myxoid liposarcoma with t(12;16) (ql3;pll) contains site-specific
TRANSCRIPT

(CANCER RESEARCH 50, 7902-7907, December 15, 1990]
Myxoid Liposarcoma with t(12;16) (ql3;pll) Contains Site-specific Differences in
Methylation Patterns Surrounding a Zinc-Finger Gene Mapped to theBreakpoint Region on Chromosome 12'
Sylvie Paulien,2 Claude Turc-Carel, Paola Dal Cin, Sheila Jani-Sait, Chandrika Sreekantaiah, Stanley P. L. Leong,3Bert Vogelstein, Kenneth W. Kinzler, Avery A. Sandberg, and Robert M. Gemmili4
Southwest Biomedicai Research Institute, Scottsdale, Arizona 85281 [S. P., P. D. C., S. J-S., C. S., A. A. S., R. M. G.J; Facultéde Médecine,Nice, France [C. T-C.J;Center of Human Genetics, University of Leuven, Leuven, Belgium [P. D. C.J; University of Arizona Cancer Center and Department of Surgery, University of Arizona,Tucson, Arizona 85724 fS. P. L. L.J; and The Johns Hopkins University, Baltimore, Maryland 21205 [B. V., K. W. K.J
ABSTRACT
The ql3 to ql5 region of human chromosome 12 is frequently andconsistently rearranged in malignant and benign adipose tissue tumors aswell as benign tumors of smooth muscle and salivary glands. A reciprocaltranslocation, (12; 16) (ql3;pll), is characteristic of the myxoid subtypeof liposarcoma, whereas translocations within 12ql3-14 are frequentlyobserved in benign lipomas. We are using pulsed-field gel electrophoresisto study the 12ql3-ql4 region in order to detect and clone the respectivetranslocation breakpoints in these tumors. The locus GLI, which encodesa zinc-finger protein, has been mapped to the same region as the myxoidliposarcoma breakpoint. Pulsed-field analysis of myxoid liposarcoma andlipoma DNA has allowed us to construct a 600-kilobase physical mapsurrounding the GLI locus, which shows that breakpoints in both typesof tumor are outside this region. However, myxoid liposarcoma DNAsamples contained altered restriction fragments detectable with GLIprobes that were highly specific and reproducible from case to case.These altered fragments are due to highly specific and reproduciblemethylation differences that are unique to myxoid liposarcoma DNA.These methylation changes may prove to be useful clinically as a diagnostic tool to differentiate subtypes of liposarcoma.
INTRODUCTION
Methylation of cytosine residues, primarily within the targetdinucleotide CpG, is the major covalent modification in vertebrate DNA (1,2). Methylation can exert a strong negative effecton expression of many genes (1-5), apparently through alteration of specific protein-DNA interactions (6). Other genes areknown for which methylation has little or no effect (7, 8).Methylation of a specific subset of CpG dinucleotides can bedetected using the restriction endonuclease isoschizomer pairHpall and Mspl (I). This assay has demonstrated that manygenes contain sites differentially methylated in a tissue-specificfashion (9-12). Methylation is widely thought to be one majorelement in the pathway controlling tissue-specific gene expression and has been implicated in processes of development anddifferentiation (2, 11, 13). Studies in the mouse have shownthat maternal and paternal genome contributions to the zygoteare not functionally equivalent, a phenomenon known as ge-nomic imprinting (14). Methylation modifications specific tooocyte and sperm nuclei may be the basis for these differences(15-17). The importance of methylation as a regulatory mechanism in these processes contrasts sharply with the lack of
Received 7/9/90; accepted 9/18/90.The costs of publication of this article were defrayed in part by the payment
of page charges. This article must therefore be hereby marked advertisement inaccordance with 18 U.S.C. Section 1734 solely to indicate this fact.
1This work was supported in part by NIH Grant CA-41183.2ARC Fellow (Association pour la Recherche contre le Cancer, Villejuif,
France). Present address: Eleanor Roosevelt Institute for Cancer Research, 1899Gaylord Street, Denver, CO 80206.
J Recipient of the American Cancer Society Clinical Oncology Career Award.4To whom requests for reprints should be addressed, at Eleanor Roosevelt
Institute for Cancer Research, 1899 Gaylord Street, Denver, CO 80206.
knowledge regarding how methylation patterns are establishedand altered. These questions will be difficult to address withoutfundamental knowledge of the patterns themselves.
Neoplastic cells are deranged in processes related to differentiation and development. All neoplastic cells have lost someor all control over cell division and many possess an alteredrepertoire of expressed genes, either making inappropriateproducts or stopping synthesis of products characteristic forthe cell type. Since cytosine methylation can affect these processes, understanding the role of methylation in neoplasia is ofgreat interest (11). Studies to date have observed generalizedhypomethylation for many solid tumors, both benign and malignant (18-20). It is important to understand how methylationpatterns are established and altered in both normal and neoplastic cells since such alterations may be related to the neoplastic phenotype.
We have been studying benign and malignant adipose tissuetumors characterized by translocations within chromosomebands 12ql3-14 (21-25). Forty % of benign lipomas, a verycommon proliferation of histologically normal adipocytes, contain rearrangements of 12ql3-14 in which the involvement of12q is consistent but the reciprocal chromosome is variable(26). MLPS,5 a rare malignant sarcoma of adipose tissue, is
similarly characterized by a translocation within 12q, t(12;16)(ql3;pll) (23-25). This translocation is always reciprocal andhas been found in nearly 100% of MLPS cases reported to date(27). In order to detect and clone these breakpoints, we havebeen using PFG electrophoresis (28, 29) to generate long-rangerestriction endonuclease cleavage maps (30) surrounding clonedDNA probes that map to 12ql3-14. One of these probes is thezinc-finger gene GLI (31, 32), which has recently been localizedto 12ql3.3-ql4.1 (33,34).
We have identified a set of very reproducible site-specificmethylation changes in MLPS DNA that occur over a 600-kilobase region surrounding the GLI locus. These changesappear to be characteristic of myxoid liposarcoma and not ofbenign lipoma or other subtypes of liposarcoma. Our observations show that tissue-specific methylation patterns can existover very large regions of the genome and be specifically alteredin neoplasia. PFG analysis provides a unique method to studymethylation patterns, both in normal and neoplastic cells, andto investigate the possible relationship between DNA methylation and cancer.
MATERIALS AND METHODS
Preparation of Tumor Specimens for PFG Analysis. Table 1 lists thespecimens that were analyzed by pulsed-field gel electrophoresis. Tumors were obtained after surgery and transported in RPMI 1640 plus
gel.' The abbreviations used are: MLPS, myxoid liposarcoma; PFG, pulsed field
7902
Research. on February 19, 2018. © 1990 American Association for Cancercancerres.aacrjournals.org Downloaded from

METHYLATION PATTERNS OF 12ql3 IN MYXOID LIPOSARCOMA
Table 1 Summary of tumor and normal tissue specimens analyzedThe proportion of cell populations that retained a cytogenetic abnormality is listed in the first column. Lymphoblastoid cell lines were established by Epstein-Barr
virus infection of peripheral blood lymphocytes. Lines TL8229, TL9542, and TL9944 were established several years before this investigation.
Tissue type abnormality Cell no. Cytogenetics
Normal fatST.88-11882
Lymphoblastoid
Fibroblast
46,XXST.88-16587nf
TL8229TL9542TL9944CL.88-11296CL.88-16940°
ST.88-16587fb"
46,XY
46.XY46,XX,t(3;8)(pl4.2;q24.1)46,XX,t(3;8)(P14.2;q24.1)46,XX46.XY
46.XY
% Cellsretaining
LiposarcomasMLPSWell-differentiatedOther
subgroupsLipomasST.88-05390ST.88-11172ST.88-13233"ST.88-18130ST.89-02510ST.88-12407ST.89-00151ST.88-04009ST.88-04019ST.88-04103ST.88-05733ST.88-06643ST.88-07389ST.88-08203ST.88-15734ST.88-21239ST.88-21338ST.89-0025146,XX,t(12;16)(ql3;pll)/46,XX46,XX,t(12;16)(ql3;pll)46,XY,t(12;16)(ql3;pll)47,XY,+8,t(12;I6)
(12/21)48,XX,+8,t(ll;17)(ql3;q21),t(12;16)(ql3;pll.2),+1947,XY,+8,t(
12; 16) (q 13;p 11.2)46,XX+ largemarkers46,X,t(l;13)(q25,ql4)46,XX,t(12;15)
(ql4;q25)/46,XX46,XX,t(7;12)(p21;ql4)/46,XX (1cell)46,XX,t(2;
12; 16) (p2 1;q 16;q24)/t(4; 17; 15;6) (q2 1;p 13,q 12;q23)/46,XX46,XX46,XX,t(2;15;12)
(q35;q21;ql6)46,XY,t(5;12)(q33;ql6)46,XX,t(6;ll)
(p23;ql3) t(ll;12)(ql3;q22)/46,XX46,XY,t(ll;12)(q23;ql3),del(13)(ql2;q22)46,XX,t(2;6)(q36;p21.3)46.XY46,XX,t(
1;9; 12)-complex/46,XXSO100100060401001010010101010109050
100100
' Those tissue samples obtained from the same patient as myxoid liposarcoma ST.88-13233.
antibiotics. Each tumor was mechanically disaggregated and treatedwith collagenase (2000 units/ml; Gibco) overnight to generate a singlecell suspension. Cells were cultured in RPMI 1640 plus 15% fetal calfserum over a period of 10 to IS days until sufficient cells were availablefor both cytogenetic and PFG analysis. Cells were typically harvestedfor cytogenetic analysis one week earlier than the balance of cells usedfor PFG analysis. Of the cells harvested for PFG analysis, a smallsample underwent repeat cytogenetic analysis to ensure that the cellsstudied via PFG retained the translocation of interest. Culture timesfor all tumor-derived cells was minimized to prevent cytogeneticchanges and overgrowth by normal fibroblasts. Nevertheless, a numberof cultures were contaminated by fibroblasts (Table 1). Lymphoblastoidand fibroblast cell lines were cultured in RPMI 1640 containing 10%fetal calf serum. Several of the lymphoblastoid cell lines used had beenextensively cultured over a period of years.
High molecular weight DNA imbedded in agarose blocks was prepared from each sample as described previously (30). Briefly, cells wereharvested, washed twice in Hanks' balanced salt solution, and resus-pended at a concentration of 4 x 107/ml. A 1% solution of InCert(FMC, Inc.) agarose in Hanks' balanced salt solution was melted, cooledto 50°C,and mixed with an equal volume of the cell suspension at42*C. The mixture was cast into blocks that were then incubated at50°Cfor 48 h in 1% Ar-laurylsarcosine-0.5 MEDTA, pH 8.0, containing
200 fig/ml of proteinase K. The blocks were washed twice in 10 mMTris HC1, l mM EDTA, pH 7.5, for 30 min at room temperature, oncefor 30 min at 50°C,then twice in the presence of 0.04 mg/ml phenyl-methylsulfonyl fluoride at 50°C.After one last Tris-EDTA wash atroom temperature, the blocks were stored at 4°C.
PFG Analysis. Digestion of agarose-imbedded DNA, separation offragments on pulsed field gels, and Southern blot analysis were performed as described previously (30) with minor modifications. ThePFG system used is based on the crossed field design of Southern et al.(35). This system uses a rotating platform to generate pulsed fields at
a 120°angle. Typical separations were performed using the following
parameters: electric field of 5 V/cm, pulse time of 60 s, agaroseconcentration of 1.5%, 0.25x TBE buffer, and a temperature of 10°C.
Molecular size standards included oligomers of X phage (rl857.S«/;;7:monomer is 48.5 kilobases), and chromosome preparations from Sac-charomyces cerevisiae strain YNN295 (Bio-Rad).
DNA was transferred to charge modified nylon membranes (BiotraceRP; Gelman) and hybridized with probes as described previously (30).Subclones of the GLI locus were labeled to high specific activity usingthe random priming method of Feinberg and Vogelstein (36).
RESULTS
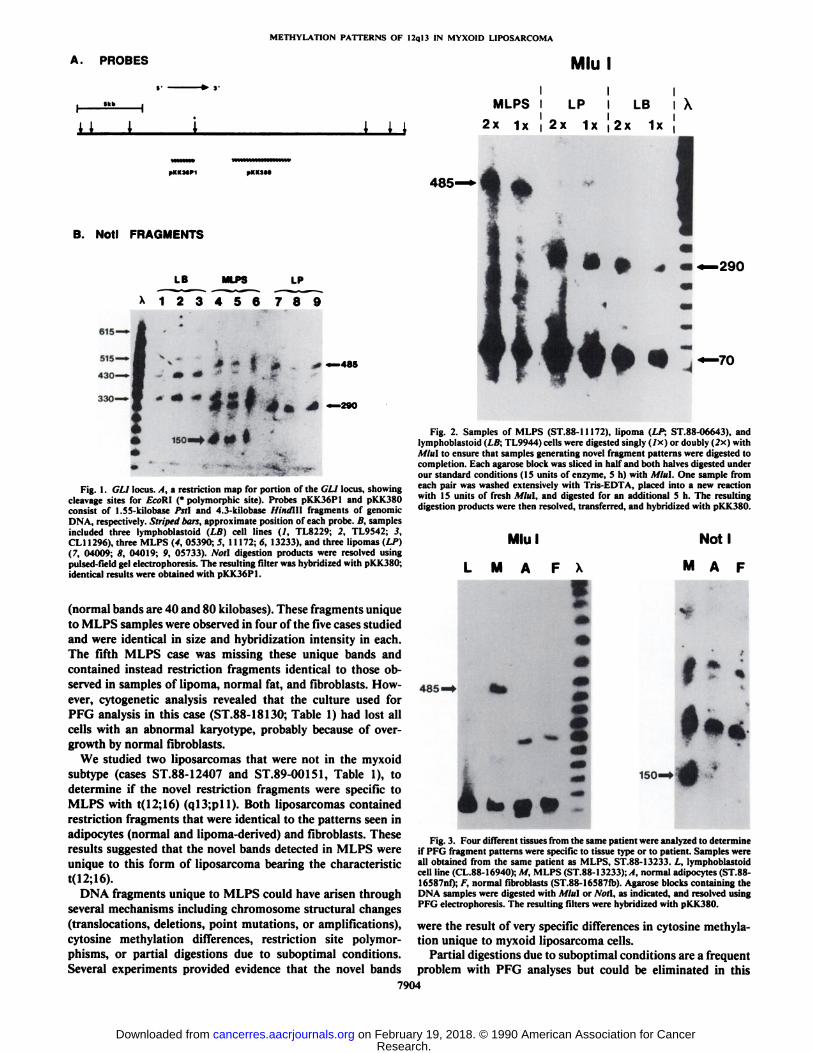
Large DNA Fragments Unique to Myxoid Liposarcoma. DNAsamples prepared from malignant and benign adipose tumors,normal fat, Epstein-Barr virus transformed lymphoblastoid celllines, and fibroblasts (Table 1) were digested to completionwith a set of eight rare-cutting restriction endonucleases. Theresulting large DNA fragments were resolved on pulsed-fieldgels, transferred to membranes, and hybridized with two probesderived from the GLI gene, pKK36Pl and pKK380 (Fig. \A).
Most endonucleases generated sets of fragments, some ofwhich appeared in tissue-specific patterns. Fig. 1A shows resultsobtained from Noti digestions of three different types of cells.The three myxoid liposarcoma samples specifically contained aunique 150-kilobase fragment in these digests, consistent withthe possibility that a translocation bearing fragment was beingdetected. Two additional endonucleases generated fragmentsunique to MLPS samples; A/M (see Figs. 2 and 3) produced a485-kilobase MLPS-specific band (normal band is 70 kilo-bases), and Sacll produced a 20-kilobase MLPS-specific band
7903
Research. on February 19, 2018. © 1990 American Association for Cancercancerres.aacrjournals.org Downloaded from
METHYLATION PATTERNS OF 12ql3 IN MYXOID L1POSARCOMA
A. PROBES Mlu I
I li
B. Not! FRAGMENTS
615—»
515—»I
430-
330—»W *
«.485
-290
150i .et
Fig. 1. GLI locus. /4, a restriction map for portion of the GLI locus, showingcleavage sites for EcoRl (* polymorphic site). Probes pKK36Pl and pKK380consist of l.SS-kilobase Psll and 4.3-kilobase Hindlll fragments of genomicDNA, respectively. Striped bars, approximate position of each probe. B, samplesincluded three lymphoblastoid (LB) cell lines (/, TL8229; 2, TL9542; 3,CL11296), three MLPS (4, 05390; 5, 11172; 6, 13233), and three lipomas (LP)(7, 04009; Ä,04019; 9, 05733). Noti digestion products were resolved usingpulsed field gel electrophoresis. The resulting filter was hybridized with pKK380;identical results were obtained with pKK36Pl.
(normal bands are 40 and 80 kilobases). These fragments uniqueto MLPS samples were observed in four of the five cases studiedand were identical in size and hybridization intensity in each.The fifth MLPS case was missing these unique bands andcontained instead restriction fragments identical to those observed in samples of lipoma, normal fat, and fibroblasts. However, cytogenetic analysis revealed that the culture used forPFG analysis in this case (ST.88-18130; Table 1) had lost allcells with an abnormal karyotype, probably because of overgrowth by normal fibroblasts.
We studied two liposarcomas that were not in the myxoidsubtype (cases ST.88-12407 and ST.89-00151, Table 1), to
determine if the novel restriction fragments were specific toMLPS with t(12;16) (ql3;pll). Both liposarcomas containedrestriction fragments that were identical to the patterns seen inadipocytes (normal and lipoma-derived) and fibroblasts. Theseresults suggested that the novel bands detected in MLPS wereunique to this form of liposarcoma bearing the characteristic
DNA fragments unique to MLPS could have arisen throughseveral mechanisms including chromosome structural changes(translocations, deletions, point mutations, or amplifications),cytosine methylation differences, restriction site polymorphisms, or partial digestions due to suboptimal conditions.Several experiments provided evidence that the novel bands
i IMLPS l LP i LB I X
2x lx|2x 1x'i2x 1x
485-
•290
•70
Fig. 2. Samples of MLPS (ST.88-11172), lipoma (LP; ST.88-06643), andlymphoblastoid (LB; TL9944) cells were digested singly (/x) or doubly (2x) withU/H! to ensure that samples generating novel fragment patterns were digested tocompletion. Each agarose block was sliced in half and both halves digested underour standard conditions (15 units of enzyme, 5 h) with Mini. One sample fromeach pair was washed extensively with Tris-EDTA, placed into a new reactionwith 15 units of fresh Mini, and digested for an additional 5 h. The resultingdigestion products were then resolved, transferred, and hybridized with pKK380.
Mlu
M A F X
Noti
MAP
f
485 • '. -1 »•«
150—»
Fig. 3. Four different tissues from the same patient were analyzed to determineif PFG fragment patterns were specific to tissue type or to patient. Samples wereall obtained from the same patient as MLPS, ST.88-13233. L, lymphoblastoidcell line (CL.88-16940); M, MLPS (ST.88-13233);/I, normal adipocytes (ST.88-16587nf); F, normal fibroblasts (ST.88-16587fb). Agarose blocks containing theDNA samples were digested with Mini or Noll, as indicated, and resolved usingPFG electrophoresis. The resulting filters were hybridized with pKK380.
were the result of very specific differences in cytosine methylation unique to myxoid liposarcoma cells.
Partial digestions due to suboptimal conditions are a frequentproblem with PFG analyses but could be eliminated in this
7904
Research. on February 19, 2018. © 1990 American Association for Cancercancerres.aacrjournals.org Downloaded from

METHYLATION PATTERNS OF 12ql3 IN MYXOID LIPOSARCOMA
analysis for several reasons: (a) the digestion patterns werecompletely reproducible from case to case and in multipleanalyses of the same case; (b) repetition of digestion on samplesresulted in no change in the pattern (Fig. 2), indicating thatcomplete digestions were being obtained under the standardconditions used; (c) the unique fragments seen with Noti andSacll were smaller than the normal fragments and thus couldnot have arisen from partial digestions.
RFLPs were eliminated by examination of four different celltypes (lymphoblasts, MLPS cells, normal adipocytes, and fibro-blasts) from the same patient (case ST.88-13233; Table 1). Allfour cell types from this case displayed primarily the fragmentpatterns expected from the tissue of origin based on our previous results (Fig. 3). The one exception is the absence of a290-kilobase Mlui fragment in the lymphoblastoid cell line(Fig. 3, Lane L) which must be due to a methylation difference.Nevertheless, the MLPS unique bands were not the result ofunusual restriction fragment length polymorphisms presentonly in sarcoma patients.
Physical Map of the GLI Locus. The possibility that thesefragments resulted from the t(12;16) (ql3;pll) was carefullyevaluated through construction of a 600-kilobase physical mapsurrounding the GLI gene. A combination of double and partialdigests was used to analyze the positions of cleavage sites inseveral types of adipocytes including those derived from normaltissue, lipoma, and myxoid liposarcoma. Both normal andlipoma-derived adipocytes generated identical maps (Fig. 4B).
Alterations specific to MLPS samples were localized by themapping process (Fig. 4C). This provided direct proof thatbands unique to MLPS samples were not the result of a trans-location or other structural aberration. Double digestions withSail showed that the Noti and MM sites on the 5' side of GLI
were identical in both normal and MLPS-derived adipocytes.This allowed the positioning of the altered sites for theseendonucleases on the 3' side of GLI. The 485-kilobase Mlul
A.
HTXOIOUPOSAHCOUASPECIFIC
Fig. 4. Physical map of GLI region. A, EcoRl cleavage map of the GLI regionis repeated here to show its relationship to the pulsed field map below. Cleavagesites within pKK.180 for A'rul and Sacll are included. B, long-range restriction
map of the 600-kilobase region surrounding GLI. This map was constructed byselected double digestions of DNA isolated from normal adipocytes and fromMLPS samples. Two large arrows, cleavage sites for A'rul and .S'«clI indicated in
A. Three or more arrows for any endonuclease indicate additional cleavage sitesfor that enzyme deduced from partial digest patterns. C, changes in long-rangemap specific for MLPS samples. Altered fragments (striped bars) and newcleavage sites (arrows) present in MLPS DNA are indicated. Two Mlul cleavagesites are missing in MLPS DNA (x). All other cleavage sites and fragments areidentical between normal adipocytes and MLPS samples and are not shown.Vertical dotted lines, region in which a hypothetical breakpoint (HB) would haveto occur.
fragment unique to MLPS samples could only be obtainedthrough either of two possibilities: (a) a translocation breakpoint occurred within the normal 70-kilobase Mlul fragment;or (b) loss of the Mlul site on the 3' side of this 70-kilobase
fragment occurred (either through point mutation or methylation) to prevent cleavage at this site and fusion of this fragmentwith the next Mlul fragment on the 3' side. Sall/Miul double
digestion yielded a single, normal, 50-kilobase product. A hypothetical breakpoint would have to be located in the 20- to 30-kilobase region (indicated by two vertical dotted lines in Fig. 4,B and C) between the 3' Sail site and the 3' Mlul site. Double
digestions between Sßland several other restriction endonucleases placed the 100-kilobase Sfil fragment as shown in Fig.4B, covering the region where a translocation breakpoint wouldhave to occur. Only a normal 100-kilobase Sfil digestion product has been observed from all the MLPS cases analyzed todate.
The novel 20-kilobase Sacll fragment was shown by the samedouble digest analysis (Fig. 4C) to be completely containedwithin the 100-kilobase Sfil fragment. Thus, two of the threeMLPS-specific fragments must have resulted from changes thatoccurred within the 100-kilobase Sfil fragment, a fragment thatdoes not vary in any of the samples studied.
Double digestion between Noti and Mlul in MLPS DNAestablished one additional important point. Two products resulted; the normal band of 70 kilobases and an additional bandof about 120 kilobases. This second band could only result fromloss of the normal Mlul site and gain of the new Noti site onthe same DNA molecule. This established that the changesobserved with Noil and Mlul occurred on the same chromosome12 homologue. Whether or not this is the translocated homologue remains to be established.
MLPS Specific Fragments Arise from Specific MethylationChanges. The changes observed in cleavage patterns specific toMLPS DNA must have resulted from alterations in restrictionenzyme cleavage sites, but the results could not distinguishbetween point mutations and changes in cytosine methylation.We hypothesized that precise changes in cytosine methylationpatterns were the most likely explanation for these results,primarily because of the great specificity required to independently convert three sequences by point mutation into specificcleavage sites in four MLPS cases. Nevertheless, it is possiblethat these precise changes are a necessary component of themalignant process.
Experimental evidence supporting the methylation hypothesis was sought in two types of experiments, (a) Cells weretreated in culture with 5'-azacytidine, an agent known to block
hemimethylation of newly replicated DNA (37, 38) and thatcan generate cell populations with reduced or altered methylation patterns. Treatment of the lymphoblastoid cell lineTL9542 with 5'-azacytidine altered the Noti fragment pattern
slightly by generation of small quantities of a 150-kilobase Notifragment that appeared similar to the 150-kilobase Noil fragment characteristic of MLPS DNA. (b) DNA isolated fromMLPS- and lipoma-derived adipocytes was tested for short-range methylation differences by cleavage with the isoschizomerpair, Hpall and Mspl, followed by Southern blot analysis usingthe two GLI probes. MLPS cells displayed the lowest methylation level, lipomas an intermediate level, while lymphoblastoidcell lines showed the highest level in the region examined.
DISCUSSION
We have identified a set of changes in restriction endonuclease cleavage sites apparently unique to myxoid liposarcoma
7905
Research. on February 19, 2018. © 1990 American Association for Cancercancerres.aacrjournals.org Downloaded from

METHYLATION PATTERNS OF 12ql3 IN MYXOID LIPOSARCOMA
cells that contain the characteristic t(12;16) (ql3;pll). Thesealterations were observed using probes for the zinc-finger protein locus, GLI, which has been mapped to 12ql3.3, very closeto the MLPS breakpoint. The 100% correlation betweenmyxoid liposarcoma and the t(12;16) strongly suggests thatalterations of one or more genes either on 12 or 16 near thesebreakpoints is a critical event in the development of this sarcoma. By analogy to the translocation t(9;22) in chronic mye-locytic leukemia (39), perhaps MLPS results from a specificrecombination between two genes, one on 12 and one on 16,both of which are required for tumor development. Although(ill was originally considered a likely candidate for one of thesegenes, no changes have been detected in either its DNA structure or transcription in liposarcomas.6 Our results excludetranslocations from a 600-kilobase region surrounding the GLIlocus in both the MLPS and lipoma samples studied here.
The changes in the GLI region specific for MLPS samplesare most readily explained as the result of differences in met h-ylation state at specific cleavage sites. The novel Noti and Sacllfragments are generated by cleavage at sites most likely exposedthrough a demethylation event, whereas the novel Mlu\ fragment is observed because normally accessible Mlul sites areblocked through de novo methylation events. Analysis of twononmyxoid liposarcomas showed that these changes were notcharacteristic of all liposarcomas, but were unique to themyxoid subtype. Myxoid tumors produce and secrete a muco-polysaccharide composed primarily of glycosaminoglycan (40).This substance is believed to be an inappropriate product forthe lipoblasts from which these liposarcomas derive.
Hypomethylation for specific genes has been observed in avariety of malignant tissues when compared with their normaltissue counterparts (11, 18-20). Such changes may be veryimportant in the malignant process because of the known effectof methylation on gene expression. Comparisons between lipoma and myxoid liposarcoma DNA reported here indicatedthat a generalized hypomethylation occurred in the immediatevicinity of GLI in MLPS samples.
Pulsed-field gel electrophoresis provides a unique opportunity to study methylation over great distances along the chromosome. Almost all restriction endonucleases that generatelarge fragments from mammalian DNA recognize and cleaveat sites that contain the rare and frequently methylated dinucle-otide, CpG (41). Since methylation of the target site inhibitscleavage for most rare-cutting enzymes, large fragments observed on PFG Southern blots represent the integration of twostates in the DNA; the presence of a cleavage site and themethylation status ofthat site. Our studies exploited this featureof PFG analysis and have allowed us to draw several importantconclusions. First, the alterations in myxoid liposarcoma DNAwere identical in four cases that retained the (12; 16) translocation. These changes could not be the result of random, generalized demethylation, but rather the product of a very specificand reproducible mechanism controlling methylation status.This is supported by the observation of both demethylation andde novo methylation within the same 600-kilobase region detectable with GLI probes. Second, the patterns of hybridizingbands revealed by our analysis occurred in a highly tissue-specific fashion. This was true for a large number of differentnormal tissue types examined at the same time as the normaland malignant adipose tissues described here.7 This suggeststhat methylation occurs in highly reproducible, tissue-specific
* B. Vogelslein, unpublished observations.1S. Paulien and R. M. Gemmili, manuscript in preparation.
patterns that can extend over at least 600 kilobases.The strong correlation between myxoid liposarcoma with the
t(12;16) and the methylation changes reported here suggest acause-and-effect relationship. Alternatively, the methylationdifferences could reflect the differentiation state of the lipoblasts present in myxoid liposarcomas. This is perhaps unlikelysince the nonmyxoid liposarcomas studied displayed methylation patterns identical to normal and lipoma-derived adipo-cytes.
A cause-and-effect relationship could work in either direction; methylation changes could precede and influence thechromosome rearrangement, or the rearrangement could influence methylation patterns. The first hypothesis was proposedby Goelz et al. (18) as a possible mechanism for the generationof chromosome aberrations in colon cancer. Methylation canapparently have a strong influence on chromosome condensation (42), particularly in centromeric heterochromatin, whichcould result in abnormal mitotic disjunction leading to multiplechromosome abnormalities characteristic of neoplasia (43). Wedo not know if such a mechanism could generate a specificchromosome translocation such as that observed in MLPS.
An intriguing possibility is suggested by the second hypothesis, that the translocation influences the methylation patternfor a large region surrounding the breakpoints. If correct, thiswould be a mechanism that could lead to altered expression fora large set of genes, both on 12 and on 16, depending on howfar these changes extended. We currently have no evidence tohelp choose between these hypotheses.
The concentration of genes in light G-bands and relativedearth of genes in dark G-bands suggests that the translocationbreakpoints in MLPS, both of which occur in light G-bands,can potentially affect many more genes than the translocationin lipoma (which usually occurs in a dark G-band). This couldpotentially explain the malignant versus benign phenotype ofthese two neoplasms.
Analysis of other myxoid sarcomas plus additional cases ofliposarcoma, both myxoid and nonmyxoid, will be necessary todetermine if the changes reported here correlate with themyxoid phenotype or with the t(12;16). These studies shouldhelp to determine if the methylation changes described herewill become a clinically useful tool for the frequently difficultpathological distinction between liposarcoma subtypes.
ACKNOWLEDGMENTS
We wish to thank Drs. David Patterson and Vince Wilson for criticalinput into this work and for helpful comments on the manuscript.
REFERENCES
1. Razin, A., and Riggs, A. D. DNA methylation and gene function. Science(Washington, DC). 210: 604-610, 1980.
2. Doerfler, W. DNA methylation and gene activity. Ann. Rev. Biochem., 32:93-124, 1983.
3. Busslinger, M., Hurst, )., and Flavell, R. A. DNA methylation and theregulation of globin gene expression. Cell, 34: 197-206, 1983.
4. Keshet, I., Yisraeli, J., and Cedar, H. Effect of regional DNA methylationon gene expression. Proc. Nati. Acad. Sci. USA, 82: 2560-2564, 1985.
5. Cedar, H. DNA methylation and gene activity. Cell, 53: 3-4, 1988.6. Dynan, W. S. Understanding the molecular mechanism by which methylation
influences gene expression. Trends in Genet., 5: 35-36, 1989.7. Bird, A. DNA methylation—how important in gene control? Nature (Lond.),
307: 503-504, 1984.8. MacLeod, D., and Bird, A. Transcription in oocytes of highly methylated
rDNA from Xenopus ¡aevissperm. Nature (Lond.), 306: 200-203, 1983.9. McGhee, J. D., and Ginder, G. D. Specific DNA methylation sites in the
vicinity of the chicken B-globin genes. Nature (Lond.), 280:419-420, 1979.10. Waalaijk, C, and Flavell, R. A. DNA Methylation at a CCGG sequence in
7906
Research. on February 19, 2018. © 1990 American Association for Cancercancerres.aacrjournals.org Downloaded from

METHYLATION PATTERNS OF 12ql3 IN MYXOID LIPOSARCOMA
the large intron of the rabbit B-globin gene: tissue specific variations. NucleicAcids Res., 5:4631-4641, 1981.
11. Jones, P. A. DNA methylation and cancer. Cancer Res., 46:461-466, 1986.12. Holliday, R. The inheritance of epigenetic defects. Science (Washington DC),
238:163-170, 1987.13. Konieczny, S. F., and Emerson, C. P. 5-Azacytidine induction of stable
mesoderma! stem cell lineages from IOTI '2 cells: evidence for regulatorygenes controlling determination. Cell, 38: 791-800, 1984.
14. Surani, M. A. H., Barton. S. C., and Morris, M. L. Influence of parentalchromosomes on spatial specificity in androgenetic-parthenogenetic chima-eras in the mouse. Nature (Lond.), 326: 395-397, 1987.
15. Reik, W., Collide, A., Norris, M. L., Barton, S. C., and Surni, A. Genomicimprinting determines methylation of parental alÃelesin transgenic mice.Nature (Lond.), 328: 248-250, 1987.
16. Sapienza, C., Peterson, A. C., Rossant, J., and Balling, R. Degree of methylation of transgenes is dependent on gamete of origin. Nature (Lond.), 328:251-254, 1987.
17. Swain, J. L., Stewart, T. A., and Leder, P. Parental legacy determinesmethylation and expression of autosomal transgene: a molecular mechanismfor parental imprinting. Cell, 50: 719-727, 1987.
18. Goelz, S., Vogelstein, B., Hamilton, S. R., and Feinberg, A. Hypomethylationof DNA from benign and malignant human colon neoplasms. Science (Washington DC), 22«:187-190, 1987.
19. Feinberg, A. P., and Vogelstein, B. Hypomethylation distinguishes genes ofsome human cancers from their normal counterparts. Nature (Lond.), 301:89-92, 1983.
20. Feinberg, A. P., Gehrke, C. W., Kuo, K. C., and Ehrlich, M. Reducedgenomic 5-methylcytosine content in human colonie neoplasia. Cancer Res.,48:1159-1161, 1988.
21. Turc-Carel, C., Dal Cin, P., Rao, U., Karakousis, C., and Sandberg, A. A.Cytogenetic studies of adipose tissue tumors. I. A benign lipoma withreciprocal translocation t(3;12) (q28;ql4). Cancer Genet. Cytogenet., 23:283-289, 1986.
22. Heim, S., Mandahl, N., Kristofferson, U., Mitelman, F., Rööser,B., Ry-dholm, A., and Willen, H. Reciprocal translocation t(3;12) (q27;ql3) inlipoma. Cancer Genet. Cytogenet., 23: 301-304, 1986.
23. Turc-Carel, C., Limon, J., Dal Cin, P., Rao, U., Karakousis, C., and Sand-berg, A. A. Cytogenetic studies of adipose tissue tumors. II. Recurrenttranslocation t(12;16) (ql3;pll) in myxoid liposarcomas. Cancer Genet.Cytogenet., 23: 291-299, 1986.
24. Smith, S., Reeves, B. V., and Wong, L. Translocation t(12;16) in a case ofmyxoid liposarcoma. Cancer Genet. Cytogenet., 26:185-186, 1987.
25. Mertens, F., Johansson, B., Mandhal, N., Heim, S., Rydholm, A., Willen,H., and Mitelman, F. Clonal chromosome abnormalities in two cases ofliposarcoma. Cancer Genet. Cytogenet., 28: 137-144, 1987.
26. Turc-Carel, C., Dal Cin, P., Boghosian, L., Leong, S. P. L., and Sandberg,A. A. Breakpoints in benign lipomas may be at 12ql3 or 12ql4. CancerGenet. Cytogenet., 36: 131-135, 1988.
27. Molenaar, W. M., Dejong, B., Buist, J., Idenburg, V. J. S., Seruca, R., Vos,A. M., and Hoekstra, H. J. Chromosomal analysis and the classification ofsoft tissue sarcomas. Lab. Invest., 60: 266-274, 1989.
28. Schwanz, C., and Cantor, R. Separation of yeast chromosome-sized DNAsby pulse field gradient gel electrophoresis. Cell, 37: 67-75, 1984.
29. Carle, G. F., and Olson, M. V. Separation of chromosomal DNA moleculesfrom yeast by orthogonal-field-alternation gel electrophoresis. Nucleic AcidsRes., 12: 5647-5664, 1984.
30. Gemmili, R. M., Coyle-Morris, J. F., McPeek, F. D., Jr., Ware-Uribe, L. F.,and Hecht, F. Construction of long-range restriction maps in human DNAusing pulsed field gel electrophoresis. Gene Anal. Tech., 4: 119-131, 1987.
31. Kinzler, K. W., Bigner, S. H., Bigner, D. D., Trent, J. M., Law, M. L.,O'Brien, S. J., Wong, A. J., and Vogelstein, B. Identification of an amplified,
highly expressed gene in a human glioma. Science (Washington DC), 236:70-73, 1987.
32. Kinzler, K. L., Ruppert, J. M., Bigner, S. H., and Vogelstein, B. The GLIgene is a member of the Krüppelfamily of zinc-finger proteins. Nature(Lond.), 332: 371-374, 1988.
33. Arheden, K., Rönne,M., Mandahl, N., Heim, S., Kinzler, K. W., Vogelstein,B., and Mitelman, F. In situ hybridization localizes the human putativeoncogene GLI to chromosome subbands 12ql3.3-14.1. Human Genet., 82:1-2, 1989.
34. Dal Cin, P., Turc-Carel, C., Sandberg, A. A., and Van Den Berghe, H. Moreprecise localization of GLI gene by in situ hybridization (Abstract 2536).Cytogenet. Cell Genet., 51: HGM10, 1989.
35. Southern, E. M., Anand, R., Brown, W. R. A., and Fletcher, D. S. A modelfor the separation of large DNA molecules by crossed field gel electrophoresis. Nucleic Acids Res., 15: 5925-5943, 1987.
36. Feinberg, A. P., and Vogelstein, B. A technique for radiolabeling DNArestriction fragments to high specific activity: addendum. Anal. Biochem.,137:266-267, 1984.
37. Jones, P. A., and Taylor, S. M. Cellular differentiation, cytidine analogs andDNA methylation. Cell, 20: 85-93, 1980.
38. Jones, P. A. Altering gene expression with 5-azacytidine. Cell, 40: 485-486,1985.
39. Kurzrock, R., Gutterman, J., and Talpaz, M. The molecular genetics ofPhiladelphia-positive leukemias. N. Engl. J. Med., 319: 990-998, 1988.
40. Enzinger, F. M., and Weiss, S. W. Soft Tissue Tumors, Ed. 2. St. Louis: TheC. V. Mosby Company, 1988.
41. Brown, W. R. A., and Bird, A. P. Long-range restriction site mapping ofmammalian genomic DNA. Nature (Lond.), 322: 477-480, 1986.
42. Schmid, M., Grunert, D., Haaf, T., and Engel, W. A direct demonstrationof somatically paired heterochromatin of human chromosomes. Cytogenet.Cell Genet., 36: 554-561, 1983.
43. Harrison, J. J., Anisowicz, A., Gadi, I. K., Raffeld, M., and Sager, R.Azacytidine-induced tumorigenesis of CHEF/18 cells: correlated DNA methylation and chromosome changes. Proc. Nati. Acad. Sci. USA, 80: 6606-6610, 1983.
7907
Research. on February 19, 2018. © 1990 American Association for Cancercancerres.aacrjournals.org Downloaded from

1990;50:7902-7907. Cancer Res Sylvie Paulien, Claude Turc-Carel, Paola Dal Cin, et al. Chromosome 12Zinc-Finger Gene Mapped to the Breakpoint Region onSite-specific Differences in Methylation Patterns Surrounding a Myxoid Liposarcoma with t(12;16) (q13;p11) Contains
Updated version
http://cancerres.aacrjournals.org/content/50/24/7902
Access the most recent version of this article at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cancerres.aacrjournals.org/content/50/24/7902To request permission to re-use all or part of this article, use this link
Research. on February 19, 2018. © 1990 American Association for Cancercancerres.aacrjournals.org Downloaded from