natural hybridization between the iberian hare (lepus ... · (i universidade do porto natural...
TRANSCRIPT

•.V \
PORTO T- FACULDADE DE CIÊNCIAS
( i UNIVERSIDADE DO PORTO
Natural hybridization between the Iberian hare (Lepus granatensis) and the brown hare (L. europaeus)
in northern Iberian Peninsula
Hélder Marques de Freitas
Porto 2006

> \ \ ^ ^
"\

' f i l l I F / ^ l Biblioteca ^BR I ■ I Faculdade da Ciências
v a u - / I y _ _ Universidade do Porto

PORTO T- FACULDADE DE CIÊNCIAS
g * UNIVERSIDADE DO PORTO
Natural hybridization between the Iberian hare (Lepus granatensis) and the brown hare (L. europaeus)
in northern Iberian Peninsula
!
l\f& jr.
Dissertação de Mestrado em Biodiversidade e Recursos Genéticos apresentada à Faculdade de Ciências da Universidade do Porto
Hélder Marques de Freitas
Porto 2006

A memória do meu Avô

Many thanks to
I am thankful to several people that contributed in various ways to this work, and I
would like to express here my gratitude.
To Dr. Pierre Boursot I would like to thank the way I was received in the GPIA,
in Montpellier and also the great help he gave me, both in some laboratorial procedures
and in more theoretical fields, with valuable discussions!
To Barbara Dod, for her sympathy, friendship and fantastic group moonlight
dinners!
To people "chez Zef', whose affection, friendship and good mood made the
adaptation to Montpellier really easy!
To Stuart Baird, for the hospitality, for the bright conversations in the varanda,
for the good coffee and, specially, for being such a good friend!
Agradecimentos
Ao longo deste trabalho, várias foram as pessoas que, directa ou indirectamente, me
ajudaram e a quem quero expressar o meu agradecimento.
Ao meu orientador, Prof. Dr. Paulo Célio Alves, quero agradecer a oportunidade
que me deu de integrar o seu grupo de investigação, tendo-me proporcionado sempre as
melhores condições possíveis. O seu apoio, a disponibilidade para ajudar e a liberdade
que me concedeu para explorar novas ideias foram muito importantes no decorrer do
trabalho desta tese.
Ao Zef, o meu muito obrigado por tudo! Pela paciência, pela ajuda, pelo apoio,
por ter sido um crítico sempre atento ao longo de todo o trabalho, por ter sido o meu
tradutor nos primeiros tempos em França.
Ao Teixas agradeço por me ter acompanhado quando cheguei ao CIBIO, por ter
sido um óptimo professor das regras e protocolos laboratoriais e pelas tardes de sábado
que dispensou para me ajudar.

Ao Esteves, pela partilha de ideias, pelas discussões sobre os mais variados
temas, por me aturar em dias de "telha", também o meu obrigado!
Ao Rui, pelas muitas e longas discussões, pela companhia em algumas noitadas
e pela grande disponibilidade para ajudar, muito obrigado!
A Catarina Pinho, minha colega de gabinete nos últimos tempos, pela partilha de
ideias, artigos e opiniões sempre válidas, obrigado!
Ao Pedro Tarroso agradeço a boa disposição com que sempre me aturou nas
várias ocasiões em que precisei da sua ajuda.
Ao Sequeira, um muito obrigado pelo apoio que me deu desde o início, pelo
acompanhamento exaustivo em determinadas fases do trabalho, pela partilha de ideias e
perspectivas diferentes e pelo imenso tempo que dispensou em sugestões e opiniões!
A Bá, pela sua aturada gestão do laboratório e pelo seu bom humor, obrigado!
A Raquel Vasconcelos agradeço a organização e asseio do laboratório, o que não
é nada fácil!
A Sara Ferreira e à Sandra Rodrigues o meu muito obrigado por manterem
sempre tudo, complexamente, na ordem!
A todas as pessoas do CIBIO, por promoverem um óptimo ambiente de trabalho,
assim como uma atmosfera científica muito saudável!
A Alexandra Dias, da Vereda das Artes, por tão bem ter tornado real a ideia para
a capa desta tese, muito obrigado!
Ao Moura agradeço a ajuda preciosa que me deu para criar algumas imagens!
Ao Ibon Telletxea agradeço a simpatia com que me recebeu em Vitoria, assim
como todas as amostras que disponibilizou e cuja organização é invejável!
Agradeço ainda ao Dr. Franz Suchentrunk, Dr. Ettore Randi, Dr. Christian
Gortazar, Dr. Rafael Villafuerte, Dr. Diego Villanua e Dr. Miguel Delibes-Mateos pela
ajuda nas campanhas de amostragem.
Finalmente, agradeço à minha família, especialmente aos meus pais, e à
Mariana, cujo apoio foi e é sempre indispensável!

Index
Resumo i
Summary iv
I. General Introduction 1 1) The occurrence of natural hybridization 1 2) The origin of hybrid zones and the influence of climate 3 3) Taxonomy of hares 6 4) Hares in the world 8 5) Hares in Europe and in the Iberian Peninsula 8 6) Hybridization in hares 9 7) Ecological requirements of the Iberian and the brown hare 12 8) Objectives and thesis organization 13
II. Paperl 15 Evidence of natural hybridization between Lepus granatensis and L. europaeus
in northern Iberian Peninsula
III. Paper2 35 Finescale analysis of a hybrid zone between Iberian and brown hare: integration
of genetical and ecological data
IV. General Discussion 51 1) Genetic structure 52 2) Natural hybridization in hares 54
2.1) Patterns revealed by microsatellites 54 2.2) Patterns revealed by mtDNA 56 2.3) Discordance between mtDNA and microsatellites 57
3) Association between genetic and ecological data 60 4) The use of new analytic methodologies 62 5) Future perspectives 63
V. Conclusion 65
VI. References 67

Resumo A hibridação interespecífica é um fenómeno há muito conhecido que se mantém no topo
do interesse em trabalhos de evolução, constituindo um importante modelo de estudo
para a compreensão de fenómenos fundamentais como, por exemplo, a especiação. As
oscilações climáticas são reconhecidas como tendo sido um importante catalisador para
a ocorrência de zonas de contacto. Na Europa, as regiões localizadas a sul (as
Penínsulas Ibérica, Itálica e os Balcãs) funcionaram como reservatórios de
biodiversidade durante as épocas menos favoráveis à ocorrência de vida no resto da
Europa e, à medida que as condições melhoraram, tornaram-se uma fonte para a
recolonização do continente europeu. A colonização para norte a partir destas
penínsulas promoveu o contacto entre espécies e, em alguns casos, conduziu à formação
de zonas híbridas.
O norte da Península Ibérica é uma área onde estão descritas zonas de contacto
secundário para diversos organismos. Entre estes, encontram-se duas espécies de lebre:
a lebre ibérica (Lepus granatensis), que representa um endemismo da Península Ibérica,
e a lebre europeia (L. europaeus), que ocupa o nordeste da Península Ibérica, bem como
uma grande extensão do restante continente europeu. Embora as rotas de expansão pós-
glacial de lebre europeia não sejam bem conhecidas, admite-se que a colonização para
Noroeste do continente europeu tenha tido origem nos Balcãs. Assim, a zona de
contacto secundário com a lebre ibérica ter-se-á formado recentemente aquando da
chegada da lebre europeia à Península Ibérica, encontrando aí a já presente lebre ibérica.
Não obstante a existência de dados que comprovam o cruzamento entre lebre variável
(L. timidus) e lebre europeia, a hibridação entre esta e a lebre ibérica nunca foi
demonstrada, apesar da existência de uma extensa zona de contacto entre estas duas
espécies no norte da Península Ibérica. Investigar se as duas espécies se cruzam com
sucesso foi o principal mote deste trabalho. Para tal, foram escolhidos marcadores
nucleares neutros (microssatélites) e o gene mitocondrial do citocromo b. Os
microssatélites utilizados provaram ser suficientemente informativos para distinguirem
as espécies em questão e permitiram a detecção de quatro indivíduos de lebre ibérica
com sinais de lebre europeia ao nível do genoma nuclear. O contrário também ocorre,
tendo-se encontrado três indivíduos de lebre europeia com introgressão nuclear de lebre
i

ibérica, embora as provas neste caso sejam menos robustas. Quanto ao DNA
mitocondrial, a análise a uma escala fina dum grande número de indivíduos das duas
espécies permitiu confirmar a já descrita forte introgressão de DNA mitocondrial de
lebre variável nas duas espécies de lebre da Península Ibérica estudadas, e ainda detectar
um considerável nível de introgressão mitocondrial de lebre ibérica para lebre europeia
(-33%). Em conjunto, os dados dos microssatélites e do DNA mitocondrial
demonstraram a existência de hibridação entre lebre ibérica e europeia. Os diferentes
padrões de introgressão observados ao nível do DNA nuclear e mitocondrial poderão
estar associados a diferenças comportamentais entre as espécies e mesmo entre os
sexos, podendo também ser explicados por hipóteses demográficas.
Os padrões de diversidade genética obtidos parecem corroborar o cenário de
uma colonização Este-Oeste por parte da lebre europeia, uma vez que é detectada uma
diminuição na diversidade genética das populações nessa direcção. Quanto à lebre
ibérica, os baixos valores de Fst observados parecem suportar a hipótese da ocorrência
de um fluxo génico elevado entre as populações, num cenário de panmixia. As
populações de lebre ibérica poderão ter-se mantido estáveis ao longo do tempo, sem
sofrerem reduções demográficas severas nem regressões para sul durante os períodos
glaciais.
Quer a lebre ibérica, quer a lebre europeia, são muito adaptáveis às condições do
habitat mas existem diferenças nos seus requisitos ecológicos que são confirmadas pelos
dados obtidos. O uso integrado de dados genéticos e ecológicos permitiu verificar que a
lebre europeia demonstra uma preferência por áreas relativamente mais húmidas e frias
em comparação com a lebre ibérica. No ecótono estudado, uma clara transição
climatológica define os limites de distribuição de ambas as espécies e é acompanhada
pelos dados do genoma nuclear. Este resultado poderá indicar um papel selectivo do
habitat na distribuição das espécies, o que torna este sistema muito interessante para
uma futura modelação recorrendo a Sistemas de Informação Geográfica. Por outro lado,
o padrão distinto encontrado para o DNA mitocondrial deverá ser estudado em maior
detalhe, nomeadamente a hipótese de selecção pelo frio para a mitocôndria da lebre
variável.
11

Este trabalho apresenta dados novos no que diz respeito à lebre ibérica e à lebre
europeia, quer no que diz respeito à sua interacção, quer no que diz respeito à sua
relação com o habitat. A mistura de genoma nuclear e uma nova introgressão
mitocondrial em lebre europeia (no norte da Península Ibérica, a lebre europeia
apresenta o seu DNA mitocondrial nativo e ainda duas linhagens introgredidas),
detectadas neste trabalho, poderão servir de base para estudos mais detalhados e para o
desenvolvimento de novas hipóteses de trabalho. O género Lepus continua a mostrar ser
um desafio para os estudos de evolução, apresentando-se como um excelente modelo de
trabalho.
111

Summary Interspecific hybridization is a long known phenomenon, and is still a matter of great
interest, constituting an important model for the understanding of fundamental
phenomena such as speciation. Climate oscillations were an important factor for the
occurrence of contact zones. In Europe, southern regions as the Iberian, Italian and
Balkans Peninsulas were biodiversity sinks during less favourable periods, and became
sources of life when conditions improved, allowing the recolonization of the European
continent. The northwards colonization routes promoted species contact and,
sometimes, hybrid zones were established.
The north of Iberian Peninsula is an area where contact zones are described for
several organisms. Among these, there are two species of hares: the Iberian hare (Lepus
granatensis), which is an endemism of Iberian Peninsula, and the brown hare (L.
europaeus), that inhabits the northeastern part of Iberia, as well as the majority of the
remaining European territory. Although post-glacial expansion routes of brown hare are
not well known, it is thought that the Northwest colonization of the European continent
was originated in the Balkans. Thus, the secondary contact with the Iberian hare
probably occurred recently, when brown hare arrived to the Iberian Peninsula. Although
it is known that the brown hare naturally hybridizes with the mountain hare (L. timidus),
hybridization between Iberian and brown hare has not been demonstrated, despite the
existence of a long contact zone between both species in northern Iberian Peninsula. To
investigate if successful interbreeding between Iberian and brown hare occurs was the
main aim of this work. For that, neutral nuclear markers (microsatellites) and the
mitochondrial gene cytochrome b were used. The chosen microsatellites set proved to
be informative and to distinguish species in a frequency base, allowing the detection of
brown hare alleles introgressed in four Iberian hare individuals. The converse also
occurs, with three brown hares displaying Iberian hare nuclear introgression, although
the evidences for these are weaker. As for the mitochondrial DNA, a thin scale analysis
of large samples of both Iberian and brown hare enabled to confirm the previously
described strong introgression from mountain hare haplotypes into the two iberian
species of hares studied, and allowed the detection of a considerable amount (-33%) of
introgression from Iberian hare into brown hare. Together, microsatellites and
IV

mitochondrial DNA data strongly support the existence of hybridization between the
two taxa. The discordance of introgression patterns between nuclear and mitochondrial
markers may be explained by behavioural differences between species and between
sexes or demographic hypothesis.
From the obtained data of genetic diversity, the scenario of East-West
colonization by the brown hare seems plausible, given the decrease in samples genetic
diversity along that axis. As for the Iberian hare, low values of pairwise Fst point to a
high gene flow between samples and the existence of a metapopulation, fitting a
panmixia scenario. Iberian hare populations may have been stable throughout time,
without severe demographic decreases or population regression to south during glacial
periods.
Iberian and brown hare are very adaptable to habitat conditions, but differences
in ecological requirements exist and are confirmed by the data obtained. The integration
of genetical and ecological data allowed to verify that the brown hare displays a
preference for relatively wetter and colder areas in comparison with Iberian hare. In the
studied ecotone, a sharp climatological transition marks species distribution limit and is
accompanied by the nuclear genome data. This result may indicate a selective role of
the habitat in species distribution, which is a very interesting system for future GIS
modelling. The distinct pattern found for mitochondrial DNA should also be further
investigated, namely the hypothesis of selection for the cold of the mountain hare
mitochondria.
This work brings new knowledge about Iberian and brown hare, the interaction
between them and the interaction of both species with the habitat. Nuclear genome
exchanges and another mitochondrial introgression into brown hare (in northern Iberia,
this species displays its native mtDNA and another two introgressed lineages) were
found and may constitute the basis for more detailed works with new working
hypothesis. The genus Lepus continuously proves to be a challenge to evolutionary
studies and much may still be learnt with it.
v

Chapter I
I. General introduction
A long way has gone since all the organisms were seen in the light of creationism, with
species regarded as static entities with no evolutionary history and no correlation
between them. With the work of geneticists as R.A. Fisher, J.B.S. Haldane and S.
Wright, the Synthetic Theory of Evolution was born around 1930, fusing Darwin's
Theory of Natural Selection with Mendel's Theory of Heredity. With the establishment
of these keystone concepts of evolution, a whole new world of ideas and experiments
were based on them. Species were no more simple objects for description, ordering and
collection, but were now dynamic units, with common features and possibly shared
evolutionary paths. Speciation left the pure taxonomical field and started to integrate
ecology, etology, physiology and genetics, for example. Nonetheless, the achieved
broadness of knowledge did not prevent the birth of controversies. Species concepts,
for instance, have been debated thoroughly (for a review, see Hey et al, 2003) but no
unifying concept has been attained. Even so, the Biological Species Concept (BSC;
Dobzhansky, 1937; Mayr, 1942) might have passed the test of time (Ridley, 1996) and
is upon it that most studies of speciation are based. This means that speciation works
are, in fact, an analysis of reproductive isolation (Coyne & Orr, 1998; Turelli et al,
2001), since the BSC advocates that "species are groups of interbreeding natural
populations that are reproductively isolated from other such groups". However, species
limits are not so well defined in natural populations, and hybridization is certainly a
challenging phenomenon to the BSC, since it blurs species boundaries.
1) The occurrence of natural hybridization
Natural hybridization has for long puzzled evolutionary biologists. Several definitions
of hybridization have been applied in scientific literature, but all of them relate to the
levels of divergence between the individuals that undergo hybridization (Arnold, 1997).
This author defines natural hybridization as a phenomenon that "involves successful
1

General Introduction
matings in nature between individuals from two populations, or groups of populations,
which are distinguishable on the basis of one or more heritable characters", where
successful matings stands for the production of some viable Fl progeny that possess
some level of fertility, even if both the number of offspring and their fertility level is
reduced. This last scenario (reduced number of offspring and level of fertility) led
several authors to minimize the importance of natural hybridization as a evolutionarily
process, but as noted by Barton & Gale (1993), "if even a small proportion of hybrids
reproduce, all the individuals in the vicinity of the hybrid zone eventually carry
introgressed alleles in some of their genes". Moreover, the occurrence of infrequent
events of hybridization may lead to new evolutionary lineages (Arnold, 1997), which
stresses the need to account for natural hybridization when studies of speciation or
population genetics are performed. This idea is strengthened by the fact that natural
hybridization occurs in a wide variety of organisms and has indeed been found to be
surprisingly more common than previously thought (Hewitt, 1988; Mallet, 2005).
Among plants, natural hybridization seems frequent: Grant (1981) suggested that the
majority of plant species were hybrids; Knobloch (1972) listed 23675 possible hybrids
among angiosperm species (although including artificial hybrids); Arnold (1994)
considered that at least 50% of the angiosperms could have a hybrid origin; and from a
review of five biosystematic floras, Ellstrand et al. (1996) calculate that there could be
27500 hybrid combinations among 250000 described plant species. So, even if
estimates of natural hybridization incidence in plants vary greatly (Otto & Whitton,
2000) and even if these phenomena are taxonomically unevenly distributed (only 16%
to 34% of plant families and 6% to 16% plant genera have one or more reported hybrid
- Rieseberg, 1997), natural hybridization seems to be of tremendous importance as a
source of variability and biodiversity.
As for animal taxa, "hybrid speciation" is not as common as seen in plants
(Turelli et al, 2001; Otto & Whitton, 2000). This may stem from the fact that
polyploidy, apparently the major characteristic driving to this kind of speciation is
much rarer in animals (Otto & Whitton, 2000). Nevertheless, natural hybridization
among animals seems to be abundant (Mallet, 2005), as attested by several works.
Species of North American freshwater fish (Hubbs, 1955) or birds (Mayr & Short,
2

Chapter I
1970) are for long known to hybridize in nature, and data from several other taxa is
accumulating. Concerning mammals, the deer (Carr et al, 1986), the house mice
(Boursot et al., 1993), the bear (Taberlet et al, 1994), the pocket gopher (Ruedi et al,
1997), the hedgehog (Santucci et al, 1998) or wild and domestic cats (Pierpaoli et al,
2003) are just some examples of the existence of natural hybridization.
Hybridization has also a geographic component. Species have a natural range of
distribution and, in some areas, transitions to another forms occur. These contact areas
define species limits where hybridization may happen, giving rise to hybrid zones.
Presently, a hybrid zone is commonly accepted as an area of interaction of genetically
distinct groups of individuals, where some admixed offspring might occur, still being
possible to find parental or pure individuals outside of that same area (Harrison, 1990).
Several models exist that predict the evolution of hybrid zones, where selection and/or
dispersal play major roles (Arnold, 1997). Among them, the Mosaic model (Rand &
Harrison, 1989) and the Tension zone model (Barton, 1979; Barton & Hewitt, 1985) are
the most commonly invoked conceptual frameworks to understand and describe hybrid
zones. Nevertheless, the Tension zone model, object of intensive theoretical work
(Barton & Hewitt, 1985; Barton & Gale, 1993; Piálek & Barton, 1997), became
predominant and has been applied throughout the years (e.g. Szymura & Barton, 1991;
Phillips et al, 2004; Bozikova et al, 2005).
2) The origin of hybrid zones and the influence of climate
The origin of hybrid zones is a major issue in evolutionary biology, with primary or
secondary contact as hypotheses. A hybrid zone of primary origin requires the
establishment of gradual clinal variation in a continuous distribution of populations,
with the maintenance of their distribution subsequent to differentiation. On the other
hand, a steep cline will arise from the contact between two allopatrically differentiated
populations, defining a hybrid zone of secondary intergradation (Hewitt, 1988;
Harrison, 1990). If some, as Mayr (1942), only considered allopatric speciation and
secondary contact as cause of hybrid zones, it has been theoretically demonstrated that
3

General Introduction
speciation in continuous populations may occur (Slatkin, 1973; Endler, 1977). The
study of the marine snail Littorina saxatilis in Galicia, Spain (RóIan-Alvarez et al.,
1997) or Sweden (Panova et al., 2006) and the Nicaraguan crater lake cichlid fish
(Barluenga et al., 2006) are some of the few examples supporting hybrid zones of
primary origin found in animals, while accumulating evidence favour secondary contact
as the path driving to the majority of the known hybrid zones (Hewitt, 2001).
Although other factors might be involved, climate has been of main importance
for the establishment of hybrid zones. Coherent explanations for genetic patterns and
species distributions come from palaeoclimatic studies (Hewitt, 2000), that unravelled a
series of major ice ages during the Quaternary period of the geological time (from 2,4
million years to present), characterized by the advancement and recession of ice sheets.
These events occurred at a global scale but were felt differently across the globe due to
regional differences in landform, ocean currents and latitude (Hewitt, 2000). In the
northern hemisphere, the impact of glaciations was so profound that almost all the
fauna and flora north of the equator was affected. Virtually no phylogeographic work of
northern hemisphere organisms ignores the effect of glaciations in the present day
distribution of species, with consequences in their genetic pool due to repeated
expansions into new habitats accompanying climate amelioration, and recessions to
glacial refugia when conditions were less favourable (Taberlet et al., 1998; Hewitt,
2000).
Europe has been mostly covered with ice during glaciations. However, a
privileged southern position of the Iberian, Italian and Balkanic Peninsulas rendered
these geographic areas reasonable conditions for the maintenance of biodiversity during
ice ages (Figure 1) and, from there, species of plants and animals recolonized Europe
when conditions improved (Taberlet et al., 1998). The colonization patterns evidenced
by many species reflect the southerly position they occupied during unfavourable
conditions, and the subsequent expansion to north, east or west they followed.
Interestingly, several species display coincident colonization routes, despite the
existence of differences in ecological requirements and dispersal.
4

Chapter I
/<7\ m /álilB Í /̂ :¾%¾¾ lJ§§ lillS;!'
ff**Ê& ̂ 1^¾¾¾.¾^%^ *^tojjrfc] jpipr
t̂fSlS™ "^^/^j / ^ / í®/i ^ ^ Î C
3i e <*»•
Figure 1: Extension of ice sheets in Europe (in blue) during the last cold period, 20000-18000 years ago. The scaled line indicates the southern limit of the permafrost. Rl, R2 and R3 correspond to the Iberian, Italian and Balkans refugia areas, respectively. Adapted from Taberlet etal., 1998.
As Europe was recolonized, closely related species that were trapped by ice
sheets in separate geographic areas became into contact, and contact zones were
created. The coincidence of species dispersal and, mainly, the existence of natural
barriers to dispersion (e.g. mountain chains, rivers) led to the overlapping of hybrid
zones of several organisms in the same area, which might still be seen. Three of the
consensually considered strips of hybridization in Europe are placed in the exit of each
of the referred southern peninsulas (Figure 2; Hewitt, 2001).
Figure 2: General position of well-known hybrid zones placed in the exit of southern refugia, adapted from Taberlet et al., 1998 and Hewitt, 2000.
5

General Introduction
These hybrid zones harbour a wide number of species and may be seen as suture
zones, described by Charles Remington (1968) as regions where many hybrid zones
from different species are located. Focusing on the Iberian Peninsula, its northern area
around the Pyrenean foothills displays a pattern that is consistent with this scenario.
Even if the Pyrenees were less of a barrier than the Alps to the northwards expansion of
species (Hewitt, 2001), they were probably enough to delay the exit of species from the
Iberian Peninsula and provided the opportunity for the contact with expanding species
from Eastern Europe. This has reportedly happened with species such as the alder
(Alnus glutinosa; King & Ferris, 1998), the crested newt (Triturus cristatus; Wallis &
Artzen, 1989) or the meadow grasshopper (Chorthippus parallelus; Szymura et al,
1996), among others. From this evidence, it is reasonable to think that more species
following similar colonization routes established contact zones, where some degree of
admixture may occur. This is the case of two hare species, Lepus granatensis and Lepus
europaeus, in Northern Iberian Peninsula.
3) Taxonomy of hares
Hares belong to the order LAGOMORPHA. This order was recognized in 1912, when a
review by Gridley separated lagomorphs from rodents (order RODENTIA), to which
they were previously allocated. The distinction is based in some morphological
characters, as the presence of a second set of incisors teeth (named peg) behind the
upper front incisors in lagomorphs. An elongated rostrum of the skull (Flux &
Angermann, 1990) and the presence of a leporine lip are other characteristic anatomical
traits of LAGOMORPHA in comparison with RODENTIA.
Within LAGOMORPHA, two families are currently recognized,
OCHOTONIDAE and LEPORIDAE (Figure 3). While the former is a monotypic
family harbouring the genus Ochotona (pikas), the latter contains eleven genera, divided
in true rabbits (ten of the eleven genera) and true hares (genus Lepus).
6

Chapter I
LAGOMORPHA
LEPORIDAE
Rabbits Brachylagus Bunolagus Caprolagus Nesolagus Oryctolagus Pentalagus Poelagus Pronolagus Romerolagus Sylvilagus
Figure 3: Families and genera of the order LAGOMORPHA (adapted from Flux & Angermann, 1990).
The species of the family LEPORIDAE have long hind legs and large movable
ears, being adapted to quick movement and flight from danger, as well as large eyes,
suited to their crepuscular and nocturnal habits (Flux & Angermann, 1990). The genus
Lepus comprises jackrabbits and hares, with some controversy in the number of species.
According to Flux & Angermann (1990), this monophyletic genus has 29 species, but
depending on the authors, the number varies between 18 and more than 30. A more
recent survey by Wilson & Reeder (2005) considers the existence of 33 hare species.
The genus Lepus is very homogeneous in cytogenetic characteristics, with every species
displaying 2n=48 chromosomes and identical G-banded karyotype (Robinson &
Matthee, 2005). Indeed, the level of variability in cytogenetic markers is generally very
low in the LEPORIDAE, suggesting a fast expansion of this family. Within
LAGOMORPHA, the most exclusive characteristic of hares is the fact that their young
are born fully furred, with their eyes open and ready to move within minutes (Corbet,
1983). Comparing with rabbits, this feature leads to a difference in behaviour, since
rabbits build nests or elaborate warrens in order to protect their young while hares will
only use a shallow depression (Flux & Angermann, 1990).
OCHOTONIDAE
_ , , Jackrabbits and Hares Ochotona Lepus
7

General Introduction
4) Hares in the world
Lepus is a widely distributed genus. In fact, hares are the lagomorphs with the most
widespread natural distribution, occurring in North and Central America, Europe, Africa
and Asia (Flux & Angermann, 1990). In general, hares are open country grazers, and so
have benefited from habitat changes caused by traditional agriculture. Hare species fit
Bergmann's rule, which states that animals tend to be larger in colder regions, i.e. in
higher latitudes, presumably for reasons of thermoregulation. Another latitude-related
characteristic is the fur coloration. The alpine or northern species of hares change from
a darker colour to white in the winter, while the rest of the species have relatively
similar "agouti" colorations in various shades of brown on the back and white or pale
buff below (Flux & Angermann, 1990).
Hares constitute a fundamental ecological resource, since they are an essential
food supply for several carnivores (e.g. the red fox - Schmidt et al., 2004), and some
species are important game animals in North America and Europe. In fact, mainly in
some European countries as France, Italy, Germany, Austria, Poland, Sweden, Spain
and Portugal, hares are a highly valuable game species, constituting an important
economic resource.
5) Hares in Europe and in the Iberian Peninsula
Five Lepus species can be naturally found in the European continent (Figure 4). Lepus
europaeus (the brown hare) and Lepus timidus (the mountain hare) are the most widely
distributed species of the genus in Europe. The former inhabits most of the continent,
from the Iberian Peninsula to Russia, while the latter occupies Northern Europe and has
isolated populations in Ireland, Scotland and in the Alps (Flux & Angermann, 1990).
With smaller distributions we can find the Italian hare, Lepus corsicanus, inhabiting
Southern Italy, the Iberian hare, Lepus granatensis, in most of the Iberian Peninsula,
and the broom hare, Lepus castroviejoi, restricted to the Cantabrian Mountains in Spain
(Figure 4).
8
Chapter I
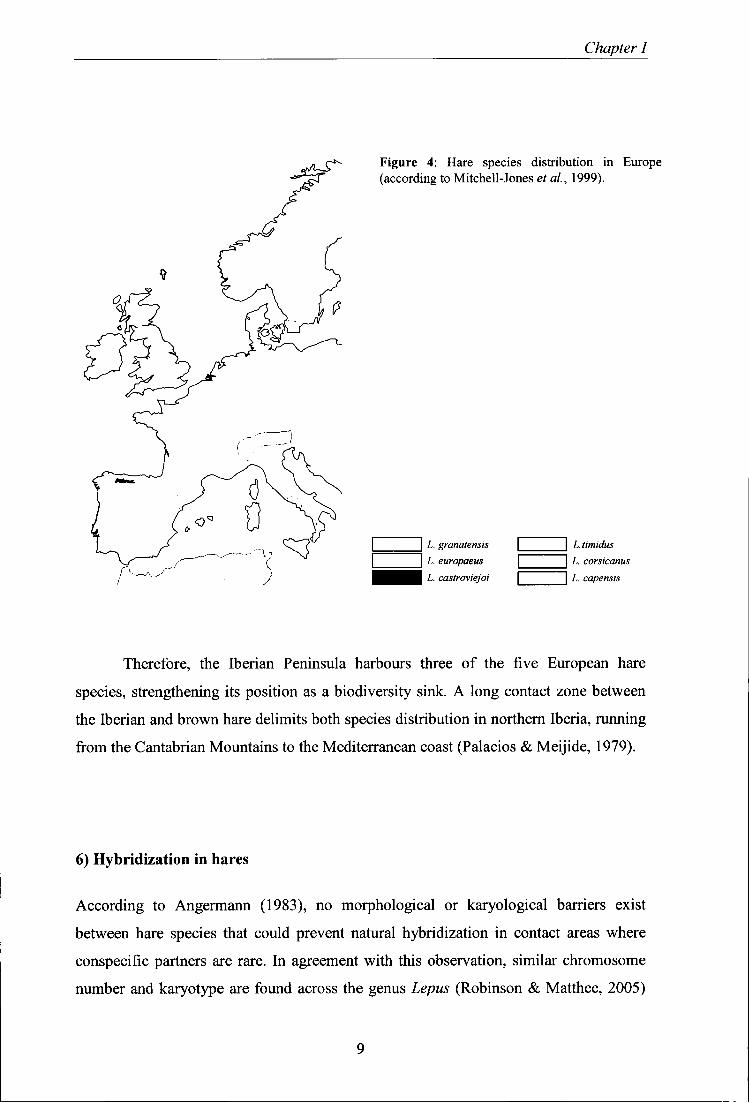
Figure 4: Hare species distribution in Europe (according to Mitchell-Jones et al, 1999).
L. granatensis L. europaeus L. castroviejoi
L.timidus ]̂ L. corsicanus
J L. capensis
Therefore, the Iberian Peninsula harbours three of the five European hare
species, strengthening its position as a biodiversity sink. A long contact zone between
the Iberian and brown hare delimits both species distribution in northern Iberia, running
from the Cantabrian Mountains to the Mediterranean coast (Palácios & Meijide, 1979).
6) Hybridization in hares
According to Angermann (1983), no morphological or karyological barriers exist
between hare species that could prevent natural hybridization in contact areas where
conspecific partners are rare. In agreement with this observation, similar chromosome
number and karyotype are found across the genus Lepus (Robinson & Matthee, 2005)
9

General Introduction
and, despite well defined morphological differences between some species (Angerbjõrn
& Flux, 1995; Palácios, 1983; Palácios, 1989), a general trend of regional variation and
phenotypic plasticity may approximate species appearance (Flux & Angerman in Flux
& Angermann, 1990).
In captivity, mountain hare females spontaneously cross with brown hare males,
whereas the converse must be performed by artificial insemination (Gustavsson &
Sundt, 1965), producing fertile offspring (Gustavsson, 1971; Schroder et al, 1987).
Using species-specific immunoglobulin markers, Schroder et al. (1987) failed to detect
natural hybridization in Finish wild populations of mountain and brown hare, but
conclusive evidence for its occurrence in Sweden has been documented by Thulin et al.
(1997). These authors described that mountain hare mtDNA is passing to brown hare
individuals by introgressive hybridization, despite mtDNA sequence divergence
between species of about 8%. In total, about 10% of the Swedish brown hares analysed
by Thulin et al. (1997) have mountain hare mtDNA, even though presently sympatric
and allopatric populations display significantly different values of introgression (~
10,4% vs. 0,6%, respectively) (Thulin & Tegelstrom, 2002). The unidirectionality of the
mtDNA transfer in Sweden is only contradicted by one individual (Thulin et al, 2006a)
but this must be a rare situation, since none had been previously found in a sample of
671 individuals of both species (Thulin et al, 1997). However, in Russia, as opposed to
the Scandinavian scenario, only introgression of brown hare mtDNA into mountain hare
has been reported (Thulin et al, 2006b), although in a small scale. Changes in
hybridization direction are possible (e.g. Bozikova et al, 2005), and in the Russia-
Sweden case, the authors argue that hybridization is frequency-dependent (Thulin et al,
2006b). Another well documented hybridization event was described by Alves et al.
(2003) and Melo-Ferreira et al. (2005) in the Iberian Peninsula. In this situation,
hybridization occurred in the past, presumably when the mountain hare was present in
Iberia, from where it is supposedly absent since the end of the last ice age. Introgressive
hybridization led to the passage of mountain hare mtDNA to the Iberian, brown and
broom hare from northern Iberian Peninsula (Alves et al, 2003; Melo-Ferreira et al,
2005). In fact, some hare populations in that region display complete replacement of the
present species mtDNA by mountain hare mitochondrial genome, configuring a
10

Chapter I
scenario of massive introgression and fixation (Melo-Ferreira et al, 2005).
Interestingly, in the Iberian Peninsula, the mountain hare mtDNA signal is currently
strong, even though the long term absence of mountain hare.
Although the brown and the mountain hare may be separated since the late
Pleistocene (Slimen et al, 2005), most of the introgression evidences among Lepus
concern this pair of species. Beside the described case of present (Thulin et al, 1997) or
past (Alves et al, 2003) transfer of mountain hare mtDNA into brown hare, there is also
data supporting the bilateral introgression between these two species of hares in natural
populations of Switzerland, at the level of allozymes and mtDNA (Suchentrunk et al,
unpubl. data). On the other hand, hybridization has been discarded between brown and
Italian hare, as well as between mountain and Italian hare in Italy, where mtDNA
control region and cytochrome b analysis clearly allocated the three species individuals
to distinct clusters (Pierpaoli et al, 1999). These authors argue for the absence of either
present or past interspecific gene flow and hypothesize completely independent
evolutionary paths for Italian, brown and mountain hares. Recently, Estonba et al
(2006) surveyed the three hare species from Iberian Peninsula with microsatellites
markers and found no evidence of hybridization. In this work, some individuals, namely
two brown hares from Bulgaria that cluster with the broom hares, are interpreted as
mislocalizations by chance, since no recent contact is reasonable due to the long
geographic distance between Iberian and Balkan Peninsulas. The microsatellites
markers allow these authors to separate all the species at the population level, which is
interpreted as compelling evidence for the existence of a barrier to neutral gene flow
between them. In contradiction, preliminary data of Melo-Ferreira et al. (2004) points to
some degree of hybridization between Iberian and brown hare, shown by the analysis of
the nuclear marker locus e. Also, one hybrid individual was obtained in captivity
resulting from the cross of an Iberian hare male with a brown hare female (P.C. Alves,
unpubl. data), although it is not known if the offspring is fertile. It does, however, show
that species barrier separating Iberian from brown hare is not sufficiently strong to
maintain total separation of species.
11

General Introduction
7) Ecological requirements of Iberian hare and brown hare
With few exceptions, hares are very flexible and adapt easily to habitat conditions. In
general, the preferred habitat of hares is open country with shrubs, bushes or rocks for
protection from predators (Flux & Angermann, 1990). Agricultural fields and wood
patches constitute a good territory for hares (Homolka, 1985), with arable areas with
intermediate-intensity farming displaying high demographic densities (Smith et al,
2005). Presently, changes in agriculture techniques, the increased mechanization, use of
pesticides and habitat fragmentation are negatively correlated with hare population
densities (Dingerkus & Montgomery, 2002; Schmidt et al, 2004; Smith et al, 2005), as
well as the extensive and monoculture farming (Smith et al, 2005).
The Iberian hare is well adapted to extremely variable habitats, occurring in
areas ranging from less than 300mm to more than 2000mm of annual rainfall (optimum
between 300mm and 600mm), and from 3°C in January to 26°C in August (Palácios &
Meijide, 1979). This species inhabits areas at the sea level or as high as about 2000m of
altitude. It occupies humid mountain areas with Atlantic type climate and agricultural
fields of rye, wheat or cork in northern Iberia; open areas of agricultural landscapes of
wheat, barley or alfalfa in continental Iberia; and dry, Mediterranean regions with olive
groves or vineyards in the south of the Iberian Peninsula (Alves & Niethammer, 2003).
The brown hare in its Iberian range is also very plastic to ecological conditions, as it
occurs in areas from the sea level up to 1700m of altitude, ranging from about 400mm
to 2000mm (optimum between 1000mm and 1500mm) and with temperatures
oscillating between 0°C in January and 18°C in August (Palácios & Meijide, 1979). It
inhabits zones of agricultural exploitation as vineyards, almond groves and carob
groves, as well as humid zones near the Cantabrian Mountains with woods of Quercus
robur, Q. pyrenaica and Fagus sylvatica, for example. It also occurs in high altitude
areas in the Pyrenees, where pine forests of Pinus unciata with underbrush of
Rhododendron ferrugineum and Vaccinium myrtillus are present (Palácios & Meijide,
1979). Some other variables as prédation (Schmidt et al, 2004), field size (Smith et al,
2005) or hunting (Marboutin et al, 2003) were assessed for the impact in brown hare
populations, but implications seem to vary regionally.
12

Chapter I
8) Objectives and thesis organization
The hare species from Iberian Peninsula have been studied mainly in the scope of
phylogenetic or phylogeographic works and taxonomic classifications (Pérez-Suarez et
al, 1994; Pierpaoli et al, 1999; Alves et al, 2003; Melo-Ferreira et al, 2005; Estonba
et al, 2006). However, other phenomena with potential evolutionary importance, such
as hybridization, may occur and this has been an overlooked question. Also, the
advancement in molecular and statistical techniques provides new tools for the
intersection of genetics and ecology (i.e., landscape genetics), which may prove
valuable to gain new insights into species dynamics. Here, both these guidelines -
hybridization and landscape genetics - were followed to study the Iberian and brown
hare in the Iberian Peninsula. Given this, this thesis has two main topics:
1) The assessment of potential admixture between Iberian and brown hare
2) The genetic characterization of a contact zone between Iberian and brown
hare and the correlation between genetic patterns, species distribution and
ecological variables.
The question of hybridization between Iberian and brown hare was tackled with
the use of microsatellites. The first task was to assemble a set of microsatellites
powerful enough to distinguish Iberian, brown and also mountain hare, since this
species molecular signature is still present in the Iberian Peninsula. These molecular
markers were then used to answer some defined objectives, namely:
1) Verify if there is any evidence for nuclear introgression of mountain
hare into brown and Iberian hare
2) Investigate the putative admixture between Iberian and brown hare
3) Describe the patterns of introgression of microsatellites markers
4) Verify the effectiveness of the hybrid zone (as indicated by the
existence of post-Fl hybrids).
13

General Introduction
All the efforts made in order to accomplish these objectives are described in Chapter II
of this thesis.
To investigate the second main topic of this work, both nuclear and
mitochondrial markers were used. Although mtDNA introgression from mountain hare
into both Iberian and brown hare was already known, a bigger sampling effort can bring
new knowledge to the subject and several objectives were pursuited:
1) Search introgression between Iberian and brown hare at the level of
mtDNA
2) Verify the concordance of nuclear and mitochondrial markers
3) Search for genetic discontinuities and correlate them with
environmental ecotones
4) Correlate species distribution with ecological variables.
This work is described in Chapter III and consisted in a fine-scale analysis of a small
study area (around 60x50 km) in Northern Iberia, using geo-referenced individuals,.
Both Chapter II and Chapter III are presented in scientific article format, and all
the results obtained in both chapters are integrated and discussed in Chapter IV, the
General Discussion. Chapters I and IV (General Introduction and General Discussion)
are structured in numbered Sections and sub-Sections. The Conclusions are presented in
Chapter V and the References used in the General Introduction and in the General
Discussion are presented in Chapter VI.
14

Chapter II
Paper 1
Evidence of natural hybridization between Lepus granatensis and L. europaeus in northern Iberian Peninsula
Abs t r ac t
Hybrid zones are a challenging subject in evolutionary biology. Climatic oscillations
provided good opportunities for allopatric speciation of organisms and for posterior
secondary contact, which is apparently the most common way of establishment of
hybrid zones. In the north of the Iberian Peninsula, two species of hares, Lepus
granatensis and L. europaeus, are in contact but hybridization has not been
demonstrated. Here, with a set of six microsatellites we aimed to assess the interaction
between both species and also to verify if the massive introgression of L. timidus
mitochondrial DNA in Iberian Peninsula hare populations was paralleled by the nuclear
genome. Our results discard a nuclear presence of L. timidus in Iberia and support the
existence of natural hybridization between L. granatensis and L. europaeus. Four
individuals of the former species and three of the latter show evidences of admixture
between both species, although introgression from L. europaeus into L. granatensis is
more strongly supported. The reciprocity of hybridization, as well as the level of
admixture or the hybrid zone structure should be reassessed. Nevertheless, Iberian and
brown hare do hybridize in northern Spain and may now be included in the list of
organisms contributing to the suture zone that exists in that region.
15

Paper 1
Introduction
Hybrid zones have for long been object of interest. Their study congregates knowledge
from multiple fields such as taxonomy, genetics, ethology or palaeoclimatology, which
renders it an intrinsic importance, but also an extra difficulty. Presently, a hybrid zone is
commonly accepted as an area of interaction between genetically distinct groups of
individuals, where some admixed offspring might occur, still being possible to find
parental or pure individuals outside of that zone (Harrison, 1990). A secondary contact
after allopatric divergence of evolutionary units is apparently the most parsimonious
explanation for the origin of many contact zones (see Hewitt 2001). In the European
Continent, which experienced several glacial and inter-glacial periods, the dynamics of
ice sheets provided good opportunities for the occurrence of speciation, colonization,
secondary contacts and hybridization phenomena (Hewitt, 2000; Taberlet et al, 1998).
Therefore, the distribution of its fauna and flora has been shaped through time by these
macroscale events coupled with a peculiar geographic context.
In Southern Europe, the Iberian, Italian and the Balkanic peninsulas constituted
biodiversity sinks, as they were the major ice age refugiai areas. So, these peninsulas
were the source of Europe recolonization by several organisms (Hewitt, 2001; Taberlet
et al, 1998) in a northwards post-glacial expansion which led to secondary contacts
between recently diverged taxa. However, in Iberian and Italian peninsulas the presence
of mountain chains (Pyrenees and Alps, respectively) strongly opposed species
northwards expansion (Hewitt, 1999). Nonetheless, after the last glacial maximum,
several contact zones have been generated (Hewitt, 1988; Hewitt, 2001). Major
biogeographic barriers as mountain chains and congruent postglacial biotic range
expansions (Hewitt, 1999; Hewitt, 2001; Swenson & Howard, 2004) led to the
coincidence of various organisms contact zones, eventually fitting Remington's concept
(1968) of a suture zone. In Europe, four such zones are usually recognized (Taberlet et
al, 1998): one corresponds to the Alpine barrier; another is located near the border
between France and Germany; the third one occurs in Scandinavia; and the fourth is
located in north-eastern Iberian Peninsula, in the Pyrenean foothills (Figure 1). In the
latter suture zone, organisms such as the meadow grasshopper Chorthippus parallelus
16

Chapter II
(Cooper et al, 1995), the alder Alnus glutinosa (King & Ferris, 1998) and the crested
newt Triturus cristatus (Arntzen & Wallis, 1991), established hybrid zones after post
glacial range expansions.
Figure 1 : Location of the four suture zones known in Europe (adapted from Taberlet et al., 1998): A) the Alps, B) France-Germany border, C) Scandinavia and D) northern Iberian Peninsula.
The brown hare (Lepus europaeus), widespread in Europe, and the Iberian hare
(Lepus granatensis), endemic to the Iberian Peninsula, also form a contact zone in
northern Spain. This contact area runs from the Mediterranean Sea to Astúrias, but the
question of hybridization is still an open issue. Within genus Lepus (LEPORIDAE;
LAGOMORPHA), some cases of hybridization have already been described. In Sweden,
unidirectional introgression of the native mountain hare {Lepus timidus) mtDNA into the
introduced brown hare was documented (Thulin et al, 1997; Thulin et al, 2006a).
Recently, hybridization between the same two species has been shown in Russia,
although the mtDNA introgression occurs in the opposite direction, L. europaeus to L.
timidus (Thulin et al, 2006b), and in the Alps where it is apparently bidirectional
(Suchentrunk et al, unpublished data). Also, although the mountain hare is presently
17

Paper 1
absent from Iberia, ancient massive introgression of its mtDNA into L. europaeus, L.
granatensis and even into the relictual population of broom hare (L. castroviejoi),
restricted to the Cantabrian Mountain (Palácios & Meijide, 1979), has been described
(Alves et al, 2003; Melo-Ferreira et al, 2005). However, despite the existence of several
contact zones between other pairs of hare species, hybridization has been discarded: for
example, no evidence for it was found between brown and Italian hare (Lepus
corsicanus) or mountain and Italian hare (Pierpaoli et al, 1999) in Italy, or between
Iberian and brown hare (Estonba et al, 2006) in the Iberian Peninsula.
Here, we present a detailed study of the contact zone between Iberian and brown
hare in the Iberian Peninsula. Given our framework, an approach with nuclear markers
that differentiate the brown, Iberian and mountain hare was needed. For that, we use
microsatellites, which are nuclear markers with a high rate of mutation, leading to good
discriminatory power in short time events. Using a set of six microsatellites markers, we
prove that Iberian and brown hare hybridize in north-eastern Iberian Peninsula and draw
plausible scenarios concerning the species dynamics.
Materials and Methods
Sampling
Ear tissue was collected from a total of 342 hare individuals (194 L. granatensis, 109 L.
europaeus and 39 L. timidus) from several localities in Europe (Figure 2 and Table 1).
Species identification in the field was done on the basis of distinctive morphological
characters (Palácios, 1989). Near the contact zone between L. granatensis and L.
europaeus in Northern Iberia, three samples of Iberian hare and three of brown hare were
obtained. Parental samples of Iberian hare were collected from broadly distributed
localities within Iberian Peninsula, while parental individuals of brown hare were taken
from Central Europe sampling localities. Also, two sampling sites were surveyed for the
mountain hare, where one (Italian Alps) represents the present southernmost distribution
of the species.
18
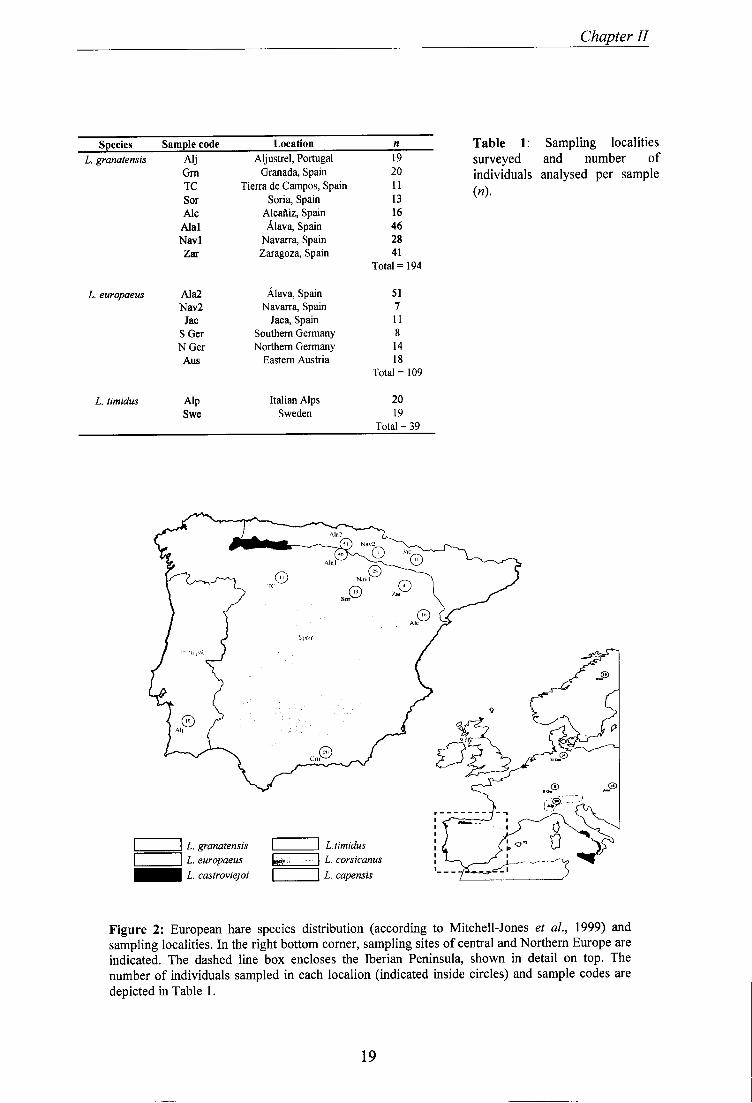
Chapter II
Species Sample code Location n L. granatensis Alj Aljustrel, Portugal 19
Grn Granada, Spain 20 TC Tierra de Campos, Spain 11 Sor Soria, Spain 13 Ale Alcafliz, Spain 16
Alal Alava, Spain 46 Navl Navarra, Spain 28 Zar Zaragoza, Spain 41
Total = 194
L. europaeus Ala2 Alava, Spain 51 Nav2 Navarra, Spain 7 Jac Jaca, Spain 11
SGer Southern Germany 8 NOer Northern Germany 14 Aus Eastern Austria 18
Total = 109
L. timidus Alp Italian Alps 20 Swe Sweden 19
Total = 39
Table 1: Sampling localities surveyed and number of individuals analysed per sample ( » ) ■
L. granatensis L. europaeus L. castroviejoi
L. timidus | L. corsicanus J L. capensis
Figure 2: European hare species distribution (according to Mitchell-Jones et al., 1999) and sampling localities. In the right bottom corner, sampling sites of central and Northern Europe are indicated. The dashed line box encloses the Iberian Peninsula, shown in detail on top. The number of individuals sampled in each localion (indicated inside circles) and sample codes are depicted in Table 1.
19

Paper 1
Microsatellites typing
Total genomic DNA was extracted from ear tissue using a standard high-salt protocol
(Sambrook et al, 1989). A set of 6 microsatellites loci was chosen from previously
published works, based on their polymorphism and allele size (Table 2). The loci Sat2,
Sat8, Satl2 and INRACCDDv358 were originally developed for the European rabbit
{Oryctolagus cuniculus), also a member of LEPORIDAE, while Lsa2 was specifically
developed for hares and Sol30Le was adapted for hares from a rabbit-specific locus. All
the used loci consist in dinucleotide repeat motifs except Sat 12, which is a
tetranucleotide microsatellites locus. The microsatellites were PCR-amplified in 15 uL
reaction volumes, with lx reaction buffer, 4 pmol of each primer, 1 pM of each dNTP,
1.5 mM MgCb and 0.2 units of Taq polymerase. For each individual, 0.25 ng of
genomic DNA was used per reaction. The thermal cycles consisted in an initial
denaturation step of 3 min at 94°C, followed by 30 cycles of 30s at 94°C, 30s at 40-58°C
and 30s at 72°C. A final 15min elongation step at 72°C was used. The microsatellites
typing was achieved using fluorescently labelled primers (Table 2) and an ABI 3100
automated sequencer (Applied Biosystems). In order to validate the allele calling results,
which was done in GeneMapper Software v3.7 (Applied Biosystems), control
individuals were included in every run and 15% of the dataset, chosen with a pseudo
random number generator, was repeated.
Table 2: Microsatellite markers identification {Locus ID and Reference), number of alleles found (Nr. Alleles), size range, fluorescent label of the primers (Label) and annealing temperature used for PCR amplification.
Locus ID Reference Nr.Alleles Size Range Label Annealing T Sat2 Mougelefa/., 1997 20 222-266 HEX 44°C Sat8 Mougel et al., 1997 7 87-101 NED 58°C Sat12 Mougelefa/., 1997 9 104-136 HEX 44°C
INRACCDv358 Chantry-Darmon ef a/., 2005 5 252-270 NED 54°C Lsa2 Kryger era/., 2002 11 237-257 NED 48°C
Sol30Le Fickelefa/., 1999 21 156-256 HEX 56°C
20

Chapter II
Data analyses Standard measures of genetic diversity as the total number of alleles per sample, mean
number of alleles per locus (MNA), expected (He) and observed (Ho) heterozygosity
were assessed using the program GENETIX version 4.05 (Belkhir et al, 2004). Linkage
disequilibrium (LD) between pairs of loci and Hardy-Weinberg Equilibrium (HWE) over
all loci across all samples were estimated using GENEPOP version 3.4 (Raymond &
Rousset, 1995). The probability tests were based on a Markov Chain Monte Carlo
(MCMC) simulation (Guo & Thompson, 1992), using GENEPOP default parameter
values and the standard Bonferroni correction for multiple test was applied. Fst was
calculated using the GENEPOP software to measure the differentiation between species
and between pairs of samples. While the latter was calculated over all the studied
samples, the former was calculated over the parental samples grouped by species.
A non-parametric 2 dimension Factorial Correspondence Analysis (FCA), as
provided by GENETIX, and a model-based Bayesian method, as implemented in
STRUCTURE (Pritchard et al, 2000), were applied to the dataset. Five replicate runs of
STRUCTURE, consisting in 500000 steps after a burnin period of 100000, were
performed for each K value, from K=2 to K=7. No prior information about the origin of
the specimens was used. Individuals were closely inspected for the possibility of
admixture when the likelihood of belonging to one species was lower than 80%.
The Iberian hare and brown hare samples were also analysed with the software
NEWHYBRIDS (Anderson & Thompon, 2002), using the same number of replicate runs
and steps as previously used in STRUCTURE. NEWHYBRIDS, as STRUCTURE, is a
model-based Bayesian software but fundamental differences stem between both models:
using multilocus genotype data, STRUCTURE infers population structure and assigns
individuals to populations, while NEWHYBRIDS assigns individuals to one of parentais
and various classes of hybrids (Fl's, F2's and various backcrosses). In the
NEWHYBRIDS analysis, prior information was used to define parental samples.
21

Paper 1
Results
A total of 73 alleles were found, ranging from 5 to 21 alleles per microsatellites locus. No
significant linkage or Hardy-Weinberg disequilibria were found. Although none of the
loci was completely diagnostic, a frequency-based species distinction was possible
(Figure 3). In fact, an almost complete concordance of microsatellites species assignment
with previous morphological identification was achieved (one possible misidentification
in 342 specimens - 99,7%).
Satl2
NOR
'z".r
Alii
Lsa2
INRACCDv358 Swt
AS S0« o KJ "fô Nav2
• % 1 I TC 1
Alj * 252 25« 2 » 258 260 262 264 266 268 270 272
N Gtr JlC
Sat8
Sol30Le
ft* 'M • • m • M iZ 0
n ^ Xiv2 0 i A1.2 0 lu
^woi. 0 Alt ^ m
All
Figure 3: Allelic frequencies found in each sample, identified in the Y-axis. Allele sizes (in bp) are shown in the X-axis. In dark blue, parental samples of Iberian hare; in light blue, Iberian hare samples from te contact zone; in dark yellow, brown hare parental samples; in light yellow, brown hare samples from the contact zone; in brown, mountain hare samples.
22

Chapter II
In general, the MNA, as well as the He and the Ho, are lower in the Iberian hare
than in brown or mountain hare, which display similar values (Table 3). The MNA tends
to increase in the samples from the contact zone when compared with the parental
samples of the respective species, being the samples from Alava (either L. granatensis or
L. europaeus) the ones that display a most conspicuous increment. The Ho also displays a
slight increase, while He suffers no modifications. However, when individuals with
suspected alien alleles (which only occur in the contact zone sampling localities) were
removed from the dataset, the recalculated values of MNA, He and Ho fitted the species
range of values or were even lower (see MNA in Alav2, Nav2 and Jac).
Table 3: Genetic diversity parameters as measured by expected heterozygosity (He), observed heterozigosity (Ho) and mean number of alleles per locus (MNA) found in each sample. For the contact zone samples, recalculated values without foreign alleles are also displayed {Recalculated). The box encloses the contact zone samples.
Sample Species He Ho MNA
Total Recalculated Total Recalculated Total Recalculated
Alj Lg 0,39 - 0,25 - 3,33 -
Grn Lg 0,39 - 0,23 - 2,67 -
TC Lg 0,36 - 0,25 - 3 -
Sor Lg 0,37 - 0,28 - 2,83 -
Ale Lg 0,32 - 0,24 - 3 -
Alal Lg 0,41 0,33 0,38 0,27 4,67 3,33
Navl Lg 0,38 0,38 0,3 0,29 3,83 3,5
Zar Lg 0,32 0,29 0,25 0,24 4,33 3,83
Ala2 Le 0,54 0,52 0,45 0,44 6,17 5
NAv2 Le 0,55 0,47 0.5 0,44 3,67 2,5
Jac Le 0,49 0,53 0,53 0,5 3,83 2,17
SGer Le 0,45 - 0,45 - 3,33 -
NGer Le 0,54 - 0,46 - 4,5 -
Aus Le 0,53 - 0,51 - 5,83 -
Alp Lt 0,51 - 0,4 - 5,67 -
Swe Lt 0,53 - 0,52 - 6,5 -
23

Paper 1
The Fst values estimated over the parental samples indicated high levels of
differentiation between species (Fst(Lg.Le) = 0,353; Fst(Lg.Lt) = 0,205; Fst(Le-Lt) = 0,208).
Accordingly, the three species are supported by the best result of STRUCTURE when
K=3 (Figure 4) and could be depicted as separate clusters in FCA (Figure 5). The K=3
value of STRUCTURE analysis was chosen since the largest log probability jump occurs
from K=2 to K=3 and, for the analysed values of K>3, we could only find the
substructuring of the Iberian hare (see K=4 in Figure 4). Moreover, K=3 fits with the
biological meaning of this work, since three recognized species were under study.
Figure 4: STRUCTURE results for K=2, K=3 and K=4. Sample codes identification can be depicted in Table 1. In K=3, blue represents the Iberian hare, light brown the brown hare and dark brown the mountain hare.
K=2
Alj
Grn TC
Sor Ale
Ala1
Nav1
Zar
Ala2
Nav2 Jac
S Ger NGer
Ans
Alp ::;f;i;;;«;i.„.,
Alp fc- "
Swe
K=3 K=4
£™WW$K§r
24

Chapter II
The tight cluster formed in FCA showed that the Iberian hare is the less diverse
species. On the other hand, the brown and the mountain hare plots displayed a bigger
dispersion. In agreement, the pairwise Fst analysis (Table 3) showed very low
differentiation between Iberian hare samples (mean 0,030; range 0 - 0,079), while brown
hare (mean 0,086; range 0,021 - 0,145) and mountain hare (0,101) showed higher
degrees of divergence between species' samples.
Table 3: Pairwise Fst between each pair of samples analysed. Samples identification is shown in Table 1. In bold are the intraspecific Fst values.
N Alj Grn TC Sor Ale Alal Navl Zar Ala2 Nav2 Jac S Ger Qer Aus Alp
Grn TC Sor Ale
Alal Navl Zar Ala2 Nav2
Jac S Ger NGer Aus Alp Swe
0,031 0,013 0,030 0,007 0,047 0,000 0,015 0,103 0,026 0,046 0,031 0,027 0,000 0,012 0,054 0,021 0,026 0,000 0,024 0,050 0,010 0,044 0,079 0,031 0,039 0,012 0,033 0,030 0,305 0,302 0,322 0,312 0,351 0,309 0,331 0,376 0,345 0,335 0,370 0,367 0,423 0,346 0,371 0,451 0,075 0,328 0,338 0,364 0,352 0,395 0,334 0,367 0,421 0,083 0,021 0,407 0,408 0,444 0,434 0,480 0,401 0,433 0,494 0,103 0,049 0,061 0,382 0,386 0,396 0,392 0,436 0,382 0,402 0,462 0,145 0,091 0,131 0,026 0,313 0,309 0,335 0,324 0,378 0,323 0,347 0,407 0,044 0,063 0,097 0,092 0,128 0,194 0,164 0,225 0,241 0,278 0,214 0,207 0,298 0,216 0,220 0,234 0,300 0,303 0,203 0,210 0,216 0,261 0,250 0,281 0,252 0,259 0,320 0,212 0,230 0,233 0,297 0,296 0,190 0,101
The FCA allowed the depiction of five individuals in an intermediate position
between the Iberian and the brown hare cluster, namely Alal 40, Alal 46, Alal 48, Alal
49 and Alal 51. As for the analysis with the software STRUCTURE, 12 individuals fall
below the defined threshold of 80% to acceptance of mixed individual ancestry. Four of
these cases (Ala2 31, Jac 4, Aus 21 and Aus 26), morphologically identified as brown
hares, assigned mostly to this species but had more than 20% of their genome attributed
to the mountain hare; one specimen (Alp 832) previously identified as a mountain hare
25

Paper 1
was mostly assigned to the brown hare cluster, although more than 20% of its genome
was assigned to the mountain hare species; and one individual (Alal 48) assigned with
similar probability to each population when K=2, 3 or 4 and randomly for higher K
values and, for this reason, was excluded from further analyses. The extant six
individuals (Alal 40, Alal 46, Alal 49, Alal 51, Alal 53 and Ala2 101) were jointly
attributed to the Iberian and brown hare cluster by the Bayesian analysis.
00
Axis 1 (7,22%)
Figure 5: FCA individuals plot. Yellow dots represent brown hares, blue dots represent Iberian hares and brown dots represent mountain hares. In green are displayed individuals morphologically identified as Iberian hares which have signs of admixture with brown hare.
Given the absence of a mountain hare signal in general and, particularly, in
Iberian hare, the mountain hare samples were removed for the analysis with the software
NEWHYBRIDS. To ensure that no signs of the mountain hare were contained in the
analysis, those individuals with some amount of potential admixture with L. timidus
were also excluded. NEWHYBRIDS results identify Alal 40, Alal 46, Alal 49, Alal
51, Ala2 56, Ala2 101 and Jac 12 as having high probabilities of being post-Fl hybrids.
26

Chapter II
Discussion
The ancient massive introgression of the mountain hare mtDNA into the Iberian hare and
the brown hare (Alves et al, 2003; Melo-Ferreira et al, 2005) complicates the
assessment of possible hybridization between the latter two species. The same event of
hybridization could have left traces on the nuclear genome of L. granatensis and L.
europaeus, although the available allozyme and molecular data do not provide evidence
of differentiation in the nuclear background between the introgressed and non-
introgressed populations of I. granatensis (Alves & Ferrand, 1999; Alves, 2002; Alves
et al. 2003). However, the genomic and geographic coverage of these data were limited,
forcing us to account for the possible contribution of L. timidus for the extant nuclear
gene pool of the Iberian species. The highly polymorphic neutral nuclear markers used in
this work allowed a clear distinction of the three studied species. Fst estimates over the
parental samples proved that species differentiation was high and FCA and
STRUCTURE both showed a good resolution, even including the contact zone samples.
Although there was a constant presence of some background noise in SRUCTURE's
analysis, no strong signs of mountain hare nuclear introgression were found. There were,
however, four brown hare individuals (Ala2 31, Jac 4, Aus 21 and Aus 26) that displayed
considerable posterior probabilities of assignment to the mountain hare species. The
admixture found in the two Austrian individuals (Aus 21 and Aus26), together with the
mountain hare individual Alp 832 from the Italian Alps, may support the occurrence of
present hybridization between brown hares from Central Europe and mountain hares
from the Alps as shown by Suchentrunk et al. (unpubl. data). The distance that separates
Aus 21 and Aus 26 from the Alps, however, may indicate that these individuals are the
evidence of past hybridization events that occurred in Eastern Austria, but such
hypothesis must be reassessed. The brown and the mountain hare, as previously shown
in Sweden (Thulin et al, 1997; Thulin et al, 2006a) and Russia (Thulin et al, 2006b),
are able to intercross and produce fertile offspring, leading to introgression events. It
should be noticed that Alp 832, although caught in the mountain hare habitat and
morphologically identified as a mountain hare, had its genome mainly assigned to the
brown hare cluster. Therefore, it constitutes the only individual with discordant
27

Paper 1
identification between morphology and microsatellites among the 342 analysed. The
mountain hare signal is also detected in two brown hare specimens from the Iberian
Peninsula (Ala2 31 and Jac 4). This may result from the referred possible hybridization
events in Central Europe between L. europaeus and L. timidus coupled with a long
distance gene flow in the brown hare, as described by Suchentrunk et al. (2000), or could
constitute the remnant signal of the ancient hybridization between L. timidus and the
Iberian species of hare, as shown to occur with mtDNA (Alves et al, 2003; Melo-
Ferreira et al, 2005). Although the absence of the same introgression signal in L.
granatensis and the short amount of time needed to erase nuclear introgression in the
presence of backcrosses (Roca et al, 2005) may argue against the latter hypothesis,
different population size (larger in Iberian hare) when hybridization occurred could
explain the detected pattern. Another possible explanation for these results is the low
number of markers used and the lack of diagnostic loci, leading to a problematic
assignment of some particular multilocus genotypes, even if our analyses showed a
complete differentiation between species using this microsatellites set.
In the analysis with the software STRUCTURE, the homogeneity of most
samples of brown hare and, specially, of Iberian hare is clear (Figure 4), which is in
conformity with Estonba et al. (2006), who recognize a compact cluster for the latter
species individuals in the NJ analysis. However, the Iberian hare sample of Alava shows
clear evidence of admixture. The mixed ancestry of four L. granatensis specimens from
Alava is supported by intersecting the three analysis softwares used. The presence of this
evidence only in one sample of Iberian hare can point to a unidirectional introgression,
with the invasion of brown hare genome into Iberian hare. The demographic dynamics of
the species can explain this unidirectionality. The Iberian hare attains higher population
densities than the brown hare (Ibón Telletxea, unpubl. data) and is progressively
replacing it in northern Iberia. This difference in demography between the species
increases the probability that hybrids backcross with Iberian hare in the contact zone
(density-dependent hybridization; Wirtz, 1999). However, this hypothesis of
unidirectional introgression is, in this particular case, contradicted by the MNA. An
increase in the MNA per locus is seen in the contact zone samples of both species, and
not only in the Iberian hare, as would be expected in such a scenario. The most sensible
28

Chapter II
increment was detected in either species samples of Alava, where robust evidence of
admixture is found. This suggests that both Iberian and brow hare gene pools are
incorporating foreign alleles (i.e., from the other species). The transferred alleles behave
as rare alleles in the new background, and that justifies the conspicuous increase of
MNA values but not of heterozygosities. Also, one brown hare individual was identified
by both Bayesian softwares as hybrid (Ala2 101) and two more (Ala2 56 and Jac 12)
were identified by NEWHYBRIDS. These results suggest that hybridization may be
bidirectional and may be detected in other populations than Alava. Nevertheless, the
signal of admixture is stronger in Iberian hare and, in this work, in the Alava sample. If
hybridization happens in low levels, as the data suggests, and given the number of
frequent private alleles of brown hare is higher than the Iberian hare ones, it is expected
that one way introgression from brown to Iberian hare is easier to detect. The low
frequency of hybridization could also mean that the admixed specimens exist only in a
narrow area where the two species meet and hybridize. Since this is the first work clearly
arguing for hybridization between Iberian and brown hare, the exact geographical
location and the width of the hybrid zone between these two species is not known. Alava
is not only our largest sample but also the most exhaustive sampling site of the contact
zone, which could explain why only there robust evidence for the existence of hybrids
were found. An experimental work using transects across the contact zone could help
solving this question. Nevertheless, the increase in the MNA per locus even in samples
where admixture is not detected (in Iberian hare samples of Navarra and Zaragoza and
brown hare samples of Navarra and Jaca) may indicate an import and posterior spread of
foreign alleles within each of the species. This scenario demands the existence of
backcrosses, which would at the same time render this hybrid zone as an effective one,
given the occurrence of fertile mixed individuals. As seen by the results obtained from
the NEWHYBRIDS analysis, all the admixed individuals were assigned to post-Fl
generations of hybrids, fitting these expectations. NEWHYBRIDS was run incorporating
prior knowledge about parental samples since these were not only morphologically
identified but strongly corroborated by STRUCTURE. In this way, using priors for the
parental samples enhanced the discriminatory power of the software. Recently, Estonba
et al. (2006), using a set of microsatellites markers as well, concluded that there is no
29

Paper 1
admixture between Iberian and brown hare. However, given the low level of
hybridization found here and the apparent narrow distribution of this event, it is
understandable that a considerably smaller and sparser sampling work did not show any
signs of genetic admixture.
Although mainly focusing on the existence of a hybrid zone between Iberian hare
and brown hare, our results can help to clarify some aspects of the past dynamics of
these two species. Apparently, the contact zone between L. granatensis and L. europaeus
resulted from a secondary contact of previously differentiated and allopatric species. The
Iberian hare has presumably been present in the Iberian Peninsula for quite some time,
since it is endemic to this region. However, during the glacial periods it could have been
pushed to lower latitudes and experienced a recent northwards post glacial expansion as
documented for other species, such as the wild rabbit (Branco et al, 2002). As for the
brown hare, even though it is still a matter of debate, several data argue for a refugiai
area in the Balkans region and subsequent expansion to west during the post-glacial
period, arriving recently to the Iberian Peninsula (Pierpaoli et al, 1999; Kasapidis et al,
2005). These historical scenarios could lead to reduction of genetic diversity from South
to North in the Iberian hare within Iberia and in brown hare from East to West across
Europe, sharpened by the crossing of the Pyrenean mountains. However, the detected
values of genetic diversity, even when calculated without foreign alleles, do not conform
to this expectation. Instead, Iberian hare samples of northern Spain display similar
(MNA and Ho) or even higher values (He) of genetic diversity parameters than the
southernmost samples. This result, coupled with the very low pairwise Fst within this
species, argues for a historical stable metapopulation of Iberian hare with high gene
flow, as also suggested by allozymes data (Alves, 2002). Concerning the brown hare,
comparing with the Balkans population (Estonba et al, 2006), the MNA decreases in a
westwards direction (Balkans samples > Central Europe samples > Northern Iberia
samples), fitting the hypothesized colonization route. As for the heterozygosities,
however, higher values are attained by the Balkans population, whereas the Central
European samples are similar to Northern Iberian ones. Such pattern could be explained
by several colonizations of northern Spain by brown hares with different genetic pools,
establishing a good subsample of the Central European genetic stock. The microsatellites
30

Chapter II
mutation rate and size-constraint may also account for the homogenization of genetic
diversity values. Together, these results join the body of evidence for a Balkan origin of
the European colonization by brown hare (Kasapidis et al, 2005), as opposed to the
possibility of the existence of an Iberian glacial refugium of this species.
Conclusion
The morphological identification of specimens was almost completely supported by
molecular data (99,7%), suggesting that these three hare species maintain enough
distinctive physical traits even when occurring in parapatry. In spite of the absence of
diagnostic loci and the possible homogenising effect of homoplasy caused by the high
mutation rate of microsatellites combined with an allele size constraint phenomenon, we
have provided unequivocal evidence of ongoing hybridization between L. granatensis
and L. europaeus in Northern Iberian Peninsula. This hybridization seems rather rare and
restricted to a narrow area of contact between the species, which has made it previously
undetected. This might also result from the very conservative conditions adopted in this
work to consider individuals as hybrids or to a low resolution power of the used
microsatellites set. The directionality of the introgression, its true level or the exact
location and width of the hybrid zone, are questions that need to be reassessed with the
use of high resolution tools such as diagnostic single nucleotide polymorphisms coupled
with thin scale sampling of the hybrid zone. Some demographic scenarios were also
tackled, with genetic diversity parameters arguing for the Balkans origin of Western
Europe colonization by the brown hare and for a stable Iberian hare metapopulation with
high gene flow within Iberian Peninsula. The Iberian and brown hare seem to be
contributing to the old Remington's concept of a suture zone, in this case located in
north-eastern Iberian Peninsula.
31

Paper 1
References
Alves PC, Ferrand N (1999) "Genetic variability in Portuguese populations of the Iberian hare, Lepus granatensis". Folia Zoológica, 48, (Suppl.l) 3-10.
Alves PC (2002) "Caracterização genética e biologia reprodutiva da lebre Ibérica, Lepus granatensis. Análise filogenética, diferenciação populacional e ciclo anual de reprodução. " PhD thesis, University of Porto.
Alves PC, Ferrand N, Suchentrunk F, Harris DJ (2003). "Ancient introgression of Lepus timidus mtDNA into L. granatensis and L. europaeus in the Iberian Peninsula." Molecular Phylogenetics and Evolution 27(1): 70-80.
Anderson EC, Thompson EA (2002) "A model-based method for identifying species hybrids using multilocus genetic data". Genetics, 160, 1217-1229.
Arntzen JW, Wallis GP (1991) "Restricted Gene Flow in a Moving Hybrid Zone of the Newts Trituras cristatus and T. marmoratus in Western France". Evolution, 45, 805-826.
Belkhir K, Borsa P, Chikhi L, Raufaste N, Bonhomme F (1996-2004) GENETIX 4.05, logiciel sous Windows TM pour la génétique des populations. Laboratoire Génome, Populations, Interactions, CNRS UMR 5171, Université de Montpellier II, Montpellier (France).
Branco M, Monnerot M, Ferrand N, Templeton AR (2002) "Postglacial dispersal of the European rabbit (Oryctolagus cuniculus) on the Iberian Peninsula reconstructed from nested clade and mismatch analysed of mitochondrial DNA genetic variation". Evolution, 56, 792-803.
Chantry-Darmon C, Urien C, Hayes H, Bertaud M, Chadi-Taourit S, Chardon P, Vaiman D, Rogel-Gaillard C (2005) "Construction of a cytogenetically anchored microsatellite map in rabbit". Mammalian Genome, 16, 442-459.
Cooper S J, Ibrahim KM, Hewitt GM (1995) "Postglacial expansion and genome subdivision in the European grasshopper Chorthippus parallelus". Molecular Ecology, 4, 49-60.
Estonba A, Solis A, Iriondo M, Sanz-Martin MJ, Pérez-Suaréz G, Markov G, Palácios F (2006). "The genetic distinctiveness of the three Iberian hare species: Lepus europaeus, L. granatensis, and L. castroviejoi." Mammalian Biology 71(1): 52-59.
Fickel J, Lieckfeldt D, Pitra C (1999) "Analysis of genetic diversity and structure in neighboring populations of the European brown hare {Lepus europaeus, Pallas 1778)". Zeitschrift fur Jagdwissenschaften, 45, 230-237.
32

Chapter II
Flux JE, Angermann R (1990) "The hares and jackrabbits". In Chapman J A & Flux JEC (Eds.), Rabbits, Hares and Pikas. Status Survey and Conservation Action plan. IUCN, Switzerland,pp. 61-94.
Guo SW, Thompson EA (1992) "Performing the exact test of Hardy-Winberg proportions for multiple alleles". Biometrics, 48, 361-372.
Harrison RG (1990) "Hybrid zones: windows on the evolutionary process". Oxford Surveys in Evolutionary Biology 7, 69-128.
Hewitt GM (1988) "Hybrid zones - natural laboratories for evolutionary studies". Trends in Ecology and Evolution, 3, 158-167.
Hewitt GM (1999) "Speciation, hybrid zones and phylogeography - or seeing genes in space and time". Molecular Ecology, 10, 537-549.
Hewitt G (2000) "The genetic legacy of the Quaternary ice ages". Nature, 405, 907-913.
Hewitt GM (2001). "Speciation, hybrid zones and phylogeography - or seeing genes in space and time." Molecular Ecology 10(3): 537-549.
Kasapidis P, Suchentrunk F, Mogoulas A, Kotoulas G (2005) "The shaping of mitochondrial DNA phylogeographic patterns of the brown hare (Lepus europaeus) under the combined influence of Late Pleistocene climatic fluctuations and anthropogenic translocations". Molecular Phylogenetics and Evolution, 34, 55-66.
King RA, Ferris C (1998) "Chloroplast DNA phylogeography of Alnus glutinosa (L.) Gaertn". Molecular Ecology, 7, 1151-1162.
Kryger U, Robinson TJ, Bloomer P (2002) "Isolation and characterization of six polymorphic microsatellite loci in South African hares {Lepus saxatilis F. Cuvier, 1823 and Lepus capensis Linnaeus, 1758)". Molecular Ecology Notes, 2, 422-424.
Melo-Ferreira J, Boursot P, Suchentrunk F, Ferrand N, Alves PC (2005). "Invasion from the cold past: extensive introgression of mountain hare (Lepus timidus) mitochondrial DNA into three other hare species in northern Iberia." Molecular Ecology 14(8): 2459-2464.
Mitchell-Jones AJ, Amori G, Bogdanowicz W (1999) Atlas of European Mammals. Academic Press, London.
Mougel F, Mounnolou JC, Monnerot M (1997) "Nine polymorphic microsatellite loci in the rabbit, Oryctolagus cuniculus". Animal Genetics, 28, 58-71.
Palácios F (1989) "Biométrie and morphologic features of the species of the genus Lepus in Spain". Mammalia, 53, 227-264.
33

Paper 1
Palácios F, Meijide M (1979) "Distribución geográfica y habitat de las liebres en la Península Ibérica". Naturalia Hispânica, 19, 1-40.
Pierpaoli M, Riga F, Trocchi V, Randi E (1999). "Species distinction and evolutionary relationships of the Italian hare (Lepus corsicanus) as described by mitochondrial DNA sequencing. " Molecular Ecology 8(11):1805-1817.
Pritchard JK, Stephens M, Donnelly P (2000) "Inference of population structure using multilocus genotype data". Genetics, 155, 945-959.
Raymond M, Rousset F (1995) "GENEPOP (version 1.2): a population genetics software for exact tests and ecumenicism". Journal of Heredity, 86, 248-249.
Remington C (1968) "Suture-zones of hybrid interaction between recently joined biotas". Evolutionary Biology, 2, 321-428.
Roca AL, Georgiadis N, O'Brien SJ (2005) "Cytonuclear genomic dissociation in African elephant species". Nature Genetics, 37, 96-100.
Sambrook E, Fritsch F, Maniatis T (1989) Molecular Cloning. Cold Spring Harbour Press, Cold Spring Harbour, New York.
Suchentrunk F, Michailov G, Markov G, Haiden A (2000). "Population genetics of Bulgarian brown hares Lepus europaeus: allozymic diversity at zoogeographical crossroads". ActaTheriologica 45, 1-12.
Swenson NG, Howard DJ (2004) "Do suture zones exist?". Evolution, 58, 2391-2397.
Taberlet P, Fumagalli L, Wust-Saucy AG, Cosson JF (1998) "Comparative phylogeography and postglacial colonization routes in Europe". Molecular Ecology, 7, 453-464.
Thulin CG, Jaarola M, Tegelstrõm H (1997) "The occurrence of mountain hare mitochondrial DNA in wild brown hares". Molecular Ecology, 6, 463-467.
Thulin CG, Stone J, Tegelstrõm H, Walker CW (2006a) "Species assignment and hybrid identification among Scandinavian hares Lepus europaeus and L. timidus". Wildlife Biology, 12, 29-38.
Thulin CG, Fang M, Averianov AO (2006b) "Introgression from Lepus europaeus to L. timidus in Russia revealed by mitochondrial single nucleotide polymorphisms and nuclear microsatellites". Hereditas, in press.
Wirtz P (1999) "Mother species-father species: unidirectional hybridization in animals with female choice". Animal.Behaviour, 58, 1-12.
34

Chapter III
Paper 2
Finescale analysis of a hybrid zone between Iberian and
brown hare: integration of genetical and ecological data
Abstract
A hybrid zone is an area of more or less pronounced transitions between states. The
widespread phenomenon of hybridization uncovers its important role in speciation
events. Distinct characteristics define hybrid zones, and even hybrid zones between the
same pair of species may differ from place to place, raising the question of what
importance has the environment and ecological conditions. In the genus Lepus, a hybrid
zone exists in northern Spain between Lepus granatensis and L. europaeus. Using
geographical information of sampled individuals, a small area was screened for six
microsatellites locus and cytochrome b mitochondrial DNA gene. Also, topographical
and meteorological conditions were integrated in the analysis, providing some insight
into both species ecological requirements. Microsatellites results were found to fit
previous morphological species identification, whereas mitochondrial DNA displayed
distinct patterns. A unidirectional introgression of Iberian hare mtDNA into seventeen
brown hare individuals was detected, strengthening the evidence that support the
existence of a hybrid zone between both species. The Iberian and brown hare
distribution in the study area (as indicated by morphological and microsatellites data)
was found to be significantly associated with some ecological variables, in contrast with
the distribution of the different mitochondrial lineages. In this work, strong evidence of
hybridization using mtDNA was found and a first approach to landscape genetics in the
area was performed.
35

Paper 2
Introduction
In order for a hybrid zone to be established and noticed, at least two evolutionary units
must come into contact. Also, these units must be to some extent genetically
differentiated, or else they would simply immediately merge into a homogeneous group.
Although the way by which differentiation is achieved might be discussed (Harrison,
1990; Barton & Hewitt, 1985), what seems clear is that in every hybrid zone some kind
of discontinuity is detected. Transitions in morphology, behaviour or molecular
information (Harrison, 1990), among others, are characteristic of these contact zones,
which have been modelled into the cline theory (Barton, 1979; Barton & Hewitt, 1985).
The maintenance and degree of concordance of clines is a result of several variables
such as the level of reproductive isolation, selection or environmental factors (Barton &
Hewitt, 1985; Hewitt, 1988; Harrison, 1990). Also, these transition zones define barriers
to gene flow, where sexual selection (McDonald et ah, 2001), prezygotic (Servedio,
2001) or poszygotic isolation (Szymura & Barton, 1991) may account for it. However,
these barriers appear to be more porous than previously thought (Rundle et ah, 2001) as
evidence of genetic exchange between species of several kinds of organisms
accumulates. Indeed, hybridization seems to be a widespread phenomenon probably
with an important role in the process of speciation and acquisition of genetic diversity
by increasing the "field of recombination" (Harrison, 1990).
The study of hybrid zones has been based in various characters and markers, as
morphology, behaviour, allozymes, mitochondrial DNA, microsatellites and non-neutral
nuclear markers, and although all have been found to overcome the genetic barrier at
some extent, discordances have been depicted in the speed, patterns and amount of
introgression in a "foreign" background. None of these parameters can be seen as
independent, and neither can be their causes. The degree of reproductive isolation of
genetically distinct organisms is certainly important for the amount of hybridization, but
ecological requirements have a significant role as well. Patterns of introgression will
characterize the genetic architecture of the barrier. As for the speed of introgression,
alleles with even a small selective advantage will penetrate relatively quickly if
compared with neutral alleles, even though phenomena as linkage disequilibrium,
36

Chapter III
hitchhiking and background selection can blur this association (Barton & Hewitt, 1985;
Pialék & Barton, 1997).
A clear discordance in introgression characteristics has long been found between
nuclear DNA (nDNA) and mitochondrial DNA (mtDNA) (Powell, 1983; Tegelstrõm,
1987). Moreover, it seems to be a common feature: fruit flies (Ballard et al, 2002),
cichlid fishes (Rognon & Guyomard, 2003), rodents (Bozíková et ai, 2005; Ruedi et
ai, 1997), newts (Babik et ai, 2003) or songbirds (Rohwer et ai, 2001) are some
examples where discordant patterns of introgression have been detected, where mtDNA
introgression is not accompanied by or at least is more extensive than nDNA.
In Northern Iberian Peninsula, two species of hare meet in a long contact zone
(Figure 1). The Europe-spread Lepus europaeus (the brown hare) and the Iberian
Peninsula endemics Lepus granatensis (the Iberian hare) hybridize to some extent in
that area, as evidenced by microsatellites markers (Chapter II) and preliminary results in
locus e (Melo-Ferreira et ai, 2004), but still maintain distinctive gene pools at this
nuclear neutral markers level. The scenario for mtDNA, however, is expected to be
complicated by the described picture of massive ancient introgression in both brown
and Iberian hare by the mountain hare {Lepus timidus) mitochondrial genome (Alves et
ai, 2003; Melo-Ferreira et ai, 2005). In fact, Melo-Ferreira et al. (2005) report the
complete replacement of the native brown hare mtDNA by the mountain hare one in
some hare populations of Northern Iberia, namely in the region of Alava, where this
study is focused (Figure 1).
Figure 1: Iberian and brown hare populations from a small study area (red square) in the province of Alava, Spain were surveyed for microsatellites and cytb. The study area is located in northern Iberian Peninsula and comprises the contact zone between L. granatensis and L. europaeus (yellow line).
37

Paper 2
We surveyed Alava's hare populations of brown and Iberian hare for six nuclear
microsatellites and mitochondrial DNA cyt b gene, and geo-referenced all the
individuals. We expect to check for the concordance between both types of markers,
hypothesizing the influence of ecological conditions, since the Cantabrian Mountain
defines an ecotone which divides both species distribution as identified in a
morphological basis. Given that our study area and experimental design are inadequate
for a cline work, we used the recently developed software GENELAND (Guillot et al.,
2005) to integrate genetical and spatial information. Also, in a geographic information
system (GIS) framework, we overlap several ecological data to draw some ideas about
species dynamics. Although the ecological requirements for the brown and Iberian hare
are not fully understood (Flux & Angermann, 1990), their allocation in this area of
abrupt transition in habitat conditions suggests some influence of the habitat in species
distribution. Our results reveal an unusual case of a species (brown hare) that harbours
three different mitochondrial genomes and may be a first approach to the modelation of
brown and Iberian hare species distribution in Iberia, leading to a finescale plot of the
hybrid zone between them.
Materials and Methods
Study area
The study area is located in the province of Alava, in the Spanish Basque Country, and
comprises around 60x50 km (Figure 1 and Figure 2). The Cantabrian Mountain
represents an ecotone, dividing this region in two main areas in a north-south axis. Both
areas differ in climatic conditions, leading to habitat heterogeneity. Hare species,
identified in a morphological basis, mimic the habitat transition, with brown hare
occupying the northern zone whereas Iberian hare inhabits the southern one.
38

Chapter III
Sampling
Ear tissue samples were obtained during the official hunting seasons between 2000 and
2005 and stored in 70% alcohol. Forty five specimens of Iberian hare and 51 of brown
hare were collected. For all individuals coordinates were taken in WGS84/UTM
coordinate system.
Figure 2: Study area (red box in Figure 1) with sampling coordinates. The size of dots is proportional to the number of individuals collected. The Cantabrian Mountain is indicated by a yellow arrow.
Microsatellites and mtDNA typing
Total genomic DNA was extracted from ear tissue using a standard high-salt protocol
(Sambrook et al, 1989). Every individual was typed for the six microsatellites set used
in Chapter II following the described conditions. As for mtDNA, a 669bp fragment of
cyt b was screened with a PCR-RFLP approach as described by Melo-Ferreira et al.
(2005).
39

Paper 2
Ecological data
Using geographic information system (GIS) software, several parameters for the study
area were obtained. From WorlClim (Hijmans et al, 2005), we use data of precipitation
(measured in millimetres), maximum temperature and minimum temperature (measured
in °C). All data was averaged to annual mean values. From a Digital Elevation Model,
elevation (measured in meters), slope (measured in degrees relative to horizontal) and
aspect (measured in degrees relative to north in clockwise direction) were calculated.
Analysis
Data was analysed in a mixed genetical and spatial framework. This was done using the
software GENELAND (Guillot et al., 2005), which tries to detect population structure
using geo-referenced individual multilocus genetic data. Microsatellites and mtDNA
data were analysed separately and results were compared. Assuming that mtDNA is
recombination-free, individual haplotypes were coded as completely species diagnostic
genotypes to fit GENELAND analysis. All the ecological parameters were integrated in
a GIS software (ArcMap9, ESRI, 2004) and their values pulled for each individual
coordinate. Significance analyses were performed by a Mann-Whitney U test for two
different groupings of individual coordinates. First, coordinates were separated by
species, i.e., coordinates where brown hare occurs were tested against coordinates
where Iberian hare is found. Second, coordinates were separated by the mtDNA
haplotype found, being one group constituted by the mountain hare-like haplotype and
the other by non-mountain hare-like haplotype.
Results
The microsatellites analysis clearly identified all the individuals with at least 96% of
posterior probability of assignment to brown or Iberian hare species.
40

Chapter III
l
I e a
I i 0 50 100
Individuals
Figure 3: Posterior probability of individual assignment to Iberian hare using microsatellites data and GENELAND software. A clear identification of Iberian hares (value 1 ) and brown hares (value 0) was obtained.
This result was concordant with the previous morphological identification of all the
specimens. There was no evidence of hybridization detected in this analysis. Moreover,
the transition in microsatellites genotypes accurately mimics the location of the ecotone
in the study area. Consequently, it fits the transition in habitat conditions and is
concordant with the species distribution (Figure 4A and 4C). However, the scenario is
very different for the mitochondrial DNA. With this marker, three groups were detected,
each corresponding to the native haplotype of brown, Iberian or mountain hare (Figure
4B). The geographic distribution of each of the haplotypes in the study area is not
concordant with the microsatellites, habitat conditions or species distribution. Instead,
mountain hare haplotypes are widespread in the study area and occur both in Iberian and
brown hares (31% and 63%, respectively), as identified by morphological traits and
microsatellites; Iberian hare haplotypes can be found in 69% of Iberian hare individuals
and in 33% of brown hares; and the native brown hare mtDNA haplotype occurs in only
4% of brown hare specimens, from the same coordinate.
41

Paper 2
~\ r 500000 510000 520000 530000 540000 550000
~~\ 1 r 500000 510000 520000 530000 540000 550000
Figure 4: Discordance between nuclear and mitochondrial markers. In (A), individual assignment as obtained with microsatellites. In (B), mtDNA haplotypes distribution. In (A) and (B) the blue area corresponds to Iberian hare, white to brown hare and yellow to mountain hare. In (C), study area showing the concordance between microsatellites discontinuity (yellow line) and the ecotone (Cantabrian Mountain).
500000 510000 520000 530000 540000 550000
As for the ecological parameters, significant differences can be found between the
brown and Iberian hare coordinates for all the studied parameters except for the slope.
In fact, the detected differences between the northern and southern regions of the study
area are highly significant, with ;?-values equal or lower than 0,054 (Table 1). The
coordinates of brown hare show higher values of precipitation (Prec), lower minimum
temperature (Tmin) and maximum temperature (Tmax), higher elevation and are placed
in colder places, since their mean Aspect is -238° (southeast), in contrast with the
south-facing Iberian hare coordinates (-181°). In general, the habitat to the north of the
Cantabrian Mountain is cooler and rainier, while to the south the habitat is hotter and
drier. When we compare points where the mountain hare mtDNA haplotype occurs with
42

Chapter III
points where we find non-mountain hare haplotypes, significant differences are found in
Tmin, Prec, Slope and Aspect. The /rvalues for these significantly different parameters
range between 0,002 and 0,035. In this comparison, Tmin, Prec and Aspect display
higher values in mountain hare haplotype coordinates.
Variable P-•value Lg vs Le /7-value Lt haplo vs non-Lt haplo Tmax 0,000 0,332 Tmin 0,054 0,002 Prec 0,000 0,035
Elevation 0,000 0,181 Slope 0,635 0,015
Aspect 0,000 0,008
Table 1 : Mann-Whitney U test p-values for each variable compared between L. granatensis and L. europaeus coordinates (p-value Lg vs Le) and compared between coordinates where L. timidus haplotype occurs and coordinates where non-Z,. timidus haplotypes are found (p-value Lt haplo vs non-Lt haplo).
Discussion
The microsatellites analysis for the hare population from Alava was totally concordant
with the previous identification, in a morphological basis, of the species to which every
individual belongs. Although a hybrid zone between Iberian and brown hare is present
in this region, as shown in Chapter II, morphological differences between both species
are maintained and allow an accurate identification. The low amount of current
hybridization may justify this, since the hybrid individuals included in the analysis are
the result of some number of generations of backcross (Chapter II). These hybrid
individuals are not detected in the present work, what may stem from the difference in
sampling between Chapter II and Chapter III and from the absence of parental
populations in the latter. Also, the presence of alien alleles in both species (Chapter II)
may contribute to mask admixed individuals, especially when using non-diagnostic loci.
Nevertheless, the microsatellites markers used have enough information to clearly
separate Iberian from brown hare. In contrast with microsatellites information,
43

Paper 2
mitochondrial DNA presents a different scenario. The finding of mountain hare
haplotypes in both Iberian and brown hares was expected, given the reported massive
ancient introgression that occurs in the north of Spain (Alves et al, 2003; Melo-Ferreira
et al, 2005). However, Melo-Ferreira and colleagues showed that the brown hare
population from Alava was fixed for the mountain hare haplotype, which is not
corroborated by this work. Our sample is more than twice the used in Melo-Ferreira et
al work (51 vs. 24) and that may account for the discrepancies. First, in our analysis
native brown hare mtDNA haplotypes are found, although in low quantity (~ 4%).
Second, we detected the presence of Iberian hare haplotypes in brown hare individuals.
This unidirectional introgression of mtDNA from Iberian to brown hare strongly
supports the hybridization scenario for this pair of species, strengthening the findings of
Chapter II. The mtDNA introgressed brown hares represent 33% of the brown hare
sample. This is a more than three-fold higher amount of introgression than the one
found from mountain to brown hare in Sweden (Thulin et al, 1997), five-fold higher
than the one from brown to mountain hare found in Russia (Thulin et al, 2006a) and
about ten times higher than the reported introgression from brown to mountain hare in
Sweden (Thulin et al, 2006b).
Another interesting issue is the discordance between the mitochondrial and
nuclear genome introgression patterns. Both markers have very distinct and
fundamental differences. The mtDNA shows a uniparental (maternal) inheritance,
haploid nature, lack of recombination (although this has been challenged by several
authors, see Rokas et al, 2003 for a review) and a smaller effective population size (1/4
of the nuclear genome), while nDNA is the complete opposite of these features. So, one
might also expect discrepant behaviour of both markers in natural populations, which
indeed happens. Commonly, mtDNA and nuclear markers are discordant in their
patterns, and this may well be considered a broadly distributed dynamics, since it is
found in a variety of organisms which display a multitude of dissimilarities in mating
behaviour, sexual selection, dispersal or ecological requirements, for example. Cichlid
fishes from West Africa (Rognon & Guyomard, 2003) show a strongly supported
separation of two species with allozymes data, but an almost complete replacement of
one species mtDNA for the other species mitochondrial genome. This has also been
44

Chapter III
found in other species of fish, as the artic char and the lake trout (Wilson and
Bernatchez, 1998) or the bull trout and the Dolly Varden (Redenbach & Taylor, 2002).
In newts, even though some nuclear introgression is detected, mtDNA introgression is
much more pronounced and bidirectional, even if a 14% excess of one of the species is
seen (Babik et al, 2003). Another noteworthy work was performed in commensal mice,
where two transects (one in Czech Republic and one in Bavaria) were analysed
(Bozikóva et al, 2005). The authors of this last work found that mtDNA clines fell into
the range of the allozymes but not with that of X markers. An interesting feature of this
study is that the mtDNA introgression occurs from one species to the other, but the
direction changes between the Czech and the Bavarian transects. The differential
introgression of mtDNA and nDNA has been reported for several other organisms, from
insects to reptiles, or molluscs to mammals (for a list, see Chan & Levin, 2005). In the
present case, microsatellites reflect the present species distribution with weak signs of
hybridization (Chapter II), while mtDNA might represent a long-lasting introgression
from Iberian to brown hare. Due to its transmission pattern, mtDNA signal will be kept
long after the nuclear genome is erased (Roca et al, 2005). The direction of the mtDNA
transfer is concordant with the principle enounced by Grant & Grant (1997), which
states that when two species meet and hybridize, it is expected that males from the
bigger species (here, the brown hare) mate with females from the smaller (Iberian hare).
As mentioned before, the genetic discontinuity found for the microsatellites is
coincident with a topographical barrier found in the field, the Cantabrian Mountain
(Figure 4A and 4C). With steep slopes and considerable elevation, this mountain poses
a very hard obstacle for some animals, as hares, to overcome. Moreover, the Cantabrian
Mountain not only rises as a physical barrier, as it interferes in the climatic conditions
of the habitat. The significant differences in precipitation, minimum and maximum
temperature, aspect and elevation between the coordinates of brown and Iberian hares
(as identified morphologically and confirmed with microsatellites) are concordant with
a broad scale description of habitat preferences for both species (Palácios & Meijide,
1979). In fact, the brown hare is apparently better adapted to colder and wetter places: it
occurs in areas with higher precipitation, with lower minimum and maximum
temperature, with higher elevation and in colder hills (higher values of aspect). As for
45

Paper 2
the comparison between coordinates registering the occurrence of mountain hare
mtDNA haplotype and coordinates with non-mountain hare haplotypes, it aimed to
check if the mountain hare haplotypes were preferably distributed in colder places,
which would be a suggestion for the positive selection of the "cold-mitochondria"
(Melo-Ferreira et al, 2005). This trend was verified by the values of precipitation and
aspect (higher values for mountain hare haplotypes) but contradicted by the mean value
of minimum temperature, which was higher for the mountain hare haplotypes
coordinates. So, no clear tendency can be inferred by these analyses. A bigger and more
homogeneous sampling of the study field, as well as thinner information of climatic
variables could be helpful to solve this question, since it appears to be a valuable
approach to the problem.
Conclusion
The widespread phenomenon of natural hybridization also occurs among Lepus species.
It has been proved to happen by the first time in Sweden by Thulin et al. (1997) and,
more recently, by Alves et al. (2003) in the Iberian Peninsula. However, in both studies
mountain hare seems to be involved in the genetic exchange. Here, robust evidence for
mtDNA unidirectional change from Iberian to brown hare was found in a considerable
amount (-33%). At the species level, the brown hare in northern Iberia attains a peculiar
situation, since it displays three different mitochondrial genomes with no apparent
decrease in fitness. Cytonuclear disequilibria studies should be performed to clarify this
question. Another interesting issue is the discordance between mtDNA and nDNA
patterns. The latter follows the habitat transition set by the Cantabrian Mountain,
whereas the former presents a more complex scenario.
This work might be the first step in the landscape genetics of Iberian and brown
hare. Overlapping the species distribution and microsatellites data with ecological
variables, models can be designed to predict the correct location of the hybrid zone
along northern Spain. A transect study should also be performed across the hybrid zone
46

Chapter III
to characterize it in a quantitative way (width, cline steepness and cline concordance),
but the first indication of markers discordance in these species is set. Also, Y and X
chromosome markers would be a natural development of such work, in order to
assemble for these species the histories of uni (maternal and paternal) and biparentaly
inherited genomes.
References
Alves PC, Ferrand N, Suchentrunk F, Harris DJ (2003). "Ancient introgression of Lepus timidus mtDNA into L. granatensis and L. europaeus in the Iberian Peninsula" Molecular Phylogenetics and Evolution 27(1): 70-80.
Babik W, Szymura JM, Rafinski J (2003) "Nuclear markers, mitochondrial DNA and male secondary sexual traits variation in a newt hybrid zone {Triturus vulgaris x T. montandoniy\ Molecular Ecology, 12, 1913-1930.
Ballard JWO, Chernoff B, James AV (2002) "Divergence of mitochondrial DNA is not corroborated by nuclear DNA, morphology, or behaviour in Drosophila simulam". Evolution, 56, 527-545.
Barton NH (1979) "The dynamics of hybrid zones". Heredity, 43, 341-359.
Barton NH, Hewitt GM (1985) "Analysis of hybrid zones". Annual Review of Ecology and Systematics, 16, 113-148.
Bozikóva E, Muclinger P, Teeter KC, Tucker PK, Macholán M, Piálek J (2005) "Mitochondrial DNA in the hybrid zone between Mus musculus musculus and Mus musculus domesticus: a comparison of two transects". Journal of the Linnean Society, 84, 363-378.
Chan KMA, Levin SA (2005) "Leaky prezygotic isolation and porous genomes: rapid introgression of maternally inherited DNA". Evolution, 59, 720-729.
Flux JE, Angermann R (1990) "The hares and jackrabbits". In Chapman JA & Flux JEC (Eds.), "Rabbits, Hares and Pikas. Status Survey and Conservation Action plan". IUCN, Switzerland,pp. 61-94.
Grant PR, Grant BR (1997) "Hybridization, Sexual Imprinting, and Mate Choice". American Naturalist, 149, 1-28.
47

Paper 2
Guillot G, Estoup A, Mortier F, Cosson JF (2005) "A Spatial Statistical Model for Landscape Genetics". Genetics, 170, 1261-1280.
Harrison RG (1990) "Hybrid zones: windows on the evolutionary process". Oxford Surveys in Evolutionary Biology 7, 69-128.
Hewitt GM (1988) "Hybrid zones - natural laboratories for evolutionary studies". Trends in Ecology and Evolution, 3, 158-167.
Hijmans RJ, Cameron SE, Parra JL, James PG, Jarvis A (2005) "Very high resolution interpolated climate surfaces for global land areas". International Journal of Clomatology, 25, 1965-1978.
McDonald DB, Clay RP, Brumfield RT, Braun MJ (2001) "Sexual selection on plumage and behaviour in an avian hybrid zone: experimental tests of male-male interactions". Evolution, 55, 1443-1451.
Melo-Ferreira J, Esteves PJ, Alves PC (2004) "First evidence of natural and artificial hybridization between Lepus granatensis and L. europaeus". 2nd World Lagomorph Conference, Vairão, Portugal.
Melo-Ferreira J, Boursot P, Suchentrunk F, Ferrand N, Alves PC (2005). "Invasion from the cold past: extensive introgression of mountain hare (Lepus timidus) mitochondrial DNA into three other hare species in northern Iberia". Molecular Ecology 14(8): 2459-2464.
Palácios F, Meijide M (1979) "Distribución geográfica y habitat de las liebres en la Península Ibérica". Naturalia Hispânica, 19, 1-40.
Piálek J, Barton NH (1997) "The Spread of an Advantageous Allele Across a Barrier: The Effects of Random Drift and Selection Against Hétérozygotes". Genetics, 145, 493-504.
Powell JR (1983) "Interspecific Cytoplasmic Gene Flow in the Absence of Nuclear Gene Flow: Evidence from Drosophila". Proceedings of the National Academy of Sciences, 80, 492-495.
Redenbach Z, Taylor EB (2002) "Evidence for historical introgression along a contact zone between two species of char (PISCES: SALMONIDAE) in Northwestern North America". Evolution, 56, 1021-1035.
Roca AL, Georgiadis N, O'Brien SJ (2005) "Cytonuclear genomic dissociation in African elephant species". Nature Genetics, 37, 96-100.
Rognon X, Guyomard R (2003) Large extent of mitochondrial DNA transfer from Oreochromis aureus to O. niloticus in West Africa. Molecular Ecology, 12, 435-445.
48

Chapter III
Rohwer S, Bermingham E, Wood C (2001) "Plumage and mitochondrial DNA haplotype variation across a moving hybrid zone". Evolution, 55, 405-422
Rokas A, Ladoukakis E, Zouros E (2003) "Animal mitochondrial DNA recombination revisited", Trends in Ecology and Evolution, 18, 411-417.
Ruedi M, Smith MF, Patton JL (1997) "Phylogenetic evidence of mitochondrial DNA introgression among pocket gophers in New Mexico (family Geomyidae) ". Molecular Ecology, 6, 453-462.
Rundle HD, Breden F, Griswold C, Mooers A0, Vos RA, Whitton J (2001) "Hybridization without guilt: gene flow and the biological species concept". Journal of Evolutionary Biology, 14, 868-869.
Sambrook E, Fritsch F, Maniatis T (1989) Molecular Cloning. Cold Spring Harbour Press, Cold Spring Harbour, New York.
Servedio MR (2001) "Beyond reinforcement: the evolution of premating isolation by direct selection on preferences and postmating, prezygotic incompatibilities". Evolution, 55, 1909-1920.
Szymura JM, Barton NH (1991) "The Genetic Structure of the Hybrid Zone between the Fire-Bellied Toads Bombina bombina and B. variegata: Comparisons between Transects and between Loci". Evolution, 45, 237-261.
Tegelstrõm H (1987) "Transfer of mitochondrial DNA from the northern red-backed vole (Clethrionomys rutilus) to the bank vole (C. glareolus)". Journal of Molecular Evolution, 24, 218-227.
Thulin CG, Jaarola M, Tegelstrõm H (1997) "The occurrence of mountain hare mitochondrial DNA in wild brown hares". Molecular Ecology, 6, 463-467.
Thulin CG, Fang M, Averianov AO (2006) "Introgression from Lepus europaeus to L. timidus in Russia revealed by mitochondrial single nucleotide polymorphisms and nuclear microsatellites". Hereditas, in press.
Thulin CG, Stone J, Tegelstrõm H, Walker CW (2006) "Species assignment and hybrid identification among Scandinavian hares Lepus europaeus and L. timidus'". Wildlife Biology, 12, 29-38.
Wilson CC, Bernatchez L (1998) "The ghost of hybrids past: fixation of arctic charr (Salvelinus alpinus) mitochondrial DNA in an introgressed population of lake trout (S. namaycushy'. Molecular Ecology, 7, 127-132.
49

Chapter IV
IV. General Discussion
Natural hybridization is an evolutionary phenomenon that contributes to the dynamics
of natural populations of organisms. Common to plants and animals, it should not be
seen as a failure to the reproductive isolation (Arnold, 1997). However, the degree of
differentiation between organisms is correlated to their ability to hybridize, although the
degree of this dependence is arguable. Climate oscillations, with the advance and retreat
of ice sheets, are a recognized fundamental process to promote divergence between
organisms in allopatric refugia and posterior secondary contacts, giving rise to several
contact or hybrid zones. Obviously, these events of population expansion and retraction
involve several stochastic (e.g., genetic drift) and/or selective processes (e.g., adaptation
to new unoccupied habitats along colonization routes), which leave a genetic signature
and shape the genetic patterns we see today.
In this work, the assessment of natural hybridization between two species of
hare, the iberian hare (L. granatensis) and the brown hare (L. europaeus), in northern
Iberian Peninsula was the main objective. Despite the existence of a long contact zone
between both species in northern Iberia, hybridization between Iberian and brown hare
has not been described and has even been discarded (Estonba et al., 2006). Here, a more
detailed analysis of the potential admixture between these species was attained by the
use of a larger sampling and by the use of new analytical methodologies. Both nuclear
and mitochondrial markers were used to obtain a more complete scenario of Iberian and
brown hare histories and interaction. The genetic structure of both species was also
described and used to infer possible phylogeographic scenarios that fit species history.
This is discussed in Section 1 of the General Discussion. Then, natural hybridization
between the analysed species of hares, as inferred from microsatellites and cytochrome
b mitochondrial gene, is discussed in Sections 2 and 3, as well as the comparison
between both markers patterns. The integration of genetic and ecological data, and the
role of habitat in species distribution, are discussed in Section 3. A critic to the
application of different methodologies is presented in Section 4 and, finally, several
possibilities to expand this work in the future are presented in Section 5.
51

General Discussion
1) Genetic structure
In this work, three hare species had to be clearly distinguished at the molecular level. A
set of six microsatellites, comprising four originally developed for the wild-rabbit, one
specifically developed for African hares and one adapted for the brown hare, proved to
be polymorphic and to separate the three studied hare species, although in a frequency
base (see Chapter II). The high amount of homoplasy could constitute a problem, as
happens in the Iberian Peninsula wild-rabbit where microsatellites do not distinguish
between two well differentiated lineages (Queney et al, 2001). This, however, does not
seem to be the case in hares. In fact, both the Factorial Correspondence Analysis (FCA)
and the Bayesian STRUCTURE analyses displayed each species as a different cluster,
which is corroborated by the high Fst estimates between species (Fst(Lg-u) = 0,353;
Fst(Lg-Lt) = 0,205; FsX^-u) = 0,208). This resolution power allowed to exclude the
possible introgression of nuclear alleles from L. timidus as was reported for the mtDNA
(Alves et al, 2003; Melo-Ferreira et al, 2005) and focus in the hybridization problem
between the Iberian and the brown hare, the main purpose of this work. In fact, two
brown hare individuals from the Iberian Peninsula display some signs of admixture with
mountain hare. We may hypothesize that this results from an ancient hybridization
event with mountain hare in the Iberian Peninsula but the long term absence of this
latter species from the region (Sesé, 1994; Alves et al, 2003; Melo-Ferreira et al,
2005), along with the fast erasing rate of nuclear markers (Roca et al, 2005) and the
non-existence of such signal in the Iberian hare (Chapter II), argue against such
hypothesis. Another possible explanation would be the import of mountain hare alleles
through present hybridization events and dispersal within brown hare. Suchentrunk et
al. (unpubl. data) described present hybridization events between both species in
Central Europe, which may be supported by the detection, in this work (Chapter II), of
two Austrian (Aus 21 and LE Aus 26) and one Italian Alps (Alp 832) specimens with
admixture signs. Also, long distance gene dispersal within brown hare is described
(Suchentrunk et al, 2000) and, together, both phenomena could explain the presence of
mountain hare signal in Iberia. Finally, we must not eliminate the possibility of lack of
resolution of the markers for some particular multilocus genotypes. The increase in
markers numbers should be enough to eradicate this problem.
52

Chapter IV
In more general terms, microsatellites reveal very interesting patterns. The
Iberian hare species displays lower values of genetic diversity, perhaps reflecting small
historical population size given its confinement to the Iberian Peninsula or the origin of
the entire current metapopulation from a small refugiai population. On the other hand,
brown and mountain hare display higher, similar genetic diversity values. The analysed
samples of this latter species represent two very different contexts: one (Sweden) was
collected within the supposedly continuous artic habitat of the species, while the other
(Italian Alps) represents the relictual southernmost distribution of L. timidus. However,
excepting the locus Sol30, no substantial differences between both samples are found.
More than one wave of colonization of the Alps by mountain hare, as suggested by
Melo-Ferreira et al. (in press) could have provided a representative sample of the
original genetic pool, overcoming the reduction in genetic diversity usually detected in
such situations. Regarding the brown hare, northern Iberia and Central Europe samples
display similar values of diversity (heterozygosities and mean number of alleles per
locus (MNA)). However, removing the putative foreign alleles (i.e., alleles that only
occur in Iberian hare parental samples) that occur in the Spanish samples leads to a
considerable decrease of their MNA values. Together with the genetic diversity analysis
performed by Estonba et al. (2006) in Bulgarian L. europaeus, a westwards decrease in
genetic diversity parameters can be observed, especially the MNA. The Iberian hare
also shows an increment in MNA values of the contact zone samples, while
heterozygosities remains stable among all samples. These results argue for a stable
metapopulation of Iberian hare with high gene flow between subpopulations within
Iberia, in agreement with the allozyme data described by Alves (2002), and for a
westwards colonization of Europe by the brown hare, with origin in a Balkans glacial
refugium, in concordance with Suchentrunk et al. (2003) and Kasapidis et al. (2005). A
recent post-glacial contact between both species apparently provided the conditions for
the occurrence of hybridization, which is supported by the high values of MNA in the
contact zone samples.
53

General Discussion
2) Natural hybridization in hares
In hares, natural hybridization has been an overlooked subject. It has been mostly
approached as a secondary issue in phylogenetic and phylogeography studies (Pérez-
Suarez et al, 1994; Pierpaoli et al, 1999; Alves et al, 2003; Melo-Ferreira et al, 2005;
Estonba et al, 2006) and not as a main question with potentially evolutionary
importance. Within the genus Lepus, only the contact zones between the brown and
mountain hare in Sweden (Thulin et al, 1997; Thulin et al, 2006a), Russia (Thulin et
al, 2006b) and Switzerland (Suchentrunk et al, unpubl. data) have been screened for
the existence of hybridization, which was found to occur. Together, these studies used
mtDNA, microsatellites and allozymes to search for admixture signs. The use of diverse
types of markers allows answering to distinct questions, since their mode of
transmission, selective pressures and intrinsic dynamics is different. In hybrid zones,
discordant introgression patterns of the markers are often detected, especially if
uniparental and biparentally inherited markers are compared (Chan & Levin, 2005). In
this work, the question of hybridization between the Iberian and brown hare in northern
Iberian Peninsula was first approached with microsatellites (Chapter II). The use of
mtDNA for this purpose was prevented by the massive ancient introgression of the
mountain hare mitochondrial genome into both Iberian and brown hare, previously
described by Alves et al (2003) and Melo-Ferreira et al (2005). Nonetheless, mtDNA
was used to check for admixture between Iberian and brown hare in a large thin scale
sample (Chapter III).
2.1) Patterns revealed by microsatellites
The microsatellites set used in this work was similar in size (six) to the one used to
distinguish the three hare species from Iberia (brown, broom and Iberian hare) by
Estonba et al (2006). Three of the microsatellites were common to both studies. Both
sets of microsatellites allowed a clear distinction between the Iberian and the brown
hare, and the one used in this thesis also identified the mountain hare as a separate
cluster (see Section 1 and Figure 1).
54

Chapter IV
Figure 1: Identification of the three species by STRUCTURE. In blue are the Iberian hare samples; in light brown the brown hare samples; in dark brown, the mountain hare samples.
\ n r | l , j p i y i V"\
JÃ &* ^0%0< ^ ^ ~ ? ^ j ^ ï ^ S ^ f e j e ^ fî»* ^V Q?
Hybridization between Iberian and brown hare was discarded by Estonba et al. (2006)
but, using a larger and more exhaustive sampling of the contact zone, robust evidence
for it was detected in the sample of Alava (Chapter II). Four specimens, previously
attributed to Iberian hares in a morphological basis, were identified by the three used
softwares as intermediate between both species. NEWHYBRIDS gives them high
probabilities of representing backcrosses with Iberian hare, which could justify their
morphological identification. If we use less restrictive conditions (i.e., not demanding
the intersection of all softwares) to accept potential hybrids, three more individuals from
Alava (one Iberian hare and two brown hares) and one from Jaca may be considered as
hybrids. These results point to a bidirectional introgression of alleles, which is in
agreement with the MNA values, that display and increase in all the contact zone
samples, either from Iberian or brown hare. Asymmetry in introgression may, however,
occur. Demographic reasons such as differences in density between species in the
contact zone may justify it (density-dependent hybridization; Wirtz, 1999), as well as
distinct behaviours (competition for females, for example, is only described for brown
hare; Flux & Angermann, 1990). Another bias in this study is the genetic diversity of
species. Brown hare has a wider range of frequent alleles, and a higher amount of those
are private to brown hare species, in comparison with the Iberian hare. This leads to an
easier detection of introgression from brown to Iberian hare, but does not exclude the
converse. More markers should be used in the future to account for this problem. From
our data, Alava seems to be the region with higher amount of hybridization. This can
however be merely due to a sampling bias. The sample of Alava is the largest of the
work and was collected exactly in the transition between both species, while the other
55

General Discussion
samples are more distantly located from the contact zone. Therefore, and since we
expect a reduction in the hybridization signal with the increasing distance from the
center of the hybrid zone, the results may simply be interpreted as a resolution problem
and not a real pattern. Increasing samples and sampling closer to the hybrid zone should
contribute for clarifying this issue.
2.2) Patterns revealed by mtDNA
At the mtDNA level, introgression of mountain hare native genome was expected in the
northern Iberia populations, but not in hare populations outside Iberia or in southern
populations within Iberia (Alves et al, 2003; Melo-Ferreira et al, 2005). This pattern
was confirmed in this work, since no mtDNA introgression was found in Northern
Germany, Southern Germany, Austria, Aljustrel or Granada samples (data not shown).
However, new knowledge was obtained from the analysis of an increased sample of
Alava (Chapter III), in comparison with the mentioned studies, since it allowed to detect
brown hare individuals with native brown hare mtDNA haplotype and, also, brown hare
specimens with introgressed Iberian hare mtDNA. None Iberian hare individual with
brown hare mtDNA was found, configuring a unidirectional transfer of mtDNA from
Iberian to brown hare (Figure 2).
Figure 2: MtDNA distribution in the study area. Blue corresponds to Iberian hare haplotypes; white corresponds to brown hare haplotypes; light brown corresponds to mountain hare haplotypes.
56

Chapter IV
Apparently, brown hare is a species that easily captures the mtDNA from related taxa.
In Sweden, unidirectional introgression from mountain to brown hare has bee reported
for the mtDNA (Thulin et al, 1997) and now a similar unidirectional transfer from
Iberian to brown hare was detected in Iberian Peninsula. The extent to which this occurs
in the analysed sample from northern Iberia (-33%) is more than three-fold higher than
the one found in Sweden (-10%). The opposite situation (brown hare mtDNA
introgressing into other species) was detected in Sweden (Thulin et al, 2006a) and
Russia (Thulin et al, 2006b), but this seems to occur to a much smaller extent (3,3%,
6,5%, respectively). In Switzerland (Suchentrunk et al, unpubl. data), bidirectional
introgression of mtDNA was found to occur, also in low amount (-6%). Thus, in the
Iberian Peninsula, the brown hare harbours three different mitochondrial genomes and,
curiously, its native mtDNA is not predominant (Melo-Ferreira et al, 2005).
2.3) Discordance between mtDNA and microsatellites
Nuclear and mitochondrial genomes have fundamental distinct characteristics, and are
differently used in population genetics studies according to them and to the proposed
objectives. Mitochondrial DNA has played and continues to play an important role in
population genetics and phylogeography studies (Moritz et al, 1987; Ballard & Rand,
2005). The ease of getting new sequence data for a set of orthologous genes and the
availability of mtDNA sequences for a wide variety of species, in part due to the
absence of recombination or selection of mtDNA (although studies exist opposing to
this strictly neutral role of mtDNA -Nachman, 1998; Rokas et al, 2003; Bazin et al,
2006), justifies the broad use of this marker. However, extreme caution must be used
when inferring population history or species relatedness solely based in mtDNA (see
Alves et al, 2006). It is known that the history of a gene might not be the history of the
species (Maddison, 1997). MtDNA is a maternally inherited genome, with none or low
levels of recombination. Therefore, it behaves as a single locus, which may bias
analysis. However, phylogenies based only in mtDNA are still being published,
sometimes with seriously flawed conclusions (e.g. Wu et al., 2005). Another potentially
biasing feature of mtDNA is its high capability of jumping species barrier and introgress
57

General Discussion
into related species (Chan & Levin, 2005). On the other hand, the diploid, biparently
inherited nuclear DNA (nDNA) presents a more complex dynamics. Portions of the
nDNA may, for example, be under strong direct selection, may be selected by hitch
hiking (Maynard-Smith & Haigh, 1974) or background selection (Charlesworth, 1994),
may suffer recombination, duplications, gene conversion or may be neutral. Again, the
history of a gene or a neutral portion of the nDNA might not be, and often is not, the
history of the species and that is the reason why biologists started to simultaneously
combine several markers and locus in their works.
In this thesis, microsatellites and mtDNA from the hybridizing hare population
of Alava are found to display discordant patterns (Figure 3 of this Chapter). Such result
was expected since a high level of introgression from the mountain hare is described,
both for the Iberian and brown hare of this area. In fact, the latter species revealed a
total replacement of its mtDNA by the mountain hare type (Melo-Ferreira et al, 2005).
However, as shown in Chapter III, native brown hare and introgressed Iberian hare
mtDNA were for the first time detected in the brown hare population from Alava, which
displays around 63% of mountain hare mtDNA, 33% of Iberian hare mtDNA and 4% of
native brown hare mtDNA. Therefore, the homogenizing mountain hare mtDNA is still
heavily present in Alava (63% in brown hare and 31% in Iberian hare) but it is now
possible to detect a unidirectional transfer of mtDNA from Iberian to brown hare, which
is not accompanied by microsatellites.
Figure 3: Distribution of microsatellites genotypes (right) and mtDNA haplotypes (left) in the study area, showing discordant patterns. Blue corresponds to Iberian hare, white to brown hare and light brown to mountain hare.
58

Chapter IV
With these markers, unidirectional introgression is seen in the opposite direction (brown
hare alleles in four Iberian hares) when using more conservative conditions to detect
hybrids. With more relaxed conditions, another Iberian hare and two brown hares are
classified as hybrids. Together with some diversity parameters that increase in the
contact zone samples of both species, bidirectional hybridization may be hypothesized.
The mtDNA pattern demands that Iberian hare females cross with brown hare males and
that the progeny backcross with brown hare. The converse (brown hare females with
Iberian hare males) is not excluded, but was not detected by the performed analysis.
Also, the backcross of hybrid individuals with Iberian hares must occur, as supported by
the detection of nuclear brown hare alleles' introgression into the Iberian hare. To
account for this mtDNA and microsatellites pattern, density-dependent hybridization
(Wirtz, 1999) can play a role, since Iberian hares seem to outnumber brown hares in the
region (Ibón Telletxea, unpubl. data). The density fluctuations suffered by hares due to
epidemics or hunting may also influence hybridization, since they can be quite severe
and so move the density trough. Also, Grant & Grant (1997) principle (when two
species meet and hybridize, crosses are usually between the females of the smaller one
with males of the larger) may be acting in hares, since Iberian hare individuals are
smaller than brown hares.
Discordant patterns between mtDNA and nDNA are commonly found in several
organisms. For example, cichlid fishes from West Africa (Rognon & Guyomard, 2003)
show a strongly supported separation of two species with allozymes data, but an almost
complete replacement of one species mtDNA for the other species mitochondrial
genome occurs. This has also been found in other species of fish, as the attic char and
the lake trout (Wilson and Bernatchez, 1998) or the bull trout and the Dolly Varden
(Redenbach & Taylor, 2002). In newts, even though some nuclear introgression is
detected, mtDNA introgression is much more pronounced and bidirectional, even if a
14% excess of one of the species is seen (Babik et al., 2003). Another noteworthy work
was performed in commensal mice, where two transects (one in Czech Republic and
one in Bavaria) were analysed (Bozikova et al., 2005). The authors of this work found
that mtDNA clines fell into the range of the allozymes but not with that of X markers.
An interesting feature of this study is that the mtDNA introgression occurs from one
59

General Discussion
species to the other, but the direction changes between the Czech and the Bavarian
transects. The hybrid zone between Iberian and brown hare in northern Spain is, thus,
another case of discordance between markers, where the introgression of a third species
mtDNA adds to the complexity of the pattern.
3) Association of genetic and ecological data
Explaining observed spatial genetic patterns with landscape variables led to the
emergence of landscape genetics (Manel et al., 2003). Its development and use was
enhanced by the improvement of molecular genetics and statistical techniques, as well
as the increase of computation power. The association between organisms distribution
and geography is for long known and constitutes the object of study of biogeography
(Crisci, 2001), whereas more recently, evolutionary biologists focused in genealogical
lineages distribution in space, constituting phylogeography (Avise, 2000). However,
landscape genetics uses typically thinner scale sampling than biogeography or
phylogeography and aims to provide insight about the interaction between landscape
features and microevolutionary processes (e.g. gene flow, genetic drift, selection; Manel
et al, 2003). The ability of unravelling population substructure and cryptic genetic
boundaries are two of the main advantages of landscape genetics. In fact, the detection
of genetic discontinuities and their correlation with environmental features (e.g.,
mountains, rivers, roads, gradient of humidity, and deforested areas) are two key steps
of this discipline (Manel et al, 2003).
In this thesis, a clear genetic discontinuity was detected in the area of Alava for
microsatellites, distinguishing two classes (Chapter III). The transition between the two
units perfectly fitted the Cantabrian Mountain, which divides the study area (Figure 4).
Species distribution, as identified in a morphological basis, was concordant with
microsatellites markers, whereas mtDNA displayed a more chaotic distribution with no
correlation with the environmental feature, due to introgression (see Chapter III and
Section 2 of this Chapter).
60

Chapter IV
Figure 4: Species distribution fits microsatellites transition (yellow line). To north of this line occurs brown hare, whereas to the south occurs Iberian hare. The microsatellites transition fits the ecotone found in the area (Cantabrian Mountain). The size of dots is proportional to the number of individuals collected in each coordinate.
The occurrence of species transition in such a sharp ecotone suggests different
specific ecological requirements. In fact, the brown hare individuals, located to the
north of the mountain, occur in significantly rainier and colder spots than the Iberian
hares, located to the south of the mountain (Chapter III). These results are in agreement
with Palácios & Meijide (1979). One must also consider another possibility: if the
hybrid zone between Iberian and brown hare is, in fact, a tension zone, it will tend to get
trapped in a density trough (Hewitt, 1988). The Cantabrian Mountain may promote such
a decrease in densities, and so the hybrid zone position may be a result of this
phenomenon and not of an active selection of the habitat by the individuals. Also
investigated was the possibility that individuals harbouring the mtDNA native of the
mountain hare would preferentially occur in colder places within the study area, since
this mitochondrial genome has been suggested to confer advantages in cold areas
(Melo-Ferreira et al, 2005). However, no evidence was found supporting this
hypothesis (Chapter III). On the other hand, the result presented here may be biased by
sampling. The number of individuals sampled is probably low for such study and
individuals were collected in a small study area (60x50 km). The resolution used is
probably inadequate to test this hypothesis and so, no conclusions can be drawn about
selection for the cold of mountain hare mtDNA type.
61

General Discussion
4) The use of new analysis methodologies
The large number of programs presently available provides researchers the ability to
tackle several questions and issues previously untreatable. However, the choice of how
to approach a problem may be a delicate theme, since it is necessary to understand the
capacity and limitations of each software to correctly apply it and, mainly, to interpret
the results.
In Chapter II, the well known and widely used individual assignment Bayesian
software STRUCTURE (Pritchard et al, 2000) was used together with the FCA
analysis of GENETIX (Belkhir et al, 2004) and the more recent NEWHYBRIDS
software (Anderson & Thompson, 2003). Although none of these analyses methods is
new, their joint use is a novelty and is justified by the following reasoning: FCA is a
canonical analysis that allowed to project individuals in microsatellites allele frequency
space with no model assumption. On the other hand, STRUCTURE implements a
model-based clustering method for inferring population structure using genotype data
consisting of unlinked markers. This program probabilistically assigns individuals in the
sample to a population or jointly to two or more populations if its genotype is found to
be admixed. Finally, NEWHYBRIDS is also a model-based software that aims to
compute the posterior distribution that individuals in a sample fall into different parental
or hybrid categories. The objective of using these three programs was i) to apply very
conservative conditions to accept the admixed ancestry of individuals and ii) to have
more resolution of the interaction between the Iberian and brown hare. The first
objective was achieved by only accepting as admixed those individuals identified as
intermediate by the three programs; the second objective was attained since hybrid
individuals were identified and also attributed to post-Fl generations of hybrids. This is
very important since it means that at least some Fl individuals are fertile and reproduce.
Such information cannot be confidently obtained from STRUCTURE'S output, given
the model underlying this software, which justifies the use of NEWHYBRIDS. The
methodology used in Chapter II was, thus, very restrictive but allowed to find robust
evidence of hybridization between Iberian and brown hare, which had been previously
discarded (see Chapter II and Section 2 of this Chapter).
62

Chapter IV
In Chapter III, the recently developed program GENELAND (Guillot et al,
2005a; Guillot et al, 2005b) was applied. It consists in a Bayesian model implemented
in a Markov chain Monte Carlo scheme that allows inference of the location of genetic
discontinuities from individual geo-referenced multilocus genotypes, without a priori
knowledge on populational units and limits (Guillot et al, 2005a). GENELAND was
used to detect genetic discontinuities for microsatellites and for mtDNA in a small study
area in Alava comprising the hybrid zone, or at least part of it. Both markers revealed
very different patterns (see Chapter III and sub-Section 2.3 of this Chapter). These
results were integrated with a GIS approach and a perfect concordance between
microsatellites transition and a landscape ecotone (the Cantabrian Mountain) was found.
Extending the GIS analyses, environmental (precipitation, minimum and maximum
annual temperature) and habitat conditions (elevation, slope and aspect) were compared
between both sides of the transition area to search for habitat preferences of both species
of hares. The use of software as GENELAND is a good tool for landscape genetics and
a valuable first approach to GIS modelling.
The results presented in this thesis using the mentioned softwares argue for the
validity of their integrated use, taking advantage of each program potential. The
assumptions underlying each of them must be taken into account in order to draw
correct inferences and to have confident results. Together, the interaction between
Iberian and brown hare, as well as some aspects of their ecology, were tackled during
this work, which must of course be further investigated.
5) Future perspectives
This work might redefine the knowledge about Iberian and brown hare interaction and
give rise to new perspectives for studying it. For the first time, evidence was found to
support hybridization between these two species. Nevertheless, several questions remain
to be answered. The amount of hybridization, its directionality, the centre and width of
the hybrid zone, the penetrance of introgressed alleles or, more generally, the type of
hybrid zone are still open issues. A well defined experimental design for a transect
study of the area is needed to answer such questions.
63

General Discussion
The number of markers should be increased to have more resolution power.
Microsatellites proved to be useful and increasing the number of microsatellites used
could render good results. Given their high mutation rate, recent genetic novelties could
be found and studied to infer some demographic parameters as dispersal. The locus
Sol30 presents a possible new allele (allele 218), which is only present in the samples of
Alava of both species. Sequencing this allele is important to understand the mechanism
by which it arose. Other rare, clustered alleles could prove valuable for microscale
demographic studies. The use of other types of markers would also help to complement
and add knowledge to what is known. Typing Y chromosome markers would input the
parental history to the scenario and diagnostic SNP's could refine the analyses and
clarify some hypothesis.
Increasingly important information to be taken is the geographic localization of
the captured individuals. A larger sampling effort in the hybrid zone and the use of
adequate resolution information layers may lead to a good modelling and extrapolation
of species habitat preferences. Such information would be useful not only for an
academic motif but also for conservation reasons. Finally, sampling all the northern
Iberia area where mountain hare mtDNA introgression occurs and using a landscape
genetics approach may lead to exciting results concerning the hypothesis of selection for
the cold of this mtDNA type.
64

Chapter V
Conclusions
The work presented in this thesis contributes to a better knowledge about the genus
Lepus by analysing a particular situation, the interaction between Iberian and brown
hare in northern Iberia. It may also prove important for the opening of new perspectives
and new working hypothesis in that context, since hybridization between both species
had been previously discarded. Some major conclusions of the performed work are:
i) The use of a microsatellites set that distinguishes Iberian, brown and
mountain hare allowed to discard a massive introgression of mountain
hare nuclear DNA into both Iberian and brown hare, as described for
mitochondrial DNA (mtDNA). The shallow evidence for such
phenomenon may stem from the used resolution power and so increasing
the number of markers should clarify this situation.
ii) Robust signs of admixture between Iberian and brown hare were
detected, both at the microsatellites and mtDNA level. The direction of
introgression is opposite for both type of markers, which can be
explained by demographic processes, but reciprocal introgression is still
a hypothesis that deserves to be further investigated. The occurrence of
introgressive hybridization demands the existence of fertile admixed
offspring, as supported by the detection of post-Fl hybrid individuals. In
this way, the hybrid zone is effective and, in this case as in several
others, Fl individuals are rarely observed.
iii) As described for several other taxa, discordance between nuclear and
mitochondrial DNA patterns was found. This stresses the need of using
different kinds of markers to prevent the bias in the analyses introduced
by differences in their dynamics, contributing to obtain a reliable history
of the species.
65

Conclusions
iv) The genetic diversity data obtained during this work joins the body of
evidence for the colonization of Western Europe by brown hare with
origin in the Balkans, having recently arrived to the Iberian Peninsula.
On the other hand, Iberian hare may fit a south to north colonization
scenario within Iberian Peninsula, but a stable metapopulation with high
gene flow between populations within this region is also plausible.
v) The genetic discontinuity found at the level of microsatellites mimics
species distribution, as identified in a morphological basis. It also
perfectly fits the ecotone found in the study area, pointing in the direction
of the important role environment may play in species geographic
allocation. In general terms, brown hare occurs in more humid and colder
areas than Iberian hare, which should be extended and tested in different
regions, namely in thin scale analyses and ecological modelling.
vi) No correlation was found between the distribution of different
mitochondrial types and ecological variables so, no evidence for the
adaptation of mountain hare mtDNA to colder regions was obtained.
66

Chapter VI
References
Alves PC (2002) "Caracterização genética e biologia reprodutiva da lebre Ibérica, Lepus granatensis. Análise filogenética, diferenciação populacional e ciclo anual de reprodução. " PhD thesis, University of Porto.
Alves PC, Ferrand N, Suchentrunk F, Harris DJ (2003). "Ancient introgression of Lepus timidus mtDNA into L. granatensis and L. europaeus in the Iberian Peninsula". Molecular Phylogenetics and Evolution, 27, 70-80.
Alves PC & Niethammer J (2003) "Die iberischen Hasen. In: Jochen Niethammer und Franz Krapp: Handbuch der Sãugetiere Europas. Band 2/II (herausgegeben von Franz Krapp). Aula-Verlag.
Anderson EC, Thompson EA (2002) "A model-based method for identifying species hybrids using multilocus genetic data". Genetics, 160, 1217-1229.
Angerbjõrn A, Flux J E C (1995) "Lepus timidus". Mammalian. Species, 495, 1-11.
Angermann R, (1983) "The taxonomy of Old World Lepus". Acta Zoológica Fennica 174, 17-21.
Arnold ML (1994) "Natural hybridization in Lousiana irises". BioScience, 44, 141-147.
Arnold ML (1997) "Natural hybridization and evolution". Oxford University Press, New York.
Avise JC (2000) "Phylogeography: the history and formation of species". Harvard University Press, Massachussets.
Ballard JWO, Rand DM (2005) "The population biology of mitochondrial DNA and its phylogenetic implications". Annual Review of Ecology and Systematics, 36, 621-642.
Barluenga M, Storting KN, Salzburger W, Muschick M, Meyer A (2006) "Sympatric speciation in Nicaraguan crater lake cichlid fish". Nature, 439, 719-723.
Barton NH (1979) "The dynamics of hybrid zones". Heredity, 43, 341-359.
Barton NH, Hewitt GM (1985) "Analysis of hybrid zones". Annual Review of Ecology and Systematics, 16, 113-148.
Barton NH, Gale KS (1993) "Genetic analysis of hybrid zones" in "Hybrid Zones and the Evolutionary Process". Oxford University Press, New York.
67

References
Bazin E, Glemin S, Galtier N (2006) "Population size does not influence mitochondrial genetic diversity in animals". Science, 312, 570-572.
Belkhir K, Borsa P, Chikhi L, Raufaste N, Bonhomme F (1996-2004) GENETIX 4.05, logiciel sous Windows TM pour la génétique des populations. Laboratoire Génome, Populations, Interactions, CNRS UMR 5171, Université de Montpellier II, Montpellier (France).
Boursot P, Auffray JC, Britton-Davidian J, Bonhomme F (1993) "The evolution of house mice". Annual Review of Ecology and Systematics, 24, 119-152.
Bozikóva E, Muclinger P, Teeter KC, Tucker PK, Macholán M, Piálek J (2005) "Mitochondrial DNA in the hybrid zone between Mus musculus musculus and Mus musculus domesticus: a comparison of two transects". Journal of the Linnean Society, 84, 363-378.
Carr SM, Ballinger SW, Derr JN, Blankenship LH, Bickham JW (1986) "Mitochondrial DNA analysis of hybridization between sympatric white-tailed deer and mule deer in west Texas". Proceedings of the National Academy of Sciences, 83, 9576-9580.
Chan KMA, Levin SA (2005) "Leaky prezygotic isolation and porous genomes: rapid introgression of maternally inherited DNA". Evolution, 59, 720-729.
Charlesworth B (1994) "The effect of background selection against deleterious mutations on weakly selected, linked variants". Genetical Research, 63, 213-27.
Corbet GB (1983) "A review of the classification in the family Leporidae". Acta Zoológica Fennica, 174, 11-15.
Coyne JA, Orr AH (1998) "The evolutionary genetics of speciation". Philosophical Transactions of the Royal Society of London, 353, 287-305.
Crisci JV (2001) "The voice of historical biogeography". Journal of Biogeography, 28, 157-168.
Dingerkus SK, Montgomery WI (2002) "A review of the status and decline in abundance of the Irish Hare (Lepus timidus hibernicus) in Northern Ireland". Mammal Review, 32, 1-11.
Dobzhansky T (1937) "Genetics and the origin of species". Columbia University Press, New York.
Ellstrand NC, Whitkus R, Rieseberg LH (1996) "Distribution of spontaneous plant hybrids". Proceedings of the National Academy of Sciences, 93, 5090-5093.
Endler JA (1977) "Geographic variation, speciation, and clines". Princeton University Press, New Jersey.
68

Chapter VI
Estonba A, Solis A, Iriondo M, Sanz-Martin MJ, Pérez-Suaréz G, Markov G, Palácios F (2006) "The genetic distinctiveness of the three Iberian hare species: Lepus europaeus, L. granatensis, and L. castroviejoi". Mammalian Biology, 71, 52-59.
Flux JE, Angermann R (1990) "The hares and jackrabbits". In Chapman JA & Flux JEC (Eds.), Rabbits, Hares and Pikas. Status Survey and Conservation Action plan. IUCN, Switzerland,pp. 61-94.
Geraldes A, Ferrand N, Nachman MW (2006) "Contrasting Patterns of Introgression at X-Linked Loci Across the Hybrid Zone Between Subspecies of the European Rabbit {Oryctolagus cuniculus)". Genetics, 143, 919-933.
Grant V (1981) "Plant speciation". Columbia University Press, New York.
Guillot G, Estoup A, Mortier F, Cosson JF (2005a) "A Spatial Statistical Model for Landscape Genetics". Genetics, 170, 1261-1280.
Guillot G, Mortier F, Estoup A (2005b) "Geneland: a computer package for landscape genetics". Molecular Ecology Notes, 5, 712-715.
Grant PR, Grant BR (1997) "Hybridization, Sexual Imprinting, and Mate Choice". American Naturalist, 149, 1-28.
Gridley J (1912) "The lagomorphs and independent order". Science, 36, 285-286.
Gustavsson, I (1971)" Mitotic and meiotic chromosomes of the variable hare (Lepus timidus L.), the common hare (Lepus europeus Pall.) and their hybrids". Hereditas 67, 27-34.
Gustavsson I, Sundt CO (1965) "Anwendung von kiinstlicher Befruchtung bei der Hybridisierung von zwei Hasenarten". Zeitschrift Fur Jagdwissenschaft, 11, 155-158.
Harrison RG (1990) "Hybrid zones: windows on the evolutionary process". Oxford Surveys in Evolutionary Biology 7, 69-128.
Hewitt GM (1988) "Hybrid zones - natural laboratories for evolutionary studies". Trends in Ecology and Evolution, 3, 158-167.
Hewitt G (2000) "The genetic legacy of the Quaternary ice ages". Nature, 405, 907-913.
Hewitt GM (2001) "Speciation, hybrid zones and phylogeography - or seeing genes in space and time". Molecular Ecology ,10, 537-549.
Hey J, Waples RS, Arnold ML, Butlin MK, Harrison RG (2003) "Understanding and confronting species uncertainty in biology and conservation". Trends in Ecology and Evolution, 18,597-603.
69

References
Homolka M (1985) "Spatial activity of hares (Lepus europaeus)". Folia Zoológica Brno, 34,217-226.
Hubbs CL (1955) "Hybridization between fish species in nature". Systematic Zoology, 4, 1-20.
Kasapidis P, Suchentrunk F, Mogoulas A, Kotoulas G (2005) "The shaping of mitochondrial DNA phylogeographic patterns of the brown hare (Lepus europaeus) under the combined influence of Late Pleistocene climatic fluctuations and anthropogenic translocations". Molecular Phylogenetics and Evolution, 34, 55-66.
King RA, Ferris C (1998) "Chloroplast DNA phylogeography of Alnus glutinosa (L.) Gaertn". Molecular Ecology, 7, 1151-1162.
Knobloch IW (1972) "Intergeneric hybridization in flowering plants". Taxon, 21, 97-103.
Maddison WP (1997) "Gene trees in species trees". Systematic Biology, 46, 523-526.
Mallet J (2005) "Hybridization as an invasion of the genome". Trends in Ecology and Evolution, 20, 229-37.
Manel S, Schwartz MK, Luikart G, Taberlet P (2003) "Landscape genetics: combining landscape ecology and population genetics". Trends in Ecology and Evolution, 18, 189-197.
Marboutin E, Bray Y, Peroux R, Mauvy B, Lartiges A (2003) "Population dynamics in European hare: breeding parameters and sustainable harvest rates". Journal of Applied Ecology, 40, 580-591.
Maynard-Smith J & Haigh J (1974) "The hitch-hiking effect of a favourable gene". Genetical Research, 23, 23-35.
Mayr E, Short LL (1970) "Species taxa of North American birds". Nuttall Ornithological Club, Publication no. 9 , Cambridge, Massachussets.
Melo-Ferreira J, Esteves PJ, Alves PC (2004) "First evidence of natural and artificial hybridization between Lepus granatensis and L. europaeus". 2nd World Lagomorph Conference, Vairão, Portugal.
Melo-Ferreira J, Boursot P, Suchentrunk F, Ferrand N, Alves PC (2005) "Invasion from the cold past: extensive introgression of mountain hare (Lepus timidus) mitochondrial DNA into three other hare species in northern Iberia". Molecular Ecology, 14, 2459-2464.
Melo-Ferreira J, Boursot P, Randi E, Kryukov A, Suchentrunk F, Ferrand N, Alves PC (2006) "The rise and fall of the mountain hare (Lepus timidus) during Pleistocene
70

Chapter VI
glaciations: expansion and retreat with hybridization in the Iberian Peninsula". Molecular Ecology, in press
Mitchell-Jones AJ, Amori G, Bogdanowicz W (1999) "Atlas of European Mammals". Academic Press, London.
Moritz C, Dowling TE, Brown WM (1987) "Evolution of Animal Mitochondrial DNA: Relevance for Population Biology and Systematics". Annual Review of Ecology and Systematics, 18, 269-292.
Nachman M, Brown WM, Stoneking M, Aquadro CF (1996) "Nonneutral Mitochondrial DNA Variation in Humans and Chimpanzees". Genetics, 142, 953-963.
Otto SP, Whitton J (2000) "Polyploid incidence and evolution". Annual Review of Genetics, 34,401-437.
Palácios F (1983) "On the taxonomic status of the genus Lepus in Spain". Acta Zoológica Fennica, 174, 27-30.
Palácios F (1989) "Biométrie and morphologic features of the species of the genus Lepus in Spain". Mammalia, 53, 227-264.
Palácios F, Meijide M (1979) "Distribución geográfica y habitat de las liebres en la Península Ibérica". Naturalia Hispânica, 19, 1-40.
Panova M, Hollander J, Johannesson K (in press) "Site-specific genetic divergence in parallel hybrid zones suggests nonallopatric evolution of reproductive barriers". Molecular Ecology.
Pérez-Suárez G, Palácios F, Boursot P (1994) "Speciation and paraphyly in western Mediterranean hares {Lepus castroviejoi, L. europaeus, L. granatensis, and L. capensis) revealed by mitochondrial DNA phylogeny". Biochemical Genetics, 32, 423-436.
Phillips BL, Baird S JE, Moritz C (2004) "When vicars meet: a narrow contact zone between morphologically cryptic phylogeographic lineages of the rainforest skink, Carlia rubrigularis".Evolution, 58, 1536-1548.
Piálek J, Barton NH (1997) "The Spread of an Advantageous Allele Across a Barrier: The Effects of Random Drift and Selection Against Hétérozygotes". Genetics, 145, 493-504.
Pierpaoli M, Riga F, Trocchi V, Randi E (1999) "Species distinction and evolutionary relationships of the Italian hare (Lepus corsicanus) as described by mitochondrial DNA sequencing". Molecular Ecology, 8, 1805-1817.
71

References
Pierpaoli M, Birò ZS, Herrmann M, Hupe K, Fernandes M, Ragni B, Szemethy L , Randi E (2003) "Genetic distinction of wildcat (Felis silvestris) populations in Europe, and hybridization with domestic cats in Hungary". Molecular Ecology, 12, 2585-2598.
Pritchard JK, Stephens M, Donnelly P (2000) "Inference of population structure using multilocus genotype data". Genetics, 155, 945-959.
Queney G, Ferrand N, Weiss S, Mougel F, Monnerot M (2000) "Stationary distributions of microsatellite loci between divergent population groups of the European rabbit {Oryctolagus cuniculus)". Molecular Biology and Evolution, 18, 2169-2178.
Rand DM, Harrison MG (1989) "Ecological Genetics of a Mosaic Hybrid Zone: Mitochondrial, Nuclear, and Reproductive Differentiation of Crickets by Soil Type". Evolution, 43, 432-449.
Redenbach Z, Taylor EB (2002) "Evidence for historical introgression along a contact zone between two species of char (PISCES: SALMONIDAE) in Northwestern North America". Evolution, 56, 1021-1035.
Remington C (1968) "Suture-zones of hybrid interaction between recently joined biotas". Evolutionary Biology, 2, 321^128.
Rieseberg LH (1997) "Hybrid Origins of Plant Species". Annual Review of Ecology and Systematics, 28, 359-389.
Ridley M (1996) "Evolution". Blackwell Science Cambridge, Massschussets.
Robinson TJ, Matthee CA (2005) "Phylogeny and evolutionary origins of the Leporidae: a review of cytogenetics, molecular analyses and a supermatrix analysis". Mammal Review, 35, 231-247.
Roca AL, Georgiadis N, O'Brien S J (2005) "Cytonuclear genomic dissociation in African elephant species". Nature Genetics, 37, 96-100.
Rognon X, Guyomard R (2003) Large extent of mitochondrial DNA transfer from Oreochromis aureus to O. niloticus in West Africa. Molecular Ecology, 12, 435-445.
Rokas A, Ladoukakis E, Zouros E (2003) "Animal mitochondrial DNA recombination revisited", Trends in Ecology and Evolution, 18, 411-417.
Rolán-Alvarez E, Johannesson K, Erlandsson J (1997) "The Maintenance of a Cline in the Marine Snail Littorina saxatilis: The Role of Home Site Advantage and Hybrid Fitness". Evolution, 51, 1838-1847.
Ruedi M, Smith MF, Patton JL (1997) "Phylogenetic evidence of mitochondrial DNA introgression among pocket gophers in New Mexico (family Geomyidae)". Molecular Ecology, 6, 453-462.
72

Chapter VI
Santucci F, Emerson B, Hewitt G (1998) "Mitochondrial DNA phylogeography of European hedgehogs". Molecular Ecology, 7, 1163-1172.
Schmidt NM, Asferg T, Forchhammer MC (2004) "Long-term patterns in European brown hare population dynamics in Denmark: effects of agriculture, prédation and climate". BMC Ecology, 4:15.
Schroder J, Soveri T, Suomalainen HA, Lindberg LA, van der Loo, W (1987) "Hybrids between Lepus timidus and Lepus europeus are rare although fertile". Hereditas 107, 185-189.
Sesé C (1994) "Paleoclimatic interpretation of the Quaternary small mammals of Spain". GEOBIOS, 27, 753-767.
Slatkin M (1973) "Gene flow and selection in a cline". Genetics, 75, 733-756.
Slimen HB, Suchentrunk F, Memmi A, Elgaaied ABA (2005) "Biochemical Genetic Relationships Among Tunisian Hares (Lepus sp.), South African Cape Hares (L. capensis), and European Brown Hares (L. europaeus)". Biochemical Genetics, 43, 577-596.
Smith R, Jennings NV, Harris S (2005) "A quantitative analysis of the abundance and demography of European hares Lepus europaeus in relation to habitat type, intensity of agriculture and climate". Mammal Review, 35, 1-24.
Suchentrunk F, Michailov G, Markov G, Haiden A (2000). "Population genetics of Bulgarian brown hares Lepus europaeus: allozymic diversity at zoogeographical crossroads". ActaTheriologica 45, 1-12.
Suchentrunk F, Mamuris Z, Sfougaris AI, Stamatis C (2003) "Biochemical genetic variability in brown hares (Lepus europaeus) from Greece". Biochemical Genetics, 41, 127-140.
Szymura JM, Barton NH (1991) "The Genetic Structure of the Hybrid Zone between the Fire-Bellied Toads Bombina bombina and B. variegata: Comparisons between Transects and between Loci". Evolution, 45, 237-261.
Szymura JM, Lunt DH, Hewitt GM (1996) "The sequence and structure of the meadow grasshopper (Chorthippus parallelus) mitochondrial srRNA, ND2, COI, COII ATPase8 and 9 tRNA genes". Insect Molecular Biology, 5, 127-139.
Taberlet P, Bouvet J (1994) "Mitochondrial DNA polymorphism, phylogeography, and conservation genetics of the brown bear (Ursus arctos) in Europe". Proceedings of the Royal Society of London B, 255, 195-200.
73

References
Taberlet P, Fumagalli L, Wust-Saucy AG, Cosson JF (1998) "Comparative phylogeography and postglacial colonization routes in Europe". Molecular Ecology, 7, 453-464.
Thulin CG, Jaarola M, Tegelstrõm H (1997) "The occurrence of mountain hare mitochondrial DNA in wild brown hares". Molecular Ecology, 6, 463-467.
Thulin, CG, Tegelstrõm H (2002) "Biased geographical distribution of mitochondrial DNA that passed the species barrier from mountain hares to brown hares (genus Lepus), an effect of genetic incompatibility and mating behaviour?". Journal of Zoology, 258, 299-306.
Thulin CG, Stone J, Tegelstrõm H, Walker CW (2006a) "Species assignment and hybrid identification among Scandinavian hares Lepus europaeus and L. timidus". Wildlife Biology, 12, 29-38.
Thulin CG, Fang M, Averianov AO (2006b) "Introgression from Lepus europaeus to L. timidus in Russia revealed by mitochondrial single nucleotide polymorphisms and nuclear microsatellites". Hereditas, in press.
Turelli M, Barton NH, Coyne JA (2001) "Theory and speciation". Trends in Ecology and Evolution, 16, 330-343.
Wallis GP, Arntzen JW (1989) "Mitochondrial-DNA Variation in the Crested Newt Superspecies: Limited Cytoplasmic Gene flow Among Species". Evolution, 43, 88-104.
Wilson CC, Bernatchez L (1998) "The ghost of hybrids past: fixation of arctic charr {Salvelinus alpinus) mitochondrial DNA in an introgressed population of lake trout (S. namaycushy\ Molecular Ecology, 7, 127-132.
Wilson DE, Reeder DM (2005) "Mammal Species of the World". Johns Hopkins University Press, Baltimore.
Wirtz P (1999) "Mother species-father species: unidirectional hybridization in animals with female choice". Animal.Behaviour, 58, 1-12.
Wu C, Wu J, Bunch TD, Li Q, Wang Y, Zhang YP (2005) "Molecular phylogenetics and biogeography of Lepus in Eastern Asia based on mitochondrial DNA sequences". Molecular Phylogenetics and Evolution, 37, 45-61.
74