microbial formation nitrosamines in vitroaem.asm.org/content/25/6/862.full.pdf · microbial...
TRANSCRIPT

APPLIED MICROBIOLOGY, June 1973, p. 862-868Copyright 0 1973 American Society for Microbiology
Vol. 25, No. 6Printed in USA.
Microbial Formation of Nitrosamines In VitroABATENI AYANABA' AND MARTIN ALEXANDER
Laboratory of Soil Microbiology, Department of Agronomy, Cornell University, Ithaca, New York 14850
Received for publication 18 December 1972
Mortierella parvispora and an unidentified bacterium converted trimethyl-amine to dimethylamine, and the bacterium (but not the fungus) formeddimethylnitrosamine in the presence of nitrite. Dimethylnitrosamine alsoappeared in cell suspensions of Escherichia coli and Streptococcus epidermidisand in hyphal mats of Aspergillus oryzae incubated with dimethylamine andnitrate. Suspensions of a number of microorganisms produced N-nitrosodi-phenylamine from diphenylamine and nitrite at pH 7.5, and soluble enzymescatalyzing the N-nitrosation of diphenylamine were obtained from two of theseorganisms. In the presence of these enzymes, several dialkylamines wereconverted to the corresponding N-nitroso compounds.
Nitrosamines have recently attracted consid-erable interest because members of this groupof compounds have been found to be carcino-genic, mutagenic, and occasionally teratogenic.Dimethylnitrosamine (DMNA), for example, isboth carcinogenic and mutagenic (7, 8, 14).Although it has been proposed that they may besignificant environmental carcinogens (13),whether they do indeed pose a hazard to humanhealth will be determined by the existence ofthe necessary precursors in nature, the occur-rence of the nitrosation reaction, and the expo-sure of humans to products of the reaction.The formation of the nitrosamine probably
requires the presence of both a secondary amineand nitrite. Secondary amines may be producedfrom tertiary amines, and nitrite is generatedmicrobiologically from ammonium and nitrate.These nitrogenous precursors are widespread.Not only are nitrate and ammonium naturallypresent in water, soil, and plants consumed byman, but large quantities of nitrate and nitriteare added to meat products as preservatives andcolor-enhancing agents (5). Secondary aminesare present in urine (3) and in many habitatswhere microorganisms proliferate (13), andtrimethylamine is found in fish (23) and algae(21).Recent evidence suggests that microorgan-
isms are involved in one or more stages ofnitrosamine synthesis. For instance, ni-trosopiperidine is formed in the rat's stomachand intestine from added nitrate and piperi-dine, and microorganisms presumably reduce
'Present address: International Institute of Tropical Agri-culture, P.M.B. 5320, Ibadan, Nigeria.
the nitrate to nitrite (2). DMNA has beenobserved in the urine of humans infected withProteus mirabilis (4). Either added trimethyla-mine or the fungicide thiram and nitrate areconverted to DMNA in water and soil, and theindigenous communities convert the tertiaryamines to dimethylamine (DMA), nitrate tonitrite, and presumably sometimes serve asnitrosating agents (Ayanaba, Verstraete, andAlexander, unpublished data). It has also beenreported that strains of several bacterial speciesform nitrosamines in vitro (10, 22).The present study was designed (i) to demon-
strate the biosynthesis of nitrosamines by cellsmaintained under conditions where nonbiologi-cal nitrosation would not confuse the interpre-tation of a presumed microbial role in thenitrosation reaction, and (ii) to obtain an activenitrosating enzyme. No such enzyme has yetbeen characterized.
MATERIALS AND METHODSSix bacterial isolates, designated strains 1 to 6, that
utilize nitrate as a nitrogen source were obtained fromgarden soil by using a medium containing 0.2%KNOs, 2.0% glucose, 0.53% KHPO4, 1.06% KHPO4,0.01% MgSO4, and micronutrients added at the ratesused by Powell and Errington (18). The solutionwithout glucose and nitrate was designated solutionS. One bacterium, strain B, which utilizes trimethyl-amine as a carbon and energy source, was isolatedfrom Beebe Lake water enriched with 0.50% of theamine as its hydrochloride, 0.53% KH2PO4, and 1.06%K2HPO4. The enriched water was incubated on arotary shaker at 30 C for 48 h, after which thesuspension was streaked on a solid medium of thesame composition. To obtain isolates that use DMAas a carbon or nitrogen source, 0.10 g of garden soil or
862
on August 30, 2018 by guest
http://aem.asm
.org/D
ownloaded from

MICROBIAL FORMATION OF NITROSAMINES
fresh cow manure or 1.0 ml of municipal sewage wasintroduced into media containing 0.50% DMA-HCl,solution S, and either 0.50% glucose, 0.20%(NH4)2SO4, or no addition. Samples from those flaskswhich became turbid were repeatedly streaked on asolid medium identical to the enrichment solution,and by this means bacterial strains 8 to 11, a fungus(strain 1), and two yeasts were obtained in axenicculture.To prepare cell suspensions of the bacteria, the
organisms were grown in the liquid isolation mediumat 30 C on a rotary shaker for 24 to 48 h. The cellswere harvested aseptically by centrifugation in thecold and washed twice with sterile 0.1 M phosphateor 0.1 M tris(hydroxymethyl)aminomethane (Tris) buf-fer, pH 7.0.To obtain large yields of bacterial cells, the cultures
were grown with constant aeration and agitation in 6liters of the appropriate medium contained in a15-liter fermentor maintained at 30 C. The cells werecollected by centrifugation during the early stationaryphase of growth (after 24 to 48 h) and washed twice inthe phosphate buffer. These cells (5 to 10 g wetweight) contained in 15 to 20 ml of Tris or phosphatebuffer were introduced into appropriate solutions.The fungi were grown at 30 C in 500-ml Fernbach
flasks containing 200 ml of the medium of Pengra,Cole, and Alexander (17), except that fungus 1 wasgrown in a medium containing 0.5% glucose and 0.5%DMA * HCl in solution S. After 7 to 10 days the culturefiltrates were carefully decanted from the flasks, andthe mycelial mats were washed twice with Tris buffer,pH 7.0.The cells were ruptured with a French pressure cell.
The microbial suspension (3 to 5 g wet weight in 30 mlof 0.1 M phosphate buffer, pH 7.5) was passed twicethrough the instrument, and a pressure of 15,000lbs/in2 was used for disruption. The cell debris andunbroken cells were removed by centrifugation at10,000 x g for 15 min, and the supernatant fluid, orcrude cell extract, was carefully removed. The crudecell extract was separated into soluble and particulatefractions by centrifugation at 144,000 x g for 60 min,and the pellet was suspended in 0.1 M phosphatebuffer, pH 7.5. The crude cell extract and the frac-tions centrifuged at 144,000 x g were used within 60min of centrifugation.To determine the nitrosating activity of the ex-
tracts, 8.0 ml of 0.1 M phosphate buffer (pH 7.5), 0.15g of NaNO,, 0.10 g of glucose, 0.05 g of diphenyl-amine, and 2.0 ml of either untreated or boiled ex-tracts were introduced into 50-ml Erlenmeyer flasks.The pH values of the reaction mixtures were main-tained at 7.5 by additions of NaOH, and the flaskswere incubated with shaking at 30 C. (Caution must beexercised in handling samples which contain or aresuspected to contain nitrosamines because of theirvolatility and toxicity.)The nitrite content of the supernatant fluid re-
maining after centrifugation at 4,600 x g for 10 minwas estimated by the method of Montgomery andDymock (16). Samples to be analyzed for DMA andnitroso compounds were steam-distilled (11), andDMA was estimated by the method of Pribyl and
Nedbalkova (20), whereas a photochemical method(6) was employed to determine nitrosamines quan-titatively.
Prior to chromatographic determinations, steamdistillates containing DMNA were concentrated indichloromethane, as described by Howard et al. (12).DMNA and N-nitrosodiphenylamine were identifiedqualitatively by thin-layer chromatography (19).
For the quantitative determination of nitrosamineson chromatography plates, the silica gel thin-layerplates were removed from the chromatography tanksand dried in air for 2 to 3 min. The areas containingthe nitrosamines, which were evident on the un-sprayed plates as dark spots under an ultraviolet (UV)lamp, were scraped into a 50-ml beaker containing 1.5ml of a 0.17% Na2CO, solution. The beakers wereplaced under a UV lamp for 15 min, and then eachreceived 1.5 ml of the sulfanilic acid and 1.5 ml of the1-naphthylethylenediamine reagents of Montgomeryand Dymock (16). After 15 min, 5.5 ml of water wasadded to each beaker, and the silica gel was removedby filtration through Whatman no. 30 filter paper.The absorbance of the colored solution was measuredat 550 nm. The limit of detection by this procedurewas 2.0 Ag of nitrosamine. Untreated areas of thesilica gel contained 1.0 to 2.0 Mg of apparent N-nitrosodiphenylamine by this method.A Varian Aerograph gas chromatograph (model
2740, Varian Associates, Walnut Creek, Calif.)equipped with a flame ionization detector was alsoused to identify DMNA. The steel column (7.62 m by4.67 mm) was packed with Porapak Q-S (100-120mesh; Waters Associates, Inc., Farmingham, Mass.),and the operating temperatures were 225, 180, and 230C for the injector, column, and detector, respectively.The carrier gas was N2 flowing at 40 mlmin. The limitof detection by this procedure was 2.0 ng of DMNA.
Authentic dimethyl-, diethyl-, and di-n-propylni-trosamine and N-nitrosodiphenylamine were ob-tained from Eastman Organic Chemicals, Rochester,N.Y.
RESULTSCell suspensions (5.0 ml) prepared from cul-
tures of 12 microorganisms were incubated for36 h in 95 ml of 0.1 M Tris buffer (pH 7.0)containing 0.34 g of trimethylamine HCl, 0.25 gof NaNO2, and 0.10 g of glucose. The pH waskept at 7.0. DMNA was present in distillates ofthe reaction mixture of bacterium B, and theconcentrations of DMNA after 0, 6, 12, 18, 24,and 36 h were 0.0, 0.0, 0.10, 0.20, 0.16, and 0.23,ug/ml, respectively. Cell suspensions of stockcultures ofAerobacter aerogenes, Proteus vulga-ris, Streptococcus epidermidis, Bacillus subti-lis, Escherichia coli, Arthrobacter sp., Nocardiasp., Aspergillus niger, Aspergillus oryzae, Mor-tierella parvispora, and Zygorhynchus moellericontained less than 0.10 ug of DMNA/ml;presumably these organisms were therefore notactive. Suspensions of bacterium B and M.
863VOL. 25, 1973
on August 30, 2018 by guest
http://aem.asm
.org/D
ownloaded from
AYANABA AND ALEXANDER
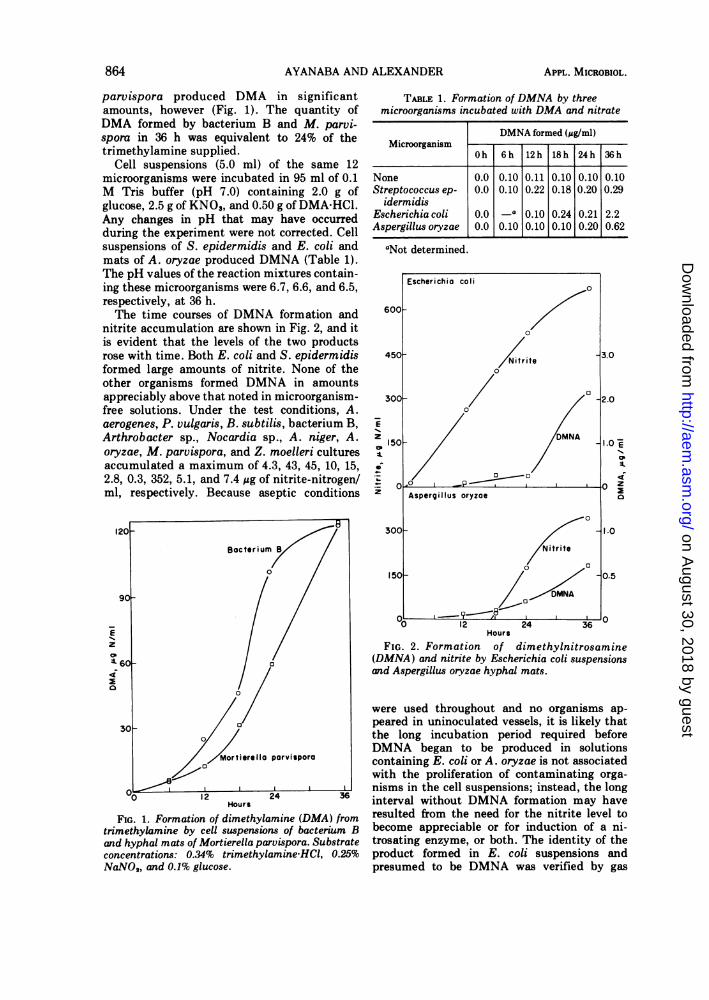
parvispora produced DMA in significantamounts, however (Fig. 1). The quantity ofDMA formed by bacterium B and M. parvi-spora in 36 h was equivalent to 24% of thetrimethylamine supplied.
Cell suspensions (5.0 ml) of the same 12microorganisms were incubated in 95 ml of 0.1M Tris buffer (pH 7.0) containing 2.0 g ofglucose, 2.5 g of KNO3, and 0.50 g of DMA-HCl.Any changes in pH that may have occurredduring the experiment were not corrected. Cellsuspensions of S. epidermidis and E. coli andmats of A. oryzae produced DMNA (Table 1).The pH values of the reaction mixtures contain-ing these microorganisms were 6.7, 6.6, and 6.5,respectively, at 36 h.The time courses of DMNA formation and
nitrite accumulation are shown in Fig. 2, and itis evident that the levels of the two productsrose with time. Both E. coli and S. epidermidisformed large amounts of nitrite. None of theother organisms formed DMNA in amountsappreciably above that noted in microorganism-free solutions. Under the test conditions, A.aerogenes, P. vulgaris, B. subtilis, bacterium B,Arthrobacter sp., Nocardia sp., A. niger, A.oryzae, M. parvispora, and Z. moelleri culturesaccumulated a maximum of 4.3, 43, 45, 10, 15,2.8, 0.3, 352, 5.1, and 7.4 ug of nitrite-nitrogen/ml, respectively. Because aseptic conditions
Hours
FIG. 1. Formation of dimethylamine (DMA) fromtrimethylamine by cell suspensions of bacterium Band hyphal mats of Mortierella parvispora. Substrateconcentrations: 0.34% trimethylamine-HCI, 0.25%NaNO2, and 0.1% glucose.
TABLE 1. Formation of DMNA by threemicroorganisms incubated with DMA and nitrate
DMNA formed (jg/ml)Microorganism
Oh 6h 12h 18h 24h 36h
None 0.0 0.10 0.11 0.10 0.10 0.10Streptococcus ep- 0.0 0.10 0.22 0.18 0.20 0.29
idermidisEscherichia coli 0.0 -a 0.10 0.24 0.21 2.2Aspergillus oryzae 0.0 0.10 0.10 0.10 0.20 0.62
aNot determined.
z
A.
z
12 24 36'Hours
FIG. 2. Formation of dimethylnitrosamine(DMNA) and nitrite by Escherichia coli suspensionsand Aspergillus oryzae hyphal mats.
were used throughout and no organisms ap-peared in uninoculated vessels, it is likely thatthe long incubation period required beforeDMNA began to be produced in solutionscontaining E. coli or A. oryzae is not associatedwith the proliferation of contaminating orga-nisms in the cell suspensions; instead, the longinterval without DMNA formation may haveresulted from the need for the nitrite level tobecome appreciable or for induction of a ni-trosating enzyme, or both. The identity of theproduct formed in E. coli suspensions andpresumed to be DMNA was verified by gas
864 APPL. MICROBIOL.
on August 30, 2018 by guest
http://aem.asm
.org/D
ownloaded from

MICROBIAL FORMATION OF NITROSAMINES
chromatography. This compound and authenticDMNA have retention times of 342 s.The presence of DMNA in test systems con-
taining several of the organisms suggested thatthey could effect the nitrosation of DMA in thepresence of nitrite. To investigate this possibil-ity, 10-ml portions of a cell suspension of E. coliwere incubated with 90 ml of 0.1 M Tris buffer(pH 7.0) containing either (i) 0.50 g of(NH,),SO, and 0.50 g of glucose; (ii)(NH)2,SO, glucose, and 0.232 g of DMA -
HCl; (iii) glucose and 0.50 g of KNO,; or (iv) ni-trate, glucose, and DMA. One-half of the ni-trate-treated vessels were shaken, and one-halfwere not. Cells incubated with ammonium andglucose or with ammonium, glucose, and DMAdid not produce nitrite or DMNA. The pHvalues in these two treatments fell graduallywith time and reached 5.7 after 5.5 h. DMNAwas formed in flasks containing DMA, nitrate,and glucose, the level reaching a maximum of2.2 Ag after 5.5 h. The yield was the samewhether the flasks were shaken or not. NoDMNA was evident in reaction mixtures notprovided with DMA. Nitrite was produced fromnitrate in the absence of DMA, and the maxi-mum yield was 350 ,g of nitrite-nitrogen.Twenty-five microorganisms were tested for
their ability to nitrosate diphenylamine in thepresence of nitrite. The bacteria and fungi weregrown for 2 to 10 days in either nutrient broth orsolution S fortified with 0.50% glucose, 0.20%KNO,, and 0.50% DMA .HCl or trimethyla-mine hydrochloride in various combinations.The cells were washed, and 3.0-ml portions ofthe resulting bacterial suspensions were addedto 7.0 ml of 0.1 M phosphate buffer, pH 7.5.The fungal mats were suspended in 100 ml ofthe same buffer contained in Fernbach flasks.The buffer was amended with diphenylamine,NaNO2, and glucose to final concentrations of0.5, 1.5, and 1.0%, respectively. The bacterialcells were incubated with shaking, whereas thehyphae were not shaken. The pH was main-tained at 7.5 ± 0.1, and the incubation wasconducted for 4 h at 30 C.Because the method for N-nitrosodiphenyl-
amine analysis was not sensitive for the detec-tion of less than about 2.0 gg of this compound,values below this amount were deemed nega-tive. The chemicals incubated together for 4 hin the buffer gave values of less than 2.0 Ag, asdid stock cultures of A. aerogenes, S. epidermi-dis, Arthrobacter sp., and Nocardia sp. grown innutrient broth, two fungal and two bacterialisolates from soil and one bacterium from sew-age grown in solution S containing 0.50% glu-cose and 0.50% DMA .HCl, five other soil bac-
teria grown in solution S containing 0.50% glu-cose and 0.20% KNO,,, and one additional soilbacterium grown in solution S containing 0.50%DMA .HC1.
However, 10 of the organisms did generateN-nitrosodiphenylamine (Table 2). All of thecultures listed contained more than 2.0 Mg of theproduct. The presence of the nitrosamine wasestablished by thin-layer chromatography. Theauthentic and biologically produced nitrosa-mine had identical R. values of 0.64 under thetest conditions.Because Pseudomonas sp. and Cryptococcus
sp. produced N-nitrosodiphenylamine spots onsilica gel plates with the largest areas andgreatest intensities, they were investigated fur-ther. The organisms were grown for 40 h insolution S amended with 0.50% DMA-HCl and0.20% of either (NH,)2S04 or KNO.. The cellswere collected and washed, and 3.0 ml of theresulting cell suspension was incubated at 30 Cwith 7.0 ml of 0.1 M phosphate buffer (pH 7.5)containing 0.1 g of glucose, 0.05 g of diphenyl-amine, and 0.15 g of NaNO,. Reaction mixturescontaining diphenylamine, glucose, and nitritewere also incubated with cells that had beenboiled for 30 min.
Suspensions of ammonium- or nitrate-growncells of the bacterium that were incubated for 4h with diphenylamine also contained less than2.0 Mg of nitrosamine. However, the yeast andthe bacterium were able to bring about a biologi-cal nitrosation of this amine (Table 3). Althoughthe nonbiological nitrosation of diphenylaminewas appreciable under the test conditions, theyield of the product after 4 h was more thantwice as high with unheated than with boiledcells. The data show that the bacterium was as
TABLE 2. Microorganisms formingN-nitrosodiphenylamine in cell suspensions or hyphal
mats
Source of Microorganism Growth mediumamicroorganism
Stock culture
Beebe LakeGarden soil
SewageCow manure
Escherichia coliBacillus subtilisBacterium BFungus 1Bacterium 8Bacterium 9Pseudomonas sp.Bacterium 10Yeast 1Cryptococcus sp.
Nutrient brothNutrient brothTrimethylamineGlucose-DMADMA-nitrateDMA-nitrateGlucose-nitrateGlucose-DMAGlucose-DMAGlucose-DMA
a All media except nutrient broth were prepared insolution S.
VOL. 25, 1973 865
on August 30, 2018 by guest
http://aem.asm
.org/D
ownloaded from

AYANABA AND ALEXANDER
active in nitrosation whether it was grown onammonium or nitrate.Crude enzyme preparations active in ni-
trosating diphenylamine were made by use ofthe French pressure cell. The crude extracts andthe particle and supernatant fractions resultingfrom a centrifugation of the extract at 144,000 xg were incubated in 0.1 M phosphate buffer (pH7.5) containing 0.5% diphenylamine, 1.5%NaNO2, and 1.0% glucose. Enzymatic nitrosa-tion was readily evident, and extracts of theyeast cells showed the greater activity (Table 4).The soluble fractions exhibited the greatest rateof nitrosation. The activity in the particle prep-arations may have resulted from soluble cellconstituents, because some of the soluble frac-tion was retained and then mixed with theparticles at the bottom of the centrifuge tube.The disappearance of the nitrosamine with timewas unexplained but may have resulted from anenzymatic cleavage of the nitrogen-nitrogenbond.A separate test of the soluble preparation
derived from the yeast confirmed that theformation of N-nitrosodiphenylamine was lin-ear with time. By using the same substrate
TABLE 3. Formation of N-nitrosodiphenylamine incell suspensions of two microorganisms incubated
with diphenylamine
NitrosamineN source formed (pg)for growth
1 h 4 h
Cryptococcus sp. Nitrate 8.5 199.0a 9.0a3.5 4.55
Pseudomonas sp. Ammonium 229.0a
Nitrate 23ioa
aCell suspension was boiled.bNo nitrite was present. All other suspensions were
incubated with nitrite.
concentrations as in the previous enzyme as-says, the nitrosamine concentration rose lin-early from 3 ,ug at zero time to 27 ,g at 120 min.Under these conditions, only 0.5 Mug of theproduct, a figure within the experimental errorof the method, appeared when an identicalenzyme preparation which was boiled for 30 minwas incubated with diphenylamine and nitritefor 120 min.To ascertain which amines could be nitro-
sated, the yeast was grown for 40 h in a mediumcontaining 0.50% DMA-HCl and 0.20% KNO3 insolution S. The soluble enzyme preparationderived from these cells was incubated at 30 Cin 0.1 M phosphate buffer (pH 7.5) containing0.50% of the amine and 1.5% of NaNO2. In a 2-hincubation period, 4 ,g of diethylnitrosamine, 3,ug of di-n-propylnitrosamine, 7 Mg of DMNA,and 24 ,g of N-nitrosodiphenylamine were pro-
duced. In each instance, the apparent nitrosa-mine present initially was subtracted to obtainthe final yield. The soluble preparation in-cubated with nitrite alone or with each of theamines alone formed <2 Mg of the nitrosamine,a quantity within the experimental error. Theyield of nitrosamine was higher with diphenyla-mine than with DMA as substrate, despite thefact that the yeast was grown on DMA.
DISCUSSIONBecause of the toxicity of nitrosamines and
the ubiquity of their precursors, information on
the mechanisms of N-nitrosation and factorsaffecting the synthesis of such toxicants undernatural conditions are of considerable signifi-cance. There is ample evidence that secondaryamines and nitrite combine nonbiologically insolutions at low pH values, as in the spontane-ous formation of di-n-propylnitrosamine (9, 22).The nonbiological production of such com-pounds in acid solutions requires that appropri-ate controls be included in any study of apossible microbial contribution to the genesis ofthese hazardous compounds. Although the
TABLE 4. Enzymatic nitrosation of diphenylamine by intracellular fractions from two microorganisms
N-nitrosodiphenylamine formed (Mg)Microorganism Fraction
0.5 h 1 h 2 h 4 h 4 ha
Pseudomonas sp. Cell extract <2 3 3 2 3Soluble 13 10 10 8 3Particulate 3 7 2 3 2
Cryptococcus sp. Cell extract 11 10 22 14 2Soluble 8 14 33 34 2Particulate 11 11 18 36 3
aFraction was boiled for 30 min before being added to reaction mixture.
866 APPL. MICROBIOL.
on August 30, 2018 by guest
http://aem.asm
.org/D
ownloaded from

MICROBIAL FORMATION OF NITROSAMINES
needed controls are not always reported upon,investigations of axenic cultures (10, 22) and ofprocesses occurring within the gastrointestinaltract of rats (1, 2) suggest a microbial contribu-tion to the formation of N-nitroso compounds.Bacterial and enzymatic C-nitrosation has alsobeen established (24).The present studies demonstrate several mi-
crobiologically catalyzed steps leading to theappearance of nitrosamines. Thus, not only domany microorganisms, including a variety ofbacteria and fungi tested here, reduce nitrate tonitrite, but some are also able to convert tri-methylamine to DMA. Both bacteria and fungilike M. parvispora are active in the dealkyla-tion. The data also show that certain micro-organisms contain enzymes that catalyze N-nitrosation; this is the first report of N-nitrosa-tion by an enzyme preparation, albeit crude,from a micro- or macroorganism.
It is possible, nevertheless, that nitrosamineformation may sometimes result from a nonen-zymatic reaction resulting from the microbialformation and concomitant increasing concen-tration of nitrite, a secondary amine, or both, orfrom a reaction effected by a cellular constitu-ent other than an enzyme. In the test systemsused here in which nitrite was present, itsconcentration decreased rather than increasedwith time, making nonbiological nitrosation lessrather than more likely. Moreover, although thepresent data do not rule out the possibility thatthe cellular constituent is not an enzyme, thefindings do show that the responsible cell com-ponent is thermolabile, a trait commonly usedto suggest, but surely not to prove, that enzymecatalysis is implicated in a particular reactionsequence.The nitrosating enzyme is particularly active
on diphenylamine, and intact cells ofCryptococcus sp. likewise formed considerablequantities of N-nitrosodiphenylamine. Whenincubated with DMA, however, yeast suspen-sions contained little DMNA. These findingsare in agreement with other reports that lessbasic amines are more easily nitrosated thanmore basic ones (10, 22).The finding that enzymes of a yeast can
catalyze the nitrosation of a secondary amine inthe presence of nitrite may be of some practicalconcern in the alcoholic fermentation. Thus,McGlashan et al. (15) found 1 to 3 ppm ofDMNA in alcoholic spirits. Although suchamounts of DMNA are small, they are possiblyof toxicological significance, and yeasts par-ticipating in the fermentation may be responsi-ble for synthesis of the toxicant.
Considerable effort is currently being ex-pended by food and residue chemists to providea definitive identification of compounds foundin natural products and presumed to be nitrosa-mines. Such firm identifications are essentialbecause the products being tested are complexand highly heterogeneous, and the precursors ofthe presumed nitrosamine are unknown. In thepresent investigation, by contrast, the amineand inorganic nitrogen precursors were added,and the test systems were simple and reasona-bly free of extraneous substances which mightnecessitate clean-up procedures to remove com-pounds which might interfere in the identifica-tion. The gas and thin-layer chromatogramsamply demonstrated the absence of an array ofpotential interfering chemicals.These studies suggest that microorganisms
may contribute to the production of nitrosa-mines in nature by synthesizing secondaryamines, by producing nitrite, or by formingenzymes which catalyze N-nitrosation. In addi-tion, microbial acidification of soil, plant tis-sues undergoing fermentation, or other sub-stances or habitats may result in conditionsfavorable to spontaneous nitrosation. In view ofthe great toxicity of N-nitroso compounds, fur-ther inquiry into the processes involved and theidentification of the types of nitrosamines thatcan be generated seem warrante.
LITERATURE CITED
1. Alam, B. S., I. B. Saporoschetz, and S. S. Epstein. 1971.Formation of N-nitrosopiperidine from piperidine andsodium nitrite in the stomach and the isolated intesti-nal loop of the rat. Nature (London) 232:116-118.
2. Alam, B. S., I. B. Saporoschetz, and S. S. Epstein. 1971.Synthesis of nitrosopiperidine from nitrate and piperi-dine in the gastrointestinal tract of the rat. Nature(London) 232:199-200.
3. Asatoor, A. M., and M. L. Simenhoff. 1965. The origin ofurinary dimethylamine. Biochim. Biophys. Acta111:384-392.
4. Brooks, J. B., W. B. Cherry, L. Thacker, and C. C. Alley.1972. Analysis by gas chromatography of amines andnitrosamines produced in vivo and in vitro by Proteusmirabilis. J. Infect. Dis. 126:143-153.
5. Committee on Nitrate Accumulation, National Academyof Sciences. 1972. Accumulation of nitrate. NationalAcademy of Sciences, Washington, D.C.
6. Daiber, D., and R. Preussmann. 1964. Quantitativecolorimetrische Bestimmung organische N-Nitroso-Verbindungen durch photochemische Spaltung derNitrosaminbindung. Z. Anal. Chem. 206:344-352.
7. DuPlessis, L. S., J. R. Nunn, and W. A. Roach. 1969.Carcinogen in a Transkeian Bantu food additive.Nature {London) 222:1198-1199.
8. Fishbein, L., W. G. Flamm, and H. L. Falk. 1970.Chemical mutagens: environmental effects on biologi-cal systems. Academic Press Inc., New York.
9. Greenblatt, M., V. Kommineni, E. Conrad, L. Wallcave,and W. Lijinsky. 1972. In vivo conversion of phenmet-
867VOL. 25, 1973
on August 30, 2018 by guest
http://aem.asm
.org/D
ownloaded from

868 AYANABA ANt
razine into its N-nitroso derivative. Nature N. Biol.236:25-26.
10. Hawksworth, G., and M. J. Hill. 1971. Bacteria and theN-nitrosation of secondary amines. Brit. J. Cancer26:520-526.
11. Heath, D. F., and J. A. E. Jarvis. 1955. The polarographicdetermination of dimethylnitrosamine in animal tis-sue. Analyst 80:613-616.
12. Howard, J. W., T. Fazio, and J. 0. Watts. 1970. Extrac-tion and gas chromatographic determination of N-nitrosodimethylamine in smoked fish: application tosmoked nitrite-treated chub. J. Ass. Offic. Anal. Chem.53:269-274.
13. Lijinsky, W., and S. S. Epstein. 1970. Nitrosamines asenvironmental carcinogens. Nature (London)225:21-23.
14. Magee, P. N. 1969. In vivo reactions of nitroso com-pounds. Ann. N. Y. Acad. Sci. 163:717-729.
15. McGlashan, N. D., C. L. Walters, and A. E. M. McLean.1968. Nitrosamines in African alcoholic spirits andoesophageal cancer. Lancet 2:1017.
16. Montgomery, H. A. C., and J. F. Dymock. 1961. Thedetermination of nitrite in water. Analyst 86:414-416.
17. Pengra, R. M., M. A. Cole, and M. Alexander. 1969. Cell
'ALEXANDER APPL. MICROBIOL.
walls and lysis of Mortierella parvispora hyphae. J.Bacteriol. 97:1056-1061.
18. Powell, E. O., and F. P. Errington. 1963. Generationtimes of individual bacteria: some corroborative meas-urements. J. Gen. Microbiol. 31:315-327.
19. Preussmann, R., D. Daiber, and H. Hengy. 1964. Asensitive colour reaction for nitrosamines on thin-layerchromatograms. Nature (London) 201:502-503.
20. Pribyl, M., and J. Nedbalkova. 1967. Spectrophotomet-rische Bestimmung von Dimethylamin in Dimethylfor-mamid. Z. Anal. Chem. 232:261-267.
21. Sakevich, A. I. 1970. Detection of methyl amines in theculture of Stephanodiscus hantzschii (in Russian).Gidrobiol. Zh. 6:98-100.
22. Sander, J. 1968. Nitrosaminsynthese durch Bakterien.Hoppe-Seyler's Z. Physiol. Chem. 349:429-432.
23. Sasajima, M. 1968. Studies on psychrotolerant bacteriain fish and shellfish. III. Reduction and decompositionof trimethylamine oxide by psychrotolerant bacteria(in Japanese). Tohoko Kaiko Suisan Kenkyujo KenkyuHokoku. 55:205-214.
24. Verstraete, W. and M. Alexander. 1972. Mechanism ofnitrification by Arthrobacter sp. J. Bacteriol.110:962-967.
D
on August 30, 2018 by guest
http://aem.asm
.org/D
ownloaded from