membranes associated with the disintegratio ofn mycetomal micro
TRANSCRIPT

J. Cell Set. 3, 65-70 (1968)
Printed in Great Britain
MEMBRANES ASSOCIATED WITH
THE DISINTEGRATION OF MYCETOMAL
MICRO-ORGANISMS IN SITOPHILUS ZEA-MAIS
(MOTS.) (COLEOPTERA)
A. J. MUSGRAVE AND I. GRINYERDepartment of Zoology andDepartment of Avian Patlwlogy and Wildlife Diseases, University of Guelph, Ontario,Canada
SUMMARY
The mycetomal micro-organisms of Sitophilus zea-mais give evidence in electron micro-graphs of having two peripheral unit membranes, an intermediate layer, intracytoplasmicmembranes and DNA-like material. They appear to disintegrate, at times, with the con-comitant formation of membrane masses. It is suggested that these are visible with the phase-microscope and that they are probably myelin figures.
INTRODUCTION
A mycetocyte is a cell that harbours micro-organisms that supposedly benefit aninsect host. In some insects mycetocytes may form a discrete structure or mycetome.In the genus Sitophilus larval mycetomes disintegrate at metamorphosis. In adultsmycetocytes with organisms are found, except in aposymbiotic strains (Musgrave,1964), in the mesentera, especially of young weevils (Musgrave, Homan & Grinyer,1964), and in the female gonads.
The morphology (and perhaps the species) of micro-organisms seems to vary withthe species of host, so that the symbiotes may be of taxonomic value (Musgrave &Homan, 1962), though there is considerable pleomorphism. In Sitophilus granarius(L.) the presence of 'nuclear equivalents' within, and of cytoplasmic membranes,and sometimes cell walls around, the typical Gram-negative organisms, and theabsence of host-provided membranes have been established (Musgrave & Singh,1965; Singh & Musgrave, 1966; Grinyer & Musgrave, 1966). Rather similar find-ings have been reported by Malke (1966) and Milburn (1966) regarding the some-what atypical Gram-positive 'bacteroid' symbiotes in certain cockroaches. Malke,however, found host-provided membranes; and Milburn found 'metamorphosedbacteroids' which were swollen and had developed 'concentric layers of strands'within them.
The mycetomal micro-organisms in Sitophilus zea-mais (Mots.) (Kuschel, 1961)differ from those of S. granarius and frequently assume C-shaped or characteristictightly spiralled forms. A study of their peripheral membranes and nuclear material
5 Cell Sci. 3

66 A. J. Musgrave and I. Grinyer
provided an insight into the origin of masses of concentric, apparently membranousmaterial that had frequently been observed in mycetomal material. These observationsare the subject of this paper.
MATERIALS AND METHODS
Weevils were reared at approximately 27 °C and 76 % relative humidity on wheat.Mycetomes and adult mesentera dissected from' the insects in drops of distilled ortap water or Bodenstein's Ringer's solution (Buck, 1953), modified Schneider's (1964)and Grace's (1962) media were examined as fresh smears by phase-contrast micro-scopy. Other smears were fixed either in the vapour of 2 % osmium tetroxide for 2 h orin Bouin's solution, and then stained in Short's modification of Heidenhain's haem-atoxylin or Twort's stain (Ollett, 1947, 1951)- Mycetomes and adult mesentera werealso dissected in Wyatt's (1956) solution without haemolymph and processed forelectron microscopy according to a method already described (Grinyer & Musgrave,1966, fixation method (b)). This material was embedded in 'Maraglas' epoxy-resin.Some of this material was sectioned at about i-o/i and examined with the lightmicroscope after staining in 1 % toluidine blue by the method of Spurlock, Kattine &Freeman (1963).
OBSERVATIONS
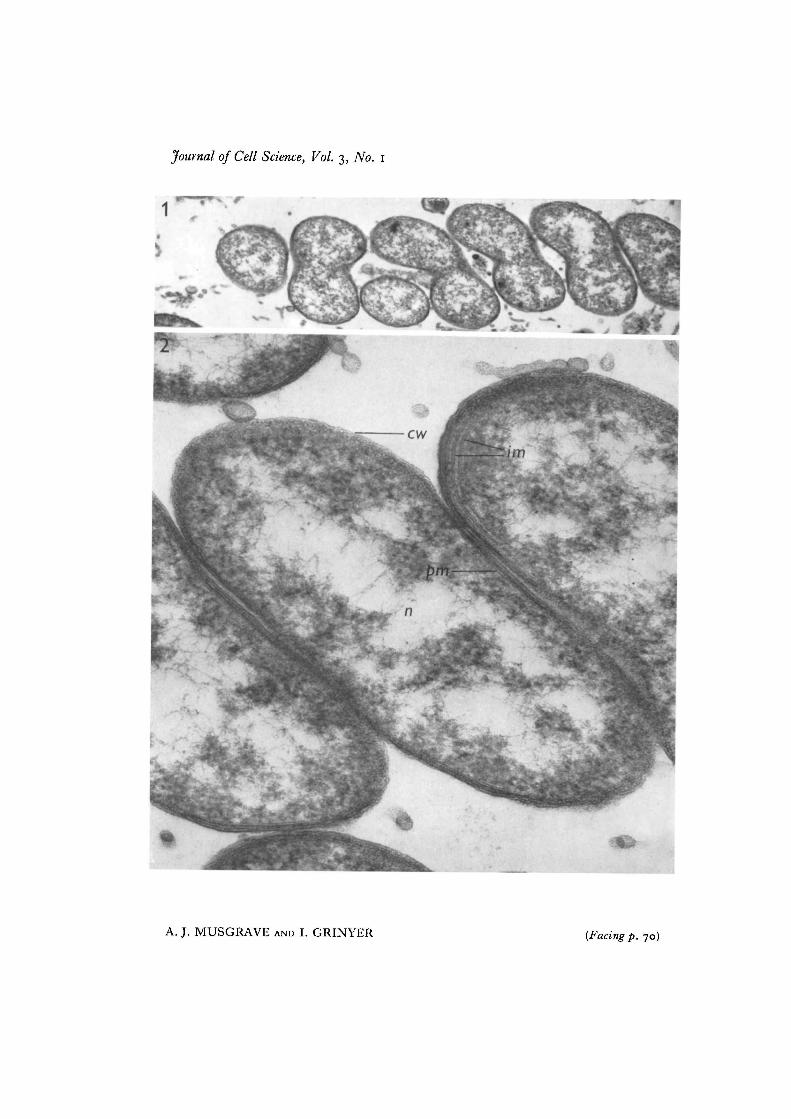
In mycetomes from larvae the electron microscope showed the tight spiralling ofsome of the organisms (Fig. 1). The presence of DNA-like material, and of two tri-laminate surface membranes with intermediate material, as well as intracytoplasmicmembranes (Fig. 2), were indications that the organisms were Gram-negative. No<*epta were seen.
Masses (presumably spheroidal) of disintegrating micro-organisms occurred associ-ated with numerous trilaminate membranes (Figs. 3-5). Apparently healthy organismswere located outside these spheroidal masses. There was evidence that as the micro-organisms disintegrated the membranes became more numerous (Fig. 4). That themembranes were a product of degeneration of the micro-organisms was suggested bythe appearance of those disintegrating individually (Fig. 6). Some of the membranemasses were larger than the mycetocyte nucleus.
A generally similar situation seemed to exist in mycetocytes in the mesentera ofadults.
There was no evidence of proliferation of the Golgi complex in those mycetocytescontaining micro-organisms, for the latter seemed to occupy most of the availablespace.
Phase-microscope studies of unfixed crushed preparations of mycetomes of larvaeand prepupae and of mesentera of adults often revealed objects conveniently referredto as 'lamellar masses' (Figs. 7, 8), of a size comparable with that of the spheroidalmasses mentioned above, associated with mycetocytes and sometimes within them.These lamellar masses seemed non-birefringent and seemed to grow to a considerable

Mycetomal micro-organisms 67
size (Fig. 11). It seemed that such lamellar masses were not simply the resultof damage to the insect cells for, in three adult weevils that were probably more than20 days old (Musgrave et al. 1964), the mesentera apparently contained neitherrecognizable micro-organisms nor lamellar masses. It seemed, too, that as the numberof lamellar masses increased, the number of spiral forms decreased. The smallerlamellar masses were not disrupted by Bouin's fixative but they appeared to be un-detectable after drying.
In an attempt to discover what configuration the spheroidal and lamellar massesmight manifest in fixed and stained smears, these were searched (using the electronmicrographs as a guide and the helical organisms as a kind of biological indicator), forevidence of spiral organisms within spheroidal objects. Although some such objectswere seen (Fig. 9) in osmium-fixed material, their true nature remained open to doubt.Spheroidal bodies with lamellar masses were clearly seen in 1 /i sections of' Maraglas '-embedded material (Fig. 10).
DISCUSSION
The various kinds of mycetomal symbiotes are generally considered to be supplyinga variety of useful metabolites to their insect hosts (see Buchner, 1966; Gilmour,1965; Henry, 1962; Musgrave, 1964; Steinhaus, 1949; and Trager, i960). Themechanism of supply is unknown. Presumably, the symbiotes pass the metabolites intothe circumambient medium or they may, additionally, perhaps, be lysed and absorbedby the host.
Although some micro-organisms were seen disintegrating in isolation, most of themseemed to disintegrate in presumptively spheroidal masses, suggesting that in myce-tocytes there may be organelles from which lysing enzymes diffuse into the surround-ing cytoplasm.
The relationship between objects seen by electron microscopy to those seen byphase-contrast and conventional light microscopy can, in the present state of ourknowledge, be established only on the basis of circumstantial evidence. The relation-ship of the membrane masses of the present electron-microscope studies to thelamellar masses of the phase-contrast studies and to spheroidal objects of the fixed andstained smears must therefore be based on this kind of evidence. The electron-microscope studies have shown clearly that the disintegration of the mycetomal micro-organisms is associated with swirls of concentric membranes which often reach a sizethat should make them visible in the light microscope; and, indeed, lamellar massesof this size were seen by phase-contrast microscopy. It is suggested, therefore, thatthe membrane masses and lamellar masses may be equated. Moreover, both kindsof masses closely resemble myelin forms or myelin figures (Freeman, 1964; Frey-Wyssling, 1953), supposedly formed as the result of cell degeneration (Cedergren, 1956;Freeman, 1964), phagocytosis (Vickerman, 1962; Freeman, 1964), or as a result ofchanges in the protein component of the lipoprotein in cellular membranes (Freeman,1964). Certain kinds of myelin forms grow by swelling and are at times birefringent(Frey-Wyssling, 1953; Mercer, 1962).
5-2

68 A. J. Musgrave and I. Grinyer
It seems justifiable then to speculate that the following is the sequence of events in themycetome of S. sea-mats. In view of Milburn's (1966) findings it is reasonable to startwith the assumption that the insect host can time its depredations on its symbioticorganisms to periods to suit its own metabolism, though it is possible that depreda-tions are continuous, even in the larval stage. Disintegration need not proceed in allmycetocytes simultaneously. Perhaps from organelles in mycetocytes lysing en-zymes diffuse into the surrounding cytoplasm, causing the disintegration of the micro-organisms with consequent formation of innumerable membranes perhaps havingthe appearance of myelin forms. These masses of membranes perhaps grow as aresult of hydration, or continuing microbial disintegration, until they are as big as, orbigger than, the nucleus of the mycetocyte. They thus become easily visible aslamellated bodies in mycetocytes in the phase-contrast microscope. As certain kindsof myelin forms (for example, those from lecithin) can grow by hydration it seems notimpossible, on the evidence given in this paper, that the very large myelin figures attimes seen in smears from S. zea-mais may be a result of symbiote degeneration.
The nature of the metabolites that its symbiotes supply to Sitophilus is unknownthough various B vitamins may be included (Musgrave, 1964). However that may be,it seems that the myelin forms described represent one way in which the host insectsderive some benefit (presumably of some generalized lipid or lipoprotein nature)from their symbiotes.
Grateful acknowledgements are made to Miss Margaret French and Mr J. Webb for technicalhelp, to Professors K. Ronald and J. Schroder, in whose Departments the work was done, andto the National Research Council of Canada for an operating grant to one of us (A. J. M.) and agrant for purchase of an electron microscope.
REFERENCES
BUCHNER, P. (1966). Endosymbiosis of Animals with Plant-like Micro-organisms. New York:John Wiley.
BUCK, J. B. (1953). Physical properties and chemical constitution of insect blood. In InsectPhysiology (ed. K. D. Roeder), pp. 147-190. New York: John Wiley.
CEDERGKEN, B. (1956). The lung tissue in mice infected by tubercle bacilli. In Electron Micro-scopy (ed. F. S. SjOstrand & J. Rhodin), pp. 248-249. New York: Academic Press.
FREEMAN, J. A. (1964). Cellular Fine Structure. London, New York, and Toronto: McGraw-Hill.FREY-WYSSLING, A. (1953). Submicroscopic Morphology of Protoplasm (transl. M. Hollander).
Amsterdam, Houston, London and New York: Elsevier.GILMOUR, D. (1965). The Metabolism of Insects. Edinburgh and London: Oliver and Boyd.GRACE, T. D. C. (1962). Establishment of four strains of cells from insect tissues grown in
vitro. Nature, Lond. 195, 788-789.GRINYER, I. & MUSGRAVE, A. J. (1966). Ultrastructure and peripheral membranes of the
mycetomal micro-organisms of Sitophilus granarius (L.) (Coleoptera). J. Cell Set. 1, 181-186.HENRY, S. M. (1962). The significance of micro-organisms in the nutrition of insects. Trans.
N.Y. Acad. Sci. (Ser. II) 24. 676-683.KUSCHEL, G. (1961). On problems of synonymy in the Sitophilus oryzae complex (30th Con-
tribution Col. Curculionidae). Ann. Mag. nat. Hist. (Ser. 13) 4, 241-245.MALKE, H. (1966). Elektronoptische Untersuchungen zur intra-cellul3ren Bakteriensymbiose
von Nauphoeta cinerea (Olivier) (Blattariae). Z. allg. Mikrobiol. 6 (3), 163-176.MERCER, E. H. (1962). The evolution of intracellular phospholipid membrane systems. In
The Interpretation of Ultrastructure (ed. R. J. C. Harris), pp. 369-384. London and NewYork: Academic Press.

Mycetomal micro-organisms 69
MILBURN, N. S. (1966). Fine structure of the pleomorphic bacteroids in the mycetocytes andovaries of several genera of cockroaches. J. Insect Physiol. 12, 1245-1254.
MUSGRAVE, A. J. (1964). Insect mycetomes. Can. Ent. 96, 377-389.MUSGRAVE, A. J. & HOMAN, R. (1962). Sitophilus sasakii (Tak.) in Canada: anatomy and myce-
tomal symbiotes as valid taxonomic characters. Can, Ent. 94, 1196-1197.MUSGRAVE, A. J., HOMAN, R. & GRINYER, I. (1964). Mycetomal and other micro-organisms in
young and aging SitopMlus (Coleoptera: Curculionidae). Can. J. Microbiol. 10, 806-808.MUSGRAVE, A. J. & SINGH, S. B. (1965). Histochemical evidence of nuclear equivalents in
mycetomal micro-organisms in Sitophtlus granarius (Linnaeus). J. Invert. Path. 7, 269-270.OLLETT, W. S. (1947). A method for staining both Gram-positive and Gram-negative bacteria in
sections, jf. Path. Bad. 59, 357-358.OLLETT, W. S. (195I). Further observations on the Gram-Twort stain. J. Path. Bact. 63, 166.SCHNEIDER, I. (1964). Differentiation of larval Z)roso/>A»7a eye-antennal discs in vitro. J. exp.Zool.
156, 91-104.SINGH, S. B. & MUSGRAVE, A. J. (1966). Some studies on the chromatin and cell wall of the
mycetomal micro-organisms of Sitophilusgranarius (L.) (Coleoptera).^. Cell Sci. 1, 175-180.SPURLOCK, B. O., KATTINE, V. C. & FREEMAN, J. A. (1963). Technical modifications in 'Mara-
glas' embedding. .7. Cell Biol. 17, 203-207.STEINHAUS, E. A. (1949). Principles of Insect Pathology. New York: McGraw Hill.TRAGER, W. (i960). Intracellular parasitism and symbiosis. In The Cell, vol. 4 (ed. J. Brachet &
A. E. Mirsky), pp. 151-213. New York and London: Academic Press.VICKERMAN, K. (1962). Patterns of cellular organization in Limax amoeba. Expl Cell Res. 26,
497-519.WYATT, S. S. (1956). Culture in vitro of tissue from the silkworm, Bombyx mori (L.). J. gen.
Physiol. 39, 841-852.
{Received 24 April 1967—Revised 5 July 1967)

70 A. J. Musgrave and I. Grinyer
Fig. i. Electron micrograph of most of a spiral organism in section. From a mycetomeof 5. zea-mais. x 32000.Fig. 2. Parts of a similar organism at greater magnification. From a mycetome (cro,cell wall; im, intracytoplasmic membranes; n, nuclear material ;pm, plasma membrane),x 150000.
Journal of Cell Science, Vol. 3, No.
A. J. MUSGRAVE AND I. GRINYER (Facing p. 70)

Fig. 3. Electron micrograph of a mass of disintegrating organisms. Normal organismsare present outside the mass. From a mycetome (dm, disintegrating micro-organism;ntn, normal organism; sm, spiral configuration of disintegrating organism), x 10000.
Fig. 4. Electron micrograph of a membrane mass. The organisms are in a moreadvanced stage of disintegration. From a mycetome (dm, disintegrating micro-organism.) x 29000.
Fig. 5. Shows the trilaminate nature of the membranes. From a mycetome. x 100000.
Fig. 6. Membranes apparently arising from an isolated, disintegrating micro-organism.From a mycetome. x 52000.

Journal of Cell Science, Vol. 3, No. 1
A. J. MUSGRAVE AND I. GRINYER

Fig. 7. Phase-contrast micrograph showing lamellar masses in an unfixed smearpreparation of a prepupal mycetome. No spiral organisms are evident, x 450.Fig. 8. As in Fig. 7, but from an adult mesenteron. x 450.Fig. 9. Evidence of spiral organism within a spheroidal object, from osmium-fixedsmear of mycetome stained in Short's modification of Heidenhain's haematoxylin.x1300.
Fig. 10. Thin (i/t) section of part of a mycetome, stained with toluidine blue. In onemycetocyte is a lamellar mass (Im). The nucleus of the mycetocyte is indicated (n).Sections of spiral organisms can be seen, x 1150.Fig. 11. Relatively large myelin figures in an unfixed crush preparation of adultmesenteron. No spiral forms are evident. Phase contrast, x 350.

Journal of Cell Science, Vol. 3, No. 1
A. J. MUSGRAVE AND I. GRINYER
