molecular cloning, functional expression, and selective regulation of ovine prostaglandin h...
TRANSCRIPT

BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS 227, 499–506 (1996)ARTICLE NO. 1536
Molecular Cloning, Functional Expression, and Selective Regulationof Ovine Prostaglandin H Synthase-21
Vivian Zhang, Mary O’Sullivan, Hameda Hussain,William T. Roswit, and Michael J. Holtzman2
Departments of Medicine and Cell Biology, Washington University School of Medicine, St. Louis, Missouri 63110
Received September 4, 1996
Structural characterization for ovine prostaglandin H synthase-1 (PGHS-1) is extensive, but the corre-sponding structure for the homologous ovine PGHS-2 isoform is undefined. Accordingly, we isolated afull-length (3.4 kb) ovine PGHS-2 cDNA from a primary-culture cell model (ovine tracheal epithelial cells)originally described as containing both PGHS isoforms. Analysis of ovine PGHS-2 cDNA sequence indi-cated conservation of critical amino acid residues, but differences in other hydrophilic regions allowed forthe development of an anti-peptide antibody highly selective for PGHS-2. Enzymatic activities of therecombinant ovine PGHS isozymes indicated significant differences in response to aspirin-acetylation consis-tent with the characteristics of endogenous cellular PGHS activities under basal and serum-induced condi-tions. The results fully account for previous evidence of two distinct PGHS activities in cultured airwayepithelial cells and provide for additional definition of PGHS structure–function relationships. q 1996
Academic Press, Inc.
Prostaglandin H synthase (PGHS) exists in two isoforms (PGHS-1 and PGHS-2) that eachconvert arachidonic acid to prostaglandin H2 (PGH2), the first committed step in the biosynthesisof biologically active prostaglandins and thromboxanes. PGHS features were defined first andremain defined most completely for ovine PGHS-1. This species was the first one cloned andsequenced (1, 2), and remains as the most extensively mutated (3-9). It is the only PGHS withthree-dimensional structure defined by x-ray crystallography (10). Together, these approacheshave defined critical structures governing PGHS-1 interaction with substrate and inhibitors.
The subsequent identification of PGHS-2 raised the possibilities that PGHS-1 and -2 maymediate constitutive versus inducible prostanoid production, respectively, and that isozyme-specific inhibitors might be designed for greater efficacy (11). Because structural features ofovine PGHS-1 are well defined, the design of inhibitors may be further facilitated by corre-sponding structural data for ovine PGHS-2. In fact, a PGHS isozyme (distinct from PGHS-1)exhibiting selective regulation and distinct pharmacology was also first identified using anovine cell culture system (12, 13), but these studies did not define ovine PGHS-2 sequenceor fully link the functional expression of 4.0 kb PGHS mRNA with the corresponding 72-kDapolypeptide. Accordingly, the present experiments were performed to (i) isolate the cDNA forovine PGHS-2 from cultured ovine tracheal epithelial cells (oTECs); (ii) compare the structurefor ovine PGHS-2 to PGHS-1; (iii) test the cDNA-encoded ovine PGHS-2 for enzymaticactivity and for reactivity with nonsteroidal anti-inflammatory drugs (especially aspirin); and
1 The nucleotide sequence described in this paper has been submitted to the GenBank database with the AccessionNo. U68486 (bankit id No. 66332).
2 To whom correspondence should be addressed at Washington University School of Medicine, 660 S. Euclid Ave.,Box 8052, St. Louis, MO 63110. Fax: (314) 362-8987; E-mail: [email protected].
Abbreviations: HETE, hydroxyeicosatetraenoic acid; oTEC, ovine tracheal epithelial cell; PG, prostaglandin; PGHS,prostaglandin H synthase.
0006-291X/96 $18.00Copyright q 1996 by Academic Press, Inc.All rights of reproduction in any form reserved.
499
AID BBRC 5523 / 690d$$$621 09-26-96 10:37:20 bbrca AP: BBRC

Vol. 227, No. 2, 1996 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
(iv) determine whether PGHS-2 is selectively regulated in (non-inflammatory) epithelial cellsthat express both isoforms.
MATERIALS AND METHODS
Materials. AMV reverse transcriptase was from Life Sciences; MMLV reverse transcriptase was from BRL; [a-32P]dCTP (3000 Ci/mmol) and [3H8]arachidonic acid (95.1 Ci/mmol) were from Dupont-New England Nuclear; PGHS-1 purified from ovine seminal vesicle was from Cayman Chemical Co., Inc.; and purified ovine placental andrecombinant chicken PGHS-2 were from Oxford Biomedical Research, Inc. Laboratory of Human Carcinogenesis(LHC) basal medium was obtained from Biofluids and was supplemented with ovine pituitary extract, epidermalgrowth factor, epinephrine, hydrocortisone, insulin, transferrin, triiodothyronine, L-glutamine, calcium chloride, traceelements, penicillin, and streptomycin (LHC-8e) as described previously for oTEC culture (12-14). Authentic referencecompounds included (15S)-hydroxy-(5Z,8Z,11Z,13E)-eicosatetraenoic acid (15-HETE) and PGB2, PGD2, PGE2, andPGF2a obtained from Biomol Research Laboratories Inc. or Cayman Chemical Co.
Antibodies. Three rabbit antisera raised against ovine seminal vesicle PGHS-1 were obtained from Oxford Biomedi-cal Research (PG-16 and PG-20) and Cayman Chemical Co. (#160103). A rabbit antisera raised against PGHS-2 N-terminus was also obtained from Oxford Biomedical Research (PG-26). In addition, rabbit antisera were raised againstdeduced amino acid sequences of murine PGHS-2 using synthetic peptides conjugated to keyhole limpet hemocyaninvia an N-terminal cysteine as antigens (15). IgG was purified from antisera by peptide-affinity chromatography usingantigenic peptide and Sulfolink coupling gel (Pierce Chemical). Peptide sequences were selected because of theirconservation across murine and chicken PGHS-2 (16-18) and their dissimilarity to ovine PGHS-1 (1, 2) as well astheir predicted hydrophilicity (19). Preliminary experiments indicated that antisera (designated MH.745/6) from bothrabbits immunized against one of these peptides exhibited selective reactivity with recombinant chicken and purifiedovine PGHS-2 at high titer (1:50,000 dilution). Neither pre-immune sera nor antisera against other peptides exhibitedPGHS reactivity.
PCR-based amplification of ovine PGHS-2 mRNA. Total RNA was isolated from cultured oTECs (12) usingguanidinium isothiocyanate lysis and centrifugation through cesium chloride (20). Poly(A)/ RNA was fractionatedwith a biotinylated oligo(dT) probe-streptavidin magnetic particle system (Promega) and was used to generate corre-sponding cDNA using MMLV reverse transcriptase. The cDNA product was amplified by PCR using an annealingcycle at 557C and Hot Tub DNA polymerase (Amersham). DNA primers were based on the sequences surroundingthe active site Tyr385 (4) and the aspirin-acetylation site Ser532 in murine PGHS-2 (17, 18) which are conserved inovine seminal vesicle PGHS-1 (1, 2) and chicken embryo fibroblast PGHS-2 (16) and were constructed to also containextensions with either XhoI (CTCGAG) or SpeI (ACTAGT) restriction sites at their 5* ends. The downstream primer5*-GATCACTCGAGATGGGATTTCCCATAAGTCCTTTCAAGGAGAA-3* contained an Xho1 site and nucleotides1668-1698 of murine PGHS-2 which included Tyr385, and the upstream primer 5*-AATGCACTAGTGAATTCAACA-CACTCTATCACTGGCACCCCCT-3* contained an SpeI site and nucleotides 1219-1251 of murine PGHS-2 whichincluded Ser532. Reverse-transcriptase PCR products were selected for further pursuit on the basis of size, Southernblotting against oligonucleotides containing internal PGHS-2 sequence, and Northern blotting against epithelial cellmRNA. PCR products were purified and ligated into XhoI/SpeI-digested pBlueScript II SK(//0) phagemid (Stratagene)with T4 DNA ligase. The resulting phagemid was used to transform E. Coli DH1 cells, and recombinant colonieswere detected by white colony formation on IPTG/XGAL plates. Phagemid insert was subjected to DNA sequencingusing the dideoxy chain termination method with the Sequenase Kit (U. S. Biochemicals) and primers based onflanking sequences of the T3 and T7 promoters in pBluescript.
cDNA library construction and screening. Poly(A)/RNA from oTECs was fractionated by oligo(dT) affinity chroma-tography and converted to double-stranded blunt-ended cDNA (21). EcoRI(NotI) linker-adapters were added usingthe reagents contained in the Copy Kit (Invitrogen Corp.). After size selection by agarose gel electrophoresis, thecDNA was ligated into a phage cloning vector (lambda ZAP II, Stratagene) and then packaged in vitro (Gigapack IIGold, Stratagene). The library was amplified in E. Coli (host strain XL1-Blue, Stratagene) and was screened with32P-labeled cDNA generated from reverse transcriptase-PCR as noted above and then radiolabeled with [a-32P]dCTPby random primer synthesis using the Klenow fragment of DNA polymerase I (Multiprime, Amersham Corp.) to aspecific activity of 109 cpm/mg. Phage from positive plaques was subjected to secondary screening and then rescuedinto pBluescript using f1 helper phage (R408, Stratagene). Insert size was determined by agarose gel eletrophoresisafter digestion with NotI. Initial sequence was obtained using primers based on flanking sequences contained inpBluescript. Both strands of a full-length cDNA for ovine PGHS-2 (designated oPGHS-2) were sequenced using alinear sequencing strategy.
Preparation of expression vectors and transfection of Cos-7 cells. For ovine PGHS-1, contiguous 0.5 kb and 1.5kb cDNAs (2) were released from M13 phage by digestion with XbaI and EcoRI and then were gel purified andsuccessively ligated into the EcoRI/XbaI-digested expression vector pECE (22) to form a 2.0 kb cDNA (oPGHS-1-coding) containing the complete open-reading frame. In addition, this insert was released with KpnI/XbaI, blunt-endedwith T4 DNA polymerase, and ligated into a new SmaI site in the expression vector pOSMLv. The parent vector
500
AID BBRC 5523 / 690d$$$621 09-26-96 10:37:20 bbrca AP: BBRC

Vol. 227, No. 2, 1996 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
(pOSML) lacking this SmaI site was obtained from W. Smith (Michigan State Univ.) and Genetics Institute, Inc (23).For the newly cloned PGHS-2, oPGHS-2 was released from pBluescript by digestion with NotI, blunt ended with theKlenow fragment of DNA polymerase I, and ligated into the SmaI site of pECE or pOSMLv. In addition, the codingregion of ovine PGHS-2 (designated oPGHS-2-coding) was expressed in pECE and pOSMLv. To generate pECE-oPGHS-2-coding, pECE was cut with BalI/XbaI (removing the 3*-UTR and a portion of the coding region of oPGHS-2) followed by PCR to restore the full coding region. To generate pOSMLv-oPGHS-2-coding, the coding region wasreleased from pECE with XbaI/HindIII, filled in with Klenow, and blunt ligated into the SmaI site in pOSMLv.Expression vector constructs (pECE- and pOSMLv-oPGHS-1-coding and pECE- and pOSMLv-oPGHS-2 and -oPGHS-2-coding) were verified by agarose gel electrophoresis after digestion with PstI or PstI/XbaI, and all inserts weresequenced to insure proper orientation and sequence reproduction. The plasmids were grown in ampicillin-containingT broth, then purified through cesium chloride, and used to transfect Cos-7 cells (ATCC CRL-1651) by lipofectin-mediated gene transfer (24).
Northern analysis of recombinant and epithelial cell PGHS. Total cellular RNA was prepared from transfectedCos-7 cells and cultured oTECs and was subjected to electrophoresis in a 1.0% agarose gel containing 1 M formalde-hyde. Equal amounts of RNA were loaded/lane on the basis of absorbance at 260 nm, and equivalency of sampleamounts was initially verified by the intensity of ethidium bromide staining of the 28S and 18S rRNA bands. RNAwas electroblotted to nylon membranes which were prehybridized for 15 min and then hybridized for 1 h at 687C in10 ml of QuikHyb solution (Stratagene) with 32P-labeled cDNAs for ovine PGHS-1 or -2 prepared as described above.32P-labeled cDNAs were added to the hybridization mixture in 300 ml of 10 mg/ml salmon sperm DNA to achieve afinal probe concentration of 1-2 1 106 cpm/ml. Blots were washed twice with 21 SSC plus 0.1% SDS for 15 minat 257C and then once in 0.11 SSC (SSC Å 0.15 M NaCl and 0.015 M sodium citrate, pH 7.0) plus 0.1% SDS for30 min at 607C and then subjected to autoradiography.
Immunoblotting of recombinant and epithelial cell PGHS. For recombinant PGHS, transfected Cos-7 cells weresonicated at 105 watts for 60 s in 50 mM Tris-HCl (pH 7.4) containing 5 mM EGTA, 1 mM EDTA, 1 mM phenol,1 mM PMSF, 1 mg/ml pepstatin, 1 mg/ml leupeptin, and 1 mM diethyldithiocarbamate (3, 12). The sonicated cellmixtures were subjected to differential centrifugation at 47C, and the 100,000 x g pellets (containing microsomes)were resuspended in sonication buffer (12). For immunoblotting, the mixture was subjected to SDS-PAGE using an8% resolving gel followed by electrophoretic transfer to PVDF membranes (Immobilon-P, Millipore Corp.) andincubation with anti-PGHS Ab and anti-rabbit IgG conjugated to horseradish peroxidase. Binding of primary Ab wasdetected using enhanced chemiluminescence (Amersham).
For epithelial PGHS, cultured oTECs were sonicated in 50 mM Tris-HCl (pH 7.4) containing 10 mM EDTA, 1%Nonidet P-40, 1 mM PMSF, 1 mg/ml pepstatin, 1 mg/ml leupeptin, and 1 mM diethyldithiocarbamate and the sonicatecentrifuged at 15,000 1 g for 30 min at 47C. The resulting supernatant was diluted in extraction buffer and thentreated with anti-PGHS Ab for 2 h followed by Protein A sepharose (Pharmacia) for 1 h at 47C. The immune complexwas washed with 50 mM Tris-HCl (pH 7.4) in 0.9% NaCl containing 5 mM EDTA, 5 mM EGTA, 1 mM PMSF, 1mg/ml pepstatin, 1 mg/ml leupeptin, 0.5% Triton X-100, and 0.1% SDS and then with the same buffer lackingdetergents. Next, the immune-complex pellet was heated at 1007C for 10 min in reducing SDS-PAGE sample buffer.The mixture was centrifuged, and the resulting supernatant was subjected to SDS-PAGE and immunoblotting asdescribed above.
Assay of recombinant and epithelial cell PGHS activities. For recombinant PGHS activity, transfected Cos-7 cells(5 1 106 cells/condition) were incubated with or without 100 mM aspirin for 30 min at 377C and then with 20 mM[3H]arachidonic acid in Hepes-buffered HBSS (pH 7.4) for 5-15 min at 377C. Cell supernatants were extracted withacidified chloroform/propanol using PGB2 as an internal standard for product recovery, and the lipid extracts wereanalyzed by reverse-phase HPLC as described previously (12). For epithelial cell PGHS activity, cultured oTECswere switched to basal culture conditions (24 h culture in LHC-basal medium) and then treated with or without serum(20% fetal bovine serum for 1-24 h) in the presence or absence of actinomycin D (5 mg/ml) or cycloheximide (5 mg/ml). Untreated and treated oTECs (1 1 106 cells/condition) were then incubated with 20 mM [3H]arachidonic acid,and cell supernatants were extracted and analyzed for arachidonate metabolites as described above.
RESULTS AND DISCUSSION
Molecular cloning and sequencing of ovine PGHS-2 cDNA. Reverse-transcriptase PCRamplification of poly(A)/RNA from cultured oTECs resulted in the generation of a 0.5 kbcDNA that selectively recognized a 4.0 kb (PGHS-2) mRNA in Northern blots of oTEC RNA.The 32P-labeled 0.5 kb cDNA identified three positive clones in a lambda ZAP II cDNA libraryprepared from cultured oTECs. Phage DNA from three plaques that strongly hybridized withthe probe were rescreened at lower plating density, then released with NotI, and the productswere analyzed by agarose gel electrophoresis. Two of these pPGHSov-2-S10 and pPGHSov-2-S24 contained overlapping sequences and a third clone PGHSov-2-S21 encoded for the entire
501
AID BBRC 5523 / 690d$$$621 09-26-96 10:37:20 bbrca AP: BBRC

Vol. 227, No. 2, 1996 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
FIG. 1. Nucleotide and deduced amino acid sequence for ovine PGHS-2. Nucleic acid and amino acid positionsare indicated on the far right. Critical residues for catalysis (Arg105, Glu509, Tyr340, Tyr370, Phe514, Ser515, Leu516, Lys517,Gly518), aspirin-acetylation (Ser515), N-glycosylation (Asn 52, 129, 395, and 579), heme coordination (His 192, 294,and 373), and inhibitor interaction (Arg120 and Val508) are each underlined and italicized. Each 3*-ATTTA motif isalso underlined.
3.4 kb mRNA (Fig. 1). This insert (designated oPGHS-2) also selectively hybridized with a4.0 kb (PGHS-2) mRNA whereas the cDNA for the coding region of PGHS-1 (oPGHS-1-coding) selectively recognized a 2.8 kb (PGHS-1) mRNA in Northern analysis of oTEC RNA(see below).
502
AID BBRC 5523 / 690d$$$621 09-26-96 10:37:20 bbrca AP: BBRC
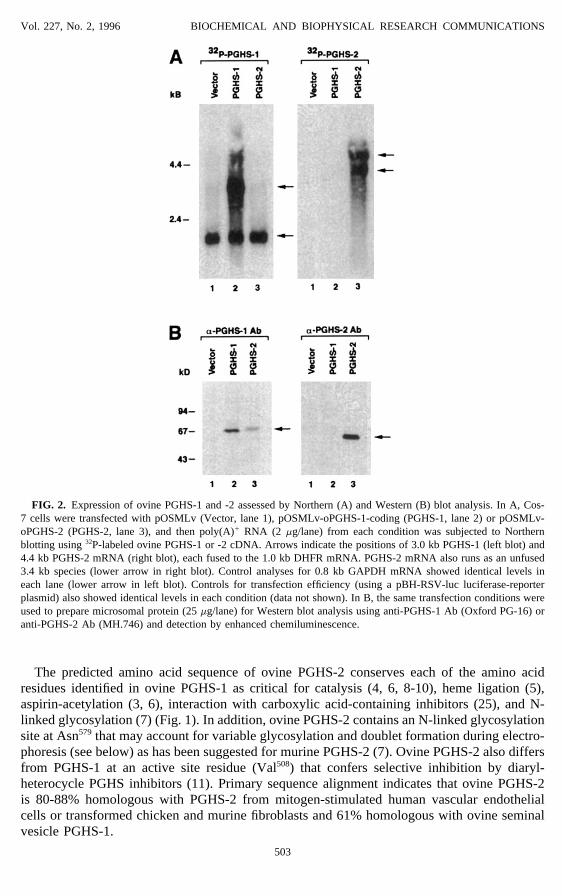
Vol. 227, No. 2, 1996 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
FIG. 2. Expression of ovine PGHS-1 and -2 assessed by Northern (A) and Western (B) blot analysis. In A, Cos-7 cells were transfected with pOSMLv (Vector, lane 1), pOSMLv-oPGHS-1-coding (PGHS-1, lane 2) or pOSMLv-oPGHS-2 (PGHS-2, lane 3), and then poly(A)/ RNA (2 mg/lane) from each condition was subjected to Northernblotting using 32P-labeled ovine PGHS-1 or -2 cDNA. Arrows indicate the positions of 3.0 kb PGHS-1 (left blot) and4.4 kb PGHS-2 mRNA (right blot), each fused to the 1.0 kb DHFR mRNA. PGHS-2 mRNA also runs as an unfused3.4 kb species (lower arrow in right blot). Control analyses for 0.8 kb GAPDH mRNA showed identical levels ineach lane (lower arrow in left blot). Controls for transfection efficiency (using a pBH-RSV-luc luciferase-reporterplasmid) also showed identical levels in each condition (data not shown). In B, the same transfection conditions wereused to prepare microsomal protein (25 mg/lane) for Western blot analysis using anti-PGHS-1 Ab (Oxford PG-16) oranti-PGHS-2 Ab (MH.746) and detection by enhanced chemiluminescence.
The predicted amino acid sequence of ovine PGHS-2 conserves each of the amino acidresidues identified in ovine PGHS-1 as critical for catalysis (4, 6, 8-10), heme ligation (5),aspirin-acetylation (3, 6), interaction with carboxylic acid-containing inhibitors (25), and N-linked glycosylation (7) (Fig. 1). In addition, ovine PGHS-2 contains an N-linked glycosylationsite at Asn579 that may account for variable glycosylation and doublet formation during electro-phoresis (see below) as has been suggested for murine PGHS-2 (7). Ovine PGHS-2 also differsfrom PGHS-1 at an active site residue (Val508) that confers selective inhibition by diaryl-heterocycle PGHS inhibitors (11). Primary sequence alignment indicates that ovine PGHS-2is 80-88% homologous with PGHS-2 from mitogen-stimulated human vascular endothelialcells or transformed chicken and murine fibroblasts and 61% homologous with ovine seminalvesicle PGHS-1.
503
AID BBRC 5523 / 690d$$$621 09-26-96 10:37:20 bbrca AP: BBRC

Vol. 227, No. 2, 1996 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
FIG. 3. Effect of aspirin on the profile of arachidonate products generated by recombinant ovine PGHS-1 (A andB) and PGHS-2 (C and D). Cos-7 cells transfected with pOSMLv-oPGHS-1-coding or pOSMLv-oPGHS-2 weretreated with vehicle (A, C) or aspirin (B, D) and then incubated with 20 mM [3H]arachidonic acid (2.2 1 105 dpm/ml) in Hepes-buffered HBSS for 15 min at 377C. Products were extracted from cell supernatants (5 1 106 cells) andanalyzed by reverse-phase HPLC. Major peaks of radioactivity coeluted with reference PGF2a (F), PGE2 (E), PGD2
(D), 15-HETE (15H), and arachidonic acid (AA). Results were identical for full-length (oPGHS-2) and coding-region(oPGHS-2-coding) constructs; control transfections with pOSMLv gave no detectable PG-forming activity (data notshown).
Expression of recombinant ovine PGH Synthases in Cos-7 cells. Expression of recombinantovine PGHS-2 (and PGHS-1) in Cos-7 cells was accomplished using pECE and pOSMLvexpression vectors (Fig. 2). Higher levels of expression attained with pOSMLv may dependon including coding sequence for murine DHFR, since this sequence appears to enhance thestability of the hybrid mRNA (26). In that context, expression of PGHS-1 and several PGHS-2 constructs (with and without the 3*-UTR) all gave similar levels of PGHS mRNA andprotein (Fig. 2) and corresponding enzymatic activity (1.3-2.2 nmol PG/mg protein/15 min).Comparisons of activity in untreated to aspirin-treated Cos-7 cells expressing ovine PGHS-1or -2 indicated that recombinant ovine PGHS-1 was completely inactivated by aspirin-acetyla-tion, but PGHS-2 still converted arachidonate to 15-HETE (Fig. 3). This profile preciselymatches PG- and 15-HETE-forming activities for endogenous PGHS isozymes in culturedovine (and human) TECs (12, 27).
Regulation of PGHS activity in cultured oTECs. Among a series of cytokines and growth
504
AID BBRC 5523 / 690d$$$621 09-26-96 10:37:20 bbrca AP: BBRC

Vol. 227, No. 2, 1996 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
FIG. 4. Selective serum-stimulation of PGHS-2 in cultured oTECs assessed at the level of mRNA by Northernblot (A), protein by Western blot (B), and enzyme activity by HPLC-based assay (C). In each case, cells were untreated(0) or treated (/) with 20% fetal bovine serum for 1 h (to detect mRNA) or 4 h (to detect protein and activity). InA, oTEC RNA (15 mg/lane) was hybridized with 32P-labeled ovine PGHS-1 or PGHS-2 cDNA under high stringencyconditions (50% formamide, 687C). Control analysis for GAPDH mRNA showed identical levels in each lane (lowerarrow). In B, oTEC protein (15 mg/lane) was immunoblotted against anti-PGHS-2 Ab (MH.746) and detected byenhanced chemiluminescence. In C, oTEC supernatants (1 1 106 cells) were extracted and analyzed by HPLC asdescribed in the Fig. 3 legend.
factors (including PMA, IL-1, TNF, and LPS), serum was the most potent stimulus of PGHS-2 mRNA levels in cultured oTECs (data not shown). Serum treatment caused little change inthe level of a 2.8 kb PGHS-1 mRNA, but a 2-4x increase in the 4.0 kb PGHS-2 mRNA (Fig.4) confirming that PGHS-2 is selectively regulated in this cell system (13). Serum treatmentcaused corresponding increases in 72-kDa PGHS-2 and a two-fold increase in PGE2-formingactivity to a final level of 1.5 nmol/mg protein/15 min (Fig. 4). Serum stimulation of PGHS-2 mRNA was completely inhibited by actinomycin D but was unchanged by cycloheximidetreatment (data not shown), indicating dependence on mRNA transcription but not proteintranslation for the rapid serum effect. Thus, PGHS-2 expression may develop more rapidly atsites of epithelial inflammation (and apoptosis) than at other sites that depend on monocyte/macrophage influx and activation (28).
In summary, we have characterized a full-length cDNA for ovine epithelial PGHS-2 thatconserves critical amino-acid residues used by ovine PGHS-1 for interaction with substrate,inhibitors, heme prosthetic groups, and carbohydrate moieties. The data provide directevidence for ovine epithelial PGHS-2 that differs from PGHS-1 in its response to aspirin-
505
AID BBRC 5523 / 690d$$$621 09-26-96 10:37:20 bbrca AP: BBRC

Vol. 227, No. 2, 1996 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
acetylation and serum-stimulation, and the results form the basis for directed analysis ofPGHS structure-function relationships in the context of mutagenesis and x-ray crystallo-graphic data for ovine PGHS-1.
ACKNOWLEDGMENTSThis research was supported by Grants HL-40078, HL-07317, DK-38111, and HL/AI-51071 from the National
Institutes of Health and the Alan A. and Edith L. Wolff Charitable Trust.
REFERENCES1. DeWitt, D. L., and Smith, W. L. (1988) Proc. Natl. Acad. Sci. U.S.A. 85, 1412–1416.2. Merlie, J. P., Fagan, D., Mudd, J., and Needleman, P. (1988) J. Biol. Chem. 263, 3550–3553.3. DeWitt, D. L., El-Harith, E. A., Kraemer, S. A., Andrews, M. J., Yao, E. F., Armstrong, R. L., and Smith, W. L.
(1990) J. Biol. Chem. 265, 5192–5198.4. Shimokawa, T., Kulmacz, R. J., DeWitt, D. L., and Smith, W. L. (1990) J. Biol. Chem. 265, 20073–20076.5. Shimokawa, T., and Smith, W. L. (1991) J. Biol. Chem. 266, 6168–6173.6. Shimokawa, T., and Smith, W. L. (1992) J. Biol. Chem. 267, 12387–12392.7. Otto, J. C., DeWitt, D. L., and Smith, W. L. (1993) J. Biol. Chem. 268, 18234–18242.8. Bhattacharyya, D. K., Lecomte, M., Rieke, C. J., Garavito, R. M., and Smith, W. L. (1996) J. Biol. Chem. 271,
2179–2184.9. Tsai, A., Hsi, L. C., Kulmacz, R. J., Palmer, G., and Smith, W. L. (1994) J. Biol. Chem. 269, 5085–5091.
10. Picot, D., Loll, P. J., and Garavito, R. M. (1994) Nature 367, 243–249.11. Gierse, J. K., McDonald, J. J., Hauser, S. D., Rangwala, S. H., Koboldt, C. M., and Siebert, K. (1996) J. Biol.
Chem. 271, 15810–15814.12. Holtzman, M. J., Turk, J., and Shornick, L. P. (1992) J. Biol. Chem. 267, 21438–21445.13. Rosen, G. D., Birkenmeier, T., Raz, A., and Holtzman, M. J. (1989) Biochem. Biophys. Res. Commun. 164, 1358–
1365.14. Shornick, L. P., and Holtzman, M. J. (1993) J. Biol. Chem. 268, 371–376.15. Bernatowicz, M. S., and Matsueda, G. R. (1986) Anal. Biochem. 155, 95–102.16. Xie, W., Chipman, J. G., Robertson, D. L., Erikson, R. L., and Simmons, D. L. (1991) Proc. Natl. Acad. Sci.
U. S. A. 88, 2692–2696.17. Kujubu, D. A., Fletcher, B. S., Varnum, B. C., Lim, R. W., and Herschman, H. R. (1991) J. Biol. Chem. 266,
12866–12872.18. O’Banion, M. K., Sadowski, H. B., Winn, V., and Young, D. A. (1991) J. Biol. Chem. 266, 23261–23267.19. Luthy, R., and Eisenberg, D. (1991) in Sequence Analysis Primer (Gribskov, M., and Devereux, J., Eds.), pp.
61–87, Stockton Press, New York.20. Chirgwin, J. M., Przybyla, A. E., MacDonald, R. J., and Rutter, W. J. (1979) Biochemistry 18, 5294–5298.21. Gubler, U., and Hoffman, B. J. (1983) Gene 25, 263–269.22. Ellis, L., Clauser, E., Morgan, D. O., Edery, M., Roth, R. A., and Rutter, W. J. (1986) Cell 45, 721–732.23. Laneuville, O., Breuer, D. K., DeWitt, D. L., Hla, T., Funk, C. D., and L., S. W. (1994) J. Pharmacol. Exp. Ther.
271, 927–934.24. Look, D. C., Pelletier, M. R., Tidwell, R. M., Roswit, W. T., and Holtzman, M. J. (1995) J. Biol. Chem. 270,
30264–30267.25. Mancini, J. A., Riendeau, D., Falgueyre, J.-P., Vickers, P. J., and O’Neill, G. P. (1995) J. Biol. Chem. 270, 29372–
29377.26. Wong, G. C., Witek, J. S., Temple, P. A., et al. (1985) Science 228,27. Holtzman, M. J. (1992) Annu. Rev. Physiol. 54, 303–329.28. DeWitt, D., and Smith, W. L. (1995) Cell 83, 345–348.
506
AID BBRC 5523 / 690d$$$621 09-26-96 10:37:20 bbrca AP: BBRC