natural imitation induced by joint attention in japanese …€¦ · · 2017-05-03eye-gaze and...
TRANSCRIPT

International Journal of Psychophysiology 50(2003) 81–99
0167-8760/03/$ - see front matter� 2003 Elsevier Science B.V. All rights reserved.doi:10.1016/S0167-8760Ž03.00126-0
Natural imitation induced by joint attention in Japanese monkeys
Mari Kumashiro *, Hidetoshi Ishibashi , Yukari Uchiyama , Shoji Itakura , Akira Murata ,a, a a b c
Atsushi Irikia
Cognitive Neurobiology Section, Department of Maxillofacial Biology, Tokyo Medical and Dental University, 1-5-45 Yushima,a
Bunkyo-ku, Tokyo 113-8549, JapanDepartment of Psychology, Graduate School of Letters, Kyoto University, Yoshida-honmachi, Sakyo-ku, Kyoto 606-8501, Japanb
Department of Physiology, School of Medicine, Kinki University, Ohno-Higashi 377-2, Osaka-sayama 589-8511, Japanc
Received 30 September 2002; accepted 29 April 2003
Abstract
To examine whether joint attention enables Japanese monkeys to imitate human actions, we presented an oralaction and manual actions directed towards targets to three monkeys who had joint attention through communicativeeye-gaze and pointing gestures and to one monkey who had incomplete joint attention and who had acquiredimperative pointing but not the use of eye-gaze gestures. Two of the monkeys who were already capable of jointattention were also able to imitate naturally, while the monkey who did not previously show joint attention was notable to imitate until acquiring joint attention capacity. We suggest that joint attention induces natural imitation duringinteraction between different species—in this case, between monkeys and humans—while individuals not showingjoint attention but only attention to a target or movement are only able to follow motion. The monkey may beendowed by nature with motion-following capacity. We speculate that motion-following capacity is developed andcontrolled through joint attention, and is connected with natural imitation.� 2003 Elsevier Science B.V. All rights reserved.
Keywords: Natural imitation; Joint attention; Eye-gaze gesture; Pointing gesture; Monkey; Bi-directional communication
1. Introduction
Bi-directional intentional communication per-mits a human to explore the internal state of otheranimals without the use of language(Kumashiroet al., 2002). Some examples of communicationmeans are:(1) communicative gazing, whichincludes gaze following;(2) eye contact;(3) gazealternation, which refers to looking alternately at
*Corresponding author. Tel.:q81-3-5803-5445; fax:q81-3-5803-0186.
E-mail address: [email protected](M. Kumashiro).
an object and at another individual’s eyes(Gomez,´
1991); and (4) communicative pointing, which isoften accompanied by gaze alternation betweenanother individual and a particular object and tendsto persist until a reward or goal is attained(Leav-ens and Hopkins, 1999; Bard, 1992). The signalsof body movement may help us communicate anintention when each individual is faced with thesame context. Several non-human species respondto another conspecific’s or human model’s atten-tional shift when indicated by cues such as eye-gaze(Anderson and Mitchell, 1999; Emery et al.,

82 M. Kumashiro et al. / International Journal of Psychophysiology 50 (2003) 81–99
Fig. 1. Schematic diagram of the representation of actions.(a) Action representation based on joint attention. Observer(O) paysattention not only to the target and hand, but also to the eye-gaze or pointing or both of the performer(P), and perceives theperformer’s target-directed action based on an understanding that the hand motion is performed by an agent of intention.(b) Actionrepresentation based on single attention, such as object-directed attention. The observer(O9) does not share attention with theperformer and perceives the motion of the hand and target regardless of an agent of intention. O, observer; P, performer; X, target.
1997; Itakura and Anderson, 1996; Kumashiro etal., 2002; Tomasello et al., 1998; Vick and Ander-son, 2000). In humans, such behavior marks apivotal change in an infant’s communicative ability(Werner and Kaplan, 1963). Above all, joint atten-tion, which is defined as an observer following thegaze of a performer and a target(Corkum andMoore, 1995; Emery, 2000), facilitates social com-munication in face-to-face interaction(Shaffer,1984) and also relates to the function of complexforms of social cognition, such as deception, empa-thy and imitation, which refers to the performanceof another individual’s action after observing itdone(Whiten, 1996). Observers who pay attentionto a performer may direct their attention to theway the performer acts on the object. It is pre-sumed that an observer represents the performer’sgaze direction and its hand manipulation and thespecific object on the basis of gaze alternation(Fig. 1a). Such an emerging triadic exchange(Fig.1a) constitutes an infant’s first attempt to simul-taneously integrate object interest and humanengagement within their focus of attention in their
first year(Bakeman and Adamson, 1984; Carpen-ter et al., 1998; Desrochers et al., 1995). As aconsequence, infants engage in a variety of behav-iors that indicate that they perceive a human’s aimin actions guided by goals(Baldwin and Moses,1994; Baron-Cohen, 1991; Tomasello, 1995). Onthe other hand, if observers pay attention only toa target without joint attention, they may representonly the target or movements(Fig. 1b).Imitation, a shared experience between the
observer and the performer, requires attention tothe performer. Although imitation occurs naturallyin human infants(Byrne, 1999; Heyes, 2001;Meltzoff, 1999; Nadel and Butterworth, 1999), itis less frequent in great apes(Bard and Russell,1999). In contrast, no evidence of imitation bymonkeys, regardless of how many years they havebeen trained(Visalberghi and Fragaszy, 1990;Whiten and Ham, 1992), has been reported. Bardand Russell(1999) suggest that apes and humanshave similarities in their development of flexiblesocial and communicative skills that could accountfor the similarities found in their early imitative

83M. Kumashiro et al. / International Journal of Psychophysiology 50 (2003) 81–99
ability, whereas monkeys appear to have moreinflexible, or hard-wired, communicative andsocial skills (Bard and Russell, 1999). If this istrue, though, monkeys who use some social com-municative gestures—e.g. the above-mentionedeye-gaze or pointing gesture—should show imita-tion ability. We previously reported that a monkeyhad the ability to recognize a human as an agentwith intention: the monkey acquired the commu-nicative pointing and gazing gestures needed torealize joint attention(Kumashiro et al., 2002).Our aim in this study was to examine whethermonkeys capable of joint attention behavior couldnaturally imitate human actions.Even if the monkey would demonstrate repro-
duction of a human’s actions, acquired imitationcan also be produced as a result of non-imitativesocial influence. For example, stimulus or localenhancement can draw the observer’s attention;the mere movement or presence of the objects canincrease the likelihood that contact is made(Vis-alberghi and Fragaszy, 1990). Thorpe (1963)defined local enhancement as apparent imitationresulting from directing the animal’s attention to aparticular object or to a particular part of theenvironment (Thorpe, 1963). This phenomenonresembles actions induced by the attention directedto an object’s character or its movements indepen-dently of another individual’s action with intention(Fig. 1b).Our principal concern was to determine whether
there is a relation between joint attention andnatural imitation in monkeys. We experimentallyraised the non-natural attention of a monkey whohad previously shown incomplete joint attentionbehavior without any eye-gaze gesture(Fig. 1b).We presented the human-motion model withoutthe use of an object, a component of joint attention,to this monkey because we expected to avoid theabove phenomenon. Furthermore, if the monkeyreproduced the human-motion after observing it, itis possible that motion reproduction and naturalimitation occurred because of a different type ofattention. We retrained the monkey to learn utili-zation and comprehension of eye-gaze, and weagain examined whether he could naturally imitatethe human’s actions in a way comparable to thatby monkeys who had demonstrated joint attention
behavior, including the eye-gaze gesture, in theprevious test.This study consisted of three parts. In theaction
test part, we investigated the monkey’s behaviorin response to several presentations of humanactions: an oral action and four manual actions. Inthe oral part, we used only tongue-protrusion(T-P) as a model because T-P is a reliable andrecognizable behavior. In themotion part, weexamined whether the monkey, who showed uni-directional use of pointing and did not show gazefollowing, could reproduce the human-motion andthen precisely control its reproduction. This partconsisted of an oral model(T-P) and five manualmodels. In theretest part, we tested the monkey’snatural imitation after the monkey acquired thejoint attention capacity. In this study, we used theterm action to mean doing something regardlessof rewards when the monkey’s attention was notexperimentally directed toward the human. Wefocused on the phased structure of the subtle andnimble motions of the monkeys in response tohuman actions or motion.
2. Materials and methods
2.1. Action test part
2.1.1. SubjectsThe subjects were four Japanese monkeys,
Macaca fuscata (Zen and Sora 5.5 kg, male; Pin6.2 kg, Ume 4.2 kg, female). The four monkeyswere separately housed and were not deprived offood or water. One monkey(Pin) showed bi-directional use of eye-gaze and pointing gestureswith humans(Kumashiro et al., 2002). Two mon-keys (Sora and Ume) were trained in the use ofthese gestures. The remaining monkey(Zen) wastrained in the same way, but showed only theunidirectional use of pointing without any eye-gaze gestures(Kumashiro et al., 2002). Three ofthe subjects had interacted with humans for severalmonths(Pin, 32 months; Zen, 21; Sora, 12). Onemonkey (Ume) began to interact with humans 1month before this study and participated in anotherexperiment. We did not mold the monkey’s handsbefore or during the study. This study wasapproved by the Animal Care and Use Committee

84 M. Kumashiro et al. / International Journal of Psychophysiology 50 (2003) 81–99
Fig. 2. Photographs of the pink-box used in the box-opening test. The box can be latched on the front side. To open the box, thelatch must be unlocked before lifting the lid.
of the Tokyo Medical and Dental University, andall husbandry and experimental procedures werein accordance with the guide for the Care and Useof Laboratory Animals of the National ResearchCouncil (1996) and the Guidelines for AnimalExperimentation at the Tokyo Medical and DentalUniversity.
2.1.2. Situation in the laboratoryThe imitation testing and training were done in
an isolated room(W=H=Ds150=185=150 or270=235=200 cm). The knob-touching test wasdone in the corridor connecting the experimentalroom to the home cages. All four walls and theceiling in the laboratory were ivory-colored. Themonkeys were seated comfortably in a primatechair (Muromachi Kikai, Co., Ltd) at a desk. Theexperimenter sat at the left or right side of thedesk because there were cameras to record themonkey’s response directly in front of the sittingmonkey. A CCD color video camera(DXC-200A,Sony) equipped with a zoom TV lens(PH6=8macro, Canon) was used to record the monkey’sbehavior. A digital video camera(DCR-PC-110,Sony) was used in its night-shot mode to recordthe eye movements. Another video camera(DCR-PC-110, Sony) was set at the side of the table tocapture the behavior. The captured images weresent to a quad switcher(YS-Q440, Sony) whichcomposed multiple images into one. The composedimage was then sent to a videotape recorder(DSR-11, Sony) for later analysis. In some experiments,
the quad switcher was bypassed and only onestream of images was fed into the videotaperecorder.
2.1.3. Procedure in the action test partEach monkey participated in the T-P and cotton-
separation action tests. Only Pin participated inthe knob-touching action test. Zen and Pin partic-ipated in the container- and box-opening tests.
2.1.3.1. Oral action. T-P action test. The exper-imenter repeatedly protruded hisyher tongue infront of the subject for approximately 5 min at 3–8 times per 10 s as one trial.
2.1.3.2. Manual actions. Cotton-separationaction test. The experimenter picked up a piece oftwo-ply white cotton (f-three, inc., 30=30=8mm), placed it on the desk, and then showed eachmonkey how to separate the cotton into two pieces.The experimenter repeated this action once, andthen presented a new piece of the cotton to themonkey.
Knob-touching action test. When the experi-menter and the monkey(Pin) were walking andstopped by a refrigerator, the experimenter pointedat a knob or vent holes on the front panel of therefrigerator(MSR-311DR(H), Sanyo) to direct themonkey’s attention toward them. The experimenterthen tapped each of them and observed the mon-key’s response.

85M. Kumashiro et al. / International Journal of Psychophysiology 50 (2003) 81–99
Opening test. The experimenter used a pink box(21=14=14 cm) that had a latch-type closure onthe front panel(Fig. 2). The gray latch was inclear contrast to the pink body of the box. Theexperimenter placed food(pieces of fresh apple)into the pink box in front of the subject andlatched it, and then presented the box to the subject4 times. The experimenter presented two kinds ofcues to help the subject discover the solution toopening the box. One cue was that the experi-menter pointed frequently to the box latch toindicate where it should be manipulated to openthe box (Experimenter pointing). The other cuewas that the experimenter actually opened the boxin full view of the monkey during the presentationof the box(Experimenter-model open).Next, we used two kinds of containers, a circular
container(7.5-cm in diameter, 5-cm high) and two50-ml polypropylene conical tubes(227 261, Grei-ner). These containers were translucent so that thecontents were visible to the monkey. The lid hadto be turned counter-clockwise relative to the bodyto be removed. The experimenter placed food(pieces of fresh apple) in the containers in fullview of the monkey, and presented each containerin turn to the monkey. The experimenter repeatedthese actions once again and then often pointed atthe lid. We conducted each presentation forapproximately 2 min.
2.1.4. Data analysisFor the T-P action test, we defined a perform-
ance as starting when the monkey moved its mouthand protruded its tongue and ending when themonkey closed its mouth. For the remaining twoaction tests, we recorded the sequential behaviorsof the monkey.
2.2. Motion part
2.2.1. Subject and situation in the laboratoryPin, Ume and Sora were engaged in another
study. Only Zen who had previously failed toacquire gazing gestures(Kumashiro et al., 2002)participated in this part of the experiment. Thelaboratory was the same as for the action test part.Zen’s head was held immobile during the initialtrials in order to direct his attention toward themodel motion experimentally.
2.2.2. Procedure for motionThis part consisted of three segments. In all
segments, a mouth model(T-P model) and fivemodels of the hands or fingers(hand–nose(H–N), hand-clap(H-clap), hand-clench(H-clench),hand–ear(H–E) and thumb-extended(T-E)) wereused. In the first segment, we performed the T-P,H–N, H-clap and H-clench models in turn for themonkey. To help in learning to imitate the subse-quent manual motions, we repeatedly presentedimitation of the T-P model. In the second segment,we repeatedly presented the finger model wherethe thumb was extended and the remaining fingersflexed. To check the influence of visual input onself-hand motion during the sessions, the monkey’shands were occluded by a rectangular sheet ofcardboard (this design allowed us to examinewhether the monkey could represent hisyher ownfingers and the human’s fingers by comparing thesimilarity between the response of the monkey andthe human’s finger model). In the third segment,we presented the H–E model to examine whetherthe repeated motion-presentation of a model simi-lar to the previous model elicited more accurateimitation. We also designed an invisible-goal sit-uation to examine how the monkey behaved whenthe target location for the human’s hand wasinvisible to the monkey. In this invisible-goalsituation, the experimenter(sitting to the right ofthe monkey) extended her right arm, moved herright hand to her right earlobe with a grasping-hand shape and gently grasped the earlobe. In thevisible situation, the experimenter sat in the sameposition and flexed her left arm and touched theleft ear.Each model was presented in the following order
on different days. A new model was started afterapproximately two sessions of the previous modeland was mixed together with previous model trials.
2.2.2.1. Oral model. T-P model. The experi-menter sat to the right of the monkey and protrudedher tongue towards the monkey 3–5 times in eachtrial which lasted for 81 s.
2.2.2.2. Manual models. H–N model. Theexperimenter sat to the right of the monkey, puther elbow on the table, and then extended her

86 M. Kumashiro et al. / International Journal of Psychophysiology 50 (2003) 81–99
fingers and touched her nose. Motion-presentationswere slowly and quickly repeated for 253 s.
H-clap model. We used two types dependingon the motion model speed. Experimenter A quick-ly and intermittently clapped hands 108 times witha clapping-noise in 88 s. Experimenter B openedboth palms and then slowly clapped hands 17times in 36 s without making a clapping-sound.
H-clench model. The experimenter moved fivefingers to make a horizontal fist with the elbowon the table or raised the clenched hand and swungit downward. The total presentation was 25 timesand lasted 33 s.
T-E model (hiding the monkey’s hands). Weattached a rectangular sheet of cardboard to themonkey’s chair in front of the monkey’s shoulderto hide the monkey’s hands from the monkey’ssight. The experimenter sat to the right of themonkey and extended hisyher thumb with the fourremaining fingers making a vertical fist. The exper-imenter slowly and intermittently presented theclench model with small finger flexure-motions inthe first period of this session for 363 s. Theexperimenter then quickly flexed four fingers withthe T-E 5 times in front of the monkey in 4 s.
H–E model. The experimenter sat to the rightof the monkey and moved her right hand to herright ear as if grasping. When the experimenterused the right arm, the monkey could not see theexperimenter’s goal because it was on the far sideof the experimenter’s body. This motion was pre-sented 4 times in 13 s. In the visible-goal situation,the experimenter sat in the same position andmoved her left hand to her left ear 8 times in 49s. When the left arm was used, the monkey couldsee the experimenter’s goal.
Use of the experimenter’s hand: During motionperiods 1, 2 and 3, use of the right or left handby the experimenter or the monkey was not con-trolled, but the experimenter generally used theright hand.
Reward: In all sessions, food, apple or sweetpotato was often given as a reward when themonkey moved the same body part as the experi-menter in accordance with each model.
Subsequent sessions after initial motion-modelpresentation: We performed the motion-presenta-tion during one session per day. Subsequent ses-
sions followed each repeated motion-presentationand proceeded in the same way. In each session,the new model was continued several times,although several models were presented in randomorder during later sessions. For the T-P models, tofocus the monkey’s attention towards the experi-menter’s mouth, the experimenter sometimesopened heryhis mouth, slowly moved the mouth,or quickly repeated this motion. We presented theopen-mouth model in the second and third ses-sions, but stopped presenting this model to themonkey since the monkey’s mouth-motion wasstrange(i.e. the monkey’s tongue became tangledor the monkey’s mouth became polygonal inshape). We stopped training when the monkeybecame fatigued during the session.(In the latersessions, Zen’s head was not held immobile.)These sessions were video-recorded through videocameras mounted on one of the walls, on the table,or both.
2.2.3. Data analysisWe performed offline analysis from the video
recording to detect any subtle behavior of themonkey. The time periods we analyzed wereapproximately 81, 253, 123, 33, 367 and 62 s forthe T-P, H-N, H-clap, H-clench, T-E and H-Emodels, respectively. We regarded one trial as aperiod from the first presentation of each model tothe reward being obtained in the T-P, H-N, H-clapand H-clench trials. We detected slight and initialbehaviors after the presentation of each model; forexample, the monkey quickly moved the samebody part as the human had, or the monkey’smotion became similar to that of the human.
2.3. Retest part
2.3.1. Subject and eye-gaze trainingThe subject was Zen. After 6 months from the
third segment in the motion part, the subject wasretrained to use an eye-gaze gesture in the sameisolated room as for the previous sessions.Eye-gaze training: The experimenter presented twocontainers that contained food. Then, if the mon-key looked at the experimenter’s eyes and aspecific object to which he was pointing alternate-ly, the experimenter gave that object to the monkey.

87M. Kumashiro et al. / International Journal of Psychophysiology 50 (2003) 81–99
This monkey acquired this communicative point-ing—that is, communicative gaze following andpointing—within several presentations of food.This training was conducted for 10 min duringordinary care twice a week. After approximately 1month from the beginning of eye-gaze training,the retest was done.
2.3.2. Procedure in the retest partThe same subject(Zen) participated in the
container- and box-manipulation test.Manualaction opening retest: We used the same procedureas for the box- and containerytube-opening tests,except that food was not placed inside the tube.We recorded the sequences of the monkey’s behav-ior (Fig. 11), at an image resolution of1280=1024 pixelsyframe and 24 framesys, witha monochrome CCD camera(CV-M4, JAI)equipped with a zoom lens(J6=11, Canon), andstored the image in our personal computer system(DiSCS, CTC).
2.3.3. Data analysisFor this test, we defined a performance as
starting when the monkey moved his hands andending when the monkey put down the box orcontainer and did not touch them further. Weinvestigated the process the monkey used to deter-mine the solution by applying time-series analysisusing the offline video recording(container retest).In the box-opening retest, we defined touching ofthe lid after the experimenter touched it as areproduced touch.
3. Results
3.1. Action part
3.1.1. Oral action
3.1.1.1. T-P test. Daily observation revealed thatbefore the test these monkeys seldom protrudedtheir tongue, except when given food. We foundthat three(Pin, Sora and Zen) of the four monkeysprotruded their tongue in the T-P test. The othermonkey(Ume) gazed at the experimenter’s mouth,but did not protrude her tongue.
3.1.2. Manual actions
3.1.2.1. Cotton-separation action test. Two mon-keys (Pin and Sora) gazed at the experimenter’saction of separating the white piece of cotton.After picking up or being given a similar piece ofcotton, these monkeys separated it in the sameway as the experimenter had(Fig. 3). Anothermonkey(Ume) gazed at the experimenter’s action,and then smelled the piece of cotton before puttingit down. The remaining monkey(Zen) did notwatch the human’s action to the end, and thensmelled and gnawed the piece of cotton after heobtained it.
3.1.2.2. Knob-touching action test. The monkey(Pin) touched the same knob and vent holes afterthe human pointed at the knob and then touchedit (Fig. 4). The monkey looked at the refrigeratorwhile standing, and lay on her stomach and lookedat the vent holes of the refrigerator when thehuman sat down. When the experimenter pointedat the holes and touched them, the monkey raisedher body and touched the vent holes as did thehuman. The experimenter next touched the knob,and the monkey touched the same position on theknob that the human had touched.
3.1.2.3. Opening test. One monkey(Zen) couldopen the box when the experimenter presented theunlatched box, but could not open the latched boxby himself and did not benefit from the model-open and the cues. Pin could open the box afterthe experimenter opened it, regardless of whichplane of the box was presented. Pin rotated thebox about various rotational planes—horizontal,coronal and saggital—until she found the latch.Zen could not open either container by using
his hands or his hands and mouth. Moreover, hethrew or bit two transparent containers in each ofthe four trials while rotating them. When the tube-container was used, Zen bit the bottom part of thetube. Zen did not obtain the food in any of thecontainers. On the other hand, Pin did not try toopen the circular container in the first trial. Instead,she gazed alternately between it and the experi-menter. When it was apparent that the experiment-er was avoiding Pin’s gaze, Pin tried to open the
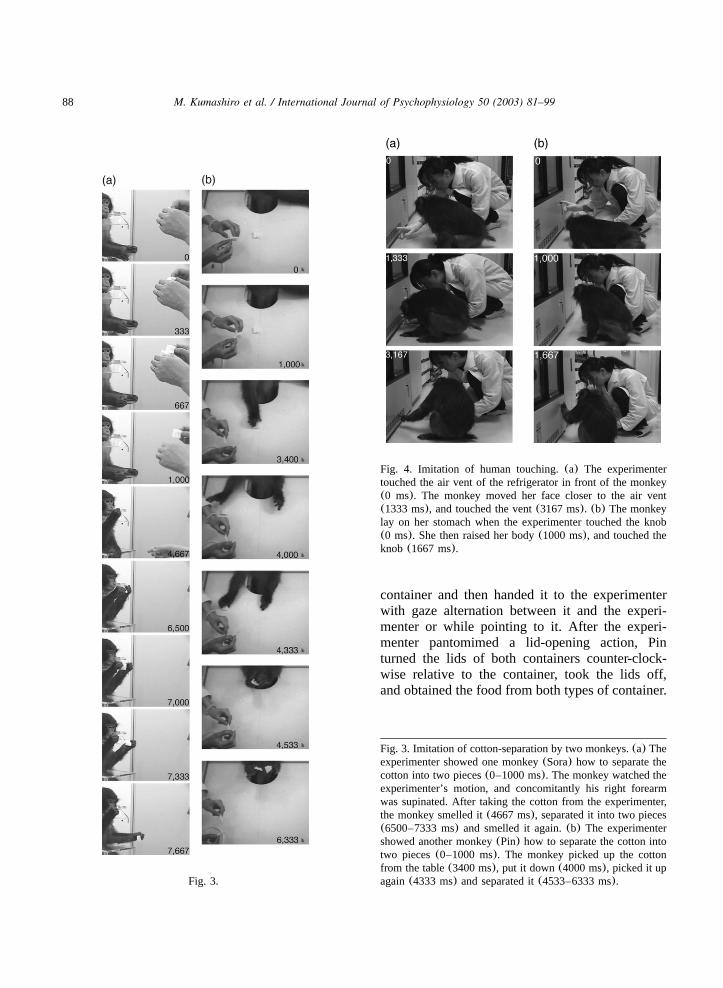
88 M. Kumashiro et al. / International Journal of Psychophysiology 50 (2003) 81–99
Fig. 4. Imitation of human touching.(a) The experimentertouched the air vent of the refrigerator in front of the monkey(0 ms). The monkey moved her face closer to the air vent(1333 ms), and touched the vent(3167 ms). (b) The monkeylay on her stomach when the experimenter touched the knob(0 ms). She then raised her body(1000 ms), and touched theknob (1667 ms).
Fig. 3.
Fig. 3. Imitation of cotton-separation by two monkeys.(a) Theexperimenter showed one monkey(Sora) how to separate thecotton into two pieces(0–1000 ms). The monkey watched theexperimenter’s motion, and concomitantly his right forearmwas supinated. After taking the cotton from the experimenter,the monkey smelled it(4667 ms), separated it into two pieces(6500–7333 ms) and smelled it again.(b) The experimentershowed another monkey(Pin) how to separate the cotton intotwo pieces(0–1000 ms). The monkey picked up the cottonfrom the table(3400 ms), put it down(4000 ms), picked it upagain(4333 ms) and separated it(4533–6333 ms).
container and then handed it to the experimenterwith gaze alternation between it and the experi-menter or while pointing to it. After the experi-menter pantomimed a lid-opening action, Pinturned the lids of both containers counter-clock-wise relative to the container, took the lids off,and obtained the food from both types of container.

89M. Kumashiro et al. / International Journal of Psychophysiology 50 (2003) 81–99
Fig. 5. Initial response to the T-P model.(a) In the first trial,the monkey thrust his lower jaw.(b) In the second trial, heopened his mouth slightly and protruded his tongue.(c) In thethird trial, he opened his mouth widely and protruded histongue.
3.2. Motion part
3.2.1. Oral motion
3.2.1.1. T-P model. The monkey moved his mouthtoward the left in the first two trials(Fig. 5a). Themonkey then protruded the tip of his tongue afterlooking at the tongue of the experimenter(Fig.5b). The monkey protruded his tongue, with thetip often directed toward the left side in the earlysessions, at approximately 81 s after the experi-menter’s first model presentation(Fig. 5c).
3.2.2. Manual motions
3.2.2.1. H–N model. For the first response(Fig.6a), the monkey looked at the experimenter’s handand faced alternately and then gazed at his ownhand. In succession, the monkey protruded histongue once, moved his hand to his mouth, andthen put his hand in his mouth. After 242 s fromthe beginning of the H–N presentation, the mon-key showed two types of behavior: slow and quickmotion, which corresponded to the two respectivetypes of H–N slow and quick presentation(Fig.6b and c). In the slow type, the monkey looked atthe experimenter and his own hand. After that, themonkey groped around the surface of his stomachfor his hair and then touched his nose when theexperimenter touched her nose while moving herarm at a moderate speed. After 1 s from this last
response, the monkey quickly moved his handtoward his mouth twice when the experimentertouched her mouth twice with a motion quickerthan that in the previous presentation. After that,the behavior of the monkey touching his noseappeared. The monkey varied the target on hisface when the speed of the experimenter’s motionschanged.
3.2.2.2. H-clap model. The monkey showed differ-ent types of clapping in response to the differentmotions of experimenters A and B. In response tothe loud, quick clapping of experimenter A, themonkey repeatedly beat the acrylic board of hischair with both hands(Fig. 7a). The monkeysometimes shut his eyes both when the experi-menter clapped and while beating the board byhimself (the fourth picture in Fig. 7a). For theslow-clapping by experimenter B, the monkeylooked at his own hand and sometimes touchedone hand with the other. The monkey did not beatthe board of the monkey’s chair during the slow-clapping model and never shut his eyes. After 254s from the first response to the quick clapping, themonkey joined both hands as if gaining impetus.The monkey beat the board 47 times in 88 s whileexperimenter A quickly presented the H-clap mod-el 108 times.(By comparison, clapping occurredtwice in 36 s when experimenter B clapped hands17 times.)
3.2.2.3. H-clench model. The monkey clenched hisright hand at approximately 33 s after the firstmodel presentation(Fig. 8). The monkey’s armoften slid from the right side of the table to thecentral position of his body when the experimenterswung down a clenched hand. The monkey thenlooked at his hand. In 33 s, he showed the hand-sliding behavior 4 times, gazed at his handyhands3 times, clapped 3 times, and showed hand-helpingbehavior(in which the monkey held his left handin his right hand) 3 times.In the second segment(hiding the monkey’s
hands), the monkey showed his representation ofthe hand posture(wrist angle) as that of the T-Emodel and his execution of the frequency inresponse to that of the human’s motions.
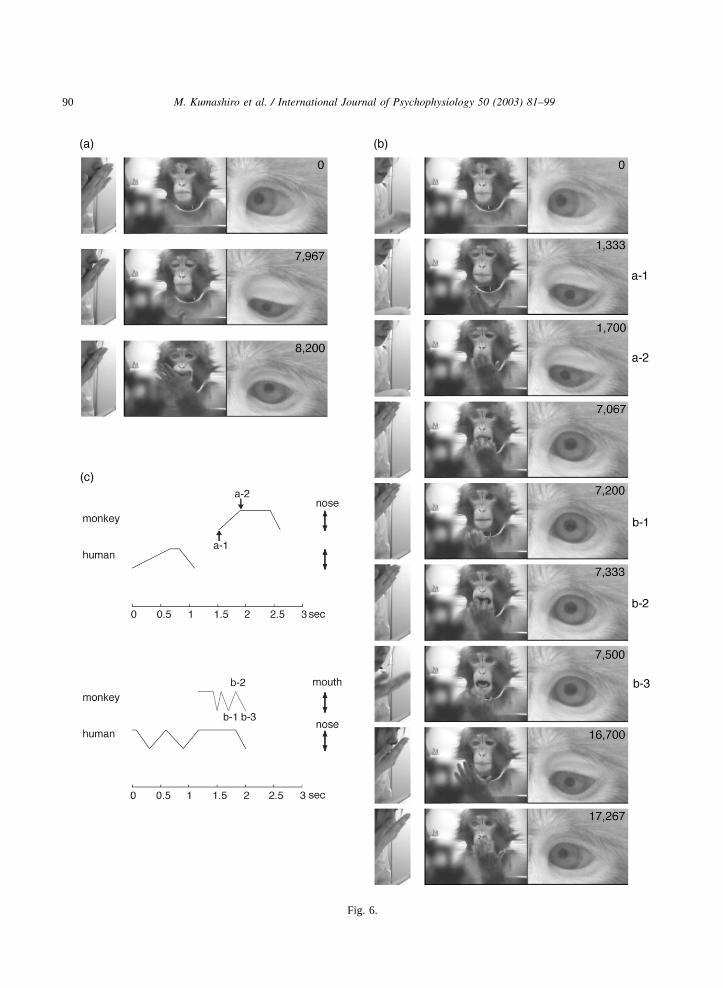
90 M. Kumashiro et al. / International Journal of Psychophysiology 50 (2003) 81–99
Fig. 6.

91M. Kumashiro et al. / International Journal of Psychophysiology 50 (2003) 81–99
Fig. 6. Initial response to the H–N model.(a) In the first trial, the money looked at the experimenter when the experimenter movedher hand to her nose(0 ms), looked at his own left hand(7967 ms) and put his left hand into his mouth(8200 ms). (b) In theeighth trial, the monkey looked at his left hand(1333–1700 ms), put his hand into his mouth(7067 ms), lowered his arm(7200ms) and put his hand into his mouth again(7333 ms), then moved his hand to his nose with the fingers extended(17 267 ms). (c)Time course of the experimenter’s and the monkey’s hand motions. The rate of the experimenter’s motions affected that of themonkey’s motions. Also, note that the number of extensions and flexions were identical between the experimenter’s and the monkey’smotions.
3.2.2.4. T-E model. When the experimenter slowlyand intermittently presented the clench model withthumbyfinger flexure-motions, the monkey verti-cally clenched the left hand once. The monkeyshowed the same hand-posture as that of the T-Emodel, but not the same finger-posture, 6 times.When the experimenter quickly flexed four fingerswith the T-E, the monkey quickly flexed all hisfingers 7 times (compared to 5 times by theexperimenter) in 4 s.In the third segment, the monkey accurately
reproduced the human’s model after exposure torepeated motion-presentations of the previousmodels.
3.2.2.5. H–E model. When the goal location ofthe model motion was hidden, the monkey oftenextended his arm to besides his body and clenchedhis right fingers in the air. When the goal locationof the model motion was visible, the monkeytouched his nose(twice) and clenched his rightfingers in the air(3 times) as if looking at hishand or the experimenter. Finally, the monkeytouched his ear at 49 s from the beginning of thevisible target. Fig. 9 shows the prominentresponses for each model.The monkey correctly produced the same
actions as the human in two sessions from thebeginning of each model. The monkey imitatedwhomever provided the model. Photographs of themonkey’s correct motions after the T-P, H–N, H-clap, H-clench and H–E sessions are shown inFig. 10.
3.3. Retest part
The monkey(Zen) showed a difference betweenbefore and after the acquisition of eye-gaze ges-tures in imitating the open-box model. After theacquisition of eye-gaze gestures, this monkey
immediately touched the latch that the human hadtouched and opened the box after demonstrationof the open-model. When the experimenter gavecues by pointing to the latch, he rotated andtouched the box, unlatched it and then opened it.The four trials proceeded as follows: Zen touchedthe latch after presentation of the model-open inthe first trial; pointed to and touched the latch inthe second trial; touched the latch and lifted it, butcould not move his arm well because of his chair,and handed the box to the experimenter in thethird trial; opened the box by moving his armposition by himself in the fourth trial.Fig. 11 shows the sequential images of this
monkey’s behavior during the container-openingtask after acquisition of the eye-gaze gestures. Themonkey’s hand-action followed the human hand-action while the monkey alternately looked at theexperimenter’s eyes and the container and touchedthe experimenter’s wrist. When the experimentertapped or pointed to the lid, the monkey touchedthe same location that the human indicated. Themonkey then rotated the lid and opened the con-tainer after watching the experimenter pantomimerotating the lid to open the container.
4. Discussion
The two monkeys who had joint attentionbehavior were able to imitate the T-P and hand-actions in the action test part of our study. Themonkey who had incomplete joint attention behav-ior—incomplete in the sense that he used impera-tive pointing and gazed at the human face, but didnot use the eye-gaze gesture in the communicativecontext—could reproduce the T-P, but not thecotton-separation. When we repeatedly exposedthis monkey to human-motions by fixing his atten-tion on the human, as we hypothesized the monkeyreproduced the same motion as that in the human

92 M. Kumashiro et al. / International Journal of Psychophysiology 50 (2003) 81–99
Fig. 7. Initial response to the H-clap model.(a) Monkey’s response after quick presentation of the model. He beat the acrylic boardof his chair with his eyes closed(4567 ms). (b and c) Monkey’s response after another experimenter’s slow and silent presentationof the model. He touched his left hand with his right hand(9100 ms), and joined two hands together(333 ms).
model. This result indicates that monkeys have thepotential to abstract the similarity between thehuman and the monkey when the monkey’s atten-tion is directed to the model. After the acquisitionof triadic gaze following(Fig. 1a), which wasaccompanied by his pointing, the monkey becameable to imitate the human’s action naturally, in amanner similar to the two previous monkeys whohad joint attention behavior. The implication ofthese findings is that there are two attention typesthat differently induce reproduction of anotherindividual’s movements. Joint attention towardbiological movement in a communicating partnerinducednatural imitation in the monkey who usedthe communicative eye-gaze and pointing gestures.The other type of attention toward the local motioninduced motion reproduction: the monkey whoexperimentally had local motion attention repro-duced the human’s motion.
4.1. Oral action and motion
The reproduction of T-P in this study allowssignificant interpretations. Both types of monkey,
either having complete or incomplete joint atten-tion, showed reproduction of the T-P in the test.Since mouth movement is related not only toeating, but also to vocalization and expressingfeelings and emotions, a monkey who is interactingwith another individual may tend to direct itsattention to the mouth movement of the partner.Furthermore, protrusion of the tongue offers furthersaliency—for monkeys, the red color of the tonguewould be in contrast to the monkey’s skin and beparticularly obvious, and the color red has intrinsicbiological significance(e.g. blood, tissue damage).Thus, monkeys may have a natural tendency todirect their attention to the mouth movements ofother individuals. This attention may make it easyfor a monkey to construct an internal representa-tion of mouth movement that can be used toinduce the same movement. In addition, the mon-keys would immediately receive multisensory inte-grated-feedback, consisting of tactile,proprioceptive and motor signals even from aslight tongue movement. Therefore, it is possiblethat mouth movement is easier to reproduce than

93M. Kumashiro et al. / International Journal of Psychophysiology 50 (2003) 81–99
Fig. 8. Initial response to the H-clench model.
the movement of other body parts. Most monkeyswho do not have joint attention with humans are,however, gaze aversive, and thus do not observethe mouth movement of another individual wellenough to reconstruct a representation of the mouthmovement, resulting in a lack of imitativebehavior.
4.2. Manual motion
In the H–N model, the monkey caught his handwith his mouth in the first response. This seemsto reflect a monkey’s conception that the mouth isa place to catch at something. In fact, monkeysfrequently catch with and place new objects intotheir mouths. Our monkey, after a while, began tolook at the human-motion and his own hand, andfinally to smell the tip of his hand. Thus, themonkey might have matched the human-motionswith the closest action he was already familiarwith, taking the speed of the motion into consid-eration. Another interpretation of his behavior isthat he correctly reproduced the model motion inthe sense that the hand was to be moved to themost protuberant part of the face. That is, themouth is the most protuberant part of a monkey’sface, while the nose is the most protuberant in ahuman face. In response to the H-clap model, themonkey behaved differently to the different rateof the model motion: the monkey beat the boardloudly in response to quick and sound-makingmodel motion and put his hands together inresponse to slow and silent model motion. Thisresult showed that the monkey abstracted themotion and repeated sound. In the third segment(H–E model), the monkey’s arm correctly movedin the air when the target was hidden, and movedtoward the ear that was the target location whenthe target was visible. There was no definedbehavior in response to the H–E model, such asthe bite- or smell-behavior that was a response tothe H–N model. The goal-directed aspects ofimitation might emerge later in human ontogeny(Bekkering et al., 2000). The monkey might havedeveloped, in a series of motion representationexperiences, the ability to reproduce the humanbody-motion. Interestingly, the number of repro-ducing motions in any model was mostly consis-
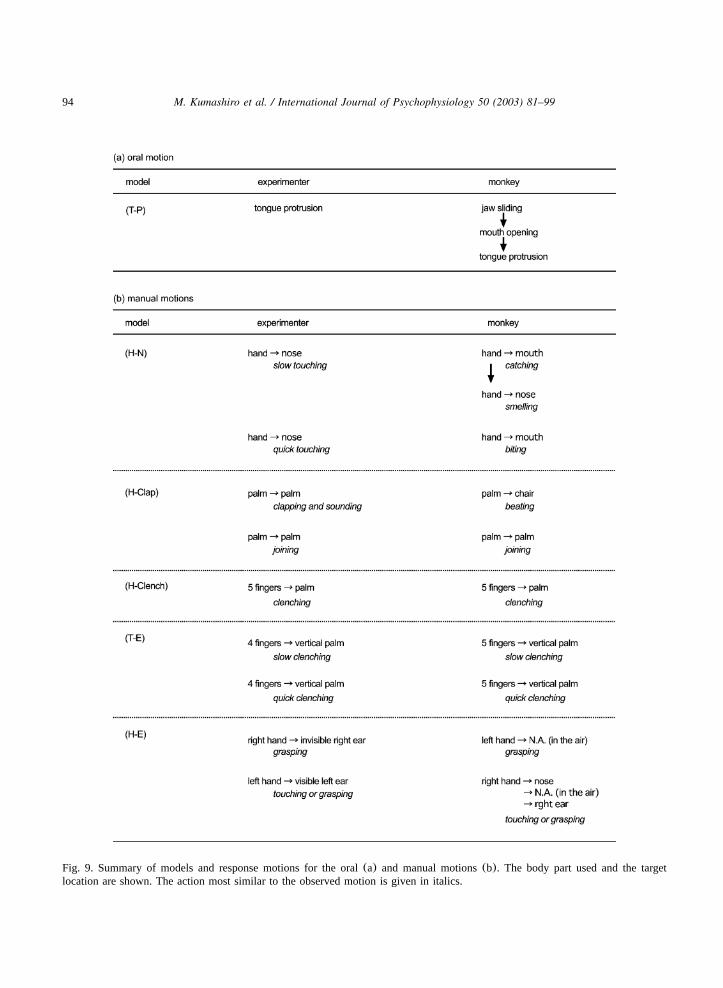
94 M. Kumashiro et al. / International Journal of Psychophysiology 50 (2003) 81–99
Fig. 9. Summary of models and response motions for the oral(a) and manual motions(b). The body part used and the targetlocation are shown. The action most similar to the observed motion is given in italics.

95M. Kumashiro et al. / International Journal of Psychophysiology 50 (2003) 81–99
Fig. 10. Reproduction of the models after repeated exposure toeach model:(a) T-P; (b) H–N; (c) clapping;(d) H-clench and(e) H–E.
tent with that of the human’s motions. It is possiblethat the monkey counted the number.During the shaping process of several motion
models, the experimenter intermittently rewardedthe monkey with food. Because of this intermittentrewarding, the monkey might have concentratedmore fully on the motion observation and itsreproduction and correction at reward intervals.Because we observed the monkey’s reproductionin the home cage after the first training of eachmotion-model, this rehearsal might also havehelped to make his reproduction more precise.
4.3. Hidden body-part motion
Our results from the second segment suggestthat the monkey could represent the motion andthe hands without visual guidance regarding hisown hands, but could not match the human’sthumb with his thumb. That is, this finding indi-cates that the monkey coded the human’s motion.We observed subtle swings of only the right thumbwhen the monkey touched his chair or desk duringthis session; however, we could not determinewhether this behavior was related to the modelpresentation. In this part, the monkey might haveabstracted the perceived motions and then inte-grated somatosensory feedback. In other words, itwas easy to code the motion, including the indi-vidual body-part motion, because of the quickfeedback signal of real-somatosensory information.
4.4. Manual action test and retest
We now have to direct our attention toward theessential mechanism ofnatural imitation from ourfindings in the Manual action and Retest parts. Wehypothesized about imitation mechanism on thebasis of several reported results and our results inthe current experiments. First, human infants from12 to 21 days old show imitation of mouth andsimple finger movements(Meltzoff and Moore,1977), and that mouth imitation emerges earlierthan hand imitation(Meltzoff and Moore, 1983,1989). Chimpanzee infants also imitate a human’smouth movements(Bard and Russell, 1999). Sec-ond, gaze shift in human infants begins to developwith rapid improvement between 9 and 16 weeks

96 M. Kumashiro et al. / International Journal of Psychophysiology 50 (2003) 81–99
Fig. 11.

97M. Kumashiro et al. / International Journal of Psychophysiology 50 (2003) 81–99
Fig. 11. Tube-opening test(a) and container-opening test(b). (a) The monkey watched the experimenter opening the cap of thetube(0 and 2750 ms). He bit the cap(15 250 ms), stopped biting and looked at the experimenter again(15 875 ms), looked at thetube he was holding(17 458 ms), bit again(20 292 ms) and pulled the cap(23 333 ms). Note that his head was leaning back dueto his pulling action. The experimenter then let the monkey see her touching the cap of the tube(25 542 ms). The monkey lightlytouched the cap as did the experimenter(38 625 ms) and held the cap with his left hand(39 292 ms and after) and the tube withhis right hand(40 375 ms and later). Note that the experimenter was holding the tube, but not the monkey’s hand.(b) The monkeypointed to the experimenter and appeared to request that the container be given to him. After presenting the container to the monkey(1417 ms), the experimenter repeatedly pointed to the monkey’s left hand that was holding the lid of the container(4375 and 7292ms). The monkey gently touched the experimenter’s wrist(6500 ms), and started holding the container until 9333 ms. He againheld the cap with his left hand and the container with his right hand(10 542 ms). He then moved his right hand closer to his bodywith his left hand fixed, resulting in the opening of the container(10 958 ms).
(Butcher et al., 2000) and infancy studies haveshown that when an adult breaks off mutual gazeand turns her head to look elsewhere, infants canfollow the adult’s gaze from the age of 12 monthsand possibly earlier(Butterworth and Jarrett, 1991;Corkum and Moore, 1995; Scaife and Bruner,1975). Children who show a deficit of socialcommunication also have a deficit of joint attentionbehavior, especially eye-gaze(Baron-Cohen et al.,2000; Carpenter et al., 2002; Sigman and Kasari,1995; Wellenius and Cullen, 2000). Third, humanshave a strong predisposition to imitate anotherindividual’s oculomotor behavior, and this is likelyto be linked to a strong tendency towards jointattention(Ricciardelli et al., 2002). Taken together,an infant’s T-P follows an adult’s T-P, an infant’sgaze follows an adult’s gaze, and self gaze followsanother’s gaze. In addition, there is evidence thatthe monkey’s gaze follows a conspecific’s gaze(Emery et al., 1997). A common feature amongthe above reported behavior isbody-motion-follow-ing. Our findings also show that the monkey’saction followed the human’s action in the test andretest parts, and the monkey’s motion followed thehuman’s motion in the motion part. We call theseaction-following andmotion-following, respective-ly. We speculate that imitation is based on afollowing mechanism, and, especially,natural imi-tation (action following) is induced by joint atten-tion (Fig. 1a).Before the acquisition of joint attention behav-
ior, the monkey reproduced the human-model inthe motion part, but this reproduction learning hadlittle effect on the monkey’s capacity fornaturalimitation (action-following) except for the mouthmovement; it does not include the object-manipu-lation. After the monkey acquired the use of triadic
gaze following and incorporated it into the inter-active behavior with humans, the monkey becameable to imitate the human’s manipulation. Themonkey could also unlatch and open the box afterobserving the human’s model actions. It may bethat the acquisition of the communicative eye-gazegesture allowed the monkey to acquire thenaturalimitation capacity. The emergence of these behav-iors supports our hypothesis. Regarding the rela-tion between eye-gaze gesture and intentionalcommunication, Gomez reported that an infant´gorilla developed eye-contact communication witha human in problem-solving situations(Gomez,´1991). To request an action or object, the gorillamade eye contact and then expressed the requestby leading the human to the target(an activityoften accompanied by eye contact while moving).Gomez suggests that the gorilla could develop the´ability to share a problem with a human owing tothe gorilla’s gazing at the human’s eyes. We alsosuggest that joint attention realized through the useof communicative gestures might pave the way fora representation of non-human primate’s intention.
4.5. Summary
In this study, we tested the imitation ability ofmonkeys in two new behavioral approaches thatthe monkeys were to reproduce of another individ-ual’s actions and motions. One wasnatural imi-tation on the basis of joint attention through useof communicative eye-gaze and pointing(Fig. 1a).It seemed probable that the observer could attendto the performer as an intentional agent manipu-lating an object or target. The other wasmotionreproduction on the basis of incomplete jointattention, near an interesting object or local motion

98 M. Kumashiro et al. / International Journal of Psychophysiology 50 (2003) 81–99
attention (Fig. 1b). This type of observer mightattend to a motion regardless of whether there wasan intentional agent. These findings suggest thatJapanese monkeys can reproduce human actionsand motion in the absence of instructional teachingthrough language. Our findings, therefore, contra-dict the belief that monkeys are unable to imitatedue to inflexible(i.e. hard-wired) communicativeand social skills. Tomasello(1999) considerschimpanzees and some other non-human primatesas able to clearly understand something of theefficacy of their own actions on the environment.Chimpanzees even engage in many kinds of inten-tional sensory-motor actions in which they usedifferent means toward the same end, removeobstacles, and use intermediaries such as tools(Tomasello, 1999). Anderson(1998) suggests thatmental condition, as well as behavior, is affectedby the degree to which an animal can express itssocial propensities(Anderson, 1998). Our findingsindicate that our monkeys could identify andabstract the body configuration common to humansand monkeys despite differences in physical prop-erties, such as size, color, or the presence of hair.We propose that monkeys might enrich the reper-toire of actions bynatural imitation, through jointattention expressed by bi-directional use of com-municative pointing and eye-gaze gestures. Hence,natural imitation may be a fundamental form ofsocial learning needed to share, use and compre-hend a common action.
Acknowledgments
This study was supported by support of youngresearchers with a term from Ministry of Educa-tion, Culture, Sports, Science and Technology ofJapan(MEXT).
References
Anderson, J.R., 1998. Social stimuli and social rewards inprimate learning and cognition. Behav. Process. 42,159–175.
Anderson, J.R., Mitchell, R.W., 1999. Macaques but not lemursco-orient visually with humans. Folia Primatol. 70, 17–22.
Bakeman, R., Adamson, L.B., 1984. Coordinating attention topeople and objects in mother–infant and peer–infant inter-action. Child Dev. 55, 1278–1289.
Baldwin, D.A., Moses, L.J., 1994. Early understanding ofreferential intent and attentional focus: evidence from lan-guage and emotion. In: Lewis, C., Mitchell, P.(Eds.),Children’s Early Understanding of Mind: Origins and Devel-opment. Lawrence Erlbaum, Hillsdale, NJ, pp. 133–156.
Bard, K.A., 1992. Intentional behavior and intentional com-munication in young free-ranging organgutans. Child Dev.62, 1186–1197.
Bard, K.A., Russell, C.L., 1999. Evolutionary foundations ofimitation: social, cognitive and developmental aspects ofimitative processes in non-human primates. In: Nadel, J.,Butterworth, G. (Eds.), Imitation in Infancy. CambridgeUniversity Press, Cambridge, pp. 89–123.
Baron-Cohen, S., 1991. Precursors to a theory of mind:understanding attention in others. In: Whiten, A.(Ed.),Natural Theories of Mind. Basil Blackwell, Oxford, pp.233–251.
Baron-Cohen, S., Tager-Flusberg, H., Cohen, D.J., 2000.Understanding Other Minds. second ed. Oxford UniversityPress, Oxford.
Bekkering, H., Wohlschlager, A., Gattis, M., 2000. Imitationof gestures in children is goal-directed. Q. J. Exp. Psychol.A 53, 153–164.
Butcher, P.R., Kalverboer, A.F., Geuze, R.H., 2000. Infants’shift of gaze from a central to a peripheral stimulus: alongitudinal study of development between 6 and 26 weeks.Infant Behav. Dev. 23, 3–21.
Butterworth, G., Jarrett, N., 1991. What minds have in com-mon is space: spatial mechanisms serving joint visualattention in infancy. Br. J. Dev. Psychol. 9, 55–72.
Byrne, R.W., 1999. Imitation without intentionality. Usingstring parsing to copy the organization of behavior. Anim.Cogn. 2, 63–72.
Carpenter, M., Nagell, K., Tomasello, M., 1998. Social cogni-tion, joint attention, and communicative competence from 9to 15 months of age. Monogr. Soc. Res. Child Dev. 63,1–143.
Carpenter, M., Pennington, B.F., Rogers, S.J., 2002. Interrela-tions among social-cognitive skills in young children withautism. J. Autism Dev. Disord. 32, 91–106.
Corkum, V., Moore, C., 1995. Joint Attention: Its Origins andRole in Development. Lawrence Erlbaum, Hillsdale, NJ.
Desrochers, S., Morissette, P., Ricard, M., 1995. Two perspec-tives on pointing in infancy. In: Moore, C., Dunham, P.J.(Eds.), Joint Attention: Its Origins and Rule in Develop-ment. Lawrence Erlbaum, Hillsdale, NJ, pp. 85–101.
Emery, N.J., 2000. The eyes have it: the neuroethology,function and evolution of social gaze. Neurosci. Biobehav.Rev. 24, 581–604.
Emery, N.J., Lorincz, E.N., Perrett, D.I., Oram, M.W., Baker,C.I., 1997. Gaze following and joint attention in rhesusmonkeys (Macaca mulatta). J. Comp. Psychol. 111,286–293.
Gomez, J.C., 1991. Visual behaviour as a window for reading´the mind of others in primates. In: Whiten, A.(Ed.), NaturalTheories of Mind: Evolution, Development and Simulation

99M. Kumashiro et al. / International Journal of Psychophysiology 50 (2003) 81–99
of Everyday Mindreading. Basil Blackwell, Oxford, pp.195–207.
Heyes, C., 2001. Causes and consequences of imitation. TrendsCogn. Sci. 5, 253–261.
Itakura, S., Anderson, J.R., 1996. Learning to use experiment-er-given cues during an object-choice task by a capuchinmonkey. Curr. Psychol. Cogn. 15, 103–112.
Kumashiro, M., Ishibashi, H., Itakura, S., Iriki, A., 2002.Bidirectional communication between a Japanese monkeyand a human through eye-gaze and pointing. Curr. Psychol.Cognit. 21, 3–32.
Leavens, D.A., Hopkins, W.D., 1999. The whole-hand point:the structure and function of pointing from a comparativeperspective. J. Comp. Psychol. 113, 417–425.
Meltzoff, A.N., 1999. Origins of theory of mind, cognitionand communication. J. Commun. Disord. 32, 251–269.
Meltzoff, A.N., Moore, M.K., 1977. Imitation of facial andmanual gestures by human neonates. Science 198, 75–78.
Meltzoff, A.N., Moore, M.K., 1983. Newborn infants imitateadult facial gestures. Child Dev. 54, 702–709.
Meltzoff, A.N., Moore, M.K., 1989. Imitation in newborninfants: exploring the range of gestures imitated and theunderlying mechanisms. Dev. Psychol. 25, 954–962.
Nadel, J., Butterworth, G., 1999. Imitation in Infancy. Cam-bridge University Press, Cambridge.
National Research Council, 1996. Guide for the Care and Useof Laboratory Animals. Institute of Laboratory AnimalResources Commission on Life Sciences. National AcademyPress, Washington D.C.
Ricciardelli, P., Bricolo, E., Aglioti, S.M., Chelazzi, L., 2002.My eyes want to look where your eyes are looking:exploring the tendency to imitate another individual’s gaze.Neuroreport 13, 2259–2264.
Scaife, M., Bruner, J.S., 1975. The capacity for joint visualattention in the infant. Nature 253, 265–266.
Shaffer, H.R., 1984. The Child’s Entry into a Social World.Academic Press, London.
Sigman, M., Kasari, C., 1995. Joint attention across contextsin normal and autistic children. In: Moore, C., Dunham, P.J.(Eds.), Joint Attention: Its Origins and Role in Develop-ment. Lawrence Erlbaum, Hillsdale, NJ, pp. 189–203.
Thorpe, W.H., 1963. Learning and Instinct in Animals. HarvardUniversity Press, Cambridge, MA.
Tomasello, M., 1995. Joint attention as social cognition. In:Moore, C., Dunham, P.J.(Eds.), Joint Attention: Its Originsand Role in Development. Lawrence Erlbaum, Hillsdale,NJ, pp. 103–130.
Tomasello, M., 1999. The Cultural Origins of Human Cogni-tion. Harvard University Press, Cambridge.
Tomasello, M., Call, J., Hare, B., 1998. Five primate speciesfollow the visual gaze of conspecifics. Anim. Behav. 55,1063–1069.
Vick, S.-J., Anderson, J.R., 2000. Learning and limits of eye-gaze by capuchion monkeys(cebus appella) in an object-choice task. J. Comp. Psychol. 114, 200–207.
Visalberghi, E., Fragaszy, D.M., 1990. Do monkeys ape? In:Parker, S.T., Gibson, K.R.(Eds.), Language and Intelligencein Monkeys and Apes. Cambridge University Press, NewYork, pp. 247–273.
Wellenius, G.A., Cullen, K.E., 2000. A comparison of head-unrestrained and head-restrained pursuit: influence of eyeposition and target velocity on latency. Exp. Brain Res. 133,139–155.
Werner, H., Kaplan, B., 1963. Symbol Formation. LawrenceErlbaum, Hillsdale, NJ.
Whiten, A., 1996. Imitation, pretence and mindreading: sec-ondary representation in comparative primatology and devel-opmental psychology. In: Russon, A.E., Bard, K.A., Parker,S.T. (Eds.), Reaching into Thought. Cambridge UniversityPress, Cambridge, pp. 300–324.
Whiten, A., Ham, R., 1992. On the nature and evolution ofimitation in the animal kingdom: reappraisal of a century ofresearch. In: Slater, P.J.B., Rosenblatt, J.S., Beer, C., Mil-inski, M. (Eds.), Advances in the Study of Behavior, vol.21. Academic Press, London, pp. 239–283.