morphogenetic fields within the human dentition: a new, clinically relevant synthesis of an old...
TRANSCRIPT

Morphogenetic fields within the human dentition: A new,clinically relevant synthesis of an old concept§
Grant Townsend a,e,*, Edward F. Harris b,e, Herve Lesot c,d,e, Francois Clauss c,d, Alan Brook e
aSchool of Dentistry, The University of Adelaide, Adelaide, South Australia 5005, AustraliabDepartment of Orthodontics, College of Dentistry, University of Tennessee, 875 Union Avenue, Memphis, TN 38163, USAc INSERM U595, Faculty of Medicine, 11, rue Humann, 67085 Strasbourg, Franced Faculty of Dentistry, Louis Pasteur University, 67085 Strasbourg Cedex, Francee International Collaborating Centre in Oro-facial Genetics and Development, The University of Liverpool, Liverpool L69 3GN, UK
a r c h i v e s o f o r a l b i o l o g y 5 4 s ( 2 0 0 9 ) s 3 4 – s 4 4
a r t i c l e i n f o
Article history:
Accepted 25 June 2008
Keywords:
Dental development
Clone model
Homeobox code
Patterning
Hypodontia
Supernumeraries
Twins
a b s t r a c t
This paper reviews the concept of morphogenetic fields within the dentition that was first
proposedbyButler (Butler PM. Studiesof themammalian dentition.Differentiation of thepost-
canine dentition. Proc Zool Soc Lond B 1939;109:1–36), then adapted for the human dentition by
Dahlberg (Dahlberg AA. The changing dentition of man. J Am Dent Assoc 1945;32:676–90;
Dahlberg AA. The dentition of the American Indian. In: Laughlin WS, editor. The Physical
Anthropology of the American Indian. New York: Viking Fund Inc.; 1951. p. 138–76). The clone
theory of dental development, proposed by Osborn (Osborn JW. Morphogenetic gradients:
fields versus clones. In: Butler PM, Joysey KA, editorsDevelopment, function and evolution of teeth.
London: Academic Press, 1978. p. 171–201), is then considered before these two important
concepts are interpreted in the light of recent findings from molecular, cellular, genetic and
theoretical and anthropological investigation. Sharpe (Sharpe PT. Homeobox genes and
orofacial development. Connect Tissue Res 1995;32:17–25) put forward the concept of an
odontogenic homeobox code to explain how different tooth classes are initiated in different
parts of the oral cavity in response to molecular cues and the expression of specific groups of
homeobox genes. Recently, Mitsiadis and Smith (Mitsiadis TA, Smith MM. How do genes make
teeth to order through development? J Exp Zool (Mol Dev Evol)2006; 306B:177–82.) proposed that
the field, clone and homeobox code models could all be incorporated into a single model to
explain dental patterning. We agree that these three models should be viewed as comple-
mentary rather than contradictory and propose that this unifying view can be extended into
the clinical setting using findings on dental patterning in individuals with missing and extra
teeth. The proposals are compatible with the unifying aetiological model developed by Brook
(Brook AH. A unifying aetiological explanation for anomalies of tooth number and size. Archs
Oral Biol 1984;29:373–78) based on human epidemiological and clinical findings. Indeed, this
new synthesis can provide a sound foundation for clinical diagnosis, counselling and manage-
ment of patients with various anomalies of dental development as well as suggesting hypoth-
eses for future studies.
# 2008 Elsevier Ltd. All rights reserved.
§ This Supplement arises from a series of papers given at an International Workshop on Oral Growth and Development held in Liverpoolon November 26–28, 2007.
* Corresponding author at: School of Dentistry, The University of Adelaide, Adelaide, South Australia 5005, Australia. Tel.: +61 8 83035968;fax: +61 8 83033444.
avai lable at www.sc iencedi rec t .com
journa l homepage: www. int l .e lsev ierhea l th .com/ journals /arob
E-mail address: [email protected] (G. Townsend).
0003–9969/$ – see front matter # 2008 Elsevier Ltd. All rights reserved.doi:10.1016/j.archoralbio.2008.06.011

a r c h i v e s o f o r a l b i o l o g y 5 4 s ( 2 0 0 9 ) s 3 4 – s 4 4 S35
1. Field and clone theories
The concept of a morphogenetic field emerged in the 19th
century, though the molecular-genetic basis for patterned
series of skeletodental elements (e.g., vertebrae, ribs, teeth)
were unknown. Ordered form was a common observation in
numerous aspects of biology,8 but the governing principles, let
alone the causes, were virtually unknown.9
The accumulation of paleontological findings, along with
information on extant species, made it clear that the
dentitions of higher vertebrates were (1) heterodont—com-
posed of multiple tooth types, and (2) organised into
morphogenetic fields. For example, Bateson,10 amassed
extensive data on meristic variation, including dental exam-
ples primarily of hypo- and hyperdontia. Bateson’s far-ranging
compilations showed that the variable dental elements
occurred at the later-forming end of a field and that these
terminal teeth were metrically and morphologically most
variable within a field.
The Zeitgeist of developmental biology in the early 20th
century was strongly influenced by Spemann’s embryological
organizing centers (Spemann11; De Robertis12). In 1934 Huxley
and De Beer combined the study of morphology with the
morphogenetic field concept in their seminal work The
Elements of Experimental Embryology, which closely preceded
Butler’s1 description of fields and morphogenetic variations in
the mammalian dentition.
Weiss13 was among the leaders to reaffirm the interpre-
tative value of the field concept. Butler1 postulated that
morphogenetic fields can account for the way in which teeth
within a particular class formed meristic series, i.e., with each
tooth displaying similarities to others nearby because of the
influence of a common field but with graded differences
because of its position. Currently these dental elements are
considered to result from the repetition of ‘developmental
modules’.14,15 Butler’s phylogenetic study of the mammalian
dentition was wholly observational, but he inferred several
features of the changes that meshed well with the biological
features of field gradients, notably (a) a size gradient within
each field, (b) a gradient of morphological complexity, from the
complex pole tooth to the simpler variable tooth, and (c) a
gradient of variability, with less size and shape variability in
the pole tooth and increasing variability away from the pole.
Documenting such field gradients was important because the
observations substantiated that teeth exist within morpho-
genetic gradients as defined by the criteria established by
Huxley and De Beer16 and Weiss.14 The phylogenetic patterns
do not always match the ontogenetic situations, however;
while incisors have lost teeth distally-to-mesially, the number
of premolars has been reduced mesially-to-distally.
Butler17 later suggested that teeth were evolving as a part of
a system rather than as individual organs. He postulated two
different effects: a ‘meristic’ effect, influencing the number
and spacing of teeth, and a ‘field’ effect produced by
substances or signals that controlled their differentiation
and final shapes. He added that the order of development of
teeth also represents a field effect.18
Dahlberg2,3 adapted Butler’s1 concepts to the human
dentition and proposed that there was a field of influence
operating on each of the tooth classes, i.e., incisors, canines,
premolars and molars. Each field was thought to have its
strongest effect on the anterior or key tooth within a class. The
more distally placed teeth, which generally develop later than
those more mesially placed, were observed to show greater
phenotypic variation. Interestingly, Dahlberg2,3 did not define
fields within the primary dentition and, without any com-
ment, he added a premolar field to Butler’s1 three-field
paradigm. In contrast, Butler19 later argues that premolars
represent modified anterior members of a permanent molar
field, with the primary second molar displaying similar
morphology to the permanent first molar located directly
distal to it.
Osborn4 proposed that a single clone of pre-programmed
cells led to the development of all the teeth within a particular
class. For example, a molar clone of cells was postulated to
induce the dental lamina to initiate molar development. As the
clone of cells grew distally, tooth buds were formed,
surrounded by zones of inhibition that prevented other teeth
developing until the migrating clone had moved on suffi-
ciently. More recent work invokes a reaction-diffusion model
in the region of a presumptive tooth, where activators induce
placode formation while negative regulators are higher in
interplacodal regions, which prevents tooth formation and,
thus, accounts for the orderly spacing of teeth.20
The demonstration that isolated presumptive first molar
tissue explants could continue to grow and form all three
molar teeth in their normal sequence, provided strong
experimental support for the clone model.21 However, the
clone model does not provide an explanation for how the
dentition develops as a whole, with different tooth classes
displaying different shapes. Furthermore, the inclusion of the
phrase ‘‘fields versus clones’’ in the title of Osborn’s original
paper4 has led many to suppose that the field and clone
models are competing, mutually exclusive concepts. Recent
findings about the roles of signalling molecules and the
expression of homeobox genes during dental development
indicate that such a distinction is unwarranted.6 Indeed, the
field and clone models can be viewed as complementary to
each other.
2. The odontogenic homeobox code
The odontogenic homeobox code model explains how dental
patterns can be generated from different domains of expression
of homeobox genes in neural crest derived ectomesench-
yme.5,22 While the code was initially developed from studies of
the mouse dentition in which there are only incisor and molar
teeth, it has been extended to explain how canines and
premolars could also be produced by overlapping domains of
gene expression.22 Experimental studies have also shown that
by modulating certain signalling molecules it is possible to alter
homeobox gene expression domains in the ectomesenchymal
tissue and to modify tooth number, size, shape and differentia-
tion.23,24 A schematic representation of how dental patterning
can be produced by an odontogenic homeobox code has been
provided recently by Cobourne and Mitsiadis.25 They describe
how an ‘‘inter-mixing’’ of genes expressed by ectomesenchyme
of the first branchial arch can lead to the establishment of
different morphogenetic fields. Patterns are established by

a r c h i v e s o f o r a l b i o l o g y 5 4 s ( 2 0 0 9 ) s 3 4 – s 4 4S36
signals from the ectoderm that induce specific domains of
homeobox gene expression in the ectomesenchyme. This
patterning is plastic initially but over time it becomes fixed
into the ‘memory’ of the ectomesenchymal cells. It is these
specific domains that are postulated to provide the molecular
information needed to specify different tooth shapes.25
There is evidence that the nature of the molecular
signalling in the upper and lower jaws may vary. The dental
formula is the same in both arches in mice and in humans, but
the shape and morphologies of the homologous teeth in the
two jaws are each distinctive one from the other.26 Biochem-
ical signalling differences have been demonstrated in the
mouse for Dlx genes27,28 and also for activin/follistatin,29
although it is not known how neural crest derived cells
migrating into the developing maxillary and mandibular
regions develop the ability to respond differently to ectoder-
mal signaling. Reports of apparently independent genetic
determination of maxillary and mandibular dentitions, based
on tooth size data derived from twins,30 are consistent with
the molecular evidence.
3. Recent perspectives of the field, clone andhomeobox models
Apart from the role of homeobox genes in determining the
location of different fields within the developing dentition, it
has been shown that expression of the signalling molecule
ectodysplasin (EDA) is important in defining the size of dental
fields. Studies using transgenic mice have shown that
alteration in tooth number occurs when EDA, its receptor
(EDAR), or the intracellular adaptor protein EDAR-binding
death domain adaptor (EDARADD) are disrupted. It has been
proposed that if there is over-production of the EDA ligand, or
the receptors for EDA are over-expressed, then the size of the
molar field in mice increases, resulting in the formation of
supernumerary teeth.31 In transgenic mice, the supernumer-
ary teeth develop mesial to the first molar, in the diastema
region that is present normally between the incisor and molar
segments. The supernumeraries usually have a reduced cusp
pattern and do not resemble normal mouse molars. We would
raise the question whether these ‘supernumeraries’ actually
arise from tooth germs that would normally undergo
apoptosis during development. In contrast to over-expression
of EDAR and its association with extra teeth, loss of EDAR
signalling leads to missing molars.32
Peterkova et al.33 have identified five different morpho-
types in the mandibular molar region of tabby/EDA mice and
reported that these result from abnormal segmentation of the
epithelial compartment of the dental lamina along a mesio-
distal axis. Increased apoptosis has been noted in the mesial
aspect of the dental epithelium of affected mice, and this
seems to be a consequence of impaired tooth development
associated with the defective segmentation process.34
In humans, apart from the X-linked form of ectodermal
dysplasia (XLHED) that involves mutation of the ectodysplasin
gene ED1 (locus Xq12-q13.1), autosomal forms of the condition
have also been described.35,36 These autosomal dominant and
recessive forms of HED are linked to mutations of EDAR at the
2q11-q13 locus or EDARADD at the 1q42.2-43 locus.
The phenotypes of autosomal dominant and recessive
forms of HED, linked to EDAR mutation, are similar to the
XLHED phenotype, including hypohidrosis, hypotrichosis and
oligodontia, which reflects the common developmental
patterns of many ectodermal derivatives.37,20 However, some
differences are evident. The skin and hair phenotypes
associated with the autosomal forms of HED seem to be
milder than those in XLHED.38 Craniofacial dysmorphic
features observed in autosomal HED also seem to be less
severe than in XLHED, with mild frontal bossing and malar
flattening.35 Moreover, dental and craniofacial phenotypic
manifestations are less severe in autosomal dominant forms
than in autosomal recessive HED.36 As for the dental
phenotype of autosomal HED, hypodontia, often severe, with
varying degrees of primary and permanent tooth agenesis is
evident, associated with anomalous conical incisors and
canines (Fig. 1).
The X-linked form of HED constitutes the majority of HED
phenotypes. Nevertheless, the ED1 mutation is not system-
atically identified in patients with HED: only 65.4% in a group
of patients exhibiting the HED phenotype,39 other loci being
mutated, such as EDAR (in 25% of non-ED1 HED) and, less
often, EDARADD.36,40
Dental phenotypic variability is observed in autosomal HED
(as shown in Fig. 1) and is probably due to genetic hetero-
geneity.38 Indeed, different non-sense mutations distributed
along the 11 exons of the EDAR gene have been identified and
are associated with varying phenotypic expression of auto-
somal HED.36 These loss-of-function mutations lead to
alteration of EDAR trimerisation and partial or total loss of
EDA-EDAR and EDAR-EDARADD molecular interactions.38 An
absence or reduced activation of NF-kB during embryogenesis
and abnormal cell signaling are the consequences of the EDAR
mutations. The same molecular mechanisms are involved in
EDARADD mutations leading to complete or partial abrogation
of NF-kB activation in both autosomal dominant and recessive
mutations.40
Line41 has emphasised that the concept of morphogenetic
fields should not be limited to the expression of a single gene
or its protein product but rather include consideration of how
various genetic influences, modulated by epigenetic effects,
affect development of the dentition. By studying variation in
the number of missing teeth in families with mutations of the
MSX1 and PAX 9 genes, Line has attempted to clarify the
relative influence of these two genes within the human
dentition, referring to their influences as ‘‘relative molecular
morphogenetic fields’’. Variations in patterns of symmetry of
missing teeth within individuals were also used to gain insight
into the effects of these two genes on the developmental
stability of dental fields. Line found that the morphogenetic
fields associated with MSX1 and PAX9 were not confined to a
single tooth class, nor did there seem to be a simple gradient
pattern. He interpreted these findings as reflecting an
interaction between different signalling molecules. Although
there was some evidence that mutations in MSX1 and PAX9
could increase the extent of asymmetry of tooth agenesis, no
consistent pattern was identified. These findings are all
consistent with the multifactorial aetiology as proposed by
Gruneberg,42 Brook,43,7 Chosack et al.,44 Brook et al.45 and
others.
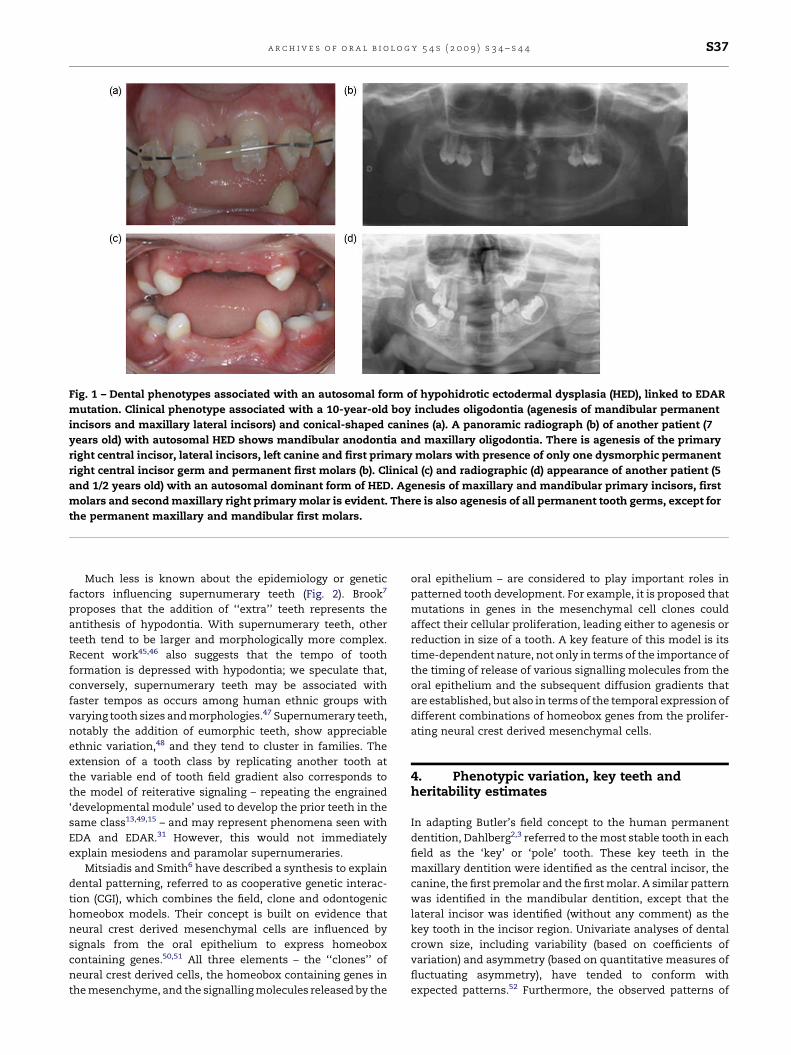
Fig. 1 – Dental phenotypes associated with an autosomal form of hypohidrotic ectodermal dysplasia (HED), linked to EDAR
mutation. Clinical phenotype associated with a 10-year-old boy includes oligodontia (agenesis of mandibular permanent
incisors and maxillary lateral incisors) and conical-shaped canines (a). A panoramic radiograph (b) of another patient (7
years old) with autosomal HED shows mandibular anodontia and maxillary oligodontia. There is agenesis of the primary
right central incisor, lateral incisors, left canine and first primary molars with presence of only one dysmorphic permanent
right central incisor germ and permanent first molars (b). Clinical (c) and radiographic (d) appearance of another patient (5
and 1/2 years old) with an autosomal dominant form of HED. Agenesis of maxillary and mandibular primary incisors, first
molars and second maxillary right primary molar is evident. There is also agenesis of all permanent tooth germs, except for
the permanent maxillary and mandibular first molars.
a r c h i v e s o f o r a l b i o l o g y 5 4 s ( 2 0 0 9 ) s 3 4 – s 4 4 S37
Much less is known about the epidemiology or genetic
factors influencing supernumerary teeth (Fig. 2). Brook7
proposes that the addition of ‘‘extra’’ teeth represents the
antithesis of hypodontia. With supernumerary teeth, other
teeth tend to be larger and morphologically more complex.
Recent work45,46 also suggests that the tempo of tooth
formation is depressed with hypodontia; we speculate that,
conversely, supernumerary teeth may be associated with
faster tempos as occurs among human ethnic groups with
varying tooth sizes and morphologies.47 Supernumerary teeth,
notably the addition of eumorphic teeth, show appreciable
ethnic variation,48 and they tend to cluster in families. The
extension of a tooth class by replicating another tooth at
the variable end of tooth field gradient also corresponds to
the model of reiterative signaling – repeating the engrained
‘developmental module’ used to develop the prior teeth in the
same class13,49,15 – and may represent phenomena seen with
EDA and EDAR.31 However, this would not immediately
explain mesiodens and paramolar supernumeraries.
Mitsiadis and Smith6 have described a synthesis to explain
dental patterning, referred to as cooperative genetic interac-
tion (CGI), which combines the field, clone and odontogenic
homeobox models. Their concept is built on evidence that
neural crest derived mesenchymal cells are influenced by
signals from the oral epithelium to express homeobox
containing genes.50,51 All three elements – the ‘‘clones’’ of
neural crest derived cells, the homeobox containing genes in
the mesenchyme, and the signalling molecules released by the
oral epithelium – are considered to play important roles in
patterned tooth development. For example, it is proposed that
mutations in genes in the mesenchymal cell clones could
affect their cellular proliferation, leading either to agenesis or
reduction in size of a tooth. A key feature of this model is its
time-dependent nature, not only in terms of the importance of
the timing of release of various signalling molecules from the
oral epithelium and the subsequent diffusion gradients that
are established, but also in terms of the temporal expression of
different combinations of homeobox genes from the prolifer-
ating neural crest derived mesenchymal cells.
4. Phenotypic variation, key teeth andheritability estimates
In adapting Butler’s field concept to the human permanent
dentition, Dahlberg2,3 referred to the most stable tooth in each
field as the ‘key’ or ‘pole’ tooth. These key teeth in the
maxillary dentition were identified as the central incisor, the
canine, the first premolar and the first molar. A similar pattern
was identified in the mandibular dentition, except that the
lateral incisor was identified (without any comment) as the
key tooth in the incisor region. Univariate analyses of dental
crown size, including variability (based on coefficients of
variation) and asymmetry (based on quantitative measures of
fluctuating asymmetry), have tended to conform with
expected patterns.52 Furthermore, the observed patterns of
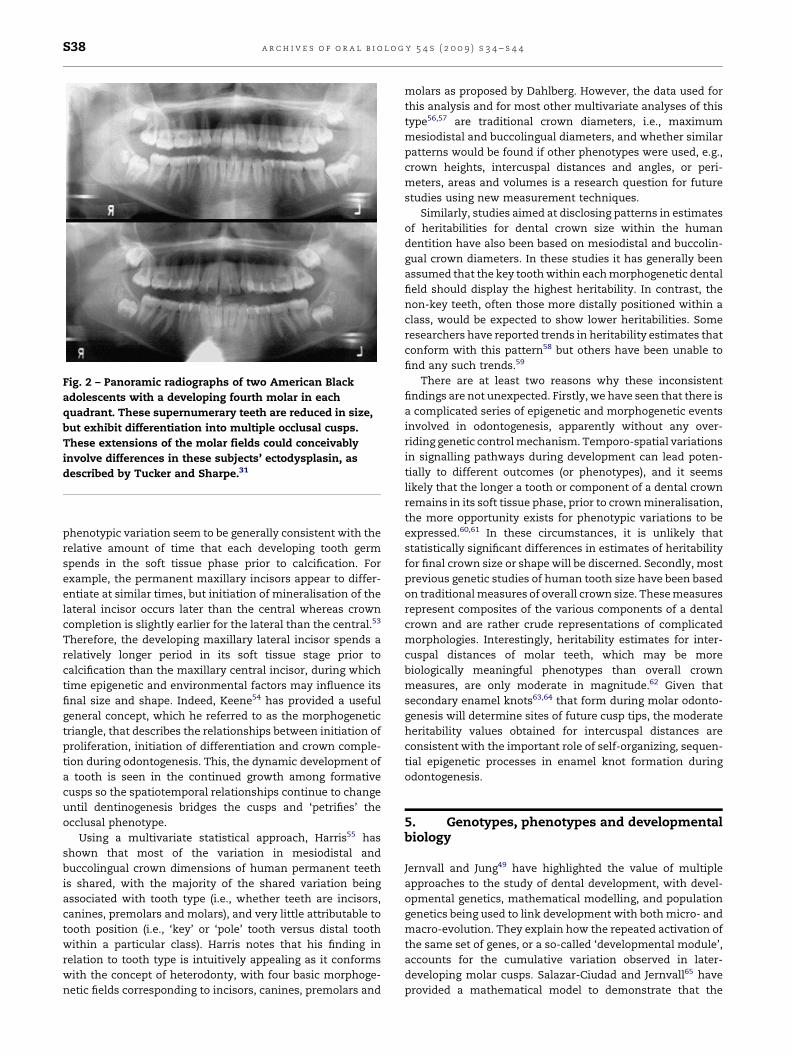
Fig. 2 – Panoramic radiographs of two American Black
adolescents with a developing fourth molar in each
quadrant. These supernumerary teeth are reduced in size,
but exhibit differentiation into multiple occlusal cusps.
These extensions of the molar fields could conceivably
involve differences in these subjects’ ectodysplasin, as
described by Tucker and Sharpe.31
a r c h i v e s o f o r a l b i o l o g y 5 4 s ( 2 0 0 9 ) s 3 4 – s 4 4S38
phenotypic variation seem to be generally consistent with the
relative amount of time that each developing tooth germ
spends in the soft tissue phase prior to calcification. For
example, the permanent maxillary incisors appear to differ-
entiate at similar times, but initiation of mineralisation of the
lateral incisor occurs later than the central whereas crown
completion is slightly earlier for the lateral than the central.53
Therefore, the developing maxillary lateral incisor spends a
relatively longer period in its soft tissue stage prior to
calcification than the maxillary central incisor, during which
time epigenetic and environmental factors may influence its
final size and shape. Indeed, Keene54 has provided a useful
general concept, which he referred to as the morphogenetic
triangle, that describes the relationships between initiation of
proliferation, initiation of differentiation and crown comple-
tion during odontogenesis. This, the dynamic development of
a tooth is seen in the continued growth among formative
cusps so the spatiotemporal relationships continue to change
until dentinogenesis bridges the cusps and ‘petrifies’ the
occlusal phenotype.
Using a multivariate statistical approach, Harris55 has
shown that most of the variation in mesiodistal and
buccolingual crown dimensions of human permanent teeth
is shared, with the majority of the shared variation being
associated with tooth type (i.e., whether teeth are incisors,
canines, premolars and molars), and very little attributable to
tooth position (i.e., ‘key’ or ‘pole’ tooth versus distal tooth
within a particular class). Harris notes that his finding in
relation to tooth type is intuitively appealing as it conforms
with the concept of heterodonty, with four basic morphoge-
netic fields corresponding to incisors, canines, premolars and
molars as proposed by Dahlberg. However, the data used for
this analysis and for most other multivariate analyses of this
type56,57 are traditional crown diameters, i.e., maximum
mesiodistal and buccolingual diameters, and whether similar
patterns would be found if other phenotypes were used, e.g.,
crown heights, intercuspal distances and angles, or peri-
meters, areas and volumes is a research question for future
studies using new measurement techniques.
Similarly, studies aimed at disclosing patterns in estimates
of heritabilities for dental crown size within the human
dentition have also been based on mesiodistal and buccolin-
gual crown diameters. In these studies it has generally been
assumed that the key tooth within each morphogenetic dental
field should display the highest heritability. In contrast, the
non-key teeth, often those more distally positioned within a
class, would be expected to show lower heritabilities. Some
researchers have reported trends in heritability estimates that
conform with this pattern58 but others have been unable to
find any such trends.59
There are at least two reasons why these inconsistent
findings are not unexpected. Firstly, we have seen that there is
a complicated series of epigenetic and morphogenetic events
involved in odontogenesis, apparently without any over-
riding genetic control mechanism. Temporo-spatial variations
in signalling pathways during development can lead poten-
tially to different outcomes (or phenotypes), and it seems
likely that the longer a tooth or component of a dental crown
remains in its soft tissue phase, prior to crown mineralisation,
the more opportunity exists for phenotypic variations to be
expressed.60,61 In these circumstances, it is unlikely that
statistically significant differences in estimates of heritability
for final crown size or shape will be discerned. Secondly, most
previous genetic studies of human tooth size have been based
on traditional measures of overall crown size. These measures
represent composites of the various components of a dental
crown and are rather crude representations of complicated
morphologies. Interestingly, heritability estimates for inter-
cuspal distances of molar teeth, which may be more
biologically meaningful phenotypes than overall crown
measures, are only moderate in magnitude.62 Given that
secondary enamel knots63,64 that form during molar odonto-
genesis will determine sites of future cusp tips, the moderate
heritability values obtained for intercuspal distances are
consistent with the important role of self-organizing, sequen-
tial epigenetic processes in enamel knot formation during
odontogenesis.
5. Genotypes, phenotypes and developmentalbiology
Jernvall and Jung49 have highlighted the value of multiple
approaches to the study of dental development, with devel-
opmental genetics, mathematical modelling, and population
genetics being used to link development with both micro- and
macro-evolution. They explain how the repeated activation of
the same set of genes, or a so-called ‘developmental module’,
accounts for the cumulative variation observed in later-
developing molar cusps. Salazar-Ciudad and Jernvall65 have
provided a mathematical model to demonstrate that the

a r c h i v e s o f o r a l b i o l o g y 5 4 s ( 2 0 0 9 ) s 3 4 – s 4 4 S39
morphology of mammalian teeth can be predicted by
integrating experimental data on gene interactions and
growth into a ‘‘morphodynamic mechanism’’, thereby linking
genotype to phenotype. Jernvall et al.66 make use of a
topographic method, Geographic Image Systems (GIS), to
show that molecular pre-patterning can predict the arrange-
ment of cusps more than a day in advance, with subtle
heterotopic shifts being postulated to play an important role
during evolution in producing new cuspal arrangements.
More recently, Salazar-Ciudad et al.67 have suggested that
developmental mechanisms used for generating patterns can
be grouped into three categories: cell autonomous mechan-
isms; inductive mechanisms; and morphogenetic mechan-
isms. The first category is postulated to include those
mechanisms by which cells become arranged into patterns
without interacting with other cells. The second category
refers to mechanisms by which cells communicate with one
another to produce patterning through reciprocal or hier-
archical alteration of phenotypes. The third category refers to
mechanisms that can lead to changes in patterns through cell
interactions that do not involve phenotypic changes in cells.
Of particular relevance to morphogenetic fields, inductive and
morphogenetic mechanisms can be combined either mor-
phostatically, in which the inductive mechanisms occur first,
or morphodynamically, in which both types of mechanisms
interact continuously. Salazar-Ciudad et al.67 suggest that the
mammalian dentition provides a good example of a develop-
mental system that employs morphodynamic mechanisms,
with induction and morphogenesis taking place concurrently
and interdependently. They also explain how morphody-
namic mechanisms use spatial epigenetic information pre-
sent in the emerging phenotype at each stage of the
developmental process to influence later development. In
this sense, morphodynamic mechanisms exhibit dependency
on the so-called ‘intermediate phenotype’.
Salazar-Ciudad et al.67 avoid using the term ‘morphoge-
netic field’ because it presupposes the concept of prospective
cell fate. They prefer to use the term ‘gene expression
territory’. They emphasise that morphodynamic mechanisms
involve interaction between cells along a developmental
trajectory that is continually interacting with a changing
molecular and geometric microenvironment. With this per-
spective of dental development, it becomes more apparent
that the field and clone models are complementary inter-
pretations of the same observations; they are mutually
supportive rather than opposing concepts. This perspective
is also entirely consistent with recent findings of discordant
dental development in monozygotic twin pairs.68,69
6. Genetics, epigenetics and environment: theMZ co-twin model
While molecular geneticists have tended to focus on methyla-
tion and acetylation of DNA when referring to epigenetics,
there is growing recognition of the need for a broader
interpretation of the term.70,71 We use the term ‘epigenetics’
in the broad sense, as proposed originally by Waddington,72,73
to refer to processes by which genotype gives rise to
phenotype. This broader view enables description of those
interactive processes that occur between cells at the local
tissue level during dental development as epigenetic events, in
addition to those that may operate directly on DNA. Molenaar
et al.74 referred to epigenetic influences as a ‘‘third source of
developmental differences’’ that can account for phenotypic
variation in development, in addition to genetic and environ-
mental factors. This third source is comprised of non-linear
epigenetic events that can cause variability at all phenotypic
levels and that these epigenetic influences result from
autonomous developmental processes with ‘‘emergent self-
organizing properties.’’ The role of epigenetic factors in the
broad sense during odontogenesis has been discussed by Lesot
et al.75 while local epigenetic influences, including cell–cell
and cell–matrix interactions and their effects on asymmetrical
growth and differential cell proliferation during dental
development have also been described.76
Interestingly, Eaves et al.77 have pointed out that Mole-
naar’s concepts can be interpreted in terms of chaos theory.
They have shown through simulations that chaotic and near-
chaotic processes may reduce the strength of correlations
between twin pairs. By simulating a simple non-linear model
behaving chaotically, Eaves et al.77 found that ‘‘small
variations in initial conditions (e.g., small quantitative
differences between MZ twins at the molecular level such
as the degree of methylation of a particular gene in a
particular tissue) will have consequences at the phenotypic
level which look like occasion-specific environmental
effects.’’ The findings of Eaves and colleagues are particularly
relevant when attempting to explain differences in expres-
sion of missing and extra teeth between monozygotic co-
twins with the same genotypes.
Eaves et al.77 provide theoretical support for our view that
minor variations in local epigenetic events during dental
development, probably relating to the spatial arrangement of
odontogenic cells and/or the timing of the signalling events
between them, can produce distinct differences in dental
features between monozygotic (MZ) twin pairs who have been
confirmed to have the same genotypes.67,68,78 These differ-
ences can include discordances in the number and/or location
of congenitally missing teeth or differences in the number of
supernumerary teeth between MZ co-twins.
7. Stepping into the clinical setting
The approaches of Line41 and Mitsiadis and Smith6 provide an
important step forward in explaining how the field, clone and
homeobox code concepts can all be used as a foundation for
understanding how phenotypic patterning is established in
the human dentition. We now provide some additional
perspectives based on our own research to position these
new concepts within a clinical context that ranges from minor
anomalies of the size and shape of teeth through to more
severe anomalies, including missing and extra teeth.
The most common clinical presentation involving missing
teeth is hypodontia of only one or a few teeth, commonly the
third molars, second premolars or maxillary lateral incisors.
On the other hand, these generalities are based on the
numerous studies of peoples of European extraction,79 and the
limited data from other ethnic groups suggest the frequencies

Fig. 3 – Dental panoramic radiographs (a and b) and
maxillary clinical views (c and d) of a pair of monozygotic
twins affected by X-linked hypohidrotic ectodermal
dysplasia (XLHED) with a mutation of the ED1 gene, coding
for ectodysplasin (EDA) morphogenetic factor.
a r c h i v e s o f o r a l b i o l o g y 5 4 s ( 2 0 0 9 ) s 3 4 – s 4 4S40
and patterns may differ.80–82 When considering supernumer-
ary teeth, the most common example is the mesiodens which
occurs in the anterior midline region of the maxilla. A
multifactorial model linking tooth size and number can
account for the different patterns of expression of tooth size,
hypodontia and supernumerary teeth observed in males and
females.7,43,45,83–85 The relatives of individuals with missing or
extra teeth have also been found to be more likely to also
display missing or extra teeth, supporting the concept of an
underlying genetic predisposition to hypodontia or super-
numerary teeth. We believe that such a multifactorial model,
with multiple genetic, epigenetic and environmental influ-
ences, provides the best explanation for our observations
involving missing and extra teeth in MZ twin pairs.
Molecular studies, primarily in mice, show that congeni-
tally missing teeth can arise from diverse developmental
causes. Tooth development can be interrupted at, at least,
three stages. First, the ectoderm may not have the capacity to
induce bud-formation in the underlying mesenchyme.20,86
Second, missing or mutant signals of a mesenchymal
transcriptional factor such as Msx1 or Pax9 can arrest
development prior to enamel knot formation.87 Third, devel-
opment can be arrested shortly after the initiation of
dentinogenesis, with subsequent apoptosis. This latter stage
corresponds to the incipient formation of lateral incisors in
mice,88,89 where these teeth never fully develop in this species.
Similarly tooth germs anterior to the first molar (i.e.,
premolars and canines) in mice undergo apoptosis.34
To illustrate the dental variability between MZ co-twins, a
pair of twins diagnosed with a mutation of the ED1 gene are
described, both showing oligodontia but with different
patterns of missing teeth (Fig. 3). One of the MZ twins (a)
showed mandibular anodontia, whereas the other exhibited a
severe mandibular hypodontia with the presence of a
mandibular left permanent molar (b). The distribution of
maxillary teeth again illustrates variability in the severity of
the phenotype. Only permanent maxillary canines and the left
first molar are evident for the first MZ twin, whereas maxillary
canines and the contra-lateral right permanent first molar are
present in the second MZ twin (b), together with the left
maxillary central incisor and a retained maxillary second right
primary molar.
The dental phenotypic variability between these XLHED co-
twins could be accounted for by different local epigenetic and
environmental factors interacting with the EDA mutation, as
described in the multifactorial theory. The dental phenotype
of the second XLHED MZ twin (b) suggests residual capacity
for initiation of odontogenesis, allowing the development of
a permanent maxillary central incisor and a permanent
mandibular molar. However, the phenotypic variability
observed shows that other epigenetic mechanisms, besides
genetic factors, represented mainly by the odontogenic
homeobox code and signaling molecules such as FGF, BMP,
Shh, and EDA-NF-kappaB,90,91 are also important for initiation
of odontogenesis and tooth morphogenesis.
The strong link between tooth number, size and shape is
illustrated by the dental phenotype of these XLHED patients.
Microdontia of canines (Fig. 3c and d), dysmorphic cone-
shaped incisors (d) and canines (c and d) and molar root fusion
and shape anomalies (a and b) are associated with the
phenotype of oligodontia and also linked to the ED1 mutation
and molecular alterations of the EDA-NF-kappaB signalling
pathway.92
By superimposing thresholds on an underlying distribution
of tooth size, it is possible to explain the relationship between
tooth size and the presence or absence of teeth. For example,
missing teeth will tend to occur below a certain threshold for
tooth size. The prevalence of missing teeth will be greater in
females who have smaller teeth, on average, than males. At
the other extreme of the distribution, where tooth size is
larger, another threshold is reached above which extra teeth
will tend to occur. The prevalence of extra teeth will tend to be
greater in males who have larger teeth, on average, than
females. These relationships are summarised in the model of
Brook (Fig. 4). It is also helpful to remember that the
‘thresholds’ actually are zones rather than well-demarcated
lines because of spatiotemporal effects of the person’s genetic
background and dynamics of his environment.93,94
Although the model is a theoretical one, it has direct
relevance to clinical practice. It emphases that common
anomalies such as missing and extra teeth represent part of a
continuous spectrum of inter-related dental phenotypes that
are influenced by a combination of genetic, epigenetic and
environmental factors. Furthermore, when linked to the

Fig. 4 – Model based on normal distributions with
superimposed thresholds to explain the relationships
between anomalies of tooth number and size after Brook.7
a r c h i v e s o f o r a l b i o l o g y 5 4 s ( 2 0 0 9 ) s 3 4 – s 4 4 S41
concept of morphogenetic fields, this conceptual model
enables clinicians to predict which teeth are most likely to
be affected, e.g., third molars, second premolars and maxillary
lateral incisors display hypodontia most commonly and are
also most often affected by certain other anomalies of
development. Such a framework provides a valuable basis
for clinical diagnosis, treatment planning and counselling.
Apart from the value to the clinician of appreciating that there
are links between tooth size, shape and number within the
dentition, reflecting underlying developmental processes,
there are many other dental phenotypes that have been
shown to be associated with congenitally missing teeth.94
These include delayed formation and eruption of other teeth,
malposition and ectopic eruption of teeth, taurodontism,
rotation of teeth, short roots and simplified morphology.95–101
Furthermore, more generalised anomalies in the craniofacial
region have also been shown to be associated with missing
teeth, including alterations in dental arch form, presence of
Class II division 2 craniofacial type and sella turcica
bridging.44,102,103 A recent report has even linked tooth
agenesis to colo-rectal cancer.104
These several associations among size and shape of
structures certainly are important clinically, but they also
reflect the integrated development of skeletodental com-
plexes, both at the molecular41 and phenotypic levels. Indeed,
Bateson10 described various examples of how nearby teeth are
themselves affected when there is hypo- or hyperdontia,
which often parallel the situations that have occurred
phylogenetically.1,105 Understanding co-variations can help
clinically as well as in the search for common developmental
etiologies.
8. Finding the genes for dental development:implications for future practice
To date, molecular-genetic studies in humans have concen-
trated mainly on locating the genes associated with missing
teeth. Mutations in three genes, MSX1, PAX9 and AXIN2104,106–
109 have been shown to be associated with familial cases of
severe hypodontia (where many teeth are missing). The
families selected for these studies had pedigrees consistent
with an autosomal dominant mode of inheritance although
variations in the number and position of teeth missing have
been noted. However, there are more than 300 genes that
appear to be involved in dental development,110 and a number
of them could be candidates for missing and extra teeth.
Furthermore, as we have indicated in the previous section of
this review, the most common clinical presentation relating to
missing teeth is simple hypodontia where only one or a few
teeth are missing rather than many, so identifying the causes
of common hypodontia is of clinical merit.
Given that there appears to be a link between the size and
shape of teeth, and hypodontia or supernumerary teeth, we
propose that there is likely to be a group of genes that exert
pleiotropic effects on multiple dental phenotypes, accounting
for their observed co-variation. Just how many genes are
involved remains to be seen, but it is possible that it may be a
relatively small number. Support for this view is provided by
Kangas et al.111 who have shown that dental characters in
mice seem to be non-independent. So too, tooth crown
dimensions as well as morphological traits are positively
intercorrelated within and among tooth types.57,112 Increasing
the levels of expression of just one gene can lead to increases
in cusp number, altered cusp shape and position, develop-
ment of longitudinal crests on teeth, and increases in tooth
number.
Rather than a simple monogenic mode of inheritance
alone, we consider that a multifactorial model7,45,83 – with
genetic, epigenetic and environmental influences – provides
the best explanation for our observations involving hypodon-
tia and supernumerary teeth in MZ twin pairs, as well as
different phenotypic expressions in MZ co-twins with ecto-
dermal dysplasia. Such a model, with superimposed thresh-
olds linking tooth size, morphology and number, provides a
plausible explanation for why MZ co-twins, who have the
same genotypes, may display different dental phenotypes.
The MZ twin pairs may have genotypes that place them near to
a threshold for either missing or extra teeth, but variations in
local epigenetic events during odontogenesis may determine
on which side of the threshold they fall.
As we have mentioned previously, and as is reported in
other papers in this special issue of Archives, measures of tooth
size that have been reported most commonly in the past have
been maximum mesiodistal and buccolingual crown dia-
meters. Often these measurements have been recorded from
dental casts with hand-held calipers that cannot provide
accuracies better than 0.1 mm. Although there have been
attempts to describe tooth size and shape using other
variables, including cusp areas, perimeter and area and
volumes,60,85,113 these types of data have rarely been used
in genetic studies. With the advances in image capturing
techniques, including 3D laser systems, we are now in a
position to define new dental phenotypes that should give
clearer insights into the nature and causes of variation in the
dentition.114
Genome-wide association studies (GWAS) are currently
being used to identify genes linked to various common
diseases, including coronary heart disease, hypertension,
diabetes and arthritis, and it is planned to use a similar
approach to identify key genes involved in dental develop-
ment using a suite of newly defined dental phenotypes in both

a r c h i v e s o f o r a l b i o l o g y 5 4 s ( 2 0 0 9 ) s 3 4 – s 4 4S42
2D and 3D.115 Although very important, the genetic aspects are
not sufficient to explain fully how various dental anomalies
arise in individuals. This is where further exploration of
epigenetic factors will be essential.
Already researchers are beginning to study epigenetic
biomarkers in an attempt to explain the reasons for observed
differences between MZ twin pairs.116 At this stage the focus is
on trying to determine the extent of differences in global
genomic DNA methylation levels but it is likely that more
specific analyses will be developed soon. Once these
approaches aimed at the level of DNA are refined further,
and our understanding of the nature of the epigenetic
influences at a local tissue level improves, we should be able
to provide a clearer picture of how genetic, epigenetic and
environmental factors influence human dental development
and lead to phenotypic patterning within the dentition.
Acknowledgements
The support of the following funding agencies is gratefully
acknowledged: National Health and Medical Research Council,
Canberra, Australian Dental Research Foundation, National
Institute of Health, USA, Wellcome Trust, UK.
Disclosures
Competing interests: None declared.
Funding: National Health and Medical Research Council of
Australia, Australian Dental Research Foundation, National
Institute of Health, USA, INSERM UMR595, Wellcome Trust,
UK, National French Reference Centre for Oral Manifestations
of Rare Diseases.
r e f e r e n c e s
1. Butler PM. Studies of the mammalian dentition.Differentiation of the post-canine dentition. Proc Zool SocLond B 1939;109:1–36.
2. Dahlberg AA. The changing dentition of man. J Am DentAssoc 1945;32:676–90.
3. Dahlberg AA. The dentition of the American Indian. In:Laughlin WS, editor. The physical anthropology ofthe American Indian. New York: Viking Fund Inc.;1951. p. 138–76.
4. Osborn JW. Morphogenetic gradients: fields versus clones.In: Butler PM, Joysey KA, editors. Development, function andevolution of teeth. London: Academic Press; 1978. p. 171–201.
5. Sharpe PT. Homeobox genes and orofacial development.Connect Tissue Res 1995;32:17–25.
6. Mitsiadis TA, Smith MM. How do genes make teeth to orderthrough development? J Exp Zool (Mol Dev Evol)2006;306B:177–82.
7. Brook AH. A unifying aetiological explanation foranomalies of tooth number and size. Archs Oral Biol1984;29:373–8.
8. Needham J. Chemical embryology. Cambridge: CambridgeUniversity Press; 1931.
9. Morgan TH. Embryology and genetics. New York: ColumbiaUniversity Press; 1934.
10. Bateson W. Materials for the study of variation: treated withspecial regard to discontinuity in the origin of species. London:Macmillan and Company; 1894.
11. Spemann H. Embryonic development and induction. NewHaven: Yale University Press; 1938.
12. De Robertis EM. Spemann’s organizer and self-regulationin amphibian embryos. Nat Rev Mol Cell Biol 2006;7:296–302.
13. Weiss P. Principles of development. New York: Holt; 1939.14. Weiss KM. Duplication with variation: metameric logic in
evolution from genes to morphology. Yrbk Phys Anthropol1990;33:1–24.
15. Jernvall J, Thesleff I. Reiterative signaling and patterningduring mammalian tooth morphogenesis. Mech Dev2000;92:19–29.
16. Huxley J, de Beer GR. The elements of experimental embryology.Cambridge: Cambridge University Press; 1934.
17. Butler PM. What happened to the field theory. In: Brook A,editor. Dental morphology. Sheffield: Sheffield AcademicPress Ltd.; 2001. p. 3–12.
18. Butler PM, Tooth morphology and primateevolution.Brothwell DR, editor. Dental anthropologySymposia of the Society for the Study of Human Biology, vol. V.New York: Pergamon Press; 1963. p. 1–13.
19. Butler PM. Comparison of the development of the seconddeciduous molar and first permanent molar in man. ArchsOral Biol 1967;12:1245–60.
20. Pispa J, Thesleff I. Mechanisms of ectodermalorganogenesis. Dev Biol 2003;262:195–205.
21. Lumsden AGS. Pattern formation in the molar dentition ofthe mouse. J Biol Buccale 1979;7:77–103.
22. McCollum MA, Sharpe PT. Developmental genetics andearly hominid craniodental evolution. Bioessays2001;23:481–93.
23. Tucker AS, Matthews K, Sharpe PT. Transformation oftooth type by inhibition of BMP signalling. Science1998;282:1136–8.
24. Plikus MV, Zeichner-David M, Mayer J-A, Reyna J, Bringas P,Thewissen JGM, et al. Morphoregulation of teeth:modulating the number, size, shape and differentiation bytuning Bnp activity. Evol Devel 2005;7:440–57.
25. Cobourne MT, Mitsiadis TA. Neural crest cells andpatterning of the mammalian dentition. J Exp Zool (Mol DevEvol) 2006;306B:251–60.
26. Cohn SA. Development of the molar teeth in the albinomouse. Am J Anat 1957;101:295–319.
27. Thomas BL, Tucker AS, Qui M, Ferguson CA, Hardcastle Z,Rubenstein JL, et al. Role of Dlx-1 and Dlx-2 genes inpatterning of the murine dentition. Development1997;124:4811–8.
28. Depew MJ, Lufkin T, Rubenstein JL. Specification of jawsubdivisions by Dlx genes. Science 2002;298:381–5.
29. Ferguson CA, Tucker AS, Heikinheimo K, Nomura M, Oh P,Li E, et al. The role of effectors of the activin signallingpathway, activin receptors IIA and IIB, and Smad2, inpatterning of tooth development. Development2001;128:4605–13.
30. Potter RH, Nance WE, Yu PL, Davis WB. A twin study ofdental dimension: II. Independent genetic determinants.Am J Phys Anthropol 1976;44:397–412.
31. Tucker A, Sharpe P. The cutting-edge of mammaliandevelopment: how the embryo makes teeth. Nature RevGenet 2004;5:499–508.
32. Ohazama A, Hu Y, Schmidt-Ullrich R, Cao Y, Scheidereit C,Karin M, et al. A dual role for Ikk alpha in toothdevelopment. Dev Cell 2004;6:219–27.
33. Peterkova R, Kristenova P, Lesot H, Lisi S, Vonesch JL,Gendrault JL, et al. Different morphotypes of thetabby (EDA) dentition in the mouse mandible result

a r c h i v e s o f o r a l b i o l o g y 5 4 s ( 2 0 0 9 ) s 3 4 – s 4 4 S43
from a defect in the mesio-distal segmentationof dental epithelium. Orthod Craniofac Res 2002;5:215–26.
34. Boran T, Lesot H, Peterka M, Peterkova R. Increasedapoptosis during morphogenesis of the lower cheek teethin tabby/EDA mice. J Dent Res 2005;84:228–33.
35. Aswegan AL, Josephson KD, Mowbray R, Pauli RM, SpritzRA, Williams MS. Autosomal dominant hypohidroticectodermal dysplasia in a large family. Am J Med Genet1997;72:462–7.
36. Chassaing N, Bourthoumieu S, Cossee M, Calvas P, VincentMC. Mutations in EDAR account for one-quarter of non-ED1-related hypohidrotic ectodermal dysplasia. Hum Mutat2006;27:255–9.
37. Hogan BL. Morphogene Cell 1999;96:225–33.38. Lind LK, Stecksen-Blicks C, Lejon K, Schmitt-Egenolf M.
EDAR mutation in autosomal dominant hypohidroticectodermal dysplasia in two Swedish families. BMC MedGenet 2006;7:80.
39. Vincent MC, Biancalana V, Ginisty D, Mandel JL, Calvas P.Mutational spectrum of the ED1 gene in X-linkedhypohidrotic ectodermal dysplasia. Eur J Hum Genet2001;9:355–63.
40. Bal E, Baala L, Cluzeau C, El Kerch F, Ouldim K, Hadj-Rabia S,et al. Autosomal dominant anhidrotic ectodermal dysplasiasat the EDARADD locus. Hum Mutat 2007;28:703–9.
41. Line SRP. Molecular morphogenetic fields in thedevelopment of human dentition. J Theor Biol 2001;211:67–75.
42. Gruneberg H. Genetical studies on the skeleton of themouse. IV. Quasi-continuous variations. J Genet 1952;51:95–114.
43. Brook AH. An epidemiological study of dental anomalies inEnglish Schoolchildren with a detailed clinical and geneticstudy of a selected group. MDS thesis, University of London1974. p. 163–73.
44. Chosack A, Eidelman E, Cohen T. Hypodontia: a polygenictrait—a family study among Israeli Jews. J Dent Res1975;54:16–9.
45. Brook AH, Lath DL, Brook BJ, Smith RN, Furtherconsideration of the aetiology of developmental anomaliesof the dentition.Zadzinska E, editor. Current trends in dentalmorphology research, Lodz: University of Lodz; 2005.p. 467–74.
46. Uslenghi S, Liversidge HM, Wong FS. A radiographic studyof tooth development in hypodontia. Archs Oral Biol2006;51:129–33.
47. Harris EF. Mineralization of the mandibular third molar: astudy of American Blacks and Whites. Am J Phys Anthropol2007;132:98–109.
48. Harris EF, Clark LL. Hypodontia: an epidemiological studyof American Blacks and whites. Am J Orthod DentofacialOrthop, in press.
49. Jernvall J, Jung HS. Genotype, phenotype anddevelopmental biology of molar tooth characters. Yrbk PhysAnthropol 2000;43:171–90.
50. Lumsden AGS. Spatial organization of the epithelium andthe role of neural crest cells in the initiation of themammalian tooth germ. Development 1988;103(Suppl.):155–69.
51. Ruch JV, Lesot H, Begue-Kirn C. Odontoblastdifferentiation. Int J Dev Biol 1995;39:51–68.
52. Townsend GC, Brown T. Morphogenetic fields within thedentition. Aust Orthod J 1981;7:3–12.
53. Reid DJ, Dean MC. Variation in modern human enamelformation times. J Hum Evol 2006;50:329–46.
54. Keene HJ. The morphogenetic triangle: a new conceptualtool for application to problems in dental morphogenesis.Am J Phys Anthropol 1982;59:281–7.
55. Harris EF. Where’s the variation? Variance components intooth sizes of the permanent dentition. Dent Anthropol2003;16:84–94.
56. Townsend GC, Brown T. Family studies of tooth size factorsin the permanent dentition. Am J Phys Anthropol1979;50:183–90.
57. Harris EF, Bailit HL. A principal components analysisof human odontometrics. Am J Phys Anthropol1988;75:87–99.
58. Alvesalo L, Tigerstedt PMA. Heritabilities of human toothdimensions. Hereditas 1974;77:311–8.
59. Dempsey PJ, Townsend GC. Genetic and environmentalcontributions to variation in human tooth size. Heredity2001;86:685–93.
60. Kondo S, Townsend G, Yamada H. Sexual dimorphism ofcusp dimensions in human maxillary molars. Am J PhysAnthropol 2005;128:870–7.
61. Takahashi M, Kondo S, Townsend G, Kanazawa E.Variability in cusp size of human maxillary molars, withparticular reference to the hypocone. Archs Oral Biol2007;52:1146–54.
62. Townsend GC, Richards LC, Hughes TE. Molar intercuspaldimensions: genetic input and phenotypic variation. J DentRes 2003;82:350–5.
63. Jernvall J, Kettunen P, Karavanova I, Martin LB, Thesleff I.Evidence for the role of the enamel knot as a control centerin mammalian tooth cusp formation: non-dividing cellsexpress growth stimulating Fgf-4 gene. Int J Dev Biol1994;38:463–9.
64. Matalova E, Antonarakis GS, Sharpe PT, Tucker AS. Celllineage of primary and secondary enamel knots. Dev Dyn2005;233:754–9.
65. Salazar-Ciudad I, Jernvall J. A gene network modelaccounting for development and evolution of mammalianteeth. PNAS 2002;99:8116–20.
66. Jernvall J, Keranen SVE, Thesleff I. Evolutionarymodification of development in mammalian teeth:quantifying gene expression patterns and topography.PNAS 2000;97:14444–8.
67. Salazar-Ciudad I, Jernvall J, Newman SA. Mechanisms ofpattern formation in development and evolution.Development 2003;130:2027–37.
68. Townsend G, Hughes T, Richards L. The dentitions ofmonozygotic twin pairs: focusing on the differences ratherthan the similarities. In: Zadzinska E, editor. Current trendsin dental morphology research. Lodz: University of Lodz Press;2005. p. 337–52.
69. Townsend G, Richards L, Hughes T, Pinkerton S,Schwerdt W. Epigenetic influences may explain dentaldifferences in monozygotic twin pairs. Aust Dent J2005;50:95–100.
70. Van de Vijver G, Van Speybroeck L, de Waele D.Epigenetics: a challenge for genetics, evolution, anddevelopment? Ann New York Acad Sci 2002;981:1–6.
71. Goldberg AD, Allis CD, Bernstein E. Epigenetics: alandscape takes shape. Cell 2007;128:635–8.
72. Waddington CH. The epigenotype. Endeavour 1942;1:18–20.73. Waddington CH. The strategy of genes: a discussion of
some aspects of theoretical biology. London: Allen & Unwin;1957.
74. Molenaar PCM, Boomsma DI, Machin G. A third sourceof developmental differences. Behav Genet 1993;23:519–24.
75. Lesot H, Lisi S, Peterkova R, Peterka M, Mitolo V, Ruch JV.Epigenetic signals during odontoblast differentiation. AdvDent Res 2001;15:8–13.
76. Obara N, Lesot H. Asymmetrical growth, differential cellproliferation, and dynamic cell rearrangement underlieepithelial morphogenesis in mouse molar development.Cell Tissue Res 2007;330:461–73.

a r c h i v e s o f o r a l b i o l o g y 5 4 s ( 2 0 0 9 ) s 3 4 – s 4 4S44
77. Eaves LJ, Kirk KM, Martin NG, Russell RJ. Some implicationsof chaos theory for the genetic analysis of humandevelopment and variation. Twin Res 1999;2:43–8.
78. Townsend GC, Brook AH. Genetic, epigenetic andenvironmental influences on dental development. OrthoTribune 2008;3:3–6.
79. Mattheeuws N, Dermaut L, Martens G. Has hypodontiaincreased in Caucasians during the 20th century? A meta-analysis. Eur J Orthod 2004;26:99–103.
80. Muller TP, Hill IN, Peterson AC, Blayney JR. A survey ofcongenitally missing permanent teeth. J Am Dent Assoc1970;81:101–7.
81. Davis PJ. Hypodontia and hyperdontia of permanent teethin Hong Kong schoolchildren. Commun Dent Oral Epidemiol1987;15:218–20.
82. Ng’ang’a RN, Ng’ang’a PM. Hypodontia of permanent teethin a Kenyan population. East Afr Med J 2001;78:200–3.
83. Brook AH, Elcock C, Al-Sharood MH, McKeown HF, KhalafK, Smith RN. Further studies of a model for the etiology ofanomalies of tooth number and size in humans. ConnectTissue Res 2002;43:289–95.
84. Khalaf K, Elcock C, Smith RN, Brook AH. Fluctuating dentalasymmetry of multiple crown variables measured by animage analysis system. Archs Oral Biol 2005;50:249–53.
85. Khalaf K, Robinson DL, Elcock C, Smith RN, Brook AH.Tooth size in patients with supernumerary teeth and acontrol group measured by image analysis system. ArchsOral Biol 2005;50:243–8.
86. Sharpe PM, Ferguson MW. Mesenchymal influences onepithelial differentiation in developing systems. J Cell SciSuppl 1988;10:195–230.
87. Peters H, Neubuser A, Kratochwil K, Balling R. Pax9-deficient mice lack pharyngeal pouch derivatives and teethand exhibit craniofacial and limb abnormalities. Genes Dev1998;12:2735–47.
88. Fitzgerald LR. Deciduous incisor teeth of the mouse (Musmusculus). Archs Oral Biol 1973;18:381–9.
89. Moss-Salentijn L. Vestigial teeth in the rabbit, rat andmouse, their relationship to the problem of lactealdentitions. In: Butler PM, Joysey KA, editors. Development,function and evolution of teeth. London: Academic Press Inc.;1978. p. 13–29.
90. Cobourne MT, Sharpe PT. Tooth and jaw: molecularmechanisms of patterning in the first branchial arch. ArchsOral Biol 2003;48:1–14.
91. Cobourne MT. Familial human hypodontia—is it all in thegenes? Br Dent J 2007;203:203–8.
92. Courtney JM, Blackburn J, Sharpe PT. The Ectodysplasinand NFkappaB signalling pathways in odontogenesis. ArchsOral Biol 2005;50:159–63.
93. Arte S, Nieminen P, Apajalahti S, Haavikko K, Thesleff I,Pirinen S. Characteristics of incisor premolar hypodontia infamilies. J Dent Res 2001;80:1445–50.
94. Harila-Kaera V, Heikkinen T, Alvesalo L, Osborne RH.Permanent tooth crown dimensions in prematurely bornchildren. Early Hum Dev 2001;62:131–47.
95. Garn SM, Lewis AB. The gradient and the pattern of crown-size reduction in simple hypodontia. Angle Orthod1970;40:51–8.
96. Brook AH, Holt RD. Taurodontism: a criterion for diagnosisand its prevalence in mandibular first permanent molars ina sample of 1115 British schoolchildren. J Int Assoc DentChild 1979;10:41–7.
97. Peck S, Peck L, Kataja M. The palatally displaced canine asa dental anomaly of genetic origin. Angle Orthod1994;64:249–56.
98. Peck S, Peck L, Kataja M. Concomitant occurrence of caninemalposition and tooth agenesis: evidence of orofacialgenetic fields. Am J Orthod Dentofacial Orthop 2002;122:657–60.
99. Kotsomitis N, Freer TJ. Inherited dental anomalies andabnormalities. ASDC J Dent Child 1997;64:405–8.
100. Seow WK, Lai PY. Association of taurodontism withhypodontia: a controlled study. Pediatr Dent 1989;11:214–9.
101. Apajalahti S, Arte S, Pirinen S. Short root anomaly infamilies and its association with other dental anomalies.Eur J Oral Sci 1999;107:97–101.
102. Basdra EK, Kiokpasoglou M, Stellzig A. The Class II division2 craniofacial type is associated with numerous congenitaltooth anomalies. Europ J Orthod 2000;22:529–35.
103. Leonardi R, Barbato E, Vichi M, Caltabiano M. A sellaturcica bridge in subjects with dental anomalies. Europ JOthod 2006;28:580–5.
104. Lammi L, Arte S, Somer M, Jarvinen H, Lahermo P, ThesleffI, et al. Mutations in AXIN2 cause familial tooth agenesisand predispose to colorectal cancer. Am J Hum Genet2004;74:1043–50.
105. Ziegler AC. A theory of the evolution of Therian dentalformulas and replacement patterns. Quart Rev Biol1971;46:226–49.
106. Nieminen P, Arte S, Tanner D, Paulin L, Alaluusua S,Thesleff I, et al. Identification of a nonsense mutation inthe PAX9 gene in molar oligodontia. Eur J Hum Genet2001;9:743–6.
107. Miletich I, Sharpe PT. Normal and abnormal dentaldevelopment. Hum Mol Genet 2003;12(Spec No 1):R69–73.
108. Kim JW, Simmer JP, Lin BP, Hu JC. Novel MSX1 frameshiftcauses autosomal-dominant oligodontia. J Dent Res2006;85:267–71.
109. Perry GH, Verrelli BC, Stone AC. Molecular evolution of theprimate developmental genes MSX1 and PAX9. Mol Biol Evol2006;23:644–54.
110. Thesleff I. The genetic basis of tooth development anddental defects. Am J Med Genet 2006;140A:2530–5.
111. Kangas AT, Evans AR, Thesleff I, Jernvall J.Nonindependence of mammalian dental characters.Nature 2004;432:211–4.
112. Scott GR, Turner CG. The anthropology of modern human teeth:dental morphology and its variation in recent humanpopulations. Cambridge: Cambridge University Press; 1997.
113. Mayhall JT, Kageyama I. A new, three-dimensional methodfor determining tooth wear. Am J Phys Anthropol1997;103:463–9.
114. Smith R, Zaitoun H, Coxon T, Karmo M, Kaur G,Townsend G, et al. Defining new dental phenotypes using3D image analysis to enhance discrimination andinsights into biological mechanisms. Archs oral Biol 2009;54:S118–25.
115. Townsend G, Hughes T, Luciano M, Brook A. Genetic andenvironmental influences on human dental variation: acritical evaluation of studies involving twins. Archs Oral Biol2009;54:S45–51.
116. Wong NC, Joo EJ, Weinrich B, Mossman D, Scott RJ, MorelyR, et al. Investigating epigenetic biomarkers underlyingphenotypic discordance in monozygotic twins. Twin ResHum Genet 2007;10(Suppl.):58.