name answer key - br.caltech.edu · name _____answer key_____ chemistry 25 (spring term 2018)...
TRANSCRIPT

1
Name ______ANSWER KEY_________
Chemistry 25 (Spring term 2018) Midterm Examination
Distributed Thursday, May 3, 2018 Due Thursday, May 10, 2018 by 1 pm in class or by 12:45 pm in 362 Broad
a drop box will be left outside 362 Broad to return midterms during the week ***late penalties enforced***
Conditions • Open this examination when you are ready to take it. • This is a 3 hour examination that must be taken in one continuous stretch. • You may use the Ch25 online lecture notes, problem sets and solutions, the course
web site and a calculator. You may use handwritten notes you have made from thermodynamics texts, including EC and KKW. You may not use any books (including EC and KKW), exams and problem sets from previous years of Ch25 (unless they are posted on the course website), other web sites, non-calculator applications of Mathematica® and related programs, discuss the exam with others, etc.
• This exam should have 19 pages total. • Show your work! All work must be completed in the provided space. Getting the right
answer is not enough – the intermediate steps are needed for credit. useful relationships 1 µm = 10-6 m; 1 nm = 10-9 m; 1 Å = 10-8 cm = 10-10 m; 1 µ = 10-6 m = 103 nm. NA = 6.022 x 1023 molecules mol-1 = Avogadro's number force/energy 1 J = 1 N m = 1 C V; 1 pN = 10-12 N; 1 pN nm = 10-21 J = 0.602 kJ mol-1; 4.184 J = 1 cal. pressure units: 1 pascal (Pa) = 1 N m-2 = 1 J m-3 1atm = 101325 Pa = 0.1013 pN nm-2 = 760 mm Hg = 14.696 lb/sq. in.
= 0.1013 J cm-3 = 101.325 kJ m-3 electrostatics F = 96.487 kJ mol-1 V-1 = 96,487 C mol-1; q = 1.602 x 10-19 Coulombs = F /NA, 1 eV = 1.602 x 10-19 J, =1.1126 x 10-10 C V-1 m-1 gas constant R 8.3144 J mol-1 K-1 = 1.986 cal mol-1 K-1 = 0.08206 liter atm mol-1 K-1 1.38066 ´ 10-23 J K-1 (= R/NA = kB, the Boltzmann constant) unless otherwise stated: you may assume T = 298 K, P = 1 atm RT = 2.48 kJ mol-1 and kBT = 4.11 x 10-21 J at T = 298 K. for water, molecular weight = 0.018 kg mol-1, liquid density = 1000 kg m-3, e = 80 liquid heat capacity = 4.18 J gm-1 K-1 = 75.4 J mol-1 K-1; ∆Hfusion = 6.01 kJ mol-1; ∆Hvap = 40.66 kJ mol-1
4πεo

2
Name________________________________ problem points 1a 2 _______ 1b 2 _______ 1c 3 _______ 1d 3 _______ 2a 10 _______ 2b 5 _______ 3a 7 _______ 3b 3 _______ 4 15 _______ 5a 10 _______ 5b 10 _______ 6a 10 _______ 6b 5 _______ 6c 5 _______ 7a 2 _______ 7b 2 _______ 7c 3 _______ 7d 3 _______ total 100 _______

3
Problem 1. (10 pts) Warming up – a few short question/short answer problems 1a. (2 pts) In 1-2 sentences, what can be concluded about the sign of the entropy change for a spontaneous endothermic reaction under constant T conditions? (ie - is ∆S always positive, always negative, or "it depends" (ie not enough information is given to draw a definite conclusion)?) _____
since ∆H > 0 for an endothermic reaction, ∆S is always > 0 in this case. 1b. (2 pts) In 1-2 sentences, what can be concluded about the sign of the entropy change for a spontaneous exothermic reaction under constant T conditions? (ie - is ∆S always positive, always negative, or "it depends" (ie not enough information is given to draw a definite conclusion)?) _____
since ∆H < 0 for an exothermic reaction, ∆S may be either positive or negative and still satisfy the ∆G < 0 requirement, so "it depends".
∆G =∆ H −T ∆ S < 0 for a spontaneous reaction⇒ ∆ S >∆ H /T
∆G =∆ H −T ∆ S < 0 for a spontaneous reaction⇒ ∆ S >∆ H /T

4
1c. (3 pts) Calculate the difference in the chemical potentials of sucrose in aqueous solution between an initial concentration of 0.01 M and a final concentration of 0.01 mM. Assume ideal solutions with T = 298 K (activity coefficients = 1), and express your answer in kJ mol-1. _____
1d. (3 pts) Calculate the difference in the chemical potentials of sodium chloride in aqueous solution between an initial concentration of 0.01 M and a final concentration of 0.01 mM. Assume ideal solutions with T = 298 K (activity coefficients = 1), and express your answer in kJ mol-1.
∆ µ = µ! + RT lncfinal − µ! − RT lncinitial = RT lncfinalcinitial
= 2.48 ln10−5
10−2 = −17.1 kJ mol−1
∆ µ = µ! + 2RT lncfinal − µ! − 2RT lncinitial = 2RT lncfinalcinitial
= 2 × 2.48 ln10−5
10−2 = −34.2 kJ mol−1

5
Problem 2. (15 pts) Double mutant cycle and salt bridge energetics In lecture 6, we analyzed the energetics of salt bridge formation in a protein using the double mutant cycle for a glutamate-lysine (EK) salt bridge in the protein ubiquitin. This salt bridge was found to stabilize the native state by ~3.6 kJ mol-1. A further test of salt bridge energetics would be to evaluate the repulsive case where two residues with the same charge are introduced into a protein. Data from one such system is summarized below, where residues 6 and 53 are substituted with either alanine (A) or glutamate (E). Tm is the temperature of heat denaturation where the free energy of protein unfolding = 0. 2a. (10 pts) For each variant, determine the change in the free energy of unfolding (∆G˚ in kJ mol-1) relative to the A6-A53 variant and complete the following table. For this calculation, it may be relevant that the enthalpy of unfolding, ∆Hm, of the A6-A53 variant at its Tm is measured to be 150 kJ mol-1. The entropy of unfolding, ∆Sm, measured for this variant may be assumed to be valid for all the variants in this temperature range. 2b. (5 pts) From the ∆G˚ values for these variants, calculate ∆∆G˚, the interaction energy in kJ mol-1 between the E6-E53 residues using a double mutant cycle. Is this value close to what you would expect for an EE interaction based on the results for the EK interaction noted above? Discuss your answer briefly.
variant Tm (˚C) ∆Tm (˚C) relative to A6-A53
∆G˚ (kJ mol-1) relative to A6-A53
A6-A53
38.2 0 0
A6-E53
40.3 +2.1 +1.0
E6-A53
39.6 +1.4 +0.7
E6-E53
32.6 -5.6 -2.7
∆∆G˚ (kJ mol-1)
∆∆G˚ (E6-E53) from double mutant
cycle
-4.4

6
∆Sm = ∆Hm/Tm = 150/(273.15+38.2) = 150/311.4 = 0.482 kJ mol-1 K-1 ∆G˚ = ∆Tm x ∆Sm ∆∆GDMC˚ = ∆G˚E6-E53 + ∆G˚A6-A53 - ∆G˚A6-E53 - ∆G˚E6-A53 = -2.7 + 0 – 1.0 – 0.7 = -4.4 kJ mol-1 A reasonable expectation is that the repulsive EE ion pair should destabilize the native state by about the same amount that the attractive EK ion pair stabilizes the native state; this is what is observed: -4.4 kJ mol-1 for EE (destabilizing) is similar to +3.5 kJ mol-1 (stabilizing) for EK.

7
Problem 3. (10 pts) Third Law Problems 3a. (7 pts) As noted in problem set 2, sulfur exists in two crystalline phases, rhombic and monoclinic. The third law of thermodynamics states that the entropy is zero for a perfectly ordered crystalline phase at 0 K. An interesting test case for the 3rd law is to establish whether or not the rhombic and monoclinic crystal forms of sulfur have the same entropies at 0 K (ie – do they both form perfectly ordered crystals at this temperature?). From the following data, evaluate the entropy change for the transition from the rhombic to the monoclinic crystal forms of sulfur at 0 K, and assess whether or not these two crystal forms have the same entropies at that temperature. The estimated uncertainties in the experimental entropies values are ~0.5 J mol-1 K-1. for the rhombic to monoclinic transition at Tm = 368.5 K, ∆Hm = 401.7 J mol-1. for the rhombic form, the entropy difference S(Tm) – S(0 K) = 36.9 J mol-1 K-1 for the monoclinic form, the entropy difference S(Tm) – S(0 K) monoclinic form = 37.8 J mol-1 K-1 (to be clear, the “S” in the last two sentences refers to “entropy” and not “sulfur”) the entropy change for the rhombic to monoclinic transition is 401.7/368.5 = 1.090 J mol-1 K-1 (see problem 4b, problem set 2)
S(0 K) monoclinic – S(0 K) rhombic = 36.9 + 1.090 – 37.8 = 0.2 J mol-1 K-1 ~ 0 within experimental error (estimated as ~0.5 J mol-1 K-1)
rhombic monoclinic
T=0 K
T = 368.5 K401.7/368.5 = 1.090 J mol-1 K-1
36.9 J mol-1 K-1 37.8 J mol-1 K-1

8
3b. (3 pts) As the preceding problem notes, calorimetric measurements can be used to calculate the entropy changes associated with a transition between two states at 0 K. Glycerol is a substance that can form two types of solid phases, a perfectly ordered crystalline phase and a disordered, metastable glass phase. The entropy difference between these two phases is 19.4 J mol-1 K-1. From the Boltzmann expression for the entropy change,
evaluate Wglass, the number of conformational states per molecule available to glycerol in the glass phase. From the structure of glycerol (HOCH2-CHOH-CH2OH) identify several factors that may contribute to the residual entropy of the glass phase of glycerol. _______________ 19.4 = R ln W W = exp(19.4/R) = 10.3 the residual entropy likely reflects the ability of glycerol molecules to exhibit alternate hydrogen bonding arrangements, alternate hydrogen positions in the hydrogen bonds (like water), and distinct torsion angles.
ΔS = Sglass − Scrystal = kB lnWglassNA − kB ln1
NA = R lnWglass

9
Problem 4. (15 pts) Completely saturated In lecture 6, we calculated that the vapor pressure of water in equilibrium with the liquid phase at 25 ˚C (298 K) is 23.6 mm Hg, from an analysis of the temperature dependence of the chemical potentials for the liquid and vapor phases of water. Since air is saturated with water vapor under these conditions, the vapor pressure of 23.6 mm Hg corresponds to 100 % relative humidity at 25 ˚C. Under these conditions of saturation, the dew point temperature, Td, the temperature below which liquid water starts to condense from the vapor phase, is also 25 ˚C. One fine spring afternoon in Pasadena, the temperature is 20 ˚C and the relative humidity is 55%, meaning the water vapor pressure is measured to be 55% of the value at saturation. What is the dew point temperature, Td, in ˚C, when this water vapor pressure corresponds to saturating conditions? For this calculation, the following thermodynamic information at 298 K may be useful – and you may also assume that the values of H and S are temperature independent and that the vapor phase obeys the ideal gas law.
phase G (kJ mol-1) H (kJ mol-1) S (J mol-1 K-1) liquid -237.2 -285.8 70.0 vapor -228.6 -241.8 189.0
hint: 0 ˚C < Td < 30 ˚C. the T dependence of µ for the gas and liquid phase are shown below, where EF corresponds to the difference in µ at 298 K, CD at 20 ˚C (293 K) and AB at Td.
The equilibrium condition , may be written
µl T( ) = µg T( )µl! T( ) = µg
! T( )+ RT ln P / P!( )µl! T( )− µg
! T( ) = RT ln P / P!( )∆ µ! T( ) = RT ln P / P!( )

10
where P˚ is the reference pressure. The difference in standard chemical potentials at a temperature T (∆µ˚(T)) may be determined from the value of ∆µ˚ at the reference temperature T (298 K):
∆S˚ is the difference in the entropies of the liquid – gas phases ( = -0.119 kJ mol-1 K-1). Hence, the vapor pressure P and temperature T are related by:
At T = 298 K, ∆µ = -8.6 kJ mol-1 (line AB), and P/P˚ = 0.0311 = 23.6 mm Hg = 3150 Pa (derived in class) At T = 20 ˚C = 293 K, ∆µ = -9.2 kJ mol-1 (line CD), and P/P˚ = 0.0229 = 17.4 mm Hg = 2325 Pa 55% relative humidity at 20 ˚C corresponds to P = 0.55x17.4 = 9.59 mm Hg and P/P˚ = 0.0126 Hence at Td (line AB):
Alternatively, the Clausius Clapeyron equation can be used to relate the change in vapor pressure P to the change in T (see PS 3, problem 3). The relevant form can be derived assuming that the volume change associated with the liquid to gas transition is given by the volume of the gas (using the ideal gas law), which can be integrated to give:
Let T1 = 293 K, and T2 = Td; P2 will equal 0.55 P1 or P2/P1 = 0.55. ∆Hvap is a function of temperature; it equals +44.0 kJ mol-1 at 298 K from the table at the start of this problem
A general rule of thumb is that for each 5% drop in relative humidity, the dew point drops by 1˚C = so 55% RH = a drop of 45% from saturation, and the dew point changes by ~9˚C. (this approximation is best when the relative humidity is above 50%)
∆ µ! T( ) =∆ µ! 298K( )+ ∂∆ µ!
∂TT − 298( )
=∆ µ! 298K( )−∆ S! 298K( ) T − 298( ) = -8.6 + 0.119 T − 298( ) kJ mol -1
RT ln P / P!( ) = −8.6 + 0.119 T − 298( )
RTd ln 0.0126( ) = -8.6 + 0.119 Td - 298( )Td =
-8.6 - 0.119 × 298R ln 0.0126( ) - 0.119= 283.6 = 10.6 ˚C
dPdT
=∆ H
vap
!
T ∆V~∆ H
vap
!
RT 2 P⇒ dPP
=∆ H
vap
!
RT 2 dT
ln P2P1
=∆ H
vap
!
R1T1
− 1T2
⎛⎝⎜
⎞⎠⎟
ln0.55 = 44.0R
1293
− 1Td
⎛⎝⎜
⎞⎠⎟
Td =44 × 293
44 − 0.0083144 × 293ln0.55= 283.6 = 10.6˚C

11
Problem 5. (20 pts) (Trying to) feel the heat A hypothetical bacteria senses temperature changes through the temperature dependent binding of a protein, P, to DNA
At T = 0 ˚C, K = 107, while at T = 37 ˚C, K = 104. 5a. (10 pts) From this information, determine ∆G˚, ∆H˚ and ∆S˚ for the binding of P to DNA at these two temperatures. Assume ∆H˚ and ∆S˚ are temperature independent (ie, ∆Cp = 0) – as we’ve seen, this is typically not a good assumption, but it simplifies this analysis.
T K ∆G˚ (kJ mol-1) ∆H˚ (kJ mol-1) ∆S˚(kJ mol-1K-1) 0 ˚C
107 -36.6
-131.9 -0.349
37 ˚C
104 -23.7 -131.9 -0.349
______________ from the K values, ∆G˚ may be calculated from the expression ∆G˚ = -RT ln K -(0.0083144)*273*ln(107) = -36.6 kJ mol-1 -(0.0083144)*310*ln(104) = -23.7 kJ mol-1 ∆S˚ may be calculated from the change in ∆G˚ with T (this is exact since ∆S is taken as temperature independent) ∆S˚ = -∆G˚/∆T = -0.349 ∆H˚ may then be calculated by ∆H˚ = ∆G˚ + T∆S˚ ∆H˚ = -36.6 + 273(-0.349) = -131.9 = -23.7 + 310(-0.349)
P + DNA!KP ·DNA, K =
P ·DNA( )P( ) DNA( )

12
5b. (10 pts) A second bacteria utilizes a different approach to thermoregulation, by fixing the binding constant K for a protein to DNA at 107, independent of temperature. For this system, determine ∆G˚, ∆H˚ and ∆S˚ for the binding of P to DNA at these two temperatures. (again, taking ∆H˚ and ∆S˚ as temperature independent).
T K ∆G˚ (kJ mol-1) ∆H˚ (kJ mol-1) ∆S˚(kJ mol-1K-
1) 0 ˚C
107 -36.6 0 0.133
37 ˚C
107 -41.5 0 0.133
from the K values, ∆G˚ may be calculated from the expression ∆G˚ = -RT ln K -(0.0083144)*273*ln(107) = -36.6 kJ mol-1 -(0.0083144)*310*ln(107) = -41.5 kJ mol-1 since K = constant, ∆H = 0 ∆S˚ may be calculated from the change in ∆G˚ with T: ∆S˚ = -∆G˚/∆T = - -4.9/37 = +0.133 or from ∆S˚ = -∆G˚/T (since ∆H=0) = 36.6/273 = 0.133

13
Problem 6. (20 pts) It’s complex 6a. (10 pts) The statistical definition of entropy as a measure of information was pioneered by Shannon in communication theory. Entropy can be used to quantitate the information of content of DNA by evaluating the number of distinct messages (strings of nucleotides) that can be encoded in a particular DNA sequence. Let’s consider a 40 base pair DNA sequence, with the following compositions: i. 40G, 0C, 0A, 0T ii. 10G, 10C, 10A, 10T iii. no sequence restriction (all possible compositions) For these three sets, calculate the total number of possible sequences and the corresponding entropies in units of kB. Sequences with restricted composition (such as (i)) are referred to as “low complexity” and they are problematic in analyses of protein and nucleic acid sequences since they will exhibit high sequence similarities to other sequences with the same restricted composition through chance and not through phylogenetic relatedness. Entropy calculations can identify these low complexity sequences and remove them from the further analysis. i. W = 40!/(40! 0! 0! 0!) = 1 S = kB ln W = 0 kB ii W = 40!/(10! 10! 10! 10!) = 4.7 x 1021 S = 49.9 kB iii W = 440 S = 55.5 kB

14

15
6b. (5 pts) The fidelity of DNA synthesis and replication is essential for storage of genetic information, but errors and mutations always take place and these randomly introduced genetic variations are crucial for evolution responding to environmental stress. A proof reading process was recruited by DNA polymerases to prevent too many mutations which lead to loss of ancestor genomic information. Suppose that upon synthesizing a molecule of DNA, on average one mutation (wrong base pair) is introduced every 1000 base pairs (Pmutation = 0.001, Pcorrect = 0.999). A 1000 base pair long DNA molecule is synthesized. What are the probabilities of finding 0, 1, or 2 mutations (wrong base pairs) in this molecule? What’s the probability of a DNA product containing more than 2 mutations? The probability of a certain number of errors may be calculated from the binomial distribution:
P 0( ) = 1000!1000!0!
0.999( )1000 = 0.368
P 1( ) = 1000!999!1!
0.999( )999 .001( ) = 0.368
P 2( ) = 1000!998!2!
0.999( )998 .001( )2 = 0.184P > 2( ) = 1.0000 − P 0( )− P 1( )− P 2( ) = 0.08

16
6c. (5 pts) The error rate for incorrect base substitution in DNA replication is measured to be ~10-8 in bacterial systems. One contribution to the incorporation of incorrect bases into DNA can arise from errors in hydrogen bond formation between the two bases in a replicated base pair. In this case, the error frequency would reflect the differences in free energies between the formation of complementary and non-complementary base pairs. In aqueous solution, these differences are estimated to be ~ 10 kJ mol-1. What would you estimate the error rate for DNA replication to be if the only mechanism was due to mistakes in base-base hydrogen bonding? This type of analysis helped highlight the importance of proof-reading mechanisms to ensure the high fidelity replication of DNA. the frequency of mis-incorporation of base pairs into DNA will be related to the difference in ∆G (10 kJ mol-1) between the incorrect and correct basepair, or
As this is ~106 x larger than the observed error frequency, base pair discrimination on the basis of hydrogen bonding energies cannot account for the observed fidelity and additional proof-reading mechanisms must be involved.
K = exp −∆G˚RT
⎡⎣⎢
⎤⎦⎥= exp − 10
2.48⎡⎣⎢
⎤⎦⎥~ 0.02

17
Problem 7. (10 pts) Cooling down – a few last short question/short answer problems 7a. (2 pts) A filled rubber balloon rapidly deflates – what happens to the temperature of the rubber during this process (does it increase, decrease or stay the same?) as the in-class demonstration demonstrated, the temperature decreases 7b. (2 pts) You measure the activity coefficient of a component in a binary mixture at a particular composition and find it to be 3.7. You report this result to professors A, B, and C, who respond A. “Just as I expected, the solution is non-ideal!” B. “Just as I expected, the solution is ideal!” C. “I don’t understand what is going on, you need to collect data at additional compositions” Briefly (1-2 sentences) explain which professor is correct, and why (and, don’t worry, we won’t tell the other 2 that you think they are wrong). C = need to know how g changes with composition – if it changes with composition, the solution is non-ideal, but if g is a constant, it could be an ideal solution, but with an unusual choice of standard state convention.
18
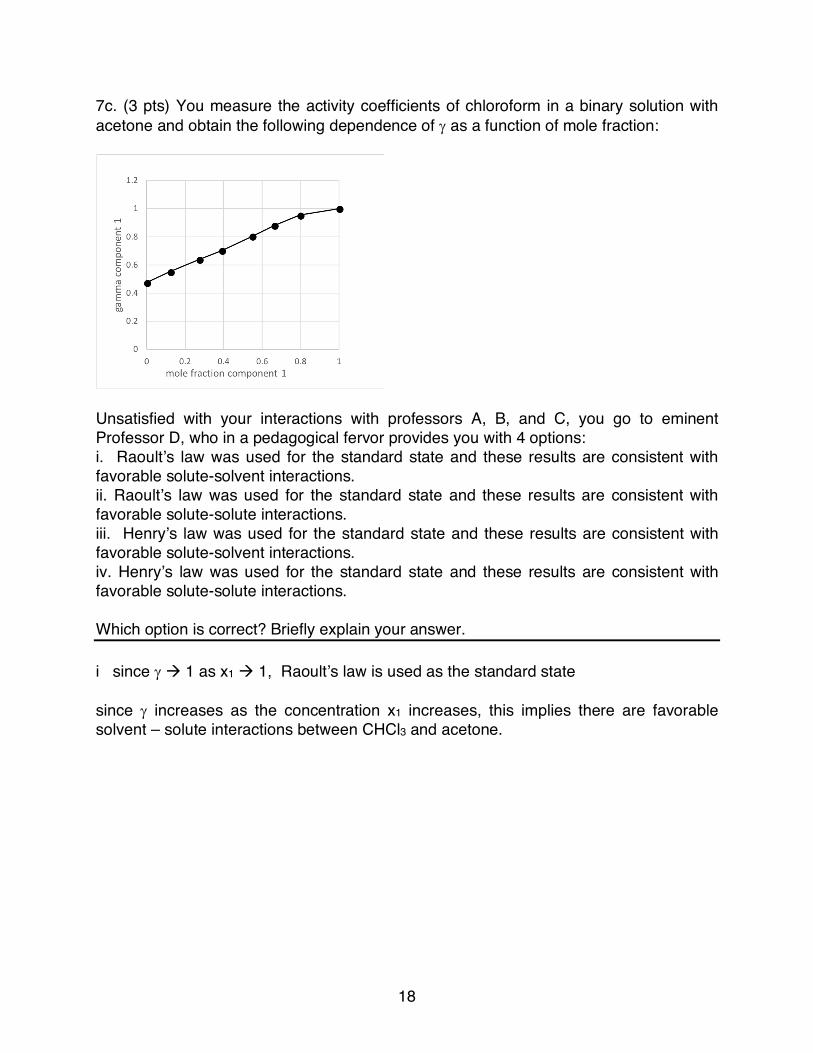
7c. (3 pts) You measure the activity coefficients of chloroform in a binary solution with acetone and obtain the following dependence of g as a function of mole fraction:
Unsatisfied with your interactions with professors A, B, and C, you go to eminent Professor D, who in a pedagogical fervor provides you with 4 options: i. Raoult’s law was used for the standard state and these results are consistent with favorable solute-solvent interactions. ii. Raoult’s law was used for the standard state and these results are consistent with favorable solute-solute interactions. iii. Henry’s law was used for the standard state and these results are consistent with favorable solute-solvent interactions. iv. Henry’s law was used for the standard state and these results are consistent with favorable solute-solute interactions. Which option is correct? Briefly explain your answer. i since g à 1 as x1 à 1, Raoult’s law is used as the standard state since g increases as the concentration x1 increases, this implies there are favorable solvent – solute interactions between CHCl3 and acetone.

19
7d. (3 pts) The pKa value for the amino acid aspartic acid is ~4 in water. A variant form of a protein is produced that has an aspartic acid side chain buried in the non-polar interior of a protein. Do you expect a change in the pKa of the aspartic acid side chain, and if so, do you expect it to increase or decrease? Explain your answer briefly. it is harder to ionize a charged group in a nonpolar environment. Therefore, the pKa of aspartic acid will increase when this residue is introduced into a buried, non-polar environment. A higher pKa means that the acid dissociation constant is smaller and it is harder to ionize the aspartic acid side chain.