neurobiologie sensorielle somesthesie 2a/2b (en ... sensorielle 2.pdf · le système...
TRANSCRIPT

www.neur-one.fr
[email protected] 1 09/09/2011
NEUROBIOLOGIE SENSORIELLE SOMESTHESIE 2A/2B
(EN PREPARATION)
LES SENSATIONS SOMESTHESIQUES
TACT (PRESSION), THERMORECEPTION, PROPRIOCEPTION CONSCIENTE, NOCICEPTION
SOMESTHESIE : GENERALITES-DEFINITIONS
Le système somatosensoriel, sensoriel somatique ou somesthésique est un système qui collecte toutes les informations sensorielles
provenant du corps.
Une classification purement anatomique permet de définir plusieurs sortes de sensibilités : la sensibilité somatique au sens strict :
o la sensibilité superficielle ou extéroceptive provenant de la peau par les EXTEROCEPTEURS,
o la sensibilité profonde ou proprioceptive qui nous renseigne sur notre position, nos sensations musculaires et
squelettiques par les PROPRIOCEPTEURS,
la sensibilité viscérale qui provient de récepteurs de l'appareil digestif, du coeur, des poumons et des glandes endocrines par
les INTEROCEPTEURS.
Fonctionnellement, le système somato-sensoriel fait partie du système nerveux sensoriel ou voie sensitive.
Diagramme général de la sensibilité somatique
Le système nerveux sensoriel contient des fibres afférentes (« apporter de », c'est-à-dire qui " viennent de“), qui conduisent les influx
nerveux provenant de tous les récepteurs sensoriels disséminés dans l'organisme vers le système nerveux central.
Ce système nerveux sensoriel peut être conscient (et c'est le cas dans la somesthésie) ou inconscient comme certains propriocepteurs, comme les récepteurs vestibulaires par exemple.
Le système somatosensoriel permet d'être conscient de ce que nous faisons et de l'impact de notre environnement sur nous.
Anatomiquement, il fait partie du système nerveux périphérique somatique et du système nerveux autonome pour les viscères.
Le système sensoriel somatique possède des récepteurs sensoriels qui nous transmettent certaines informations : Tactiles (toucher protopathique et épicritique), vibratoires (pallesthésie), de pression (baresthésie) par les mécanorécepteurs
cutanés ou proches de la peau (muqueuses),
Thermiques par les thermo-récepteurs,
Douloureuses par les nocicepteurs
Dynamiques (kinesthésie) et statiques (statesthésie) par les mécanorécepteurs propriocepteurs, contenus dans les muscles et les
articulations, les viscères,
Chimiques par les chémorécepteurs

www.neur-one.fr
[email protected] 2 09/09/2011
Le système sensoriel peut être également subdivisé en deux systèmes : un système dit " épicritique " qui véhicule les modalités tactiles fines (forme, texture, glissement, pression, vibrations) et
proprioceptives conscientes. Les fibres sont de gros calibre, c'est-à-dire, à conduction rapide, myélinisées, A et A. Leur
résolution temporelle et spatiale est élevée. Leurs récepteurs sont des mécanorécepteurs de la peau, du périoste, des articulations
et des tendons. Leur voie centrale est celle des colonnes dorsales et du lemnisque médian
un système dit " protopathique " qui véhicule les modalités thermiques, douloureuses et tactiles grossières. Les fibres sont de
petit calibre, c'est-à-dire, à conduction lente, peu ou pas myélinisées, A et C. Leur résolution temporelle et spatiale est faible.
Ce système est archaïque. Leurs récepteurs sont des mécanorécepteurs de la peau et des structures du système autonome. Leur
voie centrale est celle des voies spino-thalamiques directes et relayées.
Les deux sortes de fibres sont intimement mélangées dans les nerfs périphériques et dans les racines dorsales de la moelle.
Les sensibilités de la région céphalique sont transmises aux trois noyaux du NERF TRIJUMEAU (V).
La sensibilité proprioceptive inconsciente, en provenance des fuseaux neuro-musculaires, des récepteurs tendineux et articulaires,
empruntent des tractus spinaux qui se terminent dans le cervelet (voir chapitre suivant). Ce sont les tractus spino-cérébelleux direct (membres
inférieurs et tronc) et cunéo-cérébelleux (membre supérieur) et le tractus spino-cérébelleux croisé ou ventral (informations musculaires)
En définitive, le système somesthésique ou système sensoriel somatique, du grec « sôma », corps et « aïsthêsis », sensibilité, regroupe les mécanismes nerveux chargés de recueillir les sensations somatiques (les sensations du corps). La somesthésie est la conscience :
D’avoir un corps et
De sentir le contact qu'il a avec son environnement.
La somesthésie fait partie intégrante des différentes modalités sensorielles de perception au même titre que l’audition, la vision, la
gustation, l’olfaction, l’équilibration. C'est la sensibilité aux stimuli appliqués sur le corps. Dans ce chapitre, excluant les stimuli visuels,
auditifs, olfactifs et gustatifs, la sensibilité générale ou somesthésie correspond en fait à quatre modalités : tact, froid, chaud, douleur (la douleur sera examinée au chapitre suivant). Au sens large on peut y rattacher la kinesthésie, ainsi que la mécano-réception viscérale ou vasculaire.
Tout organisme vivant est en constante interaction avec son environnement. Ces interactions lui permettent à la fois de se déplacer ou
de réagir vis-à-vis des stimulations extérieures et de maintenir constant son milieu intérieur. Cela nécessite une prise permanente d’informations
et la circulation des messages entre les différentes cellules de l’organisme. Ces données sont centralisées par le cerveau qui préside aux modifications nerveuses et endocriniennes assurant une certaine constance des caractères physiques et chimiques du milieu intérieur de
l’organisme. Elles participent à l’HOMÉOSTASIE.
Classiquement, la somesthésie regroupe trois modalités de sensibilité somatique CONSCIENTE. La sensibilité extéroceptive:
o Tactile, ou toucher à la pression et aux vibrations, par les mécanorécepteurs ;
o Thermique, au froid et au chaud, par les mêmes récepteurs ;
o Nociceptive, à la douleur, par les mêmes récepteurs.
La sensibilité proprioceptive consciente :
o Élongation musculaire, par les fuseaux neuro-musculaires ;
o Contraction musculaire et tension tendineuse, par les récepteurs tendineux de Golgi ;
o Mouvement articulaire (kynesthésie), par les mêmes récepteurs ;
o Nociception musculo-tendino-articulaire, par les mêmes récepteurs.
La sensibilité intéroceptive consciente (essentiellement des douleurs viscérales mais aussi la sensibilité viscérale profonde comme
le rythme ventilatoire voire cardiaque, la réplétion gastrique ou vésicale etc.), par des mécanorécepteurs, des barorécepteurs etc.
La somesthésie recouvre un continuum d'activité nerveuse entre la sensation et la perception : Le niveau sensoriel
o Les informations provenant le plus directement possible des récepteurs sensoriels
Le niveau perceptif
o Intégration et interprétation des informations sensorielles
o "Construction" d’une "réalité cohérente"... d'un schéma corporel dans son environnement.
Le système somesthésique est responsable des sensations somatiques et viscérales issues de la peau, des muqueuses, des viscères et des
organes locomoteurs. Les sensibilités qui n'émergent pas à la conscience ainsi que les sensations issues d'appareils spécialisés (vision, audition,
gustation, etc.) sont donc exclues de ce chapitre, ainsi d'ailleurs que les sensibilités intéroceptives et proprioceptives inconscientes.
Les sensations somesthésiques peuvent être classées par rapport : à la modalité sensorielle : il s'agit alors de sensations tactiles, de pression, de douleur, des sensations thermiques (chaud et froid),
des sensations de mouvement ou de position d'un segment du corps ;
à la localisation des récepteurs : on parle alors d'extéroréception pour les sensations issues du revêtement tégumentaire (tact,
chaud, froid, douleur), d'intéroréception pour les sensations d'origine viscérale (douleur, soif, faim, etc.), de proprioception pour
les sensations issues de l'appareil locomoteur (sensation de position, de mouvement, douleur);

www.neur-one.fr
[email protected] 3 09/09/2011
aux caractéristiques cliniques des sensations : Head a proposé de distinguer les sensations épicritiques permettant une bonne
discrimination spatiale et temporelle et qui phylogénétiquement seraient d'apparition récente, des sensations protopathiques mal
localisables et qui phylogénétiquement seraient d'apparition ancienne.
Construction d'une réalité cohérente
Critères d’optimalité d’un système sensoriel Pour remplir sa mission, un système sensoriel doit avoir des récepteurs distincts
pour discriminer les différentes formes de stimuli
pour discriminer les différentes intensités de stimulation
pour fiabiliser la transmission du signal
pour accélérer la transmission du signal
pour filtrer les informations parasites.
C'est entre autres rôles celui des récepteurs.
LE SYSTÈME SOMESTHÉSIQUE
ou Somato-afférent général, ou Sensoriel somatique
LA SOMESTHESIE REGROUPE LES MECANISMES NERVEUX RESPONSABLES DE RECUEILLIR L'INFORMATION SENSORIELLE DU CORPS
TROIS TYPES DE SENSIBILITE CONSCIENTE
En résumé, la somesthésie regroupe le tact, la thermoréception, la proprioception et la nociception.
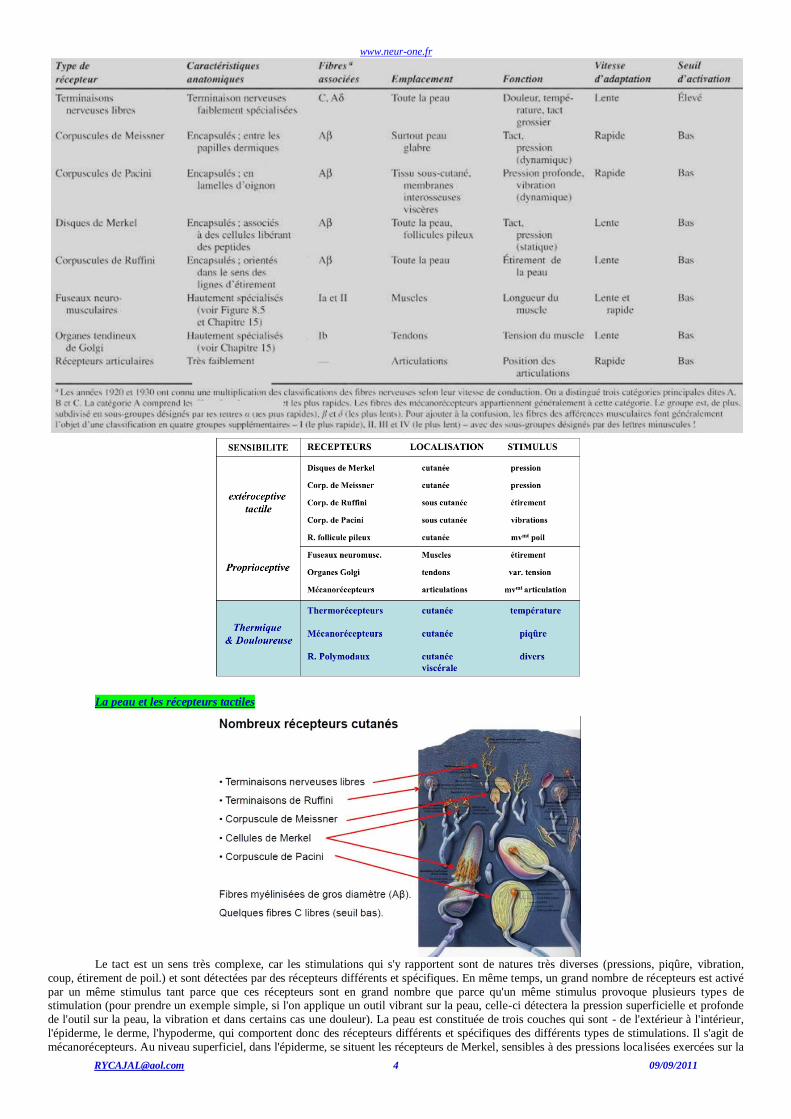
LES RECEPTEURS SOMESTHESIQUES
Catégories de récepteurs somesthésiques
Les principales catégories de récepteurs somesthésiques seront présentées dans les deux tableaux suivants.
www.neur-one.fr
[email protected] 4 09/09/2011
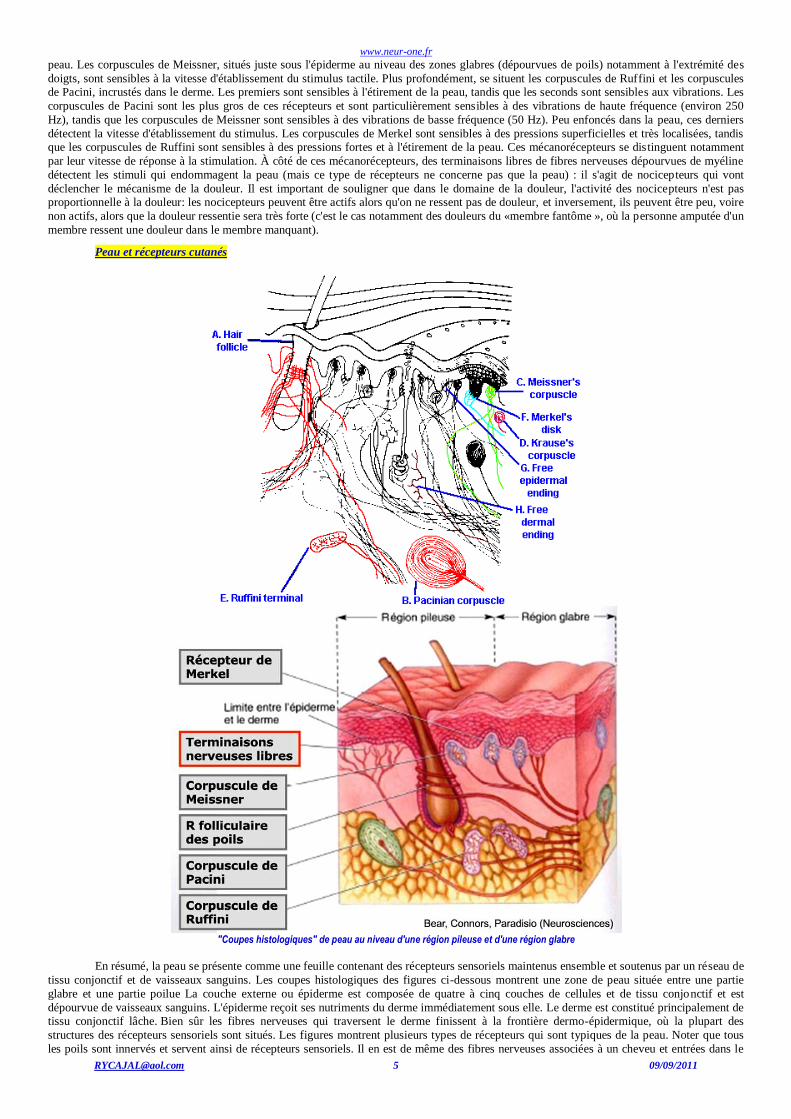
La peau et les récepteurs tactiles
Le tact est un sens très complexe, car les stimulations qui s'y rapportent sont de natures très diverses (pressions, piqûre, vibration,
coup, étirement de poil.) et sont détectées par des récepteurs différents et spécifiques. En même temps, un grand nombre de récepteurs est activé
par un même stimulus tant parce que ces récepteurs sont en grand nombre que parce qu'un même stimulus provoque plusieurs types de stimulation (pour prendre un exemple simple, si l'on applique un outil vibrant sur la peau, celle-ci détectera la pression superficielle et profonde
de l'outil sur la peau, la vibration et dans certains cas une douleur). La peau est constituée de trois couches qui sont - de l'extérieur à l'intérieur,
l'épiderme, le derme, l'hypoderme, qui comportent donc des récepteurs différents et spécifiques des différents types de stimulations. Il s'agit de
mécanorécepteurs. Au niveau superficiel, dans l'épiderme, se situent les récepteurs de Merkel, sensibles à des pressions localisées exercées sur la
www.neur-one.fr
[email protected] 5 09/09/2011
peau. Les corpuscules de Meissner, situés juste sous l'épiderme au niveau des zones glabres (dépourvues de poils) notamment à l'extrémité des
doigts, sont sensibles à la vitesse d'établissement du stimulus tactile. Plus profondément, se situent les corpuscules de Ruffini et les corpuscules de Pacini, incrustés dans le derme. Les premiers sont sensibles à l'étirement de la peau, tandis que les seconds sont sensibles aux vibrations. Les
corpuscules de Pacini sont les plus gros de ces récepteurs et sont particulièrement sensibles à des vibrations de haute fréquence (environ 250
Hz), tandis que les corpuscules de Meissner sont sensibles à des vibrations de basse fréquence (50 Hz). Peu enfoncés dans la peau, ces derniers
détectent la vitesse d'établissement du stimulus. Les corpuscules de Merkel sont sensibles à des pressions superficielles et très localisées, tandis que les corpuscules de Ruffini sont sensibles à des pressions fortes et à l'étirement de la peau. Ces mécanorécepteurs se distinguent notamment
par leur vitesse de réponse à la stimulation. À côté de ces mécanorécepteurs, des terminaisons libres de fibres nerveuses dépourvues de myéline
détectent les stimuli qui endommagent la peau (mais ce type de récepteurs ne concerne pas que la peau) : il s'agit de nocicepteurs qui vont
déclencher le mécanisme de la douleur. Il est important de souligner que dans le domaine de la douleur, l'activité des nocicepteurs n'est pas proportionnelle à la douleur: les nocicepteurs peuvent être actifs alors qu'on ne ressent pas de douleur, et inversement, ils peuvent être peu, voire
non actifs, alors que la douleur ressentie sera très forte (c'est le cas notamment des douleurs du «membre fantôme », où la personne amputée d'un
membre ressent une douleur dans le membre manquant).
Peau et récepteurs cutanés
"Coupes histologiques" de peau au niveau d'une région pileuse et d'une région glabre
En résumé, la peau se présente comme une feuille contenant des récepteurs sensoriels maintenus ensemble et soutenus par un réseau de
tissu conjonctif et de vaisseaux sanguins. Les coupes histologiques des figures ci-dessous montrent une zone de peau située entre une partie
glabre et une partie poilue La couche externe ou épiderme est composée de quatre à cinq couches de cellules et de tissu conjonctif et est
dépourvue de vaisseaux sanguins. L'épiderme reçoit ses nutriments du derme immédiatement sous elle. Le derme est constitué principalement de tissu conjonctif lâche. Bien sûr les fibres nerveuses qui traversent le derme finissent à la frontière dermo-épidermique, où la plupart des
structures des récepteurs sensoriels sont situés. Les figures montrent plusieurs types de récepteurs qui sont typiques de la peau. Noter que tous
les poils sont innervés et servent ainsi de récepteurs sensoriels. Il en est de même des fibres nerveuses associées à un cheveu et entrées dans le
www.neur-one.fr
[email protected] 6 09/09/2011
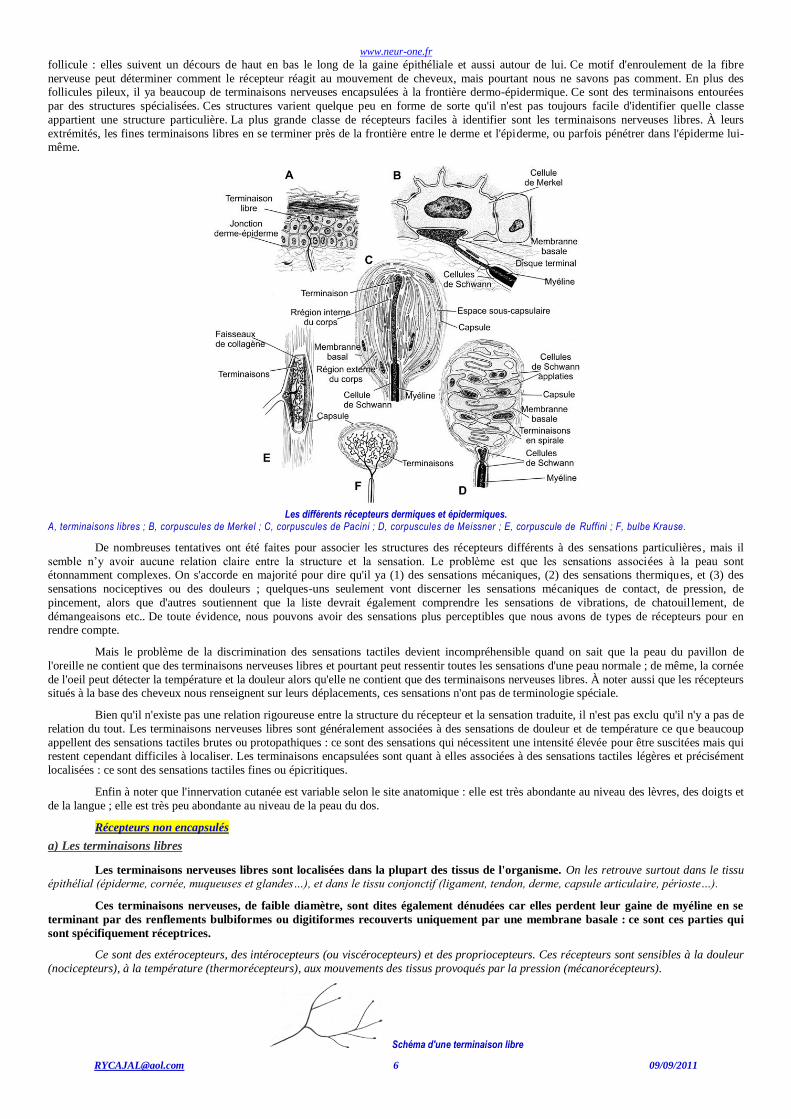
follicule : elles suivent un décours de haut en bas le long de la gaine épithéliale et aussi autour de lui. Ce motif d'enroulement de la fibre
nerveuse peut déterminer comment le récepteur réagit au mouvement de cheveux, mais pourtant nous ne savons pas comment. En plus des follicules pileux, il ya beaucoup de terminaisons nerveuses encapsulées à la frontière dermo-épidermique. Ce sont des terminaisons entourées
par des structures spécialisées. Ces structures varient quelque peu en forme de sorte qu'il n'est pas toujours facile d'identifier quelle classe
appartient une structure particulière. La plus grande classe de récepteurs faciles à identifier sont les terminaisons nerveuses libres. À leurs
extrémités, les fines terminaisons libres en se terminer près de la frontière entre le derme et l'épiderme, ou parfois pénétrer dans l'épiderme lui-même.
Les différents récepteurs dermiques et épidermiques.
A, terminaisons libres ; B, corpuscules de Merkel ; C, corpuscules de Pacini ; D, corpuscules de Meissner ; E, corpuscule de Ruffini ; F, bulbe Krause.
De nombreuses tentatives ont été faites pour associer les structures des récepteurs différents à des sensations particulières, mais il
semble n’y avoir aucune relation claire entre la structure et la sensation. Le problème est que les sensations associées à la peau sont étonnamment complexes. On s'accorde en majorité pour dire qu'il ya (1) des sensations mécaniques, (2) des sensations thermiques, et (3) des
sensations nociceptives ou des douleurs ; quelques-uns seulement vont discerner les sensations mécaniques de contact, de pression, de
pincement, alors que d'autres soutiennent que la liste devrait également comprendre les sensations de vibrations, de chatouillement, de
démangeaisons etc.. De toute évidence, nous pouvons avoir des sensations plus perceptibles que nous avons de types de récepteurs pour en rendre compte.
Mais le problème de la discrimination des sensations tactiles devient incompréhensible quand on sait que la peau du pavillon de
l'oreille ne contient que des terminaisons nerveuses libres et pourtant peut ressentir toutes les sensations d'une peau normale ; de même, la cornée
de l'oeil peut détecter la température et la douleur alors qu'elle ne contient que des terminaisons nerveuses libres. À noter aussi que les récepteurs situés à la base des cheveux nous renseignent sur leurs déplacements, ces sensations n'ont pas de terminologie spéciale.
Bien qu'il n'existe pas une relation rigoureuse entre la structure du récepteur et la sensation traduite, il n'est pas exclu qu'il n'y a pas de
relation du tout. Les terminaisons nerveuses libres sont généralement associées à des sensations de douleur et de température ce que beaucoup
appellent des sensations tactiles brutes ou protopathiques : ce sont des sensations qui nécessitent une intensité élevée pour être suscitées mais qui restent cependant difficiles à localiser. Les terminaisons encapsulées sont quant à elles associées à des sensations tactiles légères et précisément
localisées : ce sont des sensations tactiles fines ou épicritiques.
Enfin à noter que l'innervation cutanée est variable selon le site anatomique : elle est très abondante au niveau des lèvres, des doigts et
de la langue ; elle est très peu abondante au niveau de la peau du dos.
Récepteurs non encapsulés
a) Les terminaisons libres
Les terminaisons nerveuses libres sont localisées dans la plupart des tissus de l'organisme. On les retrouve surtout dans le tissu épithélial (épiderme, cornée, muqueuses et glandes…), et dans le tissu conjonctif (ligament, tendon, derme, capsule articulaire, périoste…).
Ces terminaisons nerveuses, de faible diamètre, sont dites également dénudées car elles perdent leur gaine de myéline en se
terminant par des renflements bulbiformes ou digitiformes recouverts uniquement par une membrane basale : ce sont ces parties qui
sont spécifiquement réceptrices.
Ce sont des extérocepteurs, des intérocepteurs (ou viscérocepteurs) et des propriocepteurs. Ces récepteurs sont sensibles à la douleur
(nocicepteurs), à la température (thermorécepteurs), aux mouvements des tissus provoqués par la pression (mécanorécepteurs).
Schéma d'une terminaison libre

www.neur-one.fr
[email protected] 7 09/09/2011
En fait, les terminaisons libres ne sont pas des récepteurs proprement dits. Elles portent des récepteurs de différentes natures qui sont
des capteurs moléculaires : canaux TRP, ASIC, P2X, HCN, TREK, etc (voir plus loin) : ces canaux seraient seulement responsables des sensations douloureuses voire thermiques mais il n'est pas encore prouvé qu'ils soient impliqués dans les sensations tactiles.
b) Récepteur de la racine des poils et des ongles
Chaque poil est innervé par plusieurs fibres myélinisées et non myélinisées, l'ensemble constitue un récepteur sensible à la stimulation mécanique (mécanorécepteur) constituée par un déplacement du poil. Leurs propriétés sont variables, adaptables ou non adaptables, ils peuvent
être d'une grande sensibilité et sont responsables de sensations tactiles et de déplacement. Ces récepteurs sensoriels sont des extérocepteurs et
des mécanorécepteurs. Ils comprennent : les plexus de la racine des poils
les cellules tactiles de Merkel (voir plus loin)
Les mouvements du poil engendrent une sensation tactile. La racine des ongles et le derme l'entourant contiennent de nombreuses terminaisons libres et d'autres organes encapsulés : corpuscules de Meissner et de Ruffini.
Plexus de la racine d'un poil.
Des fibres nerveuses fines s'enroulent autour des racines des follicules pileux et courent parallèlement vers le haut et le bas du poil. La fente sensorielle, formée par la perte de la gaine de myéline des terminaisons réceptrices que sur deux cotés, est stimulée par l'énergie mécanique. Les corpuscules de Meissner sont les pendants des plexus de la racine des poils. Des fibres superficielles se terminent juste sous l'épiderme. On trouve également des terminaisons nerveuses en palissades. Certaines fibres, entourant l'infundibulum dermique, sont liées à des disques de Merkel. Les mouvements du poil engendrent une déformation du follicule pileux et du derme environnant, ce qui provoque une sensation tactile.
c) Récepteur de Merkel et dôme épidermique
D'autres terminaisons nerveuses libres s'élargissent, en partie profonde de l'épiderme, au contact de 5 à 10 cellules épithéliales,
appelées cellules tactiles de Merkel (cellules sensorielles accessoires) et les kératinocytes : ce sont les disques de Merkel. Chez l'homme, les
mains et le bout des doigts, les lèvres et les organes génitaux externes en sont largement pourvus (25 % de l'innervation de la main).
Cellule de Merkel
Les cellules de Merkel possèdent des prolongements en forme de doigts qui sont déformés et provoquent une stimulation de la f ibre nerveuse : ces récepteurs sont à adaptation lente et à petit champ récepteur. Il semble qu'elles possèdent des caractéristiques neuro-endocrines : elles sécrètent des neuropeptides modulateurs (vésicules). Elles feraient partie du système endocrine diffus (système APUD).
Les disques de Merkel sont les récepteurs du toucher fin (pression légère) et discriminent les caractéristiques de l'objet : sa forme, ses bords, sa texture…Les disques de Merkel sont également associés à des fibres nerveuses du follicule pileux et réagissent aux mouvements du
poil.
Situés dans les couches superficielles de la peau (jonction derme-épiderme)
Densité élevée dans le bout des doigts, les lèvres …
Terminaisons nerveuses associées à une cellule épithéliale non neuronale formant des contacts synaptiques avec les
terminaisons nerveuses
Champ récepteur de petite taille de quelques millimètres, bien délimité
Réponse à la pression légère; permettent la discrimination statique de formes, de bords et des textures
Adaptation lente
www.neur-one.fr
[email protected] 8 09/09/2011
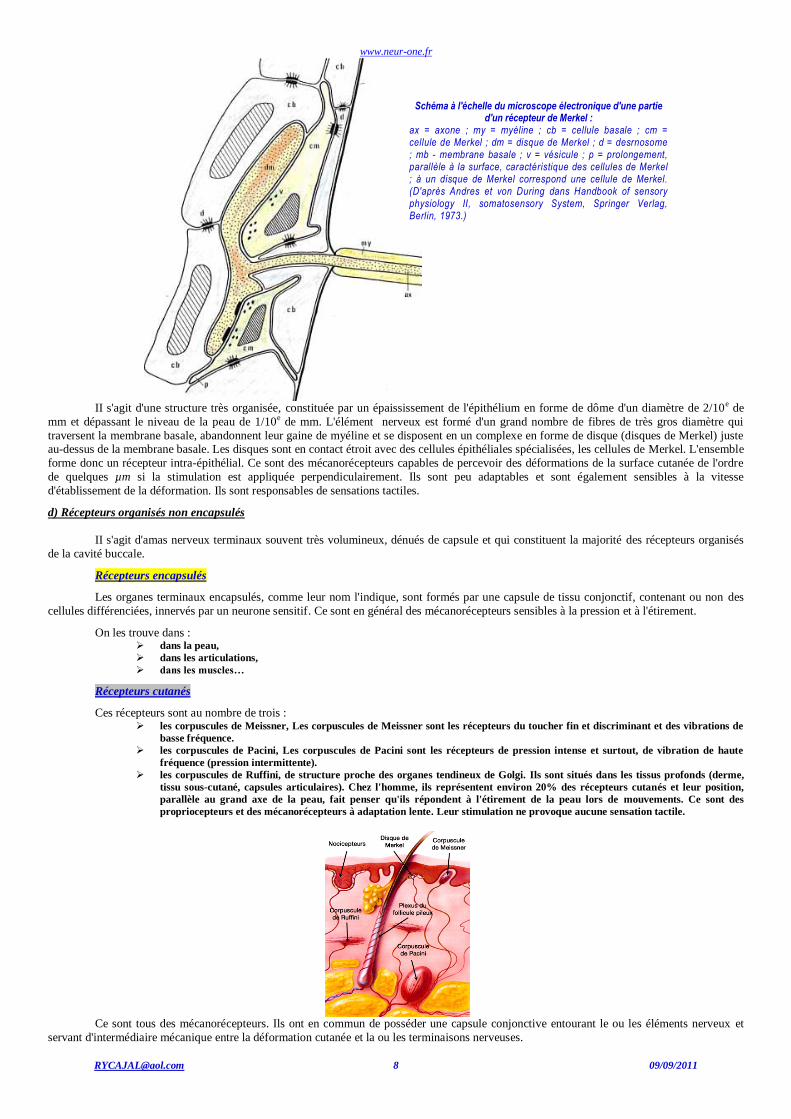
II s'agit d'une structure très organisée, constituée par un épaississement de l'épithélium en forme de dôme d'un diamètre de 2/10e de
mm et dépassant le niveau de la peau de 1/10e de mm. L'élément nerveux est formé d'un grand nombre de fibres de très gros diamètre qui
traversent la membrane basale, abandonnent leur gaine de myéline et se disposent en un complexe en forme de disque (disques de Merkel) juste au-dessus de la membrane basale. Les disques sont en contact étroit avec des cellules épithéliales spécialisées, les cellules de Merkel. L'ensemble
forme donc un récepteur intra-épithélial. Ce sont des mécanorécepteurs capables de percevoir des déformations de la surface cutanée de l'ordre
de quelques µm si la stimulation est appliquée perpendiculairement. Ils sont peu adaptables et sont également sensibles à la vitesse
d'établissement de la déformation. Ils sont responsables de sensations tactiles.
d) Récepteurs organisés non encapsulés
II s'agit d'amas nerveux terminaux souvent très volumineux, dénués de capsule et qui constituent la majorité des récepteurs organisés
de la cavité buccale.
Récepteurs encapsulés
Les organes terminaux encapsulés, comme leur nom l'indique, sont formés par une capsule de tissu conjonctif, contenant ou non des
cellules différenciées, innervés par un neurone sensitif. Ce sont en général des mécanorécepteurs sensibles à la pression et à l'étirement.
On les trouve dans : dans la peau,
dans les articulations,
dans les muscles…
Récepteurs cutanés
Ces récepteurs sont au nombre de trois : les corpuscules de Meissner, Les corpuscules de Meissner sont les récepteurs du toucher fin et discriminant et des vibrations de
basse fréquence.
les corpuscules de Pacini, Les corpuscules de Pacini sont les récepteurs de pression intense et surtout, de vibration de haute
fréquence (pression intermittente).
les corpuscules de Ruffini, de structure proche des organes tendineux de Golgi. Ils sont situés dans les tissus profonds (derme,
tissu sous-cutané, capsules articulaires). Chez l'homme, ils représentent environ 20% des récepteurs cutanés et leur position,
parallèle au grand axe de la peau, fait penser qu'ils répondent à l'étirement de la peau lors de mouvements. Ce sont des
propriocepteurs et des mécanorécepteurs à adaptation lente. Leur stimulation ne provoque aucune sensation tactile.
Ce sont tous des mécanorécepteurs. Ils ont en commun de posséder une capsule conjonctive entourant le ou les éléments nerveux et
servant d'intermédiaire mécanique entre la déformation cutanée et la ou les terminaisons nerveuses.
Schéma à l'échelle du microscope électronique d'une partie d'un récepteur de Merkel :
ax = axone ; my = myéline ; cb = cellule basale ; cm = cellule de Merkel ; dm = disque de Merkel ; d = desrnosome ; mb - membrane basale ; v = vésicule ; p = prolongement, parallèle à la surface, caractéristique des cellules de Merkel ; à un disque de Merkel correspond une cellule de Merkel. (D'après Andres et von During dans Handbook of sensory physiology II, somatosensory System, Springer Verlag, Berlin, 1973.)

www.neur-one.fr
[email protected] 9 09/09/2011
Il est probable que, sauf peut-être pour les terminaisons libres, tous les mécanorécepteurs sont en définitive excités par la déformation
mécanique de leur extrémité nerveuse. La déformation imposée par la stimulation à la peau est transmise à la fibre nerveuse par l'intermédiaire du corpuscule. Ce dernier est une structure très organisée qui modifie les caractéristiques de la stimulation de telle sorte que la déformation subie
par la terminaison libre n'a pas les mêmes propriétés spatio-temporelles que celle subie par la surface cutanée. Dans le cas du corpuscule de
Pacini, c'est la constitution du corpuscule qui entraîne cette transformation, dans le cas des corpuscules de Meissner ou de Ruffini, le mode
d'attache du corpuscule aux tissus de soutien participe à ces modifications.
a) Corpuscule de Meissner
Situés dans les couches superficielles de la peau (jonction derme-épiderme)
Densité élevée au niveau des zones glabres (doigts, lèvres…)
Capsule de tissu fibreux enfermant plusieurs lamelles de cellules de Schwann et contenant 1 ou plusieurs fibres afférentes au
centre
Champ récepteur de petite taille de quelques millimètres, bien délimité
Réponse à des dépressions minimes de la peau, des mouvements légers de surface, des vibrations lentes
Adaptation rapide
Les corpuscules de Meissner, situés dans les papilles du derme nombreux dans les parties glabres et très sensibles de la peau chez
l'homme. Les lèvres, les mamelons, les organes génitaux externes, les paupières, mais surtout les faces plantaires des pieds et des mains, en particulier chez l'homme, le bout des doigts, de l'index principalement, en sont largement pourvues (40 % de l'innervation de la main).
La racine des ongles et le derme l'entourant contiennent des corpuscules de Meissner et de Ruffini. Les corpuscules de Meissner sont
les pendants des plexus de la racine des poils.
Il est présent dans les papilles dermiques et particulièrement fréquent au niveau de l'extrémité des doigts de la main. Il est formé d'une
ou de plusieurs fibres nerveuses de gros calibre qui pénètrent une capsule ovoïde à grand axe perpendiculaire à la surface cutanée. En entrant
dans la capsule, les fibres nerveuses prennent une disposition hélicoïdale. La capsule est attachée au tissu de soutien par un système de fibres
conjonctives disposées également de façon hélicoïdale. Il est probable que cette géométrie des fibres nerveuses et des fibres élastiques joue un rôle d'amortisseur lors de stimulations mécaniques continues de la peau. Là aussi, les propriétés d'adaptation rapide de ce récepteur seraient
expliquées par la constitution de la capsule.
Ces récepteurs sont à petit champ récepteur et à adaptation rapide. Les corpuscules de Meissner, récepteurs dynamiques et des
vibrations de basse fréquence (30-50 Hz), perçoivent les mouvements d'objets sur la peau. Ils permettent, par exemple, de ressentir les vibrations des basses quand on pose la main sur une enceinte de chaîne hi-fi. Ils permettent également de saisir plus fortement un objet qui nous glisse des
mains par exemple. Les bulbes terminaux ovales de Krause, considérés comme une variante des corpuscules de Meissner, sont localisés dans les
muqueuses et les régions cutanéo-muqueuses (orifices comme les lèvres).
b) Corpuscule de Pacini
Le corpuscule de Meissner Les corpuscules de Meissner, de forme ovale, sont constitués de cellules lamellaires (lemmocytes : cellules de Schwann en piles d'assiettes, flèche 2) entre lesquelles une ou plusieurs fibres nerveuses perdent leur gaine de myéline (flèche 1). En 3, une fine capsule conjonctive emballe l'ensemble. Des fibres de collagène émergent de la capsule pour s'attacher sur les tonofibrilles de l'épiderme. Par conséquent, les déformations cutanées provoquent celle du corpuscule qui stimule la fibre sensitive.
Schéma du corpuscule de Meissner Les fibres conjonctives issues de la capsule ou de l'intérieur du corpuscule son» en continuité avec les tonofribrilles des cellules épithéliales si bien que de très faibles actions mécaniques appliquées à la surface cutanée sont répercutées au niveau de l'axone sous forme d'une torsion (flèche pleine) la flèche vide indique le mouvement consécutif de la base du corpuscule par lequel on explique que la déformation de l'axone prenne rapidement fin, ce qui rend compte des propriétés d'adaptation rapide de ce récepteur. L'intérieur du corpuscule contient, outre îles terminaisons nerveuses, des cellules de Schwann. 1.Membrane basale 2.Fibres conjonctives issues de la capsule ou de l'intérieur du corpuscule. 3.Cellule épithéliale 4.Axone dénudé 5.Capsule conjonctive 6.Papille dermique (D'après Andres et von During dans Handbook of sensory physiology II, somatosensory System, Springer Verlag, 1973.) capsule

www.neur-one.fr
[email protected] 10 09/09/2011
Situés dans les couches plus profondes de la peau (derme) et dans le tissu sous-cutané
Grande capsule en lamelle d’oignons (diamètre 1 mm) avec une terminaison nerveuse au centre
Champ récepteur large, de limites floues pouvant couvrir un doigt entier voire la moitié de la paume de la main
Discrimination de stimuli mobiles, des vibrations rapides
10- 15% des récepteurs cutanés de la main
Adaptation rapide
Les corpuscules de Pacini sont les plus grands récepteurs corpusculaires (jusqu'à 2 mm de longueur). Ils sont situés dans l'hypoderme :
doigts, plantes des pieds, organes génitaux externes, mamelons. On les retrouve également dans le périoste, à proximité des articulations et à la surface des tendons et des aponévroses, dans le mésentère et dans les intestins. Dans ce cas, ce sont des mécanorécepteurs qui sont des
intérocepteurs (ou viscérorécepteurs) et des propriocepteurs.
Dans les organes profonds, ils transmettent au squelette les vibrations du sol ou des autres éléments internes (articulation…).
Coupe transversale de deux corpuscules de Pacini
De très grande taille (de 1 à 4 mm !) il est composé de l'extrémité d'une très grosse fibre nerveuse ayant perdu sa gaine de myéline. Les corpuscules de Pacini sont formés par de nombreuses lamelles concentriques (comme la structure d'un oignon) formant trois couches distinctes : En 1, terminaison nerveuse (la massue centrale dans laquelle la fibre nerveuse pénètre en perdant sa gaine de myéline.), en 2, fibres conjonctives (la couche lamellaire externe (cellules de Schwann) dont les lamelles sont entourées par un liquide visqueux), en 3, fibroblastes.
Les corpuscules de Pacini sont les récepteurs du toucher fin (déformation de quelques microns) et des vibrations de haute fréquence
(250 à 350 Hz), à adaptation très rapide. Leur stimulation fait ressentir des chatouillements, vibrations très rapides. Leur champ récepteur est très
large : un doigt ou une partie de la paume chez l'homme. Ils permettent, par exemple, de ressentir les vibrations quand on pose la main sur une
enceinte classique de chaîne hi-fi. Les corpuscules de Golgi-Mazzoni, situés dans le derme, ont une structure quasi-identique, mais sont bien plus petits.
La déformation des corpuscules de Pacini a permis de mettre en évidence le mécanisme non-neurologique de l'adaptation neuronale.
Les corpuscules de Pacini sont formés par de nombreuses lamelles concentriques (comme la structure d'un oignon). Au centre, se trouve une
fibre sensitive. Lors de pression sur le corpuscule, l'énergie mécanique déforme la membrane neuronale de la fibre, ce qui a la propriété d'ouvrir les canaux ioniques et de provoquer l'apparition d'un potentiel récepteur. Si l'intensité du stimulus est suffisante, ce potentiel récepteur (qui est
un potentiel gradué) se transforme en potentiel d'action dans la fibre. Si la pression est continue, la fréquence des potentiels diminue.
c) Corpuscule de Ruffini
Situés dans les couches plus profondes de la peau (derme), dans le tissu sous-cutané et les capsules articulaires
Champ récepteur large, de limites floues pouvant couvrir un doigt entier
Capsule allongée en fuseau
Sensibles aux étirements persistants que produisent les mouvements des doigts et des membres
Adaptation lente
Présent dans le derme profond, il est allongé, fusiforme, la capsule est attachée au tissu de soutien par ses deux extrémités ; elle est
pénétrée par une fibre nerveuse de gros diamètre qui se ramifie et se pelotonne. Ce récepteur présente une sensibilité directionnelle puisque la
Les corpuscules de Ruffini Situés dans les couches profondes de la peau ils ont une structure et un mode de fonctionnement (sensibles à l'étirement de la peau) qui ressemblent à ceux de l'organe tendineux de Golgi : ce sont des « propriocepteurs ».

www.neur-one.fr
[email protected] 11 09/09/2011
déformation de la peau n'est efficace que si elle est appliquée selon une certaine direction. Cette propriété est sans doute liée au mode d'attache
de la capsule au tissu environnant. Il est non adaptable.
d) Corpuscules de formes intermédiaires
On a décrit d'autres corpuscules (corpuscule de Golgi, de Krause) qui ne sont que des formes intermédiaires des rnécanorécepteurs
précédemment cités.
Les thermorécepteurs
Terminaisons nerveuses libres C et Aδpour nocicepteurs et
Thermorécepteurs:
o Tact protopathique : fibre Aβet Aδ .
o Réponse au tact « grossier » avant la douleur, au médiateur de l’inflammation, a l’ischemie.
o Récepteurs polymodaux : température et douleur.
Pruricepteurs: fibres C histaminergiques (+???).
Les propriocepteurs
Les propriocepteurs musculaires
La proprioception désigne l'ensemble des récepteurs, voies et centres nerveux impliqués dans la perception, consciente ou non, de la
position relative des parties du corps. Cette définition implique :
une proprioception consciente qui est à la base de notre " schéma corporel " :
o positions et mouvements articulaires,
o sensations musculaires et tendineuses,
o perception de l'orientation du corps, des membres et de la tête,
o équilibration.
C'est le système dit " épicritique " qui véhicule les modalités proprioceptives conscientes (ainsi que les tactiles fines - forme, texture,
glissement, pression, vibrations -). Leur voie centrale est celle des colonnes dorsales et du lemnisque médian. Le schéma corporel est la représentation que chaque individu se fait de son corps afin de lui permettre de se situer dans l'espace. C'est un schéma anatomique et
fonctionnel du corps. Anatomique en ce qu'il reflète la perception qu'a un individu des rapports des différentes parties de son corps entre elles et
avec son environnement. Fonctionnel en ce qu'il exprime la perception qu'a ce même individu de la mécanique de ce corps biologique.
Les récepteurs sensoriels impliqués sont : les fuseaux neuromusculaires,
les organes tendineux de Golgi,
les récepteurs kinesthésiques articulaires,
les récepteurs cutanés (voir ci-dessus).
une proprioception inconsciente (NON TRAITÉE ICI), qui est d'adaptation très rapide, dans les phénomènes statiques ou
dynamiques comme :
o le contrôle de la contraction musculaire
o les ajustements posturaux, et chez l'homme, la station debout.
Les ajustements posturaux sont souvent préprogrammés et dépendent, en grande partie, des fuseaux neuromusculaires. La station
debout, chez l'homme, utilise des afférences visuelles, vestibulaires, proprioceptives et tactiles (récepteurs de la plante des pieds sensibles à la pression et à la température). La sensibilité prorioceptive inconsciente, en provenance des fuseaux neuro-musculaires, des récepteurs tendineux
et articulaires, empruntent des tractus spinaux qui se terminent dans le cervelet. Ce sont les tractus spino-cérébelleux direct (membres inférieurs
et tronc) et cunéo-cérébelleux (membre supérieur) et le tractus spino-cérébelleux croisé ou ventral (informations musculaires). Ces informations
sont complétées par : le système vestibulaire, le système visuel.
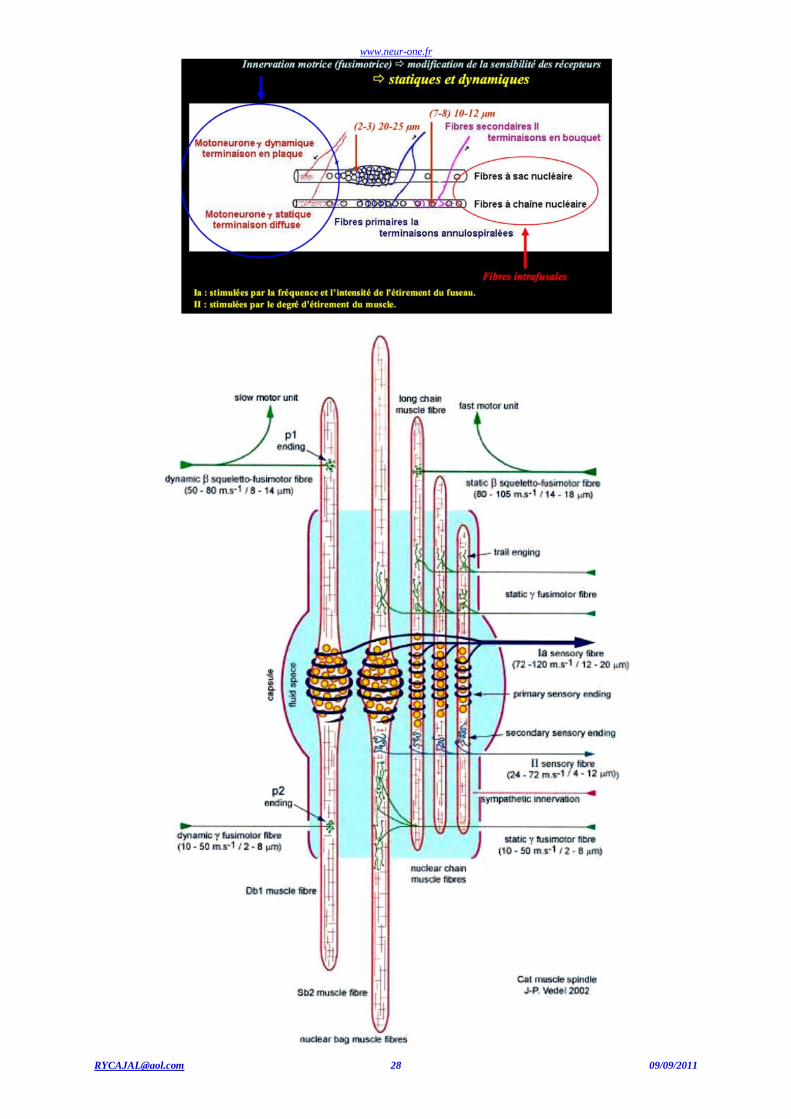
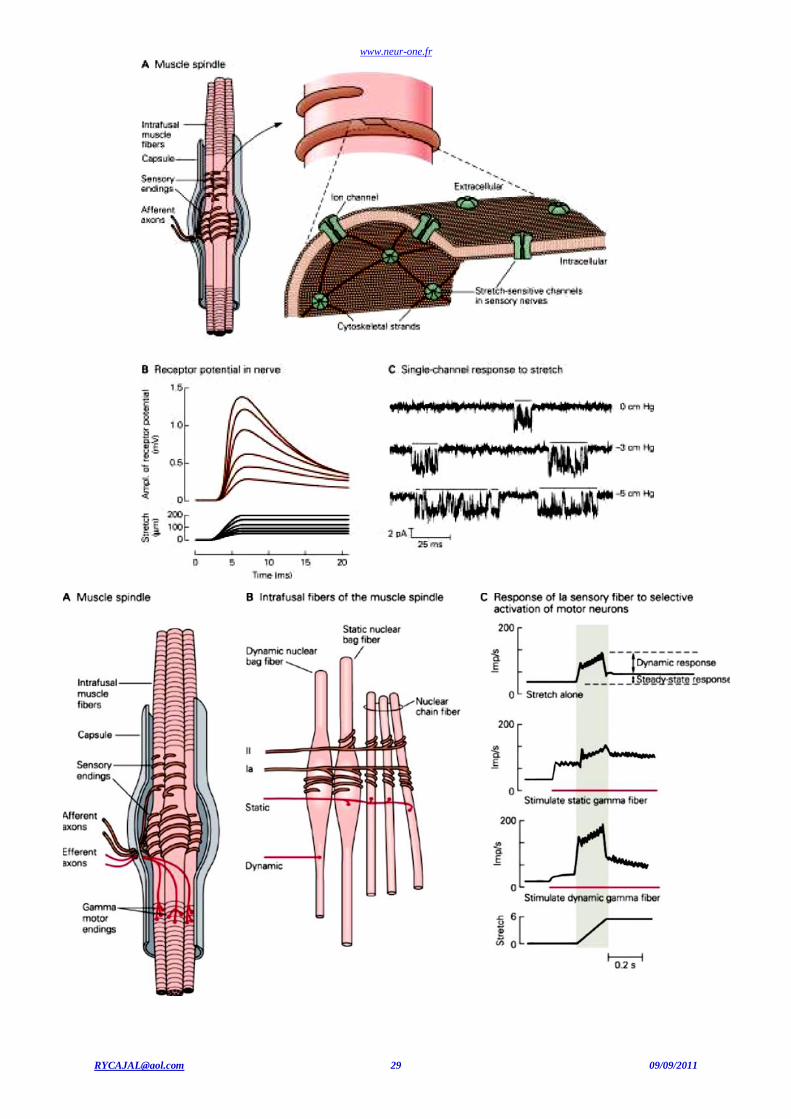
Les fuseaux neuromusculaires
Les fuseaux neuromusculaires sont situés dans les muscles striés squelettiques. Plus les mouvements sont fins, plus les fuseaux
neuromusculaires sont nombreux (par exemple, au niveau des muscles des doigts). Ces fuseaux sont formés de fibres musculaires modifiées
striées, appelées fibres intrafusoriales ou intrafusales, situées parallèlement aux fibres musculaires " normales " ou fibres extrafusoriales (en dehors du fuseau). Les fibres sensorielles s’enroulent autour des fibres d’un fuseau.
Schéma du corpuscule de Ruffini Les multiples ramifications terminales de la fibre nerveuse sont en contact étroit avec les fibres de collagène issues du tissu cutané et sous-cutané et qui pénètrent le corpuscule par ses deux pôles. Une pression exercée sur la peau se transmet aux fibres collagènes et étire le peloton nerveux central. Contrairement aux corpuscules de Pacini et de Meissner, le corpuscule de Ruffini ne présente pas de système d'amortisseur et est donc non adaptable.
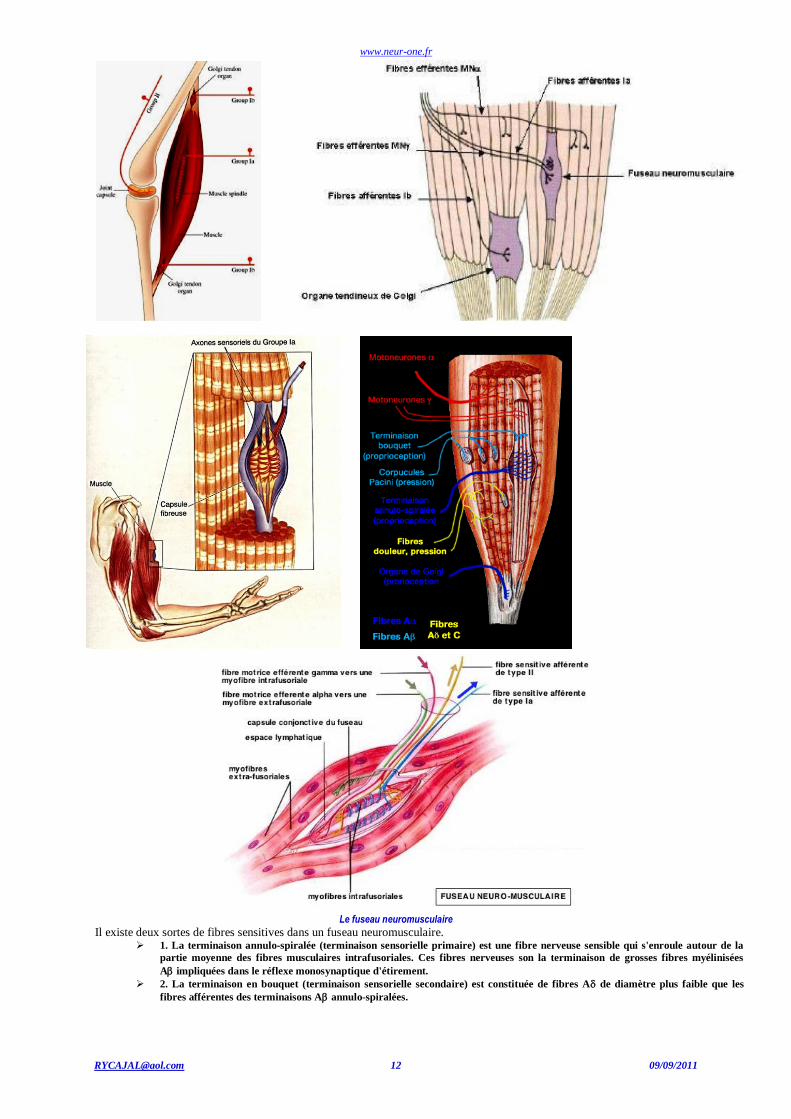
www.neur-one.fr
[email protected] 12 09/09/2011
Le fuseau neuromusculaire
Il existe deux sortes de fibres sensitives dans un fuseau neuromusculaire. 1. La terminaison annulo-spiralée (terminaison sensorielle primaire) est une fibre nerveuse sensible qui s'enroule autour de la
partie moyenne des fibres musculaires intrafusoriales. Ces fibres nerveuses son la terminaison de grosses fibres myélinisées
Aimpliquées dans le réflexe monosynaptique d'étirement.
2. La terminaison en bouquet (terminaison sensorielle secondaire) est constituée de fibres A de diamètre plus faible que les
fibres afférentes des terminaisons A annulo-spiralées.
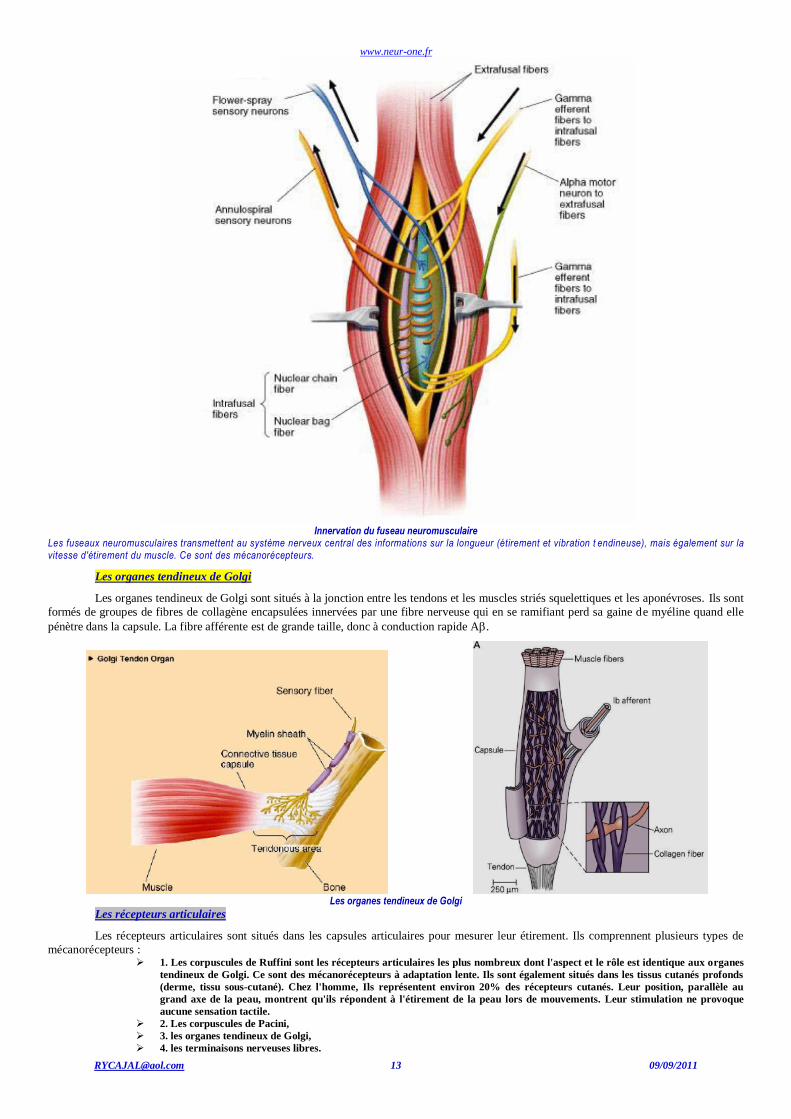
www.neur-one.fr
[email protected] 13 09/09/2011
Innervation du fuseau neuromusculaire
Les fuseaux neuromusculaires transmettent au système nerveux central des informations sur la longueur (étirement et vibration t endineuse), mais également sur la vitesse d'étirement du muscle. Ce sont des mécanorécepteurs.
Les organes tendineux de Golgi
Les organes tendineux de Golgi sont situés à la jonction entre les tendons et les muscles striés squelettiques et les aponévroses. Ils sont
formés de groupes de fibres de collagène encapsulées innervées par une fibre nerveuse qui en se ramifiant perd sa gaine de myéline quand elle
pénètre dans la capsule. La fibre afférente est de grande taille, donc à conduction rapide A.
Les organes tendineux de Golgi
Les récepteurs articulaires
Les récepteurs articulaires sont situés dans les capsules articulaires pour mesurer leur étirement. Ils comprennent plusieurs types de
mécanorécepteurs : 1. Les corpuscules de Ruffini sont les récepteurs articulaires les plus nombreux dont l'aspect et le rôle est identique aux organes
tendineux de Golgi. Ce sont des mécanorécepteurs à adaptation lente. Ils sont également situés dans les tissus cutanés profonds
(derme, tissu sous-cutané). Chez l'homme, Ils représentent environ 20% des récepteurs cutanés. Leur position, parallèle au
grand axe de la peau, montrent qu'ils répondent à l'étirement de la peau lors de mouvements. Leur stimulation ne provoque
aucune sensation tactile.
2. Les corpuscules de Pacini,
3. les organes tendineux de Golgi,
4. les terminaisons nerveuses libres.
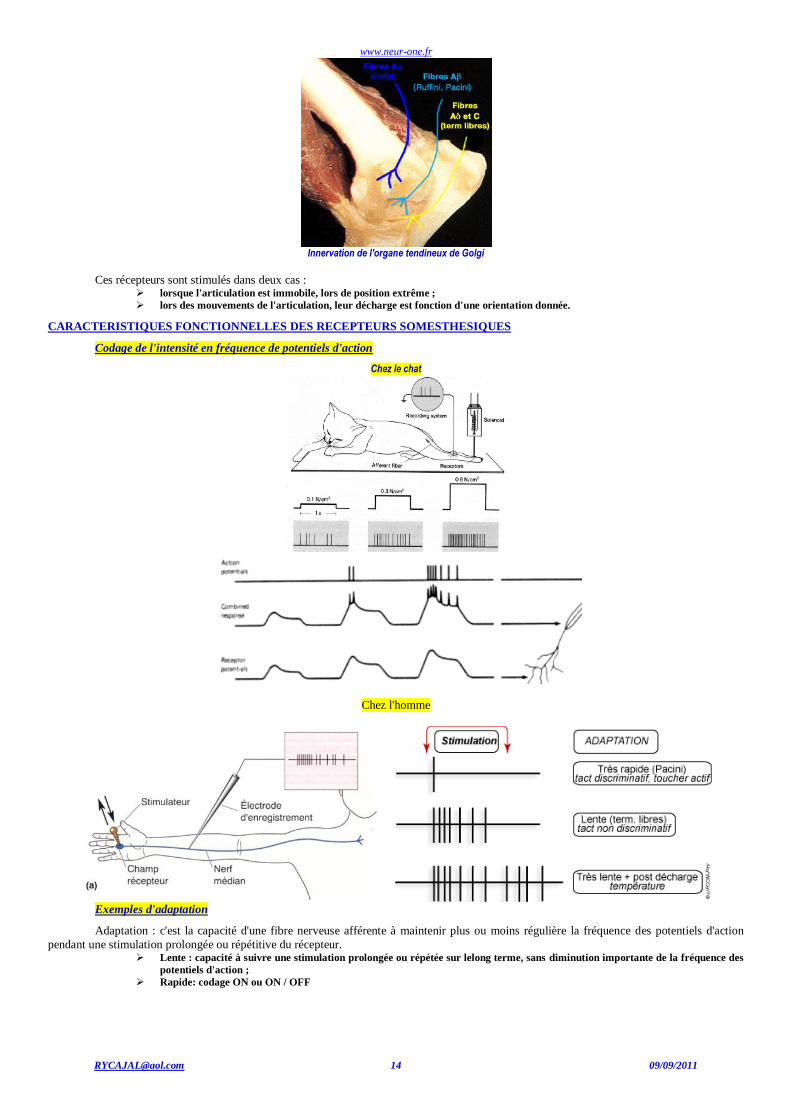
www.neur-one.fr
[email protected] 14 09/09/2011
Innervation de l'organe tendineux de Golgi
Ces récepteurs sont stimulés dans deux cas : lorsque l'articulation est immobile, lors de position extrême ;
lors des mouvements de l'articulation, leur décharge est fonction d'une orientation donnée.
CARACTERISTIQUES FONCTIONNELLES DES RECEPTEURS SOMESTHESIQUES
Codage de l'intensité en fréquence de potentiels d'action
Chez le chat
Chez l'homme
Exemples d'adaptation
Adaptation : c'est la capacité d'une fibre nerveuse afférente à maintenir plus ou moins régulière la fréquence des potentiels d'action
pendant une stimulation prolongée ou répétitive du récepteur. Lente : capacité à suivre une stimulation prolongée ou répétée sur lelong terme, sans diminution importante de la fréquence des
potentiels d'action ;
Rapide: codage ON ou ON / OFF

www.neur-one.fr
[email protected] 15 09/09/2011
Codage de la qualité, de la nature de la stimulation
Théorie spécifiste : Meissner : adaptation rapide de type I (frisson)
Pacini : adaptation rapide de type II (vibration).
Merkel : adaptation lente de type I (toucher fin, texture)
Ruffini : adaptation lente de type II (pression)
Fibres mécanoréceptrices A et C : toucher, vibration et frisson.
Théorie non spécifiste : pattern Caractéristique spatio-temporelle de la décharge.
Codage de la localisation corporelle de la stimulation
Dermatomes corporels
La sensibilité tactile n'emprunte pas les mêmes voies selon que l'information est issue de la face ou du reste du corps. Rappelons tout
d'abord que la moelle épinière est organisée de manière segmentaire. Elle comporte au niveau de chaque vertèbre (donc 31 fois) une racine dorsale et une racine ventrale par lesquelles est assurée la communication entre le SNC et la périphérie du corps. Les 31 paires de nerfs spinaux
(ou 31 segments) peuvent être regroupés en quatre groupes qui correspondent à chacun des niveaux des vertèbres: cervical, dorsal, lombaire et
sacré. La cartographie de la peau dans les différentes régions du corps correspond strictement à cette organisation: ainsi par exemple, la peau du
thorax est innervée par des nerfs spinaux pénétrant dans la moelle au niveau thoracique, la peau des mains et des bras est innervée par des nerfs spinaux pénétrant dans la moelle au niveau cervical, etc. On désigne par dermatome la région de la peau innervée par un nerf spinal. Les
ganglions des racines dorsales de la moelle épinière contiennent les corps cellulaires des neurones à l'origine des afférences primaires de la
somesthésie. Ces neurones en T ont donc un prolongement issu de la périphérie (qui pénètre dans la moelle épinière par la racine dorsale) et un
prolongement central qui atteint la corne dorsale de la moelle épinière. Les axones pénétrant dans la moelle se divisent en plusieurs branches. La branche segmentaire permet à l'information somesthésique de diffuser à travers des circuits propres de la moelle épinière impliqués dans des
réactions réflexes très rapides et non conscientes. Un exemple de ce type de réaction est le réflexe rotulien dans lequel un coup sec et bref,
appliqué sur le tendon situé sous la rotule, provoque un redressement réflexe (non volontaire) de la jambe. Ce réflexe nécessite que l'individu
soit détendu et n'oppose aucune résistance. De fait, chez l'Homme l'observation de ces réactions réflexes est difficile à mettre en évidence (alors qu'on peut observer facilement de nombreuses réactions réflexes de ce type chez l'animal) car, précisément chez l'Homme, la plupart des
réactions sont sous le contrôle volontaire: ainsi les réflexes spinaux sont-ils inhibés par le contrôle qu'exercent les centres supérieurs sur les
circuits spinaux. L'autre branche des axones pénétrant dans la moelle épinière est la branche ascendante, qui permet d'envoyer l'information vers
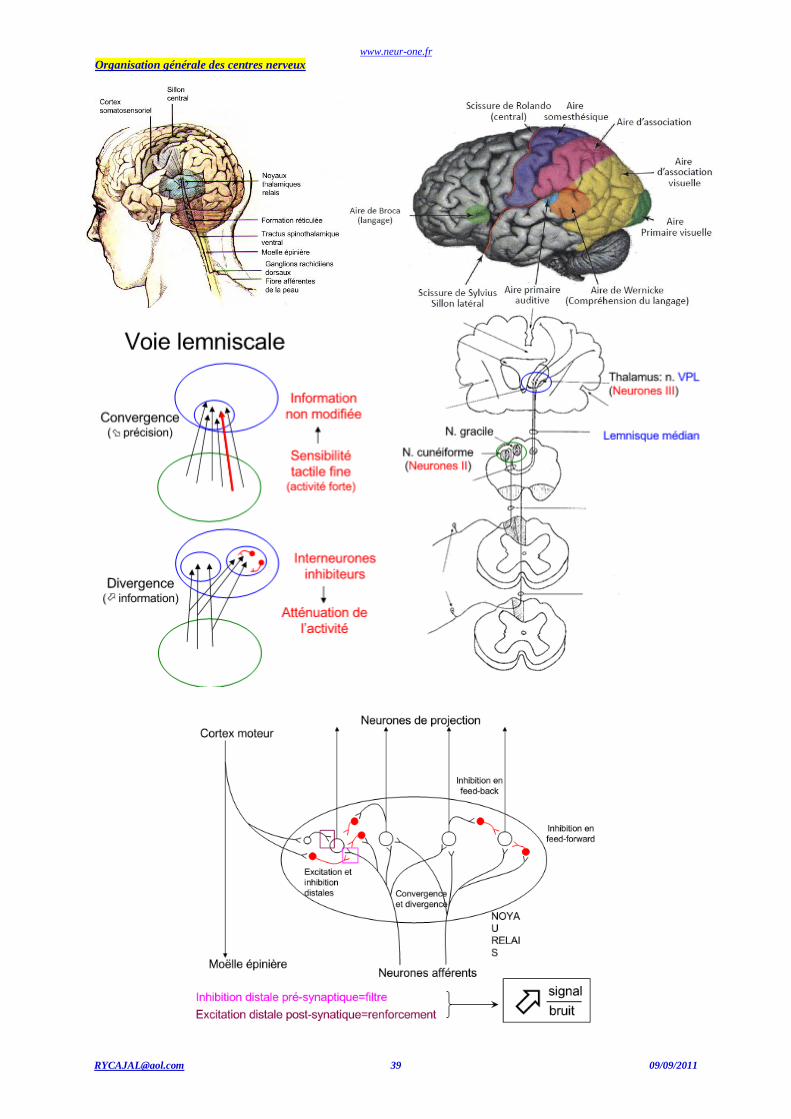
les régions supraspinales. Il existe deux grandes voies véhiculant les informations depuis la périphérie (hors visage) jusqu'au cerveau. La voie des colonnes dorsales et du lemnisque médian véhicule les informations issues des mécanorécepteurs de la sensibilité tactile et de la
proprioception, tandis que la voie spino-thalamique véhicule les informations thermiques et celles de la douleur. Seule la première de ces voies
sera décrite ici. Les axones des afférences somesthésiques ne sont pas tous de même diamètre: on distingue ainsi les fibres de très gros diamètre
Aα issues des propriocepteurs des muscles, les fibres de gros diamètres Aβ issues des mécanorécepteurs de la peau, mais aussi les fibres Aδ et C de plus petit diamètre, véhiculant les informations sur la température et la douleur. Les fibres A sont myélinisées, tandis que les fibres C sont
dépourvues de myéline. Ainsi, les fibres Aβ véhiculant les informations du toucher, de par leur diamètre et leur gaine de myéline, transmettent
les PA à la vitesse d'environ 70 m/s, tandis que les fibres C ne les transmettent qu'à environ l m/s. Les fibres Aβ pénètrent dans la colonne
dorsale et remontent vers le cerveau par la voie des colonnes dorsales et du lemnisque médian jusqu'au niveau du bulbe rachidien, sans synapse et du côté ipsilatéral. Les noyaux bulbaires constituent le premier relais sur cette. voie après lequel les neurones de second ordre décussent et
forment le lemnisque médian qui est un faisceau très dense de fibres traversant le pont et le mésencéphale et se dirigeant vers le noyau ventral
postérieur latéral (VPL) du thalamus. Les neurones de troisième ordre vont du VPL à l'aire primaire du cortex somesthésique (ou somato-
sensoriel) située dans le lobe pariétal à l'arrière de la scissure centrale (ou de Rolando). (Voir aussi la somatotopie).
Sensibilité du visage
Concernant la sensibilité du visage, l'information est véhiculée par la paire V de nerfs crâniens, les nerfs trijumeaux qui comportent
trois branches (se rapportant à la face, la bouche, la langue). D'autres régions du visage envoient leurs informations tactiles par d'autres nerfs
crâniens (par exemple le nerf IX, glossopharyngien pour la région du pharynx). Ces nerfs pénètrent dans le cerveau au niveau du pont et suivent ensuite un trajet tout à fait comparable à celui de la voie des colonnes avec un relais dans le noyau trijumeau ispsilatéral puis, après décussation,
un relais dans le noyau ventral postérieur du thalamus et enfin le cortex primaire somesthésique. Au sein de l'aire somesthésique primaire (SI),
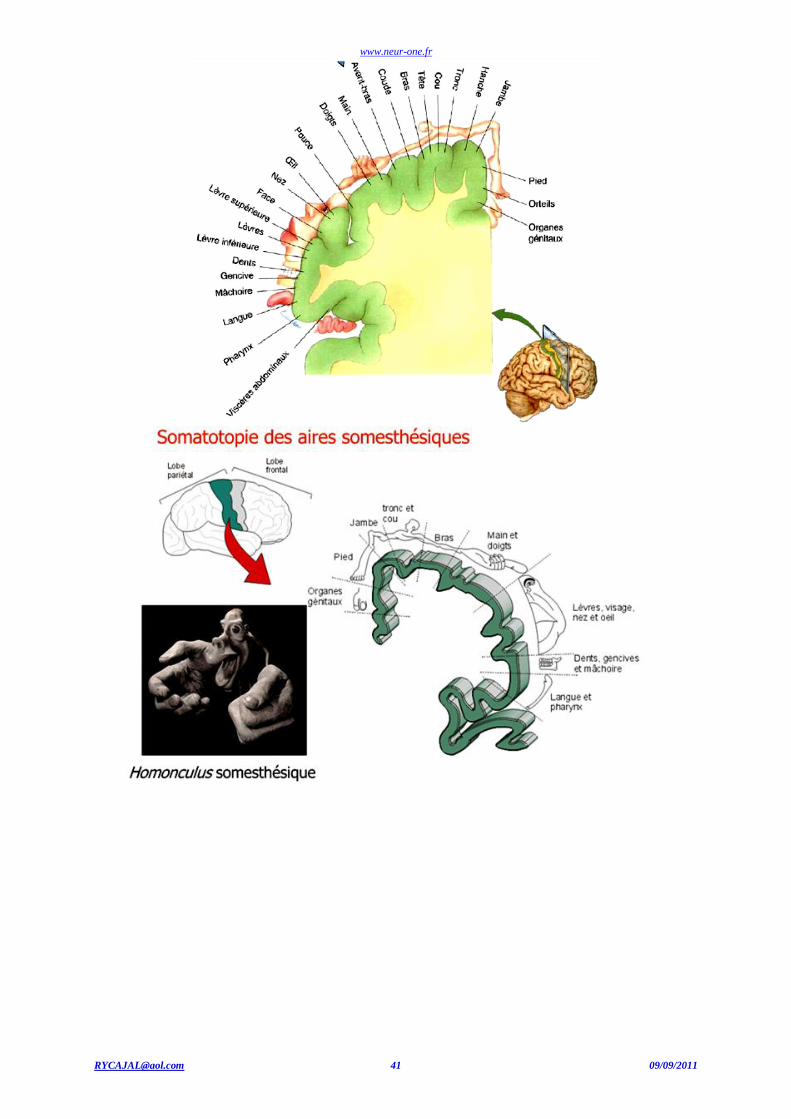
la somatotopie est strictement respectée : chacun de ses points correspond à un point précis de la périphérie et, pour une région donnée, plus il y
a de récepteurs, plus la portion de SI qui s'y rapporte est importante. La carte somatotopique du cortex somesthésique primaire, désignée sous le terme d'homunculus sensitif ou somato-sensoriel, a ainsi été établie (homunculus, i). À l'arrière de SI, le cortex pariétal postérieur comprend les
aires associatives mono sensorielles de la somesthésie et entretient des relations bidirectionnelles avec SI: c'est à ce niveau que s'opère
l'intégration en un tout cohérent des sensations tactiles et qu'émerge donc la représentation perceptive tactile. (Voir aussi la somatotopie).
www.neur-one.fr
[email protected] 16 09/09/2011
http://www.vetopsy.fr/anat/neuro/nerfs_cran/5_trijum1.php
http://www.anatomie-humaine.com/Nerf-trijumeau-V.html
www.neur-one.fr
[email protected] 17 09/09/2011
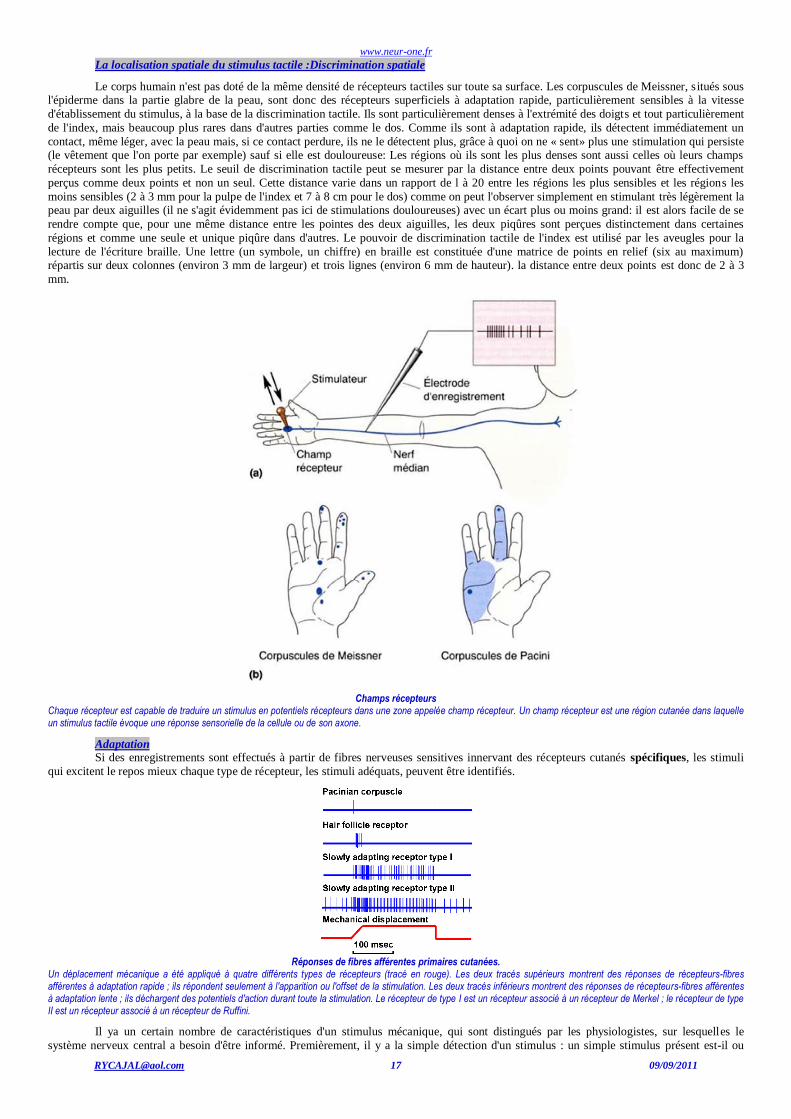
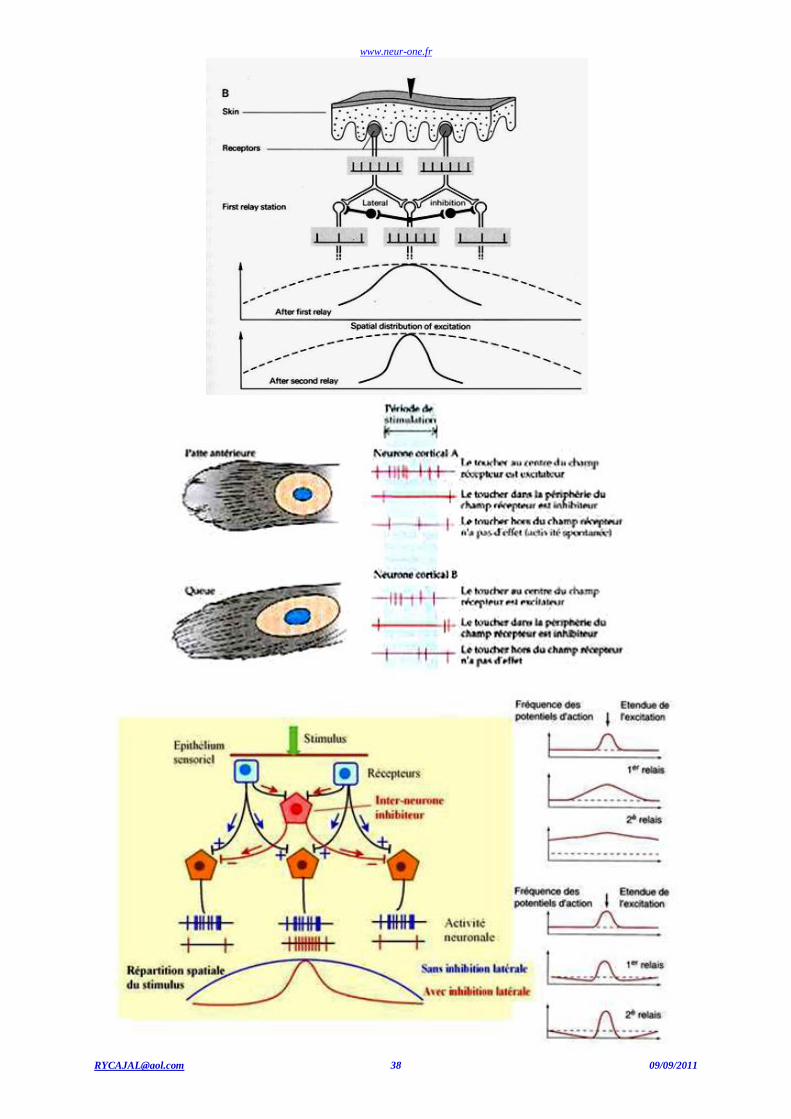
La localisation spatiale du stimulus tactile :Discrimination spatiale
Le corps humain n'est pas doté de la même densité de récepteurs tactiles sur toute sa surface. Les corpuscules de Meissner, s itués sous l'épiderme dans la partie glabre de la peau, sont donc des récepteurs superficiels à adaptation rapide, particulièrement sensibles à la vitesse
d'établissement du stimulus, à la base de la discrimination tactile. Ils sont particulièrement denses à l'extrémité des doigts et tout particulièrement
de l'index, mais beaucoup plus rares dans d'autres parties comme le dos. Comme ils sont à adaptation rapide, ils détectent immédiatement un
contact, même léger, avec la peau mais, si ce contact perdure, ils ne le détectent plus, grâce à quoi on ne « sent» plus une stimulation qui persiste (le vêtement que l'on porte par exemple) sauf si elle est douloureuse: Les régions où ils sont les plus denses sont aussi celles où leurs champs
récepteurs sont les plus petits. Le seuil de discrimination tactile peut se mesurer par la distance entre deux points pouvant être effectivement
perçus comme deux points et non un seul. Cette distance varie dans un rapport de l à 20 entre les régions les plus sensibles et les régions les
moins sensibles (2 à 3 mm pour la pulpe de l'index et 7 à 8 cm pour le dos) comme on peut l'observer simplement en stimulant très légèrement la peau par deux aiguilles (il ne s'agit évidemment pas ici de stimulations douloureuses) avec un écart plus ou moins grand: il est alors facile de se
rendre compte que, pour une même distance entre les pointes des deux aiguilles, les deux piqûres sont perçues distinctement dans certaines
régions et comme une seule et unique piqûre dans d'autres. Le pouvoir de discrimination tactile de l'index est utilisé par les aveugles pour la
lecture de l'écriture braille. Une lettre (un symbole, un chiffre) en braille est constituée d'une matrice de points en relief (six au maximum) répartis sur deux colonnes (environ 3 mm de largeur) et trois lignes (environ 6 mm de hauteur). la distance entre deux points est donc de 2 à 3
mm.
Champs récepteurs
Chaque récepteur est capable de traduire un stimulus en potentiels récepteurs dans une zone appelée champ récepteur. Un champ récepteur est une région cutanée dans laquelle un stimulus tactile évoque une réponse sensorielle de la cellule ou de son axone.
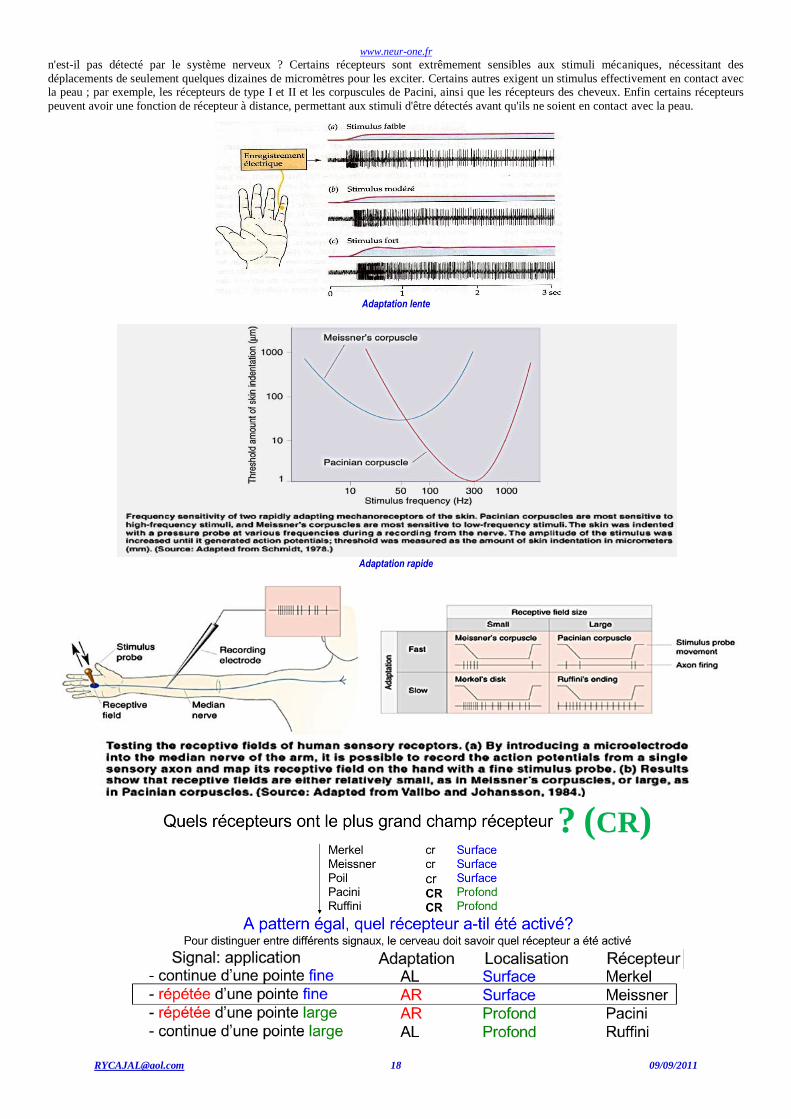
Adaptation Si des enregistrements sont effectués à partir de fibres nerveuses sensitives innervant des récepteurs cutanés spécifiques, les stimuli
qui excitent le repos mieux chaque type de récepteur, les stimuli adéquats, peuvent être identifiés.
Réponses de fibres afférentes primaires cutanées.
Un déplacement mécanique a été appliqué à quatre différents types de récepteurs (tracé en rouge). Les deux tracés supérieurs montrent des réponses de récepteurs-fibres afférentes à adaptation rapide ; ils répondent seulement à l'apparition ou l'offset de la stimulation. Les deux tracés inférieurs montrent des réponses de récepteurs-fibres afférentes à adaptation lente ; ils déchargent des potentiels d'action durant toute la stimulation. Le récepteur de type I est un récepteur associé à un récepteur de Merkel ; le récepteur de type II est un récepteur associé à un récepteur de Ruffini.
Il ya un certain nombre de caractéristiques d'un stimulus mécanique, qui sont distingués par les physiologistes, sur lesquelles le
système nerveux central a besoin d'être informé. Premièrement, il y a la simple détection d'un stimulus : un simple stimulus présent est-il ou
www.neur-one.fr
[email protected] 18 09/09/2011
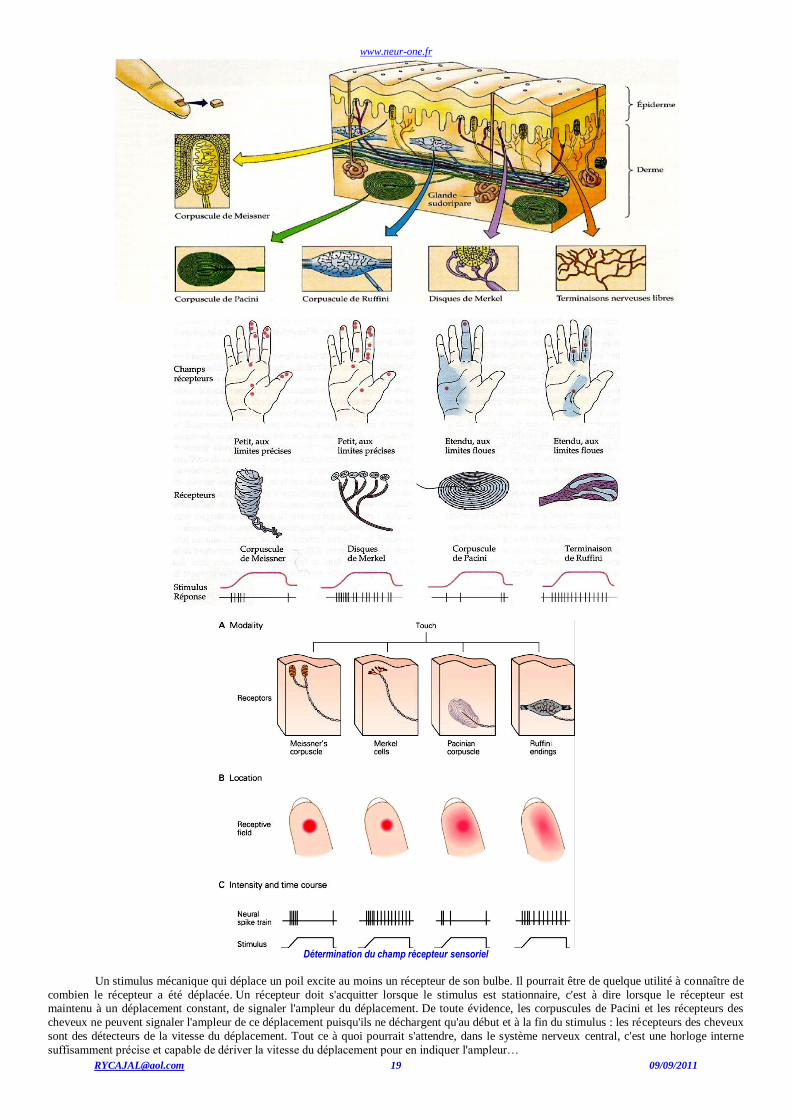
n'est-il pas détecté par le système nerveux ? Certains récepteurs sont extrêmement sensibles aux stimuli mécaniques, nécessitant des
déplacements de seulement quelques dizaines de micromètres pour les exciter. Certains autres exigent un stimulus effectivement en contact avec la peau ; par exemple, les récepteurs de type I et II et les corpuscules de Pacini, ainsi que les récepteurs des cheveux. Enfin certains récepteurs
peuvent avoir une fonction de récepteur à distance, permettant aux stimuli d'être détectés avant qu'ils ne soient en contact avec la peau.
Adaptation lente
Adaptation rapide
? (CR)
www.neur-one.fr
[email protected] 19 09/09/2011
Détermination du champ récepteur sensoriel
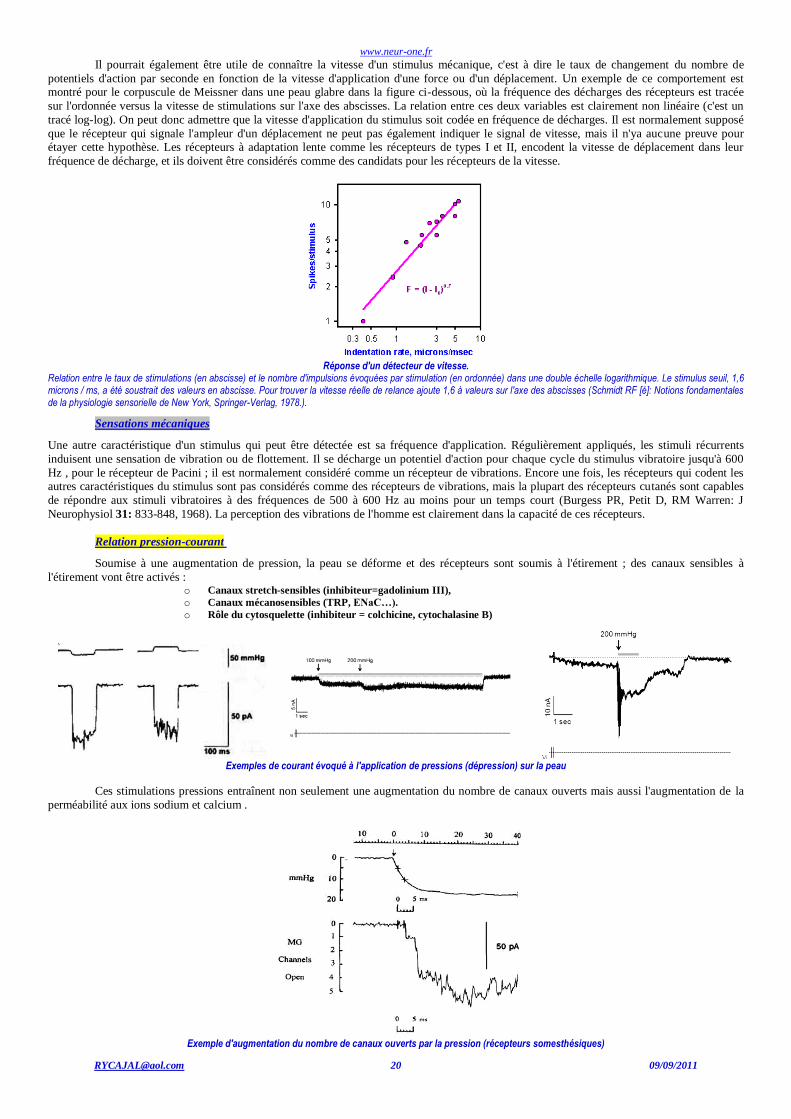
Un stimulus mécanique qui déplace un poil excite au moins un récepteur de son bulbe. Il pourrait être de quelque utilité à connaître de
combien le récepteur a été déplacée. Un récepteur doit s'acquitter lorsque le stimulus est stationnaire, c'est à dire lorsque le récepteur est maintenu à un déplacement constant, de signaler l'ampleur du déplacement. De toute évidence, les corpuscules de Pacini et les récepteurs des
cheveux ne peuvent signaler l'ampleur de ce déplacement puisqu'ils ne déchargent qu'au début et à la fin du stimulus : les récepteurs des cheveux
sont des détecteurs de la vitesse du déplacement. Tout ce à quoi pourrait s'attendre, dans le système nerveux central, c'est une horloge interne
suffisamment précise et capable de dériver la vitesse du déplacement pour en indiquer l'ampleur…
www.neur-one.fr
[email protected] 20 09/09/2011
Il pourrait également être utile de connaître la vitesse d'un stimulus mécanique, c'est à dire le taux de changement du nombre de
potentiels d'action par seconde en fonction de la vitesse d'application d'une force ou d'un déplacement. Un exemple de ce comportement est montré pour le corpuscule de Meissner dans une peau glabre dans la figure ci-dessous, où la fréquence des décharges des récepteurs est tracée
sur l'ordonnée versus la vitesse de stimulations sur l'axe des abscisses. La relation entre ces deux variables est clairement non linéaire (c'est un
tracé log-log). On peut donc admettre que la vitesse d'application du stimulus soit codée en fréquence de décharges. Il est normalement supposé
que le récepteur qui signale l'ampleur d'un déplacement ne peut pas également indiquer le signal de vitesse, mais il n'ya aucune preuve pour étayer cette hypothèse. Les récepteurs à adaptation lente comme les récepteurs de types I et II, encodent la vitesse de déplacement dans leur
fréquence de décharge, et ils doivent être considérés comme des candidats pour les récepteurs de la vitesse.
Réponse d'un détecteur de vitesse.
Relation entre le taux de stimulations (en abscisse) et le nombre d'impulsions évoquées par stimulation (en ordonnée) dans une double échelle logarithmique. Le stimulus seuil, 1,6 microns / ms, a été soustrait des valeurs en abscisse. Pour trouver la vitesse réelle de relance ajoute 1,6 à valeurs sur l'axe des abscisses (Schmidt RF [é]: Notions fondamentales de la physiologie sensorielle de New York, Springer-Verlag, 1978.).
Sensations mécaniques
Une autre caractéristique d'un stimulus qui peut être détectée est sa fréquence d'application. Régulièrement appliqués, les stimuli récurrents
induisent une sensation de vibration ou de flottement. Il se décharge un potentiel d'action pour chaque cycle du stimulus vibratoire jusqu'à 600
Hz , pour le récepteur de Pacini ; il est normalement considéré comme un récepteur de vibrations. Encore une fois, les récepteurs qui codent les autres caractéristiques du stimulus sont pas considérés comme des récepteurs de vibrations, mais la plupart des récepteurs cutanés sont capables
de répondre aux stimuli vibratoires à des fréquences de 500 à 600 Hz au moins pour un temps court (Burgess PR, Petit D, RM Warren: J
Neurophysiol 31: 833-848, 1968). La perception des vibrations de l'homme est clairement dans la capacité de ces récepteurs.
Relation pression-courant
Soumise à une augmentation de pression, la peau se déforme et des récepteurs sont soumis à l'étirement ; des canaux sensibles à
l'étirement vont être activés : o Canaux stretch-sensibles (inhibiteur=gadolinium III),
o Canaux mécanosensibles (TRP, ENaC…).
o Rôle du cytosquelette (inhibiteur = colchicine, cytochalasine B)
Exemples de courant évoqué à l'application de pressions (dépression) sur la peau
Ces stimulations pressions entraînent non seulement une augmentation du nombre de canaux ouverts mais aussi l'augmentation de la
perméabilité aux ions sodium et calcium .
Exemple d'augmentation du nombre de canaux ouverts par la pression (récepteurs somesthésiques)
www.neur-one.fr
[email protected] 21 09/09/2011
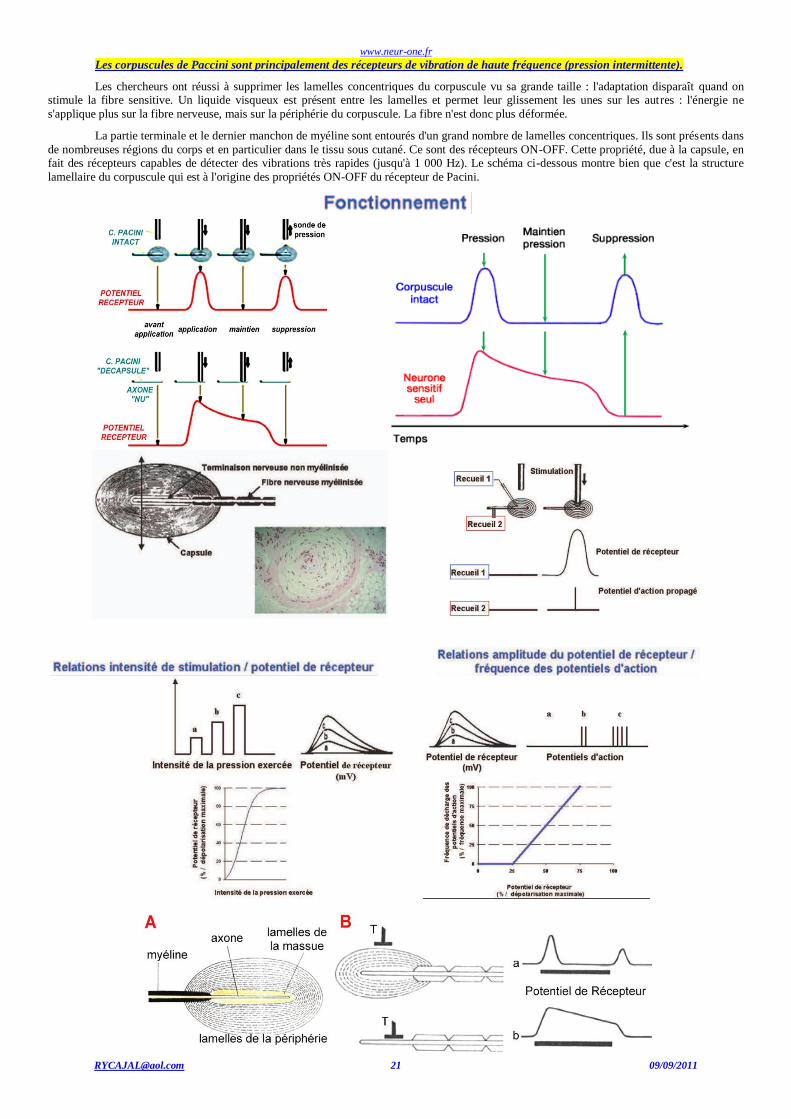
Les corpuscules de Paccini sont principalement des récepteurs de vibration de haute fréquence (pression intermittente).
Les chercheurs ont réussi à supprimer les lamelles concentriques du corpuscule vu sa grande taille : l'adaptation disparaît quand on stimule la fibre sensitive. Un liquide visqueux est présent entre les lamelles et permet leur glissement les unes sur les autres : l'énergie ne
s'applique plus sur la fibre nerveuse, mais sur la périphérie du corpuscule. La fibre n'est donc plus déformée.
La partie terminale et le dernier manchon de myéline sont entourés d'un grand nombre de lamelles concentriques. Ils sont présents dans
de nombreuses régions du corps et en particulier dans le tissu sous cutané. Ce sont des récepteurs ON-OFF. Cette propriété, due à la capsule, en fait des récepteurs capables de détecter des vibrations très rapides (jusqu'à 1 000 Hz). Le schéma ci-dessous montre bien que c'est la structure
lamellaire du corpuscule qui est à l'origine des propriétés ON-OFF du récepteur de Pacini.
www.neur-one.fr
[email protected] 22 09/09/2011
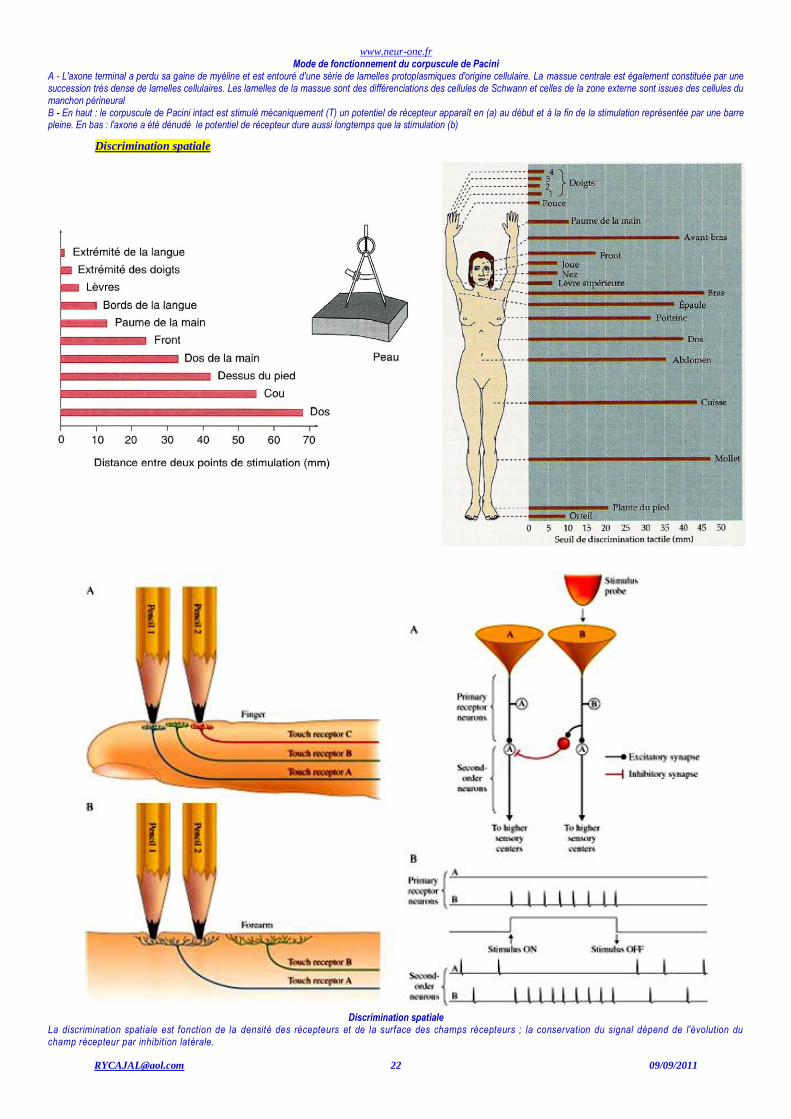
Mode de fonctionnement du corpuscule de Pacini A - L'axone terminal a perdu sa gaine de myéline et est entouré d'une série de lamelles protoplasmiques d'origine cellulaire. La massue centrale est également constituée par une succession très dense de lamelles cellulaires. Les lamelles de la massue sont des différenciations des cellules de Schwann et celles de la zone externe sont issues des cellules du manchon périneural B - En haut : le corpuscule de Pacini intact est stimulé mécaniquement (T) un potentiel de récepteur apparaît en (a) au début et à la fin de la stimulation représentée par une barre pleine. En bas : l'axone a été dénudé le potentiel de récepteur dure aussi longtemps que la stimulation (b)
Discrimination spatiale
Discrimination spatiale
La discrimination spatiale est fonction de la densité des récepteurs et de la surface des champs récepteurs ; la conservation du signal dépend de l'évolution du champ récepteur par inhibition latérale.
www.neur-one.fr
[email protected] 23 09/09/2011
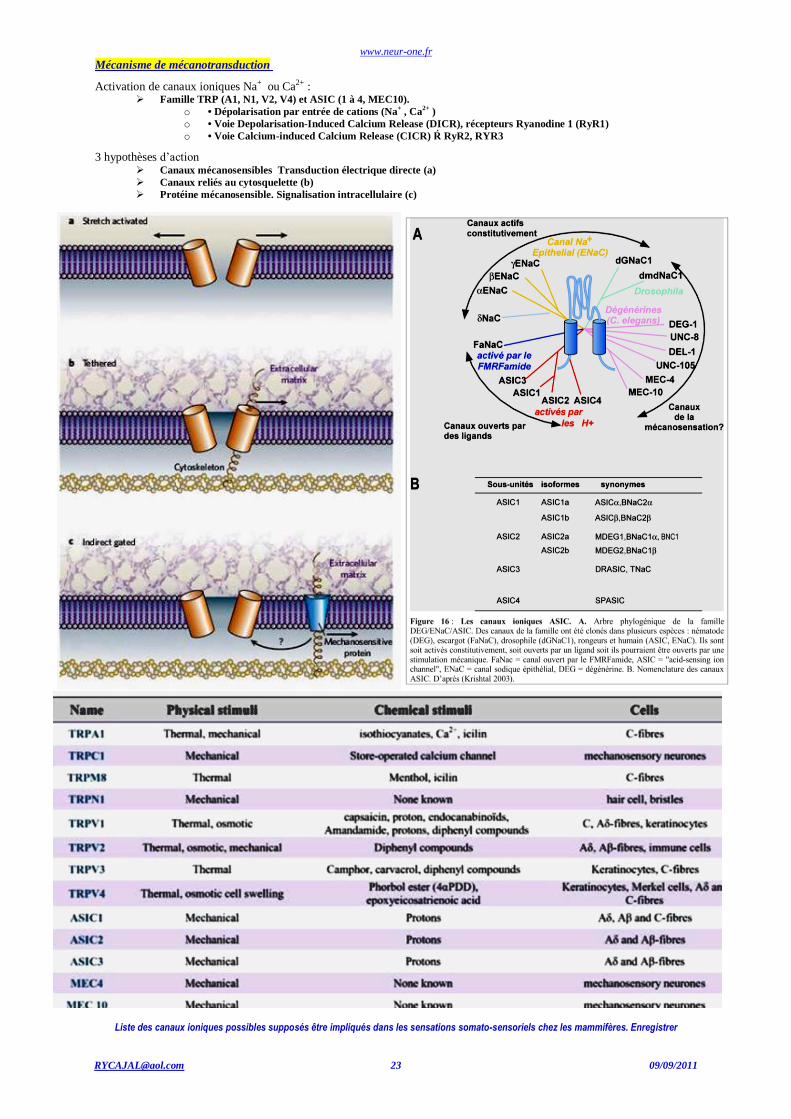
Mécanisme de mécanotransduction
Activation de canaux ioniques Na+ ou Ca2+ : Famille TRP (A1, N1, V2, V4) et ASIC (1 à 4, MEC10).
o • Dépolarisation par entrée de cations (Na+ , Ca
2+ )
o • Voie Depolarisation-Induced Calcium Release (DICR), récepteurs Ryanodine 1 (RyR1)
o • Voie Calcium-induced Calcium Release (CICR) Ŕ RyR2, RYR3
3 hypothèses d’action Canaux mécanosensibles Transduction électrique directe (a)
Canaux reliés au cytosquelette (b)
Protéine mécanosensible. Signalisation intracellulaire (c)
Liste des canaux ioniques possibles supposés être impliqués dans les sensations somato-sensoriels chez les mammifères. Enregistrer
www.neur-one.fr
[email protected] 24 09/09/2011
Thermoréception
La thermoalgie est activée lors de la thermoréception par action de la température sur des récepteurs sensibles à la chaleur. Tandis que la douleur est une sensation désagréable avec une composante affective, cognitive et culturelle, la nociception est la composante sensorielle de la
douleur.
Elle est transmise par les fibres nerveuses C, Aβ et Aα. L'extrémité des terminaisons nerveuses libres ou des récepteurs polymodaux
sensibles à la température ont un haut seuil activation.
La transmission met en jeu un relargage de substances (SP, Bradykinine, PgE2, ATP, IL …) lors d'un dommage cellulaire.
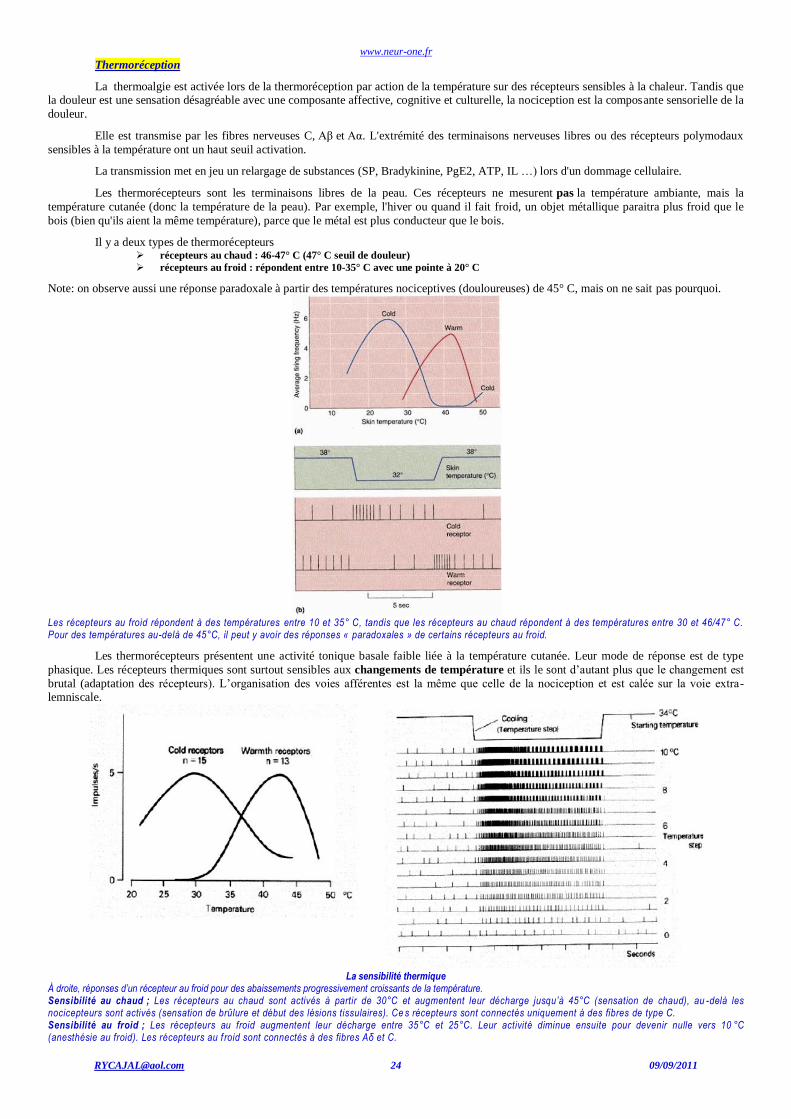
Les thermorécepteurs sont les terminaisons libres de la peau. Ces récepteurs ne mesurent pas la température ambiante, mais la
température cutanée (donc la température de la peau). Par exemple, l'hiver ou quand il fait froid, un objet métallique paraitra plus froid que le
bois (bien qu'ils aient la même température), parce que le métal est plus conducteur que le bois.
Il y a deux types de thermorécepteurs récepteurs au chaud : 46-47° C (47° C seuil de douleur)
récepteurs au froid : répondent entre 10-35° C avec une pointe à 20° C
Note: on observe aussi une réponse paradoxale à partir des températures nociceptives (douloureuses) de 45° C, mais on ne sait pas pourquoi.
Les récepteurs au froid répondent à des températures entre 10 et 35° C, tandis que les récepteurs au chaud répondent à des températures entre 30 et 46/47° C. Pour des températures au-delà de 45°C, il peut y avoir des réponses « paradoxales » de certains récepteurs au froid.
Les thermorécepteurs présentent une activité tonique basale faible liée à la température cutanée. Leur mode de réponse est de type
phasique. Les récepteurs thermiques sont surtout sensibles aux changements de température et ils le sont d’autant plus que le changement est
brutal (adaptation des récepteurs). L’organisation des voies afférentes est la même que celle de la nociception et est calée sur la voie extra-lemniscale.
La sensibilité thermique
À droite, réponses d’un récepteur au froid pour des abaissements progressivement croissants de la température. Sensibilité au chaud ; Les récepteurs au chaud sont activés à partir de 30°C et augmentent leur décharge jusqu’à 45°C (sensation de chaud), au -delà les nocicepteurs sont activés (sensation de brûlure et début des lésions tissulaires). Ce s récepteurs sont connectés uniquement à des fibres de type C. Sensibilité au froid ; Les récepteurs au froid augmentent leur décharge entre 35°C et 25°C. Leur activité diminue ensuite pour devenir nulle vers 10 °C (anesthésie au froid). Les récepteurs au f roid sont connectés à des fibres Aδ et C.

www.neur-one.fr
[email protected] 25 09/09/2011
La sensibilité thermoalgique met en jeu essentiellement des terminaisons nerveuses libres épidermiques.
Il existe plusieurs familles de récepteurs dont une qui a reçu le nom de récepteurs “TRP”. Cette famille de récepteurs est sensible aux
stimuli mécaniques, thermiques (chaud et froid) et à certaines substances chimiques et elle est divisée en six catégories (chez les mammifères),
dont 3 d’entre elles sont des récepteurs de chaleur les TRPA ,TRPM et TRPV, (transient receptor potential vanilloïde ), ces derniers récepteurs
TRPV étant sensibles à des produits chimiques de la famille des vanilloïdes mais s’activent également pour des stimuli thermiques, les TRPV1 pour des températures supérieures à 44 °C, les TRPV3 entre 32 et 38, le TRPV4 entre 25 et 30°. Les récepteurs TRPM8 sont eux sensibles entre
20 et 25 °C et les TRPA1 au dessous de 17d°C. En outre un TRP V2 est sensible au dessus de 52 °C, mais il n’est présent que dans la peau et il
y a alors souvent brûlure.
Types de canaux TRP intervenant dans la sensibilité thermique
L’agent actif contenu dans les piments est la capsaïcine. C’est un alcaloïde de la famille des vanilloïdes et son action est une façon pour la plante de se protéger contre les prédateurs. La capsaïcine active les récepteurs TRPV1 et donne donc une impression analogue à celle d’une température de 44 d°C. Elle ne pro voque donc pas de brûlure chimique mais seulement une sensation illusoire de brûlure. Le lait la transforme chimiquement et donc si vous avez la bouche en feu, parce que vous avez mâché des piments, buvez un verre de lait .
La capsaïcine est utilisée en médecine, dans des crèmes locales pour soula ger la douleur nerveuse périphérique. Le traitement consiste en l’application d’un anesthésiant jusqu’à ce que la zone soit engourdie. Ensuite la capsaïcine est placée sur la peau jusqu’à ce que le patient commence à sentir la chaleur puis est rapidement retirée. Les nerfs sont dominés par la sensation de brûlure apparente et ne peuvent communiquer la douleur pendant une longue période. Et d’autres condiments : Le thym renferme du thymol, l’origan du corvacrol, la sariette et le clou de girofle de l’eugénol, l’oignon et l’ail de l’allicine (l’oignon contient aussi d’autres produits qui font pleurer), et ces produits interfèrent aussi avec les récepteurs de la langue., mais pas avec le TRPV1 et donc ils “brûlent” moins. La plupart d’entre eux agissent sur les récepteur TRPV3 et donnent donc une sensation de chaleur très modérée. La menthe et l’ail (crus) agissent sur les récepte urs TRPA1 et produiraient donc une impression plutôt rafraichissante.
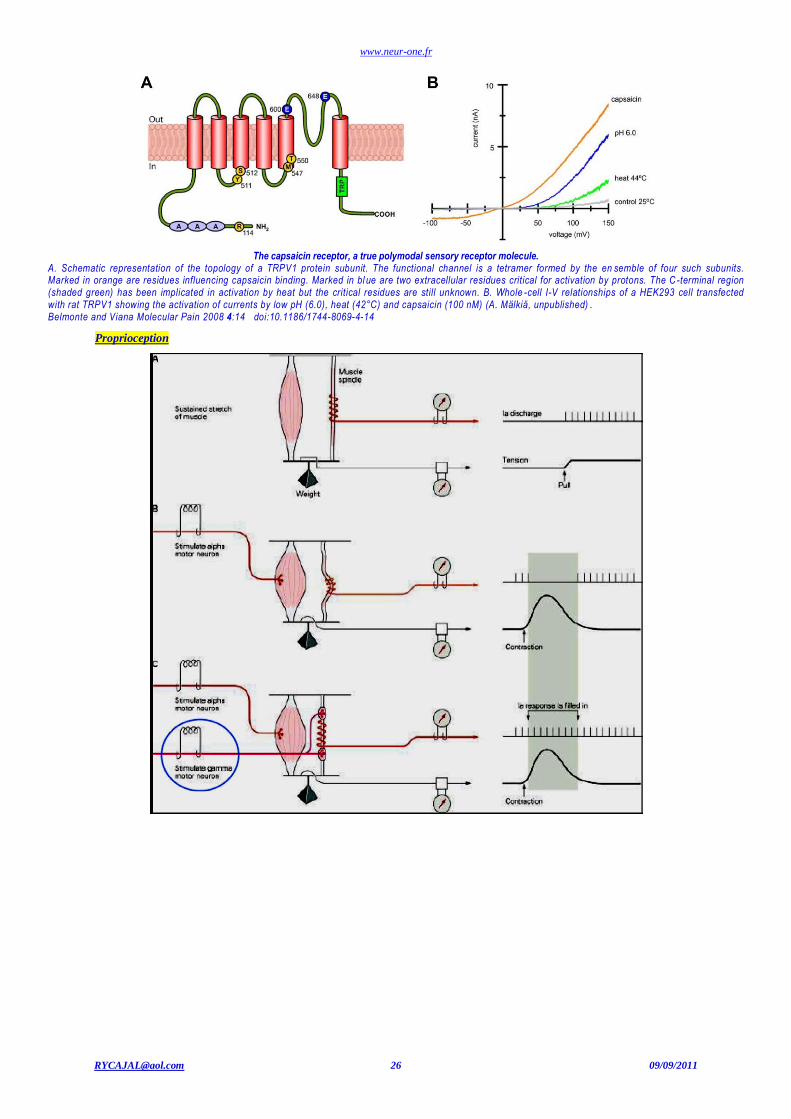
www.neur-one.fr
[email protected] 26 09/09/2011
The capsaicin receptor, a true polymodal sensory receptor molecule.
A. Schematic representation of the topology of a TRPV1 protein subunit. The functional channel is a tetramer formed by the en semble of four such subunits. Marked in orange are residues influencing capsaicin binding. Marked in bl ue are two extracellular residues critical for activation by protons. The C -terminal region (shaded green) has been implicated in activation by heat but the critical residues are still unknown. B. Whole -cell I-V relationships of a HEK293 cell transfected with rat TRPV1 showing the activation of currents by low pH (6.0), heat (42°C) and capsaicin (100 nM) (A. Mälkiä, unpublished) . Belmonte and Viana Molecular Pain 2008 4:14 doi:10.1186/1744-8069-4-14
Proprioception
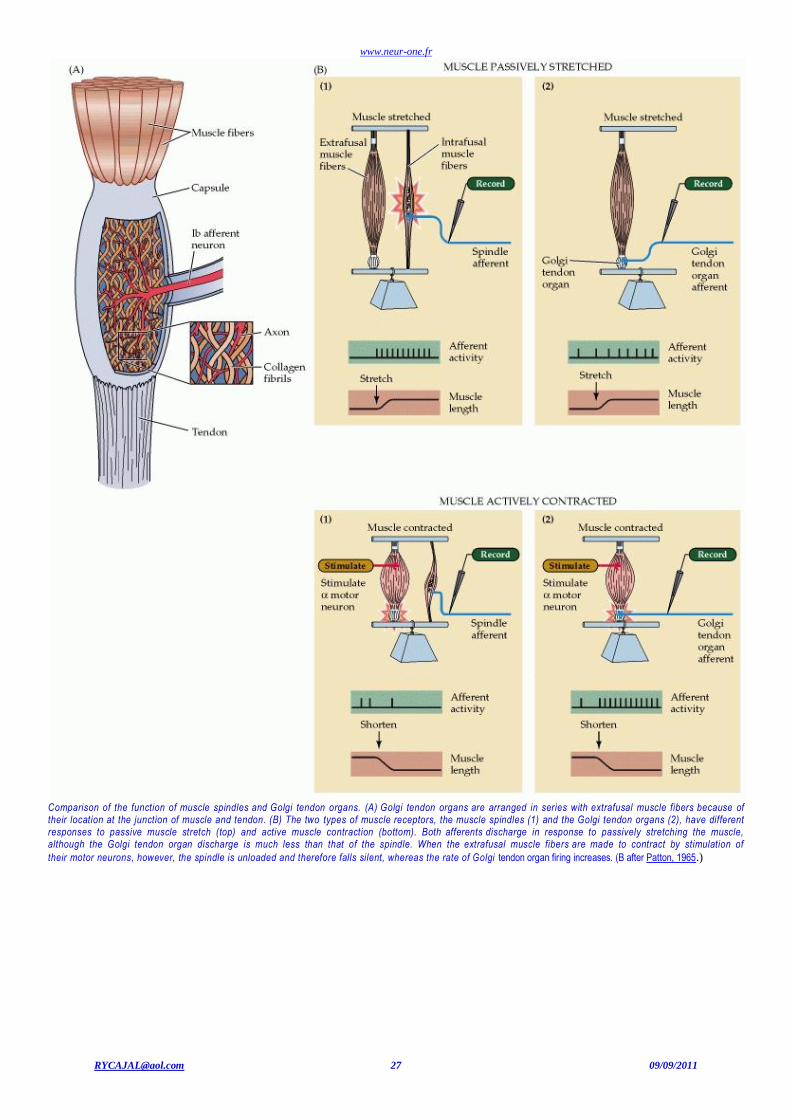
www.neur-one.fr
[email protected] 27 09/09/2011
Comparison of the function of muscle spindles and Golgi tendon organs. (A) Golgi tendon organs are arranged in series with extrafusal muscle fibers because of their location at the junction of muscle and tendon. (B) The two types of muscle receptors, the muscle spindles (1) and the Golgi tendon organs (2), have different responses to passive muscle stretch (top) and active muscle contraction (bottom). Both afferents discharge in response to passively stretching the muscle, although the Golgi tendon organ discharge is much less than that of the spindle. When the extrafusal muscle fibers are made to contract by stimulation of
their motor neurons, however, the spindle is unloaded and therefore falls silent, whereas the rate of Golgi tendon organ firing increases. (B after Patton, 1965.)
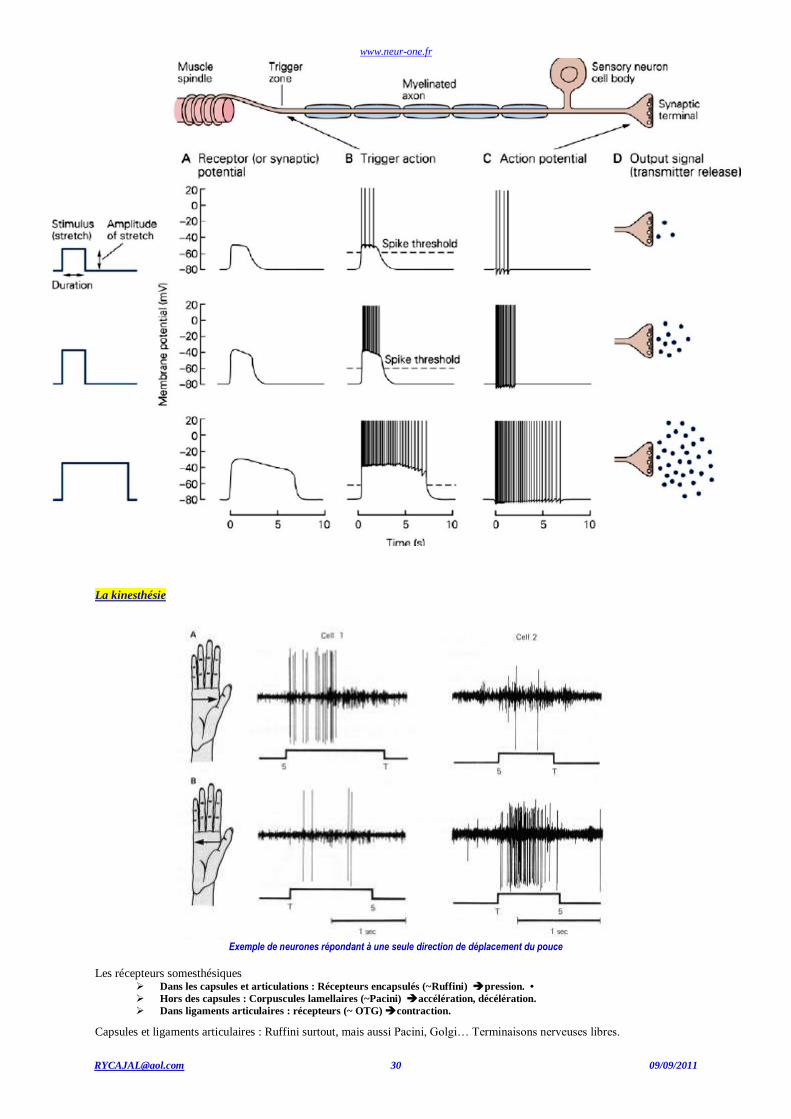
www.neur-one.fr
[email protected] 30 09/09/2011
La kinesthésie
Exemple de neurones répondant à une seule direction de déplacement du pouce
Les récepteurs somesthésiques Dans les capsules et articulations : Récepteurs encapsulés (~Ruffini) pression. •
Hors des capsules : Corpuscules lamellaires (~Pacini) accélération, décélération.
Dans ligaments articulaires : récepteurs (~ OTG) contraction.
Capsules et ligaments articulaires : Ruffini surtout, mais aussi Pacini, Golgi… Terminaisons nerveuses libres.
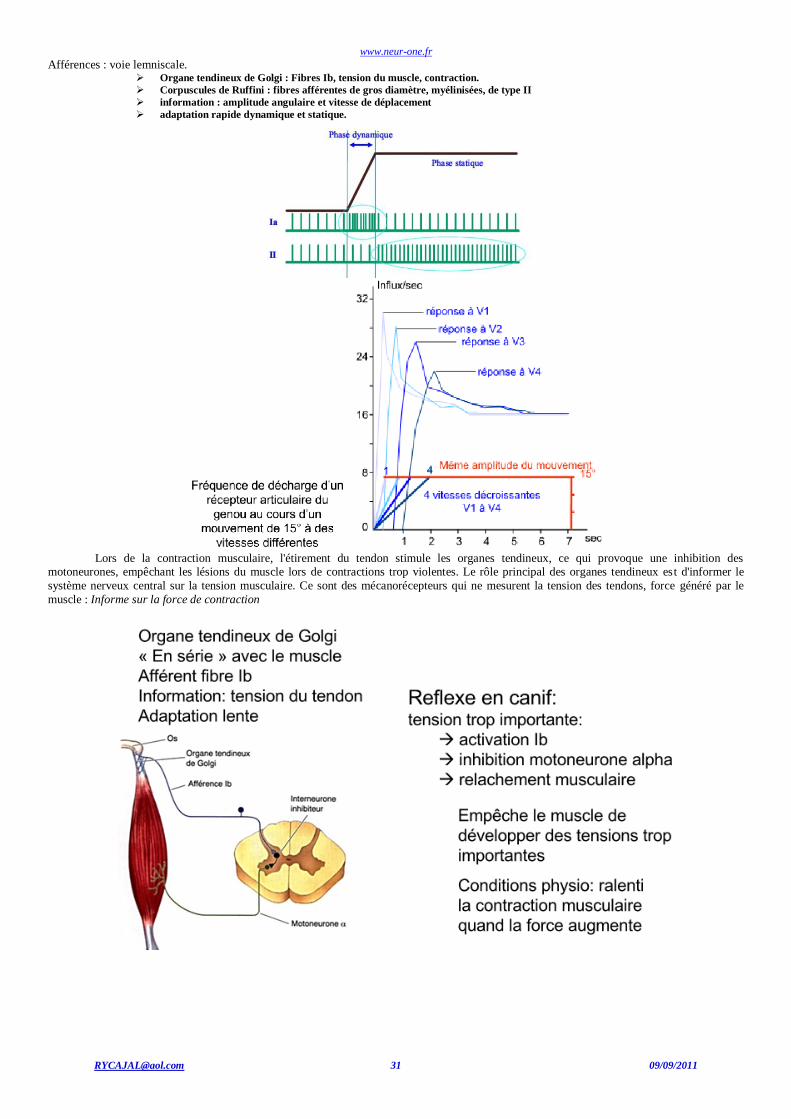
www.neur-one.fr
[email protected] 31 09/09/2011
Afférences : voie lemniscale. Organe tendineux de Golgi : Fibres Ib, tension du muscle, contraction.
Corpuscules de Ruffini : fibres afférentes de gros diamètre, myélinisées, de type II
information : amplitude angulaire et vitesse de déplacement
adaptation rapide dynamique et statique.
Lors de la contraction musculaire, l'étirement du tendon stimule les organes tendineux, ce qui provoque une inhibition des
motoneurones, empêchant les lésions du muscle lors de contractions trop violentes. Le rôle principal des organes tendineux est d'informer le
système nerveux central sur la tension musculaire. Ce sont des mécanorécepteurs qui ne mesurent la tension des tendons, force généré par le
muscle : Informe sur la force de contraction
www.neur-one.fr
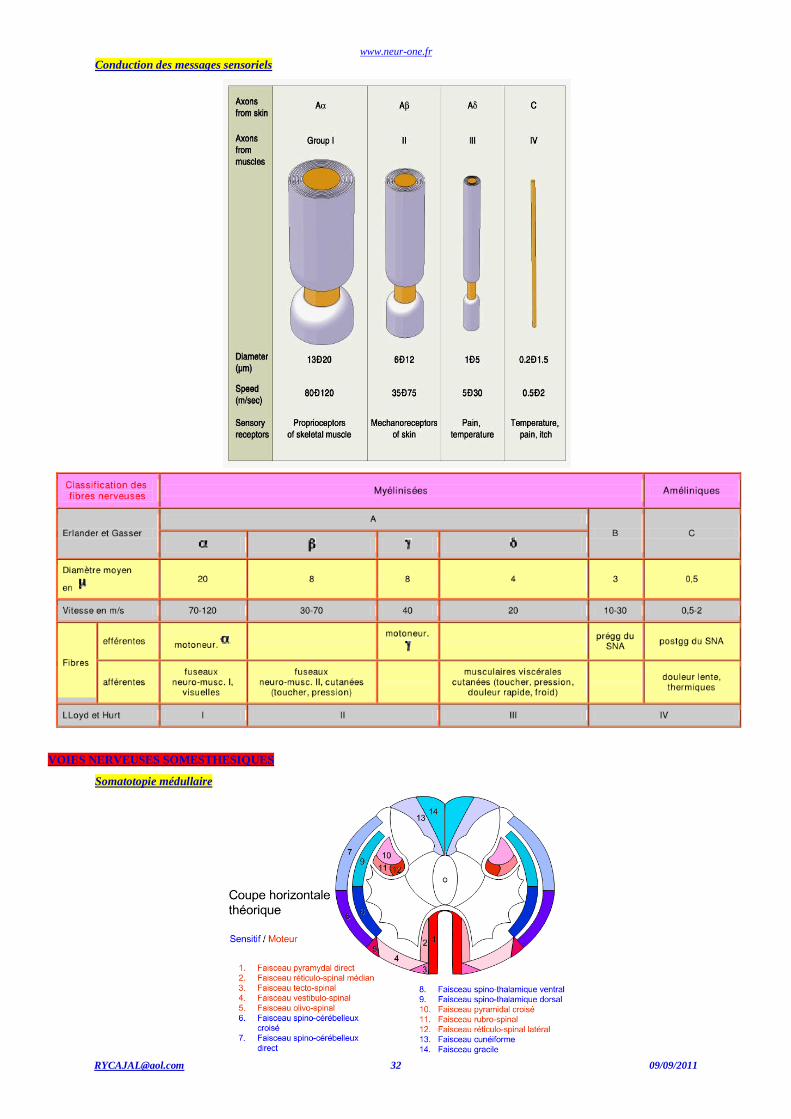
[email protected] 32 09/09/2011
Conduction des messages sensoriels
VOIES NERVEUSES SOMESTHESIQUES
Somatotopie médullaire
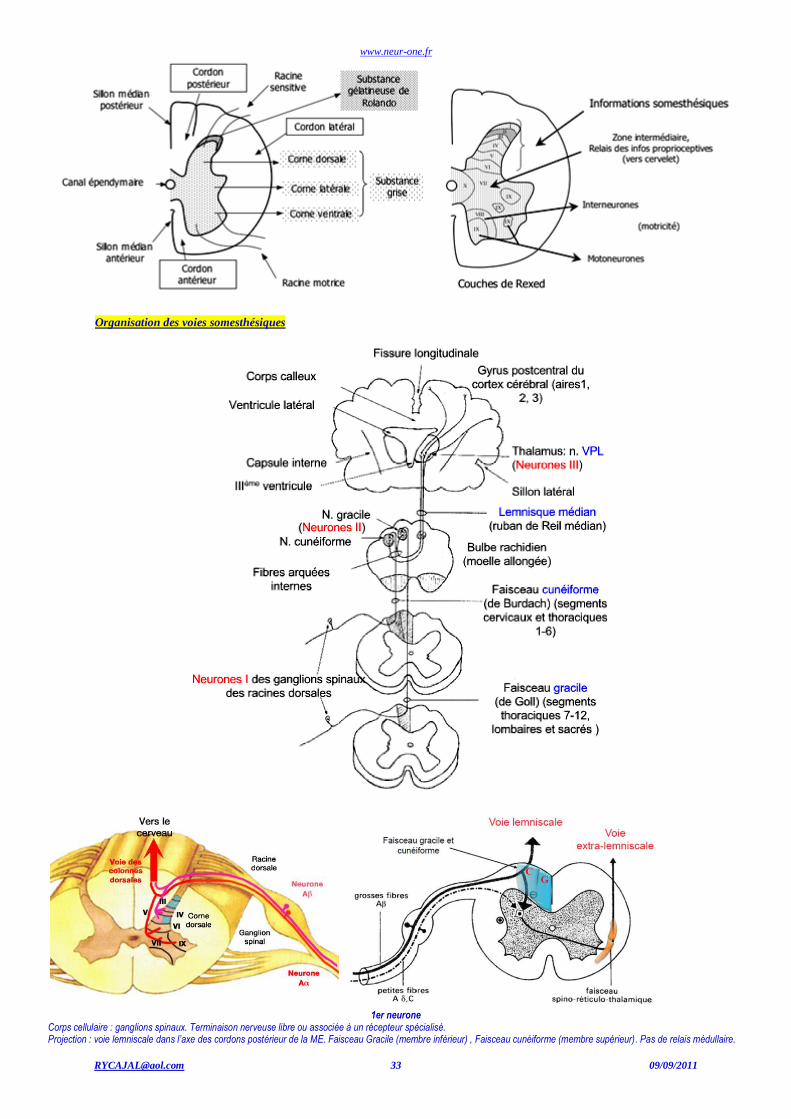
www.neur-one.fr
[email protected] 33 09/09/2011
Organisation des voies somesthésiques
1er neurone
Corps cellulaire : ganglions spinaux. Terminaison nerveuse libre ou associée à un récepteur spécialisé. Projection : voie lemniscale dans l’axe des cordons postérieur de la ME. Faisceau Gracile (membre inférieur) , Faisceau cunéiforme (membre supérieur). Pas de relais médullaire.
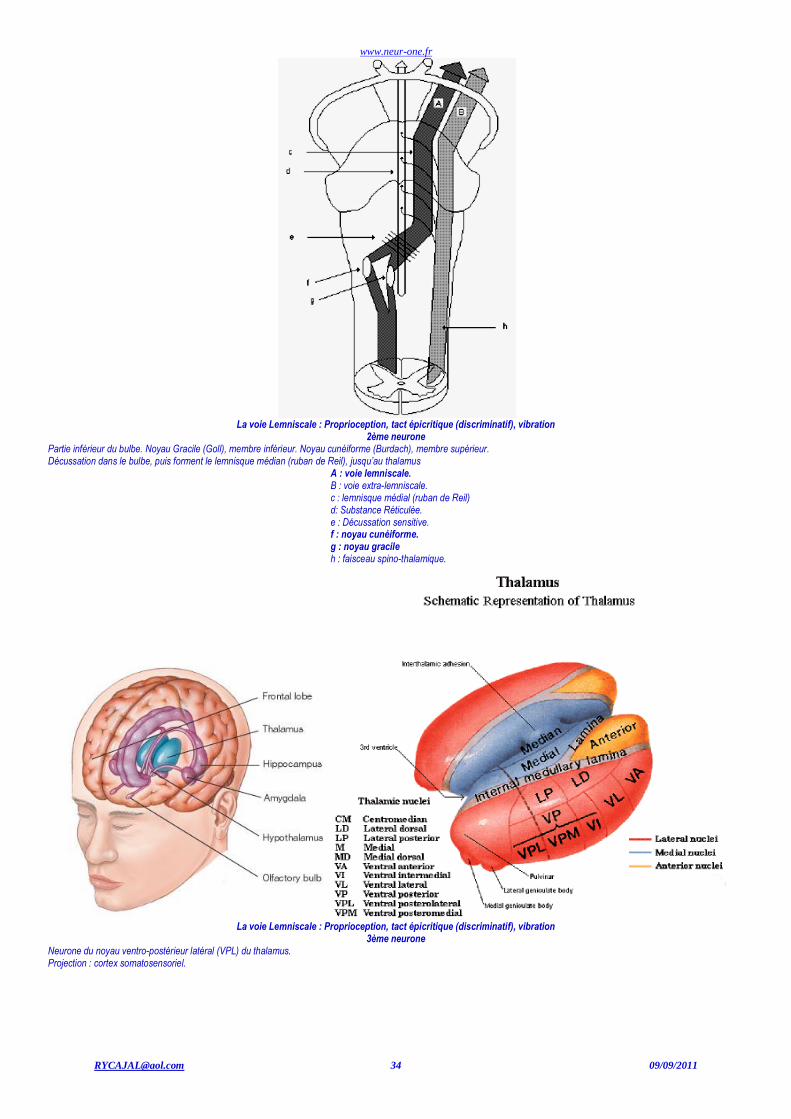
www.neur-one.fr
[email protected] 34 09/09/2011
La voie Lemniscale : Proprioception, tact épicritique (discriminatif), vibration
2ème neurone Partie inférieur du bulbe. Noyau Gracile (Goll), membre inférieur. Noyau cunéiforme (Burdach), membre supérieur. Décussation dans le bulbe, puis forment le lemnisque médian (ruban de Reil), jusqu’au thalamus
A : voie lemniscale. B : voie extra-lemniscale. c : lemnisque médial (ruban de Reil) d: Substance Réticulée. e : Décussation sensitive. f : noyau cunéiforme. g : noyau gracile h : faisceau spino-thalamique.
La voie Lemniscale : Proprioception, tact épicritique (discriminatif), vibration
3ème neurone Neurone du noyau ventro-postérieur latéral (VPL) du thalamus. Projection : cortex somatosensoriel.
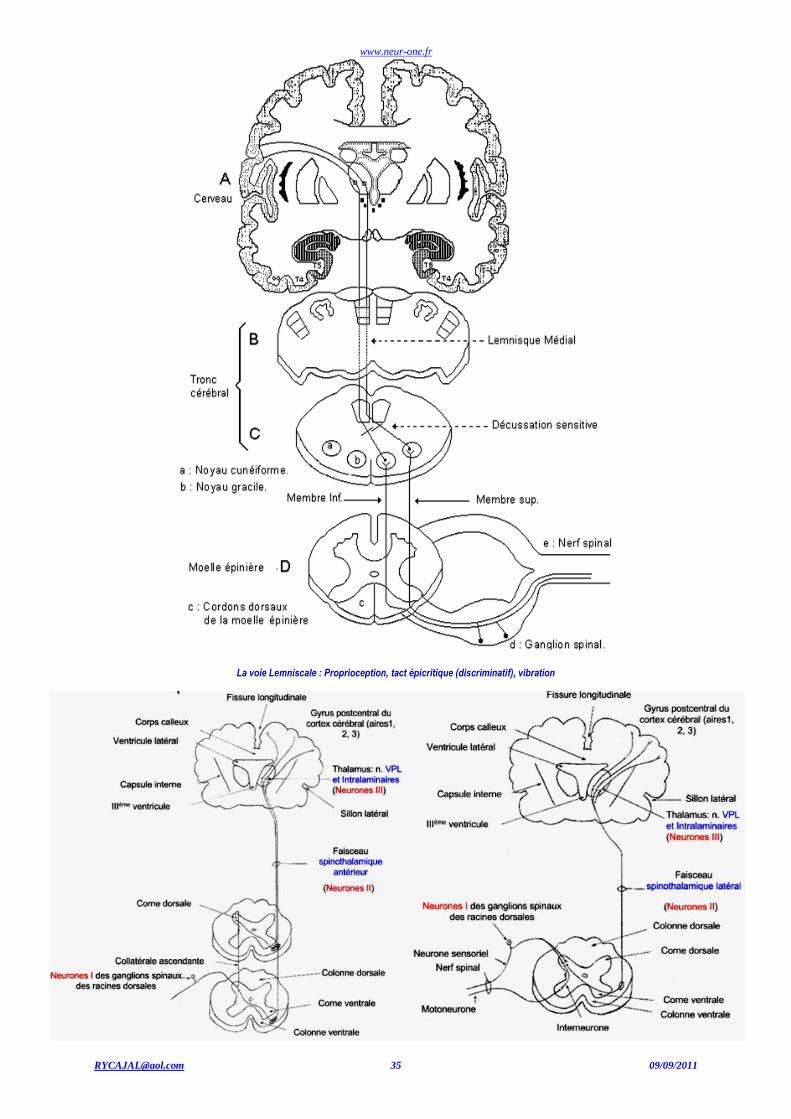
www.neur-one.fr
[email protected] 35 09/09/2011
La voie Lemniscale : Proprioception, tact épicritique (discriminatif), vibration
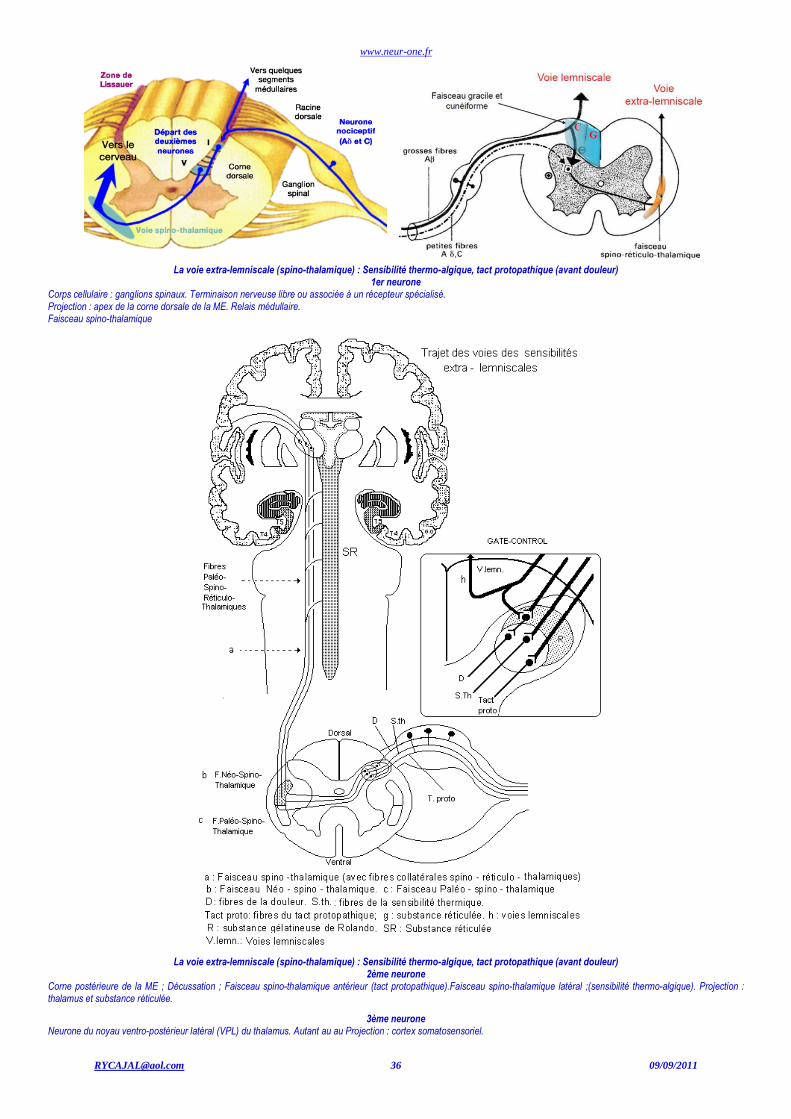
www.neur-one.fr
[email protected] 36 09/09/2011
La voie extra-lemniscale (spino-thalamique) : Sensibilité thermo-algique, tact protopathique (avant douleur)
1er neurone Corps cellulaire : ganglions spinaux. Terminaison nerveuse libre ou associée à un récepteur spécialisé. Projection : apex de la corne dorsale de la ME. Relais médullaire. Faisceau spino-thalamique
La voie extra-lemniscale (spino-thalamique) : Sensibilité thermo-algique, tact protopathique (avant douleur)
2ème neurone Corne postérieure de la ME ; Décussation ; Faisceau spino-thalamique antérieur (tact protopathique).Faisceau spino-thalamique latéral ;(sensibilité thermo-algique). Projection : thalamus et substance réticulée.
3ème neurone Neurone du noyau ventro-postérieur latéral (VPL) du thalamus. Autant au au Projection : cortex somatosensoriel.
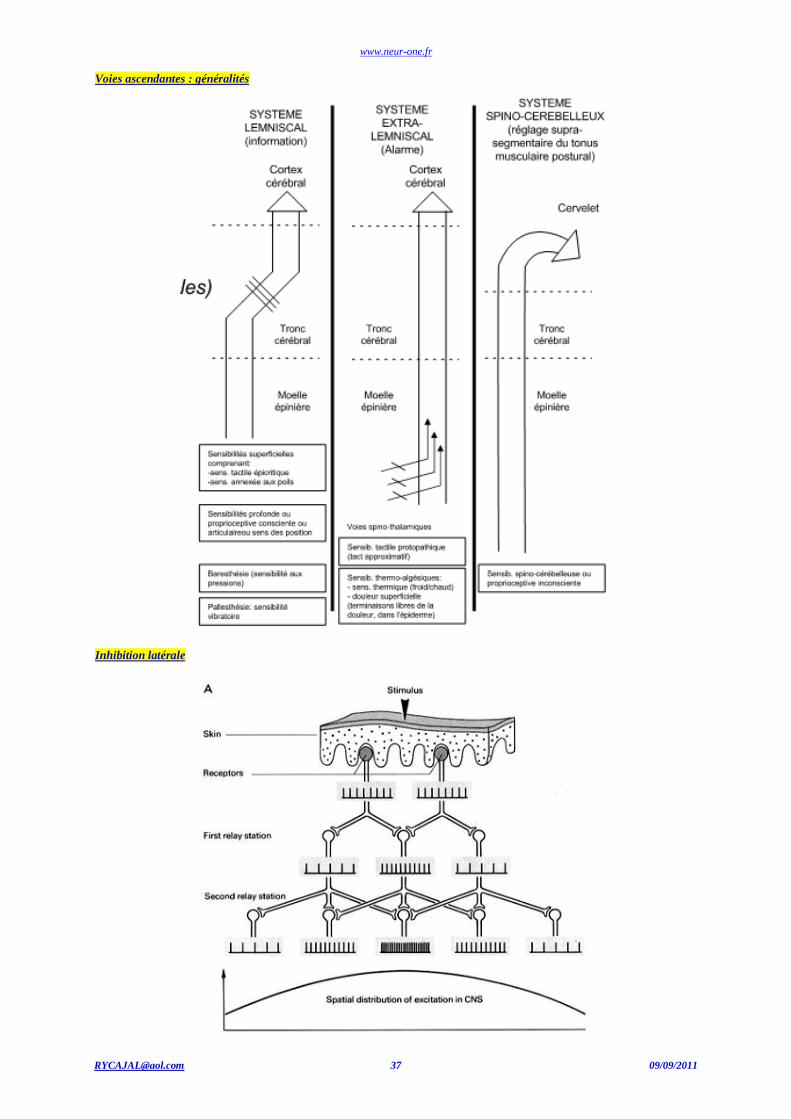
www.neur-one.fr
[email protected] 40 09/09/2011
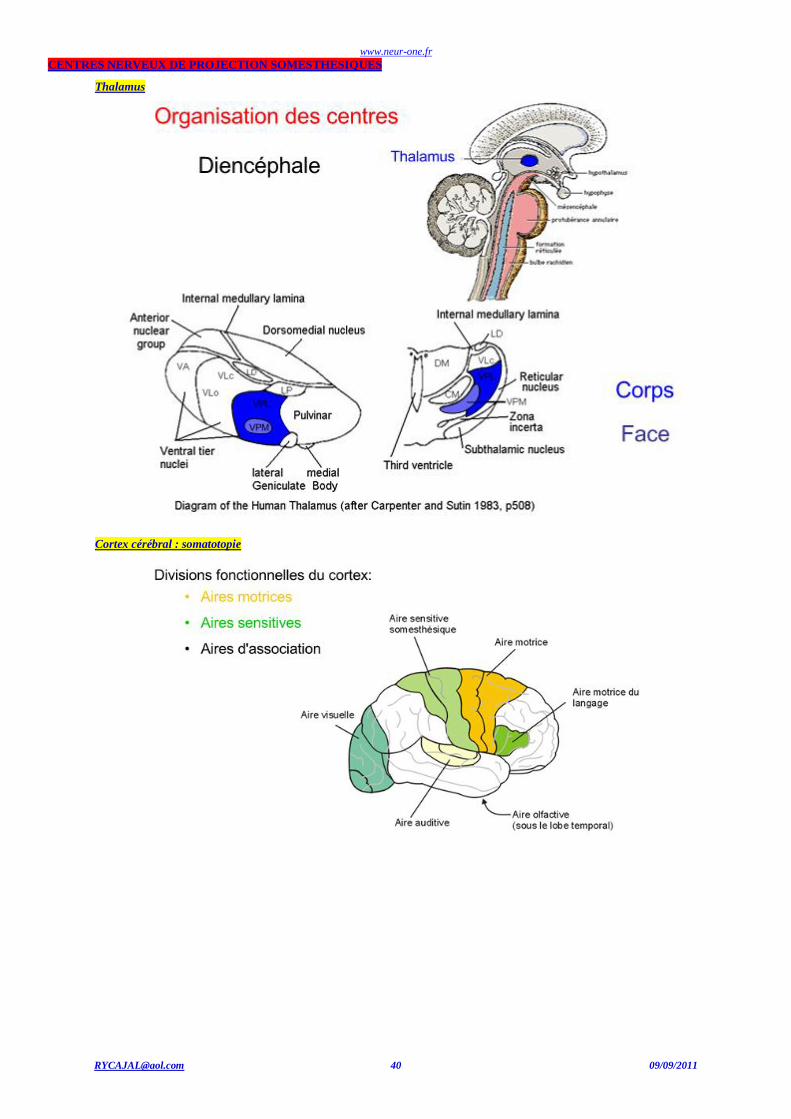
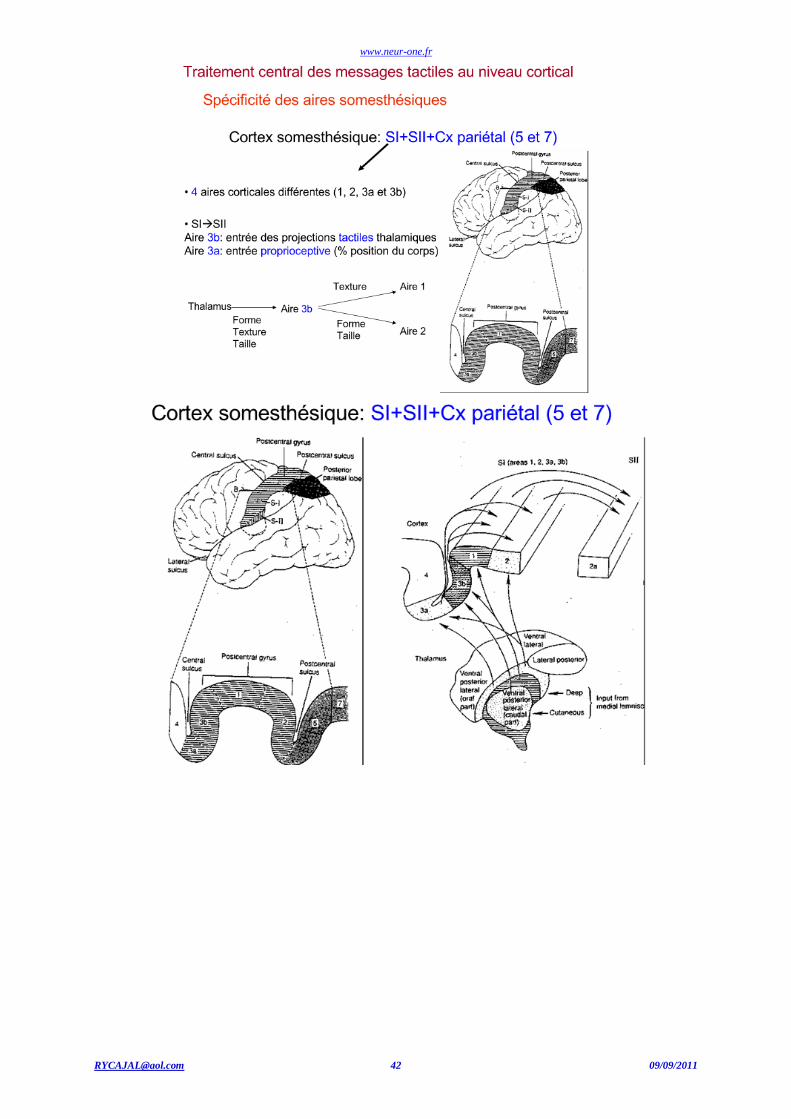
CENTRES NERVEUX DE PROJECTION SOMESTHESIQUES
Thalamus
Cortex cérébral : somatotopie

www.neur-one.fr
[email protected] 43 09/09/2011
ANNEXE
V : NERF TRIJUMEAU
Le plus volumineux
Dérive du premier arc pharyngé (arc branchial)
Mixte : branchiomoteur pour les muscles de la mastication et somatosensitif pour la face
Les neurones de la racine sensitive forment le ganglion trigéminé (ganglion de Gasser ou ganglion semilunaire) qui repose dans une dépression osseuse,
la fossette du ganglion de Gasser
Le ganglion trigéminé donne trois branches destinées aux trois étages de la face
Nerf ophtalmique de Willis
o Traverse la fissure orbitaire supérieure (fente sphénoïdale)
o Sensibilité front, paupière supérieure, cornée, fosses nasales, sinus ethmoïdal et frontal
Nerf maxillaire
o Pénètre à la base de la paroi latérale du sinus caverneux
o Sort du crâne par le foramen rotundum (trou grand rond) pour atteindre l’arrière fond de la fosse ptérygomaxillaire
o Sensibilité de la partie moyenne de la face : ailes du nez, joues, lèvre supérieure, fosses nasales, dents et palais osseux
et mou
Nerf mandibulaire (nerf maxillaire inférieur)
o Quitte le crâne par le foramen ovale (trou ovale)
o Après sa sortie donne une branche méningée récurrente réentrant par le foramen spinosum (trou petit rond)
Territoire sensitif
o sensibilité de la paroi antérieure du méat auditif externe et la face externe du tympan deux tiers antérieurs de la
langue sensibilité de la mâchoire inférieure, plancher buccal, gencives et dents sensibilité de la dure mère des étages
antérieur et moyen de la base du crâne
Territoire moteur
o Les fibres motrices du nerf trijumeau rejoignent le nerf mandibulaire elles innervent : muscles masticateurs :
temporal, masseter, ptérygoïdien latéral et ptérygoïdien médial muscle mylohyoïdien et le ventre antérieur du muscle
digastrique muscle tenseur du voile du palais muscle tenseur du tympan