murdoch research repositoryresearchrepository.murdoch.edu.au/id/eprint/31029/1/canopy decline... ·...
TRANSCRIPT

MURDOCH RESEARCH REPOSITORY
This is the author’s final version of the work, as accepted for publication following peer review but without the publisher’s layout or pagination.
The definitive version is available at http://dx.doi.org/10.1071/BT15004
Moore, T.L., Ruthrof, K.X., Craig, M.D., Valentine, L.E., Hardy, G.E.St.J. and Fleming, P.A. (2016) Living (and reproducing) on the edge: Reproductive phenology is impacted by rainfall and
canopy decline in a Mediterranean eucalypt. Australian Journal of Botany, 64 (2). pp. 129-141.
http://researchrepository.murdoch.edu.au/31029/
Copyright: © 2016 CSIRO.
It is posted here for your personal use. No further distribution is permitted.

Living (and reproducing) on the edge: reproductive phenology is impacted by rainfall and canopy decline in a Mediterranean eucalypt Moore, T. L.1, Ruthrof, K. X. 1, Craig, M. D.1, 2, Valentine, L. E.1, 2, Hardy, G. E. St J.1, Fleming, P. A.1 5
1 State Centre of Excellence for Climate Change, Woodland and Forest Health, School of Veterinary and
Life Sciences, Murdoch University. 2 ARC Centre of Excellence for Environmental Decisions, School of Plant Biology, University of Western
Australia. 10
*Corresponding author: Tracey Moore
Email: [email protected] or [email protected] 15
Address: School of Veterinary and Life Sciences,
Murdoch University,
90 South St, Murdoch, 6150,
Western Australia
Phone: 08 93607236 20
Mobile: + (614) 41789815

Abstract
Many of the worlds’ forests and woodlands are currently showing symptoms of declining condition due to a
range of factors, including changing climatic conditions, drought, and insect herbivory. Altered abiotic and 25
biotic conditions can influence the condition of trees which can, in turn, affect tree reproductive cycles.
However, the potential impact of tree decline on reproductive cycles has rarely been examined. This study
investigated the influence of canopy condition on the reproductive cycle of Eucalyptus wandoo in south-
western Australia. Canopy and seed trap monitoring were used to assess bud production, flowering, fruiting
and seed fall over 12 months at 24 sites across two locations (Dryandra Woodland and Wandoo 30
Conservation Park). Time since last fire, rainfall, ambient temperatures and the condition of individual trees
were recorded. We found that bud production, flowering and fruiting was correlated with tree condition:
healthier trees were generally associated with higher reproductive effort. Time since last fire was also
strongly related to the reproductive efforts at both locations. Declining annual rainfall and increased
temperatures also an impact on reproduction, made evident by the aborted flowering in Dryandra Woodland. 35
Decline in tree condition, coupled with changes in climate, have major implications for flowering phenology
of this species and have the potential to alter reproductive effort, recruitment, and future population
dynamics. Consideration of these issues should be incorporated into the conservation management of E.
wandoo and similar Eucalypt species.
40
Keywords: canopy condition, serotiny, weather, tree size, time since last fire, die-off, dieback, forest,
woodland, climate change, drought

Introduction
Declines in the condition of forests and woodlands have become apparent around the world (Reid and 45
Landsberg 2000; Close and Davidson 2004; Allen et al. 2010; Matusick et al. 2012; Brouwers et al. 2013).
These include Phytophthora ramorum-related oak death in Spain and Portugal (Brasier et al. 1993), drought-
induced pine forest death in the United States (Breshears et al. 2005), and drought-related Eucalyptus spp.
mortality in south-western Australia (Matusick et al. 2012; 2013). Canopy loss and death of branches are
symptoms of a decline in tree condition (Carnegie 2007; Matusick et al. 2012) and such symptoms can 50
affect the reproductive phenology of individual trees as well as the structure, functioning and composition of
an ecosystem (Allen et al. 2010). Furthermore, throughout the world, dramatic climatic changes (i.e.
increased ambient temperatures, changes in the amount of rainfall and its seasonality) are occurring
(Klausmeyer and Shaw 2009; Allen et al. 2010), potentially further altering the reproductive efforts of trees
(Ashton 1975; Pook 1986; House 1997; Law et al. 2000; Pau et al. 2011). 55
Eucalyptus wandoo ssp. wandoo (hereafter referred to as E. wandoo) is a smooth-barked serotinous species
endemic to south-west Western Australia (Yates and Hobbs 1997; Doughty 2000). Once covering most of
the Western Australian agricultural zone, much (60%) of the original 218 680 ha range of this tree species
has been cleared for agriculture. Most of the remaining extent occurs in small reserves, roadsides and as 60
isolated paddock trees (Doughty 2000; Wandoo Recovery Group 2006). Over the last few decades, patches
of E. wandoo have shown evidence of a decline in canopy condition (Whitford et al. 2008; Hooper 2009;
Dalmaris 2012; Brouwers et al. 2013). Eucalyptus wandoo canopy decline is patchy and cyclic, differing
from many other eucalypt declines in the south-west Western Australia, where large areas of canopy are lost
(Matusick et al. 2012; Matusick et al. 2013). Reproductive efforts of E. wandoo could be altered by this 65
decline in canopy condition, adding pressure to a species with an already restricted range (Mattiske & Havel
1998; Wandoo Recovery Group 2006). Furthermore, this may have effects on the wider ecosystem as
previous studies have noted that the decline in E. wandoo condition can affect woodland fauna and its
habitat (Moore et al. 2013a; Moore et al. 2013c; Moore et al. 2013b; Moore et al. 2014) and lack of
recruitment therefore can have long term effect on the overall biodiversity of an area. 70
Tree decline, weather and fire regimes have the ability to significantly affect the reproductive efforts of
eucalypts. Several studies have noted the effects of many climatic and environmental factors on the
production of flowers, fruits, and seeds over the long life spans of eucalypt species, which often exceed 200
years (Ashton 1975; House 1997; Brookshire et al. 1999). The biodiversity hotspot in south west Western 75
Australia, where E. wandoo is endemic (Klausmeyer et al. 2011), has recently been experiencing extreme
weather conditions including high temperatures and below average rainfall (Bureau of Meteorology 2010).
Additionally, prescribed burning, a biodiversity management tool, is frequently used to reduce the risk of
large scale wildfires (Burrows and Abbott 2003). The frequency or season of prescribed burns have the
potential to alter the reproductive effort of E. wandoo, as fire is a stimulant of seed germination and 80

flowering in many tree species (Pyke 1983; Burrows et al. 1990; Burrows and Abbott 2003; Lamont and
Downes 2011). Together fire and climate may influence reproductive efforts in these trees.
Current published work on the flowering phenology of E. wandoo is limited to the pollen-mediated gene
flow of the species (Byrne et al. 2008). Given the lack of information on the reproductive effort of E. 85
wandoo and how it is influenced by climatic and environmental factors, a greater understanding E. wandoo
flowering phenology is needed if remaining populations are to be effectively managed. In this study we
asked:
a. is the reproductive effort of E. wandoo related to rainfall and ambient temperature?
b. if the reproductive effort (i.e. budding, flowering, fruiting and seed fall) of E. wandoo is influenced 90
by canopy condition?
c. does time since last fire influence reproductive effort in E. wandoo?
Methods
Site description 95
Study sites were located in two locations, Dryandra Woodland (DW) (32° 48′ S, 116° 53′ E) and Wandoo
Conservation Park (WCP) (31° 54′ S, 116° 27′ E), 160km south-east and 75km east of Perth, Western
Australia, respectively. At each location, 12 sites were located in pure E. wandoo stands (total n = 24 sites).
Eucalyptus wandoo stands are generally open with an understorey comprised of Gastrolobium spp.,
Macrozamia riedlei, Xanthorrhoea preissii and a grassy herb layer (Yates and Hobbs 1997). Land clearing, 100
stock grazing, timber harvesting (logging) and prescribed burns have occurred in both locations (Department
of Conservation and Land Management 1980). Sites were initially selected using Landsat data (1999-2009)
and VegMachine (Commonwealth Scientific and Industrial Research Organisation 2010), a program that
calculates changes in vegetation reflectance over time and which can quantitatively identify healthy and
declining stands of trees. Following initial site selection, on-ground measures of tree condition were used to 105
classify sites as either healthy or declining. A healthy site was defined as one with predominately healthy
trees (trees with <50% canopy decline), whilst declining sites were composed predominately of trees in
decline (trees with >50% canopy decline). It must be noted that both healthy and declining trees were
evident at all sites, due to the spatially patchy nature of E. wandoo decline. Final site selection resulted in a
50:50 mix of healthy and declining sites. 110
Weather conditions
For comparison with the tree reproductive effort variables in this study, monthly rainfall (mm), and
minimum and maximum monthly temperature (°C) averages were downloaded from the Bureau of
Meteorology (Bureau of Meteorology 2009) database for Narrogin and York weather stations (Narrogin 115
station: 010614, 40 km from DW; York station: 010311, 40km from WCP) for the duration of this study and

to obtain long term averages (Narrogin/DW: 1913-2013, York/WCP: 1996-2013). Available total annual
rainfall and monthly minimum and maximum temperature (°C) data were also downloaded for 13 years
(1997-2010) for Narrogin/DW and 19 years (1991-2010) for York/WCP. Mean, 95% confidence intervals
(CI), and minimum and maximum values were calculated for each weather variable (Table 1). Independent 120
t-tests were performed to compare monthly rainfall, and minimum and maximum monthly temperature
between long term averages and averages during this study (IBM 2012). Statistical analysis was not
performed for summer rainfall averages as data were too sparse.
Reproductive effort 125
Two methods, canopy monitoring and seed trapping, were used to record the reproductive efforts of E.
wandoo trees over a 12-month period. Canopy monitoring involved recording the reproductive effort of 144
trees (six trees at each site, each >20cm diameter at breast height, DBH) over ten sampling events at near
monthly intervals (May 2010 - April 2011, excluding August and December), by estimating the percentage
of canopy that was: a) budding, b) flowering, and c) fruiting. Values were estimated from the same location 130
on the ground (by Tracey Moore) by visual assessment using binoculars (10 x 42, Nikon Monarch
Binoculars). Reproductive effort values measured over the year were summed to create a single value for
each of the 144 trees which was then arcsine square root-transformed to meet the assumptions of parametric
statistics (Zar 1998).
135
Seed trapping was commenced in June 2010 at each of the 24 sites when two seed traps were placed
randomly underneath the same E. wandoo tree, a tree that were also being canopy-monitored, making a total
of 48 seed traps. Seed traps were not placed under a tree if that tree had started to flower (this study
commenced just as E. wandoo was observed flowering in the area) or if its canopy overlapped with an
adjacent tree, to ensure samples were from one tree and the start of the flowering event was captured. Seed 140
traps were made from a steel ring (0.44m-diameter, 0.15m2 opening) and canvas cone (0.30m-deep) held
approximately 0.4m from the ground through attachment to three metal fence dropper posts (Yates et al.
1994). Petroleum jelly was applied to the three droppers as a deterrent to terrestrial invertebrates entering
the traps and removing seeds. Seed traps collected all leaves, sticks, buds, opercula, flowers, and fruits of E.
wandoo and were emptied nine times between July 2010 and April 2011 into labeled paper bags. The 145
biomass of each collection was dried at 40°C for 24h before being sieved and weighed. We classified the
range of reproductive tissue types collected from the seed traps into the following categories: a) failed
flowering effort; b) flowering effort; and c) aborted fruit, listed below;
a. Failed flowering effort - determined by the total mass (g) of unopened and aborted buds with 150
opercula still attached;

b. Flowering effort - represented by the total mass (g) of opercula collected (eucalyptus buds are
capped by an operculum, which is dropped as the flower opens; (opercula production is a good
indicator of the amount of flowering that occured; Semple et al. 2007); and
c. Aborted fruit - determined by the total mass (g) of unopened fruit (generally very desiccated, and 155
unlikely to hold viable seeds).
Values for each of the above reproductive effort variables were summed across the entire seed trap
monitoring period and across both seed traps to create a single value for each tree (24 trees), and arcsine
square root-transformed to meet the assumptions of parametric statistics (Zar 1998).
160 Time since last fire
Year since last fire for each site was taken from Department of Parks and Wildlife (DPaW) databases
(available from the staff at the Perth Hills and Great Southern Districts, DPaW, Western Australia). Details
on the types of fires that occurred in these areas are limited, but the majority were low, cool fires designed
reduce the fuel load in the understorey layer (prescribed burns). 165
Tree condition
A range of tree condition characteristics were estimated once for each of the 144 trees prior to the
reproductive effort monitoring (and again in 2011 on a selection of trees; Stuart 2011). USDA tree
condition assessment is a measure of tree condition characteristics originally designed and used in the USA 170
for Pinus spp. (Schomaker et al. 2007) but a range of measures can be adapted to the assessment of
eucalypts, including crown density and crown dieback. Crown density was the percent crown that contained
foliage, branches, and reproductive structures. Crown dieback was the percent crown that has undergone
recent dieback, or lost foliage. Canopy cover was estimated by taking four canopy cover measurements
1.5m from the base of each tree at north, south, east and west directions using a spherical densitometer and 175
averaged for the tree to give a single value (Lemmon 1956). Lastly, we estimated the percent of dead
branches, or the percentage of all major branches with a diameter >20 cm that are senescent, as these
characteristics have been used in other studies to investigate the relationships between tree condition and
wildlife species richness and abundance (Wentzel 2010). All these percentage values were expressed as
proportions and arcsine-square-root transformed (Zar 1998). These condition values remained constant 180
throughout the study and were also unchanged the following year (condition values on the same trees in the
following year were unchanged; Stuart 2011).
Analyses 185
Relationships between the reproductive effort of each of the 144 trees (six trees in each of the 24 sites), and
tree condition and time since last fire were analysed using generalised additive mixed models (GAMMs;
REML-restricted maximum likelihood). Dependent reproductive effort variables for canopy monitoring

were the arcsine square root-transformed values for the proportion of the canopy budding, flowering, or
fruiting. Initial analyses found location (Dryandra Woodland or Wandoo National Park) was a significant 190
variable so the two locations were analysed separately. Site (1-24) and tree (1-6) variables were nested in
each model. Fixed variables in each model included time since last fire, crown density, crown dieback,
proportion of dead branches, and canopy cover. None of these fixed independent variables were highly
correlated (r ≥ 0.40) so all variables were retained (Pearson’s r; Microsoft Excel; Appendix 1). The number
of independent variables varied between 1 and 5 in each model, but a maximum of five independent 195
variables were used in each model so as not to exceed the ratio of independent factors to replicates, to avoid
over-fitting, and strive for parsimony in terms of numbers of parameters (Burnham and Anderson 2002). A
total of 30 models were created for each dependent variable, capturing combinations of all parameters.
Seed trap monitoring was similarly analysed using GAMMs to explore the relationships between the 200
reproductive effort for each of the 24 trees, and tree condition and time since last fire. Dependent variables
were the total mass of flowering effort, failed flowering effort or aborted fruit. The two locations, Dryandra
Woodland and Wandoo National Park were again analysed separately and the five fixed variables were the
same as for the previous analysis. Data were not nested for this analysis as there was only one tree at each
of the 24 sites that underwent seed trap monitoring. The number of independent factors varied between 1 205
and 3 in each model, but a maximum of 3 independent variables were used in each model so as not to exceed
the ratio of independent factors to replicates, to avoid over-fitting, and strive for parsimony in terms of
numbers of parameters (Burnham and Anderson 2002). A total of 24 models were created for each
dependent variable, capturing combinations of all parameters.
210
An adjusted R2-value was calculated for each model as well as standardised β and P-values for each
parameter within those models. The standardised β coefficients are the regression coefficients when first
standardising all of the variables to a mean of 0 and a standard deviation of 1, and indicate the relative
contribution of each independent variable (e.g. crown density) in the prediction of the dependent variable
(e.g. proportion of canopy budding) (Statistica 9, Statsoft, Melbourne, Vic., Australia). When a 215
standardised β value was insignificant in a model, the model was compared against the same model, but
excluding that parameter. Only models with a ∆AICc of <4, models with considerable empirical support,
were considered further. The AICc model weight (ωi) was calculated for each of the models; ωi values
therefore indicate the likelihood that each model is the best model to describe the data. Model-averaged β
values (Ʃ β *wi; Burnham and Anderson 2002) were calculated to determine the importance of each habitat 220
variable in the prediction of the dependent variable. Only the most significant relationship was graphed as
scatterplots for each reproductive effort variable, using the transformed values.
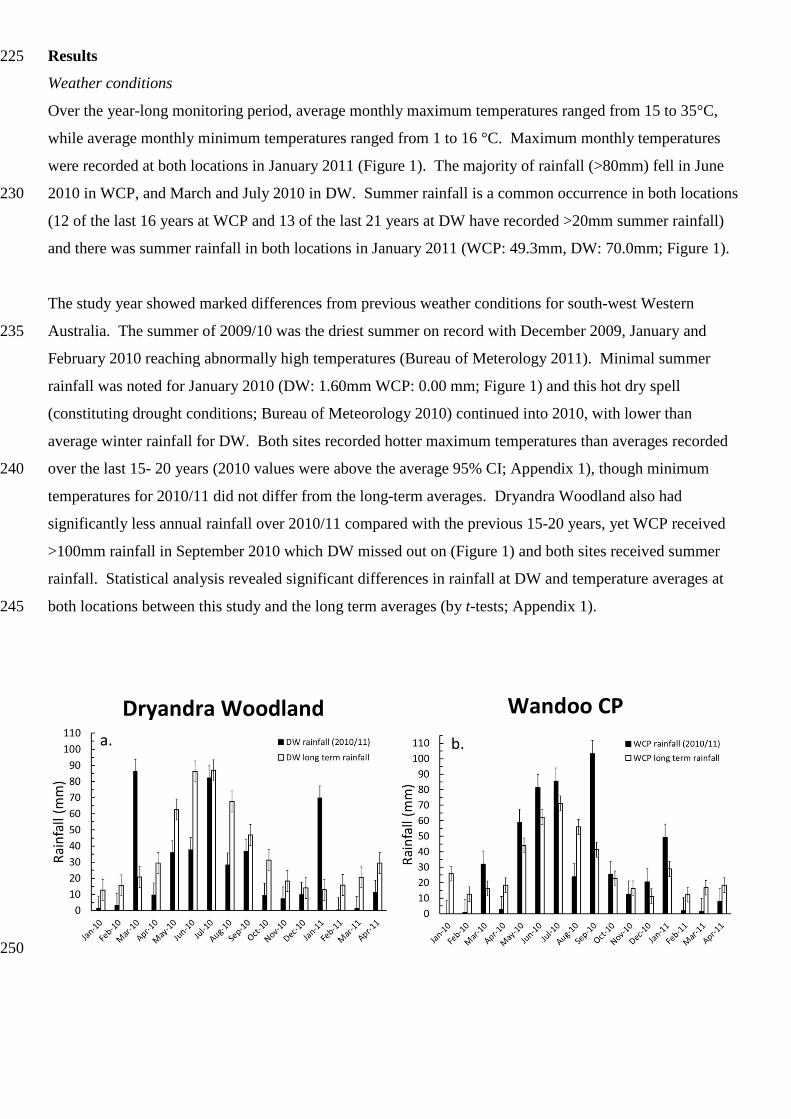
Results 225
Weather conditions
Over the year-long monitoring period, average monthly maximum temperatures ranged from 15 to 35°C,
while average monthly minimum temperatures ranged from 1 to 16 °C. Maximum monthly temperatures
were recorded at both locations in January 2011 (Figure 1). The majority of rainfall (>80mm) fell in June
2010 in WCP, and March and July 2010 in DW. Summer rainfall is a common occurrence in both locations 230
(12 of the last 16 years at WCP and 13 of the last 21 years at DW have recorded >20mm summer rainfall)
and there was summer rainfall in both locations in January 2011 (WCP: 49.3mm, DW: 70.0mm; Figure 1).
The study year showed marked differences from previous weather conditions for south-west Western
Australia. The summer of 2009/10 was the driest summer on record with December 2009, January and 235
February 2010 reaching abnormally high temperatures (Bureau of Meterology 2011). Minimal summer
rainfall was noted for January 2010 (DW: 1.60mm WCP: 0.00 mm; Figure 1) and this hot dry spell
(constituting drought conditions; Bureau of Meteorology 2010) continued into 2010, with lower than
average winter rainfall for DW. Both sites recorded hotter maximum temperatures than averages recorded
over the last 15- 20 years (2010 values were above the average 95% CI; Appendix 1), though minimum 240
temperatures for 2010/11 did not differ from the long-term averages. Dryandra Woodland also had
significantly less annual rainfall over 2010/11 compared with the previous 15-20 years, yet WCP received
>100mm rainfall in September 2010 which DW missed out on (Figure 1) and both sites received summer
rainfall. Statistical analysis revealed significant differences in rainfall at DW and temperature averages at
both locations between this study and the long term averages (by t-tests; Appendix 1). 245
250
Dryandra Woodland Wandoo CP

Fig. 1. Monthly rainfall (mm; bar graph) at Dryandra Woodland (DW; a) and Wandoo Conservation Park
(WCP; b), and maximum and minimum monthly average temperatures (°C; line graph) at Dryandra 255
Woodland (c) and Wandoo Conservation Park (d) from January 2010 to April 2011 (monitoring period was
from May 2010 to April 2011) and the respective long term averages (DW: 1913-2013, WCP: 1996-2013)
(Bureau of Meteorology 2009). All values are mean ± SE.
Reproductive effort and weather conditions 260
Our analyses indicated that the reproductive effort of E. wandoo trees was related to weather conditions in
the preceding months with the flowering of E. wandoo trees seemingly following rainfall events. Peak
flowering in WCP occurred in June and November 2010, after significant rainfall events in March (86.3mm)
and September (103.3mm), as well as the usual winter rainfall (Figure 1). Following that, trees went on to
fruit in January-April 2011. Significant flowering in DW was not observed until January 2011 after 16.8 265
mm of summer rainfall occurred in the fortnight prior (70mm of rainfall was recorded for January 2011, an
above average rainfall; Figure 1). Some trees in DW went on to fruit after the summer flowering event but
not with the same intensity as trees in WCP (Figure 2).
Monitoring period Monitoring period
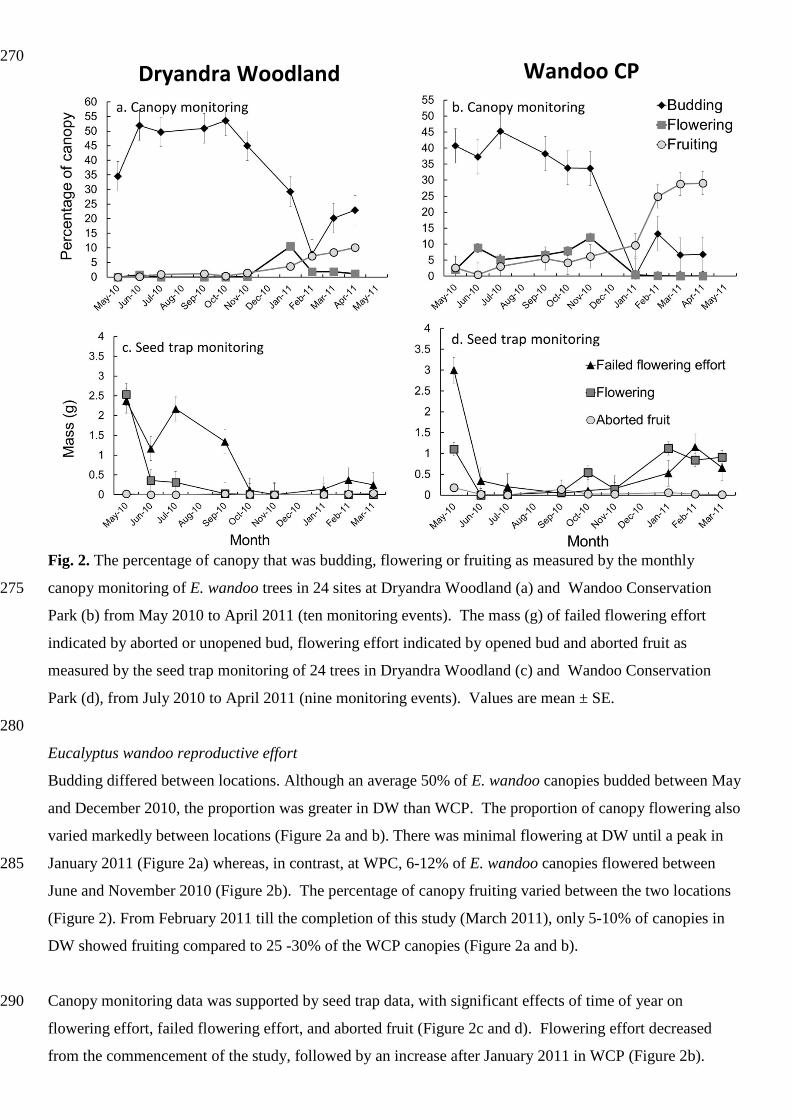
270
Fig. 2. The percentage of canopy that was budding, flowering or fruiting as measured by the monthly
canopy monitoring of E. wandoo trees in 24 sites at Dryandra Woodland (a) and Wandoo Conservation 275
Park (b) from May 2010 to April 2011 (ten monitoring events). The mass (g) of failed flowering effort
indicated by aborted or unopened bud, flowering effort indicated by opened bud and aborted fruit as
measured by the seed trap monitoring of 24 trees in Dryandra Woodland (c) and Wandoo Conservation
Park (d), from July 2010 to April 2011 (nine monitoring events). Values are mean ± SE.
280
Eucalyptus wandoo reproductive effort
Budding differed between locations. Although an average 50% of E. wandoo canopies budded between May
and December 2010, the proportion was greater in DW than WCP. The proportion of canopy flowering also
varied markedly between locations (Figure 2a and b). There was minimal flowering at DW until a peak in
January 2011 (Figure 2a) whereas, in contrast, at WPC, 6-12% of E. wandoo canopies flowered between 285
June and November 2010 (Figure 2b). The percentage of canopy fruiting varied between the two locations
(Figure 2). From February 2011 till the completion of this study (March 2011), only 5-10% of canopies in
DW showed fruiting compared to 25 -30% of the WCP canopies (Figure 2a and b).
Canopy monitoring data was supported by seed trap data, with significant effects of time of year on 290
flowering effort, failed flowering effort, and aborted fruit (Figure 2c and d). Flowering effort decreased
from the commencement of the study, followed by an increase after January 2011 in WCP (Figure 2b).
Dryandra Woodland Wandoo CP

Failed flowering efforts peaked at the commencement of the monitoring period with a continual decrease at
DW but a second peak in April 2011 at WCP (Figure 2c and d). Trees in both locations aborted fruit,
although the mass of aborted fruit was consistently < 0.2 g (Figure 2c and d). Trees aborted fruit at different 295
times (Figure 2c and d). No seeds were collected from seed traps during this study.
Reproductive effort and tree condition
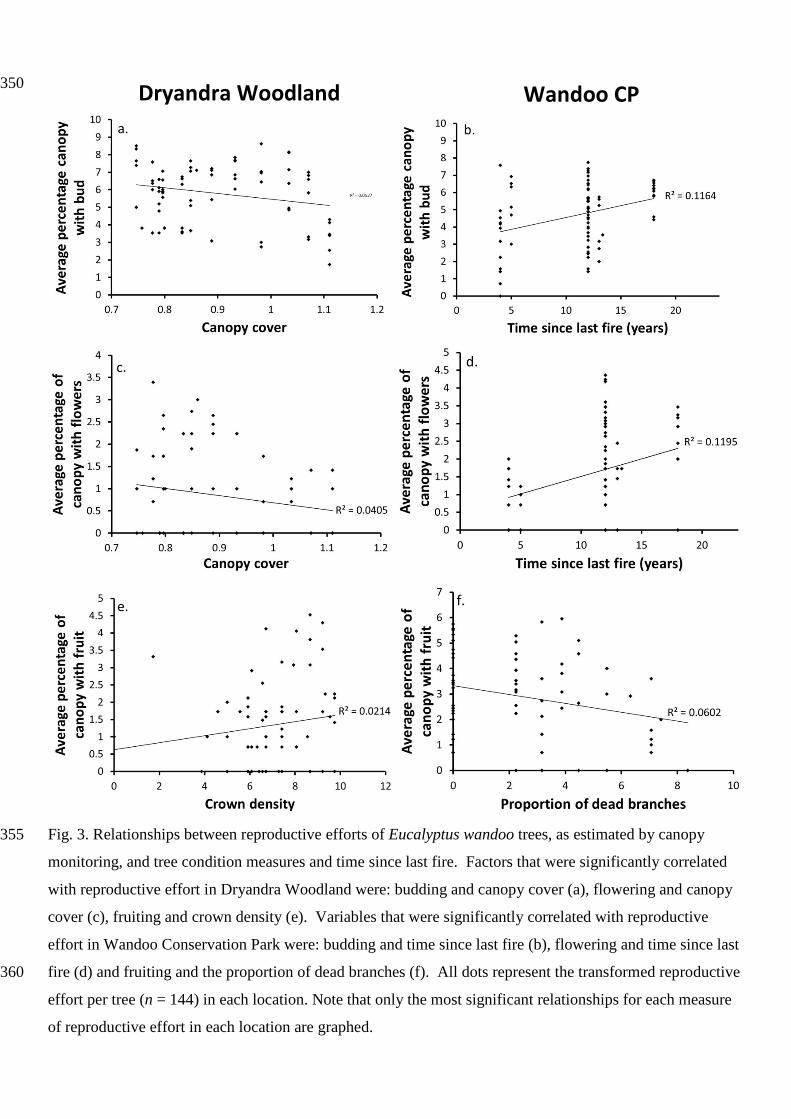
Reproductive effort in terms of budding was correlated to the condition of E. wandoo trees (Table 1 and 2)
as estimated by canopy monitoring. In DW, greater percentages of budding were seen on E. wandoo trees 300
with less canopy cover (Figure 3a), fewer dead branches and higher crown density (Table 1 and 2). In WCP,
budding was more common on trees with sparser canopies and fewer dead branches (Table 1).
Eucalyptus wandoo trees showing generally fewer signs of decline flower more often and had fewer failed
flowering events. Based on canopy monitoring, a greater proportion of flowering in the canopy in DW 305
occurred in sites with less canopy cover (Figure 3c) and fewer dead branches (Table 1). Based on seed trap
monitoring, a larger mass of flowering effort was recorded on E. wandoo trees in DW with fewer signs of
canopy loss or dead branches (Figure 4a) and full crowns (Table 2). At WCP E. wandoo trees with fewer
dead branches, or signs of crown loss, and fuller canopies showed greater flowering events (Table 1).
Similarly, as measured by the seed trap monitoring, a larger mass of flowering effort occurred in WCP in E. 310
wandoo trees with fuller canopies (Table 2). Failure to flower in DW occurred on E. wandoo trees with less
canopy cover and more crown retraction (Table 2). Results were similar in WCP with more failed attempts
to flower in E. wandoo trees with sparser canopies (Figure 4f) and more crown dieback and canopy loss
(Table 2).
315
In both locations E. wandoo trees in good condition fruited more often. Eucalyptus wandoo trees with denser
crowns (Figure 3e) and fewer signs of tree decline showed the highest fruiting values in DW (Table 1). In
WCP, trees with few dead branches (Figure 3f) and minimal crown dieback also showed the greatest
percentages of fruiting in the canopy (Table 1). For both locations, fruiting occurred primarily in trees with a
canopy cover of 40-60% which is considered of average canopy condition (Table 1). In terms of aborted 320
fruit, at DW E. wandoo trees with full canopies containing few dead branches (Figure 4c) had higher masses
of aborted fruit. Whereas in WCP E. wandoo trees with signs of canopy loss and larger percentages of dead
branches (Figure 4d; Table 2), had larger masses of aborted fruit. Summarizing that in DW fruit aborted
more often on E. wandoo trees with a full canopy but with some dead branches, whereas in WCP fruit was
aborted more commonly on E. wandoo trees in poor condition with more dead branches and crown loss. 325

Table 1: Generalised additive mixed models investigating the relationship between reproductive effort from canopy
monitoring, and tree condition, and time since last fire variables
Only those models with an Akaike information criterion adjusted for small sample size (AICc) of <∆4 of the best model are 330 shown (these models have the greatest likelihood of all model-sets to best describe the dataset). Models are ranked according to
their ∆AICc and the AICc model weight (wi); variables are listed in order of terms of decreasing absolute standardised β values.
Models for the percentage of the canopy budding, flowering or fruiting of Eucalyptus wandoo at 24 sites in Dryandra Woodland
and Wandoo Conservation Park, and tree condition, and time since last fire (fixed variables). Abbreviations for independent
variables: canopy cover (CC), time since last fire (TSLF), proportion of dead branches (PDB), crown dieback (DB) and crown 335 density (CD). Values for nested variables, site and tree (six trees per site, 144 trees in total) are not presented.
Reserve Canopy measure Model Adjusted
R2 ΔAICc w i
Dryandra Woodland
Bud CC + TSLF 0.11 0.00 0.23
CC 0.08 0.26 0.20
CC + PDB 0.11 1.23 0.12
CC + TSLF + PDB 0.13 1.91 0.09
CD + CC 0.10 2.10 0.08 Flower CC + PDB 0.12 0.00 0.31
CC 0.08 0.40 0.26
PDB 0.06 1.48 0.15
CC + PDB+ TSLF 0.12 3.47 0.06 Fruit CD 0.06 0.00 0.41
CD+ DB 0.06 3.03 0.09
CD+ CC 0.06 3.44 0.07
Wandoo Conservation Park
Bud TSLF + PDB 0.12 0.00 0.24
TSLF + CC 0.11 1.04 0.15
TSLF + PDB + CD 0.12 2.23 0.08
TSLF + CD 0.10 2.23 0.08
TSLF + PDB + CC 0.13 2.81 0.06 Flower TSLF + PDB 0.18 0.00 0.21
TSLF + CC 0.17 0.91 0.13
TSLF + DB 0.17 0.92 0.13
TSLF + CD 0.16 1.29 0.11
TSLF + PDB + DB 0.17 2.25 0.07
TSLF + PDB + CD 0.17 2.30 0.07
TSLF + CC + DB 0.18 2.73 0.05
TSLF + PDB + CC 0.18 3.03 0.05 Fruit PDB + DB 0.22 0.00 0.15
CC + PDB + DB 0.25 0.10 0.14
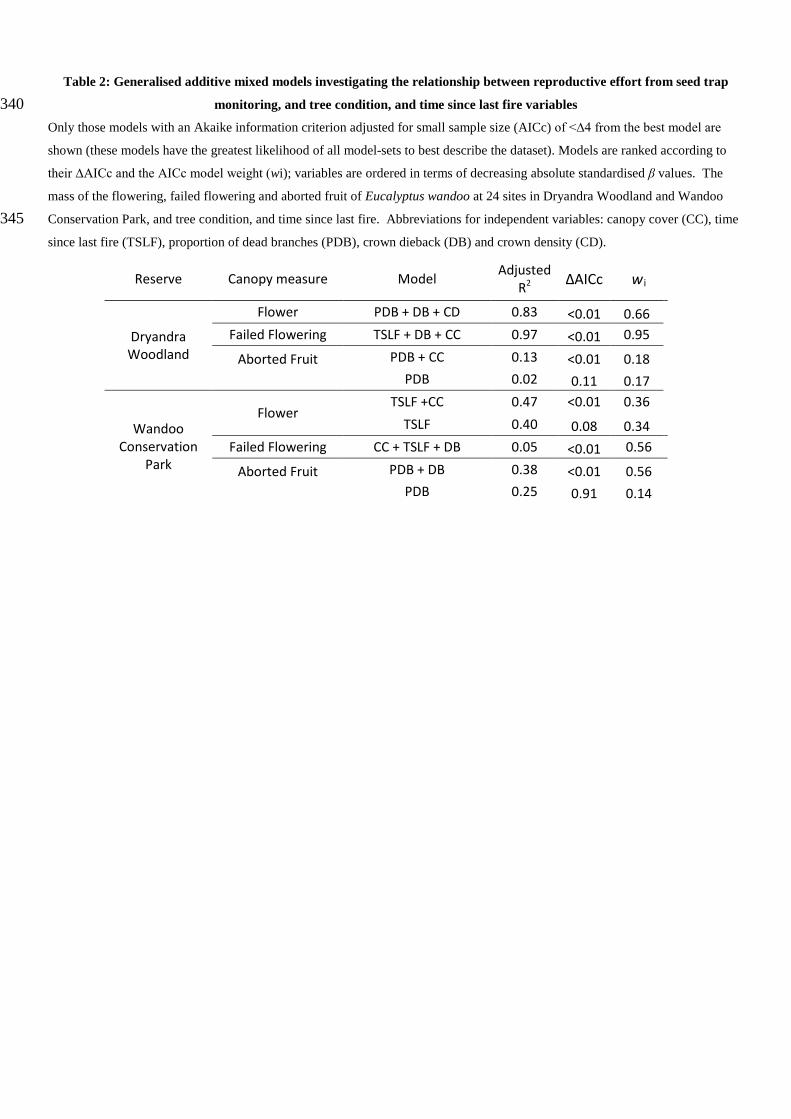
Table 2: Generalised additive mixed models investigating the relationship between reproductive effort from seed trap
monitoring, and tree condition, and time since last fire variables 340 Only those models with an Akaike information criterion adjusted for small sample size (AICc) of <∆4 from the best model are
shown (these models have the greatest likelihood of all model-sets to best describe the dataset). Models are ranked according to
their ∆AICc and the AICc model weight (wi); variables are ordered in terms of decreasing absolute standardised β values. The
mass of the flowering, failed flowering and aborted fruit of Eucalyptus wandoo at 24 sites in Dryandra Woodland and Wandoo
Conservation Park, and tree condition, and time since last fire. Abbreviations for independent variables: canopy cover (CC), time 345 since last fire (TSLF), proportion of dead branches (PDB), crown dieback (DB) and crown density (CD).
Reserve Canopy measure Model Adjusted R2 ΔAICc wi
Dryandra Woodland
Flower PDB + DB + CD 0.83 <0.01 0.66 Failed Flowering TSLF + DB + CC 0.97 <0.01 0.95
Aborted Fruit
PDB + CC 0.13 <0.01 0.18 PDB 0.02 0.11 0.17
Wandoo Conservation
Park
Flower TSLF +CC 0.47 <0.01 0.36
TSLF 0.40 0.08 0.34 Failed Flowering CC + TSLF + DB 0.05 <0.01 0.56
Aborted Fruit
PDB + DB 0.38 <0.01 0.56 PDB 0.25 0.91 0.14
350
Fig. 3. Relationships between reproductive efforts of Eucalyptus wandoo trees, as estimated by canopy 355
monitoring, and tree condition measures and time since last fire. Factors that were significantly correlated
with reproductive effort in Dryandra Woodland were: budding and canopy cover (a), flowering and canopy
cover (c), fruiting and crown density (e). Variables that were significantly correlated with reproductive
effort in Wandoo Conservation Park were: budding and time since last fire (b), flowering and time since last
fire (d) and fruiting and the proportion of dead branches (f). All dots represent the transformed reproductive 360
effort per tree (n = 144) in each location. Note that only the most significant relationships for each measure
of reproductive effort in each location are graphed.
Dryandra Woodland Wandoo CP

365
Fig. 4. Relationships between reproductive effort of Eucalyptus wandoo trees, as measured by seed trap
monitoring, and tree condition and time since last fire. Only the variables showing the most significant
relationships to reproductive effort at each location are graphed. Factors that were significantly correlated 370
with reproductive effort observed in Dryandra Woodland were flowering (a) and aborted fruit (c) and
proportion of dead branches and failed flowering and time since last fire (e). Variables that were
significantly correlated with reproductive efforts in Wandoo Conservation Park were flowering and time
since last fire (b), aborted fruit and proportion of dead branches (d) and failed flowering and canopy cover
(f). All dots represent the average transformed reproductive effort per tree (n = 24). 375
Dryandra Woodland Wandoo CP

Reproductive effort and time since last fire
Time since last fire was correlated to reproductive efforts at both locations (Table 1 and 2; Figures 3b and d,
and 4b and e). In WCP, sites with older fuel loads had higher percentages of canopy with bud and flowers
(Figure 3b and d), and a larger flowering effort as measured by the seed trap monitoring (Figure 4b). The 380
correlations between time since last fire and reproductive efforts at DW were weaker than at WCP, but less
flowering and budding in the canopy was related to older fuel loads (Table 1).
Time since last fire also influenced the failed flowering events in both locations (Table 2). In DW, sites of
older fire ages had fewer failed flowering events (Figure 4e). The inverse was true for WCP, where a larger 385
mass of failed flowering events occurred at sites of older fire age (Table 2). Fruiting or aborted fruit of E.
wandoo canopies in both locations showed no relationships with time since last fire (Tables 1 and 2).
Discussion
Budding, flowering and fruiting was variable over the months, location and sites in this study. There is 390
evidence that tree condition, climate and time since last fire affect the reproductive efforts of E. wandoo.
Reproductive effort and weather conditions
Flowering intensity and frequency in eucalypts varies from year to year (Semple et al. 2007) with rainfall
and temperature variations likely influencing reproductive efforts in many eucalypts as they do in many 395
flowering shrub species (Nagy et al. 2012). The intensity of flowering events in many trees has been shown
to be related to temperature and rainfall (Pfeifer et al. 2006; Hudson et al. 2011), such as drought during the
wet season in the Wet Tropics of Australia (Law et al. 2000). A study on temperate shrubs in Germany
noted that drought and temperature changes extended the occurrence and frequency of flowering in some
shrubs species, although this did depend on shrub or plant size (Nagy et al. 2012). Our results indicated that 400
rainfall and temperature variations likely influencing reproductive efforts in E. wandoo. Rainfall events, in
particular, appeared to be related to flowering attempts in E. wandoo, with trees from both locations
flowering after rainfall. Flowering after rainfall in 2010 was successful at WCP, as fruiting was evident
from January 2011 onwards. However, in DW, flowering was not successful over the 2010 year (with more
aborted flowers, no flowering, and little subsequent fruit), after the lower than average rainfall recorded at 405
DW in winter 2010 (May-June). However, after a summer rainfall event in DW, trees flowered in January.
A number of these trees in DW also went onto fruit after the summer flowering event but not to the same
intensity as trees in WCP; most likely due to the higher than average summer temperatures observed in DW.
It has been noted by apiarists that flowering, and therefore nectar and honey production, in E. wandoo varies
from year to year (Gaynor 2008; Law and Chidel 2008), and these annual variations could reflect rainfall 410
and, perhaps, also temperature. In particular, the flowering attempt at DW coincided with some of the
hottest summer maximum temperatures recorded for the area (up to a maximum 42.2ºC recorded), perhaps
resulting in the lower percentage of fruit seen in canopies at DW compared to WCP. Understanding the

effects of hotter summer temperatures and lower rainfall on E. wandoo flowering is therefore important to
help predict how they may respond to future climate change. 415
Drought and extreme temperatures can interfere with the reproductive cycle of eucalypts, as they do for
many tree species worldwide (Ashton 1975; Pook 1986; House 1997; Law et al. 2000; Pau et al. 2011; Nagy
et al. 2012). Furthermore, phenology is known to be influenced by weather conditions and climate change
(Cleland et al. 2007; Pau et al. 2011; Nagy et al. 2012). Continual changes in weather conditions (and 420
longer term climate change) may alter the reproductive efforts of E. wandoo in the long term (i.e. altering
the flowering time and length, changing the life cycle of pollinators, seed set and the reproduction of the
individual tree; Law and Chidel 2008; Nagy et al. 2012). Apiarists working in our locations observed that
eucalypt flowering events in recent years were diminished compared to the 1930-40s (Gaynor 2008).
Overall, it seems that E. wandoo recover and reproduce poorly from a state of decline if extreme heat and 425
drought occur regularly, leading to E. wandoo mortality and a lack of seed production in surviving trees.
Reproductive effort and tree condition
As previously mentioned, there are many factors that can influence the reproductive efforts of a eucalypt, for
example, rainfall and ambient temperatures (Ashton 1975; House 1997; Brookshire et al. 1999). This study 430
indicated that tree decline is another factor capable of altering the reproduction of eucalypts. Our study
found that reduced reproductive effort (e.g. budding, flowering, failed flowering, fruiting and aborted fruit)
in E. wandoo was strongly linked with decline symptoms (e.g. loss of canopy and more dead branches).
Decline in condition, or the loss of canopy, can reduce the reproductive capacity of eucalypts, often making
them reproductively dormant (Ferretti 1997). Generally, E. wandoo trees with few or no decline symptoms 435
(fewer dead branches and trees with healthy canopy cover) had higher percentages of budding, flowering
and fruiting, and a lower mass of aborted fruit.
Many eucalypts are serotinous, storing their seeds within the canopy for many months, seasons or even years
after a flowering event until suitable conditions for seedling establishment, such as a fire event (Burrows and 440
Abbott 2003), occur (Midgley and Enright 2000; Ruthrof et al. 2002; Cramer and Midgley 2009). No seeds
were recorded in seed traps during this study. The absence of seed fall could indicate that seed was not
formed over the previous year, which is unsurprising given the hot and dry conditions experienced over
summer 2009/10, or that the conditions were not suitable for seed release. Additionally, the lack of fire to
release the serotinous seeds could explain the lack of seeds observed (Gill and McMahon 1986; Lamont 445
1991; Burrows 2008; Lamont and Downes 2011), however, the ability of a tree to produce seeds is also
likely dependent on the canopy condition (Midgley and Enright 2000). Studies in other eucalypt species
have similarly noted that seed fall is highly variable (Semple et al. 2007) and seed fall monitoring over a
longer time frame may be required to better explore relationships between seed fall and canopy condition.
450

Failed reproductive efforts
As observed in this study, high levels of aborted buds and fruit are considered normal in many flowering
species (Stephenson 1981) as long lived plants, such as many eucalypts, have many opportunities to set seed
over the lifespan over 100 years. In many plant species not all buds go on to produce flowers and not all 455
flowers go on to produce fruit (Stephenson 1981), as some plant species reabsorb nutrients and resources
before aborting buds or fruit (Stephenson 1981; Cramer and Midgley 2009). This suggests that premature
abortion of reproductive efforts may be a survival mechanism employed when essential resources are scarce
in the environment (e.g. minimal water or nutrients in times of drought).
460
Reproductive effort and time since last fire
Fire is typically an agent of change that alters the structure of woodlands (Penn et al. 2003; Greenberg and
Waldrop 2008; Close et al. 2009), assists with the regeneration of trees and shrubs by enhancing seed
release and establishment (Burrows et al. 1990; Mercer 1994), and can stimulate flowering in many tree
species (Pyke 1983; Lamont and Downes 2011; Shepherd et al. 2011). Mass recruitment of eucalypts can 465
occur after fire events (Wellington and Noble 1985; Ruthrof et al. 2003). This study produced similar
results to the literature although the results differed between the two locations studied. Wandoo
Conservation Park generally had more budding and flowering in sites with older fire ages whereas sites in
DW with older fire ages had fewer budding and flowering events. The buildup of buds varies between E.
wandoo trees of different post-fire ages until a fire event can stimulate dehiscence of the opercula and 470
flowering. On average the post-fire age at DW is 19 years (range 9-23 years) and at WCP 11 years (range 5-
18 years), indicating that most WCP sites have undergone fires more recently than DW. As previously
stated, regular fire events can stimulate the reproduction and recruitment of many eucalypts (Burrows et al.
1990; Ruthrof et al. 2003), including E. wandoo, resulting in more budding and flowering events as is likely
occurring in WCP. In contrast, less regular fire events, like the situation at DW, may result in less regular 475
reproductive efforts. The same scenario may be relevant for the estimation of failed flowering events in
both locations as our study suggests that failed reproductive efforts may be related to certain fuel ages.
Fruiting, the precursor to seed fall (which did not occur in this study) was unrelated to time since last fire in
this study, indicating that other variables are influencing the fruiting of E. wandoo trees. Future studies
should examine factors that influence seed release from the canopy and seedling recruitment to better 480
understand the role fire can play in the management of the species.
Conclusion
Tree condition, as well as many other weather and habitat variables, can alter Eucalyptus reproduction
(Setterfield and Williams 1996; Law et al. 2000; Birtchnell and Gibson 2006; Semple et al. 2007). We have 485
shown that rainfall is a precursor to flowering events and time since last fire is related to reproductive effort.
While this study is a good starting point, its short sampling period of one year makes it difficult to

understand all the factors that influence reproductive efforts of E. wandoo in DW and WCP. Given the
dearth of studies on E. wandoo, this study, whilst only short-term, provides important insights into the
relationships between E. wandoo condition and reproductive effort. A decline in E. wandoo tree condition 490
influences reproduction and could affect future recruitment, and thus needs to be considered as part of the
conservation management of this species.
495
Acknowledgements
We thank the Holsworth Research Endowment, Wildlife Preservation Society Australia, State Centre for
Excellence of Climate Change, Woodland and Forest Health and Murdoch University for funding this
research. TLM was the recipient of a Murdoch University Research Scholarship for the duration of this
research. Field assistance was gratefully received from many volunteers, particularly Matthew Moore, 500
Bryony Palmer, Tegan Douglas and Kathryn Napier. Thanks is also given to Mike Calver and his advice on
the statistical analyses. This work was carried out with Department of Parks and Wildlife permits
(Regulation 17: SF007629).
References 505 Allen CD, Macalady AK, et al. (2010) A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. Forest Ecology and Management 259, 660-684.
Ashton DH (1975) Studies of flowering behaviour in Eucalyptus regnans F. Muell. Australian Journal of Botany 23, 399-311. 510
Birtchnell MJ, Gibson M (2006) Long-term flowering patterns of melliferous Eucalyptus (Myrtaceae) species. Australian Journal of Botany 54, 745-754.
Brasier CM, Robredo F, Ferraz JFP (1993) Evidence for Phytophthora cinnamomi involvement in Iberian 515 oak decline. Plant Pathology 42, 140-145.
Breshears DD, Cobb NS, et al. (2005) Regional vegetation die-off in response to globa-type drought. PNAS 102, 15144-15148. 520
Brookshire BL, Brosofske KD, Chen J, Crow TR, Franklin JF, Mroz GD (1999) Microclimate in forest ecosystem and landscape ecology: variations in local climate can be used to monitor and compare the effects of different management regimes Bioscience 49, 288-302.
Brouwers N, Mercer J, Lyons T, Poot P, Veneklaas E, Hardy G (2013) Climate and landscape drivers of tree 525 decline in a Mediterranean ecoregion. Ecology and Evolution 3, 67-79.

Bureau of Meteorology (2009) Climate Data Online, Bureau of Meteorology, Accessed May 19 2011, http://www.bom.gov.au/climate/data/. 530
Bureau of Meteorology (2010) A very dry year so far in southwest Western Australia, Special Climate Statement 21, Accessed October 2010, http://www.bom.gov.au/climate/current/statements/scs21.pdf.
Bureau of Meterology (2011) Perth in 2010: One of the hottest and driest years on record, Annual Report for Perth, Accessed February 535 2011, http://www.bom.gov.au/climate/current/annual/wa/archive/2010.perth.shtml.
Burnham KP, Anderson DR (2002) 'Model selection and multimodel inference: a practical information-theoretic approach.' 2nd Edition edn. (Springer-Verlag: New York, New York, USA). 540
Burrows N, Abbott I (2003) ' Fire in south-west Western Australia: synthesis of current knowledge, management implications and new research directions.' (Backhuys Publishers: Leiden, Netherlands).
Burrows N, Gardiner G, Ward B, Robinson A (1990) Regeneration of Eucalyptus wandoo following fire. Australian Forestry 53, 248-258. 545
Burrows ND (2008) Linking fire ecology and fire management in south-west Australian forest landscapes. Forest ecology and management 255, 2394-2406.
Byrne M, Elliot CP, Yates CJ, Coates DJ (2008) Maintenance of high pollen dispersal in Eucalyptus 550 wandoo, a dominant tree of the fragmented agricultural region in Western Australia. Conservation Genetics 9, 97-105.
Carnegie AJ (2007) Forest health condition in New South Wales, Australia, 1996-2005. II. Fungal damage recorded in eucalypt plantations during forest health surveys and their management. Australasian Plant 555 Pathology 36, 225-239.
Cleland EE, Chuine I, Menzel A, Mooney HA, Schwartz MD (2007) Shifting plant phenology in response to global change. The NDS in Ecology and Evolution 122, 357-365. 560
Close DC, Davidson NJ (2004) Review of rural tree decline in a changing Australian climate. Tasmanian Forests 15, 1-18.
Close DC, Davidson NJ, Johnson DW, Abrams MD, Hart SC, Lunt ID, Archibald RC, Horton B, Adams MA (2009) Premature decline of Eucalyptus and altered ecosystem processes in the absence of fire in some 565 Australian forests. Botanical Review 75, 191-205.
Commonwealth Scientific and Industrial Research Organisation (2010) Vegmachine 2.0. In. ' (Eds T Beutel, R Karfs and J Resing). (State of Queensland, Department of Employment, Economic Development and Innovation Queensland) 570

Cramer MD, Midgley JJ (2009) Maintenance costs of serotiny do not explain weak serotiny. Austral Ecology 34, 653-662.
Dalmaris E (2012) Eucalyptus wandoo: tolerance to drought and salinity in relation to provenance and 575 evolutionary history in southwestern Australia. PhD Thesis. University of Western Australia, Perth, Western Australia
Department of Conservation and Land Management (1980) Dryandra Management Plan. Department of Conservation and Land Management, Perth, Western Australia. 580
Doughty RW (2000) 'The Eucalyptus. A natural and commercial history of the gum tree.' (The John Hopkins University Press: Baltimore).
Ferretti M (1997) Forest health assessment and monitoring- issues for consideration. Environmental 585 Monitoring and Assessment 48, 45-72.
Gaynor A (2008) 'Wandoo in health and decline: a history.' (Department of Environment and Conservation.: Perth, Western Australia). 590
Gill AM, McMahon A (1986) A post-fire chronosequence of cone, follicle and seed production in Banksia ornata. Australian Journal of Botany 34, 425-433.
Greenberg CH, Waldrop TA (2008) Short-term response of reptiles and amphibians to prescribed fire and mechanical fuel reduction in a southern Appalachian upland hardwood forest Forest Ecology and 595 Management 255, 2883-2893.
Hooper RJ (2009) The role of stress and factors contributing to the decline of Eucalyptus wandoo (Blakely) in southwestern Australia. PhD thesis. University of Western Australia, Perth, Western Australia 600
House SM (1997) Reproductive biology of eucalypts. In 'Eucalypt ecology. Individuals to ecosystems. (Eds JE Williams and JCZ Woinarski) pp. 30-55. (Cambridge University Press: Cambridge).
Hudson IL, Keatley MR, Kang I (2011) Wavelet characterization of eucalypt flowering and the influence of climate. Environmental Ecological Statistics 18, 513-533. 605
IBM (2012) SPSS Statistics Software version 12.0. In. ')
Klausmeyer KR, Shaw R (2009) Climate change, habitat loss, protected areas and the climate adaptation potential of species in Mediterranean ecosystems worldwide. PLos ONE 4, e6392. 610 doi:10.1371/journal.pone.0006392.
Klausmeyer KR, Shaw R, MacKenzie JB, Cameron DR (2011) Landscape-scale indicators of biodiversity's vulnerability to climate change. Ecosphere 2, art88. doi:10.1890/ES11-00044.1. 615

Lamont BB (1991) Canopy seed storage and release: what's in a name? Oikos 60, 266-268.
Lamont BB, Downes KS (2011) Fire-stimulated flowering among resprouters and geophytes in Australia and South Africa. Plant Ecology 212, 2111-2125. 620
Law B, Mackowski C, Schoer L, Tweedie T (2000) Flowering phenology of myrtaceous trees and their relation to climatic, environmental and disturbance variables in Northern New South Wales. Austral Ecology 25, 160-178.
Law BS, Chidel M (2008) Quantifying the canopy nectar resource and the impact of logging and climate in 625 spotted gum Corymbia maculata forests. Austral Ecology 33, 999-1014.
Lemmon PE (1956) A spherical densiometer for estimation forest overstorey density. Forestry Science 2, 314-320. 630
Mattiske & Havel (1998) Vegetation mapping in the South West of Western Australia. Prepared for Environment Australia and the Department of Conservation and Land Management.
Matusick G, Ruthrof KX, Brouwers N, Dell B, Hardy GESJ (2012) Sudden forest canopy collapse corresponding with extreme drought and heat in a mediterranean-type forest in southwestern Australia. 635 Landscape Ecology 28, 69-80.
Matusick G, Ruthrof KX, Hardy GESJ (2013) Drought and heat triggers sudden and severe dieback in a dominant Mediterranean-type woodland species. Open Journal of Forestry 2, 183-186. 640
Mercer J (1994) Fire regeneration of Eucalyptus wandoo in farmland remnants. A report on results of trials. Save the Bush Program, Australian National Conservation Agency, Western Australia.
Midgley JJ, Enright NJ (2000) Serotinous species show correlation between retention time for leaves and cones. Journal of Ecology 88, 348-351. 645
Moore TL, Valentine LE, Craid MD, Hardy GESJ, Fleming PA (2014) Signs of wildlife activity and Eucalyptus wandoo condition. Australian Mammalogy Accepted.
Moore TL, Valentine LE, Craig MD, Hardy GESJ (2013a) Does woodland condition influence the diversity 650 and abundance of small mammal communities? Australian Mammalogy accepted, http://www.publish.csiro.au/paper/AM13007.htm.
Moore TL, Valentine LE, Craig MD, Hardy GESJ, Fleming PA (2013b) Do woodland birds preferentially forage in healthy Eucalyptus wandoo trees. Australian Journal of Zoology 61, 187-195. 655
Moore TL, Valentine LE, Craig MD, Hardy GESJ, Fleming PA (2013c) Is the reptile community affected by Eucalyptus wandoo tree condition? Wildlife Research http://dx.doi.org/10.1071/WR13015.

Nagy L, Kreyling J, Gellesch E, Beierkuhnlein C, Jentsch A (2012) Recurring weather extremes alter the 660 flowering phenology of two common temperate shrub. International Journal of Biometerology 27, 579-588.
Pau S, Wolkovich EM, Cook BI, Davies J, Kraft NJB, Bolmgre K., Betancourt JL, Cleland EE (2011) Predicting phenology by integrating ecology, evolution and climate science. Global Change Biology 17, 3633-3643. 665
Penn AM, Sherwin WB, Lunney D, Banks PB (2003) The effects of low-intensity fire on small mammals and lizards in a logged, burnt forest. Wildlife Research 30, 477-486.
Pfeifer M, Heinrich W, Jetschke G (2006) Climate, size and flowering history determine flowering pattern 670 of an orchid. Botanical Journal of the Linnean Society 151, 511-526.
Pook EW (1986) Canopy dynamics of Eucalyptus maculata Hook. IV* Contrasting responses to two severe droughts. Australian Journal of Botany 34, 1-14. 675
Pyke GH (1983) Relationship between time since last fire and flowering in Telopea speciosissima R. Br. and Lambertia formosa Sm. Australian Journal of Botany 31, 293-296.
Reid N, Landsberg J (2000) Tree decline in agricultural landscapes: what we stand to lose. In 'Temperate eucalypt woodlands in Australia. Biology conservation, management and restoration. (Ed. RJ Hobbs, Yates, 680 C. J.) pp. 127-166. (Surrey Beatty and Sons Pty Limited: Chipping Norton).
Ruthrof KX, Loneragan WA, Yates CJ (2003) Comparative population dynamics of Eucalyptus cladocalyx in its native habitat and as an invasive species in an urban bushland in south-western Australia. Diversity and Distributions 9, 469-484. 685
Ruthrof KX, Yates CJ, Loneragan WA (2002) The Biology of Eucalyptus gomphocephala DC. (Tuart). In 'Tuart (Eucalyptus gomphocephala) and Tuart Communities (Eds BJ Keighery and VM Longman) pp. 108-22. (Wildflower Society of Western Australia (Inc.)). 690
Schomaker ME, Zarnoch SJ, Bechtold WA, Latelle DJ, Burkman WG, Cox SM (2007) Crown-condition classification: A guide to data collection and analysis United States Department of Agriculture, Forest Service, Southern Research Station, Asheville, NC.
Semple WS, Koen T, Henderson J (2007) Seed fall and flowering in white box (Eucalyptus albens Benth.) 695 trees near Cowra, New South Wales. Australian Forestry 70, 242-252.
Setterfield SA, Williams RJ (1996) Patterns of flowering and seed production in Eucalyptus miniata and E. tetradonta in tropical savannah woodland, Northern Australia. Australian Journal of Botany 44, 335-352. 700
Shepherd BJ, Miller DL, Thetford M (2011) Fire season effects on flowering characteristics and germination of longleaf pine (Pinus palustris) savanna grasses. Restoration Ecology 20, 268-276.

Stephenson AG (1981) Flower and fruit abortion: Proximate causes and ultimate functions. Annual Review of Ecology and Systematics 12, 253-279. 705
Stuart JM (2011) Microhabitat use of small skinks in Eucalyptus wandoo woodland. Honours thesis. Murdoch University, Perth, Western Australia
Wandoo Recovery Group (2006) Wandoo crown decline. Action Plan. Department of Environment and 710 Conservation, Perth.
Wellington AB, Noble IR (1985) Post-fire recruitment and mortality in a population of the mallee Eucalyptus incrassata in semi-arid south-eastern Australia. Journal of Ecology 73, 645-656. 715
Wentzel JJ (2010) Is tuart (Eucalyptus gomphocephala) decline detrimental for fauna? PhD Thesis. Murdoch University, Perth, Western Australia
Whitford K, Manning L, Wills A (2008) Wandoo crown condition. Report of wandoo crown decline surveys, 2008. Department of Environment and Conservation and Wandoo Recovery Group. 720
Yates CJ, Hobbs RJ (1997) Temperate eucalypt woodlands: a review of their status, processes threatening their persistence and techniques for restoration. Australian Journal of Botany 45, 949-973.
Yates CJ, Hobbs RJ, Bell RW (1994) Factors limiting the recruitment of Eucalyptus salmonophloia in 725 remnant I. Patterns in flowering, seed production and seed fall. Australian Journal of Botany 42, 531-542.
Zar JH (1998) 'Biostatistical Analysis 4th Edition ' (Prentice Hall: New Jersey).
730
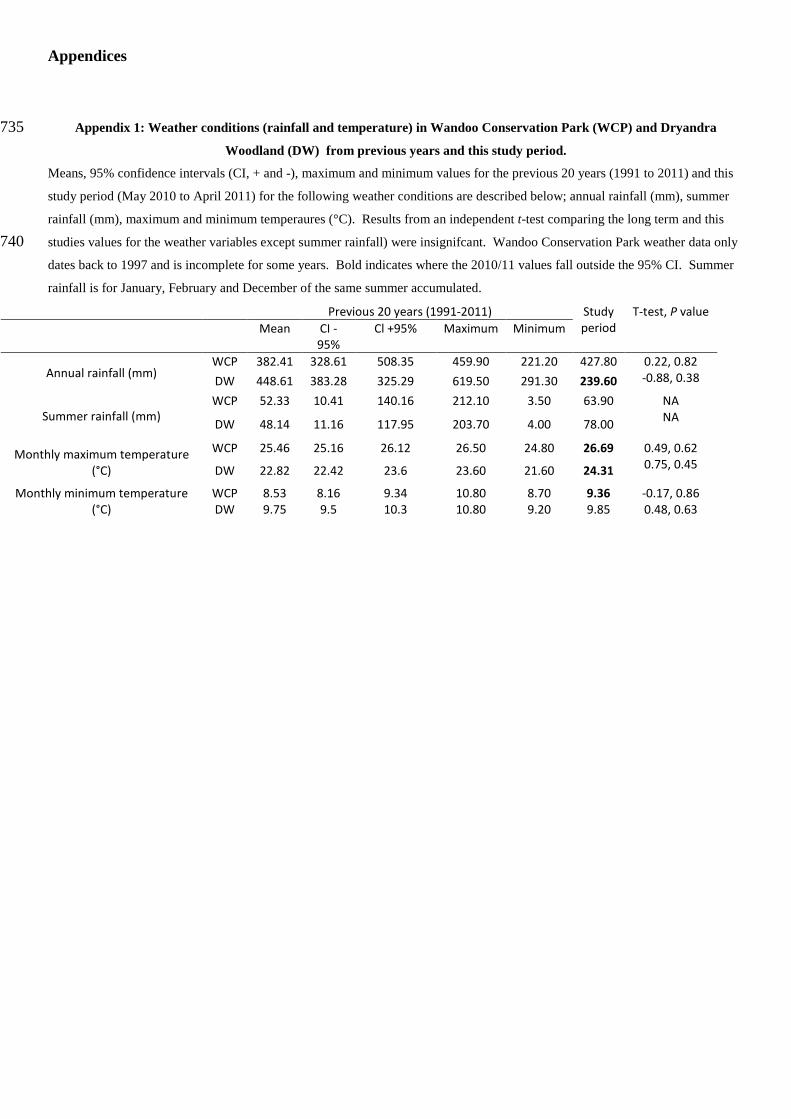
Appendices
Appendix 1: Weather conditions (rainfall and temperature) in Wandoo Conservation Park (WCP) and Dryandra 735 Woodland (DW) from previous years and this study period.
Means, 95% confidence intervals (CI, + and -), maximum and minimum values for the previous 20 years (1991 to 2011) and this
study period (May 2010 to April 2011) for the following weather conditions are described below; annual rainfall (mm), summer
rainfall (mm), maximum and minimum temperaures (°C). Results from an independent t-test comparing the long term and this
studies values for the weather variables except summer rainfall) were insignifcant. Wandoo Conservation Park weather data only 740 dates back to 1997 and is incomplete for some years. Bold indicates where the 2010/11 values fall outside the 95% CI. Summer
rainfall is for January, February and December of the same summer accumulated.
Previous 20 years (1991-2011) Study period
T-test, P value Mean CI -
95% Cl +95% Maximum Minimum
Annual rainfall (mm) WCP 382.41 328.61 508.35 459.90 221.20 427.80 0.22, 0.82 DW 448.61 383.28 325.29 619.50 291.30 239.60 -0.88, 0.38
Summer rainfall (mm) WCP 52.33 10.41 140.16 212.10 3.50 63.90 NA
DW 48.14 11.16 117.95 203.70 4.00 78.00 NA
Monthly maximum temperature (°C)
WCP 25.46 25.16 26.12 26.50 24.80 26.69 0.49, 0.62
DW 22.82 22.42 23.6 23.60 21.60 24.31 0.75, 0.45
Monthly minimum temperature (°C)
WCP 8.53 8.16 9.34 10.80 8.70 9.36 -0.17, 0.86 DW 9.75 9.5 10.3 10.80 9.20 9.85 0.48, 0.63

Appendix 2: Averaged standardised β values of the top models (with a ΔAICc <4) adjusted for small sample size, 745 explaining the percentage of the canopy budding, flowering or fruiting (from the canopy monitoring) at Dryandra
Woodland and Wandoo Conservation Park.
Abbreviations for all variables are as follows: canopy cover (CC), time since last fire (TSLF), proportion of dead branches (PDB),
crown dieback (DB) and crown density (CD).
Reserve Canopy measure CC CD DB PDB TSLF
Dryandra Woodland Bud -0.08 0.02 <0.01 -0.06 <0.01
Flower -0.14 <0.01 <0.01 -0.16 -0.02 Fruit 0.01 0.15 -0.01 <0.01 <0.01
Wandoo Conservation Park Bud -0.05 -0.02 <0.01 -0.02 0.20
Flower 0.03 0.01 -0.02 -0.08 0.25 Fruit <0.01 <0.01 -0.05 -0.07 <0.01
750

Appendix 3: Averaged standardised β values of the top models (with a ΔAICc <4) adjusted for small sample size,
explaining the mass flowering, failed flowering and aborted fruit (from the seed trap monitoring) at Dryandra Woodland
and Wandoo Conservation Park.
Abbreviations for all variables are as follows: canopy cover (CC), time since last fire (TSLF), proportion of dead branches (PDB), 755 crown dieback (DB) and crown density (CD).
Reserve Canopy measure CC CD DB PDB TSLF
Dryandra Woodland
Flower <0.01 0.01 -0.01 -0.17 <0.01 Failed Flowering -0.01 <0.01 0.10 <0.01 -0.10
Aborted Fruit 0.04 <0.01 <0.01 -0.01 <0.01
Wandoo Conservation
Park Flower 0.03 <0.01 <0.01 <0.01 0.22
Failed Flowering -0.09 <0.01 0.04 <0.01 0.08
Aborted Fruit <0.01 <0.01 0.01 0.05 <0.01