numerical taxonomy of vibrio isolated areas that are … · table 1. designation and source...
TRANSCRIPT

Vol. 17, No. 6JOURNAL OF CLINICAL MICROBIOLOGY, June 1983, p. 1102-11130095-1137/83/061102-12$02.00/0Copyright C 1983, American Society for Microbiology
Numerical Taxonomy of Vibrio cholerae and Related SpeciesIsolated from Areas That Are Endemic and Nonendemic for
CholeraLORE ANNE McNICOL, SHOUMYA P. DE,t JAMES B. KAPER,4 PAUL A. WEST, AND RITA R.
COLWELL*
Department of Microbiology, University of Maryland, College Park, Maryland 20742
Received 19 August 1982/Accepted 13 December 1982
A total of 165 strains of vibrios isolated from clinical and environmental sources
in the United States, India, and Bangladesh, 11 reference cultures, and 4duplicated cultures were compared in a numerical taxonomic study using 83 unitcharacters. Similarity between strains was computed by using the simple match-ing coefficient and the Jaccard coefficient. Strains were clustered by unweightedaverage linkage and single linkage algorithms. All methods gave similar clustercompositions. The estimated probability of error in the study was obtained from a
comparison of the results of duplicated strains and was within acceptable limits. Atotal of 174 of the 180 organisms studied were divided into eight major clusters.Two clusters were identified as Vibrio cholerae, one as Vibrio mimicus, one asVibrio parahaemolyticus, three as Vibrio species, and one as Aeromonas hydro-phila. The V. mimicus cluster could be further divided into two subclusters, andthe major V. cholerae group could be split into seven minor subclusters.Phenotypic traits routinely used to identify clinical isolates of V. cholerae can beused to identify environmental V. cholerae isolates. No distinction was foundbetween strains of V. cholerae isolatedstrains from nonendemic regions.
from regions endemic for cholera and
Vibrio cholerae is the causative agent of thedisease cholera, which is characterized by se-vere and watery diarrhea. The disease is endem-ic in Asia, and the current seventh pandemic isassociated with the El Tor biovar of V. choleraeserovar 01 (22). Antigenic structure has as-sumed fundamental importance in the identifica-tion of V. cholerae. Serovar 01 is associatedwith pandemic cholera (20), whereas strainspossessing antigens other than 01 are classifiedas non-O1 V. cholerae and are implicated insporadic cases of human disease (4).
Traditionally, the genus Vibrio has been sub-divided on the basis of sugar fermentation reac-tions. Heiberg (18) and Smith and Goodner (35)used the sugars sucrose, D-mannose, and L-arabinose to differentiate members of the genusVibrio into eight subgroups. V. cholerae 01strains fall into group I (these strains fermentsucrose and mannose, but not arabinose),whereas non-O1 V. cholerae strains are found inHeiberg groups I and II (fermenting only su-
t Present address: National Institute of Cholera and EntericDiseases, Calcutta, India.
t Present address: School of Medicine, Center for VaccineDevelopment, University of Maryland, Baltimore, MD 21201.
crose) and V (fermenting only mannose). Hei-berg group V organisms are distinct from V.cholerae in other phenotypic characters (8, 11,16) and may belong to a separate species, Vibriomimicus (12).
Since the serotaxonomic definition of V. chol-erae (20), it has become apparent that non-O1and 01 V. cholerae strains are widely distribut-ed in natural waters, even where cholera is notan endemic disease (8, 21, 24, 27). Recent taxo-nomic studies have compared the phenotypes ofclinical and environmental strains of V. choleraeisolated from areas that are nonendemic forcholera (11, 13, 14). However, an extensivetaxonomic comparison between V. choleraestrains isolated from regions that are endemicand nonendemic for cholera has not been made.In the present study we investigated whetherphenotypic differences exist between clinicaland environmental V. cholerae strains isolatedfrom countries with different disease experi-ences. Organisms isolated from ChesapeakeBay, Md., and from the Buriganga River region,Bangladesh, as well as clinical isolates from boththe United States and Bangladesh, were com-pared in an 83-character numerical taxonomicstudy. Strains of Vibrio parahaemolyticus fromenvironmental and clinical sources in India were
1102
on March 23, 2020 by guest
http://jcm.asm
.org/D
ownloaded from

COMPARATIVE STUDY OF V. CHOLERAE 1103
also examined, because this species is phenotyp-ically similar to other sucrose-negative vibriosused in the study.
MATERIALS AND METHODSStrains. All strains used are listed in Table 1.(i) Isolation and characterization of environmental
strains. The Vibrio strains from Bangladesh (BVstrains) consisted of 75 environmental isolates fromthe International Center for Diarrheal Disease Re-search study areas at Matlab and Dacca. Samples werecollected between March and May 1977 from water,plants, and sediment, and plated onto thiosulfate-citrate-bile salts-sucrose agar (Oxoid Ltd.) withoutenrichment. Pure cultures were examined by the fol-lowing preliminary identification tests: gelatinase, in-dole, and urease production; 0/129 sensitivity; motil-ity; fermentation of mannitol, sucrose, mannose, andarabinose; ornithine decarboxylase, lysine decarbox-ylase, and arginine dihydrolase; and growth in 0, 3, 8,and 10% NaCl. Cultures were stored on brain heartinfusion agar (Difco Laboratories) under oil at 20°C(31, 32).The Vibrio (V) set of 31 strains (V39 to V88) was
isolated from water, sediment, plankton, and shellfishin the Chesapeake Bay between 1976 and 1978 (7, 31).After enrichment in alkaline peptone water, sampleswere plated on thiosulfate-citrate-bile salts-sucroseagar. V. cholerae was presumptively identified by thefollowing tests: oxidase; gelatinase, H2S, and ,-galac-tosidase production; growth in 0o NaCl; glucosefermentation; lysine decarboxylase; and arginine dihy-drolase.
Strains MP18104, MP19812, and Y432 were environ-mental serovar 01 strains, provided by R. B. Sack.Most strains of V. cholerae had been previously testedfor the presence of DNA coding for cholera toxin,using the pEWD299 probe (10) in colony hybridization(J. W. Newland, M. J. Voll, and L. A. McNicol,submitted for publication).Five isolates from a Chesapeake Bay Aeromonas
hydrophila collection (strains 19, 28, 56, 75, and 85)were included as reference controls on the sensitivityof the clustering procedures (25, 31). Eighteen strainsisolated in 1976 from water, fish, seafood, and flies inthe Calcutta, India, area had been identified as V.parahaemolyticus (34).
(ii) Isolation and characterization of clinical strains.One V. cholerae serovar 01 strain (Sloccum Creek)and 12 non-O1 isolates received from the Centers forDisease Control, Atlanta, Ga., were included in thestudy. R. B. Sack kindly provided three clinicalstrains of V. cholerae (strains 30167, 30050, and62746). Seventeen strains of V. parahaemolyticus hadbeen isolated from overt cases of human gastroenteri-tis and from asymptomatic human carriers and weretested for their Kanagawa phenomenon reaction andserovar (34).
(iii) Reference strains. Eleven reference cultureswere used.Media and cultural conditions. The 180 strains were
maintained as working stocks on tryptic soy agar(Difco) slants containing a final concentration of 1%(wt/vol) NaCl under sterile mineral oil at 20°C. Unlessotherwise stated, all tests were performed at 30°C withtryptic soy agar as the basal medium, and results were
recorded after incubation for 24 and 48 h. The concen-tration of NaCI in all media was raised to 1% (wt/vol)unless otherwise stated.Taxonomic tests. Colonial morphology, pigmenta-
tion traits, motility, cell micromorphology, and theGram stain reaction were determined as described byMallory et al. (30). Resistance to the antibiotics ampi-cillin (10 ,ug), chloramphenicol (30 ,ug), kanamycin (30,ug), and polymyxin B (300 U) was determined by theagar disk diffusion method (Sensi-Discs; BBL Micro-biology Systems) (1). Growth at 5, 10, 37, and 42°C;growth in 3, 7, and 10% (wt/vol) NaCI; sensitivity to200 ,ug of 0/129 (Sigma Chemical Co.) per ml, 0.001%(wt/vol) neutral red, 0.001% (wt/vol) crystal violet,0.00125% (wt/vol) brilliant green, and 0.01% (wt/vol)sodium dodecyl sulfate; and growth at pH 4.5, 8.0, 9.0,and 10.0 were determined on tryptic soy agar medium.For the latter, the medium was adjusted to the appro-priate pH with 1 M NaOH or 1 M HCI.For the carbohydrate fermentation tests, a final
concentration of 2% (wt/vol) D-adonitol, L-arabinose,n-butanol, D-cellobiose, dextrin, dulcitol, ethanol, m-erythritol, D-fructose, D-galactose, glycerol, m-inosi-tol, D-lactose, D-mannitol, D-mannose, melezitose,melibiose, raffinose, L-rhamnose, salicin, sorbitol, su-crose, trehalose, or xylose was used. These sub-stances were either incorporated into the Hugh-Leif-son agar (Difco) basal medium and autoclaved or filtersterilized and then added to autoclaved basal medium.Butanol and ethanol were sterile as supplied. Thecarbohydrate media were dispensed into multiwelldishes (Costar) and inoculated with a multipoint inocu-lator (29).
Tests for utilization of compounds as a sole carbonsource were done with multiwell dishes and the basalyeast-nitrogen base medium described by Stevenson(38), following the procedures of Austin et al. (2). Thecompounds tested were acetate, ,-alanine, alginate, L-aspartate, citrate, formate, L-glutamate, glycine, inu-lin, DL-lactate, L-lysine, L-malonate, L-mucate, L-phenylalanine, L-proline, propanol, pyruvate, L-serine, tartrate, and L-tryptophan, at a finalconcentration of 0.2% (wt/vol or vol/vol). The mediumwas examined visually for growth at 14 days. Thepositive control was 0.2% (wt/vol) D-glucose, and thenegative control was the medium without any addi-tional carbon source.The hydrolysis of Tween 20, Tween 40, Tween 60,
Tween 80, starch, and casein was measured as de-scribed by Colwell and Wiebe (9). The methods usedto determine production of levan, catalase, gas fromglucose and lactose, indole, gelatinase, lecithinase,oxidase, H2S, phosphatase, and growth and acid oniodoacetate have been described previously (6). Argi-nine dihydrolase and lysine, ornithine, and histidinedecarboxylases were detected by using Moeller decar-boxylase base medium (Difco). Growth on thiosulfate-citrate-bile salts-sucrose agar, nitrate reduction, themethyl red test, DNase production, phenylalaninedeaminase, and urease were determined with the ap-propriate culture medium (Difco). The methods fordetecting degradation of allantoin (17), esculin hydrol-ysis (36), chitinase production (19), and P-galacto-sidase (28) have been described previously. Hemolysiswas scored after growth on 5% sheep blood agar(BBL).Computer analysis. All of the characters existed in
VOL. 17, 1983
on March 23, 2020 by guest
http://jcm.asm
.org/D
ownloaded from

TABLE 1. Designation and source of strains
Cluster Sourcea Serovarb or iden-assigned Strain tfctoSample type Location tification
1 (V. cholerae) MP19812 W B 01Y432 01MP18104 0130167ATCC Stool I, type strain 01
14035c30050V41 W Fort McHenry, CB 23C6770 Stool Alabama Non-01V72 W Magothy River, CB 106C6761 Stool Georgia Non-01C5852 Stool Texas Non-01V64 W Hawkins Point, CB 37V65 W Hawkins Point, CB 31V69 W North Point, CB 0162746BV68 W Mirpur, BV43A Duplicate
strainBV49 W Matlab, BBV40 W Meharon, BBV75 W Tejgaon, BBV19 W Meharon, BV85 W Chester River, CBBV84 W Dacca University campus
tank, BV87 W Chester River, CBBV47 W Matlab, BV88 W Chester River, CBBV23 W Meharon, BBV57 W Matlab, BBV39 W Meharon, BBV41 W Meharon, BBV56 W Matlab, BBV10 W Meharon, BBV24 W Meharon, BBV30 W Meharon, BBV29 W Meharon, BBV32 W Meharon, BBV87 W Dacca central jail, BC6487 Blood TexasV43 W Colgate Creek, CB 94V40 W Chester River, CB 94BV6 W Meharon, BV42 W Colgate Creek, CB 999V53 W Patuxent River, CB 19V56 W Bay Bush Point, CB 23BV83 W Dacca University campus
tank, BV61 W Fort McHenry, CB 18BV80 W Tejgaon, BBV81 W Tejgaon, BBV76 W Tejgaon, BBV86 W Dacca central jail, BBV88 W Husaini Dalan, BV58 W Eastern Bay, CB 999V59 W Eastern Bay, CBBV85 W Dacca central jail, BV50 W Potomac River, CB 33V47 W Choptank River, CB 999V44 W Colgate Creek, CB 31BV14 W Meharon, B
1104 McNICOL ET AL. J. CLIN. MICROBIOL.
on March 23, 2020 by guest
http://jcm.asm
.org/D
ownloaded from

COMPARATIVE STUDY OF V. CHOLERAE 1105
TABLE 1-Continued
Cluster Sourcea Serovar' or iden-assigned Strain Sample type Location tification
BV45 W Meharon, BBV79 W Begumbari, BBV28 W Meharon, BBV55 W Matlab, BV39 W Chester River, CB 343V55 W Bay Bush Point, CB 19V76 W North Point, CB 333V83 Plankton Chester River, CB 115V79 W Chester River, CB 19V80 W Chester River, CB 333V81 W Chester River, CB 19BV43 W Meharon, BBV34 W Meharon, BBV73 W Mirpur, BBV42 W Meharon, BBV46 W Meharon, BBV37 W Meharon, BBV70 W Mirpur, BBV67 W Gulshan, BBV66 W Gulshan, BBV7 W Meharon, BV65A Duplicate
strainBV27 W Meharon, BC7326 Stool California Non-01569Bc Stool India 01BV92 W Tejgaon, BC7037 Stool Arizona Non-01SLCRK Sloccum Creek 01C9414 Sputum Texas Non-01C7431 Blood North Carolina Non-01D1560 Abdominal North Carolina Non-01
fluidC4752 Gall blad- California Non-01
derV63 W Hawkins Point, CB 333BV22 W Meharon, BBV31 W Meharon, BBV35 W Meharon, BC7325 Stool California Non-01BV89 W Buriganga River at Shaddar
Ghat, B
2 (V. mimicus)2A SN72 Crab I 06:K18
BV16 W Meharon, BBV21A Duplicate
strainBV20A Duplicate
strainBV20 W Meharon, BBV21 W Meharon, BBV18 W Meharon, BBV8 W Meharon, BBV74 W Mirpur, B
2B BV82 W Dacca University campustank, B
BV52 W Matlab, BBV17 W Meharon, BBV65 W Gulshan, BBV53 W Matlab, BC6713 Blood Kentucky Non-01
VOL. 17, 1983
on March 23, 2020 by guest
http://jcm.asm
.org/D
ownloaded from

1106 McNICOL ET AL.
TABLE 1-Continued
Cluster Sourcea Serovar' or iden-Sample type Location tification
3 (V. cholerae)
4 (Vibrio spp.)
5 (V. parahaemo-lyticus)
BV51V54BV72BV93BV91
BV77BV90NIH 41cBV69
V46V45V49ATCC
33149C
W232
W225SN57SN56A348
D36
887/77
912/77
W177SN70SN36SN40SN47A271
A385
D23
SN46D17
A7
A20
W663A127
W243A148
SN67214/7A165
D51
W54
wwwww
SedimentwStoolw
wSedimentOystersEel
w
wFishFishHuman
caseHuman
caseHuman
carrierHuman
carrierwCrabPrawnPrawnFishHuman
caseHuman
caseHuman
caseFishHuman
caseHuman
caseHuman
casewHuman
casewHuman
caseCrabHuman
carrierHuman
caseHuman
casew
Matlab, BJames River, CBMirpur, BTejgaon, BTejgaon, B
Tejgaon, BTejgaon, B
Mirpur, B
Choptank River, CBFort McHenry, CBEastern Bay, CBJapan
I, KP+
I, KP-I, KP-I, KP-I, KP+
I, KP+
I, KP-
I, KP+
I, KP-I, KP-I, KP-I, KP-I, KP-I, KP+
I, KP+
I, KP+
I, KP-I, KP+
I, KP+
I, KP+
I, KP+I, KP-
I, KP-I, KP+
I, KP-I, KP+
I, KP-
I, KP+
I, KP-
14
999999999
05:K15
010:K52010:K2405:K1504:K8
03:K57
05:K17
05:K15
02:K2804:K9010:K5203:K3301:K3804:K12
08:K22
04:K55
04:K1203:K7
05:K17
04:K4
04:K1204:K9
03:K3001:K56
04:K5308:K22
010:K52
04:K49
02:K28
J. CLIN. MICROBIOL.
on March 23, 2020 by guest
http://jcm.asm
.org/D
ownloaded from

COMPARATIVE STUDY OF V. CHOLERAE 1107
TABLE 1-Continued
Cluster Source' Serovarb or iden-assigned Strain Sample type Location tification
M23/77W424M437/77915/77
912/77
ATCC17802C
BV48BV38BV15
6 (Vibrio spp.)
7 (A. hydrophila)
BV4BV11BV26BV78BV12
FlywFlyHuman
carrierHuman
carrierStool
wwPlants
wwwSedimentw
I, KP-I, KP-I, KP-I, KP-
I, KP+
Japan, KP-
Matlab, BMeharon, BMeharon, B
Meharon, BMeharon, BMeharon, BTejgaon, BMeharon, B
05:K17O10:K24O10:K24O1:K1
CF5:K15
O1:K1
CBAh 19dCBAh 56CBAh 75CBAh 28CBAh 85
8 (Vibrio spp.)
Unclustered
ATCC14181c
SAK4C
BV5ATCC14033CATCC
33148ET7717ATCC
9071cBV50
wStool
Eel
Eel
w
V. anguillarum
V. parahaemo-lyticus
Meharon, BSinai
Japan
Japan
V. cholerae
"V. anguilli-cida"
A. hydrophila
Meharon, BI Abbreviations: W, water; CB, Chesapeake Bay, Md.; I, India; B, Bangladesh; KP, Kanagawa phenomenon.b For V. cholerae, serotyping was by the Vibrio Reference Laboratory, Jefferson Medical School, Philadel-
phia, Pa., for V. parahaemolyticus, serotyping was performed with a commercial kit (Toshiba Chemical Co.,Tokyo, Japan).cType or reference strain. ATCC, American Type Culture Collection, Rockville, Md.; NIH, National
Institutes of Health, Bethesda, Md.dCBAh, Chesapeake Bay A. hydrophila collection.
one of two mutually exclusive states, scored as + or-. The test results were recorded as 0 for a negativeresult, 1 for a positive result, and 9 for noncomparableor missing data. Resistance to antibiotics and antimi-crobial agents was scored as a positive reaction. Thecombined data were analyzed for the presence ofnondiscriminatory characters and then entered into aUnivac 1108 computer. Resemblances between strainswere calculated by using simple matching and Jaccardcoefficients. The similarity matrix was transformedinto two-dimensional similarity structures (pheno-grams) by the use of single linkage and unweighted
average linkage methods on the TAXAN 6 programpackage. The IGPS3 program was used to calculatethe trait frequency results for each cluster in thephenograms.
RESULTSNon-discriminatory characters and test error.
All isolates were oxidase-positive, short, gram-negative rods with a fermentative physiology.All strains produced catalase; grew at pH 8.0,15°C, 37°C, and in the presence of 3% NaCl;
VOL. 17, 1983
on March 23, 2020 by guest
http://jcm.asm
.org/D
ownloaded from

1108 McNICOL ET AL.
TABLE 2. Frequencies of positive characters found in major clusters and subclustersa
CharacterFrequency (%) in cluster:
1 2A 2B 3 4 5 6 7(n = 96) (n = 9) (n = 11) (n = 4) (n = 4) (n = 38) (n = 5) (n = 5)
Colony morphologyWhiteYellowColorlessTranslucentCircularConvexMucoid<1-mm diameter
Growth at:50C
420CpH 4.5pH 9.0pH 10.0
Acid production from:D-AdonitolL-Arabinosen-ButanolD-CellobioseDextrinEthanolm-ErythritolD-FructoseD-Galactosem-InositolD-MannitolD-MannoseMelibioseMelezitoseRaffinoseL-RhamnoseSalicinSorbitolSucroseTrehalose
Decarboxylation of:L-ArginineL-LysineL-Ornithine
Biochemical testsIndole productionMethyl redNitrate reductionONPGbPhenylalanine deaminase
Growth in presence of:cBrilliant green (0.00125%)Crystal violet (0.001%)SDSd (0.01%)NaCl (7%)NaCl (10%)
Resistance to:Ampicillin (10 p.g)
3 0 0 0 0 5 0 200 0 0 0 50 0 0 20
97 100 100 100 50 95 100 6079 100 91 75 100 74 100 0100 100 100 100 100 92 100 10084 100 91 75 75 84 60 404 0 9 0 0 8 0 2017 11 27 25 0 5 0 20
0 0 0 0 0 0 0 4080 100 91 50 0 10 80 100100 100 100 100 25 97 80 80
0 0 0 0 0 0 0 099 100 100 100 100 100 100 10094 100 82 100 100 97 100 100
0 0 0 0 0 3 0 03 0 0 0 0 82 80 80
18 11 45 50 25 34 20 09 89 100 50 100 92 0 80
100 89 91 100 100 100 100 1001 0 0 0 0 18 0 01 0 0 0 0 0 0 0
100 100 91 100 100 100 100 10092 100 100 100 100 100 100 1000 0 0 0 0 0 0 092 100 100 75 50 97 100 10091 100 100 100 100 100 80 1000 0 0 0 25 13 0 00 0 0 0 0 0 0 00 0 0 0 0 0 0 01 11 0 0 0 3 0 01 0 0 0 100 0 80 00 0 0 0 0 0 0 0
97 0 9 75 0 8 100 60100 100 100 100 100 100 100 100
2 0 9 25 0 0 40 10099 100 91 100 100 89 20 6098 100 100 100 100 100 0 0
100 100 100 100 75 95 60 10034 22 54 25 0 3 0 0100 100 100 100 0 30 100 10056 0 65 75 75 3 100 00 0 0 25 0 3 20 20
6 022 660 00 110 11
0 25 0 0 00 25 50 0 00 0 0 0 00 0 0 100 600 0 0 42 0
00000
86 78 91 100 75
J. CLIN. MICROBIOL.
0 40 75
on March 23, 2020 by guest
http://jcm.asm
.org/D
ownloaded from

COMPARATIVE STUDY OF V. CHOLERAE 1109
TABLE 2-Continued
Character
Kanamycin (30 ,ug)Polymyxin B (300 U)
Frequency (%) in cluster:
1 2A 2B 3 4 5 6 7(n = 96) (n = 9) (n = 11) (n = 4) (n = 4) (n = 38) (n = 5) (n = 5)
98 89 81 100 100 97 100 10092 100 100 75 75 95 100 100
Sensitivity to:0/129 (200 pLg/ml)
Degradation of:EsculinCaseinChitinDNAGelatinLecithinStarchTween 40Tween 60Tween 80TyrosineUrea
Utilization of and growth on 0.2%(wt/vol):AcetateP-AlanineL-AspartateCitrateL-GlutamateGlycineInulinlodoacetateDL-LactateL-LysineMalonateMucateL-PhenylalanineL-SerineTartrateTyptophane
Miscellaneous testsBeta-hemolysisGas from glucoseLevan productionMotility
97 100 91 75 100 66 20 0
4 0 0 50 100 0 60 091 67 64 100 75 89 80 10083 44 91 100 0 5 60 100
100 100 100 100 100 100 100 10099 89 100 75 75 87 80 8099 100 100 75 100 97 100 6093 11 9 75 100 100 100 100100 100 100 100 100 100 100 100100 89 82 100 100 97 80 100100 100 82 75 100 100 80 100
1 44 36 25 0 0 0 200 0 0 0 0 5 0 0
75 100 36 75 75 89 100 10092 89 64 75 100 97 100 80100 100 100 100 100 95 100 8012 11 18 25 0 0 40 0
100 100 100 100 100 100 100 10095 89 91 75 100 97 100 100100 100 100 100 100 87 100 10098 100 100 100 100 100 100 1009 33 0 0 0 8 0 0
94 56 91 75 100 100 100 10081 79 82 25 100 97 100 10069 56 45 0 100 97 60 10011 0 0 0 25 42 60 20
100 100 100 100 50 100 100 10059 89 18 50 100 100 80 8098 100 64 75 100 100 100 100
63 11 91 0 0 0 80 401 0 0 0 0 5 0 200 0 0 0 0 0 0 0
100 100 100 100 100 100 100 100
a All strains were gram negative; formed catalase and not histidine decarboxylase; grew at pH 8.0, 15C, 37°C,and in the presence of sodium chloride (3%); produced acid from glycerol (2%, vol/vol); and were sensitive toneutral red (0.001%) and chloramphenicol (30 ,ug).
b ONPG, o-Nitrophenyl-j3-D-galactopyranoside.All percentages are expressed as wt/vol.
d SDS, Sodium dodecyl sulfate.
produced acid from glycerol; and were sensitive consistently. Other characters that were re-to neutral red and chloramphenicol. No strain moved due to poor growth compared with theproduced histidine decarboxylase. Highly sub- negative control or weak indicator changes werejective traits were also removed from the data production of acid from dulcitol, lactose, andmatrix after comparison of duplicate strains with xylose; production of gas from lactose; H2Stheir matches. production; phosphatase; degradation of Tween
Certain colonial morphology and cell micro- 20 and allantoin; and growth on alginate, for-morphology characters proved difficult to score mate, proline, propanol, and pyruvate as a sole
VOL. 17, 1983
on March 23, 2020 by guest
http://jcm.asm
.org/D
ownloaded from
1110 McNICOL ET AL.
carbon source. A total of 39 tests were removedfrom the data matrix, and the final n x t tablecontained 180 bacterial strains (n) and 83 unitcharacters (t). The inclusion of four duplicatepairs of strains measured the test error associat-ed with this study (37). The average probabilityof an erroneous two-state character result was4.1%, which does not adversely affect clusteringpatterns.
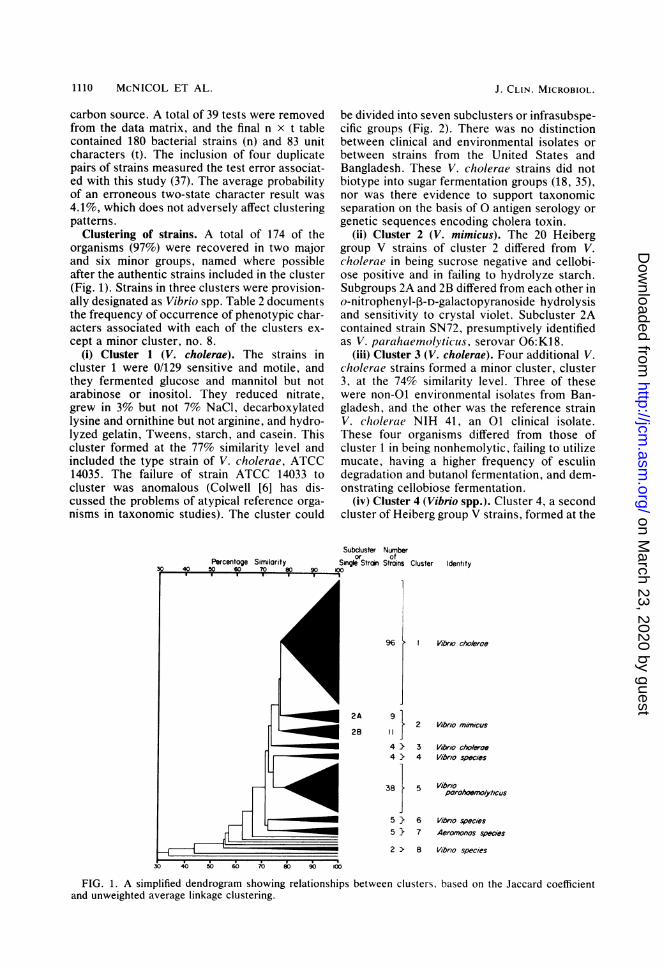
Clustering of strains. A total of 174 of theorganisms (97%) were recovered in two majorand six minor groups, named where possibleafter the authentic strains included in the cluster(Fig. 1). Strains in three clusters were provision-ally designated as Vibrio spp. Table 2 documentsthe frequency of occurrence of phenotypic char-acters associated with each of the clusters ex-cept a minor cluster, no. 8.
(i) Cluster 1 (V. cholerae). The strains incluster 1 were 0/129 sensitive and motile, andthey fermented glucose and mannitol but notarabinose or inositol. They reduced nitrate,grew in 3% but not 7% NaCl, decarboxylatedlysine and ornithine but not arginine, and hydro-lyzed gelatin, Tweens, starch, and casein. Thiscluster formed at the 77% similarity level andincluded the type strain of V. cholerae, ATCC14035. The failure of strain ATCC 14033 tocluster was anomalous (Colwell [6] has dis-cussed the problems of atypical reference orga-nisms in taxonomic studies). The cluster could
be divided into seven subclusters or infrasubspe-cific groups (Fig. 2). There was no distinctionbetween clinical and environmental isolates orbetween strains from the United States andBangladesh. These V. cholerae strains did notbiotype into sugar fermentation groups (18, 35),nor was there evidence to support taxonomicseparation on the basis of 0 antigen serology orgenetic sequences encoding cholera toxin.
(ii) Cluster 2 (V. mimicus). The 20 Heiberggroup V strains of cluster 2 differed from V.cholerae in being sucrose negative and cellobi-ose positive and in failing to hydrolyze starch.Subgroups 2A and 2B differed from each other ino-nitrophenyl-p3-D-galactopyranoside hydrolysisand sensitivity to crystal violet. Subcluster 2Acontained strain SN72, presumptively identifiedas V. parahaemolyticus, serovar 06:K18.
(iii) Cluster 3 (V. cholerae). Four additional V.cholerae strains formed a minor cluster, cluster3, at the 74% similarity level. Three of thesewere non-01 environmental isolates from Ban-gladesh, and the other was the reference strainV. cholerae NIH 41, an 01 clinical isolate.These four organisms differed from those ofcluster 1 in being nonhemolytic, failing to utilizemucate, having a higher frequency of esculindegradation and butanol fermentation, and dem-onstrating cellobiose fermentation.
(iv) Cluster 4 (Vibrio spp.). Cluster 4, a secondcluster of Heiberg group V strains, formed at the
Subcluster Numberor of
Single Strain Stroins Cluster Identity
2A
2B
96 { VibrA-o cho/eroe
i
} 2 Vlbrlo miMncus
4 }> 3 Vlbrlo cho/eroe4 } 4 Vibrlo specles
38 } 5 VbOpe7rohoon0/yt/cus
5 } 6 Vlbrio specles5 } 7 Aeromonos speaes
2 D 8 Vibrio species
FIG. 1. A simplified dendrogram showing relationships between clusters, based on the Jaccard coefficientand unweighted average linkage clustering.
J. CLIN. MICROBIOL.
on March 23, 2020 by guest
http://jcm.asm
.org/D
ownloaded from

COMPARATIVE STUDY OF V. CHOLERAE 1111
PERCENTAGE SIMILARITY STRAIN SUBCLUSTER
V432
S1040167
ATCC 140350050
V41
6770
V72
6761
5652V64
V652746
Bevs
OV40
r ~~~~~~~~~~V9
V.58V84
V878V47
vms
8V23
MV7 b
8V39
8V41
OV248V30
BV32
OW176487
V43
V40
OveV42
V53
V.6
8V76 C
V59
V50
V47
V44
.-00 ~
BV45 dOmm
V39
V76
VS3V79
514
BYe3
By7
80%simlarity evel. I incluevehe Ceapeake~~~~~ ~~~~~Baslae ndsrinAC 319
BV37"7
vmy'
7V2170379414
1560
4752
Ve3mV315135
73575 so 85 90 95 0
FIG. 2. A portio'n of the dendrogram illustrated in
Fig. 1, showing the major V. cholerae cluster in detail.
80% similarity level. It included three Chesa-
peake Bay isolates and strain ATCC 33149, a
reference culture of Vibrio vulnificus biogroup 2(39). These organisms differed from those ofcluster 2 in having a higher percentage of yellow-pigmented colonies; not growing at 10°C; fer-menting salicin; failing to reduce nitrate, de-grade chitin, or produce hemolysin; butdegrading esculin and starch. They resembled V.
parahaemolyticus, but could be distinguishedfrom strains of cluster 5 by growth at hightemperature, arabinose and salicin fermentation,o-nitrophenyl-p-D-galactopyranoside reaction,salt tolerance, and esculin degradation.
(v) Cluster 5 (V. parahaemolyticus). Cluster 5, amajor cluster, formed at the 79o similarity leveland included the V. parahaemolyticus typestrain, ATCC 17802. There was no evidence ofsubclustering of the 38 strains or of phenotypicdistinctions between clinical and environmentalisolates. Typical strains were sucrose negative,arabinose positive, able to grow on 7% NaCl,urease negative, and esculin negative.
(vi) Cluster 6 (Vibrio spp.). Cluster 6 containedfive environmental isolates from Bangladeshwhich appeared to be Vibrio spp., grouping atthe 74% similarity level. The strains were hemo-lytic and sucrose, mannose, and arabinose posi-tive. They did not ferment cellobiose, but werepositive for esculin degradation, salicin fermen-tation, and salt resistance. Some were unusual inbeing resistant to 0/129 and failing to decarbox-ylate amino acids.
(vii) Cluster 7 (A. hydrophUa). Cluster 7 formedat the 78% similarity level, comprised five orga-nisms, and was identified as A. hydrophila. Thetype strain ofA. hydrophila, ATCC 9071, did notfall into this group. The strains fermented man-nitol, but not inositol, were ornithine decarbox-ylase negative and arginine dihydrolase positive,produced opaque colonies, failed to grow on 7%NaCl, and were resistant to 0/129.
(viii) Cluster 8 (Vibrio spp.). This cluster com-prised the two reference strains V. anguillarumATCC 14181 and V. parahaemolyticus SAK4.They shared 61% similarity, indicating genericbut not species similarity.
DISCUSSIONIn regions endemic for cholera, the disease is
often initially diagnosed by dark-field microsco-py of stool samples, whereas in nonendemicareas, diagnosis generally follows isolation andidentification of the causative agent (40). Therehas been an increasing awareness of the pres-ence of V. cholerae in clinical and environmen-tal samples in the United States after the occur-rence of cases of cholera in Texas, Alabama,and Louisiana (3, 5, 41). Current identificationschemes for V. cholerae are based on the reac-tions of strains isolated from areas endemic forcholera and so far appear to be satisfactory forstrains isolated in nonendemic areas (3, 24, 27).However, there has been no overall comparisonof strains isolated from endemic and nonendem-ic areas. The results of the present taxonomicstudy show no distinction between V. choleraestrains isolated from areas endemic and nonen-demic for cholera. For example, five pairs of
VOL. 17, 1983
on March 23, 2020 by guest
http://jcm.asm
.org/D
ownloaded from

1112 McNICOL ET AL.
isolates from the Chesapeake Bay and Bangla-desh (V50/BV85, V44/BV14, V88/BV47,V87/BV84, and V85/BV19) clustered at similar-ities -greater than 94% (Fig. 2). Moreover, pairsof clinical and environmental isolates of V. chol-erae (2746/BV68 and 0050/V41) shared evenhigher similarities.The high phenotypic similarity between envi-
ronmental and clinical isolates suggests that theprevious concept that V. cholerae is highlyadapted to survival only in humans is no longertenable. Studies over the last decade on thespread of cholera, and the ecology and molecu-lar genetics of the causative organism, in regionsnonendemic for cholera have indicated a naturalaquatic habitat for toxigenic and nontoxigenicstrains of V. cholerae (3, 8, 14, 23, 27). Clearly,samples taken from diarrheal patients should beroutinely examined for the presence of V. chol-erae if there has been a recent exposure toseafood or to a brackish water environment.Infections of extraintestinal lesions due to V.cholerae after exposure to brackish waters havealso been recorded (4).
It is possible that V. cholerae may be misiden-tified as a coliform in laboratories using mediadesigned for Enterobacteriaceae species or hav-ing little experience in the identification of vib-rios. Studies have already demonstrated theefficacy of thiosulfate-citrate-bile salts-sucroseagar for the isolation of V. cholerae (42). Com-prehensive identification schemes for V. chol-erae have been published by Furniss (15) andWachsmuth et al. (40) for clinical isolates. Thepresent study demonstrates that these routinelaboratory characterization tests, originally de-veloped to identify strains of V. cholerae isolat-ed from disease cases in areas endemic forcholera, can justifiably be used for the identifica-tion of toxigenic and nontoxigenic strains of V.cholerae from both clinical and environmentalsamples. Subclustering of the major cluster of V.cholerae could not be correlated with geogra-phy, sample type, any single phenotypic charac-ter, any combination of phenotypic characters,serovar, or potential to produce cholera toxin.This study confirms the observations of Danieland Lloyd (11), who noted the failure of thecarbohydrate fermentation schemes (18, 35) tobiotype sucrose-fermenting strains of V. chol-erae when a large-scale taxonomic analysis isdone. V. cholerae clearly comprises strains fromboth clinical and environmental sources, as wellas both 01 and non-O1 serovars. In view of thepresence of the enterotoxin genes in a V. chol-erae non-O1 serovar (26), it is clear that thetraditional restriction of V. cholerae to entero-toxigenic 01 serovars is not realistic or accept-able.While the present study indicates that Heiberg
group V strains of V. cholerae are distinct fromthe major V. cholerae cluster, there was nodefinitive plexus of distinguishing phenotypiccharacters. The major differentiating tests weresucrose fermentation and starch hydrolysis. Da-vis et al. (12) placed Heiberg group V strains intothe species V. mimicus and separated the spe-cies from V. cholerae on the basis ofDNA/DNAhomology studies. The present study and that ofDavis et al. (12) indicate that there appear to befew tests which can routinely be used to distin-guish V. cholerae from V. mimicus. Furthertaxonomic studies are required since there ap-pears to be variation among strains identified asV. cholerae and V. mimicus.The results of the taxonomic analysis per-
formed in this study indicate some similaritybetween V. parahaemolyticus and V. mimicus.Neither species ferments sucrose. However, V.mimicus fails to ferment arabinose or to grow in7% NaCl, features for which V. parahaemolyti-cus frequently is positive. Differentiation be-tween these two species is important in clinicallaboratories, since both are associated with hu-man disease (4, 11).The cluster containing isolates of V. parahae-
molyticus showed no evidence of subclusters.Furthermore, the Kanagawa phenomenon, sero-var, or source of strains did not correlate withany single phenotypic character or groups ofcharacters. Thus, as was observed for V. chol-erae, there is no suggestion of separate clinicaland environmental reservoirs for this organism.
Clusters 6 and 8 contain strains which couldnot be assigned to any of the presently describedVibrio spp. Some strains failed to decarboxylatearginine, lysine, or ornithine. The taxonomicposition of some strains from these clusters iscurrently under investigation in our laboratory,since similar strains have been isolated fromhumans with pathogenic disorders (33).
ACKNOWLEDGMENTSThis work was supported by Public Health Service grant
R22 Al 14242 from the National Institutes of Health, NationalScience Foundation grants DEB-77-14646 and OCE 7682655,Department of Commerce grant NA 79AA-D-00062 and WorldHealth Organization grant C6/181/70 to R.R.C.; Department ofCommerce grant NA 79AA-D-00058 to R.R.C. and L.A.M.;and University of Maryland Biomedical Research Grant2507RR0704215 to L.A.M. Computer time was made availableby the Computer Science Center, University of Maryland,College Park.We thank K. Smith, P. R. Levine, E. R. Remmers, and
H. A. Lockman for technical assistance.LITERATURE CITED
1. Anderson, T. G. 1970. Testing of susceptibility to antimi-crobial agents and assay of antimicrobial agents in bodyfluids, p. 299-310. In J. E. Blair, E. H. Lennette, andJ. P. Truant (ed.), Manual of clinical microbiology, 1st ed.American Society for Microbiology, Washington, D.C.
2. Austin, B., S. Garges, B. Conrad, E. E. Harding, R. R.Colwell, U. Simidu, and N. Tags. 1979. Comparative studyof the aerobic, heterotrophic bacterial flora of Chesapeake
J. CLIN. MICROBIOL.
on March 23, 2020 by guest
http://jcm.asm
.org/D
ownloaded from

COMPARATIVE STUDY OF V. CHOLERAE 1113
Bay and Tokyo Bay. Appl. Environ. Microbiol. 37:704-714.
3. Blake, P. A., D. T. Allegra, J. D. Snyder, T. J. Barrett, L.McFarland, C. T. Caraway, J. C. Feeley, J. P. Craig,J. V. Lee, N. D. Puhr, and R. A. Feldman. 1980. Chol-era-a possible endemic focus in the United States. N.Engl. J. Med. 302:305-309.
4. Blake, P. A., R. E. Weaver, and D. G. Hollis. 1980. Dis-eases of humans (other than cholera) caused by vibrios.Annu. Rev. Microbiol. 34:341-367.
5. Cameron, J. M., K. Hester, W. L. Smith, E. Caviness, T.Hostz, and F. S. Wolf. 1977. Vibrio cholerae-Alabama.Morbid. Mortal. Weekly Rep. 26:159-160.
6. Colwell, R. R. 1973. Genetic and phenetic classification ofbacteria. Adv. Appl. Microbiol. 16:137-175.
7. Colwell, R. R., J. Kaper, and S. W. Joseph. 1977. Vibriocholerae, Vibrio parahaemolyticus, and other vibrios: oc-currence and distribution in Chesapeake Bay. Science198:394-396.
8. Colwell, R. R., R. J. Seidler, J. Kaper, S. W. Joseph, S.Garges, H. Lockman, D. Maneval, H. Bradford, N. Rob-erts, E. Remmers, I. Huq, and A. Huq. 1981. Occurrenceof Vibrio cholerae serotype O1 in Maryland and Louisianaestuaries. Appl. Environ. Microbiol. 41:555-558.
9. Colweil, R. R., and W. J. Wiebe. 1970. Core characteris-tics for use in classifying aerobic, heterotrophic bacteriaby numerical taxonomy. Bull. Ga. Acad. Sci. 18:165-185.
10. Dallas, W. S., D. M. Gill, and S. Falkow. 1979. Cistronsencoding Escherichia coli heat-labile toxin. J. Bacteriol.139:850-858.
11. Daniel, R. R., and B. J. Lloyd. 1981. The taxonomy ofsome cholera-like vibrios. Soc. Gen. Microbiol. Q. 8:181-182.
12. Dais, B. R., G. R. Fanning, J. M. Madden, A. G. Stei-gerwalt, H. B. Bradford, H. L. Smith, and D. J. Brenner.1981. Characterization of biochemically atypical Vibriocholerae strains and designation of a new pathogenicspecies, Vibrio mimicus. J. Clin. Microbiol. 14:631-639.
13. Dawson, C. A., and P. H. A. Sneath. 1981. An identifica-tion matrix for freshwater vibrios. Soc. Gen. Microbiol.Q. 8:181.
14. Desmarchelier, P. M., and J. L. Reichelt. 1981. Phenotyp-ic characterization of clinical and environmental isolatesof Vibrio cholerae from Australia. Curr. Microbiol. 5:123-127.
15. Furniss, A. L. 1979. Identification of human vibrios. Soc.Appl. Bacteriol. Tech. Ser. 14:143-150.
16. Furmss, A. L., J. V. Lee, and T. J. Donovan. 1978. theVibrios. Public Health Laboratory Service monographseries no. 11. Her Majesty's Stationery Office, London.
17. Gordon, R. E. 1967. The taxonomy of soil bacteria, p.293-321. In E. T. Gray and D. Parkinson (ed.), Theecology of soil bacteria. University of Liverpool Press,Liverpool.
18. Heiberg, B. 1936. The biochemical reactions of vibrios. J.Hyg. 36:114-117.
19. Hsu, S. C., and J. L. Lockwood. 1975. Powdered chitinagar as a selective medium for enumeration of actinomy-cetes in water and soil. Appl. Microbiol. 29:422-426.
20. Hugh, R., and J. C. Feeley. 1972. Report (1966-1970) ofthe Subcommittee on Taxonomy of Vibrios to the Interna-tional Committee on Nomenclature of Bacteria. Int. J.Syst. Bacteriol. 22:123.
21. Juras, H., U. Futh, D. Winkler, J. Friedmann, and T.Hfllg. 1979. Vorkommen von NAG-Vibrionen in Berlinergewassern. Z. Allg. Mikrobiol. 19:403-409.
22. Kamal, A. M. 1974. The seventh pandemic of cholera, p.1-14. In D. Barua and W. Burrows (ed.), Cholera. TheW. B. Saunders Co., Philadelphia, Pa.
23. Kaper, J. B., H. B. Bradford, N. C. Roberts, and S.Falkow. 1982. Molecular epidemiology of Vibrio choleraein the U.S. Gulf Coast. J. Clin. Microbiol. 16:129-134.
24. Kaper, J. B., H. Lockman, R. R. Colwell, and S. W.Joseph. 1979. Ecology, serology, and enterotoxin produc-tion of Vibrio cholerae in Chesapeake Bay. Appl. Envi-ron. Microbiol. 37:91-103.
25. Kaper, J. B., H. Lockman, R. R. Colwell, and S. W.Joseph. 1981. Aeromonas hydrophila: ecology and toxige-nicity of isolates from an estuary. J. Appl. Bacteriol.50:359-377.
26. Kaper, J. B., S. L. Moseley, and S. Falkow. 1981. Molecu-lar characterization of environmental and nontoxigenicstrains of Vibrio cholerae. Infect. Immun. 32:661-667.
27. Lee, J. V., D. J. Bashford, T. J. Donovan, A. L. Furniss,and P. A. West. 1982. The incidence of Vibrio cholerae inwater, animals and birds in Kent, England. J. Appl.Bacteriol. 52:281-291.
28. Le Minor, L., and F. Ben Hamida. 1962. Advantages de larecherche de la ,-galactosidase sur celle de la fermenta-tion du lactose en milieu complexe dans le diagnosticbacteriolgique en particulier des Enterobacteriaceae.Ann. Inst. Pasteur Paris 102:267-277.
29. Lovelace, T. E., and R. R. Colwell. 1968. A multipointinoculator for petri dishes. Appl. Microbiol. 16:944-945.
30. Mallory, L. M., B. Austin, and R. R. Colwell. 1977. Nu-merical taxonomy and ecology of oligotrophic bacteriaisolated from the estuarine environment. Can. J. Microbi-ol. 23:733-750.
31. McNicol, L. A., K. M. S. Aziz, I. Huq, J. B. Kaper, H. A.Lockman, E. F. Remmers, W. M. Spira, M. J. Voll, andR. R. Colwell. 1980. Isolation of drug-resistant Aeromo-nas hydrophila from aquatic environments. Antimicrob.Agents Chemother. 17:477-483.
32. McNicol, L. A., J. B. Kaper, H. A. Locnman, E. F. Rem-mers, W. M. Spira, M. J. Voll, and R. R. Colwell. 1980.R-factor carriage in a group F vibrio isolated from Bangla-desh. Antimicrob. Agents Chemother. 17:512-515.
33. Morris, J. G., H. Miller, R. Wilson, C. Tacket, D. G.Hoilis, F. Hickman, R. E. Weaver, and P. A. Blake. 1982.Illness caused by Vibrio damsela and Vibrio hollisae.Lancet 1:1294-1297.
34. Sircar, B. K., S. P. De, P. G. Sengupta, S. Mondal, D.Sen, B. C. Deb, and S. C. Pal. 1979. Studies on transmis-sion of V. parahaemolyticus infection in Calcutta commu-nities: a preliminary report. Indian J. Med. Res. 70:898-907.
35. Smith, H. L., and K. Goodner. 1965. On the classificationof vibrios. U.S. Public Health Serv. Publ. 1328:4.
36. Sneath, P. H. A. 1956. Cultural and biochemical charac-teristics of the genus Chromobacterium. J. Gen. Microbi-ol. 15:70-98.
37. Sneath, P. H. A., and R. Johnson. 1972. The influence onnumerical taxonomic similarities of errors in microbiologi-cal tests. J. Gen. Microbiol. 72:377-392.
38. Stevenson, I. L. 1967. Utilization of aromatic hydrocar-bons by Arthrobacter spp. Can. J. Microbiol. 13:205-211.
39. TIson, D. L., M. Nishibuchi, J. D. Greenwood, and R. J.Seidler. 1982. Vibrio vulnifus biogroup 2: new biogrouppathogenic for eels. Appl. Environ. Microbiol. 44:640-646.
40. Wachsmuth, I. K., G. K. Morris, and J. C. Feeley. 1980.Vibrio, p. 226-234. In E. H. Lennette, A. Balows, W. J.Hausler, Jr., and J. P. Truant (ed.), Manual of clinicalmicrobiology, 3rd ed. American Society for Microbiology,Washington, D.C.
41. Weissman, J. B., W. E. Dewitt, J. Thompson, C. N.Muchnkck, B. L. Portnoy, J. C. Feeley, and E. J. Gangar-osa. 1974. A case of cholera in Texas, 1973. Am. J.Epidemiol. 100:487-498.
42. West, P. A., E. Russek, P. R. Brayton, and R. R. Colwell.1982. Statistical evaluation of a quality control method forisolation of pathogenic Vibrio species on selected thiosul-fate-citrate-bile salts-sucrose agars. J. Clin. Microbiol.16:1110-1116.
VOL. 17, 1983
on March 23, 2020 by guest
http://jcm.asm
.org/D
ownloaded from