molecular cloning of the duck mitogen-activated protein
TRANSCRIPT

INTRODUCTION Mitogen-activated protein kinases (MAPK) are
serine/threonine-specific protein kinases that convert extracellular stimuli into a wide range of cellular re-sponses (Garcia-Rodriguez et al., 2012). Mitogen-acti-vated protein kinases are among the most ancient sig-nal transduction pathways and widely used throughout evolution in many physiological processes (Guegan et al., 2013). All eukaryotic cells possess multiple MAPK pathways, which coordinate to regulate mitosis, dif-ferentiation, metabolism, motility, apoptosis, and cell survival (Calati et al., 2013; Upadhya et al., 2013). In mammals, 14 MAPK have been characterized into 7 groups. By far the most extensively studied groups of mammalian MAPK are the ERK1/2, JNK, and p38isoforms, but recent studies have shed some light on the
function and regulation of other MAPK groups (Nix et al., 2011; Zhang et al., 2011).
Roman Dziarski had reported that soluble staphy-lococcal peptidoglycan strongly activates extracellular signal-regulated kinase (ERK) 1 and ERK2 in the mouse macrophage cell line RAW264.7, and moderately activates JNK and weakly activates p38 MAPK. In con-trast, lipopolysaccharide (LPS) strongly activates all of these kinases (Dziarski et al., 1996). Lysophosphatidic acid has significant anti-inflammatory activities involv-ing the activation of ERK 1/2, serine/threonine phos-phatases, and PI3 kinase signaling pathways (Zhang et al., 2012). Compared with the extensive research on hu-man and mammal MAPK1, our understanding of bird MAPK1 is lagging behind. There are only several re-ports on chicken MAPK1 (Xing et al., 2010; Swaggerty et al., 2011; Dupont et al., 2012; Morillo et al., 2012), and none on duck MAPK1. In our studies, the MAPK1gene was amplified from Cherry Valley duck and cloned into pMD18-T for sequence analysis. Furthermore, a quantitative real-time PCR (qRT-PCR) assay was de-veloped to detect MAPK1 expression in duck spleens following Riemerella anatipestifer yb2 infection. The current study sets the ground for further research on the role of duck MAPK1 in bacterial pathogenesis.
Molecular cloning of the duck mitogen-activated protein kinase 1 (MAPK1) gene and the development of a quantitative real-time PCR
assay to detect its expression
Shoulin Cao ,*† Xiangan Han ,* Chan Ding ,* Shaohui Wang ,* Mingxing Tian ,* Xiaolan Wang ,* Wanwan Hou ,* Jiaping yue ,* Guijun Wang ,† and Shengqing yu *1
* Shanghai Veterinary Research Institute, Chinese Academy of Agricultural Sciences, 518 Ziyue Road, Shanghai 200241, China; and † College of Animal Science and Technology, Anhui Agricultural University,
130 West Changjiang Road, Hefei, Anhui 230036, China
ABSTRACT Mitogen-activated protein kinase 1 (MAPK1) acts as an integration point for multiple bio-chemical signals, and is involved in a wide variety of biological processes such as cell proliferation and dif-ferentiation, transcription regulation, and development. Mitogen-activated protein kinase 1 plays an important role in inducing cell death in bacterial infections. In this study, the duck MAPK1 gene was cloned for the first time from the Cherry Valley duck. Sequence analy-sis showed that duck MAPK1 cDNA is 1,557 bp long, with an open reading frame of 1,107 bp. It encodes
368 amino acids, with 85.4, 84.5, and 97.3% homol-ogy with the human, mouse, and chicken MAPK1 gene, respectively. Furthermore, a SyBR Green quantita-tive real-time PCR assay was developed to detect duck MAPK1 expression. Following Riemerella anatipestiferinfection by virulent strain yb2, MAPK1 mRNA level increased more than 200-fold in the duck spleens, sug-gesting that increased duck MAPK1 expression can be used as an indicator of bacterial infection. Our results provide ground work to warrant further studies of the duck MAPK1 gene in bacterial pathogenesis.
Key words: cloning , duck mitogen-activated protein kinase 1 , quantitative real-time PCR
2014 Poultry Science 93 :2158–2167http://dx.doi.org/ 10.3382/ps.2013-03796
IMMUNOLOGY, HEALTH, AND DISEASE
Received November 28, 2013. Accepted May 10, 2014. 1 Corresponding author: [email protected]
© 2014 Poultry Science Association Inc.
2158
Dow
nloaded from https://academ
ic.oup.com/ps/article-abstract/93/9/2158/2730262 by guest on 17 D
ecember 2018

MATERIALS AND METHODS
BirdsOne-day-old Cherry Valley ducklings were obtained
from Wuxi Duck Farm (Wuxi, China) and kept under controlled temperature (28–30°C). The ducklings were housed in cages with a 12L:12D cycle and free access to food and water during the study. Care and maintenance of all animals were in accordance with the Institutional Animal Care and Use Committee guidelines set by the Shanghai Veterinary Research Institute, Chinese Acad-emy of Agricultural Sciences.
Bacterial Strains and PlasmidsEscherichia coli strain DH5α was purchased from
ComWin Biotech Co. Ltd. (Beijing, China) and cul-tured in Luria-Bertani liquid medium or on solid medi-um containing 1.5% agar at 37°C. Riemerella anatipes-tifer strain yb2 was isolated in China (Hu et al., 2011) and cultured in tryptic soy agar (Difco, Franklin Lakes, NJ) at 37°C for 24 h in 5% CO2, or in tryptic soy broth (Difco) shaking at 150 rpm for 8 to 12 h at 37°C. The pMD18-T was obtained from TaKaRa Co. (Dalian, China).
Amplification, Cloning, and Sequence Analysis of the Partial Fragment of Duck MAPK1
The partial fragment of the duck MAPK1 was ampli-fied from Cherry Valley duck spleen using primer pairs of MAPK1-AF/MAPK1-AR (Table 1), which were de-signed using Primer Premier 5.0 software according to a conserved region of chicken MAPK1 gene sequence (accession no. NM_204150). The spleen was collected
from 10-d-old Cherry Valley duck and homogenized in 10 volumes of ice-cold PBS. Total RNA was extracted using Trizol reagent (Invitrogen, Carlsbad, CA) and treated with RNase-free DNase (Promega, Fitchburg, WI) according to the manufacturer’s instructions. The cDNA was synthesized using 2 μg of total RNA, oligo d(T)15 primer, and M-MLV reverse transcriptase (Pro-mega). The partial fragment of MAPK1 was amplified from the cDNA template using primers MAPK1-AF/MAPK1-AR with the following PCR cycle: 94°C for 3 min, 25 cycles of 94°C for 30 s, 60°C for 30 s, 72°C for 60 s, followed by a 10 min final extension at 72°C. The PCR products were identified by electrophoresis and cloned into pMD18-T vector (TaKaRa Co.) for se-quencing using an automatic DNA sequencer (ABI Ap-plied Biosystems Model 377, Grand Island, Ny).
3′- and 5′- Full Rapid Amplification of cDNA Ends and Sequence Analysis
Both 3′-/5′-full rapid amplification of cDNA ends (RACE) were performed with the total RNA by using 3′-full/5′-full RACE kits (TaKaRa Co.) as described (Xiao et al., 2011; He et al., 2013). Briefly, 3′ RACE was performed using a 3′ full RACE kit. The outer PCR amplification was carried out using primers MAPK1–3GSP1 and 3′ RACE outer primer (Table 1) and 2 μL of cDNA template. Then the outer PCR product was used as a template for the inner PCR reaction using primers MAPK1–3GSP2 and 3′ RACE inner primer (Table 1). The PCR products were then subjected to gel purification, cloning, and sequence analysis.
For 5′ RACE, the total RNA was first processed us-ing a 5′-full RACE kit through dephosphorylation, 5′ cap structure removal, 5′-RACE adaptor connection, and reverse-transcription reaction steps to provide a template for the outer PCR (Xiao et al., 2011; He et
Table 1. Primers used in this study1
Primer Description Application
PCR MAPK1-AF 5′ AAGTGTTCGACGTGGGG 3′ Amplification of the internal region of
MAPK1 cDNA sequences MAPK1-AR 5′ TCCTTCGGCAAGTCATC 3′RACE-PCR MAPK1–3GSP1 5′ TTCTAAGGGTTACACCAAGTC 3′ Amplification of the 3′ and 5′ terminal of
MAPK1 cDNA 3′ RACE outer primer 5′ TACCGTCGTTCCACTAGTGATTT 3′ MAPK1–3GSP2 5′ CTATTTGCTTTCCCTACCAC 3′ 3′ RACE inner primer 5′ CGCGGATCCTCCACTAGTGATTTCACTATAGG 3′ MAPK1–5GSP1 5′ GTGGTCGTTGCTGAGGTGTTGAG 3′ 5′ RACE outer primer 5′ CATGGCTACATGCTGACAGCCTA 3′ MAPK1–5GSP2 5′ CTGGCAGTACGTCTGATGCTCAA 3′ 5′ RACE inner primer 5′ CGCGGATCCACAGCCTACTGATGATCAGTCGATG 3′Real-time PCR MAPK1-RT-F 5′ CCAGACCATGATCACACAGG 3′ Real-time quantitative PCR MAPK1-RT-R 5′ GGATCCAAGTATGCCAAGGA 3′ Arbp-RT-F 5′ CGACCTGGAAGTCCAACTAC 3′ Arbp-RT-R 5′ ATCTGCTGCATCTGCTTG 3′ MAPK1F 5′ CAACGACCACATTTGCTACTTC 3′ MAPK1R 5′ TTCAGCTACAGGCTCATCACTT 3′
1MAPK1 = mitogen-activated protein kinase 1; RACE = rapid amplification of cDNA ends; Arbp = attachment region binding protein.
2159CLONING OF DUCK MITOGEN-ACTIVATED PROTEIN KINASE 1D
ownloaded from
https://academic.oup.com
/ps/article-abstract/93/9/2158/2730262 by guest on 17 Decem
ber 2018

al., 2013). The outer and inner PCR amplifications for 5′ RACE were carried out similarly to 3′ RACE, us-ing MAPK1–5GSP1 and 5′ RACE outer primer for the outer PCR reaction and MAPK1–5GSP2 and 5′ RACE inner primer (Table 1) for the inner PCR reaction.
Sequence Assembly and Homology Analysis of the Duck MAPK1 Gene
Full-length cDNA of duck MAPK1 gene was assem-bled using Vector NTI Express Software (Life Technolo-gies, Carlsbad, CA) based on the obtained partial frag-ment and the 3′-/5′-full RACE products. Amino acid sequence and protein analysis were performed using the ExPASy proteomic tool (http://www.expasy.org/tools/) and the protein domains were predicted using SMART software (http://smart.embl-heidelberg.de/). Homologous sequences were searched using BLAST (http://ww.ncbi.nlm.nih.gov/blast/) with default set-tings on the complete nonredundant GenBank database (Wei et al., 2011; Han et al., 2012). Percent identity of deduced amino acid sequences against human, mouse, and chicken MAPK1 sequences was analyzed using the DNAStar program (DNASTAR Inc. Madison, WI).
Development of a Real-Time PCR Assay and Analysis of MAPK1 Expression in Duck Tissues
Based on the duck MAPK1 gene sequence obtained in this study, primers MAPK1-RT-F/MAPK1-RT-R (Table 1) were designed using Primer Express 3.0 to develop a real-time PCR assay for detecting duck MAPK1 expression. The target 213-bp amplicon was selected from a relatively conserved region of the gene (open reading frame 550–762 bp). The expression of attachment region binding protein (Arbp) served as an endogenous control, which was detected using prim-ers Arbp-RT-F/Arbp-RT-R (Table 1). The qPCR was performed using the SyBR Green Quantitative PCR kit (Promega). Total RNA was extracted using Trizol reagent (Invitrogen) and treated with RNase-free DN-ase (Promega) according to the manufacturer’s instruc-tions. The cDNA synthesis was performed using oligo (dT)15 primer and PrimeScript RT Master Mix (TaKa-Ra Co.). Amplifications were carried out with the fol-lowing parameters: 2 min at 95°C, followed by 40 cycles consisting of 15 s at 94°C and 60 s at 60°C. A 10-fold dilution series of plasmids containing the target gene (MAPK1) and reference gene (Arbp) fragments was used to compute the PCR efficiency. All experiments were carried out in triplicate. The changes of mRNA were expressed as fold expression and calculated using the comparative cycle threshold (2−ΔΔCt) method.
Expression of MAPK1 was analyzed in ducks in-fected with R. anatipestifer yb2. Twenty-four Cherry Valley ducks (15 d old) were divided into 2 groups of
12, injected intramuscularly with R. anatipestifer yb2 culture (0.5 mL, 106 cfu, infection group) or PBS (con-trol group) as described (Hu et al., 2011). Spleens were collected from 3 ducks in each group at 6, 12, 24, and 48 h postinjection and the respective fold expression of MAPK1 mRNA was quantitatively detected as de-scribed above. All samples were detected in triplicate. Significance was analyzed by 2-tailed Student’s t-test.
Dot Blot AssayThe primer pairs MAPK1F/MAPK1R and Arbp-RT-
F/Arbp-RT-R were designed using Primer 5.0 software and listed in Table 1. The amplified PCR products were labeled respectively with digoxigenin (DIG) us-ing the DIG Northern Starter Kit (Roche Diagnostics, Mannheim, Germany) and used as probes.
Dot blot assay was carried out using the DIG North-ern Starter Kit (Roche Diagnostics). Briefly, the total RNA was heated at 100°C for 10 min, and cooled down on ice. The denatured RNA (2 μL) was then manu-ally blotted on a positively charged nylon membrane, which was activated with 2 × SSC (0.3 M NaCl, 0.03 M sodium citrate) for 5 min. The blots were hybridized with 10 pM probes at 80°C for 2 h. After hybridization, each blot was washed twice for 5 min in 2 × SSC/0.1% SDS at room temperature with gentle agitation. Blots were then washed in DIG Wash and Block Buffer Set for 2 min and blocked in blocking solution for 30 min at room temperature. The membranes were then in-cubated with 150 mU/mL alkaline phosphatase con-jugated anti-digoxigenin antibody for another 30 min, washed twice in washing solution for 15 min, and equili-brated in 0.1 M Tris-HCl and 0.1 M NaCl (pH 9.5) for 3 min. For colorimetric detection of hybridization, substrate solution (NBT/BCIP) was used as described
Figure 1. A) PCR amplification of duck mitogen-activated protein kinase 1 (MAPK1) partial sequence M: DL2000 DNA marker. Par-tial gene sequence of duck MAPK1 was amplified from Cherry Valley duck spleen using primer pairs MAPK1-AF/MAPK1-AR, resulting in a 973-bp band. B) The 700-bp 3′ rapid amplification of cDNA ends (RACE) product of duck MAPK1 is indicated by an arrow. C) The 5′ RACE product of duck MAPK1 is 277 bp long.
2160 CAO ET AL.D
ownloaded from
https://academic.oup.com
/ps/article-abstract/93/9/2158/2730262 by guest on 17 Decem
ber 2018

in the manufacturer’s instructions. Color development was performed for 1 h and the reaction was stopped by washing the membrane with TE buffer [10 mM Tris-HCl (pH 8.0), and 1 mM EDTA] for 5 min.
RESULTS
PCR Amplification of the Duck MAPK1 Partial Sequence
The 973-bp PCR product was identified by 1% aga-rose gel electrophoresis, which is similar to the theoreti-
cal length of the gene (Figure 1A). The PCR product was then cloned into the pMD-18T vector, and trans-formed into DH5α competent cells. The obtained DNA sequence was analyzed by DNAstar software, which in-dicates that the partial sequence is 98.4% homologous to the chicken MAPK1 gene.
3′ and 5′ Full RACE of Duck MAPK1 GeneThere were 3 potential inner PCR products result-
ing from the 3′ RACE (Figure 1B). We subsequently identified the middle band as the real 3′ inner PCR
Figure 2. The full length cDNA sequence of the duck mitogen-activated protein kinase 1 (MAPK1) gene, with the 3′ and 5′ rapid amplification of cDNA ends primers marked by arrows. The coding sequence region is flanked by initiation codon ATG and termination codon TAA in bold.
2161CLONING OF DUCK MITOGEN-ACTIVATED PROTEIN KINASE 1D
ownloaded from
https://academic.oup.com
/ps/article-abstract/93/9/2158/2730262 by guest on 17 Decem
ber 2018

Figure 3. The nucleotide sequence alignment of duck, Gallus gallus (NM_204150.1), Homo sapiens (NM_002745.4), and Mus musculus (NM_011949.3) mitogen-activated protein kinase 1 (MAPK1) genes. Real-time PCR primers MAPK1-RT-F and MAPK1-RT-R were selected from a conserved region and are marked by arrows.
2162 CAO ET AL.D
ownloaded from
https://academic.oup.com
/ps/article-abstract/93/9/2158/2730262 by guest on 17 Decem
ber 2018

Figure 4. The amino acid sequence alignment of duck, Gallus gallus, Homo sapiens, and Mus musculus mitogen-activated protein kinase 1 (MAPK1) proteins. The numbers on the left indicate the residue positions. Gaps are marked by dashes, and the residues that differ from the duck MAPK1 gene are shaded with solid black.
2163CLONING OF DUCK MITOGEN-ACTIVATED PROTEIN KINASE 1D
ownloaded from
https://academic.oup.com
/ps/article-abstract/93/9/2158/2730262 by guest on 17 Decem
ber 2018

product and others as nonspecific products follow-ing cloning and sequencing of all 3 bands. A single 5′ inner PCR product was obtained (Figure 1C). The PCR products were cloned into pMD18-T vector and transformed into DH5α competent cells. The DNA se-quences were analyzed by DNAstar software, indicat-ing the partial sequences at the 3′ and 5′ ends, which were 97.7 and 97.3% homologous to chicken MAPK1, respectively.
Sequence Assembly and Homology Analysis of the Duck MAPK1 Gene
Based on the obtained partial fragment and 3′-/5′- full RACE product sequences, the full-length duck MAPK1 cDNA was assembled using the DNAStar program and submitted to GenBank (accession no: DUCK MAPK1 Duck KF482375). The duck MAPK1 full cDNA se-quence is 1,557 bp long, including a 55-bp 5′-untrans-
Figure 5. Amplification curves and standard curve. A) The melting curve analyzed the specificity of the primers MAPK1-RT-F and MAPK1-RT-R. Duck spleen total RNA was used as the template and nuclease-free water was used as a nontemplate control. The y-axis −dI/dT [also means −d(RFU)dT] represents the rate of change of the relative fluorescence units (RFU) with time (T). B) Ten-fold dilutions of the standard plasmid were used as amplification template. The x-axis represents copies of the standard plasmid, and the y-axis represents the fluorescence data used for cycle threshold (Ct) determinations. The assays were linear in the range of 107 to 102 template copies/μL, with an R2 of 0.996. The detection limit for the assay is 10 copies.
2164 CAO ET AL.D
ownloaded from
https://academic.oup.com
/ps/article-abstract/93/9/2158/2730262 by guest on 17 Decem
ber 2018
lated region, a 395-bp 3′-untranslated region, and a 1,107-bp open reading frame encoding 368 amino acids (Figure 2). The nucleotide sequences of duck, Gallus gallus (NM_204150.1), Homo sapiens (NM_002745.4), and Mus musculus (NM_011949.3) MAPK1 genes were aligned and compared using the Vector NTI Express and DNAStar programs. The alignment was performed using Clustal W v2.0. The residues that differ from duck MAPK1 were shaded with solid black (Figure 3). Duck MAPK1 gene is 85.4, 84.5, and 97.3% homolo-gous to human, rat, and chicken MAPK1, respectively. The amino acid sequence of duck MAPK1 protein was shown in Figure 4. Gaps are marked by dashes and the residues that differ from duck MAPK1 gene were shad-ed with solid black. The theoretical molecular weight of duck MAPK1 is 41.9 kDa.
Real-Time Quantitative PCR and Dot Blot Analyses of MAPK1 Expression in Ducks Infected with R. anatipestifer
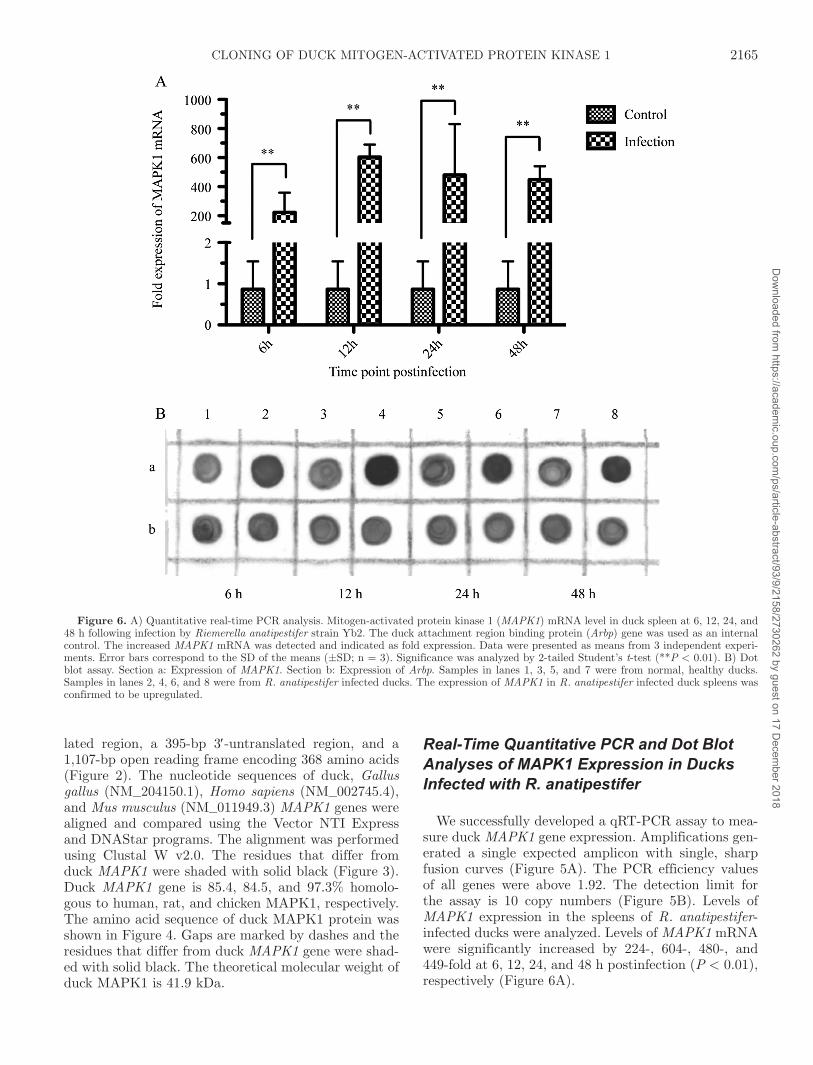
We successfully developed a qRT-PCR assay to mea-sure duck MAPK1 gene expression. Amplifications gen-erated a single expected amplicon with single, sharp fusion curves (Figure 5A). The PCR efficiency values of all genes were above 1.92. The detection limit for the assay is 10 copy numbers (Figure 5B). Levels of MAPK1 expression in the spleens of R. anatipestifer-infected ducks were analyzed. Levels of MAPK1 mRNA were significantly increased by 224-, 604-, 480-, and 449-fold at 6, 12, 24, and 48 h postinfection (P < 0.01), respectively (Figure 6A).
Figure 6. A) Quantitative real-time PCR analysis. Mitogen-activated protein kinase 1 (MAPK1) mRNA level in duck spleen at 6, 12, 24, and 48 h following infection by Riemerella anatipestifer strain yb2. The duck attachment region binding protein (Arbp) gene was used as an internal control. The increased MAPK1 mRNA was detected and indicated as fold expression. Data were presented as means from 3 independent experi-ments. Error bars correspond to the SD of the means (±SD; n = 3). Significance was analyzed by 2-tailed Student’s t-test (**P < 0.01). B) Dot blot assay. Section a: Expression of MAPK1. Section b: Expression of Arbp. Samples in lanes 1, 3, 5, and 7 were from normal, healthy ducks. Samples in lanes 2, 4, 6, and 8 were from R. anatipestifer infected ducks. The expression of MAPK1 in R. anatipestifer infected duck spleens was confirmed to be upregulated.
2165CLONING OF DUCK MITOGEN-ACTIVATED PROTEIN KINASE 1D
ownloaded from
https://academic.oup.com
/ps/article-abstract/93/9/2158/2730262 by guest on 17 Decem
ber 2018

The expression of MAPK1 in R. anatipestifer infect-ed duck spleens was also confirmed to be upregulated by dot blot assay (Figure 6B).
DISCUSSIONMitogen-activated protein kinases are a series of in-
tracellular Ser/Thr protein kinase that are highly con-served. Six identical positions of cysteine residues (C1, C3, C4, C5, C6, and C8) were found in both vertebrate and invertebrate MAPK1 (except for the Apis mellifera homolog, where C1 is replaced by S; Ponza et al., 2011). Mitogen-activated protein kinases are involved in di-recting cellular responses to a diverse array of stimuli such as mitogens, osmotic stress, heat shock, and pro-inflammatory cytokines. They regulate gene expres-sion, mitosis, proliferation, differentiation, cell survival, and apoptosis through intracellular phosphorylation of 3 consecutive enzymatic reaction cascade. The cascade consists of MAPKKK (MAPK-kinase kinase), MAPKK (MAPK kinase), and MAPK, of which MAPKK is a both Ser/Thr and tyrosine bifunctional kinase. The MAPK family has multiple subgroups, of which the ERK, JNK, p38 lightning, and ERK5 groups are most characterized at the present. The first identified MAPK family member was ERK (Jiang et al., 1996; Irving et al., 2000; Hii et al., 2004).
The orthologs of mammalian MAPK1 were identified previously in chicken cells (Duchene et al., 2008; Tum-mala et al., 2011). In this paper, we describe the clon-ing of duck MAPK1. The duck MAPK1 gene was suc-cessfully cloned from Cherry Valley ducks and shares 97.3% sequence homology with chicken MAPK1 gene.
A qRT-PCR assay was also developed in this study to measure duck MAPK1 mRNA levels. Amplifica-tion generated a single expected amplicon with single, sharp fusion curves, indicating the high specificity of the amplicons. Furthermore, MAPK1 expression was increased more than 200-fold in duck spleens follow-ing R. anatipestifer infection. This suggests that duck MAPK1 expression can be used as an indicator of bac-terial infection.
The activation of MAPK and nuclear factor κB by membrane protein in Listeria monocytogenes-infected macrophages was confirmed by the induction of ERK and IκB phosphorylation (Zhang et al., 2012). Nuclear factor κB triggers destructive inflammation accompa-nied by MAPK activation. Inflammation is driven by massive tumor necrosis factor production, which re-quires additional activation of p38 and ERK (Guma et al., 2011). Nitrous oxide production induced by bacte-rial stimulation was most effectively reduced by inhi-bition of p38 MAPK and least effectively reduced by inhibition of IκB in chicken macrophage cell line HD11 (Crippen, 2006). Brevibacillus texasporus (BT pep-tides) enhance leukocyte functional and pro-inflamma-tory cytokine and chemokine gene transcription activi-ties through MAPK pathways, and may prove useful as immune modulators in neonatal poultry (Kogut et
al., 2012). In this study, the complete sequence of duck MAPK1 gene was obtained and the increased expres-sion of duck MAPK1 was observed after R. anatipes-tifer infection. The role of duck MAPK1 in bacterial pathogenesis needs further experimental verification.
ACKNOWLEDGMENTS
This study was supported by the Chinese National High-Technology Research and Development Program (863 Program, 2011AA10A209), the Special Fund for Agro-Scientific Research in the Public Interest (grant no. 201003012), and the National Natural Science Foundation of China (31272591).
REFERENCESCalati, R., C. Crisafulli, M. Balestri, A. Serretti, E. Spina, M. Ca-
labro, A. Sidoti, D. Albani, I. Massat, P. Hofer, D. Amital, A. Juven-Wetzler, S. Kasper, J. Zohar, D. Souery, S. Montgomery, and J. Mendlewicz. 2013. Evaluation of the role of MAPK1 and CREB1 polymorphisms on treatment resistance, response and remission in mood disorder patients. Prog. Neuropsychopharma-col. Biol. Psychiatry 44:271–278. http://dx.doi.org/10.1016/j.pnpbp.2013.03.005.
Crippen, T. L. 2006. The selective inhibition of nitric oxide pro-duction in the avian macrophage cell line HD11. Vet. Immu-nol. Immunopathol. 109:127–137. http://dx.doi.org/10.1016/j.vetimm.2005.08.024.
Duchene, S., E. Audouin, S. Crochet, M. J. Duclos, J. Dupont, and S. Tesseraud. 2008. Involvement of the ERK1/2 MAPK path-way in insulin-induced S6K1 activation in avian cells. Domest. Anim. Endocrinol. 34:63–73. http://dx.doi.org/10.1016/j.domaniend.2006.11.001.
Dupont, J., S. Metayer-Coustard, B. Ji, C. Rame, C. Gespach, B. Voy, and J. Simon. 2012. Characterization of major elements of insulin signaling cascade in chicken adipose tissue: Apparent in-sulin refractoriness. Gen. Comp. Endocrinol. 176:86–93. http://dx.doi.org/10.1016/j.ygcen.2011.12.030.
Dziarski, R., y. P. Jin, and D. Gupta. 1996. Differential activation of extracellular signal-regulated kinase (ERK) 1, ERK2, p38, and c-Jun NH2-terminal kinase mitogen-activated protein kinases by bacterial peptidoglycan. J. Infect. Dis. 174:777–785.
Garcia-Rodriguez, S., J. L. Callejas-Rubio, N. Ortego-Centeno, E. Zumaquero, R. Rios-Fernandez, S. Arias-Santiago, P. Navarro, J. Sancho, and M. Zubiaur. 2012. Altered AKT1 and MAPK1 gene expression on peripheral blood mononuclear cells and cor-relation with T-helper-transcription factors in systemic lupus erythematosus patients. Mediators Inflamm. http://dx.doi.org/10.1155/2012/495934.
Guegan, J. P., F. Ezan, N. Theret, S. Langouet, and G. Baffet. 2013. MAPK signaling in cisplatin-induced death: Predominant role of ERK1 over ERK2 in human hepatocellular carcinoma cells. Car-cinogenesis 34:38–47. http://dx.doi.org/10.1093/carcin/bgs317.
Guma, M., D. Stepniak, H. Shaked, M. E. Spehlmann, S. Shenouda, H. Cheroutre, I. Vicente-Suarez, L. Eckmann, M. F. Kagnoff, and M. Karin. 2011. Constitutive intestinal NF-κB does not trigger destructive inflammation unless accompanied by MAPK activa-tion. J. Exp. Med. 208:1889–1900.
Han, X., Q. Hu, S. Ding, W. Chen, C. Ding, L. He, X. Wang, J. Ding, and S. yu. 2012. Identification and immunological charac-teristics of chaperonin GroEL in Riemerella anatipestifer. Appl. Microbiol. Biotechnol. 93:1197–1205. http://dx.doi.org/10.1007/s00253-011-3635-2.
He, S., X.-F. Liang, L. Li, W. Huang, D. Shen, and y.-X. Tao. 2013. Gene structure and expression of leptin in Chinese perch. Gen. Comp. Endocrinol. 194:183–188.
Hii, C. S., D. S. Anson, M. Costabile, V. Mukaro, K. Dunning, and A. Ferrante. 2004. Characterization of the MEK5–ERK5 module
2166 CAO ET AL.D
ownloaded from
https://academic.oup.com
/ps/article-abstract/93/9/2158/2730262 by guest on 17 Decem
ber 2018

in human neutrophils and its relationship to ERK1/ERK2 in the chemotactic response. J. Biol. Chem. 279:49825–49834. http://dx.doi.org/10.1074/jbc.M406892200.
Hu, Q., X. Han, X. Zhou, C. Ding, y. Zhu, and S. yu. 2011. OmpA is a virulence factor of Riemerella anatipestifer. Vet. Microbiol. 150:278–283. http://dx.doi.org/10.1016/j.vetmic.2011.01.022.
Irving, E. A., F. C. Barone, A. D. Reith, S. J. Hadingham, and A. A. Parsons. 2000. Differential activation of MAPK/ERK and p38/SAPK in neurones and glia following focal cerebral ischaemia in the rat. Brain Res. Mol. Brain Res. 77:65–75. http://dx.doi.org/10.1016/s0169-328x(00)00043-7.
Jiang, y., C. H. Chen, Z. J. Li, W. Guo, J. A. Gegner, S. C. Lin, and J. H. Han. 1996. Characterization of the structure and function of a new mitogen-activated protein kinase (p38 beta). J. Biol. Chem. 271:17920–17926.
Kogut, M. H., K. J. Genovese, H. He, C. L. Swaggerty, and y. W. Jiang. 2012. BT cationic peptides: Small peptides that modulate innate immune responses of chicken heterophils and monocytes. Vet. Immunol. Immunopathol. 145:151–158. http://dx.doi.org/10.1016/j.vetimm.2011.10.023.
Morillo, S. M., E. P. Abanto, M. J. Roman, and J. M. Frade. 2012. Nerve growth factor-induced cell cycle reentry in newborn neu-rons is triggered by p38(MAPK)-dependent E2F4 phosphoryla-tion. Mol. Cell. Biol. 32:2722–2737. http://dx.doi.org/10.1128/MCB.00239-12.
Nix, P., N. Hisamoto, K. Matsumoto, and M. Bastiani. 2011. Axon regeneration requires coordinate activation of p38 and JNK MAPK pathways. Proc. Natl. Acad. Sci. USA 108:10738–10743. http://dx.doi.org/10.1073/pnas.1104830108.
Ponza, P., P. yocawibun, K. Sittikankaew, R. Hiransuchalert, K. ya-mano, and S. Klinbunga. 2011. Molecular cloning and expression analysis of the Mitogen-activating protein kinase 1 (MAPK1) gene and protein during ovarian development of the giant ti-ger shrimp Penaeus monodon. Mol. Reprod. Dev. 78:347–360. http://dx.doi.org/10.1002/mrd.21310.
Swaggerty, C. L., H. He, K. J. Genovese, I. y. Pevzner, and M. H. Kogut. 2011. Protein tyrosine kinase and mitogen-activated
protein kinase signalling pathways contribute to differences in heterophil-mediated innate immune responsiveness between two lines of broilers. Avian Pathol. 40:289–297. http://dx.doi.org/10.1080/03079457.2011.565310.
Tummala, H., S. Fleming, P. M. Hocking, D. Wehner, Z. Naseem, M. Ali, C. F. Inglehearn, N. Zhelev, and D. H. Lester. 2011. The D153del mutation in gnb3 gene causes tissue specific signalling patterns and an abnormal renal morphology in Rge chickens. PLoS ONE 6. http://dx.doi.org/10.1371/journal.pone.0021156.
Upadhya, D., M. Ogata, and L. W. Reneker. 2013. MAPK1 is re-quired for establishing the pattern of cell proliferation and for cell survival during lens development. Development 140:1573–1582. http://dx.doi.org/10.1242/dev.081042.
Wei, y. C., T. S. Pan, M. X. Chang, B. Huang, Z. Xu, T. R. Luo, and P. Nie. 2011. Cloning and expression of Toll-like receptors 1 and 2 from a teleost fish, the orange-spotted grouper Epinephelus coioides. Vet. Immunol. Immunopathol. 141:173–182. http://dx.doi.org/10.1016/j.vetimm.2011.02.016.
Xiao, X., Q. Qin, and X. Chen. 2011. Molecular characterization of a Toll-like receptor 22 homologue in large yellow croaker (Pseu-dosciaena crocea) and promoter activity analysis of its 5′-flanking sequence. Fish Shellfish Immunol. 30:224–233.
Xing, Z., C. J. Cardona, J. Anunciacion, S. Adams, and N. Dao. 2010. Roles of the ERK MAPK in the regulation of proinflam-matory and apoptotic responses in chicken macrophages infected with H9N2 avian influenza virus. J. Gen. Virol. 91:343–351. http://dx.doi.org/10.1099/vir.0.015578-0.
Zhang, y., R. Zhang, H. Zhang, J. Liu, Z. yang, P. Xu, W. Cai, G. Lu, M. Cui, and R. A. Schwendener. 2012. Microparticles released by Listeria monocytogenes-infected macrophages are re-quired for dendritic cell-elicited protective immunity. Cell. Mol. Immunol. 9:489–496.
Zhang, y. y., J. W. Wu, and Z. X. Wang. 2011. Mitogen-activat-ed protein kinase (MAPK) phosphatase 3-mediated cross-talk between MAPKs ERK2 and p38 alpha. J. Biol. Chem. 286: http://dx.doi.org/10.1074/jbc.M110.203786.
2167CLONING OF DUCK MITOGEN-ACTIVATED PROTEIN KINASE 1D
ownloaded from
https://academic.oup.com
/ps/article-abstract/93/9/2158/2730262 by guest on 17 Decem
ber 2018