nonenzymic reactivation of reduced bovine pancreatic ribonuclease · pdf filenonenzymic...
TRANSCRIPT

THE ~JOURNAL OF BIOLOGKAL CHEMISTRY Vol. 250, No. 21, Issue of November 10, pp. 8477-8482, 1975
Printed in U.S.A.
Nonenzymic Reactivation of Reduced Bovine Pancreatic Ribonuclease by Air Oxidation and by Glutathione Oxidoreduction Buffers*
(Received for publication, June 17, 1974, and in revised form. March 26, 1975)
A. KARIM AHMED,$ STEPHEN W. SCHAFFER,§ AND DONALD B. WETLAUFER~!
From the Department of Biochemistry, University of Minnesota Medical School, Minneapolis, Minnesota 55455
With the glutathione system that leads to rapid regeneration of reduced lysozyme (Saxena, V. P., and Wetlaufer, D. B. (1971) Biochemistry 9, 5015), reduced pancreatic ribonuclease (RNase) regenerated activity in high yield (>90%) but at a considerably lower rate (t”, -75 min). Systematic examination of the effects upon regeneration of the concentrations and ratios of reduced and oxidized glutathione (GSH and GSSG) showed the same broad optima for RNase as were earlier found for lysozyme: [GSSG] = 5 x 10. 4 M, [GSH] = 5 x 10m3 M. Regeneration of reduced RNase by air oxidation was shown to be inhibitable by 10m4 M EDTA, whereas the glutathione regeneration was unaffected by EDTA. In addition the air-oxidative regeneration showed a strong temperature dependence, in contrast with the glutathione system. The mechanisms of these two kinds of regenerations are therefore different.
Six potentially catalytic metal ions were tested in the air-oxidative regeneration of RNase: Cu”+, Co’+, Mn*+ Fe3+ Zn’+, and Ni’+. Of these, only Cu2+ > > enhanced the rate of regeneration of RNase activity, although both CU*+ and Co’+ catalyzed thiol oxidation of reduced RNase. The rates and yields of RNase regenerations were independent of protein concentration from 3 x 10m7 M to 1.2 x 10m5 M in the glutathione system. Preincubation of freshly dissolved reduced RNase under nonoxidizing conditions before adding glutathione did not change the rate or extent of regeneration. Studies of its pH dependence showed that the glutathione regeneration depends on the deprotonation of prototropic groups with 7.5 < pK < 8.0.
The major ion exchange chromatographic peaks from glutathione and air-oxidative regenerations appeared to be identical with native RNase, by the criteria of specific activity, chromatographic mobility, and circular dichroic spectra. The glutathione system permits regeneration at much higher RNase concentration than the air regeneration, with rates and yields comparable to the greatest reported for air regeneration.
The mechanisms governing formation of three-dimensional structure in proteins continue to be a focus for investigation and speculation (l-3). Work from this laboratory has established that thiol disulfide oxidoreduction buffers will facilitate the rapid regeneration of enzymic activity from reduced hen egg lysozyme and reduced human lysozyme (4, 5). It was a matter
*This work was supported by United States Public Health Service Grants GM 10900 and 5ROl GM18814, and by the Graduate School of the University of Minnesota.
$A portion of this work was submitted by A. K. A. in partial fulfillment of the requirements for the Ph.D. degree at the LJniversity of Minnesota. Present address, Natural Resources Defense Council, Inc., 15 W. 44th St. New York, N. Y. 10036.
0 A portion of this work was submitted by S. W;. S. in partial fulfillment of the requirements for the Ph.D. degree at the University of Minnesota. Present address, Department of Chemistry, Lehigh Universitv. Bethlehem. Pa. 18015.
7 To whom inquiries concerning this paper and requests for reprints should be addressed. Present address, Department of Chemistry, University of Delaware, Newark, Del. 19711.
of considerable interest to investigate glutathione regenerations of reduced pancreatic RNase, since it was with this enzyme that Anfinsen and co-workers carried out their landmark re- generations of native protein from a reduced, disorganized precursor (1). Indeed, RNase was first used to investigate both the air-oxidative regeneration and the enzymic (“shuffle-ase”) regeneration (6, 7). Since the physiological role of the enzymic mechanism is still in doubt (3, 8), we have here chosen to com- pare the air regeneration with glutathione regeneration of reduced RNase. The present paper reports a comparison of the two kinds of regenerations, provides evidence that they are mechanistically different, and shows the results of a first round of optimization of experimental variables in glutathione regenerations.
EXPERIMENTAL. PROCEDURES
Materials-Bovine pancreatic RNase A was obtained from Sigma Chemical Co., as type XI-A, Lot 106B.8600. It was used without f’urther purification except in the experiments of Figs. 3 and 5.
8477
by guest on May 7, 2018
http://ww
w.jbc.org/
Dow
nloaded from

8478
@-Mercaptoethanol was an Eastman product and was redistilled before use. ‘I’ris(hydrox\methyl)aminomethane was obtained as Tri~ma base from Sigma. All other materials were obtained from Commercial sources.
Glass-distilled water (from a still pot containing 0.2 M H,SO,, fed with deionized water) was used in all regeneration experiments. \\‘ater was collected and stored in polyethylene containers. Glassware was cleaned by a scrupulous procedure, detailed elsewhere (9). The last two steps involve several rinses with 0.1 M HCI, followed by several rinses with glass-distilled water.
RNase Assay-RNase activity was determined by the method of Kalnitsky et al. (lo), using yeast RNA as substrate. The assay was carried out at pH 5.0, 30” for an interval of 5.0 min. Recalihrations were carried out every day the assay was used, with the calibrating digestions containing the same concentrations of reagents as were introduced from the sample aliquot (GSH. GSSG, EDTA, etc.). In a few of the later experiments a continuous recording assay method suggested by Fletcher and Hash (11) was employed. It had the advantages of improved convenience and somewhat hetter precision. Control assays were carried out to recalibrate for each experiment.
Sul)%ydry:yl Assay-The calorimetric method of Ellman (12) was used. Control experiments showed that addition of various reagents employed in the regeneration mixtures did not interfere with the assay. Protein concentration was determined by ultraviolet absorbance (c,,, = 9200 M- ’ cm ‘) for both reduced and native RNase (13).
Reduction of RNase A-The protein was reduced in 8 M urea/O.01 M
Tris buffer solution, pH 8.5. with a 100. to ~XO-fold excess of P-mercaptoethanol (13). Reduction was allowed to proceed at room temperature for 4 hours, after which the protein solution was chilled to 0” and precipitated with 10 volumes of cold acetone/l M HCl (:{9/1). The precipitate was collected by centrifugation. washed twice with acidified acetone. and washed twice with ether. It was dissolved in a small volume of water, lvophilized. and stored at 12” in a tightly stoppered bottle in a desiccator over anhydrous CaSO,. The reduced enzyme showed a content of 8.0 t 0.2 eq --SH content which proved to he stable for several weeks when stored as described.
Regeneration of Activity from Reduced RNase A-A typical regener- ation will he described. From 2.0 to 2.3 mg of lyophilized reduced RNase were quicklv dissolved in 10.0 ml of water. Within 1 min, 1.00 ml of this solution was pipetted into 9.00 ml ofTris-acetate buffer (0.10 M, pH 8.2, at 25”) in a small aperture container immersed in a thermostatted hath. For an air oxidation sy-stem. the system has been described completely. In a glutathione regeneration, the requisite amounts of GSH and GSSG were dissolved in the 9 ml of buffer just before adding the RNase solution. Control experiments consistently showed a change of pH less than 0.10 after the addition of reduced RNase and GSSG + GSH to the buffer. An aliquot of the reduced RNase solution was taken for concentration determination by means of an A,,, measurement. At suitable time intervals, O.lOO-ml aliquots were withdrawn for assay. For convenience. we refer to a regeneration solution composed of 0.09 M Tris-acetate. pH 8.2, [GSSG] 3 x 10-’ M. [GSH] 3 x 10m3 M, as an “optimal glutathione system.”
Column Chromatography of RNase-We used the system developed hy Tahorsky (14). A carhoxymethylcellulose (Whatman microgranu- lar, CM32) column (1.6 x 31 cm) was pre-equilibrated with pH 8.0 0.005 M Tris-chloride buffer at room temperature. The sample was eluted with the same buffer, on which a NaCl gradient (0 to 0.15 M) was superimposed. Fig. 5, the top curue. shows multiple components in a commercial sample of RNase, similar to Tahorsky’s findings.
Preparative Regenerations and Chromatographic Characterization of Regenerated RNase-Approximately 75 mg of purified RNase from Peak 6 of the top chromatogram of Fig. 5 were reduced with O.% M
mercaptoethanol in 8 M urea/O.10 M Tris-chloride. pH 8.5 (total volume 20 ml) for 3 lb! hours. The pH was adjusted to -3.,j and the solution was applied to a column of Sephadex G-25 coarse gel, pre-equilibrated and eluted with 0.10 M acetic acid, to separate the protein from mercapto- ethanol. Determination of’thiol titer showed 8.0 ? 0.2 -SH/RNase. To 95 ml of reduced RNase solution (0.70 mg/ml) we added 0.53 g of GSH and 0.17 g of GSSG followed by 0.01 M, pH 8.2, Tris-acetate buffer to a total volume of 500 ml. Regeneration was allowed to proceed overnight at room temperature. Activity and protein concentration measure- ments showed a regenerated product with a specific activity 93% of that of the purified fraction before reduction. The pH was adjusted to 3.5 wit,h concentrated acetic acid, and the protein was concentrated by ultrafiltration (Amicon P-10 membrane) to -25 ml. The concentrated material was passed through a Sephadex G-25 column (equilibrated and eluted with 0.05 M acetic acid) to separate the protein from
glutathione. The protein-containing solution was diluted %l’old and 0.4 M Tris base added to bring the pH to 8.0. This solution was applied to a carhoxymethylcellulose column (1.6 x 30 cm) pre-equilibrated with 0.005 M Tris-chloride buffer. The column has eluted with a buffered NaCi gradient as described above. The chromatogram obtained is shown hy the middle tracing in Fig. 5.
For characterization of an air regeneration, 48 mg of’chrornatopral)h- ically purified RNase (Fig. 5, top curve. Peak 6 material) was reduced with mercaptoethanol, acidified, and separated from mercaptoethanol as described in the preceding paragraph. The protein fractions were pooled to yield 88 ml containing 42 mg of reduced RNase. The --SH titer was found to he 7.9 + O.‘/RNase. This solution was added to 250 ml of 0.1 M ‘I‘ris-acetate buffer. pH 8.2. and the pH was readjusted to 8.2 with NaOH. Regeneration was allowed to proceed at room temperature for 2 days. After that time the specific activity was found to he 655; of that of the purified fraction before reduction. The solution was diluted 2.fold and applied to a carhox~meth~lcellulose ion exchange column prepared and operated as described in the preceding paragraph. The resulting chromatogram is seen in Fig. 5. the bottom
curue. Metal Ion Catalysis-The regenerations were carried out at :10” with
pH 8.2, 0.09 M Tris-acetate buffer, initial concentration of’ reduced RNase 10m6 M. The chloride salts oi’Cu’-, Co”, Zn”, Fez’, and Mn2+ were tested at 10 5, 1Om6, and lo-’ M. Ni2+ was used in the form of its nitrate at the same concentrations.
Circular Dichroic Spectra-Measurements were made on a Durrum- Jasco .J-10 circular dichrometer, calibrated according to the method 01 Cassim and Yang (15). Silica cells of O.lW and 1.Wcm path length were used. Frequent base-line recordings were made with the same solvent as in the protein solution of interest. RNase from the major peak of each of’ the three chromatograms of Fig. 5 was examined, each in the solvent in which the peak was eluted from the column: 0.005 M Tris-chloride. pH 8/0.1:1 M NaCl. Spectra were measured on samples at 2X-25”.
RESULTS AND DISCUSSION
Regeneration of’ activity from reduced RNase with the optimal concentrations of GSH and GSSG proceeded to a 4-hour yield of 70% in the early experiments (Fig. 1). In later experiments, yields were commonly in the range 80 to 90% (Figs. 2 and 3). The “half-time” tH, the time for regeneration of 50% of the specific activity obtained with unreduced starting material, was generally in the neighborhood of 75 min with the optimal glutathione system (Figs. 1 to 3). Results from the trials for determining optimal concentrations and ratios of GSSG and GSH are shown in Table I. Although the testing here was not as extensive as that employed for optimization of the regeneration of human and hen egg lysozyme (4, 5), the same broad optima appear to apply to RNase as were found for the lysozymes: [GSSGY[GSH] ranging from l/10 to 1, and [GSH] + [GSSG] ranging from 4 to 10 mM.
Fig. 1A shows the results of regenerations carried out by Cu+catalyzed air regeneration (trs = 220 min), by EDTA- inhibited air regeneration where the inhibition is virtually complete, and two sets of glutathione regeneration data which lie on a single curve, showing t, = 80 min. Since one of the glutathione regenerations was carried out in the presence of 10m4 M EDTA while the other regeneration contained no EDTA, it is apparent that EDTA is not an inhibitor of the glutathione regeneration.
To test whether air-oxidative regeneration may also be taking place in a glutathione regeneration we carried out the regenerations of Fig. 1B. Here three glutathione regenerations were compared, one an optimal glutathione system, the second the same as the first plus 10m7 M Cu’+, and the third the same as the first plus 10m7 M Cut+ and 1W4 M EDTA. The results of all three regenerations are seen to give the same regeneration curve. Neither deliberate “contamination” of a glutathione regeneration with cupric ion nor the addition of EDTA to such
by guest on May 7, 2018
http://ww
w.jbc.org/
Dow
nloaded from

8479
0 60 120 180 240
TIME Iminutes)
FIG. 1. Regeneration of reduced RNase by air oxidation and by glutathione systems. All experiments were carried out at 30’ in 0.09 M
Tris-acetate, pH 8.2, at 30” and the protein concentration was 0.020 mg/ml. In Panel A, X represents an “air oxidation” regeneration in which (C&O,) = lo-’ M; 0 represents an “air oxidation” regeneration to which IO-’ M Na,EDTA had been added, 0 represents a glutathi- one-facilitated regeneration system in which (GSSG) = 3 x lo-’ M,
(GSH) = 3 x lOma M; 0 represents a glutathione system in which (GSSG) = 3 x lo-’ M, (GSH) = 3 x lOWa M, and (Na,EDTA) = lo-‘M. In Panel B, 0 represents a glutathione-facilitated regeneration in which (GSSG) = 3 x lo-‘M, (GSH) = 3 x 10-a~; in A(GSSG) = 3 x lo-&~, (GSH) = 3 x 10ea M, (CuSOJ = lo-’ M; in cl (GSSG) = 3 x lo-’ M,
(GSH) = 3 x ~O-‘M, (CuSO,) = lo-‘M, and (Na,EDTA) = lo-‘M.
FIG. 2. Reactivation of reduced RNase at different temperatures by air oxidation (Panel A) and by a glutathione system (Panel B). Both sets of reactivation solutions contained 0.09 M Tris-acetate, pH 8.2 at 25”, protein concentration 0.020 mg/ml. The experiments of Panel A had no other components added, while those of Panel B contained (GSSG) = 3 x lo-’ M and (GSH) = 3 x 1O-3 M.
a “contaminated” glutathione regeneration has an effect on the regeneration. These results show that metal ion-catalyzed regeneration does not occur to any detectable extent during an optimal glutathione regeneration. It is likely that Cu2+ and other potentially catalytic metal ions are chelated either by GSH or by GSSG, or both, in a catalytically impotent form.
Fig. 2 shows the contrasting responsiveness to temperature of the air oxidation (Fig. 2A, left panel) and the glutathione system (Fig. 2B, right panel). The air-oxidative regeneration is inhibited strongly above room temperature: at 25” t, = 75 min, while at 30” t, = 190 min. These findings confirm the general trend reported (6) for air regenerations of RNase. Regeneration via the glutathione system is obviously much less temperature- sensitive: at 14’, tH = 140 min, while at 37” t,+ = 70 min. Of the temperatures examined, 37” is optimal for rapid regeneration, but the optimum is clearly broad, with only minor variations in rate between 25 and 40”. The finding of a maximum regenera- tion rate of reduced RNase at or near 37” is clearly in contrast with the inhibition of the air oxidation in this temperature
FIG. 3 (left). Glutathione-facilitated regenerations of reduced RNase at different protein concentrations. Key: A, 3.1 x lo-’ M; x, 7.6 x IO-’ M; 0, 2.6 x lo-’ M; A, 5.3 x lo-’ M; q , 1.2 x 10m5 M. (Note: 14 pg/ml = 1.0 x lOWE M RNase.) The regenerations were carried out at 30’ in pH 8.2 (0.10 M) Tris-acetate buffer, lo-’ M EDTA, [GSSG] = 3 x lo-” M, [GSH] = 3 x 1O-3 M.
FIG. 4 (right). The pH dependence of glutathione regenerations of reduced RNase. The per cent regeneration at 75 min was estimated from a smoothed plot of per cent activity versus time, employing aliquots taken at 60-, 75-, and 90-min regeneration times. The concentration of reduced RNase was 1.1 to 1.4 x lo-’ M except at pH 6.5, where it was 5.8 x 1OmB M. Regenerations were carried out at 30” in 0.10 M Tris-acetate buffers (0, l ), or in 0.10 M bis-tris acetate (0, n ). The unfilled symbols (0, Cl) represent regenerations carried out with [GSH] = 5 x lo-’ M, [GSSG] = 5 x 10e5 M; for the filled symbols ( 0, n ), [GSH] = 5 x IO-'M, [GSSG] = 5 x lo-‘M.
TABLE I
Dependence of reactivation of reduced RNase on GSSG and GSH concentrations
Regenerations were carried out in Tris-acetate buffer, 0.09 M Tris, pH 8.2, at 30”, reduced RNase concentration lo-’ M (8 x lo-’ -SH eq/liter).
lG8Hl [GSSG] Per cent activity
30 min 90 min
M M
3 x 10-s 0 m 400 0 2 1 3 x 10-Z 3x10-3 5 4000 800 9 15 3 x 10-s 3x10-’ 5 400 80 24 55 3 x 10-s 3 x 1o-a 0.5 400 800 22 53 3 x lo-’ 3x10-5 5 40 8 3 33 6 x lo-’ 3 x 10-a 0.1 80 800 16 22
0 3x10-4 0 0 80 0 3
“These ratios are expressed on the basis of equivalents of thiol and disulfide.
range, and increases the physiological plausibility of the glutathione system. The temperature dependence of RNase regeneration by the protein thiol oxidoreductase does not appear to have been determined. The strong difference ob- served in temperature dependence between air regenerations and glutathione regenerations also implies that the two regen- erations operate by different mechanisms. This conclusion was reached earlier for regenerations of hen egg lysozyme (4).
Of the six potentially catalytic metal ions examined, only Cu2+ and Co2+ showed a clear-cut acceleration of thiol oxida- tion (at concentrations between lo-’ and 1O-5 M). In a comparison of these ions’ effects on regeneration of RNase activity (Table II) we found that 10eB M Cu*+ accelerated both -SH oxidation and regeneration of RNase activity, while Coz+ catalyzed -SH oxidation without affecting regain of ac- tivity. Of the metal ions tested, it is clear that only Cu*+ is
by guest on May 7, 2018
http://ww
w.jbc.org/
Dow
nloaded from

8480
TABLE II
Comparison of effects of Cuz+ and Co2+ on air oxidation and regeneration of reduced RNase
Regeneration --SH/mol protein solution” at TO min
Control” 4.4 16 + lo-‘M CO*+ 3.9 + lom6M CO*+ 1.7 16 + lo-‘M &I’+ 1.3 44 + 1t6M cU*+ 1.0 52 + lo-55MCUZ+ 37 + lo-“MCu2+ 1.4 5
a Regeneration solutions all contained 1.4 x 10 ’ M reduced Rh‘ase in 0.10 M Tris-acetate buffer, pH 8.2; reactions were carried out at 30”. The results tabuiated above are representative values from a much larger body of data (9).
effective in catalyzing the air regeneration of RNase. Other experiments showed that the regeneration rate is invariant and reproducible over the [Cu’+] range from lo-’ to 1O-6 M, but is strongly inhibited at lo-” M.
Since catalysis of regeneration is marked at 10 ’ M, it would not be surprising to find detectable catalysis at substantially lower Cu2+ concentrations. It also seems likely that copper is the trace metal commonly responsible for adventitious cataly- sis of air regenerations of reduced proteins (6, 16, 17). Our results showing strong Cu2+ catalysis of air regeneration of RNase confirm and extend the conclusions of Takagi and Isemura (18) from studies on the air regeneration of reduced Taka-amylase A.
The effect of protein concentration on the glutathione regeneration was examined. Regenerations were carried out at reduced RNase concentrations ranging from 3 x 10m7 to 1.2 x lo-” M. The results of these experiments, shown in Fig. 3, display some scatter but show no systematic trend in the kinetics and yield of regenerated RTZase. This contrasts with the reported rate inhibition in the air regeneration at concen- trations >2 x lOwe M reduced RNase (6). We have not determined the upper limits of protein concentration for acceptable rate and yield in the glutathione regeneration, but the present work shows that a glutathione regeneration can be carried out at protein concentrations at least lo-fold higher than the air regeneration, without diminished yield or reduced rate. This offers a considerable preparative advantage.
Preincubation Experiments-These experiments were to test for the existence of conformation changes that could be part of the rate-limiting process in regeneration of activity in reduced RNase following its dissolution from a lyophilized form. Preincubation of reduced RNase, 0.20 mg/ml, was carried out for several hours at 23-25” in two different solvents: 0.10 M acetic acid containing 10m4 M EDTA, and 0.10 M
Tris-acetate buffer, pH 7.0, containing 10m4 M EDTA. Zero time was the time at which lyophilized reduced RNase was dissolved in the preincubation solvent. After varying times of preincubation, aliquots were transferred to a regeneration solution at 30”, whose composition (after aliquot addition) was 0.09 M Tris-acetate buffer, pH 8.2, [GSSG] = 3 x 10 4 M,
[GSH] = 3 x lo-’ M, lo-’ M EDTA. Enzymic activity meas- ured after 60 min of regeneration was independent of pre- incubation times from 10 min to 8 hours, for both preincuba- tion solvents. Control determinations of -SH titer on these samples showed no detectable oxidation over the &hour time interval. The results rule out rate-limiting conformation
changes following dissolution of the lyophilized reduced pro- tein.
pH Dependence-Fig. 4 shows the pH dependence of the regeneration rate of reduced RNase for two sets of regeneration experiments differing only in total glutathione concentration (same [GSSGj/[GSH]). The results obtained at the lower glutathione concentration appear to plateau at a higher rate at pH >8, but the difference is of marginal significance. Simi- larly, although the data obtained with the two buffer systems (Tsis and bis-tris’) has been represented with separate curves, there is no substantial discontinuity from one buffer to the other.
The pH dependence is consistent with rate control by ionizing groups with an apparent pK between 7.5 and 8.0. This pK could be reasonably ascribed to thiol or a-amino groups in the system, but this kind of experiment does not permit discrimination.
The pH dependence found here for glutathione regenerations is similar to that reported for air regeneration of RNase (6). This is in contrast with the oxidoreductase system for RNase regeneration, which showed (in the partly purified system) virtual pH independence from pH 7.4 to 8.2 (19). This difference suggests that the enzymic and the nonenzymic regenerations operate by different mechanisms.
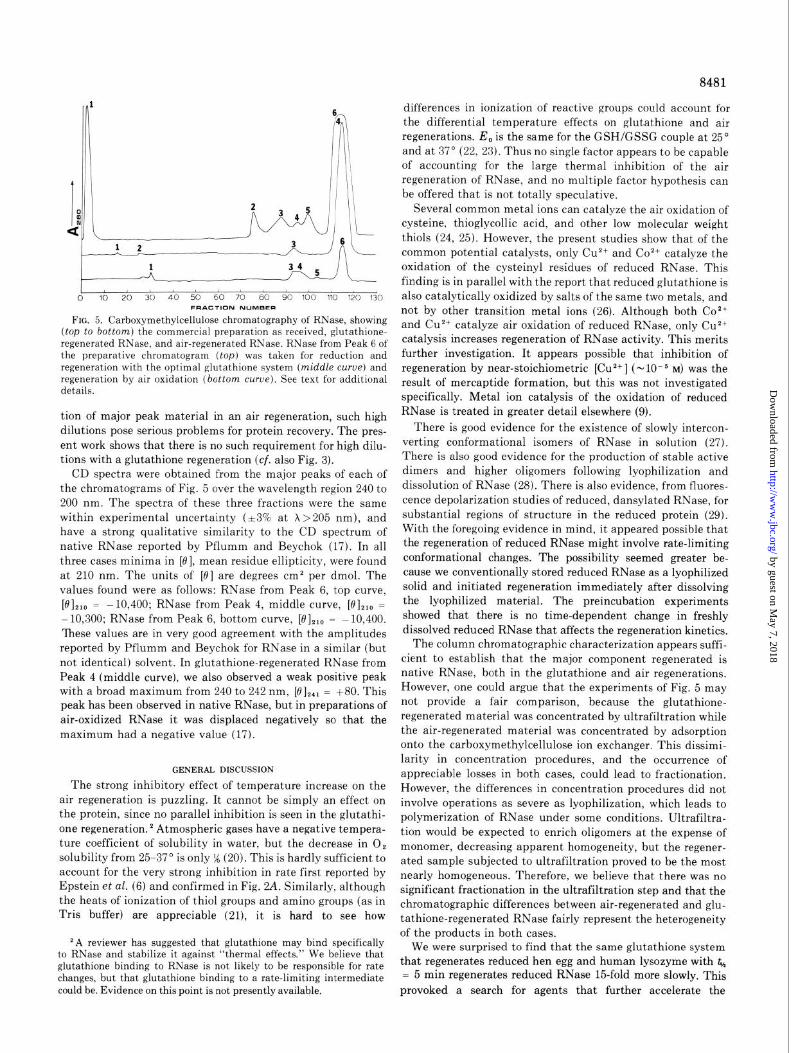
Characterization of Regenerated Products-Fig. 5 shows the column chromatographic behavior of glutathione- regenerated and air-regenerated RNase. The middle chromato- gram, obtained with glutathione-regenerated material, showed a major peak (no. 4) chromatographically identical with the purified fraction taken from chromatography of RNase ob- tained from the supplier (top curve, Peak 6). The identity is confirmed by our finding of the same specific activity in these two peaks, and by the identity of their dichroic spectra (see below). Protein from Peak 3 in the middle chromatogram showed 70% the specific activity of Peak 4 material. The unfractionated glutathione-regenerated material showed 93% the specific activity of the initial purified f’raction. The above facts are interpreted as showing that glutathione facilitation gives a high yield of native RNase.
For comparison, the bottom chromatogram of Fig. 5 was obtained from an air regeneration of reduced RNase. The specific activity of the regenerated material before fractiona- tion was 65%. Combined Peaks 3 and 4 showed 19’? specific activity, Peak 5 showed 52% specific activity, and Peak 6 showed 100% specific activity, all referred to the initial purified RNase (Peak 6, top curve). The protein of Peak 6 of the lower curve of Fig. 5 appears to be identical with that of Peak 6, upper curve, by chromatographic criteria, by specific activity, and by CD spectra (see below). We carried out the air oxidation at protein concentrations considerably above those claimed in early work to provide optimal rates and yields (6). However, Pflumm and Beychok (17) were unable to repeat those findings and indeed showed considerable chromatographic heterogene- ity in their preparations of air-regenerated RNase. The concen- tration of reduced RNase here chosen for the preparative air regeneration was a compromise. On the one hand, high dilution favors a high regeneration rate and yield. On the other hand, high dilution leads to excessive loss of S/N (signal amplitude/ noise amplitude) in the operation of the ultraviolet column monitor. Even if higher dilution should produce a higher frac-
’ The abbreviations used are: bis-tris, “,2-bis(h~drox!meth~l)- 2,2’,2”-nitriloethanol; dansyl, 5.dimethylaminonaphthalene-1-sulf’onyl.
by guest on May 7, 2018
http://ww
w.jbc.org/
Dow
nloaded from
8481
differences in ionization of reactive groups could account for the differential temperature effects on glutathione and air regenerations. E, is the same for the GSH/GSSG couple at 25” and at 37” (22, 23). Thus no single factor appears to be capable of accounting for the large thermal inhibition of the air regeneration of RNase, and no multiple factor hypothesis can be offered that is not totally speculative.
Several common metal ions can catalyze the air oxidation of cysteine, thioglycollic acid, and other low molecular weight thiols (24, 25). However, the present studies show that of the common potential catalysts, only Cu2+ and Co2+ catalyze the oxidation of the cysteinyl residues of reduced RNase. This finding is in parallel with the report that reduced glutathione is also catalytically oxidized by salts of the same two metals, and not by other transition metal ions (26). Although both Co’+ and Cu2+ catalyze air oxidation of reduced RNase, only Cu2+ catalysis increases regeneration of RNase activity. This merits further investigation. It appears possible that inhibition of regeneration by near-stoichiometric [Cu2+] (-1O-5 M) was the result of mercaptide formation, but this was not investigated specifically. Metal ion catalysis of the oxidation of reduced RNase is treated in greater detail elsewhere (9).
There is good evidence for the existence of slowly intercon- verting conformational isomers of RNase in solution (27). There is also good evidence for the production of stable active dimers and higher oligomers following lyophilization and dissolution of RNase (28). There is also evidence, from fluores- cence depolarization studies of reduced, dansylated RNase, for substantial regions of structure in the reduced protein (29). With the foregoing evidence in mind, it appeared possible that the regeneration of reduced RNase might involve rate-limiting conformational changes. The possibility seemed greater be- cause we conventionally stored reduced RNase as a lyophilized solid and initiated regeneration immediately after dissolving the lyophilized material. The preincubation experiments showed that there is no time-dependent change in freshly dissolved reduced RNase that affects the regeneration kinetics.
The column chromatographic characterization appears suffi- cient to establish that the major component regenerated is native RNase, both in the glutathione and air regenerations. However, one could argue that the experiments of Fig. 5 may not provide a fair comparison, because the glutathione- regenerated material was concentrated by ultrafiltration while the air-regenerated material was concentrated by adsorption onto the carboxymethylcellulose ion exchanger. This dissimi- larity in concentration procedures, and the occurrence of appreciable losses in both cases, could lead to fractionation. However, the differences in concentration procedures did not involve operations as severe as lyophilization, which leads to polymerization of RNase under some conditions. Ultrafiltra- tion would be expected to enrich oligomers at the expense of monomer, decreasing apparent homogeneity, but the regener- ated sample subjected to ultrafiltration proved to be the most nearly homogeneous. Therefore, we believe that there was no significant fractionation in the ultrafiltration step and that the chromatographic differences between air-regenerated and glu- tathione-regenerated RNase fairly represent the heterogeneity of the products in both cases.
We were surprised to find that the same glutathione system that regenerates reduced hen egg and human lysozyme with & = 5 min regenerates reduced RNase 1Bfold more slowly. This
provoked a search for agents that further accelerate the
.“l 6
4
B ::: 2
3 4 5
a L L
1 2 3 6
i \
1 -A
&
34 5
0 10 20 30 40 50 60 70 80 90 100 110 120 130
FF1ACTION NUMBECI
FIG. 5. Carboxymethylcellulose chromatography of RNase, showing (top to bottom) the commercial preparation as received, glutathione- regenerated RNase, and air-regenerated RNase. RNase from Peak 6 of the preparative chromatogram (top) was taken for reduction and regeneration with the optimal glutathione system (middle curue) and regeneration by air oxidation (bottom curve). See text for additional details.
tion of major peak material in an air regeneration, such high dilutions pose serious problems for protein recovery. The pres- ent work shows that there is no such requirement for high dilu- tions with a glutathione regeneration (cf. also Fig. 3).
CD spectra were obtained from the major peaks of each of the chromatograms of Fig. 5 over the wavelength region 240 to 200 nm. The spectra of these three fractions were the same within experimental uncertainty (13% at X>205 nm), and have a strong qualitative similarity to the CD spectrum of native RNase reported by Pflumm and Beychok (17). In all three cases minima in [0], mean residue ellipticity, were found at 210 nm. The units of [0] are degrees cm2 per dmol. The values found were as follows: RNase from Peak 6, top curve,
VI **a = -10,400; RNase from Peak 4, middle curve, [0],,, = -10,300; RNase from Peak 6, bottom curve, [0],,, = -10,400. ‘Ibese values are in very good agreement with the amplitudes reported by Pflumm and Beychok for RNase in a similar (but not identical) solvent. In glutathione-regenerated RNase from Peak 4 (middle curve), we also observed a weak positive peak with a broad maximum from 240 to 242 nm, [(?I,,, = +80. This peak has been observed in native RNase, but in preparations of air-oxidized RNase it was displaced negatively so that the maximum had a negative value (17).
GENERAL DISCUSSION
The strong inhibitory effect of temperature increase on the air regeneration is puzzling. It cannot be simply an effect on the protein, since no parallel inhibition is seen in the glutathi- one regeneration. * Atmospheric gases have a negative tempera- ture coefficient of solubility in water, but the decrease in O2 solubility from 25-37” is only ‘/6 (20). This is hardly sufficient to account for the very strong inhibition in rate first reported by Epstein et al. (6) and confirmed in Fig. 2A. Similarly, although the heats of ionization of thiol groups and amino groups (as in Tris buffer) are appreciable (21), it is hard to see how
*A reviewer has suggested that glutathione may bind specifically to RNase and stabilize it against “thermal effects.” We believe that glutathione binding to RNase is not likely to be responsible for rate changes, but that glutathione binding to a rate-limiting intermediate could be. Evidence on this point is not presently available.
by guest on May 7, 2018
http://ww
w.jbc.org/
Dow
nloaded from

8482
glutathione regeneration of RNase, which led to the finding of specific stimulatory and inhibitory salt effects (refer to fol- lowing paper). Returning, however, to the rate differences for regeneration of reduced lysozyme and RNase, we recall earlier evidence from another laboratory for substantial differences. The “shuffle-ase” enzyme system, complete with unidentified dialyzable cofactor, rapidly regenerates reduced RNase (30), but the enzyme regenerates lysozyme only in the absence of the cofactor. whose presence is inhibitory. In trying to relate these irz vitro regenerations to in uiuo biosynthetic mechanisms we see that the biosynthesis of nucleases and proteases poses a particular problem to the biosynthetic system. The hazard is that of nucleolytic and proteolytic digestion of the ribosome, mRNA. and tRNA, a kind of molecular suicide. A solution of this problem for proteases is their synthesis as inactive enzyme precursors. There is no evidence for the analogous solution of the problem for nucleases: inactive pro-nucleases have not been found. One may argue that premature action of nascent nucleases is avoided by an early compartmentalization of the nuclease, physically separating the enzyme from physiologi- cally inappropriate substrates. This may be so, but compelling evidence does not appear to be at hand. Our argument is, in short, that there are good reasons for expecting that the biosynthesis or folding of nucleases, or both, may involve unique components or operations, or both, not shared in the formation of other proteins. From this perspective it is less surprising that the kinetics of regeneration of reduced RNase and reduced lysozyme show substantial differences.
The results of the present investigation bear on the question of what mechanism is involved in the biosynthetic folding of RNase. but give no clear answer. While the essential compo- nents of the glutathione system are found in most tissues, including pancreas (31. 32), the rates found here with the glutathione system are slower than seems plausible for biosyn- thesis of RNase. The alternative mechanism, i.e. folding facilitated by the “shuffle-ase” enzyme, is plausibly rapid for RNase, but this enzyme could not be found in the bovine pancreas (22). Thus both candidates for physiological facilita- tion of reduced RNase have that potential, but at present both show a serious weakness.
With the present work, RNase becomes the fourth protein whose regeneration from the disulfide-reduced form is shown to be facilitated by a glutathione system. Rapid regenerations have already been shown for hen egg lysozyme (4), human lysozyme (5), and the Bowman-Birk protease inhibitor (33). Thus the nonenzymic glutathione system is gaining in func- tional generality, and in plausibility as a biosynthetic mecha- nism.
Acknowledgments-We thank Mr. G. Bratt for skilled tech- nical assistance and Mr. Peter Holmberg for assistance in
determining the pH dependence of the glutathione regenera- tion.
REFERENCES
1. Anfinsen, C. B. (1973) Science 181, 22:1-%X0 2. Scheraga, H. A. (1971) Chem. Reu. 71, 195-217 3. Wetlaufer. D. B., and Ristow, S. S. (1973) Annu. Reu. Biochem.
42,135m155 4. Saxena, V. P., and Wetlaufer, D. B. (1971) Biochemistry 9,
5015-5023 5. Wetlaufer, D. B., -Johnson. E. R., and Clauss, L. M. (1974) in
Lysozyme (Osserman, E., Canfield. R. E.. and Beychok, S.. eds) pp. 269-280, Academic Press, X. Y.
6. Epstein, C. J., Goldherger, R. F., Young, D. M., and Anfinsen, C. B. (1972) Arch. Biochem. Biophys. Suppl. I, 223-231
7. Epstein, C. ,J., and Goldberger, R. F. (1963) J. Biol. Chem. 238, 1380-1383
8. Enstein. C. ,J. (1970) in Aspects of Protein Biosynthesis (Anfinsen.
9.
10.
11.
12. 1X.
14. 15. 16. 17.
18. 19.
20. 21.
22. 23.
24.
25.
26.
27.
28
29
30
31
32 33
‘C. IX., ed) p. 367. Acaddmic Press, \J. Y. Ahmed. A. K. (1969) Ph.D. thesis, University of Minnesota,
Minneapolis Kalnitsky, G., Hummel, ,J. P., and Dierks, C. (1959) J. Biol. Chem.
234, 1512-1516 Fletcher, P. I,., Jr., and Hash, J. H. (1972) Riochemistry 11,
4274-4280 Ellman, G. L. (1959) Arch. Biochem. Biophys. 82, K-77 Anfinsen. C. B.. Haber. E., Sela, M., and \I;hite, F. H., Jr. (1961)
Proc. Natl. Acad. Sci. U. S. A. 47, 1309~1314 Taborsky, G. (1959) J. Biol. Chem. 234, 2652-2656 Cassim, J. Y.. and Yang, J. T. (1969) Biochemistry 8, 1947-1951 White, F. H., ,Jr. (1961)-J. Biol. Chem. 236, 1353-1360 Pflumm, M. N., and Beychok. S. (1969) J. Viol. &em. 244,
:3982-3989 Takagi, T., and Isemura, T. (1964) J. Rio&em. (Tokyo) 56, 344-350 Goldberger, R. F., Epstein. C. J., and Anfinsen. C. B. (1963) J.
Biol. Chem. 238, 628-635 Rattino. R.. and Clever. H. L. (1966) Chem. Reu. 66, 395-463 Bates, k. 6. (1964) De&rmination ofpH, p. 112, John Wiley and
Sons, Inc., IX. Y. .Jocelyn, P. C. (1967) Eur. J. Biochem. 2, 327-331 Gorin, G., and Doughty, G. (1968) Arch. Biochem. Biophys. 126,
547-551 < , Cecil, R. (1963) in The Proteins: Composition, Structure, and
Function (Neurath. H.. ed) 2nd Ed, Vol. 1, p. 379, Academic Press, N. Y.
Tarbell. D. S. (1961) in Organic Sulfur Compounds (Kharasch, N., ed) p. 97, Pergamon, N. Y.
Bernheim. F.. and Bernheim, M. L. C. (1939) Cold Spring Harbor Symp. Quant. Biol. 7, 174-182
Craig, L. C., King, T. P., and Crestfield. A. M. (1963) Biopolymers 1, 231-238
Crestfield, A. M., Stein, W. H., and Moore, S. (1962) Arch. Biochem. Biouhvs. Sunul. 1. 217-222
Young, D. M., and”Potts,- j. T.; Jr. (1963) J. Biol. Chem. 238, 1995 2002
Goldberger, R. F., Epstein, C. J., and Anfinsen, C. B. (1964) J. Biol. Chem. 239, 1406-1410
Long, C., King, E. J., and Sperry, W. M., eds (1961) The Biochemists’ Handbook, p. 793, Van Nostrand, N, Y.
Tietze, F. (1969) Anal. Biochem. 27, 502-522 Hogle, J. M., and Liener, I. E. (1973) Can. J. Biochem. 51, 1014-
1020
by guest on May 7, 2018
http://ww
w.jbc.org/
Dow
nloaded from

A K Ahmed, S W Schaffer and D B Wetlauferoxidation and by glutathione oxidoreduction buffers.
Nonenzymic reactivation of reduced bovine pancreatic ribonuclease by air
1975, 250:8477-8482.J. Biol. Chem.
http://www.jbc.org/content/250/21/8477Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/250/21/8477.full.html#ref-list-1
This article cites 0 references, 0 of which can be accessed free at
by guest on May 7, 2018
http://ww
w.jbc.org/
Dow
nloaded from