modeling lichen performance in relation to climate216855/fulltext01.pdf · title: modeling lichen...
TRANSCRIPT

Modeling lichen performance in relation to
climate
– scaling from thalli to landscapes
Anna Jonsson Čabrajić 2009
Department of Ecology and Environmental Sciences Umeå University SE-901 87 Umeå
Sweden
Akademisk avhandling
som med vederbörligt tillstånd av Rektorsämbetet vid Umeå universitet för avläggande av filosofie doktorsexamen i ekologi kommer att offentligen försvaras torsdagen den 4 juni, kl 10.00 i Stora Hörsalen (KB3B1), KBC.
Avhandlingen kommer att försvaras på engelska.
Examinator: Professor Lars Ericson, Umeå universitet
Fakultetsopponent: Professor Göran Ågren, Department of Ecology, Swedish University of Agricultural Sciences,
Uppsala

ORGANISATION DOCUMENT NAME Dept. of Ecology and Environmental Science Doctoral dissertation Umeå University DATE OF ISSUE SE-901 87 Umeå 14 May 2009 AUTHOR: Anna Jonsson Čabrajić TITLE: Modeling lichen performance in relation to climate – scaling from thalli to landscapes ABSTRACT Lichens can colonize nearly all terrestrial habitats on earth and are functionally important in many ecosystems. Being poikilohydric, their active growth periods are restricted to periods when the thallus is hydrated from atmospheric water sources, such as rain, fog and high relative humidity. Since lichen hydration varies greatly over time lichen growth is therefore more difficult to model compared with, for example vascular plants with more even water supply. I developed two models to predict lichen hydration under field conditions that incorporates the atmospheric water potential (Ψair), derived from air temperature and humidity, only or in combination with species-specific rehydration and desiccation rates. Using Ψair allows the prediction of hydration induced by several water sources. These models were very accurate for epiphytic lichens with a close coupling to atmospheric conditions, but they were less accurate for mat-forming lichens with substantial aerodynamic boundary layers. The hydration model was further developed to include photosynthetic activation for different species, in order to compare their performance under different micro-climatic scenarios. Water balance and activation rate had large effects on lichen activity and were positively related to habitats providing long hydration periods, for example close to streams. To study effects of climate change, a complete model for net carbon gain (photosynthesis minus respiratory losses) was developed for an epiphytic lichen with intricate responses to light, hydration and temperature. Simulation responses in different climate scenarios revealed that projected climate change on a regional scale resulted in varied local scale responses. At the lighter, exposed sites of a forest, the growth responses were positive, but were potentially negative at darker sites with closed canopy. At the local scale, fluctuating hydration, summed irradiance when wet and Chlorophyll a are variables that predict lichen growth. However, at a landscape scale, these variables may be too detailed. We tested this for two terrestrial, mat-forming lichens and developed statistical models for lichen growth in the widest possible climatic gradient in northern Scandinavia, varying in light, temperature and precipitation. Light was the most important factor for high growth at the landscape scale, reaching saturation at a site openness of 40 %, equivalent to a basal tree area of 15 m2 ha -1 in this study. Thereafter, hydration was the next limiting factor, which could be well described by precipitation for one of the species. The simplest predictor was the normal temperature in July, which was negatively correlated with growth. It was apparent that the predictive variables and their power varied at different scales. However, light and hydration are limiting at all scales, particularly by light conditions when lichens are wet. This implies that ensuring that there is sufficient light below the forest canopy is crucial for lichen growth, especially for mat-forming lichens. Hydrophilic lichens may be better preserved in open habitats with long hydration periods. It was shown that models can be powerful and “easy to use” tools to predict lichen responses in various habitats and under different climate scenarios. Models can therefore help to identify suitable habitats with optimal growth conditions, which is very important for the conservation and management of lichens and their habitats. KEYWORDS: Lichen, growth, model, micro-climate, hydration, activation, photosynthesis, respiration, landscape scale, local scale, climate change scenarios, management, mat-forming, epiphytic LANGUAGE: English ISBN: 978-91-7264-744-2 NUMBER OF PAGES: 38

Modeling lichen performance in relation to
climate
– scaling from thalli to landscapes
Anna Jonsson Čabrajić 2009
Department of Ecology and Environmental Sciences Umeå University SE-901 87 Umeå
Sweden

Detta verk är upphovsrättsskyddat enligt svensk lag ISBN: 978-91-7264-744-2 © Anna Jonsson Čabrajić 2009 Printed by VMC, KBC-HUSET, Umeå University, Umeå, 2009

“As far as the laws of mathematics refer to reality, they are not certain; and as far as they are certain, they do not refer to reality”
Albert Einstein – (1921)

“There are many methods for predicting the future. For example, you can read horoscopes, tea leaves, tarot cards, or crystal balls. Collectively, these methods are known as “nutty methods”. Or you can put well-researched facts into sophisticated computer models, more commonly referred to as - a complete waste of time.” Scott Adams

LIST OF PAPERS This thesis is based on the following papers, which will be referred to in the text by the corresponding Roman numerals. I Jonsson A.V., Moen J. & Palmqvist K. (2008). Predicting lichen hydration using biophysical models. Oecologia 156: 259-273. II Jonsson Čabrajić A.V., Lidén M., Lundmark T., Ottosson-Löfvenius M. & Palmqvist K. Modelling hydration and PSII activation in relation to in situ rain and humidity patterns- a tool to compare performance of rare and generalist epiphytic lichens. (Submitted manuscript) III Lidén M., Jonsson Čabrajić A.V, Ottosson-Löfvenius M., Palmqvist K. & Lundmark T. Species-specific activation time lags can explain habitat restrictions in hydrophilic lichens. (Submitted manuscript) IV Jonsson Čabrajić A.V., Palmqvist K. & Wennergren U. Predicting local responses of epiphytic lichen vegetation to regional climate change scenarios. (Manuscript) V Jonsson Čabrajić A.V., Moen J. & Palmqvist K. Predicting growth of mat-forming lichens on a landscape scale – comparing models with different complexities. (Submitted manuscript) Paper I is reproduced with the kind permission of Springer.
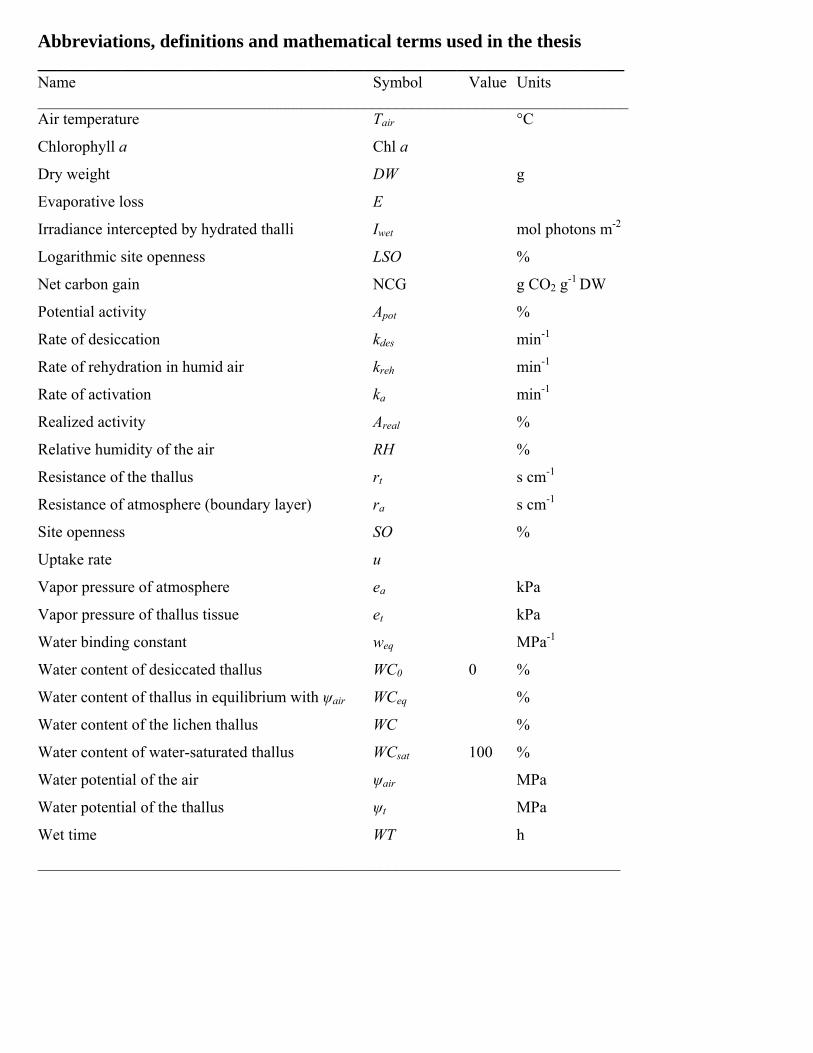
Abbreviations, definitions and mathematical terms used in the thesis _______________________________________________________________ Name Symbol Value Units __________________________________________________________________________ Air temperature Tair °C
Chlorophyll a Chl a
Dry weight DW g
Evaporative loss E
Irradiance intercepted by hydrated thalli Iwet mol photons m-2
Logarithmic site openness LSO %
Net carbon gain NCG g CO2 g-1 DW
Potential activity Apot %
Rate of desiccation kdes min-1
Rate of rehydration in humid air kreh min-1
Rate of activation ka min-1
Realized activity Areal %
Relative humidity of the air RH %
Resistance of the thallus rt s cm-1
Resistance of atmosphere (boundary layer) ra s cm-1
Site openness SO %
Uptake rate u
Vapor pressure of atmosphere ea kPa
Vapor pressure of thallus tissue et kPa
Water binding constant weq MPa-1
Water content of desiccated thallus WC0 0 %
Water content of thallus in equilibrium with ψair WCeq %
Water content of the lichen thallus WC %
Water content of water-saturated thallus WCsat 100 %
Water potential of the air ψair MPa
Water potential of the thallus ψt MPa
Wet time WT h
_________________________________________________________________________

TABLE OF CONTENTS 1. INTRODUCTION…………………………………………………………………...…………………… 1
1.1. Threats to viable lichen populations……………………………………………………………….. 1
1.2. The nature of lichens……………………………………………………………………………….. 2
1.3. Lichen physiology………………………………………………………………….......................... 3
1.3.1. Light response…………………………………………………………………....................... 4
1.3.2. Water response……………………………………………………………………………….. 5
1.3.3. Temperature response…………………………………………………………………………6
1.3.4. Nitrogen and Chlorophyll a……………………………………………………...................... 6
1.4. Growth………………………………………………………………………………....................... 6
1.5. Modeling lichen performance……………………………………………………………………… 7
1.5.1. Previous growth models…………………………………………………………………….... 7
1.5.2. Assessments of lichen hydration…………………………………………………………….. 8
2. AIMS………………………………………………………………………………………………………11
3. MODEL DEVELOPMENT……………………………………………………………............................. 11
4. MODEL SYSTEMS……………………………………………………………………………………….14
4.1. Study sites…………………………………………………………………………........................... 14
4.2. Model species………………………………………………………………………......................... 15
5. RESULTS AND DISCUSSION……………………………………………………….............................. 16
5.1. Predicting lichen wet, active periods……………………………………………………………….. 16
5.2. Modeling lichen activity……………………………………………………………………………. 17
5.3. Modeling local lichen responses in relation to regional climate change………................................ 18
5.4. Patterns and models for growth of mat-forming lichens at landscape scale……………………….. 19
5.5. Limitations to lichen growth………………………………………………………………. ………. 21
5.5.1. Micro-climatic limitations……………………………………………………........... ………. 21
5.5.2. Lichen trait limitations……………………………………………………………………….. 22
5.6. Spatial and temporal scale dependency…………………………………………............................. 23
5.6.1. Predictive variables at different scales………………………………………......................... 23
5.6.2. Predictive power of parameters................................................................................................ 24
5.7. Implications for managing lichen-rich habitats…………………………………….......................... 24
5.7.1. Generalist species…………………………………………………………………………….. 25
5.7.2. Hydrophilic lichens………………………………………………………………………….. 25
5.7.3. Climate change…………………………………………………………………..................... 26
6. CONCLUSIONS………………………………………………………………………............................. 26
7. ACKNOWLEDGEMENTS……………………………………………………………............................. 27
8. REFERENCES……………………………………………………………………………………………. 27
9. SAMMANFATTNING (SUMMARY IN SWEDISH)……………………………………………………35
TACK…………………………………………………………………………………................................... 36

1
1. INTRODUCTION
Lichens are unique as they can colonize nearly all terrestrial habitats on earth, unlike most other living
organisms. They are often found in boreal, temperate and tropic forests but are also frequently present in
deserts, arctic and alpine biomes (Kappen 1988; 2000), and have even survived exposure to outer space
conditions (Sancho et al. 2007a). In 8 % of terrestrial ecosystems, lichens are dominant, both in terms of
diversity and abundance (Larson 1987). Their appearance is extremely varied in terms of color, growth form,
growth site and position in the habitat. Terriculous lichens grow on the ground, saxicolous lichens on rocks,
boulders or cliffs, whereas epiphytic lichens grow on other plants, e.g. on trees, without directly taking up
nutrients or water from the host. Lichens are divided in three major groups based on their morphology:
crustose, foliose and fruticose. Crustose lichens adhere tightly to the substratum and cannot be removed
without destruction. The focus of this thesis, however, is on foliose and fruticose macro-lichens. Foliose
lichens are leaf-like, flat and only partially attached to the substrate (as illustrated by the two examples to the
left on the front cover). Fruticose lichens are bushy or hair-like, with either upright or pendulous morphology
(as illustrated by the four examples to the right on the front cover). Lichens are functionally important in
many ecosystems (Cornelissen et al. 2007) and significantly contribute to biomass production and nitrogen
fixation, especially in early successional ecosystems (Crews et al. 2001), and CO2 cycling (Botting &
Fredeen 2006). Others are abundant in late-successional ecosystems such as old forests. Lichens can provide
important forage for reindeer in winter (Gaare & Danell 1999; van der Wal 2006), food and shelter for
invertebrates (Pettersson et al. 1995), and are consequently important for many forest birds.
1.1. Threats to viable lichen populations Generally, lichens have relatively slow growth rates, due to their constraints being poikilohydric and
symbiotic, and therefore depend on long periods of favorable micro-climatic conditions for development. The
long-term persistence of lichen populations requires sufficient growth of the individual lichens to support a
successful life-cycle. This includes needs for successful dispersal to, and establishment in, new habitats and a
suitable micro-climate to produce sufficient new biomass to compensate for losses due to herbivory or
fragmentation (by, for example, wind or trampling). Lichens are increasingly threatened by environmental
changes, including climate change (Epstein et al. 2004), increased nitrogen deposition (Söchting 1995), air
pollution (Nash 2008), and changes in land use and forestry (Saunders et al. 1991; Esseen et al. 1996; Berg
et al. 2008).
Forestry has negatively affected lichens in several ways. Firstly, epiphytic lichens are sensitive to
logging since this will remove or reduce the amount of suitable substrate. Secondly, the short rotation periods
of commercial forestry will rarely allow lichen populations to build up large biomasses. Thirdly, many
epiphytic, pendulous lichens, especially “old forest” species such as Alectoria sarmentosa or Usnea
longissima, are sensitive to increased wind exposure at forest edges (Essen et al. 1981; Esseen & Renhorn
1998). Fourthly, most open, old-growth forests have been replaced by younger and denser forests, which

2
have changed the micro-climate adversely for both epiphytic and mat-forming lichens.
Little is known about lichen responses to climate changes, which in northern Europe are expected to
include annual warming and increased precipitation. Previous studies have mainly emphasized the general
northward movement of some lichen species (Ellis et al. 2007; van Herk et al. 2002). In addition, vegetation
studies in terrestrial systems have shown that global changes that increase vascular plant biomass will tend to
decrease the amount of lichens, mainly due to their poor competitiveness (Cornelissen et al. 2001; van Wijk
et al. 2004).
Given the contribution of lichens to biodiversity and biomass, a better understanding and ability to
predict their responses is essential for our understanding of the functioning of ecosystems and for long-term
conservation strategies. This is a particular need in view of accelerating habitat destruction (Berg et al. 2008)
and climate change (IPCC 2007).
1.2. The nature of lichens Lichens survive extreme conditions, ranging from extremely cold to hot environments and from wet to
extremely dry habitats, by an extraordinary tolerance to desiccation and extreme temperatures. The
ecological success of these organisms can be partly explained by their poikilohydric nature, since they are
inactive during the harshest conditions and active in favorable micro-climatic conditions, which are more
similar than the macro-climate (Green et al. 2007). This strategy has enabled the colonization of habitats that
are inaccessible to vascular plants. Poikilohydric organisms lack water-carrying organs and depend entirely
on the passive physical processes of water uptake and loss (Blum 1973). The water content of lichens varies
constantly via an equilibration process with the surrounding atmospheric conditions (Rundel 1982), being
metabolically metabolically activated only when hydrated. Furthermore, lichens are symbiotic associations
between nutritionally specialized fungi (mycobiont) and photosynthetic green-algae and/or cyanobacteria
(photobiont) (Honegger 1991). The fungus derive carbon and in some cases nitrogen from the photobiont,
while providing a living environment for the photobiont. The symbiotic life-style adds complexity to the
poikilohydric strategy, since poikilohydry must be favorable for both symbionts and they must manage to
instantly activate once favorable conditions appear.
Lichens are hydrated and activated almost instantaneously by rain or other sources of liquid water
(dew, stemflow and meltwater) (e.g. Coxson 1988; Lange et al. 1986) and gradually activated by high
relative humidity, although activation by this water source is restricted to green-algal lichens (Lange et al.
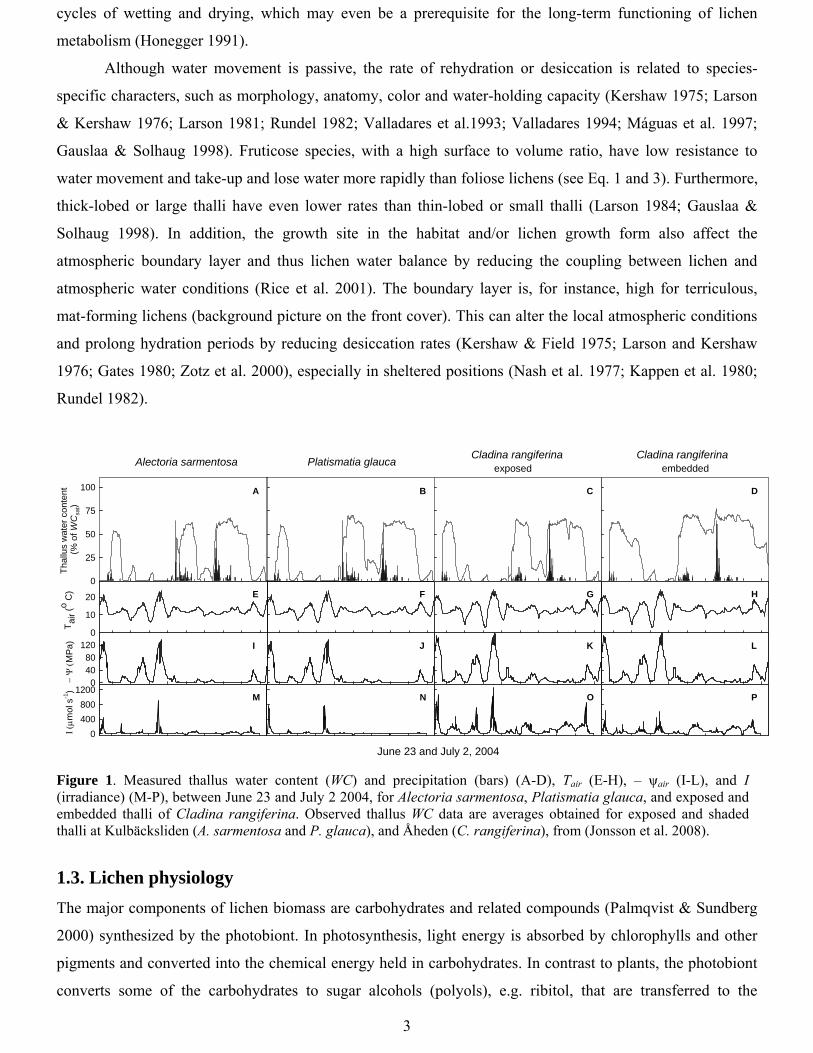
1986). Moreover, humid air seldom results in as high water contents (cf. WC Fig 1; morning June 25 and 26)
or activity, as hydration by rain (cf. WC Fig 1; June 24, 27 and 29) (Rundel 1988; Palmqvist 2000; Lange et
al. 1986; Scheidegger et al. 1995; Pintado & Sancho 2002; Lange et al. 2001). However, humid air may still
be very important for metabolic output, since humid air may be the dominant water source in dry habitats,
e.g. deserts, or it may slow down desiccation after rainfall (Kappen et al. 1980; Lange et al. 1986; 1988;
2001; Gaio-Oliveira et al. 2004; Lidén & Hilmo 2005). Lichen hydration is thus characterized by frequent
3
cycles of wetting and drying, which may even be a prerequisite for the long-term functioning of lichen
metabolism (Honegger 1991).
Although water movement is passive, the rate of rehydration or desiccation is related to species-
specific characters, such as morphology, anatomy, color and water-holding capacity (Kershaw 1975; Larson
& Kershaw 1976; Larson 1981; Rundel 1982; Valladares et al.1993; Valladares 1994; Máguas et al. 1997;
Gauslaa & Solhaug 1998). Fruticose species, with a high surface to volume ratio, have low resistance to
water movement and take-up and lose water more rapidly than foliose lichens (see Eq. 1 and 3). Furthermore,
thick-lobed or large thalli have even lower rates than thin-lobed or small thalli (Larson 1984; Gauslaa &
Solhaug 1998). In addition, the growth site in the habitat and/or lichen growth form also affect the
atmospheric boundary layer and thus lichen water balance by reducing the coupling between lichen and
atmospheric water conditions (Rice et al. 2001). The boundary layer is, for instance, high for terriculous,
mat-forming lichens (background picture on the front cover). This can alter the local atmospheric conditions
and prolong hydration periods by reducing desiccation rates (Kershaw & Field 1975; Larson and Kershaw
1976; Gates 1980; Zotz et al. 2000), especially in sheltered positions (Nash et al. 1977; Kappen et al. 1980;
Rundel 1982).
Thal
lus
wat
er c
onte
nt
(% o
f WC
sat)
0
25
50
75
100
− Ψ
( MPa
)
04080
120
T air
(o C
)
0
10
20
A B C
E F G
J KI
June 23 and July 2, 2004
Alectoria sarmentosaCladina rangiferina Cladina rangiferina
exposed embeddedPlatismatia glauca
D
H
L
Ι (μ
mol
s-1
)
0400800
1200M N O P
Figure 1. Measured thallus water content (WC) and precipitation (bars) (A-D), Tair (E-H), – ψair (I-L), and I (irradiance) (M-P), between June 23 and July 2 2004, for Alectoria sarmentosa, Platismatia glauca, and exposed and embedded thalli of Cladina rangiferina. Observed thallus WC data are averages obtained for exposed and shaded thalli at Kulbäcksliden (A. sarmentosa and P. glauca), and Åheden (C. rangiferina), from (Jonsson et al. 2008).
1.3. Lichen physiology The major components of lichen biomass are carbohydrates and related compounds (Palmqvist & Sundberg
2000) synthesized by the photobiont. In photosynthesis, light energy is absorbed by chlorophylls and other
pigments and converted into the chemical energy held in carbohydrates. In contrast to plants, the photobiont
converts some of the carbohydrates to sugar alcohols (polyols), e.g. ribitol, that are transferred to the

4
mycobiont, which rapidly and irreversibly converts them into mannitol or arabitol. It is unclear, but
suggested, that the conversion by the mycobiont generates a gradient that is the driving force of the efflux
between the symbionts (Smith 1980).
Hence, the growth of lichens is the result of the balance between carbon gain, by photosynthetic CO2
assimilation (photobiont) and losses via respiration of CO2 (by the photobiont and mycobiont), and losses
related to dispersal, fragmentation, grazing or necrosis (Palmqvist et al. 2008). Being poikilohydric, net
photosynthesis and thus carbon gain in lichens is limited to their wet active periods (Palmqvist & Sundberg
2000) and by the environmental conditions during these periods. The most important micro-climatic factors
are light intensity, thallus hydration and temperature (Nash 2008). These factors can vary greatly over small
spatial scales (Gauslaa et al. 2006; Renhorn et al. 1997), e.g. within a forest, and over temporal scales (Muir
et al. 1997; Caldiz 2004; Gaio-Oliveira et al. 2004), e.g. diurnally or seasonally.
1.3.1. Light response Lichen photosynthesis is directly regulated by available irradiance when wet (Nash 2008). The light response
curve can be described by a non-rectangular hyperbola (Lambers et al. 1998; Palmqvist 2000), since
photosynthesis is light limited at low light intensities and increases linearly until asymptotically leveling out
at light saturation (Fig 2). Lichens are generally adapted to low light environments and reach light saturation
at 100-400 μmol photons m-2 s-1 (Demmig-Adams et al. 1990). However, extreme species exist that saturate
at 20 or 1100 μmol photons m-2 s-1 (Demmig-Adams et al. 1990; Green et al. 1991). Lichens can also adapt
their photosynthesis to seasonal variations in irradiance (Kershaw 1985). However, short-term changes in
light availability, such as abrupt alterations in light exposure imposed by clear cuts at forest edges, may result
in photoinhibition that reduce their photosynthetic rate (Demmig-Adams et al. 1990; Gauslaa & Solhaug
1996). Such inhibition may be reversible in green-algal lichens, but irreversible in cyanobacterial lichens
(Demmig-Adams et al. 1990).
5
1.3.2. Water response
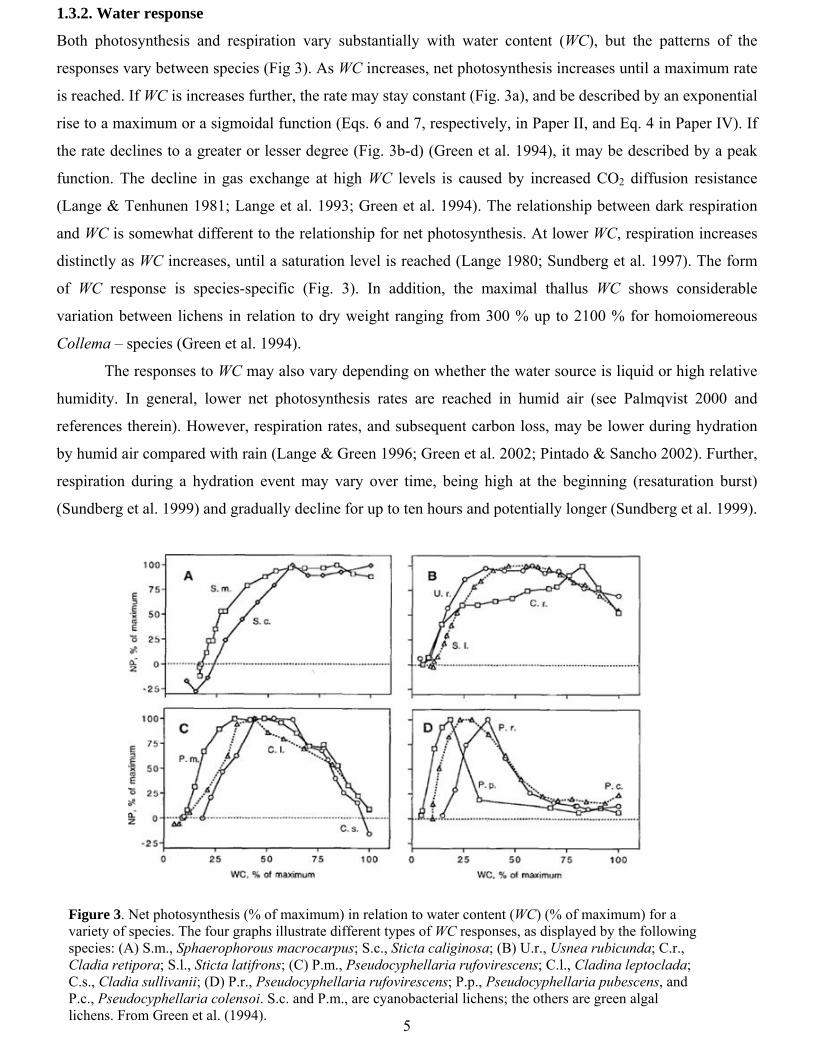
Both photosynthesis and respiration vary substantially with water content (WC), but the patterns of the
responses vary between species (Fig 3). As WC increases, net photosynthesis increases until a maximum rate
is reached. If WC is increases further, the rate may stay constant (Fig. 3a), and be described by an exponential
rise to a maximum or a sigmoidal function (Eqs. 6 and 7, respectively, in Paper II, and Eq. 4 in Paper IV). If
the rate declines to a greater or lesser degree (Fig. 3b-d) (Green et al. 1994), it may be described by a peak
function. The decline in gas exchange at high WC levels is caused by increased CO2 diffusion resistance
(Lange & Tenhunen 1981; Lange et al. 1993; Green et al. 1994). The relationship between dark respiration
and WC is somewhat different to the relationship for net photosynthesis. At lower WC, respiration increases
distinctly as WC increases, until a saturation level is reached (Lange 1980; Sundberg et al. 1997). The form
of WC response is species-specific (Fig. 3). In addition, the maximal thallus WC shows considerable
variation between lichens in relation to dry weight ranging from 300 % up to 2100 % for homoiomereous
Collema – species (Green et al. 1994).
The responses to WC may also vary depending on whether the water source is liquid or high relative
humidity. In general, lower net photosynthesis rates are reached in humid air (see Palmqvist 2000 and
references therein). However, respiration rates, and subsequent carbon loss, may be lower during hydration
by humid air compared with rain (Lange & Green 1996; Green et al. 2002; Pintado & Sancho 2002). Further,
respiration during a hydration event may vary over time, being high at the beginning (resaturation burst)
(Sundberg et al. 1999) and gradually decline for up to ten hours and potentially longer (Sundberg et al. 1999).
Figure 3. Net photosynthesis (% of maximum) in relation to water content (WC) (% of maximum) for a variety of species. The four graphs illustrate different types of WC responses, as displayed by the following species: (A) S.m., Sphaerophorous macrocarpus; S.c., Sticta caliginosa; (B) U.r., Usnea rubicunda; C.r., Cladia retipora; S.l., Sticta latifrons; (C) P.m., Pseudocyphellaria rufovirescens; C.l., Cladina leptoclada; C.s., Cladia sullivanii; (D) P.r., Pseudocyphellaria rufovirescens; P.p., Pseudocyphellaria pubescens, and P.c., Pseudocyphellaria colensoi. S.c. and P.m., are cyanobacterial lichens; the others are green algal lichens. From Green et al. (1994).

6
1.3.3. Temperature response
Rates of photosynthesis and respiration increase linearly with increases in temperature, beyond which the
processes are increasingly inhibited. Hence, net photosynthesis also increases linearly up to an optimal
temperature and then declines as temperature and respiration increases further (see Kershaw 1984; Kappen
2000). Temperature optimum varies for lichens from different habitats (Lechowitz 1982). Generally, lichens
are active at temperatures above freezing. However, lichens, especially arctic species, can also be activated at
subzero temperatures from atmospheric humidity, although gas exchange at these temperature is low (e.g.
Kappen et al. 1995; Hajek et al. 2001; Lange 2003).
1.3.4. Nitrogen and Chlorophyll a
For synthesis of new proteins, membranes and DNA, the lichen must acquire mineral nutrients, such as
nitrogen and phosphorous. The fungus absorbs most of its nutrients directly from external sources, from wet
or dry deposition, or stemflow on trees. However, 10 % of lichen species are able to fix atmospheric N2,
since they contain cyanobacteria (Ahmadijan 1967; Friedl & Büdel 1996). Two nitrogenous compounds,
chitin and chlorophylls, can be used as indirect metabolic markers. Chitin is situated in fungal cell walls and
can be used as a marker for maintenance respiration (Sundberg et al. 1999), while chlorophyll (Chl) a is well
correlated with photosynthetic capacity and RubisCO concentration, and may be useful as a marker for
photobiont concentration (Palmqvist et al. 1998; Palmqvist & Sundberg 2000; Palmqvist et al. 2002). Thallus
Chl a content is positively correlated with respiration, in addition to photosynthesis and growth (Palmqvist &
Sundberg 2000; Palmqvist et al. 2002; Palmqvist & Dahlman 2006) and increases with increases in the
nitrogen status of the thallus (Palmqvist et al. 2002; Palmqvist & Dahlman 2006). However, the specific
relationship varies among and within species (Palmqvist et al. 2002; Palmqvist et al. 1998; Palmqvist &
Dahlman 2006).
1.4. Growth Mature foliose and fruticose lichens usually have their growth zones at the apical ends of the lobes, a feature
attributed to the mycobiont tissue that dominates the lichen thallus (Wessels 1993). However, in some
pendulous species, such as Usnea longissima, incercalary growth is important (Rolstad & Rolstad 2008).
Lichen growth has frequently been expressed as a linear measure (mm y-1) of the increase in radius for
foliose and crustose lichens, or tip length for fruticose lichens. These measures are useful in lichenometry,
but have limited value when trying to understand growth responses of lichens, since the linear rate of
expansion is largely dependent on the initial thallus area and weight. A better measure is to assess area or
biomass increments in relation to the initial thallus area or biomass. Wide ranges of lichen growth have been
reported for individual thalli. For the tripartite epiphytic lichen Lobaria pulmonaria, growth ranges from 0 to
69 % biomass in a single year (Palmqvist & Sundqvist 2000; Muir et al. 1997). High growth rates (40-50 %)
have also been reported for the tripartite terriculous lichen Peltigera aphthosa (Palmqvist & Sundberg 2000;

7
Sundberg et al. 2001). For green-algal lichens, reported annual growth rates of pendulous epiphytic species
range from 3 % for Alectoria sarmentosa and Bryoria species (Renhorn & Esseen 1995; Stevensson &
Coxson 2003) to nearly 40 % for individual thalli of A. sarmentosa (Renhorn & Esseen 1995) and U.
longissima during a summer (Gauslaa et al. 2007). For mat-forming lichens, studies have reported annual
growth rates ranging from -20 – 15 % for Cladonia mitis and 8-17 % for Cladonia stellaris in northern
Finland (den Herder et al. 2003) and up to 40 % in Britain (Hyvärinen & Crittenden 1998). Hence, lichen
growth can be relatively high with similar energy use efficiency to that of vascular plants during their active
periods (Palmqvist & Sundberg 2000). Growth varies seasonally in temperate climates (Armstrong 1993;
Muir et al. 1997; Kytöviita & Crittenden 2002; Caldiz 2004) and between years (Muir et al. 1997), being
positively dependent on light availability (Renhorn et al. 1997; Gauslaa et al. 2006; 2007; Jansson et al. In
press), and precipitation (Muir et al. 1997; Gauslaa et al. 2007; Sancho et al. 2007b) or fog/dew frequency
(Rundel 1978; Lange et al. 2006; 2007).
1.5. Modeling lichen performance A mechanistic understanding of lichen performance in response to various micro-climatic conditions will
facilitate attempts to assess responses to factors such as forestry or climate change. This would also facilitate
the formulation of long-term conservation strategies of species and management of lichen-rich habitats. A
mechanistic mathematical model based on climatic conditions could potentially fulfill these needs,
particularly since growth measurements are laborious and time-consuming. However, there have been
relatively few efforts to develop such models for lichens, in contrast to the intensive modeling of vascular
plants responses.
1.5.1. Previous growth models
The poikilohydric nature of lichens limits growth to the length and frequency of their wet active periods, but
they are also affected by other prevailing climate conditions during these periods (Armstrong 1993; Muir et
al. 1997). Predicting the long-term performance of lichens is thus not straightforward, since models for
higher plants, assuming more constant water supply during the frost free period, have limited applicability
(Bergh et al. 1998; Zheng et al. 2002). Attempts to model the productivity of lichens have either been
descriptive and based on direct measurements of thallus water content (Palmqvist & Sundberg 2000; Coxson
& Coyle 2003; Gaio-Oliveira et al. 2004; 2006), or on scarce hydration modeling (cf. Kershaw & Harris
1971; Paterson et al. 1983; Lloyd 2001). Previous attempts to model lichen productivity have combined
direct measurements of thallus WC, in combination with CO2 gas exchange responses to controlled micro-
climatic conditions (e.g. Sundberg et al. 1997; Coxson & Coyle 2003; Lange et al. 2000; Lange 2003). These
attempts have shown that lichen growth is strongly correlated to the irradiation received by the wet thallus
(Iwet) (Palmqvist & Sundberg 2000, Dahlman & Palmqvist 2003; Gaio-Oliveria et al. 2004), since
photosynthesis is directly dependent on light. Lichens may thus grow better at more open, exposed forest

8
sites compared with darker, shaded sites (Renhorn et al. 1997; Gauslaa et al. 2006; 2007). A high Iwet can also
result from long wet periods under lower light conditions, which can explain the higher growth at more
humid sites or during more humid seasons (Muir et al. 1997; Kytöviita & Crittenden 2002; Caldiz 2004;
Sancho et al. 2007b). The hydration period is also influenced by the species-specific lichen morphology or
growth structure. Epiphytic lichens may experience shorter wet periods than terriculous and mat-forming
lichens, and may thus show lower growth rates (Palmqvist & Sundberg 2000). However, increased hydration
may affect lichen growth in several ways, since it may also increase respiration and decrease subsequent
growth (Gauslaa et al. 2006). This complicates attempts to determine general correlations between their
growth rates and hydration (Lange & Green 2006). Moreover, growth is also positively correlated to the
indirect marker of photosynthetic capacity, Chl a (Tretiach & Carpanelli 1992; Gaio-Oliveira et al. 2006;
Palmqvist & Dahlman 2006).
Hence, a lichen growth model must be able to identify lichen wet growth periods and account for the
micro-climatic conditions during these periods. Further, such a model must be applicable to field conditions
where mechanistic details must be balanced against information available at the scale and question
considered. It is anticipated that although lichens respond to the local micro-climatic conditions, other
measures that affect these variations may be more easily identified at larger scales.
1.5.2. Assessments of lichen hydration
Lichen water relations have been extensively studied, both in the field and the laboratory. Water content
(WC) has previously been measured gravimetrically (e.g. Larson 1979; Larson & Kershaw 1976; Lange et al.
1986; Valladares et al. 1993), or by an impedance technique (e.g. Coxson 1991; Palmqvist & Sundberg 2000;
Dahlman & Palmqvist 2003; Gaio-Oliveira et al. 2006). Since the maximum WC of a thallus varies between
species, the WC can be expressed either as a percentage in relation to dry weight (DW) (Fig. 3), or as
percentage relative to the maximum WC ranging from completely dry (0 %) to fully hydrated (100 %). The
latter approach facilitates comparisons between species of varying water holding capacities and is calculated
by Eq. 2 in Paper I. Both procedures have been adopted in this thesis, and the underlying studies.
To assess lichens’ wet and active time periods relatively easily, there is a need to develop a combined
model that mechanistically predicts the natural fluctuations of WC for longer time periods in situ, based on
quantitative meteorological variables that can be easily obtained. Further, the model should be able to handle
all the different water sources available for lichens and incorporate the species-specific characteristics
affecting the water balance. Attempts to predict lichen field water relations have previously been made
(Kershaw & Harris 1971; Pech 1989; Lloyd 2001) but without providing detailed information on the
frequency and duration of wet active periods in the field. Some models have only dealt with rain as the water
source (Kershaw & Harris 1971) and other models have overlooked the water uptake (Lloyd 2001) or diurnal
fluctuations of thallus water relations (Pech 1989).
Theoretical and mechanistic lichen water uptake (u) in humid air and loss rates (E) have been
presented by Rundel (1982) (see Eqs. 1 and 3). However, the applicability of Rundel´s theories for

9
lichens has never been tested experimentally. There are still uncertainties regarding not only how lichens
physically will equilibrate with the adjacent atmosphere, but also what measures best describe the exchange
of water.
air t
t
ur
Ψ − Ψ= Eq. 1
where u is the uptake rate, rt is the thallus diffusion resistance to water movement (s cm-1), and Ψair and Ψt
are the water potentials (MPa) of the atmosphere and thallus, respectively. Ψair, was calculated using Eq. 2
throughout this thesis, modified from Nobel (1999), by omitting the adjustment for altitude:
ln( )100
airair
RT RHV
ψ = Eq. 2
where R is the gas constant (8.31441 J mol K-1), Tair is the air temperature (K), V is the partial molar volume
of water ( 618.021 10−⋅ m3mol-1 at 10 °C), and RH is the relative humidity of the air (%). Thus, the rate of
uptake per unit thallus is positively associated with an increasing water potential gradient between the
atmosphere and the thallus, and negatively associated with thallus diffusion resistance. The above Equation 1
is valid for hydration by humid air, but not for hydration by rain when water rapidly penetrates the lichen
thallus.
According to Rundel (1982), water loss from lichens is an evaporative process, (Eq. 3):
t a
t a
e eEr r
−=
+ Eq. 3
where E is the rate of evaporative loss, et and ea are the water vapor pressures (kPa) of the thallus tissue and
the adjacent atmosphere, respectively, and ra (s cm-1) is the boundary layer resistance for diffusion of water
vapor. The vapor pressure gradient between thallus and atmosphere constitutes the driving force for the
movement of water vapor out of the lichen. The vapor pressure of the atmosphere is a function of two
physical variables, namely atmospheric humidity and temperature. The vapor pressure of the evaporating
surface of the thallus depends on thallus water potential and temperature.
The water potential of the lichen thallus is thus mainly affected by the atmospheric water potential,
Ψair, but also by the osmotic potential of the thallus (Nash et al. 1990; Hajek et al. 2006). A low osmotic
potential increases the water potential gradient between atmosphere and thallus by reducing the thallus water
potential. Coastal, maritime lichens are in this way able to utilize a hygroscopic salt film to condense liquid
water out of humid air (Rundel 1978). Thallus water potential may also be affected by thallus color in
combination with solar radiation, which in turn influences thallus temperature (Kershaw 1975), and
consequently acts to decrease the thallus water potential (i.e. more negative) (Rundel 1988). Thallus
resistance to water movement is affected by the morphology and anatomy of the thallus (Larson &

10
Kershaw 1976; Larson 1981; Valladares et al. 1993; Valladares 1994; Máguas et al. 1997; Gauslaa &
Solhaug 1998). Thin thalli with high surface to volume ratios have lower resistances, characters typical for
fruticose (cf. as illustrated by the four thalli to the right on the front cover) or thin-lobed thalli (cf. as
illustrated by the second thallus from left on the front cover). However, any reduction in thallus resistance
that increases water uptake rate will also increase the rate of water loss (Rundel 1988). The boundary layer
resistance, ra, is directly related to the square roots of the width of the thallus and inversely to the wind speed
(ref). The boundary layer is affected by lichen growth form and/or position in the habitat, e.g. terriculous,
mat-forming lichens that typically show high boundary layer resistances (Kershaw & Field 1975; Larson &
Kershaw 1976; Gates 1980; Zotz et al. 2000). Since water moves by diffusion between thallus and air
(Monteith 1965), a significant boundary layer is likely to reduce also water uptake rates, as well as water
loss. Pendulous, fruticose lichens that hang freely in the air from their substrates thus have both low thallus
and boundary layer resistances.
Despite its mechanistic advantages, the model presented by Rundel (1982) (Eqs. 1 and 3) is very
detailed and requires data on parameters that are difficult to assess for lichens, such as the thallus vapor
pressure, and time-consuming, such as the thallus water potential. Moreover, parts of the theories for lichen
water exchange are probably just transferred from theories on physical water exchange that are valid for
vascular plants. For vascular plants with a more or less constant water supply, it can be assumed that the
water potential of the evaporating surface of internal leaf surfaces is approximately close to zero (Taiz &
Zeiger 1998), since this surface is comparable to an evaporating surface of water. Hence, the vapor pressure
at the leaf may be close to the saturation vapor pressure at the specific tissue temperature. For lichens,
however, this assumption has not been tested and is likely not valid since lichens do not have a constant
water supply to the evaporating surface due to their lack of water-carrying organs. Instead, their small water
holding bodies entails rapid energy and water exchange with adjacent atmosphere. Therefore, lichen thalli
will equilibrate their water potential with the surrounding atmosphere both during water uptake (Eq. 1) and
loss and must thus clearly have lower water potentials at the surface during desiccation compared to leaves. It
is therefore probable that lichen water loss can also be described by the thallus water potential equilibrating
with the atmosphere, since this is an effective parameter for describing water movement. This parameter is
also frequently used for this purpose in studies on vascular plants and water movement between their tissues
(Marshall & Woodward 1985). However, since the thallus water potential is difficult to assess, it would be
more convenient to circumvent the thallus water potential and directly relate the thallus hydration status to
atmospheric conditions. As an effect of the rapid exchange of water by the small lichen bodies, it can be
assumed that the thallus water potential is more or less constantly the same as the continually fluctuating
atmospheric water potential. If so, it would therefore be possible to relate the hydration status of the lichen to
the atmospheric water potential directly. However, some time-lag will persist due to the inherent resistances
to water vapor diffusion of the thallus and the adjacent boundary layer, as previously discussed.

11
2. AIMS
The aim of the studies underlying this thesis was to model field performance and growth of green-algal
macro-lichens for model species varying in morphology and spatial occurrence in the habitat. The aim was
further to develop models using atmospheric parameters that can be acquired from field data and characters
accounting for the physiology of lichens. The developed models are intended to be robust and based on
realistic mechanistic assumptions, with each of them adapted for use at the spatial and temporal scale, and the
question in focus.
The specific aims were to:
• Develop and evaluate a mathematical model for lichen wet active periods in the field, based on
micro-climatic variables only, or in combination with species-specific characters.
• Develop a model of lichen activities under field conditions with contrasting micro-climates.
• Simulate carbon gain responses of an epiphytic lichen to regional climate change in contrasting
habitats.
• Study whether predictive parameters for growth at the local, habitat scale can also be used at the
landscape scale and what model complexity is required at this scale.
3. MODEL DEVELOPMENT
Any attempt to develop a mechanistic model for the field performance of poikilohydric lichens based on
atmospheric conditions, requires the key assessment of their wet and active periods in the field. To do this,
two models were developed (Paper I) to predict these periods from micro-climatic conditions at a habitat
scale. It was assumed that the equilibrium water content (WCeq) of the lichens was related to the atmospheric
humidity conditions, irrespectively of whether WCeq was attained by water loss or by uptake (see section
1.5.2.). Both models therefore related the WCeq (%) to the atmospheric humidity conditions, which was given
by the atmospheric water potential, ψair (Eq. 4) (Fig. 4):
w •eq airWC = WC eeq sat
ψ Eq. 4
Where WCeq is the equilibrium water content (%), WCsat (%) is the water content of the water-saturated
thallus (100 %), weq (MPa-1) a species-specific water-binding constant, and ψair the atmospheric water
potential that was derived from field temperature and relative humidity data (Eq. 2). Above relationship (Eq.
4) was used in a physical model, which assumes that the hydration status is constantly in equilibrium with
ψair. This model was compared with a dynamic biophysical model that, in addition to the Eq. 4, also included
the time-dependent and species-specific water uptake and loss rates that asymptotically reach WCeq (Eq. 5):

12
( )WC = k WC WCamb eqt∂
− −∂
Eq. 5
where WCamb (%) is the ambient water content and k (min-1) is the rate constant for desiccation (kdes) or
rehydration (kreh), which is equivalent to conductance or the inverse of diffusion resistance. WCeq (%) was
derived from Eq. 4. An illustration of water movement towards equilibrium is shown in Fig. 5. The models
were developed for three lichen species with varying morphology and growth position in the habitat,
parameterized to the measured lichen WC and micro-climatic conditions during one summer at a sheltered
location, and validated at a more exposed location.
Figure 4. Relationships between the negative water potential of the air (– ψair) and the apparent equilibrium water content (WCeq) for Alectoria sarmentosa (A), Platismatia glauca (B), and C. rangiferina (C). An exponential decay function of the form
eq airweq satWC = WC e ψ⋅ (Eq. 4) was fitted to the data, where the parameter weq is a species-
specific constant specifying the shape of the curve (Table 3). WCsat was set to 100 percent. From Jonsson et al. (2008).
When lichens are hydrated, there is a general conception that they are instantaneously activated
(Palmqvist & Sundberg 2000, Dahlman & Palmqvist 2003), and this has also been demonstrated by other
authors (e.g. Coxson 1988; Lange et al. 1989). However, a few studies report a delay of up to 1 h until
maximum photosynthetic activity is reached (Lange et al. 1986; Palmqvist 2000). One potential mechanism
behind the restriction of some rare hydrophilic and epiphytic lichens to humid habitats (Ahlner 1948; Thor &
Arvidsson 1999) could be their slow photosynthetic activation when hydrated. We tested this assumption in

13
Paper III and developed a photosynthetic model (Paper II) to quantitatively estimate the photosynthetic
performance of the studied lichens in different micro-climatic scenarios. The hydration model (Paper I) was
therefore used and further developed to include photosynthetic activation kinetics and maximum potential
activity in relation to lichen hydration at a habitat scale (Eq. 6):
( )reala real pot
dA k A Adt
= − − Eq. 6
where Areal (%) is the realized activity that asymptotically reaches the maximum potential activity, Apot (%),
at a given ambient WC, with the species-specific activation rate ka (min-1). To parameterize the activity
model, we used laboratory data on species-specific photosynthetic activity in relation to WC. The WC and
activity parameterization (Apot from Eqs. 6-7; Paper II) and simulations were separated between water
sources, to increase the simulated precision of WC, as suggested in Paper I. To increase WC precision further,
we used the measured WC for the model species Platismatia norvegica as WCeq, to simulate the species-
specific WC. When expressed on a relative scale, the WCeq can be more or less assumed to be independent of
the species, particularly for epiphytic species with small boundary layers (Paper I).
A complete model for carbon gain was developed for Platismatia glauca (Paper IV). In contrast to the
activity model (Paper II), both photosynthesis and respiration were parameterized to predict the net carbon
gain (NCG) (Eqs. 1-6; Paper IV) and growth. A model that takes into account the physiological responses to
the local habitat scale micro-climate was required to simulate responses of total NCG to altered climate
conditions, e.g. climate change or contrasting micro-climates within a forest. This model was developed to
assess local responses to regional climate change in relation to forest site exposure, covering seasonal
changes during an entire year. The NCG model was parameterized with data from the literature on controlled
CO2 gas exchange responses to light, WC and temperature and with observed growth responses to field
micro-climate for P. glauca collected in darker, forest interior sites (Sundberg et al. 1997). The model was
used to simulate NCG and growth at two sites with contrasting micro-climates, one in the dark, interior forest
with closed canopy and one in an open, light exposed site of the forest (Sundberg et al. 1997), and then
validated for a larger number of dark, interior and open, exposed sites (Renhorn et al. 1997). The simulated
growth exceeded the observed growth at one of the exposed sites (Sundberg et al. 1997), but was similar to
the growth at a larger number of exposed sites with similar growth conditions (Renhorn et al. 1997). These
findings implied that the developed NCG model works well for interior conditions and for lichens adapted to
light exposed sites.
Although lichens respond physiologically to the local scale micro-climatic conditions (Papers I-IV), it
is interesting from a management and conservation perspective to scale up these models and evaluate them at
a regional landscape scale (Paper V). We anticipated that the more mechanistic variables, such as wet time
(WT), Iwet, or Chl a (Palmqvist & Sundberg 2000; Tretiach & Carpanelli 1992; Dahlman & Palmqvist 2003;
Gaio-Oliverira et al. 2006), would be too detailed to be applied with the available data to larger scales.
Instead, models with more descriptive parameters, such as meteorological data on precipitation,

14
temperature and RH, and forest stand characteristics, such as site openness, which are more easily measured
or accessed for larger scales, would be more suitable to predict lichen growth. For management of lichen-rich
habitats it would further be important to identify habitats with high growth potentials. Therefore, a more
simplified model was developed for use in situations of more practical applicability, including the parameters
local basal area of the forest stand and regional scale temperature and precipitation climate normals. Growth
was measured seasonally over 16 months, covering two summer and one winter periods. The models´ ability
to predict growth at this scale was analyzed statistically, both annually and seasonally, and related to their
complexity. The best model was selected for each species with the Bayesian Information Criterion (BIC)
(Burnham & Anderson 2004), which is a tool to select the model with the highest predictive power with as
few parameters as possible.
4. MODEL SYSTEMS
4.1. Study sites All field studies were performed in the boreal forests of northern Scandinavia (Fig. 6), generally at sites
dominated by Norway spruce (Picea abies) with scattered Scots pines (Pinus sylvestris). In the landscape
scale study (Paper V), some additional sites were chosen dominated by mountain birch in the subalpine zone
in Hemavan, Västerbotten.
Habitats represented as wide a range of micro-climatic conditions as possible, to increase the
generality of the studies. In Papers I-III, we aimed to select forest sites representing a wide range of
hydration patterns. The study areas in Paper II included stream habitats with close proximity to calm flowing
streams that were compared with sites with turbulent flowing streams and more distant streams (Paper III). In
the investigations reported in Paper IV, we aimed to select habitats with contrasting micro-climates that
naturally occur within a forest stand, varying in irradiance, hydration and temperature conditions. To develop
models for lichen growth at the landscape scale (Paper V), the lichens were transplanted across northern
Scandinavia along the widest possible natural west-east gradient, varying in precipitation and temperature.
Within this climate gradient, a light exposure gradient was chosen that ranged from dense forest stands to
open clear-cuts.
The study sites were chosen either for parameterization and validation (Papers I and IV), to quantify
lichen responses in a range of micro-climatic scenarios (Papers II, III and IV), or to statistically relate local
forest stand characteristics, semi-mechanistic, and descriptive landscape-scale parameters to conditions with
high lichen growth at the landscape scale (Paper V).

15
4.2. Model species To predict the wet active periods under field conditions (Paper I), three lichen species with different
morphologies and growth positions in the habitat were chosen. These were two epiphytic species with high
coupling to atmospheric conditions – the foliose Platismatia glauca (L.) W. Culb. & C. Culb (I – V), and the
fruticose, pendulous Alectoria sarmentosa (Ach.) Ach (Paper I, V) – and one terriculous, mat-forming,
fruticose lichen, Cladina rangiferina (L.) Weber ex Wigg. To cover variations in local micro-climatic
exposure, the monitored lichen thalli were equally distributed between exposed and sheltered sites.
Four red-listed hydrophilic lichens were chosen to study their performance in relation to stream
habitats (Paper II and III): the fruticose pendulous Bryoria bicolor (Ehrh.) Brodo & D.Hawksw, the thick-
lobed foliose Lobaria amplissima (Scop.) Forssell., the foliose Platismatia norvegica (Lynge) W. L. Culb. &
C. F. Culb., and the fruticose, pendulous Usnea longissima Ach.. These were compared with a generalist
species, the thin-lobed, foliose P. glauca (Papers 1-V). We adopted the term “generalist” species for species
that are relatively abundant in a large variety of habitats.
We chose P. glauca to simulate potential climate change responses of NCG (Paper IV) partly because
extensive literature data are available on CO2 gas exchange responses for this species under a wide range of
controlled micro-climatic conditions with accompanying growth measurements (Renhorn et al. 1997;
Sundberg et al. 1997), and partly because we had acquired relevant micro-climatic data from contrasting field
habitats, also with growth measurements. The micro-climates and subsequent growth were measured over a
long reference period of 16 months. In addition, the NCG model was based on WC data that were simulated
by a lichen hydration model developed for this species (Paper I).
We chose two mat-forming terriculous lichen species, Cladina stellaris (Opiz) Brodo and Cetraria
islandica (L.) Ach., to study the patterns of growth and model requirements at the regional landscape scale
(Paper V). These species were chosen because they are relatively common and important as forage for
16
reindeer during winter (Gaare & Danell 1999). In addition, the growth of two epiphytic lichens, P. glauca
and A.sarmentosa, was also studied for limited summer periods. However, the acquired data were not
statistically processed or included in Paper V, although they are referred to in the discussion below.
5. RESULTS AND DISCUSSION
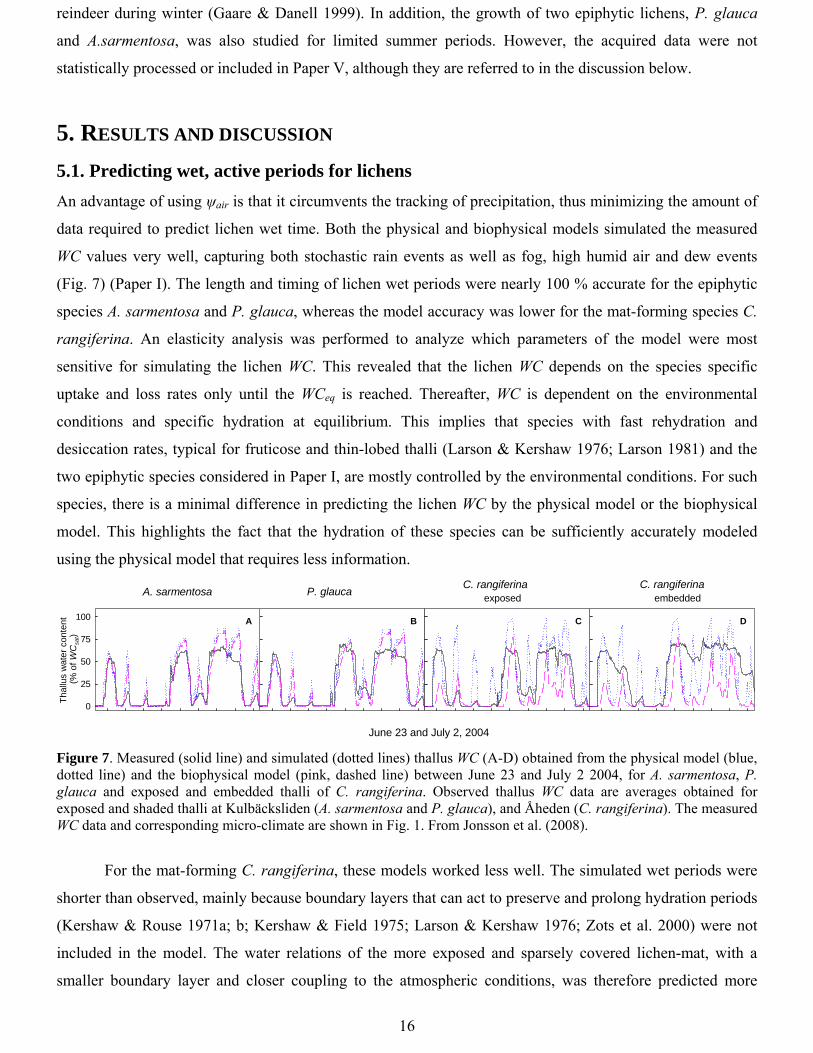
5.1. Predicting wet, active periods for lichens An advantage of using ψair is that it circumvents the tracking of precipitation, thus minimizing the amount of
data required to predict lichen wet time. Both the physical and biophysical models simulated the measured
WC values very well, capturing both stochastic rain events as well as fog, high humid air and dew events
(Fig. 7) (Paper I). The length and timing of lichen wet periods were nearly 100 % accurate for the epiphytic
species A. sarmentosa and P. glauca, whereas the model accuracy was lower for the mat-forming species C.
rangiferina. An elasticity analysis was performed to analyze which parameters of the model were most
sensitive for simulating the lichen WC. This revealed that the lichen WC depends on the species specific
uptake and loss rates only until the WCeq is reached. Thereafter, WC is dependent on the environmental
conditions and specific hydration at equilibrium. This implies that species with fast rehydration and
desiccation rates, typical for fruticose and thin-lobed thalli (Larson & Kershaw 1976; Larson 1981) and the
two epiphytic species considered in Paper I, are mostly controlled by the environmental conditions. For such
species, there is a minimal difference in predicting the lichen WC by the physical model or the biophysical
model. This highlights the fact that the hydration of these species can be sufficiently accurately modeled
using the physical model that requires less information.
0
25
50
75
100 A B C
A. sarmentosa exposed embedded
June 23 and July 2, 2004
C. rangiferina C. rangiferina
Thal
lus
wat
er c
onte
nt
(% o
f WC
sat)
P. glauca
D
Figure 7. Measured (solid line) and simulated (dotted lines) thallus WC (A-D) obtained from the physical model (blue, dotted line) and the biophysical model (pink, dashed line) between June 23 and July 2 2004, for A. sarmentosa, P. glauca and exposed and embedded thalli of C. rangiferina. Observed thallus WC data are averages obtained for exposed and shaded thalli at Kulbäcksliden (A. sarmentosa and P. glauca), and Åheden (C. rangiferina). The measured WC data and corresponding micro-climate are shown in Fig. 1. From Jonsson et al. (2008).
For the mat-forming C. rangiferina, these models worked less well. The simulated wet periods were
shorter than observed, mainly because boundary layers that can act to preserve and prolong hydration periods
(Kershaw & Rouse 1971a; b; Kershaw & Field 1975; Larson & Kershaw 1976; Zots et al. 2000) were not
included in the model. The water relations of the more exposed and sparsely covered lichen-mat, with a
smaller boundary layer and closer coupling to the atmospheric conditions, was therefore predicted more

17
accurately than those of the lichens at more sheltered sites. To better predict the water contents of lichens
with a pronounced boundary layer, a factor for net radiation should be included in the model in addition to
the ψair, since net radiation mainly drives water loss under these conditions (Monteith 1965).
Moreover, for a robust hydration model for green-algal lichens it is important to predict all wet
periods, irrespective of the water source. The physical model predicted the hydration by rain events better
than the biophysical model, since hydration by rain occurs within minutes (Blum 1973; Rundel 1988) and the
assumption of instantaneous WCeq such as in the physical model, is more appropriate for modeling water
uptake during rain. The biophysical model, on the other hand, modeled the humid air conditions more
accurately than the physical model, since the dynamic water uptake in humid air was incorporated in the
biophysical model.
The developed wet models were not adjusted to fit precise WC levels, although the simulations
apparently also captured the dynamics near maximum WC. For that purpose, the models would likely be
improved by partition between water sources in the parameterization and simulations. However, that would
entail the added complexity of tracking these water sources during collection, parameterization and
simulations. However, despite the increased complexity, a separation of water sources was applied in the
next step (Papers II and III).
The modeled wet periods of lichens can now be combined with the summed irradiance during the wet
times for correlation with lichen growth (Palmqvist & Sundberg 2000; Dahlman & Palmqvist 2003) (Paper
V). It is also possible to combine the species-specific hydration patterns with photosynthetic activity data to
predict the photosynthetic performance of lichens (Papers II and III), or to combine the simulated WC level
with the continuously varying irradiance and temperature conditions to assess instantaneous metabolic rates
(Paper IV).
5.2. Modeling lichen activity Comparison of the simulated hydration patterns with field conditions showed that the differences in species-
specific rehydration and desiccation kinetics significantly affected lichen wet time (Paper II). This confirmed
the results from Paper I and studies by other authors (Larson & Kershaw 1976; Larson 1981) that species
with thick lobes and thus slow desiccation rates can stay wet longer than species with thin lobes and faster
desiccation rates. For example, L. amplissima, stayed wet 8-26 % longer than P. glauca close to a stream.
Lichens with a higher surface to air ratio (the fruticose B. bicolor and U. longissima, and the thin-lobed P.
norvegica) had even faster desiccation rates and stayed wet for 3-13 % less time than P. glauca (Paper II-III).
Lichen activity was measured as Chl a fluorescence, which indicates photosynthetic performance.
When fully hydrated, P. glauca and L. amplissima were immediately activated, but the hydrophilic lichens
showed very slow activation rates, being fully activated after 4 h for P. norvegica, after 24 h for U.
longissima and potentially the same time for B. bicolor (Paper III). Inter-specific variations in hydration and
activity patterns had profound effects on the simulated photosynthetic performance close to the stream

18
(Papers II and III). Compared with P. glauca the realized activity was 4-38 % higher for L. amplissima, while
it was 19-22 %, 36-60 % and 52-66 % lower for P. norvegica, B. bicolor and U. longissima, respectively.
When the utilization of different water sources was compared, humid events accounted for 43-51 %
of the total wet time, but only 3-23 % of the total realized activity close to one stream site (Paper II). This
discrepancy reflects the lower photosynthetic activity induced by humid air than by rain (Paper III),
confirming earlier results (Lange et al. 1986; Scheidegger et al. 1995; Pintado & Sancho 2002). Not only
water source but also hydration patterns have an effect on photosynthetic activity. If the total wet time is
separated into many brief events, typical for humid air events, the realized activity for species with slow
activation can be further reduced. However, humid periods are important for high photosynthetic output
when they coincide with high irradiance (Brosofske et al. 1997; Danehy & Kirpes 2000), and for prolonging
wet active periods following rain events.
The differences in realized activity between species and among different water sources (Paper II)
increased with variations in micro-climatic scenarios within an examined forest stand (Paper III). The
hydration patterns vary with distance from a stream, due to the presence of a cooling water body, turbulent
water and local topography. When including stream sites varying in quality (calm and turbulent) and distance
(0 and 20 m) to streams, the simulations showed that close proximity to streams and presence of turbulent
water had a strong positive impact on realized activity among the slowly activated species (Paper III).
Compared with a distance of 20 m, the photosynthetic output of U. longissima was twice as high close to the
stream. However, the performance of L. amplissima was still three times higher than that of U. longissima
close to the stream, where L. amplissima was aided by rapid reactivation and very high water-holding
capacity. This pattern is consistent with the observed distribution patterns of hydrophilic species and could
explain their restriction to stream habitats.
5.3. Modeling local lichen responses in relation to regional climate change The modeling of lichen responses to two regional climate change scenarios of temperature, humidity
and irradiance resulted in contrasting local responses of the studied epiphytic lichen. The scenarios; IPCC
SRES A2 and B2, assume a more intense climate change or a more moderate change, respectively, in
comparison with most other SRES scenarios (IPCC 2007). Cumulative biomass growth during one year at
the darker, interior site was lower in climate change scenarios A2 and B2, 4.5 % and 4.2 %, respectively,
than in the reference climate (4.9 %). However, growth was equal or higher at the lighter, exposed site (9.7 %
and 8.2 % in the A2 and B2 scenarios, respectively, compared to 8.4 % for the reference climate). These
results are intriguing, implying that responses to general regional climate change are complex and can not
easily be applied to the local scale.
The growth response was two times higher at the exposed site compared with the closed forest
interior site, irrespective of climate scenario, which is in line with previous studies on epiphytic lichen
growth (Renhorn et al. 1997; Gauslaa et al. 2007; Jansson et al. In press). However, the response to climate

19
change was mainly due to the changed patterns of hydration and whether the concomitant light conditions
were limiting or not during the wet periods. Responses deviated compared with reference growth for three
periods: November-December, March-April and June-August. The elevated temperature increased the wet
active time for P. glauca in spring and late autumn. In November, the generally low light conditions were
limiting at the interior site with closed canopy, resulting in higher respiration and a potential weight loss
compared with the reference scenario. At the exposed site, the irradiance levels were somewhat higher,
resulting in higher photosynthesis, which offset the increased respiration. In April, the light conditions were
generally higher and apparently not limiting at either the exposed or the interior site. Growth was therefore
higher in response to altered climate change. In the projected warmer and drier summer period from June to
August, the wet time decreased, giving generally lower gross photosynthesis rates and growth compared with
the reference climate.
5.4. Patterns and models for growth of mat-forming lichens at landscape scale Irradiance was consistently the most important factor for mat-forming lichen growth at the landscape scale,
and appeared in all the best models, predicting annual growth even better than summer growth (Tables 3 and
5, Paper V). This indicates that sufficient irradiance, and thus a relatively open forest canopy, is critical for
optimal growth of this type of lichen. This was also suggested by Sulyma and Coxson (2001). Both model
species reached light saturation at a site openness value of ca 40 % (Fig. 8) (Paper V), equivalent to a forest
basal area of 15 m2 ha-1 in this study. Denser forests appeared to provide suboptimal light conditions. Optimal
light conditions could be successfully identified using site openness (SO) or basal area.
In addition, lichen growth in the most open, exposed forests was occasionally restricted, indicating
that other factors, such as water availability, were limiting. Lichens desiccate and become inactive more
rapidly at exposed sites (Gaio-Oliveira et al. 2004; Palmqvist et al. 2008). At the regional scale, the highest
growth was found at the more humid locations, compared with the more continental environments (Fig. 8., in
Paper V), as reported in other studies (cf. Armstrong 1993; Muir et al. 1997; Palmqvist & Sundberg 2000).
The combination of irradiance and hydration therefore yielded robust models, irrespective of species and
time period (Tables 3 and 5). The logarithmic SO (LSO) and precipitation explained 52 % of the variation in
annual growth, while models using LSO, WT and Iwet explained 66 % of the variation of annual growth of C.
stellaris and C. islandica. For both species, the predictive power of temperature and precipitation was greater
when only sites with optimal light conditions were considered, rather than the entire dataset (Tables 3 and 4).
Because of the negative correlation between humidity and temperature, especially for C. stellaris (Table 6),
the lichens displayed negative growth responses to increased temperatures (Tables 3 and 4).
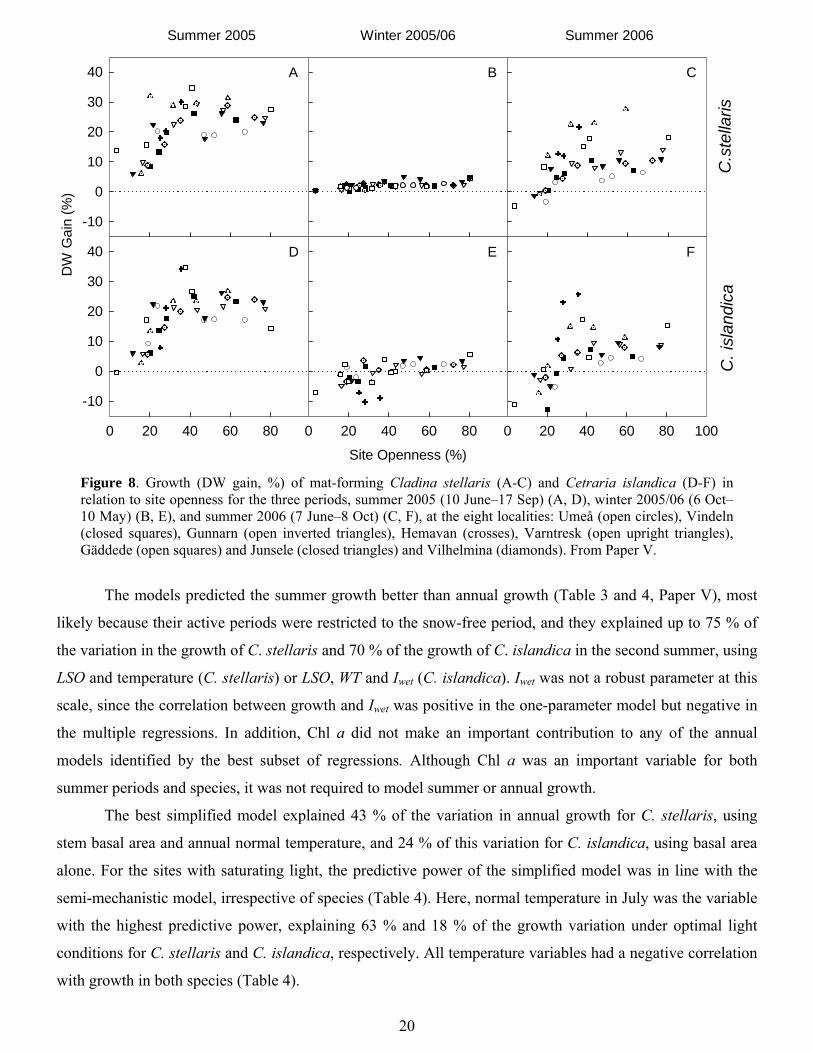
20
0 20 40 60 80 100
Vindeln
0 20 40 60 80
Vindeln
Site Openness (%)
0 20 40 60 80
DW
Gai
n (%
)
-10
0
10
20
30
40
-10
0
10
20
30
40 A B
D E F
C
C.s
tella
ris
Summer 2005
C. i
slan
dica
Winter 2005/06 Summer 2006
Figure 8. Growth (DW gain, %) of mat-forming Cladina stellaris (A-C) and Cetraria islandica (D-F) in relation to site openness for the three periods, summer 2005 (10 June–17 Sep) (A, D), winter 2005/06 (6 Oct–10 May) (B, E), and summer 2006 (7 June–8 Oct) (C, F), at the eight localities: Umeå (open circles), Vindeln (closed squares), Gunnarn (open inverted triangles), Hemavan (crosses), Varntresk (open upright triangles), Gäddede (open squares) and Junsele (closed triangles) and Vilhelmina (diamonds). From Paper V.
The models predicted the summer growth better than annual growth (Table 3 and 4, Paper V), most
likely because their active periods were restricted to the snow-free period, and they explained up to 75 % of
the variation in the growth of C. stellaris and 70 % of the growth of C. islandica in the second summer, using
LSO and temperature (C. stellaris) or LSO, WT and Iwet (C. islandica). Iwet was not a robust parameter at this
scale, since the correlation between growth and Iwet was positive in the one-parameter model but negative in
the multiple regressions. In addition, Chl a did not make an important contribution to any of the annual
models identified by the best subset of regressions. Although Chl a was an important variable for both
summer periods and species, it was not required to model summer or annual growth.
The best simplified model explained 43 % of the variation in annual growth for C. stellaris, using
stem basal area and annual normal temperature, and 24 % of this variation for C. islandica, using basal area
alone. For the sites with saturating light, the predictive power of the simplified model was in line with the
semi-mechanistic model, irrespective of species (Table 4). Here, normal temperature in July was the variable
with the highest predictive power, explaining 63 % and 18 % of the growth variation under optimal light
conditions for C. stellaris and C. islandica, respectively. All temperature variables had a negative correlation
with growth in both species (Table 4).

21
5.5. Limitations to lichen growth 5.5.1. Micro-climatic limitations
Since lichens are poikilohydric, their growth is limited to both the length and frequency of their wet active
periods and by the prevailing climate conditions during this time. The limiting factors may thus vary both
spatially and temporally. Since the simulation and/or empirical studies of lichen performance and growth
were conducted in the boreal zone, the discussion in this thesis about limitations to lichen growth is mainly
restricted to this biome.
Within a forest stand, the local variations in light exposure are larger than variations in water
availability and apparently most limiting for generalist lichen species (Fig. 8) (Papers IV and V). This was
also concluded for terriculous mat-forming lichens by Sulyma & Coxson (2001), who showed that lichens
were confined to more open habitats, whereas bryophytes were dominant at darker sites. However,
differences in water availability have large spatial effects between stands within a regional climate gradient
(Fig. 8) (Paper V) (Gauslaa et al. 2007), temporally between seasons or years (Fig. 8) (Papers IV and V)
(Armstrong 1993; Muir et al. 1997; Caldiz 2004), and between scenarios in climate change simulations
(Paper IV). At exposed sites with saturating light conditions, increased hydration generally results in
increased growth (Fig. 8) (Paper IV and V) (Gauslaa et al. 2007), since the amount of irradiance during wet
time, Iwet, increases. However, at dark interior sites of the forest, increased water availability can increase
respiration relatively more than photosynthesis and result in decreased growth (Paper IV) (Gauslaa et al.
2006). This was simulated in response to a more humid climate change scenario (Paper IV), and observed in
more humid stands in the regional climate gradient (Fig. 8) (Paper V), where the growth increment was near
zero and lowest at the landscape scale. Hence, the growth responses to a light gradient are larger and more
extreme at more humid sites, with more negative responses at the dark sites and more positive responses at
the lighter sites (Paper IV and V), as also shown by Gauslaa et al. (2007).
In the study area described in Paper V, growth and humidity was negatively correlated with
temperature (Table 6, Paper V), as also shown by Caldiz (2004). This indicates that lichen growth is more
limited by humidity than by suboptimal temperatures, as previously suggested (Sancho et al. 2007b), and that
humidity is the most important limiting factor after light for lichen growth at this landscape scale. This also
indicates that responses to increases in water availability may be stronger than responses to increases in
temperature during wet time in climate changes (Paper IV). This was also suggested by Sancho et al.
(2007b).
More specifically, lichens seem to be limited by insufficient light intensities during their wet periods
(Paper IV), rather than the total irradiance during the wet time, i.e. Iwet. This factor has a good correlation
with photosynthesis, but not to respiratory loss. Hence, although it is a simple mechanistic parameter, it is
apparently too imprecise to accurately model lichen growth at dark, interior sites.
For more sensitive hydrophilic species, such as B. bicolor and U. longissima, the water availability

22
provided by a representative forest stand in the more continental parts of the boreal region in northern
Scandinavia, may not be sufficient to provide suitable lichen habitats. These species requires more
continuous water availabilities, such as those prevailing close to streams (Papers II and III), in suboceanic
climates (Ahlner 1948; Thor & Arvidsson 1999), on north-facing slopes and ravines, adjacent to mires or in
forests with high relative humidity (Thor & Arvidsson 1999), or in habitats with high positions (altitude) in
the landscape (Esseen et al. 1981). Moreover, many of the threatened lichen species are limited by low light
availabilities in dense forests (Thor & Arvidsson 1999; Gauslaa et al. 2007; Jansson et al. In press). Other
factors, such as poor dispersal or sensitivity to wind exposure with subsequent thallus fragmentation, are also
important for occurrence, particularly for U. longissima (Esseen et al. 1981). These reasons may further
explain this species’ confinement to old-growth continuity forests that are generally more open and provide
long-term habitats for lichens to accumulate high biomass, provided they are sheltered from direct wind
(Esseen & Ericson 1982; Thor & Arvidsson 1999). However, in addition to climatic factors, the distribution
of U. longissima is also strongly influenced by logging history and occurrence of fire refugias (Esseen et al.
1981; Esseen & Ericson 1982; Josefsson et al. 2005).
5.5.2. Lichen trait limitations
Traits that affect the capacity of lichens to maintain a wet thallus under light conditions will increase carbon
assimilation. Therefore, characters such as morphological and anatomical traits that affect rehydration and
desiccation rates have an impact on lichen growth rates. This is exemplified by the thick-lobed L.
amplissima, which was found to have a substantially slower desiccation rate and significantly longer wet
active periods than the thin-lobed lichens, P. glauca and P. norvegica (Paper II), and longer wet periods than
the fruticose lichens, U. longissima and B. bicolor (Papers II and III). Moreover, thick and large foliose
lichens with slow desiccation rates have in agreement with this been shown to have higher growth rates
(Snelgar & Green 1982; Gauslaa & Solhaug 1998). However, the increased boundary layer of the mat-
forming lichen C. rangiferina also prolonged the wet time, compared with epiphytic lichens (Paper I), and
increasingly so at sheltered sites (Paper I). Since terrestrial and/or mat-forming lichens may maintain
hydration better at drier, exposed sites with high irradiance than epiphytic lichens, which are more easily
desiccated at these sites, the mat-forming lichens can show high growth rates even at exposed sites (Paper V)
(Palmqvist & Sundberg 2000). Although the growth of the mat-forming lichens was somewhat hampered at
the most open, exposed sites (Fig. 8) (Paper V), the growth depression at these sites was even more evident
for the epiphytic lichens (data not shown) (cf. Gauslaa et al. 2007). On the other hand, epiphytic lichens may
not be as adversely affected by hydration in dark, shaded conditions, due to their reduced hydration and
subsequent respiration compared with mat-forming lichens. This was indicated for the summer period in
2006, when there were some losses for the mat-forming lichens (Fig. 8) (Paper V) that were not apparent for
the epiphytic lichens (not shown). However, to gain reliable insights into this pattern, additional growth
studies (in which the epiphytic lichens are also compared) should be performed during winter.
Slow activation by some hydrophilic lichens (Papers II and III) will restrict these species to sites

23
with sufficiently long hydration periods, since photosynthetic gains may be too low during brief, intermittent
hydration periods, as discussed in the previous section.
5.6. Spatial and temporal scale dependency Due to their high dependency on atmospheric conditions, lichens respond to their local micro-climatic light,
water availability and temperature conditions (Caldiz 2004; Gauslaa et al. 2007) and the inherent Chl a
(Tretiach & Carpanelli 1992; Palmqvist & Dahlman 2006; Gauslaa et al. 2007). However, the predictive
responses and the relative importance of these variables may vary at different scales.
5.6.1. Predictive variables at different scales
The most mechanistic procedure to model lichen growth would be to use the spatially and temporally
fluctuating patterns of incident irradiation, WC, and temperature in the micro-habitat. These micro-scale
parameters were previously directly measured (Coxson 1991; Palmqvist & Sundberg 2000; Sundberg et al.
2001; Dahlman & Palmqvist 2003; Gaio-Oliveira et al. 2004; 2006).
To model WC purely mechanistically, empirical data on the water potential and/or vapor pressure of
the thallus and air would be required, in combination with thallus and boundary layer resistances (see section
1.5.2.) (Rundel 1988). Thallus water potential (ψt ) is difficult to parameterize and would require data on
osmotic potential and thallus temperature, which for example affects WC at the micro-scale via temporal sun-
flecks.
At a habitat scale (e.g. within 10 meters in a local site), the air water potential (ψair) is easy to obtain
(Eq. 2) (Papers I-IV) and is the main driver of water uptake and loss. Variations in ψt would only affect WC
at coastal, saline areas (osmotic potential), or at exposed habitats, where the thallus color in combination with
high solar radiation affect ψt more than expected by atmospheric conditions alone. This implies that the
lichen wet model (Paper I) works well in conditions where the temperatures of the thallus and the atmosphere
are similar, and in non-saline habitats. For epiphytic lichens with high exposure and rapid energy exchange
with atmospheric conditions the thallus temperature will be nearly identical to atmospheric conditions
(Coxson et al. 1984). Similarly, within sheltered forest habitats, detailed data for ψt would then not be
required to model the average WC. The epiphytic lichen WC was also accurately predicted by the lichen wet
model (Paper I), and more precisely with the species-specific rehydration and desiccation rates during humid
air events. However, for mat-forming lichens (Paper I), the model would probably be improved by including
the boundary layer resistance and a factor for radiation, especially at exposed sites where net radiation rather
than atmospheric water potential drives water loss (Monteith 1965).
At the landscape scale, detailed data on local scale ψair are not available to predict WC reliably, since
forest closure alters micro-climatic conditions at this scale (Chen et al. 1993; 1995). Instead, descriptive data
on precipitation predicted growth of C. stellaris better than the simulated WT from modeled temperature and
RH data for C. islandica (cf. Table 4, Paper V). Moreover, the temperature sum or normal temperature in

24
July (cf. Table 4, Paper V) were the single factors that best correlated with growth of C. stellaris and C.
islandica, respectively, for sites with saturating light conditions at the landscape scale. Importantly, the
correlation between temperature and growth was negative for these species.
For landscape scale assessments of lichen growth, fluctuating irradiance is not required and the light
availability at the local scale in the habitat can instead be successfully assessed indirectly by hemispherical
digital photographs to quantify site openness (or, inversely, canopy cover), as in Gauslaa et al. (2007) (Paper
V), or by basal area of trees (Paper V). If the fluctuating pattern of irradiance is still of interest, then site
openness can be combined with simulations of the fluctuating irradiance at a regional scale – as modeled in
the STRÅNG-model (Landelius et al. 2001) in Paper V – or averaged from the measured micro-climatic
variations (Paper IV).
5.6.2. Predictive power of parameters
The predictive power of a parameter may vary across temporal and spatial scales. Whilst some parameters
are valid across scales, others are affected by interacting variables at particular scales. Irradiance and
humidity are important for lichen performance at all scales (Papers II-V), although predicted using different
parameters at different scales (Papers I-V). However, Chl a may be most important at the habitat scale with
otherwise limited variation in forest openness and over a short time-period (Gaio-Oliveira et al. 2006;
Gauslaa et al. 2007), but not necessarily at a landscape scale across a wide range of light and humidity
conditions (Paper V) (Gauslaa et al. 2006). This is probably partly due to the smaller variation in Chl a
relative to light and water availability at the landscape scale and partly to growth being more limited by the
latter factors than by Chl a at this scale (Paper V). Lichen growth has also been well predicted by Iwet at a
local, habitat scale (Paper IV) (Palmqvist & Sundberg 2000; Dahlman & Palmqvist 2003), but the predictive
power was lower at the landscape scale (Paper V). The lack of predictive power is probably associated with
differences in spatial and temporal scales, since determining Iwet requires relatively precise data on both
irradiance and hydration that are not easily acquired at the landscape scale, as already discussed.
For light availability, regional scale patterns are of much less importance than the local scale patterns.
In Papers IV and V, lichen growth was shown to increase in response to higher light availabilities at a local
scale and was not reduced by lower light availabilities at the regional scale, as observed empirically close to
the mountains (Paper V), and simulated for climate scenarios with varying light intensities (Paper IV).
5.7. Implications for managing lichen-rich habitats Predictive models are potentially valuable tools that can facilitate the management of lichens that are either
rare or have important functional roles in the ecosystem, e.g. mat-forming lichens that are important for
winter grazing reindeer (Gaare & Danell 1999), or epiphytic lichens that can provide food and shelter for
invertebrates (Pettersson et al. 1995).

25
5.7.1. Generalist species
Generalist lichen species, as exemplified by P. glauca, C. stellaris and C. islandica are mostly limited by
light exposure in the habitat (Paper IV and V) (Sulyma & Coxson 2001; Gauslaa et al. 2007). Optimum
growth of mat-forming lichens occurred at sites with > 40% site openness (conversely <60 % canopy cover)
or basal tree areas < 15 m2 ha -1 (Paper V) in this study area. Ensuring sufficient irradiance below the forest
canopy is thus of crucial importance in the long-term management of mat-forming lichens (Paper V), which
was also suggested by Sulyma & Coxson (2001).
For the epiphytic lichens in this study area (Paper V), the preliminary data for short summer periods
indicate that growth is hampered at the most exposed sites. This was also found by Gauslaa et al. (2007) for
U. longissima and Lobaria pulmonaria. However, our sites represent clear-cuts where epiphytic lichens lack
natural substrates (Paper V) (Gauslaa et al. 2007). In contrast, peak growth for the epiphytic lichens at
continental sites occurred at a site openness of 40-50 % (Paper V) or 60 %, as also found by Gauslaa et al.
(2007). This site openness can be found in open old-growth forests (Gauslaa et al. 2007). At wetter sites, the
growth was hampered very little at the more exposed sites, as also shown for U. longissima (Gauslaa et al.
2007). Although the climate conditions are often favorable at exposed sites with higher irradiation levels, e.g.
at forest edges, these can induce adverse biomass loss by increased wind exposure (Esseen 2006). Open old-
growth forests would then provide favorable habitats for high growth with low wind exposure. However, to
promote high lichen biomass of epiphytic lichens at a stand-level, high growth at exposed but sparsely
forested sites must be balanced against retaining enough trees as growth substrates (Stevenson & Coxson
2007). Stevenson & Coxson (2007) concluded, in a synthesis of studies on arboreal lichens in British
Columbia, that managers should restrict basal area removal to a maximum of 30 % in partial cuts to maintain
lichen forage for the mountain caribou.
5.7.2. Hydrophilic lichens
Papers II and III emphasized the importance of extended hydration periods for lichens that are hampered by
slow activation. Both close proximity to streams and the presence of turbulent water had a consistently strong
positive impact on realized activity among these species. This pattern is consistent with the observed
distribution patterns of hydrophilic species and could hence explain their confinement to humid or sheltered
habitats, e.g. streams. To identify suitable climate conditions for hydrophilic lichens with slow activation, the
activity model (Paper II) provides an important tool in order to quantify and analyze the success of a species
in a habitat with a given water regime. Since these species are also hampered in dark forests (Thor &
Arvidsson 1999; Gauslaa et al. 2007; Jansson et al. In press), and high wind exposure (Esseen et al. 1981),
habitats that provide sheltered, humid and open conditions should be preserved, e.g. open forests close to
streams or other humid habitats.

26
5.7.3. Climate change
Understanding and predicting species-response to climate change is a key issue in conservation strategy
(Ellis et al. 2007). Climate change that causes increased growth of vascular plants tends to cause a decline in
terrestrial macrolichen biomass, due to increased levels of competition (Cornelissen et al. 2001; van Wijk et
al. 2004). However, according to (Cornelissen et al. 2001) this was not observed in the coldest high-arctic or
arctic-alpine sites, mainly due to the low competition at these sites. Lichens that are little subjected to
competition with vascular plants, i.e. saxicolous and epiphytic lichens or terrestrial lichens at dry exposed,
cold high-arctic or arctic-alpine sites, might on the other hand respond directly to climatic variables. Lichen
populations may move northward on a broad scale (Ellis et al. 2007; van Herk et al. 2002; Lättman H.,
Milberg P., Palmer M.W. & Mattsson J-E, unpublished data) following anticipated climate changes in the
boreal region, but this trend might cease in the northernmost ecosystems where epiphytic lichens will be
limited by increased respiration during warmer, dark winter periods. The climate change simulations based
on the combined effects of generally increased temperature, humidity and decreased light, indicated that
lichen populations would also be redistributed at a local scale towards open, exposed habitats (Paper IV).
More favorable growth conditions at these sites would reinforce the current pattern of increased growth at
such sites. Warmer and wetter conditions in dark, interior sites may thus promote a shift towards a more
bryophyte-dominated vegetation. These organisms are better adapted to these conditions by their low light
compensation point and water separating tissue that supports CO2 diffusion at high WCs (Proctor 1990).
Hence, with projected future climate change, exposed habitats, e.g. open forests, will be increasingly
important for maintaining high lichen biomass, preferably habitats sheltered from wind exposure; conditions
typically found in open old-growth forests.
6. CONCLUSIONS
This thesis has contributed to develop a model framework for lichen performance under field conditions.
Many of the lichen responses studied in this thesis would not have been possible without the development
and simulations of mathematical models. Models enable prediction of responses to future changes by
manipulations of the system to for instance varying micro-climate. Thus, these models provide important
tools for predicting responses of lichens in a range of habitats with varying climate conditions. This can be
used to analyze the potential success of a species in a habitat which can further help to develop optimal
management strategies for threatened or otherwise important species.
The models and results presented in this thesis can then help us to:
Predict lichen hydration in the habitat in relation to the atmospheric conditions, using micro-climatic
RH and T, and thereby accounting for several water sources with high accuracy, especially for
epiphytic lichens.
Predict photosynthetic performance of lichens in the habitat, given the species-specific hydration and

27
activity patterns. This can identify suitable habitats for lichen species, especially those that are confined
to humid habitats and more specifically to long hydration periods.
Predict responses of lichens to climate change on the local habitat scale, including their intricate
responses to projected changes in light, hydration and temperature.
Understand regional scale patterns of growth for mat-forming lichens and what predictors that can be
used to successfully predict their growth at this scale.
This thesis has also gained more knowledge about the relevant scale and factors (S&F) when studying
lichen vegetation. These must be taken into account for the specific question (Q) to be addressed:
(Q): Fine-scale lichen distribution patterns in e.g. canopy distribution, position on branch or stem.
(S&F): Reduced micro-scale; incorporating detailed, thallus-specific water relations, color, growth form,
osmotic potential and thallus-specific responses to water, light and temperature variations.
(Q): Responses to forestry, climate change and optimal micro-climatic conditions for a lichen population
within a stand
(S&F): Habitat scale; incorporating the species´ morphology, growth form, the site and position in the
habitat, species-specific activation time-lag, and their responses to water, light and temperature variations.
(Q): Regional distribution or regional patterns in lichen growth
(S&F): Landscape to regional scale; data on normal precipitation and temperature, and habitat-scale light
conditions. Species-specific growth form and position in the habitat.
7. ACKNOWLEDGEMENTS
I would like to thank Kristin Palmqvist, Erling Ögren, Per-Anders Esseen, Lars Ericson, Jon Moen and Uno
Wennergren for valuable comments on previous versions of this summary. I would like to thank SMHI,
Rossby Centre, Vindeln experimental forest, SLU, and Per-Anders Esseen for providing meteorological data,
land owners and others for providing access to study sites, Håkan Eriksson and Sonja Kivinen for preparing
Figure 6, and Ulrika Jansson and Johan Asplund for proof-reading. The work in this thesis was funded by
grants from the Swedish Research Council for Environmental, Agricultural Sciences and Spatial planning
(Formas, Sweden) and the Centre for Environmental Research (CMF, Umeå) to Kristin Palmqvist and the
Björkman Foundation to Anna Jonsson Čabrajić.
8. REFERENCES
Ahlner S. (1948) Utbredningstyper bland nordiska barrträdslavar. Acta Phytogeographica Suecica 22: 1-157.

28
Ahmadijan V. (1967) A guide to the algae occurring as lichen symbionts: isolation, culture, cultural physiology and
identification. Phycologia 6: 127.
Armstrong R.A. (1993) Seasonal growth of foliose lichens in successive years in south Gwynedd, Wales.
Environmental and Experimental Botany 33: 225-232.
Berg A., Östlund L., Moen J. & Olofsson J. (2008) A century of logging and forestry in a reindeer herding area in
northern Sweden. Forest Ecology and Management 256: 1009-1020.
Bergh J., McMurtie R.E. & Linder S. (1998) Climatic factors controlling the productivity of Norway spruce: a model
based analysis. Forest ecology and management 110: 127-139.
Blum O.B. (1973) Water relations. In: The Lichens (eds. Ahmadjian V. & Hale M.E.), pp 381-400. Academic Press,
New York.
Botting R.S. & Fredeen A.L. (2006) Net ecosystem CO2 exchange for moss and lichen dominated forest floors of old-
growth sub-boreal spruce forests in central British Columbia, Canada. Forest Ecology and Management 235: 240-
251.
Brosofske K.D., Jiquan C., Naiman R.J. & Franklin J.F. (1997) Harvesting effects on microclimatic gradients from
small streams to uplands in western Washington. Ecological Applications 7: 1188–1200.
Burnham K.P. & Anderson D.R. (2004) Multimodel interference: understanding AIC and BIC in model selection.
Sociological Methods & Research. 33: 261-305.
Caldiz M.S. (2004) Seasonal growth pattern in the lichen Pseudocyphellaria berberina in north-western Patagonia.
Lichenologist 36: 435-444.
Chen J.Q., Franklin J.F. & Spies T.A. (1993) Contrasting microclimates among clear-cut, edge, and interior of old-
growth Douglas-fir forest. Agricultural and Forest Meteorology 63: 219-237.
Chen J.Q., Franklin J.F. & Speis T.A. (1995) Growing season microclimatic gradients from clear-cut edges into old-
growth Douglas-fir forest. Ecological Applications 5: 74-86.
Cornelissen J.H.C., Callaghan T.V., Alatalo J.M., Michelsen A., Graglia E., Hartley A.E., Hik D.S., Hobbie S.E.,
Press M.C., Robinson C.H., Henry G.H.R., Shaver G.R., Phoenix G.K., Jones D.G., Jonasson S., Chapin F.S.,
Molau U., Neill C., Lee J.A., Melillo J.M., Sveinbjornsson B. & Aerts R. (2001) Global change and arctic
ecosystems: is lichen decline a function of increases in vascular plant biomass? Journal of Ecology 89: 984-994.
Cornelissen J.H.C., Lang S.I., Soudzilovskaia N.A. & During H.J. (2007) Comparative cryptogam ecology: A review
of bryophyte and lichen traits that drive biogeochemistry. Annals of Botany 99: 987-1001.
Coxson D.S. (1988) Recovery of net photosynthesis and dark respiration on rehydration of the lichen Cladina mitis,
and the influence of prior exposure to sulphur dioxide while desiccated. New Phytologist 108: 483-487.
Coxson D. (1991) Impedance measurement of thallus moisture content in lichens. Lichenologist 23: 77-84.
Coxson D.S. & Coyle M. (2003) Niche partitioning and photosynthetic response of alectorioid lichens from subalpine
spruce-fir forest in north-central British Columbia, Canada: the role of canopy microclimate gradients.
Lichenologist 35: 157-175.
Coxson D.S., Webber M.R. & Kershaw K.A. (1984) the thermal operating environment of corticolous and pendulous
tree lichens. The Bryologist 87: 197-202.
Crews T.E., Kurina L.M. & Vitousek P.M. (2001) Organic matter and nitrogen accumulation and nitrogen fixation
during early ecosystem development in Hawaii. Biogeochemistry 52: 259-279.
Dahlman L. & Palmqvist K. (2003) Growth in two foliose tripartite lichens, Nephroma arcticum and Peltigera

29
aphthosa: empirical modelling of external vs internal factors. Functional Ecology 17: 21-831.
Danehy R.J. & Kirpes B.J. (2000) Relative humidity gradients across riparian areas in eastern Oregon and Washington
forests. Northwest Science 74: 224–233.
Demmig-Adams B., Máguas C., Adams III W.W., Meyer A., Kilian E. & Lange O.L. (1990) Effect of high light on the
efficiency of photochemical energy conversion in a variety of lichen species with green and blue-green
phycobionts. Planta 180: 400-409.
den Herder M., Kytövitta M.M. & Niemela P. (2003) Growth of reindeer lichens and effects of reindeer grazing on
ground cover vegetation in a Scots pine forest and a subarctic heathland in Finnish Lapland. Ecography 26: 3-12.
Ellis C.J., Coppins B.J., Dawson T.P. & Seaward M.R.D. (2007) Response of British lichens to climate change
scenarios: Trends and uncertainties in the projected impact for contrasting biogeographic groups. Biological
Conservation 140: 217-235.
Epstein H.E., Calef M.P., Walker M.D., Chapin F.S. & Starfield A.M. (2004) Detecting changes in arctic tundra plant
communities in response to warming over decadal time scales. Global Change Biology 10: 1325-1334
Esseen P.-A. (2006) Edge influence on the old-growth forest indicator lichen Alectoria sarmentosa in natural ecotones.
Journal of Vegetation Science 17: 185-194.
Esseen P.-A. & Ericson L. (1982) Spruce forests with the lichen Usnea longissima in Sweden. Rapport PM 1513.
Swedish Environmental Protection Agency, Solna.
Esseen P.-A. & Renhorn K.E. (1998) Mass loss of epiphytic lichen litter in a boreal forest. Annales Botanici Fennici
35: 211-217.
Esseen P.-A., Ericson L., Lindström H. & Zackrisson O. (1981) Occurrence and ecology of Usnea longissima in
central Sweden. Lichenologist 13(2): 177-190.
Esseen P.-A., Renhorn K.E. & Pettersson R.B. (1996) Epiphytic lichen biomass in managed and old-growth boreal
forests: effect of branch quality. Ecological Applications 6: 228-238.
Friedl T. & Büdel B. (1996) Photobionts. In: The Biology of Lichens (ed. Nash III T.H.), pp. 8–23. Cambridge
University Press, Cambridge.
Gaare E. & Danell Ö. (1999) Bruk av betier of områder. In: Reindrift i Nordvest-Europa i 1998 – biologiske
muligheter og begrensninger (eds. Dahle H.K. Gaare Ö., & Nieminen M.). Tema Nord, 1999(510): 47-56. Nordisk
ministerråd, Kobenhavn.
Gaio-Oliveira G., Dahlman L., Máguas C. & Palmqvist, K. (2004) Growth in relation to microclimatic conditions and
physiological characteristics of four Lobaria pulmonaria populations in two contrasting habitats. Ecography 27:
13-28.
Gaio-Oliveira G., Moen J., Danell O. & Palmqvist K. (2006) Effect of simulated reindeer grazing on the re-growth
capacity of mat-forming lichens. Basic and Applied Ecology 7: 109-121.
Gates D.M. (1980) Biophysical Ecology. Springer, Berlin, Heidelberg, New York.
Gauslaa Y. & Solhaug K.A. (1996) Differences in the susceptibility to light stress between epiphytic lichens of ancient
and young boreal forest stands. Functional Ecology 10: 344-354.
Gauslaa Y. & Solhaug K.A. (1998) The significance of thallus size for the water economy of the cyanobacterial old-
forest lichen Degelia plumbea. Oecologia 116: 76-84.
Gauslaa Y., Lie M., Solhaug K. & Ohlson M. (2006) Growth and ecophysiological acclimation of the foliose lichen
Lobaria pulmonaria in forests with contrasting light climates. Oecologia 147: 406-416.

30
Gauslaa Y., Palmqvist K., Solhaug K.A., Holien H., Hilmo O., Nybakken L., Myhre L.C. & Ohlson M. (2007) Growth
of epiphytic old forest lichens across climatic and successional gradients. Canadian Journal of Forest Research
37: 1832-1845.
Green T.G.A., Kilian E. & Lange O.L. (1991) Pseudocyphellaria dissimilis: a desiccation sensitive, highly shade-
adapted lichen from New Zealand. Oecologia 85: 498-503.
Green T.G.A., Lange O.L. & Cowen I.R. (1994) Ecophysiology of lichen photosynthesis: the role of water status and
thallus diffusion resistances. Cryptogamic Botany 4: 166-178.
Green T.G.A., Schlensog M., Sancho L.G., Winkler J.B., Broom F.D. & Schroeter B. (2002) The photobiont
determines the pattern of photosynthetic activity within a single lichen thallus containing cyanobacterial and green
algal sectors (photosymbiodeme). Oecologia 130: 191-198.
Green T.G.A., Schroeter B. & Sancho L.G. (2007) Plant life in Antarctica. In: Functional Plant Ecology, 2nd edn.
(eds. Pugnaire F.I. & Valladares F.), pp. 389-433. Marcel Dekker, New York.
Hajek J., Barták M. & Gloser J. (2001) Effects of thallus temperature and hydration on photosynthetic parameters of
Cetraria islandica from contrasting habitats. Photosynthetica 39: 427-435.
Hajek J., Barták M. & Dubova J. (2006) Inhibition of photosynthetic processes in foliose lichens induced by
temperature and osmotic stress. Biologia Plantarum 50(4): 624-634
Honegger R. (1991) Functional aspects of the lichens symbiosis. Annual Review of Plant Physiology and Plant
Molecular Biology 42: 553-578.
Hyvärinen M. & Crittenden P.D. (1998) Growth of the cushion-forming lichen, Cladonia portentosa, at nitrogen-
polluted and unpolluted heathland sites. Environmental and Experimental Botany 40: 67-76.
IPCC (2007) Climate change 2007: Synthesis Report. Contribution of Working groups I, II and III to the Fourth
Assessment Report of the Intergovernmental Panel on Climate Change (eds. Core writing team, Pachuri R.K &
Reisinger A.), IPCC, Geneva, Switzerland.
Jansson U., Palmqvist K. & Esseen P.-A Growth of the old forest lichen Usnea longissima at forest edges. In press
Lichenologist
Jonsson A.V., Moen J. & Palmqvist, K. (2008) Predicting lichen hydration using biophysical models. Oecologia 156:
259-273.
Josefsson T., Hellberg E. & Östlund L. (2005) Influence of habitat history on the distribution of Usnea longissima in
boreal Scandinavia: a methodological case study. Lichenologist 37: 555-567.
Kappen L. (1988) Ecophysiological relationships in different climatic regions. In: CRC Handbook of lichenology. vol.
II. (ed. M. Galun). CRC Press, Boca Raton.
Kappen L. (2000) Some aspects of the great success of lichens in Antarctica. Antarctic Science 12: 314-324.
Kappen L., Lange O.L., Schultze E.D., Buschbom U. & Evenari M. (1980) Ecophysiological investigations on lichens
of the Negev desert VII. The influence of the habitat exposure on dew imbibition and photosynthetic productivity.
Flora 169: 216-229.
Kappen L., Sommerkorn M. & Schroeter B. (1995) Carbon acquisition and water relations of lichens in polar regions -
potentials and limitations. Lichenologist 27: 531-545.
Kershaw K.A. (1975) Studies on lichen-dominated systems. XII. The ecological significance of thallus colour.
Canadian Journal of Botany 53: 660-667.
Kershaw K.A. (1984) Seasonal photosynthetic capacity changes in lichens: a provisional mechanistic interpretation.

31
Lichenologist 16: 145-171.
Kershaw K.A. (1985) Physiological Ecology of Lichens. Cambridge University Press, Cambridge.
Kershaw K.A. & Field G.F. (1975) Studies on lichen-dominated systems. XV. The temperature and humidity profile in
a Cladina alpestris mat. Canadian Journal of Botany 53: 2614-2620.
Kershaw K.A. & Harris G.P. (1971) Simulation studies and ecology: a simple defined system model. Statistical
Ecology 3: 1-21.
Kershaw K.A. & Rouse W.R. (1971a) Studies on lichen-dominated systems. I. The water relations of Cladonia
alpestris in spruce-lichen woodland in northern Ontario. Canadian Journal of Botany 49: 1389-1399.
Kershaw K.A. & Rouse W.R. (1971b) Studies on lichen-dominated systems. II. The growth pattern of Cladonia
alpestris and Cladonia rangiferina. Canadian Journal of Botany 49: 1401-1410.
Kytöviita M.M. & Crittenden P.D. (2002). Seasonal variation in growth rate in Stereocaulon paschale. Lichenologist
34: 533-537.
Lambers H., Chapin III F.S. & Pons T.L. (1998) Photosynthesis, respiration, and long distance transport. In: Plant
Physiological Ecology (eds. Lambers H, Chapin III F.S.& Pons T.L.), pp. 10-95. Springer Verlag, Berlin.
Landelius T., Josefsson W. & Persson T. (2001) A system for modelling solar radiation parameters with mesoscale
spatial resolution. SMHI Reports Meteorology and Climatology 96.
Lange O.L. (1980) Moisture content and CO2 exchange in lichens. I. Influence of temperature on moisture dependent
net photosynthesis and dark respiration in Ramalina maciformis. Oecologia 45: 82-87.
Lange O.L. (2003) Photosynthetic productivity of the epilithic lichen Lecanora muralis: long-term field monitoring of
CO2 exchange and its physiological interpretation II. Diel and seasonal patterns of net photosynthesis and
respiration. Flora 198: 55-70.
Lange O.L. & Green T.G.A. (1996) High thallus water content severely limits photosynthetic carbon gain of central
European epilithic lichens under natural conditions. Oecologia 108: 13-20.
Lange O.L. & Green T.G.A. (2006) Nocturnal respiration of lichens in their natural habitat is not affected by
preceding diurnal net photosynthesis. Oecologia 148: 396-404.
Lange O.L. & Tenhunen J.D. (1981) Moisture content and CO2 exchange of lichens. II. Depression of net
photosynthesis in Ramalina maciformis at high water content is caused by increased thallus carbon dioxide
diffusion resistance. Oecologia 51: 426-429.
Lange O.L., Kilian E. & Ziegler H. (1986) Water vapor uptake and photosynthesis of lichens: performance differences
in species with green and blue-green algae as phycobionts. Oecologia 71: 104-110.
Lange O.L., Green T.G.A. & Ziegler H. (1988) Water status related photosynthesis and carbon isotope discrimination
in species of the genus Pseudocyphellaria with green or blue-green photobionts and in photosymbiodemes.
Oecologia 75: 494-501.
Lange O.L., Bilger W., Rimke S. & Schreiber U. (1989) Chlorophyll fluorescence of lichens containing green and
blue-green algae during hydration by water vapour uptake and by addition of liquid water. Botanica Acta 102: 306-
313.
Lange O.L., Budel B., Heber U., Meyer A., Zellner H. & Green T.G.A. (1993) Temperate rain-forest lichens in New-
Zealand - high thallus water-content can severely limit photosynthetic CO2 exchange. Oecologia 95: 303-313.
Lange O.L., Büdel B., Meyer A., Zellner H. & Zots G. (2000) Lichen carbon gain under tropical conditions: water
relations and CO2 exchange of three Leptogium species of a lower montane rainforest in Panama. Flora 195: 172-

32
190.
Lange O.L., Green T.G.A. & Heber U. (2001) Hydration-dependent photosynthetic production of lichens: what do
laboratory studies tell us about field performance? Journal of Experimental Botany 52: 2033-2042.
Lange O.L., Green T.G.A., Melzer B., Meyer A. & Zellner H. (2006) Water relations and CO2 exchange of the
terrestrial lichen Teloschistes capensis in the Namib fog desert: measurements during two seasons in the field and
under controlled conditions. Flora 201: 268-280.
Lange O.L., Green T.G.A., Meyer A. & Zellner H. (2007) Water relations and carbon dioxide exchange of epiphytic
lichens in the Namib fog desert. Flora 202: 479-487.
Larson D.W. (1979) Lichen water relations under drying conditions. New Phytologist 82: 713-731.
Larson D.W. (1981) Differential wetting in some lichens and mosses: the role of morphology. The Bryologist 84: 1-
15.
Larson D.W. (1984) Thallus size as a complicating factor in the physiological ecology of lichens. New Phytologist 97:
87-97.
Larson D.W. (1987) The absorption and release of water by lichens. Bibliotecha Lichenologia 25: 351-360.
Larson D.W. & Kershaw K.A. (1976) Studies on lichen-dominated systems. XVIII. Morphological control of
evaporation in lichens. Canadian Journal of Botany 54: 2061-2073.
Lechowicz M.J. (1982) Ecological trends in lichens photosynthesis. Oecologia 53: 330-336.
Lidén M. & Hilmo O. (2005) Population characteristics of the suboceanic lichen Platismatia norvegica in core and
fringe habitats: Relations to macroclimate, substrate, and proximity to streams. The Bryologist 108: 506-517.
Lloyd C.R. (2001) The measurement and modelling of the carbon dioxide exchange at a high Arctic site in Svalbard.
Global Change Biology 7: 405-426.
Máguas C., Valladares F. & Brugnoli E. (1997) Effects of thallus size on morphology and physiology of foliose
lichens: new findings with a new approach. Symbiosis 23: 149-164.
Marshall B. & Woodard F.I. (1985) Instrumentation for environmental physiology - Society for experimental biology
series; 22. Cambridge University Press, Cambridge.
Monteith J.L. (1965) Evaporation and environment. Symposium of the Society for experimental biology 19: 205-234.
Muir P.S., Shirazi A.M. & Patrie J. (1997) Seasonal growth dynamics in the lichen Lobaria pulmonaria. The
Bryologist 100: 458-464.
Nash III T.H. (2008) Lichen Biology, 2nd edn. Cambridge University Press. New York.
Nash III T.H., White S.L. & Marsh J.E. (1977) Lichen and moss distribution and biomass in hot desert ecosystems. The
Bryologist 80: 470-479.
Nash III T.H., Reiner A., Demmig-Adams B., Kilian E., Kaiser W.M. & Lange O.L. (1990) The effect of atmospheric
desiccation and osmotic water stress on photosynthesis and dark respiration of lichens. New Phytologist 116: 269-
276.
Nobel P.S. (1999) Physicochemical & Environmental Plant Physiology, 2nd edn. Academic Press, San Diego.
Palmqvist K. (2000) Carbon economy in lichens. New Phytologist 148: 11-36.
Palmqvist K. & Dahlman L. (2006) Responses of the green algal foliose lichen Platismatia glauca to increased
nitrogen supply. New Phytologist 171: 343-356.
Palmqvist K. & Sundberg B. (2000) Light use efficiency of dry matter gain in five macro-lichens: relative impact of
microclimate conditions and species-specific traits. Plant Cell and Environment 23: 1-14.

33
Palmqvist K., Campbell D., Ekblad A. & Johansson H. (1998) Photosynthetic capacity in relation to nitrogen content
and its partitioning in lichens with different photobionts. Plant Cell and Environment 21: 361-372.
Palmqvist K., Dahlman L., Valladares F., Tehler A., Sancho L.G. & Mattsson J.E. (2002) CO2 exchange and thallus
nitrogen across 75 contrasting lichen associations from different climate zones. Oecologia 133: 295-306.
Palmqvist K., Dahlman L., Jonsson A.V. & Nash III T.H. (2008) The carbon economy of lichens. In: Lichen Biology,
2nd edn (ed. Nash III T.H.), pp. 182-215. Cambridge University Press. New York.
Paterson D.R., Paterson E.W. & Kenworthy J.B. (1983) Physiological studies on temperate lichen species I. A
mathematical model to predict assimilation in the field, based on laboratory responses. New Phytologist 94: 605-
618.
Pech G.Y. (1989) A model to predict the moisture content of reindeer lichen. Forest Science 35: 1014-1028.
Pettersson R.B., Ball J.P., Renhorn K.-E., Esseen P.-A. & Sjöberg K. (1995) Invertebrate communities in boreal forest
canopies as influenced by forestry and lichens with implications for passerine birds. Biological Conservation 74(1):
57-63.
Pintado A. & Sancho L.G. (2002) Ecological significance of net photosynthesis activation by water vapour uptake in
Ramalina capitata from rain-protected habitats in central Spain. Lichenologist 34: 403-413.
Proctor M.C.F. (1990) The Physiological-Basis of Bryophyte Production. Botanical Journal of the Linnean Society
104: 61-77.
Renhorn K.-E. & Esseen P.-A. (1995) Biomass growth in five alectorioid lichen epiphytes. Mitt. Eidgenöss. Forsch.
Anst. Wald Scnee Landsch. 70 (1): 133-140.
Renhorn K.-E., Esseen P.-A., Palmquist K. & Sundberg B. (1997) Growth and vitality of epiphytic lichens. 1.
Responses to microclimate along a forest edge-interior gradient. Oecologia 109: 1-9.
Rice S.K., Collins D. & Anderson A.M. (2001) Functional significance of variation in bryophyte canopy structure.
American Journal of Botany 88(9): 1568-1576.
Rolstad J. & Rolstad E. (2008) Intercalary growth causes geometric length expansion in Methuselah's beard lichen
(Usnea longissima). Botany-Botanique 86: 1224-1232.
Rundel P.W. (1978) Ecological relationships of desert fog zone lichens. The Bryologist 81(2): 277-293.
Rundel P.W. (1982) The role of morphology in the water relations of desert lichens. Journal of the Hattori Botanical
Laboratory 53: 315-320.
Rundel P.W. (1988) Water relations. In Handbook of Lichenology, vol 2. (ed Galun M.), pp. 17-36. CRC Press, Boca
Raton.
Sancho L.G., de la Torre R., Horneck G., Ascaso C., de los Rios A., Pintado A., Wierzchos J. & Schuster M. (2007a)
Lichens survive in space: Results from the 2005 LICHENS experiment. Astrobiology 7: 443-454.
Sancho L.G., Green T.G.A. & Pintado A. (2007b) Slowest to fastest: Extreme range in lichen growth rates supports
their use as an indicator of climate change in Antarctica. Flora 202: 667-673.
Saunders D.A., Hobbs R.J. & Margules C.R. (1991) Biological consequences of ecosystem fragmentation: a review.
Conservation Biology 5: 18-32.
Scheidegger C., Schroeter B. & Frey B. (1995) Structural and functional processes during water vapour uptake and
desiccation in selected lichens with green algal photobionts. Planta 197: 399-409.
Smith D.C. (1980) Mechanisms of nutrient movement between lichen symbionts. In: Cellular Interactions in
Symbiosis and Parasitism (eds. Cook C.B., Pappas P.W. & Rudolph E.D.), pp. 197-227. State University Press,

34
Columbus, Ohio.
Snelgar W.P. & Green T.G.A (1982) Growth rates of Stictaceae lichens in New Zealand beech forests. The Bryologist
85: 301-306.
Stevenson S.K. & Coxson D.S. (2003) Litterfall, growth, and turnover of arboreal lichens after partial cutting in an
Engelmann spruce - subalpine fir forest in north-central British Columbia. Canadian Journal of Forest Research-
Revue Canadienne de Recherche Forestiere 33: 2306-2320.
Stevenson S.K. & Coxson D.S. (2007) Arboreal forage lichens in partial cuts - a synthesis of research results from
British Columbia, Canada. Rangifer 17: 155-165.
Sulyma R. & Coxson D.S. (2001) Microsite displacement of terrestrial lichens by feather moss mats in late seral pine-
lichen woodlands of north-central British Columbia. The Bryologist 104: 505-516.
Sundberg B., Palmquist K., Esseen P.-A. & Renhorn K.-E. (1997) Growth and vitality of epiphytic lichens. 2.
Modelling of carbon gain using field and laboratory data. Oecologia 109: 10-18.
Sundberg B., Ekblad A., Näsholm T. & Palmqvist K. (1999) Lichen respiration in relation to active time, temperature,
nitrogen and ergosterol concentrations. Functional Ecology 13: 119-125.
Sundberg B., Näsholm T. & Palmqvist K. (2001) The effect of nitrogen on growth and key thallus components in the
two tripartite lichens, Nephroma arcticum and Peltigera aphthosa. Plant, Cell and Environment 24: 517-527.
Söchting U. (1995) Lichens as monitors of nitrogen deposition. Cryptogamic botany 5: 264-269.
Taiz L. & Zeiger E. (1998) Plant physiology, 2nd ed. Sinauer associates, Massachusetts, U.S.A.
Thor G. & Arvidsson L. (1999) Rödlistade lavar i Sverige - Artfakta. ArtDatabanken, SLU, Uppsala.
Tretiach M. & Carpanelli A. (1992) Chlorophyll content and morphology as factors influencing the photosynthetic
rate of Parmelia caperata. Lichenologist 24: 81-90.
Valladares F. (1994) Texture and hygroscopic features of the upper surface of the thallus in the lichen family
Umbilicariaceae. Annals of Botany 73: 493-500.
Valladares F., Wierzchos J. & Ascaso C. (1993) Porosimetric study of the lichen family Umbilicariaceae: anatomical
interpretation and implications for water storage capacity of the thallus. American Journal of Botany 80(3): 263-
272.
van der Wal R. (2006) Do herbivores cause habitat degradation or vegetation state transition? Evidence from the
tundra. Oikos 114: 177-186.
van Herk, C.M., Aptroot A. & van Dobben H.F. (2002) Long-tern monitoring in the Netherlands suggests that lichens
respond to global warming. Lichenologist 34: 141-154.
van Wijk M.T., Clemmensen K.E., Shaver G.R., Williams M., Callaghan T.V., Chapin F.S., Cornelissen J.H.C.,
Gough L., Hobbie S.E., Jonasson S., Lee J.A., Michelsen A., Press M.C., Richardson S.J. & Rueth H. (2004)
Long-term ecosystem level experiments at Toolik Lake, Alaska, and at Abisko, Northern Sweden: generalizations
and differences in ecosystem and plant type responses to global change. Global Change Biology 10: 105-123.
Wessels J.G.H. (1993) Wall growth, protein excretion and morphogenesis in fungi. New Phytologist 123: 397-413.
Zheng D., Freeman M., Bergh J., Rosberg I. & Nilsen P. (2002) Production of Picea abies in south-east Norway in
response to climate change: A case study using process-based model simulation with field validation.
Scandinavian Journal of Forest Research 17: 35-46.
Zotz G., Schweikert A., Jetz W. & Westerman H. (2000) Water relations and carbon gain are closely related to cushion
size in the moss Grimmia pulvinata. New Phytologist 148: 59-67.

35
9. SAMMANFATTNING
Lavar kan kolonisera nästan alla terrestriska habitat i världen och är funktionellt viktiga i många ekosystem.
Eftersom lavar är poikilohydriska (växelblöta), är deras aktiva tillväxtperioder begränsade till den tid då
bålen är blöt från atmosfäriska vattenkällor, såsom regn, dimma och hög relativ fuktighet. Eftersom lavars
vatteninnehåll varierar stort över tid är lavars tillväxt svårare att modellera jämfört med till exempel
kärlväxter, med en mer jämn vattentillgång.
Jag har utvecklat två fuktmodeller som förutsäger lavars vatteninnehåll i fält. Modellerna använder
den atmosfäriska vattenpotentialen (Ψair), som erhålls från lufttemperatur och –fuktighet, antingen enbart
eller i kombination med de artspecifika uppblötnings – och uttorkningshastigheterna. Genom att använda
(Ψair) kan man förutsäga lavars vatteninnehåll från flera vattenkällor. Dessa modeller var mycket precisa för
epifytiska lavar med en nära koppling till de atmosfäriska förhållandena, men fungerade mindre väl för
mattlevande lavar med ett betydande gränsskikt. Fuktmodellen utvecklades ytterligare för att inkludera även
fotosyntetisk aktivering av olika lavar, för att kunna jämföra deras aktivitet i olika mikroklimatiska scenarior.
Vattenbalans och aktiveringshastighet hade stor effekt på på lavars aktivitet och var positivt relaterad till
habitat med tillräckligt långa fuktperioder, till exempel habitat nära strömmande vatten. För att studera
klimateffekter på lavar, utvecklade jag en total modell för nettoförvärv av kol (fotosyntes minus respiration)
för en epifytisk lav med dess intrikata förhållande mellan ljus, fukt och temperatur. Simuleringar av modellen
visade att lavens responser i förhållande till regionala klimatförändringar var kontrasterande på lokal nivå.
Vid ljusa, öppna lokaler i skogen ökade tillväxten medan de potentiellt minskade vid mörka lokaler med ett
mer slutet krontäcke.
På den lokala skalan kan fluktuerande vatteninnehåll, summerat ljus när laven är blöt, och klorofyll a-
innehåll förutsäga lavars tillväxt. Men, på en landskapsskala kan dessa variabler vara för detaljerade. Vi
testade detta för två terrestriska, mattlevande lavar och utvecklade en statistisk modell för lavars tillväxt i en
så stor klimatgradient som möjligt i norra Skandinavien genom att variera ljus, temperatur och nederbörd.
Ljus var den viktigaste faktorn för att nå hög tillväxt på landskapsnivå där en mättnad nåddes vid 40 %
öppenhet i skogen, som motsvarade en grundyta på 15 m2 ha -1 i den här studien. Fuktigheten var den näst
viktigaste begränsande faktorn och kunde beskrivas väl med nederbörd för en av arterna. Den mest
lättanvända faktorn var normaltemperaturen för juli månad, som i sin tur var negativt korrelerad till tillväxt.
Det var tydligt att de prediktiva variablerna och deras förutsägande förmåga varierade med olika
skalor. Ljus och fukt var begränsande på alla nivåer, speciellt av ljusförhållandena då lavarna är blöta.
Detta innebär att tillräckligt höga ljusnivåer under krontäcket är avgörande för lavars tillväxt, speciellt
mattlevande lavar. Hydrofila lavar torde bevaras bättre i öppna habitat med tillräckligt långa fuktperioder.
Det var tydligt att modeller kan vara betydelsefulla och lättanvända verktyg för att förutsäga lavars
responser i en bredd av habitat med olika mikroklimat. Modeller kan därför vara en hjälp för att
identifiera lämpliga habitat med optimala tillväxtförhållanden och detta är viktigt för att bevara och sköta
lavar och deras habitat.

36
Tack! Till en början var jag väl inte så imponerad av lavar, jag såg de mest som spännande modell (!) organismer.
Men under de här åren har jag fått lära mig mycket om deras värld, vad de gillar, hur de fungerar, och hur de
reagerar när grannträdet försvinner med skogsmaskinen eller om klimatet blir varmare. Jag hade en
verktygslåda för modeller (för torskar, var ni medvetna om det mina handledare ?!) som jag fått utveckla fritt
efter egen förmåga. Detta har varit otroligt utvecklande, fyllt av motgångar men också triumfer när jag
uppnått det jag velat. Jag inser nu att lavar faktiskt är märkvärdiga organismer som överlever det mesta, och
som även tål att modelleras...
För allt detta, och så mycket mer, vill jag tacka min främsta, eminenta handledare Kikki Palmqvist. Tack vare
dig var den här resan möjlig! Du har förstås lärt mig allt om lavar, men framförallt är du en bra förebild som
forskare och ledare med en skön självdistans. Det har stundtals varit ganska tufft att vara din doktorand men
jag har också tack vare det växt otroligt mycket och du har alltid lyckats ge mig en värmande knuff i rätt
riktning när jag behövt det. Du har den speciella förmågan att vara både lyhörd och vetenskapligt riktigt
skarp och våra möten har innefattat allt från en hel del fniss om livet men också invecklade resonemang.
Med dig som handledare är det svårt att misslyckas, fler borde vara som du!
Mina andra handledare vill jag tacka för ert trygga stöd i bakgrunden:
Jon – som alltid kalibrerat kompasskursen mot varför jag jobbat med det jag gjort och är en trygg vägvisare
bland luddiga och omständliga formuleringar.
P-A – som lärt mig allt om varför lavar är viktiga, egentligen. Och för att jag fick följa med dig till British
Columbia i Kanada. Det var en otrolig resa! Hoppas det går bra med dina plockhuggningsförsök, kanske det
kan bli något mer där.
Mikaell – för dina uppmuntrande kommentarer, och allt fundamentalt du lärt mig om skogsmeteorologi.
Hade inte gått utan det!
Jag vill också passa på att tacka Erling för våra diskussioner kring vattnets fysikaliska principer. Det borde vi
gjort oftare! Agneta – för stärkande input i uppföljningsgruppen. Och Uno- Tack för dina glada hejarop och
för att du alltid ser så positivt på saker. Ja, du kom ju in både först och sist kan man säga och det känns riktigt
bra att kunna knyta ihop säcken med modelleringen i Linköping. Det spelar ju ingen roll om det är torskar
eller lavar, det är det som är så kul! Så började intresset som så småningom ledde till Umeå, och för det
förtjänar också resten av modelleringsgänget där nere en speciell plats här, speciellt till Bosse! De kunskaper
och erfarenheter jag lärde mig hos er var en bra start på mitt forskningsarbete.

37
Utan mina lavbrudar hade det här arbetet inte alls varit lika roligt. Stort tack Otilia, Ullis och Marlene för ert
sällskap genom åren! Ullis, dina vänliga själ var ett bra stöd som ny (och gammal) doktorand. Marlene, vi har
haft många intressanta diskussioner kring hotade lavar och hur de kan överleva över huvudtaget. Jag är bara
så imponerad av dina verbala förmågor. Gänget utökades ju också med några lavgrabbar – Johan och Per
(kvisslingarna i Norge, vad gör man inte för att få jobba med lavar?). Det har varit riktigt skoj att få umgås
med er med mycket skratt och lustiga illustrationer. Från Ås kommer också en spesiell jente Line. Jag minns
våra strapatser i Kalifornien och i Lissabon, det var kjempegrejt (!?). Sedan finns ju ett par väldigt speciella
semi-lavare också – Torbjörn och Anna B, ni är båda härliga personer. Med er alla i lavgänget har det blivit
många intressanta diskussioner om forskning men oftast om andra mycket roligare saker i lavstugor och på
lavresor. Jag hoppas att våra vägar fortsätter att korsas många gånger framöver!
Otilia förtjänar förstås en speciell plats. Vart ska jag börja? Jag har verkligen trivts med ditt sällskap under
den här resan, för att inte tala om vilken räddare i nöden du har varit så många gånger. Tusen tack för alla
diskussioner kring lavar, forskningsvärlden och livet framförallt! För ditt glada humör, och uppmuntrande
inställning till livet efter detta (disputationen alltså). Sedan är ju våra resor tillsammans liksom svåra att
matcha. Jag menar tänk att tillsammans få möta två av lavvärldens två STORA på våra resor från
storstadspuls i Seattle, Red Wood-skog i Oregon till promenader vid Atlantens sanddyner och vinkällare i
Würzburg. Ja, vi har ju faktiskt tittat på lavar också. Och inte minst, du och Pontus har hjälpt mig så mycket
med min husering i Umeå under mina regelbundna vistelser i Umeå. Vi har i princip varit särbos. Tusen
Tack!
Tack till er jag undervisat med, det har varit väldit lärorikt och trivsamt med er! Henrik – jag uppskattar din
uppriktighet och för att du alltid tar dig tid till att kolla läget, och de Medelpadska skogarna är ju bra fina,
Johan–för dina tänkvärda reflektioner och för allt du tålmodigt lärt mig om fjällens växter. Lotta – det gick ju
faktiskt väldigt bra med vår exkursion, och det var riktigt roligt dessutom, och Melanie – lycka till i fjällen i
sommar!
Tack alla ni andra som har förgyllt min tid i Umeå; Jögga, Martin L, Jenny A, Åsa B, Victoria, Magnus,
Mikaela Carolina, Johanna T, Kristin, Pieter och alla andra doktorander. Jag blir alltid så glad när jag träffar
er! Ni nya doktorander i forskargruppen – Elina, Tim och Ed, lycka till med er forskning! Snart är ni också
här.
Det finns ju också lite annat löst folk (från SLU), som trots allt är så kul att träffa varje gång ;). Anna L och
David – det var en lyx att få låna er lägenhet. Jenny W – som föregångare har du bidragit med flera goda råd
längs vägen. Maria K och din Erik – Tack för fikaträffar, sängplats och Eriks modelleringsinsatser. Lycka till
med allt!

38
Ann – vilken klippa du är när det gäller allt praktiskt, det kan nog alla som jobbat med dig intyga. Din
lugnande atmosfär gör att allt liksom faller på plats när du är i närheten.
Jag vill också passa på att tacka alla hjälpsamma människor som hjälpt mig med fältarbetet i mitt stora
landskapsförsök Västerbotten runt. Tack markägare och nyckel(!)personer som hjälpt mig med tillgång till
alla fältförsök. Tack också till ulliga får och små barn som gärna experimenterar med de hägn som ska
utesluta just dessa orosmoment. Det piggade upp!
Ni adminsitratörer som med evigt tålamod berättar allt man behöver veta om bilnycklar, reseräkningar och
konton och som alltid ställer upp i akututryckningar. Vet ni om hur mycket ni underlättar med allt det där
andra som man inte riktigt har tid med? Tack Sussi, Ingrid, Marie, Ann-Sofie, Mattias, Birgitta och Kerstin!
Till mina vapendragare i söder, Malin, Ola, Lina, Jenny och Mia, för era uppmuntrande och värmande ord på
vägen hit. Ser fram emot våra samtal och att umgås mer med er! Också till gänget i Ludvika med Jennie,
Jocke, Bella, Emma och Anna och ni andra som följt med längs vägen.
Tack mamma och pappa för er stöttning genom alla år. Mamma – för att du höll mig sällskap på flera
Västerbotten-runt-resor. Vi får hitta andra resmål nu, som kanske inte behöver vara lika tidspressade.
Pappa – för dina frågor kring lavarna på din skog, jag tror faktiskt att jag fått dig intresserad också för de små
i skogen! Dotra era kanske faktiskt trappar ner på nomadlivet. Tack också till Bror, Veronica och Helene!
Och till Fikreta, Mika och Alisa för ert stöd, speciellt under den här sista stressigare tiden!
Till sist – min käraste Zdenko – Du är den finaste och bästaste människa jag vet, som envist uppmuntrat och
stöttat mig vid min sida under dessa år. Och så fick jag ju fick testa dina egenskaper som fältslav också, och
du höll ju måttet. Men det är slut med det nu, va? Längtar efter att bara få vara med dig!