membrane-induced folding of the plant-stress protein lti30. · 2 24 25 membrane-induced folding of...
TRANSCRIPT

1
Membrane-induced folding of the plant-stress protein Lti30. 1 2 Sylvia Eriksson1, Nadejda Eremina1, Andreas Barth1, Jens Danielsson1 and Pia 3 Harryson1* 4 5 Department of Biochemistry and Biophysics, Arrhenius Laboratories for Natural Sciences, 6 Stockholm University, 106 91 Stockholm, Sweden, Tel: + 46 8 164238 Fax: + 46 8 153679. * 7 Corresponding author: [email protected] 8 9 10 11 S.E; performed experiment, experimental design, data analysis and wrote the materials section. 12 J.D; designed and performed NMR experiments, data analysis and wrote the NMR section. 13 N.E; designed and performed the IR experiments, N.E. and A.B. 14 IR data analysis, and wrote the IR section. 15 P.H; principal investigator, conceived the project, experimental design, data analysis and wrote 16 the article with contributions from all authors. 17 18 19 Funding was from the Knut and Alice Wallenberg (J. D. and A.B) and Magnus Bergwall 20 Foundations (P.H. and J.D). 21 22 23
Plant Physiology Preview. Published on April 26, 2016, as DOI:10.1104/pp.15.01531
Copyright 2016 by the American Society of Plant Biologists
www.plantphysiol.orgon July 7, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.
www.plantphysiol.orgon July 7, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.
www.plantphysiol.orgon July 7, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.
www.plantphysiol.orgon July 7, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.
www.plantphysiol.orgon July 7, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.
www.plantphysiol.orgon July 7, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.
www.plantphysiol.orgon July 7, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.
www.plantphysiol.orgon July 7, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.
www.plantphysiol.orgon July 7, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.
www.plantphysiol.orgon July 7, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.
www.plantphysiol.orgon July 7, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.
www.plantphysiol.orgon July 7, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.
www.plantphysiol.orgon July 7, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.
www.plantphysiol.orgon July 7, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

2
24 Membrane-induced folding of the plant-stress protein Lti30. 25 26 Sylvia Eriksson1, Nadejda Eremina1, Andreas Barth1, Jens Danielsson1 and Pia 27 Harryson1* 28 29 Department of Biochemistry and Biophysics, Arrhenius Laboratories for Natural Sciences, 30 Stockholm University, 106 91 Stockholm, Sweden, Tel: + 46 8 164238 Fax: + 46 8 153679. * 31 Corresponding author: [email protected] 32 33 Dehydrins are disordered proteins, expressed in plants as a response to embryogenesis 34 and water-related stress. The molecular function and structural action of the dehydrins 35 are yet elusive, but increasing evidence point to a role in protecting the structure and 36 functional dynamics of cell membranes. An intriguing example is the cold-induced 37 dehydrin Lti30 that binds to membranes by its conserved K-segments. Moreover, this 38 binding can be regulated by pH and phosphorylation, and shifts the membrane phase 39 transition to lower temperatures, consistent with the protein’s postulated function in cold 40 stress. In this study, we reveal how the Lti30-membrane interplay works structurally at 41 atomic-level resolution. NMR analysis suggests that negatively charged lipid head 42 groups electrostatically capture the protein’s disordered K-segments, which locally fold 43 up into α-helical segments on the membrane surface. Lti30 confers thus to the general 44 theme of structure-function relationships by folding upon binding, in spite of its 45 disordered, atypically hydrophilic and repetitive sequence signatures. Moreover, the 46 fixed and well-defined structure of the membrane-bound K-segments suggest that 47 dehydrins have the molecular prerequisites for higher-level binding specificity and 48 regulation, raising new questions about the complexity of their biological function. 49 50 51 Keywords: Dehydrin, intrinsically disordered protein, Lea-proteins, membrane binding, 52 structural transition 53
www.plantphysiol.orgon July 7, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

3
INTRODUCTION 54 55 Since rooted plants cannot flee environmental changes their fitness and survival has come 56 to rely on a series of molecular defence systems, reflected in genomes that are typically 57 much larger than for animals (Rafalski 2002). One of the most basal tasks of this defence 58 is to manage cellular stress induced by desiccation, ionic strength and thermal 59 fluctuations. The molecular response includes here the production of osmo-protectants, 60 various sugars (Ingram and Bartels 1996) and up-regulated expression of dedicated stress 61 proteins like the intrinsically disordered dehydrins (Baker et al., 1988; Nylander et al., 62 2001; Bomal et al., 2002; Chakrabortee et al., 2007). Exactly how these stress proteins 63 function and interfere with the cellular processes is yet unknown, but judging by their 64 highly conserved sequence signatures their action seems under precise molecular control 65 (Close 1996; Mouillon et al., 2006; Graether and Boddington 2014). Accordingly, it is 66 speculated that the dehydrins could be evolved to selectively target and modulate the 67 properties of other proteins (Hara et al., 2001; Kovacs et al., 2008), DNA/RNA (Hara et 68 al., 2009) or membranes (Dure 1993; Danyluk et al., 1998; Koag et al., 2003). 69 Membrane targeting has come to stand out as particularly interesting, as both the 70 structure and phase properties of lipid bilayers are known to depend critically on 71 temperature and water content (Steponkus and Lynch 1989; Crowe and Crowe 1992). 72 Also, some dehydrins are found to co-localize with membrane surfaces (Danyluk et al., 73 1998; Puhakainen et al., 2004) and bind to anionic phospholipids in vitro (Koag et al., 74 2003; Kovacs et al., 2008; Koag et al., 2009; Eriksson et al., 2011). Some of these 75 interactions are driven by electrostatics and depend on positively charged residues in the 76 dehydrin sequence pairing with negatively charged lipids head-groups, following the 77 same rules as other lipid binding proteins (Lemmon 2008; Moravcevic et al., 2010; 78 Eriksson et al., 2011). A good example is the net positively charged dehydrin Lti30 from 79 Arabidopsis thaliana, which binds membranes by recognizing the negatively charged 80 head groups of phospholipids (Eriksson et al., 2011). The consequence of this membrane 81 interaction is a decrease of the main lipid phase transition by 2.5 °C in vitro (Eriksson et 82 al., 2011), intriguingly matching the decreased survival temperature of 3 °C observed of 83 upon Lti30 overexpression in Arabidopsis (Puhakainen et al., 2004). Consistent effects 84
www.plantphysiol.orgon July 7, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

4
on the membrane phase transition are reported for the dehydrin K2 (Clarke et al., 2015). 85 To achieve membrane binding, Lti30 comprises not less than 6 copies of the archetypical 86 K-segment (Figure 1). However, the K-segments are not alone accountable for the Lti30-87 membrane interaction, as binding also requires protonation of flanking pairs of histidines. 88 In essence, these co-localized pairs of histidines form a pH controlled affinity switch, 89 which can further be fine-tuned by protein phosphorylation (Eriksson et al., 2011). The 90 complexity of the Lti30-membrane interplay raises naturally the question about the 91 possible involvement of binding-induced folding (Arai et al., 2015; Wright and Dyson 92 2015). Following general structure-function relationships, such three-dimensional 93 organisation of specific structure would open up for additional layers of target specificity 94 and modes of function (Lee et al., 2010; Vuzman and Levy 2012) analogous to those 95 observed for other intrinsically disordered proteins (Kriwacki et al., 1996; Dunker and 96 Obradovic 2001; Tompa 2002; Borg et al., 2007; Sigalov and Hendricks 2009). Classical 97 examples are found among the transcription factors (Wright and Dyson 2015) but there 98 are also cases where local folding is induced by membrane binding (Jao et al., 2004; 99 Jarvet et al., 2007) resembling that of the dehydrins (Table 1) (Koag et al., 2003; Rahman 100 et al., 2010). In this study, we shed new information on this issue by identifying the 101 generic physical-chemical features of the membranes that govern Lti30 binding, its 102 dynamics, and, at atomic-level resolution, the structural changes of the K-segments that 103 accompany this membrane binding (Figure 1). The results show that, indeed, the anionic 104 charges of the lipid head groups electrostatically capture the positively charged Lti30 K-105 segments to establish a dynamic binding equilibrium. In the unbound state the K-106 segments remains fully disordered, whereas in the bound state it turns into a fixed 107 amphipathic helix that floats on the membrane surface. Such direct observation of 108 binding-mediated folding of a K-segment provides not only a strict structural base for 109 elucidating stress-defense mechanisms, but also points to the possibility that the 110 functional repertoire of the dehydrin proteins is larger than previously anticipated. 111 112 113 114 115
www.plantphysiol.orgon July 7, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

5
116 RESULTS 117 Lti30 shows electrostatic recognition of lipid head groups. Previous data examined that 118 Lti30 interact with highly negatively charged vesicles (PC:PG ratio 3:1, i.e. 25% 119 negatively charged) and assembly these into large clusters. It was further shown that the 120 binding is electrostatic and regulated by protonation of histidines in a pH dependent way 121 (Eriksson et al., 2011). Full-length Lti30 binds and aggregates lipid vesicles upon His 122 protonation, starting at pH below the pKA value of the bound state that is shifted to well 123 above 8 (Eriksson et.al., 2011). Accordingly, pH titration of Lti30 gives a binding 124 ‘midpoint’ around pH 7.5 where 50% of the vesicle-aggregation effect is manifested. 125 Because of this flexibility in achieving suitable binding regimes in vitro, we opted here 126 for pH 6.3 as ‘standard condition’. The advantage is that pH 6.3 is close to the pKA of the 127 histidines in unbound Lti30. The other pH of mechanistic interest is around the His pKA 128 in the membrane-bound state, i.e. above pH 8, but this is too high for elucidating 129 physiological action. In previous studies we examined the Lti30-lipid binding from a His 130 perspective by varying the pH (Eriksson et.al., 2011) and here we expand the analysis by 131 varying also the membrane charge. Large unilamellar vesicles (LUVs) were composed 132 of DOPG (1-40%) in the background of the neutral zwitter-ionic DOPC (99-60%) lipids. 133 The vesicles were rapidly mixed with Lti30 in a stopped flow spectrophotometer to a 134 final concentration of 0.5 mM lipids and 3.5 μM protein at pH 6.3 and the scattering 135 effect of the formed vesicle-clusters were monitored either as absorbance or fluorescence. 136 As seen in Figure 2, 2% negatively charged lipids are adequate for Lti30 to bind and 137 cluster the vesicles. The intensity of the vesicle clustering increases when the negative 138 net charge of the vesicles is elevated to 3% but seems to start to level out at 6% (Figure 139 2). Hence, below binding saturation, the binding is proportional to the acidic charges of 140 the vesicles. At high negative net charge of the vesicles (above 6%, i.e. at 25% and 40% 141 DOPG), the stopped flow absorbance/fluorescence signal changes character. This can be 142 explained by repulsion between the negative lipids head-groups that interfere with the 143 cluster reactions. The large clusters formed due to Lti30 binding are detectable with light 144 microscope and pictures of the clusters further support the stopped flow data (Figure 3). 145 The conclusion is that Lti30 has a strong affinity for low concentrations of negative lipids 146
www.plantphysiol.orgon July 7, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

6
and needs only few negative lipids to bind and cluster vesicles. This infers that Lti30 can 147 also bind to different cellular membranes in vivo, since most of these contain higher 148 concentrations of negatively charged phospholipids than tested here (Uemura et al., 1995). 149 The proposition seems even more plausible since the data presented here show vesicle 150 clustering, a reaction secondary to lipid binding, and it is not unlikely that binding of 151 Lti30 could take place at even lower vesicle charges. 152 153 Identification and His-flank regulation of the membrane-binding epitope. In previous 154 studies (Eriksson et al., 2011; Petersen et al., 2012) it have been demonstrated that 155 membrane binding of dehydrins is chiefly driven by the ubiquitous K-segments (Figure 1 156 and 4). The consensus sequence for these K-segments is EKKGIMDKIKEKLPG with 157 slight variations across the protein family (Close 1996; Graether and Boddington 2014). 158 We note here that the six K-segments of Lti30 deviate from the consensus K-segment by 159 being less charged: the Lti30 segments are close to neutral at pH 7, whereas the 160 consensus segment shows a net charge of + 2 (Figure 1 and 4). Moreover, the K-161 segments of Lti30 are all flanked by histidines. These histidine flanks occur to varied 162 extent across the dehydrin family and has previously been implicated as pH-switches in 163 regulation of membrane binding (Eriksson et al., 2011). In essence, the increased 164 positive charge from protonated histidine flanks activates binding, whereas deprotonation 165 leads to dissociation (Eriksson et al., 2011). To examine the action of such regulation in 166 Lti30 we constructed three peptide models: (i) the consensus K-segment (KC), (ii) the 167 consensus K-segment with flanking histidines (KHH-C-HH), and (iii) the K-segment of 168 Lti30 with flanking histidines (KLti30) (Figure 4). The latter occurs twice in the Lti30 169 sequence and differs only marginally from the protein’s remaining four K-segments 170 (Figure 1) which charges varies in the same range as KLti30. The results show 171 interestingly that both KHH-C-HH and KLti30 bind membranes in a pH dependent manner, 172 whereas KC that lacks the histidine flanks fails to bind in this model system (Figure 4). 173 Accordingly, the histidine flanks seem to be a functional feature of the K-segment by 174 providing extra positive charges to boost the interaction with the negatively charged 175 membrane. Sequence comparison shows, consistently, that histidine flanks are often co-176 localized with the K-segments across the dehydrin family, pointing to the possibility that 177
www.plantphysiol.orgon July 7, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

7
regulation of membrane binding by protonation is a widespread feature of the stress 178 response (Eriksson et al., 2011). For further discussion of histidine flanks and the overall 179 atypical positive net charge of dehydrins, see (Eriksson et al., 2011). From the binding 180 data in Figures 2 and 4, we conclude that the K-segments of Lti30 play a key role in the 181 vesicle binding of the full-length protein, and that this binding is modulated both by the 182 negative charge density of the membrane and the protonation state of the flanking 183 histidines. 184 Lti30 changes structure upon membrane binding as measured by FTIR spectroscopy. As 185 vesicle clustering causes high background noise in the UV region, the structural response 186 of Lti30 upon membrane association cannot be followed by conventional techniques like 187 CD. Instead we use here FTIR spectroscopy that targets conformation sensitive 188 vibrations associated with much longer wavelengths in the infrared regime. Following 189 standard procedures, we use here the second derivative of the absorption spectrum for 190 identifying peak positions as this enables easier differentiation between overlapping 191 bands. The negative bands in the second-derivative spectra correspond to the component 192 bands in the original absorption spectra. Figure 5 shows the second-derivative spectra of 193 the amide I’ vibrations of the peptide backbone, which report on secondary structure. A 194 strong band at 1644 cm-1 and a weak band at 1673 cm-1 at both pH 5 and 8.6 indicate 195 random coil structure (Figure 5A). When lipid vesicles are added, the protein amide I' 196 spectrum remains largely unchanged with both the main band near 1644 cm-1 and the 197 shoulder near 1673 cm-1 being present. However, structural changes become obvious 198 upon subtraction of the protein spectrum from the spectrum of the protein-vesicle mixture. 199 Subtractions of the second derivative spectra show negative features near 1680 and 1630 200 cm-1, indicating additional absorption when Lti30 interacts with membranes (Figures 5B 201 and C, black traces). These changes can be perceived in two ways: as (i) the appearance 202 of distinct absorption bands near 1680 and 1630 cm-1, or as (ii) a modification of the 203 structures sampled in the random coil state of Lti30, leading to a broadening of the main 204 band at 1644 cm-1 and an enhanced shoulder at 1673 cm-1. Both cases indicate that the 205 ensemble of Lti30 structures, or the range of interaction strengths, alters upon interaction 206 with vesicles. Interactions to protein carbonyls might for example strengthen due to 207 interactions with charges of the lipid head groups. Regarding the nature of the ensemble 208
www.plantphysiol.orgon July 7, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

8
alteration, the additional absorption near 1680 cm-1 is characteristic for turns, short 209 helices or the high wavenumber component of antiparallel β-sheets (Goormaghtigh et al., 210 1994; Barth and Zscherp 2002)(Nevskaya and Chirgadze 1976) and that near 1630 cm-1 211 for β-sheets or solvated α-helices (Martinez and Millhauser 1995; Reisdorf and Krimm 212 1996; Williams et al., 1996; Yoder et al., 1997; Barth and Zscherp 2002). Assignment of 213 the spectral changes to β-sheet formation is disfavoured by the generally low 214 susceptibility of monomeric Lti30 to form antiparallel β-sheets (Battaglia et al., 2008; 215 Koag et al., 2009; Thalhammer et al., 2010). At pH 5, we observe the appearance of a 216 signal at 1655 cm-1, which is characteristic of α-helices as found in globular proteins 217 (Goormaghtigh et al., 1994; Barth and Zscherp 2002). This 1655 cm-1 feature is in line 218 with the notion that Lti30 partially folds into regular, dehydrated helical segments upon 219 interaction with vesicles. Figures 5B and C show also the second-derivative spectra of 220 the vesicles alone (red traces), which are distinctly different from those of the protein-221 vesicles mixtures. In particular, differences are observed at 1750-1700 cm-1 where the 222 carbonyl stretching vibrations of the lipids absorb. These vibrations are sensitive to 223 hydrogen bonding and other parameters that affect the interfacial region of the lipids. 224 (Lewis, R. N. a H. & McElhaney, R. N. Membrane lipid phase transitions and phase 225 organization studied by Fourier transform infrared spectroscopy. Biochim. Biophys. Acta 226 1828, 2347–58 (2013). The band above 1740 cm-1 stems from non-hydrogen bonded 227 C=O groups, while the band below 1730 cm-1 comes from hydrogen bonded C=O groups. 228 When protein is present, the bands are further apart. The downshift of the band below 229 1730 cm-1 can be explained by stronger hydrogen bonded C=O groups. Moreover, the 230 Lti30 interaction decreases the amplitude of the band below 1730 cm-1. Conclusions 231 regarding the number of hydrogen bonded C=O groups, however, cannot be based on 232 amplitudes in second derivative spectra alone, as these also depend on the bandwidths of 233 the component bands. Therefore we analysed baseline corrected absorption spectra in 234 two ways: (i) we integrated the two sides of the lipid C=O band, i.e. above 1645 and 235 below 1625 cm-1 for non hydrogen bonded and hydrogen bonded groups, respectively, 236 and (ii) we fitted the lipid band with three component bands near 1723, 1736, and 1646 237 cm-1. The results show that the changes in band areas were only a few percents, 238 indicating that there is no significant change in the number of hydrogen bonded lipid 239
www.plantphysiol.orgon July 7, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

9
carbonyls upon addition of Lti30. The fits confirm also that the low wavenumber band of 240 hydrogen bonded C=O groups is downshifted in the presence of protein. Accordingly, in 241 the absence of lipid vesicles, Lti30 displays the characteristic fingerprint of a fully 242 disordered chain (Figure 5A), consistent with previous reports (Mouillon et al., 2006; 243 Mouillon et al., 2008). Then, upon interaction with DMPC:DMPG vesicles, Lti30 244 undergoes a conformational shift coupled to perturbation of the lipid bilayer (Figures 5B 245 and 5C). The nature of the latter is yet unclear but involves a reduction of the number of 246 gauche conformations in the acyl chains of the lipids, i.e. the fatty acids becomes more 247 restricted. 248 249 NMR-monitored folding of the conserved K-segment flanked by histidines. To map out at 250 atomic resolution the structural changes accompanying the membrane binding of Lti30 251 we used NMR. As a means to optimize structural resolution, we focused on the 252 interaction between the KLti30 -segment peptide with flanking histidines 253 (VHHEKKGMTEKVMEQLPGHHG) (Eriksson et al., 2011), which has previously been 254 shown to drive the Lti30-membrane association, and bicelles, i.e. mixed micelles of 255 DMPC/DMPG lipids and the detergent DHPC. Under the conditions in these 256 experiments, these bicelles form disc-shaped ellipsoids with a relatively flat lipid bilayer 257 surrounded peripherally by DHPC molecules (Andersson and Maler 2005; Lind et al., 258 2008). The free KLti30, is soluble in buffer and gives a good 1D 1H NMR spectrum, 259 typical for a fully disordered peptide (Figure 6). Upon addition of bicells, however, the 260 chemical-shift signatures of the KLti30-segment spectra undergo distinct alterations 261 consistent with binding (Figure 6). Notably, these spectra show no peaks corresponding 262 to those of the isolated K-segment, meaning either that 100 % of the peptides are bound 263 to the bicelles, or that the peptides are in fast, dynamic, exchange between free and bound 264 conformations where the observed chemical shifts are the population weighted averages. 265 In support of a scenario with dynamic exchange between free and bound peptide, the 266 NMR spectra in bicelle solution show quite narrow line width. In contrast, a 267 homogenous population of tightly bound species would display significant line 268 broadening, as the protein would here be slaved to the slower motions the large bicelles. 269 To clarify this issue we determined the translational diffusion coefficient (Dt) of the 270
www.plantphysiol.orgon July 7, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

10
isolated KLti30-segment in buffer as well as in bicellar solution. As the magnitude of Dt is 271 directly linked to the size of the diffusing object, small unbound KLti30-segments are 272 expected to diffuse faster ( ) than the much larger bicelles ( ). Thus, if all 273 KLti30-segments are firmly bound to the bicelles, i.e. co-diffuse, Dt should be the same for 274 both, i.e. approximately . In the case of dynamical exchange, on the other hand, 275 the observed Dt for the KLti30-segment ( ) is the population-weighted mean of the 276 diffusion of free peptide and the bicelle. The population of bound K-segments (pbound) at 277 every given time point can then be calculated from 278 279
bound = (Eq. 1). 280 281 In 100% D2O at 25°C, the unbound KLti30-segment yields = 1.42 ± 0.01 10-10 m2/s, 282 and the bicelles = 0.48 ± 0.01 10-10 m2/s, as determined with PFG NMR diffusion 283 experiments (Figure 5). The unbound KLti30-segment shows a diffusion coefficient that is 284 in good agreement with the expected value for an unfolded peptide in D2O (Danielsson et 285 al., 2002). In bicelle solution, however, the KLti30-segment yields = 0.97 ± 0.03 10-286 10 m2/s, suggesting a dynamic equilibrium where, at each moment, 48 % of the peptides 287 are bound to the bicelle (Eq. 1). Thus, the peptide is in unremitting exchange between 288 bound and free state, which is fast on the NMR time scale, both on the chemical shift 289 time scale and the diffusion time scale. Although the chemical shifts induced upon bicelle 290 binding are moderate at the level of population average (Figure 6), we conclude from 291 diffusion data that the chemical shift perturbations of the bound population are twice the 292 observed and, consequently, in line with folding of the KLti30-segment into ordered 293 structure. To determine at atomic resolution the structure of the membrane bound KLti30 294 we used deuterated bicelles and two-dimensional NMR. The results reveal six αHi-NHi+3 295 NOE couplings in the central part of the KLti30-segment (Figure 6C), constraining 296 residues G7 to Q15 into a well ordered α helix, when bound to bicelles (Figures 6 and 7). 297 The secondary chemical shifts that reports on secondary structure formation (Marsh et al., 298 2006), show approximately 40% fully formed helix at any given time-point, in line with a 299 dynamic equilibrium between bound and free state, in good agreement with diffusion 300
www.plantphysiol.orgon July 7, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

11
data. Interestingly, this helical structure shows archetypical amphipathic characteristics, 301 with one polar and one hydrophobic side stretching along its length. The amphipathic 302 properties of the short helix together with the fast exchanging dynamic equilibrium 303 between bound and free peptide are consistent with the structured peptide positioned on 304 the surface of the bicelle. Manually positioning the KLti30-segment structure into a lipid 305 bilayer positions the α helix parallel to the membrane plane, with the hydrophobic side 306 chains anchoring into the apolar regions of the lipids, and the polar/charged side chains 307 facing solvent and the hydrophilic environment of the lipid head groups (Figure 7). The 308 histidines of KLti30 are not part of the helix but are in good position to form electrostatic 309 contacts with the lipids. The induced folding of the conserved KLti30-segment suggests a 310 mechanism for reversible binding to negatively charged membranes. This reversible 311 binding corresponds well to that observed for several other intrinsically disordered 312 proteins upon binding specific protein targets (Jao et al., 2004; Ulmer and Bax 2005; 313 Ulmer et al., 2005). Moreover, a seemingly analogous helix induction was recently 314 observed by NMR for the dehydrin K2 upon binding to SDS micelles (Clarke et al., 2015), 315 pointing to the possibility that this mode of membrane interaction is a general feature of 316 the conserved K segments. 317 318
www.plantphysiol.orgon July 7, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

12
319 DISCUSSION. 320 Forces driving Lti30-membrane binding. To maintain in-cell solubility and freedom of 321 movement, biomolecules like proteins, membranes and DNA/RNA carry as a rule net 322 negative charge (Kurnik et al., 2012). In the few cases where proteins ‘violate’ this 323 principle of general negative repulsion by being net positive, this has been linked to 324 functional binding. Typical examples are the positively charged proteins associating with 325 DNA/RNA (Ellenberger et al., 1992; Niessing et al., 2004), and also local distributions of 326 positive side chains controlling trans-membrane protein topology (von Heijne 2006), 327 membrane binding (McLaughlin and Murray 2005; Moravcevic et al., 2010) molecular 328 dyes (Lira-De Leon et al., 2013) and protein-protein interaction (Sinha and Smith-Gill 329 2002; Muller et al., 2003; Bartels et al., 2011). In this context, it is intriguing that Lti30, 330 as well as some other members of the dehydrin family, not only stand out as ‘intrinsically 331 disordered’ but also as net positively charged (Supporting Table 1) (Eriksson et al., 2011). 332 The positive charges of these proteins are clustered in repetitive and highly conserved 333 sequence segments (Figure 1) that sometimes have the benefit of flanking histidines, 334 allowing local charge tuning (Supporting Table 1) (Eriksson et al., 2011). Together with 335 the regulation of membrane binding/net charge by phosphorylation (Eriksson et al., 2011) 336 and metal binding (Svensson et al., 2000; Alsheikh et al., 2003) this sets the stage for an 337 intricate physiological function. In this study we focus on the interaction between Lti30 338 and membranes, addressing the protein’s putative function in membrane protection 339 during environmental stress (Steponkus et al., 1998). The results show that the Lti30-340 membrane binding and macroscopic clustering of vesicles responds to the density of 341 negatively charged DOPG lipids (Figures 2 and 3). Accordingly, the interaction between 342 Lti30 and membranes is partly controlled by electrostatics, where the affinity can be 343 regulated by either protonation of the histidines flanking the protein’s K-segments 344 (Eriksson et al., 2011), or by changing the negative potential of the membrane. An 345 interesting detail is here that Lti30 manages to cluster LUVs even upon increasing their 346 DOPG content to 40 % and the inter-vesicular repulsion must be substantial (Figure 2). 347 Even though the spatial arrangement between protein and vesicles under these conditions 348 is yet unclear, the vesicular integrity seems maintained in the clusters as they do not leak 349
www.plantphysiol.orgon July 7, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

13
contained calcein (Eriksson et al., 2011) and are readily re-dispersed upon deprotonation 350 of Lti30 histidines at pH 9.0 (Supporting Figure 2). Also, Lti30-induced clustering of 351 purified thylakoids is reversed by increased ionic strength (Supporting Figure 2). Taken 352 together this points to an efficient and responsive mode of operation, as would be 353 expected if membrane binding is part of Lti30’s physiological action. 354 355 Structure of membrane-bound Lti30: the full-length protein. Following general 356 structure-function relationships, the question is then to what extent the interaction 357 between Lti30 and membranes involves specificity in terms of uniquely ordered protein 358 structure? To shed light on this issue, we used FTIR spectroscopy to probe for induction 359 of secondary structure. An advantage of this technique is that it can resolve subtle 360 rearrangements of the protein backbone in dense vesicle solutions where light scattering 361 renders conventional CD unsuitable. The results show that the Lti30 structure changes 362 from overall random coil to partly helical upon vesicle binding (Figure 5). Coil-to-helix 363 transitions, as a phenomenon, have previously been observed for dehydrins when 364 dehydration was simulated with glycerol and PEG (Mouillon et al., 2008) and LEA 365 proteins (Battaglia et al., 2008; Thalhammer et al., 2010) in desiccated samples, 366 reflecting the generic way any protein would satisfy backbone hydrogen-bonding 367 constraints in the absence of water (Tanford 1970). Similarly, dehydrins and LEA 368 proteins tend to increase their helical content when lowering the water potential by 369 osmolytic titration (Battaglia et al., 2008; Mouillon et al., 2008). In the current case, 370 however, the situation seems different as Lti30 forms helices upon binding to a putatively 371 physiological target in aqueous solution. This indicates that the structural response of 372 Lti30 yields functional specificity above that exerted by the electrostatics alone. 373 374 Structure and dynamics of membrane-bound Lti30: the KLti30. The molecular details of 375 the membrane-induced folding of Lti30 are revealed by the NMR structure of the bicelle-376 associated KLti30-segment (Figures 6 and 7). Corroborating FTIR data of the full-length 377 protein, the NMR structure of the bicelle-bound peptide shows a well-ordered α helix 378 stretching locally along residues of the KLti30-segment (i.e. G7 to Q15). Flanking this 379 helical unit, are the segments VHHEKK at N-terminal end and LPGHH at the C-terminal 380
www.plantphysiol.orgon July 7, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

14
end, none of which indicate fixed structural order (Figure 6). The helical structure 381 comprises also clear amphipathic sidedness, guiding the positioning of the K-segment to 382 the membrane surface (Figure 6 and 7). Judging by the disordered character of the 383 unbound KLti30-segment this docking could then occur either by conformational selection 384 of pre-formed helical segments or by induce folding-upon binding according the ‘fly 385 casting’ mechanism (Trizac et al., 2010). In either case, the NMR data shows that the 386 membrane-associated state is in rapid equilibrium (relaxation time faster than 387 approximate 300 s-1) with the unbound disordered state of the KLti30-segment, indicating 388 swift dynamic response. Even if the NMR analysis was done under conditions where the 389 KLti30-segment does not promote bicelle clustering, it is reasonable to assume that such 390 sandwiching of the α helical structure between membrane surfaces will significantly 391 reduce its binding dynamics. The membrane clustering in Figure 3 and Supplementary 392 figure 1 may thus be promoted by the amphipathic KLti30-segment helix binding 393 hydrophobically to one vesicle and electrostatically to the other, possibly with the aid of 394 the pH tuneable, flanking histidines (Lee et al., 2005; Eriksson et al., 2011). Although 395 this idea remains to be tested, it is clear from the ordered structure of the KLti30-segment 396 in Figures 6 and 7 that Lti30 has potential for higher degree of binding specificity and 397 diversity of targets than explored in this study. How such specificity can be achieved is 398 exemplified by the Zn fingers (Klug 2010), protein-protein interaction (Matthews et al., 399 2009) and the YscU protein (Weise et al., 2014). 400 401 402 Implications for biological function. In addition to the ubiquitous K-segment some 403 dehydrins contains additional conserved segments such as the Y and S-segments, the 404 final segment composition for each dehydrin also give the subgroup i.e K, SK, YSK, KS 405 and YK (Close 1996). In line with a broad functional repertoire of the dehydrin proteins, 406 it has been observed that different combinations of conserved segments are recruited at 407 different types of physiological stresses: lowered temperature triggers the expression of 408 K, SK and KS sub-family dehydrins (Nylander et al., 2001; Graether and Boddington 409 2014) whereas drought or increased salt exposure triggers expression of YSK dehydrins 410 (Nylander et al., 2001; Graether and Boddington 2014). Considering also the ability to 411
www.plantphysiol.orgon July 7, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

15
regulate the action of individual segments, the dehydrins may functionally be seen as a 412 permuted set of ‘Swiss-army knifes’ (Alexander et al., 2013), expressed as needed and 413 adjusted to their specific tasks by protonation (Eriksson et al., 2011), post-translational 414 phosphorylation (Alsheikh et al., 2003; Jiang and Wang 2004) and metal binding 415 (Svensson et al., 2000; Hara et al., 2005). When it comes to molecular mechanism, it is 416 suggested in this study that the K-segment fulfils a role in attaching to membranes 417 (Figure 6). Regarding the role of the other types of segments, there is yet no structural 418 information at hand. Even so, the S-segment has been identified as a key 419 phosphorylation site (Plana et al., 1991; Graether and Boddington 2014) that modulates 420 not only the proteins global charge, but also enables Ca2+ binding (Alsheikh et al., 2005). 421 Phosphorylation of S segment is further found to spatially relocate the dehydrin action by 422 moving the proteins out of the cell nucleus (Riera et al., 2004). The role of the Y 423 segment, on the other hand, is currently more difficult to envisage. From sequence blast 424 it is notable that the Y segment shows similarity with the ATP- binding motifs of the 425 chaperones GroEL and GroES (Martin et al., 1993), hinting at a role in molecular 426 recognition. The dehydrin MtCAs31, which comprises 2 Y and 4 K segments, has 427 likewise been observed to interact with the stress-associated ICE transcription factor, 428 yielding decreased number of stomata in leafs (Xie et al., 2012; Peng et al., 2014). In 429 addition to the K, S and Y segments, some dehydrins contain also H repeats found to 430 control homodimerisation (Hernandez-Sanchez et al., 2014) as well as ‘charge segments’ 431 implicated in nuclear targeting and DNA binding (Hara et al., 2005; Rosales et al., 2014). 432 Interestingly, these charge segments show further sequence similarity with a linker region 433 in HSP 90 (Mouillon et al., 2006). Several physical dimensions of the cell will be 434 affected by desiccation, such as the shape and proximity of membrane structures. As a 435 consequence, desiccation is also expected to alter the dynamics of membrane fusion, 436 budding, distort phase transitions and contribute to topological frustration. By binding to 437 membranes, the role of Lti30 may be to interfere in the progression of any these 438 processes in order to keep the membranes biologically viable. Regardless of functional 439 mode, the elements of recognition with the cellular target are likely to be the conserved 440 segments, such as the K-segment of Lti30 (Figure 7). Taken together, the diversity of 441 conserved sequence segments among the dehydrins points to a complex and versatile 442
www.plantphysiol.orgon July 7, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

16
biological function, evolvable in a simplistic modular fashion to meet new needs. A 443 pressing task is now to establish how the intriguing behaviour of dehydrins in vitro 444 translates to the crowded conditions in vivo, where other putative interaction partners will 445 likely compete with, and modulate, membrane binding. Also, it remains to find out to 446 what extent the observed behaviour in vitro reflects true functional traits, or simply 447 reports on the generic propensities of unstructured proteins (Thalhammer et al., 2014, 448 Popova et al., 2015)). In any case, the molecular clues emerging from this and other 449 studies stand out as a solid base for mapping out in detail how this unique class of 450 disordered proteins works, and can be rationally interfered with, in stressed plant cells. 451 452
www.plantphysiol.orgon July 7, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

17
MATERIAL AND METHODS 453 454 Protein production. Expression, purification and identification of the recombinant 455 Arabidopsis thaliana dehydrin Lti30 were performed as described by (Svensson et al., 456 2000), with minor changes. Glycerol stocks of the Escherichia coli strain were made and 457 150 μl was spread on Luria agar plates with 150 μg ampicillin and grown 37 °C 458 overnight. The cells were suspended and added to 2 L of Luria-Bertani medium 459 containing 50 μg/mL ampicillin and kept at 37 °C. Expression was induced at OD600 of 460 0.6-0.7 by adding 1 mM isopropyl B-D-thiogalactopyranoside and kept at 23 °C over 461 night. Cells were harvested by centrifugation at 6000 rpm for 15 min. The pellet from 1 462 L cultures was suspended in 25 mL of 20 mM Na2HPO4, pH 7.2 and 150 mM NaCl, 1 463 mM phenylmethylsulfonyl flourid and 1 tablet Complete (Roche). Cells were sonicated 464 for five 1-minute periods on ice followed by centrifugation at 18 000 rpm for 30 minutes. 465 The supernatant was placed in 80 °C water bath for 30 minutes, to precipitate heat-466 denatured proteins, and then centrifuged at 18 000 rpm for 30 minutes. Lti30 was 467 purified by metal ion affinity chromatography. The supernatant from heat precipitation 468 was diluted 1:2 with 20 mM Na2HPO4, pH 7.2, 1.88 M NaCl and 1 mM 469 phenylmethylsulfonyl fluoride. The sample was loaded on a 5-mL HiTrap IDA-Sepharos 470 column (GE Healthcare) charged with 7 mL of 3 mg/mL CuSO4. The column was 471 equilibrated with 5 volumes of 20 mM Na2HPO4, pH 7.2, and 1.0 M NaCl. The same 472 buffer (40 volumes) was used to wash off unbound sample from the column. Fractions of 473 5 mL were collected for analysis during the whole run. Elution was performed with 2 M 474 NH4Cl in 20 mM Na2HPO4, pH 7.2, and 1.0 M NaCl in one step. The column was then 475 equilibrated with 10 volumes of 20 mM Na2HPO4, pH 7.2, followed by elution of the 476 copper with 10 mM EDTA in 20 mM Na2HPO4, pH 7.2. Precipitation of protein was 477 performed with 80% (NH4)2SO4 over night and protein was collected by centrifugation at 478 18 000 rpm for 45 minutes. Lti30 pellet was suspended with 2.5 mL of 5 mM MES, pH 479 6.3, and in case of FTIR 2.5 mL dH2O. The protein was desalted on 2 PD-10 columns 480 (GE Healthcare). Purity was analyzed by Ready gel SDS-PAGE system (Biorad). Protein 481 quantification was measured with bicinchoninic acid assay (Sigma-Aldrich). 482 483
www.plantphysiol.orgon July 7, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

18
Preparation of LUVs and bicells. All lipids were purchased from Avanti Polar Lipids. 484 LUVs of DOPC:DOPG, were prepared with an extrusion method. The lipids were 485 dissolved in chloroform, and lipid mixtures were dried under a gentle flow of fluid 486 nitrogen for 3-4 hours. The lipid film was solved in 5 mM MES pH 6.3 and then vortexed 487 for 10 minutes, for FTIR experiments the lipids were solved in 10 mM of potassium-488 phosphate in D2O at pH 5.0 and 8.6. 5 freeze thaw cycles in liquid nitrogen to reduce 489 lamellarity followed and then the lipid solution was extruded 20 times through an hand-490 driven extruder with 0.1 μm pore-size polycarbonate filter. The measurements of size 491 distribution of produced LUVs were done on a ALV/CGS-3 compact goniometer system 492 (ALV-GmbH, Langen, Germany) with a HeNe laser (632.8 nm), with a scattering angle 493 of 150 degrees. Bicells for NMR-measurements were prepared with 1,2-dihexanoyl-sn-494 glycero-3-phosphocoline (DHPC) as a detergent. DMPC:DMPG (4:1) lipids were used to 495 provide the bilayer of the bicells. Bicells with q=0.3 (q=([DMPC]+[DMPG])/[DHPC], q 496 equals the molar ratio of lipids and detergents) were prepared by mixing DMPC:DMPG 497 (4:1) with 20 mM of potassium-phosphate pH 6.3. The mixture was vortexed followed 498 by centrifugation 13 300 rpm for 1 minute, this cycle was repeated until the solution 499 looked homogenous. DHPC was added and the vortex-centrifuged cycle was repeated 500 until a clear non-viscous lipid-solution was obtained. 501 502 Thylakoid Membrane Preparation. Forty grams of spinach (Spinacia oleracea) and 100 503 mL of 0.3 M Sucrose, 5 mM MgCl2 and 50 mM Na-phosphate, pH 7.4 was placed in a 504 cold mixer and mixed for 5 x 10 s. The solution was filtered and centrifuged at 3000 rpm 505 for 3 min, 4 °C. The pellet was suspended in 30 mL of 0.3 M sucrose, 5 mM MgCl2 and 506 50 mM Na-phosphate pH 7.4 and centrifuged at 4500 for 5 min, 4 °C. The solution was 507 homogenized in 30 mL 5 mM MgCl2, 5 mM NaCl and 10 mM Na-phosphate, pH 7.4 and 508 centrifuged at 4500 rpm for 5 min, 4 °C. The pellet was homogenized in 12 mL of 0.1 M 509 sucrose, 5 mM MgCl2, 5 mM NaCl and 10 mM phosphate, pH 7.4. 510 511 Stopped flow measurements. Scattering of protein-lipid cluster formation were monitored 512 on SX.18-MV stopped flow fluorimeter (Applied Photophysics). Excitation was at 400 513
www.plantphysiol.orgon July 7, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

19
nm, and all measurements were done at room temperature in 5 mM MES pH 6.3. Protein 514 concentration was after mixing 3.5 μM and lipid concentration was 0.5 mM. 515 Absorbance measurements. Absorbance measurements were performed on an Ultrospec 516 3300 pro (Amersham) at 400 nm and at 25 °C. 517 518 Light microscopic pictures. To examine light microscopic pictures an inverted Zeiss 519 Axiovert 40 CFL microscope equipped with a digital Aciocam ICc1 camera at 10 times 520 magnification was used. Solution of Lti30 (14 μM) together with vesicles (1.4 mM) was 521 added on a glass slide and three images per droplet were examined. 522 523 FTIR. All samples were prepared with 10 mM potassium-phosphate in D2O at pH 5.0 or 524 8.6. Samples containing Lti30 had a concentration of 0.2 mM and samples containing 525 lipids of DOPC:DOPG (3:1) had a concentration of 8 mM. Samples of 3 μL were 526 deposited on a CaF2 window with a trough of 55 μm, covered with a flat CaF2 window 527 and placed in the spectrometer. Infrared spectra were recorded with a Tensor 37 Fourier-528 transform spectrometer (Bruker Optics, Germany), equipped with a liquid N2-cooled 529 HgCdTe detector and continuously purged with CO2-free, dry air. Samples were 530 mounted on a two-position sample shuttle, which allowed for interleaved acquisition of 531 sample and background spectra. A wait time of 15 min was allowed after inserting the 532 sample to ensure complete purging. All measurements were made at room temperature. 533 Interferograms were recorded at a resolution of 2 cm−1, apodised using a 3-term 534 Blackman–Harris apodisation function and Fourier-transformed with a zero-filling factor 535 of 2. Sixteen consecutive interferograms were averaged to obtain a single sample or 536 background spectrum. Infrared spectra were recorded and analyzed using the OPUS 537 software from the instrument manufacturer. Second derivative spectra were calculated 538 using a smoothing length of 13 data points (approx. 13 cm−1). 539 540 NMR. In all NMR experiments we used a un-labelled synthesized peptide, and the 541 experiments where performed at 298K, on a Bruker Avance 600MHz spectrometer 542 equipped with a room temperature probe with z-gradients. To obtain proton assignment 543 we recorded two homonuclear TOCSY experiments with 80 and 100 ms Hartman-Hahn 544
www.plantphysiol.orgon July 7, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

20
mixing times (Bax and Davis 1985), two homonuclear NOESY experiments with 150 and 545 250 ms mixing times (Bax and Davis 1985; Hwang and Shaka 1995). The NOESY 546 experiments were also used for identification of long range contacts. All 2D experiments 547 were recorded with 512 increments in the indirect detected dimension, and used 548 excitation sculpting to reduce the water signal. Processing was performed in nmrPipe 549 (Delaglio et al., 1995) and analysis was performed using Sparky (T. D. Goddard and D. G. 550 Kneller, SPARKY 3, University of California, San Francisco). 551 The structure was generated by using PEP-fold (Maupetit et al., 2009) and using 6 552 experimentally determined (i, i+3) HαHN distances as constraints when generating the 553 structure. The obtained coarse grained structure was further energy-minimized using the 554 Yasara force field (Krieger et al., 2009) and the best 5 structures were selected as 555 representative models for KLti30 peptide structure in membrane mimetica. The PDB 556 representation of the double-layered lipid membrane was generated using the CHARMM-557 GUI membrane builder (Jo et al., 2007; Jo et al., 2008; Jo et al., 2009; Wu et al., 2014). 558 Diffusion experiments were performed in 100% D2O at 298K. A list of 16 linearly spaced 559 gradient strengths was used. Gradient pulse length was set to 5ms and diffusion time was 560 set to 100ms. Diffusion coefficients were obtained by integration of the aromatic side-561 chain protons. 562 563 Estimation of the end-to-end distance of the vesicle-bound Lti30. The size dimension of 564 Lti30 located on a vesicle surface was estimated as the end-to-end distance of a self-565 avoiding random walk of a bead-on-a-string Cα model on a two-dimensional surface. 566 The overall dimensions (e.g. radius of gyration) scales similarly as the end-to-end 567 distance. Here we model the protein by a chain of N = 193 spheres with 3.5 Å diameter, 568 linked by 3.88 Å long bonds. No overlap of the spheres is allowed. As Lti30 is an 569 intrinsically disordered protein the size is a distribution of sizes, rather than a well-570 defined value. To estimate the distribution we calculated 104 chains, using an in-house 571 developed script, and found that the end-to-end distance, as expected, follow a skewed 572 Gaussian distribution where the mean end-to-end distance is 7.5 ± 3.5 nm. In the vesicle 573 bound state Lti30 coordinates the charged lipid head groups, and as these are sparsely and 574 evenly distributed over the vesicle surface, we assume Lti30 to be significantly more 575
www.plantphysiol.orgon July 7, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

21
extended than a random Gaussian chain. Therefore, to estimate the length distribution on 576 such vesicles we only included the 2% most extended conformations of the ensemble. 577 This sub-ensemble has a mean end-to end-distance of 20.4 ± 2.5 nm. 578 579
www.plantphysiol.orgon July 7, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

22
580 FIGURE LEGENDS 581 Figure 1. Schematic picture of disordered Lti30 binding to lipid (PC:PG) 582 vesicles. Blue is positive charge and red is negative charge. 583 (A) Sequence of Lti30 with K-segments (bold) and the positions of the K 584 segments matching the KLti30 peptide (gray boxes). (B) Sequence of the KLti30 585 peptide. (C) Binding equilibrium between Lti30 and lipid vesicles where 586 attraction is influenced by charge. Binding is promoted by protonation of the 587 flanking His pairs (Eriksson et al., 2011), and down regulated by phosphorylation 588 of the K segments Treonines (Eriksson et al., 2011). Upon Lti30 binding, the 589 membrane phase decreases 2.5 °C and the vesicles cluster also into 590 macroscopic aggregates (Eriksson et al., 2011). For size reference, Lti30 has an 591 extended length of 20 nm and the diameter of the vesicles is 100 nm. 592 593 Figure 2. Stopped-flow data of Lti30 binding and clustering PC:PG lipid vesicles 594 (LUVs). 595 3.5 μM Lti30 was rapidly mixed with 0.5 mM PC:PG vesicles at 1:142 protein:lipid 596 ratio. Binding of positively charged Lti30 promotes vesicle clustering monitored 597 by scattering at 400 nm. The process depends on the LUV net negative charge, 598 which was varied between PG (1-40%) and PC (99-60%). (A) Scattering at 400 599 nm. (B) Absorbance at 400 nm. (C) Scattering amplitude at 400 nm at time = 600 0.5 seconds indicating two binding modes. 601 602 Figure 3. Light-microscope analysis of the Lti30-induced clustering of PC:PG 603 LUVs . 604 Lti30 (14 μM) in the presence of LUVs (1.4 mM) with 1, 3 or 25 % PG at pH 6.3. 605 Protein-to-lipid ratio is 1:100. The schematic cartoons show a reductionist 606 interpretation of data: Lti30 (blue) and LUVs with increasing content of negatively 607 charged PG (red). (A) Lti30 and PC:PG (99:1). No visible clusters. (B) Lti30 and 608 PC:PG (97:3). Small dispersed clusters less than 5μm. (C) Lti30 and PC:PG 609 (75:25). Clear macroscopic clusters, larger than 5μm (reduced magnification, 610 size bar 50 μm). 611 612 613 614 Figure 4. Vesicle clustering by versions of K-segments correlates to peptide net 615 charge. 616
www.plantphysiol.orgon July 7, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

23
Titration of different versions of the K-segment into 0.5mM PC:PG vesicles (1:3) 617 at pH 6.3 (blue lines) or pH 4.3 (red lines). Vesicle clustering by K-peptides 618 correlates to peptide net charge. KLti30 represent one type of K- segment in Lti30 619 and KC is the generic K-segment and KHHCHH is with the addition of histidine 620 flanks. KHHCHH bind regardless of pH, i.e. the pH tuned binding as with KLti30, is 621 lost. Clustering of lipid vesicles measured by absorbance at 400nm. The net 622 charge of the peptides are calculated by http://protcalc.sourceforge.net. 623 Figure 5. FTIR spectra of Lti30 only and mixed with PC:PG vesicles. 624 (A) Second derivative IR absorption spectra of Lti30 alone in the amide I region 625 at pH 5 (black) and pH 8.6 (red). (B) and (C) Corresponding spectra of vesicles 626 alone (red line) and the Lti30-vesicle mix (black line) at pH 5 and pH 8.6, 627 respectively. 628 629 Figure 6. Structural analysis of membrane bound KLti30 peptide by NMR. 630 (A) The five best structures of bicelle-bound KLti30 peptide from 1H-NMR 631 constraints, where the central nine residues of the peptide adopt a fixed α-helix, 632 whereas the N and C termini are more disordered. (B) Secondary-structure 633 propensity (SSP) from 1Ha, 1HN and 1Hb chemical shifts, where positive values 634 show the induced α-helix in the central part of the peptide. Negative SSP values 635 indicate extended conformation in the N-terminal region of the peptide. (C) 636 Determined NOE connectivity between peptide residues along the KLti30. The 637 ordered n+3 couplings within the residue segment G7 to Q15 are the hallmark of 638 α-helical structure. 639 640 Figure 7. The atomic-resolution structure of the membrane binding domain of 641 Lti30. 642 (A) (B) (C) Atomic model of the membrane-binding KLti30-segment of Lti30, in 643 side, diagonal and front views, respectively. Positively charged residues are in 644 blue, negatively charged residues in red, and the hydrophobic side of the 645 amphipathic α-helix faces downwards. (D) NMR diffusion data of KLti30 alone, 646 bicelles alone and KLti30 mixed with bicelles, verifying KLti30-micelle binding. (E) 647 Modeling of the positioning of the histidine flanked KLti30-segment into a lipid 648 bilayer (PC:PG 98:2 mix), using periodic boundary conditions. Consistent with 649 the binding data in Figures 2-5, the positive side chains H2, H3, K5 and K6 650 coordinate the negative charge of a PG head group. 651 652 List of supplemental materials. 653 Supplementary Table 1. Summary of positive net charge among LEA proteins. 654 Sequence annotations from data in Battaglia et al., 2008. 655
www.plantphysiol.orgon July 7, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

24
Supplementary Figure 1. Light-microscope analysis of the K-segment induced 656 clustering of PC:PG LUVs 657 Light-microscopy analysis showing that vesicle clustering by different K-segments 658 follows pH and thus peptide net charge. Peptide concentration 0.7 M in 1.4 mM PC:PG 659 vesicles at 1:3 ratio. 660 Supplementary Figure 2. Effects of NaCl on thylakoid and LUV clustering byLti30 and 661 KLti30. 662 (A) Light microscopy image of isolated thylakoid membranes (0.2mg/ml) from spinach. 663 (B) Thylakoid membranes (0.2mg/ml) in Lti30 (0.2mg/ml). (C) Thylakoid membranes 664 (0.2mg/ml) and Lti30 (0.2mg/ml), showing that clustering of Membranes can be reversed 665 by 20 mM NaCl. (D) Clustering of LUVs (1.4 mM PC:PG 3:1 molar ratio in the presence 666 of KLti30 peptide (0.3 mg/ml) at pH 4.3 titrated with NaCl. Light scattering measured by 667 absorbance at 400nm. 668 Supplementary Figure 3. FTIR spectra of Lti30, lipid vesicles or a mix of Lti30 and 669 vesicles. 670 The figure shows the absorption spectra of the pH 5 experiment. Spectra are shifted 671 vertically for a clearer presentation. The blue spectrum is for the protein vesicle mixture, 672 the red spectrum for the protein sample, the green spectrum for the vesicle sample and the 673 black spectrum is a buffer spectrum. The spectral range shown is between the strong 674 absorption bands of the stretching and bending vibrations of D2O. The spectra are 675 largely dominated by the absorption of HOD and D2O. Note that the buffer spectra is not 676 subtracted form the other spectra before we calculated the second derivatives, shown in 677 the other figures. Since the buffer spectrum contains only broad bands, its second 678 derivative is featureless above 1600 cm-1. 679 680 681
www.plantphysiol.orgon July 7, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

25
682 683 Table 1. Summary of dehydrins previously observed to bind lipid vesicles and reported 684 corresponding structural transitions. 685 686
Protein Plant Type of lipid Gain of structure References
DHN1 Zea PA (100 %) α-helix
(CD)
Koag et al., 2003
Lti30 Arabidopsis PC:PG (3:1) α-helix
(MNR, FTIR)
ERD10 Arabidopsis PC:PS (1:1) Disordered
(CD)
Kovacs et al., 2008
ERD14 Arabidopsis PC:PS (1:1) Disordered
(CD)
Kovacs et al., 2008
TsDHN1 Thellungiella PC:PS:PI
(33:47:20) β-strand, α-helix
(FTIR, CD)
Rahman et al.,
2010
TsDHN2 Thellungiella PC:PS:PI
(33:47:20) β-strand, α-helix
(FTIR, CD)
Rahman et al.,
2010
K2 Vitis riparia PC:PA (1:1) α-helix
(CD)
Clarke et al., 2015
687 688
www.plantphysiol.orgon July 7, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

26
Acknowledgements. We are grateful to Prof. Mikael Oliveberg for helpful discussions. 689 690 691 692 REFERENCES 693 694 Alexander CG, Jurgens MC, Shepherd DA, Freund SM, Ashcroft AE, Ferguson N 695 (2013) Thermodynamic origins of protein folding, allostery, and capsid formation in 696 the human hepatitis B virus core protein. Proc Natl Acad Sci U S A 110(30): E2782-697 2791 698 Alsheikh MK, Heyen BJ, Randall SK (2003) Ion binding properties of the dehydrin 699 ERD14 are dependent upon phosphorylation. J Biol Chem 278(42): 40882-40889 700 Alsheikh MK, Svensson JT, Randall SK (2005) Phosphorylation regulated ion-701 binding is a property shared by the acidic subclass dehydrins. Plant Cell and 702 Environment 28(9): 1114-1122 703 Andersson A, Maler L (2005) Magnetic resonance investigations of lipid motion in 704 isotropic bicelles. Langmuir 21(17): 7702-7709 705 Arai M, Sugase K, Dyson HJ, Wright PE (2015) Conformational propensities of 706 intrinsically disordered proteins influence the mechanism of binding and folding. 707 Proc Natl Acad Sci U S A 112(31): 9614-9619 708 Baker J, Steele C, Dure L (1988) Sequence and Characterization of 6 Lea Proteins 709 and Their Genes from Cotton. Plant Molecular Biology 11(3): 277-291 710 Bartels T, Choi JG, Selkoe DJ (2011) alpha-Synuclein occurs physiologically as a 711 helically folded tetramer that resists aggregation. Nature 477(7362): 107-110 712 Barth A, Zscherp C (2002) What vibrations tell us about proteins. Q Rev Biophys 713 35(4): 369-430 714 Battaglia M, Olvera-Carrillo Y, Garciarrubio A, Campos F, Covarrubias AA 715 (2008) The enigmatic LEA proteins and other hydrophilins. Plant Physiol 148(1): 6-716 24 717 Bomal C, Le VQ, Tremblay FM (2002) Induction of tolerance to fast desiccation in 718 black spruce (Picea mariana) somatic embryos: relationship between partial water 719 loss, sugars, and dehydrins. Physiol Plant 115(4): 523-530 720 Borg M, Mittag T, Pawson T, Tyers M, Forman-Kay JD, Chan HS (2007) 721 Polyelectrostatic interactions of disordered ligands suggest a physical basis for 722 ultrasensitivity. Proc Natl Acad Sci U S A 104(23): 9650-9655 723 Candat A, Paszkiewicz G, Neveu M, Gautier R, Logan DC, Avelange-Macherel MH, 724 Macherel D (2014) The ubiquitous distribution of late embryogenesis abundant 725 proteins across cell compartments in Arabidopsis offers tailored protection against 726 abiotic stress. Plant Cell 26(7): 3148-3166 727 Chakrabortee S, Boschetti C, Walton LJ, Sarkar S, Rubinsztein DC, Tunnacliffe A 728 (2007) Hydrophilic protein associated with desiccation tolerance exhibits broad 729 protein stabilization function. Proc Natl Acad Sci U S A 104(46): 18073-18078 730 Clarke MW, Boddington KF, Warnica JM, Atkinson J, McKenna S, Madge J, 731 Barker CH, Graether SP (2015) Structural and Functional Insights into the 732 Cryoprotection of Membranes by the Intrinsically Disordered Dehydrins. J Biol 733 Chem 290(45): 26900-26913 734
www.plantphysiol.orgon July 7, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

27
Close TJ (1996) Dehydrins: Emergence of a biochemical role of a family of plant 735 dehydration proteins. Physiologia Plantarum 97(4): 795-803 736 Crowe LM, Crowe JH (1992) Anhydrobiosis: a strategy for survival. Adv Space Res 737 12(4): 239-247 738 Danielsson J, Jarvet J, Damberg P, Graslund A (2002) Translational diffusion 739 measured by PFG-NMR on full length and fragments of the Alzheimer A beta(1-40) 740 peptide. Determination of hydrodynamic radii of random coil peptides of varying 741 length. Magnetic Resonance in Chemistry 40: S89-S97 742 Danyluk J, Perron A, Houde M, Limin A, Fowler B, Benhamou N, Sarhan F 743 (1998) Accumulation of an acidic dehydrin in the vicinity of the plasma membrane 744 during cold acclimation of wheat. Plant Cell 10(4): 623-638 745 Delaglio F, Grzesiek S, Vuister GW, Zhu G, Pfeifer J, Bax A (1995) Nmrpipe - a 746 Multidimensional Spectral Processing System Based on Unix Pipes. Journal of 747 Biomolecular Nmr 6(3): 277-293 748 Dunker AK, Obradovic Z (2001) The protein trinity--linking function and disorder. 749 Nat Biotechnol 19(9): 805-806 750 Dure L, 3rd (1993) A repeating 11-mer amino acid motif and plant desiccation. 751 Plant J 3(3): 363-369 752 Ellenberger TE, Brandl CJ, Struhl K, Harrison SC (1992) The GCN4 basic region 753 leucine zipper binds DNA as a dimer of uninterrupted alpha helices: crystal 754 structure of the protein-DNA complex. Cell 71(7): 1223-1237 755 Eriksson SK, Kutzer M, Procek J, Grobner G, Harryson P (2011) Tunable 756 membrane binding of the intrinsically disordered dehydrin Lti30, a cold-induced 757 plant stress protein. Plant Cell 23(6): 2391-2404 758 Goormaghtigh E, Cabiaux V, Ruysschaert JM (1994) Determination of soluble and 759 membrane protein structure by Fourier transform infrared spectroscopy. I. 760 Assignments and model compounds. Subcell Biochem 23: 329-362 761 Graether SP, Boddington KF (2014) Disorder and function: a review of the 762 dehydrin protein family. Front Plant Sci 5: 576 763 Hara M, Fujinaga M, Kuboi T (2005) Metal binding by citrus dehydrin with 764 histidine-rich domains. J Exp Bot 56(420): 2695-2703 765 Hara M, Shinoda Y, Tanaka Y, Kuboi T (2009) DNA binding of citrus dehydrin 766 promoted by zinc ion. Plant Cell Environ 32(5): 532-541 767 Hara M, Terashima S, Kuboi T (2001) Characterization and cryoprotective activity 768 of cold-responsive dehydrin from Citrus unshiu. Journal of Plant Physiology 769 158(10): 1333-1339 770 Hernandez-Sanchez IE, Martynowicz DM, Rodriguez-Hernandez AA, Perez-771 Morales MB, Graether SP, Jimenez-Bremont JF (2014) A dehydrin-dehydrin 772 interaction: the case of SK3 from Opuntia streptacantha. Front Plant Sci 5: 520 773 Ingram J, Bartels D (1996) The Molecular Basis of Dehydration Tolerance in Plants. 774 Annu Rev Plant Physiol Plant Mol Biol 47: 377-403 775 Jao CC, Der-Sarkissian A, Chen J, Langen R (2004) Structure of membrane-bound 776 alpha-synuclein studied by site-directed spin labeling. Proc Natl Acad Sci U S A 777 101(22): 8331-8336 778
www.plantphysiol.orgon July 7, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

28
Jarvet J, Danielsson J, Damberg P, Oleszczuk M, Graslund A (2007) Positioning of 779 the Alzheimer Abeta(1-40) peptide in SDS micelles using NMR and paramagnetic 780 probes. J Biomol NMR 39(1): 63-72 781 Jiang X, Wang Y (2004) Beta-elimination coupled with tandem mass spectrometry 782 for the identification of in vivo and in vitro phosphorylation sites in maize dehydrin 783 DHN1 protein. Biochemistry 43(49): 15567-15576 784 Jo S, Kim T, Im W (2007) Automated builder and database of protein/membrane 785 complexes for molecular dynamics simulations. PLoS One 2(9): e880 786 Jo S, Kim T, Iyer VG, Im W (2008) CHARMM-GUI: a web-based graphical user 787 interface for CHARMM. J Comput Chem 29(11): 1859-1865 788 Jo S, Lim JB, Klauda JB, Im W (2009) CHARMM-GUI Membrane Builder for mixed 789 bilayers and its application to yeast membranes. Biophys J 97(1): 50-58 790 Klug A (2010) The discovery of zinc fingers and their development for practical 791 applications in gene regulation and genome manipulation. Q Rev Biophys 43(1): 1-792 21 793 Koag MC, Fenton RD, Wilkens S, Close TJ (2003) The binding of maize DHN1 to 794 lipid vesicles. Gain of structure and lipid specificity. Plant Physiol 131(1): 309-316 795 Koag MC, Wilkens S, Fenton RD, Resnik J, Vo E, Close TJ (2009) The K-segment of 796 maize DHN1 mediates binding to anionic phospholipid vesicles and concomitant 797 structural changes. Plant Physiol 150(3): 1503-1514 798 Kovacs D, Kalmar E, Torok Z, Tompa P (2008) Chaperone activity of ERD10 and 799 ERD14, two disordered stress-related plant proteins. Plant Physiol 147(1): 381-390 800 Krieger E, Joo K, Lee J, Lee J, Raman S, Thompson J, Tyka M, Baker D, Karplus K 801 (2009) Improving physical realism, stereochemistry, and side-chain accuracy in 802 homology modeling: Four approaches that performed well in CASP8. Proteins-803 Structure Function and Bioinformatics 77: 114-122 804 Kriwacki RW, Hengst L, Tennant L, Reed SI, Wright PE (1996) Structural studies 805 of p21Waf1/Cip1/Sdi1 in the free and Cdk2-bound state: conformational disorder 806 mediates binding diversity. Proc Natl Acad Sci U S A 93(21): 11504-11509 807 Kurnik M, Hedberg L, Danielsson J, Oliveberg M (2012) Folding without charges. 808 Proc Natl Acad Sci U S A 109(15): 5705-5710 809 Lee CW, Martinez-Yamout MA, Dyson HJ, Wright PE (2010) Structure of the p53 810 transactivation domain in complex with the nuclear receptor coactivator binding 811 domain of CREB binding protein. Biochemistry 49(46): 9964-9971 812 Lee SA, Eyeson R, Cheever ML, Geng J, Verkhusha VV, Burd C, Overduin M, 813 Kutateladze TG (2005) Targeting of the FYVE domain to endosomal membranes is 814 regulated by a histidine switch. Proc Natl Acad Sci U S A 102(37): 13052-13057 815 Lemmon MA (2008) Membrane recognition by phospholipid-binding domains. Nat 816 Rev Mol Cell Biol 9(2): 99-111 817 Lind J, Nordin J, Maler L (2008) Lipid dynamics in fast-tumbling bicelles with 818 varying bilayer thickness: effect of model transmembrane peptides. Biochim 819 Biophys Acta 1778(11): 2526-2534 820 Lira-De Leon KI, Garcia-Gutierrez P, Serratos IN, Palomera-Cardenas M, 821 Figueroa-Corona Mdel P, Campos-Pena V, Meraz-Rios MA (2013) Molecular 822 mechanism of tau aggregation induced by anionic and cationic dyes. J Alzheimers 823 Dis 35(2): 319-334 824
www.plantphysiol.orgon July 7, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

29
Marsh JA, Singh VK, Jia Z, Forman-Kay JD (2006) Sensitivity of secondary 825 structure propensities to sequence differences between alpha- and gamma-826 synuclein: implications for fibrillation. Protein Sci 15(12): 2795-2804 827 Martin J, Geromanos S, Tempst P, Hartl FU (1993) Identification of nucleotide-828 binding regions in the chaperonin proteins GroEL and GroES. Nature 366(6452): 829 279-282 830 Martinez G, Millhauser G (1995) FTIR spectroscopy of alanine-based peptides: 831 assignment of the amide I' modes for random coil and helix. J Struct Biol 114(1): 23-832 27 833 Matthews JM, Bhati M, Lehtomaki E, Mansfield RE, Cubeddu L, Mackay JP 834 (2009) It takes two to tango: the structure and function of LIM, RING, PHD and 835 MYND domains. Curr Pharm Des 15(31): 3681-3696 836 Maupetit J, Derreumaux P, Tuffery P (2009) PEP-FOLD: an online resource for de 837 novo peptide structure prediction. Nucleic Acids Research 37: W498-W503 838 McLaughlin S, Murray D (2005) Plasma membrane phosphoinositide organization 839 by protein electrostatics. Nature 438(7068): 605-611 840 Moravcevic K, Mendrola JM, Schmitz KR, Wang YH, Slochower D, Janmey PA, 841 Lemmon MA (2010) Kinase associated-1 domains drive MARK/PAR1 kinases to 842 membrane targets by binding acidic phospholipids. Cell 143(6): 966-977 843 Mouillon JM, Eriksson SK, Harryson P (2008) Mimicking the plant cell interior 844 under water stress by macromolecular crowding: disordered dehydrin proteins are 845 highly resistant to structural collapse. Plant Physiol 148(4): 1925-1937 846 Mouillon JM, Gustafsson P, Harryson P (2006) Structural investigation of 847 disordered stress proteins. Comparison of full-length dehydrins with isolated 848 peptides of their conserved segments. Plant Physiol 141(2): 638-650 849 Muller JJ, Lapko A, Ruckpaul K, Heinemann U (2003) Modeling of electrostatic 850 recognition processes in the mammalian mitochondrial steroid hydroxylase system. 851 Biophys Chem 100(1-3): 281-292 852 Nevskaya NA, Chirgadze YN (1976) Infrared spectra and resonance interactions of 853 amide-I and II vibration of alpha-helix. Biopolymers 15(4): 637-648 854 Niessing D, Huttelmaier S, Zenklusen D, Singer RH, Burley SK (2004) She2p is a 855 novel RNA binding protein with a basic helical hairpin motif. Cell 119(4): 491-502 856 Nylander M, Svensson J, Palva ET, Welin BV (2001) Stress-induced accumulation 857 and tissue-specific localization of dehydrins in Arabidopsis thaliana. Plant Mol Biol 858 45(3): 263-279 859 Peng PH, Lin CH, Tsai HW, Lin TY (2014) Cold response in Phalaenopsis aphrodite 860 and characterization of PaCBF1 and PaICE1. Plant Cell Physiol 55(9): 1623-1635 861 Petersen J, Eriksson SK, Harryson P, Pierog S, Colby T, Bartels D, Rohrig H 862 (2012) The lysine-rich motif of intrinsically disordered stress protein CDeT11-24 863 from Craterostigma plantagineum is responsible for phosphatidic acid binding and 864 protection of enzymes from damaging effects caused by desiccation. J Exp Bot 865 63(13): 4919-4929 866 Plana M, Itarte E, Eritja R, Goday A, Pages M, Martinez MC (1991) 867 Phosphorylation of maize RAB-17 protein by casein kinase 2. J Biol Chem 266(33): 868 22510-22514 869
www.plantphysiol.orgon July 7, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

30
Popova AV, Rausch S, Hundertmark M, Gibon Y, Hincha DK (2015) The 870 intrinsically disordered protein LEA7 from Arabidopsis thaliana protects the 871 isolated enzyme lactate dehydrogenase and enzymes in a soluble leaf proteome 872 during freezing and drying. Biochim Biophys Acta 1854(10 Pt A): 1517-1525 873 Puhakainen T, Hess MW, Makela P, Svensson J, Heino P, Palva ET (2004) 874 Overexpression of multiple dehydrin genes enhances tolerance to freezing stress in 875 Arabidopsis. Plant Mol Biol 54(5): 743-753 876 Rafalski JA (2002) Plant genomics: present state and a perspective on future 877 developments. Brief Funct Genomic Proteomic 1(1): 80-94 878 Rahman LN, Chen L, Nazim S, Bamm VV, Yaish MW, Moffatt BA, Dutcher JR, 879 Harauz G (2010) Interactions of intrinsically disordered Thellungiella salsuginea 880 dehydrins TsDHN-1 and TsDHN-2 with membranes - synergistic effects of lipid 881 composition and temperature on secondary structure. Biochem Cell Biol 88(5): 791-882 807 883 Reisdorf WC, Jr., Krimm S (1996) Infrared amide I' band of the coiled coil. 884 Biochemistry 35(5): 1383-1386 885 Riera M, Figueras M, Lopez C, Goday A, Pages M (2004) Protein kinase CK2 886 modulates developmental functions of the abscisic acid responsive protein Rab17 887 from maize. Proc Natl Acad Sci U S A 101(26): 9879-9884 888 Rosales R, Romero I, Escribano MI, Merodio C, Sanchez-Ballesta MT (2014) The 889 crucial role of Phi- and K-segments in the in vitro functionality of Vitis vinifera 890 dehydrin DHN1a. Phytochemistry 108: 17-25 891 Sigalov AB, Hendricks GM (2009) Membrane binding mode of intrinsically 892 disordered cytoplasmic domains of T cell receptor signaling subunits depends on 893 lipid composition. Biochem Biophys Res Commun 389(2): 388-393 894 Sinha N, Smith-Gill SJ (2002) Electrostatics in protein binding and function. Curr 895 Protein Pept Sci 3(6): 601-614 896 Steponkus PL, Lynch DV (1989) Freeze/thaw-induced destabilization of the 897 plasma membrane and the effects of cold acclimation. J Bioenerg Biomembr 21(1): 898 21-41 899 Steponkus PL, Uemura M, Joseph RA, Gilmour SJ, Thomashow MF (1998) Mode 900 of action of the COR15a gene on the freezing tolerance of Arabidopsis thaliana. Proc 901 Natl Acad Sci U S A 95(24): 14570-14575 902 Svensson J, Palva ET, Welin B (2000) Purification of recombinant Arabidopsis 903 thaliana dehydrins by metal ion affinity chromatography. Protein Expr Purif 20(2): 904 169-178 905 Tanford C (1970) Protein denaturation. C. Theoretical models for the mechanism of 906 denaturation. Adv Protein Chem 24: 1-95 907 Thalhammer A, Bryant G, Sulpice R, Hincha DK (2014) Disordered cold 908 regulated15 proteins protect chloroplast membranes during freezing through 909 binding and folding, but do not stabilize chloroplast enzymes in vivo. Plant Physiol 910 166(1): 190-201 911 Thalhammer A, Hundertmark M, Popova AV, Seckler R, Hincha DK (2010) 912 Interaction of two intrinsically disordered plant stress proteins (COR15A and 913 COR15B) with lipid membranes in the dry state. Biochim Biophys Acta 1798(9): 914 1812-1820 915
www.plantphysiol.orgon July 7, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

31
Tompa P (2002) Intrinsically unstructured proteins. Trends Biochem Sci 27(10): 916 527-533 917 Trizac E, Levy Y, Wolynes PG (2010) Capillarity theory for the fly-casting 918 mechanism. Proc Natl Acad Sci U S A 107(7): 2746-2750 919 Uemura M, Joseph RA, Steponkus PL (1995) Cold Acclimation of Arabidopsis 920 thaliana (Effect on Plasma Membrane Lipid Composition and Freeze-Induced 921 Lesions). Plant Physiol 109(1): 15-30 922 Ulmer TS, Bax A (2005) Comparison of structure and dynamics of micelle-bound 923 human alpha-synuclein and Parkinson disease variants. J Biol Chem 280(52): 924 43179-43187 925 Ulmer TS, Bax A, Cole NB, Nussbaum RL (2005) Structure and dynamics of 926 micelle-bound human alpha-synuclein. J Biol Chem 280(10): 9595-9603 927 von Heijne G (2006) Membrane-protein topology. Nat Rev Mol Cell Biol 7(12): 909-928 918 929 Vuzman D, Levy Y (2012) Intrinsically disordered regions as affinity tuners in 930 protein-DNA interactions. Mol Biosyst 8(1): 47-57 931 Weise CF, Login FH, Ho O, Grobner G, Wolf-Watz H, Wolf-Watz M (2014) 932 Negatively charged lipid membranes promote a disorder-order transition in the 933 Yersinia YscU protein. Biophys J 107(8): 1950-1961 934 Williams S, Causgrove TP, Gilmanshin R, Fang KS, Callender RH, Woodruff WH, 935 Dyer RB (1996) Fast events in protein folding: helix melting and formation in a 936 small peptide. Biochemistry 35(3): 691-697 937 Wright PE, Dyson HJ (2015) Intrinsically disordered proteins in cellular signalling 938 and regulation. Nat Rev Mol Cell Biol 16(1): 18-29 939 Wu EL, Cheng X, Jo S, Rui H, Song KC, Davila-Contreras EM, Qi Y, Lee J, Monje-940 Galvan V, Venable RM, Klauda JB, Im W (2014) CHARMM-GUI Membrane Builder 941 toward realistic biological membrane simulations. J Comput Chem 35(27): 1997-942 2004 943 Xie C, Zhang R, Qu Y, Miao Z, Zhang Y, Shen X, Wang T, Dong J (2012) 944 Overexpression of MtCAS31 enhances drought tolerance in transgenic Arabidopsis 945 by reducing stomatal density. New Phytol 195(1): 124-135 946 Yoder G, Pancoska P, Keiderling TA (1997) Characterization of alanine-rich 947 peptides, Ac-(AAKAA)n-GY-NH2 (n = 1-4), using vibrational circular dichroism and 948 Fourier transform infrared. Conformational determination and thermal unfolding. 949 Biochemistry 36(49): 15123-15133 950 951 952
www.plantphysiol.orgon July 7, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

Figure 1. Schematic picture of disordered Lti30 binding to lipid (PC:PG) vesicles. Blue is positive charge and red is negative charge. (A) Sequence of Lti30 with K-segments (bold) and the positions of the K segments matching the KLti30 peptide (gray boxes). (B) Sequence of the KLti30 peptide. (C) Binding equilibrium between Lti30 and lipid vesicles where attraction is influenced by charge. Binding is promoted by protonation of the flanking His pairs and down regulated by phosphorylation of the K segments threonines (Eriksson et al. 2011 ). Upon Lti30 binding, the membrane phase decreases 2.5 °C and the vesicles cluster also into macroscopic aggregates (Eriksson et al. 2011 ). For size reference, Lti30 has an extended length of 20 nm and the diameter of the vesicles is 100 nm.
A.! MNSHQNQTGVQKKGITEKIMEKLPGHHGPTNTGVVHHEKKGMTEKVMEQLPGHHGATGTGGVHHEKKGMTEKVMEQLPGHHG SHQTGTNTTYGTTNTGGVHHEKKSVTEKVMEKLPGHHGSHQTGTNTAYGTNTNVVHHEKKGIAEKIKEQLPGHHGTHKTGTTTSYGNTGVVHHENKSTMDKIKEKLPGGHH!
Lti30 !(+13 to +20)!
PC:PG vesicle !(– 630 to – 25400)!
HH—KK—T—K——HH!!
+!+! +!
HH—KK—T—K——HH!!
+!+! +!
P -
+!+!+!+!
HH—KK—T—K——HH!!
+!+! +! +!+!+!+!
protonation of H-switch
phosphorylation
+!
- P
- PG
- PG
- PG
- PG
- PG
- PG
vesicle clustering
Lti30-induced shift of membrane phase transition
regulation!
C.!
VHHEKKGMTEKVMEQLPGHHG!KLti30 peptide :!B.!

Figure 2. Stopped-flow data of Lti30 binding and clustering PC:PG lipid vesicles (LUVs). 3.5 µM Lti30 was rapidly mixed with 0.5 mM PC:PG vesicles at 1:142 protein:lipid ratio. Binding of positively charged Lti30 promotes vesicle clustering monitored by scattering at 400 nm. The process depends on the LUV net negative charge, which was varied between PG (1-40%) and PC (99-60%). (A) Scattering at 400 nm. (B) Absorbance at 400 nm. (C) Scattering amplitude at 400 nm at time = 0.5 seconds indicating two binding modes.
C.!
0.0!
1.0!
2.0!
3.0!
4.0!
time (s)!0.0! 1.0! 2.0! 3.0! 4.0! 5.0!
1 %!2 % !in
tens
ity a
t 400
nm!
3 % !6 % !
25 %!
A.!
% PG !10.0! 20.0! 30.0! 40.0!1.0!
abso
rban
ce a
t 400
nm!
0.00!
0.02!
0.04!
0.06!
0.08!
0.10!
0.12!
0.0! 1.0! 2.0! 3.0! 4.0! 5.0!
1 %!
2 %!
6 %!
25 %!
B.!
3 %!
time (s)!
inte
nsity
at 4
00 n
m a
t 0.5
sec!
0.0!
1.0!
2.0!
3.0!
4.0!
40%!
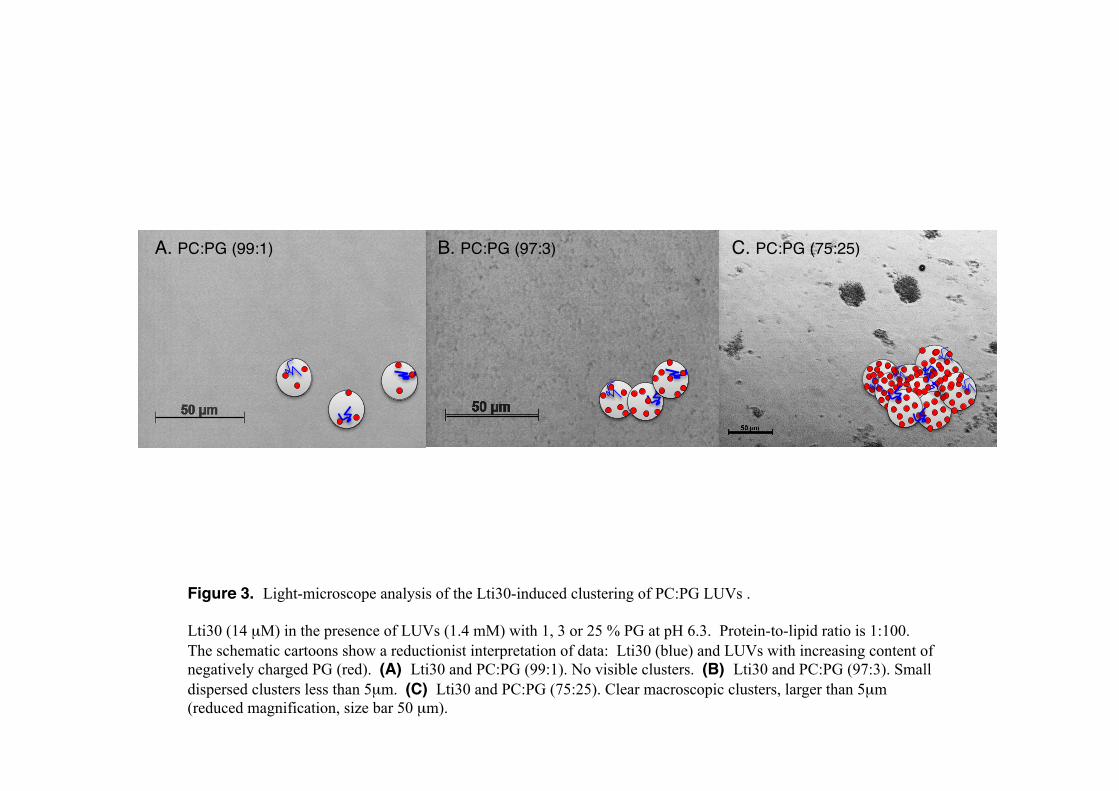
Figure 3. Light-microscope analysis of the Lti30-induced clustering of PC:PG LUVs . Lti30 (14 µM) in the presence of LUVs (1.4 mM) with 1, 3 or 25 % PG at pH 6.3. Protein-to-lipid ratio is 1:100. The schematic cartoons show a reductionist interpretation of data: Lti30 (blue) and LUVs with increasing content of negatively charged PG (red). (A) Lti30 and PC:PG (99:1). No visible clusters. (B) Lti30 and PC:PG (97:3). Small dispersed clusters less than 5µm. (C) Lti30 and PC:PG (75:25). Clear macroscopic clusters, larger than 5µm (reduced magnification, size bar 50 µm).
B. PC:PG (97:3)!A. PC:PG (99:1)! C. PC:PG (75:25)!

Figure 4. Vesicle clustering by titration of various K-segments correlates to peptide net charge. Clustering of lipid vesicles was measured by absorbance at 400nm and peptide net charge was calculated at http://protcalc.sourceforge.net. (A) Titration of different K-segments into 0.5 mM PC:PG vesicles (1:3) at pH 6.3 (blue lines) and pH 4.3 (red lines). The vesicle clustering follows peptide net charge. KLti30 represent the dominating type of K-segment in Lti30, KC is the consensus K-segment and KHHCHH is KC with the addition of histidine flanks. KC fail to bind vesicles regardless of pH, showing the regulatory impact of positively charged histidine flanks. (B) Peptide sequences, with positively and negative residues in blue and red, respectively. !
!KLti30 VHHEKKGMTEKVMEQLPGHHG!KHHCHH HHEKKGIMDKIKEKLPGHH!KC EKKGIMDKIKEKLPG!
abso
rban
ce a
t 400
nm!
[K-segment] (mg/ml)!
1.0!
0.3!0.2!0.1!0!
0.5!
0!
KLti30 !
(+5.7, pH 4.3) !KHHCHH !
(+7.7, pH 4.3) !KHHCHH !(+4.5, pH 6.3) !
KLti30 (+2.5, pH 6.3) !KC (+2.0, pH 6.3) !
KC (+3.5, pH 4.3) !
A.!
B.!

Figure 5. FTIR spectra of Lti30 only and mixed with PC:PG vesicles.
(A) Second derivative IR absorption spectra of Lti30 alone in the amide I region at pH 5 (black) and pH 8.6 (red).
(B) and (C) Corresponding spectra of vesicles alone (red line) and the Lti30-vesicle mix (black line) at pH 5 and
pH 8.6, respectively.
A. Lti30 only
pH 5.0
wavenumber (cm-1) wavenumber (cm-1)
second d
erivative x
1000
second d
erivative x
1000
B.
Lti30-vesicle mix
vesicles only
vesicle binding at pH 5.0 C.
wavenumber (cm-1)
second d
erivative x
1000
Lti30-vesicle mix
vesicles only
vesicle binding pH 8.6
pH 8.6

Figure 6. Structural analysis of membrane bound KLti30 peptide by NMR. (A) The five best structures of bicelle-bound KLti30 peptide from 1H-NMR constraints, where the central nine residues of the peptide adopt a fixed α-helix, whereas the N and C termini are more disordered. (B) Secondary-structure propensity (SSP) from 1Ha, 1HN and 1Hb chemical shifts, where positive values show the induced α-helix in the central part of the peptide. Negative SSP values indicate extended conformation in the N-terminal region of the peptide. (C) Determined NOE connectivity between peptide residues along the KLti30. The ordered n+3 couplings within the residue segment G7 to Q15 are the hallmark of α-helical structure.
A.! B.!
V1 H2 H3 E4 K5 K6 G7 M8 T9 E10 K11 V12 M13 E14 Q15 L16 P17 G18 H19 H20 G21 !
C. NOE connectivity!
KLti30 sequence !
KLti30 sequence !
SSP!
2 6 10 14 18!
0.6!
0.4!
0.2!
0!
-0.2!
-0.4!
-0.6!
n!
c!

Figure 7. The atomic-resolution structure of the membrane binding domain of Lti30. (A) (B) (C) Atomic model of the membrane-binding KLti30-segment of Lti30, in side, diagonal and front views, respectively. Positively charged residues are in blue, negatively charged residues in red, and the hydrophobic side of the amphipathic α-helix faces downwards. (D) NMR diffusion data of KLti30 alone, bicelles alone and KLti30 mixed with bicelles, verifying KLti30-micelle binding. (E) Molecular dynamics modeling of the histidine flanked KLti30-segment into a lipid bilayer (PC:PG 98:2 mix), using periodic boundary conditions. Consistent with the binding data in Figures 2-5, the positive side chains H2, H3, K5 and K6 coordinate the negative charge of a PG head group.
gradient strength (a.u.)!10! 30!
rela
tive
sign
al in
tens
ity !
1!
0.6!
0.2!
D.! KLti30 !bicelles !complex!
E.!
H2!
H20! H3!
H19!
PG!
E14!PG!
A.! V1!
G21!
K6!E10!
K11!E14!
H19!
H3!
H2!
K5!
H20!E4!
H19!
G21!
E10!
K11!
E14!H20!
H3!
E4!
V1!H2 !H3!
G21!
E4!
K6!E10!
K11!E14!
H19!
H20!
K5!
B.! C.!

Parsed CitationsAlexander CG, Jurgens MC, Shepherd DA, Freund SM, Ashcroft AE, Ferguson N (2013) Thermodynamic origins of protein folding,allostery, and capsid formation in the human hepatitis B virus core protein. Proc Natl Acad Sci U S A 110(30): E2782-2791
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Alsheikh MK, Heyen BJ, Randall SK (2003) Ion binding properties of the dehydrin ERD14 are dependent upon phosphorylation. JBiol Chem 278(42): 40882-40889
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Alsheikh MK, Svensson JT, Randall SK (2005) Phosphorylation regulated ion-binding is a property shared by the acidic subclassdehydrins. Plant Cell and Environment 28(9): 1114-1122
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Andersson A, Maler L (2005) Magnetic resonance investigations of lipid motion in isotropic bicelles. Langmuir 21(17): 7702-7709Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Arai M, Sugase K, Dyson HJ, Wright PE (2015) Conformational propensities of intrinsically disordered proteins influence themechanism of binding and folding. Proc Natl Acad Sci U S A 112(31): 9614-9619
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Baker J, Steele C, Dure L (1988) Sequence and Characterization of 6 Lea Proteins and Their Genes from Cotton. Plant MolecularBiology 11(3): 277-291
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Bartels T, Choi JG, Selkoe DJ (2011) alpha-Synuclein occurs physiologically as a helically folded tetramer that resists aggregation.Nature 477(7362): 107-110
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Barth A, Zscherp C (2002) What vibrations tell us about proteins. Q Rev Biophys 35(4): 369-430Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Battaglia M, Olvera-Carrillo Y, Garciarrubio A, Campos F, Covarrubias AA (2008) The enigmatic LEA proteins and otherhydrophilins. Plant Physiol 148(1): 6-24
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Bomal C, Le VQ, Tremblay FM (2002) Induction of tolerance to fast desiccation in black spruce (Picea mariana) somatic embryos:relationship between partial water loss, sugars, and dehydrins. Physiol Plant 115(4): 523-530
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Borg M, Mittag T, Pawson T, Tyers M, Forman-Kay JD, Chan HS (2007) Polyelectrostatic interactions of disordered ligands suggesta physical basis for ultrasensitivity. Proc Natl Acad Sci U S A 104(23): 9650-9655
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Candat A, Paszkiewicz G, Neveu M, Gautier R, Logan DC, Avelange-Macherel MH, Macherel D (2014) The ubiquitous distributionof late embryogenesis abundant proteins across cell compartments in Arabidopsis offers tailored protection against abiotic stress.Plant Cell 26(7): 3148-3166
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Chakrabortee S, Boschetti C, Walton LJ, Sarkar S, Rubinsztein DC, Tunnacliffe A (2007) Hydrophilic protein associated withdesiccation tolerance exhibits broad protein stabilization function. Proc Natl Acad Sci U S A 104(46): 18073-18078
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Clarke MW, Boddington KF, Warnica JM, Atkinson J, McKenna S, Madge J, Barker CH, Graether SP (2015) Structural and

Functional Insights into the Cryoprotection of Membranes by the Intrinsically Disordered Dehydrins. J Biol Chem 290(45): 26900-26913
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Close TJ (1996) Dehydrins: Emergence of a biochemical role of a family of plant dehydration proteins. Physiologia Plantarum 97(4):795-803
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Crowe LM, Crowe JH (1992) Anhydrobiosis: a strategy for survival. Adv Space Res 12(4): 239-247Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Danielsson J, Jarvet J, Damberg P, Graslund A (2002) Translational diffusion measured by PFG-NMR on full length and fragmentsof the Alzheimer A beta(1-40) peptide. Determination of hydrodynamic radii of random coil peptides of varying length. MagneticResonance in Chemistry 40: S89-S97
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Danyluk J, Perron A, Houde M, Limin A, Fowler B, Benhamou N, Sarhan F (1998) Accumulation of an acidic dehydrin in the vicinityof the plasma membrane during cold acclimation of wheat. Plant Cell 10(4): 623-638
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Delaglio F, Grzesiek S, Vuister GW, Zhu G, Pfeifer J, Bax A (1995) Nmrpipe - a Multidimensional Spectral Processing System Basedon Unix Pipes. Journal of Biomolecular Nmr 6(3): 277-293
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Dunker AK, Obradovic Z (2001) The protein trinity--linking function and disorder. Nat Biotechnol 19(9): 805-806Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Dure L, 3rd (1993) A repeating 11-mer amino acid motif and plant desiccation. Plant J 3(3): 363-369Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Ellenberger TE, Brandl CJ, Struhl K, Harrison SC (1992) The GCN4 basic region leucine zipper binds DNA as a dimer ofuninterrupted alpha helices: crystal structure of the protein-DNA complex. Cell 71(7): 1223-1237
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Eriksson SK, Kutzer M, Procek J, Grobner G, Harryson P (2011) Tunable membrane binding of the intrinsically disordereddehydrin Lti30, a cold-induced plant stress protein. Plant Cell 23(6): 2391-2404
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Goormaghtigh E, Cabiaux V, Ruysschaert JM (1994) Determination of soluble and membrane protein structure by Fourier transforminfrared spectroscopy. I. Assignments and model compounds. Subcell Biochem 23: 329-362
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Graether SP, Boddington KF (2014) Disorder and function: a review of the dehydrin protein family. Front Plant Sci 5: 576Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Hara M, Fujinaga M, Kuboi T (2005) Metal binding by citrus dehydrin with histidine-rich domains. J Exp Bot 56(420): 2695-2703Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Hara M, Shinoda Y, Tanaka Y, Kuboi T (2009) DNA binding of citrus dehydrin promoted by zinc ion. Plant Cell Environ 32(5): 532-541
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Hara M, Terashima S, Kuboi T (2001) Characterization and cryoprotective activity of cold-responsive dehydrin from Citrus unshiu.

Journal of Plant Physiology 158(10): 1333-1339Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Hernandez-Sanchez IE, Martynowicz DM, Rodriguez-Hernandez AA, Perez-Morales MB, Graether SP, Jimenez-Bremont JF (2014)A dehydrin-dehydrin interaction: the case of SK3 from Opuntia streptacantha. Front Plant Sci 5: 520
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Ingram J, Bartels D (1996) The Molecular Basis of Dehydration Tolerance in Plants. Annu Rev Plant Physiol Plant Mol Biol 47: 377-403
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Jao CC, Der-Sarkissian A, Chen J, Langen R (2004) Structure of membrane-bound alpha-synuclein studied by site-directed spinlabeling. Proc Natl Acad Sci U S A 101(22): 8331-8336
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Jarvet J, Danielsson J, Damberg P, Oleszczuk M, Graslund A (2007) Positioning of the Alzheimer Abeta(1-40) peptide in SDSmicelles using NMR and paramagnetic probes. J Biomol NMR 39(1): 63-72
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Jiang X, Wang Y (2004) Beta-elimination coupled with tandem mass spectrometry for the identification of in vivo and in vitrophosphorylation sites in maize dehydrin DHN1 protein. Biochemistry 43(49): 15567-15576
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Jo S, Kim T, Im W (2007) Automated builder and database of protein/membrane complexes for molecular dynamics simulations.PLoS One 2(9): e880
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Jo S, Kim T, Iyer VG, Im W (2008) CHARMM-GUI: a web-based graphical user interface for CHARMM. J Comput Chem 29(11): 1859-1865
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Jo S, Lim JB, Klauda JB, Im W (2009) CHARMM-GUI Membrane Builder for mixed bilayers and its application to yeast membranes.Biophys J 97(1): 50-58
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Klug A (2010) The discovery of zinc fingers and their development for practical applications in gene regulation and genomemanipulation. Q Rev Biophys 43(1): 1-21
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Koag MC, Fenton RD, Wilkens S, Close TJ (2003) The binding of maize DHN1 to lipid vesicles. Gain of structure and lipidspecificity. Plant Physiol 131(1): 309-316
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Koag MC, Wilkens S, Fenton RD, Resnik J, Vo E, Close TJ (2009) The K-segment of maize DHN1 mediates binding to anionicphospholipid vesicles and concomitant structural changes. Plant Physiol 150(3): 1503-1514
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Kovacs D, Kalmar E, Torok Z, Tompa P (2008) Chaperone activity of ERD10 and ERD14, two disordered stress-related plantproteins. Plant Physiol 147(1): 381-390
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Krieger E, Joo K, Lee J, Lee J, Raman S, Thompson J, Tyka M, Baker D, Karplus K (2009) Improving physical realism,stereochemistry, and side-chain accuracy in homology modeling: Four approaches that performed well in CASP8. Proteins-Structure Function and Bioinformatics 77: 114-122

Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Kriwacki RW, Hengst L, Tennant L, Reed SI, Wright PE (1996) Structural studies of p21Waf1/Cip1/Sdi1 in the free and Cdk2-boundstate: conformational disorder mediates binding diversity. Proc Natl Acad Sci U S A 93(21): 11504-11509
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Kurnik M, Hedberg L, Danielsson J, Oliveberg M (2012) Folding without charges. Proc Natl Acad Sci U S A 109(15): 5705-5710Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Lee CW, Martinez-Yamout MA, Dyson HJ, Wright PE (2010) Structure of the p53 transactivation domain in complex with the nuclearreceptor coactivator binding domain of CREB binding protein. Biochemistry 49(46): 9964-9971
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Lee SA, Eyeson R, Cheever ML, Geng J, Verkhusha VV, Burd C, Overduin M, Kutateladze TG (2005) Targeting of the FYVE domainto endosomal membranes is regulated by a histidine switch. Proc Natl Acad Sci U S A 102(37): 13052-13057
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Lemmon MA (2008) Membrane recognition by phospholipid-binding domains. Nat Rev Mol Cell Biol 9(2): 99-111Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Lind J, Nordin J, Maler L (2008) Lipid dynamics in fast-tumbling bicelles with varying bilayer thickness: effect of modeltransmembrane peptides. Biochim Biophys Acta 1778(11): 2526-2534
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Lira-De Leon KI, Garcia-Gutierrez P, Serratos IN, Palomera-Cardenas M, Figueroa-Corona Mdel P, Campos-Pena V, Meraz-RiosMA (2013) Molecular mechanism of tau aggregation induced by anionic and cationic dyes. J Alzheimers Dis 35(2): 319-334
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Marsh JA, Singh VK, Jia Z, Forman-Kay JD (2006) Sensitivity of secondary structure propensities to sequence differences betweenalpha- and gamma-synuclein: implications for fibrillation. Protein Sci 15(12): 2795-2804
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Martin J, Geromanos S, Tempst P, Hartl FU (1993) Identification of nucleotide-binding regions in the chaperonin proteins GroELand GroES. Nature 366(6452): 279-282
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Martinez G, Millhauser G (1995) FTIR spectroscopy of alanine-based peptides: assignment of the amide I' modes for random coiland helix. J Struct Biol 114(1): 23-27
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Matthews JM, Bhati M, Lehtomaki E, Mansfield RE, Cubeddu L, Mackay JP (2009) It takes two to tango: the structure and functionof LIM, RING, PHD and MYND domains. Curr Pharm Des 15(31): 3681-3696
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Maupetit J, Derreumaux P, Tuffery P (2009) PEP-FOLD: an online resource for de novo peptide structure prediction. Nucleic AcidsResearch 37: W498-W503
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
McLaughlin S, Murray D (2005) Plasma membrane phosphoinositide organization by protein electrostatics. Nature 438(7068): 605-611
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Moravcevic K, Mendrola JM, Schmitz KR, Wang YH, Slochower D, Janmey PA, Lemmon MA (2010) Kinase associated-1 domains

drive MARK/PAR1 kinases to membrane targets by binding acidic phospholipids. Cell 143(6): 966-977Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Mouillon JM, Eriksson SK, Harryson P (2008) Mimicking the plant cell interior under water stress by macromolecular crowding:disordered dehydrin proteins are highly resistant to structural collapse. Plant Physiol 148(4): 1925-1937
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Mouillon JM, Gustafsson P, Harryson P (2006) Structural investigation of disordered stress proteins. Comparison of full-lengthdehydrins with isolated peptides of their conserved segments. Plant Physiol 141(2): 638-650
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Muller JJ, Lapko A, Ruckpaul K, Heinemann U (2003) Modeling of electrostatic recognition processes in the mammalianmitochondrial steroid hydroxylase system. Biophys Chem 100(1-3): 281-292
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Nevskaya NA, Chirgadze YN (1976) Infrared spectra and resonance interactions of amide-I and II vibration of alpha-helix.Biopolymers 15(4): 637-648
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Niessing D, Huttelmaier S, Zenklusen D, Singer RH, Burley SK (2004) She2p is a novel RNA binding protein with a basic helicalhairpin motif. Cell 119(4): 491-502
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Nylander M, Svensson J, Palva ET, Welin BV (2001) Stress-induced accumulation and tissue-specific localization of dehydrins inArabidopsis thaliana. Plant Mol Biol 45(3): 263-279
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Peng PH, Lin CH, Tsai HW, Lin TY (2014) Cold response in Phalaenopsis aphrodite and characterization of PaCBF1 and PaICE1.Plant Cell Physiol 55(9): 1623-1635
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Petersen J, Eriksson SK, Harryson P, Pierog S, Colby T, Bartels D, Rohrig H (2012) The lysine-rich motif of intrinsically disorderedstress protein CDeT11-24 from Craterostigma plantagineum is responsible for phosphatidic acid binding and protection ofenzymes from damaging effects caused by desiccation. J Exp Bot 63(13): 4919-4929
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Plana M, Itarte E, Eritja R, Goday A, Pages M, Martinez MC (1991) Phosphorylation of maize RAB-17 protein by casein kinase 2. JBiol Chem 266(33): 22510-22514
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Popova AV, Rausch S, Hundertmark M, Gibon Y, Hincha DK (2015) The intrinsically disordered protein LEA7 from Arabidopsisthaliana protects the isolated enzyme lactate dehydrogenase and enzymes in a soluble leaf proteome during freezing and drying.Biochim Biophys Acta 1854(10 Pt A): 1517-1525
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Puhakainen T, Hess MW, Makela P, Svensson J, Heino P, Palva ET (2004) Overexpression of multiple dehydrin genes enhancestolerance to freezing stress in Arabidopsis. Plant Mol Biol 54(5): 743-753
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Rafalski JA (2002) Plant genomics: present state and a perspective on future developments. Brief Funct Genomic Proteomic 1(1):80-94
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Rahman LN, Chen L, Nazim S, Bamm VV, Yaish MW, Moffatt BA, Dutcher JR, Harauz G (2010) Interactions of intrinsically disordered

Thellungiella salsuginea dehydrins TsDHN-1 and TsDHN-2 with membranes - synergistic effects of lipid composition andtemperature on secondary structure. Biochem Cell Biol 88(5): 791-807
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Reisdorf WC, Jr., Krimm S (1996) Infrared amide I' band of the coiled coil. Biochemistry 35(5): 1383-1386Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Riera M, Figueras M, Lopez C, Goday A, Pages M (2004) Protein kinase CK2 modulates developmental functions of the abscisicacid responsive protein Rab17 from maize. Proc Natl Acad Sci U S A 101(26): 9879-9884
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Rosales R, Romero I, Escribano MI, Merodio C, Sanchez-Ballesta MT (2014) The crucial role of Phi- and K-segments in the in vitrofunctionality of Vitis vinifera dehydrin DHN1a. Phytochemistry 108: 17-25
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Sigalov AB, Hendricks GM (2009) Membrane binding mode of intrinsically disordered cytoplasmic domains of T cell receptorsignaling subunits depends on lipid composition. Biochem Biophys Res Commun 389(2): 388-393
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Sinha N, Smith-Gill SJ (2002) Electrostatics in protein binding and function. Curr Protein Pept Sci 3(6): 601-614Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Steponkus PL, Lynch DV (1989) Freeze/thaw-induced destabilization of the plasma membrane and the effects of cold acclimation. JBioenerg Biomembr 21(1): 21-41
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Steponkus PL, Uemura M, Joseph RA, Gilmour SJ, Thomashow MF (1998) Mode of action of the COR15a gene on the freezingtolerance of Arabidopsis thaliana. Proc Natl Acad Sci U S A 95(24): 14570-14575
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Svensson J, Palva ET, Welin B (2000) Purification of recombinant Arabidopsis thaliana dehydrins by metal ion affinitychromatography. Protein Expr Purif 20(2): 169-178
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Tanford C (1970) Protein denaturation. C. Theoretical models for the mechanism of denaturation. Adv Protein Chem 24: 1-95Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Thalhammer A, Bryant G, Sulpice R, Hincha DK (2014) Disordered cold regulated15 proteins protect chloroplast membranes duringfreezing through binding and folding, but do not stabilize chloroplast enzymes in vivo. Plant Physiol 166(1): 190-201
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Thalhammer A, Hundertmark M, Popova AV, Seckler R, Hincha DK (2010) Interaction of two intrinsically disordered plant stressproteins (COR15A and COR15B) with lipid membranes in the dry state. Biochim Biophys Acta 1798(9): 1812-1820
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Tompa P (2002) Intrinsically unstructured proteins. Trends Biochem Sci 27(10): 527-533Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Trizac E, Levy Y, Wolynes PG (2010) Capillarity theory for the fly-casting mechanism. Proc Natl Acad Sci U S A 107(7): 2746-2750Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Uemura M, Joseph RA, Steponkus PL (1995) Cold Acclimation of Arabidopsis thaliana (Effect on Plasma Membrane LipidComposition and Freeze-Induced Lesions). Plant Physiol 109(1): 15-30

Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Ulmer TS, Bax A (2005) Comparison of structure and dynamics of micelle-bound human alpha-synuclein and Parkinson diseasevariants. J Biol Chem 280(52): 43179-43187
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Ulmer TS, Bax A, Cole NB, Nussbaum RL (2005) Structure and dynamics of micelle-bound human alpha-synuclein. J Biol Chem280(10): 9595-9603
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
von Heijne G (2006) Membrane-protein topology. Nat Rev Mol Cell Biol 7(12): 909-918Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Vuzman D, Levy Y (2012) Intrinsically disordered regions as affinity tuners in protein-DNA interactions. Mol Biosyst 8(1): 47-57Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Weise CF, Login FH, Ho O, Grobner G, Wolf-Watz H, Wolf-Watz M (2014) Negatively charged lipid membranes promote a disorder-order transition in the Yersinia YscU protein. Biophys J 107(8): 1950-1961
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Williams S, Causgrove TP, Gilmanshin R, Fang KS, Callender RH, Woodruff WH, Dyer RB (1996) Fast events in protein folding:helix melting and formation in a small peptide. Biochemistry 35(3): 691-697
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Wright PE, Dyson HJ (2015) Intrinsically disordered proteins in cellular signalling and regulation. Nat Rev Mol Cell Biol 16(1): 18-29Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Wu EL, Cheng X, Jo S, Rui H, Song KC, Davila-Contreras EM, Qi Y, Lee J, Monje-Galvan V, Venable RM, Klauda JB, Im W (2014)CHARMM-GUI Membrane Builder toward realistic biological membrane simulations. J Comput Chem 35(27): 1997-2004
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Xie C, Zhang R, Qu Y, Miao Z, Zhang Y, Shen X, Wang T, Dong J (2012) Overexpression of MtCAS31 enhances drought tolerance intransgenic Arabidopsis by reducing stomatal density. New Phytol 195(1): 124-135
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Yoder G, Pancoska P, Keiderling TA (1997) Characterization of alanine-rich peptides, Ac-(AAKAA)n-GY-NH2 (n = 1-4), usingvibrational circular dichroism and Fourier transform infrared. Conformational determination and thermal unfolding. Biochemistry36(49): 15123-15133
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title