nucleolar chromosome variation and evolution in trie · pdf filenucleolar chromosome variation...
TRANSCRIPT

CARYOLOGIA Vol. 51, n. 2: 133-147, 1998
Nucleolar chromosome variation and evolution intrie genus Vicia
M.S. BISHT, K. KESAVACHARYULU and S.N. RAINA *Laboratory of Cellular and Molecular Cytogenetics, Department of Botany, University of Delhi, Delhi -110 007, India.
SUMMARY — The nucleolar chromosomes in the genus Vicia usually comprise one orrarely two pairs of the complement in the diploid (2n= 10, 12, 14; x = 5, 6, 7) taxa.Barring V. anatolica nucleolar chromosomes in complements of various species have onesecondary constriction and one satellite each. In V. anatolica (2n = 2x— 10) one of thetwo nucleolar chromosome pairs contained two secondary constrictions and twosatellites. The nucleolar chromosomes in Vicia species show substantial variation, insatellite size from very minute to highly conspicuous, in the location of primary andsecondary constrictions from terminal to mid region, and in the size of mid arm fromvery small to about 90 per cent of the total chromosome size. On the basis ofproportionate size of the three stainable components, the nucleolar chromosomes in Viciaspecies have been classified into eighteen distinct types. The comparative morphology ofthe nucleolar chromosomes shows that interchanges including Robert-sonian fusion,deletion and/or inversion have mainly contributed to their divergence and evolution.Overall, the nucleolar chromosome variation showed a good correlation with thetaxonomic classification. The present results provide probably the first example of anastonishing array of diversity in the form and shape of nucleolar chromosomes within agenus.
INTRODUCTION
The genus Vicia, an important member of legume family, includes about166 species (ALLKIN et al. 1986) out of which 40 species are of considerableeconomic importance (HARLAN 1956; KUPICHA 1976; HANELT and METTIN 1989).Unlike many plant genera (STEBBINS 1950, 1971; VED BRAT 1965; JOHN and LEWIS 1968;RAINA and KHOSHOO 1971; RAINA et al. 1989) the genus Vicia carries structuralalterations in the chromosomes on a very large scale (YAMA-MOTO 1973; RAINA andREES 1983a; RAINA et al. 1989; RAINA 1990; MAXTED et
* Corresponding author.

134 BISHT, KESAVACHARYULU and RAINA
al. 1991). There are several species that have complements of median andsubmedian chromosomes alone while in others all chromosomes are subtelocen-tric. The size differences between individual chromosomes of the set are small inmany species and they grade into one another in an almost a continuous series,while in some species the arm length asymmetry and drastic change in size offew chromosomes within the complement have resulted in most aberrantkaryotypes. Apart from chromosomal repatterning, there is large scale variationin chromosome size, 2C nuclear DNA content, repetitive and non-repetitive DNAsequences, chromosome number (2n = 10, 12, 14, 24, 28, 36, 42; x = 5, 6, 7),euchromatin (75-88%) and heterochromatin (11-25%), base compositionalheterogeneity (17.08-19.52), satellite DNA sequences, G + C content (34.89-38.06%) ribosomal DNA repeat unit and DNA density (0.096-0.263) per unitarea (Cnooi 1971; RAINA and REES 1983^, b; RAINA and NARAYAN 1984; NARAYANet al. 1985; RAINA and BISHT 1988; RAINA 1990; RAINA and OGIHARA 1995).
In view of numerous and divergent changes in the evolution of Viciagenomes including that of detailed karyotypic rearrangements, the present studywas undertaken to establish the nature and extent of variation in NOR location inthe nucleolar chromosomes in 67 Vicia species/subspecies. This region of rRNAgenes (rDNA) stains poorly with Feulgen, Orcein or Giemsa and is visible atmetaphase as a faintly stained secondary constriction. These particular markerchromosomes are powerful experimental vehicles in the light of generalagreement that the evolutionary change(s) due to chromosomal mutation, if any,can be more easily identified and measured in them.
MATERIALS AND METHODS
Plant material. — The seed samples of various species/subspecies were obtainedfrom Zentralinstitut fur Genetik und Kulturpflanzenforschung der Akedemie derWissenschaften, Gatersleben, Germany; Department of Biology, The University ofSouthampton, U.K.; C.N.R. Istituto del Germoplasma, Bari, Italy; United StatesDepartment of Agriculture (USDA), Beltsville, USA and International Centre forAgricultural Research in the Dry Areas (ICARDA), Aleppo, Syria to whom we aregrateful. We have adopted KUPICHA'S (1976) classification.
Karyotypes. — Actively growing root tips of about 1-2 cm length were immersed indistilled water and cold treated at 4°C for 24 h. They were subsequently washed and fixedin freshly prepared 1:3 acetic alcohol for a minimum period of 24 h. Fixed root tipswashed in distilled water were hydrolysed in 5N HC1 for 1 h at room temperature andstained in 0.5% Feulgen solution (SIGMA) for 1 h before squashing. The nucleolarchromosomes were analyzed from a minimum of seven cells for each taxon for arm ratioand other morphological details.

NUCLEOLAR CHROMOSOME VARIATION IN VICIA 135
Cluster analysis. — Pair wise comparisons, based on arm ratio of the nucleolarchromosome, were done to calculate percentage dissimilarity values. Cluster analysis wasconducted using Unweighted Centroid method, and the dendrogram was constructed on thebasis of resulting clusters. The analysis was undertaken using the software MVSP.
OBSERVATIONS
The chromosome numbers, 2C DNA amounts and arm ratio of the nucleolarchromosomes determined in 67 species/subspecies are given in Table 1.

136 BISHT, KESAVACHARYULU and RAINA

The number of nucleolar chromosomes.
In most of the diploid (2n=2x=1O, 12, 14) species the chromosomecomplement contained only one pair of nucleolar chromosomes but there were somespecies/subspecies with more than one pair (Table 2).
Nucleolar chromosome types.
It was amazing to find that in the genus Vicia as many as 18 distinct types ofnucleolar chromosomes could be described. These chromosomes, on the basis ofrelative position of primary and secondary constrictions, are extremely diverse(Figs. 1 and 2, Table 1) and distinctive in form and structure. Hetero-morphy inthe pair(s) of nucleolar chromosomes within the complement was nonexistent inthe species investigated. The different types of nucleolar chromosomes have beennamed according to the type species and they are as follows.
NUCLEOLAR CHROMOSOME VARIATION IN VICIA ' 137

138 BISHT, KESAVACHARYULU and RAINA
TABLE 2- Vicia species with two types of nucleolar chromosomes.
Nucleolar chromosomes with one secondary constriction and one satellite.
Villosa. — This is characteristic of relatively primitive section Cracca (see KUPICHA1976; HANELT and METTIN 1989; MAXTED et al. 1991), and has been observed in six specieswith most primitive chromosome number (2n = 2x=14 ) (see RAINA and REES 1983(2).The present authors suggest that the submedian chromosome with a prominent satellite onthe long arm constitutes the basic type of nucleolar chromosome in the genus. The middlesegment, flanked by primary and secondary constrictions, is smallest in size. The tworemaining parts of the chromosome are of similar size (Fig. 1A).
Sativa and Macrocarpa. — The section Vicia is characterized by the nucleolarchromosomes of both Sativa and Macrocarpa types. Barring V. incisa all the speciesbelonging to the V. sativa aggregate (HANELT and METTIN 1989) have nucleolarchromosomes of Sativa or Macrocarpa types. The middle segment in the subterminalchromosome is very prominent in both cases but the clear difference between the twotypes lies in the satellite size which almost equals the size of short arm in Macrocarpatype (Fig. 1B-C). The Macrocarpa type is commonly found in the species having 2n=12and never in species with 2 n = l 0 or 14, whereas the Sativa type has been observed inspecies with 2n= 10, 12, 14.
Fig. 1. — Nucleolar chromosome types. (A) Villosa - V. pulchella, V. dasycarpa, V. eriocarpa, V. villosa, V.benghalemis, V. vicioides; (B) Sativa - V, cordata, V. lathyroides, V. sativa, V. pilosa, V. striata; (C)Macrocarpa - V. nigra, V. melanosperma, V. angustijolia ssp. angustifolia, V. segetalis, V. macrocarpa; (D)Monantha - V. monantha, V. cirrhosa, V. disperma, V. ludovkiana; (E) Grandiflora - V. gmndiflom var.ketaibeliana, V. cassubica, V. incisa, V. pyrenaica, V. orobus, V. tenuissima; (F) Biennis - V. palaestina, V.biennis, V. picta; (G) Tetrasperma - V. tetrasperma, V. bithynica, V. articulata; (H) Hybrida - V. hybrida, V.sericocarpa; (I) Peregrina - V. peregrina, V. michauxii, V. persepolitana, V. pannonica ssp. purpura-scens.The lower constriction represents the secondary constriction.


140 ' BISHT, KESAVACHARYULU and RAINA
Monantha. — This type is characteristic also for some species of section Cracca.The centromere position is submedian. The prominent feature of this type of nucleolarchromosome is the equal size of the satellite and the middle segment, and in theformer, the stalk is, more often than not, highly extended displaying sometimes achromomere along its length (Fig. ID). It is found in species with 2n= 14 and not in thespecies with 2n — 10 or 12.
Grandzflora. — This is subtelocentric chromosome with the secondary constrictionbisecting the short arm into two equal halves. The other arm is more than four timeslonger than the short arm or the satellite (Fig. IE).
Eiennis. — The nucleolar chromosome in this type has a submedian centromereand a satellite on the short arm. It has been found only in species with 2n= 14 (Fig. IF).
Tetrasperma. — It is marked by a "compound constriction" which appears to beattenuated owing to the close proximity of the centromere (Fig. 1G) to the nucleolarconstriction (LA COUR 1950). It is present in species with 2n= 14.
Hybrida. — The difference between Sativa, Macrocarpa and Hybrida types lies inthe arm ratios of the three components of the NOR chromosome. All the three types,otherwise have large middle segment and two unequal - equal arms (Fig. 1H). It ischaracteristic of the species (2n= 12) belonging to section Hypechusa.
Peregrina. — It has a subterminal centromere and a very large satellite on the longarm (Fig. II). It is found in species with 2n = 14 or 10.
Eristalioides. — This type is characterized by the close proximity of primary andsecondary constrictions (Fig. 2A). The satellite in the short arm in the submedianchromosome is very large as in the Peregrina type.
Minutiflora. — This submedian nucleolar chromosome with a minute satellite onthe long arm occurs in two species with 2n= 14 (Fig. 2B).
Cretica. — The satellite body size approximates to that of the short arm, and themid segment is larger in size than the one present in the Villosa type (Fig. 2C).
Amurensis. — The secondary constriction in this type bisects the long arm intotwo equal halves (Fig. 2D). In proportional terms the short arm is smaller in size thanthe Cretica type.
Fig. 2. — Nucleolar chromosome types. (A) Eristalioides - V. peregrina, V. michauxii, V. eristalioides; (B)Minutiflora - V. altissima, V. minutiflora; (C) Cretica - V. cretica ssp. cretica, V. vicioides, V. pannonicassp. pannonica; (D) Amurensis - V. amurensis, V. unijuga, V. pannonica ssp. purpurascem, V. anatolica, V,hajastana; (E) Hyrcanica - V. unijuga, V. noeana var. megalodonta, V. sericocarpa, V. pannonica ssp.pannonica, V. hyrcanica, V. hybrida, V. lutea; (F) Pisiformis - V. elegans, V. incana, V. neglecta, V.pisiformis, V. dumetorum; (G) Faba - V. galeata, V. esdmelonensis, V. bajastana, V. melanops, V, faba; (H)Anatolica - V. anatolica. The lower constriction represents the secondary constriction.
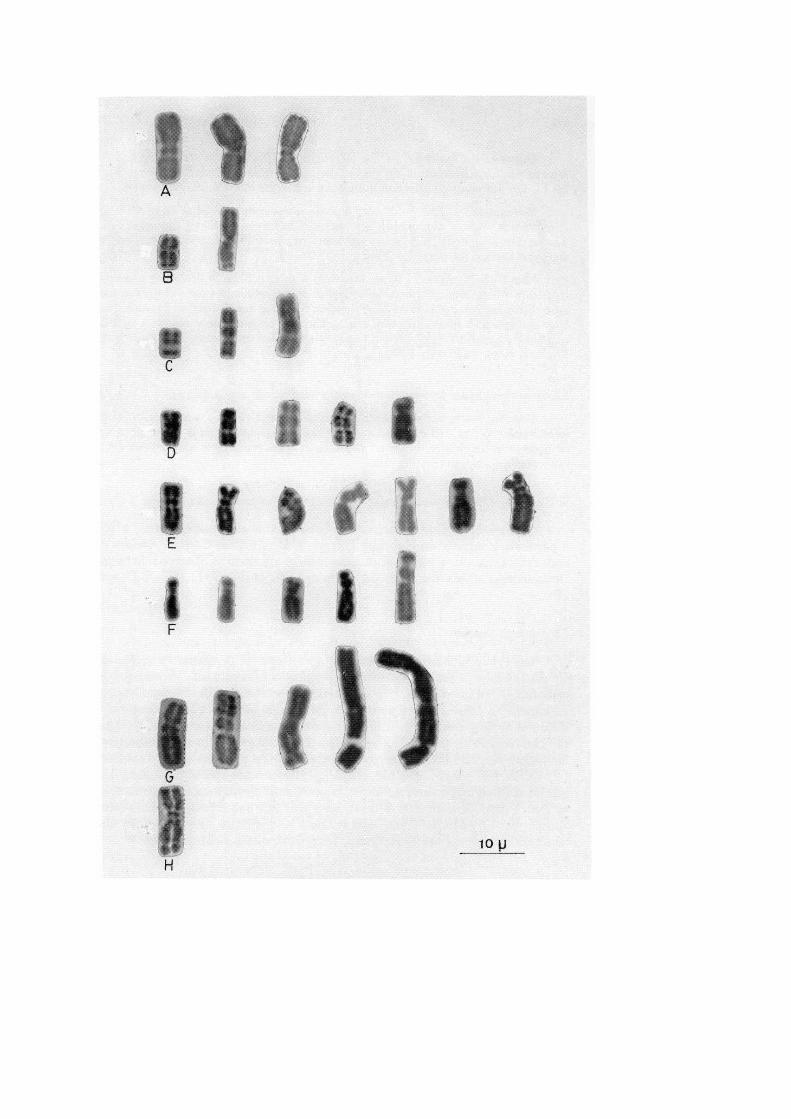

142 BISHT, KESAVACHARYULU and RAINA
Hyrcanica and Pisiformis. — The Hyrcanica and Pisiformis types are one and thesame except for differences in the sizes of the short arm and the middle segment (Figs.2E-F). The secondary constriction in the long arm is present near the centromere.
Narbonensis. — This NOR chromosome with mean ratio of 2.79:1.00:1.83 ischaracteristic of the V. narbonensis aggregate of eight species belonging to section Faba.The satellite component is bigger in size than the remaining two components, middle andshort arms of the nucleolar chromosome.
Faba. — It is commonly found in species with 2 n = 10 or 12 but never in specieshaving 2 n = 14. Although this group of nucleolar chromosomes from different specieshave dissimilar arm ratios (Fig. 2G), they have been grouped together because these largechromosomes have a common origin. The long arms of nonhomologous subtelo-centricsare united by Robertsonian fusion to a common centromere (RAINA and REES 1983a, b; RAINAand BISHT 1988).
Nucleolar chromosome with two secondary constrictions and two satellites.
Anatolica. — It has a submedian centromere, and two secondary constrictionsintersecting short as well as long arms. The nucleolar constriction is proximal to thecentromere in the former and distal in the latter, thus resulting in a very large and a smallsatellite, respectively (Fig. 2H).
Size variation between nucleolar chromosomes within same nucleolar type.
Another major kind of variation in the nucleolar chromosome form, which isparticularly striking in Hybrida, Peregrina, Minutiflora, Cretica, Amurensis andPisiformis types, is that of size which are clearly observed between chromosomeswithin the nucleolar types as a whole (Figs. 1H, 1I; 2B-D, and2F).
Phylogenetic analysis.
Ten (Vicia vicioides, V. hybrida, V. sericocarpa, V. peregrina, V. michauxii, V.pannonica ssp. purpurascens, V. pannonica ssp. pannonica, V. unijuga, V. anatolica,V. hajastana] and five (V. galeata, V. esdraelonensis, V. hajastana, V. melanops, V.faba) species were separated from others because they form different groups,possessing either two pairs of nucleolar chromosomes within the complement orthe nucleolar chromosome type arisen through Robertsonian fusion, respectively,and an analysis was made on the remaining 53 species.
With few exceptions, the nucleolar chromosome variation among the speciesshowed a good correlation with taxonomic classification. For example, sectionCracca, except for V. palaestina and V. minutijlora, section Vicia, except for V.grandiflora var. kitaebeliana, V. pyrenaica and V. incisa, and section Faba, exceptfor V. bithynica, form three distinct clusters (Fig. 4).

Fig. 3. — Classification and evolution of nucleolar chromosomes in the genus Vicia. The line across theopen space marks secondary constriction.
DISCUSSION
The present comparative data on 67 Vicia species/subspecies have revealedgreat variety in the nucleolus organizing chromosomes. It is a general belief thatin comparison to many detailed accounts given in literature of this type of variationbetween species (VED BRAT 1965; RAINA and KHOSHOO 1971; SHARMA et al. 1986; KOULand WAFAI 1980; LANGER and KOUL 1983a, b), the variation affecting these markerchromosomes in Vicia species is much more substantial. There are two categoriesof diversity in the nucleolar chromosomes in the Vicia species. First, thenumerical nucleolar chromosome changes range from one occasionally to twodissimilar pairs. The occurrence of more than one pair of nucleolar chromosomesis not uncommon, and it has been reported in a number of diploid plant species (VEDBRAT 1965; MAGGINI and GARBARI 1977; MAGGINI and DE DOMINICIS 1977; RAJHATHY1983; SHARMA et al. 1986). According to MAGGINI and GARBARI (1977) theinterspecific differences in the
NUCLEOLAR CHROMOSOME VARIATION IN VICIA 143

144 BISHT, KESAVACHARYULU and RAINA
number of nucleolar chromosomes in Allium species is related to the amount ofrDNA, breeding system and ecogeographic distribution of species. MENSINKAI(1940) considered it as the secondary balanced condition. In Vicia there isevidence to suggest that major cytogenetic differentiation and/or structuraldivergence of karyotypes is partly due to hybridization, and segmental inter-changes (SCHAFER 1973; ROLLINGS and STAGE 1974; LADIZINSKY and TEMKIN 1978;LADIZINSKY 1981; YAMAMOTO 1986; LIU et al. 1988; RAINA et al 1989; HANELT andMETTIN 1989) by the complete elimination of heterozygotes and establishment ofhomozygotes. As a consequence of such event(s) it is therefore, not surprising tofind numerical variations in the nucleolar chromosomes of Vicia species.
The second category of diversity lies in the existence of great variety in thestructure of nucleolar chromosomes. It reveals a wealth of interspecific variationand suggests that karyotype evolution involving these marker chromosomes isvery active. There is substantial variation, in the satellite size from virtuallyabsent to highly conspicuous, location of primary and secondary constrictionsfrom terminal to mid region, size of middle arm flanked by two constrictionsfrom very small to about 90 per cent of the total size, position of nucleolusorganizer in either of the two arms and finally number of satellites from one totwo per chromosome. Barring the two subspecies of V. pannonica, comparisons ofthe nucleolar chromosomes observed in the present study with those presented byother workers show a general agreement in the occurrence of uniform type(s) ofnucleolar chromosomes within a species.
Fig. 3 provides a diagrammatic representation of scheme which illustratesand summarizes the evolutionary pathways that might have occurred in thesemarker chromosomes through inversion, deletion and interchange events. Thenucleolar chromosome morphology of Villosa type, as mentioned before, possiblyrepresents a state more primitive than that of types Sativa to Anatolica. NICOLOFFet al. (1977a, b), ANASTASSOVA-KRISTEVA et al. (1977), MICHAELIS and RIEGER (1971) andMATSUURA (1942) have given experimental proof that the morphological changeswhich occur in the reconstructed nucleolar chromosomes result from reciprocaltranslocation and inversion. In the case of Vicia species the morphology of NORchromosomes in the Hybrida, Cretica, Hyr-canica and Faba types appears similarto what should be expected from a reciprocal or interstitial translocation orRobertsonian fusion in the chromosomes which are similar to the Villosa type.Similarly the Sativa type could arise through pericentric inversion, as inCichorieae (STEBBINS 1958), of the Villosa type. The Monantha type has ashorter satellite than the Villosa type and the secondary constriction divides thearm into two equal halves.
Presumably it has arisen by means of either deletion or reciprocal translo-cation, more probably the latter. Subsequent to the above changes in the Villosatype there have been further alterations in the derived nucleolar chromosomes asdepicted in Fig. 3. As is clear from Fig. 3, inversion in the

Fig. 4. — Dendrogram showing relationships among 53 Vicia species based on nucleolar chromosome armratio, and costructed by Unweighted Centroid method.
nucleolar arm also seems to have contributed to the structural changes. Whiledetails about the chromosome complements of the various species are not j relevanthere, mention may be made that chromosomal repatterning especially pericentricinversion and translocation have as a whole played a very important role in thedivergence and evolution of the Vicia species (RAINA and REES 1983a, b; RAINA etal. 1989; RAINA 1990; HANELT and METTIN 1989; LADI-ZINSKY and TEMKIN 1978;LADIZINSKY 1978; ROLLINGS and STAGE 1974).
The Anatolica type unusually possesses four NORs and four satellites. Sucha type might have arisen as a result of Robertsonian fusion between twodissimilar nucleolar chromosomes, presumably of Macrocarpa and Eristalioidestypes.
NUCLEOLAR CHROMOSOME VARIATION IN VICIA 145

146 BISHT, KESAVACHARYULU and RAINA
REFERENCES
ALLKIN R., GOYDER D.J., BISBY F.A. and WHITE R.J., 1986. — Names and synonyms of species andsubspecies in the Vicieae. Vicieae Database Project, Publication 7, Issue 3.
ANASTASSOVA-KRISTEVA M., NICOLOFF H., KUNZEL G. and RIEGER R., 1977. — Nucleolus formation in structurallyreconstructed barley karyotypes with six satellite chromosomes. Chromosoma, 62: 111-119.
CHOOI W.Y., 1971. — Variation in nuclear DNA content in the genus Vicia. Genetics, 68: 195-211.HANELT P. and METTIN D., 1989. — Biosystematics of the genus Vicia L. (Leguminosae). Annu. Rev.
Ecol. System., 20: 199-223.HARLAN J.R., 1956. — Theory and dynamics of grassland griculture. D. Van. Noztrand Co. Inc.,
Princeton.HOLLINGS E. and STAGE C.A., 1974. — Karyotype variation and evolution in the Vicia saliva aggregate.
New Phytol., 73: 195-208.JOHN B. and LEWIS K.R., 1968. — The chromosome complement. Protoplasmatologia, VI A. Springer -
Verlag, Wien.KOUL A.K. and WAFAI B.A., 1980. — Chromosome polymorphism and nucleolar organization in some
species of Fritillaria Linn. Cytologia, 45: 675-682.KUPICHA F.K., 1976. — The infrageneric structure of Vicia. Notes Roy. Bot. Gard. Edinburgh, 34: 287-
326.LA COUR L.F., 1950. — Compound constriction. Appendix to hybridity selection in Campanula.
Heredity, 4: 217-248.LADIZINSKY G., 1978. — Chromosomal polymorphism in wild populations of Vicia saliva L. Caryologia,
31: 233-241.—, 1981. — Consequences of hybridization in Vicia saliva aggregate. Heredity, 47: 431-438.LADIZINSKY G. and TEMKIN R., 1978. — The cytogenetic structure of Vicia sativa aggregate. Theor. Appl.
Genet., 53: 33-42.LANGER A. and KOUL A.K., 1983a. — Studies on nucleolus and nucleolar chromosomes in angiosperms.
VII. Nature of nucleolar chromosome polymorphism in Allium cepa var. viviparum (Metzg.) Alef.Cytologia, 48: 323-332.
—, 1983b. — Studies on nucleolus and nucleolar chromosomes in angiosperms. IX. Lilium Linn.Cytologia, 48: 519-526.
Liu Y.A., YAMAMOTO K. and RAINA S.N., 1988. — Chromosome constitution of two karyotypes in Viciamacrocarpa. Japan. Jour. Breed., 38: 35-42.
MAGGINI F. and DE DOMINICIS R.I., 1977. — The ribosomal RNA gene number and the length of thenucleolar secondary costrictions in Bellevalia romana and B. dubia (Liliaceae). A possible correlation.Caryologia, 30: 97-103
MAGGINI F. and GARBARI F., 1977. — Amounts of ribosomalDNA in Allium (Liliaceae). Plant System.Evol.,128: 201-208.
MATSUURA H., 1942. — Chromosome studies on Trillium kamtschaticum Pall. XVI. Alternations ofthenucleolus chromosome system due to irradiation. Cytologia, 12: 271-288.
MAXTED N., CALLIMASSIA M.A. and BENNETT M.D., 1991. — Cytotaxonomic studies of East Mediterranean Viciaspecies (Leguminosae). Plant System. Evol., 177: 221-234.
MENSINKAI S.W., 1940. — Cytogenetic studies in genus Allium. Jour. Genet., 39: 1-45MICHAELIS A. and RIEGER R., 1971. — New karyotypes of Vicia faba L. Chromosoma, 35: 1-8.NARAYAN R.K.J., RAMACHANDRAN C. and RAINA S.N., 1985. — The distribution of satellite DNA in thechromosome
complements of Vicia species (Leguminosae). Genetica, 66: 115-121NICOLOFF H., ANNASTASSOVA-KRISTEVA M. and KUNZEL G., I977a. — Changes in nucleolar organizer
activity due to segmental interchanges between satellite chromosomes in -barley. Biol. Zbl., 96: 223-227.
NICOLOFF H., ANNASTASSOVA-KRISTEVA M., KUNZEL G. and RIEGER R., 1977b. — The behaviour ofnucleolus organizers in structurally changed karyotypes of barley. Chromosoma, 62: 103-109.
RAINA S.N., 1990. — Genome organisation and evolution in the genus Vicia. In: S. Kawano (ed.)."Biological approaches and evolutionary trends in plants", pp. 183-201, Academic Press,London.

NUCLEOLAR CHROMOSOME VARIATION IN VICIA 147
RAINA S.N. and BISHT M.S., 1988. — DAM amounts and chromatin compactness in Vicia. Genetica, 77:65-77.
RAINA S.N. and KHOSHOO T.N., 1971. — Cytogenetics of tropical bulbous ornamentals. II. Variation inmitotic complement in Crinum. Nucleus, 14: 23-39
. RAINA S.N. and OGIHARA Y., 1995. — Ribosomal DNA repeat unit polymorphism in 49 Vicia species.Theor. Appl. Genet., 90: 477-486.
RAINA S.N. and NARAYAN R.K.J., 1984. — Changes in DNA composition in the evolution of Viciaspecies. Theor. Appl. Genet. 68: 187-192.
RAINA S.N. and REES H., 1983a. — DNA variation between and within chromosome complements ofVicia species. Heredity, 51: 335-346.
—, 1983b. — Variation in chromosomal DNA associated with the evolution of Vicia species. In: P.E.Brandham and M.D. Bennett (eds), "Kew chromosome conference. II", pp. 360, George Alienand Unwin, London.
RAINA S.N., YAMAMOTO K. and MURAKAMI M., 1989. — Intraspecific hybridization and its bearing onchromosome evolution in Vicia narbonensis (Fabaceae). Plant System. Evol., 167: 201-217.
RAJHATHY T., 1983. — The chromosomal architecture of Avena. In: M.S. Swaminathan, P.K. Guptaand U. Sinha (eds), "Cytogenetics of Crop Plants", pp. 283-307, Macmillan India Limited,Delhi.
SCHAFER H.I., 1973. — Zur Taxonomie der Vicia narbonensis Gruppe. Kulturpflanze, 21: 211-273. SHARMA P.K., LANGER A. and KOUL A.K., 1986. — Studies on nucleolus and nucleolar chromosomes in
angiosperms. XL Plantago L. Ann. Bot., 57: 145-154. STEBBINS G.L., 1950. — Variation and evolution in plants. Columbia Univ. Press, New York.—, 1958. — Longevity, habitat and release of variability in the higher plants. Cold Spring Harbour Symp.
Quater. Biol., 23: 365-378.—, 1971. — Chromosomal evolution in higher plants. Addison - Wesley Publishing Co., London.VED BRAT S., 1965. — Genetic system in Allium. I. Chromosome variation. Chromosoma, 16: 486-499. YAMAMOTO K., 1973. — Karyotaxonomical studies on Vicia L. On the karyotype and character of someannual species of Vicia. Japan Jour. Genet., 48: 315-327.—, 1986. — Interspecific hybridization among Vicia narbonensis and its wild relatives. Biol. Zentralbl.,
105: 181-197.
Received 10 February 1998; accepted 17 March 1998