mesenchymal stem cells maintain the microenvironment of
TRANSCRIPT

Full Terms & Conditions of access and use can be found athttp://www.tandfonline.com/action/journalInformation?journalCode=ines20
Download by: [Chao Xu] Date: 19 May 2017, At: 01:52
International Journal of Neuroscience
ISSN: 0020-7454 (Print) 1543-5245 (Online) Journal homepage: http://www.tandfonline.com/loi/ines20
Mesenchymal stem cells maintain themicroenvironment of central nervous systemby regulating the polarization of macrophages/microglia after traumatic brain injury
Chao Xu, Feng Fu, Xiaohong Li & Sai Zhang
To cite this article: Chao Xu, Feng Fu, Xiaohong Li & Sai Zhang (2017): Mesenchymal stemcells maintain the microenvironment of central nervous system by regulating the polarization ofmacrophages/microglia after traumatic brain injury, International Journal of Neuroscience, DOI:10.1080/00207454.2017.1325884
To link to this article: http://dx.doi.org/10.1080/00207454.2017.1325884
Accepted author version posted online: 03May 2017.Published online: 19 May 2017.
Submit your article to this journal
Article views: 33
View related articles
View Crossmark data

Mesenchymal stem cells maintain the microenvironment of central nervoussystem by regulating the polarization of macrophages/microglia after traumaticbrain injury
Chao Xu*, Feng Fu*, Xiaohong Li and Sai Zhang
Institute of Traumatic Brain Injury and Neurology, Pingjin Hospital, Logistics University of Chinese People’s Armed Police Forces, Tianjin 300162,China
ARTICLE HISTORYReceived 22 November 2016Revised 19 April 2017Accepted 27 April 2017Published online 19 May2017
ABSTRACTMesenchymal stem cells (MSCs), which are regarded as promising candidates for cell replacementtherapies, are able to regulate immune responses after traumatic brain injury (TBI). Secondaryimmune response following the mechanical injury is the essential factor leading to the necrosisand apoptosis of neural cells during and after the cerebral edema has subsided and there is lack ofefficient agent that can mitigate such neuroinflammation in the clinical application. By means ofthree molecular pathways (prostaglandin E2 (PGE2), tumor-necrosis-factor-inducible gene 6 protein(TSG-6), and progesterone receptor (PR) and glucocorticoid receptors (GR)), MSCs induce theactivation of macrophages/microglia and drive them polarize into the M2 phenotypes, whichinhibits the release of pro-inflammatory cytokines and promotes tissue repair and nerveregeneration. The regulation of MSCs and the polarization of macrophages/microglia aredynamically changing based on the inflammatory environment. Under the stimulation of plateletlysate (PL), MSCs also promote the release of pro-inflammatory cytokines. Meanwhile, the statue ofmacrophages/microglia exerts significant effects on the survival, proliferation, differentiation andactivation of MSCs by changing the niche of cells. They form positive feedback loops inmaintaining the homeostasis after TBI to relieving the secondary injury and promoting tissuerepair. MSC therapies have obtained great achievements in several central nervous system diseaseclinical trials, which will accelerate the application of MSCs in TBI treatment.
KEYWORDSMesenchymal stem cells;macrophage/microgliapolarization; traumatic braininjury; cell-based therapy;neuroinflammation
Introduction
Mesenchymal stem cells (MSCs), a type of multipotentstem cells, are capable of regulating the immune system[1] and are widely distributed in the connective tissues oforgans. It has been confirmed that MSCs can differentiateinto effector cells, such as neurons [2,3] and glial cells [4],to replace and restore damaged tissues [5]. MSCs can evenimprove the short-term and long-term behavior [6], andthus have been regarded as one of the most promisingtherapies for central nervous system (CNS) diseases to pro-mote neuroprotection, regeneration and repair [6–8]. Aftertraumatic brain injury (TBI), stress reactions are switchedon, which begins the protection, scavenger and repair pro-grams by driving inflammatory events and changing thelocal immune environment [9]. Stem cells such as neuralstem cells (NSCs) and neural progenitor cells (NPCs) derivedfrom the subventricular zone (SVZ) of lateral ventricles andthe subgranular zone (SGZ) of dentate gyrus are activatedto differentiate into mature neurons and glial cells [10].
However, MSCs are not able to replace the lost neural cellsas expected [11], even though they are migrating to sitesof lesions [12,13]. Instead, they modulate the immunemicroenvironment of the injury sites or lesions reducingneurotoxicity, protecting nervous cells, accelerating self-repair [14,15], improving impaired motor-sensory functionsfollowed by secondary inflammation of TBI and by acceler-ating the proliferation and differentiation of NSCs andNPCs [16]. Neuroinflammation induced by various harmfulfactors, including TBI, is reduced after transplanting MSCsinto the brains of animal models [17] because of their abil-ity to modulate focal immune components especially byregulating the polarization of macrophages or microglialcells via the release of biomolecules [18]. This mechanismhas been highlighted as a delivery platform in cell thera-pies [19]. Macrophages and microglial cells, which are themacrophages of the CNS, are the pivotal targets of immu-noregulation and the primary sites of inflammatory activi-ties. MSCs drive macrophages/microglia to polarize into
CONTACT Sai Zhang [email protected]*Xu Chao and Fu Feng contributed equally to this article.
© 2017 Informa UK Limited, trading as Taylor & Francis Group
INTERNATIONAL JOURNAL OF NEUROSCIENCE, 2017https://doi.org/10.1080/00207454.2017.1325884

M2-like cells and inhibit the secretion of pro-inflammatorycytokines of the M1-like cells [20] that promote immuno-pathologic reactions. However, these cytokines play animportant role in the MSC niche where the MSCs survive,proliferate, differentiate and interact with the surroundings[1]. Therefore, MSCs are activated by inflammatory media-tors related to macrophages/microglia when injuries occuror the homeostasis is shifted [21,22].
The use of therapeutic MSC transplantation is stillunder clinical trials of CNS diseases (stroke [23,24], multi-ple sclerosis [25], and spinal cord injury [26]) and graftversus host disease (GVHD) [27], and although the safetyof allogeneic MSCs has been confirmed [28], they havenot been approved by health care departments aroundthe world [29]. In addition, there is lack of evidence rec-ommending cell transplantation as an established ther-apy that can be generalized widely for inflammatorydiseases. Additionally, despite intensive studies at themolecular level, the mechanism of interaction betweenMSCs and macrophages/microglia has only been par-tially identified. In this review, we have summarized theresults from current studies to map out the mechanismof interaction between MSCs and macrophages/micro-glia, thereby demonstrating the detrimental or beneficialimpacts of MSCs on damaged tissues and their functionin regulating the immune system to maintain the CNS ina steady inflammatory state after TBI.
Mechanism of MSCs regulating macrophagepolarization
MSCs change their immune properties and functionalcharacteristics according to the culture condition or sur-roundings [30]. They respond by changing the immunemicroenvironment by releasing various paracrine cyto-kines [31]. The cytokines released by MSCs can inhibit ormitigate inflammation (prostaglandin E2 (PGE2) andtumor necrosis factor-stimulated gene 6 protein (TSG-6)[32]), enhance vascularization (vascular endothelialgrowth factor (VEGF)) [33] and enhance cells proliferation(interleukin (IL)-6 [34] and transforming growth factor(TGF)-b1 [35]). MSCs also produce chemokines, such asmonocyte chemotactic protein (MCP)-1 and C-X-C motifligand (CXCL)-4, to activate Notch or CD95/Fas signalingpathways [36], which modulate proliferation, activationand effector functions of T lymphocytes [37], dendriticcells (DC) [38] and natural killer (NK) cells [39] by directcell-to-cell contacts and indirect connections. Further-more, MSCs also affect the cellular components of theCNS, e.g. by regulating survival, metabolic activities andcell cycling of astrocytes through various signaling path-ways [40]. However, most importantly, MSCs regulate thepolarization of macrophages and microglial cells, which
are the pivotal for carrying out immunoreactions, and bydoing so, they control the process of inflammation, limitexcessive immunological defense mechanism [41,42],attenuate tissue damage caused by inflammation, andeven enhance the capability of self-repair [43] in order tomaintain the stability of the lesion.
Macrophages/microglia polarize into pro-inflammatoryM1 phenotypes and anti-inflammatory M2 phenotypes
Macrophages are located in the peripheral organs, such aslymph nodes, blood and perivascular tissues, while micro-glia are located in the CNS [44]. Macrophages can travelthrough the blood vessel and adjacent tissues and theirtropism is dictated by molecules of the injured cells [45].Microglial aggregations are serve to eliminate necroticneurons [46] and activating the immune system [47]. Mac-rophages/microglia are also the essential cells that exertsignificant effects on the regulation of the immune sys-tem and the activation of further responses against infec-tions as well as injury during inflammatory processes [48].They control the “on–off” mechanism of microglia-medi-ated inflammation [47] when detrimental factors invadeinto healthy tissues, and they also activate and managethe progression of the endogenous tissue repair program[49]. When inflammation develops, pro- and anti-inflam-matory mechanisms take effects on secondary injury andtissue repair through macrophages/microglias [50]. Theconcept of macrophage polarization was first reported in1992, when IL-4 was found to significantly enhance thefunction of mannose receptor, CD206, in rodent macro-phages [41]. Activated macrophages are divided into twomajor phenotypes according to the different inflamma-tory microenvironments – classic M1 and alternative (orprotective) M2 [51]. The M1/M2 balance is one of themost important mechanisms of injury/repair regulation.Following a myocardial infarction, the quantities of M1phenotypes and M2 phenotypes change dynamically andthey are able to transform into one another in the ische-mic tissues [13,21]. The classic M1 phenotype is inducedby lipopolysaccharide (LPS) and interferon (IFN)-g , and itmainly produces pro-inflammatory cytokines IL-1b, IL-6,tumor necrosis factor (TNF)-a, inducible nitric oxide syn-thase (iNOS), IL-12p40, IL-23 and IFN-g . These cytokinesrecruit peripheral inflammatory cells and activate inflam-matory cascades. The alternative M2 phenotype isinduced by IL-4 and IL-13, and it secretes IL-10, IL-1RA,TGF-b1, TGF-b3 and expresses CD206 and arginase (Arg)-1 [21,52,53]. Recently, some studies have shown that theprocess of macrophage polarization in vivo is more com-plex than the co-culture conditions in vitro [54,55].According to the gene analysis, activated resident macro-phages are showing a complicated plasticity that
2 C. XU ET AL.
responds to particular pathological process [56]. Two hun-dred ninety-nine macrophage transcriptomes were identi-fied under different kinds of stimulation and 49 distinctco-expression clusters were found in transcriptionalresponses between M1 and M2 [57,58], which suggestedthat every macrophage bore its own specific characteris-tics following the environmental signals. However, only asmall amount of macrophage populations among thespectrum were comprehensively recognized. Previousstudies have illustrated that the M2 phenotypes aredivided into M2a [59], M2b [60], M2c [14] and M2r [61].The M2a subtype promotes tissue repair while the M2b
and M2c subtypes have increased phagocytosis capacityand regulate immune responses [62]. M2r cells producedprogrammed cell death 1 ligand-2, IL-10 and TGF-b, andwere able to deactivate M1 phenotype macrophages andsuppress T cell proliferation [61]. Another subtype, theMox subtype, activated by oxidized low-density lipopro-teins (ox-LDLs), exerts potent effects on atherosclerosis[63]. The finding of rapid invasion (wormhole travel) ofmature macrophages that contained special functionsfrom cavity [64] released novel inflammatory cell recruit-ment and tissue repair [65] which might generate newdiscussions about the inflammatory response of CNS.
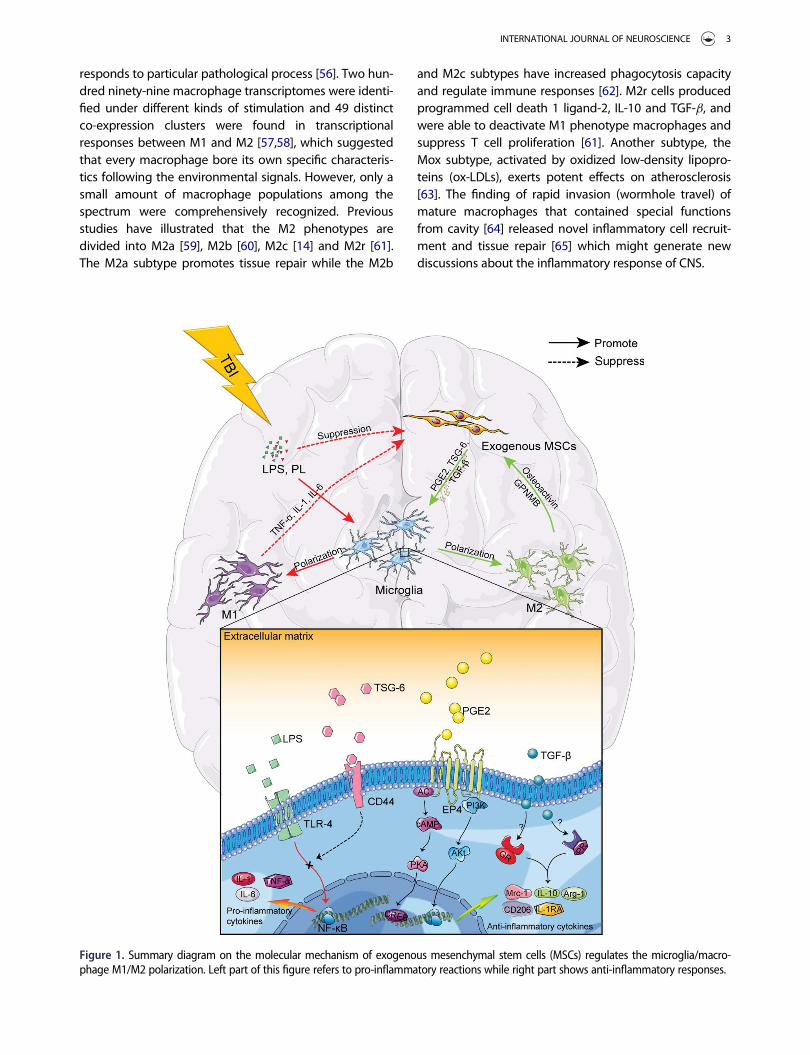
Figure 1. Summary diagram on the molecular mechanism of exogenous mesenchymal stem cells (MSCs) regulates the microglia/macro-phage M1/M2 polarization. Left part of this figure refers to pro-inflammatory reactions while right part shows anti-inflammatory responses.
INTERNATIONAL JOURNAL OF NEUROSCIENCE 3

Vitality of MSCs in regulating macrophage/microgliapolarization
MSCs regulate the activation of macrophages/microglia,transforming the classic M1 phenotype into alternativeM2 phenotype, expressing Arg-1, CD206, IL-10, andPGE2, which strengthens phototrophic activity [52], miti-gates the release of pro-inflammation cytokines andmodulates inflammatory activity [66]. These changes sta-bilize the environment of the injury site and make regen-erative stable niche for ischemic tissue [67]. Experimentalstudies of bone marrow derived macrophages and MSCco-cultures showed the markers of M1 phenotype, suchas IL-6, IL-1b, MCP-1 and iNOS, were decreased signifi-cantly while the markers of M2 phenotype, such as IL-10,IL-4, CD206 and Arg-1 [14], were increased indicatingthat the immunoregulation property of MSCs exerts theessential effect on the transformation of M1 to M2 phe-notypes [52].
Mechanism of MSCs regulating macrophage/microglia polarization
After wounding or injury, a variety of effector moleculesand mechanisms, particularly reactive oxygen species(ROS) [68] and pro-inflammatory cytokines, change thebalance of M1/M2 and their activated states [69]. Nuclearfactor kappa-light-chain-enhancer of activated B cells(NF-kB), the pivotal factor controlling DNA transcriptionand cytokine production, serves a critical role in macro-phage/microglia polarization and is the regulatory site ofmost intervention mechanisms [68,70]. It is believed thatMSCs regulate the activation and polarization of macro-phages/microglia mainly through three pathways: PGE2pathway, TSG-6 pathway [71] and glucocorticoid recep-tors (GR)/progesterone receptors (PR) (Figure 1) [72]. ThePGE2 pathway depends on activated MSCs releasingPGE2, which drives the transformation of M1 phenotypecells mainly colonized in the peripheral tissues into theprotective M2 phenotype [1]. The TSG-6 pathway takeseffect by activating the reaction of TSG-6 and CD44,which blocks the Toll-like receptor (TLR)-4/NF-kB signal-ing pathway of macrophages. This pathway reduces theproduction of pro-inflammatory cytokines and controlsthe effect of pro-inflammatory M1 phenotype [73]. Thethird pathway is linked by macrophage surface recep-tors, GR and PR, which determine macrophage differenti-ation [20].
MSCs drive macrophages to polarize into M2phenotype through PGE2 releasingTo increase the therapeutic potentials [74] and enhancethe anti-inflammatory properties [75], researchers co-
cultured MSCs and human adult dermal fibroblasts as 3Dspheroids and collected conditioned medium. Then,macrophages that had been induced by LPS (M1 pheno-type) were cultured with the collected conditionedmedium. The result demonstrated that TNF-a leveldeclined noticeably, while IL-10, IL-1RA level and CD206positive cells increased [53]. Furthermore, pro-inflamma-tory genes, Tnf and Csf2, were downregulated and theanti-inflammatory gene, Tgm, was upregulated. This sug-gested that the MSCs drove LPS-induced macrophagesinto the protective M2 phenotype. Moreover, theresearchers found that PGE2 levels from microarray anal-yses were much higher than before and when inhibitorsof cyclooxygenase (COX)-2, which is an essential enzymein the PGE2 synthesis, were added into the conditionedmedium, the anti-inflammatory effect of the M2 pheno-type macrophages was immediately weakened, and thesecretion of IL-10 and IL-1RA was reduced. Thus, it wasconfirmed that PGE2 is one of the mechanisms that con-nects MSCs to macrophage/microglia polarization [53].Another study has found that during inflammationcaused by injuries, platelet lysate (PL) activates NF-kB inMSCs. NF-kB promotes mPGE expression, thus producingmore PGE2 [76].
Among the four receptors of PGE2, only EP4 can stopthe reduction in TNF-a levels and increase in IL-10 levels[77], which indicates that EP4 on the surface of macro-phages modulate the same anti-inflammatory effect thatPGE2 mediates. According to the results of a formerstudy, PGE2 released by MSCs in 3D spheroids rely ontheir own cysteine aspartic protease (Caspase) and theactivation of NF-kB signaling pathway [53].
By activating Caspase [32], NF-kB [70] and cAMP [78]signaling pathways, the endogenous or exogenous stim-uli (such as mechanical damage [79] and IL-1b [80])imposed on MSCs increase the production and secretionof PGE2. Serving as the direct contact between MSCsand macrophages/microglia, PGE2 can attach to EP4 onthe surface of macrophages, thereby phosphorylatingintracellular cAMP-response element binding protein(CREB). Thus the expression of transcription factor C/EBP-b is upregulated [81]. Finally, with the expression ofArg-1, IL-10 and Mrc-1 being increased [82], the transfor-mation from M1 phenotype to M2 phenotype inducedby the PGE2 pathway is complete.
MSCs inhibit M1 phenotypes producing pro-inflammatory cytokines by releasing TSG-6TSG-6 is able to inhibit the release of pro-inflammatorycytokines [73] and enhances the tissue repair [83]. LPSand IFN-g are capable of inducing subsequent intracellu-lar signaling pathways and motivate the degradation ofIkB, the inhibitory protein bound to NF-kB [70]. When
4 C. XU ET AL.

NF-kB is free from the inhibitory protein, it translocatesinto the nucleus and binds to the promoters of varioustarget genes to initiate the expression of pro-inflamma-tory cytokines and chemokines. High levels of TNF-a, IL-1b, IL-6, MIP-1a and MCP-1 are produced quickly, andthe inflammation process becomes aggressive [84]. Itwas found that, compared to the control group, theTSG-6 level of experimental rats that were intravenouslyadministered MSCs was significantly increased in 12–72 hours after cortex injury while NF-kB and various pro-inflammatory cytokines were significantly decreased[17]. Besides, TSG-6 released by MSCs inhibit NF-kB sig-naling pathway and block blood–brain barrier (BBB) dis-ruption in ICH [85]. These results suggest that TSG-6works through NF-kB signaling pathway.
When TSG-6 expression was suppressed with siRNA,the regulation microglia polarization by MSCs was weak-ened [73]. Compared to the control group, the phos-phorylation of p38, c-Jun N-terminal kinase (JNK),extracellular regulated protein kinases (ERK) and mito-gen-activated protein kinase (MAPK) were reduced inthe BV2 microglia that had been induced by TSG-6 afterLPS stimulation. Meanwhile, TSG-6 interferes with theactivation of NF-kB signaling pathway which is modu-lated by LPS/TLR-2 [86], suggesting that MSCs restrainNF-kB and MAPK signaling pathways within the micro-glia by releasing TSG-6 in order to block the secretion ofpro-inflammatory factors. Another study showed thatthe interaction between TSG-6 and CD44 on the surfaceof macrophages reduced the nuclear translocation ofNF-kB which was regulated by zymosan/TLR-2 [86].Therefore, the TSG-6 pathway serves its function throughsurface molecule CD44, subsequently stopping TLR-2from activating the NF-kB signaling pathway and block-ing the production of pro-inflammatory cytokines.
MSCs modulate the process of macrophage/microgliapolarization through GR and PRProgesterone was previously regarded as a neuroprotec-tive reagent in animal models of TBI [87] and was shownto improve the recovery of neural function by reducingthe production of inflammatory cytokines and inhibitingthe activation of microglia via PR and g-aminobutyricacid type A (GABA (A)) receptors [72]. PR is widely distrib-uted in the neural cells, and its expression is indepen-dent of the level of steroid, particularly membraneprogesterone receptor (mPR)-a, which is expressed inthe activated microglia, astrocytes and oligodendrocytesafter TBI [88]. However, it did not show any advantage inclinical trials and recent clinical trial results have indi-cated that progesterone did not improve the outcomeof TBI [89]. These clinical trials almost always focus onthe early stages of moderate or severe TBI and their
short-term outcome without long-term follow-up andlack of standardized treatment protocol. Thus, therehave been a lack of prediction and evaluation of progno-sis for the survivors.
We believe that PR and GR play a more important role inregulating inflammation and neuroprotection and in modu-lating the microglia because they really mitigate the pro-duction of inflammatory molecules [90]. When mifepristone(inhibitor of GR and PR) was added into the culture systemof human umbilical derived MSCs, the released factors byMSCs to modulate macrophage differentiation was partlyblocked [20]. We can, therefore, infer that MSCs modulatethe process of M1 cells polarization to M2 cells through theeffect of GR and PR on macrophages. But there is little evi-dence indicating that MSCs are capable of secreting gluco-corticoid or progesterone [91]. Instead, they regulate thefunction of GR through TGF-b. Unfortunately, there is littleevidence demonstrating the potential interactions betweenTGF-b and PR [92]. Eventually, NF-kB can be activated andMSCs achieve regulation of the polarization process [93].
Recent studies have revealed that, rather than beingsimply stagnant, the immunomodulation properties andbiological functions of MSCs in regulating macrophagesare continuously adjusting depending on the immunemicroenvironment and the period of inflammation pro-gression [30,94,95], which differs cell therapies from phar-macotherapy. In early stages, under PL activation, MSCsproduce granulocyte colony-stimulating factor (G-CSF)driving monocytes to differentiate toward pro-inflamma-tory M1 macrophages that release IL-6 and TNF-a, whichpromote the expansion of inflammation and tissue repair[76]. These findings have indicated that MSCs are notonly exerting effects on blocking the inflammatoryresponse, but also promoting early immune response,and might be dynamically keeping the microenvironmentrelatively stable. Nevertheless, more experimental dataare needed to demonstrate the potential mechanism ofhomeostatic regulation by MSCs.
Macrophage polarizing to M2 phenotype assistsMSCs survival, proliferation and migration
During the study of MSCs in cell therapies and immunehomeostasis, it was discovered that MSCs could adapt tovarious extreme physical conditions, such as hypoxiaand starving, and immune environments [96]. Especiallyin the inflammatory niche with TNF-a and IL-1, the MSCsare activated to function in immune suppression [97].When MSCs regulate macrophage/microglia differentia-tion, their own growth and effector functions alsobecome influenced by the polarized macrophages/microglia [45]. An in vitro study showed that the growthof MSCs was restricted under the condition that contains
INTERNATIONAL JOURNAL OF NEUROSCIENCE 5

M1 phenotype but was promoted under the influence ofthe M2 phenotype and its cytokines [21]. TNF-a, IL-1band IL-6 released by macrophage enhance the ability ofMSCs to produce inflammatory cytokines [20] and theirability to migrate [95,97]. It has been described inanother article that compared to the classic M1 pheno-type, the M2 phenotype expresses more osteoactivin/glycoprotein non-metastatic melanoma protein B(GPNMB), which activates the ERK/JNK signaling path-way and assists in MSC survival, proliferation and migra-tion [98]. According to these findings, we hypothesizedthat there might be a feedback loop between endoge-nous MSCs and the regulation of their immune functionby macrophages/microglia. An immune microenviron-ment that maintains the injured tissue in a comparativelystable inflammatory condition is built by MSCs, macro-phages/microglia and their downstream components,especially T lymphocytes and B lymphocytes. This inter-active network balances pro-inflammatory and anti-inflammatory effects, tissue damage and repair. There-fore, it provides us with a totally new prospect when weare trying to improve the immune microenvironmentand reduce inflammatory injury through autologous orallogeneic transplantation of exogenous MSCs.
Role of MSC-induced macrophage polarizationin TBI
Focal immune environment caused by TBI
TBI is a complicated socioeconomic disease of which theincidence and outcome are tightly dependent on theincome and the medical system of countries andregions, and the epidemiological and demographicalfeatures are changing gradually [99]. According to theCenters for Disease Control (CDC), 1.7 million people suf-fer from TBI each year in the USA and the major cause ofTBI falls rather than traffic accidents [100,101]. CNS dam-age caused by TBI can be divided into two phases. Theearly phase is the immediate effect of trauma, whichleads to BBB rupture, cerebral edema, and intracranialhemorrhage [9]. With the development of severe dam-age, increasing intracranial pressure (ICP) and neural cellnecrosis are the major pathophysiologic causes of deathand disability in the acute stage [102]. Current treat-ments, such as decompressive craniectomy, prophylactichypothermia, hyperosmolar therapy and cerebrospinalfluid drainage, are mainly focusing on this phase andmanaging ICP, cerebral perfusion pressure (CPP) andvital signs based on a large amount of random clinicaltrials according to the novel edition guidelines [103].
The next phase is the secondary injury caused by neu-roinflammation [9,104]. Immune cells located in both
CNS and peripheral tissues respond to the damage andare recruited to the injury sites, releasing pro-inflamma-tory mediators [105], impairing neural cells and inducingdiffused neuroinflammation [106]. During this time, apo-ptosis of neural cells occur continuously and this processcan last more than 12 months after TBI. Novel guidelinesinhibit the use of steroids to control the increase of ICPand inflammatory responses because of the high ste-roid-related death rate [103]. However, the inflammatoryenvironment indirectly created by TBI turns the mononu-clear phagocyte system into a more aggressive and over-driving state, which promotes the phagocytic andantigen-presenting ability of macrophages, causingthem to produce more pro-inflammatory molecules andsignaling proteins, and activating functional downstreamcells [107]. A variety of T lymphocytes, B lymphocytesand complements are activated, and cytolysis is under-way [108]. In the meantime, the inflammatory environ-ment also blocks the recruitment of endogenous stemcells (NSCs and NPCs) and neural regeneration [10]. CNSinflammation in rats with TBI has been found to takeplace in two stages as well [109]. One stage is acute andmild neural inflammation where the immune responselasts at least 24 hours. The other stage is subacute oreven chronic state with more severe inflammatory activi-ties, which peak at 3 days after TBI. Researchers havefound that in female rat brains injured by a controlledcortical impact, microglial immune activity peak at 5–7 days post TBI and M2 phenotype macrophages relatedmarkers also peaked at 5 days post TBI [51].
Nevertheless, the mechanism of occurrence of neuro-inflammation following TBI is complicated and potentneuroprotective agents aimed at specific targets havenot been found until now [110]. A series of clinical trialshave provided powerful evidence that a variety of drugs,such as erythropoietin [111], steroids [112], non-steroidalanti-inflammatory drugs (NSAIDs) [113], minocycline[114] and BBB permeability drugs that could inhibit neu-roinflammation did not improve the outcome of TBI sig-nificantly [110,115], which has challenged thebreakthrough we had achieved in cellular biology andanimal models. Because of the complicated connectivenetworks, pointing to a certain single site cannot easilylimit the development of the inflammatory response[116]. Therefore, cell-based therapies, including MSCtransplantation may provide more advantages in meet-ing the physiological needs.
The effect of MSC-induced macrophage polarizationin neuroinflammation after TBI
Transplantation of MSCs to the injured sites of rodentbrains after TBI induced macrophages/microglia to
6 C. XU ET AL.

polarize into alternative M2 phenotype [14,17,51]. Thegene expression signature of M2 cells was upregulatedat 3–7 days post injury [14]. In addition, the inflammatoryactivity was relieved simultaneously, and cerebral edemawas mitigated and the damage range was limited. Com-pared to the control group, rats transplanted with MSCsgot significantly higher neuroscores at every time pointsfor 7–35 days post TBI. This indicated that the MSCswere being beneficial by regulating the inflammatoryenvironment, protecting neural function and promotingtissue repair and neural regeneration. Besides, clinical tri-als have also confirmed that MSCs are able to promotepositive outcomes in patients with TBI [117]. However,studies focusing on the mechanism of MSCs after TBI inmaintaining stable immune system in vivo by regulatingmacrophage/microglia polarization and controllingdownstream functional inflammatory cells are quite rare.The improvement of the focal immune environmentby MSCs is a pivotal process. The improved inflammatoryenvironment promotes the release of anti-inflammatorycytokines and further creates a beneficial niche for theproliferation and function of MSCs, which forms a posi-tive feedback loop. By modulating macrophage polariza-tion, the MSCs reduce secondary damage and activatetissue repair after TBI [10]. Furthermore, it is obvious thatthe polarized M2 cells improve the environment of oligo-dendrocyte regeneration and nerve fiber remyelination[90].
Previously, we regarded the CNS as a completely iso-lated structure lacking of “classic” lymphatic drainagesystems except blood vessels linked to the peripheral cir-culation [118]. And the BBB thus separates the immunesystem into two independent parts: the CNS and theperipheral immune systems. In 2015, functional lym-phatic vessels arranged in dural sinuses were finallyfound within the meningeal compartment [119]. Theselymphatic vessels are able to transport both fluid andimmune cells from the cerebrospinal fluid to the deepcervical lymph nodes. This finding disproved our tradi-tional concept about the CNS immune system andshowed that the whole immune system is integrated.Hence, components located in the peripheral tissue orthose migrating in vessels are able to function in theCNS. Thus, CNS diseases could be treated using periph-eral pathways and therapeutic cell transplantation couldbe a promising method to reduce CNS inflammation byin situ regulation of the pro-inflammatory activities ofmigratory immune cells after TBI.
Discussion and perspective
TBI has become a worldwide disease with irreversiblehigh morbidity and disability [100] because of the lack of
effective methods to improve the poor outcome andlong-term impairment of behavioral function due tospontaneous self-protective inflammatory responses [9].The balance of damage/repair is controlled by the prog-ress of neuroinflammation [120]. Inhibiting inflammationmay not be the best choice for TBI patients with severesecondary injury, because of the risk of the reduction inneural regeneration [121]. Compared to regular thera-pies, which target components of the inflammatory reac-tion chains, cell-based therapies have more advantagesin modulating the excessive inflammation and enhanc-ing tissue repair [10,110]. This is due to their special biol-ogy and immune properties acting through differentkinds of pathways, which exert comprehensive and inte-grated impacts on the injured sites. Concepts like thesethat demonstrate autogenous regulation have obviouspotential for benefiting patients.
In recent years, studies concerning the ability of MSCsto improve neural function after TBI are drawing plentyof attention. Starting with transplantation of primaryMSCs, engineered MSCs have been designed to enhancecellular function to improve outcome of various indica-tions, especially TBI [122]. For example, the secretion ofimmunoregulatory cytokines by MSCs was enhanced orinhibited to limit CNS autoimmune diseases [66], andtemperature-sensitive MSCs were established to treatTBI combined with mild hypothermia [123]. Additionally,in order to simulate the niche of the grafted cells, 3Dspheroids [53], alginate micro-encapsulation [124], andco-graft system of Schwann cells and MSC scaffolds[125] were developed to enhance the modulation ofneuroinflammation.
It has been accepted that the macrophage/microgliapolarization is one of that pathways regulated by MSCsto maintain the stability of the immune microenviron-ment [1]. With the progression of studies focused on themechanism of interaction between MSCs and macro-phages and their downstream immune components,transplantation of therapeutic MSCs is going to be thenew prospect of clinical practice, despite the controver-sies regarding treatment opportunity, duration and path-way of transplantation. Actually, researchers haveconfirmed that MSCs are very biocompatible whichmakes them suitable to be used in common deliveryconditions [126]. Many physicians are cautious about celltherapies, limited by their practical resource [127],because of the safety and feasibility of such therapies.This has been preventing the application of MSCs incomprehensive clinical treatments even though manyclinical trials are ongoing. However, with the latestbreakthrough in the benefits of MSC transplantation instroke patients (Unique identifier: NCT01287936) [24], itis likely that the safety and efficiency of cell therapies
INTERNATIONAL JOURNAL OF NEUROSCIENCE 7

will be accepted and related studies regarding CNS dis-eases, especially TBI, will be developed rapidly.
Neuroinflammation is induced by an intricate networkof activating immune cells in which MSCs are involved inimmunoregulation [108]. A part of this conclusion hasbeen based on in vitro experiments or simple animalmodels. Whether MSCs regulate the immune microenvi-ronment in the same pathways in TBI is still unclear. Weare not able to fully declare that MSCs are capable ofserving as a mature and safe immunoregulator used inclinical treatments after TBI yet. Unlike animal models,there are still some unanswered questions that are inter-fering with the clinical application of MSCs. First, thecomplex ethics of stem cell clinical use is widely dis-cussed [128]. The International Society for Stem CellResearch updated guidelines for the translation of stemcells into clinical use on 12 May 2016 [29]. This standardemphasized the ethics of such application. Second, thesafety of cell-based therapies is controversial. In the newguidelines, the responsibilities of regulator and supervi-sors are emphasized to guarantee the health of stemcells and the transparency of clinical trials. Someresearchers reported that MSCs enhance the progressionof tumor in animal models [129,130], which is one of themost threatening features. Meanwhile, potential organdamage also puzzles therapists. Then, clinical trials lackof standardized “patients” and treatment protocols andwhich influences the favorable results of preclinical stud-ies. Besides, it is difficult to detect the survival, prolifera-tion and function of MSCs that are implanted intopatients by a non-invasive and safe methods and long-term follow-ups. In addition, economic and commercialpressure is a practical hamper of translation of MSCtransplantation [131].
More intensive studies are needed in the future todetermine if there are other mechanisms operating in thepolarization of macrophages/microglia and their down-stream effectors while more rigorous clinical trials withlong-term follow-up are needed to clarify the exact effectsand unfavorable adverse effects of MSC transplantation.
Disclosure statement
The authors report no conflicts of interest. The authors aloneare responsible for the content and writing of the paper.
Funding
This work was financially supported by National Natural Sci-ence Foundation of China [grant number 81271392], [grantnumber 81301050], [grant number 81401067], [grant number81471275], [grant number 81541034]; Natural Science Founda-tion of Tianjin City [grant number 14JCQNJC10200], [grantnumber 15JCQNJC11100].
References
1. Liu J, Kuwabara A, Kamio Y, et al. Human mesenchymalstem cell-derived microvesicles prevent the rupture ofintracranial aneurysm in part by suppression of mast cellactivation via a PGE2-dependent mechanism. Stem Cells2016;34:2943–55.
2. Zhao E, Jia Y, Wang D, et al. Effect of p65 gene inhibitedby siRNA on differentiation of rat marrow mesenchymalstem cells into neurons. Chinese J Appl Physiol2015;31:254–8.
3. Zeng X, Qiu X-C, Ma Y-H, et al. Integration of donor mes-enchymal stem cell-derived neuron-like cells into hostneural network after rat spinal cord transection. Biomate-rials 2015;53:184–201.
4. Pirzad Jahromi G, Shabanzadeh Pirsaraei A, Sadr SS, et al.Multipotent bone marrow stromal cell therapy promotesendogenous cell proliferation following ischemic stroke.Clin Exp Pharmacol Physiol 2015;42:1158–67.
5. Deng K, Lin DL, Hanzlicek B, et al. Mesenchymal stem cellsand their secretome partially restore nerve and urethralfunction in a dual muscle and nerve injury stress urinaryincontinence model. Am J Physiol Renal Physiol 2015;308:F92–f100. Epub 2014/11/08.
6. Kota DJ, Prabhakara KS, van Brummen AJ, et al. Proprano-lol and mesenchymal stromal cells combine to treat trau-matic brain injury. Stem Cells Trans Med 2016;5:33–44.Epub 2015/11/21.
7. Tang G, Liu Y, Zhang Z, et al. Mesenchymal stem cellsmaintain blood�brain barrier integrity by inhibitingAquaporin�4 upregulation after cerebral ischemia. StemCells 2014;32:3150–62.
8. Cerri S, Greco R, Levandis G, et al. Intracarotid infusion ofmesenchymal stem cells in an animal model of Parkin-son’s disease, focusing on cell distribution and neuropro-tective and behavioral effects. Stem Cells Trans Med2015;4:1073–85.
9. Chiu CC, Liao YE, Yang LY, et al. Neuroinflammation in ani-mal models of traumatic brain injury. J Neurosci Methods2016;272:38–49. Epub 2016/07/07.
10. Gennai S, Monsel A, Hao Q, et al. Cell-based therapy fortraumatic brain injury. Br J Anaesthesia 2015;115:203–12.
11. Neirinckx V, Marquet A, Coste C, et al. Adult bone marrowneural crest stem cells and mesenchymal stem cells arenot able to replace lost neurons in acute MPTP-lesionedmice. PloS One 2013;8:e64723. Epub 2013/06/07.
12. Chen A, Siow B, Blamire AM, et al. Transplantation of mag-netically labeled mesenchymal stem cells in a model ofperinatal brain injury. Stem Cell Res 2010;5:255–66. Epub2010/09/30.
13. Nakajima H, Uchida K, Guerrero AR, et al. Transplantationof mesenchymal stem cells promotes an alternative path-way of macrophage activation and functional recoveryafter spinal cord injury. J Neurotrauma 2012;29:1614–25.
14. Zanier ER, Pischiutta F, Riganti L, et al. Bone marrow mes-enchymal stromal cells drive protective M2 microgliapolarization after brain trauma. Neurotherapeutics2014;11:679–95.
15. Donega V, Nijboer CH, van Tilborg G, et al. Intranasallyadministered mesenchymal stem cells promote a regen-erative niche for repair of neonatal ischemic brain injury.Exp Neurol 2014;261:53–64. Epub 2014/06/20.
8 C. XU ET AL.

16. Zhao Y, Gibb SL, Zhao J, et al. Wnt3a, a protein secretedby mesenchymal stem cells is neuroprotective and pro-motes neurocognitive recovery following traumatic braininjury. Stem Cells 2016;34:1263–72. Epub 2016/02/04.
17. Zhang R, Liu Y, Yan K, et al. Anti-inflammatory and immu-nomodulatory mechanisms of mesenchymal stem celltransplantation in experimental traumatic brain injury. JNeuroinflam 2013;10:106. Epub 2013/08/27.
18. Park HJ, Oh SH, Kim HN, et al. Mesenchymal stem cellsenhance alpha-synuclein clearance via M2 microgliapolarization in experimental and human parkinsonian dis-order. Acta Neuropathol 2016;132:685–701. Epub 2016/10/22.
19. D’souza N, Rossignoli F, Golinelli G, et al. Mesenchymalstem/stromal cells as a delivery platform in cell and genetherapies. BMC Med 2015;13:186.
20. Abumaree MH, Al Jumah MA, Kalionis B, et al. Human pla-cental mesenchymal stem cells (pMSCs) play a role asimmune suppressive cells by shifting macrophage differ-entiation from inflammatory M1 to anti-inflammatory M2macrophages. Stem Cell Rev 2013;9:620–41.
21. Freytes DO, Kang JW, Marcos-Campos I, Vunjak-NovakovicG. Macrophages modulate the viability and growth ofhuman mesenchymal stem cells. J Cell Biochem2013;114:220–9.
22. Chen H, Min XH, Wang QY, et al. Pre-activation of mesen-chymal stem cells with TNF-alpha, IL-1beta and nitricoxide enhances its paracrine effects on radiation-inducedintestinal injury. Sci Rep 2015;5:8718.
23. Diez-Tejedor E, Gutierrez-Fernandez M, Martinez-SanchezP, et al. Reparative therapy for acute ischemic stroke withallogeneic mesenchymal stem cells from adipose tissue: asafety assessment: a phase II randomized, double-blind,placebo-controlled, single-center, pilot clinical trial. J StrokeCerebrovasc Dis 2014;23:2694–700. Epub 2014/10/12.
24. Steinberg GK, Kondziolka D, Wechsler LR, et al. Clinicaloutcomes of transplanted modified bone marrow-derivedmesenchymal stem cells in stroke: a phase 1/2a study.Stroke 2016;47(7):1817–24. Epub 2016/06/04.
25. Lublin FD, Bowen JD, Huddlestone J, et al. Human pla-centa-derived cells (PDA-001) for the treatment of adultswith multiple sclerosis: a randomized, placebo-controlled,multiple-dose study. Mult Scler Relat Disord 2014;3:696–704. Epub 2015/04/22.
26. Pal R, Venkataramana NK, Bansal A, et al. Ex vivo-expanded autologous bone marrow-derived mesenchy-mal stromal cells in human spinal cord injury/paraplegia:a pilot clinical study. Cytotherapy 2009;11:897–911. Epub2009/11/12.
27. Introna M, Lucchini G, Dander E, et al. Treatment of graftversus host disease with mesenchymal stromal cells: aphase I study on 40 adult and pediatric patients. BiolBlood Marrow Transplant 2014;20:375–81. Epub 2013/12/11.
28. Reinders ME, Dreyer GJ, Bank JR, et al. Safety of allogeneicbone marrow derived mesenchymal stromal cell therapyin renal transplant recipients: the Neptune study. J TranslMed 2015;13:344. Epub 2015/11/06.
29. Kimmelman J, Hyun I, Benvenisty N, et al. Policy: globalstandards for stem-cell research. Nature 2016;533:311–3.Epub 2016/05/20.
30. Mounayar M, Kefaloyianni E, Smith B, et al. PI3kalpha andSTAT1 interplay regulates human mesenchymal stem cellimmune polarization. Stem Cells 2015;33:1892–901.
31. Pati S, Gerber MH, Menge TD, et al. Bone marrow derivedmesenchymal stem cells inhibit inflammation and pre-serve vascular endothelial integrity in the lungs afterhemorrhagic shock. PloS One 2011;6:e25171. Epub 2011/10/08.
32. Bartosh TJ, Ylostalo JH, Bazhanov N, et al. Dynamic com-paction of human mesenchymal stem/precursor cells intospheres self-activates caspase-dependent IL1 signaling toenhance secretion of modulators of inflammation andimmunity (PGE2, TSG6, and STC1). Stem Cells2013;31:2443–56. Epub 2013/08/08.
33. Arutyunyan I, Fatkhudinov T, Kananykhina E, et al. Role ofVEGF-A in angiogenesis promoted by umbilical cord-derived mesenchymal stromal/stem cells: in vitro study.Stem Cell Res Therapy 2016;7:46. Epub 2016/03/24.
34. Gu Y, He M, Zhou X, et al. Endogenous IL-6 of mesenchy-mal stem cell improves behavioral outcome of hypoxic-ischemic brain damage neonatal rats by supressing apo-ptosis in astrocyte. Sci Rep 2016;6:18587. Epub 2016/01/15.
35. Noh MY, Lim SM, Oh KW, et al. Mesenchymal stem cellsmodulate the functional properties of microglia via TGF-beta secretion. Stem Cells Transl Med 2016;5:1538–49.Epub 2016/07/13.
36. Griffin MD, Elliman SJ, Cahill E, et al. Concise review: adultmesenchymal stromal cell therapy for inflammatory dis-eases: how well are we joining the dots? Stem Cells2013;31:2033–41.
37. Giuliani M, Fleury M, Vernochet A, et al. Long-lastinginhibitory effects of fetal liver mesenchymal stem cells onT-lymphocyte proliferation. PloS One 2011;6:e19988.Epub 2011/06/01.
38. Abomaray FM, Al Jumah MA, Kalionis B, et al. Human cho-rionic villous mesenchymal stem cells modify the func-tions of human dendritic cells, and induce an anti-inflammatory phenotype in CD1+ dendritic cells. StemCell Rev 2015;11:423–41.
39. Cui R, Rekasi H, Hepner-Schefczyk M, et al. Human mesen-chymal stromal/stem cells acquire immunostimulatorycapacity upon cross-talk with natural killer cells and mightimprove the NK cell function of immunocompromisedpatients. Stem Cell Res Therapy 2016;7:88. Epub 2016/07/09.
40. Huang W, Lv B, Zeng H, et al. Paracrine factors secreted byMSCs promote astrocyte survival associated with GFAPdownregulation after ischemic stroke via p38 MAPK andJNK. J Cell Physiol 2015;230:2461–75.
41. Zheng G, Ge M, Qiu G, et al. Mesenchymal stromal cellsaffect disease outcomes via macrophage polarization.Stem Cells Int 2015;2015:989473.
42. Hattori H, Ishihara M. Altered protein secretions duringinteractions between adipose tissue-or bone marrow-derived stromal cells and inflammatory cells. Stem CellRes Therapy 2015;6:70.
43. Prockop DJ, Oh JY. Medical therapies with adult stem/pro-genitor cells (MSCs): a backward journey from dramaticresults in vivo to the cellular and molecular explanations.J Cell Biochem 2012;113:1460–9.
INTERNATIONAL JOURNAL OF NEUROSCIENCE 9

44. Gomez Perdiguero E, Schulz C, Geissmann F. Developmentand homeostasis of "resident" myeloid cells: the case ofthe microglia. Glia 2013;61:112–20. Epub 2012/08/01.
45. Anton K, Banerjee D, Glod J. Macrophage-associated mes-enchymal stem cells assume an activated, migratory, pro-inflammatory phenotype with increased IL-6 and CXCL10secretion. PloS One 2012;7:e35036. Epub 2012/04/13.
46. Brown GC, Vilalta A. How microglia kill neurons. Brain Res2015;1628:288–97. Epub 2015/09/06.
47. Baby N, Patnala R, Ling EA, Dheen ST. Nanomedicine andits application in treatment of microglia-mediated neuro-inflammation. Curr Med Chem 2014;21:4215–26. Epub2014/07/22.
48. Bedi SS, Smith P, Hetz RA, et al. Immunomagnetic enrich-ment and flow cytometric characterization of mousemicroglia. J Neurosci Methods 2013;219:176–82. Epub2013/08/10.
49. Ariel A, Timor O. Hanging in the balance: endogenousanti-inflammatory mechanisms in tissue repair and fibro-sis. J Pathol 2013;229:250–63. Epub 2012/09/26.
50. Su P, Zhang J, Wang D, et al. The role of autophagy inmodulation of neuroinflammation in microglia. Neurosci-ence 2016;319:155–67. Epub 2016/02/02.
51. Turtzo LC, Lescher J, Janes L, et al. Macrophagic andmicroglial responses after focal traumatic brain injury inthe female rat. J Neuroinflam 2014;11:82.
52. Cho DI, Kim MR, Jeong HY, et al. Mesenchymal stem cellsreciprocally regulate the M1/M2 balance in mouse bonemarrow-derived macrophages. Exp Mol Med 2014;46:e70.
53. Ylostalo JH, Bartosh TJ, Coble K, Prockop DJ. Human mes-enchymal stem/stromal cells cultured as spheroids areself-activated to produce prostaglandin E2 that directsstimulated macrophages into an anti-inflammatory phe-notype. Stem Cells 2012;30:2283–96.
54. Arnold CE, Whyte CS, Gordon P, et al. A critical role forsuppressor of cytokine signalling 3 in promoting M1 mac-rophage activation and function in vitro and in vivo.Immunology 2014;141:96–110. Epub 2013/10/04.
55. Hu X, Leak RK, Shi Y, et al. Microglial and macrophagepolarization-new prospects for brain repair. Nat Rev Neu-rol 2015;11:56–64.
56. Mosser DM, Edwards JP. Exploring the full spectrum ofmacrophage activation. Nat Rev Immunol 2008;8:958–69.Epub 2008/11/26.
57. Xue J, Schmidt SV, Sander J, et al. Transcriptome-basednetwork analysis reveals a spectrum model of humanmacrophage activation. Immunity 2014;40:274–88. Epub2014/02/18.
58. Hume DA. The many alternative faces of macrophageactivation. Front Immunol 2015;6:370. Epub 2015/08/11.
59. Pepe G, Calderazzi G, De Maglie M, et al. Heterogeneousinduction of microglia M2a phenotype by central admin-istration of interleukin-4. J Neuroinflam 2014;11:211.Epub 2015/01/01.
60. Kudlik G, Hegyi B, Czibula A, et al. Mesenchymal stem cellspromote macrophage polarization toward M2b-like cells.Exp Cell Res 2016;348:36–45. Epub 2016/09/01.
61. Parsa R, Andresen P, Gillett A, et al. Adoptive transfer ofimmunomodulatory M2 macrophages prevents type 1diabetes in NOD mice. Diabetes 2012;61:2881–92.
62. Ohlsson SM, Linge CP, Gullstrand B, et al. Serum frompatients with systemic vasculitis induces alternatively
activated macrophage M2c polarization. Clin Immunol2014;152:10–9. Epub 2014/03/19.
63. Moore KJ, Sheedy FJ, Fisher EA. Macrophages in athero-sclerosis: a dynamic balance. Nat Rev Immunol2013;13:709–21.
64. Wang J, Kubes P. A reservoir of mature cavity macro-phages that can rapidly invade visceral organs to affecttissue repair. Cell 2016;165:668–78. Epub 2016/04/12.
65. Okabe Y, Medzhitov R. Wormhole travel for macrophages.Cell 2016;165:518–9. Epub 2016/04/23.
66. Payne NL, Sun G, McDonald C, et al. Human adipose-derived mesenchymal stem cells engineered to secreteIL-10 inhibit APC function and limit CNS autoimmunity.Brain Behav Immun 2013;30:103–14. Epub 2013/02/02.
67. Hegyi B, Kornyei Z, Ferenczi S, et al. Regulation of mousemicroglia activation and effector functions by bone mar-row-derived mesenchymal stem cells. Stem Cells Dev2014;23:2600–12.
68. Taetzsch T, Levesque S, McGraw C, et al. Redox regulationof NF-kappaB p50 and M1 polarization in microglia. Glia2015;63:423–40. Epub 2014/10/22.
69. Hu X, Leak RK, Shi Y, et al. Microglial and macrophagepolarization-new prospects for brain repair. Nat Rev Neu-rol 2015;11:56–64.
70. Huang C, Wang J, Lu X, et al. Z-guggulsterone negativelycontrols microglia-mediated neuroinflammation viablocking IkappaB-alpha-NF-kappaB signals. Neurosci Lett2016;619:34–42. Epub 2016/02/18.
71. Prockop DJ. Concise review: two negative feedback loopsplace mesenchymal stem/stromal cells at the center of earlyregulators of inflammation. Stem Cells 2013;31:2042–6.
72. Schumacher M, Mattern C, Ghoumari A, et al. Revisitingthe roles of progesterone and allopregnanolone in thenervous system: resurgence of the progesterone recep-tors. Prog Neurobiol 2014;113:6–39.
73. Liu Y, Zhang R, Yan K, et al. Mesenchymal stem cells inhibitlipopolysaccharide-induced inflammatory responses of BV2microglial cells through TSG-6. J Neuroinflam 2014;11:135.
74. Xu Y, Shi T, Xu A, Zhang L. 3D spheroid culture enhancessurvival and therapeutic capacities of MSCs injected intoischemic kidney. J Cell Mole Med 2016;20:1203–13. Epub2016/02/26.
75. Xie L, Mao M, Zhou L, Zhang L. Signal factors secreted by2D and spheroid mesenchymal stem cells and by cocul-tures of mesenchymal stem cells derived microvesiclesand retinal photoreceptor neurons. Stem Cells Int2017;2017:2730472.
76. Ulivi V, Tasso R, Cancedda R, Descalzi F. Mesenchymalstem cell paracrine activity is modulated by plateletlysate: induction of an inflammatory response and secre-tion of factors maintaining macrophages in a proinflam-matory phenotype. Stem Cells Dev 2014;23:1858–69.
77. Yasui M, Tamura Y, Minami M, et al. The prostaglandin E2receptor EP4 regulates obesity-related inflammation andinsulin sensitivity. PloS One 2015;10:e0136304.
78. Luan B, Yoon YS, Le Lay J, et al. CREB pathway links PGE2signaling with macrophage polarization. Proc Natl AcadSci U S A 2015;112:15642–7. Epub 2015/12/09.
79. Diaz MF, Vaidya AB, Evans SM, et al. Biomechanical forcespromote immune regulatory function of bone marrowmesenchymal stromal cells. Stem Cells 2017;35(5):1259–72. Epub 2017/03/05.
10 C. XU ET AL.

80. Gray A, Maguire T, Schloss R, Yarmush ML. Identificationof IL-1beta and LPS as optimal activators of monolayerand alginate-encapsulated mesenchymal stromal cellimmunomodulation using design of experiments and sta-tistical methods. Biotechnol Progr 2015;31:1058–70. Epub2015/05/12.
81. Barminko JA, Nativ NI, Schloss R, Yarmush ML. Fractionalfactorial design to investigate stromal cell regulation ofmacrophage plasticity. Biotechnol Bioeng 2014;111:2239–51. Epub 2014/06/04.
82. Na YR, Jung D, Yoon BR, et al. Endogenous prostaglandinE2 potentiates anti-inflammatory phenotype of macro-phage through the CREB-C/EBP-beta cascade. Eur JImmunol 2015;45:2661–71.
83. Qi Y, Jiang D, Sindrilaru A, et al. TSG-6 released from intra-dermally injected mesenchymal stem cells accelerateswound healing and reduces tissue fibrosis in murine full-thickness skin wounds. J Invest Dermatol 2014;134:526–37. Epub 2013/08/08.
84. Park S, Choi JJ, Park BK, et al. Pheophytin a and chloro-phyll a suppress neuroinflammatory responses in lipo-polysaccharide and interferon-gamma-stimulated BV2microglia. Life Sci 2014;103:59–67.
85. Chen M, Li X, Zhang X, et al. The inhibitory effect of mes-enchymal stem cell on blood-brain barrier disruption fol-lowing intracerebral hemorrhage in rats: contribution ofTSG-6. J Neuroinflam 2015;12:61. Epub 2015/04/19.
86. Choi H, Lee RH, Bazhanov N, et al. Anti-inflammatory pro-tein TSG-6 secreted by activated MSCs attenuates zymo-san-induced mouse peritonitis by decreasing TLR2/NF-kappaB signaling in resident macrophages. Blood2011;118:330–8. Epub 2011/05/10.
87. Lopez-Rodriguez AB, Acaz-Fonseca E, Giatti S, et al. Corre-lation of brain levels of progesterone and dehydroepian-drosterone with neurological recovery after traumaticbrain injury in female mice. Psychoneuroendocrinology2015;56:1–11.
88. Meffre D, Labombarda F, Delespierre B, et al. Distributionof membrane progesterone receptor alpha in the malemouse and rat brain and its regulation after traumaticbrain injury. Neuroscience 2013;231:111–24. Epub 2012/12/06.
89. Goldstein FC, Caveney AF, Hertzberg VS, et al. Very earlyadministration of progesterone does not improve neuro-psychological outcomes in subjects with moderate tosevere traumatic brain injury. J Neurotraum 2016;34(1):115–20. Epub 2016/03/15.
90. Labombarda F, Jure I, Gonzalez S, et al. A functional pro-gesterone receptor is required for immunomodulation,reduction of reactive gliosis and survival of oligodendro-cyte precursors in the injured spinal cord. J Steroid Bio-chem Mol Biol 2015;154:274–84. Epub 2015/09/16.
91. Lavoie JR, Rosu-Myles M. Uncovering the secretes of mes-enchymal stem cells. Biochimie 2013;95:2212–21. Epub2013/07/03.
92. Aversa S, Ondolo C, Abbadessa G, et al. Steroid resistancein nasal polyposis: role of glucocorticoid receptor andTGF-beta1. Rhinology 2012;50:427–35. Epub 2012/11/30.
93. Chantong B, Kratschmar DV, Nashev LG, et al. Mineralo-corticoid and glucocorticoid receptors differentially regu-late NF-kappaB activity and pro-inflammatory cytokine
production in murine BV-2 microglial cells. J Neuroinflam2012;9:260. Epub 2012/11/30.
94. Barrachina L, Remacha AR, Romero A, et al. Primingequine bone marrow-derived mesenchymal stem cellswith proinflammatory cytokines: implications in immuno-modulation-immunogenicity balance, cell viability, anddifferentiation potential. Stem Cells Dev 2017;26:15–24.Epub 2016/11/05.
95. Spaeth EL, Kidd S, Marini FC. Tracking inflammation-induced mobilization of mesenchymal stem cells. MethMole Biol. 2012;904:173–90. Epub 2012/08/15.
96. Ciavarella C, Fittipaldi S, Pedrini S, et al. In vitro alterationof physiological parameters do not hamper the growth ofhuman multipotent vascular wall-mesenchymal stemcells. Front Cell Dev Biol 2015;3:36.
97. Toupet K, Maumus M, Luz-Crawford P, et al. Survival andbiodistribution of xenogenic adipose mesenchymal stemcells is not affected by the degree of inflammation inarthritis. PloS One 2015;10:e0114962.
98. Wu B, Sondag G, Malcuit C, et al. Macrophage-associatedosteoactivin/gpnmb mediates mesenchymal stem cellsurvival, proliferation, and migration via a CD44-depen-dent mechanism. J Cell Biochem 2015;117(7):1511–21.
99. Andriessen TM, Horn J, Franschman G, et al. Epidemiol-ogy, severity classification, and outcome of moderateand severe traumatic brain injury: a prospective multicen-ter study. J Neurotraum 2011;28:2019–31. Epub 2011/07/27.
100. Roozenbeek B, Maas AI, Menon DK. Changing patterns inthe epidemiology of traumatic brain injury. Nat Rev Neu-rol 2013;9:231–6.
101. Maas AI, Stocchetti N, Bullock R. Moderate and severetraumatic brain injury in adults. Lancet Neurol2008;7:728–41. Epub 2008/07/19.
102. Olivecrona M, Rodling-Wahlstrom M, Naredi S, KoskinenLO. Effective ICP reduction by decompressive craniec-tomy in patients with severe traumatic brain injurytreated by an ICP-targeted therapy. J Neurotraum2007;24:927–35. Epub 2007/06/30.
103. Carney N, Totten AM, O’Reilly C, et al. Guidelines for themanagement of severe traumatic brain injury, fourth edi-tion. Neurosurgery 2017;80:6–15. Epub 2016/09/23.
104. Simon DW, McGeachy MJ, Bayir H, et al. The far-reachingscope of neuroinflammation after traumatic brain injury.Nat Rev Neurol 2017;13(3):171–91. Epub 2017/02/12.
105. Balu R. Inflammation and immune system activation aftertraumatic brain injury. Curr Neurol Neurosci Rep2014;14:484. Epub 2014/08/21.
106. Hinson HE, Rowell S, Schreiber M. Clinical evidence ofinflammation driving secondary brain injury: a systematicreview. J Trauma Acute Care Surg 2015;78:184–91. Epub2014/12/30.
107. Benarroch EE. Microglia: multiple roles in surveillance, cir-cuit shaping, and response to injury. Neurology2013;81:1079–88. Epub 2013/08/16.
108. Czigner A, Mihaly A, Farkas O, et al. Kinetics of the cellularimmune response following closed head injury. Acta Neu-rochir 2007;149:281–9. Epub 2007/02/09.
109. Watanabe J, Shetty AK, Hattiangady B, et al. Administra-tion of TSG-6 improves memory after traumatic braininjury in mice. Neurobiol Dis 2013;59:86–99.
INTERNATIONAL JOURNAL OF NEUROSCIENCE 11

110. Hellewell S, Semple BD, Morganti-Kossmann MC. Thera-pies negating neuroinflammation after brain trauma.Brain Res 2015.
111. Schober ME, Requena DF, Block B, et al. Erythropoietinimproved cognitive function and decreased hippocampalcaspase activity in rat pups after traumatic brain injury. JNeurotraum 2014;31(4):358–69. Epub 2013/08/27.
112. Webster KM, Wright DK, Sun M, et al. Progesterone treat-ment reduces neuroinflammation, oxidative stress andbrain damage and improves long-term outcomes in a ratmodel of repeated mild traumatic brain injury. J Neuroin-flam 2015;12:238. Epub 2015/12/20.
113. Lee M, Sparatore A, Del Soldato P, et al. Hydrogen sulfide-releasing NSAIDs attenuate neuroinflammation inducedby microglial and astrocytic activation. Glia 2010;58:103–13. Epub 2009/06/23.
114. Ng SY, Semple BD, Morganti-Kossmann MC, Bye N. Atten-uation of microglial activation with minocycline is notassociated with changes in neurogenesis after focal trau-matic brain injury in adult mice. J Neurotraum2012;29:1410–25. Epub 2012/01/21.
115. Liu WC, Wen L, Xie T, et al. Therapeutic effect of erythro-poietin in patients with traumatic brain injury: a meta-analysis of randomized controlled trials. J Neurosurg.Published online 2016 July 1:1–8. Epub 2016/07/02.
116. Margulies SS, Hicks R. Combination therapies for trau-matic brain injury: prospective considerations. J Neuro-traum 2009;26:925–39.
117. Tian C, Wang X, Wang X, et al. Autologous bone marrowmesenchymal stem cell therapy in the subacute stage oftraumatic brain injury by lumbar puncture. Exp Clin Trans-plant 2013;11:176–81.
118. Hart BA, den Dunnen WF. Commentary on special issue:CNS diseases and the immune system. J NeuroimmunePharmacol 2013;8:757–9. Epub 2013/06/12.
119. Louveau A, Smirnov I, Keyes TJ, et al. Structural and func-tional features of central nervous system lymphatic ves-sels. Nature 2015;523:337–41.
120. Woodcock T, Morganti-Kossmann MC. The role of markersof inflammation in traumatic brain injury. Front Neurol2013;4:18. Epub 2013/03/06.
121. Marin MA, de Lima S, Gilbert HY, et al. Reassembly ofexcitable domains after CNS Axon regeneration. J Neuro-sci 2016;36(35):9148–60.
122. Wang Z, Wang Y, Wang Z, et al. Engineered mesenchymalstem cells with enhanced tropism and paracrine secretionof cytokines and growth factors to treat traumatic braininjury. Stem Cells 2015;33:456–67. Epub 2014/10/28.
123. Tu Y, Chen C, Sun HT, et al. Combination of temperature-sensitive stem cells and mild hypothermia: a new poten-tial therapy for severe traumatic brain injury. J Neuro-traum 2012;29:2393–403. Epub 2012/06/05.
124. Stucky EC, Schloss RS, Yarmush ML, Shreiber DI. Alginatemicro-encapsulation of mesenchymal stromal cells enhan-ces modulation of the neuro-inflammatory response.Cytotherapy 2015;17:1353–64. Epub 2015/07/27.
125. Zhou L-N, Zhang J-W, Liu X-L, Zhou L-H. Co-graft of bonemarrow stromal cells and Schwann cells into acellularnerve scaffold for sciatic nerve regeneration in rats. J OralMaxil Surg 2015;73:1651–60.
126. Walker PA, Jimenez F, Gerber MH, et al. Effect of needlediameter and flow rate on rat and human mesenchymalstromal cell characterization and viability. Tissue Eng PartC 2010;16:989–97. Epub 2009/12/17.
127. Moll G, Geissler S, Catar R, et al. Cryopreserved or freshmesenchymal stromal cells: only a matter of taste or keyto unleash the full clinical potential of MSC therapy? AdvExp Med Biol 2016;951:77–98. Epub 2016/11/12.
128. Yarborough M, Tempkin T, Nolta J, Joyce N. The complexethics of first in human stem cell clinical trials. AJOB Neu-rosci 2012;3(2):14–23. Epub 2012/01/01.
129. Ke CC, Liu RS, Suetsugu A, et al. In vivo fluorescence imag-ing reveals the promotion of mammary tumorigenesis bymesenchymal stromal cells. PloS One 2013;8:e69658.Epub 2013/08/13.
130. Zhu W, Xu W, Jiang R, et al. Mesenchymal stem cellsderived from bone marrow favor tumor cell growth invivo. Exp Mol Pathol 2006;80:267–74. Epub 2005/10/11.
131. Dunnett SB, Rosser AE. Challenges for taking primary andstem cells into clinical neurotransplantation trials for neu-rodegenerative disease. Neurobiol Dis 2014;61:79–89.Epub 2013/05/22.
12 C. XU ET AL.