microglia activation in basal ganglia is a late event in
TRANSCRIPT

ARTICLE OPEN ACCESS
Microglia Activation in Basal Ganglia Is a LateEvent in Huntington Disease PathophysiologyNatalia P Rocha PhD Odelin Charron MSc Leigh B Latham PhD Gabriela D Colpo PhD
Paolo Zanotti-Fregonara MD PhD Meixiang Yu PhD Leorah Freeman MD PhD Erin Furr Stimming MD
and Antonio L Teixeira MD PhD
Neurol Neuroimmunol Neuroinflamm 20218e984 doi101212NXI0000000000000984
Correspondence
Dr Rocha
npessoarochagmailcom
AbstractObjectiveTo define the role played by microglia in different stages of Huntington disease (HD) we usedthe TSPO radioligand [11C]-ER176 and PET to evaluate microglial activation in relation toneurodegeneration and in relation to the clinical features seen at premanifest and manifeststages of the disease
MethodsThis is a cross-sectional study in which 18 subjects (6 controls 6 premanifest and 6 manifestHD gene carriers) underwent a [11C]-ER176 PET scan and an MRI for anatomic localizationSegmentation of regions of interest (ROIs) was performed and group differences in [11C]-ER176 binding (used to evaluate the extent of microglial activation) were assessed by thestandardized uptake value ratio (SUVR) Microglial activation was correlated with ROIs vol-umes disease burden and the scores obtained in the clinical scales As an exploratory aim weevaluated the dynamic functions of microglia in vitro by using induced microglia-like (iMG)cells from peripheral blood monocytes
ResultsIndividuals with manifest HD present higher [11C]-ER176 SUVR in both globi pallidi andputamina in comparison with controls No differences were observed when we comparedpremanifest HD with controls or with manifest HD We also found a significant correlationbetween increased microglial activation and cumulative disease burden and with reducedvolumes iMG from controls premanifest HD and manifest HD patients showed similarphagocytic capacity
ConclusionsAltogether our data demonstrate that microglial activation is involved in HD pathophysiologyand is associated with disease progression
These authors contributed equally to this work as condashsenior authors
From the Mitchell Center for Alzheimerrsquos Disease and Related Brain Disorders (NPR) Department of Neurology McGovern Medical School The University of Texas Health ScienceCenter Houston Department of Neurology (OC LF) The University of Texas at Austin School of Medicine (LBL) University of Washington Seattle Neuropsychiatry Program(GDC ALT) Department of Psychiatry and Behavioral Sciences McGovern Medical School University of Texas Houston Houston Methodist Research Institute and Weill CornellMedicine (PZ-F MY) TX and HDSA Center of Excellence at University of Texas Health Science Center at Houston (EFS)
Go to NeurologyorgNN for full disclosures Funding information is provided at the end of the article
The Article Processing Charge was funded by the Department of Psychiatry and Behavioral Sciences UTHealth
This is an open access article distributed under the terms of the Creative Commons Attribution-NonCommercial-NoDerivatives License 40 (CC BY-NC-ND) which permits downloadingand sharing the work provided it is properly cited The work cannot be changed in any way or used commercially without permission from the journal
Copyright copy 2021 The Author(s) Published by Wolters Kluwer Health Inc on behalf of the American Academy of Neurology 1
Huntington disease (HD) is an autosomal dominant diseasecaused by a trinucleotide (CAG) repeat expansion in thehuntingtin gene (HTT)1 The toxic gain of function of theexpanded mutant huntingtin protein (HTT) seems to be thekey event involved in the progressive neuronal dysfunctionand neuronal loss particularly in the striatum and cortex As aresult individuals with HD classically present with motordysfunction cognitive decline and behavioral disorders2 Al-though the cause of HD is well established the mechanismsunderlying neuronal dysfunction and death are not com-pletely understood Mutant HTT seems to trigger a patho-genic cascade that includes oxidative stress and immuneinflammatory mechanisms which are further stimulated byneuronal dysfunctiondeath Together these mechanismshave been regarded as key contributors to the pathophysiol-ogy of HD3
Neuroinflammation in HD has been reported since earlypostmortem studies which described reactive astrocytes45
and microglial activation6 in brain regions associated with HDpathogenesis Inflammatory mechanisms in the CNS arethought to be initiated as compensatory responses againstmisfolded protein oligomers andor deposits7 On the otherhand inflammatory responses can trigger neuronal damageand thus contribute to disease progression8 Noteworthy theabove-mentioned postmortem studies showed that bothmicroglial activation and reactive astrocytosis grades correlatewith disease severity4-6 These results motivated studies toevaluate neuroinflammation in HD in vivo In this regardPET neuroimaging studies have focused on evaluatingmicroglial activation through specific molecular targets invivo Microglial activation imaging is currently performed byusing PET tracers binding to the 18-kDa translocator protein(TSPO) a mitochondrial protein that is highly expressed inphagocytic inflammatory cells including activated microgliain the brain and macrophages in the periphery910 TSPO isexpressed at very low levels in the CNS but its levels areupregulated by activated microglia10
The prototype isoquinoline ligand for TSPO [11C]-PK11195 has been introduced in the field of molecularbrain imaging of microglial activation more than 20 yearsago9 However PK11195 shows low specific signal-to-noiseratios even in clinical conditions in which microglial activa-tion is a prominent and well-established feature11 This limi-tation motivated the development of alternative TSPO PETligands with better signal-to-noise ratios such as [11C]-
PBR28 and other second-generation ligands11 Howeverother technical issues became evident mainly the aberrantlylow TSPO binding in some individuals due to the single nu-cleotide polymorphism rs6971 More recently a series of4-phenylquinazoline-2-carboxamides lacking allelic sensitivityto the human single nucleotide polymorphism rs6971 hasbeen developed Noteworthy [11C]-ER176 (11C-(R)-N-sec-butyl-4-(2-chlorophenyl)-N-methylquinazoline-2-carboxamide) a new analog of [11C]-PK1119512 presentshigher specific binding when compared with most of theavailable TSPO binders13 Although still sensitive to TSPOpolymorphism ER176 allows the inclusion of low-affinitybinders because of its high in vivo binding potential14 Ofnote [11C]-ER176 has a higher specific binding and a smallerintersubject variability compared with [11C]-PBR28 thusresulting in higher statistical power and requiring fewer sub-jects for clinical studies Therefore [11C]-ER176 should bepreferred over [11C]-PBR28 for TSPO studies in humans15
To date 6 studies have used PET neuroimaging with TSPObinders to evaluate microglial activation in HD Five studiesused the first-generation TSPO radiotracer [11C]-PK1119516-20 and one used the second-generation TSPObinder [11C]-PBR2821 These studies revealed increasedmicroglial activation in HD gene carriers which correlatedwith disease stage andor severity of symptoms The availablefindings indicate that microglial activation is involved in HDpathophysiology and TSPO PET imaging may be a valuabletool to monitor HD progression and therapeutic efficacy ofdrugs targeting neuroinflammation However these indica-tions have yet to be explored So far no study has evaluatedthe use of the novel TSPO tracer [11C]-ER176 as a markerof microglial activation in HD gene carriers Therefore thecurrent study was designed to define the role played bymicroglia in different stages of HD by using [11C]-ER176PET We evaluated microglial activation in HD gene carriersin relation to neurodegeneration and in relation to the clinicalfeatures seen at premanifest andmanifest stages of the diseaseAs an exploratory aim we evaluated the dynamic functions ofmicroglia in vitro by using induced microglia-like (iMG) cellsfrom human peripheral blood cells
MethodsSubjects and Clinical EvaluationThis cross-sectional study included 18 subjects 6 controls 6premanifest and 6 manifest HD gene carriers The HTT
Glossary3D = 3 dimensional CAP = CAG age product HD = Huntington disease IL = interleukin iMG = induced microglia-likeIND = Investigational NewDrug ApplicationMFI =median fluorescence intensityMMSE =MinindashMental State ExaminationPBA-s = short version of the Problem Behaviors Assessment PE = phycoerythrin ROI = region of interest SDMT = SymbolDigit Modalities Test SUVR = standardized uptake value ratio TNF = tumor necrosis factor UHDRS = Unified HD RatingScale
2 Neurology Neuroimmunology amp Neuroinflammation | Volume 8 Number 3 | May 2021 NeurologyorgNN
expansion was confirmed by a genotype larger CAG allelege36 A movement disorder specialist evaluated all HD genecarriers and the clinical diagnosis of HD was based on themotor signs certainty ie a Diagnostic Confidence Level of 4in the Unified HD Rating Scale (UHDRS)222 Patients wererecruited from the Huntington Disease Society of AmericaCenter of Excellence at UTHealth Controls were recruitedfrom the local community comprising a group of people withno history of neurologic or psychiatric disorder We excludedindividuals who have had infectious or autoimmune diseasesin activity or who have used anti-inflammatories in the 4weeks before the study pregnant or breastfeeding womenand participants with claustrophobia metal implants in-compatible with MRI previous neurosurgery or current se-rious comorbidity (eg cancer) Participantsrsquo recruitment andstudy procedures were performed from July 2018 to January2019
HD gene carriers completed the UHDRS22 including thetotal motor scale the total functional capacity and in-dependence scales The cognitive evaluation included theMini-Mental State Examination (MMSE)23 the SymbolDigit Modalities Test (SDMT) the Stroop InterferenceTest and the Verbal fluency test22 Behavioral symptomswere assessed with the short version of the Problem Be-haviors Assessment (PBA-s)24
The CAG age product (CAP) score was used to estimate theprogression of HD pathology as a function of CAG repeatlength and time of exposure to the effects of the expansion2
The CAP has been defined as
CAP = AGE times (CAG minus L)K
where AGE is the current age of the individual CAG is therepeat length and L and K are constants L is an estimate ofthe lower limit of the CAG expansion at which phenotypicexpression of the effects of mutant HTT could be observedand K is a normalizing constant We used the CAP proposedby Warner and Sampaio25 in which L = 30 and K = 627 andthe CAP will be equal to 100 at the subjectrsquos expected age ofmotor symptoms onset
Assessment of Microglial Activation In VivoUsing PETParticipants received 1 [11C]-ER176 PET scan on a GE Dis-covery RX PETCT scanner Subjects were scanned at rest AnIV line was placed in the antecubital fossa of the arm Subjectswere placed supine on the camera table with their head firmlysecured using a thermoplastic facemask Attenuation correctionCT was performed before PET acquisition Vital signs wereobtained before administration of [11C]-ER176 and at the endof the scan All subjects received a single IV bolus of about 20mCi of [11C]-ER176 in 10 mL (injected mass dose lt10 μg)and PET data were acquired for 90 minutes Subjects wereobserved during scanning and no earlier determination waswarranted Bladder voiding was encouraged to reduce bladderradiation exposure after PET scanning
For the purpose of anatomic localization the participants alsounderwent a whole-brain MRI scan on a 3T Phillips Ingeniascanner using an 8-channel head coil TheMRI protocol includeda 3-dimensional (3D) T1-weighted sequence magnetization-prepared gradient echo repetition time = 8100 ms time to echo= 37milliseconds voxel size 094times 094 times 15mm and 170 slicesPET frames were aligned for motion correction and the regionaltime-activity curves were obtained after segmentation with theautomated anatomic labelingndashmerged atlas implemented in thePNEURO module of PMOD 38 (PMOD Technologies)15
Time-activity curves were averaged from 60 to 90 minutes andnormalized over that of the whole cerebellum (standardizeduptake value ratio [SUVR]) following the methodology de-scribed by Lyoo et al26
In terms of analysis cortical (frontal cingulate occipitaltemporal parietal and insular cortices) and subcortical(thalamus caudate putamen pallidum hippocampus andamygdala) regions of interest (ROIs) were selected Corticalreconstruction and segmentation of the selected ROIs wereperformed with the recon-all pipeline from FreeSurfer v530image analysis suite27 on the 3DT1 image The PET para-metric map was then registered onto the FreeSurfer processedT1 image using ITK-SNAP28 in a semiautomatic way Aver-aged [11C]-ER176 SUVR and the associated ROI volumeswere then extracted from each ROI using FreeSurfer
In Vitro Evaluation of the Dynamic Functionsof MicrogliaThe same individuals subjected to the PETMRI scans (N =18 N = 6 individuals per group) were subjected to a blooddrawn Tenmilliliters of blood were collected by venipuncturein vacuum tubes containing heparin on the same day of thePET scan Monocytes were used to generate iMG cells aspreviously described29 Briefly peripheral blood mononuclearcells were isolated by Ficoll gradient centrifugation and platedat a density of 4 times 105 cellsmL After overnight incubationnonadherent cells were removed and the adherent cells(mainly monocytes) were cultured with granulocyte-macrophage colony-stimulating factor (10 ngmL) and in-terleukin (IL)-34 (100 ngmL) for 14 days to develop iMGcells Microglial characterization was evaluated throughmorphologic and phenotypical changes observed in thefluorescence microscopy using CX3CR1CCR2 doublestaining
Phagocytosis was examined by fluorescent microscopy andflow cytometry using the Phagocytosis Assay Kit (CaymanChemical Ann Arbor MI) At day 14 phycoerythrin (PE)-conjugated beads were added to the media (1250) for 24hours Cells were observed under the fluorescence micro-scope and then harvested by nonenzymatic cell dissociationsolution (Sigma-Aldrich St Louis MO) and cell lifter Thecells were washed and acquired using a BD FACSJazzcytometer (BD Biosciences San Jose CA) Flow cytometrydata were analyzed using FlowJo V10 software (Tree StarAshland OR)
NeurologyorgNN Neurology Neuroimmunology amp Neuroinflammation | Volume 8 Number 3 | May 2021 3
Secretion of inflammatory cytokines (tumor necrosis factor[TNF]-α IL-6 and IL-1β) during phagocytosis was mea-sured from culture supernatant using a Multiplex Immuno-assay following the manufacturer instructions (Bio-RadLaboratories Hercules CA) Briefly the samples and stan-dards were incubated with magnetic microspheres co-valently coupled with antibodies directed against the desired
biomarker (ie TNFα IL-1β and IL-6) After a series ofwashes to remove unbound protein a biotinylated detectionantibody was added to create a sandwich complex (similar tothat of a sandwich ELISA) The final detection complex wasformed with the addition of streptavidin-PE conjugate PEserved as a fluorescent indicator Data from standards andsamples and a standard curve were acquired in a Bio-Plex 200
Table Demographic and Clinical Characteristics of Premanifest HD Gene Carriers and Manifest Patients With HDIncluded in This Study
VariableControls(N = 6)
HD gene carriers
pValue
Premanifest HD(N = 6)
Manifest HD(N = 6)
Age y mean plusmn SD (median) 4328 plusmn 879(43)
3815 plusmn 718 (35) 5075 plusmn 1155(51)
0121a
Sex female n () 5 (83) 4 (67) 6 (100) 0301b
Educational level y mean plusmn SD (median) 1642 plusmn 432(175)
1583 plusmn 445 (145) 1467 plusmn 333(135)
0598a
TSPO genotype n ()
Thr147Thr147 0 0 0
Ala147Thr147 2 (33) 3 (50) 2 (33) 0792b
Ala147Ala147 4 (67) 3 (50) 4 (67)
CAP score mdash 7488 plusmn 1027 11509 plusmn 888 0002c
CAG repeats (larger) mean plusmn SD (median) mdash 4250 plusmn 187 (425) 4467 plusmn 242 (44) 0180c
UHDRS-TMS mean plusmn SD (median) mdash 367 plusmn 356 (3) 3083 plusmn 231 (31) 0002c
UHDRS-TFC mean plusmn SD (median) mdash 1250 plusmn 123 (13) 783 plusmn 184 (7) 0004c
Independence scale mdash 9750 plusmn 612 (100) 8417 plusmn 861 (85) 0015c
SDMT (total correct) mean plusmn SD (median) 5450 plusmn 784(54)
5417 plusmn 915 (575) 3067 plusmn 1019(335)
0004d
Verbal fluency test (category) no of correct responses in 1 min mean plusmn SD(median)
2383 plusmn 392(24)
2167 plusmn 403 (21) 1100 plusmn 525(105)
0008e
Stroop Interference Test no of correct responses mean plusmn SD (median) 4383 plusmn 1329(445)
4050 plusmn 599 (405) 2867 plusmn 1626(255)
0091a
MMSE mean plusmn SD (median) 2867 plusmn 186(29)
2883 plusmn 098 (285) 2660 plusmn 321 (28) 0138a
PBA-s domain scores mean plusmn SD (median)
Depression 400 plusmn 555 (2) 617 plusmn 546 (65) 717 plusmn 1096 (25) 0871a
Irritabilityaggression 133 plusmn 234 (05) 283 plusmn 325 (2) 233 plusmn 266 (2) 0845a
Psychosis 000 plusmn 000 (0) 017 plusmn 041 (0) 000 plusmn 000 (0) 0368a
Apathy 100 plusmn 089 (1) 250 plusmn 356 (05) 383 plusmn 440 (35) 0546a
Executive function 000 plusmn 000 (0) 450 plusmn 446 (4) 467 plusmn 350 (5) 0024f
Abbreviations HD = Huntington disease MMSE = Mini-Mental State Examination PBA-s = short version of the Problem Behaviors Assessment SDMT =Symbol Digit Modalities Test TFC = total functional capacity TMS = total motor score UHDRS = Unified Huntingtonrsquos Disease Rating ScaleSignificant values are highlighted in bolda Kruskal-Wallis testb Pearson χ2 testc Mann-Whitney testd Kruskal-Wallis followed by Dunn multiple comparisons test (controls = premanifest HD) lt manifest HDe Kruskal-Wallis followed by Dunn multiple comparisons test post hoc (controls = premanifest HD) gt manifest HDf Kruskal-Wallis followed by Dunn multiple comparisons test post hoc (controls = premanifest HD) gt manifest HD
4 Neurology Neuroimmunology amp Neuroinflammation | Volume 8 Number 3 | May 2021 NeurologyorgNN
Reader and analyzed in a Bio-Plex Manager Software whichpresented data as median fluorescence intensity (MFI) Thecytokine concentrations in the samples were calculatedbased on a standard curve in which the MFI values obtainedfrom the standard were plotted against the known concen-trations (in picograms per milliliter) Cytokine levels (inpicograms per milliliter) were measured in the supernatant24 hours after bead exposure or 24 hours of culture withoutany stimulus (baseline condition) and the values were cor-rected by protein levels (in milligrams per milliliter) in thecell lysates using the Pierce BCA Protein Assay Kit (Ther-moFisher Scientific Waltham MA) The final results areprovided in pgmg
DNA Polymorphism GenotypingPeripheral blood mononuclear cells were used for DNAextraction using the QIAamp DNA Blood Mini Kit
(Qiagen Hilden Germany) and subsequent genotyping forthe rs6971 polymorphism (Ala147Thr) in the TSPO gene(TaqMan assay ID C_2512465_20 Thermo Fisher Sci-entific) Allele Ala147 was linked to Vic and allele Thr147was linked to FAM PCRs were performed in QuantStudio7 Flex Real-Time PCR system (Life TechnologiesThermoFisher Scientific)
Statistical AnalysisAssociation between dichotomous variables was assessedwith the χ2 test Our limited sample size made it difficult toascertain data distribution and therefore nonparametrictests were used Comparisons between the 3 groups weremade by the Kruskal-Wallis test When appropriate posthoc analyses were used to determine significant differencesbetween pairs of groups (Dunn Multiple ComparisonTest) The Mann-Whitney test was used for 2-group
Figure 1 Microglial Activation in HD
(A) PET image showing increased [11C]-ER176 standardized uptake value ratio (SUVR) in bilateral putamina and pallidi of a patient with manifest HD incomparison with a control Note that there is no visible difference in [11C]-ER176 SUVR for the premanifest HD gene carrier (central image) (B) Groupcomparisons in left (L) and right (R) putamina and pallidi Significant differences between groups are indicated by different letters (Kruskal-Wallis testfollowed by Dunn multiple comparisons test) Horizontal bars show the mean and the standard error of the mean Data from other regions of interest areshown in figure e-1 (linkslwwcomNXIA459) HD = Huntington disease
NeurologyorgNN Neurology Neuroimmunology amp Neuroinflammation | Volume 8 Number 3 | May 2021 5
comparisons (premanifest HD vs manifest HD) Spearmancorrelations were performed to examine the relationshipbetween [11]C-ER176 SUVR in the ROIs significantlydifferent between groups and (1) CAP score (2) volume inthe same ROIs and (3) scores in the clinical scales Allstatistical tests were 2 tailed and were performed using asignificance level of α = 005 Statistical analyses wereperformed using SPSS software version 260 and GraphPadPrism version 50
Standard Protocol Approvals Registrationsand Patient ConsentsAll subjects provided written informed consent before ad-mission to the study The Research Ethics Committees ofUTHealth and Houston Methodist Research Institute ap-proved this study
An Investigational New Drug Application (IND) was sub-mitted and authorization was obtained from the Food andDrug Administration to use [11C]-ER176 as a PET Imagingagent in this study (IND 139220)
Data AvailabilityThe raw data not provided in the article will be made availableby the authors at the request of other investigators for pur-poses of replicating procedures and results
ResultsClinical Evaluation and TSPO GenotypingDemographic and clinical characteristics of participants included inthis study are shown in the table There was no significant differ-ence between controls premanifest HD gene carriers and patientswith HD regarding age sex and educational level As expectedpatients with manifest HD presented a higher CAP score andworse scores in the scales that evaluatedmotor symptoms severityfunctional capacity and independence in comparison with pre-manifest HD (table) The age at clinical diagnosis was 4767 plusmn1035 years for the patients with manifest HD Premanifest HDgene carriers had amean of 1298 plusmn 617 years before the predictedclinical diagnosis of HD The 3 groups presented similar perfor-mance on the MMSE Patients with the clinical diagnosis of HDpresented worse performance on the SDMT and verbal fluencytest in comparisonwith premanifest and control subjects Controlsindividuals with premanifest HD and patients with HD presentedsimilar scores in the PBA-s subscales except for executive functionin which HD gene carriers (both premanifest and manifest HD)scored worse than controls evidencing that those problems arepresent before clinical diagnosis of HD (table)
The groups presented similar distribution of TSPO genotype(table) Individuals with genotype Ala147Ala147 were clas-sified as high-affinity binders and those with genotype
Figure 2 Microglial Activation in HD Is Associated With Disease Burden
Among HD gene carriers (premanifest andmanifest HD) a higher CAP score (a proxy for cumulative disease burden) was significantly associated with higher[11C]-ER176 standardized uptake value ratio (SUVR) in the (A) left putamen (ρ = 0664 p = 0018) (B) left pallidum (ρ = 0657 p = 0020) (C) right putamen (ρ =0671 p = 0017) and (D) right pallidum (ρ = 0734 p = 0007) Note the 2 clusters in [11C]-ER176 SUVR composed of premanifest HD gene carriers (CAP lt100)and manifest patients with HD (CAP gt100) CAP = CAG age product HD = Huntingtonrsquos disease
6 Neurology Neuroimmunology amp Neuroinflammation | Volume 8 Number 3 | May 2021 NeurologyorgNN
Ala147Thr147 as mixed affinity None of our patients pre-sented the genotype Thr147Thr147 (low-affinity binders)Because the groups presented comparable distribution ofTSPO genotype (table) and the outcome parameter wasSUVR which minimizes genotype differences no correctionfor genotype-related binding affinity was performed
In Vivo Assessment of Microglial ActivationPatients with HD demonstrate higher [11C]-ER176 SUVR inboth the left and right globi pallidi and putamina in comparisonwith controls These differences were not significant when wecompared premanifest HD gene carriers with patients withHDorcontrols (figure 1 and figure e-1 linkslwwcomNXIA459)These results indicate increased microglial activation in the puta-mina and globi pallidi of patients with manifest HDWe found nodifferences between different TSPO genotypes on [11C]-ER176SUVR when controlling for HD diagnosis (data not shown)
Among HD gene carriers (premanifest and manifest HD) ahigher CAP score was significantly associated with higher[11C]-ER176 SUVR in the left putamen (ρ = 0664 p =0018) left pallidum (ρ = 0657 p = 0020) right putamen(ρ = 0671 p = 0017) and right pallidum (ρ = 0734 p =0007) (figure 2) Corroborating these findings increased[11C]-ER176 SUVRwas associated with decreased volume inthe same ROIs (figure 3)
In Vitro Evaluation of the Dynamic Functionsof MicrogliaWe confirmed that the iMG cells have the immunophenotypeof microglial cells such as CX3CR1highCCR2low and atypical (ie ramified) morphology (figure 4) The iMG cellsexpress dynamic functions such as phagocytosis and secretionof inflammatory cytokines making them a valuable tool toassess microglial functioning30 The phagocytosis assaydemonstrated that iMG cells obtained from controls pre-manifest HD and HD patients (figure 5 AndashC) showedsimilar phagocytic capacity (figure 5E) Secretion of in-flammatory cytokines (TNFα IL-6 and IL-1β) duringphagocytosis was measured from culture supernatant Asshown in figure 5F although iMG cells from controls pre-manifest HD and patients with HD increased the productionof the evaluated cytokines during phagocytosis the valueswere significant only for TNFα in controls and manifest HD(figure 5F)
DiscussionBy using [11C]-ER176 PET we verified that patients withHD have increased microglial activation in basal gangliastructures that are involved in HD pathophysiologyie putamina and globi pallidi In addition we found that
Figure 3 Microglial Activation Is Associated With Brain Atrophy
Increased microglial activation is associated with decreased volume in putamen and pallidus Controls are displayed in red dots premanifest HD genecarriers in green dots andmanifest HD in black dots A higher [11C]-ER176 standardized uptake value ratio (SUVR) was associatedwith a decreased volume inthe (A) left putamen (ρ = minus0569 p = 0014) (B) left pallidum (ρ = minus0591 p = 0010) (C) right putamen (ρ = minus0601 p = 0008) and (D) right pallidum (ρ = minus0686 p= 0002) ICV = intracranial volume The dashed line was placed to emphasize the 2 clusters composed of controls + premanifest HD and manifest HD genecarriers HD = Huntington disease
NeurologyorgNN Neurology Neuroimmunology amp Neuroinflammation | Volume 8 Number 3 | May 2021 7
premanifest HD gene carriers demonstrate [11C]-ER176SUVRs comparable to controls suggesting that microglialactivation is a later process in HD pathophysiology becomingevident only after the onset of motor symptoms (figure 1)Supporting these findings we reported a significant correla-tion between increased microglial activation and higher CAPscores a proxy for cumulative disease burden (figure 2)Moreover increased microglial activation in putamina andpallidi was significantly associated with reduced volumes inthe same ROIs (figure 3) Individuals with manifest HDpresented a significant decrease in the cortical volume incomparison with controls (data not shown) Despite alreadypresenting cortical atrophy manifest HD individuals do nothave increased microglial activation in the cortex (figure 1)The only areas displaying increased microglia activation arethose that are known to be affected early in HD pathophysi-ology Because PET scans require individuals to be still for 90minutes our sample did not include patients in moderatelatestages of HD Whether increased microglia activation willoccur in the cortex later in the HD course is yet to be de-termined Altogether our data demonstrate that microglialactivation is involved in HD pathophysiology and is associ-ated with disease progression (as demonstrated by the CAPscore and atrophy in basal ganglia structures) Our findingssupport the conclusions of previous research that reportedthat microglial activation correlates with disease severitycontributing to the ongoing neuronal degeneration in HD16
HD has been regarded as an important model to study neu-rodegenerative diseases as it is caused by a single geneticmutation and is amenable to predictive genetic testing en-abling the study of patients before clinical diagnosis2 In this
study we assessed microglial activation not only in patientswith HD but also in premanifest HD gene carriers with amean of 13 years before the predicted onset of clinicalsymptoms Our results corroborate the view that microglialactivation is unlikely to be the initiating event in HD but maycontribute to neuronal death through multiple pathways31
Similar to other neurodegenerative diseases the mecha-nisms underlying selective neuronal dysfunction and deathin HD are still uncertain In addition to impairment in sys-tems for handling abnormal proteins other metabolicpathways and mechanisms might contribute to neuro-degeneration and progression of HD such as mitochondrialdysfunction impaired neurotrophic support oxidativestress and inflammatoryimmune mechanisms83233 In thecurrent study we described a significant increase in micro-glial activation in regions of neurodegeneration in HDsuggesting that this process is related to the disease patho-physiology and not to random premorbid events Accord-ingly postmortem studies reported significant microglialactivation in regions implicated in HD pathogenesis46 Thefact that we found significant microglial activation in patientswith manifest HD (but not in premanifest HD gene carriers)in comparison with controls suggests that this event occursin response or concomitantly to neuronal death This as-sumption is further corroborated by the finding of significantassociations between microglial activation and ROI volumesas well as CAP scores Here again our data are corroboratedby postmortem findings in which the number of activatedmicroglia in the striatum and cortex correlated with neuronalloss confirming the hypothesis that neuroinflammation maybe elicited by degenerating neurons6
Figure 4 Inducing Microglia-Like Cells From Human Peripheral Blood Monocytes
Peripheral blood monocytes were cultured for 14 days in conventional conditions (A) or with GM-CSF and IL-34 resulting in induced microglia like cells thatexhibit microglia-like morphology and increased CX3CR1CCR2 ratio (B)
8 Neurology Neuroimmunology amp Neuroinflammation | Volume 8 Number 3 | May 2021 NeurologyorgNN
Neuroinflammation elicited by neuronal death can be at firstregarded as a positive event as it promotes clearance of celldebris However inflammatory mechanisms contribute toneurodegeneration and dying neurons further activate in-flammatory responses resulting in a vicious cycle of in-flammation and neuronal death34 Therefore inflammatoryresponses although essential for tissue homeostasis cancontribute to neuronal injury As neural tissues have a
restricted cell renewal and regenerative capacity the CNS isextremely vulnerable to uncontrolled andor chronic immuneand inflammatory processes34 Interestingly enough our datacorroborate this hypothesis as we observed increased micro-glial activation later in the HD course (ie manifest HD)reinforcing the idea that neuroinflammation contributes todisease progression rather than to disease onset Activatedmicroglia releases proinflammatory mediators including
Figure 5 Phagocytosis by Induced Microglia-Like (iMG) Cells
Peripheral blood monocytes (day 1 panel A) were cultured for 14 days with GM-CSF and IL-34 resulting in iMG cells (day 14 panel B) Phagocytosis wasassessed by exposing iMG cells to PE-conjugated (1250) beads (panel C) for 24 hours The iMG cells showed the ability of phagocytosis with morphologicalchanges (C) The read arrows show PE-conjugated beads phagocytosed by iMG cells After washing beads out cells were acquired in a flow cytometer andanalyzed iMG were first gated by forwardside scatter (not shown) PE uptake was analyzed as shown in (D) orange (left) histogram shows the cells withoutbeads (negative control) red (right) histogram shows the cells that phagocytosed to the PE-conjugated beads Quantification is shown in (E) iMG fromcontrols premanifest HD and HD patients showed similar phagocytic capacity (no significant difference found at the Kruskal-Wallis test) (F) Cytokineproduction during phagocytosis iMG cells were incubated with PE-conjugated beads (1250) for 24 hours Supernatant was collected and cytokines wereanalyzed by multiparametric assay Cytokine levels (in picograms per milliliter) were corrected by protein levels (in milligrams per milliliter) Although iMGcells from controls premanifest HD and patients with HD increased the production of the evaluated cytokines during phagocytosis the values weresignificant only for TNFα in controls andmanifest HD (p values are provided in the figure Wilcoxonmatched-pairs signed-rank test) Scale bar (AndashC) = 10mmHorizontal bars (E and F) show the mean and the standard error of the mean HD = Huntington disease PE = phycoerythrin TNF = tumor necrosis factor
NeurologyorgNN Neurology Neuroimmunology amp Neuroinflammation | Volume 8 Number 3 | May 2021 9
cytokines and reactive oxygen species which in turn will alsocontribute to neuronal death and further microglial activa-tion34We observed an increase in cytokine release (especiallyTNFα) following a phagocytic stimulus in our in vitro ex-periments In line with our findings postmortem studies havealso reported increased levels of inflammatory cytokines inbrain samples of patients with HD in comparison withcontrols3536 It is worth mentioning that cytokine release is amicroglial reaction to a proinflammatory challenge and undercontrolled conditions (such as the iMG cultures) phagocy-tosis is a noninflammatory function of microglia A proin-flammatory stimulus would be needed to appropriately assessdifferences in cytokine release between controls premanifestHD and manifest HD iMG cultures
The current study used a novel PET tracer for TSPO [11C]-ER176 to assess microglial activation in HD By using [11C]-PK11195 PET studies have shown a significant increase inmicroglial activation in the striatum of patients with HD16 aswell as in premanifest HD gene carriers18 compared withcontrols In both studies [11C]-PK11195 binding significantlycorrelated with disease severity as measured by decreased[11C]-raclopride (a marker of dopamine D2 and D3 receptoravailability) binding in the striatum1618 Similar results weredescribed in the hypothalamus17 Later the same work groupused a multimodal imaging approach combiningMRI and PETanalyses and worsening in atrophy evaluated by MRI was ac-companied by a reduction in [11C]-raclopride and an increasein [11C]-PK11195 bindings in patients with HD In pre-manifest HD increased level of microglial activation in theassociative striatum and in the regional network associated withcognition correlated with 5-year probability of HD onset19
More recently increased microglial activation in the somato-sensory cortex (as evaluated through [11C]-PK11195 binding)was associatedwith higher levels of IL-1β IL-6 IL-8 and TNFαproduced by stimulated peripheral blood monocytes frompremanifest HD gene carriers2037 Although these studiesprovided valuable information about neuroinflammation inHD indicating microglial activation even in premanifest sub-jects their results should be interpreted with caution The poorsignal-to-background ratio of first-generation TSPO radio-tracers poses important limitations to the analyses The ratio ofspecific to nonspecific binding of [11C]-PK11195 in humanbrain has been reported to be as low as 023839 As an attempt tomitigate this problem a recent study used the second-generation TSPO tracer [11C]-PBR28 to evaluate micro-glial activation in HD21 Of interest the results were similar toours ie increased levels of [11C]-PBR28 binding in theputamina and pallidi of HD gene carriers (which included 7manifest patients with HD and only 1 person with premanifestHD) in comparison with controls Of note the results of thesingle premanifest subject were midway between controls andthe remaining patients suggesting that microglial activationwas associated with disease progression21 The authors didconfirm that [11C]-PBR28 provides a high signal-to-background ratio an important feature required for the anal-ysis at the individual level and within individual brain regions
An important limitation of [11C]-PBR28 and other second-generation TSPO radioligands is that they have varying degreesof sensitivity to the single nucleotide polymorphism rs6971 inthe TSPO gene40 [11C]-ER176 has unique advantages overthe other TSPO binders being the only known TSPO radio-ligand that allows the inclusion of low-affinity binders Thebinding of ER176 in low-affinity binders was found to becomparable to that for PBR28 in high-affinity binders More-over ER176 was proven to be the only radioligand not con-taminated by radiometabolites accumulating in the brain14
Our findings must be interpreted while taking into accountthe limitations of the study which include the relatively smallsample size and the cross-sectional design The cross-sectionaldesign prevented the assessment of microglial activation inthe context of disease progression and development of motorand nonmotor symptoms In addition the use of an objectivemethod to measure striatal degeneration (eg [11C]-raclopride PET) would provide more accurate informationabout striatal neuron dysfunctiondeath Regarding the invitro experiments the use of a proinflammatory challenge(other than the beads added for the phagocytosis assay) in ourexperiments would provide valuable information on cytokinesrelease by iMG from the different groups It is worth men-tioning the strengths of the study that include the use of aunique TSPO ligand [11C]-ER176 the enrollment of bothpremanifest and manifest subjects and the exploratory in-vestigation of microglia activity
In conclusion our findings strengthen the evidence thatmicroglial activation is associated with HD pathophysiologyMicroglial activation is unlikely to be the initiating event inHD Yet it may exacerbate neuronal dysfunction and neu-ronal death thus contributing to disease progression It isimportant to refine our understanding of the more specificimmuneinflammatory mechanisms that are involved in HDand in neurodegeneration This may foster the developmentof new therapeutic interventions to halt the progression ofHD and other neurodegenerative diseases
AcknowledgmentThe authors acknowledge and thank all of the volunteers thatparticipated in this study and are indebted to their families fortheir magnificent support They also thank the staff from theUTHealth McGovern Medical School 3T MRI Center andfrom the HoustonMethodist Research Institute TranslationalImaging Core Facilities especially Vips Patel and Kim MDoan for their skilled technical support Dr Gabriel Fries forhis assistance with the genotyping analyses and the teammembers of the HDSA Center of Excellence at UTHealth
Study FundingThis study was funded by the HD Human BiologyProjectmdashHuntingtonrsquos Disease Society of America (HDSA)E Furr Stimming receives research funding from RocheGenetech Vaccinex Cures Within Reach HDSA Uniqureand CHDI Foundation
10 Neurology Neuroimmunology amp Neuroinflammation | Volume 8 Number 3 | May 2021 NeurologyorgNN
DisclosureThe authors report no disclosures relevant to the manuscriptGo to NeurologyorgNN for full disclosures
Publication HistoryReceived by Neurology Neuroimmunology amp NeuroinflammationSeptember 9 2020 Accepted in final form January 20 2021
References1 The Huntingtonrsquos Disease Collaborative Research Group A novel gene containing a
trinucleotide repeat that is expanded and unstable on Huntingtonrsquos disease chro-mosomes Cell 199372971ndash983
2 Ross CA Aylward EH Wild EJ et al Huntington disease natural history biomarkersand prospects for therapeutics Nat Rev Neurol 201410204ndash216
3 Soulet D Cicchetti F The role of immunity in Huntingtonrsquos disease Mol Psychiatry201116889ndash902
4 Myers RH Vonsattel JP Paskevich PA et al Decreased neuronal and increasedoligodendroglial densities in Huntingtonrsquos disease caudate nucleus J NeuropatholExp Neurol 199150729ndash742
5 Vonsattel JP Myers RH Stevens TJ Ferrante RJ Bird ED Richardson EP JrNeuropathological classification of Huntingtonrsquos disease J Neuropathol Exp Neurol198544559ndash577
6 Sapp E Kegel KB Aronin N et al Early and progressive accumulation of reactivemicroglia in the Huntington disease brain J Neuropathol Exp Neurol 200160161ndash172
7 Ciccocioppo F Bologna G Ercolino E et al Neurodegenerative diseases asproteinopathies-driven immune disorders Neural Regen Res 202015850ndash856
8 Rocha NP Ribeiro FM Furr-Stimming E Teixeira AL Neuroimmunology of Hun-tingtonrsquos disease revisiting evidence from human studies Mediators Inflamm 201620168653132
9 Vowinckel E Reutens D Becher B et al PK11195 binding to the peripheral ben-zodiazepine receptor as a marker of microglia activation in multiple sclerosis andexperimental autoimmune encephalomyelitis J Neurosci Res 199750345ndash353
10 Papadopoulos V Baraldi M Guilarte TR et al Translocator protein (18kDa) newnomenclature for the peripheral-type benzodiazepine receptor based on its structureand molecular function Trends Pharmacol Sci 200627402ndash409
11 Cumming P Burgher B Patkar O et al Sifting through the surfeit of neuro-inflammation tracers J Cereb Blood Flow Metab 201838204ndash224
12 Zanotti-Fregonara P Zhang Y Jenko KJ et al Synthesis and evaluation of translocator18 kDa protein (TSPO) positron emission tomography (PET) radioligands with lowbinding sensitivity to human single nucleotide polymorphism rs6971 ACS ChemNeurosci 20145963ndash971
13 Ikawa M Lohith TG Shrestha S et al 11C-ER176 a radioligand for 18-kDa trans-locator protein has adequate sensitivity to robustly image all three affinity genotypesin human brain J Nucl Med 201758320ndash325
14 Fujita M Kobayashi M Ikawa M et al Comparison of four (11)C-labeled PETligands to quantify translocator protein 18 kDa (TSPO) in human brain (R)-PK11195 PBR28 DPA-713 and ER176-based on recent publications that measuredspecific-to-non-displaceable ratios EJNMMI Res 2017784
15 Zanotti-Fregonara P Pascual B Veronese M et al Head-to-head comparison of (11)C-PBR28 and (11)C-ER176 for quantification of the translocator protein in thehuman brain Eur J Nucl Med Mol Imaging 2019461822ndash1829
16 Pavese N Gerhard A Tai YF et al Microglial activation correlates with severity inHuntington disease a clinical and PET study Neurology 2006661638ndash1643
17 Politis M Pavese N Tai YF Tabrizi SJ Barker RA Piccini P Hypothalamic in-volvement in Huntingtonrsquos disease an in vivo PET study Brain 20081312860ndash2869
18 Tai YF Pavese N Gerhard A et al Microglial activation in presymptomatic Hun-tingtonrsquos disease gene carriers Brain 20071301759ndash1766
19 Politis M Pavese N Tai YF et al Microglial activation in regions related to cognitivefunction predicts disease onset in Huntingtonrsquos disease a multimodal imaging studyHum Brain Mapp 201132258ndash270
20 Politis M Lahiri N Niccolini F et al Increased central microglial activation associatedwith peripheral cytokine levels in premanifest Huntingtonrsquos disease gene carriersNeurobiol Dis 201583115ndash121
21 Lois C Gonzalez I Izquierdo-Garcia D et al Neuroinflammation in Huntingtonrsquosdisease new insights with (11)C-PBR28 PETMRI ACS Chem Neurosci 201892563ndash2571
22 Huntington Study Group Unified Huntingtonrsquos Disease Rating Scale reliability andconsistency Mov Disord 199611136ndash142
23 Folstein MF Folstein SE McHugh PR ldquoMini-mental staterdquo A practical method forgrading the cognitive state of patients for the clinician J Psychiatr Res 197512189ndash198
24 McNally G Rickards H Horton M Craufurd D Exploring the validity of the shortversion of the Problem Behaviours Assessment (PBA-s) for Huntingtonrsquos disease arasch analysis J Huntingtons Dis 20154347ndash369
25 Warner JH Sampaio C Modeling variability in the progression of Huntingtonrsquosdisease a novel modeling approach applied to structural imaging markers fromTRACK-HD CPT Pharmacometrics Syst Pharmacol 20165437ndash445
26 Lyoo CH Ikawa M Liow JS et al Cerebellum can serve as a pseudo-reference regionin Alzheimer disease to detect neuroinflammation measured with PET radioligandbinding to translocator protein J Nucl Med 201556701ndash706
27 Reuter M Schmansky NJ Rosas HD Fischl B Within-subject template estimation forunbiased longitudinal image analysis Neuroimage 2012611402ndash1418
28 Yushkevich PA Piven J Hazlett HC et al User-guided 3D active contour segmen-tation of anatomical structures significantly improved efficiency and reliabilityNeuroimage 2006311116ndash1128
29 Ohgidani M Kato TA Setoyama D et al Direct induction of ramified microglia-likecells from human monocytes dynamic microglial dysfunction in Nasu-Hakola dis-ease Sci Rep 201444957
30 Ohgidani M Kato TA Kanba S Introducing directly induced microglia-like (iMG)cells from fresh human monocytes a novel translational research tool for psychiatricdisorders Front Cell Neurosci 20159184
31 Kim SU de Vellis J Microglia in health and disease J Neurosci Res 200581302-31332 Ross CA Tabrizi SJ Huntingtonrsquos disease from molecular pathogenesis to clinical
treatment Lancet Neurol 20111083ndash9833 Chitnis TWeiner HL CNS inflammation and neurodegeneration J Clin Invest 2017
1273577ndash3587
Appendix Authors
Name Location Contribution
Natalia PRocha PhD
The Mitchell Center forAlzheimerrsquos Disease andRelated Brain DisordersDepartment of NeurologyMcGovern Medical SchoolThe University of TexasHealth Science CenterHouston
Designed andconceptualized the studymajor role in theacquisition of dataanalyzed the data anddrafted the manuscript forintellectual content
OdelinCharron MS
Department of NeurologyThe University of Texas atAustin
Major role in data analysis
Leigh BLathamPhD
School of MedicineUniversity of WashingtonSeattle
Major role in theacquisition of data
Gabriela DColpo PhD
Neuropsychiatry ProgramDepartment of Psychiatryand Behavioral SciencesMcGovern Medical SchoolUniversity of TexasHouston
Major role in theacquisition of data
PaoloZanotti-FregonaraMD PhD
Houston MethodistResearch Institute andWeill Cornell Medicine TX
Major role in the studydesign and the acquisitionof data
MeixiangYu PhD
Houston MethodistResearch Institute andWeill Cornell Medicine TX
Major role in theacquisition of data
LeorahFreemanMD PhD
Department of NeurologyThe University of Texas atAustin
Major role in the studydesign data analysis andinterpretation and theacquisition of data
Erin FurrStimmingMD
HDSA Center of Excellenceat University of TexasHealth Science Center atHouston
Major role in the studydesign and acquisition ofdata interpreted the dataand revised themanuscript for intellectualcontent
Antonio LTeixeiraMD PhD
Neuropsychiatry ProgramDepartment of Psychiatryand Behavioral SciencesMcGovern Medical SchoolUniversity of TexasHouston
Major role in the studydesign and acquisition ofdata interpreted the dataand revised themanuscript for intellectualcontent
NeurologyorgNN Neurology Neuroimmunology amp Neuroinflammation | Volume 8 Number 3 | May 2021 11
34 Rocha NP de Miranda AS Teixeira AL Insights into neuroinflammation in Parkin-sonrsquos disease from biomarkers to anti-inflammatory based therapies Biomed Res Int20152015628192
35 Silvestroni A Faull RL Strand AD Moller T Distinct neuroinflammatory profile inpost-mortem human Huntingtonrsquos disease Neuroreport 2009201098ndash1103
36 Bjorkqvist M Wild EJ Thiele J et al A novel pathogenic pathway of immune acti-vation detectable before clinical onset in Huntingtonrsquos disease J Exp Med 20082051869ndash1877
37 Politis M Lahiri N Niccolini F et al Corrigendum to ldquoIncreased central microglialactivation associated with peripheral cytokine levels in premanifest Huntingtonrsquos
disease gene carriersrdquo [Neurobiol Dis 83 (2015) 115-121] Neurobiol Dis 201798162
38 Kropholler MA Boellaard R Schuitemaker A Folkersma H van Berckel BN Lam-mertsma AA Evaluation of reference tissue models for the analysis of [11C](R)-PK11195 studies J Cereb Blood Flow Metab 2006261431ndash1441
39 Turkheimer FE Edison P Pavese N et al Reference and target region modeling of[11C]-(R)-PK11195 brain studies J Nucl Med 200748158ndash167
40 Kreisl WC Jenko KJ Hines CS et al A genetic polymorphism for translocator protein18 kDa affects both in vitro and in vivo radioligand binding in human brain to thisputative biomarker of neuroinflammation J Cereb Blood FlowMetab 20133353ndash58
12 Neurology Neuroimmunology amp Neuroinflammation | Volume 8 Number 3 | May 2021 NeurologyorgNN
DOI 101212NXI000000000000098420218 Neurol Neuroimmunol Neuroinflamm
Natalia P Rocha Odelin Charron Leigh B Latham et al Pathophysiology
Microglia Activation in Basal Ganglia Is a Late Event in Huntington Disease
This information is current as of April 1 2021
ServicesUpdated Information amp
httpnnneurologyorgcontent83e984fullhtmlincluding high resolution figures can be found at
References httpnnneurologyorgcontent83e984fullhtmlref-list-1
This article cites 40 articles 4 of which you can access for free at
Subspecialty Collections
httpnnneurologyorgcgicollectionpetPET
httpnnneurologyorgcgicollectionhuntingtons_diseaseHuntingtons diseasefollowing collection(s) This article along with others on similar topics appears in the
Permissions amp Licensing
httpnnneurologyorgmiscaboutxhtmlpermissionsits entirety can be found online atInformation about reproducing this article in parts (figurestables) or in
Reprints
httpnnneurologyorgmiscaddirxhtmlreprintsusInformation about ordering reprints can be found online
Academy of Neurology All rights reserved Online ISSN 2332-7812Copyright copy 2021 The Author(s) Published by Wolters Kluwer Health Inc on behalf of the AmericanPublished since April 2014 it is an open-access online-only continuous publication journal Copyright
is an official journal of the American Academy of NeurologyNeurol Neuroimmunol Neuroinflamm

Huntington disease (HD) is an autosomal dominant diseasecaused by a trinucleotide (CAG) repeat expansion in thehuntingtin gene (HTT)1 The toxic gain of function of theexpanded mutant huntingtin protein (HTT) seems to be thekey event involved in the progressive neuronal dysfunctionand neuronal loss particularly in the striatum and cortex As aresult individuals with HD classically present with motordysfunction cognitive decline and behavioral disorders2 Al-though the cause of HD is well established the mechanismsunderlying neuronal dysfunction and death are not com-pletely understood Mutant HTT seems to trigger a patho-genic cascade that includes oxidative stress and immuneinflammatory mechanisms which are further stimulated byneuronal dysfunctiondeath Together these mechanismshave been regarded as key contributors to the pathophysiol-ogy of HD3
Neuroinflammation in HD has been reported since earlypostmortem studies which described reactive astrocytes45
and microglial activation6 in brain regions associated with HDpathogenesis Inflammatory mechanisms in the CNS arethought to be initiated as compensatory responses againstmisfolded protein oligomers andor deposits7 On the otherhand inflammatory responses can trigger neuronal damageand thus contribute to disease progression8 Noteworthy theabove-mentioned postmortem studies showed that bothmicroglial activation and reactive astrocytosis grades correlatewith disease severity4-6 These results motivated studies toevaluate neuroinflammation in HD in vivo In this regardPET neuroimaging studies have focused on evaluatingmicroglial activation through specific molecular targets invivo Microglial activation imaging is currently performed byusing PET tracers binding to the 18-kDa translocator protein(TSPO) a mitochondrial protein that is highly expressed inphagocytic inflammatory cells including activated microgliain the brain and macrophages in the periphery910 TSPO isexpressed at very low levels in the CNS but its levels areupregulated by activated microglia10
The prototype isoquinoline ligand for TSPO [11C]-PK11195 has been introduced in the field of molecularbrain imaging of microglial activation more than 20 yearsago9 However PK11195 shows low specific signal-to-noiseratios even in clinical conditions in which microglial activa-tion is a prominent and well-established feature11 This limi-tation motivated the development of alternative TSPO PETligands with better signal-to-noise ratios such as [11C]-
PBR28 and other second-generation ligands11 Howeverother technical issues became evident mainly the aberrantlylow TSPO binding in some individuals due to the single nu-cleotide polymorphism rs6971 More recently a series of4-phenylquinazoline-2-carboxamides lacking allelic sensitivityto the human single nucleotide polymorphism rs6971 hasbeen developed Noteworthy [11C]-ER176 (11C-(R)-N-sec-butyl-4-(2-chlorophenyl)-N-methylquinazoline-2-carboxamide) a new analog of [11C]-PK1119512 presentshigher specific binding when compared with most of theavailable TSPO binders13 Although still sensitive to TSPOpolymorphism ER176 allows the inclusion of low-affinitybinders because of its high in vivo binding potential14 Ofnote [11C]-ER176 has a higher specific binding and a smallerintersubject variability compared with [11C]-PBR28 thusresulting in higher statistical power and requiring fewer sub-jects for clinical studies Therefore [11C]-ER176 should bepreferred over [11C]-PBR28 for TSPO studies in humans15
To date 6 studies have used PET neuroimaging with TSPObinders to evaluate microglial activation in HD Five studiesused the first-generation TSPO radiotracer [11C]-PK1119516-20 and one used the second-generation TSPObinder [11C]-PBR2821 These studies revealed increasedmicroglial activation in HD gene carriers which correlatedwith disease stage andor severity of symptoms The availablefindings indicate that microglial activation is involved in HDpathophysiology and TSPO PET imaging may be a valuabletool to monitor HD progression and therapeutic efficacy ofdrugs targeting neuroinflammation However these indica-tions have yet to be explored So far no study has evaluatedthe use of the novel TSPO tracer [11C]-ER176 as a markerof microglial activation in HD gene carriers Therefore thecurrent study was designed to define the role played bymicroglia in different stages of HD by using [11C]-ER176PET We evaluated microglial activation in HD gene carriersin relation to neurodegeneration and in relation to the clinicalfeatures seen at premanifest andmanifest stages of the diseaseAs an exploratory aim we evaluated the dynamic functions ofmicroglia in vitro by using induced microglia-like (iMG) cellsfrom human peripheral blood cells
MethodsSubjects and Clinical EvaluationThis cross-sectional study included 18 subjects 6 controls 6premanifest and 6 manifest HD gene carriers The HTT
Glossary3D = 3 dimensional CAP = CAG age product HD = Huntington disease IL = interleukin iMG = induced microglia-likeIND = Investigational NewDrug ApplicationMFI =median fluorescence intensityMMSE =MinindashMental State ExaminationPBA-s = short version of the Problem Behaviors Assessment PE = phycoerythrin ROI = region of interest SDMT = SymbolDigit Modalities Test SUVR = standardized uptake value ratio TNF = tumor necrosis factor UHDRS = Unified HD RatingScale
2 Neurology Neuroimmunology amp Neuroinflammation | Volume 8 Number 3 | May 2021 NeurologyorgNN
expansion was confirmed by a genotype larger CAG allelege36 A movement disorder specialist evaluated all HD genecarriers and the clinical diagnosis of HD was based on themotor signs certainty ie a Diagnostic Confidence Level of 4in the Unified HD Rating Scale (UHDRS)222 Patients wererecruited from the Huntington Disease Society of AmericaCenter of Excellence at UTHealth Controls were recruitedfrom the local community comprising a group of people withno history of neurologic or psychiatric disorder We excludedindividuals who have had infectious or autoimmune diseasesin activity or who have used anti-inflammatories in the 4weeks before the study pregnant or breastfeeding womenand participants with claustrophobia metal implants in-compatible with MRI previous neurosurgery or current se-rious comorbidity (eg cancer) Participantsrsquo recruitment andstudy procedures were performed from July 2018 to January2019
HD gene carriers completed the UHDRS22 including thetotal motor scale the total functional capacity and in-dependence scales The cognitive evaluation included theMini-Mental State Examination (MMSE)23 the SymbolDigit Modalities Test (SDMT) the Stroop InterferenceTest and the Verbal fluency test22 Behavioral symptomswere assessed with the short version of the Problem Be-haviors Assessment (PBA-s)24
The CAG age product (CAP) score was used to estimate theprogression of HD pathology as a function of CAG repeatlength and time of exposure to the effects of the expansion2
The CAP has been defined as
CAP = AGE times (CAG minus L)K
where AGE is the current age of the individual CAG is therepeat length and L and K are constants L is an estimate ofthe lower limit of the CAG expansion at which phenotypicexpression of the effects of mutant HTT could be observedand K is a normalizing constant We used the CAP proposedby Warner and Sampaio25 in which L = 30 and K = 627 andthe CAP will be equal to 100 at the subjectrsquos expected age ofmotor symptoms onset
Assessment of Microglial Activation In VivoUsing PETParticipants received 1 [11C]-ER176 PET scan on a GE Dis-covery RX PETCT scanner Subjects were scanned at rest AnIV line was placed in the antecubital fossa of the arm Subjectswere placed supine on the camera table with their head firmlysecured using a thermoplastic facemask Attenuation correctionCT was performed before PET acquisition Vital signs wereobtained before administration of [11C]-ER176 and at the endof the scan All subjects received a single IV bolus of about 20mCi of [11C]-ER176 in 10 mL (injected mass dose lt10 μg)and PET data were acquired for 90 minutes Subjects wereobserved during scanning and no earlier determination waswarranted Bladder voiding was encouraged to reduce bladderradiation exposure after PET scanning
For the purpose of anatomic localization the participants alsounderwent a whole-brain MRI scan on a 3T Phillips Ingeniascanner using an 8-channel head coil TheMRI protocol includeda 3-dimensional (3D) T1-weighted sequence magnetization-prepared gradient echo repetition time = 8100 ms time to echo= 37milliseconds voxel size 094times 094 times 15mm and 170 slicesPET frames were aligned for motion correction and the regionaltime-activity curves were obtained after segmentation with theautomated anatomic labelingndashmerged atlas implemented in thePNEURO module of PMOD 38 (PMOD Technologies)15
Time-activity curves were averaged from 60 to 90 minutes andnormalized over that of the whole cerebellum (standardizeduptake value ratio [SUVR]) following the methodology de-scribed by Lyoo et al26
In terms of analysis cortical (frontal cingulate occipitaltemporal parietal and insular cortices) and subcortical(thalamus caudate putamen pallidum hippocampus andamygdala) regions of interest (ROIs) were selected Corticalreconstruction and segmentation of the selected ROIs wereperformed with the recon-all pipeline from FreeSurfer v530image analysis suite27 on the 3DT1 image The PET para-metric map was then registered onto the FreeSurfer processedT1 image using ITK-SNAP28 in a semiautomatic way Aver-aged [11C]-ER176 SUVR and the associated ROI volumeswere then extracted from each ROI using FreeSurfer
In Vitro Evaluation of the Dynamic Functionsof MicrogliaThe same individuals subjected to the PETMRI scans (N =18 N = 6 individuals per group) were subjected to a blooddrawn Tenmilliliters of blood were collected by venipuncturein vacuum tubes containing heparin on the same day of thePET scan Monocytes were used to generate iMG cells aspreviously described29 Briefly peripheral blood mononuclearcells were isolated by Ficoll gradient centrifugation and platedat a density of 4 times 105 cellsmL After overnight incubationnonadherent cells were removed and the adherent cells(mainly monocytes) were cultured with granulocyte-macrophage colony-stimulating factor (10 ngmL) and in-terleukin (IL)-34 (100 ngmL) for 14 days to develop iMGcells Microglial characterization was evaluated throughmorphologic and phenotypical changes observed in thefluorescence microscopy using CX3CR1CCR2 doublestaining
Phagocytosis was examined by fluorescent microscopy andflow cytometry using the Phagocytosis Assay Kit (CaymanChemical Ann Arbor MI) At day 14 phycoerythrin (PE)-conjugated beads were added to the media (1250) for 24hours Cells were observed under the fluorescence micro-scope and then harvested by nonenzymatic cell dissociationsolution (Sigma-Aldrich St Louis MO) and cell lifter Thecells were washed and acquired using a BD FACSJazzcytometer (BD Biosciences San Jose CA) Flow cytometrydata were analyzed using FlowJo V10 software (Tree StarAshland OR)
NeurologyorgNN Neurology Neuroimmunology amp Neuroinflammation | Volume 8 Number 3 | May 2021 3
Secretion of inflammatory cytokines (tumor necrosis factor[TNF]-α IL-6 and IL-1β) during phagocytosis was mea-sured from culture supernatant using a Multiplex Immuno-assay following the manufacturer instructions (Bio-RadLaboratories Hercules CA) Briefly the samples and stan-dards were incubated with magnetic microspheres co-valently coupled with antibodies directed against the desired
biomarker (ie TNFα IL-1β and IL-6) After a series ofwashes to remove unbound protein a biotinylated detectionantibody was added to create a sandwich complex (similar tothat of a sandwich ELISA) The final detection complex wasformed with the addition of streptavidin-PE conjugate PEserved as a fluorescent indicator Data from standards andsamples and a standard curve were acquired in a Bio-Plex 200
Table Demographic and Clinical Characteristics of Premanifest HD Gene Carriers and Manifest Patients With HDIncluded in This Study
VariableControls(N = 6)
HD gene carriers
pValue
Premanifest HD(N = 6)
Manifest HD(N = 6)
Age y mean plusmn SD (median) 4328 plusmn 879(43)
3815 plusmn 718 (35) 5075 plusmn 1155(51)
0121a
Sex female n () 5 (83) 4 (67) 6 (100) 0301b
Educational level y mean plusmn SD (median) 1642 plusmn 432(175)
1583 plusmn 445 (145) 1467 plusmn 333(135)
0598a
TSPO genotype n ()
Thr147Thr147 0 0 0
Ala147Thr147 2 (33) 3 (50) 2 (33) 0792b
Ala147Ala147 4 (67) 3 (50) 4 (67)
CAP score mdash 7488 plusmn 1027 11509 plusmn 888 0002c
CAG repeats (larger) mean plusmn SD (median) mdash 4250 plusmn 187 (425) 4467 plusmn 242 (44) 0180c
UHDRS-TMS mean plusmn SD (median) mdash 367 plusmn 356 (3) 3083 plusmn 231 (31) 0002c
UHDRS-TFC mean plusmn SD (median) mdash 1250 plusmn 123 (13) 783 plusmn 184 (7) 0004c
Independence scale mdash 9750 plusmn 612 (100) 8417 plusmn 861 (85) 0015c
SDMT (total correct) mean plusmn SD (median) 5450 plusmn 784(54)
5417 plusmn 915 (575) 3067 plusmn 1019(335)
0004d
Verbal fluency test (category) no of correct responses in 1 min mean plusmn SD(median)
2383 plusmn 392(24)
2167 plusmn 403 (21) 1100 plusmn 525(105)
0008e
Stroop Interference Test no of correct responses mean plusmn SD (median) 4383 plusmn 1329(445)
4050 plusmn 599 (405) 2867 plusmn 1626(255)
0091a
MMSE mean plusmn SD (median) 2867 plusmn 186(29)
2883 plusmn 098 (285) 2660 plusmn 321 (28) 0138a
PBA-s domain scores mean plusmn SD (median)
Depression 400 plusmn 555 (2) 617 plusmn 546 (65) 717 plusmn 1096 (25) 0871a
Irritabilityaggression 133 plusmn 234 (05) 283 plusmn 325 (2) 233 plusmn 266 (2) 0845a
Psychosis 000 plusmn 000 (0) 017 plusmn 041 (0) 000 plusmn 000 (0) 0368a
Apathy 100 plusmn 089 (1) 250 plusmn 356 (05) 383 plusmn 440 (35) 0546a
Executive function 000 plusmn 000 (0) 450 plusmn 446 (4) 467 plusmn 350 (5) 0024f
Abbreviations HD = Huntington disease MMSE = Mini-Mental State Examination PBA-s = short version of the Problem Behaviors Assessment SDMT =Symbol Digit Modalities Test TFC = total functional capacity TMS = total motor score UHDRS = Unified Huntingtonrsquos Disease Rating ScaleSignificant values are highlighted in bolda Kruskal-Wallis testb Pearson χ2 testc Mann-Whitney testd Kruskal-Wallis followed by Dunn multiple comparisons test (controls = premanifest HD) lt manifest HDe Kruskal-Wallis followed by Dunn multiple comparisons test post hoc (controls = premanifest HD) gt manifest HDf Kruskal-Wallis followed by Dunn multiple comparisons test post hoc (controls = premanifest HD) gt manifest HD
4 Neurology Neuroimmunology amp Neuroinflammation | Volume 8 Number 3 | May 2021 NeurologyorgNN
Reader and analyzed in a Bio-Plex Manager Software whichpresented data as median fluorescence intensity (MFI) Thecytokine concentrations in the samples were calculatedbased on a standard curve in which the MFI values obtainedfrom the standard were plotted against the known concen-trations (in picograms per milliliter) Cytokine levels (inpicograms per milliliter) were measured in the supernatant24 hours after bead exposure or 24 hours of culture withoutany stimulus (baseline condition) and the values were cor-rected by protein levels (in milligrams per milliliter) in thecell lysates using the Pierce BCA Protein Assay Kit (Ther-moFisher Scientific Waltham MA) The final results areprovided in pgmg
DNA Polymorphism GenotypingPeripheral blood mononuclear cells were used for DNAextraction using the QIAamp DNA Blood Mini Kit
(Qiagen Hilden Germany) and subsequent genotyping forthe rs6971 polymorphism (Ala147Thr) in the TSPO gene(TaqMan assay ID C_2512465_20 Thermo Fisher Sci-entific) Allele Ala147 was linked to Vic and allele Thr147was linked to FAM PCRs were performed in QuantStudio7 Flex Real-Time PCR system (Life TechnologiesThermoFisher Scientific)
Statistical AnalysisAssociation between dichotomous variables was assessedwith the χ2 test Our limited sample size made it difficult toascertain data distribution and therefore nonparametrictests were used Comparisons between the 3 groups weremade by the Kruskal-Wallis test When appropriate posthoc analyses were used to determine significant differencesbetween pairs of groups (Dunn Multiple ComparisonTest) The Mann-Whitney test was used for 2-group
Figure 1 Microglial Activation in HD
(A) PET image showing increased [11C]-ER176 standardized uptake value ratio (SUVR) in bilateral putamina and pallidi of a patient with manifest HD incomparison with a control Note that there is no visible difference in [11C]-ER176 SUVR for the premanifest HD gene carrier (central image) (B) Groupcomparisons in left (L) and right (R) putamina and pallidi Significant differences between groups are indicated by different letters (Kruskal-Wallis testfollowed by Dunn multiple comparisons test) Horizontal bars show the mean and the standard error of the mean Data from other regions of interest areshown in figure e-1 (linkslwwcomNXIA459) HD = Huntington disease
NeurologyorgNN Neurology Neuroimmunology amp Neuroinflammation | Volume 8 Number 3 | May 2021 5
comparisons (premanifest HD vs manifest HD) Spearmancorrelations were performed to examine the relationshipbetween [11]C-ER176 SUVR in the ROIs significantlydifferent between groups and (1) CAP score (2) volume inthe same ROIs and (3) scores in the clinical scales Allstatistical tests were 2 tailed and were performed using asignificance level of α = 005 Statistical analyses wereperformed using SPSS software version 260 and GraphPadPrism version 50
Standard Protocol Approvals Registrationsand Patient ConsentsAll subjects provided written informed consent before ad-mission to the study The Research Ethics Committees ofUTHealth and Houston Methodist Research Institute ap-proved this study
An Investigational New Drug Application (IND) was sub-mitted and authorization was obtained from the Food andDrug Administration to use [11C]-ER176 as a PET Imagingagent in this study (IND 139220)
Data AvailabilityThe raw data not provided in the article will be made availableby the authors at the request of other investigators for pur-poses of replicating procedures and results
ResultsClinical Evaluation and TSPO GenotypingDemographic and clinical characteristics of participants included inthis study are shown in the table There was no significant differ-ence between controls premanifest HD gene carriers and patientswith HD regarding age sex and educational level As expectedpatients with manifest HD presented a higher CAP score andworse scores in the scales that evaluatedmotor symptoms severityfunctional capacity and independence in comparison with pre-manifest HD (table) The age at clinical diagnosis was 4767 plusmn1035 years for the patients with manifest HD Premanifest HDgene carriers had amean of 1298 plusmn 617 years before the predictedclinical diagnosis of HD The 3 groups presented similar perfor-mance on the MMSE Patients with the clinical diagnosis of HDpresented worse performance on the SDMT and verbal fluencytest in comparisonwith premanifest and control subjects Controlsindividuals with premanifest HD and patients with HD presentedsimilar scores in the PBA-s subscales except for executive functionin which HD gene carriers (both premanifest and manifest HD)scored worse than controls evidencing that those problems arepresent before clinical diagnosis of HD (table)
The groups presented similar distribution of TSPO genotype(table) Individuals with genotype Ala147Ala147 were clas-sified as high-affinity binders and those with genotype
Figure 2 Microglial Activation in HD Is Associated With Disease Burden
Among HD gene carriers (premanifest andmanifest HD) a higher CAP score (a proxy for cumulative disease burden) was significantly associated with higher[11C]-ER176 standardized uptake value ratio (SUVR) in the (A) left putamen (ρ = 0664 p = 0018) (B) left pallidum (ρ = 0657 p = 0020) (C) right putamen (ρ =0671 p = 0017) and (D) right pallidum (ρ = 0734 p = 0007) Note the 2 clusters in [11C]-ER176 SUVR composed of premanifest HD gene carriers (CAP lt100)and manifest patients with HD (CAP gt100) CAP = CAG age product HD = Huntingtonrsquos disease
6 Neurology Neuroimmunology amp Neuroinflammation | Volume 8 Number 3 | May 2021 NeurologyorgNN
Ala147Thr147 as mixed affinity None of our patients pre-sented the genotype Thr147Thr147 (low-affinity binders)Because the groups presented comparable distribution ofTSPO genotype (table) and the outcome parameter wasSUVR which minimizes genotype differences no correctionfor genotype-related binding affinity was performed
In Vivo Assessment of Microglial ActivationPatients with HD demonstrate higher [11C]-ER176 SUVR inboth the left and right globi pallidi and putamina in comparisonwith controls These differences were not significant when wecompared premanifest HD gene carriers with patients withHDorcontrols (figure 1 and figure e-1 linkslwwcomNXIA459)These results indicate increased microglial activation in the puta-mina and globi pallidi of patients with manifest HDWe found nodifferences between different TSPO genotypes on [11C]-ER176SUVR when controlling for HD diagnosis (data not shown)
Among HD gene carriers (premanifest and manifest HD) ahigher CAP score was significantly associated with higher[11C]-ER176 SUVR in the left putamen (ρ = 0664 p =0018) left pallidum (ρ = 0657 p = 0020) right putamen(ρ = 0671 p = 0017) and right pallidum (ρ = 0734 p =0007) (figure 2) Corroborating these findings increased[11C]-ER176 SUVRwas associated with decreased volume inthe same ROIs (figure 3)
In Vitro Evaluation of the Dynamic Functionsof MicrogliaWe confirmed that the iMG cells have the immunophenotypeof microglial cells such as CX3CR1highCCR2low and atypical (ie ramified) morphology (figure 4) The iMG cellsexpress dynamic functions such as phagocytosis and secretionof inflammatory cytokines making them a valuable tool toassess microglial functioning30 The phagocytosis assaydemonstrated that iMG cells obtained from controls pre-manifest HD and HD patients (figure 5 AndashC) showedsimilar phagocytic capacity (figure 5E) Secretion of in-flammatory cytokines (TNFα IL-6 and IL-1β) duringphagocytosis was measured from culture supernatant Asshown in figure 5F although iMG cells from controls pre-manifest HD and patients with HD increased the productionof the evaluated cytokines during phagocytosis the valueswere significant only for TNFα in controls and manifest HD(figure 5F)
DiscussionBy using [11C]-ER176 PET we verified that patients withHD have increased microglial activation in basal gangliastructures that are involved in HD pathophysiologyie putamina and globi pallidi In addition we found that
Figure 3 Microglial Activation Is Associated With Brain Atrophy
Increased microglial activation is associated with decreased volume in putamen and pallidus Controls are displayed in red dots premanifest HD genecarriers in green dots andmanifest HD in black dots A higher [11C]-ER176 standardized uptake value ratio (SUVR) was associatedwith a decreased volume inthe (A) left putamen (ρ = minus0569 p = 0014) (B) left pallidum (ρ = minus0591 p = 0010) (C) right putamen (ρ = minus0601 p = 0008) and (D) right pallidum (ρ = minus0686 p= 0002) ICV = intracranial volume The dashed line was placed to emphasize the 2 clusters composed of controls + premanifest HD and manifest HD genecarriers HD = Huntington disease
NeurologyorgNN Neurology Neuroimmunology amp Neuroinflammation | Volume 8 Number 3 | May 2021 7
premanifest HD gene carriers demonstrate [11C]-ER176SUVRs comparable to controls suggesting that microglialactivation is a later process in HD pathophysiology becomingevident only after the onset of motor symptoms (figure 1)Supporting these findings we reported a significant correla-tion between increased microglial activation and higher CAPscores a proxy for cumulative disease burden (figure 2)Moreover increased microglial activation in putamina andpallidi was significantly associated with reduced volumes inthe same ROIs (figure 3) Individuals with manifest HDpresented a significant decrease in the cortical volume incomparison with controls (data not shown) Despite alreadypresenting cortical atrophy manifest HD individuals do nothave increased microglial activation in the cortex (figure 1)The only areas displaying increased microglia activation arethose that are known to be affected early in HD pathophysi-ology Because PET scans require individuals to be still for 90minutes our sample did not include patients in moderatelatestages of HD Whether increased microglia activation willoccur in the cortex later in the HD course is yet to be de-termined Altogether our data demonstrate that microglialactivation is involved in HD pathophysiology and is associ-ated with disease progression (as demonstrated by the CAPscore and atrophy in basal ganglia structures) Our findingssupport the conclusions of previous research that reportedthat microglial activation correlates with disease severitycontributing to the ongoing neuronal degeneration in HD16
HD has been regarded as an important model to study neu-rodegenerative diseases as it is caused by a single geneticmutation and is amenable to predictive genetic testing en-abling the study of patients before clinical diagnosis2 In this
study we assessed microglial activation not only in patientswith HD but also in premanifest HD gene carriers with amean of 13 years before the predicted onset of clinicalsymptoms Our results corroborate the view that microglialactivation is unlikely to be the initiating event in HD but maycontribute to neuronal death through multiple pathways31
Similar to other neurodegenerative diseases the mecha-nisms underlying selective neuronal dysfunction and deathin HD are still uncertain In addition to impairment in sys-tems for handling abnormal proteins other metabolicpathways and mechanisms might contribute to neuro-degeneration and progression of HD such as mitochondrialdysfunction impaired neurotrophic support oxidativestress and inflammatoryimmune mechanisms83233 In thecurrent study we described a significant increase in micro-glial activation in regions of neurodegeneration in HDsuggesting that this process is related to the disease patho-physiology and not to random premorbid events Accord-ingly postmortem studies reported significant microglialactivation in regions implicated in HD pathogenesis46 Thefact that we found significant microglial activation in patientswith manifest HD (but not in premanifest HD gene carriers)in comparison with controls suggests that this event occursin response or concomitantly to neuronal death This as-sumption is further corroborated by the finding of significantassociations between microglial activation and ROI volumesas well as CAP scores Here again our data are corroboratedby postmortem findings in which the number of activatedmicroglia in the striatum and cortex correlated with neuronalloss confirming the hypothesis that neuroinflammation maybe elicited by degenerating neurons6
Figure 4 Inducing Microglia-Like Cells From Human Peripheral Blood Monocytes
Peripheral blood monocytes were cultured for 14 days in conventional conditions (A) or with GM-CSF and IL-34 resulting in induced microglia like cells thatexhibit microglia-like morphology and increased CX3CR1CCR2 ratio (B)
8 Neurology Neuroimmunology amp Neuroinflammation | Volume 8 Number 3 | May 2021 NeurologyorgNN
Neuroinflammation elicited by neuronal death can be at firstregarded as a positive event as it promotes clearance of celldebris However inflammatory mechanisms contribute toneurodegeneration and dying neurons further activate in-flammatory responses resulting in a vicious cycle of in-flammation and neuronal death34 Therefore inflammatoryresponses although essential for tissue homeostasis cancontribute to neuronal injury As neural tissues have a
restricted cell renewal and regenerative capacity the CNS isextremely vulnerable to uncontrolled andor chronic immuneand inflammatory processes34 Interestingly enough our datacorroborate this hypothesis as we observed increased micro-glial activation later in the HD course (ie manifest HD)reinforcing the idea that neuroinflammation contributes todisease progression rather than to disease onset Activatedmicroglia releases proinflammatory mediators including
Figure 5 Phagocytosis by Induced Microglia-Like (iMG) Cells
Peripheral blood monocytes (day 1 panel A) were cultured for 14 days with GM-CSF and IL-34 resulting in iMG cells (day 14 panel B) Phagocytosis wasassessed by exposing iMG cells to PE-conjugated (1250) beads (panel C) for 24 hours The iMG cells showed the ability of phagocytosis with morphologicalchanges (C) The read arrows show PE-conjugated beads phagocytosed by iMG cells After washing beads out cells were acquired in a flow cytometer andanalyzed iMG were first gated by forwardside scatter (not shown) PE uptake was analyzed as shown in (D) orange (left) histogram shows the cells withoutbeads (negative control) red (right) histogram shows the cells that phagocytosed to the PE-conjugated beads Quantification is shown in (E) iMG fromcontrols premanifest HD and HD patients showed similar phagocytic capacity (no significant difference found at the Kruskal-Wallis test) (F) Cytokineproduction during phagocytosis iMG cells were incubated with PE-conjugated beads (1250) for 24 hours Supernatant was collected and cytokines wereanalyzed by multiparametric assay Cytokine levels (in picograms per milliliter) were corrected by protein levels (in milligrams per milliliter) Although iMGcells from controls premanifest HD and patients with HD increased the production of the evaluated cytokines during phagocytosis the values weresignificant only for TNFα in controls andmanifest HD (p values are provided in the figure Wilcoxonmatched-pairs signed-rank test) Scale bar (AndashC) = 10mmHorizontal bars (E and F) show the mean and the standard error of the mean HD = Huntington disease PE = phycoerythrin TNF = tumor necrosis factor
NeurologyorgNN Neurology Neuroimmunology amp Neuroinflammation | Volume 8 Number 3 | May 2021 9
cytokines and reactive oxygen species which in turn will alsocontribute to neuronal death and further microglial activa-tion34We observed an increase in cytokine release (especiallyTNFα) following a phagocytic stimulus in our in vitro ex-periments In line with our findings postmortem studies havealso reported increased levels of inflammatory cytokines inbrain samples of patients with HD in comparison withcontrols3536 It is worth mentioning that cytokine release is amicroglial reaction to a proinflammatory challenge and undercontrolled conditions (such as the iMG cultures) phagocy-tosis is a noninflammatory function of microglia A proin-flammatory stimulus would be needed to appropriately assessdifferences in cytokine release between controls premanifestHD and manifest HD iMG cultures
The current study used a novel PET tracer for TSPO [11C]-ER176 to assess microglial activation in HD By using [11C]-PK11195 PET studies have shown a significant increase inmicroglial activation in the striatum of patients with HD16 aswell as in premanifest HD gene carriers18 compared withcontrols In both studies [11C]-PK11195 binding significantlycorrelated with disease severity as measured by decreased[11C]-raclopride (a marker of dopamine D2 and D3 receptoravailability) binding in the striatum1618 Similar results weredescribed in the hypothalamus17 Later the same work groupused a multimodal imaging approach combiningMRI and PETanalyses and worsening in atrophy evaluated by MRI was ac-companied by a reduction in [11C]-raclopride and an increasein [11C]-PK11195 bindings in patients with HD In pre-manifest HD increased level of microglial activation in theassociative striatum and in the regional network associated withcognition correlated with 5-year probability of HD onset19
More recently increased microglial activation in the somato-sensory cortex (as evaluated through [11C]-PK11195 binding)was associatedwith higher levels of IL-1β IL-6 IL-8 and TNFαproduced by stimulated peripheral blood monocytes frompremanifest HD gene carriers2037 Although these studiesprovided valuable information about neuroinflammation inHD indicating microglial activation even in premanifest sub-jects their results should be interpreted with caution The poorsignal-to-background ratio of first-generation TSPO radio-tracers poses important limitations to the analyses The ratio ofspecific to nonspecific binding of [11C]-PK11195 in humanbrain has been reported to be as low as 023839 As an attempt tomitigate this problem a recent study used the second-generation TSPO tracer [11C]-PBR28 to evaluate micro-glial activation in HD21 Of interest the results were similar toours ie increased levels of [11C]-PBR28 binding in theputamina and pallidi of HD gene carriers (which included 7manifest patients with HD and only 1 person with premanifestHD) in comparison with controls Of note the results of thesingle premanifest subject were midway between controls andthe remaining patients suggesting that microglial activationwas associated with disease progression21 The authors didconfirm that [11C]-PBR28 provides a high signal-to-background ratio an important feature required for the anal-ysis at the individual level and within individual brain regions
An important limitation of [11C]-PBR28 and other second-generation TSPO radioligands is that they have varying degreesof sensitivity to the single nucleotide polymorphism rs6971 inthe TSPO gene40 [11C]-ER176 has unique advantages overthe other TSPO binders being the only known TSPO radio-ligand that allows the inclusion of low-affinity binders Thebinding of ER176 in low-affinity binders was found to becomparable to that for PBR28 in high-affinity binders More-over ER176 was proven to be the only radioligand not con-taminated by radiometabolites accumulating in the brain14
Our findings must be interpreted while taking into accountthe limitations of the study which include the relatively smallsample size and the cross-sectional design The cross-sectionaldesign prevented the assessment of microglial activation inthe context of disease progression and development of motorand nonmotor symptoms In addition the use of an objectivemethod to measure striatal degeneration (eg [11C]-raclopride PET) would provide more accurate informationabout striatal neuron dysfunctiondeath Regarding the invitro experiments the use of a proinflammatory challenge(other than the beads added for the phagocytosis assay) in ourexperiments would provide valuable information on cytokinesrelease by iMG from the different groups It is worth men-tioning the strengths of the study that include the use of aunique TSPO ligand [11C]-ER176 the enrollment of bothpremanifest and manifest subjects and the exploratory in-vestigation of microglia activity
In conclusion our findings strengthen the evidence thatmicroglial activation is associated with HD pathophysiologyMicroglial activation is unlikely to be the initiating event inHD Yet it may exacerbate neuronal dysfunction and neu-ronal death thus contributing to disease progression It isimportant to refine our understanding of the more specificimmuneinflammatory mechanisms that are involved in HDand in neurodegeneration This may foster the developmentof new therapeutic interventions to halt the progression ofHD and other neurodegenerative diseases
AcknowledgmentThe authors acknowledge and thank all of the volunteers thatparticipated in this study and are indebted to their families fortheir magnificent support They also thank the staff from theUTHealth McGovern Medical School 3T MRI Center andfrom the HoustonMethodist Research Institute TranslationalImaging Core Facilities especially Vips Patel and Kim MDoan for their skilled technical support Dr Gabriel Fries forhis assistance with the genotyping analyses and the teammembers of the HDSA Center of Excellence at UTHealth
Study FundingThis study was funded by the HD Human BiologyProjectmdashHuntingtonrsquos Disease Society of America (HDSA)E Furr Stimming receives research funding from RocheGenetech Vaccinex Cures Within Reach HDSA Uniqureand CHDI Foundation
10 Neurology Neuroimmunology amp Neuroinflammation | Volume 8 Number 3 | May 2021 NeurologyorgNN
DisclosureThe authors report no disclosures relevant to the manuscriptGo to NeurologyorgNN for full disclosures
Publication HistoryReceived by Neurology Neuroimmunology amp NeuroinflammationSeptember 9 2020 Accepted in final form January 20 2021
References1 The Huntingtonrsquos Disease Collaborative Research Group A novel gene containing a
trinucleotide repeat that is expanded and unstable on Huntingtonrsquos disease chro-mosomes Cell 199372971ndash983
2 Ross CA Aylward EH Wild EJ et al Huntington disease natural history biomarkersand prospects for therapeutics Nat Rev Neurol 201410204ndash216
3 Soulet D Cicchetti F The role of immunity in Huntingtonrsquos disease Mol Psychiatry201116889ndash902
4 Myers RH Vonsattel JP Paskevich PA et al Decreased neuronal and increasedoligodendroglial densities in Huntingtonrsquos disease caudate nucleus J NeuropatholExp Neurol 199150729ndash742
5 Vonsattel JP Myers RH Stevens TJ Ferrante RJ Bird ED Richardson EP JrNeuropathological classification of Huntingtonrsquos disease J Neuropathol Exp Neurol198544559ndash577
6 Sapp E Kegel KB Aronin N et al Early and progressive accumulation of reactivemicroglia in the Huntington disease brain J Neuropathol Exp Neurol 200160161ndash172
7 Ciccocioppo F Bologna G Ercolino E et al Neurodegenerative diseases asproteinopathies-driven immune disorders Neural Regen Res 202015850ndash856
8 Rocha NP Ribeiro FM Furr-Stimming E Teixeira AL Neuroimmunology of Hun-tingtonrsquos disease revisiting evidence from human studies Mediators Inflamm 201620168653132
9 Vowinckel E Reutens D Becher B et al PK11195 binding to the peripheral ben-zodiazepine receptor as a marker of microglia activation in multiple sclerosis andexperimental autoimmune encephalomyelitis J Neurosci Res 199750345ndash353
10 Papadopoulos V Baraldi M Guilarte TR et al Translocator protein (18kDa) newnomenclature for the peripheral-type benzodiazepine receptor based on its structureand molecular function Trends Pharmacol Sci 200627402ndash409
11 Cumming P Burgher B Patkar O et al Sifting through the surfeit of neuro-inflammation tracers J Cereb Blood Flow Metab 201838204ndash224
12 Zanotti-Fregonara P Zhang Y Jenko KJ et al Synthesis and evaluation of translocator18 kDa protein (TSPO) positron emission tomography (PET) radioligands with lowbinding sensitivity to human single nucleotide polymorphism rs6971 ACS ChemNeurosci 20145963ndash971
13 Ikawa M Lohith TG Shrestha S et al 11C-ER176 a radioligand for 18-kDa trans-locator protein has adequate sensitivity to robustly image all three affinity genotypesin human brain J Nucl Med 201758320ndash325
14 Fujita M Kobayashi M Ikawa M et al Comparison of four (11)C-labeled PETligands to quantify translocator protein 18 kDa (TSPO) in human brain (R)-PK11195 PBR28 DPA-713 and ER176-based on recent publications that measuredspecific-to-non-displaceable ratios EJNMMI Res 2017784
15 Zanotti-Fregonara P Pascual B Veronese M et al Head-to-head comparison of (11)C-PBR28 and (11)C-ER176 for quantification of the translocator protein in thehuman brain Eur J Nucl Med Mol Imaging 2019461822ndash1829
16 Pavese N Gerhard A Tai YF et al Microglial activation correlates with severity inHuntington disease a clinical and PET study Neurology 2006661638ndash1643
17 Politis M Pavese N Tai YF Tabrizi SJ Barker RA Piccini P Hypothalamic in-volvement in Huntingtonrsquos disease an in vivo PET study Brain 20081312860ndash2869
18 Tai YF Pavese N Gerhard A et al Microglial activation in presymptomatic Hun-tingtonrsquos disease gene carriers Brain 20071301759ndash1766
19 Politis M Pavese N Tai YF et al Microglial activation in regions related to cognitivefunction predicts disease onset in Huntingtonrsquos disease a multimodal imaging studyHum Brain Mapp 201132258ndash270
20 Politis M Lahiri N Niccolini F et al Increased central microglial activation associatedwith peripheral cytokine levels in premanifest Huntingtonrsquos disease gene carriersNeurobiol Dis 201583115ndash121
21 Lois C Gonzalez I Izquierdo-Garcia D et al Neuroinflammation in Huntingtonrsquosdisease new insights with (11)C-PBR28 PETMRI ACS Chem Neurosci 201892563ndash2571
22 Huntington Study Group Unified Huntingtonrsquos Disease Rating Scale reliability andconsistency Mov Disord 199611136ndash142
23 Folstein MF Folstein SE McHugh PR ldquoMini-mental staterdquo A practical method forgrading the cognitive state of patients for the clinician J Psychiatr Res 197512189ndash198
24 McNally G Rickards H Horton M Craufurd D Exploring the validity of the shortversion of the Problem Behaviours Assessment (PBA-s) for Huntingtonrsquos disease arasch analysis J Huntingtons Dis 20154347ndash369
25 Warner JH Sampaio C Modeling variability in the progression of Huntingtonrsquosdisease a novel modeling approach applied to structural imaging markers fromTRACK-HD CPT Pharmacometrics Syst Pharmacol 20165437ndash445
26 Lyoo CH Ikawa M Liow JS et al Cerebellum can serve as a pseudo-reference regionin Alzheimer disease to detect neuroinflammation measured with PET radioligandbinding to translocator protein J Nucl Med 201556701ndash706
27 Reuter M Schmansky NJ Rosas HD Fischl B Within-subject template estimation forunbiased longitudinal image analysis Neuroimage 2012611402ndash1418
28 Yushkevich PA Piven J Hazlett HC et al User-guided 3D active contour segmen-tation of anatomical structures significantly improved efficiency and reliabilityNeuroimage 2006311116ndash1128
29 Ohgidani M Kato TA Setoyama D et al Direct induction of ramified microglia-likecells from human monocytes dynamic microglial dysfunction in Nasu-Hakola dis-ease Sci Rep 201444957
30 Ohgidani M Kato TA Kanba S Introducing directly induced microglia-like (iMG)cells from fresh human monocytes a novel translational research tool for psychiatricdisorders Front Cell Neurosci 20159184
31 Kim SU de Vellis J Microglia in health and disease J Neurosci Res 200581302-31332 Ross CA Tabrizi SJ Huntingtonrsquos disease from molecular pathogenesis to clinical
treatment Lancet Neurol 20111083ndash9833 Chitnis TWeiner HL CNS inflammation and neurodegeneration J Clin Invest 2017
1273577ndash3587
Appendix Authors
Name Location Contribution
Natalia PRocha PhD
The Mitchell Center forAlzheimerrsquos Disease andRelated Brain DisordersDepartment of NeurologyMcGovern Medical SchoolThe University of TexasHealth Science CenterHouston
Designed andconceptualized the studymajor role in theacquisition of dataanalyzed the data anddrafted the manuscript forintellectual content
OdelinCharron MS
Department of NeurologyThe University of Texas atAustin
Major role in data analysis
Leigh BLathamPhD
School of MedicineUniversity of WashingtonSeattle
Major role in theacquisition of data
Gabriela DColpo PhD
Neuropsychiatry ProgramDepartment of Psychiatryand Behavioral SciencesMcGovern Medical SchoolUniversity of TexasHouston
Major role in theacquisition of data
PaoloZanotti-FregonaraMD PhD
Houston MethodistResearch Institute andWeill Cornell Medicine TX
Major role in the studydesign and the acquisitionof data
MeixiangYu PhD
Houston MethodistResearch Institute andWeill Cornell Medicine TX
Major role in theacquisition of data
LeorahFreemanMD PhD
Department of NeurologyThe University of Texas atAustin
Major role in the studydesign data analysis andinterpretation and theacquisition of data
Erin FurrStimmingMD
HDSA Center of Excellenceat University of TexasHealth Science Center atHouston
Major role in the studydesign and acquisition ofdata interpreted the dataand revised themanuscript for intellectualcontent
Antonio LTeixeiraMD PhD
Neuropsychiatry ProgramDepartment of Psychiatryand Behavioral SciencesMcGovern Medical SchoolUniversity of TexasHouston
Major role in the studydesign and acquisition ofdata interpreted the dataand revised themanuscript for intellectualcontent
NeurologyorgNN Neurology Neuroimmunology amp Neuroinflammation | Volume 8 Number 3 | May 2021 11
34 Rocha NP de Miranda AS Teixeira AL Insights into neuroinflammation in Parkin-sonrsquos disease from biomarkers to anti-inflammatory based therapies Biomed Res Int20152015628192
35 Silvestroni A Faull RL Strand AD Moller T Distinct neuroinflammatory profile inpost-mortem human Huntingtonrsquos disease Neuroreport 2009201098ndash1103
36 Bjorkqvist M Wild EJ Thiele J et al A novel pathogenic pathway of immune acti-vation detectable before clinical onset in Huntingtonrsquos disease J Exp Med 20082051869ndash1877
37 Politis M Lahiri N Niccolini F et al Corrigendum to ldquoIncreased central microglialactivation associated with peripheral cytokine levels in premanifest Huntingtonrsquos
disease gene carriersrdquo [Neurobiol Dis 83 (2015) 115-121] Neurobiol Dis 201798162
38 Kropholler MA Boellaard R Schuitemaker A Folkersma H van Berckel BN Lam-mertsma AA Evaluation of reference tissue models for the analysis of [11C](R)-PK11195 studies J Cereb Blood Flow Metab 2006261431ndash1441
39 Turkheimer FE Edison P Pavese N et al Reference and target region modeling of[11C]-(R)-PK11195 brain studies J Nucl Med 200748158ndash167
40 Kreisl WC Jenko KJ Hines CS et al A genetic polymorphism for translocator protein18 kDa affects both in vitro and in vivo radioligand binding in human brain to thisputative biomarker of neuroinflammation J Cereb Blood FlowMetab 20133353ndash58
12 Neurology Neuroimmunology amp Neuroinflammation | Volume 8 Number 3 | May 2021 NeurologyorgNN
DOI 101212NXI000000000000098420218 Neurol Neuroimmunol Neuroinflamm
Natalia P Rocha Odelin Charron Leigh B Latham et al Pathophysiology
Microglia Activation in Basal Ganglia Is a Late Event in Huntington Disease
This information is current as of April 1 2021
ServicesUpdated Information amp
httpnnneurologyorgcontent83e984fullhtmlincluding high resolution figures can be found at
References httpnnneurologyorgcontent83e984fullhtmlref-list-1
This article cites 40 articles 4 of which you can access for free at
Subspecialty Collections
httpnnneurologyorgcgicollectionpetPET
httpnnneurologyorgcgicollectionhuntingtons_diseaseHuntingtons diseasefollowing collection(s) This article along with others on similar topics appears in the
Permissions amp Licensing
httpnnneurologyorgmiscaboutxhtmlpermissionsits entirety can be found online atInformation about reproducing this article in parts (figurestables) or in
Reprints
httpnnneurologyorgmiscaddirxhtmlreprintsusInformation about ordering reprints can be found online
Academy of Neurology All rights reserved Online ISSN 2332-7812Copyright copy 2021 The Author(s) Published by Wolters Kluwer Health Inc on behalf of the AmericanPublished since April 2014 it is an open-access online-only continuous publication journal Copyright
is an official journal of the American Academy of NeurologyNeurol Neuroimmunol Neuroinflamm

expansion was confirmed by a genotype larger CAG allelege36 A movement disorder specialist evaluated all HD genecarriers and the clinical diagnosis of HD was based on themotor signs certainty ie a Diagnostic Confidence Level of 4in the Unified HD Rating Scale (UHDRS)222 Patients wererecruited from the Huntington Disease Society of AmericaCenter of Excellence at UTHealth Controls were recruitedfrom the local community comprising a group of people withno history of neurologic or psychiatric disorder We excludedindividuals who have had infectious or autoimmune diseasesin activity or who have used anti-inflammatories in the 4weeks before the study pregnant or breastfeeding womenand participants with claustrophobia metal implants in-compatible with MRI previous neurosurgery or current se-rious comorbidity (eg cancer) Participantsrsquo recruitment andstudy procedures were performed from July 2018 to January2019
HD gene carriers completed the UHDRS22 including thetotal motor scale the total functional capacity and in-dependence scales The cognitive evaluation included theMini-Mental State Examination (MMSE)23 the SymbolDigit Modalities Test (SDMT) the Stroop InterferenceTest and the Verbal fluency test22 Behavioral symptomswere assessed with the short version of the Problem Be-haviors Assessment (PBA-s)24
The CAG age product (CAP) score was used to estimate theprogression of HD pathology as a function of CAG repeatlength and time of exposure to the effects of the expansion2
The CAP has been defined as
CAP = AGE times (CAG minus L)K
where AGE is the current age of the individual CAG is therepeat length and L and K are constants L is an estimate ofthe lower limit of the CAG expansion at which phenotypicexpression of the effects of mutant HTT could be observedand K is a normalizing constant We used the CAP proposedby Warner and Sampaio25 in which L = 30 and K = 627 andthe CAP will be equal to 100 at the subjectrsquos expected age ofmotor symptoms onset
Assessment of Microglial Activation In VivoUsing PETParticipants received 1 [11C]-ER176 PET scan on a GE Dis-covery RX PETCT scanner Subjects were scanned at rest AnIV line was placed in the antecubital fossa of the arm Subjectswere placed supine on the camera table with their head firmlysecured using a thermoplastic facemask Attenuation correctionCT was performed before PET acquisition Vital signs wereobtained before administration of [11C]-ER176 and at the endof the scan All subjects received a single IV bolus of about 20mCi of [11C]-ER176 in 10 mL (injected mass dose lt10 μg)and PET data were acquired for 90 minutes Subjects wereobserved during scanning and no earlier determination waswarranted Bladder voiding was encouraged to reduce bladderradiation exposure after PET scanning
For the purpose of anatomic localization the participants alsounderwent a whole-brain MRI scan on a 3T Phillips Ingeniascanner using an 8-channel head coil TheMRI protocol includeda 3-dimensional (3D) T1-weighted sequence magnetization-prepared gradient echo repetition time = 8100 ms time to echo= 37milliseconds voxel size 094times 094 times 15mm and 170 slicesPET frames were aligned for motion correction and the regionaltime-activity curves were obtained after segmentation with theautomated anatomic labelingndashmerged atlas implemented in thePNEURO module of PMOD 38 (PMOD Technologies)15
Time-activity curves were averaged from 60 to 90 minutes andnormalized over that of the whole cerebellum (standardizeduptake value ratio [SUVR]) following the methodology de-scribed by Lyoo et al26
In terms of analysis cortical (frontal cingulate occipitaltemporal parietal and insular cortices) and subcortical(thalamus caudate putamen pallidum hippocampus andamygdala) regions of interest (ROIs) were selected Corticalreconstruction and segmentation of the selected ROIs wereperformed with the recon-all pipeline from FreeSurfer v530image analysis suite27 on the 3DT1 image The PET para-metric map was then registered onto the FreeSurfer processedT1 image using ITK-SNAP28 in a semiautomatic way Aver-aged [11C]-ER176 SUVR and the associated ROI volumeswere then extracted from each ROI using FreeSurfer
In Vitro Evaluation of the Dynamic Functionsof MicrogliaThe same individuals subjected to the PETMRI scans (N =18 N = 6 individuals per group) were subjected to a blooddrawn Tenmilliliters of blood were collected by venipuncturein vacuum tubes containing heparin on the same day of thePET scan Monocytes were used to generate iMG cells aspreviously described29 Briefly peripheral blood mononuclearcells were isolated by Ficoll gradient centrifugation and platedat a density of 4 times 105 cellsmL After overnight incubationnonadherent cells were removed and the adherent cells(mainly monocytes) were cultured with granulocyte-macrophage colony-stimulating factor (10 ngmL) and in-terleukin (IL)-34 (100 ngmL) for 14 days to develop iMGcells Microglial characterization was evaluated throughmorphologic and phenotypical changes observed in thefluorescence microscopy using CX3CR1CCR2 doublestaining
Phagocytosis was examined by fluorescent microscopy andflow cytometry using the Phagocytosis Assay Kit (CaymanChemical Ann Arbor MI) At day 14 phycoerythrin (PE)-conjugated beads were added to the media (1250) for 24hours Cells were observed under the fluorescence micro-scope and then harvested by nonenzymatic cell dissociationsolution (Sigma-Aldrich St Louis MO) and cell lifter Thecells were washed and acquired using a BD FACSJazzcytometer (BD Biosciences San Jose CA) Flow cytometrydata were analyzed using FlowJo V10 software (Tree StarAshland OR)
NeurologyorgNN Neurology Neuroimmunology amp Neuroinflammation | Volume 8 Number 3 | May 2021 3
Secretion of inflammatory cytokines (tumor necrosis factor[TNF]-α IL-6 and IL-1β) during phagocytosis was mea-sured from culture supernatant using a Multiplex Immuno-assay following the manufacturer instructions (Bio-RadLaboratories Hercules CA) Briefly the samples and stan-dards were incubated with magnetic microspheres co-valently coupled with antibodies directed against the desired
biomarker (ie TNFα IL-1β and IL-6) After a series ofwashes to remove unbound protein a biotinylated detectionantibody was added to create a sandwich complex (similar tothat of a sandwich ELISA) The final detection complex wasformed with the addition of streptavidin-PE conjugate PEserved as a fluorescent indicator Data from standards andsamples and a standard curve were acquired in a Bio-Plex 200
Table Demographic and Clinical Characteristics of Premanifest HD Gene Carriers and Manifest Patients With HDIncluded in This Study
VariableControls(N = 6)
HD gene carriers
pValue
Premanifest HD(N = 6)
Manifest HD(N = 6)
Age y mean plusmn SD (median) 4328 plusmn 879(43)
3815 plusmn 718 (35) 5075 plusmn 1155(51)
0121a
Sex female n () 5 (83) 4 (67) 6 (100) 0301b
Educational level y mean plusmn SD (median) 1642 plusmn 432(175)
1583 plusmn 445 (145) 1467 plusmn 333(135)
0598a
TSPO genotype n ()
Thr147Thr147 0 0 0
Ala147Thr147 2 (33) 3 (50) 2 (33) 0792b
Ala147Ala147 4 (67) 3 (50) 4 (67)
CAP score mdash 7488 plusmn 1027 11509 plusmn 888 0002c
CAG repeats (larger) mean plusmn SD (median) mdash 4250 plusmn 187 (425) 4467 plusmn 242 (44) 0180c
UHDRS-TMS mean plusmn SD (median) mdash 367 plusmn 356 (3) 3083 plusmn 231 (31) 0002c
UHDRS-TFC mean plusmn SD (median) mdash 1250 plusmn 123 (13) 783 plusmn 184 (7) 0004c
Independence scale mdash 9750 plusmn 612 (100) 8417 plusmn 861 (85) 0015c
SDMT (total correct) mean plusmn SD (median) 5450 plusmn 784(54)
5417 plusmn 915 (575) 3067 plusmn 1019(335)
0004d
Verbal fluency test (category) no of correct responses in 1 min mean plusmn SD(median)
2383 plusmn 392(24)
2167 plusmn 403 (21) 1100 plusmn 525(105)
0008e
Stroop Interference Test no of correct responses mean plusmn SD (median) 4383 plusmn 1329(445)
4050 plusmn 599 (405) 2867 plusmn 1626(255)
0091a
MMSE mean plusmn SD (median) 2867 plusmn 186(29)
2883 plusmn 098 (285) 2660 plusmn 321 (28) 0138a
PBA-s domain scores mean plusmn SD (median)
Depression 400 plusmn 555 (2) 617 plusmn 546 (65) 717 plusmn 1096 (25) 0871a
Irritabilityaggression 133 plusmn 234 (05) 283 plusmn 325 (2) 233 plusmn 266 (2) 0845a
Psychosis 000 plusmn 000 (0) 017 plusmn 041 (0) 000 plusmn 000 (0) 0368a
Apathy 100 plusmn 089 (1) 250 plusmn 356 (05) 383 plusmn 440 (35) 0546a
Executive function 000 plusmn 000 (0) 450 plusmn 446 (4) 467 plusmn 350 (5) 0024f
Abbreviations HD = Huntington disease MMSE = Mini-Mental State Examination PBA-s = short version of the Problem Behaviors Assessment SDMT =Symbol Digit Modalities Test TFC = total functional capacity TMS = total motor score UHDRS = Unified Huntingtonrsquos Disease Rating ScaleSignificant values are highlighted in bolda Kruskal-Wallis testb Pearson χ2 testc Mann-Whitney testd Kruskal-Wallis followed by Dunn multiple comparisons test (controls = premanifest HD) lt manifest HDe Kruskal-Wallis followed by Dunn multiple comparisons test post hoc (controls = premanifest HD) gt manifest HDf Kruskal-Wallis followed by Dunn multiple comparisons test post hoc (controls = premanifest HD) gt manifest HD
4 Neurology Neuroimmunology amp Neuroinflammation | Volume 8 Number 3 | May 2021 NeurologyorgNN
Reader and analyzed in a Bio-Plex Manager Software whichpresented data as median fluorescence intensity (MFI) Thecytokine concentrations in the samples were calculatedbased on a standard curve in which the MFI values obtainedfrom the standard were plotted against the known concen-trations (in picograms per milliliter) Cytokine levels (inpicograms per milliliter) were measured in the supernatant24 hours after bead exposure or 24 hours of culture withoutany stimulus (baseline condition) and the values were cor-rected by protein levels (in milligrams per milliliter) in thecell lysates using the Pierce BCA Protein Assay Kit (Ther-moFisher Scientific Waltham MA) The final results areprovided in pgmg
DNA Polymorphism GenotypingPeripheral blood mononuclear cells were used for DNAextraction using the QIAamp DNA Blood Mini Kit
(Qiagen Hilden Germany) and subsequent genotyping forthe rs6971 polymorphism (Ala147Thr) in the TSPO gene(TaqMan assay ID C_2512465_20 Thermo Fisher Sci-entific) Allele Ala147 was linked to Vic and allele Thr147was linked to FAM PCRs were performed in QuantStudio7 Flex Real-Time PCR system (Life TechnologiesThermoFisher Scientific)
Statistical AnalysisAssociation between dichotomous variables was assessedwith the χ2 test Our limited sample size made it difficult toascertain data distribution and therefore nonparametrictests were used Comparisons between the 3 groups weremade by the Kruskal-Wallis test When appropriate posthoc analyses were used to determine significant differencesbetween pairs of groups (Dunn Multiple ComparisonTest) The Mann-Whitney test was used for 2-group
Figure 1 Microglial Activation in HD
(A) PET image showing increased [11C]-ER176 standardized uptake value ratio (SUVR) in bilateral putamina and pallidi of a patient with manifest HD incomparison with a control Note that there is no visible difference in [11C]-ER176 SUVR for the premanifest HD gene carrier (central image) (B) Groupcomparisons in left (L) and right (R) putamina and pallidi Significant differences between groups are indicated by different letters (Kruskal-Wallis testfollowed by Dunn multiple comparisons test) Horizontal bars show the mean and the standard error of the mean Data from other regions of interest areshown in figure e-1 (linkslwwcomNXIA459) HD = Huntington disease
NeurologyorgNN Neurology Neuroimmunology amp Neuroinflammation | Volume 8 Number 3 | May 2021 5
comparisons (premanifest HD vs manifest HD) Spearmancorrelations were performed to examine the relationshipbetween [11]C-ER176 SUVR in the ROIs significantlydifferent between groups and (1) CAP score (2) volume inthe same ROIs and (3) scores in the clinical scales Allstatistical tests were 2 tailed and were performed using asignificance level of α = 005 Statistical analyses wereperformed using SPSS software version 260 and GraphPadPrism version 50
Standard Protocol Approvals Registrationsand Patient ConsentsAll subjects provided written informed consent before ad-mission to the study The Research Ethics Committees ofUTHealth and Houston Methodist Research Institute ap-proved this study
An Investigational New Drug Application (IND) was sub-mitted and authorization was obtained from the Food andDrug Administration to use [11C]-ER176 as a PET Imagingagent in this study (IND 139220)
Data AvailabilityThe raw data not provided in the article will be made availableby the authors at the request of other investigators for pur-poses of replicating procedures and results
ResultsClinical Evaluation and TSPO GenotypingDemographic and clinical characteristics of participants included inthis study are shown in the table There was no significant differ-ence between controls premanifest HD gene carriers and patientswith HD regarding age sex and educational level As expectedpatients with manifest HD presented a higher CAP score andworse scores in the scales that evaluatedmotor symptoms severityfunctional capacity and independence in comparison with pre-manifest HD (table) The age at clinical diagnosis was 4767 plusmn1035 years for the patients with manifest HD Premanifest HDgene carriers had amean of 1298 plusmn 617 years before the predictedclinical diagnosis of HD The 3 groups presented similar perfor-mance on the MMSE Patients with the clinical diagnosis of HDpresented worse performance on the SDMT and verbal fluencytest in comparisonwith premanifest and control subjects Controlsindividuals with premanifest HD and patients with HD presentedsimilar scores in the PBA-s subscales except for executive functionin which HD gene carriers (both premanifest and manifest HD)scored worse than controls evidencing that those problems arepresent before clinical diagnosis of HD (table)
The groups presented similar distribution of TSPO genotype(table) Individuals with genotype Ala147Ala147 were clas-sified as high-affinity binders and those with genotype
Figure 2 Microglial Activation in HD Is Associated With Disease Burden
Among HD gene carriers (premanifest andmanifest HD) a higher CAP score (a proxy for cumulative disease burden) was significantly associated with higher[11C]-ER176 standardized uptake value ratio (SUVR) in the (A) left putamen (ρ = 0664 p = 0018) (B) left pallidum (ρ = 0657 p = 0020) (C) right putamen (ρ =0671 p = 0017) and (D) right pallidum (ρ = 0734 p = 0007) Note the 2 clusters in [11C]-ER176 SUVR composed of premanifest HD gene carriers (CAP lt100)and manifest patients with HD (CAP gt100) CAP = CAG age product HD = Huntingtonrsquos disease
6 Neurology Neuroimmunology amp Neuroinflammation | Volume 8 Number 3 | May 2021 NeurologyorgNN
Ala147Thr147 as mixed affinity None of our patients pre-sented the genotype Thr147Thr147 (low-affinity binders)Because the groups presented comparable distribution ofTSPO genotype (table) and the outcome parameter wasSUVR which minimizes genotype differences no correctionfor genotype-related binding affinity was performed
In Vivo Assessment of Microglial ActivationPatients with HD demonstrate higher [11C]-ER176 SUVR inboth the left and right globi pallidi and putamina in comparisonwith controls These differences were not significant when wecompared premanifest HD gene carriers with patients withHDorcontrols (figure 1 and figure e-1 linkslwwcomNXIA459)These results indicate increased microglial activation in the puta-mina and globi pallidi of patients with manifest HDWe found nodifferences between different TSPO genotypes on [11C]-ER176SUVR when controlling for HD diagnosis (data not shown)
Among HD gene carriers (premanifest and manifest HD) ahigher CAP score was significantly associated with higher[11C]-ER176 SUVR in the left putamen (ρ = 0664 p =0018) left pallidum (ρ = 0657 p = 0020) right putamen(ρ = 0671 p = 0017) and right pallidum (ρ = 0734 p =0007) (figure 2) Corroborating these findings increased[11C]-ER176 SUVRwas associated with decreased volume inthe same ROIs (figure 3)
In Vitro Evaluation of the Dynamic Functionsof MicrogliaWe confirmed that the iMG cells have the immunophenotypeof microglial cells such as CX3CR1highCCR2low and atypical (ie ramified) morphology (figure 4) The iMG cellsexpress dynamic functions such as phagocytosis and secretionof inflammatory cytokines making them a valuable tool toassess microglial functioning30 The phagocytosis assaydemonstrated that iMG cells obtained from controls pre-manifest HD and HD patients (figure 5 AndashC) showedsimilar phagocytic capacity (figure 5E) Secretion of in-flammatory cytokines (TNFα IL-6 and IL-1β) duringphagocytosis was measured from culture supernatant Asshown in figure 5F although iMG cells from controls pre-manifest HD and patients with HD increased the productionof the evaluated cytokines during phagocytosis the valueswere significant only for TNFα in controls and manifest HD(figure 5F)
DiscussionBy using [11C]-ER176 PET we verified that patients withHD have increased microglial activation in basal gangliastructures that are involved in HD pathophysiologyie putamina and globi pallidi In addition we found that
Figure 3 Microglial Activation Is Associated With Brain Atrophy
Increased microglial activation is associated with decreased volume in putamen and pallidus Controls are displayed in red dots premanifest HD genecarriers in green dots andmanifest HD in black dots A higher [11C]-ER176 standardized uptake value ratio (SUVR) was associatedwith a decreased volume inthe (A) left putamen (ρ = minus0569 p = 0014) (B) left pallidum (ρ = minus0591 p = 0010) (C) right putamen (ρ = minus0601 p = 0008) and (D) right pallidum (ρ = minus0686 p= 0002) ICV = intracranial volume The dashed line was placed to emphasize the 2 clusters composed of controls + premanifest HD and manifest HD genecarriers HD = Huntington disease
NeurologyorgNN Neurology Neuroimmunology amp Neuroinflammation | Volume 8 Number 3 | May 2021 7
premanifest HD gene carriers demonstrate [11C]-ER176SUVRs comparable to controls suggesting that microglialactivation is a later process in HD pathophysiology becomingevident only after the onset of motor symptoms (figure 1)Supporting these findings we reported a significant correla-tion between increased microglial activation and higher CAPscores a proxy for cumulative disease burden (figure 2)Moreover increased microglial activation in putamina andpallidi was significantly associated with reduced volumes inthe same ROIs (figure 3) Individuals with manifest HDpresented a significant decrease in the cortical volume incomparison with controls (data not shown) Despite alreadypresenting cortical atrophy manifest HD individuals do nothave increased microglial activation in the cortex (figure 1)The only areas displaying increased microglia activation arethose that are known to be affected early in HD pathophysi-ology Because PET scans require individuals to be still for 90minutes our sample did not include patients in moderatelatestages of HD Whether increased microglia activation willoccur in the cortex later in the HD course is yet to be de-termined Altogether our data demonstrate that microglialactivation is involved in HD pathophysiology and is associ-ated with disease progression (as demonstrated by the CAPscore and atrophy in basal ganglia structures) Our findingssupport the conclusions of previous research that reportedthat microglial activation correlates with disease severitycontributing to the ongoing neuronal degeneration in HD16
HD has been regarded as an important model to study neu-rodegenerative diseases as it is caused by a single geneticmutation and is amenable to predictive genetic testing en-abling the study of patients before clinical diagnosis2 In this
study we assessed microglial activation not only in patientswith HD but also in premanifest HD gene carriers with amean of 13 years before the predicted onset of clinicalsymptoms Our results corroborate the view that microglialactivation is unlikely to be the initiating event in HD but maycontribute to neuronal death through multiple pathways31
Similar to other neurodegenerative diseases the mecha-nisms underlying selective neuronal dysfunction and deathin HD are still uncertain In addition to impairment in sys-tems for handling abnormal proteins other metabolicpathways and mechanisms might contribute to neuro-degeneration and progression of HD such as mitochondrialdysfunction impaired neurotrophic support oxidativestress and inflammatoryimmune mechanisms83233 In thecurrent study we described a significant increase in micro-glial activation in regions of neurodegeneration in HDsuggesting that this process is related to the disease patho-physiology and not to random premorbid events Accord-ingly postmortem studies reported significant microglialactivation in regions implicated in HD pathogenesis46 Thefact that we found significant microglial activation in patientswith manifest HD (but not in premanifest HD gene carriers)in comparison with controls suggests that this event occursin response or concomitantly to neuronal death This as-sumption is further corroborated by the finding of significantassociations between microglial activation and ROI volumesas well as CAP scores Here again our data are corroboratedby postmortem findings in which the number of activatedmicroglia in the striatum and cortex correlated with neuronalloss confirming the hypothesis that neuroinflammation maybe elicited by degenerating neurons6
Figure 4 Inducing Microglia-Like Cells From Human Peripheral Blood Monocytes
Peripheral blood monocytes were cultured for 14 days in conventional conditions (A) or with GM-CSF and IL-34 resulting in induced microglia like cells thatexhibit microglia-like morphology and increased CX3CR1CCR2 ratio (B)
8 Neurology Neuroimmunology amp Neuroinflammation | Volume 8 Number 3 | May 2021 NeurologyorgNN
Neuroinflammation elicited by neuronal death can be at firstregarded as a positive event as it promotes clearance of celldebris However inflammatory mechanisms contribute toneurodegeneration and dying neurons further activate in-flammatory responses resulting in a vicious cycle of in-flammation and neuronal death34 Therefore inflammatoryresponses although essential for tissue homeostasis cancontribute to neuronal injury As neural tissues have a
restricted cell renewal and regenerative capacity the CNS isextremely vulnerable to uncontrolled andor chronic immuneand inflammatory processes34 Interestingly enough our datacorroborate this hypothesis as we observed increased micro-glial activation later in the HD course (ie manifest HD)reinforcing the idea that neuroinflammation contributes todisease progression rather than to disease onset Activatedmicroglia releases proinflammatory mediators including
Figure 5 Phagocytosis by Induced Microglia-Like (iMG) Cells
Peripheral blood monocytes (day 1 panel A) were cultured for 14 days with GM-CSF and IL-34 resulting in iMG cells (day 14 panel B) Phagocytosis wasassessed by exposing iMG cells to PE-conjugated (1250) beads (panel C) for 24 hours The iMG cells showed the ability of phagocytosis with morphologicalchanges (C) The read arrows show PE-conjugated beads phagocytosed by iMG cells After washing beads out cells were acquired in a flow cytometer andanalyzed iMG were first gated by forwardside scatter (not shown) PE uptake was analyzed as shown in (D) orange (left) histogram shows the cells withoutbeads (negative control) red (right) histogram shows the cells that phagocytosed to the PE-conjugated beads Quantification is shown in (E) iMG fromcontrols premanifest HD and HD patients showed similar phagocytic capacity (no significant difference found at the Kruskal-Wallis test) (F) Cytokineproduction during phagocytosis iMG cells were incubated with PE-conjugated beads (1250) for 24 hours Supernatant was collected and cytokines wereanalyzed by multiparametric assay Cytokine levels (in picograms per milliliter) were corrected by protein levels (in milligrams per milliliter) Although iMGcells from controls premanifest HD and patients with HD increased the production of the evaluated cytokines during phagocytosis the values weresignificant only for TNFα in controls andmanifest HD (p values are provided in the figure Wilcoxonmatched-pairs signed-rank test) Scale bar (AndashC) = 10mmHorizontal bars (E and F) show the mean and the standard error of the mean HD = Huntington disease PE = phycoerythrin TNF = tumor necrosis factor
NeurologyorgNN Neurology Neuroimmunology amp Neuroinflammation | Volume 8 Number 3 | May 2021 9
cytokines and reactive oxygen species which in turn will alsocontribute to neuronal death and further microglial activa-tion34We observed an increase in cytokine release (especiallyTNFα) following a phagocytic stimulus in our in vitro ex-periments In line with our findings postmortem studies havealso reported increased levels of inflammatory cytokines inbrain samples of patients with HD in comparison withcontrols3536 It is worth mentioning that cytokine release is amicroglial reaction to a proinflammatory challenge and undercontrolled conditions (such as the iMG cultures) phagocy-tosis is a noninflammatory function of microglia A proin-flammatory stimulus would be needed to appropriately assessdifferences in cytokine release between controls premanifestHD and manifest HD iMG cultures
The current study used a novel PET tracer for TSPO [11C]-ER176 to assess microglial activation in HD By using [11C]-PK11195 PET studies have shown a significant increase inmicroglial activation in the striatum of patients with HD16 aswell as in premanifest HD gene carriers18 compared withcontrols In both studies [11C]-PK11195 binding significantlycorrelated with disease severity as measured by decreased[11C]-raclopride (a marker of dopamine D2 and D3 receptoravailability) binding in the striatum1618 Similar results weredescribed in the hypothalamus17 Later the same work groupused a multimodal imaging approach combiningMRI and PETanalyses and worsening in atrophy evaluated by MRI was ac-companied by a reduction in [11C]-raclopride and an increasein [11C]-PK11195 bindings in patients with HD In pre-manifest HD increased level of microglial activation in theassociative striatum and in the regional network associated withcognition correlated with 5-year probability of HD onset19
More recently increased microglial activation in the somato-sensory cortex (as evaluated through [11C]-PK11195 binding)was associatedwith higher levels of IL-1β IL-6 IL-8 and TNFαproduced by stimulated peripheral blood monocytes frompremanifest HD gene carriers2037 Although these studiesprovided valuable information about neuroinflammation inHD indicating microglial activation even in premanifest sub-jects their results should be interpreted with caution The poorsignal-to-background ratio of first-generation TSPO radio-tracers poses important limitations to the analyses The ratio ofspecific to nonspecific binding of [11C]-PK11195 in humanbrain has been reported to be as low as 023839 As an attempt tomitigate this problem a recent study used the second-generation TSPO tracer [11C]-PBR28 to evaluate micro-glial activation in HD21 Of interest the results were similar toours ie increased levels of [11C]-PBR28 binding in theputamina and pallidi of HD gene carriers (which included 7manifest patients with HD and only 1 person with premanifestHD) in comparison with controls Of note the results of thesingle premanifest subject were midway between controls andthe remaining patients suggesting that microglial activationwas associated with disease progression21 The authors didconfirm that [11C]-PBR28 provides a high signal-to-background ratio an important feature required for the anal-ysis at the individual level and within individual brain regions
An important limitation of [11C]-PBR28 and other second-generation TSPO radioligands is that they have varying degreesof sensitivity to the single nucleotide polymorphism rs6971 inthe TSPO gene40 [11C]-ER176 has unique advantages overthe other TSPO binders being the only known TSPO radio-ligand that allows the inclusion of low-affinity binders Thebinding of ER176 in low-affinity binders was found to becomparable to that for PBR28 in high-affinity binders More-over ER176 was proven to be the only radioligand not con-taminated by radiometabolites accumulating in the brain14
Our findings must be interpreted while taking into accountthe limitations of the study which include the relatively smallsample size and the cross-sectional design The cross-sectionaldesign prevented the assessment of microglial activation inthe context of disease progression and development of motorand nonmotor symptoms In addition the use of an objectivemethod to measure striatal degeneration (eg [11C]-raclopride PET) would provide more accurate informationabout striatal neuron dysfunctiondeath Regarding the invitro experiments the use of a proinflammatory challenge(other than the beads added for the phagocytosis assay) in ourexperiments would provide valuable information on cytokinesrelease by iMG from the different groups It is worth men-tioning the strengths of the study that include the use of aunique TSPO ligand [11C]-ER176 the enrollment of bothpremanifest and manifest subjects and the exploratory in-vestigation of microglia activity
In conclusion our findings strengthen the evidence thatmicroglial activation is associated with HD pathophysiologyMicroglial activation is unlikely to be the initiating event inHD Yet it may exacerbate neuronal dysfunction and neu-ronal death thus contributing to disease progression It isimportant to refine our understanding of the more specificimmuneinflammatory mechanisms that are involved in HDand in neurodegeneration This may foster the developmentof new therapeutic interventions to halt the progression ofHD and other neurodegenerative diseases
AcknowledgmentThe authors acknowledge and thank all of the volunteers thatparticipated in this study and are indebted to their families fortheir magnificent support They also thank the staff from theUTHealth McGovern Medical School 3T MRI Center andfrom the HoustonMethodist Research Institute TranslationalImaging Core Facilities especially Vips Patel and Kim MDoan for their skilled technical support Dr Gabriel Fries forhis assistance with the genotyping analyses and the teammembers of the HDSA Center of Excellence at UTHealth
Study FundingThis study was funded by the HD Human BiologyProjectmdashHuntingtonrsquos Disease Society of America (HDSA)E Furr Stimming receives research funding from RocheGenetech Vaccinex Cures Within Reach HDSA Uniqureand CHDI Foundation
10 Neurology Neuroimmunology amp Neuroinflammation | Volume 8 Number 3 | May 2021 NeurologyorgNN
DisclosureThe authors report no disclosures relevant to the manuscriptGo to NeurologyorgNN for full disclosures
Publication HistoryReceived by Neurology Neuroimmunology amp NeuroinflammationSeptember 9 2020 Accepted in final form January 20 2021
References1 The Huntingtonrsquos Disease Collaborative Research Group A novel gene containing a
trinucleotide repeat that is expanded and unstable on Huntingtonrsquos disease chro-mosomes Cell 199372971ndash983
2 Ross CA Aylward EH Wild EJ et al Huntington disease natural history biomarkersand prospects for therapeutics Nat Rev Neurol 201410204ndash216
3 Soulet D Cicchetti F The role of immunity in Huntingtonrsquos disease Mol Psychiatry201116889ndash902
4 Myers RH Vonsattel JP Paskevich PA et al Decreased neuronal and increasedoligodendroglial densities in Huntingtonrsquos disease caudate nucleus J NeuropatholExp Neurol 199150729ndash742
5 Vonsattel JP Myers RH Stevens TJ Ferrante RJ Bird ED Richardson EP JrNeuropathological classification of Huntingtonrsquos disease J Neuropathol Exp Neurol198544559ndash577
6 Sapp E Kegel KB Aronin N et al Early and progressive accumulation of reactivemicroglia in the Huntington disease brain J Neuropathol Exp Neurol 200160161ndash172
7 Ciccocioppo F Bologna G Ercolino E et al Neurodegenerative diseases asproteinopathies-driven immune disorders Neural Regen Res 202015850ndash856
8 Rocha NP Ribeiro FM Furr-Stimming E Teixeira AL Neuroimmunology of Hun-tingtonrsquos disease revisiting evidence from human studies Mediators Inflamm 201620168653132
9 Vowinckel E Reutens D Becher B et al PK11195 binding to the peripheral ben-zodiazepine receptor as a marker of microglia activation in multiple sclerosis andexperimental autoimmune encephalomyelitis J Neurosci Res 199750345ndash353
10 Papadopoulos V Baraldi M Guilarte TR et al Translocator protein (18kDa) newnomenclature for the peripheral-type benzodiazepine receptor based on its structureand molecular function Trends Pharmacol Sci 200627402ndash409
11 Cumming P Burgher B Patkar O et al Sifting through the surfeit of neuro-inflammation tracers J Cereb Blood Flow Metab 201838204ndash224
12 Zanotti-Fregonara P Zhang Y Jenko KJ et al Synthesis and evaluation of translocator18 kDa protein (TSPO) positron emission tomography (PET) radioligands with lowbinding sensitivity to human single nucleotide polymorphism rs6971 ACS ChemNeurosci 20145963ndash971
13 Ikawa M Lohith TG Shrestha S et al 11C-ER176 a radioligand for 18-kDa trans-locator protein has adequate sensitivity to robustly image all three affinity genotypesin human brain J Nucl Med 201758320ndash325
14 Fujita M Kobayashi M Ikawa M et al Comparison of four (11)C-labeled PETligands to quantify translocator protein 18 kDa (TSPO) in human brain (R)-PK11195 PBR28 DPA-713 and ER176-based on recent publications that measuredspecific-to-non-displaceable ratios EJNMMI Res 2017784
15 Zanotti-Fregonara P Pascual B Veronese M et al Head-to-head comparison of (11)C-PBR28 and (11)C-ER176 for quantification of the translocator protein in thehuman brain Eur J Nucl Med Mol Imaging 2019461822ndash1829
16 Pavese N Gerhard A Tai YF et al Microglial activation correlates with severity inHuntington disease a clinical and PET study Neurology 2006661638ndash1643
17 Politis M Pavese N Tai YF Tabrizi SJ Barker RA Piccini P Hypothalamic in-volvement in Huntingtonrsquos disease an in vivo PET study Brain 20081312860ndash2869
18 Tai YF Pavese N Gerhard A et al Microglial activation in presymptomatic Hun-tingtonrsquos disease gene carriers Brain 20071301759ndash1766
19 Politis M Pavese N Tai YF et al Microglial activation in regions related to cognitivefunction predicts disease onset in Huntingtonrsquos disease a multimodal imaging studyHum Brain Mapp 201132258ndash270
20 Politis M Lahiri N Niccolini F et al Increased central microglial activation associatedwith peripheral cytokine levels in premanifest Huntingtonrsquos disease gene carriersNeurobiol Dis 201583115ndash121
21 Lois C Gonzalez I Izquierdo-Garcia D et al Neuroinflammation in Huntingtonrsquosdisease new insights with (11)C-PBR28 PETMRI ACS Chem Neurosci 201892563ndash2571
22 Huntington Study Group Unified Huntingtonrsquos Disease Rating Scale reliability andconsistency Mov Disord 199611136ndash142
23 Folstein MF Folstein SE McHugh PR ldquoMini-mental staterdquo A practical method forgrading the cognitive state of patients for the clinician J Psychiatr Res 197512189ndash198
24 McNally G Rickards H Horton M Craufurd D Exploring the validity of the shortversion of the Problem Behaviours Assessment (PBA-s) for Huntingtonrsquos disease arasch analysis J Huntingtons Dis 20154347ndash369
25 Warner JH Sampaio C Modeling variability in the progression of Huntingtonrsquosdisease a novel modeling approach applied to structural imaging markers fromTRACK-HD CPT Pharmacometrics Syst Pharmacol 20165437ndash445
26 Lyoo CH Ikawa M Liow JS et al Cerebellum can serve as a pseudo-reference regionin Alzheimer disease to detect neuroinflammation measured with PET radioligandbinding to translocator protein J Nucl Med 201556701ndash706
27 Reuter M Schmansky NJ Rosas HD Fischl B Within-subject template estimation forunbiased longitudinal image analysis Neuroimage 2012611402ndash1418
28 Yushkevich PA Piven J Hazlett HC et al User-guided 3D active contour segmen-tation of anatomical structures significantly improved efficiency and reliabilityNeuroimage 2006311116ndash1128
29 Ohgidani M Kato TA Setoyama D et al Direct induction of ramified microglia-likecells from human monocytes dynamic microglial dysfunction in Nasu-Hakola dis-ease Sci Rep 201444957
30 Ohgidani M Kato TA Kanba S Introducing directly induced microglia-like (iMG)cells from fresh human monocytes a novel translational research tool for psychiatricdisorders Front Cell Neurosci 20159184
31 Kim SU de Vellis J Microglia in health and disease J Neurosci Res 200581302-31332 Ross CA Tabrizi SJ Huntingtonrsquos disease from molecular pathogenesis to clinical
treatment Lancet Neurol 20111083ndash9833 Chitnis TWeiner HL CNS inflammation and neurodegeneration J Clin Invest 2017
1273577ndash3587
Appendix Authors
Name Location Contribution
Natalia PRocha PhD
The Mitchell Center forAlzheimerrsquos Disease andRelated Brain DisordersDepartment of NeurologyMcGovern Medical SchoolThe University of TexasHealth Science CenterHouston
Designed andconceptualized the studymajor role in theacquisition of dataanalyzed the data anddrafted the manuscript forintellectual content
OdelinCharron MS
Department of NeurologyThe University of Texas atAustin
Major role in data analysis
Leigh BLathamPhD
School of MedicineUniversity of WashingtonSeattle
Major role in theacquisition of data
Gabriela DColpo PhD
Neuropsychiatry ProgramDepartment of Psychiatryand Behavioral SciencesMcGovern Medical SchoolUniversity of TexasHouston
Major role in theacquisition of data
PaoloZanotti-FregonaraMD PhD
Houston MethodistResearch Institute andWeill Cornell Medicine TX
Major role in the studydesign and the acquisitionof data
MeixiangYu PhD
Houston MethodistResearch Institute andWeill Cornell Medicine TX
Major role in theacquisition of data
LeorahFreemanMD PhD
Department of NeurologyThe University of Texas atAustin
Major role in the studydesign data analysis andinterpretation and theacquisition of data
Erin FurrStimmingMD
HDSA Center of Excellenceat University of TexasHealth Science Center atHouston
Major role in the studydesign and acquisition ofdata interpreted the dataand revised themanuscript for intellectualcontent
Antonio LTeixeiraMD PhD
Neuropsychiatry ProgramDepartment of Psychiatryand Behavioral SciencesMcGovern Medical SchoolUniversity of TexasHouston
Major role in the studydesign and acquisition ofdata interpreted the dataand revised themanuscript for intellectualcontent
NeurologyorgNN Neurology Neuroimmunology amp Neuroinflammation | Volume 8 Number 3 | May 2021 11
34 Rocha NP de Miranda AS Teixeira AL Insights into neuroinflammation in Parkin-sonrsquos disease from biomarkers to anti-inflammatory based therapies Biomed Res Int20152015628192
35 Silvestroni A Faull RL Strand AD Moller T Distinct neuroinflammatory profile inpost-mortem human Huntingtonrsquos disease Neuroreport 2009201098ndash1103
36 Bjorkqvist M Wild EJ Thiele J et al A novel pathogenic pathway of immune acti-vation detectable before clinical onset in Huntingtonrsquos disease J Exp Med 20082051869ndash1877
37 Politis M Lahiri N Niccolini F et al Corrigendum to ldquoIncreased central microglialactivation associated with peripheral cytokine levels in premanifest Huntingtonrsquos
disease gene carriersrdquo [Neurobiol Dis 83 (2015) 115-121] Neurobiol Dis 201798162
38 Kropholler MA Boellaard R Schuitemaker A Folkersma H van Berckel BN Lam-mertsma AA Evaluation of reference tissue models for the analysis of [11C](R)-PK11195 studies J Cereb Blood Flow Metab 2006261431ndash1441
39 Turkheimer FE Edison P Pavese N et al Reference and target region modeling of[11C]-(R)-PK11195 brain studies J Nucl Med 200748158ndash167
40 Kreisl WC Jenko KJ Hines CS et al A genetic polymorphism for translocator protein18 kDa affects both in vitro and in vivo radioligand binding in human brain to thisputative biomarker of neuroinflammation J Cereb Blood FlowMetab 20133353ndash58
12 Neurology Neuroimmunology amp Neuroinflammation | Volume 8 Number 3 | May 2021 NeurologyorgNN
DOI 101212NXI000000000000098420218 Neurol Neuroimmunol Neuroinflamm
Natalia P Rocha Odelin Charron Leigh B Latham et al Pathophysiology
Microglia Activation in Basal Ganglia Is a Late Event in Huntington Disease
This information is current as of April 1 2021
ServicesUpdated Information amp
httpnnneurologyorgcontent83e984fullhtmlincluding high resolution figures can be found at
References httpnnneurologyorgcontent83e984fullhtmlref-list-1
This article cites 40 articles 4 of which you can access for free at
Subspecialty Collections
httpnnneurologyorgcgicollectionpetPET
httpnnneurologyorgcgicollectionhuntingtons_diseaseHuntingtons diseasefollowing collection(s) This article along with others on similar topics appears in the
Permissions amp Licensing
httpnnneurologyorgmiscaboutxhtmlpermissionsits entirety can be found online atInformation about reproducing this article in parts (figurestables) or in
Reprints
httpnnneurologyorgmiscaddirxhtmlreprintsusInformation about ordering reprints can be found online
Academy of Neurology All rights reserved Online ISSN 2332-7812Copyright copy 2021 The Author(s) Published by Wolters Kluwer Health Inc on behalf of the AmericanPublished since April 2014 it is an open-access online-only continuous publication journal Copyright
is an official journal of the American Academy of NeurologyNeurol Neuroimmunol Neuroinflamm
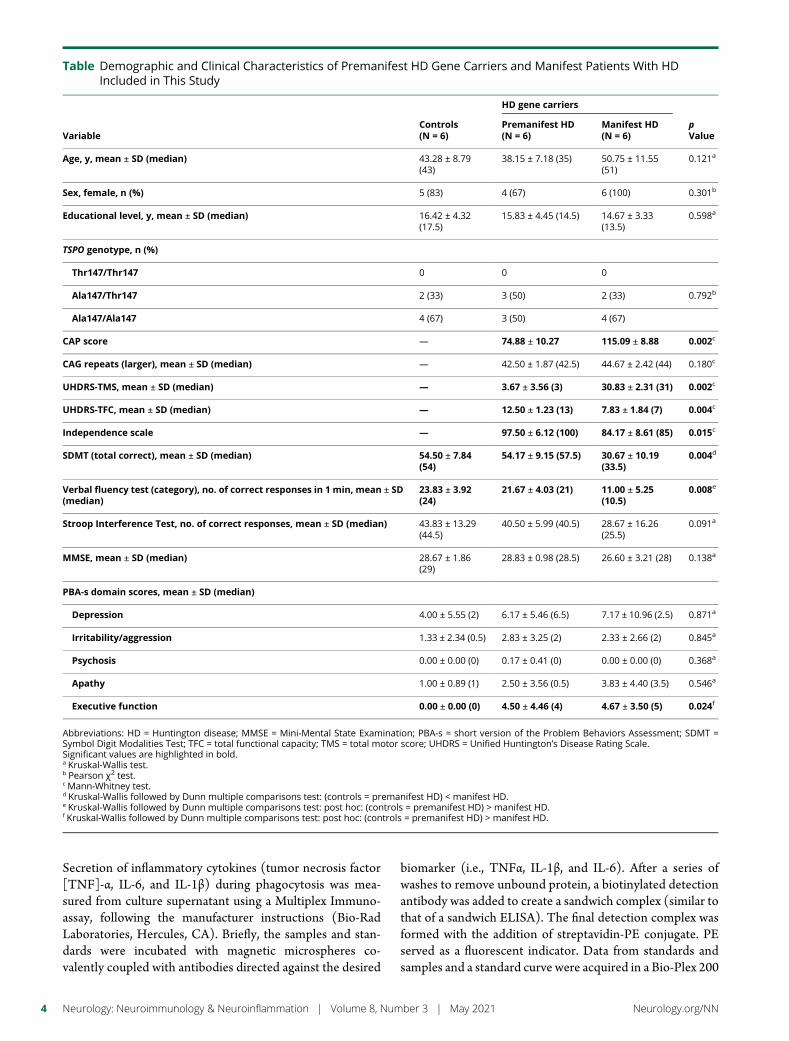
Secretion of inflammatory cytokines (tumor necrosis factor[TNF]-α IL-6 and IL-1β) during phagocytosis was mea-sured from culture supernatant using a Multiplex Immuno-assay following the manufacturer instructions (Bio-RadLaboratories Hercules CA) Briefly the samples and stan-dards were incubated with magnetic microspheres co-valently coupled with antibodies directed against the desired
biomarker (ie TNFα IL-1β and IL-6) After a series ofwashes to remove unbound protein a biotinylated detectionantibody was added to create a sandwich complex (similar tothat of a sandwich ELISA) The final detection complex wasformed with the addition of streptavidin-PE conjugate PEserved as a fluorescent indicator Data from standards andsamples and a standard curve were acquired in a Bio-Plex 200
Table Demographic and Clinical Characteristics of Premanifest HD Gene Carriers and Manifest Patients With HDIncluded in This Study
VariableControls(N = 6)
HD gene carriers
pValue
Premanifest HD(N = 6)
Manifest HD(N = 6)
Age y mean plusmn SD (median) 4328 plusmn 879(43)
3815 plusmn 718 (35) 5075 plusmn 1155(51)
0121a
Sex female n () 5 (83) 4 (67) 6 (100) 0301b
Educational level y mean plusmn SD (median) 1642 plusmn 432(175)
1583 plusmn 445 (145) 1467 plusmn 333(135)
0598a
TSPO genotype n ()
Thr147Thr147 0 0 0
Ala147Thr147 2 (33) 3 (50) 2 (33) 0792b
Ala147Ala147 4 (67) 3 (50) 4 (67)
CAP score mdash 7488 plusmn 1027 11509 plusmn 888 0002c
CAG repeats (larger) mean plusmn SD (median) mdash 4250 plusmn 187 (425) 4467 plusmn 242 (44) 0180c
UHDRS-TMS mean plusmn SD (median) mdash 367 plusmn 356 (3) 3083 plusmn 231 (31) 0002c
UHDRS-TFC mean plusmn SD (median) mdash 1250 plusmn 123 (13) 783 plusmn 184 (7) 0004c
Independence scale mdash 9750 plusmn 612 (100) 8417 plusmn 861 (85) 0015c
SDMT (total correct) mean plusmn SD (median) 5450 plusmn 784(54)
5417 plusmn 915 (575) 3067 plusmn 1019(335)
0004d
Verbal fluency test (category) no of correct responses in 1 min mean plusmn SD(median)
2383 plusmn 392(24)
2167 plusmn 403 (21) 1100 plusmn 525(105)
0008e
Stroop Interference Test no of correct responses mean plusmn SD (median) 4383 plusmn 1329(445)
4050 plusmn 599 (405) 2867 plusmn 1626(255)
0091a
MMSE mean plusmn SD (median) 2867 plusmn 186(29)
2883 plusmn 098 (285) 2660 plusmn 321 (28) 0138a
PBA-s domain scores mean plusmn SD (median)
Depression 400 plusmn 555 (2) 617 plusmn 546 (65) 717 plusmn 1096 (25) 0871a
Irritabilityaggression 133 plusmn 234 (05) 283 plusmn 325 (2) 233 plusmn 266 (2) 0845a
Psychosis 000 plusmn 000 (0) 017 plusmn 041 (0) 000 plusmn 000 (0) 0368a
Apathy 100 plusmn 089 (1) 250 plusmn 356 (05) 383 plusmn 440 (35) 0546a
Executive function 000 plusmn 000 (0) 450 plusmn 446 (4) 467 plusmn 350 (5) 0024f
Abbreviations HD = Huntington disease MMSE = Mini-Mental State Examination PBA-s = short version of the Problem Behaviors Assessment SDMT =Symbol Digit Modalities Test TFC = total functional capacity TMS = total motor score UHDRS = Unified Huntingtonrsquos Disease Rating ScaleSignificant values are highlighted in bolda Kruskal-Wallis testb Pearson χ2 testc Mann-Whitney testd Kruskal-Wallis followed by Dunn multiple comparisons test (controls = premanifest HD) lt manifest HDe Kruskal-Wallis followed by Dunn multiple comparisons test post hoc (controls = premanifest HD) gt manifest HDf Kruskal-Wallis followed by Dunn multiple comparisons test post hoc (controls = premanifest HD) gt manifest HD
4 Neurology Neuroimmunology amp Neuroinflammation | Volume 8 Number 3 | May 2021 NeurologyorgNN
Reader and analyzed in a Bio-Plex Manager Software whichpresented data as median fluorescence intensity (MFI) Thecytokine concentrations in the samples were calculatedbased on a standard curve in which the MFI values obtainedfrom the standard were plotted against the known concen-trations (in picograms per milliliter) Cytokine levels (inpicograms per milliliter) were measured in the supernatant24 hours after bead exposure or 24 hours of culture withoutany stimulus (baseline condition) and the values were cor-rected by protein levels (in milligrams per milliliter) in thecell lysates using the Pierce BCA Protein Assay Kit (Ther-moFisher Scientific Waltham MA) The final results areprovided in pgmg
DNA Polymorphism GenotypingPeripheral blood mononuclear cells were used for DNAextraction using the QIAamp DNA Blood Mini Kit
(Qiagen Hilden Germany) and subsequent genotyping forthe rs6971 polymorphism (Ala147Thr) in the TSPO gene(TaqMan assay ID C_2512465_20 Thermo Fisher Sci-entific) Allele Ala147 was linked to Vic and allele Thr147was linked to FAM PCRs were performed in QuantStudio7 Flex Real-Time PCR system (Life TechnologiesThermoFisher Scientific)
Statistical AnalysisAssociation between dichotomous variables was assessedwith the χ2 test Our limited sample size made it difficult toascertain data distribution and therefore nonparametrictests were used Comparisons between the 3 groups weremade by the Kruskal-Wallis test When appropriate posthoc analyses were used to determine significant differencesbetween pairs of groups (Dunn Multiple ComparisonTest) The Mann-Whitney test was used for 2-group
Figure 1 Microglial Activation in HD
(A) PET image showing increased [11C]-ER176 standardized uptake value ratio (SUVR) in bilateral putamina and pallidi of a patient with manifest HD incomparison with a control Note that there is no visible difference in [11C]-ER176 SUVR for the premanifest HD gene carrier (central image) (B) Groupcomparisons in left (L) and right (R) putamina and pallidi Significant differences between groups are indicated by different letters (Kruskal-Wallis testfollowed by Dunn multiple comparisons test) Horizontal bars show the mean and the standard error of the mean Data from other regions of interest areshown in figure e-1 (linkslwwcomNXIA459) HD = Huntington disease
NeurologyorgNN Neurology Neuroimmunology amp Neuroinflammation | Volume 8 Number 3 | May 2021 5
comparisons (premanifest HD vs manifest HD) Spearmancorrelations were performed to examine the relationshipbetween [11]C-ER176 SUVR in the ROIs significantlydifferent between groups and (1) CAP score (2) volume inthe same ROIs and (3) scores in the clinical scales Allstatistical tests were 2 tailed and were performed using asignificance level of α = 005 Statistical analyses wereperformed using SPSS software version 260 and GraphPadPrism version 50
Standard Protocol Approvals Registrationsand Patient ConsentsAll subjects provided written informed consent before ad-mission to the study The Research Ethics Committees ofUTHealth and Houston Methodist Research Institute ap-proved this study
An Investigational New Drug Application (IND) was sub-mitted and authorization was obtained from the Food andDrug Administration to use [11C]-ER176 as a PET Imagingagent in this study (IND 139220)
Data AvailabilityThe raw data not provided in the article will be made availableby the authors at the request of other investigators for pur-poses of replicating procedures and results
ResultsClinical Evaluation and TSPO GenotypingDemographic and clinical characteristics of participants included inthis study are shown in the table There was no significant differ-ence between controls premanifest HD gene carriers and patientswith HD regarding age sex and educational level As expectedpatients with manifest HD presented a higher CAP score andworse scores in the scales that evaluatedmotor symptoms severityfunctional capacity and independence in comparison with pre-manifest HD (table) The age at clinical diagnosis was 4767 plusmn1035 years for the patients with manifest HD Premanifest HDgene carriers had amean of 1298 plusmn 617 years before the predictedclinical diagnosis of HD The 3 groups presented similar perfor-mance on the MMSE Patients with the clinical diagnosis of HDpresented worse performance on the SDMT and verbal fluencytest in comparisonwith premanifest and control subjects Controlsindividuals with premanifest HD and patients with HD presentedsimilar scores in the PBA-s subscales except for executive functionin which HD gene carriers (both premanifest and manifest HD)scored worse than controls evidencing that those problems arepresent before clinical diagnosis of HD (table)
The groups presented similar distribution of TSPO genotype(table) Individuals with genotype Ala147Ala147 were clas-sified as high-affinity binders and those with genotype
Figure 2 Microglial Activation in HD Is Associated With Disease Burden
Among HD gene carriers (premanifest andmanifest HD) a higher CAP score (a proxy for cumulative disease burden) was significantly associated with higher[11C]-ER176 standardized uptake value ratio (SUVR) in the (A) left putamen (ρ = 0664 p = 0018) (B) left pallidum (ρ = 0657 p = 0020) (C) right putamen (ρ =0671 p = 0017) and (D) right pallidum (ρ = 0734 p = 0007) Note the 2 clusters in [11C]-ER176 SUVR composed of premanifest HD gene carriers (CAP lt100)and manifest patients with HD (CAP gt100) CAP = CAG age product HD = Huntingtonrsquos disease
6 Neurology Neuroimmunology amp Neuroinflammation | Volume 8 Number 3 | May 2021 NeurologyorgNN
Ala147Thr147 as mixed affinity None of our patients pre-sented the genotype Thr147Thr147 (low-affinity binders)Because the groups presented comparable distribution ofTSPO genotype (table) and the outcome parameter wasSUVR which minimizes genotype differences no correctionfor genotype-related binding affinity was performed
In Vivo Assessment of Microglial ActivationPatients with HD demonstrate higher [11C]-ER176 SUVR inboth the left and right globi pallidi and putamina in comparisonwith controls These differences were not significant when wecompared premanifest HD gene carriers with patients withHDorcontrols (figure 1 and figure e-1 linkslwwcomNXIA459)These results indicate increased microglial activation in the puta-mina and globi pallidi of patients with manifest HDWe found nodifferences between different TSPO genotypes on [11C]-ER176SUVR when controlling for HD diagnosis (data not shown)
Among HD gene carriers (premanifest and manifest HD) ahigher CAP score was significantly associated with higher[11C]-ER176 SUVR in the left putamen (ρ = 0664 p =0018) left pallidum (ρ = 0657 p = 0020) right putamen(ρ = 0671 p = 0017) and right pallidum (ρ = 0734 p =0007) (figure 2) Corroborating these findings increased[11C]-ER176 SUVRwas associated with decreased volume inthe same ROIs (figure 3)
In Vitro Evaluation of the Dynamic Functionsof MicrogliaWe confirmed that the iMG cells have the immunophenotypeof microglial cells such as CX3CR1highCCR2low and atypical (ie ramified) morphology (figure 4) The iMG cellsexpress dynamic functions such as phagocytosis and secretionof inflammatory cytokines making them a valuable tool toassess microglial functioning30 The phagocytosis assaydemonstrated that iMG cells obtained from controls pre-manifest HD and HD patients (figure 5 AndashC) showedsimilar phagocytic capacity (figure 5E) Secretion of in-flammatory cytokines (TNFα IL-6 and IL-1β) duringphagocytosis was measured from culture supernatant Asshown in figure 5F although iMG cells from controls pre-manifest HD and patients with HD increased the productionof the evaluated cytokines during phagocytosis the valueswere significant only for TNFα in controls and manifest HD(figure 5F)
DiscussionBy using [11C]-ER176 PET we verified that patients withHD have increased microglial activation in basal gangliastructures that are involved in HD pathophysiologyie putamina and globi pallidi In addition we found that
Figure 3 Microglial Activation Is Associated With Brain Atrophy
Increased microglial activation is associated with decreased volume in putamen and pallidus Controls are displayed in red dots premanifest HD genecarriers in green dots andmanifest HD in black dots A higher [11C]-ER176 standardized uptake value ratio (SUVR) was associatedwith a decreased volume inthe (A) left putamen (ρ = minus0569 p = 0014) (B) left pallidum (ρ = minus0591 p = 0010) (C) right putamen (ρ = minus0601 p = 0008) and (D) right pallidum (ρ = minus0686 p= 0002) ICV = intracranial volume The dashed line was placed to emphasize the 2 clusters composed of controls + premanifest HD and manifest HD genecarriers HD = Huntington disease
NeurologyorgNN Neurology Neuroimmunology amp Neuroinflammation | Volume 8 Number 3 | May 2021 7
premanifest HD gene carriers demonstrate [11C]-ER176SUVRs comparable to controls suggesting that microglialactivation is a later process in HD pathophysiology becomingevident only after the onset of motor symptoms (figure 1)Supporting these findings we reported a significant correla-tion between increased microglial activation and higher CAPscores a proxy for cumulative disease burden (figure 2)Moreover increased microglial activation in putamina andpallidi was significantly associated with reduced volumes inthe same ROIs (figure 3) Individuals with manifest HDpresented a significant decrease in the cortical volume incomparison with controls (data not shown) Despite alreadypresenting cortical atrophy manifest HD individuals do nothave increased microglial activation in the cortex (figure 1)The only areas displaying increased microglia activation arethose that are known to be affected early in HD pathophysi-ology Because PET scans require individuals to be still for 90minutes our sample did not include patients in moderatelatestages of HD Whether increased microglia activation willoccur in the cortex later in the HD course is yet to be de-termined Altogether our data demonstrate that microglialactivation is involved in HD pathophysiology and is associ-ated with disease progression (as demonstrated by the CAPscore and atrophy in basal ganglia structures) Our findingssupport the conclusions of previous research that reportedthat microglial activation correlates with disease severitycontributing to the ongoing neuronal degeneration in HD16
HD has been regarded as an important model to study neu-rodegenerative diseases as it is caused by a single geneticmutation and is amenable to predictive genetic testing en-abling the study of patients before clinical diagnosis2 In this
study we assessed microglial activation not only in patientswith HD but also in premanifest HD gene carriers with amean of 13 years before the predicted onset of clinicalsymptoms Our results corroborate the view that microglialactivation is unlikely to be the initiating event in HD but maycontribute to neuronal death through multiple pathways31
Similar to other neurodegenerative diseases the mecha-nisms underlying selective neuronal dysfunction and deathin HD are still uncertain In addition to impairment in sys-tems for handling abnormal proteins other metabolicpathways and mechanisms might contribute to neuro-degeneration and progression of HD such as mitochondrialdysfunction impaired neurotrophic support oxidativestress and inflammatoryimmune mechanisms83233 In thecurrent study we described a significant increase in micro-glial activation in regions of neurodegeneration in HDsuggesting that this process is related to the disease patho-physiology and not to random premorbid events Accord-ingly postmortem studies reported significant microglialactivation in regions implicated in HD pathogenesis46 Thefact that we found significant microglial activation in patientswith manifest HD (but not in premanifest HD gene carriers)in comparison with controls suggests that this event occursin response or concomitantly to neuronal death This as-sumption is further corroborated by the finding of significantassociations between microglial activation and ROI volumesas well as CAP scores Here again our data are corroboratedby postmortem findings in which the number of activatedmicroglia in the striatum and cortex correlated with neuronalloss confirming the hypothesis that neuroinflammation maybe elicited by degenerating neurons6
Figure 4 Inducing Microglia-Like Cells From Human Peripheral Blood Monocytes
Peripheral blood monocytes were cultured for 14 days in conventional conditions (A) or with GM-CSF and IL-34 resulting in induced microglia like cells thatexhibit microglia-like morphology and increased CX3CR1CCR2 ratio (B)
8 Neurology Neuroimmunology amp Neuroinflammation | Volume 8 Number 3 | May 2021 NeurologyorgNN
Neuroinflammation elicited by neuronal death can be at firstregarded as a positive event as it promotes clearance of celldebris However inflammatory mechanisms contribute toneurodegeneration and dying neurons further activate in-flammatory responses resulting in a vicious cycle of in-flammation and neuronal death34 Therefore inflammatoryresponses although essential for tissue homeostasis cancontribute to neuronal injury As neural tissues have a
restricted cell renewal and regenerative capacity the CNS isextremely vulnerable to uncontrolled andor chronic immuneand inflammatory processes34 Interestingly enough our datacorroborate this hypothesis as we observed increased micro-glial activation later in the HD course (ie manifest HD)reinforcing the idea that neuroinflammation contributes todisease progression rather than to disease onset Activatedmicroglia releases proinflammatory mediators including
Figure 5 Phagocytosis by Induced Microglia-Like (iMG) Cells
Peripheral blood monocytes (day 1 panel A) were cultured for 14 days with GM-CSF and IL-34 resulting in iMG cells (day 14 panel B) Phagocytosis wasassessed by exposing iMG cells to PE-conjugated (1250) beads (panel C) for 24 hours The iMG cells showed the ability of phagocytosis with morphologicalchanges (C) The read arrows show PE-conjugated beads phagocytosed by iMG cells After washing beads out cells were acquired in a flow cytometer andanalyzed iMG were first gated by forwardside scatter (not shown) PE uptake was analyzed as shown in (D) orange (left) histogram shows the cells withoutbeads (negative control) red (right) histogram shows the cells that phagocytosed to the PE-conjugated beads Quantification is shown in (E) iMG fromcontrols premanifest HD and HD patients showed similar phagocytic capacity (no significant difference found at the Kruskal-Wallis test) (F) Cytokineproduction during phagocytosis iMG cells were incubated with PE-conjugated beads (1250) for 24 hours Supernatant was collected and cytokines wereanalyzed by multiparametric assay Cytokine levels (in picograms per milliliter) were corrected by protein levels (in milligrams per milliliter) Although iMGcells from controls premanifest HD and patients with HD increased the production of the evaluated cytokines during phagocytosis the values weresignificant only for TNFα in controls andmanifest HD (p values are provided in the figure Wilcoxonmatched-pairs signed-rank test) Scale bar (AndashC) = 10mmHorizontal bars (E and F) show the mean and the standard error of the mean HD = Huntington disease PE = phycoerythrin TNF = tumor necrosis factor
NeurologyorgNN Neurology Neuroimmunology amp Neuroinflammation | Volume 8 Number 3 | May 2021 9
cytokines and reactive oxygen species which in turn will alsocontribute to neuronal death and further microglial activa-tion34We observed an increase in cytokine release (especiallyTNFα) following a phagocytic stimulus in our in vitro ex-periments In line with our findings postmortem studies havealso reported increased levels of inflammatory cytokines inbrain samples of patients with HD in comparison withcontrols3536 It is worth mentioning that cytokine release is amicroglial reaction to a proinflammatory challenge and undercontrolled conditions (such as the iMG cultures) phagocy-tosis is a noninflammatory function of microglia A proin-flammatory stimulus would be needed to appropriately assessdifferences in cytokine release between controls premanifestHD and manifest HD iMG cultures
The current study used a novel PET tracer for TSPO [11C]-ER176 to assess microglial activation in HD By using [11C]-PK11195 PET studies have shown a significant increase inmicroglial activation in the striatum of patients with HD16 aswell as in premanifest HD gene carriers18 compared withcontrols In both studies [11C]-PK11195 binding significantlycorrelated with disease severity as measured by decreased[11C]-raclopride (a marker of dopamine D2 and D3 receptoravailability) binding in the striatum1618 Similar results weredescribed in the hypothalamus17 Later the same work groupused a multimodal imaging approach combiningMRI and PETanalyses and worsening in atrophy evaluated by MRI was ac-companied by a reduction in [11C]-raclopride and an increasein [11C]-PK11195 bindings in patients with HD In pre-manifest HD increased level of microglial activation in theassociative striatum and in the regional network associated withcognition correlated with 5-year probability of HD onset19
More recently increased microglial activation in the somato-sensory cortex (as evaluated through [11C]-PK11195 binding)was associatedwith higher levels of IL-1β IL-6 IL-8 and TNFαproduced by stimulated peripheral blood monocytes frompremanifest HD gene carriers2037 Although these studiesprovided valuable information about neuroinflammation inHD indicating microglial activation even in premanifest sub-jects their results should be interpreted with caution The poorsignal-to-background ratio of first-generation TSPO radio-tracers poses important limitations to the analyses The ratio ofspecific to nonspecific binding of [11C]-PK11195 in humanbrain has been reported to be as low as 023839 As an attempt tomitigate this problem a recent study used the second-generation TSPO tracer [11C]-PBR28 to evaluate micro-glial activation in HD21 Of interest the results were similar toours ie increased levels of [11C]-PBR28 binding in theputamina and pallidi of HD gene carriers (which included 7manifest patients with HD and only 1 person with premanifestHD) in comparison with controls Of note the results of thesingle premanifest subject were midway between controls andthe remaining patients suggesting that microglial activationwas associated with disease progression21 The authors didconfirm that [11C]-PBR28 provides a high signal-to-background ratio an important feature required for the anal-ysis at the individual level and within individual brain regions
An important limitation of [11C]-PBR28 and other second-generation TSPO radioligands is that they have varying degreesof sensitivity to the single nucleotide polymorphism rs6971 inthe TSPO gene40 [11C]-ER176 has unique advantages overthe other TSPO binders being the only known TSPO radio-ligand that allows the inclusion of low-affinity binders Thebinding of ER176 in low-affinity binders was found to becomparable to that for PBR28 in high-affinity binders More-over ER176 was proven to be the only radioligand not con-taminated by radiometabolites accumulating in the brain14
Our findings must be interpreted while taking into accountthe limitations of the study which include the relatively smallsample size and the cross-sectional design The cross-sectionaldesign prevented the assessment of microglial activation inthe context of disease progression and development of motorand nonmotor symptoms In addition the use of an objectivemethod to measure striatal degeneration (eg [11C]-raclopride PET) would provide more accurate informationabout striatal neuron dysfunctiondeath Regarding the invitro experiments the use of a proinflammatory challenge(other than the beads added for the phagocytosis assay) in ourexperiments would provide valuable information on cytokinesrelease by iMG from the different groups It is worth men-tioning the strengths of the study that include the use of aunique TSPO ligand [11C]-ER176 the enrollment of bothpremanifest and manifest subjects and the exploratory in-vestigation of microglia activity
In conclusion our findings strengthen the evidence thatmicroglial activation is associated with HD pathophysiologyMicroglial activation is unlikely to be the initiating event inHD Yet it may exacerbate neuronal dysfunction and neu-ronal death thus contributing to disease progression It isimportant to refine our understanding of the more specificimmuneinflammatory mechanisms that are involved in HDand in neurodegeneration This may foster the developmentof new therapeutic interventions to halt the progression ofHD and other neurodegenerative diseases
AcknowledgmentThe authors acknowledge and thank all of the volunteers thatparticipated in this study and are indebted to their families fortheir magnificent support They also thank the staff from theUTHealth McGovern Medical School 3T MRI Center andfrom the HoustonMethodist Research Institute TranslationalImaging Core Facilities especially Vips Patel and Kim MDoan for their skilled technical support Dr Gabriel Fries forhis assistance with the genotyping analyses and the teammembers of the HDSA Center of Excellence at UTHealth
Study FundingThis study was funded by the HD Human BiologyProjectmdashHuntingtonrsquos Disease Society of America (HDSA)E Furr Stimming receives research funding from RocheGenetech Vaccinex Cures Within Reach HDSA Uniqureand CHDI Foundation
10 Neurology Neuroimmunology amp Neuroinflammation | Volume 8 Number 3 | May 2021 NeurologyorgNN
DisclosureThe authors report no disclosures relevant to the manuscriptGo to NeurologyorgNN for full disclosures
Publication HistoryReceived by Neurology Neuroimmunology amp NeuroinflammationSeptember 9 2020 Accepted in final form January 20 2021
References1 The Huntingtonrsquos Disease Collaborative Research Group A novel gene containing a
trinucleotide repeat that is expanded and unstable on Huntingtonrsquos disease chro-mosomes Cell 199372971ndash983
2 Ross CA Aylward EH Wild EJ et al Huntington disease natural history biomarkersand prospects for therapeutics Nat Rev Neurol 201410204ndash216
3 Soulet D Cicchetti F The role of immunity in Huntingtonrsquos disease Mol Psychiatry201116889ndash902
4 Myers RH Vonsattel JP Paskevich PA et al Decreased neuronal and increasedoligodendroglial densities in Huntingtonrsquos disease caudate nucleus J NeuropatholExp Neurol 199150729ndash742
5 Vonsattel JP Myers RH Stevens TJ Ferrante RJ Bird ED Richardson EP JrNeuropathological classification of Huntingtonrsquos disease J Neuropathol Exp Neurol198544559ndash577
6 Sapp E Kegel KB Aronin N et al Early and progressive accumulation of reactivemicroglia in the Huntington disease brain J Neuropathol Exp Neurol 200160161ndash172
7 Ciccocioppo F Bologna G Ercolino E et al Neurodegenerative diseases asproteinopathies-driven immune disorders Neural Regen Res 202015850ndash856
8 Rocha NP Ribeiro FM Furr-Stimming E Teixeira AL Neuroimmunology of Hun-tingtonrsquos disease revisiting evidence from human studies Mediators Inflamm 201620168653132
9 Vowinckel E Reutens D Becher B et al PK11195 binding to the peripheral ben-zodiazepine receptor as a marker of microglia activation in multiple sclerosis andexperimental autoimmune encephalomyelitis J Neurosci Res 199750345ndash353
10 Papadopoulos V Baraldi M Guilarte TR et al Translocator protein (18kDa) newnomenclature for the peripheral-type benzodiazepine receptor based on its structureand molecular function Trends Pharmacol Sci 200627402ndash409
11 Cumming P Burgher B Patkar O et al Sifting through the surfeit of neuro-inflammation tracers J Cereb Blood Flow Metab 201838204ndash224
12 Zanotti-Fregonara P Zhang Y Jenko KJ et al Synthesis and evaluation of translocator18 kDa protein (TSPO) positron emission tomography (PET) radioligands with lowbinding sensitivity to human single nucleotide polymorphism rs6971 ACS ChemNeurosci 20145963ndash971
13 Ikawa M Lohith TG Shrestha S et al 11C-ER176 a radioligand for 18-kDa trans-locator protein has adequate sensitivity to robustly image all three affinity genotypesin human brain J Nucl Med 201758320ndash325
14 Fujita M Kobayashi M Ikawa M et al Comparison of four (11)C-labeled PETligands to quantify translocator protein 18 kDa (TSPO) in human brain (R)-PK11195 PBR28 DPA-713 and ER176-based on recent publications that measuredspecific-to-non-displaceable ratios EJNMMI Res 2017784
15 Zanotti-Fregonara P Pascual B Veronese M et al Head-to-head comparison of (11)C-PBR28 and (11)C-ER176 for quantification of the translocator protein in thehuman brain Eur J Nucl Med Mol Imaging 2019461822ndash1829
16 Pavese N Gerhard A Tai YF et al Microglial activation correlates with severity inHuntington disease a clinical and PET study Neurology 2006661638ndash1643
17 Politis M Pavese N Tai YF Tabrizi SJ Barker RA Piccini P Hypothalamic in-volvement in Huntingtonrsquos disease an in vivo PET study Brain 20081312860ndash2869
18 Tai YF Pavese N Gerhard A et al Microglial activation in presymptomatic Hun-tingtonrsquos disease gene carriers Brain 20071301759ndash1766
19 Politis M Pavese N Tai YF et al Microglial activation in regions related to cognitivefunction predicts disease onset in Huntingtonrsquos disease a multimodal imaging studyHum Brain Mapp 201132258ndash270
20 Politis M Lahiri N Niccolini F et al Increased central microglial activation associatedwith peripheral cytokine levels in premanifest Huntingtonrsquos disease gene carriersNeurobiol Dis 201583115ndash121
21 Lois C Gonzalez I Izquierdo-Garcia D et al Neuroinflammation in Huntingtonrsquosdisease new insights with (11)C-PBR28 PETMRI ACS Chem Neurosci 201892563ndash2571
22 Huntington Study Group Unified Huntingtonrsquos Disease Rating Scale reliability andconsistency Mov Disord 199611136ndash142
23 Folstein MF Folstein SE McHugh PR ldquoMini-mental staterdquo A practical method forgrading the cognitive state of patients for the clinician J Psychiatr Res 197512189ndash198
24 McNally G Rickards H Horton M Craufurd D Exploring the validity of the shortversion of the Problem Behaviours Assessment (PBA-s) for Huntingtonrsquos disease arasch analysis J Huntingtons Dis 20154347ndash369
25 Warner JH Sampaio C Modeling variability in the progression of Huntingtonrsquosdisease a novel modeling approach applied to structural imaging markers fromTRACK-HD CPT Pharmacometrics Syst Pharmacol 20165437ndash445
26 Lyoo CH Ikawa M Liow JS et al Cerebellum can serve as a pseudo-reference regionin Alzheimer disease to detect neuroinflammation measured with PET radioligandbinding to translocator protein J Nucl Med 201556701ndash706
27 Reuter M Schmansky NJ Rosas HD Fischl B Within-subject template estimation forunbiased longitudinal image analysis Neuroimage 2012611402ndash1418
28 Yushkevich PA Piven J Hazlett HC et al User-guided 3D active contour segmen-tation of anatomical structures significantly improved efficiency and reliabilityNeuroimage 2006311116ndash1128
29 Ohgidani M Kato TA Setoyama D et al Direct induction of ramified microglia-likecells from human monocytes dynamic microglial dysfunction in Nasu-Hakola dis-ease Sci Rep 201444957
30 Ohgidani M Kato TA Kanba S Introducing directly induced microglia-like (iMG)cells from fresh human monocytes a novel translational research tool for psychiatricdisorders Front Cell Neurosci 20159184
31 Kim SU de Vellis J Microglia in health and disease J Neurosci Res 200581302-31332 Ross CA Tabrizi SJ Huntingtonrsquos disease from molecular pathogenesis to clinical
treatment Lancet Neurol 20111083ndash9833 Chitnis TWeiner HL CNS inflammation and neurodegeneration J Clin Invest 2017
1273577ndash3587
Appendix Authors
Name Location Contribution
Natalia PRocha PhD
The Mitchell Center forAlzheimerrsquos Disease andRelated Brain DisordersDepartment of NeurologyMcGovern Medical SchoolThe University of TexasHealth Science CenterHouston
Designed andconceptualized the studymajor role in theacquisition of dataanalyzed the data anddrafted the manuscript forintellectual content
OdelinCharron MS
Department of NeurologyThe University of Texas atAustin
Major role in data analysis
Leigh BLathamPhD
School of MedicineUniversity of WashingtonSeattle
Major role in theacquisition of data
Gabriela DColpo PhD
Neuropsychiatry ProgramDepartment of Psychiatryand Behavioral SciencesMcGovern Medical SchoolUniversity of TexasHouston
Major role in theacquisition of data
PaoloZanotti-FregonaraMD PhD
Houston MethodistResearch Institute andWeill Cornell Medicine TX
Major role in the studydesign and the acquisitionof data
MeixiangYu PhD
Houston MethodistResearch Institute andWeill Cornell Medicine TX
Major role in theacquisition of data
LeorahFreemanMD PhD
Department of NeurologyThe University of Texas atAustin
Major role in the studydesign data analysis andinterpretation and theacquisition of data
Erin FurrStimmingMD
HDSA Center of Excellenceat University of TexasHealth Science Center atHouston
Major role in the studydesign and acquisition ofdata interpreted the dataand revised themanuscript for intellectualcontent
Antonio LTeixeiraMD PhD
Neuropsychiatry ProgramDepartment of Psychiatryand Behavioral SciencesMcGovern Medical SchoolUniversity of TexasHouston
Major role in the studydesign and acquisition ofdata interpreted the dataand revised themanuscript for intellectualcontent
NeurologyorgNN Neurology Neuroimmunology amp Neuroinflammation | Volume 8 Number 3 | May 2021 11
34 Rocha NP de Miranda AS Teixeira AL Insights into neuroinflammation in Parkin-sonrsquos disease from biomarkers to anti-inflammatory based therapies Biomed Res Int20152015628192
35 Silvestroni A Faull RL Strand AD Moller T Distinct neuroinflammatory profile inpost-mortem human Huntingtonrsquos disease Neuroreport 2009201098ndash1103
36 Bjorkqvist M Wild EJ Thiele J et al A novel pathogenic pathway of immune acti-vation detectable before clinical onset in Huntingtonrsquos disease J Exp Med 20082051869ndash1877
37 Politis M Lahiri N Niccolini F et al Corrigendum to ldquoIncreased central microglialactivation associated with peripheral cytokine levels in premanifest Huntingtonrsquos
disease gene carriersrdquo [Neurobiol Dis 83 (2015) 115-121] Neurobiol Dis 201798162
38 Kropholler MA Boellaard R Schuitemaker A Folkersma H van Berckel BN Lam-mertsma AA Evaluation of reference tissue models for the analysis of [11C](R)-PK11195 studies J Cereb Blood Flow Metab 2006261431ndash1441
39 Turkheimer FE Edison P Pavese N et al Reference and target region modeling of[11C]-(R)-PK11195 brain studies J Nucl Med 200748158ndash167
40 Kreisl WC Jenko KJ Hines CS et al A genetic polymorphism for translocator protein18 kDa affects both in vitro and in vivo radioligand binding in human brain to thisputative biomarker of neuroinflammation J Cereb Blood FlowMetab 20133353ndash58
12 Neurology Neuroimmunology amp Neuroinflammation | Volume 8 Number 3 | May 2021 NeurologyorgNN
DOI 101212NXI000000000000098420218 Neurol Neuroimmunol Neuroinflamm
Natalia P Rocha Odelin Charron Leigh B Latham et al Pathophysiology
Microglia Activation in Basal Ganglia Is a Late Event in Huntington Disease
This information is current as of April 1 2021
ServicesUpdated Information amp
httpnnneurologyorgcontent83e984fullhtmlincluding high resolution figures can be found at
References httpnnneurologyorgcontent83e984fullhtmlref-list-1
This article cites 40 articles 4 of which you can access for free at
Subspecialty Collections
httpnnneurologyorgcgicollectionpetPET
httpnnneurologyorgcgicollectionhuntingtons_diseaseHuntingtons diseasefollowing collection(s) This article along with others on similar topics appears in the
Permissions amp Licensing
httpnnneurologyorgmiscaboutxhtmlpermissionsits entirety can be found online atInformation about reproducing this article in parts (figurestables) or in
Reprints
httpnnneurologyorgmiscaddirxhtmlreprintsusInformation about ordering reprints can be found online
Academy of Neurology All rights reserved Online ISSN 2332-7812Copyright copy 2021 The Author(s) Published by Wolters Kluwer Health Inc on behalf of the AmericanPublished since April 2014 it is an open-access online-only continuous publication journal Copyright
is an official journal of the American Academy of NeurologyNeurol Neuroimmunol Neuroinflamm

Reader and analyzed in a Bio-Plex Manager Software whichpresented data as median fluorescence intensity (MFI) Thecytokine concentrations in the samples were calculatedbased on a standard curve in which the MFI values obtainedfrom the standard were plotted against the known concen-trations (in picograms per milliliter) Cytokine levels (inpicograms per milliliter) were measured in the supernatant24 hours after bead exposure or 24 hours of culture withoutany stimulus (baseline condition) and the values were cor-rected by protein levels (in milligrams per milliliter) in thecell lysates using the Pierce BCA Protein Assay Kit (Ther-moFisher Scientific Waltham MA) The final results areprovided in pgmg
DNA Polymorphism GenotypingPeripheral blood mononuclear cells were used for DNAextraction using the QIAamp DNA Blood Mini Kit
(Qiagen Hilden Germany) and subsequent genotyping forthe rs6971 polymorphism (Ala147Thr) in the TSPO gene(TaqMan assay ID C_2512465_20 Thermo Fisher Sci-entific) Allele Ala147 was linked to Vic and allele Thr147was linked to FAM PCRs were performed in QuantStudio7 Flex Real-Time PCR system (Life TechnologiesThermoFisher Scientific)
Statistical AnalysisAssociation between dichotomous variables was assessedwith the χ2 test Our limited sample size made it difficult toascertain data distribution and therefore nonparametrictests were used Comparisons between the 3 groups weremade by the Kruskal-Wallis test When appropriate posthoc analyses were used to determine significant differencesbetween pairs of groups (Dunn Multiple ComparisonTest) The Mann-Whitney test was used for 2-group
Figure 1 Microglial Activation in HD
(A) PET image showing increased [11C]-ER176 standardized uptake value ratio (SUVR) in bilateral putamina and pallidi of a patient with manifest HD incomparison with a control Note that there is no visible difference in [11C]-ER176 SUVR for the premanifest HD gene carrier (central image) (B) Groupcomparisons in left (L) and right (R) putamina and pallidi Significant differences between groups are indicated by different letters (Kruskal-Wallis testfollowed by Dunn multiple comparisons test) Horizontal bars show the mean and the standard error of the mean Data from other regions of interest areshown in figure e-1 (linkslwwcomNXIA459) HD = Huntington disease
NeurologyorgNN Neurology Neuroimmunology amp Neuroinflammation | Volume 8 Number 3 | May 2021 5
comparisons (premanifest HD vs manifest HD) Spearmancorrelations were performed to examine the relationshipbetween [11]C-ER176 SUVR in the ROIs significantlydifferent between groups and (1) CAP score (2) volume inthe same ROIs and (3) scores in the clinical scales Allstatistical tests were 2 tailed and were performed using asignificance level of α = 005 Statistical analyses wereperformed using SPSS software version 260 and GraphPadPrism version 50
Standard Protocol Approvals Registrationsand Patient ConsentsAll subjects provided written informed consent before ad-mission to the study The Research Ethics Committees ofUTHealth and Houston Methodist Research Institute ap-proved this study
An Investigational New Drug Application (IND) was sub-mitted and authorization was obtained from the Food andDrug Administration to use [11C]-ER176 as a PET Imagingagent in this study (IND 139220)
Data AvailabilityThe raw data not provided in the article will be made availableby the authors at the request of other investigators for pur-poses of replicating procedures and results
ResultsClinical Evaluation and TSPO GenotypingDemographic and clinical characteristics of participants included inthis study are shown in the table There was no significant differ-ence between controls premanifest HD gene carriers and patientswith HD regarding age sex and educational level As expectedpatients with manifest HD presented a higher CAP score andworse scores in the scales that evaluatedmotor symptoms severityfunctional capacity and independence in comparison with pre-manifest HD (table) The age at clinical diagnosis was 4767 plusmn1035 years for the patients with manifest HD Premanifest HDgene carriers had amean of 1298 plusmn 617 years before the predictedclinical diagnosis of HD The 3 groups presented similar perfor-mance on the MMSE Patients with the clinical diagnosis of HDpresented worse performance on the SDMT and verbal fluencytest in comparisonwith premanifest and control subjects Controlsindividuals with premanifest HD and patients with HD presentedsimilar scores in the PBA-s subscales except for executive functionin which HD gene carriers (both premanifest and manifest HD)scored worse than controls evidencing that those problems arepresent before clinical diagnosis of HD (table)
The groups presented similar distribution of TSPO genotype(table) Individuals with genotype Ala147Ala147 were clas-sified as high-affinity binders and those with genotype
Figure 2 Microglial Activation in HD Is Associated With Disease Burden
Among HD gene carriers (premanifest andmanifest HD) a higher CAP score (a proxy for cumulative disease burden) was significantly associated with higher[11C]-ER176 standardized uptake value ratio (SUVR) in the (A) left putamen (ρ = 0664 p = 0018) (B) left pallidum (ρ = 0657 p = 0020) (C) right putamen (ρ =0671 p = 0017) and (D) right pallidum (ρ = 0734 p = 0007) Note the 2 clusters in [11C]-ER176 SUVR composed of premanifest HD gene carriers (CAP lt100)and manifest patients with HD (CAP gt100) CAP = CAG age product HD = Huntingtonrsquos disease
6 Neurology Neuroimmunology amp Neuroinflammation | Volume 8 Number 3 | May 2021 NeurologyorgNN
Ala147Thr147 as mixed affinity None of our patients pre-sented the genotype Thr147Thr147 (low-affinity binders)Because the groups presented comparable distribution ofTSPO genotype (table) and the outcome parameter wasSUVR which minimizes genotype differences no correctionfor genotype-related binding affinity was performed
In Vivo Assessment of Microglial ActivationPatients with HD demonstrate higher [11C]-ER176 SUVR inboth the left and right globi pallidi and putamina in comparisonwith controls These differences were not significant when wecompared premanifest HD gene carriers with patients withHDorcontrols (figure 1 and figure e-1 linkslwwcomNXIA459)These results indicate increased microglial activation in the puta-mina and globi pallidi of patients with manifest HDWe found nodifferences between different TSPO genotypes on [11C]-ER176SUVR when controlling for HD diagnosis (data not shown)
Among HD gene carriers (premanifest and manifest HD) ahigher CAP score was significantly associated with higher[11C]-ER176 SUVR in the left putamen (ρ = 0664 p =0018) left pallidum (ρ = 0657 p = 0020) right putamen(ρ = 0671 p = 0017) and right pallidum (ρ = 0734 p =0007) (figure 2) Corroborating these findings increased[11C]-ER176 SUVRwas associated with decreased volume inthe same ROIs (figure 3)
In Vitro Evaluation of the Dynamic Functionsof MicrogliaWe confirmed that the iMG cells have the immunophenotypeof microglial cells such as CX3CR1highCCR2low and atypical (ie ramified) morphology (figure 4) The iMG cellsexpress dynamic functions such as phagocytosis and secretionof inflammatory cytokines making them a valuable tool toassess microglial functioning30 The phagocytosis assaydemonstrated that iMG cells obtained from controls pre-manifest HD and HD patients (figure 5 AndashC) showedsimilar phagocytic capacity (figure 5E) Secretion of in-flammatory cytokines (TNFα IL-6 and IL-1β) duringphagocytosis was measured from culture supernatant Asshown in figure 5F although iMG cells from controls pre-manifest HD and patients with HD increased the productionof the evaluated cytokines during phagocytosis the valueswere significant only for TNFα in controls and manifest HD(figure 5F)
DiscussionBy using [11C]-ER176 PET we verified that patients withHD have increased microglial activation in basal gangliastructures that are involved in HD pathophysiologyie putamina and globi pallidi In addition we found that
Figure 3 Microglial Activation Is Associated With Brain Atrophy
Increased microglial activation is associated with decreased volume in putamen and pallidus Controls are displayed in red dots premanifest HD genecarriers in green dots andmanifest HD in black dots A higher [11C]-ER176 standardized uptake value ratio (SUVR) was associatedwith a decreased volume inthe (A) left putamen (ρ = minus0569 p = 0014) (B) left pallidum (ρ = minus0591 p = 0010) (C) right putamen (ρ = minus0601 p = 0008) and (D) right pallidum (ρ = minus0686 p= 0002) ICV = intracranial volume The dashed line was placed to emphasize the 2 clusters composed of controls + premanifest HD and manifest HD genecarriers HD = Huntington disease
NeurologyorgNN Neurology Neuroimmunology amp Neuroinflammation | Volume 8 Number 3 | May 2021 7
premanifest HD gene carriers demonstrate [11C]-ER176SUVRs comparable to controls suggesting that microglialactivation is a later process in HD pathophysiology becomingevident only after the onset of motor symptoms (figure 1)Supporting these findings we reported a significant correla-tion between increased microglial activation and higher CAPscores a proxy for cumulative disease burden (figure 2)Moreover increased microglial activation in putamina andpallidi was significantly associated with reduced volumes inthe same ROIs (figure 3) Individuals with manifest HDpresented a significant decrease in the cortical volume incomparison with controls (data not shown) Despite alreadypresenting cortical atrophy manifest HD individuals do nothave increased microglial activation in the cortex (figure 1)The only areas displaying increased microglia activation arethose that are known to be affected early in HD pathophysi-ology Because PET scans require individuals to be still for 90minutes our sample did not include patients in moderatelatestages of HD Whether increased microglia activation willoccur in the cortex later in the HD course is yet to be de-termined Altogether our data demonstrate that microglialactivation is involved in HD pathophysiology and is associ-ated with disease progression (as demonstrated by the CAPscore and atrophy in basal ganglia structures) Our findingssupport the conclusions of previous research that reportedthat microglial activation correlates with disease severitycontributing to the ongoing neuronal degeneration in HD16
HD has been regarded as an important model to study neu-rodegenerative diseases as it is caused by a single geneticmutation and is amenable to predictive genetic testing en-abling the study of patients before clinical diagnosis2 In this
study we assessed microglial activation not only in patientswith HD but also in premanifest HD gene carriers with amean of 13 years before the predicted onset of clinicalsymptoms Our results corroborate the view that microglialactivation is unlikely to be the initiating event in HD but maycontribute to neuronal death through multiple pathways31
Similar to other neurodegenerative diseases the mecha-nisms underlying selective neuronal dysfunction and deathin HD are still uncertain In addition to impairment in sys-tems for handling abnormal proteins other metabolicpathways and mechanisms might contribute to neuro-degeneration and progression of HD such as mitochondrialdysfunction impaired neurotrophic support oxidativestress and inflammatoryimmune mechanisms83233 In thecurrent study we described a significant increase in micro-glial activation in regions of neurodegeneration in HDsuggesting that this process is related to the disease patho-physiology and not to random premorbid events Accord-ingly postmortem studies reported significant microglialactivation in regions implicated in HD pathogenesis46 Thefact that we found significant microglial activation in patientswith manifest HD (but not in premanifest HD gene carriers)in comparison with controls suggests that this event occursin response or concomitantly to neuronal death This as-sumption is further corroborated by the finding of significantassociations between microglial activation and ROI volumesas well as CAP scores Here again our data are corroboratedby postmortem findings in which the number of activatedmicroglia in the striatum and cortex correlated with neuronalloss confirming the hypothesis that neuroinflammation maybe elicited by degenerating neurons6
Figure 4 Inducing Microglia-Like Cells From Human Peripheral Blood Monocytes
Peripheral blood monocytes were cultured for 14 days in conventional conditions (A) or with GM-CSF and IL-34 resulting in induced microglia like cells thatexhibit microglia-like morphology and increased CX3CR1CCR2 ratio (B)
8 Neurology Neuroimmunology amp Neuroinflammation | Volume 8 Number 3 | May 2021 NeurologyorgNN
Neuroinflammation elicited by neuronal death can be at firstregarded as a positive event as it promotes clearance of celldebris However inflammatory mechanisms contribute toneurodegeneration and dying neurons further activate in-flammatory responses resulting in a vicious cycle of in-flammation and neuronal death34 Therefore inflammatoryresponses although essential for tissue homeostasis cancontribute to neuronal injury As neural tissues have a
restricted cell renewal and regenerative capacity the CNS isextremely vulnerable to uncontrolled andor chronic immuneand inflammatory processes34 Interestingly enough our datacorroborate this hypothesis as we observed increased micro-glial activation later in the HD course (ie manifest HD)reinforcing the idea that neuroinflammation contributes todisease progression rather than to disease onset Activatedmicroglia releases proinflammatory mediators including
Figure 5 Phagocytosis by Induced Microglia-Like (iMG) Cells
Peripheral blood monocytes (day 1 panel A) were cultured for 14 days with GM-CSF and IL-34 resulting in iMG cells (day 14 panel B) Phagocytosis wasassessed by exposing iMG cells to PE-conjugated (1250) beads (panel C) for 24 hours The iMG cells showed the ability of phagocytosis with morphologicalchanges (C) The read arrows show PE-conjugated beads phagocytosed by iMG cells After washing beads out cells were acquired in a flow cytometer andanalyzed iMG were first gated by forwardside scatter (not shown) PE uptake was analyzed as shown in (D) orange (left) histogram shows the cells withoutbeads (negative control) red (right) histogram shows the cells that phagocytosed to the PE-conjugated beads Quantification is shown in (E) iMG fromcontrols premanifest HD and HD patients showed similar phagocytic capacity (no significant difference found at the Kruskal-Wallis test) (F) Cytokineproduction during phagocytosis iMG cells were incubated with PE-conjugated beads (1250) for 24 hours Supernatant was collected and cytokines wereanalyzed by multiparametric assay Cytokine levels (in picograms per milliliter) were corrected by protein levels (in milligrams per milliliter) Although iMGcells from controls premanifest HD and patients with HD increased the production of the evaluated cytokines during phagocytosis the values weresignificant only for TNFα in controls andmanifest HD (p values are provided in the figure Wilcoxonmatched-pairs signed-rank test) Scale bar (AndashC) = 10mmHorizontal bars (E and F) show the mean and the standard error of the mean HD = Huntington disease PE = phycoerythrin TNF = tumor necrosis factor
NeurologyorgNN Neurology Neuroimmunology amp Neuroinflammation | Volume 8 Number 3 | May 2021 9
cytokines and reactive oxygen species which in turn will alsocontribute to neuronal death and further microglial activa-tion34We observed an increase in cytokine release (especiallyTNFα) following a phagocytic stimulus in our in vitro ex-periments In line with our findings postmortem studies havealso reported increased levels of inflammatory cytokines inbrain samples of patients with HD in comparison withcontrols3536 It is worth mentioning that cytokine release is amicroglial reaction to a proinflammatory challenge and undercontrolled conditions (such as the iMG cultures) phagocy-tosis is a noninflammatory function of microglia A proin-flammatory stimulus would be needed to appropriately assessdifferences in cytokine release between controls premanifestHD and manifest HD iMG cultures
The current study used a novel PET tracer for TSPO [11C]-ER176 to assess microglial activation in HD By using [11C]-PK11195 PET studies have shown a significant increase inmicroglial activation in the striatum of patients with HD16 aswell as in premanifest HD gene carriers18 compared withcontrols In both studies [11C]-PK11195 binding significantlycorrelated with disease severity as measured by decreased[11C]-raclopride (a marker of dopamine D2 and D3 receptoravailability) binding in the striatum1618 Similar results weredescribed in the hypothalamus17 Later the same work groupused a multimodal imaging approach combiningMRI and PETanalyses and worsening in atrophy evaluated by MRI was ac-companied by a reduction in [11C]-raclopride and an increasein [11C]-PK11195 bindings in patients with HD In pre-manifest HD increased level of microglial activation in theassociative striatum and in the regional network associated withcognition correlated with 5-year probability of HD onset19
More recently increased microglial activation in the somato-sensory cortex (as evaluated through [11C]-PK11195 binding)was associatedwith higher levels of IL-1β IL-6 IL-8 and TNFαproduced by stimulated peripheral blood monocytes frompremanifest HD gene carriers2037 Although these studiesprovided valuable information about neuroinflammation inHD indicating microglial activation even in premanifest sub-jects their results should be interpreted with caution The poorsignal-to-background ratio of first-generation TSPO radio-tracers poses important limitations to the analyses The ratio ofspecific to nonspecific binding of [11C]-PK11195 in humanbrain has been reported to be as low as 023839 As an attempt tomitigate this problem a recent study used the second-generation TSPO tracer [11C]-PBR28 to evaluate micro-glial activation in HD21 Of interest the results were similar toours ie increased levels of [11C]-PBR28 binding in theputamina and pallidi of HD gene carriers (which included 7manifest patients with HD and only 1 person with premanifestHD) in comparison with controls Of note the results of thesingle premanifest subject were midway between controls andthe remaining patients suggesting that microglial activationwas associated with disease progression21 The authors didconfirm that [11C]-PBR28 provides a high signal-to-background ratio an important feature required for the anal-ysis at the individual level and within individual brain regions
An important limitation of [11C]-PBR28 and other second-generation TSPO radioligands is that they have varying degreesof sensitivity to the single nucleotide polymorphism rs6971 inthe TSPO gene40 [11C]-ER176 has unique advantages overthe other TSPO binders being the only known TSPO radio-ligand that allows the inclusion of low-affinity binders Thebinding of ER176 in low-affinity binders was found to becomparable to that for PBR28 in high-affinity binders More-over ER176 was proven to be the only radioligand not con-taminated by radiometabolites accumulating in the brain14
Our findings must be interpreted while taking into accountthe limitations of the study which include the relatively smallsample size and the cross-sectional design The cross-sectionaldesign prevented the assessment of microglial activation inthe context of disease progression and development of motorand nonmotor symptoms In addition the use of an objectivemethod to measure striatal degeneration (eg [11C]-raclopride PET) would provide more accurate informationabout striatal neuron dysfunctiondeath Regarding the invitro experiments the use of a proinflammatory challenge(other than the beads added for the phagocytosis assay) in ourexperiments would provide valuable information on cytokinesrelease by iMG from the different groups It is worth men-tioning the strengths of the study that include the use of aunique TSPO ligand [11C]-ER176 the enrollment of bothpremanifest and manifest subjects and the exploratory in-vestigation of microglia activity
In conclusion our findings strengthen the evidence thatmicroglial activation is associated with HD pathophysiologyMicroglial activation is unlikely to be the initiating event inHD Yet it may exacerbate neuronal dysfunction and neu-ronal death thus contributing to disease progression It isimportant to refine our understanding of the more specificimmuneinflammatory mechanisms that are involved in HDand in neurodegeneration This may foster the developmentof new therapeutic interventions to halt the progression ofHD and other neurodegenerative diseases
AcknowledgmentThe authors acknowledge and thank all of the volunteers thatparticipated in this study and are indebted to their families fortheir magnificent support They also thank the staff from theUTHealth McGovern Medical School 3T MRI Center andfrom the HoustonMethodist Research Institute TranslationalImaging Core Facilities especially Vips Patel and Kim MDoan for their skilled technical support Dr Gabriel Fries forhis assistance with the genotyping analyses and the teammembers of the HDSA Center of Excellence at UTHealth
Study FundingThis study was funded by the HD Human BiologyProjectmdashHuntingtonrsquos Disease Society of America (HDSA)E Furr Stimming receives research funding from RocheGenetech Vaccinex Cures Within Reach HDSA Uniqureand CHDI Foundation
10 Neurology Neuroimmunology amp Neuroinflammation | Volume 8 Number 3 | May 2021 NeurologyorgNN
DisclosureThe authors report no disclosures relevant to the manuscriptGo to NeurologyorgNN for full disclosures
Publication HistoryReceived by Neurology Neuroimmunology amp NeuroinflammationSeptember 9 2020 Accepted in final form January 20 2021
References1 The Huntingtonrsquos Disease Collaborative Research Group A novel gene containing a
trinucleotide repeat that is expanded and unstable on Huntingtonrsquos disease chro-mosomes Cell 199372971ndash983
2 Ross CA Aylward EH Wild EJ et al Huntington disease natural history biomarkersand prospects for therapeutics Nat Rev Neurol 201410204ndash216
3 Soulet D Cicchetti F The role of immunity in Huntingtonrsquos disease Mol Psychiatry201116889ndash902
4 Myers RH Vonsattel JP Paskevich PA et al Decreased neuronal and increasedoligodendroglial densities in Huntingtonrsquos disease caudate nucleus J NeuropatholExp Neurol 199150729ndash742
5 Vonsattel JP Myers RH Stevens TJ Ferrante RJ Bird ED Richardson EP JrNeuropathological classification of Huntingtonrsquos disease J Neuropathol Exp Neurol198544559ndash577
6 Sapp E Kegel KB Aronin N et al Early and progressive accumulation of reactivemicroglia in the Huntington disease brain J Neuropathol Exp Neurol 200160161ndash172
7 Ciccocioppo F Bologna G Ercolino E et al Neurodegenerative diseases asproteinopathies-driven immune disorders Neural Regen Res 202015850ndash856
8 Rocha NP Ribeiro FM Furr-Stimming E Teixeira AL Neuroimmunology of Hun-tingtonrsquos disease revisiting evidence from human studies Mediators Inflamm 201620168653132
9 Vowinckel E Reutens D Becher B et al PK11195 binding to the peripheral ben-zodiazepine receptor as a marker of microglia activation in multiple sclerosis andexperimental autoimmune encephalomyelitis J Neurosci Res 199750345ndash353
10 Papadopoulos V Baraldi M Guilarte TR et al Translocator protein (18kDa) newnomenclature for the peripheral-type benzodiazepine receptor based on its structureand molecular function Trends Pharmacol Sci 200627402ndash409
11 Cumming P Burgher B Patkar O et al Sifting through the surfeit of neuro-inflammation tracers J Cereb Blood Flow Metab 201838204ndash224
12 Zanotti-Fregonara P Zhang Y Jenko KJ et al Synthesis and evaluation of translocator18 kDa protein (TSPO) positron emission tomography (PET) radioligands with lowbinding sensitivity to human single nucleotide polymorphism rs6971 ACS ChemNeurosci 20145963ndash971
13 Ikawa M Lohith TG Shrestha S et al 11C-ER176 a radioligand for 18-kDa trans-locator protein has adequate sensitivity to robustly image all three affinity genotypesin human brain J Nucl Med 201758320ndash325
14 Fujita M Kobayashi M Ikawa M et al Comparison of four (11)C-labeled PETligands to quantify translocator protein 18 kDa (TSPO) in human brain (R)-PK11195 PBR28 DPA-713 and ER176-based on recent publications that measuredspecific-to-non-displaceable ratios EJNMMI Res 2017784
15 Zanotti-Fregonara P Pascual B Veronese M et al Head-to-head comparison of (11)C-PBR28 and (11)C-ER176 for quantification of the translocator protein in thehuman brain Eur J Nucl Med Mol Imaging 2019461822ndash1829
16 Pavese N Gerhard A Tai YF et al Microglial activation correlates with severity inHuntington disease a clinical and PET study Neurology 2006661638ndash1643
17 Politis M Pavese N Tai YF Tabrizi SJ Barker RA Piccini P Hypothalamic in-volvement in Huntingtonrsquos disease an in vivo PET study Brain 20081312860ndash2869
18 Tai YF Pavese N Gerhard A et al Microglial activation in presymptomatic Hun-tingtonrsquos disease gene carriers Brain 20071301759ndash1766
19 Politis M Pavese N Tai YF et al Microglial activation in regions related to cognitivefunction predicts disease onset in Huntingtonrsquos disease a multimodal imaging studyHum Brain Mapp 201132258ndash270
20 Politis M Lahiri N Niccolini F et al Increased central microglial activation associatedwith peripheral cytokine levels in premanifest Huntingtonrsquos disease gene carriersNeurobiol Dis 201583115ndash121
21 Lois C Gonzalez I Izquierdo-Garcia D et al Neuroinflammation in Huntingtonrsquosdisease new insights with (11)C-PBR28 PETMRI ACS Chem Neurosci 201892563ndash2571
22 Huntington Study Group Unified Huntingtonrsquos Disease Rating Scale reliability andconsistency Mov Disord 199611136ndash142
23 Folstein MF Folstein SE McHugh PR ldquoMini-mental staterdquo A practical method forgrading the cognitive state of patients for the clinician J Psychiatr Res 197512189ndash198
24 McNally G Rickards H Horton M Craufurd D Exploring the validity of the shortversion of the Problem Behaviours Assessment (PBA-s) for Huntingtonrsquos disease arasch analysis J Huntingtons Dis 20154347ndash369
25 Warner JH Sampaio C Modeling variability in the progression of Huntingtonrsquosdisease a novel modeling approach applied to structural imaging markers fromTRACK-HD CPT Pharmacometrics Syst Pharmacol 20165437ndash445
26 Lyoo CH Ikawa M Liow JS et al Cerebellum can serve as a pseudo-reference regionin Alzheimer disease to detect neuroinflammation measured with PET radioligandbinding to translocator protein J Nucl Med 201556701ndash706
27 Reuter M Schmansky NJ Rosas HD Fischl B Within-subject template estimation forunbiased longitudinal image analysis Neuroimage 2012611402ndash1418
28 Yushkevich PA Piven J Hazlett HC et al User-guided 3D active contour segmen-tation of anatomical structures significantly improved efficiency and reliabilityNeuroimage 2006311116ndash1128
29 Ohgidani M Kato TA Setoyama D et al Direct induction of ramified microglia-likecells from human monocytes dynamic microglial dysfunction in Nasu-Hakola dis-ease Sci Rep 201444957
30 Ohgidani M Kato TA Kanba S Introducing directly induced microglia-like (iMG)cells from fresh human monocytes a novel translational research tool for psychiatricdisorders Front Cell Neurosci 20159184
31 Kim SU de Vellis J Microglia in health and disease J Neurosci Res 200581302-31332 Ross CA Tabrizi SJ Huntingtonrsquos disease from molecular pathogenesis to clinical
treatment Lancet Neurol 20111083ndash9833 Chitnis TWeiner HL CNS inflammation and neurodegeneration J Clin Invest 2017
1273577ndash3587
Appendix Authors
Name Location Contribution
Natalia PRocha PhD
The Mitchell Center forAlzheimerrsquos Disease andRelated Brain DisordersDepartment of NeurologyMcGovern Medical SchoolThe University of TexasHealth Science CenterHouston
Designed andconceptualized the studymajor role in theacquisition of dataanalyzed the data anddrafted the manuscript forintellectual content
OdelinCharron MS
Department of NeurologyThe University of Texas atAustin
Major role in data analysis
Leigh BLathamPhD
School of MedicineUniversity of WashingtonSeattle
Major role in theacquisition of data
Gabriela DColpo PhD
Neuropsychiatry ProgramDepartment of Psychiatryand Behavioral SciencesMcGovern Medical SchoolUniversity of TexasHouston
Major role in theacquisition of data
PaoloZanotti-FregonaraMD PhD
Houston MethodistResearch Institute andWeill Cornell Medicine TX
Major role in the studydesign and the acquisitionof data
MeixiangYu PhD
Houston MethodistResearch Institute andWeill Cornell Medicine TX
Major role in theacquisition of data
LeorahFreemanMD PhD
Department of NeurologyThe University of Texas atAustin
Major role in the studydesign data analysis andinterpretation and theacquisition of data
Erin FurrStimmingMD
HDSA Center of Excellenceat University of TexasHealth Science Center atHouston
Major role in the studydesign and acquisition ofdata interpreted the dataand revised themanuscript for intellectualcontent
Antonio LTeixeiraMD PhD
Neuropsychiatry ProgramDepartment of Psychiatryand Behavioral SciencesMcGovern Medical SchoolUniversity of TexasHouston
Major role in the studydesign and acquisition ofdata interpreted the dataand revised themanuscript for intellectualcontent
NeurologyorgNN Neurology Neuroimmunology amp Neuroinflammation | Volume 8 Number 3 | May 2021 11
34 Rocha NP de Miranda AS Teixeira AL Insights into neuroinflammation in Parkin-sonrsquos disease from biomarkers to anti-inflammatory based therapies Biomed Res Int20152015628192
35 Silvestroni A Faull RL Strand AD Moller T Distinct neuroinflammatory profile inpost-mortem human Huntingtonrsquos disease Neuroreport 2009201098ndash1103
36 Bjorkqvist M Wild EJ Thiele J et al A novel pathogenic pathway of immune acti-vation detectable before clinical onset in Huntingtonrsquos disease J Exp Med 20082051869ndash1877
37 Politis M Lahiri N Niccolini F et al Corrigendum to ldquoIncreased central microglialactivation associated with peripheral cytokine levels in premanifest Huntingtonrsquos
disease gene carriersrdquo [Neurobiol Dis 83 (2015) 115-121] Neurobiol Dis 201798162
38 Kropholler MA Boellaard R Schuitemaker A Folkersma H van Berckel BN Lam-mertsma AA Evaluation of reference tissue models for the analysis of [11C](R)-PK11195 studies J Cereb Blood Flow Metab 2006261431ndash1441
39 Turkheimer FE Edison P Pavese N et al Reference and target region modeling of[11C]-(R)-PK11195 brain studies J Nucl Med 200748158ndash167
40 Kreisl WC Jenko KJ Hines CS et al A genetic polymorphism for translocator protein18 kDa affects both in vitro and in vivo radioligand binding in human brain to thisputative biomarker of neuroinflammation J Cereb Blood FlowMetab 20133353ndash58
12 Neurology Neuroimmunology amp Neuroinflammation | Volume 8 Number 3 | May 2021 NeurologyorgNN
DOI 101212NXI000000000000098420218 Neurol Neuroimmunol Neuroinflamm
Natalia P Rocha Odelin Charron Leigh B Latham et al Pathophysiology
Microglia Activation in Basal Ganglia Is a Late Event in Huntington Disease
This information is current as of April 1 2021
ServicesUpdated Information amp
httpnnneurologyorgcontent83e984fullhtmlincluding high resolution figures can be found at
References httpnnneurologyorgcontent83e984fullhtmlref-list-1
This article cites 40 articles 4 of which you can access for free at
Subspecialty Collections
httpnnneurologyorgcgicollectionpetPET
httpnnneurologyorgcgicollectionhuntingtons_diseaseHuntingtons diseasefollowing collection(s) This article along with others on similar topics appears in the
Permissions amp Licensing
httpnnneurologyorgmiscaboutxhtmlpermissionsits entirety can be found online atInformation about reproducing this article in parts (figurestables) or in
Reprints
httpnnneurologyorgmiscaddirxhtmlreprintsusInformation about ordering reprints can be found online
Academy of Neurology All rights reserved Online ISSN 2332-7812Copyright copy 2021 The Author(s) Published by Wolters Kluwer Health Inc on behalf of the AmericanPublished since April 2014 it is an open-access online-only continuous publication journal Copyright
is an official journal of the American Academy of NeurologyNeurol Neuroimmunol Neuroinflamm
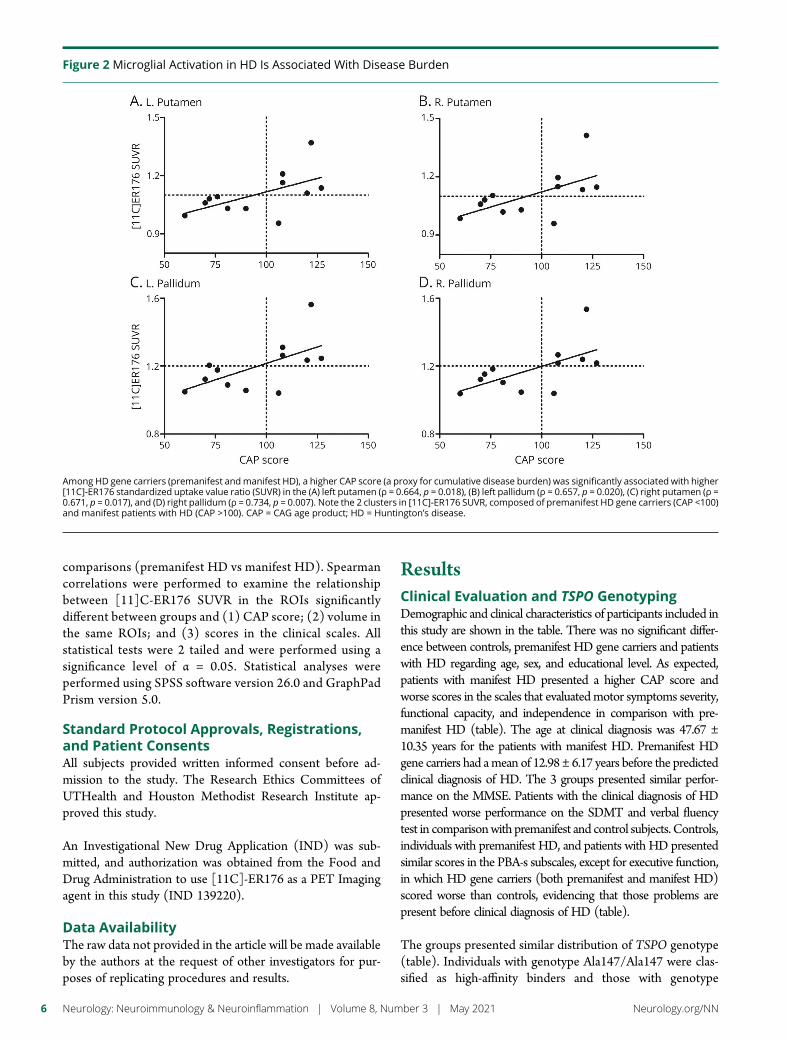
comparisons (premanifest HD vs manifest HD) Spearmancorrelations were performed to examine the relationshipbetween [11]C-ER176 SUVR in the ROIs significantlydifferent between groups and (1) CAP score (2) volume inthe same ROIs and (3) scores in the clinical scales Allstatistical tests were 2 tailed and were performed using asignificance level of α = 005 Statistical analyses wereperformed using SPSS software version 260 and GraphPadPrism version 50
Standard Protocol Approvals Registrationsand Patient ConsentsAll subjects provided written informed consent before ad-mission to the study The Research Ethics Committees ofUTHealth and Houston Methodist Research Institute ap-proved this study
An Investigational New Drug Application (IND) was sub-mitted and authorization was obtained from the Food andDrug Administration to use [11C]-ER176 as a PET Imagingagent in this study (IND 139220)
Data AvailabilityThe raw data not provided in the article will be made availableby the authors at the request of other investigators for pur-poses of replicating procedures and results
ResultsClinical Evaluation and TSPO GenotypingDemographic and clinical characteristics of participants included inthis study are shown in the table There was no significant differ-ence between controls premanifest HD gene carriers and patientswith HD regarding age sex and educational level As expectedpatients with manifest HD presented a higher CAP score andworse scores in the scales that evaluatedmotor symptoms severityfunctional capacity and independence in comparison with pre-manifest HD (table) The age at clinical diagnosis was 4767 plusmn1035 years for the patients with manifest HD Premanifest HDgene carriers had amean of 1298 plusmn 617 years before the predictedclinical diagnosis of HD The 3 groups presented similar perfor-mance on the MMSE Patients with the clinical diagnosis of HDpresented worse performance on the SDMT and verbal fluencytest in comparisonwith premanifest and control subjects Controlsindividuals with premanifest HD and patients with HD presentedsimilar scores in the PBA-s subscales except for executive functionin which HD gene carriers (both premanifest and manifest HD)scored worse than controls evidencing that those problems arepresent before clinical diagnosis of HD (table)
The groups presented similar distribution of TSPO genotype(table) Individuals with genotype Ala147Ala147 were clas-sified as high-affinity binders and those with genotype
Figure 2 Microglial Activation in HD Is Associated With Disease Burden
Among HD gene carriers (premanifest andmanifest HD) a higher CAP score (a proxy for cumulative disease burden) was significantly associated with higher[11C]-ER176 standardized uptake value ratio (SUVR) in the (A) left putamen (ρ = 0664 p = 0018) (B) left pallidum (ρ = 0657 p = 0020) (C) right putamen (ρ =0671 p = 0017) and (D) right pallidum (ρ = 0734 p = 0007) Note the 2 clusters in [11C]-ER176 SUVR composed of premanifest HD gene carriers (CAP lt100)and manifest patients with HD (CAP gt100) CAP = CAG age product HD = Huntingtonrsquos disease
6 Neurology Neuroimmunology amp Neuroinflammation | Volume 8 Number 3 | May 2021 NeurologyorgNN
Ala147Thr147 as mixed affinity None of our patients pre-sented the genotype Thr147Thr147 (low-affinity binders)Because the groups presented comparable distribution ofTSPO genotype (table) and the outcome parameter wasSUVR which minimizes genotype differences no correctionfor genotype-related binding affinity was performed
In Vivo Assessment of Microglial ActivationPatients with HD demonstrate higher [11C]-ER176 SUVR inboth the left and right globi pallidi and putamina in comparisonwith controls These differences were not significant when wecompared premanifest HD gene carriers with patients withHDorcontrols (figure 1 and figure e-1 linkslwwcomNXIA459)These results indicate increased microglial activation in the puta-mina and globi pallidi of patients with manifest HDWe found nodifferences between different TSPO genotypes on [11C]-ER176SUVR when controlling for HD diagnosis (data not shown)
Among HD gene carriers (premanifest and manifest HD) ahigher CAP score was significantly associated with higher[11C]-ER176 SUVR in the left putamen (ρ = 0664 p =0018) left pallidum (ρ = 0657 p = 0020) right putamen(ρ = 0671 p = 0017) and right pallidum (ρ = 0734 p =0007) (figure 2) Corroborating these findings increased[11C]-ER176 SUVRwas associated with decreased volume inthe same ROIs (figure 3)
In Vitro Evaluation of the Dynamic Functionsof MicrogliaWe confirmed that the iMG cells have the immunophenotypeof microglial cells such as CX3CR1highCCR2low and atypical (ie ramified) morphology (figure 4) The iMG cellsexpress dynamic functions such as phagocytosis and secretionof inflammatory cytokines making them a valuable tool toassess microglial functioning30 The phagocytosis assaydemonstrated that iMG cells obtained from controls pre-manifest HD and HD patients (figure 5 AndashC) showedsimilar phagocytic capacity (figure 5E) Secretion of in-flammatory cytokines (TNFα IL-6 and IL-1β) duringphagocytosis was measured from culture supernatant Asshown in figure 5F although iMG cells from controls pre-manifest HD and patients with HD increased the productionof the evaluated cytokines during phagocytosis the valueswere significant only for TNFα in controls and manifest HD(figure 5F)
DiscussionBy using [11C]-ER176 PET we verified that patients withHD have increased microglial activation in basal gangliastructures that are involved in HD pathophysiologyie putamina and globi pallidi In addition we found that
Figure 3 Microglial Activation Is Associated With Brain Atrophy
Increased microglial activation is associated with decreased volume in putamen and pallidus Controls are displayed in red dots premanifest HD genecarriers in green dots andmanifest HD in black dots A higher [11C]-ER176 standardized uptake value ratio (SUVR) was associatedwith a decreased volume inthe (A) left putamen (ρ = minus0569 p = 0014) (B) left pallidum (ρ = minus0591 p = 0010) (C) right putamen (ρ = minus0601 p = 0008) and (D) right pallidum (ρ = minus0686 p= 0002) ICV = intracranial volume The dashed line was placed to emphasize the 2 clusters composed of controls + premanifest HD and manifest HD genecarriers HD = Huntington disease
NeurologyorgNN Neurology Neuroimmunology amp Neuroinflammation | Volume 8 Number 3 | May 2021 7
premanifest HD gene carriers demonstrate [11C]-ER176SUVRs comparable to controls suggesting that microglialactivation is a later process in HD pathophysiology becomingevident only after the onset of motor symptoms (figure 1)Supporting these findings we reported a significant correla-tion between increased microglial activation and higher CAPscores a proxy for cumulative disease burden (figure 2)Moreover increased microglial activation in putamina andpallidi was significantly associated with reduced volumes inthe same ROIs (figure 3) Individuals with manifest HDpresented a significant decrease in the cortical volume incomparison with controls (data not shown) Despite alreadypresenting cortical atrophy manifest HD individuals do nothave increased microglial activation in the cortex (figure 1)The only areas displaying increased microglia activation arethose that are known to be affected early in HD pathophysi-ology Because PET scans require individuals to be still for 90minutes our sample did not include patients in moderatelatestages of HD Whether increased microglia activation willoccur in the cortex later in the HD course is yet to be de-termined Altogether our data demonstrate that microglialactivation is involved in HD pathophysiology and is associ-ated with disease progression (as demonstrated by the CAPscore and atrophy in basal ganglia structures) Our findingssupport the conclusions of previous research that reportedthat microglial activation correlates with disease severitycontributing to the ongoing neuronal degeneration in HD16
HD has been regarded as an important model to study neu-rodegenerative diseases as it is caused by a single geneticmutation and is amenable to predictive genetic testing en-abling the study of patients before clinical diagnosis2 In this
study we assessed microglial activation not only in patientswith HD but also in premanifest HD gene carriers with amean of 13 years before the predicted onset of clinicalsymptoms Our results corroborate the view that microglialactivation is unlikely to be the initiating event in HD but maycontribute to neuronal death through multiple pathways31
Similar to other neurodegenerative diseases the mecha-nisms underlying selective neuronal dysfunction and deathin HD are still uncertain In addition to impairment in sys-tems for handling abnormal proteins other metabolicpathways and mechanisms might contribute to neuro-degeneration and progression of HD such as mitochondrialdysfunction impaired neurotrophic support oxidativestress and inflammatoryimmune mechanisms83233 In thecurrent study we described a significant increase in micro-glial activation in regions of neurodegeneration in HDsuggesting that this process is related to the disease patho-physiology and not to random premorbid events Accord-ingly postmortem studies reported significant microglialactivation in regions implicated in HD pathogenesis46 Thefact that we found significant microglial activation in patientswith manifest HD (but not in premanifest HD gene carriers)in comparison with controls suggests that this event occursin response or concomitantly to neuronal death This as-sumption is further corroborated by the finding of significantassociations between microglial activation and ROI volumesas well as CAP scores Here again our data are corroboratedby postmortem findings in which the number of activatedmicroglia in the striatum and cortex correlated with neuronalloss confirming the hypothesis that neuroinflammation maybe elicited by degenerating neurons6
Figure 4 Inducing Microglia-Like Cells From Human Peripheral Blood Monocytes
Peripheral blood monocytes were cultured for 14 days in conventional conditions (A) or with GM-CSF and IL-34 resulting in induced microglia like cells thatexhibit microglia-like morphology and increased CX3CR1CCR2 ratio (B)
8 Neurology Neuroimmunology amp Neuroinflammation | Volume 8 Number 3 | May 2021 NeurologyorgNN
Neuroinflammation elicited by neuronal death can be at firstregarded as a positive event as it promotes clearance of celldebris However inflammatory mechanisms contribute toneurodegeneration and dying neurons further activate in-flammatory responses resulting in a vicious cycle of in-flammation and neuronal death34 Therefore inflammatoryresponses although essential for tissue homeostasis cancontribute to neuronal injury As neural tissues have a
restricted cell renewal and regenerative capacity the CNS isextremely vulnerable to uncontrolled andor chronic immuneand inflammatory processes34 Interestingly enough our datacorroborate this hypothesis as we observed increased micro-glial activation later in the HD course (ie manifest HD)reinforcing the idea that neuroinflammation contributes todisease progression rather than to disease onset Activatedmicroglia releases proinflammatory mediators including
Figure 5 Phagocytosis by Induced Microglia-Like (iMG) Cells
Peripheral blood monocytes (day 1 panel A) were cultured for 14 days with GM-CSF and IL-34 resulting in iMG cells (day 14 panel B) Phagocytosis wasassessed by exposing iMG cells to PE-conjugated (1250) beads (panel C) for 24 hours The iMG cells showed the ability of phagocytosis with morphologicalchanges (C) The read arrows show PE-conjugated beads phagocytosed by iMG cells After washing beads out cells were acquired in a flow cytometer andanalyzed iMG were first gated by forwardside scatter (not shown) PE uptake was analyzed as shown in (D) orange (left) histogram shows the cells withoutbeads (negative control) red (right) histogram shows the cells that phagocytosed to the PE-conjugated beads Quantification is shown in (E) iMG fromcontrols premanifest HD and HD patients showed similar phagocytic capacity (no significant difference found at the Kruskal-Wallis test) (F) Cytokineproduction during phagocytosis iMG cells were incubated with PE-conjugated beads (1250) for 24 hours Supernatant was collected and cytokines wereanalyzed by multiparametric assay Cytokine levels (in picograms per milliliter) were corrected by protein levels (in milligrams per milliliter) Although iMGcells from controls premanifest HD and patients with HD increased the production of the evaluated cytokines during phagocytosis the values weresignificant only for TNFα in controls andmanifest HD (p values are provided in the figure Wilcoxonmatched-pairs signed-rank test) Scale bar (AndashC) = 10mmHorizontal bars (E and F) show the mean and the standard error of the mean HD = Huntington disease PE = phycoerythrin TNF = tumor necrosis factor
NeurologyorgNN Neurology Neuroimmunology amp Neuroinflammation | Volume 8 Number 3 | May 2021 9
cytokines and reactive oxygen species which in turn will alsocontribute to neuronal death and further microglial activa-tion34We observed an increase in cytokine release (especiallyTNFα) following a phagocytic stimulus in our in vitro ex-periments In line with our findings postmortem studies havealso reported increased levels of inflammatory cytokines inbrain samples of patients with HD in comparison withcontrols3536 It is worth mentioning that cytokine release is amicroglial reaction to a proinflammatory challenge and undercontrolled conditions (such as the iMG cultures) phagocy-tosis is a noninflammatory function of microglia A proin-flammatory stimulus would be needed to appropriately assessdifferences in cytokine release between controls premanifestHD and manifest HD iMG cultures
The current study used a novel PET tracer for TSPO [11C]-ER176 to assess microglial activation in HD By using [11C]-PK11195 PET studies have shown a significant increase inmicroglial activation in the striatum of patients with HD16 aswell as in premanifest HD gene carriers18 compared withcontrols In both studies [11C]-PK11195 binding significantlycorrelated with disease severity as measured by decreased[11C]-raclopride (a marker of dopamine D2 and D3 receptoravailability) binding in the striatum1618 Similar results weredescribed in the hypothalamus17 Later the same work groupused a multimodal imaging approach combiningMRI and PETanalyses and worsening in atrophy evaluated by MRI was ac-companied by a reduction in [11C]-raclopride and an increasein [11C]-PK11195 bindings in patients with HD In pre-manifest HD increased level of microglial activation in theassociative striatum and in the regional network associated withcognition correlated with 5-year probability of HD onset19
More recently increased microglial activation in the somato-sensory cortex (as evaluated through [11C]-PK11195 binding)was associatedwith higher levels of IL-1β IL-6 IL-8 and TNFαproduced by stimulated peripheral blood monocytes frompremanifest HD gene carriers2037 Although these studiesprovided valuable information about neuroinflammation inHD indicating microglial activation even in premanifest sub-jects their results should be interpreted with caution The poorsignal-to-background ratio of first-generation TSPO radio-tracers poses important limitations to the analyses The ratio ofspecific to nonspecific binding of [11C]-PK11195 in humanbrain has been reported to be as low as 023839 As an attempt tomitigate this problem a recent study used the second-generation TSPO tracer [11C]-PBR28 to evaluate micro-glial activation in HD21 Of interest the results were similar toours ie increased levels of [11C]-PBR28 binding in theputamina and pallidi of HD gene carriers (which included 7manifest patients with HD and only 1 person with premanifestHD) in comparison with controls Of note the results of thesingle premanifest subject were midway between controls andthe remaining patients suggesting that microglial activationwas associated with disease progression21 The authors didconfirm that [11C]-PBR28 provides a high signal-to-background ratio an important feature required for the anal-ysis at the individual level and within individual brain regions
An important limitation of [11C]-PBR28 and other second-generation TSPO radioligands is that they have varying degreesof sensitivity to the single nucleotide polymorphism rs6971 inthe TSPO gene40 [11C]-ER176 has unique advantages overthe other TSPO binders being the only known TSPO radio-ligand that allows the inclusion of low-affinity binders Thebinding of ER176 in low-affinity binders was found to becomparable to that for PBR28 in high-affinity binders More-over ER176 was proven to be the only radioligand not con-taminated by radiometabolites accumulating in the brain14
Our findings must be interpreted while taking into accountthe limitations of the study which include the relatively smallsample size and the cross-sectional design The cross-sectionaldesign prevented the assessment of microglial activation inthe context of disease progression and development of motorand nonmotor symptoms In addition the use of an objectivemethod to measure striatal degeneration (eg [11C]-raclopride PET) would provide more accurate informationabout striatal neuron dysfunctiondeath Regarding the invitro experiments the use of a proinflammatory challenge(other than the beads added for the phagocytosis assay) in ourexperiments would provide valuable information on cytokinesrelease by iMG from the different groups It is worth men-tioning the strengths of the study that include the use of aunique TSPO ligand [11C]-ER176 the enrollment of bothpremanifest and manifest subjects and the exploratory in-vestigation of microglia activity
In conclusion our findings strengthen the evidence thatmicroglial activation is associated with HD pathophysiologyMicroglial activation is unlikely to be the initiating event inHD Yet it may exacerbate neuronal dysfunction and neu-ronal death thus contributing to disease progression It isimportant to refine our understanding of the more specificimmuneinflammatory mechanisms that are involved in HDand in neurodegeneration This may foster the developmentof new therapeutic interventions to halt the progression ofHD and other neurodegenerative diseases
AcknowledgmentThe authors acknowledge and thank all of the volunteers thatparticipated in this study and are indebted to their families fortheir magnificent support They also thank the staff from theUTHealth McGovern Medical School 3T MRI Center andfrom the HoustonMethodist Research Institute TranslationalImaging Core Facilities especially Vips Patel and Kim MDoan for their skilled technical support Dr Gabriel Fries forhis assistance with the genotyping analyses and the teammembers of the HDSA Center of Excellence at UTHealth
Study FundingThis study was funded by the HD Human BiologyProjectmdashHuntingtonrsquos Disease Society of America (HDSA)E Furr Stimming receives research funding from RocheGenetech Vaccinex Cures Within Reach HDSA Uniqureand CHDI Foundation
10 Neurology Neuroimmunology amp Neuroinflammation | Volume 8 Number 3 | May 2021 NeurologyorgNN
DisclosureThe authors report no disclosures relevant to the manuscriptGo to NeurologyorgNN for full disclosures
Publication HistoryReceived by Neurology Neuroimmunology amp NeuroinflammationSeptember 9 2020 Accepted in final form January 20 2021
References1 The Huntingtonrsquos Disease Collaborative Research Group A novel gene containing a
trinucleotide repeat that is expanded and unstable on Huntingtonrsquos disease chro-mosomes Cell 199372971ndash983
2 Ross CA Aylward EH Wild EJ et al Huntington disease natural history biomarkersand prospects for therapeutics Nat Rev Neurol 201410204ndash216
3 Soulet D Cicchetti F The role of immunity in Huntingtonrsquos disease Mol Psychiatry201116889ndash902
4 Myers RH Vonsattel JP Paskevich PA et al Decreased neuronal and increasedoligodendroglial densities in Huntingtonrsquos disease caudate nucleus J NeuropatholExp Neurol 199150729ndash742
5 Vonsattel JP Myers RH Stevens TJ Ferrante RJ Bird ED Richardson EP JrNeuropathological classification of Huntingtonrsquos disease J Neuropathol Exp Neurol198544559ndash577
6 Sapp E Kegel KB Aronin N et al Early and progressive accumulation of reactivemicroglia in the Huntington disease brain J Neuropathol Exp Neurol 200160161ndash172
7 Ciccocioppo F Bologna G Ercolino E et al Neurodegenerative diseases asproteinopathies-driven immune disorders Neural Regen Res 202015850ndash856
8 Rocha NP Ribeiro FM Furr-Stimming E Teixeira AL Neuroimmunology of Hun-tingtonrsquos disease revisiting evidence from human studies Mediators Inflamm 201620168653132
9 Vowinckel E Reutens D Becher B et al PK11195 binding to the peripheral ben-zodiazepine receptor as a marker of microglia activation in multiple sclerosis andexperimental autoimmune encephalomyelitis J Neurosci Res 199750345ndash353
10 Papadopoulos V Baraldi M Guilarte TR et al Translocator protein (18kDa) newnomenclature for the peripheral-type benzodiazepine receptor based on its structureand molecular function Trends Pharmacol Sci 200627402ndash409
11 Cumming P Burgher B Patkar O et al Sifting through the surfeit of neuro-inflammation tracers J Cereb Blood Flow Metab 201838204ndash224
12 Zanotti-Fregonara P Zhang Y Jenko KJ et al Synthesis and evaluation of translocator18 kDa protein (TSPO) positron emission tomography (PET) radioligands with lowbinding sensitivity to human single nucleotide polymorphism rs6971 ACS ChemNeurosci 20145963ndash971
13 Ikawa M Lohith TG Shrestha S et al 11C-ER176 a radioligand for 18-kDa trans-locator protein has adequate sensitivity to robustly image all three affinity genotypesin human brain J Nucl Med 201758320ndash325
14 Fujita M Kobayashi M Ikawa M et al Comparison of four (11)C-labeled PETligands to quantify translocator protein 18 kDa (TSPO) in human brain (R)-PK11195 PBR28 DPA-713 and ER176-based on recent publications that measuredspecific-to-non-displaceable ratios EJNMMI Res 2017784
15 Zanotti-Fregonara P Pascual B Veronese M et al Head-to-head comparison of (11)C-PBR28 and (11)C-ER176 for quantification of the translocator protein in thehuman brain Eur J Nucl Med Mol Imaging 2019461822ndash1829
16 Pavese N Gerhard A Tai YF et al Microglial activation correlates with severity inHuntington disease a clinical and PET study Neurology 2006661638ndash1643
17 Politis M Pavese N Tai YF Tabrizi SJ Barker RA Piccini P Hypothalamic in-volvement in Huntingtonrsquos disease an in vivo PET study Brain 20081312860ndash2869
18 Tai YF Pavese N Gerhard A et al Microglial activation in presymptomatic Hun-tingtonrsquos disease gene carriers Brain 20071301759ndash1766
19 Politis M Pavese N Tai YF et al Microglial activation in regions related to cognitivefunction predicts disease onset in Huntingtonrsquos disease a multimodal imaging studyHum Brain Mapp 201132258ndash270
20 Politis M Lahiri N Niccolini F et al Increased central microglial activation associatedwith peripheral cytokine levels in premanifest Huntingtonrsquos disease gene carriersNeurobiol Dis 201583115ndash121
21 Lois C Gonzalez I Izquierdo-Garcia D et al Neuroinflammation in Huntingtonrsquosdisease new insights with (11)C-PBR28 PETMRI ACS Chem Neurosci 201892563ndash2571
22 Huntington Study Group Unified Huntingtonrsquos Disease Rating Scale reliability andconsistency Mov Disord 199611136ndash142
23 Folstein MF Folstein SE McHugh PR ldquoMini-mental staterdquo A practical method forgrading the cognitive state of patients for the clinician J Psychiatr Res 197512189ndash198
24 McNally G Rickards H Horton M Craufurd D Exploring the validity of the shortversion of the Problem Behaviours Assessment (PBA-s) for Huntingtonrsquos disease arasch analysis J Huntingtons Dis 20154347ndash369
25 Warner JH Sampaio C Modeling variability in the progression of Huntingtonrsquosdisease a novel modeling approach applied to structural imaging markers fromTRACK-HD CPT Pharmacometrics Syst Pharmacol 20165437ndash445
26 Lyoo CH Ikawa M Liow JS et al Cerebellum can serve as a pseudo-reference regionin Alzheimer disease to detect neuroinflammation measured with PET radioligandbinding to translocator protein J Nucl Med 201556701ndash706
27 Reuter M Schmansky NJ Rosas HD Fischl B Within-subject template estimation forunbiased longitudinal image analysis Neuroimage 2012611402ndash1418
28 Yushkevich PA Piven J Hazlett HC et al User-guided 3D active contour segmen-tation of anatomical structures significantly improved efficiency and reliabilityNeuroimage 2006311116ndash1128
29 Ohgidani M Kato TA Setoyama D et al Direct induction of ramified microglia-likecells from human monocytes dynamic microglial dysfunction in Nasu-Hakola dis-ease Sci Rep 201444957
30 Ohgidani M Kato TA Kanba S Introducing directly induced microglia-like (iMG)cells from fresh human monocytes a novel translational research tool for psychiatricdisorders Front Cell Neurosci 20159184
31 Kim SU de Vellis J Microglia in health and disease J Neurosci Res 200581302-31332 Ross CA Tabrizi SJ Huntingtonrsquos disease from molecular pathogenesis to clinical
treatment Lancet Neurol 20111083ndash9833 Chitnis TWeiner HL CNS inflammation and neurodegeneration J Clin Invest 2017
1273577ndash3587
Appendix Authors
Name Location Contribution
Natalia PRocha PhD
The Mitchell Center forAlzheimerrsquos Disease andRelated Brain DisordersDepartment of NeurologyMcGovern Medical SchoolThe University of TexasHealth Science CenterHouston
Designed andconceptualized the studymajor role in theacquisition of dataanalyzed the data anddrafted the manuscript forintellectual content
OdelinCharron MS
Department of NeurologyThe University of Texas atAustin
Major role in data analysis
Leigh BLathamPhD
School of MedicineUniversity of WashingtonSeattle
Major role in theacquisition of data
Gabriela DColpo PhD
Neuropsychiatry ProgramDepartment of Psychiatryand Behavioral SciencesMcGovern Medical SchoolUniversity of TexasHouston
Major role in theacquisition of data
PaoloZanotti-FregonaraMD PhD
Houston MethodistResearch Institute andWeill Cornell Medicine TX
Major role in the studydesign and the acquisitionof data
MeixiangYu PhD
Houston MethodistResearch Institute andWeill Cornell Medicine TX
Major role in theacquisition of data
LeorahFreemanMD PhD
Department of NeurologyThe University of Texas atAustin
Major role in the studydesign data analysis andinterpretation and theacquisition of data
Erin FurrStimmingMD
HDSA Center of Excellenceat University of TexasHealth Science Center atHouston
Major role in the studydesign and acquisition ofdata interpreted the dataand revised themanuscript for intellectualcontent
Antonio LTeixeiraMD PhD
Neuropsychiatry ProgramDepartment of Psychiatryand Behavioral SciencesMcGovern Medical SchoolUniversity of TexasHouston
Major role in the studydesign and acquisition ofdata interpreted the dataand revised themanuscript for intellectualcontent
NeurologyorgNN Neurology Neuroimmunology amp Neuroinflammation | Volume 8 Number 3 | May 2021 11
34 Rocha NP de Miranda AS Teixeira AL Insights into neuroinflammation in Parkin-sonrsquos disease from biomarkers to anti-inflammatory based therapies Biomed Res Int20152015628192
35 Silvestroni A Faull RL Strand AD Moller T Distinct neuroinflammatory profile inpost-mortem human Huntingtonrsquos disease Neuroreport 2009201098ndash1103
36 Bjorkqvist M Wild EJ Thiele J et al A novel pathogenic pathway of immune acti-vation detectable before clinical onset in Huntingtonrsquos disease J Exp Med 20082051869ndash1877
37 Politis M Lahiri N Niccolini F et al Corrigendum to ldquoIncreased central microglialactivation associated with peripheral cytokine levels in premanifest Huntingtonrsquos
disease gene carriersrdquo [Neurobiol Dis 83 (2015) 115-121] Neurobiol Dis 201798162
38 Kropholler MA Boellaard R Schuitemaker A Folkersma H van Berckel BN Lam-mertsma AA Evaluation of reference tissue models for the analysis of [11C](R)-PK11195 studies J Cereb Blood Flow Metab 2006261431ndash1441
39 Turkheimer FE Edison P Pavese N et al Reference and target region modeling of[11C]-(R)-PK11195 brain studies J Nucl Med 200748158ndash167
40 Kreisl WC Jenko KJ Hines CS et al A genetic polymorphism for translocator protein18 kDa affects both in vitro and in vivo radioligand binding in human brain to thisputative biomarker of neuroinflammation J Cereb Blood FlowMetab 20133353ndash58
12 Neurology Neuroimmunology amp Neuroinflammation | Volume 8 Number 3 | May 2021 NeurologyorgNN
DOI 101212NXI000000000000098420218 Neurol Neuroimmunol Neuroinflamm
Natalia P Rocha Odelin Charron Leigh B Latham et al Pathophysiology
Microglia Activation in Basal Ganglia Is a Late Event in Huntington Disease
This information is current as of April 1 2021
ServicesUpdated Information amp
httpnnneurologyorgcontent83e984fullhtmlincluding high resolution figures can be found at
References httpnnneurologyorgcontent83e984fullhtmlref-list-1
This article cites 40 articles 4 of which you can access for free at
Subspecialty Collections
httpnnneurologyorgcgicollectionpetPET
httpnnneurologyorgcgicollectionhuntingtons_diseaseHuntingtons diseasefollowing collection(s) This article along with others on similar topics appears in the
Permissions amp Licensing
httpnnneurologyorgmiscaboutxhtmlpermissionsits entirety can be found online atInformation about reproducing this article in parts (figurestables) or in
Reprints
httpnnneurologyorgmiscaddirxhtmlreprintsusInformation about ordering reprints can be found online
Academy of Neurology All rights reserved Online ISSN 2332-7812Copyright copy 2021 The Author(s) Published by Wolters Kluwer Health Inc on behalf of the AmericanPublished since April 2014 it is an open-access online-only continuous publication journal Copyright
is an official journal of the American Academy of NeurologyNeurol Neuroimmunol Neuroinflamm

Ala147Thr147 as mixed affinity None of our patients pre-sented the genotype Thr147Thr147 (low-affinity binders)Because the groups presented comparable distribution ofTSPO genotype (table) and the outcome parameter wasSUVR which minimizes genotype differences no correctionfor genotype-related binding affinity was performed
In Vivo Assessment of Microglial ActivationPatients with HD demonstrate higher [11C]-ER176 SUVR inboth the left and right globi pallidi and putamina in comparisonwith controls These differences were not significant when wecompared premanifest HD gene carriers with patients withHDorcontrols (figure 1 and figure e-1 linkslwwcomNXIA459)These results indicate increased microglial activation in the puta-mina and globi pallidi of patients with manifest HDWe found nodifferences between different TSPO genotypes on [11C]-ER176SUVR when controlling for HD diagnosis (data not shown)
Among HD gene carriers (premanifest and manifest HD) ahigher CAP score was significantly associated with higher[11C]-ER176 SUVR in the left putamen (ρ = 0664 p =0018) left pallidum (ρ = 0657 p = 0020) right putamen(ρ = 0671 p = 0017) and right pallidum (ρ = 0734 p =0007) (figure 2) Corroborating these findings increased[11C]-ER176 SUVRwas associated with decreased volume inthe same ROIs (figure 3)
In Vitro Evaluation of the Dynamic Functionsof MicrogliaWe confirmed that the iMG cells have the immunophenotypeof microglial cells such as CX3CR1highCCR2low and atypical (ie ramified) morphology (figure 4) The iMG cellsexpress dynamic functions such as phagocytosis and secretionof inflammatory cytokines making them a valuable tool toassess microglial functioning30 The phagocytosis assaydemonstrated that iMG cells obtained from controls pre-manifest HD and HD patients (figure 5 AndashC) showedsimilar phagocytic capacity (figure 5E) Secretion of in-flammatory cytokines (TNFα IL-6 and IL-1β) duringphagocytosis was measured from culture supernatant Asshown in figure 5F although iMG cells from controls pre-manifest HD and patients with HD increased the productionof the evaluated cytokines during phagocytosis the valueswere significant only for TNFα in controls and manifest HD(figure 5F)
DiscussionBy using [11C]-ER176 PET we verified that patients withHD have increased microglial activation in basal gangliastructures that are involved in HD pathophysiologyie putamina and globi pallidi In addition we found that
Figure 3 Microglial Activation Is Associated With Brain Atrophy
Increased microglial activation is associated with decreased volume in putamen and pallidus Controls are displayed in red dots premanifest HD genecarriers in green dots andmanifest HD in black dots A higher [11C]-ER176 standardized uptake value ratio (SUVR) was associatedwith a decreased volume inthe (A) left putamen (ρ = minus0569 p = 0014) (B) left pallidum (ρ = minus0591 p = 0010) (C) right putamen (ρ = minus0601 p = 0008) and (D) right pallidum (ρ = minus0686 p= 0002) ICV = intracranial volume The dashed line was placed to emphasize the 2 clusters composed of controls + premanifest HD and manifest HD genecarriers HD = Huntington disease
NeurologyorgNN Neurology Neuroimmunology amp Neuroinflammation | Volume 8 Number 3 | May 2021 7
premanifest HD gene carriers demonstrate [11C]-ER176SUVRs comparable to controls suggesting that microglialactivation is a later process in HD pathophysiology becomingevident only after the onset of motor symptoms (figure 1)Supporting these findings we reported a significant correla-tion between increased microglial activation and higher CAPscores a proxy for cumulative disease burden (figure 2)Moreover increased microglial activation in putamina andpallidi was significantly associated with reduced volumes inthe same ROIs (figure 3) Individuals with manifest HDpresented a significant decrease in the cortical volume incomparison with controls (data not shown) Despite alreadypresenting cortical atrophy manifest HD individuals do nothave increased microglial activation in the cortex (figure 1)The only areas displaying increased microglia activation arethose that are known to be affected early in HD pathophysi-ology Because PET scans require individuals to be still for 90minutes our sample did not include patients in moderatelatestages of HD Whether increased microglia activation willoccur in the cortex later in the HD course is yet to be de-termined Altogether our data demonstrate that microglialactivation is involved in HD pathophysiology and is associ-ated with disease progression (as demonstrated by the CAPscore and atrophy in basal ganglia structures) Our findingssupport the conclusions of previous research that reportedthat microglial activation correlates with disease severitycontributing to the ongoing neuronal degeneration in HD16
HD has been regarded as an important model to study neu-rodegenerative diseases as it is caused by a single geneticmutation and is amenable to predictive genetic testing en-abling the study of patients before clinical diagnosis2 In this
study we assessed microglial activation not only in patientswith HD but also in premanifest HD gene carriers with amean of 13 years before the predicted onset of clinicalsymptoms Our results corroborate the view that microglialactivation is unlikely to be the initiating event in HD but maycontribute to neuronal death through multiple pathways31
Similar to other neurodegenerative diseases the mecha-nisms underlying selective neuronal dysfunction and deathin HD are still uncertain In addition to impairment in sys-tems for handling abnormal proteins other metabolicpathways and mechanisms might contribute to neuro-degeneration and progression of HD such as mitochondrialdysfunction impaired neurotrophic support oxidativestress and inflammatoryimmune mechanisms83233 In thecurrent study we described a significant increase in micro-glial activation in regions of neurodegeneration in HDsuggesting that this process is related to the disease patho-physiology and not to random premorbid events Accord-ingly postmortem studies reported significant microglialactivation in regions implicated in HD pathogenesis46 Thefact that we found significant microglial activation in patientswith manifest HD (but not in premanifest HD gene carriers)in comparison with controls suggests that this event occursin response or concomitantly to neuronal death This as-sumption is further corroborated by the finding of significantassociations between microglial activation and ROI volumesas well as CAP scores Here again our data are corroboratedby postmortem findings in which the number of activatedmicroglia in the striatum and cortex correlated with neuronalloss confirming the hypothesis that neuroinflammation maybe elicited by degenerating neurons6
Figure 4 Inducing Microglia-Like Cells From Human Peripheral Blood Monocytes
Peripheral blood monocytes were cultured for 14 days in conventional conditions (A) or with GM-CSF and IL-34 resulting in induced microglia like cells thatexhibit microglia-like morphology and increased CX3CR1CCR2 ratio (B)
8 Neurology Neuroimmunology amp Neuroinflammation | Volume 8 Number 3 | May 2021 NeurologyorgNN
Neuroinflammation elicited by neuronal death can be at firstregarded as a positive event as it promotes clearance of celldebris However inflammatory mechanisms contribute toneurodegeneration and dying neurons further activate in-flammatory responses resulting in a vicious cycle of in-flammation and neuronal death34 Therefore inflammatoryresponses although essential for tissue homeostasis cancontribute to neuronal injury As neural tissues have a
restricted cell renewal and regenerative capacity the CNS isextremely vulnerable to uncontrolled andor chronic immuneand inflammatory processes34 Interestingly enough our datacorroborate this hypothesis as we observed increased micro-glial activation later in the HD course (ie manifest HD)reinforcing the idea that neuroinflammation contributes todisease progression rather than to disease onset Activatedmicroglia releases proinflammatory mediators including
Figure 5 Phagocytosis by Induced Microglia-Like (iMG) Cells
Peripheral blood monocytes (day 1 panel A) were cultured for 14 days with GM-CSF and IL-34 resulting in iMG cells (day 14 panel B) Phagocytosis wasassessed by exposing iMG cells to PE-conjugated (1250) beads (panel C) for 24 hours The iMG cells showed the ability of phagocytosis with morphologicalchanges (C) The read arrows show PE-conjugated beads phagocytosed by iMG cells After washing beads out cells were acquired in a flow cytometer andanalyzed iMG were first gated by forwardside scatter (not shown) PE uptake was analyzed as shown in (D) orange (left) histogram shows the cells withoutbeads (negative control) red (right) histogram shows the cells that phagocytosed to the PE-conjugated beads Quantification is shown in (E) iMG fromcontrols premanifest HD and HD patients showed similar phagocytic capacity (no significant difference found at the Kruskal-Wallis test) (F) Cytokineproduction during phagocytosis iMG cells were incubated with PE-conjugated beads (1250) for 24 hours Supernatant was collected and cytokines wereanalyzed by multiparametric assay Cytokine levels (in picograms per milliliter) were corrected by protein levels (in milligrams per milliliter) Although iMGcells from controls premanifest HD and patients with HD increased the production of the evaluated cytokines during phagocytosis the values weresignificant only for TNFα in controls andmanifest HD (p values are provided in the figure Wilcoxonmatched-pairs signed-rank test) Scale bar (AndashC) = 10mmHorizontal bars (E and F) show the mean and the standard error of the mean HD = Huntington disease PE = phycoerythrin TNF = tumor necrosis factor
NeurologyorgNN Neurology Neuroimmunology amp Neuroinflammation | Volume 8 Number 3 | May 2021 9
cytokines and reactive oxygen species which in turn will alsocontribute to neuronal death and further microglial activa-tion34We observed an increase in cytokine release (especiallyTNFα) following a phagocytic stimulus in our in vitro ex-periments In line with our findings postmortem studies havealso reported increased levels of inflammatory cytokines inbrain samples of patients with HD in comparison withcontrols3536 It is worth mentioning that cytokine release is amicroglial reaction to a proinflammatory challenge and undercontrolled conditions (such as the iMG cultures) phagocy-tosis is a noninflammatory function of microglia A proin-flammatory stimulus would be needed to appropriately assessdifferences in cytokine release between controls premanifestHD and manifest HD iMG cultures
The current study used a novel PET tracer for TSPO [11C]-ER176 to assess microglial activation in HD By using [11C]-PK11195 PET studies have shown a significant increase inmicroglial activation in the striatum of patients with HD16 aswell as in premanifest HD gene carriers18 compared withcontrols In both studies [11C]-PK11195 binding significantlycorrelated with disease severity as measured by decreased[11C]-raclopride (a marker of dopamine D2 and D3 receptoravailability) binding in the striatum1618 Similar results weredescribed in the hypothalamus17 Later the same work groupused a multimodal imaging approach combiningMRI and PETanalyses and worsening in atrophy evaluated by MRI was ac-companied by a reduction in [11C]-raclopride and an increasein [11C]-PK11195 bindings in patients with HD In pre-manifest HD increased level of microglial activation in theassociative striatum and in the regional network associated withcognition correlated with 5-year probability of HD onset19
More recently increased microglial activation in the somato-sensory cortex (as evaluated through [11C]-PK11195 binding)was associatedwith higher levels of IL-1β IL-6 IL-8 and TNFαproduced by stimulated peripheral blood monocytes frompremanifest HD gene carriers2037 Although these studiesprovided valuable information about neuroinflammation inHD indicating microglial activation even in premanifest sub-jects their results should be interpreted with caution The poorsignal-to-background ratio of first-generation TSPO radio-tracers poses important limitations to the analyses The ratio ofspecific to nonspecific binding of [11C]-PK11195 in humanbrain has been reported to be as low as 023839 As an attempt tomitigate this problem a recent study used the second-generation TSPO tracer [11C]-PBR28 to evaluate micro-glial activation in HD21 Of interest the results were similar toours ie increased levels of [11C]-PBR28 binding in theputamina and pallidi of HD gene carriers (which included 7manifest patients with HD and only 1 person with premanifestHD) in comparison with controls Of note the results of thesingle premanifest subject were midway between controls andthe remaining patients suggesting that microglial activationwas associated with disease progression21 The authors didconfirm that [11C]-PBR28 provides a high signal-to-background ratio an important feature required for the anal-ysis at the individual level and within individual brain regions
An important limitation of [11C]-PBR28 and other second-generation TSPO radioligands is that they have varying degreesof sensitivity to the single nucleotide polymorphism rs6971 inthe TSPO gene40 [11C]-ER176 has unique advantages overthe other TSPO binders being the only known TSPO radio-ligand that allows the inclusion of low-affinity binders Thebinding of ER176 in low-affinity binders was found to becomparable to that for PBR28 in high-affinity binders More-over ER176 was proven to be the only radioligand not con-taminated by radiometabolites accumulating in the brain14
Our findings must be interpreted while taking into accountthe limitations of the study which include the relatively smallsample size and the cross-sectional design The cross-sectionaldesign prevented the assessment of microglial activation inthe context of disease progression and development of motorand nonmotor symptoms In addition the use of an objectivemethod to measure striatal degeneration (eg [11C]-raclopride PET) would provide more accurate informationabout striatal neuron dysfunctiondeath Regarding the invitro experiments the use of a proinflammatory challenge(other than the beads added for the phagocytosis assay) in ourexperiments would provide valuable information on cytokinesrelease by iMG from the different groups It is worth men-tioning the strengths of the study that include the use of aunique TSPO ligand [11C]-ER176 the enrollment of bothpremanifest and manifest subjects and the exploratory in-vestigation of microglia activity
In conclusion our findings strengthen the evidence thatmicroglial activation is associated with HD pathophysiologyMicroglial activation is unlikely to be the initiating event inHD Yet it may exacerbate neuronal dysfunction and neu-ronal death thus contributing to disease progression It isimportant to refine our understanding of the more specificimmuneinflammatory mechanisms that are involved in HDand in neurodegeneration This may foster the developmentof new therapeutic interventions to halt the progression ofHD and other neurodegenerative diseases
AcknowledgmentThe authors acknowledge and thank all of the volunteers thatparticipated in this study and are indebted to their families fortheir magnificent support They also thank the staff from theUTHealth McGovern Medical School 3T MRI Center andfrom the HoustonMethodist Research Institute TranslationalImaging Core Facilities especially Vips Patel and Kim MDoan for their skilled technical support Dr Gabriel Fries forhis assistance with the genotyping analyses and the teammembers of the HDSA Center of Excellence at UTHealth
Study FundingThis study was funded by the HD Human BiologyProjectmdashHuntingtonrsquos Disease Society of America (HDSA)E Furr Stimming receives research funding from RocheGenetech Vaccinex Cures Within Reach HDSA Uniqureand CHDI Foundation
10 Neurology Neuroimmunology amp Neuroinflammation | Volume 8 Number 3 | May 2021 NeurologyorgNN
DisclosureThe authors report no disclosures relevant to the manuscriptGo to NeurologyorgNN for full disclosures
Publication HistoryReceived by Neurology Neuroimmunology amp NeuroinflammationSeptember 9 2020 Accepted in final form January 20 2021
References1 The Huntingtonrsquos Disease Collaborative Research Group A novel gene containing a
trinucleotide repeat that is expanded and unstable on Huntingtonrsquos disease chro-mosomes Cell 199372971ndash983
2 Ross CA Aylward EH Wild EJ et al Huntington disease natural history biomarkersand prospects for therapeutics Nat Rev Neurol 201410204ndash216
3 Soulet D Cicchetti F The role of immunity in Huntingtonrsquos disease Mol Psychiatry201116889ndash902
4 Myers RH Vonsattel JP Paskevich PA et al Decreased neuronal and increasedoligodendroglial densities in Huntingtonrsquos disease caudate nucleus J NeuropatholExp Neurol 199150729ndash742
5 Vonsattel JP Myers RH Stevens TJ Ferrante RJ Bird ED Richardson EP JrNeuropathological classification of Huntingtonrsquos disease J Neuropathol Exp Neurol198544559ndash577
6 Sapp E Kegel KB Aronin N et al Early and progressive accumulation of reactivemicroglia in the Huntington disease brain J Neuropathol Exp Neurol 200160161ndash172
7 Ciccocioppo F Bologna G Ercolino E et al Neurodegenerative diseases asproteinopathies-driven immune disorders Neural Regen Res 202015850ndash856
8 Rocha NP Ribeiro FM Furr-Stimming E Teixeira AL Neuroimmunology of Hun-tingtonrsquos disease revisiting evidence from human studies Mediators Inflamm 201620168653132
9 Vowinckel E Reutens D Becher B et al PK11195 binding to the peripheral ben-zodiazepine receptor as a marker of microglia activation in multiple sclerosis andexperimental autoimmune encephalomyelitis J Neurosci Res 199750345ndash353
10 Papadopoulos V Baraldi M Guilarte TR et al Translocator protein (18kDa) newnomenclature for the peripheral-type benzodiazepine receptor based on its structureand molecular function Trends Pharmacol Sci 200627402ndash409
11 Cumming P Burgher B Patkar O et al Sifting through the surfeit of neuro-inflammation tracers J Cereb Blood Flow Metab 201838204ndash224
12 Zanotti-Fregonara P Zhang Y Jenko KJ et al Synthesis and evaluation of translocator18 kDa protein (TSPO) positron emission tomography (PET) radioligands with lowbinding sensitivity to human single nucleotide polymorphism rs6971 ACS ChemNeurosci 20145963ndash971
13 Ikawa M Lohith TG Shrestha S et al 11C-ER176 a radioligand for 18-kDa trans-locator protein has adequate sensitivity to robustly image all three affinity genotypesin human brain J Nucl Med 201758320ndash325
14 Fujita M Kobayashi M Ikawa M et al Comparison of four (11)C-labeled PETligands to quantify translocator protein 18 kDa (TSPO) in human brain (R)-PK11195 PBR28 DPA-713 and ER176-based on recent publications that measuredspecific-to-non-displaceable ratios EJNMMI Res 2017784
15 Zanotti-Fregonara P Pascual B Veronese M et al Head-to-head comparison of (11)C-PBR28 and (11)C-ER176 for quantification of the translocator protein in thehuman brain Eur J Nucl Med Mol Imaging 2019461822ndash1829
16 Pavese N Gerhard A Tai YF et al Microglial activation correlates with severity inHuntington disease a clinical and PET study Neurology 2006661638ndash1643
17 Politis M Pavese N Tai YF Tabrizi SJ Barker RA Piccini P Hypothalamic in-volvement in Huntingtonrsquos disease an in vivo PET study Brain 20081312860ndash2869
18 Tai YF Pavese N Gerhard A et al Microglial activation in presymptomatic Hun-tingtonrsquos disease gene carriers Brain 20071301759ndash1766
19 Politis M Pavese N Tai YF et al Microglial activation in regions related to cognitivefunction predicts disease onset in Huntingtonrsquos disease a multimodal imaging studyHum Brain Mapp 201132258ndash270
20 Politis M Lahiri N Niccolini F et al Increased central microglial activation associatedwith peripheral cytokine levels in premanifest Huntingtonrsquos disease gene carriersNeurobiol Dis 201583115ndash121
21 Lois C Gonzalez I Izquierdo-Garcia D et al Neuroinflammation in Huntingtonrsquosdisease new insights with (11)C-PBR28 PETMRI ACS Chem Neurosci 201892563ndash2571
22 Huntington Study Group Unified Huntingtonrsquos Disease Rating Scale reliability andconsistency Mov Disord 199611136ndash142
23 Folstein MF Folstein SE McHugh PR ldquoMini-mental staterdquo A practical method forgrading the cognitive state of patients for the clinician J Psychiatr Res 197512189ndash198
24 McNally G Rickards H Horton M Craufurd D Exploring the validity of the shortversion of the Problem Behaviours Assessment (PBA-s) for Huntingtonrsquos disease arasch analysis J Huntingtons Dis 20154347ndash369
25 Warner JH Sampaio C Modeling variability in the progression of Huntingtonrsquosdisease a novel modeling approach applied to structural imaging markers fromTRACK-HD CPT Pharmacometrics Syst Pharmacol 20165437ndash445
26 Lyoo CH Ikawa M Liow JS et al Cerebellum can serve as a pseudo-reference regionin Alzheimer disease to detect neuroinflammation measured with PET radioligandbinding to translocator protein J Nucl Med 201556701ndash706
27 Reuter M Schmansky NJ Rosas HD Fischl B Within-subject template estimation forunbiased longitudinal image analysis Neuroimage 2012611402ndash1418
28 Yushkevich PA Piven J Hazlett HC et al User-guided 3D active contour segmen-tation of anatomical structures significantly improved efficiency and reliabilityNeuroimage 2006311116ndash1128
29 Ohgidani M Kato TA Setoyama D et al Direct induction of ramified microglia-likecells from human monocytes dynamic microglial dysfunction in Nasu-Hakola dis-ease Sci Rep 201444957
30 Ohgidani M Kato TA Kanba S Introducing directly induced microglia-like (iMG)cells from fresh human monocytes a novel translational research tool for psychiatricdisorders Front Cell Neurosci 20159184
31 Kim SU de Vellis J Microglia in health and disease J Neurosci Res 200581302-31332 Ross CA Tabrizi SJ Huntingtonrsquos disease from molecular pathogenesis to clinical
treatment Lancet Neurol 20111083ndash9833 Chitnis TWeiner HL CNS inflammation and neurodegeneration J Clin Invest 2017
1273577ndash3587
Appendix Authors
Name Location Contribution
Natalia PRocha PhD
The Mitchell Center forAlzheimerrsquos Disease andRelated Brain DisordersDepartment of NeurologyMcGovern Medical SchoolThe University of TexasHealth Science CenterHouston
Designed andconceptualized the studymajor role in theacquisition of dataanalyzed the data anddrafted the manuscript forintellectual content
OdelinCharron MS
Department of NeurologyThe University of Texas atAustin
Major role in data analysis
Leigh BLathamPhD
School of MedicineUniversity of WashingtonSeattle
Major role in theacquisition of data
Gabriela DColpo PhD
Neuropsychiatry ProgramDepartment of Psychiatryand Behavioral SciencesMcGovern Medical SchoolUniversity of TexasHouston
Major role in theacquisition of data
PaoloZanotti-FregonaraMD PhD
Houston MethodistResearch Institute andWeill Cornell Medicine TX
Major role in the studydesign and the acquisitionof data
MeixiangYu PhD
Houston MethodistResearch Institute andWeill Cornell Medicine TX
Major role in theacquisition of data
LeorahFreemanMD PhD
Department of NeurologyThe University of Texas atAustin
Major role in the studydesign data analysis andinterpretation and theacquisition of data
Erin FurrStimmingMD
HDSA Center of Excellenceat University of TexasHealth Science Center atHouston
Major role in the studydesign and acquisition ofdata interpreted the dataand revised themanuscript for intellectualcontent
Antonio LTeixeiraMD PhD
Neuropsychiatry ProgramDepartment of Psychiatryand Behavioral SciencesMcGovern Medical SchoolUniversity of TexasHouston
Major role in the studydesign and acquisition ofdata interpreted the dataand revised themanuscript for intellectualcontent
NeurologyorgNN Neurology Neuroimmunology amp Neuroinflammation | Volume 8 Number 3 | May 2021 11
34 Rocha NP de Miranda AS Teixeira AL Insights into neuroinflammation in Parkin-sonrsquos disease from biomarkers to anti-inflammatory based therapies Biomed Res Int20152015628192
35 Silvestroni A Faull RL Strand AD Moller T Distinct neuroinflammatory profile inpost-mortem human Huntingtonrsquos disease Neuroreport 2009201098ndash1103
36 Bjorkqvist M Wild EJ Thiele J et al A novel pathogenic pathway of immune acti-vation detectable before clinical onset in Huntingtonrsquos disease J Exp Med 20082051869ndash1877
37 Politis M Lahiri N Niccolini F et al Corrigendum to ldquoIncreased central microglialactivation associated with peripheral cytokine levels in premanifest Huntingtonrsquos
disease gene carriersrdquo [Neurobiol Dis 83 (2015) 115-121] Neurobiol Dis 201798162
38 Kropholler MA Boellaard R Schuitemaker A Folkersma H van Berckel BN Lam-mertsma AA Evaluation of reference tissue models for the analysis of [11C](R)-PK11195 studies J Cereb Blood Flow Metab 2006261431ndash1441
39 Turkheimer FE Edison P Pavese N et al Reference and target region modeling of[11C]-(R)-PK11195 brain studies J Nucl Med 200748158ndash167
40 Kreisl WC Jenko KJ Hines CS et al A genetic polymorphism for translocator protein18 kDa affects both in vitro and in vivo radioligand binding in human brain to thisputative biomarker of neuroinflammation J Cereb Blood FlowMetab 20133353ndash58
12 Neurology Neuroimmunology amp Neuroinflammation | Volume 8 Number 3 | May 2021 NeurologyorgNN
DOI 101212NXI000000000000098420218 Neurol Neuroimmunol Neuroinflamm
Natalia P Rocha Odelin Charron Leigh B Latham et al Pathophysiology
Microglia Activation in Basal Ganglia Is a Late Event in Huntington Disease
This information is current as of April 1 2021
ServicesUpdated Information amp
httpnnneurologyorgcontent83e984fullhtmlincluding high resolution figures can be found at
References httpnnneurologyorgcontent83e984fullhtmlref-list-1
This article cites 40 articles 4 of which you can access for free at
Subspecialty Collections
httpnnneurologyorgcgicollectionpetPET
httpnnneurologyorgcgicollectionhuntingtons_diseaseHuntingtons diseasefollowing collection(s) This article along with others on similar topics appears in the
Permissions amp Licensing
httpnnneurologyorgmiscaboutxhtmlpermissionsits entirety can be found online atInformation about reproducing this article in parts (figurestables) or in
Reprints
httpnnneurologyorgmiscaddirxhtmlreprintsusInformation about ordering reprints can be found online
Academy of Neurology All rights reserved Online ISSN 2332-7812Copyright copy 2021 The Author(s) Published by Wolters Kluwer Health Inc on behalf of the AmericanPublished since April 2014 it is an open-access online-only continuous publication journal Copyright
is an official journal of the American Academy of NeurologyNeurol Neuroimmunol Neuroinflamm
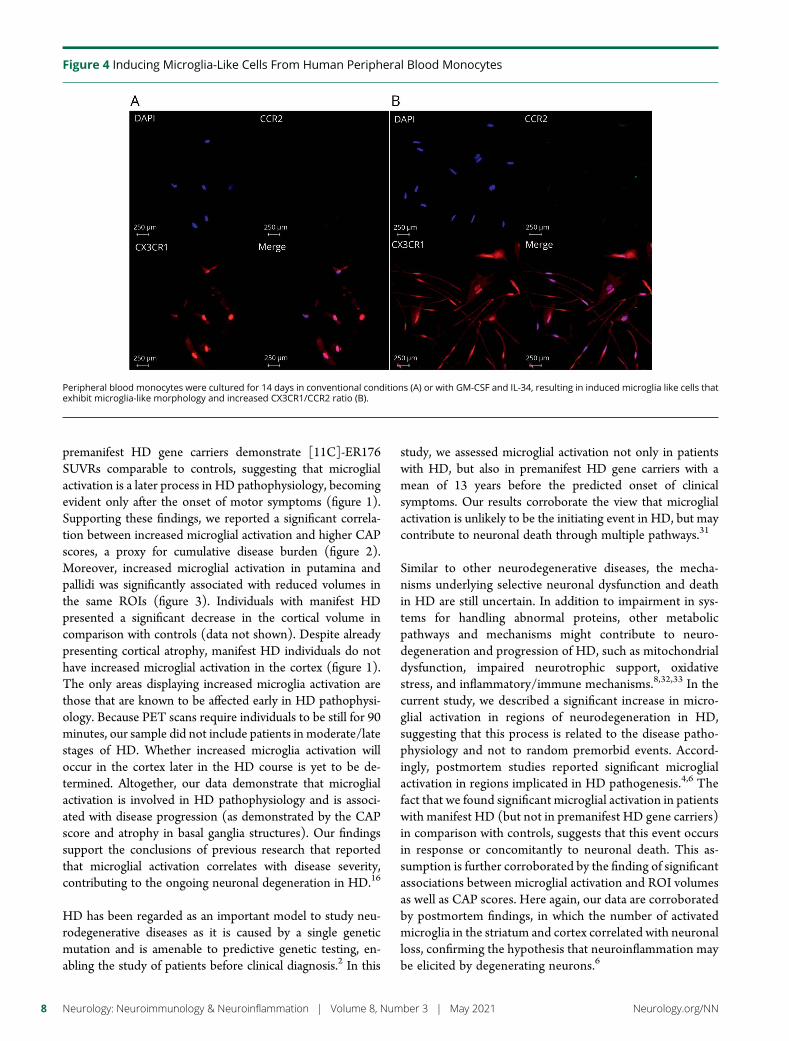
premanifest HD gene carriers demonstrate [11C]-ER176SUVRs comparable to controls suggesting that microglialactivation is a later process in HD pathophysiology becomingevident only after the onset of motor symptoms (figure 1)Supporting these findings we reported a significant correla-tion between increased microglial activation and higher CAPscores a proxy for cumulative disease burden (figure 2)Moreover increased microglial activation in putamina andpallidi was significantly associated with reduced volumes inthe same ROIs (figure 3) Individuals with manifest HDpresented a significant decrease in the cortical volume incomparison with controls (data not shown) Despite alreadypresenting cortical atrophy manifest HD individuals do nothave increased microglial activation in the cortex (figure 1)The only areas displaying increased microglia activation arethose that are known to be affected early in HD pathophysi-ology Because PET scans require individuals to be still for 90minutes our sample did not include patients in moderatelatestages of HD Whether increased microglia activation willoccur in the cortex later in the HD course is yet to be de-termined Altogether our data demonstrate that microglialactivation is involved in HD pathophysiology and is associ-ated with disease progression (as demonstrated by the CAPscore and atrophy in basal ganglia structures) Our findingssupport the conclusions of previous research that reportedthat microglial activation correlates with disease severitycontributing to the ongoing neuronal degeneration in HD16
HD has been regarded as an important model to study neu-rodegenerative diseases as it is caused by a single geneticmutation and is amenable to predictive genetic testing en-abling the study of patients before clinical diagnosis2 In this
study we assessed microglial activation not only in patientswith HD but also in premanifest HD gene carriers with amean of 13 years before the predicted onset of clinicalsymptoms Our results corroborate the view that microglialactivation is unlikely to be the initiating event in HD but maycontribute to neuronal death through multiple pathways31
Similar to other neurodegenerative diseases the mecha-nisms underlying selective neuronal dysfunction and deathin HD are still uncertain In addition to impairment in sys-tems for handling abnormal proteins other metabolicpathways and mechanisms might contribute to neuro-degeneration and progression of HD such as mitochondrialdysfunction impaired neurotrophic support oxidativestress and inflammatoryimmune mechanisms83233 In thecurrent study we described a significant increase in micro-glial activation in regions of neurodegeneration in HDsuggesting that this process is related to the disease patho-physiology and not to random premorbid events Accord-ingly postmortem studies reported significant microglialactivation in regions implicated in HD pathogenesis46 Thefact that we found significant microglial activation in patientswith manifest HD (but not in premanifest HD gene carriers)in comparison with controls suggests that this event occursin response or concomitantly to neuronal death This as-sumption is further corroborated by the finding of significantassociations between microglial activation and ROI volumesas well as CAP scores Here again our data are corroboratedby postmortem findings in which the number of activatedmicroglia in the striatum and cortex correlated with neuronalloss confirming the hypothesis that neuroinflammation maybe elicited by degenerating neurons6
Figure 4 Inducing Microglia-Like Cells From Human Peripheral Blood Monocytes
Peripheral blood monocytes were cultured for 14 days in conventional conditions (A) or with GM-CSF and IL-34 resulting in induced microglia like cells thatexhibit microglia-like morphology and increased CX3CR1CCR2 ratio (B)
8 Neurology Neuroimmunology amp Neuroinflammation | Volume 8 Number 3 | May 2021 NeurologyorgNN
Neuroinflammation elicited by neuronal death can be at firstregarded as a positive event as it promotes clearance of celldebris However inflammatory mechanisms contribute toneurodegeneration and dying neurons further activate in-flammatory responses resulting in a vicious cycle of in-flammation and neuronal death34 Therefore inflammatoryresponses although essential for tissue homeostasis cancontribute to neuronal injury As neural tissues have a
restricted cell renewal and regenerative capacity the CNS isextremely vulnerable to uncontrolled andor chronic immuneand inflammatory processes34 Interestingly enough our datacorroborate this hypothesis as we observed increased micro-glial activation later in the HD course (ie manifest HD)reinforcing the idea that neuroinflammation contributes todisease progression rather than to disease onset Activatedmicroglia releases proinflammatory mediators including
Figure 5 Phagocytosis by Induced Microglia-Like (iMG) Cells
Peripheral blood monocytes (day 1 panel A) were cultured for 14 days with GM-CSF and IL-34 resulting in iMG cells (day 14 panel B) Phagocytosis wasassessed by exposing iMG cells to PE-conjugated (1250) beads (panel C) for 24 hours The iMG cells showed the ability of phagocytosis with morphologicalchanges (C) The read arrows show PE-conjugated beads phagocytosed by iMG cells After washing beads out cells were acquired in a flow cytometer andanalyzed iMG were first gated by forwardside scatter (not shown) PE uptake was analyzed as shown in (D) orange (left) histogram shows the cells withoutbeads (negative control) red (right) histogram shows the cells that phagocytosed to the PE-conjugated beads Quantification is shown in (E) iMG fromcontrols premanifest HD and HD patients showed similar phagocytic capacity (no significant difference found at the Kruskal-Wallis test) (F) Cytokineproduction during phagocytosis iMG cells were incubated with PE-conjugated beads (1250) for 24 hours Supernatant was collected and cytokines wereanalyzed by multiparametric assay Cytokine levels (in picograms per milliliter) were corrected by protein levels (in milligrams per milliliter) Although iMGcells from controls premanifest HD and patients with HD increased the production of the evaluated cytokines during phagocytosis the values weresignificant only for TNFα in controls andmanifest HD (p values are provided in the figure Wilcoxonmatched-pairs signed-rank test) Scale bar (AndashC) = 10mmHorizontal bars (E and F) show the mean and the standard error of the mean HD = Huntington disease PE = phycoerythrin TNF = tumor necrosis factor
NeurologyorgNN Neurology Neuroimmunology amp Neuroinflammation | Volume 8 Number 3 | May 2021 9
cytokines and reactive oxygen species which in turn will alsocontribute to neuronal death and further microglial activa-tion34We observed an increase in cytokine release (especiallyTNFα) following a phagocytic stimulus in our in vitro ex-periments In line with our findings postmortem studies havealso reported increased levels of inflammatory cytokines inbrain samples of patients with HD in comparison withcontrols3536 It is worth mentioning that cytokine release is amicroglial reaction to a proinflammatory challenge and undercontrolled conditions (such as the iMG cultures) phagocy-tosis is a noninflammatory function of microglia A proin-flammatory stimulus would be needed to appropriately assessdifferences in cytokine release between controls premanifestHD and manifest HD iMG cultures
The current study used a novel PET tracer for TSPO [11C]-ER176 to assess microglial activation in HD By using [11C]-PK11195 PET studies have shown a significant increase inmicroglial activation in the striatum of patients with HD16 aswell as in premanifest HD gene carriers18 compared withcontrols In both studies [11C]-PK11195 binding significantlycorrelated with disease severity as measured by decreased[11C]-raclopride (a marker of dopamine D2 and D3 receptoravailability) binding in the striatum1618 Similar results weredescribed in the hypothalamus17 Later the same work groupused a multimodal imaging approach combiningMRI and PETanalyses and worsening in atrophy evaluated by MRI was ac-companied by a reduction in [11C]-raclopride and an increasein [11C]-PK11195 bindings in patients with HD In pre-manifest HD increased level of microglial activation in theassociative striatum and in the regional network associated withcognition correlated with 5-year probability of HD onset19
More recently increased microglial activation in the somato-sensory cortex (as evaluated through [11C]-PK11195 binding)was associatedwith higher levels of IL-1β IL-6 IL-8 and TNFαproduced by stimulated peripheral blood monocytes frompremanifest HD gene carriers2037 Although these studiesprovided valuable information about neuroinflammation inHD indicating microglial activation even in premanifest sub-jects their results should be interpreted with caution The poorsignal-to-background ratio of first-generation TSPO radio-tracers poses important limitations to the analyses The ratio ofspecific to nonspecific binding of [11C]-PK11195 in humanbrain has been reported to be as low as 023839 As an attempt tomitigate this problem a recent study used the second-generation TSPO tracer [11C]-PBR28 to evaluate micro-glial activation in HD21 Of interest the results were similar toours ie increased levels of [11C]-PBR28 binding in theputamina and pallidi of HD gene carriers (which included 7manifest patients with HD and only 1 person with premanifestHD) in comparison with controls Of note the results of thesingle premanifest subject were midway between controls andthe remaining patients suggesting that microglial activationwas associated with disease progression21 The authors didconfirm that [11C]-PBR28 provides a high signal-to-background ratio an important feature required for the anal-ysis at the individual level and within individual brain regions
An important limitation of [11C]-PBR28 and other second-generation TSPO radioligands is that they have varying degreesof sensitivity to the single nucleotide polymorphism rs6971 inthe TSPO gene40 [11C]-ER176 has unique advantages overthe other TSPO binders being the only known TSPO radio-ligand that allows the inclusion of low-affinity binders Thebinding of ER176 in low-affinity binders was found to becomparable to that for PBR28 in high-affinity binders More-over ER176 was proven to be the only radioligand not con-taminated by radiometabolites accumulating in the brain14
Our findings must be interpreted while taking into accountthe limitations of the study which include the relatively smallsample size and the cross-sectional design The cross-sectionaldesign prevented the assessment of microglial activation inthe context of disease progression and development of motorand nonmotor symptoms In addition the use of an objectivemethod to measure striatal degeneration (eg [11C]-raclopride PET) would provide more accurate informationabout striatal neuron dysfunctiondeath Regarding the invitro experiments the use of a proinflammatory challenge(other than the beads added for the phagocytosis assay) in ourexperiments would provide valuable information on cytokinesrelease by iMG from the different groups It is worth men-tioning the strengths of the study that include the use of aunique TSPO ligand [11C]-ER176 the enrollment of bothpremanifest and manifest subjects and the exploratory in-vestigation of microglia activity
In conclusion our findings strengthen the evidence thatmicroglial activation is associated with HD pathophysiologyMicroglial activation is unlikely to be the initiating event inHD Yet it may exacerbate neuronal dysfunction and neu-ronal death thus contributing to disease progression It isimportant to refine our understanding of the more specificimmuneinflammatory mechanisms that are involved in HDand in neurodegeneration This may foster the developmentof new therapeutic interventions to halt the progression ofHD and other neurodegenerative diseases
AcknowledgmentThe authors acknowledge and thank all of the volunteers thatparticipated in this study and are indebted to their families fortheir magnificent support They also thank the staff from theUTHealth McGovern Medical School 3T MRI Center andfrom the HoustonMethodist Research Institute TranslationalImaging Core Facilities especially Vips Patel and Kim MDoan for their skilled technical support Dr Gabriel Fries forhis assistance with the genotyping analyses and the teammembers of the HDSA Center of Excellence at UTHealth
Study FundingThis study was funded by the HD Human BiologyProjectmdashHuntingtonrsquos Disease Society of America (HDSA)E Furr Stimming receives research funding from RocheGenetech Vaccinex Cures Within Reach HDSA Uniqureand CHDI Foundation
10 Neurology Neuroimmunology amp Neuroinflammation | Volume 8 Number 3 | May 2021 NeurologyorgNN
DisclosureThe authors report no disclosures relevant to the manuscriptGo to NeurologyorgNN for full disclosures
Publication HistoryReceived by Neurology Neuroimmunology amp NeuroinflammationSeptember 9 2020 Accepted in final form January 20 2021
References1 The Huntingtonrsquos Disease Collaborative Research Group A novel gene containing a
trinucleotide repeat that is expanded and unstable on Huntingtonrsquos disease chro-mosomes Cell 199372971ndash983
2 Ross CA Aylward EH Wild EJ et al Huntington disease natural history biomarkersand prospects for therapeutics Nat Rev Neurol 201410204ndash216
3 Soulet D Cicchetti F The role of immunity in Huntingtonrsquos disease Mol Psychiatry201116889ndash902
4 Myers RH Vonsattel JP Paskevich PA et al Decreased neuronal and increasedoligodendroglial densities in Huntingtonrsquos disease caudate nucleus J NeuropatholExp Neurol 199150729ndash742
5 Vonsattel JP Myers RH Stevens TJ Ferrante RJ Bird ED Richardson EP JrNeuropathological classification of Huntingtonrsquos disease J Neuropathol Exp Neurol198544559ndash577
6 Sapp E Kegel KB Aronin N et al Early and progressive accumulation of reactivemicroglia in the Huntington disease brain J Neuropathol Exp Neurol 200160161ndash172
7 Ciccocioppo F Bologna G Ercolino E et al Neurodegenerative diseases asproteinopathies-driven immune disorders Neural Regen Res 202015850ndash856
8 Rocha NP Ribeiro FM Furr-Stimming E Teixeira AL Neuroimmunology of Hun-tingtonrsquos disease revisiting evidence from human studies Mediators Inflamm 201620168653132
9 Vowinckel E Reutens D Becher B et al PK11195 binding to the peripheral ben-zodiazepine receptor as a marker of microglia activation in multiple sclerosis andexperimental autoimmune encephalomyelitis J Neurosci Res 199750345ndash353
10 Papadopoulos V Baraldi M Guilarte TR et al Translocator protein (18kDa) newnomenclature for the peripheral-type benzodiazepine receptor based on its structureand molecular function Trends Pharmacol Sci 200627402ndash409
11 Cumming P Burgher B Patkar O et al Sifting through the surfeit of neuro-inflammation tracers J Cereb Blood Flow Metab 201838204ndash224
12 Zanotti-Fregonara P Zhang Y Jenko KJ et al Synthesis and evaluation of translocator18 kDa protein (TSPO) positron emission tomography (PET) radioligands with lowbinding sensitivity to human single nucleotide polymorphism rs6971 ACS ChemNeurosci 20145963ndash971
13 Ikawa M Lohith TG Shrestha S et al 11C-ER176 a radioligand for 18-kDa trans-locator protein has adequate sensitivity to robustly image all three affinity genotypesin human brain J Nucl Med 201758320ndash325
14 Fujita M Kobayashi M Ikawa M et al Comparison of four (11)C-labeled PETligands to quantify translocator protein 18 kDa (TSPO) in human brain (R)-PK11195 PBR28 DPA-713 and ER176-based on recent publications that measuredspecific-to-non-displaceable ratios EJNMMI Res 2017784
15 Zanotti-Fregonara P Pascual B Veronese M et al Head-to-head comparison of (11)C-PBR28 and (11)C-ER176 for quantification of the translocator protein in thehuman brain Eur J Nucl Med Mol Imaging 2019461822ndash1829
16 Pavese N Gerhard A Tai YF et al Microglial activation correlates with severity inHuntington disease a clinical and PET study Neurology 2006661638ndash1643
17 Politis M Pavese N Tai YF Tabrizi SJ Barker RA Piccini P Hypothalamic in-volvement in Huntingtonrsquos disease an in vivo PET study Brain 20081312860ndash2869
18 Tai YF Pavese N Gerhard A et al Microglial activation in presymptomatic Hun-tingtonrsquos disease gene carriers Brain 20071301759ndash1766
19 Politis M Pavese N Tai YF et al Microglial activation in regions related to cognitivefunction predicts disease onset in Huntingtonrsquos disease a multimodal imaging studyHum Brain Mapp 201132258ndash270
20 Politis M Lahiri N Niccolini F et al Increased central microglial activation associatedwith peripheral cytokine levels in premanifest Huntingtonrsquos disease gene carriersNeurobiol Dis 201583115ndash121
21 Lois C Gonzalez I Izquierdo-Garcia D et al Neuroinflammation in Huntingtonrsquosdisease new insights with (11)C-PBR28 PETMRI ACS Chem Neurosci 201892563ndash2571
22 Huntington Study Group Unified Huntingtonrsquos Disease Rating Scale reliability andconsistency Mov Disord 199611136ndash142
23 Folstein MF Folstein SE McHugh PR ldquoMini-mental staterdquo A practical method forgrading the cognitive state of patients for the clinician J Psychiatr Res 197512189ndash198
24 McNally G Rickards H Horton M Craufurd D Exploring the validity of the shortversion of the Problem Behaviours Assessment (PBA-s) for Huntingtonrsquos disease arasch analysis J Huntingtons Dis 20154347ndash369
25 Warner JH Sampaio C Modeling variability in the progression of Huntingtonrsquosdisease a novel modeling approach applied to structural imaging markers fromTRACK-HD CPT Pharmacometrics Syst Pharmacol 20165437ndash445
26 Lyoo CH Ikawa M Liow JS et al Cerebellum can serve as a pseudo-reference regionin Alzheimer disease to detect neuroinflammation measured with PET radioligandbinding to translocator protein J Nucl Med 201556701ndash706
27 Reuter M Schmansky NJ Rosas HD Fischl B Within-subject template estimation forunbiased longitudinal image analysis Neuroimage 2012611402ndash1418
28 Yushkevich PA Piven J Hazlett HC et al User-guided 3D active contour segmen-tation of anatomical structures significantly improved efficiency and reliabilityNeuroimage 2006311116ndash1128
29 Ohgidani M Kato TA Setoyama D et al Direct induction of ramified microglia-likecells from human monocytes dynamic microglial dysfunction in Nasu-Hakola dis-ease Sci Rep 201444957
30 Ohgidani M Kato TA Kanba S Introducing directly induced microglia-like (iMG)cells from fresh human monocytes a novel translational research tool for psychiatricdisorders Front Cell Neurosci 20159184
31 Kim SU de Vellis J Microglia in health and disease J Neurosci Res 200581302-31332 Ross CA Tabrizi SJ Huntingtonrsquos disease from molecular pathogenesis to clinical
treatment Lancet Neurol 20111083ndash9833 Chitnis TWeiner HL CNS inflammation and neurodegeneration J Clin Invest 2017
1273577ndash3587
Appendix Authors
Name Location Contribution
Natalia PRocha PhD
The Mitchell Center forAlzheimerrsquos Disease andRelated Brain DisordersDepartment of NeurologyMcGovern Medical SchoolThe University of TexasHealth Science CenterHouston
Designed andconceptualized the studymajor role in theacquisition of dataanalyzed the data anddrafted the manuscript forintellectual content
OdelinCharron MS
Department of NeurologyThe University of Texas atAustin
Major role in data analysis
Leigh BLathamPhD
School of MedicineUniversity of WashingtonSeattle
Major role in theacquisition of data
Gabriela DColpo PhD
Neuropsychiatry ProgramDepartment of Psychiatryand Behavioral SciencesMcGovern Medical SchoolUniversity of TexasHouston
Major role in theacquisition of data
PaoloZanotti-FregonaraMD PhD
Houston MethodistResearch Institute andWeill Cornell Medicine TX
Major role in the studydesign and the acquisitionof data
MeixiangYu PhD
Houston MethodistResearch Institute andWeill Cornell Medicine TX
Major role in theacquisition of data
LeorahFreemanMD PhD
Department of NeurologyThe University of Texas atAustin
Major role in the studydesign data analysis andinterpretation and theacquisition of data
Erin FurrStimmingMD
HDSA Center of Excellenceat University of TexasHealth Science Center atHouston
Major role in the studydesign and acquisition ofdata interpreted the dataand revised themanuscript for intellectualcontent
Antonio LTeixeiraMD PhD
Neuropsychiatry ProgramDepartment of Psychiatryand Behavioral SciencesMcGovern Medical SchoolUniversity of TexasHouston
Major role in the studydesign and acquisition ofdata interpreted the dataand revised themanuscript for intellectualcontent
NeurologyorgNN Neurology Neuroimmunology amp Neuroinflammation | Volume 8 Number 3 | May 2021 11
34 Rocha NP de Miranda AS Teixeira AL Insights into neuroinflammation in Parkin-sonrsquos disease from biomarkers to anti-inflammatory based therapies Biomed Res Int20152015628192
35 Silvestroni A Faull RL Strand AD Moller T Distinct neuroinflammatory profile inpost-mortem human Huntingtonrsquos disease Neuroreport 2009201098ndash1103
36 Bjorkqvist M Wild EJ Thiele J et al A novel pathogenic pathway of immune acti-vation detectable before clinical onset in Huntingtonrsquos disease J Exp Med 20082051869ndash1877
37 Politis M Lahiri N Niccolini F et al Corrigendum to ldquoIncreased central microglialactivation associated with peripheral cytokine levels in premanifest Huntingtonrsquos
disease gene carriersrdquo [Neurobiol Dis 83 (2015) 115-121] Neurobiol Dis 201798162
38 Kropholler MA Boellaard R Schuitemaker A Folkersma H van Berckel BN Lam-mertsma AA Evaluation of reference tissue models for the analysis of [11C](R)-PK11195 studies J Cereb Blood Flow Metab 2006261431ndash1441
39 Turkheimer FE Edison P Pavese N et al Reference and target region modeling of[11C]-(R)-PK11195 brain studies J Nucl Med 200748158ndash167
40 Kreisl WC Jenko KJ Hines CS et al A genetic polymorphism for translocator protein18 kDa affects both in vitro and in vivo radioligand binding in human brain to thisputative biomarker of neuroinflammation J Cereb Blood FlowMetab 20133353ndash58
12 Neurology Neuroimmunology amp Neuroinflammation | Volume 8 Number 3 | May 2021 NeurologyorgNN
DOI 101212NXI000000000000098420218 Neurol Neuroimmunol Neuroinflamm
Natalia P Rocha Odelin Charron Leigh B Latham et al Pathophysiology
Microglia Activation in Basal Ganglia Is a Late Event in Huntington Disease
This information is current as of April 1 2021
ServicesUpdated Information amp
httpnnneurologyorgcontent83e984fullhtmlincluding high resolution figures can be found at
References httpnnneurologyorgcontent83e984fullhtmlref-list-1
This article cites 40 articles 4 of which you can access for free at
Subspecialty Collections
httpnnneurologyorgcgicollectionpetPET
httpnnneurologyorgcgicollectionhuntingtons_diseaseHuntingtons diseasefollowing collection(s) This article along with others on similar topics appears in the
Permissions amp Licensing
httpnnneurologyorgmiscaboutxhtmlpermissionsits entirety can be found online atInformation about reproducing this article in parts (figurestables) or in
Reprints
httpnnneurologyorgmiscaddirxhtmlreprintsusInformation about ordering reprints can be found online
Academy of Neurology All rights reserved Online ISSN 2332-7812Copyright copy 2021 The Author(s) Published by Wolters Kluwer Health Inc on behalf of the AmericanPublished since April 2014 it is an open-access online-only continuous publication journal Copyright
is an official journal of the American Academy of NeurologyNeurol Neuroimmunol Neuroinflamm

Neuroinflammation elicited by neuronal death can be at firstregarded as a positive event as it promotes clearance of celldebris However inflammatory mechanisms contribute toneurodegeneration and dying neurons further activate in-flammatory responses resulting in a vicious cycle of in-flammation and neuronal death34 Therefore inflammatoryresponses although essential for tissue homeostasis cancontribute to neuronal injury As neural tissues have a
restricted cell renewal and regenerative capacity the CNS isextremely vulnerable to uncontrolled andor chronic immuneand inflammatory processes34 Interestingly enough our datacorroborate this hypothesis as we observed increased micro-glial activation later in the HD course (ie manifest HD)reinforcing the idea that neuroinflammation contributes todisease progression rather than to disease onset Activatedmicroglia releases proinflammatory mediators including
Figure 5 Phagocytosis by Induced Microglia-Like (iMG) Cells
Peripheral blood monocytes (day 1 panel A) were cultured for 14 days with GM-CSF and IL-34 resulting in iMG cells (day 14 panel B) Phagocytosis wasassessed by exposing iMG cells to PE-conjugated (1250) beads (panel C) for 24 hours The iMG cells showed the ability of phagocytosis with morphologicalchanges (C) The read arrows show PE-conjugated beads phagocytosed by iMG cells After washing beads out cells were acquired in a flow cytometer andanalyzed iMG were first gated by forwardside scatter (not shown) PE uptake was analyzed as shown in (D) orange (left) histogram shows the cells withoutbeads (negative control) red (right) histogram shows the cells that phagocytosed to the PE-conjugated beads Quantification is shown in (E) iMG fromcontrols premanifest HD and HD patients showed similar phagocytic capacity (no significant difference found at the Kruskal-Wallis test) (F) Cytokineproduction during phagocytosis iMG cells were incubated with PE-conjugated beads (1250) for 24 hours Supernatant was collected and cytokines wereanalyzed by multiparametric assay Cytokine levels (in picograms per milliliter) were corrected by protein levels (in milligrams per milliliter) Although iMGcells from controls premanifest HD and patients with HD increased the production of the evaluated cytokines during phagocytosis the values weresignificant only for TNFα in controls andmanifest HD (p values are provided in the figure Wilcoxonmatched-pairs signed-rank test) Scale bar (AndashC) = 10mmHorizontal bars (E and F) show the mean and the standard error of the mean HD = Huntington disease PE = phycoerythrin TNF = tumor necrosis factor
NeurologyorgNN Neurology Neuroimmunology amp Neuroinflammation | Volume 8 Number 3 | May 2021 9
cytokines and reactive oxygen species which in turn will alsocontribute to neuronal death and further microglial activa-tion34We observed an increase in cytokine release (especiallyTNFα) following a phagocytic stimulus in our in vitro ex-periments In line with our findings postmortem studies havealso reported increased levels of inflammatory cytokines inbrain samples of patients with HD in comparison withcontrols3536 It is worth mentioning that cytokine release is amicroglial reaction to a proinflammatory challenge and undercontrolled conditions (such as the iMG cultures) phagocy-tosis is a noninflammatory function of microglia A proin-flammatory stimulus would be needed to appropriately assessdifferences in cytokine release between controls premanifestHD and manifest HD iMG cultures
The current study used a novel PET tracer for TSPO [11C]-ER176 to assess microglial activation in HD By using [11C]-PK11195 PET studies have shown a significant increase inmicroglial activation in the striatum of patients with HD16 aswell as in premanifest HD gene carriers18 compared withcontrols In both studies [11C]-PK11195 binding significantlycorrelated with disease severity as measured by decreased[11C]-raclopride (a marker of dopamine D2 and D3 receptoravailability) binding in the striatum1618 Similar results weredescribed in the hypothalamus17 Later the same work groupused a multimodal imaging approach combiningMRI and PETanalyses and worsening in atrophy evaluated by MRI was ac-companied by a reduction in [11C]-raclopride and an increasein [11C]-PK11195 bindings in patients with HD In pre-manifest HD increased level of microglial activation in theassociative striatum and in the regional network associated withcognition correlated with 5-year probability of HD onset19
More recently increased microglial activation in the somato-sensory cortex (as evaluated through [11C]-PK11195 binding)was associatedwith higher levels of IL-1β IL-6 IL-8 and TNFαproduced by stimulated peripheral blood monocytes frompremanifest HD gene carriers2037 Although these studiesprovided valuable information about neuroinflammation inHD indicating microglial activation even in premanifest sub-jects their results should be interpreted with caution The poorsignal-to-background ratio of first-generation TSPO radio-tracers poses important limitations to the analyses The ratio ofspecific to nonspecific binding of [11C]-PK11195 in humanbrain has been reported to be as low as 023839 As an attempt tomitigate this problem a recent study used the second-generation TSPO tracer [11C]-PBR28 to evaluate micro-glial activation in HD21 Of interest the results were similar toours ie increased levels of [11C]-PBR28 binding in theputamina and pallidi of HD gene carriers (which included 7manifest patients with HD and only 1 person with premanifestHD) in comparison with controls Of note the results of thesingle premanifest subject were midway between controls andthe remaining patients suggesting that microglial activationwas associated with disease progression21 The authors didconfirm that [11C]-PBR28 provides a high signal-to-background ratio an important feature required for the anal-ysis at the individual level and within individual brain regions
An important limitation of [11C]-PBR28 and other second-generation TSPO radioligands is that they have varying degreesof sensitivity to the single nucleotide polymorphism rs6971 inthe TSPO gene40 [11C]-ER176 has unique advantages overthe other TSPO binders being the only known TSPO radio-ligand that allows the inclusion of low-affinity binders Thebinding of ER176 in low-affinity binders was found to becomparable to that for PBR28 in high-affinity binders More-over ER176 was proven to be the only radioligand not con-taminated by radiometabolites accumulating in the brain14
Our findings must be interpreted while taking into accountthe limitations of the study which include the relatively smallsample size and the cross-sectional design The cross-sectionaldesign prevented the assessment of microglial activation inthe context of disease progression and development of motorand nonmotor symptoms In addition the use of an objectivemethod to measure striatal degeneration (eg [11C]-raclopride PET) would provide more accurate informationabout striatal neuron dysfunctiondeath Regarding the invitro experiments the use of a proinflammatory challenge(other than the beads added for the phagocytosis assay) in ourexperiments would provide valuable information on cytokinesrelease by iMG from the different groups It is worth men-tioning the strengths of the study that include the use of aunique TSPO ligand [11C]-ER176 the enrollment of bothpremanifest and manifest subjects and the exploratory in-vestigation of microglia activity
In conclusion our findings strengthen the evidence thatmicroglial activation is associated with HD pathophysiologyMicroglial activation is unlikely to be the initiating event inHD Yet it may exacerbate neuronal dysfunction and neu-ronal death thus contributing to disease progression It isimportant to refine our understanding of the more specificimmuneinflammatory mechanisms that are involved in HDand in neurodegeneration This may foster the developmentof new therapeutic interventions to halt the progression ofHD and other neurodegenerative diseases
AcknowledgmentThe authors acknowledge and thank all of the volunteers thatparticipated in this study and are indebted to their families fortheir magnificent support They also thank the staff from theUTHealth McGovern Medical School 3T MRI Center andfrom the HoustonMethodist Research Institute TranslationalImaging Core Facilities especially Vips Patel and Kim MDoan for their skilled technical support Dr Gabriel Fries forhis assistance with the genotyping analyses and the teammembers of the HDSA Center of Excellence at UTHealth
Study FundingThis study was funded by the HD Human BiologyProjectmdashHuntingtonrsquos Disease Society of America (HDSA)E Furr Stimming receives research funding from RocheGenetech Vaccinex Cures Within Reach HDSA Uniqureand CHDI Foundation
10 Neurology Neuroimmunology amp Neuroinflammation | Volume 8 Number 3 | May 2021 NeurologyorgNN
DisclosureThe authors report no disclosures relevant to the manuscriptGo to NeurologyorgNN for full disclosures
Publication HistoryReceived by Neurology Neuroimmunology amp NeuroinflammationSeptember 9 2020 Accepted in final form January 20 2021
References1 The Huntingtonrsquos Disease Collaborative Research Group A novel gene containing a
trinucleotide repeat that is expanded and unstable on Huntingtonrsquos disease chro-mosomes Cell 199372971ndash983
2 Ross CA Aylward EH Wild EJ et al Huntington disease natural history biomarkersand prospects for therapeutics Nat Rev Neurol 201410204ndash216
3 Soulet D Cicchetti F The role of immunity in Huntingtonrsquos disease Mol Psychiatry201116889ndash902
4 Myers RH Vonsattel JP Paskevich PA et al Decreased neuronal and increasedoligodendroglial densities in Huntingtonrsquos disease caudate nucleus J NeuropatholExp Neurol 199150729ndash742
5 Vonsattel JP Myers RH Stevens TJ Ferrante RJ Bird ED Richardson EP JrNeuropathological classification of Huntingtonrsquos disease J Neuropathol Exp Neurol198544559ndash577
6 Sapp E Kegel KB Aronin N et al Early and progressive accumulation of reactivemicroglia in the Huntington disease brain J Neuropathol Exp Neurol 200160161ndash172
7 Ciccocioppo F Bologna G Ercolino E et al Neurodegenerative diseases asproteinopathies-driven immune disorders Neural Regen Res 202015850ndash856
8 Rocha NP Ribeiro FM Furr-Stimming E Teixeira AL Neuroimmunology of Hun-tingtonrsquos disease revisiting evidence from human studies Mediators Inflamm 201620168653132
9 Vowinckel E Reutens D Becher B et al PK11195 binding to the peripheral ben-zodiazepine receptor as a marker of microglia activation in multiple sclerosis andexperimental autoimmune encephalomyelitis J Neurosci Res 199750345ndash353
10 Papadopoulos V Baraldi M Guilarte TR et al Translocator protein (18kDa) newnomenclature for the peripheral-type benzodiazepine receptor based on its structureand molecular function Trends Pharmacol Sci 200627402ndash409
11 Cumming P Burgher B Patkar O et al Sifting through the surfeit of neuro-inflammation tracers J Cereb Blood Flow Metab 201838204ndash224
12 Zanotti-Fregonara P Zhang Y Jenko KJ et al Synthesis and evaluation of translocator18 kDa protein (TSPO) positron emission tomography (PET) radioligands with lowbinding sensitivity to human single nucleotide polymorphism rs6971 ACS ChemNeurosci 20145963ndash971
13 Ikawa M Lohith TG Shrestha S et al 11C-ER176 a radioligand for 18-kDa trans-locator protein has adequate sensitivity to robustly image all three affinity genotypesin human brain J Nucl Med 201758320ndash325
14 Fujita M Kobayashi M Ikawa M et al Comparison of four (11)C-labeled PETligands to quantify translocator protein 18 kDa (TSPO) in human brain (R)-PK11195 PBR28 DPA-713 and ER176-based on recent publications that measuredspecific-to-non-displaceable ratios EJNMMI Res 2017784
15 Zanotti-Fregonara P Pascual B Veronese M et al Head-to-head comparison of (11)C-PBR28 and (11)C-ER176 for quantification of the translocator protein in thehuman brain Eur J Nucl Med Mol Imaging 2019461822ndash1829
16 Pavese N Gerhard A Tai YF et al Microglial activation correlates with severity inHuntington disease a clinical and PET study Neurology 2006661638ndash1643
17 Politis M Pavese N Tai YF Tabrizi SJ Barker RA Piccini P Hypothalamic in-volvement in Huntingtonrsquos disease an in vivo PET study Brain 20081312860ndash2869
18 Tai YF Pavese N Gerhard A et al Microglial activation in presymptomatic Hun-tingtonrsquos disease gene carriers Brain 20071301759ndash1766
19 Politis M Pavese N Tai YF et al Microglial activation in regions related to cognitivefunction predicts disease onset in Huntingtonrsquos disease a multimodal imaging studyHum Brain Mapp 201132258ndash270
20 Politis M Lahiri N Niccolini F et al Increased central microglial activation associatedwith peripheral cytokine levels in premanifest Huntingtonrsquos disease gene carriersNeurobiol Dis 201583115ndash121
21 Lois C Gonzalez I Izquierdo-Garcia D et al Neuroinflammation in Huntingtonrsquosdisease new insights with (11)C-PBR28 PETMRI ACS Chem Neurosci 201892563ndash2571
22 Huntington Study Group Unified Huntingtonrsquos Disease Rating Scale reliability andconsistency Mov Disord 199611136ndash142
23 Folstein MF Folstein SE McHugh PR ldquoMini-mental staterdquo A practical method forgrading the cognitive state of patients for the clinician J Psychiatr Res 197512189ndash198
24 McNally G Rickards H Horton M Craufurd D Exploring the validity of the shortversion of the Problem Behaviours Assessment (PBA-s) for Huntingtonrsquos disease arasch analysis J Huntingtons Dis 20154347ndash369
25 Warner JH Sampaio C Modeling variability in the progression of Huntingtonrsquosdisease a novel modeling approach applied to structural imaging markers fromTRACK-HD CPT Pharmacometrics Syst Pharmacol 20165437ndash445
26 Lyoo CH Ikawa M Liow JS et al Cerebellum can serve as a pseudo-reference regionin Alzheimer disease to detect neuroinflammation measured with PET radioligandbinding to translocator protein J Nucl Med 201556701ndash706
27 Reuter M Schmansky NJ Rosas HD Fischl B Within-subject template estimation forunbiased longitudinal image analysis Neuroimage 2012611402ndash1418
28 Yushkevich PA Piven J Hazlett HC et al User-guided 3D active contour segmen-tation of anatomical structures significantly improved efficiency and reliabilityNeuroimage 2006311116ndash1128
29 Ohgidani M Kato TA Setoyama D et al Direct induction of ramified microglia-likecells from human monocytes dynamic microglial dysfunction in Nasu-Hakola dis-ease Sci Rep 201444957
30 Ohgidani M Kato TA Kanba S Introducing directly induced microglia-like (iMG)cells from fresh human monocytes a novel translational research tool for psychiatricdisorders Front Cell Neurosci 20159184
31 Kim SU de Vellis J Microglia in health and disease J Neurosci Res 200581302-31332 Ross CA Tabrizi SJ Huntingtonrsquos disease from molecular pathogenesis to clinical
treatment Lancet Neurol 20111083ndash9833 Chitnis TWeiner HL CNS inflammation and neurodegeneration J Clin Invest 2017
1273577ndash3587
Appendix Authors
Name Location Contribution
Natalia PRocha PhD
The Mitchell Center forAlzheimerrsquos Disease andRelated Brain DisordersDepartment of NeurologyMcGovern Medical SchoolThe University of TexasHealth Science CenterHouston
Designed andconceptualized the studymajor role in theacquisition of dataanalyzed the data anddrafted the manuscript forintellectual content
OdelinCharron MS
Department of NeurologyThe University of Texas atAustin
Major role in data analysis
Leigh BLathamPhD
School of MedicineUniversity of WashingtonSeattle
Major role in theacquisition of data
Gabriela DColpo PhD
Neuropsychiatry ProgramDepartment of Psychiatryand Behavioral SciencesMcGovern Medical SchoolUniversity of TexasHouston
Major role in theacquisition of data
PaoloZanotti-FregonaraMD PhD
Houston MethodistResearch Institute andWeill Cornell Medicine TX
Major role in the studydesign and the acquisitionof data
MeixiangYu PhD
Houston MethodistResearch Institute andWeill Cornell Medicine TX
Major role in theacquisition of data
LeorahFreemanMD PhD
Department of NeurologyThe University of Texas atAustin
Major role in the studydesign data analysis andinterpretation and theacquisition of data
Erin FurrStimmingMD
HDSA Center of Excellenceat University of TexasHealth Science Center atHouston
Major role in the studydesign and acquisition ofdata interpreted the dataand revised themanuscript for intellectualcontent
Antonio LTeixeiraMD PhD
Neuropsychiatry ProgramDepartment of Psychiatryand Behavioral SciencesMcGovern Medical SchoolUniversity of TexasHouston
Major role in the studydesign and acquisition ofdata interpreted the dataand revised themanuscript for intellectualcontent
NeurologyorgNN Neurology Neuroimmunology amp Neuroinflammation | Volume 8 Number 3 | May 2021 11
34 Rocha NP de Miranda AS Teixeira AL Insights into neuroinflammation in Parkin-sonrsquos disease from biomarkers to anti-inflammatory based therapies Biomed Res Int20152015628192
35 Silvestroni A Faull RL Strand AD Moller T Distinct neuroinflammatory profile inpost-mortem human Huntingtonrsquos disease Neuroreport 2009201098ndash1103
36 Bjorkqvist M Wild EJ Thiele J et al A novel pathogenic pathway of immune acti-vation detectable before clinical onset in Huntingtonrsquos disease J Exp Med 20082051869ndash1877
37 Politis M Lahiri N Niccolini F et al Corrigendum to ldquoIncreased central microglialactivation associated with peripheral cytokine levels in premanifest Huntingtonrsquos
disease gene carriersrdquo [Neurobiol Dis 83 (2015) 115-121] Neurobiol Dis 201798162
38 Kropholler MA Boellaard R Schuitemaker A Folkersma H van Berckel BN Lam-mertsma AA Evaluation of reference tissue models for the analysis of [11C](R)-PK11195 studies J Cereb Blood Flow Metab 2006261431ndash1441
39 Turkheimer FE Edison P Pavese N et al Reference and target region modeling of[11C]-(R)-PK11195 brain studies J Nucl Med 200748158ndash167
40 Kreisl WC Jenko KJ Hines CS et al A genetic polymorphism for translocator protein18 kDa affects both in vitro and in vivo radioligand binding in human brain to thisputative biomarker of neuroinflammation J Cereb Blood FlowMetab 20133353ndash58
12 Neurology Neuroimmunology amp Neuroinflammation | Volume 8 Number 3 | May 2021 NeurologyorgNN
DOI 101212NXI000000000000098420218 Neurol Neuroimmunol Neuroinflamm
Natalia P Rocha Odelin Charron Leigh B Latham et al Pathophysiology
Microglia Activation in Basal Ganglia Is a Late Event in Huntington Disease
This information is current as of April 1 2021
ServicesUpdated Information amp
httpnnneurologyorgcontent83e984fullhtmlincluding high resolution figures can be found at
References httpnnneurologyorgcontent83e984fullhtmlref-list-1
This article cites 40 articles 4 of which you can access for free at
Subspecialty Collections
httpnnneurologyorgcgicollectionpetPET
httpnnneurologyorgcgicollectionhuntingtons_diseaseHuntingtons diseasefollowing collection(s) This article along with others on similar topics appears in the
Permissions amp Licensing
httpnnneurologyorgmiscaboutxhtmlpermissionsits entirety can be found online atInformation about reproducing this article in parts (figurestables) or in
Reprints
httpnnneurologyorgmiscaddirxhtmlreprintsusInformation about ordering reprints can be found online
Academy of Neurology All rights reserved Online ISSN 2332-7812Copyright copy 2021 The Author(s) Published by Wolters Kluwer Health Inc on behalf of the AmericanPublished since April 2014 it is an open-access online-only continuous publication journal Copyright
is an official journal of the American Academy of NeurologyNeurol Neuroimmunol Neuroinflamm

cytokines and reactive oxygen species which in turn will alsocontribute to neuronal death and further microglial activa-tion34We observed an increase in cytokine release (especiallyTNFα) following a phagocytic stimulus in our in vitro ex-periments In line with our findings postmortem studies havealso reported increased levels of inflammatory cytokines inbrain samples of patients with HD in comparison withcontrols3536 It is worth mentioning that cytokine release is amicroglial reaction to a proinflammatory challenge and undercontrolled conditions (such as the iMG cultures) phagocy-tosis is a noninflammatory function of microglia A proin-flammatory stimulus would be needed to appropriately assessdifferences in cytokine release between controls premanifestHD and manifest HD iMG cultures
The current study used a novel PET tracer for TSPO [11C]-ER176 to assess microglial activation in HD By using [11C]-PK11195 PET studies have shown a significant increase inmicroglial activation in the striatum of patients with HD16 aswell as in premanifest HD gene carriers18 compared withcontrols In both studies [11C]-PK11195 binding significantlycorrelated with disease severity as measured by decreased[11C]-raclopride (a marker of dopamine D2 and D3 receptoravailability) binding in the striatum1618 Similar results weredescribed in the hypothalamus17 Later the same work groupused a multimodal imaging approach combiningMRI and PETanalyses and worsening in atrophy evaluated by MRI was ac-companied by a reduction in [11C]-raclopride and an increasein [11C]-PK11195 bindings in patients with HD In pre-manifest HD increased level of microglial activation in theassociative striatum and in the regional network associated withcognition correlated with 5-year probability of HD onset19
More recently increased microglial activation in the somato-sensory cortex (as evaluated through [11C]-PK11195 binding)was associatedwith higher levels of IL-1β IL-6 IL-8 and TNFαproduced by stimulated peripheral blood monocytes frompremanifest HD gene carriers2037 Although these studiesprovided valuable information about neuroinflammation inHD indicating microglial activation even in premanifest sub-jects their results should be interpreted with caution The poorsignal-to-background ratio of first-generation TSPO radio-tracers poses important limitations to the analyses The ratio ofspecific to nonspecific binding of [11C]-PK11195 in humanbrain has been reported to be as low as 023839 As an attempt tomitigate this problem a recent study used the second-generation TSPO tracer [11C]-PBR28 to evaluate micro-glial activation in HD21 Of interest the results were similar toours ie increased levels of [11C]-PBR28 binding in theputamina and pallidi of HD gene carriers (which included 7manifest patients with HD and only 1 person with premanifestHD) in comparison with controls Of note the results of thesingle premanifest subject were midway between controls andthe remaining patients suggesting that microglial activationwas associated with disease progression21 The authors didconfirm that [11C]-PBR28 provides a high signal-to-background ratio an important feature required for the anal-ysis at the individual level and within individual brain regions
An important limitation of [11C]-PBR28 and other second-generation TSPO radioligands is that they have varying degreesof sensitivity to the single nucleotide polymorphism rs6971 inthe TSPO gene40 [11C]-ER176 has unique advantages overthe other TSPO binders being the only known TSPO radio-ligand that allows the inclusion of low-affinity binders Thebinding of ER176 in low-affinity binders was found to becomparable to that for PBR28 in high-affinity binders More-over ER176 was proven to be the only radioligand not con-taminated by radiometabolites accumulating in the brain14
Our findings must be interpreted while taking into accountthe limitations of the study which include the relatively smallsample size and the cross-sectional design The cross-sectionaldesign prevented the assessment of microglial activation inthe context of disease progression and development of motorand nonmotor symptoms In addition the use of an objectivemethod to measure striatal degeneration (eg [11C]-raclopride PET) would provide more accurate informationabout striatal neuron dysfunctiondeath Regarding the invitro experiments the use of a proinflammatory challenge(other than the beads added for the phagocytosis assay) in ourexperiments would provide valuable information on cytokinesrelease by iMG from the different groups It is worth men-tioning the strengths of the study that include the use of aunique TSPO ligand [11C]-ER176 the enrollment of bothpremanifest and manifest subjects and the exploratory in-vestigation of microglia activity
In conclusion our findings strengthen the evidence thatmicroglial activation is associated with HD pathophysiologyMicroglial activation is unlikely to be the initiating event inHD Yet it may exacerbate neuronal dysfunction and neu-ronal death thus contributing to disease progression It isimportant to refine our understanding of the more specificimmuneinflammatory mechanisms that are involved in HDand in neurodegeneration This may foster the developmentof new therapeutic interventions to halt the progression ofHD and other neurodegenerative diseases
AcknowledgmentThe authors acknowledge and thank all of the volunteers thatparticipated in this study and are indebted to their families fortheir magnificent support They also thank the staff from theUTHealth McGovern Medical School 3T MRI Center andfrom the HoustonMethodist Research Institute TranslationalImaging Core Facilities especially Vips Patel and Kim MDoan for their skilled technical support Dr Gabriel Fries forhis assistance with the genotyping analyses and the teammembers of the HDSA Center of Excellence at UTHealth
Study FundingThis study was funded by the HD Human BiologyProjectmdashHuntingtonrsquos Disease Society of America (HDSA)E Furr Stimming receives research funding from RocheGenetech Vaccinex Cures Within Reach HDSA Uniqureand CHDI Foundation
10 Neurology Neuroimmunology amp Neuroinflammation | Volume 8 Number 3 | May 2021 NeurologyorgNN
DisclosureThe authors report no disclosures relevant to the manuscriptGo to NeurologyorgNN for full disclosures
Publication HistoryReceived by Neurology Neuroimmunology amp NeuroinflammationSeptember 9 2020 Accepted in final form January 20 2021
References1 The Huntingtonrsquos Disease Collaborative Research Group A novel gene containing a
trinucleotide repeat that is expanded and unstable on Huntingtonrsquos disease chro-mosomes Cell 199372971ndash983
2 Ross CA Aylward EH Wild EJ et al Huntington disease natural history biomarkersand prospects for therapeutics Nat Rev Neurol 201410204ndash216
3 Soulet D Cicchetti F The role of immunity in Huntingtonrsquos disease Mol Psychiatry201116889ndash902
4 Myers RH Vonsattel JP Paskevich PA et al Decreased neuronal and increasedoligodendroglial densities in Huntingtonrsquos disease caudate nucleus J NeuropatholExp Neurol 199150729ndash742
5 Vonsattel JP Myers RH Stevens TJ Ferrante RJ Bird ED Richardson EP JrNeuropathological classification of Huntingtonrsquos disease J Neuropathol Exp Neurol198544559ndash577
6 Sapp E Kegel KB Aronin N et al Early and progressive accumulation of reactivemicroglia in the Huntington disease brain J Neuropathol Exp Neurol 200160161ndash172
7 Ciccocioppo F Bologna G Ercolino E et al Neurodegenerative diseases asproteinopathies-driven immune disorders Neural Regen Res 202015850ndash856
8 Rocha NP Ribeiro FM Furr-Stimming E Teixeira AL Neuroimmunology of Hun-tingtonrsquos disease revisiting evidence from human studies Mediators Inflamm 201620168653132
9 Vowinckel E Reutens D Becher B et al PK11195 binding to the peripheral ben-zodiazepine receptor as a marker of microglia activation in multiple sclerosis andexperimental autoimmune encephalomyelitis J Neurosci Res 199750345ndash353
10 Papadopoulos V Baraldi M Guilarte TR et al Translocator protein (18kDa) newnomenclature for the peripheral-type benzodiazepine receptor based on its structureand molecular function Trends Pharmacol Sci 200627402ndash409
11 Cumming P Burgher B Patkar O et al Sifting through the surfeit of neuro-inflammation tracers J Cereb Blood Flow Metab 201838204ndash224
12 Zanotti-Fregonara P Zhang Y Jenko KJ et al Synthesis and evaluation of translocator18 kDa protein (TSPO) positron emission tomography (PET) radioligands with lowbinding sensitivity to human single nucleotide polymorphism rs6971 ACS ChemNeurosci 20145963ndash971
13 Ikawa M Lohith TG Shrestha S et al 11C-ER176 a radioligand for 18-kDa trans-locator protein has adequate sensitivity to robustly image all three affinity genotypesin human brain J Nucl Med 201758320ndash325
14 Fujita M Kobayashi M Ikawa M et al Comparison of four (11)C-labeled PETligands to quantify translocator protein 18 kDa (TSPO) in human brain (R)-PK11195 PBR28 DPA-713 and ER176-based on recent publications that measuredspecific-to-non-displaceable ratios EJNMMI Res 2017784
15 Zanotti-Fregonara P Pascual B Veronese M et al Head-to-head comparison of (11)C-PBR28 and (11)C-ER176 for quantification of the translocator protein in thehuman brain Eur J Nucl Med Mol Imaging 2019461822ndash1829
16 Pavese N Gerhard A Tai YF et al Microglial activation correlates with severity inHuntington disease a clinical and PET study Neurology 2006661638ndash1643
17 Politis M Pavese N Tai YF Tabrizi SJ Barker RA Piccini P Hypothalamic in-volvement in Huntingtonrsquos disease an in vivo PET study Brain 20081312860ndash2869
18 Tai YF Pavese N Gerhard A et al Microglial activation in presymptomatic Hun-tingtonrsquos disease gene carriers Brain 20071301759ndash1766
19 Politis M Pavese N Tai YF et al Microglial activation in regions related to cognitivefunction predicts disease onset in Huntingtonrsquos disease a multimodal imaging studyHum Brain Mapp 201132258ndash270
20 Politis M Lahiri N Niccolini F et al Increased central microglial activation associatedwith peripheral cytokine levels in premanifest Huntingtonrsquos disease gene carriersNeurobiol Dis 201583115ndash121
21 Lois C Gonzalez I Izquierdo-Garcia D et al Neuroinflammation in Huntingtonrsquosdisease new insights with (11)C-PBR28 PETMRI ACS Chem Neurosci 201892563ndash2571
22 Huntington Study Group Unified Huntingtonrsquos Disease Rating Scale reliability andconsistency Mov Disord 199611136ndash142
23 Folstein MF Folstein SE McHugh PR ldquoMini-mental staterdquo A practical method forgrading the cognitive state of patients for the clinician J Psychiatr Res 197512189ndash198
24 McNally G Rickards H Horton M Craufurd D Exploring the validity of the shortversion of the Problem Behaviours Assessment (PBA-s) for Huntingtonrsquos disease arasch analysis J Huntingtons Dis 20154347ndash369
25 Warner JH Sampaio C Modeling variability in the progression of Huntingtonrsquosdisease a novel modeling approach applied to structural imaging markers fromTRACK-HD CPT Pharmacometrics Syst Pharmacol 20165437ndash445
26 Lyoo CH Ikawa M Liow JS et al Cerebellum can serve as a pseudo-reference regionin Alzheimer disease to detect neuroinflammation measured with PET radioligandbinding to translocator protein J Nucl Med 201556701ndash706
27 Reuter M Schmansky NJ Rosas HD Fischl B Within-subject template estimation forunbiased longitudinal image analysis Neuroimage 2012611402ndash1418
28 Yushkevich PA Piven J Hazlett HC et al User-guided 3D active contour segmen-tation of anatomical structures significantly improved efficiency and reliabilityNeuroimage 2006311116ndash1128
29 Ohgidani M Kato TA Setoyama D et al Direct induction of ramified microglia-likecells from human monocytes dynamic microglial dysfunction in Nasu-Hakola dis-ease Sci Rep 201444957
30 Ohgidani M Kato TA Kanba S Introducing directly induced microglia-like (iMG)cells from fresh human monocytes a novel translational research tool for psychiatricdisorders Front Cell Neurosci 20159184
31 Kim SU de Vellis J Microglia in health and disease J Neurosci Res 200581302-31332 Ross CA Tabrizi SJ Huntingtonrsquos disease from molecular pathogenesis to clinical
treatment Lancet Neurol 20111083ndash9833 Chitnis TWeiner HL CNS inflammation and neurodegeneration J Clin Invest 2017
1273577ndash3587
Appendix Authors
Name Location Contribution
Natalia PRocha PhD
The Mitchell Center forAlzheimerrsquos Disease andRelated Brain DisordersDepartment of NeurologyMcGovern Medical SchoolThe University of TexasHealth Science CenterHouston
Designed andconceptualized the studymajor role in theacquisition of dataanalyzed the data anddrafted the manuscript forintellectual content
OdelinCharron MS
Department of NeurologyThe University of Texas atAustin
Major role in data analysis
Leigh BLathamPhD
School of MedicineUniversity of WashingtonSeattle
Major role in theacquisition of data
Gabriela DColpo PhD
Neuropsychiatry ProgramDepartment of Psychiatryand Behavioral SciencesMcGovern Medical SchoolUniversity of TexasHouston
Major role in theacquisition of data
PaoloZanotti-FregonaraMD PhD
Houston MethodistResearch Institute andWeill Cornell Medicine TX
Major role in the studydesign and the acquisitionof data
MeixiangYu PhD
Houston MethodistResearch Institute andWeill Cornell Medicine TX
Major role in theacquisition of data
LeorahFreemanMD PhD
Department of NeurologyThe University of Texas atAustin
Major role in the studydesign data analysis andinterpretation and theacquisition of data
Erin FurrStimmingMD
HDSA Center of Excellenceat University of TexasHealth Science Center atHouston
Major role in the studydesign and acquisition ofdata interpreted the dataand revised themanuscript for intellectualcontent
Antonio LTeixeiraMD PhD
Neuropsychiatry ProgramDepartment of Psychiatryand Behavioral SciencesMcGovern Medical SchoolUniversity of TexasHouston
Major role in the studydesign and acquisition ofdata interpreted the dataand revised themanuscript for intellectualcontent
NeurologyorgNN Neurology Neuroimmunology amp Neuroinflammation | Volume 8 Number 3 | May 2021 11
34 Rocha NP de Miranda AS Teixeira AL Insights into neuroinflammation in Parkin-sonrsquos disease from biomarkers to anti-inflammatory based therapies Biomed Res Int20152015628192
35 Silvestroni A Faull RL Strand AD Moller T Distinct neuroinflammatory profile inpost-mortem human Huntingtonrsquos disease Neuroreport 2009201098ndash1103
36 Bjorkqvist M Wild EJ Thiele J et al A novel pathogenic pathway of immune acti-vation detectable before clinical onset in Huntingtonrsquos disease J Exp Med 20082051869ndash1877
37 Politis M Lahiri N Niccolini F et al Corrigendum to ldquoIncreased central microglialactivation associated with peripheral cytokine levels in premanifest Huntingtonrsquos
disease gene carriersrdquo [Neurobiol Dis 83 (2015) 115-121] Neurobiol Dis 201798162
38 Kropholler MA Boellaard R Schuitemaker A Folkersma H van Berckel BN Lam-mertsma AA Evaluation of reference tissue models for the analysis of [11C](R)-PK11195 studies J Cereb Blood Flow Metab 2006261431ndash1441
39 Turkheimer FE Edison P Pavese N et al Reference and target region modeling of[11C]-(R)-PK11195 brain studies J Nucl Med 200748158ndash167
40 Kreisl WC Jenko KJ Hines CS et al A genetic polymorphism for translocator protein18 kDa affects both in vitro and in vivo radioligand binding in human brain to thisputative biomarker of neuroinflammation J Cereb Blood FlowMetab 20133353ndash58
12 Neurology Neuroimmunology amp Neuroinflammation | Volume 8 Number 3 | May 2021 NeurologyorgNN
DOI 101212NXI000000000000098420218 Neurol Neuroimmunol Neuroinflamm
Natalia P Rocha Odelin Charron Leigh B Latham et al Pathophysiology
Microglia Activation in Basal Ganglia Is a Late Event in Huntington Disease
This information is current as of April 1 2021
ServicesUpdated Information amp
httpnnneurologyorgcontent83e984fullhtmlincluding high resolution figures can be found at
References httpnnneurologyorgcontent83e984fullhtmlref-list-1
This article cites 40 articles 4 of which you can access for free at
Subspecialty Collections
httpnnneurologyorgcgicollectionpetPET
httpnnneurologyorgcgicollectionhuntingtons_diseaseHuntingtons diseasefollowing collection(s) This article along with others on similar topics appears in the
Permissions amp Licensing
httpnnneurologyorgmiscaboutxhtmlpermissionsits entirety can be found online atInformation about reproducing this article in parts (figurestables) or in
Reprints
httpnnneurologyorgmiscaddirxhtmlreprintsusInformation about ordering reprints can be found online
Academy of Neurology All rights reserved Online ISSN 2332-7812Copyright copy 2021 The Author(s) Published by Wolters Kluwer Health Inc on behalf of the AmericanPublished since April 2014 it is an open-access online-only continuous publication journal Copyright
is an official journal of the American Academy of NeurologyNeurol Neuroimmunol Neuroinflamm

DisclosureThe authors report no disclosures relevant to the manuscriptGo to NeurologyorgNN for full disclosures
Publication HistoryReceived by Neurology Neuroimmunology amp NeuroinflammationSeptember 9 2020 Accepted in final form January 20 2021
References1 The Huntingtonrsquos Disease Collaborative Research Group A novel gene containing a
trinucleotide repeat that is expanded and unstable on Huntingtonrsquos disease chro-mosomes Cell 199372971ndash983
2 Ross CA Aylward EH Wild EJ et al Huntington disease natural history biomarkersand prospects for therapeutics Nat Rev Neurol 201410204ndash216
3 Soulet D Cicchetti F The role of immunity in Huntingtonrsquos disease Mol Psychiatry201116889ndash902
4 Myers RH Vonsattel JP Paskevich PA et al Decreased neuronal and increasedoligodendroglial densities in Huntingtonrsquos disease caudate nucleus J NeuropatholExp Neurol 199150729ndash742
5 Vonsattel JP Myers RH Stevens TJ Ferrante RJ Bird ED Richardson EP JrNeuropathological classification of Huntingtonrsquos disease J Neuropathol Exp Neurol198544559ndash577
6 Sapp E Kegel KB Aronin N et al Early and progressive accumulation of reactivemicroglia in the Huntington disease brain J Neuropathol Exp Neurol 200160161ndash172
7 Ciccocioppo F Bologna G Ercolino E et al Neurodegenerative diseases asproteinopathies-driven immune disorders Neural Regen Res 202015850ndash856
8 Rocha NP Ribeiro FM Furr-Stimming E Teixeira AL Neuroimmunology of Hun-tingtonrsquos disease revisiting evidence from human studies Mediators Inflamm 201620168653132
9 Vowinckel E Reutens D Becher B et al PK11195 binding to the peripheral ben-zodiazepine receptor as a marker of microglia activation in multiple sclerosis andexperimental autoimmune encephalomyelitis J Neurosci Res 199750345ndash353
10 Papadopoulos V Baraldi M Guilarte TR et al Translocator protein (18kDa) newnomenclature for the peripheral-type benzodiazepine receptor based on its structureand molecular function Trends Pharmacol Sci 200627402ndash409
11 Cumming P Burgher B Patkar O et al Sifting through the surfeit of neuro-inflammation tracers J Cereb Blood Flow Metab 201838204ndash224
12 Zanotti-Fregonara P Zhang Y Jenko KJ et al Synthesis and evaluation of translocator18 kDa protein (TSPO) positron emission tomography (PET) radioligands with lowbinding sensitivity to human single nucleotide polymorphism rs6971 ACS ChemNeurosci 20145963ndash971
13 Ikawa M Lohith TG Shrestha S et al 11C-ER176 a radioligand for 18-kDa trans-locator protein has adequate sensitivity to robustly image all three affinity genotypesin human brain J Nucl Med 201758320ndash325
14 Fujita M Kobayashi M Ikawa M et al Comparison of four (11)C-labeled PETligands to quantify translocator protein 18 kDa (TSPO) in human brain (R)-PK11195 PBR28 DPA-713 and ER176-based on recent publications that measuredspecific-to-non-displaceable ratios EJNMMI Res 2017784
15 Zanotti-Fregonara P Pascual B Veronese M et al Head-to-head comparison of (11)C-PBR28 and (11)C-ER176 for quantification of the translocator protein in thehuman brain Eur J Nucl Med Mol Imaging 2019461822ndash1829
16 Pavese N Gerhard A Tai YF et al Microglial activation correlates with severity inHuntington disease a clinical and PET study Neurology 2006661638ndash1643
17 Politis M Pavese N Tai YF Tabrizi SJ Barker RA Piccini P Hypothalamic in-volvement in Huntingtonrsquos disease an in vivo PET study Brain 20081312860ndash2869
18 Tai YF Pavese N Gerhard A et al Microglial activation in presymptomatic Hun-tingtonrsquos disease gene carriers Brain 20071301759ndash1766
19 Politis M Pavese N Tai YF et al Microglial activation in regions related to cognitivefunction predicts disease onset in Huntingtonrsquos disease a multimodal imaging studyHum Brain Mapp 201132258ndash270
20 Politis M Lahiri N Niccolini F et al Increased central microglial activation associatedwith peripheral cytokine levels in premanifest Huntingtonrsquos disease gene carriersNeurobiol Dis 201583115ndash121
21 Lois C Gonzalez I Izquierdo-Garcia D et al Neuroinflammation in Huntingtonrsquosdisease new insights with (11)C-PBR28 PETMRI ACS Chem Neurosci 201892563ndash2571
22 Huntington Study Group Unified Huntingtonrsquos Disease Rating Scale reliability andconsistency Mov Disord 199611136ndash142
23 Folstein MF Folstein SE McHugh PR ldquoMini-mental staterdquo A practical method forgrading the cognitive state of patients for the clinician J Psychiatr Res 197512189ndash198
24 McNally G Rickards H Horton M Craufurd D Exploring the validity of the shortversion of the Problem Behaviours Assessment (PBA-s) for Huntingtonrsquos disease arasch analysis J Huntingtons Dis 20154347ndash369
25 Warner JH Sampaio C Modeling variability in the progression of Huntingtonrsquosdisease a novel modeling approach applied to structural imaging markers fromTRACK-HD CPT Pharmacometrics Syst Pharmacol 20165437ndash445
26 Lyoo CH Ikawa M Liow JS et al Cerebellum can serve as a pseudo-reference regionin Alzheimer disease to detect neuroinflammation measured with PET radioligandbinding to translocator protein J Nucl Med 201556701ndash706
27 Reuter M Schmansky NJ Rosas HD Fischl B Within-subject template estimation forunbiased longitudinal image analysis Neuroimage 2012611402ndash1418
28 Yushkevich PA Piven J Hazlett HC et al User-guided 3D active contour segmen-tation of anatomical structures significantly improved efficiency and reliabilityNeuroimage 2006311116ndash1128
29 Ohgidani M Kato TA Setoyama D et al Direct induction of ramified microglia-likecells from human monocytes dynamic microglial dysfunction in Nasu-Hakola dis-ease Sci Rep 201444957
30 Ohgidani M Kato TA Kanba S Introducing directly induced microglia-like (iMG)cells from fresh human monocytes a novel translational research tool for psychiatricdisorders Front Cell Neurosci 20159184
31 Kim SU de Vellis J Microglia in health and disease J Neurosci Res 200581302-31332 Ross CA Tabrizi SJ Huntingtonrsquos disease from molecular pathogenesis to clinical
treatment Lancet Neurol 20111083ndash9833 Chitnis TWeiner HL CNS inflammation and neurodegeneration J Clin Invest 2017
1273577ndash3587
Appendix Authors
Name Location Contribution
Natalia PRocha PhD
The Mitchell Center forAlzheimerrsquos Disease andRelated Brain DisordersDepartment of NeurologyMcGovern Medical SchoolThe University of TexasHealth Science CenterHouston
Designed andconceptualized the studymajor role in theacquisition of dataanalyzed the data anddrafted the manuscript forintellectual content
OdelinCharron MS
Department of NeurologyThe University of Texas atAustin
Major role in data analysis
Leigh BLathamPhD
School of MedicineUniversity of WashingtonSeattle
Major role in theacquisition of data
Gabriela DColpo PhD
Neuropsychiatry ProgramDepartment of Psychiatryand Behavioral SciencesMcGovern Medical SchoolUniversity of TexasHouston
Major role in theacquisition of data
PaoloZanotti-FregonaraMD PhD
Houston MethodistResearch Institute andWeill Cornell Medicine TX
Major role in the studydesign and the acquisitionof data
MeixiangYu PhD
Houston MethodistResearch Institute andWeill Cornell Medicine TX
Major role in theacquisition of data
LeorahFreemanMD PhD
Department of NeurologyThe University of Texas atAustin
Major role in the studydesign data analysis andinterpretation and theacquisition of data
Erin FurrStimmingMD
HDSA Center of Excellenceat University of TexasHealth Science Center atHouston
Major role in the studydesign and acquisition ofdata interpreted the dataand revised themanuscript for intellectualcontent
Antonio LTeixeiraMD PhD
Neuropsychiatry ProgramDepartment of Psychiatryand Behavioral SciencesMcGovern Medical SchoolUniversity of TexasHouston
Major role in the studydesign and acquisition ofdata interpreted the dataand revised themanuscript for intellectualcontent
NeurologyorgNN Neurology Neuroimmunology amp Neuroinflammation | Volume 8 Number 3 | May 2021 11
34 Rocha NP de Miranda AS Teixeira AL Insights into neuroinflammation in Parkin-sonrsquos disease from biomarkers to anti-inflammatory based therapies Biomed Res Int20152015628192
35 Silvestroni A Faull RL Strand AD Moller T Distinct neuroinflammatory profile inpost-mortem human Huntingtonrsquos disease Neuroreport 2009201098ndash1103
36 Bjorkqvist M Wild EJ Thiele J et al A novel pathogenic pathway of immune acti-vation detectable before clinical onset in Huntingtonrsquos disease J Exp Med 20082051869ndash1877
37 Politis M Lahiri N Niccolini F et al Corrigendum to ldquoIncreased central microglialactivation associated with peripheral cytokine levels in premanifest Huntingtonrsquos
disease gene carriersrdquo [Neurobiol Dis 83 (2015) 115-121] Neurobiol Dis 201798162
38 Kropholler MA Boellaard R Schuitemaker A Folkersma H van Berckel BN Lam-mertsma AA Evaluation of reference tissue models for the analysis of [11C](R)-PK11195 studies J Cereb Blood Flow Metab 2006261431ndash1441
39 Turkheimer FE Edison P Pavese N et al Reference and target region modeling of[11C]-(R)-PK11195 brain studies J Nucl Med 200748158ndash167
40 Kreisl WC Jenko KJ Hines CS et al A genetic polymorphism for translocator protein18 kDa affects both in vitro and in vivo radioligand binding in human brain to thisputative biomarker of neuroinflammation J Cereb Blood FlowMetab 20133353ndash58
12 Neurology Neuroimmunology amp Neuroinflammation | Volume 8 Number 3 | May 2021 NeurologyorgNN
DOI 101212NXI000000000000098420218 Neurol Neuroimmunol Neuroinflamm
Natalia P Rocha Odelin Charron Leigh B Latham et al Pathophysiology
Microglia Activation in Basal Ganglia Is a Late Event in Huntington Disease
This information is current as of April 1 2021
ServicesUpdated Information amp
httpnnneurologyorgcontent83e984fullhtmlincluding high resolution figures can be found at
References httpnnneurologyorgcontent83e984fullhtmlref-list-1
This article cites 40 articles 4 of which you can access for free at
Subspecialty Collections
httpnnneurologyorgcgicollectionpetPET
httpnnneurologyorgcgicollectionhuntingtons_diseaseHuntingtons diseasefollowing collection(s) This article along with others on similar topics appears in the
Permissions amp Licensing
httpnnneurologyorgmiscaboutxhtmlpermissionsits entirety can be found online atInformation about reproducing this article in parts (figurestables) or in
Reprints
httpnnneurologyorgmiscaddirxhtmlreprintsusInformation about ordering reprints can be found online
Academy of Neurology All rights reserved Online ISSN 2332-7812Copyright copy 2021 The Author(s) Published by Wolters Kluwer Health Inc on behalf of the AmericanPublished since April 2014 it is an open-access online-only continuous publication journal Copyright
is an official journal of the American Academy of NeurologyNeurol Neuroimmunol Neuroinflamm

34 Rocha NP de Miranda AS Teixeira AL Insights into neuroinflammation in Parkin-sonrsquos disease from biomarkers to anti-inflammatory based therapies Biomed Res Int20152015628192
35 Silvestroni A Faull RL Strand AD Moller T Distinct neuroinflammatory profile inpost-mortem human Huntingtonrsquos disease Neuroreport 2009201098ndash1103
36 Bjorkqvist M Wild EJ Thiele J et al A novel pathogenic pathway of immune acti-vation detectable before clinical onset in Huntingtonrsquos disease J Exp Med 20082051869ndash1877
37 Politis M Lahiri N Niccolini F et al Corrigendum to ldquoIncreased central microglialactivation associated with peripheral cytokine levels in premanifest Huntingtonrsquos
disease gene carriersrdquo [Neurobiol Dis 83 (2015) 115-121] Neurobiol Dis 201798162
38 Kropholler MA Boellaard R Schuitemaker A Folkersma H van Berckel BN Lam-mertsma AA Evaluation of reference tissue models for the analysis of [11C](R)-PK11195 studies J Cereb Blood Flow Metab 2006261431ndash1441
39 Turkheimer FE Edison P Pavese N et al Reference and target region modeling of[11C]-(R)-PK11195 brain studies J Nucl Med 200748158ndash167
40 Kreisl WC Jenko KJ Hines CS et al A genetic polymorphism for translocator protein18 kDa affects both in vitro and in vivo radioligand binding in human brain to thisputative biomarker of neuroinflammation J Cereb Blood FlowMetab 20133353ndash58
12 Neurology Neuroimmunology amp Neuroinflammation | Volume 8 Number 3 | May 2021 NeurologyorgNN
DOI 101212NXI000000000000098420218 Neurol Neuroimmunol Neuroinflamm
Natalia P Rocha Odelin Charron Leigh B Latham et al Pathophysiology
Microglia Activation in Basal Ganglia Is a Late Event in Huntington Disease
This information is current as of April 1 2021
ServicesUpdated Information amp
httpnnneurologyorgcontent83e984fullhtmlincluding high resolution figures can be found at
References httpnnneurologyorgcontent83e984fullhtmlref-list-1
This article cites 40 articles 4 of which you can access for free at
Subspecialty Collections
httpnnneurologyorgcgicollectionpetPET
httpnnneurologyorgcgicollectionhuntingtons_diseaseHuntingtons diseasefollowing collection(s) This article along with others on similar topics appears in the
Permissions amp Licensing
httpnnneurologyorgmiscaboutxhtmlpermissionsits entirety can be found online atInformation about reproducing this article in parts (figurestables) or in
Reprints
httpnnneurologyorgmiscaddirxhtmlreprintsusInformation about ordering reprints can be found online
Academy of Neurology All rights reserved Online ISSN 2332-7812Copyright copy 2021 The Author(s) Published by Wolters Kluwer Health Inc on behalf of the AmericanPublished since April 2014 it is an open-access online-only continuous publication journal Copyright
is an official journal of the American Academy of NeurologyNeurol Neuroimmunol Neuroinflamm

DOI 101212NXI000000000000098420218 Neurol Neuroimmunol Neuroinflamm
Natalia P Rocha Odelin Charron Leigh B Latham et al Pathophysiology
Microglia Activation in Basal Ganglia Is a Late Event in Huntington Disease
This information is current as of April 1 2021
ServicesUpdated Information amp
httpnnneurologyorgcontent83e984fullhtmlincluding high resolution figures can be found at
References httpnnneurologyorgcontent83e984fullhtmlref-list-1
This article cites 40 articles 4 of which you can access for free at
Subspecialty Collections
httpnnneurologyorgcgicollectionpetPET
httpnnneurologyorgcgicollectionhuntingtons_diseaseHuntingtons diseasefollowing collection(s) This article along with others on similar topics appears in the
Permissions amp Licensing
httpnnneurologyorgmiscaboutxhtmlpermissionsits entirety can be found online atInformation about reproducing this article in parts (figurestables) or in
Reprints
httpnnneurologyorgmiscaddirxhtmlreprintsusInformation about ordering reprints can be found online
Academy of Neurology All rights reserved Online ISSN 2332-7812Copyright copy 2021 The Author(s) Published by Wolters Kluwer Health Inc on behalf of the AmericanPublished since April 2014 it is an open-access online-only continuous publication journal Copyright
is an official journal of the American Academy of NeurologyNeurol Neuroimmunol Neuroinflamm