
anodes in the MESs The average current density produced with Fe nanoparticle
decorated anodes increased up to 59-fold higher than plain graphite anodes A whole
genome microarray is utilized to analyze the possible mechanism of enhanced current
density as responded to nanoparticle decorated anodes
The fourth paper describes the effects of carbon nanotube (CNT) modified anode on
the performance S oneidensis MR-1 in MESs Results demonstrate that current densities
produced with CNT decorated anodes are up to 56-fold higher than plain graphite anodes
The possible mechanisms of enhanced current density by CNT decorated are explored
13
Chapter 2
Bacterial community analysis of mixed consortium in higher
power density MESs
Shoutao Xu and Hong Liu
1 Introduction
The improvement in the performance of mixed culture MFCs requires an
understanding of the ecology in microbial communities of MFCs Many researchers have
attempted to characterize microbial populations and activities to elucidate the behaviors
and ecology of microorganisms in MFCs (Wrighton et al 2008 Fedorovich et al 2009)
In order to study the microbial ecology of the mixed culture in a MFC and select the
appropriate isolation medium for dominant bacterial species in the mixed culture the
fundamental steps are to analyze the bacterial diversity in the mixed culture of MFCs and
identity the dominant bacterial species in bacteria communities in MFCs
14
As for the identification of bacterial communities typically there are two general
methods The first method for identification of bacterial community is the traditional
cultivation processes using selective nutrients to promote the growth of different types of
bacteria within the samples (Amman et al 2000) The community structure can then be
assessed by identifying the isolates from the dominant colonies that were cultured This
can often be costly and laborious as each isolate has to be further studied by examining
its physiology taxonomy and reactivity to stains (Adwards et al 1989)
The second method relies on utilizing molecular techniques to analyze bacterial
community DNA Several molecular methods involving the extraction and analysis of
DNA from entire bacterial communities are used to identify genetic fingerprints of
bacteria These methods including the cloning and sequencing of 16S rDNA automated
ribosomal intergenic spacer analysis (ARISA) terminal restriction fragment length
polymorphism (T-RFLP) and denaturing gradient gel electrophoresis (DGGE) generate
profiles of bacterial community structures They can rapidly assess complex communities
from various environments (Amman et al 2000)
Due to the conservative characteristic of 16S rDNA in bacteria during the process
of evolution 16S rDNA sequencing can be used to identify different species of bacteria
Therefore the molecular techniques of denaturing gradient gel electrophoresis (DGGE)
with PCR and 16S rDNA clone library are used for analysis of the microbial diversity
These methods are more convenient and save time compared to traditional
isolationcultivation methods for microorganism analysis
In this work a biofilm bacterial community from an anode of a MFCs wase studied
by using denaturing gradient gel electrophoresis (DGGE) of PCR-amplified partial 16S
15
rRNA genes followed by cloning and sequencing of 16S rDNA The results provided
essential information for dominant bacterial isolation in mixed culture MFCs
2 Materials and methods
21 High power generation of mixed culture MFCs
Mixed cultures were originally inoculated from domestic wastewater (Corvallis
Wastewater Treatment Plant Corvallis OR) to MFCs after operating for 2 years using a
defined medium solution (Lovely 2002) with sodium acetate as the carbon source A
new MFCs was inoculated from the operating MFCs The polarization curves were
performed to measure the power generation when maximal stable power were established
(normally 3-5 batches post-inoculation) and the genomic DNA from the microbial
biofilm on the anode was extracted under the sterile conditions for further bacterial
community analysis
22 Denaturing gradient gel electrophoresis (DGGE)
DGGE with PCR is a method of analysis of bacterial community composition
based on different GC percentage of 16s rDNA in different bacterial species PCR-DGGE
comprises three steps (i) extraction of total community DNA from the sample (ii) PCR-
controlled amplification using specific oligonucleotide primers and (iii) separation of the
amplicons using DGGE For this purpose a reproducible and efficient method for total
DNA extraction is indispensable and needs to be evaluated and optimized depending on
the nature of the sample In the subsequent PCR step multiple PCR primer sets with
different resolution can be used In most PCR-DGGE applications on bacteria universal
or specific primers are targeting the 16S rRNA gene Upon electrophoresis of the PCR
16
amplicons and gel staining (using ethidium bromide silver staining or SYBR green)
DGGE gels are digitally captured and further analyzed using computer software packages
The richness of DNA fragment bands in DGGE indicates the bacterial diversity of the
sample
The detailed steps are as followed Biofilms were scratched from the anodes of high
power generated MFCs fed with sodium acetate Bacterial genomic DNA was extracted
from the biofilm samples using the DNeasy blood and tissue kit (Qiagen) according to the
manufacturerrsquos instructions The universal primer set was used to amplify the 16S rDNA
from the extracted genomic DNA then the V3 region of bacterial 16S ribosomal DNA
(rDNA) was amplified with GC-clamp primer (Muyzer et al 1993) for DGGE PCR
amplification was performed in a thermocycler DGGE of the PCR products was carried
out in a DcodeTM
Universal Mutation Detection System) The 8 (wv) polyacrylamide
gels contains from 30 to 55 denaturing gradients Electrophoresis was conducted
using a 1timesTAE (Tris-Acetate-EDTA) buffer at 130V and 60degC for 5 hours After
electrophoresis the gel was stained with ethidium bromide in 1timesTAE buffer for 15
minutes and washed in 1timesTAE buffer for 10 minutes The fragments were visualized
under a UV transilluminator The richness of single band in DGGE gel picture
preliminarily showed that the bacterial diversity in mixed culture since the single band in
gel represents one bacterial species
23 16S rDNA clone library construction method
16S rDNA-DGGE fingerprinting has proven to be particularly useful as an initial
investigation into bacterial communities and is suitable for identifying the predominant
bacteria that comprise 1 or more of cells within a given sample In the 16s rDNA clone
17
library construction process the first step is the extratction of the total genomic DNA
then the genomic DNA of the mixed culture biofilm was used as template for PCR
amplification of approximately 1500 bp of 16S rDNA with universal primers The PCR
products of 16S rDNA were purified and inverted into pGEM-T Easy vector system
before they were transformed into competent E coli The transformed cells were spread
on Luria-Bertani (LB) plates containing the selective antibiotic ampicillin and X-Gal and
incubated overnight at 37 OC after the more than 80 inverted 16s DNA fragment E coli
colonies were picked up and inoculated in the 1-3 ml liquid LB medium and left to grow
for 16 hours The plasmid DNA were extracted and sequenced The sequences were
compared directly to all known sequences deposited in GenBank databases using the
basic local alignment search tool (BLAST)
24 Bioinformatics Analysis
16S rDNA sequencing results of more than 80 colones were queried against the
GenBank and Ribosomal Database Project (RDP) databases using BLAST and
SEQUATCH algorithms (Cole et al 2007 Wang et al 2007 Cole et al 2009) The
neighbor-joining trees were constructed with the Molecular Evolutionary Genetics
Analysis package (MEGA Version 4) based on 1000 bootstrap analysis (Tamura et al
2007)
25 Dominant bacterial species band in DGGE identification
The DGGE results gave the information of dominant bacterial richness of bacterial
species in mixed culture based on the theory that single band represents one bacterial
species which showed the diversity of bacterial community in the mixed culture of a
MFC 16S rDNA clone library results provided the whole picture of bacterial community
18
including the bacterial species name and bacterial species percentage in the mixed culture
The known pure bacterial species which have been sequenced can be used as markers to
identify dominant bacterial species in DGGE The whole experimental procedure is
similar to the procedure of the mixed culture DGGE steps The first step is to extract the
collect the mixed culture biofilm genomic DNA and pure bacterial species marker
genomic DNA Then the universal primer set was used to amplify the 16S rDNA from
the extracted genomic DNA After that the V3 region of bacterial 16S ribosomal DNA
(rDNA) was amplified with GC-clamp primer (Muyzer et al 1993) for DGGE The
subsequent steps were similar to the mixed culture DGGE experimental procedure The
dominant bacterial bands in DGGE were determined by comparisons of the single band
position in the mixed culture DGGE with single pure bacterial species location in DGGE
26 Dominant bacterial species isolation and power generation
The bacterial community of mixed culture in MFCs were predominantly composed
of Geobacter sp Arcobacter sp and Citrobacter sp Two dominant bacterial species
have been isolated
Citrobacter sp SX-1 was isolated from the anode biofilm by using a solid agar
medium containing NH4Cl 6mM KCl 2mM CH3COONa (electron donor) 30 mM
FeC6H5O7 (electron acceptor) 20mM minerals and vitamins (Lovley and Phillips 1988)
Single colonies were picked up after culturing on the plate for 36 hours at 30 degC and
transferred two times on the agar plate for purification Then the isolates grew in a liquid
medium solution in anaerobic tubes containing the same constituents as the solid
medium All isolation process was operated in a glove box anaerobic chamber (Coy
Laboratory Products Grass Lake MI)
19
Arcobacter sp SX-2 was isolated from the anode biofilm of a MFC fed with sodium
acetate by using the same method of isolation of pure bacterial species Citrobacter XS-1
except using Trypticase Soy Agar with 5 Sheep Blood solid (from American type
culture collection (ATCC)) agar medium instead of Citrobacter sp medium Single
colonies were picked up after culturing on the plate for 36 hours at 30 degC and transferred
twice on the agar plate for purification Then the isolates grew in a liquid medium
solution microaerobically
Geobacter sp have been trying to isolate by using three different methods They are
the colony pickup after different condition enrichment Goebacter medium isolation and
dilution to extinction by multiple channel mini MFCs isolation however the Geobacter
sp has not been isolated yet due to some limits so far But they provide valuable
information for further isolation of Geobacter sp
Single chamber MFCs were used to evaluate power generation by different isolates
The MFCs were constructed as described previously (Liu and Logan 2004) and modified
with 3 cm2 carbon cloth anodes and 7 cm
2 carbon clothPt cathodes The total liquid
volume of each MFCs was about 15 ml with electrode spacing about 17 cm All MFCs
were operated in an autoclaved closed plastic box and sterile cotton was attached to the
outer surface of the air cathodes to prevent contamination A MFCs without bacterial
culture was used as control MFCs were inoculated with 3 ml late exponential phase
cultures of SX-1 in the medium solution reported previously (Liu and Logan 2004)
3 Results and discussion
31 Power production by mixed culture MFCs
20
A polarization curve was used to characterize current as a function of voltage in the
MFCs The polarization curves are performed by varying a series of external resistances
The results are shown in Figure 21 With MFCs acclimated to 25 Ω external resistance
the maximum power density was achieved at 65Wm2 based on the polarization data At
this point the current density is 21mAcm2 The power density was three times higher
than previous report (20 Wm2) using phosphate buffer (Fan et al 2007) Power density
was also 2 times higher than (28 m Wm2) using a bicarbonate buffer ( Fan et al 2007)
32 Bacterial community structure assessed by PCR-DGGE
Bacterial DGGE profiles for higher power generating MFCs mixed culture setups
were relatively complex (more than 10 bands) However there are three brightly stained
bands and other less intense bands (Fig 2-2) These results indicated there were probably
three dominant bacterial species in mixed culture MFCs
010
020
030
040
050
060
070
2
3
4
5
6
7
04 05 05 06 07 08 10 11 13 15 21 22 23 24
Volta
ge (
V)
Pow
er
densi
ty (
Wm
2)
Current density (mAcm2)
Power
Voltage
power density 65wm2 at current
density208 mAcm2
Figure 2-1 Polarization curves of high power mixed culture MFCs
21
33 16S rDNA clone library results
A total of 83 colonies were sequenced for 16S rDNA clone library construction They
represented the bacterial community structure in mixed culture in MFCs The results
revealed that the predominant bacterial species in mixed culture are Geobacter sp (66
of the mixed culture) Arcobacter sp (12 of the mixed culture) and Citrobacter sp (11
of the mixed culture) Those three genuses comprised 88 of the total bacterial species
(Fig 23) This result is consistent with the DGGE result which showed that three bright
bands in the DGGE gel represented three dominant bacterial species in the mixed culture
The phylogenetic analysis of the colonies (83 colony sequences) is shown in Figure 24
Figure 22 DGGE result of high power generation mixed culture MFC (1 mixed
culture 2 pure culture control)
1 2
22
34 Dominant bacterial band in DGGE identification
Geobacter
66
Arcobacter
12
others
6
Pseudomonas
1
Citrobacter
11
Clostridium
2
Anaerovorax
2 Geobacter
Arcobacter
Citrobacter
Clostridium
Anaerovorax
Pseudomonas
others
Figure 23 Bacterial species and percentage of bacterial community of mixed
culture MFC lt 1 Alcaligensgt
lt1 Commamonasgt
lt1 Pseudomonas aeruginosagt
lt1 Klebsiellagt
lt11 Citrobactergt
lt1 Wolinellagt
lt12 Arobactergt
lt2 Anaeroboraxgt
lt2 Clostridium gt
lt1 unclear bacteriagt
lt66Geobactergt
100
100
85
100
100
82
99
87
100
002
Figure 24 Phylogenetic trees of bacterial species from mixed culture MFC
23
The dominant bacterial species including Goebacter sp Citrobacter sp and Arcobacter
sp in the mixed culture are shown in Figure 25 The upper band is Arcobacter sp and
Citrobacter sp is located in the bottom band of DGGE Geobacter sp is the middle one
The analysis of the bacterial community using denaturing gradient gel electrophoresis
(DGGE) of PCR-amplified 16S rRNA gene fragments and 16S rDNA clone library
construction method showed great phylogenetic diversity of mixed culture in MFCs with
the identification of sequences derived from bacteria of the taxa deltaproteobacteria
(Geobacter sp) gammaproteobacteria (Citrobacter sp) and epsilonproteobacteria
(Arcobacter sp) The result is consistent with the conclusion that among the isolated
exoelectrogens gammaproteobacteria and deltaprotebactia appear to be dominant among
others (Parot et al 2009 Liu et al 2010)
35 The dominant bacterial species isolation and power generation
Figure 25 Dominant bacterial species identification in DGGE
A B C D
A Citrobacter sp
B Geobacter sp
C Arobacter butzmeri
D mixed culture control
24
Two dominant bacterial species Citrobacter sp XS-1 and Arcobacter sp XS-2 have been
isolated and tested for power generation The power densities generated by the two
isolates are shown in the Fig 26 Current densities generated by isolate Citrobacter sp
and Arcobacter sp were 98 mAm2 and 20 mAm
2 respectively which were much less
than that (21Am2) generated by the mixed culture from which they were isolated
Figure 26 Current density generated by different dominant isolates
The maximum current density generated by mixed culture is much higher than the
one produced by pure bacterial species Citrobacter sp XS-1 and Arcobacter sp XS-2 in
the same structure MFCs Although the Geobacter sp in our mixed culture has not been
isolated a previous study showed that maximum current density generated by Geobacter
sp was 080 Am2
(Bond and Lovley 2003) The current density results seem partially
supporting the conclusion that mixed-culture generated higher power densities than their
pure-culture counterparts in MFCs (Jung and Regan 2007) This result suggests there
may be synergistic interactions within the anode bacterial communities
0
500
1000
1500
2000
2500
Mixed culture Citrobacter sp Arcobacter Sp
Cu
rre
nt
de
nsi
ty (
mA
m2)
25
Geobacter sp are strictly anaerobic bacteria which were enriched in most of the anode
biofilms (George 2005) Citrobacter species are facultative anaerobic bacteria (George
2005) which can be found in a wide variety of habitats including in soil water and
wastewater Arcobacter sp are micro-aerobic bacteria including both environmental
nonpathogens and opportunistic human pathogens They are able to grow in aerobic
conditions but in the optimal growth under micro-aerobic conditions (George 2005)
Arcobacter sp has been discovered in the MFCs by Fedorovich (Fedorovich et al 2009)
It is interesting that the mixed culture bacterial community composed of primarily these
three bacterial species has demonstrated to generate high power at non-strictly anaerobic
condition while leading dominant bacterial species Geobacter sp (66) is strict
anaerobic bacteria and generated powder in the MFCs only under strict anaerobic
condition (George 2005) Therefore it is possible that microaerobic bacteria such as
Arcobacter sp and facultative anaerobic bacteria Citrobacter sp can facilitate to create
anaerobic condition for Geobacter sp when they utilized oxygen for growing in the
mixed culture Syntrophic communities study have showed that anaerobic bacteria and
methanogenic archaea form compact microbial structures that operate like an organ rather
than a set of microorganisms functioning independently (Stams and Plugge 2009) Some
substrates have been degradeted within these communities while they are not able to be
fermented by individual species alone interspecies electron transfer also have been
presented in these communities (Stams and Plugge 2009) Summers et al (2010) also
discovered that direct exchange of electrons happened within coculture of Geobacter
metallireducens and Geobacter sulfurreducens Therefore we believe that there are
26
more complex synergistic interactions between different bacteria species in mixed culture
in the MFCs
Acknowledgements
The authors acknowledge support from the US National Science Foundation
(CBET 0955124) We also thank Keaton Lesnik for the review of this manuscript
27
Chapter 3
New Exoelectrogen Citrobacter sp SX-1 Isolated from a Microbial Fuel Cell
Shoutao Xu and Hong Liu
Published in
Journal of Applied Microbiology 111(5)1108-1115 (2011)
ABSTRACT
Exoelectrogenic bacterial strain SX-1 was isolated from a mediator-less microbial fuel
cell by conventional plating techniques with ferric citrate as electron acceptor under
anaerobic condition Phylogenetic analysis of the 16S rDNA sequence revealed that it
28
was related to the members of Citrobacter genus with Citrobacter sp sdy-48 being the
most closely related species The bacterial strain SX-1 produced electricity from citrate
acetate glucose sucrose glycerol and lactose in MFCs with the highest current density
of 205 mA m-2
generated from citrate Cyclic voltammetry analysis indicated that
membrane associated proteins may play an important role in facilitating the electrons
transferring from bacteria to electrode This is the first study that demonstrates that
Citrobacter species can transfer electrons to extracellular electron acceptors Citrobacter
strain SX-1 is capable of generating electricity from a wide range of substrates in MFCs
This finding increases the known diversity of power generating exoelectrogens and
provided a new strain to explore the mechanisms of extracellular electron transfer from
bacteria to electrode The wide range of substrate utilization by SX-1 increases the
application potential of MFCs in renewable energy generation and waste treatment
Keywords Microbial fuel cell Exoelectrogen Citrobacter sp SX-1 Extracellular
electron transfer
29
Introduction
Microbial Fuel Cells (MFCs) technology has become an active research area
recently as a promising approach for renewable energy generation wastewater treatment
and bioremediation (Rabaey and Verstraete 2005 Lovley 2006 Fan et al 2008 Logan
2009 Liu et al 2010) The key feature of MFCs system is the microbe-catalyzed electron
transfer from organic matter to anodes Many studies have shown the presence of diverse
bacterial communities on MFCs anodes (Reimers et al 2001 Liu et al 2004b Kim et al
2005) Over 20 exoelectrogens ie microbes that can transfer electrons exocellularly to
electrodes have been reported in the past 10 years (Logan 2009 Liu et al 2010) The
exoelectrogens so far belong to diverse genetic groups including alphaproteobacteria
(Rhodopseudomonas Ochrobactrum and Acidiphilium) (Malki et al 2008 Xing et al
2008 Zuo et al 2008) betaproteobacteria (Rhodoferax) (Chaudhuri et al 2003)
gammaproteobacteria (Shewanella Pseudomonas Klebsiella Enterobacter and
Aeronomas) (Kim et al 2002 Pham et al 2003 Rabaey et al 2005 Bretschger et al
2007 Zhang et al 2008 Chung et al 2009 Rezaei et al 2009) deltaproteobacteria
(Geobacter Geopsychrobacter Desulfuromonas and Desulfobulbus) (Bond et al 2002
Bond and Lovley 2003 Holmes et al 2004ab 2006) Epsilonproteobacteria (Arcobacter)
(Fedorovich et al 2009) Firmicutes (Clostridium and Thermincola) (Park et al 2001
Wrighton et al 2008) Acidobacteria (Geothrix) (Bond and Lovley 2005) and
actinobacteria (Propionibacterium) (Wang et al 2008) A wider range of
exoelectrogenic species are expected to be discovered
Three mechanisms have been proposed for exocellular transport of electrons by
exoelectrogens without artificial mediators Some exoelectrogenic bacteria can transfer
30
electrons to electrodes through soluble redox compounds excreted by microorganisms
(Bond and Lovley 2005 Rabaey et al 2005) and others can directly transfer electrons to
anodes via outer cell membrane proteins (Myers and Myers 1992) Recently more and
more evidence supports the involvement of bacterial nanowires in extracellular electron
transport (Reguera et al 2005 2006 Gorby et al 2006) In spite of the discovery of
many bacterial species that can transfer the electrons to electrode without the need of
artificial mediators the investigation of extracellular electron transfer mechanisms was
mainly focused on a few species such as those from Geobacter and Shewanella genera
(Kim et al 1999 2002 Reguera et al 2005 2006 Gorby et al 2006) The electron
transfer mechanisms for many of the isolated exoelectrogens species are still not well
studied
In the present study we isolated a new exoelectrogen strain SX-1 from a MFC a
strain phylogenetically related to Citrobacter sp Power generation from various carbon
sources by this strain was evaluated using single chamber MFCs Plausible extracellular
electron transfer mechanisms were also discussed based on the characterization of anodic
biofilms by cyclic voltammetry (CV)
Materials and methods
Bacterial strain SX-1 isolation
Bacterial strain SX-1 was isolated from the anode biofilm of a MFC fed with sodium
acetate operated in fed-batch mode over a period of six months The original source of
the inoculum is wastewater from a local waste water treatment plant Bacterial cells were
released from the carbon cloth anode by shaking the cloth in a sterile flask with 15 mL
sterile 1 NaCl solution and glass beads (2 mm diameter) The suspension was then
31
serially diluted from 10 times to 105
times and plated on a petri dish with a solid agar
medium containing NH4Cl 6mM KCl 2mM CH3COONa (electron donor) 30 mM
FeC6H5O7 (electron acceptor) 20mM minerals and vitamins (Lovley and Phillips 1988)
Single colony was randomly selected after culturing on the plate for 36 hours at 30 degC
and purified on a new agar plate following a procedure reported previously (Chung and
Okabe 2009) Then the isolate was picked up and grew in a liquid medium solution in
anaerobic tubes containing the same constituents as the solid medium for further analysis
All isolation process was operated in a glove box anaerobic chamber (Coy Laboratory
Products Grass Lake MI)
16S rDNA sequencing and phylogenetic analysis
Bacterial genomic DNA was extracted from the isolated strain SX-1 using the DNeasy
tissue Kits (Qiagen Valencia CA) according to the manufacturerrsquos instructions The 16S
rDNA was amplified by PCR using a pair of universal primers 27F (5rsquo-
AGAGTTTGATCMTGGCTCAG-3rsquo) and 1492R (5rsquo-GGTTACCTTTGTTACGACTT-
3rsquo) (Park et al 2001) The conditions used for PCR were described previously
(Ferna ndez et al 1999) PCR products were purified ligated and cloned according to a
procedure described in a previous report (Xing et al 2008) Plasmids were isolated from
randomly selected clone colonies with a QIAprep Spin Miniprep kit (Qiagen Valencia
CA) and three plasmid inserts were then sequenced in both directions using an ABI 3730
DNA sequencer (Applied Biosystems Carlsbad CA)United States) Due to the identical
16S rDNA sequence of the three colones only one was queried against the GenBank and
Ribosomal Database Project (RDP) databases using BLAST and SEQUATCH algorithms
(Cole et al 2007 Wang et al 2007 Cole et al 2009) The 16S rDNA sequence of
32
strain SX-1 and closely related type strains were aligned using CLUSTALX software
(Thompson et al 1997) The neighbor-joining trees were constructed with the Molecular
Evolutionary Genetics Analysis package (MEGA Version 4) based on 1000 bootstrap
analysis (Tamura et al 2007)
SEM
The morphologies of SX-1 cells growing overnight in anaerobic tubes were examined
with a scanning electron microscope (FEI Quanta 600 FEG FEI Company Hillsboro
OR) The suspended cells were fixed in a 25 glutaraldehyde and 01M phosphate
buffer solution and dehydrated with a graded ethanol series from 30 to 100 After
dehydration the samples were dried in a critical point dryer and then sputter-coated with
AuPd for SEM examination (Liu and Logan 2004)
Evaluation of power generation by SX-1 in MFCs
Single chamber MFCs were used to evaluate power generation by SX-1 using various
substrates The MFCs were constructed as described previously (Liu and Logan 2004)
and modified with 3 cm2 carbon cloth anodes and 7cm
2 carbon clothPt cathodes The
total liquid volume of each MFCs was about 15 ml with electrode spacing about 17 cm
All MFCs were operated in an autoclaved closed plastic box and sterile cotton was
attached to the outer surface of the air cathodes to prevent contamination A MFC
without bacterial culture was used as control MFCs were inoculated with 3 ml late
exponential phase cultures of SX-1 in the medium solution reported previously (Liu and
Logan 2004) Seven substrates including citrate acetate glucose sucrose glycerol and
lactose were evaluated individually for power generation in a fed-batch mode in a
temperature control chamber (30 plusmn2degC) For each substrate at least 8 batches were run to
33
investigate the effect of biofilm formation on current output at fixed external resistance of
1kΩ and the same initial substrate concentration of 30 mM Phosphate buffer (100mM)
was used to maintain the solution pH and conductivity The MFCs with sodium citrate as
electron donors were also examined for maximum power output by varying the external
resistance from 20 kΩ to 4 kΩ A data acquisition system was used to record voltage data
during the MFCs operation (Keithley 2700 Keithley Instruments Inc Cleveland OH) It
took about 15 to 30 minutes for the MFCs to stabilize depending on the external
resistance At each resistance we collected at least five data at the steady condition to
make the polarization curves Averaged voltages were used to calculate the power density
(mW m-2
) according to P=IVA where I was the current V was voltage and A was cross-
sectional area of the anode
CV analysis
CV was used to characterize the oxidation and reduction reactions on the anodic surface
of the MFCs fed with citrate and operated at 7 KΩ This external resistance was selected
because the maximum power density was obtained at this resistance based on the
polarization experiment We assumed the biofilm on the anode of MFCs were well-
developed when stable power output was obtained after 3 batches of operation The
MFCs were then used directly for CV analysis at four current generating stages of the
fourth batch (1) initial exponential current increasing stage (middle point of stage about
4 h after media change) (2) current plateau stage (middle point of stage about 8 h after
media change) and (3) current decreasing stage (middle point of stage about 16h after
media change) (4) right after the complete replacement of medium solution (Figure 6)
The anode was used as working electrode the cathode as counter electrode and an
34
AgAgCl electrode was selected as the reference The CV curves were scanned from 200
to -600 mV at a rate of 5 mV s-1
using a potentiostat (G300 Gamry Instrument
IncWarminster PA) Control experiment was also conducted using new anode (without
biofilm) and new medium solution
Nucleotide sequence accession number
The 16S r DNA sequence determined in this study has been deposited in the GenBank
database under accession number HQ845373
Results
Identification of the strain SX-1
An almost-complete 16S rDNA sequence (1475 bp) of strain SX-1 was obtained and
subjected to comparative analysis with the 16S rDNA of closely related reference strains
A phylogenetic analysis revealed that strain SX-1 should be assigned to the genus
Citrobacter and was most closely related to Citrobacter sp sdy-48 EJ463782 (990
sequence similarity) and Citrobacter sp yy-21 FJ463779 (975 sequence similarity)
These three strains formed a distinct sub cluster in the neighbor-joining in which the new
isolate and Citrobacter sp sdy-48 EJ463782 formed a distinct subline (Figure 31)
The SEM image illustrated that the strain SX-1 is a rod-shaped bacterium 03-06 microm
wide and 12-17 microm long (Figure 32) Strain SX-1 colonies on nutrient agar appeared 2-
3 mm in diameter smooth moist opaque with a shiny surface after 5 days incubation
under anaerobic condition The fact that strain SX-1 grew both aerobically and
anaerobically using FeC6H5O7 as electron acceptor suggests that the strain is facultatively
anaerobic Strain SX-1 showed robust growth on citrate as a carbon source These D
35
properties were similar to those of genus Citrobacter described in Bergeyrsquos Manual of
Systematic Bacteriology (George 2005)
Figure 31 phylogenetic tree of strain SX-1 and closely related species based on 16S
rDNA sequences The tree was constructed using the neighbor-joining method
Figure 32 SEM image of planktonic cells of Citrobacter sp SX-1
Strain SX-1
Citrobacter sp sdy-48 (FJ463782)
Citrobacter sp yy-21 (FJ463779)
Citrobacter koseri E639 (ATCC 25408)
Citrobater koseri CDC 3613-63(AF025372)
Citrobacter koseri CDC 8132-86(AF025366)
Salmonella enterica SL483 (CP001138)
Salmonella enterica AKU12601(AY696668)
Shigella sonnei Ss046(CP000038)
E coli C2 (AF403733)
E coli E24377A(CP000800)88
100
100
76
44
92
53
36
0001
1 microm
36
Electricity production by strain SX-1 in MFCs
The isolated strain SX-1 was first tested for its ability to generate current from sodium
citrate at 1kΩ Current gradually increased to a maximum 50 mA m-2
after cell
inoculation and then decreased (Figure 33) After the MFCs was refilled with new
substrate solution the current recovered rapidly and reached a higher level than the first
batch After 4 batches operation the maximum current output of each batch became
stable The highest current density achieved in the MFCs with Citrobacter sp SX-1 at 1
KΩ was 98 mA m-2
(Figure 33)
Figure 33 Electricity generation by Citrobacter sp SX-1 in a single chamber MFCs
with sodium citrate (30 mM) as substrate at 1 KΩ
0
20
40
60
80
100
120
0 50 100 150 200
Curr
ent
den
sity
(m
A m
-2)
Time (h)
37
Current generation by SX-1 from other substrates including glucose lactose sodium
acetate glycerol and sucrose were also assessed at an external resistance of 1KΩ
(Figure 34) When repeatable cycles of current output were obtained for these substrates
glycerol generated the highest maximum current density of 58 mA m-2
followed by
lactose and sucrose with 29 mA m-2
and 27 mA m-2
respectively Glucose and acetate
produced the lowest maximum current density of 96 and 43 mA m-2
respectively These
results indicated that strain SX-1 can utilize a wide range of substrates for electricity
generation in MFCs but with different power generation potentials
Figure 34 Electricity generation of Citrobacter sp SX-1 using different substrates in
single chamber air-cathode MFCs at 1 KΩ (The error bars stand for the standard
deviation of 3 replicates)
0
10
20
30
40
50
60
70
Glucose Lactose Glycerol Acetate Sucrose
Curr
ent
den
sity
(m
A m
-2)
38
Polarization experiment was further conducted to determine the maximum power
density generated by strain SX-1 in the single chamber MFCs with citrate as substrate A
maximum power density of 881 mW m-2
was obtained at current density of 205 mA m-2
at an external resistance of 7 KΩ (Figure 35)
Figure 35 Power and voltage generation by Citrobacter sp SX-1 as a function of
current density using sodium citrate (30 mM) as substrate The error bars stand for the
standard deviation of the 3 voltagepower outputs obtained in three MFCs
20
30
40
50
60
70
80
90
100
01
02
03
04
05
06
07
80 110 140 170 200 230
Po
wer
den
sity
(m
W m
-2)
Vo
ltag
e (V
)
Current density (mA m-2)
Voltage
Power
39
Figure 36 Four current generating stages for CV analysis current increasing stage (stage
1) current plateau stage (stage 2) current decreasing stage (stage 3) and right after the
replacement of medium solution (stage 4) The MFCs was operated at 7 KΩ with sodium
citrate (30 mM) as substrate
Cyclic Voltammetry
To determine the presence of redox active compounds produced by SX-1 and the
location of these compounds CV scan of the anodic biofilms at four current output stages
and the supernatant of MFCs medium solution at the end of the batch experiment were
performed Reduction peaks at range of -310 ndash -350 mV and oxidation peaks at range -
100 ndash -150 mV were observed at all current generating stages (Figure 36 Figure 37A)
suggesting the presence of redox active compounds may involve in extracellular electron
transfer by Citrobacter sp SX-1 The amplitude of the peaks increased according to the
growth stage of the batch and the highest peaks were present after the current plateau
stage which indicated the redox active compounds mainly were secreted in the current
plateau stage While the current density at stage 3 (deceasing stage) was lower than that
at stage 2 (current plateau stage) the redox peak at stage 3 was higher than that in stage 2
0
50
100
150
200
250
50 55 60 65 70 75 80 85
Curr
ent
den
sity
(m
A m
-2)
Time (h)
A
A
Stage 1
Stage 2 Stage 3
Stage 4
40
A plausible explanation for this pattern was that redox active compounds were
continuously being secreted and accumulated after the current peak was reached
resulting in more redox compounds present after the plateau stage But since most of the
carbon source had already been being used up many of the redox compounds at this
stage had not been as active as those at current plateau stage due to much less electrons
were available to be transferred The bigger peak amplitude in stage 4 than in stage 1
indicates that the redox active compounds were continuously being secreted and
accumulated in the biofilm even after stable power was generated at 7 kΩ The slightly
change of the peak locations at different stages was possible due to the solution chemistry
change during the current generation process or the slightly location change of reference
electrode during the measurement (Figure 37B) When the medium solution in the MFCs
was replaced by fresh medium at the end of the batch the oxidization and reduction
peaks were still presented in the CV (Figure 37B) but no peaks were observed when the
supernatant and fresh mediums were tested in a MFCs with no biofilm (Figure 37C)
These results suggest the compounds involved in the electron transfer were located not in
the supernatant (solution) but in the biofilm
41
Figure 37 Cyclic Voltammetry analysis of Citrobacter sp SX-1 biofilms at (A) current
increasing stage current plateau stage and decreasing stage (end of the batch) (B) end of
the batch and replaced with fresh medium and (C) controls
-40E-04
-30E-04
-20E-04
-10E-04
00E+00
10E-04
20E-04
30E-04
-060 -050 -040 -030 -020 -010 000 010 020
Curr
ent
(A)
Potential (V) vs AgAgCl
Current plateau stage
Current increasing stage
Current decreasing stage
-40E-04
-30E-04
-20E-04
-10E-04
00E+00
10E-04
20E-04
30E-04
-060 -050 -040 -030 -020 -010 000 010 020
Curr
ent
(A)
Potential (V) vs AgAgCl
Current decreasing stage
Biofilm with fresh medium
-40E-04
-35E-04
-30E-04
-25E-04
-20E-04
-15E-04
-10E-04
-50E-05
00E+00
-060 -050 -040 -030 -020 -010 000 010 020
Curr
ent(
A)
Potential (V) vs AgAgCl
Supernatant
Fresh medium
A
B
C
B C
42
Discussion
Citrobacter species belonging to Gammaproteobacteria Enterobacteriales
Enterobacteriaceae are facultative anaerobic bacteria (George 2005) They can grow with
oxygen as an electron acceptor but also with a variety of electron accepters such as Fe (III)
in the absence of oxygen (George 2005) Citrobacter species can be found in a wide
variety of habitats including in soil water and wastewater Several Citrobacter species
have been studied as important bioremediation bacterium for heavy metal removals
sulfate reduction phenol degradation and chlorophenol degradation (Macaskie et al 1995
Narde et al 2004 Qiu et al 2009) Citrobacter species have also been found on the
anodic biofilm of a diesel-degrading MFCs (Morris et al 2009) However there was no
study reported that Citrobacter species can transfer electrons to extracellular electron
acceptors This finding that Citrobacter strain SX-1 can generate electricity in MFCs
increased the diversity of power generating exoelectrogens and provided a new strain to
explore the mechanisms of extracellular electron transfer from bacteria to electrode
Among the isolated exoelectrogens gammaproteobacteria and deltaprotebactia appear to
be dominant among others (Parot et al 2009 Liu et al 2010) The isolated strain
Citrobacter sp SX-1 phylogenetically belongs to gammaproteobacteria which provides
another evidence to support this observation
Most exoelectrogens utilize a limited range of substrates for power generation For
example Shewanella oniedensis MR-1 can oxidize lactate under anaerobic condition but
cannot utilize acetate for electricity generation (Kim et al 2002) Geobacter
sulfurreducens can completely oxidize acetate for power generation but it cannot utilize
simple sugars (Bond and Lovley 2003 Caccavo et al 1994)) In comparison strain SX-1
43
not only can oxidize citrate for power generation but also utilize a wide range of
substrates for power generation including glycerol glucose lactose sucrose and acetate
Interestingly the higher current density generated by strain SX-1 from glycerol a main
by-product of biodiesel production process suggests that strain SX-1 may be potentially
used for harvesting energy from biodiesel wastes using MFCs However the current
density generated by strain SX-1 (205 mA m-2
) is lower than that (805 mA m-2
at 1 kΩ)
generated by the mixed culture from which the SX-1 was isolated indicating the
existence of other higher power generating bacteria andor complex ecology in the mixed
culture community
Understanding mechanisms of microbial extracellular electron transfer is critical for
enhancing the electron transfer rate from bacteria to electrode through metabolic or
genetic engineering of exoelectrogens Since the spent supernatant solution of MFCs run
with SX-1 showed no redox properties the redox compounds produced by SX-1 may
have been retained in the biofilm Alternatively SX-1 may utilize one or both of the
other two known extracellular electron transfer mechanisms For example it is possible
that strain SX-1 transfers electrons to the anode via cell-bound outer membrane proteins
The two distinct oxidation and reduction peaks observed at -100mv- -150 mV and -310--
350 mV of the SX-1 biofilm have a midpoint potential range from -205mV to -250mV
which is close to the redox potential of some c-type cytochromes (Field et al 2000 Meitl
et al 2009) which are well-known to play an important role in extracellular electron
trasnfer (Eggleston et al 2008 Field et al 2000 Meitl et al 2009) Field et al (2000)
reported that the c-type cytochromes CymA had the midpoint potentials of -229 mV
Meitl et al (2009) and Eggleston et al (2008) reported that the c- type cytochromes
44
OmcA in Shewanella oneidensis MR-1 had the midpoint potentials of -201 mV and -208
mV
While itrsquos possible that SX-1 may also have conductive appendages to enhance the
extracellular transfer electron similar to the nanowires discovered in some Geobacter
and Shewanella species (Reguera et al 2005 2006 Gorby et al 2006) further
investigation on the presence of these appendages on the cell surface of SX-1 is needed
Acknowledgements
The authors acknowledge support from the US National Science Foundation
(CBET 0955124) We also thank Teresa Sawyer for her helps with the SEM analysis and
Jeremy Chignell and Yanzhen Fan for their review of this manuscript
45
Chapter 4
Enhanced Performance and Mechanism Study of Microbial Electrolysis Cells
Using Fe Nanoparticles Decorated Anodes
Shoutao Xu Hong LiuYanzhen Fan Rebecca Schaller Jun Jiao Frank Chaplen
Published in
Applied Microbiology and biotechnology 93(2)871-880 (2012)
46
ABSTRACT
Anode properties are critical for performance of microbial electrolysis cells (MECs) In
the present study Fe nanoparticle modified graphite disks were used as anodes to
investigate the effects of nanoparticles on the performance of Shewanella oneidensis MR-
1 in MECs Results demonstrated that average current densities produced with Fe
nanoparticle decorated anodes were up to 56-fold higher than plain graphite anodes
Whole genome microarray analysis of the gene expression showed that genes encoding
biofilm formation were significantly up-regulated as response to nanoparticle decorated
anodes Increased expression of genes related to nanowires flavins and c-type
cytochromes indicate that enhanced mechanisms of electron transfer to the anode may
also have contributed to the observed increases in current density The majority of the
remaining differentially expressed genes were associated with electron transport and
anaerobic metabolism demonstrating a systemic response to increased power loads
Keywords Microbial electrochemical system microbial fuel cell microbial electrolysis
cell nanotechnology differential gene expression DNA microarray
47
Introduction
Microbial electrochemical systems (MESs) have been intensively studied since Lewis
achieved practical advances in this field (Logan 2007) however they attracted much
research attention in recent years due to their promising applications in renewable energy
generation bioremediation and wastewater treatment In a MES microorganisms
interact with electrodes via electrons catalyzing oxidation and reduction reactions at the
anode and the cathode The most-described type of MESs is microbial fuel cells (MFCs)
in which useful power is generated from electron donors typically biodegradable organic
materials (Logan et al 2006) Various novel MESs have recently been developed to
produce hydrogen (microbial electrolysis cells (MECs)) (Liu et al 2005 Logan et al
2008 Rozendal et al 2006 Schaller et al 2009) to power an autonomous sensor or
sensor network (Ringeisen et al 2006) to reduce bioremediation targets (Lovley 2006
Shea et al 2008) Sukkasem et al 2008) and to desalinate water (Cao et al 2009) The
key feature shared by these systems is the microbe-catalyzed electron transfer from
organic matter to electrodes (anodes) (Rabaey et al 2004) Enhancing the anodic current
output which highly depends on the performance of the electrodes is critical for the
successful application of all these processes (Logan et al 2007 Park and Zeilus 2002
2003)
Nanomaterials have received much attention from researchers in the context of
microbiology due to their unique physical electrical and chemical properties which
facilitate the study of interactions between bacteria and surfaces (Liu 2006) Previous
studies have demonstrated that electrodes decorated with different nanostructures such
as carbon nanotubes nanostructured polyanilinetitanium dioxide composite and titania
48
nanotubes have the potential to enhance power generation in MFCs (Morozan et al 2007
Qiao et al 2008 Quan et al 2005) Furthermore our recent study shows nanoparticle
(NP) decorated anodes greatly increased the electrochemical electron transfer rate in
MFCs (Fan et al 2011) However Fe NPs as an ideal candidate for decorating electrodes
because of respectively low price and high conductivity compared to other materials has
not been focused to study in MECs
S oneidensis MR-1 an important electrochemically active bacterial strain has been
exploited for the electricity generation in MFCs (Logan et al 2007 Logan 2009 Park
and Zeikus 2002) The availability of genome sequence for this strain makes it possible to
use transcriptome assays to globally measure the responses to different growth conditions
and environmental stresses (Murray et al 2001) S oneidensis MR-1 gene expression
response to heat shock cold shock Cr (VI) and U (VI) reduction chromate stress and
iron and acid tolerance have been studied in the past several years (Bencheikh-Latmani et
al 2005 Brown et al 2006 Gao et al2006 Gao et al 2004 Yang et al 2008) However
no studies have been focused on the Shewanella gene expression response to NPs in
MESs
The mechanism of increased electron transfer rate exhibited by nanoparticle decorated
electrodes is not well understood yet In the present study graphite disks decorated with
Fe NPs were used as anodes to explore the effects of nanostructures on current generation
in a multi-anode MECs DNA microarrays were utilized to investigate differences in the
global gene expression profile of S oneidensis MR-1 grown on plain versus Fe NPs
decorated anodes
Materials and methods
49
Bacterial cultures
S oneidensis MR-1 was purchased from American Type Culture collection (ATCC
700550) and stored as freezer stocks at -80degC Stock cells of S oneidensis MR-1 was
grown aerobically using a medium containing 30 gL Trypticase Soy Broth (BD 211825
Becton Dickinson Company Franklin lakes NJ) at 30degC Cells in early stationary
growth phase (optical density of 15-16 at 600nm (Spectrophotometer UV-1700
Shimadzu Columbia MD) were washed two times then injected into the chamber of the
MECs for current production Trypticase Soy Broth medium with 30 mM sodium lactate
as electron donor was used in MECs Phosphate buffer (100mM) was used to maintain
the solution pH 7 and solution conductivity at 15 mScm
Characterization of nanostructured anodes
Superfine isomolded graphite disks (16 cm x 05 cm GraphiteStorecom Inc) were
polished with ultrafine sand paper (2000 grit 3M Company) as the base for the NP
decorated and control anodes Fe NPs decorated anodes were fabricated by thermal
annealing of metal thin film coated graphite disks Thin films of Fe were firstly deposited
on the polished graphite disk by using sputter coating for 95 min and the samples then
were annealed in the Chemical vapor deposition (CVD) at 750degC for 20 min to form Fe
NPs Fe-NPs were produced with a diameter of 200plusmn100 nm and a density range
(distance between NPs) of 200plusmn100 nm with a coating time of 95 min SEM images of
Fe NPs decorated anode and control surfaces were shown in the Figure 41
50
Figure 41 SEM images of Fe NPs decorated anodes (A) Graphite disk control (B) Fe
Nanoparticle decorated
Multiple channel MECs construction and operation
A MECs with removable multiple anodes with each effective anode area 07 cm2
was
constructed and used to evaluate the effects of nanostructure on current density of
according to a previous report (Fan et al 2011) The cathode was made of wet-proof
(30) carbon cloth (type B E-TEK Division Inc USA) coated with PtC (20 E-TEK
Division Inc USA) following a previously reported procedure (Liu et al 2005) The
final platinum loading was 05 mgcm2 per projected cathode area The size of cathode
(150 cm2) was 25 times larger than the total effective area of the 8 graphite disk anodes
(56 cm2) to prevent cathode limitations on the performance of the MECs system All
MECs with Fe-NP decorated anodes and control anodes were tested for current
generation for 24 h with medium (without bacteria) before injecting the bacterial cells in
order to determine whether the Fe-NP decorated anodes demonstrate chemical current
generation compared to control anodes All the testing anodes were set up in the MECs
B
500 nm
A
500 nm
51
at once so that the S oneidensis MR-1 culture was applied to all of them simultaneously
Short electrode spacing (17 cm) was used in the MECs design to reduce the internal
resistance A voltage of 06 V was applied the MECs for current generation after the cells
of S oneidensis MR-1 were inoculated into MECs at early stationary phase A multimeter
with a data acquisition system (2700 Keithly USA) was used to monitor the current
change by measuring the voltage (every 300 seconds) drop through a resistor at 300 ohm
Fig 4 2 Effects of Fe-NP decorated anodes on the current density in MECs Control is
plain graphite disk anode Fe-NP is the anode with Fe-NP decoration
Microarray analysis
Biofilms for whole gene microarray analysis were aseptically removed from the plain
and Fe-NP decorated anodes of MECs after 110 hours further incubation at 30oC when
current density obviously started to decease Total RNA was extracted using Trizol
(Ambion inc AustinTX) following the manufacturerrsquos instructions Integrity of the
52
RNA samples was confirmed using the Agilent 2100 Bioanalyzer (Agilent Technology
Inc Palo Alto CA) The same amount of RNA (200 ng) from each sample was used for
further analysis after amplification by using the MessageAmpTM
II-Bacteria Prokaryotic
RNA Amplification kit (Ambion AustinTX) according to the manufacturerrsquos
instructions All of the 4295 genes (resource from NCBI) represented on the S oneidensis
MR-1 whole-gene microarrays were ordered from Roche NimbleGen Inc (Madison WI)
Biological triplicates of biofilms on the Fe-NP decorated anodes and control were
analyzed respectively cDNA synthesis labeling and hybridization were carried out by
the CGRB Core Laboratories at Oregon State University DNASTAR ArrayStarTM
3
software was used to identify genes that were up- or down-regulated more than 2-fold
when grown on the nanoparticle modified anodes using the unpaired two sample t-test
with a cutoff p-value of 005 The complete microarray data set generated in this study is
deposited for public access in the Gene Expression Omnibus
(httpwwwncbinlmnihgovgeo) under accession number GSE31535
Results
Enhancement of current generation by using NP decorated anodes in MECs
The current density generated by Fe-NP decorated anode MECs with S onidensis MR-1
increased to approximately 43 microAcm2 20 h following inoculation and then slowly
decreased Figure 42 shows that the maximum current density achieved by NP
decorated anodes was 83 times higher than that (51 microAcm2) generated by the control
(plain graphite disk) The average current density improvement of 110 hours was 59
times of that generated by the control The current densities generated in MECs in the
53
absence of bacteria were negligible (03 microAcm2) which demonstrated that the current
enhancement observed with the Fe-NP decorated anodes was biologically-derived
Global transcriptome analysis
Whole-genome DNA microarrays were used to attain a comprehensive general
overview
of the transcriptional response to Fe-NP decorated anodes by S oneidensis MR-1 in MEC
Of a total 4295 genes assayed 392 (188 induced and 204 repressed) exhibited significant
(P lt 005) differential expression at a 2-fold (log (expression ratio 1) level in 3
replicates in response to Fe-NPs These
total gene numbers present 9 of the 4295 open
reading frames (ORFs) presented on the array
Fig 43 Differentially expressed genes grouped by functional classification according to
the TIGR S oneidensis genome database (wwwtigrorg) Columns 1 amino acid
biosynthesis 2 biosynthesis of cofactors prosthetic groups and carriers 3 cell
envelope 4 cellular processes 5 central intermediary metabolism 6 DNA metabolism
7 energy metabolism 8 fatty acid and phospholipid metabolism 9 other categories 10
protein fate 11 protein synthesis 12 purines pyrimidines nucleosides and nucleotides
13 regulatory functions 14 signal transduction 15 transcription 16 transport and
binding proteins 17 unknown function 18 hypothetical proteins
Up-Regulated
-75
-50
-25
0
25
50
75
100
125
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18
Down-Regulated
Num
ber
of
Gen
es
54
Figure 43 summarizes the overall genomic response of S oneidensis MR-1 to Fe-NP
decorated anodes by grouping the differentially expressed genes into their functional role
categories based on The Institute for Genomic Researchrsquos annotation (Version 8) of the
MR-1 genome sequence The wide distribution of putative functional roles attributed to
the differentially expressed genes indicated the extent of the molecular response of
Soniedenis MR-1 to the NP decorated anodes Among genes with defined function a
large number of up-regulated genes were associated with cellular processes (group 4)
energy metabolism (group 7) other categories (group 9) and hypothetical proteins (group
18) While those genes encoding proteins involved in amino acid biosynthesis (group 1)
cellular processes (group 4) substrate transport (group 16) and hypothetical proteins
(group 18) were among the most down-regulated genes Two groups of genes that were
the most noticeable among the functional gene groups genes related to energy
metabolism (group 7) and genes related to amino acid biosynthesis (group 1) The ratios
of up-regulated genes to down-regulated genes in these functional groupings were much
higher than other functional gene groupings there were 21 up-regulated genes and 7
down-regulated genes in energy metabolism gene group and 2 up-regulated and 22 down
regulated genes in the amino acid biosynthesis group
Genes related to biofilm formation
In order to determine whether there was a correlation between the observed biofilm
enhancements by NP decorated anodes and the genes related to biofilm formation the
significant modified genes related to biofilm formation were examined and summarized
in the Table 41 These included SO_3223 and SO_3228 genes encoding the flagellum
proteins (Thormann et al 2004) which has critical impacts on initial attachment to the
55
surface as swimming motility functions were up-regulated over 10 and 09 fold
respectively The gene clusters SO_4178-SO_4180 which are thought to be essential for
formation of a three-dimensional biofilm were up-regulated from 12 to 13 fold
(Thormann et al 2004) In addition the genes SO_4105 encoding IV pilus which is
known to represent a critical factor for biofilm formation on a biotic surfaces (Thormann
et al 2004) were also induced 08 fold (Table 41) The genes related to flagellum
motility (SO_4286 SO_1529 and SO_1530) were also induced slightly (Thormann et al
2004)
Table 41 Expression levels of genes related to biofilm formation
Locus Tag Gene product descriptions Fold change
SO_3228 flagellar basal-body MS-ring and collar protein FliF 10
SO_3223 flagellar hook-length control protein FliK 09
SO_4103 MshA minor pilin protein MshD 05
SO_4105 MSHA major pilin protein MshA 08
SO_4178 expressed protein of unknown function MxdC 12
SO_4179 inner membrane family 2 glycosyltransferase MxdB 13
SO_4180 diguanylate cyclase-like protein MxdA 10
a fold changes were presented after log2
b positive number represented up-regulated and negative represented down-regulated
Genes related to energy metabolism
A closer consideration of energy metabolism genes was undertaken as anaerobic
metabolism for electron generation and electron transport functions may play critical
roles in enhanced current density generation in MECs Genes associated with energy
56
metabolism and with significantly modified gene expression levels are summarized in the
Table 42 Several of the up-regulated genes were related to formate dehydrogenase
proteins Most notably four genes related to formate dehydrogenase were up-regulated
more than 2-fold SO_0102 SO_0103 SO_0104 and SO_0107 The other genes
expression related to formate dehydrogenase proteins SO_4513 SO_4514 SO_4515 also
significantly increased Another interesting gene is that encoding the cytochrome c
oxidase protein gene SO_4609 which was up-regulated 11 fold while the gene
SO_0479 related periplasmic octaheme cytochrome c MccA was down-regulated
However SO_1519 the gene encoding lactate dehydrogenase was down-regulated 12
fold despite lactate being the primary carbon source in the media
Table 4 2 Genes related to anaerobic growth and electron transfer with significantly change expression level
Locus Tag Gene product descriptions Fold
change
SO_0101 nitrate-inducible formate dehydrogenase molybdopterin-binding
subunit FdnG
18
SO_0102 formate dehydrogenase nitrate-inducible iron-sulfur subunit 21
SO_0103 formate dehydrogenase nitrate-inducible cytochrome b556 subunit 24
SO_0104 formate dehydrogenase accessory protein FdhE 22
SO_0107 formate dehydrogenase accessory protein fdhD 24
SO_0397 quinolfumarate reductase menaquinol-oxidizing subunit FrdC_2 12
SO_0452 thioredoxin 2 15
SO_0903 Na-translocating NADH-quinone reductase subunit B NqrB_1 12
SO_1012 NADH-ubiquinone oxidoreductase subunit K NuoK 12
SO_1013 NADH-ubiquinone oxidoreductase subunit J NuoJ 12
SO_1363 hydroxylamine reductase 13
57
SO_2417 ferredoxin cofactor maintenance protein YfaE 10
SO_3922 formate dehydrogenase cytochrome b Fdh -12
SO_4513 formate dehydrogenase molybdopterin-binding subunit _2 15
SO_4514 formate dehydrogenase FeS subunit FdhB_2 12
SO_4515 formate dehydrogenase cytochrome b subunit FdhC_2 14
SO_4609 aa3 type cytochrome c oxidase subunit III CoxC 11
SO_0479 periplasmic octaheme cytochrome c MccA -12
SO_0847 periplasmic nitrate reductase ferredoxin component NapG -15
SO_1519 L-lactate dehydrogenase iron-sulfur cluster-binding protein LldF -10
SO_1251 ferredoxin 4Fe-4S -11
SO_37411 hypothetical inner membrane protein -11
a fold changes were presented after log2
b positive number represented up-regulated and negative represented down-regulated
Flavin and cytochrome related genes
Flavins can be secreted by Shewanella species as electron shuttle to facilitate
extracellular electron transfer (Cansterin et al 2008) C-type cytochromes play the
important roles on the process of extracellular electron transfer (Shi et al 2007) The
genes related to flavins synthesis and the genes encoding the cytochromes electron
transport proteins were shown in the table 43 the genes SO_1414 and SO_3058 related
to flavins synthesis was up-regulated to 03 and 01 folds respectively The genes related
to s C-type cytochromes SO_0135 SO_4105 SO_0169 SO_1778 are increased to 10
08 05 04 01 folds respectively However most of genes have shown no significant
changes (less one-fold change) as response to nano particle decorated anode in MECs
58
Table 43 Expression level of flavin and cytochrome genes
Locus Tag Gene product descriptions Fold
change
SO_1414 flavocytochrome c flavin subunit putative 03
SO_3468 riboflavin synthase subunit alpha -02
SO_3058 flavocytochrome c flavin subunit 01
SO_4105 MSHA pilin protein MshA 08
SO_0169 general secretion pathway protein GspG 05
SO_1778 outer membrane decaheme cytochrome c lipoprotein MtrC 04
SO_1779 outer membrane decaheme cytochrome c OmcA 01
SO_0135 lipoprotein of unknown function DUF333 10
SO_0136 conserved hypothetical inner membrane protein 01
a fold changes were presented after log2
b positive number represented up-regulated and negative represented down-regulated
Other genes with significantly modified expression levels
These genes over 15 fold change and possibly related to current enhancements were
shown in Table 44 SO_3104 (expressed inner membrane protein) was over expressed
and the gene SO_2194 which encodes outer membrane protein (OmpA family protein)
was repressed as response to nanoparticle decorated anode however it is unclear the
relationships of this modified membrane protein genes with the current enhancement
Another interesting phenomenon was possible co-regulation of several gene clusters
indicating possible operon associations This includes three sets of genes that were
induced more than 15 fold SO_0108SO_0109 and SO_1042SO_1043SO_1044 The
59
consistency of expression of these genes under the NP conditions provides basic evidence
to support operon structure However the correlation of these significant changed gene
with current density enhance are unclear which showed there are unknown multiple and
complex responses of S oneidensis MR-1 to nanoparticle anode of MECs
Table 44 Other genes with significantly changed expression levels
Locus Tag Gene product descriptions Fold change
SO_0101 nitrate-inducible formate dehydrogenase molybdopterin-binding
subunit FdnG
18
SO_0108 integral membrane protein of unknown function DUF39 YedE 17
SO_0109 SirA family protein YedF 17
SO_0275 N-acetyl-gamma-glutamyl-phosphate reductase -31
SO_0277 ornithine carbamoyltransferase -21
SO_0279 argininosuccinate lyase -24
SO_0404 zinc dependent metalloprotease domain lipoprotein 21
SO_0956 alkyl hydroperoxide reductase F subunit -20
SO_1042 amino acid ABC transporter ATP-binding protein -18
SO_1043 amino acid ABC transporter permease protein -27
SO_1044 amino acid ABC transporter periplasmic amino acid-binding protein -27
SO_1072 chitin-binding protein putative 21
SO_1405 transglutaminase family protein 20
SO_1822 TonB-dependent receptor putative -22
SO_2069
1-(5-phosphoribosyl)-5-[(5-phosphoribosylamino)
methylideneamino] imidazole-4-carboxamide isomerase
-19
SO_2070 amidotransferase HisH -26
SO_2071 imidazole glycerol-phosphate dehydratasehistidinol phosphatase -27
60
SO_2072 histidinol-phosphate aminotransferase -28
SO_2073 histidinol dehydrogenase -30
SO_2194 OmpA family protein -36
SO_2195 inter-alpha-trypsin inhibitor domain protein -27
SO_2767 asparagine synthetase B -26
SO_2945 prophage LambdaSo tail fiber protein 20
SO_2963 prophage LambdaSo major capsid protein HK97 family 20
SO_2987 prophage LambdaSo hypothetical protein 19
SO_2988 prophage LambdaSo expressed protein of unknown function 18
SO_3104 expressed inner membrane protein 131
SO_3408 conserved hypothetical inner membrane protein -18
SO_3585 azoreductase putative -18
SO_3586 glyoxalase family protein -17
SO_3687 curli production assemblytransport component CsgE putative -18
SO_38191 hypothetical ammonia permease -19
SO_4014 AcrBAcrDAcrF family protein 18
SO_40151 type I secretion system membrane fusion protein RND family 18
SO_4054 510-methylenetetrahydrofolate reductase 18
SO_4245 N-acetylglutamate synthase -22
SO_45251 hypothetical transcriptional regulator LysR family -17
SO_4527 integral membrane domain protein -19
SO_4705 transcriptional regulator putative -19
a fold changes were presented after log2
b positive number represented up-regulated and negative represented down-regulated
Discussion
61
The addition of NP to anodes significantly impacts current densities with the elemental
composition being a critical factor The chemical composition of NPs significantly affects
the current enhancement in MECs (Fan et al 2011) because the chemical composition of
NP properties is a critical factor in determining the conductivity which significantly
affects the efficiency of electron transfer from bacteria to electrode Fe is an ideal
candidate for NP formation for current enhancement in MESs because of high
conductivity and low price compared to gold and other conductive materials Gold NPs
has also been studies for enhancement of electricity conductivity (Bao et al 2008)
However the disadvantages of the high price of gold limit their practical application in
MECs (Fan et al 2011) The different size and density of the same composition of NP
also has essential effects on the current density enhancement (Fan et al 2011) In this
study the results showed the current density enhancement with Fe-NP size range of
200plusmn100 nm and density range of 200plusmn100 nm is considerable enhancement consequence
There are a number of reports of studies of current density enhancement in MFCs
by nanostructured anodes (Fan et al 2011 Qiao et al 2008 Sharma et al 2008 Tsai et al
2009) Based on bacterial behavior on nanostructured anode surface Qiao et al (2008)
believes that Escherichia coli cells on the nanostructured electrode surface produce hair-
like structures similar to pili that could facilitate the electron transfer between the cells
and electrode The production of hair-like structures is believed to be stimulated by
nanostructures and could play the key role on current density enhancement in pili-
producing bacteria in MFCs (Qiao et al 2008) S oneidensis can produce electrically
conductive bacterial nanowires which have similar structure to the hair-like structures in
Ecoli (Gorby et al 2006 Reguera et al 2005) A number of possible genes related
62
nanowires of S oneidensis have been identified including mshA (SO_4105) and gspG
(SO_0169) (Konstantinidis et al 2009) They are up-regulated 08 and 05 fold in the
current study respectively even though they are not significant induced This result
supported the increased nanowire genes had contributions to enhance current density in
MFCs by NPs decorated anodes
Based on the previous research on electron transfer mechanisms for S oneidensis it is
known that self-produced mediators (flavins) and multihaem c-type cytochromes (c-Cyts)
may play critical roles in the electron transfer of S oneidensis to extracellular insoluble
electrode (Canstein et al 2008 Myers and Myers 1992 Scott and Nealson 1994 Shi et al
2007) The flavins secreted by S oneidensis can facilitate to transfer electrons from
bacteria to solid anode electrodes by acting as extracellular electron shuttles (Canstein et
al 2008) In this study the genes encoding flavins (SO_1414 SO_3468 and SO_3058)
had no significant increase (less than 1 fold change) as a response to nanoparticle
decorated anode in MECs which suggests that increased flavin gene expression does not
significantly contribute to the current density enhancements in nano-particle decorated
anode The genes encoding cytochromes electron transport proteins (SO_4104 SO_0417
SO_177879 SO_013536) had no significant increase (less than 1-fold change)
However those genes were up-regulated slightly These results suggest the increased
expression flavin and c type cytochromes genes had partial contributions even not
significantly to enhance current density in MFCs by NPs decorated anodes
The thickness of bacterial biofilm on the anode can affect the power generation in
MFCs (Biffinger et al 2007 Heydorn et al 2000) since increased biofilm thickness on
the anode represents more active bacteria involving in the process of electron generation
63
and transfer in MFCs Biofilm formation related genes included flagellar genes IV pilus
genes and other genes The flagellar related genes SO_3228 SO_3229 (fliB and fliK) and
IV pilus genes SO_4105 (mshA) were induced from 19 to 09 fold the genes (SO_4178
-4180) related to formation of a three-dimensional biofilm were up-regulated from 10 to
13 folds even though the genes related bacterial motility protein (SO_ 4286 SO_4287
SO_1529 and SO_1530) are induced slightly (less than 0 5 fold) However the motility
genes most probably play a role for initial biofilm attachment on the anode surfaces
(Thormann et al 2004 Thormann et al 2006) with the rest of biofilm genes being more
essential for biofilm maintenance in mature cultures as were presented at harvest in this
study This enhancement of biofilm density under conditions of increase current density
has been observed for gold NP decorated anodes using confocal light microscopy (data
not shown) but has not yet been confirmed for Fe NP decorated anodes In any event an
increase in the expression level of genes related biofilm formation therefore also
supported the possibility of enhanced biofilm formation on NPs decorated anodes thus
facilitating electron transfer from bacteria to anodes
Beliaev et al described a model of the Fe reduction pathway in S oneidensis MR-1 using
solid substrates as the electron acceptor (Beliaev et al 2001) This model proposes that
electrons are generated and released in cytoplasm then transferred to the quinone pool
eg menaquinone in the cytoplasmic membrane The reduced menaquinone in turn
reduces CymA and from there electrons are then transferred to a periplasmic shuttle (eg
CctA) and then to MtrA a periplasmic soluble decaheme cytochrome c located in
associated with the outer membrane via interaction with MtrB Finally surface displayed
outer membrane proteins decaheme cytochromes (OmcA MtrC or MtrF) can transfer
64
electron to Fe or other solid substrates (Beliaev et al 2001)) Bencheikh-Latmani et al
demonstrated that the same electron transport pathway may be used for more than one
electron acceptor through studies of the response of S oneidensis MR-1 to U(VI) and
Cr(VI) under anaerobic conditions particularly focused on the critical genes
(SO_1777mtrA SO_1776mtrB SO_1778mtrC) (Bencheikh-Latmani et al 2005)
However our studies showed that these genes had no significant expression changes in
response to nanostructured anodes Contrastingly one of expressed inner membrane
protein genes SO_3104 and one of cytochromes genes SO_4609 encoding cytochrome c
oxidase subunit III demonstrated a 13-fold and 11-fold up-regulation respectively in this
study while the gene SO_2194 which encodes outer membrane protein (OmpA family
protein) was down regulated 36 fold Our results therefore suggest that S oneidensis
MR-1 does not utilize the same set of core critical genes (SO_1777 mtrA SO_1776
mtrB SO_1778 mtrC) products to transfer electrons in all instances This is the similar
result with the study of Bretschger et al (2007) which indicated that the electron
transport system in S oneidensis MR-1 is complex with several different proteins able to
participate in electron transfer to the anode of MECs
It should also be mentioned that the gene (SO_1519) encoding lactate
dehydrogenase repressed (-12 fold) here suggesting that lactate was depleted at the time
of cell harvest from the MECs and that other pathways of anaerobiosis had been activated
including those associated with amino acid uptake and consumption the growth media
was a complex mixture including yeast extract In particular the catabolic pathway for
histidine of S oneidensis MR-1 a pathway that may formate as an intermediate is up-
regulated with the genes for histidine ammonia lyase SO_0098 SO_3057 and SO_4374
65
up-regulated 13 11 and 11-fold respectively The possible production of formate
during histidine degradation may in turn explain the high levels of formate
dehydrogenates as evidenced by upregulation of SO_0102 SO_0103 SO_0104 and
SO_0107 annotated as formate dehydrogenase proteins These genes had the highest
levels of up-regulated expression as response to Fe NP decorated anodes (greater than 2-
fold)
Acknowledgements
This research was partially supported by the US National Science Foundation CBET
0828544 and the funds from ONAMIDOD (ARL-DOD Cooperative Agreement
W911NF-07-2-0083) We thank Barbara Gvakharia and Caprice Rosato for helpful
suggestions We also thank Margaret Romine of Pacific Northwest Laboratories for
valuable comments on manuscripts We also thank anonymous reviewers for significant
suggested improvements imparted as part of prior review of this manuscript
66
Chapter 5
Global Transcriptome Analysis of the Response of Shewanella oneidensis MR-1 to
Carbon Nanotube Decorated Anodes in Microbial Electrochemical Systems
Shoutao Xu Yanzhen Fan Rebecca Schaller Frank Chaplen Jun Jiao Hong Liu
67
Abstract
Shewanella oneidensis MR-1 is an important model microorganism for metabolic studies
on the effects of different environmental factors because of its diverse respiratory
capabilities Carbon nanotube (CNT) modified graphite disks were used as anodes to
investigate the effects of nanomatrials on the performance of S oneidensis MR-1 in
microbial electrolysis cells (MECs) The current densities produced with CNT decorated
anodes were on average 56-fold higher than plain graphite anodes Whole genome
microarray analysis of gene expression showed that up-regulation of cytochromes c genes
associated with extracellular electron transfer are strongly correlated to current increases
in CNT decorated anodes Genes involved in flavin biosynthesis also contribute to
current increase in CNT decorated anode MECs
Keywords Microbial electrochemical system microbial fuel cell microbial
electrolysis cell carbon nanotube gene expression DNA microarray
68
Introduction
Shewanella oneidensis MR-1 is an important model microorganism for metabolic
studies of the effects of different environmental factors because of its diverse respiratory
capabilities It has been used for transcriptome analysis to investigate the responses to
different growth conditions and environmental stresses (Murray et al 2001) Gene
expression patterns under different conditions such as heat shock cold shock Cr (VI) U
(VI) reduction chromate stress iron and acid tolerance have been studied previously
(Bencheikh-Latmani et al 2005 Brown et al 2006 Gao et al2006 Gao et al 2004
Yang et al 2008) More recently it has been exploited as a model species for power
generation in microbial electrochemical systems (MESs)which have potential
applications in renewable energy generation bioremediation and wastewater treatment
(Logan et al 2007 Logan 2009 Park and Zeikus 2002 Fan et al 2011 Xu et al 2012)
In a MES electrochemically active microorganisms oxidize organic matter in the an
ode chamber to release electrons Electrons are then transferred to the anode electrode thr
ough various mechanisms (Logan et al 2006 Lovley 2006 Rabaey et al 2003) and fina
lly travel to the cathode electrode and combine with the terminal electron acceptor The
key feature of MESs is the microbe-catalyzed electron transfer from the organic matter to
the anode (Rabaey et al 2004) Enhancing the current output which highly depends on
the performance of the anode electrode is critical for the successful application of MESs
(Logan et al 2007 Park and Zeilus 2002 2003) Several nanomaterials including TiO2
gold (Au) and palladium (Pd) nanoparticles have been utilized to decorate electrodes to
enhance power density in MESs (Qiao et al 2008 Quan et al 2005 Fan et al 2011) The
possible mechanisms for increased current densities have been studied by using Fe
69
nanoparticle-decorated anodes (Xu et al 2012)
Carbon nanotubes (CNT) are cylindrically shaped nanomaterials with extremely
high surface area excellent electrical conductivity and chemical inertness (He et al
2005a Serp et al 2003) These unique properties make CNT a promising electrode
material (Liang et al 2008) The biocompatibility of microorganisms and carbon
nanostructures has been evaluated (Morozan et al 2007) The effects of CNT on anodic
biofilms have been tested in MFCs (Liang et al 2011) However no studies have been re
ported on global transcriptome analysis for the response of S oneidensis MR-1 to CNT
decorated anode in MECs
In the present study the graphite disks decorated with CNT were used as anodes to
investigate the effects of nanomaterials on current generation in multi-anode MECs
DNA microarrays were used to analyze differences in the global gene expression profile
of S oneidensis MR-1 grown on plain versus CNT decorated anodes
Materials and methods
Bacterial cultures S oneidensis MR-1 was purchased from American Type Culture
collection (ATCC 700550) and stored as freezer stocks at -80degC Stock S oneidensis
MR-1 cells were grown aerobically using M1 medium at 30degC Cells in early stationary
growth phase (optical density of 15-16 at 600nm (Spectrophotometer UV-1700
Shimadzu Columbia MD) were injected into the chamber of the MECs for current
production Sodium lactate (final concentration 30mM) was added as the additional
electron donor
Fabrication and characterization of nanostructured anodes Superfine isomolded
graphite disks (16 cm x 05 cm GraphiteStorecom Inc) were polished with ultrafine
70
sand paper (2000 grit 3M Company) as the base for the CNT decorated and control
anodes Aligned multi-walled carbon nanotube samples were synthesized via plasma
enhanced chemical vapor deposition (PECVD) Samples superfine isomolded graphite
pillars with a 16 cm diameter and 05 cm thickness were sputtered with a thin film of Ni
in a CRC 300 magnetron sputter coater for 1 to 2 min They were then placed in the
PECVD reactor where the Ni was annealed into small catalyst islands C2H2 was bled
into the chamber for growth and an NH3 plasma was used to vertically align the growth
of the multi-walled carbon nanotubes (MWCNTs) The temperature and time were varied
between 725degC and 775oC and 15 to 60 min to control the length of the tubes and density
of samples Samples were then analyzed in a FEI Sirion field emission scanning electron
microscope (FESEM) SEM images of CNT decorated anode and control surfaces were
shown in the Figure 51
Figure 51 SEM images of CNT decorated anodes (A) Graphite disk control (B) CNT
decorated anode
Multiple channel MECs construction and operation A MECs with multiple
removable anodes each with an effective anode area of 07 cm2
was constructed and used
to evaluate the effects of nanostructures on the current density of according to a previous
B A
71
study (Fan et al 2011) The cathode was made of wet-proof (30) carbon cloth (type B
E-TEK Division Inc USA) coated with PtC (20 E-TEK Division Inc USA)
following a previously reported procedure (Liu et al 2005) The final platinum loading
was 05 mgcm2 per projected cathode area The size of the cathode (150 cm
2) was 25
times larger than the total effective area of the 8 graphite disk anodes (56 cm2) to prevent
cathode limitations on the performance of the MECs system All MECs with CNT
decorated anodes and control anodes were tested for current generation for 24 h with
sterile a medium (without bacteria) before injecting the bacterial cells in order to
determine whether the CNT decorated anodes demonstrate chemical current generation
compared to control anodes All the testing anodes were set up in the MECs
simultaneously so that the S oneidensis MR-1 culture was applied to all of them
simultaneously Short electrode spacing (17 cm) was used in the MECs design to reduce
the internal resistance A voltage of 06 V was applied the MECs for current generation
after the cells of S oneidensis MR-1 were inoculated into the MECs in the early
stationary phase A multimeter with a data acquisition system (2700 Keithly USA) was
used to monitor the current change by measuring the voltage drop through a resistor
Microarray analysis Biofilms for whole gene microarray analysis were aseptically
removed from the plain and CNT decorated anodes of MECs after 80 hours further
incubation at 30oC when current density obviously reached to a stable phase Total RNA
was extracted using Trizol (Ambion inc AustinTX) following the manufacturerrsquos
instructions Integrity of the RNA samples was confirmed using the Agilent 2100
Bioanalyzer (Agilent Technology Inc Palo Alto CA) Total RNA (200 ng) from each
sample was amplified using the MessageAmpTM
II-Bacteria Prokaryotic RNA
72
Amplification kit (Ambion AustinTX) according to the manufacturerrsquos instructions The
S oneidensis MR-1 whole-gene microarrays representing all of the 4295 genes (resource
from NCBI) were ordered from Roche NimbleGen Inc (Madison WI) Biological
triplicates of biofilms on the CNT decorated anodes and control were analyzed cDNA
synthesis labeling and hybridization were carried out by the CGRB Core Laboratories at
Oregon State University DNASTAR ArrayStarTM
3 software was used to identify genes
that were up- or down-regulated more than 2-fold when grown on the nanoparticle
modified anodes using the unpaired two sample t-test with a cutoff p-value of 005 with
bonferroni correction
Results and discussion
Enhancement of current generation using CNT decorated anodes in MECs The
current density generated by CNT decorated anode MECs with S onidensis MR-1
increased to 36 microAcm2 10 hours post-inoculation and reached a maximum of 40 microAcm
2
at 29 hours inoculation then gradually decreased The current density stabilized at
30microAcm2 after 78 h operation Figure 52 shows that the current density curve generated
by CNT decorated anode in MECs The average current density generated by CNT
decorated anodes was 56 times that of the control The current density generated in the
MECs without bacteria was negligible (data not shown) This result demonstrates that the
current enhancement observed with the CNT decorated anodes was biologically-derived
A
73
Figure 52 Effects of CNT decorated anodes on the current density in MECs (3 replicates)
Control is plain graphite disk anode CNT is the anode with CNT decoration (Error bars
represent standard deviation)
It has been reported that nanostructured decorated anodes have significant impacts on
current densities (Fan et al 2011 Qiao et al 2006 Sharma 2008 Tsai et al 2009) The
CNT decorated anodes had significantly enhanced current densities in MECs in this
study This result is consistent with previous studies Interestingly the current density
curve generated by CNT decorated anode MECs is quite different from the current
density curve generated by Au NP and Pd NP decorated anodes using same bacterial
species (Fan et al 2011) In the Au and Pd NP decorated anode MECs the current
density curve was not significantly increased in the beginning compared with the control
but gradually increased after the inoculation the current density reached the maximum
current density more than 50 hours after the inoculation However the current density
generated by CNT decorated anode MECs started to increase significantly immediately
after the inoculation It reached a maximum current density 29 hours post-inoculation
-1
6
13
20
27
34
41
48
0 10 20 30 40 50 60 70 80
Curr
ent
den
sity
(micro
Ac
m2)
Time (Hours)
Control
CNT
74
These results indicate that there might be different current density enhancement
mechanisms between CNT with other metal NP decorated anodes in MECs
Global transcriptome analysis Whole-genome DNA microarrays were used to attain a
comprehensive general overview of the transcriptional response of S oneidensis
MR-1 of
S oneidensis MR-1 to CNT decorated anodes in MECs In a total of 4295 genes analyzed
457 genes (250 genes up-regulated and 207 genes down-regulated) exhibited significantly
(P lt 005) changed expression at a 2-fold level in at least 3 replicates in the response to
CNT The total number of the regulated genes present 11 of the 4295 open reading
frames (ORFs) presented on the microarray The total number of genes at significant
expression levels in response to CNT decorated anodes is close to that to Fe NP decorated
anodes in MECs (392 9) ( Xu et al 2012)
Fig 53 Differentially expressed genes grouped by functional classification according to
the TIGR S oneidensis genome database (wwwtigrorg) Columns 1 amino acid
biosynthesis 2 biosynthesis of cofactors prosthetic groups and carriers 3 cell
envelope 4 cellular processes 5 central intermediary metabolism 6 DNA metabolism
7 energy metabolism 8 fatty acid and phospholipid metabolism 9 other categories 10
protein fate 11 protein synthesis 12 purines pyrimidines nucleosides and nucleotides
-80
-60
-40
-20
0
20
40
60
80
100
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18
Nu
mb
er
of
Ge
ne
s
Up-regulated
Down-regulated
75
13 regulatory functions 14 signal transduction 15 transcription 16 transport and
binding proteins 17 unknown function 18 hypothetical proteins
The overall transcript genomic response of S oneidensis MR-1 to CNT decorated
anodes was summarized in Figure 53 by grouping the differentially expressed genes into
their functional role categories based on The Institute for Genomic Researchrsquos annotation
(Version 8) of the MR-1 genome sequence The wide distribution of putative functional
roles attributed to the differentially expressed genes indicated the extent of the molecular
response of Soniedenis MR-1 to the CNT decorated anodes A large number of up-
regulated genes were presented in genes associated with biosynthesis of cofactors
prosthetic groups and carriers (group 2) cell envelope (group 3) protein fate (group 10)
protein synthesis (group 11) and purines pyrimidines nucleosides and nucleotides
(group 12) in this study The number of up-regulated genes was two times more than that
of down-regulated genes in each functional group Contrastingly in the genomic response
S oneidensis MR-1 to Fe NP decorated anodes (chapter 4) most of up-regulated genes
were associated with cellular processes (group 4) energy metabolism (group 7) other
categories (group 9) and hypothetical proteins (group 18) The down-regulated genes
presented in the amino acid biosynthesis (group 1) cellular processes (group 4) substrate
transport (group 16) and hypothetical proteins (group 18) While those genes encoding
proteins involved in amino acid biosynthesis (group 1) central intermediary metabolism
( group 5) DNA metabolism( group 6) fatty acid and phospholipid metabolism (group
8) and other categories (group 9) were among the most down-regulated genes in this
study However the two groups of genes that were the most noticeable between the
genomic response S oneidensis MR-1 to CNT and Fe NP decorated anodes which they
are the genes related to energy metabolism (group 7) and hypothetical proteins (group
76
18) These results indicated there are different and common genomic responses to S
oneidensis MR-1 on CNT and Fe NP decorated anodes in MECs
Electron transfer related genes Based on previous electron transfer mechanism
studies on Shewanella a serial of group proteins collectively described as the Mtr
pathway play the role to transfer electron rom the inner bacterial body to the outer
membrane (Beliaev et al 2001 Shi et al 2007 Brutinel and Gralnick 2012) In the Mtr
pathway firstly electrons generated by bacteria received by CymA a tetraheme c-type
cytochrome anchored in the inner membrane then electrons were transferred to MtrA a
periplasmic decaheme c-type cytochrome After that electrons were transferred from
MtrA to MtrC an outer-membrane decaheme c-type cytochrome is facilitated by MtrB a
non-heme containing integral outer-membrane proteinOuter-membrane decaheme c-type
cytochrome MtrC and OmcA are in change of electron transfer to exreacellular electron
acceptor (Shi et al 2007 Brutinel and Gralnick 2012 ) These genes encoding electron
transport proteins are shown in the table 51 The critical genes SO_1777 mtrA SO_1776
mtrB SO_1778 mtrC are unregulated 41 to 80 fold in response to CNT decorated
anodes other cytochrome c genes related to electron transfer in MtrA pathway also are
significantly increased in response to CNT decorated anodes These genes encoding outer
membrane proteins have different level increases supporting that the increase of
cytochrome c gene expression contributes to the current enhancements of MECs as
response to CNT decorated anodes
77
Table 51 Expression level of cytochrome c as response to CNT decorated anodes
Locus Tag Gene product descriptions Fold change CNT
SO_0165 general secretion pathway protein GspC 31
SO_0167 general secretion pathway protein GspE 45
SO_1776 outer membrane protein precursor MtrB 41
SO_1777 outer membrane decaheme cytochrome c
lipoprotein MtrA
80
SO_1778 outer membrane decaheme cytochrome c
lipoprotein MtrC
43
SO_1779 outer membrane decaheme cytochrome c
OmcA
63
SO_0135 lipoprotein of unknown function DUF333 22
SO_0136 conserved hypothetical inner membrane protein 24
Flavin related genes It is known that self-produced mediators play critical roles in the
electron transfer of S oneidensis to extracellular insoluble electrode (Canstein von et al 2
008 Myers et al 1992 Scott et al 1994 Shi et al 2007) It has been proved that the
flavins secreted by S oneidensis can facilitate electron transfer from bacteria to solid
anode electrodes by acting as extracellular electron shuttles (Cansterin von et al 2008) T
wo critical genes (SO_1414 and SO_3468) related to flavin synthesis were shown increa
sed significantly (more than 2 folds) increase in this study (Table 52) This result support
s the hypothesis that flavin production increases as the response to CNT decorated anodes
in MECs have significant contribution to the current density enhancements
78
Table 52 Expression levels of genes related to flavin synthesis as response to CNT
decorated anode
Locus Tag Gene product descriptions Fold change CNT
SO_1414 flavocytochrome c flavin subunit putative 22
SO_3468 riboflavin synthase subunit alpha 20
The amount of bacterial biofilm biomass on the anode can affect the power
generation in MFCs (Biffinger et al 2007 Heydorn et al 2000) Biofilm formation
related genes include flagellar gene IV pilus genes and other genes the flagellar related
genes (fliB and fliK) and IV pilus genes (mshA) were induced only from 105 to 134
fold as response of S oneidensis MR-1 to CNT decorated anodes in this study No
biofilm-associated genes presented significant changes in expression levels in this study
indicating there is no direct connections between biofilm enhancements to CNT
decorated anodes in this study This result is consistent with the Liang et al (2011)
conclusion that using CNT enhanced the electrochemical activity of anodic biofilms but
did not result in a significant increase of biomass in the anodic biofilms
Our microarray results showed two significantly up-regulated gene groups the
genes encoding proteins localized on the outer membrane and the genes involved in
flavin biosynthesis contributed to current density enhancement by CNT decorated anodes
Among 457 significantly changed genes to CNT in this study there are also a relatively
large number of genes encoding proteins with unknown functions which are either up- or
down-regulated in the response to CNT decorated anodes which indicated more
79
complicated responses of S oneidensis MR-1 to CNT decorated anodes in MECs and
further study is needed
80
Chapter 6
Summary
Microbial electrochemical systems (MESs) with a mixed culture initially inoculated
from Corvallis wastewater treatment plant have been studied for more than 6 years for
varying purposes including power generation hydrogen production heavy metal
removal and wastewater treatment Experiment results have shown that the mixed culture
is quite stable with excellent performance in MESs Our community analysis using
denaturing gradient gel electrophoresis (DGGE) and 16S rDNA clone library construction
suggests that the mixed culture is composed predominantly of Geobacter sp (66)
Arcobacter sp (12) and Citrobacter sp (11) These results not only improved our
understanding of the mixed culture community but also guided our studies on the
cultivation and isolation of the different bacterial species
Exoelectrogenic Citrobacter sp SX-1 was first isolated and characterized from a
MFC operated with the mixed culture Citrobacter sp SX-1 has been demonstrated to
produce electricity from wide range of different substrates including citrate acetate
glucose sucrose glycerol and lactosein MFCs Cyclic voltammetry analysis indicated
that membrane associated proteins may play an important role in facilitating the electrons
transferring from bacteria to electrode The strain SX-1 increased the known diversity of
power generating exoelectrogens and provided a unique bacterial species for study in
renewable energy generation and waste treatment
81
To enhance the electron transfer from bacteria to anode nano decorated anodes
including Fe NP and CNT were developed and characterized and evaluated in MECs
using Shewanella oneidensis MR-1 as a model species Both nanostructures have
significantly increased current density compared with the control Whole genome
microarray analysis elucidated the possible mechanisms of power enhancement in
response to these nano-decorated anodes
These results benefit to understanding of physiology and ecology of mixed
cultures in MFCs and improve the efficiency of current generation in MESs which will
facilitate the viability of niche applications for MESs in near future
82
Bibliography
1 Amman R Ludwig W (2000) Ribosomal RNA-targeted nucleic acid probes for
studies in microbial ecology FEMS Microbiol Ecol 24 555ndash565
1 Bao SJ Li CM Zang JF Cui XQ Qiao Y Guo J (2008) Novel Nanoporous TiO2
Electrode for Directly Electrochemistry of glucose oxidase Adv Funct Mater
18(4) 591-599
2 Beliaev AS Saffarini DA McLaughlin JL Hunnicutt D (2001) MtrC an outer
membrane decahaem c cytochrome required for metal reduction in Shewanella
putrefaciens MR-1 Mol Microbiol 39722-30
3 Bencheikh-Latmani R Williams SM Haucke L Criddle CS Wu L Criddle CS
Zhou J Tebo BM (2005) Global transcriptional profiling of Shewanella oneidensis
MR-1 during Cr(VI) and U(VI) reduction Appl Environ Microbiol 71 7453ndash7460
4 Biffinger JC Pietron J Ray R Little B and Ringeisen BR (2007) A biofilm
enhanced miniature microbial fuel cell using Shewanella oneidensis DSP10 and
oxygen reduction cathodes Biosens Bioelectron 221672ndash1679
5 Bodelier PLE Roslev P Henckel T FenzelP (2000) Stimulation by ammonium
based fertilizers of methane oxidation in soil around rice root Nature 40 421ndash424
6 Bond DR Holmes DE Tender LM Lovley DR (2002) Electrode-reducing
microorganisms that harvest energy from marine sediments Science 295 483-485
7 Bond DR Lovley DR (2003) Electricity production by Geobacter sulfurreducens
attached to electrodes Appl Environ Microbiol 69 1548-1555
8 Bond DR Lovley DR (2005) Evidence for involvement of an electron shuttle in
electricity generation by Geothrix fermentans Appl Environ Microbiol 71 2186-
2189
9 Bretschger O Obraztsova A Sturm CA Chang IS Gorby YA Reed SB Culley
DE Reardon CL Barua S Romine MF Zhou J Beliaev AS Bouhenni R
Saffarini D Mansfeld F Kim BH Fredrickson JK Nealson KH (2007) Current
production and metal oxide reduction by Shewanella oneidensis MR-1 wildtype and
mutants Appl Environ Microbiol 73 7003ndash7012
10 Brown SD Thompson MR Verberkmoes NC Chourey K Shah M Zhou J
Hettich RL and Thompson DK (2006) Molecular dynamics of the Shewanella
oneidensis response to chromate stress Mol Cell Proteomics 51054-1071
11 Brutinel ED Gralnick JA (2012) Shuttling happens soluble flavin mediators of
extracellular electron transfer in Shewanella Appl Microbiol Biotechnol 93(1) 41-8
12 Butler JE Kaufmann F Coppi MV Nuacutentildeez C Lovley DR (2004) MacA a diheme
c-type cytochrome involved in Fe(III) reduction by Geobacter sulfurreducens J
Bacteriol 186(12) 4042ndash4045
83
13 Caccavo F Lonergan DJ Lovley DR Davis M Stolz JF McInerney MJ (1994)
Geobacter sulfurreducens sp nov a hydrogen- and acetate oxidizing dissimilatory
metal-reducing microorganism Appl Environ Microbiol 60 3752ndash3759
14 Canstein von H Ogawa J Shimizu S Lloyd JR (2008) Secretion of flavins by
Shewanella species and their role in extracellular electron transfer Appl Environ
Microbiol 74 615ndash623
15 Cao X Huang X Liang P Xiao K Zhou Y Zhang X Logan BE (2009) A New
Method for Water Desalination Using Microbial Desalination Cells Environ Sci
Technol 43(18)7148-7152
16 Chaudhuri SK Lovley DR (2003) Electricity generation by direct oxidation of
glucose in mediatorless microbial fuel cells Nat Biotechnol 21 1229-1232
17 Cheng S Liu H Logan B E (2006) Increased power generation in a continuous
flow MFC with advective flow through the porous anode and reduced electrode
spacing Environ Sci Technol 402426-2432
18 Cheng S Logan B E (2007) Sustainable and efficient biohydrogen production via
electrohydrogenesis PNAS 104 18871-18873
19 Chignell J (2010) Energy Recovery from Biodiesel Waste Performance of
Microbial Electrochemical Systems on Glycerol Thesis
20 Chung K Okabe S (2009) Characterization of electrochemical activity of a strain
ISO2-3 phylogenetically related to Aeromonas sp isolated from a glucose-fed
microbial fuel cell Biotechnol Bioeng 104(5) 901-910
21 Cole JR Chai B Farris RJ Wang Q Kulam-Syed-Mohideen AS McGarrell
DM Bandela AM Cardenas E Garrity GM Tiedje JM (2007) The ribosomal
database project (RDP-II) introducing myRDP space and quality controlled public
data Nucleic Acids Res 35 (Database issue) D169-D172 doi 101093nargkl889
22 Cole JR Wang Q Cardena E Fish J Chai B Farris RJ Kulam-Syed-Mohideen
AS McGarrell DM Marsh T Garrity GM Tiedje JM (2009) The Ribosomal
Database Project improved alignments and new tools for rRNA analysis Nucleic
Acids Res 37 (Database issue) D141-D145 doi 101093nargkn879
23 Dey A De S De A De SK (2004) Characterization and dielectric properties of
polyaniline-TiO2 nanocomposites Nanotechnol 15 1277-1283
24 Edwards U Rogall T Bloumlcker H Emde M Boumlttger EC (1989) Isolation and
direct complete nucleotide determination of entire genesmdashcharacterization of a gene
coding for 16S-ribosomal RNA NucleicAcids Res 17 7843ndash7853
25 Eggleston CM Voumlroumls J Shi L Lower BH Droubay TC Colberg PJS (2008)
Binding and direct electrochemistry of OmcA an outer-membrane cytochrome from
an iron reducing bacterium with oxide electrodes a candidate biofuel cell system
Inorg Chim Acta 361769ndash777
26 Fan Y Sharbrough E Liu H (2008) Quantification of the Internal Resistance
Distribution of Microbial Fuel Cells Environ Sci Technol 42 8101ndash8107
84
27 Fan Y Xu S Schaller R Jiao J Chaplen F Liu H (2011) Nanoparticle decorated
anodes for enhanced current generation in microbial electrochemical
cells Biosensors and Bioelectronics 26 (5)1908ndash1912
28 Fan Y Hu H Liu H (2007)Sustainable power generation in microbial fuel cells
using bicarbonate buffer and proton transfer mechanisms Environ Sci
Technol 41(23) 8154-8
29 Fedorovich V Knighton MC Pagaling E Ward FB Free A Goryanin I (2009)
Novel electrochemically active bacterium phylogenetically related to Arcobacter
butzleri isolated from a microbial fuel cell Appl Environ Microbiol 75(23) 7326-
7334
30 Ferna ndez A Huang S Seston S Xing J Hickey R Criddle C Tiedje J (1999)
How stable is stable Function versus community composition Appl Environ
Microbiol 65 3697ndash3704
31 Field SJ Dobbin PS Cheesman MR Watmough NJ Thomson AJ Richardson
DJ (2000) Purification and magneto-optical spectroscopic characterization of
cytoplasmic membrane and outer membrane multiheme c-type cytochromes from
Shewanella frigidimarina NCIMB400 J Biol Chem 275 8515ndash8522
32 Freguia S Masuda M Tsujimura S Kano K (2009) Lactococcus lactis catalyses
electricity generation at microbial fuel cell anodes via excretion of a soluble quinone
Bioelectrochemistry 7614ndash18
33 Fricke K Harnisch F and Schroumlder U (2008) On the use of cyclic voltammetry for
the study of the anodic electron transfer in microbial fuel cells Energy Environ
Sci 1 144ndash147
34 Gao H Wang Y Liu X Yan T Wu L Alm E Arkin A Thompson DK and
Zhou J (2004) Global transcriptome analysis of the heat shock response of
Shewanella oneidensis J Bacteriol 1867796-7803
35 Gao H Yang ZK Wu L Thompson DK Zhou J (2006) Global transcriptome
analysis of the cold shock response of Shewanella oneidensis MR-1 and mutational
analysis of its classical cold shock proteins J Bacteriol 1884560-4569
36 George MG (2005) Bergeyrsquos Manual of Systematic Bacteriology Volume Two The
Proteobacteria Part B The Gammaproteobacteria Second Edition New York
Springer 651p
37 Gonzaacutelez-Toril E Llobet-Brossa E Casamayor EO Amann R Amils R (2003)
Microbial ecology of an extreme acidic environment the Tinto river Appl Environ
Microbiol 69(8) 4853ndash4865
38 Gorby YA Yanina S Mclean JS Rosso KM Moyles D Dohnalkova A
Beveridge TJ Chang IS Kim BH Kim KS Culley DE Reed SB Romine M
F Saffarini DA Hill EA Shi L Elias DA Kennedy DW Pinchuk G
Watanabe K Ishii S Logan B Nealson KH Fredrickson JK (2006) Electrically
conductive bacterial nanowires produced by Shewanella oneidensis strain MR-1 and
other microorganisms Proc Natl Acad Sci USA 10311358-11363
85
39 He Z Wagner N Minteer SD Angenent LT (2006) An upflow microbial fuel cell
with an interior cathodes assessment of the internal resistance by impedance
spectroscopy Environ Sci Technol 40 5212-5217
40 Heydorn A Nielsen AT Hentzer M Sternberg C Givskov M Ersboslashll BK Molin
S (2000) Quantification of biofilm structures by the novel computer program
COMSTAT Microbiol 1462395-2407
41 Holmes DE Bond DR Lovley DR (2004a) Electron transfer by Desulfobulbus
propionicus to Fe(III) and graphite electrodes Appl Environ Microbiol 701234ndash
1237
42 Holmes DE Chaudhuri SK Nevin KP Mehta T Methe BA Liu A Ward JE
Woodard TL Webster J Lovley DR (2006) Microarray and genetic analysis of
electron transfer to electrodes in Geobacter sulfurreducens Environ Microbiol
81805-1815
43 Holmes DE Nicoll JS Bond DR Lovley DR (2004b) Potential role of a novel
psychrotolerant member of the family Geobacteraceae Geopsychrobacter
electrodiphilus gen nov sp nov in electricity production by a marine sediment
fuel cell Appl Environ Microbiol 70 6023ndash6030
44 Holmes DE Chaudhuri SK Nevin KP Mehta T Metheacute BA Liu A Ward JE
Woodard TL Webster J Lovley DR (2006) microarray and genetic analysis of
electron transfer to electrodes in Geobacter sulfurreducens Environ Microbial
8(10)1805-1815
45 Hu H (2009) Enhancing Hydrogen Production in Microbial Electrolysis Cells
through Development of Platinum-free Cathode and Improvement of Reactor
Dissertation
46 Ince BK Ayman O N Turker G Ccedilelikkol S Ince O (2010) Microbial ecology of
anaerobic reactors for treatment of alcohol industry wastewaters a review Current
research technology and education topics in applied microbiology and microbial
ecology 988-999
47 Jung S Regan JR (2007) Comparison of anode bacterial communities and
performance in microbial fuel cells with different electron donors Appl Microbiol
Biotechnol 77393ndash 402
48 Kim BH Kim HJ Hyun MS Park DH (1999) Direct electrode reaction of Fe(III)
reducing bacterium Shwwanella putrefaciens J Microbial Biotechnol 9127-131
49 Kim GT Hyun MS Chang IS Kim HJ Park HS Kim BH Kim SD Wimpenny
JW Weightman AJ (2005) Dissimilatory Fe(III) reduction by an electrochemically
active lactic acid bacterium phylogenetically related to Enterococcus gallinarum
isolated from submerged soil J Appl Microbiol 99(4) 978ndash987
50 Kim GT Webster G Wimpenny JW Kim BH Kim HJ Weightman AJ (2006)
Bacterial community structure compartmentalization and activity in a microbial fuel
cell J Appl Microbiol 101(3) 698ndash710
86
51 Kim HJ Park HS Hyun MS Chang IS Kim M Kim BH (2002) A mediator-
less microbial fuel cell using a metal reducing bacterium Shewanella putrefaciens
Enzyme Microb Technol 30 145-152
52 Kim JR Min B Logan BE (2005) Evaluation of procedures to acclimate a
microbial fuel cell for electricity production Appl Microbiol Biotechnol 68(1)23-30
53 Kim JR Cheng S Oh SE Logan BE (2007) Power generation using different
cation anion and ultrafiltration membranes in microbial fuel cells Environ Sci
Technol 41 1004-1009
54 Konstantinidis KT Serres MH Romine MF Rodrigues JL Auchtung J McCue
LA Lipton MS Obraztsova A Giometti CS Nealson KH Fredrickson JK
Tiedje JM (2009) Comparative systems biology across an evolutionary gradient
within the Shewanella genus Proc Natl Acad Sci U S A 106(37)15909-15914
55 Lee J Phung NT Chang IS Kim BH Sung HC (2003) Use of acetate for
enrichment of electrochemically active microorganisms and their 16S rDNA
analyses FEMS Microbiol Lett 223185ndash191
56 Liang P Wang H Xia X Huang X Mo Y Cao X Fan M(2011)Carbon
nanotube powders as electrode modifier to enhance the activity of anodic biofilm
in microbial fuel cells Biosens Bioelectron 26(6)3000-4
57 Lies DP Hernandez ME Kappler A Mielke RE Gralnick JA Newman DK
(2005) Shewanaella oneidensis MR-1 uses overlapping pathways for iron reduction
at a distance and by direct contact under conditions relevant for biofilms Appl
Environ Microbiol 71 4414-4426
58 Liu H and Logan B E (2004) Electricity generation using an air-cathode single
chamber microbial fuel cell in the presence and absence of a proton exchange
membrane Environ Sci Technol 38 4040-4046
59 Liu H Grot S Logan BE (2005) Electrochemically assisted microbial production
of hydrogen from acetate Environ Sci Technol 39 4317 ndash 320
60 Liu H Hu H Chignell J Fan Y (2010) Microbial Electrolysis Novel Technology
for Hydrogen Production from Biomass Biofuels 1(1) 129ndash142
61 Liu H Ramnarayanan R Logan BE (2004) Production of electricity during
wastewater treatment using a single chamber microbial fuel cell Environ Sci Technol
382281-2285
62 Liu WT Marsh TL Cheng H Forney LJ (1997) Characterization of microbial
diversity by determining terminal restriction fragment length polymorphisms of gene
encoding 16S rRNA Appl Environ Microbiol 63 4516ndash4522
63 Logan B E Hamelers B Rozendal R Schroder U Keller J Freguia S Aelterman
P Verstraete W Rabaey K (2006) Microbial fuel cells methodology and
technology Environ Sci Technol 40 (17) 5181-5192
64 Logan B E and Regan JM (2006) Electricity-producing bacterial communities in
microbial fuel cells Trends Microbiol 14512ndash518
87
65 Logan B Cheng S Watson V Estadt G (2007) Graphite fiber brush anodes for
increased power production in air-cathode microbial fuel cells Environ Sci Technol
41(9)3341-3346
66 Logan BE (2009) Exoelectrogenic bacteria that power microbial fuel cells Nat Rev
Microbiol 5 375-381
67 Logan BE Call D Cheng S Hamelers HV Sleutels TH Jeremiasse AW
Rozendal RA (2008) Microbial electrolysis cells for high yield hydrogen gas
production from organic matter Environ Sci Technol 42 8630-8640
68 Logan BE Hamelers B Rozendal R Schroder U Keller J Freguia S Aelterman
P Verstraete W Rabaey K (2006) Microbial fuel cells methodology and
technology Environ SciTechnol 40 5181-5192
69 Lovely DR (2006) Bug juice harvesting electricity with microorganisms Nature
Review| Microbiology 4 497-508
70 Lovley DR Phillips EJ (1988) Novel Mode of Microbial Energy Metabolism
Organic Carbon Oxidation Coupled to Dissimilatory Reduction of Iron or
Manganese Appl Environ Microbiol 54(6) 1472-1480
71 Ly HK Sezer M Wisitruangsakul N Feng JJ Kranich A Millo D Weidinger
IM Zebger I Murgida DH Hildebrandt P (2011) Surface-enhanced vibrational
spectroscopy for probing transient interactions of proteins with biomimetic interfaces
electric field effects on structure dynamics and function of cytochrome c FEBS J
278(9)1382-90
72 Macaskie LE Hewitt CJ Shearer JA Kent CA (1995) Biomass production for
the removal of heavy metals from aqueous solutions at low pH using growth-
decoupled cells of a Citrobacter sp Int Biodeter Biodegr 3573ndash92
73 Malki M De Lacey AL Rodriguez N Amils R Fernandez VM (2008)
Preferential use of an anode as an electron acceptor by an acidophilic bacterium in
the presence of oxygen Appl Environ Microbiol 74 4472ndash4476
74 Meitl LA Eggleston CM Colberg PJS Khare N Reardon CL Shi L (2009)
Electrochemical interaction of Shewanella oneidensis MR-1 and its outer membrane
cytochromes OmcA and MtrC with hematite electrodes Geochim Cosmochim
Ac 73(18) 5292-5307
75 Morozan A Stamatin L and Nastase
F (2007) The biocompatibility
microorganisms-carbon nanostructures for applications in microbial fuel cells Phys
Stat Sol 6 1797-1803
76 Morris JM Jin S Crimi B Pruden A (2009) Microbial fuel cell in enhancing
anaerobic biodegradation of diesel Chem Eng J 146 161ndash167
77 Murray AE Lies D Li G Nealson K Zhou J Tiedje JM (2001) DNADNA
hybridization to microarrays reveals gene-specific differences between closely
related microbial genomes Proc Natl Acad Sci U S A 98 9853-9858
78 Muyzer G de Waal EC Uitterlinden AG (1993) Profiling of complex microbial
communities by denaturing gradient gel electrophoresis analysis of polymerase chain
88
reaction amplified genes coding for 16S rRNA Appl Environ Microbiol 59 695ndash
700
79 Myers CR Myers JM (1992) Localization of cytochromes to the outer membrane
of anaerobically grown Shewanella putrefacians MR-1 J Bacteriol 174 3429ndash3438
80 Narde GK Kapley A Purohit HJ (2004) Isolation and characterization of
Citrobacter strain HPC255 for broad-range substrate specificity for chlorophenols
Curr Microbiol 48(6) 419-423
81 Okabe S Ito T Sugita K Satoh H (2005) Succession of internal sulfur cycles and
sulfur-oxidizing bacterial communities in microaerophilic wastewater biofilms Appl
Environ Microbiol 71(5) 2520-2529
82 Park DH and Zeikus JG (2000) Electricity generation in microbial fuel cells using
neutral red as an electronophore Appl Environ Microbial 66 292-1297
83 Park DH and Zeikus JG (2002) Impact of electrode composition on electricity
generation in a single-compartment fuel cell using Shewanella putrefaciens Appl
Microbiol Biotechnol 5958ndash56
84 Park DH and Zeikus JG (2003) Improved fuel cell and electrode designs for
producing electricity from microbial degradation Biotechnol Bioeng 81 348ndash355
85 Park HS Kim BH Kim HS Kim HJ Kim GT Kim M Chang IS Park Y
K Chang HI (2001) A novel electrochemically active and Fe(III)-reducing
bacterium phylogenetically related to Clostridium butyricum isolated from a
microbial fuel cell Anaerobe 7 297-306
86 Parot S Nercessian O Delia ML Achouak W Bergel A (2009) Electrochemical
checking of aerobic isolates from electrochemically active biofilms formed in
compost J Appl Microbiol 106(4) 1350-1359
87 Pham CA Jung SJ Phung NT Lee J Chang IS Kim BH Yi H Chun J
(2003) A novel electrochemically active and Fe(III)-reducing bacterium
phylogenetically related to Aeromonas hydrophila isolated from a microbial fuel cell
FEMS Microbiol Lett 223 129-134
88 Qiao Y Bao SJ Li CM Cui XQ Lu ZS Guo J (2008) Nanostructured
polyanilinetitanium dioxide composite anode for microbial fuel cells ACS Nano 2
113-119
89 Qiu R Zhao B Liu J Huang X Li Q Brewer E Wang S Shi N (2009) Sulfate
reduction and copper precipitation by a Citrobacter sp isolated from a mining area J
Hazard Mater 1641310-1315
90 Quan X Yang SG Ruan XL and Zhao HM (2005) Preparation of titania
nanotubes and their environmental applications as electrode Environ Sci Technol 39
3770ndash3775
91 Rabaey K Boon N Hofte M Verstraete W (2005) Microbial phenazine
production enhances electron transfer in biofuel cells Environ Sci Technol 393401ndash
3408
89
92 Rabaey K Boon N Siciliano S D Verhaege M Verstraete W (2004) Biofuel cells
select for microbial consortia that self-mediate electron transfer Appl Environ
Microbiol 70(9) 5373ndash5382
93 Rabaey K Lissens G Siciliano S D Verstraete W (2003) Amicrobial fuel cell
capable of converting glucose to electricity at high rate and efficiency Biotechnol
Lett 25 1531-15
94 Rabaey K Verstraete W (2005) Microbial fuel cells novel biotechnology for
energy generation Trends Biotechnol 23(6) 291-298
95 Reguera G McCarthy KD Mehta T Nicoll JS Tuominen MT Lovley DR
(2005) Extracellular electron transfer via microbial nanowires Nature 435 1098-
1101
96 Reguera G Nevin KP Nicoll JS Covalla SF Woodard TL Lovley DR (2006)
Biofilm and nanowire production leads to increased current in Geobacter
sulfurreducens fuel cells Appl Environ Microbiol 72 7345-7348
97 Reimers CE Tender LM Fertig S Wang W (2001) Harvesting energy from the
marine sediment-water interface Environ Sci Technol 35192-195
98 Rezaei F Xing D Wagner R Regan JM Richard TM Logan BE (2009)
Simultaneous cellulose degradation and electricity production by Enterobacter
cloacae in a microbial fuel cell Appl Microbiol Biotechnol 75 3673ndash3678
99 Richter H Nevin KP Jia H Lowy DA Lovley DR Tender LM (2009) Cyclic
voltammetry of biofilms of wild type and mutant Geobacter sulfurreducens on fuel
cell anodes indicates possible roles of OmcB OmcZ type IV pili and protons in
extracellular electron transfer Energy Environ Sci 2 506ndash516
100 Riesenfeld SC Schloss DP Handelsman J(2004) Metagenomics Genomic
Analysis of Microbial Communities Annual Review Genetics 38525-552
101 Rozendal RA Hamelers HV Buisman CJ (2006) Effects of membrane cation
transport on pH and microbial fuel cell performance Environ Sci Technol 40
5206-5211
102 Rozendal RA Hamelers HVM Euverink GJW Metz SJ Buisman CJN (2006)
Principle and perspectives of hydrogen production through biocatalyzed electrolysis
Int J Hydrogen Energ 311632-1640
103 Rozendal RA Hamelers HV Molenkamp RJ Buisman CJ (2007) Performance of
single chamber biocatalyzed electrolysis with different types of ion exchange
membrances Water Res 41 1984-1994
104 Schluumlter A Bekel T Diaz NN Dondrup M Eichenlaub R Gartemann
KH Krahn I Krause L Kroumlmeke H Kruse O Mussgnug JH Neuweger
H Niehaus K Puumlhler A Runte KJ Szczepanowski R Tauch A Tilker
A Viehoumlver P Goesmann A (2008) The metagenome of a biogas-producing
microbial community of a production-scale biogas plant fermenter analyzed by the
454-pyrosequencing technology J Biotechnol136 77-90
90
105 Scott J H and Nealson KH (1994) A biochemical study of the intermediary carbon
metabolism of Shewanella putrefaciens J Bacteriol 176(11) 3408ndash3411
106 Sekiguchi H Tomioka H Nakahara T and Uchiyama H (2001) A single band
does not always represent single bacterial strains in denaturing gradient gel
electrophoresis analysis Biotechnology Letters23 1205ndash1208
107 Sharma T Leela M Reddy A Chandra TS Ramaprabhu S (2008) Development
of carbon nanotubes and nanofluids based microbial fuel cell Int J Hydrogen Energ
336749ndash6754
108 Shea C Clauwaert P Verstraete W Nerenberg R (2008) Adapting a denitrifying
biocathode for perchlorate reduction Water Sci Technol 58 (10) 1941-1946
109 Shi L Squier TC Zachara JM Fredrickson JK (2007) Respiration of metal (hydr)
oxides by Shewanella and Geobacter a key role for multihaem c-type cytochromes
Mol Microbiol 6512-20
110 Stams AJ and Plugge CM(2009)Electron transfer in syntrophic communities
of anaerobic bacteria and archaea Nat Rev Microbiol7(8)568-77
111 Sukkasem C Xu S Park S Boonsawang P Liu H (2008) Effect of Nitrate on the
Performance of Single Chamber Air Cathode Microbial Fuel Cells Water Res 42(19)
4743-4750
112 Summers ZM Fogarty HE Leang C Franks AE Malvankar NS Lovley DR
(2010)Direct exchange of electrons within aggregates of an evolved syntrophic
coculture of anaerobic bacteria Science 330 (6009)1413-5
113 Szczepanowski R Bekel T Goesmann A Krause L Kroumlmeke H Kaiser
O Eichler W Puumlhler A Schluumlter A (2008) Insight into the plasmid metagenome
of wastewater treatment plant bacteria showing reduced susceptibility to
antimicrobial drugs analysed by the 454-pyrosequencing technology J
Biotechnol 136 54-64
114 Tamura K Dudley J Nei M Kumar S (2007) MEGA4 Molecular Evolutionary
Genetics Analysis (MEGA) software version 40 MolBiol Evol 24(8) 1596-1599
115 Thompson JD Gibson TJ Plewniak F Jeanmougin F Higgins DG (1997) The
CLUSTAL_X windows interface flexible strategies for multiple sequence alignment
aided by quality analysis tools Nucleic Acids Res 25 4876-4882
116 Thormann KM Duttler S Saville RM Hyodo M Shukla S Hayakawa Y
Spormann AM (2006) Control of formation and cellular detachment from
Shewanella oneidensis MR-1 biofilms by cyclic di-GMP J Bacteriol 1882681-2691
117 Thormann KM Saville RM Shukla S Pelletier DA Spormann AM (2004)
Initial Phases of biofilm formation in Shewanella oneidensis MR-1 J Bacteriol
186(23) 8096-104
118 Torres CI Kato Marcus A Rittmann BE (2008b) Proton transport inside the
biofilm limits electrical current generation by anode-respiring bacteria Biotechnol
Bioeng 100 872ndash881
91
119 Torres CI Marcus AK Lee HS Parameswaran P Krajmalnik-Brown R Rittmann
BE (2010) A kinetic perspective on extracellular electron transfer by anode-
respiring bacteria FEMS Microbiol Rev 234(1)3-17
120 Torres CI Marcus AK Parameswaran P Rittmann BE (2008a) Kinetic
experiments for evaluating the NernstndashMonod model for anode-respiring bacteria
(ARB) in a biofilm anode Environ Sci Technol 42 6593ndash6597
121 Tsai HY Wu Ch-Ch Lee Ch-Y Shih EP (2009) Microbial fuel cell performance
of multiwall carbon nanotubes on carbon cloth as electrodes J Power Sources
194199-205
122 Tyson GW Chapman J Hugenholtz P Allen EE Ram RJ Richardson
PM Solovyev VV Rubin EM Rokhsar DS Banfield JF(2004) Community
structure and metabolism through reconstruction of microbial genomes from the
environment Nature42837-43
123 Venkateswaran K Moser DP Dollhopf ME Lies DP Saffarini D A MacGregor
BJ Ringelberg DB White DC Nishijima M Sano H Burghardt J Stackebrandt
E Nealson KH (1999) Polyphasic taxonomy of the genus Shewanella and
description of Shewanella oneidensis sp Int J Syst Bacteriol 49705-724
124 von Canstein H Ogawa J Shimizu S Lloyd JR (2008) secretion of Flavins
by Shewanella Species and Their Role in Extracellular Electron Transfer Appl
Environ Microbiol 74 3615-623
125 Wang A Liu W Cheng Sh Xing D Zhou J Logan BE (2009) Source of methane
and methods to control its formation in single chamber microbial electrolysis cells
Int J Hydrogen Energ 34 3653-3658
126 Wang J Ma T Zhao L Lv J Li G Liang F Liu R (2008) PCRndashDGGE method
for analyzing the bacterial community in a high temperature petroleum reservoir
World J Microbiol Biotechnol 2 9694-9702
127 Wang Q Garrity GM Tiedje JM Cole JR (2007) Naiumlve Bayesian Classifier for
Rapid Assignment of rRNA Sequences into the New Bacterial Taxonomy Appl
Environ Microbiol 73(16) 5261-5267
128 Wang YF Masuda M Tsulimura S Kano K (2008) Electrochemical regulation of
the end-product profile in Propionibacterium freudenreichii ET-3 with an
endogenous mediator Biotechnol Bioeng 101(3) 579ndash586
129 Wrighton KC Agbo P Warnecke F Weber KA Brodie EL DeSantis TZ
Hugenholtz P Andersen GL Coates JD (2008) A novel ecological role of the
Firmicutes identified in thermophilic microbial fuel cells ISME J 2 1146ndash1156
130 Xing D Zuo Y Cheng S Regan JM Logan BE (2008) Electricity generation by
Rhodopseudomonas palustris DX-1 Environ Sci Technol 42 4146-4151
131 Xu S Liu H Fan Y Schaller R Jiao J Chaplen F (2012) Enhanced performance
and mechanism study of microbial electrolysis cells using Fenanoparticle-decorated
anodes Appl Microbiol Biotechnol 93(2)871-880
92
132 Yang Y Harris DP Luo F Wu L Parsons AB Palumbo AV Zhou J (2008)
Characterization of the Shewanella oneidensis Fur gene roles in iron and acid
tolerance response BMC Genomics 9 Suppl 1S11
133 Zhang L Zhou S Zhuang L Li W Zhang J Lu N Deng L (2008) Microbial
fuel cell based on Klebsiella pneumoniae biofilm Electrochem Commun10 1641ndash
1643
134 Zuo Y Xing DF Regan JM Logan BE (2008) Isolation of the exoelectrogenic
bacterium Ochrobactrum anthropi YZ-1 by using a U-tube microbial fuel cell Appl
Environ Microbiol 74 3130-3137
93
APPENDICES
List of published papers during PhD study
1 Shoutao Xu Hong Liu Yanzhen Fan Rebecca Schaller Jun Jiao and Frank
Chaplen (2012) Enhanced performance and mechanism study
of microbial electrolysis cells using Fe nanoparticle-decorated anodes Applied
Microbiology and Biotechnology 93(2)871-880
2 Shoutao Xu Hong Liu (2011) New exoelectrogen Citrobacter sp SX-1 isolated
from a microbial fuel cell Journal of Applied Microbiology111(5)1108-1115
3 Yanzhen Fan Shoutao Xu Rebecca Schaller Jun Jiao Frank Chaplen Hong Liu
(2011) Nanoparticle decorated anodes for enhanced current generation in microbial
electrochemical cells Biosensors and Bioelectronics 26(5) 1908-1912
4 Tunc Catal Shoutao Xu Kaichang Li Hakan Bermek Hong Liu
(2008)
Electricity generation from polyalcohols in single-chamber microbial fuel
cells Biosensors and Bioelectronics 24(4)849-854
5 Chontisa Sukkasema Shoutao Xu Sunhwa Park Piyarat Boonsawang Hong Liu
(2008) Effect of nitrate on the performance of single chamber air cathode
microbial fuel cells Water research 424743-4750
6 Rebecca Schaller Yanzhen Fan Shoutao Xu Alan Fern Frank Chaplen Hong
Liu and Jun Jiao (2009) Vertically Aligned Multi-walled Carbon Nanotube
Decorated Anodes for Microbial Fuel Cells Proceedings of Materials Research
Society 2009 1170 R05-13

produced with Fe nanoparticle decorated anodes were up to 59-fold higher than plain
graphite anodes Whole genome microarray analysis of the gene expression showed that
genes encoding biofilm formation were significantly up-regulated as a response to
nanoparticle decorated anodes Increased expression of genes related to nanowires
flavins and c-type cytochromes indicate that enhanced mechanisms of electron transfer
to the anode may also have contributed to the observed increases in current density The
majority of the remaining differentially expressed genes were associated with electron
transport and anaerobic metabolism demonstrating a systemic response to increased
power loads
The carbon nanotube (CNT) is another form of nano materials Carbon nanotube
(CNT) modified graphite disks were used as anodes to investigate the effects of
nanostructures on the performance S oneidensis MR-1 in microbial electrolysis cells
(MECs) The current densities produced with CNT decorated anodes were up to 56-fold
higher than plain graphite anodes Global transcriptome analysis showed that cytochrome
c genes associated with extracellular electron transfer are up-expressed by CNT
decorated anodes which is the leading factor to contribute current increase in CNT
decorated anode MECs The up regulated genes encoded to flavin also contribute to
current enhancement in CNT decorated anode MECs
copyCopyright by Shoutao Xu
June 15 2012
All Rights Reserved
Bacterial Community Analysis New Exoelectrogen Isolation and Enhanced Performance
of Microbial Electrochemical Systems Using Nano-Decorated Anodes
by
Shoutao Xu
A DISSERTATION
Submitted to
Oregon State University
in partial fulfillment of
the requirements for the
degree of
Doctor of Philosophy
Presented June 15 2012
Commencement June 2013
Doctor of Philosophy dissertation of Shoutao Xu presented on June 15 2012
APPROVED
Co-Major Professor representing Biological and Ecological Engineering
Co-Major Professor representing Biological and Ecological Engineering
Head of the Department of Biological and Ecological Engineering
Dean of the Graduate School
I understand that my dissertation will become part of the permanent collection of
Oregon State University libraries My signature below authorizes release of my
dissertation to any reader upon request
Shoutao Xu Author
ACKNOWLEDGEMENTS
I would like to thank all people who have helped and inspired me during my
doctoral study Foremost I would like to express my sincere gratitude to my advisors Dr
Hong Liu and Dr Frank Chaplen for their continuous support of my PhD study and the
research of Microbial Fuel Cells at Oregon State University Hong inspired me to devote
myself to the field of Bio-energy with her great patience and enthusiasm towards
scientific educations Frank was always available and willing to help me with my study
especially during the period of Hongrsquos sabbatical leave Thanks to his kindness and
assistance my study at OSU became smooth and rewarding
Besides my advisors I would like to thank everyone in my dissertation committee Dr
Martin Schuster Dr Clare Reimers and Dr Mark Dolan Due to their encouragement
and insightful comments on my research I could always have the courage and knowledge
to overcome difficulties in my research I benefited greatly from Martinrsquos valuable
suggestions on my writing skills and his generous help regarding microarray data analysis
Clarersquos advice helped me build a solid foundation of Electrochemistry Also it was a
great honor to have Mark as my committee member His teaching gave me an insight into
the world of environmental engineering and enhanced the depth and width of my research
I thanked Dr Yanzhen Fan for always giving me guidance about the designs of the
reactors in my experiments
It was also important for me to say thanks to my current and previous lab-mates
Keaton Lesnik Kuhuan Chien Cheng Li Corale Abourached Anthony Janicek
Hongqiang Hu Jeremy Chignell Yudith Nieto and Wengguo Wu I had a wonderful
time enjoying doing research with them Their friendship and help made me confident of
my ability to do research as a scientist In addition I cherished and appreciated the
friendly environment in our BEE department Faculty members especially Dr John Bolte
Dr John Selker Dr Roger Ely Dr Ganti Murthy and Desiree Tullos made BEE an
excellent department for our students
My deepest gratitude went to my father Fuqin Wang and my mother Jinzhi Wang
for their endless care love and support throughout my life Also I would like to say
thanks to my best-loved wife Songhua Zhu She always supported me and never
complained that I could spend so little time accompanying with her This dissertation was
also for my lovely son Gabriel Hong-Yi Xu
TABLE OF CONTENTS
Page
1 General Introductionhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 1
2 Bacterial Community Analysis of Mixed Consortium in Microbial
Electrochemical Systemshelliphelliphelliphellip 13
3 New Exoelectrogen Citrobacter sp SX-1 Isolation and
Characterizationhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 27
4
Enhanced Performance and Mechanism Study of Microbial Electrolysis
Cells using Fe Nanoparticles Decorated Anodeshelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 45
5 Global Transcriptome Analysis of Response of Shewanella oneidensis
MR-1 to CNT Nanostructure Decorated Anodes in Microbial
Electrochemical System
66
6 Summaryhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 80
7 Bibliography helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 82
8 Appendices helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 93
LIST OF FIGURES
Figure Page
1-1 Generalized schematics (a) Microbial fuel cell (MFCs)(b) Microbial
electrolysis cell (MECs) helliphelliphelliphelliphelliphelliphelliphelliphelliphellip helliphelliphelliphellip
2
1-2 Mechanisms for extracellular electron transport in a MFCs anode 7
2-1 Polarization curves of high power mixed culture MFCshelliphelliphelliphelliphelliphellip 20
2-2 DGGE result of high power generation mixed culture MFCshelliphelliphellip 21
2-3 Bacterial species and percentage of bacterial community of mixed
culture MFCs helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip
22
2-4 Phylogenetic trees of bacterial species from mixed culture MFCs 22
2-5 Dominant bacterial species identification in DGGEhelliphelliphelliphelliphelliphelliphelliphellip 23
2-6 Current density generated by different dominant isolateshelliphelliphelliphelliphelliphellip 24
3-1 Phylogenetic tree of strain SX-1 and closely related species based on
16S rDNA sequenceshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip
35
3-2 SEM image of planktonic cells of Citrobacter sp SX-1helliphelliphelliphelliphelliphellip 35
3-3 Electricity generation by Citrobacter sp SX-1 in a single chamber
MFCs with sodium citrate helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip
36
3-4 Electricity generation of Citrobacter sp SX-1 using different
substrates in single chamber air-cathode MFCs at 1 KΩhelliphelliphellip
37
3-5 Power and voltage generation by Citrobacter sp SX-1 as a function
of current density helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip
38
3-6 Four current generating stages for CV analysis helliphelliphelliphelliphelliphelliphelliphellip 39
3-7 Cyclic Voltammetry analysis of of Citrobacter sp SX-1 biofilms helliphellip 41
4-1 SEM images of Fe NPs decorated anodeshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 50
4-2 Effects of Fe-NP decorated anodes on the current density in MECs helliphellip 51
4-3 Differentially expressed genes grouped by functional classification in
Fe NP decorated anodeshelliphelliphelliphelliphelliphellip helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip
53
5-1 SEM images of CNT decorated anodeshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 70
5-2 Effects of CNT decorated anodes on the current density in MECshelliphellip 73
5-3 Differentially expressed genes grouped by functional classification in
CNT decorated anodes helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip
74
LIST OF TABLES
Table
1-1
4-1
4-2
4-3
Taxa of bacteria current density generated and reactor configuration
used in microbial electrochemical cell experimentshelliphelliphelliphellip
Expression levels of genes related to biofilm formationhelliphelliphelliphelliphellip
Genes related to anaerobic growth and electron transfer with
significantly change expression levelhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip
Expression level of flavin and cytochrome geneshelliphelliphelliphelliphelliphelliphelliphellip
Page
4
55
56
58
4-4
5-1
5-2
Other genes with significantly changed expression level
Expression level of cytochrome c as response to CNT decorated anodes
Expression level of genes related to flavin synthesis as response to CNT
decorated anode in CNT decorated anodeshelliphellip
59
76
78
1
1
Bacterial Community Analysis New Exoelectrogen Isolation and Enhanced Performance
of Microbial Electrochemical Systems Using Nano-Decorated Anodes
Chapter 1
General Introduction
11 Microbial Fuel Cells and Microbial Electrolysis Cells
Microbial electrochemical systems (MESs) have drawn the attention of researchers
in recent years due to their promising applications in a variety of scientific fields such as
renewable energy generation bioremediation and wastewater treatment In a MES
microorganisms interact with electrodes catalyzing oxidation and reduction reactions at
the anode and the cathode
The most-described type of MESs is the microbial fuel cells (MFCs) in which
useful power is generated directly using the catalytic action of active microorganisms
(Kim 2002 Cheng and Logan 2007 Cheng et al 2006) In a typical two-chamber
MFCs (Figure 1a) organic matter is oxidized by electrochemically active
microorganisms in the anode chamber to release protons and electrons Protons diffuse
into the cathode chamber through a proton exchange membrane Meanwhile electrons
are transferred to the anode through various mechanisms (Logan et al 2006 Lovley
2
2006 Rabaey et al 2003) and then travel to the cathode where normally they combine
with oxygen and protons to form water
Another common used type of MES is the microbial electrolysis cell (MECs) in
which hydrogen is produced instead of electricity by applying a circuit voltage to the
MFCs system and maintaining anaerobic conditions in the cathode chamber (Liu et al
2005 Logan et al 2008 Rozendal et al 2006 Schaller et al 2009) (Figure 1b) A
cathode potential of at least - 410 mV (vs standard hydrogen electrode at pH 70) is
required to produce hydrogen at the cathode Therefore a theoretical potential of 110 mV
(ie 410-300 mV) is needed but voltages of gt02 V are normally applied due to various
overpotentials
Fig 1-1 Generalized schematics (a) Microbial fuel cells (MFCs) where electricity is
captured through the resistance R (b) Microbial electrolysis cells (MECs) where
hydrogen gas is produced in the cathode chamber R for resistor PS for power supply and
PEM for proton exchange membrane
The fundamental feature shared by microbial electrochemical systems (MESs)
(MECs and MFCs) is the microbe-catalyzed electron transfer from organic matter to
electrodes (anodes) (Rebaey et al 2004) Therefore different types of MESs can be
PEM
R
H+
e- e-
Anode
Bacte
ria
a
Cath
ode
H2O
PEM
PS
H+
e- e-
Anode
Bacte
ria
b
Cath
ode
H2
3
utilized to investigate the phenomena and mechanisms of interactions between microbes
and electrodes
12 Bacteria and Electron Transfer Mechanisms in MESs
The electrochemically active bacteria in MFCs are thought to be iron-reducing
bacteria such as Shewanella and Geobacter species (Gorby et al 2006) They have great
importance in the natural environment principally in metal oxidation and reduction
However recent studies have shown that the diversity of bacterial communities is much
greater than these model iron reducers on MFCs anodes (Reimers et al 2001 Liu et al
2004b Kim et al 2005) Over 20 electrochemically active bacterial species those can
transfer electrons exocellularly to electrodes have been reported in the past 10 years
(Logan 2009 Liu et al 2010) These bacteria have been categorized into diverse genetic
groups so far including alphaproteobacteria (Rhodopseudomonas Ochrobactrum and
Acidiphilium) (Malki et al 2008 Xing et al 2008 Zuo et al 2008) betaproteobacteria
(Rhodoferax) (Chaudhuri et al 2003) gammaproteobacteria (Shewanella Pseudomonas
Klebsiella Enterobacter and Aeronomas) (Kim et al 2002 Pham et al 2003 Rabaey et
al 2005 Bretschger et al 2007 Zhang et al 2008 Chung et al 2009 Rezaei et al 2009)
deltaproteobacteria (Geobacter Geopsychrobacter Desulfuromonas and Desulfobulbus)
(Bond et al 2002 Bond and Lovley 2003 Holmes et al 2004ab 2006)
Epsilonproteobacteria (Arcobacter) (Fedorovich et al 2009) Firmicutes (Clostridium and
Thermincola) (Park et al 2001 Wrighton et al 2008) Acidobacteria (Geothrix) (Bond
and Lovley 2005) and actinobacteria (Propionibacterium) (Wang et al 2008) A wider
range of electrochemically active bacteria are expected to be discovered
4
Table 1 Taxa of bacteria current density generated and reactor configuration used in
microbial electrochemical cell experiments (Liu et al 2010)
The electrochemically active bacterial species that possess the ability to transfer
electrons outside of the cell are called exoelectrogens in the MESs research field The
different exoelectrogens have demonstrated a wide-ranging power generation ability in
MFCs For example Shewanella oneidensis MR-1 has demonstrated the ability to
generate 0018 Am2 current density in single chamber MFCs while the Geobacter
Taxon Microorganisms Current Density (Am2
)
α-proteobacteria
Rhodopseudomonas palustris DX-1 003
Ochrobactrum anthropi YZ-1 071
Acidiphilium sp 32sup5 300
β-proteobacteria Rhodoferax ferrireducens 0031
γ-proteobacteria
Shewanella putrefaciens IR-1 0016
Shewanella oneidensis DSP10 0013
Shewanella oneidensis MR-1 0018
Pseudomonas aeruginosa KRA3 0017
Escherichia coli K12 HB101 100
Klebsiella pneumoniae L17 120
Enterobacter cloacae 013
Aeromonas hydrophila PA3 030
δ-proteobacteria
Geobacter metallireducens 065
Geobacter sulfurreducens 800
Desulfuromonas acetoxidans 0005
Geopsychrobacter
Electrodiphilus strain A2
0066
Desulfobulbus propionicus 003
Firmicutes
Lactococcus lactis 003
Thermincola sp strain Jr 020
Clostridium butyricum EG3 022
Thermincola ferriacetica Z-0001 040
Brevibacillus spp PTH1 0009
Desulfitobacterium hafniense DCB2 110
Acidobacteria Geothrix fermentans 0097
Actinobacteria Propionibacterium freudenreichiiET-3 120
5
sulfurreducens species has generated current densities as high as 80 Am2 which is 400
times higher than the one generated by Shewanella oneidensis MR-1 However mixed-
culture communities have generated much higher power densities than their pure-culture
counterparts in MFCs perhaps due to synergistic interactions within the anode bacterial
communities and the participation of currently unknown bacteria species and mechanisms
(Jung and Regan 2007) The dominant bacteria species vary in mixed cultures bacteria
communities in MFCs due to the enrichment of different substrates
Traditional methods of extroelectrogen study depend on cultivation hampered novel
exoelectrogens discovery because the inadequacy of defined media underestimates the
actual microbial diversity in MFCs Slow growth rate and unknown growth requirements
of anaerobic microorganisms make some anaerobic extroelectrogens cultivation difficult
However these limitations have been overcome by using molecular biological methods
based on DNARNA analysis Molecular biological techniques are now widely applied to
assess the diversity of microbial communities by analyzing the 16S rDNA sequence The
most commonly used molecular biological techniques for bacterial community analysis
include denaturing gradient gel electrophoresis (DGGE) restriction fragment length
polymorphism (RFLP) and 16S rDNA sequencing These methods are much less time
consuming than traditional isolation and cultivation methods
Isolated exoelectrogens were utilized to explore the mechanism of electron transfer
to the anode However the mechanisms of electron transfer to extracellular electron
acceptors are not well understood Three mechanisms have been proposed for exocellular
transport of electrons by exoelectrogens to anodes in MFCs so far Some exoelectrogenic
bacteria can transfer electrons to electrodes through soluble redox compounds (Bond and
6
Lovley 2005) These compounds include artificial mediators and mediators secreted by
exoelectrogenic bacteria A variety of chemicals have been used to facilitate the shuttling
of electrons from inside of cell to electrodes outside the cell These exogenous mediators
include neutral red (Park et al 1999) anthraquinone-2-6 disulfonate (AQDS) thionin
potassium ferricyanide (Bond et al 2002) methyl viologen and others (Logan 2004
Rabaey and Verstraete 2005) Some bacteria can secrete their own chemical mediator
for example Pseudomonas aeruginosa secretes pyocyanin and phenazine-1-carboxamide
to transfer electrons toward the MFCs anode (Rabaey et al 2005) Another evidence for
mediator production by bacteria is Geothrix fermentans When the medium was replaced
in a MFCs that had stable power generation with this bacteria power dropped by 50
and required 10 days to resume the original level
Some bacteria can directly transfer electrons to anodes via outer cell membrane
proteins (Myers and Myers 1992) The outer cell membrane protein cytochrome c is
thought to play a critical role in to transferring electrons to anodes Ly et al (2011)
isolated the haem protein cytochrome c and demonstrated that electric field effects may
be functional for the natural redox processes of cytochrome c in the respiratory chain
Shewanella oneidensis is the example of a bacterium that is capable of transfer electron to
anode via outer cell membrane proteins When Shewanella oneidensis adhered to an iron
surface the greater force has showed benefits to grow cells because closer contact
required for electron transfer from cell bound cytochromes (Lower et al 2001)
7
Fig1-2 Mechanisms for extracellular electron transport in an MFCs anode (1) direct
contact (top in green) (2) by nanowires (middle in purple) and (3) self-produced
mediators (bottom in blue) (Logan 2009)
More and more evidence supports the involvement of bacterial nanowires in
extracellular electron transport (Reguera et al 2005 2006 Gorby et al 2006) Nanowires
are conductive appendages produced by both Geobacter and Shewanella species (Gorby
and Beveridge 2005) The conductivity of the appendages was examined and confirmed
by using conductive scanning tunneling microscopy (STM) (Gorby et al 2006)
Nanowires can carry electrons from the cell to the anode surface of MFCs
The solid component of the extracellular biofilm matrix has high efficiency on
extracellular electron transfer compared with other extracellular electron transfer
8
mechanisms and recently Torres et al (2010) hypothesized that the solid component of
the extracellular biofilm matrix formed by exoelectogens is another way bacteria transfer
electrons to electrodes This hypothesis was based on kinetic analysis of each EET
mechanism reported in available literature (Torres et al 2010)
13 Anode electrodes
In MFCsMECs anode electrodes are a critical component because exoelectrogens
adhere to the surface of anodes to transfer electrons to the electrode The characteristics
of anodes have significant effects on electron transfer rate from bacteria to anode
electrodes in MFCs The requirements of an anode material are it should be highly
conductive non-corrosive have a high specific surface area (area per volume) high
porosity be non-fouling inexpensive and easily scaled to larger sizes Of these
properties the most important one that is different from other biofilm reactors is that the
material must be electrically conductive Normally they are made of various carbon
materials including carbon fiber carbon clothe and carbon paper due to their stability
high conductivity and high specific surface-area Nevertheless they have little
electrocatalytic activity for the anode microbial reactions and thus a modification of the
carbon materials is the main approach for improving their performance Consequently
there is a great need to develop a new type of anode material for MFCsMECs
It is a great challenge to develop a new anode material to further increase the power
density of a MFCs The nature of the catalytic mechanism of a MFCrsquos anode involves not
only a biological but also an electrocatalytic process An optimal nanostructure with a
high specific surface area favorable for both catalytic processes could play a critical role
in improving the power density of the MFCs such a structure needs to host the bacteria
9
with high bioactivity while enhancing the electron-transfer rate Schroumlder et al (2008)
employed PANI to modify a platinum anode for MFCs and achieved a current density 1
order of magnitude higher than the previously reported value PANIinorganic
composites are also reported to have better conductivity Qiao et al also applied a new
mesoporous TiO2 electrode material with uniform nanopore distribution and a high
specific surface area to anode in comparison to previously reported work with E coli
MFCs the composite anode delivers 2-fold higher power density (Qiao et al 2008)
Thus it has great potential for use as the anode in a high-power MFCs and may be a new
approach for improving performance of MFCs
14 Other parts of MESs
141 Membranes and ion transport
The ion exchange membrane is another one of the critical components in two-
chamber MESs systems It separates anode and cathode chambers and at the same time
maintains the electron neutrality of the system ie transport of electrons to the cathode
needs to be compensated by transport of an equal amount of positive charge to the
cathode chamber A proton exchange membrane (PEM) such as Nafion a per-fluorinated
sulphonic acid membrane (PFSAs) consisting of a hydrophobic fluorocarbon backbone to
which hydrophilic sulfonate groups (-SO3-) are attached is commonly used in chemical
fuel cell systems For MFCs systems however mainly cation species like Na+ and K
+
other than proton are often responsible for the dominant transport of positive charge
through the cation exchange membrane (CEM) to maintain electroneutrality due to the
low proton concentration in any aqueous medium with near neutral pH (Rozendal et al
2006) Consequently the pH increases in the cathode chamber due to the consumption of
10
protons and decreases in the anode chamber because of the accumulation of protons
(Rozendal et al 2007) This pH change in two-chamber systems results in a decrease of
the cathode potential and performance The application of anion exchange membrane
(AEM) has also been explored in MFCs systems (Cheng and Logan 2007 Kim and
Logan 2007) where it has been proposed that protons are transferred via pH buffers like
phosphate anions
142 Cathodes and Catalysts
The cathode is another challenge for making MFCs commercially available
technology because the chemical reaction that occurs at the cathode is difficult to
engineer as the electrons protons and oxygen must all meet at a catalyst in a tri-phase
reaction (solid catalyst air and water) The catalyst must be on a conductive surface and
must be exposed to both water and air so that protons and electrons in these different
phases can reach the same point The most commonly used material for a cathode is
commercially available carbon paper pre-loaded with a Pt catalyst on one side When it is
used in a MFC the side that contains the catalyst faces the water and the uncoated side
faces air To reduce the high cathode cost associated with platinum catalyst other precious-
metal-free cathodes such as (NiMo and NiW) were developed by electrodepositing on a
carbon fiber They have achieved comparable performance with Pt catalyst with same
loading at a much lower cathode fabrication cost (Hu 2010)
The requirements of cathode for MECs are quite similar to the requirements of
cathode for MFCs but easier than cathode for MFCs for the manufacturing process
because the cathode in MECs is not necessarily to exposed to air Therefore it can be
made of the exact same materials of cathode in MFCs except a waterproof layer
11
Recently some researchers have attempted to apply microorganisms as a biocatalyst to
precede the combination of electron with oxygen in the cathodes
15 Dissertation overview
Low power densities in MESs limit practical applications The improvement of
MESs performance requires a detailed understanding of the physiology and ecology of
microorganisms in MESs including the mechanism of electron transfer to the anode from
the microorganism
This dissertation focuses on the problem of the low power density of MESs The
bacterial community structure of a high power generated mixed culture communities in
MFCs will be identified firstly and then one of major exoelectrogens will be isolated and
characterized This information will be helpful to understand the physiology and ecology
of exoelectrogens in MESs Consequently they will be beneficial to improve power
density of MFCs Lastly the nanostructure decorated anodes in MECs will be utilized to
improve the power density The power enhancement mechanism will be explored by
using a whole genome microarray They are presented here as four papers
In the first paper the cultivation independent molecular biological techniques
denaturing gradient gel electrophoresis (DGGE) and 16S rDNA clone library are utilized
to analyze the bacterial community structure of a higher power mixed culture MFCs The
analyzed results provide fundamental information for isolating the dominant bacteria in
mixed culture MFCs Two of dominant bacterial species has been isolated and used aone
to test power generation in MFCs The possible interaction among different bacterial
species in mixed culture is discussed
12
In the second paper one isolated exoelectrogenic bacterial strain SX-1 is
characterized It is identified as a member of the Citrobacter genus and power generation
is tested ultilizing a wide range of different substrates The electron transfer mechanism is
explored using Cyclic Voltammetry (CV) This study increases the known diversity of
power generating exoelectrogens and provides a new strain to explore the mechanisms of
extracellular electron transfer from bacteria to electrodes
The third paper shows effects on MESs performance by Fe nanoparticle decorated
anodes in the MESs The average current density produced with Fe nanoparticle
decorated anodes increased up to 59-fold higher than plain graphite anodes A whole
genome microarray is utilized to analyze the possible mechanism of enhanced current
density as responded to nanoparticle decorated anodes
The fourth paper describes the effects of carbon nanotube (CNT) modified anode on
the performance S oneidensis MR-1 in MESs Results demonstrate that current densities
produced with CNT decorated anodes are up to 56-fold higher than plain graphite anodes
The possible mechanisms of enhanced current density by CNT decorated are explored
13
Chapter 2
Bacterial community analysis of mixed consortium in higher
power density MESs
Shoutao Xu and Hong Liu
1 Introduction
The improvement in the performance of mixed culture MFCs requires an
understanding of the ecology in microbial communities of MFCs Many researchers have
attempted to characterize microbial populations and activities to elucidate the behaviors
and ecology of microorganisms in MFCs (Wrighton et al 2008 Fedorovich et al 2009)
In order to study the microbial ecology of the mixed culture in a MFC and select the
appropriate isolation medium for dominant bacterial species in the mixed culture the
fundamental steps are to analyze the bacterial diversity in the mixed culture of MFCs and
identity the dominant bacterial species in bacteria communities in MFCs
14
As for the identification of bacterial communities typically there are two general
methods The first method for identification of bacterial community is the traditional
cultivation processes using selective nutrients to promote the growth of different types of
bacteria within the samples (Amman et al 2000) The community structure can then be
assessed by identifying the isolates from the dominant colonies that were cultured This
can often be costly and laborious as each isolate has to be further studied by examining
its physiology taxonomy and reactivity to stains (Adwards et al 1989)
The second method relies on utilizing molecular techniques to analyze bacterial
community DNA Several molecular methods involving the extraction and analysis of
DNA from entire bacterial communities are used to identify genetic fingerprints of
bacteria These methods including the cloning and sequencing of 16S rDNA automated
ribosomal intergenic spacer analysis (ARISA) terminal restriction fragment length
polymorphism (T-RFLP) and denaturing gradient gel electrophoresis (DGGE) generate
profiles of bacterial community structures They can rapidly assess complex communities
from various environments (Amman et al 2000)
Due to the conservative characteristic of 16S rDNA in bacteria during the process
of evolution 16S rDNA sequencing can be used to identify different species of bacteria
Therefore the molecular techniques of denaturing gradient gel electrophoresis (DGGE)
with PCR and 16S rDNA clone library are used for analysis of the microbial diversity
These methods are more convenient and save time compared to traditional
isolationcultivation methods for microorganism analysis
In this work a biofilm bacterial community from an anode of a MFCs wase studied
by using denaturing gradient gel electrophoresis (DGGE) of PCR-amplified partial 16S
15
rRNA genes followed by cloning and sequencing of 16S rDNA The results provided
essential information for dominant bacterial isolation in mixed culture MFCs
2 Materials and methods
21 High power generation of mixed culture MFCs
Mixed cultures were originally inoculated from domestic wastewater (Corvallis
Wastewater Treatment Plant Corvallis OR) to MFCs after operating for 2 years using a
defined medium solution (Lovely 2002) with sodium acetate as the carbon source A
new MFCs was inoculated from the operating MFCs The polarization curves were
performed to measure the power generation when maximal stable power were established
(normally 3-5 batches post-inoculation) and the genomic DNA from the microbial
biofilm on the anode was extracted under the sterile conditions for further bacterial
community analysis
22 Denaturing gradient gel electrophoresis (DGGE)
DGGE with PCR is a method of analysis of bacterial community composition
based on different GC percentage of 16s rDNA in different bacterial species PCR-DGGE
comprises three steps (i) extraction of total community DNA from the sample (ii) PCR-
controlled amplification using specific oligonucleotide primers and (iii) separation of the
amplicons using DGGE For this purpose a reproducible and efficient method for total
DNA extraction is indispensable and needs to be evaluated and optimized depending on
the nature of the sample In the subsequent PCR step multiple PCR primer sets with
different resolution can be used In most PCR-DGGE applications on bacteria universal
or specific primers are targeting the 16S rRNA gene Upon electrophoresis of the PCR
16
amplicons and gel staining (using ethidium bromide silver staining or SYBR green)
DGGE gels are digitally captured and further analyzed using computer software packages
The richness of DNA fragment bands in DGGE indicates the bacterial diversity of the
sample
The detailed steps are as followed Biofilms were scratched from the anodes of high
power generated MFCs fed with sodium acetate Bacterial genomic DNA was extracted
from the biofilm samples using the DNeasy blood and tissue kit (Qiagen) according to the
manufacturerrsquos instructions The universal primer set was used to amplify the 16S rDNA
from the extracted genomic DNA then the V3 region of bacterial 16S ribosomal DNA
(rDNA) was amplified with GC-clamp primer (Muyzer et al 1993) for DGGE PCR
amplification was performed in a thermocycler DGGE of the PCR products was carried
out in a DcodeTM
Universal Mutation Detection System) The 8 (wv) polyacrylamide
gels contains from 30 to 55 denaturing gradients Electrophoresis was conducted
using a 1timesTAE (Tris-Acetate-EDTA) buffer at 130V and 60degC for 5 hours After
electrophoresis the gel was stained with ethidium bromide in 1timesTAE buffer for 15
minutes and washed in 1timesTAE buffer for 10 minutes The fragments were visualized
under a UV transilluminator The richness of single band in DGGE gel picture
preliminarily showed that the bacterial diversity in mixed culture since the single band in
gel represents one bacterial species
23 16S rDNA clone library construction method
16S rDNA-DGGE fingerprinting has proven to be particularly useful as an initial
investigation into bacterial communities and is suitable for identifying the predominant
bacteria that comprise 1 or more of cells within a given sample In the 16s rDNA clone
17
library construction process the first step is the extratction of the total genomic DNA
then the genomic DNA of the mixed culture biofilm was used as template for PCR
amplification of approximately 1500 bp of 16S rDNA with universal primers The PCR
products of 16S rDNA were purified and inverted into pGEM-T Easy vector system
before they were transformed into competent E coli The transformed cells were spread
on Luria-Bertani (LB) plates containing the selective antibiotic ampicillin and X-Gal and
incubated overnight at 37 OC after the more than 80 inverted 16s DNA fragment E coli
colonies were picked up and inoculated in the 1-3 ml liquid LB medium and left to grow
for 16 hours The plasmid DNA were extracted and sequenced The sequences were
compared directly to all known sequences deposited in GenBank databases using the
basic local alignment search tool (BLAST)
24 Bioinformatics Analysis
16S rDNA sequencing results of more than 80 colones were queried against the
GenBank and Ribosomal Database Project (RDP) databases using BLAST and
SEQUATCH algorithms (Cole et al 2007 Wang et al 2007 Cole et al 2009) The
neighbor-joining trees were constructed with the Molecular Evolutionary Genetics
Analysis package (MEGA Version 4) based on 1000 bootstrap analysis (Tamura et al
2007)
25 Dominant bacterial species band in DGGE identification
The DGGE results gave the information of dominant bacterial richness of bacterial
species in mixed culture based on the theory that single band represents one bacterial
species which showed the diversity of bacterial community in the mixed culture of a
MFC 16S rDNA clone library results provided the whole picture of bacterial community
18
including the bacterial species name and bacterial species percentage in the mixed culture
The known pure bacterial species which have been sequenced can be used as markers to
identify dominant bacterial species in DGGE The whole experimental procedure is
similar to the procedure of the mixed culture DGGE steps The first step is to extract the
collect the mixed culture biofilm genomic DNA and pure bacterial species marker
genomic DNA Then the universal primer set was used to amplify the 16S rDNA from
the extracted genomic DNA After that the V3 region of bacterial 16S ribosomal DNA
(rDNA) was amplified with GC-clamp primer (Muyzer et al 1993) for DGGE The
subsequent steps were similar to the mixed culture DGGE experimental procedure The
dominant bacterial bands in DGGE were determined by comparisons of the single band
position in the mixed culture DGGE with single pure bacterial species location in DGGE
26 Dominant bacterial species isolation and power generation
The bacterial community of mixed culture in MFCs were predominantly composed
of Geobacter sp Arcobacter sp and Citrobacter sp Two dominant bacterial species
have been isolated
Citrobacter sp SX-1 was isolated from the anode biofilm by using a solid agar
medium containing NH4Cl 6mM KCl 2mM CH3COONa (electron donor) 30 mM
FeC6H5O7 (electron acceptor) 20mM minerals and vitamins (Lovley and Phillips 1988)
Single colonies were picked up after culturing on the plate for 36 hours at 30 degC and
transferred two times on the agar plate for purification Then the isolates grew in a liquid
medium solution in anaerobic tubes containing the same constituents as the solid
medium All isolation process was operated in a glove box anaerobic chamber (Coy
Laboratory Products Grass Lake MI)
19
Arcobacter sp SX-2 was isolated from the anode biofilm of a MFC fed with sodium
acetate by using the same method of isolation of pure bacterial species Citrobacter XS-1
except using Trypticase Soy Agar with 5 Sheep Blood solid (from American type
culture collection (ATCC)) agar medium instead of Citrobacter sp medium Single
colonies were picked up after culturing on the plate for 36 hours at 30 degC and transferred
twice on the agar plate for purification Then the isolates grew in a liquid medium
solution microaerobically
Geobacter sp have been trying to isolate by using three different methods They are
the colony pickup after different condition enrichment Goebacter medium isolation and
dilution to extinction by multiple channel mini MFCs isolation however the Geobacter
sp has not been isolated yet due to some limits so far But they provide valuable
information for further isolation of Geobacter sp
Single chamber MFCs were used to evaluate power generation by different isolates
The MFCs were constructed as described previously (Liu and Logan 2004) and modified
with 3 cm2 carbon cloth anodes and 7 cm
2 carbon clothPt cathodes The total liquid
volume of each MFCs was about 15 ml with electrode spacing about 17 cm All MFCs
were operated in an autoclaved closed plastic box and sterile cotton was attached to the
outer surface of the air cathodes to prevent contamination A MFCs without bacterial
culture was used as control MFCs were inoculated with 3 ml late exponential phase
cultures of SX-1 in the medium solution reported previously (Liu and Logan 2004)
3 Results and discussion
31 Power production by mixed culture MFCs
20
A polarization curve was used to characterize current as a function of voltage in the
MFCs The polarization curves are performed by varying a series of external resistances
The results are shown in Figure 21 With MFCs acclimated to 25 Ω external resistance
the maximum power density was achieved at 65Wm2 based on the polarization data At
this point the current density is 21mAcm2 The power density was three times higher
than previous report (20 Wm2) using phosphate buffer (Fan et al 2007) Power density
was also 2 times higher than (28 m Wm2) using a bicarbonate buffer ( Fan et al 2007)
32 Bacterial community structure assessed by PCR-DGGE
Bacterial DGGE profiles for higher power generating MFCs mixed culture setups
were relatively complex (more than 10 bands) However there are three brightly stained
bands and other less intense bands (Fig 2-2) These results indicated there were probably
three dominant bacterial species in mixed culture MFCs
010
020
030
040
050
060
070
2
3
4
5
6
7
04 05 05 06 07 08 10 11 13 15 21 22 23 24
Volta
ge (
V)
Pow
er
densi
ty (
Wm
2)
Current density (mAcm2)
Power
Voltage
power density 65wm2 at current
density208 mAcm2
Figure 2-1 Polarization curves of high power mixed culture MFCs
21
33 16S rDNA clone library results
A total of 83 colonies were sequenced for 16S rDNA clone library construction They
represented the bacterial community structure in mixed culture in MFCs The results
revealed that the predominant bacterial species in mixed culture are Geobacter sp (66
of the mixed culture) Arcobacter sp (12 of the mixed culture) and Citrobacter sp (11
of the mixed culture) Those three genuses comprised 88 of the total bacterial species
(Fig 23) This result is consistent with the DGGE result which showed that three bright
bands in the DGGE gel represented three dominant bacterial species in the mixed culture
The phylogenetic analysis of the colonies (83 colony sequences) is shown in Figure 24
Figure 22 DGGE result of high power generation mixed culture MFC (1 mixed
culture 2 pure culture control)
1 2
22
34 Dominant bacterial band in DGGE identification
Geobacter
66
Arcobacter
12
others
6
Pseudomonas
1
Citrobacter
11
Clostridium
2
Anaerovorax
2 Geobacter
Arcobacter
Citrobacter
Clostridium
Anaerovorax
Pseudomonas
others
Figure 23 Bacterial species and percentage of bacterial community of mixed
culture MFC lt 1 Alcaligensgt
lt1 Commamonasgt
lt1 Pseudomonas aeruginosagt
lt1 Klebsiellagt
lt11 Citrobactergt
lt1 Wolinellagt
lt12 Arobactergt
lt2 Anaeroboraxgt
lt2 Clostridium gt
lt1 unclear bacteriagt
lt66Geobactergt
100
100
85
100
100
82
99
87
100
002
Figure 24 Phylogenetic trees of bacterial species from mixed culture MFC
23
The dominant bacterial species including Goebacter sp Citrobacter sp and Arcobacter
sp in the mixed culture are shown in Figure 25 The upper band is Arcobacter sp and
Citrobacter sp is located in the bottom band of DGGE Geobacter sp is the middle one
The analysis of the bacterial community using denaturing gradient gel electrophoresis
(DGGE) of PCR-amplified 16S rRNA gene fragments and 16S rDNA clone library
construction method showed great phylogenetic diversity of mixed culture in MFCs with
the identification of sequences derived from bacteria of the taxa deltaproteobacteria
(Geobacter sp) gammaproteobacteria (Citrobacter sp) and epsilonproteobacteria
(Arcobacter sp) The result is consistent with the conclusion that among the isolated
exoelectrogens gammaproteobacteria and deltaprotebactia appear to be dominant among
others (Parot et al 2009 Liu et al 2010)
35 The dominant bacterial species isolation and power generation
Figure 25 Dominant bacterial species identification in DGGE
A B C D
A Citrobacter sp
B Geobacter sp
C Arobacter butzmeri
D mixed culture control
24
Two dominant bacterial species Citrobacter sp XS-1 and Arcobacter sp XS-2 have been
isolated and tested for power generation The power densities generated by the two
isolates are shown in the Fig 26 Current densities generated by isolate Citrobacter sp
and Arcobacter sp were 98 mAm2 and 20 mAm
2 respectively which were much less
than that (21Am2) generated by the mixed culture from which they were isolated
Figure 26 Current density generated by different dominant isolates
The maximum current density generated by mixed culture is much higher than the
one produced by pure bacterial species Citrobacter sp XS-1 and Arcobacter sp XS-2 in
the same structure MFCs Although the Geobacter sp in our mixed culture has not been
isolated a previous study showed that maximum current density generated by Geobacter
sp was 080 Am2
(Bond and Lovley 2003) The current density results seem partially
supporting the conclusion that mixed-culture generated higher power densities than their
pure-culture counterparts in MFCs (Jung and Regan 2007) This result suggests there
may be synergistic interactions within the anode bacterial communities
0
500
1000
1500
2000
2500
Mixed culture Citrobacter sp Arcobacter Sp
Cu
rre
nt
de
nsi
ty (
mA
m2)
25
Geobacter sp are strictly anaerobic bacteria which were enriched in most of the anode
biofilms (George 2005) Citrobacter species are facultative anaerobic bacteria (George
2005) which can be found in a wide variety of habitats including in soil water and
wastewater Arcobacter sp are micro-aerobic bacteria including both environmental
nonpathogens and opportunistic human pathogens They are able to grow in aerobic
conditions but in the optimal growth under micro-aerobic conditions (George 2005)
Arcobacter sp has been discovered in the MFCs by Fedorovich (Fedorovich et al 2009)
It is interesting that the mixed culture bacterial community composed of primarily these
three bacterial species has demonstrated to generate high power at non-strictly anaerobic
condition while leading dominant bacterial species Geobacter sp (66) is strict
anaerobic bacteria and generated powder in the MFCs only under strict anaerobic
condition (George 2005) Therefore it is possible that microaerobic bacteria such as
Arcobacter sp and facultative anaerobic bacteria Citrobacter sp can facilitate to create
anaerobic condition for Geobacter sp when they utilized oxygen for growing in the
mixed culture Syntrophic communities study have showed that anaerobic bacteria and
methanogenic archaea form compact microbial structures that operate like an organ rather
than a set of microorganisms functioning independently (Stams and Plugge 2009) Some
substrates have been degradeted within these communities while they are not able to be
fermented by individual species alone interspecies electron transfer also have been
presented in these communities (Stams and Plugge 2009) Summers et al (2010) also
discovered that direct exchange of electrons happened within coculture of Geobacter
metallireducens and Geobacter sulfurreducens Therefore we believe that there are
26
more complex synergistic interactions between different bacteria species in mixed culture
in the MFCs
Acknowledgements
The authors acknowledge support from the US National Science Foundation
(CBET 0955124) We also thank Keaton Lesnik for the review of this manuscript
27
Chapter 3
New Exoelectrogen Citrobacter sp SX-1 Isolated from a Microbial Fuel Cell
Shoutao Xu and Hong Liu
Published in
Journal of Applied Microbiology 111(5)1108-1115 (2011)
ABSTRACT
Exoelectrogenic bacterial strain SX-1 was isolated from a mediator-less microbial fuel
cell by conventional plating techniques with ferric citrate as electron acceptor under
anaerobic condition Phylogenetic analysis of the 16S rDNA sequence revealed that it
28
was related to the members of Citrobacter genus with Citrobacter sp sdy-48 being the
most closely related species The bacterial strain SX-1 produced electricity from citrate
acetate glucose sucrose glycerol and lactose in MFCs with the highest current density
of 205 mA m-2
generated from citrate Cyclic voltammetry analysis indicated that
membrane associated proteins may play an important role in facilitating the electrons
transferring from bacteria to electrode This is the first study that demonstrates that
Citrobacter species can transfer electrons to extracellular electron acceptors Citrobacter
strain SX-1 is capable of generating electricity from a wide range of substrates in MFCs
This finding increases the known diversity of power generating exoelectrogens and
provided a new strain to explore the mechanisms of extracellular electron transfer from
bacteria to electrode The wide range of substrate utilization by SX-1 increases the
application potential of MFCs in renewable energy generation and waste treatment
Keywords Microbial fuel cell Exoelectrogen Citrobacter sp SX-1 Extracellular
electron transfer
29
Introduction
Microbial Fuel Cells (MFCs) technology has become an active research area
recently as a promising approach for renewable energy generation wastewater treatment
and bioremediation (Rabaey and Verstraete 2005 Lovley 2006 Fan et al 2008 Logan
2009 Liu et al 2010) The key feature of MFCs system is the microbe-catalyzed electron
transfer from organic matter to anodes Many studies have shown the presence of diverse
bacterial communities on MFCs anodes (Reimers et al 2001 Liu et al 2004b Kim et al
2005) Over 20 exoelectrogens ie microbes that can transfer electrons exocellularly to
electrodes have been reported in the past 10 years (Logan 2009 Liu et al 2010) The
exoelectrogens so far belong to diverse genetic groups including alphaproteobacteria
(Rhodopseudomonas Ochrobactrum and Acidiphilium) (Malki et al 2008 Xing et al
2008 Zuo et al 2008) betaproteobacteria (Rhodoferax) (Chaudhuri et al 2003)
gammaproteobacteria (Shewanella Pseudomonas Klebsiella Enterobacter and
Aeronomas) (Kim et al 2002 Pham et al 2003 Rabaey et al 2005 Bretschger et al
2007 Zhang et al 2008 Chung et al 2009 Rezaei et al 2009) deltaproteobacteria
(Geobacter Geopsychrobacter Desulfuromonas and Desulfobulbus) (Bond et al 2002
Bond and Lovley 2003 Holmes et al 2004ab 2006) Epsilonproteobacteria (Arcobacter)
(Fedorovich et al 2009) Firmicutes (Clostridium and Thermincola) (Park et al 2001
Wrighton et al 2008) Acidobacteria (Geothrix) (Bond and Lovley 2005) and
actinobacteria (Propionibacterium) (Wang et al 2008) A wider range of
exoelectrogenic species are expected to be discovered
Three mechanisms have been proposed for exocellular transport of electrons by
exoelectrogens without artificial mediators Some exoelectrogenic bacteria can transfer
30
electrons to electrodes through soluble redox compounds excreted by microorganisms
(Bond and Lovley 2005 Rabaey et al 2005) and others can directly transfer electrons to
anodes via outer cell membrane proteins (Myers and Myers 1992) Recently more and
more evidence supports the involvement of bacterial nanowires in extracellular electron
transport (Reguera et al 2005 2006 Gorby et al 2006) In spite of the discovery of
many bacterial species that can transfer the electrons to electrode without the need of
artificial mediators the investigation of extracellular electron transfer mechanisms was
mainly focused on a few species such as those from Geobacter and Shewanella genera
(Kim et al 1999 2002 Reguera et al 2005 2006 Gorby et al 2006) The electron
transfer mechanisms for many of the isolated exoelectrogens species are still not well
studied
In the present study we isolated a new exoelectrogen strain SX-1 from a MFC a
strain phylogenetically related to Citrobacter sp Power generation from various carbon
sources by this strain was evaluated using single chamber MFCs Plausible extracellular
electron transfer mechanisms were also discussed based on the characterization of anodic
biofilms by cyclic voltammetry (CV)
Materials and methods
Bacterial strain SX-1 isolation
Bacterial strain SX-1 was isolated from the anode biofilm of a MFC fed with sodium
acetate operated in fed-batch mode over a period of six months The original source of
the inoculum is wastewater from a local waste water treatment plant Bacterial cells were
released from the carbon cloth anode by shaking the cloth in a sterile flask with 15 mL
sterile 1 NaCl solution and glass beads (2 mm diameter) The suspension was then
31
serially diluted from 10 times to 105
times and plated on a petri dish with a solid agar
medium containing NH4Cl 6mM KCl 2mM CH3COONa (electron donor) 30 mM
FeC6H5O7 (electron acceptor) 20mM minerals and vitamins (Lovley and Phillips 1988)
Single colony was randomly selected after culturing on the plate for 36 hours at 30 degC
and purified on a new agar plate following a procedure reported previously (Chung and
Okabe 2009) Then the isolate was picked up and grew in a liquid medium solution in
anaerobic tubes containing the same constituents as the solid medium for further analysis
All isolation process was operated in a glove box anaerobic chamber (Coy Laboratory
Products Grass Lake MI)
16S rDNA sequencing and phylogenetic analysis
Bacterial genomic DNA was extracted from the isolated strain SX-1 using the DNeasy
tissue Kits (Qiagen Valencia CA) according to the manufacturerrsquos instructions The 16S
rDNA was amplified by PCR using a pair of universal primers 27F (5rsquo-
AGAGTTTGATCMTGGCTCAG-3rsquo) and 1492R (5rsquo-GGTTACCTTTGTTACGACTT-
3rsquo) (Park et al 2001) The conditions used for PCR were described previously
(Ferna ndez et al 1999) PCR products were purified ligated and cloned according to a
procedure described in a previous report (Xing et al 2008) Plasmids were isolated from
randomly selected clone colonies with a QIAprep Spin Miniprep kit (Qiagen Valencia
CA) and three plasmid inserts were then sequenced in both directions using an ABI 3730
DNA sequencer (Applied Biosystems Carlsbad CA)United States) Due to the identical
16S rDNA sequence of the three colones only one was queried against the GenBank and
Ribosomal Database Project (RDP) databases using BLAST and SEQUATCH algorithms
(Cole et al 2007 Wang et al 2007 Cole et al 2009) The 16S rDNA sequence of
32
strain SX-1 and closely related type strains were aligned using CLUSTALX software
(Thompson et al 1997) The neighbor-joining trees were constructed with the Molecular
Evolutionary Genetics Analysis package (MEGA Version 4) based on 1000 bootstrap
analysis (Tamura et al 2007)
SEM
The morphologies of SX-1 cells growing overnight in anaerobic tubes were examined
with a scanning electron microscope (FEI Quanta 600 FEG FEI Company Hillsboro
OR) The suspended cells were fixed in a 25 glutaraldehyde and 01M phosphate
buffer solution and dehydrated with a graded ethanol series from 30 to 100 After
dehydration the samples were dried in a critical point dryer and then sputter-coated with
AuPd for SEM examination (Liu and Logan 2004)
Evaluation of power generation by SX-1 in MFCs
Single chamber MFCs were used to evaluate power generation by SX-1 using various
substrates The MFCs were constructed as described previously (Liu and Logan 2004)
and modified with 3 cm2 carbon cloth anodes and 7cm
2 carbon clothPt cathodes The
total liquid volume of each MFCs was about 15 ml with electrode spacing about 17 cm
All MFCs were operated in an autoclaved closed plastic box and sterile cotton was
attached to the outer surface of the air cathodes to prevent contamination A MFC
without bacterial culture was used as control MFCs were inoculated with 3 ml late
exponential phase cultures of SX-1 in the medium solution reported previously (Liu and
Logan 2004) Seven substrates including citrate acetate glucose sucrose glycerol and
lactose were evaluated individually for power generation in a fed-batch mode in a
temperature control chamber (30 plusmn2degC) For each substrate at least 8 batches were run to
33
investigate the effect of biofilm formation on current output at fixed external resistance of
1kΩ and the same initial substrate concentration of 30 mM Phosphate buffer (100mM)
was used to maintain the solution pH and conductivity The MFCs with sodium citrate as
electron donors were also examined for maximum power output by varying the external
resistance from 20 kΩ to 4 kΩ A data acquisition system was used to record voltage data
during the MFCs operation (Keithley 2700 Keithley Instruments Inc Cleveland OH) It
took about 15 to 30 minutes for the MFCs to stabilize depending on the external
resistance At each resistance we collected at least five data at the steady condition to
make the polarization curves Averaged voltages were used to calculate the power density
(mW m-2
) according to P=IVA where I was the current V was voltage and A was cross-
sectional area of the anode
CV analysis
CV was used to characterize the oxidation and reduction reactions on the anodic surface
of the MFCs fed with citrate and operated at 7 KΩ This external resistance was selected
because the maximum power density was obtained at this resistance based on the
polarization experiment We assumed the biofilm on the anode of MFCs were well-
developed when stable power output was obtained after 3 batches of operation The
MFCs were then used directly for CV analysis at four current generating stages of the
fourth batch (1) initial exponential current increasing stage (middle point of stage about
4 h after media change) (2) current plateau stage (middle point of stage about 8 h after
media change) and (3) current decreasing stage (middle point of stage about 16h after
media change) (4) right after the complete replacement of medium solution (Figure 6)
The anode was used as working electrode the cathode as counter electrode and an
34
AgAgCl electrode was selected as the reference The CV curves were scanned from 200
to -600 mV at a rate of 5 mV s-1
using a potentiostat (G300 Gamry Instrument
IncWarminster PA) Control experiment was also conducted using new anode (without
biofilm) and new medium solution
Nucleotide sequence accession number
The 16S r DNA sequence determined in this study has been deposited in the GenBank
database under accession number HQ845373
Results
Identification of the strain SX-1
An almost-complete 16S rDNA sequence (1475 bp) of strain SX-1 was obtained and
subjected to comparative analysis with the 16S rDNA of closely related reference strains
A phylogenetic analysis revealed that strain SX-1 should be assigned to the genus
Citrobacter and was most closely related to Citrobacter sp sdy-48 EJ463782 (990
sequence similarity) and Citrobacter sp yy-21 FJ463779 (975 sequence similarity)
These three strains formed a distinct sub cluster in the neighbor-joining in which the new
isolate and Citrobacter sp sdy-48 EJ463782 formed a distinct subline (Figure 31)
The SEM image illustrated that the strain SX-1 is a rod-shaped bacterium 03-06 microm
wide and 12-17 microm long (Figure 32) Strain SX-1 colonies on nutrient agar appeared 2-
3 mm in diameter smooth moist opaque with a shiny surface after 5 days incubation
under anaerobic condition The fact that strain SX-1 grew both aerobically and
anaerobically using FeC6H5O7 as electron acceptor suggests that the strain is facultatively
anaerobic Strain SX-1 showed robust growth on citrate as a carbon source These D
35
properties were similar to those of genus Citrobacter described in Bergeyrsquos Manual of
Systematic Bacteriology (George 2005)
Figure 31 phylogenetic tree of strain SX-1 and closely related species based on 16S
rDNA sequences The tree was constructed using the neighbor-joining method
Figure 32 SEM image of planktonic cells of Citrobacter sp SX-1
Strain SX-1
Citrobacter sp sdy-48 (FJ463782)
Citrobacter sp yy-21 (FJ463779)
Citrobacter koseri E639 (ATCC 25408)
Citrobater koseri CDC 3613-63(AF025372)
Citrobacter koseri CDC 8132-86(AF025366)
Salmonella enterica SL483 (CP001138)
Salmonella enterica AKU12601(AY696668)
Shigella sonnei Ss046(CP000038)
E coli C2 (AF403733)
E coli E24377A(CP000800)88
100
100
76
44
92
53
36
0001
1 microm
36
Electricity production by strain SX-1 in MFCs
The isolated strain SX-1 was first tested for its ability to generate current from sodium
citrate at 1kΩ Current gradually increased to a maximum 50 mA m-2
after cell
inoculation and then decreased (Figure 33) After the MFCs was refilled with new
substrate solution the current recovered rapidly and reached a higher level than the first
batch After 4 batches operation the maximum current output of each batch became
stable The highest current density achieved in the MFCs with Citrobacter sp SX-1 at 1
KΩ was 98 mA m-2
(Figure 33)
Figure 33 Electricity generation by Citrobacter sp SX-1 in a single chamber MFCs
with sodium citrate (30 mM) as substrate at 1 KΩ
0
20
40
60
80
100
120
0 50 100 150 200
Curr
ent
den
sity
(m
A m
-2)
Time (h)
37
Current generation by SX-1 from other substrates including glucose lactose sodium
acetate glycerol and sucrose were also assessed at an external resistance of 1KΩ
(Figure 34) When repeatable cycles of current output were obtained for these substrates
glycerol generated the highest maximum current density of 58 mA m-2
followed by
lactose and sucrose with 29 mA m-2
and 27 mA m-2
respectively Glucose and acetate
produced the lowest maximum current density of 96 and 43 mA m-2
respectively These
results indicated that strain SX-1 can utilize a wide range of substrates for electricity
generation in MFCs but with different power generation potentials
Figure 34 Electricity generation of Citrobacter sp SX-1 using different substrates in
single chamber air-cathode MFCs at 1 KΩ (The error bars stand for the standard
deviation of 3 replicates)
0
10
20
30
40
50
60
70
Glucose Lactose Glycerol Acetate Sucrose
Curr
ent
den
sity
(m
A m
-2)
38
Polarization experiment was further conducted to determine the maximum power
density generated by strain SX-1 in the single chamber MFCs with citrate as substrate A
maximum power density of 881 mW m-2
was obtained at current density of 205 mA m-2
at an external resistance of 7 KΩ (Figure 35)
Figure 35 Power and voltage generation by Citrobacter sp SX-1 as a function of
current density using sodium citrate (30 mM) as substrate The error bars stand for the
standard deviation of the 3 voltagepower outputs obtained in three MFCs
20
30
40
50
60
70
80
90
100
01
02
03
04
05
06
07
80 110 140 170 200 230
Po
wer
den
sity
(m
W m
-2)
Vo
ltag
e (V
)
Current density (mA m-2)
Voltage
Power
39
Figure 36 Four current generating stages for CV analysis current increasing stage (stage
1) current plateau stage (stage 2) current decreasing stage (stage 3) and right after the
replacement of medium solution (stage 4) The MFCs was operated at 7 KΩ with sodium
citrate (30 mM) as substrate
Cyclic Voltammetry
To determine the presence of redox active compounds produced by SX-1 and the
location of these compounds CV scan of the anodic biofilms at four current output stages
and the supernatant of MFCs medium solution at the end of the batch experiment were
performed Reduction peaks at range of -310 ndash -350 mV and oxidation peaks at range -
100 ndash -150 mV were observed at all current generating stages (Figure 36 Figure 37A)
suggesting the presence of redox active compounds may involve in extracellular electron
transfer by Citrobacter sp SX-1 The amplitude of the peaks increased according to the
growth stage of the batch and the highest peaks were present after the current plateau
stage which indicated the redox active compounds mainly were secreted in the current
plateau stage While the current density at stage 3 (deceasing stage) was lower than that
at stage 2 (current plateau stage) the redox peak at stage 3 was higher than that in stage 2
0
50
100
150
200
250
50 55 60 65 70 75 80 85
Curr
ent
den
sity
(m
A m
-2)
Time (h)
A
A
Stage 1
Stage 2 Stage 3
Stage 4
40
A plausible explanation for this pattern was that redox active compounds were
continuously being secreted and accumulated after the current peak was reached
resulting in more redox compounds present after the plateau stage But since most of the
carbon source had already been being used up many of the redox compounds at this
stage had not been as active as those at current plateau stage due to much less electrons
were available to be transferred The bigger peak amplitude in stage 4 than in stage 1
indicates that the redox active compounds were continuously being secreted and
accumulated in the biofilm even after stable power was generated at 7 kΩ The slightly
change of the peak locations at different stages was possible due to the solution chemistry
change during the current generation process or the slightly location change of reference
electrode during the measurement (Figure 37B) When the medium solution in the MFCs
was replaced by fresh medium at the end of the batch the oxidization and reduction
peaks were still presented in the CV (Figure 37B) but no peaks were observed when the
supernatant and fresh mediums were tested in a MFCs with no biofilm (Figure 37C)
These results suggest the compounds involved in the electron transfer were located not in
the supernatant (solution) but in the biofilm
41
Figure 37 Cyclic Voltammetry analysis of Citrobacter sp SX-1 biofilms at (A) current
increasing stage current plateau stage and decreasing stage (end of the batch) (B) end of
the batch and replaced with fresh medium and (C) controls
-40E-04
-30E-04
-20E-04
-10E-04
00E+00
10E-04
20E-04
30E-04
-060 -050 -040 -030 -020 -010 000 010 020
Curr
ent
(A)
Potential (V) vs AgAgCl
Current plateau stage
Current increasing stage
Current decreasing stage
-40E-04
-30E-04
-20E-04
-10E-04
00E+00
10E-04
20E-04
30E-04
-060 -050 -040 -030 -020 -010 000 010 020
Curr
ent
(A)
Potential (V) vs AgAgCl
Current decreasing stage
Biofilm with fresh medium
-40E-04
-35E-04
-30E-04
-25E-04
-20E-04
-15E-04
-10E-04
-50E-05
00E+00
-060 -050 -040 -030 -020 -010 000 010 020
Curr
ent(
A)
Potential (V) vs AgAgCl
Supernatant
Fresh medium
A
B
C
B C
42
Discussion
Citrobacter species belonging to Gammaproteobacteria Enterobacteriales
Enterobacteriaceae are facultative anaerobic bacteria (George 2005) They can grow with
oxygen as an electron acceptor but also with a variety of electron accepters such as Fe (III)
in the absence of oxygen (George 2005) Citrobacter species can be found in a wide
variety of habitats including in soil water and wastewater Several Citrobacter species
have been studied as important bioremediation bacterium for heavy metal removals
sulfate reduction phenol degradation and chlorophenol degradation (Macaskie et al 1995
Narde et al 2004 Qiu et al 2009) Citrobacter species have also been found on the
anodic biofilm of a diesel-degrading MFCs (Morris et al 2009) However there was no
study reported that Citrobacter species can transfer electrons to extracellular electron
acceptors This finding that Citrobacter strain SX-1 can generate electricity in MFCs
increased the diversity of power generating exoelectrogens and provided a new strain to
explore the mechanisms of extracellular electron transfer from bacteria to electrode
Among the isolated exoelectrogens gammaproteobacteria and deltaprotebactia appear to
be dominant among others (Parot et al 2009 Liu et al 2010) The isolated strain
Citrobacter sp SX-1 phylogenetically belongs to gammaproteobacteria which provides
another evidence to support this observation
Most exoelectrogens utilize a limited range of substrates for power generation For
example Shewanella oniedensis MR-1 can oxidize lactate under anaerobic condition but
cannot utilize acetate for electricity generation (Kim et al 2002) Geobacter
sulfurreducens can completely oxidize acetate for power generation but it cannot utilize
simple sugars (Bond and Lovley 2003 Caccavo et al 1994)) In comparison strain SX-1
43
not only can oxidize citrate for power generation but also utilize a wide range of
substrates for power generation including glycerol glucose lactose sucrose and acetate
Interestingly the higher current density generated by strain SX-1 from glycerol a main
by-product of biodiesel production process suggests that strain SX-1 may be potentially
used for harvesting energy from biodiesel wastes using MFCs However the current
density generated by strain SX-1 (205 mA m-2
) is lower than that (805 mA m-2
at 1 kΩ)
generated by the mixed culture from which the SX-1 was isolated indicating the
existence of other higher power generating bacteria andor complex ecology in the mixed
culture community
Understanding mechanisms of microbial extracellular electron transfer is critical for
enhancing the electron transfer rate from bacteria to electrode through metabolic or
genetic engineering of exoelectrogens Since the spent supernatant solution of MFCs run
with SX-1 showed no redox properties the redox compounds produced by SX-1 may
have been retained in the biofilm Alternatively SX-1 may utilize one or both of the
other two known extracellular electron transfer mechanisms For example it is possible
that strain SX-1 transfers electrons to the anode via cell-bound outer membrane proteins
The two distinct oxidation and reduction peaks observed at -100mv- -150 mV and -310--
350 mV of the SX-1 biofilm have a midpoint potential range from -205mV to -250mV
which is close to the redox potential of some c-type cytochromes (Field et al 2000 Meitl
et al 2009) which are well-known to play an important role in extracellular electron
trasnfer (Eggleston et al 2008 Field et al 2000 Meitl et al 2009) Field et al (2000)
reported that the c-type cytochromes CymA had the midpoint potentials of -229 mV
Meitl et al (2009) and Eggleston et al (2008) reported that the c- type cytochromes
44
OmcA in Shewanella oneidensis MR-1 had the midpoint potentials of -201 mV and -208
mV
While itrsquos possible that SX-1 may also have conductive appendages to enhance the
extracellular transfer electron similar to the nanowires discovered in some Geobacter
and Shewanella species (Reguera et al 2005 2006 Gorby et al 2006) further
investigation on the presence of these appendages on the cell surface of SX-1 is needed
Acknowledgements
The authors acknowledge support from the US National Science Foundation
(CBET 0955124) We also thank Teresa Sawyer for her helps with the SEM analysis and
Jeremy Chignell and Yanzhen Fan for their review of this manuscript
45
Chapter 4
Enhanced Performance and Mechanism Study of Microbial Electrolysis Cells
Using Fe Nanoparticles Decorated Anodes
Shoutao Xu Hong LiuYanzhen Fan Rebecca Schaller Jun Jiao Frank Chaplen
Published in
Applied Microbiology and biotechnology 93(2)871-880 (2012)
46
ABSTRACT
Anode properties are critical for performance of microbial electrolysis cells (MECs) In
the present study Fe nanoparticle modified graphite disks were used as anodes to
investigate the effects of nanoparticles on the performance of Shewanella oneidensis MR-
1 in MECs Results demonstrated that average current densities produced with Fe
nanoparticle decorated anodes were up to 56-fold higher than plain graphite anodes
Whole genome microarray analysis of the gene expression showed that genes encoding
biofilm formation were significantly up-regulated as response to nanoparticle decorated
anodes Increased expression of genes related to nanowires flavins and c-type
cytochromes indicate that enhanced mechanisms of electron transfer to the anode may
also have contributed to the observed increases in current density The majority of the
remaining differentially expressed genes were associated with electron transport and
anaerobic metabolism demonstrating a systemic response to increased power loads
Keywords Microbial electrochemical system microbial fuel cell microbial electrolysis
cell nanotechnology differential gene expression DNA microarray
47
Introduction
Microbial electrochemical systems (MESs) have been intensively studied since Lewis
achieved practical advances in this field (Logan 2007) however they attracted much
research attention in recent years due to their promising applications in renewable energy
generation bioremediation and wastewater treatment In a MES microorganisms
interact with electrodes via electrons catalyzing oxidation and reduction reactions at the
anode and the cathode The most-described type of MESs is microbial fuel cells (MFCs)
in which useful power is generated from electron donors typically biodegradable organic
materials (Logan et al 2006) Various novel MESs have recently been developed to
produce hydrogen (microbial electrolysis cells (MECs)) (Liu et al 2005 Logan et al
2008 Rozendal et al 2006 Schaller et al 2009) to power an autonomous sensor or
sensor network (Ringeisen et al 2006) to reduce bioremediation targets (Lovley 2006
Shea et al 2008) Sukkasem et al 2008) and to desalinate water (Cao et al 2009) The
key feature shared by these systems is the microbe-catalyzed electron transfer from
organic matter to electrodes (anodes) (Rabaey et al 2004) Enhancing the anodic current
output which highly depends on the performance of the electrodes is critical for the
successful application of all these processes (Logan et al 2007 Park and Zeilus 2002
2003)
Nanomaterials have received much attention from researchers in the context of
microbiology due to their unique physical electrical and chemical properties which
facilitate the study of interactions between bacteria and surfaces (Liu 2006) Previous
studies have demonstrated that electrodes decorated with different nanostructures such
as carbon nanotubes nanostructured polyanilinetitanium dioxide composite and titania
48
nanotubes have the potential to enhance power generation in MFCs (Morozan et al 2007
Qiao et al 2008 Quan et al 2005) Furthermore our recent study shows nanoparticle
(NP) decorated anodes greatly increased the electrochemical electron transfer rate in
MFCs (Fan et al 2011) However Fe NPs as an ideal candidate for decorating electrodes
because of respectively low price and high conductivity compared to other materials has
not been focused to study in MECs
S oneidensis MR-1 an important electrochemically active bacterial strain has been
exploited for the electricity generation in MFCs (Logan et al 2007 Logan 2009 Park
and Zeikus 2002) The availability of genome sequence for this strain makes it possible to
use transcriptome assays to globally measure the responses to different growth conditions
and environmental stresses (Murray et al 2001) S oneidensis MR-1 gene expression
response to heat shock cold shock Cr (VI) and U (VI) reduction chromate stress and
iron and acid tolerance have been studied in the past several years (Bencheikh-Latmani et
al 2005 Brown et al 2006 Gao et al2006 Gao et al 2004 Yang et al 2008) However
no studies have been focused on the Shewanella gene expression response to NPs in
MESs
The mechanism of increased electron transfer rate exhibited by nanoparticle decorated
electrodes is not well understood yet In the present study graphite disks decorated with
Fe NPs were used as anodes to explore the effects of nanostructures on current generation
in a multi-anode MECs DNA microarrays were utilized to investigate differences in the
global gene expression profile of S oneidensis MR-1 grown on plain versus Fe NPs
decorated anodes
Materials and methods
49
Bacterial cultures
S oneidensis MR-1 was purchased from American Type Culture collection (ATCC
700550) and stored as freezer stocks at -80degC Stock cells of S oneidensis MR-1 was
grown aerobically using a medium containing 30 gL Trypticase Soy Broth (BD 211825
Becton Dickinson Company Franklin lakes NJ) at 30degC Cells in early stationary
growth phase (optical density of 15-16 at 600nm (Spectrophotometer UV-1700
Shimadzu Columbia MD) were washed two times then injected into the chamber of the
MECs for current production Trypticase Soy Broth medium with 30 mM sodium lactate
as electron donor was used in MECs Phosphate buffer (100mM) was used to maintain
the solution pH 7 and solution conductivity at 15 mScm
Characterization of nanostructured anodes
Superfine isomolded graphite disks (16 cm x 05 cm GraphiteStorecom Inc) were
polished with ultrafine sand paper (2000 grit 3M Company) as the base for the NP
decorated and control anodes Fe NPs decorated anodes were fabricated by thermal
annealing of metal thin film coated graphite disks Thin films of Fe were firstly deposited
on the polished graphite disk by using sputter coating for 95 min and the samples then
were annealed in the Chemical vapor deposition (CVD) at 750degC for 20 min to form Fe
NPs Fe-NPs were produced with a diameter of 200plusmn100 nm and a density range
(distance between NPs) of 200plusmn100 nm with a coating time of 95 min SEM images of
Fe NPs decorated anode and control surfaces were shown in the Figure 41
50
Figure 41 SEM images of Fe NPs decorated anodes (A) Graphite disk control (B) Fe
Nanoparticle decorated
Multiple channel MECs construction and operation
A MECs with removable multiple anodes with each effective anode area 07 cm2
was
constructed and used to evaluate the effects of nanostructure on current density of
according to a previous report (Fan et al 2011) The cathode was made of wet-proof
(30) carbon cloth (type B E-TEK Division Inc USA) coated with PtC (20 E-TEK
Division Inc USA) following a previously reported procedure (Liu et al 2005) The
final platinum loading was 05 mgcm2 per projected cathode area The size of cathode
(150 cm2) was 25 times larger than the total effective area of the 8 graphite disk anodes
(56 cm2) to prevent cathode limitations on the performance of the MECs system All
MECs with Fe-NP decorated anodes and control anodes were tested for current
generation for 24 h with medium (without bacteria) before injecting the bacterial cells in
order to determine whether the Fe-NP decorated anodes demonstrate chemical current
generation compared to control anodes All the testing anodes were set up in the MECs
B
500 nm
A
500 nm
51
at once so that the S oneidensis MR-1 culture was applied to all of them simultaneously
Short electrode spacing (17 cm) was used in the MECs design to reduce the internal
resistance A voltage of 06 V was applied the MECs for current generation after the cells
of S oneidensis MR-1 were inoculated into MECs at early stationary phase A multimeter
with a data acquisition system (2700 Keithly USA) was used to monitor the current
change by measuring the voltage (every 300 seconds) drop through a resistor at 300 ohm
Fig 4 2 Effects of Fe-NP decorated anodes on the current density in MECs Control is
plain graphite disk anode Fe-NP is the anode with Fe-NP decoration
Microarray analysis
Biofilms for whole gene microarray analysis were aseptically removed from the plain
and Fe-NP decorated anodes of MECs after 110 hours further incubation at 30oC when
current density obviously started to decease Total RNA was extracted using Trizol
(Ambion inc AustinTX) following the manufacturerrsquos instructions Integrity of the
52
RNA samples was confirmed using the Agilent 2100 Bioanalyzer (Agilent Technology
Inc Palo Alto CA) The same amount of RNA (200 ng) from each sample was used for
further analysis after amplification by using the MessageAmpTM
II-Bacteria Prokaryotic
RNA Amplification kit (Ambion AustinTX) according to the manufacturerrsquos
instructions All of the 4295 genes (resource from NCBI) represented on the S oneidensis
MR-1 whole-gene microarrays were ordered from Roche NimbleGen Inc (Madison WI)
Biological triplicates of biofilms on the Fe-NP decorated anodes and control were
analyzed respectively cDNA synthesis labeling and hybridization were carried out by
the CGRB Core Laboratories at Oregon State University DNASTAR ArrayStarTM
3
software was used to identify genes that were up- or down-regulated more than 2-fold
when grown on the nanoparticle modified anodes using the unpaired two sample t-test
with a cutoff p-value of 005 The complete microarray data set generated in this study is
deposited for public access in the Gene Expression Omnibus
(httpwwwncbinlmnihgovgeo) under accession number GSE31535
Results
Enhancement of current generation by using NP decorated anodes in MECs
The current density generated by Fe-NP decorated anode MECs with S onidensis MR-1
increased to approximately 43 microAcm2 20 h following inoculation and then slowly
decreased Figure 42 shows that the maximum current density achieved by NP
decorated anodes was 83 times higher than that (51 microAcm2) generated by the control
(plain graphite disk) The average current density improvement of 110 hours was 59
times of that generated by the control The current densities generated in MECs in the
53
absence of bacteria were negligible (03 microAcm2) which demonstrated that the current
enhancement observed with the Fe-NP decorated anodes was biologically-derived
Global transcriptome analysis
Whole-genome DNA microarrays were used to attain a comprehensive general
overview
of the transcriptional response to Fe-NP decorated anodes by S oneidensis MR-1 in MEC
Of a total 4295 genes assayed 392 (188 induced and 204 repressed) exhibited significant
(P lt 005) differential expression at a 2-fold (log (expression ratio 1) level in 3
replicates in response to Fe-NPs These
total gene numbers present 9 of the 4295 open
reading frames (ORFs) presented on the array
Fig 43 Differentially expressed genes grouped by functional classification according to
the TIGR S oneidensis genome database (wwwtigrorg) Columns 1 amino acid
biosynthesis 2 biosynthesis of cofactors prosthetic groups and carriers 3 cell
envelope 4 cellular processes 5 central intermediary metabolism 6 DNA metabolism
7 energy metabolism 8 fatty acid and phospholipid metabolism 9 other categories 10
protein fate 11 protein synthesis 12 purines pyrimidines nucleosides and nucleotides
13 regulatory functions 14 signal transduction 15 transcription 16 transport and
binding proteins 17 unknown function 18 hypothetical proteins
Up-Regulated
-75
-50
-25
0
25
50
75
100
125
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18
Down-Regulated
Num
ber
of
Gen
es
54
Figure 43 summarizes the overall genomic response of S oneidensis MR-1 to Fe-NP
decorated anodes by grouping the differentially expressed genes into their functional role
categories based on The Institute for Genomic Researchrsquos annotation (Version 8) of the
MR-1 genome sequence The wide distribution of putative functional roles attributed to
the differentially expressed genes indicated the extent of the molecular response of
Soniedenis MR-1 to the NP decorated anodes Among genes with defined function a
large number of up-regulated genes were associated with cellular processes (group 4)
energy metabolism (group 7) other categories (group 9) and hypothetical proteins (group
18) While those genes encoding proteins involved in amino acid biosynthesis (group 1)
cellular processes (group 4) substrate transport (group 16) and hypothetical proteins
(group 18) were among the most down-regulated genes Two groups of genes that were
the most noticeable among the functional gene groups genes related to energy
metabolism (group 7) and genes related to amino acid biosynthesis (group 1) The ratios
of up-regulated genes to down-regulated genes in these functional groupings were much
higher than other functional gene groupings there were 21 up-regulated genes and 7
down-regulated genes in energy metabolism gene group and 2 up-regulated and 22 down
regulated genes in the amino acid biosynthesis group
Genes related to biofilm formation
In order to determine whether there was a correlation between the observed biofilm
enhancements by NP decorated anodes and the genes related to biofilm formation the
significant modified genes related to biofilm formation were examined and summarized
in the Table 41 These included SO_3223 and SO_3228 genes encoding the flagellum
proteins (Thormann et al 2004) which has critical impacts on initial attachment to the
55
surface as swimming motility functions were up-regulated over 10 and 09 fold
respectively The gene clusters SO_4178-SO_4180 which are thought to be essential for
formation of a three-dimensional biofilm were up-regulated from 12 to 13 fold
(Thormann et al 2004) In addition the genes SO_4105 encoding IV pilus which is
known to represent a critical factor for biofilm formation on a biotic surfaces (Thormann
et al 2004) were also induced 08 fold (Table 41) The genes related to flagellum
motility (SO_4286 SO_1529 and SO_1530) were also induced slightly (Thormann et al
2004)
Table 41 Expression levels of genes related to biofilm formation
Locus Tag Gene product descriptions Fold change
SO_3228 flagellar basal-body MS-ring and collar protein FliF 10
SO_3223 flagellar hook-length control protein FliK 09
SO_4103 MshA minor pilin protein MshD 05
SO_4105 MSHA major pilin protein MshA 08
SO_4178 expressed protein of unknown function MxdC 12
SO_4179 inner membrane family 2 glycosyltransferase MxdB 13
SO_4180 diguanylate cyclase-like protein MxdA 10
a fold changes were presented after log2
b positive number represented up-regulated and negative represented down-regulated
Genes related to energy metabolism
A closer consideration of energy metabolism genes was undertaken as anaerobic
metabolism for electron generation and electron transport functions may play critical
roles in enhanced current density generation in MECs Genes associated with energy
56
metabolism and with significantly modified gene expression levels are summarized in the
Table 42 Several of the up-regulated genes were related to formate dehydrogenase
proteins Most notably four genes related to formate dehydrogenase were up-regulated
more than 2-fold SO_0102 SO_0103 SO_0104 and SO_0107 The other genes
expression related to formate dehydrogenase proteins SO_4513 SO_4514 SO_4515 also
significantly increased Another interesting gene is that encoding the cytochrome c
oxidase protein gene SO_4609 which was up-regulated 11 fold while the gene
SO_0479 related periplasmic octaheme cytochrome c MccA was down-regulated
However SO_1519 the gene encoding lactate dehydrogenase was down-regulated 12
fold despite lactate being the primary carbon source in the media
Table 4 2 Genes related to anaerobic growth and electron transfer with significantly change expression level
Locus Tag Gene product descriptions Fold
change
SO_0101 nitrate-inducible formate dehydrogenase molybdopterin-binding
subunit FdnG
18
SO_0102 formate dehydrogenase nitrate-inducible iron-sulfur subunit 21
SO_0103 formate dehydrogenase nitrate-inducible cytochrome b556 subunit 24
SO_0104 formate dehydrogenase accessory protein FdhE 22
SO_0107 formate dehydrogenase accessory protein fdhD 24
SO_0397 quinolfumarate reductase menaquinol-oxidizing subunit FrdC_2 12
SO_0452 thioredoxin 2 15
SO_0903 Na-translocating NADH-quinone reductase subunit B NqrB_1 12
SO_1012 NADH-ubiquinone oxidoreductase subunit K NuoK 12
SO_1013 NADH-ubiquinone oxidoreductase subunit J NuoJ 12
SO_1363 hydroxylamine reductase 13
57
SO_2417 ferredoxin cofactor maintenance protein YfaE 10
SO_3922 formate dehydrogenase cytochrome b Fdh -12
SO_4513 formate dehydrogenase molybdopterin-binding subunit _2 15
SO_4514 formate dehydrogenase FeS subunit FdhB_2 12
SO_4515 formate dehydrogenase cytochrome b subunit FdhC_2 14
SO_4609 aa3 type cytochrome c oxidase subunit III CoxC 11
SO_0479 periplasmic octaheme cytochrome c MccA -12
SO_0847 periplasmic nitrate reductase ferredoxin component NapG -15
SO_1519 L-lactate dehydrogenase iron-sulfur cluster-binding protein LldF -10
SO_1251 ferredoxin 4Fe-4S -11
SO_37411 hypothetical inner membrane protein -11
a fold changes were presented after log2
b positive number represented up-regulated and negative represented down-regulated
Flavin and cytochrome related genes
Flavins can be secreted by Shewanella species as electron shuttle to facilitate
extracellular electron transfer (Cansterin et al 2008) C-type cytochromes play the
important roles on the process of extracellular electron transfer (Shi et al 2007) The
genes related to flavins synthesis and the genes encoding the cytochromes electron
transport proteins were shown in the table 43 the genes SO_1414 and SO_3058 related
to flavins synthesis was up-regulated to 03 and 01 folds respectively The genes related
to s C-type cytochromes SO_0135 SO_4105 SO_0169 SO_1778 are increased to 10
08 05 04 01 folds respectively However most of genes have shown no significant
changes (less one-fold change) as response to nano particle decorated anode in MECs
58
Table 43 Expression level of flavin and cytochrome genes
Locus Tag Gene product descriptions Fold
change
SO_1414 flavocytochrome c flavin subunit putative 03
SO_3468 riboflavin synthase subunit alpha -02
SO_3058 flavocytochrome c flavin subunit 01
SO_4105 MSHA pilin protein MshA 08
SO_0169 general secretion pathway protein GspG 05
SO_1778 outer membrane decaheme cytochrome c lipoprotein MtrC 04
SO_1779 outer membrane decaheme cytochrome c OmcA 01
SO_0135 lipoprotein of unknown function DUF333 10
SO_0136 conserved hypothetical inner membrane protein 01
a fold changes were presented after log2
b positive number represented up-regulated and negative represented down-regulated
Other genes with significantly modified expression levels
These genes over 15 fold change and possibly related to current enhancements were
shown in Table 44 SO_3104 (expressed inner membrane protein) was over expressed
and the gene SO_2194 which encodes outer membrane protein (OmpA family protein)
was repressed as response to nanoparticle decorated anode however it is unclear the
relationships of this modified membrane protein genes with the current enhancement
Another interesting phenomenon was possible co-regulation of several gene clusters
indicating possible operon associations This includes three sets of genes that were
induced more than 15 fold SO_0108SO_0109 and SO_1042SO_1043SO_1044 The
59
consistency of expression of these genes under the NP conditions provides basic evidence
to support operon structure However the correlation of these significant changed gene
with current density enhance are unclear which showed there are unknown multiple and
complex responses of S oneidensis MR-1 to nanoparticle anode of MECs
Table 44 Other genes with significantly changed expression levels
Locus Tag Gene product descriptions Fold change
SO_0101 nitrate-inducible formate dehydrogenase molybdopterin-binding
subunit FdnG
18
SO_0108 integral membrane protein of unknown function DUF39 YedE 17
SO_0109 SirA family protein YedF 17
SO_0275 N-acetyl-gamma-glutamyl-phosphate reductase -31
SO_0277 ornithine carbamoyltransferase -21
SO_0279 argininosuccinate lyase -24
SO_0404 zinc dependent metalloprotease domain lipoprotein 21
SO_0956 alkyl hydroperoxide reductase F subunit -20
SO_1042 amino acid ABC transporter ATP-binding protein -18
SO_1043 amino acid ABC transporter permease protein -27
SO_1044 amino acid ABC transporter periplasmic amino acid-binding protein -27
SO_1072 chitin-binding protein putative 21
SO_1405 transglutaminase family protein 20
SO_1822 TonB-dependent receptor putative -22
SO_2069
1-(5-phosphoribosyl)-5-[(5-phosphoribosylamino)
methylideneamino] imidazole-4-carboxamide isomerase
-19
SO_2070 amidotransferase HisH -26
SO_2071 imidazole glycerol-phosphate dehydratasehistidinol phosphatase -27
60
SO_2072 histidinol-phosphate aminotransferase -28
SO_2073 histidinol dehydrogenase -30
SO_2194 OmpA family protein -36
SO_2195 inter-alpha-trypsin inhibitor domain protein -27
SO_2767 asparagine synthetase B -26
SO_2945 prophage LambdaSo tail fiber protein 20
SO_2963 prophage LambdaSo major capsid protein HK97 family 20
SO_2987 prophage LambdaSo hypothetical protein 19
SO_2988 prophage LambdaSo expressed protein of unknown function 18
SO_3104 expressed inner membrane protein 131
SO_3408 conserved hypothetical inner membrane protein -18
SO_3585 azoreductase putative -18
SO_3586 glyoxalase family protein -17
SO_3687 curli production assemblytransport component CsgE putative -18
SO_38191 hypothetical ammonia permease -19
SO_4014 AcrBAcrDAcrF family protein 18
SO_40151 type I secretion system membrane fusion protein RND family 18
SO_4054 510-methylenetetrahydrofolate reductase 18
SO_4245 N-acetylglutamate synthase -22
SO_45251 hypothetical transcriptional regulator LysR family -17
SO_4527 integral membrane domain protein -19
SO_4705 transcriptional regulator putative -19
a fold changes were presented after log2
b positive number represented up-regulated and negative represented down-regulated
Discussion
61
The addition of NP to anodes significantly impacts current densities with the elemental
composition being a critical factor The chemical composition of NPs significantly affects
the current enhancement in MECs (Fan et al 2011) because the chemical composition of
NP properties is a critical factor in determining the conductivity which significantly
affects the efficiency of electron transfer from bacteria to electrode Fe is an ideal
candidate for NP formation for current enhancement in MESs because of high
conductivity and low price compared to gold and other conductive materials Gold NPs
has also been studies for enhancement of electricity conductivity (Bao et al 2008)
However the disadvantages of the high price of gold limit their practical application in
MECs (Fan et al 2011) The different size and density of the same composition of NP
also has essential effects on the current density enhancement (Fan et al 2011) In this
study the results showed the current density enhancement with Fe-NP size range of
200plusmn100 nm and density range of 200plusmn100 nm is considerable enhancement consequence
There are a number of reports of studies of current density enhancement in MFCs
by nanostructured anodes (Fan et al 2011 Qiao et al 2008 Sharma et al 2008 Tsai et al
2009) Based on bacterial behavior on nanostructured anode surface Qiao et al (2008)
believes that Escherichia coli cells on the nanostructured electrode surface produce hair-
like structures similar to pili that could facilitate the electron transfer between the cells
and electrode The production of hair-like structures is believed to be stimulated by
nanostructures and could play the key role on current density enhancement in pili-
producing bacteria in MFCs (Qiao et al 2008) S oneidensis can produce electrically
conductive bacterial nanowires which have similar structure to the hair-like structures in
Ecoli (Gorby et al 2006 Reguera et al 2005) A number of possible genes related
62
nanowires of S oneidensis have been identified including mshA (SO_4105) and gspG
(SO_0169) (Konstantinidis et al 2009) They are up-regulated 08 and 05 fold in the
current study respectively even though they are not significant induced This result
supported the increased nanowire genes had contributions to enhance current density in
MFCs by NPs decorated anodes
Based on the previous research on electron transfer mechanisms for S oneidensis it is
known that self-produced mediators (flavins) and multihaem c-type cytochromes (c-Cyts)
may play critical roles in the electron transfer of S oneidensis to extracellular insoluble
electrode (Canstein et al 2008 Myers and Myers 1992 Scott and Nealson 1994 Shi et al
2007) The flavins secreted by S oneidensis can facilitate to transfer electrons from
bacteria to solid anode electrodes by acting as extracellular electron shuttles (Canstein et
al 2008) In this study the genes encoding flavins (SO_1414 SO_3468 and SO_3058)
had no significant increase (less than 1 fold change) as a response to nanoparticle
decorated anode in MECs which suggests that increased flavin gene expression does not
significantly contribute to the current density enhancements in nano-particle decorated
anode The genes encoding cytochromes electron transport proteins (SO_4104 SO_0417
SO_177879 SO_013536) had no significant increase (less than 1-fold change)
However those genes were up-regulated slightly These results suggest the increased
expression flavin and c type cytochromes genes had partial contributions even not
significantly to enhance current density in MFCs by NPs decorated anodes
The thickness of bacterial biofilm on the anode can affect the power generation in
MFCs (Biffinger et al 2007 Heydorn et al 2000) since increased biofilm thickness on
the anode represents more active bacteria involving in the process of electron generation
63
and transfer in MFCs Biofilm formation related genes included flagellar genes IV pilus
genes and other genes The flagellar related genes SO_3228 SO_3229 (fliB and fliK) and
IV pilus genes SO_4105 (mshA) were induced from 19 to 09 fold the genes (SO_4178
-4180) related to formation of a three-dimensional biofilm were up-regulated from 10 to
13 folds even though the genes related bacterial motility protein (SO_ 4286 SO_4287
SO_1529 and SO_1530) are induced slightly (less than 0 5 fold) However the motility
genes most probably play a role for initial biofilm attachment on the anode surfaces
(Thormann et al 2004 Thormann et al 2006) with the rest of biofilm genes being more
essential for biofilm maintenance in mature cultures as were presented at harvest in this
study This enhancement of biofilm density under conditions of increase current density
has been observed for gold NP decorated anodes using confocal light microscopy (data
not shown) but has not yet been confirmed for Fe NP decorated anodes In any event an
increase in the expression level of genes related biofilm formation therefore also
supported the possibility of enhanced biofilm formation on NPs decorated anodes thus
facilitating electron transfer from bacteria to anodes
Beliaev et al described a model of the Fe reduction pathway in S oneidensis MR-1 using
solid substrates as the electron acceptor (Beliaev et al 2001) This model proposes that
electrons are generated and released in cytoplasm then transferred to the quinone pool
eg menaquinone in the cytoplasmic membrane The reduced menaquinone in turn
reduces CymA and from there electrons are then transferred to a periplasmic shuttle (eg
CctA) and then to MtrA a periplasmic soluble decaheme cytochrome c located in
associated with the outer membrane via interaction with MtrB Finally surface displayed
outer membrane proteins decaheme cytochromes (OmcA MtrC or MtrF) can transfer
64
electron to Fe or other solid substrates (Beliaev et al 2001)) Bencheikh-Latmani et al
demonstrated that the same electron transport pathway may be used for more than one
electron acceptor through studies of the response of S oneidensis MR-1 to U(VI) and
Cr(VI) under anaerobic conditions particularly focused on the critical genes
(SO_1777mtrA SO_1776mtrB SO_1778mtrC) (Bencheikh-Latmani et al 2005)
However our studies showed that these genes had no significant expression changes in
response to nanostructured anodes Contrastingly one of expressed inner membrane
protein genes SO_3104 and one of cytochromes genes SO_4609 encoding cytochrome c
oxidase subunit III demonstrated a 13-fold and 11-fold up-regulation respectively in this
study while the gene SO_2194 which encodes outer membrane protein (OmpA family
protein) was down regulated 36 fold Our results therefore suggest that S oneidensis
MR-1 does not utilize the same set of core critical genes (SO_1777 mtrA SO_1776
mtrB SO_1778 mtrC) products to transfer electrons in all instances This is the similar
result with the study of Bretschger et al (2007) which indicated that the electron
transport system in S oneidensis MR-1 is complex with several different proteins able to
participate in electron transfer to the anode of MECs
It should also be mentioned that the gene (SO_1519) encoding lactate
dehydrogenase repressed (-12 fold) here suggesting that lactate was depleted at the time
of cell harvest from the MECs and that other pathways of anaerobiosis had been activated
including those associated with amino acid uptake and consumption the growth media
was a complex mixture including yeast extract In particular the catabolic pathway for
histidine of S oneidensis MR-1 a pathway that may formate as an intermediate is up-
regulated with the genes for histidine ammonia lyase SO_0098 SO_3057 and SO_4374
65
up-regulated 13 11 and 11-fold respectively The possible production of formate
during histidine degradation may in turn explain the high levels of formate
dehydrogenates as evidenced by upregulation of SO_0102 SO_0103 SO_0104 and
SO_0107 annotated as formate dehydrogenase proteins These genes had the highest
levels of up-regulated expression as response to Fe NP decorated anodes (greater than 2-
fold)
Acknowledgements
This research was partially supported by the US National Science Foundation CBET
0828544 and the funds from ONAMIDOD (ARL-DOD Cooperative Agreement
W911NF-07-2-0083) We thank Barbara Gvakharia and Caprice Rosato for helpful
suggestions We also thank Margaret Romine of Pacific Northwest Laboratories for
valuable comments on manuscripts We also thank anonymous reviewers for significant
suggested improvements imparted as part of prior review of this manuscript
66
Chapter 5
Global Transcriptome Analysis of the Response of Shewanella oneidensis MR-1 to
Carbon Nanotube Decorated Anodes in Microbial Electrochemical Systems
Shoutao Xu Yanzhen Fan Rebecca Schaller Frank Chaplen Jun Jiao Hong Liu
67
Abstract
Shewanella oneidensis MR-1 is an important model microorganism for metabolic studies
on the effects of different environmental factors because of its diverse respiratory
capabilities Carbon nanotube (CNT) modified graphite disks were used as anodes to
investigate the effects of nanomatrials on the performance of S oneidensis MR-1 in
microbial electrolysis cells (MECs) The current densities produced with CNT decorated
anodes were on average 56-fold higher than plain graphite anodes Whole genome
microarray analysis of gene expression showed that up-regulation of cytochromes c genes
associated with extracellular electron transfer are strongly correlated to current increases
in CNT decorated anodes Genes involved in flavin biosynthesis also contribute to
current increase in CNT decorated anode MECs
Keywords Microbial electrochemical system microbial fuel cell microbial
electrolysis cell carbon nanotube gene expression DNA microarray
68
Introduction
Shewanella oneidensis MR-1 is an important model microorganism for metabolic
studies of the effects of different environmental factors because of its diverse respiratory
capabilities It has been used for transcriptome analysis to investigate the responses to
different growth conditions and environmental stresses (Murray et al 2001) Gene
expression patterns under different conditions such as heat shock cold shock Cr (VI) U
(VI) reduction chromate stress iron and acid tolerance have been studied previously
(Bencheikh-Latmani et al 2005 Brown et al 2006 Gao et al2006 Gao et al 2004
Yang et al 2008) More recently it has been exploited as a model species for power
generation in microbial electrochemical systems (MESs)which have potential
applications in renewable energy generation bioremediation and wastewater treatment
(Logan et al 2007 Logan 2009 Park and Zeikus 2002 Fan et al 2011 Xu et al 2012)
In a MES electrochemically active microorganisms oxidize organic matter in the an
ode chamber to release electrons Electrons are then transferred to the anode electrode thr
ough various mechanisms (Logan et al 2006 Lovley 2006 Rabaey et al 2003) and fina
lly travel to the cathode electrode and combine with the terminal electron acceptor The
key feature of MESs is the microbe-catalyzed electron transfer from the organic matter to
the anode (Rabaey et al 2004) Enhancing the current output which highly depends on
the performance of the anode electrode is critical for the successful application of MESs
(Logan et al 2007 Park and Zeilus 2002 2003) Several nanomaterials including TiO2
gold (Au) and palladium (Pd) nanoparticles have been utilized to decorate electrodes to
enhance power density in MESs (Qiao et al 2008 Quan et al 2005 Fan et al 2011) The
possible mechanisms for increased current densities have been studied by using Fe
69
nanoparticle-decorated anodes (Xu et al 2012)
Carbon nanotubes (CNT) are cylindrically shaped nanomaterials with extremely
high surface area excellent electrical conductivity and chemical inertness (He et al
2005a Serp et al 2003) These unique properties make CNT a promising electrode
material (Liang et al 2008) The biocompatibility of microorganisms and carbon
nanostructures has been evaluated (Morozan et al 2007) The effects of CNT on anodic
biofilms have been tested in MFCs (Liang et al 2011) However no studies have been re
ported on global transcriptome analysis for the response of S oneidensis MR-1 to CNT
decorated anode in MECs
In the present study the graphite disks decorated with CNT were used as anodes to
investigate the effects of nanomaterials on current generation in multi-anode MECs
DNA microarrays were used to analyze differences in the global gene expression profile
of S oneidensis MR-1 grown on plain versus CNT decorated anodes
Materials and methods
Bacterial cultures S oneidensis MR-1 was purchased from American Type Culture
collection (ATCC 700550) and stored as freezer stocks at -80degC Stock S oneidensis
MR-1 cells were grown aerobically using M1 medium at 30degC Cells in early stationary
growth phase (optical density of 15-16 at 600nm (Spectrophotometer UV-1700
Shimadzu Columbia MD) were injected into the chamber of the MECs for current
production Sodium lactate (final concentration 30mM) was added as the additional
electron donor
Fabrication and characterization of nanostructured anodes Superfine isomolded
graphite disks (16 cm x 05 cm GraphiteStorecom Inc) were polished with ultrafine
70
sand paper (2000 grit 3M Company) as the base for the CNT decorated and control
anodes Aligned multi-walled carbon nanotube samples were synthesized via plasma
enhanced chemical vapor deposition (PECVD) Samples superfine isomolded graphite
pillars with a 16 cm diameter and 05 cm thickness were sputtered with a thin film of Ni
in a CRC 300 magnetron sputter coater for 1 to 2 min They were then placed in the
PECVD reactor where the Ni was annealed into small catalyst islands C2H2 was bled
into the chamber for growth and an NH3 plasma was used to vertically align the growth
of the multi-walled carbon nanotubes (MWCNTs) The temperature and time were varied
between 725degC and 775oC and 15 to 60 min to control the length of the tubes and density
of samples Samples were then analyzed in a FEI Sirion field emission scanning electron
microscope (FESEM) SEM images of CNT decorated anode and control surfaces were
shown in the Figure 51
Figure 51 SEM images of CNT decorated anodes (A) Graphite disk control (B) CNT
decorated anode
Multiple channel MECs construction and operation A MECs with multiple
removable anodes each with an effective anode area of 07 cm2
was constructed and used
to evaluate the effects of nanostructures on the current density of according to a previous
B A
71
study (Fan et al 2011) The cathode was made of wet-proof (30) carbon cloth (type B
E-TEK Division Inc USA) coated with PtC (20 E-TEK Division Inc USA)
following a previously reported procedure (Liu et al 2005) The final platinum loading
was 05 mgcm2 per projected cathode area The size of the cathode (150 cm
2) was 25
times larger than the total effective area of the 8 graphite disk anodes (56 cm2) to prevent
cathode limitations on the performance of the MECs system All MECs with CNT
decorated anodes and control anodes were tested for current generation for 24 h with
sterile a medium (without bacteria) before injecting the bacterial cells in order to
determine whether the CNT decorated anodes demonstrate chemical current generation
compared to control anodes All the testing anodes were set up in the MECs
simultaneously so that the S oneidensis MR-1 culture was applied to all of them
simultaneously Short electrode spacing (17 cm) was used in the MECs design to reduce
the internal resistance A voltage of 06 V was applied the MECs for current generation
after the cells of S oneidensis MR-1 were inoculated into the MECs in the early
stationary phase A multimeter with a data acquisition system (2700 Keithly USA) was
used to monitor the current change by measuring the voltage drop through a resistor
Microarray analysis Biofilms for whole gene microarray analysis were aseptically
removed from the plain and CNT decorated anodes of MECs after 80 hours further
incubation at 30oC when current density obviously reached to a stable phase Total RNA
was extracted using Trizol (Ambion inc AustinTX) following the manufacturerrsquos
instructions Integrity of the RNA samples was confirmed using the Agilent 2100
Bioanalyzer (Agilent Technology Inc Palo Alto CA) Total RNA (200 ng) from each
sample was amplified using the MessageAmpTM
II-Bacteria Prokaryotic RNA
72
Amplification kit (Ambion AustinTX) according to the manufacturerrsquos instructions The
S oneidensis MR-1 whole-gene microarrays representing all of the 4295 genes (resource
from NCBI) were ordered from Roche NimbleGen Inc (Madison WI) Biological
triplicates of biofilms on the CNT decorated anodes and control were analyzed cDNA
synthesis labeling and hybridization were carried out by the CGRB Core Laboratories at
Oregon State University DNASTAR ArrayStarTM
3 software was used to identify genes
that were up- or down-regulated more than 2-fold when grown on the nanoparticle
modified anodes using the unpaired two sample t-test with a cutoff p-value of 005 with
bonferroni correction
Results and discussion
Enhancement of current generation using CNT decorated anodes in MECs The
current density generated by CNT decorated anode MECs with S onidensis MR-1
increased to 36 microAcm2 10 hours post-inoculation and reached a maximum of 40 microAcm
2
at 29 hours inoculation then gradually decreased The current density stabilized at
30microAcm2 after 78 h operation Figure 52 shows that the current density curve generated
by CNT decorated anode in MECs The average current density generated by CNT
decorated anodes was 56 times that of the control The current density generated in the
MECs without bacteria was negligible (data not shown) This result demonstrates that the
current enhancement observed with the CNT decorated anodes was biologically-derived
A
73
Figure 52 Effects of CNT decorated anodes on the current density in MECs (3 replicates)
Control is plain graphite disk anode CNT is the anode with CNT decoration (Error bars
represent standard deviation)
It has been reported that nanostructured decorated anodes have significant impacts on
current densities (Fan et al 2011 Qiao et al 2006 Sharma 2008 Tsai et al 2009) The
CNT decorated anodes had significantly enhanced current densities in MECs in this
study This result is consistent with previous studies Interestingly the current density
curve generated by CNT decorated anode MECs is quite different from the current
density curve generated by Au NP and Pd NP decorated anodes using same bacterial
species (Fan et al 2011) In the Au and Pd NP decorated anode MECs the current
density curve was not significantly increased in the beginning compared with the control
but gradually increased after the inoculation the current density reached the maximum
current density more than 50 hours after the inoculation However the current density
generated by CNT decorated anode MECs started to increase significantly immediately
after the inoculation It reached a maximum current density 29 hours post-inoculation
-1
6
13
20
27
34
41
48
0 10 20 30 40 50 60 70 80
Curr
ent
den
sity
(micro
Ac
m2)
Time (Hours)
Control
CNT
74
These results indicate that there might be different current density enhancement
mechanisms between CNT with other metal NP decorated anodes in MECs
Global transcriptome analysis Whole-genome DNA microarrays were used to attain a
comprehensive general overview of the transcriptional response of S oneidensis
MR-1 of
S oneidensis MR-1 to CNT decorated anodes in MECs In a total of 4295 genes analyzed
457 genes (250 genes up-regulated and 207 genes down-regulated) exhibited significantly
(P lt 005) changed expression at a 2-fold level in at least 3 replicates in the response to
CNT The total number of the regulated genes present 11 of the 4295 open reading
frames (ORFs) presented on the microarray The total number of genes at significant
expression levels in response to CNT decorated anodes is close to that to Fe NP decorated
anodes in MECs (392 9) ( Xu et al 2012)
Fig 53 Differentially expressed genes grouped by functional classification according to
the TIGR S oneidensis genome database (wwwtigrorg) Columns 1 amino acid
biosynthesis 2 biosynthesis of cofactors prosthetic groups and carriers 3 cell
envelope 4 cellular processes 5 central intermediary metabolism 6 DNA metabolism
7 energy metabolism 8 fatty acid and phospholipid metabolism 9 other categories 10
protein fate 11 protein synthesis 12 purines pyrimidines nucleosides and nucleotides
-80
-60
-40
-20
0
20
40
60
80
100
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18
Nu
mb
er
of
Ge
ne
s
Up-regulated
Down-regulated
75
13 regulatory functions 14 signal transduction 15 transcription 16 transport and
binding proteins 17 unknown function 18 hypothetical proteins
The overall transcript genomic response of S oneidensis MR-1 to CNT decorated
anodes was summarized in Figure 53 by grouping the differentially expressed genes into
their functional role categories based on The Institute for Genomic Researchrsquos annotation
(Version 8) of the MR-1 genome sequence The wide distribution of putative functional
roles attributed to the differentially expressed genes indicated the extent of the molecular
response of Soniedenis MR-1 to the CNT decorated anodes A large number of up-
regulated genes were presented in genes associated with biosynthesis of cofactors
prosthetic groups and carriers (group 2) cell envelope (group 3) protein fate (group 10)
protein synthesis (group 11) and purines pyrimidines nucleosides and nucleotides
(group 12) in this study The number of up-regulated genes was two times more than that
of down-regulated genes in each functional group Contrastingly in the genomic response
S oneidensis MR-1 to Fe NP decorated anodes (chapter 4) most of up-regulated genes
were associated with cellular processes (group 4) energy metabolism (group 7) other
categories (group 9) and hypothetical proteins (group 18) The down-regulated genes
presented in the amino acid biosynthesis (group 1) cellular processes (group 4) substrate
transport (group 16) and hypothetical proteins (group 18) While those genes encoding
proteins involved in amino acid biosynthesis (group 1) central intermediary metabolism
( group 5) DNA metabolism( group 6) fatty acid and phospholipid metabolism (group
8) and other categories (group 9) were among the most down-regulated genes in this
study However the two groups of genes that were the most noticeable between the
genomic response S oneidensis MR-1 to CNT and Fe NP decorated anodes which they
are the genes related to energy metabolism (group 7) and hypothetical proteins (group
76
18) These results indicated there are different and common genomic responses to S
oneidensis MR-1 on CNT and Fe NP decorated anodes in MECs
Electron transfer related genes Based on previous electron transfer mechanism
studies on Shewanella a serial of group proteins collectively described as the Mtr
pathway play the role to transfer electron rom the inner bacterial body to the outer
membrane (Beliaev et al 2001 Shi et al 2007 Brutinel and Gralnick 2012) In the Mtr
pathway firstly electrons generated by bacteria received by CymA a tetraheme c-type
cytochrome anchored in the inner membrane then electrons were transferred to MtrA a
periplasmic decaheme c-type cytochrome After that electrons were transferred from
MtrA to MtrC an outer-membrane decaheme c-type cytochrome is facilitated by MtrB a
non-heme containing integral outer-membrane proteinOuter-membrane decaheme c-type
cytochrome MtrC and OmcA are in change of electron transfer to exreacellular electron
acceptor (Shi et al 2007 Brutinel and Gralnick 2012 ) These genes encoding electron
transport proteins are shown in the table 51 The critical genes SO_1777 mtrA SO_1776
mtrB SO_1778 mtrC are unregulated 41 to 80 fold in response to CNT decorated
anodes other cytochrome c genes related to electron transfer in MtrA pathway also are
significantly increased in response to CNT decorated anodes These genes encoding outer
membrane proteins have different level increases supporting that the increase of
cytochrome c gene expression contributes to the current enhancements of MECs as
response to CNT decorated anodes
77
Table 51 Expression level of cytochrome c as response to CNT decorated anodes
Locus Tag Gene product descriptions Fold change CNT
SO_0165 general secretion pathway protein GspC 31
SO_0167 general secretion pathway protein GspE 45
SO_1776 outer membrane protein precursor MtrB 41
SO_1777 outer membrane decaheme cytochrome c
lipoprotein MtrA
80
SO_1778 outer membrane decaheme cytochrome c
lipoprotein MtrC
43
SO_1779 outer membrane decaheme cytochrome c
OmcA
63
SO_0135 lipoprotein of unknown function DUF333 22
SO_0136 conserved hypothetical inner membrane protein 24
Flavin related genes It is known that self-produced mediators play critical roles in the
electron transfer of S oneidensis to extracellular insoluble electrode (Canstein von et al 2
008 Myers et al 1992 Scott et al 1994 Shi et al 2007) It has been proved that the
flavins secreted by S oneidensis can facilitate electron transfer from bacteria to solid
anode electrodes by acting as extracellular electron shuttles (Cansterin von et al 2008) T
wo critical genes (SO_1414 and SO_3468) related to flavin synthesis were shown increa
sed significantly (more than 2 folds) increase in this study (Table 52) This result support
s the hypothesis that flavin production increases as the response to CNT decorated anodes
in MECs have significant contribution to the current density enhancements
78
Table 52 Expression levels of genes related to flavin synthesis as response to CNT
decorated anode
Locus Tag Gene product descriptions Fold change CNT
SO_1414 flavocytochrome c flavin subunit putative 22
SO_3468 riboflavin synthase subunit alpha 20
The amount of bacterial biofilm biomass on the anode can affect the power
generation in MFCs (Biffinger et al 2007 Heydorn et al 2000) Biofilm formation
related genes include flagellar gene IV pilus genes and other genes the flagellar related
genes (fliB and fliK) and IV pilus genes (mshA) were induced only from 105 to 134
fold as response of S oneidensis MR-1 to CNT decorated anodes in this study No
biofilm-associated genes presented significant changes in expression levels in this study
indicating there is no direct connections between biofilm enhancements to CNT
decorated anodes in this study This result is consistent with the Liang et al (2011)
conclusion that using CNT enhanced the electrochemical activity of anodic biofilms but
did not result in a significant increase of biomass in the anodic biofilms
Our microarray results showed two significantly up-regulated gene groups the
genes encoding proteins localized on the outer membrane and the genes involved in
flavin biosynthesis contributed to current density enhancement by CNT decorated anodes
Among 457 significantly changed genes to CNT in this study there are also a relatively
large number of genes encoding proteins with unknown functions which are either up- or
down-regulated in the response to CNT decorated anodes which indicated more
79
complicated responses of S oneidensis MR-1 to CNT decorated anodes in MECs and
further study is needed
80
Chapter 6
Summary
Microbial electrochemical systems (MESs) with a mixed culture initially inoculated
from Corvallis wastewater treatment plant have been studied for more than 6 years for
varying purposes including power generation hydrogen production heavy metal
removal and wastewater treatment Experiment results have shown that the mixed culture
is quite stable with excellent performance in MESs Our community analysis using
denaturing gradient gel electrophoresis (DGGE) and 16S rDNA clone library construction
suggests that the mixed culture is composed predominantly of Geobacter sp (66)
Arcobacter sp (12) and Citrobacter sp (11) These results not only improved our
understanding of the mixed culture community but also guided our studies on the
cultivation and isolation of the different bacterial species
Exoelectrogenic Citrobacter sp SX-1 was first isolated and characterized from a
MFC operated with the mixed culture Citrobacter sp SX-1 has been demonstrated to
produce electricity from wide range of different substrates including citrate acetate
glucose sucrose glycerol and lactosein MFCs Cyclic voltammetry analysis indicated
that membrane associated proteins may play an important role in facilitating the electrons
transferring from bacteria to electrode The strain SX-1 increased the known diversity of
power generating exoelectrogens and provided a unique bacterial species for study in
renewable energy generation and waste treatment
81
To enhance the electron transfer from bacteria to anode nano decorated anodes
including Fe NP and CNT were developed and characterized and evaluated in MECs
using Shewanella oneidensis MR-1 as a model species Both nanostructures have
significantly increased current density compared with the control Whole genome
microarray analysis elucidated the possible mechanisms of power enhancement in
response to these nano-decorated anodes
These results benefit to understanding of physiology and ecology of mixed
cultures in MFCs and improve the efficiency of current generation in MESs which will
facilitate the viability of niche applications for MESs in near future
82
Bibliography
1 Amman R Ludwig W (2000) Ribosomal RNA-targeted nucleic acid probes for
studies in microbial ecology FEMS Microbiol Ecol 24 555ndash565
1 Bao SJ Li CM Zang JF Cui XQ Qiao Y Guo J (2008) Novel Nanoporous TiO2
Electrode for Directly Electrochemistry of glucose oxidase Adv Funct Mater
18(4) 591-599
2 Beliaev AS Saffarini DA McLaughlin JL Hunnicutt D (2001) MtrC an outer
membrane decahaem c cytochrome required for metal reduction in Shewanella
putrefaciens MR-1 Mol Microbiol 39722-30
3 Bencheikh-Latmani R Williams SM Haucke L Criddle CS Wu L Criddle CS
Zhou J Tebo BM (2005) Global transcriptional profiling of Shewanella oneidensis
MR-1 during Cr(VI) and U(VI) reduction Appl Environ Microbiol 71 7453ndash7460
4 Biffinger JC Pietron J Ray R Little B and Ringeisen BR (2007) A biofilm
enhanced miniature microbial fuel cell using Shewanella oneidensis DSP10 and
oxygen reduction cathodes Biosens Bioelectron 221672ndash1679
5 Bodelier PLE Roslev P Henckel T FenzelP (2000) Stimulation by ammonium
based fertilizers of methane oxidation in soil around rice root Nature 40 421ndash424
6 Bond DR Holmes DE Tender LM Lovley DR (2002) Electrode-reducing
microorganisms that harvest energy from marine sediments Science 295 483-485
7 Bond DR Lovley DR (2003) Electricity production by Geobacter sulfurreducens
attached to electrodes Appl Environ Microbiol 69 1548-1555
8 Bond DR Lovley DR (2005) Evidence for involvement of an electron shuttle in
electricity generation by Geothrix fermentans Appl Environ Microbiol 71 2186-
2189
9 Bretschger O Obraztsova A Sturm CA Chang IS Gorby YA Reed SB Culley
DE Reardon CL Barua S Romine MF Zhou J Beliaev AS Bouhenni R
Saffarini D Mansfeld F Kim BH Fredrickson JK Nealson KH (2007) Current
production and metal oxide reduction by Shewanella oneidensis MR-1 wildtype and
mutants Appl Environ Microbiol 73 7003ndash7012
10 Brown SD Thompson MR Verberkmoes NC Chourey K Shah M Zhou J
Hettich RL and Thompson DK (2006) Molecular dynamics of the Shewanella
oneidensis response to chromate stress Mol Cell Proteomics 51054-1071
11 Brutinel ED Gralnick JA (2012) Shuttling happens soluble flavin mediators of
extracellular electron transfer in Shewanella Appl Microbiol Biotechnol 93(1) 41-8
12 Butler JE Kaufmann F Coppi MV Nuacutentildeez C Lovley DR (2004) MacA a diheme
c-type cytochrome involved in Fe(III) reduction by Geobacter sulfurreducens J
Bacteriol 186(12) 4042ndash4045
83
13 Caccavo F Lonergan DJ Lovley DR Davis M Stolz JF McInerney MJ (1994)
Geobacter sulfurreducens sp nov a hydrogen- and acetate oxidizing dissimilatory
metal-reducing microorganism Appl Environ Microbiol 60 3752ndash3759
14 Canstein von H Ogawa J Shimizu S Lloyd JR (2008) Secretion of flavins by
Shewanella species and their role in extracellular electron transfer Appl Environ
Microbiol 74 615ndash623
15 Cao X Huang X Liang P Xiao K Zhou Y Zhang X Logan BE (2009) A New
Method for Water Desalination Using Microbial Desalination Cells Environ Sci
Technol 43(18)7148-7152
16 Chaudhuri SK Lovley DR (2003) Electricity generation by direct oxidation of
glucose in mediatorless microbial fuel cells Nat Biotechnol 21 1229-1232
17 Cheng S Liu H Logan B E (2006) Increased power generation in a continuous
flow MFC with advective flow through the porous anode and reduced electrode
spacing Environ Sci Technol 402426-2432
18 Cheng S Logan B E (2007) Sustainable and efficient biohydrogen production via
electrohydrogenesis PNAS 104 18871-18873
19 Chignell J (2010) Energy Recovery from Biodiesel Waste Performance of
Microbial Electrochemical Systems on Glycerol Thesis
20 Chung K Okabe S (2009) Characterization of electrochemical activity of a strain
ISO2-3 phylogenetically related to Aeromonas sp isolated from a glucose-fed
microbial fuel cell Biotechnol Bioeng 104(5) 901-910
21 Cole JR Chai B Farris RJ Wang Q Kulam-Syed-Mohideen AS McGarrell
DM Bandela AM Cardenas E Garrity GM Tiedje JM (2007) The ribosomal
database project (RDP-II) introducing myRDP space and quality controlled public
data Nucleic Acids Res 35 (Database issue) D169-D172 doi 101093nargkl889
22 Cole JR Wang Q Cardena E Fish J Chai B Farris RJ Kulam-Syed-Mohideen
AS McGarrell DM Marsh T Garrity GM Tiedje JM (2009) The Ribosomal
Database Project improved alignments and new tools for rRNA analysis Nucleic
Acids Res 37 (Database issue) D141-D145 doi 101093nargkn879
23 Dey A De S De A De SK (2004) Characterization and dielectric properties of
polyaniline-TiO2 nanocomposites Nanotechnol 15 1277-1283
24 Edwards U Rogall T Bloumlcker H Emde M Boumlttger EC (1989) Isolation and
direct complete nucleotide determination of entire genesmdashcharacterization of a gene
coding for 16S-ribosomal RNA NucleicAcids Res 17 7843ndash7853
25 Eggleston CM Voumlroumls J Shi L Lower BH Droubay TC Colberg PJS (2008)
Binding and direct electrochemistry of OmcA an outer-membrane cytochrome from
an iron reducing bacterium with oxide electrodes a candidate biofuel cell system
Inorg Chim Acta 361769ndash777
26 Fan Y Sharbrough E Liu H (2008) Quantification of the Internal Resistance
Distribution of Microbial Fuel Cells Environ Sci Technol 42 8101ndash8107
84
27 Fan Y Xu S Schaller R Jiao J Chaplen F Liu H (2011) Nanoparticle decorated
anodes for enhanced current generation in microbial electrochemical
cells Biosensors and Bioelectronics 26 (5)1908ndash1912
28 Fan Y Hu H Liu H (2007)Sustainable power generation in microbial fuel cells
using bicarbonate buffer and proton transfer mechanisms Environ Sci
Technol 41(23) 8154-8
29 Fedorovich V Knighton MC Pagaling E Ward FB Free A Goryanin I (2009)
Novel electrochemically active bacterium phylogenetically related to Arcobacter
butzleri isolated from a microbial fuel cell Appl Environ Microbiol 75(23) 7326-
7334
30 Ferna ndez A Huang S Seston S Xing J Hickey R Criddle C Tiedje J (1999)
How stable is stable Function versus community composition Appl Environ
Microbiol 65 3697ndash3704
31 Field SJ Dobbin PS Cheesman MR Watmough NJ Thomson AJ Richardson
DJ (2000) Purification and magneto-optical spectroscopic characterization of
cytoplasmic membrane and outer membrane multiheme c-type cytochromes from
Shewanella frigidimarina NCIMB400 J Biol Chem 275 8515ndash8522
32 Freguia S Masuda M Tsujimura S Kano K (2009) Lactococcus lactis catalyses
electricity generation at microbial fuel cell anodes via excretion of a soluble quinone
Bioelectrochemistry 7614ndash18
33 Fricke K Harnisch F and Schroumlder U (2008) On the use of cyclic voltammetry for
the study of the anodic electron transfer in microbial fuel cells Energy Environ
Sci 1 144ndash147
34 Gao H Wang Y Liu X Yan T Wu L Alm E Arkin A Thompson DK and
Zhou J (2004) Global transcriptome analysis of the heat shock response of
Shewanella oneidensis J Bacteriol 1867796-7803
35 Gao H Yang ZK Wu L Thompson DK Zhou J (2006) Global transcriptome
analysis of the cold shock response of Shewanella oneidensis MR-1 and mutational
analysis of its classical cold shock proteins J Bacteriol 1884560-4569
36 George MG (2005) Bergeyrsquos Manual of Systematic Bacteriology Volume Two The
Proteobacteria Part B The Gammaproteobacteria Second Edition New York
Springer 651p
37 Gonzaacutelez-Toril E Llobet-Brossa E Casamayor EO Amann R Amils R (2003)
Microbial ecology of an extreme acidic environment the Tinto river Appl Environ
Microbiol 69(8) 4853ndash4865
38 Gorby YA Yanina S Mclean JS Rosso KM Moyles D Dohnalkova A
Beveridge TJ Chang IS Kim BH Kim KS Culley DE Reed SB Romine M
F Saffarini DA Hill EA Shi L Elias DA Kennedy DW Pinchuk G
Watanabe K Ishii S Logan B Nealson KH Fredrickson JK (2006) Electrically
conductive bacterial nanowires produced by Shewanella oneidensis strain MR-1 and
other microorganisms Proc Natl Acad Sci USA 10311358-11363
85
39 He Z Wagner N Minteer SD Angenent LT (2006) An upflow microbial fuel cell
with an interior cathodes assessment of the internal resistance by impedance
spectroscopy Environ Sci Technol 40 5212-5217
40 Heydorn A Nielsen AT Hentzer M Sternberg C Givskov M Ersboslashll BK Molin
S (2000) Quantification of biofilm structures by the novel computer program
COMSTAT Microbiol 1462395-2407
41 Holmes DE Bond DR Lovley DR (2004a) Electron transfer by Desulfobulbus
propionicus to Fe(III) and graphite electrodes Appl Environ Microbiol 701234ndash
1237
42 Holmes DE Chaudhuri SK Nevin KP Mehta T Methe BA Liu A Ward JE
Woodard TL Webster J Lovley DR (2006) Microarray and genetic analysis of
electron transfer to electrodes in Geobacter sulfurreducens Environ Microbiol
81805-1815
43 Holmes DE Nicoll JS Bond DR Lovley DR (2004b) Potential role of a novel
psychrotolerant member of the family Geobacteraceae Geopsychrobacter
electrodiphilus gen nov sp nov in electricity production by a marine sediment
fuel cell Appl Environ Microbiol 70 6023ndash6030
44 Holmes DE Chaudhuri SK Nevin KP Mehta T Metheacute BA Liu A Ward JE
Woodard TL Webster J Lovley DR (2006) microarray and genetic analysis of
electron transfer to electrodes in Geobacter sulfurreducens Environ Microbial
8(10)1805-1815
45 Hu H (2009) Enhancing Hydrogen Production in Microbial Electrolysis Cells
through Development of Platinum-free Cathode and Improvement of Reactor
Dissertation
46 Ince BK Ayman O N Turker G Ccedilelikkol S Ince O (2010) Microbial ecology of
anaerobic reactors for treatment of alcohol industry wastewaters a review Current
research technology and education topics in applied microbiology and microbial
ecology 988-999
47 Jung S Regan JR (2007) Comparison of anode bacterial communities and
performance in microbial fuel cells with different electron donors Appl Microbiol
Biotechnol 77393ndash 402
48 Kim BH Kim HJ Hyun MS Park DH (1999) Direct electrode reaction of Fe(III)
reducing bacterium Shwwanella putrefaciens J Microbial Biotechnol 9127-131
49 Kim GT Hyun MS Chang IS Kim HJ Park HS Kim BH Kim SD Wimpenny
JW Weightman AJ (2005) Dissimilatory Fe(III) reduction by an electrochemically
active lactic acid bacterium phylogenetically related to Enterococcus gallinarum
isolated from submerged soil J Appl Microbiol 99(4) 978ndash987
50 Kim GT Webster G Wimpenny JW Kim BH Kim HJ Weightman AJ (2006)
Bacterial community structure compartmentalization and activity in a microbial fuel
cell J Appl Microbiol 101(3) 698ndash710
86
51 Kim HJ Park HS Hyun MS Chang IS Kim M Kim BH (2002) A mediator-
less microbial fuel cell using a metal reducing bacterium Shewanella putrefaciens
Enzyme Microb Technol 30 145-152
52 Kim JR Min B Logan BE (2005) Evaluation of procedures to acclimate a
microbial fuel cell for electricity production Appl Microbiol Biotechnol 68(1)23-30
53 Kim JR Cheng S Oh SE Logan BE (2007) Power generation using different
cation anion and ultrafiltration membranes in microbial fuel cells Environ Sci
Technol 41 1004-1009
54 Konstantinidis KT Serres MH Romine MF Rodrigues JL Auchtung J McCue
LA Lipton MS Obraztsova A Giometti CS Nealson KH Fredrickson JK
Tiedje JM (2009) Comparative systems biology across an evolutionary gradient
within the Shewanella genus Proc Natl Acad Sci U S A 106(37)15909-15914
55 Lee J Phung NT Chang IS Kim BH Sung HC (2003) Use of acetate for
enrichment of electrochemically active microorganisms and their 16S rDNA
analyses FEMS Microbiol Lett 223185ndash191
56 Liang P Wang H Xia X Huang X Mo Y Cao X Fan M(2011)Carbon
nanotube powders as electrode modifier to enhance the activity of anodic biofilm
in microbial fuel cells Biosens Bioelectron 26(6)3000-4
57 Lies DP Hernandez ME Kappler A Mielke RE Gralnick JA Newman DK
(2005) Shewanaella oneidensis MR-1 uses overlapping pathways for iron reduction
at a distance and by direct contact under conditions relevant for biofilms Appl
Environ Microbiol 71 4414-4426
58 Liu H and Logan B E (2004) Electricity generation using an air-cathode single
chamber microbial fuel cell in the presence and absence of a proton exchange
membrane Environ Sci Technol 38 4040-4046
59 Liu H Grot S Logan BE (2005) Electrochemically assisted microbial production
of hydrogen from acetate Environ Sci Technol 39 4317 ndash 320
60 Liu H Hu H Chignell J Fan Y (2010) Microbial Electrolysis Novel Technology
for Hydrogen Production from Biomass Biofuels 1(1) 129ndash142
61 Liu H Ramnarayanan R Logan BE (2004) Production of electricity during
wastewater treatment using a single chamber microbial fuel cell Environ Sci Technol
382281-2285
62 Liu WT Marsh TL Cheng H Forney LJ (1997) Characterization of microbial
diversity by determining terminal restriction fragment length polymorphisms of gene
encoding 16S rRNA Appl Environ Microbiol 63 4516ndash4522
63 Logan B E Hamelers B Rozendal R Schroder U Keller J Freguia S Aelterman
P Verstraete W Rabaey K (2006) Microbial fuel cells methodology and
technology Environ Sci Technol 40 (17) 5181-5192
64 Logan B E and Regan JM (2006) Electricity-producing bacterial communities in
microbial fuel cells Trends Microbiol 14512ndash518
87
65 Logan B Cheng S Watson V Estadt G (2007) Graphite fiber brush anodes for
increased power production in air-cathode microbial fuel cells Environ Sci Technol
41(9)3341-3346
66 Logan BE (2009) Exoelectrogenic bacteria that power microbial fuel cells Nat Rev
Microbiol 5 375-381
67 Logan BE Call D Cheng S Hamelers HV Sleutels TH Jeremiasse AW
Rozendal RA (2008) Microbial electrolysis cells for high yield hydrogen gas
production from organic matter Environ Sci Technol 42 8630-8640
68 Logan BE Hamelers B Rozendal R Schroder U Keller J Freguia S Aelterman
P Verstraete W Rabaey K (2006) Microbial fuel cells methodology and
technology Environ SciTechnol 40 5181-5192
69 Lovely DR (2006) Bug juice harvesting electricity with microorganisms Nature
Review| Microbiology 4 497-508
70 Lovley DR Phillips EJ (1988) Novel Mode of Microbial Energy Metabolism
Organic Carbon Oxidation Coupled to Dissimilatory Reduction of Iron or
Manganese Appl Environ Microbiol 54(6) 1472-1480
71 Ly HK Sezer M Wisitruangsakul N Feng JJ Kranich A Millo D Weidinger
IM Zebger I Murgida DH Hildebrandt P (2011) Surface-enhanced vibrational
spectroscopy for probing transient interactions of proteins with biomimetic interfaces
electric field effects on structure dynamics and function of cytochrome c FEBS J
278(9)1382-90
72 Macaskie LE Hewitt CJ Shearer JA Kent CA (1995) Biomass production for
the removal of heavy metals from aqueous solutions at low pH using growth-
decoupled cells of a Citrobacter sp Int Biodeter Biodegr 3573ndash92
73 Malki M De Lacey AL Rodriguez N Amils R Fernandez VM (2008)
Preferential use of an anode as an electron acceptor by an acidophilic bacterium in
the presence of oxygen Appl Environ Microbiol 74 4472ndash4476
74 Meitl LA Eggleston CM Colberg PJS Khare N Reardon CL Shi L (2009)
Electrochemical interaction of Shewanella oneidensis MR-1 and its outer membrane
cytochromes OmcA and MtrC with hematite electrodes Geochim Cosmochim
Ac 73(18) 5292-5307
75 Morozan A Stamatin L and Nastase
F (2007) The biocompatibility
microorganisms-carbon nanostructures for applications in microbial fuel cells Phys
Stat Sol 6 1797-1803
76 Morris JM Jin S Crimi B Pruden A (2009) Microbial fuel cell in enhancing
anaerobic biodegradation of diesel Chem Eng J 146 161ndash167
77 Murray AE Lies D Li G Nealson K Zhou J Tiedje JM (2001) DNADNA
hybridization to microarrays reveals gene-specific differences between closely
related microbial genomes Proc Natl Acad Sci U S A 98 9853-9858
78 Muyzer G de Waal EC Uitterlinden AG (1993) Profiling of complex microbial
communities by denaturing gradient gel electrophoresis analysis of polymerase chain
88
reaction amplified genes coding for 16S rRNA Appl Environ Microbiol 59 695ndash
700
79 Myers CR Myers JM (1992) Localization of cytochromes to the outer membrane
of anaerobically grown Shewanella putrefacians MR-1 J Bacteriol 174 3429ndash3438
80 Narde GK Kapley A Purohit HJ (2004) Isolation and characterization of
Citrobacter strain HPC255 for broad-range substrate specificity for chlorophenols
Curr Microbiol 48(6) 419-423
81 Okabe S Ito T Sugita K Satoh H (2005) Succession of internal sulfur cycles and
sulfur-oxidizing bacterial communities in microaerophilic wastewater biofilms Appl
Environ Microbiol 71(5) 2520-2529
82 Park DH and Zeikus JG (2000) Electricity generation in microbial fuel cells using
neutral red as an electronophore Appl Environ Microbial 66 292-1297
83 Park DH and Zeikus JG (2002) Impact of electrode composition on electricity
generation in a single-compartment fuel cell using Shewanella putrefaciens Appl
Microbiol Biotechnol 5958ndash56
84 Park DH and Zeikus JG (2003) Improved fuel cell and electrode designs for
producing electricity from microbial degradation Biotechnol Bioeng 81 348ndash355
85 Park HS Kim BH Kim HS Kim HJ Kim GT Kim M Chang IS Park Y
K Chang HI (2001) A novel electrochemically active and Fe(III)-reducing
bacterium phylogenetically related to Clostridium butyricum isolated from a
microbial fuel cell Anaerobe 7 297-306
86 Parot S Nercessian O Delia ML Achouak W Bergel A (2009) Electrochemical
checking of aerobic isolates from electrochemically active biofilms formed in
compost J Appl Microbiol 106(4) 1350-1359
87 Pham CA Jung SJ Phung NT Lee J Chang IS Kim BH Yi H Chun J
(2003) A novel electrochemically active and Fe(III)-reducing bacterium
phylogenetically related to Aeromonas hydrophila isolated from a microbial fuel cell
FEMS Microbiol Lett 223 129-134
88 Qiao Y Bao SJ Li CM Cui XQ Lu ZS Guo J (2008) Nanostructured
polyanilinetitanium dioxide composite anode for microbial fuel cells ACS Nano 2
113-119
89 Qiu R Zhao B Liu J Huang X Li Q Brewer E Wang S Shi N (2009) Sulfate
reduction and copper precipitation by a Citrobacter sp isolated from a mining area J
Hazard Mater 1641310-1315
90 Quan X Yang SG Ruan XL and Zhao HM (2005) Preparation of titania
nanotubes and their environmental applications as electrode Environ Sci Technol 39
3770ndash3775
91 Rabaey K Boon N Hofte M Verstraete W (2005) Microbial phenazine
production enhances electron transfer in biofuel cells Environ Sci Technol 393401ndash
3408
89
92 Rabaey K Boon N Siciliano S D Verhaege M Verstraete W (2004) Biofuel cells
select for microbial consortia that self-mediate electron transfer Appl Environ
Microbiol 70(9) 5373ndash5382
93 Rabaey K Lissens G Siciliano S D Verstraete W (2003) Amicrobial fuel cell
capable of converting glucose to electricity at high rate and efficiency Biotechnol
Lett 25 1531-15
94 Rabaey K Verstraete W (2005) Microbial fuel cells novel biotechnology for
energy generation Trends Biotechnol 23(6) 291-298
95 Reguera G McCarthy KD Mehta T Nicoll JS Tuominen MT Lovley DR
(2005) Extracellular electron transfer via microbial nanowires Nature 435 1098-
1101
96 Reguera G Nevin KP Nicoll JS Covalla SF Woodard TL Lovley DR (2006)
Biofilm and nanowire production leads to increased current in Geobacter
sulfurreducens fuel cells Appl Environ Microbiol 72 7345-7348
97 Reimers CE Tender LM Fertig S Wang W (2001) Harvesting energy from the
marine sediment-water interface Environ Sci Technol 35192-195
98 Rezaei F Xing D Wagner R Regan JM Richard TM Logan BE (2009)
Simultaneous cellulose degradation and electricity production by Enterobacter
cloacae in a microbial fuel cell Appl Microbiol Biotechnol 75 3673ndash3678
99 Richter H Nevin KP Jia H Lowy DA Lovley DR Tender LM (2009) Cyclic
voltammetry of biofilms of wild type and mutant Geobacter sulfurreducens on fuel
cell anodes indicates possible roles of OmcB OmcZ type IV pili and protons in
extracellular electron transfer Energy Environ Sci 2 506ndash516
100 Riesenfeld SC Schloss DP Handelsman J(2004) Metagenomics Genomic
Analysis of Microbial Communities Annual Review Genetics 38525-552
101 Rozendal RA Hamelers HV Buisman CJ (2006) Effects of membrane cation
transport on pH and microbial fuel cell performance Environ Sci Technol 40
5206-5211
102 Rozendal RA Hamelers HVM Euverink GJW Metz SJ Buisman CJN (2006)
Principle and perspectives of hydrogen production through biocatalyzed electrolysis
Int J Hydrogen Energ 311632-1640
103 Rozendal RA Hamelers HV Molenkamp RJ Buisman CJ (2007) Performance of
single chamber biocatalyzed electrolysis with different types of ion exchange
membrances Water Res 41 1984-1994
104 Schluumlter A Bekel T Diaz NN Dondrup M Eichenlaub R Gartemann
KH Krahn I Krause L Kroumlmeke H Kruse O Mussgnug JH Neuweger
H Niehaus K Puumlhler A Runte KJ Szczepanowski R Tauch A Tilker
A Viehoumlver P Goesmann A (2008) The metagenome of a biogas-producing
microbial community of a production-scale biogas plant fermenter analyzed by the
454-pyrosequencing technology J Biotechnol136 77-90
90
105 Scott J H and Nealson KH (1994) A biochemical study of the intermediary carbon
metabolism of Shewanella putrefaciens J Bacteriol 176(11) 3408ndash3411
106 Sekiguchi H Tomioka H Nakahara T and Uchiyama H (2001) A single band
does not always represent single bacterial strains in denaturing gradient gel
electrophoresis analysis Biotechnology Letters23 1205ndash1208
107 Sharma T Leela M Reddy A Chandra TS Ramaprabhu S (2008) Development
of carbon nanotubes and nanofluids based microbial fuel cell Int J Hydrogen Energ
336749ndash6754
108 Shea C Clauwaert P Verstraete W Nerenberg R (2008) Adapting a denitrifying
biocathode for perchlorate reduction Water Sci Technol 58 (10) 1941-1946
109 Shi L Squier TC Zachara JM Fredrickson JK (2007) Respiration of metal (hydr)
oxides by Shewanella and Geobacter a key role for multihaem c-type cytochromes
Mol Microbiol 6512-20
110 Stams AJ and Plugge CM(2009)Electron transfer in syntrophic communities
of anaerobic bacteria and archaea Nat Rev Microbiol7(8)568-77
111 Sukkasem C Xu S Park S Boonsawang P Liu H (2008) Effect of Nitrate on the
Performance of Single Chamber Air Cathode Microbial Fuel Cells Water Res 42(19)
4743-4750
112 Summers ZM Fogarty HE Leang C Franks AE Malvankar NS Lovley DR
(2010)Direct exchange of electrons within aggregates of an evolved syntrophic
coculture of anaerobic bacteria Science 330 (6009)1413-5
113 Szczepanowski R Bekel T Goesmann A Krause L Kroumlmeke H Kaiser
O Eichler W Puumlhler A Schluumlter A (2008) Insight into the plasmid metagenome
of wastewater treatment plant bacteria showing reduced susceptibility to
antimicrobial drugs analysed by the 454-pyrosequencing technology J
Biotechnol 136 54-64
114 Tamura K Dudley J Nei M Kumar S (2007) MEGA4 Molecular Evolutionary
Genetics Analysis (MEGA) software version 40 MolBiol Evol 24(8) 1596-1599
115 Thompson JD Gibson TJ Plewniak F Jeanmougin F Higgins DG (1997) The
CLUSTAL_X windows interface flexible strategies for multiple sequence alignment
aided by quality analysis tools Nucleic Acids Res 25 4876-4882
116 Thormann KM Duttler S Saville RM Hyodo M Shukla S Hayakawa Y
Spormann AM (2006) Control of formation and cellular detachment from
Shewanella oneidensis MR-1 biofilms by cyclic di-GMP J Bacteriol 1882681-2691
117 Thormann KM Saville RM Shukla S Pelletier DA Spormann AM (2004)
Initial Phases of biofilm formation in Shewanella oneidensis MR-1 J Bacteriol
186(23) 8096-104
118 Torres CI Kato Marcus A Rittmann BE (2008b) Proton transport inside the
biofilm limits electrical current generation by anode-respiring bacteria Biotechnol
Bioeng 100 872ndash881
91
119 Torres CI Marcus AK Lee HS Parameswaran P Krajmalnik-Brown R Rittmann
BE (2010) A kinetic perspective on extracellular electron transfer by anode-
respiring bacteria FEMS Microbiol Rev 234(1)3-17
120 Torres CI Marcus AK Parameswaran P Rittmann BE (2008a) Kinetic
experiments for evaluating the NernstndashMonod model for anode-respiring bacteria
(ARB) in a biofilm anode Environ Sci Technol 42 6593ndash6597
121 Tsai HY Wu Ch-Ch Lee Ch-Y Shih EP (2009) Microbial fuel cell performance
of multiwall carbon nanotubes on carbon cloth as electrodes J Power Sources
194199-205
122 Tyson GW Chapman J Hugenholtz P Allen EE Ram RJ Richardson
PM Solovyev VV Rubin EM Rokhsar DS Banfield JF(2004) Community
structure and metabolism through reconstruction of microbial genomes from the
environment Nature42837-43
123 Venkateswaran K Moser DP Dollhopf ME Lies DP Saffarini D A MacGregor
BJ Ringelberg DB White DC Nishijima M Sano H Burghardt J Stackebrandt
E Nealson KH (1999) Polyphasic taxonomy of the genus Shewanella and
description of Shewanella oneidensis sp Int J Syst Bacteriol 49705-724
124 von Canstein H Ogawa J Shimizu S Lloyd JR (2008) secretion of Flavins
by Shewanella Species and Their Role in Extracellular Electron Transfer Appl
Environ Microbiol 74 3615-623
125 Wang A Liu W Cheng Sh Xing D Zhou J Logan BE (2009) Source of methane
and methods to control its formation in single chamber microbial electrolysis cells
Int J Hydrogen Energ 34 3653-3658
126 Wang J Ma T Zhao L Lv J Li G Liang F Liu R (2008) PCRndashDGGE method
for analyzing the bacterial community in a high temperature petroleum reservoir
World J Microbiol Biotechnol 2 9694-9702
127 Wang Q Garrity GM Tiedje JM Cole JR (2007) Naiumlve Bayesian Classifier for
Rapid Assignment of rRNA Sequences into the New Bacterial Taxonomy Appl
Environ Microbiol 73(16) 5261-5267
128 Wang YF Masuda M Tsulimura S Kano K (2008) Electrochemical regulation of
the end-product profile in Propionibacterium freudenreichii ET-3 with an
endogenous mediator Biotechnol Bioeng 101(3) 579ndash586
129 Wrighton KC Agbo P Warnecke F Weber KA Brodie EL DeSantis TZ
Hugenholtz P Andersen GL Coates JD (2008) A novel ecological role of the
Firmicutes identified in thermophilic microbial fuel cells ISME J 2 1146ndash1156
130 Xing D Zuo Y Cheng S Regan JM Logan BE (2008) Electricity generation by
Rhodopseudomonas palustris DX-1 Environ Sci Technol 42 4146-4151
131 Xu S Liu H Fan Y Schaller R Jiao J Chaplen F (2012) Enhanced performance
and mechanism study of microbial electrolysis cells using Fenanoparticle-decorated
anodes Appl Microbiol Biotechnol 93(2)871-880
92
132 Yang Y Harris DP Luo F Wu L Parsons AB Palumbo AV Zhou J (2008)
Characterization of the Shewanella oneidensis Fur gene roles in iron and acid
tolerance response BMC Genomics 9 Suppl 1S11
133 Zhang L Zhou S Zhuang L Li W Zhang J Lu N Deng L (2008) Microbial
fuel cell based on Klebsiella pneumoniae biofilm Electrochem Commun10 1641ndash
1643
134 Zuo Y Xing DF Regan JM Logan BE (2008) Isolation of the exoelectrogenic
bacterium Ochrobactrum anthropi YZ-1 by using a U-tube microbial fuel cell Appl
Environ Microbiol 74 3130-3137
93
APPENDICES
List of published papers during PhD study
1 Shoutao Xu Hong Liu Yanzhen Fan Rebecca Schaller Jun Jiao and Frank
Chaplen (2012) Enhanced performance and mechanism study
of microbial electrolysis cells using Fe nanoparticle-decorated anodes Applied
Microbiology and Biotechnology 93(2)871-880
2 Shoutao Xu Hong Liu (2011) New exoelectrogen Citrobacter sp SX-1 isolated
from a microbial fuel cell Journal of Applied Microbiology111(5)1108-1115
3 Yanzhen Fan Shoutao Xu Rebecca Schaller Jun Jiao Frank Chaplen Hong Liu
(2011) Nanoparticle decorated anodes for enhanced current generation in microbial
electrochemical cells Biosensors and Bioelectronics 26(5) 1908-1912
4 Tunc Catal Shoutao Xu Kaichang Li Hakan Bermek Hong Liu
(2008)
Electricity generation from polyalcohols in single-chamber microbial fuel
cells Biosensors and Bioelectronics 24(4)849-854
5 Chontisa Sukkasema Shoutao Xu Sunhwa Park Piyarat Boonsawang Hong Liu
(2008) Effect of nitrate on the performance of single chamber air cathode
microbial fuel cells Water research 424743-4750
6 Rebecca Schaller Yanzhen Fan Shoutao Xu Alan Fern Frank Chaplen Hong
Liu and Jun Jiao (2009) Vertically Aligned Multi-walled Carbon Nanotube
Decorated Anodes for Microbial Fuel Cells Proceedings of Materials Research
Society 2009 1170 R05-13

copyCopyright by Shoutao Xu
June 15 2012
All Rights Reserved
Bacterial Community Analysis New Exoelectrogen Isolation and Enhanced Performance
of Microbial Electrochemical Systems Using Nano-Decorated Anodes
by
Shoutao Xu
A DISSERTATION
Submitted to
Oregon State University
in partial fulfillment of
the requirements for the
degree of
Doctor of Philosophy
Presented June 15 2012
Commencement June 2013
Doctor of Philosophy dissertation of Shoutao Xu presented on June 15 2012
APPROVED
Co-Major Professor representing Biological and Ecological Engineering
Co-Major Professor representing Biological and Ecological Engineering
Head of the Department of Biological and Ecological Engineering
Dean of the Graduate School
I understand that my dissertation will become part of the permanent collection of
Oregon State University libraries My signature below authorizes release of my
dissertation to any reader upon request
Shoutao Xu Author
ACKNOWLEDGEMENTS
I would like to thank all people who have helped and inspired me during my
doctoral study Foremost I would like to express my sincere gratitude to my advisors Dr
Hong Liu and Dr Frank Chaplen for their continuous support of my PhD study and the
research of Microbial Fuel Cells at Oregon State University Hong inspired me to devote
myself to the field of Bio-energy with her great patience and enthusiasm towards
scientific educations Frank was always available and willing to help me with my study
especially during the period of Hongrsquos sabbatical leave Thanks to his kindness and
assistance my study at OSU became smooth and rewarding
Besides my advisors I would like to thank everyone in my dissertation committee Dr
Martin Schuster Dr Clare Reimers and Dr Mark Dolan Due to their encouragement
and insightful comments on my research I could always have the courage and knowledge
to overcome difficulties in my research I benefited greatly from Martinrsquos valuable
suggestions on my writing skills and his generous help regarding microarray data analysis
Clarersquos advice helped me build a solid foundation of Electrochemistry Also it was a
great honor to have Mark as my committee member His teaching gave me an insight into
the world of environmental engineering and enhanced the depth and width of my research
I thanked Dr Yanzhen Fan for always giving me guidance about the designs of the
reactors in my experiments
It was also important for me to say thanks to my current and previous lab-mates
Keaton Lesnik Kuhuan Chien Cheng Li Corale Abourached Anthony Janicek
Hongqiang Hu Jeremy Chignell Yudith Nieto and Wengguo Wu I had a wonderful
time enjoying doing research with them Their friendship and help made me confident of
my ability to do research as a scientist In addition I cherished and appreciated the
friendly environment in our BEE department Faculty members especially Dr John Bolte
Dr John Selker Dr Roger Ely Dr Ganti Murthy and Desiree Tullos made BEE an
excellent department for our students
My deepest gratitude went to my father Fuqin Wang and my mother Jinzhi Wang
for their endless care love and support throughout my life Also I would like to say
thanks to my best-loved wife Songhua Zhu She always supported me and never
complained that I could spend so little time accompanying with her This dissertation was
also for my lovely son Gabriel Hong-Yi Xu
TABLE OF CONTENTS
Page
1 General Introductionhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 1
2 Bacterial Community Analysis of Mixed Consortium in Microbial
Electrochemical Systemshelliphelliphelliphellip 13
3 New Exoelectrogen Citrobacter sp SX-1 Isolation and
Characterizationhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 27
4
Enhanced Performance and Mechanism Study of Microbial Electrolysis
Cells using Fe Nanoparticles Decorated Anodeshelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 45
5 Global Transcriptome Analysis of Response of Shewanella oneidensis
MR-1 to CNT Nanostructure Decorated Anodes in Microbial
Electrochemical System
66
6 Summaryhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 80
7 Bibliography helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 82
8 Appendices helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 93
LIST OF FIGURES
Figure Page
1-1 Generalized schematics (a) Microbial fuel cell (MFCs)(b) Microbial
electrolysis cell (MECs) helliphelliphelliphelliphelliphelliphelliphelliphelliphellip helliphelliphelliphellip
2
1-2 Mechanisms for extracellular electron transport in a MFCs anode 7
2-1 Polarization curves of high power mixed culture MFCshelliphelliphelliphelliphelliphellip 20
2-2 DGGE result of high power generation mixed culture MFCshelliphelliphellip 21
2-3 Bacterial species and percentage of bacterial community of mixed
culture MFCs helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip
22
2-4 Phylogenetic trees of bacterial species from mixed culture MFCs 22
2-5 Dominant bacterial species identification in DGGEhelliphelliphelliphelliphelliphelliphelliphellip 23
2-6 Current density generated by different dominant isolateshelliphelliphelliphelliphelliphellip 24
3-1 Phylogenetic tree of strain SX-1 and closely related species based on
16S rDNA sequenceshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip
35
3-2 SEM image of planktonic cells of Citrobacter sp SX-1helliphelliphelliphelliphelliphellip 35
3-3 Electricity generation by Citrobacter sp SX-1 in a single chamber
MFCs with sodium citrate helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip
36
3-4 Electricity generation of Citrobacter sp SX-1 using different
substrates in single chamber air-cathode MFCs at 1 KΩhelliphelliphellip
37
3-5 Power and voltage generation by Citrobacter sp SX-1 as a function
of current density helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip
38
3-6 Four current generating stages for CV analysis helliphelliphelliphelliphelliphelliphelliphellip 39
3-7 Cyclic Voltammetry analysis of of Citrobacter sp SX-1 biofilms helliphellip 41
4-1 SEM images of Fe NPs decorated anodeshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 50
4-2 Effects of Fe-NP decorated anodes on the current density in MECs helliphellip 51
4-3 Differentially expressed genes grouped by functional classification in
Fe NP decorated anodeshelliphelliphelliphelliphelliphellip helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip
53
5-1 SEM images of CNT decorated anodeshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 70
5-2 Effects of CNT decorated anodes on the current density in MECshelliphellip 73
5-3 Differentially expressed genes grouped by functional classification in
CNT decorated anodes helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip
74
LIST OF TABLES
Table
1-1
4-1
4-2
4-3
Taxa of bacteria current density generated and reactor configuration
used in microbial electrochemical cell experimentshelliphelliphelliphellip
Expression levels of genes related to biofilm formationhelliphelliphelliphelliphellip
Genes related to anaerobic growth and electron transfer with
significantly change expression levelhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip
Expression level of flavin and cytochrome geneshelliphelliphelliphelliphelliphelliphelliphellip
Page
4
55
56
58
4-4
5-1
5-2
Other genes with significantly changed expression level
Expression level of cytochrome c as response to CNT decorated anodes
Expression level of genes related to flavin synthesis as response to CNT
decorated anode in CNT decorated anodeshelliphellip
59
76
78
1
1
Bacterial Community Analysis New Exoelectrogen Isolation and Enhanced Performance
of Microbial Electrochemical Systems Using Nano-Decorated Anodes
Chapter 1
General Introduction
11 Microbial Fuel Cells and Microbial Electrolysis Cells
Microbial electrochemical systems (MESs) have drawn the attention of researchers
in recent years due to their promising applications in a variety of scientific fields such as
renewable energy generation bioremediation and wastewater treatment In a MES
microorganisms interact with electrodes catalyzing oxidation and reduction reactions at
the anode and the cathode
The most-described type of MESs is the microbial fuel cells (MFCs) in which
useful power is generated directly using the catalytic action of active microorganisms
(Kim 2002 Cheng and Logan 2007 Cheng et al 2006) In a typical two-chamber
MFCs (Figure 1a) organic matter is oxidized by electrochemically active
microorganisms in the anode chamber to release protons and electrons Protons diffuse
into the cathode chamber through a proton exchange membrane Meanwhile electrons
are transferred to the anode through various mechanisms (Logan et al 2006 Lovley
2
2006 Rabaey et al 2003) and then travel to the cathode where normally they combine
with oxygen and protons to form water
Another common used type of MES is the microbial electrolysis cell (MECs) in
which hydrogen is produced instead of electricity by applying a circuit voltage to the
MFCs system and maintaining anaerobic conditions in the cathode chamber (Liu et al
2005 Logan et al 2008 Rozendal et al 2006 Schaller et al 2009) (Figure 1b) A
cathode potential of at least - 410 mV (vs standard hydrogen electrode at pH 70) is
required to produce hydrogen at the cathode Therefore a theoretical potential of 110 mV
(ie 410-300 mV) is needed but voltages of gt02 V are normally applied due to various
overpotentials
Fig 1-1 Generalized schematics (a) Microbial fuel cells (MFCs) where electricity is
captured through the resistance R (b) Microbial electrolysis cells (MECs) where
hydrogen gas is produced in the cathode chamber R for resistor PS for power supply and
PEM for proton exchange membrane
The fundamental feature shared by microbial electrochemical systems (MESs)
(MECs and MFCs) is the microbe-catalyzed electron transfer from organic matter to
electrodes (anodes) (Rebaey et al 2004) Therefore different types of MESs can be
PEM
R
H+
e- e-
Anode
Bacte
ria
a
Cath
ode
H2O
PEM
PS
H+
e- e-
Anode
Bacte
ria
b
Cath
ode
H2
3
utilized to investigate the phenomena and mechanisms of interactions between microbes
and electrodes
12 Bacteria and Electron Transfer Mechanisms in MESs
The electrochemically active bacteria in MFCs are thought to be iron-reducing
bacteria such as Shewanella and Geobacter species (Gorby et al 2006) They have great
importance in the natural environment principally in metal oxidation and reduction
However recent studies have shown that the diversity of bacterial communities is much
greater than these model iron reducers on MFCs anodes (Reimers et al 2001 Liu et al
2004b Kim et al 2005) Over 20 electrochemically active bacterial species those can
transfer electrons exocellularly to electrodes have been reported in the past 10 years
(Logan 2009 Liu et al 2010) These bacteria have been categorized into diverse genetic
groups so far including alphaproteobacteria (Rhodopseudomonas Ochrobactrum and
Acidiphilium) (Malki et al 2008 Xing et al 2008 Zuo et al 2008) betaproteobacteria
(Rhodoferax) (Chaudhuri et al 2003) gammaproteobacteria (Shewanella Pseudomonas
Klebsiella Enterobacter and Aeronomas) (Kim et al 2002 Pham et al 2003 Rabaey et
al 2005 Bretschger et al 2007 Zhang et al 2008 Chung et al 2009 Rezaei et al 2009)
deltaproteobacteria (Geobacter Geopsychrobacter Desulfuromonas and Desulfobulbus)
(Bond et al 2002 Bond and Lovley 2003 Holmes et al 2004ab 2006)
Epsilonproteobacteria (Arcobacter) (Fedorovich et al 2009) Firmicutes (Clostridium and
Thermincola) (Park et al 2001 Wrighton et al 2008) Acidobacteria (Geothrix) (Bond
and Lovley 2005) and actinobacteria (Propionibacterium) (Wang et al 2008) A wider
range of electrochemically active bacteria are expected to be discovered
4
Table 1 Taxa of bacteria current density generated and reactor configuration used in
microbial electrochemical cell experiments (Liu et al 2010)
The electrochemically active bacterial species that possess the ability to transfer
electrons outside of the cell are called exoelectrogens in the MESs research field The
different exoelectrogens have demonstrated a wide-ranging power generation ability in
MFCs For example Shewanella oneidensis MR-1 has demonstrated the ability to
generate 0018 Am2 current density in single chamber MFCs while the Geobacter
Taxon Microorganisms Current Density (Am2
)
α-proteobacteria
Rhodopseudomonas palustris DX-1 003
Ochrobactrum anthropi YZ-1 071
Acidiphilium sp 32sup5 300
β-proteobacteria Rhodoferax ferrireducens 0031
γ-proteobacteria
Shewanella putrefaciens IR-1 0016
Shewanella oneidensis DSP10 0013
Shewanella oneidensis MR-1 0018
Pseudomonas aeruginosa KRA3 0017
Escherichia coli K12 HB101 100
Klebsiella pneumoniae L17 120
Enterobacter cloacae 013
Aeromonas hydrophila PA3 030
δ-proteobacteria
Geobacter metallireducens 065
Geobacter sulfurreducens 800
Desulfuromonas acetoxidans 0005
Geopsychrobacter
Electrodiphilus strain A2
0066
Desulfobulbus propionicus 003
Firmicutes
Lactococcus lactis 003
Thermincola sp strain Jr 020
Clostridium butyricum EG3 022
Thermincola ferriacetica Z-0001 040
Brevibacillus spp PTH1 0009
Desulfitobacterium hafniense DCB2 110
Acidobacteria Geothrix fermentans 0097
Actinobacteria Propionibacterium freudenreichiiET-3 120
5
sulfurreducens species has generated current densities as high as 80 Am2 which is 400
times higher than the one generated by Shewanella oneidensis MR-1 However mixed-
culture communities have generated much higher power densities than their pure-culture
counterparts in MFCs perhaps due to synergistic interactions within the anode bacterial
communities and the participation of currently unknown bacteria species and mechanisms
(Jung and Regan 2007) The dominant bacteria species vary in mixed cultures bacteria
communities in MFCs due to the enrichment of different substrates
Traditional methods of extroelectrogen study depend on cultivation hampered novel
exoelectrogens discovery because the inadequacy of defined media underestimates the
actual microbial diversity in MFCs Slow growth rate and unknown growth requirements
of anaerobic microorganisms make some anaerobic extroelectrogens cultivation difficult
However these limitations have been overcome by using molecular biological methods
based on DNARNA analysis Molecular biological techniques are now widely applied to
assess the diversity of microbial communities by analyzing the 16S rDNA sequence The
most commonly used molecular biological techniques for bacterial community analysis
include denaturing gradient gel electrophoresis (DGGE) restriction fragment length
polymorphism (RFLP) and 16S rDNA sequencing These methods are much less time
consuming than traditional isolation and cultivation methods
Isolated exoelectrogens were utilized to explore the mechanism of electron transfer
to the anode However the mechanisms of electron transfer to extracellular electron
acceptors are not well understood Three mechanisms have been proposed for exocellular
transport of electrons by exoelectrogens to anodes in MFCs so far Some exoelectrogenic
bacteria can transfer electrons to electrodes through soluble redox compounds (Bond and
6
Lovley 2005) These compounds include artificial mediators and mediators secreted by
exoelectrogenic bacteria A variety of chemicals have been used to facilitate the shuttling
of electrons from inside of cell to electrodes outside the cell These exogenous mediators
include neutral red (Park et al 1999) anthraquinone-2-6 disulfonate (AQDS) thionin
potassium ferricyanide (Bond et al 2002) methyl viologen and others (Logan 2004
Rabaey and Verstraete 2005) Some bacteria can secrete their own chemical mediator
for example Pseudomonas aeruginosa secretes pyocyanin and phenazine-1-carboxamide
to transfer electrons toward the MFCs anode (Rabaey et al 2005) Another evidence for
mediator production by bacteria is Geothrix fermentans When the medium was replaced
in a MFCs that had stable power generation with this bacteria power dropped by 50
and required 10 days to resume the original level
Some bacteria can directly transfer electrons to anodes via outer cell membrane
proteins (Myers and Myers 1992) The outer cell membrane protein cytochrome c is
thought to play a critical role in to transferring electrons to anodes Ly et al (2011)
isolated the haem protein cytochrome c and demonstrated that electric field effects may
be functional for the natural redox processes of cytochrome c in the respiratory chain
Shewanella oneidensis is the example of a bacterium that is capable of transfer electron to
anode via outer cell membrane proteins When Shewanella oneidensis adhered to an iron
surface the greater force has showed benefits to grow cells because closer contact
required for electron transfer from cell bound cytochromes (Lower et al 2001)
7
Fig1-2 Mechanisms for extracellular electron transport in an MFCs anode (1) direct
contact (top in green) (2) by nanowires (middle in purple) and (3) self-produced
mediators (bottom in blue) (Logan 2009)
More and more evidence supports the involvement of bacterial nanowires in
extracellular electron transport (Reguera et al 2005 2006 Gorby et al 2006) Nanowires
are conductive appendages produced by both Geobacter and Shewanella species (Gorby
and Beveridge 2005) The conductivity of the appendages was examined and confirmed
by using conductive scanning tunneling microscopy (STM) (Gorby et al 2006)
Nanowires can carry electrons from the cell to the anode surface of MFCs
The solid component of the extracellular biofilm matrix has high efficiency on
extracellular electron transfer compared with other extracellular electron transfer
8
mechanisms and recently Torres et al (2010) hypothesized that the solid component of
the extracellular biofilm matrix formed by exoelectogens is another way bacteria transfer
electrons to electrodes This hypothesis was based on kinetic analysis of each EET
mechanism reported in available literature (Torres et al 2010)
13 Anode electrodes
In MFCsMECs anode electrodes are a critical component because exoelectrogens
adhere to the surface of anodes to transfer electrons to the electrode The characteristics
of anodes have significant effects on electron transfer rate from bacteria to anode
electrodes in MFCs The requirements of an anode material are it should be highly
conductive non-corrosive have a high specific surface area (area per volume) high
porosity be non-fouling inexpensive and easily scaled to larger sizes Of these
properties the most important one that is different from other biofilm reactors is that the
material must be electrically conductive Normally they are made of various carbon
materials including carbon fiber carbon clothe and carbon paper due to their stability
high conductivity and high specific surface-area Nevertheless they have little
electrocatalytic activity for the anode microbial reactions and thus a modification of the
carbon materials is the main approach for improving their performance Consequently
there is a great need to develop a new type of anode material for MFCsMECs
It is a great challenge to develop a new anode material to further increase the power
density of a MFCs The nature of the catalytic mechanism of a MFCrsquos anode involves not
only a biological but also an electrocatalytic process An optimal nanostructure with a
high specific surface area favorable for both catalytic processes could play a critical role
in improving the power density of the MFCs such a structure needs to host the bacteria
9
with high bioactivity while enhancing the electron-transfer rate Schroumlder et al (2008)
employed PANI to modify a platinum anode for MFCs and achieved a current density 1
order of magnitude higher than the previously reported value PANIinorganic
composites are also reported to have better conductivity Qiao et al also applied a new
mesoporous TiO2 electrode material with uniform nanopore distribution and a high
specific surface area to anode in comparison to previously reported work with E coli
MFCs the composite anode delivers 2-fold higher power density (Qiao et al 2008)
Thus it has great potential for use as the anode in a high-power MFCs and may be a new
approach for improving performance of MFCs
14 Other parts of MESs
141 Membranes and ion transport
The ion exchange membrane is another one of the critical components in two-
chamber MESs systems It separates anode and cathode chambers and at the same time
maintains the electron neutrality of the system ie transport of electrons to the cathode
needs to be compensated by transport of an equal amount of positive charge to the
cathode chamber A proton exchange membrane (PEM) such as Nafion a per-fluorinated
sulphonic acid membrane (PFSAs) consisting of a hydrophobic fluorocarbon backbone to
which hydrophilic sulfonate groups (-SO3-) are attached is commonly used in chemical
fuel cell systems For MFCs systems however mainly cation species like Na+ and K
+
other than proton are often responsible for the dominant transport of positive charge
through the cation exchange membrane (CEM) to maintain electroneutrality due to the
low proton concentration in any aqueous medium with near neutral pH (Rozendal et al
2006) Consequently the pH increases in the cathode chamber due to the consumption of
10
protons and decreases in the anode chamber because of the accumulation of protons
(Rozendal et al 2007) This pH change in two-chamber systems results in a decrease of
the cathode potential and performance The application of anion exchange membrane
(AEM) has also been explored in MFCs systems (Cheng and Logan 2007 Kim and
Logan 2007) where it has been proposed that protons are transferred via pH buffers like
phosphate anions
142 Cathodes and Catalysts
The cathode is another challenge for making MFCs commercially available
technology because the chemical reaction that occurs at the cathode is difficult to
engineer as the electrons protons and oxygen must all meet at a catalyst in a tri-phase
reaction (solid catalyst air and water) The catalyst must be on a conductive surface and
must be exposed to both water and air so that protons and electrons in these different
phases can reach the same point The most commonly used material for a cathode is
commercially available carbon paper pre-loaded with a Pt catalyst on one side When it is
used in a MFC the side that contains the catalyst faces the water and the uncoated side
faces air To reduce the high cathode cost associated with platinum catalyst other precious-
metal-free cathodes such as (NiMo and NiW) were developed by electrodepositing on a
carbon fiber They have achieved comparable performance with Pt catalyst with same
loading at a much lower cathode fabrication cost (Hu 2010)
The requirements of cathode for MECs are quite similar to the requirements of
cathode for MFCs but easier than cathode for MFCs for the manufacturing process
because the cathode in MECs is not necessarily to exposed to air Therefore it can be
made of the exact same materials of cathode in MFCs except a waterproof layer
11
Recently some researchers have attempted to apply microorganisms as a biocatalyst to
precede the combination of electron with oxygen in the cathodes
15 Dissertation overview
Low power densities in MESs limit practical applications The improvement of
MESs performance requires a detailed understanding of the physiology and ecology of
microorganisms in MESs including the mechanism of electron transfer to the anode from
the microorganism
This dissertation focuses on the problem of the low power density of MESs The
bacterial community structure of a high power generated mixed culture communities in
MFCs will be identified firstly and then one of major exoelectrogens will be isolated and
characterized This information will be helpful to understand the physiology and ecology
of exoelectrogens in MESs Consequently they will be beneficial to improve power
density of MFCs Lastly the nanostructure decorated anodes in MECs will be utilized to
improve the power density The power enhancement mechanism will be explored by
using a whole genome microarray They are presented here as four papers
In the first paper the cultivation independent molecular biological techniques
denaturing gradient gel electrophoresis (DGGE) and 16S rDNA clone library are utilized
to analyze the bacterial community structure of a higher power mixed culture MFCs The
analyzed results provide fundamental information for isolating the dominant bacteria in
mixed culture MFCs Two of dominant bacterial species has been isolated and used aone
to test power generation in MFCs The possible interaction among different bacterial
species in mixed culture is discussed
12
In the second paper one isolated exoelectrogenic bacterial strain SX-1 is
characterized It is identified as a member of the Citrobacter genus and power generation
is tested ultilizing a wide range of different substrates The electron transfer mechanism is
explored using Cyclic Voltammetry (CV) This study increases the known diversity of
power generating exoelectrogens and provides a new strain to explore the mechanisms of
extracellular electron transfer from bacteria to electrodes
The third paper shows effects on MESs performance by Fe nanoparticle decorated
anodes in the MESs The average current density produced with Fe nanoparticle
decorated anodes increased up to 59-fold higher than plain graphite anodes A whole
genome microarray is utilized to analyze the possible mechanism of enhanced current
density as responded to nanoparticle decorated anodes
The fourth paper describes the effects of carbon nanotube (CNT) modified anode on
the performance S oneidensis MR-1 in MESs Results demonstrate that current densities
produced with CNT decorated anodes are up to 56-fold higher than plain graphite anodes
The possible mechanisms of enhanced current density by CNT decorated are explored
13
Chapter 2
Bacterial community analysis of mixed consortium in higher
power density MESs
Shoutao Xu and Hong Liu
1 Introduction
The improvement in the performance of mixed culture MFCs requires an
understanding of the ecology in microbial communities of MFCs Many researchers have
attempted to characterize microbial populations and activities to elucidate the behaviors
and ecology of microorganisms in MFCs (Wrighton et al 2008 Fedorovich et al 2009)
In order to study the microbial ecology of the mixed culture in a MFC and select the
appropriate isolation medium for dominant bacterial species in the mixed culture the
fundamental steps are to analyze the bacterial diversity in the mixed culture of MFCs and
identity the dominant bacterial species in bacteria communities in MFCs
14
As for the identification of bacterial communities typically there are two general
methods The first method for identification of bacterial community is the traditional
cultivation processes using selective nutrients to promote the growth of different types of
bacteria within the samples (Amman et al 2000) The community structure can then be
assessed by identifying the isolates from the dominant colonies that were cultured This
can often be costly and laborious as each isolate has to be further studied by examining
its physiology taxonomy and reactivity to stains (Adwards et al 1989)
The second method relies on utilizing molecular techniques to analyze bacterial
community DNA Several molecular methods involving the extraction and analysis of
DNA from entire bacterial communities are used to identify genetic fingerprints of
bacteria These methods including the cloning and sequencing of 16S rDNA automated
ribosomal intergenic spacer analysis (ARISA) terminal restriction fragment length
polymorphism (T-RFLP) and denaturing gradient gel electrophoresis (DGGE) generate
profiles of bacterial community structures They can rapidly assess complex communities
from various environments (Amman et al 2000)
Due to the conservative characteristic of 16S rDNA in bacteria during the process
of evolution 16S rDNA sequencing can be used to identify different species of bacteria
Therefore the molecular techniques of denaturing gradient gel electrophoresis (DGGE)
with PCR and 16S rDNA clone library are used for analysis of the microbial diversity
These methods are more convenient and save time compared to traditional
isolationcultivation methods for microorganism analysis
In this work a biofilm bacterial community from an anode of a MFCs wase studied
by using denaturing gradient gel electrophoresis (DGGE) of PCR-amplified partial 16S
15
rRNA genes followed by cloning and sequencing of 16S rDNA The results provided
essential information for dominant bacterial isolation in mixed culture MFCs
2 Materials and methods
21 High power generation of mixed culture MFCs
Mixed cultures were originally inoculated from domestic wastewater (Corvallis
Wastewater Treatment Plant Corvallis OR) to MFCs after operating for 2 years using a
defined medium solution (Lovely 2002) with sodium acetate as the carbon source A
new MFCs was inoculated from the operating MFCs The polarization curves were
performed to measure the power generation when maximal stable power were established
(normally 3-5 batches post-inoculation) and the genomic DNA from the microbial
biofilm on the anode was extracted under the sterile conditions for further bacterial
community analysis
22 Denaturing gradient gel electrophoresis (DGGE)
DGGE with PCR is a method of analysis of bacterial community composition
based on different GC percentage of 16s rDNA in different bacterial species PCR-DGGE
comprises three steps (i) extraction of total community DNA from the sample (ii) PCR-
controlled amplification using specific oligonucleotide primers and (iii) separation of the
amplicons using DGGE For this purpose a reproducible and efficient method for total
DNA extraction is indispensable and needs to be evaluated and optimized depending on
the nature of the sample In the subsequent PCR step multiple PCR primer sets with
different resolution can be used In most PCR-DGGE applications on bacteria universal
or specific primers are targeting the 16S rRNA gene Upon electrophoresis of the PCR
16
amplicons and gel staining (using ethidium bromide silver staining or SYBR green)
DGGE gels are digitally captured and further analyzed using computer software packages
The richness of DNA fragment bands in DGGE indicates the bacterial diversity of the
sample
The detailed steps are as followed Biofilms were scratched from the anodes of high
power generated MFCs fed with sodium acetate Bacterial genomic DNA was extracted
from the biofilm samples using the DNeasy blood and tissue kit (Qiagen) according to the
manufacturerrsquos instructions The universal primer set was used to amplify the 16S rDNA
from the extracted genomic DNA then the V3 region of bacterial 16S ribosomal DNA
(rDNA) was amplified with GC-clamp primer (Muyzer et al 1993) for DGGE PCR
amplification was performed in a thermocycler DGGE of the PCR products was carried
out in a DcodeTM
Universal Mutation Detection System) The 8 (wv) polyacrylamide
gels contains from 30 to 55 denaturing gradients Electrophoresis was conducted
using a 1timesTAE (Tris-Acetate-EDTA) buffer at 130V and 60degC for 5 hours After
electrophoresis the gel was stained with ethidium bromide in 1timesTAE buffer for 15
minutes and washed in 1timesTAE buffer for 10 minutes The fragments were visualized
under a UV transilluminator The richness of single band in DGGE gel picture
preliminarily showed that the bacterial diversity in mixed culture since the single band in
gel represents one bacterial species
23 16S rDNA clone library construction method
16S rDNA-DGGE fingerprinting has proven to be particularly useful as an initial
investigation into bacterial communities and is suitable for identifying the predominant
bacteria that comprise 1 or more of cells within a given sample In the 16s rDNA clone
17
library construction process the first step is the extratction of the total genomic DNA
then the genomic DNA of the mixed culture biofilm was used as template for PCR
amplification of approximately 1500 bp of 16S rDNA with universal primers The PCR
products of 16S rDNA were purified and inverted into pGEM-T Easy vector system
before they were transformed into competent E coli The transformed cells were spread
on Luria-Bertani (LB) plates containing the selective antibiotic ampicillin and X-Gal and
incubated overnight at 37 OC after the more than 80 inverted 16s DNA fragment E coli
colonies were picked up and inoculated in the 1-3 ml liquid LB medium and left to grow
for 16 hours The plasmid DNA were extracted and sequenced The sequences were
compared directly to all known sequences deposited in GenBank databases using the
basic local alignment search tool (BLAST)
24 Bioinformatics Analysis
16S rDNA sequencing results of more than 80 colones were queried against the
GenBank and Ribosomal Database Project (RDP) databases using BLAST and
SEQUATCH algorithms (Cole et al 2007 Wang et al 2007 Cole et al 2009) The
neighbor-joining trees were constructed with the Molecular Evolutionary Genetics
Analysis package (MEGA Version 4) based on 1000 bootstrap analysis (Tamura et al
2007)
25 Dominant bacterial species band in DGGE identification
The DGGE results gave the information of dominant bacterial richness of bacterial
species in mixed culture based on the theory that single band represents one bacterial
species which showed the diversity of bacterial community in the mixed culture of a
MFC 16S rDNA clone library results provided the whole picture of bacterial community
18
including the bacterial species name and bacterial species percentage in the mixed culture
The known pure bacterial species which have been sequenced can be used as markers to
identify dominant bacterial species in DGGE The whole experimental procedure is
similar to the procedure of the mixed culture DGGE steps The first step is to extract the
collect the mixed culture biofilm genomic DNA and pure bacterial species marker
genomic DNA Then the universal primer set was used to amplify the 16S rDNA from
the extracted genomic DNA After that the V3 region of bacterial 16S ribosomal DNA
(rDNA) was amplified with GC-clamp primer (Muyzer et al 1993) for DGGE The
subsequent steps were similar to the mixed culture DGGE experimental procedure The
dominant bacterial bands in DGGE were determined by comparisons of the single band
position in the mixed culture DGGE with single pure bacterial species location in DGGE
26 Dominant bacterial species isolation and power generation
The bacterial community of mixed culture in MFCs were predominantly composed
of Geobacter sp Arcobacter sp and Citrobacter sp Two dominant bacterial species
have been isolated
Citrobacter sp SX-1 was isolated from the anode biofilm by using a solid agar
medium containing NH4Cl 6mM KCl 2mM CH3COONa (electron donor) 30 mM
FeC6H5O7 (electron acceptor) 20mM minerals and vitamins (Lovley and Phillips 1988)
Single colonies were picked up after culturing on the plate for 36 hours at 30 degC and
transferred two times on the agar plate for purification Then the isolates grew in a liquid
medium solution in anaerobic tubes containing the same constituents as the solid
medium All isolation process was operated in a glove box anaerobic chamber (Coy
Laboratory Products Grass Lake MI)
19
Arcobacter sp SX-2 was isolated from the anode biofilm of a MFC fed with sodium
acetate by using the same method of isolation of pure bacterial species Citrobacter XS-1
except using Trypticase Soy Agar with 5 Sheep Blood solid (from American type
culture collection (ATCC)) agar medium instead of Citrobacter sp medium Single
colonies were picked up after culturing on the plate for 36 hours at 30 degC and transferred
twice on the agar plate for purification Then the isolates grew in a liquid medium
solution microaerobically
Geobacter sp have been trying to isolate by using three different methods They are
the colony pickup after different condition enrichment Goebacter medium isolation and
dilution to extinction by multiple channel mini MFCs isolation however the Geobacter
sp has not been isolated yet due to some limits so far But they provide valuable
information for further isolation of Geobacter sp
Single chamber MFCs were used to evaluate power generation by different isolates
The MFCs were constructed as described previously (Liu and Logan 2004) and modified
with 3 cm2 carbon cloth anodes and 7 cm
2 carbon clothPt cathodes The total liquid
volume of each MFCs was about 15 ml with electrode spacing about 17 cm All MFCs
were operated in an autoclaved closed plastic box and sterile cotton was attached to the
outer surface of the air cathodes to prevent contamination A MFCs without bacterial
culture was used as control MFCs were inoculated with 3 ml late exponential phase
cultures of SX-1 in the medium solution reported previously (Liu and Logan 2004)
3 Results and discussion
31 Power production by mixed culture MFCs
20
A polarization curve was used to characterize current as a function of voltage in the
MFCs The polarization curves are performed by varying a series of external resistances
The results are shown in Figure 21 With MFCs acclimated to 25 Ω external resistance
the maximum power density was achieved at 65Wm2 based on the polarization data At
this point the current density is 21mAcm2 The power density was three times higher
than previous report (20 Wm2) using phosphate buffer (Fan et al 2007) Power density
was also 2 times higher than (28 m Wm2) using a bicarbonate buffer ( Fan et al 2007)
32 Bacterial community structure assessed by PCR-DGGE
Bacterial DGGE profiles for higher power generating MFCs mixed culture setups
were relatively complex (more than 10 bands) However there are three brightly stained
bands and other less intense bands (Fig 2-2) These results indicated there were probably
three dominant bacterial species in mixed culture MFCs
010
020
030
040
050
060
070
2
3
4
5
6
7
04 05 05 06 07 08 10 11 13 15 21 22 23 24
Volta
ge (
V)
Pow
er
densi
ty (
Wm
2)
Current density (mAcm2)
Power
Voltage
power density 65wm2 at current
density208 mAcm2
Figure 2-1 Polarization curves of high power mixed culture MFCs
21
33 16S rDNA clone library results
A total of 83 colonies were sequenced for 16S rDNA clone library construction They
represented the bacterial community structure in mixed culture in MFCs The results
revealed that the predominant bacterial species in mixed culture are Geobacter sp (66
of the mixed culture) Arcobacter sp (12 of the mixed culture) and Citrobacter sp (11
of the mixed culture) Those three genuses comprised 88 of the total bacterial species
(Fig 23) This result is consistent with the DGGE result which showed that three bright
bands in the DGGE gel represented three dominant bacterial species in the mixed culture
The phylogenetic analysis of the colonies (83 colony sequences) is shown in Figure 24
Figure 22 DGGE result of high power generation mixed culture MFC (1 mixed
culture 2 pure culture control)
1 2
22
34 Dominant bacterial band in DGGE identification
Geobacter
66
Arcobacter
12
others
6
Pseudomonas
1
Citrobacter
11
Clostridium
2
Anaerovorax
2 Geobacter
Arcobacter
Citrobacter
Clostridium
Anaerovorax
Pseudomonas
others
Figure 23 Bacterial species and percentage of bacterial community of mixed
culture MFC lt 1 Alcaligensgt
lt1 Commamonasgt
lt1 Pseudomonas aeruginosagt
lt1 Klebsiellagt
lt11 Citrobactergt
lt1 Wolinellagt
lt12 Arobactergt
lt2 Anaeroboraxgt
lt2 Clostridium gt
lt1 unclear bacteriagt
lt66Geobactergt
100
100
85
100
100
82
99
87
100
002
Figure 24 Phylogenetic trees of bacterial species from mixed culture MFC
23
The dominant bacterial species including Goebacter sp Citrobacter sp and Arcobacter
sp in the mixed culture are shown in Figure 25 The upper band is Arcobacter sp and
Citrobacter sp is located in the bottom band of DGGE Geobacter sp is the middle one
The analysis of the bacterial community using denaturing gradient gel electrophoresis
(DGGE) of PCR-amplified 16S rRNA gene fragments and 16S rDNA clone library
construction method showed great phylogenetic diversity of mixed culture in MFCs with
the identification of sequences derived from bacteria of the taxa deltaproteobacteria
(Geobacter sp) gammaproteobacteria (Citrobacter sp) and epsilonproteobacteria
(Arcobacter sp) The result is consistent with the conclusion that among the isolated
exoelectrogens gammaproteobacteria and deltaprotebactia appear to be dominant among
others (Parot et al 2009 Liu et al 2010)
35 The dominant bacterial species isolation and power generation
Figure 25 Dominant bacterial species identification in DGGE
A B C D
A Citrobacter sp
B Geobacter sp
C Arobacter butzmeri
D mixed culture control
24
Two dominant bacterial species Citrobacter sp XS-1 and Arcobacter sp XS-2 have been
isolated and tested for power generation The power densities generated by the two
isolates are shown in the Fig 26 Current densities generated by isolate Citrobacter sp
and Arcobacter sp were 98 mAm2 and 20 mAm
2 respectively which were much less
than that (21Am2) generated by the mixed culture from which they were isolated
Figure 26 Current density generated by different dominant isolates
The maximum current density generated by mixed culture is much higher than the
one produced by pure bacterial species Citrobacter sp XS-1 and Arcobacter sp XS-2 in
the same structure MFCs Although the Geobacter sp in our mixed culture has not been
isolated a previous study showed that maximum current density generated by Geobacter
sp was 080 Am2
(Bond and Lovley 2003) The current density results seem partially
supporting the conclusion that mixed-culture generated higher power densities than their
pure-culture counterparts in MFCs (Jung and Regan 2007) This result suggests there
may be synergistic interactions within the anode bacterial communities
0
500
1000
1500
2000
2500
Mixed culture Citrobacter sp Arcobacter Sp
Cu
rre
nt
de
nsi
ty (
mA
m2)
25
Geobacter sp are strictly anaerobic bacteria which were enriched in most of the anode
biofilms (George 2005) Citrobacter species are facultative anaerobic bacteria (George
2005) which can be found in a wide variety of habitats including in soil water and
wastewater Arcobacter sp are micro-aerobic bacteria including both environmental
nonpathogens and opportunistic human pathogens They are able to grow in aerobic
conditions but in the optimal growth under micro-aerobic conditions (George 2005)
Arcobacter sp has been discovered in the MFCs by Fedorovich (Fedorovich et al 2009)
It is interesting that the mixed culture bacterial community composed of primarily these
three bacterial species has demonstrated to generate high power at non-strictly anaerobic
condition while leading dominant bacterial species Geobacter sp (66) is strict
anaerobic bacteria and generated powder in the MFCs only under strict anaerobic
condition (George 2005) Therefore it is possible that microaerobic bacteria such as
Arcobacter sp and facultative anaerobic bacteria Citrobacter sp can facilitate to create
anaerobic condition for Geobacter sp when they utilized oxygen for growing in the
mixed culture Syntrophic communities study have showed that anaerobic bacteria and
methanogenic archaea form compact microbial structures that operate like an organ rather
than a set of microorganisms functioning independently (Stams and Plugge 2009) Some
substrates have been degradeted within these communities while they are not able to be
fermented by individual species alone interspecies electron transfer also have been
presented in these communities (Stams and Plugge 2009) Summers et al (2010) also
discovered that direct exchange of electrons happened within coculture of Geobacter
metallireducens and Geobacter sulfurreducens Therefore we believe that there are
26
more complex synergistic interactions between different bacteria species in mixed culture
in the MFCs
Acknowledgements
The authors acknowledge support from the US National Science Foundation
(CBET 0955124) We also thank Keaton Lesnik for the review of this manuscript
27
Chapter 3
New Exoelectrogen Citrobacter sp SX-1 Isolated from a Microbial Fuel Cell
Shoutao Xu and Hong Liu
Published in
Journal of Applied Microbiology 111(5)1108-1115 (2011)
ABSTRACT
Exoelectrogenic bacterial strain SX-1 was isolated from a mediator-less microbial fuel
cell by conventional plating techniques with ferric citrate as electron acceptor under
anaerobic condition Phylogenetic analysis of the 16S rDNA sequence revealed that it
28
was related to the members of Citrobacter genus with Citrobacter sp sdy-48 being the
most closely related species The bacterial strain SX-1 produced electricity from citrate
acetate glucose sucrose glycerol and lactose in MFCs with the highest current density
of 205 mA m-2
generated from citrate Cyclic voltammetry analysis indicated that
membrane associated proteins may play an important role in facilitating the electrons
transferring from bacteria to electrode This is the first study that demonstrates that
Citrobacter species can transfer electrons to extracellular electron acceptors Citrobacter
strain SX-1 is capable of generating electricity from a wide range of substrates in MFCs
This finding increases the known diversity of power generating exoelectrogens and
provided a new strain to explore the mechanisms of extracellular electron transfer from
bacteria to electrode The wide range of substrate utilization by SX-1 increases the
application potential of MFCs in renewable energy generation and waste treatment
Keywords Microbial fuel cell Exoelectrogen Citrobacter sp SX-1 Extracellular
electron transfer
29
Introduction
Microbial Fuel Cells (MFCs) technology has become an active research area
recently as a promising approach for renewable energy generation wastewater treatment
and bioremediation (Rabaey and Verstraete 2005 Lovley 2006 Fan et al 2008 Logan
2009 Liu et al 2010) The key feature of MFCs system is the microbe-catalyzed electron
transfer from organic matter to anodes Many studies have shown the presence of diverse
bacterial communities on MFCs anodes (Reimers et al 2001 Liu et al 2004b Kim et al
2005) Over 20 exoelectrogens ie microbes that can transfer electrons exocellularly to
electrodes have been reported in the past 10 years (Logan 2009 Liu et al 2010) The
exoelectrogens so far belong to diverse genetic groups including alphaproteobacteria
(Rhodopseudomonas Ochrobactrum and Acidiphilium) (Malki et al 2008 Xing et al
2008 Zuo et al 2008) betaproteobacteria (Rhodoferax) (Chaudhuri et al 2003)
gammaproteobacteria (Shewanella Pseudomonas Klebsiella Enterobacter and
Aeronomas) (Kim et al 2002 Pham et al 2003 Rabaey et al 2005 Bretschger et al
2007 Zhang et al 2008 Chung et al 2009 Rezaei et al 2009) deltaproteobacteria
(Geobacter Geopsychrobacter Desulfuromonas and Desulfobulbus) (Bond et al 2002
Bond and Lovley 2003 Holmes et al 2004ab 2006) Epsilonproteobacteria (Arcobacter)
(Fedorovich et al 2009) Firmicutes (Clostridium and Thermincola) (Park et al 2001
Wrighton et al 2008) Acidobacteria (Geothrix) (Bond and Lovley 2005) and
actinobacteria (Propionibacterium) (Wang et al 2008) A wider range of
exoelectrogenic species are expected to be discovered
Three mechanisms have been proposed for exocellular transport of electrons by
exoelectrogens without artificial mediators Some exoelectrogenic bacteria can transfer
30
electrons to electrodes through soluble redox compounds excreted by microorganisms
(Bond and Lovley 2005 Rabaey et al 2005) and others can directly transfer electrons to
anodes via outer cell membrane proteins (Myers and Myers 1992) Recently more and
more evidence supports the involvement of bacterial nanowires in extracellular electron
transport (Reguera et al 2005 2006 Gorby et al 2006) In spite of the discovery of
many bacterial species that can transfer the electrons to electrode without the need of
artificial mediators the investigation of extracellular electron transfer mechanisms was
mainly focused on a few species such as those from Geobacter and Shewanella genera
(Kim et al 1999 2002 Reguera et al 2005 2006 Gorby et al 2006) The electron
transfer mechanisms for many of the isolated exoelectrogens species are still not well
studied
In the present study we isolated a new exoelectrogen strain SX-1 from a MFC a
strain phylogenetically related to Citrobacter sp Power generation from various carbon
sources by this strain was evaluated using single chamber MFCs Plausible extracellular
electron transfer mechanisms were also discussed based on the characterization of anodic
biofilms by cyclic voltammetry (CV)
Materials and methods
Bacterial strain SX-1 isolation
Bacterial strain SX-1 was isolated from the anode biofilm of a MFC fed with sodium
acetate operated in fed-batch mode over a period of six months The original source of
the inoculum is wastewater from a local waste water treatment plant Bacterial cells were
released from the carbon cloth anode by shaking the cloth in a sterile flask with 15 mL
sterile 1 NaCl solution and glass beads (2 mm diameter) The suspension was then
31
serially diluted from 10 times to 105
times and plated on a petri dish with a solid agar
medium containing NH4Cl 6mM KCl 2mM CH3COONa (electron donor) 30 mM
FeC6H5O7 (electron acceptor) 20mM minerals and vitamins (Lovley and Phillips 1988)
Single colony was randomly selected after culturing on the plate for 36 hours at 30 degC
and purified on a new agar plate following a procedure reported previously (Chung and
Okabe 2009) Then the isolate was picked up and grew in a liquid medium solution in
anaerobic tubes containing the same constituents as the solid medium for further analysis
All isolation process was operated in a glove box anaerobic chamber (Coy Laboratory
Products Grass Lake MI)
16S rDNA sequencing and phylogenetic analysis
Bacterial genomic DNA was extracted from the isolated strain SX-1 using the DNeasy
tissue Kits (Qiagen Valencia CA) according to the manufacturerrsquos instructions The 16S
rDNA was amplified by PCR using a pair of universal primers 27F (5rsquo-
AGAGTTTGATCMTGGCTCAG-3rsquo) and 1492R (5rsquo-GGTTACCTTTGTTACGACTT-
3rsquo) (Park et al 2001) The conditions used for PCR were described previously
(Ferna ndez et al 1999) PCR products were purified ligated and cloned according to a
procedure described in a previous report (Xing et al 2008) Plasmids were isolated from
randomly selected clone colonies with a QIAprep Spin Miniprep kit (Qiagen Valencia
CA) and three plasmid inserts were then sequenced in both directions using an ABI 3730
DNA sequencer (Applied Biosystems Carlsbad CA)United States) Due to the identical
16S rDNA sequence of the three colones only one was queried against the GenBank and
Ribosomal Database Project (RDP) databases using BLAST and SEQUATCH algorithms
(Cole et al 2007 Wang et al 2007 Cole et al 2009) The 16S rDNA sequence of
32
strain SX-1 and closely related type strains were aligned using CLUSTALX software
(Thompson et al 1997) The neighbor-joining trees were constructed with the Molecular
Evolutionary Genetics Analysis package (MEGA Version 4) based on 1000 bootstrap
analysis (Tamura et al 2007)
SEM
The morphologies of SX-1 cells growing overnight in anaerobic tubes were examined
with a scanning electron microscope (FEI Quanta 600 FEG FEI Company Hillsboro
OR) The suspended cells were fixed in a 25 glutaraldehyde and 01M phosphate
buffer solution and dehydrated with a graded ethanol series from 30 to 100 After
dehydration the samples were dried in a critical point dryer and then sputter-coated with
AuPd for SEM examination (Liu and Logan 2004)
Evaluation of power generation by SX-1 in MFCs
Single chamber MFCs were used to evaluate power generation by SX-1 using various
substrates The MFCs were constructed as described previously (Liu and Logan 2004)
and modified with 3 cm2 carbon cloth anodes and 7cm
2 carbon clothPt cathodes The
total liquid volume of each MFCs was about 15 ml with electrode spacing about 17 cm
All MFCs were operated in an autoclaved closed plastic box and sterile cotton was
attached to the outer surface of the air cathodes to prevent contamination A MFC
without bacterial culture was used as control MFCs were inoculated with 3 ml late
exponential phase cultures of SX-1 in the medium solution reported previously (Liu and
Logan 2004) Seven substrates including citrate acetate glucose sucrose glycerol and
lactose were evaluated individually for power generation in a fed-batch mode in a
temperature control chamber (30 plusmn2degC) For each substrate at least 8 batches were run to
33
investigate the effect of biofilm formation on current output at fixed external resistance of
1kΩ and the same initial substrate concentration of 30 mM Phosphate buffer (100mM)
was used to maintain the solution pH and conductivity The MFCs with sodium citrate as
electron donors were also examined for maximum power output by varying the external
resistance from 20 kΩ to 4 kΩ A data acquisition system was used to record voltage data
during the MFCs operation (Keithley 2700 Keithley Instruments Inc Cleveland OH) It
took about 15 to 30 minutes for the MFCs to stabilize depending on the external
resistance At each resistance we collected at least five data at the steady condition to
make the polarization curves Averaged voltages were used to calculate the power density
(mW m-2
) according to P=IVA where I was the current V was voltage and A was cross-
sectional area of the anode
CV analysis
CV was used to characterize the oxidation and reduction reactions on the anodic surface
of the MFCs fed with citrate and operated at 7 KΩ This external resistance was selected
because the maximum power density was obtained at this resistance based on the
polarization experiment We assumed the biofilm on the anode of MFCs were well-
developed when stable power output was obtained after 3 batches of operation The
MFCs were then used directly for CV analysis at four current generating stages of the
fourth batch (1) initial exponential current increasing stage (middle point of stage about
4 h after media change) (2) current plateau stage (middle point of stage about 8 h after
media change) and (3) current decreasing stage (middle point of stage about 16h after
media change) (4) right after the complete replacement of medium solution (Figure 6)
The anode was used as working electrode the cathode as counter electrode and an
34
AgAgCl electrode was selected as the reference The CV curves were scanned from 200
to -600 mV at a rate of 5 mV s-1
using a potentiostat (G300 Gamry Instrument
IncWarminster PA) Control experiment was also conducted using new anode (without
biofilm) and new medium solution
Nucleotide sequence accession number
The 16S r DNA sequence determined in this study has been deposited in the GenBank
database under accession number HQ845373
Results
Identification of the strain SX-1
An almost-complete 16S rDNA sequence (1475 bp) of strain SX-1 was obtained and
subjected to comparative analysis with the 16S rDNA of closely related reference strains
A phylogenetic analysis revealed that strain SX-1 should be assigned to the genus
Citrobacter and was most closely related to Citrobacter sp sdy-48 EJ463782 (990
sequence similarity) and Citrobacter sp yy-21 FJ463779 (975 sequence similarity)
These three strains formed a distinct sub cluster in the neighbor-joining in which the new
isolate and Citrobacter sp sdy-48 EJ463782 formed a distinct subline (Figure 31)
The SEM image illustrated that the strain SX-1 is a rod-shaped bacterium 03-06 microm
wide and 12-17 microm long (Figure 32) Strain SX-1 colonies on nutrient agar appeared 2-
3 mm in diameter smooth moist opaque with a shiny surface after 5 days incubation
under anaerobic condition The fact that strain SX-1 grew both aerobically and
anaerobically using FeC6H5O7 as electron acceptor suggests that the strain is facultatively
anaerobic Strain SX-1 showed robust growth on citrate as a carbon source These D
35
properties were similar to those of genus Citrobacter described in Bergeyrsquos Manual of
Systematic Bacteriology (George 2005)
Figure 31 phylogenetic tree of strain SX-1 and closely related species based on 16S
rDNA sequences The tree was constructed using the neighbor-joining method
Figure 32 SEM image of planktonic cells of Citrobacter sp SX-1
Strain SX-1
Citrobacter sp sdy-48 (FJ463782)
Citrobacter sp yy-21 (FJ463779)
Citrobacter koseri E639 (ATCC 25408)
Citrobater koseri CDC 3613-63(AF025372)
Citrobacter koseri CDC 8132-86(AF025366)
Salmonella enterica SL483 (CP001138)
Salmonella enterica AKU12601(AY696668)
Shigella sonnei Ss046(CP000038)
E coli C2 (AF403733)
E coli E24377A(CP000800)88
100
100
76
44
92
53
36
0001
1 microm
36
Electricity production by strain SX-1 in MFCs
The isolated strain SX-1 was first tested for its ability to generate current from sodium
citrate at 1kΩ Current gradually increased to a maximum 50 mA m-2
after cell
inoculation and then decreased (Figure 33) After the MFCs was refilled with new
substrate solution the current recovered rapidly and reached a higher level than the first
batch After 4 batches operation the maximum current output of each batch became
stable The highest current density achieved in the MFCs with Citrobacter sp SX-1 at 1
KΩ was 98 mA m-2
(Figure 33)
Figure 33 Electricity generation by Citrobacter sp SX-1 in a single chamber MFCs
with sodium citrate (30 mM) as substrate at 1 KΩ
0
20
40
60
80
100
120
0 50 100 150 200
Curr
ent
den
sity
(m
A m
-2)
Time (h)
37
Current generation by SX-1 from other substrates including glucose lactose sodium
acetate glycerol and sucrose were also assessed at an external resistance of 1KΩ
(Figure 34) When repeatable cycles of current output were obtained for these substrates
glycerol generated the highest maximum current density of 58 mA m-2
followed by
lactose and sucrose with 29 mA m-2
and 27 mA m-2
respectively Glucose and acetate
produced the lowest maximum current density of 96 and 43 mA m-2
respectively These
results indicated that strain SX-1 can utilize a wide range of substrates for electricity
generation in MFCs but with different power generation potentials
Figure 34 Electricity generation of Citrobacter sp SX-1 using different substrates in
single chamber air-cathode MFCs at 1 KΩ (The error bars stand for the standard
deviation of 3 replicates)
0
10
20
30
40
50
60
70
Glucose Lactose Glycerol Acetate Sucrose
Curr
ent
den
sity
(m
A m
-2)
38
Polarization experiment was further conducted to determine the maximum power
density generated by strain SX-1 in the single chamber MFCs with citrate as substrate A
maximum power density of 881 mW m-2
was obtained at current density of 205 mA m-2
at an external resistance of 7 KΩ (Figure 35)
Figure 35 Power and voltage generation by Citrobacter sp SX-1 as a function of
current density using sodium citrate (30 mM) as substrate The error bars stand for the
standard deviation of the 3 voltagepower outputs obtained in three MFCs
20
30
40
50
60
70
80
90
100
01
02
03
04
05
06
07
80 110 140 170 200 230
Po
wer
den
sity
(m
W m
-2)
Vo
ltag
e (V
)
Current density (mA m-2)
Voltage
Power
39
Figure 36 Four current generating stages for CV analysis current increasing stage (stage
1) current plateau stage (stage 2) current decreasing stage (stage 3) and right after the
replacement of medium solution (stage 4) The MFCs was operated at 7 KΩ with sodium
citrate (30 mM) as substrate
Cyclic Voltammetry
To determine the presence of redox active compounds produced by SX-1 and the
location of these compounds CV scan of the anodic biofilms at four current output stages
and the supernatant of MFCs medium solution at the end of the batch experiment were
performed Reduction peaks at range of -310 ndash -350 mV and oxidation peaks at range -
100 ndash -150 mV were observed at all current generating stages (Figure 36 Figure 37A)
suggesting the presence of redox active compounds may involve in extracellular electron
transfer by Citrobacter sp SX-1 The amplitude of the peaks increased according to the
growth stage of the batch and the highest peaks were present after the current plateau
stage which indicated the redox active compounds mainly were secreted in the current
plateau stage While the current density at stage 3 (deceasing stage) was lower than that
at stage 2 (current plateau stage) the redox peak at stage 3 was higher than that in stage 2
0
50
100
150
200
250
50 55 60 65 70 75 80 85
Curr
ent
den
sity
(m
A m
-2)
Time (h)
A
A
Stage 1
Stage 2 Stage 3
Stage 4
40
A plausible explanation for this pattern was that redox active compounds were
continuously being secreted and accumulated after the current peak was reached
resulting in more redox compounds present after the plateau stage But since most of the
carbon source had already been being used up many of the redox compounds at this
stage had not been as active as those at current plateau stage due to much less electrons
were available to be transferred The bigger peak amplitude in stage 4 than in stage 1
indicates that the redox active compounds were continuously being secreted and
accumulated in the biofilm even after stable power was generated at 7 kΩ The slightly
change of the peak locations at different stages was possible due to the solution chemistry
change during the current generation process or the slightly location change of reference
electrode during the measurement (Figure 37B) When the medium solution in the MFCs
was replaced by fresh medium at the end of the batch the oxidization and reduction
peaks were still presented in the CV (Figure 37B) but no peaks were observed when the
supernatant and fresh mediums were tested in a MFCs with no biofilm (Figure 37C)
These results suggest the compounds involved in the electron transfer were located not in
the supernatant (solution) but in the biofilm
41
Figure 37 Cyclic Voltammetry analysis of Citrobacter sp SX-1 biofilms at (A) current
increasing stage current plateau stage and decreasing stage (end of the batch) (B) end of
the batch and replaced with fresh medium and (C) controls
-40E-04
-30E-04
-20E-04
-10E-04
00E+00
10E-04
20E-04
30E-04
-060 -050 -040 -030 -020 -010 000 010 020
Curr
ent
(A)
Potential (V) vs AgAgCl
Current plateau stage
Current increasing stage
Current decreasing stage
-40E-04
-30E-04
-20E-04
-10E-04
00E+00
10E-04
20E-04
30E-04
-060 -050 -040 -030 -020 -010 000 010 020
Curr
ent
(A)
Potential (V) vs AgAgCl
Current decreasing stage
Biofilm with fresh medium
-40E-04
-35E-04
-30E-04
-25E-04
-20E-04
-15E-04
-10E-04
-50E-05
00E+00
-060 -050 -040 -030 -020 -010 000 010 020
Curr
ent(
A)
Potential (V) vs AgAgCl
Supernatant
Fresh medium
A
B
C
B C
42
Discussion
Citrobacter species belonging to Gammaproteobacteria Enterobacteriales
Enterobacteriaceae are facultative anaerobic bacteria (George 2005) They can grow with
oxygen as an electron acceptor but also with a variety of electron accepters such as Fe (III)
in the absence of oxygen (George 2005) Citrobacter species can be found in a wide
variety of habitats including in soil water and wastewater Several Citrobacter species
have been studied as important bioremediation bacterium for heavy metal removals
sulfate reduction phenol degradation and chlorophenol degradation (Macaskie et al 1995
Narde et al 2004 Qiu et al 2009) Citrobacter species have also been found on the
anodic biofilm of a diesel-degrading MFCs (Morris et al 2009) However there was no
study reported that Citrobacter species can transfer electrons to extracellular electron
acceptors This finding that Citrobacter strain SX-1 can generate electricity in MFCs
increased the diversity of power generating exoelectrogens and provided a new strain to
explore the mechanisms of extracellular electron transfer from bacteria to electrode
Among the isolated exoelectrogens gammaproteobacteria and deltaprotebactia appear to
be dominant among others (Parot et al 2009 Liu et al 2010) The isolated strain
Citrobacter sp SX-1 phylogenetically belongs to gammaproteobacteria which provides
another evidence to support this observation
Most exoelectrogens utilize a limited range of substrates for power generation For
example Shewanella oniedensis MR-1 can oxidize lactate under anaerobic condition but
cannot utilize acetate for electricity generation (Kim et al 2002) Geobacter
sulfurreducens can completely oxidize acetate for power generation but it cannot utilize
simple sugars (Bond and Lovley 2003 Caccavo et al 1994)) In comparison strain SX-1
43
not only can oxidize citrate for power generation but also utilize a wide range of
substrates for power generation including glycerol glucose lactose sucrose and acetate
Interestingly the higher current density generated by strain SX-1 from glycerol a main
by-product of biodiesel production process suggests that strain SX-1 may be potentially
used for harvesting energy from biodiesel wastes using MFCs However the current
density generated by strain SX-1 (205 mA m-2
) is lower than that (805 mA m-2
at 1 kΩ)
generated by the mixed culture from which the SX-1 was isolated indicating the
existence of other higher power generating bacteria andor complex ecology in the mixed
culture community
Understanding mechanisms of microbial extracellular electron transfer is critical for
enhancing the electron transfer rate from bacteria to electrode through metabolic or
genetic engineering of exoelectrogens Since the spent supernatant solution of MFCs run
with SX-1 showed no redox properties the redox compounds produced by SX-1 may
have been retained in the biofilm Alternatively SX-1 may utilize one or both of the
other two known extracellular electron transfer mechanisms For example it is possible
that strain SX-1 transfers electrons to the anode via cell-bound outer membrane proteins
The two distinct oxidation and reduction peaks observed at -100mv- -150 mV and -310--
350 mV of the SX-1 biofilm have a midpoint potential range from -205mV to -250mV
which is close to the redox potential of some c-type cytochromes (Field et al 2000 Meitl
et al 2009) which are well-known to play an important role in extracellular electron
trasnfer (Eggleston et al 2008 Field et al 2000 Meitl et al 2009) Field et al (2000)
reported that the c-type cytochromes CymA had the midpoint potentials of -229 mV
Meitl et al (2009) and Eggleston et al (2008) reported that the c- type cytochromes
44
OmcA in Shewanella oneidensis MR-1 had the midpoint potentials of -201 mV and -208
mV
While itrsquos possible that SX-1 may also have conductive appendages to enhance the
extracellular transfer electron similar to the nanowires discovered in some Geobacter
and Shewanella species (Reguera et al 2005 2006 Gorby et al 2006) further
investigation on the presence of these appendages on the cell surface of SX-1 is needed
Acknowledgements
The authors acknowledge support from the US National Science Foundation
(CBET 0955124) We also thank Teresa Sawyer for her helps with the SEM analysis and
Jeremy Chignell and Yanzhen Fan for their review of this manuscript
45
Chapter 4
Enhanced Performance and Mechanism Study of Microbial Electrolysis Cells
Using Fe Nanoparticles Decorated Anodes
Shoutao Xu Hong LiuYanzhen Fan Rebecca Schaller Jun Jiao Frank Chaplen
Published in
Applied Microbiology and biotechnology 93(2)871-880 (2012)
46
ABSTRACT
Anode properties are critical for performance of microbial electrolysis cells (MECs) In
the present study Fe nanoparticle modified graphite disks were used as anodes to
investigate the effects of nanoparticles on the performance of Shewanella oneidensis MR-
1 in MECs Results demonstrated that average current densities produced with Fe
nanoparticle decorated anodes were up to 56-fold higher than plain graphite anodes
Whole genome microarray analysis of the gene expression showed that genes encoding
biofilm formation were significantly up-regulated as response to nanoparticle decorated
anodes Increased expression of genes related to nanowires flavins and c-type
cytochromes indicate that enhanced mechanisms of electron transfer to the anode may
also have contributed to the observed increases in current density The majority of the
remaining differentially expressed genes were associated with electron transport and
anaerobic metabolism demonstrating a systemic response to increased power loads
Keywords Microbial electrochemical system microbial fuel cell microbial electrolysis
cell nanotechnology differential gene expression DNA microarray
47
Introduction
Microbial electrochemical systems (MESs) have been intensively studied since Lewis
achieved practical advances in this field (Logan 2007) however they attracted much
research attention in recent years due to their promising applications in renewable energy
generation bioremediation and wastewater treatment In a MES microorganisms
interact with electrodes via electrons catalyzing oxidation and reduction reactions at the
anode and the cathode The most-described type of MESs is microbial fuel cells (MFCs)
in which useful power is generated from electron donors typically biodegradable organic
materials (Logan et al 2006) Various novel MESs have recently been developed to
produce hydrogen (microbial electrolysis cells (MECs)) (Liu et al 2005 Logan et al
2008 Rozendal et al 2006 Schaller et al 2009) to power an autonomous sensor or
sensor network (Ringeisen et al 2006) to reduce bioremediation targets (Lovley 2006
Shea et al 2008) Sukkasem et al 2008) and to desalinate water (Cao et al 2009) The
key feature shared by these systems is the microbe-catalyzed electron transfer from
organic matter to electrodes (anodes) (Rabaey et al 2004) Enhancing the anodic current
output which highly depends on the performance of the electrodes is critical for the
successful application of all these processes (Logan et al 2007 Park and Zeilus 2002
2003)
Nanomaterials have received much attention from researchers in the context of
microbiology due to their unique physical electrical and chemical properties which
facilitate the study of interactions between bacteria and surfaces (Liu 2006) Previous
studies have demonstrated that electrodes decorated with different nanostructures such
as carbon nanotubes nanostructured polyanilinetitanium dioxide composite and titania
48
nanotubes have the potential to enhance power generation in MFCs (Morozan et al 2007
Qiao et al 2008 Quan et al 2005) Furthermore our recent study shows nanoparticle
(NP) decorated anodes greatly increased the electrochemical electron transfer rate in
MFCs (Fan et al 2011) However Fe NPs as an ideal candidate for decorating electrodes
because of respectively low price and high conductivity compared to other materials has
not been focused to study in MECs
S oneidensis MR-1 an important electrochemically active bacterial strain has been
exploited for the electricity generation in MFCs (Logan et al 2007 Logan 2009 Park
and Zeikus 2002) The availability of genome sequence for this strain makes it possible to
use transcriptome assays to globally measure the responses to different growth conditions
and environmental stresses (Murray et al 2001) S oneidensis MR-1 gene expression
response to heat shock cold shock Cr (VI) and U (VI) reduction chromate stress and
iron and acid tolerance have been studied in the past several years (Bencheikh-Latmani et
al 2005 Brown et al 2006 Gao et al2006 Gao et al 2004 Yang et al 2008) However
no studies have been focused on the Shewanella gene expression response to NPs in
MESs
The mechanism of increased electron transfer rate exhibited by nanoparticle decorated
electrodes is not well understood yet In the present study graphite disks decorated with
Fe NPs were used as anodes to explore the effects of nanostructures on current generation
in a multi-anode MECs DNA microarrays were utilized to investigate differences in the
global gene expression profile of S oneidensis MR-1 grown on plain versus Fe NPs
decorated anodes
Materials and methods
49
Bacterial cultures
S oneidensis MR-1 was purchased from American Type Culture collection (ATCC
700550) and stored as freezer stocks at -80degC Stock cells of S oneidensis MR-1 was
grown aerobically using a medium containing 30 gL Trypticase Soy Broth (BD 211825
Becton Dickinson Company Franklin lakes NJ) at 30degC Cells in early stationary
growth phase (optical density of 15-16 at 600nm (Spectrophotometer UV-1700
Shimadzu Columbia MD) were washed two times then injected into the chamber of the
MECs for current production Trypticase Soy Broth medium with 30 mM sodium lactate
as electron donor was used in MECs Phosphate buffer (100mM) was used to maintain
the solution pH 7 and solution conductivity at 15 mScm
Characterization of nanostructured anodes
Superfine isomolded graphite disks (16 cm x 05 cm GraphiteStorecom Inc) were
polished with ultrafine sand paper (2000 grit 3M Company) as the base for the NP
decorated and control anodes Fe NPs decorated anodes were fabricated by thermal
annealing of metal thin film coated graphite disks Thin films of Fe were firstly deposited
on the polished graphite disk by using sputter coating for 95 min and the samples then
were annealed in the Chemical vapor deposition (CVD) at 750degC for 20 min to form Fe
NPs Fe-NPs were produced with a diameter of 200plusmn100 nm and a density range
(distance between NPs) of 200plusmn100 nm with a coating time of 95 min SEM images of
Fe NPs decorated anode and control surfaces were shown in the Figure 41
50
Figure 41 SEM images of Fe NPs decorated anodes (A) Graphite disk control (B) Fe
Nanoparticle decorated
Multiple channel MECs construction and operation
A MECs with removable multiple anodes with each effective anode area 07 cm2
was
constructed and used to evaluate the effects of nanostructure on current density of
according to a previous report (Fan et al 2011) The cathode was made of wet-proof
(30) carbon cloth (type B E-TEK Division Inc USA) coated with PtC (20 E-TEK
Division Inc USA) following a previously reported procedure (Liu et al 2005) The
final platinum loading was 05 mgcm2 per projected cathode area The size of cathode
(150 cm2) was 25 times larger than the total effective area of the 8 graphite disk anodes
(56 cm2) to prevent cathode limitations on the performance of the MECs system All
MECs with Fe-NP decorated anodes and control anodes were tested for current
generation for 24 h with medium (without bacteria) before injecting the bacterial cells in
order to determine whether the Fe-NP decorated anodes demonstrate chemical current
generation compared to control anodes All the testing anodes were set up in the MECs
B
500 nm
A
500 nm
51
at once so that the S oneidensis MR-1 culture was applied to all of them simultaneously
Short electrode spacing (17 cm) was used in the MECs design to reduce the internal
resistance A voltage of 06 V was applied the MECs for current generation after the cells
of S oneidensis MR-1 were inoculated into MECs at early stationary phase A multimeter
with a data acquisition system (2700 Keithly USA) was used to monitor the current
change by measuring the voltage (every 300 seconds) drop through a resistor at 300 ohm
Fig 4 2 Effects of Fe-NP decorated anodes on the current density in MECs Control is
plain graphite disk anode Fe-NP is the anode with Fe-NP decoration
Microarray analysis
Biofilms for whole gene microarray analysis were aseptically removed from the plain
and Fe-NP decorated anodes of MECs after 110 hours further incubation at 30oC when
current density obviously started to decease Total RNA was extracted using Trizol
(Ambion inc AustinTX) following the manufacturerrsquos instructions Integrity of the
52
RNA samples was confirmed using the Agilent 2100 Bioanalyzer (Agilent Technology
Inc Palo Alto CA) The same amount of RNA (200 ng) from each sample was used for
further analysis after amplification by using the MessageAmpTM
II-Bacteria Prokaryotic
RNA Amplification kit (Ambion AustinTX) according to the manufacturerrsquos
instructions All of the 4295 genes (resource from NCBI) represented on the S oneidensis
MR-1 whole-gene microarrays were ordered from Roche NimbleGen Inc (Madison WI)
Biological triplicates of biofilms on the Fe-NP decorated anodes and control were
analyzed respectively cDNA synthesis labeling and hybridization were carried out by
the CGRB Core Laboratories at Oregon State University DNASTAR ArrayStarTM
3
software was used to identify genes that were up- or down-regulated more than 2-fold
when grown on the nanoparticle modified anodes using the unpaired two sample t-test
with a cutoff p-value of 005 The complete microarray data set generated in this study is
deposited for public access in the Gene Expression Omnibus
(httpwwwncbinlmnihgovgeo) under accession number GSE31535
Results
Enhancement of current generation by using NP decorated anodes in MECs
The current density generated by Fe-NP decorated anode MECs with S onidensis MR-1
increased to approximately 43 microAcm2 20 h following inoculation and then slowly
decreased Figure 42 shows that the maximum current density achieved by NP
decorated anodes was 83 times higher than that (51 microAcm2) generated by the control
(plain graphite disk) The average current density improvement of 110 hours was 59
times of that generated by the control The current densities generated in MECs in the
53
absence of bacteria were negligible (03 microAcm2) which demonstrated that the current
enhancement observed with the Fe-NP decorated anodes was biologically-derived
Global transcriptome analysis
Whole-genome DNA microarrays were used to attain a comprehensive general
overview
of the transcriptional response to Fe-NP decorated anodes by S oneidensis MR-1 in MEC
Of a total 4295 genes assayed 392 (188 induced and 204 repressed) exhibited significant
(P lt 005) differential expression at a 2-fold (log (expression ratio 1) level in 3
replicates in response to Fe-NPs These
total gene numbers present 9 of the 4295 open
reading frames (ORFs) presented on the array
Fig 43 Differentially expressed genes grouped by functional classification according to
the TIGR S oneidensis genome database (wwwtigrorg) Columns 1 amino acid
biosynthesis 2 biosynthesis of cofactors prosthetic groups and carriers 3 cell
envelope 4 cellular processes 5 central intermediary metabolism 6 DNA metabolism
7 energy metabolism 8 fatty acid and phospholipid metabolism 9 other categories 10
protein fate 11 protein synthesis 12 purines pyrimidines nucleosides and nucleotides
13 regulatory functions 14 signal transduction 15 transcription 16 transport and
binding proteins 17 unknown function 18 hypothetical proteins
Up-Regulated
-75
-50
-25
0
25
50
75
100
125
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18
Down-Regulated
Num
ber
of
Gen
es
54
Figure 43 summarizes the overall genomic response of S oneidensis MR-1 to Fe-NP
decorated anodes by grouping the differentially expressed genes into their functional role
categories based on The Institute for Genomic Researchrsquos annotation (Version 8) of the
MR-1 genome sequence The wide distribution of putative functional roles attributed to
the differentially expressed genes indicated the extent of the molecular response of
Soniedenis MR-1 to the NP decorated anodes Among genes with defined function a
large number of up-regulated genes were associated with cellular processes (group 4)
energy metabolism (group 7) other categories (group 9) and hypothetical proteins (group
18) While those genes encoding proteins involved in amino acid biosynthesis (group 1)
cellular processes (group 4) substrate transport (group 16) and hypothetical proteins
(group 18) were among the most down-regulated genes Two groups of genes that were
the most noticeable among the functional gene groups genes related to energy
metabolism (group 7) and genes related to amino acid biosynthesis (group 1) The ratios
of up-regulated genes to down-regulated genes in these functional groupings were much
higher than other functional gene groupings there were 21 up-regulated genes and 7
down-regulated genes in energy metabolism gene group and 2 up-regulated and 22 down
regulated genes in the amino acid biosynthesis group
Genes related to biofilm formation
In order to determine whether there was a correlation between the observed biofilm
enhancements by NP decorated anodes and the genes related to biofilm formation the
significant modified genes related to biofilm formation were examined and summarized
in the Table 41 These included SO_3223 and SO_3228 genes encoding the flagellum
proteins (Thormann et al 2004) which has critical impacts on initial attachment to the
55
surface as swimming motility functions were up-regulated over 10 and 09 fold
respectively The gene clusters SO_4178-SO_4180 which are thought to be essential for
formation of a three-dimensional biofilm were up-regulated from 12 to 13 fold
(Thormann et al 2004) In addition the genes SO_4105 encoding IV pilus which is
known to represent a critical factor for biofilm formation on a biotic surfaces (Thormann
et al 2004) were also induced 08 fold (Table 41) The genes related to flagellum
motility (SO_4286 SO_1529 and SO_1530) were also induced slightly (Thormann et al
2004)
Table 41 Expression levels of genes related to biofilm formation
Locus Tag Gene product descriptions Fold change
SO_3228 flagellar basal-body MS-ring and collar protein FliF 10
SO_3223 flagellar hook-length control protein FliK 09
SO_4103 MshA minor pilin protein MshD 05
SO_4105 MSHA major pilin protein MshA 08
SO_4178 expressed protein of unknown function MxdC 12
SO_4179 inner membrane family 2 glycosyltransferase MxdB 13
SO_4180 diguanylate cyclase-like protein MxdA 10
a fold changes were presented after log2
b positive number represented up-regulated and negative represented down-regulated
Genes related to energy metabolism
A closer consideration of energy metabolism genes was undertaken as anaerobic
metabolism for electron generation and electron transport functions may play critical
roles in enhanced current density generation in MECs Genes associated with energy
56
metabolism and with significantly modified gene expression levels are summarized in the
Table 42 Several of the up-regulated genes were related to formate dehydrogenase
proteins Most notably four genes related to formate dehydrogenase were up-regulated
more than 2-fold SO_0102 SO_0103 SO_0104 and SO_0107 The other genes
expression related to formate dehydrogenase proteins SO_4513 SO_4514 SO_4515 also
significantly increased Another interesting gene is that encoding the cytochrome c
oxidase protein gene SO_4609 which was up-regulated 11 fold while the gene
SO_0479 related periplasmic octaheme cytochrome c MccA was down-regulated
However SO_1519 the gene encoding lactate dehydrogenase was down-regulated 12
fold despite lactate being the primary carbon source in the media
Table 4 2 Genes related to anaerobic growth and electron transfer with significantly change expression level
Locus Tag Gene product descriptions Fold
change
SO_0101 nitrate-inducible formate dehydrogenase molybdopterin-binding
subunit FdnG
18
SO_0102 formate dehydrogenase nitrate-inducible iron-sulfur subunit 21
SO_0103 formate dehydrogenase nitrate-inducible cytochrome b556 subunit 24
SO_0104 formate dehydrogenase accessory protein FdhE 22
SO_0107 formate dehydrogenase accessory protein fdhD 24
SO_0397 quinolfumarate reductase menaquinol-oxidizing subunit FrdC_2 12
SO_0452 thioredoxin 2 15
SO_0903 Na-translocating NADH-quinone reductase subunit B NqrB_1 12
SO_1012 NADH-ubiquinone oxidoreductase subunit K NuoK 12
SO_1013 NADH-ubiquinone oxidoreductase subunit J NuoJ 12
SO_1363 hydroxylamine reductase 13
57
SO_2417 ferredoxin cofactor maintenance protein YfaE 10
SO_3922 formate dehydrogenase cytochrome b Fdh -12
SO_4513 formate dehydrogenase molybdopterin-binding subunit _2 15
SO_4514 formate dehydrogenase FeS subunit FdhB_2 12
SO_4515 formate dehydrogenase cytochrome b subunit FdhC_2 14
SO_4609 aa3 type cytochrome c oxidase subunit III CoxC 11
SO_0479 periplasmic octaheme cytochrome c MccA -12
SO_0847 periplasmic nitrate reductase ferredoxin component NapG -15
SO_1519 L-lactate dehydrogenase iron-sulfur cluster-binding protein LldF -10
SO_1251 ferredoxin 4Fe-4S -11
SO_37411 hypothetical inner membrane protein -11
a fold changes were presented after log2
b positive number represented up-regulated and negative represented down-regulated
Flavin and cytochrome related genes
Flavins can be secreted by Shewanella species as electron shuttle to facilitate
extracellular electron transfer (Cansterin et al 2008) C-type cytochromes play the
important roles on the process of extracellular electron transfer (Shi et al 2007) The
genes related to flavins synthesis and the genes encoding the cytochromes electron
transport proteins were shown in the table 43 the genes SO_1414 and SO_3058 related
to flavins synthesis was up-regulated to 03 and 01 folds respectively The genes related
to s C-type cytochromes SO_0135 SO_4105 SO_0169 SO_1778 are increased to 10
08 05 04 01 folds respectively However most of genes have shown no significant
changes (less one-fold change) as response to nano particle decorated anode in MECs
58
Table 43 Expression level of flavin and cytochrome genes
Locus Tag Gene product descriptions Fold
change
SO_1414 flavocytochrome c flavin subunit putative 03
SO_3468 riboflavin synthase subunit alpha -02
SO_3058 flavocytochrome c flavin subunit 01
SO_4105 MSHA pilin protein MshA 08
SO_0169 general secretion pathway protein GspG 05
SO_1778 outer membrane decaheme cytochrome c lipoprotein MtrC 04
SO_1779 outer membrane decaheme cytochrome c OmcA 01
SO_0135 lipoprotein of unknown function DUF333 10
SO_0136 conserved hypothetical inner membrane protein 01
a fold changes were presented after log2
b positive number represented up-regulated and negative represented down-regulated
Other genes with significantly modified expression levels
These genes over 15 fold change and possibly related to current enhancements were
shown in Table 44 SO_3104 (expressed inner membrane protein) was over expressed
and the gene SO_2194 which encodes outer membrane protein (OmpA family protein)
was repressed as response to nanoparticle decorated anode however it is unclear the
relationships of this modified membrane protein genes with the current enhancement
Another interesting phenomenon was possible co-regulation of several gene clusters
indicating possible operon associations This includes three sets of genes that were
induced more than 15 fold SO_0108SO_0109 and SO_1042SO_1043SO_1044 The
59
consistency of expression of these genes under the NP conditions provides basic evidence
to support operon structure However the correlation of these significant changed gene
with current density enhance are unclear which showed there are unknown multiple and
complex responses of S oneidensis MR-1 to nanoparticle anode of MECs
Table 44 Other genes with significantly changed expression levels
Locus Tag Gene product descriptions Fold change
SO_0101 nitrate-inducible formate dehydrogenase molybdopterin-binding
subunit FdnG
18
SO_0108 integral membrane protein of unknown function DUF39 YedE 17
SO_0109 SirA family protein YedF 17
SO_0275 N-acetyl-gamma-glutamyl-phosphate reductase -31
SO_0277 ornithine carbamoyltransferase -21
SO_0279 argininosuccinate lyase -24
SO_0404 zinc dependent metalloprotease domain lipoprotein 21
SO_0956 alkyl hydroperoxide reductase F subunit -20
SO_1042 amino acid ABC transporter ATP-binding protein -18
SO_1043 amino acid ABC transporter permease protein -27
SO_1044 amino acid ABC transporter periplasmic amino acid-binding protein -27
SO_1072 chitin-binding protein putative 21
SO_1405 transglutaminase family protein 20
SO_1822 TonB-dependent receptor putative -22
SO_2069
1-(5-phosphoribosyl)-5-[(5-phosphoribosylamino)
methylideneamino] imidazole-4-carboxamide isomerase
-19
SO_2070 amidotransferase HisH -26
SO_2071 imidazole glycerol-phosphate dehydratasehistidinol phosphatase -27
60
SO_2072 histidinol-phosphate aminotransferase -28
SO_2073 histidinol dehydrogenase -30
SO_2194 OmpA family protein -36
SO_2195 inter-alpha-trypsin inhibitor domain protein -27
SO_2767 asparagine synthetase B -26
SO_2945 prophage LambdaSo tail fiber protein 20
SO_2963 prophage LambdaSo major capsid protein HK97 family 20
SO_2987 prophage LambdaSo hypothetical protein 19
SO_2988 prophage LambdaSo expressed protein of unknown function 18
SO_3104 expressed inner membrane protein 131
SO_3408 conserved hypothetical inner membrane protein -18
SO_3585 azoreductase putative -18
SO_3586 glyoxalase family protein -17
SO_3687 curli production assemblytransport component CsgE putative -18
SO_38191 hypothetical ammonia permease -19
SO_4014 AcrBAcrDAcrF family protein 18
SO_40151 type I secretion system membrane fusion protein RND family 18
SO_4054 510-methylenetetrahydrofolate reductase 18
SO_4245 N-acetylglutamate synthase -22
SO_45251 hypothetical transcriptional regulator LysR family -17
SO_4527 integral membrane domain protein -19
SO_4705 transcriptional regulator putative -19
a fold changes were presented after log2
b positive number represented up-regulated and negative represented down-regulated
Discussion
61
The addition of NP to anodes significantly impacts current densities with the elemental
composition being a critical factor The chemical composition of NPs significantly affects
the current enhancement in MECs (Fan et al 2011) because the chemical composition of
NP properties is a critical factor in determining the conductivity which significantly
affects the efficiency of electron transfer from bacteria to electrode Fe is an ideal
candidate for NP formation for current enhancement in MESs because of high
conductivity and low price compared to gold and other conductive materials Gold NPs
has also been studies for enhancement of electricity conductivity (Bao et al 2008)
However the disadvantages of the high price of gold limit their practical application in
MECs (Fan et al 2011) The different size and density of the same composition of NP
also has essential effects on the current density enhancement (Fan et al 2011) In this
study the results showed the current density enhancement with Fe-NP size range of
200plusmn100 nm and density range of 200plusmn100 nm is considerable enhancement consequence
There are a number of reports of studies of current density enhancement in MFCs
by nanostructured anodes (Fan et al 2011 Qiao et al 2008 Sharma et al 2008 Tsai et al
2009) Based on bacterial behavior on nanostructured anode surface Qiao et al (2008)
believes that Escherichia coli cells on the nanostructured electrode surface produce hair-
like structures similar to pili that could facilitate the electron transfer between the cells
and electrode The production of hair-like structures is believed to be stimulated by
nanostructures and could play the key role on current density enhancement in pili-
producing bacteria in MFCs (Qiao et al 2008) S oneidensis can produce electrically
conductive bacterial nanowires which have similar structure to the hair-like structures in
Ecoli (Gorby et al 2006 Reguera et al 2005) A number of possible genes related
62
nanowires of S oneidensis have been identified including mshA (SO_4105) and gspG
(SO_0169) (Konstantinidis et al 2009) They are up-regulated 08 and 05 fold in the
current study respectively even though they are not significant induced This result
supported the increased nanowire genes had contributions to enhance current density in
MFCs by NPs decorated anodes
Based on the previous research on electron transfer mechanisms for S oneidensis it is
known that self-produced mediators (flavins) and multihaem c-type cytochromes (c-Cyts)
may play critical roles in the electron transfer of S oneidensis to extracellular insoluble
electrode (Canstein et al 2008 Myers and Myers 1992 Scott and Nealson 1994 Shi et al
2007) The flavins secreted by S oneidensis can facilitate to transfer electrons from
bacteria to solid anode electrodes by acting as extracellular electron shuttles (Canstein et
al 2008) In this study the genes encoding flavins (SO_1414 SO_3468 and SO_3058)
had no significant increase (less than 1 fold change) as a response to nanoparticle
decorated anode in MECs which suggests that increased flavin gene expression does not
significantly contribute to the current density enhancements in nano-particle decorated
anode The genes encoding cytochromes electron transport proteins (SO_4104 SO_0417
SO_177879 SO_013536) had no significant increase (less than 1-fold change)
However those genes were up-regulated slightly These results suggest the increased
expression flavin and c type cytochromes genes had partial contributions even not
significantly to enhance current density in MFCs by NPs decorated anodes
The thickness of bacterial biofilm on the anode can affect the power generation in
MFCs (Biffinger et al 2007 Heydorn et al 2000) since increased biofilm thickness on
the anode represents more active bacteria involving in the process of electron generation
63
and transfer in MFCs Biofilm formation related genes included flagellar genes IV pilus
genes and other genes The flagellar related genes SO_3228 SO_3229 (fliB and fliK) and
IV pilus genes SO_4105 (mshA) were induced from 19 to 09 fold the genes (SO_4178
-4180) related to formation of a three-dimensional biofilm were up-regulated from 10 to
13 folds even though the genes related bacterial motility protein (SO_ 4286 SO_4287
SO_1529 and SO_1530) are induced slightly (less than 0 5 fold) However the motility
genes most probably play a role for initial biofilm attachment on the anode surfaces
(Thormann et al 2004 Thormann et al 2006) with the rest of biofilm genes being more
essential for biofilm maintenance in mature cultures as were presented at harvest in this
study This enhancement of biofilm density under conditions of increase current density
has been observed for gold NP decorated anodes using confocal light microscopy (data
not shown) but has not yet been confirmed for Fe NP decorated anodes In any event an
increase in the expression level of genes related biofilm formation therefore also
supported the possibility of enhanced biofilm formation on NPs decorated anodes thus
facilitating electron transfer from bacteria to anodes
Beliaev et al described a model of the Fe reduction pathway in S oneidensis MR-1 using
solid substrates as the electron acceptor (Beliaev et al 2001) This model proposes that
electrons are generated and released in cytoplasm then transferred to the quinone pool
eg menaquinone in the cytoplasmic membrane The reduced menaquinone in turn
reduces CymA and from there electrons are then transferred to a periplasmic shuttle (eg
CctA) and then to MtrA a periplasmic soluble decaheme cytochrome c located in
associated with the outer membrane via interaction with MtrB Finally surface displayed
outer membrane proteins decaheme cytochromes (OmcA MtrC or MtrF) can transfer
64
electron to Fe or other solid substrates (Beliaev et al 2001)) Bencheikh-Latmani et al
demonstrated that the same electron transport pathway may be used for more than one
electron acceptor through studies of the response of S oneidensis MR-1 to U(VI) and
Cr(VI) under anaerobic conditions particularly focused on the critical genes
(SO_1777mtrA SO_1776mtrB SO_1778mtrC) (Bencheikh-Latmani et al 2005)
However our studies showed that these genes had no significant expression changes in
response to nanostructured anodes Contrastingly one of expressed inner membrane
protein genes SO_3104 and one of cytochromes genes SO_4609 encoding cytochrome c
oxidase subunit III demonstrated a 13-fold and 11-fold up-regulation respectively in this
study while the gene SO_2194 which encodes outer membrane protein (OmpA family
protein) was down regulated 36 fold Our results therefore suggest that S oneidensis
MR-1 does not utilize the same set of core critical genes (SO_1777 mtrA SO_1776
mtrB SO_1778 mtrC) products to transfer electrons in all instances This is the similar
result with the study of Bretschger et al (2007) which indicated that the electron
transport system in S oneidensis MR-1 is complex with several different proteins able to
participate in electron transfer to the anode of MECs
It should also be mentioned that the gene (SO_1519) encoding lactate
dehydrogenase repressed (-12 fold) here suggesting that lactate was depleted at the time
of cell harvest from the MECs and that other pathways of anaerobiosis had been activated
including those associated with amino acid uptake and consumption the growth media
was a complex mixture including yeast extract In particular the catabolic pathway for
histidine of S oneidensis MR-1 a pathway that may formate as an intermediate is up-
regulated with the genes for histidine ammonia lyase SO_0098 SO_3057 and SO_4374
65
up-regulated 13 11 and 11-fold respectively The possible production of formate
during histidine degradation may in turn explain the high levels of formate
dehydrogenates as evidenced by upregulation of SO_0102 SO_0103 SO_0104 and
SO_0107 annotated as formate dehydrogenase proteins These genes had the highest
levels of up-regulated expression as response to Fe NP decorated anodes (greater than 2-
fold)
Acknowledgements
This research was partially supported by the US National Science Foundation CBET
0828544 and the funds from ONAMIDOD (ARL-DOD Cooperative Agreement
W911NF-07-2-0083) We thank Barbara Gvakharia and Caprice Rosato for helpful
suggestions We also thank Margaret Romine of Pacific Northwest Laboratories for
valuable comments on manuscripts We also thank anonymous reviewers for significant
suggested improvements imparted as part of prior review of this manuscript
66
Chapter 5
Global Transcriptome Analysis of the Response of Shewanella oneidensis MR-1 to
Carbon Nanotube Decorated Anodes in Microbial Electrochemical Systems
Shoutao Xu Yanzhen Fan Rebecca Schaller Frank Chaplen Jun Jiao Hong Liu
67
Abstract
Shewanella oneidensis MR-1 is an important model microorganism for metabolic studies
on the effects of different environmental factors because of its diverse respiratory
capabilities Carbon nanotube (CNT) modified graphite disks were used as anodes to
investigate the effects of nanomatrials on the performance of S oneidensis MR-1 in
microbial electrolysis cells (MECs) The current densities produced with CNT decorated
anodes were on average 56-fold higher than plain graphite anodes Whole genome
microarray analysis of gene expression showed that up-regulation of cytochromes c genes
associated with extracellular electron transfer are strongly correlated to current increases
in CNT decorated anodes Genes involved in flavin biosynthesis also contribute to
current increase in CNT decorated anode MECs
Keywords Microbial electrochemical system microbial fuel cell microbial
electrolysis cell carbon nanotube gene expression DNA microarray
68
Introduction
Shewanella oneidensis MR-1 is an important model microorganism for metabolic
studies of the effects of different environmental factors because of its diverse respiratory
capabilities It has been used for transcriptome analysis to investigate the responses to
different growth conditions and environmental stresses (Murray et al 2001) Gene
expression patterns under different conditions such as heat shock cold shock Cr (VI) U
(VI) reduction chromate stress iron and acid tolerance have been studied previously
(Bencheikh-Latmani et al 2005 Brown et al 2006 Gao et al2006 Gao et al 2004
Yang et al 2008) More recently it has been exploited as a model species for power
generation in microbial electrochemical systems (MESs)which have potential
applications in renewable energy generation bioremediation and wastewater treatment
(Logan et al 2007 Logan 2009 Park and Zeikus 2002 Fan et al 2011 Xu et al 2012)
In a MES electrochemically active microorganisms oxidize organic matter in the an
ode chamber to release electrons Electrons are then transferred to the anode electrode thr
ough various mechanisms (Logan et al 2006 Lovley 2006 Rabaey et al 2003) and fina
lly travel to the cathode electrode and combine with the terminal electron acceptor The
key feature of MESs is the microbe-catalyzed electron transfer from the organic matter to
the anode (Rabaey et al 2004) Enhancing the current output which highly depends on
the performance of the anode electrode is critical for the successful application of MESs
(Logan et al 2007 Park and Zeilus 2002 2003) Several nanomaterials including TiO2
gold (Au) and palladium (Pd) nanoparticles have been utilized to decorate electrodes to
enhance power density in MESs (Qiao et al 2008 Quan et al 2005 Fan et al 2011) The
possible mechanisms for increased current densities have been studied by using Fe
69
nanoparticle-decorated anodes (Xu et al 2012)
Carbon nanotubes (CNT) are cylindrically shaped nanomaterials with extremely
high surface area excellent electrical conductivity and chemical inertness (He et al
2005a Serp et al 2003) These unique properties make CNT a promising electrode
material (Liang et al 2008) The biocompatibility of microorganisms and carbon
nanostructures has been evaluated (Morozan et al 2007) The effects of CNT on anodic
biofilms have been tested in MFCs (Liang et al 2011) However no studies have been re
ported on global transcriptome analysis for the response of S oneidensis MR-1 to CNT
decorated anode in MECs
In the present study the graphite disks decorated with CNT were used as anodes to
investigate the effects of nanomaterials on current generation in multi-anode MECs
DNA microarrays were used to analyze differences in the global gene expression profile
of S oneidensis MR-1 grown on plain versus CNT decorated anodes
Materials and methods
Bacterial cultures S oneidensis MR-1 was purchased from American Type Culture
collection (ATCC 700550) and stored as freezer stocks at -80degC Stock S oneidensis
MR-1 cells were grown aerobically using M1 medium at 30degC Cells in early stationary
growth phase (optical density of 15-16 at 600nm (Spectrophotometer UV-1700
Shimadzu Columbia MD) were injected into the chamber of the MECs for current
production Sodium lactate (final concentration 30mM) was added as the additional
electron donor
Fabrication and characterization of nanostructured anodes Superfine isomolded
graphite disks (16 cm x 05 cm GraphiteStorecom Inc) were polished with ultrafine
70
sand paper (2000 grit 3M Company) as the base for the CNT decorated and control
anodes Aligned multi-walled carbon nanotube samples were synthesized via plasma
enhanced chemical vapor deposition (PECVD) Samples superfine isomolded graphite
pillars with a 16 cm diameter and 05 cm thickness were sputtered with a thin film of Ni
in a CRC 300 magnetron sputter coater for 1 to 2 min They were then placed in the
PECVD reactor where the Ni was annealed into small catalyst islands C2H2 was bled
into the chamber for growth and an NH3 plasma was used to vertically align the growth
of the multi-walled carbon nanotubes (MWCNTs) The temperature and time were varied
between 725degC and 775oC and 15 to 60 min to control the length of the tubes and density
of samples Samples were then analyzed in a FEI Sirion field emission scanning electron
microscope (FESEM) SEM images of CNT decorated anode and control surfaces were
shown in the Figure 51
Figure 51 SEM images of CNT decorated anodes (A) Graphite disk control (B) CNT
decorated anode
Multiple channel MECs construction and operation A MECs with multiple
removable anodes each with an effective anode area of 07 cm2
was constructed and used
to evaluate the effects of nanostructures on the current density of according to a previous
B A
71
study (Fan et al 2011) The cathode was made of wet-proof (30) carbon cloth (type B
E-TEK Division Inc USA) coated with PtC (20 E-TEK Division Inc USA)
following a previously reported procedure (Liu et al 2005) The final platinum loading
was 05 mgcm2 per projected cathode area The size of the cathode (150 cm
2) was 25
times larger than the total effective area of the 8 graphite disk anodes (56 cm2) to prevent
cathode limitations on the performance of the MECs system All MECs with CNT
decorated anodes and control anodes were tested for current generation for 24 h with
sterile a medium (without bacteria) before injecting the bacterial cells in order to
determine whether the CNT decorated anodes demonstrate chemical current generation
compared to control anodes All the testing anodes were set up in the MECs
simultaneously so that the S oneidensis MR-1 culture was applied to all of them
simultaneously Short electrode spacing (17 cm) was used in the MECs design to reduce
the internal resistance A voltage of 06 V was applied the MECs for current generation
after the cells of S oneidensis MR-1 were inoculated into the MECs in the early
stationary phase A multimeter with a data acquisition system (2700 Keithly USA) was
used to monitor the current change by measuring the voltage drop through a resistor
Microarray analysis Biofilms for whole gene microarray analysis were aseptically
removed from the plain and CNT decorated anodes of MECs after 80 hours further
incubation at 30oC when current density obviously reached to a stable phase Total RNA
was extracted using Trizol (Ambion inc AustinTX) following the manufacturerrsquos
instructions Integrity of the RNA samples was confirmed using the Agilent 2100
Bioanalyzer (Agilent Technology Inc Palo Alto CA) Total RNA (200 ng) from each
sample was amplified using the MessageAmpTM
II-Bacteria Prokaryotic RNA
72
Amplification kit (Ambion AustinTX) according to the manufacturerrsquos instructions The
S oneidensis MR-1 whole-gene microarrays representing all of the 4295 genes (resource
from NCBI) were ordered from Roche NimbleGen Inc (Madison WI) Biological
triplicates of biofilms on the CNT decorated anodes and control were analyzed cDNA
synthesis labeling and hybridization were carried out by the CGRB Core Laboratories at
Oregon State University DNASTAR ArrayStarTM
3 software was used to identify genes
that were up- or down-regulated more than 2-fold when grown on the nanoparticle
modified anodes using the unpaired two sample t-test with a cutoff p-value of 005 with
bonferroni correction
Results and discussion
Enhancement of current generation using CNT decorated anodes in MECs The
current density generated by CNT decorated anode MECs with S onidensis MR-1
increased to 36 microAcm2 10 hours post-inoculation and reached a maximum of 40 microAcm
2
at 29 hours inoculation then gradually decreased The current density stabilized at
30microAcm2 after 78 h operation Figure 52 shows that the current density curve generated
by CNT decorated anode in MECs The average current density generated by CNT
decorated anodes was 56 times that of the control The current density generated in the
MECs without bacteria was negligible (data not shown) This result demonstrates that the
current enhancement observed with the CNT decorated anodes was biologically-derived
A
73
Figure 52 Effects of CNT decorated anodes on the current density in MECs (3 replicates)
Control is plain graphite disk anode CNT is the anode with CNT decoration (Error bars
represent standard deviation)
It has been reported that nanostructured decorated anodes have significant impacts on
current densities (Fan et al 2011 Qiao et al 2006 Sharma 2008 Tsai et al 2009) The
CNT decorated anodes had significantly enhanced current densities in MECs in this
study This result is consistent with previous studies Interestingly the current density
curve generated by CNT decorated anode MECs is quite different from the current
density curve generated by Au NP and Pd NP decorated anodes using same bacterial
species (Fan et al 2011) In the Au and Pd NP decorated anode MECs the current
density curve was not significantly increased in the beginning compared with the control
but gradually increased after the inoculation the current density reached the maximum
current density more than 50 hours after the inoculation However the current density
generated by CNT decorated anode MECs started to increase significantly immediately
after the inoculation It reached a maximum current density 29 hours post-inoculation
-1
6
13
20
27
34
41
48
0 10 20 30 40 50 60 70 80
Curr
ent
den
sity
(micro
Ac
m2)
Time (Hours)
Control
CNT
74
These results indicate that there might be different current density enhancement
mechanisms between CNT with other metal NP decorated anodes in MECs
Global transcriptome analysis Whole-genome DNA microarrays were used to attain a
comprehensive general overview of the transcriptional response of S oneidensis
MR-1 of
S oneidensis MR-1 to CNT decorated anodes in MECs In a total of 4295 genes analyzed
457 genes (250 genes up-regulated and 207 genes down-regulated) exhibited significantly
(P lt 005) changed expression at a 2-fold level in at least 3 replicates in the response to
CNT The total number of the regulated genes present 11 of the 4295 open reading
frames (ORFs) presented on the microarray The total number of genes at significant
expression levels in response to CNT decorated anodes is close to that to Fe NP decorated
anodes in MECs (392 9) ( Xu et al 2012)
Fig 53 Differentially expressed genes grouped by functional classification according to
the TIGR S oneidensis genome database (wwwtigrorg) Columns 1 amino acid
biosynthesis 2 biosynthesis of cofactors prosthetic groups and carriers 3 cell
envelope 4 cellular processes 5 central intermediary metabolism 6 DNA metabolism
7 energy metabolism 8 fatty acid and phospholipid metabolism 9 other categories 10
protein fate 11 protein synthesis 12 purines pyrimidines nucleosides and nucleotides
-80
-60
-40
-20
0
20
40
60
80
100
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18
Nu
mb
er
of
Ge
ne
s
Up-regulated
Down-regulated
75
13 regulatory functions 14 signal transduction 15 transcription 16 transport and
binding proteins 17 unknown function 18 hypothetical proteins
The overall transcript genomic response of S oneidensis MR-1 to CNT decorated
anodes was summarized in Figure 53 by grouping the differentially expressed genes into
their functional role categories based on The Institute for Genomic Researchrsquos annotation
(Version 8) of the MR-1 genome sequence The wide distribution of putative functional
roles attributed to the differentially expressed genes indicated the extent of the molecular
response of Soniedenis MR-1 to the CNT decorated anodes A large number of up-
regulated genes were presented in genes associated with biosynthesis of cofactors
prosthetic groups and carriers (group 2) cell envelope (group 3) protein fate (group 10)
protein synthesis (group 11) and purines pyrimidines nucleosides and nucleotides
(group 12) in this study The number of up-regulated genes was two times more than that
of down-regulated genes in each functional group Contrastingly in the genomic response
S oneidensis MR-1 to Fe NP decorated anodes (chapter 4) most of up-regulated genes
were associated with cellular processes (group 4) energy metabolism (group 7) other
categories (group 9) and hypothetical proteins (group 18) The down-regulated genes
presented in the amino acid biosynthesis (group 1) cellular processes (group 4) substrate
transport (group 16) and hypothetical proteins (group 18) While those genes encoding
proteins involved in amino acid biosynthesis (group 1) central intermediary metabolism
( group 5) DNA metabolism( group 6) fatty acid and phospholipid metabolism (group
8) and other categories (group 9) were among the most down-regulated genes in this
study However the two groups of genes that were the most noticeable between the
genomic response S oneidensis MR-1 to CNT and Fe NP decorated anodes which they
are the genes related to energy metabolism (group 7) and hypothetical proteins (group
76
18) These results indicated there are different and common genomic responses to S
oneidensis MR-1 on CNT and Fe NP decorated anodes in MECs
Electron transfer related genes Based on previous electron transfer mechanism
studies on Shewanella a serial of group proteins collectively described as the Mtr
pathway play the role to transfer electron rom the inner bacterial body to the outer
membrane (Beliaev et al 2001 Shi et al 2007 Brutinel and Gralnick 2012) In the Mtr
pathway firstly electrons generated by bacteria received by CymA a tetraheme c-type
cytochrome anchored in the inner membrane then electrons were transferred to MtrA a
periplasmic decaheme c-type cytochrome After that electrons were transferred from
MtrA to MtrC an outer-membrane decaheme c-type cytochrome is facilitated by MtrB a
non-heme containing integral outer-membrane proteinOuter-membrane decaheme c-type
cytochrome MtrC and OmcA are in change of electron transfer to exreacellular electron
acceptor (Shi et al 2007 Brutinel and Gralnick 2012 ) These genes encoding electron
transport proteins are shown in the table 51 The critical genes SO_1777 mtrA SO_1776
mtrB SO_1778 mtrC are unregulated 41 to 80 fold in response to CNT decorated
anodes other cytochrome c genes related to electron transfer in MtrA pathway also are
significantly increased in response to CNT decorated anodes These genes encoding outer
membrane proteins have different level increases supporting that the increase of
cytochrome c gene expression contributes to the current enhancements of MECs as
response to CNT decorated anodes
77
Table 51 Expression level of cytochrome c as response to CNT decorated anodes
Locus Tag Gene product descriptions Fold change CNT
SO_0165 general secretion pathway protein GspC 31
SO_0167 general secretion pathway protein GspE 45
SO_1776 outer membrane protein precursor MtrB 41
SO_1777 outer membrane decaheme cytochrome c
lipoprotein MtrA
80
SO_1778 outer membrane decaheme cytochrome c
lipoprotein MtrC
43
SO_1779 outer membrane decaheme cytochrome c
OmcA
63
SO_0135 lipoprotein of unknown function DUF333 22
SO_0136 conserved hypothetical inner membrane protein 24
Flavin related genes It is known that self-produced mediators play critical roles in the
electron transfer of S oneidensis to extracellular insoluble electrode (Canstein von et al 2
008 Myers et al 1992 Scott et al 1994 Shi et al 2007) It has been proved that the
flavins secreted by S oneidensis can facilitate electron transfer from bacteria to solid
anode electrodes by acting as extracellular electron shuttles (Cansterin von et al 2008) T
wo critical genes (SO_1414 and SO_3468) related to flavin synthesis were shown increa
sed significantly (more than 2 folds) increase in this study (Table 52) This result support
s the hypothesis that flavin production increases as the response to CNT decorated anodes
in MECs have significant contribution to the current density enhancements
78
Table 52 Expression levels of genes related to flavin synthesis as response to CNT
decorated anode
Locus Tag Gene product descriptions Fold change CNT
SO_1414 flavocytochrome c flavin subunit putative 22
SO_3468 riboflavin synthase subunit alpha 20
The amount of bacterial biofilm biomass on the anode can affect the power
generation in MFCs (Biffinger et al 2007 Heydorn et al 2000) Biofilm formation
related genes include flagellar gene IV pilus genes and other genes the flagellar related
genes (fliB and fliK) and IV pilus genes (mshA) were induced only from 105 to 134
fold as response of S oneidensis MR-1 to CNT decorated anodes in this study No
biofilm-associated genes presented significant changes in expression levels in this study
indicating there is no direct connections between biofilm enhancements to CNT
decorated anodes in this study This result is consistent with the Liang et al (2011)
conclusion that using CNT enhanced the electrochemical activity of anodic biofilms but
did not result in a significant increase of biomass in the anodic biofilms
Our microarray results showed two significantly up-regulated gene groups the
genes encoding proteins localized on the outer membrane and the genes involved in
flavin biosynthesis contributed to current density enhancement by CNT decorated anodes
Among 457 significantly changed genes to CNT in this study there are also a relatively
large number of genes encoding proteins with unknown functions which are either up- or
down-regulated in the response to CNT decorated anodes which indicated more
79
complicated responses of S oneidensis MR-1 to CNT decorated anodes in MECs and
further study is needed
80
Chapter 6
Summary
Microbial electrochemical systems (MESs) with a mixed culture initially inoculated
from Corvallis wastewater treatment plant have been studied for more than 6 years for
varying purposes including power generation hydrogen production heavy metal
removal and wastewater treatment Experiment results have shown that the mixed culture
is quite stable with excellent performance in MESs Our community analysis using
denaturing gradient gel electrophoresis (DGGE) and 16S rDNA clone library construction
suggests that the mixed culture is composed predominantly of Geobacter sp (66)
Arcobacter sp (12) and Citrobacter sp (11) These results not only improved our
understanding of the mixed culture community but also guided our studies on the
cultivation and isolation of the different bacterial species
Exoelectrogenic Citrobacter sp SX-1 was first isolated and characterized from a
MFC operated with the mixed culture Citrobacter sp SX-1 has been demonstrated to
produce electricity from wide range of different substrates including citrate acetate
glucose sucrose glycerol and lactosein MFCs Cyclic voltammetry analysis indicated
that membrane associated proteins may play an important role in facilitating the electrons
transferring from bacteria to electrode The strain SX-1 increased the known diversity of
power generating exoelectrogens and provided a unique bacterial species for study in
renewable energy generation and waste treatment
81
To enhance the electron transfer from bacteria to anode nano decorated anodes
including Fe NP and CNT were developed and characterized and evaluated in MECs
using Shewanella oneidensis MR-1 as a model species Both nanostructures have
significantly increased current density compared with the control Whole genome
microarray analysis elucidated the possible mechanisms of power enhancement in
response to these nano-decorated anodes
These results benefit to understanding of physiology and ecology of mixed
cultures in MFCs and improve the efficiency of current generation in MESs which will
facilitate the viability of niche applications for MESs in near future
82
Bibliography
1 Amman R Ludwig W (2000) Ribosomal RNA-targeted nucleic acid probes for
studies in microbial ecology FEMS Microbiol Ecol 24 555ndash565
1 Bao SJ Li CM Zang JF Cui XQ Qiao Y Guo J (2008) Novel Nanoporous TiO2
Electrode for Directly Electrochemistry of glucose oxidase Adv Funct Mater
18(4) 591-599
2 Beliaev AS Saffarini DA McLaughlin JL Hunnicutt D (2001) MtrC an outer
membrane decahaem c cytochrome required for metal reduction in Shewanella
putrefaciens MR-1 Mol Microbiol 39722-30
3 Bencheikh-Latmani R Williams SM Haucke L Criddle CS Wu L Criddle CS
Zhou J Tebo BM (2005) Global transcriptional profiling of Shewanella oneidensis
MR-1 during Cr(VI) and U(VI) reduction Appl Environ Microbiol 71 7453ndash7460
4 Biffinger JC Pietron J Ray R Little B and Ringeisen BR (2007) A biofilm
enhanced miniature microbial fuel cell using Shewanella oneidensis DSP10 and
oxygen reduction cathodes Biosens Bioelectron 221672ndash1679
5 Bodelier PLE Roslev P Henckel T FenzelP (2000) Stimulation by ammonium
based fertilizers of methane oxidation in soil around rice root Nature 40 421ndash424
6 Bond DR Holmes DE Tender LM Lovley DR (2002) Electrode-reducing
microorganisms that harvest energy from marine sediments Science 295 483-485
7 Bond DR Lovley DR (2003) Electricity production by Geobacter sulfurreducens
attached to electrodes Appl Environ Microbiol 69 1548-1555
8 Bond DR Lovley DR (2005) Evidence for involvement of an electron shuttle in
electricity generation by Geothrix fermentans Appl Environ Microbiol 71 2186-
2189
9 Bretschger O Obraztsova A Sturm CA Chang IS Gorby YA Reed SB Culley
DE Reardon CL Barua S Romine MF Zhou J Beliaev AS Bouhenni R
Saffarini D Mansfeld F Kim BH Fredrickson JK Nealson KH (2007) Current
production and metal oxide reduction by Shewanella oneidensis MR-1 wildtype and
mutants Appl Environ Microbiol 73 7003ndash7012
10 Brown SD Thompson MR Verberkmoes NC Chourey K Shah M Zhou J
Hettich RL and Thompson DK (2006) Molecular dynamics of the Shewanella
oneidensis response to chromate stress Mol Cell Proteomics 51054-1071
11 Brutinel ED Gralnick JA (2012) Shuttling happens soluble flavin mediators of
extracellular electron transfer in Shewanella Appl Microbiol Biotechnol 93(1) 41-8
12 Butler JE Kaufmann F Coppi MV Nuacutentildeez C Lovley DR (2004) MacA a diheme
c-type cytochrome involved in Fe(III) reduction by Geobacter sulfurreducens J
Bacteriol 186(12) 4042ndash4045
83
13 Caccavo F Lonergan DJ Lovley DR Davis M Stolz JF McInerney MJ (1994)
Geobacter sulfurreducens sp nov a hydrogen- and acetate oxidizing dissimilatory
metal-reducing microorganism Appl Environ Microbiol 60 3752ndash3759
14 Canstein von H Ogawa J Shimizu S Lloyd JR (2008) Secretion of flavins by
Shewanella species and their role in extracellular electron transfer Appl Environ
Microbiol 74 615ndash623
15 Cao X Huang X Liang P Xiao K Zhou Y Zhang X Logan BE (2009) A New
Method for Water Desalination Using Microbial Desalination Cells Environ Sci
Technol 43(18)7148-7152
16 Chaudhuri SK Lovley DR (2003) Electricity generation by direct oxidation of
glucose in mediatorless microbial fuel cells Nat Biotechnol 21 1229-1232
17 Cheng S Liu H Logan B E (2006) Increased power generation in a continuous
flow MFC with advective flow through the porous anode and reduced electrode
spacing Environ Sci Technol 402426-2432
18 Cheng S Logan B E (2007) Sustainable and efficient biohydrogen production via
electrohydrogenesis PNAS 104 18871-18873
19 Chignell J (2010) Energy Recovery from Biodiesel Waste Performance of
Microbial Electrochemical Systems on Glycerol Thesis
20 Chung K Okabe S (2009) Characterization of electrochemical activity of a strain
ISO2-3 phylogenetically related to Aeromonas sp isolated from a glucose-fed
microbial fuel cell Biotechnol Bioeng 104(5) 901-910
21 Cole JR Chai B Farris RJ Wang Q Kulam-Syed-Mohideen AS McGarrell
DM Bandela AM Cardenas E Garrity GM Tiedje JM (2007) The ribosomal
database project (RDP-II) introducing myRDP space and quality controlled public
data Nucleic Acids Res 35 (Database issue) D169-D172 doi 101093nargkl889
22 Cole JR Wang Q Cardena E Fish J Chai B Farris RJ Kulam-Syed-Mohideen
AS McGarrell DM Marsh T Garrity GM Tiedje JM (2009) The Ribosomal
Database Project improved alignments and new tools for rRNA analysis Nucleic
Acids Res 37 (Database issue) D141-D145 doi 101093nargkn879
23 Dey A De S De A De SK (2004) Characterization and dielectric properties of
polyaniline-TiO2 nanocomposites Nanotechnol 15 1277-1283
24 Edwards U Rogall T Bloumlcker H Emde M Boumlttger EC (1989) Isolation and
direct complete nucleotide determination of entire genesmdashcharacterization of a gene
coding for 16S-ribosomal RNA NucleicAcids Res 17 7843ndash7853
25 Eggleston CM Voumlroumls J Shi L Lower BH Droubay TC Colberg PJS (2008)
Binding and direct electrochemistry of OmcA an outer-membrane cytochrome from
an iron reducing bacterium with oxide electrodes a candidate biofuel cell system
Inorg Chim Acta 361769ndash777
26 Fan Y Sharbrough E Liu H (2008) Quantification of the Internal Resistance
Distribution of Microbial Fuel Cells Environ Sci Technol 42 8101ndash8107
84
27 Fan Y Xu S Schaller R Jiao J Chaplen F Liu H (2011) Nanoparticle decorated
anodes for enhanced current generation in microbial electrochemical
cells Biosensors and Bioelectronics 26 (5)1908ndash1912
28 Fan Y Hu H Liu H (2007)Sustainable power generation in microbial fuel cells
using bicarbonate buffer and proton transfer mechanisms Environ Sci
Technol 41(23) 8154-8
29 Fedorovich V Knighton MC Pagaling E Ward FB Free A Goryanin I (2009)
Novel electrochemically active bacterium phylogenetically related to Arcobacter
butzleri isolated from a microbial fuel cell Appl Environ Microbiol 75(23) 7326-
7334
30 Ferna ndez A Huang S Seston S Xing J Hickey R Criddle C Tiedje J (1999)
How stable is stable Function versus community composition Appl Environ
Microbiol 65 3697ndash3704
31 Field SJ Dobbin PS Cheesman MR Watmough NJ Thomson AJ Richardson
DJ (2000) Purification and magneto-optical spectroscopic characterization of
cytoplasmic membrane and outer membrane multiheme c-type cytochromes from
Shewanella frigidimarina NCIMB400 J Biol Chem 275 8515ndash8522
32 Freguia S Masuda M Tsujimura S Kano K (2009) Lactococcus lactis catalyses
electricity generation at microbial fuel cell anodes via excretion of a soluble quinone
Bioelectrochemistry 7614ndash18
33 Fricke K Harnisch F and Schroumlder U (2008) On the use of cyclic voltammetry for
the study of the anodic electron transfer in microbial fuel cells Energy Environ
Sci 1 144ndash147
34 Gao H Wang Y Liu X Yan T Wu L Alm E Arkin A Thompson DK and
Zhou J (2004) Global transcriptome analysis of the heat shock response of
Shewanella oneidensis J Bacteriol 1867796-7803
35 Gao H Yang ZK Wu L Thompson DK Zhou J (2006) Global transcriptome
analysis of the cold shock response of Shewanella oneidensis MR-1 and mutational
analysis of its classical cold shock proteins J Bacteriol 1884560-4569
36 George MG (2005) Bergeyrsquos Manual of Systematic Bacteriology Volume Two The
Proteobacteria Part B The Gammaproteobacteria Second Edition New York
Springer 651p
37 Gonzaacutelez-Toril E Llobet-Brossa E Casamayor EO Amann R Amils R (2003)
Microbial ecology of an extreme acidic environment the Tinto river Appl Environ
Microbiol 69(8) 4853ndash4865
38 Gorby YA Yanina S Mclean JS Rosso KM Moyles D Dohnalkova A
Beveridge TJ Chang IS Kim BH Kim KS Culley DE Reed SB Romine M
F Saffarini DA Hill EA Shi L Elias DA Kennedy DW Pinchuk G
Watanabe K Ishii S Logan B Nealson KH Fredrickson JK (2006) Electrically
conductive bacterial nanowires produced by Shewanella oneidensis strain MR-1 and
other microorganisms Proc Natl Acad Sci USA 10311358-11363
85
39 He Z Wagner N Minteer SD Angenent LT (2006) An upflow microbial fuel cell
with an interior cathodes assessment of the internal resistance by impedance
spectroscopy Environ Sci Technol 40 5212-5217
40 Heydorn A Nielsen AT Hentzer M Sternberg C Givskov M Ersboslashll BK Molin
S (2000) Quantification of biofilm structures by the novel computer program
COMSTAT Microbiol 1462395-2407
41 Holmes DE Bond DR Lovley DR (2004a) Electron transfer by Desulfobulbus
propionicus to Fe(III) and graphite electrodes Appl Environ Microbiol 701234ndash
1237
42 Holmes DE Chaudhuri SK Nevin KP Mehta T Methe BA Liu A Ward JE
Woodard TL Webster J Lovley DR (2006) Microarray and genetic analysis of
electron transfer to electrodes in Geobacter sulfurreducens Environ Microbiol
81805-1815
43 Holmes DE Nicoll JS Bond DR Lovley DR (2004b) Potential role of a novel
psychrotolerant member of the family Geobacteraceae Geopsychrobacter
electrodiphilus gen nov sp nov in electricity production by a marine sediment
fuel cell Appl Environ Microbiol 70 6023ndash6030
44 Holmes DE Chaudhuri SK Nevin KP Mehta T Metheacute BA Liu A Ward JE
Woodard TL Webster J Lovley DR (2006) microarray and genetic analysis of
electron transfer to electrodes in Geobacter sulfurreducens Environ Microbial
8(10)1805-1815
45 Hu H (2009) Enhancing Hydrogen Production in Microbial Electrolysis Cells
through Development of Platinum-free Cathode and Improvement of Reactor
Dissertation
46 Ince BK Ayman O N Turker G Ccedilelikkol S Ince O (2010) Microbial ecology of
anaerobic reactors for treatment of alcohol industry wastewaters a review Current
research technology and education topics in applied microbiology and microbial
ecology 988-999
47 Jung S Regan JR (2007) Comparison of anode bacterial communities and
performance in microbial fuel cells with different electron donors Appl Microbiol
Biotechnol 77393ndash 402
48 Kim BH Kim HJ Hyun MS Park DH (1999) Direct electrode reaction of Fe(III)
reducing bacterium Shwwanella putrefaciens J Microbial Biotechnol 9127-131
49 Kim GT Hyun MS Chang IS Kim HJ Park HS Kim BH Kim SD Wimpenny
JW Weightman AJ (2005) Dissimilatory Fe(III) reduction by an electrochemically
active lactic acid bacterium phylogenetically related to Enterococcus gallinarum
isolated from submerged soil J Appl Microbiol 99(4) 978ndash987
50 Kim GT Webster G Wimpenny JW Kim BH Kim HJ Weightman AJ (2006)
Bacterial community structure compartmentalization and activity in a microbial fuel
cell J Appl Microbiol 101(3) 698ndash710
86
51 Kim HJ Park HS Hyun MS Chang IS Kim M Kim BH (2002) A mediator-
less microbial fuel cell using a metal reducing bacterium Shewanella putrefaciens
Enzyme Microb Technol 30 145-152
52 Kim JR Min B Logan BE (2005) Evaluation of procedures to acclimate a
microbial fuel cell for electricity production Appl Microbiol Biotechnol 68(1)23-30
53 Kim JR Cheng S Oh SE Logan BE (2007) Power generation using different
cation anion and ultrafiltration membranes in microbial fuel cells Environ Sci
Technol 41 1004-1009
54 Konstantinidis KT Serres MH Romine MF Rodrigues JL Auchtung J McCue
LA Lipton MS Obraztsova A Giometti CS Nealson KH Fredrickson JK
Tiedje JM (2009) Comparative systems biology across an evolutionary gradient
within the Shewanella genus Proc Natl Acad Sci U S A 106(37)15909-15914
55 Lee J Phung NT Chang IS Kim BH Sung HC (2003) Use of acetate for
enrichment of electrochemically active microorganisms and their 16S rDNA
analyses FEMS Microbiol Lett 223185ndash191
56 Liang P Wang H Xia X Huang X Mo Y Cao X Fan M(2011)Carbon
nanotube powders as electrode modifier to enhance the activity of anodic biofilm
in microbial fuel cells Biosens Bioelectron 26(6)3000-4
57 Lies DP Hernandez ME Kappler A Mielke RE Gralnick JA Newman DK
(2005) Shewanaella oneidensis MR-1 uses overlapping pathways for iron reduction
at a distance and by direct contact under conditions relevant for biofilms Appl
Environ Microbiol 71 4414-4426
58 Liu H and Logan B E (2004) Electricity generation using an air-cathode single
chamber microbial fuel cell in the presence and absence of a proton exchange
membrane Environ Sci Technol 38 4040-4046
59 Liu H Grot S Logan BE (2005) Electrochemically assisted microbial production
of hydrogen from acetate Environ Sci Technol 39 4317 ndash 320
60 Liu H Hu H Chignell J Fan Y (2010) Microbial Electrolysis Novel Technology
for Hydrogen Production from Biomass Biofuels 1(1) 129ndash142
61 Liu H Ramnarayanan R Logan BE (2004) Production of electricity during
wastewater treatment using a single chamber microbial fuel cell Environ Sci Technol
382281-2285
62 Liu WT Marsh TL Cheng H Forney LJ (1997) Characterization of microbial
diversity by determining terminal restriction fragment length polymorphisms of gene
encoding 16S rRNA Appl Environ Microbiol 63 4516ndash4522
63 Logan B E Hamelers B Rozendal R Schroder U Keller J Freguia S Aelterman
P Verstraete W Rabaey K (2006) Microbial fuel cells methodology and
technology Environ Sci Technol 40 (17) 5181-5192
64 Logan B E and Regan JM (2006) Electricity-producing bacterial communities in
microbial fuel cells Trends Microbiol 14512ndash518
87
65 Logan B Cheng S Watson V Estadt G (2007) Graphite fiber brush anodes for
increased power production in air-cathode microbial fuel cells Environ Sci Technol
41(9)3341-3346
66 Logan BE (2009) Exoelectrogenic bacteria that power microbial fuel cells Nat Rev
Microbiol 5 375-381
67 Logan BE Call D Cheng S Hamelers HV Sleutels TH Jeremiasse AW
Rozendal RA (2008) Microbial electrolysis cells for high yield hydrogen gas
production from organic matter Environ Sci Technol 42 8630-8640
68 Logan BE Hamelers B Rozendal R Schroder U Keller J Freguia S Aelterman
P Verstraete W Rabaey K (2006) Microbial fuel cells methodology and
technology Environ SciTechnol 40 5181-5192
69 Lovely DR (2006) Bug juice harvesting electricity with microorganisms Nature
Review| Microbiology 4 497-508
70 Lovley DR Phillips EJ (1988) Novel Mode of Microbial Energy Metabolism
Organic Carbon Oxidation Coupled to Dissimilatory Reduction of Iron or
Manganese Appl Environ Microbiol 54(6) 1472-1480
71 Ly HK Sezer M Wisitruangsakul N Feng JJ Kranich A Millo D Weidinger
IM Zebger I Murgida DH Hildebrandt P (2011) Surface-enhanced vibrational
spectroscopy for probing transient interactions of proteins with biomimetic interfaces
electric field effects on structure dynamics and function of cytochrome c FEBS J
278(9)1382-90
72 Macaskie LE Hewitt CJ Shearer JA Kent CA (1995) Biomass production for
the removal of heavy metals from aqueous solutions at low pH using growth-
decoupled cells of a Citrobacter sp Int Biodeter Biodegr 3573ndash92
73 Malki M De Lacey AL Rodriguez N Amils R Fernandez VM (2008)
Preferential use of an anode as an electron acceptor by an acidophilic bacterium in
the presence of oxygen Appl Environ Microbiol 74 4472ndash4476
74 Meitl LA Eggleston CM Colberg PJS Khare N Reardon CL Shi L (2009)
Electrochemical interaction of Shewanella oneidensis MR-1 and its outer membrane
cytochromes OmcA and MtrC with hematite electrodes Geochim Cosmochim
Ac 73(18) 5292-5307
75 Morozan A Stamatin L and Nastase
F (2007) The biocompatibility
microorganisms-carbon nanostructures for applications in microbial fuel cells Phys
Stat Sol 6 1797-1803
76 Morris JM Jin S Crimi B Pruden A (2009) Microbial fuel cell in enhancing
anaerobic biodegradation of diesel Chem Eng J 146 161ndash167
77 Murray AE Lies D Li G Nealson K Zhou J Tiedje JM (2001) DNADNA
hybridization to microarrays reveals gene-specific differences between closely
related microbial genomes Proc Natl Acad Sci U S A 98 9853-9858
78 Muyzer G de Waal EC Uitterlinden AG (1993) Profiling of complex microbial
communities by denaturing gradient gel electrophoresis analysis of polymerase chain
88
reaction amplified genes coding for 16S rRNA Appl Environ Microbiol 59 695ndash
700
79 Myers CR Myers JM (1992) Localization of cytochromes to the outer membrane
of anaerobically grown Shewanella putrefacians MR-1 J Bacteriol 174 3429ndash3438
80 Narde GK Kapley A Purohit HJ (2004) Isolation and characterization of
Citrobacter strain HPC255 for broad-range substrate specificity for chlorophenols
Curr Microbiol 48(6) 419-423
81 Okabe S Ito T Sugita K Satoh H (2005) Succession of internal sulfur cycles and
sulfur-oxidizing bacterial communities in microaerophilic wastewater biofilms Appl
Environ Microbiol 71(5) 2520-2529
82 Park DH and Zeikus JG (2000) Electricity generation in microbial fuel cells using
neutral red as an electronophore Appl Environ Microbial 66 292-1297
83 Park DH and Zeikus JG (2002) Impact of electrode composition on electricity
generation in a single-compartment fuel cell using Shewanella putrefaciens Appl
Microbiol Biotechnol 5958ndash56
84 Park DH and Zeikus JG (2003) Improved fuel cell and electrode designs for
producing electricity from microbial degradation Biotechnol Bioeng 81 348ndash355
85 Park HS Kim BH Kim HS Kim HJ Kim GT Kim M Chang IS Park Y
K Chang HI (2001) A novel electrochemically active and Fe(III)-reducing
bacterium phylogenetically related to Clostridium butyricum isolated from a
microbial fuel cell Anaerobe 7 297-306
86 Parot S Nercessian O Delia ML Achouak W Bergel A (2009) Electrochemical
checking of aerobic isolates from electrochemically active biofilms formed in
compost J Appl Microbiol 106(4) 1350-1359
87 Pham CA Jung SJ Phung NT Lee J Chang IS Kim BH Yi H Chun J
(2003) A novel electrochemically active and Fe(III)-reducing bacterium
phylogenetically related to Aeromonas hydrophila isolated from a microbial fuel cell
FEMS Microbiol Lett 223 129-134
88 Qiao Y Bao SJ Li CM Cui XQ Lu ZS Guo J (2008) Nanostructured
polyanilinetitanium dioxide composite anode for microbial fuel cells ACS Nano 2
113-119
89 Qiu R Zhao B Liu J Huang X Li Q Brewer E Wang S Shi N (2009) Sulfate
reduction and copper precipitation by a Citrobacter sp isolated from a mining area J
Hazard Mater 1641310-1315
90 Quan X Yang SG Ruan XL and Zhao HM (2005) Preparation of titania
nanotubes and their environmental applications as electrode Environ Sci Technol 39
3770ndash3775
91 Rabaey K Boon N Hofte M Verstraete W (2005) Microbial phenazine
production enhances electron transfer in biofuel cells Environ Sci Technol 393401ndash
3408
89
92 Rabaey K Boon N Siciliano S D Verhaege M Verstraete W (2004) Biofuel cells
select for microbial consortia that self-mediate electron transfer Appl Environ
Microbiol 70(9) 5373ndash5382
93 Rabaey K Lissens G Siciliano S D Verstraete W (2003) Amicrobial fuel cell
capable of converting glucose to electricity at high rate and efficiency Biotechnol
Lett 25 1531-15
94 Rabaey K Verstraete W (2005) Microbial fuel cells novel biotechnology for
energy generation Trends Biotechnol 23(6) 291-298
95 Reguera G McCarthy KD Mehta T Nicoll JS Tuominen MT Lovley DR
(2005) Extracellular electron transfer via microbial nanowires Nature 435 1098-
1101
96 Reguera G Nevin KP Nicoll JS Covalla SF Woodard TL Lovley DR (2006)
Biofilm and nanowire production leads to increased current in Geobacter
sulfurreducens fuel cells Appl Environ Microbiol 72 7345-7348
97 Reimers CE Tender LM Fertig S Wang W (2001) Harvesting energy from the
marine sediment-water interface Environ Sci Technol 35192-195
98 Rezaei F Xing D Wagner R Regan JM Richard TM Logan BE (2009)
Simultaneous cellulose degradation and electricity production by Enterobacter
cloacae in a microbial fuel cell Appl Microbiol Biotechnol 75 3673ndash3678
99 Richter H Nevin KP Jia H Lowy DA Lovley DR Tender LM (2009) Cyclic
voltammetry of biofilms of wild type and mutant Geobacter sulfurreducens on fuel
cell anodes indicates possible roles of OmcB OmcZ type IV pili and protons in
extracellular electron transfer Energy Environ Sci 2 506ndash516
100 Riesenfeld SC Schloss DP Handelsman J(2004) Metagenomics Genomic
Analysis of Microbial Communities Annual Review Genetics 38525-552
101 Rozendal RA Hamelers HV Buisman CJ (2006) Effects of membrane cation
transport on pH and microbial fuel cell performance Environ Sci Technol 40
5206-5211
102 Rozendal RA Hamelers HVM Euverink GJW Metz SJ Buisman CJN (2006)
Principle and perspectives of hydrogen production through biocatalyzed electrolysis
Int J Hydrogen Energ 311632-1640
103 Rozendal RA Hamelers HV Molenkamp RJ Buisman CJ (2007) Performance of
single chamber biocatalyzed electrolysis with different types of ion exchange
membrances Water Res 41 1984-1994
104 Schluumlter A Bekel T Diaz NN Dondrup M Eichenlaub R Gartemann
KH Krahn I Krause L Kroumlmeke H Kruse O Mussgnug JH Neuweger
H Niehaus K Puumlhler A Runte KJ Szczepanowski R Tauch A Tilker
A Viehoumlver P Goesmann A (2008) The metagenome of a biogas-producing
microbial community of a production-scale biogas plant fermenter analyzed by the
454-pyrosequencing technology J Biotechnol136 77-90
90
105 Scott J H and Nealson KH (1994) A biochemical study of the intermediary carbon
metabolism of Shewanella putrefaciens J Bacteriol 176(11) 3408ndash3411
106 Sekiguchi H Tomioka H Nakahara T and Uchiyama H (2001) A single band
does not always represent single bacterial strains in denaturing gradient gel
electrophoresis analysis Biotechnology Letters23 1205ndash1208
107 Sharma T Leela M Reddy A Chandra TS Ramaprabhu S (2008) Development
of carbon nanotubes and nanofluids based microbial fuel cell Int J Hydrogen Energ
336749ndash6754
108 Shea C Clauwaert P Verstraete W Nerenberg R (2008) Adapting a denitrifying
biocathode for perchlorate reduction Water Sci Technol 58 (10) 1941-1946
109 Shi L Squier TC Zachara JM Fredrickson JK (2007) Respiration of metal (hydr)
oxides by Shewanella and Geobacter a key role for multihaem c-type cytochromes
Mol Microbiol 6512-20
110 Stams AJ and Plugge CM(2009)Electron transfer in syntrophic communities
of anaerobic bacteria and archaea Nat Rev Microbiol7(8)568-77
111 Sukkasem C Xu S Park S Boonsawang P Liu H (2008) Effect of Nitrate on the
Performance of Single Chamber Air Cathode Microbial Fuel Cells Water Res 42(19)
4743-4750
112 Summers ZM Fogarty HE Leang C Franks AE Malvankar NS Lovley DR
(2010)Direct exchange of electrons within aggregates of an evolved syntrophic
coculture of anaerobic bacteria Science 330 (6009)1413-5
113 Szczepanowski R Bekel T Goesmann A Krause L Kroumlmeke H Kaiser
O Eichler W Puumlhler A Schluumlter A (2008) Insight into the plasmid metagenome
of wastewater treatment plant bacteria showing reduced susceptibility to
antimicrobial drugs analysed by the 454-pyrosequencing technology J
Biotechnol 136 54-64
114 Tamura K Dudley J Nei M Kumar S (2007) MEGA4 Molecular Evolutionary
Genetics Analysis (MEGA) software version 40 MolBiol Evol 24(8) 1596-1599
115 Thompson JD Gibson TJ Plewniak F Jeanmougin F Higgins DG (1997) The
CLUSTAL_X windows interface flexible strategies for multiple sequence alignment
aided by quality analysis tools Nucleic Acids Res 25 4876-4882
116 Thormann KM Duttler S Saville RM Hyodo M Shukla S Hayakawa Y
Spormann AM (2006) Control of formation and cellular detachment from
Shewanella oneidensis MR-1 biofilms by cyclic di-GMP J Bacteriol 1882681-2691
117 Thormann KM Saville RM Shukla S Pelletier DA Spormann AM (2004)
Initial Phases of biofilm formation in Shewanella oneidensis MR-1 J Bacteriol
186(23) 8096-104
118 Torres CI Kato Marcus A Rittmann BE (2008b) Proton transport inside the
biofilm limits electrical current generation by anode-respiring bacteria Biotechnol
Bioeng 100 872ndash881
91
119 Torres CI Marcus AK Lee HS Parameswaran P Krajmalnik-Brown R Rittmann
BE (2010) A kinetic perspective on extracellular electron transfer by anode-
respiring bacteria FEMS Microbiol Rev 234(1)3-17
120 Torres CI Marcus AK Parameswaran P Rittmann BE (2008a) Kinetic
experiments for evaluating the NernstndashMonod model for anode-respiring bacteria
(ARB) in a biofilm anode Environ Sci Technol 42 6593ndash6597
121 Tsai HY Wu Ch-Ch Lee Ch-Y Shih EP (2009) Microbial fuel cell performance
of multiwall carbon nanotubes on carbon cloth as electrodes J Power Sources
194199-205
122 Tyson GW Chapman J Hugenholtz P Allen EE Ram RJ Richardson
PM Solovyev VV Rubin EM Rokhsar DS Banfield JF(2004) Community
structure and metabolism through reconstruction of microbial genomes from the
environment Nature42837-43
123 Venkateswaran K Moser DP Dollhopf ME Lies DP Saffarini D A MacGregor
BJ Ringelberg DB White DC Nishijima M Sano H Burghardt J Stackebrandt
E Nealson KH (1999) Polyphasic taxonomy of the genus Shewanella and
description of Shewanella oneidensis sp Int J Syst Bacteriol 49705-724
124 von Canstein H Ogawa J Shimizu S Lloyd JR (2008) secretion of Flavins
by Shewanella Species and Their Role in Extracellular Electron Transfer Appl
Environ Microbiol 74 3615-623
125 Wang A Liu W Cheng Sh Xing D Zhou J Logan BE (2009) Source of methane
and methods to control its formation in single chamber microbial electrolysis cells
Int J Hydrogen Energ 34 3653-3658
126 Wang J Ma T Zhao L Lv J Li G Liang F Liu R (2008) PCRndashDGGE method
for analyzing the bacterial community in a high temperature petroleum reservoir
World J Microbiol Biotechnol 2 9694-9702
127 Wang Q Garrity GM Tiedje JM Cole JR (2007) Naiumlve Bayesian Classifier for
Rapid Assignment of rRNA Sequences into the New Bacterial Taxonomy Appl
Environ Microbiol 73(16) 5261-5267
128 Wang YF Masuda M Tsulimura S Kano K (2008) Electrochemical regulation of
the end-product profile in Propionibacterium freudenreichii ET-3 with an
endogenous mediator Biotechnol Bioeng 101(3) 579ndash586
129 Wrighton KC Agbo P Warnecke F Weber KA Brodie EL DeSantis TZ
Hugenholtz P Andersen GL Coates JD (2008) A novel ecological role of the
Firmicutes identified in thermophilic microbial fuel cells ISME J 2 1146ndash1156
130 Xing D Zuo Y Cheng S Regan JM Logan BE (2008) Electricity generation by
Rhodopseudomonas palustris DX-1 Environ Sci Technol 42 4146-4151
131 Xu S Liu H Fan Y Schaller R Jiao J Chaplen F (2012) Enhanced performance
and mechanism study of microbial electrolysis cells using Fenanoparticle-decorated
anodes Appl Microbiol Biotechnol 93(2)871-880
92
132 Yang Y Harris DP Luo F Wu L Parsons AB Palumbo AV Zhou J (2008)
Characterization of the Shewanella oneidensis Fur gene roles in iron and acid
tolerance response BMC Genomics 9 Suppl 1S11
133 Zhang L Zhou S Zhuang L Li W Zhang J Lu N Deng L (2008) Microbial
fuel cell based on Klebsiella pneumoniae biofilm Electrochem Commun10 1641ndash
1643
134 Zuo Y Xing DF Regan JM Logan BE (2008) Isolation of the exoelectrogenic
bacterium Ochrobactrum anthropi YZ-1 by using a U-tube microbial fuel cell Appl
Environ Microbiol 74 3130-3137
93
APPENDICES
List of published papers during PhD study
1 Shoutao Xu Hong Liu Yanzhen Fan Rebecca Schaller Jun Jiao and Frank
Chaplen (2012) Enhanced performance and mechanism study
of microbial electrolysis cells using Fe nanoparticle-decorated anodes Applied
Microbiology and Biotechnology 93(2)871-880
2 Shoutao Xu Hong Liu (2011) New exoelectrogen Citrobacter sp SX-1 isolated
from a microbial fuel cell Journal of Applied Microbiology111(5)1108-1115
3 Yanzhen Fan Shoutao Xu Rebecca Schaller Jun Jiao Frank Chaplen Hong Liu
(2011) Nanoparticle decorated anodes for enhanced current generation in microbial
electrochemical cells Biosensors and Bioelectronics 26(5) 1908-1912
4 Tunc Catal Shoutao Xu Kaichang Li Hakan Bermek Hong Liu
(2008)
Electricity generation from polyalcohols in single-chamber microbial fuel
cells Biosensors and Bioelectronics 24(4)849-854
5 Chontisa Sukkasema Shoutao Xu Sunhwa Park Piyarat Boonsawang Hong Liu
(2008) Effect of nitrate on the performance of single chamber air cathode
microbial fuel cells Water research 424743-4750
6 Rebecca Schaller Yanzhen Fan Shoutao Xu Alan Fern Frank Chaplen Hong
Liu and Jun Jiao (2009) Vertically Aligned Multi-walled Carbon Nanotube
Decorated Anodes for Microbial Fuel Cells Proceedings of Materials Research
Society 2009 1170 R05-13

Bacterial Community Analysis New Exoelectrogen Isolation and Enhanced Performance
of Microbial Electrochemical Systems Using Nano-Decorated Anodes
by
Shoutao Xu
A DISSERTATION
Submitted to
Oregon State University
in partial fulfillment of
the requirements for the
degree of
Doctor of Philosophy
Presented June 15 2012
Commencement June 2013
Doctor of Philosophy dissertation of Shoutao Xu presented on June 15 2012
APPROVED
Co-Major Professor representing Biological and Ecological Engineering
Co-Major Professor representing Biological and Ecological Engineering
Head of the Department of Biological and Ecological Engineering
Dean of the Graduate School
I understand that my dissertation will become part of the permanent collection of
Oregon State University libraries My signature below authorizes release of my
dissertation to any reader upon request
Shoutao Xu Author
ACKNOWLEDGEMENTS
I would like to thank all people who have helped and inspired me during my
doctoral study Foremost I would like to express my sincere gratitude to my advisors Dr
Hong Liu and Dr Frank Chaplen for their continuous support of my PhD study and the
research of Microbial Fuel Cells at Oregon State University Hong inspired me to devote
myself to the field of Bio-energy with her great patience and enthusiasm towards
scientific educations Frank was always available and willing to help me with my study
especially during the period of Hongrsquos sabbatical leave Thanks to his kindness and
assistance my study at OSU became smooth and rewarding
Besides my advisors I would like to thank everyone in my dissertation committee Dr
Martin Schuster Dr Clare Reimers and Dr Mark Dolan Due to their encouragement
and insightful comments on my research I could always have the courage and knowledge
to overcome difficulties in my research I benefited greatly from Martinrsquos valuable
suggestions on my writing skills and his generous help regarding microarray data analysis
Clarersquos advice helped me build a solid foundation of Electrochemistry Also it was a
great honor to have Mark as my committee member His teaching gave me an insight into
the world of environmental engineering and enhanced the depth and width of my research
I thanked Dr Yanzhen Fan for always giving me guidance about the designs of the
reactors in my experiments
It was also important for me to say thanks to my current and previous lab-mates
Keaton Lesnik Kuhuan Chien Cheng Li Corale Abourached Anthony Janicek
Hongqiang Hu Jeremy Chignell Yudith Nieto and Wengguo Wu I had a wonderful
time enjoying doing research with them Their friendship and help made me confident of
my ability to do research as a scientist In addition I cherished and appreciated the
friendly environment in our BEE department Faculty members especially Dr John Bolte
Dr John Selker Dr Roger Ely Dr Ganti Murthy and Desiree Tullos made BEE an
excellent department for our students
My deepest gratitude went to my father Fuqin Wang and my mother Jinzhi Wang
for their endless care love and support throughout my life Also I would like to say
thanks to my best-loved wife Songhua Zhu She always supported me and never
complained that I could spend so little time accompanying with her This dissertation was
also for my lovely son Gabriel Hong-Yi Xu
TABLE OF CONTENTS
Page
1 General Introductionhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 1
2 Bacterial Community Analysis of Mixed Consortium in Microbial
Electrochemical Systemshelliphelliphelliphellip 13
3 New Exoelectrogen Citrobacter sp SX-1 Isolation and
Characterizationhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 27
4
Enhanced Performance and Mechanism Study of Microbial Electrolysis
Cells using Fe Nanoparticles Decorated Anodeshelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 45
5 Global Transcriptome Analysis of Response of Shewanella oneidensis
MR-1 to CNT Nanostructure Decorated Anodes in Microbial
Electrochemical System
66
6 Summaryhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 80
7 Bibliography helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 82
8 Appendices helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 93
LIST OF FIGURES
Figure Page
1-1 Generalized schematics (a) Microbial fuel cell (MFCs)(b) Microbial
electrolysis cell (MECs) helliphelliphelliphelliphelliphelliphelliphelliphelliphellip helliphelliphelliphellip
2
1-2 Mechanisms for extracellular electron transport in a MFCs anode 7
2-1 Polarization curves of high power mixed culture MFCshelliphelliphelliphelliphelliphellip 20
2-2 DGGE result of high power generation mixed culture MFCshelliphelliphellip 21
2-3 Bacterial species and percentage of bacterial community of mixed
culture MFCs helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip
22
2-4 Phylogenetic trees of bacterial species from mixed culture MFCs 22
2-5 Dominant bacterial species identification in DGGEhelliphelliphelliphelliphelliphelliphelliphellip 23
2-6 Current density generated by different dominant isolateshelliphelliphelliphelliphelliphellip 24
3-1 Phylogenetic tree of strain SX-1 and closely related species based on
16S rDNA sequenceshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip
35
3-2 SEM image of planktonic cells of Citrobacter sp SX-1helliphelliphelliphelliphelliphellip 35
3-3 Electricity generation by Citrobacter sp SX-1 in a single chamber
MFCs with sodium citrate helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip
36
3-4 Electricity generation of Citrobacter sp SX-1 using different
substrates in single chamber air-cathode MFCs at 1 KΩhelliphelliphellip
37
3-5 Power and voltage generation by Citrobacter sp SX-1 as a function
of current density helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip
38
3-6 Four current generating stages for CV analysis helliphelliphelliphelliphelliphelliphelliphellip 39
3-7 Cyclic Voltammetry analysis of of Citrobacter sp SX-1 biofilms helliphellip 41
4-1 SEM images of Fe NPs decorated anodeshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 50
4-2 Effects of Fe-NP decorated anodes on the current density in MECs helliphellip 51
4-3 Differentially expressed genes grouped by functional classification in
Fe NP decorated anodeshelliphelliphelliphelliphelliphellip helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip
53
5-1 SEM images of CNT decorated anodeshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 70
5-2 Effects of CNT decorated anodes on the current density in MECshelliphellip 73
5-3 Differentially expressed genes grouped by functional classification in
CNT decorated anodes helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip
74
LIST OF TABLES
Table
1-1
4-1
4-2
4-3
Taxa of bacteria current density generated and reactor configuration
used in microbial electrochemical cell experimentshelliphelliphelliphellip
Expression levels of genes related to biofilm formationhelliphelliphelliphelliphellip
Genes related to anaerobic growth and electron transfer with
significantly change expression levelhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip
Expression level of flavin and cytochrome geneshelliphelliphelliphelliphelliphelliphelliphellip
Page
4
55
56
58
4-4
5-1
5-2
Other genes with significantly changed expression level
Expression level of cytochrome c as response to CNT decorated anodes
Expression level of genes related to flavin synthesis as response to CNT
decorated anode in CNT decorated anodeshelliphellip
59
76
78
1
1
Bacterial Community Analysis New Exoelectrogen Isolation and Enhanced Performance
of Microbial Electrochemical Systems Using Nano-Decorated Anodes
Chapter 1
General Introduction
11 Microbial Fuel Cells and Microbial Electrolysis Cells
Microbial electrochemical systems (MESs) have drawn the attention of researchers
in recent years due to their promising applications in a variety of scientific fields such as
renewable energy generation bioremediation and wastewater treatment In a MES
microorganisms interact with electrodes catalyzing oxidation and reduction reactions at
the anode and the cathode
The most-described type of MESs is the microbial fuel cells (MFCs) in which
useful power is generated directly using the catalytic action of active microorganisms
(Kim 2002 Cheng and Logan 2007 Cheng et al 2006) In a typical two-chamber
MFCs (Figure 1a) organic matter is oxidized by electrochemically active
microorganisms in the anode chamber to release protons and electrons Protons diffuse
into the cathode chamber through a proton exchange membrane Meanwhile electrons
are transferred to the anode through various mechanisms (Logan et al 2006 Lovley
2
2006 Rabaey et al 2003) and then travel to the cathode where normally they combine
with oxygen and protons to form water
Another common used type of MES is the microbial electrolysis cell (MECs) in
which hydrogen is produced instead of electricity by applying a circuit voltage to the
MFCs system and maintaining anaerobic conditions in the cathode chamber (Liu et al
2005 Logan et al 2008 Rozendal et al 2006 Schaller et al 2009) (Figure 1b) A
cathode potential of at least - 410 mV (vs standard hydrogen electrode at pH 70) is
required to produce hydrogen at the cathode Therefore a theoretical potential of 110 mV
(ie 410-300 mV) is needed but voltages of gt02 V are normally applied due to various
overpotentials
Fig 1-1 Generalized schematics (a) Microbial fuel cells (MFCs) where electricity is
captured through the resistance R (b) Microbial electrolysis cells (MECs) where
hydrogen gas is produced in the cathode chamber R for resistor PS for power supply and
PEM for proton exchange membrane
The fundamental feature shared by microbial electrochemical systems (MESs)
(MECs and MFCs) is the microbe-catalyzed electron transfer from organic matter to
electrodes (anodes) (Rebaey et al 2004) Therefore different types of MESs can be
PEM
R
H+
e- e-
Anode
Bacte
ria
a
Cath
ode
H2O
PEM
PS
H+
e- e-
Anode
Bacte
ria
b
Cath
ode
H2
3
utilized to investigate the phenomena and mechanisms of interactions between microbes
and electrodes
12 Bacteria and Electron Transfer Mechanisms in MESs
The electrochemically active bacteria in MFCs are thought to be iron-reducing
bacteria such as Shewanella and Geobacter species (Gorby et al 2006) They have great
importance in the natural environment principally in metal oxidation and reduction
However recent studies have shown that the diversity of bacterial communities is much
greater than these model iron reducers on MFCs anodes (Reimers et al 2001 Liu et al
2004b Kim et al 2005) Over 20 electrochemically active bacterial species those can
transfer electrons exocellularly to electrodes have been reported in the past 10 years
(Logan 2009 Liu et al 2010) These bacteria have been categorized into diverse genetic
groups so far including alphaproteobacteria (Rhodopseudomonas Ochrobactrum and
Acidiphilium) (Malki et al 2008 Xing et al 2008 Zuo et al 2008) betaproteobacteria
(Rhodoferax) (Chaudhuri et al 2003) gammaproteobacteria (Shewanella Pseudomonas
Klebsiella Enterobacter and Aeronomas) (Kim et al 2002 Pham et al 2003 Rabaey et
al 2005 Bretschger et al 2007 Zhang et al 2008 Chung et al 2009 Rezaei et al 2009)
deltaproteobacteria (Geobacter Geopsychrobacter Desulfuromonas and Desulfobulbus)
(Bond et al 2002 Bond and Lovley 2003 Holmes et al 2004ab 2006)
Epsilonproteobacteria (Arcobacter) (Fedorovich et al 2009) Firmicutes (Clostridium and
Thermincola) (Park et al 2001 Wrighton et al 2008) Acidobacteria (Geothrix) (Bond
and Lovley 2005) and actinobacteria (Propionibacterium) (Wang et al 2008) A wider
range of electrochemically active bacteria are expected to be discovered
4
Table 1 Taxa of bacteria current density generated and reactor configuration used in
microbial electrochemical cell experiments (Liu et al 2010)
The electrochemically active bacterial species that possess the ability to transfer
electrons outside of the cell are called exoelectrogens in the MESs research field The
different exoelectrogens have demonstrated a wide-ranging power generation ability in
MFCs For example Shewanella oneidensis MR-1 has demonstrated the ability to
generate 0018 Am2 current density in single chamber MFCs while the Geobacter
Taxon Microorganisms Current Density (Am2
)
α-proteobacteria
Rhodopseudomonas palustris DX-1 003
Ochrobactrum anthropi YZ-1 071
Acidiphilium sp 32sup5 300
β-proteobacteria Rhodoferax ferrireducens 0031
γ-proteobacteria
Shewanella putrefaciens IR-1 0016
Shewanella oneidensis DSP10 0013
Shewanella oneidensis MR-1 0018
Pseudomonas aeruginosa KRA3 0017
Escherichia coli K12 HB101 100
Klebsiella pneumoniae L17 120
Enterobacter cloacae 013
Aeromonas hydrophila PA3 030
δ-proteobacteria
Geobacter metallireducens 065
Geobacter sulfurreducens 800
Desulfuromonas acetoxidans 0005
Geopsychrobacter
Electrodiphilus strain A2
0066
Desulfobulbus propionicus 003
Firmicutes
Lactococcus lactis 003
Thermincola sp strain Jr 020
Clostridium butyricum EG3 022
Thermincola ferriacetica Z-0001 040
Brevibacillus spp PTH1 0009
Desulfitobacterium hafniense DCB2 110
Acidobacteria Geothrix fermentans 0097
Actinobacteria Propionibacterium freudenreichiiET-3 120
5
sulfurreducens species has generated current densities as high as 80 Am2 which is 400
times higher than the one generated by Shewanella oneidensis MR-1 However mixed-
culture communities have generated much higher power densities than their pure-culture
counterparts in MFCs perhaps due to synergistic interactions within the anode bacterial
communities and the participation of currently unknown bacteria species and mechanisms
(Jung and Regan 2007) The dominant bacteria species vary in mixed cultures bacteria
communities in MFCs due to the enrichment of different substrates
Traditional methods of extroelectrogen study depend on cultivation hampered novel
exoelectrogens discovery because the inadequacy of defined media underestimates the
actual microbial diversity in MFCs Slow growth rate and unknown growth requirements
of anaerobic microorganisms make some anaerobic extroelectrogens cultivation difficult
However these limitations have been overcome by using molecular biological methods
based on DNARNA analysis Molecular biological techniques are now widely applied to
assess the diversity of microbial communities by analyzing the 16S rDNA sequence The
most commonly used molecular biological techniques for bacterial community analysis
include denaturing gradient gel electrophoresis (DGGE) restriction fragment length
polymorphism (RFLP) and 16S rDNA sequencing These methods are much less time
consuming than traditional isolation and cultivation methods
Isolated exoelectrogens were utilized to explore the mechanism of electron transfer
to the anode However the mechanisms of electron transfer to extracellular electron
acceptors are not well understood Three mechanisms have been proposed for exocellular
transport of electrons by exoelectrogens to anodes in MFCs so far Some exoelectrogenic
bacteria can transfer electrons to electrodes through soluble redox compounds (Bond and
6
Lovley 2005) These compounds include artificial mediators and mediators secreted by
exoelectrogenic bacteria A variety of chemicals have been used to facilitate the shuttling
of electrons from inside of cell to electrodes outside the cell These exogenous mediators
include neutral red (Park et al 1999) anthraquinone-2-6 disulfonate (AQDS) thionin
potassium ferricyanide (Bond et al 2002) methyl viologen and others (Logan 2004
Rabaey and Verstraete 2005) Some bacteria can secrete their own chemical mediator
for example Pseudomonas aeruginosa secretes pyocyanin and phenazine-1-carboxamide
to transfer electrons toward the MFCs anode (Rabaey et al 2005) Another evidence for
mediator production by bacteria is Geothrix fermentans When the medium was replaced
in a MFCs that had stable power generation with this bacteria power dropped by 50
and required 10 days to resume the original level
Some bacteria can directly transfer electrons to anodes via outer cell membrane
proteins (Myers and Myers 1992) The outer cell membrane protein cytochrome c is
thought to play a critical role in to transferring electrons to anodes Ly et al (2011)
isolated the haem protein cytochrome c and demonstrated that electric field effects may
be functional for the natural redox processes of cytochrome c in the respiratory chain
Shewanella oneidensis is the example of a bacterium that is capable of transfer electron to
anode via outer cell membrane proteins When Shewanella oneidensis adhered to an iron
surface the greater force has showed benefits to grow cells because closer contact
required for electron transfer from cell bound cytochromes (Lower et al 2001)
7
Fig1-2 Mechanisms for extracellular electron transport in an MFCs anode (1) direct
contact (top in green) (2) by nanowires (middle in purple) and (3) self-produced
mediators (bottom in blue) (Logan 2009)
More and more evidence supports the involvement of bacterial nanowires in
extracellular electron transport (Reguera et al 2005 2006 Gorby et al 2006) Nanowires
are conductive appendages produced by both Geobacter and Shewanella species (Gorby
and Beveridge 2005) The conductivity of the appendages was examined and confirmed
by using conductive scanning tunneling microscopy (STM) (Gorby et al 2006)
Nanowires can carry electrons from the cell to the anode surface of MFCs
The solid component of the extracellular biofilm matrix has high efficiency on
extracellular electron transfer compared with other extracellular electron transfer
8
mechanisms and recently Torres et al (2010) hypothesized that the solid component of
the extracellular biofilm matrix formed by exoelectogens is another way bacteria transfer
electrons to electrodes This hypothesis was based on kinetic analysis of each EET
mechanism reported in available literature (Torres et al 2010)
13 Anode electrodes
In MFCsMECs anode electrodes are a critical component because exoelectrogens
adhere to the surface of anodes to transfer electrons to the electrode The characteristics
of anodes have significant effects on electron transfer rate from bacteria to anode
electrodes in MFCs The requirements of an anode material are it should be highly
conductive non-corrosive have a high specific surface area (area per volume) high
porosity be non-fouling inexpensive and easily scaled to larger sizes Of these
properties the most important one that is different from other biofilm reactors is that the
material must be electrically conductive Normally they are made of various carbon
materials including carbon fiber carbon clothe and carbon paper due to their stability
high conductivity and high specific surface-area Nevertheless they have little
electrocatalytic activity for the anode microbial reactions and thus a modification of the
carbon materials is the main approach for improving their performance Consequently
there is a great need to develop a new type of anode material for MFCsMECs
It is a great challenge to develop a new anode material to further increase the power
density of a MFCs The nature of the catalytic mechanism of a MFCrsquos anode involves not
only a biological but also an electrocatalytic process An optimal nanostructure with a
high specific surface area favorable for both catalytic processes could play a critical role
in improving the power density of the MFCs such a structure needs to host the bacteria
9
with high bioactivity while enhancing the electron-transfer rate Schroumlder et al (2008)
employed PANI to modify a platinum anode for MFCs and achieved a current density 1
order of magnitude higher than the previously reported value PANIinorganic
composites are also reported to have better conductivity Qiao et al also applied a new
mesoporous TiO2 electrode material with uniform nanopore distribution and a high
specific surface area to anode in comparison to previously reported work with E coli
MFCs the composite anode delivers 2-fold higher power density (Qiao et al 2008)
Thus it has great potential for use as the anode in a high-power MFCs and may be a new
approach for improving performance of MFCs
14 Other parts of MESs
141 Membranes and ion transport
The ion exchange membrane is another one of the critical components in two-
chamber MESs systems It separates anode and cathode chambers and at the same time
maintains the electron neutrality of the system ie transport of electrons to the cathode
needs to be compensated by transport of an equal amount of positive charge to the
cathode chamber A proton exchange membrane (PEM) such as Nafion a per-fluorinated
sulphonic acid membrane (PFSAs) consisting of a hydrophobic fluorocarbon backbone to
which hydrophilic sulfonate groups (-SO3-) are attached is commonly used in chemical
fuel cell systems For MFCs systems however mainly cation species like Na+ and K
+
other than proton are often responsible for the dominant transport of positive charge
through the cation exchange membrane (CEM) to maintain electroneutrality due to the
low proton concentration in any aqueous medium with near neutral pH (Rozendal et al
2006) Consequently the pH increases in the cathode chamber due to the consumption of
10
protons and decreases in the anode chamber because of the accumulation of protons
(Rozendal et al 2007) This pH change in two-chamber systems results in a decrease of
the cathode potential and performance The application of anion exchange membrane
(AEM) has also been explored in MFCs systems (Cheng and Logan 2007 Kim and
Logan 2007) where it has been proposed that protons are transferred via pH buffers like
phosphate anions
142 Cathodes and Catalysts
The cathode is another challenge for making MFCs commercially available
technology because the chemical reaction that occurs at the cathode is difficult to
engineer as the electrons protons and oxygen must all meet at a catalyst in a tri-phase
reaction (solid catalyst air and water) The catalyst must be on a conductive surface and
must be exposed to both water and air so that protons and electrons in these different
phases can reach the same point The most commonly used material for a cathode is
commercially available carbon paper pre-loaded with a Pt catalyst on one side When it is
used in a MFC the side that contains the catalyst faces the water and the uncoated side
faces air To reduce the high cathode cost associated with platinum catalyst other precious-
metal-free cathodes such as (NiMo and NiW) were developed by electrodepositing on a
carbon fiber They have achieved comparable performance with Pt catalyst with same
loading at a much lower cathode fabrication cost (Hu 2010)
The requirements of cathode for MECs are quite similar to the requirements of
cathode for MFCs but easier than cathode for MFCs for the manufacturing process
because the cathode in MECs is not necessarily to exposed to air Therefore it can be
made of the exact same materials of cathode in MFCs except a waterproof layer
11
Recently some researchers have attempted to apply microorganisms as a biocatalyst to
precede the combination of electron with oxygen in the cathodes
15 Dissertation overview
Low power densities in MESs limit practical applications The improvement of
MESs performance requires a detailed understanding of the physiology and ecology of
microorganisms in MESs including the mechanism of electron transfer to the anode from
the microorganism
This dissertation focuses on the problem of the low power density of MESs The
bacterial community structure of a high power generated mixed culture communities in
MFCs will be identified firstly and then one of major exoelectrogens will be isolated and
characterized This information will be helpful to understand the physiology and ecology
of exoelectrogens in MESs Consequently they will be beneficial to improve power
density of MFCs Lastly the nanostructure decorated anodes in MECs will be utilized to
improve the power density The power enhancement mechanism will be explored by
using a whole genome microarray They are presented here as four papers
In the first paper the cultivation independent molecular biological techniques
denaturing gradient gel electrophoresis (DGGE) and 16S rDNA clone library are utilized
to analyze the bacterial community structure of a higher power mixed culture MFCs The
analyzed results provide fundamental information for isolating the dominant bacteria in
mixed culture MFCs Two of dominant bacterial species has been isolated and used aone
to test power generation in MFCs The possible interaction among different bacterial
species in mixed culture is discussed
12
In the second paper one isolated exoelectrogenic bacterial strain SX-1 is
characterized It is identified as a member of the Citrobacter genus and power generation
is tested ultilizing a wide range of different substrates The electron transfer mechanism is
explored using Cyclic Voltammetry (CV) This study increases the known diversity of
power generating exoelectrogens and provides a new strain to explore the mechanisms of
extracellular electron transfer from bacteria to electrodes
The third paper shows effects on MESs performance by Fe nanoparticle decorated
anodes in the MESs The average current density produced with Fe nanoparticle
decorated anodes increased up to 59-fold higher than plain graphite anodes A whole
genome microarray is utilized to analyze the possible mechanism of enhanced current
density as responded to nanoparticle decorated anodes
The fourth paper describes the effects of carbon nanotube (CNT) modified anode on
the performance S oneidensis MR-1 in MESs Results demonstrate that current densities
produced with CNT decorated anodes are up to 56-fold higher than plain graphite anodes
The possible mechanisms of enhanced current density by CNT decorated are explored
13
Chapter 2
Bacterial community analysis of mixed consortium in higher
power density MESs
Shoutao Xu and Hong Liu
1 Introduction
The improvement in the performance of mixed culture MFCs requires an
understanding of the ecology in microbial communities of MFCs Many researchers have
attempted to characterize microbial populations and activities to elucidate the behaviors
and ecology of microorganisms in MFCs (Wrighton et al 2008 Fedorovich et al 2009)
In order to study the microbial ecology of the mixed culture in a MFC and select the
appropriate isolation medium for dominant bacterial species in the mixed culture the
fundamental steps are to analyze the bacterial diversity in the mixed culture of MFCs and
identity the dominant bacterial species in bacteria communities in MFCs
14
As for the identification of bacterial communities typically there are two general
methods The first method for identification of bacterial community is the traditional
cultivation processes using selective nutrients to promote the growth of different types of
bacteria within the samples (Amman et al 2000) The community structure can then be
assessed by identifying the isolates from the dominant colonies that were cultured This
can often be costly and laborious as each isolate has to be further studied by examining
its physiology taxonomy and reactivity to stains (Adwards et al 1989)
The second method relies on utilizing molecular techniques to analyze bacterial
community DNA Several molecular methods involving the extraction and analysis of
DNA from entire bacterial communities are used to identify genetic fingerprints of
bacteria These methods including the cloning and sequencing of 16S rDNA automated
ribosomal intergenic spacer analysis (ARISA) terminal restriction fragment length
polymorphism (T-RFLP) and denaturing gradient gel electrophoresis (DGGE) generate
profiles of bacterial community structures They can rapidly assess complex communities
from various environments (Amman et al 2000)
Due to the conservative characteristic of 16S rDNA in bacteria during the process
of evolution 16S rDNA sequencing can be used to identify different species of bacteria
Therefore the molecular techniques of denaturing gradient gel electrophoresis (DGGE)
with PCR and 16S rDNA clone library are used for analysis of the microbial diversity
These methods are more convenient and save time compared to traditional
isolationcultivation methods for microorganism analysis
In this work a biofilm bacterial community from an anode of a MFCs wase studied
by using denaturing gradient gel electrophoresis (DGGE) of PCR-amplified partial 16S
15
rRNA genes followed by cloning and sequencing of 16S rDNA The results provided
essential information for dominant bacterial isolation in mixed culture MFCs
2 Materials and methods
21 High power generation of mixed culture MFCs
Mixed cultures were originally inoculated from domestic wastewater (Corvallis
Wastewater Treatment Plant Corvallis OR) to MFCs after operating for 2 years using a
defined medium solution (Lovely 2002) with sodium acetate as the carbon source A
new MFCs was inoculated from the operating MFCs The polarization curves were
performed to measure the power generation when maximal stable power were established
(normally 3-5 batches post-inoculation) and the genomic DNA from the microbial
biofilm on the anode was extracted under the sterile conditions for further bacterial
community analysis
22 Denaturing gradient gel electrophoresis (DGGE)
DGGE with PCR is a method of analysis of bacterial community composition
based on different GC percentage of 16s rDNA in different bacterial species PCR-DGGE
comprises three steps (i) extraction of total community DNA from the sample (ii) PCR-
controlled amplification using specific oligonucleotide primers and (iii) separation of the
amplicons using DGGE For this purpose a reproducible and efficient method for total
DNA extraction is indispensable and needs to be evaluated and optimized depending on
the nature of the sample In the subsequent PCR step multiple PCR primer sets with
different resolution can be used In most PCR-DGGE applications on bacteria universal
or specific primers are targeting the 16S rRNA gene Upon electrophoresis of the PCR
16
amplicons and gel staining (using ethidium bromide silver staining or SYBR green)
DGGE gels are digitally captured and further analyzed using computer software packages
The richness of DNA fragment bands in DGGE indicates the bacterial diversity of the
sample
The detailed steps are as followed Biofilms were scratched from the anodes of high
power generated MFCs fed with sodium acetate Bacterial genomic DNA was extracted
from the biofilm samples using the DNeasy blood and tissue kit (Qiagen) according to the
manufacturerrsquos instructions The universal primer set was used to amplify the 16S rDNA
from the extracted genomic DNA then the V3 region of bacterial 16S ribosomal DNA
(rDNA) was amplified with GC-clamp primer (Muyzer et al 1993) for DGGE PCR
amplification was performed in a thermocycler DGGE of the PCR products was carried
out in a DcodeTM
Universal Mutation Detection System) The 8 (wv) polyacrylamide
gels contains from 30 to 55 denaturing gradients Electrophoresis was conducted
using a 1timesTAE (Tris-Acetate-EDTA) buffer at 130V and 60degC for 5 hours After
electrophoresis the gel was stained with ethidium bromide in 1timesTAE buffer for 15
minutes and washed in 1timesTAE buffer for 10 minutes The fragments were visualized
under a UV transilluminator The richness of single band in DGGE gel picture
preliminarily showed that the bacterial diversity in mixed culture since the single band in
gel represents one bacterial species
23 16S rDNA clone library construction method
16S rDNA-DGGE fingerprinting has proven to be particularly useful as an initial
investigation into bacterial communities and is suitable for identifying the predominant
bacteria that comprise 1 or more of cells within a given sample In the 16s rDNA clone
17
library construction process the first step is the extratction of the total genomic DNA
then the genomic DNA of the mixed culture biofilm was used as template for PCR
amplification of approximately 1500 bp of 16S rDNA with universal primers The PCR
products of 16S rDNA were purified and inverted into pGEM-T Easy vector system
before they were transformed into competent E coli The transformed cells were spread
on Luria-Bertani (LB) plates containing the selective antibiotic ampicillin and X-Gal and
incubated overnight at 37 OC after the more than 80 inverted 16s DNA fragment E coli
colonies were picked up and inoculated in the 1-3 ml liquid LB medium and left to grow
for 16 hours The plasmid DNA were extracted and sequenced The sequences were
compared directly to all known sequences deposited in GenBank databases using the
basic local alignment search tool (BLAST)
24 Bioinformatics Analysis
16S rDNA sequencing results of more than 80 colones were queried against the
GenBank and Ribosomal Database Project (RDP) databases using BLAST and
SEQUATCH algorithms (Cole et al 2007 Wang et al 2007 Cole et al 2009) The
neighbor-joining trees were constructed with the Molecular Evolutionary Genetics
Analysis package (MEGA Version 4) based on 1000 bootstrap analysis (Tamura et al
2007)
25 Dominant bacterial species band in DGGE identification
The DGGE results gave the information of dominant bacterial richness of bacterial
species in mixed culture based on the theory that single band represents one bacterial
species which showed the diversity of bacterial community in the mixed culture of a
MFC 16S rDNA clone library results provided the whole picture of bacterial community
18
including the bacterial species name and bacterial species percentage in the mixed culture
The known pure bacterial species which have been sequenced can be used as markers to
identify dominant bacterial species in DGGE The whole experimental procedure is
similar to the procedure of the mixed culture DGGE steps The first step is to extract the
collect the mixed culture biofilm genomic DNA and pure bacterial species marker
genomic DNA Then the universal primer set was used to amplify the 16S rDNA from
the extracted genomic DNA After that the V3 region of bacterial 16S ribosomal DNA
(rDNA) was amplified with GC-clamp primer (Muyzer et al 1993) for DGGE The
subsequent steps were similar to the mixed culture DGGE experimental procedure The
dominant bacterial bands in DGGE were determined by comparisons of the single band
position in the mixed culture DGGE with single pure bacterial species location in DGGE
26 Dominant bacterial species isolation and power generation
The bacterial community of mixed culture in MFCs were predominantly composed
of Geobacter sp Arcobacter sp and Citrobacter sp Two dominant bacterial species
have been isolated
Citrobacter sp SX-1 was isolated from the anode biofilm by using a solid agar
medium containing NH4Cl 6mM KCl 2mM CH3COONa (electron donor) 30 mM
FeC6H5O7 (electron acceptor) 20mM minerals and vitamins (Lovley and Phillips 1988)
Single colonies were picked up after culturing on the plate for 36 hours at 30 degC and
transferred two times on the agar plate for purification Then the isolates grew in a liquid
medium solution in anaerobic tubes containing the same constituents as the solid
medium All isolation process was operated in a glove box anaerobic chamber (Coy
Laboratory Products Grass Lake MI)
19
Arcobacter sp SX-2 was isolated from the anode biofilm of a MFC fed with sodium
acetate by using the same method of isolation of pure bacterial species Citrobacter XS-1
except using Trypticase Soy Agar with 5 Sheep Blood solid (from American type
culture collection (ATCC)) agar medium instead of Citrobacter sp medium Single
colonies were picked up after culturing on the plate for 36 hours at 30 degC and transferred
twice on the agar plate for purification Then the isolates grew in a liquid medium
solution microaerobically
Geobacter sp have been trying to isolate by using three different methods They are
the colony pickup after different condition enrichment Goebacter medium isolation and
dilution to extinction by multiple channel mini MFCs isolation however the Geobacter
sp has not been isolated yet due to some limits so far But they provide valuable
information for further isolation of Geobacter sp
Single chamber MFCs were used to evaluate power generation by different isolates
The MFCs were constructed as described previously (Liu and Logan 2004) and modified
with 3 cm2 carbon cloth anodes and 7 cm
2 carbon clothPt cathodes The total liquid
volume of each MFCs was about 15 ml with electrode spacing about 17 cm All MFCs
were operated in an autoclaved closed plastic box and sterile cotton was attached to the
outer surface of the air cathodes to prevent contamination A MFCs without bacterial
culture was used as control MFCs were inoculated with 3 ml late exponential phase
cultures of SX-1 in the medium solution reported previously (Liu and Logan 2004)
3 Results and discussion
31 Power production by mixed culture MFCs
20
A polarization curve was used to characterize current as a function of voltage in the
MFCs The polarization curves are performed by varying a series of external resistances
The results are shown in Figure 21 With MFCs acclimated to 25 Ω external resistance
the maximum power density was achieved at 65Wm2 based on the polarization data At
this point the current density is 21mAcm2 The power density was three times higher
than previous report (20 Wm2) using phosphate buffer (Fan et al 2007) Power density
was also 2 times higher than (28 m Wm2) using a bicarbonate buffer ( Fan et al 2007)
32 Bacterial community structure assessed by PCR-DGGE
Bacterial DGGE profiles for higher power generating MFCs mixed culture setups
were relatively complex (more than 10 bands) However there are three brightly stained
bands and other less intense bands (Fig 2-2) These results indicated there were probably
three dominant bacterial species in mixed culture MFCs
010
020
030
040
050
060
070
2
3
4
5
6
7
04 05 05 06 07 08 10 11 13 15 21 22 23 24
Volta
ge (
V)
Pow
er
densi
ty (
Wm
2)
Current density (mAcm2)
Power
Voltage
power density 65wm2 at current
density208 mAcm2
Figure 2-1 Polarization curves of high power mixed culture MFCs
21
33 16S rDNA clone library results
A total of 83 colonies were sequenced for 16S rDNA clone library construction They
represented the bacterial community structure in mixed culture in MFCs The results
revealed that the predominant bacterial species in mixed culture are Geobacter sp (66
of the mixed culture) Arcobacter sp (12 of the mixed culture) and Citrobacter sp (11
of the mixed culture) Those three genuses comprised 88 of the total bacterial species
(Fig 23) This result is consistent with the DGGE result which showed that three bright
bands in the DGGE gel represented three dominant bacterial species in the mixed culture
The phylogenetic analysis of the colonies (83 colony sequences) is shown in Figure 24
Figure 22 DGGE result of high power generation mixed culture MFC (1 mixed
culture 2 pure culture control)
1 2
22
34 Dominant bacterial band in DGGE identification
Geobacter
66
Arcobacter
12
others
6
Pseudomonas
1
Citrobacter
11
Clostridium
2
Anaerovorax
2 Geobacter
Arcobacter
Citrobacter
Clostridium
Anaerovorax
Pseudomonas
others
Figure 23 Bacterial species and percentage of bacterial community of mixed
culture MFC lt 1 Alcaligensgt
lt1 Commamonasgt
lt1 Pseudomonas aeruginosagt
lt1 Klebsiellagt
lt11 Citrobactergt
lt1 Wolinellagt
lt12 Arobactergt
lt2 Anaeroboraxgt
lt2 Clostridium gt
lt1 unclear bacteriagt
lt66Geobactergt
100
100
85
100
100
82
99
87
100
002
Figure 24 Phylogenetic trees of bacterial species from mixed culture MFC
23
The dominant bacterial species including Goebacter sp Citrobacter sp and Arcobacter
sp in the mixed culture are shown in Figure 25 The upper band is Arcobacter sp and
Citrobacter sp is located in the bottom band of DGGE Geobacter sp is the middle one
The analysis of the bacterial community using denaturing gradient gel electrophoresis
(DGGE) of PCR-amplified 16S rRNA gene fragments and 16S rDNA clone library
construction method showed great phylogenetic diversity of mixed culture in MFCs with
the identification of sequences derived from bacteria of the taxa deltaproteobacteria
(Geobacter sp) gammaproteobacteria (Citrobacter sp) and epsilonproteobacteria
(Arcobacter sp) The result is consistent with the conclusion that among the isolated
exoelectrogens gammaproteobacteria and deltaprotebactia appear to be dominant among
others (Parot et al 2009 Liu et al 2010)
35 The dominant bacterial species isolation and power generation
Figure 25 Dominant bacterial species identification in DGGE
A B C D
A Citrobacter sp
B Geobacter sp
C Arobacter butzmeri
D mixed culture control
24
Two dominant bacterial species Citrobacter sp XS-1 and Arcobacter sp XS-2 have been
isolated and tested for power generation The power densities generated by the two
isolates are shown in the Fig 26 Current densities generated by isolate Citrobacter sp
and Arcobacter sp were 98 mAm2 and 20 mAm
2 respectively which were much less
than that (21Am2) generated by the mixed culture from which they were isolated
Figure 26 Current density generated by different dominant isolates
The maximum current density generated by mixed culture is much higher than the
one produced by pure bacterial species Citrobacter sp XS-1 and Arcobacter sp XS-2 in
the same structure MFCs Although the Geobacter sp in our mixed culture has not been
isolated a previous study showed that maximum current density generated by Geobacter
sp was 080 Am2
(Bond and Lovley 2003) The current density results seem partially
supporting the conclusion that mixed-culture generated higher power densities than their
pure-culture counterparts in MFCs (Jung and Regan 2007) This result suggests there
may be synergistic interactions within the anode bacterial communities
0
500
1000
1500
2000
2500
Mixed culture Citrobacter sp Arcobacter Sp
Cu
rre
nt
de
nsi
ty (
mA
m2)
25
Geobacter sp are strictly anaerobic bacteria which were enriched in most of the anode
biofilms (George 2005) Citrobacter species are facultative anaerobic bacteria (George
2005) which can be found in a wide variety of habitats including in soil water and
wastewater Arcobacter sp are micro-aerobic bacteria including both environmental
nonpathogens and opportunistic human pathogens They are able to grow in aerobic
conditions but in the optimal growth under micro-aerobic conditions (George 2005)
Arcobacter sp has been discovered in the MFCs by Fedorovich (Fedorovich et al 2009)
It is interesting that the mixed culture bacterial community composed of primarily these
three bacterial species has demonstrated to generate high power at non-strictly anaerobic
condition while leading dominant bacterial species Geobacter sp (66) is strict
anaerobic bacteria and generated powder in the MFCs only under strict anaerobic
condition (George 2005) Therefore it is possible that microaerobic bacteria such as
Arcobacter sp and facultative anaerobic bacteria Citrobacter sp can facilitate to create
anaerobic condition for Geobacter sp when they utilized oxygen for growing in the
mixed culture Syntrophic communities study have showed that anaerobic bacteria and
methanogenic archaea form compact microbial structures that operate like an organ rather
than a set of microorganisms functioning independently (Stams and Plugge 2009) Some
substrates have been degradeted within these communities while they are not able to be
fermented by individual species alone interspecies electron transfer also have been
presented in these communities (Stams and Plugge 2009) Summers et al (2010) also
discovered that direct exchange of electrons happened within coculture of Geobacter
metallireducens and Geobacter sulfurreducens Therefore we believe that there are
26
more complex synergistic interactions between different bacteria species in mixed culture
in the MFCs
Acknowledgements
The authors acknowledge support from the US National Science Foundation
(CBET 0955124) We also thank Keaton Lesnik for the review of this manuscript
27
Chapter 3
New Exoelectrogen Citrobacter sp SX-1 Isolated from a Microbial Fuel Cell
Shoutao Xu and Hong Liu
Published in
Journal of Applied Microbiology 111(5)1108-1115 (2011)
ABSTRACT
Exoelectrogenic bacterial strain SX-1 was isolated from a mediator-less microbial fuel
cell by conventional plating techniques with ferric citrate as electron acceptor under
anaerobic condition Phylogenetic analysis of the 16S rDNA sequence revealed that it
28
was related to the members of Citrobacter genus with Citrobacter sp sdy-48 being the
most closely related species The bacterial strain SX-1 produced electricity from citrate
acetate glucose sucrose glycerol and lactose in MFCs with the highest current density
of 205 mA m-2
generated from citrate Cyclic voltammetry analysis indicated that
membrane associated proteins may play an important role in facilitating the electrons
transferring from bacteria to electrode This is the first study that demonstrates that
Citrobacter species can transfer electrons to extracellular electron acceptors Citrobacter
strain SX-1 is capable of generating electricity from a wide range of substrates in MFCs
This finding increases the known diversity of power generating exoelectrogens and
provided a new strain to explore the mechanisms of extracellular electron transfer from
bacteria to electrode The wide range of substrate utilization by SX-1 increases the
application potential of MFCs in renewable energy generation and waste treatment
Keywords Microbial fuel cell Exoelectrogen Citrobacter sp SX-1 Extracellular
electron transfer
29
Introduction
Microbial Fuel Cells (MFCs) technology has become an active research area
recently as a promising approach for renewable energy generation wastewater treatment
and bioremediation (Rabaey and Verstraete 2005 Lovley 2006 Fan et al 2008 Logan
2009 Liu et al 2010) The key feature of MFCs system is the microbe-catalyzed electron
transfer from organic matter to anodes Many studies have shown the presence of diverse
bacterial communities on MFCs anodes (Reimers et al 2001 Liu et al 2004b Kim et al
2005) Over 20 exoelectrogens ie microbes that can transfer electrons exocellularly to
electrodes have been reported in the past 10 years (Logan 2009 Liu et al 2010) The
exoelectrogens so far belong to diverse genetic groups including alphaproteobacteria
(Rhodopseudomonas Ochrobactrum and Acidiphilium) (Malki et al 2008 Xing et al
2008 Zuo et al 2008) betaproteobacteria (Rhodoferax) (Chaudhuri et al 2003)
gammaproteobacteria (Shewanella Pseudomonas Klebsiella Enterobacter and
Aeronomas) (Kim et al 2002 Pham et al 2003 Rabaey et al 2005 Bretschger et al
2007 Zhang et al 2008 Chung et al 2009 Rezaei et al 2009) deltaproteobacteria
(Geobacter Geopsychrobacter Desulfuromonas and Desulfobulbus) (Bond et al 2002
Bond and Lovley 2003 Holmes et al 2004ab 2006) Epsilonproteobacteria (Arcobacter)
(Fedorovich et al 2009) Firmicutes (Clostridium and Thermincola) (Park et al 2001
Wrighton et al 2008) Acidobacteria (Geothrix) (Bond and Lovley 2005) and
actinobacteria (Propionibacterium) (Wang et al 2008) A wider range of
exoelectrogenic species are expected to be discovered
Three mechanisms have been proposed for exocellular transport of electrons by
exoelectrogens without artificial mediators Some exoelectrogenic bacteria can transfer
30
electrons to electrodes through soluble redox compounds excreted by microorganisms
(Bond and Lovley 2005 Rabaey et al 2005) and others can directly transfer electrons to
anodes via outer cell membrane proteins (Myers and Myers 1992) Recently more and
more evidence supports the involvement of bacterial nanowires in extracellular electron
transport (Reguera et al 2005 2006 Gorby et al 2006) In spite of the discovery of
many bacterial species that can transfer the electrons to electrode without the need of
artificial mediators the investigation of extracellular electron transfer mechanisms was
mainly focused on a few species such as those from Geobacter and Shewanella genera
(Kim et al 1999 2002 Reguera et al 2005 2006 Gorby et al 2006) The electron
transfer mechanisms for many of the isolated exoelectrogens species are still not well
studied
In the present study we isolated a new exoelectrogen strain SX-1 from a MFC a
strain phylogenetically related to Citrobacter sp Power generation from various carbon
sources by this strain was evaluated using single chamber MFCs Plausible extracellular
electron transfer mechanisms were also discussed based on the characterization of anodic
biofilms by cyclic voltammetry (CV)
Materials and methods
Bacterial strain SX-1 isolation
Bacterial strain SX-1 was isolated from the anode biofilm of a MFC fed with sodium
acetate operated in fed-batch mode over a period of six months The original source of
the inoculum is wastewater from a local waste water treatment plant Bacterial cells were
released from the carbon cloth anode by shaking the cloth in a sterile flask with 15 mL
sterile 1 NaCl solution and glass beads (2 mm diameter) The suspension was then
31
serially diluted from 10 times to 105
times and plated on a petri dish with a solid agar
medium containing NH4Cl 6mM KCl 2mM CH3COONa (electron donor) 30 mM
FeC6H5O7 (electron acceptor) 20mM minerals and vitamins (Lovley and Phillips 1988)
Single colony was randomly selected after culturing on the plate for 36 hours at 30 degC
and purified on a new agar plate following a procedure reported previously (Chung and
Okabe 2009) Then the isolate was picked up and grew in a liquid medium solution in
anaerobic tubes containing the same constituents as the solid medium for further analysis
All isolation process was operated in a glove box anaerobic chamber (Coy Laboratory
Products Grass Lake MI)
16S rDNA sequencing and phylogenetic analysis
Bacterial genomic DNA was extracted from the isolated strain SX-1 using the DNeasy
tissue Kits (Qiagen Valencia CA) according to the manufacturerrsquos instructions The 16S
rDNA was amplified by PCR using a pair of universal primers 27F (5rsquo-
AGAGTTTGATCMTGGCTCAG-3rsquo) and 1492R (5rsquo-GGTTACCTTTGTTACGACTT-
3rsquo) (Park et al 2001) The conditions used for PCR were described previously
(Ferna ndez et al 1999) PCR products were purified ligated and cloned according to a
procedure described in a previous report (Xing et al 2008) Plasmids were isolated from
randomly selected clone colonies with a QIAprep Spin Miniprep kit (Qiagen Valencia
CA) and three plasmid inserts were then sequenced in both directions using an ABI 3730
DNA sequencer (Applied Biosystems Carlsbad CA)United States) Due to the identical
16S rDNA sequence of the three colones only one was queried against the GenBank and
Ribosomal Database Project (RDP) databases using BLAST and SEQUATCH algorithms
(Cole et al 2007 Wang et al 2007 Cole et al 2009) The 16S rDNA sequence of
32
strain SX-1 and closely related type strains were aligned using CLUSTALX software
(Thompson et al 1997) The neighbor-joining trees were constructed with the Molecular
Evolutionary Genetics Analysis package (MEGA Version 4) based on 1000 bootstrap
analysis (Tamura et al 2007)
SEM
The morphologies of SX-1 cells growing overnight in anaerobic tubes were examined
with a scanning electron microscope (FEI Quanta 600 FEG FEI Company Hillsboro
OR) The suspended cells were fixed in a 25 glutaraldehyde and 01M phosphate
buffer solution and dehydrated with a graded ethanol series from 30 to 100 After
dehydration the samples were dried in a critical point dryer and then sputter-coated with
AuPd for SEM examination (Liu and Logan 2004)
Evaluation of power generation by SX-1 in MFCs
Single chamber MFCs were used to evaluate power generation by SX-1 using various
substrates The MFCs were constructed as described previously (Liu and Logan 2004)
and modified with 3 cm2 carbon cloth anodes and 7cm
2 carbon clothPt cathodes The
total liquid volume of each MFCs was about 15 ml with electrode spacing about 17 cm
All MFCs were operated in an autoclaved closed plastic box and sterile cotton was
attached to the outer surface of the air cathodes to prevent contamination A MFC
without bacterial culture was used as control MFCs were inoculated with 3 ml late
exponential phase cultures of SX-1 in the medium solution reported previously (Liu and
Logan 2004) Seven substrates including citrate acetate glucose sucrose glycerol and
lactose were evaluated individually for power generation in a fed-batch mode in a
temperature control chamber (30 plusmn2degC) For each substrate at least 8 batches were run to
33
investigate the effect of biofilm formation on current output at fixed external resistance of
1kΩ and the same initial substrate concentration of 30 mM Phosphate buffer (100mM)
was used to maintain the solution pH and conductivity The MFCs with sodium citrate as
electron donors were also examined for maximum power output by varying the external
resistance from 20 kΩ to 4 kΩ A data acquisition system was used to record voltage data
during the MFCs operation (Keithley 2700 Keithley Instruments Inc Cleveland OH) It
took about 15 to 30 minutes for the MFCs to stabilize depending on the external
resistance At each resistance we collected at least five data at the steady condition to
make the polarization curves Averaged voltages were used to calculate the power density
(mW m-2
) according to P=IVA where I was the current V was voltage and A was cross-
sectional area of the anode
CV analysis
CV was used to characterize the oxidation and reduction reactions on the anodic surface
of the MFCs fed with citrate and operated at 7 KΩ This external resistance was selected
because the maximum power density was obtained at this resistance based on the
polarization experiment We assumed the biofilm on the anode of MFCs were well-
developed when stable power output was obtained after 3 batches of operation The
MFCs were then used directly for CV analysis at four current generating stages of the
fourth batch (1) initial exponential current increasing stage (middle point of stage about
4 h after media change) (2) current plateau stage (middle point of stage about 8 h after
media change) and (3) current decreasing stage (middle point of stage about 16h after
media change) (4) right after the complete replacement of medium solution (Figure 6)
The anode was used as working electrode the cathode as counter electrode and an
34
AgAgCl electrode was selected as the reference The CV curves were scanned from 200
to -600 mV at a rate of 5 mV s-1
using a potentiostat (G300 Gamry Instrument
IncWarminster PA) Control experiment was also conducted using new anode (without
biofilm) and new medium solution
Nucleotide sequence accession number
The 16S r DNA sequence determined in this study has been deposited in the GenBank
database under accession number HQ845373
Results
Identification of the strain SX-1
An almost-complete 16S rDNA sequence (1475 bp) of strain SX-1 was obtained and
subjected to comparative analysis with the 16S rDNA of closely related reference strains
A phylogenetic analysis revealed that strain SX-1 should be assigned to the genus
Citrobacter and was most closely related to Citrobacter sp sdy-48 EJ463782 (990
sequence similarity) and Citrobacter sp yy-21 FJ463779 (975 sequence similarity)
These three strains formed a distinct sub cluster in the neighbor-joining in which the new
isolate and Citrobacter sp sdy-48 EJ463782 formed a distinct subline (Figure 31)
The SEM image illustrated that the strain SX-1 is a rod-shaped bacterium 03-06 microm
wide and 12-17 microm long (Figure 32) Strain SX-1 colonies on nutrient agar appeared 2-
3 mm in diameter smooth moist opaque with a shiny surface after 5 days incubation
under anaerobic condition The fact that strain SX-1 grew both aerobically and
anaerobically using FeC6H5O7 as electron acceptor suggests that the strain is facultatively
anaerobic Strain SX-1 showed robust growth on citrate as a carbon source These D
35
properties were similar to those of genus Citrobacter described in Bergeyrsquos Manual of
Systematic Bacteriology (George 2005)
Figure 31 phylogenetic tree of strain SX-1 and closely related species based on 16S
rDNA sequences The tree was constructed using the neighbor-joining method
Figure 32 SEM image of planktonic cells of Citrobacter sp SX-1
Strain SX-1
Citrobacter sp sdy-48 (FJ463782)
Citrobacter sp yy-21 (FJ463779)
Citrobacter koseri E639 (ATCC 25408)
Citrobater koseri CDC 3613-63(AF025372)
Citrobacter koseri CDC 8132-86(AF025366)
Salmonella enterica SL483 (CP001138)
Salmonella enterica AKU12601(AY696668)
Shigella sonnei Ss046(CP000038)
E coli C2 (AF403733)
E coli E24377A(CP000800)88
100
100
76
44
92
53
36
0001
1 microm
36
Electricity production by strain SX-1 in MFCs
The isolated strain SX-1 was first tested for its ability to generate current from sodium
citrate at 1kΩ Current gradually increased to a maximum 50 mA m-2
after cell
inoculation and then decreased (Figure 33) After the MFCs was refilled with new
substrate solution the current recovered rapidly and reached a higher level than the first
batch After 4 batches operation the maximum current output of each batch became
stable The highest current density achieved in the MFCs with Citrobacter sp SX-1 at 1
KΩ was 98 mA m-2
(Figure 33)
Figure 33 Electricity generation by Citrobacter sp SX-1 in a single chamber MFCs
with sodium citrate (30 mM) as substrate at 1 KΩ
0
20
40
60
80
100
120
0 50 100 150 200
Curr
ent
den
sity
(m
A m
-2)
Time (h)
37
Current generation by SX-1 from other substrates including glucose lactose sodium
acetate glycerol and sucrose were also assessed at an external resistance of 1KΩ
(Figure 34) When repeatable cycles of current output were obtained for these substrates
glycerol generated the highest maximum current density of 58 mA m-2
followed by
lactose and sucrose with 29 mA m-2
and 27 mA m-2
respectively Glucose and acetate
produced the lowest maximum current density of 96 and 43 mA m-2
respectively These
results indicated that strain SX-1 can utilize a wide range of substrates for electricity
generation in MFCs but with different power generation potentials
Figure 34 Electricity generation of Citrobacter sp SX-1 using different substrates in
single chamber air-cathode MFCs at 1 KΩ (The error bars stand for the standard
deviation of 3 replicates)
0
10
20
30
40
50
60
70
Glucose Lactose Glycerol Acetate Sucrose
Curr
ent
den
sity
(m
A m
-2)
38
Polarization experiment was further conducted to determine the maximum power
density generated by strain SX-1 in the single chamber MFCs with citrate as substrate A
maximum power density of 881 mW m-2
was obtained at current density of 205 mA m-2
at an external resistance of 7 KΩ (Figure 35)
Figure 35 Power and voltage generation by Citrobacter sp SX-1 as a function of
current density using sodium citrate (30 mM) as substrate The error bars stand for the
standard deviation of the 3 voltagepower outputs obtained in three MFCs
20
30
40
50
60
70
80
90
100
01
02
03
04
05
06
07
80 110 140 170 200 230
Po
wer
den
sity
(m
W m
-2)
Vo
ltag
e (V
)
Current density (mA m-2)
Voltage
Power
39
Figure 36 Four current generating stages for CV analysis current increasing stage (stage
1) current plateau stage (stage 2) current decreasing stage (stage 3) and right after the
replacement of medium solution (stage 4) The MFCs was operated at 7 KΩ with sodium
citrate (30 mM) as substrate
Cyclic Voltammetry
To determine the presence of redox active compounds produced by SX-1 and the
location of these compounds CV scan of the anodic biofilms at four current output stages
and the supernatant of MFCs medium solution at the end of the batch experiment were
performed Reduction peaks at range of -310 ndash -350 mV and oxidation peaks at range -
100 ndash -150 mV were observed at all current generating stages (Figure 36 Figure 37A)
suggesting the presence of redox active compounds may involve in extracellular electron
transfer by Citrobacter sp SX-1 The amplitude of the peaks increased according to the
growth stage of the batch and the highest peaks were present after the current plateau
stage which indicated the redox active compounds mainly were secreted in the current
plateau stage While the current density at stage 3 (deceasing stage) was lower than that
at stage 2 (current plateau stage) the redox peak at stage 3 was higher than that in stage 2
0
50
100
150
200
250
50 55 60 65 70 75 80 85
Curr
ent
den
sity
(m
A m
-2)
Time (h)
A
A
Stage 1
Stage 2 Stage 3
Stage 4
40
A plausible explanation for this pattern was that redox active compounds were
continuously being secreted and accumulated after the current peak was reached
resulting in more redox compounds present after the plateau stage But since most of the
carbon source had already been being used up many of the redox compounds at this
stage had not been as active as those at current plateau stage due to much less electrons
were available to be transferred The bigger peak amplitude in stage 4 than in stage 1
indicates that the redox active compounds were continuously being secreted and
accumulated in the biofilm even after stable power was generated at 7 kΩ The slightly
change of the peak locations at different stages was possible due to the solution chemistry
change during the current generation process or the slightly location change of reference
electrode during the measurement (Figure 37B) When the medium solution in the MFCs
was replaced by fresh medium at the end of the batch the oxidization and reduction
peaks were still presented in the CV (Figure 37B) but no peaks were observed when the
supernatant and fresh mediums were tested in a MFCs with no biofilm (Figure 37C)
These results suggest the compounds involved in the electron transfer were located not in
the supernatant (solution) but in the biofilm
41
Figure 37 Cyclic Voltammetry analysis of Citrobacter sp SX-1 biofilms at (A) current
increasing stage current plateau stage and decreasing stage (end of the batch) (B) end of
the batch and replaced with fresh medium and (C) controls
-40E-04
-30E-04
-20E-04
-10E-04
00E+00
10E-04
20E-04
30E-04
-060 -050 -040 -030 -020 -010 000 010 020
Curr
ent
(A)
Potential (V) vs AgAgCl
Current plateau stage
Current increasing stage
Current decreasing stage
-40E-04
-30E-04
-20E-04
-10E-04
00E+00
10E-04
20E-04
30E-04
-060 -050 -040 -030 -020 -010 000 010 020
Curr
ent
(A)
Potential (V) vs AgAgCl
Current decreasing stage
Biofilm with fresh medium
-40E-04
-35E-04
-30E-04
-25E-04
-20E-04
-15E-04
-10E-04
-50E-05
00E+00
-060 -050 -040 -030 -020 -010 000 010 020
Curr
ent(
A)
Potential (V) vs AgAgCl
Supernatant
Fresh medium
A
B
C
B C
42
Discussion
Citrobacter species belonging to Gammaproteobacteria Enterobacteriales
Enterobacteriaceae are facultative anaerobic bacteria (George 2005) They can grow with
oxygen as an electron acceptor but also with a variety of electron accepters such as Fe (III)
in the absence of oxygen (George 2005) Citrobacter species can be found in a wide
variety of habitats including in soil water and wastewater Several Citrobacter species
have been studied as important bioremediation bacterium for heavy metal removals
sulfate reduction phenol degradation and chlorophenol degradation (Macaskie et al 1995
Narde et al 2004 Qiu et al 2009) Citrobacter species have also been found on the
anodic biofilm of a diesel-degrading MFCs (Morris et al 2009) However there was no
study reported that Citrobacter species can transfer electrons to extracellular electron
acceptors This finding that Citrobacter strain SX-1 can generate electricity in MFCs
increased the diversity of power generating exoelectrogens and provided a new strain to
explore the mechanisms of extracellular electron transfer from bacteria to electrode
Among the isolated exoelectrogens gammaproteobacteria and deltaprotebactia appear to
be dominant among others (Parot et al 2009 Liu et al 2010) The isolated strain
Citrobacter sp SX-1 phylogenetically belongs to gammaproteobacteria which provides
another evidence to support this observation
Most exoelectrogens utilize a limited range of substrates for power generation For
example Shewanella oniedensis MR-1 can oxidize lactate under anaerobic condition but
cannot utilize acetate for electricity generation (Kim et al 2002) Geobacter
sulfurreducens can completely oxidize acetate for power generation but it cannot utilize
simple sugars (Bond and Lovley 2003 Caccavo et al 1994)) In comparison strain SX-1
43
not only can oxidize citrate for power generation but also utilize a wide range of
substrates for power generation including glycerol glucose lactose sucrose and acetate
Interestingly the higher current density generated by strain SX-1 from glycerol a main
by-product of biodiesel production process suggests that strain SX-1 may be potentially
used for harvesting energy from biodiesel wastes using MFCs However the current
density generated by strain SX-1 (205 mA m-2
) is lower than that (805 mA m-2
at 1 kΩ)
generated by the mixed culture from which the SX-1 was isolated indicating the
existence of other higher power generating bacteria andor complex ecology in the mixed
culture community
Understanding mechanisms of microbial extracellular electron transfer is critical for
enhancing the electron transfer rate from bacteria to electrode through metabolic or
genetic engineering of exoelectrogens Since the spent supernatant solution of MFCs run
with SX-1 showed no redox properties the redox compounds produced by SX-1 may
have been retained in the biofilm Alternatively SX-1 may utilize one or both of the
other two known extracellular electron transfer mechanisms For example it is possible
that strain SX-1 transfers electrons to the anode via cell-bound outer membrane proteins
The two distinct oxidation and reduction peaks observed at -100mv- -150 mV and -310--
350 mV of the SX-1 biofilm have a midpoint potential range from -205mV to -250mV
which is close to the redox potential of some c-type cytochromes (Field et al 2000 Meitl
et al 2009) which are well-known to play an important role in extracellular electron
trasnfer (Eggleston et al 2008 Field et al 2000 Meitl et al 2009) Field et al (2000)
reported that the c-type cytochromes CymA had the midpoint potentials of -229 mV
Meitl et al (2009) and Eggleston et al (2008) reported that the c- type cytochromes
44
OmcA in Shewanella oneidensis MR-1 had the midpoint potentials of -201 mV and -208
mV
While itrsquos possible that SX-1 may also have conductive appendages to enhance the
extracellular transfer electron similar to the nanowires discovered in some Geobacter
and Shewanella species (Reguera et al 2005 2006 Gorby et al 2006) further
investigation on the presence of these appendages on the cell surface of SX-1 is needed
Acknowledgements
The authors acknowledge support from the US National Science Foundation
(CBET 0955124) We also thank Teresa Sawyer for her helps with the SEM analysis and
Jeremy Chignell and Yanzhen Fan for their review of this manuscript
45
Chapter 4
Enhanced Performance and Mechanism Study of Microbial Electrolysis Cells
Using Fe Nanoparticles Decorated Anodes
Shoutao Xu Hong LiuYanzhen Fan Rebecca Schaller Jun Jiao Frank Chaplen
Published in
Applied Microbiology and biotechnology 93(2)871-880 (2012)
46
ABSTRACT
Anode properties are critical for performance of microbial electrolysis cells (MECs) In
the present study Fe nanoparticle modified graphite disks were used as anodes to
investigate the effects of nanoparticles on the performance of Shewanella oneidensis MR-
1 in MECs Results demonstrated that average current densities produced with Fe
nanoparticle decorated anodes were up to 56-fold higher than plain graphite anodes
Whole genome microarray analysis of the gene expression showed that genes encoding
biofilm formation were significantly up-regulated as response to nanoparticle decorated
anodes Increased expression of genes related to nanowires flavins and c-type
cytochromes indicate that enhanced mechanisms of electron transfer to the anode may
also have contributed to the observed increases in current density The majority of the
remaining differentially expressed genes were associated with electron transport and
anaerobic metabolism demonstrating a systemic response to increased power loads
Keywords Microbial electrochemical system microbial fuel cell microbial electrolysis
cell nanotechnology differential gene expression DNA microarray
47
Introduction
Microbial electrochemical systems (MESs) have been intensively studied since Lewis
achieved practical advances in this field (Logan 2007) however they attracted much
research attention in recent years due to their promising applications in renewable energy
generation bioremediation and wastewater treatment In a MES microorganisms
interact with electrodes via electrons catalyzing oxidation and reduction reactions at the
anode and the cathode The most-described type of MESs is microbial fuel cells (MFCs)
in which useful power is generated from electron donors typically biodegradable organic
materials (Logan et al 2006) Various novel MESs have recently been developed to
produce hydrogen (microbial electrolysis cells (MECs)) (Liu et al 2005 Logan et al
2008 Rozendal et al 2006 Schaller et al 2009) to power an autonomous sensor or
sensor network (Ringeisen et al 2006) to reduce bioremediation targets (Lovley 2006
Shea et al 2008) Sukkasem et al 2008) and to desalinate water (Cao et al 2009) The
key feature shared by these systems is the microbe-catalyzed electron transfer from
organic matter to electrodes (anodes) (Rabaey et al 2004) Enhancing the anodic current
output which highly depends on the performance of the electrodes is critical for the
successful application of all these processes (Logan et al 2007 Park and Zeilus 2002
2003)
Nanomaterials have received much attention from researchers in the context of
microbiology due to their unique physical electrical and chemical properties which
facilitate the study of interactions between bacteria and surfaces (Liu 2006) Previous
studies have demonstrated that electrodes decorated with different nanostructures such
as carbon nanotubes nanostructured polyanilinetitanium dioxide composite and titania
48
nanotubes have the potential to enhance power generation in MFCs (Morozan et al 2007
Qiao et al 2008 Quan et al 2005) Furthermore our recent study shows nanoparticle
(NP) decorated anodes greatly increased the electrochemical electron transfer rate in
MFCs (Fan et al 2011) However Fe NPs as an ideal candidate for decorating electrodes
because of respectively low price and high conductivity compared to other materials has
not been focused to study in MECs
S oneidensis MR-1 an important electrochemically active bacterial strain has been
exploited for the electricity generation in MFCs (Logan et al 2007 Logan 2009 Park
and Zeikus 2002) The availability of genome sequence for this strain makes it possible to
use transcriptome assays to globally measure the responses to different growth conditions
and environmental stresses (Murray et al 2001) S oneidensis MR-1 gene expression
response to heat shock cold shock Cr (VI) and U (VI) reduction chromate stress and
iron and acid tolerance have been studied in the past several years (Bencheikh-Latmani et
al 2005 Brown et al 2006 Gao et al2006 Gao et al 2004 Yang et al 2008) However
no studies have been focused on the Shewanella gene expression response to NPs in
MESs
The mechanism of increased electron transfer rate exhibited by nanoparticle decorated
electrodes is not well understood yet In the present study graphite disks decorated with
Fe NPs were used as anodes to explore the effects of nanostructures on current generation
in a multi-anode MECs DNA microarrays were utilized to investigate differences in the
global gene expression profile of S oneidensis MR-1 grown on plain versus Fe NPs
decorated anodes
Materials and methods
49
Bacterial cultures
S oneidensis MR-1 was purchased from American Type Culture collection (ATCC
700550) and stored as freezer stocks at -80degC Stock cells of S oneidensis MR-1 was
grown aerobically using a medium containing 30 gL Trypticase Soy Broth (BD 211825
Becton Dickinson Company Franklin lakes NJ) at 30degC Cells in early stationary
growth phase (optical density of 15-16 at 600nm (Spectrophotometer UV-1700
Shimadzu Columbia MD) were washed two times then injected into the chamber of the
MECs for current production Trypticase Soy Broth medium with 30 mM sodium lactate
as electron donor was used in MECs Phosphate buffer (100mM) was used to maintain
the solution pH 7 and solution conductivity at 15 mScm
Characterization of nanostructured anodes
Superfine isomolded graphite disks (16 cm x 05 cm GraphiteStorecom Inc) were
polished with ultrafine sand paper (2000 grit 3M Company) as the base for the NP
decorated and control anodes Fe NPs decorated anodes were fabricated by thermal
annealing of metal thin film coated graphite disks Thin films of Fe were firstly deposited
on the polished graphite disk by using sputter coating for 95 min and the samples then
were annealed in the Chemical vapor deposition (CVD) at 750degC for 20 min to form Fe
NPs Fe-NPs were produced with a diameter of 200plusmn100 nm and a density range
(distance between NPs) of 200plusmn100 nm with a coating time of 95 min SEM images of
Fe NPs decorated anode and control surfaces were shown in the Figure 41
50
Figure 41 SEM images of Fe NPs decorated anodes (A) Graphite disk control (B) Fe
Nanoparticle decorated
Multiple channel MECs construction and operation
A MECs with removable multiple anodes with each effective anode area 07 cm2
was
constructed and used to evaluate the effects of nanostructure on current density of
according to a previous report (Fan et al 2011) The cathode was made of wet-proof
(30) carbon cloth (type B E-TEK Division Inc USA) coated with PtC (20 E-TEK
Division Inc USA) following a previously reported procedure (Liu et al 2005) The
final platinum loading was 05 mgcm2 per projected cathode area The size of cathode
(150 cm2) was 25 times larger than the total effective area of the 8 graphite disk anodes
(56 cm2) to prevent cathode limitations on the performance of the MECs system All
MECs with Fe-NP decorated anodes and control anodes were tested for current
generation for 24 h with medium (without bacteria) before injecting the bacterial cells in
order to determine whether the Fe-NP decorated anodes demonstrate chemical current
generation compared to control anodes All the testing anodes were set up in the MECs
B
500 nm
A
500 nm
51
at once so that the S oneidensis MR-1 culture was applied to all of them simultaneously
Short electrode spacing (17 cm) was used in the MECs design to reduce the internal
resistance A voltage of 06 V was applied the MECs for current generation after the cells
of S oneidensis MR-1 were inoculated into MECs at early stationary phase A multimeter
with a data acquisition system (2700 Keithly USA) was used to monitor the current
change by measuring the voltage (every 300 seconds) drop through a resistor at 300 ohm
Fig 4 2 Effects of Fe-NP decorated anodes on the current density in MECs Control is
plain graphite disk anode Fe-NP is the anode with Fe-NP decoration
Microarray analysis
Biofilms for whole gene microarray analysis were aseptically removed from the plain
and Fe-NP decorated anodes of MECs after 110 hours further incubation at 30oC when
current density obviously started to decease Total RNA was extracted using Trizol
(Ambion inc AustinTX) following the manufacturerrsquos instructions Integrity of the
52
RNA samples was confirmed using the Agilent 2100 Bioanalyzer (Agilent Technology
Inc Palo Alto CA) The same amount of RNA (200 ng) from each sample was used for
further analysis after amplification by using the MessageAmpTM
II-Bacteria Prokaryotic
RNA Amplification kit (Ambion AustinTX) according to the manufacturerrsquos
instructions All of the 4295 genes (resource from NCBI) represented on the S oneidensis
MR-1 whole-gene microarrays were ordered from Roche NimbleGen Inc (Madison WI)
Biological triplicates of biofilms on the Fe-NP decorated anodes and control were
analyzed respectively cDNA synthesis labeling and hybridization were carried out by
the CGRB Core Laboratories at Oregon State University DNASTAR ArrayStarTM
3
software was used to identify genes that were up- or down-regulated more than 2-fold
when grown on the nanoparticle modified anodes using the unpaired two sample t-test
with a cutoff p-value of 005 The complete microarray data set generated in this study is
deposited for public access in the Gene Expression Omnibus
(httpwwwncbinlmnihgovgeo) under accession number GSE31535
Results
Enhancement of current generation by using NP decorated anodes in MECs
The current density generated by Fe-NP decorated anode MECs with S onidensis MR-1
increased to approximately 43 microAcm2 20 h following inoculation and then slowly
decreased Figure 42 shows that the maximum current density achieved by NP
decorated anodes was 83 times higher than that (51 microAcm2) generated by the control
(plain graphite disk) The average current density improvement of 110 hours was 59
times of that generated by the control The current densities generated in MECs in the
53
absence of bacteria were negligible (03 microAcm2) which demonstrated that the current
enhancement observed with the Fe-NP decorated anodes was biologically-derived
Global transcriptome analysis
Whole-genome DNA microarrays were used to attain a comprehensive general
overview
of the transcriptional response to Fe-NP decorated anodes by S oneidensis MR-1 in MEC
Of a total 4295 genes assayed 392 (188 induced and 204 repressed) exhibited significant
(P lt 005) differential expression at a 2-fold (log (expression ratio 1) level in 3
replicates in response to Fe-NPs These
total gene numbers present 9 of the 4295 open
reading frames (ORFs) presented on the array
Fig 43 Differentially expressed genes grouped by functional classification according to
the TIGR S oneidensis genome database (wwwtigrorg) Columns 1 amino acid
biosynthesis 2 biosynthesis of cofactors prosthetic groups and carriers 3 cell
envelope 4 cellular processes 5 central intermediary metabolism 6 DNA metabolism
7 energy metabolism 8 fatty acid and phospholipid metabolism 9 other categories 10
protein fate 11 protein synthesis 12 purines pyrimidines nucleosides and nucleotides
13 regulatory functions 14 signal transduction 15 transcription 16 transport and
binding proteins 17 unknown function 18 hypothetical proteins
Up-Regulated
-75
-50
-25
0
25
50
75
100
125
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18
Down-Regulated
Num
ber
of
Gen
es
54
Figure 43 summarizes the overall genomic response of S oneidensis MR-1 to Fe-NP
decorated anodes by grouping the differentially expressed genes into their functional role
categories based on The Institute for Genomic Researchrsquos annotation (Version 8) of the
MR-1 genome sequence The wide distribution of putative functional roles attributed to
the differentially expressed genes indicated the extent of the molecular response of
Soniedenis MR-1 to the NP decorated anodes Among genes with defined function a
large number of up-regulated genes were associated with cellular processes (group 4)
energy metabolism (group 7) other categories (group 9) and hypothetical proteins (group
18) While those genes encoding proteins involved in amino acid biosynthesis (group 1)
cellular processes (group 4) substrate transport (group 16) and hypothetical proteins
(group 18) were among the most down-regulated genes Two groups of genes that were
the most noticeable among the functional gene groups genes related to energy
metabolism (group 7) and genes related to amino acid biosynthesis (group 1) The ratios
of up-regulated genes to down-regulated genes in these functional groupings were much
higher than other functional gene groupings there were 21 up-regulated genes and 7
down-regulated genes in energy metabolism gene group and 2 up-regulated and 22 down
regulated genes in the amino acid biosynthesis group
Genes related to biofilm formation
In order to determine whether there was a correlation between the observed biofilm
enhancements by NP decorated anodes and the genes related to biofilm formation the
significant modified genes related to biofilm formation were examined and summarized
in the Table 41 These included SO_3223 and SO_3228 genes encoding the flagellum
proteins (Thormann et al 2004) which has critical impacts on initial attachment to the
55
surface as swimming motility functions were up-regulated over 10 and 09 fold
respectively The gene clusters SO_4178-SO_4180 which are thought to be essential for
formation of a three-dimensional biofilm were up-regulated from 12 to 13 fold
(Thormann et al 2004) In addition the genes SO_4105 encoding IV pilus which is
known to represent a critical factor for biofilm formation on a biotic surfaces (Thormann
et al 2004) were also induced 08 fold (Table 41) The genes related to flagellum
motility (SO_4286 SO_1529 and SO_1530) were also induced slightly (Thormann et al
2004)
Table 41 Expression levels of genes related to biofilm formation
Locus Tag Gene product descriptions Fold change
SO_3228 flagellar basal-body MS-ring and collar protein FliF 10
SO_3223 flagellar hook-length control protein FliK 09
SO_4103 MshA minor pilin protein MshD 05
SO_4105 MSHA major pilin protein MshA 08
SO_4178 expressed protein of unknown function MxdC 12
SO_4179 inner membrane family 2 glycosyltransferase MxdB 13
SO_4180 diguanylate cyclase-like protein MxdA 10
a fold changes were presented after log2
b positive number represented up-regulated and negative represented down-regulated
Genes related to energy metabolism
A closer consideration of energy metabolism genes was undertaken as anaerobic
metabolism for electron generation and electron transport functions may play critical
roles in enhanced current density generation in MECs Genes associated with energy
56
metabolism and with significantly modified gene expression levels are summarized in the
Table 42 Several of the up-regulated genes were related to formate dehydrogenase
proteins Most notably four genes related to formate dehydrogenase were up-regulated
more than 2-fold SO_0102 SO_0103 SO_0104 and SO_0107 The other genes
expression related to formate dehydrogenase proteins SO_4513 SO_4514 SO_4515 also
significantly increased Another interesting gene is that encoding the cytochrome c
oxidase protein gene SO_4609 which was up-regulated 11 fold while the gene
SO_0479 related periplasmic octaheme cytochrome c MccA was down-regulated
However SO_1519 the gene encoding lactate dehydrogenase was down-regulated 12
fold despite lactate being the primary carbon source in the media
Table 4 2 Genes related to anaerobic growth and electron transfer with significantly change expression level
Locus Tag Gene product descriptions Fold
change
SO_0101 nitrate-inducible formate dehydrogenase molybdopterin-binding
subunit FdnG
18
SO_0102 formate dehydrogenase nitrate-inducible iron-sulfur subunit 21
SO_0103 formate dehydrogenase nitrate-inducible cytochrome b556 subunit 24
SO_0104 formate dehydrogenase accessory protein FdhE 22
SO_0107 formate dehydrogenase accessory protein fdhD 24
SO_0397 quinolfumarate reductase menaquinol-oxidizing subunit FrdC_2 12
SO_0452 thioredoxin 2 15
SO_0903 Na-translocating NADH-quinone reductase subunit B NqrB_1 12
SO_1012 NADH-ubiquinone oxidoreductase subunit K NuoK 12
SO_1013 NADH-ubiquinone oxidoreductase subunit J NuoJ 12
SO_1363 hydroxylamine reductase 13
57
SO_2417 ferredoxin cofactor maintenance protein YfaE 10
SO_3922 formate dehydrogenase cytochrome b Fdh -12
SO_4513 formate dehydrogenase molybdopterin-binding subunit _2 15
SO_4514 formate dehydrogenase FeS subunit FdhB_2 12
SO_4515 formate dehydrogenase cytochrome b subunit FdhC_2 14
SO_4609 aa3 type cytochrome c oxidase subunit III CoxC 11
SO_0479 periplasmic octaheme cytochrome c MccA -12
SO_0847 periplasmic nitrate reductase ferredoxin component NapG -15
SO_1519 L-lactate dehydrogenase iron-sulfur cluster-binding protein LldF -10
SO_1251 ferredoxin 4Fe-4S -11
SO_37411 hypothetical inner membrane protein -11
a fold changes were presented after log2
b positive number represented up-regulated and negative represented down-regulated
Flavin and cytochrome related genes
Flavins can be secreted by Shewanella species as electron shuttle to facilitate
extracellular electron transfer (Cansterin et al 2008) C-type cytochromes play the
important roles on the process of extracellular electron transfer (Shi et al 2007) The
genes related to flavins synthesis and the genes encoding the cytochromes electron
transport proteins were shown in the table 43 the genes SO_1414 and SO_3058 related
to flavins synthesis was up-regulated to 03 and 01 folds respectively The genes related
to s C-type cytochromes SO_0135 SO_4105 SO_0169 SO_1778 are increased to 10
08 05 04 01 folds respectively However most of genes have shown no significant
changes (less one-fold change) as response to nano particle decorated anode in MECs
58
Table 43 Expression level of flavin and cytochrome genes
Locus Tag Gene product descriptions Fold
change
SO_1414 flavocytochrome c flavin subunit putative 03
SO_3468 riboflavin synthase subunit alpha -02
SO_3058 flavocytochrome c flavin subunit 01
SO_4105 MSHA pilin protein MshA 08
SO_0169 general secretion pathway protein GspG 05
SO_1778 outer membrane decaheme cytochrome c lipoprotein MtrC 04
SO_1779 outer membrane decaheme cytochrome c OmcA 01
SO_0135 lipoprotein of unknown function DUF333 10
SO_0136 conserved hypothetical inner membrane protein 01
a fold changes were presented after log2
b positive number represented up-regulated and negative represented down-regulated
Other genes with significantly modified expression levels
These genes over 15 fold change and possibly related to current enhancements were
shown in Table 44 SO_3104 (expressed inner membrane protein) was over expressed
and the gene SO_2194 which encodes outer membrane protein (OmpA family protein)
was repressed as response to nanoparticle decorated anode however it is unclear the
relationships of this modified membrane protein genes with the current enhancement
Another interesting phenomenon was possible co-regulation of several gene clusters
indicating possible operon associations This includes three sets of genes that were
induced more than 15 fold SO_0108SO_0109 and SO_1042SO_1043SO_1044 The
59
consistency of expression of these genes under the NP conditions provides basic evidence
to support operon structure However the correlation of these significant changed gene
with current density enhance are unclear which showed there are unknown multiple and
complex responses of S oneidensis MR-1 to nanoparticle anode of MECs
Table 44 Other genes with significantly changed expression levels
Locus Tag Gene product descriptions Fold change
SO_0101 nitrate-inducible formate dehydrogenase molybdopterin-binding
subunit FdnG
18
SO_0108 integral membrane protein of unknown function DUF39 YedE 17
SO_0109 SirA family protein YedF 17
SO_0275 N-acetyl-gamma-glutamyl-phosphate reductase -31
SO_0277 ornithine carbamoyltransferase -21
SO_0279 argininosuccinate lyase -24
SO_0404 zinc dependent metalloprotease domain lipoprotein 21
SO_0956 alkyl hydroperoxide reductase F subunit -20
SO_1042 amino acid ABC transporter ATP-binding protein -18
SO_1043 amino acid ABC transporter permease protein -27
SO_1044 amino acid ABC transporter periplasmic amino acid-binding protein -27
SO_1072 chitin-binding protein putative 21
SO_1405 transglutaminase family protein 20
SO_1822 TonB-dependent receptor putative -22
SO_2069
1-(5-phosphoribosyl)-5-[(5-phosphoribosylamino)
methylideneamino] imidazole-4-carboxamide isomerase
-19
SO_2070 amidotransferase HisH -26
SO_2071 imidazole glycerol-phosphate dehydratasehistidinol phosphatase -27
60
SO_2072 histidinol-phosphate aminotransferase -28
SO_2073 histidinol dehydrogenase -30
SO_2194 OmpA family protein -36
SO_2195 inter-alpha-trypsin inhibitor domain protein -27
SO_2767 asparagine synthetase B -26
SO_2945 prophage LambdaSo tail fiber protein 20
SO_2963 prophage LambdaSo major capsid protein HK97 family 20
SO_2987 prophage LambdaSo hypothetical protein 19
SO_2988 prophage LambdaSo expressed protein of unknown function 18
SO_3104 expressed inner membrane protein 131
SO_3408 conserved hypothetical inner membrane protein -18
SO_3585 azoreductase putative -18
SO_3586 glyoxalase family protein -17
SO_3687 curli production assemblytransport component CsgE putative -18
SO_38191 hypothetical ammonia permease -19
SO_4014 AcrBAcrDAcrF family protein 18
SO_40151 type I secretion system membrane fusion protein RND family 18
SO_4054 510-methylenetetrahydrofolate reductase 18
SO_4245 N-acetylglutamate synthase -22
SO_45251 hypothetical transcriptional regulator LysR family -17
SO_4527 integral membrane domain protein -19
SO_4705 transcriptional regulator putative -19
a fold changes were presented after log2
b positive number represented up-regulated and negative represented down-regulated
Discussion
61
The addition of NP to anodes significantly impacts current densities with the elemental
composition being a critical factor The chemical composition of NPs significantly affects
the current enhancement in MECs (Fan et al 2011) because the chemical composition of
NP properties is a critical factor in determining the conductivity which significantly
affects the efficiency of electron transfer from bacteria to electrode Fe is an ideal
candidate for NP formation for current enhancement in MESs because of high
conductivity and low price compared to gold and other conductive materials Gold NPs
has also been studies for enhancement of electricity conductivity (Bao et al 2008)
However the disadvantages of the high price of gold limit their practical application in
MECs (Fan et al 2011) The different size and density of the same composition of NP
also has essential effects on the current density enhancement (Fan et al 2011) In this
study the results showed the current density enhancement with Fe-NP size range of
200plusmn100 nm and density range of 200plusmn100 nm is considerable enhancement consequence
There are a number of reports of studies of current density enhancement in MFCs
by nanostructured anodes (Fan et al 2011 Qiao et al 2008 Sharma et al 2008 Tsai et al
2009) Based on bacterial behavior on nanostructured anode surface Qiao et al (2008)
believes that Escherichia coli cells on the nanostructured electrode surface produce hair-
like structures similar to pili that could facilitate the electron transfer between the cells
and electrode The production of hair-like structures is believed to be stimulated by
nanostructures and could play the key role on current density enhancement in pili-
producing bacteria in MFCs (Qiao et al 2008) S oneidensis can produce electrically
conductive bacterial nanowires which have similar structure to the hair-like structures in
Ecoli (Gorby et al 2006 Reguera et al 2005) A number of possible genes related
62
nanowires of S oneidensis have been identified including mshA (SO_4105) and gspG
(SO_0169) (Konstantinidis et al 2009) They are up-regulated 08 and 05 fold in the
current study respectively even though they are not significant induced This result
supported the increased nanowire genes had contributions to enhance current density in
MFCs by NPs decorated anodes
Based on the previous research on electron transfer mechanisms for S oneidensis it is
known that self-produced mediators (flavins) and multihaem c-type cytochromes (c-Cyts)
may play critical roles in the electron transfer of S oneidensis to extracellular insoluble
electrode (Canstein et al 2008 Myers and Myers 1992 Scott and Nealson 1994 Shi et al
2007) The flavins secreted by S oneidensis can facilitate to transfer electrons from
bacteria to solid anode electrodes by acting as extracellular electron shuttles (Canstein et
al 2008) In this study the genes encoding flavins (SO_1414 SO_3468 and SO_3058)
had no significant increase (less than 1 fold change) as a response to nanoparticle
decorated anode in MECs which suggests that increased flavin gene expression does not
significantly contribute to the current density enhancements in nano-particle decorated
anode The genes encoding cytochromes electron transport proteins (SO_4104 SO_0417
SO_177879 SO_013536) had no significant increase (less than 1-fold change)
However those genes were up-regulated slightly These results suggest the increased
expression flavin and c type cytochromes genes had partial contributions even not
significantly to enhance current density in MFCs by NPs decorated anodes
The thickness of bacterial biofilm on the anode can affect the power generation in
MFCs (Biffinger et al 2007 Heydorn et al 2000) since increased biofilm thickness on
the anode represents more active bacteria involving in the process of electron generation
63
and transfer in MFCs Biofilm formation related genes included flagellar genes IV pilus
genes and other genes The flagellar related genes SO_3228 SO_3229 (fliB and fliK) and
IV pilus genes SO_4105 (mshA) were induced from 19 to 09 fold the genes (SO_4178
-4180) related to formation of a three-dimensional biofilm were up-regulated from 10 to
13 folds even though the genes related bacterial motility protein (SO_ 4286 SO_4287
SO_1529 and SO_1530) are induced slightly (less than 0 5 fold) However the motility
genes most probably play a role for initial biofilm attachment on the anode surfaces
(Thormann et al 2004 Thormann et al 2006) with the rest of biofilm genes being more
essential for biofilm maintenance in mature cultures as were presented at harvest in this
study This enhancement of biofilm density under conditions of increase current density
has been observed for gold NP decorated anodes using confocal light microscopy (data
not shown) but has not yet been confirmed for Fe NP decorated anodes In any event an
increase in the expression level of genes related biofilm formation therefore also
supported the possibility of enhanced biofilm formation on NPs decorated anodes thus
facilitating electron transfer from bacteria to anodes
Beliaev et al described a model of the Fe reduction pathway in S oneidensis MR-1 using
solid substrates as the electron acceptor (Beliaev et al 2001) This model proposes that
electrons are generated and released in cytoplasm then transferred to the quinone pool
eg menaquinone in the cytoplasmic membrane The reduced menaquinone in turn
reduces CymA and from there electrons are then transferred to a periplasmic shuttle (eg
CctA) and then to MtrA a periplasmic soluble decaheme cytochrome c located in
associated with the outer membrane via interaction with MtrB Finally surface displayed
outer membrane proteins decaheme cytochromes (OmcA MtrC or MtrF) can transfer
64
electron to Fe or other solid substrates (Beliaev et al 2001)) Bencheikh-Latmani et al
demonstrated that the same electron transport pathway may be used for more than one
electron acceptor through studies of the response of S oneidensis MR-1 to U(VI) and
Cr(VI) under anaerobic conditions particularly focused on the critical genes
(SO_1777mtrA SO_1776mtrB SO_1778mtrC) (Bencheikh-Latmani et al 2005)
However our studies showed that these genes had no significant expression changes in
response to nanostructured anodes Contrastingly one of expressed inner membrane
protein genes SO_3104 and one of cytochromes genes SO_4609 encoding cytochrome c
oxidase subunit III demonstrated a 13-fold and 11-fold up-regulation respectively in this
study while the gene SO_2194 which encodes outer membrane protein (OmpA family
protein) was down regulated 36 fold Our results therefore suggest that S oneidensis
MR-1 does not utilize the same set of core critical genes (SO_1777 mtrA SO_1776
mtrB SO_1778 mtrC) products to transfer electrons in all instances This is the similar
result with the study of Bretschger et al (2007) which indicated that the electron
transport system in S oneidensis MR-1 is complex with several different proteins able to
participate in electron transfer to the anode of MECs
It should also be mentioned that the gene (SO_1519) encoding lactate
dehydrogenase repressed (-12 fold) here suggesting that lactate was depleted at the time
of cell harvest from the MECs and that other pathways of anaerobiosis had been activated
including those associated with amino acid uptake and consumption the growth media
was a complex mixture including yeast extract In particular the catabolic pathway for
histidine of S oneidensis MR-1 a pathway that may formate as an intermediate is up-
regulated with the genes for histidine ammonia lyase SO_0098 SO_3057 and SO_4374
65
up-regulated 13 11 and 11-fold respectively The possible production of formate
during histidine degradation may in turn explain the high levels of formate
dehydrogenates as evidenced by upregulation of SO_0102 SO_0103 SO_0104 and
SO_0107 annotated as formate dehydrogenase proteins These genes had the highest
levels of up-regulated expression as response to Fe NP decorated anodes (greater than 2-
fold)
Acknowledgements
This research was partially supported by the US National Science Foundation CBET
0828544 and the funds from ONAMIDOD (ARL-DOD Cooperative Agreement
W911NF-07-2-0083) We thank Barbara Gvakharia and Caprice Rosato for helpful
suggestions We also thank Margaret Romine of Pacific Northwest Laboratories for
valuable comments on manuscripts We also thank anonymous reviewers for significant
suggested improvements imparted as part of prior review of this manuscript
66
Chapter 5
Global Transcriptome Analysis of the Response of Shewanella oneidensis MR-1 to
Carbon Nanotube Decorated Anodes in Microbial Electrochemical Systems
Shoutao Xu Yanzhen Fan Rebecca Schaller Frank Chaplen Jun Jiao Hong Liu
67
Abstract
Shewanella oneidensis MR-1 is an important model microorganism for metabolic studies
on the effects of different environmental factors because of its diverse respiratory
capabilities Carbon nanotube (CNT) modified graphite disks were used as anodes to
investigate the effects of nanomatrials on the performance of S oneidensis MR-1 in
microbial electrolysis cells (MECs) The current densities produced with CNT decorated
anodes were on average 56-fold higher than plain graphite anodes Whole genome
microarray analysis of gene expression showed that up-regulation of cytochromes c genes
associated with extracellular electron transfer are strongly correlated to current increases
in CNT decorated anodes Genes involved in flavin biosynthesis also contribute to
current increase in CNT decorated anode MECs
Keywords Microbial electrochemical system microbial fuel cell microbial
electrolysis cell carbon nanotube gene expression DNA microarray
68
Introduction
Shewanella oneidensis MR-1 is an important model microorganism for metabolic
studies of the effects of different environmental factors because of its diverse respiratory
capabilities It has been used for transcriptome analysis to investigate the responses to
different growth conditions and environmental stresses (Murray et al 2001) Gene
expression patterns under different conditions such as heat shock cold shock Cr (VI) U
(VI) reduction chromate stress iron and acid tolerance have been studied previously
(Bencheikh-Latmani et al 2005 Brown et al 2006 Gao et al2006 Gao et al 2004
Yang et al 2008) More recently it has been exploited as a model species for power
generation in microbial electrochemical systems (MESs)which have potential
applications in renewable energy generation bioremediation and wastewater treatment
(Logan et al 2007 Logan 2009 Park and Zeikus 2002 Fan et al 2011 Xu et al 2012)
In a MES electrochemically active microorganisms oxidize organic matter in the an
ode chamber to release electrons Electrons are then transferred to the anode electrode thr
ough various mechanisms (Logan et al 2006 Lovley 2006 Rabaey et al 2003) and fina
lly travel to the cathode electrode and combine with the terminal electron acceptor The
key feature of MESs is the microbe-catalyzed electron transfer from the organic matter to
the anode (Rabaey et al 2004) Enhancing the current output which highly depends on
the performance of the anode electrode is critical for the successful application of MESs
(Logan et al 2007 Park and Zeilus 2002 2003) Several nanomaterials including TiO2
gold (Au) and palladium (Pd) nanoparticles have been utilized to decorate electrodes to
enhance power density in MESs (Qiao et al 2008 Quan et al 2005 Fan et al 2011) The
possible mechanisms for increased current densities have been studied by using Fe
69
nanoparticle-decorated anodes (Xu et al 2012)
Carbon nanotubes (CNT) are cylindrically shaped nanomaterials with extremely
high surface area excellent electrical conductivity and chemical inertness (He et al
2005a Serp et al 2003) These unique properties make CNT a promising electrode
material (Liang et al 2008) The biocompatibility of microorganisms and carbon
nanostructures has been evaluated (Morozan et al 2007) The effects of CNT on anodic
biofilms have been tested in MFCs (Liang et al 2011) However no studies have been re
ported on global transcriptome analysis for the response of S oneidensis MR-1 to CNT
decorated anode in MECs
In the present study the graphite disks decorated with CNT were used as anodes to
investigate the effects of nanomaterials on current generation in multi-anode MECs
DNA microarrays were used to analyze differences in the global gene expression profile
of S oneidensis MR-1 grown on plain versus CNT decorated anodes
Materials and methods
Bacterial cultures S oneidensis MR-1 was purchased from American Type Culture
collection (ATCC 700550) and stored as freezer stocks at -80degC Stock S oneidensis
MR-1 cells were grown aerobically using M1 medium at 30degC Cells in early stationary
growth phase (optical density of 15-16 at 600nm (Spectrophotometer UV-1700
Shimadzu Columbia MD) were injected into the chamber of the MECs for current
production Sodium lactate (final concentration 30mM) was added as the additional
electron donor
Fabrication and characterization of nanostructured anodes Superfine isomolded
graphite disks (16 cm x 05 cm GraphiteStorecom Inc) were polished with ultrafine
70
sand paper (2000 grit 3M Company) as the base for the CNT decorated and control
anodes Aligned multi-walled carbon nanotube samples were synthesized via plasma
enhanced chemical vapor deposition (PECVD) Samples superfine isomolded graphite
pillars with a 16 cm diameter and 05 cm thickness were sputtered with a thin film of Ni
in a CRC 300 magnetron sputter coater for 1 to 2 min They were then placed in the
PECVD reactor where the Ni was annealed into small catalyst islands C2H2 was bled
into the chamber for growth and an NH3 plasma was used to vertically align the growth
of the multi-walled carbon nanotubes (MWCNTs) The temperature and time were varied
between 725degC and 775oC and 15 to 60 min to control the length of the tubes and density
of samples Samples were then analyzed in a FEI Sirion field emission scanning electron
microscope (FESEM) SEM images of CNT decorated anode and control surfaces were
shown in the Figure 51
Figure 51 SEM images of CNT decorated anodes (A) Graphite disk control (B) CNT
decorated anode
Multiple channel MECs construction and operation A MECs with multiple
removable anodes each with an effective anode area of 07 cm2
was constructed and used
to evaluate the effects of nanostructures on the current density of according to a previous
B A
71
study (Fan et al 2011) The cathode was made of wet-proof (30) carbon cloth (type B
E-TEK Division Inc USA) coated with PtC (20 E-TEK Division Inc USA)
following a previously reported procedure (Liu et al 2005) The final platinum loading
was 05 mgcm2 per projected cathode area The size of the cathode (150 cm
2) was 25
times larger than the total effective area of the 8 graphite disk anodes (56 cm2) to prevent
cathode limitations on the performance of the MECs system All MECs with CNT
decorated anodes and control anodes were tested for current generation for 24 h with
sterile a medium (without bacteria) before injecting the bacterial cells in order to
determine whether the CNT decorated anodes demonstrate chemical current generation
compared to control anodes All the testing anodes were set up in the MECs
simultaneously so that the S oneidensis MR-1 culture was applied to all of them
simultaneously Short electrode spacing (17 cm) was used in the MECs design to reduce
the internal resistance A voltage of 06 V was applied the MECs for current generation
after the cells of S oneidensis MR-1 were inoculated into the MECs in the early
stationary phase A multimeter with a data acquisition system (2700 Keithly USA) was
used to monitor the current change by measuring the voltage drop through a resistor
Microarray analysis Biofilms for whole gene microarray analysis were aseptically
removed from the plain and CNT decorated anodes of MECs after 80 hours further
incubation at 30oC when current density obviously reached to a stable phase Total RNA
was extracted using Trizol (Ambion inc AustinTX) following the manufacturerrsquos
instructions Integrity of the RNA samples was confirmed using the Agilent 2100
Bioanalyzer (Agilent Technology Inc Palo Alto CA) Total RNA (200 ng) from each
sample was amplified using the MessageAmpTM
II-Bacteria Prokaryotic RNA
72
Amplification kit (Ambion AustinTX) according to the manufacturerrsquos instructions The
S oneidensis MR-1 whole-gene microarrays representing all of the 4295 genes (resource
from NCBI) were ordered from Roche NimbleGen Inc (Madison WI) Biological
triplicates of biofilms on the CNT decorated anodes and control were analyzed cDNA
synthesis labeling and hybridization were carried out by the CGRB Core Laboratories at
Oregon State University DNASTAR ArrayStarTM
3 software was used to identify genes
that were up- or down-regulated more than 2-fold when grown on the nanoparticle
modified anodes using the unpaired two sample t-test with a cutoff p-value of 005 with
bonferroni correction
Results and discussion
Enhancement of current generation using CNT decorated anodes in MECs The
current density generated by CNT decorated anode MECs with S onidensis MR-1
increased to 36 microAcm2 10 hours post-inoculation and reached a maximum of 40 microAcm
2
at 29 hours inoculation then gradually decreased The current density stabilized at
30microAcm2 after 78 h operation Figure 52 shows that the current density curve generated
by CNT decorated anode in MECs The average current density generated by CNT
decorated anodes was 56 times that of the control The current density generated in the
MECs without bacteria was negligible (data not shown) This result demonstrates that the
current enhancement observed with the CNT decorated anodes was biologically-derived
A
73
Figure 52 Effects of CNT decorated anodes on the current density in MECs (3 replicates)
Control is plain graphite disk anode CNT is the anode with CNT decoration (Error bars
represent standard deviation)
It has been reported that nanostructured decorated anodes have significant impacts on
current densities (Fan et al 2011 Qiao et al 2006 Sharma 2008 Tsai et al 2009) The
CNT decorated anodes had significantly enhanced current densities in MECs in this
study This result is consistent with previous studies Interestingly the current density
curve generated by CNT decorated anode MECs is quite different from the current
density curve generated by Au NP and Pd NP decorated anodes using same bacterial
species (Fan et al 2011) In the Au and Pd NP decorated anode MECs the current
density curve was not significantly increased in the beginning compared with the control
but gradually increased after the inoculation the current density reached the maximum
current density more than 50 hours after the inoculation However the current density
generated by CNT decorated anode MECs started to increase significantly immediately
after the inoculation It reached a maximum current density 29 hours post-inoculation
-1
6
13
20
27
34
41
48
0 10 20 30 40 50 60 70 80
Curr
ent
den
sity
(micro
Ac
m2)
Time (Hours)
Control
CNT
74
These results indicate that there might be different current density enhancement
mechanisms between CNT with other metal NP decorated anodes in MECs
Global transcriptome analysis Whole-genome DNA microarrays were used to attain a
comprehensive general overview of the transcriptional response of S oneidensis
MR-1 of
S oneidensis MR-1 to CNT decorated anodes in MECs In a total of 4295 genes analyzed
457 genes (250 genes up-regulated and 207 genes down-regulated) exhibited significantly
(P lt 005) changed expression at a 2-fold level in at least 3 replicates in the response to
CNT The total number of the regulated genes present 11 of the 4295 open reading
frames (ORFs) presented on the microarray The total number of genes at significant
expression levels in response to CNT decorated anodes is close to that to Fe NP decorated
anodes in MECs (392 9) ( Xu et al 2012)
Fig 53 Differentially expressed genes grouped by functional classification according to
the TIGR S oneidensis genome database (wwwtigrorg) Columns 1 amino acid
biosynthesis 2 biosynthesis of cofactors prosthetic groups and carriers 3 cell
envelope 4 cellular processes 5 central intermediary metabolism 6 DNA metabolism
7 energy metabolism 8 fatty acid and phospholipid metabolism 9 other categories 10
protein fate 11 protein synthesis 12 purines pyrimidines nucleosides and nucleotides
-80
-60
-40
-20
0
20
40
60
80
100
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18
Nu
mb
er
of
Ge
ne
s
Up-regulated
Down-regulated
75
13 regulatory functions 14 signal transduction 15 transcription 16 transport and
binding proteins 17 unknown function 18 hypothetical proteins
The overall transcript genomic response of S oneidensis MR-1 to CNT decorated
anodes was summarized in Figure 53 by grouping the differentially expressed genes into
their functional role categories based on The Institute for Genomic Researchrsquos annotation
(Version 8) of the MR-1 genome sequence The wide distribution of putative functional
roles attributed to the differentially expressed genes indicated the extent of the molecular
response of Soniedenis MR-1 to the CNT decorated anodes A large number of up-
regulated genes were presented in genes associated with biosynthesis of cofactors
prosthetic groups and carriers (group 2) cell envelope (group 3) protein fate (group 10)
protein synthesis (group 11) and purines pyrimidines nucleosides and nucleotides
(group 12) in this study The number of up-regulated genes was two times more than that
of down-regulated genes in each functional group Contrastingly in the genomic response
S oneidensis MR-1 to Fe NP decorated anodes (chapter 4) most of up-regulated genes
were associated with cellular processes (group 4) energy metabolism (group 7) other
categories (group 9) and hypothetical proteins (group 18) The down-regulated genes
presented in the amino acid biosynthesis (group 1) cellular processes (group 4) substrate
transport (group 16) and hypothetical proteins (group 18) While those genes encoding
proteins involved in amino acid biosynthesis (group 1) central intermediary metabolism
( group 5) DNA metabolism( group 6) fatty acid and phospholipid metabolism (group
8) and other categories (group 9) were among the most down-regulated genes in this
study However the two groups of genes that were the most noticeable between the
genomic response S oneidensis MR-1 to CNT and Fe NP decorated anodes which they
are the genes related to energy metabolism (group 7) and hypothetical proteins (group
76
18) These results indicated there are different and common genomic responses to S
oneidensis MR-1 on CNT and Fe NP decorated anodes in MECs
Electron transfer related genes Based on previous electron transfer mechanism
studies on Shewanella a serial of group proteins collectively described as the Mtr
pathway play the role to transfer electron rom the inner bacterial body to the outer
membrane (Beliaev et al 2001 Shi et al 2007 Brutinel and Gralnick 2012) In the Mtr
pathway firstly electrons generated by bacteria received by CymA a tetraheme c-type
cytochrome anchored in the inner membrane then electrons were transferred to MtrA a
periplasmic decaheme c-type cytochrome After that electrons were transferred from
MtrA to MtrC an outer-membrane decaheme c-type cytochrome is facilitated by MtrB a
non-heme containing integral outer-membrane proteinOuter-membrane decaheme c-type
cytochrome MtrC and OmcA are in change of electron transfer to exreacellular electron
acceptor (Shi et al 2007 Brutinel and Gralnick 2012 ) These genes encoding electron
transport proteins are shown in the table 51 The critical genes SO_1777 mtrA SO_1776
mtrB SO_1778 mtrC are unregulated 41 to 80 fold in response to CNT decorated
anodes other cytochrome c genes related to electron transfer in MtrA pathway also are
significantly increased in response to CNT decorated anodes These genes encoding outer
membrane proteins have different level increases supporting that the increase of
cytochrome c gene expression contributes to the current enhancements of MECs as
response to CNT decorated anodes
77
Table 51 Expression level of cytochrome c as response to CNT decorated anodes
Locus Tag Gene product descriptions Fold change CNT
SO_0165 general secretion pathway protein GspC 31
SO_0167 general secretion pathway protein GspE 45
SO_1776 outer membrane protein precursor MtrB 41
SO_1777 outer membrane decaheme cytochrome c
lipoprotein MtrA
80
SO_1778 outer membrane decaheme cytochrome c
lipoprotein MtrC
43
SO_1779 outer membrane decaheme cytochrome c
OmcA
63
SO_0135 lipoprotein of unknown function DUF333 22
SO_0136 conserved hypothetical inner membrane protein 24
Flavin related genes It is known that self-produced mediators play critical roles in the
electron transfer of S oneidensis to extracellular insoluble electrode (Canstein von et al 2
008 Myers et al 1992 Scott et al 1994 Shi et al 2007) It has been proved that the
flavins secreted by S oneidensis can facilitate electron transfer from bacteria to solid
anode electrodes by acting as extracellular electron shuttles (Cansterin von et al 2008) T
wo critical genes (SO_1414 and SO_3468) related to flavin synthesis were shown increa
sed significantly (more than 2 folds) increase in this study (Table 52) This result support
s the hypothesis that flavin production increases as the response to CNT decorated anodes
in MECs have significant contribution to the current density enhancements
78
Table 52 Expression levels of genes related to flavin synthesis as response to CNT
decorated anode
Locus Tag Gene product descriptions Fold change CNT
SO_1414 flavocytochrome c flavin subunit putative 22
SO_3468 riboflavin synthase subunit alpha 20
The amount of bacterial biofilm biomass on the anode can affect the power
generation in MFCs (Biffinger et al 2007 Heydorn et al 2000) Biofilm formation
related genes include flagellar gene IV pilus genes and other genes the flagellar related
genes (fliB and fliK) and IV pilus genes (mshA) were induced only from 105 to 134
fold as response of S oneidensis MR-1 to CNT decorated anodes in this study No
biofilm-associated genes presented significant changes in expression levels in this study
indicating there is no direct connections between biofilm enhancements to CNT
decorated anodes in this study This result is consistent with the Liang et al (2011)
conclusion that using CNT enhanced the electrochemical activity of anodic biofilms but
did not result in a significant increase of biomass in the anodic biofilms
Our microarray results showed two significantly up-regulated gene groups the
genes encoding proteins localized on the outer membrane and the genes involved in
flavin biosynthesis contributed to current density enhancement by CNT decorated anodes
Among 457 significantly changed genes to CNT in this study there are also a relatively
large number of genes encoding proteins with unknown functions which are either up- or
down-regulated in the response to CNT decorated anodes which indicated more
79
complicated responses of S oneidensis MR-1 to CNT decorated anodes in MECs and
further study is needed
80
Chapter 6
Summary
Microbial electrochemical systems (MESs) with a mixed culture initially inoculated
from Corvallis wastewater treatment plant have been studied for more than 6 years for
varying purposes including power generation hydrogen production heavy metal
removal and wastewater treatment Experiment results have shown that the mixed culture
is quite stable with excellent performance in MESs Our community analysis using
denaturing gradient gel electrophoresis (DGGE) and 16S rDNA clone library construction
suggests that the mixed culture is composed predominantly of Geobacter sp (66)
Arcobacter sp (12) and Citrobacter sp (11) These results not only improved our
understanding of the mixed culture community but also guided our studies on the
cultivation and isolation of the different bacterial species
Exoelectrogenic Citrobacter sp SX-1 was first isolated and characterized from a
MFC operated with the mixed culture Citrobacter sp SX-1 has been demonstrated to
produce electricity from wide range of different substrates including citrate acetate
glucose sucrose glycerol and lactosein MFCs Cyclic voltammetry analysis indicated
that membrane associated proteins may play an important role in facilitating the electrons
transferring from bacteria to electrode The strain SX-1 increased the known diversity of
power generating exoelectrogens and provided a unique bacterial species for study in
renewable energy generation and waste treatment
81
To enhance the electron transfer from bacteria to anode nano decorated anodes
including Fe NP and CNT were developed and characterized and evaluated in MECs
using Shewanella oneidensis MR-1 as a model species Both nanostructures have
significantly increased current density compared with the control Whole genome
microarray analysis elucidated the possible mechanisms of power enhancement in
response to these nano-decorated anodes
These results benefit to understanding of physiology and ecology of mixed
cultures in MFCs and improve the efficiency of current generation in MESs which will
facilitate the viability of niche applications for MESs in near future
82
Bibliography
1 Amman R Ludwig W (2000) Ribosomal RNA-targeted nucleic acid probes for
studies in microbial ecology FEMS Microbiol Ecol 24 555ndash565
1 Bao SJ Li CM Zang JF Cui XQ Qiao Y Guo J (2008) Novel Nanoporous TiO2
Electrode for Directly Electrochemistry of glucose oxidase Adv Funct Mater
18(4) 591-599
2 Beliaev AS Saffarini DA McLaughlin JL Hunnicutt D (2001) MtrC an outer
membrane decahaem c cytochrome required for metal reduction in Shewanella
putrefaciens MR-1 Mol Microbiol 39722-30
3 Bencheikh-Latmani R Williams SM Haucke L Criddle CS Wu L Criddle CS
Zhou J Tebo BM (2005) Global transcriptional profiling of Shewanella oneidensis
MR-1 during Cr(VI) and U(VI) reduction Appl Environ Microbiol 71 7453ndash7460
4 Biffinger JC Pietron J Ray R Little B and Ringeisen BR (2007) A biofilm
enhanced miniature microbial fuel cell using Shewanella oneidensis DSP10 and
oxygen reduction cathodes Biosens Bioelectron 221672ndash1679
5 Bodelier PLE Roslev P Henckel T FenzelP (2000) Stimulation by ammonium
based fertilizers of methane oxidation in soil around rice root Nature 40 421ndash424
6 Bond DR Holmes DE Tender LM Lovley DR (2002) Electrode-reducing
microorganisms that harvest energy from marine sediments Science 295 483-485
7 Bond DR Lovley DR (2003) Electricity production by Geobacter sulfurreducens
attached to electrodes Appl Environ Microbiol 69 1548-1555
8 Bond DR Lovley DR (2005) Evidence for involvement of an electron shuttle in
electricity generation by Geothrix fermentans Appl Environ Microbiol 71 2186-
2189
9 Bretschger O Obraztsova A Sturm CA Chang IS Gorby YA Reed SB Culley
DE Reardon CL Barua S Romine MF Zhou J Beliaev AS Bouhenni R
Saffarini D Mansfeld F Kim BH Fredrickson JK Nealson KH (2007) Current
production and metal oxide reduction by Shewanella oneidensis MR-1 wildtype and
mutants Appl Environ Microbiol 73 7003ndash7012
10 Brown SD Thompson MR Verberkmoes NC Chourey K Shah M Zhou J
Hettich RL and Thompson DK (2006) Molecular dynamics of the Shewanella
oneidensis response to chromate stress Mol Cell Proteomics 51054-1071
11 Brutinel ED Gralnick JA (2012) Shuttling happens soluble flavin mediators of
extracellular electron transfer in Shewanella Appl Microbiol Biotechnol 93(1) 41-8
12 Butler JE Kaufmann F Coppi MV Nuacutentildeez C Lovley DR (2004) MacA a diheme
c-type cytochrome involved in Fe(III) reduction by Geobacter sulfurreducens J
Bacteriol 186(12) 4042ndash4045
83
13 Caccavo F Lonergan DJ Lovley DR Davis M Stolz JF McInerney MJ (1994)
Geobacter sulfurreducens sp nov a hydrogen- and acetate oxidizing dissimilatory
metal-reducing microorganism Appl Environ Microbiol 60 3752ndash3759
14 Canstein von H Ogawa J Shimizu S Lloyd JR (2008) Secretion of flavins by
Shewanella species and their role in extracellular electron transfer Appl Environ
Microbiol 74 615ndash623
15 Cao X Huang X Liang P Xiao K Zhou Y Zhang X Logan BE (2009) A New
Method for Water Desalination Using Microbial Desalination Cells Environ Sci
Technol 43(18)7148-7152
16 Chaudhuri SK Lovley DR (2003) Electricity generation by direct oxidation of
glucose in mediatorless microbial fuel cells Nat Biotechnol 21 1229-1232
17 Cheng S Liu H Logan B E (2006) Increased power generation in a continuous
flow MFC with advective flow through the porous anode and reduced electrode
spacing Environ Sci Technol 402426-2432
18 Cheng S Logan B E (2007) Sustainable and efficient biohydrogen production via
electrohydrogenesis PNAS 104 18871-18873
19 Chignell J (2010) Energy Recovery from Biodiesel Waste Performance of
Microbial Electrochemical Systems on Glycerol Thesis
20 Chung K Okabe S (2009) Characterization of electrochemical activity of a strain
ISO2-3 phylogenetically related to Aeromonas sp isolated from a glucose-fed
microbial fuel cell Biotechnol Bioeng 104(5) 901-910
21 Cole JR Chai B Farris RJ Wang Q Kulam-Syed-Mohideen AS McGarrell
DM Bandela AM Cardenas E Garrity GM Tiedje JM (2007) The ribosomal
database project (RDP-II) introducing myRDP space and quality controlled public
data Nucleic Acids Res 35 (Database issue) D169-D172 doi 101093nargkl889
22 Cole JR Wang Q Cardena E Fish J Chai B Farris RJ Kulam-Syed-Mohideen
AS McGarrell DM Marsh T Garrity GM Tiedje JM (2009) The Ribosomal
Database Project improved alignments and new tools for rRNA analysis Nucleic
Acids Res 37 (Database issue) D141-D145 doi 101093nargkn879
23 Dey A De S De A De SK (2004) Characterization and dielectric properties of
polyaniline-TiO2 nanocomposites Nanotechnol 15 1277-1283
24 Edwards U Rogall T Bloumlcker H Emde M Boumlttger EC (1989) Isolation and
direct complete nucleotide determination of entire genesmdashcharacterization of a gene
coding for 16S-ribosomal RNA NucleicAcids Res 17 7843ndash7853
25 Eggleston CM Voumlroumls J Shi L Lower BH Droubay TC Colberg PJS (2008)
Binding and direct electrochemistry of OmcA an outer-membrane cytochrome from
an iron reducing bacterium with oxide electrodes a candidate biofuel cell system
Inorg Chim Acta 361769ndash777
26 Fan Y Sharbrough E Liu H (2008) Quantification of the Internal Resistance
Distribution of Microbial Fuel Cells Environ Sci Technol 42 8101ndash8107
84
27 Fan Y Xu S Schaller R Jiao J Chaplen F Liu H (2011) Nanoparticle decorated
anodes for enhanced current generation in microbial electrochemical
cells Biosensors and Bioelectronics 26 (5)1908ndash1912
28 Fan Y Hu H Liu H (2007)Sustainable power generation in microbial fuel cells
using bicarbonate buffer and proton transfer mechanisms Environ Sci
Technol 41(23) 8154-8
29 Fedorovich V Knighton MC Pagaling E Ward FB Free A Goryanin I (2009)
Novel electrochemically active bacterium phylogenetically related to Arcobacter
butzleri isolated from a microbial fuel cell Appl Environ Microbiol 75(23) 7326-
7334
30 Ferna ndez A Huang S Seston S Xing J Hickey R Criddle C Tiedje J (1999)
How stable is stable Function versus community composition Appl Environ
Microbiol 65 3697ndash3704
31 Field SJ Dobbin PS Cheesman MR Watmough NJ Thomson AJ Richardson
DJ (2000) Purification and magneto-optical spectroscopic characterization of
cytoplasmic membrane and outer membrane multiheme c-type cytochromes from
Shewanella frigidimarina NCIMB400 J Biol Chem 275 8515ndash8522
32 Freguia S Masuda M Tsujimura S Kano K (2009) Lactococcus lactis catalyses
electricity generation at microbial fuel cell anodes via excretion of a soluble quinone
Bioelectrochemistry 7614ndash18
33 Fricke K Harnisch F and Schroumlder U (2008) On the use of cyclic voltammetry for
the study of the anodic electron transfer in microbial fuel cells Energy Environ
Sci 1 144ndash147
34 Gao H Wang Y Liu X Yan T Wu L Alm E Arkin A Thompson DK and
Zhou J (2004) Global transcriptome analysis of the heat shock response of
Shewanella oneidensis J Bacteriol 1867796-7803
35 Gao H Yang ZK Wu L Thompson DK Zhou J (2006) Global transcriptome
analysis of the cold shock response of Shewanella oneidensis MR-1 and mutational
analysis of its classical cold shock proteins J Bacteriol 1884560-4569
36 George MG (2005) Bergeyrsquos Manual of Systematic Bacteriology Volume Two The
Proteobacteria Part B The Gammaproteobacteria Second Edition New York
Springer 651p
37 Gonzaacutelez-Toril E Llobet-Brossa E Casamayor EO Amann R Amils R (2003)
Microbial ecology of an extreme acidic environment the Tinto river Appl Environ
Microbiol 69(8) 4853ndash4865
38 Gorby YA Yanina S Mclean JS Rosso KM Moyles D Dohnalkova A
Beveridge TJ Chang IS Kim BH Kim KS Culley DE Reed SB Romine M
F Saffarini DA Hill EA Shi L Elias DA Kennedy DW Pinchuk G
Watanabe K Ishii S Logan B Nealson KH Fredrickson JK (2006) Electrically
conductive bacterial nanowires produced by Shewanella oneidensis strain MR-1 and
other microorganisms Proc Natl Acad Sci USA 10311358-11363
85
39 He Z Wagner N Minteer SD Angenent LT (2006) An upflow microbial fuel cell
with an interior cathodes assessment of the internal resistance by impedance
spectroscopy Environ Sci Technol 40 5212-5217
40 Heydorn A Nielsen AT Hentzer M Sternberg C Givskov M Ersboslashll BK Molin
S (2000) Quantification of biofilm structures by the novel computer program
COMSTAT Microbiol 1462395-2407
41 Holmes DE Bond DR Lovley DR (2004a) Electron transfer by Desulfobulbus
propionicus to Fe(III) and graphite electrodes Appl Environ Microbiol 701234ndash
1237
42 Holmes DE Chaudhuri SK Nevin KP Mehta T Methe BA Liu A Ward JE
Woodard TL Webster J Lovley DR (2006) Microarray and genetic analysis of
electron transfer to electrodes in Geobacter sulfurreducens Environ Microbiol
81805-1815
43 Holmes DE Nicoll JS Bond DR Lovley DR (2004b) Potential role of a novel
psychrotolerant member of the family Geobacteraceae Geopsychrobacter
electrodiphilus gen nov sp nov in electricity production by a marine sediment
fuel cell Appl Environ Microbiol 70 6023ndash6030
44 Holmes DE Chaudhuri SK Nevin KP Mehta T Metheacute BA Liu A Ward JE
Woodard TL Webster J Lovley DR (2006) microarray and genetic analysis of
electron transfer to electrodes in Geobacter sulfurreducens Environ Microbial
8(10)1805-1815
45 Hu H (2009) Enhancing Hydrogen Production in Microbial Electrolysis Cells
through Development of Platinum-free Cathode and Improvement of Reactor
Dissertation
46 Ince BK Ayman O N Turker G Ccedilelikkol S Ince O (2010) Microbial ecology of
anaerobic reactors for treatment of alcohol industry wastewaters a review Current
research technology and education topics in applied microbiology and microbial
ecology 988-999
47 Jung S Regan JR (2007) Comparison of anode bacterial communities and
performance in microbial fuel cells with different electron donors Appl Microbiol
Biotechnol 77393ndash 402
48 Kim BH Kim HJ Hyun MS Park DH (1999) Direct electrode reaction of Fe(III)
reducing bacterium Shwwanella putrefaciens J Microbial Biotechnol 9127-131
49 Kim GT Hyun MS Chang IS Kim HJ Park HS Kim BH Kim SD Wimpenny
JW Weightman AJ (2005) Dissimilatory Fe(III) reduction by an electrochemically
active lactic acid bacterium phylogenetically related to Enterococcus gallinarum
isolated from submerged soil J Appl Microbiol 99(4) 978ndash987
50 Kim GT Webster G Wimpenny JW Kim BH Kim HJ Weightman AJ (2006)
Bacterial community structure compartmentalization and activity in a microbial fuel
cell J Appl Microbiol 101(3) 698ndash710
86
51 Kim HJ Park HS Hyun MS Chang IS Kim M Kim BH (2002) A mediator-
less microbial fuel cell using a metal reducing bacterium Shewanella putrefaciens
Enzyme Microb Technol 30 145-152
52 Kim JR Min B Logan BE (2005) Evaluation of procedures to acclimate a
microbial fuel cell for electricity production Appl Microbiol Biotechnol 68(1)23-30
53 Kim JR Cheng S Oh SE Logan BE (2007) Power generation using different
cation anion and ultrafiltration membranes in microbial fuel cells Environ Sci
Technol 41 1004-1009
54 Konstantinidis KT Serres MH Romine MF Rodrigues JL Auchtung J McCue
LA Lipton MS Obraztsova A Giometti CS Nealson KH Fredrickson JK
Tiedje JM (2009) Comparative systems biology across an evolutionary gradient
within the Shewanella genus Proc Natl Acad Sci U S A 106(37)15909-15914
55 Lee J Phung NT Chang IS Kim BH Sung HC (2003) Use of acetate for
enrichment of electrochemically active microorganisms and their 16S rDNA
analyses FEMS Microbiol Lett 223185ndash191
56 Liang P Wang H Xia X Huang X Mo Y Cao X Fan M(2011)Carbon
nanotube powders as electrode modifier to enhance the activity of anodic biofilm
in microbial fuel cells Biosens Bioelectron 26(6)3000-4
57 Lies DP Hernandez ME Kappler A Mielke RE Gralnick JA Newman DK
(2005) Shewanaella oneidensis MR-1 uses overlapping pathways for iron reduction
at a distance and by direct contact under conditions relevant for biofilms Appl
Environ Microbiol 71 4414-4426
58 Liu H and Logan B E (2004) Electricity generation using an air-cathode single
chamber microbial fuel cell in the presence and absence of a proton exchange
membrane Environ Sci Technol 38 4040-4046
59 Liu H Grot S Logan BE (2005) Electrochemically assisted microbial production
of hydrogen from acetate Environ Sci Technol 39 4317 ndash 320
60 Liu H Hu H Chignell J Fan Y (2010) Microbial Electrolysis Novel Technology
for Hydrogen Production from Biomass Biofuels 1(1) 129ndash142
61 Liu H Ramnarayanan R Logan BE (2004) Production of electricity during
wastewater treatment using a single chamber microbial fuel cell Environ Sci Technol
382281-2285
62 Liu WT Marsh TL Cheng H Forney LJ (1997) Characterization of microbial
diversity by determining terminal restriction fragment length polymorphisms of gene
encoding 16S rRNA Appl Environ Microbiol 63 4516ndash4522
63 Logan B E Hamelers B Rozendal R Schroder U Keller J Freguia S Aelterman
P Verstraete W Rabaey K (2006) Microbial fuel cells methodology and
technology Environ Sci Technol 40 (17) 5181-5192
64 Logan B E and Regan JM (2006) Electricity-producing bacterial communities in
microbial fuel cells Trends Microbiol 14512ndash518
87
65 Logan B Cheng S Watson V Estadt G (2007) Graphite fiber brush anodes for
increased power production in air-cathode microbial fuel cells Environ Sci Technol
41(9)3341-3346
66 Logan BE (2009) Exoelectrogenic bacteria that power microbial fuel cells Nat Rev
Microbiol 5 375-381
67 Logan BE Call D Cheng S Hamelers HV Sleutels TH Jeremiasse AW
Rozendal RA (2008) Microbial electrolysis cells for high yield hydrogen gas
production from organic matter Environ Sci Technol 42 8630-8640
68 Logan BE Hamelers B Rozendal R Schroder U Keller J Freguia S Aelterman
P Verstraete W Rabaey K (2006) Microbial fuel cells methodology and
technology Environ SciTechnol 40 5181-5192
69 Lovely DR (2006) Bug juice harvesting electricity with microorganisms Nature
Review| Microbiology 4 497-508
70 Lovley DR Phillips EJ (1988) Novel Mode of Microbial Energy Metabolism
Organic Carbon Oxidation Coupled to Dissimilatory Reduction of Iron or
Manganese Appl Environ Microbiol 54(6) 1472-1480
71 Ly HK Sezer M Wisitruangsakul N Feng JJ Kranich A Millo D Weidinger
IM Zebger I Murgida DH Hildebrandt P (2011) Surface-enhanced vibrational
spectroscopy for probing transient interactions of proteins with biomimetic interfaces
electric field effects on structure dynamics and function of cytochrome c FEBS J
278(9)1382-90
72 Macaskie LE Hewitt CJ Shearer JA Kent CA (1995) Biomass production for
the removal of heavy metals from aqueous solutions at low pH using growth-
decoupled cells of a Citrobacter sp Int Biodeter Biodegr 3573ndash92
73 Malki M De Lacey AL Rodriguez N Amils R Fernandez VM (2008)
Preferential use of an anode as an electron acceptor by an acidophilic bacterium in
the presence of oxygen Appl Environ Microbiol 74 4472ndash4476
74 Meitl LA Eggleston CM Colberg PJS Khare N Reardon CL Shi L (2009)
Electrochemical interaction of Shewanella oneidensis MR-1 and its outer membrane
cytochromes OmcA and MtrC with hematite electrodes Geochim Cosmochim
Ac 73(18) 5292-5307
75 Morozan A Stamatin L and Nastase
F (2007) The biocompatibility
microorganisms-carbon nanostructures for applications in microbial fuel cells Phys
Stat Sol 6 1797-1803
76 Morris JM Jin S Crimi B Pruden A (2009) Microbial fuel cell in enhancing
anaerobic biodegradation of diesel Chem Eng J 146 161ndash167
77 Murray AE Lies D Li G Nealson K Zhou J Tiedje JM (2001) DNADNA
hybridization to microarrays reveals gene-specific differences between closely
related microbial genomes Proc Natl Acad Sci U S A 98 9853-9858
78 Muyzer G de Waal EC Uitterlinden AG (1993) Profiling of complex microbial
communities by denaturing gradient gel electrophoresis analysis of polymerase chain
88
reaction amplified genes coding for 16S rRNA Appl Environ Microbiol 59 695ndash
700
79 Myers CR Myers JM (1992) Localization of cytochromes to the outer membrane
of anaerobically grown Shewanella putrefacians MR-1 J Bacteriol 174 3429ndash3438
80 Narde GK Kapley A Purohit HJ (2004) Isolation and characterization of
Citrobacter strain HPC255 for broad-range substrate specificity for chlorophenols
Curr Microbiol 48(6) 419-423
81 Okabe S Ito T Sugita K Satoh H (2005) Succession of internal sulfur cycles and
sulfur-oxidizing bacterial communities in microaerophilic wastewater biofilms Appl
Environ Microbiol 71(5) 2520-2529
82 Park DH and Zeikus JG (2000) Electricity generation in microbial fuel cells using
neutral red as an electronophore Appl Environ Microbial 66 292-1297
83 Park DH and Zeikus JG (2002) Impact of electrode composition on electricity
generation in a single-compartment fuel cell using Shewanella putrefaciens Appl
Microbiol Biotechnol 5958ndash56
84 Park DH and Zeikus JG (2003) Improved fuel cell and electrode designs for
producing electricity from microbial degradation Biotechnol Bioeng 81 348ndash355
85 Park HS Kim BH Kim HS Kim HJ Kim GT Kim M Chang IS Park Y
K Chang HI (2001) A novel electrochemically active and Fe(III)-reducing
bacterium phylogenetically related to Clostridium butyricum isolated from a
microbial fuel cell Anaerobe 7 297-306
86 Parot S Nercessian O Delia ML Achouak W Bergel A (2009) Electrochemical
checking of aerobic isolates from electrochemically active biofilms formed in
compost J Appl Microbiol 106(4) 1350-1359
87 Pham CA Jung SJ Phung NT Lee J Chang IS Kim BH Yi H Chun J
(2003) A novel electrochemically active and Fe(III)-reducing bacterium
phylogenetically related to Aeromonas hydrophila isolated from a microbial fuel cell
FEMS Microbiol Lett 223 129-134
88 Qiao Y Bao SJ Li CM Cui XQ Lu ZS Guo J (2008) Nanostructured
polyanilinetitanium dioxide composite anode for microbial fuel cells ACS Nano 2
113-119
89 Qiu R Zhao B Liu J Huang X Li Q Brewer E Wang S Shi N (2009) Sulfate
reduction and copper precipitation by a Citrobacter sp isolated from a mining area J
Hazard Mater 1641310-1315
90 Quan X Yang SG Ruan XL and Zhao HM (2005) Preparation of titania
nanotubes and their environmental applications as electrode Environ Sci Technol 39
3770ndash3775
91 Rabaey K Boon N Hofte M Verstraete W (2005) Microbial phenazine
production enhances electron transfer in biofuel cells Environ Sci Technol 393401ndash
3408
89
92 Rabaey K Boon N Siciliano S D Verhaege M Verstraete W (2004) Biofuel cells
select for microbial consortia that self-mediate electron transfer Appl Environ
Microbiol 70(9) 5373ndash5382
93 Rabaey K Lissens G Siciliano S D Verstraete W (2003) Amicrobial fuel cell
capable of converting glucose to electricity at high rate and efficiency Biotechnol
Lett 25 1531-15
94 Rabaey K Verstraete W (2005) Microbial fuel cells novel biotechnology for
energy generation Trends Biotechnol 23(6) 291-298
95 Reguera G McCarthy KD Mehta T Nicoll JS Tuominen MT Lovley DR
(2005) Extracellular electron transfer via microbial nanowires Nature 435 1098-
1101
96 Reguera G Nevin KP Nicoll JS Covalla SF Woodard TL Lovley DR (2006)
Biofilm and nanowire production leads to increased current in Geobacter
sulfurreducens fuel cells Appl Environ Microbiol 72 7345-7348
97 Reimers CE Tender LM Fertig S Wang W (2001) Harvesting energy from the
marine sediment-water interface Environ Sci Technol 35192-195
98 Rezaei F Xing D Wagner R Regan JM Richard TM Logan BE (2009)
Simultaneous cellulose degradation and electricity production by Enterobacter
cloacae in a microbial fuel cell Appl Microbiol Biotechnol 75 3673ndash3678
99 Richter H Nevin KP Jia H Lowy DA Lovley DR Tender LM (2009) Cyclic
voltammetry of biofilms of wild type and mutant Geobacter sulfurreducens on fuel
cell anodes indicates possible roles of OmcB OmcZ type IV pili and protons in
extracellular electron transfer Energy Environ Sci 2 506ndash516
100 Riesenfeld SC Schloss DP Handelsman J(2004) Metagenomics Genomic
Analysis of Microbial Communities Annual Review Genetics 38525-552
101 Rozendal RA Hamelers HV Buisman CJ (2006) Effects of membrane cation
transport on pH and microbial fuel cell performance Environ Sci Technol 40
5206-5211
102 Rozendal RA Hamelers HVM Euverink GJW Metz SJ Buisman CJN (2006)
Principle and perspectives of hydrogen production through biocatalyzed electrolysis
Int J Hydrogen Energ 311632-1640
103 Rozendal RA Hamelers HV Molenkamp RJ Buisman CJ (2007) Performance of
single chamber biocatalyzed electrolysis with different types of ion exchange
membrances Water Res 41 1984-1994
104 Schluumlter A Bekel T Diaz NN Dondrup M Eichenlaub R Gartemann
KH Krahn I Krause L Kroumlmeke H Kruse O Mussgnug JH Neuweger
H Niehaus K Puumlhler A Runte KJ Szczepanowski R Tauch A Tilker
A Viehoumlver P Goesmann A (2008) The metagenome of a biogas-producing
microbial community of a production-scale biogas plant fermenter analyzed by the
454-pyrosequencing technology J Biotechnol136 77-90
90
105 Scott J H and Nealson KH (1994) A biochemical study of the intermediary carbon
metabolism of Shewanella putrefaciens J Bacteriol 176(11) 3408ndash3411
106 Sekiguchi H Tomioka H Nakahara T and Uchiyama H (2001) A single band
does not always represent single bacterial strains in denaturing gradient gel
electrophoresis analysis Biotechnology Letters23 1205ndash1208
107 Sharma T Leela M Reddy A Chandra TS Ramaprabhu S (2008) Development
of carbon nanotubes and nanofluids based microbial fuel cell Int J Hydrogen Energ
336749ndash6754
108 Shea C Clauwaert P Verstraete W Nerenberg R (2008) Adapting a denitrifying
biocathode for perchlorate reduction Water Sci Technol 58 (10) 1941-1946
109 Shi L Squier TC Zachara JM Fredrickson JK (2007) Respiration of metal (hydr)
oxides by Shewanella and Geobacter a key role for multihaem c-type cytochromes
Mol Microbiol 6512-20
110 Stams AJ and Plugge CM(2009)Electron transfer in syntrophic communities
of anaerobic bacteria and archaea Nat Rev Microbiol7(8)568-77
111 Sukkasem C Xu S Park S Boonsawang P Liu H (2008) Effect of Nitrate on the
Performance of Single Chamber Air Cathode Microbial Fuel Cells Water Res 42(19)
4743-4750
112 Summers ZM Fogarty HE Leang C Franks AE Malvankar NS Lovley DR
(2010)Direct exchange of electrons within aggregates of an evolved syntrophic
coculture of anaerobic bacteria Science 330 (6009)1413-5
113 Szczepanowski R Bekel T Goesmann A Krause L Kroumlmeke H Kaiser
O Eichler W Puumlhler A Schluumlter A (2008) Insight into the plasmid metagenome
of wastewater treatment plant bacteria showing reduced susceptibility to
antimicrobial drugs analysed by the 454-pyrosequencing technology J
Biotechnol 136 54-64
114 Tamura K Dudley J Nei M Kumar S (2007) MEGA4 Molecular Evolutionary
Genetics Analysis (MEGA) software version 40 MolBiol Evol 24(8) 1596-1599
115 Thompson JD Gibson TJ Plewniak F Jeanmougin F Higgins DG (1997) The
CLUSTAL_X windows interface flexible strategies for multiple sequence alignment
aided by quality analysis tools Nucleic Acids Res 25 4876-4882
116 Thormann KM Duttler S Saville RM Hyodo M Shukla S Hayakawa Y
Spormann AM (2006) Control of formation and cellular detachment from
Shewanella oneidensis MR-1 biofilms by cyclic di-GMP J Bacteriol 1882681-2691
117 Thormann KM Saville RM Shukla S Pelletier DA Spormann AM (2004)
Initial Phases of biofilm formation in Shewanella oneidensis MR-1 J Bacteriol
186(23) 8096-104
118 Torres CI Kato Marcus A Rittmann BE (2008b) Proton transport inside the
biofilm limits electrical current generation by anode-respiring bacteria Biotechnol
Bioeng 100 872ndash881
91
119 Torres CI Marcus AK Lee HS Parameswaran P Krajmalnik-Brown R Rittmann
BE (2010) A kinetic perspective on extracellular electron transfer by anode-
respiring bacteria FEMS Microbiol Rev 234(1)3-17
120 Torres CI Marcus AK Parameswaran P Rittmann BE (2008a) Kinetic
experiments for evaluating the NernstndashMonod model for anode-respiring bacteria
(ARB) in a biofilm anode Environ Sci Technol 42 6593ndash6597
121 Tsai HY Wu Ch-Ch Lee Ch-Y Shih EP (2009) Microbial fuel cell performance
of multiwall carbon nanotubes on carbon cloth as electrodes J Power Sources
194199-205
122 Tyson GW Chapman J Hugenholtz P Allen EE Ram RJ Richardson
PM Solovyev VV Rubin EM Rokhsar DS Banfield JF(2004) Community
structure and metabolism through reconstruction of microbial genomes from the
environment Nature42837-43
123 Venkateswaran K Moser DP Dollhopf ME Lies DP Saffarini D A MacGregor
BJ Ringelberg DB White DC Nishijima M Sano H Burghardt J Stackebrandt
E Nealson KH (1999) Polyphasic taxonomy of the genus Shewanella and
description of Shewanella oneidensis sp Int J Syst Bacteriol 49705-724
124 von Canstein H Ogawa J Shimizu S Lloyd JR (2008) secretion of Flavins
by Shewanella Species and Their Role in Extracellular Electron Transfer Appl
Environ Microbiol 74 3615-623
125 Wang A Liu W Cheng Sh Xing D Zhou J Logan BE (2009) Source of methane
and methods to control its formation in single chamber microbial electrolysis cells
Int J Hydrogen Energ 34 3653-3658
126 Wang J Ma T Zhao L Lv J Li G Liang F Liu R (2008) PCRndashDGGE method
for analyzing the bacterial community in a high temperature petroleum reservoir
World J Microbiol Biotechnol 2 9694-9702
127 Wang Q Garrity GM Tiedje JM Cole JR (2007) Naiumlve Bayesian Classifier for
Rapid Assignment of rRNA Sequences into the New Bacterial Taxonomy Appl
Environ Microbiol 73(16) 5261-5267
128 Wang YF Masuda M Tsulimura S Kano K (2008) Electrochemical regulation of
the end-product profile in Propionibacterium freudenreichii ET-3 with an
endogenous mediator Biotechnol Bioeng 101(3) 579ndash586
129 Wrighton KC Agbo P Warnecke F Weber KA Brodie EL DeSantis TZ
Hugenholtz P Andersen GL Coates JD (2008) A novel ecological role of the
Firmicutes identified in thermophilic microbial fuel cells ISME J 2 1146ndash1156
130 Xing D Zuo Y Cheng S Regan JM Logan BE (2008) Electricity generation by
Rhodopseudomonas palustris DX-1 Environ Sci Technol 42 4146-4151
131 Xu S Liu H Fan Y Schaller R Jiao J Chaplen F (2012) Enhanced performance
and mechanism study of microbial electrolysis cells using Fenanoparticle-decorated
anodes Appl Microbiol Biotechnol 93(2)871-880
92
132 Yang Y Harris DP Luo F Wu L Parsons AB Palumbo AV Zhou J (2008)
Characterization of the Shewanella oneidensis Fur gene roles in iron and acid
tolerance response BMC Genomics 9 Suppl 1S11
133 Zhang L Zhou S Zhuang L Li W Zhang J Lu N Deng L (2008) Microbial
fuel cell based on Klebsiella pneumoniae biofilm Electrochem Commun10 1641ndash
1643
134 Zuo Y Xing DF Regan JM Logan BE (2008) Isolation of the exoelectrogenic
bacterium Ochrobactrum anthropi YZ-1 by using a U-tube microbial fuel cell Appl
Environ Microbiol 74 3130-3137
93
APPENDICES
List of published papers during PhD study
1 Shoutao Xu Hong Liu Yanzhen Fan Rebecca Schaller Jun Jiao and Frank
Chaplen (2012) Enhanced performance and mechanism study
of microbial electrolysis cells using Fe nanoparticle-decorated anodes Applied
Microbiology and Biotechnology 93(2)871-880
2 Shoutao Xu Hong Liu (2011) New exoelectrogen Citrobacter sp SX-1 isolated
from a microbial fuel cell Journal of Applied Microbiology111(5)1108-1115
3 Yanzhen Fan Shoutao Xu Rebecca Schaller Jun Jiao Frank Chaplen Hong Liu
(2011) Nanoparticle decorated anodes for enhanced current generation in microbial
electrochemical cells Biosensors and Bioelectronics 26(5) 1908-1912
4 Tunc Catal Shoutao Xu Kaichang Li Hakan Bermek Hong Liu
(2008)
Electricity generation from polyalcohols in single-chamber microbial fuel
cells Biosensors and Bioelectronics 24(4)849-854
5 Chontisa Sukkasema Shoutao Xu Sunhwa Park Piyarat Boonsawang Hong Liu
(2008) Effect of nitrate on the performance of single chamber air cathode
microbial fuel cells Water research 424743-4750
6 Rebecca Schaller Yanzhen Fan Shoutao Xu Alan Fern Frank Chaplen Hong
Liu and Jun Jiao (2009) Vertically Aligned Multi-walled Carbon Nanotube
Decorated Anodes for Microbial Fuel Cells Proceedings of Materials Research
Society 2009 1170 R05-13

Doctor of Philosophy dissertation of Shoutao Xu presented on June 15 2012
APPROVED
Co-Major Professor representing Biological and Ecological Engineering
Co-Major Professor representing Biological and Ecological Engineering
Head of the Department of Biological and Ecological Engineering
Dean of the Graduate School
I understand that my dissertation will become part of the permanent collection of
Oregon State University libraries My signature below authorizes release of my
dissertation to any reader upon request
Shoutao Xu Author
ACKNOWLEDGEMENTS
I would like to thank all people who have helped and inspired me during my
doctoral study Foremost I would like to express my sincere gratitude to my advisors Dr
Hong Liu and Dr Frank Chaplen for their continuous support of my PhD study and the
research of Microbial Fuel Cells at Oregon State University Hong inspired me to devote
myself to the field of Bio-energy with her great patience and enthusiasm towards
scientific educations Frank was always available and willing to help me with my study
especially during the period of Hongrsquos sabbatical leave Thanks to his kindness and
assistance my study at OSU became smooth and rewarding
Besides my advisors I would like to thank everyone in my dissertation committee Dr
Martin Schuster Dr Clare Reimers and Dr Mark Dolan Due to their encouragement
and insightful comments on my research I could always have the courage and knowledge
to overcome difficulties in my research I benefited greatly from Martinrsquos valuable
suggestions on my writing skills and his generous help regarding microarray data analysis
Clarersquos advice helped me build a solid foundation of Electrochemistry Also it was a
great honor to have Mark as my committee member His teaching gave me an insight into
the world of environmental engineering and enhanced the depth and width of my research
I thanked Dr Yanzhen Fan for always giving me guidance about the designs of the
reactors in my experiments
It was also important for me to say thanks to my current and previous lab-mates
Keaton Lesnik Kuhuan Chien Cheng Li Corale Abourached Anthony Janicek
Hongqiang Hu Jeremy Chignell Yudith Nieto and Wengguo Wu I had a wonderful
time enjoying doing research with them Their friendship and help made me confident of
my ability to do research as a scientist In addition I cherished and appreciated the
friendly environment in our BEE department Faculty members especially Dr John Bolte
Dr John Selker Dr Roger Ely Dr Ganti Murthy and Desiree Tullos made BEE an
excellent department for our students
My deepest gratitude went to my father Fuqin Wang and my mother Jinzhi Wang
for their endless care love and support throughout my life Also I would like to say
thanks to my best-loved wife Songhua Zhu She always supported me and never
complained that I could spend so little time accompanying with her This dissertation was
also for my lovely son Gabriel Hong-Yi Xu
TABLE OF CONTENTS
Page
1 General Introductionhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 1
2 Bacterial Community Analysis of Mixed Consortium in Microbial
Electrochemical Systemshelliphelliphelliphellip 13
3 New Exoelectrogen Citrobacter sp SX-1 Isolation and
Characterizationhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 27
4
Enhanced Performance and Mechanism Study of Microbial Electrolysis
Cells using Fe Nanoparticles Decorated Anodeshelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 45
5 Global Transcriptome Analysis of Response of Shewanella oneidensis
MR-1 to CNT Nanostructure Decorated Anodes in Microbial
Electrochemical System
66
6 Summaryhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 80
7 Bibliography helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 82
8 Appendices helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 93
LIST OF FIGURES
Figure Page
1-1 Generalized schematics (a) Microbial fuel cell (MFCs)(b) Microbial
electrolysis cell (MECs) helliphelliphelliphelliphelliphelliphelliphelliphelliphellip helliphelliphelliphellip
2
1-2 Mechanisms for extracellular electron transport in a MFCs anode 7
2-1 Polarization curves of high power mixed culture MFCshelliphelliphelliphelliphelliphellip 20
2-2 DGGE result of high power generation mixed culture MFCshelliphelliphellip 21
2-3 Bacterial species and percentage of bacterial community of mixed
culture MFCs helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip
22
2-4 Phylogenetic trees of bacterial species from mixed culture MFCs 22
2-5 Dominant bacterial species identification in DGGEhelliphelliphelliphelliphelliphelliphelliphellip 23
2-6 Current density generated by different dominant isolateshelliphelliphelliphelliphelliphellip 24
3-1 Phylogenetic tree of strain SX-1 and closely related species based on
16S rDNA sequenceshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip
35
3-2 SEM image of planktonic cells of Citrobacter sp SX-1helliphelliphelliphelliphelliphellip 35
3-3 Electricity generation by Citrobacter sp SX-1 in a single chamber
MFCs with sodium citrate helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip
36
3-4 Electricity generation of Citrobacter sp SX-1 using different
substrates in single chamber air-cathode MFCs at 1 KΩhelliphelliphellip
37
3-5 Power and voltage generation by Citrobacter sp SX-1 as a function
of current density helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip
38
3-6 Four current generating stages for CV analysis helliphelliphelliphelliphelliphelliphelliphellip 39
3-7 Cyclic Voltammetry analysis of of Citrobacter sp SX-1 biofilms helliphellip 41
4-1 SEM images of Fe NPs decorated anodeshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 50
4-2 Effects of Fe-NP decorated anodes on the current density in MECs helliphellip 51
4-3 Differentially expressed genes grouped by functional classification in
Fe NP decorated anodeshelliphelliphelliphelliphelliphellip helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip
53
5-1 SEM images of CNT decorated anodeshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 70
5-2 Effects of CNT decorated anodes on the current density in MECshelliphellip 73
5-3 Differentially expressed genes grouped by functional classification in
CNT decorated anodes helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip
74
LIST OF TABLES
Table
1-1
4-1
4-2
4-3
Taxa of bacteria current density generated and reactor configuration
used in microbial electrochemical cell experimentshelliphelliphelliphellip
Expression levels of genes related to biofilm formationhelliphelliphelliphelliphellip
Genes related to anaerobic growth and electron transfer with
significantly change expression levelhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip
Expression level of flavin and cytochrome geneshelliphelliphelliphelliphelliphelliphelliphellip
Page
4
55
56
58
4-4
5-1
5-2
Other genes with significantly changed expression level
Expression level of cytochrome c as response to CNT decorated anodes
Expression level of genes related to flavin synthesis as response to CNT
decorated anode in CNT decorated anodeshelliphellip
59
76
78
1
1
Bacterial Community Analysis New Exoelectrogen Isolation and Enhanced Performance
of Microbial Electrochemical Systems Using Nano-Decorated Anodes
Chapter 1
General Introduction
11 Microbial Fuel Cells and Microbial Electrolysis Cells
Microbial electrochemical systems (MESs) have drawn the attention of researchers
in recent years due to their promising applications in a variety of scientific fields such as
renewable energy generation bioremediation and wastewater treatment In a MES
microorganisms interact with electrodes catalyzing oxidation and reduction reactions at
the anode and the cathode
The most-described type of MESs is the microbial fuel cells (MFCs) in which
useful power is generated directly using the catalytic action of active microorganisms
(Kim 2002 Cheng and Logan 2007 Cheng et al 2006) In a typical two-chamber
MFCs (Figure 1a) organic matter is oxidized by electrochemically active
microorganisms in the anode chamber to release protons and electrons Protons diffuse
into the cathode chamber through a proton exchange membrane Meanwhile electrons
are transferred to the anode through various mechanisms (Logan et al 2006 Lovley
2
2006 Rabaey et al 2003) and then travel to the cathode where normally they combine
with oxygen and protons to form water
Another common used type of MES is the microbial electrolysis cell (MECs) in
which hydrogen is produced instead of electricity by applying a circuit voltage to the
MFCs system and maintaining anaerobic conditions in the cathode chamber (Liu et al
2005 Logan et al 2008 Rozendal et al 2006 Schaller et al 2009) (Figure 1b) A
cathode potential of at least - 410 mV (vs standard hydrogen electrode at pH 70) is
required to produce hydrogen at the cathode Therefore a theoretical potential of 110 mV
(ie 410-300 mV) is needed but voltages of gt02 V are normally applied due to various
overpotentials
Fig 1-1 Generalized schematics (a) Microbial fuel cells (MFCs) where electricity is
captured through the resistance R (b) Microbial electrolysis cells (MECs) where
hydrogen gas is produced in the cathode chamber R for resistor PS for power supply and
PEM for proton exchange membrane
The fundamental feature shared by microbial electrochemical systems (MESs)
(MECs and MFCs) is the microbe-catalyzed electron transfer from organic matter to
electrodes (anodes) (Rebaey et al 2004) Therefore different types of MESs can be
PEM
R
H+
e- e-
Anode
Bacte
ria
a
Cath
ode
H2O
PEM
PS
H+
e- e-
Anode
Bacte
ria
b
Cath
ode
H2
3
utilized to investigate the phenomena and mechanisms of interactions between microbes
and electrodes
12 Bacteria and Electron Transfer Mechanisms in MESs
The electrochemically active bacteria in MFCs are thought to be iron-reducing
bacteria such as Shewanella and Geobacter species (Gorby et al 2006) They have great
importance in the natural environment principally in metal oxidation and reduction
However recent studies have shown that the diversity of bacterial communities is much
greater than these model iron reducers on MFCs anodes (Reimers et al 2001 Liu et al
2004b Kim et al 2005) Over 20 electrochemically active bacterial species those can
transfer electrons exocellularly to electrodes have been reported in the past 10 years
(Logan 2009 Liu et al 2010) These bacteria have been categorized into diverse genetic
groups so far including alphaproteobacteria (Rhodopseudomonas Ochrobactrum and
Acidiphilium) (Malki et al 2008 Xing et al 2008 Zuo et al 2008) betaproteobacteria
(Rhodoferax) (Chaudhuri et al 2003) gammaproteobacteria (Shewanella Pseudomonas
Klebsiella Enterobacter and Aeronomas) (Kim et al 2002 Pham et al 2003 Rabaey et
al 2005 Bretschger et al 2007 Zhang et al 2008 Chung et al 2009 Rezaei et al 2009)
deltaproteobacteria (Geobacter Geopsychrobacter Desulfuromonas and Desulfobulbus)
(Bond et al 2002 Bond and Lovley 2003 Holmes et al 2004ab 2006)
Epsilonproteobacteria (Arcobacter) (Fedorovich et al 2009) Firmicutes (Clostridium and
Thermincola) (Park et al 2001 Wrighton et al 2008) Acidobacteria (Geothrix) (Bond
and Lovley 2005) and actinobacteria (Propionibacterium) (Wang et al 2008) A wider
range of electrochemically active bacteria are expected to be discovered
4
Table 1 Taxa of bacteria current density generated and reactor configuration used in
microbial electrochemical cell experiments (Liu et al 2010)
The electrochemically active bacterial species that possess the ability to transfer
electrons outside of the cell are called exoelectrogens in the MESs research field The
different exoelectrogens have demonstrated a wide-ranging power generation ability in
MFCs For example Shewanella oneidensis MR-1 has demonstrated the ability to
generate 0018 Am2 current density in single chamber MFCs while the Geobacter
Taxon Microorganisms Current Density (Am2
)
α-proteobacteria
Rhodopseudomonas palustris DX-1 003
Ochrobactrum anthropi YZ-1 071
Acidiphilium sp 32sup5 300
β-proteobacteria Rhodoferax ferrireducens 0031
γ-proteobacteria
Shewanella putrefaciens IR-1 0016
Shewanella oneidensis DSP10 0013
Shewanella oneidensis MR-1 0018
Pseudomonas aeruginosa KRA3 0017
Escherichia coli K12 HB101 100
Klebsiella pneumoniae L17 120
Enterobacter cloacae 013
Aeromonas hydrophila PA3 030
δ-proteobacteria
Geobacter metallireducens 065
Geobacter sulfurreducens 800
Desulfuromonas acetoxidans 0005
Geopsychrobacter
Electrodiphilus strain A2
0066
Desulfobulbus propionicus 003
Firmicutes
Lactococcus lactis 003
Thermincola sp strain Jr 020
Clostridium butyricum EG3 022
Thermincola ferriacetica Z-0001 040
Brevibacillus spp PTH1 0009
Desulfitobacterium hafniense DCB2 110
Acidobacteria Geothrix fermentans 0097
Actinobacteria Propionibacterium freudenreichiiET-3 120
5
sulfurreducens species has generated current densities as high as 80 Am2 which is 400
times higher than the one generated by Shewanella oneidensis MR-1 However mixed-
culture communities have generated much higher power densities than their pure-culture
counterparts in MFCs perhaps due to synergistic interactions within the anode bacterial
communities and the participation of currently unknown bacteria species and mechanisms
(Jung and Regan 2007) The dominant bacteria species vary in mixed cultures bacteria
communities in MFCs due to the enrichment of different substrates
Traditional methods of extroelectrogen study depend on cultivation hampered novel
exoelectrogens discovery because the inadequacy of defined media underestimates the
actual microbial diversity in MFCs Slow growth rate and unknown growth requirements
of anaerobic microorganisms make some anaerobic extroelectrogens cultivation difficult
However these limitations have been overcome by using molecular biological methods
based on DNARNA analysis Molecular biological techniques are now widely applied to
assess the diversity of microbial communities by analyzing the 16S rDNA sequence The
most commonly used molecular biological techniques for bacterial community analysis
include denaturing gradient gel electrophoresis (DGGE) restriction fragment length
polymorphism (RFLP) and 16S rDNA sequencing These methods are much less time
consuming than traditional isolation and cultivation methods
Isolated exoelectrogens were utilized to explore the mechanism of electron transfer
to the anode However the mechanisms of electron transfer to extracellular electron
acceptors are not well understood Three mechanisms have been proposed for exocellular
transport of electrons by exoelectrogens to anodes in MFCs so far Some exoelectrogenic
bacteria can transfer electrons to electrodes through soluble redox compounds (Bond and
6
Lovley 2005) These compounds include artificial mediators and mediators secreted by
exoelectrogenic bacteria A variety of chemicals have been used to facilitate the shuttling
of electrons from inside of cell to electrodes outside the cell These exogenous mediators
include neutral red (Park et al 1999) anthraquinone-2-6 disulfonate (AQDS) thionin
potassium ferricyanide (Bond et al 2002) methyl viologen and others (Logan 2004
Rabaey and Verstraete 2005) Some bacteria can secrete their own chemical mediator
for example Pseudomonas aeruginosa secretes pyocyanin and phenazine-1-carboxamide
to transfer electrons toward the MFCs anode (Rabaey et al 2005) Another evidence for
mediator production by bacteria is Geothrix fermentans When the medium was replaced
in a MFCs that had stable power generation with this bacteria power dropped by 50
and required 10 days to resume the original level
Some bacteria can directly transfer electrons to anodes via outer cell membrane
proteins (Myers and Myers 1992) The outer cell membrane protein cytochrome c is
thought to play a critical role in to transferring electrons to anodes Ly et al (2011)
isolated the haem protein cytochrome c and demonstrated that electric field effects may
be functional for the natural redox processes of cytochrome c in the respiratory chain
Shewanella oneidensis is the example of a bacterium that is capable of transfer electron to
anode via outer cell membrane proteins When Shewanella oneidensis adhered to an iron
surface the greater force has showed benefits to grow cells because closer contact
required for electron transfer from cell bound cytochromes (Lower et al 2001)
7
Fig1-2 Mechanisms for extracellular electron transport in an MFCs anode (1) direct
contact (top in green) (2) by nanowires (middle in purple) and (3) self-produced
mediators (bottom in blue) (Logan 2009)
More and more evidence supports the involvement of bacterial nanowires in
extracellular electron transport (Reguera et al 2005 2006 Gorby et al 2006) Nanowires
are conductive appendages produced by both Geobacter and Shewanella species (Gorby
and Beveridge 2005) The conductivity of the appendages was examined and confirmed
by using conductive scanning tunneling microscopy (STM) (Gorby et al 2006)
Nanowires can carry electrons from the cell to the anode surface of MFCs
The solid component of the extracellular biofilm matrix has high efficiency on
extracellular electron transfer compared with other extracellular electron transfer
8
mechanisms and recently Torres et al (2010) hypothesized that the solid component of
the extracellular biofilm matrix formed by exoelectogens is another way bacteria transfer
electrons to electrodes This hypothesis was based on kinetic analysis of each EET
mechanism reported in available literature (Torres et al 2010)
13 Anode electrodes
In MFCsMECs anode electrodes are a critical component because exoelectrogens
adhere to the surface of anodes to transfer electrons to the electrode The characteristics
of anodes have significant effects on electron transfer rate from bacteria to anode
electrodes in MFCs The requirements of an anode material are it should be highly
conductive non-corrosive have a high specific surface area (area per volume) high
porosity be non-fouling inexpensive and easily scaled to larger sizes Of these
properties the most important one that is different from other biofilm reactors is that the
material must be electrically conductive Normally they are made of various carbon
materials including carbon fiber carbon clothe and carbon paper due to their stability
high conductivity and high specific surface-area Nevertheless they have little
electrocatalytic activity for the anode microbial reactions and thus a modification of the
carbon materials is the main approach for improving their performance Consequently
there is a great need to develop a new type of anode material for MFCsMECs
It is a great challenge to develop a new anode material to further increase the power
density of a MFCs The nature of the catalytic mechanism of a MFCrsquos anode involves not
only a biological but also an electrocatalytic process An optimal nanostructure with a
high specific surface area favorable for both catalytic processes could play a critical role
in improving the power density of the MFCs such a structure needs to host the bacteria
9
with high bioactivity while enhancing the electron-transfer rate Schroumlder et al (2008)
employed PANI to modify a platinum anode for MFCs and achieved a current density 1
order of magnitude higher than the previously reported value PANIinorganic
composites are also reported to have better conductivity Qiao et al also applied a new
mesoporous TiO2 electrode material with uniform nanopore distribution and a high
specific surface area to anode in comparison to previously reported work with E coli
MFCs the composite anode delivers 2-fold higher power density (Qiao et al 2008)
Thus it has great potential for use as the anode in a high-power MFCs and may be a new
approach for improving performance of MFCs
14 Other parts of MESs
141 Membranes and ion transport
The ion exchange membrane is another one of the critical components in two-
chamber MESs systems It separates anode and cathode chambers and at the same time
maintains the electron neutrality of the system ie transport of electrons to the cathode
needs to be compensated by transport of an equal amount of positive charge to the
cathode chamber A proton exchange membrane (PEM) such as Nafion a per-fluorinated
sulphonic acid membrane (PFSAs) consisting of a hydrophobic fluorocarbon backbone to
which hydrophilic sulfonate groups (-SO3-) are attached is commonly used in chemical
fuel cell systems For MFCs systems however mainly cation species like Na+ and K
+
other than proton are often responsible for the dominant transport of positive charge
through the cation exchange membrane (CEM) to maintain electroneutrality due to the
low proton concentration in any aqueous medium with near neutral pH (Rozendal et al
2006) Consequently the pH increases in the cathode chamber due to the consumption of
10
protons and decreases in the anode chamber because of the accumulation of protons
(Rozendal et al 2007) This pH change in two-chamber systems results in a decrease of
the cathode potential and performance The application of anion exchange membrane
(AEM) has also been explored in MFCs systems (Cheng and Logan 2007 Kim and
Logan 2007) where it has been proposed that protons are transferred via pH buffers like
phosphate anions
142 Cathodes and Catalysts
The cathode is another challenge for making MFCs commercially available
technology because the chemical reaction that occurs at the cathode is difficult to
engineer as the electrons protons and oxygen must all meet at a catalyst in a tri-phase
reaction (solid catalyst air and water) The catalyst must be on a conductive surface and
must be exposed to both water and air so that protons and electrons in these different
phases can reach the same point The most commonly used material for a cathode is
commercially available carbon paper pre-loaded with a Pt catalyst on one side When it is
used in a MFC the side that contains the catalyst faces the water and the uncoated side
faces air To reduce the high cathode cost associated with platinum catalyst other precious-
metal-free cathodes such as (NiMo and NiW) were developed by electrodepositing on a
carbon fiber They have achieved comparable performance with Pt catalyst with same
loading at a much lower cathode fabrication cost (Hu 2010)
The requirements of cathode for MECs are quite similar to the requirements of
cathode for MFCs but easier than cathode for MFCs for the manufacturing process
because the cathode in MECs is not necessarily to exposed to air Therefore it can be
made of the exact same materials of cathode in MFCs except a waterproof layer
11
Recently some researchers have attempted to apply microorganisms as a biocatalyst to
precede the combination of electron with oxygen in the cathodes
15 Dissertation overview
Low power densities in MESs limit practical applications The improvement of
MESs performance requires a detailed understanding of the physiology and ecology of
microorganisms in MESs including the mechanism of electron transfer to the anode from
the microorganism
This dissertation focuses on the problem of the low power density of MESs The
bacterial community structure of a high power generated mixed culture communities in
MFCs will be identified firstly and then one of major exoelectrogens will be isolated and
characterized This information will be helpful to understand the physiology and ecology
of exoelectrogens in MESs Consequently they will be beneficial to improve power
density of MFCs Lastly the nanostructure decorated anodes in MECs will be utilized to
improve the power density The power enhancement mechanism will be explored by
using a whole genome microarray They are presented here as four papers
In the first paper the cultivation independent molecular biological techniques
denaturing gradient gel electrophoresis (DGGE) and 16S rDNA clone library are utilized
to analyze the bacterial community structure of a higher power mixed culture MFCs The
analyzed results provide fundamental information for isolating the dominant bacteria in
mixed culture MFCs Two of dominant bacterial species has been isolated and used aone
to test power generation in MFCs The possible interaction among different bacterial
species in mixed culture is discussed
12
In the second paper one isolated exoelectrogenic bacterial strain SX-1 is
characterized It is identified as a member of the Citrobacter genus and power generation
is tested ultilizing a wide range of different substrates The electron transfer mechanism is
explored using Cyclic Voltammetry (CV) This study increases the known diversity of
power generating exoelectrogens and provides a new strain to explore the mechanisms of
extracellular electron transfer from bacteria to electrodes
The third paper shows effects on MESs performance by Fe nanoparticle decorated
anodes in the MESs The average current density produced with Fe nanoparticle
decorated anodes increased up to 59-fold higher than plain graphite anodes A whole
genome microarray is utilized to analyze the possible mechanism of enhanced current
density as responded to nanoparticle decorated anodes
The fourth paper describes the effects of carbon nanotube (CNT) modified anode on
the performance S oneidensis MR-1 in MESs Results demonstrate that current densities
produced with CNT decorated anodes are up to 56-fold higher than plain graphite anodes
The possible mechanisms of enhanced current density by CNT decorated are explored
13
Chapter 2
Bacterial community analysis of mixed consortium in higher
power density MESs
Shoutao Xu and Hong Liu
1 Introduction
The improvement in the performance of mixed culture MFCs requires an
understanding of the ecology in microbial communities of MFCs Many researchers have
attempted to characterize microbial populations and activities to elucidate the behaviors
and ecology of microorganisms in MFCs (Wrighton et al 2008 Fedorovich et al 2009)
In order to study the microbial ecology of the mixed culture in a MFC and select the
appropriate isolation medium for dominant bacterial species in the mixed culture the
fundamental steps are to analyze the bacterial diversity in the mixed culture of MFCs and
identity the dominant bacterial species in bacteria communities in MFCs
14
As for the identification of bacterial communities typically there are two general
methods The first method for identification of bacterial community is the traditional
cultivation processes using selective nutrients to promote the growth of different types of
bacteria within the samples (Amman et al 2000) The community structure can then be
assessed by identifying the isolates from the dominant colonies that were cultured This
can often be costly and laborious as each isolate has to be further studied by examining
its physiology taxonomy and reactivity to stains (Adwards et al 1989)
The second method relies on utilizing molecular techniques to analyze bacterial
community DNA Several molecular methods involving the extraction and analysis of
DNA from entire bacterial communities are used to identify genetic fingerprints of
bacteria These methods including the cloning and sequencing of 16S rDNA automated
ribosomal intergenic spacer analysis (ARISA) terminal restriction fragment length
polymorphism (T-RFLP) and denaturing gradient gel electrophoresis (DGGE) generate
profiles of bacterial community structures They can rapidly assess complex communities
from various environments (Amman et al 2000)
Due to the conservative characteristic of 16S rDNA in bacteria during the process
of evolution 16S rDNA sequencing can be used to identify different species of bacteria
Therefore the molecular techniques of denaturing gradient gel electrophoresis (DGGE)
with PCR and 16S rDNA clone library are used for analysis of the microbial diversity
These methods are more convenient and save time compared to traditional
isolationcultivation methods for microorganism analysis
In this work a biofilm bacterial community from an anode of a MFCs wase studied
by using denaturing gradient gel electrophoresis (DGGE) of PCR-amplified partial 16S
15
rRNA genes followed by cloning and sequencing of 16S rDNA The results provided
essential information for dominant bacterial isolation in mixed culture MFCs
2 Materials and methods
21 High power generation of mixed culture MFCs
Mixed cultures were originally inoculated from domestic wastewater (Corvallis
Wastewater Treatment Plant Corvallis OR) to MFCs after operating for 2 years using a
defined medium solution (Lovely 2002) with sodium acetate as the carbon source A
new MFCs was inoculated from the operating MFCs The polarization curves were
performed to measure the power generation when maximal stable power were established
(normally 3-5 batches post-inoculation) and the genomic DNA from the microbial
biofilm on the anode was extracted under the sterile conditions for further bacterial
community analysis
22 Denaturing gradient gel electrophoresis (DGGE)
DGGE with PCR is a method of analysis of bacterial community composition
based on different GC percentage of 16s rDNA in different bacterial species PCR-DGGE
comprises three steps (i) extraction of total community DNA from the sample (ii) PCR-
controlled amplification using specific oligonucleotide primers and (iii) separation of the
amplicons using DGGE For this purpose a reproducible and efficient method for total
DNA extraction is indispensable and needs to be evaluated and optimized depending on
the nature of the sample In the subsequent PCR step multiple PCR primer sets with
different resolution can be used In most PCR-DGGE applications on bacteria universal
or specific primers are targeting the 16S rRNA gene Upon electrophoresis of the PCR
16
amplicons and gel staining (using ethidium bromide silver staining or SYBR green)
DGGE gels are digitally captured and further analyzed using computer software packages
The richness of DNA fragment bands in DGGE indicates the bacterial diversity of the
sample
The detailed steps are as followed Biofilms were scratched from the anodes of high
power generated MFCs fed with sodium acetate Bacterial genomic DNA was extracted
from the biofilm samples using the DNeasy blood and tissue kit (Qiagen) according to the
manufacturerrsquos instructions The universal primer set was used to amplify the 16S rDNA
from the extracted genomic DNA then the V3 region of bacterial 16S ribosomal DNA
(rDNA) was amplified with GC-clamp primer (Muyzer et al 1993) for DGGE PCR
amplification was performed in a thermocycler DGGE of the PCR products was carried
out in a DcodeTM
Universal Mutation Detection System) The 8 (wv) polyacrylamide
gels contains from 30 to 55 denaturing gradients Electrophoresis was conducted
using a 1timesTAE (Tris-Acetate-EDTA) buffer at 130V and 60degC for 5 hours After
electrophoresis the gel was stained with ethidium bromide in 1timesTAE buffer for 15
minutes and washed in 1timesTAE buffer for 10 minutes The fragments were visualized
under a UV transilluminator The richness of single band in DGGE gel picture
preliminarily showed that the bacterial diversity in mixed culture since the single band in
gel represents one bacterial species
23 16S rDNA clone library construction method
16S rDNA-DGGE fingerprinting has proven to be particularly useful as an initial
investigation into bacterial communities and is suitable for identifying the predominant
bacteria that comprise 1 or more of cells within a given sample In the 16s rDNA clone
17
library construction process the first step is the extratction of the total genomic DNA
then the genomic DNA of the mixed culture biofilm was used as template for PCR
amplification of approximately 1500 bp of 16S rDNA with universal primers The PCR
products of 16S rDNA were purified and inverted into pGEM-T Easy vector system
before they were transformed into competent E coli The transformed cells were spread
on Luria-Bertani (LB) plates containing the selective antibiotic ampicillin and X-Gal and
incubated overnight at 37 OC after the more than 80 inverted 16s DNA fragment E coli
colonies were picked up and inoculated in the 1-3 ml liquid LB medium and left to grow
for 16 hours The plasmid DNA were extracted and sequenced The sequences were
compared directly to all known sequences deposited in GenBank databases using the
basic local alignment search tool (BLAST)
24 Bioinformatics Analysis
16S rDNA sequencing results of more than 80 colones were queried against the
GenBank and Ribosomal Database Project (RDP) databases using BLAST and
SEQUATCH algorithms (Cole et al 2007 Wang et al 2007 Cole et al 2009) The
neighbor-joining trees were constructed with the Molecular Evolutionary Genetics
Analysis package (MEGA Version 4) based on 1000 bootstrap analysis (Tamura et al
2007)
25 Dominant bacterial species band in DGGE identification
The DGGE results gave the information of dominant bacterial richness of bacterial
species in mixed culture based on the theory that single band represents one bacterial
species which showed the diversity of bacterial community in the mixed culture of a
MFC 16S rDNA clone library results provided the whole picture of bacterial community
18
including the bacterial species name and bacterial species percentage in the mixed culture
The known pure bacterial species which have been sequenced can be used as markers to
identify dominant bacterial species in DGGE The whole experimental procedure is
similar to the procedure of the mixed culture DGGE steps The first step is to extract the
collect the mixed culture biofilm genomic DNA and pure bacterial species marker
genomic DNA Then the universal primer set was used to amplify the 16S rDNA from
the extracted genomic DNA After that the V3 region of bacterial 16S ribosomal DNA
(rDNA) was amplified with GC-clamp primer (Muyzer et al 1993) for DGGE The
subsequent steps were similar to the mixed culture DGGE experimental procedure The
dominant bacterial bands in DGGE were determined by comparisons of the single band
position in the mixed culture DGGE with single pure bacterial species location in DGGE
26 Dominant bacterial species isolation and power generation
The bacterial community of mixed culture in MFCs were predominantly composed
of Geobacter sp Arcobacter sp and Citrobacter sp Two dominant bacterial species
have been isolated
Citrobacter sp SX-1 was isolated from the anode biofilm by using a solid agar
medium containing NH4Cl 6mM KCl 2mM CH3COONa (electron donor) 30 mM
FeC6H5O7 (electron acceptor) 20mM minerals and vitamins (Lovley and Phillips 1988)
Single colonies were picked up after culturing on the plate for 36 hours at 30 degC and
transferred two times on the agar plate for purification Then the isolates grew in a liquid
medium solution in anaerobic tubes containing the same constituents as the solid
medium All isolation process was operated in a glove box anaerobic chamber (Coy
Laboratory Products Grass Lake MI)
19
Arcobacter sp SX-2 was isolated from the anode biofilm of a MFC fed with sodium
acetate by using the same method of isolation of pure bacterial species Citrobacter XS-1
except using Trypticase Soy Agar with 5 Sheep Blood solid (from American type
culture collection (ATCC)) agar medium instead of Citrobacter sp medium Single
colonies were picked up after culturing on the plate for 36 hours at 30 degC and transferred
twice on the agar plate for purification Then the isolates grew in a liquid medium
solution microaerobically
Geobacter sp have been trying to isolate by using three different methods They are
the colony pickup after different condition enrichment Goebacter medium isolation and
dilution to extinction by multiple channel mini MFCs isolation however the Geobacter
sp has not been isolated yet due to some limits so far But they provide valuable
information for further isolation of Geobacter sp
Single chamber MFCs were used to evaluate power generation by different isolates
The MFCs were constructed as described previously (Liu and Logan 2004) and modified
with 3 cm2 carbon cloth anodes and 7 cm
2 carbon clothPt cathodes The total liquid
volume of each MFCs was about 15 ml with electrode spacing about 17 cm All MFCs
were operated in an autoclaved closed plastic box and sterile cotton was attached to the
outer surface of the air cathodes to prevent contamination A MFCs without bacterial
culture was used as control MFCs were inoculated with 3 ml late exponential phase
cultures of SX-1 in the medium solution reported previously (Liu and Logan 2004)
3 Results and discussion
31 Power production by mixed culture MFCs
20
A polarization curve was used to characterize current as a function of voltage in the
MFCs The polarization curves are performed by varying a series of external resistances
The results are shown in Figure 21 With MFCs acclimated to 25 Ω external resistance
the maximum power density was achieved at 65Wm2 based on the polarization data At
this point the current density is 21mAcm2 The power density was three times higher
than previous report (20 Wm2) using phosphate buffer (Fan et al 2007) Power density
was also 2 times higher than (28 m Wm2) using a bicarbonate buffer ( Fan et al 2007)
32 Bacterial community structure assessed by PCR-DGGE
Bacterial DGGE profiles for higher power generating MFCs mixed culture setups
were relatively complex (more than 10 bands) However there are three brightly stained
bands and other less intense bands (Fig 2-2) These results indicated there were probably
three dominant bacterial species in mixed culture MFCs
010
020
030
040
050
060
070
2
3
4
5
6
7
04 05 05 06 07 08 10 11 13 15 21 22 23 24
Volta
ge (
V)
Pow
er
densi
ty (
Wm
2)
Current density (mAcm2)
Power
Voltage
power density 65wm2 at current
density208 mAcm2
Figure 2-1 Polarization curves of high power mixed culture MFCs
21
33 16S rDNA clone library results
A total of 83 colonies were sequenced for 16S rDNA clone library construction They
represented the bacterial community structure in mixed culture in MFCs The results
revealed that the predominant bacterial species in mixed culture are Geobacter sp (66
of the mixed culture) Arcobacter sp (12 of the mixed culture) and Citrobacter sp (11
of the mixed culture) Those three genuses comprised 88 of the total bacterial species
(Fig 23) This result is consistent with the DGGE result which showed that three bright
bands in the DGGE gel represented three dominant bacterial species in the mixed culture
The phylogenetic analysis of the colonies (83 colony sequences) is shown in Figure 24
Figure 22 DGGE result of high power generation mixed culture MFC (1 mixed
culture 2 pure culture control)
1 2
22
34 Dominant bacterial band in DGGE identification
Geobacter
66
Arcobacter
12
others
6
Pseudomonas
1
Citrobacter
11
Clostridium
2
Anaerovorax
2 Geobacter
Arcobacter
Citrobacter
Clostridium
Anaerovorax
Pseudomonas
others
Figure 23 Bacterial species and percentage of bacterial community of mixed
culture MFC lt 1 Alcaligensgt
lt1 Commamonasgt
lt1 Pseudomonas aeruginosagt
lt1 Klebsiellagt
lt11 Citrobactergt
lt1 Wolinellagt
lt12 Arobactergt
lt2 Anaeroboraxgt
lt2 Clostridium gt
lt1 unclear bacteriagt
lt66Geobactergt
100
100
85
100
100
82
99
87
100
002
Figure 24 Phylogenetic trees of bacterial species from mixed culture MFC
23
The dominant bacterial species including Goebacter sp Citrobacter sp and Arcobacter
sp in the mixed culture are shown in Figure 25 The upper band is Arcobacter sp and
Citrobacter sp is located in the bottom band of DGGE Geobacter sp is the middle one
The analysis of the bacterial community using denaturing gradient gel electrophoresis
(DGGE) of PCR-amplified 16S rRNA gene fragments and 16S rDNA clone library
construction method showed great phylogenetic diversity of mixed culture in MFCs with
the identification of sequences derived from bacteria of the taxa deltaproteobacteria
(Geobacter sp) gammaproteobacteria (Citrobacter sp) and epsilonproteobacteria
(Arcobacter sp) The result is consistent with the conclusion that among the isolated
exoelectrogens gammaproteobacteria and deltaprotebactia appear to be dominant among
others (Parot et al 2009 Liu et al 2010)
35 The dominant bacterial species isolation and power generation
Figure 25 Dominant bacterial species identification in DGGE
A B C D
A Citrobacter sp
B Geobacter sp
C Arobacter butzmeri
D mixed culture control
24
Two dominant bacterial species Citrobacter sp XS-1 and Arcobacter sp XS-2 have been
isolated and tested for power generation The power densities generated by the two
isolates are shown in the Fig 26 Current densities generated by isolate Citrobacter sp
and Arcobacter sp were 98 mAm2 and 20 mAm
2 respectively which were much less
than that (21Am2) generated by the mixed culture from which they were isolated
Figure 26 Current density generated by different dominant isolates
The maximum current density generated by mixed culture is much higher than the
one produced by pure bacterial species Citrobacter sp XS-1 and Arcobacter sp XS-2 in
the same structure MFCs Although the Geobacter sp in our mixed culture has not been
isolated a previous study showed that maximum current density generated by Geobacter
sp was 080 Am2
(Bond and Lovley 2003) The current density results seem partially
supporting the conclusion that mixed-culture generated higher power densities than their
pure-culture counterparts in MFCs (Jung and Regan 2007) This result suggests there
may be synergistic interactions within the anode bacterial communities
0
500
1000
1500
2000
2500
Mixed culture Citrobacter sp Arcobacter Sp
Cu
rre
nt
de
nsi
ty (
mA
m2)
25
Geobacter sp are strictly anaerobic bacteria which were enriched in most of the anode
biofilms (George 2005) Citrobacter species are facultative anaerobic bacteria (George
2005) which can be found in a wide variety of habitats including in soil water and
wastewater Arcobacter sp are micro-aerobic bacteria including both environmental
nonpathogens and opportunistic human pathogens They are able to grow in aerobic
conditions but in the optimal growth under micro-aerobic conditions (George 2005)
Arcobacter sp has been discovered in the MFCs by Fedorovich (Fedorovich et al 2009)
It is interesting that the mixed culture bacterial community composed of primarily these
three bacterial species has demonstrated to generate high power at non-strictly anaerobic
condition while leading dominant bacterial species Geobacter sp (66) is strict
anaerobic bacteria and generated powder in the MFCs only under strict anaerobic
condition (George 2005) Therefore it is possible that microaerobic bacteria such as
Arcobacter sp and facultative anaerobic bacteria Citrobacter sp can facilitate to create
anaerobic condition for Geobacter sp when they utilized oxygen for growing in the
mixed culture Syntrophic communities study have showed that anaerobic bacteria and
methanogenic archaea form compact microbial structures that operate like an organ rather
than a set of microorganisms functioning independently (Stams and Plugge 2009) Some
substrates have been degradeted within these communities while they are not able to be
fermented by individual species alone interspecies electron transfer also have been
presented in these communities (Stams and Plugge 2009) Summers et al (2010) also
discovered that direct exchange of electrons happened within coculture of Geobacter
metallireducens and Geobacter sulfurreducens Therefore we believe that there are
26
more complex synergistic interactions between different bacteria species in mixed culture
in the MFCs
Acknowledgements
The authors acknowledge support from the US National Science Foundation
(CBET 0955124) We also thank Keaton Lesnik for the review of this manuscript
27
Chapter 3
New Exoelectrogen Citrobacter sp SX-1 Isolated from a Microbial Fuel Cell
Shoutao Xu and Hong Liu
Published in
Journal of Applied Microbiology 111(5)1108-1115 (2011)
ABSTRACT
Exoelectrogenic bacterial strain SX-1 was isolated from a mediator-less microbial fuel
cell by conventional plating techniques with ferric citrate as electron acceptor under
anaerobic condition Phylogenetic analysis of the 16S rDNA sequence revealed that it
28
was related to the members of Citrobacter genus with Citrobacter sp sdy-48 being the
most closely related species The bacterial strain SX-1 produced electricity from citrate
acetate glucose sucrose glycerol and lactose in MFCs with the highest current density
of 205 mA m-2
generated from citrate Cyclic voltammetry analysis indicated that
membrane associated proteins may play an important role in facilitating the electrons
transferring from bacteria to electrode This is the first study that demonstrates that
Citrobacter species can transfer electrons to extracellular electron acceptors Citrobacter
strain SX-1 is capable of generating electricity from a wide range of substrates in MFCs
This finding increases the known diversity of power generating exoelectrogens and
provided a new strain to explore the mechanisms of extracellular electron transfer from
bacteria to electrode The wide range of substrate utilization by SX-1 increases the
application potential of MFCs in renewable energy generation and waste treatment
Keywords Microbial fuel cell Exoelectrogen Citrobacter sp SX-1 Extracellular
electron transfer
29
Introduction
Microbial Fuel Cells (MFCs) technology has become an active research area
recently as a promising approach for renewable energy generation wastewater treatment
and bioremediation (Rabaey and Verstraete 2005 Lovley 2006 Fan et al 2008 Logan
2009 Liu et al 2010) The key feature of MFCs system is the microbe-catalyzed electron
transfer from organic matter to anodes Many studies have shown the presence of diverse
bacterial communities on MFCs anodes (Reimers et al 2001 Liu et al 2004b Kim et al
2005) Over 20 exoelectrogens ie microbes that can transfer electrons exocellularly to
electrodes have been reported in the past 10 years (Logan 2009 Liu et al 2010) The
exoelectrogens so far belong to diverse genetic groups including alphaproteobacteria
(Rhodopseudomonas Ochrobactrum and Acidiphilium) (Malki et al 2008 Xing et al
2008 Zuo et al 2008) betaproteobacteria (Rhodoferax) (Chaudhuri et al 2003)
gammaproteobacteria (Shewanella Pseudomonas Klebsiella Enterobacter and
Aeronomas) (Kim et al 2002 Pham et al 2003 Rabaey et al 2005 Bretschger et al
2007 Zhang et al 2008 Chung et al 2009 Rezaei et al 2009) deltaproteobacteria
(Geobacter Geopsychrobacter Desulfuromonas and Desulfobulbus) (Bond et al 2002
Bond and Lovley 2003 Holmes et al 2004ab 2006) Epsilonproteobacteria (Arcobacter)
(Fedorovich et al 2009) Firmicutes (Clostridium and Thermincola) (Park et al 2001
Wrighton et al 2008) Acidobacteria (Geothrix) (Bond and Lovley 2005) and
actinobacteria (Propionibacterium) (Wang et al 2008) A wider range of
exoelectrogenic species are expected to be discovered
Three mechanisms have been proposed for exocellular transport of electrons by
exoelectrogens without artificial mediators Some exoelectrogenic bacteria can transfer
30
electrons to electrodes through soluble redox compounds excreted by microorganisms
(Bond and Lovley 2005 Rabaey et al 2005) and others can directly transfer electrons to
anodes via outer cell membrane proteins (Myers and Myers 1992) Recently more and
more evidence supports the involvement of bacterial nanowires in extracellular electron
transport (Reguera et al 2005 2006 Gorby et al 2006) In spite of the discovery of
many bacterial species that can transfer the electrons to electrode without the need of
artificial mediators the investigation of extracellular electron transfer mechanisms was
mainly focused on a few species such as those from Geobacter and Shewanella genera
(Kim et al 1999 2002 Reguera et al 2005 2006 Gorby et al 2006) The electron
transfer mechanisms for many of the isolated exoelectrogens species are still not well
studied
In the present study we isolated a new exoelectrogen strain SX-1 from a MFC a
strain phylogenetically related to Citrobacter sp Power generation from various carbon
sources by this strain was evaluated using single chamber MFCs Plausible extracellular
electron transfer mechanisms were also discussed based on the characterization of anodic
biofilms by cyclic voltammetry (CV)
Materials and methods
Bacterial strain SX-1 isolation
Bacterial strain SX-1 was isolated from the anode biofilm of a MFC fed with sodium
acetate operated in fed-batch mode over a period of six months The original source of
the inoculum is wastewater from a local waste water treatment plant Bacterial cells were
released from the carbon cloth anode by shaking the cloth in a sterile flask with 15 mL
sterile 1 NaCl solution and glass beads (2 mm diameter) The suspension was then
31
serially diluted from 10 times to 105
times and plated on a petri dish with a solid agar
medium containing NH4Cl 6mM KCl 2mM CH3COONa (electron donor) 30 mM
FeC6H5O7 (electron acceptor) 20mM minerals and vitamins (Lovley and Phillips 1988)
Single colony was randomly selected after culturing on the plate for 36 hours at 30 degC
and purified on a new agar plate following a procedure reported previously (Chung and
Okabe 2009) Then the isolate was picked up and grew in a liquid medium solution in
anaerobic tubes containing the same constituents as the solid medium for further analysis
All isolation process was operated in a glove box anaerobic chamber (Coy Laboratory
Products Grass Lake MI)
16S rDNA sequencing and phylogenetic analysis
Bacterial genomic DNA was extracted from the isolated strain SX-1 using the DNeasy
tissue Kits (Qiagen Valencia CA) according to the manufacturerrsquos instructions The 16S
rDNA was amplified by PCR using a pair of universal primers 27F (5rsquo-
AGAGTTTGATCMTGGCTCAG-3rsquo) and 1492R (5rsquo-GGTTACCTTTGTTACGACTT-
3rsquo) (Park et al 2001) The conditions used for PCR were described previously
(Ferna ndez et al 1999) PCR products were purified ligated and cloned according to a
procedure described in a previous report (Xing et al 2008) Plasmids were isolated from
randomly selected clone colonies with a QIAprep Spin Miniprep kit (Qiagen Valencia
CA) and three plasmid inserts were then sequenced in both directions using an ABI 3730
DNA sequencer (Applied Biosystems Carlsbad CA)United States) Due to the identical
16S rDNA sequence of the three colones only one was queried against the GenBank and
Ribosomal Database Project (RDP) databases using BLAST and SEQUATCH algorithms
(Cole et al 2007 Wang et al 2007 Cole et al 2009) The 16S rDNA sequence of
32
strain SX-1 and closely related type strains were aligned using CLUSTALX software
(Thompson et al 1997) The neighbor-joining trees were constructed with the Molecular
Evolutionary Genetics Analysis package (MEGA Version 4) based on 1000 bootstrap
analysis (Tamura et al 2007)
SEM
The morphologies of SX-1 cells growing overnight in anaerobic tubes were examined
with a scanning electron microscope (FEI Quanta 600 FEG FEI Company Hillsboro
OR) The suspended cells were fixed in a 25 glutaraldehyde and 01M phosphate
buffer solution and dehydrated with a graded ethanol series from 30 to 100 After
dehydration the samples were dried in a critical point dryer and then sputter-coated with
AuPd for SEM examination (Liu and Logan 2004)
Evaluation of power generation by SX-1 in MFCs
Single chamber MFCs were used to evaluate power generation by SX-1 using various
substrates The MFCs were constructed as described previously (Liu and Logan 2004)
and modified with 3 cm2 carbon cloth anodes and 7cm
2 carbon clothPt cathodes The
total liquid volume of each MFCs was about 15 ml with electrode spacing about 17 cm
All MFCs were operated in an autoclaved closed plastic box and sterile cotton was
attached to the outer surface of the air cathodes to prevent contamination A MFC
without bacterial culture was used as control MFCs were inoculated with 3 ml late
exponential phase cultures of SX-1 in the medium solution reported previously (Liu and
Logan 2004) Seven substrates including citrate acetate glucose sucrose glycerol and
lactose were evaluated individually for power generation in a fed-batch mode in a
temperature control chamber (30 plusmn2degC) For each substrate at least 8 batches were run to
33
investigate the effect of biofilm formation on current output at fixed external resistance of
1kΩ and the same initial substrate concentration of 30 mM Phosphate buffer (100mM)
was used to maintain the solution pH and conductivity The MFCs with sodium citrate as
electron donors were also examined for maximum power output by varying the external
resistance from 20 kΩ to 4 kΩ A data acquisition system was used to record voltage data
during the MFCs operation (Keithley 2700 Keithley Instruments Inc Cleveland OH) It
took about 15 to 30 minutes for the MFCs to stabilize depending on the external
resistance At each resistance we collected at least five data at the steady condition to
make the polarization curves Averaged voltages were used to calculate the power density
(mW m-2
) according to P=IVA where I was the current V was voltage and A was cross-
sectional area of the anode
CV analysis
CV was used to characterize the oxidation and reduction reactions on the anodic surface
of the MFCs fed with citrate and operated at 7 KΩ This external resistance was selected
because the maximum power density was obtained at this resistance based on the
polarization experiment We assumed the biofilm on the anode of MFCs were well-
developed when stable power output was obtained after 3 batches of operation The
MFCs were then used directly for CV analysis at four current generating stages of the
fourth batch (1) initial exponential current increasing stage (middle point of stage about
4 h after media change) (2) current plateau stage (middle point of stage about 8 h after
media change) and (3) current decreasing stage (middle point of stage about 16h after
media change) (4) right after the complete replacement of medium solution (Figure 6)
The anode was used as working electrode the cathode as counter electrode and an
34
AgAgCl electrode was selected as the reference The CV curves were scanned from 200
to -600 mV at a rate of 5 mV s-1
using a potentiostat (G300 Gamry Instrument
IncWarminster PA) Control experiment was also conducted using new anode (without
biofilm) and new medium solution
Nucleotide sequence accession number
The 16S r DNA sequence determined in this study has been deposited in the GenBank
database under accession number HQ845373
Results
Identification of the strain SX-1
An almost-complete 16S rDNA sequence (1475 bp) of strain SX-1 was obtained and
subjected to comparative analysis with the 16S rDNA of closely related reference strains
A phylogenetic analysis revealed that strain SX-1 should be assigned to the genus
Citrobacter and was most closely related to Citrobacter sp sdy-48 EJ463782 (990
sequence similarity) and Citrobacter sp yy-21 FJ463779 (975 sequence similarity)
These three strains formed a distinct sub cluster in the neighbor-joining in which the new
isolate and Citrobacter sp sdy-48 EJ463782 formed a distinct subline (Figure 31)
The SEM image illustrated that the strain SX-1 is a rod-shaped bacterium 03-06 microm
wide and 12-17 microm long (Figure 32) Strain SX-1 colonies on nutrient agar appeared 2-
3 mm in diameter smooth moist opaque with a shiny surface after 5 days incubation
under anaerobic condition The fact that strain SX-1 grew both aerobically and
anaerobically using FeC6H5O7 as electron acceptor suggests that the strain is facultatively
anaerobic Strain SX-1 showed robust growth on citrate as a carbon source These D
35
properties were similar to those of genus Citrobacter described in Bergeyrsquos Manual of
Systematic Bacteriology (George 2005)
Figure 31 phylogenetic tree of strain SX-1 and closely related species based on 16S
rDNA sequences The tree was constructed using the neighbor-joining method
Figure 32 SEM image of planktonic cells of Citrobacter sp SX-1
Strain SX-1
Citrobacter sp sdy-48 (FJ463782)
Citrobacter sp yy-21 (FJ463779)
Citrobacter koseri E639 (ATCC 25408)
Citrobater koseri CDC 3613-63(AF025372)
Citrobacter koseri CDC 8132-86(AF025366)
Salmonella enterica SL483 (CP001138)
Salmonella enterica AKU12601(AY696668)
Shigella sonnei Ss046(CP000038)
E coli C2 (AF403733)
E coli E24377A(CP000800)88
100
100
76
44
92
53
36
0001
1 microm
36
Electricity production by strain SX-1 in MFCs
The isolated strain SX-1 was first tested for its ability to generate current from sodium
citrate at 1kΩ Current gradually increased to a maximum 50 mA m-2
after cell
inoculation and then decreased (Figure 33) After the MFCs was refilled with new
substrate solution the current recovered rapidly and reached a higher level than the first
batch After 4 batches operation the maximum current output of each batch became
stable The highest current density achieved in the MFCs with Citrobacter sp SX-1 at 1
KΩ was 98 mA m-2
(Figure 33)
Figure 33 Electricity generation by Citrobacter sp SX-1 in a single chamber MFCs
with sodium citrate (30 mM) as substrate at 1 KΩ
0
20
40
60
80
100
120
0 50 100 150 200
Curr
ent
den
sity
(m
A m
-2)
Time (h)
37
Current generation by SX-1 from other substrates including glucose lactose sodium
acetate glycerol and sucrose were also assessed at an external resistance of 1KΩ
(Figure 34) When repeatable cycles of current output were obtained for these substrates
glycerol generated the highest maximum current density of 58 mA m-2
followed by
lactose and sucrose with 29 mA m-2
and 27 mA m-2
respectively Glucose and acetate
produced the lowest maximum current density of 96 and 43 mA m-2
respectively These
results indicated that strain SX-1 can utilize a wide range of substrates for electricity
generation in MFCs but with different power generation potentials
Figure 34 Electricity generation of Citrobacter sp SX-1 using different substrates in
single chamber air-cathode MFCs at 1 KΩ (The error bars stand for the standard
deviation of 3 replicates)
0
10
20
30
40
50
60
70
Glucose Lactose Glycerol Acetate Sucrose
Curr
ent
den
sity
(m
A m
-2)
38
Polarization experiment was further conducted to determine the maximum power
density generated by strain SX-1 in the single chamber MFCs with citrate as substrate A
maximum power density of 881 mW m-2
was obtained at current density of 205 mA m-2
at an external resistance of 7 KΩ (Figure 35)
Figure 35 Power and voltage generation by Citrobacter sp SX-1 as a function of
current density using sodium citrate (30 mM) as substrate The error bars stand for the
standard deviation of the 3 voltagepower outputs obtained in three MFCs
20
30
40
50
60
70
80
90
100
01
02
03
04
05
06
07
80 110 140 170 200 230
Po
wer
den
sity
(m
W m
-2)
Vo
ltag
e (V
)
Current density (mA m-2)
Voltage
Power
39
Figure 36 Four current generating stages for CV analysis current increasing stage (stage
1) current plateau stage (stage 2) current decreasing stage (stage 3) and right after the
replacement of medium solution (stage 4) The MFCs was operated at 7 KΩ with sodium
citrate (30 mM) as substrate
Cyclic Voltammetry
To determine the presence of redox active compounds produced by SX-1 and the
location of these compounds CV scan of the anodic biofilms at four current output stages
and the supernatant of MFCs medium solution at the end of the batch experiment were
performed Reduction peaks at range of -310 ndash -350 mV and oxidation peaks at range -
100 ndash -150 mV were observed at all current generating stages (Figure 36 Figure 37A)
suggesting the presence of redox active compounds may involve in extracellular electron
transfer by Citrobacter sp SX-1 The amplitude of the peaks increased according to the
growth stage of the batch and the highest peaks were present after the current plateau
stage which indicated the redox active compounds mainly were secreted in the current
plateau stage While the current density at stage 3 (deceasing stage) was lower than that
at stage 2 (current plateau stage) the redox peak at stage 3 was higher than that in stage 2
0
50
100
150
200
250
50 55 60 65 70 75 80 85
Curr
ent
den
sity
(m
A m
-2)
Time (h)
A
A
Stage 1
Stage 2 Stage 3
Stage 4
40
A plausible explanation for this pattern was that redox active compounds were
continuously being secreted and accumulated after the current peak was reached
resulting in more redox compounds present after the plateau stage But since most of the
carbon source had already been being used up many of the redox compounds at this
stage had not been as active as those at current plateau stage due to much less electrons
were available to be transferred The bigger peak amplitude in stage 4 than in stage 1
indicates that the redox active compounds were continuously being secreted and
accumulated in the biofilm even after stable power was generated at 7 kΩ The slightly
change of the peak locations at different stages was possible due to the solution chemistry
change during the current generation process or the slightly location change of reference
electrode during the measurement (Figure 37B) When the medium solution in the MFCs
was replaced by fresh medium at the end of the batch the oxidization and reduction
peaks were still presented in the CV (Figure 37B) but no peaks were observed when the
supernatant and fresh mediums were tested in a MFCs with no biofilm (Figure 37C)
These results suggest the compounds involved in the electron transfer were located not in
the supernatant (solution) but in the biofilm
41
Figure 37 Cyclic Voltammetry analysis of Citrobacter sp SX-1 biofilms at (A) current
increasing stage current plateau stage and decreasing stage (end of the batch) (B) end of
the batch and replaced with fresh medium and (C) controls
-40E-04
-30E-04
-20E-04
-10E-04
00E+00
10E-04
20E-04
30E-04
-060 -050 -040 -030 -020 -010 000 010 020
Curr
ent
(A)
Potential (V) vs AgAgCl
Current plateau stage
Current increasing stage
Current decreasing stage
-40E-04
-30E-04
-20E-04
-10E-04
00E+00
10E-04
20E-04
30E-04
-060 -050 -040 -030 -020 -010 000 010 020
Curr
ent
(A)
Potential (V) vs AgAgCl
Current decreasing stage
Biofilm with fresh medium
-40E-04
-35E-04
-30E-04
-25E-04
-20E-04
-15E-04
-10E-04
-50E-05
00E+00
-060 -050 -040 -030 -020 -010 000 010 020
Curr
ent(
A)
Potential (V) vs AgAgCl
Supernatant
Fresh medium
A
B
C
B C
42
Discussion
Citrobacter species belonging to Gammaproteobacteria Enterobacteriales
Enterobacteriaceae are facultative anaerobic bacteria (George 2005) They can grow with
oxygen as an electron acceptor but also with a variety of electron accepters such as Fe (III)
in the absence of oxygen (George 2005) Citrobacter species can be found in a wide
variety of habitats including in soil water and wastewater Several Citrobacter species
have been studied as important bioremediation bacterium for heavy metal removals
sulfate reduction phenol degradation and chlorophenol degradation (Macaskie et al 1995
Narde et al 2004 Qiu et al 2009) Citrobacter species have also been found on the
anodic biofilm of a diesel-degrading MFCs (Morris et al 2009) However there was no
study reported that Citrobacter species can transfer electrons to extracellular electron
acceptors This finding that Citrobacter strain SX-1 can generate electricity in MFCs
increased the diversity of power generating exoelectrogens and provided a new strain to
explore the mechanisms of extracellular electron transfer from bacteria to electrode
Among the isolated exoelectrogens gammaproteobacteria and deltaprotebactia appear to
be dominant among others (Parot et al 2009 Liu et al 2010) The isolated strain
Citrobacter sp SX-1 phylogenetically belongs to gammaproteobacteria which provides
another evidence to support this observation
Most exoelectrogens utilize a limited range of substrates for power generation For
example Shewanella oniedensis MR-1 can oxidize lactate under anaerobic condition but
cannot utilize acetate for electricity generation (Kim et al 2002) Geobacter
sulfurreducens can completely oxidize acetate for power generation but it cannot utilize
simple sugars (Bond and Lovley 2003 Caccavo et al 1994)) In comparison strain SX-1
43
not only can oxidize citrate for power generation but also utilize a wide range of
substrates for power generation including glycerol glucose lactose sucrose and acetate
Interestingly the higher current density generated by strain SX-1 from glycerol a main
by-product of biodiesel production process suggests that strain SX-1 may be potentially
used for harvesting energy from biodiesel wastes using MFCs However the current
density generated by strain SX-1 (205 mA m-2
) is lower than that (805 mA m-2
at 1 kΩ)
generated by the mixed culture from which the SX-1 was isolated indicating the
existence of other higher power generating bacteria andor complex ecology in the mixed
culture community
Understanding mechanisms of microbial extracellular electron transfer is critical for
enhancing the electron transfer rate from bacteria to electrode through metabolic or
genetic engineering of exoelectrogens Since the spent supernatant solution of MFCs run
with SX-1 showed no redox properties the redox compounds produced by SX-1 may
have been retained in the biofilm Alternatively SX-1 may utilize one or both of the
other two known extracellular electron transfer mechanisms For example it is possible
that strain SX-1 transfers electrons to the anode via cell-bound outer membrane proteins
The two distinct oxidation and reduction peaks observed at -100mv- -150 mV and -310--
350 mV of the SX-1 biofilm have a midpoint potential range from -205mV to -250mV
which is close to the redox potential of some c-type cytochromes (Field et al 2000 Meitl
et al 2009) which are well-known to play an important role in extracellular electron
trasnfer (Eggleston et al 2008 Field et al 2000 Meitl et al 2009) Field et al (2000)
reported that the c-type cytochromes CymA had the midpoint potentials of -229 mV
Meitl et al (2009) and Eggleston et al (2008) reported that the c- type cytochromes
44
OmcA in Shewanella oneidensis MR-1 had the midpoint potentials of -201 mV and -208
mV
While itrsquos possible that SX-1 may also have conductive appendages to enhance the
extracellular transfer electron similar to the nanowires discovered in some Geobacter
and Shewanella species (Reguera et al 2005 2006 Gorby et al 2006) further
investigation on the presence of these appendages on the cell surface of SX-1 is needed
Acknowledgements
The authors acknowledge support from the US National Science Foundation
(CBET 0955124) We also thank Teresa Sawyer for her helps with the SEM analysis and
Jeremy Chignell and Yanzhen Fan for their review of this manuscript
45
Chapter 4
Enhanced Performance and Mechanism Study of Microbial Electrolysis Cells
Using Fe Nanoparticles Decorated Anodes
Shoutao Xu Hong LiuYanzhen Fan Rebecca Schaller Jun Jiao Frank Chaplen
Published in
Applied Microbiology and biotechnology 93(2)871-880 (2012)
46
ABSTRACT
Anode properties are critical for performance of microbial electrolysis cells (MECs) In
the present study Fe nanoparticle modified graphite disks were used as anodes to
investigate the effects of nanoparticles on the performance of Shewanella oneidensis MR-
1 in MECs Results demonstrated that average current densities produced with Fe
nanoparticle decorated anodes were up to 56-fold higher than plain graphite anodes
Whole genome microarray analysis of the gene expression showed that genes encoding
biofilm formation were significantly up-regulated as response to nanoparticle decorated
anodes Increased expression of genes related to nanowires flavins and c-type
cytochromes indicate that enhanced mechanisms of electron transfer to the anode may
also have contributed to the observed increases in current density The majority of the
remaining differentially expressed genes were associated with electron transport and
anaerobic metabolism demonstrating a systemic response to increased power loads
Keywords Microbial electrochemical system microbial fuel cell microbial electrolysis
cell nanotechnology differential gene expression DNA microarray
47
Introduction
Microbial electrochemical systems (MESs) have been intensively studied since Lewis
achieved practical advances in this field (Logan 2007) however they attracted much
research attention in recent years due to their promising applications in renewable energy
generation bioremediation and wastewater treatment In a MES microorganisms
interact with electrodes via electrons catalyzing oxidation and reduction reactions at the
anode and the cathode The most-described type of MESs is microbial fuel cells (MFCs)
in which useful power is generated from electron donors typically biodegradable organic
materials (Logan et al 2006) Various novel MESs have recently been developed to
produce hydrogen (microbial electrolysis cells (MECs)) (Liu et al 2005 Logan et al
2008 Rozendal et al 2006 Schaller et al 2009) to power an autonomous sensor or
sensor network (Ringeisen et al 2006) to reduce bioremediation targets (Lovley 2006
Shea et al 2008) Sukkasem et al 2008) and to desalinate water (Cao et al 2009) The
key feature shared by these systems is the microbe-catalyzed electron transfer from
organic matter to electrodes (anodes) (Rabaey et al 2004) Enhancing the anodic current
output which highly depends on the performance of the electrodes is critical for the
successful application of all these processes (Logan et al 2007 Park and Zeilus 2002
2003)
Nanomaterials have received much attention from researchers in the context of
microbiology due to their unique physical electrical and chemical properties which
facilitate the study of interactions between bacteria and surfaces (Liu 2006) Previous
studies have demonstrated that electrodes decorated with different nanostructures such
as carbon nanotubes nanostructured polyanilinetitanium dioxide composite and titania
48
nanotubes have the potential to enhance power generation in MFCs (Morozan et al 2007
Qiao et al 2008 Quan et al 2005) Furthermore our recent study shows nanoparticle
(NP) decorated anodes greatly increased the electrochemical electron transfer rate in
MFCs (Fan et al 2011) However Fe NPs as an ideal candidate for decorating electrodes
because of respectively low price and high conductivity compared to other materials has
not been focused to study in MECs
S oneidensis MR-1 an important electrochemically active bacterial strain has been
exploited for the electricity generation in MFCs (Logan et al 2007 Logan 2009 Park
and Zeikus 2002) The availability of genome sequence for this strain makes it possible to
use transcriptome assays to globally measure the responses to different growth conditions
and environmental stresses (Murray et al 2001) S oneidensis MR-1 gene expression
response to heat shock cold shock Cr (VI) and U (VI) reduction chromate stress and
iron and acid tolerance have been studied in the past several years (Bencheikh-Latmani et
al 2005 Brown et al 2006 Gao et al2006 Gao et al 2004 Yang et al 2008) However
no studies have been focused on the Shewanella gene expression response to NPs in
MESs
The mechanism of increased electron transfer rate exhibited by nanoparticle decorated
electrodes is not well understood yet In the present study graphite disks decorated with
Fe NPs were used as anodes to explore the effects of nanostructures on current generation
in a multi-anode MECs DNA microarrays were utilized to investigate differences in the
global gene expression profile of S oneidensis MR-1 grown on plain versus Fe NPs
decorated anodes
Materials and methods
49
Bacterial cultures
S oneidensis MR-1 was purchased from American Type Culture collection (ATCC
700550) and stored as freezer stocks at -80degC Stock cells of S oneidensis MR-1 was
grown aerobically using a medium containing 30 gL Trypticase Soy Broth (BD 211825
Becton Dickinson Company Franklin lakes NJ) at 30degC Cells in early stationary
growth phase (optical density of 15-16 at 600nm (Spectrophotometer UV-1700
Shimadzu Columbia MD) were washed two times then injected into the chamber of the
MECs for current production Trypticase Soy Broth medium with 30 mM sodium lactate
as electron donor was used in MECs Phosphate buffer (100mM) was used to maintain
the solution pH 7 and solution conductivity at 15 mScm
Characterization of nanostructured anodes
Superfine isomolded graphite disks (16 cm x 05 cm GraphiteStorecom Inc) were
polished with ultrafine sand paper (2000 grit 3M Company) as the base for the NP
decorated and control anodes Fe NPs decorated anodes were fabricated by thermal
annealing of metal thin film coated graphite disks Thin films of Fe were firstly deposited
on the polished graphite disk by using sputter coating for 95 min and the samples then
were annealed in the Chemical vapor deposition (CVD) at 750degC for 20 min to form Fe
NPs Fe-NPs were produced with a diameter of 200plusmn100 nm and a density range
(distance between NPs) of 200plusmn100 nm with a coating time of 95 min SEM images of
Fe NPs decorated anode and control surfaces were shown in the Figure 41
50
Figure 41 SEM images of Fe NPs decorated anodes (A) Graphite disk control (B) Fe
Nanoparticle decorated
Multiple channel MECs construction and operation
A MECs with removable multiple anodes with each effective anode area 07 cm2
was
constructed and used to evaluate the effects of nanostructure on current density of
according to a previous report (Fan et al 2011) The cathode was made of wet-proof
(30) carbon cloth (type B E-TEK Division Inc USA) coated with PtC (20 E-TEK
Division Inc USA) following a previously reported procedure (Liu et al 2005) The
final platinum loading was 05 mgcm2 per projected cathode area The size of cathode
(150 cm2) was 25 times larger than the total effective area of the 8 graphite disk anodes
(56 cm2) to prevent cathode limitations on the performance of the MECs system All
MECs with Fe-NP decorated anodes and control anodes were tested for current
generation for 24 h with medium (without bacteria) before injecting the bacterial cells in
order to determine whether the Fe-NP decorated anodes demonstrate chemical current
generation compared to control anodes All the testing anodes were set up in the MECs
B
500 nm
A
500 nm
51
at once so that the S oneidensis MR-1 culture was applied to all of them simultaneously
Short electrode spacing (17 cm) was used in the MECs design to reduce the internal
resistance A voltage of 06 V was applied the MECs for current generation after the cells
of S oneidensis MR-1 were inoculated into MECs at early stationary phase A multimeter
with a data acquisition system (2700 Keithly USA) was used to monitor the current
change by measuring the voltage (every 300 seconds) drop through a resistor at 300 ohm
Fig 4 2 Effects of Fe-NP decorated anodes on the current density in MECs Control is
plain graphite disk anode Fe-NP is the anode with Fe-NP decoration
Microarray analysis
Biofilms for whole gene microarray analysis were aseptically removed from the plain
and Fe-NP decorated anodes of MECs after 110 hours further incubation at 30oC when
current density obviously started to decease Total RNA was extracted using Trizol
(Ambion inc AustinTX) following the manufacturerrsquos instructions Integrity of the
52
RNA samples was confirmed using the Agilent 2100 Bioanalyzer (Agilent Technology
Inc Palo Alto CA) The same amount of RNA (200 ng) from each sample was used for
further analysis after amplification by using the MessageAmpTM
II-Bacteria Prokaryotic
RNA Amplification kit (Ambion AustinTX) according to the manufacturerrsquos
instructions All of the 4295 genes (resource from NCBI) represented on the S oneidensis
MR-1 whole-gene microarrays were ordered from Roche NimbleGen Inc (Madison WI)
Biological triplicates of biofilms on the Fe-NP decorated anodes and control were
analyzed respectively cDNA synthesis labeling and hybridization were carried out by
the CGRB Core Laboratories at Oregon State University DNASTAR ArrayStarTM
3
software was used to identify genes that were up- or down-regulated more than 2-fold
when grown on the nanoparticle modified anodes using the unpaired two sample t-test
with a cutoff p-value of 005 The complete microarray data set generated in this study is
deposited for public access in the Gene Expression Omnibus
(httpwwwncbinlmnihgovgeo) under accession number GSE31535
Results
Enhancement of current generation by using NP decorated anodes in MECs
The current density generated by Fe-NP decorated anode MECs with S onidensis MR-1
increased to approximately 43 microAcm2 20 h following inoculation and then slowly
decreased Figure 42 shows that the maximum current density achieved by NP
decorated anodes was 83 times higher than that (51 microAcm2) generated by the control
(plain graphite disk) The average current density improvement of 110 hours was 59
times of that generated by the control The current densities generated in MECs in the
53
absence of bacteria were negligible (03 microAcm2) which demonstrated that the current
enhancement observed with the Fe-NP decorated anodes was biologically-derived
Global transcriptome analysis
Whole-genome DNA microarrays were used to attain a comprehensive general
overview
of the transcriptional response to Fe-NP decorated anodes by S oneidensis MR-1 in MEC
Of a total 4295 genes assayed 392 (188 induced and 204 repressed) exhibited significant
(P lt 005) differential expression at a 2-fold (log (expression ratio 1) level in 3
replicates in response to Fe-NPs These
total gene numbers present 9 of the 4295 open
reading frames (ORFs) presented on the array
Fig 43 Differentially expressed genes grouped by functional classification according to
the TIGR S oneidensis genome database (wwwtigrorg) Columns 1 amino acid
biosynthesis 2 biosynthesis of cofactors prosthetic groups and carriers 3 cell
envelope 4 cellular processes 5 central intermediary metabolism 6 DNA metabolism
7 energy metabolism 8 fatty acid and phospholipid metabolism 9 other categories 10
protein fate 11 protein synthesis 12 purines pyrimidines nucleosides and nucleotides
13 regulatory functions 14 signal transduction 15 transcription 16 transport and
binding proteins 17 unknown function 18 hypothetical proteins
Up-Regulated
-75
-50
-25
0
25
50
75
100
125
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18
Down-Regulated
Num
ber
of
Gen
es
54
Figure 43 summarizes the overall genomic response of S oneidensis MR-1 to Fe-NP
decorated anodes by grouping the differentially expressed genes into their functional role
categories based on The Institute for Genomic Researchrsquos annotation (Version 8) of the
MR-1 genome sequence The wide distribution of putative functional roles attributed to
the differentially expressed genes indicated the extent of the molecular response of
Soniedenis MR-1 to the NP decorated anodes Among genes with defined function a
large number of up-regulated genes were associated with cellular processes (group 4)
energy metabolism (group 7) other categories (group 9) and hypothetical proteins (group
18) While those genes encoding proteins involved in amino acid biosynthesis (group 1)
cellular processes (group 4) substrate transport (group 16) and hypothetical proteins
(group 18) were among the most down-regulated genes Two groups of genes that were
the most noticeable among the functional gene groups genes related to energy
metabolism (group 7) and genes related to amino acid biosynthesis (group 1) The ratios
of up-regulated genes to down-regulated genes in these functional groupings were much
higher than other functional gene groupings there were 21 up-regulated genes and 7
down-regulated genes in energy metabolism gene group and 2 up-regulated and 22 down
regulated genes in the amino acid biosynthesis group
Genes related to biofilm formation
In order to determine whether there was a correlation between the observed biofilm
enhancements by NP decorated anodes and the genes related to biofilm formation the
significant modified genes related to biofilm formation were examined and summarized
in the Table 41 These included SO_3223 and SO_3228 genes encoding the flagellum
proteins (Thormann et al 2004) which has critical impacts on initial attachment to the
55
surface as swimming motility functions were up-regulated over 10 and 09 fold
respectively The gene clusters SO_4178-SO_4180 which are thought to be essential for
formation of a three-dimensional biofilm were up-regulated from 12 to 13 fold
(Thormann et al 2004) In addition the genes SO_4105 encoding IV pilus which is
known to represent a critical factor for biofilm formation on a biotic surfaces (Thormann
et al 2004) were also induced 08 fold (Table 41) The genes related to flagellum
motility (SO_4286 SO_1529 and SO_1530) were also induced slightly (Thormann et al
2004)
Table 41 Expression levels of genes related to biofilm formation
Locus Tag Gene product descriptions Fold change
SO_3228 flagellar basal-body MS-ring and collar protein FliF 10
SO_3223 flagellar hook-length control protein FliK 09
SO_4103 MshA minor pilin protein MshD 05
SO_4105 MSHA major pilin protein MshA 08
SO_4178 expressed protein of unknown function MxdC 12
SO_4179 inner membrane family 2 glycosyltransferase MxdB 13
SO_4180 diguanylate cyclase-like protein MxdA 10
a fold changes were presented after log2
b positive number represented up-regulated and negative represented down-regulated
Genes related to energy metabolism
A closer consideration of energy metabolism genes was undertaken as anaerobic
metabolism for electron generation and electron transport functions may play critical
roles in enhanced current density generation in MECs Genes associated with energy
56
metabolism and with significantly modified gene expression levels are summarized in the
Table 42 Several of the up-regulated genes were related to formate dehydrogenase
proteins Most notably four genes related to formate dehydrogenase were up-regulated
more than 2-fold SO_0102 SO_0103 SO_0104 and SO_0107 The other genes
expression related to formate dehydrogenase proteins SO_4513 SO_4514 SO_4515 also
significantly increased Another interesting gene is that encoding the cytochrome c
oxidase protein gene SO_4609 which was up-regulated 11 fold while the gene
SO_0479 related periplasmic octaheme cytochrome c MccA was down-regulated
However SO_1519 the gene encoding lactate dehydrogenase was down-regulated 12
fold despite lactate being the primary carbon source in the media
Table 4 2 Genes related to anaerobic growth and electron transfer with significantly change expression level
Locus Tag Gene product descriptions Fold
change
SO_0101 nitrate-inducible formate dehydrogenase molybdopterin-binding
subunit FdnG
18
SO_0102 formate dehydrogenase nitrate-inducible iron-sulfur subunit 21
SO_0103 formate dehydrogenase nitrate-inducible cytochrome b556 subunit 24
SO_0104 formate dehydrogenase accessory protein FdhE 22
SO_0107 formate dehydrogenase accessory protein fdhD 24
SO_0397 quinolfumarate reductase menaquinol-oxidizing subunit FrdC_2 12
SO_0452 thioredoxin 2 15
SO_0903 Na-translocating NADH-quinone reductase subunit B NqrB_1 12
SO_1012 NADH-ubiquinone oxidoreductase subunit K NuoK 12
SO_1013 NADH-ubiquinone oxidoreductase subunit J NuoJ 12
SO_1363 hydroxylamine reductase 13
57
SO_2417 ferredoxin cofactor maintenance protein YfaE 10
SO_3922 formate dehydrogenase cytochrome b Fdh -12
SO_4513 formate dehydrogenase molybdopterin-binding subunit _2 15
SO_4514 formate dehydrogenase FeS subunit FdhB_2 12
SO_4515 formate dehydrogenase cytochrome b subunit FdhC_2 14
SO_4609 aa3 type cytochrome c oxidase subunit III CoxC 11
SO_0479 periplasmic octaheme cytochrome c MccA -12
SO_0847 periplasmic nitrate reductase ferredoxin component NapG -15
SO_1519 L-lactate dehydrogenase iron-sulfur cluster-binding protein LldF -10
SO_1251 ferredoxin 4Fe-4S -11
SO_37411 hypothetical inner membrane protein -11
a fold changes were presented after log2
b positive number represented up-regulated and negative represented down-regulated
Flavin and cytochrome related genes
Flavins can be secreted by Shewanella species as electron shuttle to facilitate
extracellular electron transfer (Cansterin et al 2008) C-type cytochromes play the
important roles on the process of extracellular electron transfer (Shi et al 2007) The
genes related to flavins synthesis and the genes encoding the cytochromes electron
transport proteins were shown in the table 43 the genes SO_1414 and SO_3058 related
to flavins synthesis was up-regulated to 03 and 01 folds respectively The genes related
to s C-type cytochromes SO_0135 SO_4105 SO_0169 SO_1778 are increased to 10
08 05 04 01 folds respectively However most of genes have shown no significant
changes (less one-fold change) as response to nano particle decorated anode in MECs
58
Table 43 Expression level of flavin and cytochrome genes
Locus Tag Gene product descriptions Fold
change
SO_1414 flavocytochrome c flavin subunit putative 03
SO_3468 riboflavin synthase subunit alpha -02
SO_3058 flavocytochrome c flavin subunit 01
SO_4105 MSHA pilin protein MshA 08
SO_0169 general secretion pathway protein GspG 05
SO_1778 outer membrane decaheme cytochrome c lipoprotein MtrC 04
SO_1779 outer membrane decaheme cytochrome c OmcA 01
SO_0135 lipoprotein of unknown function DUF333 10
SO_0136 conserved hypothetical inner membrane protein 01
a fold changes were presented after log2
b positive number represented up-regulated and negative represented down-regulated
Other genes with significantly modified expression levels
These genes over 15 fold change and possibly related to current enhancements were
shown in Table 44 SO_3104 (expressed inner membrane protein) was over expressed
and the gene SO_2194 which encodes outer membrane protein (OmpA family protein)
was repressed as response to nanoparticle decorated anode however it is unclear the
relationships of this modified membrane protein genes with the current enhancement
Another interesting phenomenon was possible co-regulation of several gene clusters
indicating possible operon associations This includes three sets of genes that were
induced more than 15 fold SO_0108SO_0109 and SO_1042SO_1043SO_1044 The
59
consistency of expression of these genes under the NP conditions provides basic evidence
to support operon structure However the correlation of these significant changed gene
with current density enhance are unclear which showed there are unknown multiple and
complex responses of S oneidensis MR-1 to nanoparticle anode of MECs
Table 44 Other genes with significantly changed expression levels
Locus Tag Gene product descriptions Fold change
SO_0101 nitrate-inducible formate dehydrogenase molybdopterin-binding
subunit FdnG
18
SO_0108 integral membrane protein of unknown function DUF39 YedE 17
SO_0109 SirA family protein YedF 17
SO_0275 N-acetyl-gamma-glutamyl-phosphate reductase -31
SO_0277 ornithine carbamoyltransferase -21
SO_0279 argininosuccinate lyase -24
SO_0404 zinc dependent metalloprotease domain lipoprotein 21
SO_0956 alkyl hydroperoxide reductase F subunit -20
SO_1042 amino acid ABC transporter ATP-binding protein -18
SO_1043 amino acid ABC transporter permease protein -27
SO_1044 amino acid ABC transporter periplasmic amino acid-binding protein -27
SO_1072 chitin-binding protein putative 21
SO_1405 transglutaminase family protein 20
SO_1822 TonB-dependent receptor putative -22
SO_2069
1-(5-phosphoribosyl)-5-[(5-phosphoribosylamino)
methylideneamino] imidazole-4-carboxamide isomerase
-19
SO_2070 amidotransferase HisH -26
SO_2071 imidazole glycerol-phosphate dehydratasehistidinol phosphatase -27
60
SO_2072 histidinol-phosphate aminotransferase -28
SO_2073 histidinol dehydrogenase -30
SO_2194 OmpA family protein -36
SO_2195 inter-alpha-trypsin inhibitor domain protein -27
SO_2767 asparagine synthetase B -26
SO_2945 prophage LambdaSo tail fiber protein 20
SO_2963 prophage LambdaSo major capsid protein HK97 family 20
SO_2987 prophage LambdaSo hypothetical protein 19
SO_2988 prophage LambdaSo expressed protein of unknown function 18
SO_3104 expressed inner membrane protein 131
SO_3408 conserved hypothetical inner membrane protein -18
SO_3585 azoreductase putative -18
SO_3586 glyoxalase family protein -17
SO_3687 curli production assemblytransport component CsgE putative -18
SO_38191 hypothetical ammonia permease -19
SO_4014 AcrBAcrDAcrF family protein 18
SO_40151 type I secretion system membrane fusion protein RND family 18
SO_4054 510-methylenetetrahydrofolate reductase 18
SO_4245 N-acetylglutamate synthase -22
SO_45251 hypothetical transcriptional regulator LysR family -17
SO_4527 integral membrane domain protein -19
SO_4705 transcriptional regulator putative -19
a fold changes were presented after log2
b positive number represented up-regulated and negative represented down-regulated
Discussion
61
The addition of NP to anodes significantly impacts current densities with the elemental
composition being a critical factor The chemical composition of NPs significantly affects
the current enhancement in MECs (Fan et al 2011) because the chemical composition of
NP properties is a critical factor in determining the conductivity which significantly
affects the efficiency of electron transfer from bacteria to electrode Fe is an ideal
candidate for NP formation for current enhancement in MESs because of high
conductivity and low price compared to gold and other conductive materials Gold NPs
has also been studies for enhancement of electricity conductivity (Bao et al 2008)
However the disadvantages of the high price of gold limit their practical application in
MECs (Fan et al 2011) The different size and density of the same composition of NP
also has essential effects on the current density enhancement (Fan et al 2011) In this
study the results showed the current density enhancement with Fe-NP size range of
200plusmn100 nm and density range of 200plusmn100 nm is considerable enhancement consequence
There are a number of reports of studies of current density enhancement in MFCs
by nanostructured anodes (Fan et al 2011 Qiao et al 2008 Sharma et al 2008 Tsai et al
2009) Based on bacterial behavior on nanostructured anode surface Qiao et al (2008)
believes that Escherichia coli cells on the nanostructured electrode surface produce hair-
like structures similar to pili that could facilitate the electron transfer between the cells
and electrode The production of hair-like structures is believed to be stimulated by
nanostructures and could play the key role on current density enhancement in pili-
producing bacteria in MFCs (Qiao et al 2008) S oneidensis can produce electrically
conductive bacterial nanowires which have similar structure to the hair-like structures in
Ecoli (Gorby et al 2006 Reguera et al 2005) A number of possible genes related
62
nanowires of S oneidensis have been identified including mshA (SO_4105) and gspG
(SO_0169) (Konstantinidis et al 2009) They are up-regulated 08 and 05 fold in the
current study respectively even though they are not significant induced This result
supported the increased nanowire genes had contributions to enhance current density in
MFCs by NPs decorated anodes
Based on the previous research on electron transfer mechanisms for S oneidensis it is
known that self-produced mediators (flavins) and multihaem c-type cytochromes (c-Cyts)
may play critical roles in the electron transfer of S oneidensis to extracellular insoluble
electrode (Canstein et al 2008 Myers and Myers 1992 Scott and Nealson 1994 Shi et al
2007) The flavins secreted by S oneidensis can facilitate to transfer electrons from
bacteria to solid anode electrodes by acting as extracellular electron shuttles (Canstein et
al 2008) In this study the genes encoding flavins (SO_1414 SO_3468 and SO_3058)
had no significant increase (less than 1 fold change) as a response to nanoparticle
decorated anode in MECs which suggests that increased flavin gene expression does not
significantly contribute to the current density enhancements in nano-particle decorated
anode The genes encoding cytochromes electron transport proteins (SO_4104 SO_0417
SO_177879 SO_013536) had no significant increase (less than 1-fold change)
However those genes were up-regulated slightly These results suggest the increased
expression flavin and c type cytochromes genes had partial contributions even not
significantly to enhance current density in MFCs by NPs decorated anodes
The thickness of bacterial biofilm on the anode can affect the power generation in
MFCs (Biffinger et al 2007 Heydorn et al 2000) since increased biofilm thickness on
the anode represents more active bacteria involving in the process of electron generation
63
and transfer in MFCs Biofilm formation related genes included flagellar genes IV pilus
genes and other genes The flagellar related genes SO_3228 SO_3229 (fliB and fliK) and
IV pilus genes SO_4105 (mshA) were induced from 19 to 09 fold the genes (SO_4178
-4180) related to formation of a three-dimensional biofilm were up-regulated from 10 to
13 folds even though the genes related bacterial motility protein (SO_ 4286 SO_4287
SO_1529 and SO_1530) are induced slightly (less than 0 5 fold) However the motility
genes most probably play a role for initial biofilm attachment on the anode surfaces
(Thormann et al 2004 Thormann et al 2006) with the rest of biofilm genes being more
essential for biofilm maintenance in mature cultures as were presented at harvest in this
study This enhancement of biofilm density under conditions of increase current density
has been observed for gold NP decorated anodes using confocal light microscopy (data
not shown) but has not yet been confirmed for Fe NP decorated anodes In any event an
increase in the expression level of genes related biofilm formation therefore also
supported the possibility of enhanced biofilm formation on NPs decorated anodes thus
facilitating electron transfer from bacteria to anodes
Beliaev et al described a model of the Fe reduction pathway in S oneidensis MR-1 using
solid substrates as the electron acceptor (Beliaev et al 2001) This model proposes that
electrons are generated and released in cytoplasm then transferred to the quinone pool
eg menaquinone in the cytoplasmic membrane The reduced menaquinone in turn
reduces CymA and from there electrons are then transferred to a periplasmic shuttle (eg
CctA) and then to MtrA a periplasmic soluble decaheme cytochrome c located in
associated with the outer membrane via interaction with MtrB Finally surface displayed
outer membrane proteins decaheme cytochromes (OmcA MtrC or MtrF) can transfer
64
electron to Fe or other solid substrates (Beliaev et al 2001)) Bencheikh-Latmani et al
demonstrated that the same electron transport pathway may be used for more than one
electron acceptor through studies of the response of S oneidensis MR-1 to U(VI) and
Cr(VI) under anaerobic conditions particularly focused on the critical genes
(SO_1777mtrA SO_1776mtrB SO_1778mtrC) (Bencheikh-Latmani et al 2005)
However our studies showed that these genes had no significant expression changes in
response to nanostructured anodes Contrastingly one of expressed inner membrane
protein genes SO_3104 and one of cytochromes genes SO_4609 encoding cytochrome c
oxidase subunit III demonstrated a 13-fold and 11-fold up-regulation respectively in this
study while the gene SO_2194 which encodes outer membrane protein (OmpA family
protein) was down regulated 36 fold Our results therefore suggest that S oneidensis
MR-1 does not utilize the same set of core critical genes (SO_1777 mtrA SO_1776
mtrB SO_1778 mtrC) products to transfer electrons in all instances This is the similar
result with the study of Bretschger et al (2007) which indicated that the electron
transport system in S oneidensis MR-1 is complex with several different proteins able to
participate in electron transfer to the anode of MECs
It should also be mentioned that the gene (SO_1519) encoding lactate
dehydrogenase repressed (-12 fold) here suggesting that lactate was depleted at the time
of cell harvest from the MECs and that other pathways of anaerobiosis had been activated
including those associated with amino acid uptake and consumption the growth media
was a complex mixture including yeast extract In particular the catabolic pathway for
histidine of S oneidensis MR-1 a pathway that may formate as an intermediate is up-
regulated with the genes for histidine ammonia lyase SO_0098 SO_3057 and SO_4374
65
up-regulated 13 11 and 11-fold respectively The possible production of formate
during histidine degradation may in turn explain the high levels of formate
dehydrogenates as evidenced by upregulation of SO_0102 SO_0103 SO_0104 and
SO_0107 annotated as formate dehydrogenase proteins These genes had the highest
levels of up-regulated expression as response to Fe NP decorated anodes (greater than 2-
fold)
Acknowledgements
This research was partially supported by the US National Science Foundation CBET
0828544 and the funds from ONAMIDOD (ARL-DOD Cooperative Agreement
W911NF-07-2-0083) We thank Barbara Gvakharia and Caprice Rosato for helpful
suggestions We also thank Margaret Romine of Pacific Northwest Laboratories for
valuable comments on manuscripts We also thank anonymous reviewers for significant
suggested improvements imparted as part of prior review of this manuscript
66
Chapter 5
Global Transcriptome Analysis of the Response of Shewanella oneidensis MR-1 to
Carbon Nanotube Decorated Anodes in Microbial Electrochemical Systems
Shoutao Xu Yanzhen Fan Rebecca Schaller Frank Chaplen Jun Jiao Hong Liu
67
Abstract
Shewanella oneidensis MR-1 is an important model microorganism for metabolic studies
on the effects of different environmental factors because of its diverse respiratory
capabilities Carbon nanotube (CNT) modified graphite disks were used as anodes to
investigate the effects of nanomatrials on the performance of S oneidensis MR-1 in
microbial electrolysis cells (MECs) The current densities produced with CNT decorated
anodes were on average 56-fold higher than plain graphite anodes Whole genome
microarray analysis of gene expression showed that up-regulation of cytochromes c genes
associated with extracellular electron transfer are strongly correlated to current increases
in CNT decorated anodes Genes involved in flavin biosynthesis also contribute to
current increase in CNT decorated anode MECs
Keywords Microbial electrochemical system microbial fuel cell microbial
electrolysis cell carbon nanotube gene expression DNA microarray
68
Introduction
Shewanella oneidensis MR-1 is an important model microorganism for metabolic
studies of the effects of different environmental factors because of its diverse respiratory
capabilities It has been used for transcriptome analysis to investigate the responses to
different growth conditions and environmental stresses (Murray et al 2001) Gene
expression patterns under different conditions such as heat shock cold shock Cr (VI) U
(VI) reduction chromate stress iron and acid tolerance have been studied previously
(Bencheikh-Latmani et al 2005 Brown et al 2006 Gao et al2006 Gao et al 2004
Yang et al 2008) More recently it has been exploited as a model species for power
generation in microbial electrochemical systems (MESs)which have potential
applications in renewable energy generation bioremediation and wastewater treatment
(Logan et al 2007 Logan 2009 Park and Zeikus 2002 Fan et al 2011 Xu et al 2012)
In a MES electrochemically active microorganisms oxidize organic matter in the an
ode chamber to release electrons Electrons are then transferred to the anode electrode thr
ough various mechanisms (Logan et al 2006 Lovley 2006 Rabaey et al 2003) and fina
lly travel to the cathode electrode and combine with the terminal electron acceptor The
key feature of MESs is the microbe-catalyzed electron transfer from the organic matter to
the anode (Rabaey et al 2004) Enhancing the current output which highly depends on
the performance of the anode electrode is critical for the successful application of MESs
(Logan et al 2007 Park and Zeilus 2002 2003) Several nanomaterials including TiO2
gold (Au) and palladium (Pd) nanoparticles have been utilized to decorate electrodes to
enhance power density in MESs (Qiao et al 2008 Quan et al 2005 Fan et al 2011) The
possible mechanisms for increased current densities have been studied by using Fe
69
nanoparticle-decorated anodes (Xu et al 2012)
Carbon nanotubes (CNT) are cylindrically shaped nanomaterials with extremely
high surface area excellent electrical conductivity and chemical inertness (He et al
2005a Serp et al 2003) These unique properties make CNT a promising electrode
material (Liang et al 2008) The biocompatibility of microorganisms and carbon
nanostructures has been evaluated (Morozan et al 2007) The effects of CNT on anodic
biofilms have been tested in MFCs (Liang et al 2011) However no studies have been re
ported on global transcriptome analysis for the response of S oneidensis MR-1 to CNT
decorated anode in MECs
In the present study the graphite disks decorated with CNT were used as anodes to
investigate the effects of nanomaterials on current generation in multi-anode MECs
DNA microarrays were used to analyze differences in the global gene expression profile
of S oneidensis MR-1 grown on plain versus CNT decorated anodes
Materials and methods
Bacterial cultures S oneidensis MR-1 was purchased from American Type Culture
collection (ATCC 700550) and stored as freezer stocks at -80degC Stock S oneidensis
MR-1 cells were grown aerobically using M1 medium at 30degC Cells in early stationary
growth phase (optical density of 15-16 at 600nm (Spectrophotometer UV-1700
Shimadzu Columbia MD) were injected into the chamber of the MECs for current
production Sodium lactate (final concentration 30mM) was added as the additional
electron donor
Fabrication and characterization of nanostructured anodes Superfine isomolded
graphite disks (16 cm x 05 cm GraphiteStorecom Inc) were polished with ultrafine
70
sand paper (2000 grit 3M Company) as the base for the CNT decorated and control
anodes Aligned multi-walled carbon nanotube samples were synthesized via plasma
enhanced chemical vapor deposition (PECVD) Samples superfine isomolded graphite
pillars with a 16 cm diameter and 05 cm thickness were sputtered with a thin film of Ni
in a CRC 300 magnetron sputter coater for 1 to 2 min They were then placed in the
PECVD reactor where the Ni was annealed into small catalyst islands C2H2 was bled
into the chamber for growth and an NH3 plasma was used to vertically align the growth
of the multi-walled carbon nanotubes (MWCNTs) The temperature and time were varied
between 725degC and 775oC and 15 to 60 min to control the length of the tubes and density
of samples Samples were then analyzed in a FEI Sirion field emission scanning electron
microscope (FESEM) SEM images of CNT decorated anode and control surfaces were
shown in the Figure 51
Figure 51 SEM images of CNT decorated anodes (A) Graphite disk control (B) CNT
decorated anode
Multiple channel MECs construction and operation A MECs with multiple
removable anodes each with an effective anode area of 07 cm2
was constructed and used
to evaluate the effects of nanostructures on the current density of according to a previous
B A
71
study (Fan et al 2011) The cathode was made of wet-proof (30) carbon cloth (type B
E-TEK Division Inc USA) coated with PtC (20 E-TEK Division Inc USA)
following a previously reported procedure (Liu et al 2005) The final platinum loading
was 05 mgcm2 per projected cathode area The size of the cathode (150 cm
2) was 25
times larger than the total effective area of the 8 graphite disk anodes (56 cm2) to prevent
cathode limitations on the performance of the MECs system All MECs with CNT
decorated anodes and control anodes were tested for current generation for 24 h with
sterile a medium (without bacteria) before injecting the bacterial cells in order to
determine whether the CNT decorated anodes demonstrate chemical current generation
compared to control anodes All the testing anodes were set up in the MECs
simultaneously so that the S oneidensis MR-1 culture was applied to all of them
simultaneously Short electrode spacing (17 cm) was used in the MECs design to reduce
the internal resistance A voltage of 06 V was applied the MECs for current generation
after the cells of S oneidensis MR-1 were inoculated into the MECs in the early
stationary phase A multimeter with a data acquisition system (2700 Keithly USA) was
used to monitor the current change by measuring the voltage drop through a resistor
Microarray analysis Biofilms for whole gene microarray analysis were aseptically
removed from the plain and CNT decorated anodes of MECs after 80 hours further
incubation at 30oC when current density obviously reached to a stable phase Total RNA
was extracted using Trizol (Ambion inc AustinTX) following the manufacturerrsquos
instructions Integrity of the RNA samples was confirmed using the Agilent 2100
Bioanalyzer (Agilent Technology Inc Palo Alto CA) Total RNA (200 ng) from each
sample was amplified using the MessageAmpTM
II-Bacteria Prokaryotic RNA
72
Amplification kit (Ambion AustinTX) according to the manufacturerrsquos instructions The
S oneidensis MR-1 whole-gene microarrays representing all of the 4295 genes (resource
from NCBI) were ordered from Roche NimbleGen Inc (Madison WI) Biological
triplicates of biofilms on the CNT decorated anodes and control were analyzed cDNA
synthesis labeling and hybridization were carried out by the CGRB Core Laboratories at
Oregon State University DNASTAR ArrayStarTM
3 software was used to identify genes
that were up- or down-regulated more than 2-fold when grown on the nanoparticle
modified anodes using the unpaired two sample t-test with a cutoff p-value of 005 with
bonferroni correction
Results and discussion
Enhancement of current generation using CNT decorated anodes in MECs The
current density generated by CNT decorated anode MECs with S onidensis MR-1
increased to 36 microAcm2 10 hours post-inoculation and reached a maximum of 40 microAcm
2
at 29 hours inoculation then gradually decreased The current density stabilized at
30microAcm2 after 78 h operation Figure 52 shows that the current density curve generated
by CNT decorated anode in MECs The average current density generated by CNT
decorated anodes was 56 times that of the control The current density generated in the
MECs without bacteria was negligible (data not shown) This result demonstrates that the
current enhancement observed with the CNT decorated anodes was biologically-derived
A
73
Figure 52 Effects of CNT decorated anodes on the current density in MECs (3 replicates)
Control is plain graphite disk anode CNT is the anode with CNT decoration (Error bars
represent standard deviation)
It has been reported that nanostructured decorated anodes have significant impacts on
current densities (Fan et al 2011 Qiao et al 2006 Sharma 2008 Tsai et al 2009) The
CNT decorated anodes had significantly enhanced current densities in MECs in this
study This result is consistent with previous studies Interestingly the current density
curve generated by CNT decorated anode MECs is quite different from the current
density curve generated by Au NP and Pd NP decorated anodes using same bacterial
species (Fan et al 2011) In the Au and Pd NP decorated anode MECs the current
density curve was not significantly increased in the beginning compared with the control
but gradually increased after the inoculation the current density reached the maximum
current density more than 50 hours after the inoculation However the current density
generated by CNT decorated anode MECs started to increase significantly immediately
after the inoculation It reached a maximum current density 29 hours post-inoculation
-1
6
13
20
27
34
41
48
0 10 20 30 40 50 60 70 80
Curr
ent
den
sity
(micro
Ac
m2)
Time (Hours)
Control
CNT
74
These results indicate that there might be different current density enhancement
mechanisms between CNT with other metal NP decorated anodes in MECs
Global transcriptome analysis Whole-genome DNA microarrays were used to attain a
comprehensive general overview of the transcriptional response of S oneidensis
MR-1 of
S oneidensis MR-1 to CNT decorated anodes in MECs In a total of 4295 genes analyzed
457 genes (250 genes up-regulated and 207 genes down-regulated) exhibited significantly
(P lt 005) changed expression at a 2-fold level in at least 3 replicates in the response to
CNT The total number of the regulated genes present 11 of the 4295 open reading
frames (ORFs) presented on the microarray The total number of genes at significant
expression levels in response to CNT decorated anodes is close to that to Fe NP decorated
anodes in MECs (392 9) ( Xu et al 2012)
Fig 53 Differentially expressed genes grouped by functional classification according to
the TIGR S oneidensis genome database (wwwtigrorg) Columns 1 amino acid
biosynthesis 2 biosynthesis of cofactors prosthetic groups and carriers 3 cell
envelope 4 cellular processes 5 central intermediary metabolism 6 DNA metabolism
7 energy metabolism 8 fatty acid and phospholipid metabolism 9 other categories 10
protein fate 11 protein synthesis 12 purines pyrimidines nucleosides and nucleotides
-80
-60
-40
-20
0
20
40
60
80
100
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18
Nu
mb
er
of
Ge
ne
s
Up-regulated
Down-regulated
75
13 regulatory functions 14 signal transduction 15 transcription 16 transport and
binding proteins 17 unknown function 18 hypothetical proteins
The overall transcript genomic response of S oneidensis MR-1 to CNT decorated
anodes was summarized in Figure 53 by grouping the differentially expressed genes into
their functional role categories based on The Institute for Genomic Researchrsquos annotation
(Version 8) of the MR-1 genome sequence The wide distribution of putative functional
roles attributed to the differentially expressed genes indicated the extent of the molecular
response of Soniedenis MR-1 to the CNT decorated anodes A large number of up-
regulated genes were presented in genes associated with biosynthesis of cofactors
prosthetic groups and carriers (group 2) cell envelope (group 3) protein fate (group 10)
protein synthesis (group 11) and purines pyrimidines nucleosides and nucleotides
(group 12) in this study The number of up-regulated genes was two times more than that
of down-regulated genes in each functional group Contrastingly in the genomic response
S oneidensis MR-1 to Fe NP decorated anodes (chapter 4) most of up-regulated genes
were associated with cellular processes (group 4) energy metabolism (group 7) other
categories (group 9) and hypothetical proteins (group 18) The down-regulated genes
presented in the amino acid biosynthesis (group 1) cellular processes (group 4) substrate
transport (group 16) and hypothetical proteins (group 18) While those genes encoding
proteins involved in amino acid biosynthesis (group 1) central intermediary metabolism
( group 5) DNA metabolism( group 6) fatty acid and phospholipid metabolism (group
8) and other categories (group 9) were among the most down-regulated genes in this
study However the two groups of genes that were the most noticeable between the
genomic response S oneidensis MR-1 to CNT and Fe NP decorated anodes which they
are the genes related to energy metabolism (group 7) and hypothetical proteins (group
76
18) These results indicated there are different and common genomic responses to S
oneidensis MR-1 on CNT and Fe NP decorated anodes in MECs
Electron transfer related genes Based on previous electron transfer mechanism
studies on Shewanella a serial of group proteins collectively described as the Mtr
pathway play the role to transfer electron rom the inner bacterial body to the outer
membrane (Beliaev et al 2001 Shi et al 2007 Brutinel and Gralnick 2012) In the Mtr
pathway firstly electrons generated by bacteria received by CymA a tetraheme c-type
cytochrome anchored in the inner membrane then electrons were transferred to MtrA a
periplasmic decaheme c-type cytochrome After that electrons were transferred from
MtrA to MtrC an outer-membrane decaheme c-type cytochrome is facilitated by MtrB a
non-heme containing integral outer-membrane proteinOuter-membrane decaheme c-type
cytochrome MtrC and OmcA are in change of electron transfer to exreacellular electron
acceptor (Shi et al 2007 Brutinel and Gralnick 2012 ) These genes encoding electron
transport proteins are shown in the table 51 The critical genes SO_1777 mtrA SO_1776
mtrB SO_1778 mtrC are unregulated 41 to 80 fold in response to CNT decorated
anodes other cytochrome c genes related to electron transfer in MtrA pathway also are
significantly increased in response to CNT decorated anodes These genes encoding outer
membrane proteins have different level increases supporting that the increase of
cytochrome c gene expression contributes to the current enhancements of MECs as
response to CNT decorated anodes
77
Table 51 Expression level of cytochrome c as response to CNT decorated anodes
Locus Tag Gene product descriptions Fold change CNT
SO_0165 general secretion pathway protein GspC 31
SO_0167 general secretion pathway protein GspE 45
SO_1776 outer membrane protein precursor MtrB 41
SO_1777 outer membrane decaheme cytochrome c
lipoprotein MtrA
80
SO_1778 outer membrane decaheme cytochrome c
lipoprotein MtrC
43
SO_1779 outer membrane decaheme cytochrome c
OmcA
63
SO_0135 lipoprotein of unknown function DUF333 22
SO_0136 conserved hypothetical inner membrane protein 24
Flavin related genes It is known that self-produced mediators play critical roles in the
electron transfer of S oneidensis to extracellular insoluble electrode (Canstein von et al 2
008 Myers et al 1992 Scott et al 1994 Shi et al 2007) It has been proved that the
flavins secreted by S oneidensis can facilitate electron transfer from bacteria to solid
anode electrodes by acting as extracellular electron shuttles (Cansterin von et al 2008) T
wo critical genes (SO_1414 and SO_3468) related to flavin synthesis were shown increa
sed significantly (more than 2 folds) increase in this study (Table 52) This result support
s the hypothesis that flavin production increases as the response to CNT decorated anodes
in MECs have significant contribution to the current density enhancements
78
Table 52 Expression levels of genes related to flavin synthesis as response to CNT
decorated anode
Locus Tag Gene product descriptions Fold change CNT
SO_1414 flavocytochrome c flavin subunit putative 22
SO_3468 riboflavin synthase subunit alpha 20
The amount of bacterial biofilm biomass on the anode can affect the power
generation in MFCs (Biffinger et al 2007 Heydorn et al 2000) Biofilm formation
related genes include flagellar gene IV pilus genes and other genes the flagellar related
genes (fliB and fliK) and IV pilus genes (mshA) were induced only from 105 to 134
fold as response of S oneidensis MR-1 to CNT decorated anodes in this study No
biofilm-associated genes presented significant changes in expression levels in this study
indicating there is no direct connections between biofilm enhancements to CNT
decorated anodes in this study This result is consistent with the Liang et al (2011)
conclusion that using CNT enhanced the electrochemical activity of anodic biofilms but
did not result in a significant increase of biomass in the anodic biofilms
Our microarray results showed two significantly up-regulated gene groups the
genes encoding proteins localized on the outer membrane and the genes involved in
flavin biosynthesis contributed to current density enhancement by CNT decorated anodes
Among 457 significantly changed genes to CNT in this study there are also a relatively
large number of genes encoding proteins with unknown functions which are either up- or
down-regulated in the response to CNT decorated anodes which indicated more
79
complicated responses of S oneidensis MR-1 to CNT decorated anodes in MECs and
further study is needed
80
Chapter 6
Summary
Microbial electrochemical systems (MESs) with a mixed culture initially inoculated
from Corvallis wastewater treatment plant have been studied for more than 6 years for
varying purposes including power generation hydrogen production heavy metal
removal and wastewater treatment Experiment results have shown that the mixed culture
is quite stable with excellent performance in MESs Our community analysis using
denaturing gradient gel electrophoresis (DGGE) and 16S rDNA clone library construction
suggests that the mixed culture is composed predominantly of Geobacter sp (66)
Arcobacter sp (12) and Citrobacter sp (11) These results not only improved our
understanding of the mixed culture community but also guided our studies on the
cultivation and isolation of the different bacterial species
Exoelectrogenic Citrobacter sp SX-1 was first isolated and characterized from a
MFC operated with the mixed culture Citrobacter sp SX-1 has been demonstrated to
produce electricity from wide range of different substrates including citrate acetate
glucose sucrose glycerol and lactosein MFCs Cyclic voltammetry analysis indicated
that membrane associated proteins may play an important role in facilitating the electrons
transferring from bacteria to electrode The strain SX-1 increased the known diversity of
power generating exoelectrogens and provided a unique bacterial species for study in
renewable energy generation and waste treatment
81
To enhance the electron transfer from bacteria to anode nano decorated anodes
including Fe NP and CNT were developed and characterized and evaluated in MECs
using Shewanella oneidensis MR-1 as a model species Both nanostructures have
significantly increased current density compared with the control Whole genome
microarray analysis elucidated the possible mechanisms of power enhancement in
response to these nano-decorated anodes
These results benefit to understanding of physiology and ecology of mixed
cultures in MFCs and improve the efficiency of current generation in MESs which will
facilitate the viability of niche applications for MESs in near future
82
Bibliography
1 Amman R Ludwig W (2000) Ribosomal RNA-targeted nucleic acid probes for
studies in microbial ecology FEMS Microbiol Ecol 24 555ndash565
1 Bao SJ Li CM Zang JF Cui XQ Qiao Y Guo J (2008) Novel Nanoporous TiO2
Electrode for Directly Electrochemistry of glucose oxidase Adv Funct Mater
18(4) 591-599
2 Beliaev AS Saffarini DA McLaughlin JL Hunnicutt D (2001) MtrC an outer
membrane decahaem c cytochrome required for metal reduction in Shewanella
putrefaciens MR-1 Mol Microbiol 39722-30
3 Bencheikh-Latmani R Williams SM Haucke L Criddle CS Wu L Criddle CS
Zhou J Tebo BM (2005) Global transcriptional profiling of Shewanella oneidensis
MR-1 during Cr(VI) and U(VI) reduction Appl Environ Microbiol 71 7453ndash7460
4 Biffinger JC Pietron J Ray R Little B and Ringeisen BR (2007) A biofilm
enhanced miniature microbial fuel cell using Shewanella oneidensis DSP10 and
oxygen reduction cathodes Biosens Bioelectron 221672ndash1679
5 Bodelier PLE Roslev P Henckel T FenzelP (2000) Stimulation by ammonium
based fertilizers of methane oxidation in soil around rice root Nature 40 421ndash424
6 Bond DR Holmes DE Tender LM Lovley DR (2002) Electrode-reducing
microorganisms that harvest energy from marine sediments Science 295 483-485
7 Bond DR Lovley DR (2003) Electricity production by Geobacter sulfurreducens
attached to electrodes Appl Environ Microbiol 69 1548-1555
8 Bond DR Lovley DR (2005) Evidence for involvement of an electron shuttle in
electricity generation by Geothrix fermentans Appl Environ Microbiol 71 2186-
2189
9 Bretschger O Obraztsova A Sturm CA Chang IS Gorby YA Reed SB Culley
DE Reardon CL Barua S Romine MF Zhou J Beliaev AS Bouhenni R
Saffarini D Mansfeld F Kim BH Fredrickson JK Nealson KH (2007) Current
production and metal oxide reduction by Shewanella oneidensis MR-1 wildtype and
mutants Appl Environ Microbiol 73 7003ndash7012
10 Brown SD Thompson MR Verberkmoes NC Chourey K Shah M Zhou J
Hettich RL and Thompson DK (2006) Molecular dynamics of the Shewanella
oneidensis response to chromate stress Mol Cell Proteomics 51054-1071
11 Brutinel ED Gralnick JA (2012) Shuttling happens soluble flavin mediators of
extracellular electron transfer in Shewanella Appl Microbiol Biotechnol 93(1) 41-8
12 Butler JE Kaufmann F Coppi MV Nuacutentildeez C Lovley DR (2004) MacA a diheme
c-type cytochrome involved in Fe(III) reduction by Geobacter sulfurreducens J
Bacteriol 186(12) 4042ndash4045
83
13 Caccavo F Lonergan DJ Lovley DR Davis M Stolz JF McInerney MJ (1994)
Geobacter sulfurreducens sp nov a hydrogen- and acetate oxidizing dissimilatory
metal-reducing microorganism Appl Environ Microbiol 60 3752ndash3759
14 Canstein von H Ogawa J Shimizu S Lloyd JR (2008) Secretion of flavins by
Shewanella species and their role in extracellular electron transfer Appl Environ
Microbiol 74 615ndash623
15 Cao X Huang X Liang P Xiao K Zhou Y Zhang X Logan BE (2009) A New
Method for Water Desalination Using Microbial Desalination Cells Environ Sci
Technol 43(18)7148-7152
16 Chaudhuri SK Lovley DR (2003) Electricity generation by direct oxidation of
glucose in mediatorless microbial fuel cells Nat Biotechnol 21 1229-1232
17 Cheng S Liu H Logan B E (2006) Increased power generation in a continuous
flow MFC with advective flow through the porous anode and reduced electrode
spacing Environ Sci Technol 402426-2432
18 Cheng S Logan B E (2007) Sustainable and efficient biohydrogen production via
electrohydrogenesis PNAS 104 18871-18873
19 Chignell J (2010) Energy Recovery from Biodiesel Waste Performance of
Microbial Electrochemical Systems on Glycerol Thesis
20 Chung K Okabe S (2009) Characterization of electrochemical activity of a strain
ISO2-3 phylogenetically related to Aeromonas sp isolated from a glucose-fed
microbial fuel cell Biotechnol Bioeng 104(5) 901-910
21 Cole JR Chai B Farris RJ Wang Q Kulam-Syed-Mohideen AS McGarrell
DM Bandela AM Cardenas E Garrity GM Tiedje JM (2007) The ribosomal
database project (RDP-II) introducing myRDP space and quality controlled public
data Nucleic Acids Res 35 (Database issue) D169-D172 doi 101093nargkl889
22 Cole JR Wang Q Cardena E Fish J Chai B Farris RJ Kulam-Syed-Mohideen
AS McGarrell DM Marsh T Garrity GM Tiedje JM (2009) The Ribosomal
Database Project improved alignments and new tools for rRNA analysis Nucleic
Acids Res 37 (Database issue) D141-D145 doi 101093nargkn879
23 Dey A De S De A De SK (2004) Characterization and dielectric properties of
polyaniline-TiO2 nanocomposites Nanotechnol 15 1277-1283
24 Edwards U Rogall T Bloumlcker H Emde M Boumlttger EC (1989) Isolation and
direct complete nucleotide determination of entire genesmdashcharacterization of a gene
coding for 16S-ribosomal RNA NucleicAcids Res 17 7843ndash7853
25 Eggleston CM Voumlroumls J Shi L Lower BH Droubay TC Colberg PJS (2008)
Binding and direct electrochemistry of OmcA an outer-membrane cytochrome from
an iron reducing bacterium with oxide electrodes a candidate biofuel cell system
Inorg Chim Acta 361769ndash777
26 Fan Y Sharbrough E Liu H (2008) Quantification of the Internal Resistance
Distribution of Microbial Fuel Cells Environ Sci Technol 42 8101ndash8107
84
27 Fan Y Xu S Schaller R Jiao J Chaplen F Liu H (2011) Nanoparticle decorated
anodes for enhanced current generation in microbial electrochemical
cells Biosensors and Bioelectronics 26 (5)1908ndash1912
28 Fan Y Hu H Liu H (2007)Sustainable power generation in microbial fuel cells
using bicarbonate buffer and proton transfer mechanisms Environ Sci
Technol 41(23) 8154-8
29 Fedorovich V Knighton MC Pagaling E Ward FB Free A Goryanin I (2009)
Novel electrochemically active bacterium phylogenetically related to Arcobacter
butzleri isolated from a microbial fuel cell Appl Environ Microbiol 75(23) 7326-
7334
30 Ferna ndez A Huang S Seston S Xing J Hickey R Criddle C Tiedje J (1999)
How stable is stable Function versus community composition Appl Environ
Microbiol 65 3697ndash3704
31 Field SJ Dobbin PS Cheesman MR Watmough NJ Thomson AJ Richardson
DJ (2000) Purification and magneto-optical spectroscopic characterization of
cytoplasmic membrane and outer membrane multiheme c-type cytochromes from
Shewanella frigidimarina NCIMB400 J Biol Chem 275 8515ndash8522
32 Freguia S Masuda M Tsujimura S Kano K (2009) Lactococcus lactis catalyses
electricity generation at microbial fuel cell anodes via excretion of a soluble quinone
Bioelectrochemistry 7614ndash18
33 Fricke K Harnisch F and Schroumlder U (2008) On the use of cyclic voltammetry for
the study of the anodic electron transfer in microbial fuel cells Energy Environ
Sci 1 144ndash147
34 Gao H Wang Y Liu X Yan T Wu L Alm E Arkin A Thompson DK and
Zhou J (2004) Global transcriptome analysis of the heat shock response of
Shewanella oneidensis J Bacteriol 1867796-7803
35 Gao H Yang ZK Wu L Thompson DK Zhou J (2006) Global transcriptome
analysis of the cold shock response of Shewanella oneidensis MR-1 and mutational
analysis of its classical cold shock proteins J Bacteriol 1884560-4569
36 George MG (2005) Bergeyrsquos Manual of Systematic Bacteriology Volume Two The
Proteobacteria Part B The Gammaproteobacteria Second Edition New York
Springer 651p
37 Gonzaacutelez-Toril E Llobet-Brossa E Casamayor EO Amann R Amils R (2003)
Microbial ecology of an extreme acidic environment the Tinto river Appl Environ
Microbiol 69(8) 4853ndash4865
38 Gorby YA Yanina S Mclean JS Rosso KM Moyles D Dohnalkova A
Beveridge TJ Chang IS Kim BH Kim KS Culley DE Reed SB Romine M
F Saffarini DA Hill EA Shi L Elias DA Kennedy DW Pinchuk G
Watanabe K Ishii S Logan B Nealson KH Fredrickson JK (2006) Electrically
conductive bacterial nanowires produced by Shewanella oneidensis strain MR-1 and
other microorganisms Proc Natl Acad Sci USA 10311358-11363
85
39 He Z Wagner N Minteer SD Angenent LT (2006) An upflow microbial fuel cell
with an interior cathodes assessment of the internal resistance by impedance
spectroscopy Environ Sci Technol 40 5212-5217
40 Heydorn A Nielsen AT Hentzer M Sternberg C Givskov M Ersboslashll BK Molin
S (2000) Quantification of biofilm structures by the novel computer program
COMSTAT Microbiol 1462395-2407
41 Holmes DE Bond DR Lovley DR (2004a) Electron transfer by Desulfobulbus
propionicus to Fe(III) and graphite electrodes Appl Environ Microbiol 701234ndash
1237
42 Holmes DE Chaudhuri SK Nevin KP Mehta T Methe BA Liu A Ward JE
Woodard TL Webster J Lovley DR (2006) Microarray and genetic analysis of
electron transfer to electrodes in Geobacter sulfurreducens Environ Microbiol
81805-1815
43 Holmes DE Nicoll JS Bond DR Lovley DR (2004b) Potential role of a novel
psychrotolerant member of the family Geobacteraceae Geopsychrobacter
electrodiphilus gen nov sp nov in electricity production by a marine sediment
fuel cell Appl Environ Microbiol 70 6023ndash6030
44 Holmes DE Chaudhuri SK Nevin KP Mehta T Metheacute BA Liu A Ward JE
Woodard TL Webster J Lovley DR (2006) microarray and genetic analysis of
electron transfer to electrodes in Geobacter sulfurreducens Environ Microbial
8(10)1805-1815
45 Hu H (2009) Enhancing Hydrogen Production in Microbial Electrolysis Cells
through Development of Platinum-free Cathode and Improvement of Reactor
Dissertation
46 Ince BK Ayman O N Turker G Ccedilelikkol S Ince O (2010) Microbial ecology of
anaerobic reactors for treatment of alcohol industry wastewaters a review Current
research technology and education topics in applied microbiology and microbial
ecology 988-999
47 Jung S Regan JR (2007) Comparison of anode bacterial communities and
performance in microbial fuel cells with different electron donors Appl Microbiol
Biotechnol 77393ndash 402
48 Kim BH Kim HJ Hyun MS Park DH (1999) Direct electrode reaction of Fe(III)
reducing bacterium Shwwanella putrefaciens J Microbial Biotechnol 9127-131
49 Kim GT Hyun MS Chang IS Kim HJ Park HS Kim BH Kim SD Wimpenny
JW Weightman AJ (2005) Dissimilatory Fe(III) reduction by an electrochemically
active lactic acid bacterium phylogenetically related to Enterococcus gallinarum
isolated from submerged soil J Appl Microbiol 99(4) 978ndash987
50 Kim GT Webster G Wimpenny JW Kim BH Kim HJ Weightman AJ (2006)
Bacterial community structure compartmentalization and activity in a microbial fuel
cell J Appl Microbiol 101(3) 698ndash710
86
51 Kim HJ Park HS Hyun MS Chang IS Kim M Kim BH (2002) A mediator-
less microbial fuel cell using a metal reducing bacterium Shewanella putrefaciens
Enzyme Microb Technol 30 145-152
52 Kim JR Min B Logan BE (2005) Evaluation of procedures to acclimate a
microbial fuel cell for electricity production Appl Microbiol Biotechnol 68(1)23-30
53 Kim JR Cheng S Oh SE Logan BE (2007) Power generation using different
cation anion and ultrafiltration membranes in microbial fuel cells Environ Sci
Technol 41 1004-1009
54 Konstantinidis KT Serres MH Romine MF Rodrigues JL Auchtung J McCue
LA Lipton MS Obraztsova A Giometti CS Nealson KH Fredrickson JK
Tiedje JM (2009) Comparative systems biology across an evolutionary gradient
within the Shewanella genus Proc Natl Acad Sci U S A 106(37)15909-15914
55 Lee J Phung NT Chang IS Kim BH Sung HC (2003) Use of acetate for
enrichment of electrochemically active microorganisms and their 16S rDNA
analyses FEMS Microbiol Lett 223185ndash191
56 Liang P Wang H Xia X Huang X Mo Y Cao X Fan M(2011)Carbon
nanotube powders as electrode modifier to enhance the activity of anodic biofilm
in microbial fuel cells Biosens Bioelectron 26(6)3000-4
57 Lies DP Hernandez ME Kappler A Mielke RE Gralnick JA Newman DK
(2005) Shewanaella oneidensis MR-1 uses overlapping pathways for iron reduction
at a distance and by direct contact under conditions relevant for biofilms Appl
Environ Microbiol 71 4414-4426
58 Liu H and Logan B E (2004) Electricity generation using an air-cathode single
chamber microbial fuel cell in the presence and absence of a proton exchange
membrane Environ Sci Technol 38 4040-4046
59 Liu H Grot S Logan BE (2005) Electrochemically assisted microbial production
of hydrogen from acetate Environ Sci Technol 39 4317 ndash 320
60 Liu H Hu H Chignell J Fan Y (2010) Microbial Electrolysis Novel Technology
for Hydrogen Production from Biomass Biofuels 1(1) 129ndash142
61 Liu H Ramnarayanan R Logan BE (2004) Production of electricity during
wastewater treatment using a single chamber microbial fuel cell Environ Sci Technol
382281-2285
62 Liu WT Marsh TL Cheng H Forney LJ (1997) Characterization of microbial
diversity by determining terminal restriction fragment length polymorphisms of gene
encoding 16S rRNA Appl Environ Microbiol 63 4516ndash4522
63 Logan B E Hamelers B Rozendal R Schroder U Keller J Freguia S Aelterman
P Verstraete W Rabaey K (2006) Microbial fuel cells methodology and
technology Environ Sci Technol 40 (17) 5181-5192
64 Logan B E and Regan JM (2006) Electricity-producing bacterial communities in
microbial fuel cells Trends Microbiol 14512ndash518
87
65 Logan B Cheng S Watson V Estadt G (2007) Graphite fiber brush anodes for
increased power production in air-cathode microbial fuel cells Environ Sci Technol
41(9)3341-3346
66 Logan BE (2009) Exoelectrogenic bacteria that power microbial fuel cells Nat Rev
Microbiol 5 375-381
67 Logan BE Call D Cheng S Hamelers HV Sleutels TH Jeremiasse AW
Rozendal RA (2008) Microbial electrolysis cells for high yield hydrogen gas
production from organic matter Environ Sci Technol 42 8630-8640
68 Logan BE Hamelers B Rozendal R Schroder U Keller J Freguia S Aelterman
P Verstraete W Rabaey K (2006) Microbial fuel cells methodology and
technology Environ SciTechnol 40 5181-5192
69 Lovely DR (2006) Bug juice harvesting electricity with microorganisms Nature
Review| Microbiology 4 497-508
70 Lovley DR Phillips EJ (1988) Novel Mode of Microbial Energy Metabolism
Organic Carbon Oxidation Coupled to Dissimilatory Reduction of Iron or
Manganese Appl Environ Microbiol 54(6) 1472-1480
71 Ly HK Sezer M Wisitruangsakul N Feng JJ Kranich A Millo D Weidinger
IM Zebger I Murgida DH Hildebrandt P (2011) Surface-enhanced vibrational
spectroscopy for probing transient interactions of proteins with biomimetic interfaces
electric field effects on structure dynamics and function of cytochrome c FEBS J
278(9)1382-90
72 Macaskie LE Hewitt CJ Shearer JA Kent CA (1995) Biomass production for
the removal of heavy metals from aqueous solutions at low pH using growth-
decoupled cells of a Citrobacter sp Int Biodeter Biodegr 3573ndash92
73 Malki M De Lacey AL Rodriguez N Amils R Fernandez VM (2008)
Preferential use of an anode as an electron acceptor by an acidophilic bacterium in
the presence of oxygen Appl Environ Microbiol 74 4472ndash4476
74 Meitl LA Eggleston CM Colberg PJS Khare N Reardon CL Shi L (2009)
Electrochemical interaction of Shewanella oneidensis MR-1 and its outer membrane
cytochromes OmcA and MtrC with hematite electrodes Geochim Cosmochim
Ac 73(18) 5292-5307
75 Morozan A Stamatin L and Nastase
F (2007) The biocompatibility
microorganisms-carbon nanostructures for applications in microbial fuel cells Phys
Stat Sol 6 1797-1803
76 Morris JM Jin S Crimi B Pruden A (2009) Microbial fuel cell in enhancing
anaerobic biodegradation of diesel Chem Eng J 146 161ndash167
77 Murray AE Lies D Li G Nealson K Zhou J Tiedje JM (2001) DNADNA
hybridization to microarrays reveals gene-specific differences between closely
related microbial genomes Proc Natl Acad Sci U S A 98 9853-9858
78 Muyzer G de Waal EC Uitterlinden AG (1993) Profiling of complex microbial
communities by denaturing gradient gel electrophoresis analysis of polymerase chain
88
reaction amplified genes coding for 16S rRNA Appl Environ Microbiol 59 695ndash
700
79 Myers CR Myers JM (1992) Localization of cytochromes to the outer membrane
of anaerobically grown Shewanella putrefacians MR-1 J Bacteriol 174 3429ndash3438
80 Narde GK Kapley A Purohit HJ (2004) Isolation and characterization of
Citrobacter strain HPC255 for broad-range substrate specificity for chlorophenols
Curr Microbiol 48(6) 419-423
81 Okabe S Ito T Sugita K Satoh H (2005) Succession of internal sulfur cycles and
sulfur-oxidizing bacterial communities in microaerophilic wastewater biofilms Appl
Environ Microbiol 71(5) 2520-2529
82 Park DH and Zeikus JG (2000) Electricity generation in microbial fuel cells using
neutral red as an electronophore Appl Environ Microbial 66 292-1297
83 Park DH and Zeikus JG (2002) Impact of electrode composition on electricity
generation in a single-compartment fuel cell using Shewanella putrefaciens Appl
Microbiol Biotechnol 5958ndash56
84 Park DH and Zeikus JG (2003) Improved fuel cell and electrode designs for
producing electricity from microbial degradation Biotechnol Bioeng 81 348ndash355
85 Park HS Kim BH Kim HS Kim HJ Kim GT Kim M Chang IS Park Y
K Chang HI (2001) A novel electrochemically active and Fe(III)-reducing
bacterium phylogenetically related to Clostridium butyricum isolated from a
microbial fuel cell Anaerobe 7 297-306
86 Parot S Nercessian O Delia ML Achouak W Bergel A (2009) Electrochemical
checking of aerobic isolates from electrochemically active biofilms formed in
compost J Appl Microbiol 106(4) 1350-1359
87 Pham CA Jung SJ Phung NT Lee J Chang IS Kim BH Yi H Chun J
(2003) A novel electrochemically active and Fe(III)-reducing bacterium
phylogenetically related to Aeromonas hydrophila isolated from a microbial fuel cell
FEMS Microbiol Lett 223 129-134
88 Qiao Y Bao SJ Li CM Cui XQ Lu ZS Guo J (2008) Nanostructured
polyanilinetitanium dioxide composite anode for microbial fuel cells ACS Nano 2
113-119
89 Qiu R Zhao B Liu J Huang X Li Q Brewer E Wang S Shi N (2009) Sulfate
reduction and copper precipitation by a Citrobacter sp isolated from a mining area J
Hazard Mater 1641310-1315
90 Quan X Yang SG Ruan XL and Zhao HM (2005) Preparation of titania
nanotubes and their environmental applications as electrode Environ Sci Technol 39
3770ndash3775
91 Rabaey K Boon N Hofte M Verstraete W (2005) Microbial phenazine
production enhances electron transfer in biofuel cells Environ Sci Technol 393401ndash
3408
89
92 Rabaey K Boon N Siciliano S D Verhaege M Verstraete W (2004) Biofuel cells
select for microbial consortia that self-mediate electron transfer Appl Environ
Microbiol 70(9) 5373ndash5382
93 Rabaey K Lissens G Siciliano S D Verstraete W (2003) Amicrobial fuel cell
capable of converting glucose to electricity at high rate and efficiency Biotechnol
Lett 25 1531-15
94 Rabaey K Verstraete W (2005) Microbial fuel cells novel biotechnology for
energy generation Trends Biotechnol 23(6) 291-298
95 Reguera G McCarthy KD Mehta T Nicoll JS Tuominen MT Lovley DR
(2005) Extracellular electron transfer via microbial nanowires Nature 435 1098-
1101
96 Reguera G Nevin KP Nicoll JS Covalla SF Woodard TL Lovley DR (2006)
Biofilm and nanowire production leads to increased current in Geobacter
sulfurreducens fuel cells Appl Environ Microbiol 72 7345-7348
97 Reimers CE Tender LM Fertig S Wang W (2001) Harvesting energy from the
marine sediment-water interface Environ Sci Technol 35192-195
98 Rezaei F Xing D Wagner R Regan JM Richard TM Logan BE (2009)
Simultaneous cellulose degradation and electricity production by Enterobacter
cloacae in a microbial fuel cell Appl Microbiol Biotechnol 75 3673ndash3678
99 Richter H Nevin KP Jia H Lowy DA Lovley DR Tender LM (2009) Cyclic
voltammetry of biofilms of wild type and mutant Geobacter sulfurreducens on fuel
cell anodes indicates possible roles of OmcB OmcZ type IV pili and protons in
extracellular electron transfer Energy Environ Sci 2 506ndash516
100 Riesenfeld SC Schloss DP Handelsman J(2004) Metagenomics Genomic
Analysis of Microbial Communities Annual Review Genetics 38525-552
101 Rozendal RA Hamelers HV Buisman CJ (2006) Effects of membrane cation
transport on pH and microbial fuel cell performance Environ Sci Technol 40
5206-5211
102 Rozendal RA Hamelers HVM Euverink GJW Metz SJ Buisman CJN (2006)
Principle and perspectives of hydrogen production through biocatalyzed electrolysis
Int J Hydrogen Energ 311632-1640
103 Rozendal RA Hamelers HV Molenkamp RJ Buisman CJ (2007) Performance of
single chamber biocatalyzed electrolysis with different types of ion exchange
membrances Water Res 41 1984-1994
104 Schluumlter A Bekel T Diaz NN Dondrup M Eichenlaub R Gartemann
KH Krahn I Krause L Kroumlmeke H Kruse O Mussgnug JH Neuweger
H Niehaus K Puumlhler A Runte KJ Szczepanowski R Tauch A Tilker
A Viehoumlver P Goesmann A (2008) The metagenome of a biogas-producing
microbial community of a production-scale biogas plant fermenter analyzed by the
454-pyrosequencing technology J Biotechnol136 77-90
90
105 Scott J H and Nealson KH (1994) A biochemical study of the intermediary carbon
metabolism of Shewanella putrefaciens J Bacteriol 176(11) 3408ndash3411
106 Sekiguchi H Tomioka H Nakahara T and Uchiyama H (2001) A single band
does not always represent single bacterial strains in denaturing gradient gel
electrophoresis analysis Biotechnology Letters23 1205ndash1208
107 Sharma T Leela M Reddy A Chandra TS Ramaprabhu S (2008) Development
of carbon nanotubes and nanofluids based microbial fuel cell Int J Hydrogen Energ
336749ndash6754
108 Shea C Clauwaert P Verstraete W Nerenberg R (2008) Adapting a denitrifying
biocathode for perchlorate reduction Water Sci Technol 58 (10) 1941-1946
109 Shi L Squier TC Zachara JM Fredrickson JK (2007) Respiration of metal (hydr)
oxides by Shewanella and Geobacter a key role for multihaem c-type cytochromes
Mol Microbiol 6512-20
110 Stams AJ and Plugge CM(2009)Electron transfer in syntrophic communities
of anaerobic bacteria and archaea Nat Rev Microbiol7(8)568-77
111 Sukkasem C Xu S Park S Boonsawang P Liu H (2008) Effect of Nitrate on the
Performance of Single Chamber Air Cathode Microbial Fuel Cells Water Res 42(19)
4743-4750
112 Summers ZM Fogarty HE Leang C Franks AE Malvankar NS Lovley DR
(2010)Direct exchange of electrons within aggregates of an evolved syntrophic
coculture of anaerobic bacteria Science 330 (6009)1413-5
113 Szczepanowski R Bekel T Goesmann A Krause L Kroumlmeke H Kaiser
O Eichler W Puumlhler A Schluumlter A (2008) Insight into the plasmid metagenome
of wastewater treatment plant bacteria showing reduced susceptibility to
antimicrobial drugs analysed by the 454-pyrosequencing technology J
Biotechnol 136 54-64
114 Tamura K Dudley J Nei M Kumar S (2007) MEGA4 Molecular Evolutionary
Genetics Analysis (MEGA) software version 40 MolBiol Evol 24(8) 1596-1599
115 Thompson JD Gibson TJ Plewniak F Jeanmougin F Higgins DG (1997) The
CLUSTAL_X windows interface flexible strategies for multiple sequence alignment
aided by quality analysis tools Nucleic Acids Res 25 4876-4882
116 Thormann KM Duttler S Saville RM Hyodo M Shukla S Hayakawa Y
Spormann AM (2006) Control of formation and cellular detachment from
Shewanella oneidensis MR-1 biofilms by cyclic di-GMP J Bacteriol 1882681-2691
117 Thormann KM Saville RM Shukla S Pelletier DA Spormann AM (2004)
Initial Phases of biofilm formation in Shewanella oneidensis MR-1 J Bacteriol
186(23) 8096-104
118 Torres CI Kato Marcus A Rittmann BE (2008b) Proton transport inside the
biofilm limits electrical current generation by anode-respiring bacteria Biotechnol
Bioeng 100 872ndash881
91
119 Torres CI Marcus AK Lee HS Parameswaran P Krajmalnik-Brown R Rittmann
BE (2010) A kinetic perspective on extracellular electron transfer by anode-
respiring bacteria FEMS Microbiol Rev 234(1)3-17
120 Torres CI Marcus AK Parameswaran P Rittmann BE (2008a) Kinetic
experiments for evaluating the NernstndashMonod model for anode-respiring bacteria
(ARB) in a biofilm anode Environ Sci Technol 42 6593ndash6597
121 Tsai HY Wu Ch-Ch Lee Ch-Y Shih EP (2009) Microbial fuel cell performance
of multiwall carbon nanotubes on carbon cloth as electrodes J Power Sources
194199-205
122 Tyson GW Chapman J Hugenholtz P Allen EE Ram RJ Richardson
PM Solovyev VV Rubin EM Rokhsar DS Banfield JF(2004) Community
structure and metabolism through reconstruction of microbial genomes from the
environment Nature42837-43
123 Venkateswaran K Moser DP Dollhopf ME Lies DP Saffarini D A MacGregor
BJ Ringelberg DB White DC Nishijima M Sano H Burghardt J Stackebrandt
E Nealson KH (1999) Polyphasic taxonomy of the genus Shewanella and
description of Shewanella oneidensis sp Int J Syst Bacteriol 49705-724
124 von Canstein H Ogawa J Shimizu S Lloyd JR (2008) secretion of Flavins
by Shewanella Species and Their Role in Extracellular Electron Transfer Appl
Environ Microbiol 74 3615-623
125 Wang A Liu W Cheng Sh Xing D Zhou J Logan BE (2009) Source of methane
and methods to control its formation in single chamber microbial electrolysis cells
Int J Hydrogen Energ 34 3653-3658
126 Wang J Ma T Zhao L Lv J Li G Liang F Liu R (2008) PCRndashDGGE method
for analyzing the bacterial community in a high temperature petroleum reservoir
World J Microbiol Biotechnol 2 9694-9702
127 Wang Q Garrity GM Tiedje JM Cole JR (2007) Naiumlve Bayesian Classifier for
Rapid Assignment of rRNA Sequences into the New Bacterial Taxonomy Appl
Environ Microbiol 73(16) 5261-5267
128 Wang YF Masuda M Tsulimura S Kano K (2008) Electrochemical regulation of
the end-product profile in Propionibacterium freudenreichii ET-3 with an
endogenous mediator Biotechnol Bioeng 101(3) 579ndash586
129 Wrighton KC Agbo P Warnecke F Weber KA Brodie EL DeSantis TZ
Hugenholtz P Andersen GL Coates JD (2008) A novel ecological role of the
Firmicutes identified in thermophilic microbial fuel cells ISME J 2 1146ndash1156
130 Xing D Zuo Y Cheng S Regan JM Logan BE (2008) Electricity generation by
Rhodopseudomonas palustris DX-1 Environ Sci Technol 42 4146-4151
131 Xu S Liu H Fan Y Schaller R Jiao J Chaplen F (2012) Enhanced performance
and mechanism study of microbial electrolysis cells using Fenanoparticle-decorated
anodes Appl Microbiol Biotechnol 93(2)871-880
92
132 Yang Y Harris DP Luo F Wu L Parsons AB Palumbo AV Zhou J (2008)
Characterization of the Shewanella oneidensis Fur gene roles in iron and acid
tolerance response BMC Genomics 9 Suppl 1S11
133 Zhang L Zhou S Zhuang L Li W Zhang J Lu N Deng L (2008) Microbial
fuel cell based on Klebsiella pneumoniae biofilm Electrochem Commun10 1641ndash
1643
134 Zuo Y Xing DF Regan JM Logan BE (2008) Isolation of the exoelectrogenic
bacterium Ochrobactrum anthropi YZ-1 by using a U-tube microbial fuel cell Appl
Environ Microbiol 74 3130-3137
93
APPENDICES
List of published papers during PhD study
1 Shoutao Xu Hong Liu Yanzhen Fan Rebecca Schaller Jun Jiao and Frank
Chaplen (2012) Enhanced performance and mechanism study
of microbial electrolysis cells using Fe nanoparticle-decorated anodes Applied
Microbiology and Biotechnology 93(2)871-880
2 Shoutao Xu Hong Liu (2011) New exoelectrogen Citrobacter sp SX-1 isolated
from a microbial fuel cell Journal of Applied Microbiology111(5)1108-1115
3 Yanzhen Fan Shoutao Xu Rebecca Schaller Jun Jiao Frank Chaplen Hong Liu
(2011) Nanoparticle decorated anodes for enhanced current generation in microbial
electrochemical cells Biosensors and Bioelectronics 26(5) 1908-1912
4 Tunc Catal Shoutao Xu Kaichang Li Hakan Bermek Hong Liu
(2008)
Electricity generation from polyalcohols in single-chamber microbial fuel
cells Biosensors and Bioelectronics 24(4)849-854
5 Chontisa Sukkasema Shoutao Xu Sunhwa Park Piyarat Boonsawang Hong Liu
(2008) Effect of nitrate on the performance of single chamber air cathode
microbial fuel cells Water research 424743-4750
6 Rebecca Schaller Yanzhen Fan Shoutao Xu Alan Fern Frank Chaplen Hong
Liu and Jun Jiao (2009) Vertically Aligned Multi-walled Carbon Nanotube
Decorated Anodes for Microbial Fuel Cells Proceedings of Materials Research
Society 2009 1170 R05-13

ACKNOWLEDGEMENTS
I would like to thank all people who have helped and inspired me during my
doctoral study Foremost I would like to express my sincere gratitude to my advisors Dr
Hong Liu and Dr Frank Chaplen for their continuous support of my PhD study and the
research of Microbial Fuel Cells at Oregon State University Hong inspired me to devote
myself to the field of Bio-energy with her great patience and enthusiasm towards
scientific educations Frank was always available and willing to help me with my study
especially during the period of Hongrsquos sabbatical leave Thanks to his kindness and
assistance my study at OSU became smooth and rewarding
Besides my advisors I would like to thank everyone in my dissertation committee Dr
Martin Schuster Dr Clare Reimers and Dr Mark Dolan Due to their encouragement
and insightful comments on my research I could always have the courage and knowledge
to overcome difficulties in my research I benefited greatly from Martinrsquos valuable
suggestions on my writing skills and his generous help regarding microarray data analysis
Clarersquos advice helped me build a solid foundation of Electrochemistry Also it was a
great honor to have Mark as my committee member His teaching gave me an insight into
the world of environmental engineering and enhanced the depth and width of my research
I thanked Dr Yanzhen Fan for always giving me guidance about the designs of the
reactors in my experiments
It was also important for me to say thanks to my current and previous lab-mates
Keaton Lesnik Kuhuan Chien Cheng Li Corale Abourached Anthony Janicek
Hongqiang Hu Jeremy Chignell Yudith Nieto and Wengguo Wu I had a wonderful
time enjoying doing research with them Their friendship and help made me confident of
my ability to do research as a scientist In addition I cherished and appreciated the
friendly environment in our BEE department Faculty members especially Dr John Bolte
Dr John Selker Dr Roger Ely Dr Ganti Murthy and Desiree Tullos made BEE an
excellent department for our students
My deepest gratitude went to my father Fuqin Wang and my mother Jinzhi Wang
for their endless care love and support throughout my life Also I would like to say
thanks to my best-loved wife Songhua Zhu She always supported me and never
complained that I could spend so little time accompanying with her This dissertation was
also for my lovely son Gabriel Hong-Yi Xu
TABLE OF CONTENTS
Page
1 General Introductionhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 1
2 Bacterial Community Analysis of Mixed Consortium in Microbial
Electrochemical Systemshelliphelliphelliphellip 13
3 New Exoelectrogen Citrobacter sp SX-1 Isolation and
Characterizationhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 27
4
Enhanced Performance and Mechanism Study of Microbial Electrolysis
Cells using Fe Nanoparticles Decorated Anodeshelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 45
5 Global Transcriptome Analysis of Response of Shewanella oneidensis
MR-1 to CNT Nanostructure Decorated Anodes in Microbial
Electrochemical System
66
6 Summaryhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 80
7 Bibliography helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 82
8 Appendices helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 93
LIST OF FIGURES
Figure Page
1-1 Generalized schematics (a) Microbial fuel cell (MFCs)(b) Microbial
electrolysis cell (MECs) helliphelliphelliphelliphelliphelliphelliphelliphelliphellip helliphelliphelliphellip
2
1-2 Mechanisms for extracellular electron transport in a MFCs anode 7
2-1 Polarization curves of high power mixed culture MFCshelliphelliphelliphelliphelliphellip 20
2-2 DGGE result of high power generation mixed culture MFCshelliphelliphellip 21
2-3 Bacterial species and percentage of bacterial community of mixed
culture MFCs helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip
22
2-4 Phylogenetic trees of bacterial species from mixed culture MFCs 22
2-5 Dominant bacterial species identification in DGGEhelliphelliphelliphelliphelliphelliphelliphellip 23
2-6 Current density generated by different dominant isolateshelliphelliphelliphelliphelliphellip 24
3-1 Phylogenetic tree of strain SX-1 and closely related species based on
16S rDNA sequenceshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip
35
3-2 SEM image of planktonic cells of Citrobacter sp SX-1helliphelliphelliphelliphelliphellip 35
3-3 Electricity generation by Citrobacter sp SX-1 in a single chamber
MFCs with sodium citrate helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip
36
3-4 Electricity generation of Citrobacter sp SX-1 using different
substrates in single chamber air-cathode MFCs at 1 KΩhelliphelliphellip
37
3-5 Power and voltage generation by Citrobacter sp SX-1 as a function
of current density helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip
38
3-6 Four current generating stages for CV analysis helliphelliphelliphelliphelliphelliphelliphellip 39
3-7 Cyclic Voltammetry analysis of of Citrobacter sp SX-1 biofilms helliphellip 41
4-1 SEM images of Fe NPs decorated anodeshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 50
4-2 Effects of Fe-NP decorated anodes on the current density in MECs helliphellip 51
4-3 Differentially expressed genes grouped by functional classification in
Fe NP decorated anodeshelliphelliphelliphelliphelliphellip helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip
53
5-1 SEM images of CNT decorated anodeshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 70
5-2 Effects of CNT decorated anodes on the current density in MECshelliphellip 73
5-3 Differentially expressed genes grouped by functional classification in
CNT decorated anodes helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip
74
LIST OF TABLES
Table
1-1
4-1
4-2
4-3
Taxa of bacteria current density generated and reactor configuration
used in microbial electrochemical cell experimentshelliphelliphelliphellip
Expression levels of genes related to biofilm formationhelliphelliphelliphelliphellip
Genes related to anaerobic growth and electron transfer with
significantly change expression levelhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip
Expression level of flavin and cytochrome geneshelliphelliphelliphelliphelliphelliphelliphellip
Page
4
55
56
58
4-4
5-1
5-2
Other genes with significantly changed expression level
Expression level of cytochrome c as response to CNT decorated anodes
Expression level of genes related to flavin synthesis as response to CNT
decorated anode in CNT decorated anodeshelliphellip
59
76
78
1
1
Bacterial Community Analysis New Exoelectrogen Isolation and Enhanced Performance
of Microbial Electrochemical Systems Using Nano-Decorated Anodes
Chapter 1
General Introduction
11 Microbial Fuel Cells and Microbial Electrolysis Cells
Microbial electrochemical systems (MESs) have drawn the attention of researchers
in recent years due to their promising applications in a variety of scientific fields such as
renewable energy generation bioremediation and wastewater treatment In a MES
microorganisms interact with electrodes catalyzing oxidation and reduction reactions at
the anode and the cathode
The most-described type of MESs is the microbial fuel cells (MFCs) in which
useful power is generated directly using the catalytic action of active microorganisms
(Kim 2002 Cheng and Logan 2007 Cheng et al 2006) In a typical two-chamber
MFCs (Figure 1a) organic matter is oxidized by electrochemically active
microorganisms in the anode chamber to release protons and electrons Protons diffuse
into the cathode chamber through a proton exchange membrane Meanwhile electrons
are transferred to the anode through various mechanisms (Logan et al 2006 Lovley
2
2006 Rabaey et al 2003) and then travel to the cathode where normally they combine
with oxygen and protons to form water
Another common used type of MES is the microbial electrolysis cell (MECs) in
which hydrogen is produced instead of electricity by applying a circuit voltage to the
MFCs system and maintaining anaerobic conditions in the cathode chamber (Liu et al
2005 Logan et al 2008 Rozendal et al 2006 Schaller et al 2009) (Figure 1b) A
cathode potential of at least - 410 mV (vs standard hydrogen electrode at pH 70) is
required to produce hydrogen at the cathode Therefore a theoretical potential of 110 mV
(ie 410-300 mV) is needed but voltages of gt02 V are normally applied due to various
overpotentials
Fig 1-1 Generalized schematics (a) Microbial fuel cells (MFCs) where electricity is
captured through the resistance R (b) Microbial electrolysis cells (MECs) where
hydrogen gas is produced in the cathode chamber R for resistor PS for power supply and
PEM for proton exchange membrane
The fundamental feature shared by microbial electrochemical systems (MESs)
(MECs and MFCs) is the microbe-catalyzed electron transfer from organic matter to
electrodes (anodes) (Rebaey et al 2004) Therefore different types of MESs can be
PEM
R
H+
e- e-
Anode
Bacte
ria
a
Cath
ode
H2O
PEM
PS
H+
e- e-
Anode
Bacte
ria
b
Cath
ode
H2
3
utilized to investigate the phenomena and mechanisms of interactions between microbes
and electrodes
12 Bacteria and Electron Transfer Mechanisms in MESs
The electrochemically active bacteria in MFCs are thought to be iron-reducing
bacteria such as Shewanella and Geobacter species (Gorby et al 2006) They have great
importance in the natural environment principally in metal oxidation and reduction
However recent studies have shown that the diversity of bacterial communities is much
greater than these model iron reducers on MFCs anodes (Reimers et al 2001 Liu et al
2004b Kim et al 2005) Over 20 electrochemically active bacterial species those can
transfer electrons exocellularly to electrodes have been reported in the past 10 years
(Logan 2009 Liu et al 2010) These bacteria have been categorized into diverse genetic
groups so far including alphaproteobacteria (Rhodopseudomonas Ochrobactrum and
Acidiphilium) (Malki et al 2008 Xing et al 2008 Zuo et al 2008) betaproteobacteria
(Rhodoferax) (Chaudhuri et al 2003) gammaproteobacteria (Shewanella Pseudomonas
Klebsiella Enterobacter and Aeronomas) (Kim et al 2002 Pham et al 2003 Rabaey et
al 2005 Bretschger et al 2007 Zhang et al 2008 Chung et al 2009 Rezaei et al 2009)
deltaproteobacteria (Geobacter Geopsychrobacter Desulfuromonas and Desulfobulbus)
(Bond et al 2002 Bond and Lovley 2003 Holmes et al 2004ab 2006)
Epsilonproteobacteria (Arcobacter) (Fedorovich et al 2009) Firmicutes (Clostridium and
Thermincola) (Park et al 2001 Wrighton et al 2008) Acidobacteria (Geothrix) (Bond
and Lovley 2005) and actinobacteria (Propionibacterium) (Wang et al 2008) A wider
range of electrochemically active bacteria are expected to be discovered
4
Table 1 Taxa of bacteria current density generated and reactor configuration used in
microbial electrochemical cell experiments (Liu et al 2010)
The electrochemically active bacterial species that possess the ability to transfer
electrons outside of the cell are called exoelectrogens in the MESs research field The
different exoelectrogens have demonstrated a wide-ranging power generation ability in
MFCs For example Shewanella oneidensis MR-1 has demonstrated the ability to
generate 0018 Am2 current density in single chamber MFCs while the Geobacter
Taxon Microorganisms Current Density (Am2
)
α-proteobacteria
Rhodopseudomonas palustris DX-1 003
Ochrobactrum anthropi YZ-1 071
Acidiphilium sp 32sup5 300
β-proteobacteria Rhodoferax ferrireducens 0031
γ-proteobacteria
Shewanella putrefaciens IR-1 0016
Shewanella oneidensis DSP10 0013
Shewanella oneidensis MR-1 0018
Pseudomonas aeruginosa KRA3 0017
Escherichia coli K12 HB101 100
Klebsiella pneumoniae L17 120
Enterobacter cloacae 013
Aeromonas hydrophila PA3 030
δ-proteobacteria
Geobacter metallireducens 065
Geobacter sulfurreducens 800
Desulfuromonas acetoxidans 0005
Geopsychrobacter
Electrodiphilus strain A2
0066
Desulfobulbus propionicus 003
Firmicutes
Lactococcus lactis 003
Thermincola sp strain Jr 020
Clostridium butyricum EG3 022
Thermincola ferriacetica Z-0001 040
Brevibacillus spp PTH1 0009
Desulfitobacterium hafniense DCB2 110
Acidobacteria Geothrix fermentans 0097
Actinobacteria Propionibacterium freudenreichiiET-3 120
5
sulfurreducens species has generated current densities as high as 80 Am2 which is 400
times higher than the one generated by Shewanella oneidensis MR-1 However mixed-
culture communities have generated much higher power densities than their pure-culture
counterparts in MFCs perhaps due to synergistic interactions within the anode bacterial
communities and the participation of currently unknown bacteria species and mechanisms
(Jung and Regan 2007) The dominant bacteria species vary in mixed cultures bacteria
communities in MFCs due to the enrichment of different substrates
Traditional methods of extroelectrogen study depend on cultivation hampered novel
exoelectrogens discovery because the inadequacy of defined media underestimates the
actual microbial diversity in MFCs Slow growth rate and unknown growth requirements
of anaerobic microorganisms make some anaerobic extroelectrogens cultivation difficult
However these limitations have been overcome by using molecular biological methods
based on DNARNA analysis Molecular biological techniques are now widely applied to
assess the diversity of microbial communities by analyzing the 16S rDNA sequence The
most commonly used molecular biological techniques for bacterial community analysis
include denaturing gradient gel electrophoresis (DGGE) restriction fragment length
polymorphism (RFLP) and 16S rDNA sequencing These methods are much less time
consuming than traditional isolation and cultivation methods
Isolated exoelectrogens were utilized to explore the mechanism of electron transfer
to the anode However the mechanisms of electron transfer to extracellular electron
acceptors are not well understood Three mechanisms have been proposed for exocellular
transport of electrons by exoelectrogens to anodes in MFCs so far Some exoelectrogenic
bacteria can transfer electrons to electrodes through soluble redox compounds (Bond and
6
Lovley 2005) These compounds include artificial mediators and mediators secreted by
exoelectrogenic bacteria A variety of chemicals have been used to facilitate the shuttling
of electrons from inside of cell to electrodes outside the cell These exogenous mediators
include neutral red (Park et al 1999) anthraquinone-2-6 disulfonate (AQDS) thionin
potassium ferricyanide (Bond et al 2002) methyl viologen and others (Logan 2004
Rabaey and Verstraete 2005) Some bacteria can secrete their own chemical mediator
for example Pseudomonas aeruginosa secretes pyocyanin and phenazine-1-carboxamide
to transfer electrons toward the MFCs anode (Rabaey et al 2005) Another evidence for
mediator production by bacteria is Geothrix fermentans When the medium was replaced
in a MFCs that had stable power generation with this bacteria power dropped by 50
and required 10 days to resume the original level
Some bacteria can directly transfer electrons to anodes via outer cell membrane
proteins (Myers and Myers 1992) The outer cell membrane protein cytochrome c is
thought to play a critical role in to transferring electrons to anodes Ly et al (2011)
isolated the haem protein cytochrome c and demonstrated that electric field effects may
be functional for the natural redox processes of cytochrome c in the respiratory chain
Shewanella oneidensis is the example of a bacterium that is capable of transfer electron to
anode via outer cell membrane proteins When Shewanella oneidensis adhered to an iron
surface the greater force has showed benefits to grow cells because closer contact
required for electron transfer from cell bound cytochromes (Lower et al 2001)
7
Fig1-2 Mechanisms for extracellular electron transport in an MFCs anode (1) direct
contact (top in green) (2) by nanowires (middle in purple) and (3) self-produced
mediators (bottom in blue) (Logan 2009)
More and more evidence supports the involvement of bacterial nanowires in
extracellular electron transport (Reguera et al 2005 2006 Gorby et al 2006) Nanowires
are conductive appendages produced by both Geobacter and Shewanella species (Gorby
and Beveridge 2005) The conductivity of the appendages was examined and confirmed
by using conductive scanning tunneling microscopy (STM) (Gorby et al 2006)
Nanowires can carry electrons from the cell to the anode surface of MFCs
The solid component of the extracellular biofilm matrix has high efficiency on
extracellular electron transfer compared with other extracellular electron transfer
8
mechanisms and recently Torres et al (2010) hypothesized that the solid component of
the extracellular biofilm matrix formed by exoelectogens is another way bacteria transfer
electrons to electrodes This hypothesis was based on kinetic analysis of each EET
mechanism reported in available literature (Torres et al 2010)
13 Anode electrodes
In MFCsMECs anode electrodes are a critical component because exoelectrogens
adhere to the surface of anodes to transfer electrons to the electrode The characteristics
of anodes have significant effects on electron transfer rate from bacteria to anode
electrodes in MFCs The requirements of an anode material are it should be highly
conductive non-corrosive have a high specific surface area (area per volume) high
porosity be non-fouling inexpensive and easily scaled to larger sizes Of these
properties the most important one that is different from other biofilm reactors is that the
material must be electrically conductive Normally they are made of various carbon
materials including carbon fiber carbon clothe and carbon paper due to their stability
high conductivity and high specific surface-area Nevertheless they have little
electrocatalytic activity for the anode microbial reactions and thus a modification of the
carbon materials is the main approach for improving their performance Consequently
there is a great need to develop a new type of anode material for MFCsMECs
It is a great challenge to develop a new anode material to further increase the power
density of a MFCs The nature of the catalytic mechanism of a MFCrsquos anode involves not
only a biological but also an electrocatalytic process An optimal nanostructure with a
high specific surface area favorable for both catalytic processes could play a critical role
in improving the power density of the MFCs such a structure needs to host the bacteria
9
with high bioactivity while enhancing the electron-transfer rate Schroumlder et al (2008)
employed PANI to modify a platinum anode for MFCs and achieved a current density 1
order of magnitude higher than the previously reported value PANIinorganic
composites are also reported to have better conductivity Qiao et al also applied a new
mesoporous TiO2 electrode material with uniform nanopore distribution and a high
specific surface area to anode in comparison to previously reported work with E coli
MFCs the composite anode delivers 2-fold higher power density (Qiao et al 2008)
Thus it has great potential for use as the anode in a high-power MFCs and may be a new
approach for improving performance of MFCs
14 Other parts of MESs
141 Membranes and ion transport
The ion exchange membrane is another one of the critical components in two-
chamber MESs systems It separates anode and cathode chambers and at the same time
maintains the electron neutrality of the system ie transport of electrons to the cathode
needs to be compensated by transport of an equal amount of positive charge to the
cathode chamber A proton exchange membrane (PEM) such as Nafion a per-fluorinated
sulphonic acid membrane (PFSAs) consisting of a hydrophobic fluorocarbon backbone to
which hydrophilic sulfonate groups (-SO3-) are attached is commonly used in chemical
fuel cell systems For MFCs systems however mainly cation species like Na+ and K
+
other than proton are often responsible for the dominant transport of positive charge
through the cation exchange membrane (CEM) to maintain electroneutrality due to the
low proton concentration in any aqueous medium with near neutral pH (Rozendal et al
2006) Consequently the pH increases in the cathode chamber due to the consumption of
10
protons and decreases in the anode chamber because of the accumulation of protons
(Rozendal et al 2007) This pH change in two-chamber systems results in a decrease of
the cathode potential and performance The application of anion exchange membrane
(AEM) has also been explored in MFCs systems (Cheng and Logan 2007 Kim and
Logan 2007) where it has been proposed that protons are transferred via pH buffers like
phosphate anions
142 Cathodes and Catalysts
The cathode is another challenge for making MFCs commercially available
technology because the chemical reaction that occurs at the cathode is difficult to
engineer as the electrons protons and oxygen must all meet at a catalyst in a tri-phase
reaction (solid catalyst air and water) The catalyst must be on a conductive surface and
must be exposed to both water and air so that protons and electrons in these different
phases can reach the same point The most commonly used material for a cathode is
commercially available carbon paper pre-loaded with a Pt catalyst on one side When it is
used in a MFC the side that contains the catalyst faces the water and the uncoated side
faces air To reduce the high cathode cost associated with platinum catalyst other precious-
metal-free cathodes such as (NiMo and NiW) were developed by electrodepositing on a
carbon fiber They have achieved comparable performance with Pt catalyst with same
loading at a much lower cathode fabrication cost (Hu 2010)
The requirements of cathode for MECs are quite similar to the requirements of
cathode for MFCs but easier than cathode for MFCs for the manufacturing process
because the cathode in MECs is not necessarily to exposed to air Therefore it can be
made of the exact same materials of cathode in MFCs except a waterproof layer
11
Recently some researchers have attempted to apply microorganisms as a biocatalyst to
precede the combination of electron with oxygen in the cathodes
15 Dissertation overview
Low power densities in MESs limit practical applications The improvement of
MESs performance requires a detailed understanding of the physiology and ecology of
microorganisms in MESs including the mechanism of electron transfer to the anode from
the microorganism
This dissertation focuses on the problem of the low power density of MESs The
bacterial community structure of a high power generated mixed culture communities in
MFCs will be identified firstly and then one of major exoelectrogens will be isolated and
characterized This information will be helpful to understand the physiology and ecology
of exoelectrogens in MESs Consequently they will be beneficial to improve power
density of MFCs Lastly the nanostructure decorated anodes in MECs will be utilized to
improve the power density The power enhancement mechanism will be explored by
using a whole genome microarray They are presented here as four papers
In the first paper the cultivation independent molecular biological techniques
denaturing gradient gel electrophoresis (DGGE) and 16S rDNA clone library are utilized
to analyze the bacterial community structure of a higher power mixed culture MFCs The
analyzed results provide fundamental information for isolating the dominant bacteria in
mixed culture MFCs Two of dominant bacterial species has been isolated and used aone
to test power generation in MFCs The possible interaction among different bacterial
species in mixed culture is discussed
12
In the second paper one isolated exoelectrogenic bacterial strain SX-1 is
characterized It is identified as a member of the Citrobacter genus and power generation
is tested ultilizing a wide range of different substrates The electron transfer mechanism is
explored using Cyclic Voltammetry (CV) This study increases the known diversity of
power generating exoelectrogens and provides a new strain to explore the mechanisms of
extracellular electron transfer from bacteria to electrodes
The third paper shows effects on MESs performance by Fe nanoparticle decorated
anodes in the MESs The average current density produced with Fe nanoparticle
decorated anodes increased up to 59-fold higher than plain graphite anodes A whole
genome microarray is utilized to analyze the possible mechanism of enhanced current
density as responded to nanoparticle decorated anodes
The fourth paper describes the effects of carbon nanotube (CNT) modified anode on
the performance S oneidensis MR-1 in MESs Results demonstrate that current densities
produced with CNT decorated anodes are up to 56-fold higher than plain graphite anodes
The possible mechanisms of enhanced current density by CNT decorated are explored
13
Chapter 2
Bacterial community analysis of mixed consortium in higher
power density MESs
Shoutao Xu and Hong Liu
1 Introduction
The improvement in the performance of mixed culture MFCs requires an
understanding of the ecology in microbial communities of MFCs Many researchers have
attempted to characterize microbial populations and activities to elucidate the behaviors
and ecology of microorganisms in MFCs (Wrighton et al 2008 Fedorovich et al 2009)
In order to study the microbial ecology of the mixed culture in a MFC and select the
appropriate isolation medium for dominant bacterial species in the mixed culture the
fundamental steps are to analyze the bacterial diversity in the mixed culture of MFCs and
identity the dominant bacterial species in bacteria communities in MFCs
14
As for the identification of bacterial communities typically there are two general
methods The first method for identification of bacterial community is the traditional
cultivation processes using selective nutrients to promote the growth of different types of
bacteria within the samples (Amman et al 2000) The community structure can then be
assessed by identifying the isolates from the dominant colonies that were cultured This
can often be costly and laborious as each isolate has to be further studied by examining
its physiology taxonomy and reactivity to stains (Adwards et al 1989)
The second method relies on utilizing molecular techniques to analyze bacterial
community DNA Several molecular methods involving the extraction and analysis of
DNA from entire bacterial communities are used to identify genetic fingerprints of
bacteria These methods including the cloning and sequencing of 16S rDNA automated
ribosomal intergenic spacer analysis (ARISA) terminal restriction fragment length
polymorphism (T-RFLP) and denaturing gradient gel electrophoresis (DGGE) generate
profiles of bacterial community structures They can rapidly assess complex communities
from various environments (Amman et al 2000)
Due to the conservative characteristic of 16S rDNA in bacteria during the process
of evolution 16S rDNA sequencing can be used to identify different species of bacteria
Therefore the molecular techniques of denaturing gradient gel electrophoresis (DGGE)
with PCR and 16S rDNA clone library are used for analysis of the microbial diversity
These methods are more convenient and save time compared to traditional
isolationcultivation methods for microorganism analysis
In this work a biofilm bacterial community from an anode of a MFCs wase studied
by using denaturing gradient gel electrophoresis (DGGE) of PCR-amplified partial 16S
15
rRNA genes followed by cloning and sequencing of 16S rDNA The results provided
essential information for dominant bacterial isolation in mixed culture MFCs
2 Materials and methods
21 High power generation of mixed culture MFCs
Mixed cultures were originally inoculated from domestic wastewater (Corvallis
Wastewater Treatment Plant Corvallis OR) to MFCs after operating for 2 years using a
defined medium solution (Lovely 2002) with sodium acetate as the carbon source A
new MFCs was inoculated from the operating MFCs The polarization curves were
performed to measure the power generation when maximal stable power were established
(normally 3-5 batches post-inoculation) and the genomic DNA from the microbial
biofilm on the anode was extracted under the sterile conditions for further bacterial
community analysis
22 Denaturing gradient gel electrophoresis (DGGE)
DGGE with PCR is a method of analysis of bacterial community composition
based on different GC percentage of 16s rDNA in different bacterial species PCR-DGGE
comprises three steps (i) extraction of total community DNA from the sample (ii) PCR-
controlled amplification using specific oligonucleotide primers and (iii) separation of the
amplicons using DGGE For this purpose a reproducible and efficient method for total
DNA extraction is indispensable and needs to be evaluated and optimized depending on
the nature of the sample In the subsequent PCR step multiple PCR primer sets with
different resolution can be used In most PCR-DGGE applications on bacteria universal
or specific primers are targeting the 16S rRNA gene Upon electrophoresis of the PCR
16
amplicons and gel staining (using ethidium bromide silver staining or SYBR green)
DGGE gels are digitally captured and further analyzed using computer software packages
The richness of DNA fragment bands in DGGE indicates the bacterial diversity of the
sample
The detailed steps are as followed Biofilms were scratched from the anodes of high
power generated MFCs fed with sodium acetate Bacterial genomic DNA was extracted
from the biofilm samples using the DNeasy blood and tissue kit (Qiagen) according to the
manufacturerrsquos instructions The universal primer set was used to amplify the 16S rDNA
from the extracted genomic DNA then the V3 region of bacterial 16S ribosomal DNA
(rDNA) was amplified with GC-clamp primer (Muyzer et al 1993) for DGGE PCR
amplification was performed in a thermocycler DGGE of the PCR products was carried
out in a DcodeTM
Universal Mutation Detection System) The 8 (wv) polyacrylamide
gels contains from 30 to 55 denaturing gradients Electrophoresis was conducted
using a 1timesTAE (Tris-Acetate-EDTA) buffer at 130V and 60degC for 5 hours After
electrophoresis the gel was stained with ethidium bromide in 1timesTAE buffer for 15
minutes and washed in 1timesTAE buffer for 10 minutes The fragments were visualized
under a UV transilluminator The richness of single band in DGGE gel picture
preliminarily showed that the bacterial diversity in mixed culture since the single band in
gel represents one bacterial species
23 16S rDNA clone library construction method
16S rDNA-DGGE fingerprinting has proven to be particularly useful as an initial
investigation into bacterial communities and is suitable for identifying the predominant
bacteria that comprise 1 or more of cells within a given sample In the 16s rDNA clone
17
library construction process the first step is the extratction of the total genomic DNA
then the genomic DNA of the mixed culture biofilm was used as template for PCR
amplification of approximately 1500 bp of 16S rDNA with universal primers The PCR
products of 16S rDNA were purified and inverted into pGEM-T Easy vector system
before they were transformed into competent E coli The transformed cells were spread
on Luria-Bertani (LB) plates containing the selective antibiotic ampicillin and X-Gal and
incubated overnight at 37 OC after the more than 80 inverted 16s DNA fragment E coli
colonies were picked up and inoculated in the 1-3 ml liquid LB medium and left to grow
for 16 hours The plasmid DNA were extracted and sequenced The sequences were
compared directly to all known sequences deposited in GenBank databases using the
basic local alignment search tool (BLAST)
24 Bioinformatics Analysis
16S rDNA sequencing results of more than 80 colones were queried against the
GenBank and Ribosomal Database Project (RDP) databases using BLAST and
SEQUATCH algorithms (Cole et al 2007 Wang et al 2007 Cole et al 2009) The
neighbor-joining trees were constructed with the Molecular Evolutionary Genetics
Analysis package (MEGA Version 4) based on 1000 bootstrap analysis (Tamura et al
2007)
25 Dominant bacterial species band in DGGE identification
The DGGE results gave the information of dominant bacterial richness of bacterial
species in mixed culture based on the theory that single band represents one bacterial
species which showed the diversity of bacterial community in the mixed culture of a
MFC 16S rDNA clone library results provided the whole picture of bacterial community
18
including the bacterial species name and bacterial species percentage in the mixed culture
The known pure bacterial species which have been sequenced can be used as markers to
identify dominant bacterial species in DGGE The whole experimental procedure is
similar to the procedure of the mixed culture DGGE steps The first step is to extract the
collect the mixed culture biofilm genomic DNA and pure bacterial species marker
genomic DNA Then the universal primer set was used to amplify the 16S rDNA from
the extracted genomic DNA After that the V3 region of bacterial 16S ribosomal DNA
(rDNA) was amplified with GC-clamp primer (Muyzer et al 1993) for DGGE The
subsequent steps were similar to the mixed culture DGGE experimental procedure The
dominant bacterial bands in DGGE were determined by comparisons of the single band
position in the mixed culture DGGE with single pure bacterial species location in DGGE
26 Dominant bacterial species isolation and power generation
The bacterial community of mixed culture in MFCs were predominantly composed
of Geobacter sp Arcobacter sp and Citrobacter sp Two dominant bacterial species
have been isolated
Citrobacter sp SX-1 was isolated from the anode biofilm by using a solid agar
medium containing NH4Cl 6mM KCl 2mM CH3COONa (electron donor) 30 mM
FeC6H5O7 (electron acceptor) 20mM minerals and vitamins (Lovley and Phillips 1988)
Single colonies were picked up after culturing on the plate for 36 hours at 30 degC and
transferred two times on the agar plate for purification Then the isolates grew in a liquid
medium solution in anaerobic tubes containing the same constituents as the solid
medium All isolation process was operated in a glove box anaerobic chamber (Coy
Laboratory Products Grass Lake MI)
19
Arcobacter sp SX-2 was isolated from the anode biofilm of a MFC fed with sodium
acetate by using the same method of isolation of pure bacterial species Citrobacter XS-1
except using Trypticase Soy Agar with 5 Sheep Blood solid (from American type
culture collection (ATCC)) agar medium instead of Citrobacter sp medium Single
colonies were picked up after culturing on the plate for 36 hours at 30 degC and transferred
twice on the agar plate for purification Then the isolates grew in a liquid medium
solution microaerobically
Geobacter sp have been trying to isolate by using three different methods They are
the colony pickup after different condition enrichment Goebacter medium isolation and
dilution to extinction by multiple channel mini MFCs isolation however the Geobacter
sp has not been isolated yet due to some limits so far But they provide valuable
information for further isolation of Geobacter sp
Single chamber MFCs were used to evaluate power generation by different isolates
The MFCs were constructed as described previously (Liu and Logan 2004) and modified
with 3 cm2 carbon cloth anodes and 7 cm
2 carbon clothPt cathodes The total liquid
volume of each MFCs was about 15 ml with electrode spacing about 17 cm All MFCs
were operated in an autoclaved closed plastic box and sterile cotton was attached to the
outer surface of the air cathodes to prevent contamination A MFCs without bacterial
culture was used as control MFCs were inoculated with 3 ml late exponential phase
cultures of SX-1 in the medium solution reported previously (Liu and Logan 2004)
3 Results and discussion
31 Power production by mixed culture MFCs
20
A polarization curve was used to characterize current as a function of voltage in the
MFCs The polarization curves are performed by varying a series of external resistances
The results are shown in Figure 21 With MFCs acclimated to 25 Ω external resistance
the maximum power density was achieved at 65Wm2 based on the polarization data At
this point the current density is 21mAcm2 The power density was three times higher
than previous report (20 Wm2) using phosphate buffer (Fan et al 2007) Power density
was also 2 times higher than (28 m Wm2) using a bicarbonate buffer ( Fan et al 2007)
32 Bacterial community structure assessed by PCR-DGGE
Bacterial DGGE profiles for higher power generating MFCs mixed culture setups
were relatively complex (more than 10 bands) However there are three brightly stained
bands and other less intense bands (Fig 2-2) These results indicated there were probably
three dominant bacterial species in mixed culture MFCs
010
020
030
040
050
060
070
2
3
4
5
6
7
04 05 05 06 07 08 10 11 13 15 21 22 23 24
Volta
ge (
V)
Pow
er
densi
ty (
Wm
2)
Current density (mAcm2)
Power
Voltage
power density 65wm2 at current
density208 mAcm2
Figure 2-1 Polarization curves of high power mixed culture MFCs
21
33 16S rDNA clone library results
A total of 83 colonies were sequenced for 16S rDNA clone library construction They
represented the bacterial community structure in mixed culture in MFCs The results
revealed that the predominant bacterial species in mixed culture are Geobacter sp (66
of the mixed culture) Arcobacter sp (12 of the mixed culture) and Citrobacter sp (11
of the mixed culture) Those three genuses comprised 88 of the total bacterial species
(Fig 23) This result is consistent with the DGGE result which showed that three bright
bands in the DGGE gel represented three dominant bacterial species in the mixed culture
The phylogenetic analysis of the colonies (83 colony sequences) is shown in Figure 24
Figure 22 DGGE result of high power generation mixed culture MFC (1 mixed
culture 2 pure culture control)
1 2
22
34 Dominant bacterial band in DGGE identification
Geobacter
66
Arcobacter
12
others
6
Pseudomonas
1
Citrobacter
11
Clostridium
2
Anaerovorax
2 Geobacter
Arcobacter
Citrobacter
Clostridium
Anaerovorax
Pseudomonas
others
Figure 23 Bacterial species and percentage of bacterial community of mixed
culture MFC lt 1 Alcaligensgt
lt1 Commamonasgt
lt1 Pseudomonas aeruginosagt
lt1 Klebsiellagt
lt11 Citrobactergt
lt1 Wolinellagt
lt12 Arobactergt
lt2 Anaeroboraxgt
lt2 Clostridium gt
lt1 unclear bacteriagt
lt66Geobactergt
100
100
85
100
100
82
99
87
100
002
Figure 24 Phylogenetic trees of bacterial species from mixed culture MFC
23
The dominant bacterial species including Goebacter sp Citrobacter sp and Arcobacter
sp in the mixed culture are shown in Figure 25 The upper band is Arcobacter sp and
Citrobacter sp is located in the bottom band of DGGE Geobacter sp is the middle one
The analysis of the bacterial community using denaturing gradient gel electrophoresis
(DGGE) of PCR-amplified 16S rRNA gene fragments and 16S rDNA clone library
construction method showed great phylogenetic diversity of mixed culture in MFCs with
the identification of sequences derived from bacteria of the taxa deltaproteobacteria
(Geobacter sp) gammaproteobacteria (Citrobacter sp) and epsilonproteobacteria
(Arcobacter sp) The result is consistent with the conclusion that among the isolated
exoelectrogens gammaproteobacteria and deltaprotebactia appear to be dominant among
others (Parot et al 2009 Liu et al 2010)
35 The dominant bacterial species isolation and power generation
Figure 25 Dominant bacterial species identification in DGGE
A B C D
A Citrobacter sp
B Geobacter sp
C Arobacter butzmeri
D mixed culture control
24
Two dominant bacterial species Citrobacter sp XS-1 and Arcobacter sp XS-2 have been
isolated and tested for power generation The power densities generated by the two
isolates are shown in the Fig 26 Current densities generated by isolate Citrobacter sp
and Arcobacter sp were 98 mAm2 and 20 mAm
2 respectively which were much less
than that (21Am2) generated by the mixed culture from which they were isolated
Figure 26 Current density generated by different dominant isolates
The maximum current density generated by mixed culture is much higher than the
one produced by pure bacterial species Citrobacter sp XS-1 and Arcobacter sp XS-2 in
the same structure MFCs Although the Geobacter sp in our mixed culture has not been
isolated a previous study showed that maximum current density generated by Geobacter
sp was 080 Am2
(Bond and Lovley 2003) The current density results seem partially
supporting the conclusion that mixed-culture generated higher power densities than their
pure-culture counterparts in MFCs (Jung and Regan 2007) This result suggests there
may be synergistic interactions within the anode bacterial communities
0
500
1000
1500
2000
2500
Mixed culture Citrobacter sp Arcobacter Sp
Cu
rre
nt
de
nsi
ty (
mA
m2)
25
Geobacter sp are strictly anaerobic bacteria which were enriched in most of the anode
biofilms (George 2005) Citrobacter species are facultative anaerobic bacteria (George
2005) which can be found in a wide variety of habitats including in soil water and
wastewater Arcobacter sp are micro-aerobic bacteria including both environmental
nonpathogens and opportunistic human pathogens They are able to grow in aerobic
conditions but in the optimal growth under micro-aerobic conditions (George 2005)
Arcobacter sp has been discovered in the MFCs by Fedorovich (Fedorovich et al 2009)
It is interesting that the mixed culture bacterial community composed of primarily these
three bacterial species has demonstrated to generate high power at non-strictly anaerobic
condition while leading dominant bacterial species Geobacter sp (66) is strict
anaerobic bacteria and generated powder in the MFCs only under strict anaerobic
condition (George 2005) Therefore it is possible that microaerobic bacteria such as
Arcobacter sp and facultative anaerobic bacteria Citrobacter sp can facilitate to create
anaerobic condition for Geobacter sp when they utilized oxygen for growing in the
mixed culture Syntrophic communities study have showed that anaerobic bacteria and
methanogenic archaea form compact microbial structures that operate like an organ rather
than a set of microorganisms functioning independently (Stams and Plugge 2009) Some
substrates have been degradeted within these communities while they are not able to be
fermented by individual species alone interspecies electron transfer also have been
presented in these communities (Stams and Plugge 2009) Summers et al (2010) also
discovered that direct exchange of electrons happened within coculture of Geobacter
metallireducens and Geobacter sulfurreducens Therefore we believe that there are
26
more complex synergistic interactions between different bacteria species in mixed culture
in the MFCs
Acknowledgements
The authors acknowledge support from the US National Science Foundation
(CBET 0955124) We also thank Keaton Lesnik for the review of this manuscript
27
Chapter 3
New Exoelectrogen Citrobacter sp SX-1 Isolated from a Microbial Fuel Cell
Shoutao Xu and Hong Liu
Published in
Journal of Applied Microbiology 111(5)1108-1115 (2011)
ABSTRACT
Exoelectrogenic bacterial strain SX-1 was isolated from a mediator-less microbial fuel
cell by conventional plating techniques with ferric citrate as electron acceptor under
anaerobic condition Phylogenetic analysis of the 16S rDNA sequence revealed that it
28
was related to the members of Citrobacter genus with Citrobacter sp sdy-48 being the
most closely related species The bacterial strain SX-1 produced electricity from citrate
acetate glucose sucrose glycerol and lactose in MFCs with the highest current density
of 205 mA m-2
generated from citrate Cyclic voltammetry analysis indicated that
membrane associated proteins may play an important role in facilitating the electrons
transferring from bacteria to electrode This is the first study that demonstrates that
Citrobacter species can transfer electrons to extracellular electron acceptors Citrobacter
strain SX-1 is capable of generating electricity from a wide range of substrates in MFCs
This finding increases the known diversity of power generating exoelectrogens and
provided a new strain to explore the mechanisms of extracellular electron transfer from
bacteria to electrode The wide range of substrate utilization by SX-1 increases the
application potential of MFCs in renewable energy generation and waste treatment
Keywords Microbial fuel cell Exoelectrogen Citrobacter sp SX-1 Extracellular
electron transfer
29
Introduction
Microbial Fuel Cells (MFCs) technology has become an active research area
recently as a promising approach for renewable energy generation wastewater treatment
and bioremediation (Rabaey and Verstraete 2005 Lovley 2006 Fan et al 2008 Logan
2009 Liu et al 2010) The key feature of MFCs system is the microbe-catalyzed electron
transfer from organic matter to anodes Many studies have shown the presence of diverse
bacterial communities on MFCs anodes (Reimers et al 2001 Liu et al 2004b Kim et al
2005) Over 20 exoelectrogens ie microbes that can transfer electrons exocellularly to
electrodes have been reported in the past 10 years (Logan 2009 Liu et al 2010) The
exoelectrogens so far belong to diverse genetic groups including alphaproteobacteria
(Rhodopseudomonas Ochrobactrum and Acidiphilium) (Malki et al 2008 Xing et al
2008 Zuo et al 2008) betaproteobacteria (Rhodoferax) (Chaudhuri et al 2003)
gammaproteobacteria (Shewanella Pseudomonas Klebsiella Enterobacter and
Aeronomas) (Kim et al 2002 Pham et al 2003 Rabaey et al 2005 Bretschger et al
2007 Zhang et al 2008 Chung et al 2009 Rezaei et al 2009) deltaproteobacteria
(Geobacter Geopsychrobacter Desulfuromonas and Desulfobulbus) (Bond et al 2002
Bond and Lovley 2003 Holmes et al 2004ab 2006) Epsilonproteobacteria (Arcobacter)
(Fedorovich et al 2009) Firmicutes (Clostridium and Thermincola) (Park et al 2001
Wrighton et al 2008) Acidobacteria (Geothrix) (Bond and Lovley 2005) and
actinobacteria (Propionibacterium) (Wang et al 2008) A wider range of
exoelectrogenic species are expected to be discovered
Three mechanisms have been proposed for exocellular transport of electrons by
exoelectrogens without artificial mediators Some exoelectrogenic bacteria can transfer
30
electrons to electrodes through soluble redox compounds excreted by microorganisms
(Bond and Lovley 2005 Rabaey et al 2005) and others can directly transfer electrons to
anodes via outer cell membrane proteins (Myers and Myers 1992) Recently more and
more evidence supports the involvement of bacterial nanowires in extracellular electron
transport (Reguera et al 2005 2006 Gorby et al 2006) In spite of the discovery of
many bacterial species that can transfer the electrons to electrode without the need of
artificial mediators the investigation of extracellular electron transfer mechanisms was
mainly focused on a few species such as those from Geobacter and Shewanella genera
(Kim et al 1999 2002 Reguera et al 2005 2006 Gorby et al 2006) The electron
transfer mechanisms for many of the isolated exoelectrogens species are still not well
studied
In the present study we isolated a new exoelectrogen strain SX-1 from a MFC a
strain phylogenetically related to Citrobacter sp Power generation from various carbon
sources by this strain was evaluated using single chamber MFCs Plausible extracellular
electron transfer mechanisms were also discussed based on the characterization of anodic
biofilms by cyclic voltammetry (CV)
Materials and methods
Bacterial strain SX-1 isolation
Bacterial strain SX-1 was isolated from the anode biofilm of a MFC fed with sodium
acetate operated in fed-batch mode over a period of six months The original source of
the inoculum is wastewater from a local waste water treatment plant Bacterial cells were
released from the carbon cloth anode by shaking the cloth in a sterile flask with 15 mL
sterile 1 NaCl solution and glass beads (2 mm diameter) The suspension was then
31
serially diluted from 10 times to 105
times and plated on a petri dish with a solid agar
medium containing NH4Cl 6mM KCl 2mM CH3COONa (electron donor) 30 mM
FeC6H5O7 (electron acceptor) 20mM minerals and vitamins (Lovley and Phillips 1988)
Single colony was randomly selected after culturing on the plate for 36 hours at 30 degC
and purified on a new agar plate following a procedure reported previously (Chung and
Okabe 2009) Then the isolate was picked up and grew in a liquid medium solution in
anaerobic tubes containing the same constituents as the solid medium for further analysis
All isolation process was operated in a glove box anaerobic chamber (Coy Laboratory
Products Grass Lake MI)
16S rDNA sequencing and phylogenetic analysis
Bacterial genomic DNA was extracted from the isolated strain SX-1 using the DNeasy
tissue Kits (Qiagen Valencia CA) according to the manufacturerrsquos instructions The 16S
rDNA was amplified by PCR using a pair of universal primers 27F (5rsquo-
AGAGTTTGATCMTGGCTCAG-3rsquo) and 1492R (5rsquo-GGTTACCTTTGTTACGACTT-
3rsquo) (Park et al 2001) The conditions used for PCR were described previously
(Ferna ndez et al 1999) PCR products were purified ligated and cloned according to a
procedure described in a previous report (Xing et al 2008) Plasmids were isolated from
randomly selected clone colonies with a QIAprep Spin Miniprep kit (Qiagen Valencia
CA) and three plasmid inserts were then sequenced in both directions using an ABI 3730
DNA sequencer (Applied Biosystems Carlsbad CA)United States) Due to the identical
16S rDNA sequence of the three colones only one was queried against the GenBank and
Ribosomal Database Project (RDP) databases using BLAST and SEQUATCH algorithms
(Cole et al 2007 Wang et al 2007 Cole et al 2009) The 16S rDNA sequence of
32
strain SX-1 and closely related type strains were aligned using CLUSTALX software
(Thompson et al 1997) The neighbor-joining trees were constructed with the Molecular
Evolutionary Genetics Analysis package (MEGA Version 4) based on 1000 bootstrap
analysis (Tamura et al 2007)
SEM
The morphologies of SX-1 cells growing overnight in anaerobic tubes were examined
with a scanning electron microscope (FEI Quanta 600 FEG FEI Company Hillsboro
OR) The suspended cells were fixed in a 25 glutaraldehyde and 01M phosphate
buffer solution and dehydrated with a graded ethanol series from 30 to 100 After
dehydration the samples were dried in a critical point dryer and then sputter-coated with
AuPd for SEM examination (Liu and Logan 2004)
Evaluation of power generation by SX-1 in MFCs
Single chamber MFCs were used to evaluate power generation by SX-1 using various
substrates The MFCs were constructed as described previously (Liu and Logan 2004)
and modified with 3 cm2 carbon cloth anodes and 7cm
2 carbon clothPt cathodes The
total liquid volume of each MFCs was about 15 ml with electrode spacing about 17 cm
All MFCs were operated in an autoclaved closed plastic box and sterile cotton was
attached to the outer surface of the air cathodes to prevent contamination A MFC
without bacterial culture was used as control MFCs were inoculated with 3 ml late
exponential phase cultures of SX-1 in the medium solution reported previously (Liu and
Logan 2004) Seven substrates including citrate acetate glucose sucrose glycerol and
lactose were evaluated individually for power generation in a fed-batch mode in a
temperature control chamber (30 plusmn2degC) For each substrate at least 8 batches were run to
33
investigate the effect of biofilm formation on current output at fixed external resistance of
1kΩ and the same initial substrate concentration of 30 mM Phosphate buffer (100mM)
was used to maintain the solution pH and conductivity The MFCs with sodium citrate as
electron donors were also examined for maximum power output by varying the external
resistance from 20 kΩ to 4 kΩ A data acquisition system was used to record voltage data
during the MFCs operation (Keithley 2700 Keithley Instruments Inc Cleveland OH) It
took about 15 to 30 minutes for the MFCs to stabilize depending on the external
resistance At each resistance we collected at least five data at the steady condition to
make the polarization curves Averaged voltages were used to calculate the power density
(mW m-2
) according to P=IVA where I was the current V was voltage and A was cross-
sectional area of the anode
CV analysis
CV was used to characterize the oxidation and reduction reactions on the anodic surface
of the MFCs fed with citrate and operated at 7 KΩ This external resistance was selected
because the maximum power density was obtained at this resistance based on the
polarization experiment We assumed the biofilm on the anode of MFCs were well-
developed when stable power output was obtained after 3 batches of operation The
MFCs were then used directly for CV analysis at four current generating stages of the
fourth batch (1) initial exponential current increasing stage (middle point of stage about
4 h after media change) (2) current plateau stage (middle point of stage about 8 h after
media change) and (3) current decreasing stage (middle point of stage about 16h after
media change) (4) right after the complete replacement of medium solution (Figure 6)
The anode was used as working electrode the cathode as counter electrode and an
34
AgAgCl electrode was selected as the reference The CV curves were scanned from 200
to -600 mV at a rate of 5 mV s-1
using a potentiostat (G300 Gamry Instrument
IncWarminster PA) Control experiment was also conducted using new anode (without
biofilm) and new medium solution
Nucleotide sequence accession number
The 16S r DNA sequence determined in this study has been deposited in the GenBank
database under accession number HQ845373
Results
Identification of the strain SX-1
An almost-complete 16S rDNA sequence (1475 bp) of strain SX-1 was obtained and
subjected to comparative analysis with the 16S rDNA of closely related reference strains
A phylogenetic analysis revealed that strain SX-1 should be assigned to the genus
Citrobacter and was most closely related to Citrobacter sp sdy-48 EJ463782 (990
sequence similarity) and Citrobacter sp yy-21 FJ463779 (975 sequence similarity)
These three strains formed a distinct sub cluster in the neighbor-joining in which the new
isolate and Citrobacter sp sdy-48 EJ463782 formed a distinct subline (Figure 31)
The SEM image illustrated that the strain SX-1 is a rod-shaped bacterium 03-06 microm
wide and 12-17 microm long (Figure 32) Strain SX-1 colonies on nutrient agar appeared 2-
3 mm in diameter smooth moist opaque with a shiny surface after 5 days incubation
under anaerobic condition The fact that strain SX-1 grew both aerobically and
anaerobically using FeC6H5O7 as electron acceptor suggests that the strain is facultatively
anaerobic Strain SX-1 showed robust growth on citrate as a carbon source These D
35
properties were similar to those of genus Citrobacter described in Bergeyrsquos Manual of
Systematic Bacteriology (George 2005)
Figure 31 phylogenetic tree of strain SX-1 and closely related species based on 16S
rDNA sequences The tree was constructed using the neighbor-joining method
Figure 32 SEM image of planktonic cells of Citrobacter sp SX-1
Strain SX-1
Citrobacter sp sdy-48 (FJ463782)
Citrobacter sp yy-21 (FJ463779)
Citrobacter koseri E639 (ATCC 25408)
Citrobater koseri CDC 3613-63(AF025372)
Citrobacter koseri CDC 8132-86(AF025366)
Salmonella enterica SL483 (CP001138)
Salmonella enterica AKU12601(AY696668)
Shigella sonnei Ss046(CP000038)
E coli C2 (AF403733)
E coli E24377A(CP000800)88
100
100
76
44
92
53
36
0001
1 microm
36
Electricity production by strain SX-1 in MFCs
The isolated strain SX-1 was first tested for its ability to generate current from sodium
citrate at 1kΩ Current gradually increased to a maximum 50 mA m-2
after cell
inoculation and then decreased (Figure 33) After the MFCs was refilled with new
substrate solution the current recovered rapidly and reached a higher level than the first
batch After 4 batches operation the maximum current output of each batch became
stable The highest current density achieved in the MFCs with Citrobacter sp SX-1 at 1
KΩ was 98 mA m-2
(Figure 33)
Figure 33 Electricity generation by Citrobacter sp SX-1 in a single chamber MFCs
with sodium citrate (30 mM) as substrate at 1 KΩ
0
20
40
60
80
100
120
0 50 100 150 200
Curr
ent
den
sity
(m
A m
-2)
Time (h)
37
Current generation by SX-1 from other substrates including glucose lactose sodium
acetate glycerol and sucrose were also assessed at an external resistance of 1KΩ
(Figure 34) When repeatable cycles of current output were obtained for these substrates
glycerol generated the highest maximum current density of 58 mA m-2
followed by
lactose and sucrose with 29 mA m-2
and 27 mA m-2
respectively Glucose and acetate
produced the lowest maximum current density of 96 and 43 mA m-2
respectively These
results indicated that strain SX-1 can utilize a wide range of substrates for electricity
generation in MFCs but with different power generation potentials
Figure 34 Electricity generation of Citrobacter sp SX-1 using different substrates in
single chamber air-cathode MFCs at 1 KΩ (The error bars stand for the standard
deviation of 3 replicates)
0
10
20
30
40
50
60
70
Glucose Lactose Glycerol Acetate Sucrose
Curr
ent
den
sity
(m
A m
-2)
38
Polarization experiment was further conducted to determine the maximum power
density generated by strain SX-1 in the single chamber MFCs with citrate as substrate A
maximum power density of 881 mW m-2
was obtained at current density of 205 mA m-2
at an external resistance of 7 KΩ (Figure 35)
Figure 35 Power and voltage generation by Citrobacter sp SX-1 as a function of
current density using sodium citrate (30 mM) as substrate The error bars stand for the
standard deviation of the 3 voltagepower outputs obtained in three MFCs
20
30
40
50
60
70
80
90
100
01
02
03
04
05
06
07
80 110 140 170 200 230
Po
wer
den
sity
(m
W m
-2)
Vo
ltag
e (V
)
Current density (mA m-2)
Voltage
Power
39
Figure 36 Four current generating stages for CV analysis current increasing stage (stage
1) current plateau stage (stage 2) current decreasing stage (stage 3) and right after the
replacement of medium solution (stage 4) The MFCs was operated at 7 KΩ with sodium
citrate (30 mM) as substrate
Cyclic Voltammetry
To determine the presence of redox active compounds produced by SX-1 and the
location of these compounds CV scan of the anodic biofilms at four current output stages
and the supernatant of MFCs medium solution at the end of the batch experiment were
performed Reduction peaks at range of -310 ndash -350 mV and oxidation peaks at range -
100 ndash -150 mV were observed at all current generating stages (Figure 36 Figure 37A)
suggesting the presence of redox active compounds may involve in extracellular electron
transfer by Citrobacter sp SX-1 The amplitude of the peaks increased according to the
growth stage of the batch and the highest peaks were present after the current plateau
stage which indicated the redox active compounds mainly were secreted in the current
plateau stage While the current density at stage 3 (deceasing stage) was lower than that
at stage 2 (current plateau stage) the redox peak at stage 3 was higher than that in stage 2
0
50
100
150
200
250
50 55 60 65 70 75 80 85
Curr
ent
den
sity
(m
A m
-2)
Time (h)
A
A
Stage 1
Stage 2 Stage 3
Stage 4
40
A plausible explanation for this pattern was that redox active compounds were
continuously being secreted and accumulated after the current peak was reached
resulting in more redox compounds present after the plateau stage But since most of the
carbon source had already been being used up many of the redox compounds at this
stage had not been as active as those at current plateau stage due to much less electrons
were available to be transferred The bigger peak amplitude in stage 4 than in stage 1
indicates that the redox active compounds were continuously being secreted and
accumulated in the biofilm even after stable power was generated at 7 kΩ The slightly
change of the peak locations at different stages was possible due to the solution chemistry
change during the current generation process or the slightly location change of reference
electrode during the measurement (Figure 37B) When the medium solution in the MFCs
was replaced by fresh medium at the end of the batch the oxidization and reduction
peaks were still presented in the CV (Figure 37B) but no peaks were observed when the
supernatant and fresh mediums were tested in a MFCs with no biofilm (Figure 37C)
These results suggest the compounds involved in the electron transfer were located not in
the supernatant (solution) but in the biofilm
41
Figure 37 Cyclic Voltammetry analysis of Citrobacter sp SX-1 biofilms at (A) current
increasing stage current plateau stage and decreasing stage (end of the batch) (B) end of
the batch and replaced with fresh medium and (C) controls
-40E-04
-30E-04
-20E-04
-10E-04
00E+00
10E-04
20E-04
30E-04
-060 -050 -040 -030 -020 -010 000 010 020
Curr
ent
(A)
Potential (V) vs AgAgCl
Current plateau stage
Current increasing stage
Current decreasing stage
-40E-04
-30E-04
-20E-04
-10E-04
00E+00
10E-04
20E-04
30E-04
-060 -050 -040 -030 -020 -010 000 010 020
Curr
ent
(A)
Potential (V) vs AgAgCl
Current decreasing stage
Biofilm with fresh medium
-40E-04
-35E-04
-30E-04
-25E-04
-20E-04
-15E-04
-10E-04
-50E-05
00E+00
-060 -050 -040 -030 -020 -010 000 010 020
Curr
ent(
A)
Potential (V) vs AgAgCl
Supernatant
Fresh medium
A
B
C
B C
42
Discussion
Citrobacter species belonging to Gammaproteobacteria Enterobacteriales
Enterobacteriaceae are facultative anaerobic bacteria (George 2005) They can grow with
oxygen as an electron acceptor but also with a variety of electron accepters such as Fe (III)
in the absence of oxygen (George 2005) Citrobacter species can be found in a wide
variety of habitats including in soil water and wastewater Several Citrobacter species
have been studied as important bioremediation bacterium for heavy metal removals
sulfate reduction phenol degradation and chlorophenol degradation (Macaskie et al 1995
Narde et al 2004 Qiu et al 2009) Citrobacter species have also been found on the
anodic biofilm of a diesel-degrading MFCs (Morris et al 2009) However there was no
study reported that Citrobacter species can transfer electrons to extracellular electron
acceptors This finding that Citrobacter strain SX-1 can generate electricity in MFCs
increased the diversity of power generating exoelectrogens and provided a new strain to
explore the mechanisms of extracellular electron transfer from bacteria to electrode
Among the isolated exoelectrogens gammaproteobacteria and deltaprotebactia appear to
be dominant among others (Parot et al 2009 Liu et al 2010) The isolated strain
Citrobacter sp SX-1 phylogenetically belongs to gammaproteobacteria which provides
another evidence to support this observation
Most exoelectrogens utilize a limited range of substrates for power generation For
example Shewanella oniedensis MR-1 can oxidize lactate under anaerobic condition but
cannot utilize acetate for electricity generation (Kim et al 2002) Geobacter
sulfurreducens can completely oxidize acetate for power generation but it cannot utilize
simple sugars (Bond and Lovley 2003 Caccavo et al 1994)) In comparison strain SX-1
43
not only can oxidize citrate for power generation but also utilize a wide range of
substrates for power generation including glycerol glucose lactose sucrose and acetate
Interestingly the higher current density generated by strain SX-1 from glycerol a main
by-product of biodiesel production process suggests that strain SX-1 may be potentially
used for harvesting energy from biodiesel wastes using MFCs However the current
density generated by strain SX-1 (205 mA m-2
) is lower than that (805 mA m-2
at 1 kΩ)
generated by the mixed culture from which the SX-1 was isolated indicating the
existence of other higher power generating bacteria andor complex ecology in the mixed
culture community
Understanding mechanisms of microbial extracellular electron transfer is critical for
enhancing the electron transfer rate from bacteria to electrode through metabolic or
genetic engineering of exoelectrogens Since the spent supernatant solution of MFCs run
with SX-1 showed no redox properties the redox compounds produced by SX-1 may
have been retained in the biofilm Alternatively SX-1 may utilize one or both of the
other two known extracellular electron transfer mechanisms For example it is possible
that strain SX-1 transfers electrons to the anode via cell-bound outer membrane proteins
The two distinct oxidation and reduction peaks observed at -100mv- -150 mV and -310--
350 mV of the SX-1 biofilm have a midpoint potential range from -205mV to -250mV
which is close to the redox potential of some c-type cytochromes (Field et al 2000 Meitl
et al 2009) which are well-known to play an important role in extracellular electron
trasnfer (Eggleston et al 2008 Field et al 2000 Meitl et al 2009) Field et al (2000)
reported that the c-type cytochromes CymA had the midpoint potentials of -229 mV
Meitl et al (2009) and Eggleston et al (2008) reported that the c- type cytochromes
44
OmcA in Shewanella oneidensis MR-1 had the midpoint potentials of -201 mV and -208
mV
While itrsquos possible that SX-1 may also have conductive appendages to enhance the
extracellular transfer electron similar to the nanowires discovered in some Geobacter
and Shewanella species (Reguera et al 2005 2006 Gorby et al 2006) further
investigation on the presence of these appendages on the cell surface of SX-1 is needed
Acknowledgements
The authors acknowledge support from the US National Science Foundation
(CBET 0955124) We also thank Teresa Sawyer for her helps with the SEM analysis and
Jeremy Chignell and Yanzhen Fan for their review of this manuscript
45
Chapter 4
Enhanced Performance and Mechanism Study of Microbial Electrolysis Cells
Using Fe Nanoparticles Decorated Anodes
Shoutao Xu Hong LiuYanzhen Fan Rebecca Schaller Jun Jiao Frank Chaplen
Published in
Applied Microbiology and biotechnology 93(2)871-880 (2012)
46
ABSTRACT
Anode properties are critical for performance of microbial electrolysis cells (MECs) In
the present study Fe nanoparticle modified graphite disks were used as anodes to
investigate the effects of nanoparticles on the performance of Shewanella oneidensis MR-
1 in MECs Results demonstrated that average current densities produced with Fe
nanoparticle decorated anodes were up to 56-fold higher than plain graphite anodes
Whole genome microarray analysis of the gene expression showed that genes encoding
biofilm formation were significantly up-regulated as response to nanoparticle decorated
anodes Increased expression of genes related to nanowires flavins and c-type
cytochromes indicate that enhanced mechanisms of electron transfer to the anode may
also have contributed to the observed increases in current density The majority of the
remaining differentially expressed genes were associated with electron transport and
anaerobic metabolism demonstrating a systemic response to increased power loads
Keywords Microbial electrochemical system microbial fuel cell microbial electrolysis
cell nanotechnology differential gene expression DNA microarray
47
Introduction
Microbial electrochemical systems (MESs) have been intensively studied since Lewis
achieved practical advances in this field (Logan 2007) however they attracted much
research attention in recent years due to their promising applications in renewable energy
generation bioremediation and wastewater treatment In a MES microorganisms
interact with electrodes via electrons catalyzing oxidation and reduction reactions at the
anode and the cathode The most-described type of MESs is microbial fuel cells (MFCs)
in which useful power is generated from electron donors typically biodegradable organic
materials (Logan et al 2006) Various novel MESs have recently been developed to
produce hydrogen (microbial electrolysis cells (MECs)) (Liu et al 2005 Logan et al
2008 Rozendal et al 2006 Schaller et al 2009) to power an autonomous sensor or
sensor network (Ringeisen et al 2006) to reduce bioremediation targets (Lovley 2006
Shea et al 2008) Sukkasem et al 2008) and to desalinate water (Cao et al 2009) The
key feature shared by these systems is the microbe-catalyzed electron transfer from
organic matter to electrodes (anodes) (Rabaey et al 2004) Enhancing the anodic current
output which highly depends on the performance of the electrodes is critical for the
successful application of all these processes (Logan et al 2007 Park and Zeilus 2002
2003)
Nanomaterials have received much attention from researchers in the context of
microbiology due to their unique physical electrical and chemical properties which
facilitate the study of interactions between bacteria and surfaces (Liu 2006) Previous
studies have demonstrated that electrodes decorated with different nanostructures such
as carbon nanotubes nanostructured polyanilinetitanium dioxide composite and titania
48
nanotubes have the potential to enhance power generation in MFCs (Morozan et al 2007
Qiao et al 2008 Quan et al 2005) Furthermore our recent study shows nanoparticle
(NP) decorated anodes greatly increased the electrochemical electron transfer rate in
MFCs (Fan et al 2011) However Fe NPs as an ideal candidate for decorating electrodes
because of respectively low price and high conductivity compared to other materials has
not been focused to study in MECs
S oneidensis MR-1 an important electrochemically active bacterial strain has been
exploited for the electricity generation in MFCs (Logan et al 2007 Logan 2009 Park
and Zeikus 2002) The availability of genome sequence for this strain makes it possible to
use transcriptome assays to globally measure the responses to different growth conditions
and environmental stresses (Murray et al 2001) S oneidensis MR-1 gene expression
response to heat shock cold shock Cr (VI) and U (VI) reduction chromate stress and
iron and acid tolerance have been studied in the past several years (Bencheikh-Latmani et
al 2005 Brown et al 2006 Gao et al2006 Gao et al 2004 Yang et al 2008) However
no studies have been focused on the Shewanella gene expression response to NPs in
MESs
The mechanism of increased electron transfer rate exhibited by nanoparticle decorated
electrodes is not well understood yet In the present study graphite disks decorated with
Fe NPs were used as anodes to explore the effects of nanostructures on current generation
in a multi-anode MECs DNA microarrays were utilized to investigate differences in the
global gene expression profile of S oneidensis MR-1 grown on plain versus Fe NPs
decorated anodes
Materials and methods
49
Bacterial cultures
S oneidensis MR-1 was purchased from American Type Culture collection (ATCC
700550) and stored as freezer stocks at -80degC Stock cells of S oneidensis MR-1 was
grown aerobically using a medium containing 30 gL Trypticase Soy Broth (BD 211825
Becton Dickinson Company Franklin lakes NJ) at 30degC Cells in early stationary
growth phase (optical density of 15-16 at 600nm (Spectrophotometer UV-1700
Shimadzu Columbia MD) were washed two times then injected into the chamber of the
MECs for current production Trypticase Soy Broth medium with 30 mM sodium lactate
as electron donor was used in MECs Phosphate buffer (100mM) was used to maintain
the solution pH 7 and solution conductivity at 15 mScm
Characterization of nanostructured anodes
Superfine isomolded graphite disks (16 cm x 05 cm GraphiteStorecom Inc) were
polished with ultrafine sand paper (2000 grit 3M Company) as the base for the NP
decorated and control anodes Fe NPs decorated anodes were fabricated by thermal
annealing of metal thin film coated graphite disks Thin films of Fe were firstly deposited
on the polished graphite disk by using sputter coating for 95 min and the samples then
were annealed in the Chemical vapor deposition (CVD) at 750degC for 20 min to form Fe
NPs Fe-NPs were produced with a diameter of 200plusmn100 nm and a density range
(distance between NPs) of 200plusmn100 nm with a coating time of 95 min SEM images of
Fe NPs decorated anode and control surfaces were shown in the Figure 41
50
Figure 41 SEM images of Fe NPs decorated anodes (A) Graphite disk control (B) Fe
Nanoparticle decorated
Multiple channel MECs construction and operation
A MECs with removable multiple anodes with each effective anode area 07 cm2
was
constructed and used to evaluate the effects of nanostructure on current density of
according to a previous report (Fan et al 2011) The cathode was made of wet-proof
(30) carbon cloth (type B E-TEK Division Inc USA) coated with PtC (20 E-TEK
Division Inc USA) following a previously reported procedure (Liu et al 2005) The
final platinum loading was 05 mgcm2 per projected cathode area The size of cathode
(150 cm2) was 25 times larger than the total effective area of the 8 graphite disk anodes
(56 cm2) to prevent cathode limitations on the performance of the MECs system All
MECs with Fe-NP decorated anodes and control anodes were tested for current
generation for 24 h with medium (without bacteria) before injecting the bacterial cells in
order to determine whether the Fe-NP decorated anodes demonstrate chemical current
generation compared to control anodes All the testing anodes were set up in the MECs
B
500 nm
A
500 nm
51
at once so that the S oneidensis MR-1 culture was applied to all of them simultaneously
Short electrode spacing (17 cm) was used in the MECs design to reduce the internal
resistance A voltage of 06 V was applied the MECs for current generation after the cells
of S oneidensis MR-1 were inoculated into MECs at early stationary phase A multimeter
with a data acquisition system (2700 Keithly USA) was used to monitor the current
change by measuring the voltage (every 300 seconds) drop through a resistor at 300 ohm
Fig 4 2 Effects of Fe-NP decorated anodes on the current density in MECs Control is
plain graphite disk anode Fe-NP is the anode with Fe-NP decoration
Microarray analysis
Biofilms for whole gene microarray analysis were aseptically removed from the plain
and Fe-NP decorated anodes of MECs after 110 hours further incubation at 30oC when
current density obviously started to decease Total RNA was extracted using Trizol
(Ambion inc AustinTX) following the manufacturerrsquos instructions Integrity of the
52
RNA samples was confirmed using the Agilent 2100 Bioanalyzer (Agilent Technology
Inc Palo Alto CA) The same amount of RNA (200 ng) from each sample was used for
further analysis after amplification by using the MessageAmpTM
II-Bacteria Prokaryotic
RNA Amplification kit (Ambion AustinTX) according to the manufacturerrsquos
instructions All of the 4295 genes (resource from NCBI) represented on the S oneidensis
MR-1 whole-gene microarrays were ordered from Roche NimbleGen Inc (Madison WI)
Biological triplicates of biofilms on the Fe-NP decorated anodes and control were
analyzed respectively cDNA synthesis labeling and hybridization were carried out by
the CGRB Core Laboratories at Oregon State University DNASTAR ArrayStarTM
3
software was used to identify genes that were up- or down-regulated more than 2-fold
when grown on the nanoparticle modified anodes using the unpaired two sample t-test
with a cutoff p-value of 005 The complete microarray data set generated in this study is
deposited for public access in the Gene Expression Omnibus
(httpwwwncbinlmnihgovgeo) under accession number GSE31535
Results
Enhancement of current generation by using NP decorated anodes in MECs
The current density generated by Fe-NP decorated anode MECs with S onidensis MR-1
increased to approximately 43 microAcm2 20 h following inoculation and then slowly
decreased Figure 42 shows that the maximum current density achieved by NP
decorated anodes was 83 times higher than that (51 microAcm2) generated by the control
(plain graphite disk) The average current density improvement of 110 hours was 59
times of that generated by the control The current densities generated in MECs in the
53
absence of bacteria were negligible (03 microAcm2) which demonstrated that the current
enhancement observed with the Fe-NP decorated anodes was biologically-derived
Global transcriptome analysis
Whole-genome DNA microarrays were used to attain a comprehensive general
overview
of the transcriptional response to Fe-NP decorated anodes by S oneidensis MR-1 in MEC
Of a total 4295 genes assayed 392 (188 induced and 204 repressed) exhibited significant
(P lt 005) differential expression at a 2-fold (log (expression ratio 1) level in 3
replicates in response to Fe-NPs These
total gene numbers present 9 of the 4295 open
reading frames (ORFs) presented on the array
Fig 43 Differentially expressed genes grouped by functional classification according to
the TIGR S oneidensis genome database (wwwtigrorg) Columns 1 amino acid
biosynthesis 2 biosynthesis of cofactors prosthetic groups and carriers 3 cell
envelope 4 cellular processes 5 central intermediary metabolism 6 DNA metabolism
7 energy metabolism 8 fatty acid and phospholipid metabolism 9 other categories 10
protein fate 11 protein synthesis 12 purines pyrimidines nucleosides and nucleotides
13 regulatory functions 14 signal transduction 15 transcription 16 transport and
binding proteins 17 unknown function 18 hypothetical proteins
Up-Regulated
-75
-50
-25
0
25
50
75
100
125
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18
Down-Regulated
Num
ber
of
Gen
es
54
Figure 43 summarizes the overall genomic response of S oneidensis MR-1 to Fe-NP
decorated anodes by grouping the differentially expressed genes into their functional role
categories based on The Institute for Genomic Researchrsquos annotation (Version 8) of the
MR-1 genome sequence The wide distribution of putative functional roles attributed to
the differentially expressed genes indicated the extent of the molecular response of
Soniedenis MR-1 to the NP decorated anodes Among genes with defined function a
large number of up-regulated genes were associated with cellular processes (group 4)
energy metabolism (group 7) other categories (group 9) and hypothetical proteins (group
18) While those genes encoding proteins involved in amino acid biosynthesis (group 1)
cellular processes (group 4) substrate transport (group 16) and hypothetical proteins
(group 18) were among the most down-regulated genes Two groups of genes that were
the most noticeable among the functional gene groups genes related to energy
metabolism (group 7) and genes related to amino acid biosynthesis (group 1) The ratios
of up-regulated genes to down-regulated genes in these functional groupings were much
higher than other functional gene groupings there were 21 up-regulated genes and 7
down-regulated genes in energy metabolism gene group and 2 up-regulated and 22 down
regulated genes in the amino acid biosynthesis group
Genes related to biofilm formation
In order to determine whether there was a correlation between the observed biofilm
enhancements by NP decorated anodes and the genes related to biofilm formation the
significant modified genes related to biofilm formation were examined and summarized
in the Table 41 These included SO_3223 and SO_3228 genes encoding the flagellum
proteins (Thormann et al 2004) which has critical impacts on initial attachment to the
55
surface as swimming motility functions were up-regulated over 10 and 09 fold
respectively The gene clusters SO_4178-SO_4180 which are thought to be essential for
formation of a three-dimensional biofilm were up-regulated from 12 to 13 fold
(Thormann et al 2004) In addition the genes SO_4105 encoding IV pilus which is
known to represent a critical factor for biofilm formation on a biotic surfaces (Thormann
et al 2004) were also induced 08 fold (Table 41) The genes related to flagellum
motility (SO_4286 SO_1529 and SO_1530) were also induced slightly (Thormann et al
2004)
Table 41 Expression levels of genes related to biofilm formation
Locus Tag Gene product descriptions Fold change
SO_3228 flagellar basal-body MS-ring and collar protein FliF 10
SO_3223 flagellar hook-length control protein FliK 09
SO_4103 MshA minor pilin protein MshD 05
SO_4105 MSHA major pilin protein MshA 08
SO_4178 expressed protein of unknown function MxdC 12
SO_4179 inner membrane family 2 glycosyltransferase MxdB 13
SO_4180 diguanylate cyclase-like protein MxdA 10
a fold changes were presented after log2
b positive number represented up-regulated and negative represented down-regulated
Genes related to energy metabolism
A closer consideration of energy metabolism genes was undertaken as anaerobic
metabolism for electron generation and electron transport functions may play critical
roles in enhanced current density generation in MECs Genes associated with energy
56
metabolism and with significantly modified gene expression levels are summarized in the
Table 42 Several of the up-regulated genes were related to formate dehydrogenase
proteins Most notably four genes related to formate dehydrogenase were up-regulated
more than 2-fold SO_0102 SO_0103 SO_0104 and SO_0107 The other genes
expression related to formate dehydrogenase proteins SO_4513 SO_4514 SO_4515 also
significantly increased Another interesting gene is that encoding the cytochrome c
oxidase protein gene SO_4609 which was up-regulated 11 fold while the gene
SO_0479 related periplasmic octaheme cytochrome c MccA was down-regulated
However SO_1519 the gene encoding lactate dehydrogenase was down-regulated 12
fold despite lactate being the primary carbon source in the media
Table 4 2 Genes related to anaerobic growth and electron transfer with significantly change expression level
Locus Tag Gene product descriptions Fold
change
SO_0101 nitrate-inducible formate dehydrogenase molybdopterin-binding
subunit FdnG
18
SO_0102 formate dehydrogenase nitrate-inducible iron-sulfur subunit 21
SO_0103 formate dehydrogenase nitrate-inducible cytochrome b556 subunit 24
SO_0104 formate dehydrogenase accessory protein FdhE 22
SO_0107 formate dehydrogenase accessory protein fdhD 24
SO_0397 quinolfumarate reductase menaquinol-oxidizing subunit FrdC_2 12
SO_0452 thioredoxin 2 15
SO_0903 Na-translocating NADH-quinone reductase subunit B NqrB_1 12
SO_1012 NADH-ubiquinone oxidoreductase subunit K NuoK 12
SO_1013 NADH-ubiquinone oxidoreductase subunit J NuoJ 12
SO_1363 hydroxylamine reductase 13
57
SO_2417 ferredoxin cofactor maintenance protein YfaE 10
SO_3922 formate dehydrogenase cytochrome b Fdh -12
SO_4513 formate dehydrogenase molybdopterin-binding subunit _2 15
SO_4514 formate dehydrogenase FeS subunit FdhB_2 12
SO_4515 formate dehydrogenase cytochrome b subunit FdhC_2 14
SO_4609 aa3 type cytochrome c oxidase subunit III CoxC 11
SO_0479 periplasmic octaheme cytochrome c MccA -12
SO_0847 periplasmic nitrate reductase ferredoxin component NapG -15
SO_1519 L-lactate dehydrogenase iron-sulfur cluster-binding protein LldF -10
SO_1251 ferredoxin 4Fe-4S -11
SO_37411 hypothetical inner membrane protein -11
a fold changes were presented after log2
b positive number represented up-regulated and negative represented down-regulated
Flavin and cytochrome related genes
Flavins can be secreted by Shewanella species as electron shuttle to facilitate
extracellular electron transfer (Cansterin et al 2008) C-type cytochromes play the
important roles on the process of extracellular electron transfer (Shi et al 2007) The
genes related to flavins synthesis and the genes encoding the cytochromes electron
transport proteins were shown in the table 43 the genes SO_1414 and SO_3058 related
to flavins synthesis was up-regulated to 03 and 01 folds respectively The genes related
to s C-type cytochromes SO_0135 SO_4105 SO_0169 SO_1778 are increased to 10
08 05 04 01 folds respectively However most of genes have shown no significant
changes (less one-fold change) as response to nano particle decorated anode in MECs
58
Table 43 Expression level of flavin and cytochrome genes
Locus Tag Gene product descriptions Fold
change
SO_1414 flavocytochrome c flavin subunit putative 03
SO_3468 riboflavin synthase subunit alpha -02
SO_3058 flavocytochrome c flavin subunit 01
SO_4105 MSHA pilin protein MshA 08
SO_0169 general secretion pathway protein GspG 05
SO_1778 outer membrane decaheme cytochrome c lipoprotein MtrC 04
SO_1779 outer membrane decaheme cytochrome c OmcA 01
SO_0135 lipoprotein of unknown function DUF333 10
SO_0136 conserved hypothetical inner membrane protein 01
a fold changes were presented after log2
b positive number represented up-regulated and negative represented down-regulated
Other genes with significantly modified expression levels
These genes over 15 fold change and possibly related to current enhancements were
shown in Table 44 SO_3104 (expressed inner membrane protein) was over expressed
and the gene SO_2194 which encodes outer membrane protein (OmpA family protein)
was repressed as response to nanoparticle decorated anode however it is unclear the
relationships of this modified membrane protein genes with the current enhancement
Another interesting phenomenon was possible co-regulation of several gene clusters
indicating possible operon associations This includes three sets of genes that were
induced more than 15 fold SO_0108SO_0109 and SO_1042SO_1043SO_1044 The
59
consistency of expression of these genes under the NP conditions provides basic evidence
to support operon structure However the correlation of these significant changed gene
with current density enhance are unclear which showed there are unknown multiple and
complex responses of S oneidensis MR-1 to nanoparticle anode of MECs
Table 44 Other genes with significantly changed expression levels
Locus Tag Gene product descriptions Fold change
SO_0101 nitrate-inducible formate dehydrogenase molybdopterin-binding
subunit FdnG
18
SO_0108 integral membrane protein of unknown function DUF39 YedE 17
SO_0109 SirA family protein YedF 17
SO_0275 N-acetyl-gamma-glutamyl-phosphate reductase -31
SO_0277 ornithine carbamoyltransferase -21
SO_0279 argininosuccinate lyase -24
SO_0404 zinc dependent metalloprotease domain lipoprotein 21
SO_0956 alkyl hydroperoxide reductase F subunit -20
SO_1042 amino acid ABC transporter ATP-binding protein -18
SO_1043 amino acid ABC transporter permease protein -27
SO_1044 amino acid ABC transporter periplasmic amino acid-binding protein -27
SO_1072 chitin-binding protein putative 21
SO_1405 transglutaminase family protein 20
SO_1822 TonB-dependent receptor putative -22
SO_2069
1-(5-phosphoribosyl)-5-[(5-phosphoribosylamino)
methylideneamino] imidazole-4-carboxamide isomerase
-19
SO_2070 amidotransferase HisH -26
SO_2071 imidazole glycerol-phosphate dehydratasehistidinol phosphatase -27
60
SO_2072 histidinol-phosphate aminotransferase -28
SO_2073 histidinol dehydrogenase -30
SO_2194 OmpA family protein -36
SO_2195 inter-alpha-trypsin inhibitor domain protein -27
SO_2767 asparagine synthetase B -26
SO_2945 prophage LambdaSo tail fiber protein 20
SO_2963 prophage LambdaSo major capsid protein HK97 family 20
SO_2987 prophage LambdaSo hypothetical protein 19
SO_2988 prophage LambdaSo expressed protein of unknown function 18
SO_3104 expressed inner membrane protein 131
SO_3408 conserved hypothetical inner membrane protein -18
SO_3585 azoreductase putative -18
SO_3586 glyoxalase family protein -17
SO_3687 curli production assemblytransport component CsgE putative -18
SO_38191 hypothetical ammonia permease -19
SO_4014 AcrBAcrDAcrF family protein 18
SO_40151 type I secretion system membrane fusion protein RND family 18
SO_4054 510-methylenetetrahydrofolate reductase 18
SO_4245 N-acetylglutamate synthase -22
SO_45251 hypothetical transcriptional regulator LysR family -17
SO_4527 integral membrane domain protein -19
SO_4705 transcriptional regulator putative -19
a fold changes were presented after log2
b positive number represented up-regulated and negative represented down-regulated
Discussion
61
The addition of NP to anodes significantly impacts current densities with the elemental
composition being a critical factor The chemical composition of NPs significantly affects
the current enhancement in MECs (Fan et al 2011) because the chemical composition of
NP properties is a critical factor in determining the conductivity which significantly
affects the efficiency of electron transfer from bacteria to electrode Fe is an ideal
candidate for NP formation for current enhancement in MESs because of high
conductivity and low price compared to gold and other conductive materials Gold NPs
has also been studies for enhancement of electricity conductivity (Bao et al 2008)
However the disadvantages of the high price of gold limit their practical application in
MECs (Fan et al 2011) The different size and density of the same composition of NP
also has essential effects on the current density enhancement (Fan et al 2011) In this
study the results showed the current density enhancement with Fe-NP size range of
200plusmn100 nm and density range of 200plusmn100 nm is considerable enhancement consequence
There are a number of reports of studies of current density enhancement in MFCs
by nanostructured anodes (Fan et al 2011 Qiao et al 2008 Sharma et al 2008 Tsai et al
2009) Based on bacterial behavior on nanostructured anode surface Qiao et al (2008)
believes that Escherichia coli cells on the nanostructured electrode surface produce hair-
like structures similar to pili that could facilitate the electron transfer between the cells
and electrode The production of hair-like structures is believed to be stimulated by
nanostructures and could play the key role on current density enhancement in pili-
producing bacteria in MFCs (Qiao et al 2008) S oneidensis can produce electrically
conductive bacterial nanowires which have similar structure to the hair-like structures in
Ecoli (Gorby et al 2006 Reguera et al 2005) A number of possible genes related
62
nanowires of S oneidensis have been identified including mshA (SO_4105) and gspG
(SO_0169) (Konstantinidis et al 2009) They are up-regulated 08 and 05 fold in the
current study respectively even though they are not significant induced This result
supported the increased nanowire genes had contributions to enhance current density in
MFCs by NPs decorated anodes
Based on the previous research on electron transfer mechanisms for S oneidensis it is
known that self-produced mediators (flavins) and multihaem c-type cytochromes (c-Cyts)
may play critical roles in the electron transfer of S oneidensis to extracellular insoluble
electrode (Canstein et al 2008 Myers and Myers 1992 Scott and Nealson 1994 Shi et al
2007) The flavins secreted by S oneidensis can facilitate to transfer electrons from
bacteria to solid anode electrodes by acting as extracellular electron shuttles (Canstein et
al 2008) In this study the genes encoding flavins (SO_1414 SO_3468 and SO_3058)
had no significant increase (less than 1 fold change) as a response to nanoparticle
decorated anode in MECs which suggests that increased flavin gene expression does not
significantly contribute to the current density enhancements in nano-particle decorated
anode The genes encoding cytochromes electron transport proteins (SO_4104 SO_0417
SO_177879 SO_013536) had no significant increase (less than 1-fold change)
However those genes were up-regulated slightly These results suggest the increased
expression flavin and c type cytochromes genes had partial contributions even not
significantly to enhance current density in MFCs by NPs decorated anodes
The thickness of bacterial biofilm on the anode can affect the power generation in
MFCs (Biffinger et al 2007 Heydorn et al 2000) since increased biofilm thickness on
the anode represents more active bacteria involving in the process of electron generation
63
and transfer in MFCs Biofilm formation related genes included flagellar genes IV pilus
genes and other genes The flagellar related genes SO_3228 SO_3229 (fliB and fliK) and
IV pilus genes SO_4105 (mshA) were induced from 19 to 09 fold the genes (SO_4178
-4180) related to formation of a three-dimensional biofilm were up-regulated from 10 to
13 folds even though the genes related bacterial motility protein (SO_ 4286 SO_4287
SO_1529 and SO_1530) are induced slightly (less than 0 5 fold) However the motility
genes most probably play a role for initial biofilm attachment on the anode surfaces
(Thormann et al 2004 Thormann et al 2006) with the rest of biofilm genes being more
essential for biofilm maintenance in mature cultures as were presented at harvest in this
study This enhancement of biofilm density under conditions of increase current density
has been observed for gold NP decorated anodes using confocal light microscopy (data
not shown) but has not yet been confirmed for Fe NP decorated anodes In any event an
increase in the expression level of genes related biofilm formation therefore also
supported the possibility of enhanced biofilm formation on NPs decorated anodes thus
facilitating electron transfer from bacteria to anodes
Beliaev et al described a model of the Fe reduction pathway in S oneidensis MR-1 using
solid substrates as the electron acceptor (Beliaev et al 2001) This model proposes that
electrons are generated and released in cytoplasm then transferred to the quinone pool
eg menaquinone in the cytoplasmic membrane The reduced menaquinone in turn
reduces CymA and from there electrons are then transferred to a periplasmic shuttle (eg
CctA) and then to MtrA a periplasmic soluble decaheme cytochrome c located in
associated with the outer membrane via interaction with MtrB Finally surface displayed
outer membrane proteins decaheme cytochromes (OmcA MtrC or MtrF) can transfer
64
electron to Fe or other solid substrates (Beliaev et al 2001)) Bencheikh-Latmani et al
demonstrated that the same electron transport pathway may be used for more than one
electron acceptor through studies of the response of S oneidensis MR-1 to U(VI) and
Cr(VI) under anaerobic conditions particularly focused on the critical genes
(SO_1777mtrA SO_1776mtrB SO_1778mtrC) (Bencheikh-Latmani et al 2005)
However our studies showed that these genes had no significant expression changes in
response to nanostructured anodes Contrastingly one of expressed inner membrane
protein genes SO_3104 and one of cytochromes genes SO_4609 encoding cytochrome c
oxidase subunit III demonstrated a 13-fold and 11-fold up-regulation respectively in this
study while the gene SO_2194 which encodes outer membrane protein (OmpA family
protein) was down regulated 36 fold Our results therefore suggest that S oneidensis
MR-1 does not utilize the same set of core critical genes (SO_1777 mtrA SO_1776
mtrB SO_1778 mtrC) products to transfer electrons in all instances This is the similar
result with the study of Bretschger et al (2007) which indicated that the electron
transport system in S oneidensis MR-1 is complex with several different proteins able to
participate in electron transfer to the anode of MECs
It should also be mentioned that the gene (SO_1519) encoding lactate
dehydrogenase repressed (-12 fold) here suggesting that lactate was depleted at the time
of cell harvest from the MECs and that other pathways of anaerobiosis had been activated
including those associated with amino acid uptake and consumption the growth media
was a complex mixture including yeast extract In particular the catabolic pathway for
histidine of S oneidensis MR-1 a pathway that may formate as an intermediate is up-
regulated with the genes for histidine ammonia lyase SO_0098 SO_3057 and SO_4374
65
up-regulated 13 11 and 11-fold respectively The possible production of formate
during histidine degradation may in turn explain the high levels of formate
dehydrogenates as evidenced by upregulation of SO_0102 SO_0103 SO_0104 and
SO_0107 annotated as formate dehydrogenase proteins These genes had the highest
levels of up-regulated expression as response to Fe NP decorated anodes (greater than 2-
fold)
Acknowledgements
This research was partially supported by the US National Science Foundation CBET
0828544 and the funds from ONAMIDOD (ARL-DOD Cooperative Agreement
W911NF-07-2-0083) We thank Barbara Gvakharia and Caprice Rosato for helpful
suggestions We also thank Margaret Romine of Pacific Northwest Laboratories for
valuable comments on manuscripts We also thank anonymous reviewers for significant
suggested improvements imparted as part of prior review of this manuscript
66
Chapter 5
Global Transcriptome Analysis of the Response of Shewanella oneidensis MR-1 to
Carbon Nanotube Decorated Anodes in Microbial Electrochemical Systems
Shoutao Xu Yanzhen Fan Rebecca Schaller Frank Chaplen Jun Jiao Hong Liu
67
Abstract
Shewanella oneidensis MR-1 is an important model microorganism for metabolic studies
on the effects of different environmental factors because of its diverse respiratory
capabilities Carbon nanotube (CNT) modified graphite disks were used as anodes to
investigate the effects of nanomatrials on the performance of S oneidensis MR-1 in
microbial electrolysis cells (MECs) The current densities produced with CNT decorated
anodes were on average 56-fold higher than plain graphite anodes Whole genome
microarray analysis of gene expression showed that up-regulation of cytochromes c genes
associated with extracellular electron transfer are strongly correlated to current increases
in CNT decorated anodes Genes involved in flavin biosynthesis also contribute to
current increase in CNT decorated anode MECs
Keywords Microbial electrochemical system microbial fuel cell microbial
electrolysis cell carbon nanotube gene expression DNA microarray
68
Introduction
Shewanella oneidensis MR-1 is an important model microorganism for metabolic
studies of the effects of different environmental factors because of its diverse respiratory
capabilities It has been used for transcriptome analysis to investigate the responses to
different growth conditions and environmental stresses (Murray et al 2001) Gene
expression patterns under different conditions such as heat shock cold shock Cr (VI) U
(VI) reduction chromate stress iron and acid tolerance have been studied previously
(Bencheikh-Latmani et al 2005 Brown et al 2006 Gao et al2006 Gao et al 2004
Yang et al 2008) More recently it has been exploited as a model species for power
generation in microbial electrochemical systems (MESs)which have potential
applications in renewable energy generation bioremediation and wastewater treatment
(Logan et al 2007 Logan 2009 Park and Zeikus 2002 Fan et al 2011 Xu et al 2012)
In a MES electrochemically active microorganisms oxidize organic matter in the an
ode chamber to release electrons Electrons are then transferred to the anode electrode thr
ough various mechanisms (Logan et al 2006 Lovley 2006 Rabaey et al 2003) and fina
lly travel to the cathode electrode and combine with the terminal electron acceptor The
key feature of MESs is the microbe-catalyzed electron transfer from the organic matter to
the anode (Rabaey et al 2004) Enhancing the current output which highly depends on
the performance of the anode electrode is critical for the successful application of MESs
(Logan et al 2007 Park and Zeilus 2002 2003) Several nanomaterials including TiO2
gold (Au) and palladium (Pd) nanoparticles have been utilized to decorate electrodes to
enhance power density in MESs (Qiao et al 2008 Quan et al 2005 Fan et al 2011) The
possible mechanisms for increased current densities have been studied by using Fe
69
nanoparticle-decorated anodes (Xu et al 2012)
Carbon nanotubes (CNT) are cylindrically shaped nanomaterials with extremely
high surface area excellent electrical conductivity and chemical inertness (He et al
2005a Serp et al 2003) These unique properties make CNT a promising electrode
material (Liang et al 2008) The biocompatibility of microorganisms and carbon
nanostructures has been evaluated (Morozan et al 2007) The effects of CNT on anodic
biofilms have been tested in MFCs (Liang et al 2011) However no studies have been re
ported on global transcriptome analysis for the response of S oneidensis MR-1 to CNT
decorated anode in MECs
In the present study the graphite disks decorated with CNT were used as anodes to
investigate the effects of nanomaterials on current generation in multi-anode MECs
DNA microarrays were used to analyze differences in the global gene expression profile
of S oneidensis MR-1 grown on plain versus CNT decorated anodes
Materials and methods
Bacterial cultures S oneidensis MR-1 was purchased from American Type Culture
collection (ATCC 700550) and stored as freezer stocks at -80degC Stock S oneidensis
MR-1 cells were grown aerobically using M1 medium at 30degC Cells in early stationary
growth phase (optical density of 15-16 at 600nm (Spectrophotometer UV-1700
Shimadzu Columbia MD) were injected into the chamber of the MECs for current
production Sodium lactate (final concentration 30mM) was added as the additional
electron donor
Fabrication and characterization of nanostructured anodes Superfine isomolded
graphite disks (16 cm x 05 cm GraphiteStorecom Inc) were polished with ultrafine
70
sand paper (2000 grit 3M Company) as the base for the CNT decorated and control
anodes Aligned multi-walled carbon nanotube samples were synthesized via plasma
enhanced chemical vapor deposition (PECVD) Samples superfine isomolded graphite
pillars with a 16 cm diameter and 05 cm thickness were sputtered with a thin film of Ni
in a CRC 300 magnetron sputter coater for 1 to 2 min They were then placed in the
PECVD reactor where the Ni was annealed into small catalyst islands C2H2 was bled
into the chamber for growth and an NH3 plasma was used to vertically align the growth
of the multi-walled carbon nanotubes (MWCNTs) The temperature and time were varied
between 725degC and 775oC and 15 to 60 min to control the length of the tubes and density
of samples Samples were then analyzed in a FEI Sirion field emission scanning electron
microscope (FESEM) SEM images of CNT decorated anode and control surfaces were
shown in the Figure 51
Figure 51 SEM images of CNT decorated anodes (A) Graphite disk control (B) CNT
decorated anode
Multiple channel MECs construction and operation A MECs with multiple
removable anodes each with an effective anode area of 07 cm2
was constructed and used
to evaluate the effects of nanostructures on the current density of according to a previous
B A
71
study (Fan et al 2011) The cathode was made of wet-proof (30) carbon cloth (type B
E-TEK Division Inc USA) coated with PtC (20 E-TEK Division Inc USA)
following a previously reported procedure (Liu et al 2005) The final platinum loading
was 05 mgcm2 per projected cathode area The size of the cathode (150 cm
2) was 25
times larger than the total effective area of the 8 graphite disk anodes (56 cm2) to prevent
cathode limitations on the performance of the MECs system All MECs with CNT
decorated anodes and control anodes were tested for current generation for 24 h with
sterile a medium (without bacteria) before injecting the bacterial cells in order to
determine whether the CNT decorated anodes demonstrate chemical current generation
compared to control anodes All the testing anodes were set up in the MECs
simultaneously so that the S oneidensis MR-1 culture was applied to all of them
simultaneously Short electrode spacing (17 cm) was used in the MECs design to reduce
the internal resistance A voltage of 06 V was applied the MECs for current generation
after the cells of S oneidensis MR-1 were inoculated into the MECs in the early
stationary phase A multimeter with a data acquisition system (2700 Keithly USA) was
used to monitor the current change by measuring the voltage drop through a resistor
Microarray analysis Biofilms for whole gene microarray analysis were aseptically
removed from the plain and CNT decorated anodes of MECs after 80 hours further
incubation at 30oC when current density obviously reached to a stable phase Total RNA
was extracted using Trizol (Ambion inc AustinTX) following the manufacturerrsquos
instructions Integrity of the RNA samples was confirmed using the Agilent 2100
Bioanalyzer (Agilent Technology Inc Palo Alto CA) Total RNA (200 ng) from each
sample was amplified using the MessageAmpTM
II-Bacteria Prokaryotic RNA
72
Amplification kit (Ambion AustinTX) according to the manufacturerrsquos instructions The
S oneidensis MR-1 whole-gene microarrays representing all of the 4295 genes (resource
from NCBI) were ordered from Roche NimbleGen Inc (Madison WI) Biological
triplicates of biofilms on the CNT decorated anodes and control were analyzed cDNA
synthesis labeling and hybridization were carried out by the CGRB Core Laboratories at
Oregon State University DNASTAR ArrayStarTM
3 software was used to identify genes
that were up- or down-regulated more than 2-fold when grown on the nanoparticle
modified anodes using the unpaired two sample t-test with a cutoff p-value of 005 with
bonferroni correction
Results and discussion
Enhancement of current generation using CNT decorated anodes in MECs The
current density generated by CNT decorated anode MECs with S onidensis MR-1
increased to 36 microAcm2 10 hours post-inoculation and reached a maximum of 40 microAcm
2
at 29 hours inoculation then gradually decreased The current density stabilized at
30microAcm2 after 78 h operation Figure 52 shows that the current density curve generated
by CNT decorated anode in MECs The average current density generated by CNT
decorated anodes was 56 times that of the control The current density generated in the
MECs without bacteria was negligible (data not shown) This result demonstrates that the
current enhancement observed with the CNT decorated anodes was biologically-derived
A
73
Figure 52 Effects of CNT decorated anodes on the current density in MECs (3 replicates)
Control is plain graphite disk anode CNT is the anode with CNT decoration (Error bars
represent standard deviation)
It has been reported that nanostructured decorated anodes have significant impacts on
current densities (Fan et al 2011 Qiao et al 2006 Sharma 2008 Tsai et al 2009) The
CNT decorated anodes had significantly enhanced current densities in MECs in this
study This result is consistent with previous studies Interestingly the current density
curve generated by CNT decorated anode MECs is quite different from the current
density curve generated by Au NP and Pd NP decorated anodes using same bacterial
species (Fan et al 2011) In the Au and Pd NP decorated anode MECs the current
density curve was not significantly increased in the beginning compared with the control
but gradually increased after the inoculation the current density reached the maximum
current density more than 50 hours after the inoculation However the current density
generated by CNT decorated anode MECs started to increase significantly immediately
after the inoculation It reached a maximum current density 29 hours post-inoculation
-1
6
13
20
27
34
41
48
0 10 20 30 40 50 60 70 80
Curr
ent
den
sity
(micro
Ac
m2)
Time (Hours)
Control
CNT
74
These results indicate that there might be different current density enhancement
mechanisms between CNT with other metal NP decorated anodes in MECs
Global transcriptome analysis Whole-genome DNA microarrays were used to attain a
comprehensive general overview of the transcriptional response of S oneidensis
MR-1 of
S oneidensis MR-1 to CNT decorated anodes in MECs In a total of 4295 genes analyzed
457 genes (250 genes up-regulated and 207 genes down-regulated) exhibited significantly
(P lt 005) changed expression at a 2-fold level in at least 3 replicates in the response to
CNT The total number of the regulated genes present 11 of the 4295 open reading
frames (ORFs) presented on the microarray The total number of genes at significant
expression levels in response to CNT decorated anodes is close to that to Fe NP decorated
anodes in MECs (392 9) ( Xu et al 2012)
Fig 53 Differentially expressed genes grouped by functional classification according to
the TIGR S oneidensis genome database (wwwtigrorg) Columns 1 amino acid
biosynthesis 2 biosynthesis of cofactors prosthetic groups and carriers 3 cell
envelope 4 cellular processes 5 central intermediary metabolism 6 DNA metabolism
7 energy metabolism 8 fatty acid and phospholipid metabolism 9 other categories 10
protein fate 11 protein synthesis 12 purines pyrimidines nucleosides and nucleotides
-80
-60
-40
-20
0
20
40
60
80
100
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18
Nu
mb
er
of
Ge
ne
s
Up-regulated
Down-regulated
75
13 regulatory functions 14 signal transduction 15 transcription 16 transport and
binding proteins 17 unknown function 18 hypothetical proteins
The overall transcript genomic response of S oneidensis MR-1 to CNT decorated
anodes was summarized in Figure 53 by grouping the differentially expressed genes into
their functional role categories based on The Institute for Genomic Researchrsquos annotation
(Version 8) of the MR-1 genome sequence The wide distribution of putative functional
roles attributed to the differentially expressed genes indicated the extent of the molecular
response of Soniedenis MR-1 to the CNT decorated anodes A large number of up-
regulated genes were presented in genes associated with biosynthesis of cofactors
prosthetic groups and carriers (group 2) cell envelope (group 3) protein fate (group 10)
protein synthesis (group 11) and purines pyrimidines nucleosides and nucleotides
(group 12) in this study The number of up-regulated genes was two times more than that
of down-regulated genes in each functional group Contrastingly in the genomic response
S oneidensis MR-1 to Fe NP decorated anodes (chapter 4) most of up-regulated genes
were associated with cellular processes (group 4) energy metabolism (group 7) other
categories (group 9) and hypothetical proteins (group 18) The down-regulated genes
presented in the amino acid biosynthesis (group 1) cellular processes (group 4) substrate
transport (group 16) and hypothetical proteins (group 18) While those genes encoding
proteins involved in amino acid biosynthesis (group 1) central intermediary metabolism
( group 5) DNA metabolism( group 6) fatty acid and phospholipid metabolism (group
8) and other categories (group 9) were among the most down-regulated genes in this
study However the two groups of genes that were the most noticeable between the
genomic response S oneidensis MR-1 to CNT and Fe NP decorated anodes which they
are the genes related to energy metabolism (group 7) and hypothetical proteins (group
76
18) These results indicated there are different and common genomic responses to S
oneidensis MR-1 on CNT and Fe NP decorated anodes in MECs
Electron transfer related genes Based on previous electron transfer mechanism
studies on Shewanella a serial of group proteins collectively described as the Mtr
pathway play the role to transfer electron rom the inner bacterial body to the outer
membrane (Beliaev et al 2001 Shi et al 2007 Brutinel and Gralnick 2012) In the Mtr
pathway firstly electrons generated by bacteria received by CymA a tetraheme c-type
cytochrome anchored in the inner membrane then electrons were transferred to MtrA a
periplasmic decaheme c-type cytochrome After that electrons were transferred from
MtrA to MtrC an outer-membrane decaheme c-type cytochrome is facilitated by MtrB a
non-heme containing integral outer-membrane proteinOuter-membrane decaheme c-type
cytochrome MtrC and OmcA are in change of electron transfer to exreacellular electron
acceptor (Shi et al 2007 Brutinel and Gralnick 2012 ) These genes encoding electron
transport proteins are shown in the table 51 The critical genes SO_1777 mtrA SO_1776
mtrB SO_1778 mtrC are unregulated 41 to 80 fold in response to CNT decorated
anodes other cytochrome c genes related to electron transfer in MtrA pathway also are
significantly increased in response to CNT decorated anodes These genes encoding outer
membrane proteins have different level increases supporting that the increase of
cytochrome c gene expression contributes to the current enhancements of MECs as
response to CNT decorated anodes
77
Table 51 Expression level of cytochrome c as response to CNT decorated anodes
Locus Tag Gene product descriptions Fold change CNT
SO_0165 general secretion pathway protein GspC 31
SO_0167 general secretion pathway protein GspE 45
SO_1776 outer membrane protein precursor MtrB 41
SO_1777 outer membrane decaheme cytochrome c
lipoprotein MtrA
80
SO_1778 outer membrane decaheme cytochrome c
lipoprotein MtrC
43
SO_1779 outer membrane decaheme cytochrome c
OmcA
63
SO_0135 lipoprotein of unknown function DUF333 22
SO_0136 conserved hypothetical inner membrane protein 24
Flavin related genes It is known that self-produced mediators play critical roles in the
electron transfer of S oneidensis to extracellular insoluble electrode (Canstein von et al 2
008 Myers et al 1992 Scott et al 1994 Shi et al 2007) It has been proved that the
flavins secreted by S oneidensis can facilitate electron transfer from bacteria to solid
anode electrodes by acting as extracellular electron shuttles (Cansterin von et al 2008) T
wo critical genes (SO_1414 and SO_3468) related to flavin synthesis were shown increa
sed significantly (more than 2 folds) increase in this study (Table 52) This result support
s the hypothesis that flavin production increases as the response to CNT decorated anodes
in MECs have significant contribution to the current density enhancements
78
Table 52 Expression levels of genes related to flavin synthesis as response to CNT
decorated anode
Locus Tag Gene product descriptions Fold change CNT
SO_1414 flavocytochrome c flavin subunit putative 22
SO_3468 riboflavin synthase subunit alpha 20
The amount of bacterial biofilm biomass on the anode can affect the power
generation in MFCs (Biffinger et al 2007 Heydorn et al 2000) Biofilm formation
related genes include flagellar gene IV pilus genes and other genes the flagellar related
genes (fliB and fliK) and IV pilus genes (mshA) were induced only from 105 to 134
fold as response of S oneidensis MR-1 to CNT decorated anodes in this study No
biofilm-associated genes presented significant changes in expression levels in this study
indicating there is no direct connections between biofilm enhancements to CNT
decorated anodes in this study This result is consistent with the Liang et al (2011)
conclusion that using CNT enhanced the electrochemical activity of anodic biofilms but
did not result in a significant increase of biomass in the anodic biofilms
Our microarray results showed two significantly up-regulated gene groups the
genes encoding proteins localized on the outer membrane and the genes involved in
flavin biosynthesis contributed to current density enhancement by CNT decorated anodes
Among 457 significantly changed genes to CNT in this study there are also a relatively
large number of genes encoding proteins with unknown functions which are either up- or
down-regulated in the response to CNT decorated anodes which indicated more
79
complicated responses of S oneidensis MR-1 to CNT decorated anodes in MECs and
further study is needed
80
Chapter 6
Summary
Microbial electrochemical systems (MESs) with a mixed culture initially inoculated
from Corvallis wastewater treatment plant have been studied for more than 6 years for
varying purposes including power generation hydrogen production heavy metal
removal and wastewater treatment Experiment results have shown that the mixed culture
is quite stable with excellent performance in MESs Our community analysis using
denaturing gradient gel electrophoresis (DGGE) and 16S rDNA clone library construction
suggests that the mixed culture is composed predominantly of Geobacter sp (66)
Arcobacter sp (12) and Citrobacter sp (11) These results not only improved our
understanding of the mixed culture community but also guided our studies on the
cultivation and isolation of the different bacterial species
Exoelectrogenic Citrobacter sp SX-1 was first isolated and characterized from a
MFC operated with the mixed culture Citrobacter sp SX-1 has been demonstrated to
produce electricity from wide range of different substrates including citrate acetate
glucose sucrose glycerol and lactosein MFCs Cyclic voltammetry analysis indicated
that membrane associated proteins may play an important role in facilitating the electrons
transferring from bacteria to electrode The strain SX-1 increased the known diversity of
power generating exoelectrogens and provided a unique bacterial species for study in
renewable energy generation and waste treatment
81
To enhance the electron transfer from bacteria to anode nano decorated anodes
including Fe NP and CNT were developed and characterized and evaluated in MECs
using Shewanella oneidensis MR-1 as a model species Both nanostructures have
significantly increased current density compared with the control Whole genome
microarray analysis elucidated the possible mechanisms of power enhancement in
response to these nano-decorated anodes
These results benefit to understanding of physiology and ecology of mixed
cultures in MFCs and improve the efficiency of current generation in MESs which will
facilitate the viability of niche applications for MESs in near future
82
Bibliography
1 Amman R Ludwig W (2000) Ribosomal RNA-targeted nucleic acid probes for
studies in microbial ecology FEMS Microbiol Ecol 24 555ndash565
1 Bao SJ Li CM Zang JF Cui XQ Qiao Y Guo J (2008) Novel Nanoporous TiO2
Electrode for Directly Electrochemistry of glucose oxidase Adv Funct Mater
18(4) 591-599
2 Beliaev AS Saffarini DA McLaughlin JL Hunnicutt D (2001) MtrC an outer
membrane decahaem c cytochrome required for metal reduction in Shewanella
putrefaciens MR-1 Mol Microbiol 39722-30
3 Bencheikh-Latmani R Williams SM Haucke L Criddle CS Wu L Criddle CS
Zhou J Tebo BM (2005) Global transcriptional profiling of Shewanella oneidensis
MR-1 during Cr(VI) and U(VI) reduction Appl Environ Microbiol 71 7453ndash7460
4 Biffinger JC Pietron J Ray R Little B and Ringeisen BR (2007) A biofilm
enhanced miniature microbial fuel cell using Shewanella oneidensis DSP10 and
oxygen reduction cathodes Biosens Bioelectron 221672ndash1679
5 Bodelier PLE Roslev P Henckel T FenzelP (2000) Stimulation by ammonium
based fertilizers of methane oxidation in soil around rice root Nature 40 421ndash424
6 Bond DR Holmes DE Tender LM Lovley DR (2002) Electrode-reducing
microorganisms that harvest energy from marine sediments Science 295 483-485
7 Bond DR Lovley DR (2003) Electricity production by Geobacter sulfurreducens
attached to electrodes Appl Environ Microbiol 69 1548-1555
8 Bond DR Lovley DR (2005) Evidence for involvement of an electron shuttle in
electricity generation by Geothrix fermentans Appl Environ Microbiol 71 2186-
2189
9 Bretschger O Obraztsova A Sturm CA Chang IS Gorby YA Reed SB Culley
DE Reardon CL Barua S Romine MF Zhou J Beliaev AS Bouhenni R
Saffarini D Mansfeld F Kim BH Fredrickson JK Nealson KH (2007) Current
production and metal oxide reduction by Shewanella oneidensis MR-1 wildtype and
mutants Appl Environ Microbiol 73 7003ndash7012
10 Brown SD Thompson MR Verberkmoes NC Chourey K Shah M Zhou J
Hettich RL and Thompson DK (2006) Molecular dynamics of the Shewanella
oneidensis response to chromate stress Mol Cell Proteomics 51054-1071
11 Brutinel ED Gralnick JA (2012) Shuttling happens soluble flavin mediators of
extracellular electron transfer in Shewanella Appl Microbiol Biotechnol 93(1) 41-8
12 Butler JE Kaufmann F Coppi MV Nuacutentildeez C Lovley DR (2004) MacA a diheme
c-type cytochrome involved in Fe(III) reduction by Geobacter sulfurreducens J
Bacteriol 186(12) 4042ndash4045
83
13 Caccavo F Lonergan DJ Lovley DR Davis M Stolz JF McInerney MJ (1994)
Geobacter sulfurreducens sp nov a hydrogen- and acetate oxidizing dissimilatory
metal-reducing microorganism Appl Environ Microbiol 60 3752ndash3759
14 Canstein von H Ogawa J Shimizu S Lloyd JR (2008) Secretion of flavins by
Shewanella species and their role in extracellular electron transfer Appl Environ
Microbiol 74 615ndash623
15 Cao X Huang X Liang P Xiao K Zhou Y Zhang X Logan BE (2009) A New
Method for Water Desalination Using Microbial Desalination Cells Environ Sci
Technol 43(18)7148-7152
16 Chaudhuri SK Lovley DR (2003) Electricity generation by direct oxidation of
glucose in mediatorless microbial fuel cells Nat Biotechnol 21 1229-1232
17 Cheng S Liu H Logan B E (2006) Increased power generation in a continuous
flow MFC with advective flow through the porous anode and reduced electrode
spacing Environ Sci Technol 402426-2432
18 Cheng S Logan B E (2007) Sustainable and efficient biohydrogen production via
electrohydrogenesis PNAS 104 18871-18873
19 Chignell J (2010) Energy Recovery from Biodiesel Waste Performance of
Microbial Electrochemical Systems on Glycerol Thesis
20 Chung K Okabe S (2009) Characterization of electrochemical activity of a strain
ISO2-3 phylogenetically related to Aeromonas sp isolated from a glucose-fed
microbial fuel cell Biotechnol Bioeng 104(5) 901-910
21 Cole JR Chai B Farris RJ Wang Q Kulam-Syed-Mohideen AS McGarrell
DM Bandela AM Cardenas E Garrity GM Tiedje JM (2007) The ribosomal
database project (RDP-II) introducing myRDP space and quality controlled public
data Nucleic Acids Res 35 (Database issue) D169-D172 doi 101093nargkl889
22 Cole JR Wang Q Cardena E Fish J Chai B Farris RJ Kulam-Syed-Mohideen
AS McGarrell DM Marsh T Garrity GM Tiedje JM (2009) The Ribosomal
Database Project improved alignments and new tools for rRNA analysis Nucleic
Acids Res 37 (Database issue) D141-D145 doi 101093nargkn879
23 Dey A De S De A De SK (2004) Characterization and dielectric properties of
polyaniline-TiO2 nanocomposites Nanotechnol 15 1277-1283
24 Edwards U Rogall T Bloumlcker H Emde M Boumlttger EC (1989) Isolation and
direct complete nucleotide determination of entire genesmdashcharacterization of a gene
coding for 16S-ribosomal RNA NucleicAcids Res 17 7843ndash7853
25 Eggleston CM Voumlroumls J Shi L Lower BH Droubay TC Colberg PJS (2008)
Binding and direct electrochemistry of OmcA an outer-membrane cytochrome from
an iron reducing bacterium with oxide electrodes a candidate biofuel cell system
Inorg Chim Acta 361769ndash777
26 Fan Y Sharbrough E Liu H (2008) Quantification of the Internal Resistance
Distribution of Microbial Fuel Cells Environ Sci Technol 42 8101ndash8107
84
27 Fan Y Xu S Schaller R Jiao J Chaplen F Liu H (2011) Nanoparticle decorated
anodes for enhanced current generation in microbial electrochemical
cells Biosensors and Bioelectronics 26 (5)1908ndash1912
28 Fan Y Hu H Liu H (2007)Sustainable power generation in microbial fuel cells
using bicarbonate buffer and proton transfer mechanisms Environ Sci
Technol 41(23) 8154-8
29 Fedorovich V Knighton MC Pagaling E Ward FB Free A Goryanin I (2009)
Novel electrochemically active bacterium phylogenetically related to Arcobacter
butzleri isolated from a microbial fuel cell Appl Environ Microbiol 75(23) 7326-
7334
30 Ferna ndez A Huang S Seston S Xing J Hickey R Criddle C Tiedje J (1999)
How stable is stable Function versus community composition Appl Environ
Microbiol 65 3697ndash3704
31 Field SJ Dobbin PS Cheesman MR Watmough NJ Thomson AJ Richardson
DJ (2000) Purification and magneto-optical spectroscopic characterization of
cytoplasmic membrane and outer membrane multiheme c-type cytochromes from
Shewanella frigidimarina NCIMB400 J Biol Chem 275 8515ndash8522
32 Freguia S Masuda M Tsujimura S Kano K (2009) Lactococcus lactis catalyses
electricity generation at microbial fuel cell anodes via excretion of a soluble quinone
Bioelectrochemistry 7614ndash18
33 Fricke K Harnisch F and Schroumlder U (2008) On the use of cyclic voltammetry for
the study of the anodic electron transfer in microbial fuel cells Energy Environ
Sci 1 144ndash147
34 Gao H Wang Y Liu X Yan T Wu L Alm E Arkin A Thompson DK and
Zhou J (2004) Global transcriptome analysis of the heat shock response of
Shewanella oneidensis J Bacteriol 1867796-7803
35 Gao H Yang ZK Wu L Thompson DK Zhou J (2006) Global transcriptome
analysis of the cold shock response of Shewanella oneidensis MR-1 and mutational
analysis of its classical cold shock proteins J Bacteriol 1884560-4569
36 George MG (2005) Bergeyrsquos Manual of Systematic Bacteriology Volume Two The
Proteobacteria Part B The Gammaproteobacteria Second Edition New York
Springer 651p
37 Gonzaacutelez-Toril E Llobet-Brossa E Casamayor EO Amann R Amils R (2003)
Microbial ecology of an extreme acidic environment the Tinto river Appl Environ
Microbiol 69(8) 4853ndash4865
38 Gorby YA Yanina S Mclean JS Rosso KM Moyles D Dohnalkova A
Beveridge TJ Chang IS Kim BH Kim KS Culley DE Reed SB Romine M
F Saffarini DA Hill EA Shi L Elias DA Kennedy DW Pinchuk G
Watanabe K Ishii S Logan B Nealson KH Fredrickson JK (2006) Electrically
conductive bacterial nanowires produced by Shewanella oneidensis strain MR-1 and
other microorganisms Proc Natl Acad Sci USA 10311358-11363
85
39 He Z Wagner N Minteer SD Angenent LT (2006) An upflow microbial fuel cell
with an interior cathodes assessment of the internal resistance by impedance
spectroscopy Environ Sci Technol 40 5212-5217
40 Heydorn A Nielsen AT Hentzer M Sternberg C Givskov M Ersboslashll BK Molin
S (2000) Quantification of biofilm structures by the novel computer program
COMSTAT Microbiol 1462395-2407
41 Holmes DE Bond DR Lovley DR (2004a) Electron transfer by Desulfobulbus
propionicus to Fe(III) and graphite electrodes Appl Environ Microbiol 701234ndash
1237
42 Holmes DE Chaudhuri SK Nevin KP Mehta T Methe BA Liu A Ward JE
Woodard TL Webster J Lovley DR (2006) Microarray and genetic analysis of
electron transfer to electrodes in Geobacter sulfurreducens Environ Microbiol
81805-1815
43 Holmes DE Nicoll JS Bond DR Lovley DR (2004b) Potential role of a novel
psychrotolerant member of the family Geobacteraceae Geopsychrobacter
electrodiphilus gen nov sp nov in electricity production by a marine sediment
fuel cell Appl Environ Microbiol 70 6023ndash6030
44 Holmes DE Chaudhuri SK Nevin KP Mehta T Metheacute BA Liu A Ward JE
Woodard TL Webster J Lovley DR (2006) microarray and genetic analysis of
electron transfer to electrodes in Geobacter sulfurreducens Environ Microbial
8(10)1805-1815
45 Hu H (2009) Enhancing Hydrogen Production in Microbial Electrolysis Cells
through Development of Platinum-free Cathode and Improvement of Reactor
Dissertation
46 Ince BK Ayman O N Turker G Ccedilelikkol S Ince O (2010) Microbial ecology of
anaerobic reactors for treatment of alcohol industry wastewaters a review Current
research technology and education topics in applied microbiology and microbial
ecology 988-999
47 Jung S Regan JR (2007) Comparison of anode bacterial communities and
performance in microbial fuel cells with different electron donors Appl Microbiol
Biotechnol 77393ndash 402
48 Kim BH Kim HJ Hyun MS Park DH (1999) Direct electrode reaction of Fe(III)
reducing bacterium Shwwanella putrefaciens J Microbial Biotechnol 9127-131
49 Kim GT Hyun MS Chang IS Kim HJ Park HS Kim BH Kim SD Wimpenny
JW Weightman AJ (2005) Dissimilatory Fe(III) reduction by an electrochemically
active lactic acid bacterium phylogenetically related to Enterococcus gallinarum
isolated from submerged soil J Appl Microbiol 99(4) 978ndash987
50 Kim GT Webster G Wimpenny JW Kim BH Kim HJ Weightman AJ (2006)
Bacterial community structure compartmentalization and activity in a microbial fuel
cell J Appl Microbiol 101(3) 698ndash710
86
51 Kim HJ Park HS Hyun MS Chang IS Kim M Kim BH (2002) A mediator-
less microbial fuel cell using a metal reducing bacterium Shewanella putrefaciens
Enzyme Microb Technol 30 145-152
52 Kim JR Min B Logan BE (2005) Evaluation of procedures to acclimate a
microbial fuel cell for electricity production Appl Microbiol Biotechnol 68(1)23-30
53 Kim JR Cheng S Oh SE Logan BE (2007) Power generation using different
cation anion and ultrafiltration membranes in microbial fuel cells Environ Sci
Technol 41 1004-1009
54 Konstantinidis KT Serres MH Romine MF Rodrigues JL Auchtung J McCue
LA Lipton MS Obraztsova A Giometti CS Nealson KH Fredrickson JK
Tiedje JM (2009) Comparative systems biology across an evolutionary gradient
within the Shewanella genus Proc Natl Acad Sci U S A 106(37)15909-15914
55 Lee J Phung NT Chang IS Kim BH Sung HC (2003) Use of acetate for
enrichment of electrochemically active microorganisms and their 16S rDNA
analyses FEMS Microbiol Lett 223185ndash191
56 Liang P Wang H Xia X Huang X Mo Y Cao X Fan M(2011)Carbon
nanotube powders as electrode modifier to enhance the activity of anodic biofilm
in microbial fuel cells Biosens Bioelectron 26(6)3000-4
57 Lies DP Hernandez ME Kappler A Mielke RE Gralnick JA Newman DK
(2005) Shewanaella oneidensis MR-1 uses overlapping pathways for iron reduction
at a distance and by direct contact under conditions relevant for biofilms Appl
Environ Microbiol 71 4414-4426
58 Liu H and Logan B E (2004) Electricity generation using an air-cathode single
chamber microbial fuel cell in the presence and absence of a proton exchange
membrane Environ Sci Technol 38 4040-4046
59 Liu H Grot S Logan BE (2005) Electrochemically assisted microbial production
of hydrogen from acetate Environ Sci Technol 39 4317 ndash 320
60 Liu H Hu H Chignell J Fan Y (2010) Microbial Electrolysis Novel Technology
for Hydrogen Production from Biomass Biofuels 1(1) 129ndash142
61 Liu H Ramnarayanan R Logan BE (2004) Production of electricity during
wastewater treatment using a single chamber microbial fuel cell Environ Sci Technol
382281-2285
62 Liu WT Marsh TL Cheng H Forney LJ (1997) Characterization of microbial
diversity by determining terminal restriction fragment length polymorphisms of gene
encoding 16S rRNA Appl Environ Microbiol 63 4516ndash4522
63 Logan B E Hamelers B Rozendal R Schroder U Keller J Freguia S Aelterman
P Verstraete W Rabaey K (2006) Microbial fuel cells methodology and
technology Environ Sci Technol 40 (17) 5181-5192
64 Logan B E and Regan JM (2006) Electricity-producing bacterial communities in
microbial fuel cells Trends Microbiol 14512ndash518
87
65 Logan B Cheng S Watson V Estadt G (2007) Graphite fiber brush anodes for
increased power production in air-cathode microbial fuel cells Environ Sci Technol
41(9)3341-3346
66 Logan BE (2009) Exoelectrogenic bacteria that power microbial fuel cells Nat Rev
Microbiol 5 375-381
67 Logan BE Call D Cheng S Hamelers HV Sleutels TH Jeremiasse AW
Rozendal RA (2008) Microbial electrolysis cells for high yield hydrogen gas
production from organic matter Environ Sci Technol 42 8630-8640
68 Logan BE Hamelers B Rozendal R Schroder U Keller J Freguia S Aelterman
P Verstraete W Rabaey K (2006) Microbial fuel cells methodology and
technology Environ SciTechnol 40 5181-5192
69 Lovely DR (2006) Bug juice harvesting electricity with microorganisms Nature
Review| Microbiology 4 497-508
70 Lovley DR Phillips EJ (1988) Novel Mode of Microbial Energy Metabolism
Organic Carbon Oxidation Coupled to Dissimilatory Reduction of Iron or
Manganese Appl Environ Microbiol 54(6) 1472-1480
71 Ly HK Sezer M Wisitruangsakul N Feng JJ Kranich A Millo D Weidinger
IM Zebger I Murgida DH Hildebrandt P (2011) Surface-enhanced vibrational
spectroscopy for probing transient interactions of proteins with biomimetic interfaces
electric field effects on structure dynamics and function of cytochrome c FEBS J
278(9)1382-90
72 Macaskie LE Hewitt CJ Shearer JA Kent CA (1995) Biomass production for
the removal of heavy metals from aqueous solutions at low pH using growth-
decoupled cells of a Citrobacter sp Int Biodeter Biodegr 3573ndash92
73 Malki M De Lacey AL Rodriguez N Amils R Fernandez VM (2008)
Preferential use of an anode as an electron acceptor by an acidophilic bacterium in
the presence of oxygen Appl Environ Microbiol 74 4472ndash4476
74 Meitl LA Eggleston CM Colberg PJS Khare N Reardon CL Shi L (2009)
Electrochemical interaction of Shewanella oneidensis MR-1 and its outer membrane
cytochromes OmcA and MtrC with hematite electrodes Geochim Cosmochim
Ac 73(18) 5292-5307
75 Morozan A Stamatin L and Nastase
F (2007) The biocompatibility
microorganisms-carbon nanostructures for applications in microbial fuel cells Phys
Stat Sol 6 1797-1803
76 Morris JM Jin S Crimi B Pruden A (2009) Microbial fuel cell in enhancing
anaerobic biodegradation of diesel Chem Eng J 146 161ndash167
77 Murray AE Lies D Li G Nealson K Zhou J Tiedje JM (2001) DNADNA
hybridization to microarrays reveals gene-specific differences between closely
related microbial genomes Proc Natl Acad Sci U S A 98 9853-9858
78 Muyzer G de Waal EC Uitterlinden AG (1993) Profiling of complex microbial
communities by denaturing gradient gel electrophoresis analysis of polymerase chain
88
reaction amplified genes coding for 16S rRNA Appl Environ Microbiol 59 695ndash
700
79 Myers CR Myers JM (1992) Localization of cytochromes to the outer membrane
of anaerobically grown Shewanella putrefacians MR-1 J Bacteriol 174 3429ndash3438
80 Narde GK Kapley A Purohit HJ (2004) Isolation and characterization of
Citrobacter strain HPC255 for broad-range substrate specificity for chlorophenols
Curr Microbiol 48(6) 419-423
81 Okabe S Ito T Sugita K Satoh H (2005) Succession of internal sulfur cycles and
sulfur-oxidizing bacterial communities in microaerophilic wastewater biofilms Appl
Environ Microbiol 71(5) 2520-2529
82 Park DH and Zeikus JG (2000) Electricity generation in microbial fuel cells using
neutral red as an electronophore Appl Environ Microbial 66 292-1297
83 Park DH and Zeikus JG (2002) Impact of electrode composition on electricity
generation in a single-compartment fuel cell using Shewanella putrefaciens Appl
Microbiol Biotechnol 5958ndash56
84 Park DH and Zeikus JG (2003) Improved fuel cell and electrode designs for
producing electricity from microbial degradation Biotechnol Bioeng 81 348ndash355
85 Park HS Kim BH Kim HS Kim HJ Kim GT Kim M Chang IS Park Y
K Chang HI (2001) A novel electrochemically active and Fe(III)-reducing
bacterium phylogenetically related to Clostridium butyricum isolated from a
microbial fuel cell Anaerobe 7 297-306
86 Parot S Nercessian O Delia ML Achouak W Bergel A (2009) Electrochemical
checking of aerobic isolates from electrochemically active biofilms formed in
compost J Appl Microbiol 106(4) 1350-1359
87 Pham CA Jung SJ Phung NT Lee J Chang IS Kim BH Yi H Chun J
(2003) A novel electrochemically active and Fe(III)-reducing bacterium
phylogenetically related to Aeromonas hydrophila isolated from a microbial fuel cell
FEMS Microbiol Lett 223 129-134
88 Qiao Y Bao SJ Li CM Cui XQ Lu ZS Guo J (2008) Nanostructured
polyanilinetitanium dioxide composite anode for microbial fuel cells ACS Nano 2
113-119
89 Qiu R Zhao B Liu J Huang X Li Q Brewer E Wang S Shi N (2009) Sulfate
reduction and copper precipitation by a Citrobacter sp isolated from a mining area J
Hazard Mater 1641310-1315
90 Quan X Yang SG Ruan XL and Zhao HM (2005) Preparation of titania
nanotubes and their environmental applications as electrode Environ Sci Technol 39
3770ndash3775
91 Rabaey K Boon N Hofte M Verstraete W (2005) Microbial phenazine
production enhances electron transfer in biofuel cells Environ Sci Technol 393401ndash
3408
89
92 Rabaey K Boon N Siciliano S D Verhaege M Verstraete W (2004) Biofuel cells
select for microbial consortia that self-mediate electron transfer Appl Environ
Microbiol 70(9) 5373ndash5382
93 Rabaey K Lissens G Siciliano S D Verstraete W (2003) Amicrobial fuel cell
capable of converting glucose to electricity at high rate and efficiency Biotechnol
Lett 25 1531-15
94 Rabaey K Verstraete W (2005) Microbial fuel cells novel biotechnology for
energy generation Trends Biotechnol 23(6) 291-298
95 Reguera G McCarthy KD Mehta T Nicoll JS Tuominen MT Lovley DR
(2005) Extracellular electron transfer via microbial nanowires Nature 435 1098-
1101
96 Reguera G Nevin KP Nicoll JS Covalla SF Woodard TL Lovley DR (2006)
Biofilm and nanowire production leads to increased current in Geobacter
sulfurreducens fuel cells Appl Environ Microbiol 72 7345-7348
97 Reimers CE Tender LM Fertig S Wang W (2001) Harvesting energy from the
marine sediment-water interface Environ Sci Technol 35192-195
98 Rezaei F Xing D Wagner R Regan JM Richard TM Logan BE (2009)
Simultaneous cellulose degradation and electricity production by Enterobacter
cloacae in a microbial fuel cell Appl Microbiol Biotechnol 75 3673ndash3678
99 Richter H Nevin KP Jia H Lowy DA Lovley DR Tender LM (2009) Cyclic
voltammetry of biofilms of wild type and mutant Geobacter sulfurreducens on fuel
cell anodes indicates possible roles of OmcB OmcZ type IV pili and protons in
extracellular electron transfer Energy Environ Sci 2 506ndash516
100 Riesenfeld SC Schloss DP Handelsman J(2004) Metagenomics Genomic
Analysis of Microbial Communities Annual Review Genetics 38525-552
101 Rozendal RA Hamelers HV Buisman CJ (2006) Effects of membrane cation
transport on pH and microbial fuel cell performance Environ Sci Technol 40
5206-5211
102 Rozendal RA Hamelers HVM Euverink GJW Metz SJ Buisman CJN (2006)
Principle and perspectives of hydrogen production through biocatalyzed electrolysis
Int J Hydrogen Energ 311632-1640
103 Rozendal RA Hamelers HV Molenkamp RJ Buisman CJ (2007) Performance of
single chamber biocatalyzed electrolysis with different types of ion exchange
membrances Water Res 41 1984-1994
104 Schluumlter A Bekel T Diaz NN Dondrup M Eichenlaub R Gartemann
KH Krahn I Krause L Kroumlmeke H Kruse O Mussgnug JH Neuweger
H Niehaus K Puumlhler A Runte KJ Szczepanowski R Tauch A Tilker
A Viehoumlver P Goesmann A (2008) The metagenome of a biogas-producing
microbial community of a production-scale biogas plant fermenter analyzed by the
454-pyrosequencing technology J Biotechnol136 77-90
90
105 Scott J H and Nealson KH (1994) A biochemical study of the intermediary carbon
metabolism of Shewanella putrefaciens J Bacteriol 176(11) 3408ndash3411
106 Sekiguchi H Tomioka H Nakahara T and Uchiyama H (2001) A single band
does not always represent single bacterial strains in denaturing gradient gel
electrophoresis analysis Biotechnology Letters23 1205ndash1208
107 Sharma T Leela M Reddy A Chandra TS Ramaprabhu S (2008) Development
of carbon nanotubes and nanofluids based microbial fuel cell Int J Hydrogen Energ
336749ndash6754
108 Shea C Clauwaert P Verstraete W Nerenberg R (2008) Adapting a denitrifying
biocathode for perchlorate reduction Water Sci Technol 58 (10) 1941-1946
109 Shi L Squier TC Zachara JM Fredrickson JK (2007) Respiration of metal (hydr)
oxides by Shewanella and Geobacter a key role for multihaem c-type cytochromes
Mol Microbiol 6512-20
110 Stams AJ and Plugge CM(2009)Electron transfer in syntrophic communities
of anaerobic bacteria and archaea Nat Rev Microbiol7(8)568-77
111 Sukkasem C Xu S Park S Boonsawang P Liu H (2008) Effect of Nitrate on the
Performance of Single Chamber Air Cathode Microbial Fuel Cells Water Res 42(19)
4743-4750
112 Summers ZM Fogarty HE Leang C Franks AE Malvankar NS Lovley DR
(2010)Direct exchange of electrons within aggregates of an evolved syntrophic
coculture of anaerobic bacteria Science 330 (6009)1413-5
113 Szczepanowski R Bekel T Goesmann A Krause L Kroumlmeke H Kaiser
O Eichler W Puumlhler A Schluumlter A (2008) Insight into the plasmid metagenome
of wastewater treatment plant bacteria showing reduced susceptibility to
antimicrobial drugs analysed by the 454-pyrosequencing technology J
Biotechnol 136 54-64
114 Tamura K Dudley J Nei M Kumar S (2007) MEGA4 Molecular Evolutionary
Genetics Analysis (MEGA) software version 40 MolBiol Evol 24(8) 1596-1599
115 Thompson JD Gibson TJ Plewniak F Jeanmougin F Higgins DG (1997) The
CLUSTAL_X windows interface flexible strategies for multiple sequence alignment
aided by quality analysis tools Nucleic Acids Res 25 4876-4882
116 Thormann KM Duttler S Saville RM Hyodo M Shukla S Hayakawa Y
Spormann AM (2006) Control of formation and cellular detachment from
Shewanella oneidensis MR-1 biofilms by cyclic di-GMP J Bacteriol 1882681-2691
117 Thormann KM Saville RM Shukla S Pelletier DA Spormann AM (2004)
Initial Phases of biofilm formation in Shewanella oneidensis MR-1 J Bacteriol
186(23) 8096-104
118 Torres CI Kato Marcus A Rittmann BE (2008b) Proton transport inside the
biofilm limits electrical current generation by anode-respiring bacteria Biotechnol
Bioeng 100 872ndash881
91
119 Torres CI Marcus AK Lee HS Parameswaran P Krajmalnik-Brown R Rittmann
BE (2010) A kinetic perspective on extracellular electron transfer by anode-
respiring bacteria FEMS Microbiol Rev 234(1)3-17
120 Torres CI Marcus AK Parameswaran P Rittmann BE (2008a) Kinetic
experiments for evaluating the NernstndashMonod model for anode-respiring bacteria
(ARB) in a biofilm anode Environ Sci Technol 42 6593ndash6597
121 Tsai HY Wu Ch-Ch Lee Ch-Y Shih EP (2009) Microbial fuel cell performance
of multiwall carbon nanotubes on carbon cloth as electrodes J Power Sources
194199-205
122 Tyson GW Chapman J Hugenholtz P Allen EE Ram RJ Richardson
PM Solovyev VV Rubin EM Rokhsar DS Banfield JF(2004) Community
structure and metabolism through reconstruction of microbial genomes from the
environment Nature42837-43
123 Venkateswaran K Moser DP Dollhopf ME Lies DP Saffarini D A MacGregor
BJ Ringelberg DB White DC Nishijima M Sano H Burghardt J Stackebrandt
E Nealson KH (1999) Polyphasic taxonomy of the genus Shewanella and
description of Shewanella oneidensis sp Int J Syst Bacteriol 49705-724
124 von Canstein H Ogawa J Shimizu S Lloyd JR (2008) secretion of Flavins
by Shewanella Species and Their Role in Extracellular Electron Transfer Appl
Environ Microbiol 74 3615-623
125 Wang A Liu W Cheng Sh Xing D Zhou J Logan BE (2009) Source of methane
and methods to control its formation in single chamber microbial electrolysis cells
Int J Hydrogen Energ 34 3653-3658
126 Wang J Ma T Zhao L Lv J Li G Liang F Liu R (2008) PCRndashDGGE method
for analyzing the bacterial community in a high temperature petroleum reservoir
World J Microbiol Biotechnol 2 9694-9702
127 Wang Q Garrity GM Tiedje JM Cole JR (2007) Naiumlve Bayesian Classifier for
Rapid Assignment of rRNA Sequences into the New Bacterial Taxonomy Appl
Environ Microbiol 73(16) 5261-5267
128 Wang YF Masuda M Tsulimura S Kano K (2008) Electrochemical regulation of
the end-product profile in Propionibacterium freudenreichii ET-3 with an
endogenous mediator Biotechnol Bioeng 101(3) 579ndash586
129 Wrighton KC Agbo P Warnecke F Weber KA Brodie EL DeSantis TZ
Hugenholtz P Andersen GL Coates JD (2008) A novel ecological role of the
Firmicutes identified in thermophilic microbial fuel cells ISME J 2 1146ndash1156
130 Xing D Zuo Y Cheng S Regan JM Logan BE (2008) Electricity generation by
Rhodopseudomonas palustris DX-1 Environ Sci Technol 42 4146-4151
131 Xu S Liu H Fan Y Schaller R Jiao J Chaplen F (2012) Enhanced performance
and mechanism study of microbial electrolysis cells using Fenanoparticle-decorated
anodes Appl Microbiol Biotechnol 93(2)871-880
92
132 Yang Y Harris DP Luo F Wu L Parsons AB Palumbo AV Zhou J (2008)
Characterization of the Shewanella oneidensis Fur gene roles in iron and acid
tolerance response BMC Genomics 9 Suppl 1S11
133 Zhang L Zhou S Zhuang L Li W Zhang J Lu N Deng L (2008) Microbial
fuel cell based on Klebsiella pneumoniae biofilm Electrochem Commun10 1641ndash
1643
134 Zuo Y Xing DF Regan JM Logan BE (2008) Isolation of the exoelectrogenic
bacterium Ochrobactrum anthropi YZ-1 by using a U-tube microbial fuel cell Appl
Environ Microbiol 74 3130-3137
93
APPENDICES
List of published papers during PhD study
1 Shoutao Xu Hong Liu Yanzhen Fan Rebecca Schaller Jun Jiao and Frank
Chaplen (2012) Enhanced performance and mechanism study
of microbial electrolysis cells using Fe nanoparticle-decorated anodes Applied
Microbiology and Biotechnology 93(2)871-880
2 Shoutao Xu Hong Liu (2011) New exoelectrogen Citrobacter sp SX-1 isolated
from a microbial fuel cell Journal of Applied Microbiology111(5)1108-1115
3 Yanzhen Fan Shoutao Xu Rebecca Schaller Jun Jiao Frank Chaplen Hong Liu
(2011) Nanoparticle decorated anodes for enhanced current generation in microbial
electrochemical cells Biosensors and Bioelectronics 26(5) 1908-1912
4 Tunc Catal Shoutao Xu Kaichang Li Hakan Bermek Hong Liu
(2008)
Electricity generation from polyalcohols in single-chamber microbial fuel
cells Biosensors and Bioelectronics 24(4)849-854
5 Chontisa Sukkasema Shoutao Xu Sunhwa Park Piyarat Boonsawang Hong Liu
(2008) Effect of nitrate on the performance of single chamber air cathode
microbial fuel cells Water research 424743-4750
6 Rebecca Schaller Yanzhen Fan Shoutao Xu Alan Fern Frank Chaplen Hong
Liu and Jun Jiao (2009) Vertically Aligned Multi-walled Carbon Nanotube
Decorated Anodes for Microbial Fuel Cells Proceedings of Materials Research
Society 2009 1170 R05-13

time enjoying doing research with them Their friendship and help made me confident of
my ability to do research as a scientist In addition I cherished and appreciated the
friendly environment in our BEE department Faculty members especially Dr John Bolte
Dr John Selker Dr Roger Ely Dr Ganti Murthy and Desiree Tullos made BEE an
excellent department for our students
My deepest gratitude went to my father Fuqin Wang and my mother Jinzhi Wang
for their endless care love and support throughout my life Also I would like to say
thanks to my best-loved wife Songhua Zhu She always supported me and never
complained that I could spend so little time accompanying with her This dissertation was
also for my lovely son Gabriel Hong-Yi Xu
TABLE OF CONTENTS
Page
1 General Introductionhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 1
2 Bacterial Community Analysis of Mixed Consortium in Microbial
Electrochemical Systemshelliphelliphelliphellip 13
3 New Exoelectrogen Citrobacter sp SX-1 Isolation and
Characterizationhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 27
4
Enhanced Performance and Mechanism Study of Microbial Electrolysis
Cells using Fe Nanoparticles Decorated Anodeshelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 45
5 Global Transcriptome Analysis of Response of Shewanella oneidensis
MR-1 to CNT Nanostructure Decorated Anodes in Microbial
Electrochemical System
66
6 Summaryhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 80
7 Bibliography helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 82
8 Appendices helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 93
LIST OF FIGURES
Figure Page
1-1 Generalized schematics (a) Microbial fuel cell (MFCs)(b) Microbial
electrolysis cell (MECs) helliphelliphelliphelliphelliphelliphelliphelliphelliphellip helliphelliphelliphellip
2
1-2 Mechanisms for extracellular electron transport in a MFCs anode 7
2-1 Polarization curves of high power mixed culture MFCshelliphelliphelliphelliphelliphellip 20
2-2 DGGE result of high power generation mixed culture MFCshelliphelliphellip 21
2-3 Bacterial species and percentage of bacterial community of mixed
culture MFCs helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip
22
2-4 Phylogenetic trees of bacterial species from mixed culture MFCs 22
2-5 Dominant bacterial species identification in DGGEhelliphelliphelliphelliphelliphelliphelliphellip 23
2-6 Current density generated by different dominant isolateshelliphelliphelliphelliphelliphellip 24
3-1 Phylogenetic tree of strain SX-1 and closely related species based on
16S rDNA sequenceshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip
35
3-2 SEM image of planktonic cells of Citrobacter sp SX-1helliphelliphelliphelliphelliphellip 35
3-3 Electricity generation by Citrobacter sp SX-1 in a single chamber
MFCs with sodium citrate helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip
36
3-4 Electricity generation of Citrobacter sp SX-1 using different
substrates in single chamber air-cathode MFCs at 1 KΩhelliphelliphellip
37
3-5 Power and voltage generation by Citrobacter sp SX-1 as a function
of current density helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip
38
3-6 Four current generating stages for CV analysis helliphelliphelliphelliphelliphelliphelliphellip 39
3-7 Cyclic Voltammetry analysis of of Citrobacter sp SX-1 biofilms helliphellip 41
4-1 SEM images of Fe NPs decorated anodeshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 50
4-2 Effects of Fe-NP decorated anodes on the current density in MECs helliphellip 51
4-3 Differentially expressed genes grouped by functional classification in
Fe NP decorated anodeshelliphelliphelliphelliphelliphellip helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip
53
5-1 SEM images of CNT decorated anodeshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 70
5-2 Effects of CNT decorated anodes on the current density in MECshelliphellip 73
5-3 Differentially expressed genes grouped by functional classification in
CNT decorated anodes helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip
74
LIST OF TABLES
Table
1-1
4-1
4-2
4-3
Taxa of bacteria current density generated and reactor configuration
used in microbial electrochemical cell experimentshelliphelliphelliphellip
Expression levels of genes related to biofilm formationhelliphelliphelliphelliphellip
Genes related to anaerobic growth and electron transfer with
significantly change expression levelhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip
Expression level of flavin and cytochrome geneshelliphelliphelliphelliphelliphelliphelliphellip
Page
4
55
56
58
4-4
5-1
5-2
Other genes with significantly changed expression level
Expression level of cytochrome c as response to CNT decorated anodes
Expression level of genes related to flavin synthesis as response to CNT
decorated anode in CNT decorated anodeshelliphellip
59
76
78
1
1
Bacterial Community Analysis New Exoelectrogen Isolation and Enhanced Performance
of Microbial Electrochemical Systems Using Nano-Decorated Anodes
Chapter 1
General Introduction
11 Microbial Fuel Cells and Microbial Electrolysis Cells
Microbial electrochemical systems (MESs) have drawn the attention of researchers
in recent years due to their promising applications in a variety of scientific fields such as
renewable energy generation bioremediation and wastewater treatment In a MES
microorganisms interact with electrodes catalyzing oxidation and reduction reactions at
the anode and the cathode
The most-described type of MESs is the microbial fuel cells (MFCs) in which
useful power is generated directly using the catalytic action of active microorganisms
(Kim 2002 Cheng and Logan 2007 Cheng et al 2006) In a typical two-chamber
MFCs (Figure 1a) organic matter is oxidized by electrochemically active
microorganisms in the anode chamber to release protons and electrons Protons diffuse
into the cathode chamber through a proton exchange membrane Meanwhile electrons
are transferred to the anode through various mechanisms (Logan et al 2006 Lovley
2
2006 Rabaey et al 2003) and then travel to the cathode where normally they combine
with oxygen and protons to form water
Another common used type of MES is the microbial electrolysis cell (MECs) in
which hydrogen is produced instead of electricity by applying a circuit voltage to the
MFCs system and maintaining anaerobic conditions in the cathode chamber (Liu et al
2005 Logan et al 2008 Rozendal et al 2006 Schaller et al 2009) (Figure 1b) A
cathode potential of at least - 410 mV (vs standard hydrogen electrode at pH 70) is
required to produce hydrogen at the cathode Therefore a theoretical potential of 110 mV
(ie 410-300 mV) is needed but voltages of gt02 V are normally applied due to various
overpotentials
Fig 1-1 Generalized schematics (a) Microbial fuel cells (MFCs) where electricity is
captured through the resistance R (b) Microbial electrolysis cells (MECs) where
hydrogen gas is produced in the cathode chamber R for resistor PS for power supply and
PEM for proton exchange membrane
The fundamental feature shared by microbial electrochemical systems (MESs)
(MECs and MFCs) is the microbe-catalyzed electron transfer from organic matter to
electrodes (anodes) (Rebaey et al 2004) Therefore different types of MESs can be
PEM
R
H+
e- e-
Anode
Bacte
ria
a
Cath
ode
H2O
PEM
PS
H+
e- e-
Anode
Bacte
ria
b
Cath
ode
H2
3
utilized to investigate the phenomena and mechanisms of interactions between microbes
and electrodes
12 Bacteria and Electron Transfer Mechanisms in MESs
The electrochemically active bacteria in MFCs are thought to be iron-reducing
bacteria such as Shewanella and Geobacter species (Gorby et al 2006) They have great
importance in the natural environment principally in metal oxidation and reduction
However recent studies have shown that the diversity of bacterial communities is much
greater than these model iron reducers on MFCs anodes (Reimers et al 2001 Liu et al
2004b Kim et al 2005) Over 20 electrochemically active bacterial species those can
transfer electrons exocellularly to electrodes have been reported in the past 10 years
(Logan 2009 Liu et al 2010) These bacteria have been categorized into diverse genetic
groups so far including alphaproteobacteria (Rhodopseudomonas Ochrobactrum and
Acidiphilium) (Malki et al 2008 Xing et al 2008 Zuo et al 2008) betaproteobacteria
(Rhodoferax) (Chaudhuri et al 2003) gammaproteobacteria (Shewanella Pseudomonas
Klebsiella Enterobacter and Aeronomas) (Kim et al 2002 Pham et al 2003 Rabaey et
al 2005 Bretschger et al 2007 Zhang et al 2008 Chung et al 2009 Rezaei et al 2009)
deltaproteobacteria (Geobacter Geopsychrobacter Desulfuromonas and Desulfobulbus)
(Bond et al 2002 Bond and Lovley 2003 Holmes et al 2004ab 2006)
Epsilonproteobacteria (Arcobacter) (Fedorovich et al 2009) Firmicutes (Clostridium and
Thermincola) (Park et al 2001 Wrighton et al 2008) Acidobacteria (Geothrix) (Bond
and Lovley 2005) and actinobacteria (Propionibacterium) (Wang et al 2008) A wider
range of electrochemically active bacteria are expected to be discovered
4
Table 1 Taxa of bacteria current density generated and reactor configuration used in
microbial electrochemical cell experiments (Liu et al 2010)
The electrochemically active bacterial species that possess the ability to transfer
electrons outside of the cell are called exoelectrogens in the MESs research field The
different exoelectrogens have demonstrated a wide-ranging power generation ability in
MFCs For example Shewanella oneidensis MR-1 has demonstrated the ability to
generate 0018 Am2 current density in single chamber MFCs while the Geobacter
Taxon Microorganisms Current Density (Am2
)
α-proteobacteria
Rhodopseudomonas palustris DX-1 003
Ochrobactrum anthropi YZ-1 071
Acidiphilium sp 32sup5 300
β-proteobacteria Rhodoferax ferrireducens 0031
γ-proteobacteria
Shewanella putrefaciens IR-1 0016
Shewanella oneidensis DSP10 0013
Shewanella oneidensis MR-1 0018
Pseudomonas aeruginosa KRA3 0017
Escherichia coli K12 HB101 100
Klebsiella pneumoniae L17 120
Enterobacter cloacae 013
Aeromonas hydrophila PA3 030
δ-proteobacteria
Geobacter metallireducens 065
Geobacter sulfurreducens 800
Desulfuromonas acetoxidans 0005
Geopsychrobacter
Electrodiphilus strain A2
0066
Desulfobulbus propionicus 003
Firmicutes
Lactococcus lactis 003
Thermincola sp strain Jr 020
Clostridium butyricum EG3 022
Thermincola ferriacetica Z-0001 040
Brevibacillus spp PTH1 0009
Desulfitobacterium hafniense DCB2 110
Acidobacteria Geothrix fermentans 0097
Actinobacteria Propionibacterium freudenreichiiET-3 120
5
sulfurreducens species has generated current densities as high as 80 Am2 which is 400
times higher than the one generated by Shewanella oneidensis MR-1 However mixed-
culture communities have generated much higher power densities than their pure-culture
counterparts in MFCs perhaps due to synergistic interactions within the anode bacterial
communities and the participation of currently unknown bacteria species and mechanisms
(Jung and Regan 2007) The dominant bacteria species vary in mixed cultures bacteria
communities in MFCs due to the enrichment of different substrates
Traditional methods of extroelectrogen study depend on cultivation hampered novel
exoelectrogens discovery because the inadequacy of defined media underestimates the
actual microbial diversity in MFCs Slow growth rate and unknown growth requirements
of anaerobic microorganisms make some anaerobic extroelectrogens cultivation difficult
However these limitations have been overcome by using molecular biological methods
based on DNARNA analysis Molecular biological techniques are now widely applied to
assess the diversity of microbial communities by analyzing the 16S rDNA sequence The
most commonly used molecular biological techniques for bacterial community analysis
include denaturing gradient gel electrophoresis (DGGE) restriction fragment length
polymorphism (RFLP) and 16S rDNA sequencing These methods are much less time
consuming than traditional isolation and cultivation methods
Isolated exoelectrogens were utilized to explore the mechanism of electron transfer
to the anode However the mechanisms of electron transfer to extracellular electron
acceptors are not well understood Three mechanisms have been proposed for exocellular
transport of electrons by exoelectrogens to anodes in MFCs so far Some exoelectrogenic
bacteria can transfer electrons to electrodes through soluble redox compounds (Bond and
6
Lovley 2005) These compounds include artificial mediators and mediators secreted by
exoelectrogenic bacteria A variety of chemicals have been used to facilitate the shuttling
of electrons from inside of cell to electrodes outside the cell These exogenous mediators
include neutral red (Park et al 1999) anthraquinone-2-6 disulfonate (AQDS) thionin
potassium ferricyanide (Bond et al 2002) methyl viologen and others (Logan 2004
Rabaey and Verstraete 2005) Some bacteria can secrete their own chemical mediator
for example Pseudomonas aeruginosa secretes pyocyanin and phenazine-1-carboxamide
to transfer electrons toward the MFCs anode (Rabaey et al 2005) Another evidence for
mediator production by bacteria is Geothrix fermentans When the medium was replaced
in a MFCs that had stable power generation with this bacteria power dropped by 50
and required 10 days to resume the original level
Some bacteria can directly transfer electrons to anodes via outer cell membrane
proteins (Myers and Myers 1992) The outer cell membrane protein cytochrome c is
thought to play a critical role in to transferring electrons to anodes Ly et al (2011)
isolated the haem protein cytochrome c and demonstrated that electric field effects may
be functional for the natural redox processes of cytochrome c in the respiratory chain
Shewanella oneidensis is the example of a bacterium that is capable of transfer electron to
anode via outer cell membrane proteins When Shewanella oneidensis adhered to an iron
surface the greater force has showed benefits to grow cells because closer contact
required for electron transfer from cell bound cytochromes (Lower et al 2001)
7
Fig1-2 Mechanisms for extracellular electron transport in an MFCs anode (1) direct
contact (top in green) (2) by nanowires (middle in purple) and (3) self-produced
mediators (bottom in blue) (Logan 2009)
More and more evidence supports the involvement of bacterial nanowires in
extracellular electron transport (Reguera et al 2005 2006 Gorby et al 2006) Nanowires
are conductive appendages produced by both Geobacter and Shewanella species (Gorby
and Beveridge 2005) The conductivity of the appendages was examined and confirmed
by using conductive scanning tunneling microscopy (STM) (Gorby et al 2006)
Nanowires can carry electrons from the cell to the anode surface of MFCs
The solid component of the extracellular biofilm matrix has high efficiency on
extracellular electron transfer compared with other extracellular electron transfer
8
mechanisms and recently Torres et al (2010) hypothesized that the solid component of
the extracellular biofilm matrix formed by exoelectogens is another way bacteria transfer
electrons to electrodes This hypothesis was based on kinetic analysis of each EET
mechanism reported in available literature (Torres et al 2010)
13 Anode electrodes
In MFCsMECs anode electrodes are a critical component because exoelectrogens
adhere to the surface of anodes to transfer electrons to the electrode The characteristics
of anodes have significant effects on electron transfer rate from bacteria to anode
electrodes in MFCs The requirements of an anode material are it should be highly
conductive non-corrosive have a high specific surface area (area per volume) high
porosity be non-fouling inexpensive and easily scaled to larger sizes Of these
properties the most important one that is different from other biofilm reactors is that the
material must be electrically conductive Normally they are made of various carbon
materials including carbon fiber carbon clothe and carbon paper due to their stability
high conductivity and high specific surface-area Nevertheless they have little
electrocatalytic activity for the anode microbial reactions and thus a modification of the
carbon materials is the main approach for improving their performance Consequently
there is a great need to develop a new type of anode material for MFCsMECs
It is a great challenge to develop a new anode material to further increase the power
density of a MFCs The nature of the catalytic mechanism of a MFCrsquos anode involves not
only a biological but also an electrocatalytic process An optimal nanostructure with a
high specific surface area favorable for both catalytic processes could play a critical role
in improving the power density of the MFCs such a structure needs to host the bacteria
9
with high bioactivity while enhancing the electron-transfer rate Schroumlder et al (2008)
employed PANI to modify a platinum anode for MFCs and achieved a current density 1
order of magnitude higher than the previously reported value PANIinorganic
composites are also reported to have better conductivity Qiao et al also applied a new
mesoporous TiO2 electrode material with uniform nanopore distribution and a high
specific surface area to anode in comparison to previously reported work with E coli
MFCs the composite anode delivers 2-fold higher power density (Qiao et al 2008)
Thus it has great potential for use as the anode in a high-power MFCs and may be a new
approach for improving performance of MFCs
14 Other parts of MESs
141 Membranes and ion transport
The ion exchange membrane is another one of the critical components in two-
chamber MESs systems It separates anode and cathode chambers and at the same time
maintains the electron neutrality of the system ie transport of electrons to the cathode
needs to be compensated by transport of an equal amount of positive charge to the
cathode chamber A proton exchange membrane (PEM) such as Nafion a per-fluorinated
sulphonic acid membrane (PFSAs) consisting of a hydrophobic fluorocarbon backbone to
which hydrophilic sulfonate groups (-SO3-) are attached is commonly used in chemical
fuel cell systems For MFCs systems however mainly cation species like Na+ and K
+
other than proton are often responsible for the dominant transport of positive charge
through the cation exchange membrane (CEM) to maintain electroneutrality due to the
low proton concentration in any aqueous medium with near neutral pH (Rozendal et al
2006) Consequently the pH increases in the cathode chamber due to the consumption of
10
protons and decreases in the anode chamber because of the accumulation of protons
(Rozendal et al 2007) This pH change in two-chamber systems results in a decrease of
the cathode potential and performance The application of anion exchange membrane
(AEM) has also been explored in MFCs systems (Cheng and Logan 2007 Kim and
Logan 2007) where it has been proposed that protons are transferred via pH buffers like
phosphate anions
142 Cathodes and Catalysts
The cathode is another challenge for making MFCs commercially available
technology because the chemical reaction that occurs at the cathode is difficult to
engineer as the electrons protons and oxygen must all meet at a catalyst in a tri-phase
reaction (solid catalyst air and water) The catalyst must be on a conductive surface and
must be exposed to both water and air so that protons and electrons in these different
phases can reach the same point The most commonly used material for a cathode is
commercially available carbon paper pre-loaded with a Pt catalyst on one side When it is
used in a MFC the side that contains the catalyst faces the water and the uncoated side
faces air To reduce the high cathode cost associated with platinum catalyst other precious-
metal-free cathodes such as (NiMo and NiW) were developed by electrodepositing on a
carbon fiber They have achieved comparable performance with Pt catalyst with same
loading at a much lower cathode fabrication cost (Hu 2010)
The requirements of cathode for MECs are quite similar to the requirements of
cathode for MFCs but easier than cathode for MFCs for the manufacturing process
because the cathode in MECs is not necessarily to exposed to air Therefore it can be
made of the exact same materials of cathode in MFCs except a waterproof layer
11
Recently some researchers have attempted to apply microorganisms as a biocatalyst to
precede the combination of electron with oxygen in the cathodes
15 Dissertation overview
Low power densities in MESs limit practical applications The improvement of
MESs performance requires a detailed understanding of the physiology and ecology of
microorganisms in MESs including the mechanism of electron transfer to the anode from
the microorganism
This dissertation focuses on the problem of the low power density of MESs The
bacterial community structure of a high power generated mixed culture communities in
MFCs will be identified firstly and then one of major exoelectrogens will be isolated and
characterized This information will be helpful to understand the physiology and ecology
of exoelectrogens in MESs Consequently they will be beneficial to improve power
density of MFCs Lastly the nanostructure decorated anodes in MECs will be utilized to
improve the power density The power enhancement mechanism will be explored by
using a whole genome microarray They are presented here as four papers
In the first paper the cultivation independent molecular biological techniques
denaturing gradient gel electrophoresis (DGGE) and 16S rDNA clone library are utilized
to analyze the bacterial community structure of a higher power mixed culture MFCs The
analyzed results provide fundamental information for isolating the dominant bacteria in
mixed culture MFCs Two of dominant bacterial species has been isolated and used aone
to test power generation in MFCs The possible interaction among different bacterial
species in mixed culture is discussed
12
In the second paper one isolated exoelectrogenic bacterial strain SX-1 is
characterized It is identified as a member of the Citrobacter genus and power generation
is tested ultilizing a wide range of different substrates The electron transfer mechanism is
explored using Cyclic Voltammetry (CV) This study increases the known diversity of
power generating exoelectrogens and provides a new strain to explore the mechanisms of
extracellular electron transfer from bacteria to electrodes
The third paper shows effects on MESs performance by Fe nanoparticle decorated
anodes in the MESs The average current density produced with Fe nanoparticle
decorated anodes increased up to 59-fold higher than plain graphite anodes A whole
genome microarray is utilized to analyze the possible mechanism of enhanced current
density as responded to nanoparticle decorated anodes
The fourth paper describes the effects of carbon nanotube (CNT) modified anode on
the performance S oneidensis MR-1 in MESs Results demonstrate that current densities
produced with CNT decorated anodes are up to 56-fold higher than plain graphite anodes
The possible mechanisms of enhanced current density by CNT decorated are explored
13
Chapter 2
Bacterial community analysis of mixed consortium in higher
power density MESs
Shoutao Xu and Hong Liu
1 Introduction
The improvement in the performance of mixed culture MFCs requires an
understanding of the ecology in microbial communities of MFCs Many researchers have
attempted to characterize microbial populations and activities to elucidate the behaviors
and ecology of microorganisms in MFCs (Wrighton et al 2008 Fedorovich et al 2009)
In order to study the microbial ecology of the mixed culture in a MFC and select the
appropriate isolation medium for dominant bacterial species in the mixed culture the
fundamental steps are to analyze the bacterial diversity in the mixed culture of MFCs and
identity the dominant bacterial species in bacteria communities in MFCs
14
As for the identification of bacterial communities typically there are two general
methods The first method for identification of bacterial community is the traditional
cultivation processes using selective nutrients to promote the growth of different types of
bacteria within the samples (Amman et al 2000) The community structure can then be
assessed by identifying the isolates from the dominant colonies that were cultured This
can often be costly and laborious as each isolate has to be further studied by examining
its physiology taxonomy and reactivity to stains (Adwards et al 1989)
The second method relies on utilizing molecular techniques to analyze bacterial
community DNA Several molecular methods involving the extraction and analysis of
DNA from entire bacterial communities are used to identify genetic fingerprints of
bacteria These methods including the cloning and sequencing of 16S rDNA automated
ribosomal intergenic spacer analysis (ARISA) terminal restriction fragment length
polymorphism (T-RFLP) and denaturing gradient gel electrophoresis (DGGE) generate
profiles of bacterial community structures They can rapidly assess complex communities
from various environments (Amman et al 2000)
Due to the conservative characteristic of 16S rDNA in bacteria during the process
of evolution 16S rDNA sequencing can be used to identify different species of bacteria
Therefore the molecular techniques of denaturing gradient gel electrophoresis (DGGE)
with PCR and 16S rDNA clone library are used for analysis of the microbial diversity
These methods are more convenient and save time compared to traditional
isolationcultivation methods for microorganism analysis
In this work a biofilm bacterial community from an anode of a MFCs wase studied
by using denaturing gradient gel electrophoresis (DGGE) of PCR-amplified partial 16S
15
rRNA genes followed by cloning and sequencing of 16S rDNA The results provided
essential information for dominant bacterial isolation in mixed culture MFCs
2 Materials and methods
21 High power generation of mixed culture MFCs
Mixed cultures were originally inoculated from domestic wastewater (Corvallis
Wastewater Treatment Plant Corvallis OR) to MFCs after operating for 2 years using a
defined medium solution (Lovely 2002) with sodium acetate as the carbon source A
new MFCs was inoculated from the operating MFCs The polarization curves were
performed to measure the power generation when maximal stable power were established
(normally 3-5 batches post-inoculation) and the genomic DNA from the microbial
biofilm on the anode was extracted under the sterile conditions for further bacterial
community analysis
22 Denaturing gradient gel electrophoresis (DGGE)
DGGE with PCR is a method of analysis of bacterial community composition
based on different GC percentage of 16s rDNA in different bacterial species PCR-DGGE
comprises three steps (i) extraction of total community DNA from the sample (ii) PCR-
controlled amplification using specific oligonucleotide primers and (iii) separation of the
amplicons using DGGE For this purpose a reproducible and efficient method for total
DNA extraction is indispensable and needs to be evaluated and optimized depending on
the nature of the sample In the subsequent PCR step multiple PCR primer sets with
different resolution can be used In most PCR-DGGE applications on bacteria universal
or specific primers are targeting the 16S rRNA gene Upon electrophoresis of the PCR
16
amplicons and gel staining (using ethidium bromide silver staining or SYBR green)
DGGE gels are digitally captured and further analyzed using computer software packages
The richness of DNA fragment bands in DGGE indicates the bacterial diversity of the
sample
The detailed steps are as followed Biofilms were scratched from the anodes of high
power generated MFCs fed with sodium acetate Bacterial genomic DNA was extracted
from the biofilm samples using the DNeasy blood and tissue kit (Qiagen) according to the
manufacturerrsquos instructions The universal primer set was used to amplify the 16S rDNA
from the extracted genomic DNA then the V3 region of bacterial 16S ribosomal DNA
(rDNA) was amplified with GC-clamp primer (Muyzer et al 1993) for DGGE PCR
amplification was performed in a thermocycler DGGE of the PCR products was carried
out in a DcodeTM
Universal Mutation Detection System) The 8 (wv) polyacrylamide
gels contains from 30 to 55 denaturing gradients Electrophoresis was conducted
using a 1timesTAE (Tris-Acetate-EDTA) buffer at 130V and 60degC for 5 hours After
electrophoresis the gel was stained with ethidium bromide in 1timesTAE buffer for 15
minutes and washed in 1timesTAE buffer for 10 minutes The fragments were visualized
under a UV transilluminator The richness of single band in DGGE gel picture
preliminarily showed that the bacterial diversity in mixed culture since the single band in
gel represents one bacterial species
23 16S rDNA clone library construction method
16S rDNA-DGGE fingerprinting has proven to be particularly useful as an initial
investigation into bacterial communities and is suitable for identifying the predominant
bacteria that comprise 1 or more of cells within a given sample In the 16s rDNA clone
17
library construction process the first step is the extratction of the total genomic DNA
then the genomic DNA of the mixed culture biofilm was used as template for PCR
amplification of approximately 1500 bp of 16S rDNA with universal primers The PCR
products of 16S rDNA were purified and inverted into pGEM-T Easy vector system
before they were transformed into competent E coli The transformed cells were spread
on Luria-Bertani (LB) plates containing the selective antibiotic ampicillin and X-Gal and
incubated overnight at 37 OC after the more than 80 inverted 16s DNA fragment E coli
colonies were picked up and inoculated in the 1-3 ml liquid LB medium and left to grow
for 16 hours The plasmid DNA were extracted and sequenced The sequences were
compared directly to all known sequences deposited in GenBank databases using the
basic local alignment search tool (BLAST)
24 Bioinformatics Analysis
16S rDNA sequencing results of more than 80 colones were queried against the
GenBank and Ribosomal Database Project (RDP) databases using BLAST and
SEQUATCH algorithms (Cole et al 2007 Wang et al 2007 Cole et al 2009) The
neighbor-joining trees were constructed with the Molecular Evolutionary Genetics
Analysis package (MEGA Version 4) based on 1000 bootstrap analysis (Tamura et al
2007)
25 Dominant bacterial species band in DGGE identification
The DGGE results gave the information of dominant bacterial richness of bacterial
species in mixed culture based on the theory that single band represents one bacterial
species which showed the diversity of bacterial community in the mixed culture of a
MFC 16S rDNA clone library results provided the whole picture of bacterial community
18
including the bacterial species name and bacterial species percentage in the mixed culture
The known pure bacterial species which have been sequenced can be used as markers to
identify dominant bacterial species in DGGE The whole experimental procedure is
similar to the procedure of the mixed culture DGGE steps The first step is to extract the
collect the mixed culture biofilm genomic DNA and pure bacterial species marker
genomic DNA Then the universal primer set was used to amplify the 16S rDNA from
the extracted genomic DNA After that the V3 region of bacterial 16S ribosomal DNA
(rDNA) was amplified with GC-clamp primer (Muyzer et al 1993) for DGGE The
subsequent steps were similar to the mixed culture DGGE experimental procedure The
dominant bacterial bands in DGGE were determined by comparisons of the single band
position in the mixed culture DGGE with single pure bacterial species location in DGGE
26 Dominant bacterial species isolation and power generation
The bacterial community of mixed culture in MFCs were predominantly composed
of Geobacter sp Arcobacter sp and Citrobacter sp Two dominant bacterial species
have been isolated
Citrobacter sp SX-1 was isolated from the anode biofilm by using a solid agar
medium containing NH4Cl 6mM KCl 2mM CH3COONa (electron donor) 30 mM
FeC6H5O7 (electron acceptor) 20mM minerals and vitamins (Lovley and Phillips 1988)
Single colonies were picked up after culturing on the plate for 36 hours at 30 degC and
transferred two times on the agar plate for purification Then the isolates grew in a liquid
medium solution in anaerobic tubes containing the same constituents as the solid
medium All isolation process was operated in a glove box anaerobic chamber (Coy
Laboratory Products Grass Lake MI)
19
Arcobacter sp SX-2 was isolated from the anode biofilm of a MFC fed with sodium
acetate by using the same method of isolation of pure bacterial species Citrobacter XS-1
except using Trypticase Soy Agar with 5 Sheep Blood solid (from American type
culture collection (ATCC)) agar medium instead of Citrobacter sp medium Single
colonies were picked up after culturing on the plate for 36 hours at 30 degC and transferred
twice on the agar plate for purification Then the isolates grew in a liquid medium
solution microaerobically
Geobacter sp have been trying to isolate by using three different methods They are
the colony pickup after different condition enrichment Goebacter medium isolation and
dilution to extinction by multiple channel mini MFCs isolation however the Geobacter
sp has not been isolated yet due to some limits so far But they provide valuable
information for further isolation of Geobacter sp
Single chamber MFCs were used to evaluate power generation by different isolates
The MFCs were constructed as described previously (Liu and Logan 2004) and modified
with 3 cm2 carbon cloth anodes and 7 cm
2 carbon clothPt cathodes The total liquid
volume of each MFCs was about 15 ml with electrode spacing about 17 cm All MFCs
were operated in an autoclaved closed plastic box and sterile cotton was attached to the
outer surface of the air cathodes to prevent contamination A MFCs without bacterial
culture was used as control MFCs were inoculated with 3 ml late exponential phase
cultures of SX-1 in the medium solution reported previously (Liu and Logan 2004)
3 Results and discussion
31 Power production by mixed culture MFCs
20
A polarization curve was used to characterize current as a function of voltage in the
MFCs The polarization curves are performed by varying a series of external resistances
The results are shown in Figure 21 With MFCs acclimated to 25 Ω external resistance
the maximum power density was achieved at 65Wm2 based on the polarization data At
this point the current density is 21mAcm2 The power density was three times higher
than previous report (20 Wm2) using phosphate buffer (Fan et al 2007) Power density
was also 2 times higher than (28 m Wm2) using a bicarbonate buffer ( Fan et al 2007)
32 Bacterial community structure assessed by PCR-DGGE
Bacterial DGGE profiles for higher power generating MFCs mixed culture setups
were relatively complex (more than 10 bands) However there are three brightly stained
bands and other less intense bands (Fig 2-2) These results indicated there were probably
three dominant bacterial species in mixed culture MFCs
010
020
030
040
050
060
070
2
3
4
5
6
7
04 05 05 06 07 08 10 11 13 15 21 22 23 24
Volta
ge (
V)
Pow
er
densi
ty (
Wm
2)
Current density (mAcm2)
Power
Voltage
power density 65wm2 at current
density208 mAcm2
Figure 2-1 Polarization curves of high power mixed culture MFCs
21
33 16S rDNA clone library results
A total of 83 colonies were sequenced for 16S rDNA clone library construction They
represented the bacterial community structure in mixed culture in MFCs The results
revealed that the predominant bacterial species in mixed culture are Geobacter sp (66
of the mixed culture) Arcobacter sp (12 of the mixed culture) and Citrobacter sp (11
of the mixed culture) Those three genuses comprised 88 of the total bacterial species
(Fig 23) This result is consistent with the DGGE result which showed that three bright
bands in the DGGE gel represented three dominant bacterial species in the mixed culture
The phylogenetic analysis of the colonies (83 colony sequences) is shown in Figure 24
Figure 22 DGGE result of high power generation mixed culture MFC (1 mixed
culture 2 pure culture control)
1 2
22
34 Dominant bacterial band in DGGE identification
Geobacter
66
Arcobacter
12
others
6
Pseudomonas
1
Citrobacter
11
Clostridium
2
Anaerovorax
2 Geobacter
Arcobacter
Citrobacter
Clostridium
Anaerovorax
Pseudomonas
others
Figure 23 Bacterial species and percentage of bacterial community of mixed
culture MFC lt 1 Alcaligensgt
lt1 Commamonasgt
lt1 Pseudomonas aeruginosagt
lt1 Klebsiellagt
lt11 Citrobactergt
lt1 Wolinellagt
lt12 Arobactergt
lt2 Anaeroboraxgt
lt2 Clostridium gt
lt1 unclear bacteriagt
lt66Geobactergt
100
100
85
100
100
82
99
87
100
002
Figure 24 Phylogenetic trees of bacterial species from mixed culture MFC
23
The dominant bacterial species including Goebacter sp Citrobacter sp and Arcobacter
sp in the mixed culture are shown in Figure 25 The upper band is Arcobacter sp and
Citrobacter sp is located in the bottom band of DGGE Geobacter sp is the middle one
The analysis of the bacterial community using denaturing gradient gel electrophoresis
(DGGE) of PCR-amplified 16S rRNA gene fragments and 16S rDNA clone library
construction method showed great phylogenetic diversity of mixed culture in MFCs with
the identification of sequences derived from bacteria of the taxa deltaproteobacteria
(Geobacter sp) gammaproteobacteria (Citrobacter sp) and epsilonproteobacteria
(Arcobacter sp) The result is consistent with the conclusion that among the isolated
exoelectrogens gammaproteobacteria and deltaprotebactia appear to be dominant among
others (Parot et al 2009 Liu et al 2010)
35 The dominant bacterial species isolation and power generation
Figure 25 Dominant bacterial species identification in DGGE
A B C D
A Citrobacter sp
B Geobacter sp
C Arobacter butzmeri
D mixed culture control
24
Two dominant bacterial species Citrobacter sp XS-1 and Arcobacter sp XS-2 have been
isolated and tested for power generation The power densities generated by the two
isolates are shown in the Fig 26 Current densities generated by isolate Citrobacter sp
and Arcobacter sp were 98 mAm2 and 20 mAm
2 respectively which were much less
than that (21Am2) generated by the mixed culture from which they were isolated
Figure 26 Current density generated by different dominant isolates
The maximum current density generated by mixed culture is much higher than the
one produced by pure bacterial species Citrobacter sp XS-1 and Arcobacter sp XS-2 in
the same structure MFCs Although the Geobacter sp in our mixed culture has not been
isolated a previous study showed that maximum current density generated by Geobacter
sp was 080 Am2
(Bond and Lovley 2003) The current density results seem partially
supporting the conclusion that mixed-culture generated higher power densities than their
pure-culture counterparts in MFCs (Jung and Regan 2007) This result suggests there
may be synergistic interactions within the anode bacterial communities
0
500
1000
1500
2000
2500
Mixed culture Citrobacter sp Arcobacter Sp
Cu
rre
nt
de
nsi
ty (
mA
m2)
25
Geobacter sp are strictly anaerobic bacteria which were enriched in most of the anode
biofilms (George 2005) Citrobacter species are facultative anaerobic bacteria (George
2005) which can be found in a wide variety of habitats including in soil water and
wastewater Arcobacter sp are micro-aerobic bacteria including both environmental
nonpathogens and opportunistic human pathogens They are able to grow in aerobic
conditions but in the optimal growth under micro-aerobic conditions (George 2005)
Arcobacter sp has been discovered in the MFCs by Fedorovich (Fedorovich et al 2009)
It is interesting that the mixed culture bacterial community composed of primarily these
three bacterial species has demonstrated to generate high power at non-strictly anaerobic
condition while leading dominant bacterial species Geobacter sp (66) is strict
anaerobic bacteria and generated powder in the MFCs only under strict anaerobic
condition (George 2005) Therefore it is possible that microaerobic bacteria such as
Arcobacter sp and facultative anaerobic bacteria Citrobacter sp can facilitate to create
anaerobic condition for Geobacter sp when they utilized oxygen for growing in the
mixed culture Syntrophic communities study have showed that anaerobic bacteria and
methanogenic archaea form compact microbial structures that operate like an organ rather
than a set of microorganisms functioning independently (Stams and Plugge 2009) Some
substrates have been degradeted within these communities while they are not able to be
fermented by individual species alone interspecies electron transfer also have been
presented in these communities (Stams and Plugge 2009) Summers et al (2010) also
discovered that direct exchange of electrons happened within coculture of Geobacter
metallireducens and Geobacter sulfurreducens Therefore we believe that there are
26
more complex synergistic interactions between different bacteria species in mixed culture
in the MFCs
Acknowledgements
The authors acknowledge support from the US National Science Foundation
(CBET 0955124) We also thank Keaton Lesnik for the review of this manuscript
27
Chapter 3
New Exoelectrogen Citrobacter sp SX-1 Isolated from a Microbial Fuel Cell
Shoutao Xu and Hong Liu
Published in
Journal of Applied Microbiology 111(5)1108-1115 (2011)
ABSTRACT
Exoelectrogenic bacterial strain SX-1 was isolated from a mediator-less microbial fuel
cell by conventional plating techniques with ferric citrate as electron acceptor under
anaerobic condition Phylogenetic analysis of the 16S rDNA sequence revealed that it
28
was related to the members of Citrobacter genus with Citrobacter sp sdy-48 being the
most closely related species The bacterial strain SX-1 produced electricity from citrate
acetate glucose sucrose glycerol and lactose in MFCs with the highest current density
of 205 mA m-2
generated from citrate Cyclic voltammetry analysis indicated that
membrane associated proteins may play an important role in facilitating the electrons
transferring from bacteria to electrode This is the first study that demonstrates that
Citrobacter species can transfer electrons to extracellular electron acceptors Citrobacter
strain SX-1 is capable of generating electricity from a wide range of substrates in MFCs
This finding increases the known diversity of power generating exoelectrogens and
provided a new strain to explore the mechanisms of extracellular electron transfer from
bacteria to electrode The wide range of substrate utilization by SX-1 increases the
application potential of MFCs in renewable energy generation and waste treatment
Keywords Microbial fuel cell Exoelectrogen Citrobacter sp SX-1 Extracellular
electron transfer
29
Introduction
Microbial Fuel Cells (MFCs) technology has become an active research area
recently as a promising approach for renewable energy generation wastewater treatment
and bioremediation (Rabaey and Verstraete 2005 Lovley 2006 Fan et al 2008 Logan
2009 Liu et al 2010) The key feature of MFCs system is the microbe-catalyzed electron
transfer from organic matter to anodes Many studies have shown the presence of diverse
bacterial communities on MFCs anodes (Reimers et al 2001 Liu et al 2004b Kim et al
2005) Over 20 exoelectrogens ie microbes that can transfer electrons exocellularly to
electrodes have been reported in the past 10 years (Logan 2009 Liu et al 2010) The
exoelectrogens so far belong to diverse genetic groups including alphaproteobacteria
(Rhodopseudomonas Ochrobactrum and Acidiphilium) (Malki et al 2008 Xing et al
2008 Zuo et al 2008) betaproteobacteria (Rhodoferax) (Chaudhuri et al 2003)
gammaproteobacteria (Shewanella Pseudomonas Klebsiella Enterobacter and
Aeronomas) (Kim et al 2002 Pham et al 2003 Rabaey et al 2005 Bretschger et al
2007 Zhang et al 2008 Chung et al 2009 Rezaei et al 2009) deltaproteobacteria
(Geobacter Geopsychrobacter Desulfuromonas and Desulfobulbus) (Bond et al 2002
Bond and Lovley 2003 Holmes et al 2004ab 2006) Epsilonproteobacteria (Arcobacter)
(Fedorovich et al 2009) Firmicutes (Clostridium and Thermincola) (Park et al 2001
Wrighton et al 2008) Acidobacteria (Geothrix) (Bond and Lovley 2005) and
actinobacteria (Propionibacterium) (Wang et al 2008) A wider range of
exoelectrogenic species are expected to be discovered
Three mechanisms have been proposed for exocellular transport of electrons by
exoelectrogens without artificial mediators Some exoelectrogenic bacteria can transfer
30
electrons to electrodes through soluble redox compounds excreted by microorganisms
(Bond and Lovley 2005 Rabaey et al 2005) and others can directly transfer electrons to
anodes via outer cell membrane proteins (Myers and Myers 1992) Recently more and
more evidence supports the involvement of bacterial nanowires in extracellular electron
transport (Reguera et al 2005 2006 Gorby et al 2006) In spite of the discovery of
many bacterial species that can transfer the electrons to electrode without the need of
artificial mediators the investigation of extracellular electron transfer mechanisms was
mainly focused on a few species such as those from Geobacter and Shewanella genera
(Kim et al 1999 2002 Reguera et al 2005 2006 Gorby et al 2006) The electron
transfer mechanisms for many of the isolated exoelectrogens species are still not well
studied
In the present study we isolated a new exoelectrogen strain SX-1 from a MFC a
strain phylogenetically related to Citrobacter sp Power generation from various carbon
sources by this strain was evaluated using single chamber MFCs Plausible extracellular
electron transfer mechanisms were also discussed based on the characterization of anodic
biofilms by cyclic voltammetry (CV)
Materials and methods
Bacterial strain SX-1 isolation
Bacterial strain SX-1 was isolated from the anode biofilm of a MFC fed with sodium
acetate operated in fed-batch mode over a period of six months The original source of
the inoculum is wastewater from a local waste water treatment plant Bacterial cells were
released from the carbon cloth anode by shaking the cloth in a sterile flask with 15 mL
sterile 1 NaCl solution and glass beads (2 mm diameter) The suspension was then
31
serially diluted from 10 times to 105
times and plated on a petri dish with a solid agar
medium containing NH4Cl 6mM KCl 2mM CH3COONa (electron donor) 30 mM
FeC6H5O7 (electron acceptor) 20mM minerals and vitamins (Lovley and Phillips 1988)
Single colony was randomly selected after culturing on the plate for 36 hours at 30 degC
and purified on a new agar plate following a procedure reported previously (Chung and
Okabe 2009) Then the isolate was picked up and grew in a liquid medium solution in
anaerobic tubes containing the same constituents as the solid medium for further analysis
All isolation process was operated in a glove box anaerobic chamber (Coy Laboratory
Products Grass Lake MI)
16S rDNA sequencing and phylogenetic analysis
Bacterial genomic DNA was extracted from the isolated strain SX-1 using the DNeasy
tissue Kits (Qiagen Valencia CA) according to the manufacturerrsquos instructions The 16S
rDNA was amplified by PCR using a pair of universal primers 27F (5rsquo-
AGAGTTTGATCMTGGCTCAG-3rsquo) and 1492R (5rsquo-GGTTACCTTTGTTACGACTT-
3rsquo) (Park et al 2001) The conditions used for PCR were described previously
(Ferna ndez et al 1999) PCR products were purified ligated and cloned according to a
procedure described in a previous report (Xing et al 2008) Plasmids were isolated from
randomly selected clone colonies with a QIAprep Spin Miniprep kit (Qiagen Valencia
CA) and three plasmid inserts were then sequenced in both directions using an ABI 3730
DNA sequencer (Applied Biosystems Carlsbad CA)United States) Due to the identical
16S rDNA sequence of the three colones only one was queried against the GenBank and
Ribosomal Database Project (RDP) databases using BLAST and SEQUATCH algorithms
(Cole et al 2007 Wang et al 2007 Cole et al 2009) The 16S rDNA sequence of
32
strain SX-1 and closely related type strains were aligned using CLUSTALX software
(Thompson et al 1997) The neighbor-joining trees were constructed with the Molecular
Evolutionary Genetics Analysis package (MEGA Version 4) based on 1000 bootstrap
analysis (Tamura et al 2007)
SEM
The morphologies of SX-1 cells growing overnight in anaerobic tubes were examined
with a scanning electron microscope (FEI Quanta 600 FEG FEI Company Hillsboro
OR) The suspended cells were fixed in a 25 glutaraldehyde and 01M phosphate
buffer solution and dehydrated with a graded ethanol series from 30 to 100 After
dehydration the samples were dried in a critical point dryer and then sputter-coated with
AuPd for SEM examination (Liu and Logan 2004)
Evaluation of power generation by SX-1 in MFCs
Single chamber MFCs were used to evaluate power generation by SX-1 using various
substrates The MFCs were constructed as described previously (Liu and Logan 2004)
and modified with 3 cm2 carbon cloth anodes and 7cm
2 carbon clothPt cathodes The
total liquid volume of each MFCs was about 15 ml with electrode spacing about 17 cm
All MFCs were operated in an autoclaved closed plastic box and sterile cotton was
attached to the outer surface of the air cathodes to prevent contamination A MFC
without bacterial culture was used as control MFCs were inoculated with 3 ml late
exponential phase cultures of SX-1 in the medium solution reported previously (Liu and
Logan 2004) Seven substrates including citrate acetate glucose sucrose glycerol and
lactose were evaluated individually for power generation in a fed-batch mode in a
temperature control chamber (30 plusmn2degC) For each substrate at least 8 batches were run to
33
investigate the effect of biofilm formation on current output at fixed external resistance of
1kΩ and the same initial substrate concentration of 30 mM Phosphate buffer (100mM)
was used to maintain the solution pH and conductivity The MFCs with sodium citrate as
electron donors were also examined for maximum power output by varying the external
resistance from 20 kΩ to 4 kΩ A data acquisition system was used to record voltage data
during the MFCs operation (Keithley 2700 Keithley Instruments Inc Cleveland OH) It
took about 15 to 30 minutes for the MFCs to stabilize depending on the external
resistance At each resistance we collected at least five data at the steady condition to
make the polarization curves Averaged voltages were used to calculate the power density
(mW m-2
) according to P=IVA where I was the current V was voltage and A was cross-
sectional area of the anode
CV analysis
CV was used to characterize the oxidation and reduction reactions on the anodic surface
of the MFCs fed with citrate and operated at 7 KΩ This external resistance was selected
because the maximum power density was obtained at this resistance based on the
polarization experiment We assumed the biofilm on the anode of MFCs were well-
developed when stable power output was obtained after 3 batches of operation The
MFCs were then used directly for CV analysis at four current generating stages of the
fourth batch (1) initial exponential current increasing stage (middle point of stage about
4 h after media change) (2) current plateau stage (middle point of stage about 8 h after
media change) and (3) current decreasing stage (middle point of stage about 16h after
media change) (4) right after the complete replacement of medium solution (Figure 6)
The anode was used as working electrode the cathode as counter electrode and an
34
AgAgCl electrode was selected as the reference The CV curves were scanned from 200
to -600 mV at a rate of 5 mV s-1
using a potentiostat (G300 Gamry Instrument
IncWarminster PA) Control experiment was also conducted using new anode (without
biofilm) and new medium solution
Nucleotide sequence accession number
The 16S r DNA sequence determined in this study has been deposited in the GenBank
database under accession number HQ845373
Results
Identification of the strain SX-1
An almost-complete 16S rDNA sequence (1475 bp) of strain SX-1 was obtained and
subjected to comparative analysis with the 16S rDNA of closely related reference strains
A phylogenetic analysis revealed that strain SX-1 should be assigned to the genus
Citrobacter and was most closely related to Citrobacter sp sdy-48 EJ463782 (990
sequence similarity) and Citrobacter sp yy-21 FJ463779 (975 sequence similarity)
These three strains formed a distinct sub cluster in the neighbor-joining in which the new
isolate and Citrobacter sp sdy-48 EJ463782 formed a distinct subline (Figure 31)
The SEM image illustrated that the strain SX-1 is a rod-shaped bacterium 03-06 microm
wide and 12-17 microm long (Figure 32) Strain SX-1 colonies on nutrient agar appeared 2-
3 mm in diameter smooth moist opaque with a shiny surface after 5 days incubation
under anaerobic condition The fact that strain SX-1 grew both aerobically and
anaerobically using FeC6H5O7 as electron acceptor suggests that the strain is facultatively
anaerobic Strain SX-1 showed robust growth on citrate as a carbon source These D
35
properties were similar to those of genus Citrobacter described in Bergeyrsquos Manual of
Systematic Bacteriology (George 2005)
Figure 31 phylogenetic tree of strain SX-1 and closely related species based on 16S
rDNA sequences The tree was constructed using the neighbor-joining method
Figure 32 SEM image of planktonic cells of Citrobacter sp SX-1
Strain SX-1
Citrobacter sp sdy-48 (FJ463782)
Citrobacter sp yy-21 (FJ463779)
Citrobacter koseri E639 (ATCC 25408)
Citrobater koseri CDC 3613-63(AF025372)
Citrobacter koseri CDC 8132-86(AF025366)
Salmonella enterica SL483 (CP001138)
Salmonella enterica AKU12601(AY696668)
Shigella sonnei Ss046(CP000038)
E coli C2 (AF403733)
E coli E24377A(CP000800)88
100
100
76
44
92
53
36
0001
1 microm
36
Electricity production by strain SX-1 in MFCs
The isolated strain SX-1 was first tested for its ability to generate current from sodium
citrate at 1kΩ Current gradually increased to a maximum 50 mA m-2
after cell
inoculation and then decreased (Figure 33) After the MFCs was refilled with new
substrate solution the current recovered rapidly and reached a higher level than the first
batch After 4 batches operation the maximum current output of each batch became
stable The highest current density achieved in the MFCs with Citrobacter sp SX-1 at 1
KΩ was 98 mA m-2
(Figure 33)
Figure 33 Electricity generation by Citrobacter sp SX-1 in a single chamber MFCs
with sodium citrate (30 mM) as substrate at 1 KΩ
0
20
40
60
80
100
120
0 50 100 150 200
Curr
ent
den
sity
(m
A m
-2)
Time (h)
37
Current generation by SX-1 from other substrates including glucose lactose sodium
acetate glycerol and sucrose were also assessed at an external resistance of 1KΩ
(Figure 34) When repeatable cycles of current output were obtained for these substrates
glycerol generated the highest maximum current density of 58 mA m-2
followed by
lactose and sucrose with 29 mA m-2
and 27 mA m-2
respectively Glucose and acetate
produced the lowest maximum current density of 96 and 43 mA m-2
respectively These
results indicated that strain SX-1 can utilize a wide range of substrates for electricity
generation in MFCs but with different power generation potentials
Figure 34 Electricity generation of Citrobacter sp SX-1 using different substrates in
single chamber air-cathode MFCs at 1 KΩ (The error bars stand for the standard
deviation of 3 replicates)
0
10
20
30
40
50
60
70
Glucose Lactose Glycerol Acetate Sucrose
Curr
ent
den
sity
(m
A m
-2)
38
Polarization experiment was further conducted to determine the maximum power
density generated by strain SX-1 in the single chamber MFCs with citrate as substrate A
maximum power density of 881 mW m-2
was obtained at current density of 205 mA m-2
at an external resistance of 7 KΩ (Figure 35)
Figure 35 Power and voltage generation by Citrobacter sp SX-1 as a function of
current density using sodium citrate (30 mM) as substrate The error bars stand for the
standard deviation of the 3 voltagepower outputs obtained in three MFCs
20
30
40
50
60
70
80
90
100
01
02
03
04
05
06
07
80 110 140 170 200 230
Po
wer
den
sity
(m
W m
-2)
Vo
ltag
e (V
)
Current density (mA m-2)
Voltage
Power
39
Figure 36 Four current generating stages for CV analysis current increasing stage (stage
1) current plateau stage (stage 2) current decreasing stage (stage 3) and right after the
replacement of medium solution (stage 4) The MFCs was operated at 7 KΩ with sodium
citrate (30 mM) as substrate
Cyclic Voltammetry
To determine the presence of redox active compounds produced by SX-1 and the
location of these compounds CV scan of the anodic biofilms at four current output stages
and the supernatant of MFCs medium solution at the end of the batch experiment were
performed Reduction peaks at range of -310 ndash -350 mV and oxidation peaks at range -
100 ndash -150 mV were observed at all current generating stages (Figure 36 Figure 37A)
suggesting the presence of redox active compounds may involve in extracellular electron
transfer by Citrobacter sp SX-1 The amplitude of the peaks increased according to the
growth stage of the batch and the highest peaks were present after the current plateau
stage which indicated the redox active compounds mainly were secreted in the current
plateau stage While the current density at stage 3 (deceasing stage) was lower than that
at stage 2 (current plateau stage) the redox peak at stage 3 was higher than that in stage 2
0
50
100
150
200
250
50 55 60 65 70 75 80 85
Curr
ent
den
sity
(m
A m
-2)
Time (h)
A
A
Stage 1
Stage 2 Stage 3
Stage 4
40
A plausible explanation for this pattern was that redox active compounds were
continuously being secreted and accumulated after the current peak was reached
resulting in more redox compounds present after the plateau stage But since most of the
carbon source had already been being used up many of the redox compounds at this
stage had not been as active as those at current plateau stage due to much less electrons
were available to be transferred The bigger peak amplitude in stage 4 than in stage 1
indicates that the redox active compounds were continuously being secreted and
accumulated in the biofilm even after stable power was generated at 7 kΩ The slightly
change of the peak locations at different stages was possible due to the solution chemistry
change during the current generation process or the slightly location change of reference
electrode during the measurement (Figure 37B) When the medium solution in the MFCs
was replaced by fresh medium at the end of the batch the oxidization and reduction
peaks were still presented in the CV (Figure 37B) but no peaks were observed when the
supernatant and fresh mediums were tested in a MFCs with no biofilm (Figure 37C)
These results suggest the compounds involved in the electron transfer were located not in
the supernatant (solution) but in the biofilm
41
Figure 37 Cyclic Voltammetry analysis of Citrobacter sp SX-1 biofilms at (A) current
increasing stage current plateau stage and decreasing stage (end of the batch) (B) end of
the batch and replaced with fresh medium and (C) controls
-40E-04
-30E-04
-20E-04
-10E-04
00E+00
10E-04
20E-04
30E-04
-060 -050 -040 -030 -020 -010 000 010 020
Curr
ent
(A)
Potential (V) vs AgAgCl
Current plateau stage
Current increasing stage
Current decreasing stage
-40E-04
-30E-04
-20E-04
-10E-04
00E+00
10E-04
20E-04
30E-04
-060 -050 -040 -030 -020 -010 000 010 020
Curr
ent
(A)
Potential (V) vs AgAgCl
Current decreasing stage
Biofilm with fresh medium
-40E-04
-35E-04
-30E-04
-25E-04
-20E-04
-15E-04
-10E-04
-50E-05
00E+00
-060 -050 -040 -030 -020 -010 000 010 020
Curr
ent(
A)
Potential (V) vs AgAgCl
Supernatant
Fresh medium
A
B
C
B C
42
Discussion
Citrobacter species belonging to Gammaproteobacteria Enterobacteriales
Enterobacteriaceae are facultative anaerobic bacteria (George 2005) They can grow with
oxygen as an electron acceptor but also with a variety of electron accepters such as Fe (III)
in the absence of oxygen (George 2005) Citrobacter species can be found in a wide
variety of habitats including in soil water and wastewater Several Citrobacter species
have been studied as important bioremediation bacterium for heavy metal removals
sulfate reduction phenol degradation and chlorophenol degradation (Macaskie et al 1995
Narde et al 2004 Qiu et al 2009) Citrobacter species have also been found on the
anodic biofilm of a diesel-degrading MFCs (Morris et al 2009) However there was no
study reported that Citrobacter species can transfer electrons to extracellular electron
acceptors This finding that Citrobacter strain SX-1 can generate electricity in MFCs
increased the diversity of power generating exoelectrogens and provided a new strain to
explore the mechanisms of extracellular electron transfer from bacteria to electrode
Among the isolated exoelectrogens gammaproteobacteria and deltaprotebactia appear to
be dominant among others (Parot et al 2009 Liu et al 2010) The isolated strain
Citrobacter sp SX-1 phylogenetically belongs to gammaproteobacteria which provides
another evidence to support this observation
Most exoelectrogens utilize a limited range of substrates for power generation For
example Shewanella oniedensis MR-1 can oxidize lactate under anaerobic condition but
cannot utilize acetate for electricity generation (Kim et al 2002) Geobacter
sulfurreducens can completely oxidize acetate for power generation but it cannot utilize
simple sugars (Bond and Lovley 2003 Caccavo et al 1994)) In comparison strain SX-1
43
not only can oxidize citrate for power generation but also utilize a wide range of
substrates for power generation including glycerol glucose lactose sucrose and acetate
Interestingly the higher current density generated by strain SX-1 from glycerol a main
by-product of biodiesel production process suggests that strain SX-1 may be potentially
used for harvesting energy from biodiesel wastes using MFCs However the current
density generated by strain SX-1 (205 mA m-2
) is lower than that (805 mA m-2
at 1 kΩ)
generated by the mixed culture from which the SX-1 was isolated indicating the
existence of other higher power generating bacteria andor complex ecology in the mixed
culture community
Understanding mechanisms of microbial extracellular electron transfer is critical for
enhancing the electron transfer rate from bacteria to electrode through metabolic or
genetic engineering of exoelectrogens Since the spent supernatant solution of MFCs run
with SX-1 showed no redox properties the redox compounds produced by SX-1 may
have been retained in the biofilm Alternatively SX-1 may utilize one or both of the
other two known extracellular electron transfer mechanisms For example it is possible
that strain SX-1 transfers electrons to the anode via cell-bound outer membrane proteins
The two distinct oxidation and reduction peaks observed at -100mv- -150 mV and -310--
350 mV of the SX-1 biofilm have a midpoint potential range from -205mV to -250mV
which is close to the redox potential of some c-type cytochromes (Field et al 2000 Meitl
et al 2009) which are well-known to play an important role in extracellular electron
trasnfer (Eggleston et al 2008 Field et al 2000 Meitl et al 2009) Field et al (2000)
reported that the c-type cytochromes CymA had the midpoint potentials of -229 mV
Meitl et al (2009) and Eggleston et al (2008) reported that the c- type cytochromes
44
OmcA in Shewanella oneidensis MR-1 had the midpoint potentials of -201 mV and -208
mV
While itrsquos possible that SX-1 may also have conductive appendages to enhance the
extracellular transfer electron similar to the nanowires discovered in some Geobacter
and Shewanella species (Reguera et al 2005 2006 Gorby et al 2006) further
investigation on the presence of these appendages on the cell surface of SX-1 is needed
Acknowledgements
The authors acknowledge support from the US National Science Foundation
(CBET 0955124) We also thank Teresa Sawyer for her helps with the SEM analysis and
Jeremy Chignell and Yanzhen Fan for their review of this manuscript
45
Chapter 4
Enhanced Performance and Mechanism Study of Microbial Electrolysis Cells
Using Fe Nanoparticles Decorated Anodes
Shoutao Xu Hong LiuYanzhen Fan Rebecca Schaller Jun Jiao Frank Chaplen
Published in
Applied Microbiology and biotechnology 93(2)871-880 (2012)
46
ABSTRACT
Anode properties are critical for performance of microbial electrolysis cells (MECs) In
the present study Fe nanoparticle modified graphite disks were used as anodes to
investigate the effects of nanoparticles on the performance of Shewanella oneidensis MR-
1 in MECs Results demonstrated that average current densities produced with Fe
nanoparticle decorated anodes were up to 56-fold higher than plain graphite anodes
Whole genome microarray analysis of the gene expression showed that genes encoding
biofilm formation were significantly up-regulated as response to nanoparticle decorated
anodes Increased expression of genes related to nanowires flavins and c-type
cytochromes indicate that enhanced mechanisms of electron transfer to the anode may
also have contributed to the observed increases in current density The majority of the
remaining differentially expressed genes were associated with electron transport and
anaerobic metabolism demonstrating a systemic response to increased power loads
Keywords Microbial electrochemical system microbial fuel cell microbial electrolysis
cell nanotechnology differential gene expression DNA microarray
47
Introduction
Microbial electrochemical systems (MESs) have been intensively studied since Lewis
achieved practical advances in this field (Logan 2007) however they attracted much
research attention in recent years due to their promising applications in renewable energy
generation bioremediation and wastewater treatment In a MES microorganisms
interact with electrodes via electrons catalyzing oxidation and reduction reactions at the
anode and the cathode The most-described type of MESs is microbial fuel cells (MFCs)
in which useful power is generated from electron donors typically biodegradable organic
materials (Logan et al 2006) Various novel MESs have recently been developed to
produce hydrogen (microbial electrolysis cells (MECs)) (Liu et al 2005 Logan et al
2008 Rozendal et al 2006 Schaller et al 2009) to power an autonomous sensor or
sensor network (Ringeisen et al 2006) to reduce bioremediation targets (Lovley 2006
Shea et al 2008) Sukkasem et al 2008) and to desalinate water (Cao et al 2009) The
key feature shared by these systems is the microbe-catalyzed electron transfer from
organic matter to electrodes (anodes) (Rabaey et al 2004) Enhancing the anodic current
output which highly depends on the performance of the electrodes is critical for the
successful application of all these processes (Logan et al 2007 Park and Zeilus 2002
2003)
Nanomaterials have received much attention from researchers in the context of
microbiology due to their unique physical electrical and chemical properties which
facilitate the study of interactions between bacteria and surfaces (Liu 2006) Previous
studies have demonstrated that electrodes decorated with different nanostructures such
as carbon nanotubes nanostructured polyanilinetitanium dioxide composite and titania
48
nanotubes have the potential to enhance power generation in MFCs (Morozan et al 2007
Qiao et al 2008 Quan et al 2005) Furthermore our recent study shows nanoparticle
(NP) decorated anodes greatly increased the electrochemical electron transfer rate in
MFCs (Fan et al 2011) However Fe NPs as an ideal candidate for decorating electrodes
because of respectively low price and high conductivity compared to other materials has
not been focused to study in MECs
S oneidensis MR-1 an important electrochemically active bacterial strain has been
exploited for the electricity generation in MFCs (Logan et al 2007 Logan 2009 Park
and Zeikus 2002) The availability of genome sequence for this strain makes it possible to
use transcriptome assays to globally measure the responses to different growth conditions
and environmental stresses (Murray et al 2001) S oneidensis MR-1 gene expression
response to heat shock cold shock Cr (VI) and U (VI) reduction chromate stress and
iron and acid tolerance have been studied in the past several years (Bencheikh-Latmani et
al 2005 Brown et al 2006 Gao et al2006 Gao et al 2004 Yang et al 2008) However
no studies have been focused on the Shewanella gene expression response to NPs in
MESs
The mechanism of increased electron transfer rate exhibited by nanoparticle decorated
electrodes is not well understood yet In the present study graphite disks decorated with
Fe NPs were used as anodes to explore the effects of nanostructures on current generation
in a multi-anode MECs DNA microarrays were utilized to investigate differences in the
global gene expression profile of S oneidensis MR-1 grown on plain versus Fe NPs
decorated anodes
Materials and methods
49
Bacterial cultures
S oneidensis MR-1 was purchased from American Type Culture collection (ATCC
700550) and stored as freezer stocks at -80degC Stock cells of S oneidensis MR-1 was
grown aerobically using a medium containing 30 gL Trypticase Soy Broth (BD 211825
Becton Dickinson Company Franklin lakes NJ) at 30degC Cells in early stationary
growth phase (optical density of 15-16 at 600nm (Spectrophotometer UV-1700
Shimadzu Columbia MD) were washed two times then injected into the chamber of the
MECs for current production Trypticase Soy Broth medium with 30 mM sodium lactate
as electron donor was used in MECs Phosphate buffer (100mM) was used to maintain
the solution pH 7 and solution conductivity at 15 mScm
Characterization of nanostructured anodes
Superfine isomolded graphite disks (16 cm x 05 cm GraphiteStorecom Inc) were
polished with ultrafine sand paper (2000 grit 3M Company) as the base for the NP
decorated and control anodes Fe NPs decorated anodes were fabricated by thermal
annealing of metal thin film coated graphite disks Thin films of Fe were firstly deposited
on the polished graphite disk by using sputter coating for 95 min and the samples then
were annealed in the Chemical vapor deposition (CVD) at 750degC for 20 min to form Fe
NPs Fe-NPs were produced with a diameter of 200plusmn100 nm and a density range
(distance between NPs) of 200plusmn100 nm with a coating time of 95 min SEM images of
Fe NPs decorated anode and control surfaces were shown in the Figure 41
50
Figure 41 SEM images of Fe NPs decorated anodes (A) Graphite disk control (B) Fe
Nanoparticle decorated
Multiple channel MECs construction and operation
A MECs with removable multiple anodes with each effective anode area 07 cm2
was
constructed and used to evaluate the effects of nanostructure on current density of
according to a previous report (Fan et al 2011) The cathode was made of wet-proof
(30) carbon cloth (type B E-TEK Division Inc USA) coated with PtC (20 E-TEK
Division Inc USA) following a previously reported procedure (Liu et al 2005) The
final platinum loading was 05 mgcm2 per projected cathode area The size of cathode
(150 cm2) was 25 times larger than the total effective area of the 8 graphite disk anodes
(56 cm2) to prevent cathode limitations on the performance of the MECs system All
MECs with Fe-NP decorated anodes and control anodes were tested for current
generation for 24 h with medium (without bacteria) before injecting the bacterial cells in
order to determine whether the Fe-NP decorated anodes demonstrate chemical current
generation compared to control anodes All the testing anodes were set up in the MECs
B
500 nm
A
500 nm
51
at once so that the S oneidensis MR-1 culture was applied to all of them simultaneously
Short electrode spacing (17 cm) was used in the MECs design to reduce the internal
resistance A voltage of 06 V was applied the MECs for current generation after the cells
of S oneidensis MR-1 were inoculated into MECs at early stationary phase A multimeter
with a data acquisition system (2700 Keithly USA) was used to monitor the current
change by measuring the voltage (every 300 seconds) drop through a resistor at 300 ohm
Fig 4 2 Effects of Fe-NP decorated anodes on the current density in MECs Control is
plain graphite disk anode Fe-NP is the anode with Fe-NP decoration
Microarray analysis
Biofilms for whole gene microarray analysis were aseptically removed from the plain
and Fe-NP decorated anodes of MECs after 110 hours further incubation at 30oC when
current density obviously started to decease Total RNA was extracted using Trizol
(Ambion inc AustinTX) following the manufacturerrsquos instructions Integrity of the
52
RNA samples was confirmed using the Agilent 2100 Bioanalyzer (Agilent Technology
Inc Palo Alto CA) The same amount of RNA (200 ng) from each sample was used for
further analysis after amplification by using the MessageAmpTM
II-Bacteria Prokaryotic
RNA Amplification kit (Ambion AustinTX) according to the manufacturerrsquos
instructions All of the 4295 genes (resource from NCBI) represented on the S oneidensis
MR-1 whole-gene microarrays were ordered from Roche NimbleGen Inc (Madison WI)
Biological triplicates of biofilms on the Fe-NP decorated anodes and control were
analyzed respectively cDNA synthesis labeling and hybridization were carried out by
the CGRB Core Laboratories at Oregon State University DNASTAR ArrayStarTM
3
software was used to identify genes that were up- or down-regulated more than 2-fold
when grown on the nanoparticle modified anodes using the unpaired two sample t-test
with a cutoff p-value of 005 The complete microarray data set generated in this study is
deposited for public access in the Gene Expression Omnibus
(httpwwwncbinlmnihgovgeo) under accession number GSE31535
Results
Enhancement of current generation by using NP decorated anodes in MECs
The current density generated by Fe-NP decorated anode MECs with S onidensis MR-1
increased to approximately 43 microAcm2 20 h following inoculation and then slowly
decreased Figure 42 shows that the maximum current density achieved by NP
decorated anodes was 83 times higher than that (51 microAcm2) generated by the control
(plain graphite disk) The average current density improvement of 110 hours was 59
times of that generated by the control The current densities generated in MECs in the
53
absence of bacteria were negligible (03 microAcm2) which demonstrated that the current
enhancement observed with the Fe-NP decorated anodes was biologically-derived
Global transcriptome analysis
Whole-genome DNA microarrays were used to attain a comprehensive general
overview
of the transcriptional response to Fe-NP decorated anodes by S oneidensis MR-1 in MEC
Of a total 4295 genes assayed 392 (188 induced and 204 repressed) exhibited significant
(P lt 005) differential expression at a 2-fold (log (expression ratio 1) level in 3
replicates in response to Fe-NPs These
total gene numbers present 9 of the 4295 open
reading frames (ORFs) presented on the array
Fig 43 Differentially expressed genes grouped by functional classification according to
the TIGR S oneidensis genome database (wwwtigrorg) Columns 1 amino acid
biosynthesis 2 biosynthesis of cofactors prosthetic groups and carriers 3 cell
envelope 4 cellular processes 5 central intermediary metabolism 6 DNA metabolism
7 energy metabolism 8 fatty acid and phospholipid metabolism 9 other categories 10
protein fate 11 protein synthesis 12 purines pyrimidines nucleosides and nucleotides
13 regulatory functions 14 signal transduction 15 transcription 16 transport and
binding proteins 17 unknown function 18 hypothetical proteins
Up-Regulated
-75
-50
-25
0
25
50
75
100
125
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18
Down-Regulated
Num
ber
of
Gen
es
54
Figure 43 summarizes the overall genomic response of S oneidensis MR-1 to Fe-NP
decorated anodes by grouping the differentially expressed genes into their functional role
categories based on The Institute for Genomic Researchrsquos annotation (Version 8) of the
MR-1 genome sequence The wide distribution of putative functional roles attributed to
the differentially expressed genes indicated the extent of the molecular response of
Soniedenis MR-1 to the NP decorated anodes Among genes with defined function a
large number of up-regulated genes were associated with cellular processes (group 4)
energy metabolism (group 7) other categories (group 9) and hypothetical proteins (group
18) While those genes encoding proteins involved in amino acid biosynthesis (group 1)
cellular processes (group 4) substrate transport (group 16) and hypothetical proteins
(group 18) were among the most down-regulated genes Two groups of genes that were
the most noticeable among the functional gene groups genes related to energy
metabolism (group 7) and genes related to amino acid biosynthesis (group 1) The ratios
of up-regulated genes to down-regulated genes in these functional groupings were much
higher than other functional gene groupings there were 21 up-regulated genes and 7
down-regulated genes in energy metabolism gene group and 2 up-regulated and 22 down
regulated genes in the amino acid biosynthesis group
Genes related to biofilm formation
In order to determine whether there was a correlation between the observed biofilm
enhancements by NP decorated anodes and the genes related to biofilm formation the
significant modified genes related to biofilm formation were examined and summarized
in the Table 41 These included SO_3223 and SO_3228 genes encoding the flagellum
proteins (Thormann et al 2004) which has critical impacts on initial attachment to the
55
surface as swimming motility functions were up-regulated over 10 and 09 fold
respectively The gene clusters SO_4178-SO_4180 which are thought to be essential for
formation of a three-dimensional biofilm were up-regulated from 12 to 13 fold
(Thormann et al 2004) In addition the genes SO_4105 encoding IV pilus which is
known to represent a critical factor for biofilm formation on a biotic surfaces (Thormann
et al 2004) were also induced 08 fold (Table 41) The genes related to flagellum
motility (SO_4286 SO_1529 and SO_1530) were also induced slightly (Thormann et al
2004)
Table 41 Expression levels of genes related to biofilm formation
Locus Tag Gene product descriptions Fold change
SO_3228 flagellar basal-body MS-ring and collar protein FliF 10
SO_3223 flagellar hook-length control protein FliK 09
SO_4103 MshA minor pilin protein MshD 05
SO_4105 MSHA major pilin protein MshA 08
SO_4178 expressed protein of unknown function MxdC 12
SO_4179 inner membrane family 2 glycosyltransferase MxdB 13
SO_4180 diguanylate cyclase-like protein MxdA 10
a fold changes were presented after log2
b positive number represented up-regulated and negative represented down-regulated
Genes related to energy metabolism
A closer consideration of energy metabolism genes was undertaken as anaerobic
metabolism for electron generation and electron transport functions may play critical
roles in enhanced current density generation in MECs Genes associated with energy
56
metabolism and with significantly modified gene expression levels are summarized in the
Table 42 Several of the up-regulated genes were related to formate dehydrogenase
proteins Most notably four genes related to formate dehydrogenase were up-regulated
more than 2-fold SO_0102 SO_0103 SO_0104 and SO_0107 The other genes
expression related to formate dehydrogenase proteins SO_4513 SO_4514 SO_4515 also
significantly increased Another interesting gene is that encoding the cytochrome c
oxidase protein gene SO_4609 which was up-regulated 11 fold while the gene
SO_0479 related periplasmic octaheme cytochrome c MccA was down-regulated
However SO_1519 the gene encoding lactate dehydrogenase was down-regulated 12
fold despite lactate being the primary carbon source in the media
Table 4 2 Genes related to anaerobic growth and electron transfer with significantly change expression level
Locus Tag Gene product descriptions Fold
change
SO_0101 nitrate-inducible formate dehydrogenase molybdopterin-binding
subunit FdnG
18
SO_0102 formate dehydrogenase nitrate-inducible iron-sulfur subunit 21
SO_0103 formate dehydrogenase nitrate-inducible cytochrome b556 subunit 24
SO_0104 formate dehydrogenase accessory protein FdhE 22
SO_0107 formate dehydrogenase accessory protein fdhD 24
SO_0397 quinolfumarate reductase menaquinol-oxidizing subunit FrdC_2 12
SO_0452 thioredoxin 2 15
SO_0903 Na-translocating NADH-quinone reductase subunit B NqrB_1 12
SO_1012 NADH-ubiquinone oxidoreductase subunit K NuoK 12
SO_1013 NADH-ubiquinone oxidoreductase subunit J NuoJ 12
SO_1363 hydroxylamine reductase 13
57
SO_2417 ferredoxin cofactor maintenance protein YfaE 10
SO_3922 formate dehydrogenase cytochrome b Fdh -12
SO_4513 formate dehydrogenase molybdopterin-binding subunit _2 15
SO_4514 formate dehydrogenase FeS subunit FdhB_2 12
SO_4515 formate dehydrogenase cytochrome b subunit FdhC_2 14
SO_4609 aa3 type cytochrome c oxidase subunit III CoxC 11
SO_0479 periplasmic octaheme cytochrome c MccA -12
SO_0847 periplasmic nitrate reductase ferredoxin component NapG -15
SO_1519 L-lactate dehydrogenase iron-sulfur cluster-binding protein LldF -10
SO_1251 ferredoxin 4Fe-4S -11
SO_37411 hypothetical inner membrane protein -11
a fold changes were presented after log2
b positive number represented up-regulated and negative represented down-regulated
Flavin and cytochrome related genes
Flavins can be secreted by Shewanella species as electron shuttle to facilitate
extracellular electron transfer (Cansterin et al 2008) C-type cytochromes play the
important roles on the process of extracellular electron transfer (Shi et al 2007) The
genes related to flavins synthesis and the genes encoding the cytochromes electron
transport proteins were shown in the table 43 the genes SO_1414 and SO_3058 related
to flavins synthesis was up-regulated to 03 and 01 folds respectively The genes related
to s C-type cytochromes SO_0135 SO_4105 SO_0169 SO_1778 are increased to 10
08 05 04 01 folds respectively However most of genes have shown no significant
changes (less one-fold change) as response to nano particle decorated anode in MECs
58
Table 43 Expression level of flavin and cytochrome genes
Locus Tag Gene product descriptions Fold
change
SO_1414 flavocytochrome c flavin subunit putative 03
SO_3468 riboflavin synthase subunit alpha -02
SO_3058 flavocytochrome c flavin subunit 01
SO_4105 MSHA pilin protein MshA 08
SO_0169 general secretion pathway protein GspG 05
SO_1778 outer membrane decaheme cytochrome c lipoprotein MtrC 04
SO_1779 outer membrane decaheme cytochrome c OmcA 01
SO_0135 lipoprotein of unknown function DUF333 10
SO_0136 conserved hypothetical inner membrane protein 01
a fold changes were presented after log2
b positive number represented up-regulated and negative represented down-regulated
Other genes with significantly modified expression levels
These genes over 15 fold change and possibly related to current enhancements were
shown in Table 44 SO_3104 (expressed inner membrane protein) was over expressed
and the gene SO_2194 which encodes outer membrane protein (OmpA family protein)
was repressed as response to nanoparticle decorated anode however it is unclear the
relationships of this modified membrane protein genes with the current enhancement
Another interesting phenomenon was possible co-regulation of several gene clusters
indicating possible operon associations This includes three sets of genes that were
induced more than 15 fold SO_0108SO_0109 and SO_1042SO_1043SO_1044 The
59
consistency of expression of these genes under the NP conditions provides basic evidence
to support operon structure However the correlation of these significant changed gene
with current density enhance are unclear which showed there are unknown multiple and
complex responses of S oneidensis MR-1 to nanoparticle anode of MECs
Table 44 Other genes with significantly changed expression levels
Locus Tag Gene product descriptions Fold change
SO_0101 nitrate-inducible formate dehydrogenase molybdopterin-binding
subunit FdnG
18
SO_0108 integral membrane protein of unknown function DUF39 YedE 17
SO_0109 SirA family protein YedF 17
SO_0275 N-acetyl-gamma-glutamyl-phosphate reductase -31
SO_0277 ornithine carbamoyltransferase -21
SO_0279 argininosuccinate lyase -24
SO_0404 zinc dependent metalloprotease domain lipoprotein 21
SO_0956 alkyl hydroperoxide reductase F subunit -20
SO_1042 amino acid ABC transporter ATP-binding protein -18
SO_1043 amino acid ABC transporter permease protein -27
SO_1044 amino acid ABC transporter periplasmic amino acid-binding protein -27
SO_1072 chitin-binding protein putative 21
SO_1405 transglutaminase family protein 20
SO_1822 TonB-dependent receptor putative -22
SO_2069
1-(5-phosphoribosyl)-5-[(5-phosphoribosylamino)
methylideneamino] imidazole-4-carboxamide isomerase
-19
SO_2070 amidotransferase HisH -26
SO_2071 imidazole glycerol-phosphate dehydratasehistidinol phosphatase -27
60
SO_2072 histidinol-phosphate aminotransferase -28
SO_2073 histidinol dehydrogenase -30
SO_2194 OmpA family protein -36
SO_2195 inter-alpha-trypsin inhibitor domain protein -27
SO_2767 asparagine synthetase B -26
SO_2945 prophage LambdaSo tail fiber protein 20
SO_2963 prophage LambdaSo major capsid protein HK97 family 20
SO_2987 prophage LambdaSo hypothetical protein 19
SO_2988 prophage LambdaSo expressed protein of unknown function 18
SO_3104 expressed inner membrane protein 131
SO_3408 conserved hypothetical inner membrane protein -18
SO_3585 azoreductase putative -18
SO_3586 glyoxalase family protein -17
SO_3687 curli production assemblytransport component CsgE putative -18
SO_38191 hypothetical ammonia permease -19
SO_4014 AcrBAcrDAcrF family protein 18
SO_40151 type I secretion system membrane fusion protein RND family 18
SO_4054 510-methylenetetrahydrofolate reductase 18
SO_4245 N-acetylglutamate synthase -22
SO_45251 hypothetical transcriptional regulator LysR family -17
SO_4527 integral membrane domain protein -19
SO_4705 transcriptional regulator putative -19
a fold changes were presented after log2
b positive number represented up-regulated and negative represented down-regulated
Discussion
61
The addition of NP to anodes significantly impacts current densities with the elemental
composition being a critical factor The chemical composition of NPs significantly affects
the current enhancement in MECs (Fan et al 2011) because the chemical composition of
NP properties is a critical factor in determining the conductivity which significantly
affects the efficiency of electron transfer from bacteria to electrode Fe is an ideal
candidate for NP formation for current enhancement in MESs because of high
conductivity and low price compared to gold and other conductive materials Gold NPs
has also been studies for enhancement of electricity conductivity (Bao et al 2008)
However the disadvantages of the high price of gold limit their practical application in
MECs (Fan et al 2011) The different size and density of the same composition of NP
also has essential effects on the current density enhancement (Fan et al 2011) In this
study the results showed the current density enhancement with Fe-NP size range of
200plusmn100 nm and density range of 200plusmn100 nm is considerable enhancement consequence
There are a number of reports of studies of current density enhancement in MFCs
by nanostructured anodes (Fan et al 2011 Qiao et al 2008 Sharma et al 2008 Tsai et al
2009) Based on bacterial behavior on nanostructured anode surface Qiao et al (2008)
believes that Escherichia coli cells on the nanostructured electrode surface produce hair-
like structures similar to pili that could facilitate the electron transfer between the cells
and electrode The production of hair-like structures is believed to be stimulated by
nanostructures and could play the key role on current density enhancement in pili-
producing bacteria in MFCs (Qiao et al 2008) S oneidensis can produce electrically
conductive bacterial nanowires which have similar structure to the hair-like structures in
Ecoli (Gorby et al 2006 Reguera et al 2005) A number of possible genes related
62
nanowires of S oneidensis have been identified including mshA (SO_4105) and gspG
(SO_0169) (Konstantinidis et al 2009) They are up-regulated 08 and 05 fold in the
current study respectively even though they are not significant induced This result
supported the increased nanowire genes had contributions to enhance current density in
MFCs by NPs decorated anodes
Based on the previous research on electron transfer mechanisms for S oneidensis it is
known that self-produced mediators (flavins) and multihaem c-type cytochromes (c-Cyts)
may play critical roles in the electron transfer of S oneidensis to extracellular insoluble
electrode (Canstein et al 2008 Myers and Myers 1992 Scott and Nealson 1994 Shi et al
2007) The flavins secreted by S oneidensis can facilitate to transfer electrons from
bacteria to solid anode electrodes by acting as extracellular electron shuttles (Canstein et
al 2008) In this study the genes encoding flavins (SO_1414 SO_3468 and SO_3058)
had no significant increase (less than 1 fold change) as a response to nanoparticle
decorated anode in MECs which suggests that increased flavin gene expression does not
significantly contribute to the current density enhancements in nano-particle decorated
anode The genes encoding cytochromes electron transport proteins (SO_4104 SO_0417
SO_177879 SO_013536) had no significant increase (less than 1-fold change)
However those genes were up-regulated slightly These results suggest the increased
expression flavin and c type cytochromes genes had partial contributions even not
significantly to enhance current density in MFCs by NPs decorated anodes
The thickness of bacterial biofilm on the anode can affect the power generation in
MFCs (Biffinger et al 2007 Heydorn et al 2000) since increased biofilm thickness on
the anode represents more active bacteria involving in the process of electron generation
63
and transfer in MFCs Biofilm formation related genes included flagellar genes IV pilus
genes and other genes The flagellar related genes SO_3228 SO_3229 (fliB and fliK) and
IV pilus genes SO_4105 (mshA) were induced from 19 to 09 fold the genes (SO_4178
-4180) related to formation of a three-dimensional biofilm were up-regulated from 10 to
13 folds even though the genes related bacterial motility protein (SO_ 4286 SO_4287
SO_1529 and SO_1530) are induced slightly (less than 0 5 fold) However the motility
genes most probably play a role for initial biofilm attachment on the anode surfaces
(Thormann et al 2004 Thormann et al 2006) with the rest of biofilm genes being more
essential for biofilm maintenance in mature cultures as were presented at harvest in this
study This enhancement of biofilm density under conditions of increase current density
has been observed for gold NP decorated anodes using confocal light microscopy (data
not shown) but has not yet been confirmed for Fe NP decorated anodes In any event an
increase in the expression level of genes related biofilm formation therefore also
supported the possibility of enhanced biofilm formation on NPs decorated anodes thus
facilitating electron transfer from bacteria to anodes
Beliaev et al described a model of the Fe reduction pathway in S oneidensis MR-1 using
solid substrates as the electron acceptor (Beliaev et al 2001) This model proposes that
electrons are generated and released in cytoplasm then transferred to the quinone pool
eg menaquinone in the cytoplasmic membrane The reduced menaquinone in turn
reduces CymA and from there electrons are then transferred to a periplasmic shuttle (eg
CctA) and then to MtrA a periplasmic soluble decaheme cytochrome c located in
associated with the outer membrane via interaction with MtrB Finally surface displayed
outer membrane proteins decaheme cytochromes (OmcA MtrC or MtrF) can transfer
64
electron to Fe or other solid substrates (Beliaev et al 2001)) Bencheikh-Latmani et al
demonstrated that the same electron transport pathway may be used for more than one
electron acceptor through studies of the response of S oneidensis MR-1 to U(VI) and
Cr(VI) under anaerobic conditions particularly focused on the critical genes
(SO_1777mtrA SO_1776mtrB SO_1778mtrC) (Bencheikh-Latmani et al 2005)
However our studies showed that these genes had no significant expression changes in
response to nanostructured anodes Contrastingly one of expressed inner membrane
protein genes SO_3104 and one of cytochromes genes SO_4609 encoding cytochrome c
oxidase subunit III demonstrated a 13-fold and 11-fold up-regulation respectively in this
study while the gene SO_2194 which encodes outer membrane protein (OmpA family
protein) was down regulated 36 fold Our results therefore suggest that S oneidensis
MR-1 does not utilize the same set of core critical genes (SO_1777 mtrA SO_1776
mtrB SO_1778 mtrC) products to transfer electrons in all instances This is the similar
result with the study of Bretschger et al (2007) which indicated that the electron
transport system in S oneidensis MR-1 is complex with several different proteins able to
participate in electron transfer to the anode of MECs
It should also be mentioned that the gene (SO_1519) encoding lactate
dehydrogenase repressed (-12 fold) here suggesting that lactate was depleted at the time
of cell harvest from the MECs and that other pathways of anaerobiosis had been activated
including those associated with amino acid uptake and consumption the growth media
was a complex mixture including yeast extract In particular the catabolic pathway for
histidine of S oneidensis MR-1 a pathway that may formate as an intermediate is up-
regulated with the genes for histidine ammonia lyase SO_0098 SO_3057 and SO_4374
65
up-regulated 13 11 and 11-fold respectively The possible production of formate
during histidine degradation may in turn explain the high levels of formate
dehydrogenates as evidenced by upregulation of SO_0102 SO_0103 SO_0104 and
SO_0107 annotated as formate dehydrogenase proteins These genes had the highest
levels of up-regulated expression as response to Fe NP decorated anodes (greater than 2-
fold)
Acknowledgements
This research was partially supported by the US National Science Foundation CBET
0828544 and the funds from ONAMIDOD (ARL-DOD Cooperative Agreement
W911NF-07-2-0083) We thank Barbara Gvakharia and Caprice Rosato for helpful
suggestions We also thank Margaret Romine of Pacific Northwest Laboratories for
valuable comments on manuscripts We also thank anonymous reviewers for significant
suggested improvements imparted as part of prior review of this manuscript
66
Chapter 5
Global Transcriptome Analysis of the Response of Shewanella oneidensis MR-1 to
Carbon Nanotube Decorated Anodes in Microbial Electrochemical Systems
Shoutao Xu Yanzhen Fan Rebecca Schaller Frank Chaplen Jun Jiao Hong Liu
67
Abstract
Shewanella oneidensis MR-1 is an important model microorganism for metabolic studies
on the effects of different environmental factors because of its diverse respiratory
capabilities Carbon nanotube (CNT) modified graphite disks were used as anodes to
investigate the effects of nanomatrials on the performance of S oneidensis MR-1 in
microbial electrolysis cells (MECs) The current densities produced with CNT decorated
anodes were on average 56-fold higher than plain graphite anodes Whole genome
microarray analysis of gene expression showed that up-regulation of cytochromes c genes
associated with extracellular electron transfer are strongly correlated to current increases
in CNT decorated anodes Genes involved in flavin biosynthesis also contribute to
current increase in CNT decorated anode MECs
Keywords Microbial electrochemical system microbial fuel cell microbial
electrolysis cell carbon nanotube gene expression DNA microarray
68
Introduction
Shewanella oneidensis MR-1 is an important model microorganism for metabolic
studies of the effects of different environmental factors because of its diverse respiratory
capabilities It has been used for transcriptome analysis to investigate the responses to
different growth conditions and environmental stresses (Murray et al 2001) Gene
expression patterns under different conditions such as heat shock cold shock Cr (VI) U
(VI) reduction chromate stress iron and acid tolerance have been studied previously
(Bencheikh-Latmani et al 2005 Brown et al 2006 Gao et al2006 Gao et al 2004
Yang et al 2008) More recently it has been exploited as a model species for power
generation in microbial electrochemical systems (MESs)which have potential
applications in renewable energy generation bioremediation and wastewater treatment
(Logan et al 2007 Logan 2009 Park and Zeikus 2002 Fan et al 2011 Xu et al 2012)
In a MES electrochemically active microorganisms oxidize organic matter in the an
ode chamber to release electrons Electrons are then transferred to the anode electrode thr
ough various mechanisms (Logan et al 2006 Lovley 2006 Rabaey et al 2003) and fina
lly travel to the cathode electrode and combine with the terminal electron acceptor The
key feature of MESs is the microbe-catalyzed electron transfer from the organic matter to
the anode (Rabaey et al 2004) Enhancing the current output which highly depends on
the performance of the anode electrode is critical for the successful application of MESs
(Logan et al 2007 Park and Zeilus 2002 2003) Several nanomaterials including TiO2
gold (Au) and palladium (Pd) nanoparticles have been utilized to decorate electrodes to
enhance power density in MESs (Qiao et al 2008 Quan et al 2005 Fan et al 2011) The
possible mechanisms for increased current densities have been studied by using Fe
69
nanoparticle-decorated anodes (Xu et al 2012)
Carbon nanotubes (CNT) are cylindrically shaped nanomaterials with extremely
high surface area excellent electrical conductivity and chemical inertness (He et al
2005a Serp et al 2003) These unique properties make CNT a promising electrode
material (Liang et al 2008) The biocompatibility of microorganisms and carbon
nanostructures has been evaluated (Morozan et al 2007) The effects of CNT on anodic
biofilms have been tested in MFCs (Liang et al 2011) However no studies have been re
ported on global transcriptome analysis for the response of S oneidensis MR-1 to CNT
decorated anode in MECs
In the present study the graphite disks decorated with CNT were used as anodes to
investigate the effects of nanomaterials on current generation in multi-anode MECs
DNA microarrays were used to analyze differences in the global gene expression profile
of S oneidensis MR-1 grown on plain versus CNT decorated anodes
Materials and methods
Bacterial cultures S oneidensis MR-1 was purchased from American Type Culture
collection (ATCC 700550) and stored as freezer stocks at -80degC Stock S oneidensis
MR-1 cells were grown aerobically using M1 medium at 30degC Cells in early stationary
growth phase (optical density of 15-16 at 600nm (Spectrophotometer UV-1700
Shimadzu Columbia MD) were injected into the chamber of the MECs for current
production Sodium lactate (final concentration 30mM) was added as the additional
electron donor
Fabrication and characterization of nanostructured anodes Superfine isomolded
graphite disks (16 cm x 05 cm GraphiteStorecom Inc) were polished with ultrafine
70
sand paper (2000 grit 3M Company) as the base for the CNT decorated and control
anodes Aligned multi-walled carbon nanotube samples were synthesized via plasma
enhanced chemical vapor deposition (PECVD) Samples superfine isomolded graphite
pillars with a 16 cm diameter and 05 cm thickness were sputtered with a thin film of Ni
in a CRC 300 magnetron sputter coater for 1 to 2 min They were then placed in the
PECVD reactor where the Ni was annealed into small catalyst islands C2H2 was bled
into the chamber for growth and an NH3 plasma was used to vertically align the growth
of the multi-walled carbon nanotubes (MWCNTs) The temperature and time were varied
between 725degC and 775oC and 15 to 60 min to control the length of the tubes and density
of samples Samples were then analyzed in a FEI Sirion field emission scanning electron
microscope (FESEM) SEM images of CNT decorated anode and control surfaces were
shown in the Figure 51
Figure 51 SEM images of CNT decorated anodes (A) Graphite disk control (B) CNT
decorated anode
Multiple channel MECs construction and operation A MECs with multiple
removable anodes each with an effective anode area of 07 cm2
was constructed and used
to evaluate the effects of nanostructures on the current density of according to a previous
B A
71
study (Fan et al 2011) The cathode was made of wet-proof (30) carbon cloth (type B
E-TEK Division Inc USA) coated with PtC (20 E-TEK Division Inc USA)
following a previously reported procedure (Liu et al 2005) The final platinum loading
was 05 mgcm2 per projected cathode area The size of the cathode (150 cm
2) was 25
times larger than the total effective area of the 8 graphite disk anodes (56 cm2) to prevent
cathode limitations on the performance of the MECs system All MECs with CNT
decorated anodes and control anodes were tested for current generation for 24 h with
sterile a medium (without bacteria) before injecting the bacterial cells in order to
determine whether the CNT decorated anodes demonstrate chemical current generation
compared to control anodes All the testing anodes were set up in the MECs
simultaneously so that the S oneidensis MR-1 culture was applied to all of them
simultaneously Short electrode spacing (17 cm) was used in the MECs design to reduce
the internal resistance A voltage of 06 V was applied the MECs for current generation
after the cells of S oneidensis MR-1 were inoculated into the MECs in the early
stationary phase A multimeter with a data acquisition system (2700 Keithly USA) was
used to monitor the current change by measuring the voltage drop through a resistor
Microarray analysis Biofilms for whole gene microarray analysis were aseptically
removed from the plain and CNT decorated anodes of MECs after 80 hours further
incubation at 30oC when current density obviously reached to a stable phase Total RNA
was extracted using Trizol (Ambion inc AustinTX) following the manufacturerrsquos
instructions Integrity of the RNA samples was confirmed using the Agilent 2100
Bioanalyzer (Agilent Technology Inc Palo Alto CA) Total RNA (200 ng) from each
sample was amplified using the MessageAmpTM
II-Bacteria Prokaryotic RNA
72
Amplification kit (Ambion AustinTX) according to the manufacturerrsquos instructions The
S oneidensis MR-1 whole-gene microarrays representing all of the 4295 genes (resource
from NCBI) were ordered from Roche NimbleGen Inc (Madison WI) Biological
triplicates of biofilms on the CNT decorated anodes and control were analyzed cDNA
synthesis labeling and hybridization were carried out by the CGRB Core Laboratories at
Oregon State University DNASTAR ArrayStarTM
3 software was used to identify genes
that were up- or down-regulated more than 2-fold when grown on the nanoparticle
modified anodes using the unpaired two sample t-test with a cutoff p-value of 005 with
bonferroni correction
Results and discussion
Enhancement of current generation using CNT decorated anodes in MECs The
current density generated by CNT decorated anode MECs with S onidensis MR-1
increased to 36 microAcm2 10 hours post-inoculation and reached a maximum of 40 microAcm
2
at 29 hours inoculation then gradually decreased The current density stabilized at
30microAcm2 after 78 h operation Figure 52 shows that the current density curve generated
by CNT decorated anode in MECs The average current density generated by CNT
decorated anodes was 56 times that of the control The current density generated in the
MECs without bacteria was negligible (data not shown) This result demonstrates that the
current enhancement observed with the CNT decorated anodes was biologically-derived
A
73
Figure 52 Effects of CNT decorated anodes on the current density in MECs (3 replicates)
Control is plain graphite disk anode CNT is the anode with CNT decoration (Error bars
represent standard deviation)
It has been reported that nanostructured decorated anodes have significant impacts on
current densities (Fan et al 2011 Qiao et al 2006 Sharma 2008 Tsai et al 2009) The
CNT decorated anodes had significantly enhanced current densities in MECs in this
study This result is consistent with previous studies Interestingly the current density
curve generated by CNT decorated anode MECs is quite different from the current
density curve generated by Au NP and Pd NP decorated anodes using same bacterial
species (Fan et al 2011) In the Au and Pd NP decorated anode MECs the current
density curve was not significantly increased in the beginning compared with the control
but gradually increased after the inoculation the current density reached the maximum
current density more than 50 hours after the inoculation However the current density
generated by CNT decorated anode MECs started to increase significantly immediately
after the inoculation It reached a maximum current density 29 hours post-inoculation
-1
6
13
20
27
34
41
48
0 10 20 30 40 50 60 70 80
Curr
ent
den
sity
(micro
Ac
m2)
Time (Hours)
Control
CNT
74
These results indicate that there might be different current density enhancement
mechanisms between CNT with other metal NP decorated anodes in MECs
Global transcriptome analysis Whole-genome DNA microarrays were used to attain a
comprehensive general overview of the transcriptional response of S oneidensis
MR-1 of
S oneidensis MR-1 to CNT decorated anodes in MECs In a total of 4295 genes analyzed
457 genes (250 genes up-regulated and 207 genes down-regulated) exhibited significantly
(P lt 005) changed expression at a 2-fold level in at least 3 replicates in the response to
CNT The total number of the regulated genes present 11 of the 4295 open reading
frames (ORFs) presented on the microarray The total number of genes at significant
expression levels in response to CNT decorated anodes is close to that to Fe NP decorated
anodes in MECs (392 9) ( Xu et al 2012)
Fig 53 Differentially expressed genes grouped by functional classification according to
the TIGR S oneidensis genome database (wwwtigrorg) Columns 1 amino acid
biosynthesis 2 biosynthesis of cofactors prosthetic groups and carriers 3 cell
envelope 4 cellular processes 5 central intermediary metabolism 6 DNA metabolism
7 energy metabolism 8 fatty acid and phospholipid metabolism 9 other categories 10
protein fate 11 protein synthesis 12 purines pyrimidines nucleosides and nucleotides
-80
-60
-40
-20
0
20
40
60
80
100
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18
Nu
mb
er
of
Ge
ne
s
Up-regulated
Down-regulated
75
13 regulatory functions 14 signal transduction 15 transcription 16 transport and
binding proteins 17 unknown function 18 hypothetical proteins
The overall transcript genomic response of S oneidensis MR-1 to CNT decorated
anodes was summarized in Figure 53 by grouping the differentially expressed genes into
their functional role categories based on The Institute for Genomic Researchrsquos annotation
(Version 8) of the MR-1 genome sequence The wide distribution of putative functional
roles attributed to the differentially expressed genes indicated the extent of the molecular
response of Soniedenis MR-1 to the CNT decorated anodes A large number of up-
regulated genes were presented in genes associated with biosynthesis of cofactors
prosthetic groups and carriers (group 2) cell envelope (group 3) protein fate (group 10)
protein synthesis (group 11) and purines pyrimidines nucleosides and nucleotides
(group 12) in this study The number of up-regulated genes was two times more than that
of down-regulated genes in each functional group Contrastingly in the genomic response
S oneidensis MR-1 to Fe NP decorated anodes (chapter 4) most of up-regulated genes
were associated with cellular processes (group 4) energy metabolism (group 7) other
categories (group 9) and hypothetical proteins (group 18) The down-regulated genes
presented in the amino acid biosynthesis (group 1) cellular processes (group 4) substrate
transport (group 16) and hypothetical proteins (group 18) While those genes encoding
proteins involved in amino acid biosynthesis (group 1) central intermediary metabolism
( group 5) DNA metabolism( group 6) fatty acid and phospholipid metabolism (group
8) and other categories (group 9) were among the most down-regulated genes in this
study However the two groups of genes that were the most noticeable between the
genomic response S oneidensis MR-1 to CNT and Fe NP decorated anodes which they
are the genes related to energy metabolism (group 7) and hypothetical proteins (group
76
18) These results indicated there are different and common genomic responses to S
oneidensis MR-1 on CNT and Fe NP decorated anodes in MECs
Electron transfer related genes Based on previous electron transfer mechanism
studies on Shewanella a serial of group proteins collectively described as the Mtr
pathway play the role to transfer electron rom the inner bacterial body to the outer
membrane (Beliaev et al 2001 Shi et al 2007 Brutinel and Gralnick 2012) In the Mtr
pathway firstly electrons generated by bacteria received by CymA a tetraheme c-type
cytochrome anchored in the inner membrane then electrons were transferred to MtrA a
periplasmic decaheme c-type cytochrome After that electrons were transferred from
MtrA to MtrC an outer-membrane decaheme c-type cytochrome is facilitated by MtrB a
non-heme containing integral outer-membrane proteinOuter-membrane decaheme c-type
cytochrome MtrC and OmcA are in change of electron transfer to exreacellular electron
acceptor (Shi et al 2007 Brutinel and Gralnick 2012 ) These genes encoding electron
transport proteins are shown in the table 51 The critical genes SO_1777 mtrA SO_1776
mtrB SO_1778 mtrC are unregulated 41 to 80 fold in response to CNT decorated
anodes other cytochrome c genes related to electron transfer in MtrA pathway also are
significantly increased in response to CNT decorated anodes These genes encoding outer
membrane proteins have different level increases supporting that the increase of
cytochrome c gene expression contributes to the current enhancements of MECs as
response to CNT decorated anodes
77
Table 51 Expression level of cytochrome c as response to CNT decorated anodes
Locus Tag Gene product descriptions Fold change CNT
SO_0165 general secretion pathway protein GspC 31
SO_0167 general secretion pathway protein GspE 45
SO_1776 outer membrane protein precursor MtrB 41
SO_1777 outer membrane decaheme cytochrome c
lipoprotein MtrA
80
SO_1778 outer membrane decaheme cytochrome c
lipoprotein MtrC
43
SO_1779 outer membrane decaheme cytochrome c
OmcA
63
SO_0135 lipoprotein of unknown function DUF333 22
SO_0136 conserved hypothetical inner membrane protein 24
Flavin related genes It is known that self-produced mediators play critical roles in the
electron transfer of S oneidensis to extracellular insoluble electrode (Canstein von et al 2
008 Myers et al 1992 Scott et al 1994 Shi et al 2007) It has been proved that the
flavins secreted by S oneidensis can facilitate electron transfer from bacteria to solid
anode electrodes by acting as extracellular electron shuttles (Cansterin von et al 2008) T
wo critical genes (SO_1414 and SO_3468) related to flavin synthesis were shown increa
sed significantly (more than 2 folds) increase in this study (Table 52) This result support
s the hypothesis that flavin production increases as the response to CNT decorated anodes
in MECs have significant contribution to the current density enhancements
78
Table 52 Expression levels of genes related to flavin synthesis as response to CNT
decorated anode
Locus Tag Gene product descriptions Fold change CNT
SO_1414 flavocytochrome c flavin subunit putative 22
SO_3468 riboflavin synthase subunit alpha 20
The amount of bacterial biofilm biomass on the anode can affect the power
generation in MFCs (Biffinger et al 2007 Heydorn et al 2000) Biofilm formation
related genes include flagellar gene IV pilus genes and other genes the flagellar related
genes (fliB and fliK) and IV pilus genes (mshA) were induced only from 105 to 134
fold as response of S oneidensis MR-1 to CNT decorated anodes in this study No
biofilm-associated genes presented significant changes in expression levels in this study
indicating there is no direct connections between biofilm enhancements to CNT
decorated anodes in this study This result is consistent with the Liang et al (2011)
conclusion that using CNT enhanced the electrochemical activity of anodic biofilms but
did not result in a significant increase of biomass in the anodic biofilms
Our microarray results showed two significantly up-regulated gene groups the
genes encoding proteins localized on the outer membrane and the genes involved in
flavin biosynthesis contributed to current density enhancement by CNT decorated anodes
Among 457 significantly changed genes to CNT in this study there are also a relatively
large number of genes encoding proteins with unknown functions which are either up- or
down-regulated in the response to CNT decorated anodes which indicated more
79
complicated responses of S oneidensis MR-1 to CNT decorated anodes in MECs and
further study is needed
80
Chapter 6
Summary
Microbial electrochemical systems (MESs) with a mixed culture initially inoculated
from Corvallis wastewater treatment plant have been studied for more than 6 years for
varying purposes including power generation hydrogen production heavy metal
removal and wastewater treatment Experiment results have shown that the mixed culture
is quite stable with excellent performance in MESs Our community analysis using
denaturing gradient gel electrophoresis (DGGE) and 16S rDNA clone library construction
suggests that the mixed culture is composed predominantly of Geobacter sp (66)
Arcobacter sp (12) and Citrobacter sp (11) These results not only improved our
understanding of the mixed culture community but also guided our studies on the
cultivation and isolation of the different bacterial species
Exoelectrogenic Citrobacter sp SX-1 was first isolated and characterized from a
MFC operated with the mixed culture Citrobacter sp SX-1 has been demonstrated to
produce electricity from wide range of different substrates including citrate acetate
glucose sucrose glycerol and lactosein MFCs Cyclic voltammetry analysis indicated
that membrane associated proteins may play an important role in facilitating the electrons
transferring from bacteria to electrode The strain SX-1 increased the known diversity of
power generating exoelectrogens and provided a unique bacterial species for study in
renewable energy generation and waste treatment
81
To enhance the electron transfer from bacteria to anode nano decorated anodes
including Fe NP and CNT were developed and characterized and evaluated in MECs
using Shewanella oneidensis MR-1 as a model species Both nanostructures have
significantly increased current density compared with the control Whole genome
microarray analysis elucidated the possible mechanisms of power enhancement in
response to these nano-decorated anodes
These results benefit to understanding of physiology and ecology of mixed
cultures in MFCs and improve the efficiency of current generation in MESs which will
facilitate the viability of niche applications for MESs in near future
82
Bibliography
1 Amman R Ludwig W (2000) Ribosomal RNA-targeted nucleic acid probes for
studies in microbial ecology FEMS Microbiol Ecol 24 555ndash565
1 Bao SJ Li CM Zang JF Cui XQ Qiao Y Guo J (2008) Novel Nanoporous TiO2
Electrode for Directly Electrochemistry of glucose oxidase Adv Funct Mater
18(4) 591-599
2 Beliaev AS Saffarini DA McLaughlin JL Hunnicutt D (2001) MtrC an outer
membrane decahaem c cytochrome required for metal reduction in Shewanella
putrefaciens MR-1 Mol Microbiol 39722-30
3 Bencheikh-Latmani R Williams SM Haucke L Criddle CS Wu L Criddle CS
Zhou J Tebo BM (2005) Global transcriptional profiling of Shewanella oneidensis
MR-1 during Cr(VI) and U(VI) reduction Appl Environ Microbiol 71 7453ndash7460
4 Biffinger JC Pietron J Ray R Little B and Ringeisen BR (2007) A biofilm
enhanced miniature microbial fuel cell using Shewanella oneidensis DSP10 and
oxygen reduction cathodes Biosens Bioelectron 221672ndash1679
5 Bodelier PLE Roslev P Henckel T FenzelP (2000) Stimulation by ammonium
based fertilizers of methane oxidation in soil around rice root Nature 40 421ndash424
6 Bond DR Holmes DE Tender LM Lovley DR (2002) Electrode-reducing
microorganisms that harvest energy from marine sediments Science 295 483-485
7 Bond DR Lovley DR (2003) Electricity production by Geobacter sulfurreducens
attached to electrodes Appl Environ Microbiol 69 1548-1555
8 Bond DR Lovley DR (2005) Evidence for involvement of an electron shuttle in
electricity generation by Geothrix fermentans Appl Environ Microbiol 71 2186-
2189
9 Bretschger O Obraztsova A Sturm CA Chang IS Gorby YA Reed SB Culley
DE Reardon CL Barua S Romine MF Zhou J Beliaev AS Bouhenni R
Saffarini D Mansfeld F Kim BH Fredrickson JK Nealson KH (2007) Current
production and metal oxide reduction by Shewanella oneidensis MR-1 wildtype and
mutants Appl Environ Microbiol 73 7003ndash7012
10 Brown SD Thompson MR Verberkmoes NC Chourey K Shah M Zhou J
Hettich RL and Thompson DK (2006) Molecular dynamics of the Shewanella
oneidensis response to chromate stress Mol Cell Proteomics 51054-1071
11 Brutinel ED Gralnick JA (2012) Shuttling happens soluble flavin mediators of
extracellular electron transfer in Shewanella Appl Microbiol Biotechnol 93(1) 41-8
12 Butler JE Kaufmann F Coppi MV Nuacutentildeez C Lovley DR (2004) MacA a diheme
c-type cytochrome involved in Fe(III) reduction by Geobacter sulfurreducens J
Bacteriol 186(12) 4042ndash4045
83
13 Caccavo F Lonergan DJ Lovley DR Davis M Stolz JF McInerney MJ (1994)
Geobacter sulfurreducens sp nov a hydrogen- and acetate oxidizing dissimilatory
metal-reducing microorganism Appl Environ Microbiol 60 3752ndash3759
14 Canstein von H Ogawa J Shimizu S Lloyd JR (2008) Secretion of flavins by
Shewanella species and their role in extracellular electron transfer Appl Environ
Microbiol 74 615ndash623
15 Cao X Huang X Liang P Xiao K Zhou Y Zhang X Logan BE (2009) A New
Method for Water Desalination Using Microbial Desalination Cells Environ Sci
Technol 43(18)7148-7152
16 Chaudhuri SK Lovley DR (2003) Electricity generation by direct oxidation of
glucose in mediatorless microbial fuel cells Nat Biotechnol 21 1229-1232
17 Cheng S Liu H Logan B E (2006) Increased power generation in a continuous
flow MFC with advective flow through the porous anode and reduced electrode
spacing Environ Sci Technol 402426-2432
18 Cheng S Logan B E (2007) Sustainable and efficient biohydrogen production via
electrohydrogenesis PNAS 104 18871-18873
19 Chignell J (2010) Energy Recovery from Biodiesel Waste Performance of
Microbial Electrochemical Systems on Glycerol Thesis
20 Chung K Okabe S (2009) Characterization of electrochemical activity of a strain
ISO2-3 phylogenetically related to Aeromonas sp isolated from a glucose-fed
microbial fuel cell Biotechnol Bioeng 104(5) 901-910
21 Cole JR Chai B Farris RJ Wang Q Kulam-Syed-Mohideen AS McGarrell
DM Bandela AM Cardenas E Garrity GM Tiedje JM (2007) The ribosomal
database project (RDP-II) introducing myRDP space and quality controlled public
data Nucleic Acids Res 35 (Database issue) D169-D172 doi 101093nargkl889
22 Cole JR Wang Q Cardena E Fish J Chai B Farris RJ Kulam-Syed-Mohideen
AS McGarrell DM Marsh T Garrity GM Tiedje JM (2009) The Ribosomal
Database Project improved alignments and new tools for rRNA analysis Nucleic
Acids Res 37 (Database issue) D141-D145 doi 101093nargkn879
23 Dey A De S De A De SK (2004) Characterization and dielectric properties of
polyaniline-TiO2 nanocomposites Nanotechnol 15 1277-1283
24 Edwards U Rogall T Bloumlcker H Emde M Boumlttger EC (1989) Isolation and
direct complete nucleotide determination of entire genesmdashcharacterization of a gene
coding for 16S-ribosomal RNA NucleicAcids Res 17 7843ndash7853
25 Eggleston CM Voumlroumls J Shi L Lower BH Droubay TC Colberg PJS (2008)
Binding and direct electrochemistry of OmcA an outer-membrane cytochrome from
an iron reducing bacterium with oxide electrodes a candidate biofuel cell system
Inorg Chim Acta 361769ndash777
26 Fan Y Sharbrough E Liu H (2008) Quantification of the Internal Resistance
Distribution of Microbial Fuel Cells Environ Sci Technol 42 8101ndash8107
84
27 Fan Y Xu S Schaller R Jiao J Chaplen F Liu H (2011) Nanoparticle decorated
anodes for enhanced current generation in microbial electrochemical
cells Biosensors and Bioelectronics 26 (5)1908ndash1912
28 Fan Y Hu H Liu H (2007)Sustainable power generation in microbial fuel cells
using bicarbonate buffer and proton transfer mechanisms Environ Sci
Technol 41(23) 8154-8
29 Fedorovich V Knighton MC Pagaling E Ward FB Free A Goryanin I (2009)
Novel electrochemically active bacterium phylogenetically related to Arcobacter
butzleri isolated from a microbial fuel cell Appl Environ Microbiol 75(23) 7326-
7334
30 Ferna ndez A Huang S Seston S Xing J Hickey R Criddle C Tiedje J (1999)
How stable is stable Function versus community composition Appl Environ
Microbiol 65 3697ndash3704
31 Field SJ Dobbin PS Cheesman MR Watmough NJ Thomson AJ Richardson
DJ (2000) Purification and magneto-optical spectroscopic characterization of
cytoplasmic membrane and outer membrane multiheme c-type cytochromes from
Shewanella frigidimarina NCIMB400 J Biol Chem 275 8515ndash8522
32 Freguia S Masuda M Tsujimura S Kano K (2009) Lactococcus lactis catalyses
electricity generation at microbial fuel cell anodes via excretion of a soluble quinone
Bioelectrochemistry 7614ndash18
33 Fricke K Harnisch F and Schroumlder U (2008) On the use of cyclic voltammetry for
the study of the anodic electron transfer in microbial fuel cells Energy Environ
Sci 1 144ndash147
34 Gao H Wang Y Liu X Yan T Wu L Alm E Arkin A Thompson DK and
Zhou J (2004) Global transcriptome analysis of the heat shock response of
Shewanella oneidensis J Bacteriol 1867796-7803
35 Gao H Yang ZK Wu L Thompson DK Zhou J (2006) Global transcriptome
analysis of the cold shock response of Shewanella oneidensis MR-1 and mutational
analysis of its classical cold shock proteins J Bacteriol 1884560-4569
36 George MG (2005) Bergeyrsquos Manual of Systematic Bacteriology Volume Two The
Proteobacteria Part B The Gammaproteobacteria Second Edition New York
Springer 651p
37 Gonzaacutelez-Toril E Llobet-Brossa E Casamayor EO Amann R Amils R (2003)
Microbial ecology of an extreme acidic environment the Tinto river Appl Environ
Microbiol 69(8) 4853ndash4865
38 Gorby YA Yanina S Mclean JS Rosso KM Moyles D Dohnalkova A
Beveridge TJ Chang IS Kim BH Kim KS Culley DE Reed SB Romine M
F Saffarini DA Hill EA Shi L Elias DA Kennedy DW Pinchuk G
Watanabe K Ishii S Logan B Nealson KH Fredrickson JK (2006) Electrically
conductive bacterial nanowires produced by Shewanella oneidensis strain MR-1 and
other microorganisms Proc Natl Acad Sci USA 10311358-11363
85
39 He Z Wagner N Minteer SD Angenent LT (2006) An upflow microbial fuel cell
with an interior cathodes assessment of the internal resistance by impedance
spectroscopy Environ Sci Technol 40 5212-5217
40 Heydorn A Nielsen AT Hentzer M Sternberg C Givskov M Ersboslashll BK Molin
S (2000) Quantification of biofilm structures by the novel computer program
COMSTAT Microbiol 1462395-2407
41 Holmes DE Bond DR Lovley DR (2004a) Electron transfer by Desulfobulbus
propionicus to Fe(III) and graphite electrodes Appl Environ Microbiol 701234ndash
1237
42 Holmes DE Chaudhuri SK Nevin KP Mehta T Methe BA Liu A Ward JE
Woodard TL Webster J Lovley DR (2006) Microarray and genetic analysis of
electron transfer to electrodes in Geobacter sulfurreducens Environ Microbiol
81805-1815
43 Holmes DE Nicoll JS Bond DR Lovley DR (2004b) Potential role of a novel
psychrotolerant member of the family Geobacteraceae Geopsychrobacter
electrodiphilus gen nov sp nov in electricity production by a marine sediment
fuel cell Appl Environ Microbiol 70 6023ndash6030
44 Holmes DE Chaudhuri SK Nevin KP Mehta T Metheacute BA Liu A Ward JE
Woodard TL Webster J Lovley DR (2006) microarray and genetic analysis of
electron transfer to electrodes in Geobacter sulfurreducens Environ Microbial
8(10)1805-1815
45 Hu H (2009) Enhancing Hydrogen Production in Microbial Electrolysis Cells
through Development of Platinum-free Cathode and Improvement of Reactor
Dissertation
46 Ince BK Ayman O N Turker G Ccedilelikkol S Ince O (2010) Microbial ecology of
anaerobic reactors for treatment of alcohol industry wastewaters a review Current
research technology and education topics in applied microbiology and microbial
ecology 988-999
47 Jung S Regan JR (2007) Comparison of anode bacterial communities and
performance in microbial fuel cells with different electron donors Appl Microbiol
Biotechnol 77393ndash 402
48 Kim BH Kim HJ Hyun MS Park DH (1999) Direct electrode reaction of Fe(III)
reducing bacterium Shwwanella putrefaciens J Microbial Biotechnol 9127-131
49 Kim GT Hyun MS Chang IS Kim HJ Park HS Kim BH Kim SD Wimpenny
JW Weightman AJ (2005) Dissimilatory Fe(III) reduction by an electrochemically
active lactic acid bacterium phylogenetically related to Enterococcus gallinarum
isolated from submerged soil J Appl Microbiol 99(4) 978ndash987
50 Kim GT Webster G Wimpenny JW Kim BH Kim HJ Weightman AJ (2006)
Bacterial community structure compartmentalization and activity in a microbial fuel
cell J Appl Microbiol 101(3) 698ndash710
86
51 Kim HJ Park HS Hyun MS Chang IS Kim M Kim BH (2002) A mediator-
less microbial fuel cell using a metal reducing bacterium Shewanella putrefaciens
Enzyme Microb Technol 30 145-152
52 Kim JR Min B Logan BE (2005) Evaluation of procedures to acclimate a
microbial fuel cell for electricity production Appl Microbiol Biotechnol 68(1)23-30
53 Kim JR Cheng S Oh SE Logan BE (2007) Power generation using different
cation anion and ultrafiltration membranes in microbial fuel cells Environ Sci
Technol 41 1004-1009
54 Konstantinidis KT Serres MH Romine MF Rodrigues JL Auchtung J McCue
LA Lipton MS Obraztsova A Giometti CS Nealson KH Fredrickson JK
Tiedje JM (2009) Comparative systems biology across an evolutionary gradient
within the Shewanella genus Proc Natl Acad Sci U S A 106(37)15909-15914
55 Lee J Phung NT Chang IS Kim BH Sung HC (2003) Use of acetate for
enrichment of electrochemically active microorganisms and their 16S rDNA
analyses FEMS Microbiol Lett 223185ndash191
56 Liang P Wang H Xia X Huang X Mo Y Cao X Fan M(2011)Carbon
nanotube powders as electrode modifier to enhance the activity of anodic biofilm
in microbial fuel cells Biosens Bioelectron 26(6)3000-4
57 Lies DP Hernandez ME Kappler A Mielke RE Gralnick JA Newman DK
(2005) Shewanaella oneidensis MR-1 uses overlapping pathways for iron reduction
at a distance and by direct contact under conditions relevant for biofilms Appl
Environ Microbiol 71 4414-4426
58 Liu H and Logan B E (2004) Electricity generation using an air-cathode single
chamber microbial fuel cell in the presence and absence of a proton exchange
membrane Environ Sci Technol 38 4040-4046
59 Liu H Grot S Logan BE (2005) Electrochemically assisted microbial production
of hydrogen from acetate Environ Sci Technol 39 4317 ndash 320
60 Liu H Hu H Chignell J Fan Y (2010) Microbial Electrolysis Novel Technology
for Hydrogen Production from Biomass Biofuels 1(1) 129ndash142
61 Liu H Ramnarayanan R Logan BE (2004) Production of electricity during
wastewater treatment using a single chamber microbial fuel cell Environ Sci Technol
382281-2285
62 Liu WT Marsh TL Cheng H Forney LJ (1997) Characterization of microbial
diversity by determining terminal restriction fragment length polymorphisms of gene
encoding 16S rRNA Appl Environ Microbiol 63 4516ndash4522
63 Logan B E Hamelers B Rozendal R Schroder U Keller J Freguia S Aelterman
P Verstraete W Rabaey K (2006) Microbial fuel cells methodology and
technology Environ Sci Technol 40 (17) 5181-5192
64 Logan B E and Regan JM (2006) Electricity-producing bacterial communities in
microbial fuel cells Trends Microbiol 14512ndash518
87
65 Logan B Cheng S Watson V Estadt G (2007) Graphite fiber brush anodes for
increased power production in air-cathode microbial fuel cells Environ Sci Technol
41(9)3341-3346
66 Logan BE (2009) Exoelectrogenic bacteria that power microbial fuel cells Nat Rev
Microbiol 5 375-381
67 Logan BE Call D Cheng S Hamelers HV Sleutels TH Jeremiasse AW
Rozendal RA (2008) Microbial electrolysis cells for high yield hydrogen gas
production from organic matter Environ Sci Technol 42 8630-8640
68 Logan BE Hamelers B Rozendal R Schroder U Keller J Freguia S Aelterman
P Verstraete W Rabaey K (2006) Microbial fuel cells methodology and
technology Environ SciTechnol 40 5181-5192
69 Lovely DR (2006) Bug juice harvesting electricity with microorganisms Nature
Review| Microbiology 4 497-508
70 Lovley DR Phillips EJ (1988) Novel Mode of Microbial Energy Metabolism
Organic Carbon Oxidation Coupled to Dissimilatory Reduction of Iron or
Manganese Appl Environ Microbiol 54(6) 1472-1480
71 Ly HK Sezer M Wisitruangsakul N Feng JJ Kranich A Millo D Weidinger
IM Zebger I Murgida DH Hildebrandt P (2011) Surface-enhanced vibrational
spectroscopy for probing transient interactions of proteins with biomimetic interfaces
electric field effects on structure dynamics and function of cytochrome c FEBS J
278(9)1382-90
72 Macaskie LE Hewitt CJ Shearer JA Kent CA (1995) Biomass production for
the removal of heavy metals from aqueous solutions at low pH using growth-
decoupled cells of a Citrobacter sp Int Biodeter Biodegr 3573ndash92
73 Malki M De Lacey AL Rodriguez N Amils R Fernandez VM (2008)
Preferential use of an anode as an electron acceptor by an acidophilic bacterium in
the presence of oxygen Appl Environ Microbiol 74 4472ndash4476
74 Meitl LA Eggleston CM Colberg PJS Khare N Reardon CL Shi L (2009)
Electrochemical interaction of Shewanella oneidensis MR-1 and its outer membrane
cytochromes OmcA and MtrC with hematite electrodes Geochim Cosmochim
Ac 73(18) 5292-5307
75 Morozan A Stamatin L and Nastase
F (2007) The biocompatibility
microorganisms-carbon nanostructures for applications in microbial fuel cells Phys
Stat Sol 6 1797-1803
76 Morris JM Jin S Crimi B Pruden A (2009) Microbial fuel cell in enhancing
anaerobic biodegradation of diesel Chem Eng J 146 161ndash167
77 Murray AE Lies D Li G Nealson K Zhou J Tiedje JM (2001) DNADNA
hybridization to microarrays reveals gene-specific differences between closely
related microbial genomes Proc Natl Acad Sci U S A 98 9853-9858
78 Muyzer G de Waal EC Uitterlinden AG (1993) Profiling of complex microbial
communities by denaturing gradient gel electrophoresis analysis of polymerase chain
88
reaction amplified genes coding for 16S rRNA Appl Environ Microbiol 59 695ndash
700
79 Myers CR Myers JM (1992) Localization of cytochromes to the outer membrane
of anaerobically grown Shewanella putrefacians MR-1 J Bacteriol 174 3429ndash3438
80 Narde GK Kapley A Purohit HJ (2004) Isolation and characterization of
Citrobacter strain HPC255 for broad-range substrate specificity for chlorophenols
Curr Microbiol 48(6) 419-423
81 Okabe S Ito T Sugita K Satoh H (2005) Succession of internal sulfur cycles and
sulfur-oxidizing bacterial communities in microaerophilic wastewater biofilms Appl
Environ Microbiol 71(5) 2520-2529
82 Park DH and Zeikus JG (2000) Electricity generation in microbial fuel cells using
neutral red as an electronophore Appl Environ Microbial 66 292-1297
83 Park DH and Zeikus JG (2002) Impact of electrode composition on electricity
generation in a single-compartment fuel cell using Shewanella putrefaciens Appl
Microbiol Biotechnol 5958ndash56
84 Park DH and Zeikus JG (2003) Improved fuel cell and electrode designs for
producing electricity from microbial degradation Biotechnol Bioeng 81 348ndash355
85 Park HS Kim BH Kim HS Kim HJ Kim GT Kim M Chang IS Park Y
K Chang HI (2001) A novel electrochemically active and Fe(III)-reducing
bacterium phylogenetically related to Clostridium butyricum isolated from a
microbial fuel cell Anaerobe 7 297-306
86 Parot S Nercessian O Delia ML Achouak W Bergel A (2009) Electrochemical
checking of aerobic isolates from electrochemically active biofilms formed in
compost J Appl Microbiol 106(4) 1350-1359
87 Pham CA Jung SJ Phung NT Lee J Chang IS Kim BH Yi H Chun J
(2003) A novel electrochemically active and Fe(III)-reducing bacterium
phylogenetically related to Aeromonas hydrophila isolated from a microbial fuel cell
FEMS Microbiol Lett 223 129-134
88 Qiao Y Bao SJ Li CM Cui XQ Lu ZS Guo J (2008) Nanostructured
polyanilinetitanium dioxide composite anode for microbial fuel cells ACS Nano 2
113-119
89 Qiu R Zhao B Liu J Huang X Li Q Brewer E Wang S Shi N (2009) Sulfate
reduction and copper precipitation by a Citrobacter sp isolated from a mining area J
Hazard Mater 1641310-1315
90 Quan X Yang SG Ruan XL and Zhao HM (2005) Preparation of titania
nanotubes and their environmental applications as electrode Environ Sci Technol 39
3770ndash3775
91 Rabaey K Boon N Hofte M Verstraete W (2005) Microbial phenazine
production enhances electron transfer in biofuel cells Environ Sci Technol 393401ndash
3408
89
92 Rabaey K Boon N Siciliano S D Verhaege M Verstraete W (2004) Biofuel cells
select for microbial consortia that self-mediate electron transfer Appl Environ
Microbiol 70(9) 5373ndash5382
93 Rabaey K Lissens G Siciliano S D Verstraete W (2003) Amicrobial fuel cell
capable of converting glucose to electricity at high rate and efficiency Biotechnol
Lett 25 1531-15
94 Rabaey K Verstraete W (2005) Microbial fuel cells novel biotechnology for
energy generation Trends Biotechnol 23(6) 291-298
95 Reguera G McCarthy KD Mehta T Nicoll JS Tuominen MT Lovley DR
(2005) Extracellular electron transfer via microbial nanowires Nature 435 1098-
1101
96 Reguera G Nevin KP Nicoll JS Covalla SF Woodard TL Lovley DR (2006)
Biofilm and nanowire production leads to increased current in Geobacter
sulfurreducens fuel cells Appl Environ Microbiol 72 7345-7348
97 Reimers CE Tender LM Fertig S Wang W (2001) Harvesting energy from the
marine sediment-water interface Environ Sci Technol 35192-195
98 Rezaei F Xing D Wagner R Regan JM Richard TM Logan BE (2009)
Simultaneous cellulose degradation and electricity production by Enterobacter
cloacae in a microbial fuel cell Appl Microbiol Biotechnol 75 3673ndash3678
99 Richter H Nevin KP Jia H Lowy DA Lovley DR Tender LM (2009) Cyclic
voltammetry of biofilms of wild type and mutant Geobacter sulfurreducens on fuel
cell anodes indicates possible roles of OmcB OmcZ type IV pili and protons in
extracellular electron transfer Energy Environ Sci 2 506ndash516
100 Riesenfeld SC Schloss DP Handelsman J(2004) Metagenomics Genomic
Analysis of Microbial Communities Annual Review Genetics 38525-552
101 Rozendal RA Hamelers HV Buisman CJ (2006) Effects of membrane cation
transport on pH and microbial fuel cell performance Environ Sci Technol 40
5206-5211
102 Rozendal RA Hamelers HVM Euverink GJW Metz SJ Buisman CJN (2006)
Principle and perspectives of hydrogen production through biocatalyzed electrolysis
Int J Hydrogen Energ 311632-1640
103 Rozendal RA Hamelers HV Molenkamp RJ Buisman CJ (2007) Performance of
single chamber biocatalyzed electrolysis with different types of ion exchange
membrances Water Res 41 1984-1994
104 Schluumlter A Bekel T Diaz NN Dondrup M Eichenlaub R Gartemann
KH Krahn I Krause L Kroumlmeke H Kruse O Mussgnug JH Neuweger
H Niehaus K Puumlhler A Runte KJ Szczepanowski R Tauch A Tilker
A Viehoumlver P Goesmann A (2008) The metagenome of a biogas-producing
microbial community of a production-scale biogas plant fermenter analyzed by the
454-pyrosequencing technology J Biotechnol136 77-90
90
105 Scott J H and Nealson KH (1994) A biochemical study of the intermediary carbon
metabolism of Shewanella putrefaciens J Bacteriol 176(11) 3408ndash3411
106 Sekiguchi H Tomioka H Nakahara T and Uchiyama H (2001) A single band
does not always represent single bacterial strains in denaturing gradient gel
electrophoresis analysis Biotechnology Letters23 1205ndash1208
107 Sharma T Leela M Reddy A Chandra TS Ramaprabhu S (2008) Development
of carbon nanotubes and nanofluids based microbial fuel cell Int J Hydrogen Energ
336749ndash6754
108 Shea C Clauwaert P Verstraete W Nerenberg R (2008) Adapting a denitrifying
biocathode for perchlorate reduction Water Sci Technol 58 (10) 1941-1946
109 Shi L Squier TC Zachara JM Fredrickson JK (2007) Respiration of metal (hydr)
oxides by Shewanella and Geobacter a key role for multihaem c-type cytochromes
Mol Microbiol 6512-20
110 Stams AJ and Plugge CM(2009)Electron transfer in syntrophic communities
of anaerobic bacteria and archaea Nat Rev Microbiol7(8)568-77
111 Sukkasem C Xu S Park S Boonsawang P Liu H (2008) Effect of Nitrate on the
Performance of Single Chamber Air Cathode Microbial Fuel Cells Water Res 42(19)
4743-4750
112 Summers ZM Fogarty HE Leang C Franks AE Malvankar NS Lovley DR
(2010)Direct exchange of electrons within aggregates of an evolved syntrophic
coculture of anaerobic bacteria Science 330 (6009)1413-5
113 Szczepanowski R Bekel T Goesmann A Krause L Kroumlmeke H Kaiser
O Eichler W Puumlhler A Schluumlter A (2008) Insight into the plasmid metagenome
of wastewater treatment plant bacteria showing reduced susceptibility to
antimicrobial drugs analysed by the 454-pyrosequencing technology J
Biotechnol 136 54-64
114 Tamura K Dudley J Nei M Kumar S (2007) MEGA4 Molecular Evolutionary
Genetics Analysis (MEGA) software version 40 MolBiol Evol 24(8) 1596-1599
115 Thompson JD Gibson TJ Plewniak F Jeanmougin F Higgins DG (1997) The
CLUSTAL_X windows interface flexible strategies for multiple sequence alignment
aided by quality analysis tools Nucleic Acids Res 25 4876-4882
116 Thormann KM Duttler S Saville RM Hyodo M Shukla S Hayakawa Y
Spormann AM (2006) Control of formation and cellular detachment from
Shewanella oneidensis MR-1 biofilms by cyclic di-GMP J Bacteriol 1882681-2691
117 Thormann KM Saville RM Shukla S Pelletier DA Spormann AM (2004)
Initial Phases of biofilm formation in Shewanella oneidensis MR-1 J Bacteriol
186(23) 8096-104
118 Torres CI Kato Marcus A Rittmann BE (2008b) Proton transport inside the
biofilm limits electrical current generation by anode-respiring bacteria Biotechnol
Bioeng 100 872ndash881
91
119 Torres CI Marcus AK Lee HS Parameswaran P Krajmalnik-Brown R Rittmann
BE (2010) A kinetic perspective on extracellular electron transfer by anode-
respiring bacteria FEMS Microbiol Rev 234(1)3-17
120 Torres CI Marcus AK Parameswaran P Rittmann BE (2008a) Kinetic
experiments for evaluating the NernstndashMonod model for anode-respiring bacteria
(ARB) in a biofilm anode Environ Sci Technol 42 6593ndash6597
121 Tsai HY Wu Ch-Ch Lee Ch-Y Shih EP (2009) Microbial fuel cell performance
of multiwall carbon nanotubes on carbon cloth as electrodes J Power Sources
194199-205
122 Tyson GW Chapman J Hugenholtz P Allen EE Ram RJ Richardson
PM Solovyev VV Rubin EM Rokhsar DS Banfield JF(2004) Community
structure and metabolism through reconstruction of microbial genomes from the
environment Nature42837-43
123 Venkateswaran K Moser DP Dollhopf ME Lies DP Saffarini D A MacGregor
BJ Ringelberg DB White DC Nishijima M Sano H Burghardt J Stackebrandt
E Nealson KH (1999) Polyphasic taxonomy of the genus Shewanella and
description of Shewanella oneidensis sp Int J Syst Bacteriol 49705-724
124 von Canstein H Ogawa J Shimizu S Lloyd JR (2008) secretion of Flavins
by Shewanella Species and Their Role in Extracellular Electron Transfer Appl
Environ Microbiol 74 3615-623
125 Wang A Liu W Cheng Sh Xing D Zhou J Logan BE (2009) Source of methane
and methods to control its formation in single chamber microbial electrolysis cells
Int J Hydrogen Energ 34 3653-3658
126 Wang J Ma T Zhao L Lv J Li G Liang F Liu R (2008) PCRndashDGGE method
for analyzing the bacterial community in a high temperature petroleum reservoir
World J Microbiol Biotechnol 2 9694-9702
127 Wang Q Garrity GM Tiedje JM Cole JR (2007) Naiumlve Bayesian Classifier for
Rapid Assignment of rRNA Sequences into the New Bacterial Taxonomy Appl
Environ Microbiol 73(16) 5261-5267
128 Wang YF Masuda M Tsulimura S Kano K (2008) Electrochemical regulation of
the end-product profile in Propionibacterium freudenreichii ET-3 with an
endogenous mediator Biotechnol Bioeng 101(3) 579ndash586
129 Wrighton KC Agbo P Warnecke F Weber KA Brodie EL DeSantis TZ
Hugenholtz P Andersen GL Coates JD (2008) A novel ecological role of the
Firmicutes identified in thermophilic microbial fuel cells ISME J 2 1146ndash1156
130 Xing D Zuo Y Cheng S Regan JM Logan BE (2008) Electricity generation by
Rhodopseudomonas palustris DX-1 Environ Sci Technol 42 4146-4151
131 Xu S Liu H Fan Y Schaller R Jiao J Chaplen F (2012) Enhanced performance
and mechanism study of microbial electrolysis cells using Fenanoparticle-decorated
anodes Appl Microbiol Biotechnol 93(2)871-880
92
132 Yang Y Harris DP Luo F Wu L Parsons AB Palumbo AV Zhou J (2008)
Characterization of the Shewanella oneidensis Fur gene roles in iron and acid
tolerance response BMC Genomics 9 Suppl 1S11
133 Zhang L Zhou S Zhuang L Li W Zhang J Lu N Deng L (2008) Microbial
fuel cell based on Klebsiella pneumoniae biofilm Electrochem Commun10 1641ndash
1643
134 Zuo Y Xing DF Regan JM Logan BE (2008) Isolation of the exoelectrogenic
bacterium Ochrobactrum anthropi YZ-1 by using a U-tube microbial fuel cell Appl
Environ Microbiol 74 3130-3137
93
APPENDICES
List of published papers during PhD study
1 Shoutao Xu Hong Liu Yanzhen Fan Rebecca Schaller Jun Jiao and Frank
Chaplen (2012) Enhanced performance and mechanism study
of microbial electrolysis cells using Fe nanoparticle-decorated anodes Applied
Microbiology and Biotechnology 93(2)871-880
2 Shoutao Xu Hong Liu (2011) New exoelectrogen Citrobacter sp SX-1 isolated
from a microbial fuel cell Journal of Applied Microbiology111(5)1108-1115
3 Yanzhen Fan Shoutao Xu Rebecca Schaller Jun Jiao Frank Chaplen Hong Liu
(2011) Nanoparticle decorated anodes for enhanced current generation in microbial
electrochemical cells Biosensors and Bioelectronics 26(5) 1908-1912
4 Tunc Catal Shoutao Xu Kaichang Li Hakan Bermek Hong Liu
(2008)
Electricity generation from polyalcohols in single-chamber microbial fuel
cells Biosensors and Bioelectronics 24(4)849-854
5 Chontisa Sukkasema Shoutao Xu Sunhwa Park Piyarat Boonsawang Hong Liu
(2008) Effect of nitrate on the performance of single chamber air cathode
microbial fuel cells Water research 424743-4750
6 Rebecca Schaller Yanzhen Fan Shoutao Xu Alan Fern Frank Chaplen Hong
Liu and Jun Jiao (2009) Vertically Aligned Multi-walled Carbon Nanotube
Decorated Anodes for Microbial Fuel Cells Proceedings of Materials Research
Society 2009 1170 R05-13

TABLE OF CONTENTS
Page
1 General Introductionhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 1
2 Bacterial Community Analysis of Mixed Consortium in Microbial
Electrochemical Systemshelliphelliphelliphellip 13
3 New Exoelectrogen Citrobacter sp SX-1 Isolation and
Characterizationhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 27
4
Enhanced Performance and Mechanism Study of Microbial Electrolysis
Cells using Fe Nanoparticles Decorated Anodeshelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 45
5 Global Transcriptome Analysis of Response of Shewanella oneidensis
MR-1 to CNT Nanostructure Decorated Anodes in Microbial
Electrochemical System
66
6 Summaryhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 80
7 Bibliography helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 82
8 Appendices helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 93
LIST OF FIGURES
Figure Page
1-1 Generalized schematics (a) Microbial fuel cell (MFCs)(b) Microbial
electrolysis cell (MECs) helliphelliphelliphelliphelliphelliphelliphelliphelliphellip helliphelliphelliphellip
2
1-2 Mechanisms for extracellular electron transport in a MFCs anode 7
2-1 Polarization curves of high power mixed culture MFCshelliphelliphelliphelliphelliphellip 20
2-2 DGGE result of high power generation mixed culture MFCshelliphelliphellip 21
2-3 Bacterial species and percentage of bacterial community of mixed
culture MFCs helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip
22
2-4 Phylogenetic trees of bacterial species from mixed culture MFCs 22
2-5 Dominant bacterial species identification in DGGEhelliphelliphelliphelliphelliphelliphelliphellip 23
2-6 Current density generated by different dominant isolateshelliphelliphelliphelliphelliphellip 24
3-1 Phylogenetic tree of strain SX-1 and closely related species based on
16S rDNA sequenceshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip
35
3-2 SEM image of planktonic cells of Citrobacter sp SX-1helliphelliphelliphelliphelliphellip 35
3-3 Electricity generation by Citrobacter sp SX-1 in a single chamber
MFCs with sodium citrate helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip
36
3-4 Electricity generation of Citrobacter sp SX-1 using different
substrates in single chamber air-cathode MFCs at 1 KΩhelliphelliphellip
37
3-5 Power and voltage generation by Citrobacter sp SX-1 as a function
of current density helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip
38
3-6 Four current generating stages for CV analysis helliphelliphelliphelliphelliphelliphelliphellip 39
3-7 Cyclic Voltammetry analysis of of Citrobacter sp SX-1 biofilms helliphellip 41
4-1 SEM images of Fe NPs decorated anodeshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 50
4-2 Effects of Fe-NP decorated anodes on the current density in MECs helliphellip 51
4-3 Differentially expressed genes grouped by functional classification in
Fe NP decorated anodeshelliphelliphelliphelliphelliphellip helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip
53
5-1 SEM images of CNT decorated anodeshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 70
5-2 Effects of CNT decorated anodes on the current density in MECshelliphellip 73
5-3 Differentially expressed genes grouped by functional classification in
CNT decorated anodes helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip
74
LIST OF TABLES
Table
1-1
4-1
4-2
4-3
Taxa of bacteria current density generated and reactor configuration
used in microbial electrochemical cell experimentshelliphelliphelliphellip
Expression levels of genes related to biofilm formationhelliphelliphelliphelliphellip
Genes related to anaerobic growth and electron transfer with
significantly change expression levelhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip
Expression level of flavin and cytochrome geneshelliphelliphelliphelliphelliphelliphelliphellip
Page
4
55
56
58
4-4
5-1
5-2
Other genes with significantly changed expression level
Expression level of cytochrome c as response to CNT decorated anodes
Expression level of genes related to flavin synthesis as response to CNT
decorated anode in CNT decorated anodeshelliphellip
59
76
78
1
1
Bacterial Community Analysis New Exoelectrogen Isolation and Enhanced Performance
of Microbial Electrochemical Systems Using Nano-Decorated Anodes
Chapter 1
General Introduction
11 Microbial Fuel Cells and Microbial Electrolysis Cells
Microbial electrochemical systems (MESs) have drawn the attention of researchers
in recent years due to their promising applications in a variety of scientific fields such as
renewable energy generation bioremediation and wastewater treatment In a MES
microorganisms interact with electrodes catalyzing oxidation and reduction reactions at
the anode and the cathode
The most-described type of MESs is the microbial fuel cells (MFCs) in which
useful power is generated directly using the catalytic action of active microorganisms
(Kim 2002 Cheng and Logan 2007 Cheng et al 2006) In a typical two-chamber
MFCs (Figure 1a) organic matter is oxidized by electrochemically active
microorganisms in the anode chamber to release protons and electrons Protons diffuse
into the cathode chamber through a proton exchange membrane Meanwhile electrons
are transferred to the anode through various mechanisms (Logan et al 2006 Lovley
2
2006 Rabaey et al 2003) and then travel to the cathode where normally they combine
with oxygen and protons to form water
Another common used type of MES is the microbial electrolysis cell (MECs) in
which hydrogen is produced instead of electricity by applying a circuit voltage to the
MFCs system and maintaining anaerobic conditions in the cathode chamber (Liu et al
2005 Logan et al 2008 Rozendal et al 2006 Schaller et al 2009) (Figure 1b) A
cathode potential of at least - 410 mV (vs standard hydrogen electrode at pH 70) is
required to produce hydrogen at the cathode Therefore a theoretical potential of 110 mV
(ie 410-300 mV) is needed but voltages of gt02 V are normally applied due to various
overpotentials
Fig 1-1 Generalized schematics (a) Microbial fuel cells (MFCs) where electricity is
captured through the resistance R (b) Microbial electrolysis cells (MECs) where
hydrogen gas is produced in the cathode chamber R for resistor PS for power supply and
PEM for proton exchange membrane
The fundamental feature shared by microbial electrochemical systems (MESs)
(MECs and MFCs) is the microbe-catalyzed electron transfer from organic matter to
electrodes (anodes) (Rebaey et al 2004) Therefore different types of MESs can be
PEM
R
H+
e- e-
Anode
Bacte
ria
a
Cath
ode
H2O
PEM
PS
H+
e- e-
Anode
Bacte
ria
b
Cath
ode
H2
3
utilized to investigate the phenomena and mechanisms of interactions between microbes
and electrodes
12 Bacteria and Electron Transfer Mechanisms in MESs
The electrochemically active bacteria in MFCs are thought to be iron-reducing
bacteria such as Shewanella and Geobacter species (Gorby et al 2006) They have great
importance in the natural environment principally in metal oxidation and reduction
However recent studies have shown that the diversity of bacterial communities is much
greater than these model iron reducers on MFCs anodes (Reimers et al 2001 Liu et al
2004b Kim et al 2005) Over 20 electrochemically active bacterial species those can
transfer electrons exocellularly to electrodes have been reported in the past 10 years
(Logan 2009 Liu et al 2010) These bacteria have been categorized into diverse genetic
groups so far including alphaproteobacteria (Rhodopseudomonas Ochrobactrum and
Acidiphilium) (Malki et al 2008 Xing et al 2008 Zuo et al 2008) betaproteobacteria
(Rhodoferax) (Chaudhuri et al 2003) gammaproteobacteria (Shewanella Pseudomonas
Klebsiella Enterobacter and Aeronomas) (Kim et al 2002 Pham et al 2003 Rabaey et
al 2005 Bretschger et al 2007 Zhang et al 2008 Chung et al 2009 Rezaei et al 2009)
deltaproteobacteria (Geobacter Geopsychrobacter Desulfuromonas and Desulfobulbus)
(Bond et al 2002 Bond and Lovley 2003 Holmes et al 2004ab 2006)
Epsilonproteobacteria (Arcobacter) (Fedorovich et al 2009) Firmicutes (Clostridium and
Thermincola) (Park et al 2001 Wrighton et al 2008) Acidobacteria (Geothrix) (Bond
and Lovley 2005) and actinobacteria (Propionibacterium) (Wang et al 2008) A wider
range of electrochemically active bacteria are expected to be discovered
4
Table 1 Taxa of bacteria current density generated and reactor configuration used in
microbial electrochemical cell experiments (Liu et al 2010)
The electrochemically active bacterial species that possess the ability to transfer
electrons outside of the cell are called exoelectrogens in the MESs research field The
different exoelectrogens have demonstrated a wide-ranging power generation ability in
MFCs For example Shewanella oneidensis MR-1 has demonstrated the ability to
generate 0018 Am2 current density in single chamber MFCs while the Geobacter
Taxon Microorganisms Current Density (Am2
)
α-proteobacteria
Rhodopseudomonas palustris DX-1 003
Ochrobactrum anthropi YZ-1 071
Acidiphilium sp 32sup5 300
β-proteobacteria Rhodoferax ferrireducens 0031
γ-proteobacteria
Shewanella putrefaciens IR-1 0016
Shewanella oneidensis DSP10 0013
Shewanella oneidensis MR-1 0018
Pseudomonas aeruginosa KRA3 0017
Escherichia coli K12 HB101 100
Klebsiella pneumoniae L17 120
Enterobacter cloacae 013
Aeromonas hydrophila PA3 030
δ-proteobacteria
Geobacter metallireducens 065
Geobacter sulfurreducens 800
Desulfuromonas acetoxidans 0005
Geopsychrobacter
Electrodiphilus strain A2
0066
Desulfobulbus propionicus 003
Firmicutes
Lactococcus lactis 003
Thermincola sp strain Jr 020
Clostridium butyricum EG3 022
Thermincola ferriacetica Z-0001 040
Brevibacillus spp PTH1 0009
Desulfitobacterium hafniense DCB2 110
Acidobacteria Geothrix fermentans 0097
Actinobacteria Propionibacterium freudenreichiiET-3 120
5
sulfurreducens species has generated current densities as high as 80 Am2 which is 400
times higher than the one generated by Shewanella oneidensis MR-1 However mixed-
culture communities have generated much higher power densities than their pure-culture
counterparts in MFCs perhaps due to synergistic interactions within the anode bacterial
communities and the participation of currently unknown bacteria species and mechanisms
(Jung and Regan 2007) The dominant bacteria species vary in mixed cultures bacteria
communities in MFCs due to the enrichment of different substrates
Traditional methods of extroelectrogen study depend on cultivation hampered novel
exoelectrogens discovery because the inadequacy of defined media underestimates the
actual microbial diversity in MFCs Slow growth rate and unknown growth requirements
of anaerobic microorganisms make some anaerobic extroelectrogens cultivation difficult
However these limitations have been overcome by using molecular biological methods
based on DNARNA analysis Molecular biological techniques are now widely applied to
assess the diversity of microbial communities by analyzing the 16S rDNA sequence The
most commonly used molecular biological techniques for bacterial community analysis
include denaturing gradient gel electrophoresis (DGGE) restriction fragment length
polymorphism (RFLP) and 16S rDNA sequencing These methods are much less time
consuming than traditional isolation and cultivation methods
Isolated exoelectrogens were utilized to explore the mechanism of electron transfer
to the anode However the mechanisms of electron transfer to extracellular electron
acceptors are not well understood Three mechanisms have been proposed for exocellular
transport of electrons by exoelectrogens to anodes in MFCs so far Some exoelectrogenic
bacteria can transfer electrons to electrodes through soluble redox compounds (Bond and
6
Lovley 2005) These compounds include artificial mediators and mediators secreted by
exoelectrogenic bacteria A variety of chemicals have been used to facilitate the shuttling
of electrons from inside of cell to electrodes outside the cell These exogenous mediators
include neutral red (Park et al 1999) anthraquinone-2-6 disulfonate (AQDS) thionin
potassium ferricyanide (Bond et al 2002) methyl viologen and others (Logan 2004
Rabaey and Verstraete 2005) Some bacteria can secrete their own chemical mediator
for example Pseudomonas aeruginosa secretes pyocyanin and phenazine-1-carboxamide
to transfer electrons toward the MFCs anode (Rabaey et al 2005) Another evidence for
mediator production by bacteria is Geothrix fermentans When the medium was replaced
in a MFCs that had stable power generation with this bacteria power dropped by 50
and required 10 days to resume the original level
Some bacteria can directly transfer electrons to anodes via outer cell membrane
proteins (Myers and Myers 1992) The outer cell membrane protein cytochrome c is
thought to play a critical role in to transferring electrons to anodes Ly et al (2011)
isolated the haem protein cytochrome c and demonstrated that electric field effects may
be functional for the natural redox processes of cytochrome c in the respiratory chain
Shewanella oneidensis is the example of a bacterium that is capable of transfer electron to
anode via outer cell membrane proteins When Shewanella oneidensis adhered to an iron
surface the greater force has showed benefits to grow cells because closer contact
required for electron transfer from cell bound cytochromes (Lower et al 2001)
7
Fig1-2 Mechanisms for extracellular electron transport in an MFCs anode (1) direct
contact (top in green) (2) by nanowires (middle in purple) and (3) self-produced
mediators (bottom in blue) (Logan 2009)
More and more evidence supports the involvement of bacterial nanowires in
extracellular electron transport (Reguera et al 2005 2006 Gorby et al 2006) Nanowires
are conductive appendages produced by both Geobacter and Shewanella species (Gorby
and Beveridge 2005) The conductivity of the appendages was examined and confirmed
by using conductive scanning tunneling microscopy (STM) (Gorby et al 2006)
Nanowires can carry electrons from the cell to the anode surface of MFCs
The solid component of the extracellular biofilm matrix has high efficiency on
extracellular electron transfer compared with other extracellular electron transfer
8
mechanisms and recently Torres et al (2010) hypothesized that the solid component of
the extracellular biofilm matrix formed by exoelectogens is another way bacteria transfer
electrons to electrodes This hypothesis was based on kinetic analysis of each EET
mechanism reported in available literature (Torres et al 2010)
13 Anode electrodes
In MFCsMECs anode electrodes are a critical component because exoelectrogens
adhere to the surface of anodes to transfer electrons to the electrode The characteristics
of anodes have significant effects on electron transfer rate from bacteria to anode
electrodes in MFCs The requirements of an anode material are it should be highly
conductive non-corrosive have a high specific surface area (area per volume) high
porosity be non-fouling inexpensive and easily scaled to larger sizes Of these
properties the most important one that is different from other biofilm reactors is that the
material must be electrically conductive Normally they are made of various carbon
materials including carbon fiber carbon clothe and carbon paper due to their stability
high conductivity and high specific surface-area Nevertheless they have little
electrocatalytic activity for the anode microbial reactions and thus a modification of the
carbon materials is the main approach for improving their performance Consequently
there is a great need to develop a new type of anode material for MFCsMECs
It is a great challenge to develop a new anode material to further increase the power
density of a MFCs The nature of the catalytic mechanism of a MFCrsquos anode involves not
only a biological but also an electrocatalytic process An optimal nanostructure with a
high specific surface area favorable for both catalytic processes could play a critical role
in improving the power density of the MFCs such a structure needs to host the bacteria
9
with high bioactivity while enhancing the electron-transfer rate Schroumlder et al (2008)
employed PANI to modify a platinum anode for MFCs and achieved a current density 1
order of magnitude higher than the previously reported value PANIinorganic
composites are also reported to have better conductivity Qiao et al also applied a new
mesoporous TiO2 electrode material with uniform nanopore distribution and a high
specific surface area to anode in comparison to previously reported work with E coli
MFCs the composite anode delivers 2-fold higher power density (Qiao et al 2008)
Thus it has great potential for use as the anode in a high-power MFCs and may be a new
approach for improving performance of MFCs
14 Other parts of MESs
141 Membranes and ion transport
The ion exchange membrane is another one of the critical components in two-
chamber MESs systems It separates anode and cathode chambers and at the same time
maintains the electron neutrality of the system ie transport of electrons to the cathode
needs to be compensated by transport of an equal amount of positive charge to the
cathode chamber A proton exchange membrane (PEM) such as Nafion a per-fluorinated
sulphonic acid membrane (PFSAs) consisting of a hydrophobic fluorocarbon backbone to
which hydrophilic sulfonate groups (-SO3-) are attached is commonly used in chemical
fuel cell systems For MFCs systems however mainly cation species like Na+ and K
+
other than proton are often responsible for the dominant transport of positive charge
through the cation exchange membrane (CEM) to maintain electroneutrality due to the
low proton concentration in any aqueous medium with near neutral pH (Rozendal et al
2006) Consequently the pH increases in the cathode chamber due to the consumption of
10
protons and decreases in the anode chamber because of the accumulation of protons
(Rozendal et al 2007) This pH change in two-chamber systems results in a decrease of
the cathode potential and performance The application of anion exchange membrane
(AEM) has also been explored in MFCs systems (Cheng and Logan 2007 Kim and
Logan 2007) where it has been proposed that protons are transferred via pH buffers like
phosphate anions
142 Cathodes and Catalysts
The cathode is another challenge for making MFCs commercially available
technology because the chemical reaction that occurs at the cathode is difficult to
engineer as the electrons protons and oxygen must all meet at a catalyst in a tri-phase
reaction (solid catalyst air and water) The catalyst must be on a conductive surface and
must be exposed to both water and air so that protons and electrons in these different
phases can reach the same point The most commonly used material for a cathode is
commercially available carbon paper pre-loaded with a Pt catalyst on one side When it is
used in a MFC the side that contains the catalyst faces the water and the uncoated side
faces air To reduce the high cathode cost associated with platinum catalyst other precious-
metal-free cathodes such as (NiMo and NiW) were developed by electrodepositing on a
carbon fiber They have achieved comparable performance with Pt catalyst with same
loading at a much lower cathode fabrication cost (Hu 2010)
The requirements of cathode for MECs are quite similar to the requirements of
cathode for MFCs but easier than cathode for MFCs for the manufacturing process
because the cathode in MECs is not necessarily to exposed to air Therefore it can be
made of the exact same materials of cathode in MFCs except a waterproof layer
11
Recently some researchers have attempted to apply microorganisms as a biocatalyst to
precede the combination of electron with oxygen in the cathodes
15 Dissertation overview
Low power densities in MESs limit practical applications The improvement of
MESs performance requires a detailed understanding of the physiology and ecology of
microorganisms in MESs including the mechanism of electron transfer to the anode from
the microorganism
This dissertation focuses on the problem of the low power density of MESs The
bacterial community structure of a high power generated mixed culture communities in
MFCs will be identified firstly and then one of major exoelectrogens will be isolated and
characterized This information will be helpful to understand the physiology and ecology
of exoelectrogens in MESs Consequently they will be beneficial to improve power
density of MFCs Lastly the nanostructure decorated anodes in MECs will be utilized to
improve the power density The power enhancement mechanism will be explored by
using a whole genome microarray They are presented here as four papers
In the first paper the cultivation independent molecular biological techniques
denaturing gradient gel electrophoresis (DGGE) and 16S rDNA clone library are utilized
to analyze the bacterial community structure of a higher power mixed culture MFCs The
analyzed results provide fundamental information for isolating the dominant bacteria in
mixed culture MFCs Two of dominant bacterial species has been isolated and used aone
to test power generation in MFCs The possible interaction among different bacterial
species in mixed culture is discussed
12
In the second paper one isolated exoelectrogenic bacterial strain SX-1 is
characterized It is identified as a member of the Citrobacter genus and power generation
is tested ultilizing a wide range of different substrates The electron transfer mechanism is
explored using Cyclic Voltammetry (CV) This study increases the known diversity of
power generating exoelectrogens and provides a new strain to explore the mechanisms of
extracellular electron transfer from bacteria to electrodes
The third paper shows effects on MESs performance by Fe nanoparticle decorated
anodes in the MESs The average current density produced with Fe nanoparticle
decorated anodes increased up to 59-fold higher than plain graphite anodes A whole
genome microarray is utilized to analyze the possible mechanism of enhanced current
density as responded to nanoparticle decorated anodes
The fourth paper describes the effects of carbon nanotube (CNT) modified anode on
the performance S oneidensis MR-1 in MESs Results demonstrate that current densities
produced with CNT decorated anodes are up to 56-fold higher than plain graphite anodes
The possible mechanisms of enhanced current density by CNT decorated are explored
13
Chapter 2
Bacterial community analysis of mixed consortium in higher
power density MESs
Shoutao Xu and Hong Liu
1 Introduction
The improvement in the performance of mixed culture MFCs requires an
understanding of the ecology in microbial communities of MFCs Many researchers have
attempted to characterize microbial populations and activities to elucidate the behaviors
and ecology of microorganisms in MFCs (Wrighton et al 2008 Fedorovich et al 2009)
In order to study the microbial ecology of the mixed culture in a MFC and select the
appropriate isolation medium for dominant bacterial species in the mixed culture the
fundamental steps are to analyze the bacterial diversity in the mixed culture of MFCs and
identity the dominant bacterial species in bacteria communities in MFCs
14
As for the identification of bacterial communities typically there are two general
methods The first method for identification of bacterial community is the traditional
cultivation processes using selective nutrients to promote the growth of different types of
bacteria within the samples (Amman et al 2000) The community structure can then be
assessed by identifying the isolates from the dominant colonies that were cultured This
can often be costly and laborious as each isolate has to be further studied by examining
its physiology taxonomy and reactivity to stains (Adwards et al 1989)
The second method relies on utilizing molecular techniques to analyze bacterial
community DNA Several molecular methods involving the extraction and analysis of
DNA from entire bacterial communities are used to identify genetic fingerprints of
bacteria These methods including the cloning and sequencing of 16S rDNA automated
ribosomal intergenic spacer analysis (ARISA) terminal restriction fragment length
polymorphism (T-RFLP) and denaturing gradient gel electrophoresis (DGGE) generate
profiles of bacterial community structures They can rapidly assess complex communities
from various environments (Amman et al 2000)
Due to the conservative characteristic of 16S rDNA in bacteria during the process
of evolution 16S rDNA sequencing can be used to identify different species of bacteria
Therefore the molecular techniques of denaturing gradient gel electrophoresis (DGGE)
with PCR and 16S rDNA clone library are used for analysis of the microbial diversity
These methods are more convenient and save time compared to traditional
isolationcultivation methods for microorganism analysis
In this work a biofilm bacterial community from an anode of a MFCs wase studied
by using denaturing gradient gel electrophoresis (DGGE) of PCR-amplified partial 16S
15
rRNA genes followed by cloning and sequencing of 16S rDNA The results provided
essential information for dominant bacterial isolation in mixed culture MFCs
2 Materials and methods
21 High power generation of mixed culture MFCs
Mixed cultures were originally inoculated from domestic wastewater (Corvallis
Wastewater Treatment Plant Corvallis OR) to MFCs after operating for 2 years using a
defined medium solution (Lovely 2002) with sodium acetate as the carbon source A
new MFCs was inoculated from the operating MFCs The polarization curves were
performed to measure the power generation when maximal stable power were established
(normally 3-5 batches post-inoculation) and the genomic DNA from the microbial
biofilm on the anode was extracted under the sterile conditions for further bacterial
community analysis
22 Denaturing gradient gel electrophoresis (DGGE)
DGGE with PCR is a method of analysis of bacterial community composition
based on different GC percentage of 16s rDNA in different bacterial species PCR-DGGE
comprises three steps (i) extraction of total community DNA from the sample (ii) PCR-
controlled amplification using specific oligonucleotide primers and (iii) separation of the
amplicons using DGGE For this purpose a reproducible and efficient method for total
DNA extraction is indispensable and needs to be evaluated and optimized depending on
the nature of the sample In the subsequent PCR step multiple PCR primer sets with
different resolution can be used In most PCR-DGGE applications on bacteria universal
or specific primers are targeting the 16S rRNA gene Upon electrophoresis of the PCR
16
amplicons and gel staining (using ethidium bromide silver staining or SYBR green)
DGGE gels are digitally captured and further analyzed using computer software packages
The richness of DNA fragment bands in DGGE indicates the bacterial diversity of the
sample
The detailed steps are as followed Biofilms were scratched from the anodes of high
power generated MFCs fed with sodium acetate Bacterial genomic DNA was extracted
from the biofilm samples using the DNeasy blood and tissue kit (Qiagen) according to the
manufacturerrsquos instructions The universal primer set was used to amplify the 16S rDNA
from the extracted genomic DNA then the V3 region of bacterial 16S ribosomal DNA
(rDNA) was amplified with GC-clamp primer (Muyzer et al 1993) for DGGE PCR
amplification was performed in a thermocycler DGGE of the PCR products was carried
out in a DcodeTM
Universal Mutation Detection System) The 8 (wv) polyacrylamide
gels contains from 30 to 55 denaturing gradients Electrophoresis was conducted
using a 1timesTAE (Tris-Acetate-EDTA) buffer at 130V and 60degC for 5 hours After
electrophoresis the gel was stained with ethidium bromide in 1timesTAE buffer for 15
minutes and washed in 1timesTAE buffer for 10 minutes The fragments were visualized
under a UV transilluminator The richness of single band in DGGE gel picture
preliminarily showed that the bacterial diversity in mixed culture since the single band in
gel represents one bacterial species
23 16S rDNA clone library construction method
16S rDNA-DGGE fingerprinting has proven to be particularly useful as an initial
investigation into bacterial communities and is suitable for identifying the predominant
bacteria that comprise 1 or more of cells within a given sample In the 16s rDNA clone
17
library construction process the first step is the extratction of the total genomic DNA
then the genomic DNA of the mixed culture biofilm was used as template for PCR
amplification of approximately 1500 bp of 16S rDNA with universal primers The PCR
products of 16S rDNA were purified and inverted into pGEM-T Easy vector system
before they were transformed into competent E coli The transformed cells were spread
on Luria-Bertani (LB) plates containing the selective antibiotic ampicillin and X-Gal and
incubated overnight at 37 OC after the more than 80 inverted 16s DNA fragment E coli
colonies were picked up and inoculated in the 1-3 ml liquid LB medium and left to grow
for 16 hours The plasmid DNA were extracted and sequenced The sequences were
compared directly to all known sequences deposited in GenBank databases using the
basic local alignment search tool (BLAST)
24 Bioinformatics Analysis
16S rDNA sequencing results of more than 80 colones were queried against the
GenBank and Ribosomal Database Project (RDP) databases using BLAST and
SEQUATCH algorithms (Cole et al 2007 Wang et al 2007 Cole et al 2009) The
neighbor-joining trees were constructed with the Molecular Evolutionary Genetics
Analysis package (MEGA Version 4) based on 1000 bootstrap analysis (Tamura et al
2007)
25 Dominant bacterial species band in DGGE identification
The DGGE results gave the information of dominant bacterial richness of bacterial
species in mixed culture based on the theory that single band represents one bacterial
species which showed the diversity of bacterial community in the mixed culture of a
MFC 16S rDNA clone library results provided the whole picture of bacterial community
18
including the bacterial species name and bacterial species percentage in the mixed culture
The known pure bacterial species which have been sequenced can be used as markers to
identify dominant bacterial species in DGGE The whole experimental procedure is
similar to the procedure of the mixed culture DGGE steps The first step is to extract the
collect the mixed culture biofilm genomic DNA and pure bacterial species marker
genomic DNA Then the universal primer set was used to amplify the 16S rDNA from
the extracted genomic DNA After that the V3 region of bacterial 16S ribosomal DNA
(rDNA) was amplified with GC-clamp primer (Muyzer et al 1993) for DGGE The
subsequent steps were similar to the mixed culture DGGE experimental procedure The
dominant bacterial bands in DGGE were determined by comparisons of the single band
position in the mixed culture DGGE with single pure bacterial species location in DGGE
26 Dominant bacterial species isolation and power generation
The bacterial community of mixed culture in MFCs were predominantly composed
of Geobacter sp Arcobacter sp and Citrobacter sp Two dominant bacterial species
have been isolated
Citrobacter sp SX-1 was isolated from the anode biofilm by using a solid agar
medium containing NH4Cl 6mM KCl 2mM CH3COONa (electron donor) 30 mM
FeC6H5O7 (electron acceptor) 20mM minerals and vitamins (Lovley and Phillips 1988)
Single colonies were picked up after culturing on the plate for 36 hours at 30 degC and
transferred two times on the agar plate for purification Then the isolates grew in a liquid
medium solution in anaerobic tubes containing the same constituents as the solid
medium All isolation process was operated in a glove box anaerobic chamber (Coy
Laboratory Products Grass Lake MI)
19
Arcobacter sp SX-2 was isolated from the anode biofilm of a MFC fed with sodium
acetate by using the same method of isolation of pure bacterial species Citrobacter XS-1
except using Trypticase Soy Agar with 5 Sheep Blood solid (from American type
culture collection (ATCC)) agar medium instead of Citrobacter sp medium Single
colonies were picked up after culturing on the plate for 36 hours at 30 degC and transferred
twice on the agar plate for purification Then the isolates grew in a liquid medium
solution microaerobically
Geobacter sp have been trying to isolate by using three different methods They are
the colony pickup after different condition enrichment Goebacter medium isolation and
dilution to extinction by multiple channel mini MFCs isolation however the Geobacter
sp has not been isolated yet due to some limits so far But they provide valuable
information for further isolation of Geobacter sp
Single chamber MFCs were used to evaluate power generation by different isolates
The MFCs were constructed as described previously (Liu and Logan 2004) and modified
with 3 cm2 carbon cloth anodes and 7 cm
2 carbon clothPt cathodes The total liquid
volume of each MFCs was about 15 ml with electrode spacing about 17 cm All MFCs
were operated in an autoclaved closed plastic box and sterile cotton was attached to the
outer surface of the air cathodes to prevent contamination A MFCs without bacterial
culture was used as control MFCs were inoculated with 3 ml late exponential phase
cultures of SX-1 in the medium solution reported previously (Liu and Logan 2004)
3 Results and discussion
31 Power production by mixed culture MFCs
20
A polarization curve was used to characterize current as a function of voltage in the
MFCs The polarization curves are performed by varying a series of external resistances
The results are shown in Figure 21 With MFCs acclimated to 25 Ω external resistance
the maximum power density was achieved at 65Wm2 based on the polarization data At
this point the current density is 21mAcm2 The power density was three times higher
than previous report (20 Wm2) using phosphate buffer (Fan et al 2007) Power density
was also 2 times higher than (28 m Wm2) using a bicarbonate buffer ( Fan et al 2007)
32 Bacterial community structure assessed by PCR-DGGE
Bacterial DGGE profiles for higher power generating MFCs mixed culture setups
were relatively complex (more than 10 bands) However there are three brightly stained
bands and other less intense bands (Fig 2-2) These results indicated there were probably
three dominant bacterial species in mixed culture MFCs
010
020
030
040
050
060
070
2
3
4
5
6
7
04 05 05 06 07 08 10 11 13 15 21 22 23 24
Volta
ge (
V)
Pow
er
densi
ty (
Wm
2)
Current density (mAcm2)
Power
Voltage
power density 65wm2 at current
density208 mAcm2
Figure 2-1 Polarization curves of high power mixed culture MFCs
21
33 16S rDNA clone library results
A total of 83 colonies were sequenced for 16S rDNA clone library construction They
represented the bacterial community structure in mixed culture in MFCs The results
revealed that the predominant bacterial species in mixed culture are Geobacter sp (66
of the mixed culture) Arcobacter sp (12 of the mixed culture) and Citrobacter sp (11
of the mixed culture) Those three genuses comprised 88 of the total bacterial species
(Fig 23) This result is consistent with the DGGE result which showed that three bright
bands in the DGGE gel represented three dominant bacterial species in the mixed culture
The phylogenetic analysis of the colonies (83 colony sequences) is shown in Figure 24
Figure 22 DGGE result of high power generation mixed culture MFC (1 mixed
culture 2 pure culture control)
1 2
22
34 Dominant bacterial band in DGGE identification
Geobacter
66
Arcobacter
12
others
6
Pseudomonas
1
Citrobacter
11
Clostridium
2
Anaerovorax
2 Geobacter
Arcobacter
Citrobacter
Clostridium
Anaerovorax
Pseudomonas
others
Figure 23 Bacterial species and percentage of bacterial community of mixed
culture MFC lt 1 Alcaligensgt
lt1 Commamonasgt
lt1 Pseudomonas aeruginosagt
lt1 Klebsiellagt
lt11 Citrobactergt
lt1 Wolinellagt
lt12 Arobactergt
lt2 Anaeroboraxgt
lt2 Clostridium gt
lt1 unclear bacteriagt
lt66Geobactergt
100
100
85
100
100
82
99
87
100
002
Figure 24 Phylogenetic trees of bacterial species from mixed culture MFC
23
The dominant bacterial species including Goebacter sp Citrobacter sp and Arcobacter
sp in the mixed culture are shown in Figure 25 The upper band is Arcobacter sp and
Citrobacter sp is located in the bottom band of DGGE Geobacter sp is the middle one
The analysis of the bacterial community using denaturing gradient gel electrophoresis
(DGGE) of PCR-amplified 16S rRNA gene fragments and 16S rDNA clone library
construction method showed great phylogenetic diversity of mixed culture in MFCs with
the identification of sequences derived from bacteria of the taxa deltaproteobacteria
(Geobacter sp) gammaproteobacteria (Citrobacter sp) and epsilonproteobacteria
(Arcobacter sp) The result is consistent with the conclusion that among the isolated
exoelectrogens gammaproteobacteria and deltaprotebactia appear to be dominant among
others (Parot et al 2009 Liu et al 2010)
35 The dominant bacterial species isolation and power generation
Figure 25 Dominant bacterial species identification in DGGE
A B C D
A Citrobacter sp
B Geobacter sp
C Arobacter butzmeri
D mixed culture control
24
Two dominant bacterial species Citrobacter sp XS-1 and Arcobacter sp XS-2 have been
isolated and tested for power generation The power densities generated by the two
isolates are shown in the Fig 26 Current densities generated by isolate Citrobacter sp
and Arcobacter sp were 98 mAm2 and 20 mAm
2 respectively which were much less
than that (21Am2) generated by the mixed culture from which they were isolated
Figure 26 Current density generated by different dominant isolates
The maximum current density generated by mixed culture is much higher than the
one produced by pure bacterial species Citrobacter sp XS-1 and Arcobacter sp XS-2 in
the same structure MFCs Although the Geobacter sp in our mixed culture has not been
isolated a previous study showed that maximum current density generated by Geobacter
sp was 080 Am2
(Bond and Lovley 2003) The current density results seem partially
supporting the conclusion that mixed-culture generated higher power densities than their
pure-culture counterparts in MFCs (Jung and Regan 2007) This result suggests there
may be synergistic interactions within the anode bacterial communities
0
500
1000
1500
2000
2500
Mixed culture Citrobacter sp Arcobacter Sp
Cu
rre
nt
de
nsi
ty (
mA
m2)
25
Geobacter sp are strictly anaerobic bacteria which were enriched in most of the anode
biofilms (George 2005) Citrobacter species are facultative anaerobic bacteria (George
2005) which can be found in a wide variety of habitats including in soil water and
wastewater Arcobacter sp are micro-aerobic bacteria including both environmental
nonpathogens and opportunistic human pathogens They are able to grow in aerobic
conditions but in the optimal growth under micro-aerobic conditions (George 2005)
Arcobacter sp has been discovered in the MFCs by Fedorovich (Fedorovich et al 2009)
It is interesting that the mixed culture bacterial community composed of primarily these
three bacterial species has demonstrated to generate high power at non-strictly anaerobic
condition while leading dominant bacterial species Geobacter sp (66) is strict
anaerobic bacteria and generated powder in the MFCs only under strict anaerobic
condition (George 2005) Therefore it is possible that microaerobic bacteria such as
Arcobacter sp and facultative anaerobic bacteria Citrobacter sp can facilitate to create
anaerobic condition for Geobacter sp when they utilized oxygen for growing in the
mixed culture Syntrophic communities study have showed that anaerobic bacteria and
methanogenic archaea form compact microbial structures that operate like an organ rather
than a set of microorganisms functioning independently (Stams and Plugge 2009) Some
substrates have been degradeted within these communities while they are not able to be
fermented by individual species alone interspecies electron transfer also have been
presented in these communities (Stams and Plugge 2009) Summers et al (2010) also
discovered that direct exchange of electrons happened within coculture of Geobacter
metallireducens and Geobacter sulfurreducens Therefore we believe that there are
26
more complex synergistic interactions between different bacteria species in mixed culture
in the MFCs
Acknowledgements
The authors acknowledge support from the US National Science Foundation
(CBET 0955124) We also thank Keaton Lesnik for the review of this manuscript
27
Chapter 3
New Exoelectrogen Citrobacter sp SX-1 Isolated from a Microbial Fuel Cell
Shoutao Xu and Hong Liu
Published in
Journal of Applied Microbiology 111(5)1108-1115 (2011)
ABSTRACT
Exoelectrogenic bacterial strain SX-1 was isolated from a mediator-less microbial fuel
cell by conventional plating techniques with ferric citrate as electron acceptor under
anaerobic condition Phylogenetic analysis of the 16S rDNA sequence revealed that it
28
was related to the members of Citrobacter genus with Citrobacter sp sdy-48 being the
most closely related species The bacterial strain SX-1 produced electricity from citrate
acetate glucose sucrose glycerol and lactose in MFCs with the highest current density
of 205 mA m-2
generated from citrate Cyclic voltammetry analysis indicated that
membrane associated proteins may play an important role in facilitating the electrons
transferring from bacteria to electrode This is the first study that demonstrates that
Citrobacter species can transfer electrons to extracellular electron acceptors Citrobacter
strain SX-1 is capable of generating electricity from a wide range of substrates in MFCs
This finding increases the known diversity of power generating exoelectrogens and
provided a new strain to explore the mechanisms of extracellular electron transfer from
bacteria to electrode The wide range of substrate utilization by SX-1 increases the
application potential of MFCs in renewable energy generation and waste treatment
Keywords Microbial fuel cell Exoelectrogen Citrobacter sp SX-1 Extracellular
electron transfer
29
Introduction
Microbial Fuel Cells (MFCs) technology has become an active research area
recently as a promising approach for renewable energy generation wastewater treatment
and bioremediation (Rabaey and Verstraete 2005 Lovley 2006 Fan et al 2008 Logan
2009 Liu et al 2010) The key feature of MFCs system is the microbe-catalyzed electron
transfer from organic matter to anodes Many studies have shown the presence of diverse
bacterial communities on MFCs anodes (Reimers et al 2001 Liu et al 2004b Kim et al
2005) Over 20 exoelectrogens ie microbes that can transfer electrons exocellularly to
electrodes have been reported in the past 10 years (Logan 2009 Liu et al 2010) The
exoelectrogens so far belong to diverse genetic groups including alphaproteobacteria
(Rhodopseudomonas Ochrobactrum and Acidiphilium) (Malki et al 2008 Xing et al
2008 Zuo et al 2008) betaproteobacteria (Rhodoferax) (Chaudhuri et al 2003)
gammaproteobacteria (Shewanella Pseudomonas Klebsiella Enterobacter and
Aeronomas) (Kim et al 2002 Pham et al 2003 Rabaey et al 2005 Bretschger et al
2007 Zhang et al 2008 Chung et al 2009 Rezaei et al 2009) deltaproteobacteria
(Geobacter Geopsychrobacter Desulfuromonas and Desulfobulbus) (Bond et al 2002
Bond and Lovley 2003 Holmes et al 2004ab 2006) Epsilonproteobacteria (Arcobacter)
(Fedorovich et al 2009) Firmicutes (Clostridium and Thermincola) (Park et al 2001
Wrighton et al 2008) Acidobacteria (Geothrix) (Bond and Lovley 2005) and
actinobacteria (Propionibacterium) (Wang et al 2008) A wider range of
exoelectrogenic species are expected to be discovered
Three mechanisms have been proposed for exocellular transport of electrons by
exoelectrogens without artificial mediators Some exoelectrogenic bacteria can transfer
30
electrons to electrodes through soluble redox compounds excreted by microorganisms
(Bond and Lovley 2005 Rabaey et al 2005) and others can directly transfer electrons to
anodes via outer cell membrane proteins (Myers and Myers 1992) Recently more and
more evidence supports the involvement of bacterial nanowires in extracellular electron
transport (Reguera et al 2005 2006 Gorby et al 2006) In spite of the discovery of
many bacterial species that can transfer the electrons to electrode without the need of
artificial mediators the investigation of extracellular electron transfer mechanisms was
mainly focused on a few species such as those from Geobacter and Shewanella genera
(Kim et al 1999 2002 Reguera et al 2005 2006 Gorby et al 2006) The electron
transfer mechanisms for many of the isolated exoelectrogens species are still not well
studied
In the present study we isolated a new exoelectrogen strain SX-1 from a MFC a
strain phylogenetically related to Citrobacter sp Power generation from various carbon
sources by this strain was evaluated using single chamber MFCs Plausible extracellular
electron transfer mechanisms were also discussed based on the characterization of anodic
biofilms by cyclic voltammetry (CV)
Materials and methods
Bacterial strain SX-1 isolation
Bacterial strain SX-1 was isolated from the anode biofilm of a MFC fed with sodium
acetate operated in fed-batch mode over a period of six months The original source of
the inoculum is wastewater from a local waste water treatment plant Bacterial cells were
released from the carbon cloth anode by shaking the cloth in a sterile flask with 15 mL
sterile 1 NaCl solution and glass beads (2 mm diameter) The suspension was then
31
serially diluted from 10 times to 105
times and plated on a petri dish with a solid agar
medium containing NH4Cl 6mM KCl 2mM CH3COONa (electron donor) 30 mM
FeC6H5O7 (electron acceptor) 20mM minerals and vitamins (Lovley and Phillips 1988)
Single colony was randomly selected after culturing on the plate for 36 hours at 30 degC
and purified on a new agar plate following a procedure reported previously (Chung and
Okabe 2009) Then the isolate was picked up and grew in a liquid medium solution in
anaerobic tubes containing the same constituents as the solid medium for further analysis
All isolation process was operated in a glove box anaerobic chamber (Coy Laboratory
Products Grass Lake MI)
16S rDNA sequencing and phylogenetic analysis
Bacterial genomic DNA was extracted from the isolated strain SX-1 using the DNeasy
tissue Kits (Qiagen Valencia CA) according to the manufacturerrsquos instructions The 16S
rDNA was amplified by PCR using a pair of universal primers 27F (5rsquo-
AGAGTTTGATCMTGGCTCAG-3rsquo) and 1492R (5rsquo-GGTTACCTTTGTTACGACTT-
3rsquo) (Park et al 2001) The conditions used for PCR were described previously
(Ferna ndez et al 1999) PCR products were purified ligated and cloned according to a
procedure described in a previous report (Xing et al 2008) Plasmids were isolated from
randomly selected clone colonies with a QIAprep Spin Miniprep kit (Qiagen Valencia
CA) and three plasmid inserts were then sequenced in both directions using an ABI 3730
DNA sequencer (Applied Biosystems Carlsbad CA)United States) Due to the identical
16S rDNA sequence of the three colones only one was queried against the GenBank and
Ribosomal Database Project (RDP) databases using BLAST and SEQUATCH algorithms
(Cole et al 2007 Wang et al 2007 Cole et al 2009) The 16S rDNA sequence of
32
strain SX-1 and closely related type strains were aligned using CLUSTALX software
(Thompson et al 1997) The neighbor-joining trees were constructed with the Molecular
Evolutionary Genetics Analysis package (MEGA Version 4) based on 1000 bootstrap
analysis (Tamura et al 2007)
SEM
The morphologies of SX-1 cells growing overnight in anaerobic tubes were examined
with a scanning electron microscope (FEI Quanta 600 FEG FEI Company Hillsboro
OR) The suspended cells were fixed in a 25 glutaraldehyde and 01M phosphate
buffer solution and dehydrated with a graded ethanol series from 30 to 100 After
dehydration the samples were dried in a critical point dryer and then sputter-coated with
AuPd for SEM examination (Liu and Logan 2004)
Evaluation of power generation by SX-1 in MFCs
Single chamber MFCs were used to evaluate power generation by SX-1 using various
substrates The MFCs were constructed as described previously (Liu and Logan 2004)
and modified with 3 cm2 carbon cloth anodes and 7cm
2 carbon clothPt cathodes The
total liquid volume of each MFCs was about 15 ml with electrode spacing about 17 cm
All MFCs were operated in an autoclaved closed plastic box and sterile cotton was
attached to the outer surface of the air cathodes to prevent contamination A MFC
without bacterial culture was used as control MFCs were inoculated with 3 ml late
exponential phase cultures of SX-1 in the medium solution reported previously (Liu and
Logan 2004) Seven substrates including citrate acetate glucose sucrose glycerol and
lactose were evaluated individually for power generation in a fed-batch mode in a
temperature control chamber (30 plusmn2degC) For each substrate at least 8 batches were run to
33
investigate the effect of biofilm formation on current output at fixed external resistance of
1kΩ and the same initial substrate concentration of 30 mM Phosphate buffer (100mM)
was used to maintain the solution pH and conductivity The MFCs with sodium citrate as
electron donors were also examined for maximum power output by varying the external
resistance from 20 kΩ to 4 kΩ A data acquisition system was used to record voltage data
during the MFCs operation (Keithley 2700 Keithley Instruments Inc Cleveland OH) It
took about 15 to 30 minutes for the MFCs to stabilize depending on the external
resistance At each resistance we collected at least five data at the steady condition to
make the polarization curves Averaged voltages were used to calculate the power density
(mW m-2
) according to P=IVA where I was the current V was voltage and A was cross-
sectional area of the anode
CV analysis
CV was used to characterize the oxidation and reduction reactions on the anodic surface
of the MFCs fed with citrate and operated at 7 KΩ This external resistance was selected
because the maximum power density was obtained at this resistance based on the
polarization experiment We assumed the biofilm on the anode of MFCs were well-
developed when stable power output was obtained after 3 batches of operation The
MFCs were then used directly for CV analysis at four current generating stages of the
fourth batch (1) initial exponential current increasing stage (middle point of stage about
4 h after media change) (2) current plateau stage (middle point of stage about 8 h after
media change) and (3) current decreasing stage (middle point of stage about 16h after
media change) (4) right after the complete replacement of medium solution (Figure 6)
The anode was used as working electrode the cathode as counter electrode and an
34
AgAgCl electrode was selected as the reference The CV curves were scanned from 200
to -600 mV at a rate of 5 mV s-1
using a potentiostat (G300 Gamry Instrument
IncWarminster PA) Control experiment was also conducted using new anode (without
biofilm) and new medium solution
Nucleotide sequence accession number
The 16S r DNA sequence determined in this study has been deposited in the GenBank
database under accession number HQ845373
Results
Identification of the strain SX-1
An almost-complete 16S rDNA sequence (1475 bp) of strain SX-1 was obtained and
subjected to comparative analysis with the 16S rDNA of closely related reference strains
A phylogenetic analysis revealed that strain SX-1 should be assigned to the genus
Citrobacter and was most closely related to Citrobacter sp sdy-48 EJ463782 (990
sequence similarity) and Citrobacter sp yy-21 FJ463779 (975 sequence similarity)
These three strains formed a distinct sub cluster in the neighbor-joining in which the new
isolate and Citrobacter sp sdy-48 EJ463782 formed a distinct subline (Figure 31)
The SEM image illustrated that the strain SX-1 is a rod-shaped bacterium 03-06 microm
wide and 12-17 microm long (Figure 32) Strain SX-1 colonies on nutrient agar appeared 2-
3 mm in diameter smooth moist opaque with a shiny surface after 5 days incubation
under anaerobic condition The fact that strain SX-1 grew both aerobically and
anaerobically using FeC6H5O7 as electron acceptor suggests that the strain is facultatively
anaerobic Strain SX-1 showed robust growth on citrate as a carbon source These D
35
properties were similar to those of genus Citrobacter described in Bergeyrsquos Manual of
Systematic Bacteriology (George 2005)
Figure 31 phylogenetic tree of strain SX-1 and closely related species based on 16S
rDNA sequences The tree was constructed using the neighbor-joining method
Figure 32 SEM image of planktonic cells of Citrobacter sp SX-1
Strain SX-1
Citrobacter sp sdy-48 (FJ463782)
Citrobacter sp yy-21 (FJ463779)
Citrobacter koseri E639 (ATCC 25408)
Citrobater koseri CDC 3613-63(AF025372)
Citrobacter koseri CDC 8132-86(AF025366)
Salmonella enterica SL483 (CP001138)
Salmonella enterica AKU12601(AY696668)
Shigella sonnei Ss046(CP000038)
E coli C2 (AF403733)
E coli E24377A(CP000800)88
100
100
76
44
92
53
36
0001
1 microm
36
Electricity production by strain SX-1 in MFCs
The isolated strain SX-1 was first tested for its ability to generate current from sodium
citrate at 1kΩ Current gradually increased to a maximum 50 mA m-2
after cell
inoculation and then decreased (Figure 33) After the MFCs was refilled with new
substrate solution the current recovered rapidly and reached a higher level than the first
batch After 4 batches operation the maximum current output of each batch became
stable The highest current density achieved in the MFCs with Citrobacter sp SX-1 at 1
KΩ was 98 mA m-2
(Figure 33)
Figure 33 Electricity generation by Citrobacter sp SX-1 in a single chamber MFCs
with sodium citrate (30 mM) as substrate at 1 KΩ
0
20
40
60
80
100
120
0 50 100 150 200
Curr
ent
den
sity
(m
A m
-2)
Time (h)
37
Current generation by SX-1 from other substrates including glucose lactose sodium
acetate glycerol and sucrose were also assessed at an external resistance of 1KΩ
(Figure 34) When repeatable cycles of current output were obtained for these substrates
glycerol generated the highest maximum current density of 58 mA m-2
followed by
lactose and sucrose with 29 mA m-2
and 27 mA m-2
respectively Glucose and acetate
produced the lowest maximum current density of 96 and 43 mA m-2
respectively These
results indicated that strain SX-1 can utilize a wide range of substrates for electricity
generation in MFCs but with different power generation potentials
Figure 34 Electricity generation of Citrobacter sp SX-1 using different substrates in
single chamber air-cathode MFCs at 1 KΩ (The error bars stand for the standard
deviation of 3 replicates)
0
10
20
30
40
50
60
70
Glucose Lactose Glycerol Acetate Sucrose
Curr
ent
den
sity
(m
A m
-2)
38
Polarization experiment was further conducted to determine the maximum power
density generated by strain SX-1 in the single chamber MFCs with citrate as substrate A
maximum power density of 881 mW m-2
was obtained at current density of 205 mA m-2
at an external resistance of 7 KΩ (Figure 35)
Figure 35 Power and voltage generation by Citrobacter sp SX-1 as a function of
current density using sodium citrate (30 mM) as substrate The error bars stand for the
standard deviation of the 3 voltagepower outputs obtained in three MFCs
20
30
40
50
60
70
80
90
100
01
02
03
04
05
06
07
80 110 140 170 200 230
Po
wer
den
sity
(m
W m
-2)
Vo
ltag
e (V
)
Current density (mA m-2)
Voltage
Power
39
Figure 36 Four current generating stages for CV analysis current increasing stage (stage
1) current plateau stage (stage 2) current decreasing stage (stage 3) and right after the
replacement of medium solution (stage 4) The MFCs was operated at 7 KΩ with sodium
citrate (30 mM) as substrate
Cyclic Voltammetry
To determine the presence of redox active compounds produced by SX-1 and the
location of these compounds CV scan of the anodic biofilms at four current output stages
and the supernatant of MFCs medium solution at the end of the batch experiment were
performed Reduction peaks at range of -310 ndash -350 mV and oxidation peaks at range -
100 ndash -150 mV were observed at all current generating stages (Figure 36 Figure 37A)
suggesting the presence of redox active compounds may involve in extracellular electron
transfer by Citrobacter sp SX-1 The amplitude of the peaks increased according to the
growth stage of the batch and the highest peaks were present after the current plateau
stage which indicated the redox active compounds mainly were secreted in the current
plateau stage While the current density at stage 3 (deceasing stage) was lower than that
at stage 2 (current plateau stage) the redox peak at stage 3 was higher than that in stage 2
0
50
100
150
200
250
50 55 60 65 70 75 80 85
Curr
ent
den
sity
(m
A m
-2)
Time (h)
A
A
Stage 1
Stage 2 Stage 3
Stage 4
40
A plausible explanation for this pattern was that redox active compounds were
continuously being secreted and accumulated after the current peak was reached
resulting in more redox compounds present after the plateau stage But since most of the
carbon source had already been being used up many of the redox compounds at this
stage had not been as active as those at current plateau stage due to much less electrons
were available to be transferred The bigger peak amplitude in stage 4 than in stage 1
indicates that the redox active compounds were continuously being secreted and
accumulated in the biofilm even after stable power was generated at 7 kΩ The slightly
change of the peak locations at different stages was possible due to the solution chemistry
change during the current generation process or the slightly location change of reference
electrode during the measurement (Figure 37B) When the medium solution in the MFCs
was replaced by fresh medium at the end of the batch the oxidization and reduction
peaks were still presented in the CV (Figure 37B) but no peaks were observed when the
supernatant and fresh mediums were tested in a MFCs with no biofilm (Figure 37C)
These results suggest the compounds involved in the electron transfer were located not in
the supernatant (solution) but in the biofilm
41
Figure 37 Cyclic Voltammetry analysis of Citrobacter sp SX-1 biofilms at (A) current
increasing stage current plateau stage and decreasing stage (end of the batch) (B) end of
the batch and replaced with fresh medium and (C) controls
-40E-04
-30E-04
-20E-04
-10E-04
00E+00
10E-04
20E-04
30E-04
-060 -050 -040 -030 -020 -010 000 010 020
Curr
ent
(A)
Potential (V) vs AgAgCl
Current plateau stage
Current increasing stage
Current decreasing stage
-40E-04
-30E-04
-20E-04
-10E-04
00E+00
10E-04
20E-04
30E-04
-060 -050 -040 -030 -020 -010 000 010 020
Curr
ent
(A)
Potential (V) vs AgAgCl
Current decreasing stage
Biofilm with fresh medium
-40E-04
-35E-04
-30E-04
-25E-04
-20E-04
-15E-04
-10E-04
-50E-05
00E+00
-060 -050 -040 -030 -020 -010 000 010 020
Curr
ent(
A)
Potential (V) vs AgAgCl
Supernatant
Fresh medium
A
B
C
B C
42
Discussion
Citrobacter species belonging to Gammaproteobacteria Enterobacteriales
Enterobacteriaceae are facultative anaerobic bacteria (George 2005) They can grow with
oxygen as an electron acceptor but also with a variety of electron accepters such as Fe (III)
in the absence of oxygen (George 2005) Citrobacter species can be found in a wide
variety of habitats including in soil water and wastewater Several Citrobacter species
have been studied as important bioremediation bacterium for heavy metal removals
sulfate reduction phenol degradation and chlorophenol degradation (Macaskie et al 1995
Narde et al 2004 Qiu et al 2009) Citrobacter species have also been found on the
anodic biofilm of a diesel-degrading MFCs (Morris et al 2009) However there was no
study reported that Citrobacter species can transfer electrons to extracellular electron
acceptors This finding that Citrobacter strain SX-1 can generate electricity in MFCs
increased the diversity of power generating exoelectrogens and provided a new strain to
explore the mechanisms of extracellular electron transfer from bacteria to electrode
Among the isolated exoelectrogens gammaproteobacteria and deltaprotebactia appear to
be dominant among others (Parot et al 2009 Liu et al 2010) The isolated strain
Citrobacter sp SX-1 phylogenetically belongs to gammaproteobacteria which provides
another evidence to support this observation
Most exoelectrogens utilize a limited range of substrates for power generation For
example Shewanella oniedensis MR-1 can oxidize lactate under anaerobic condition but
cannot utilize acetate for electricity generation (Kim et al 2002) Geobacter
sulfurreducens can completely oxidize acetate for power generation but it cannot utilize
simple sugars (Bond and Lovley 2003 Caccavo et al 1994)) In comparison strain SX-1
43
not only can oxidize citrate for power generation but also utilize a wide range of
substrates for power generation including glycerol glucose lactose sucrose and acetate
Interestingly the higher current density generated by strain SX-1 from glycerol a main
by-product of biodiesel production process suggests that strain SX-1 may be potentially
used for harvesting energy from biodiesel wastes using MFCs However the current
density generated by strain SX-1 (205 mA m-2
) is lower than that (805 mA m-2
at 1 kΩ)
generated by the mixed culture from which the SX-1 was isolated indicating the
existence of other higher power generating bacteria andor complex ecology in the mixed
culture community
Understanding mechanisms of microbial extracellular electron transfer is critical for
enhancing the electron transfer rate from bacteria to electrode through metabolic or
genetic engineering of exoelectrogens Since the spent supernatant solution of MFCs run
with SX-1 showed no redox properties the redox compounds produced by SX-1 may
have been retained in the biofilm Alternatively SX-1 may utilize one or both of the
other two known extracellular electron transfer mechanisms For example it is possible
that strain SX-1 transfers electrons to the anode via cell-bound outer membrane proteins
The two distinct oxidation and reduction peaks observed at -100mv- -150 mV and -310--
350 mV of the SX-1 biofilm have a midpoint potential range from -205mV to -250mV
which is close to the redox potential of some c-type cytochromes (Field et al 2000 Meitl
et al 2009) which are well-known to play an important role in extracellular electron
trasnfer (Eggleston et al 2008 Field et al 2000 Meitl et al 2009) Field et al (2000)
reported that the c-type cytochromes CymA had the midpoint potentials of -229 mV
Meitl et al (2009) and Eggleston et al (2008) reported that the c- type cytochromes
44
OmcA in Shewanella oneidensis MR-1 had the midpoint potentials of -201 mV and -208
mV
While itrsquos possible that SX-1 may also have conductive appendages to enhance the
extracellular transfer electron similar to the nanowires discovered in some Geobacter
and Shewanella species (Reguera et al 2005 2006 Gorby et al 2006) further
investigation on the presence of these appendages on the cell surface of SX-1 is needed
Acknowledgements
The authors acknowledge support from the US National Science Foundation
(CBET 0955124) We also thank Teresa Sawyer for her helps with the SEM analysis and
Jeremy Chignell and Yanzhen Fan for their review of this manuscript
45
Chapter 4
Enhanced Performance and Mechanism Study of Microbial Electrolysis Cells
Using Fe Nanoparticles Decorated Anodes
Shoutao Xu Hong LiuYanzhen Fan Rebecca Schaller Jun Jiao Frank Chaplen
Published in
Applied Microbiology and biotechnology 93(2)871-880 (2012)
46
ABSTRACT
Anode properties are critical for performance of microbial electrolysis cells (MECs) In
the present study Fe nanoparticle modified graphite disks were used as anodes to
investigate the effects of nanoparticles on the performance of Shewanella oneidensis MR-
1 in MECs Results demonstrated that average current densities produced with Fe
nanoparticle decorated anodes were up to 56-fold higher than plain graphite anodes
Whole genome microarray analysis of the gene expression showed that genes encoding
biofilm formation were significantly up-regulated as response to nanoparticle decorated
anodes Increased expression of genes related to nanowires flavins and c-type
cytochromes indicate that enhanced mechanisms of electron transfer to the anode may
also have contributed to the observed increases in current density The majority of the
remaining differentially expressed genes were associated with electron transport and
anaerobic metabolism demonstrating a systemic response to increased power loads
Keywords Microbial electrochemical system microbial fuel cell microbial electrolysis
cell nanotechnology differential gene expression DNA microarray
47
Introduction
Microbial electrochemical systems (MESs) have been intensively studied since Lewis
achieved practical advances in this field (Logan 2007) however they attracted much
research attention in recent years due to their promising applications in renewable energy
generation bioremediation and wastewater treatment In a MES microorganisms
interact with electrodes via electrons catalyzing oxidation and reduction reactions at the
anode and the cathode The most-described type of MESs is microbial fuel cells (MFCs)
in which useful power is generated from electron donors typically biodegradable organic
materials (Logan et al 2006) Various novel MESs have recently been developed to
produce hydrogen (microbial electrolysis cells (MECs)) (Liu et al 2005 Logan et al
2008 Rozendal et al 2006 Schaller et al 2009) to power an autonomous sensor or
sensor network (Ringeisen et al 2006) to reduce bioremediation targets (Lovley 2006
Shea et al 2008) Sukkasem et al 2008) and to desalinate water (Cao et al 2009) The
key feature shared by these systems is the microbe-catalyzed electron transfer from
organic matter to electrodes (anodes) (Rabaey et al 2004) Enhancing the anodic current
output which highly depends on the performance of the electrodes is critical for the
successful application of all these processes (Logan et al 2007 Park and Zeilus 2002
2003)
Nanomaterials have received much attention from researchers in the context of
microbiology due to their unique physical electrical and chemical properties which
facilitate the study of interactions between bacteria and surfaces (Liu 2006) Previous
studies have demonstrated that electrodes decorated with different nanostructures such
as carbon nanotubes nanostructured polyanilinetitanium dioxide composite and titania
48
nanotubes have the potential to enhance power generation in MFCs (Morozan et al 2007
Qiao et al 2008 Quan et al 2005) Furthermore our recent study shows nanoparticle
(NP) decorated anodes greatly increased the electrochemical electron transfer rate in
MFCs (Fan et al 2011) However Fe NPs as an ideal candidate for decorating electrodes
because of respectively low price and high conductivity compared to other materials has
not been focused to study in MECs
S oneidensis MR-1 an important electrochemically active bacterial strain has been
exploited for the electricity generation in MFCs (Logan et al 2007 Logan 2009 Park
and Zeikus 2002) The availability of genome sequence for this strain makes it possible to
use transcriptome assays to globally measure the responses to different growth conditions
and environmental stresses (Murray et al 2001) S oneidensis MR-1 gene expression
response to heat shock cold shock Cr (VI) and U (VI) reduction chromate stress and
iron and acid tolerance have been studied in the past several years (Bencheikh-Latmani et
al 2005 Brown et al 2006 Gao et al2006 Gao et al 2004 Yang et al 2008) However
no studies have been focused on the Shewanella gene expression response to NPs in
MESs
The mechanism of increased electron transfer rate exhibited by nanoparticle decorated
electrodes is not well understood yet In the present study graphite disks decorated with
Fe NPs were used as anodes to explore the effects of nanostructures on current generation
in a multi-anode MECs DNA microarrays were utilized to investigate differences in the
global gene expression profile of S oneidensis MR-1 grown on plain versus Fe NPs
decorated anodes
Materials and methods
49
Bacterial cultures
S oneidensis MR-1 was purchased from American Type Culture collection (ATCC
700550) and stored as freezer stocks at -80degC Stock cells of S oneidensis MR-1 was
grown aerobically using a medium containing 30 gL Trypticase Soy Broth (BD 211825
Becton Dickinson Company Franklin lakes NJ) at 30degC Cells in early stationary
growth phase (optical density of 15-16 at 600nm (Spectrophotometer UV-1700
Shimadzu Columbia MD) were washed two times then injected into the chamber of the
MECs for current production Trypticase Soy Broth medium with 30 mM sodium lactate
as electron donor was used in MECs Phosphate buffer (100mM) was used to maintain
the solution pH 7 and solution conductivity at 15 mScm
Characterization of nanostructured anodes
Superfine isomolded graphite disks (16 cm x 05 cm GraphiteStorecom Inc) were
polished with ultrafine sand paper (2000 grit 3M Company) as the base for the NP
decorated and control anodes Fe NPs decorated anodes were fabricated by thermal
annealing of metal thin film coated graphite disks Thin films of Fe were firstly deposited
on the polished graphite disk by using sputter coating for 95 min and the samples then
were annealed in the Chemical vapor deposition (CVD) at 750degC for 20 min to form Fe
NPs Fe-NPs were produced with a diameter of 200plusmn100 nm and a density range
(distance between NPs) of 200plusmn100 nm with a coating time of 95 min SEM images of
Fe NPs decorated anode and control surfaces were shown in the Figure 41
50
Figure 41 SEM images of Fe NPs decorated anodes (A) Graphite disk control (B) Fe
Nanoparticle decorated
Multiple channel MECs construction and operation
A MECs with removable multiple anodes with each effective anode area 07 cm2
was
constructed and used to evaluate the effects of nanostructure on current density of
according to a previous report (Fan et al 2011) The cathode was made of wet-proof
(30) carbon cloth (type B E-TEK Division Inc USA) coated with PtC (20 E-TEK
Division Inc USA) following a previously reported procedure (Liu et al 2005) The
final platinum loading was 05 mgcm2 per projected cathode area The size of cathode
(150 cm2) was 25 times larger than the total effective area of the 8 graphite disk anodes
(56 cm2) to prevent cathode limitations on the performance of the MECs system All
MECs with Fe-NP decorated anodes and control anodes were tested for current
generation for 24 h with medium (without bacteria) before injecting the bacterial cells in
order to determine whether the Fe-NP decorated anodes demonstrate chemical current
generation compared to control anodes All the testing anodes were set up in the MECs
B
500 nm
A
500 nm
51
at once so that the S oneidensis MR-1 culture was applied to all of them simultaneously
Short electrode spacing (17 cm) was used in the MECs design to reduce the internal
resistance A voltage of 06 V was applied the MECs for current generation after the cells
of S oneidensis MR-1 were inoculated into MECs at early stationary phase A multimeter
with a data acquisition system (2700 Keithly USA) was used to monitor the current
change by measuring the voltage (every 300 seconds) drop through a resistor at 300 ohm
Fig 4 2 Effects of Fe-NP decorated anodes on the current density in MECs Control is
plain graphite disk anode Fe-NP is the anode with Fe-NP decoration
Microarray analysis
Biofilms for whole gene microarray analysis were aseptically removed from the plain
and Fe-NP decorated anodes of MECs after 110 hours further incubation at 30oC when
current density obviously started to decease Total RNA was extracted using Trizol
(Ambion inc AustinTX) following the manufacturerrsquos instructions Integrity of the
52
RNA samples was confirmed using the Agilent 2100 Bioanalyzer (Agilent Technology
Inc Palo Alto CA) The same amount of RNA (200 ng) from each sample was used for
further analysis after amplification by using the MessageAmpTM
II-Bacteria Prokaryotic
RNA Amplification kit (Ambion AustinTX) according to the manufacturerrsquos
instructions All of the 4295 genes (resource from NCBI) represented on the S oneidensis
MR-1 whole-gene microarrays were ordered from Roche NimbleGen Inc (Madison WI)
Biological triplicates of biofilms on the Fe-NP decorated anodes and control were
analyzed respectively cDNA synthesis labeling and hybridization were carried out by
the CGRB Core Laboratories at Oregon State University DNASTAR ArrayStarTM
3
software was used to identify genes that were up- or down-regulated more than 2-fold
when grown on the nanoparticle modified anodes using the unpaired two sample t-test
with a cutoff p-value of 005 The complete microarray data set generated in this study is
deposited for public access in the Gene Expression Omnibus
(httpwwwncbinlmnihgovgeo) under accession number GSE31535
Results
Enhancement of current generation by using NP decorated anodes in MECs
The current density generated by Fe-NP decorated anode MECs with S onidensis MR-1
increased to approximately 43 microAcm2 20 h following inoculation and then slowly
decreased Figure 42 shows that the maximum current density achieved by NP
decorated anodes was 83 times higher than that (51 microAcm2) generated by the control
(plain graphite disk) The average current density improvement of 110 hours was 59
times of that generated by the control The current densities generated in MECs in the
53
absence of bacteria were negligible (03 microAcm2) which demonstrated that the current
enhancement observed with the Fe-NP decorated anodes was biologically-derived
Global transcriptome analysis
Whole-genome DNA microarrays were used to attain a comprehensive general
overview
of the transcriptional response to Fe-NP decorated anodes by S oneidensis MR-1 in MEC
Of a total 4295 genes assayed 392 (188 induced and 204 repressed) exhibited significant
(P lt 005) differential expression at a 2-fold (log (expression ratio 1) level in 3
replicates in response to Fe-NPs These
total gene numbers present 9 of the 4295 open
reading frames (ORFs) presented on the array
Fig 43 Differentially expressed genes grouped by functional classification according to
the TIGR S oneidensis genome database (wwwtigrorg) Columns 1 amino acid
biosynthesis 2 biosynthesis of cofactors prosthetic groups and carriers 3 cell
envelope 4 cellular processes 5 central intermediary metabolism 6 DNA metabolism
7 energy metabolism 8 fatty acid and phospholipid metabolism 9 other categories 10
protein fate 11 protein synthesis 12 purines pyrimidines nucleosides and nucleotides
13 regulatory functions 14 signal transduction 15 transcription 16 transport and
binding proteins 17 unknown function 18 hypothetical proteins
Up-Regulated
-75
-50
-25
0
25
50
75
100
125
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18
Down-Regulated
Num
ber
of
Gen
es
54
Figure 43 summarizes the overall genomic response of S oneidensis MR-1 to Fe-NP
decorated anodes by grouping the differentially expressed genes into their functional role
categories based on The Institute for Genomic Researchrsquos annotation (Version 8) of the
MR-1 genome sequence The wide distribution of putative functional roles attributed to
the differentially expressed genes indicated the extent of the molecular response of
Soniedenis MR-1 to the NP decorated anodes Among genes with defined function a
large number of up-regulated genes were associated with cellular processes (group 4)
energy metabolism (group 7) other categories (group 9) and hypothetical proteins (group
18) While those genes encoding proteins involved in amino acid biosynthesis (group 1)
cellular processes (group 4) substrate transport (group 16) and hypothetical proteins
(group 18) were among the most down-regulated genes Two groups of genes that were
the most noticeable among the functional gene groups genes related to energy
metabolism (group 7) and genes related to amino acid biosynthesis (group 1) The ratios
of up-regulated genes to down-regulated genes in these functional groupings were much
higher than other functional gene groupings there were 21 up-regulated genes and 7
down-regulated genes in energy metabolism gene group and 2 up-regulated and 22 down
regulated genes in the amino acid biosynthesis group
Genes related to biofilm formation
In order to determine whether there was a correlation between the observed biofilm
enhancements by NP decorated anodes and the genes related to biofilm formation the
significant modified genes related to biofilm formation were examined and summarized
in the Table 41 These included SO_3223 and SO_3228 genes encoding the flagellum
proteins (Thormann et al 2004) which has critical impacts on initial attachment to the
55
surface as swimming motility functions were up-regulated over 10 and 09 fold
respectively The gene clusters SO_4178-SO_4180 which are thought to be essential for
formation of a three-dimensional biofilm were up-regulated from 12 to 13 fold
(Thormann et al 2004) In addition the genes SO_4105 encoding IV pilus which is
known to represent a critical factor for biofilm formation on a biotic surfaces (Thormann
et al 2004) were also induced 08 fold (Table 41) The genes related to flagellum
motility (SO_4286 SO_1529 and SO_1530) were also induced slightly (Thormann et al
2004)
Table 41 Expression levels of genes related to biofilm formation
Locus Tag Gene product descriptions Fold change
SO_3228 flagellar basal-body MS-ring and collar protein FliF 10
SO_3223 flagellar hook-length control protein FliK 09
SO_4103 MshA minor pilin protein MshD 05
SO_4105 MSHA major pilin protein MshA 08
SO_4178 expressed protein of unknown function MxdC 12
SO_4179 inner membrane family 2 glycosyltransferase MxdB 13
SO_4180 diguanylate cyclase-like protein MxdA 10
a fold changes were presented after log2
b positive number represented up-regulated and negative represented down-regulated
Genes related to energy metabolism
A closer consideration of energy metabolism genes was undertaken as anaerobic
metabolism for electron generation and electron transport functions may play critical
roles in enhanced current density generation in MECs Genes associated with energy
56
metabolism and with significantly modified gene expression levels are summarized in the
Table 42 Several of the up-regulated genes were related to formate dehydrogenase
proteins Most notably four genes related to formate dehydrogenase were up-regulated
more than 2-fold SO_0102 SO_0103 SO_0104 and SO_0107 The other genes
expression related to formate dehydrogenase proteins SO_4513 SO_4514 SO_4515 also
significantly increased Another interesting gene is that encoding the cytochrome c
oxidase protein gene SO_4609 which was up-regulated 11 fold while the gene
SO_0479 related periplasmic octaheme cytochrome c MccA was down-regulated
However SO_1519 the gene encoding lactate dehydrogenase was down-regulated 12
fold despite lactate being the primary carbon source in the media
Table 4 2 Genes related to anaerobic growth and electron transfer with significantly change expression level
Locus Tag Gene product descriptions Fold
change
SO_0101 nitrate-inducible formate dehydrogenase molybdopterin-binding
subunit FdnG
18
SO_0102 formate dehydrogenase nitrate-inducible iron-sulfur subunit 21
SO_0103 formate dehydrogenase nitrate-inducible cytochrome b556 subunit 24
SO_0104 formate dehydrogenase accessory protein FdhE 22
SO_0107 formate dehydrogenase accessory protein fdhD 24
SO_0397 quinolfumarate reductase menaquinol-oxidizing subunit FrdC_2 12
SO_0452 thioredoxin 2 15
SO_0903 Na-translocating NADH-quinone reductase subunit B NqrB_1 12
SO_1012 NADH-ubiquinone oxidoreductase subunit K NuoK 12
SO_1013 NADH-ubiquinone oxidoreductase subunit J NuoJ 12
SO_1363 hydroxylamine reductase 13
57
SO_2417 ferredoxin cofactor maintenance protein YfaE 10
SO_3922 formate dehydrogenase cytochrome b Fdh -12
SO_4513 formate dehydrogenase molybdopterin-binding subunit _2 15
SO_4514 formate dehydrogenase FeS subunit FdhB_2 12
SO_4515 formate dehydrogenase cytochrome b subunit FdhC_2 14
SO_4609 aa3 type cytochrome c oxidase subunit III CoxC 11
SO_0479 periplasmic octaheme cytochrome c MccA -12
SO_0847 periplasmic nitrate reductase ferredoxin component NapG -15
SO_1519 L-lactate dehydrogenase iron-sulfur cluster-binding protein LldF -10
SO_1251 ferredoxin 4Fe-4S -11
SO_37411 hypothetical inner membrane protein -11
a fold changes were presented after log2
b positive number represented up-regulated and negative represented down-regulated
Flavin and cytochrome related genes
Flavins can be secreted by Shewanella species as electron shuttle to facilitate
extracellular electron transfer (Cansterin et al 2008) C-type cytochromes play the
important roles on the process of extracellular electron transfer (Shi et al 2007) The
genes related to flavins synthesis and the genes encoding the cytochromes electron
transport proteins were shown in the table 43 the genes SO_1414 and SO_3058 related
to flavins synthesis was up-regulated to 03 and 01 folds respectively The genes related
to s C-type cytochromes SO_0135 SO_4105 SO_0169 SO_1778 are increased to 10
08 05 04 01 folds respectively However most of genes have shown no significant
changes (less one-fold change) as response to nano particle decorated anode in MECs
58
Table 43 Expression level of flavin and cytochrome genes
Locus Tag Gene product descriptions Fold
change
SO_1414 flavocytochrome c flavin subunit putative 03
SO_3468 riboflavin synthase subunit alpha -02
SO_3058 flavocytochrome c flavin subunit 01
SO_4105 MSHA pilin protein MshA 08
SO_0169 general secretion pathway protein GspG 05
SO_1778 outer membrane decaheme cytochrome c lipoprotein MtrC 04
SO_1779 outer membrane decaheme cytochrome c OmcA 01
SO_0135 lipoprotein of unknown function DUF333 10
SO_0136 conserved hypothetical inner membrane protein 01
a fold changes were presented after log2
b positive number represented up-regulated and negative represented down-regulated
Other genes with significantly modified expression levels
These genes over 15 fold change and possibly related to current enhancements were
shown in Table 44 SO_3104 (expressed inner membrane protein) was over expressed
and the gene SO_2194 which encodes outer membrane protein (OmpA family protein)
was repressed as response to nanoparticle decorated anode however it is unclear the
relationships of this modified membrane protein genes with the current enhancement
Another interesting phenomenon was possible co-regulation of several gene clusters
indicating possible operon associations This includes three sets of genes that were
induced more than 15 fold SO_0108SO_0109 and SO_1042SO_1043SO_1044 The
59
consistency of expression of these genes under the NP conditions provides basic evidence
to support operon structure However the correlation of these significant changed gene
with current density enhance are unclear which showed there are unknown multiple and
complex responses of S oneidensis MR-1 to nanoparticle anode of MECs
Table 44 Other genes with significantly changed expression levels
Locus Tag Gene product descriptions Fold change
SO_0101 nitrate-inducible formate dehydrogenase molybdopterin-binding
subunit FdnG
18
SO_0108 integral membrane protein of unknown function DUF39 YedE 17
SO_0109 SirA family protein YedF 17
SO_0275 N-acetyl-gamma-glutamyl-phosphate reductase -31
SO_0277 ornithine carbamoyltransferase -21
SO_0279 argininosuccinate lyase -24
SO_0404 zinc dependent metalloprotease domain lipoprotein 21
SO_0956 alkyl hydroperoxide reductase F subunit -20
SO_1042 amino acid ABC transporter ATP-binding protein -18
SO_1043 amino acid ABC transporter permease protein -27
SO_1044 amino acid ABC transporter periplasmic amino acid-binding protein -27
SO_1072 chitin-binding protein putative 21
SO_1405 transglutaminase family protein 20
SO_1822 TonB-dependent receptor putative -22
SO_2069
1-(5-phosphoribosyl)-5-[(5-phosphoribosylamino)
methylideneamino] imidazole-4-carboxamide isomerase
-19
SO_2070 amidotransferase HisH -26
SO_2071 imidazole glycerol-phosphate dehydratasehistidinol phosphatase -27
60
SO_2072 histidinol-phosphate aminotransferase -28
SO_2073 histidinol dehydrogenase -30
SO_2194 OmpA family protein -36
SO_2195 inter-alpha-trypsin inhibitor domain protein -27
SO_2767 asparagine synthetase B -26
SO_2945 prophage LambdaSo tail fiber protein 20
SO_2963 prophage LambdaSo major capsid protein HK97 family 20
SO_2987 prophage LambdaSo hypothetical protein 19
SO_2988 prophage LambdaSo expressed protein of unknown function 18
SO_3104 expressed inner membrane protein 131
SO_3408 conserved hypothetical inner membrane protein -18
SO_3585 azoreductase putative -18
SO_3586 glyoxalase family protein -17
SO_3687 curli production assemblytransport component CsgE putative -18
SO_38191 hypothetical ammonia permease -19
SO_4014 AcrBAcrDAcrF family protein 18
SO_40151 type I secretion system membrane fusion protein RND family 18
SO_4054 510-methylenetetrahydrofolate reductase 18
SO_4245 N-acetylglutamate synthase -22
SO_45251 hypothetical transcriptional regulator LysR family -17
SO_4527 integral membrane domain protein -19
SO_4705 transcriptional regulator putative -19
a fold changes were presented after log2
b positive number represented up-regulated and negative represented down-regulated
Discussion
61
The addition of NP to anodes significantly impacts current densities with the elemental
composition being a critical factor The chemical composition of NPs significantly affects
the current enhancement in MECs (Fan et al 2011) because the chemical composition of
NP properties is a critical factor in determining the conductivity which significantly
affects the efficiency of electron transfer from bacteria to electrode Fe is an ideal
candidate for NP formation for current enhancement in MESs because of high
conductivity and low price compared to gold and other conductive materials Gold NPs
has also been studies for enhancement of electricity conductivity (Bao et al 2008)
However the disadvantages of the high price of gold limit their practical application in
MECs (Fan et al 2011) The different size and density of the same composition of NP
also has essential effects on the current density enhancement (Fan et al 2011) In this
study the results showed the current density enhancement with Fe-NP size range of
200plusmn100 nm and density range of 200plusmn100 nm is considerable enhancement consequence
There are a number of reports of studies of current density enhancement in MFCs
by nanostructured anodes (Fan et al 2011 Qiao et al 2008 Sharma et al 2008 Tsai et al
2009) Based on bacterial behavior on nanostructured anode surface Qiao et al (2008)
believes that Escherichia coli cells on the nanostructured electrode surface produce hair-
like structures similar to pili that could facilitate the electron transfer between the cells
and electrode The production of hair-like structures is believed to be stimulated by
nanostructures and could play the key role on current density enhancement in pili-
producing bacteria in MFCs (Qiao et al 2008) S oneidensis can produce electrically
conductive bacterial nanowires which have similar structure to the hair-like structures in
Ecoli (Gorby et al 2006 Reguera et al 2005) A number of possible genes related
62
nanowires of S oneidensis have been identified including mshA (SO_4105) and gspG
(SO_0169) (Konstantinidis et al 2009) They are up-regulated 08 and 05 fold in the
current study respectively even though they are not significant induced This result
supported the increased nanowire genes had contributions to enhance current density in
MFCs by NPs decorated anodes
Based on the previous research on electron transfer mechanisms for S oneidensis it is
known that self-produced mediators (flavins) and multihaem c-type cytochromes (c-Cyts)
may play critical roles in the electron transfer of S oneidensis to extracellular insoluble
electrode (Canstein et al 2008 Myers and Myers 1992 Scott and Nealson 1994 Shi et al
2007) The flavins secreted by S oneidensis can facilitate to transfer electrons from
bacteria to solid anode electrodes by acting as extracellular electron shuttles (Canstein et
al 2008) In this study the genes encoding flavins (SO_1414 SO_3468 and SO_3058)
had no significant increase (less than 1 fold change) as a response to nanoparticle
decorated anode in MECs which suggests that increased flavin gene expression does not
significantly contribute to the current density enhancements in nano-particle decorated
anode The genes encoding cytochromes electron transport proteins (SO_4104 SO_0417
SO_177879 SO_013536) had no significant increase (less than 1-fold change)
However those genes were up-regulated slightly These results suggest the increased
expression flavin and c type cytochromes genes had partial contributions even not
significantly to enhance current density in MFCs by NPs decorated anodes
The thickness of bacterial biofilm on the anode can affect the power generation in
MFCs (Biffinger et al 2007 Heydorn et al 2000) since increased biofilm thickness on
the anode represents more active bacteria involving in the process of electron generation
63
and transfer in MFCs Biofilm formation related genes included flagellar genes IV pilus
genes and other genes The flagellar related genes SO_3228 SO_3229 (fliB and fliK) and
IV pilus genes SO_4105 (mshA) were induced from 19 to 09 fold the genes (SO_4178
-4180) related to formation of a three-dimensional biofilm were up-regulated from 10 to
13 folds even though the genes related bacterial motility protein (SO_ 4286 SO_4287
SO_1529 and SO_1530) are induced slightly (less than 0 5 fold) However the motility
genes most probably play a role for initial biofilm attachment on the anode surfaces
(Thormann et al 2004 Thormann et al 2006) with the rest of biofilm genes being more
essential for biofilm maintenance in mature cultures as were presented at harvest in this
study This enhancement of biofilm density under conditions of increase current density
has been observed for gold NP decorated anodes using confocal light microscopy (data
not shown) but has not yet been confirmed for Fe NP decorated anodes In any event an
increase in the expression level of genes related biofilm formation therefore also
supported the possibility of enhanced biofilm formation on NPs decorated anodes thus
facilitating electron transfer from bacteria to anodes
Beliaev et al described a model of the Fe reduction pathway in S oneidensis MR-1 using
solid substrates as the electron acceptor (Beliaev et al 2001) This model proposes that
electrons are generated and released in cytoplasm then transferred to the quinone pool
eg menaquinone in the cytoplasmic membrane The reduced menaquinone in turn
reduces CymA and from there electrons are then transferred to a periplasmic shuttle (eg
CctA) and then to MtrA a periplasmic soluble decaheme cytochrome c located in
associated with the outer membrane via interaction with MtrB Finally surface displayed
outer membrane proteins decaheme cytochromes (OmcA MtrC or MtrF) can transfer
64
electron to Fe or other solid substrates (Beliaev et al 2001)) Bencheikh-Latmani et al
demonstrated that the same electron transport pathway may be used for more than one
electron acceptor through studies of the response of S oneidensis MR-1 to U(VI) and
Cr(VI) under anaerobic conditions particularly focused on the critical genes
(SO_1777mtrA SO_1776mtrB SO_1778mtrC) (Bencheikh-Latmani et al 2005)
However our studies showed that these genes had no significant expression changes in
response to nanostructured anodes Contrastingly one of expressed inner membrane
protein genes SO_3104 and one of cytochromes genes SO_4609 encoding cytochrome c
oxidase subunit III demonstrated a 13-fold and 11-fold up-regulation respectively in this
study while the gene SO_2194 which encodes outer membrane protein (OmpA family
protein) was down regulated 36 fold Our results therefore suggest that S oneidensis
MR-1 does not utilize the same set of core critical genes (SO_1777 mtrA SO_1776
mtrB SO_1778 mtrC) products to transfer electrons in all instances This is the similar
result with the study of Bretschger et al (2007) which indicated that the electron
transport system in S oneidensis MR-1 is complex with several different proteins able to
participate in electron transfer to the anode of MECs
It should also be mentioned that the gene (SO_1519) encoding lactate
dehydrogenase repressed (-12 fold) here suggesting that lactate was depleted at the time
of cell harvest from the MECs and that other pathways of anaerobiosis had been activated
including those associated with amino acid uptake and consumption the growth media
was a complex mixture including yeast extract In particular the catabolic pathway for
histidine of S oneidensis MR-1 a pathway that may formate as an intermediate is up-
regulated with the genes for histidine ammonia lyase SO_0098 SO_3057 and SO_4374
65
up-regulated 13 11 and 11-fold respectively The possible production of formate
during histidine degradation may in turn explain the high levels of formate
dehydrogenates as evidenced by upregulation of SO_0102 SO_0103 SO_0104 and
SO_0107 annotated as formate dehydrogenase proteins These genes had the highest
levels of up-regulated expression as response to Fe NP decorated anodes (greater than 2-
fold)
Acknowledgements
This research was partially supported by the US National Science Foundation CBET
0828544 and the funds from ONAMIDOD (ARL-DOD Cooperative Agreement
W911NF-07-2-0083) We thank Barbara Gvakharia and Caprice Rosato for helpful
suggestions We also thank Margaret Romine of Pacific Northwest Laboratories for
valuable comments on manuscripts We also thank anonymous reviewers for significant
suggested improvements imparted as part of prior review of this manuscript
66
Chapter 5
Global Transcriptome Analysis of the Response of Shewanella oneidensis MR-1 to
Carbon Nanotube Decorated Anodes in Microbial Electrochemical Systems
Shoutao Xu Yanzhen Fan Rebecca Schaller Frank Chaplen Jun Jiao Hong Liu
67
Abstract
Shewanella oneidensis MR-1 is an important model microorganism for metabolic studies
on the effects of different environmental factors because of its diverse respiratory
capabilities Carbon nanotube (CNT) modified graphite disks were used as anodes to
investigate the effects of nanomatrials on the performance of S oneidensis MR-1 in
microbial electrolysis cells (MECs) The current densities produced with CNT decorated
anodes were on average 56-fold higher than plain graphite anodes Whole genome
microarray analysis of gene expression showed that up-regulation of cytochromes c genes
associated with extracellular electron transfer are strongly correlated to current increases
in CNT decorated anodes Genes involved in flavin biosynthesis also contribute to
current increase in CNT decorated anode MECs
Keywords Microbial electrochemical system microbial fuel cell microbial
electrolysis cell carbon nanotube gene expression DNA microarray
68
Introduction
Shewanella oneidensis MR-1 is an important model microorganism for metabolic
studies of the effects of different environmental factors because of its diverse respiratory
capabilities It has been used for transcriptome analysis to investigate the responses to
different growth conditions and environmental stresses (Murray et al 2001) Gene
expression patterns under different conditions such as heat shock cold shock Cr (VI) U
(VI) reduction chromate stress iron and acid tolerance have been studied previously
(Bencheikh-Latmani et al 2005 Brown et al 2006 Gao et al2006 Gao et al 2004
Yang et al 2008) More recently it has been exploited as a model species for power
generation in microbial electrochemical systems (MESs)which have potential
applications in renewable energy generation bioremediation and wastewater treatment
(Logan et al 2007 Logan 2009 Park and Zeikus 2002 Fan et al 2011 Xu et al 2012)
In a MES electrochemically active microorganisms oxidize organic matter in the an
ode chamber to release electrons Electrons are then transferred to the anode electrode thr
ough various mechanisms (Logan et al 2006 Lovley 2006 Rabaey et al 2003) and fina
lly travel to the cathode electrode and combine with the terminal electron acceptor The
key feature of MESs is the microbe-catalyzed electron transfer from the organic matter to
the anode (Rabaey et al 2004) Enhancing the current output which highly depends on
the performance of the anode electrode is critical for the successful application of MESs
(Logan et al 2007 Park and Zeilus 2002 2003) Several nanomaterials including TiO2
gold (Au) and palladium (Pd) nanoparticles have been utilized to decorate electrodes to
enhance power density in MESs (Qiao et al 2008 Quan et al 2005 Fan et al 2011) The
possible mechanisms for increased current densities have been studied by using Fe
69
nanoparticle-decorated anodes (Xu et al 2012)
Carbon nanotubes (CNT) are cylindrically shaped nanomaterials with extremely
high surface area excellent electrical conductivity and chemical inertness (He et al
2005a Serp et al 2003) These unique properties make CNT a promising electrode
material (Liang et al 2008) The biocompatibility of microorganisms and carbon
nanostructures has been evaluated (Morozan et al 2007) The effects of CNT on anodic
biofilms have been tested in MFCs (Liang et al 2011) However no studies have been re
ported on global transcriptome analysis for the response of S oneidensis MR-1 to CNT
decorated anode in MECs
In the present study the graphite disks decorated with CNT were used as anodes to
investigate the effects of nanomaterials on current generation in multi-anode MECs
DNA microarrays were used to analyze differences in the global gene expression profile
of S oneidensis MR-1 grown on plain versus CNT decorated anodes
Materials and methods
Bacterial cultures S oneidensis MR-1 was purchased from American Type Culture
collection (ATCC 700550) and stored as freezer stocks at -80degC Stock S oneidensis
MR-1 cells were grown aerobically using M1 medium at 30degC Cells in early stationary
growth phase (optical density of 15-16 at 600nm (Spectrophotometer UV-1700
Shimadzu Columbia MD) were injected into the chamber of the MECs for current
production Sodium lactate (final concentration 30mM) was added as the additional
electron donor
Fabrication and characterization of nanostructured anodes Superfine isomolded
graphite disks (16 cm x 05 cm GraphiteStorecom Inc) were polished with ultrafine
70
sand paper (2000 grit 3M Company) as the base for the CNT decorated and control
anodes Aligned multi-walled carbon nanotube samples were synthesized via plasma
enhanced chemical vapor deposition (PECVD) Samples superfine isomolded graphite
pillars with a 16 cm diameter and 05 cm thickness were sputtered with a thin film of Ni
in a CRC 300 magnetron sputter coater for 1 to 2 min They were then placed in the
PECVD reactor where the Ni was annealed into small catalyst islands C2H2 was bled
into the chamber for growth and an NH3 plasma was used to vertically align the growth
of the multi-walled carbon nanotubes (MWCNTs) The temperature and time were varied
between 725degC and 775oC and 15 to 60 min to control the length of the tubes and density
of samples Samples were then analyzed in a FEI Sirion field emission scanning electron
microscope (FESEM) SEM images of CNT decorated anode and control surfaces were
shown in the Figure 51
Figure 51 SEM images of CNT decorated anodes (A) Graphite disk control (B) CNT
decorated anode
Multiple channel MECs construction and operation A MECs with multiple
removable anodes each with an effective anode area of 07 cm2
was constructed and used
to evaluate the effects of nanostructures on the current density of according to a previous
B A
71
study (Fan et al 2011) The cathode was made of wet-proof (30) carbon cloth (type B
E-TEK Division Inc USA) coated with PtC (20 E-TEK Division Inc USA)
following a previously reported procedure (Liu et al 2005) The final platinum loading
was 05 mgcm2 per projected cathode area The size of the cathode (150 cm
2) was 25
times larger than the total effective area of the 8 graphite disk anodes (56 cm2) to prevent
cathode limitations on the performance of the MECs system All MECs with CNT
decorated anodes and control anodes were tested for current generation for 24 h with
sterile a medium (without bacteria) before injecting the bacterial cells in order to
determine whether the CNT decorated anodes demonstrate chemical current generation
compared to control anodes All the testing anodes were set up in the MECs
simultaneously so that the S oneidensis MR-1 culture was applied to all of them
simultaneously Short electrode spacing (17 cm) was used in the MECs design to reduce
the internal resistance A voltage of 06 V was applied the MECs for current generation
after the cells of S oneidensis MR-1 were inoculated into the MECs in the early
stationary phase A multimeter with a data acquisition system (2700 Keithly USA) was
used to monitor the current change by measuring the voltage drop through a resistor
Microarray analysis Biofilms for whole gene microarray analysis were aseptically
removed from the plain and CNT decorated anodes of MECs after 80 hours further
incubation at 30oC when current density obviously reached to a stable phase Total RNA
was extracted using Trizol (Ambion inc AustinTX) following the manufacturerrsquos
instructions Integrity of the RNA samples was confirmed using the Agilent 2100
Bioanalyzer (Agilent Technology Inc Palo Alto CA) Total RNA (200 ng) from each
sample was amplified using the MessageAmpTM
II-Bacteria Prokaryotic RNA
72
Amplification kit (Ambion AustinTX) according to the manufacturerrsquos instructions The
S oneidensis MR-1 whole-gene microarrays representing all of the 4295 genes (resource
from NCBI) were ordered from Roche NimbleGen Inc (Madison WI) Biological
triplicates of biofilms on the CNT decorated anodes and control were analyzed cDNA
synthesis labeling and hybridization were carried out by the CGRB Core Laboratories at
Oregon State University DNASTAR ArrayStarTM
3 software was used to identify genes
that were up- or down-regulated more than 2-fold when grown on the nanoparticle
modified anodes using the unpaired two sample t-test with a cutoff p-value of 005 with
bonferroni correction
Results and discussion
Enhancement of current generation using CNT decorated anodes in MECs The
current density generated by CNT decorated anode MECs with S onidensis MR-1
increased to 36 microAcm2 10 hours post-inoculation and reached a maximum of 40 microAcm
2
at 29 hours inoculation then gradually decreased The current density stabilized at
30microAcm2 after 78 h operation Figure 52 shows that the current density curve generated
by CNT decorated anode in MECs The average current density generated by CNT
decorated anodes was 56 times that of the control The current density generated in the
MECs without bacteria was negligible (data not shown) This result demonstrates that the
current enhancement observed with the CNT decorated anodes was biologically-derived
A
73
Figure 52 Effects of CNT decorated anodes on the current density in MECs (3 replicates)
Control is plain graphite disk anode CNT is the anode with CNT decoration (Error bars
represent standard deviation)
It has been reported that nanostructured decorated anodes have significant impacts on
current densities (Fan et al 2011 Qiao et al 2006 Sharma 2008 Tsai et al 2009) The
CNT decorated anodes had significantly enhanced current densities in MECs in this
study This result is consistent with previous studies Interestingly the current density
curve generated by CNT decorated anode MECs is quite different from the current
density curve generated by Au NP and Pd NP decorated anodes using same bacterial
species (Fan et al 2011) In the Au and Pd NP decorated anode MECs the current
density curve was not significantly increased in the beginning compared with the control
but gradually increased after the inoculation the current density reached the maximum
current density more than 50 hours after the inoculation However the current density
generated by CNT decorated anode MECs started to increase significantly immediately
after the inoculation It reached a maximum current density 29 hours post-inoculation
-1
6
13
20
27
34
41
48
0 10 20 30 40 50 60 70 80
Curr
ent
den
sity
(micro
Ac
m2)
Time (Hours)
Control
CNT
74
These results indicate that there might be different current density enhancement
mechanisms between CNT with other metal NP decorated anodes in MECs
Global transcriptome analysis Whole-genome DNA microarrays were used to attain a
comprehensive general overview of the transcriptional response of S oneidensis
MR-1 of
S oneidensis MR-1 to CNT decorated anodes in MECs In a total of 4295 genes analyzed
457 genes (250 genes up-regulated and 207 genes down-regulated) exhibited significantly
(P lt 005) changed expression at a 2-fold level in at least 3 replicates in the response to
CNT The total number of the regulated genes present 11 of the 4295 open reading
frames (ORFs) presented on the microarray The total number of genes at significant
expression levels in response to CNT decorated anodes is close to that to Fe NP decorated
anodes in MECs (392 9) ( Xu et al 2012)
Fig 53 Differentially expressed genes grouped by functional classification according to
the TIGR S oneidensis genome database (wwwtigrorg) Columns 1 amino acid
biosynthesis 2 biosynthesis of cofactors prosthetic groups and carriers 3 cell
envelope 4 cellular processes 5 central intermediary metabolism 6 DNA metabolism
7 energy metabolism 8 fatty acid and phospholipid metabolism 9 other categories 10
protein fate 11 protein synthesis 12 purines pyrimidines nucleosides and nucleotides
-80
-60
-40
-20
0
20
40
60
80
100
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18
Nu
mb
er
of
Ge
ne
s
Up-regulated
Down-regulated
75
13 regulatory functions 14 signal transduction 15 transcription 16 transport and
binding proteins 17 unknown function 18 hypothetical proteins
The overall transcript genomic response of S oneidensis MR-1 to CNT decorated
anodes was summarized in Figure 53 by grouping the differentially expressed genes into
their functional role categories based on The Institute for Genomic Researchrsquos annotation
(Version 8) of the MR-1 genome sequence The wide distribution of putative functional
roles attributed to the differentially expressed genes indicated the extent of the molecular
response of Soniedenis MR-1 to the CNT decorated anodes A large number of up-
regulated genes were presented in genes associated with biosynthesis of cofactors
prosthetic groups and carriers (group 2) cell envelope (group 3) protein fate (group 10)
protein synthesis (group 11) and purines pyrimidines nucleosides and nucleotides
(group 12) in this study The number of up-regulated genes was two times more than that
of down-regulated genes in each functional group Contrastingly in the genomic response
S oneidensis MR-1 to Fe NP decorated anodes (chapter 4) most of up-regulated genes
were associated with cellular processes (group 4) energy metabolism (group 7) other
categories (group 9) and hypothetical proteins (group 18) The down-regulated genes
presented in the amino acid biosynthesis (group 1) cellular processes (group 4) substrate
transport (group 16) and hypothetical proteins (group 18) While those genes encoding
proteins involved in amino acid biosynthesis (group 1) central intermediary metabolism
( group 5) DNA metabolism( group 6) fatty acid and phospholipid metabolism (group
8) and other categories (group 9) were among the most down-regulated genes in this
study However the two groups of genes that were the most noticeable between the
genomic response S oneidensis MR-1 to CNT and Fe NP decorated anodes which they
are the genes related to energy metabolism (group 7) and hypothetical proteins (group
76
18) These results indicated there are different and common genomic responses to S
oneidensis MR-1 on CNT and Fe NP decorated anodes in MECs
Electron transfer related genes Based on previous electron transfer mechanism
studies on Shewanella a serial of group proteins collectively described as the Mtr
pathway play the role to transfer electron rom the inner bacterial body to the outer
membrane (Beliaev et al 2001 Shi et al 2007 Brutinel and Gralnick 2012) In the Mtr
pathway firstly electrons generated by bacteria received by CymA a tetraheme c-type
cytochrome anchored in the inner membrane then electrons were transferred to MtrA a
periplasmic decaheme c-type cytochrome After that electrons were transferred from
MtrA to MtrC an outer-membrane decaheme c-type cytochrome is facilitated by MtrB a
non-heme containing integral outer-membrane proteinOuter-membrane decaheme c-type
cytochrome MtrC and OmcA are in change of electron transfer to exreacellular electron
acceptor (Shi et al 2007 Brutinel and Gralnick 2012 ) These genes encoding electron
transport proteins are shown in the table 51 The critical genes SO_1777 mtrA SO_1776
mtrB SO_1778 mtrC are unregulated 41 to 80 fold in response to CNT decorated
anodes other cytochrome c genes related to electron transfer in MtrA pathway also are
significantly increased in response to CNT decorated anodes These genes encoding outer
membrane proteins have different level increases supporting that the increase of
cytochrome c gene expression contributes to the current enhancements of MECs as
response to CNT decorated anodes
77
Table 51 Expression level of cytochrome c as response to CNT decorated anodes
Locus Tag Gene product descriptions Fold change CNT
SO_0165 general secretion pathway protein GspC 31
SO_0167 general secretion pathway protein GspE 45
SO_1776 outer membrane protein precursor MtrB 41
SO_1777 outer membrane decaheme cytochrome c
lipoprotein MtrA
80
SO_1778 outer membrane decaheme cytochrome c
lipoprotein MtrC
43
SO_1779 outer membrane decaheme cytochrome c
OmcA
63
SO_0135 lipoprotein of unknown function DUF333 22
SO_0136 conserved hypothetical inner membrane protein 24
Flavin related genes It is known that self-produced mediators play critical roles in the
electron transfer of S oneidensis to extracellular insoluble electrode (Canstein von et al 2
008 Myers et al 1992 Scott et al 1994 Shi et al 2007) It has been proved that the
flavins secreted by S oneidensis can facilitate electron transfer from bacteria to solid
anode electrodes by acting as extracellular electron shuttles (Cansterin von et al 2008) T
wo critical genes (SO_1414 and SO_3468) related to flavin synthesis were shown increa
sed significantly (more than 2 folds) increase in this study (Table 52) This result support
s the hypothesis that flavin production increases as the response to CNT decorated anodes
in MECs have significant contribution to the current density enhancements
78
Table 52 Expression levels of genes related to flavin synthesis as response to CNT
decorated anode
Locus Tag Gene product descriptions Fold change CNT
SO_1414 flavocytochrome c flavin subunit putative 22
SO_3468 riboflavin synthase subunit alpha 20
The amount of bacterial biofilm biomass on the anode can affect the power
generation in MFCs (Biffinger et al 2007 Heydorn et al 2000) Biofilm formation
related genes include flagellar gene IV pilus genes and other genes the flagellar related
genes (fliB and fliK) and IV pilus genes (mshA) were induced only from 105 to 134
fold as response of S oneidensis MR-1 to CNT decorated anodes in this study No
biofilm-associated genes presented significant changes in expression levels in this study
indicating there is no direct connections between biofilm enhancements to CNT
decorated anodes in this study This result is consistent with the Liang et al (2011)
conclusion that using CNT enhanced the electrochemical activity of anodic biofilms but
did not result in a significant increase of biomass in the anodic biofilms
Our microarray results showed two significantly up-regulated gene groups the
genes encoding proteins localized on the outer membrane and the genes involved in
flavin biosynthesis contributed to current density enhancement by CNT decorated anodes
Among 457 significantly changed genes to CNT in this study there are also a relatively
large number of genes encoding proteins with unknown functions which are either up- or
down-regulated in the response to CNT decorated anodes which indicated more
79
complicated responses of S oneidensis MR-1 to CNT decorated anodes in MECs and
further study is needed
80
Chapter 6
Summary
Microbial electrochemical systems (MESs) with a mixed culture initially inoculated
from Corvallis wastewater treatment plant have been studied for more than 6 years for
varying purposes including power generation hydrogen production heavy metal
removal and wastewater treatment Experiment results have shown that the mixed culture
is quite stable with excellent performance in MESs Our community analysis using
denaturing gradient gel electrophoresis (DGGE) and 16S rDNA clone library construction
suggests that the mixed culture is composed predominantly of Geobacter sp (66)
Arcobacter sp (12) and Citrobacter sp (11) These results not only improved our
understanding of the mixed culture community but also guided our studies on the
cultivation and isolation of the different bacterial species
Exoelectrogenic Citrobacter sp SX-1 was first isolated and characterized from a
MFC operated with the mixed culture Citrobacter sp SX-1 has been demonstrated to
produce electricity from wide range of different substrates including citrate acetate
glucose sucrose glycerol and lactosein MFCs Cyclic voltammetry analysis indicated
that membrane associated proteins may play an important role in facilitating the electrons
transferring from bacteria to electrode The strain SX-1 increased the known diversity of
power generating exoelectrogens and provided a unique bacterial species for study in
renewable energy generation and waste treatment
81
To enhance the electron transfer from bacteria to anode nano decorated anodes
including Fe NP and CNT were developed and characterized and evaluated in MECs
using Shewanella oneidensis MR-1 as a model species Both nanostructures have
significantly increased current density compared with the control Whole genome
microarray analysis elucidated the possible mechanisms of power enhancement in
response to these nano-decorated anodes
These results benefit to understanding of physiology and ecology of mixed
cultures in MFCs and improve the efficiency of current generation in MESs which will
facilitate the viability of niche applications for MESs in near future
82
Bibliography
1 Amman R Ludwig W (2000) Ribosomal RNA-targeted nucleic acid probes for
studies in microbial ecology FEMS Microbiol Ecol 24 555ndash565
1 Bao SJ Li CM Zang JF Cui XQ Qiao Y Guo J (2008) Novel Nanoporous TiO2
Electrode for Directly Electrochemistry of glucose oxidase Adv Funct Mater
18(4) 591-599
2 Beliaev AS Saffarini DA McLaughlin JL Hunnicutt D (2001) MtrC an outer
membrane decahaem c cytochrome required for metal reduction in Shewanella
putrefaciens MR-1 Mol Microbiol 39722-30
3 Bencheikh-Latmani R Williams SM Haucke L Criddle CS Wu L Criddle CS
Zhou J Tebo BM (2005) Global transcriptional profiling of Shewanella oneidensis
MR-1 during Cr(VI) and U(VI) reduction Appl Environ Microbiol 71 7453ndash7460
4 Biffinger JC Pietron J Ray R Little B and Ringeisen BR (2007) A biofilm
enhanced miniature microbial fuel cell using Shewanella oneidensis DSP10 and
oxygen reduction cathodes Biosens Bioelectron 221672ndash1679
5 Bodelier PLE Roslev P Henckel T FenzelP (2000) Stimulation by ammonium
based fertilizers of methane oxidation in soil around rice root Nature 40 421ndash424
6 Bond DR Holmes DE Tender LM Lovley DR (2002) Electrode-reducing
microorganisms that harvest energy from marine sediments Science 295 483-485
7 Bond DR Lovley DR (2003) Electricity production by Geobacter sulfurreducens
attached to electrodes Appl Environ Microbiol 69 1548-1555
8 Bond DR Lovley DR (2005) Evidence for involvement of an electron shuttle in
electricity generation by Geothrix fermentans Appl Environ Microbiol 71 2186-
2189
9 Bretschger O Obraztsova A Sturm CA Chang IS Gorby YA Reed SB Culley
DE Reardon CL Barua S Romine MF Zhou J Beliaev AS Bouhenni R
Saffarini D Mansfeld F Kim BH Fredrickson JK Nealson KH (2007) Current
production and metal oxide reduction by Shewanella oneidensis MR-1 wildtype and
mutants Appl Environ Microbiol 73 7003ndash7012
10 Brown SD Thompson MR Verberkmoes NC Chourey K Shah M Zhou J
Hettich RL and Thompson DK (2006) Molecular dynamics of the Shewanella
oneidensis response to chromate stress Mol Cell Proteomics 51054-1071
11 Brutinel ED Gralnick JA (2012) Shuttling happens soluble flavin mediators of
extracellular electron transfer in Shewanella Appl Microbiol Biotechnol 93(1) 41-8
12 Butler JE Kaufmann F Coppi MV Nuacutentildeez C Lovley DR (2004) MacA a diheme
c-type cytochrome involved in Fe(III) reduction by Geobacter sulfurreducens J
Bacteriol 186(12) 4042ndash4045
83
13 Caccavo F Lonergan DJ Lovley DR Davis M Stolz JF McInerney MJ (1994)
Geobacter sulfurreducens sp nov a hydrogen- and acetate oxidizing dissimilatory
metal-reducing microorganism Appl Environ Microbiol 60 3752ndash3759
14 Canstein von H Ogawa J Shimizu S Lloyd JR (2008) Secretion of flavins by
Shewanella species and their role in extracellular electron transfer Appl Environ
Microbiol 74 615ndash623
15 Cao X Huang X Liang P Xiao K Zhou Y Zhang X Logan BE (2009) A New
Method for Water Desalination Using Microbial Desalination Cells Environ Sci
Technol 43(18)7148-7152
16 Chaudhuri SK Lovley DR (2003) Electricity generation by direct oxidation of
glucose in mediatorless microbial fuel cells Nat Biotechnol 21 1229-1232
17 Cheng S Liu H Logan B E (2006) Increased power generation in a continuous
flow MFC with advective flow through the porous anode and reduced electrode
spacing Environ Sci Technol 402426-2432
18 Cheng S Logan B E (2007) Sustainable and efficient biohydrogen production via
electrohydrogenesis PNAS 104 18871-18873
19 Chignell J (2010) Energy Recovery from Biodiesel Waste Performance of
Microbial Electrochemical Systems on Glycerol Thesis
20 Chung K Okabe S (2009) Characterization of electrochemical activity of a strain
ISO2-3 phylogenetically related to Aeromonas sp isolated from a glucose-fed
microbial fuel cell Biotechnol Bioeng 104(5) 901-910
21 Cole JR Chai B Farris RJ Wang Q Kulam-Syed-Mohideen AS McGarrell
DM Bandela AM Cardenas E Garrity GM Tiedje JM (2007) The ribosomal
database project (RDP-II) introducing myRDP space and quality controlled public
data Nucleic Acids Res 35 (Database issue) D169-D172 doi 101093nargkl889
22 Cole JR Wang Q Cardena E Fish J Chai B Farris RJ Kulam-Syed-Mohideen
AS McGarrell DM Marsh T Garrity GM Tiedje JM (2009) The Ribosomal
Database Project improved alignments and new tools for rRNA analysis Nucleic
Acids Res 37 (Database issue) D141-D145 doi 101093nargkn879
23 Dey A De S De A De SK (2004) Characterization and dielectric properties of
polyaniline-TiO2 nanocomposites Nanotechnol 15 1277-1283
24 Edwards U Rogall T Bloumlcker H Emde M Boumlttger EC (1989) Isolation and
direct complete nucleotide determination of entire genesmdashcharacterization of a gene
coding for 16S-ribosomal RNA NucleicAcids Res 17 7843ndash7853
25 Eggleston CM Voumlroumls J Shi L Lower BH Droubay TC Colberg PJS (2008)
Binding and direct electrochemistry of OmcA an outer-membrane cytochrome from
an iron reducing bacterium with oxide electrodes a candidate biofuel cell system
Inorg Chim Acta 361769ndash777
26 Fan Y Sharbrough E Liu H (2008) Quantification of the Internal Resistance
Distribution of Microbial Fuel Cells Environ Sci Technol 42 8101ndash8107
84
27 Fan Y Xu S Schaller R Jiao J Chaplen F Liu H (2011) Nanoparticle decorated
anodes for enhanced current generation in microbial electrochemical
cells Biosensors and Bioelectronics 26 (5)1908ndash1912
28 Fan Y Hu H Liu H (2007)Sustainable power generation in microbial fuel cells
using bicarbonate buffer and proton transfer mechanisms Environ Sci
Technol 41(23) 8154-8
29 Fedorovich V Knighton MC Pagaling E Ward FB Free A Goryanin I (2009)
Novel electrochemically active bacterium phylogenetically related to Arcobacter
butzleri isolated from a microbial fuel cell Appl Environ Microbiol 75(23) 7326-
7334
30 Ferna ndez A Huang S Seston S Xing J Hickey R Criddle C Tiedje J (1999)
How stable is stable Function versus community composition Appl Environ
Microbiol 65 3697ndash3704
31 Field SJ Dobbin PS Cheesman MR Watmough NJ Thomson AJ Richardson
DJ (2000) Purification and magneto-optical spectroscopic characterization of
cytoplasmic membrane and outer membrane multiheme c-type cytochromes from
Shewanella frigidimarina NCIMB400 J Biol Chem 275 8515ndash8522
32 Freguia S Masuda M Tsujimura S Kano K (2009) Lactococcus lactis catalyses
electricity generation at microbial fuel cell anodes via excretion of a soluble quinone
Bioelectrochemistry 7614ndash18
33 Fricke K Harnisch F and Schroumlder U (2008) On the use of cyclic voltammetry for
the study of the anodic electron transfer in microbial fuel cells Energy Environ
Sci 1 144ndash147
34 Gao H Wang Y Liu X Yan T Wu L Alm E Arkin A Thompson DK and
Zhou J (2004) Global transcriptome analysis of the heat shock response of
Shewanella oneidensis J Bacteriol 1867796-7803
35 Gao H Yang ZK Wu L Thompson DK Zhou J (2006) Global transcriptome
analysis of the cold shock response of Shewanella oneidensis MR-1 and mutational
analysis of its classical cold shock proteins J Bacteriol 1884560-4569
36 George MG (2005) Bergeyrsquos Manual of Systematic Bacteriology Volume Two The
Proteobacteria Part B The Gammaproteobacteria Second Edition New York
Springer 651p
37 Gonzaacutelez-Toril E Llobet-Brossa E Casamayor EO Amann R Amils R (2003)
Microbial ecology of an extreme acidic environment the Tinto river Appl Environ
Microbiol 69(8) 4853ndash4865
38 Gorby YA Yanina S Mclean JS Rosso KM Moyles D Dohnalkova A
Beveridge TJ Chang IS Kim BH Kim KS Culley DE Reed SB Romine M
F Saffarini DA Hill EA Shi L Elias DA Kennedy DW Pinchuk G
Watanabe K Ishii S Logan B Nealson KH Fredrickson JK (2006) Electrically
conductive bacterial nanowires produced by Shewanella oneidensis strain MR-1 and
other microorganisms Proc Natl Acad Sci USA 10311358-11363
85
39 He Z Wagner N Minteer SD Angenent LT (2006) An upflow microbial fuel cell
with an interior cathodes assessment of the internal resistance by impedance
spectroscopy Environ Sci Technol 40 5212-5217
40 Heydorn A Nielsen AT Hentzer M Sternberg C Givskov M Ersboslashll BK Molin
S (2000) Quantification of biofilm structures by the novel computer program
COMSTAT Microbiol 1462395-2407
41 Holmes DE Bond DR Lovley DR (2004a) Electron transfer by Desulfobulbus
propionicus to Fe(III) and graphite electrodes Appl Environ Microbiol 701234ndash
1237
42 Holmes DE Chaudhuri SK Nevin KP Mehta T Methe BA Liu A Ward JE
Woodard TL Webster J Lovley DR (2006) Microarray and genetic analysis of
electron transfer to electrodes in Geobacter sulfurreducens Environ Microbiol
81805-1815
43 Holmes DE Nicoll JS Bond DR Lovley DR (2004b) Potential role of a novel
psychrotolerant member of the family Geobacteraceae Geopsychrobacter
electrodiphilus gen nov sp nov in electricity production by a marine sediment
fuel cell Appl Environ Microbiol 70 6023ndash6030
44 Holmes DE Chaudhuri SK Nevin KP Mehta T Metheacute BA Liu A Ward JE
Woodard TL Webster J Lovley DR (2006) microarray and genetic analysis of
electron transfer to electrodes in Geobacter sulfurreducens Environ Microbial
8(10)1805-1815
45 Hu H (2009) Enhancing Hydrogen Production in Microbial Electrolysis Cells
through Development of Platinum-free Cathode and Improvement of Reactor
Dissertation
46 Ince BK Ayman O N Turker G Ccedilelikkol S Ince O (2010) Microbial ecology of
anaerobic reactors for treatment of alcohol industry wastewaters a review Current
research technology and education topics in applied microbiology and microbial
ecology 988-999
47 Jung S Regan JR (2007) Comparison of anode bacterial communities and
performance in microbial fuel cells with different electron donors Appl Microbiol
Biotechnol 77393ndash 402
48 Kim BH Kim HJ Hyun MS Park DH (1999) Direct electrode reaction of Fe(III)
reducing bacterium Shwwanella putrefaciens J Microbial Biotechnol 9127-131
49 Kim GT Hyun MS Chang IS Kim HJ Park HS Kim BH Kim SD Wimpenny
JW Weightman AJ (2005) Dissimilatory Fe(III) reduction by an electrochemically
active lactic acid bacterium phylogenetically related to Enterococcus gallinarum
isolated from submerged soil J Appl Microbiol 99(4) 978ndash987
50 Kim GT Webster G Wimpenny JW Kim BH Kim HJ Weightman AJ (2006)
Bacterial community structure compartmentalization and activity in a microbial fuel
cell J Appl Microbiol 101(3) 698ndash710
86
51 Kim HJ Park HS Hyun MS Chang IS Kim M Kim BH (2002) A mediator-
less microbial fuel cell using a metal reducing bacterium Shewanella putrefaciens
Enzyme Microb Technol 30 145-152
52 Kim JR Min B Logan BE (2005) Evaluation of procedures to acclimate a
microbial fuel cell for electricity production Appl Microbiol Biotechnol 68(1)23-30
53 Kim JR Cheng S Oh SE Logan BE (2007) Power generation using different
cation anion and ultrafiltration membranes in microbial fuel cells Environ Sci
Technol 41 1004-1009
54 Konstantinidis KT Serres MH Romine MF Rodrigues JL Auchtung J McCue
LA Lipton MS Obraztsova A Giometti CS Nealson KH Fredrickson JK
Tiedje JM (2009) Comparative systems biology across an evolutionary gradient
within the Shewanella genus Proc Natl Acad Sci U S A 106(37)15909-15914
55 Lee J Phung NT Chang IS Kim BH Sung HC (2003) Use of acetate for
enrichment of electrochemically active microorganisms and their 16S rDNA
analyses FEMS Microbiol Lett 223185ndash191
56 Liang P Wang H Xia X Huang X Mo Y Cao X Fan M(2011)Carbon
nanotube powders as electrode modifier to enhance the activity of anodic biofilm
in microbial fuel cells Biosens Bioelectron 26(6)3000-4
57 Lies DP Hernandez ME Kappler A Mielke RE Gralnick JA Newman DK
(2005) Shewanaella oneidensis MR-1 uses overlapping pathways for iron reduction
at a distance and by direct contact under conditions relevant for biofilms Appl
Environ Microbiol 71 4414-4426
58 Liu H and Logan B E (2004) Electricity generation using an air-cathode single
chamber microbial fuel cell in the presence and absence of a proton exchange
membrane Environ Sci Technol 38 4040-4046
59 Liu H Grot S Logan BE (2005) Electrochemically assisted microbial production
of hydrogen from acetate Environ Sci Technol 39 4317 ndash 320
60 Liu H Hu H Chignell J Fan Y (2010) Microbial Electrolysis Novel Technology
for Hydrogen Production from Biomass Biofuels 1(1) 129ndash142
61 Liu H Ramnarayanan R Logan BE (2004) Production of electricity during
wastewater treatment using a single chamber microbial fuel cell Environ Sci Technol
382281-2285
62 Liu WT Marsh TL Cheng H Forney LJ (1997) Characterization of microbial
diversity by determining terminal restriction fragment length polymorphisms of gene
encoding 16S rRNA Appl Environ Microbiol 63 4516ndash4522
63 Logan B E Hamelers B Rozendal R Schroder U Keller J Freguia S Aelterman
P Verstraete W Rabaey K (2006) Microbial fuel cells methodology and
technology Environ Sci Technol 40 (17) 5181-5192
64 Logan B E and Regan JM (2006) Electricity-producing bacterial communities in
microbial fuel cells Trends Microbiol 14512ndash518
87
65 Logan B Cheng S Watson V Estadt G (2007) Graphite fiber brush anodes for
increased power production in air-cathode microbial fuel cells Environ Sci Technol
41(9)3341-3346
66 Logan BE (2009) Exoelectrogenic bacteria that power microbial fuel cells Nat Rev
Microbiol 5 375-381
67 Logan BE Call D Cheng S Hamelers HV Sleutels TH Jeremiasse AW
Rozendal RA (2008) Microbial electrolysis cells for high yield hydrogen gas
production from organic matter Environ Sci Technol 42 8630-8640
68 Logan BE Hamelers B Rozendal R Schroder U Keller J Freguia S Aelterman
P Verstraete W Rabaey K (2006) Microbial fuel cells methodology and
technology Environ SciTechnol 40 5181-5192
69 Lovely DR (2006) Bug juice harvesting electricity with microorganisms Nature
Review| Microbiology 4 497-508
70 Lovley DR Phillips EJ (1988) Novel Mode of Microbial Energy Metabolism
Organic Carbon Oxidation Coupled to Dissimilatory Reduction of Iron or
Manganese Appl Environ Microbiol 54(6) 1472-1480
71 Ly HK Sezer M Wisitruangsakul N Feng JJ Kranich A Millo D Weidinger
IM Zebger I Murgida DH Hildebrandt P (2011) Surface-enhanced vibrational
spectroscopy for probing transient interactions of proteins with biomimetic interfaces
electric field effects on structure dynamics and function of cytochrome c FEBS J
278(9)1382-90
72 Macaskie LE Hewitt CJ Shearer JA Kent CA (1995) Biomass production for
the removal of heavy metals from aqueous solutions at low pH using growth-
decoupled cells of a Citrobacter sp Int Biodeter Biodegr 3573ndash92
73 Malki M De Lacey AL Rodriguez N Amils R Fernandez VM (2008)
Preferential use of an anode as an electron acceptor by an acidophilic bacterium in
the presence of oxygen Appl Environ Microbiol 74 4472ndash4476
74 Meitl LA Eggleston CM Colberg PJS Khare N Reardon CL Shi L (2009)
Electrochemical interaction of Shewanella oneidensis MR-1 and its outer membrane
cytochromes OmcA and MtrC with hematite electrodes Geochim Cosmochim
Ac 73(18) 5292-5307
75 Morozan A Stamatin L and Nastase
F (2007) The biocompatibility
microorganisms-carbon nanostructures for applications in microbial fuel cells Phys
Stat Sol 6 1797-1803
76 Morris JM Jin S Crimi B Pruden A (2009) Microbial fuel cell in enhancing
anaerobic biodegradation of diesel Chem Eng J 146 161ndash167
77 Murray AE Lies D Li G Nealson K Zhou J Tiedje JM (2001) DNADNA
hybridization to microarrays reveals gene-specific differences between closely
related microbial genomes Proc Natl Acad Sci U S A 98 9853-9858
78 Muyzer G de Waal EC Uitterlinden AG (1993) Profiling of complex microbial
communities by denaturing gradient gel electrophoresis analysis of polymerase chain
88
reaction amplified genes coding for 16S rRNA Appl Environ Microbiol 59 695ndash
700
79 Myers CR Myers JM (1992) Localization of cytochromes to the outer membrane
of anaerobically grown Shewanella putrefacians MR-1 J Bacteriol 174 3429ndash3438
80 Narde GK Kapley A Purohit HJ (2004) Isolation and characterization of
Citrobacter strain HPC255 for broad-range substrate specificity for chlorophenols
Curr Microbiol 48(6) 419-423
81 Okabe S Ito T Sugita K Satoh H (2005) Succession of internal sulfur cycles and
sulfur-oxidizing bacterial communities in microaerophilic wastewater biofilms Appl
Environ Microbiol 71(5) 2520-2529
82 Park DH and Zeikus JG (2000) Electricity generation in microbial fuel cells using
neutral red as an electronophore Appl Environ Microbial 66 292-1297
83 Park DH and Zeikus JG (2002) Impact of electrode composition on electricity
generation in a single-compartment fuel cell using Shewanella putrefaciens Appl
Microbiol Biotechnol 5958ndash56
84 Park DH and Zeikus JG (2003) Improved fuel cell and electrode designs for
producing electricity from microbial degradation Biotechnol Bioeng 81 348ndash355
85 Park HS Kim BH Kim HS Kim HJ Kim GT Kim M Chang IS Park Y
K Chang HI (2001) A novel electrochemically active and Fe(III)-reducing
bacterium phylogenetically related to Clostridium butyricum isolated from a
microbial fuel cell Anaerobe 7 297-306
86 Parot S Nercessian O Delia ML Achouak W Bergel A (2009) Electrochemical
checking of aerobic isolates from electrochemically active biofilms formed in
compost J Appl Microbiol 106(4) 1350-1359
87 Pham CA Jung SJ Phung NT Lee J Chang IS Kim BH Yi H Chun J
(2003) A novel electrochemically active and Fe(III)-reducing bacterium
phylogenetically related to Aeromonas hydrophila isolated from a microbial fuel cell
FEMS Microbiol Lett 223 129-134
88 Qiao Y Bao SJ Li CM Cui XQ Lu ZS Guo J (2008) Nanostructured
polyanilinetitanium dioxide composite anode for microbial fuel cells ACS Nano 2
113-119
89 Qiu R Zhao B Liu J Huang X Li Q Brewer E Wang S Shi N (2009) Sulfate
reduction and copper precipitation by a Citrobacter sp isolated from a mining area J
Hazard Mater 1641310-1315
90 Quan X Yang SG Ruan XL and Zhao HM (2005) Preparation of titania
nanotubes and their environmental applications as electrode Environ Sci Technol 39
3770ndash3775
91 Rabaey K Boon N Hofte M Verstraete W (2005) Microbial phenazine
production enhances electron transfer in biofuel cells Environ Sci Technol 393401ndash
3408
89
92 Rabaey K Boon N Siciliano S D Verhaege M Verstraete W (2004) Biofuel cells
select for microbial consortia that self-mediate electron transfer Appl Environ
Microbiol 70(9) 5373ndash5382
93 Rabaey K Lissens G Siciliano S D Verstraete W (2003) Amicrobial fuel cell
capable of converting glucose to electricity at high rate and efficiency Biotechnol
Lett 25 1531-15
94 Rabaey K Verstraete W (2005) Microbial fuel cells novel biotechnology for
energy generation Trends Biotechnol 23(6) 291-298
95 Reguera G McCarthy KD Mehta T Nicoll JS Tuominen MT Lovley DR
(2005) Extracellular electron transfer via microbial nanowires Nature 435 1098-
1101
96 Reguera G Nevin KP Nicoll JS Covalla SF Woodard TL Lovley DR (2006)
Biofilm and nanowire production leads to increased current in Geobacter
sulfurreducens fuel cells Appl Environ Microbiol 72 7345-7348
97 Reimers CE Tender LM Fertig S Wang W (2001) Harvesting energy from the
marine sediment-water interface Environ Sci Technol 35192-195
98 Rezaei F Xing D Wagner R Regan JM Richard TM Logan BE (2009)
Simultaneous cellulose degradation and electricity production by Enterobacter
cloacae in a microbial fuel cell Appl Microbiol Biotechnol 75 3673ndash3678
99 Richter H Nevin KP Jia H Lowy DA Lovley DR Tender LM (2009) Cyclic
voltammetry of biofilms of wild type and mutant Geobacter sulfurreducens on fuel
cell anodes indicates possible roles of OmcB OmcZ type IV pili and protons in
extracellular electron transfer Energy Environ Sci 2 506ndash516
100 Riesenfeld SC Schloss DP Handelsman J(2004) Metagenomics Genomic
Analysis of Microbial Communities Annual Review Genetics 38525-552
101 Rozendal RA Hamelers HV Buisman CJ (2006) Effects of membrane cation
transport on pH and microbial fuel cell performance Environ Sci Technol 40
5206-5211
102 Rozendal RA Hamelers HVM Euverink GJW Metz SJ Buisman CJN (2006)
Principle and perspectives of hydrogen production through biocatalyzed electrolysis
Int J Hydrogen Energ 311632-1640
103 Rozendal RA Hamelers HV Molenkamp RJ Buisman CJ (2007) Performance of
single chamber biocatalyzed electrolysis with different types of ion exchange
membrances Water Res 41 1984-1994
104 Schluumlter A Bekel T Diaz NN Dondrup M Eichenlaub R Gartemann
KH Krahn I Krause L Kroumlmeke H Kruse O Mussgnug JH Neuweger
H Niehaus K Puumlhler A Runte KJ Szczepanowski R Tauch A Tilker
A Viehoumlver P Goesmann A (2008) The metagenome of a biogas-producing
microbial community of a production-scale biogas plant fermenter analyzed by the
454-pyrosequencing technology J Biotechnol136 77-90
90
105 Scott J H and Nealson KH (1994) A biochemical study of the intermediary carbon
metabolism of Shewanella putrefaciens J Bacteriol 176(11) 3408ndash3411
106 Sekiguchi H Tomioka H Nakahara T and Uchiyama H (2001) A single band
does not always represent single bacterial strains in denaturing gradient gel
electrophoresis analysis Biotechnology Letters23 1205ndash1208
107 Sharma T Leela M Reddy A Chandra TS Ramaprabhu S (2008) Development
of carbon nanotubes and nanofluids based microbial fuel cell Int J Hydrogen Energ
336749ndash6754
108 Shea C Clauwaert P Verstraete W Nerenberg R (2008) Adapting a denitrifying
biocathode for perchlorate reduction Water Sci Technol 58 (10) 1941-1946
109 Shi L Squier TC Zachara JM Fredrickson JK (2007) Respiration of metal (hydr)
oxides by Shewanella and Geobacter a key role for multihaem c-type cytochromes
Mol Microbiol 6512-20
110 Stams AJ and Plugge CM(2009)Electron transfer in syntrophic communities
of anaerobic bacteria and archaea Nat Rev Microbiol7(8)568-77
111 Sukkasem C Xu S Park S Boonsawang P Liu H (2008) Effect of Nitrate on the
Performance of Single Chamber Air Cathode Microbial Fuel Cells Water Res 42(19)
4743-4750
112 Summers ZM Fogarty HE Leang C Franks AE Malvankar NS Lovley DR
(2010)Direct exchange of electrons within aggregates of an evolved syntrophic
coculture of anaerobic bacteria Science 330 (6009)1413-5
113 Szczepanowski R Bekel T Goesmann A Krause L Kroumlmeke H Kaiser
O Eichler W Puumlhler A Schluumlter A (2008) Insight into the plasmid metagenome
of wastewater treatment plant bacteria showing reduced susceptibility to
antimicrobial drugs analysed by the 454-pyrosequencing technology J
Biotechnol 136 54-64
114 Tamura K Dudley J Nei M Kumar S (2007) MEGA4 Molecular Evolutionary
Genetics Analysis (MEGA) software version 40 MolBiol Evol 24(8) 1596-1599
115 Thompson JD Gibson TJ Plewniak F Jeanmougin F Higgins DG (1997) The
CLUSTAL_X windows interface flexible strategies for multiple sequence alignment
aided by quality analysis tools Nucleic Acids Res 25 4876-4882
116 Thormann KM Duttler S Saville RM Hyodo M Shukla S Hayakawa Y
Spormann AM (2006) Control of formation and cellular detachment from
Shewanella oneidensis MR-1 biofilms by cyclic di-GMP J Bacteriol 1882681-2691
117 Thormann KM Saville RM Shukla S Pelletier DA Spormann AM (2004)
Initial Phases of biofilm formation in Shewanella oneidensis MR-1 J Bacteriol
186(23) 8096-104
118 Torres CI Kato Marcus A Rittmann BE (2008b) Proton transport inside the
biofilm limits electrical current generation by anode-respiring bacteria Biotechnol
Bioeng 100 872ndash881
91
119 Torres CI Marcus AK Lee HS Parameswaran P Krajmalnik-Brown R Rittmann
BE (2010) A kinetic perspective on extracellular electron transfer by anode-
respiring bacteria FEMS Microbiol Rev 234(1)3-17
120 Torres CI Marcus AK Parameswaran P Rittmann BE (2008a) Kinetic
experiments for evaluating the NernstndashMonod model for anode-respiring bacteria
(ARB) in a biofilm anode Environ Sci Technol 42 6593ndash6597
121 Tsai HY Wu Ch-Ch Lee Ch-Y Shih EP (2009) Microbial fuel cell performance
of multiwall carbon nanotubes on carbon cloth as electrodes J Power Sources
194199-205
122 Tyson GW Chapman J Hugenholtz P Allen EE Ram RJ Richardson
PM Solovyev VV Rubin EM Rokhsar DS Banfield JF(2004) Community
structure and metabolism through reconstruction of microbial genomes from the
environment Nature42837-43
123 Venkateswaran K Moser DP Dollhopf ME Lies DP Saffarini D A MacGregor
BJ Ringelberg DB White DC Nishijima M Sano H Burghardt J Stackebrandt
E Nealson KH (1999) Polyphasic taxonomy of the genus Shewanella and
description of Shewanella oneidensis sp Int J Syst Bacteriol 49705-724
124 von Canstein H Ogawa J Shimizu S Lloyd JR (2008) secretion of Flavins
by Shewanella Species and Their Role in Extracellular Electron Transfer Appl
Environ Microbiol 74 3615-623
125 Wang A Liu W Cheng Sh Xing D Zhou J Logan BE (2009) Source of methane
and methods to control its formation in single chamber microbial electrolysis cells
Int J Hydrogen Energ 34 3653-3658
126 Wang J Ma T Zhao L Lv J Li G Liang F Liu R (2008) PCRndashDGGE method
for analyzing the bacterial community in a high temperature petroleum reservoir
World J Microbiol Biotechnol 2 9694-9702
127 Wang Q Garrity GM Tiedje JM Cole JR (2007) Naiumlve Bayesian Classifier for
Rapid Assignment of rRNA Sequences into the New Bacterial Taxonomy Appl
Environ Microbiol 73(16) 5261-5267
128 Wang YF Masuda M Tsulimura S Kano K (2008) Electrochemical regulation of
the end-product profile in Propionibacterium freudenreichii ET-3 with an
endogenous mediator Biotechnol Bioeng 101(3) 579ndash586
129 Wrighton KC Agbo P Warnecke F Weber KA Brodie EL DeSantis TZ
Hugenholtz P Andersen GL Coates JD (2008) A novel ecological role of the
Firmicutes identified in thermophilic microbial fuel cells ISME J 2 1146ndash1156
130 Xing D Zuo Y Cheng S Regan JM Logan BE (2008) Electricity generation by
Rhodopseudomonas palustris DX-1 Environ Sci Technol 42 4146-4151
131 Xu S Liu H Fan Y Schaller R Jiao J Chaplen F (2012) Enhanced performance
and mechanism study of microbial electrolysis cells using Fenanoparticle-decorated
anodes Appl Microbiol Biotechnol 93(2)871-880
92
132 Yang Y Harris DP Luo F Wu L Parsons AB Palumbo AV Zhou J (2008)
Characterization of the Shewanella oneidensis Fur gene roles in iron and acid
tolerance response BMC Genomics 9 Suppl 1S11
133 Zhang L Zhou S Zhuang L Li W Zhang J Lu N Deng L (2008) Microbial
fuel cell based on Klebsiella pneumoniae biofilm Electrochem Commun10 1641ndash
1643
134 Zuo Y Xing DF Regan JM Logan BE (2008) Isolation of the exoelectrogenic
bacterium Ochrobactrum anthropi YZ-1 by using a U-tube microbial fuel cell Appl
Environ Microbiol 74 3130-3137
93
APPENDICES
List of published papers during PhD study
1 Shoutao Xu Hong Liu Yanzhen Fan Rebecca Schaller Jun Jiao and Frank
Chaplen (2012) Enhanced performance and mechanism study
of microbial electrolysis cells using Fe nanoparticle-decorated anodes Applied
Microbiology and Biotechnology 93(2)871-880
2 Shoutao Xu Hong Liu (2011) New exoelectrogen Citrobacter sp SX-1 isolated
from a microbial fuel cell Journal of Applied Microbiology111(5)1108-1115
3 Yanzhen Fan Shoutao Xu Rebecca Schaller Jun Jiao Frank Chaplen Hong Liu
(2011) Nanoparticle decorated anodes for enhanced current generation in microbial
electrochemical cells Biosensors and Bioelectronics 26(5) 1908-1912
4 Tunc Catal Shoutao Xu Kaichang Li Hakan Bermek Hong Liu
(2008)
Electricity generation from polyalcohols in single-chamber microbial fuel
cells Biosensors and Bioelectronics 24(4)849-854
5 Chontisa Sukkasema Shoutao Xu Sunhwa Park Piyarat Boonsawang Hong Liu
(2008) Effect of nitrate on the performance of single chamber air cathode
microbial fuel cells Water research 424743-4750
6 Rebecca Schaller Yanzhen Fan Shoutao Xu Alan Fern Frank Chaplen Hong
Liu and Jun Jiao (2009) Vertically Aligned Multi-walled Carbon Nanotube
Decorated Anodes for Microbial Fuel Cells Proceedings of Materials Research
Society 2009 1170 R05-13

LIST OF FIGURES
Figure Page
1-1 Generalized schematics (a) Microbial fuel cell (MFCs)(b) Microbial
electrolysis cell (MECs) helliphelliphelliphelliphelliphelliphelliphelliphelliphellip helliphelliphelliphellip
2
1-2 Mechanisms for extracellular electron transport in a MFCs anode 7
2-1 Polarization curves of high power mixed culture MFCshelliphelliphelliphelliphelliphellip 20
2-2 DGGE result of high power generation mixed culture MFCshelliphelliphellip 21
2-3 Bacterial species and percentage of bacterial community of mixed
culture MFCs helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip
22
2-4 Phylogenetic trees of bacterial species from mixed culture MFCs 22
2-5 Dominant bacterial species identification in DGGEhelliphelliphelliphelliphelliphelliphelliphellip 23
2-6 Current density generated by different dominant isolateshelliphelliphelliphelliphelliphellip 24
3-1 Phylogenetic tree of strain SX-1 and closely related species based on
16S rDNA sequenceshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip
35
3-2 SEM image of planktonic cells of Citrobacter sp SX-1helliphelliphelliphelliphelliphellip 35
3-3 Electricity generation by Citrobacter sp SX-1 in a single chamber
MFCs with sodium citrate helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip
36
3-4 Electricity generation of Citrobacter sp SX-1 using different
substrates in single chamber air-cathode MFCs at 1 KΩhelliphelliphellip
37
3-5 Power and voltage generation by Citrobacter sp SX-1 as a function
of current density helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip
38
3-6 Four current generating stages for CV analysis helliphelliphelliphelliphelliphelliphelliphellip 39
3-7 Cyclic Voltammetry analysis of of Citrobacter sp SX-1 biofilms helliphellip 41
4-1 SEM images of Fe NPs decorated anodeshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 50
4-2 Effects of Fe-NP decorated anodes on the current density in MECs helliphellip 51
4-3 Differentially expressed genes grouped by functional classification in
Fe NP decorated anodeshelliphelliphelliphelliphelliphellip helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip
53
5-1 SEM images of CNT decorated anodeshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 70
5-2 Effects of CNT decorated anodes on the current density in MECshelliphellip 73
5-3 Differentially expressed genes grouped by functional classification in
CNT decorated anodes helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip
74
LIST OF TABLES
Table
1-1
4-1
4-2
4-3
Taxa of bacteria current density generated and reactor configuration
used in microbial electrochemical cell experimentshelliphelliphelliphellip
Expression levels of genes related to biofilm formationhelliphelliphelliphelliphellip
Genes related to anaerobic growth and electron transfer with
significantly change expression levelhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip
Expression level of flavin and cytochrome geneshelliphelliphelliphelliphelliphelliphelliphellip
Page
4
55
56
58
4-4
5-1
5-2
Other genes with significantly changed expression level
Expression level of cytochrome c as response to CNT decorated anodes
Expression level of genes related to flavin synthesis as response to CNT
decorated anode in CNT decorated anodeshelliphellip
59
76
78
1
1
Bacterial Community Analysis New Exoelectrogen Isolation and Enhanced Performance
of Microbial Electrochemical Systems Using Nano-Decorated Anodes
Chapter 1
General Introduction
11 Microbial Fuel Cells and Microbial Electrolysis Cells
Microbial electrochemical systems (MESs) have drawn the attention of researchers
in recent years due to their promising applications in a variety of scientific fields such as
renewable energy generation bioremediation and wastewater treatment In a MES
microorganisms interact with electrodes catalyzing oxidation and reduction reactions at
the anode and the cathode
The most-described type of MESs is the microbial fuel cells (MFCs) in which
useful power is generated directly using the catalytic action of active microorganisms
(Kim 2002 Cheng and Logan 2007 Cheng et al 2006) In a typical two-chamber
MFCs (Figure 1a) organic matter is oxidized by electrochemically active
microorganisms in the anode chamber to release protons and electrons Protons diffuse
into the cathode chamber through a proton exchange membrane Meanwhile electrons
are transferred to the anode through various mechanisms (Logan et al 2006 Lovley
2
2006 Rabaey et al 2003) and then travel to the cathode where normally they combine
with oxygen and protons to form water
Another common used type of MES is the microbial electrolysis cell (MECs) in
which hydrogen is produced instead of electricity by applying a circuit voltage to the
MFCs system and maintaining anaerobic conditions in the cathode chamber (Liu et al
2005 Logan et al 2008 Rozendal et al 2006 Schaller et al 2009) (Figure 1b) A
cathode potential of at least - 410 mV (vs standard hydrogen electrode at pH 70) is
required to produce hydrogen at the cathode Therefore a theoretical potential of 110 mV
(ie 410-300 mV) is needed but voltages of gt02 V are normally applied due to various
overpotentials
Fig 1-1 Generalized schematics (a) Microbial fuel cells (MFCs) where electricity is
captured through the resistance R (b) Microbial electrolysis cells (MECs) where
hydrogen gas is produced in the cathode chamber R for resistor PS for power supply and
PEM for proton exchange membrane
The fundamental feature shared by microbial electrochemical systems (MESs)
(MECs and MFCs) is the microbe-catalyzed electron transfer from organic matter to
electrodes (anodes) (Rebaey et al 2004) Therefore different types of MESs can be
PEM
R
H+
e- e-
Anode
Bacte
ria
a
Cath
ode
H2O
PEM
PS
H+
e- e-
Anode
Bacte
ria
b
Cath
ode
H2
3
utilized to investigate the phenomena and mechanisms of interactions between microbes
and electrodes
12 Bacteria and Electron Transfer Mechanisms in MESs
The electrochemically active bacteria in MFCs are thought to be iron-reducing
bacteria such as Shewanella and Geobacter species (Gorby et al 2006) They have great
importance in the natural environment principally in metal oxidation and reduction
However recent studies have shown that the diversity of bacterial communities is much
greater than these model iron reducers on MFCs anodes (Reimers et al 2001 Liu et al
2004b Kim et al 2005) Over 20 electrochemically active bacterial species those can
transfer electrons exocellularly to electrodes have been reported in the past 10 years
(Logan 2009 Liu et al 2010) These bacteria have been categorized into diverse genetic
groups so far including alphaproteobacteria (Rhodopseudomonas Ochrobactrum and
Acidiphilium) (Malki et al 2008 Xing et al 2008 Zuo et al 2008) betaproteobacteria
(Rhodoferax) (Chaudhuri et al 2003) gammaproteobacteria (Shewanella Pseudomonas
Klebsiella Enterobacter and Aeronomas) (Kim et al 2002 Pham et al 2003 Rabaey et
al 2005 Bretschger et al 2007 Zhang et al 2008 Chung et al 2009 Rezaei et al 2009)
deltaproteobacteria (Geobacter Geopsychrobacter Desulfuromonas and Desulfobulbus)
(Bond et al 2002 Bond and Lovley 2003 Holmes et al 2004ab 2006)
Epsilonproteobacteria (Arcobacter) (Fedorovich et al 2009) Firmicutes (Clostridium and
Thermincola) (Park et al 2001 Wrighton et al 2008) Acidobacteria (Geothrix) (Bond
and Lovley 2005) and actinobacteria (Propionibacterium) (Wang et al 2008) A wider
range of electrochemically active bacteria are expected to be discovered
4
Table 1 Taxa of bacteria current density generated and reactor configuration used in
microbial electrochemical cell experiments (Liu et al 2010)
The electrochemically active bacterial species that possess the ability to transfer
electrons outside of the cell are called exoelectrogens in the MESs research field The
different exoelectrogens have demonstrated a wide-ranging power generation ability in
MFCs For example Shewanella oneidensis MR-1 has demonstrated the ability to
generate 0018 Am2 current density in single chamber MFCs while the Geobacter
Taxon Microorganisms Current Density (Am2
)
α-proteobacteria
Rhodopseudomonas palustris DX-1 003
Ochrobactrum anthropi YZ-1 071
Acidiphilium sp 32sup5 300
β-proteobacteria Rhodoferax ferrireducens 0031
γ-proteobacteria
Shewanella putrefaciens IR-1 0016
Shewanella oneidensis DSP10 0013
Shewanella oneidensis MR-1 0018
Pseudomonas aeruginosa KRA3 0017
Escherichia coli K12 HB101 100
Klebsiella pneumoniae L17 120
Enterobacter cloacae 013
Aeromonas hydrophila PA3 030
δ-proteobacteria
Geobacter metallireducens 065
Geobacter sulfurreducens 800
Desulfuromonas acetoxidans 0005
Geopsychrobacter
Electrodiphilus strain A2
0066
Desulfobulbus propionicus 003
Firmicutes
Lactococcus lactis 003
Thermincola sp strain Jr 020
Clostridium butyricum EG3 022
Thermincola ferriacetica Z-0001 040
Brevibacillus spp PTH1 0009
Desulfitobacterium hafniense DCB2 110
Acidobacteria Geothrix fermentans 0097
Actinobacteria Propionibacterium freudenreichiiET-3 120
5
sulfurreducens species has generated current densities as high as 80 Am2 which is 400
times higher than the one generated by Shewanella oneidensis MR-1 However mixed-
culture communities have generated much higher power densities than their pure-culture
counterparts in MFCs perhaps due to synergistic interactions within the anode bacterial
communities and the participation of currently unknown bacteria species and mechanisms
(Jung and Regan 2007) The dominant bacteria species vary in mixed cultures bacteria
communities in MFCs due to the enrichment of different substrates
Traditional methods of extroelectrogen study depend on cultivation hampered novel
exoelectrogens discovery because the inadequacy of defined media underestimates the
actual microbial diversity in MFCs Slow growth rate and unknown growth requirements
of anaerobic microorganisms make some anaerobic extroelectrogens cultivation difficult
However these limitations have been overcome by using molecular biological methods
based on DNARNA analysis Molecular biological techniques are now widely applied to
assess the diversity of microbial communities by analyzing the 16S rDNA sequence The
most commonly used molecular biological techniques for bacterial community analysis
include denaturing gradient gel electrophoresis (DGGE) restriction fragment length
polymorphism (RFLP) and 16S rDNA sequencing These methods are much less time
consuming than traditional isolation and cultivation methods
Isolated exoelectrogens were utilized to explore the mechanism of electron transfer
to the anode However the mechanisms of electron transfer to extracellular electron
acceptors are not well understood Three mechanisms have been proposed for exocellular
transport of electrons by exoelectrogens to anodes in MFCs so far Some exoelectrogenic
bacteria can transfer electrons to electrodes through soluble redox compounds (Bond and
6
Lovley 2005) These compounds include artificial mediators and mediators secreted by
exoelectrogenic bacteria A variety of chemicals have been used to facilitate the shuttling
of electrons from inside of cell to electrodes outside the cell These exogenous mediators
include neutral red (Park et al 1999) anthraquinone-2-6 disulfonate (AQDS) thionin
potassium ferricyanide (Bond et al 2002) methyl viologen and others (Logan 2004
Rabaey and Verstraete 2005) Some bacteria can secrete their own chemical mediator
for example Pseudomonas aeruginosa secretes pyocyanin and phenazine-1-carboxamide
to transfer electrons toward the MFCs anode (Rabaey et al 2005) Another evidence for
mediator production by bacteria is Geothrix fermentans When the medium was replaced
in a MFCs that had stable power generation with this bacteria power dropped by 50
and required 10 days to resume the original level
Some bacteria can directly transfer electrons to anodes via outer cell membrane
proteins (Myers and Myers 1992) The outer cell membrane protein cytochrome c is
thought to play a critical role in to transferring electrons to anodes Ly et al (2011)
isolated the haem protein cytochrome c and demonstrated that electric field effects may
be functional for the natural redox processes of cytochrome c in the respiratory chain
Shewanella oneidensis is the example of a bacterium that is capable of transfer electron to
anode via outer cell membrane proteins When Shewanella oneidensis adhered to an iron
surface the greater force has showed benefits to grow cells because closer contact
required for electron transfer from cell bound cytochromes (Lower et al 2001)
7
Fig1-2 Mechanisms for extracellular electron transport in an MFCs anode (1) direct
contact (top in green) (2) by nanowires (middle in purple) and (3) self-produced
mediators (bottom in blue) (Logan 2009)
More and more evidence supports the involvement of bacterial nanowires in
extracellular electron transport (Reguera et al 2005 2006 Gorby et al 2006) Nanowires
are conductive appendages produced by both Geobacter and Shewanella species (Gorby
and Beveridge 2005) The conductivity of the appendages was examined and confirmed
by using conductive scanning tunneling microscopy (STM) (Gorby et al 2006)
Nanowires can carry electrons from the cell to the anode surface of MFCs
The solid component of the extracellular biofilm matrix has high efficiency on
extracellular electron transfer compared with other extracellular electron transfer
8
mechanisms and recently Torres et al (2010) hypothesized that the solid component of
the extracellular biofilm matrix formed by exoelectogens is another way bacteria transfer
electrons to electrodes This hypothesis was based on kinetic analysis of each EET
mechanism reported in available literature (Torres et al 2010)
13 Anode electrodes
In MFCsMECs anode electrodes are a critical component because exoelectrogens
adhere to the surface of anodes to transfer electrons to the electrode The characteristics
of anodes have significant effects on electron transfer rate from bacteria to anode
electrodes in MFCs The requirements of an anode material are it should be highly
conductive non-corrosive have a high specific surface area (area per volume) high
porosity be non-fouling inexpensive and easily scaled to larger sizes Of these
properties the most important one that is different from other biofilm reactors is that the
material must be electrically conductive Normally they are made of various carbon
materials including carbon fiber carbon clothe and carbon paper due to their stability
high conductivity and high specific surface-area Nevertheless they have little
electrocatalytic activity for the anode microbial reactions and thus a modification of the
carbon materials is the main approach for improving their performance Consequently
there is a great need to develop a new type of anode material for MFCsMECs
It is a great challenge to develop a new anode material to further increase the power
density of a MFCs The nature of the catalytic mechanism of a MFCrsquos anode involves not
only a biological but also an electrocatalytic process An optimal nanostructure with a
high specific surface area favorable for both catalytic processes could play a critical role
in improving the power density of the MFCs such a structure needs to host the bacteria
9
with high bioactivity while enhancing the electron-transfer rate Schroumlder et al (2008)
employed PANI to modify a platinum anode for MFCs and achieved a current density 1
order of magnitude higher than the previously reported value PANIinorganic
composites are also reported to have better conductivity Qiao et al also applied a new
mesoporous TiO2 electrode material with uniform nanopore distribution and a high
specific surface area to anode in comparison to previously reported work with E coli
MFCs the composite anode delivers 2-fold higher power density (Qiao et al 2008)
Thus it has great potential for use as the anode in a high-power MFCs and may be a new
approach for improving performance of MFCs
14 Other parts of MESs
141 Membranes and ion transport
The ion exchange membrane is another one of the critical components in two-
chamber MESs systems It separates anode and cathode chambers and at the same time
maintains the electron neutrality of the system ie transport of electrons to the cathode
needs to be compensated by transport of an equal amount of positive charge to the
cathode chamber A proton exchange membrane (PEM) such as Nafion a per-fluorinated
sulphonic acid membrane (PFSAs) consisting of a hydrophobic fluorocarbon backbone to
which hydrophilic sulfonate groups (-SO3-) are attached is commonly used in chemical
fuel cell systems For MFCs systems however mainly cation species like Na+ and K
+
other than proton are often responsible for the dominant transport of positive charge
through the cation exchange membrane (CEM) to maintain electroneutrality due to the
low proton concentration in any aqueous medium with near neutral pH (Rozendal et al
2006) Consequently the pH increases in the cathode chamber due to the consumption of
10
protons and decreases in the anode chamber because of the accumulation of protons
(Rozendal et al 2007) This pH change in two-chamber systems results in a decrease of
the cathode potential and performance The application of anion exchange membrane
(AEM) has also been explored in MFCs systems (Cheng and Logan 2007 Kim and
Logan 2007) where it has been proposed that protons are transferred via pH buffers like
phosphate anions
142 Cathodes and Catalysts
The cathode is another challenge for making MFCs commercially available
technology because the chemical reaction that occurs at the cathode is difficult to
engineer as the electrons protons and oxygen must all meet at a catalyst in a tri-phase
reaction (solid catalyst air and water) The catalyst must be on a conductive surface and
must be exposed to both water and air so that protons and electrons in these different
phases can reach the same point The most commonly used material for a cathode is
commercially available carbon paper pre-loaded with a Pt catalyst on one side When it is
used in a MFC the side that contains the catalyst faces the water and the uncoated side
faces air To reduce the high cathode cost associated with platinum catalyst other precious-
metal-free cathodes such as (NiMo and NiW) were developed by electrodepositing on a
carbon fiber They have achieved comparable performance with Pt catalyst with same
loading at a much lower cathode fabrication cost (Hu 2010)
The requirements of cathode for MECs are quite similar to the requirements of
cathode for MFCs but easier than cathode for MFCs for the manufacturing process
because the cathode in MECs is not necessarily to exposed to air Therefore it can be
made of the exact same materials of cathode in MFCs except a waterproof layer
11
Recently some researchers have attempted to apply microorganisms as a biocatalyst to
precede the combination of electron with oxygen in the cathodes
15 Dissertation overview
Low power densities in MESs limit practical applications The improvement of
MESs performance requires a detailed understanding of the physiology and ecology of
microorganisms in MESs including the mechanism of electron transfer to the anode from
the microorganism
This dissertation focuses on the problem of the low power density of MESs The
bacterial community structure of a high power generated mixed culture communities in
MFCs will be identified firstly and then one of major exoelectrogens will be isolated and
characterized This information will be helpful to understand the physiology and ecology
of exoelectrogens in MESs Consequently they will be beneficial to improve power
density of MFCs Lastly the nanostructure decorated anodes in MECs will be utilized to
improve the power density The power enhancement mechanism will be explored by
using a whole genome microarray They are presented here as four papers
In the first paper the cultivation independent molecular biological techniques
denaturing gradient gel electrophoresis (DGGE) and 16S rDNA clone library are utilized
to analyze the bacterial community structure of a higher power mixed culture MFCs The
analyzed results provide fundamental information for isolating the dominant bacteria in
mixed culture MFCs Two of dominant bacterial species has been isolated and used aone
to test power generation in MFCs The possible interaction among different bacterial
species in mixed culture is discussed
12
In the second paper one isolated exoelectrogenic bacterial strain SX-1 is
characterized It is identified as a member of the Citrobacter genus and power generation
is tested ultilizing a wide range of different substrates The electron transfer mechanism is
explored using Cyclic Voltammetry (CV) This study increases the known diversity of
power generating exoelectrogens and provides a new strain to explore the mechanisms of
extracellular electron transfer from bacteria to electrodes
The third paper shows effects on MESs performance by Fe nanoparticle decorated
anodes in the MESs The average current density produced with Fe nanoparticle
decorated anodes increased up to 59-fold higher than plain graphite anodes A whole
genome microarray is utilized to analyze the possible mechanism of enhanced current
density as responded to nanoparticle decorated anodes
The fourth paper describes the effects of carbon nanotube (CNT) modified anode on
the performance S oneidensis MR-1 in MESs Results demonstrate that current densities
produced with CNT decorated anodes are up to 56-fold higher than plain graphite anodes
The possible mechanisms of enhanced current density by CNT decorated are explored
13
Chapter 2
Bacterial community analysis of mixed consortium in higher
power density MESs
Shoutao Xu and Hong Liu
1 Introduction
The improvement in the performance of mixed culture MFCs requires an
understanding of the ecology in microbial communities of MFCs Many researchers have
attempted to characterize microbial populations and activities to elucidate the behaviors
and ecology of microorganisms in MFCs (Wrighton et al 2008 Fedorovich et al 2009)
In order to study the microbial ecology of the mixed culture in a MFC and select the
appropriate isolation medium for dominant bacterial species in the mixed culture the
fundamental steps are to analyze the bacterial diversity in the mixed culture of MFCs and
identity the dominant bacterial species in bacteria communities in MFCs
14
As for the identification of bacterial communities typically there are two general
methods The first method for identification of bacterial community is the traditional
cultivation processes using selective nutrients to promote the growth of different types of
bacteria within the samples (Amman et al 2000) The community structure can then be
assessed by identifying the isolates from the dominant colonies that were cultured This
can often be costly and laborious as each isolate has to be further studied by examining
its physiology taxonomy and reactivity to stains (Adwards et al 1989)
The second method relies on utilizing molecular techniques to analyze bacterial
community DNA Several molecular methods involving the extraction and analysis of
DNA from entire bacterial communities are used to identify genetic fingerprints of
bacteria These methods including the cloning and sequencing of 16S rDNA automated
ribosomal intergenic spacer analysis (ARISA) terminal restriction fragment length
polymorphism (T-RFLP) and denaturing gradient gel electrophoresis (DGGE) generate
profiles of bacterial community structures They can rapidly assess complex communities
from various environments (Amman et al 2000)
Due to the conservative characteristic of 16S rDNA in bacteria during the process
of evolution 16S rDNA sequencing can be used to identify different species of bacteria
Therefore the molecular techniques of denaturing gradient gel electrophoresis (DGGE)
with PCR and 16S rDNA clone library are used for analysis of the microbial diversity
These methods are more convenient and save time compared to traditional
isolationcultivation methods for microorganism analysis
In this work a biofilm bacterial community from an anode of a MFCs wase studied
by using denaturing gradient gel electrophoresis (DGGE) of PCR-amplified partial 16S
15
rRNA genes followed by cloning and sequencing of 16S rDNA The results provided
essential information for dominant bacterial isolation in mixed culture MFCs
2 Materials and methods
21 High power generation of mixed culture MFCs
Mixed cultures were originally inoculated from domestic wastewater (Corvallis
Wastewater Treatment Plant Corvallis OR) to MFCs after operating for 2 years using a
defined medium solution (Lovely 2002) with sodium acetate as the carbon source A
new MFCs was inoculated from the operating MFCs The polarization curves were
performed to measure the power generation when maximal stable power were established
(normally 3-5 batches post-inoculation) and the genomic DNA from the microbial
biofilm on the anode was extracted under the sterile conditions for further bacterial
community analysis
22 Denaturing gradient gel electrophoresis (DGGE)
DGGE with PCR is a method of analysis of bacterial community composition
based on different GC percentage of 16s rDNA in different bacterial species PCR-DGGE
comprises three steps (i) extraction of total community DNA from the sample (ii) PCR-
controlled amplification using specific oligonucleotide primers and (iii) separation of the
amplicons using DGGE For this purpose a reproducible and efficient method for total
DNA extraction is indispensable and needs to be evaluated and optimized depending on
the nature of the sample In the subsequent PCR step multiple PCR primer sets with
different resolution can be used In most PCR-DGGE applications on bacteria universal
or specific primers are targeting the 16S rRNA gene Upon electrophoresis of the PCR
16
amplicons and gel staining (using ethidium bromide silver staining or SYBR green)
DGGE gels are digitally captured and further analyzed using computer software packages
The richness of DNA fragment bands in DGGE indicates the bacterial diversity of the
sample
The detailed steps are as followed Biofilms were scratched from the anodes of high
power generated MFCs fed with sodium acetate Bacterial genomic DNA was extracted
from the biofilm samples using the DNeasy blood and tissue kit (Qiagen) according to the
manufacturerrsquos instructions The universal primer set was used to amplify the 16S rDNA
from the extracted genomic DNA then the V3 region of bacterial 16S ribosomal DNA
(rDNA) was amplified with GC-clamp primer (Muyzer et al 1993) for DGGE PCR
amplification was performed in a thermocycler DGGE of the PCR products was carried
out in a DcodeTM
Universal Mutation Detection System) The 8 (wv) polyacrylamide
gels contains from 30 to 55 denaturing gradients Electrophoresis was conducted
using a 1timesTAE (Tris-Acetate-EDTA) buffer at 130V and 60degC for 5 hours After
electrophoresis the gel was stained with ethidium bromide in 1timesTAE buffer for 15
minutes and washed in 1timesTAE buffer for 10 minutes The fragments were visualized
under a UV transilluminator The richness of single band in DGGE gel picture
preliminarily showed that the bacterial diversity in mixed culture since the single band in
gel represents one bacterial species
23 16S rDNA clone library construction method
16S rDNA-DGGE fingerprinting has proven to be particularly useful as an initial
investigation into bacterial communities and is suitable for identifying the predominant
bacteria that comprise 1 or more of cells within a given sample In the 16s rDNA clone
17
library construction process the first step is the extratction of the total genomic DNA
then the genomic DNA of the mixed culture biofilm was used as template for PCR
amplification of approximately 1500 bp of 16S rDNA with universal primers The PCR
products of 16S rDNA were purified and inverted into pGEM-T Easy vector system
before they were transformed into competent E coli The transformed cells were spread
on Luria-Bertani (LB) plates containing the selective antibiotic ampicillin and X-Gal and
incubated overnight at 37 OC after the more than 80 inverted 16s DNA fragment E coli
colonies were picked up and inoculated in the 1-3 ml liquid LB medium and left to grow
for 16 hours The plasmid DNA were extracted and sequenced The sequences were
compared directly to all known sequences deposited in GenBank databases using the
basic local alignment search tool (BLAST)
24 Bioinformatics Analysis
16S rDNA sequencing results of more than 80 colones were queried against the
GenBank and Ribosomal Database Project (RDP) databases using BLAST and
SEQUATCH algorithms (Cole et al 2007 Wang et al 2007 Cole et al 2009) The
neighbor-joining trees were constructed with the Molecular Evolutionary Genetics
Analysis package (MEGA Version 4) based on 1000 bootstrap analysis (Tamura et al
2007)
25 Dominant bacterial species band in DGGE identification
The DGGE results gave the information of dominant bacterial richness of bacterial
species in mixed culture based on the theory that single band represents one bacterial
species which showed the diversity of bacterial community in the mixed culture of a
MFC 16S rDNA clone library results provided the whole picture of bacterial community
18
including the bacterial species name and bacterial species percentage in the mixed culture
The known pure bacterial species which have been sequenced can be used as markers to
identify dominant bacterial species in DGGE The whole experimental procedure is
similar to the procedure of the mixed culture DGGE steps The first step is to extract the
collect the mixed culture biofilm genomic DNA and pure bacterial species marker
genomic DNA Then the universal primer set was used to amplify the 16S rDNA from
the extracted genomic DNA After that the V3 region of bacterial 16S ribosomal DNA
(rDNA) was amplified with GC-clamp primer (Muyzer et al 1993) for DGGE The
subsequent steps were similar to the mixed culture DGGE experimental procedure The
dominant bacterial bands in DGGE were determined by comparisons of the single band
position in the mixed culture DGGE with single pure bacterial species location in DGGE
26 Dominant bacterial species isolation and power generation
The bacterial community of mixed culture in MFCs were predominantly composed
of Geobacter sp Arcobacter sp and Citrobacter sp Two dominant bacterial species
have been isolated
Citrobacter sp SX-1 was isolated from the anode biofilm by using a solid agar
medium containing NH4Cl 6mM KCl 2mM CH3COONa (electron donor) 30 mM
FeC6H5O7 (electron acceptor) 20mM minerals and vitamins (Lovley and Phillips 1988)
Single colonies were picked up after culturing on the plate for 36 hours at 30 degC and
transferred two times on the agar plate for purification Then the isolates grew in a liquid
medium solution in anaerobic tubes containing the same constituents as the solid
medium All isolation process was operated in a glove box anaerobic chamber (Coy
Laboratory Products Grass Lake MI)
19
Arcobacter sp SX-2 was isolated from the anode biofilm of a MFC fed with sodium
acetate by using the same method of isolation of pure bacterial species Citrobacter XS-1
except using Trypticase Soy Agar with 5 Sheep Blood solid (from American type
culture collection (ATCC)) agar medium instead of Citrobacter sp medium Single
colonies were picked up after culturing on the plate for 36 hours at 30 degC and transferred
twice on the agar plate for purification Then the isolates grew in a liquid medium
solution microaerobically
Geobacter sp have been trying to isolate by using three different methods They are
the colony pickup after different condition enrichment Goebacter medium isolation and
dilution to extinction by multiple channel mini MFCs isolation however the Geobacter
sp has not been isolated yet due to some limits so far But they provide valuable
information for further isolation of Geobacter sp
Single chamber MFCs were used to evaluate power generation by different isolates
The MFCs were constructed as described previously (Liu and Logan 2004) and modified
with 3 cm2 carbon cloth anodes and 7 cm
2 carbon clothPt cathodes The total liquid
volume of each MFCs was about 15 ml with electrode spacing about 17 cm All MFCs
were operated in an autoclaved closed plastic box and sterile cotton was attached to the
outer surface of the air cathodes to prevent contamination A MFCs without bacterial
culture was used as control MFCs were inoculated with 3 ml late exponential phase
cultures of SX-1 in the medium solution reported previously (Liu and Logan 2004)
3 Results and discussion
31 Power production by mixed culture MFCs
20
A polarization curve was used to characterize current as a function of voltage in the
MFCs The polarization curves are performed by varying a series of external resistances
The results are shown in Figure 21 With MFCs acclimated to 25 Ω external resistance
the maximum power density was achieved at 65Wm2 based on the polarization data At
this point the current density is 21mAcm2 The power density was three times higher
than previous report (20 Wm2) using phosphate buffer (Fan et al 2007) Power density
was also 2 times higher than (28 m Wm2) using a bicarbonate buffer ( Fan et al 2007)
32 Bacterial community structure assessed by PCR-DGGE
Bacterial DGGE profiles for higher power generating MFCs mixed culture setups
were relatively complex (more than 10 bands) However there are three brightly stained
bands and other less intense bands (Fig 2-2) These results indicated there were probably
three dominant bacterial species in mixed culture MFCs
010
020
030
040
050
060
070
2
3
4
5
6
7
04 05 05 06 07 08 10 11 13 15 21 22 23 24
Volta
ge (
V)
Pow
er
densi
ty (
Wm
2)
Current density (mAcm2)
Power
Voltage
power density 65wm2 at current
density208 mAcm2
Figure 2-1 Polarization curves of high power mixed culture MFCs
21
33 16S rDNA clone library results
A total of 83 colonies were sequenced for 16S rDNA clone library construction They
represented the bacterial community structure in mixed culture in MFCs The results
revealed that the predominant bacterial species in mixed culture are Geobacter sp (66
of the mixed culture) Arcobacter sp (12 of the mixed culture) and Citrobacter sp (11
of the mixed culture) Those three genuses comprised 88 of the total bacterial species
(Fig 23) This result is consistent with the DGGE result which showed that three bright
bands in the DGGE gel represented three dominant bacterial species in the mixed culture
The phylogenetic analysis of the colonies (83 colony sequences) is shown in Figure 24
Figure 22 DGGE result of high power generation mixed culture MFC (1 mixed
culture 2 pure culture control)
1 2
22
34 Dominant bacterial band in DGGE identification
Geobacter
66
Arcobacter
12
others
6
Pseudomonas
1
Citrobacter
11
Clostridium
2
Anaerovorax
2 Geobacter
Arcobacter
Citrobacter
Clostridium
Anaerovorax
Pseudomonas
others
Figure 23 Bacterial species and percentage of bacterial community of mixed
culture MFC lt 1 Alcaligensgt
lt1 Commamonasgt
lt1 Pseudomonas aeruginosagt
lt1 Klebsiellagt
lt11 Citrobactergt
lt1 Wolinellagt
lt12 Arobactergt
lt2 Anaeroboraxgt
lt2 Clostridium gt
lt1 unclear bacteriagt
lt66Geobactergt
100
100
85
100
100
82
99
87
100
002
Figure 24 Phylogenetic trees of bacterial species from mixed culture MFC
23
The dominant bacterial species including Goebacter sp Citrobacter sp and Arcobacter
sp in the mixed culture are shown in Figure 25 The upper band is Arcobacter sp and
Citrobacter sp is located in the bottom band of DGGE Geobacter sp is the middle one
The analysis of the bacterial community using denaturing gradient gel electrophoresis
(DGGE) of PCR-amplified 16S rRNA gene fragments and 16S rDNA clone library
construction method showed great phylogenetic diversity of mixed culture in MFCs with
the identification of sequences derived from bacteria of the taxa deltaproteobacteria
(Geobacter sp) gammaproteobacteria (Citrobacter sp) and epsilonproteobacteria
(Arcobacter sp) The result is consistent with the conclusion that among the isolated
exoelectrogens gammaproteobacteria and deltaprotebactia appear to be dominant among
others (Parot et al 2009 Liu et al 2010)
35 The dominant bacterial species isolation and power generation
Figure 25 Dominant bacterial species identification in DGGE
A B C D
A Citrobacter sp
B Geobacter sp
C Arobacter butzmeri
D mixed culture control
24
Two dominant bacterial species Citrobacter sp XS-1 and Arcobacter sp XS-2 have been
isolated and tested for power generation The power densities generated by the two
isolates are shown in the Fig 26 Current densities generated by isolate Citrobacter sp
and Arcobacter sp were 98 mAm2 and 20 mAm
2 respectively which were much less
than that (21Am2) generated by the mixed culture from which they were isolated
Figure 26 Current density generated by different dominant isolates
The maximum current density generated by mixed culture is much higher than the
one produced by pure bacterial species Citrobacter sp XS-1 and Arcobacter sp XS-2 in
the same structure MFCs Although the Geobacter sp in our mixed culture has not been
isolated a previous study showed that maximum current density generated by Geobacter
sp was 080 Am2
(Bond and Lovley 2003) The current density results seem partially
supporting the conclusion that mixed-culture generated higher power densities than their
pure-culture counterparts in MFCs (Jung and Regan 2007) This result suggests there
may be synergistic interactions within the anode bacterial communities
0
500
1000
1500
2000
2500
Mixed culture Citrobacter sp Arcobacter Sp
Cu
rre
nt
de
nsi
ty (
mA
m2)
25
Geobacter sp are strictly anaerobic bacteria which were enriched in most of the anode
biofilms (George 2005) Citrobacter species are facultative anaerobic bacteria (George
2005) which can be found in a wide variety of habitats including in soil water and
wastewater Arcobacter sp are micro-aerobic bacteria including both environmental
nonpathogens and opportunistic human pathogens They are able to grow in aerobic
conditions but in the optimal growth under micro-aerobic conditions (George 2005)
Arcobacter sp has been discovered in the MFCs by Fedorovich (Fedorovich et al 2009)
It is interesting that the mixed culture bacterial community composed of primarily these
three bacterial species has demonstrated to generate high power at non-strictly anaerobic
condition while leading dominant bacterial species Geobacter sp (66) is strict
anaerobic bacteria and generated powder in the MFCs only under strict anaerobic
condition (George 2005) Therefore it is possible that microaerobic bacteria such as
Arcobacter sp and facultative anaerobic bacteria Citrobacter sp can facilitate to create
anaerobic condition for Geobacter sp when they utilized oxygen for growing in the
mixed culture Syntrophic communities study have showed that anaerobic bacteria and
methanogenic archaea form compact microbial structures that operate like an organ rather
than a set of microorganisms functioning independently (Stams and Plugge 2009) Some
substrates have been degradeted within these communities while they are not able to be
fermented by individual species alone interspecies electron transfer also have been
presented in these communities (Stams and Plugge 2009) Summers et al (2010) also
discovered that direct exchange of electrons happened within coculture of Geobacter
metallireducens and Geobacter sulfurreducens Therefore we believe that there are
26
more complex synergistic interactions between different bacteria species in mixed culture
in the MFCs
Acknowledgements
The authors acknowledge support from the US National Science Foundation
(CBET 0955124) We also thank Keaton Lesnik for the review of this manuscript
27
Chapter 3
New Exoelectrogen Citrobacter sp SX-1 Isolated from a Microbial Fuel Cell
Shoutao Xu and Hong Liu
Published in
Journal of Applied Microbiology 111(5)1108-1115 (2011)
ABSTRACT
Exoelectrogenic bacterial strain SX-1 was isolated from a mediator-less microbial fuel
cell by conventional plating techniques with ferric citrate as electron acceptor under
anaerobic condition Phylogenetic analysis of the 16S rDNA sequence revealed that it
28
was related to the members of Citrobacter genus with Citrobacter sp sdy-48 being the
most closely related species The bacterial strain SX-1 produced electricity from citrate
acetate glucose sucrose glycerol and lactose in MFCs with the highest current density
of 205 mA m-2
generated from citrate Cyclic voltammetry analysis indicated that
membrane associated proteins may play an important role in facilitating the electrons
transferring from bacteria to electrode This is the first study that demonstrates that
Citrobacter species can transfer electrons to extracellular electron acceptors Citrobacter
strain SX-1 is capable of generating electricity from a wide range of substrates in MFCs
This finding increases the known diversity of power generating exoelectrogens and
provided a new strain to explore the mechanisms of extracellular electron transfer from
bacteria to electrode The wide range of substrate utilization by SX-1 increases the
application potential of MFCs in renewable energy generation and waste treatment
Keywords Microbial fuel cell Exoelectrogen Citrobacter sp SX-1 Extracellular
electron transfer
29
Introduction
Microbial Fuel Cells (MFCs) technology has become an active research area
recently as a promising approach for renewable energy generation wastewater treatment
and bioremediation (Rabaey and Verstraete 2005 Lovley 2006 Fan et al 2008 Logan
2009 Liu et al 2010) The key feature of MFCs system is the microbe-catalyzed electron
transfer from organic matter to anodes Many studies have shown the presence of diverse
bacterial communities on MFCs anodes (Reimers et al 2001 Liu et al 2004b Kim et al
2005) Over 20 exoelectrogens ie microbes that can transfer electrons exocellularly to
electrodes have been reported in the past 10 years (Logan 2009 Liu et al 2010) The
exoelectrogens so far belong to diverse genetic groups including alphaproteobacteria
(Rhodopseudomonas Ochrobactrum and Acidiphilium) (Malki et al 2008 Xing et al
2008 Zuo et al 2008) betaproteobacteria (Rhodoferax) (Chaudhuri et al 2003)
gammaproteobacteria (Shewanella Pseudomonas Klebsiella Enterobacter and
Aeronomas) (Kim et al 2002 Pham et al 2003 Rabaey et al 2005 Bretschger et al
2007 Zhang et al 2008 Chung et al 2009 Rezaei et al 2009) deltaproteobacteria
(Geobacter Geopsychrobacter Desulfuromonas and Desulfobulbus) (Bond et al 2002
Bond and Lovley 2003 Holmes et al 2004ab 2006) Epsilonproteobacteria (Arcobacter)
(Fedorovich et al 2009) Firmicutes (Clostridium and Thermincola) (Park et al 2001
Wrighton et al 2008) Acidobacteria (Geothrix) (Bond and Lovley 2005) and
actinobacteria (Propionibacterium) (Wang et al 2008) A wider range of
exoelectrogenic species are expected to be discovered
Three mechanisms have been proposed for exocellular transport of electrons by
exoelectrogens without artificial mediators Some exoelectrogenic bacteria can transfer
30
electrons to electrodes through soluble redox compounds excreted by microorganisms
(Bond and Lovley 2005 Rabaey et al 2005) and others can directly transfer electrons to
anodes via outer cell membrane proteins (Myers and Myers 1992) Recently more and
more evidence supports the involvement of bacterial nanowires in extracellular electron
transport (Reguera et al 2005 2006 Gorby et al 2006) In spite of the discovery of
many bacterial species that can transfer the electrons to electrode without the need of
artificial mediators the investigation of extracellular electron transfer mechanisms was
mainly focused on a few species such as those from Geobacter and Shewanella genera
(Kim et al 1999 2002 Reguera et al 2005 2006 Gorby et al 2006) The electron
transfer mechanisms for many of the isolated exoelectrogens species are still not well
studied
In the present study we isolated a new exoelectrogen strain SX-1 from a MFC a
strain phylogenetically related to Citrobacter sp Power generation from various carbon
sources by this strain was evaluated using single chamber MFCs Plausible extracellular
electron transfer mechanisms were also discussed based on the characterization of anodic
biofilms by cyclic voltammetry (CV)
Materials and methods
Bacterial strain SX-1 isolation
Bacterial strain SX-1 was isolated from the anode biofilm of a MFC fed with sodium
acetate operated in fed-batch mode over a period of six months The original source of
the inoculum is wastewater from a local waste water treatment plant Bacterial cells were
released from the carbon cloth anode by shaking the cloth in a sterile flask with 15 mL
sterile 1 NaCl solution and glass beads (2 mm diameter) The suspension was then
31
serially diluted from 10 times to 105
times and plated on a petri dish with a solid agar
medium containing NH4Cl 6mM KCl 2mM CH3COONa (electron donor) 30 mM
FeC6H5O7 (electron acceptor) 20mM minerals and vitamins (Lovley and Phillips 1988)
Single colony was randomly selected after culturing on the plate for 36 hours at 30 degC
and purified on a new agar plate following a procedure reported previously (Chung and
Okabe 2009) Then the isolate was picked up and grew in a liquid medium solution in
anaerobic tubes containing the same constituents as the solid medium for further analysis
All isolation process was operated in a glove box anaerobic chamber (Coy Laboratory
Products Grass Lake MI)
16S rDNA sequencing and phylogenetic analysis
Bacterial genomic DNA was extracted from the isolated strain SX-1 using the DNeasy
tissue Kits (Qiagen Valencia CA) according to the manufacturerrsquos instructions The 16S
rDNA was amplified by PCR using a pair of universal primers 27F (5rsquo-
AGAGTTTGATCMTGGCTCAG-3rsquo) and 1492R (5rsquo-GGTTACCTTTGTTACGACTT-
3rsquo) (Park et al 2001) The conditions used for PCR were described previously
(Ferna ndez et al 1999) PCR products were purified ligated and cloned according to a
procedure described in a previous report (Xing et al 2008) Plasmids were isolated from
randomly selected clone colonies with a QIAprep Spin Miniprep kit (Qiagen Valencia
CA) and three plasmid inserts were then sequenced in both directions using an ABI 3730
DNA sequencer (Applied Biosystems Carlsbad CA)United States) Due to the identical
16S rDNA sequence of the three colones only one was queried against the GenBank and
Ribosomal Database Project (RDP) databases using BLAST and SEQUATCH algorithms
(Cole et al 2007 Wang et al 2007 Cole et al 2009) The 16S rDNA sequence of
32
strain SX-1 and closely related type strains were aligned using CLUSTALX software
(Thompson et al 1997) The neighbor-joining trees were constructed with the Molecular
Evolutionary Genetics Analysis package (MEGA Version 4) based on 1000 bootstrap
analysis (Tamura et al 2007)
SEM
The morphologies of SX-1 cells growing overnight in anaerobic tubes were examined
with a scanning electron microscope (FEI Quanta 600 FEG FEI Company Hillsboro
OR) The suspended cells were fixed in a 25 glutaraldehyde and 01M phosphate
buffer solution and dehydrated with a graded ethanol series from 30 to 100 After
dehydration the samples were dried in a critical point dryer and then sputter-coated with
AuPd for SEM examination (Liu and Logan 2004)
Evaluation of power generation by SX-1 in MFCs
Single chamber MFCs were used to evaluate power generation by SX-1 using various
substrates The MFCs were constructed as described previously (Liu and Logan 2004)
and modified with 3 cm2 carbon cloth anodes and 7cm
2 carbon clothPt cathodes The
total liquid volume of each MFCs was about 15 ml with electrode spacing about 17 cm
All MFCs were operated in an autoclaved closed plastic box and sterile cotton was
attached to the outer surface of the air cathodes to prevent contamination A MFC
without bacterial culture was used as control MFCs were inoculated with 3 ml late
exponential phase cultures of SX-1 in the medium solution reported previously (Liu and
Logan 2004) Seven substrates including citrate acetate glucose sucrose glycerol and
lactose were evaluated individually for power generation in a fed-batch mode in a
temperature control chamber (30 plusmn2degC) For each substrate at least 8 batches were run to
33
investigate the effect of biofilm formation on current output at fixed external resistance of
1kΩ and the same initial substrate concentration of 30 mM Phosphate buffer (100mM)
was used to maintain the solution pH and conductivity The MFCs with sodium citrate as
electron donors were also examined for maximum power output by varying the external
resistance from 20 kΩ to 4 kΩ A data acquisition system was used to record voltage data
during the MFCs operation (Keithley 2700 Keithley Instruments Inc Cleveland OH) It
took about 15 to 30 minutes for the MFCs to stabilize depending on the external
resistance At each resistance we collected at least five data at the steady condition to
make the polarization curves Averaged voltages were used to calculate the power density
(mW m-2
) according to P=IVA where I was the current V was voltage and A was cross-
sectional area of the anode
CV analysis
CV was used to characterize the oxidation and reduction reactions on the anodic surface
of the MFCs fed with citrate and operated at 7 KΩ This external resistance was selected
because the maximum power density was obtained at this resistance based on the
polarization experiment We assumed the biofilm on the anode of MFCs were well-
developed when stable power output was obtained after 3 batches of operation The
MFCs were then used directly for CV analysis at four current generating stages of the
fourth batch (1) initial exponential current increasing stage (middle point of stage about
4 h after media change) (2) current plateau stage (middle point of stage about 8 h after
media change) and (3) current decreasing stage (middle point of stage about 16h after
media change) (4) right after the complete replacement of medium solution (Figure 6)
The anode was used as working electrode the cathode as counter electrode and an
34
AgAgCl electrode was selected as the reference The CV curves were scanned from 200
to -600 mV at a rate of 5 mV s-1
using a potentiostat (G300 Gamry Instrument
IncWarminster PA) Control experiment was also conducted using new anode (without
biofilm) and new medium solution
Nucleotide sequence accession number
The 16S r DNA sequence determined in this study has been deposited in the GenBank
database under accession number HQ845373
Results
Identification of the strain SX-1
An almost-complete 16S rDNA sequence (1475 bp) of strain SX-1 was obtained and
subjected to comparative analysis with the 16S rDNA of closely related reference strains
A phylogenetic analysis revealed that strain SX-1 should be assigned to the genus
Citrobacter and was most closely related to Citrobacter sp sdy-48 EJ463782 (990
sequence similarity) and Citrobacter sp yy-21 FJ463779 (975 sequence similarity)
These three strains formed a distinct sub cluster in the neighbor-joining in which the new
isolate and Citrobacter sp sdy-48 EJ463782 formed a distinct subline (Figure 31)
The SEM image illustrated that the strain SX-1 is a rod-shaped bacterium 03-06 microm
wide and 12-17 microm long (Figure 32) Strain SX-1 colonies on nutrient agar appeared 2-
3 mm in diameter smooth moist opaque with a shiny surface after 5 days incubation
under anaerobic condition The fact that strain SX-1 grew both aerobically and
anaerobically using FeC6H5O7 as electron acceptor suggests that the strain is facultatively
anaerobic Strain SX-1 showed robust growth on citrate as a carbon source These D
35
properties were similar to those of genus Citrobacter described in Bergeyrsquos Manual of
Systematic Bacteriology (George 2005)
Figure 31 phylogenetic tree of strain SX-1 and closely related species based on 16S
rDNA sequences The tree was constructed using the neighbor-joining method
Figure 32 SEM image of planktonic cells of Citrobacter sp SX-1
Strain SX-1
Citrobacter sp sdy-48 (FJ463782)
Citrobacter sp yy-21 (FJ463779)
Citrobacter koseri E639 (ATCC 25408)
Citrobater koseri CDC 3613-63(AF025372)
Citrobacter koseri CDC 8132-86(AF025366)
Salmonella enterica SL483 (CP001138)
Salmonella enterica AKU12601(AY696668)
Shigella sonnei Ss046(CP000038)
E coli C2 (AF403733)
E coli E24377A(CP000800)88
100
100
76
44
92
53
36
0001
1 microm
36
Electricity production by strain SX-1 in MFCs
The isolated strain SX-1 was first tested for its ability to generate current from sodium
citrate at 1kΩ Current gradually increased to a maximum 50 mA m-2
after cell
inoculation and then decreased (Figure 33) After the MFCs was refilled with new
substrate solution the current recovered rapidly and reached a higher level than the first
batch After 4 batches operation the maximum current output of each batch became
stable The highest current density achieved in the MFCs with Citrobacter sp SX-1 at 1
KΩ was 98 mA m-2
(Figure 33)
Figure 33 Electricity generation by Citrobacter sp SX-1 in a single chamber MFCs
with sodium citrate (30 mM) as substrate at 1 KΩ
0
20
40
60
80
100
120
0 50 100 150 200
Curr
ent
den
sity
(m
A m
-2)
Time (h)
37
Current generation by SX-1 from other substrates including glucose lactose sodium
acetate glycerol and sucrose were also assessed at an external resistance of 1KΩ
(Figure 34) When repeatable cycles of current output were obtained for these substrates
glycerol generated the highest maximum current density of 58 mA m-2
followed by
lactose and sucrose with 29 mA m-2
and 27 mA m-2
respectively Glucose and acetate
produced the lowest maximum current density of 96 and 43 mA m-2
respectively These
results indicated that strain SX-1 can utilize a wide range of substrates for electricity
generation in MFCs but with different power generation potentials
Figure 34 Electricity generation of Citrobacter sp SX-1 using different substrates in
single chamber air-cathode MFCs at 1 KΩ (The error bars stand for the standard
deviation of 3 replicates)
0
10
20
30
40
50
60
70
Glucose Lactose Glycerol Acetate Sucrose
Curr
ent
den
sity
(m
A m
-2)
38
Polarization experiment was further conducted to determine the maximum power
density generated by strain SX-1 in the single chamber MFCs with citrate as substrate A
maximum power density of 881 mW m-2
was obtained at current density of 205 mA m-2
at an external resistance of 7 KΩ (Figure 35)
Figure 35 Power and voltage generation by Citrobacter sp SX-1 as a function of
current density using sodium citrate (30 mM) as substrate The error bars stand for the
standard deviation of the 3 voltagepower outputs obtained in three MFCs
20
30
40
50
60
70
80
90
100
01
02
03
04
05
06
07
80 110 140 170 200 230
Po
wer
den
sity
(m
W m
-2)
Vo
ltag
e (V
)
Current density (mA m-2)
Voltage
Power
39
Figure 36 Four current generating stages for CV analysis current increasing stage (stage
1) current plateau stage (stage 2) current decreasing stage (stage 3) and right after the
replacement of medium solution (stage 4) The MFCs was operated at 7 KΩ with sodium
citrate (30 mM) as substrate
Cyclic Voltammetry
To determine the presence of redox active compounds produced by SX-1 and the
location of these compounds CV scan of the anodic biofilms at four current output stages
and the supernatant of MFCs medium solution at the end of the batch experiment were
performed Reduction peaks at range of -310 ndash -350 mV and oxidation peaks at range -
100 ndash -150 mV were observed at all current generating stages (Figure 36 Figure 37A)
suggesting the presence of redox active compounds may involve in extracellular electron
transfer by Citrobacter sp SX-1 The amplitude of the peaks increased according to the
growth stage of the batch and the highest peaks were present after the current plateau
stage which indicated the redox active compounds mainly were secreted in the current
plateau stage While the current density at stage 3 (deceasing stage) was lower than that
at stage 2 (current plateau stage) the redox peak at stage 3 was higher than that in stage 2
0
50
100
150
200
250
50 55 60 65 70 75 80 85
Curr
ent
den
sity
(m
A m
-2)
Time (h)
A
A
Stage 1
Stage 2 Stage 3
Stage 4
40
A plausible explanation for this pattern was that redox active compounds were
continuously being secreted and accumulated after the current peak was reached
resulting in more redox compounds present after the plateau stage But since most of the
carbon source had already been being used up many of the redox compounds at this
stage had not been as active as those at current plateau stage due to much less electrons
were available to be transferred The bigger peak amplitude in stage 4 than in stage 1
indicates that the redox active compounds were continuously being secreted and
accumulated in the biofilm even after stable power was generated at 7 kΩ The slightly
change of the peak locations at different stages was possible due to the solution chemistry
change during the current generation process or the slightly location change of reference
electrode during the measurement (Figure 37B) When the medium solution in the MFCs
was replaced by fresh medium at the end of the batch the oxidization and reduction
peaks were still presented in the CV (Figure 37B) but no peaks were observed when the
supernatant and fresh mediums were tested in a MFCs with no biofilm (Figure 37C)
These results suggest the compounds involved in the electron transfer were located not in
the supernatant (solution) but in the biofilm
41
Figure 37 Cyclic Voltammetry analysis of Citrobacter sp SX-1 biofilms at (A) current
increasing stage current plateau stage and decreasing stage (end of the batch) (B) end of
the batch and replaced with fresh medium and (C) controls
-40E-04
-30E-04
-20E-04
-10E-04
00E+00
10E-04
20E-04
30E-04
-060 -050 -040 -030 -020 -010 000 010 020
Curr
ent
(A)
Potential (V) vs AgAgCl
Current plateau stage
Current increasing stage
Current decreasing stage
-40E-04
-30E-04
-20E-04
-10E-04
00E+00
10E-04
20E-04
30E-04
-060 -050 -040 -030 -020 -010 000 010 020
Curr
ent
(A)
Potential (V) vs AgAgCl
Current decreasing stage
Biofilm with fresh medium
-40E-04
-35E-04
-30E-04
-25E-04
-20E-04
-15E-04
-10E-04
-50E-05
00E+00
-060 -050 -040 -030 -020 -010 000 010 020
Curr
ent(
A)
Potential (V) vs AgAgCl
Supernatant
Fresh medium
A
B
C
B C
42
Discussion
Citrobacter species belonging to Gammaproteobacteria Enterobacteriales
Enterobacteriaceae are facultative anaerobic bacteria (George 2005) They can grow with
oxygen as an electron acceptor but also with a variety of electron accepters such as Fe (III)
in the absence of oxygen (George 2005) Citrobacter species can be found in a wide
variety of habitats including in soil water and wastewater Several Citrobacter species
have been studied as important bioremediation bacterium for heavy metal removals
sulfate reduction phenol degradation and chlorophenol degradation (Macaskie et al 1995
Narde et al 2004 Qiu et al 2009) Citrobacter species have also been found on the
anodic biofilm of a diesel-degrading MFCs (Morris et al 2009) However there was no
study reported that Citrobacter species can transfer electrons to extracellular electron
acceptors This finding that Citrobacter strain SX-1 can generate electricity in MFCs
increased the diversity of power generating exoelectrogens and provided a new strain to
explore the mechanisms of extracellular electron transfer from bacteria to electrode
Among the isolated exoelectrogens gammaproteobacteria and deltaprotebactia appear to
be dominant among others (Parot et al 2009 Liu et al 2010) The isolated strain
Citrobacter sp SX-1 phylogenetically belongs to gammaproteobacteria which provides
another evidence to support this observation
Most exoelectrogens utilize a limited range of substrates for power generation For
example Shewanella oniedensis MR-1 can oxidize lactate under anaerobic condition but
cannot utilize acetate for electricity generation (Kim et al 2002) Geobacter
sulfurreducens can completely oxidize acetate for power generation but it cannot utilize
simple sugars (Bond and Lovley 2003 Caccavo et al 1994)) In comparison strain SX-1
43
not only can oxidize citrate for power generation but also utilize a wide range of
substrates for power generation including glycerol glucose lactose sucrose and acetate
Interestingly the higher current density generated by strain SX-1 from glycerol a main
by-product of biodiesel production process suggests that strain SX-1 may be potentially
used for harvesting energy from biodiesel wastes using MFCs However the current
density generated by strain SX-1 (205 mA m-2
) is lower than that (805 mA m-2
at 1 kΩ)
generated by the mixed culture from which the SX-1 was isolated indicating the
existence of other higher power generating bacteria andor complex ecology in the mixed
culture community
Understanding mechanisms of microbial extracellular electron transfer is critical for
enhancing the electron transfer rate from bacteria to electrode through metabolic or
genetic engineering of exoelectrogens Since the spent supernatant solution of MFCs run
with SX-1 showed no redox properties the redox compounds produced by SX-1 may
have been retained in the biofilm Alternatively SX-1 may utilize one or both of the
other two known extracellular electron transfer mechanisms For example it is possible
that strain SX-1 transfers electrons to the anode via cell-bound outer membrane proteins
The two distinct oxidation and reduction peaks observed at -100mv- -150 mV and -310--
350 mV of the SX-1 biofilm have a midpoint potential range from -205mV to -250mV
which is close to the redox potential of some c-type cytochromes (Field et al 2000 Meitl
et al 2009) which are well-known to play an important role in extracellular electron
trasnfer (Eggleston et al 2008 Field et al 2000 Meitl et al 2009) Field et al (2000)
reported that the c-type cytochromes CymA had the midpoint potentials of -229 mV
Meitl et al (2009) and Eggleston et al (2008) reported that the c- type cytochromes
44
OmcA in Shewanella oneidensis MR-1 had the midpoint potentials of -201 mV and -208
mV
While itrsquos possible that SX-1 may also have conductive appendages to enhance the
extracellular transfer electron similar to the nanowires discovered in some Geobacter
and Shewanella species (Reguera et al 2005 2006 Gorby et al 2006) further
investigation on the presence of these appendages on the cell surface of SX-1 is needed
Acknowledgements
The authors acknowledge support from the US National Science Foundation
(CBET 0955124) We also thank Teresa Sawyer for her helps with the SEM analysis and
Jeremy Chignell and Yanzhen Fan for their review of this manuscript
45
Chapter 4
Enhanced Performance and Mechanism Study of Microbial Electrolysis Cells
Using Fe Nanoparticles Decorated Anodes
Shoutao Xu Hong LiuYanzhen Fan Rebecca Schaller Jun Jiao Frank Chaplen
Published in
Applied Microbiology and biotechnology 93(2)871-880 (2012)
46
ABSTRACT
Anode properties are critical for performance of microbial electrolysis cells (MECs) In
the present study Fe nanoparticle modified graphite disks were used as anodes to
investigate the effects of nanoparticles on the performance of Shewanella oneidensis MR-
1 in MECs Results demonstrated that average current densities produced with Fe
nanoparticle decorated anodes were up to 56-fold higher than plain graphite anodes
Whole genome microarray analysis of the gene expression showed that genes encoding
biofilm formation were significantly up-regulated as response to nanoparticle decorated
anodes Increased expression of genes related to nanowires flavins and c-type
cytochromes indicate that enhanced mechanisms of electron transfer to the anode may
also have contributed to the observed increases in current density The majority of the
remaining differentially expressed genes were associated with electron transport and
anaerobic metabolism demonstrating a systemic response to increased power loads
Keywords Microbial electrochemical system microbial fuel cell microbial electrolysis
cell nanotechnology differential gene expression DNA microarray
47
Introduction
Microbial electrochemical systems (MESs) have been intensively studied since Lewis
achieved practical advances in this field (Logan 2007) however they attracted much
research attention in recent years due to their promising applications in renewable energy
generation bioremediation and wastewater treatment In a MES microorganisms
interact with electrodes via electrons catalyzing oxidation and reduction reactions at the
anode and the cathode The most-described type of MESs is microbial fuel cells (MFCs)
in which useful power is generated from electron donors typically biodegradable organic
materials (Logan et al 2006) Various novel MESs have recently been developed to
produce hydrogen (microbial electrolysis cells (MECs)) (Liu et al 2005 Logan et al
2008 Rozendal et al 2006 Schaller et al 2009) to power an autonomous sensor or
sensor network (Ringeisen et al 2006) to reduce bioremediation targets (Lovley 2006
Shea et al 2008) Sukkasem et al 2008) and to desalinate water (Cao et al 2009) The
key feature shared by these systems is the microbe-catalyzed electron transfer from
organic matter to electrodes (anodes) (Rabaey et al 2004) Enhancing the anodic current
output which highly depends on the performance of the electrodes is critical for the
successful application of all these processes (Logan et al 2007 Park and Zeilus 2002
2003)
Nanomaterials have received much attention from researchers in the context of
microbiology due to their unique physical electrical and chemical properties which
facilitate the study of interactions between bacteria and surfaces (Liu 2006) Previous
studies have demonstrated that electrodes decorated with different nanostructures such
as carbon nanotubes nanostructured polyanilinetitanium dioxide composite and titania
48
nanotubes have the potential to enhance power generation in MFCs (Morozan et al 2007
Qiao et al 2008 Quan et al 2005) Furthermore our recent study shows nanoparticle
(NP) decorated anodes greatly increased the electrochemical electron transfer rate in
MFCs (Fan et al 2011) However Fe NPs as an ideal candidate for decorating electrodes
because of respectively low price and high conductivity compared to other materials has
not been focused to study in MECs
S oneidensis MR-1 an important electrochemically active bacterial strain has been
exploited for the electricity generation in MFCs (Logan et al 2007 Logan 2009 Park
and Zeikus 2002) The availability of genome sequence for this strain makes it possible to
use transcriptome assays to globally measure the responses to different growth conditions
and environmental stresses (Murray et al 2001) S oneidensis MR-1 gene expression
response to heat shock cold shock Cr (VI) and U (VI) reduction chromate stress and
iron and acid tolerance have been studied in the past several years (Bencheikh-Latmani et
al 2005 Brown et al 2006 Gao et al2006 Gao et al 2004 Yang et al 2008) However
no studies have been focused on the Shewanella gene expression response to NPs in
MESs
The mechanism of increased electron transfer rate exhibited by nanoparticle decorated
electrodes is not well understood yet In the present study graphite disks decorated with
Fe NPs were used as anodes to explore the effects of nanostructures on current generation
in a multi-anode MECs DNA microarrays were utilized to investigate differences in the
global gene expression profile of S oneidensis MR-1 grown on plain versus Fe NPs
decorated anodes
Materials and methods
49
Bacterial cultures
S oneidensis MR-1 was purchased from American Type Culture collection (ATCC
700550) and stored as freezer stocks at -80degC Stock cells of S oneidensis MR-1 was
grown aerobically using a medium containing 30 gL Trypticase Soy Broth (BD 211825
Becton Dickinson Company Franklin lakes NJ) at 30degC Cells in early stationary
growth phase (optical density of 15-16 at 600nm (Spectrophotometer UV-1700
Shimadzu Columbia MD) were washed two times then injected into the chamber of the
MECs for current production Trypticase Soy Broth medium with 30 mM sodium lactate
as electron donor was used in MECs Phosphate buffer (100mM) was used to maintain
the solution pH 7 and solution conductivity at 15 mScm
Characterization of nanostructured anodes
Superfine isomolded graphite disks (16 cm x 05 cm GraphiteStorecom Inc) were
polished with ultrafine sand paper (2000 grit 3M Company) as the base for the NP
decorated and control anodes Fe NPs decorated anodes were fabricated by thermal
annealing of metal thin film coated graphite disks Thin films of Fe were firstly deposited
on the polished graphite disk by using sputter coating for 95 min and the samples then
were annealed in the Chemical vapor deposition (CVD) at 750degC for 20 min to form Fe
NPs Fe-NPs were produced with a diameter of 200plusmn100 nm and a density range
(distance between NPs) of 200plusmn100 nm with a coating time of 95 min SEM images of
Fe NPs decorated anode and control surfaces were shown in the Figure 41
50
Figure 41 SEM images of Fe NPs decorated anodes (A) Graphite disk control (B) Fe
Nanoparticle decorated
Multiple channel MECs construction and operation
A MECs with removable multiple anodes with each effective anode area 07 cm2
was
constructed and used to evaluate the effects of nanostructure on current density of
according to a previous report (Fan et al 2011) The cathode was made of wet-proof
(30) carbon cloth (type B E-TEK Division Inc USA) coated with PtC (20 E-TEK
Division Inc USA) following a previously reported procedure (Liu et al 2005) The
final platinum loading was 05 mgcm2 per projected cathode area The size of cathode
(150 cm2) was 25 times larger than the total effective area of the 8 graphite disk anodes
(56 cm2) to prevent cathode limitations on the performance of the MECs system All
MECs with Fe-NP decorated anodes and control anodes were tested for current
generation for 24 h with medium (without bacteria) before injecting the bacterial cells in
order to determine whether the Fe-NP decorated anodes demonstrate chemical current
generation compared to control anodes All the testing anodes were set up in the MECs
B
500 nm
A
500 nm
51
at once so that the S oneidensis MR-1 culture was applied to all of them simultaneously
Short electrode spacing (17 cm) was used in the MECs design to reduce the internal
resistance A voltage of 06 V was applied the MECs for current generation after the cells
of S oneidensis MR-1 were inoculated into MECs at early stationary phase A multimeter
with a data acquisition system (2700 Keithly USA) was used to monitor the current
change by measuring the voltage (every 300 seconds) drop through a resistor at 300 ohm
Fig 4 2 Effects of Fe-NP decorated anodes on the current density in MECs Control is
plain graphite disk anode Fe-NP is the anode with Fe-NP decoration
Microarray analysis
Biofilms for whole gene microarray analysis were aseptically removed from the plain
and Fe-NP decorated anodes of MECs after 110 hours further incubation at 30oC when
current density obviously started to decease Total RNA was extracted using Trizol
(Ambion inc AustinTX) following the manufacturerrsquos instructions Integrity of the
52
RNA samples was confirmed using the Agilent 2100 Bioanalyzer (Agilent Technology
Inc Palo Alto CA) The same amount of RNA (200 ng) from each sample was used for
further analysis after amplification by using the MessageAmpTM
II-Bacteria Prokaryotic
RNA Amplification kit (Ambion AustinTX) according to the manufacturerrsquos
instructions All of the 4295 genes (resource from NCBI) represented on the S oneidensis
MR-1 whole-gene microarrays were ordered from Roche NimbleGen Inc (Madison WI)
Biological triplicates of biofilms on the Fe-NP decorated anodes and control were
analyzed respectively cDNA synthesis labeling and hybridization were carried out by
the CGRB Core Laboratories at Oregon State University DNASTAR ArrayStarTM
3
software was used to identify genes that were up- or down-regulated more than 2-fold
when grown on the nanoparticle modified anodes using the unpaired two sample t-test
with a cutoff p-value of 005 The complete microarray data set generated in this study is
deposited for public access in the Gene Expression Omnibus
(httpwwwncbinlmnihgovgeo) under accession number GSE31535
Results
Enhancement of current generation by using NP decorated anodes in MECs
The current density generated by Fe-NP decorated anode MECs with S onidensis MR-1
increased to approximately 43 microAcm2 20 h following inoculation and then slowly
decreased Figure 42 shows that the maximum current density achieved by NP
decorated anodes was 83 times higher than that (51 microAcm2) generated by the control
(plain graphite disk) The average current density improvement of 110 hours was 59
times of that generated by the control The current densities generated in MECs in the
53
absence of bacteria were negligible (03 microAcm2) which demonstrated that the current
enhancement observed with the Fe-NP decorated anodes was biologically-derived
Global transcriptome analysis
Whole-genome DNA microarrays were used to attain a comprehensive general
overview
of the transcriptional response to Fe-NP decorated anodes by S oneidensis MR-1 in MEC
Of a total 4295 genes assayed 392 (188 induced and 204 repressed) exhibited significant
(P lt 005) differential expression at a 2-fold (log (expression ratio 1) level in 3
replicates in response to Fe-NPs These
total gene numbers present 9 of the 4295 open
reading frames (ORFs) presented on the array
Fig 43 Differentially expressed genes grouped by functional classification according to
the TIGR S oneidensis genome database (wwwtigrorg) Columns 1 amino acid
biosynthesis 2 biosynthesis of cofactors prosthetic groups and carriers 3 cell
envelope 4 cellular processes 5 central intermediary metabolism 6 DNA metabolism
7 energy metabolism 8 fatty acid and phospholipid metabolism 9 other categories 10
protein fate 11 protein synthesis 12 purines pyrimidines nucleosides and nucleotides
13 regulatory functions 14 signal transduction 15 transcription 16 transport and
binding proteins 17 unknown function 18 hypothetical proteins
Up-Regulated
-75
-50
-25
0
25
50
75
100
125
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18
Down-Regulated
Num
ber
of
Gen
es
54
Figure 43 summarizes the overall genomic response of S oneidensis MR-1 to Fe-NP
decorated anodes by grouping the differentially expressed genes into their functional role
categories based on The Institute for Genomic Researchrsquos annotation (Version 8) of the
MR-1 genome sequence The wide distribution of putative functional roles attributed to
the differentially expressed genes indicated the extent of the molecular response of
Soniedenis MR-1 to the NP decorated anodes Among genes with defined function a
large number of up-regulated genes were associated with cellular processes (group 4)
energy metabolism (group 7) other categories (group 9) and hypothetical proteins (group
18) While those genes encoding proteins involved in amino acid biosynthesis (group 1)
cellular processes (group 4) substrate transport (group 16) and hypothetical proteins
(group 18) were among the most down-regulated genes Two groups of genes that were
the most noticeable among the functional gene groups genes related to energy
metabolism (group 7) and genes related to amino acid biosynthesis (group 1) The ratios
of up-regulated genes to down-regulated genes in these functional groupings were much
higher than other functional gene groupings there were 21 up-regulated genes and 7
down-regulated genes in energy metabolism gene group and 2 up-regulated and 22 down
regulated genes in the amino acid biosynthesis group
Genes related to biofilm formation
In order to determine whether there was a correlation between the observed biofilm
enhancements by NP decorated anodes and the genes related to biofilm formation the
significant modified genes related to biofilm formation were examined and summarized
in the Table 41 These included SO_3223 and SO_3228 genes encoding the flagellum
proteins (Thormann et al 2004) which has critical impacts on initial attachment to the
55
surface as swimming motility functions were up-regulated over 10 and 09 fold
respectively The gene clusters SO_4178-SO_4180 which are thought to be essential for
formation of a three-dimensional biofilm were up-regulated from 12 to 13 fold
(Thormann et al 2004) In addition the genes SO_4105 encoding IV pilus which is
known to represent a critical factor for biofilm formation on a biotic surfaces (Thormann
et al 2004) were also induced 08 fold (Table 41) The genes related to flagellum
motility (SO_4286 SO_1529 and SO_1530) were also induced slightly (Thormann et al
2004)
Table 41 Expression levels of genes related to biofilm formation
Locus Tag Gene product descriptions Fold change
SO_3228 flagellar basal-body MS-ring and collar protein FliF 10
SO_3223 flagellar hook-length control protein FliK 09
SO_4103 MshA minor pilin protein MshD 05
SO_4105 MSHA major pilin protein MshA 08
SO_4178 expressed protein of unknown function MxdC 12
SO_4179 inner membrane family 2 glycosyltransferase MxdB 13
SO_4180 diguanylate cyclase-like protein MxdA 10
a fold changes were presented after log2
b positive number represented up-regulated and negative represented down-regulated
Genes related to energy metabolism
A closer consideration of energy metabolism genes was undertaken as anaerobic
metabolism for electron generation and electron transport functions may play critical
roles in enhanced current density generation in MECs Genes associated with energy
56
metabolism and with significantly modified gene expression levels are summarized in the
Table 42 Several of the up-regulated genes were related to formate dehydrogenase
proteins Most notably four genes related to formate dehydrogenase were up-regulated
more than 2-fold SO_0102 SO_0103 SO_0104 and SO_0107 The other genes
expression related to formate dehydrogenase proteins SO_4513 SO_4514 SO_4515 also
significantly increased Another interesting gene is that encoding the cytochrome c
oxidase protein gene SO_4609 which was up-regulated 11 fold while the gene
SO_0479 related periplasmic octaheme cytochrome c MccA was down-regulated
However SO_1519 the gene encoding lactate dehydrogenase was down-regulated 12
fold despite lactate being the primary carbon source in the media
Table 4 2 Genes related to anaerobic growth and electron transfer with significantly change expression level
Locus Tag Gene product descriptions Fold
change
SO_0101 nitrate-inducible formate dehydrogenase molybdopterin-binding
subunit FdnG
18
SO_0102 formate dehydrogenase nitrate-inducible iron-sulfur subunit 21
SO_0103 formate dehydrogenase nitrate-inducible cytochrome b556 subunit 24
SO_0104 formate dehydrogenase accessory protein FdhE 22
SO_0107 formate dehydrogenase accessory protein fdhD 24
SO_0397 quinolfumarate reductase menaquinol-oxidizing subunit FrdC_2 12
SO_0452 thioredoxin 2 15
SO_0903 Na-translocating NADH-quinone reductase subunit B NqrB_1 12
SO_1012 NADH-ubiquinone oxidoreductase subunit K NuoK 12
SO_1013 NADH-ubiquinone oxidoreductase subunit J NuoJ 12
SO_1363 hydroxylamine reductase 13
57
SO_2417 ferredoxin cofactor maintenance protein YfaE 10
SO_3922 formate dehydrogenase cytochrome b Fdh -12
SO_4513 formate dehydrogenase molybdopterin-binding subunit _2 15
SO_4514 formate dehydrogenase FeS subunit FdhB_2 12
SO_4515 formate dehydrogenase cytochrome b subunit FdhC_2 14
SO_4609 aa3 type cytochrome c oxidase subunit III CoxC 11
SO_0479 periplasmic octaheme cytochrome c MccA -12
SO_0847 periplasmic nitrate reductase ferredoxin component NapG -15
SO_1519 L-lactate dehydrogenase iron-sulfur cluster-binding protein LldF -10
SO_1251 ferredoxin 4Fe-4S -11
SO_37411 hypothetical inner membrane protein -11
a fold changes were presented after log2
b positive number represented up-regulated and negative represented down-regulated
Flavin and cytochrome related genes
Flavins can be secreted by Shewanella species as electron shuttle to facilitate
extracellular electron transfer (Cansterin et al 2008) C-type cytochromes play the
important roles on the process of extracellular electron transfer (Shi et al 2007) The
genes related to flavins synthesis and the genes encoding the cytochromes electron
transport proteins were shown in the table 43 the genes SO_1414 and SO_3058 related
to flavins synthesis was up-regulated to 03 and 01 folds respectively The genes related
to s C-type cytochromes SO_0135 SO_4105 SO_0169 SO_1778 are increased to 10
08 05 04 01 folds respectively However most of genes have shown no significant
changes (less one-fold change) as response to nano particle decorated anode in MECs
58
Table 43 Expression level of flavin and cytochrome genes
Locus Tag Gene product descriptions Fold
change
SO_1414 flavocytochrome c flavin subunit putative 03
SO_3468 riboflavin synthase subunit alpha -02
SO_3058 flavocytochrome c flavin subunit 01
SO_4105 MSHA pilin protein MshA 08
SO_0169 general secretion pathway protein GspG 05
SO_1778 outer membrane decaheme cytochrome c lipoprotein MtrC 04
SO_1779 outer membrane decaheme cytochrome c OmcA 01
SO_0135 lipoprotein of unknown function DUF333 10
SO_0136 conserved hypothetical inner membrane protein 01
a fold changes were presented after log2
b positive number represented up-regulated and negative represented down-regulated
Other genes with significantly modified expression levels
These genes over 15 fold change and possibly related to current enhancements were
shown in Table 44 SO_3104 (expressed inner membrane protein) was over expressed
and the gene SO_2194 which encodes outer membrane protein (OmpA family protein)
was repressed as response to nanoparticle decorated anode however it is unclear the
relationships of this modified membrane protein genes with the current enhancement
Another interesting phenomenon was possible co-regulation of several gene clusters
indicating possible operon associations This includes three sets of genes that were
induced more than 15 fold SO_0108SO_0109 and SO_1042SO_1043SO_1044 The
59
consistency of expression of these genes under the NP conditions provides basic evidence
to support operon structure However the correlation of these significant changed gene
with current density enhance are unclear which showed there are unknown multiple and
complex responses of S oneidensis MR-1 to nanoparticle anode of MECs
Table 44 Other genes with significantly changed expression levels
Locus Tag Gene product descriptions Fold change
SO_0101 nitrate-inducible formate dehydrogenase molybdopterin-binding
subunit FdnG
18
SO_0108 integral membrane protein of unknown function DUF39 YedE 17
SO_0109 SirA family protein YedF 17
SO_0275 N-acetyl-gamma-glutamyl-phosphate reductase -31
SO_0277 ornithine carbamoyltransferase -21
SO_0279 argininosuccinate lyase -24
SO_0404 zinc dependent metalloprotease domain lipoprotein 21
SO_0956 alkyl hydroperoxide reductase F subunit -20
SO_1042 amino acid ABC transporter ATP-binding protein -18
SO_1043 amino acid ABC transporter permease protein -27
SO_1044 amino acid ABC transporter periplasmic amino acid-binding protein -27
SO_1072 chitin-binding protein putative 21
SO_1405 transglutaminase family protein 20
SO_1822 TonB-dependent receptor putative -22
SO_2069
1-(5-phosphoribosyl)-5-[(5-phosphoribosylamino)
methylideneamino] imidazole-4-carboxamide isomerase
-19
SO_2070 amidotransferase HisH -26
SO_2071 imidazole glycerol-phosphate dehydratasehistidinol phosphatase -27
60
SO_2072 histidinol-phosphate aminotransferase -28
SO_2073 histidinol dehydrogenase -30
SO_2194 OmpA family protein -36
SO_2195 inter-alpha-trypsin inhibitor domain protein -27
SO_2767 asparagine synthetase B -26
SO_2945 prophage LambdaSo tail fiber protein 20
SO_2963 prophage LambdaSo major capsid protein HK97 family 20
SO_2987 prophage LambdaSo hypothetical protein 19
SO_2988 prophage LambdaSo expressed protein of unknown function 18
SO_3104 expressed inner membrane protein 131
SO_3408 conserved hypothetical inner membrane protein -18
SO_3585 azoreductase putative -18
SO_3586 glyoxalase family protein -17
SO_3687 curli production assemblytransport component CsgE putative -18
SO_38191 hypothetical ammonia permease -19
SO_4014 AcrBAcrDAcrF family protein 18
SO_40151 type I secretion system membrane fusion protein RND family 18
SO_4054 510-methylenetetrahydrofolate reductase 18
SO_4245 N-acetylglutamate synthase -22
SO_45251 hypothetical transcriptional regulator LysR family -17
SO_4527 integral membrane domain protein -19
SO_4705 transcriptional regulator putative -19
a fold changes were presented after log2
b positive number represented up-regulated and negative represented down-regulated
Discussion
61
The addition of NP to anodes significantly impacts current densities with the elemental
composition being a critical factor The chemical composition of NPs significantly affects
the current enhancement in MECs (Fan et al 2011) because the chemical composition of
NP properties is a critical factor in determining the conductivity which significantly
affects the efficiency of electron transfer from bacteria to electrode Fe is an ideal
candidate for NP formation for current enhancement in MESs because of high
conductivity and low price compared to gold and other conductive materials Gold NPs
has also been studies for enhancement of electricity conductivity (Bao et al 2008)
However the disadvantages of the high price of gold limit their practical application in
MECs (Fan et al 2011) The different size and density of the same composition of NP
also has essential effects on the current density enhancement (Fan et al 2011) In this
study the results showed the current density enhancement with Fe-NP size range of
200plusmn100 nm and density range of 200plusmn100 nm is considerable enhancement consequence
There are a number of reports of studies of current density enhancement in MFCs
by nanostructured anodes (Fan et al 2011 Qiao et al 2008 Sharma et al 2008 Tsai et al
2009) Based on bacterial behavior on nanostructured anode surface Qiao et al (2008)
believes that Escherichia coli cells on the nanostructured electrode surface produce hair-
like structures similar to pili that could facilitate the electron transfer between the cells
and electrode The production of hair-like structures is believed to be stimulated by
nanostructures and could play the key role on current density enhancement in pili-
producing bacteria in MFCs (Qiao et al 2008) S oneidensis can produce electrically
conductive bacterial nanowires which have similar structure to the hair-like structures in
Ecoli (Gorby et al 2006 Reguera et al 2005) A number of possible genes related
62
nanowires of S oneidensis have been identified including mshA (SO_4105) and gspG
(SO_0169) (Konstantinidis et al 2009) They are up-regulated 08 and 05 fold in the
current study respectively even though they are not significant induced This result
supported the increased nanowire genes had contributions to enhance current density in
MFCs by NPs decorated anodes
Based on the previous research on electron transfer mechanisms for S oneidensis it is
known that self-produced mediators (flavins) and multihaem c-type cytochromes (c-Cyts)
may play critical roles in the electron transfer of S oneidensis to extracellular insoluble
electrode (Canstein et al 2008 Myers and Myers 1992 Scott and Nealson 1994 Shi et al
2007) The flavins secreted by S oneidensis can facilitate to transfer electrons from
bacteria to solid anode electrodes by acting as extracellular electron shuttles (Canstein et
al 2008) In this study the genes encoding flavins (SO_1414 SO_3468 and SO_3058)
had no significant increase (less than 1 fold change) as a response to nanoparticle
decorated anode in MECs which suggests that increased flavin gene expression does not
significantly contribute to the current density enhancements in nano-particle decorated
anode The genes encoding cytochromes electron transport proteins (SO_4104 SO_0417
SO_177879 SO_013536) had no significant increase (less than 1-fold change)
However those genes were up-regulated slightly These results suggest the increased
expression flavin and c type cytochromes genes had partial contributions even not
significantly to enhance current density in MFCs by NPs decorated anodes
The thickness of bacterial biofilm on the anode can affect the power generation in
MFCs (Biffinger et al 2007 Heydorn et al 2000) since increased biofilm thickness on
the anode represents more active bacteria involving in the process of electron generation
63
and transfer in MFCs Biofilm formation related genes included flagellar genes IV pilus
genes and other genes The flagellar related genes SO_3228 SO_3229 (fliB and fliK) and
IV pilus genes SO_4105 (mshA) were induced from 19 to 09 fold the genes (SO_4178
-4180) related to formation of a three-dimensional biofilm were up-regulated from 10 to
13 folds even though the genes related bacterial motility protein (SO_ 4286 SO_4287
SO_1529 and SO_1530) are induced slightly (less than 0 5 fold) However the motility
genes most probably play a role for initial biofilm attachment on the anode surfaces
(Thormann et al 2004 Thormann et al 2006) with the rest of biofilm genes being more
essential for biofilm maintenance in mature cultures as were presented at harvest in this
study This enhancement of biofilm density under conditions of increase current density
has been observed for gold NP decorated anodes using confocal light microscopy (data
not shown) but has not yet been confirmed for Fe NP decorated anodes In any event an
increase in the expression level of genes related biofilm formation therefore also
supported the possibility of enhanced biofilm formation on NPs decorated anodes thus
facilitating electron transfer from bacteria to anodes
Beliaev et al described a model of the Fe reduction pathway in S oneidensis MR-1 using
solid substrates as the electron acceptor (Beliaev et al 2001) This model proposes that
electrons are generated and released in cytoplasm then transferred to the quinone pool
eg menaquinone in the cytoplasmic membrane The reduced menaquinone in turn
reduces CymA and from there electrons are then transferred to a periplasmic shuttle (eg
CctA) and then to MtrA a periplasmic soluble decaheme cytochrome c located in
associated with the outer membrane via interaction with MtrB Finally surface displayed
outer membrane proteins decaheme cytochromes (OmcA MtrC or MtrF) can transfer
64
electron to Fe or other solid substrates (Beliaev et al 2001)) Bencheikh-Latmani et al
demonstrated that the same electron transport pathway may be used for more than one
electron acceptor through studies of the response of S oneidensis MR-1 to U(VI) and
Cr(VI) under anaerobic conditions particularly focused on the critical genes
(SO_1777mtrA SO_1776mtrB SO_1778mtrC) (Bencheikh-Latmani et al 2005)
However our studies showed that these genes had no significant expression changes in
response to nanostructured anodes Contrastingly one of expressed inner membrane
protein genes SO_3104 and one of cytochromes genes SO_4609 encoding cytochrome c
oxidase subunit III demonstrated a 13-fold and 11-fold up-regulation respectively in this
study while the gene SO_2194 which encodes outer membrane protein (OmpA family
protein) was down regulated 36 fold Our results therefore suggest that S oneidensis
MR-1 does not utilize the same set of core critical genes (SO_1777 mtrA SO_1776
mtrB SO_1778 mtrC) products to transfer electrons in all instances This is the similar
result with the study of Bretschger et al (2007) which indicated that the electron
transport system in S oneidensis MR-1 is complex with several different proteins able to
participate in electron transfer to the anode of MECs
It should also be mentioned that the gene (SO_1519) encoding lactate
dehydrogenase repressed (-12 fold) here suggesting that lactate was depleted at the time
of cell harvest from the MECs and that other pathways of anaerobiosis had been activated
including those associated with amino acid uptake and consumption the growth media
was a complex mixture including yeast extract In particular the catabolic pathway for
histidine of S oneidensis MR-1 a pathway that may formate as an intermediate is up-
regulated with the genes for histidine ammonia lyase SO_0098 SO_3057 and SO_4374
65
up-regulated 13 11 and 11-fold respectively The possible production of formate
during histidine degradation may in turn explain the high levels of formate
dehydrogenates as evidenced by upregulation of SO_0102 SO_0103 SO_0104 and
SO_0107 annotated as formate dehydrogenase proteins These genes had the highest
levels of up-regulated expression as response to Fe NP decorated anodes (greater than 2-
fold)
Acknowledgements
This research was partially supported by the US National Science Foundation CBET
0828544 and the funds from ONAMIDOD (ARL-DOD Cooperative Agreement
W911NF-07-2-0083) We thank Barbara Gvakharia and Caprice Rosato for helpful
suggestions We also thank Margaret Romine of Pacific Northwest Laboratories for
valuable comments on manuscripts We also thank anonymous reviewers for significant
suggested improvements imparted as part of prior review of this manuscript
66
Chapter 5
Global Transcriptome Analysis of the Response of Shewanella oneidensis MR-1 to
Carbon Nanotube Decorated Anodes in Microbial Electrochemical Systems
Shoutao Xu Yanzhen Fan Rebecca Schaller Frank Chaplen Jun Jiao Hong Liu
67
Abstract
Shewanella oneidensis MR-1 is an important model microorganism for metabolic studies
on the effects of different environmental factors because of its diverse respiratory
capabilities Carbon nanotube (CNT) modified graphite disks were used as anodes to
investigate the effects of nanomatrials on the performance of S oneidensis MR-1 in
microbial electrolysis cells (MECs) The current densities produced with CNT decorated
anodes were on average 56-fold higher than plain graphite anodes Whole genome
microarray analysis of gene expression showed that up-regulation of cytochromes c genes
associated with extracellular electron transfer are strongly correlated to current increases
in CNT decorated anodes Genes involved in flavin biosynthesis also contribute to
current increase in CNT decorated anode MECs
Keywords Microbial electrochemical system microbial fuel cell microbial
electrolysis cell carbon nanotube gene expression DNA microarray
68
Introduction
Shewanella oneidensis MR-1 is an important model microorganism for metabolic
studies of the effects of different environmental factors because of its diverse respiratory
capabilities It has been used for transcriptome analysis to investigate the responses to
different growth conditions and environmental stresses (Murray et al 2001) Gene
expression patterns under different conditions such as heat shock cold shock Cr (VI) U
(VI) reduction chromate stress iron and acid tolerance have been studied previously
(Bencheikh-Latmani et al 2005 Brown et al 2006 Gao et al2006 Gao et al 2004
Yang et al 2008) More recently it has been exploited as a model species for power
generation in microbial electrochemical systems (MESs)which have potential
applications in renewable energy generation bioremediation and wastewater treatment
(Logan et al 2007 Logan 2009 Park and Zeikus 2002 Fan et al 2011 Xu et al 2012)
In a MES electrochemically active microorganisms oxidize organic matter in the an
ode chamber to release electrons Electrons are then transferred to the anode electrode thr
ough various mechanisms (Logan et al 2006 Lovley 2006 Rabaey et al 2003) and fina
lly travel to the cathode electrode and combine with the terminal electron acceptor The
key feature of MESs is the microbe-catalyzed electron transfer from the organic matter to
the anode (Rabaey et al 2004) Enhancing the current output which highly depends on
the performance of the anode electrode is critical for the successful application of MESs
(Logan et al 2007 Park and Zeilus 2002 2003) Several nanomaterials including TiO2
gold (Au) and palladium (Pd) nanoparticles have been utilized to decorate electrodes to
enhance power density in MESs (Qiao et al 2008 Quan et al 2005 Fan et al 2011) The
possible mechanisms for increased current densities have been studied by using Fe
69
nanoparticle-decorated anodes (Xu et al 2012)
Carbon nanotubes (CNT) are cylindrically shaped nanomaterials with extremely
high surface area excellent electrical conductivity and chemical inertness (He et al
2005a Serp et al 2003) These unique properties make CNT a promising electrode
material (Liang et al 2008) The biocompatibility of microorganisms and carbon
nanostructures has been evaluated (Morozan et al 2007) The effects of CNT on anodic
biofilms have been tested in MFCs (Liang et al 2011) However no studies have been re
ported on global transcriptome analysis for the response of S oneidensis MR-1 to CNT
decorated anode in MECs
In the present study the graphite disks decorated with CNT were used as anodes to
investigate the effects of nanomaterials on current generation in multi-anode MECs
DNA microarrays were used to analyze differences in the global gene expression profile
of S oneidensis MR-1 grown on plain versus CNT decorated anodes
Materials and methods
Bacterial cultures S oneidensis MR-1 was purchased from American Type Culture
collection (ATCC 700550) and stored as freezer stocks at -80degC Stock S oneidensis
MR-1 cells were grown aerobically using M1 medium at 30degC Cells in early stationary
growth phase (optical density of 15-16 at 600nm (Spectrophotometer UV-1700
Shimadzu Columbia MD) were injected into the chamber of the MECs for current
production Sodium lactate (final concentration 30mM) was added as the additional
electron donor
Fabrication and characterization of nanostructured anodes Superfine isomolded
graphite disks (16 cm x 05 cm GraphiteStorecom Inc) were polished with ultrafine
70
sand paper (2000 grit 3M Company) as the base for the CNT decorated and control
anodes Aligned multi-walled carbon nanotube samples were synthesized via plasma
enhanced chemical vapor deposition (PECVD) Samples superfine isomolded graphite
pillars with a 16 cm diameter and 05 cm thickness were sputtered with a thin film of Ni
in a CRC 300 magnetron sputter coater for 1 to 2 min They were then placed in the
PECVD reactor where the Ni was annealed into small catalyst islands C2H2 was bled
into the chamber for growth and an NH3 plasma was used to vertically align the growth
of the multi-walled carbon nanotubes (MWCNTs) The temperature and time were varied
between 725degC and 775oC and 15 to 60 min to control the length of the tubes and density
of samples Samples were then analyzed in a FEI Sirion field emission scanning electron
microscope (FESEM) SEM images of CNT decorated anode and control surfaces were
shown in the Figure 51
Figure 51 SEM images of CNT decorated anodes (A) Graphite disk control (B) CNT
decorated anode
Multiple channel MECs construction and operation A MECs with multiple
removable anodes each with an effective anode area of 07 cm2
was constructed and used
to evaluate the effects of nanostructures on the current density of according to a previous
B A
71
study (Fan et al 2011) The cathode was made of wet-proof (30) carbon cloth (type B
E-TEK Division Inc USA) coated with PtC (20 E-TEK Division Inc USA)
following a previously reported procedure (Liu et al 2005) The final platinum loading
was 05 mgcm2 per projected cathode area The size of the cathode (150 cm
2) was 25
times larger than the total effective area of the 8 graphite disk anodes (56 cm2) to prevent
cathode limitations on the performance of the MECs system All MECs with CNT
decorated anodes and control anodes were tested for current generation for 24 h with
sterile a medium (without bacteria) before injecting the bacterial cells in order to
determine whether the CNT decorated anodes demonstrate chemical current generation
compared to control anodes All the testing anodes were set up in the MECs
simultaneously so that the S oneidensis MR-1 culture was applied to all of them
simultaneously Short electrode spacing (17 cm) was used in the MECs design to reduce
the internal resistance A voltage of 06 V was applied the MECs for current generation
after the cells of S oneidensis MR-1 were inoculated into the MECs in the early
stationary phase A multimeter with a data acquisition system (2700 Keithly USA) was
used to monitor the current change by measuring the voltage drop through a resistor
Microarray analysis Biofilms for whole gene microarray analysis were aseptically
removed from the plain and CNT decorated anodes of MECs after 80 hours further
incubation at 30oC when current density obviously reached to a stable phase Total RNA
was extracted using Trizol (Ambion inc AustinTX) following the manufacturerrsquos
instructions Integrity of the RNA samples was confirmed using the Agilent 2100
Bioanalyzer (Agilent Technology Inc Palo Alto CA) Total RNA (200 ng) from each
sample was amplified using the MessageAmpTM
II-Bacteria Prokaryotic RNA
72
Amplification kit (Ambion AustinTX) according to the manufacturerrsquos instructions The
S oneidensis MR-1 whole-gene microarrays representing all of the 4295 genes (resource
from NCBI) were ordered from Roche NimbleGen Inc (Madison WI) Biological
triplicates of biofilms on the CNT decorated anodes and control were analyzed cDNA
synthesis labeling and hybridization were carried out by the CGRB Core Laboratories at
Oregon State University DNASTAR ArrayStarTM
3 software was used to identify genes
that were up- or down-regulated more than 2-fold when grown on the nanoparticle
modified anodes using the unpaired two sample t-test with a cutoff p-value of 005 with
bonferroni correction
Results and discussion
Enhancement of current generation using CNT decorated anodes in MECs The
current density generated by CNT decorated anode MECs with S onidensis MR-1
increased to 36 microAcm2 10 hours post-inoculation and reached a maximum of 40 microAcm
2
at 29 hours inoculation then gradually decreased The current density stabilized at
30microAcm2 after 78 h operation Figure 52 shows that the current density curve generated
by CNT decorated anode in MECs The average current density generated by CNT
decorated anodes was 56 times that of the control The current density generated in the
MECs without bacteria was negligible (data not shown) This result demonstrates that the
current enhancement observed with the CNT decorated anodes was biologically-derived
A
73
Figure 52 Effects of CNT decorated anodes on the current density in MECs (3 replicates)
Control is plain graphite disk anode CNT is the anode with CNT decoration (Error bars
represent standard deviation)
It has been reported that nanostructured decorated anodes have significant impacts on
current densities (Fan et al 2011 Qiao et al 2006 Sharma 2008 Tsai et al 2009) The
CNT decorated anodes had significantly enhanced current densities in MECs in this
study This result is consistent with previous studies Interestingly the current density
curve generated by CNT decorated anode MECs is quite different from the current
density curve generated by Au NP and Pd NP decorated anodes using same bacterial
species (Fan et al 2011) In the Au and Pd NP decorated anode MECs the current
density curve was not significantly increased in the beginning compared with the control
but gradually increased after the inoculation the current density reached the maximum
current density more than 50 hours after the inoculation However the current density
generated by CNT decorated anode MECs started to increase significantly immediately
after the inoculation It reached a maximum current density 29 hours post-inoculation
-1
6
13
20
27
34
41
48
0 10 20 30 40 50 60 70 80
Curr
ent
den
sity
(micro
Ac
m2)
Time (Hours)
Control
CNT
74
These results indicate that there might be different current density enhancement
mechanisms between CNT with other metal NP decorated anodes in MECs
Global transcriptome analysis Whole-genome DNA microarrays were used to attain a
comprehensive general overview of the transcriptional response of S oneidensis
MR-1 of
S oneidensis MR-1 to CNT decorated anodes in MECs In a total of 4295 genes analyzed
457 genes (250 genes up-regulated and 207 genes down-regulated) exhibited significantly
(P lt 005) changed expression at a 2-fold level in at least 3 replicates in the response to
CNT The total number of the regulated genes present 11 of the 4295 open reading
frames (ORFs) presented on the microarray The total number of genes at significant
expression levels in response to CNT decorated anodes is close to that to Fe NP decorated
anodes in MECs (392 9) ( Xu et al 2012)
Fig 53 Differentially expressed genes grouped by functional classification according to
the TIGR S oneidensis genome database (wwwtigrorg) Columns 1 amino acid
biosynthesis 2 biosynthesis of cofactors prosthetic groups and carriers 3 cell
envelope 4 cellular processes 5 central intermediary metabolism 6 DNA metabolism
7 energy metabolism 8 fatty acid and phospholipid metabolism 9 other categories 10
protein fate 11 protein synthesis 12 purines pyrimidines nucleosides and nucleotides
-80
-60
-40
-20
0
20
40
60
80
100
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18
Nu
mb
er
of
Ge
ne
s
Up-regulated
Down-regulated
75
13 regulatory functions 14 signal transduction 15 transcription 16 transport and
binding proteins 17 unknown function 18 hypothetical proteins
The overall transcript genomic response of S oneidensis MR-1 to CNT decorated
anodes was summarized in Figure 53 by grouping the differentially expressed genes into
their functional role categories based on The Institute for Genomic Researchrsquos annotation
(Version 8) of the MR-1 genome sequence The wide distribution of putative functional
roles attributed to the differentially expressed genes indicated the extent of the molecular
response of Soniedenis MR-1 to the CNT decorated anodes A large number of up-
regulated genes were presented in genes associated with biosynthesis of cofactors
prosthetic groups and carriers (group 2) cell envelope (group 3) protein fate (group 10)
protein synthesis (group 11) and purines pyrimidines nucleosides and nucleotides
(group 12) in this study The number of up-regulated genes was two times more than that
of down-regulated genes in each functional group Contrastingly in the genomic response
S oneidensis MR-1 to Fe NP decorated anodes (chapter 4) most of up-regulated genes
were associated with cellular processes (group 4) energy metabolism (group 7) other
categories (group 9) and hypothetical proteins (group 18) The down-regulated genes
presented in the amino acid biosynthesis (group 1) cellular processes (group 4) substrate
transport (group 16) and hypothetical proteins (group 18) While those genes encoding
proteins involved in amino acid biosynthesis (group 1) central intermediary metabolism
( group 5) DNA metabolism( group 6) fatty acid and phospholipid metabolism (group
8) and other categories (group 9) were among the most down-regulated genes in this
study However the two groups of genes that were the most noticeable between the
genomic response S oneidensis MR-1 to CNT and Fe NP decorated anodes which they
are the genes related to energy metabolism (group 7) and hypothetical proteins (group
76
18) These results indicated there are different and common genomic responses to S
oneidensis MR-1 on CNT and Fe NP decorated anodes in MECs
Electron transfer related genes Based on previous electron transfer mechanism
studies on Shewanella a serial of group proteins collectively described as the Mtr
pathway play the role to transfer electron rom the inner bacterial body to the outer
membrane (Beliaev et al 2001 Shi et al 2007 Brutinel and Gralnick 2012) In the Mtr
pathway firstly electrons generated by bacteria received by CymA a tetraheme c-type
cytochrome anchored in the inner membrane then electrons were transferred to MtrA a
periplasmic decaheme c-type cytochrome After that electrons were transferred from
MtrA to MtrC an outer-membrane decaheme c-type cytochrome is facilitated by MtrB a
non-heme containing integral outer-membrane proteinOuter-membrane decaheme c-type
cytochrome MtrC and OmcA are in change of electron transfer to exreacellular electron
acceptor (Shi et al 2007 Brutinel and Gralnick 2012 ) These genes encoding electron
transport proteins are shown in the table 51 The critical genes SO_1777 mtrA SO_1776
mtrB SO_1778 mtrC are unregulated 41 to 80 fold in response to CNT decorated
anodes other cytochrome c genes related to electron transfer in MtrA pathway also are
significantly increased in response to CNT decorated anodes These genes encoding outer
membrane proteins have different level increases supporting that the increase of
cytochrome c gene expression contributes to the current enhancements of MECs as
response to CNT decorated anodes
77
Table 51 Expression level of cytochrome c as response to CNT decorated anodes
Locus Tag Gene product descriptions Fold change CNT
SO_0165 general secretion pathway protein GspC 31
SO_0167 general secretion pathway protein GspE 45
SO_1776 outer membrane protein precursor MtrB 41
SO_1777 outer membrane decaheme cytochrome c
lipoprotein MtrA
80
SO_1778 outer membrane decaheme cytochrome c
lipoprotein MtrC
43
SO_1779 outer membrane decaheme cytochrome c
OmcA
63
SO_0135 lipoprotein of unknown function DUF333 22
SO_0136 conserved hypothetical inner membrane protein 24
Flavin related genes It is known that self-produced mediators play critical roles in the
electron transfer of S oneidensis to extracellular insoluble electrode (Canstein von et al 2
008 Myers et al 1992 Scott et al 1994 Shi et al 2007) It has been proved that the
flavins secreted by S oneidensis can facilitate electron transfer from bacteria to solid
anode electrodes by acting as extracellular electron shuttles (Cansterin von et al 2008) T
wo critical genes (SO_1414 and SO_3468) related to flavin synthesis were shown increa
sed significantly (more than 2 folds) increase in this study (Table 52) This result support
s the hypothesis that flavin production increases as the response to CNT decorated anodes
in MECs have significant contribution to the current density enhancements
78
Table 52 Expression levels of genes related to flavin synthesis as response to CNT
decorated anode
Locus Tag Gene product descriptions Fold change CNT
SO_1414 flavocytochrome c flavin subunit putative 22
SO_3468 riboflavin synthase subunit alpha 20
The amount of bacterial biofilm biomass on the anode can affect the power
generation in MFCs (Biffinger et al 2007 Heydorn et al 2000) Biofilm formation
related genes include flagellar gene IV pilus genes and other genes the flagellar related
genes (fliB and fliK) and IV pilus genes (mshA) were induced only from 105 to 134
fold as response of S oneidensis MR-1 to CNT decorated anodes in this study No
biofilm-associated genes presented significant changes in expression levels in this study
indicating there is no direct connections between biofilm enhancements to CNT
decorated anodes in this study This result is consistent with the Liang et al (2011)
conclusion that using CNT enhanced the electrochemical activity of anodic biofilms but
did not result in a significant increase of biomass in the anodic biofilms
Our microarray results showed two significantly up-regulated gene groups the
genes encoding proteins localized on the outer membrane and the genes involved in
flavin biosynthesis contributed to current density enhancement by CNT decorated anodes
Among 457 significantly changed genes to CNT in this study there are also a relatively
large number of genes encoding proteins with unknown functions which are either up- or
down-regulated in the response to CNT decorated anodes which indicated more
79
complicated responses of S oneidensis MR-1 to CNT decorated anodes in MECs and
further study is needed
80
Chapter 6
Summary
Microbial electrochemical systems (MESs) with a mixed culture initially inoculated
from Corvallis wastewater treatment plant have been studied for more than 6 years for
varying purposes including power generation hydrogen production heavy metal
removal and wastewater treatment Experiment results have shown that the mixed culture
is quite stable with excellent performance in MESs Our community analysis using
denaturing gradient gel electrophoresis (DGGE) and 16S rDNA clone library construction
suggests that the mixed culture is composed predominantly of Geobacter sp (66)
Arcobacter sp (12) and Citrobacter sp (11) These results not only improved our
understanding of the mixed culture community but also guided our studies on the
cultivation and isolation of the different bacterial species
Exoelectrogenic Citrobacter sp SX-1 was first isolated and characterized from a
MFC operated with the mixed culture Citrobacter sp SX-1 has been demonstrated to
produce electricity from wide range of different substrates including citrate acetate
glucose sucrose glycerol and lactosein MFCs Cyclic voltammetry analysis indicated
that membrane associated proteins may play an important role in facilitating the electrons
transferring from bacteria to electrode The strain SX-1 increased the known diversity of
power generating exoelectrogens and provided a unique bacterial species for study in
renewable energy generation and waste treatment
81
To enhance the electron transfer from bacteria to anode nano decorated anodes
including Fe NP and CNT were developed and characterized and evaluated in MECs
using Shewanella oneidensis MR-1 as a model species Both nanostructures have
significantly increased current density compared with the control Whole genome
microarray analysis elucidated the possible mechanisms of power enhancement in
response to these nano-decorated anodes
These results benefit to understanding of physiology and ecology of mixed
cultures in MFCs and improve the efficiency of current generation in MESs which will
facilitate the viability of niche applications for MESs in near future
82
Bibliography
1 Amman R Ludwig W (2000) Ribosomal RNA-targeted nucleic acid probes for
studies in microbial ecology FEMS Microbiol Ecol 24 555ndash565
1 Bao SJ Li CM Zang JF Cui XQ Qiao Y Guo J (2008) Novel Nanoporous TiO2
Electrode for Directly Electrochemistry of glucose oxidase Adv Funct Mater
18(4) 591-599
2 Beliaev AS Saffarini DA McLaughlin JL Hunnicutt D (2001) MtrC an outer
membrane decahaem c cytochrome required for metal reduction in Shewanella
putrefaciens MR-1 Mol Microbiol 39722-30
3 Bencheikh-Latmani R Williams SM Haucke L Criddle CS Wu L Criddle CS
Zhou J Tebo BM (2005) Global transcriptional profiling of Shewanella oneidensis
MR-1 during Cr(VI) and U(VI) reduction Appl Environ Microbiol 71 7453ndash7460
4 Biffinger JC Pietron J Ray R Little B and Ringeisen BR (2007) A biofilm
enhanced miniature microbial fuel cell using Shewanella oneidensis DSP10 and
oxygen reduction cathodes Biosens Bioelectron 221672ndash1679
5 Bodelier PLE Roslev P Henckel T FenzelP (2000) Stimulation by ammonium
based fertilizers of methane oxidation in soil around rice root Nature 40 421ndash424
6 Bond DR Holmes DE Tender LM Lovley DR (2002) Electrode-reducing
microorganisms that harvest energy from marine sediments Science 295 483-485
7 Bond DR Lovley DR (2003) Electricity production by Geobacter sulfurreducens
attached to electrodes Appl Environ Microbiol 69 1548-1555
8 Bond DR Lovley DR (2005) Evidence for involvement of an electron shuttle in
electricity generation by Geothrix fermentans Appl Environ Microbiol 71 2186-
2189
9 Bretschger O Obraztsova A Sturm CA Chang IS Gorby YA Reed SB Culley
DE Reardon CL Barua S Romine MF Zhou J Beliaev AS Bouhenni R
Saffarini D Mansfeld F Kim BH Fredrickson JK Nealson KH (2007) Current
production and metal oxide reduction by Shewanella oneidensis MR-1 wildtype and
mutants Appl Environ Microbiol 73 7003ndash7012
10 Brown SD Thompson MR Verberkmoes NC Chourey K Shah M Zhou J
Hettich RL and Thompson DK (2006) Molecular dynamics of the Shewanella
oneidensis response to chromate stress Mol Cell Proteomics 51054-1071
11 Brutinel ED Gralnick JA (2012) Shuttling happens soluble flavin mediators of
extracellular electron transfer in Shewanella Appl Microbiol Biotechnol 93(1) 41-8
12 Butler JE Kaufmann F Coppi MV Nuacutentildeez C Lovley DR (2004) MacA a diheme
c-type cytochrome involved in Fe(III) reduction by Geobacter sulfurreducens J
Bacteriol 186(12) 4042ndash4045
83
13 Caccavo F Lonergan DJ Lovley DR Davis M Stolz JF McInerney MJ (1994)
Geobacter sulfurreducens sp nov a hydrogen- and acetate oxidizing dissimilatory
metal-reducing microorganism Appl Environ Microbiol 60 3752ndash3759
14 Canstein von H Ogawa J Shimizu S Lloyd JR (2008) Secretion of flavins by
Shewanella species and their role in extracellular electron transfer Appl Environ
Microbiol 74 615ndash623
15 Cao X Huang X Liang P Xiao K Zhou Y Zhang X Logan BE (2009) A New
Method for Water Desalination Using Microbial Desalination Cells Environ Sci
Technol 43(18)7148-7152
16 Chaudhuri SK Lovley DR (2003) Electricity generation by direct oxidation of
glucose in mediatorless microbial fuel cells Nat Biotechnol 21 1229-1232
17 Cheng S Liu H Logan B E (2006) Increased power generation in a continuous
flow MFC with advective flow through the porous anode and reduced electrode
spacing Environ Sci Technol 402426-2432
18 Cheng S Logan B E (2007) Sustainable and efficient biohydrogen production via
electrohydrogenesis PNAS 104 18871-18873
19 Chignell J (2010) Energy Recovery from Biodiesel Waste Performance of
Microbial Electrochemical Systems on Glycerol Thesis
20 Chung K Okabe S (2009) Characterization of electrochemical activity of a strain
ISO2-3 phylogenetically related to Aeromonas sp isolated from a glucose-fed
microbial fuel cell Biotechnol Bioeng 104(5) 901-910
21 Cole JR Chai B Farris RJ Wang Q Kulam-Syed-Mohideen AS McGarrell
DM Bandela AM Cardenas E Garrity GM Tiedje JM (2007) The ribosomal
database project (RDP-II) introducing myRDP space and quality controlled public
data Nucleic Acids Res 35 (Database issue) D169-D172 doi 101093nargkl889
22 Cole JR Wang Q Cardena E Fish J Chai B Farris RJ Kulam-Syed-Mohideen
AS McGarrell DM Marsh T Garrity GM Tiedje JM (2009) The Ribosomal
Database Project improved alignments and new tools for rRNA analysis Nucleic
Acids Res 37 (Database issue) D141-D145 doi 101093nargkn879
23 Dey A De S De A De SK (2004) Characterization and dielectric properties of
polyaniline-TiO2 nanocomposites Nanotechnol 15 1277-1283
24 Edwards U Rogall T Bloumlcker H Emde M Boumlttger EC (1989) Isolation and
direct complete nucleotide determination of entire genesmdashcharacterization of a gene
coding for 16S-ribosomal RNA NucleicAcids Res 17 7843ndash7853
25 Eggleston CM Voumlroumls J Shi L Lower BH Droubay TC Colberg PJS (2008)
Binding and direct electrochemistry of OmcA an outer-membrane cytochrome from
an iron reducing bacterium with oxide electrodes a candidate biofuel cell system
Inorg Chim Acta 361769ndash777
26 Fan Y Sharbrough E Liu H (2008) Quantification of the Internal Resistance
Distribution of Microbial Fuel Cells Environ Sci Technol 42 8101ndash8107
84
27 Fan Y Xu S Schaller R Jiao J Chaplen F Liu H (2011) Nanoparticle decorated
anodes for enhanced current generation in microbial electrochemical
cells Biosensors and Bioelectronics 26 (5)1908ndash1912
28 Fan Y Hu H Liu H (2007)Sustainable power generation in microbial fuel cells
using bicarbonate buffer and proton transfer mechanisms Environ Sci
Technol 41(23) 8154-8
29 Fedorovich V Knighton MC Pagaling E Ward FB Free A Goryanin I (2009)
Novel electrochemically active bacterium phylogenetically related to Arcobacter
butzleri isolated from a microbial fuel cell Appl Environ Microbiol 75(23) 7326-
7334
30 Ferna ndez A Huang S Seston S Xing J Hickey R Criddle C Tiedje J (1999)
How stable is stable Function versus community composition Appl Environ
Microbiol 65 3697ndash3704
31 Field SJ Dobbin PS Cheesman MR Watmough NJ Thomson AJ Richardson
DJ (2000) Purification and magneto-optical spectroscopic characterization of
cytoplasmic membrane and outer membrane multiheme c-type cytochromes from
Shewanella frigidimarina NCIMB400 J Biol Chem 275 8515ndash8522
32 Freguia S Masuda M Tsujimura S Kano K (2009) Lactococcus lactis catalyses
electricity generation at microbial fuel cell anodes via excretion of a soluble quinone
Bioelectrochemistry 7614ndash18
33 Fricke K Harnisch F and Schroumlder U (2008) On the use of cyclic voltammetry for
the study of the anodic electron transfer in microbial fuel cells Energy Environ
Sci 1 144ndash147
34 Gao H Wang Y Liu X Yan T Wu L Alm E Arkin A Thompson DK and
Zhou J (2004) Global transcriptome analysis of the heat shock response of
Shewanella oneidensis J Bacteriol 1867796-7803
35 Gao H Yang ZK Wu L Thompson DK Zhou J (2006) Global transcriptome
analysis of the cold shock response of Shewanella oneidensis MR-1 and mutational
analysis of its classical cold shock proteins J Bacteriol 1884560-4569
36 George MG (2005) Bergeyrsquos Manual of Systematic Bacteriology Volume Two The
Proteobacteria Part B The Gammaproteobacteria Second Edition New York
Springer 651p
37 Gonzaacutelez-Toril E Llobet-Brossa E Casamayor EO Amann R Amils R (2003)
Microbial ecology of an extreme acidic environment the Tinto river Appl Environ
Microbiol 69(8) 4853ndash4865
38 Gorby YA Yanina S Mclean JS Rosso KM Moyles D Dohnalkova A
Beveridge TJ Chang IS Kim BH Kim KS Culley DE Reed SB Romine M
F Saffarini DA Hill EA Shi L Elias DA Kennedy DW Pinchuk G
Watanabe K Ishii S Logan B Nealson KH Fredrickson JK (2006) Electrically
conductive bacterial nanowires produced by Shewanella oneidensis strain MR-1 and
other microorganisms Proc Natl Acad Sci USA 10311358-11363
85
39 He Z Wagner N Minteer SD Angenent LT (2006) An upflow microbial fuel cell
with an interior cathodes assessment of the internal resistance by impedance
spectroscopy Environ Sci Technol 40 5212-5217
40 Heydorn A Nielsen AT Hentzer M Sternberg C Givskov M Ersboslashll BK Molin
S (2000) Quantification of biofilm structures by the novel computer program
COMSTAT Microbiol 1462395-2407
41 Holmes DE Bond DR Lovley DR (2004a) Electron transfer by Desulfobulbus
propionicus to Fe(III) and graphite electrodes Appl Environ Microbiol 701234ndash
1237
42 Holmes DE Chaudhuri SK Nevin KP Mehta T Methe BA Liu A Ward JE
Woodard TL Webster J Lovley DR (2006) Microarray and genetic analysis of
electron transfer to electrodes in Geobacter sulfurreducens Environ Microbiol
81805-1815
43 Holmes DE Nicoll JS Bond DR Lovley DR (2004b) Potential role of a novel
psychrotolerant member of the family Geobacteraceae Geopsychrobacter
electrodiphilus gen nov sp nov in electricity production by a marine sediment
fuel cell Appl Environ Microbiol 70 6023ndash6030
44 Holmes DE Chaudhuri SK Nevin KP Mehta T Metheacute BA Liu A Ward JE
Woodard TL Webster J Lovley DR (2006) microarray and genetic analysis of
electron transfer to electrodes in Geobacter sulfurreducens Environ Microbial
8(10)1805-1815
45 Hu H (2009) Enhancing Hydrogen Production in Microbial Electrolysis Cells
through Development of Platinum-free Cathode and Improvement of Reactor
Dissertation
46 Ince BK Ayman O N Turker G Ccedilelikkol S Ince O (2010) Microbial ecology of
anaerobic reactors for treatment of alcohol industry wastewaters a review Current
research technology and education topics in applied microbiology and microbial
ecology 988-999
47 Jung S Regan JR (2007) Comparison of anode bacterial communities and
performance in microbial fuel cells with different electron donors Appl Microbiol
Biotechnol 77393ndash 402
48 Kim BH Kim HJ Hyun MS Park DH (1999) Direct electrode reaction of Fe(III)
reducing bacterium Shwwanella putrefaciens J Microbial Biotechnol 9127-131
49 Kim GT Hyun MS Chang IS Kim HJ Park HS Kim BH Kim SD Wimpenny
JW Weightman AJ (2005) Dissimilatory Fe(III) reduction by an electrochemically
active lactic acid bacterium phylogenetically related to Enterococcus gallinarum
isolated from submerged soil J Appl Microbiol 99(4) 978ndash987
50 Kim GT Webster G Wimpenny JW Kim BH Kim HJ Weightman AJ (2006)
Bacterial community structure compartmentalization and activity in a microbial fuel
cell J Appl Microbiol 101(3) 698ndash710
86
51 Kim HJ Park HS Hyun MS Chang IS Kim M Kim BH (2002) A mediator-
less microbial fuel cell using a metal reducing bacterium Shewanella putrefaciens
Enzyme Microb Technol 30 145-152
52 Kim JR Min B Logan BE (2005) Evaluation of procedures to acclimate a
microbial fuel cell for electricity production Appl Microbiol Biotechnol 68(1)23-30
53 Kim JR Cheng S Oh SE Logan BE (2007) Power generation using different
cation anion and ultrafiltration membranes in microbial fuel cells Environ Sci
Technol 41 1004-1009
54 Konstantinidis KT Serres MH Romine MF Rodrigues JL Auchtung J McCue
LA Lipton MS Obraztsova A Giometti CS Nealson KH Fredrickson JK
Tiedje JM (2009) Comparative systems biology across an evolutionary gradient
within the Shewanella genus Proc Natl Acad Sci U S A 106(37)15909-15914
55 Lee J Phung NT Chang IS Kim BH Sung HC (2003) Use of acetate for
enrichment of electrochemically active microorganisms and their 16S rDNA
analyses FEMS Microbiol Lett 223185ndash191
56 Liang P Wang H Xia X Huang X Mo Y Cao X Fan M(2011)Carbon
nanotube powders as electrode modifier to enhance the activity of anodic biofilm
in microbial fuel cells Biosens Bioelectron 26(6)3000-4
57 Lies DP Hernandez ME Kappler A Mielke RE Gralnick JA Newman DK
(2005) Shewanaella oneidensis MR-1 uses overlapping pathways for iron reduction
at a distance and by direct contact under conditions relevant for biofilms Appl
Environ Microbiol 71 4414-4426
58 Liu H and Logan B E (2004) Electricity generation using an air-cathode single
chamber microbial fuel cell in the presence and absence of a proton exchange
membrane Environ Sci Technol 38 4040-4046
59 Liu H Grot S Logan BE (2005) Electrochemically assisted microbial production
of hydrogen from acetate Environ Sci Technol 39 4317 ndash 320
60 Liu H Hu H Chignell J Fan Y (2010) Microbial Electrolysis Novel Technology
for Hydrogen Production from Biomass Biofuels 1(1) 129ndash142
61 Liu H Ramnarayanan R Logan BE (2004) Production of electricity during
wastewater treatment using a single chamber microbial fuel cell Environ Sci Technol
382281-2285
62 Liu WT Marsh TL Cheng H Forney LJ (1997) Characterization of microbial
diversity by determining terminal restriction fragment length polymorphisms of gene
encoding 16S rRNA Appl Environ Microbiol 63 4516ndash4522
63 Logan B E Hamelers B Rozendal R Schroder U Keller J Freguia S Aelterman
P Verstraete W Rabaey K (2006) Microbial fuel cells methodology and
technology Environ Sci Technol 40 (17) 5181-5192
64 Logan B E and Regan JM (2006) Electricity-producing bacterial communities in
microbial fuel cells Trends Microbiol 14512ndash518
87
65 Logan B Cheng S Watson V Estadt G (2007) Graphite fiber brush anodes for
increased power production in air-cathode microbial fuel cells Environ Sci Technol
41(9)3341-3346
66 Logan BE (2009) Exoelectrogenic bacteria that power microbial fuel cells Nat Rev
Microbiol 5 375-381
67 Logan BE Call D Cheng S Hamelers HV Sleutels TH Jeremiasse AW
Rozendal RA (2008) Microbial electrolysis cells for high yield hydrogen gas
production from organic matter Environ Sci Technol 42 8630-8640
68 Logan BE Hamelers B Rozendal R Schroder U Keller J Freguia S Aelterman
P Verstraete W Rabaey K (2006) Microbial fuel cells methodology and
technology Environ SciTechnol 40 5181-5192
69 Lovely DR (2006) Bug juice harvesting electricity with microorganisms Nature
Review| Microbiology 4 497-508
70 Lovley DR Phillips EJ (1988) Novel Mode of Microbial Energy Metabolism
Organic Carbon Oxidation Coupled to Dissimilatory Reduction of Iron or
Manganese Appl Environ Microbiol 54(6) 1472-1480
71 Ly HK Sezer M Wisitruangsakul N Feng JJ Kranich A Millo D Weidinger
IM Zebger I Murgida DH Hildebrandt P (2011) Surface-enhanced vibrational
spectroscopy for probing transient interactions of proteins with biomimetic interfaces
electric field effects on structure dynamics and function of cytochrome c FEBS J
278(9)1382-90
72 Macaskie LE Hewitt CJ Shearer JA Kent CA (1995) Biomass production for
the removal of heavy metals from aqueous solutions at low pH using growth-
decoupled cells of a Citrobacter sp Int Biodeter Biodegr 3573ndash92
73 Malki M De Lacey AL Rodriguez N Amils R Fernandez VM (2008)
Preferential use of an anode as an electron acceptor by an acidophilic bacterium in
the presence of oxygen Appl Environ Microbiol 74 4472ndash4476
74 Meitl LA Eggleston CM Colberg PJS Khare N Reardon CL Shi L (2009)
Electrochemical interaction of Shewanella oneidensis MR-1 and its outer membrane
cytochromes OmcA and MtrC with hematite electrodes Geochim Cosmochim
Ac 73(18) 5292-5307
75 Morozan A Stamatin L and Nastase
F (2007) The biocompatibility
microorganisms-carbon nanostructures for applications in microbial fuel cells Phys
Stat Sol 6 1797-1803
76 Morris JM Jin S Crimi B Pruden A (2009) Microbial fuel cell in enhancing
anaerobic biodegradation of diesel Chem Eng J 146 161ndash167
77 Murray AE Lies D Li G Nealson K Zhou J Tiedje JM (2001) DNADNA
hybridization to microarrays reveals gene-specific differences between closely
related microbial genomes Proc Natl Acad Sci U S A 98 9853-9858
78 Muyzer G de Waal EC Uitterlinden AG (1993) Profiling of complex microbial
communities by denaturing gradient gel electrophoresis analysis of polymerase chain
88
reaction amplified genes coding for 16S rRNA Appl Environ Microbiol 59 695ndash
700
79 Myers CR Myers JM (1992) Localization of cytochromes to the outer membrane
of anaerobically grown Shewanella putrefacians MR-1 J Bacteriol 174 3429ndash3438
80 Narde GK Kapley A Purohit HJ (2004) Isolation and characterization of
Citrobacter strain HPC255 for broad-range substrate specificity for chlorophenols
Curr Microbiol 48(6) 419-423
81 Okabe S Ito T Sugita K Satoh H (2005) Succession of internal sulfur cycles and
sulfur-oxidizing bacterial communities in microaerophilic wastewater biofilms Appl
Environ Microbiol 71(5) 2520-2529
82 Park DH and Zeikus JG (2000) Electricity generation in microbial fuel cells using
neutral red as an electronophore Appl Environ Microbial 66 292-1297
83 Park DH and Zeikus JG (2002) Impact of electrode composition on electricity
generation in a single-compartment fuel cell using Shewanella putrefaciens Appl
Microbiol Biotechnol 5958ndash56
84 Park DH and Zeikus JG (2003) Improved fuel cell and electrode designs for
producing electricity from microbial degradation Biotechnol Bioeng 81 348ndash355
85 Park HS Kim BH Kim HS Kim HJ Kim GT Kim M Chang IS Park Y
K Chang HI (2001) A novel electrochemically active and Fe(III)-reducing
bacterium phylogenetically related to Clostridium butyricum isolated from a
microbial fuel cell Anaerobe 7 297-306
86 Parot S Nercessian O Delia ML Achouak W Bergel A (2009) Electrochemical
checking of aerobic isolates from electrochemically active biofilms formed in
compost J Appl Microbiol 106(4) 1350-1359
87 Pham CA Jung SJ Phung NT Lee J Chang IS Kim BH Yi H Chun J
(2003) A novel electrochemically active and Fe(III)-reducing bacterium
phylogenetically related to Aeromonas hydrophila isolated from a microbial fuel cell
FEMS Microbiol Lett 223 129-134
88 Qiao Y Bao SJ Li CM Cui XQ Lu ZS Guo J (2008) Nanostructured
polyanilinetitanium dioxide composite anode for microbial fuel cells ACS Nano 2
113-119
89 Qiu R Zhao B Liu J Huang X Li Q Brewer E Wang S Shi N (2009) Sulfate
reduction and copper precipitation by a Citrobacter sp isolated from a mining area J
Hazard Mater 1641310-1315
90 Quan X Yang SG Ruan XL and Zhao HM (2005) Preparation of titania
nanotubes and their environmental applications as electrode Environ Sci Technol 39
3770ndash3775
91 Rabaey K Boon N Hofte M Verstraete W (2005) Microbial phenazine
production enhances electron transfer in biofuel cells Environ Sci Technol 393401ndash
3408
89
92 Rabaey K Boon N Siciliano S D Verhaege M Verstraete W (2004) Biofuel cells
select for microbial consortia that self-mediate electron transfer Appl Environ
Microbiol 70(9) 5373ndash5382
93 Rabaey K Lissens G Siciliano S D Verstraete W (2003) Amicrobial fuel cell
capable of converting glucose to electricity at high rate and efficiency Biotechnol
Lett 25 1531-15
94 Rabaey K Verstraete W (2005) Microbial fuel cells novel biotechnology for
energy generation Trends Biotechnol 23(6) 291-298
95 Reguera G McCarthy KD Mehta T Nicoll JS Tuominen MT Lovley DR
(2005) Extracellular electron transfer via microbial nanowires Nature 435 1098-
1101
96 Reguera G Nevin KP Nicoll JS Covalla SF Woodard TL Lovley DR (2006)
Biofilm and nanowire production leads to increased current in Geobacter
sulfurreducens fuel cells Appl Environ Microbiol 72 7345-7348
97 Reimers CE Tender LM Fertig S Wang W (2001) Harvesting energy from the
marine sediment-water interface Environ Sci Technol 35192-195
98 Rezaei F Xing D Wagner R Regan JM Richard TM Logan BE (2009)
Simultaneous cellulose degradation and electricity production by Enterobacter
cloacae in a microbial fuel cell Appl Microbiol Biotechnol 75 3673ndash3678
99 Richter H Nevin KP Jia H Lowy DA Lovley DR Tender LM (2009) Cyclic
voltammetry of biofilms of wild type and mutant Geobacter sulfurreducens on fuel
cell anodes indicates possible roles of OmcB OmcZ type IV pili and protons in
extracellular electron transfer Energy Environ Sci 2 506ndash516
100 Riesenfeld SC Schloss DP Handelsman J(2004) Metagenomics Genomic
Analysis of Microbial Communities Annual Review Genetics 38525-552
101 Rozendal RA Hamelers HV Buisman CJ (2006) Effects of membrane cation
transport on pH and microbial fuel cell performance Environ Sci Technol 40
5206-5211
102 Rozendal RA Hamelers HVM Euverink GJW Metz SJ Buisman CJN (2006)
Principle and perspectives of hydrogen production through biocatalyzed electrolysis
Int J Hydrogen Energ 311632-1640
103 Rozendal RA Hamelers HV Molenkamp RJ Buisman CJ (2007) Performance of
single chamber biocatalyzed electrolysis with different types of ion exchange
membrances Water Res 41 1984-1994
104 Schluumlter A Bekel T Diaz NN Dondrup M Eichenlaub R Gartemann
KH Krahn I Krause L Kroumlmeke H Kruse O Mussgnug JH Neuweger
H Niehaus K Puumlhler A Runte KJ Szczepanowski R Tauch A Tilker
A Viehoumlver P Goesmann A (2008) The metagenome of a biogas-producing
microbial community of a production-scale biogas plant fermenter analyzed by the
454-pyrosequencing technology J Biotechnol136 77-90
90
105 Scott J H and Nealson KH (1994) A biochemical study of the intermediary carbon
metabolism of Shewanella putrefaciens J Bacteriol 176(11) 3408ndash3411
106 Sekiguchi H Tomioka H Nakahara T and Uchiyama H (2001) A single band
does not always represent single bacterial strains in denaturing gradient gel
electrophoresis analysis Biotechnology Letters23 1205ndash1208
107 Sharma T Leela M Reddy A Chandra TS Ramaprabhu S (2008) Development
of carbon nanotubes and nanofluids based microbial fuel cell Int J Hydrogen Energ
336749ndash6754
108 Shea C Clauwaert P Verstraete W Nerenberg R (2008) Adapting a denitrifying
biocathode for perchlorate reduction Water Sci Technol 58 (10) 1941-1946
109 Shi L Squier TC Zachara JM Fredrickson JK (2007) Respiration of metal (hydr)
oxides by Shewanella and Geobacter a key role for multihaem c-type cytochromes
Mol Microbiol 6512-20
110 Stams AJ and Plugge CM(2009)Electron transfer in syntrophic communities
of anaerobic bacteria and archaea Nat Rev Microbiol7(8)568-77
111 Sukkasem C Xu S Park S Boonsawang P Liu H (2008) Effect of Nitrate on the
Performance of Single Chamber Air Cathode Microbial Fuel Cells Water Res 42(19)
4743-4750
112 Summers ZM Fogarty HE Leang C Franks AE Malvankar NS Lovley DR
(2010)Direct exchange of electrons within aggregates of an evolved syntrophic
coculture of anaerobic bacteria Science 330 (6009)1413-5
113 Szczepanowski R Bekel T Goesmann A Krause L Kroumlmeke H Kaiser
O Eichler W Puumlhler A Schluumlter A (2008) Insight into the plasmid metagenome
of wastewater treatment plant bacteria showing reduced susceptibility to
antimicrobial drugs analysed by the 454-pyrosequencing technology J
Biotechnol 136 54-64
114 Tamura K Dudley J Nei M Kumar S (2007) MEGA4 Molecular Evolutionary
Genetics Analysis (MEGA) software version 40 MolBiol Evol 24(8) 1596-1599
115 Thompson JD Gibson TJ Plewniak F Jeanmougin F Higgins DG (1997) The
CLUSTAL_X windows interface flexible strategies for multiple sequence alignment
aided by quality analysis tools Nucleic Acids Res 25 4876-4882
116 Thormann KM Duttler S Saville RM Hyodo M Shukla S Hayakawa Y
Spormann AM (2006) Control of formation and cellular detachment from
Shewanella oneidensis MR-1 biofilms by cyclic di-GMP J Bacteriol 1882681-2691
117 Thormann KM Saville RM Shukla S Pelletier DA Spormann AM (2004)
Initial Phases of biofilm formation in Shewanella oneidensis MR-1 J Bacteriol
186(23) 8096-104
118 Torres CI Kato Marcus A Rittmann BE (2008b) Proton transport inside the
biofilm limits electrical current generation by anode-respiring bacteria Biotechnol
Bioeng 100 872ndash881
91
119 Torres CI Marcus AK Lee HS Parameswaran P Krajmalnik-Brown R Rittmann
BE (2010) A kinetic perspective on extracellular electron transfer by anode-
respiring bacteria FEMS Microbiol Rev 234(1)3-17
120 Torres CI Marcus AK Parameswaran P Rittmann BE (2008a) Kinetic
experiments for evaluating the NernstndashMonod model for anode-respiring bacteria
(ARB) in a biofilm anode Environ Sci Technol 42 6593ndash6597
121 Tsai HY Wu Ch-Ch Lee Ch-Y Shih EP (2009) Microbial fuel cell performance
of multiwall carbon nanotubes on carbon cloth as electrodes J Power Sources
194199-205
122 Tyson GW Chapman J Hugenholtz P Allen EE Ram RJ Richardson
PM Solovyev VV Rubin EM Rokhsar DS Banfield JF(2004) Community
structure and metabolism through reconstruction of microbial genomes from the
environment Nature42837-43
123 Venkateswaran K Moser DP Dollhopf ME Lies DP Saffarini D A MacGregor
BJ Ringelberg DB White DC Nishijima M Sano H Burghardt J Stackebrandt
E Nealson KH (1999) Polyphasic taxonomy of the genus Shewanella and
description of Shewanella oneidensis sp Int J Syst Bacteriol 49705-724
124 von Canstein H Ogawa J Shimizu S Lloyd JR (2008) secretion of Flavins
by Shewanella Species and Their Role in Extracellular Electron Transfer Appl
Environ Microbiol 74 3615-623
125 Wang A Liu W Cheng Sh Xing D Zhou J Logan BE (2009) Source of methane
and methods to control its formation in single chamber microbial electrolysis cells
Int J Hydrogen Energ 34 3653-3658
126 Wang J Ma T Zhao L Lv J Li G Liang F Liu R (2008) PCRndashDGGE method
for analyzing the bacterial community in a high temperature petroleum reservoir
World J Microbiol Biotechnol 2 9694-9702
127 Wang Q Garrity GM Tiedje JM Cole JR (2007) Naiumlve Bayesian Classifier for
Rapid Assignment of rRNA Sequences into the New Bacterial Taxonomy Appl
Environ Microbiol 73(16) 5261-5267
128 Wang YF Masuda M Tsulimura S Kano K (2008) Electrochemical regulation of
the end-product profile in Propionibacterium freudenreichii ET-3 with an
endogenous mediator Biotechnol Bioeng 101(3) 579ndash586
129 Wrighton KC Agbo P Warnecke F Weber KA Brodie EL DeSantis TZ
Hugenholtz P Andersen GL Coates JD (2008) A novel ecological role of the
Firmicutes identified in thermophilic microbial fuel cells ISME J 2 1146ndash1156
130 Xing D Zuo Y Cheng S Regan JM Logan BE (2008) Electricity generation by
Rhodopseudomonas palustris DX-1 Environ Sci Technol 42 4146-4151
131 Xu S Liu H Fan Y Schaller R Jiao J Chaplen F (2012) Enhanced performance
and mechanism study of microbial electrolysis cells using Fenanoparticle-decorated
anodes Appl Microbiol Biotechnol 93(2)871-880
92
132 Yang Y Harris DP Luo F Wu L Parsons AB Palumbo AV Zhou J (2008)
Characterization of the Shewanella oneidensis Fur gene roles in iron and acid
tolerance response BMC Genomics 9 Suppl 1S11
133 Zhang L Zhou S Zhuang L Li W Zhang J Lu N Deng L (2008) Microbial
fuel cell based on Klebsiella pneumoniae biofilm Electrochem Commun10 1641ndash
1643
134 Zuo Y Xing DF Regan JM Logan BE (2008) Isolation of the exoelectrogenic
bacterium Ochrobactrum anthropi YZ-1 by using a U-tube microbial fuel cell Appl
Environ Microbiol 74 3130-3137
93
APPENDICES
List of published papers during PhD study
1 Shoutao Xu Hong Liu Yanzhen Fan Rebecca Schaller Jun Jiao and Frank
Chaplen (2012) Enhanced performance and mechanism study
of microbial electrolysis cells using Fe nanoparticle-decorated anodes Applied
Microbiology and Biotechnology 93(2)871-880
2 Shoutao Xu Hong Liu (2011) New exoelectrogen Citrobacter sp SX-1 isolated
from a microbial fuel cell Journal of Applied Microbiology111(5)1108-1115
3 Yanzhen Fan Shoutao Xu Rebecca Schaller Jun Jiao Frank Chaplen Hong Liu
(2011) Nanoparticle decorated anodes for enhanced current generation in microbial
electrochemical cells Biosensors and Bioelectronics 26(5) 1908-1912
4 Tunc Catal Shoutao Xu Kaichang Li Hakan Bermek Hong Liu
(2008)
Electricity generation from polyalcohols in single-chamber microbial fuel
cells Biosensors and Bioelectronics 24(4)849-854
5 Chontisa Sukkasema Shoutao Xu Sunhwa Park Piyarat Boonsawang Hong Liu
(2008) Effect of nitrate on the performance of single chamber air cathode
microbial fuel cells Water research 424743-4750
6 Rebecca Schaller Yanzhen Fan Shoutao Xu Alan Fern Frank Chaplen Hong
Liu and Jun Jiao (2009) Vertically Aligned Multi-walled Carbon Nanotube
Decorated Anodes for Microbial Fuel Cells Proceedings of Materials Research
Society 2009 1170 R05-13

LIST OF TABLES
Table
1-1
4-1
4-2
4-3
Taxa of bacteria current density generated and reactor configuration
used in microbial electrochemical cell experimentshelliphelliphelliphellip
Expression levels of genes related to biofilm formationhelliphelliphelliphelliphellip
Genes related to anaerobic growth and electron transfer with
significantly change expression levelhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip
Expression level of flavin and cytochrome geneshelliphelliphelliphelliphelliphelliphelliphellip
Page
4
55
56
58
4-4
5-1
5-2
Other genes with significantly changed expression level
Expression level of cytochrome c as response to CNT decorated anodes
Expression level of genes related to flavin synthesis as response to CNT
decorated anode in CNT decorated anodeshelliphellip
59
76
78
1
1
Bacterial Community Analysis New Exoelectrogen Isolation and Enhanced Performance
of Microbial Electrochemical Systems Using Nano-Decorated Anodes
Chapter 1
General Introduction
11 Microbial Fuel Cells and Microbial Electrolysis Cells
Microbial electrochemical systems (MESs) have drawn the attention of researchers
in recent years due to their promising applications in a variety of scientific fields such as
renewable energy generation bioremediation and wastewater treatment In a MES
microorganisms interact with electrodes catalyzing oxidation and reduction reactions at
the anode and the cathode
The most-described type of MESs is the microbial fuel cells (MFCs) in which
useful power is generated directly using the catalytic action of active microorganisms
(Kim 2002 Cheng and Logan 2007 Cheng et al 2006) In a typical two-chamber
MFCs (Figure 1a) organic matter is oxidized by electrochemically active
microorganisms in the anode chamber to release protons and electrons Protons diffuse
into the cathode chamber through a proton exchange membrane Meanwhile electrons
are transferred to the anode through various mechanisms (Logan et al 2006 Lovley
2
2006 Rabaey et al 2003) and then travel to the cathode where normally they combine
with oxygen and protons to form water
Another common used type of MES is the microbial electrolysis cell (MECs) in
which hydrogen is produced instead of electricity by applying a circuit voltage to the
MFCs system and maintaining anaerobic conditions in the cathode chamber (Liu et al
2005 Logan et al 2008 Rozendal et al 2006 Schaller et al 2009) (Figure 1b) A
cathode potential of at least - 410 mV (vs standard hydrogen electrode at pH 70) is
required to produce hydrogen at the cathode Therefore a theoretical potential of 110 mV
(ie 410-300 mV) is needed but voltages of gt02 V are normally applied due to various
overpotentials
Fig 1-1 Generalized schematics (a) Microbial fuel cells (MFCs) where electricity is
captured through the resistance R (b) Microbial electrolysis cells (MECs) where
hydrogen gas is produced in the cathode chamber R for resistor PS for power supply and
PEM for proton exchange membrane
The fundamental feature shared by microbial electrochemical systems (MESs)
(MECs and MFCs) is the microbe-catalyzed electron transfer from organic matter to
electrodes (anodes) (Rebaey et al 2004) Therefore different types of MESs can be
PEM
R
H+
e- e-
Anode
Bacte
ria
a
Cath
ode
H2O
PEM
PS
H+
e- e-
Anode
Bacte
ria
b
Cath
ode
H2
3
utilized to investigate the phenomena and mechanisms of interactions between microbes
and electrodes
12 Bacteria and Electron Transfer Mechanisms in MESs
The electrochemically active bacteria in MFCs are thought to be iron-reducing
bacteria such as Shewanella and Geobacter species (Gorby et al 2006) They have great
importance in the natural environment principally in metal oxidation and reduction
However recent studies have shown that the diversity of bacterial communities is much
greater than these model iron reducers on MFCs anodes (Reimers et al 2001 Liu et al
2004b Kim et al 2005) Over 20 electrochemically active bacterial species those can
transfer electrons exocellularly to electrodes have been reported in the past 10 years
(Logan 2009 Liu et al 2010) These bacteria have been categorized into diverse genetic
groups so far including alphaproteobacteria (Rhodopseudomonas Ochrobactrum and
Acidiphilium) (Malki et al 2008 Xing et al 2008 Zuo et al 2008) betaproteobacteria
(Rhodoferax) (Chaudhuri et al 2003) gammaproteobacteria (Shewanella Pseudomonas
Klebsiella Enterobacter and Aeronomas) (Kim et al 2002 Pham et al 2003 Rabaey et
al 2005 Bretschger et al 2007 Zhang et al 2008 Chung et al 2009 Rezaei et al 2009)
deltaproteobacteria (Geobacter Geopsychrobacter Desulfuromonas and Desulfobulbus)
(Bond et al 2002 Bond and Lovley 2003 Holmes et al 2004ab 2006)
Epsilonproteobacteria (Arcobacter) (Fedorovich et al 2009) Firmicutes (Clostridium and
Thermincola) (Park et al 2001 Wrighton et al 2008) Acidobacteria (Geothrix) (Bond
and Lovley 2005) and actinobacteria (Propionibacterium) (Wang et al 2008) A wider
range of electrochemically active bacteria are expected to be discovered
4
Table 1 Taxa of bacteria current density generated and reactor configuration used in
microbial electrochemical cell experiments (Liu et al 2010)
The electrochemically active bacterial species that possess the ability to transfer
electrons outside of the cell are called exoelectrogens in the MESs research field The
different exoelectrogens have demonstrated a wide-ranging power generation ability in
MFCs For example Shewanella oneidensis MR-1 has demonstrated the ability to
generate 0018 Am2 current density in single chamber MFCs while the Geobacter
Taxon Microorganisms Current Density (Am2
)
α-proteobacteria
Rhodopseudomonas palustris DX-1 003
Ochrobactrum anthropi YZ-1 071
Acidiphilium sp 32sup5 300
β-proteobacteria Rhodoferax ferrireducens 0031
γ-proteobacteria
Shewanella putrefaciens IR-1 0016
Shewanella oneidensis DSP10 0013
Shewanella oneidensis MR-1 0018
Pseudomonas aeruginosa KRA3 0017
Escherichia coli K12 HB101 100
Klebsiella pneumoniae L17 120
Enterobacter cloacae 013
Aeromonas hydrophila PA3 030
δ-proteobacteria
Geobacter metallireducens 065
Geobacter sulfurreducens 800
Desulfuromonas acetoxidans 0005
Geopsychrobacter
Electrodiphilus strain A2
0066
Desulfobulbus propionicus 003
Firmicutes
Lactococcus lactis 003
Thermincola sp strain Jr 020
Clostridium butyricum EG3 022
Thermincola ferriacetica Z-0001 040
Brevibacillus spp PTH1 0009
Desulfitobacterium hafniense DCB2 110
Acidobacteria Geothrix fermentans 0097
Actinobacteria Propionibacterium freudenreichiiET-3 120
5
sulfurreducens species has generated current densities as high as 80 Am2 which is 400
times higher than the one generated by Shewanella oneidensis MR-1 However mixed-
culture communities have generated much higher power densities than their pure-culture
counterparts in MFCs perhaps due to synergistic interactions within the anode bacterial
communities and the participation of currently unknown bacteria species and mechanisms
(Jung and Regan 2007) The dominant bacteria species vary in mixed cultures bacteria
communities in MFCs due to the enrichment of different substrates
Traditional methods of extroelectrogen study depend on cultivation hampered novel
exoelectrogens discovery because the inadequacy of defined media underestimates the
actual microbial diversity in MFCs Slow growth rate and unknown growth requirements
of anaerobic microorganisms make some anaerobic extroelectrogens cultivation difficult
However these limitations have been overcome by using molecular biological methods
based on DNARNA analysis Molecular biological techniques are now widely applied to
assess the diversity of microbial communities by analyzing the 16S rDNA sequence The
most commonly used molecular biological techniques for bacterial community analysis
include denaturing gradient gel electrophoresis (DGGE) restriction fragment length
polymorphism (RFLP) and 16S rDNA sequencing These methods are much less time
consuming than traditional isolation and cultivation methods
Isolated exoelectrogens were utilized to explore the mechanism of electron transfer
to the anode However the mechanisms of electron transfer to extracellular electron
acceptors are not well understood Three mechanisms have been proposed for exocellular
transport of electrons by exoelectrogens to anodes in MFCs so far Some exoelectrogenic
bacteria can transfer electrons to electrodes through soluble redox compounds (Bond and
6
Lovley 2005) These compounds include artificial mediators and mediators secreted by
exoelectrogenic bacteria A variety of chemicals have been used to facilitate the shuttling
of electrons from inside of cell to electrodes outside the cell These exogenous mediators
include neutral red (Park et al 1999) anthraquinone-2-6 disulfonate (AQDS) thionin
potassium ferricyanide (Bond et al 2002) methyl viologen and others (Logan 2004
Rabaey and Verstraete 2005) Some bacteria can secrete their own chemical mediator
for example Pseudomonas aeruginosa secretes pyocyanin and phenazine-1-carboxamide
to transfer electrons toward the MFCs anode (Rabaey et al 2005) Another evidence for
mediator production by bacteria is Geothrix fermentans When the medium was replaced
in a MFCs that had stable power generation with this bacteria power dropped by 50
and required 10 days to resume the original level
Some bacteria can directly transfer electrons to anodes via outer cell membrane
proteins (Myers and Myers 1992) The outer cell membrane protein cytochrome c is
thought to play a critical role in to transferring electrons to anodes Ly et al (2011)
isolated the haem protein cytochrome c and demonstrated that electric field effects may
be functional for the natural redox processes of cytochrome c in the respiratory chain
Shewanella oneidensis is the example of a bacterium that is capable of transfer electron to
anode via outer cell membrane proteins When Shewanella oneidensis adhered to an iron
surface the greater force has showed benefits to grow cells because closer contact
required for electron transfer from cell bound cytochromes (Lower et al 2001)
7
Fig1-2 Mechanisms for extracellular electron transport in an MFCs anode (1) direct
contact (top in green) (2) by nanowires (middle in purple) and (3) self-produced
mediators (bottom in blue) (Logan 2009)
More and more evidence supports the involvement of bacterial nanowires in
extracellular electron transport (Reguera et al 2005 2006 Gorby et al 2006) Nanowires
are conductive appendages produced by both Geobacter and Shewanella species (Gorby
and Beveridge 2005) The conductivity of the appendages was examined and confirmed
by using conductive scanning tunneling microscopy (STM) (Gorby et al 2006)
Nanowires can carry electrons from the cell to the anode surface of MFCs
The solid component of the extracellular biofilm matrix has high efficiency on
extracellular electron transfer compared with other extracellular electron transfer
8
mechanisms and recently Torres et al (2010) hypothesized that the solid component of
the extracellular biofilm matrix formed by exoelectogens is another way bacteria transfer
electrons to electrodes This hypothesis was based on kinetic analysis of each EET
mechanism reported in available literature (Torres et al 2010)
13 Anode electrodes
In MFCsMECs anode electrodes are a critical component because exoelectrogens
adhere to the surface of anodes to transfer electrons to the electrode The characteristics
of anodes have significant effects on electron transfer rate from bacteria to anode
electrodes in MFCs The requirements of an anode material are it should be highly
conductive non-corrosive have a high specific surface area (area per volume) high
porosity be non-fouling inexpensive and easily scaled to larger sizes Of these
properties the most important one that is different from other biofilm reactors is that the
material must be electrically conductive Normally they are made of various carbon
materials including carbon fiber carbon clothe and carbon paper due to their stability
high conductivity and high specific surface-area Nevertheless they have little
electrocatalytic activity for the anode microbial reactions and thus a modification of the
carbon materials is the main approach for improving their performance Consequently
there is a great need to develop a new type of anode material for MFCsMECs
It is a great challenge to develop a new anode material to further increase the power
density of a MFCs The nature of the catalytic mechanism of a MFCrsquos anode involves not
only a biological but also an electrocatalytic process An optimal nanostructure with a
high specific surface area favorable for both catalytic processes could play a critical role
in improving the power density of the MFCs such a structure needs to host the bacteria
9
with high bioactivity while enhancing the electron-transfer rate Schroumlder et al (2008)
employed PANI to modify a platinum anode for MFCs and achieved a current density 1
order of magnitude higher than the previously reported value PANIinorganic
composites are also reported to have better conductivity Qiao et al also applied a new
mesoporous TiO2 electrode material with uniform nanopore distribution and a high
specific surface area to anode in comparison to previously reported work with E coli
MFCs the composite anode delivers 2-fold higher power density (Qiao et al 2008)
Thus it has great potential for use as the anode in a high-power MFCs and may be a new
approach for improving performance of MFCs
14 Other parts of MESs
141 Membranes and ion transport
The ion exchange membrane is another one of the critical components in two-
chamber MESs systems It separates anode and cathode chambers and at the same time
maintains the electron neutrality of the system ie transport of electrons to the cathode
needs to be compensated by transport of an equal amount of positive charge to the
cathode chamber A proton exchange membrane (PEM) such as Nafion a per-fluorinated
sulphonic acid membrane (PFSAs) consisting of a hydrophobic fluorocarbon backbone to
which hydrophilic sulfonate groups (-SO3-) are attached is commonly used in chemical
fuel cell systems For MFCs systems however mainly cation species like Na+ and K
+
other than proton are often responsible for the dominant transport of positive charge
through the cation exchange membrane (CEM) to maintain electroneutrality due to the
low proton concentration in any aqueous medium with near neutral pH (Rozendal et al
2006) Consequently the pH increases in the cathode chamber due to the consumption of
10
protons and decreases in the anode chamber because of the accumulation of protons
(Rozendal et al 2007) This pH change in two-chamber systems results in a decrease of
the cathode potential and performance The application of anion exchange membrane
(AEM) has also been explored in MFCs systems (Cheng and Logan 2007 Kim and
Logan 2007) where it has been proposed that protons are transferred via pH buffers like
phosphate anions
142 Cathodes and Catalysts
The cathode is another challenge for making MFCs commercially available
technology because the chemical reaction that occurs at the cathode is difficult to
engineer as the electrons protons and oxygen must all meet at a catalyst in a tri-phase
reaction (solid catalyst air and water) The catalyst must be on a conductive surface and
must be exposed to both water and air so that protons and electrons in these different
phases can reach the same point The most commonly used material for a cathode is
commercially available carbon paper pre-loaded with a Pt catalyst on one side When it is
used in a MFC the side that contains the catalyst faces the water and the uncoated side
faces air To reduce the high cathode cost associated with platinum catalyst other precious-
metal-free cathodes such as (NiMo and NiW) were developed by electrodepositing on a
carbon fiber They have achieved comparable performance with Pt catalyst with same
loading at a much lower cathode fabrication cost (Hu 2010)
The requirements of cathode for MECs are quite similar to the requirements of
cathode for MFCs but easier than cathode for MFCs for the manufacturing process
because the cathode in MECs is not necessarily to exposed to air Therefore it can be
made of the exact same materials of cathode in MFCs except a waterproof layer
11
Recently some researchers have attempted to apply microorganisms as a biocatalyst to
precede the combination of electron with oxygen in the cathodes
15 Dissertation overview
Low power densities in MESs limit practical applications The improvement of
MESs performance requires a detailed understanding of the physiology and ecology of
microorganisms in MESs including the mechanism of electron transfer to the anode from
the microorganism
This dissertation focuses on the problem of the low power density of MESs The
bacterial community structure of a high power generated mixed culture communities in
MFCs will be identified firstly and then one of major exoelectrogens will be isolated and
characterized This information will be helpful to understand the physiology and ecology
of exoelectrogens in MESs Consequently they will be beneficial to improve power
density of MFCs Lastly the nanostructure decorated anodes in MECs will be utilized to
improve the power density The power enhancement mechanism will be explored by
using a whole genome microarray They are presented here as four papers
In the first paper the cultivation independent molecular biological techniques
denaturing gradient gel electrophoresis (DGGE) and 16S rDNA clone library are utilized
to analyze the bacterial community structure of a higher power mixed culture MFCs The
analyzed results provide fundamental information for isolating the dominant bacteria in
mixed culture MFCs Two of dominant bacterial species has been isolated and used aone
to test power generation in MFCs The possible interaction among different bacterial
species in mixed culture is discussed
12
In the second paper one isolated exoelectrogenic bacterial strain SX-1 is
characterized It is identified as a member of the Citrobacter genus and power generation
is tested ultilizing a wide range of different substrates The electron transfer mechanism is
explored using Cyclic Voltammetry (CV) This study increases the known diversity of
power generating exoelectrogens and provides a new strain to explore the mechanisms of
extracellular electron transfer from bacteria to electrodes
The third paper shows effects on MESs performance by Fe nanoparticle decorated
anodes in the MESs The average current density produced with Fe nanoparticle
decorated anodes increased up to 59-fold higher than plain graphite anodes A whole
genome microarray is utilized to analyze the possible mechanism of enhanced current
density as responded to nanoparticle decorated anodes
The fourth paper describes the effects of carbon nanotube (CNT) modified anode on
the performance S oneidensis MR-1 in MESs Results demonstrate that current densities
produced with CNT decorated anodes are up to 56-fold higher than plain graphite anodes
The possible mechanisms of enhanced current density by CNT decorated are explored
13
Chapter 2
Bacterial community analysis of mixed consortium in higher
power density MESs
Shoutao Xu and Hong Liu
1 Introduction
The improvement in the performance of mixed culture MFCs requires an
understanding of the ecology in microbial communities of MFCs Many researchers have
attempted to characterize microbial populations and activities to elucidate the behaviors
and ecology of microorganisms in MFCs (Wrighton et al 2008 Fedorovich et al 2009)
In order to study the microbial ecology of the mixed culture in a MFC and select the
appropriate isolation medium for dominant bacterial species in the mixed culture the
fundamental steps are to analyze the bacterial diversity in the mixed culture of MFCs and
identity the dominant bacterial species in bacteria communities in MFCs
14
As for the identification of bacterial communities typically there are two general
methods The first method for identification of bacterial community is the traditional
cultivation processes using selective nutrients to promote the growth of different types of
bacteria within the samples (Amman et al 2000) The community structure can then be
assessed by identifying the isolates from the dominant colonies that were cultured This
can often be costly and laborious as each isolate has to be further studied by examining
its physiology taxonomy and reactivity to stains (Adwards et al 1989)
The second method relies on utilizing molecular techniques to analyze bacterial
community DNA Several molecular methods involving the extraction and analysis of
DNA from entire bacterial communities are used to identify genetic fingerprints of
bacteria These methods including the cloning and sequencing of 16S rDNA automated
ribosomal intergenic spacer analysis (ARISA) terminal restriction fragment length
polymorphism (T-RFLP) and denaturing gradient gel electrophoresis (DGGE) generate
profiles of bacterial community structures They can rapidly assess complex communities
from various environments (Amman et al 2000)
Due to the conservative characteristic of 16S rDNA in bacteria during the process
of evolution 16S rDNA sequencing can be used to identify different species of bacteria
Therefore the molecular techniques of denaturing gradient gel electrophoresis (DGGE)
with PCR and 16S rDNA clone library are used for analysis of the microbial diversity
These methods are more convenient and save time compared to traditional
isolationcultivation methods for microorganism analysis
In this work a biofilm bacterial community from an anode of a MFCs wase studied
by using denaturing gradient gel electrophoresis (DGGE) of PCR-amplified partial 16S
15
rRNA genes followed by cloning and sequencing of 16S rDNA The results provided
essential information for dominant bacterial isolation in mixed culture MFCs
2 Materials and methods
21 High power generation of mixed culture MFCs
Mixed cultures were originally inoculated from domestic wastewater (Corvallis
Wastewater Treatment Plant Corvallis OR) to MFCs after operating for 2 years using a
defined medium solution (Lovely 2002) with sodium acetate as the carbon source A
new MFCs was inoculated from the operating MFCs The polarization curves were
performed to measure the power generation when maximal stable power were established
(normally 3-5 batches post-inoculation) and the genomic DNA from the microbial
biofilm on the anode was extracted under the sterile conditions for further bacterial
community analysis
22 Denaturing gradient gel electrophoresis (DGGE)
DGGE with PCR is a method of analysis of bacterial community composition
based on different GC percentage of 16s rDNA in different bacterial species PCR-DGGE
comprises three steps (i) extraction of total community DNA from the sample (ii) PCR-
controlled amplification using specific oligonucleotide primers and (iii) separation of the
amplicons using DGGE For this purpose a reproducible and efficient method for total
DNA extraction is indispensable and needs to be evaluated and optimized depending on
the nature of the sample In the subsequent PCR step multiple PCR primer sets with
different resolution can be used In most PCR-DGGE applications on bacteria universal
or specific primers are targeting the 16S rRNA gene Upon electrophoresis of the PCR
16
amplicons and gel staining (using ethidium bromide silver staining or SYBR green)
DGGE gels are digitally captured and further analyzed using computer software packages
The richness of DNA fragment bands in DGGE indicates the bacterial diversity of the
sample
The detailed steps are as followed Biofilms were scratched from the anodes of high
power generated MFCs fed with sodium acetate Bacterial genomic DNA was extracted
from the biofilm samples using the DNeasy blood and tissue kit (Qiagen) according to the
manufacturerrsquos instructions The universal primer set was used to amplify the 16S rDNA
from the extracted genomic DNA then the V3 region of bacterial 16S ribosomal DNA
(rDNA) was amplified with GC-clamp primer (Muyzer et al 1993) for DGGE PCR
amplification was performed in a thermocycler DGGE of the PCR products was carried
out in a DcodeTM
Universal Mutation Detection System) The 8 (wv) polyacrylamide
gels contains from 30 to 55 denaturing gradients Electrophoresis was conducted
using a 1timesTAE (Tris-Acetate-EDTA) buffer at 130V and 60degC for 5 hours After
electrophoresis the gel was stained with ethidium bromide in 1timesTAE buffer for 15
minutes and washed in 1timesTAE buffer for 10 minutes The fragments were visualized
under a UV transilluminator The richness of single band in DGGE gel picture
preliminarily showed that the bacterial diversity in mixed culture since the single band in
gel represents one bacterial species
23 16S rDNA clone library construction method
16S rDNA-DGGE fingerprinting has proven to be particularly useful as an initial
investigation into bacterial communities and is suitable for identifying the predominant
bacteria that comprise 1 or more of cells within a given sample In the 16s rDNA clone
17
library construction process the first step is the extratction of the total genomic DNA
then the genomic DNA of the mixed culture biofilm was used as template for PCR
amplification of approximately 1500 bp of 16S rDNA with universal primers The PCR
products of 16S rDNA were purified and inverted into pGEM-T Easy vector system
before they were transformed into competent E coli The transformed cells were spread
on Luria-Bertani (LB) plates containing the selective antibiotic ampicillin and X-Gal and
incubated overnight at 37 OC after the more than 80 inverted 16s DNA fragment E coli
colonies were picked up and inoculated in the 1-3 ml liquid LB medium and left to grow
for 16 hours The plasmid DNA were extracted and sequenced The sequences were
compared directly to all known sequences deposited in GenBank databases using the
basic local alignment search tool (BLAST)
24 Bioinformatics Analysis
16S rDNA sequencing results of more than 80 colones were queried against the
GenBank and Ribosomal Database Project (RDP) databases using BLAST and
SEQUATCH algorithms (Cole et al 2007 Wang et al 2007 Cole et al 2009) The
neighbor-joining trees were constructed with the Molecular Evolutionary Genetics
Analysis package (MEGA Version 4) based on 1000 bootstrap analysis (Tamura et al
2007)
25 Dominant bacterial species band in DGGE identification
The DGGE results gave the information of dominant bacterial richness of bacterial
species in mixed culture based on the theory that single band represents one bacterial
species which showed the diversity of bacterial community in the mixed culture of a
MFC 16S rDNA clone library results provided the whole picture of bacterial community
18
including the bacterial species name and bacterial species percentage in the mixed culture
The known pure bacterial species which have been sequenced can be used as markers to
identify dominant bacterial species in DGGE The whole experimental procedure is
similar to the procedure of the mixed culture DGGE steps The first step is to extract the
collect the mixed culture biofilm genomic DNA and pure bacterial species marker
genomic DNA Then the universal primer set was used to amplify the 16S rDNA from
the extracted genomic DNA After that the V3 region of bacterial 16S ribosomal DNA
(rDNA) was amplified with GC-clamp primer (Muyzer et al 1993) for DGGE The
subsequent steps were similar to the mixed culture DGGE experimental procedure The
dominant bacterial bands in DGGE were determined by comparisons of the single band
position in the mixed culture DGGE with single pure bacterial species location in DGGE
26 Dominant bacterial species isolation and power generation
The bacterial community of mixed culture in MFCs were predominantly composed
of Geobacter sp Arcobacter sp and Citrobacter sp Two dominant bacterial species
have been isolated
Citrobacter sp SX-1 was isolated from the anode biofilm by using a solid agar
medium containing NH4Cl 6mM KCl 2mM CH3COONa (electron donor) 30 mM
FeC6H5O7 (electron acceptor) 20mM minerals and vitamins (Lovley and Phillips 1988)
Single colonies were picked up after culturing on the plate for 36 hours at 30 degC and
transferred two times on the agar plate for purification Then the isolates grew in a liquid
medium solution in anaerobic tubes containing the same constituents as the solid
medium All isolation process was operated in a glove box anaerobic chamber (Coy
Laboratory Products Grass Lake MI)
19
Arcobacter sp SX-2 was isolated from the anode biofilm of a MFC fed with sodium
acetate by using the same method of isolation of pure bacterial species Citrobacter XS-1
except using Trypticase Soy Agar with 5 Sheep Blood solid (from American type
culture collection (ATCC)) agar medium instead of Citrobacter sp medium Single
colonies were picked up after culturing on the plate for 36 hours at 30 degC and transferred
twice on the agar plate for purification Then the isolates grew in a liquid medium
solution microaerobically
Geobacter sp have been trying to isolate by using three different methods They are
the colony pickup after different condition enrichment Goebacter medium isolation and
dilution to extinction by multiple channel mini MFCs isolation however the Geobacter
sp has not been isolated yet due to some limits so far But they provide valuable
information for further isolation of Geobacter sp
Single chamber MFCs were used to evaluate power generation by different isolates
The MFCs were constructed as described previously (Liu and Logan 2004) and modified
with 3 cm2 carbon cloth anodes and 7 cm
2 carbon clothPt cathodes The total liquid
volume of each MFCs was about 15 ml with electrode spacing about 17 cm All MFCs
were operated in an autoclaved closed plastic box and sterile cotton was attached to the
outer surface of the air cathodes to prevent contamination A MFCs without bacterial
culture was used as control MFCs were inoculated with 3 ml late exponential phase
cultures of SX-1 in the medium solution reported previously (Liu and Logan 2004)
3 Results and discussion
31 Power production by mixed culture MFCs
20
A polarization curve was used to characterize current as a function of voltage in the
MFCs The polarization curves are performed by varying a series of external resistances
The results are shown in Figure 21 With MFCs acclimated to 25 Ω external resistance
the maximum power density was achieved at 65Wm2 based on the polarization data At
this point the current density is 21mAcm2 The power density was three times higher
than previous report (20 Wm2) using phosphate buffer (Fan et al 2007) Power density
was also 2 times higher than (28 m Wm2) using a bicarbonate buffer ( Fan et al 2007)
32 Bacterial community structure assessed by PCR-DGGE
Bacterial DGGE profiles for higher power generating MFCs mixed culture setups
were relatively complex (more than 10 bands) However there are three brightly stained
bands and other less intense bands (Fig 2-2) These results indicated there were probably
three dominant bacterial species in mixed culture MFCs
010
020
030
040
050
060
070
2
3
4
5
6
7
04 05 05 06 07 08 10 11 13 15 21 22 23 24
Volta
ge (
V)
Pow
er
densi
ty (
Wm
2)
Current density (mAcm2)
Power
Voltage
power density 65wm2 at current
density208 mAcm2
Figure 2-1 Polarization curves of high power mixed culture MFCs
21
33 16S rDNA clone library results
A total of 83 colonies were sequenced for 16S rDNA clone library construction They
represented the bacterial community structure in mixed culture in MFCs The results
revealed that the predominant bacterial species in mixed culture are Geobacter sp (66
of the mixed culture) Arcobacter sp (12 of the mixed culture) and Citrobacter sp (11
of the mixed culture) Those three genuses comprised 88 of the total bacterial species
(Fig 23) This result is consistent with the DGGE result which showed that three bright
bands in the DGGE gel represented three dominant bacterial species in the mixed culture
The phylogenetic analysis of the colonies (83 colony sequences) is shown in Figure 24
Figure 22 DGGE result of high power generation mixed culture MFC (1 mixed
culture 2 pure culture control)
1 2
22
34 Dominant bacterial band in DGGE identification
Geobacter
66
Arcobacter
12
others
6
Pseudomonas
1
Citrobacter
11
Clostridium
2
Anaerovorax
2 Geobacter
Arcobacter
Citrobacter
Clostridium
Anaerovorax
Pseudomonas
others
Figure 23 Bacterial species and percentage of bacterial community of mixed
culture MFC lt 1 Alcaligensgt
lt1 Commamonasgt
lt1 Pseudomonas aeruginosagt
lt1 Klebsiellagt
lt11 Citrobactergt
lt1 Wolinellagt
lt12 Arobactergt
lt2 Anaeroboraxgt
lt2 Clostridium gt
lt1 unclear bacteriagt
lt66Geobactergt
100
100
85
100
100
82
99
87
100
002
Figure 24 Phylogenetic trees of bacterial species from mixed culture MFC
23
The dominant bacterial species including Goebacter sp Citrobacter sp and Arcobacter
sp in the mixed culture are shown in Figure 25 The upper band is Arcobacter sp and
Citrobacter sp is located in the bottom band of DGGE Geobacter sp is the middle one
The analysis of the bacterial community using denaturing gradient gel electrophoresis
(DGGE) of PCR-amplified 16S rRNA gene fragments and 16S rDNA clone library
construction method showed great phylogenetic diversity of mixed culture in MFCs with
the identification of sequences derived from bacteria of the taxa deltaproteobacteria
(Geobacter sp) gammaproteobacteria (Citrobacter sp) and epsilonproteobacteria
(Arcobacter sp) The result is consistent with the conclusion that among the isolated
exoelectrogens gammaproteobacteria and deltaprotebactia appear to be dominant among
others (Parot et al 2009 Liu et al 2010)
35 The dominant bacterial species isolation and power generation
Figure 25 Dominant bacterial species identification in DGGE
A B C D
A Citrobacter sp
B Geobacter sp
C Arobacter butzmeri
D mixed culture control
24
Two dominant bacterial species Citrobacter sp XS-1 and Arcobacter sp XS-2 have been
isolated and tested for power generation The power densities generated by the two
isolates are shown in the Fig 26 Current densities generated by isolate Citrobacter sp
and Arcobacter sp were 98 mAm2 and 20 mAm
2 respectively which were much less
than that (21Am2) generated by the mixed culture from which they were isolated
Figure 26 Current density generated by different dominant isolates
The maximum current density generated by mixed culture is much higher than the
one produced by pure bacterial species Citrobacter sp XS-1 and Arcobacter sp XS-2 in
the same structure MFCs Although the Geobacter sp in our mixed culture has not been
isolated a previous study showed that maximum current density generated by Geobacter
sp was 080 Am2
(Bond and Lovley 2003) The current density results seem partially
supporting the conclusion that mixed-culture generated higher power densities than their
pure-culture counterparts in MFCs (Jung and Regan 2007) This result suggests there
may be synergistic interactions within the anode bacterial communities
0
500
1000
1500
2000
2500
Mixed culture Citrobacter sp Arcobacter Sp
Cu
rre
nt
de
nsi
ty (
mA
m2)
25
Geobacter sp are strictly anaerobic bacteria which were enriched in most of the anode
biofilms (George 2005) Citrobacter species are facultative anaerobic bacteria (George
2005) which can be found in a wide variety of habitats including in soil water and
wastewater Arcobacter sp are micro-aerobic bacteria including both environmental
nonpathogens and opportunistic human pathogens They are able to grow in aerobic
conditions but in the optimal growth under micro-aerobic conditions (George 2005)
Arcobacter sp has been discovered in the MFCs by Fedorovich (Fedorovich et al 2009)
It is interesting that the mixed culture bacterial community composed of primarily these
three bacterial species has demonstrated to generate high power at non-strictly anaerobic
condition while leading dominant bacterial species Geobacter sp (66) is strict
anaerobic bacteria and generated powder in the MFCs only under strict anaerobic
condition (George 2005) Therefore it is possible that microaerobic bacteria such as
Arcobacter sp and facultative anaerobic bacteria Citrobacter sp can facilitate to create
anaerobic condition for Geobacter sp when they utilized oxygen for growing in the
mixed culture Syntrophic communities study have showed that anaerobic bacteria and
methanogenic archaea form compact microbial structures that operate like an organ rather
than a set of microorganisms functioning independently (Stams and Plugge 2009) Some
substrates have been degradeted within these communities while they are not able to be
fermented by individual species alone interspecies electron transfer also have been
presented in these communities (Stams and Plugge 2009) Summers et al (2010) also
discovered that direct exchange of electrons happened within coculture of Geobacter
metallireducens and Geobacter sulfurreducens Therefore we believe that there are
26
more complex synergistic interactions between different bacteria species in mixed culture
in the MFCs
Acknowledgements
The authors acknowledge support from the US National Science Foundation
(CBET 0955124) We also thank Keaton Lesnik for the review of this manuscript
27
Chapter 3
New Exoelectrogen Citrobacter sp SX-1 Isolated from a Microbial Fuel Cell
Shoutao Xu and Hong Liu
Published in
Journal of Applied Microbiology 111(5)1108-1115 (2011)
ABSTRACT
Exoelectrogenic bacterial strain SX-1 was isolated from a mediator-less microbial fuel
cell by conventional plating techniques with ferric citrate as electron acceptor under
anaerobic condition Phylogenetic analysis of the 16S rDNA sequence revealed that it
28
was related to the members of Citrobacter genus with Citrobacter sp sdy-48 being the
most closely related species The bacterial strain SX-1 produced electricity from citrate
acetate glucose sucrose glycerol and lactose in MFCs with the highest current density
of 205 mA m-2
generated from citrate Cyclic voltammetry analysis indicated that
membrane associated proteins may play an important role in facilitating the electrons
transferring from bacteria to electrode This is the first study that demonstrates that
Citrobacter species can transfer electrons to extracellular electron acceptors Citrobacter
strain SX-1 is capable of generating electricity from a wide range of substrates in MFCs
This finding increases the known diversity of power generating exoelectrogens and
provided a new strain to explore the mechanisms of extracellular electron transfer from
bacteria to electrode The wide range of substrate utilization by SX-1 increases the
application potential of MFCs in renewable energy generation and waste treatment
Keywords Microbial fuel cell Exoelectrogen Citrobacter sp SX-1 Extracellular
electron transfer
29
Introduction
Microbial Fuel Cells (MFCs) technology has become an active research area
recently as a promising approach for renewable energy generation wastewater treatment
and bioremediation (Rabaey and Verstraete 2005 Lovley 2006 Fan et al 2008 Logan
2009 Liu et al 2010) The key feature of MFCs system is the microbe-catalyzed electron
transfer from organic matter to anodes Many studies have shown the presence of diverse
bacterial communities on MFCs anodes (Reimers et al 2001 Liu et al 2004b Kim et al
2005) Over 20 exoelectrogens ie microbes that can transfer electrons exocellularly to
electrodes have been reported in the past 10 years (Logan 2009 Liu et al 2010) The
exoelectrogens so far belong to diverse genetic groups including alphaproteobacteria
(Rhodopseudomonas Ochrobactrum and Acidiphilium) (Malki et al 2008 Xing et al
2008 Zuo et al 2008) betaproteobacteria (Rhodoferax) (Chaudhuri et al 2003)
gammaproteobacteria (Shewanella Pseudomonas Klebsiella Enterobacter and
Aeronomas) (Kim et al 2002 Pham et al 2003 Rabaey et al 2005 Bretschger et al
2007 Zhang et al 2008 Chung et al 2009 Rezaei et al 2009) deltaproteobacteria
(Geobacter Geopsychrobacter Desulfuromonas and Desulfobulbus) (Bond et al 2002
Bond and Lovley 2003 Holmes et al 2004ab 2006) Epsilonproteobacteria (Arcobacter)
(Fedorovich et al 2009) Firmicutes (Clostridium and Thermincola) (Park et al 2001
Wrighton et al 2008) Acidobacteria (Geothrix) (Bond and Lovley 2005) and
actinobacteria (Propionibacterium) (Wang et al 2008) A wider range of
exoelectrogenic species are expected to be discovered
Three mechanisms have been proposed for exocellular transport of electrons by
exoelectrogens without artificial mediators Some exoelectrogenic bacteria can transfer
30
electrons to electrodes through soluble redox compounds excreted by microorganisms
(Bond and Lovley 2005 Rabaey et al 2005) and others can directly transfer electrons to
anodes via outer cell membrane proteins (Myers and Myers 1992) Recently more and
more evidence supports the involvement of bacterial nanowires in extracellular electron
transport (Reguera et al 2005 2006 Gorby et al 2006) In spite of the discovery of
many bacterial species that can transfer the electrons to electrode without the need of
artificial mediators the investigation of extracellular electron transfer mechanisms was
mainly focused on a few species such as those from Geobacter and Shewanella genera
(Kim et al 1999 2002 Reguera et al 2005 2006 Gorby et al 2006) The electron
transfer mechanisms for many of the isolated exoelectrogens species are still not well
studied
In the present study we isolated a new exoelectrogen strain SX-1 from a MFC a
strain phylogenetically related to Citrobacter sp Power generation from various carbon
sources by this strain was evaluated using single chamber MFCs Plausible extracellular
electron transfer mechanisms were also discussed based on the characterization of anodic
biofilms by cyclic voltammetry (CV)
Materials and methods
Bacterial strain SX-1 isolation
Bacterial strain SX-1 was isolated from the anode biofilm of a MFC fed with sodium
acetate operated in fed-batch mode over a period of six months The original source of
the inoculum is wastewater from a local waste water treatment plant Bacterial cells were
released from the carbon cloth anode by shaking the cloth in a sterile flask with 15 mL
sterile 1 NaCl solution and glass beads (2 mm diameter) The suspension was then
31
serially diluted from 10 times to 105
times and plated on a petri dish with a solid agar
medium containing NH4Cl 6mM KCl 2mM CH3COONa (electron donor) 30 mM
FeC6H5O7 (electron acceptor) 20mM minerals and vitamins (Lovley and Phillips 1988)
Single colony was randomly selected after culturing on the plate for 36 hours at 30 degC
and purified on a new agar plate following a procedure reported previously (Chung and
Okabe 2009) Then the isolate was picked up and grew in a liquid medium solution in
anaerobic tubes containing the same constituents as the solid medium for further analysis
All isolation process was operated in a glove box anaerobic chamber (Coy Laboratory
Products Grass Lake MI)
16S rDNA sequencing and phylogenetic analysis
Bacterial genomic DNA was extracted from the isolated strain SX-1 using the DNeasy
tissue Kits (Qiagen Valencia CA) according to the manufacturerrsquos instructions The 16S
rDNA was amplified by PCR using a pair of universal primers 27F (5rsquo-
AGAGTTTGATCMTGGCTCAG-3rsquo) and 1492R (5rsquo-GGTTACCTTTGTTACGACTT-
3rsquo) (Park et al 2001) The conditions used for PCR were described previously
(Ferna ndez et al 1999) PCR products were purified ligated and cloned according to a
procedure described in a previous report (Xing et al 2008) Plasmids were isolated from
randomly selected clone colonies with a QIAprep Spin Miniprep kit (Qiagen Valencia
CA) and three plasmid inserts were then sequenced in both directions using an ABI 3730
DNA sequencer (Applied Biosystems Carlsbad CA)United States) Due to the identical
16S rDNA sequence of the three colones only one was queried against the GenBank and
Ribosomal Database Project (RDP) databases using BLAST and SEQUATCH algorithms
(Cole et al 2007 Wang et al 2007 Cole et al 2009) The 16S rDNA sequence of
32
strain SX-1 and closely related type strains were aligned using CLUSTALX software
(Thompson et al 1997) The neighbor-joining trees were constructed with the Molecular
Evolutionary Genetics Analysis package (MEGA Version 4) based on 1000 bootstrap
analysis (Tamura et al 2007)
SEM
The morphologies of SX-1 cells growing overnight in anaerobic tubes were examined
with a scanning electron microscope (FEI Quanta 600 FEG FEI Company Hillsboro
OR) The suspended cells were fixed in a 25 glutaraldehyde and 01M phosphate
buffer solution and dehydrated with a graded ethanol series from 30 to 100 After
dehydration the samples were dried in a critical point dryer and then sputter-coated with
AuPd for SEM examination (Liu and Logan 2004)
Evaluation of power generation by SX-1 in MFCs
Single chamber MFCs were used to evaluate power generation by SX-1 using various
substrates The MFCs were constructed as described previously (Liu and Logan 2004)
and modified with 3 cm2 carbon cloth anodes and 7cm
2 carbon clothPt cathodes The
total liquid volume of each MFCs was about 15 ml with electrode spacing about 17 cm
All MFCs were operated in an autoclaved closed plastic box and sterile cotton was
attached to the outer surface of the air cathodes to prevent contamination A MFC
without bacterial culture was used as control MFCs were inoculated with 3 ml late
exponential phase cultures of SX-1 in the medium solution reported previously (Liu and
Logan 2004) Seven substrates including citrate acetate glucose sucrose glycerol and
lactose were evaluated individually for power generation in a fed-batch mode in a
temperature control chamber (30 plusmn2degC) For each substrate at least 8 batches were run to
33
investigate the effect of biofilm formation on current output at fixed external resistance of
1kΩ and the same initial substrate concentration of 30 mM Phosphate buffer (100mM)
was used to maintain the solution pH and conductivity The MFCs with sodium citrate as
electron donors were also examined for maximum power output by varying the external
resistance from 20 kΩ to 4 kΩ A data acquisition system was used to record voltage data
during the MFCs operation (Keithley 2700 Keithley Instruments Inc Cleveland OH) It
took about 15 to 30 minutes for the MFCs to stabilize depending on the external
resistance At each resistance we collected at least five data at the steady condition to
make the polarization curves Averaged voltages were used to calculate the power density
(mW m-2
) according to P=IVA where I was the current V was voltage and A was cross-
sectional area of the anode
CV analysis
CV was used to characterize the oxidation and reduction reactions on the anodic surface
of the MFCs fed with citrate and operated at 7 KΩ This external resistance was selected
because the maximum power density was obtained at this resistance based on the
polarization experiment We assumed the biofilm on the anode of MFCs were well-
developed when stable power output was obtained after 3 batches of operation The
MFCs were then used directly for CV analysis at four current generating stages of the
fourth batch (1) initial exponential current increasing stage (middle point of stage about
4 h after media change) (2) current plateau stage (middle point of stage about 8 h after
media change) and (3) current decreasing stage (middle point of stage about 16h after
media change) (4) right after the complete replacement of medium solution (Figure 6)
The anode was used as working electrode the cathode as counter electrode and an
34
AgAgCl electrode was selected as the reference The CV curves were scanned from 200
to -600 mV at a rate of 5 mV s-1
using a potentiostat (G300 Gamry Instrument
IncWarminster PA) Control experiment was also conducted using new anode (without
biofilm) and new medium solution
Nucleotide sequence accession number
The 16S r DNA sequence determined in this study has been deposited in the GenBank
database under accession number HQ845373
Results
Identification of the strain SX-1
An almost-complete 16S rDNA sequence (1475 bp) of strain SX-1 was obtained and
subjected to comparative analysis with the 16S rDNA of closely related reference strains
A phylogenetic analysis revealed that strain SX-1 should be assigned to the genus
Citrobacter and was most closely related to Citrobacter sp sdy-48 EJ463782 (990
sequence similarity) and Citrobacter sp yy-21 FJ463779 (975 sequence similarity)
These three strains formed a distinct sub cluster in the neighbor-joining in which the new
isolate and Citrobacter sp sdy-48 EJ463782 formed a distinct subline (Figure 31)
The SEM image illustrated that the strain SX-1 is a rod-shaped bacterium 03-06 microm
wide and 12-17 microm long (Figure 32) Strain SX-1 colonies on nutrient agar appeared 2-
3 mm in diameter smooth moist opaque with a shiny surface after 5 days incubation
under anaerobic condition The fact that strain SX-1 grew both aerobically and
anaerobically using FeC6H5O7 as electron acceptor suggests that the strain is facultatively
anaerobic Strain SX-1 showed robust growth on citrate as a carbon source These D
35
properties were similar to those of genus Citrobacter described in Bergeyrsquos Manual of
Systematic Bacteriology (George 2005)
Figure 31 phylogenetic tree of strain SX-1 and closely related species based on 16S
rDNA sequences The tree was constructed using the neighbor-joining method
Figure 32 SEM image of planktonic cells of Citrobacter sp SX-1
Strain SX-1
Citrobacter sp sdy-48 (FJ463782)
Citrobacter sp yy-21 (FJ463779)
Citrobacter koseri E639 (ATCC 25408)
Citrobater koseri CDC 3613-63(AF025372)
Citrobacter koseri CDC 8132-86(AF025366)
Salmonella enterica SL483 (CP001138)
Salmonella enterica AKU12601(AY696668)
Shigella sonnei Ss046(CP000038)
E coli C2 (AF403733)
E coli E24377A(CP000800)88
100
100
76
44
92
53
36
0001
1 microm
36
Electricity production by strain SX-1 in MFCs
The isolated strain SX-1 was first tested for its ability to generate current from sodium
citrate at 1kΩ Current gradually increased to a maximum 50 mA m-2
after cell
inoculation and then decreased (Figure 33) After the MFCs was refilled with new
substrate solution the current recovered rapidly and reached a higher level than the first
batch After 4 batches operation the maximum current output of each batch became
stable The highest current density achieved in the MFCs with Citrobacter sp SX-1 at 1
KΩ was 98 mA m-2
(Figure 33)
Figure 33 Electricity generation by Citrobacter sp SX-1 in a single chamber MFCs
with sodium citrate (30 mM) as substrate at 1 KΩ
0
20
40
60
80
100
120
0 50 100 150 200
Curr
ent
den
sity
(m
A m
-2)
Time (h)
37
Current generation by SX-1 from other substrates including glucose lactose sodium
acetate glycerol and sucrose were also assessed at an external resistance of 1KΩ
(Figure 34) When repeatable cycles of current output were obtained for these substrates
glycerol generated the highest maximum current density of 58 mA m-2
followed by
lactose and sucrose with 29 mA m-2
and 27 mA m-2
respectively Glucose and acetate
produced the lowest maximum current density of 96 and 43 mA m-2
respectively These
results indicated that strain SX-1 can utilize a wide range of substrates for electricity
generation in MFCs but with different power generation potentials
Figure 34 Electricity generation of Citrobacter sp SX-1 using different substrates in
single chamber air-cathode MFCs at 1 KΩ (The error bars stand for the standard
deviation of 3 replicates)
0
10
20
30
40
50
60
70
Glucose Lactose Glycerol Acetate Sucrose
Curr
ent
den
sity
(m
A m
-2)
38
Polarization experiment was further conducted to determine the maximum power
density generated by strain SX-1 in the single chamber MFCs with citrate as substrate A
maximum power density of 881 mW m-2
was obtained at current density of 205 mA m-2
at an external resistance of 7 KΩ (Figure 35)
Figure 35 Power and voltage generation by Citrobacter sp SX-1 as a function of
current density using sodium citrate (30 mM) as substrate The error bars stand for the
standard deviation of the 3 voltagepower outputs obtained in three MFCs
20
30
40
50
60
70
80
90
100
01
02
03
04
05
06
07
80 110 140 170 200 230
Po
wer
den
sity
(m
W m
-2)
Vo
ltag
e (V
)
Current density (mA m-2)
Voltage
Power
39
Figure 36 Four current generating stages for CV analysis current increasing stage (stage
1) current plateau stage (stage 2) current decreasing stage (stage 3) and right after the
replacement of medium solution (stage 4) The MFCs was operated at 7 KΩ with sodium
citrate (30 mM) as substrate
Cyclic Voltammetry
To determine the presence of redox active compounds produced by SX-1 and the
location of these compounds CV scan of the anodic biofilms at four current output stages
and the supernatant of MFCs medium solution at the end of the batch experiment were
performed Reduction peaks at range of -310 ndash -350 mV and oxidation peaks at range -
100 ndash -150 mV were observed at all current generating stages (Figure 36 Figure 37A)
suggesting the presence of redox active compounds may involve in extracellular electron
transfer by Citrobacter sp SX-1 The amplitude of the peaks increased according to the
growth stage of the batch and the highest peaks were present after the current plateau
stage which indicated the redox active compounds mainly were secreted in the current
plateau stage While the current density at stage 3 (deceasing stage) was lower than that
at stage 2 (current plateau stage) the redox peak at stage 3 was higher than that in stage 2
0
50
100
150
200
250
50 55 60 65 70 75 80 85
Curr
ent
den
sity
(m
A m
-2)
Time (h)
A
A
Stage 1
Stage 2 Stage 3
Stage 4
40
A plausible explanation for this pattern was that redox active compounds were
continuously being secreted and accumulated after the current peak was reached
resulting in more redox compounds present after the plateau stage But since most of the
carbon source had already been being used up many of the redox compounds at this
stage had not been as active as those at current plateau stage due to much less electrons
were available to be transferred The bigger peak amplitude in stage 4 than in stage 1
indicates that the redox active compounds were continuously being secreted and
accumulated in the biofilm even after stable power was generated at 7 kΩ The slightly
change of the peak locations at different stages was possible due to the solution chemistry
change during the current generation process or the slightly location change of reference
electrode during the measurement (Figure 37B) When the medium solution in the MFCs
was replaced by fresh medium at the end of the batch the oxidization and reduction
peaks were still presented in the CV (Figure 37B) but no peaks were observed when the
supernatant and fresh mediums were tested in a MFCs with no biofilm (Figure 37C)
These results suggest the compounds involved in the electron transfer were located not in
the supernatant (solution) but in the biofilm
41
Figure 37 Cyclic Voltammetry analysis of Citrobacter sp SX-1 biofilms at (A) current
increasing stage current plateau stage and decreasing stage (end of the batch) (B) end of
the batch and replaced with fresh medium and (C) controls
-40E-04
-30E-04
-20E-04
-10E-04
00E+00
10E-04
20E-04
30E-04
-060 -050 -040 -030 -020 -010 000 010 020
Curr
ent
(A)
Potential (V) vs AgAgCl
Current plateau stage
Current increasing stage
Current decreasing stage
-40E-04
-30E-04
-20E-04
-10E-04
00E+00
10E-04
20E-04
30E-04
-060 -050 -040 -030 -020 -010 000 010 020
Curr
ent
(A)
Potential (V) vs AgAgCl
Current decreasing stage
Biofilm with fresh medium
-40E-04
-35E-04
-30E-04
-25E-04
-20E-04
-15E-04
-10E-04
-50E-05
00E+00
-060 -050 -040 -030 -020 -010 000 010 020
Curr
ent(
A)
Potential (V) vs AgAgCl
Supernatant
Fresh medium
A
B
C
B C
42
Discussion
Citrobacter species belonging to Gammaproteobacteria Enterobacteriales
Enterobacteriaceae are facultative anaerobic bacteria (George 2005) They can grow with
oxygen as an electron acceptor but also with a variety of electron accepters such as Fe (III)
in the absence of oxygen (George 2005) Citrobacter species can be found in a wide
variety of habitats including in soil water and wastewater Several Citrobacter species
have been studied as important bioremediation bacterium for heavy metal removals
sulfate reduction phenol degradation and chlorophenol degradation (Macaskie et al 1995
Narde et al 2004 Qiu et al 2009) Citrobacter species have also been found on the
anodic biofilm of a diesel-degrading MFCs (Morris et al 2009) However there was no
study reported that Citrobacter species can transfer electrons to extracellular electron
acceptors This finding that Citrobacter strain SX-1 can generate electricity in MFCs
increased the diversity of power generating exoelectrogens and provided a new strain to
explore the mechanisms of extracellular electron transfer from bacteria to electrode
Among the isolated exoelectrogens gammaproteobacteria and deltaprotebactia appear to
be dominant among others (Parot et al 2009 Liu et al 2010) The isolated strain
Citrobacter sp SX-1 phylogenetically belongs to gammaproteobacteria which provides
another evidence to support this observation
Most exoelectrogens utilize a limited range of substrates for power generation For
example Shewanella oniedensis MR-1 can oxidize lactate under anaerobic condition but
cannot utilize acetate for electricity generation (Kim et al 2002) Geobacter
sulfurreducens can completely oxidize acetate for power generation but it cannot utilize
simple sugars (Bond and Lovley 2003 Caccavo et al 1994)) In comparison strain SX-1
43
not only can oxidize citrate for power generation but also utilize a wide range of
substrates for power generation including glycerol glucose lactose sucrose and acetate
Interestingly the higher current density generated by strain SX-1 from glycerol a main
by-product of biodiesel production process suggests that strain SX-1 may be potentially
used for harvesting energy from biodiesel wastes using MFCs However the current
density generated by strain SX-1 (205 mA m-2
) is lower than that (805 mA m-2
at 1 kΩ)
generated by the mixed culture from which the SX-1 was isolated indicating the
existence of other higher power generating bacteria andor complex ecology in the mixed
culture community
Understanding mechanisms of microbial extracellular electron transfer is critical for
enhancing the electron transfer rate from bacteria to electrode through metabolic or
genetic engineering of exoelectrogens Since the spent supernatant solution of MFCs run
with SX-1 showed no redox properties the redox compounds produced by SX-1 may
have been retained in the biofilm Alternatively SX-1 may utilize one or both of the
other two known extracellular electron transfer mechanisms For example it is possible
that strain SX-1 transfers electrons to the anode via cell-bound outer membrane proteins
The two distinct oxidation and reduction peaks observed at -100mv- -150 mV and -310--
350 mV of the SX-1 biofilm have a midpoint potential range from -205mV to -250mV
which is close to the redox potential of some c-type cytochromes (Field et al 2000 Meitl
et al 2009) which are well-known to play an important role in extracellular electron
trasnfer (Eggleston et al 2008 Field et al 2000 Meitl et al 2009) Field et al (2000)
reported that the c-type cytochromes CymA had the midpoint potentials of -229 mV
Meitl et al (2009) and Eggleston et al (2008) reported that the c- type cytochromes
44
OmcA in Shewanella oneidensis MR-1 had the midpoint potentials of -201 mV and -208
mV
While itrsquos possible that SX-1 may also have conductive appendages to enhance the
extracellular transfer electron similar to the nanowires discovered in some Geobacter
and Shewanella species (Reguera et al 2005 2006 Gorby et al 2006) further
investigation on the presence of these appendages on the cell surface of SX-1 is needed
Acknowledgements
The authors acknowledge support from the US National Science Foundation
(CBET 0955124) We also thank Teresa Sawyer for her helps with the SEM analysis and
Jeremy Chignell and Yanzhen Fan for their review of this manuscript
45
Chapter 4
Enhanced Performance and Mechanism Study of Microbial Electrolysis Cells
Using Fe Nanoparticles Decorated Anodes
Shoutao Xu Hong LiuYanzhen Fan Rebecca Schaller Jun Jiao Frank Chaplen
Published in
Applied Microbiology and biotechnology 93(2)871-880 (2012)
46
ABSTRACT
Anode properties are critical for performance of microbial electrolysis cells (MECs) In
the present study Fe nanoparticle modified graphite disks were used as anodes to
investigate the effects of nanoparticles on the performance of Shewanella oneidensis MR-
1 in MECs Results demonstrated that average current densities produced with Fe
nanoparticle decorated anodes were up to 56-fold higher than plain graphite anodes
Whole genome microarray analysis of the gene expression showed that genes encoding
biofilm formation were significantly up-regulated as response to nanoparticle decorated
anodes Increased expression of genes related to nanowires flavins and c-type
cytochromes indicate that enhanced mechanisms of electron transfer to the anode may
also have contributed to the observed increases in current density The majority of the
remaining differentially expressed genes were associated with electron transport and
anaerobic metabolism demonstrating a systemic response to increased power loads
Keywords Microbial electrochemical system microbial fuel cell microbial electrolysis
cell nanotechnology differential gene expression DNA microarray
47
Introduction
Microbial electrochemical systems (MESs) have been intensively studied since Lewis
achieved practical advances in this field (Logan 2007) however they attracted much
research attention in recent years due to their promising applications in renewable energy
generation bioremediation and wastewater treatment In a MES microorganisms
interact with electrodes via electrons catalyzing oxidation and reduction reactions at the
anode and the cathode The most-described type of MESs is microbial fuel cells (MFCs)
in which useful power is generated from electron donors typically biodegradable organic
materials (Logan et al 2006) Various novel MESs have recently been developed to
produce hydrogen (microbial electrolysis cells (MECs)) (Liu et al 2005 Logan et al
2008 Rozendal et al 2006 Schaller et al 2009) to power an autonomous sensor or
sensor network (Ringeisen et al 2006) to reduce bioremediation targets (Lovley 2006
Shea et al 2008) Sukkasem et al 2008) and to desalinate water (Cao et al 2009) The
key feature shared by these systems is the microbe-catalyzed electron transfer from
organic matter to electrodes (anodes) (Rabaey et al 2004) Enhancing the anodic current
output which highly depends on the performance of the electrodes is critical for the
successful application of all these processes (Logan et al 2007 Park and Zeilus 2002
2003)
Nanomaterials have received much attention from researchers in the context of
microbiology due to their unique physical electrical and chemical properties which
facilitate the study of interactions between bacteria and surfaces (Liu 2006) Previous
studies have demonstrated that electrodes decorated with different nanostructures such
as carbon nanotubes nanostructured polyanilinetitanium dioxide composite and titania
48
nanotubes have the potential to enhance power generation in MFCs (Morozan et al 2007
Qiao et al 2008 Quan et al 2005) Furthermore our recent study shows nanoparticle
(NP) decorated anodes greatly increased the electrochemical electron transfer rate in
MFCs (Fan et al 2011) However Fe NPs as an ideal candidate for decorating electrodes
because of respectively low price and high conductivity compared to other materials has
not been focused to study in MECs
S oneidensis MR-1 an important electrochemically active bacterial strain has been
exploited for the electricity generation in MFCs (Logan et al 2007 Logan 2009 Park
and Zeikus 2002) The availability of genome sequence for this strain makes it possible to
use transcriptome assays to globally measure the responses to different growth conditions
and environmental stresses (Murray et al 2001) S oneidensis MR-1 gene expression
response to heat shock cold shock Cr (VI) and U (VI) reduction chromate stress and
iron and acid tolerance have been studied in the past several years (Bencheikh-Latmani et
al 2005 Brown et al 2006 Gao et al2006 Gao et al 2004 Yang et al 2008) However
no studies have been focused on the Shewanella gene expression response to NPs in
MESs
The mechanism of increased electron transfer rate exhibited by nanoparticle decorated
electrodes is not well understood yet In the present study graphite disks decorated with
Fe NPs were used as anodes to explore the effects of nanostructures on current generation
in a multi-anode MECs DNA microarrays were utilized to investigate differences in the
global gene expression profile of S oneidensis MR-1 grown on plain versus Fe NPs
decorated anodes
Materials and methods
49
Bacterial cultures
S oneidensis MR-1 was purchased from American Type Culture collection (ATCC
700550) and stored as freezer stocks at -80degC Stock cells of S oneidensis MR-1 was
grown aerobically using a medium containing 30 gL Trypticase Soy Broth (BD 211825
Becton Dickinson Company Franklin lakes NJ) at 30degC Cells in early stationary
growth phase (optical density of 15-16 at 600nm (Spectrophotometer UV-1700
Shimadzu Columbia MD) were washed two times then injected into the chamber of the
MECs for current production Trypticase Soy Broth medium with 30 mM sodium lactate
as electron donor was used in MECs Phosphate buffer (100mM) was used to maintain
the solution pH 7 and solution conductivity at 15 mScm
Characterization of nanostructured anodes
Superfine isomolded graphite disks (16 cm x 05 cm GraphiteStorecom Inc) were
polished with ultrafine sand paper (2000 grit 3M Company) as the base for the NP
decorated and control anodes Fe NPs decorated anodes were fabricated by thermal
annealing of metal thin film coated graphite disks Thin films of Fe were firstly deposited
on the polished graphite disk by using sputter coating for 95 min and the samples then
were annealed in the Chemical vapor deposition (CVD) at 750degC for 20 min to form Fe
NPs Fe-NPs were produced with a diameter of 200plusmn100 nm and a density range
(distance between NPs) of 200plusmn100 nm with a coating time of 95 min SEM images of
Fe NPs decorated anode and control surfaces were shown in the Figure 41
50
Figure 41 SEM images of Fe NPs decorated anodes (A) Graphite disk control (B) Fe
Nanoparticle decorated
Multiple channel MECs construction and operation
A MECs with removable multiple anodes with each effective anode area 07 cm2
was
constructed and used to evaluate the effects of nanostructure on current density of
according to a previous report (Fan et al 2011) The cathode was made of wet-proof
(30) carbon cloth (type B E-TEK Division Inc USA) coated with PtC (20 E-TEK
Division Inc USA) following a previously reported procedure (Liu et al 2005) The
final platinum loading was 05 mgcm2 per projected cathode area The size of cathode
(150 cm2) was 25 times larger than the total effective area of the 8 graphite disk anodes
(56 cm2) to prevent cathode limitations on the performance of the MECs system All
MECs with Fe-NP decorated anodes and control anodes were tested for current
generation for 24 h with medium (without bacteria) before injecting the bacterial cells in
order to determine whether the Fe-NP decorated anodes demonstrate chemical current
generation compared to control anodes All the testing anodes were set up in the MECs
B
500 nm
A
500 nm
51
at once so that the S oneidensis MR-1 culture was applied to all of them simultaneously
Short electrode spacing (17 cm) was used in the MECs design to reduce the internal
resistance A voltage of 06 V was applied the MECs for current generation after the cells
of S oneidensis MR-1 were inoculated into MECs at early stationary phase A multimeter
with a data acquisition system (2700 Keithly USA) was used to monitor the current
change by measuring the voltage (every 300 seconds) drop through a resistor at 300 ohm
Fig 4 2 Effects of Fe-NP decorated anodes on the current density in MECs Control is
plain graphite disk anode Fe-NP is the anode with Fe-NP decoration
Microarray analysis
Biofilms for whole gene microarray analysis were aseptically removed from the plain
and Fe-NP decorated anodes of MECs after 110 hours further incubation at 30oC when
current density obviously started to decease Total RNA was extracted using Trizol
(Ambion inc AustinTX) following the manufacturerrsquos instructions Integrity of the
52
RNA samples was confirmed using the Agilent 2100 Bioanalyzer (Agilent Technology
Inc Palo Alto CA) The same amount of RNA (200 ng) from each sample was used for
further analysis after amplification by using the MessageAmpTM
II-Bacteria Prokaryotic
RNA Amplification kit (Ambion AustinTX) according to the manufacturerrsquos
instructions All of the 4295 genes (resource from NCBI) represented on the S oneidensis
MR-1 whole-gene microarrays were ordered from Roche NimbleGen Inc (Madison WI)
Biological triplicates of biofilms on the Fe-NP decorated anodes and control were
analyzed respectively cDNA synthesis labeling and hybridization were carried out by
the CGRB Core Laboratories at Oregon State University DNASTAR ArrayStarTM
3
software was used to identify genes that were up- or down-regulated more than 2-fold
when grown on the nanoparticle modified anodes using the unpaired two sample t-test
with a cutoff p-value of 005 The complete microarray data set generated in this study is
deposited for public access in the Gene Expression Omnibus
(httpwwwncbinlmnihgovgeo) under accession number GSE31535
Results
Enhancement of current generation by using NP decorated anodes in MECs
The current density generated by Fe-NP decorated anode MECs with S onidensis MR-1
increased to approximately 43 microAcm2 20 h following inoculation and then slowly
decreased Figure 42 shows that the maximum current density achieved by NP
decorated anodes was 83 times higher than that (51 microAcm2) generated by the control
(plain graphite disk) The average current density improvement of 110 hours was 59
times of that generated by the control The current densities generated in MECs in the
53
absence of bacteria were negligible (03 microAcm2) which demonstrated that the current
enhancement observed with the Fe-NP decorated anodes was biologically-derived
Global transcriptome analysis
Whole-genome DNA microarrays were used to attain a comprehensive general
overview
of the transcriptional response to Fe-NP decorated anodes by S oneidensis MR-1 in MEC
Of a total 4295 genes assayed 392 (188 induced and 204 repressed) exhibited significant
(P lt 005) differential expression at a 2-fold (log (expression ratio 1) level in 3
replicates in response to Fe-NPs These
total gene numbers present 9 of the 4295 open
reading frames (ORFs) presented on the array
Fig 43 Differentially expressed genes grouped by functional classification according to
the TIGR S oneidensis genome database (wwwtigrorg) Columns 1 amino acid
biosynthesis 2 biosynthesis of cofactors prosthetic groups and carriers 3 cell
envelope 4 cellular processes 5 central intermediary metabolism 6 DNA metabolism
7 energy metabolism 8 fatty acid and phospholipid metabolism 9 other categories 10
protein fate 11 protein synthesis 12 purines pyrimidines nucleosides and nucleotides
13 regulatory functions 14 signal transduction 15 transcription 16 transport and
binding proteins 17 unknown function 18 hypothetical proteins
Up-Regulated
-75
-50
-25
0
25
50
75
100
125
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18
Down-Regulated
Num
ber
of
Gen
es
54
Figure 43 summarizes the overall genomic response of S oneidensis MR-1 to Fe-NP
decorated anodes by grouping the differentially expressed genes into their functional role
categories based on The Institute for Genomic Researchrsquos annotation (Version 8) of the
MR-1 genome sequence The wide distribution of putative functional roles attributed to
the differentially expressed genes indicated the extent of the molecular response of
Soniedenis MR-1 to the NP decorated anodes Among genes with defined function a
large number of up-regulated genes were associated with cellular processes (group 4)
energy metabolism (group 7) other categories (group 9) and hypothetical proteins (group
18) While those genes encoding proteins involved in amino acid biosynthesis (group 1)
cellular processes (group 4) substrate transport (group 16) and hypothetical proteins
(group 18) were among the most down-regulated genes Two groups of genes that were
the most noticeable among the functional gene groups genes related to energy
metabolism (group 7) and genes related to amino acid biosynthesis (group 1) The ratios
of up-regulated genes to down-regulated genes in these functional groupings were much
higher than other functional gene groupings there were 21 up-regulated genes and 7
down-regulated genes in energy metabolism gene group and 2 up-regulated and 22 down
regulated genes in the amino acid biosynthesis group
Genes related to biofilm formation
In order to determine whether there was a correlation between the observed biofilm
enhancements by NP decorated anodes and the genes related to biofilm formation the
significant modified genes related to biofilm formation were examined and summarized
in the Table 41 These included SO_3223 and SO_3228 genes encoding the flagellum
proteins (Thormann et al 2004) which has critical impacts on initial attachment to the
55
surface as swimming motility functions were up-regulated over 10 and 09 fold
respectively The gene clusters SO_4178-SO_4180 which are thought to be essential for
formation of a three-dimensional biofilm were up-regulated from 12 to 13 fold
(Thormann et al 2004) In addition the genes SO_4105 encoding IV pilus which is
known to represent a critical factor for biofilm formation on a biotic surfaces (Thormann
et al 2004) were also induced 08 fold (Table 41) The genes related to flagellum
motility (SO_4286 SO_1529 and SO_1530) were also induced slightly (Thormann et al
2004)
Table 41 Expression levels of genes related to biofilm formation
Locus Tag Gene product descriptions Fold change
SO_3228 flagellar basal-body MS-ring and collar protein FliF 10
SO_3223 flagellar hook-length control protein FliK 09
SO_4103 MshA minor pilin protein MshD 05
SO_4105 MSHA major pilin protein MshA 08
SO_4178 expressed protein of unknown function MxdC 12
SO_4179 inner membrane family 2 glycosyltransferase MxdB 13
SO_4180 diguanylate cyclase-like protein MxdA 10
a fold changes were presented after log2
b positive number represented up-regulated and negative represented down-regulated
Genes related to energy metabolism
A closer consideration of energy metabolism genes was undertaken as anaerobic
metabolism for electron generation and electron transport functions may play critical
roles in enhanced current density generation in MECs Genes associated with energy
56
metabolism and with significantly modified gene expression levels are summarized in the
Table 42 Several of the up-regulated genes were related to formate dehydrogenase
proteins Most notably four genes related to formate dehydrogenase were up-regulated
more than 2-fold SO_0102 SO_0103 SO_0104 and SO_0107 The other genes
expression related to formate dehydrogenase proteins SO_4513 SO_4514 SO_4515 also
significantly increased Another interesting gene is that encoding the cytochrome c
oxidase protein gene SO_4609 which was up-regulated 11 fold while the gene
SO_0479 related periplasmic octaheme cytochrome c MccA was down-regulated
However SO_1519 the gene encoding lactate dehydrogenase was down-regulated 12
fold despite lactate being the primary carbon source in the media
Table 4 2 Genes related to anaerobic growth and electron transfer with significantly change expression level
Locus Tag Gene product descriptions Fold
change
SO_0101 nitrate-inducible formate dehydrogenase molybdopterin-binding
subunit FdnG
18
SO_0102 formate dehydrogenase nitrate-inducible iron-sulfur subunit 21
SO_0103 formate dehydrogenase nitrate-inducible cytochrome b556 subunit 24
SO_0104 formate dehydrogenase accessory protein FdhE 22
SO_0107 formate dehydrogenase accessory protein fdhD 24
SO_0397 quinolfumarate reductase menaquinol-oxidizing subunit FrdC_2 12
SO_0452 thioredoxin 2 15
SO_0903 Na-translocating NADH-quinone reductase subunit B NqrB_1 12
SO_1012 NADH-ubiquinone oxidoreductase subunit K NuoK 12
SO_1013 NADH-ubiquinone oxidoreductase subunit J NuoJ 12
SO_1363 hydroxylamine reductase 13
57
SO_2417 ferredoxin cofactor maintenance protein YfaE 10
SO_3922 formate dehydrogenase cytochrome b Fdh -12
SO_4513 formate dehydrogenase molybdopterin-binding subunit _2 15
SO_4514 formate dehydrogenase FeS subunit FdhB_2 12
SO_4515 formate dehydrogenase cytochrome b subunit FdhC_2 14
SO_4609 aa3 type cytochrome c oxidase subunit III CoxC 11
SO_0479 periplasmic octaheme cytochrome c MccA -12
SO_0847 periplasmic nitrate reductase ferredoxin component NapG -15
SO_1519 L-lactate dehydrogenase iron-sulfur cluster-binding protein LldF -10
SO_1251 ferredoxin 4Fe-4S -11
SO_37411 hypothetical inner membrane protein -11
a fold changes were presented after log2
b positive number represented up-regulated and negative represented down-regulated
Flavin and cytochrome related genes
Flavins can be secreted by Shewanella species as electron shuttle to facilitate
extracellular electron transfer (Cansterin et al 2008) C-type cytochromes play the
important roles on the process of extracellular electron transfer (Shi et al 2007) The
genes related to flavins synthesis and the genes encoding the cytochromes electron
transport proteins were shown in the table 43 the genes SO_1414 and SO_3058 related
to flavins synthesis was up-regulated to 03 and 01 folds respectively The genes related
to s C-type cytochromes SO_0135 SO_4105 SO_0169 SO_1778 are increased to 10
08 05 04 01 folds respectively However most of genes have shown no significant
changes (less one-fold change) as response to nano particle decorated anode in MECs
58
Table 43 Expression level of flavin and cytochrome genes
Locus Tag Gene product descriptions Fold
change
SO_1414 flavocytochrome c flavin subunit putative 03
SO_3468 riboflavin synthase subunit alpha -02
SO_3058 flavocytochrome c flavin subunit 01
SO_4105 MSHA pilin protein MshA 08
SO_0169 general secretion pathway protein GspG 05
SO_1778 outer membrane decaheme cytochrome c lipoprotein MtrC 04
SO_1779 outer membrane decaheme cytochrome c OmcA 01
SO_0135 lipoprotein of unknown function DUF333 10
SO_0136 conserved hypothetical inner membrane protein 01
a fold changes were presented after log2
b positive number represented up-regulated and negative represented down-regulated
Other genes with significantly modified expression levels
These genes over 15 fold change and possibly related to current enhancements were
shown in Table 44 SO_3104 (expressed inner membrane protein) was over expressed
and the gene SO_2194 which encodes outer membrane protein (OmpA family protein)
was repressed as response to nanoparticle decorated anode however it is unclear the
relationships of this modified membrane protein genes with the current enhancement
Another interesting phenomenon was possible co-regulation of several gene clusters
indicating possible operon associations This includes three sets of genes that were
induced more than 15 fold SO_0108SO_0109 and SO_1042SO_1043SO_1044 The
59
consistency of expression of these genes under the NP conditions provides basic evidence
to support operon structure However the correlation of these significant changed gene
with current density enhance are unclear which showed there are unknown multiple and
complex responses of S oneidensis MR-1 to nanoparticle anode of MECs
Table 44 Other genes with significantly changed expression levels
Locus Tag Gene product descriptions Fold change
SO_0101 nitrate-inducible formate dehydrogenase molybdopterin-binding
subunit FdnG
18
SO_0108 integral membrane protein of unknown function DUF39 YedE 17
SO_0109 SirA family protein YedF 17
SO_0275 N-acetyl-gamma-glutamyl-phosphate reductase -31
SO_0277 ornithine carbamoyltransferase -21
SO_0279 argininosuccinate lyase -24
SO_0404 zinc dependent metalloprotease domain lipoprotein 21
SO_0956 alkyl hydroperoxide reductase F subunit -20
SO_1042 amino acid ABC transporter ATP-binding protein -18
SO_1043 amino acid ABC transporter permease protein -27
SO_1044 amino acid ABC transporter periplasmic amino acid-binding protein -27
SO_1072 chitin-binding protein putative 21
SO_1405 transglutaminase family protein 20
SO_1822 TonB-dependent receptor putative -22
SO_2069
1-(5-phosphoribosyl)-5-[(5-phosphoribosylamino)
methylideneamino] imidazole-4-carboxamide isomerase
-19
SO_2070 amidotransferase HisH -26
SO_2071 imidazole glycerol-phosphate dehydratasehistidinol phosphatase -27
60
SO_2072 histidinol-phosphate aminotransferase -28
SO_2073 histidinol dehydrogenase -30
SO_2194 OmpA family protein -36
SO_2195 inter-alpha-trypsin inhibitor domain protein -27
SO_2767 asparagine synthetase B -26
SO_2945 prophage LambdaSo tail fiber protein 20
SO_2963 prophage LambdaSo major capsid protein HK97 family 20
SO_2987 prophage LambdaSo hypothetical protein 19
SO_2988 prophage LambdaSo expressed protein of unknown function 18
SO_3104 expressed inner membrane protein 131
SO_3408 conserved hypothetical inner membrane protein -18
SO_3585 azoreductase putative -18
SO_3586 glyoxalase family protein -17
SO_3687 curli production assemblytransport component CsgE putative -18
SO_38191 hypothetical ammonia permease -19
SO_4014 AcrBAcrDAcrF family protein 18
SO_40151 type I secretion system membrane fusion protein RND family 18
SO_4054 510-methylenetetrahydrofolate reductase 18
SO_4245 N-acetylglutamate synthase -22
SO_45251 hypothetical transcriptional regulator LysR family -17
SO_4527 integral membrane domain protein -19
SO_4705 transcriptional regulator putative -19
a fold changes were presented after log2
b positive number represented up-regulated and negative represented down-regulated
Discussion
61
The addition of NP to anodes significantly impacts current densities with the elemental
composition being a critical factor The chemical composition of NPs significantly affects
the current enhancement in MECs (Fan et al 2011) because the chemical composition of
NP properties is a critical factor in determining the conductivity which significantly
affects the efficiency of electron transfer from bacteria to electrode Fe is an ideal
candidate for NP formation for current enhancement in MESs because of high
conductivity and low price compared to gold and other conductive materials Gold NPs
has also been studies for enhancement of electricity conductivity (Bao et al 2008)
However the disadvantages of the high price of gold limit their practical application in
MECs (Fan et al 2011) The different size and density of the same composition of NP
also has essential effects on the current density enhancement (Fan et al 2011) In this
study the results showed the current density enhancement with Fe-NP size range of
200plusmn100 nm and density range of 200plusmn100 nm is considerable enhancement consequence
There are a number of reports of studies of current density enhancement in MFCs
by nanostructured anodes (Fan et al 2011 Qiao et al 2008 Sharma et al 2008 Tsai et al
2009) Based on bacterial behavior on nanostructured anode surface Qiao et al (2008)
believes that Escherichia coli cells on the nanostructured electrode surface produce hair-
like structures similar to pili that could facilitate the electron transfer between the cells
and electrode The production of hair-like structures is believed to be stimulated by
nanostructures and could play the key role on current density enhancement in pili-
producing bacteria in MFCs (Qiao et al 2008) S oneidensis can produce electrically
conductive bacterial nanowires which have similar structure to the hair-like structures in
Ecoli (Gorby et al 2006 Reguera et al 2005) A number of possible genes related
62
nanowires of S oneidensis have been identified including mshA (SO_4105) and gspG
(SO_0169) (Konstantinidis et al 2009) They are up-regulated 08 and 05 fold in the
current study respectively even though they are not significant induced This result
supported the increased nanowire genes had contributions to enhance current density in
MFCs by NPs decorated anodes
Based on the previous research on electron transfer mechanisms for S oneidensis it is
known that self-produced mediators (flavins) and multihaem c-type cytochromes (c-Cyts)
may play critical roles in the electron transfer of S oneidensis to extracellular insoluble
electrode (Canstein et al 2008 Myers and Myers 1992 Scott and Nealson 1994 Shi et al
2007) The flavins secreted by S oneidensis can facilitate to transfer electrons from
bacteria to solid anode electrodes by acting as extracellular electron shuttles (Canstein et
al 2008) In this study the genes encoding flavins (SO_1414 SO_3468 and SO_3058)
had no significant increase (less than 1 fold change) as a response to nanoparticle
decorated anode in MECs which suggests that increased flavin gene expression does not
significantly contribute to the current density enhancements in nano-particle decorated
anode The genes encoding cytochromes electron transport proteins (SO_4104 SO_0417
SO_177879 SO_013536) had no significant increase (less than 1-fold change)
However those genes were up-regulated slightly These results suggest the increased
expression flavin and c type cytochromes genes had partial contributions even not
significantly to enhance current density in MFCs by NPs decorated anodes
The thickness of bacterial biofilm on the anode can affect the power generation in
MFCs (Biffinger et al 2007 Heydorn et al 2000) since increased biofilm thickness on
the anode represents more active bacteria involving in the process of electron generation
63
and transfer in MFCs Biofilm formation related genes included flagellar genes IV pilus
genes and other genes The flagellar related genes SO_3228 SO_3229 (fliB and fliK) and
IV pilus genes SO_4105 (mshA) were induced from 19 to 09 fold the genes (SO_4178
-4180) related to formation of a three-dimensional biofilm were up-regulated from 10 to
13 folds even though the genes related bacterial motility protein (SO_ 4286 SO_4287
SO_1529 and SO_1530) are induced slightly (less than 0 5 fold) However the motility
genes most probably play a role for initial biofilm attachment on the anode surfaces
(Thormann et al 2004 Thormann et al 2006) with the rest of biofilm genes being more
essential for biofilm maintenance in mature cultures as were presented at harvest in this
study This enhancement of biofilm density under conditions of increase current density
has been observed for gold NP decorated anodes using confocal light microscopy (data
not shown) but has not yet been confirmed for Fe NP decorated anodes In any event an
increase in the expression level of genes related biofilm formation therefore also
supported the possibility of enhanced biofilm formation on NPs decorated anodes thus
facilitating electron transfer from bacteria to anodes
Beliaev et al described a model of the Fe reduction pathway in S oneidensis MR-1 using
solid substrates as the electron acceptor (Beliaev et al 2001) This model proposes that
electrons are generated and released in cytoplasm then transferred to the quinone pool
eg menaquinone in the cytoplasmic membrane The reduced menaquinone in turn
reduces CymA and from there electrons are then transferred to a periplasmic shuttle (eg
CctA) and then to MtrA a periplasmic soluble decaheme cytochrome c located in
associated with the outer membrane via interaction with MtrB Finally surface displayed
outer membrane proteins decaheme cytochromes (OmcA MtrC or MtrF) can transfer
64
electron to Fe or other solid substrates (Beliaev et al 2001)) Bencheikh-Latmani et al
demonstrated that the same electron transport pathway may be used for more than one
electron acceptor through studies of the response of S oneidensis MR-1 to U(VI) and
Cr(VI) under anaerobic conditions particularly focused on the critical genes
(SO_1777mtrA SO_1776mtrB SO_1778mtrC) (Bencheikh-Latmani et al 2005)
However our studies showed that these genes had no significant expression changes in
response to nanostructured anodes Contrastingly one of expressed inner membrane
protein genes SO_3104 and one of cytochromes genes SO_4609 encoding cytochrome c
oxidase subunit III demonstrated a 13-fold and 11-fold up-regulation respectively in this
study while the gene SO_2194 which encodes outer membrane protein (OmpA family
protein) was down regulated 36 fold Our results therefore suggest that S oneidensis
MR-1 does not utilize the same set of core critical genes (SO_1777 mtrA SO_1776
mtrB SO_1778 mtrC) products to transfer electrons in all instances This is the similar
result with the study of Bretschger et al (2007) which indicated that the electron
transport system in S oneidensis MR-1 is complex with several different proteins able to
participate in electron transfer to the anode of MECs
It should also be mentioned that the gene (SO_1519) encoding lactate
dehydrogenase repressed (-12 fold) here suggesting that lactate was depleted at the time
of cell harvest from the MECs and that other pathways of anaerobiosis had been activated
including those associated with amino acid uptake and consumption the growth media
was a complex mixture including yeast extract In particular the catabolic pathway for
histidine of S oneidensis MR-1 a pathway that may formate as an intermediate is up-
regulated with the genes for histidine ammonia lyase SO_0098 SO_3057 and SO_4374
65
up-regulated 13 11 and 11-fold respectively The possible production of formate
during histidine degradation may in turn explain the high levels of formate
dehydrogenates as evidenced by upregulation of SO_0102 SO_0103 SO_0104 and
SO_0107 annotated as formate dehydrogenase proteins These genes had the highest
levels of up-regulated expression as response to Fe NP decorated anodes (greater than 2-
fold)
Acknowledgements
This research was partially supported by the US National Science Foundation CBET
0828544 and the funds from ONAMIDOD (ARL-DOD Cooperative Agreement
W911NF-07-2-0083) We thank Barbara Gvakharia and Caprice Rosato for helpful
suggestions We also thank Margaret Romine of Pacific Northwest Laboratories for
valuable comments on manuscripts We also thank anonymous reviewers for significant
suggested improvements imparted as part of prior review of this manuscript
66
Chapter 5
Global Transcriptome Analysis of the Response of Shewanella oneidensis MR-1 to
Carbon Nanotube Decorated Anodes in Microbial Electrochemical Systems
Shoutao Xu Yanzhen Fan Rebecca Schaller Frank Chaplen Jun Jiao Hong Liu
67
Abstract
Shewanella oneidensis MR-1 is an important model microorganism for metabolic studies
on the effects of different environmental factors because of its diverse respiratory
capabilities Carbon nanotube (CNT) modified graphite disks were used as anodes to
investigate the effects of nanomatrials on the performance of S oneidensis MR-1 in
microbial electrolysis cells (MECs) The current densities produced with CNT decorated
anodes were on average 56-fold higher than plain graphite anodes Whole genome
microarray analysis of gene expression showed that up-regulation of cytochromes c genes
associated with extracellular electron transfer are strongly correlated to current increases
in CNT decorated anodes Genes involved in flavin biosynthesis also contribute to
current increase in CNT decorated anode MECs
Keywords Microbial electrochemical system microbial fuel cell microbial
electrolysis cell carbon nanotube gene expression DNA microarray
68
Introduction
Shewanella oneidensis MR-1 is an important model microorganism for metabolic
studies of the effects of different environmental factors because of its diverse respiratory
capabilities It has been used for transcriptome analysis to investigate the responses to
different growth conditions and environmental stresses (Murray et al 2001) Gene
expression patterns under different conditions such as heat shock cold shock Cr (VI) U
(VI) reduction chromate stress iron and acid tolerance have been studied previously
(Bencheikh-Latmani et al 2005 Brown et al 2006 Gao et al2006 Gao et al 2004
Yang et al 2008) More recently it has been exploited as a model species for power
generation in microbial electrochemical systems (MESs)which have potential
applications in renewable energy generation bioremediation and wastewater treatment
(Logan et al 2007 Logan 2009 Park and Zeikus 2002 Fan et al 2011 Xu et al 2012)
In a MES electrochemically active microorganisms oxidize organic matter in the an
ode chamber to release electrons Electrons are then transferred to the anode electrode thr
ough various mechanisms (Logan et al 2006 Lovley 2006 Rabaey et al 2003) and fina
lly travel to the cathode electrode and combine with the terminal electron acceptor The
key feature of MESs is the microbe-catalyzed electron transfer from the organic matter to
the anode (Rabaey et al 2004) Enhancing the current output which highly depends on
the performance of the anode electrode is critical for the successful application of MESs
(Logan et al 2007 Park and Zeilus 2002 2003) Several nanomaterials including TiO2
gold (Au) and palladium (Pd) nanoparticles have been utilized to decorate electrodes to
enhance power density in MESs (Qiao et al 2008 Quan et al 2005 Fan et al 2011) The
possible mechanisms for increased current densities have been studied by using Fe
69
nanoparticle-decorated anodes (Xu et al 2012)
Carbon nanotubes (CNT) are cylindrically shaped nanomaterials with extremely
high surface area excellent electrical conductivity and chemical inertness (He et al
2005a Serp et al 2003) These unique properties make CNT a promising electrode
material (Liang et al 2008) The biocompatibility of microorganisms and carbon
nanostructures has been evaluated (Morozan et al 2007) The effects of CNT on anodic
biofilms have been tested in MFCs (Liang et al 2011) However no studies have been re
ported on global transcriptome analysis for the response of S oneidensis MR-1 to CNT
decorated anode in MECs
In the present study the graphite disks decorated with CNT were used as anodes to
investigate the effects of nanomaterials on current generation in multi-anode MECs
DNA microarrays were used to analyze differences in the global gene expression profile
of S oneidensis MR-1 grown on plain versus CNT decorated anodes
Materials and methods
Bacterial cultures S oneidensis MR-1 was purchased from American Type Culture
collection (ATCC 700550) and stored as freezer stocks at -80degC Stock S oneidensis
MR-1 cells were grown aerobically using M1 medium at 30degC Cells in early stationary
growth phase (optical density of 15-16 at 600nm (Spectrophotometer UV-1700
Shimadzu Columbia MD) were injected into the chamber of the MECs for current
production Sodium lactate (final concentration 30mM) was added as the additional
electron donor
Fabrication and characterization of nanostructured anodes Superfine isomolded
graphite disks (16 cm x 05 cm GraphiteStorecom Inc) were polished with ultrafine
70
sand paper (2000 grit 3M Company) as the base for the CNT decorated and control
anodes Aligned multi-walled carbon nanotube samples were synthesized via plasma
enhanced chemical vapor deposition (PECVD) Samples superfine isomolded graphite
pillars with a 16 cm diameter and 05 cm thickness were sputtered with a thin film of Ni
in a CRC 300 magnetron sputter coater for 1 to 2 min They were then placed in the
PECVD reactor where the Ni was annealed into small catalyst islands C2H2 was bled
into the chamber for growth and an NH3 plasma was used to vertically align the growth
of the multi-walled carbon nanotubes (MWCNTs) The temperature and time were varied
between 725degC and 775oC and 15 to 60 min to control the length of the tubes and density
of samples Samples were then analyzed in a FEI Sirion field emission scanning electron
microscope (FESEM) SEM images of CNT decorated anode and control surfaces were
shown in the Figure 51
Figure 51 SEM images of CNT decorated anodes (A) Graphite disk control (B) CNT
decorated anode
Multiple channel MECs construction and operation A MECs with multiple
removable anodes each with an effective anode area of 07 cm2
was constructed and used
to evaluate the effects of nanostructures on the current density of according to a previous
B A
71
study (Fan et al 2011) The cathode was made of wet-proof (30) carbon cloth (type B
E-TEK Division Inc USA) coated with PtC (20 E-TEK Division Inc USA)
following a previously reported procedure (Liu et al 2005) The final platinum loading
was 05 mgcm2 per projected cathode area The size of the cathode (150 cm
2) was 25
times larger than the total effective area of the 8 graphite disk anodes (56 cm2) to prevent
cathode limitations on the performance of the MECs system All MECs with CNT
decorated anodes and control anodes were tested for current generation for 24 h with
sterile a medium (without bacteria) before injecting the bacterial cells in order to
determine whether the CNT decorated anodes demonstrate chemical current generation
compared to control anodes All the testing anodes were set up in the MECs
simultaneously so that the S oneidensis MR-1 culture was applied to all of them
simultaneously Short electrode spacing (17 cm) was used in the MECs design to reduce
the internal resistance A voltage of 06 V was applied the MECs for current generation
after the cells of S oneidensis MR-1 were inoculated into the MECs in the early
stationary phase A multimeter with a data acquisition system (2700 Keithly USA) was
used to monitor the current change by measuring the voltage drop through a resistor
Microarray analysis Biofilms for whole gene microarray analysis were aseptically
removed from the plain and CNT decorated anodes of MECs after 80 hours further
incubation at 30oC when current density obviously reached to a stable phase Total RNA
was extracted using Trizol (Ambion inc AustinTX) following the manufacturerrsquos
instructions Integrity of the RNA samples was confirmed using the Agilent 2100
Bioanalyzer (Agilent Technology Inc Palo Alto CA) Total RNA (200 ng) from each
sample was amplified using the MessageAmpTM
II-Bacteria Prokaryotic RNA
72
Amplification kit (Ambion AustinTX) according to the manufacturerrsquos instructions The
S oneidensis MR-1 whole-gene microarrays representing all of the 4295 genes (resource
from NCBI) were ordered from Roche NimbleGen Inc (Madison WI) Biological
triplicates of biofilms on the CNT decorated anodes and control were analyzed cDNA
synthesis labeling and hybridization were carried out by the CGRB Core Laboratories at
Oregon State University DNASTAR ArrayStarTM
3 software was used to identify genes
that were up- or down-regulated more than 2-fold when grown on the nanoparticle
modified anodes using the unpaired two sample t-test with a cutoff p-value of 005 with
bonferroni correction
Results and discussion
Enhancement of current generation using CNT decorated anodes in MECs The
current density generated by CNT decorated anode MECs with S onidensis MR-1
increased to 36 microAcm2 10 hours post-inoculation and reached a maximum of 40 microAcm
2
at 29 hours inoculation then gradually decreased The current density stabilized at
30microAcm2 after 78 h operation Figure 52 shows that the current density curve generated
by CNT decorated anode in MECs The average current density generated by CNT
decorated anodes was 56 times that of the control The current density generated in the
MECs without bacteria was negligible (data not shown) This result demonstrates that the
current enhancement observed with the CNT decorated anodes was biologically-derived
A
73
Figure 52 Effects of CNT decorated anodes on the current density in MECs (3 replicates)
Control is plain graphite disk anode CNT is the anode with CNT decoration (Error bars
represent standard deviation)
It has been reported that nanostructured decorated anodes have significant impacts on
current densities (Fan et al 2011 Qiao et al 2006 Sharma 2008 Tsai et al 2009) The
CNT decorated anodes had significantly enhanced current densities in MECs in this
study This result is consistent with previous studies Interestingly the current density
curve generated by CNT decorated anode MECs is quite different from the current
density curve generated by Au NP and Pd NP decorated anodes using same bacterial
species (Fan et al 2011) In the Au and Pd NP decorated anode MECs the current
density curve was not significantly increased in the beginning compared with the control
but gradually increased after the inoculation the current density reached the maximum
current density more than 50 hours after the inoculation However the current density
generated by CNT decorated anode MECs started to increase significantly immediately
after the inoculation It reached a maximum current density 29 hours post-inoculation
-1
6
13
20
27
34
41
48
0 10 20 30 40 50 60 70 80
Curr
ent
den
sity
(micro
Ac
m2)
Time (Hours)
Control
CNT
74
These results indicate that there might be different current density enhancement
mechanisms between CNT with other metal NP decorated anodes in MECs
Global transcriptome analysis Whole-genome DNA microarrays were used to attain a
comprehensive general overview of the transcriptional response of S oneidensis
MR-1 of
S oneidensis MR-1 to CNT decorated anodes in MECs In a total of 4295 genes analyzed
457 genes (250 genes up-regulated and 207 genes down-regulated) exhibited significantly
(P lt 005) changed expression at a 2-fold level in at least 3 replicates in the response to
CNT The total number of the regulated genes present 11 of the 4295 open reading
frames (ORFs) presented on the microarray The total number of genes at significant
expression levels in response to CNT decorated anodes is close to that to Fe NP decorated
anodes in MECs (392 9) ( Xu et al 2012)
Fig 53 Differentially expressed genes grouped by functional classification according to
the TIGR S oneidensis genome database (wwwtigrorg) Columns 1 amino acid
biosynthesis 2 biosynthesis of cofactors prosthetic groups and carriers 3 cell
envelope 4 cellular processes 5 central intermediary metabolism 6 DNA metabolism
7 energy metabolism 8 fatty acid and phospholipid metabolism 9 other categories 10
protein fate 11 protein synthesis 12 purines pyrimidines nucleosides and nucleotides
-80
-60
-40
-20
0
20
40
60
80
100
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18
Nu
mb
er
of
Ge
ne
s
Up-regulated
Down-regulated
75
13 regulatory functions 14 signal transduction 15 transcription 16 transport and
binding proteins 17 unknown function 18 hypothetical proteins
The overall transcript genomic response of S oneidensis MR-1 to CNT decorated
anodes was summarized in Figure 53 by grouping the differentially expressed genes into
their functional role categories based on The Institute for Genomic Researchrsquos annotation
(Version 8) of the MR-1 genome sequence The wide distribution of putative functional
roles attributed to the differentially expressed genes indicated the extent of the molecular
response of Soniedenis MR-1 to the CNT decorated anodes A large number of up-
regulated genes were presented in genes associated with biosynthesis of cofactors
prosthetic groups and carriers (group 2) cell envelope (group 3) protein fate (group 10)
protein synthesis (group 11) and purines pyrimidines nucleosides and nucleotides
(group 12) in this study The number of up-regulated genes was two times more than that
of down-regulated genes in each functional group Contrastingly in the genomic response
S oneidensis MR-1 to Fe NP decorated anodes (chapter 4) most of up-regulated genes
were associated with cellular processes (group 4) energy metabolism (group 7) other
categories (group 9) and hypothetical proteins (group 18) The down-regulated genes
presented in the amino acid biosynthesis (group 1) cellular processes (group 4) substrate
transport (group 16) and hypothetical proteins (group 18) While those genes encoding
proteins involved in amino acid biosynthesis (group 1) central intermediary metabolism
( group 5) DNA metabolism( group 6) fatty acid and phospholipid metabolism (group
8) and other categories (group 9) were among the most down-regulated genes in this
study However the two groups of genes that were the most noticeable between the
genomic response S oneidensis MR-1 to CNT and Fe NP decorated anodes which they
are the genes related to energy metabolism (group 7) and hypothetical proteins (group
76
18) These results indicated there are different and common genomic responses to S
oneidensis MR-1 on CNT and Fe NP decorated anodes in MECs
Electron transfer related genes Based on previous electron transfer mechanism
studies on Shewanella a serial of group proteins collectively described as the Mtr
pathway play the role to transfer electron rom the inner bacterial body to the outer
membrane (Beliaev et al 2001 Shi et al 2007 Brutinel and Gralnick 2012) In the Mtr
pathway firstly electrons generated by bacteria received by CymA a tetraheme c-type
cytochrome anchored in the inner membrane then electrons were transferred to MtrA a
periplasmic decaheme c-type cytochrome After that electrons were transferred from
MtrA to MtrC an outer-membrane decaheme c-type cytochrome is facilitated by MtrB a
non-heme containing integral outer-membrane proteinOuter-membrane decaheme c-type
cytochrome MtrC and OmcA are in change of electron transfer to exreacellular electron
acceptor (Shi et al 2007 Brutinel and Gralnick 2012 ) These genes encoding electron
transport proteins are shown in the table 51 The critical genes SO_1777 mtrA SO_1776
mtrB SO_1778 mtrC are unregulated 41 to 80 fold in response to CNT decorated
anodes other cytochrome c genes related to electron transfer in MtrA pathway also are
significantly increased in response to CNT decorated anodes These genes encoding outer
membrane proteins have different level increases supporting that the increase of
cytochrome c gene expression contributes to the current enhancements of MECs as
response to CNT decorated anodes
77
Table 51 Expression level of cytochrome c as response to CNT decorated anodes
Locus Tag Gene product descriptions Fold change CNT
SO_0165 general secretion pathway protein GspC 31
SO_0167 general secretion pathway protein GspE 45
SO_1776 outer membrane protein precursor MtrB 41
SO_1777 outer membrane decaheme cytochrome c
lipoprotein MtrA
80
SO_1778 outer membrane decaheme cytochrome c
lipoprotein MtrC
43
SO_1779 outer membrane decaheme cytochrome c
OmcA
63
SO_0135 lipoprotein of unknown function DUF333 22
SO_0136 conserved hypothetical inner membrane protein 24
Flavin related genes It is known that self-produced mediators play critical roles in the
electron transfer of S oneidensis to extracellular insoluble electrode (Canstein von et al 2
008 Myers et al 1992 Scott et al 1994 Shi et al 2007) It has been proved that the
flavins secreted by S oneidensis can facilitate electron transfer from bacteria to solid
anode electrodes by acting as extracellular electron shuttles (Cansterin von et al 2008) T
wo critical genes (SO_1414 and SO_3468) related to flavin synthesis were shown increa
sed significantly (more than 2 folds) increase in this study (Table 52) This result support
s the hypothesis that flavin production increases as the response to CNT decorated anodes
in MECs have significant contribution to the current density enhancements
78
Table 52 Expression levels of genes related to flavin synthesis as response to CNT
decorated anode
Locus Tag Gene product descriptions Fold change CNT
SO_1414 flavocytochrome c flavin subunit putative 22
SO_3468 riboflavin synthase subunit alpha 20
The amount of bacterial biofilm biomass on the anode can affect the power
generation in MFCs (Biffinger et al 2007 Heydorn et al 2000) Biofilm formation
related genes include flagellar gene IV pilus genes and other genes the flagellar related
genes (fliB and fliK) and IV pilus genes (mshA) were induced only from 105 to 134
fold as response of S oneidensis MR-1 to CNT decorated anodes in this study No
biofilm-associated genes presented significant changes in expression levels in this study
indicating there is no direct connections between biofilm enhancements to CNT
decorated anodes in this study This result is consistent with the Liang et al (2011)
conclusion that using CNT enhanced the electrochemical activity of anodic biofilms but
did not result in a significant increase of biomass in the anodic biofilms
Our microarray results showed two significantly up-regulated gene groups the
genes encoding proteins localized on the outer membrane and the genes involved in
flavin biosynthesis contributed to current density enhancement by CNT decorated anodes
Among 457 significantly changed genes to CNT in this study there are also a relatively
large number of genes encoding proteins with unknown functions which are either up- or
down-regulated in the response to CNT decorated anodes which indicated more
79
complicated responses of S oneidensis MR-1 to CNT decorated anodes in MECs and
further study is needed
80
Chapter 6
Summary
Microbial electrochemical systems (MESs) with a mixed culture initially inoculated
from Corvallis wastewater treatment plant have been studied for more than 6 years for
varying purposes including power generation hydrogen production heavy metal
removal and wastewater treatment Experiment results have shown that the mixed culture
is quite stable with excellent performance in MESs Our community analysis using
denaturing gradient gel electrophoresis (DGGE) and 16S rDNA clone library construction
suggests that the mixed culture is composed predominantly of Geobacter sp (66)
Arcobacter sp (12) and Citrobacter sp (11) These results not only improved our
understanding of the mixed culture community but also guided our studies on the
cultivation and isolation of the different bacterial species
Exoelectrogenic Citrobacter sp SX-1 was first isolated and characterized from a
MFC operated with the mixed culture Citrobacter sp SX-1 has been demonstrated to
produce electricity from wide range of different substrates including citrate acetate
glucose sucrose glycerol and lactosein MFCs Cyclic voltammetry analysis indicated
that membrane associated proteins may play an important role in facilitating the electrons
transferring from bacteria to electrode The strain SX-1 increased the known diversity of
power generating exoelectrogens and provided a unique bacterial species for study in
renewable energy generation and waste treatment
81
To enhance the electron transfer from bacteria to anode nano decorated anodes
including Fe NP and CNT were developed and characterized and evaluated in MECs
using Shewanella oneidensis MR-1 as a model species Both nanostructures have
significantly increased current density compared with the control Whole genome
microarray analysis elucidated the possible mechanisms of power enhancement in
response to these nano-decorated anodes
These results benefit to understanding of physiology and ecology of mixed
cultures in MFCs and improve the efficiency of current generation in MESs which will
facilitate the viability of niche applications for MESs in near future
82
Bibliography
1 Amman R Ludwig W (2000) Ribosomal RNA-targeted nucleic acid probes for
studies in microbial ecology FEMS Microbiol Ecol 24 555ndash565
1 Bao SJ Li CM Zang JF Cui XQ Qiao Y Guo J (2008) Novel Nanoporous TiO2
Electrode for Directly Electrochemistry of glucose oxidase Adv Funct Mater
18(4) 591-599
2 Beliaev AS Saffarini DA McLaughlin JL Hunnicutt D (2001) MtrC an outer
membrane decahaem c cytochrome required for metal reduction in Shewanella
putrefaciens MR-1 Mol Microbiol 39722-30
3 Bencheikh-Latmani R Williams SM Haucke L Criddle CS Wu L Criddle CS
Zhou J Tebo BM (2005) Global transcriptional profiling of Shewanella oneidensis
MR-1 during Cr(VI) and U(VI) reduction Appl Environ Microbiol 71 7453ndash7460
4 Biffinger JC Pietron J Ray R Little B and Ringeisen BR (2007) A biofilm
enhanced miniature microbial fuel cell using Shewanella oneidensis DSP10 and
oxygen reduction cathodes Biosens Bioelectron 221672ndash1679
5 Bodelier PLE Roslev P Henckel T FenzelP (2000) Stimulation by ammonium
based fertilizers of methane oxidation in soil around rice root Nature 40 421ndash424
6 Bond DR Holmes DE Tender LM Lovley DR (2002) Electrode-reducing
microorganisms that harvest energy from marine sediments Science 295 483-485
7 Bond DR Lovley DR (2003) Electricity production by Geobacter sulfurreducens
attached to electrodes Appl Environ Microbiol 69 1548-1555
8 Bond DR Lovley DR (2005) Evidence for involvement of an electron shuttle in
electricity generation by Geothrix fermentans Appl Environ Microbiol 71 2186-
2189
9 Bretschger O Obraztsova A Sturm CA Chang IS Gorby YA Reed SB Culley
DE Reardon CL Barua S Romine MF Zhou J Beliaev AS Bouhenni R
Saffarini D Mansfeld F Kim BH Fredrickson JK Nealson KH (2007) Current
production and metal oxide reduction by Shewanella oneidensis MR-1 wildtype and
mutants Appl Environ Microbiol 73 7003ndash7012
10 Brown SD Thompson MR Verberkmoes NC Chourey K Shah M Zhou J
Hettich RL and Thompson DK (2006) Molecular dynamics of the Shewanella
oneidensis response to chromate stress Mol Cell Proteomics 51054-1071
11 Brutinel ED Gralnick JA (2012) Shuttling happens soluble flavin mediators of
extracellular electron transfer in Shewanella Appl Microbiol Biotechnol 93(1) 41-8
12 Butler JE Kaufmann F Coppi MV Nuacutentildeez C Lovley DR (2004) MacA a diheme
c-type cytochrome involved in Fe(III) reduction by Geobacter sulfurreducens J
Bacteriol 186(12) 4042ndash4045
83
13 Caccavo F Lonergan DJ Lovley DR Davis M Stolz JF McInerney MJ (1994)
Geobacter sulfurreducens sp nov a hydrogen- and acetate oxidizing dissimilatory
metal-reducing microorganism Appl Environ Microbiol 60 3752ndash3759
14 Canstein von H Ogawa J Shimizu S Lloyd JR (2008) Secretion of flavins by
Shewanella species and their role in extracellular electron transfer Appl Environ
Microbiol 74 615ndash623
15 Cao X Huang X Liang P Xiao K Zhou Y Zhang X Logan BE (2009) A New
Method for Water Desalination Using Microbial Desalination Cells Environ Sci
Technol 43(18)7148-7152
16 Chaudhuri SK Lovley DR (2003) Electricity generation by direct oxidation of
glucose in mediatorless microbial fuel cells Nat Biotechnol 21 1229-1232
17 Cheng S Liu H Logan B E (2006) Increased power generation in a continuous
flow MFC with advective flow through the porous anode and reduced electrode
spacing Environ Sci Technol 402426-2432
18 Cheng S Logan B E (2007) Sustainable and efficient biohydrogen production via
electrohydrogenesis PNAS 104 18871-18873
19 Chignell J (2010) Energy Recovery from Biodiesel Waste Performance of
Microbial Electrochemical Systems on Glycerol Thesis
20 Chung K Okabe S (2009) Characterization of electrochemical activity of a strain
ISO2-3 phylogenetically related to Aeromonas sp isolated from a glucose-fed
microbial fuel cell Biotechnol Bioeng 104(5) 901-910
21 Cole JR Chai B Farris RJ Wang Q Kulam-Syed-Mohideen AS McGarrell
DM Bandela AM Cardenas E Garrity GM Tiedje JM (2007) The ribosomal
database project (RDP-II) introducing myRDP space and quality controlled public
data Nucleic Acids Res 35 (Database issue) D169-D172 doi 101093nargkl889
22 Cole JR Wang Q Cardena E Fish J Chai B Farris RJ Kulam-Syed-Mohideen
AS McGarrell DM Marsh T Garrity GM Tiedje JM (2009) The Ribosomal
Database Project improved alignments and new tools for rRNA analysis Nucleic
Acids Res 37 (Database issue) D141-D145 doi 101093nargkn879
23 Dey A De S De A De SK (2004) Characterization and dielectric properties of
polyaniline-TiO2 nanocomposites Nanotechnol 15 1277-1283
24 Edwards U Rogall T Bloumlcker H Emde M Boumlttger EC (1989) Isolation and
direct complete nucleotide determination of entire genesmdashcharacterization of a gene
coding for 16S-ribosomal RNA NucleicAcids Res 17 7843ndash7853
25 Eggleston CM Voumlroumls J Shi L Lower BH Droubay TC Colberg PJS (2008)
Binding and direct electrochemistry of OmcA an outer-membrane cytochrome from
an iron reducing bacterium with oxide electrodes a candidate biofuel cell system
Inorg Chim Acta 361769ndash777
26 Fan Y Sharbrough E Liu H (2008) Quantification of the Internal Resistance
Distribution of Microbial Fuel Cells Environ Sci Technol 42 8101ndash8107
84
27 Fan Y Xu S Schaller R Jiao J Chaplen F Liu H (2011) Nanoparticle decorated
anodes for enhanced current generation in microbial electrochemical
cells Biosensors and Bioelectronics 26 (5)1908ndash1912
28 Fan Y Hu H Liu H (2007)Sustainable power generation in microbial fuel cells
using bicarbonate buffer and proton transfer mechanisms Environ Sci
Technol 41(23) 8154-8
29 Fedorovich V Knighton MC Pagaling E Ward FB Free A Goryanin I (2009)
Novel electrochemically active bacterium phylogenetically related to Arcobacter
butzleri isolated from a microbial fuel cell Appl Environ Microbiol 75(23) 7326-
7334
30 Ferna ndez A Huang S Seston S Xing J Hickey R Criddle C Tiedje J (1999)
How stable is stable Function versus community composition Appl Environ
Microbiol 65 3697ndash3704
31 Field SJ Dobbin PS Cheesman MR Watmough NJ Thomson AJ Richardson
DJ (2000) Purification and magneto-optical spectroscopic characterization of
cytoplasmic membrane and outer membrane multiheme c-type cytochromes from
Shewanella frigidimarina NCIMB400 J Biol Chem 275 8515ndash8522
32 Freguia S Masuda M Tsujimura S Kano K (2009) Lactococcus lactis catalyses
electricity generation at microbial fuel cell anodes via excretion of a soluble quinone
Bioelectrochemistry 7614ndash18
33 Fricke K Harnisch F and Schroumlder U (2008) On the use of cyclic voltammetry for
the study of the anodic electron transfer in microbial fuel cells Energy Environ
Sci 1 144ndash147
34 Gao H Wang Y Liu X Yan T Wu L Alm E Arkin A Thompson DK and
Zhou J (2004) Global transcriptome analysis of the heat shock response of
Shewanella oneidensis J Bacteriol 1867796-7803
35 Gao H Yang ZK Wu L Thompson DK Zhou J (2006) Global transcriptome
analysis of the cold shock response of Shewanella oneidensis MR-1 and mutational
analysis of its classical cold shock proteins J Bacteriol 1884560-4569
36 George MG (2005) Bergeyrsquos Manual of Systematic Bacteriology Volume Two The
Proteobacteria Part B The Gammaproteobacteria Second Edition New York
Springer 651p
37 Gonzaacutelez-Toril E Llobet-Brossa E Casamayor EO Amann R Amils R (2003)
Microbial ecology of an extreme acidic environment the Tinto river Appl Environ
Microbiol 69(8) 4853ndash4865
38 Gorby YA Yanina S Mclean JS Rosso KM Moyles D Dohnalkova A
Beveridge TJ Chang IS Kim BH Kim KS Culley DE Reed SB Romine M
F Saffarini DA Hill EA Shi L Elias DA Kennedy DW Pinchuk G
Watanabe K Ishii S Logan B Nealson KH Fredrickson JK (2006) Electrically
conductive bacterial nanowires produced by Shewanella oneidensis strain MR-1 and
other microorganisms Proc Natl Acad Sci USA 10311358-11363
85
39 He Z Wagner N Minteer SD Angenent LT (2006) An upflow microbial fuel cell
with an interior cathodes assessment of the internal resistance by impedance
spectroscopy Environ Sci Technol 40 5212-5217
40 Heydorn A Nielsen AT Hentzer M Sternberg C Givskov M Ersboslashll BK Molin
S (2000) Quantification of biofilm structures by the novel computer program
COMSTAT Microbiol 1462395-2407
41 Holmes DE Bond DR Lovley DR (2004a) Electron transfer by Desulfobulbus
propionicus to Fe(III) and graphite electrodes Appl Environ Microbiol 701234ndash
1237
42 Holmes DE Chaudhuri SK Nevin KP Mehta T Methe BA Liu A Ward JE
Woodard TL Webster J Lovley DR (2006) Microarray and genetic analysis of
electron transfer to electrodes in Geobacter sulfurreducens Environ Microbiol
81805-1815
43 Holmes DE Nicoll JS Bond DR Lovley DR (2004b) Potential role of a novel
psychrotolerant member of the family Geobacteraceae Geopsychrobacter
electrodiphilus gen nov sp nov in electricity production by a marine sediment
fuel cell Appl Environ Microbiol 70 6023ndash6030
44 Holmes DE Chaudhuri SK Nevin KP Mehta T Metheacute BA Liu A Ward JE
Woodard TL Webster J Lovley DR (2006) microarray and genetic analysis of
electron transfer to electrodes in Geobacter sulfurreducens Environ Microbial
8(10)1805-1815
45 Hu H (2009) Enhancing Hydrogen Production in Microbial Electrolysis Cells
through Development of Platinum-free Cathode and Improvement of Reactor
Dissertation
46 Ince BK Ayman O N Turker G Ccedilelikkol S Ince O (2010) Microbial ecology of
anaerobic reactors for treatment of alcohol industry wastewaters a review Current
research technology and education topics in applied microbiology and microbial
ecology 988-999
47 Jung S Regan JR (2007) Comparison of anode bacterial communities and
performance in microbial fuel cells with different electron donors Appl Microbiol
Biotechnol 77393ndash 402
48 Kim BH Kim HJ Hyun MS Park DH (1999) Direct electrode reaction of Fe(III)
reducing bacterium Shwwanella putrefaciens J Microbial Biotechnol 9127-131
49 Kim GT Hyun MS Chang IS Kim HJ Park HS Kim BH Kim SD Wimpenny
JW Weightman AJ (2005) Dissimilatory Fe(III) reduction by an electrochemically
active lactic acid bacterium phylogenetically related to Enterococcus gallinarum
isolated from submerged soil J Appl Microbiol 99(4) 978ndash987
50 Kim GT Webster G Wimpenny JW Kim BH Kim HJ Weightman AJ (2006)
Bacterial community structure compartmentalization and activity in a microbial fuel
cell J Appl Microbiol 101(3) 698ndash710
86
51 Kim HJ Park HS Hyun MS Chang IS Kim M Kim BH (2002) A mediator-
less microbial fuel cell using a metal reducing bacterium Shewanella putrefaciens
Enzyme Microb Technol 30 145-152
52 Kim JR Min B Logan BE (2005) Evaluation of procedures to acclimate a
microbial fuel cell for electricity production Appl Microbiol Biotechnol 68(1)23-30
53 Kim JR Cheng S Oh SE Logan BE (2007) Power generation using different
cation anion and ultrafiltration membranes in microbial fuel cells Environ Sci
Technol 41 1004-1009
54 Konstantinidis KT Serres MH Romine MF Rodrigues JL Auchtung J McCue
LA Lipton MS Obraztsova A Giometti CS Nealson KH Fredrickson JK
Tiedje JM (2009) Comparative systems biology across an evolutionary gradient
within the Shewanella genus Proc Natl Acad Sci U S A 106(37)15909-15914
55 Lee J Phung NT Chang IS Kim BH Sung HC (2003) Use of acetate for
enrichment of electrochemically active microorganisms and their 16S rDNA
analyses FEMS Microbiol Lett 223185ndash191
56 Liang P Wang H Xia X Huang X Mo Y Cao X Fan M(2011)Carbon
nanotube powders as electrode modifier to enhance the activity of anodic biofilm
in microbial fuel cells Biosens Bioelectron 26(6)3000-4
57 Lies DP Hernandez ME Kappler A Mielke RE Gralnick JA Newman DK
(2005) Shewanaella oneidensis MR-1 uses overlapping pathways for iron reduction
at a distance and by direct contact under conditions relevant for biofilms Appl
Environ Microbiol 71 4414-4426
58 Liu H and Logan B E (2004) Electricity generation using an air-cathode single
chamber microbial fuel cell in the presence and absence of a proton exchange
membrane Environ Sci Technol 38 4040-4046
59 Liu H Grot S Logan BE (2005) Electrochemically assisted microbial production
of hydrogen from acetate Environ Sci Technol 39 4317 ndash 320
60 Liu H Hu H Chignell J Fan Y (2010) Microbial Electrolysis Novel Technology
for Hydrogen Production from Biomass Biofuels 1(1) 129ndash142
61 Liu H Ramnarayanan R Logan BE (2004) Production of electricity during
wastewater treatment using a single chamber microbial fuel cell Environ Sci Technol
382281-2285
62 Liu WT Marsh TL Cheng H Forney LJ (1997) Characterization of microbial
diversity by determining terminal restriction fragment length polymorphisms of gene
encoding 16S rRNA Appl Environ Microbiol 63 4516ndash4522
63 Logan B E Hamelers B Rozendal R Schroder U Keller J Freguia S Aelterman
P Verstraete W Rabaey K (2006) Microbial fuel cells methodology and
technology Environ Sci Technol 40 (17) 5181-5192
64 Logan B E and Regan JM (2006) Electricity-producing bacterial communities in
microbial fuel cells Trends Microbiol 14512ndash518
87
65 Logan B Cheng S Watson V Estadt G (2007) Graphite fiber brush anodes for
increased power production in air-cathode microbial fuel cells Environ Sci Technol
41(9)3341-3346
66 Logan BE (2009) Exoelectrogenic bacteria that power microbial fuel cells Nat Rev
Microbiol 5 375-381
67 Logan BE Call D Cheng S Hamelers HV Sleutels TH Jeremiasse AW
Rozendal RA (2008) Microbial electrolysis cells for high yield hydrogen gas
production from organic matter Environ Sci Technol 42 8630-8640
68 Logan BE Hamelers B Rozendal R Schroder U Keller J Freguia S Aelterman
P Verstraete W Rabaey K (2006) Microbial fuel cells methodology and
technology Environ SciTechnol 40 5181-5192
69 Lovely DR (2006) Bug juice harvesting electricity with microorganisms Nature
Review| Microbiology 4 497-508
70 Lovley DR Phillips EJ (1988) Novel Mode of Microbial Energy Metabolism
Organic Carbon Oxidation Coupled to Dissimilatory Reduction of Iron or
Manganese Appl Environ Microbiol 54(6) 1472-1480
71 Ly HK Sezer M Wisitruangsakul N Feng JJ Kranich A Millo D Weidinger
IM Zebger I Murgida DH Hildebrandt P (2011) Surface-enhanced vibrational
spectroscopy for probing transient interactions of proteins with biomimetic interfaces
electric field effects on structure dynamics and function of cytochrome c FEBS J
278(9)1382-90
72 Macaskie LE Hewitt CJ Shearer JA Kent CA (1995) Biomass production for
the removal of heavy metals from aqueous solutions at low pH using growth-
decoupled cells of a Citrobacter sp Int Biodeter Biodegr 3573ndash92
73 Malki M De Lacey AL Rodriguez N Amils R Fernandez VM (2008)
Preferential use of an anode as an electron acceptor by an acidophilic bacterium in
the presence of oxygen Appl Environ Microbiol 74 4472ndash4476
74 Meitl LA Eggleston CM Colberg PJS Khare N Reardon CL Shi L (2009)
Electrochemical interaction of Shewanella oneidensis MR-1 and its outer membrane
cytochromes OmcA and MtrC with hematite electrodes Geochim Cosmochim
Ac 73(18) 5292-5307
75 Morozan A Stamatin L and Nastase
F (2007) The biocompatibility
microorganisms-carbon nanostructures for applications in microbial fuel cells Phys
Stat Sol 6 1797-1803
76 Morris JM Jin S Crimi B Pruden A (2009) Microbial fuel cell in enhancing
anaerobic biodegradation of diesel Chem Eng J 146 161ndash167
77 Murray AE Lies D Li G Nealson K Zhou J Tiedje JM (2001) DNADNA
hybridization to microarrays reveals gene-specific differences between closely
related microbial genomes Proc Natl Acad Sci U S A 98 9853-9858
78 Muyzer G de Waal EC Uitterlinden AG (1993) Profiling of complex microbial
communities by denaturing gradient gel electrophoresis analysis of polymerase chain
88
reaction amplified genes coding for 16S rRNA Appl Environ Microbiol 59 695ndash
700
79 Myers CR Myers JM (1992) Localization of cytochromes to the outer membrane
of anaerobically grown Shewanella putrefacians MR-1 J Bacteriol 174 3429ndash3438
80 Narde GK Kapley A Purohit HJ (2004) Isolation and characterization of
Citrobacter strain HPC255 for broad-range substrate specificity for chlorophenols
Curr Microbiol 48(6) 419-423
81 Okabe S Ito T Sugita K Satoh H (2005) Succession of internal sulfur cycles and
sulfur-oxidizing bacterial communities in microaerophilic wastewater biofilms Appl
Environ Microbiol 71(5) 2520-2529
82 Park DH and Zeikus JG (2000) Electricity generation in microbial fuel cells using
neutral red as an electronophore Appl Environ Microbial 66 292-1297
83 Park DH and Zeikus JG (2002) Impact of electrode composition on electricity
generation in a single-compartment fuel cell using Shewanella putrefaciens Appl
Microbiol Biotechnol 5958ndash56
84 Park DH and Zeikus JG (2003) Improved fuel cell and electrode designs for
producing electricity from microbial degradation Biotechnol Bioeng 81 348ndash355
85 Park HS Kim BH Kim HS Kim HJ Kim GT Kim M Chang IS Park Y
K Chang HI (2001) A novel electrochemically active and Fe(III)-reducing
bacterium phylogenetically related to Clostridium butyricum isolated from a
microbial fuel cell Anaerobe 7 297-306
86 Parot S Nercessian O Delia ML Achouak W Bergel A (2009) Electrochemical
checking of aerobic isolates from electrochemically active biofilms formed in
compost J Appl Microbiol 106(4) 1350-1359
87 Pham CA Jung SJ Phung NT Lee J Chang IS Kim BH Yi H Chun J
(2003) A novel electrochemically active and Fe(III)-reducing bacterium
phylogenetically related to Aeromonas hydrophila isolated from a microbial fuel cell
FEMS Microbiol Lett 223 129-134
88 Qiao Y Bao SJ Li CM Cui XQ Lu ZS Guo J (2008) Nanostructured
polyanilinetitanium dioxide composite anode for microbial fuel cells ACS Nano 2
113-119
89 Qiu R Zhao B Liu J Huang X Li Q Brewer E Wang S Shi N (2009) Sulfate
reduction and copper precipitation by a Citrobacter sp isolated from a mining area J
Hazard Mater 1641310-1315
90 Quan X Yang SG Ruan XL and Zhao HM (2005) Preparation of titania
nanotubes and their environmental applications as electrode Environ Sci Technol 39
3770ndash3775
91 Rabaey K Boon N Hofte M Verstraete W (2005) Microbial phenazine
production enhances electron transfer in biofuel cells Environ Sci Technol 393401ndash
3408
89
92 Rabaey K Boon N Siciliano S D Verhaege M Verstraete W (2004) Biofuel cells
select for microbial consortia that self-mediate electron transfer Appl Environ
Microbiol 70(9) 5373ndash5382
93 Rabaey K Lissens G Siciliano S D Verstraete W (2003) Amicrobial fuel cell
capable of converting glucose to electricity at high rate and efficiency Biotechnol
Lett 25 1531-15
94 Rabaey K Verstraete W (2005) Microbial fuel cells novel biotechnology for
energy generation Trends Biotechnol 23(6) 291-298
95 Reguera G McCarthy KD Mehta T Nicoll JS Tuominen MT Lovley DR
(2005) Extracellular electron transfer via microbial nanowires Nature 435 1098-
1101
96 Reguera G Nevin KP Nicoll JS Covalla SF Woodard TL Lovley DR (2006)
Biofilm and nanowire production leads to increased current in Geobacter
sulfurreducens fuel cells Appl Environ Microbiol 72 7345-7348
97 Reimers CE Tender LM Fertig S Wang W (2001) Harvesting energy from the
marine sediment-water interface Environ Sci Technol 35192-195
98 Rezaei F Xing D Wagner R Regan JM Richard TM Logan BE (2009)
Simultaneous cellulose degradation and electricity production by Enterobacter
cloacae in a microbial fuel cell Appl Microbiol Biotechnol 75 3673ndash3678
99 Richter H Nevin KP Jia H Lowy DA Lovley DR Tender LM (2009) Cyclic
voltammetry of biofilms of wild type and mutant Geobacter sulfurreducens on fuel
cell anodes indicates possible roles of OmcB OmcZ type IV pili and protons in
extracellular electron transfer Energy Environ Sci 2 506ndash516
100 Riesenfeld SC Schloss DP Handelsman J(2004) Metagenomics Genomic
Analysis of Microbial Communities Annual Review Genetics 38525-552
101 Rozendal RA Hamelers HV Buisman CJ (2006) Effects of membrane cation
transport on pH and microbial fuel cell performance Environ Sci Technol 40
5206-5211
102 Rozendal RA Hamelers HVM Euverink GJW Metz SJ Buisman CJN (2006)
Principle and perspectives of hydrogen production through biocatalyzed electrolysis
Int J Hydrogen Energ 311632-1640
103 Rozendal RA Hamelers HV Molenkamp RJ Buisman CJ (2007) Performance of
single chamber biocatalyzed electrolysis with different types of ion exchange
membrances Water Res 41 1984-1994
104 Schluumlter A Bekel T Diaz NN Dondrup M Eichenlaub R Gartemann
KH Krahn I Krause L Kroumlmeke H Kruse O Mussgnug JH Neuweger
H Niehaus K Puumlhler A Runte KJ Szczepanowski R Tauch A Tilker
A Viehoumlver P Goesmann A (2008) The metagenome of a biogas-producing
microbial community of a production-scale biogas plant fermenter analyzed by the
454-pyrosequencing technology J Biotechnol136 77-90
90
105 Scott J H and Nealson KH (1994) A biochemical study of the intermediary carbon
metabolism of Shewanella putrefaciens J Bacteriol 176(11) 3408ndash3411
106 Sekiguchi H Tomioka H Nakahara T and Uchiyama H (2001) A single band
does not always represent single bacterial strains in denaturing gradient gel
electrophoresis analysis Biotechnology Letters23 1205ndash1208
107 Sharma T Leela M Reddy A Chandra TS Ramaprabhu S (2008) Development
of carbon nanotubes and nanofluids based microbial fuel cell Int J Hydrogen Energ
336749ndash6754
108 Shea C Clauwaert P Verstraete W Nerenberg R (2008) Adapting a denitrifying
biocathode for perchlorate reduction Water Sci Technol 58 (10) 1941-1946
109 Shi L Squier TC Zachara JM Fredrickson JK (2007) Respiration of metal (hydr)
oxides by Shewanella and Geobacter a key role for multihaem c-type cytochromes
Mol Microbiol 6512-20
110 Stams AJ and Plugge CM(2009)Electron transfer in syntrophic communities
of anaerobic bacteria and archaea Nat Rev Microbiol7(8)568-77
111 Sukkasem C Xu S Park S Boonsawang P Liu H (2008) Effect of Nitrate on the
Performance of Single Chamber Air Cathode Microbial Fuel Cells Water Res 42(19)
4743-4750
112 Summers ZM Fogarty HE Leang C Franks AE Malvankar NS Lovley DR
(2010)Direct exchange of electrons within aggregates of an evolved syntrophic
coculture of anaerobic bacteria Science 330 (6009)1413-5
113 Szczepanowski R Bekel T Goesmann A Krause L Kroumlmeke H Kaiser
O Eichler W Puumlhler A Schluumlter A (2008) Insight into the plasmid metagenome
of wastewater treatment plant bacteria showing reduced susceptibility to
antimicrobial drugs analysed by the 454-pyrosequencing technology J
Biotechnol 136 54-64
114 Tamura K Dudley J Nei M Kumar S (2007) MEGA4 Molecular Evolutionary
Genetics Analysis (MEGA) software version 40 MolBiol Evol 24(8) 1596-1599
115 Thompson JD Gibson TJ Plewniak F Jeanmougin F Higgins DG (1997) The
CLUSTAL_X windows interface flexible strategies for multiple sequence alignment
aided by quality analysis tools Nucleic Acids Res 25 4876-4882
116 Thormann KM Duttler S Saville RM Hyodo M Shukla S Hayakawa Y
Spormann AM (2006) Control of formation and cellular detachment from
Shewanella oneidensis MR-1 biofilms by cyclic di-GMP J Bacteriol 1882681-2691
117 Thormann KM Saville RM Shukla S Pelletier DA Spormann AM (2004)
Initial Phases of biofilm formation in Shewanella oneidensis MR-1 J Bacteriol
186(23) 8096-104
118 Torres CI Kato Marcus A Rittmann BE (2008b) Proton transport inside the
biofilm limits electrical current generation by anode-respiring bacteria Biotechnol
Bioeng 100 872ndash881
91
119 Torres CI Marcus AK Lee HS Parameswaran P Krajmalnik-Brown R Rittmann
BE (2010) A kinetic perspective on extracellular electron transfer by anode-
respiring bacteria FEMS Microbiol Rev 234(1)3-17
120 Torres CI Marcus AK Parameswaran P Rittmann BE (2008a) Kinetic
experiments for evaluating the NernstndashMonod model for anode-respiring bacteria
(ARB) in a biofilm anode Environ Sci Technol 42 6593ndash6597
121 Tsai HY Wu Ch-Ch Lee Ch-Y Shih EP (2009) Microbial fuel cell performance
of multiwall carbon nanotubes on carbon cloth as electrodes J Power Sources
194199-205
122 Tyson GW Chapman J Hugenholtz P Allen EE Ram RJ Richardson
PM Solovyev VV Rubin EM Rokhsar DS Banfield JF(2004) Community
structure and metabolism through reconstruction of microbial genomes from the
environment Nature42837-43
123 Venkateswaran K Moser DP Dollhopf ME Lies DP Saffarini D A MacGregor
BJ Ringelberg DB White DC Nishijima M Sano H Burghardt J Stackebrandt
E Nealson KH (1999) Polyphasic taxonomy of the genus Shewanella and
description of Shewanella oneidensis sp Int J Syst Bacteriol 49705-724
124 von Canstein H Ogawa J Shimizu S Lloyd JR (2008) secretion of Flavins
by Shewanella Species and Their Role in Extracellular Electron Transfer Appl
Environ Microbiol 74 3615-623
125 Wang A Liu W Cheng Sh Xing D Zhou J Logan BE (2009) Source of methane
and methods to control its formation in single chamber microbial electrolysis cells
Int J Hydrogen Energ 34 3653-3658
126 Wang J Ma T Zhao L Lv J Li G Liang F Liu R (2008) PCRndashDGGE method
for analyzing the bacterial community in a high temperature petroleum reservoir
World J Microbiol Biotechnol 2 9694-9702
127 Wang Q Garrity GM Tiedje JM Cole JR (2007) Naiumlve Bayesian Classifier for
Rapid Assignment of rRNA Sequences into the New Bacterial Taxonomy Appl
Environ Microbiol 73(16) 5261-5267
128 Wang YF Masuda M Tsulimura S Kano K (2008) Electrochemical regulation of
the end-product profile in Propionibacterium freudenreichii ET-3 with an
endogenous mediator Biotechnol Bioeng 101(3) 579ndash586
129 Wrighton KC Agbo P Warnecke F Weber KA Brodie EL DeSantis TZ
Hugenholtz P Andersen GL Coates JD (2008) A novel ecological role of the
Firmicutes identified in thermophilic microbial fuel cells ISME J 2 1146ndash1156
130 Xing D Zuo Y Cheng S Regan JM Logan BE (2008) Electricity generation by
Rhodopseudomonas palustris DX-1 Environ Sci Technol 42 4146-4151
131 Xu S Liu H Fan Y Schaller R Jiao J Chaplen F (2012) Enhanced performance
and mechanism study of microbial electrolysis cells using Fenanoparticle-decorated
anodes Appl Microbiol Biotechnol 93(2)871-880
92
132 Yang Y Harris DP Luo F Wu L Parsons AB Palumbo AV Zhou J (2008)
Characterization of the Shewanella oneidensis Fur gene roles in iron and acid
tolerance response BMC Genomics 9 Suppl 1S11
133 Zhang L Zhou S Zhuang L Li W Zhang J Lu N Deng L (2008) Microbial
fuel cell based on Klebsiella pneumoniae biofilm Electrochem Commun10 1641ndash
1643
134 Zuo Y Xing DF Regan JM Logan BE (2008) Isolation of the exoelectrogenic
bacterium Ochrobactrum anthropi YZ-1 by using a U-tube microbial fuel cell Appl
Environ Microbiol 74 3130-3137
93
APPENDICES
List of published papers during PhD study
1 Shoutao Xu Hong Liu Yanzhen Fan Rebecca Schaller Jun Jiao and Frank
Chaplen (2012) Enhanced performance and mechanism study
of microbial electrolysis cells using Fe nanoparticle-decorated anodes Applied
Microbiology and Biotechnology 93(2)871-880
2 Shoutao Xu Hong Liu (2011) New exoelectrogen Citrobacter sp SX-1 isolated
from a microbial fuel cell Journal of Applied Microbiology111(5)1108-1115
3 Yanzhen Fan Shoutao Xu Rebecca Schaller Jun Jiao Frank Chaplen Hong Liu
(2011) Nanoparticle decorated anodes for enhanced current generation in microbial
electrochemical cells Biosensors and Bioelectronics 26(5) 1908-1912
4 Tunc Catal Shoutao Xu Kaichang Li Hakan Bermek Hong Liu
(2008)
Electricity generation from polyalcohols in single-chamber microbial fuel
cells Biosensors and Bioelectronics 24(4)849-854
5 Chontisa Sukkasema Shoutao Xu Sunhwa Park Piyarat Boonsawang Hong Liu
(2008) Effect of nitrate on the performance of single chamber air cathode
microbial fuel cells Water research 424743-4750
6 Rebecca Schaller Yanzhen Fan Shoutao Xu Alan Fern Frank Chaplen Hong
Liu and Jun Jiao (2009) Vertically Aligned Multi-walled Carbon Nanotube
Decorated Anodes for Microbial Fuel Cells Proceedings of Materials Research
Society 2009 1170 R05-13

1
1
Bacterial Community Analysis New Exoelectrogen Isolation and Enhanced Performance
of Microbial Electrochemical Systems Using Nano-Decorated Anodes
Chapter 1
General Introduction
11 Microbial Fuel Cells and Microbial Electrolysis Cells
Microbial electrochemical systems (MESs) have drawn the attention of researchers
in recent years due to their promising applications in a variety of scientific fields such as
renewable energy generation bioremediation and wastewater treatment In a MES
microorganisms interact with electrodes catalyzing oxidation and reduction reactions at
the anode and the cathode
The most-described type of MESs is the microbial fuel cells (MFCs) in which
useful power is generated directly using the catalytic action of active microorganisms
(Kim 2002 Cheng and Logan 2007 Cheng et al 2006) In a typical two-chamber
MFCs (Figure 1a) organic matter is oxidized by electrochemically active
microorganisms in the anode chamber to release protons and electrons Protons diffuse
into the cathode chamber through a proton exchange membrane Meanwhile electrons
are transferred to the anode through various mechanisms (Logan et al 2006 Lovley
2
2006 Rabaey et al 2003) and then travel to the cathode where normally they combine
with oxygen and protons to form water
Another common used type of MES is the microbial electrolysis cell (MECs) in
which hydrogen is produced instead of electricity by applying a circuit voltage to the
MFCs system and maintaining anaerobic conditions in the cathode chamber (Liu et al
2005 Logan et al 2008 Rozendal et al 2006 Schaller et al 2009) (Figure 1b) A
cathode potential of at least - 410 mV (vs standard hydrogen electrode at pH 70) is
required to produce hydrogen at the cathode Therefore a theoretical potential of 110 mV
(ie 410-300 mV) is needed but voltages of gt02 V are normally applied due to various
overpotentials
Fig 1-1 Generalized schematics (a) Microbial fuel cells (MFCs) where electricity is
captured through the resistance R (b) Microbial electrolysis cells (MECs) where
hydrogen gas is produced in the cathode chamber R for resistor PS for power supply and
PEM for proton exchange membrane
The fundamental feature shared by microbial electrochemical systems (MESs)
(MECs and MFCs) is the microbe-catalyzed electron transfer from organic matter to
electrodes (anodes) (Rebaey et al 2004) Therefore different types of MESs can be
PEM
R
H+
e- e-
Anode
Bacte
ria
a
Cath
ode
H2O
PEM
PS
H+
e- e-
Anode
Bacte
ria
b
Cath
ode
H2
3
utilized to investigate the phenomena and mechanisms of interactions between microbes
and electrodes
12 Bacteria and Electron Transfer Mechanisms in MESs
The electrochemically active bacteria in MFCs are thought to be iron-reducing
bacteria such as Shewanella and Geobacter species (Gorby et al 2006) They have great
importance in the natural environment principally in metal oxidation and reduction
However recent studies have shown that the diversity of bacterial communities is much
greater than these model iron reducers on MFCs anodes (Reimers et al 2001 Liu et al
2004b Kim et al 2005) Over 20 electrochemically active bacterial species those can
transfer electrons exocellularly to electrodes have been reported in the past 10 years
(Logan 2009 Liu et al 2010) These bacteria have been categorized into diverse genetic
groups so far including alphaproteobacteria (Rhodopseudomonas Ochrobactrum and
Acidiphilium) (Malki et al 2008 Xing et al 2008 Zuo et al 2008) betaproteobacteria
(Rhodoferax) (Chaudhuri et al 2003) gammaproteobacteria (Shewanella Pseudomonas
Klebsiella Enterobacter and Aeronomas) (Kim et al 2002 Pham et al 2003 Rabaey et
al 2005 Bretschger et al 2007 Zhang et al 2008 Chung et al 2009 Rezaei et al 2009)
deltaproteobacteria (Geobacter Geopsychrobacter Desulfuromonas and Desulfobulbus)
(Bond et al 2002 Bond and Lovley 2003 Holmes et al 2004ab 2006)
Epsilonproteobacteria (Arcobacter) (Fedorovich et al 2009) Firmicutes (Clostridium and
Thermincola) (Park et al 2001 Wrighton et al 2008) Acidobacteria (Geothrix) (Bond
and Lovley 2005) and actinobacteria (Propionibacterium) (Wang et al 2008) A wider
range of electrochemically active bacteria are expected to be discovered
4
Table 1 Taxa of bacteria current density generated and reactor configuration used in
microbial electrochemical cell experiments (Liu et al 2010)
The electrochemically active bacterial species that possess the ability to transfer
electrons outside of the cell are called exoelectrogens in the MESs research field The
different exoelectrogens have demonstrated a wide-ranging power generation ability in
MFCs For example Shewanella oneidensis MR-1 has demonstrated the ability to
generate 0018 Am2 current density in single chamber MFCs while the Geobacter
Taxon Microorganisms Current Density (Am2
)
α-proteobacteria
Rhodopseudomonas palustris DX-1 003
Ochrobactrum anthropi YZ-1 071
Acidiphilium sp 32sup5 300
β-proteobacteria Rhodoferax ferrireducens 0031
γ-proteobacteria
Shewanella putrefaciens IR-1 0016
Shewanella oneidensis DSP10 0013
Shewanella oneidensis MR-1 0018
Pseudomonas aeruginosa KRA3 0017
Escherichia coli K12 HB101 100
Klebsiella pneumoniae L17 120
Enterobacter cloacae 013
Aeromonas hydrophila PA3 030
δ-proteobacteria
Geobacter metallireducens 065
Geobacter sulfurreducens 800
Desulfuromonas acetoxidans 0005
Geopsychrobacter
Electrodiphilus strain A2
0066
Desulfobulbus propionicus 003
Firmicutes
Lactococcus lactis 003
Thermincola sp strain Jr 020
Clostridium butyricum EG3 022
Thermincola ferriacetica Z-0001 040
Brevibacillus spp PTH1 0009
Desulfitobacterium hafniense DCB2 110
Acidobacteria Geothrix fermentans 0097
Actinobacteria Propionibacterium freudenreichiiET-3 120
5
sulfurreducens species has generated current densities as high as 80 Am2 which is 400
times higher than the one generated by Shewanella oneidensis MR-1 However mixed-
culture communities have generated much higher power densities than their pure-culture
counterparts in MFCs perhaps due to synergistic interactions within the anode bacterial
communities and the participation of currently unknown bacteria species and mechanisms
(Jung and Regan 2007) The dominant bacteria species vary in mixed cultures bacteria
communities in MFCs due to the enrichment of different substrates
Traditional methods of extroelectrogen study depend on cultivation hampered novel
exoelectrogens discovery because the inadequacy of defined media underestimates the
actual microbial diversity in MFCs Slow growth rate and unknown growth requirements
of anaerobic microorganisms make some anaerobic extroelectrogens cultivation difficult
However these limitations have been overcome by using molecular biological methods
based on DNARNA analysis Molecular biological techniques are now widely applied to
assess the diversity of microbial communities by analyzing the 16S rDNA sequence The
most commonly used molecular biological techniques for bacterial community analysis
include denaturing gradient gel electrophoresis (DGGE) restriction fragment length
polymorphism (RFLP) and 16S rDNA sequencing These methods are much less time
consuming than traditional isolation and cultivation methods
Isolated exoelectrogens were utilized to explore the mechanism of electron transfer
to the anode However the mechanisms of electron transfer to extracellular electron
acceptors are not well understood Three mechanisms have been proposed for exocellular
transport of electrons by exoelectrogens to anodes in MFCs so far Some exoelectrogenic
bacteria can transfer electrons to electrodes through soluble redox compounds (Bond and
6
Lovley 2005) These compounds include artificial mediators and mediators secreted by
exoelectrogenic bacteria A variety of chemicals have been used to facilitate the shuttling
of electrons from inside of cell to electrodes outside the cell These exogenous mediators
include neutral red (Park et al 1999) anthraquinone-2-6 disulfonate (AQDS) thionin
potassium ferricyanide (Bond et al 2002) methyl viologen and others (Logan 2004
Rabaey and Verstraete 2005) Some bacteria can secrete their own chemical mediator
for example Pseudomonas aeruginosa secretes pyocyanin and phenazine-1-carboxamide
to transfer electrons toward the MFCs anode (Rabaey et al 2005) Another evidence for
mediator production by bacteria is Geothrix fermentans When the medium was replaced
in a MFCs that had stable power generation with this bacteria power dropped by 50
and required 10 days to resume the original level
Some bacteria can directly transfer electrons to anodes via outer cell membrane
proteins (Myers and Myers 1992) The outer cell membrane protein cytochrome c is
thought to play a critical role in to transferring electrons to anodes Ly et al (2011)
isolated the haem protein cytochrome c and demonstrated that electric field effects may
be functional for the natural redox processes of cytochrome c in the respiratory chain
Shewanella oneidensis is the example of a bacterium that is capable of transfer electron to
anode via outer cell membrane proteins When Shewanella oneidensis adhered to an iron
surface the greater force has showed benefits to grow cells because closer contact
required for electron transfer from cell bound cytochromes (Lower et al 2001)
7
Fig1-2 Mechanisms for extracellular electron transport in an MFCs anode (1) direct
contact (top in green) (2) by nanowires (middle in purple) and (3) self-produced
mediators (bottom in blue) (Logan 2009)
More and more evidence supports the involvement of bacterial nanowires in
extracellular electron transport (Reguera et al 2005 2006 Gorby et al 2006) Nanowires
are conductive appendages produced by both Geobacter and Shewanella species (Gorby
and Beveridge 2005) The conductivity of the appendages was examined and confirmed
by using conductive scanning tunneling microscopy (STM) (Gorby et al 2006)
Nanowires can carry electrons from the cell to the anode surface of MFCs
The solid component of the extracellular biofilm matrix has high efficiency on
extracellular electron transfer compared with other extracellular electron transfer
8
mechanisms and recently Torres et al (2010) hypothesized that the solid component of
the extracellular biofilm matrix formed by exoelectogens is another way bacteria transfer
electrons to electrodes This hypothesis was based on kinetic analysis of each EET
mechanism reported in available literature (Torres et al 2010)
13 Anode electrodes
In MFCsMECs anode electrodes are a critical component because exoelectrogens
adhere to the surface of anodes to transfer electrons to the electrode The characteristics
of anodes have significant effects on electron transfer rate from bacteria to anode
electrodes in MFCs The requirements of an anode material are it should be highly
conductive non-corrosive have a high specific surface area (area per volume) high
porosity be non-fouling inexpensive and easily scaled to larger sizes Of these
properties the most important one that is different from other biofilm reactors is that the
material must be electrically conductive Normally they are made of various carbon
materials including carbon fiber carbon clothe and carbon paper due to their stability
high conductivity and high specific surface-area Nevertheless they have little
electrocatalytic activity for the anode microbial reactions and thus a modification of the
carbon materials is the main approach for improving their performance Consequently
there is a great need to develop a new type of anode material for MFCsMECs
It is a great challenge to develop a new anode material to further increase the power
density of a MFCs The nature of the catalytic mechanism of a MFCrsquos anode involves not
only a biological but also an electrocatalytic process An optimal nanostructure with a
high specific surface area favorable for both catalytic processes could play a critical role
in improving the power density of the MFCs such a structure needs to host the bacteria
9
with high bioactivity while enhancing the electron-transfer rate Schroumlder et al (2008)
employed PANI to modify a platinum anode for MFCs and achieved a current density 1
order of magnitude higher than the previously reported value PANIinorganic
composites are also reported to have better conductivity Qiao et al also applied a new
mesoporous TiO2 electrode material with uniform nanopore distribution and a high
specific surface area to anode in comparison to previously reported work with E coli
MFCs the composite anode delivers 2-fold higher power density (Qiao et al 2008)
Thus it has great potential for use as the anode in a high-power MFCs and may be a new
approach for improving performance of MFCs
14 Other parts of MESs
141 Membranes and ion transport
The ion exchange membrane is another one of the critical components in two-
chamber MESs systems It separates anode and cathode chambers and at the same time
maintains the electron neutrality of the system ie transport of electrons to the cathode
needs to be compensated by transport of an equal amount of positive charge to the
cathode chamber A proton exchange membrane (PEM) such as Nafion a per-fluorinated
sulphonic acid membrane (PFSAs) consisting of a hydrophobic fluorocarbon backbone to
which hydrophilic sulfonate groups (-SO3-) are attached is commonly used in chemical
fuel cell systems For MFCs systems however mainly cation species like Na+ and K
+
other than proton are often responsible for the dominant transport of positive charge
through the cation exchange membrane (CEM) to maintain electroneutrality due to the
low proton concentration in any aqueous medium with near neutral pH (Rozendal et al
2006) Consequently the pH increases in the cathode chamber due to the consumption of
10
protons and decreases in the anode chamber because of the accumulation of protons
(Rozendal et al 2007) This pH change in two-chamber systems results in a decrease of
the cathode potential and performance The application of anion exchange membrane
(AEM) has also been explored in MFCs systems (Cheng and Logan 2007 Kim and
Logan 2007) where it has been proposed that protons are transferred via pH buffers like
phosphate anions
142 Cathodes and Catalysts
The cathode is another challenge for making MFCs commercially available
technology because the chemical reaction that occurs at the cathode is difficult to
engineer as the electrons protons and oxygen must all meet at a catalyst in a tri-phase
reaction (solid catalyst air and water) The catalyst must be on a conductive surface and
must be exposed to both water and air so that protons and electrons in these different
phases can reach the same point The most commonly used material for a cathode is
commercially available carbon paper pre-loaded with a Pt catalyst on one side When it is
used in a MFC the side that contains the catalyst faces the water and the uncoated side
faces air To reduce the high cathode cost associated with platinum catalyst other precious-
metal-free cathodes such as (NiMo and NiW) were developed by electrodepositing on a
carbon fiber They have achieved comparable performance with Pt catalyst with same
loading at a much lower cathode fabrication cost (Hu 2010)
The requirements of cathode for MECs are quite similar to the requirements of
cathode for MFCs but easier than cathode for MFCs for the manufacturing process
because the cathode in MECs is not necessarily to exposed to air Therefore it can be
made of the exact same materials of cathode in MFCs except a waterproof layer
11
Recently some researchers have attempted to apply microorganisms as a biocatalyst to
precede the combination of electron with oxygen in the cathodes
15 Dissertation overview
Low power densities in MESs limit practical applications The improvement of
MESs performance requires a detailed understanding of the physiology and ecology of
microorganisms in MESs including the mechanism of electron transfer to the anode from
the microorganism
This dissertation focuses on the problem of the low power density of MESs The
bacterial community structure of a high power generated mixed culture communities in
MFCs will be identified firstly and then one of major exoelectrogens will be isolated and
characterized This information will be helpful to understand the physiology and ecology
of exoelectrogens in MESs Consequently they will be beneficial to improve power
density of MFCs Lastly the nanostructure decorated anodes in MECs will be utilized to
improve the power density The power enhancement mechanism will be explored by
using a whole genome microarray They are presented here as four papers
In the first paper the cultivation independent molecular biological techniques
denaturing gradient gel electrophoresis (DGGE) and 16S rDNA clone library are utilized
to analyze the bacterial community structure of a higher power mixed culture MFCs The
analyzed results provide fundamental information for isolating the dominant bacteria in
mixed culture MFCs Two of dominant bacterial species has been isolated and used aone
to test power generation in MFCs The possible interaction among different bacterial
species in mixed culture is discussed
12
In the second paper one isolated exoelectrogenic bacterial strain SX-1 is
characterized It is identified as a member of the Citrobacter genus and power generation
is tested ultilizing a wide range of different substrates The electron transfer mechanism is
explored using Cyclic Voltammetry (CV) This study increases the known diversity of
power generating exoelectrogens and provides a new strain to explore the mechanisms of
extracellular electron transfer from bacteria to electrodes
The third paper shows effects on MESs performance by Fe nanoparticle decorated
anodes in the MESs The average current density produced with Fe nanoparticle
decorated anodes increased up to 59-fold higher than plain graphite anodes A whole
genome microarray is utilized to analyze the possible mechanism of enhanced current
density as responded to nanoparticle decorated anodes
The fourth paper describes the effects of carbon nanotube (CNT) modified anode on
the performance S oneidensis MR-1 in MESs Results demonstrate that current densities
produced with CNT decorated anodes are up to 56-fold higher than plain graphite anodes
The possible mechanisms of enhanced current density by CNT decorated are explored
13
Chapter 2
Bacterial community analysis of mixed consortium in higher
power density MESs
Shoutao Xu and Hong Liu
1 Introduction
The improvement in the performance of mixed culture MFCs requires an
understanding of the ecology in microbial communities of MFCs Many researchers have
attempted to characterize microbial populations and activities to elucidate the behaviors
and ecology of microorganisms in MFCs (Wrighton et al 2008 Fedorovich et al 2009)
In order to study the microbial ecology of the mixed culture in a MFC and select the
appropriate isolation medium for dominant bacterial species in the mixed culture the
fundamental steps are to analyze the bacterial diversity in the mixed culture of MFCs and
identity the dominant bacterial species in bacteria communities in MFCs
14
As for the identification of bacterial communities typically there are two general
methods The first method for identification of bacterial community is the traditional
cultivation processes using selective nutrients to promote the growth of different types of
bacteria within the samples (Amman et al 2000) The community structure can then be
assessed by identifying the isolates from the dominant colonies that were cultured This
can often be costly and laborious as each isolate has to be further studied by examining
its physiology taxonomy and reactivity to stains (Adwards et al 1989)
The second method relies on utilizing molecular techniques to analyze bacterial
community DNA Several molecular methods involving the extraction and analysis of
DNA from entire bacterial communities are used to identify genetic fingerprints of
bacteria These methods including the cloning and sequencing of 16S rDNA automated
ribosomal intergenic spacer analysis (ARISA) terminal restriction fragment length
polymorphism (T-RFLP) and denaturing gradient gel electrophoresis (DGGE) generate
profiles of bacterial community structures They can rapidly assess complex communities
from various environments (Amman et al 2000)
Due to the conservative characteristic of 16S rDNA in bacteria during the process
of evolution 16S rDNA sequencing can be used to identify different species of bacteria
Therefore the molecular techniques of denaturing gradient gel electrophoresis (DGGE)
with PCR and 16S rDNA clone library are used for analysis of the microbial diversity
These methods are more convenient and save time compared to traditional
isolationcultivation methods for microorganism analysis
In this work a biofilm bacterial community from an anode of a MFCs wase studied
by using denaturing gradient gel electrophoresis (DGGE) of PCR-amplified partial 16S
15
rRNA genes followed by cloning and sequencing of 16S rDNA The results provided
essential information for dominant bacterial isolation in mixed culture MFCs
2 Materials and methods
21 High power generation of mixed culture MFCs
Mixed cultures were originally inoculated from domestic wastewater (Corvallis
Wastewater Treatment Plant Corvallis OR) to MFCs after operating for 2 years using a
defined medium solution (Lovely 2002) with sodium acetate as the carbon source A
new MFCs was inoculated from the operating MFCs The polarization curves were
performed to measure the power generation when maximal stable power were established
(normally 3-5 batches post-inoculation) and the genomic DNA from the microbial
biofilm on the anode was extracted under the sterile conditions for further bacterial
community analysis
22 Denaturing gradient gel electrophoresis (DGGE)
DGGE with PCR is a method of analysis of bacterial community composition
based on different GC percentage of 16s rDNA in different bacterial species PCR-DGGE
comprises three steps (i) extraction of total community DNA from the sample (ii) PCR-
controlled amplification using specific oligonucleotide primers and (iii) separation of the
amplicons using DGGE For this purpose a reproducible and efficient method for total
DNA extraction is indispensable and needs to be evaluated and optimized depending on
the nature of the sample In the subsequent PCR step multiple PCR primer sets with
different resolution can be used In most PCR-DGGE applications on bacteria universal
or specific primers are targeting the 16S rRNA gene Upon electrophoresis of the PCR
16
amplicons and gel staining (using ethidium bromide silver staining or SYBR green)
DGGE gels are digitally captured and further analyzed using computer software packages
The richness of DNA fragment bands in DGGE indicates the bacterial diversity of the
sample
The detailed steps are as followed Biofilms were scratched from the anodes of high
power generated MFCs fed with sodium acetate Bacterial genomic DNA was extracted
from the biofilm samples using the DNeasy blood and tissue kit (Qiagen) according to the
manufacturerrsquos instructions The universal primer set was used to amplify the 16S rDNA
from the extracted genomic DNA then the V3 region of bacterial 16S ribosomal DNA
(rDNA) was amplified with GC-clamp primer (Muyzer et al 1993) for DGGE PCR
amplification was performed in a thermocycler DGGE of the PCR products was carried
out in a DcodeTM
Universal Mutation Detection System) The 8 (wv) polyacrylamide
gels contains from 30 to 55 denaturing gradients Electrophoresis was conducted
using a 1timesTAE (Tris-Acetate-EDTA) buffer at 130V and 60degC for 5 hours After
electrophoresis the gel was stained with ethidium bromide in 1timesTAE buffer for 15
minutes and washed in 1timesTAE buffer for 10 minutes The fragments were visualized
under a UV transilluminator The richness of single band in DGGE gel picture
preliminarily showed that the bacterial diversity in mixed culture since the single band in
gel represents one bacterial species
23 16S rDNA clone library construction method
16S rDNA-DGGE fingerprinting has proven to be particularly useful as an initial
investigation into bacterial communities and is suitable for identifying the predominant
bacteria that comprise 1 or more of cells within a given sample In the 16s rDNA clone
17
library construction process the first step is the extratction of the total genomic DNA
then the genomic DNA of the mixed culture biofilm was used as template for PCR
amplification of approximately 1500 bp of 16S rDNA with universal primers The PCR
products of 16S rDNA were purified and inverted into pGEM-T Easy vector system
before they were transformed into competent E coli The transformed cells were spread
on Luria-Bertani (LB) plates containing the selective antibiotic ampicillin and X-Gal and
incubated overnight at 37 OC after the more than 80 inverted 16s DNA fragment E coli
colonies were picked up and inoculated in the 1-3 ml liquid LB medium and left to grow
for 16 hours The plasmid DNA were extracted and sequenced The sequences were
compared directly to all known sequences deposited in GenBank databases using the
basic local alignment search tool (BLAST)
24 Bioinformatics Analysis
16S rDNA sequencing results of more than 80 colones were queried against the
GenBank and Ribosomal Database Project (RDP) databases using BLAST and
SEQUATCH algorithms (Cole et al 2007 Wang et al 2007 Cole et al 2009) The
neighbor-joining trees were constructed with the Molecular Evolutionary Genetics
Analysis package (MEGA Version 4) based on 1000 bootstrap analysis (Tamura et al
2007)
25 Dominant bacterial species band in DGGE identification
The DGGE results gave the information of dominant bacterial richness of bacterial
species in mixed culture based on the theory that single band represents one bacterial
species which showed the diversity of bacterial community in the mixed culture of a
MFC 16S rDNA clone library results provided the whole picture of bacterial community
18
including the bacterial species name and bacterial species percentage in the mixed culture
The known pure bacterial species which have been sequenced can be used as markers to
identify dominant bacterial species in DGGE The whole experimental procedure is
similar to the procedure of the mixed culture DGGE steps The first step is to extract the
collect the mixed culture biofilm genomic DNA and pure bacterial species marker
genomic DNA Then the universal primer set was used to amplify the 16S rDNA from
the extracted genomic DNA After that the V3 region of bacterial 16S ribosomal DNA
(rDNA) was amplified with GC-clamp primer (Muyzer et al 1993) for DGGE The
subsequent steps were similar to the mixed culture DGGE experimental procedure The
dominant bacterial bands in DGGE were determined by comparisons of the single band
position in the mixed culture DGGE with single pure bacterial species location in DGGE
26 Dominant bacterial species isolation and power generation
The bacterial community of mixed culture in MFCs were predominantly composed
of Geobacter sp Arcobacter sp and Citrobacter sp Two dominant bacterial species
have been isolated
Citrobacter sp SX-1 was isolated from the anode biofilm by using a solid agar
medium containing NH4Cl 6mM KCl 2mM CH3COONa (electron donor) 30 mM
FeC6H5O7 (electron acceptor) 20mM minerals and vitamins (Lovley and Phillips 1988)
Single colonies were picked up after culturing on the plate for 36 hours at 30 degC and
transferred two times on the agar plate for purification Then the isolates grew in a liquid
medium solution in anaerobic tubes containing the same constituents as the solid
medium All isolation process was operated in a glove box anaerobic chamber (Coy
Laboratory Products Grass Lake MI)
19
Arcobacter sp SX-2 was isolated from the anode biofilm of a MFC fed with sodium
acetate by using the same method of isolation of pure bacterial species Citrobacter XS-1
except using Trypticase Soy Agar with 5 Sheep Blood solid (from American type
culture collection (ATCC)) agar medium instead of Citrobacter sp medium Single
colonies were picked up after culturing on the plate for 36 hours at 30 degC and transferred
twice on the agar plate for purification Then the isolates grew in a liquid medium
solution microaerobically
Geobacter sp have been trying to isolate by using three different methods They are
the colony pickup after different condition enrichment Goebacter medium isolation and
dilution to extinction by multiple channel mini MFCs isolation however the Geobacter
sp has not been isolated yet due to some limits so far But they provide valuable
information for further isolation of Geobacter sp
Single chamber MFCs were used to evaluate power generation by different isolates
The MFCs were constructed as described previously (Liu and Logan 2004) and modified
with 3 cm2 carbon cloth anodes and 7 cm
2 carbon clothPt cathodes The total liquid
volume of each MFCs was about 15 ml with electrode spacing about 17 cm All MFCs
were operated in an autoclaved closed plastic box and sterile cotton was attached to the
outer surface of the air cathodes to prevent contamination A MFCs without bacterial
culture was used as control MFCs were inoculated with 3 ml late exponential phase
cultures of SX-1 in the medium solution reported previously (Liu and Logan 2004)
3 Results and discussion
31 Power production by mixed culture MFCs
20
A polarization curve was used to characterize current as a function of voltage in the
MFCs The polarization curves are performed by varying a series of external resistances
The results are shown in Figure 21 With MFCs acclimated to 25 Ω external resistance
the maximum power density was achieved at 65Wm2 based on the polarization data At
this point the current density is 21mAcm2 The power density was three times higher
than previous report (20 Wm2) using phosphate buffer (Fan et al 2007) Power density
was also 2 times higher than (28 m Wm2) using a bicarbonate buffer ( Fan et al 2007)
32 Bacterial community structure assessed by PCR-DGGE
Bacterial DGGE profiles for higher power generating MFCs mixed culture setups
were relatively complex (more than 10 bands) However there are three brightly stained
bands and other less intense bands (Fig 2-2) These results indicated there were probably
three dominant bacterial species in mixed culture MFCs
010
020
030
040
050
060
070
2
3
4
5
6
7
04 05 05 06 07 08 10 11 13 15 21 22 23 24
Volta
ge (
V)
Pow
er
densi
ty (
Wm
2)
Current density (mAcm2)
Power
Voltage
power density 65wm2 at current
density208 mAcm2
Figure 2-1 Polarization curves of high power mixed culture MFCs
21
33 16S rDNA clone library results
A total of 83 colonies were sequenced for 16S rDNA clone library construction They
represented the bacterial community structure in mixed culture in MFCs The results
revealed that the predominant bacterial species in mixed culture are Geobacter sp (66
of the mixed culture) Arcobacter sp (12 of the mixed culture) and Citrobacter sp (11
of the mixed culture) Those three genuses comprised 88 of the total bacterial species
(Fig 23) This result is consistent with the DGGE result which showed that three bright
bands in the DGGE gel represented three dominant bacterial species in the mixed culture
The phylogenetic analysis of the colonies (83 colony sequences) is shown in Figure 24
Figure 22 DGGE result of high power generation mixed culture MFC (1 mixed
culture 2 pure culture control)
1 2
22
34 Dominant bacterial band in DGGE identification
Geobacter
66
Arcobacter
12
others
6
Pseudomonas
1
Citrobacter
11
Clostridium
2
Anaerovorax
2 Geobacter
Arcobacter
Citrobacter
Clostridium
Anaerovorax
Pseudomonas
others
Figure 23 Bacterial species and percentage of bacterial community of mixed
culture MFC lt 1 Alcaligensgt
lt1 Commamonasgt
lt1 Pseudomonas aeruginosagt
lt1 Klebsiellagt
lt11 Citrobactergt
lt1 Wolinellagt
lt12 Arobactergt
lt2 Anaeroboraxgt
lt2 Clostridium gt
lt1 unclear bacteriagt
lt66Geobactergt
100
100
85
100
100
82
99
87
100
002
Figure 24 Phylogenetic trees of bacterial species from mixed culture MFC
23
The dominant bacterial species including Goebacter sp Citrobacter sp and Arcobacter
sp in the mixed culture are shown in Figure 25 The upper band is Arcobacter sp and
Citrobacter sp is located in the bottom band of DGGE Geobacter sp is the middle one
The analysis of the bacterial community using denaturing gradient gel electrophoresis
(DGGE) of PCR-amplified 16S rRNA gene fragments and 16S rDNA clone library
construction method showed great phylogenetic diversity of mixed culture in MFCs with
the identification of sequences derived from bacteria of the taxa deltaproteobacteria
(Geobacter sp) gammaproteobacteria (Citrobacter sp) and epsilonproteobacteria
(Arcobacter sp) The result is consistent with the conclusion that among the isolated
exoelectrogens gammaproteobacteria and deltaprotebactia appear to be dominant among
others (Parot et al 2009 Liu et al 2010)
35 The dominant bacterial species isolation and power generation
Figure 25 Dominant bacterial species identification in DGGE
A B C D
A Citrobacter sp
B Geobacter sp
C Arobacter butzmeri
D mixed culture control
24
Two dominant bacterial species Citrobacter sp XS-1 and Arcobacter sp XS-2 have been
isolated and tested for power generation The power densities generated by the two
isolates are shown in the Fig 26 Current densities generated by isolate Citrobacter sp
and Arcobacter sp were 98 mAm2 and 20 mAm
2 respectively which were much less
than that (21Am2) generated by the mixed culture from which they were isolated
Figure 26 Current density generated by different dominant isolates
The maximum current density generated by mixed culture is much higher than the
one produced by pure bacterial species Citrobacter sp XS-1 and Arcobacter sp XS-2 in
the same structure MFCs Although the Geobacter sp in our mixed culture has not been
isolated a previous study showed that maximum current density generated by Geobacter
sp was 080 Am2
(Bond and Lovley 2003) The current density results seem partially
supporting the conclusion that mixed-culture generated higher power densities than their
pure-culture counterparts in MFCs (Jung and Regan 2007) This result suggests there
may be synergistic interactions within the anode bacterial communities
0
500
1000
1500
2000
2500
Mixed culture Citrobacter sp Arcobacter Sp
Cu
rre
nt
de
nsi
ty (
mA
m2)
25
Geobacter sp are strictly anaerobic bacteria which were enriched in most of the anode
biofilms (George 2005) Citrobacter species are facultative anaerobic bacteria (George
2005) which can be found in a wide variety of habitats including in soil water and
wastewater Arcobacter sp are micro-aerobic bacteria including both environmental
nonpathogens and opportunistic human pathogens They are able to grow in aerobic
conditions but in the optimal growth under micro-aerobic conditions (George 2005)
Arcobacter sp has been discovered in the MFCs by Fedorovich (Fedorovich et al 2009)
It is interesting that the mixed culture bacterial community composed of primarily these
three bacterial species has demonstrated to generate high power at non-strictly anaerobic
condition while leading dominant bacterial species Geobacter sp (66) is strict
anaerobic bacteria and generated powder in the MFCs only under strict anaerobic
condition (George 2005) Therefore it is possible that microaerobic bacteria such as
Arcobacter sp and facultative anaerobic bacteria Citrobacter sp can facilitate to create
anaerobic condition for Geobacter sp when they utilized oxygen for growing in the
mixed culture Syntrophic communities study have showed that anaerobic bacteria and
methanogenic archaea form compact microbial structures that operate like an organ rather
than a set of microorganisms functioning independently (Stams and Plugge 2009) Some
substrates have been degradeted within these communities while they are not able to be
fermented by individual species alone interspecies electron transfer also have been
presented in these communities (Stams and Plugge 2009) Summers et al (2010) also
discovered that direct exchange of electrons happened within coculture of Geobacter
metallireducens and Geobacter sulfurreducens Therefore we believe that there are
26
more complex synergistic interactions between different bacteria species in mixed culture
in the MFCs
Acknowledgements
The authors acknowledge support from the US National Science Foundation
(CBET 0955124) We also thank Keaton Lesnik for the review of this manuscript
27
Chapter 3
New Exoelectrogen Citrobacter sp SX-1 Isolated from a Microbial Fuel Cell
Shoutao Xu and Hong Liu
Published in
Journal of Applied Microbiology 111(5)1108-1115 (2011)
ABSTRACT
Exoelectrogenic bacterial strain SX-1 was isolated from a mediator-less microbial fuel
cell by conventional plating techniques with ferric citrate as electron acceptor under
anaerobic condition Phylogenetic analysis of the 16S rDNA sequence revealed that it
28
was related to the members of Citrobacter genus with Citrobacter sp sdy-48 being the
most closely related species The bacterial strain SX-1 produced electricity from citrate
acetate glucose sucrose glycerol and lactose in MFCs with the highest current density
of 205 mA m-2
generated from citrate Cyclic voltammetry analysis indicated that
membrane associated proteins may play an important role in facilitating the electrons
transferring from bacteria to electrode This is the first study that demonstrates that
Citrobacter species can transfer electrons to extracellular electron acceptors Citrobacter
strain SX-1 is capable of generating electricity from a wide range of substrates in MFCs
This finding increases the known diversity of power generating exoelectrogens and
provided a new strain to explore the mechanisms of extracellular electron transfer from
bacteria to electrode The wide range of substrate utilization by SX-1 increases the
application potential of MFCs in renewable energy generation and waste treatment
Keywords Microbial fuel cell Exoelectrogen Citrobacter sp SX-1 Extracellular
electron transfer
29
Introduction
Microbial Fuel Cells (MFCs) technology has become an active research area
recently as a promising approach for renewable energy generation wastewater treatment
and bioremediation (Rabaey and Verstraete 2005 Lovley 2006 Fan et al 2008 Logan
2009 Liu et al 2010) The key feature of MFCs system is the microbe-catalyzed electron
transfer from organic matter to anodes Many studies have shown the presence of diverse
bacterial communities on MFCs anodes (Reimers et al 2001 Liu et al 2004b Kim et al
2005) Over 20 exoelectrogens ie microbes that can transfer electrons exocellularly to
electrodes have been reported in the past 10 years (Logan 2009 Liu et al 2010) The
exoelectrogens so far belong to diverse genetic groups including alphaproteobacteria
(Rhodopseudomonas Ochrobactrum and Acidiphilium) (Malki et al 2008 Xing et al
2008 Zuo et al 2008) betaproteobacteria (Rhodoferax) (Chaudhuri et al 2003)
gammaproteobacteria (Shewanella Pseudomonas Klebsiella Enterobacter and
Aeronomas) (Kim et al 2002 Pham et al 2003 Rabaey et al 2005 Bretschger et al
2007 Zhang et al 2008 Chung et al 2009 Rezaei et al 2009) deltaproteobacteria
(Geobacter Geopsychrobacter Desulfuromonas and Desulfobulbus) (Bond et al 2002
Bond and Lovley 2003 Holmes et al 2004ab 2006) Epsilonproteobacteria (Arcobacter)
(Fedorovich et al 2009) Firmicutes (Clostridium and Thermincola) (Park et al 2001
Wrighton et al 2008) Acidobacteria (Geothrix) (Bond and Lovley 2005) and
actinobacteria (Propionibacterium) (Wang et al 2008) A wider range of
exoelectrogenic species are expected to be discovered
Three mechanisms have been proposed for exocellular transport of electrons by
exoelectrogens without artificial mediators Some exoelectrogenic bacteria can transfer
30
electrons to electrodes through soluble redox compounds excreted by microorganisms
(Bond and Lovley 2005 Rabaey et al 2005) and others can directly transfer electrons to
anodes via outer cell membrane proteins (Myers and Myers 1992) Recently more and
more evidence supports the involvement of bacterial nanowires in extracellular electron
transport (Reguera et al 2005 2006 Gorby et al 2006) In spite of the discovery of
many bacterial species that can transfer the electrons to electrode without the need of
artificial mediators the investigation of extracellular electron transfer mechanisms was
mainly focused on a few species such as those from Geobacter and Shewanella genera
(Kim et al 1999 2002 Reguera et al 2005 2006 Gorby et al 2006) The electron
transfer mechanisms for many of the isolated exoelectrogens species are still not well
studied
In the present study we isolated a new exoelectrogen strain SX-1 from a MFC a
strain phylogenetically related to Citrobacter sp Power generation from various carbon
sources by this strain was evaluated using single chamber MFCs Plausible extracellular
electron transfer mechanisms were also discussed based on the characterization of anodic
biofilms by cyclic voltammetry (CV)
Materials and methods
Bacterial strain SX-1 isolation
Bacterial strain SX-1 was isolated from the anode biofilm of a MFC fed with sodium
acetate operated in fed-batch mode over a period of six months The original source of
the inoculum is wastewater from a local waste water treatment plant Bacterial cells were
released from the carbon cloth anode by shaking the cloth in a sterile flask with 15 mL
sterile 1 NaCl solution and glass beads (2 mm diameter) The suspension was then
31
serially diluted from 10 times to 105
times and plated on a petri dish with a solid agar
medium containing NH4Cl 6mM KCl 2mM CH3COONa (electron donor) 30 mM
FeC6H5O7 (electron acceptor) 20mM minerals and vitamins (Lovley and Phillips 1988)
Single colony was randomly selected after culturing on the plate for 36 hours at 30 degC
and purified on a new agar plate following a procedure reported previously (Chung and
Okabe 2009) Then the isolate was picked up and grew in a liquid medium solution in
anaerobic tubes containing the same constituents as the solid medium for further analysis
All isolation process was operated in a glove box anaerobic chamber (Coy Laboratory
Products Grass Lake MI)
16S rDNA sequencing and phylogenetic analysis
Bacterial genomic DNA was extracted from the isolated strain SX-1 using the DNeasy
tissue Kits (Qiagen Valencia CA) according to the manufacturerrsquos instructions The 16S
rDNA was amplified by PCR using a pair of universal primers 27F (5rsquo-
AGAGTTTGATCMTGGCTCAG-3rsquo) and 1492R (5rsquo-GGTTACCTTTGTTACGACTT-
3rsquo) (Park et al 2001) The conditions used for PCR were described previously
(Ferna ndez et al 1999) PCR products were purified ligated and cloned according to a
procedure described in a previous report (Xing et al 2008) Plasmids were isolated from
randomly selected clone colonies with a QIAprep Spin Miniprep kit (Qiagen Valencia
CA) and three plasmid inserts were then sequenced in both directions using an ABI 3730
DNA sequencer (Applied Biosystems Carlsbad CA)United States) Due to the identical
16S rDNA sequence of the three colones only one was queried against the GenBank and
Ribosomal Database Project (RDP) databases using BLAST and SEQUATCH algorithms
(Cole et al 2007 Wang et al 2007 Cole et al 2009) The 16S rDNA sequence of
32
strain SX-1 and closely related type strains were aligned using CLUSTALX software
(Thompson et al 1997) The neighbor-joining trees were constructed with the Molecular
Evolutionary Genetics Analysis package (MEGA Version 4) based on 1000 bootstrap
analysis (Tamura et al 2007)
SEM
The morphologies of SX-1 cells growing overnight in anaerobic tubes were examined
with a scanning electron microscope (FEI Quanta 600 FEG FEI Company Hillsboro
OR) The suspended cells were fixed in a 25 glutaraldehyde and 01M phosphate
buffer solution and dehydrated with a graded ethanol series from 30 to 100 After
dehydration the samples were dried in a critical point dryer and then sputter-coated with
AuPd for SEM examination (Liu and Logan 2004)
Evaluation of power generation by SX-1 in MFCs
Single chamber MFCs were used to evaluate power generation by SX-1 using various
substrates The MFCs were constructed as described previously (Liu and Logan 2004)
and modified with 3 cm2 carbon cloth anodes and 7cm
2 carbon clothPt cathodes The
total liquid volume of each MFCs was about 15 ml with electrode spacing about 17 cm
All MFCs were operated in an autoclaved closed plastic box and sterile cotton was
attached to the outer surface of the air cathodes to prevent contamination A MFC
without bacterial culture was used as control MFCs were inoculated with 3 ml late
exponential phase cultures of SX-1 in the medium solution reported previously (Liu and
Logan 2004) Seven substrates including citrate acetate glucose sucrose glycerol and
lactose were evaluated individually for power generation in a fed-batch mode in a
temperature control chamber (30 plusmn2degC) For each substrate at least 8 batches were run to
33
investigate the effect of biofilm formation on current output at fixed external resistance of
1kΩ and the same initial substrate concentration of 30 mM Phosphate buffer (100mM)
was used to maintain the solution pH and conductivity The MFCs with sodium citrate as
electron donors were also examined for maximum power output by varying the external
resistance from 20 kΩ to 4 kΩ A data acquisition system was used to record voltage data
during the MFCs operation (Keithley 2700 Keithley Instruments Inc Cleveland OH) It
took about 15 to 30 minutes for the MFCs to stabilize depending on the external
resistance At each resistance we collected at least five data at the steady condition to
make the polarization curves Averaged voltages were used to calculate the power density
(mW m-2
) according to P=IVA where I was the current V was voltage and A was cross-
sectional area of the anode
CV analysis
CV was used to characterize the oxidation and reduction reactions on the anodic surface
of the MFCs fed with citrate and operated at 7 KΩ This external resistance was selected
because the maximum power density was obtained at this resistance based on the
polarization experiment We assumed the biofilm on the anode of MFCs were well-
developed when stable power output was obtained after 3 batches of operation The
MFCs were then used directly for CV analysis at four current generating stages of the
fourth batch (1) initial exponential current increasing stage (middle point of stage about
4 h after media change) (2) current plateau stage (middle point of stage about 8 h after
media change) and (3) current decreasing stage (middle point of stage about 16h after
media change) (4) right after the complete replacement of medium solution (Figure 6)
The anode was used as working electrode the cathode as counter electrode and an
34
AgAgCl electrode was selected as the reference The CV curves were scanned from 200
to -600 mV at a rate of 5 mV s-1
using a potentiostat (G300 Gamry Instrument
IncWarminster PA) Control experiment was also conducted using new anode (without
biofilm) and new medium solution
Nucleotide sequence accession number
The 16S r DNA sequence determined in this study has been deposited in the GenBank
database under accession number HQ845373
Results
Identification of the strain SX-1
An almost-complete 16S rDNA sequence (1475 bp) of strain SX-1 was obtained and
subjected to comparative analysis with the 16S rDNA of closely related reference strains
A phylogenetic analysis revealed that strain SX-1 should be assigned to the genus
Citrobacter and was most closely related to Citrobacter sp sdy-48 EJ463782 (990
sequence similarity) and Citrobacter sp yy-21 FJ463779 (975 sequence similarity)
These three strains formed a distinct sub cluster in the neighbor-joining in which the new
isolate and Citrobacter sp sdy-48 EJ463782 formed a distinct subline (Figure 31)
The SEM image illustrated that the strain SX-1 is a rod-shaped bacterium 03-06 microm
wide and 12-17 microm long (Figure 32) Strain SX-1 colonies on nutrient agar appeared 2-
3 mm in diameter smooth moist opaque with a shiny surface after 5 days incubation
under anaerobic condition The fact that strain SX-1 grew both aerobically and
anaerobically using FeC6H5O7 as electron acceptor suggests that the strain is facultatively
anaerobic Strain SX-1 showed robust growth on citrate as a carbon source These D
35
properties were similar to those of genus Citrobacter described in Bergeyrsquos Manual of
Systematic Bacteriology (George 2005)
Figure 31 phylogenetic tree of strain SX-1 and closely related species based on 16S
rDNA sequences The tree was constructed using the neighbor-joining method
Figure 32 SEM image of planktonic cells of Citrobacter sp SX-1
Strain SX-1
Citrobacter sp sdy-48 (FJ463782)
Citrobacter sp yy-21 (FJ463779)
Citrobacter koseri E639 (ATCC 25408)
Citrobater koseri CDC 3613-63(AF025372)
Citrobacter koseri CDC 8132-86(AF025366)
Salmonella enterica SL483 (CP001138)
Salmonella enterica AKU12601(AY696668)
Shigella sonnei Ss046(CP000038)
E coli C2 (AF403733)
E coli E24377A(CP000800)88
100
100
76
44
92
53
36
0001
1 microm
36
Electricity production by strain SX-1 in MFCs
The isolated strain SX-1 was first tested for its ability to generate current from sodium
citrate at 1kΩ Current gradually increased to a maximum 50 mA m-2
after cell
inoculation and then decreased (Figure 33) After the MFCs was refilled with new
substrate solution the current recovered rapidly and reached a higher level than the first
batch After 4 batches operation the maximum current output of each batch became
stable The highest current density achieved in the MFCs with Citrobacter sp SX-1 at 1
KΩ was 98 mA m-2
(Figure 33)
Figure 33 Electricity generation by Citrobacter sp SX-1 in a single chamber MFCs
with sodium citrate (30 mM) as substrate at 1 KΩ
0
20
40
60
80
100
120
0 50 100 150 200
Curr
ent
den
sity
(m
A m
-2)
Time (h)
37
Current generation by SX-1 from other substrates including glucose lactose sodium
acetate glycerol and sucrose were also assessed at an external resistance of 1KΩ
(Figure 34) When repeatable cycles of current output were obtained for these substrates
glycerol generated the highest maximum current density of 58 mA m-2
followed by
lactose and sucrose with 29 mA m-2
and 27 mA m-2
respectively Glucose and acetate
produced the lowest maximum current density of 96 and 43 mA m-2
respectively These
results indicated that strain SX-1 can utilize a wide range of substrates for electricity
generation in MFCs but with different power generation potentials
Figure 34 Electricity generation of Citrobacter sp SX-1 using different substrates in
single chamber air-cathode MFCs at 1 KΩ (The error bars stand for the standard
deviation of 3 replicates)
0
10
20
30
40
50
60
70
Glucose Lactose Glycerol Acetate Sucrose
Curr
ent
den
sity
(m
A m
-2)
38
Polarization experiment was further conducted to determine the maximum power
density generated by strain SX-1 in the single chamber MFCs with citrate as substrate A
maximum power density of 881 mW m-2
was obtained at current density of 205 mA m-2
at an external resistance of 7 KΩ (Figure 35)
Figure 35 Power and voltage generation by Citrobacter sp SX-1 as a function of
current density using sodium citrate (30 mM) as substrate The error bars stand for the
standard deviation of the 3 voltagepower outputs obtained in three MFCs
20
30
40
50
60
70
80
90
100
01
02
03
04
05
06
07
80 110 140 170 200 230
Po
wer
den
sity
(m
W m
-2)
Vo
ltag
e (V
)
Current density (mA m-2)
Voltage
Power
39
Figure 36 Four current generating stages for CV analysis current increasing stage (stage
1) current plateau stage (stage 2) current decreasing stage (stage 3) and right after the
replacement of medium solution (stage 4) The MFCs was operated at 7 KΩ with sodium
citrate (30 mM) as substrate
Cyclic Voltammetry
To determine the presence of redox active compounds produced by SX-1 and the
location of these compounds CV scan of the anodic biofilms at four current output stages
and the supernatant of MFCs medium solution at the end of the batch experiment were
performed Reduction peaks at range of -310 ndash -350 mV and oxidation peaks at range -
100 ndash -150 mV were observed at all current generating stages (Figure 36 Figure 37A)
suggesting the presence of redox active compounds may involve in extracellular electron
transfer by Citrobacter sp SX-1 The amplitude of the peaks increased according to the
growth stage of the batch and the highest peaks were present after the current plateau
stage which indicated the redox active compounds mainly were secreted in the current
plateau stage While the current density at stage 3 (deceasing stage) was lower than that
at stage 2 (current plateau stage) the redox peak at stage 3 was higher than that in stage 2
0
50
100
150
200
250
50 55 60 65 70 75 80 85
Curr
ent
den
sity
(m
A m
-2)
Time (h)
A
A
Stage 1
Stage 2 Stage 3
Stage 4
40
A plausible explanation for this pattern was that redox active compounds were
continuously being secreted and accumulated after the current peak was reached
resulting in more redox compounds present after the plateau stage But since most of the
carbon source had already been being used up many of the redox compounds at this
stage had not been as active as those at current plateau stage due to much less electrons
were available to be transferred The bigger peak amplitude in stage 4 than in stage 1
indicates that the redox active compounds were continuously being secreted and
accumulated in the biofilm even after stable power was generated at 7 kΩ The slightly
change of the peak locations at different stages was possible due to the solution chemistry
change during the current generation process or the slightly location change of reference
electrode during the measurement (Figure 37B) When the medium solution in the MFCs
was replaced by fresh medium at the end of the batch the oxidization and reduction
peaks were still presented in the CV (Figure 37B) but no peaks were observed when the
supernatant and fresh mediums were tested in a MFCs with no biofilm (Figure 37C)
These results suggest the compounds involved in the electron transfer were located not in
the supernatant (solution) but in the biofilm
41
Figure 37 Cyclic Voltammetry analysis of Citrobacter sp SX-1 biofilms at (A) current
increasing stage current plateau stage and decreasing stage (end of the batch) (B) end of
the batch and replaced with fresh medium and (C) controls
-40E-04
-30E-04
-20E-04
-10E-04
00E+00
10E-04
20E-04
30E-04
-060 -050 -040 -030 -020 -010 000 010 020
Curr
ent
(A)
Potential (V) vs AgAgCl
Current plateau stage
Current increasing stage
Current decreasing stage
-40E-04
-30E-04
-20E-04
-10E-04
00E+00
10E-04
20E-04
30E-04
-060 -050 -040 -030 -020 -010 000 010 020
Curr
ent
(A)
Potential (V) vs AgAgCl
Current decreasing stage
Biofilm with fresh medium
-40E-04
-35E-04
-30E-04
-25E-04
-20E-04
-15E-04
-10E-04
-50E-05
00E+00
-060 -050 -040 -030 -020 -010 000 010 020
Curr
ent(
A)
Potential (V) vs AgAgCl
Supernatant
Fresh medium
A
B
C
B C
42
Discussion
Citrobacter species belonging to Gammaproteobacteria Enterobacteriales
Enterobacteriaceae are facultative anaerobic bacteria (George 2005) They can grow with
oxygen as an electron acceptor but also with a variety of electron accepters such as Fe (III)
in the absence of oxygen (George 2005) Citrobacter species can be found in a wide
variety of habitats including in soil water and wastewater Several Citrobacter species
have been studied as important bioremediation bacterium for heavy metal removals
sulfate reduction phenol degradation and chlorophenol degradation (Macaskie et al 1995
Narde et al 2004 Qiu et al 2009) Citrobacter species have also been found on the
anodic biofilm of a diesel-degrading MFCs (Morris et al 2009) However there was no
study reported that Citrobacter species can transfer electrons to extracellular electron
acceptors This finding that Citrobacter strain SX-1 can generate electricity in MFCs
increased the diversity of power generating exoelectrogens and provided a new strain to
explore the mechanisms of extracellular electron transfer from bacteria to electrode
Among the isolated exoelectrogens gammaproteobacteria and deltaprotebactia appear to
be dominant among others (Parot et al 2009 Liu et al 2010) The isolated strain
Citrobacter sp SX-1 phylogenetically belongs to gammaproteobacteria which provides
another evidence to support this observation
Most exoelectrogens utilize a limited range of substrates for power generation For
example Shewanella oniedensis MR-1 can oxidize lactate under anaerobic condition but
cannot utilize acetate for electricity generation (Kim et al 2002) Geobacter
sulfurreducens can completely oxidize acetate for power generation but it cannot utilize
simple sugars (Bond and Lovley 2003 Caccavo et al 1994)) In comparison strain SX-1
43
not only can oxidize citrate for power generation but also utilize a wide range of
substrates for power generation including glycerol glucose lactose sucrose and acetate
Interestingly the higher current density generated by strain SX-1 from glycerol a main
by-product of biodiesel production process suggests that strain SX-1 may be potentially
used for harvesting energy from biodiesel wastes using MFCs However the current
density generated by strain SX-1 (205 mA m-2
) is lower than that (805 mA m-2
at 1 kΩ)
generated by the mixed culture from which the SX-1 was isolated indicating the
existence of other higher power generating bacteria andor complex ecology in the mixed
culture community
Understanding mechanisms of microbial extracellular electron transfer is critical for
enhancing the electron transfer rate from bacteria to electrode through metabolic or
genetic engineering of exoelectrogens Since the spent supernatant solution of MFCs run
with SX-1 showed no redox properties the redox compounds produced by SX-1 may
have been retained in the biofilm Alternatively SX-1 may utilize one or both of the
other two known extracellular electron transfer mechanisms For example it is possible
that strain SX-1 transfers electrons to the anode via cell-bound outer membrane proteins
The two distinct oxidation and reduction peaks observed at -100mv- -150 mV and -310--
350 mV of the SX-1 biofilm have a midpoint potential range from -205mV to -250mV
which is close to the redox potential of some c-type cytochromes (Field et al 2000 Meitl
et al 2009) which are well-known to play an important role in extracellular electron
trasnfer (Eggleston et al 2008 Field et al 2000 Meitl et al 2009) Field et al (2000)
reported that the c-type cytochromes CymA had the midpoint potentials of -229 mV
Meitl et al (2009) and Eggleston et al (2008) reported that the c- type cytochromes
44
OmcA in Shewanella oneidensis MR-1 had the midpoint potentials of -201 mV and -208
mV
While itrsquos possible that SX-1 may also have conductive appendages to enhance the
extracellular transfer electron similar to the nanowires discovered in some Geobacter
and Shewanella species (Reguera et al 2005 2006 Gorby et al 2006) further
investigation on the presence of these appendages on the cell surface of SX-1 is needed
Acknowledgements
The authors acknowledge support from the US National Science Foundation
(CBET 0955124) We also thank Teresa Sawyer for her helps with the SEM analysis and
Jeremy Chignell and Yanzhen Fan for their review of this manuscript
45
Chapter 4
Enhanced Performance and Mechanism Study of Microbial Electrolysis Cells
Using Fe Nanoparticles Decorated Anodes
Shoutao Xu Hong LiuYanzhen Fan Rebecca Schaller Jun Jiao Frank Chaplen
Published in
Applied Microbiology and biotechnology 93(2)871-880 (2012)
46
ABSTRACT
Anode properties are critical for performance of microbial electrolysis cells (MECs) In
the present study Fe nanoparticle modified graphite disks were used as anodes to
investigate the effects of nanoparticles on the performance of Shewanella oneidensis MR-
1 in MECs Results demonstrated that average current densities produced with Fe
nanoparticle decorated anodes were up to 56-fold higher than plain graphite anodes
Whole genome microarray analysis of the gene expression showed that genes encoding
biofilm formation were significantly up-regulated as response to nanoparticle decorated
anodes Increased expression of genes related to nanowires flavins and c-type
cytochromes indicate that enhanced mechanisms of electron transfer to the anode may
also have contributed to the observed increases in current density The majority of the
remaining differentially expressed genes were associated with electron transport and
anaerobic metabolism demonstrating a systemic response to increased power loads
Keywords Microbial electrochemical system microbial fuel cell microbial electrolysis
cell nanotechnology differential gene expression DNA microarray
47
Introduction
Microbial electrochemical systems (MESs) have been intensively studied since Lewis
achieved practical advances in this field (Logan 2007) however they attracted much
research attention in recent years due to their promising applications in renewable energy
generation bioremediation and wastewater treatment In a MES microorganisms
interact with electrodes via electrons catalyzing oxidation and reduction reactions at the
anode and the cathode The most-described type of MESs is microbial fuel cells (MFCs)
in which useful power is generated from electron donors typically biodegradable organic
materials (Logan et al 2006) Various novel MESs have recently been developed to
produce hydrogen (microbial electrolysis cells (MECs)) (Liu et al 2005 Logan et al
2008 Rozendal et al 2006 Schaller et al 2009) to power an autonomous sensor or
sensor network (Ringeisen et al 2006) to reduce bioremediation targets (Lovley 2006
Shea et al 2008) Sukkasem et al 2008) and to desalinate water (Cao et al 2009) The
key feature shared by these systems is the microbe-catalyzed electron transfer from
organic matter to electrodes (anodes) (Rabaey et al 2004) Enhancing the anodic current
output which highly depends on the performance of the electrodes is critical for the
successful application of all these processes (Logan et al 2007 Park and Zeilus 2002
2003)
Nanomaterials have received much attention from researchers in the context of
microbiology due to their unique physical electrical and chemical properties which
facilitate the study of interactions between bacteria and surfaces (Liu 2006) Previous
studies have demonstrated that electrodes decorated with different nanostructures such
as carbon nanotubes nanostructured polyanilinetitanium dioxide composite and titania
48
nanotubes have the potential to enhance power generation in MFCs (Morozan et al 2007
Qiao et al 2008 Quan et al 2005) Furthermore our recent study shows nanoparticle
(NP) decorated anodes greatly increased the electrochemical electron transfer rate in
MFCs (Fan et al 2011) However Fe NPs as an ideal candidate for decorating electrodes
because of respectively low price and high conductivity compared to other materials has
not been focused to study in MECs
S oneidensis MR-1 an important electrochemically active bacterial strain has been
exploited for the electricity generation in MFCs (Logan et al 2007 Logan 2009 Park
and Zeikus 2002) The availability of genome sequence for this strain makes it possible to
use transcriptome assays to globally measure the responses to different growth conditions
and environmental stresses (Murray et al 2001) S oneidensis MR-1 gene expression
response to heat shock cold shock Cr (VI) and U (VI) reduction chromate stress and
iron and acid tolerance have been studied in the past several years (Bencheikh-Latmani et
al 2005 Brown et al 2006 Gao et al2006 Gao et al 2004 Yang et al 2008) However
no studies have been focused on the Shewanella gene expression response to NPs in
MESs
The mechanism of increased electron transfer rate exhibited by nanoparticle decorated
electrodes is not well understood yet In the present study graphite disks decorated with
Fe NPs were used as anodes to explore the effects of nanostructures on current generation
in a multi-anode MECs DNA microarrays were utilized to investigate differences in the
global gene expression profile of S oneidensis MR-1 grown on plain versus Fe NPs
decorated anodes
Materials and methods
49
Bacterial cultures
S oneidensis MR-1 was purchased from American Type Culture collection (ATCC
700550) and stored as freezer stocks at -80degC Stock cells of S oneidensis MR-1 was
grown aerobically using a medium containing 30 gL Trypticase Soy Broth (BD 211825
Becton Dickinson Company Franklin lakes NJ) at 30degC Cells in early stationary
growth phase (optical density of 15-16 at 600nm (Spectrophotometer UV-1700
Shimadzu Columbia MD) were washed two times then injected into the chamber of the
MECs for current production Trypticase Soy Broth medium with 30 mM sodium lactate
as electron donor was used in MECs Phosphate buffer (100mM) was used to maintain
the solution pH 7 and solution conductivity at 15 mScm
Characterization of nanostructured anodes
Superfine isomolded graphite disks (16 cm x 05 cm GraphiteStorecom Inc) were
polished with ultrafine sand paper (2000 grit 3M Company) as the base for the NP
decorated and control anodes Fe NPs decorated anodes were fabricated by thermal
annealing of metal thin film coated graphite disks Thin films of Fe were firstly deposited
on the polished graphite disk by using sputter coating for 95 min and the samples then
were annealed in the Chemical vapor deposition (CVD) at 750degC for 20 min to form Fe
NPs Fe-NPs were produced with a diameter of 200plusmn100 nm and a density range
(distance between NPs) of 200plusmn100 nm with a coating time of 95 min SEM images of
Fe NPs decorated anode and control surfaces were shown in the Figure 41
50
Figure 41 SEM images of Fe NPs decorated anodes (A) Graphite disk control (B) Fe
Nanoparticle decorated
Multiple channel MECs construction and operation
A MECs with removable multiple anodes with each effective anode area 07 cm2
was
constructed and used to evaluate the effects of nanostructure on current density of
according to a previous report (Fan et al 2011) The cathode was made of wet-proof
(30) carbon cloth (type B E-TEK Division Inc USA) coated with PtC (20 E-TEK
Division Inc USA) following a previously reported procedure (Liu et al 2005) The
final platinum loading was 05 mgcm2 per projected cathode area The size of cathode
(150 cm2) was 25 times larger than the total effective area of the 8 graphite disk anodes
(56 cm2) to prevent cathode limitations on the performance of the MECs system All
MECs with Fe-NP decorated anodes and control anodes were tested for current
generation for 24 h with medium (without bacteria) before injecting the bacterial cells in
order to determine whether the Fe-NP decorated anodes demonstrate chemical current
generation compared to control anodes All the testing anodes were set up in the MECs
B
500 nm
A
500 nm
51
at once so that the S oneidensis MR-1 culture was applied to all of them simultaneously
Short electrode spacing (17 cm) was used in the MECs design to reduce the internal
resistance A voltage of 06 V was applied the MECs for current generation after the cells
of S oneidensis MR-1 were inoculated into MECs at early stationary phase A multimeter
with a data acquisition system (2700 Keithly USA) was used to monitor the current
change by measuring the voltage (every 300 seconds) drop through a resistor at 300 ohm
Fig 4 2 Effects of Fe-NP decorated anodes on the current density in MECs Control is
plain graphite disk anode Fe-NP is the anode with Fe-NP decoration
Microarray analysis
Biofilms for whole gene microarray analysis were aseptically removed from the plain
and Fe-NP decorated anodes of MECs after 110 hours further incubation at 30oC when
current density obviously started to decease Total RNA was extracted using Trizol
(Ambion inc AustinTX) following the manufacturerrsquos instructions Integrity of the
52
RNA samples was confirmed using the Agilent 2100 Bioanalyzer (Agilent Technology
Inc Palo Alto CA) The same amount of RNA (200 ng) from each sample was used for
further analysis after amplification by using the MessageAmpTM
II-Bacteria Prokaryotic
RNA Amplification kit (Ambion AustinTX) according to the manufacturerrsquos
instructions All of the 4295 genes (resource from NCBI) represented on the S oneidensis
MR-1 whole-gene microarrays were ordered from Roche NimbleGen Inc (Madison WI)
Biological triplicates of biofilms on the Fe-NP decorated anodes and control were
analyzed respectively cDNA synthesis labeling and hybridization were carried out by
the CGRB Core Laboratories at Oregon State University DNASTAR ArrayStarTM
3
software was used to identify genes that were up- or down-regulated more than 2-fold
when grown on the nanoparticle modified anodes using the unpaired two sample t-test
with a cutoff p-value of 005 The complete microarray data set generated in this study is
deposited for public access in the Gene Expression Omnibus
(httpwwwncbinlmnihgovgeo) under accession number GSE31535
Results
Enhancement of current generation by using NP decorated anodes in MECs
The current density generated by Fe-NP decorated anode MECs with S onidensis MR-1
increased to approximately 43 microAcm2 20 h following inoculation and then slowly
decreased Figure 42 shows that the maximum current density achieved by NP
decorated anodes was 83 times higher than that (51 microAcm2) generated by the control
(plain graphite disk) The average current density improvement of 110 hours was 59
times of that generated by the control The current densities generated in MECs in the
53
absence of bacteria were negligible (03 microAcm2) which demonstrated that the current
enhancement observed with the Fe-NP decorated anodes was biologically-derived
Global transcriptome analysis
Whole-genome DNA microarrays were used to attain a comprehensive general
overview
of the transcriptional response to Fe-NP decorated anodes by S oneidensis MR-1 in MEC
Of a total 4295 genes assayed 392 (188 induced and 204 repressed) exhibited significant
(P lt 005) differential expression at a 2-fold (log (expression ratio 1) level in 3
replicates in response to Fe-NPs These
total gene numbers present 9 of the 4295 open
reading frames (ORFs) presented on the array
Fig 43 Differentially expressed genes grouped by functional classification according to
the TIGR S oneidensis genome database (wwwtigrorg) Columns 1 amino acid
biosynthesis 2 biosynthesis of cofactors prosthetic groups and carriers 3 cell
envelope 4 cellular processes 5 central intermediary metabolism 6 DNA metabolism
7 energy metabolism 8 fatty acid and phospholipid metabolism 9 other categories 10
protein fate 11 protein synthesis 12 purines pyrimidines nucleosides and nucleotides
13 regulatory functions 14 signal transduction 15 transcription 16 transport and
binding proteins 17 unknown function 18 hypothetical proteins
Up-Regulated
-75
-50
-25
0
25
50
75
100
125
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18
Down-Regulated
Num
ber
of
Gen
es
54
Figure 43 summarizes the overall genomic response of S oneidensis MR-1 to Fe-NP
decorated anodes by grouping the differentially expressed genes into their functional role
categories based on The Institute for Genomic Researchrsquos annotation (Version 8) of the
MR-1 genome sequence The wide distribution of putative functional roles attributed to
the differentially expressed genes indicated the extent of the molecular response of
Soniedenis MR-1 to the NP decorated anodes Among genes with defined function a
large number of up-regulated genes were associated with cellular processes (group 4)
energy metabolism (group 7) other categories (group 9) and hypothetical proteins (group
18) While those genes encoding proteins involved in amino acid biosynthesis (group 1)
cellular processes (group 4) substrate transport (group 16) and hypothetical proteins
(group 18) were among the most down-regulated genes Two groups of genes that were
the most noticeable among the functional gene groups genes related to energy
metabolism (group 7) and genes related to amino acid biosynthesis (group 1) The ratios
of up-regulated genes to down-regulated genes in these functional groupings were much
higher than other functional gene groupings there were 21 up-regulated genes and 7
down-regulated genes in energy metabolism gene group and 2 up-regulated and 22 down
regulated genes in the amino acid biosynthesis group
Genes related to biofilm formation
In order to determine whether there was a correlation between the observed biofilm
enhancements by NP decorated anodes and the genes related to biofilm formation the
significant modified genes related to biofilm formation were examined and summarized
in the Table 41 These included SO_3223 and SO_3228 genes encoding the flagellum
proteins (Thormann et al 2004) which has critical impacts on initial attachment to the
55
surface as swimming motility functions were up-regulated over 10 and 09 fold
respectively The gene clusters SO_4178-SO_4180 which are thought to be essential for
formation of a three-dimensional biofilm were up-regulated from 12 to 13 fold
(Thormann et al 2004) In addition the genes SO_4105 encoding IV pilus which is
known to represent a critical factor for biofilm formation on a biotic surfaces (Thormann
et al 2004) were also induced 08 fold (Table 41) The genes related to flagellum
motility (SO_4286 SO_1529 and SO_1530) were also induced slightly (Thormann et al
2004)
Table 41 Expression levels of genes related to biofilm formation
Locus Tag Gene product descriptions Fold change
SO_3228 flagellar basal-body MS-ring and collar protein FliF 10
SO_3223 flagellar hook-length control protein FliK 09
SO_4103 MshA minor pilin protein MshD 05
SO_4105 MSHA major pilin protein MshA 08
SO_4178 expressed protein of unknown function MxdC 12
SO_4179 inner membrane family 2 glycosyltransferase MxdB 13
SO_4180 diguanylate cyclase-like protein MxdA 10
a fold changes were presented after log2
b positive number represented up-regulated and negative represented down-regulated
Genes related to energy metabolism
A closer consideration of energy metabolism genes was undertaken as anaerobic
metabolism for electron generation and electron transport functions may play critical
roles in enhanced current density generation in MECs Genes associated with energy
56
metabolism and with significantly modified gene expression levels are summarized in the
Table 42 Several of the up-regulated genes were related to formate dehydrogenase
proteins Most notably four genes related to formate dehydrogenase were up-regulated
more than 2-fold SO_0102 SO_0103 SO_0104 and SO_0107 The other genes
expression related to formate dehydrogenase proteins SO_4513 SO_4514 SO_4515 also
significantly increased Another interesting gene is that encoding the cytochrome c
oxidase protein gene SO_4609 which was up-regulated 11 fold while the gene
SO_0479 related periplasmic octaheme cytochrome c MccA was down-regulated
However SO_1519 the gene encoding lactate dehydrogenase was down-regulated 12
fold despite lactate being the primary carbon source in the media
Table 4 2 Genes related to anaerobic growth and electron transfer with significantly change expression level
Locus Tag Gene product descriptions Fold
change
SO_0101 nitrate-inducible formate dehydrogenase molybdopterin-binding
subunit FdnG
18
SO_0102 formate dehydrogenase nitrate-inducible iron-sulfur subunit 21
SO_0103 formate dehydrogenase nitrate-inducible cytochrome b556 subunit 24
SO_0104 formate dehydrogenase accessory protein FdhE 22
SO_0107 formate dehydrogenase accessory protein fdhD 24
SO_0397 quinolfumarate reductase menaquinol-oxidizing subunit FrdC_2 12
SO_0452 thioredoxin 2 15
SO_0903 Na-translocating NADH-quinone reductase subunit B NqrB_1 12
SO_1012 NADH-ubiquinone oxidoreductase subunit K NuoK 12
SO_1013 NADH-ubiquinone oxidoreductase subunit J NuoJ 12
SO_1363 hydroxylamine reductase 13
57
SO_2417 ferredoxin cofactor maintenance protein YfaE 10
SO_3922 formate dehydrogenase cytochrome b Fdh -12
SO_4513 formate dehydrogenase molybdopterin-binding subunit _2 15
SO_4514 formate dehydrogenase FeS subunit FdhB_2 12
SO_4515 formate dehydrogenase cytochrome b subunit FdhC_2 14
SO_4609 aa3 type cytochrome c oxidase subunit III CoxC 11
SO_0479 periplasmic octaheme cytochrome c MccA -12
SO_0847 periplasmic nitrate reductase ferredoxin component NapG -15
SO_1519 L-lactate dehydrogenase iron-sulfur cluster-binding protein LldF -10
SO_1251 ferredoxin 4Fe-4S -11
SO_37411 hypothetical inner membrane protein -11
a fold changes were presented after log2
b positive number represented up-regulated and negative represented down-regulated
Flavin and cytochrome related genes
Flavins can be secreted by Shewanella species as electron shuttle to facilitate
extracellular electron transfer (Cansterin et al 2008) C-type cytochromes play the
important roles on the process of extracellular electron transfer (Shi et al 2007) The
genes related to flavins synthesis and the genes encoding the cytochromes electron
transport proteins were shown in the table 43 the genes SO_1414 and SO_3058 related
to flavins synthesis was up-regulated to 03 and 01 folds respectively The genes related
to s C-type cytochromes SO_0135 SO_4105 SO_0169 SO_1778 are increased to 10
08 05 04 01 folds respectively However most of genes have shown no significant
changes (less one-fold change) as response to nano particle decorated anode in MECs
58
Table 43 Expression level of flavin and cytochrome genes
Locus Tag Gene product descriptions Fold
change
SO_1414 flavocytochrome c flavin subunit putative 03
SO_3468 riboflavin synthase subunit alpha -02
SO_3058 flavocytochrome c flavin subunit 01
SO_4105 MSHA pilin protein MshA 08
SO_0169 general secretion pathway protein GspG 05
SO_1778 outer membrane decaheme cytochrome c lipoprotein MtrC 04
SO_1779 outer membrane decaheme cytochrome c OmcA 01
SO_0135 lipoprotein of unknown function DUF333 10
SO_0136 conserved hypothetical inner membrane protein 01
a fold changes were presented after log2
b positive number represented up-regulated and negative represented down-regulated
Other genes with significantly modified expression levels
These genes over 15 fold change and possibly related to current enhancements were
shown in Table 44 SO_3104 (expressed inner membrane protein) was over expressed
and the gene SO_2194 which encodes outer membrane protein (OmpA family protein)
was repressed as response to nanoparticle decorated anode however it is unclear the
relationships of this modified membrane protein genes with the current enhancement
Another interesting phenomenon was possible co-regulation of several gene clusters
indicating possible operon associations This includes three sets of genes that were
induced more than 15 fold SO_0108SO_0109 and SO_1042SO_1043SO_1044 The
59
consistency of expression of these genes under the NP conditions provides basic evidence
to support operon structure However the correlation of these significant changed gene
with current density enhance are unclear which showed there are unknown multiple and
complex responses of S oneidensis MR-1 to nanoparticle anode of MECs
Table 44 Other genes with significantly changed expression levels
Locus Tag Gene product descriptions Fold change
SO_0101 nitrate-inducible formate dehydrogenase molybdopterin-binding
subunit FdnG
18
SO_0108 integral membrane protein of unknown function DUF39 YedE 17
SO_0109 SirA family protein YedF 17
SO_0275 N-acetyl-gamma-glutamyl-phosphate reductase -31
SO_0277 ornithine carbamoyltransferase -21
SO_0279 argininosuccinate lyase -24
SO_0404 zinc dependent metalloprotease domain lipoprotein 21
SO_0956 alkyl hydroperoxide reductase F subunit -20
SO_1042 amino acid ABC transporter ATP-binding protein -18
SO_1043 amino acid ABC transporter permease protein -27
SO_1044 amino acid ABC transporter periplasmic amino acid-binding protein -27
SO_1072 chitin-binding protein putative 21
SO_1405 transglutaminase family protein 20
SO_1822 TonB-dependent receptor putative -22
SO_2069
1-(5-phosphoribosyl)-5-[(5-phosphoribosylamino)
methylideneamino] imidazole-4-carboxamide isomerase
-19
SO_2070 amidotransferase HisH -26
SO_2071 imidazole glycerol-phosphate dehydratasehistidinol phosphatase -27
60
SO_2072 histidinol-phosphate aminotransferase -28
SO_2073 histidinol dehydrogenase -30
SO_2194 OmpA family protein -36
SO_2195 inter-alpha-trypsin inhibitor domain protein -27
SO_2767 asparagine synthetase B -26
SO_2945 prophage LambdaSo tail fiber protein 20
SO_2963 prophage LambdaSo major capsid protein HK97 family 20
SO_2987 prophage LambdaSo hypothetical protein 19
SO_2988 prophage LambdaSo expressed protein of unknown function 18
SO_3104 expressed inner membrane protein 131
SO_3408 conserved hypothetical inner membrane protein -18
SO_3585 azoreductase putative -18
SO_3586 glyoxalase family protein -17
SO_3687 curli production assemblytransport component CsgE putative -18
SO_38191 hypothetical ammonia permease -19
SO_4014 AcrBAcrDAcrF family protein 18
SO_40151 type I secretion system membrane fusion protein RND family 18
SO_4054 510-methylenetetrahydrofolate reductase 18
SO_4245 N-acetylglutamate synthase -22
SO_45251 hypothetical transcriptional regulator LysR family -17
SO_4527 integral membrane domain protein -19
SO_4705 transcriptional regulator putative -19
a fold changes were presented after log2
b positive number represented up-regulated and negative represented down-regulated
Discussion
61
The addition of NP to anodes significantly impacts current densities with the elemental
composition being a critical factor The chemical composition of NPs significantly affects
the current enhancement in MECs (Fan et al 2011) because the chemical composition of
NP properties is a critical factor in determining the conductivity which significantly
affects the efficiency of electron transfer from bacteria to electrode Fe is an ideal
candidate for NP formation for current enhancement in MESs because of high
conductivity and low price compared to gold and other conductive materials Gold NPs
has also been studies for enhancement of electricity conductivity (Bao et al 2008)
However the disadvantages of the high price of gold limit their practical application in
MECs (Fan et al 2011) The different size and density of the same composition of NP
also has essential effects on the current density enhancement (Fan et al 2011) In this
study the results showed the current density enhancement with Fe-NP size range of
200plusmn100 nm and density range of 200plusmn100 nm is considerable enhancement consequence
There are a number of reports of studies of current density enhancement in MFCs
by nanostructured anodes (Fan et al 2011 Qiao et al 2008 Sharma et al 2008 Tsai et al
2009) Based on bacterial behavior on nanostructured anode surface Qiao et al (2008)
believes that Escherichia coli cells on the nanostructured electrode surface produce hair-
like structures similar to pili that could facilitate the electron transfer between the cells
and electrode The production of hair-like structures is believed to be stimulated by
nanostructures and could play the key role on current density enhancement in pili-
producing bacteria in MFCs (Qiao et al 2008) S oneidensis can produce electrically
conductive bacterial nanowires which have similar structure to the hair-like structures in
Ecoli (Gorby et al 2006 Reguera et al 2005) A number of possible genes related
62
nanowires of S oneidensis have been identified including mshA (SO_4105) and gspG
(SO_0169) (Konstantinidis et al 2009) They are up-regulated 08 and 05 fold in the
current study respectively even though they are not significant induced This result
supported the increased nanowire genes had contributions to enhance current density in
MFCs by NPs decorated anodes
Based on the previous research on electron transfer mechanisms for S oneidensis it is
known that self-produced mediators (flavins) and multihaem c-type cytochromes (c-Cyts)
may play critical roles in the electron transfer of S oneidensis to extracellular insoluble
electrode (Canstein et al 2008 Myers and Myers 1992 Scott and Nealson 1994 Shi et al
2007) The flavins secreted by S oneidensis can facilitate to transfer electrons from
bacteria to solid anode electrodes by acting as extracellular electron shuttles (Canstein et
al 2008) In this study the genes encoding flavins (SO_1414 SO_3468 and SO_3058)
had no significant increase (less than 1 fold change) as a response to nanoparticle
decorated anode in MECs which suggests that increased flavin gene expression does not
significantly contribute to the current density enhancements in nano-particle decorated
anode The genes encoding cytochromes electron transport proteins (SO_4104 SO_0417
SO_177879 SO_013536) had no significant increase (less than 1-fold change)
However those genes were up-regulated slightly These results suggest the increased
expression flavin and c type cytochromes genes had partial contributions even not
significantly to enhance current density in MFCs by NPs decorated anodes
The thickness of bacterial biofilm on the anode can affect the power generation in
MFCs (Biffinger et al 2007 Heydorn et al 2000) since increased biofilm thickness on
the anode represents more active bacteria involving in the process of electron generation
63
and transfer in MFCs Biofilm formation related genes included flagellar genes IV pilus
genes and other genes The flagellar related genes SO_3228 SO_3229 (fliB and fliK) and
IV pilus genes SO_4105 (mshA) were induced from 19 to 09 fold the genes (SO_4178
-4180) related to formation of a three-dimensional biofilm were up-regulated from 10 to
13 folds even though the genes related bacterial motility protein (SO_ 4286 SO_4287
SO_1529 and SO_1530) are induced slightly (less than 0 5 fold) However the motility
genes most probably play a role for initial biofilm attachment on the anode surfaces
(Thormann et al 2004 Thormann et al 2006) with the rest of biofilm genes being more
essential for biofilm maintenance in mature cultures as were presented at harvest in this
study This enhancement of biofilm density under conditions of increase current density
has been observed for gold NP decorated anodes using confocal light microscopy (data
not shown) but has not yet been confirmed for Fe NP decorated anodes In any event an
increase in the expression level of genes related biofilm formation therefore also
supported the possibility of enhanced biofilm formation on NPs decorated anodes thus
facilitating electron transfer from bacteria to anodes
Beliaev et al described a model of the Fe reduction pathway in S oneidensis MR-1 using
solid substrates as the electron acceptor (Beliaev et al 2001) This model proposes that
electrons are generated and released in cytoplasm then transferred to the quinone pool
eg menaquinone in the cytoplasmic membrane The reduced menaquinone in turn
reduces CymA and from there electrons are then transferred to a periplasmic shuttle (eg
CctA) and then to MtrA a periplasmic soluble decaheme cytochrome c located in
associated with the outer membrane via interaction with MtrB Finally surface displayed
outer membrane proteins decaheme cytochromes (OmcA MtrC or MtrF) can transfer
64
electron to Fe or other solid substrates (Beliaev et al 2001)) Bencheikh-Latmani et al
demonstrated that the same electron transport pathway may be used for more than one
electron acceptor through studies of the response of S oneidensis MR-1 to U(VI) and
Cr(VI) under anaerobic conditions particularly focused on the critical genes
(SO_1777mtrA SO_1776mtrB SO_1778mtrC) (Bencheikh-Latmani et al 2005)
However our studies showed that these genes had no significant expression changes in
response to nanostructured anodes Contrastingly one of expressed inner membrane
protein genes SO_3104 and one of cytochromes genes SO_4609 encoding cytochrome c
oxidase subunit III demonstrated a 13-fold and 11-fold up-regulation respectively in this
study while the gene SO_2194 which encodes outer membrane protein (OmpA family
protein) was down regulated 36 fold Our results therefore suggest that S oneidensis
MR-1 does not utilize the same set of core critical genes (SO_1777 mtrA SO_1776
mtrB SO_1778 mtrC) products to transfer electrons in all instances This is the similar
result with the study of Bretschger et al (2007) which indicated that the electron
transport system in S oneidensis MR-1 is complex with several different proteins able to
participate in electron transfer to the anode of MECs
It should also be mentioned that the gene (SO_1519) encoding lactate
dehydrogenase repressed (-12 fold) here suggesting that lactate was depleted at the time
of cell harvest from the MECs and that other pathways of anaerobiosis had been activated
including those associated with amino acid uptake and consumption the growth media
was a complex mixture including yeast extract In particular the catabolic pathway for
histidine of S oneidensis MR-1 a pathway that may formate as an intermediate is up-
regulated with the genes for histidine ammonia lyase SO_0098 SO_3057 and SO_4374
65
up-regulated 13 11 and 11-fold respectively The possible production of formate
during histidine degradation may in turn explain the high levels of formate
dehydrogenates as evidenced by upregulation of SO_0102 SO_0103 SO_0104 and
SO_0107 annotated as formate dehydrogenase proteins These genes had the highest
levels of up-regulated expression as response to Fe NP decorated anodes (greater than 2-
fold)
Acknowledgements
This research was partially supported by the US National Science Foundation CBET
0828544 and the funds from ONAMIDOD (ARL-DOD Cooperative Agreement
W911NF-07-2-0083) We thank Barbara Gvakharia and Caprice Rosato for helpful
suggestions We also thank Margaret Romine of Pacific Northwest Laboratories for
valuable comments on manuscripts We also thank anonymous reviewers for significant
suggested improvements imparted as part of prior review of this manuscript
66
Chapter 5
Global Transcriptome Analysis of the Response of Shewanella oneidensis MR-1 to
Carbon Nanotube Decorated Anodes in Microbial Electrochemical Systems
Shoutao Xu Yanzhen Fan Rebecca Schaller Frank Chaplen Jun Jiao Hong Liu
67
Abstract
Shewanella oneidensis MR-1 is an important model microorganism for metabolic studies
on the effects of different environmental factors because of its diverse respiratory
capabilities Carbon nanotube (CNT) modified graphite disks were used as anodes to
investigate the effects of nanomatrials on the performance of S oneidensis MR-1 in
microbial electrolysis cells (MECs) The current densities produced with CNT decorated
anodes were on average 56-fold higher than plain graphite anodes Whole genome
microarray analysis of gene expression showed that up-regulation of cytochromes c genes
associated with extracellular electron transfer are strongly correlated to current increases
in CNT decorated anodes Genes involved in flavin biosynthesis also contribute to
current increase in CNT decorated anode MECs
Keywords Microbial electrochemical system microbial fuel cell microbial
electrolysis cell carbon nanotube gene expression DNA microarray
68
Introduction
Shewanella oneidensis MR-1 is an important model microorganism for metabolic
studies of the effects of different environmental factors because of its diverse respiratory
capabilities It has been used for transcriptome analysis to investigate the responses to
different growth conditions and environmental stresses (Murray et al 2001) Gene
expression patterns under different conditions such as heat shock cold shock Cr (VI) U
(VI) reduction chromate stress iron and acid tolerance have been studied previously
(Bencheikh-Latmani et al 2005 Brown et al 2006 Gao et al2006 Gao et al 2004
Yang et al 2008) More recently it has been exploited as a model species for power
generation in microbial electrochemical systems (MESs)which have potential
applications in renewable energy generation bioremediation and wastewater treatment
(Logan et al 2007 Logan 2009 Park and Zeikus 2002 Fan et al 2011 Xu et al 2012)
In a MES electrochemically active microorganisms oxidize organic matter in the an
ode chamber to release electrons Electrons are then transferred to the anode electrode thr
ough various mechanisms (Logan et al 2006 Lovley 2006 Rabaey et al 2003) and fina
lly travel to the cathode electrode and combine with the terminal electron acceptor The
key feature of MESs is the microbe-catalyzed electron transfer from the organic matter to
the anode (Rabaey et al 2004) Enhancing the current output which highly depends on
the performance of the anode electrode is critical for the successful application of MESs
(Logan et al 2007 Park and Zeilus 2002 2003) Several nanomaterials including TiO2
gold (Au) and palladium (Pd) nanoparticles have been utilized to decorate electrodes to
enhance power density in MESs (Qiao et al 2008 Quan et al 2005 Fan et al 2011) The
possible mechanisms for increased current densities have been studied by using Fe
69
nanoparticle-decorated anodes (Xu et al 2012)
Carbon nanotubes (CNT) are cylindrically shaped nanomaterials with extremely
high surface area excellent electrical conductivity and chemical inertness (He et al
2005a Serp et al 2003) These unique properties make CNT a promising electrode
material (Liang et al 2008) The biocompatibility of microorganisms and carbon
nanostructures has been evaluated (Morozan et al 2007) The effects of CNT on anodic
biofilms have been tested in MFCs (Liang et al 2011) However no studies have been re
ported on global transcriptome analysis for the response of S oneidensis MR-1 to CNT
decorated anode in MECs
In the present study the graphite disks decorated with CNT were used as anodes to
investigate the effects of nanomaterials on current generation in multi-anode MECs
DNA microarrays were used to analyze differences in the global gene expression profile
of S oneidensis MR-1 grown on plain versus CNT decorated anodes
Materials and methods
Bacterial cultures S oneidensis MR-1 was purchased from American Type Culture
collection (ATCC 700550) and stored as freezer stocks at -80degC Stock S oneidensis
MR-1 cells were grown aerobically using M1 medium at 30degC Cells in early stationary
growth phase (optical density of 15-16 at 600nm (Spectrophotometer UV-1700
Shimadzu Columbia MD) were injected into the chamber of the MECs for current
production Sodium lactate (final concentration 30mM) was added as the additional
electron donor
Fabrication and characterization of nanostructured anodes Superfine isomolded
graphite disks (16 cm x 05 cm GraphiteStorecom Inc) were polished with ultrafine
70
sand paper (2000 grit 3M Company) as the base for the CNT decorated and control
anodes Aligned multi-walled carbon nanotube samples were synthesized via plasma
enhanced chemical vapor deposition (PECVD) Samples superfine isomolded graphite
pillars with a 16 cm diameter and 05 cm thickness were sputtered with a thin film of Ni
in a CRC 300 magnetron sputter coater for 1 to 2 min They were then placed in the
PECVD reactor where the Ni was annealed into small catalyst islands C2H2 was bled
into the chamber for growth and an NH3 plasma was used to vertically align the growth
of the multi-walled carbon nanotubes (MWCNTs) The temperature and time were varied
between 725degC and 775oC and 15 to 60 min to control the length of the tubes and density
of samples Samples were then analyzed in a FEI Sirion field emission scanning electron
microscope (FESEM) SEM images of CNT decorated anode and control surfaces were
shown in the Figure 51
Figure 51 SEM images of CNT decorated anodes (A) Graphite disk control (B) CNT
decorated anode
Multiple channel MECs construction and operation A MECs with multiple
removable anodes each with an effective anode area of 07 cm2
was constructed and used
to evaluate the effects of nanostructures on the current density of according to a previous
B A
71
study (Fan et al 2011) The cathode was made of wet-proof (30) carbon cloth (type B
E-TEK Division Inc USA) coated with PtC (20 E-TEK Division Inc USA)
following a previously reported procedure (Liu et al 2005) The final platinum loading
was 05 mgcm2 per projected cathode area The size of the cathode (150 cm
2) was 25
times larger than the total effective area of the 8 graphite disk anodes (56 cm2) to prevent
cathode limitations on the performance of the MECs system All MECs with CNT
decorated anodes and control anodes were tested for current generation for 24 h with
sterile a medium (without bacteria) before injecting the bacterial cells in order to
determine whether the CNT decorated anodes demonstrate chemical current generation
compared to control anodes All the testing anodes were set up in the MECs
simultaneously so that the S oneidensis MR-1 culture was applied to all of them
simultaneously Short electrode spacing (17 cm) was used in the MECs design to reduce
the internal resistance A voltage of 06 V was applied the MECs for current generation
after the cells of S oneidensis MR-1 were inoculated into the MECs in the early
stationary phase A multimeter with a data acquisition system (2700 Keithly USA) was
used to monitor the current change by measuring the voltage drop through a resistor
Microarray analysis Biofilms for whole gene microarray analysis were aseptically
removed from the plain and CNT decorated anodes of MECs after 80 hours further
incubation at 30oC when current density obviously reached to a stable phase Total RNA
was extracted using Trizol (Ambion inc AustinTX) following the manufacturerrsquos
instructions Integrity of the RNA samples was confirmed using the Agilent 2100
Bioanalyzer (Agilent Technology Inc Palo Alto CA) Total RNA (200 ng) from each
sample was amplified using the MessageAmpTM
II-Bacteria Prokaryotic RNA
72
Amplification kit (Ambion AustinTX) according to the manufacturerrsquos instructions The
S oneidensis MR-1 whole-gene microarrays representing all of the 4295 genes (resource
from NCBI) were ordered from Roche NimbleGen Inc (Madison WI) Biological
triplicates of biofilms on the CNT decorated anodes and control were analyzed cDNA
synthesis labeling and hybridization were carried out by the CGRB Core Laboratories at
Oregon State University DNASTAR ArrayStarTM
3 software was used to identify genes
that were up- or down-regulated more than 2-fold when grown on the nanoparticle
modified anodes using the unpaired two sample t-test with a cutoff p-value of 005 with
bonferroni correction
Results and discussion
Enhancement of current generation using CNT decorated anodes in MECs The
current density generated by CNT decorated anode MECs with S onidensis MR-1
increased to 36 microAcm2 10 hours post-inoculation and reached a maximum of 40 microAcm
2
at 29 hours inoculation then gradually decreased The current density stabilized at
30microAcm2 after 78 h operation Figure 52 shows that the current density curve generated
by CNT decorated anode in MECs The average current density generated by CNT
decorated anodes was 56 times that of the control The current density generated in the
MECs without bacteria was negligible (data not shown) This result demonstrates that the
current enhancement observed with the CNT decorated anodes was biologically-derived
A
73
Figure 52 Effects of CNT decorated anodes on the current density in MECs (3 replicates)
Control is plain graphite disk anode CNT is the anode with CNT decoration (Error bars
represent standard deviation)
It has been reported that nanostructured decorated anodes have significant impacts on
current densities (Fan et al 2011 Qiao et al 2006 Sharma 2008 Tsai et al 2009) The
CNT decorated anodes had significantly enhanced current densities in MECs in this
study This result is consistent with previous studies Interestingly the current density
curve generated by CNT decorated anode MECs is quite different from the current
density curve generated by Au NP and Pd NP decorated anodes using same bacterial
species (Fan et al 2011) In the Au and Pd NP decorated anode MECs the current
density curve was not significantly increased in the beginning compared with the control
but gradually increased after the inoculation the current density reached the maximum
current density more than 50 hours after the inoculation However the current density
generated by CNT decorated anode MECs started to increase significantly immediately
after the inoculation It reached a maximum current density 29 hours post-inoculation
-1
6
13
20
27
34
41
48
0 10 20 30 40 50 60 70 80
Curr
ent
den
sity
(micro
Ac
m2)
Time (Hours)
Control
CNT
74
These results indicate that there might be different current density enhancement
mechanisms between CNT with other metal NP decorated anodes in MECs
Global transcriptome analysis Whole-genome DNA microarrays were used to attain a
comprehensive general overview of the transcriptional response of S oneidensis
MR-1 of
S oneidensis MR-1 to CNT decorated anodes in MECs In a total of 4295 genes analyzed
457 genes (250 genes up-regulated and 207 genes down-regulated) exhibited significantly
(P lt 005) changed expression at a 2-fold level in at least 3 replicates in the response to
CNT The total number of the regulated genes present 11 of the 4295 open reading
frames (ORFs) presented on the microarray The total number of genes at significant
expression levels in response to CNT decorated anodes is close to that to Fe NP decorated
anodes in MECs (392 9) ( Xu et al 2012)
Fig 53 Differentially expressed genes grouped by functional classification according to
the TIGR S oneidensis genome database (wwwtigrorg) Columns 1 amino acid
biosynthesis 2 biosynthesis of cofactors prosthetic groups and carriers 3 cell
envelope 4 cellular processes 5 central intermediary metabolism 6 DNA metabolism
7 energy metabolism 8 fatty acid and phospholipid metabolism 9 other categories 10
protein fate 11 protein synthesis 12 purines pyrimidines nucleosides and nucleotides
-80
-60
-40
-20
0
20
40
60
80
100
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18
Nu
mb
er
of
Ge
ne
s
Up-regulated
Down-regulated
75
13 regulatory functions 14 signal transduction 15 transcription 16 transport and
binding proteins 17 unknown function 18 hypothetical proteins
The overall transcript genomic response of S oneidensis MR-1 to CNT decorated
anodes was summarized in Figure 53 by grouping the differentially expressed genes into
their functional role categories based on The Institute for Genomic Researchrsquos annotation
(Version 8) of the MR-1 genome sequence The wide distribution of putative functional
roles attributed to the differentially expressed genes indicated the extent of the molecular
response of Soniedenis MR-1 to the CNT decorated anodes A large number of up-
regulated genes were presented in genes associated with biosynthesis of cofactors
prosthetic groups and carriers (group 2) cell envelope (group 3) protein fate (group 10)
protein synthesis (group 11) and purines pyrimidines nucleosides and nucleotides
(group 12) in this study The number of up-regulated genes was two times more than that
of down-regulated genes in each functional group Contrastingly in the genomic response
S oneidensis MR-1 to Fe NP decorated anodes (chapter 4) most of up-regulated genes
were associated with cellular processes (group 4) energy metabolism (group 7) other
categories (group 9) and hypothetical proteins (group 18) The down-regulated genes
presented in the amino acid biosynthesis (group 1) cellular processes (group 4) substrate
transport (group 16) and hypothetical proteins (group 18) While those genes encoding
proteins involved in amino acid biosynthesis (group 1) central intermediary metabolism
( group 5) DNA metabolism( group 6) fatty acid and phospholipid metabolism (group
8) and other categories (group 9) were among the most down-regulated genes in this
study However the two groups of genes that were the most noticeable between the
genomic response S oneidensis MR-1 to CNT and Fe NP decorated anodes which they
are the genes related to energy metabolism (group 7) and hypothetical proteins (group
76
18) These results indicated there are different and common genomic responses to S
oneidensis MR-1 on CNT and Fe NP decorated anodes in MECs
Electron transfer related genes Based on previous electron transfer mechanism
studies on Shewanella a serial of group proteins collectively described as the Mtr
pathway play the role to transfer electron rom the inner bacterial body to the outer
membrane (Beliaev et al 2001 Shi et al 2007 Brutinel and Gralnick 2012) In the Mtr
pathway firstly electrons generated by bacteria received by CymA a tetraheme c-type
cytochrome anchored in the inner membrane then electrons were transferred to MtrA a
periplasmic decaheme c-type cytochrome After that electrons were transferred from
MtrA to MtrC an outer-membrane decaheme c-type cytochrome is facilitated by MtrB a
non-heme containing integral outer-membrane proteinOuter-membrane decaheme c-type
cytochrome MtrC and OmcA are in change of electron transfer to exreacellular electron
acceptor (Shi et al 2007 Brutinel and Gralnick 2012 ) These genes encoding electron
transport proteins are shown in the table 51 The critical genes SO_1777 mtrA SO_1776
mtrB SO_1778 mtrC are unregulated 41 to 80 fold in response to CNT decorated
anodes other cytochrome c genes related to electron transfer in MtrA pathway also are
significantly increased in response to CNT decorated anodes These genes encoding outer
membrane proteins have different level increases supporting that the increase of
cytochrome c gene expression contributes to the current enhancements of MECs as
response to CNT decorated anodes
77
Table 51 Expression level of cytochrome c as response to CNT decorated anodes
Locus Tag Gene product descriptions Fold change CNT
SO_0165 general secretion pathway protein GspC 31
SO_0167 general secretion pathway protein GspE 45
SO_1776 outer membrane protein precursor MtrB 41
SO_1777 outer membrane decaheme cytochrome c
lipoprotein MtrA
80
SO_1778 outer membrane decaheme cytochrome c
lipoprotein MtrC
43
SO_1779 outer membrane decaheme cytochrome c
OmcA
63
SO_0135 lipoprotein of unknown function DUF333 22
SO_0136 conserved hypothetical inner membrane protein 24
Flavin related genes It is known that self-produced mediators play critical roles in the
electron transfer of S oneidensis to extracellular insoluble electrode (Canstein von et al 2
008 Myers et al 1992 Scott et al 1994 Shi et al 2007) It has been proved that the
flavins secreted by S oneidensis can facilitate electron transfer from bacteria to solid
anode electrodes by acting as extracellular electron shuttles (Cansterin von et al 2008) T
wo critical genes (SO_1414 and SO_3468) related to flavin synthesis were shown increa
sed significantly (more than 2 folds) increase in this study (Table 52) This result support
s the hypothesis that flavin production increases as the response to CNT decorated anodes
in MECs have significant contribution to the current density enhancements
78
Table 52 Expression levels of genes related to flavin synthesis as response to CNT
decorated anode
Locus Tag Gene product descriptions Fold change CNT
SO_1414 flavocytochrome c flavin subunit putative 22
SO_3468 riboflavin synthase subunit alpha 20
The amount of bacterial biofilm biomass on the anode can affect the power
generation in MFCs (Biffinger et al 2007 Heydorn et al 2000) Biofilm formation
related genes include flagellar gene IV pilus genes and other genes the flagellar related
genes (fliB and fliK) and IV pilus genes (mshA) were induced only from 105 to 134
fold as response of S oneidensis MR-1 to CNT decorated anodes in this study No
biofilm-associated genes presented significant changes in expression levels in this study
indicating there is no direct connections between biofilm enhancements to CNT
decorated anodes in this study This result is consistent with the Liang et al (2011)
conclusion that using CNT enhanced the electrochemical activity of anodic biofilms but
did not result in a significant increase of biomass in the anodic biofilms
Our microarray results showed two significantly up-regulated gene groups the
genes encoding proteins localized on the outer membrane and the genes involved in
flavin biosynthesis contributed to current density enhancement by CNT decorated anodes
Among 457 significantly changed genes to CNT in this study there are also a relatively
large number of genes encoding proteins with unknown functions which are either up- or
down-regulated in the response to CNT decorated anodes which indicated more
79
complicated responses of S oneidensis MR-1 to CNT decorated anodes in MECs and
further study is needed
80
Chapter 6
Summary
Microbial electrochemical systems (MESs) with a mixed culture initially inoculated
from Corvallis wastewater treatment plant have been studied for more than 6 years for
varying purposes including power generation hydrogen production heavy metal
removal and wastewater treatment Experiment results have shown that the mixed culture
is quite stable with excellent performance in MESs Our community analysis using
denaturing gradient gel electrophoresis (DGGE) and 16S rDNA clone library construction
suggests that the mixed culture is composed predominantly of Geobacter sp (66)
Arcobacter sp (12) and Citrobacter sp (11) These results not only improved our
understanding of the mixed culture community but also guided our studies on the
cultivation and isolation of the different bacterial species
Exoelectrogenic Citrobacter sp SX-1 was first isolated and characterized from a
MFC operated with the mixed culture Citrobacter sp SX-1 has been demonstrated to
produce electricity from wide range of different substrates including citrate acetate
glucose sucrose glycerol and lactosein MFCs Cyclic voltammetry analysis indicated
that membrane associated proteins may play an important role in facilitating the electrons
transferring from bacteria to electrode The strain SX-1 increased the known diversity of
power generating exoelectrogens and provided a unique bacterial species for study in
renewable energy generation and waste treatment
81
To enhance the electron transfer from bacteria to anode nano decorated anodes
including Fe NP and CNT were developed and characterized and evaluated in MECs
using Shewanella oneidensis MR-1 as a model species Both nanostructures have
significantly increased current density compared with the control Whole genome
microarray analysis elucidated the possible mechanisms of power enhancement in
response to these nano-decorated anodes
These results benefit to understanding of physiology and ecology of mixed
cultures in MFCs and improve the efficiency of current generation in MESs which will
facilitate the viability of niche applications for MESs in near future
82
Bibliography
1 Amman R Ludwig W (2000) Ribosomal RNA-targeted nucleic acid probes for
studies in microbial ecology FEMS Microbiol Ecol 24 555ndash565
1 Bao SJ Li CM Zang JF Cui XQ Qiao Y Guo J (2008) Novel Nanoporous TiO2
Electrode for Directly Electrochemistry of glucose oxidase Adv Funct Mater
18(4) 591-599
2 Beliaev AS Saffarini DA McLaughlin JL Hunnicutt D (2001) MtrC an outer
membrane decahaem c cytochrome required for metal reduction in Shewanella
putrefaciens MR-1 Mol Microbiol 39722-30
3 Bencheikh-Latmani R Williams SM Haucke L Criddle CS Wu L Criddle CS
Zhou J Tebo BM (2005) Global transcriptional profiling of Shewanella oneidensis
MR-1 during Cr(VI) and U(VI) reduction Appl Environ Microbiol 71 7453ndash7460
4 Biffinger JC Pietron J Ray R Little B and Ringeisen BR (2007) A biofilm
enhanced miniature microbial fuel cell using Shewanella oneidensis DSP10 and
oxygen reduction cathodes Biosens Bioelectron 221672ndash1679
5 Bodelier PLE Roslev P Henckel T FenzelP (2000) Stimulation by ammonium
based fertilizers of methane oxidation in soil around rice root Nature 40 421ndash424
6 Bond DR Holmes DE Tender LM Lovley DR (2002) Electrode-reducing
microorganisms that harvest energy from marine sediments Science 295 483-485
7 Bond DR Lovley DR (2003) Electricity production by Geobacter sulfurreducens
attached to electrodes Appl Environ Microbiol 69 1548-1555
8 Bond DR Lovley DR (2005) Evidence for involvement of an electron shuttle in
electricity generation by Geothrix fermentans Appl Environ Microbiol 71 2186-
2189
9 Bretschger O Obraztsova A Sturm CA Chang IS Gorby YA Reed SB Culley
DE Reardon CL Barua S Romine MF Zhou J Beliaev AS Bouhenni R
Saffarini D Mansfeld F Kim BH Fredrickson JK Nealson KH (2007) Current
production and metal oxide reduction by Shewanella oneidensis MR-1 wildtype and
mutants Appl Environ Microbiol 73 7003ndash7012
10 Brown SD Thompson MR Verberkmoes NC Chourey K Shah M Zhou J
Hettich RL and Thompson DK (2006) Molecular dynamics of the Shewanella
oneidensis response to chromate stress Mol Cell Proteomics 51054-1071
11 Brutinel ED Gralnick JA (2012) Shuttling happens soluble flavin mediators of
extracellular electron transfer in Shewanella Appl Microbiol Biotechnol 93(1) 41-8
12 Butler JE Kaufmann F Coppi MV Nuacutentildeez C Lovley DR (2004) MacA a diheme
c-type cytochrome involved in Fe(III) reduction by Geobacter sulfurreducens J
Bacteriol 186(12) 4042ndash4045
83
13 Caccavo F Lonergan DJ Lovley DR Davis M Stolz JF McInerney MJ (1994)
Geobacter sulfurreducens sp nov a hydrogen- and acetate oxidizing dissimilatory
metal-reducing microorganism Appl Environ Microbiol 60 3752ndash3759
14 Canstein von H Ogawa J Shimizu S Lloyd JR (2008) Secretion of flavins by
Shewanella species and their role in extracellular electron transfer Appl Environ
Microbiol 74 615ndash623
15 Cao X Huang X Liang P Xiao K Zhou Y Zhang X Logan BE (2009) A New
Method for Water Desalination Using Microbial Desalination Cells Environ Sci
Technol 43(18)7148-7152
16 Chaudhuri SK Lovley DR (2003) Electricity generation by direct oxidation of
glucose in mediatorless microbial fuel cells Nat Biotechnol 21 1229-1232
17 Cheng S Liu H Logan B E (2006) Increased power generation in a continuous
flow MFC with advective flow through the porous anode and reduced electrode
spacing Environ Sci Technol 402426-2432
18 Cheng S Logan B E (2007) Sustainable and efficient biohydrogen production via
electrohydrogenesis PNAS 104 18871-18873
19 Chignell J (2010) Energy Recovery from Biodiesel Waste Performance of
Microbial Electrochemical Systems on Glycerol Thesis
20 Chung K Okabe S (2009) Characterization of electrochemical activity of a strain
ISO2-3 phylogenetically related to Aeromonas sp isolated from a glucose-fed
microbial fuel cell Biotechnol Bioeng 104(5) 901-910
21 Cole JR Chai B Farris RJ Wang Q Kulam-Syed-Mohideen AS McGarrell
DM Bandela AM Cardenas E Garrity GM Tiedje JM (2007) The ribosomal
database project (RDP-II) introducing myRDP space and quality controlled public
data Nucleic Acids Res 35 (Database issue) D169-D172 doi 101093nargkl889
22 Cole JR Wang Q Cardena E Fish J Chai B Farris RJ Kulam-Syed-Mohideen
AS McGarrell DM Marsh T Garrity GM Tiedje JM (2009) The Ribosomal
Database Project improved alignments and new tools for rRNA analysis Nucleic
Acids Res 37 (Database issue) D141-D145 doi 101093nargkn879
23 Dey A De S De A De SK (2004) Characterization and dielectric properties of
polyaniline-TiO2 nanocomposites Nanotechnol 15 1277-1283
24 Edwards U Rogall T Bloumlcker H Emde M Boumlttger EC (1989) Isolation and
direct complete nucleotide determination of entire genesmdashcharacterization of a gene
coding for 16S-ribosomal RNA NucleicAcids Res 17 7843ndash7853
25 Eggleston CM Voumlroumls J Shi L Lower BH Droubay TC Colberg PJS (2008)
Binding and direct electrochemistry of OmcA an outer-membrane cytochrome from
an iron reducing bacterium with oxide electrodes a candidate biofuel cell system
Inorg Chim Acta 361769ndash777
26 Fan Y Sharbrough E Liu H (2008) Quantification of the Internal Resistance
Distribution of Microbial Fuel Cells Environ Sci Technol 42 8101ndash8107
84
27 Fan Y Xu S Schaller R Jiao J Chaplen F Liu H (2011) Nanoparticle decorated
anodes for enhanced current generation in microbial electrochemical
cells Biosensors and Bioelectronics 26 (5)1908ndash1912
28 Fan Y Hu H Liu H (2007)Sustainable power generation in microbial fuel cells
using bicarbonate buffer and proton transfer mechanisms Environ Sci
Technol 41(23) 8154-8
29 Fedorovich V Knighton MC Pagaling E Ward FB Free A Goryanin I (2009)
Novel electrochemically active bacterium phylogenetically related to Arcobacter
butzleri isolated from a microbial fuel cell Appl Environ Microbiol 75(23) 7326-
7334
30 Ferna ndez A Huang S Seston S Xing J Hickey R Criddle C Tiedje J (1999)
How stable is stable Function versus community composition Appl Environ
Microbiol 65 3697ndash3704
31 Field SJ Dobbin PS Cheesman MR Watmough NJ Thomson AJ Richardson
DJ (2000) Purification and magneto-optical spectroscopic characterization of
cytoplasmic membrane and outer membrane multiheme c-type cytochromes from
Shewanella frigidimarina NCIMB400 J Biol Chem 275 8515ndash8522
32 Freguia S Masuda M Tsujimura S Kano K (2009) Lactococcus lactis catalyses
electricity generation at microbial fuel cell anodes via excretion of a soluble quinone
Bioelectrochemistry 7614ndash18
33 Fricke K Harnisch F and Schroumlder U (2008) On the use of cyclic voltammetry for
the study of the anodic electron transfer in microbial fuel cells Energy Environ
Sci 1 144ndash147
34 Gao H Wang Y Liu X Yan T Wu L Alm E Arkin A Thompson DK and
Zhou J (2004) Global transcriptome analysis of the heat shock response of
Shewanella oneidensis J Bacteriol 1867796-7803
35 Gao H Yang ZK Wu L Thompson DK Zhou J (2006) Global transcriptome
analysis of the cold shock response of Shewanella oneidensis MR-1 and mutational
analysis of its classical cold shock proteins J Bacteriol 1884560-4569
36 George MG (2005) Bergeyrsquos Manual of Systematic Bacteriology Volume Two The
Proteobacteria Part B The Gammaproteobacteria Second Edition New York
Springer 651p
37 Gonzaacutelez-Toril E Llobet-Brossa E Casamayor EO Amann R Amils R (2003)
Microbial ecology of an extreme acidic environment the Tinto river Appl Environ
Microbiol 69(8) 4853ndash4865
38 Gorby YA Yanina S Mclean JS Rosso KM Moyles D Dohnalkova A
Beveridge TJ Chang IS Kim BH Kim KS Culley DE Reed SB Romine M
F Saffarini DA Hill EA Shi L Elias DA Kennedy DW Pinchuk G
Watanabe K Ishii S Logan B Nealson KH Fredrickson JK (2006) Electrically
conductive bacterial nanowires produced by Shewanella oneidensis strain MR-1 and
other microorganisms Proc Natl Acad Sci USA 10311358-11363
85
39 He Z Wagner N Minteer SD Angenent LT (2006) An upflow microbial fuel cell
with an interior cathodes assessment of the internal resistance by impedance
spectroscopy Environ Sci Technol 40 5212-5217
40 Heydorn A Nielsen AT Hentzer M Sternberg C Givskov M Ersboslashll BK Molin
S (2000) Quantification of biofilm structures by the novel computer program
COMSTAT Microbiol 1462395-2407
41 Holmes DE Bond DR Lovley DR (2004a) Electron transfer by Desulfobulbus
propionicus to Fe(III) and graphite electrodes Appl Environ Microbiol 701234ndash
1237
42 Holmes DE Chaudhuri SK Nevin KP Mehta T Methe BA Liu A Ward JE
Woodard TL Webster J Lovley DR (2006) Microarray and genetic analysis of
electron transfer to electrodes in Geobacter sulfurreducens Environ Microbiol
81805-1815
43 Holmes DE Nicoll JS Bond DR Lovley DR (2004b) Potential role of a novel
psychrotolerant member of the family Geobacteraceae Geopsychrobacter
electrodiphilus gen nov sp nov in electricity production by a marine sediment
fuel cell Appl Environ Microbiol 70 6023ndash6030
44 Holmes DE Chaudhuri SK Nevin KP Mehta T Metheacute BA Liu A Ward JE
Woodard TL Webster J Lovley DR (2006) microarray and genetic analysis of
electron transfer to electrodes in Geobacter sulfurreducens Environ Microbial
8(10)1805-1815
45 Hu H (2009) Enhancing Hydrogen Production in Microbial Electrolysis Cells
through Development of Platinum-free Cathode and Improvement of Reactor
Dissertation
46 Ince BK Ayman O N Turker G Ccedilelikkol S Ince O (2010) Microbial ecology of
anaerobic reactors for treatment of alcohol industry wastewaters a review Current
research technology and education topics in applied microbiology and microbial
ecology 988-999
47 Jung S Regan JR (2007) Comparison of anode bacterial communities and
performance in microbial fuel cells with different electron donors Appl Microbiol
Biotechnol 77393ndash 402
48 Kim BH Kim HJ Hyun MS Park DH (1999) Direct electrode reaction of Fe(III)
reducing bacterium Shwwanella putrefaciens J Microbial Biotechnol 9127-131
49 Kim GT Hyun MS Chang IS Kim HJ Park HS Kim BH Kim SD Wimpenny
JW Weightman AJ (2005) Dissimilatory Fe(III) reduction by an electrochemically
active lactic acid bacterium phylogenetically related to Enterococcus gallinarum
isolated from submerged soil J Appl Microbiol 99(4) 978ndash987
50 Kim GT Webster G Wimpenny JW Kim BH Kim HJ Weightman AJ (2006)
Bacterial community structure compartmentalization and activity in a microbial fuel
cell J Appl Microbiol 101(3) 698ndash710
86
51 Kim HJ Park HS Hyun MS Chang IS Kim M Kim BH (2002) A mediator-
less microbial fuel cell using a metal reducing bacterium Shewanella putrefaciens
Enzyme Microb Technol 30 145-152
52 Kim JR Min B Logan BE (2005) Evaluation of procedures to acclimate a
microbial fuel cell for electricity production Appl Microbiol Biotechnol 68(1)23-30
53 Kim JR Cheng S Oh SE Logan BE (2007) Power generation using different
cation anion and ultrafiltration membranes in microbial fuel cells Environ Sci
Technol 41 1004-1009
54 Konstantinidis KT Serres MH Romine MF Rodrigues JL Auchtung J McCue
LA Lipton MS Obraztsova A Giometti CS Nealson KH Fredrickson JK
Tiedje JM (2009) Comparative systems biology across an evolutionary gradient
within the Shewanella genus Proc Natl Acad Sci U S A 106(37)15909-15914
55 Lee J Phung NT Chang IS Kim BH Sung HC (2003) Use of acetate for
enrichment of electrochemically active microorganisms and their 16S rDNA
analyses FEMS Microbiol Lett 223185ndash191
56 Liang P Wang H Xia X Huang X Mo Y Cao X Fan M(2011)Carbon
nanotube powders as electrode modifier to enhance the activity of anodic biofilm
in microbial fuel cells Biosens Bioelectron 26(6)3000-4
57 Lies DP Hernandez ME Kappler A Mielke RE Gralnick JA Newman DK
(2005) Shewanaella oneidensis MR-1 uses overlapping pathways for iron reduction
at a distance and by direct contact under conditions relevant for biofilms Appl
Environ Microbiol 71 4414-4426
58 Liu H and Logan B E (2004) Electricity generation using an air-cathode single
chamber microbial fuel cell in the presence and absence of a proton exchange
membrane Environ Sci Technol 38 4040-4046
59 Liu H Grot S Logan BE (2005) Electrochemically assisted microbial production
of hydrogen from acetate Environ Sci Technol 39 4317 ndash 320
60 Liu H Hu H Chignell J Fan Y (2010) Microbial Electrolysis Novel Technology
for Hydrogen Production from Biomass Biofuels 1(1) 129ndash142
61 Liu H Ramnarayanan R Logan BE (2004) Production of electricity during
wastewater treatment using a single chamber microbial fuel cell Environ Sci Technol
382281-2285
62 Liu WT Marsh TL Cheng H Forney LJ (1997) Characterization of microbial
diversity by determining terminal restriction fragment length polymorphisms of gene
encoding 16S rRNA Appl Environ Microbiol 63 4516ndash4522
63 Logan B E Hamelers B Rozendal R Schroder U Keller J Freguia S Aelterman
P Verstraete W Rabaey K (2006) Microbial fuel cells methodology and
technology Environ Sci Technol 40 (17) 5181-5192
64 Logan B E and Regan JM (2006) Electricity-producing bacterial communities in
microbial fuel cells Trends Microbiol 14512ndash518
87
65 Logan B Cheng S Watson V Estadt G (2007) Graphite fiber brush anodes for
increased power production in air-cathode microbial fuel cells Environ Sci Technol
41(9)3341-3346
66 Logan BE (2009) Exoelectrogenic bacteria that power microbial fuel cells Nat Rev
Microbiol 5 375-381
67 Logan BE Call D Cheng S Hamelers HV Sleutels TH Jeremiasse AW
Rozendal RA (2008) Microbial electrolysis cells for high yield hydrogen gas
production from organic matter Environ Sci Technol 42 8630-8640
68 Logan BE Hamelers B Rozendal R Schroder U Keller J Freguia S Aelterman
P Verstraete W Rabaey K (2006) Microbial fuel cells methodology and
technology Environ SciTechnol 40 5181-5192
69 Lovely DR (2006) Bug juice harvesting electricity with microorganisms Nature
Review| Microbiology 4 497-508
70 Lovley DR Phillips EJ (1988) Novel Mode of Microbial Energy Metabolism
Organic Carbon Oxidation Coupled to Dissimilatory Reduction of Iron or
Manganese Appl Environ Microbiol 54(6) 1472-1480
71 Ly HK Sezer M Wisitruangsakul N Feng JJ Kranich A Millo D Weidinger
IM Zebger I Murgida DH Hildebrandt P (2011) Surface-enhanced vibrational
spectroscopy for probing transient interactions of proteins with biomimetic interfaces
electric field effects on structure dynamics and function of cytochrome c FEBS J
278(9)1382-90
72 Macaskie LE Hewitt CJ Shearer JA Kent CA (1995) Biomass production for
the removal of heavy metals from aqueous solutions at low pH using growth-
decoupled cells of a Citrobacter sp Int Biodeter Biodegr 3573ndash92
73 Malki M De Lacey AL Rodriguez N Amils R Fernandez VM (2008)
Preferential use of an anode as an electron acceptor by an acidophilic bacterium in
the presence of oxygen Appl Environ Microbiol 74 4472ndash4476
74 Meitl LA Eggleston CM Colberg PJS Khare N Reardon CL Shi L (2009)
Electrochemical interaction of Shewanella oneidensis MR-1 and its outer membrane
cytochromes OmcA and MtrC with hematite electrodes Geochim Cosmochim
Ac 73(18) 5292-5307
75 Morozan A Stamatin L and Nastase
F (2007) The biocompatibility
microorganisms-carbon nanostructures for applications in microbial fuel cells Phys
Stat Sol 6 1797-1803
76 Morris JM Jin S Crimi B Pruden A (2009) Microbial fuel cell in enhancing
anaerobic biodegradation of diesel Chem Eng J 146 161ndash167
77 Murray AE Lies D Li G Nealson K Zhou J Tiedje JM (2001) DNADNA
hybridization to microarrays reveals gene-specific differences between closely
related microbial genomes Proc Natl Acad Sci U S A 98 9853-9858
78 Muyzer G de Waal EC Uitterlinden AG (1993) Profiling of complex microbial
communities by denaturing gradient gel electrophoresis analysis of polymerase chain
88
reaction amplified genes coding for 16S rRNA Appl Environ Microbiol 59 695ndash
700
79 Myers CR Myers JM (1992) Localization of cytochromes to the outer membrane
of anaerobically grown Shewanella putrefacians MR-1 J Bacteriol 174 3429ndash3438
80 Narde GK Kapley A Purohit HJ (2004) Isolation and characterization of
Citrobacter strain HPC255 for broad-range substrate specificity for chlorophenols
Curr Microbiol 48(6) 419-423
81 Okabe S Ito T Sugita K Satoh H (2005) Succession of internal sulfur cycles and
sulfur-oxidizing bacterial communities in microaerophilic wastewater biofilms Appl
Environ Microbiol 71(5) 2520-2529
82 Park DH and Zeikus JG (2000) Electricity generation in microbial fuel cells using
neutral red as an electronophore Appl Environ Microbial 66 292-1297
83 Park DH and Zeikus JG (2002) Impact of electrode composition on electricity
generation in a single-compartment fuel cell using Shewanella putrefaciens Appl
Microbiol Biotechnol 5958ndash56
84 Park DH and Zeikus JG (2003) Improved fuel cell and electrode designs for
producing electricity from microbial degradation Biotechnol Bioeng 81 348ndash355
85 Park HS Kim BH Kim HS Kim HJ Kim GT Kim M Chang IS Park Y
K Chang HI (2001) A novel electrochemically active and Fe(III)-reducing
bacterium phylogenetically related to Clostridium butyricum isolated from a
microbial fuel cell Anaerobe 7 297-306
86 Parot S Nercessian O Delia ML Achouak W Bergel A (2009) Electrochemical
checking of aerobic isolates from electrochemically active biofilms formed in
compost J Appl Microbiol 106(4) 1350-1359
87 Pham CA Jung SJ Phung NT Lee J Chang IS Kim BH Yi H Chun J
(2003) A novel electrochemically active and Fe(III)-reducing bacterium
phylogenetically related to Aeromonas hydrophila isolated from a microbial fuel cell
FEMS Microbiol Lett 223 129-134
88 Qiao Y Bao SJ Li CM Cui XQ Lu ZS Guo J (2008) Nanostructured
polyanilinetitanium dioxide composite anode for microbial fuel cells ACS Nano 2
113-119
89 Qiu R Zhao B Liu J Huang X Li Q Brewer E Wang S Shi N (2009) Sulfate
reduction and copper precipitation by a Citrobacter sp isolated from a mining area J
Hazard Mater 1641310-1315
90 Quan X Yang SG Ruan XL and Zhao HM (2005) Preparation of titania
nanotubes and their environmental applications as electrode Environ Sci Technol 39
3770ndash3775
91 Rabaey K Boon N Hofte M Verstraete W (2005) Microbial phenazine
production enhances electron transfer in biofuel cells Environ Sci Technol 393401ndash
3408
89
92 Rabaey K Boon N Siciliano S D Verhaege M Verstraete W (2004) Biofuel cells
select for microbial consortia that self-mediate electron transfer Appl Environ
Microbiol 70(9) 5373ndash5382
93 Rabaey K Lissens G Siciliano S D Verstraete W (2003) Amicrobial fuel cell
capable of converting glucose to electricity at high rate and efficiency Biotechnol
Lett 25 1531-15
94 Rabaey K Verstraete W (2005) Microbial fuel cells novel biotechnology for
energy generation Trends Biotechnol 23(6) 291-298
95 Reguera G McCarthy KD Mehta T Nicoll JS Tuominen MT Lovley DR
(2005) Extracellular electron transfer via microbial nanowires Nature 435 1098-
1101
96 Reguera G Nevin KP Nicoll JS Covalla SF Woodard TL Lovley DR (2006)
Biofilm and nanowire production leads to increased current in Geobacter
sulfurreducens fuel cells Appl Environ Microbiol 72 7345-7348
97 Reimers CE Tender LM Fertig S Wang W (2001) Harvesting energy from the
marine sediment-water interface Environ Sci Technol 35192-195
98 Rezaei F Xing D Wagner R Regan JM Richard TM Logan BE (2009)
Simultaneous cellulose degradation and electricity production by Enterobacter
cloacae in a microbial fuel cell Appl Microbiol Biotechnol 75 3673ndash3678
99 Richter H Nevin KP Jia H Lowy DA Lovley DR Tender LM (2009) Cyclic
voltammetry of biofilms of wild type and mutant Geobacter sulfurreducens on fuel
cell anodes indicates possible roles of OmcB OmcZ type IV pili and protons in
extracellular electron transfer Energy Environ Sci 2 506ndash516
100 Riesenfeld SC Schloss DP Handelsman J(2004) Metagenomics Genomic
Analysis of Microbial Communities Annual Review Genetics 38525-552
101 Rozendal RA Hamelers HV Buisman CJ (2006) Effects of membrane cation
transport on pH and microbial fuel cell performance Environ Sci Technol 40
5206-5211
102 Rozendal RA Hamelers HVM Euverink GJW Metz SJ Buisman CJN (2006)
Principle and perspectives of hydrogen production through biocatalyzed electrolysis
Int J Hydrogen Energ 311632-1640
103 Rozendal RA Hamelers HV Molenkamp RJ Buisman CJ (2007) Performance of
single chamber biocatalyzed electrolysis with different types of ion exchange
membrances Water Res 41 1984-1994
104 Schluumlter A Bekel T Diaz NN Dondrup M Eichenlaub R Gartemann
KH Krahn I Krause L Kroumlmeke H Kruse O Mussgnug JH Neuweger
H Niehaus K Puumlhler A Runte KJ Szczepanowski R Tauch A Tilker
A Viehoumlver P Goesmann A (2008) The metagenome of a biogas-producing
microbial community of a production-scale biogas plant fermenter analyzed by the
454-pyrosequencing technology J Biotechnol136 77-90
90
105 Scott J H and Nealson KH (1994) A biochemical study of the intermediary carbon
metabolism of Shewanella putrefaciens J Bacteriol 176(11) 3408ndash3411
106 Sekiguchi H Tomioka H Nakahara T and Uchiyama H (2001) A single band
does not always represent single bacterial strains in denaturing gradient gel
electrophoresis analysis Biotechnology Letters23 1205ndash1208
107 Sharma T Leela M Reddy A Chandra TS Ramaprabhu S (2008) Development
of carbon nanotubes and nanofluids based microbial fuel cell Int J Hydrogen Energ
336749ndash6754
108 Shea C Clauwaert P Verstraete W Nerenberg R (2008) Adapting a denitrifying
biocathode for perchlorate reduction Water Sci Technol 58 (10) 1941-1946
109 Shi L Squier TC Zachara JM Fredrickson JK (2007) Respiration of metal (hydr)
oxides by Shewanella and Geobacter a key role for multihaem c-type cytochromes
Mol Microbiol 6512-20
110 Stams AJ and Plugge CM(2009)Electron transfer in syntrophic communities
of anaerobic bacteria and archaea Nat Rev Microbiol7(8)568-77
111 Sukkasem C Xu S Park S Boonsawang P Liu H (2008) Effect of Nitrate on the
Performance of Single Chamber Air Cathode Microbial Fuel Cells Water Res 42(19)
4743-4750
112 Summers ZM Fogarty HE Leang C Franks AE Malvankar NS Lovley DR
(2010)Direct exchange of electrons within aggregates of an evolved syntrophic
coculture of anaerobic bacteria Science 330 (6009)1413-5
113 Szczepanowski R Bekel T Goesmann A Krause L Kroumlmeke H Kaiser
O Eichler W Puumlhler A Schluumlter A (2008) Insight into the plasmid metagenome
of wastewater treatment plant bacteria showing reduced susceptibility to
antimicrobial drugs analysed by the 454-pyrosequencing technology J
Biotechnol 136 54-64
114 Tamura K Dudley J Nei M Kumar S (2007) MEGA4 Molecular Evolutionary
Genetics Analysis (MEGA) software version 40 MolBiol Evol 24(8) 1596-1599
115 Thompson JD Gibson TJ Plewniak F Jeanmougin F Higgins DG (1997) The
CLUSTAL_X windows interface flexible strategies for multiple sequence alignment
aided by quality analysis tools Nucleic Acids Res 25 4876-4882
116 Thormann KM Duttler S Saville RM Hyodo M Shukla S Hayakawa Y
Spormann AM (2006) Control of formation and cellular detachment from
Shewanella oneidensis MR-1 biofilms by cyclic di-GMP J Bacteriol 1882681-2691
117 Thormann KM Saville RM Shukla S Pelletier DA Spormann AM (2004)
Initial Phases of biofilm formation in Shewanella oneidensis MR-1 J Bacteriol
186(23) 8096-104
118 Torres CI Kato Marcus A Rittmann BE (2008b) Proton transport inside the
biofilm limits electrical current generation by anode-respiring bacteria Biotechnol
Bioeng 100 872ndash881
91
119 Torres CI Marcus AK Lee HS Parameswaran P Krajmalnik-Brown R Rittmann
BE (2010) A kinetic perspective on extracellular electron transfer by anode-
respiring bacteria FEMS Microbiol Rev 234(1)3-17
120 Torres CI Marcus AK Parameswaran P Rittmann BE (2008a) Kinetic
experiments for evaluating the NernstndashMonod model for anode-respiring bacteria
(ARB) in a biofilm anode Environ Sci Technol 42 6593ndash6597
121 Tsai HY Wu Ch-Ch Lee Ch-Y Shih EP (2009) Microbial fuel cell performance
of multiwall carbon nanotubes on carbon cloth as electrodes J Power Sources
194199-205
122 Tyson GW Chapman J Hugenholtz P Allen EE Ram RJ Richardson
PM Solovyev VV Rubin EM Rokhsar DS Banfield JF(2004) Community
structure and metabolism through reconstruction of microbial genomes from the
environment Nature42837-43
123 Venkateswaran K Moser DP Dollhopf ME Lies DP Saffarini D A MacGregor
BJ Ringelberg DB White DC Nishijima M Sano H Burghardt J Stackebrandt
E Nealson KH (1999) Polyphasic taxonomy of the genus Shewanella and
description of Shewanella oneidensis sp Int J Syst Bacteriol 49705-724
124 von Canstein H Ogawa J Shimizu S Lloyd JR (2008) secretion of Flavins
by Shewanella Species and Their Role in Extracellular Electron Transfer Appl
Environ Microbiol 74 3615-623
125 Wang A Liu W Cheng Sh Xing D Zhou J Logan BE (2009) Source of methane
and methods to control its formation in single chamber microbial electrolysis cells
Int J Hydrogen Energ 34 3653-3658
126 Wang J Ma T Zhao L Lv J Li G Liang F Liu R (2008) PCRndashDGGE method
for analyzing the bacterial community in a high temperature petroleum reservoir
World J Microbiol Biotechnol 2 9694-9702
127 Wang Q Garrity GM Tiedje JM Cole JR (2007) Naiumlve Bayesian Classifier for
Rapid Assignment of rRNA Sequences into the New Bacterial Taxonomy Appl
Environ Microbiol 73(16) 5261-5267
128 Wang YF Masuda M Tsulimura S Kano K (2008) Electrochemical regulation of
the end-product profile in Propionibacterium freudenreichii ET-3 with an
endogenous mediator Biotechnol Bioeng 101(3) 579ndash586
129 Wrighton KC Agbo P Warnecke F Weber KA Brodie EL DeSantis TZ
Hugenholtz P Andersen GL Coates JD (2008) A novel ecological role of the
Firmicutes identified in thermophilic microbial fuel cells ISME J 2 1146ndash1156
130 Xing D Zuo Y Cheng S Regan JM Logan BE (2008) Electricity generation by
Rhodopseudomonas palustris DX-1 Environ Sci Technol 42 4146-4151
131 Xu S Liu H Fan Y Schaller R Jiao J Chaplen F (2012) Enhanced performance
and mechanism study of microbial electrolysis cells using Fenanoparticle-decorated
anodes Appl Microbiol Biotechnol 93(2)871-880
92
132 Yang Y Harris DP Luo F Wu L Parsons AB Palumbo AV Zhou J (2008)
Characterization of the Shewanella oneidensis Fur gene roles in iron and acid
tolerance response BMC Genomics 9 Suppl 1S11
133 Zhang L Zhou S Zhuang L Li W Zhang J Lu N Deng L (2008) Microbial
fuel cell based on Klebsiella pneumoniae biofilm Electrochem Commun10 1641ndash
1643
134 Zuo Y Xing DF Regan JM Logan BE (2008) Isolation of the exoelectrogenic
bacterium Ochrobactrum anthropi YZ-1 by using a U-tube microbial fuel cell Appl
Environ Microbiol 74 3130-3137
93
APPENDICES
List of published papers during PhD study
1 Shoutao Xu Hong Liu Yanzhen Fan Rebecca Schaller Jun Jiao and Frank
Chaplen (2012) Enhanced performance and mechanism study
of microbial electrolysis cells using Fe nanoparticle-decorated anodes Applied
Microbiology and Biotechnology 93(2)871-880
2 Shoutao Xu Hong Liu (2011) New exoelectrogen Citrobacter sp SX-1 isolated
from a microbial fuel cell Journal of Applied Microbiology111(5)1108-1115
3 Yanzhen Fan Shoutao Xu Rebecca Schaller Jun Jiao Frank Chaplen Hong Liu
(2011) Nanoparticle decorated anodes for enhanced current generation in microbial
electrochemical cells Biosensors and Bioelectronics 26(5) 1908-1912
4 Tunc Catal Shoutao Xu Kaichang Li Hakan Bermek Hong Liu
(2008)
Electricity generation from polyalcohols in single-chamber microbial fuel
cells Biosensors and Bioelectronics 24(4)849-854
5 Chontisa Sukkasema Shoutao Xu Sunhwa Park Piyarat Boonsawang Hong Liu
(2008) Effect of nitrate on the performance of single chamber air cathode
microbial fuel cells Water research 424743-4750
6 Rebecca Schaller Yanzhen Fan Shoutao Xu Alan Fern Frank Chaplen Hong
Liu and Jun Jiao (2009) Vertically Aligned Multi-walled Carbon Nanotube
Decorated Anodes for Microbial Fuel Cells Proceedings of Materials Research
Society 2009 1170 R05-13