ministry of agriculture, fisheries and...
TRANSCRIPT

MINISTRY OF AGRICULTURE, FISHERIES AND FOOD CSG15Research and Development
Final Project Report(Not to be used for LINK projects)
Project title
MAFF project code
Contractor organisationAnd location
Total MAFF project costs
Project start date Project end date
This work addresses the DEFRA objectives of ensuring the reliable supply of high quality produce from competitive UK sources by putting in place underlying knowledge on the post harvest deterioration of ethylene-insensitive cut flowers. Cut flowers are sold on appearance and the ability to retain high quality appearance in the vase adds value to the product. The processes that occur during ageing of the flowers that lead to loss of quality are known as senescence. The senescence of flowers determines the time at which consumers will discard cut flowers and thus affects the value and profitability of the flowers to the grower, wholesaler and retailer. Ethylene is a growth substance i.e. a compound produced naturally in plants that controls specific aspects of their development. Many cut flowers respond to ethylene, by accelerated senescence or petal drop. For these, production of ethylene by the plant, and sensitivity to the plant growth regulator ethylene are the main factors linked to reduced longevity and shelf life and a number of treatments, including chemical and genetic approaches, can be used to increase floral longevity by inactivating the ethylene system. However, for many other commercially important flower species, senescence is not sensitive to ethylene and the techniques presently available are ineffective at prolonging their vase life. Alternative techniques are needed for these ethylene-insensitive plants. Design of these requires detailed knowledge of the physiological, biochemical and molecular biological changes associated with their senescence. With this in mind, the objective of this project was to establish the biochemical and molecular basis of petal deterioration in Alstroemeria. This species was chosen because it is both an important UK crop and, according to the literature, does not show ethylene sensitivity. The project involved collaboration between laboratories at the Universities of Cardiff and Royal Holloway and HRI,
Two hard copies of this form should be returned to:Research Policy and International Division, Final Reports UnitMAFF, Area 6/011A Page Street, London SW1P 4PQ
An electronic version should be e-mailed to [email protected]
The biochemical basis of post-harvest quality in cut flowers
HH2122TOF
Cardiff School of Biosciences, Cardiff UniversityP.O. Box 915, Cardiff CF10 3TL
£ 257 515
01/04/98 30/06/01
Executive summary ( maximum 2 sides A4)

The biochemical basis of post-harvest quality in cut flowers HH2122TOFProject MAFFTitle project codeWellesbourne. Three key areas of the biology of petal deterioration were investigated; protein breakdown, cell membrane biochemistry and cell death. Proteins are important because they direct the biochemistry of the cell, they have structural roles within the cell and they act as a store of nitrogen. Cell membranes are needed to maintain the integrity of the cell, which is lost during senescence. Senescence leads to cell death, which may be a pre-programmed stage of the life cycle as found in some other biological situations. Initial physiological studies identified seven stages in petal deterioration in Alstroemeria
designated numerically from S0 to S6 and representing 12 days in time. Briefly, S0 and S1 are the stages of floral development when the buds are opening and by S2 the flowers were fully open with the sepals reflexed. At S3 the top three anthers anthesed and two days, later at S4, the petals showed initial signs of in-rolling and discoloration (visible senescence) while the bottom three anthers had also anthesed. Stage 5 (S5) was defined by the separation of the stigmatic lobes and further signs of petal discoloration and in-rolling. Abscission of the perianth occurred at S6. These stages were used to provide a common framework to co-ordinate investigations in the three laboratories involved in the project.
Protein Breakdown studies: Changes in protein levels and the activity of proteases, the enzymes that cause protein breakdown, were investigated over the duration of the flower vase life. Protein levels fell continuously from the unopened bud, throughout development, until petal abscission while total protease activity in the petals increased more than tenfold between flower opening and S5. Three distinct protease activities could be distinguished and on the basis of studies with specific inhibitors all three were shown to be of the cysteine proteases class. This may be interesting, as cysteine proteases have been found to be associated with cell death in other species. The levels of these activities changed during flower development and senescence with one of the three activities showing an increase. Studies on the expression of an Alstroemeria cysteine protease gene sequence revealed that the gene was expressed from the earliest stages analysed but a dramatic increase took place as the petals aged. A well-known mechanism for protein degradation involves attaching ubiquitin, itself a protein, to the protein destined for breakdown. Ubiquitin acts as a label and targets the degradation process to the attached protein. In Alstroemeria, no important changes were detected in the expression of the ubiquitin gene or ubiquinated proteins throughout petal development and senescence. These results suggest that, in contrast to other lilies such as Hemerocallis, ubiquitination may not be an important mechanism controlling protein degradation in this species.
Membrane integrity and lipid metabolism: The project has provided new insights into lipid and membrane breakdown during flower ageing. During senescence, most of the lipids in the membrane were lost through metabolism. As the membranes aged and were depleted, their integrity was progressively lost, as measured by leakage of ions. Lipid peroxidation had previously been hypothesised as the decisive factor in membrane deterioration, but although it was detected, its importance was less than in some other flower species and lipoyxgenases also had a lesser role. Oxidative damage may have played a part in flower deterioration as the levels of carotenoids, which are antioxidants were higher in petals than sepals and this was correlated with reduced damage in petal tissues.
Cell death: The involvement of programmed cell death in Alstroemeria petals was tested for by three standard methods. DNA laddering was used to reveal patterns of DNA fragmentation which have been found in other systems to be a hallmark of programmed cell death. Laddering was evident in Alstroemeria petals even in young petals although more extensive laddering was detectable during petal deterioration. The second approach was to look for genes that act in other systems that act as markers of cell death. One of these, DAD1 (defender against apoptotic death) is down-regulated as cells enter cell death in most systems studied and a homologue of DAD1 was isolated from Alstroemeria as a partial petal cDNA clone. The expression pattern of this gene showed a sharp decrease at stage 4 and hence may provide a useful marker for this stage in petal deterioration. A further method, the TUNEL assay for DNA breakdown was unsuccessful in this project.
Major conclusions and suggestions for further work: The major overall research finding is that there is no simple triggering event for petal senescence in Alstroemeria, as in ethylene sensitive species, but petal ageing represents a gradual deterioration. A number of genes and processes were identified as changing concurrently in ageing flowers. Diagnostics for post harvest quality therefore will need to be based on simultaneous monitoring of multiple components. An additional finding is that some of the processes activated during petal senescence may already be in place while the petals are still developing. This is significant as it indicates that it may be possible to develop diagnostic approaches for flowers at the stage of picking as well as later in their shelf life. It also suggests that it will CSG15 (1/00) 2

The biochemical basis of post-harvest quality in cut flowers HH2122TOFProject MAFFTitle project codebe essential to understand the effect of treatments during packaging and shipping of cut flowers on the senescence process.Besides contributing to DEFRA objectives of ensuring the reliable supply of high quality produce from competitive UK sources these results provide the foundation and guidance to their further achievement through the application of appropriate (post-genomic) technologies. We propose the next steps to build on the existing work.1 Develop DNA microarray chips for diagnostic studies on flower deterioration 2 Use these diagnostics to identify sensitive points in the supply, storage and distribution chain.
This project has provided important new information and understanding of the factors involved in deterioration of ethylene insensitive cut flowers. This will underpin the competitive position of UK Horticulture in the growing market for cut flowers. The information has been disseminated through peer reviewed and horticultural trade publications and at national and international conferences. UK industry has confirmed the relevance of the work by supporting proposals to follow the work up via BBSRC and DEFRA.
Objectives and relative milestones which were changed during the course of the project and Reasons for making the changes:
Milestone number
Original milestone New milestone Reason for change
02/05 Northern analysis of delta-9 desaturase and LOX genes
Northern analysis of Alstroemeria LOX genes
Work on rose was not continued and delta-9-desaturase was less relevant to Alstroemeria
S03/02/01 Preparation of probes to genes coding for enzymes involved in the ligation of ubiquitin to proteins
comparison of methods of lipid hydroperoxide quantification
Results from transcriptional and western analysis of ubiquitin expression suggests that ubiquitination may not be a major player in this system
S02/04/02 Assess GST levels using CDNB substrate
Antioxidant status of sepals and petals Antioxidant studies were considered to be
S02/04/03 Determination of further substrates for GST assay
Petal and sepal pigment levels, including total carotenoids, chlorophyll and anthocyanins
more relevant to the emerging picture
S02/05/01 Assay delta9 desaturase using radiolabelling
RemovedWork on rose was not continued and delta-
S02/05/02 Preparation of probes for analysis of delta 9 desaturase and LOX
Preparation of probes for analysis of LOX expression
9-desaturase was less relevant to Alstroemeria
At an early stage in the project it was agreed with the PMO that efforts would be concentrated on Alstroemeria and thus work on rose varieties would not be undertaken. This decision was based on difficulties in finding a suitable variety for the work, and an appreciation that petal senescence in lileaceous species is of greater scientific and commercial relevance.
CSG15 (1/00) 3
Scientific report (maximum 20 sides A4)

The biochemical basis of post-harvest quality in cut flowers HH2122TOFProject MAFFTitle project code
CSG7 objectives and to what extent they were metObjective Title Objective fully met?01 Establish a common set of parameters for assessment of wilting in the two species.
Including fresh weight, surface area appearance etc. yes02 Investigate membrane integrity during petal senescence:02/01 Undertake measurements of conductivity and/or osmolarity
and factors affecting cell membrane permeability (Y1)yes
02/02 Quantification and identification of membrane lipids (Y1) yes 02/03 Activity of enzymes involved in lipid peroxidation (Y2) yes 02/04 [Activity of GSTs using a range of suitable substrates
substituted with analysis of antioxidant status ] (Y2)abandoned in favour of important studies on petal antioxidant status
02/05 Northern analysis for timing expression of related genes: LOX and [(in roses) delta-9-desaturase removed 31/08/99.(Y1-2)]
yes for LOX expression in Alstroemeria, using alternative method
03 Protein degradation during petal senescence:03/01 Western and northern analysis for protein degradation via the
ubiquitin pathway and ubiquitin gene expression(Y1)yes, using alternative method, although the results were not very informative
S03/01/02 Determine overall protein changes using SDS PAGE (Y1) yes03/02 [Using probes to genes encoding enzymes in the ubiquitin
pathway, follow their expression:substituted with comparison of methods of lipid hydroperoxide quantification] (Y3)
abandoned due to the nature of the results from 03/01. New objective was met in full
03/03 Using probes to cysteine proteases, determine the timing of expression of these genes in relation to loss of protein. (Y1-2)
Yes, using an alternative method for expression analysis
04 Charting cell death during petal senescence:04/01 Using TUNEL, investigate levels of DNA fragmentation (Y3) Attempted but not successful04/02 Assess levels of DNA laddering (Y3) yes04/03 Use northern analysis to investigate expression of the dad1
gene (a key control gene for cell death) (Y3)Yes, using an alternative method for expression analysis
05 Technology transfer and communication of results:05/01 Preparation of papers for publication in refereed journals in
the three major areas of research (Y2-3)Yes (see Annex)
05/02 Preparation of article for horticultural trade press (Y3) Yes (see Annex)05/03 Presentation at Grower Conference(s) (Y2-3) Yes (see Annex)
IntroductionThe senescence of flowers determines the time at which consumers will discard cut flowers and, in consequence, affects the value and profitability of the flowers to the grower, wholesaler and retailer. The production of cut flowers in the UK is worth in excess of £100m pa compared with an import value of over £220m (MAFF statistics, 1995 data). Increased quality and shelf-life of flowers would be of considerable economic benefit to producers (increased crop value) and retailers (increased shelf life) as well as the consumer (increased vase life). In order to bring about such improvements a thorough understanding of the biochemical and physiological processes which lead to the deterioration of the products is required. Research to date has identified that the plant growth regulator ethylene is intimately linked with the reduced longevity and shelf life of some flowers. For these species, treatments, including chemical and molecular means, can be used to increase floral longevity. However, senescence of many economically important species is not ethylene sensitive and thus the techniques presently available are ineffectual at prolonging their vase life. As yet, there is little or no information on either the controlling mechanisms which initiate the senescence of these tissues (van Doorn and Stead, 1995), or the biochemical events which occur during the senescence process. Understanding the molecular and physiological changes associated with senescence will allow a targeted approach to this problem. Alstroemeria was chosen for the subject of this study as it is both an important UK crop (sales increased by 35% in 1998 relative to 1997, Flower
CSG15 (1/00) 4

The biochemical basis of post-harvest quality in cut flowers HH2122TOFProject MAFFTitle project codeBusiness International, Feb, 1999) and, according to the literature (Woltering and van Doorn, 1988), shows little or no ethylene sensitivity.The primary purpose of this work, therefore, was to gain an understanding of the processes occurring during the degeneration of ethylene-insensitive flowers. Three key areas were chosen for study: changes in membrane composition, protein degradation and processes associated with cell death. These processes were chosen as all three have been associated with senescence and were likely to provide information on the timing of biochemical and molecular changes occurring in Alstroemeria petal development and senescence. Protein degradation is important both globally in the re-mobilisation of nutrients from the senescing tissue, but also degradation of specific proteins and activation of specific proteases are key control elements in other systems (Glotzer et al, 1991; Lam et al, 1999). Membrane integrity is an essential component of living cells and is lost during senescence, thus investigation of this process was also considered to be important in charting petal wilting in Alstroemeria. Cell death is the final
process in terminal senescence and the timing of its onset indicates that the final phase in this developmental process has been reached.
(1) Membrane damageThe process of lipid peroxidation (the addition of oxygen to membrane unsaturated fatty acids) is an important event associated with senescence (Lesham, 1992). In some tissues this appears to be a late phenomenon occurring coincidentally with cellular death and
may, therefore, be a consequence rather than a cause of the loss of membrane integrity. However, in other cases, lipid peroxidation precedes any obvious signs of senescence and therefore appears to be a primary event associated with the onset of senescence (Zhuang et al, 1995). In either case, the initial changes in lipid composition are likely to be associated with the de -acylation of the membrane lipid components through the action of phospholipases. The released fatty acids then serve as substrates for lipoxygenase (LOX) mediated oxygenation and/or autoxidation (ie non-enzyme catalysed reactions) processes.
The role of enzymes that protect against membrane damage such as glutathione-S transferases (GSTs) was also investigated Understanding the role of GSTs and timing of their expression during petal senescence may be important in understanding the processes leading to irreversible membrane damage. GST encoding genes have been associated with petal senescence in other species (Itzhaki and Woodson, 1993) and may be important in delaying membrane damage while remobilisation of nutrients takes place. This is achieved by conjugating glutathione to cytotoxic compounds produced during membrane breakdown, resulting in their detoxification (Marrs, 1996).
(2) Protein breakdownThe ability to selectively degrade proteins is essential in all tissues, therefore changes in the pattern of protein degradation would be expected during both flower development and senescence. Moreover, the loss of membrane permeability seen during petal senescence may be the result of the selective degradation of integral membrane proteins. One way in which this may be achieved is to target specific proteins by conjugating ubiquitin moieties to the target protein, the ubiquitinated protein is then degraded by an ATP-dependent protease (Callis, 1995).In the ethylene-insensitive flowers of Hemerocallis patterns of protein ubiquitination clearly alter suggesting that specific proteins are targeted for degradation in this way. Furthermore, treatment with cylcoheximide (CHI - an inhibitor of translation) delayed flower senescence in daylilies (Courtney et al., 1994) and iris (van Doorn et al, 1995), whilst senescence was also delayed after CHI treatment in the ethylene-sensitive petal of carnation (Wulster et al, 1982a,b). In daylily several different ubiquitin transcripts were detected by northern analysis which were differentially expressed during flower development and senescence and CHI treatment resulted in the accumulation of several of these, however one transcript was down regulated (Courtney et al, 1994). The identification of specific proteins which become ubiquitinated prior to visual signs of petal wilting will assist in the characterisation of critical events controlling the wilting process.Specific protein degradation can also occur as a result of the action of targeted proteases. This is a well-understood process resulting in proteolytic cascades in a wide range of systems. A class of cysteine proteases (caspases), in particular, have been identified in a complex proteolytic network controlling programmed cell death in animal systems. To date although cysteine proteases are commonly found in senescent plant tissues, a caspase controlled regulatory system has not (Lam et al, 1999). However it is possible that a parallel system does exist and that other proteases are performing an analogous function to the animal caspases.
(3) Cell death processesPetal senescence is a clearly defined process involving re-mobilisation of resources into the rest of the plant and, ultimately cell death. A number of genes have been identified in animals which are switched CSG15 (1/00) 5

The biochemical basis of post-harvest quality in cut flowers HH2122TOFProject MAFFTitle project codeon or off during cell death, however homologues for very few of these genes have been identified in plants. A number of genes have also been identified in plants, associated either broadly with senescence or more specifically with pathogen interactions. At the time when this project was initiated few of these had been isolated in more than one plant species and or had been shown to be active in developmental cell death. One plant homologue dad1 had been recently been identified in several plant species, and studies of pea flowers demonstrated that the expression of this gene declines dramatically after anthesis and is undetectable in senescent petals (Orzaez and Granell, 1997). We therefore chose to use dad1 as a marker for cell death in our system. Another valuable marker for cell death, identified first in animal systems, and recently shown to be important in plant cell death processes is genomic DNA fragmentation. Two tools are available for detecting fragmentation, DNA laddering and TUNEL. DNA laddering can be easily detected by separation of genomic DNA on agarose gels, while the TUNEL assay, developed for use in mammalian systems measures DNA fragmentation by labelling DNA ends, and has been adapted for use with plants (Mittler and Lam, 1995). TUNEL has the advantage that spatial information on the distribution of cells undergoing cell death is obtainable, however it is technically more demanding. Studies on pea petals show that both DNA laddering and fragmentation using the TUNEL assay can be detected in senescent pea petals, indicating that these are useful markers for petal senescence.
Materials and MethodsPlant materialSeven stages of Alstroemeria peruviensis var. Samora petal development and senescence were used (Wagstaff et al., 2001). Flowers were removed from the plant (at stage 1) 2 days before flower opening (outer sepals fully pigmented) and transported back to the laboratory dry. Individual cymes were then removed from each inflorescence and placed into dH2O. The flowers were fully open at stage 2 (day 0) when the sepals were reflexed. At stage 3 (day +2) the top three anthers had anthesed and two days later at stage 4 (day +4) the bottom three anthers had done likewise. Stage 5 (day +6) was defined by the separation of the stigmatic lobes and by stage 6 (day +8) the petals were showing signs of discoloration and in-rolling (Fig 1). Abscission of the perianth occurred at stage 7 (day +10). Petals from each stage were used for protein and RNA extraction.
Fresh Weight and Dry Weight DeterminationsTwo flowers (5 replicates), were harvested from each developmental stage, and their fresh weights determined. They were then dried at 80oC for three days to measure dry weight. To convert fresh weight measurements to equivalent dry weight, the fresh weight for each stage was divided by the dry weight to yield a conversion factor.
Electrolyte Leakage10 discs, 7mm in diameter, were cut from both sepals and petals using a cork borer, avoiding the midrib, and directly weighed to determine their fresh weight. The tissue discs were washed with water for 10 min with constant agitation. The wastewater was then removed and an additional 10ml of fresh water added. Conductivity was determined at the start (background) and following 2h incubation. All measurements were made in triplicate, and the experiment repeated twice and the results averaged (n=6). Measurements were made on an 800 conductivity meter, with a cell constant of K = 1, (EDT, UK). Measurements are expressed as the percent of total membrane leakage relative to electrolyte leakage determined by slowly freezing, 10, S1 discs of floral tissue for 24 h, at –20oC
Protein extractionProtein was extracted from each stage of development by grinding 400mg tissue in liquid nitrogen (number of petals/sepals in each sample was also recorded) in 1.5ml buffer containing 50mM NaPO4, 2mM EDTA, 5mM PCMPS, 1mM PMSF, 5mM iodoacetamide, 20M leupeptin. The contents of the mortar were transferred to an eppendorf tube and placed on ice for 10min. CSG15 (1/00) 6
Fig 1: Stages of floral development and senescence in Alstroemeria
1 Coloured bud2 Sepals reflexed3 Fully open, no anthesis4 Fully open, anthesis
Time course of Alstroemeria pruviana CV Samora Senescence.
1 2 3 4 5 6 7
5 In-rolling, discolouration of petals6 Translucence of sepals7 Abscission of corolla

The biochemical basis of post-harvest quality in cut flowers HH2122TOFProject MAFFTitle project code500mM DTT was added to bring the final concentration to 10mM and the samples were then incubated on ice for a further 10min prior to microcentrifugation (4C, x12000g for 10min). The supernatant was split into 200l aliquots and 1.2ml ice-cold acetone was added to each aliquot. Samples were incubated on ice for 10min followed by centrifugation as before. The supernatant was discarded and the pellet resuspended in 200l 1% SDS and stored at -20C. Total protein concentration was determined by a dye binding assay (Smith et al., 1985).
Total protease activityTotal protease activity of the petals was determined by using a method based on that of Neiri (1998) using azocasein as a synthetic substrate with additional modifications. A crude extract was prepared by weighing and then grinding individual petals from all seven stages in 500l ice-cold 50mM Tris-HCl pH 7.4. The samples were transferred to microcentrifuge tubes and centrifuged for 3min at x 10000g, 4C. The supernatant was retained and stored at -80C until required. Protease activity of the crude extract was determined by adding 100l from each extract to 200l 50mM Na-acetate pH 5.0 containing 0.5% (w/v) azocasein. The samples were incubated at 37C for 24h and the reactions were then terminated by the addition of 50l 50% TCA and incubation on ice for 1h. A duplicate reaction for each sample was prepared with the TCA added at the start to act as a blank. The samples were then centrifuged as before and 100l aliquots were placed on a microtitre plate and then alkalised with 15l 10M NaOH per well prior to reading at 492nm. Sample activity was related to a standard curve consisting of 1x10 -5 – 10mg/ml solutions of crude protease type I (Sigma) in 50mM Tris-HCl pH 7.4.
ZymographyProteins were extracted for zymography from petal tissue using the method of Eason and de Vre (1995) and immediately stored at -80C. The protein content of the samples was determined using the method above. Zymograms were prepared using 11% polyacrylamide gels containing 0.1% gelatin (type I from porcine skin (Sigma)) substrate with a 4% stacking gel overlaid. 5g protein was loaded in each lane in 2 x volumes zymogram loading buffer (50mM Tris-HCl pH 6.8, 10% (v/v) glycerol, 1% (w/v) SDS, 0.01% (w/v) bromophenol blue). The proteins were run under denaturing conditions at 180V for 30min against a molecular weight marker. The gels were then renatured for 1h in 2.5% Triton X-100 and then developed overnight (15-18h) at 37C in buffer containing 50mM sodium citrate, 5mM DTT, 5mM CaCl2, 1mM ZnCl2, pH 5.0. The following day the gels were stained in 0.05% Coomassie R-250 and destained in 10% acetic acid, 40% methanol solution. Areas of protease activity were revealed as cleared bands on a blue background where the gelatin had been digested. Protease inhibitors were used to determine the specificity of the bands visualised by zymography. Proteases were inhibited by including either 2M leupeptin in dH2O (inhibits serine and cysteine proteases) or 100M L-trans -epoxysuccinyl-leucylamido (4-guanidino) butane (E-64, Sigma) in 50% ethanol (inhibits cysteine proteases) in the overnight development buffer.
Western blottingProteins were separated on a 10% gel by SDS-PAGE (Biorad mini-gel sysytem) and transferred to nitrocellulose membranes (Sartorius, Germany) and detected by chemiluminescence (ECL + Plus Western blotting, AmershamPharmacia, UK). Lipoxygenase antidody raised to recombinant cucumber oil body lox was used as the primary antibody. Ubiquitin antibody was raised in half-English lop rabbits using a modification of the procedure of Haas and Bright (1985) with additional modifications according to Hershko et al. (1982) to produce antisera with higher titres for conjugated ubiquitin as opposed to free ubiquitin.
RNA extractionTwo batches of 1g petals were extracted from each stage using the method of Wilkins and Smart (1996) including the amendments suggested by Hunter and Reid (2001). Total RNA (2.5g) from each stage was DNAse treated (Promega) and then used to make cDNA using Superscript II MMLV reverse transcriptase (Gibco). Controls lacking reverse transcriptase were included in the cDNA synthesis to verify the complete removal of genomic DNA in the DNAase treatment step.
Cloning of partial cDNAs from Alstroemeria:Degenerate primers to important genes involved in protein degradation, lipid metabolism and cell death were designed from a comparison of conserved regions of the genes from monocotyledonous species available in the database (Table 1). Partial cDNAs were amplified from Alstroemeria cDNA using 0.625 units of Quiagen Taq polymerase, Quiagen buffer, 125ng of cDNA from petals of flowers at stages 1, 3 and 5, 1.5mM MgCl2, 0.2 mM dNTPs and 1g of each PCR primer. Reactions were cycled in a Hybaid OMNE machine for 35 cycles of {94oC 1min, 60oC 1min, 72oC 1min}. PCR products were cloned into pGEMEasy T (Promega) and sequenced using an ABI 377 automated sequencer.
Table 1: Degenerate primers designed to amplify important genes in Alstroemeriagene Orientation/
nameTm Product
sizePrimer sequence
Cysteine protease CYPFON 62/60 GACATGASCWRSSAGGARTTCYPR1N 62/56 333 GAAYTSGAAYGCATARTCCAT
CSG15 (1/00) 7

The biochemical basis of post-harvest quality in cut flowers HH2122TOFProject MAFFTitle project codeUbiquitin UQF GCNAAGATCCAGGACAA
UQR 278 ATSARVCGCTGCTGGTGlutathione-S-transferase
GSTPOS3 62/58 SGTGTGGMYGGAGGTGGA
GSTNEG4 60/56 126 CTCGTASTYSYCSAGCAClipoxygenase LOXFN 64/60 GCVGTCAACTTYGGSCAGTA
LOXRA 62/58 465 GAKRTGCTGTTGGGGATGCLOXR414 64/60 414 GTCGGAGGTRTTNGGRTA
Defender against apoptotic death (dad-1)
DADF00 58/50 GGGTCRTTYCCHTTYAAC
DADR00 58/54 147 CAYAGRACGAAAWCWGCAAA
Semi-quantitative RT-PCRSpecific Alstroemeria PCR primers were designed from the partial cDNA clones (Table 2). An initial PCR reaction was conducted using the Alstroemeria specific primers on 100ng of the cloned partial cDNA to estimate the optimal cycle number for exponential amplification. PCR conditions were as described above except that the appropriate Tm was used for each primer set. (Table 2). A cycle number producing a barely visible band was subsequently used for semi-quantitative RT-PCR (Table 2) in which 125ng of cDNA from the seven defined stages of Alstroemeria petals was used as a template under the conditions described above.
Spectrophotometry and ethidium bromide stained gels were used to establish that equal amounts of RNA were used for the RT-PCR. PCR products were separated on 1.5% agarose gels and Southern blotted onto nylon membrane to enhance the signal for quantitation. Pre-hybridisation and hybridisation were performed at 60oC in a solution containing 5 x Denharts, 6 x SSC, 0.1% SDS, 5% PEG, 0.1% tetrasodium pyrophosphate and 100 gml-1 denatured herring sperm DNA. Random primed probes were prepared as described by Feinberg and Vogelstein (1983). The probe used was the respective PCR product. Washes were in 2 x SSC 0.1 x SDS at 60oC. Blots were exposed to phosphorimaging film (Kodak) and images were analysed in a Bio-Rad phosphorimager using a Quantity One (Bio-Rad) image analysis package.
Table 2: Specific Alstroemeria primers and PCR conditionss used for expression analyses.gene Orientation/name Tm used Product size/bp Cycle no. used Primer sequenceCYP CYPAF 54 AGGAGTTCAGAACCCATTA
CYPAR 54 327 22 TCGAATGCATAGTCCATGAGST ALSGSTF 62 TGGTGTGGATGGAGGTGGA
ALSGSTR 58 126 28 TCTGGTAGACGCCGAGCALOX LOXAF 56 GCGGTCAACTTCGGGCAG
LOXAR 54 442 24 GAGGTGCTGTTGGGGATLOXA414R 58 414 24 GTCGGAGGTGTTCGGGTA
DAD DADAF 54 GGGTCGTTTCCATTCAACDADAR 54 151 28 CATAGGACGAAATCTGCAA
DNA laddering assayGenomic DNA was extracted from 2g petals at each stage of development. Petals were ground in liquid nitrogen before adding to 15ml extraction buffer (100mM Tris-HCl pH 8.0, 1.4M NaCl;, 20mM EDTA, 2% CTAB (hexadecyltrimethylammonium bromide)) containing 0.4% -mercaptoethanol prewarmed to 60C and then mixed gently and incubate at 60C for 20min. An equal volume of chloroform: isoamyl alcohol (24:1) was added and the solution was rocked vigorously for a further 30min. Cellular material was spun down at 4C in a Sorvall SS34 rotor for 10min at 12000g. The supernatant was removed and ethanol precipitated (2.5 x volume 100% EtOH, 0.1 x 3M NaOAc pH 5.2) overnight at -20 C. The following day the solution was centrifuged at 6000g for 30min at 4C and the supernatant was dicarded prior to washing the pellet with 75% EtOH before drying. The pellet was resuspended in Tris-EDTA (TE, pH 8.0) and an RNAse treatment was performed at 37 C for 1h. A phenol:chloroform:isoamylalcohol extraction (25:24:1) was then followed by a second ethanol precipitation as before. The final pellet was resuspended in 50l TE. Equal concentrations (approx 30g) of DNA were run on a 1.5% agarose gel stained with ethidium bromide and then Southern blotted onto nylon membrane. Prehybridisation and probe preparation were as above. A probe was made using 5g genomic DNA cut with the Sau3A restriction enzyme.
Extraction of lipids Total lipids were extracted from tissues by a modification of the method of Bligh and Dyer (1959) according Griffiths et al. (2000). All procedures were performed in dim light at 4 oC using chilled solvents (containing BHT, 0.01% w/v) and glassware. Tissues (approx. 0.5g fresh weight) were homogenised with a pestle and mortar containing 0.15M acetic acid (1ml) and chloroform/methanol (1:2 v/v; 7.5ml) for approx. 2min and transferred to culture tubes (Pyrex, UK). The pestle and mortar were
CSG15 (1/00) 8

The biochemical basis of post-harvest quality in cut flowers HH2122TOFProject MAFFTitle project coderinsed with chloroform (2.25ml) and combined with the extract to which was added dH2O (2.25ml). Phase separation was facilitated by low speed centrifugation, the lower chloroform phase containing the lipids was removed and stored at –20C.
Spectrophotometric determination of lipid hydroperoxidesPreparation of FOX ReagentFOX reagent was prepared according to Nourooz-Zadeh et al. (1995) by dissolving xylenol orange (Sigma, UK) and ammonium ferrous sulphate in 250mM H2SO4 to final concentrations of 1 and 2.5mM, respectively. One volume of this concentrated reagent was added to 9 volumes of HPLC grade methanol containing 4.4mM BHT to make the working reagent which comprised 250µM ammonium ferrous sulphate, 100µM xylenol orange, 25mM H2SO4 and 4mM BHT in 90% v/v methanol.
Analytical proceduresLipid hydroperoxides were determined in chloroform extracts immediately following extraction and aliquots were dispensed into amber vials (Hewlett Packard, USA) and evaporated under N2. Samples were resuspended in HPLC grade methanol either in l for samples without TPP or in 90l methanol to which were added 10l TPP (25mM in methanol). Samples were incubated at room temperature for 30min in the dark and then for a further 30min following the addition of the working FOX reagent. The concentration of LHPOs was determined spectrophotometrically at 560nm using a molar absorption coefficient derived for standard linoleate hydroperoxide (=6.0 x 104 M-1 cm-1 , Gay et al., 1999). Standard curves were constructed using linoleate hydroperoxide as standard synthesized by the method of Gardiner (1997). 13S-hydroperoxy-9Z, 11E-octadecadienoic acid (HPODE) was enzymatically synthesised using soybean lipoxygenase (Sigma, lipoxidase type 1-S). Product formation was monitored at 234nm and the purity of the eluting fractions determined at the same wavelength by HPLC (Hewlett-Packard 1100
series chromatograph) on a 5m Partisil column (Fisons chromatography, Leicester, UK) using isocratic elution in 0.75% ethanol in n-hexane at a flow rate of 4ml min (Chan and Levett, 1977).
Lipid purification and quantitationLipids were purified by thin layer chromatography (TLC) on pre-coated silica gel plates (Merck, silica gel 60). Neutral lipids were separated using hexane/diethylether/acetic acid/ (70:30:1, by vol.) and polar lipids using chloroform/methanol/acetic acid/water (170: 30: 20: 7, by vol.) (Griffiths et al., 1997). Lipids were located by lightly staining with iodine vapour and then removed from the plates for transmethylation for the analysis of fatty acids. Fatty acid methyl esters were prepared in 2.5% (by vol.) sulphuric acid in anhydrous methanol (2ml) and quantified by gas liquid chromatography using heptadecanoic acid as the internal standard and on a 10% SP-2330 100/120 Chromosorb W AW (Supelco) column at 135 oC using a 6890 gas liquid chromatograph equipped with a flame ionization detector (FID) and a mass selective detector (Agilent Technologies).
Extraction and Detection of lox products by HPLCOxidised fatty acids were extracted according to the method of Kohlmann et al. (1999). One gram of frozen floral tissue was added to 20ml of extraction medium [hexane/isopropanol, 3/2 by vol with 0.0025% wt/vol. BHT) and immediately homogenised with an ultra Turax for 30s under a stream of argon on ice. The extract was centrifuged for 15min at 4500 g at 4 oC. The clear upper phase was collected and the pellet extracted three times with 3ml each of extraction medium. To the combined organic phases a 6.7% (wt/vol.) solution of potassium sulfate was added to a volume of 47ml. After vigourous shaking the upper hexane-rich layer was removed. The upper organic phase containing the hydroperoxy fatty acid derivatives were dried under nitrogen and redissolved in chloroform and stored under argon at –80oC until use. HPLC analysis was carried out on a Beckman HPLC system coupled to a diode array detector (Feussner et al., 1995) Straight phase (SP)-HPLC of the hydroperoxy fatty acids was carried out on a Zorbax SIL column (DuPont, 250 x 4.6 mm, 5m particle size) with a solvent system of n-hexane/2-propanol/acetic acid 100/2/0.1 by volume. and a flow rate of 1mlmin-1 . Absorbance at 234nm was monitored.
Thiobarbituric acid reactive substance determination (TBARS)Malondialdehyde was synthesised as described by Gutteridge (1982) and the endogenous levels present in the tissues were determined by a modified version of the thiobarbituric reactive substance assay recently described by Hodges et al. (1999). MDA was extracted by grinding 0.5g floral tissue, in 5ml 80% ethanol containing 0.01% BHT at 4 oC. The homogenate was adjusted to a tissue:solvent ratio of 1:25, mixed and centrifuged at 2000g for 10min at 4oC. 1ml aliquots of the resulting supernatant were added to 1ml of reagent containing 20% trichloroacteic acid, 0.01% BHT, 0.13mM disodium EDTA, ,+ or – TBA 0.65% then incubated at 95oC for 25min. The samples were cooled on ice for 5min and centrifuged again at 2000g for 10min at 4 oC. Absorbances were measured at 440, 532 and 600nm and the concentration of the MDA equivalents calculated as described by Hodges et al. (1999).
Lipoxygenase assays Total proteins were extracted from approximately 1g of floral tissue in a pestle and mortar in 2 vols of 50mM potassium phosphate, pH7.0, containing: 1% PVP (polyvinylpolypyrrolidone), 0.1% Triton-X 100 (t-Octylphenoxypolyethoxyethanol), 0.04% Potasium bisulphite and 1mM DTT (DL-Dithiothreitol) (modified from Koch et al., 1992) at 4 oC. The homogenate was centrifuged at 13,000g, for 2min and the resulting supernatant was passed through a PD-10 column (Amersham Pharmacia Biotech UK), pre-equilibrated with 50mM potassium phosphate buffer, pH 7.0. The protein eluate was resuspended in glycerol (20
CSG15 (1/00) 9

The biochemical basis of post-harvest quality in cut flowers HH2122TOFProject MAFFTitle project code% v/v final concentration) and snap frozen in liquid nitrogen and stored at –80 oC until required. Lox activity was measured spectrophotometrically by following the increase in A234 (formation of conjugated dienes) from added fatty acid substrate (10-100nmol per assay in ethanol) in a 1ml reaction volume at 25C with 0.005%triton X-100 (Pierce ultrapure) in the buffer. Oleic acid (C18:1 cis was used as a control fatty acid and the rate minus background (lacking addition of unsaturated fatty acids) was used to correct for activity determination.
TUNELPetals cut to approximately 2mm square pieces from all developmental stages were fixed in xylene for 15min, followed by a wash in 100% EtOH and then incubated for 2h in a fresh solution of PBS pH 7.4, 70mM EGTA, 4% paraformaldehyde, 0.25%glutaldehyde, 0.1% TWEEN, 10% DMSO. Fixed tissue was then stored in 70% EtOH at -20C. Proteins were removed using a 20min incubation at 37C in 20gml-1 proteinase K solution in PBST. The specimens were then washed twice in PBST for 5min. The remaining procedure was completed according to that given in association with the Apoptag Fluorescein In Situ Apoptosis Detection Kit (Intergen, Oxford, UK). Samples were mounted on slides and dual stained with 80gml-1 propidium iodide and anti-digoxigenin conjugate and then viewed on an Olympus BH2 fluorescent light microscope with a FITC filter (excitation 490nm, emission 520nm).
Results1. PhysiologyFresh and dry massInitially a distinction was made between sepals and petals and between the upper two petals and the lower one petal as they are visually quite distinct. For example, the average fresh weight of each of the upper petals at the time of flower opening was 218.4mg whilst the lower petal averaged 79.2mg and each of the sepals was 415.1mg. However the changes in fresh and dry weight were similar in all petals but somewhat different to the changes that occurred in the sepals (Fig. 2). The dry mass of the petals remained reasonably constant during flower opening and senescence (Fig. 2). At the time of tepal abscission the dry mass of the sepals was 63% of the maximum recorded mass and for the petals the corresponding value was at least 90%. Compared to many species this loss of dry mass is minimal, for example in daylily petal dry mass at the end of vase life is only 33% of the maximum recorded value (Lay-Yee et al., 1992). However, in other species where petal abscission occurs the loss may be considerably smaller, for example in Digitalis the loss of dry weight is less than 15% (Stead & Moore, 1977). Fresh mass reached a maximum at the time of flower opening (Fig. 2) and the sepals lost about 50% of their fresh weight by the time that they abscised. Such a loss of fresh weight suggests that Alstroemeria flowers show a wilting like response as seen in flowers such as Petunia (Lovell et al., 1987) and carnation (Woltering and van Doorn, 1988).
Figure 2: Fresh weight and dry weight of petals and sepals throughout flower development and senescence (n>10sd)
2. Protein degradationTotal protein concentration and protease activityChanges in protein levels and protease activity were investigated over the duration of flower vase life. The petals lost fresh weight rapidly towards the end of their vase life that resulted in an apparent increase in protein levels at stage 6 when calculations were made per g FW (Fig. 3) but fell overall during vase-life from 11 to 8g.mg-1 FW. Total protease activity increased more than 200 fold in the last two stages when expressed in g.g-1 FW (equivalent to more than 100 fold on a dry weight basis) relative to the TCA soluble peptides released from bovine pancreas protease standards.
CSG15 (1/00) 10
Fresh weight change of H2O treated cymes
050
100150200250300350400450500
day -
1
day +
1
day +
3
day +
5
day +
7
Fres
h w
eigh
t (m
g)
upper petalslower petalsepals
Dry weight change of H2O treated cymes
0
5
10
15
20
25
30
35
40
day -
1
day +
1
day +
3
day +
5
day +
7
Dry
wei
ght (
mg)
upper petalslower petalsepals

The biochemical basis of post-harvest quality in cut flowers HH2122TOFProject MAFFTitle project code
Figure 3: Protein content and protease activity of Alstroemeria petals during senescence. Stage 1 (day –2), stage 2 (day 0), stage 3 (day +2), stage 4 (day +4), stage 5 (day +6), stage 6 (day +8), stage 7 (day +10). Bars = range calculated from a mean of n=3 (protein) independent tissue extractions and n=10 (protease). Protein content was determined using a BCA dye-binding assay, protease content was determined by comparison to the liberation of soluble peptides from azocasein by bovine pancreas crude protease extract over 24h.
Characterisation of the protease activity by zymographyZymography revealed protease activity in three distinct bands visible in the lower part of the gel apparently ranging from 12 to 26KDa (Fig. 4A). No protease activity was visible prior to stage 3 (day +2). Band I (26KDal)of protease activity showed down-regulation from stage 3 (day +2) onwards, activity of band II (16 Kdal) fluctuated during flower development and band III (12 Kdal) was up-regulated over the same period. Zymograms were conducted with specific inhibitors to identify the classes of proteases represented by these bands of activity. Leupeptin inhibited both serine and cysteine proteases (Fig. 4B), whereas E64
inhibited cysteine proteases specifically (Fig. 4C). Both treatments eliminated all three bands of protease activity leading to the conclusion that all represented cysteine proteases.
Figure 4: Protease activity in Alstroemeria petals determined by zymography. Extracts of Alstroemeria petals corresponding to 5g protein were subjected to SDS-PAGE in an 11% polyacrylamide gel containing 0.1% gelatin substrate. (representative data are presented from three separate experiments). Gels were post-stained with 0.5% Coomassie blue. Protease activity was revealed as cleared bands within the background of the gel. stage 1 (day –2), stage 2 (day 0), stage 3 (day +2), stage 4 (day +4), stage 5 (day +6), stage 6 (day +8), stage 7 (day +10).A: Control samples with no protease inhibitors.B: Samples treated with 2M leupeptin in dH2O (inhibits serine and cysteine proteases) present in the overnight development buffer. C:Samples treated with 100M L-trans–epoxysuccinyl-leucylamido (4-guanidino) butane (E-64) (inhibits cysteine proteases) in the overnight development buffer.Control gels stained with Coomassie brilliant blue confirmed equal loading and integrity of the protein samples.
Isolation and characterisation of a partial Alstroemeria cysteine protease and ubiquitin cDNAs.Using degenerate PCR primers (Table 1), a 333bp partial cDNA (ALSCYP) and a 278bp ubiquitin partial cDNA (ALSUQ) were obtained from Alstroemeria petals. (The nucleotide sequences will appear in the EMBL, Genbank and DDBJ Nucleotide Sequence Databases under Accession numbers AY034030 and AY034031 respectively.) Analysis of the putative cysteine protease sequence suggests that this gene is a papain-like cysteine protease. The predicted amino acid sequence of ALSCYP begins approximately 390bp from the N terminus of other plant cysteine proteases and contains several of the elements characteristic of the structure of papain (Light et al., 1964; Mitchel et al., 1970). The three highly conserved amino acids (Cys, His and Asn) that make up the catalytic triad characteristic of the active site of cysteine proteases (Storer and Menard, 1994) are present. In addition the sequence obtained contains four conserved Gly residues that are involved in maintaining an active enzyme conformation (Kamphuis, 1985), two conserved Cys residues that form disulphide bridges (Koehler and Ho, 1990) and a Gln active site residue (Kambouris et al., 1992). Alignment of the putative protein sequence deduced from the partial cDNA with cysteine proteases identified in floral tissue shows 53-65% homology at the amino acid level with the same regions of SEN102 (Hemerocallis), SEN11 (Hemerocallis), DCCP1 (Dianthus) and cysteine proteases from Phalaenopsis and Sandersonia (Fig. 5). CSG15 (1/00) 11
6
7
8
9
10
11
12
13
14
1 2 3 4 5 6 7
Stages
Prot
ein
(g.m
g FW
-1)
0
50
100
150
200
250
300
Prot
ease
(g.
g FW
-1)
ProteinProtease
1 2 3 4 5 6 726KDa16KDa12KDa
1 2 3 4 5 6 726KDa16KDa12KDa
1 2 3 4 5 6 7
26KDa16KDa12KDa
A
B
(stages)
(stages)
(stages)C

The biochemical basis of post-harvest quality in cut flowers HH2122TOFProject MAFFTitle project code
Figure 5: Alignment of predicted amino acid sequence of ALSCYP cysteine protease cloned from Alstroemeria with other floral papain-class cysteine proteases namely SEN102 (Hemerocallis), SEN11 (Hemerocallis), DCCP1 (Dianthus) and cysteine proteases from Phalaenopsis and Sandersonia (Accession numbers P43156, U12637, U17135, U34747 and AAD28477 respectively). Alignment of the same region of the cDNA as ALSCYP was conducted using Clustal method analysis in Megalign (DNAStar Inc.) Amino acids matching the consensus are shaded. Cys residue forming a disulphide bridge; Gly residues maintaining an active enzyme conformation; * Gln active site residue; catalytic Cys, His and Asn triad of the active site.
ALSCYP 1 MSCEEFRTHYAGSKIDHHRARRGSPRGN-AG-EFMYERAT--APP--AVDWRQKGAVADVKDQGQCGSCWAFSTIAAVEGINQIKSGDLVSLSEQELVDCDTKGENQGCNGGLMDYAF DIANTHUS 1 --NDEFRRIYFG--VKRPEKAESVKSD---R--YAVKEGD--ELP-ESVDWRKKGAVSHVKDQGQCGSCWAFSAIGAVEGINKIVTGDLITLSEQELVDCDTSY-NSGCDGGLMDY-- PHAL 1 --NHEFRSTYAGSRINHHRSLRGSRRGGATN-SFMYQSLDSRSLP-ASIDWRQKGAVTAVKDQGQCGSCWAFSTVAAVEGINQIKTKKLLSLSEQELIDCDTDE-NNGCNGGLMDY-- SEN11 1 --NQEFRSTYAGSKIDHHMTLRGV-KD--AG-EFSYEKFH--DLP-TSVDWREKGAVTGVKDQGQCGSCWAFSTVVAVEGINQIKTNELVSLSEQQLVDCDTK--NSGCNGGLMDY-- SEN102 1 --NQEFRSKYAGSKIQHHRSQRGIQKN--TG-SFMYENVG--SLPAASIDWRAKGAVTGVKDQGQCGSCWAFSTIASVEGINQIKTGELVSLSEQELVDCDTSY-NEGCNGGLMDY-- SAND 1 --NQEFRAKYAGSKVHHHRTMKGSRHGSGSGAKFMYENAV--APP--SIDWRERGAVAAVKNQGQCGSCWAFSAIAAVEGINQIVTKELVPLSEQELIDCDTDQ-NQGCSGGLMDY-- Consensus 1 --N EFR YAGSKI HHR RG - G- FMYE -- LP- S DWR KGAV VKDQGQCGSCWAFSTIAAVEGINQIKT LVSLSEQELVDCDT -N GCNGGLMDY--
*
The partial ubiquitin cDNA (ALSUQ), cloned using degenerate primers from Alstroemeria petals (Table 1), spans the repeat sequence that is conserved among eukaryotic organisms in poly-ubiquitin and mono-ubiquitin genes (Fig 6A). The repeat is 228bp in length and comparison of the sequence to poly-ubiquitin genes from other monocots (Fig 6B) shows the high levels of homology of these genes across species even at the nucleotide level, the highest homology being to rice (91%). Specific primers for Alstroemeria were designed, and were used in subsequent semi-quantitative RT-PCR experiments to detect expression both of mono-ubiquitin and poly-ubiquitin transcripts.
Figure 6A Alstroemeria ALSUQ sequence showing repeat unit (two shaded regions) and primers used for amplification of the cDNA portion (underlined). Due to the repetitive nature of the gene, primers will amplify a 50bp PCR product from mono-ubiquitin as well as the 278 bp fragment from poly-ubiquitin transcripts using RT-PCR.
GCCAAGATCCAGGACAAGGAGGGCATCCCCCCTGACCAGCAGCGTCTCATCTTTGCCGGCAAGCAGCTTGAGGATGGGCGTACCCTGGCTGATTACAACATCCAGAAGGAGTCCACCCTCCACTCGGTGCTCCGTCTCCGTGGTGGTATGCAGATCTTTGTTAAGACCCTCACTGGCAAGACCATCACCCTTGAGGTTGAGAGCTCTGACACCATTGACAATGTCAAGGCGAAGATCCAGGACAAGGAGGGCATCCCCCCTGACCAGCAGCGTCTGAT
Figure 6B Alignment of nucleotide sequence of ALSUQ cloned from Alstroemeria, with other poly-ubiquitin genes from monocotyledonous species namely: RUBQ2 (rice), HvB074 (barley) and MubC5 (maize) (Accession numbers: AF184280, AJ228945 and U29158). Alignment of the same region of the cDNA as ALSUQ was conducted using Clustal method analysis in BioEdit. Nucleotides matching the consensus are shaded.
Ubiquitination of proteins during petal development and senescenceWestern blotting was used to determine levels of ubiquitinated proteins in petals throughout development and senescence, and to investigate whether ubiquitination of specific proteins changed. Little overall ubiquitination was detectable compared to other systems (e.g. Hemerocallis), and there was no clear evidence of changes in patterns of ubiquitination ( data not shown)
Expression programme of ALSCYP and ALSUQ during petal development and senescenceSemi quantative RT-PCR was used to investigate the expression programme of ALSCYP and ALSUQ in petals. Note that since a PCR method was used, it important to treat the reults as semi-quantitative rather than an absolute quantification of transcript levels. A continuous increase in ALSCYP mRNA levels during flower vase life was found (Fig. 7). As described in the Materials and Methods section, spectrophotometry and ethidium bromide gels were used to equalise the RNA amounts used for the RT-PCR. This is important: as internal controls for this type of developmental series are likely to be unreliable due to the expected global changes in gene expression. Tubulin primers spanning an intron were used to check for the presence of genomic DNA. This
CSG15 (1/00) 12
ALSUQ 1 GCCAAGATCCAGGACAAGGAGGGCATCCCCCCTGACCAGCAGCGTCTCATCTTTGCCGGCAAGCAGCTTGAGGATGGGCGTACCCTGGCTGATTACAACATCCAGAAGGAGTCCACCCTC maize 1 GCCAAGATCCAGGACAAGGAAGGCATTCCCCCGGACCAGCAGCGGCTCATCTTCGCTGGCAAGCAGCTCGAGGACGGGCGCACACTTGCCGACTACAACATCCAGAAGGAGAGCACCCTC barley 1 GCGAAGATCCAGGACAAGGAGGGCATCCCTCCGGACCAGCAGCGCCTCATCTTTGCTGGCAAGCAGCTCGAGGATGGCCGCACCCTGGCTGACTATAACATCCAGAAGGAGTCCACTCTC rice 1 GCCAAGATCCAGGACAAGGAGGGCATTCCCCCGGACCAGCAGCGCCTCATCTTTGCTGGCAAGCAGCTGGAGGATGGCAGGACCCTTGCTGACTACAACATCCAGAAGGAGTCCACCCTC Consensus 1 GCCAAGATCCAGGACAAGGAAGGCATTCCCCCGGACCAGCAGCGGCTCATCTTCGCTGGCAAGCAGCTCGAGGACGGGCGCACACTTGCCGACTACAACATCCAGAAGGAGAGCACCCTC
ALSUQ 121 CACTCGGTGCTCCGTCTCCGTGGTGGTATGCAGATCTTTGTTAAGACCCTCACTGGCAAGACCATCACCCTTGAGGTTGAGAGCTCTGACACCATTGACAATGTCAAGGCGAAGATCCAG maize 121 CACCTTGTTCTCCGCCTCAGGGGAGGCATGCAGATCTTCGTCAAGACCCTCACTGGAAAGACTATCACCCTCGAGGTGGAGTCTTCTGACACCATCGACAACGTCAAGGCCAAGATCCAG barley 121 CACCTGGTGCTCAGGCTTCGTGGAGGCATGCAGATTTTCGTGAAGACCCTCACCGGCAAGACCATCACACTGGAGGTGGAGTCCTCTGACACCATCGACAACGTCAAGGCGAAGATCCAG rice 121 CACCTTGTGCTCCGCCTTCGTGGTGGTATGCAGATCTTTGTCAAGACCCTCACAGGCAAGACCATCACCCTGGAGGTTGAGAGCTCGGACACCATCGACAACGTCAAGGCCAAGATCCAG Consensus 121 CACCTTGTTCTCCGCCTCAGGGGAGGCATGCAGATCTTCGTCAAGACCCTCACTGGAAAGACTATCACCCTCGAGGTGGAGTCTTCTGACACCATCGACAACGTCAAGGCCAAGATCCAG
ALSUQ 241 GACAAGGAGGGCATCCCCCCTGACCAGCAGCGTCTGAT maize 241 GACAAGGAGGGGATTCCTCCGGACCAGCAGCGCCTGAT barley 241 GACAAGGAGGGCATCCCTCCGGACCAGCAGCGCCTCAT rice 241 GACAAGGAGGGCATCCCCCCAGACCAGCAGCGTCTCAT Consensus 241 GACAAGGAGGGGATTCCTCCGGACCAGCAGCGCCTGAT

The biochemical basis of post-harvest quality in cut flowers HH2122TOFProject MAFFTitle project codewas clearly absent from the RT controls. The tubulin expression levels fluctuated during the developmental programme: this is not unexpected given that the primers used will detect multiple members of the tubulin gene family. Evidence from other species (e.g. maize, Rogers et al, 1993) indicates that each tubulin gene has its own characteristic expression pattern. The pattern seen is therefore a composite of this underlying expression of total tubulin. Faint signals are visible in the RT-control reactions possibly due to very low levels of genomic DNA or plasmid contamination. These were subtracted prior to quantification of the expression levels.
Figure 7: Semi-quantitative expression of Alstroemeria petal transcripts isolated as cDNA clones Stage 1 (day –2), stage 2 (day 0), stage 3 (day +2), stage 4 (day +4), stage 5 (day +6), stage 6 (day +8), stage 7 (day +10). Autoradiograph of gene expression following limited PCR cycles. The products were Southern blotted and a probe made from a similar reaction was hybridised to the membrane overnight. At least two separate PCR amplifications were compared for each transcript, and similar patterns of expression obtained. The left hand side shows products from a reaction containing 125ng cDNA from each stage, the right hand side shows negative controls without reverse transcriptase.The gene target is indicated to the right of each panel. he primers used for the tubulin PCR span an intron. The extreme right hand lane is the larger size PCR product obtained from using these primers with genomic DNA as a control and acts as further confirmation of the absence of genomic DNA contamination from the cDNA samples.
Quantitation of the RT-PCR signal (Fig 8) revealed an approximately 30 fold increase in signal from stage 1 (day-2) to stage 7 (day +10) indicating strong up-regulation of this gene during petal senescence. This is in contrast to the levels of ubiquitin expression determined using the same method from the identical batch of cDNA. Ubiquitin expression remained more or less constant throughout vase life (Fig. 7) with the
maximal variation in signal less than 1.5 times that of the minimum signal recorded during quantification. The appearance of multiple bands on the autoradiograph is presumably due to amplification of multiples of the repeat sequence that is characteristic of the ubiquitin gene ( see Fig6A above).
CSG15 (1/00) 13
0
1000
2000
3000
4000
5000
6000
7000
1 2 3 4 5 6 7
stage
peak
are
a
0100
200300400
500600
1 2 3 4 5 6 7
stage
peak
are
a
ALSCYP ALSUQ
ALSCYP
DAD1
cDNA -RT
Glutathione-S-Transferase
ALSUQ
1 2 3 4 5 6 7 1 2 3 4 5 6 7
LOX1
LOX2
tubulin
327 bp
151 bp
126 bp
442 bp
414 bp
230 bp
278 bp550 bp

The biochemical basis of post-harvest quality in cut flowers HH2122TOFProject MAFFTitle project code
Fig 8: Quantitation of the signal from a representative autoradiographs showing ALSCYP and ALSUQ expression using a phosphorimager screen and Quantity One image analysis (BioRad). The area of the peak was calculated for each stage (arbitrary units) and the corresponding background region in the –RT controls was subtracted for each stage.
3) Membrane integrity and lipid metabolisme maintenance of cellular integrity and subcellular compartmentation is integral to cell function. However, during senescence of both ethylene sensitive and ethylene insensitive flowers, marked changes occur in the biochemical and biophysical properties of the cell membranes. These result from losses of membrane phospholipids, increases in neutral lipids, increases in sterol/phospholipid ratio and increases in the saturation:unsaturation index of fatty acids (Lesham, 1992, Thompson et al., 1998). Membrane polyunsaturated fatty acids are prone to oxidation either by enzymatic means (lipoxygenase, lox) or through autoxidative events (non-enzyme catalyzed). In a number of floral tissues such as carnation (Fobel et al., 1987), daylily (Panavas and Rubinstein, 1998) and rose (Fukuchi-Mizutani et al., 2000), lox activity increases prior to the onset of electrolyte leakage (a marker of loss of membrane semi-permeability). Increase in lipid peroxidation, usually estimated as thiobarbituric acid reactive
substances (TBARS), accompanies the increase in lox activity and the products of peroxidation are considered to perturb membrane function, partly at least, by causing increased membrane rigidification (Thompson et al., 1998).
Electrolyte leakageThere was little loss of membrane integrity, determined by measurements of electrolyte leakage from petal and sepal discs, prior to
tepal abscission. There was, however, a small increase in leakage from old flowers which was detected about 6 days after flower opening (Fig 9). This increase in leakage suggested that the membrane integrity is compromised as the flowers age, possibly due to changes in the composition of the membranes, which were therefore investigated.
Fig 9 Electrolyte leakage throughout senescence expressed as the percent of total leakage (n=6 )
Changes in lipid composition during senescenceTotal lipid content (as fatty acid methyl esters relative to heptadecanoic acid) showed a marked decline in both sepals and petals after S2 (Fig. 10), representing a loss of 56 and 68% of total lipid respectively. Of the major fatty acid constituents to be degraded, linoleic acid (C18:2 cis 9, 12) showed the largest decline, losing almost 80% of the initial content. Decreases in palmitic acid
(C16:0) and 18:3 (linolenic acid, C18:3 all cis 9,12,15) were also observed. Similar changes in sepal fatty acid constituents were observed.
Figure 10: Total fatty acid content of sepals and petals during senescence. Fatty acids were extracted in a chloroform:methanol based solvent system, separated by gas chromatography as their methyl esters and quantified using heptadecanoic acid as an internal standard. Data points represent the mean of n=3±SD.
The complex lipid profile was almost identical for both sepal and petal and the same changes observed. Therefore only data for petal tissues are shown (Fig. 11).
CSG15 (1/00) 14Stage
S1 S2 S3 S4 S5 S6 S720
40
60
80
100
120 SepalPetal
mol/gDW

The biochemical basis of post-harvest quality in cut flowers HH2122TOFProject MAFFTitle project code
Figure 11: Changes in the major complex lipids during petal senescence. Lipids were purified by thin layer chromatography and analysed as their fatty acid methyl esters, by gas chromatography. Lipids purified consisted of, phosphatidylcholine (PC), phosphatidylethanolamine (PE), monogalactosyldiacylglycerol (MGDG), digalactosyldiacylglycerol (DGDG). Results are representative of analyses repeated twice.
In stages from S1-S5 phospholipids were the major components representing almost 60% of the total lipids of which phosphatidylcholine (PC) was 35±3% with phosphatidylethanolamine (PE) being the next most abundant lipid over this time period (22±2%). The chloroplast galactolipids, monogalactosyldiaglycerol (MGDG) and digalactosyldiacylglycerol (DGDG), were present in similar amounts and jointly constituted around
25% of the total tissue lipids at S1. Phospshatidylinositol, phosphatidic acid and phosphatidylserine were detected in low amounts (<5% of the total lipids combined, data not shown). Neutral lipids were present in young S1 tissues with triacylglycerol (4%), diacylglycerol (3%) unesterified fatty acids (4%) and sterol esters (4%) detected (data not shown). During senescence phospholipids and galactolipids were degraded and by S7, 87% of PC, 83% of PE, 95% of MGDG, and 89% of DGDG had been utilised (Fig.11). The levels of neutral lipids, by contrast, remained similar throughout senescence although some increase in the UFA level was detected (data not shown).
(a) Lipoxygenase gene expressionUsing two overlapping sets of degenerate primers (Table 1), two LOX partial petal cDNAs were isolated (ALSLOX1 and ALSLOX2) which are distinct from each other sharing only 43% homology at the amino acid level suggesting that they represent
independent genes (Fig. 12). Figure 12: Phylogenetic tree using Neighbour-joining method showing relationship of ALSLOX1 and ALSLOX2 from Alstroemeria to other species over the same region as the partial cDNA isolated. An alignment of the Alstroemeria partial clones with lipoxygenase genes from other monocotyledonous species reveals that LOX2 is more similar to the major group of monocot LOX genes over this region of the protein, whereas LOX1 is more divergent. Sequence analysis at the amino acid level (Fig. 13) over
* ++ + + + + +ALSLOX1 1 AVNFGQYPFGGYPPNRPTLLKRLIPEEDEPE---YESFLSN---PQHTFLTSLPSQLQATQIMAVQDTLSTHSPDEEYLGQVYESHSHWIDDSQVLRSFDKFSARLEEIEVAINRRNKDL ALSLOX2 1 AVNFGQYAYAGYMPNRPTISRRFMPEPGTEE---YEELRRN---PERVFLKTITSQLLTILGVALIEVLSRHSSDEVYLGQ--RDSPEWTSNRAALEAFRRFGERLVEIEDRINVMNNDE BLYLOXA 710 AVNFGQYPYAGFLPNRPTVSRRRMPEPGTEE---YAELERD---PERAFIHTITSQIQTIIGVSLLEVLSKHSSDELYLGQ--RDTPEWTSDPKALEVFKRFSDRLVEIESKVVGMNHDP BLYLOXB 720 AVNFGQYPYAGYLPNRPTVSRRPMPKEGDEE---YEQLKEGGEAADMVFIHTITSQFQTILGITLIEILSKHSSDEVYLGQ--RDTPEWTSDAKALEAFKRFGSRLVEIEKRILDMNKDP BLYLOXC 712 AVNFGQYPYSGYHPNKPSASRRPMPVQGSEE---YAELERD---PEKAFIRTITSQFHALVGISLMEILSKHSSDEVYLGQ--HDTPAWTSDAKALEAFKRFGAKLEGIEKQVVAMNSDP RICECL2 774 AVNFGQYDFGGYFPNRPSIARTVMPVEEPVDGAAMERFLDN---PDQALRECFPSQVQATVVMAVLDVLSSHSTDEEYLGG--EQTRPGTATRRLQAAYDGFAARLKEIEGVIDGRNKDR RICECML1 670 AVNFGQYDFGGYFPNRPSIARTVMPVEEPVDGAAMERFLDN---PDQRLRECFPSQVQATVVMAVLDVLSSHSTDEEYLGG--EQTRPWNSDAAVQAAYDGFAARLKEIEGVIDGRNKDR RICECML2 533 AVNFGQYDFGGYFPNRPSIARTVMPVEEPVDGAAMERFLDN---PDQALRECFPSQVQATVVMAVLDVLSTHSTDEEYLGG--EQTRPWNSDAAVQAAYAGFTARLKEIEGVIDGRNKDR RICEL2 713 AVNFGQYPYAGYLPNRPSVSRRAMPEPGTKE---YDELARD---PEKVFVRTITKQMQAIVGISLLEILSKHSSDEVYLGQ--RDTPEWTSDAKALEAFKRFGARLTEIESRVVAMNKDP RICEL3 713 AVNFGQYPYAGYLPNRPSVSRRAMPEPGTKE---YDELARD---PEKVFVRTITKQMQAIVGISLLEILSKHSSDEVYLGQ--RDTPEWTSDAKALEAFKRFGARLTEIESRVVAMNKDP WHTLOX1T 364 AVNFGQYSYAGYLPNRPTVSRRQMPEPGTKE---YKEVETD---PDLAFIHTITSQLQSIIGVSVIEVLSNHSSDEVYLGQ--RDEPKWTSDVKAKKAFEDFSQKLIDIEKRIMNMNANR ZEALOXL2 535 AVNFGQYPYAGYLPNRPTVSRRRMPEPGSKE---YEELERD---PERAFIHTITSQIQTIIGISLIEILSKHSSDEVYLGQ--RDTPEWTSDARALAAFKRFSDALVKIEGKVVGENRDP ZEALOX 712 AVNFGQYPYAGYLPNRPTVSRRRMPEPGSKE---YEELERD---PERGFIHTITSQIQTIIGISLIEILSKHSSDEVYLGQ--RDTPEWTSDARALAAFKRFSDALVKIEGKVVGENRDP ZEALOXJA 721 AVNFGQYPYAGYLPNRPSVSRKPMPAPGSDE---YAELERK---PEKVFVRTITSQFQALVGISLLEILSSHSSDEVYLGQ--RDTKEWTSDAKAQEAFKRFGARLTEIEKRVVTMNADP Consensus AVNFGQYPYAGY PNRP SRR MP G E---Y EL ---P F TITSQ Q G L E LS HSSDEVYLGQ--RDTP WTSDA AL AF RF RL EIE N D
*ALSLOX1 115 RLKNRSAAGIPPYELLLP-SS--GP----GVTGRGIPNSTSI ALSLOX2 113 GLKNRNGPVRVPYTLLYPNTSD-------------------- BLYLOXA 823 ELKNRNGPAKFPYMLLYPNTSDHKG-AAAGLTAKGIPNSISI BLYLOXB 836 ALKNRNGPVKMPYMLLYPNTSDANGEKALGLTAMGIPNSVSI BLYLOXC 825 QLKNRTGPAKFPYMLLYPNTSDHTG-QAEGLTARGIPNSISI RICECL2 890 KLKNRCGAGILPYQLMKP-FS--DS----GVTGMGIPNSTSI RICECML1 786 KLKNRCGAGILPYQLMKP-FS--DS----GVTGMGIPNSTSI RICECML2 649 KLKNRCGAGILPYQLMKP-FS--DA----GVTGMGIPNSTSI RICEL2 826 HRKNRVGPTNFPYTLLYPNTSDLKG-DAAGLSARGIPNSISI RICEL3 826 HRKNRVGPTNFPYTLLYPNTSDLKG-DAAGLSARGIPNSISI WHTLOX1T 477 QLKNRNGPAKFPYMLLYPNTSDIDGESATGITAKGIPNSISI ZEALOXL2 648 QLRNRNGPAEFPYMLLYPNTSDHSG-AAAGLTAKGIPNSISI ZEALOX 825 QLRNRNGPAEFPYMLLYPNTSDHSG-AAAGLTAKGIPNSISI ZEALOXJA 834 RLKNRNGPAEFPYTLLYPNTSDTKG-DAAGITAKGIPNSISI
CSG15 (1/00) 15
PC PE MGDG DGDG0
5
10
15
20S1S3S5S7
mol/gDW
ALSLOX1WHEATLOX
ALSLOX2
BLYLOXB
BLYLOXC
RICECL2
BLYLOXA
ZEALOXJA
RICEL3RICEL2
ZEALOXZEALOX2
RICECML2RICECML1
Fig 13 Sequence alignment of predicted amino acid sequences of ALSLOX1 and ALSLOX2 with similar regions of monocotyledonous lipoxygenase genes from barley: BLYLOXA (L35931), BLYLOXB (L37359), BLYLOXC (L37358), rice: RICECL2 (D14000), RICECML1 (AF09585), RICECML2 (AF095896), RICEL2 (X64396), RICEL3 (E03480), wheat: WHTLOX1T (U32428)and maize: ZEALOXL2 (AF149803), ZEALOX (AF329371), ZEALOXJA (AF271894). Accession numbers are shown in brackets. Symbols – star (*) = conserved residue believed to be involved in binding iron atom, plus sign (+) = highly conserved residue.

The biochemical basis of post-harvest quality in cut flowers HH2122TOFProject MAFFTitle project codeConsensus LKNR GP PY LLYPNTSD G- A G TA GIPNS SI
the same region of the gene in other species shows that the two Alstroemeria cDNAs share several highly conserved residues present in this region among plant lipoxygenases (Royo et al., 1996) that are believed to form part of the internal cavity of the enzyme that is likely to accommodate the fatty acid substrate. In addition an isoleucine residue and an asparagine residue are present within this region that are associated with binding of the iron atom essential for enzymatic activity of lipoxygenases.
Expression patterns of the two LOX genes from Alstroemeria were determined by semi-quantitative RT-PCR (Fig. 7 and Fig 14). LOX1 decreased markedly from S1 and by S3 contained over fourfold less transcript. LOX2 transcript accumulation peaked at S1 and again had decreased over four fold by S6 although some enhanced expression was evident at S7. In contrast tubulin and ubiquitin levels remained relatively constant during senescence with fluctuation being less than two fold over this period for either gene.
Fig 14: Quantitation of the signal from representative autoradiographs showing ALSLOX1 and ALSLOX2 expression using a phosphorimager screen and Quantity One image analysis (BioRad). The area of the peak (volume) was calculated for each stage (arbitrary units) and the corresponding background region in the –RT controls was subtracted for each stage.
(b) Lipoxygenase activity
The floral lox had a pH optima between 5.5-6.0 (Fig. 15a) and was inhibited, in a dose dependent manner, by esculetin (50% inhibition with 100uM) (Fig. 15b). The lox showed activity to a range unsaturated fatty acids although a preference for 18:2 over 18:3 was observed. The enzyme also readily used linolenic acid (C18:3 all cis 6,9,12) and arachidonic acid (C20:4 all cis 5,8,11,14) which are not endogenous fatty acids in the floral tissues (Fig. 15c). During the course of senescence the activity of lox declined rapidly by 50% from S1 to S3 and continued to decline to S7 (Fig. 16). This decline in activity was paralleled by a decrease in the level of lox protein detected in western blots (Fig. 16 insert).The lox antibody was raised in cucumber to a conserved region of the 9- and 13-lox and therefore detects both types of lox protein. The lox assay also measures total lox activity since it is based on conjugated diene formation (measured at 234nm) and does not distinguish between 9 and 13-lox activity either. In effect the total lox activity profile and the total protein profile are in agreement but do not indicate the specificity of the oxygenation site. We know from stereo and regio chemical analyses that both lox isoforms are present in Alstroemeria floral tissues Stereochemical profile of LOX derived hydroperoxy fatty acidsFatty acids either present in complex lipids or released as UFA by acyl hydrolase activity are prone to peroxidation by either chemical (autoxidation) or enzymatic (lox) processes. These two routes can be distinguished by the positional or stereochemical addition of oxygen to the acyl chain. Enzymatically, lox will catalyse the addition of oxygen to a specific C-atom of the unsaturation centres of the polyunsaturated fatty acids. For C18:2 (cis 9,12) this will be at either the C9 or C13 position and the enantiomeric form will be of the S-type. In autoxidation, molecular oxygen will add to both C9 and C13 positions and generate an almost equimolar ratio of R:S enantiomers. The major lipid oxidation products detected were derived from 13-lox activity acting on linoleic acid and linolenic acid esterified in complex lipids ie of enzymatic origin. Fatty acids, released from complex lipids by acyl hydrolase activity were also oxygenated again predominately at the 13C position although unlike complex lipids 9-lox products were also detected in this CSG15 (1/00) 16
lox2 volumes
800000
1000000
1200000
1400000
1600000
1800000
2000000
1 2 3 4 5 6 7
Stages
Volu
mes
lox1 volumes
0
50000
100000
150000
200000
250000
300000
1 2 3 4 5 6 7
stages
volu
mes
LALSLOX1 LALSLOX2

The biochemical basis of post-harvest quality in cut flowers HH2122TOFProject MAFFTitle project codefraction. This suggested that the 13-lox acts specifically on lipid linked fatty acid substrates whereas 9-lox activity is directed towards fatty acids released by acyl hydrolases. Thus two types of enzymatic activity were detected acting on different substrates. Interestingly high levels of keto fatty acid products were also detected (30% of total lox products). These are usually minor secondary products of lox activity accounting for 5% lox activity. In Alstroemeria tissues however their levels are exceptionally high. Whether this is a specific property of the lox requires further characterisation as does their potential biological role.
In the esterified lipid hydroperoxide fraction, the predominant positional isomers were the 13-oxy derivatives of C18:3 (all cis 9,12,15; 13-HOTE) and C18:2 (cis 9,12;13-HODE) and the enantiomeric form was >94% in the S-form indicating an enzymaticorigin. Small amounts of 16-HOTE, 9-HOTE and 9-HODE were also detected and were racemic indicating an autoxidative origin. In the unesterified fatty acid hydroperoxide pool the predominant positional isomer at S0, S2 and S4 were 13-HOTE (84 ± 2%) and 13-HODE (74 ± 4%) while their corresponding 9-oxy derivatives constituted 16 ± 2% (9-HOTE) and 26 ± 4 % (9-HODE), respectively. The 9-oxy derivatives were predominantly of the S-type (86-97 %) again indicating an enzymatic origin. Thus the majority of peroxidation in both tissues was of enzymatic origin, predominantly due to 13-lox activity. However some 9-lox activity (20% of total lox activity) was detected and some autoxidation was also detected in both sepals and petals. Autoxidation was higher in sepals than petals probably due in part to chemical photo-oxidation.
Figure 15: Characterisation of Alstroemeria lox activity, (A) pH optima, (B) inhibition of activity by esculetin and (C) fatty acid substrate specificity. Lox activity was monitored in 13,000g supernatants of petal extract by measuring the increase at A234, following the formation of conjugated dienes from fatty acid substrate. Both the pH optima and escultein inhibition analyses were performed using 18:2 as a substrate, all data points represent n=9±SD.
Figure 16: Total lox activity in petal tissue throughout senescence.
Lox activity was determined in 13,000g supernatants by following conjugated diene formation at 234nm, using
18:2 as a substrate. Data points represent n=9±SD. The insert shows a western blot probed with antibodies raised against a recombinant cucumber lipid body lox, results are representative of analyses repeated twice.
Endogenous oxidative damage in freshly harvested floral tissuesTo determine changes in hydroperoxy fatty acid content we developed a rapid and sensitive spectrophotometric method for detecting plant lipid hydroperoxides based upon hydroperoxide-mediated oxidation of ferrous to ferric ions under acidic conditions (Griffiths et al., 2000). Triphenylphosphine is use to authenticate the signal generated. In sepals lipid hydroperoxides accumulated but declined in petals (Fig. 17). Lipid hydroperoxide levels, detected using this technique, were substantially higher (2-3 fold) in sepals compared with petals.
Figure 17: Assessment of lipid peroxidation in senescing floral tissues. Lipid hydroperoxide content determined by the FOX assay.
CSG15 (1/00) 17
C
Substrate
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
1.6
B
Esculetin Concentration
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
LOX Control
Ethanol Control10 M
50 M
100 M200 M
A
pH5.0 5.5 6.0 6.5 7.0 7.5 8.0
mol
LH
PO fo
rmed
/min
/mg
prot
ien
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
mol
LH
PO fo
rmed
/min
/mg
prot
ien
mol
LH
PO fo
rmed
/min
/mg
prot
ien
18:2 18:3 18:3 20:4
S2 S3 S4 S5 S6 S7S1
S1 S2 S3 S4 S5 S6 S7

The biochemical basis of post-harvest quality in cut flowers HH2122TOFProject MAFFTitle project code
Antioxidant status and pigment content of floral tissues The differences in lipid oxidation between petals and sepals could be due to their differing total aqueous soluble antioxidants (TASA). 2,2-azino-di [3-ethylbenzthiazoline sulphonate]) (ABTS) was incubated with peroxidase (metmyoglobin) and hydrogen peroxide to generate the radical cation ABTS+, addition of tissue extracts to this mixture causes suppression of cation formation proportional to their concentration. No clear difference in the TASA content between the two tissue types was evident in stages S1 to S4. However, from S5 to S7, the level of antioxidants actually increased in sepals yet remained relatively constant in petals (data not shown). Thus increased lipid oxidation in sepals can not be explained by reduced levels of antioxidants.
The increase in sepal TASA is not due to an increase in anthocyanidins as the levels in petals is more than double that in the sepals at S3 and S4 and similarly carotenoids are more abundant in petals than sepals (Fig 18). In both tissues, chlorophyll content declines as the buds expand and open but the rate of loss is greater from the petals. The loss is over 50% of the maximum recorded by S4 in petals but in sepals over 70% still remains at S6 (Fig. 18). Thus marked differences exist in the content of the major pigments in the different floral tissues of Alstroemeria.
Figure 18. Antioxidant status and pigment content of senescing floral tissues. (A) carotenoid content and B) chlorophyll content throughout senescence. n=9±SD.
Petal extracts inhibit in vitro lipid peroxidationTo confirm that sepals have a lower potential to prevent lipid peroxidation tissue extracts were added to 13-HOTE or 13-HODE and the production of TBARS determined. 13-HOTE generated more TBARS than 13-HODE and showed a linear increase with concentration and was thus chosen for further investigation on in vitro lipid peroxidation using tissue extracts. In the absence of exogenous substrate, sepals generated over twice as much TBARS as petals (Fig. 19). Addition of increasing
CSG15 (1/00) 18
S2 S3 S4 S5 S6 S7S1
A
B

The biochemical basis of post-harvest quality in cut flowers HH2122TOFProject MAFFTitle project codeamounts of 13-HOTE resulted in an increase in TBAR production in sepal extracts but not in petals. These results indicate that extracts of petals are inherently less prone to lipid peroxidation than those prepared from sepals.
Figure 19: In vitro generation of TBARS. Generation of TBARS in whole tissue extracts by exogenously added HPOTE. Samples, (4 to 5 mg protein) , were spiked with 5-25 nmol/ml of HPODE just prior to grinding. The recovery of TBARS product was determined as described in materials and methods. Representative data are shown for experiments performed twice.
Isolation and characterisation of a partial Alstroemeria GST cDNAs and its expression during petal senescence.Degenerate primers were also designed to conserved regions of glutathione-S transferases (GSTs)and used to isolate a partial cDNA from Alstroemeria petals, ALSGST. Alignment of the sequence with plant GSTs confirmed the putative identity of the cDNA clone, which shows highest homology to an Arabidopsis GST (Fig. 20). Expression analysis using RT-PCR revealed some expression throughout the petal life span with some fluctuation (Fig. 7 and Fig 21). There was no clear trend although perhaps expression was higher in younger petals compared to old. A low level of PCR product was also detected in the -RT controls for this experiment, suggesting low levels of genomic DNA or plasmid contamination.
Figure 20: Alignment of predicted amino acid sequence of Alstroemeria GST with GST genes from other species: ATGST (Arabidopsis), EUGST (Euphorbia), ALSGST (Alstroemeria) NICGST (Nicotiana tabacum), ORYZAGST (rice), SIGGST (Silene), ZMGST (maize)
Figure 21: Quantitation of the signal from representative autoradiograph showing ALSGST using a phosphorimager screen and Quantity One image analysis (BioRad). The area of the peak was calculated for each stage (arbitrary units) and the corresponding background region in the –RT controls was subtracted for each stage.
4) Cell deathOne of the hall marks of programmed cell death is DNA fragmentation. This was investigated in two ways, by direct observation of DNA laddering and via the TUNEL assay that reveals cells in which DNA fragmentation has occurred via an enzyme-linked immunofluorescence assay, another indication of programmed cell death.
DNA laddering CSG15 (1/00) 19
V W M E V E A Q Q F D P V A S K L A F E L V F K P M - F G M T T D E A V V E E N E A K L A K V L D V Maj or i t y
10 20 30 40 50I G M Q V E D H Q F D P V A S K L A F E Q I F K S I - Y G L T T D E A V V A E E E A K L A K V L D V1 ATGST. PROV W L E V E A Q K F E P V A S K L N W E I V F K P M - F G M Q T D P A A V E E N E T K L A G V L D V1 EUGST. PROV W M E V E S Q Q F N P P I Y H I A Y Q T I F A S M - L G A S T D E A V V A T N A A K L G A V L G V1 GSTEF. PROV W M E V E G Q K F D P P A S K L T F E L G I K P I - I G M T T D D A A V K E S E E Q L S K V L D I1 NI CGST. PROV W L E V E S N Q Y T P A L N P I L F Q C L I R P M M F G A P P D E K V V E E N L E K L K K V L E V1 ORYZAGST. PROV W E E V E A H Q F D P V A S K L A W E L V F K G I - F G M Q T D T T V V E E N E A K L A K V L D V1 SI GST. PROV W I E V E A N Q Y T A A L N P I L F Q V L I S P M L - G G T T D Q K V V D E N L E K L K K V L E V1 ZMGST. PRO
Y E Maj or i t y
Y E 50 ATGST. PROY E 50 EUGST. PROY E 50 GSTEF. PROY E 50 NI CGST. PROY E 51 ORYZAGST. PROY E 50 SI GST. PROY E 50 ZMGST. PRO
Decor at i on ' Decor at i on #1' : Shade ( wi t h bl ack at 40% fi l l ) r esi dues t hat mat ch t he Consensus exact l y.
ALSGST.PRO
ALSGST.PRO

The biochemical basis of post-harvest quality in cut flowers HH2122TOFProject MAFFTitle project code
Genomic DNA was extracted from petals throughout their life span and separated by electrophoresis. To enhance the visualisation of any laddering, the electrophoresed DNA was transferred to a nylon membrane by Southern blotting and probed using a digest of the genomic DNA itself. Clear evidence of DNA laddering was obtained even by visualisation with ethidium bromide, but was clearer after the Southern blotting (Fig. 22).
Figure 22: DNA laddering. Genomic DNA extracted from petals throughout senescence, separated by agarose gel electrophoresis Southern blotted and probed with genomic DNA probe to enhance visualisation of laddering. Arrows show bands of DNA indicative of DNA laddering. This experiment was repeated twice with very similar results
TUNEL assaySeveral attempts were made to use this assay to detect DNA fragmentation however none was apparent. The nuclei were clearly labelled with propidium iodide so it would appear that the stain was able to permeate the cells. This assay has previously been shown to label fragmenting nuclei in a tobacco BY2 cell population in which cell death was induced by ethylene treatment in our hands so it would appear that in Alstroemeria petals this phenomenon is not present, or the technique required further optimisation for this species.
Isolation and characterisation of a partial Alstroemeria DAD-1 cDNA and its expression during petal senescence.Degenerate primers were designed by comparison to plant DAD-1 genes in the databases (Table 1) and used to isolate a partial DAD-1 cDNA from Alstroemeria petal tissue. Comparison of the sequence to DAD-1 genes from other species (Fig.23) confirms the putative identity of this cDNA. All the plant DAD-1 genes show very close homology at the amino acid level (Fig. 23).
Figure 23: Alignment of predicted amino acid sequence of ALSDAD1 with DAD1 genes from other plant speciesTOMDAD1 (tomato), ATHDAD1(Arabidopsis) , HORDDAD1 (barley), MAIZEDAD1 (maize) showing high degree of conservation across plant species
TOMDAD1 1 MAKSSATKDAQALFHSLRSAYAATPTNLKIIDLYVIFAISTALIQVVYMAIVGSFPFNSFLSGVLSCIGTAVLAVCLRIQVNKENKEFKDLPPERAFADFVLCNLVLHLVIMNFLG ATHDAD1 1 MVKSTS-KDAQDLFRSLRSAYSATPTNLKIIDLYVVFAVFTALIQVVYMALVGSFPFNSFLSGVLSCIGTAVLAVCLRIQVNKENKEFKDLAPERAFADFVLYNLVLHLVIINFLG HORDDAD1 1 MPKAAG--DAKLLIQSLNKAYAATPTNLKIIDLYVVFAVVTALLQVVYMGIVGSFPFNSFLSGVLSCIGTAVLAVCHRIQVNKDNKEFKDLAPERAFADFVLCSLVLHLVIMNFLG MAIZEDAD1 1 -------------------------------------AVATALIQVAYMGLVGSFPFNSFLSGVLSCIGTAVLAVCLRIQVNKDNKEFKDLPPERAFADFVLCNLVLHLVIMNFLG ALSDAD1 1 ---------------------------------------------------IGSFPFNSFLSGVLSCVGTAILAVCLRIQVNKENKDFKDLAPERAFAVSSYESL----------- CONSENSUS 1 M K --DA L SL AY ATPTNLKIIDLYV FAV TALIQVVYM VGSFPFNSFLSGVLSCIGTAVLAVCLRIQVNKENKEFKDLAPERAFADFVLCNLVLHLVIMNFLG
Expression analysis, performed as above, revealed an interesting pattern of expression. Early in petal development (stages 0-4) there was significant expression which then tailed off as the petals began to senesce with expression falling almost ten-fold by stage 6 (Fig. 7 and Fig 24) although there is a small rise at stage 7 which may indicate levels fluctuate towards the end of senescence.
Figure 24 Quantitation of the signal from representative autoradiograph showing ALSDAD1 expression using a phosphorimager screen and Quantity One image analysis (BioRad). The area of the peak (volume) was calculated for each stage (arbitrary units) and the corresponding background region in the –RT controls was subtracted for each stage.
Discussion and implicationsPhysiological markers of senescence in AlstroemeriaOur initial choice of species was based on reports that Alstroemeria is insensitive to the plant growth regulator ethylene (Woltering and Van Doorn, 1988). However little was known about the precise timing of senescence indicators or how these relate to biochemical and molecular events. Initial studies have defined seven stages in the development and senscence of Alstroemeria
CSG15 (1/00) 20
Days relative to flower opening +2 +5 +7 +9 +11 +13
ALSDAD1DAD1 28B
0
500
1000
1500
2000
2500
3000
1 2 3 4 5 6 7
Stage
peak
are
a
ALSDAD1

The biochemical basis of post-harvest quality in cut flowers HH2122TOFProject MAFFTitle project code(Wagstaff et al., 2001) based on morphological features. During this detailed work we also re-evaluated the role of ethylene and concluded that the initial reports were not entirely accurate. Our results (See Annex IV) suggest that although this species produces very little ethylene and its senescence can only be delayed relatively little by ethylene inhibitors, abscission of the petals is accelerated by exogenous ethylene and in fact Alstroemeria is very sensitive to applications of exogenous ethylene. This is an important finding in practical terms as Alstroemeria is often used in mixed bouquets. It would thus be desirable to limit the production of ethylene by other components of bouquets for achieving the longest vase life of Alstroemeria flowers. However it does not in our view affect the use of this species for the study of ethylene-independent senescence: our findings confirm reports in the literature that ethylene is not involved in endogenously controlling senescence in this species. Thus Alstroemeria still provides a good model in which to study the control and progression of senescence which is not coordinated by ethylene.
Another interesting finding, from work carried out as part of a parallel independently funded project, was that the lifespan could be increased significantly by pre-treating with STS and combinations of trimming and/or supplying sugars. (Full details are available in Chasanut, 2001. The findings are summarised in Annex VI and mentioned here only as they complement some of the physiological studies performed in the MAFF-funded project). Some of these procedures are already used in the industry for other species, however, these simple treatments are not used universally for improving vase life of Alstroemeria in the UK. As predicted from studies in other species, the petal fresh weight declined gradually over the lifespan of the flower, and no clear diagnostic or predictive features could be derived from such simple measurements.
Proteolysis during senescenceProtein degradation in Alstroemeria petals was measured throughout their lifespan. We were interested to find that proteolytic activity was rising well before any visible signs of petal deterioration. This was paralleled by a steady increase in the expression of at least one cysteine protease gene. This suggests to us that the machinery for executing the high levels of protein degradation that are seen at the end of the vase life are already in place very early. Activity of the proteases may be controlled during early floral development via compartmentalisation, post-transcriptional or post-translational regulatory systems. Thus genes controlling proteolysis and potential predictors of lifespan may need to be sought much earlier than one might have anticipated from the visible physiological changes taking place. Another important finding was that the majority of the proteolytic activity appears to be due to the action of cysteine proteases. Zymography in the presence of specific inhibitors of different protease classes did not reveal any other types of proteases,
although these might be present at levels below the sensitivity of this system or require a different assay system for their detection. Cysteine proteases have been implicated in senescence processes in a number of plant species both in leaves and flowers (Buchanan-Wollaston, 1997; Valpuesta et al., 1995). A major interest in this class of proteases derives also from the extensive work on cell death in animals which has revealed a complex cascade of proteases known as caspases which represent a particular type of cysteine protease. Homologues of caspases have so far not been found in plants thus there is speculation that if a similar proteolytic cascade control mechanism operates in plants other forms of cysteine protease may be fulfilling this role. Attempts were made to obtain full-length clones for the Alstroemeria cysteine protease which we isolated. This would be a necessary step for further experiments to confirm proteolytic activity and potential substrates, however, time restrictions did not allow us to pursue this further.
The role of ubiquitin seemed another interesting area to investigate as ubiquitination is known to be a mechanism for targeting specific proteins for degradation (Wilkinson, 1995) and thus has the potential for acting as a regulatory mechanism. It also has the potential for providing useful markers of the progress of senescence. However, our preliminary investigations of gene expression and levels of protein ubiquitination lead us to conclude that perhaps ubiquitination is not a major player in Alstroemeria petal senescence. This is in contrast to results from related species (e.g. Hemerocallis) where ubiquitination of specific proteins was seen to change during senescence (Courtney et al., 1994). The methods generally used for this type of assay are of course relatively crude and it is still possible that regulatory proteins, which are present at levels below the detection of western blotting, might be regulated in turn by ubiquitination and be important in the control of senescence in Alstroemeria. A more detailed study was, however, beyond the scope of this project.
Membrane integrity and lipid metabolism Lipid catabolism was extensive in Alstroemeria floral tissues resulting in over 80% depletion of phospholipids and galactolipids during senescence, one possibility is that these are mobilised and utilized in the development of other tissues. Although complex lipids were degraded, the level of neutral lipids remained fairly constant throughout senescence, unlike in other senescing tissues where increases in neutral lipid biosynthesis have been reported with consequential damage to the biophysical properties of membranes (Borochov et al., 1997). Lipid peroxidation is linked with senescence although whether it is a primary event that initiates senescence or is a consequence of senescence remains unclear. Lipid peroxidation is usually measured by determining MDA, hydrocarbon gases or decrease in PUFA, the most widely used is the TBAR assay which measures malondialdehydes (3 carbon fragments) derived from C18-PUFA. However, MDA can only be formed from fatty acids with three or more double bonds (Halliwell and Gutteridge, 1989) but plant tissues often contain high levels of 18:2 ( cis ,9,12), therefore the TBA assay may underestimate the actual extent of CSG15 (1/00) 21

The biochemical basis of post-harvest quality in cut flowers HH2122TOFProject MAFFTitle project codeperoxidation. In Alstroemeria floral tissues, 18:2 ( cis,9,12) is the predominant acyl constituent and to accurately estimate the extent of peroxidation we developed a new method to detect primary oxygenated fatty acid metabolites, moreover, the levels of these compounds are more likely to be physiologically relevant than MDAs (Griffiths et al., 2000). The LHPO accumulation pattern differed between sepals and petals even though they had similar fatty acid and complex lipid profiles. This difference was not related to differences in lox enzymatic activity, which decreased from S1 in both tissue types. Stereochemical analysis of lipid oxidation products revealed that 13-oxy fatty acid isomers were exclusively detected in complex lipids. Whether 13-lox acts directly on complex lipids in situ or whether UFAs were oxygenated and selectively transferred to the complex lipids by acyltransferase activity was not investigated. Analysis of the oxygenated UFA, however, revealed that a 9-lox was also present which accounted for some 20% of the total lox activity present in both tissues. These 9-lox products were not detected in complex lipids. The detection of 2 lox activities in the tissues agrees with detection of two lox gene products during senescence namely LOX 1 and LOX 2. Although no specific assignment of enzymatic activity can be given to these gene products, LOX 1 expression would be consistent with the marked decrease in 13-lox activity (the major activity in the floral tissue). LOX 2 expression, although not abolished at S6, could be related to a minor lox activity which can not be detected in extracts from that stage of senescence in activity determinations.
Perhaps the most surprising result was the difference in the pattern of LHPOs in the two tissues yet despite this both tissues exhibited increased electrolyte leakage at the same developmental time point. In sepals and petals the basal level of oxidation of the cell membrane was 1% of total fatty acids similar to that recently calculated for a wide range of plant tissues (Griffiths et al.,
2000). In sepals this level rose to 3% during the course of senescence whereas it remained constant at <1% in petal tissues. The degradation of LHPO by tissue extracts of Alstroemeria petals and sepals was similar and the activity was maintained throughout senescence (data not shown). The observations that lox activity declines but LHPO degradation is maintained in both sepal and petal tissues indicates that the increase LHPO in sepals is the result of non-enzyme catalyzed reactions relating to autoxidation.
The products of lipid autoxidation are complex and include a range of aldehydes (Kohlmann et al., 1999) which are strongly inhibitory to various SH dependent enzyme reactions, inhibit DNA, RNA and protein synthesis and are acutely cytotoxic (Esterbauer and Zollner 1989). In this regard, suppression of hydroperoxide lyase activity in transgenic potato resulted in increased generation of autoxidation volatile products (Vancanneyt et al., 2001) but did not lead to induction of visible leaf senescence symptoms despite an almost 2-fold increase in endogenous LHPO levels (Griffiths et al., 2000).
In non-senescent tissues autoxidation is low and controlled by antioxidant components within the cells. These include the enzymes of the ascorbic acid cycle and other detoxifying enzymes that remove potentially harmful reactive oxygen species (ROS) such as SOD and catalase. Pigments such as anthocyanins and carotenioids are also free radical scavengers and limit ROS propagation within plant cells. The level of total aqueous-soluble antioxidants (TASA) increased in sepals coincident with the increase in LHPO levels. Thus, TASA (including anthocyanidins) appear ineffective in preventing oxidative damage to the lipid components. However, the level of carotenoids was almost double in the petals and during the course of senescence the level of chlorophyll also declined more rapidly in petals. Thus during the course of senescence the ratio of chlorophyll/carotenoid increased in sepals and decreased in petals. Light absorption by chlorophyll can initiate the formation of free radicals in sensitised photooxidation (Chan and Cotton, 1987). The ratio of chlorophyll/carotenoid would favour this in sepal tissues and could therefore be a contributory factor in generating the higher levels of LHPO seen in sepal tissues.
A survey of the different types of floral senescence patterns observed in the plant kingdom indicate that two categories can be defined on the basis ethylene sensitivity or insensitivity. In the ethylene sensitive category, lox activity may play a positive role in promoting senescence through oxidative membrane damage as seen in rose, carnation, petunia and Ipomea. In rose petals (ethylene sensitive), a lipoxygenase has been isolated using tomato lox cDNA fragments as probes. Northern blot analysis showed that the transcript was dramatically increased during senescence and its expression was stimulated by ethylene (Fukuchi-Mizutani et al., 2000). On the other hand, ethylene sensitive plants such as Phalaenopsis and Dendrobium (Orchidaceae) have been characterised in which lox does not play any apparent role in this process (Porat et al., 1995). In the ethylene insensitive category, lox promotion of senescence has been identified in daylily. Petals showed a 5-fold increase in TBAR from the time of opening to the point of electrolyte leakage and the activity of lox, which increased 2-fold, mirrored the increase in TBAR production. mRNA levels of AOS were also upregulated suggesting the possible involvement of downstream oxylipins (including JA) in the promotion of senescence in this species. In Alstroemeria, floral tissues however, lox activity had declined by some 80% at the point of electrolyte leakage initiation and the accumulation of oxidised lipids (LHPOs and TBARS) was not coincident with electrolyte leakage indicating an unlikely role for lox in perturbing membrane function in this species. The loss of membrane semi-permeability most likely results from the loss of bulk lipids and proteins (Wagstaff et al., 2001) which occurs during senescence. Thus, Alstroemeria represents a distinct pattern of floral senescence which is both ethylene and lox-independent.
Cell death processesThe end point of senescence is clearly the death of cells and since senescence itself is a tightly regulated process we were interested to investigate when cell death is initiated in Alstroemeria petals and whether this could be linked to other physiological biochemical and molecular markers. We chose three markers of cell death. Two of these are linked to DNA degradation: TUNEL
CSG15 (1/00) 22

The biochemical basis of post-harvest quality in cut flowers HH2122TOFProject MAFFTitle project codeand DNA laddering and both measure the extent to which DNA degradation has occurred. Although TUNEL has been used successfully in our laboratory in other systems, we were unable to obtain convincing data using this technique with Alstroemeria. However, DNA laddering was very evident even from early stages in floral development and became increasingly apparent with petal age. This finding is in agreement with our results from the protein degradation suggesting that senescence processes may be initiated considerably earlier than we anticipated. We also isolated the Alstroemeria homologue to a gene, which has been identified as a marker of programmed cell death (PCD) processes in both plant and animal systems (Nakashima et al., 1993; Orzaez and Granell, 1997): DAD1. Although recent literature (Kelleher and Gilmore, 1997) concludes that this gene is probably not part of the cell death machinery itself, it is down-regulated in cells which have entered into PCD. The expression pattern of this gene in Alstroemeria petals was consistent with this role. Moreover, this gene appears to decrease sharply in expression in stage 4 petals rather than changing expression gradually as was seen in some of the other genes we isolated. This is significant as this may provide a useful marker for flowers that have reached this terminal stage in the senescence process.
Future workDuring this project we chose to target a small number of processes which were already known to be implicated in floral and or leaf senescence. Our approach was focussed on understanding the underlying scientific principles behind floral senescence in Alstroemeria. We are now well equipped with detailed physiological data on the progression of senescence and a much better understanding of the role of ethylene and simple treatments on floral longevity in this species. Three approaches to future studies on ethylene insensitive cut flowers are evident to us:
1. With recent advances in genomic technologies including specifically the microarray technology, opportunities now arise to take a much broader approach to the molecular events accompanying and controlling floral senescence. This may lead to fundamental insights into how senescence is controlled in species where ethylene appears not to be a major regulator. It also has potential for the provision of a suite of molecular markers for predicting the progression of senescence. Many of the genes we isolated showed a gradual change in expression through development and senescence, although for example DAD1 showed a sharper change. We suggest that diagnostics of practical use to the industry may consist of a comparative approach using a suite of genes and assessing threshold levels of each gene to determine the senescence status of the flower. This might be possible through future developments of microarray technology to make it more accessible to the industry.
2. One of the major findings of our work was that some of the processes activated during petal senescence may already be in place much earlier while the petals are still developing. This is significant as it indicates that it may be possible to develop diagnostic approaches for flowers at the stage of picking as well as later in their shelf life. It also suggests to us that it may be important to study the effect of treatments during packaging and shipping of cut flowers and the effect these have on gene expression. If senescence is already initiated very early, then sub-optimal storage prior to sale may have an important effect in accelerating the onset of physiological changes, which result in poor quality for the consumer. Thus a detailed study of the effects on storage and handling, ideally making use of post-genomic technologies to survey a broad range of genes, may provide further important information both on key genes affecting the process and also on the effects of treatments cut-flower quality.
3. Although the focus of our work has been on Alstroemeria this is just one of a number of important cut flower species which are classed as ethylene insensitive. An important focus of future work will be to assess the extent to which parallels can be drawn between the biochemical and molecular events associated with petal senescence in Alstroemeria and processes in related species such as other lilies. Again a broader approach may be useful in providing a suite of genes which can be compared. Broader comparisons to ethylene dependent species may also reveal common features to all types of floral senescence, which would be both of fundamental and practical significance. Such comparisons are facilitated e.g. by work in other laboratories producing micoraarrays for ethylene sensitive species such as petunia and carnation.
ReferencesBligh EG and Dyer WJ. 1959. A rapid method of total lipid extraction and purification. Can J Biochem and Physiol. 37: 911-917Borochov A, Spielgelstein H, Philosoph-Hada S. 1997. Ethylene and flower petal senescence: Interrelationship with membrane lipid catabolism. Physiol Plant. 100: 606-612Buchanan-Wollaston V. 1997. The molecular biology of leaf senescence. Journal of Experimental Botany. 48: 181-199.Callis J. 1995. Regulation of protein degradation. The Plant Cell. 7: 845-857.Chan HW-S, Levett G. 1977. Autoxidation of methyl linoleate. Separation and analysis of isomeric mixtures of methyl linoleate hydroperoxides and methyl hydroxylinoleates. Lipids 12: 99-104Chan HW-S, Cotton DT. 1987. Lipid hydroperoxides. In: Chan H W-S, ed. Autoxidation of unsaturated lipids. Academic Press, UK, 17-50.Courtney SE, Rider CC and Stead AD. 1994. Changes in protein ubiquitination and the expression of ubiquitin-encoding transcripts in daylily petals during floral development and senescence. Physiologia Plantarum. 91: 196-204.Eason JR and de Vre L. 1995. Ethylene-insensitive floral senescence in Sandersonia aurantiaca (Hook). New Zealand Journal of Crop and Horticultural Science. 23: 447-454.Esterbauer H, Zollner H. 1989. Methods for determination of aldehydic lipid peroxidation products. Free Radical Biol Med. 7: 197-203.Feinberg AP and Vogelstein B. 1983. A technique for radiolabeling DNA restriction endonuclease fragments to high specific activity. Analytical Biochemistry. 132: 6-13.Fobel M, Lynch DV, Thompson JE 1987 Membrane deterioration in senescing carnation flowers. Plant Physiol 85: 204-211CSG15 (1/00) 23

The biochemical basis of post-harvest quality in cut flowers HH2122TOFProject MAFFTitle project codeFukuchi-Mizutani M, Ishiguro K, Nakayama T, Utsunomiya Y, Tanaka Y, Kusumi T, Ueda T (2000) Molecular and functional characterization of a rose lipoxygenase cDNA related to flower senescence. Plant Sci 160: 129-137Gardner HW. 1997. Analysis of plant lipoxygenase metabolites. In: Christie, WW, ed. Advances in lipid methodology-Four. The Oily Press, UK, 1-43. Griffiths G, Jones HE, Eaton CL, Stobart AK. 1997. Effect of n-6 polyunsaturated fatty acids on growth and lipid composition of neoplastic and non-neoplastic canine prostate epithelial cell cultures. The Prostate 31, 29-36.Griffiths G, Leverentz M, Silkowski, H, Gill, N, Sanchez-Serrano JJ. 2000. Lipid hydroperoxide levels in plant tissues. J Exp Bot. 51: 1363-1370Guerrero C, de la Calle M, Reid MS and Valpuesta V. 1998. Analysis of the expression of two thiolprotease genes from daylily (Hemerocallis spp.) during flower senescence. Plant Molecular Biology. 36: 565-571.Gutteridge JMC. 1982. Free-radical damage to lipids, amino acids, carbohydrates and nucleic acids determined by thiobarbituric acid reactivity. Int. J. Biochem 14:649-563.Halliwell B, Gutteridge JMC. 1989. Free radicals in Biology and Medicine, Clarendon Press, Oxford: 188-275Haas AL and Bright PM. 1985. The immunochemical detection and quantitation of intracellular ubiquitin-protein conjugates. J. Biol. Chem. 260: 12464-12473. Hershko A, Eytan E, Ciechanover A and Haas AL. 1982. Immunochemical analysis of the turnover of ubiquitin-protein conjugates in intact cells. Relationship to the breakdown of abnormal proteins. J. Biol. Chem. 257: 13964-13970.Hodges DM, Delong JM, Forney CF, Prange RK. 1999. Improving the thiobarbituric acid-reactive-substances assay for estimating lipid peroxidation in plant tissues containing anthocyanin and other interfering compounds. Planta 207: 604-611
Itzhaki, H. and Woodson, W.R. 1993. Characterization of an ethylene-responsive glutathione-s-transferase gene-cluster in carnation. Plant Mol. Biol. 22: 43-58.Jones ML, Larsen PB and Woodson WR. 1995. Ethylene-regulated expression of a carnation cysteine proteinase during flower petal senescence. Plant Molecular Biology. 28: 505-512.Kambouris NG, Burke DJ and Creutz CE. 1992. Cloning and characterisation of a cysteine proteinase from Saccharomyces cerevisiae. Journal of Biological Chemistry. 267: 21570-21576.Kamphuis IG, Drenth J and Baker EN. 1985. Thiolproteases. Comparitive studies based on the high-resolution structures of papain and actinidin, and on amino acid sequence from cathepsins B and H, and stem bromelain. Journal of Molecular Biology. 182: 317-329.Kelleher DJand Gilmore R, 1997 DAD1 the defender against apoptotic cell death, is a subunit of the mammalian oligosaccaryltransferase. Proc. Natl. Acad. Sci. USA 94: 4994-4999.Koehler S and Ho T-HD. 1990. A major gibberellic acid-induced barley aleurone cysteine proteinase which digests hordein. Purification and characterization. Plant Physiology. 94: 251-258.Koch E, Meier BM, Eiben, HG, Sulsarenko A. 1992. A Lipoxygenase from leaves of tomato (Lycopersicon esculentum Mill.) is induced in response to plant pathogenic pseudomonads. Plant Physiol 99: 571-576Kohlmann M, Bachmann A, Weichert H, Kolbe A, Balkenhohl, Wasternack C, Feussner I 1999. Formation of lipoxygenase-pathway-derived aldehydes in barley leaves upon methyl jasmonate treatment. Eur J Biochem 260: 885-895Lay-Yee M, Reid MS and Stead AD. 1992. Flower senescence in daylily (Hemerocallis). Physiologia Plantarum. 86: 308-314.Lesham, YY. 1992. Plant Membranes: A biophysical approach to structure, development and senescence. Kluwer Academic Publishers.Light A, Frater R, Kimmel JR and Smith EL. 1964. Current status of the structure of papain: the linear sequence, active sulfhydryl group, and the disulfide bridges. Proceedings of the National Academy of Science USA. 52: 1276-1283.Lovell PJ, Lovell PH, Nichols R 1987 The control of flower senescence in petunia (Petunia-hybrida) Annals of Botany 60: 49-59.Marrs, K.A. 1996. The functions and regulation of glutathione S-transferases in plants. Ann. Rev. Plant Physiol. Plant Mol. Biol. 47: 127-58.Mitchel REJ, Chaiken IM and Smith EM. 1970. The complete amino acid sequence of papain. Journal of Biological Chemistry. 245: 3485-3492.Mittler, R. and Lam, E. 1995. In-situ detection of nDNA fragmentation during the differentiation of tracheary elements in higher-plants. Plant Physiol., 108: 489-493Nakashima T, Sekiguchi T, Kuraoka A, Fukushima K, Shibata Y, Komiyama S, Nishimoto T. 1993. Molecular-cloning of a human cDNA-encoding a novel protein, DAD1, whose defect causes apoptotic cell-death in hamster BHK-21-cells. Molecular and Cellular Biology 13: 6367-6374 Nieri B, Canino S, Versace R and Alpi A. 1998. Purification and characterisation of an endoprotease from alfalfa senescent leaves. Phytochemistry. 49: 643-649.Nourooz-Zadeh J, Tajaddini-Sarmadi J, Birlouez-Aragon I, Wolff SP 1995. Measurement of hydroperoxides in edible oils using the ferrous oxidation in xylenol orange assay. J Agric Food Chem 43: 17-21Orzaez, D. and Granell, A. 1997. The plant homologue of the defender against apoptotic death gene is down-regulated during senescence of flower petals. FEBS Letters, 404: 275-278.Panavas T, Rubinstein B 1998 Oxidative events during programmed cell death of daylily (Hemerocallis Hybrid) petals. Plant Sci 133: 125-138Porat R, Reiss N, Atzorn, Halevy AH, Borochov A. 1995. Examination of the possible involvement of lipoxygenase and jasmonates in pollintion-induced senescence of Phalaenopsis and Dendrobium orchid flowers. Physiol Plant 94: 205-210Rogers H.J., Greenland A., Hussey P.J. 1993. Four members of the maize ß-tubulin gene family are expressed in the male gametophyte. The Plant Journal 4: 875-882.Royo J, Vancanneyt G, Perez AG, Sanz C, Stormann K, Rosahl S and Sanchez-Serrano JJ. 1996. Characterisation of three potato lipoxygenases with distinct enzymatic activities and different organ-specific and wound-regulated expression patterns. J. Biol. Chem. 271: 21012-21019.Smith PK, Krohn RI, Hermanson GT, Mallia AK, Gartner FH, Provenzano MD, Fujimoto EK, Goeke NM, Olson BJ, Klenk DC. 1985. Measurement of protein using bicinchoninic acid. Analytical Biochemistry. 150: 76-85.Stead AD and Moore KG. 1979. Flower development and senescence in Digitalis purpurea L cv. Foxy. Annals of Botany. 41: 283-292.
CSG15 (1/00) 24

The biochemical basis of post-harvest quality in cut flowers HH2122TOFProject MAFFTitle project codeStephenson P and Rubinstein B. 1998. Characterisation of proteolytic activity during senescence in daylilies. Physiologia Plantarum. 104: 463-473.Storer AC and Menard R. 1994. Catalytic mechanism in papain family of cysteine peptidases. Methods in Enzymology. 244: 486-511.Thompson JE, Froese CD, Madey E, Smith MD, Hong YW. 1998. Lipid metabolism during plant senescence. Prog Lipid Res 37: 119-141.Valpuesta V, Lange NE, Guerrero C and Reid MS. 1995. Up-regulation of a cysteine protease accompanies the ethylene-insensitive senescence of daylily (Hemerocallis) flowers. Plant Molecular Biology. 28: 575-582.Vancanneyt G, Sanz C, Farmaki T, Paneque M, Ortego F, Castanera P, Sanchez-Serrano JJ. 2001. Proc Natl Acad Sci USA (in press)Wagstaff C, Rogers HJ, Leverentz MK, Thomas B, Chanasut U and Stead AD. 2001. Characterisation of Alstroemeria flower vase life. Acta Horticulturae. 543: 161-175.Wilkinson KD. 1995. Roles of ubiquitination in proteolysis and cellular regulation. Annual Review of Nutrition. 15: 161-189.Woltering EJ and van Doorn WG. 1988. Regulation of ethylene in senescence of petals – morphological and taxonomic relationships. Journal of Experimental Botany. 39: 1605-1616.Zhuang,H., Hildebrand, D.F. and Barth, M.M. 1995. Senescence of Broccoli buds is related to changes in lipid peroxidation. J. Agr. Food Chem. 43: 2585-2591.
Action resulting from research (e.g. IP)
The ability to assess floral quality and predict floral longevity has long been a goal of the floral industry, the realisation that the expression patterns of some genes may be indicative of both these important commercial parameters offers the chance to develop diagnostic tools of great interest to the industry.
The development of these techniques is the subject of a follow on project funded by DEFRA and is also addressed in a further grant applications (BBSRC – ‘Exploiting Genomics)
Annex of Published papers/presentations
Papers (full text and figures attached):Characterisation of Alstroemeria flower vase life C. Wagstaff, H.J. Rogers, M. K. Leverentz, G. Griffiths, B. Thomas, U. Chanasut A.D. Stead (2001) Acta Horticulturae 543: 161-175 (Annex I)
Cysteine protease gene expression and proteolytic activity during senescence of Alstroemeria petals.Carol Wagstaff, Michael K. Leverentz, Gareth Griffiths, Brian Thomas, Usawadee Chanasut, Anthony D. Stead and Hilary J. Rogers Journal of Experimental Botany (in press) (Annex II)
Characterisation of a novel lipoxygenase-independent senescence mechanism in ethylene insensitive Alstroemeria floral tissueMichael K Leverentz, Carol Wagstaff, Hilary J Rogers, Antony D Stead, Helena Silkowski, Usawadee Chanasut Brian Thomas, Ivo Feussner and Gareth Griffiths (To be submitted to Plant Physiology ) (Annex III)
Presentations:Characterisation of Alstroemeria vase lifeC. Wagstaff , H.J. Rogers, U. Chanasut and A.D. SteadPoster presented at the Seventh International Symposium on Postharvest Physiology of Ornamental Plants, Ft. Lauderdale, Florida 13-18 Nov 1999
Changes in protease activity and gene expression during Alstroemeria petal senescenceC. Wagstaff, H.J. Rogers, U. Chanasut and A.D. Stead
Poster and short talk presented at the Gordon Research Conference on Plant Senescence, Abscission and Programmed Cell Death, Plymouth New Hampshire, 16-21 July 2000
Lipid peroxidation and lipoxygenase activity during floral senescence in Peruvian lily (Alstroemeria peruviana), M. K. Leverentz, H. Rogers, C. Wagstaff, H. Silkowski, A Stead, B. Thomas & G. Griffiths
CSG15 (1/00) 25
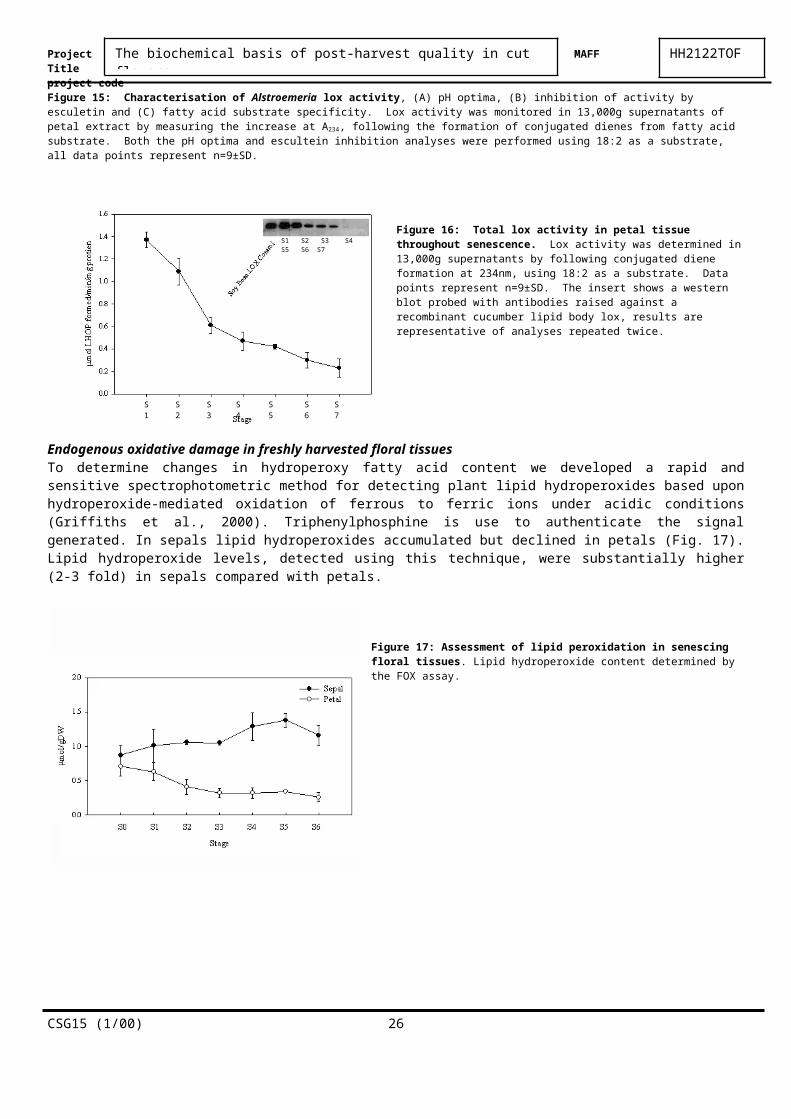
The biochemical basis of post-harvest quality in cut flowers HH2122TOFProject MAFFTitle project codePoster and short talk presented at the Gordon Research Conference on Plant Senescence, Abscission and Programmed Cell Death, Plymouth New Hampshire, 16-21 July 2000
Reports on Allied workReport on: Determination of ethylene production from cut Alstroemeria flowers (Annex IV)Report on: Determination of ethane production from cut Alstroemeria flowers (Annex V)Abstract of thesis: Post-Harvest Physiology of Alstroemeria Var. Rebecca Cut Flowers. Usawadee Chanasut.PhD Thesis University of London, 2001. (Annex VI)
CSG15 (1/00) 26