mitochondrial-dependent apoptosis in experimental rodent abdominal aortic aneurysms
TRANSCRIPT

8
Mitochondrial-dependent apoptosisin experimental rodent abdominalaortic aneurysmsIndranil Sinha, BA,a Amiya P. Sinha-Hikim, PhD,b Kevin K. Hannawa, BS,a Peter K. Henke, MD,a
Matthew J. Eagleton, MD,a James C. Stanley, MD,a and Gilbert R. Upchurch Jr, MD,a Ann Arbor, Mich,and Torrance, Calif
Objectives. While extrinsic mechanisms of apoptosis in abdominal aortic aneurysms (AAAs) arerecognized, this project hypothesizes that an intrinsic, mitochondrial-dependent, mechanism of apoptosisalso contributes to experimental AAA formation.Methods. Rat aortas were perfused with either saline or elastase (N = 5 per group) and harvested 7 dayspostperfusion. The aortas were placed in gluteraldehyde for subsequent transmission electron microscopy,Bouin’s solution for TUNEL, or paraformaldehyde for immunohistochemical staining for caspase-9,caspase-3, and Bid.Results. Abdominal aortic diameters increased 168 ± 25% (mean ± SEM) after elastase perfusion.compared with 30 ± 5% after saline perfusion (P < .001). Apoptosis of aortic smooth muscle cells,macrophages, and neutrophils was evidenced by transmission electron microscopy and TUNEL in theelastase-perfused aneurysmal aortas. Quantitative analysis of the apoptotic cells revealed a significant(P < .01) increase in the number of total apoptotic cells in the elastase-perfused aortas (12 ± 3 cells perhigh-power field), compared with that of saline-infused controls (1.3 ± 0.2). Caspase-9, the key initiatorin the mitochondrial-dependent apoptotic pathway, stained positively in only elastase-perfused aortas.Bid staining was not detected in either the elastase-perfused aortas or the saline controls.Conclusions. Apoptosis is evident in multiple cell lines in elastase-perfused aneurysmal aortas, but rarelyobserved in control aortas. Caspase-9, the key initiator of intrinsic apoptosis, was documented only inelastase-perfused aortas. These results suggest that mitochondrial-dependent apoptosis is associated withabdominal aortic aneurysm formation. (Surgery 2005;138:806-11.)
From the Jobst Vascular Research Laboratories, Section of Vascular Surgery, Department of Surgery, Universityof Michigan,a Ann Arbor, and the Los Angeles Biomedical Institute at Harbor-UCLA,b Torrance, Calif
ABDOMINAL AORTIC ANEURYSM (AAA) pathogenesis iscomplex and multifactorial. It is characterized bydestruction of the extracellular matrix (ECM), pri-marily the structural proteins elastin and colla-gen.1 The destruction of the ECM, primarily inthe media and adventitia, leads to subsequent aor-tic dilatation and life-threatening complications,
Presented at the 66th Annual Meeting of the Society of Univer-sity Surgeons, Nashville, Tennessee, February 9-12, 2005.
Supported by grant KO8 (HL67885-02) from the National Insti-tutes of Health (G.R.U.), the VonLeibig Award-Lifeline Founda-tion (G.R.U.), the Lifeline Medical Student Research Award(I.S. and K.H.), the Griswold and Margery H. Ruth AlphaOmega Alpha Medical Student Research Fellowship (I.S.), andthe Jobst Foundation.
Reprint requests: Gilbert R. Upchurch Jr, MD, University ofMichigan Hospital, 2210 THCC, 1500 East Medical Center Dr,Ann Arbor, MI 48109-0329. E-mail: [email protected].
0039-6060/$ - see front matter
� 2005 Mosby, Inc. All rights reserved.
doi:10.1016/j.surg.2005.07.011
06 SURGERY
such as aortic rupture. One mechanism for the de-crease in ECM stability is an increase in collagenand elastin proteolysis mediated by matrix-metallo-proteinase expression, a family of proteolytic en-zymes.2,3 A second mechanism, not exclusive ofthe first, is decreased elastin and collagen produc-tion by aortic smooth muscle cells (SMCs).4 SMCsare involved in the secretion of elastin, collagen,and other structural ECM proteins both directlyand indirectly.5 Thereby, depletion of SMCs, aknown component of AAA pathogenesis, may con-tribute to the structural degeneration accompany-ing aneurysm development.6 Recently, apoptosishas been identified as a cause of decreased SMCnumbers in aneurysmal aortas.7-9
Apoptosis can be initiated by either an extrinsicand/or intrinsic pathway, both leading to a finalcommon pathway in which activated downstreamcaspases are involved in DNA degradation.10 In-flammation, involving macrophages or lympho-cytes and death ligands, are recognized as a part

SurgeryVolume 138, Number 4
Sinha et al 807
of the extrinsic pathway, whereas the intrinsic path-way is instead secondary to DNA instability andoxidative stress.11-13
A study by Henderson et al9 previously demon-strated inflammation-mediated apoptosis in humanAAAs via the Fas/Fas ligand (FasL) pathway. Apo-ptosis also has been linked to oxidative stress inAAA tissue, although an association was establishedand no mechanism proposed.14 Oxidative stressinitiates apoptosis through the intrinsic pathway ofapoptosis, which is initiated by mitochondrial cyto-chrome c translocation into the cytoplasmic space,rather than by the Fas/FasL–activating system.11-13
An intracellular mechanism of apoptosis has notbeen described previously in AAAs, but may serveas another cause of decreased SMC numbers andan explanation for oxidative stress–induced SMCdepletion. This present study examines the intracel-lularmechanismof apoptosis in elastase-inducedex-perimental abdominal aortic aneurysms in rodents.
MATERIAL AND METHODS
Rats were obtained from Charles River Labora-tories (Wilmington, Mass). All experiments andprocedures were approved by the University ofMichigan Universal Committee on the Use andCare of Animals (UCUCA #8566).
Elastase perfusion. Pancreatic porcine elastaseperfusion was performed as previously described.15
Briefly, male Sprague-Dawley rats (200-250 g) wereanesthetized under 2% isoflurane inhalation, andthe infrarenal aorta was isolated. Digital imagesof the aorta were obtained with the use of SpotInsight Color Optic Camera (Diagnostic Instru-ments, Sterling Heights, Mich) attached to an op-erating microscope (Nikon, Melville, NY). Aorticdiameter was measured with the use of ImagePro Express (Media Cybernetics, LP, Silver Springs,Md). Temporary control of the aorta was obtained,and an aortotomy was made near the bifurcationwith a 30-gauge needle. The aorta was then cannu-lated with PE-10 tubing and perfused with either2 mL isotonic saline (N = 5) or 12 units total ofporcine pancreatic elastase in 2 mL isotonic saline(N = 5) for a total of 30 minutes. The aortotomywas repaired with 10-0 suture. Aortic diametermeasurements were repeated at the time of har-vest. Rodent aortas were harvested 7 days postoper-atively and placed in 5% glutaraldehyde. Aortaswere processed for routine paraffin embeddingfor either in situ detection of apoptosis orimmunohistochemistry.
Assessment of apoptosis. In situ detection ofcells with DNA strand breaks was performed in
glutaraldehyde-fixed, paraffin-embedded aorticsections by TUNEL with the use of an ApopTagPeroxidase Kit, as described previously (Integren,Purchase, NY).16 Arteries were examined with anAmerican Optical microscope with 340 objectiveand a 310 eyepiece. A square grid fitted within 1eyepiece provided a reference area of 62,500 lm2.TUNEL-positive cells (SMCs, macrophages, andneutrophils) within the frame were counted. Foreach aorta, at least 10 grid fields were examined.17
Transmission electron microscopy. Portions ofglutaraldehyde-fixed aortic tissue were diced intosmaller pieces, postfixed into 1% osmium tetrox-ide, and embedded in Epon 812. Thin selectionsfrom selected tissue blocks were cut with an LKBultramicrotome, stained with uranyl acetate andlead citrate, and examined with a Hitachi 600electron microscope (Hialeah, Fla).
Immunohistochemical analysis. Hematoxylin-eosin (H&E) staining was performed on aorticsections. Bouin’s-fixed, paraffin-embedded aorticsections were immunostained as described previ-ously.17 Primary antibodies included (1) a rabbitpolyclonal–cleaved caspase-9 (1:50; this antibodydetects only the cleaved product p38 and p17 of ac-tivated caspase-9), (2) a rabbit polyclonal caspase-3antibody (1:1000; this antibody recognizes the p18and p12 subunits of activated caspase-3, but notthe inactive zymogen), and (3) a rabbit polyclonalBid antibody (1:50; Novus Biologicals Inc, Little-ton, Colo) that recognizes only cleaved, activated,murine Bid.18,19 Immunoreactivity was detectedwith the use of biotinylated goat antirabbit IgG sec-ondary antibody followed by avadin-biotinylatedhorseradish peroxidase complex visualized withdiaminobenzadine tetrahydrochloride accordingto the manufacturer’s protocol (Unitect DetectionSystem, Oncogene Science, San Diego, Calif).
Negative and positive controls were run for allassays. Negative controls were processed in anidentical manner, except the primary antibodieswere substituted by the same volume of buffer.Testicular sections from a rat who had beenexposed to short-term local testicular heatingwere used as positive controls for caspase-9 andcaspase 3.16 Bouin’s-fixed, paraffin-embedded sec-tions of mice 6 hours after treatment with a singleintraperitoneal injection of lipopolysaccharide ata dose of 25 mg/kg body weight were used as a pos-itive control for Bid.19
Statistical analysis. Data analysis was performedby using nonpaired t test with statistical signifi-cance assigned as P < .05. Analysis was performedon GraphPad Prism 3.0 (GraphPad Software, Inc,San Diego, Calif).

SurgeryOctober 2005
808 Sinha et al
RESULTS
Abdominal aortic diameters increased by amean (± SEM) of 168% ± 25% 7 days after elastaseperfusion, compared with 30% ± 5% in the saline-perfused group (P < .001). Of the elastase-treatedaortas, 80% (4/5) formed aneurysms by 7 days asdefined by a 100% growth in aortic diameter. Incontrast, none of the aortas (0/5) was aneurysmalby day 7 in the saline-perfused group.
Elastase exposure is associated with increasedapoptosis. Histologic changes byH&E in the aorta 7days after saline or elastase perfusion is shown in Fig-ure 1. Saline-perfused aortas showminimal architec-tural changes with an intact internal elastic lamellae(Fig 1, A). The elastase-perfused aortas, in contrast,exhibit marked medial thickening, destruction ofthe internal elastic lamellae, and infiltration of leu-kocytes (Fig 1, B). Using a modified TUNEL assay,apoptotic cells were rarely observed in control aor-tas (Fig 2, A), but clear evidence of apoptotic cells
Fig 1. H & E staining at 320 of saline-perfused controlaortas (A) demonstrates minimal histologic changes,whereas staining of elastase-perfused aortas (B) demon-strates marked histologic changes, including intimalthickening, destruction of internal elastic lamellae, andinfiltrating leukocytes. L, Lumen.
were seen in elastase-perfused aortas. Quantitativeanalysis of the apoptotic cells revealed a significant(P < .01) increase in the number of total apoptoticcells in elastase-perfused aortas (12.13 ± 2.76cells/hpf) compared with saline-perfused aortas(1.34 ± 0.24 cells/hpf). Electronmicroscopy charac-terized these cells as neutrophils, macrophages, andSMCs in the elastase-perfused aortas (Fig 3, A-D).
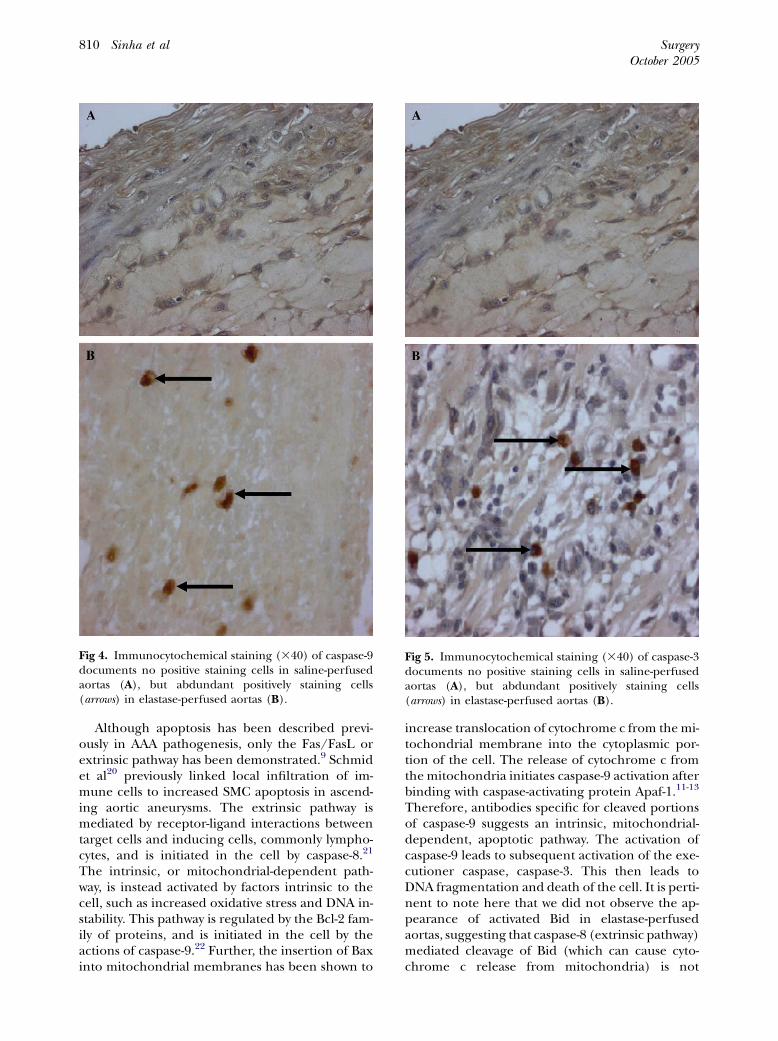
Activation of the initiator caspase-9 and theexecutioner caspase-3 is associated with AAAformation. Since the principal intracellular enzymesin apoptosis are caspases, we further examined theactivation of the initiator caspase-9 and the execu-tioner caspase-3 in saline- and elastase-perfusedaortas. Activation of both caspase-9 (Fig 4, A and
Fig 2. Representative sections demonstrating no apopto-tic cells detected in saline-perfused control aortas (A),with abundant apoptotic cells (arrows) in elastase-perfused aortas (B) by TUNEL. L, Lumen.

SurgeryVolume 138, Number 4
Sinha et al 809
Fig 3. Multiple apoptotic cell types are visualized in the elastase-perfused aortas by TEM. Arrows are point-ing towards chromatin. A, An apoptotic macrophage shows chromatin fragmentation characteristic ofapoptosis. B, An apoptotic neutrophil also undergoing chromatic fragmentation and cytoplasmiccondensation. C, A representative SMC from control aorta shows normal ultrastructural morphology.D, In contrast, a SMC from an elastase-perfused aorta demonstrates apoptotic ultrastructural changes,including chromatin fragmentation and cytoplasmic condensation.
B), the key initiator in the intrinsic (mitochondria-dependent) pathway and caspase-3 (Fig 5, A andB)was evidenced by immunocytochemical staining inonly elastase-perfused aortas. There was no evidenceof Bid activation from immunocytochemical stain-ing for cleaved Bid in either the elastase- orsaline-perfused aortas (data not shown).
DISCUSSION
This is the first study to suggest that an intrinsic,mitochondrial-dependent, mechanism of apoptosismay be relevant in AAA pathogenesis. TUNELstaining demonstrated apoptosis of cells in the
aortic wall in rodent experimental AAAs, whereasfew positive cells were noted in saline-perfusedaortas. Specific apoptotic cells, as shown by TEM,include macrophages, neutrophils, and SMCs. Tothe best of our knowledge, this is the first time TEMhas demonstrated definitively apoptosis in leuko-cytes in aortic tissue. Importantly, immunohisto-chemical staining of activated caspase-9 and caspase-3 was detectable only in elastase-perfused aortas.Caspase-9 is specific for the intrinsic pathway, whichsuggests that apoptosis in experimental AAAs maybe triggeredbymitochondrial-dependent apoptoticsignaling.
SurgeryOctober 2005
810 Sinha et al
Although apoptosis has been described previ-ously in AAA pathogenesis, only the Fas/FasL orextrinsic pathway has been demonstrated.9 Schmidet al20 previously linked local infiltration of im-mune cells to increased SMC apoptosis in ascend-ing aortic aneurysms. The extrinsic pathway ismediated by receptor-ligand interactions betweentarget cells and inducing cells, commonly lympho-cytes, and is initiated in the cell by caspase-8.21
The intrinsic, or mitochondrial-dependent path-way, is instead activated by factors intrinsic to thecell, such as increased oxidative stress and DNA in-stability. This pathway is regulated by the Bcl-2 fam-ily of proteins, and is initiated in the cell by theactions of caspase-9.22 Further, the insertion of Baxinto mitochondrial membranes has been shown to
Fig 4. Immunocytochemical staining (340) of caspase-9documents no positive staining cells in saline-perfusedaortas (A), but abdundant positively staining cells(arrows) in elastase-perfused aortas (B).
increase translocation of cytochrome c from themi-tochondrial membrane into the cytoplasmic por-tion of the cell. The release of cytochrome c fromthe mitochondria initiates caspase-9 activation afterbinding with caspase-activating protein Apaf-1.11-13
Therefore, antibodies specific for cleaved portionsof caspase-9 suggests an intrinsic, mitochondrial-dependent, apoptotic pathway. The activation ofcaspase-9 leads to subsequent activation of the exe-cutioner caspase, caspase-3. This then leads toDNA fragmentation and death of the cell. It is perti-nent to note here that we did not observe the ap-pearance of activated Bid in elastase-perfusedaortas, suggesting that caspase-8 (extrinsic pathway)mediated cleavage of Bid (which can cause cyto-chrome c release from mitochondria) is not
Fig 5. Immunocytochemical staining (340) of caspase-3documents no positive staining cells in saline-perfusedaortas (A), but abdundant positively staining cells(arrows) in elastase-perfused aortas (B).

SurgeryVolume 138, Number 4
Sinha et al 811
responsible for the observed caspase-9 activation inapoptosis-induced cell death.12,13,19
The present study, although primarily observa-tional andbasedonhistology, suggests that caspase-9is increasedduringAAA formation and that intrinsicmechanisms of apoptosis, perhaps mediated by re-active oxygen species (ROS), may be important inAAA pathogenesis. Studies indirectly supportingthis mechanism as being important in the patho-genesis of AAAs have been performed.23-26 Zhanget al23 demonstrated increased apoptosis in conjunc-tion with high levels of inducible nitric oxide syn-thase and speculated that increased oxidative stresswas responsible for increased apoptosis in humanAAAs. Others have shown that increased levels ofangiotensin II also have a proapoptotic effect onvascular tissue, specifically endothelial cells, asmediated by ROS.24,25
The relative importance of the intrinsic andextrinsic pathways of apoptosis in the aneurysmalwall remains unclear. The relevance in humanAAA disease also is not addressed in the presentstudy; thus, studies using human aortic tissue toconfirm that both pathways occur and are impor-tant in human AAA development are needed. Inaddition, although TUNEL, TEM, and immuno-histochemical staining of activated caspase-3 allprovide evidence that apoptosis is occurring, im-munohistochemical staining of caspase-9 is theonly evidence of the intrinsic mechanism of apo-ptosis. Experiments are necessary to demonstratecytoplasmic translocation of cytochrome c, a keyevent in intrinsic apoptotic signaling.
REFERENCES
1. Wills A, Thompson MM, Crowther M, et al. Pathogenesisof abdominal aortic aneurysms cellular and biomechanicalmechanisms. Eur J Vasc Endovasc Surg 1996;12:391-400.
2. Shah PK. Inflammation, metalloproteinases, and increasedproteolysis: an emerging pathophysiological paradigm inaortic aneurysm. Circulation 1997;96:2115-7.
3. Thompson RW, Liao S, Curci JA. Vascular smooth musclecell apoptosis in abdominal aortic aneurysms. Coron ArteryDis 1997;8:623-31.
4. Ailawadi G, Eliason JL, Upchurch GR. Current concepts inthe pathogenesis of abdominal aortic aneurysm. J Vasc Surg2003;38:584-8.
5. Rasmusses LM, Wolf YG, Ruoslahti E. Vascular smoothmuscle cells from injured rat aortas display elevated matrixproduction associated with transforming growth factor-bactivity. Am J Pathol 1995;147:1041-8.
6. Thompson RW. Reflections on the pathogenesis of abdom-inal aortic aneurysms. Cadiovasc Surg 2002;10:389-94.
7. Walton LJ, Franklin IJ, Bayston T, et al. Inhibitionof prostaglandin E2 synthesis in abdominal aortic aneu-rysms: implications for smooth muscle cell viability, in-flammatory processes, and the expansion of abdominalaortic aneurysms. Circulation 1999;100:48-54.
8. Lopez-Candales A, Holmes DR, Liao S, et al. Decreased vas-cular smooth muscle cell density in medial degeneration ofhuman abdominal aortic aneurysms. Am J Pathol 1997;150:993-1007.
9. Henderson EL, Geng Y-J, Sukhova GK, et al. Death ofsmooth muscle cells and expression of mediators of apopto-sis by T lymphocytes in human abdominal aortic aneurysms.Circulation 1999;99:96-104.
10. Leszczynski D, Zhao Y, Luokkamaki M, et al. Apoptosis ofvascular smooth muscle cells: protein kinase C and onco-protein Bcl-2 are involved in apoptosis in non-transformedrat vascular smooth muscle cells. Am J Pathol 1994;145:1265-70.
11. Green DR. Apoptotic pathways: paper wraps stone bluntsscissors. Cell 2000;102:104.
12. Hengartner MO. The biochemistry of apoptosis. Nature2000;407:770-6.
13. Reed JC. Mechanisms of apoptosis. Am J Pathol 2000;157:1415-30.
14. Miller FJ, Sharp WJ, Xiang F, et al. Oxidative stress inhuman abdominal aortic aneurysms: a potential mediatorin aneurysmal remodeling. Arterioscler Thromb Vasc Biol2002;22:560-5.
15. Anidjar S, Salzmann JL, Gentric D, et al. Elastase-inducedexperimental aneurysms in rats. Circulation 1990;82:973-81.
16. Sinha-Hikim AP, Lue H, Yamamoto CM, et al. Key apoptoticpathways for heat-induced programmed germ cell death inthe testis. Endocrinology 2003;144:3167-75.
17. Yamamoto CM, Sinha-Hikim AP, Huynh PN, et al. Redistri-bution of Bax is an early step in an apoptotic pathway lead-ing to germ cell death in rats, triggered by mild testicularhyperthermia. Biol Reprod 2003;63:1683-90.
18. Srinivasan A, Roth KA, Sayers RO, et al. In situ immunode-tection of activated caspase-3 in apoptotic neurons in thedeveloping nervous system. Cell Death Differ 1998;5:1004-6.
19. Kang SJ, Wang S, Kuida K, Yuan J. Distinct downstreampathways of caspase-11 in regulating apoptosis and cytokinematuration during septic shock. Cell Death Differ 2002;9:1115-25.
20. Schmid FX, Bielenberg K, Schneider A, et al. Ascendingaortic aneurysm associated with bicuspid and tricuspid aor-tic valve: involvement and clinical relevance of smooth mus-cle cell apoptosis and expression of cell death-initiatingproteins. Euro J Cardio-Thoracic Surg 2003;23:537-43.
21. Nagata S, Golstein P. The Fas death factor. Science 1995;267:1449-55.
22. Adams JM, Cory S. The bcl-2 protein family: arbiters of cellsurvival. Science. 281:1305-8.
23. Zhang J, Schmidt J, Ryschich E, et al. Inducible nitric oxidesynthase is present in human abdominal aortic aneurysmand promotes oxidative vascular injury. J Vasc Surg 2003;38:360-7.
24. Griendling KK, Minieri CA, Ollerenshaw JD, et al. Angi-otensin II stimulated NADH and NADPH oxidase activ-ity in cultured smooth muscle cells. Circ Res 1994;74:1141-8.
25. Dimmeler S, Rippmann V, Weiland U, et al. Angiotensin IIinduces apoptosis of human endothelial cells. Circ Res1997;81:970-6.
26. Grigoryants V, Hannawa KK, Pearce CG, et al. Tamoxifenup-regulates catalase production, inhibits vessel wall neutro-phil infiltration, and attenuates development of abdominalaortic aneurysms. J Vasc Surg 2005;41:108-14.