milkweed bug hox rnai - home |...
TRANSCRIPT

INTRODUCTION
Our current understanding of insect development is deep, butnarrow. While important insights have come from the extensivedevelopmental genetic studies in the fruit fly Drosophilamelanogaster, we have little idea of how generally applicablethese mechanisms and pathways are to other insects. Especiallyin terms of head development, it is very difficult to generalizefrom flies to other orders of insects, since both the Drosophilalarva and adult have very specialized head appendages. Workin Tribolium is helping to remedy this situation. Triboliumcastaneum, the red flour beetle, has mandibulate mouthparts,which are both ancestral and typical of most insects. Geneticanalysis of head development in Tribolium provides a usefulfoil for comparison to Drosophila, but these two species alonecannot represent the diversity of morphology within theinsects. To understand the morphological and developmentalvariability of the insects, we need to study representatives ofadditional orders.
The Hemiptera represent a basal outgroup to both flies andbeetles, and as hemimetabolous insects they exhibit a very
different developmental pattern, which is well suited tophenotypic studies, since most adult structures are present athatching (Fig. 1A,B). They are particularly interesting in termsof head development. In the Hemiptera, the mandibular andmaxillary segments give rise to two similar pairs of long, thinstylets (Fig. 1D). The paired maxillary appendages formchannels for liquid flow and the piercing mandibles lie oneither side. These four interlocked stylets run down a groovein the long, fused labium, which provides support. Althoughin typical mandibulate insects the maxillary and labialappendages are very similar (Fig. 1C), in the Hemiptera, it isthe mandibular and maxillary appendages that share a highlyunusual morphology very different from the labium (Fig. 1D).These specialized mouthparts represent an importantevolutionary innovation that allowed this order of insects tofeed by extracting fluids from other organisms. The questionthen arises: what developmental mechanisms changed topromote such novel morphology?
Based on work in Drosophilaand other animals, the anteriorHox genes are undoubtedly important regulators of identity forthe head appendages. Each Hox gene controls the
3683Development 127, 3683-3694 (2000)Printed in Great Britain © The Company of Biologists Limited 2000DEV8752
Insects have evolved a large variety of specialized feedingstrategies, with a corresponding variability in mouthpartmorphology. We have, however, little understanding of thedevelopmental mechanisms that underlie this diversity.Until recently it was difficult to perform any analysis ofgene function outside of the genetic model insectsDrosophila melanogasterand Tribolium castaneum. In thispaper, we report the use of dsRNA-mediated interference(RNAi) to dissect gene function in the development of themilkweed bug Oncopeltus fasciatus, which has specializedsuctorial mouthparts. The Hox genes Deformed (Dfd),proboscipedia (pb) and Sex combs reduced(Scr) havepreviously been shown to be expressed in the gnathalappendages of this species. Strikingly, the milkweed bugwas found to have an unusual expression pattern of pb.Here, by analyzing single and combination RNAidepletions, we find that Dfd, pb and Scr are used in themilkweed bug to specify the identity of the mouthparts. The
exact roles of the genes, however, are different from whatis known in the two genetic model insects. The maxillaryappendages in the bug are determined by the activities ofthe genes Dfd and Scr, rather than Dfd and pbas in the flyand beetle. The mandibular appendages are specified byDfd, but their unique morphology in Oncopeltussuggeststhat Dfd’s target genes are different. As in flies and beetles,the labium is specified by the combined activities of pb andScr, but again, the function of pb appears to be different.Additionally, the regulatory control of pb by the other twogenes seems to be different in the bug than in either of theother species. These novelties in Hox function, expressionpattern and regulatory relationships may have beenimportant for the evolution of the unique Hemipteran head.
Key words: RNAi, Hemiptera, Milkweed bug, Oncopeltus, Head,Stylate-haustellate mouthpart, Mandible, Maxilla, Labium, Hox,Deformed, Dfd, proboscipedia, pb, Sex combs reduced, Scr
SUMMARY
RNAi analysis of Deformed , proboscipedia and Sex combs reduced in the
milkweed bug Oncopeltus fasciatus : novel roles for Hox genes in the
Hemipteran head
Cynthia L. Hughes and Thomas C. Kaufman*
Howard Hughes Medical Institute, Department of Biology, Indiana University, Bloomington, IN 47405, USA*Author for correspondence (e-mail: [email protected])
Accepted 20 June; published on WWW 9 August 2000

3684
developmental fate of one or more segments, to the extent thatremoval of that gene creates a homeotic transformation to thefate of another segment. For instance, the proboscipedia(pb)mutation in flies causes transformation of the proboscis (thefused labial appendages) to a pair of legs (Kaufman, 1978). Wecan think of a Hox gene mutant phenotype as reflecting theremoval of an entire developmental module. This is becauseHox genes are transcription factors that specify segmentalidentity by controlling perhaps hundreds of target genessimultaneously (Botas and Auwers, 1996; Mastick et al.,1995). In addition, since Hox genes also regulate each other,mutant phenotypes are often complicated by a concomitantshift in expression of another Hox gene which is normallyexcluded from or promoted in a given segment. Analysis of themutant phenotype of a Hox gene can thus provide informationabout the domain of activity, the type of identity conferredthrough targeted effector genes and regulatory interactionswith other Hox genes.
Because of their importance in development, the Hox genesare thought to be important for evolutionary change indevelopment as well. Since the DNA-binding homeodomain ishighly conserved among animals and orthologs from verydistant species can rescue fly mutations (e.g., Lutz et al., 1996),it is thought that the influence of Hox genes on evolution ofthe arthropods is generally not due to changes in biochemicalfunction (for an exception, see Falciani et al., 1996). Rather,the function of a Hox gene within the arthropods is likely tochange primarily via three possible mechanisms: changingexpression pattern, changing regulatory relationships, orchanging suites of target genes. A shift in the expressionpattern of a Hox gene, for instance, would carry a wholemodule of developmental instructions into a new tissue. Achange in regulatory interactions between Hox genes wouldhave a similar effect. These shifts in Hox expression couldresult in either dramatic or relatively subtle morphologicalchanges in evolution (Averof and Patel, 1997; Stern, 1998).Finally, changing suites of target loci activated or repressed bya given Hox gene would change the effective function of thatHox gene in terms of developmental fate. Because of the‘relaxed’ requirement for binding, whereby the homeodomaincan bind many variations of sites that include the core TAAT,it is relatively easy for the Hox genes to acquire new target loci(Hayashi and Scott, 1990).
Previous work has revealed the expression patterns of Dfd,pb and Scr in Oncopeltus, which have some interestingdifferences compared to Drosophilaand other insects (Rogersand Kaufman, 1997; Rogers et al., 1997). For instance, unlikethe broad bands of prothoracic (T1) expression in theDrosophila embryo, in Oncopeltus, the T1 ectodermalexpression of Sex combs reducedis limited to a dorsal patch,which is presumed to suppress wing formation, and a spot onthe leg, which is thought to control comb development (Rogerset al., 1997). The expression pattern of proboscipediaisespecially unusual in the bug. Unlike other insects, pbis notexpressed in the maxillary appendage, but only in a smalldorsal maxillary spot. Interestingly, this lack of pb expressionin the maxillary appendage is correlated with the Hemipteranstylate-haustellate mouthparts (Rogers and Kaufman, 1997).
While these differences in Hox expression patterns betweenflies and bugs are provocative, they also raise additionalquestions, since we cannot know the roles of these genes
without functional testing. A particular domain of expressionmay not necessarily reveal a gene’s function. For instance,although pbhas a discrete expression pattern in the fly embryo,due in part to regulation by Dfdand Scr, loss-of-functionmutations in pb do not affect fly embryonic development(Rusch and Kaufman, 2000; Pultz et al., 1988). Therefore thebest way to infer gene function is to analyze the phenotyperesulting from a disruption of gene activity. Until recently, theability to knockout gene function in non-model insects seemedto be a distant possibility. With development of the dsRNA-mediated inhibition technique (RNAi), however, we canpotentially bring loss-of-function studies to additionalarthropods (Fire et al, 1998; Kennerdell and Carthew, 1998;Brown et al., 1999).
To examine the role of Hox genes in Hemipteran mouthpartspecification, we used the RNAi technique to create loss-of-function phenocopies of the genes Dfd, pb and Scrin themilkweed bug. Analysis of the resulting morphologicaltransformations provides insight into the roles of these Hoxgenes in the Hemipteran head and how they differ from otherinsects. Despite the reputation of Hox loci as some of the mosthighly conserved developmental genes, we find notabledifferences in the roles, functional domains and regulatoryinteractions of Deformed, proboscipediaand Sex combsreducedin the milkweed bug as compared to DrosophilaandTribolium.
MATERIALS AND METHODS
HusbandryA strain of milkweed bugs (Oncopeltus fasciatus) adapted to feed onsunflower seeds was supplied by Carolina Biological Supply. Bugswere reared at room temperature, fed on organically grown sunflowerseeds and provided with sterilized cotton for laying eggs.
CloningEggs of mixed stages were collected and RNA was prepared usingTrizol (GibcoBRL/Life Technologies). Poly(A) RNA was selectedusing the Oligotex mRNA minikit (Qiagen). cDNA was preparedusing the 5′/3′RACE kit (Boehringer Mannheim). Primers to DfdandScr were designed using homeobox sequences cloned previously(Rogers and Kaufman, 1997; Rogers et al., 1997). 3′ RACE was doneusing the RACE kit in combination with either Taq Polymerase(Fisher) or a blend of Taq and Pfu polymerases from the TaqPlus LongPCR System kit (Stratagene). PCR products were viewed on standardagarose gels, gel-extracted (Qiagen), blunt-ligated into Bluescript andsequenced. For specificity, the homeobox and poly(A) region weretrimmed off and the remaining portion of the 3′end was subclonedinto Bluescript. Clones with identical sequences were isolated fromindependent PCRs. For pb, a previously isolated clone was used(Rogers and Kaufman, 1997). Although it contains the homeoboxregion, the sequence identity with Scris only 63% with, at most, 24contiguous identical nucleotides. The sequence identity between theDfd and Scr3′ end clones is negligible. Final clones consisted of 358bp (Dfd), 262 bp (Scr) and 258 bp (pb). GenBank accession numbersare AF279336, AF279337 and AF279338.
Preparation of dsRNARNA was prepared using the MEGAscript kit (Ambion). Plasmidswere linearized by restriction digestion, and sense and antisensestrands were transcribed in two separate reactions. The RNA wasphenol-chloroform extracted and isopropanol precipitated. Then theresuspended RNA strands were combined and annealed in a thermal
C. L. Hughes and T. C. Kaufman

3685Milkweed bug Hox RNAi
cycler (Perkin Elmer) using the following program: 85°C 3 minutes,20 minute ramp down to 55°C, hold 10 minutes, 10 minute ramp downto 40°C, hold 20 minutes, 5 minute ramp down to 30°C, hold 10minutes, and hold at 4°C.
RNA was resuspended in injection buffer (0.1 mM NaPO4, 5 mMKCl, pH 6.8), and the inert dye tetramethylrhodamine dextran(Molecular Probes) was added to the RNA at 2 mg/ml to aid indetection of the injected liquid (under normal illumination the dye isbright pink). RNA was injected at the following concentrations: Dfd,11 and 20 µM; pb, 20 µM; Scr, 8 and 20 µM; Dfd pb, 20 µM each;Dfd Scr, 9 µM each; pb Scr, 18 µM each; and Dfd pb Scr, 19 µMeach.
InjectionEggs were always less than 4 hours old at the time of injection. Twoinjection methods were used. For anterior injection, eggs were linedup on Scotch Doublestick™ tape on a microscope slide. A coating ofoil or other liquid was not necessary. For lateral injections, eggs werelined up against the edge of a second microscope slide. Injectionmethod did not affect the resulting phenotypes, except that lateralinjections occasionally led to more asymmetrical effects.
The RNA was backloaded into glass needles (pulled on a Model P-87 micropipette puller, Sutter). RNA was injected into each egg withpressures between 40 and 80 psi using a Narishige model IM 300microinjector. The injection amount was necessarily variable anddifficult to quantify due to needle and egg variability; it ranged fromabout 0.05% to 5% of overall egg volume.
Phenocopy analysisAfter injection, the embryos were allowed to develop at roomtemperature in a humid Petri dish. After approximately 9 days, whenthe unaffected cohort had hatched, unhatched embryos were dissectedand stored in 75% ethanol. Embryo morphology was analyzed usinga dissection microscope (Nikon), a transmission microsope (Zeiss)and by scanning electron microscopy (Jeol, model #JSM-5800LV).For this study, only external morphology was analyzed, focusing onthe appendages.
RESULTS
Injection of dsRNA creates loss-of-functionphenocopies in the milkweed bugDouble-stranded RNAs corresponding to portions of the genesDeformed(Dfd), proboscipedia(pb) and Sex combs reduced(Scr) were produced and injected into freshly laid Oncopeltusfasciatuseggs. Injections included individual genes as well asall possible combinations. Eggs were injected either anteriorlyor laterally; point of injection did not affect the resultingphenotypes, except that laterally injected embryos showslightly more asymmetrical defects (data not shown). Negativecontrols were injected with buffer and dye only. The negativecontrols resulted in a set of defects present in all experiments,which are presumed to be caused by the injection process itself.These include developmental arrest (50% of injected negativecontrols, 54-91% of injected experimental animals) as well asnon-specific developmental defects, e.g., undifferentiated headappendages (17% of developed negative controls, 6-17%of developed experimental animals). No appendagetransformations were seen in the negative controls (Table 1).
For the experimental animals, RNA injection results in ahigh penetrance of phenocopies among those embryoscompleting development (42-88%; Table 1). The phenocopieswere somewhat variable, but fell into a range representing a
clear suite of transformations. Left and right appendages weregenerally affected similarly and were scored as pairs. Thespectrum of transformation from mild to severe is probably dueto differences in the volume of RNA injected, which isnecessarily variable due to inconsistencies in needle size andegg turgor.
Double or triple depletion analysis was possible by injectinga mixture of dsRNAs corresponding to the different genes.These multiple depletions are extremely effective; in the DfdScr double, for example, just 8% of the developing animalshave a phenocopy of only one of the genes, while 42% exhibitthe double phenocopy. The facility of the double depletion inOncopeltusprovides a great advantage for the analysis ofgenetic interactions.
Except for the aforementioned non-specific effects, thephenotypes produced by each injected RNA were unique.Moreover, based on previously determined expression patterns,they primarily affected the expected appendages (Rogers andKaufman, 1997; Rogers et al., 1997). Finally, each RNAidepletion resulted in homeotic transformations, the hallmark ofHox gene mutations. These results give us confidence that the
Fig. 1. Wild-type Oncopeltusmorphology and mouthparts. (A,B) Two wild-type embryos from negative control injections, athatching. Stylets are visible in B at the top of the labium. (C) Typicalmandibulate insect mouthparts versus (D) Hemipteran stylate-haustellate mouthparts. In the Hemiptera, the mandibular andmaxillary segments form long thin stylets, which run down a groovein the fused labium (the haustellum). Mn, mandibles (red); Mx,maxillae (blue); Lb, labium (green). SEMs by Rudi Turner. (C,D) Adapted from Daly et al. (1998), with permission.

3686
RNAi technique produces true loss-of-function phenocopies ofthe targeted gene.
Deformed RNAi transforms the mandibular andmaxillary appendages Injection of dsRNA corresponding to a portion of the Deformedgene resulted in transformation of the mandibular andmaxillary appendages (penetrance=62% of developed animals;Fig. 2). The mandibular appendage takes on distal antennalidentity, with a short fused base (98% of affected animals). Theresulting short, plump, bristled appendages are quite distinctfrom the thin, wire-like stylets usually formed from thissegment (Fig. 1). The form of the appendage, as well as thetype of bristle, is very similar to the most distal segment of theantenna, and is clearly recognizable as such in the vast majorityof affected embryos. Moreover, 12% of these transformedmandibular appendages are partially fused to the genuineantennae rather than attached directly to the head capsule (Fig.2C). Of the majority that emerge directly from the head, it isnot clear precisely which segment(s) are associated with thebase of the appendage, as the more proximal portion isdistorted. Based on the relative position of this antenna-likeappendage, however, we conclude that it is derived from themandibular segment. Interestingly, despite the consistent andobvious distal transformation, the proximal antennal segmentsare missing and a full-length mandibular antenna is never seenin the Dfddepletions.
The transformed fate of the maxillary appendages in the Dfddepletions is much less clear. They range from short, bristledappendages (93% of affected animals) to larger fusedstructures with short, bristled lumps at the tip (7%). In bothcases, the appendages resemble the distal antenna, but theoverall shape is curved and distorted, generally curvingtowards the posterior. Overall, in the Dfd depletions themaxillary appendages seem to take on a unique form, butwhich is nevertheless largely antennal in identity. The labiumis not affected by DfdRNAi depletion.
proboscipedia RNAi transforms the labium to legidentityInjection of dsRNA corresponding to a portion of theproboscipediagene results in transformation of the distallabium to prothoracic legs (penetrance=71% of developedanimals; Fig. 3). The mandibular and maxillary appendages areunaffected. Injection of 20 µM RNA results in a highpenetrance of an extreme tuning-fork-like phenotype (52% ofdeveloped animals, 73% of affected animals), in which theproximal first two sections of the labium are wild type, whilethe third section is split midway into a pair of tibia, tarsi andpretarsi, complete with T1 combs, claws and pulvilli (Fig.3A,B). Transformation of the proximal labium is never seen.
C. L. Hughes and T. C. Kaufman
Fig. 2.The Dfd RNAi phenotype. (A,C) Two typical individuals,exhibiting the transformation of the mandibular and maxillary styletsto antenna-like appendages. (B) The mandibular and maxillaryappendages, removed from a third individual. The mandibularappendages (Mn) resemble the distal antenna. The mandibularappendages usually originate from the head capsule (A,B), but aresometimes fused basally to the genuine antennae (Ant) (C, asterisks).The maxillary appendages (Mx) are short and generally curlposteriorly (A,C). The labium (Lb) is wild type in theDfd depletionanimals.
Fig. 3.The pbRNAi phenotype. (A) A light micrograph and (B) SEM of typical individuals exhibiting the tuning-fork-likephenotype. While the first two segments of the labium are wild type(I, II), the third (III) splits into a pair of distal legs composed of tibia(ti), tarsus (tr) and pretarsus (pt), complete with claws and pulvilli.The mandibular and maxillary stylets are wild type (B, arrow).Compare (C) a wild-type distal labium and (E) a wild-type distal leg,with (D) the distal labial appendage of a pbdepletion animal. Notethe claws (arrowheads) and pulvilli (asterisks) present in D,E. Scalebar, 10 µm.
3687Milkweed bug Hox RNAi
Occasionally, only one side of the labium produces aprothoracic leg (19% of developed animals); this mosaic effectis seen with lateral injections but not anterior injections (datanot shown). The mandibular and maxillary stylets are presentand properly extended in the majority of the affected animals,even in conjunction with a strongly transformed labium (87%of affected animals; Fig. 3B). The strong pb phenotype isassociated with surprisingly high viability: 80% of affectedanimals hatch and tan, although they are unable to feed.
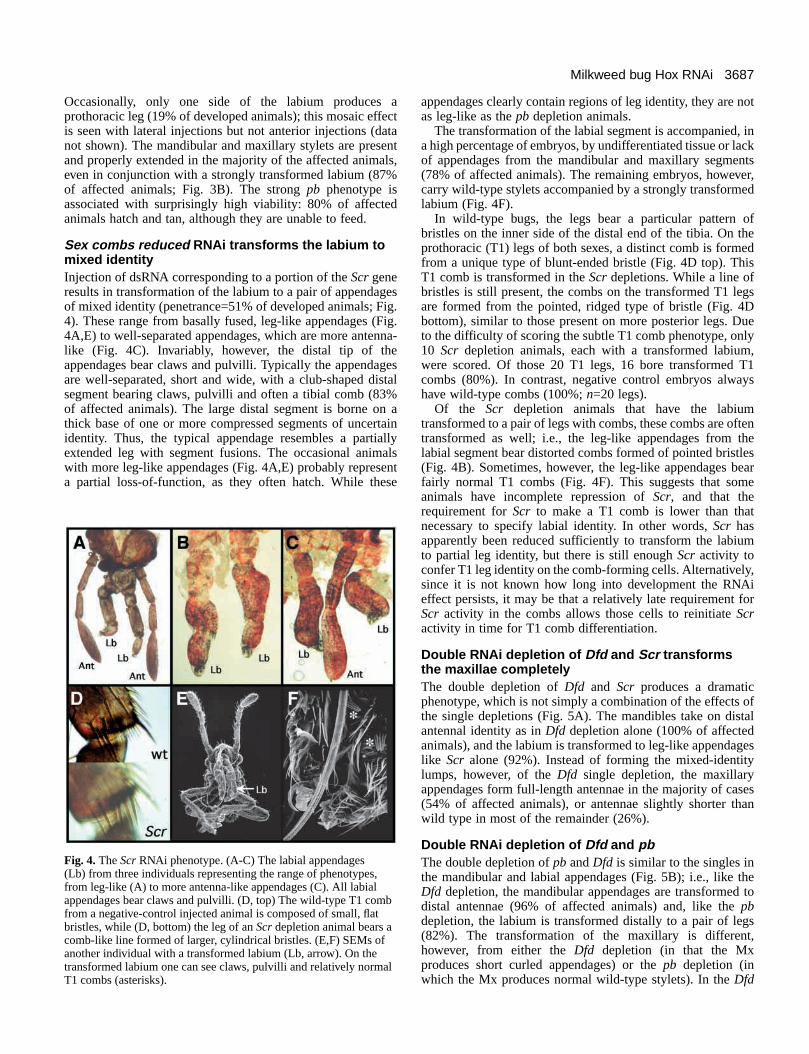
Sex combs reduced RNAi transforms the labium tomixed identityInjection of dsRNA corresponding to a portion of the Scrgeneresults in transformation of the labium to a pair of appendagesof mixed identity (penetrance=51% of developed animals; Fig.4). These range from basally fused, leg-like appendages (Fig.4A,E) to well-separated appendages, which are more antenna-like (Fig. 4C). Invariably, however, the distal tip of theappendages bear claws and pulvilli. Typically the appendagesare well-separated, short and wide, with a club-shaped distalsegment bearing claws, pulvilli and often a tibial comb (83%of affected animals). The large distal segment is borne on athick base of one or more compressed segments of uncertainidentity. Thus, the typical appendage resembles a partiallyextended leg with segment fusions. The occasional animalswith more leg-like appendages (Fig. 4A,E) probably representa partial loss-of-function, as they often hatch. While these
appendages clearly contain regions of leg identity, they are notas leg-like as the pb depletion animals.
The transformation of the labial segment is accompanied, ina high percentage of embryos, by undifferentiated tissue or lackof appendages from the mandibular and maxillary segments(78% of affected animals). The remaining embryos, however,carry wild-type stylets accompanied by a strongly transformedlabium (Fig. 4F).
In wild-type bugs, the legs bear a particular pattern ofbristles on the inner side of the distal end of the tibia. On theprothoracic (T1) legs of both sexes, a distinct comb is formedfrom a unique type of blunt-ended bristle (Fig. 4D top). ThisT1 comb is transformed in the Scrdepletions. While a line ofbristles is still present, the combs on the transformed T1 legsare formed from the pointed, ridged type of bristle (Fig. 4Dbottom), similar to those present on more posterior legs. Dueto the difficulty of scoring the subtle T1 comb phenotype, only10 Scr depletion animals, each with a transformed labium,were scored. Of those 20 T1 legs, 16 bore transformed T1combs (80%). In contrast, negative control embryos alwayshave wild-type combs (100%; n=20 legs).
Of the Scr depletion animals that have the labiumtransformed to a pair of legs with combs, these combs are oftentransformed as well; i.e., the leg-like appendages from thelabial segment bear distorted combs formed of pointed bristles(Fig. 4B). Sometimes, however, the leg-like appendages bearfairly normal T1 combs (Fig. 4F). This suggests that someanimals have incomplete repression of Scr, and that therequirement for Scrto make a T1 comb is lower than thatnecessary to specify labial identity. In other words, Scr hasapparently been reduced sufficiently to transform the labiumto partial leg identity, but there is still enough Scr activity toconfer T1 leg identity on the comb-forming cells. Alternatively,since it is not known how long into development the RNAieffect persists, it may be that a relatively late requirement forScr activity in the combs allows those cells to reinitiate Scractivity in time for T1 comb differentiation.
Double RNAi depletion of Dfd and Scr transformsthe maxillae completelyThe double depletion of Dfdand Scrproduces a dramaticphenotype, which is not simply a combination of the effects ofthe single depletions (Fig. 5A). The mandibles take on distalantennal identity as in Dfd depletion alone (100% of affectedanimals), and the labium is transformed to leg-like appendageslike Scr alone (92%). Instead of forming the mixed-identitylumps, however, of the Dfd single depletion, the maxillaryappendages form full-length antennae in the majority of cases(54% of affected animals), or antennae slightly shorter thanwild type in most of the remainder (26%).
Double RNAi depletion of Dfd and pbThe double depletion of pb and Dfdis similar to the singles inthe mandibular and labial appendages (Fig. 5B); i.e., like theDfd depletion, the mandibular appendages are transformed todistal antennae (96% of affected animals) and, like the pbdepletion, the labium is transformed distally to a pair of legs(82%). The transformation of the maxillary is different,however, from either the Dfd depletion (in that the Mxproduces short curled appendages) or the pb depletion (inwhich the Mx produces normal wild-type stylets). In the Dfd
Fig. 4. The ScrRNAi phenotype. (A-C) The labial appendages (Lb) from three individuals representing the range of phenotypes,from leg-like (A) to more antenna-like appendages (C). All labialappendages bear claws and pulvilli. (D, top) The wild-type T1 combfrom a negative-control injected animal is composed of small, flatbristles, while (D, bottom) the leg of an Scrdepletion animal bears acomb-like line formed of larger, cylindrical bristles. (E,F) SEMs ofanother individual with a transformed labium (Lb, arrow). On thetransformed labium one can see claws, pulvilli and relatively normalT1 combs (asterisks).

3688
pb double depletion, the maxillary segment produced short,straight appendages which resemble distal antennae, similar tothe transformed mandibles (67% of affected animals). Thedistal tips of these transformed limbs, however, often bearclaws as well (75%). Thus the transformed maxillaryappendage seems to have mixed leg and antennal identity.
Double RNAi depletion of pb and Scr transformslabium to antennaeInjection of dsRNAs corresponding to pb and Scr results in anextreme transformation of the labium to a pair of antennae (Fig.5C), which is unlike the effects of either pbor Scralone. Whilethe affected appendages fall into a range between leg andantennal identity; the majority are clearly antenna-like and lackclaws (65% of affected animals). Very few of the appendagesare overall truly leg-like (5%), and the remainder are unclearor have mixed identity between leg and antenna (29%).
The labial transformation is nearly always accompanied byundifferentiated or unelongated mandibular and maxillaryappendages (97% of affected animals). In addition, the T1 legssometimes exhibit a knobby appearance, due to a thickenedtibia (20%). Finally the genuine antennae are usuallyundifferentiated on their distal ends, often with their tipspinched off or fused to each other (79% of affected animals;Fig. 5C). Rather than being a direct effect, this phenotypelikely represents an indirect, physical effect of the transformedlabium on the antennae, since in the egg these appendages liein close proximity.
Triple RNAi depletion transforms all gnathalappendages to antennaeBy combining preparations of dsRNA corresponding to Dfd,pb and Scr, all three genes were depleted simultaneously. Thisis helpful for the analysis, as it produces a ‘default state’ forthe fate of the appendages in the absence of all three genes. Afew animals (6%) were affected in only a subset of thethree appendages, but 67% of developed animals hadtransformations of the Mn, Mx and Lb appendages (Fig. 5D,E).
Like theDfd depletion alone, the mandibular appendages areconsistently transformed to distal antennae (94% of affectedanimals; Fig. 5D,E). Like the Dfd Scr double depletion, themaxillary appendages are generally transformed to full-lengthor mid-length antennae (59% and 28% of affected animalsrespectively; Fig. 5D,E). The phenotype of the labialappendages, however, falls into a range that probablyrepresents the extent of repression of gene activity. The labiumis transformed to a pair of appendages ranging from leg toantennal identity: leg-like with claws (31% of affectedanimals), antenna-like with claws (25%; Fig. 5E), antenna-like(28%), or complete antennal transformation (13%; Fig. 5D).We conclude that, as in the pb Scr double, the full-lengthantenna represents the phenotype of the most complete loss-of-function.
DISCUSSION
RNAi allows functional analysis in the bugThe molecular mechanism of RNAi is still somewhatmysterious, but C. elegans researchers have characterizedseveral of the necessary cellular components (Hunter, 2000),
and embraced the technique as a quick method for producingloss-of-function phenocopies, which can guide subsequentdetailed genetic studies (e.g. Boxem et al., 1999; Fay et al.,1999; Hong et al., 1998).
Without the balancers, markers and the short generation timeof models like Drosophila, traditional genetics is oftenimpractical in other arthropods. Up to the present, researchersinterested in comparative arthropod development have had tobe content with inferences based on gene expression patterns,by analogy to what is known in Drosophila. This approach hasbeen fruitful but also frustrating, since expression patterns areoften misleading and novel expression can be difficult tointerpret. RNAi, however, provides a potential tool forfunctional analysis in non-model organisms.
Unfortunately, however, the success of RNAi has thus farbeen limited in the arthropods. It seems to work well inDrosophila, Tribolium and Oncopeltus, but results in moredistant taxa have been discouraging (Kennerdell and Carthew,
C. L. Hughes and T. C. Kaufman
Fig. 5. Combination RNAi phenotypes. (A) The Dfd Scr doubledepletion. The mandibular appendages (Mn) form short distalantennae, the maxillary appendages (Mx) form full-length antennaeand the labium (Lb) forms leg-like appendages. (B) The Dfd pbdouble depletion. The mandibular appendages form short distalantennae. The maxillary appendages form similar short appendages,often with claws and pulvilli, as here. The distal labial appendagesare transformed to legs. (C) The pb Scrdouble depletion. Themandibular and maxillary appendages are undeveloped. The labialappendages form full-length antennae. The genuine antennae (Ant)are under-differentiated and pinched-off at the distal tip. (D,E) TheDfd pb Scrtriple depletion. (D) The extreme phenotype, in which allgnathal appendages are antenna-like. The mandibular appendagesform distal antennae; the maxillary and labial appendages form full-length antennae. (E) The typical phenotype, in which all appendagesare very antenna-like, but the labial appendages bear claws andpulvilli at the tip (arrowheads).

3689Milkweed bug Hox RNAi
1998; Brown et al., 1999). This is surprising, since thenecessary components for RNAi seem to exist phylogeneticallyas far away as C. elegansand even fungi (Tabara et al., 1999;Catalanotto et al., 2000). We conclude, then, that RNAi mayyet be effective in other arthropods, but that for technicalreasons it may be more difficult than in flies, beetles and bugs.For instance, the nuclei may be less exposed to the diffusionof dsRNA before cellularization than in the higher insects.Alternatively, there may simply be higher levels of nativeRNases in some insects that destroy the injected dsRNA.Techniques for targeting or protecting the dsRNA may proveeffective in these organisms. It is hoped an improved
understanding of the RNAi mechanism will expand thetechnique’s usefulness into more taxa in the future.
Even when effective, RNAi phenocopies may be somewhatdifficult to interpret. When using RNAi, we do knowimmediately that we have some sort of loss-of-function of thetargeted gene, rather than the hypermorphs or neomorphspossible with mutagenesis. Moreover, we tend to get a rangeof phenotypes, similar to an allelic series. This can be a helpfulfeature, especially in Oncopeltuswhere the range ofphenotypes seems to result from the amount of dsRNAinjected, and is not merely a mosaic effect caused by limiteddiffusion. There is, however, difficulty in knowing if and when
From left to right: negative control injection (NC), single, double and triple gene depletions. Typical phenotype, the typical resulting phenotypes are illustrated diagrammatically for the mandibular (Mn), maxillary (Mx) and labial
(Lb) appendages. No. injected, the number of eggs either mock-injected with buffer (NC) or injected with dsRNA. No. developed, the number of embryos that completed development.Penetrance, the number of embryos exhibiting the typical phenotypes. Non-specific, the number of embryos exhibiting undifferentiated-appendage phenotypes due to the injection process itself. Other, additional phenotypes seen, either instead of the typical phenotype, or in addition to the typical phenotype (+). All percentages refer to the number of embryos with a given phenotype divided by the number of embryos that completed development.
NA, not applicable.*The Lb-to-antenna transformation illustrated here is the extreme phenotype, rather than the typical phenotype for the triple depletion.
The typical phenotype is an antenna-like appendage with claws.**These embryos were scored from a separate batch of injections.
No. injected 392 675 230 667 291 279 670 223
No. developed 196 311 21 196 70 92 151 48
Penetrance NA 193 15 99 51 39 133 3262% 71% 51% 73% 42% 88% 67%
Non-specific 34 53 2 13 12 11 16 317% 17% 10% 7% 17% 12% 11% 6%
Other TI comb wt + Ant undiff Dfd alone 3% Scralone 1% + Ant undiff Ant undiff100% 21% pbalone 1% Dfd alone 7% 79% 19%
(n=20 legs) +T1 comb txd Mx leg-like 5% + T1 leg80% (16/20 legs)** distorted 20%
Table 1. Summary of RNAi depletion experimentsNC Dfd pb Scr Dfd pb Dfd Scr pb Scr Dfd pb Scr
Typi
cal p
heno
type
Mn
Mx
Lb

3690
we have truly mimicked a complete null for the targeted gene.Assuming that there are multiple RNAi complexes present ineach cell, it is likely that many cells may be incompletelysaturated with dsRNA, and merely have lowered transcriptlevels of the target gene. Thus presently we can only proposethat the most extreme phenotypes are probably closest torepresenting a complete null phenocopy.
Dfd specifies the mandibleFrom the phenotypes produced by the various individualdepletions and combinations, we can infer that, of the Hoxgenes tested, Dfd is the sole gene responsible for mandibularidentity. This conforms with the expression of Dfdin themandibular segment (Rogers and Kaufman, 1997). The singledepletion of Dfd transforms the mandibular appendages todistal antennal identity, rather than the long thin styletsnormally formed (Fig. 2; Table 1). Thus Dfd is necessary forproper mandibular development.
The depletion of pb, on the other hand, leaves themandibular appendages untouched (Fig. 3; Table 1). Thus pbis not necessary for mandibular development. The phenotypeof the Scr depletion is more difficult to interpret, as themandibular stylets generally fail to grow, and are merely shortbristles in a mass of undifferentiated tissue. Since Scris notexpressed in the mandibular segment, we suspect that this isan indirect effect of the Scr phenotype on other segments ofthe head (Rogers et al., 1997). Because head development isintegrated to some degree, non-local, indirect effects are oftenseen in Drosophila Hox mutants, particularly labial andDeformed(Merrill et al., 1987, 1989). In the case of milkweedbug Scr, disruption of the proximal labium may be interferingwith the normal development of the adjacent mandibular andmaxillary appendages.
The Dfd Scrdouble depletion corroborates this view (Fig. 5;Table 1). The mandibles in the double form distal antennae,indistinguishable from either the Dfd depletion alone or thetriple. So we can infer that, when present, Scr is not acting in
the mandibular appendage to directly specify any identity overthat of the default state.
The inferred activity of Dfd, but not pb and Scr, in themandibular appendages matches the predictions based onexpression patterns. While Dfd is expressed strongly in themandibular appendages, pb and Scr are not (Rogers andKaufman, 1997; Rogers et al., 1997). The role of Dfd as thesole Hox gene regulating mandibular identity also matches thesituation in Tribolium, where Dfd mutants transform Mnstructures, while maxillopedia(mxp, the pbhomologue) andCephalothorax(Cx, the Scrhomologue) mutants leave the Mnunaffected (Brown et al., 2000, Shippy et al., 2000; Beemanet al., 1993). The effect of Hox mutations on the reducedmandibular structures of Drosophila, however, is more difficultto determine. In the embryo, Dfd mutations disrupt thedorsolateral papillae of the terminal sense organ, which arethought to derive from the Mn segment (Merrill et al., 1987).In the Drosophilaadult, Dfd mutations disrupt parts of the headcapsule (Merrill et al., 1987). Mutations in pb and Scrdo notappear to affect these presumed mandibular structures (Pultz etal., 1988; Pattatucci et al., 1991).
Although both beetles and milkweed bugs use Dfd to specifythe mandibular segment, the resulting appendages are verydifferent. In contrast to the chewing mandibles of the beetle,the bug mandibles are very thin stylets nearly as long as thebody (Fig. 1B,D). Again, these are very different from themandibular appendages in Drosophila, which are internalstructures in the embryo, and are either missing or incorporatedinto the head capsule of the adult. Thus we conclude that whileDfd’s basic role in the mandible may be conserved, thedevelopmental module driven by Dfd is extremely labile.
Dfd and Scr specify the maxillaeIn the maxillary segment, Dfd depletion results in only a partialtransformation of the maxilla to antennal identity (Fig. 2; Table1). The Dfd Scrdouble depletion (Fig. 5A; Table 1), however,results in complete transformation of the maxillae to antennae.Thus we can conclude that Dfd acts in concert with Scr in themaxillary segment. The curled phenotype of the Dfd depletionmaxillary appendage (Fig. 2; Table 1) suggests that Scr maybe repressing growth of the transformed limb in its posteriordomain; this repression is released in the Dfd Scrdouble (Fig.5A; Table 1).
The phenotypes in the maxillary segment are somewhatconflicting, however, regarding the role of pb. Two resultssuggest that pbis not acting to specify maxillary identity.Depletion of pbalone leaves the maxillary segment unaffected,therefore pb is not necessary for wild-type maxillarydevelopment (Fig. 3; Table 1). Secondly, the phenotype of theDfd Scr double is the same as the triple depletion (i.e.,antennae; Fig. 5A,D; Table 1), so we can infer that pb is notacting to confer any identity over that of the default (seebelow). There is, however, a subtle difference between thephenotypes of the Dfdsingle and Dfd pb double depletion(compare Fig. 2 with Fig. 5B). While the Dfd depletionproduces short, curled antenna-like appendages, the Dfd pbdouble produces straight antenna-like appendages, often withpretarsal claws (Fig. 5B). This suggests that in the absence ofDfd, pb can affect the maxillary appendages. Whether thisactivity derives from its small dorsal maxillary domain ofaccumulation, or from an expanded domain of pb expression
C. L. Hughes and T. C. Kaufman
Fig. 6. Differences in expression, regulation and function of thegnathal Hox genes in Oncopeltus, Drosophilaand Triboliummouthparts. The three insects control the development of themaxillary appendages using different combinations of Hox genes. Inthe maxillary and labial appendages, there are different regulatoryrelationships between the three genes. The functions of these Hoxgenes also differ between the three species. E, embryonic; A, adult.(Unpublished Tribolium data courtesy of S. Brown.)

3691Milkweed bug Hox RNAi
in the Dfddepletion, is not yet clear. Nevertheless, based onthe pb depletion, we conclude that, in wild-type embryos, pbis not active in the specification of the maxillary stylet.
This lack of pbfunction in determining the maxillary styletmatches the prediction made based on the expression patternof pb in the bug. Thus far this is the only insect known not touse pb to determine maxillary identity (Fig. 6). In Drosophilaand Tribolium, pb/mxpmutations cause transformation of themaxillary palps, which become reduced in Drosophila, andtransform to legs in Tribolium(Pultz et al., 1988; Beeman etal., 1993; Shippy et al., 2000). In the bug, it would appear thatScrassumes much of that role. This is in contrast to Drosophilaor Tribolium, where the Scr/Cx mutation does not affect themaxillary appendage (Pattatucci et al., 1991; Beeman et al.,1993).
While Dfd and Scr work together to specify maxillaryidentity in the bug, it seems that Dfdhas the principal role.Perhaps by activating a similar set of target genes as in themandible, Dfd may induce a similar stylet identity in themaxillary appendage. This contrasts with mandibulate insects,in which the maxillary morphology is most similar to thelabium, probably due to the activity of pb in both of thoseappendages.
Scr and pb specify the labiumBy analyzing the depletion phenotypes, we can infer that pband Scr cooperate to pattern the labium. Dfd depletion leavesthe labium unaffected, so we can conclude that Dfd is notnecessary for wild-type labial development (Fig. 2; Table 1).Depletion of pbresults in a labium that is fused and wild-typebasally, but splits distally into a pair of normal T1 legs (Fig.3). From this, we infer that pb is necessary for distal, but notproximal, labial development. Scr depletion results intransformation of the entire appendage to a mixed identitybetween leg and antenna (Fig. 4). From this we infer that Scris necessary for development of both proximal and distallabium. The pb Scrdouble depletion, as well as the triple,transforms the labium to a pair of full-length antennae (Fig. 5C,D; Table 1), which is very different from either singledepletion alone. Therefore in the bug as well as the fly andbeetle, pb and Scrspecify the labium (Fig. 6).
The Scr depletion phenotype in the labium supports thehypothesis that Scr normally functions to fuse the two labialappendages (Rogers et al., 1997). This conclusion is derivedfrom the observation that, in wild-type and the pb depletionanimals, the transformed labial appendage is basally fused,while the Scrdepletion limbs are often well-separated (Fig. 4).
By comparing the pb depletion to the pb Scrdouble, we caninfer that the function of Scr in the absence of pbis to induceT1 legs. In light of the Drosophilamodel, this may seemreasonable, since Scris also expressed in T1 but, in the caseof the milkweed bug, it is actually a bit surprising. Based onits ectodermal pattern of accumulation in a single discrete spoton the T1 leg, Scrwould not appear to be capable of conferringoverall leg identity to this appendage. Rather, in ventral T1 Scrappears to function solely in the specification of the T1 legcomb (see below).
The role of pb in the gnathal appendages is provided by acomparison of the Scr single depletion animals to the pb Scrdoubles. The Scrdepletion labium is transformed to a pair ofappendages with features of both antenna and leg. This
suggests that the activity of pbalone is conferring some legidentity over the default antennal state. This contrasts stronglywith the function of pbin flies. In two situations in Drosophila,the presence of pb alone appears to induce maxillary identity:first, in Scrhypomorphic mutant adults, the labium is partiallytransformed to maxillae; second, when pb+ is ectopicallyexpressed in the antennae, they are transformed to maxillarypalps (Pattatucci et al., 1991; Cribbs et al., 1995). Since in thebug pb is inducing partial leg identity rather than maxillaryidentity, this suggests that pb may have a very differentfunction than in the fly.
Since both pb and Scr, when acting alone, each seem to bespecifying some leg identity, this suggests that pband Scrhaveoverlapping functions. We know, however, that the two genesare not entirely redundant, because their depletion phenotypesare different and because when both are present the result islabial identity. Thus the activity of wild-type pb or Scr aloneinduces leg identity, but the combined synergistic activity ofthe two together creates a new overriding labium identity.Whether the combinatorial effect of pb and Scris at the levelof transcription of their target genes, or in the activity of theircombined target gene products, is not yet clear.
One aspect of the Scrdepletion phenotype deserves specialmention. Although expression studies have shown that Scr isnot expressed in the milkweed bug labium distally (Rogers etal., 1997), the Scrdepletion results in a change of identity forthe distal labium. Instead of the sensory structures normallyformed there (Fig. 3C), pretarsal claws and pulvilli appear (Fig.4F). This suggests that the presence of Scr elsewhere in theappendage normally acts non-cell autonomously to inducelabial identity. Non-cell-autonomous functions for Scr andother Hox genes have been suggested for other tissues inDrosophila (D. F. B. Miller, S. S. Holtzman, A. Kalkbrennerand T. C. K., unpublished data; Percival-Smith et al., 1997).
Scr function in the thoraxUnlike the broad band of Screxpression across the firstthoracic segment in fly embryos, in milkweed bugs and otherinsects, the pattern of Scr is restricted to three thoracicdomains: a T1 leg patch, a dorsal T1 patch and the mesodermof all legs (Rogers et al., 1997). We have been able to addressa hypothesis about the function of Scr in one domain, butcannot yet describe the functions of the other thoracic domainsin the milkweed bug.
As noted above, Scr is expressed in the T1 limb in ananterior patch midway down the leg. This accumulationcorrelates with the position of a comb on the distal tibia of thisleg and it has been proposed that Screxpression is necessaryfor the development of that structure (Rogers et al., 1997). TheScrdepletions reported here support this hypothesis. Althoughcomb transformation was not invariably associated with labiumtransformation, of the animals that also had a transformedlabium, 80% of the legs had a transformed comb, with bristlesresembling those found in a comparable position of the moreposterior legs (Fig. 4D bottom). This suggests that, in wild-type, the patch of Screxpression is promoting the formation ofthe T1 comb from a row of bristles on the distal tibia, ascompared to the serially homologous structures found on thesecond and third legs.
The second domain of Scr expression is a patch on dorsalT1. Tribolium and Drosophila Cx/Scrmutants develop ectopic

3692
wings on dorsal T1; thus it has been proposed that the dorsalpatch of Scr functions to repress wing development in thatsegment in the other insects as well (Beeman, 1987; Rogers etal., 1997). Unfortunately, this could not be tested with the Scrdepletions reported here since, in the bug, the wings do notdevelop until several instars after hatching. Because depletionsof Scralso disrupt the mouthparts, affected animals could notbe raised to determine if wing rudiments developed on the firstthoracic segment.
The third thoracic domain of Scr is in the mesoderm of allthree segments. This expression of Scr has been proposed tonon-cell autonomously direct tarsus development in the fly(Percival-Smith et al., 1997). In the Scr depletions, noconsistent effect on the tarsi was observed. It would bedifficult, however, to ensure that the depletion had completelyremoved Scr function from all cells of the mesoderm; activityin just a few cells might be sufficient to create enoughdownstream diffusible signal. Thus determining the generalityof this proposed function for Scr requires furtherexperimentation.
Antennae are specified in the absence of HoxSince the triple depletion animals should lack all Hox activityin the gnathal segments, we can infer the default state of theseappendages in the absence of Hox function. Antennae havebeen thought to represent the default appendage state in theabsence of Hox activity, since a large deletion of much of theHom-C in the beetle was found to produce a larva bearing aseries of antennae instead of mouthparts and legs (Stuart et al.,1991). Likewise, in the adult fly, the pband Scrdouble mutantcombination in the adult labium gives rise to antennae(Percival-Smith et al., 1997).
Our triple depletion indicates that, in the milkweed bug aswell, antennae represent the no-Hox state for the gnathalappendages (Fig. 5D; Table 1). In the triple depletion, themaxillary and labial segments form full-length antennae. Sincewild-type antennae are the only paired appendages that lackHox expression, it is reasonable that this morphology isproduced in the absence of Hox function. Furthermore, thisresult indicates that the genetic program that specifies thepresence of an appendage is independent of Hox expression,and that the function of the Hox gene products is to confer aspecific morphology onto a generic appendage program. In theabsence of Hox activity, this generic appendage turns on theantennal differentiation pathway.
In the absence of Hox activity, the mandibular appendages,however, each form only a distal antenna on a short base. Thissuggests that a portion of the generic appendage is missing.Previous work has suggested that insect mandibles, which lackDistal-less (Dll) expression, are gnathobasic (lack distalportions of the appendage) (Scholtz et al., 1998; Popadic et al.,1998). In Drosophilaand Tribolium, Dll is repressed by Dfd inthe mandibular segment (O’Hara et al., 1993; Brown et al.,2000). In the milkweed bug, therefore, depletion of Dfd islikely associated with Dll derepression. Consistent with thishypothesis, the morphology of the transformed mandibularappendage is primarily to distal antenna. Interestingly, as notedthe basal portions of the appendage are distorted or missing,compared to the maxillary or labial default appendages. Thissuggests that some aspects of the reduced structure of the insectmandible are independent of the Dfd/Dll pathway.
Novel regulatory relationshipsIn Drosophila, the expression pattern of pb in the maxillarysegment of the embryo is dependent on the activity of Dfd (D.F. B. Miller, B. T. Rogers, A. Kalkbrenner and T. C. K.,unpublished data; Rusch and Kaufman, 2000). In Tribolium,however, Dfd is not required to activate mxpin the maxillarypalp (S. Brown, personal communication). Therefore, it isinteresting to analyze the regulatory relationship between Dfdand pb in the milkweed bug. The lack of pb expression andfunction in the maxillary appendage suggests that Dfdis notactivating pb. It is possible, however, that Dfd represses pb. Ifthis were the case Dfddepletion might be expected to derepresspb in the maxillary segment. Moreover, since in the absence ofScr activity the labium is leg-like, one would expect to seesome leg-like identity in the transformed maxillary appendagein Dfd Scrdouble depletion animals. This treatment, however,produces an antennal appendage on the maxillary segment.Thus it would appear that the expression of pb is largelyindependent of Dfd (Fig. 6).
In the Drosophila and Tribolium embryo, Scr/Cx isnecessary to activate the expression of pb/mxp(Fig. 6). InScr/Cxmutants, the expression of pb/mxpis greatly reduced inthe labial segment (D. F. B. Miller, B. T. Rogers, A.Kalkbrenner and T. C. K., unpublished data; S. Brown,personal communication). If Scr were acting similarly in thebug, we would expect that the Scrsingle depletion wouldresemble the pb Scr double depletion; i.e., if Scr werenecessary for the activity of pb, then in the Scr depletionanimals pb would be inactive as well. This is not the case.While the pb Scrdouble depletion transforms the labium to apair of antennae, in the Scr depletion, the labium has muchmore leg-like identity, indicating the activity of pb. Thereforethe regulation of pbin the milkweed bug appears to beindependent of either Dfdor Scr, and thus is different fromboth Drosophilaand Tribolium (Fig. 6). Likewise, the activityof Dfd and Scr seem to be independent of pb or each others’activity. There may, however, be subtle regulatory interactionsbetween the three genes that would not be detectable in thisanalysis; these interactions await further study.
Evolutionary innovation and Hox genesHox genes have been studied both as indicators and possiblemediators of evolutionary change. In this study, we findevidence of remodeling in the role of Hox genes in headappendage development. This change in the role of the Hoxgenes is due to three mechanisms: (1) change in expressionpatterns, (2) change in regulatory relationships and (3) changein function.
Previous work had shown that pbpossesses a uniqueexpression pattern in the milkweed bug (Rogers and Kaufman,1997). The RNAi analysis confirms that the unusual expressionpattern does in fact reflect an unusual domain of activity.Unlike Drosophila, Triboliumand probably most other insects,pb is not necessary for development of the maxillaryappendages in the milkweed bug.
The pb gene is also regulated differently in the bug, whereit appears to be independent of the activity of Dfd and Scr. Thiscontrasts strongly with the regulatory interactions describedfor the fly and the beetle (D. F. B. Miller, B. T. Rogers,A. Kalkbrenner and T. C. K., unpublished data; Rusch andKaufman, 2000; S. Brown, personal communication). Studies
C. L. Hughes and T. C. Kaufman

3693Milkweed bug Hox RNAi
of more distant insects are needed, however, before we canconfidently infer the ancestral state and the direction of changefor these regulatory interactions.
Lastly, we find evidence of novel functions in the bug, whichprobably reflect differences in the suite of target genesactivated by the Hox genes. For example, the function of pb isdifferent in the bug, where it induces a mixed leg-like identityrather than a maxillary palp. Even in the mandibular segment,where Dfd is the conserved controller of identity, the resultingmandibular appendage is strikingly different from themandibles of other insects, indicating a very differentdevelopmental module at work downstream of the Hox signal.We cannot tell, however, whether the repertoire of target genesis different, whether the orthologous target genes themselveshave different functions or if both possibilities are correct.
These differences in Hox function are provocative, but weare still some way from describing a likely scenario for theevolution of the Hemipteran mouthparts. The exclusion of pbfrom the maxillary segment may have caused the loss of themaxillary palp – or the expression of pb may be the result ofloss of the palp, which would remove selection to maintain pbin an inactive tissue. Moreover, the evolution of Hemipteranstylate-haustellate mouthparts was clearly a multistep processwithin the Hemipteroid group: the Psocoptera (booklice) andsome Phthiraptera (chewing lice) have pronounced lacinia; theThysanoptera (thrips), some Phthiraptera (sucking lice) and theHemiptera have long stylets; and finally, the Hemipterans havelost the maxillary palps entirely (Daly et al., 1998).Understanding the changes that accompanied the evolution ofthese mouthparts will require additional study of theembryology, homologies and gene expression of these variousinsect orders.
We wish to thank Paul Z. Liu for innumerable helpful discussionsand development of the lateral injection technique. Special thanks toRudi Turner for preparing the SEMs. We also thank Sue Brown,Teresa Shippy, Kathy Matthews, Paul Liu and two anonymousreviewers for careful reading of the manuscript. C. L. H. is supportedby a Howard Hughes Medical Institute predoctoral fellowship. T. C.K. is an investigator of the Howard Hughes Medical Institute.
REFERENCES
Averof, M. and Patel, N. H. (1997). Crustacean appendage evolutionassociated with changes in Hox gene expression. Nature, 388, 682-686.
Beeman, R. W.(1987). A homeotic gene cluster in the red flour beetle. Nature327, 247-249.
Beeman, R. W., Stuart, J. J., Brown, S. J. and Denell, R. E.(1993). Structureand function of the Homeotic Gene Complex (HOM-C) in the beetle,Tribolium castaneum. Bioessays, 15, 439-444.
Botas, J. and Auwers, L.(1996). Chromosomal binding sites of Ultrabithoraxhomeotic proteins. Mech. Dev. 56, 129-138.
Boxem, M., Srinivasan, D. G. and van den Heuvel, S.(1999). TheCaenorhabditis elegansgene ncc-1encodes a cdc2-related kinase requiredfor M phase in meiotic and mitotic cell divisions, but not for S phase.Development126, 2227-2239.
Brown, S. J., Mahaffey, J. P., Lorenzen, M. D., Denell, R. E. and Mahaffey,J. W. (1999). Using RNAi to investigate orthologous homeotic genefunction during development of distantly related insects. Evol. Dev.1, 11-15.
Brown, S., DeCamillis, M., Gonzalez-Charneco, K., Denell, M., Haas, S.,Beeman, R., Nie, W. and Denell, R.(2000). Implications of the TriboliumDeformedmutant phenotype for the evolution of Hox gene function. Proc.Natn. Acad. Sci., USA97, 4510-4514.
Catalanotto, C., Azzalin, G., Macino, G. and Cogoni, C.(2000). Genesilencing in worms and fungi. Nature404, 245.
Cribbs, D. L., Benassayag, C., Randazzo, F. M. and Kaufman, T. C.(1995).Levels of homeotic protein function can determine developmental identity:Evidence from low-level expression of the Drosophila homeotic geneproboscipediaunder Hsp70 control. EMBO J. 14, 767-778.
Daly, H. V., Doyen, J. T. and Purcell, A. H.(1998). Introduction to InsectBiology and Diversity. Oxford: Oxford University Press.
Falciani, F., Hausdorf, B., Schröder, R., Akam, M., Tautz, D., Denell, R.and Brown, S.(1996). Class 3 Hox genes in insects and the origin of zen.Proc. Natn. Acad. Sci., USA93, 8479-8484.
Fay, D. S., Stanley, H. M., Han, M. and Wood, W. B.(1999). ACaenorhabditis eleganshomologue of hunchbackis required for late stagesof development but not early embryonic patterning. Dev. Biol. 205, 240-253.
Fire, A., Xu, S., Montgomery, M. K., Kostas, S. A., Driver, S. E.; andMello, C. C. (1998). Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans. Nature391, 806-811.
Hayashi, S. and Scott, M.(1990). What determines the specificity of actionof Drosophilahomeodomain proteins? Cell63, 883-894.
Hong, Y., Roy, R. and Ambros, V.(1998). Developmental regulation of acyclin-dependent kinase inhibitor controls postembryonic cell cycleprogression in Caenorhabditis elegans. Development125, 3585-3597.
Hunter, C. P. (2000). Gene silencing: shrinking the black box of RNAi.Curr.Biol. 10, R137-R140.
Kaufman, T. C. (1978). Cytogenetic analysis of chromosome 3 in Drosophilamelanogatsre: mapping of the proximal portion of the right arm. Genetics80, 733-752.
Kennerdell, J. R. and Carthew, R. W. (1998). Use of dsRNA-mediatedgenetic interference to demonstate that frizzled and frizzled 2act in theWingless pathway. Cell 95, 1017-1026.
Lutz, B., Lu, H., Eichele, G., Miller, D., Kaufman, T. C.(1996). Rescue ofDrosophila labialnull mutant by the chicken ortholog Hoxb-1demonstratesthat the function of Hox genes is phylogenetically conserved. Genes Dev.10, 176-184.
Mastick, G. S., McKay, R., Oligino, T., Donovan, K. and Lopez, A. J.(1995). Identification of target genes regulated by homeotic proteins inDrosophila melanogasterthrough genetic selection of Ultrabithorax protein-binding sites in yeast. Genetics139, 349-363.
Merrill, V. K., Turner, F. R. and Kaufman, T. C. (1987). A genetic anddevelopmental analysis of mutations in the Deformedlocus in Drosophilamelanogaster. Dev. Biol. 122, 379-395.
Merrill, V. K., Diederich, R. J., Turner, F. R. and Kaufman, T. C. (1989).A genetic and developmental analysis of mutations in labial, a genenecessary for proper head formation in Drosophila melanogaster. Dev. Biol.135, 376-391.
O’Hara, E., Cohen, B., Cohen, S. M. and McGinnis, W. (1993). Distal-lessis a downstream gene of Deformed required for ventral maxillary identity.Development 117, 847-856.
Pattatucci, A. M., Otteson D. C. and Kaufman, T. C.(1991). A functionaland structural analysis of the Sex combs reducedlocus of Drosophilamelanogaster. Genetics2, 423-441.
Percival-Smith, A., Weber, J., Gilfoyle, E. and Wilson, P.(1997). Geneticcharacterization of the role of the two HOX proteins, Proboscipedia and SexCombs Reduced, in determination of adult antennal, tarsal, maxillary palpand proboscis identities in Drosophila melanogaster. Development124,5049-5062.
Popadic, A., Panganiban, G., Rusch, D., Shear, B. and Kaufman, T. C.(1998). Molecular evidence for the gnathobasic derivation of arthropodmandibles and for the appendicular origin of the labrum and other structures.Dev. Genes Evol. 208, 142-150
Pultz, M. A., Diederich, R. J., Cribbs, D. L. and Kaufman T. C.(1988).The proboscipedialocus of the Antennapedia Complex: a molecular andgenetic analysis. Genes Dev. 2, 901-920.
Rogers, B. T. and Kaufman, T. C.(1997). Structure of the insect head inontogeny and phylogeny: A view from Drosophila. Int. Rev. Cytol.174, 1-84.
Rogers, B. T., Peterson, M. D. and Kaufman, T. C.(1997). Evolution of theinsect body plan as revealed by the Sex combs reducedexpression pattern.Development124, 149-157.
Rusch, D. B. and Kaufman, T. C. (2000). Regulation of proboscipediabyhomeotic selector genes. Genetics(in press).
Scholtz, G., Mittmann, B. and Gerberding, M.(1998). The pattern of Distal-lessexpression in the mouthparts of crustaceans, myriapods and insects:New evidence for a gnathobasic mandible and the common origin ofmandibulata. Int. J. Dev. Biol. 42, 801-810.

3694
Shippy, T. D., Guo, J., Brown, S. J., Beeman, R. W., Haas, M. S. andDenell, R. E. (2000). Analysis of maxillopediaexpression pattern and larvalcuticular phenotype in wild-type and mutant Tribolium. Genetics155, 721-731.
Stern, D. L. (1998). A role of Ultrabithorax in morphological differencesbetween Drosophilaspecies. Nature396, 463-466.
Stuart, J. J., Brown, S., J., Beeman, R. W. and Denell, R. E.(1991). Adeficiency of the homeotic complex of the beetle Tribolium. Nature350, 72-74.
Tabara, H., Sarkissian, M., Kelly, W. G., Fleenor, J., Grishok, A.,Timmons, L., Fire, A. and Mello, C. C. (1999). The rde-1 gene, RNAinterference, and transposon silencing in C. elegans. Cell99, 123-132.
C. L. Hughes and T. C. Kaufman