mouthparts of male aedes (stegomta) mosquttoes · aedes, inchtding 5 strains of aedes aegypti and 6...
TRANSCRIPT

Journal of the American Mosquito Control Association, 18(2):73-80,2OO2Copyright @ 2OO2 by the American Mosquito Control Association, Inc.
MOUTHPARTS OF MALE AEDES (STEGOMTA) MOSQUTTOESISRA WAHID, TOSHIHIKO SUNAHARA rNo MOTOyOSHI MOGI
Division of Parasitoktgy, Department of Microbiology, Saga Medical SchoolNabeshima 5-1-1, Saga 849-8501, Japan
ABSTRACT- Mouthparts of adult males of l7 strains of 8 species from the subgenus Stegomyia of the genusAedes, inchtding 5 strains of Aedes aegypti and 6 strains of Aedes albopictus, were examined. Lengths ofmaxillae, mandibles, maxillary palpi, and proboscises were measured under light microscopy and their detailedstructures were examined by scanning electron microscopy. Lengths were presented as ratios to proboscis lengths.In contrast to previous reports, mandibles were found in all 5 strains of male Ae, aegypti examined. Variationsin maxillary and mandibular lengths were significant among strains, even within,4e. aegypti and Ae. albopictus.High variation of these structures among and within species indicates that the average length of these structuresin only 1 species may not be a reliable representative of a subgenus, and those of I strain may not be reliablefbr a species. However, their range in length (maxillae O.f3 {.50, mandibles O.04-{.17 length of the proboscis)may be regarded as a subgeneric attribute. Maxillae and mandibles distinctly shorter than the proboscis, togetherwith their delicate structures and the large coefficient of variation, suggest that they exist only as vestigialstructures. A positive correlation was found between lengths of maxillae and those of mandibles, but mandiblesare usually shorter than maxillae. The hypopharynx is discernible from the labium wall by its texture and border,and this suggests that it was a free stylet in the past.
KEY WORDS Aedes, Stegomyfui, maxllla, mandible. hypopharynx. male mouthparts
INTRODUCTION
Because most female mosquitoes feed on blood,their mouthparts are highly specialized for piercingthe host skin and sucking blood. The piercing-sucking tools, known collectively as the fascicle,contain I pair of teeth-bearing maxillary stylets, Ipair of mandibular stylets, a labrum, and a hypo-pharynx with its salivary canal. All these structuresare packed in the gutterlike labium and constitutea proboscis.
The labrum forms a food canal and provides ri-gidity for the fascicle, whereas the maxillae anchorthemselves to the host skin by their teeth. Whetherthe mandibles play an active role during piercingas in other bloodsucking nematoceran flies(Downes 1970), or simply serve as a closure of thedistal opening of the labral food canal (Clements1992) is not clear. The hypopharynx, which par-tially makes the ventral closure of the labral foodcanal, enters the host skin together with other partsand releases saliva from a gutter on its surface (thesalivary canal) during feeding.
Male mosquitoes do not take blood. Their foodsources are mainly floral and extrafloral nectaries,honeydew (Foster 1995), or even plant tissue(Schlein and Muller 1995). Their mouthparts arenot developed for piercing. The maxillae and man-dibles are much shorter than the proboscis (Vizzi1953, Snodgrass 1959, Downes 1970, Clements1992) and are considered to be functionless (Vizzi1953, Snodgrass 1959), as in nonbloodsucking ne-matocerans (Downes l97O). On the other hand, thelabrum and the hypopharynx do reach the tip of theproboscis. The hypopharynx, with its salivary ca-nal, fuses with the inner wall of the labium and mayform the ventral closure of the labral food canal(Clements 1992).
As a result of the elevation of the subgenus Och-lerotatus to generic rank by Reinert (2000), thesubgenera Stegomyia and Aedes remain in the ge-nus Aedes, whereas subgenera Ochlerotatus andFinlaya belong to the genus Ochlerotatus. To avoidconfusion, references to previous comparative mor-phological studies of subgenera will be on the basisof the older classification. The implications of theReinert classification on these studies will be dis-cussed later.
Marshall and Staley (1935) reported variations ofmaxillae and mandibles among male mosquitoesfrom the United Kingdom. They found maxillarystylets to be present in 8 subgenera examined, butthe lengths of the stylets were found to be highlyvariable arnong the subgenera, whereas mandibleswere absent in subgenera Aedes and Ochlerotatusof the genus Aedes. Marshall and Staley regardedthe lengths of these structures as characteristic toeach subgenus. However, because they examinedonly l-4 species from the United Kingdom foreach subgenus, whether these attributes are com-mon throughout each subgenus is not clear. Vizzi(1953) reported a wide range of individual variationin mouthpart lengths of male Anopheles quadri-maculatus Say, even for specimens taken from Ilaboratory colony.
The subgenus Stegomyia of the genus Aedes in-cludes vectors of human filariasis and a number ofviral diseases and is a dominant subgenus in theOriental Region (Huang 1979). However, reportson mouthparts of male Stegomyia are limited tothose dealing with Aedes aegypti (L.). Christophers(1960) and Lee (1974) described the strucrures ofmouthparts of both males and females under lightmicroscopy and scanning electron microscopy, re-spectively. Both of these authors stated that man-dibles are absent in males of Ae. aegypti.
73

JounNel op rnr AunnrcAN Moseurro CoNrnol AssocrartoN V o r . 1 8 , N o . 2
Table l. Aedes (Stegomyta) species and strainsexamined.
Species Strain
Aedes aegypti Makassat Indonesia 20Liverpoolr 18Jakarta, Indonesia 13Timor, Indonesia 15Polewali, Indonesia 20Matsumoto, Japan 18Polewali, Indonesia 12Tanegashima, Japan 15Makassar, Indonesia I IOkinawa, Japan 11Mambi, Indonesia l0Seram, Indonesia 20Seram, Indonesia 20Kabeshima, Japan l7Seburi, Japan 10Kyongi-do, Korea 18Mambi, Indonesia 9
Aedes albopictus
Aedes paullusiAedes scutellarisAedes riversiAedes flavopictusAedes galloisiAede s pseudoalbolineatus
' The Liverpool strain originally was collected in Thailand.
Our preliminary observation suggested the pres-ence of maxillae and mandibles of males in somespecies of the subgents Stegomyia. Thus, our ob-jectives were to describe the mouthparts of maleAedes (Stegonyla) species, with confirmation of themorphological status of mandibles of Ae. aegyptimales; and to examine whether the lengths of max-illae and mandibles are common attributes throush-out species belonging to this subgenus.
MATERIALS AND METHODS
Specimens included 17 strains of 8 species from3 species groups of Stegomyia: Ae. aegypti repre-sented the aegypti group; the scutellaris group in-cluded Aedes alboplcrzs (Skuse), Aedes paullusiStone and Farner, Aedes scutellaris (Walker), Aedesriversi Bohart and Ingram, Aetles flavopictus Ya-mada, and Aedes galloisi Yamada; and Aedes pseu-doalbolineatus Brug represented the albolineatusgroup. For Ae. aegypti and Ae. albopictus,5 and 6strains were examined, respectively (Table 1). lden-tifications were based on keys of Wepster (1954),Huang (1979), and Tanaka et al. (1979). Numbersof specimens varied from 9 to 20 individuals, de-pending on availability. Specimens were rearedfrom larvae collected in the field or obtained fromlaboratory colonies, and kept alive for at least 24 hafter emergence. Until examination, specimenswere kept in 7O7o ethanol.
Because the labium, labrum, and hypopharynx inmale Stegomyia arc almost the same length andreach the tip of the proboscis, we measured onlythe lengths of maxillae, mandibles, maxillary palpi,and the proboscis (the length of the labium fromthe base of the prementum to the tip of the labella)(Fig. 1). All measurements were converted to mil-limeters and then presented as ratios of proboscislengths (Marshall and Staley 1935). This enabled
Fig. 1. Head and mouthparts of a male of Aedes(Stegomyia).
us to compare the relative lengths of the structures,because the absolute values are affected by bodysize. We measured the lengths of maxillary palpi,because they are a well-developed functional organin males (Clements 1992) and are connected di-rectly to the base of maxillary stylets. The lengthsof maxillary palpi can be used to compare the ex-tent of variation between structures with functionsand those without function.
Specimens in 7O7o ethanol were transferred to2%o KOH solution and heated at 50'C for 2 h. Theywere then dehydrated by passage through 7O7o,9OVo. and 99Vo ethanol in this order, and stainedwith acid fuchsin. Specimens were kept in the stain-ing solution at least for 2 h. Just before examina-tion, the specimens were put on tissue paper for afew seconds to absorb the stain, and then put on amicroscope slide. One or 2 drops of Methyl Cel-losolve (Nakarai Chemicals, Ltd., Kyoto, Japan)were placed on the specimen, which was then ex-amined under both dissecting and compound mi-croscopes.
First, the head was separated from the body by
Maxilla
Maxillary
El
(s
o$3occ



JuNe 2002 Mournpanrs or Mala AEDES (SrEGoMya) MoseurroEs
I' 6
-8P o
3
' I
3 o.:6.
;6
o 0.2
i s € ! ! ;F E " : - i . :* - = 9 \ S €
\ !" \ €{
E € ? E E E ' P € g e, E A 7 ; i E f : : E B- E : € : E S $ e o E$ i i i $ i r € s F P: $ $ $ E . $ i i $ $ E* ; ; ; = : € i i i s
r < = s s' \i
s B $ € ; ; ; € i € € i i i i $ tFseIFF$$$s,$
r !
Fig. 5. Average lengths of maxillary palpi of maleStegomyia. Proboscis length : l.
riversi, Ae. flavopictus, and Ae. galloisi. However,in Ae. pseudoalbolineatus, lengths of maxillary pal-pi are about one half of the proboscis length (Fig.5). Statistical tests showed significant differencesamong all the strains examined (Kruskal*Wallistest, 11 : 203.7, df : 16, P < 0.001). Signif icantdifferences were found even within the same spe-cies (Ae. aegypti , H: 11.6, df : 4, P < O.021;Ae. albopictus, H : 23.3, dt : 5, P < 0.OOl).
Maxillae are much shorter than proboscises, witha range of 0.13-0.5 of the proboscis lengths. TheKruskal-Wallis test showed signiflcant differencesamong all the strains examined (H : 140.6, df :16, P < 0.001), within Ae. aegypri (H : 36.1, df: 4, P < 0.001), and within Ae. albopictus (H :2 4 . 6 , d t : 5 , P < 0 . 0 0 1 ) .
Figure 6 shows that a maxilla of Ae. pseudoal-bolineatus is the shortest and it is significantly dif-f-erent from those of all the other species. Withinthe scutellaris group (except Ae. albopictus), themaxillae of Ae. paullusi are significantly longerthan those of the other species. Intraspecific varia-tion within Ae. albopictus and Ae. aegypti is con-siderable, and some strains of Ae. albopictus, aswell as some strains of Ae. aegyptl, are significantlydifl'erent from the other species, but some are not.Among strains within the same species, at least theMatsumoto strain of Ae. albopictu.r apparently isdiff'erent fiom the Mambi strain, and the Jakartaand Polewali strains of Ae. aegypti are differentfrom the Timor strain.
The mandibles are shorter than maxillae, with arange of O.O44.ll of the proboscis lengths, excepttbr 2 individuals mentioned below- Mandibles, asmaxillae, vary among all the strains examined (ll
Fig. 6. Variation of maxillary lengths of male Stego-myia. The box indicates the interquartile range with themedian as a transversal line; upper and lower whiskers aremaximum and minimum values within 1.5 of the inter-quartile range. Outliers (more than 1.5 times of interquar-tile ranges) are plotted separately and marked with densecircles. Vertical gray bars across the medians are957o con-fidence intervals. Horizontal lines at the bottom of thegraph are (A) strains of Aedes aegypti, (B) strains ofAe.albopictus, and (C) scutellaris group. The aegypti groupand, albolineatus group are represented by Ae. aegypti andAe. pse udoalbo I ineatus, respectively.
: 148.5, df : 16. P < 0.001). and also wi th inAe.aegypti (H : 23.7, dt : 4, P < 0.001) anrJ Ae.albopictus (H : 13.7, df : 5, P < 0.05).
Figure 7 shows that within the scutellaris group(except Ae. albopiuus), lengths of mandibles of Ae.paullusi and Ae. galloisi are different from that ofAe. scutellaris, as well as from those of Ae. riversiand Ae. flavopictus. The latter 2 species are differ-ent also from Ae. pseudoalbolineatus (the alboli-neatus group). Within Ae. aegypti, the Makassarstrain is clearly different from other strains, where-as strains of Ae. albopictus do not vary appreciably(conf,dence interval bars almost overlap).
Maxillae and mandibles show wide variation intheir lengths even within each strain. Examples arethe maxillae of Ae. aegypri from Timor and themandibles of Ae. albopictus tiom Makassar. Notethat in the Makassar strain of Ae. albopictus, 2 un-usual mandible lengths occur: 0.41 and 0.45 of theproboscis lengths, which exceed the length of max-illae (0.25 of the proboscis lengths in both cases)in their pairs. These are unique because in all otherspecimens, maxillae are longer than mandibles. Weeliminated the possibility of confusing maxillae asmandibles and vice versa by careful examination oftheir bases.
JounNnr- oF THE AMERTcAN Moseurro CoNrr.or- AssocrnrroN Vor . 18 , No.2
o
TT I* T r l r l
Itryf;[fi*;;+**o+*flRE 0 r
EEE€€g€g3aEiisiSsi i S " � $ : i ; : E ? . = ; Es g Y . * g i $ g 5 5 i
; =
o 0.4Jo 0 4 1
Fig. 7. Variation of mandibular lengths of male .Srega-myia. For explanation of symbols, see Figure 6.
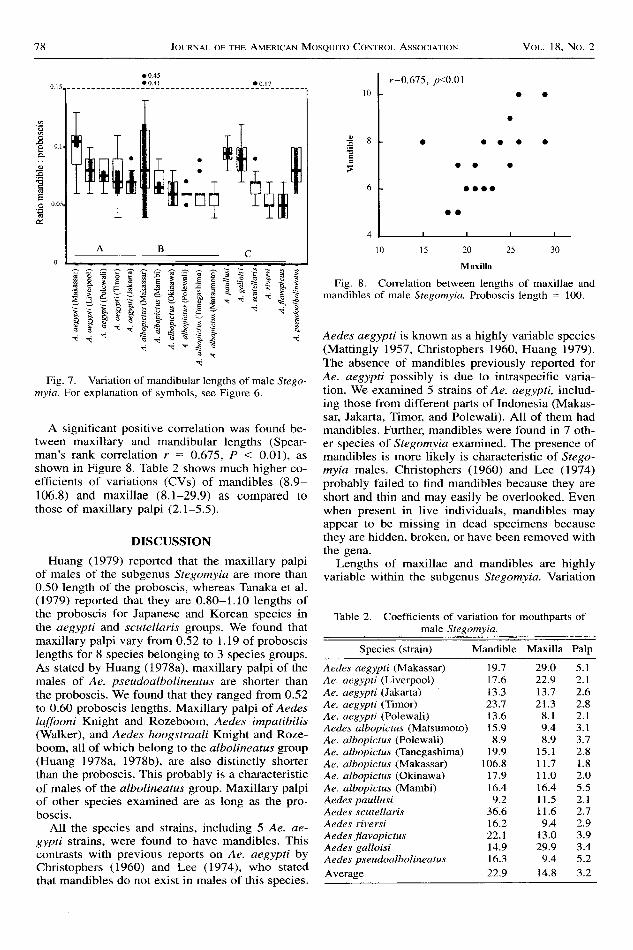
A significant positive correlation was found be-tween maxillary and mandibular lengths (Spear-man's rank correlation r = 0.675, P < 0.01), asshown in Figure 8. Table 2 shows much higher co-efficients of variations (CVs) of mandibles (8.9-106.8) and maxillae (8.1-29.9) as compared tothose of maxillary palpi (2.1-5.5).
DISCUSSION
Huang (1979) reported that the maxillary palpiof males of the subgenus Stegomyia are more than0.50 length of the proboscis, whereas Tanaka et al.(1979) reported that they are 0.80-1.10 lengths ofthe proboscis for Japanese and Korean species inthe aegypti and scutellarls groups. We found thatmaxillary palpi vary from 0.52 to I . 19 of proboscislengths for 8 species belonging to 3 species groups.As stated by Huang (1978a), maxillary palpi of themales of Ae. pseudoalbolineatus are shorter thanthe proboscis. We found that they ranged from 0.52to 0.60 proboscis lengths. Maxillary palpi of Aedeslaffooni Knight and Rozeboom, Aedes impatibilis(Walker), and Aedes hoogstraali Knight and Roze-boom, all of which belong rc rhe albolineatus gro.op(Huang 1978a, 1978b), are also distinctly shorterthan the proboscis. This probably is a characteristicof males of the albolineatus grolup. Maxillary palpiof other species examined are as long as the pro-boscis.
A1l the species and strains, including 5 Ae. ae-gypti strains, were found to have mandibles. Thiscontrasts with previous reports on Ae. aegypti byChristophers (1960) and Lee (1974), who statedthat mandibles do not exist in males of this species.
Maxilla
Fig. 8. Correlation between lengths of maxillae andmanditrles of male Stegomyia. Proboscis length : 1O0.
Aedes aegypti is known as a highly variable species(Mattingly 1957, Christophers 1960, Huang 1979).The absence of mandibles previously reported forAe. aegypti possibly is due to intraspecific varia-tion. We examined 5 strains of Ae. aegypti, includ-ing those from different parts of Indonesia (Makas-sar, Jakarta, Timor, and Polewali). All of them hadmandibles. Furthet mandibles were found in 7 oth-er species of Stegomyia examined. The presence ofmandibles is more likely is characteristic of Stego-myia maIes. Christophers (1960) and Lee (1974)probably failed to flnd mandibles because they areshort and thin and may easily be overlooked. Evenwhen present in live individuals, mandibles mayappear to be missing in dead specimens becausethey are hidden, broken, or have been removed withthe gena.
Lengths of maxillae and mandibles are highlyvariable within the subgenus Stegomyia. Variation
Table 2. Coefficients of variation for mouthparts ofmale Stegomyia.
Species (strain) Mandible Maxilla Palp
Aedes aegypti (Makassar) 19.7Ae. aegypti (Liverpool) 1'1.6Ae. aegypti (Jakarta) 13.3Ae. aegypti (Timor) 23.7Ae. aegypti (Polewali) 13.6Aedes albopictus (Matsumoto) 15.9Ae. albopictus (Polewali) 8.9Ae. albopictus (Tanegashima) 19.9Ae. albopictus (Makassar) 106.8Ae. albopictus (Okinawa) 17.9Ae. albopictus (Mambi) 16.4Aedes paullusi 9.2Aedes scutellaris 36.6Aedes riversi 16.2Aedes frdvopictus 22.1Aedes galloisi 14.9Aedes pseudoalbolineatus 16.3
Average 22.9
29.O22.913.721.3
8 . 19.48.9
1 5 . 1r1.71 1 . 016.4I 1 . 5I 1 . 69.4
13.029.99.4
14.8
5 . 12 .12.62.82 . 13 . 1J . t
2.81.82.O5.52 . 12.72.93.93.45 .23 .2
r:0.675, p<0.01

JUNB 2002 MOUTHPARTS or Mer-r AEDES (STEGoMra) MoseurroEs 79
exists even among strains within the same species(Ae. aegypti and Ae. albopictus). If multiple strainsare examined for other species, similar intraspecificvariation, as it Ae. aegypti and Ae. albopictus,might also be found. Despite this variation, thelengths of maxillae and mandibles of the subgenusStegomyia have distinct ranges (maxillae: 0. 13-0.50, mandibles: 0.04-0.17, excluding 2 unusualvalues). These ranges may be regarded as subge-neric attributes, although species of the edwardsiand w-albus groups in the Oriental Region (Huang1979) and species groups of Africa (Huang 1990)have not been examined.
Compared with other subgenera of the genus Ae-des examined by Marshall and Staley (1935), thesubgenus Stegomstia has, on average, longer max-illae with a greater range of length (0. 13-{.50) thanthose of the subgenera Aedes (0.06-0.09), Finlaya(0.20-0.28), and Ochlerotatus (O.2O4.28). Man-dibles of males of the subgenus Stegomyia (0.04-0.17) are more variable than those of the subgenusFinlaya (0.09-0.14), whereas mandibles were notfound by these authors for subgenera Aedes andOchlerotatus.
According to the recent new classification of thegents Aedes, subgenera Stegomyia and Aede,s re-main in the genus Aedes, whereas subgenera Oc&-lerotatus and Finlaya belong to the genus Ochler-otatus (Reinert 2000). Following this new system,maxillae and mandibles of males appea,r to be at-tributes that do not distinguish these genera, as isalso the case of maxillary palpi of males (Reinert2000).
Marshall and Staley (1935) reported that maxil-lae of species of genera Orthopodomyia (O.28-0.37) and Culex (0.09-O.19), as well as their man-dlbles (Orthopodomyia: 0.05-0.07, Culex: O.O5-0.07), are also short. On the other hand, themaxillae of species of the genera Culiseta (0.58-O.94) and Anopheles (0.44-0.68), as well as theirmandibles (Culiseta; O.O74.28, Anopheles: O.38-0.46) were distinctly longer. Other subgenera of Ae-des and other genera in the Oriental Region arebeing examined to determine the taxonomic andphylogenetic significance of the mouthparts inmales.
As stated by Yizzi ( 1953) for males of An. quad-rimaculatus, and by Snodgrass (1959) for femalesof Toxorhynchites, delicate, short, and irregularlyshaped maxillae and mandibles suggest the lack offunction, at least for piercing. Much higher CV val-ues of maxillae (14.8) and mandibles (22.9\ tnanthose of maxillary palpi (3.21 strengthen this view.Structures with function naturally are less variablethan those without function, because they need tohold a flxed shape or size to carry out their func-tion. For example, CV values calculated from thedata of wing, hind femur, and scutum lengths offemale Ae. albopictus in t}le laboratory (Mori 1929,Thbles 5-7, 12-14) remain usually less than 5.0,although the absolute lengths vary greatly depend-
ing on nutrition and density conditions. Even forwild specimens exposed to diverse nutrition, den-sity, and temperature conditions during larval de-velopment, CV values for body size were usuallyless than 20.0 for mosquitoes breeding in groundpools (Fish 1985). Thus, high variability of maxil-lae and mandibles of male mosquitoes supports anexplanation that they existjust as functionless, ves-tigial structures (Downes 1970).
The positive correlation that occurs betweenmaxillary and mandibular lengths indicates the si-multaneous reduction of both organs in males. Thefact that mandibles are usually shorter than maxil-Iae might suggest that mandibles lost their functionearlier than did the maxillae.
Snodgrass (1959) stated that the hypopharynx inmale mosquitoes is completely fused with the la-bium and forms the labiohypopharynx. Christo-phers (1960) recognized the hypopharynx only asa ridge on the labium. On the other hand, Vizzi(1953) found the hypopharynx of An. quadrima-culatus to be a sclerotic plate on the labial gutter.We found the hypopharynx of males of Stegomyia,although it is not a free stylet, to be a structureclearly discernible from the inner wall of the labi-um, thus retaining evidence of its free origin. Thisstructure is also found in the genera Anopheles, Cu-lex, and Tripteroides (unpublished data), and isprobably cofilmon to other mosquitoes as well.
Because the hypopharynx in female mosquitoes,together with other stylets, enters the host skin dur-ing bloodfeeding, the free hypopharynx clearly iscorrelated with their ability to suck blood. Silva andGrunewald (2000) reported that the hypopharynx ofmale LutTomyia migonei Franca (Psychodidae) is afree stylet. Probably the earlier form of the hypo-pharynx of male nematoceran flies was a free stylet.Fusion of the hypopharynx of the male mosquitowith the labium may have taken place after the an-cestor of the mosquito separated from the sandfly'sancestor.
ACKNOWLEDGMENTS
We gratefully thank T. Takahashi and T. Tabata,who helped in preparing scanning electron micro-scope images; I. Miyagi, who made many sugges-tions on staining and dissection methods; Y. Egu-chi, who provided some of the specimens fromlaboratory colonies; and the 2 anonymous review-ers, who gave useful comments to improve themanuscript. Collection of Indonesian strains wassupported by Grant-in-Aid for Monbusho Interna-tional Scientific Research Program (Field Research)(1004 1200).
REFERENCES CTTED
Benjamini Y. 1988. Opening the box of a boxplot. AzSnr 42:257-262.
Christophers SR. 1960. Aedes aegypti (L.) the yellow fe-

80 JounNlr- oF THE AMERICIN Moseurro CoNtnol Assocr,qrror Vor-. 18, No. 2
ver mosquitoes Cambridge, United Kingdom: Cam-bridge University Press.
Cfements AN. 1992. The biology of mosquitoes Volume1. London, United Kingdom: Chapman & Hall.
Downes JA. 1970. The ecology of blood sucking Diptera:an evolutionary perspective. In: Fallis AM, ed. Ecologyand physiology of parasites Toronto, Ontario, Canada:University of Toronto Press. p 232-238.
Fish D. 1985. An analysis of adult size variation withinnatural mosquito populations. In: Lounibus LP, Rey JR,Frank JH, eds. Ecology of mosquitoes Vero Beach, FL:Florida Medical Entomology Laboratory. p 419-43O.
Foster WA. 1995. Mosquito sugar feeding and reproduc-tive energetics. Annu Rev Entumol 4O:443-474.
Harbach RE, Knight KL. 1980. Taxonomist's gktssary ofmosquito anatomy Marlton, NJ: Plexus Publishing, Inc.
Huang YM. 1978a. Taxonomic status of Aedes (Stego-myia) laJfooni Knight and Rozeboom with redescriptionof Aedes (Stegomyia) pseudoalbolineatus Brug (Dip-tera: Culicidae). Mosq Syst 10:351-364.
Huang YM. 1978b. The identity of two species of Stego-nyia belonging to the Aedes albolineatus group (Dip-tera: Culicidae). Mosq Sy.st l0:197-210.
Huang YM. 1979. Medical entomology studies-Xl. Thesubgenus Stegomyia of Aedes in the Oriental Regionwith keys to the species (Diptera: Culicidae). ContribAm Entomol Inst (Ann Arbor) 15(.6):l-79.
Huang YM. 1990. The subgenus Stegomyia of Aedes rnthe Afiotropical Region l. The africanus group of spe-cies (Diptera: Culicidae). Contrib Am Entomol Inst(Ann Arbor) 26( l ) : l -90.
Jobling B. 198'/. Anatomical drawing of biting.flies Lon-don, United Kingdom: British Museum (Natural His-tory).
Lee R. I974. Structure and function of the fascicular sty-lets and the labral and cibarial sense organ of male andf'emale of Aedes aegypti (L.) (Diptera: Culicidae).
Quaest Entomol 10:187-215.Marshall JE Staley J. 1935. Generic and subgeneric dif-
ferences in the mouthparts of male mosquitoes. BzllEntomol Res 26:53 l-532.
Mattingly PE 1957. Genetical aspects oftheAedes aegyptiproblem. Ann Trop Med Parasitol 51:392-408.
McGill R, Larsen WA, Tukey JW. 1978. Variations of boxplots. Arn Stat 32:12-16.
Mori A. 1979. Effects of larval density and nutrition onsome attributes of immature and adrlJt Aedes albopictus.Trop Med 21:85-103.
Reinert JE 2000. New classification for the composite ge-us Aed.es (Diptera: Culicidae: Aedini), elevation ofsubgenus Ochlerotatus to generic rank, reclassificationof the other subgenera, and notes on certain subgeneraand species. J Am Mosq Control Assoc l6:175-188.
Schlein X Muller G. 1995. Assessment of plant tissuefeeding by sand flies (Diptera: Psychodidae) and mos-quitoes (Diptera: Culicidae). J Med Entomol 32:882-887.
Silva OS, Grunewald J. 2000. Comparative study of themouthparts of males and females of Lutlomyia migonei(Diptera: Psychodidae) by scanning electron microsco-py. J Med Entomol 37:748-753.
Snodgrass RE. 1959. The anatomical li.fe of the mosqui-loes Smithsonian Miscellaneous Collections 139 No 8.Washington, DC: Smithsonian Institution.
Sokal RR, Rohlf FJ. 1995. Biometry: the principles andpractice of stcttistics in biological research 3rd ed. NewYork, NY: WH Freeman and Co.
Tanaka K, Mizusawa K, Saugstad ES. 1979. A revisionof the adult and larval mosquitoes in Japan (includingthe Ryukyu Archipelago and the Ogasawara Islands)and Korea (Diptera: Culicidae). Contrib Am EntomolInst (Ann Arbor) 16:.l-987.
Yizzi FF. 1953. The mouthparts of the male mosquitoesAnopheles quadrimaculatus Say. Ann Entomol Soc Am46:496-5O4.
Wepster JB. 1954. Synopsis of a hundred common nonanopheline mosquitoes oJ the Greater and Lesser Sun-das, the Moluccas and New Guinea. Royal Tropical In-stitute of Amsterdam Special Publication CXI. Amster-dam, The Netherlands: Elsevier Publishing Co.