(nutritionally variant streptococcus serotype ii) group antigen
TRANSCRIPT

INFECTION AND IMMUNITY, Feb. 1991, p. 592-599 Vol. 59, No. 20019-9567/91/020592-08$02.00/0Copyright © 1991, American Society for Microbiology
Purification and Characterization of Streptococcus adjacens(Nutritionally Variant Streptococcus Serotype II) Group Antigen
P. A. SIELING AND I. VAN DE RIJN*Wake Forest University Medical Center, 300 S. Hawthorne Road,
Winston-Salem, North Carolina 27103
Received 26 September 1990/Accepted 19 November 1990
Nutritionally variant streptococci (NVS) possess amphiphiles which are serologically distinct from lipote-ichoic acid and which serve as group-specific antigens for NVS. The objective of this study was to purifyand characterize the NVS serotype II (Streptococcus adjacens) amphiphile. Amphiphile was isolated fromstationary-phase culture supernatants of NVS strain 81 (NVS serotype II). Phenol-water extracts of culturesupernatants were subjected to hydrophobic interaction chromatography and gel filtration chromatography. Ahomogeneous preparation of amphiphile (22 mg; 8.5 x 106 hemagglutination units) was recovered, and itsapproximate molecular size (23,000 to 24,000 Da) and chemical composition were determined. Purified S.adjacens amphiphile contained phosphorus, ribitol, galactose, galactosamine, alanine, and fatty acids in molarratios of 1.00:0.88:1.39:1.10:0.08:0.24. Since ribitol, galactose, and galactosamine were the primary carbo-hydrate components, the amphiphile may exist as a polyribitol phosphate with galactose and galactosaminesubstituents. Preliminary structural analysis demonstrated the presence of phosphodiester bonds within theamphiphile structure. Finally, the amphiphile serves as the S. adjacens group antigen.
Amphiphiles of gram-positive bacteria have various chem-ical compositions, though each maintains a common struc-tural pattern of hydrophilic and hydrophobic domains (43).Lipoteichoic acids (LTAs) of many streptococci are com-posed of a 1-3-linked poly(glycerol phosphate) backbonesubstituted periodically at the C-2 position of the glycerolmolecule with glucose, galactose, or D-alanine in esterlinkage. Covalently attached to a terminal glycerol residue inphosphodiester linkage is either a glycolipid, phosphatidyl-glycolipid, or a fatty acid (8, 42). Streptococcus mitis is oneexample of a number of gram-positive bacteria lacking thepoly(glycerol phosphate) of LTA (29). Nutritionally variantstreptococci (NVS) possess amphiphiles with serologicalproperties distinct from those of LTA (3). The serotype Iamphiphile from NVS was previously purified and charac-terized as a lipid-substituted poly(ribitol phosphate) (10).
Presumably, the amphiphile's hydrophobic portion an-chors in the cytoplasmic membrane while the hydrophilicregion extends through the cell wall and beyond, givingamphiphiles a cell surface location (38). Amphiphiles havealso been found in the culture supernatants of gram-positivebacteria (17, 25). Their heterogeneous chemical nature andsurface location make amphiphiles useful serological mark-ers. Indeed, the group antigens of Streptococcus groups Dand N and Lactobacillus group F are LTAs (6, 20, 41).NVS have been classified serologically by van de Rijn and
George, with serotypes I and II comprising 97% of isolates(37). These serological differences corresponded to immuno-logical differences when examined in an experimental animalmodel of bacterial endocarditis (35). Also, serotypes I and IIwere distinguishable by the ability of serotype I to produce,-galactosidase and serotype II to produce 0-glucuronidase(4). Further evidence for differentiation of the two NVStypes was recently published by Bouvet et al. (2). In theirinvestigation, serotypes I and II were compared by chromo-somal DNA hybridization and found to share less than 10%
* Corresponding author.
identity. These immunological, biochemical, and geneticdifferences indicate that NVS types I and II are distinctorganisms. On the basis of these differences and theirdistinction from other streptococci, Bouvet et al. (2) havedesignated serotype I NVS as S. defectivus and serotype IIas S. adjacens. Therefore, the serotype antigens now serveas group antigens for the two streptococcal species.NVS are important human pathogens, as evidenced by
their occurrence in 5% of streptococcal endocarditis infec-tions (27) and their presence in numerous other bacterialinfections (12). Identification of NVS from endocarditispatients can be hindered because they may exist in so-calledculture-negative isolates (27). Appropriate antimicrobialtherapy may be less effective in the event of delayeddiagnosis of NVS infection. Precise identification of NVSwould be enhanced through the purification and characteri-zation of group-specific antigens.
This report describes the purification and characterizationof the S. adjacens amphiphile from NVS. S. adjacens strainswere compared for production of both intracellular andextracellular amphiphiles. Maximal amphiphile productionduring the growth phase was also examined. The S. adjacensamphiphile was purified by hydrophobic interaction chroma-tography (HIC) and gel filtration chromatography. The pu-rified amphiphile was demonstrated to contain phosphorus,ribitol, galactose, galactosamine, alanine, and fatty acids in a1.00:0.88:1.39:1.10:0.08:0.24 molar ratio.
MATERIALS AND METHODS
Bacterial strains. Stock cultures of NVS strains were keptfrozen at -70°C in a 50% (vol/vol) solution of chemicallydefined medium supplemented with Todd-Hewitt dialysate(CDMT) (3) and glycerol. The identity of NVS isolates wasdetermined by the ability of the strains to form satellitecolonies around colonies of Staphylococcus epidermidis andby the presence of a pH-dependent chromophore (36).
Bacterial maintenance. NVS strains were grown in CDMTat 37°C without shaking. The growth of streptococcal cul-
592
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 29
Nov
embe
r 20
21 b
y 21
1.22
9.62
.37.

S. ADJACENS GROUP ANTIGEN 593
tures was measured in 18-mm tubes by using a Spectronic 20spectrophotometer at a wavelength of 650 nm.
Purification of amphiphile. Amphiphile purification wasbased on the method used to purify the NVS serotype Iantigen (now S. defectivus group antigen) (11). Bacteria (360liters) were grown to stationary phase (30 h) at 37°C andwere sedimented in a refrigerated high-speed centrifuge(Sharples-Stokes Div., Pennwalt Corp., Warminster, Pa.).The culture supernatant was then concentrated (with aDC101 concentrator and a HlOP30-20 hollow fiber cartridge;Amicon Corp., Lexington, Mass.) and dialyzed againstdeionized water. The concentrated supernatant was centri-fuged (7,000 x g, 20 min), filtered through a 0.45-,um-pore-size filter, resedimented to remove any remaining bacteria,and then lyophilized. Lyophilized culture supernatant wasrehydrated in 800 ml of sodium phosphate buffer, pH 7.0.DNase and RNase (40 mg each; Sigma Chemical Co., St.Louis, Mo.) were added, and the solution was incubated for2 h at 37°C with stirring, followed by an identical 2-htreatment with 40 mg of trypsin (Sigma). Enzyme-treatedculture supernatant was then extracted with an equal volumeof 90% phenol at 68°C by the method of Kessler andShockman (18).
Lyophilized phenol-water extracts of amphiphile wererehydrated in 0.1 M sodium acetate-15% 1-propanol, pH 6.0,at a concentration of 10 mg/ml and loaded onto a column (2.5by 34 cm) of octyl-Sepharose (CL-4B; Sigma) for HIC. Afterloading the column with extract (10 ml/h), unbound materialwas removed with two column volumes of the column bufferand eluted at a flow rate of 20 ml/h with two column volumesof 0.1 M sodium acetate-50% 1-propanol, pH 6.0. Fractions(5 ml) were collected throughout the chromatographic pro-cess and analyzed for sheep erythrocyte (SRBC) sensitiza-tion activity, phosphorus, and hexose.Column fractions containing SRBC sensitization activity
were pooled and diluted with 0.1 M sodium acetate buffer(pH 6.0) to restore the propanol concentration to 15%. HICwas repeated on the pooled fractions under the conditionsdescribed above until phosphorus and hexose were no longerdetected in fall-through fractions (three times total). Frac-tions with SRBC sensitization activity were pooled, dialyzedagainst deionized water (48 h at 4°C with six changes), andlyophilized.Amphiphile preparation isolated via HIC was rehydrated
in 0.1 M sodium acetate-0.5% Chapso, pH 6.5, (4 mg/ml) andloaded onto a Sephadex G-75 (Sigma) gel filtration column(2.5 by 39 cm). Amphiphile was eluted with the same buffer(10 mlIh), and fractions (5 ml) were collected. SRBC sensi-tizing fractions were pooled, dialyzed against water, andlyophilized. Amphiphile was rechromatographed on octyl-Sepharose as described above to remove detergent from thepreparation.
Determination of amphiphile homogeneity. (i) RIE and CIE.Rocket immunoelectrophoresis (RIE) and crossed immuno-electrophoresis (CIE) of S. adjacens amphiphile were per-formed with sodium barbital buffer (ionic strength, 0.02; pH8.8) in the presence or absence of 1% (vol/vol) Triton X-100(19). The immunoprecipitates were visualized by stainingwith 0.5% (wt/vol) Coomassie brilliant blue in 45% ethanol-10% acetic acid (vol/vol).
(ii) SDS-PAGE. S. adjacens amphiphile was analyzed bysodium dodecyl sulfate-polyacrylamide gel electrophoresis(SDS-PAGE) by the system of Laemmli (21). Resolving gels(12.5% acrylamide) were prepared in Laemmli buffers con-taining 0.002 M EDTA. Standards (lipopolysaccharide [LPS]and LTA) and samples mixed with sample buffer (0.15 M
Tris [pH 6.8], 5% SDS, 25% glycerol, bromphenol blue)were boiled for 3 min before loading onto the gel. Sampleswere electrophoresed at 40 mA and silver stained (26).Amphiphile molecular size determination. Purified NVS-81
amphiphile (53 ,ug) was dissolved in 0.1 M sodium acetate-0.5% Chapso, pH 6.5, and loaded onto a column of Sepha-cryl S-200 (0.7 by 28.5 cm) equilibrated in the same buffer.Amphiphile was eluted from the column with equilibrationbuffer at a flow rate of 6 ml/h. Fractions (0.5 ml) wereanalyzed for antigenic activity by RIE and SRBC sensitiza-tion activity. Molecular size determination was performedby chromatographing dextrans (average molecular weights,66,300, 40,200, and 10,000 Da) on the S-200 column underthe same conditions as the amphiphile. Peak elution volumeswere converted to Kav, and a standard curve was generatedfrom these values.
Amphiphile chemical composition. (i) Hexose. Hexose wasdetermined by the phenol-sulfuric acid method of Dubois etal. (5) by using a glucose standard. The standards andsamples were measured in triplicate and read against a bufferblank.
(ii) Phosphorus. The acid-soluble and inorganic-plus-labilephosphorus content of the samples was determined by themethod of Lowry et al. (23) with a KH2PO4 standard. Thestandards and samples were measured in triplicate.
(iii) Protein. Protein quantity was determined by theLowry protein assay (24) with bovine serum albumin as astandard.
(iv) Nucleic acid. Nucleic acid content was measured byscanning the A850 to A200 of amphiphile preparations. Strep-tococcal DNA was used as a standard for quantitationpurposes.
(v) Glycosides and glycitols. Neutral sugars were identifiedand quantitated as their alditol acetates (13) after hydrolysisof the amphiphile in 2 N trifluoroacetic acid for 90 min at120°C. Fatty acids were extracted with hexane before de-rivatization of the sample. To ensure complete quantitationof polyalcohols and sugars, the hydrolyzed samples weretreated with phosphomonoesterase (see below). Ribitol wasthe only new peak detected after this treatment. Monosac-charides released by trifluoroacetic acid hydrolysis werereduced with 0.6 N NaBH4 in 1 N NH40H (60 min, roomtemperature). Excess borohydride was removed with glacialacetic acid, and the resulting borate was removed by dis-solving in 1 ml of methanol followed by evaporation (repeat-ed five times). Acetylation was performed by incubatingsamples in the presence of acetic anhydride (250 ,lI) for 30min at 120°C. Derivatized sugars were then chromato-graphed on a 15-m fused silica capillary column (SP2330;Supelco, Inc., Bellefonte, Pa.) in a Carlo Erba Strumentazi-one gas chromatograph with helium as the carrier gas (2.7ml/min). The chromatograph was equipped with a flameionization detector set at 295°C and a manual injection portset at 245°C. Oven temperatures were programmed as gra-dients from 100 to 245°C (30°C/min) with a 1-min initial holdand an 18-min final hold. The identification of sugars was
based on comparison of retention times of known standards.Amphiphile components were quantitated with the Maxima820 software system (Waters Associates, Inc., Milford,Mass.) with xylose as the internal standard.
(vi) Fatty acids. Fatty acids were detected and quantitatedas their methyl esters by a modification of the method ofRogozinski (28). Samples were chromatographed on a 30-mfused silica column coated with DB-225 (J and W Scientific,Inc., Folsom, Calif.) in a Hewlett-Packard 5890A gas chro-matograph with helium as the carrier gas (1 ml/min). The
VOL. 59, 1991
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 29
Nov
embe
r 20
21 b
y 21
1.22
9.62
.37.
594 SIELING AND VAN DE RIJN
chromatograph was equipped with a flame ionization detec-tor at 280°C and a manual injection port at 230°C. Oventemperatures were programmed as gradients from 165 to226°C (3°C/min) with an initial hold of 2.0 min. Unknownswere identified by comparisons with reference standards.Heptadecanoic acid, added as an internal standard, allowedfor quantitation of identified fatty acids.
(vii) Amino acids and amino sugars. Amino acids andamino sugars were identified as their phenylisothiocarbamyl(PITC) derivatives and quantitated by high-performanceliquid chromatography (HPLC) by using the PicoTag method(1). Amphiphile (100 pug) was hydrolyzed in 6 N HCI (vaporphase) for 18 to 24 h at 110°C. Hydrolysates were labeledwith PITC and chromatographed on an RCM column (8 by10) (Waters) with a Nova-Pak C18 packing at 42°C accordingto the procedure of Bidlingmeyer et al. (1). PITC derivativeswere detected at 254 nm. All samples were assayed intriplicate and identified and quantitated on the basis ofretention times and peak areas of reference standards. Thelimit of quantitation for amino acids was 25 pmol. A standardcurve for quantitation of galactosamine was derived bytreating known amounts of galactosamine under the sameconditions as amphiphile.Phosphomonoesterase treatment of S. adjacens amphiphile.
S. adjacens amphiphile (100 nmol of phosphorus) was dis-solved in a buffer of 0.1 M glycine, 1 mM MgCl2, and 1 mMZnCl2 (pH 10.4). Phosphomonoesterase (0.5 U) (bovinealkaline phosphatase; Sigma) was added, and the mixture(500 ,ul) was incubated at 37°C for 60 min. Phosphatase-treated amphiphile was examined for the release of inorganicphosphate, as described above, compared with the amountof inorganic phosphate present in untreated amphiphile(incubated with buffer alone).
Preparation of antisera. (i) Streptococcal whole cell sera.Bacterial strains to be used as inocula for immunization weregrown to stationary phase and killed by exposure to a UVlight source for 30 min. Rabbits were immunized by theprotocol described by Rotta et al. (30).
(ii) NVS culture supernatant antisera. Animals were vac-cinated with 1.0 ml (administered in four aliquots at fourdifferent sites) of an emulsion of the culture supernatant (5mg/ml in water) and Freund complete (first vaccination) orincomplete (subsequent vaccinations) adjuvant. Immuniza-tion was performed weekly for 3 weeks. At week 3, therabbits were bled and immunized for 6 to 9 additional weeks.Culture supernatant antisera were also produced by thedouble adjuvant method described below.
(iii) Antisera against partially purified amphiphile. Am-phiphile (1 mg/ml) isolated by HIC was mixed with methyl-ated bovine serum albumin (1 mglml). The conjugate wasthen mixed with Freund incomplete adjuvant (1:1, vol/vol),followed by the addition of 2% Tween 80 in saline (1:1 ratio,vol/vol) (7). The double adjuvant mixture (1 ml) was admin-istered to rabbits intravenously once a week for the first 3weeks. Rabbits were bled at week 3 and bled and immunizedfor 6 to 9 additional weeks.HA assays. Hemagglutination (HA) titers of antiam-
phiphile antisera were determined in duplicate by previouslypublished methods (16). Crude amphiphile preparations (10,ug/ml, final concentration) were added to a 2% solution ofSRBCs. Sensitization of the SRBC solution was performedat 37°C for 60 min, followed by 4°C for 16 h. SensitizedSRBCs were then washed with saline to remove unboundamphiphile and were resuspended at a 1% concentration.Anti-S. adjacens antiserum titers were determined by serialdilution of antisera in microtiter plates followed by addition
-
o
0 40 80 120 160
Fraction No.
0
I-
0-
0
a0
a' o200
1~~~J40
9
6
20
3310
0 G1haaaa10 &&*00 40 80 120 160 200
2
4c
007S
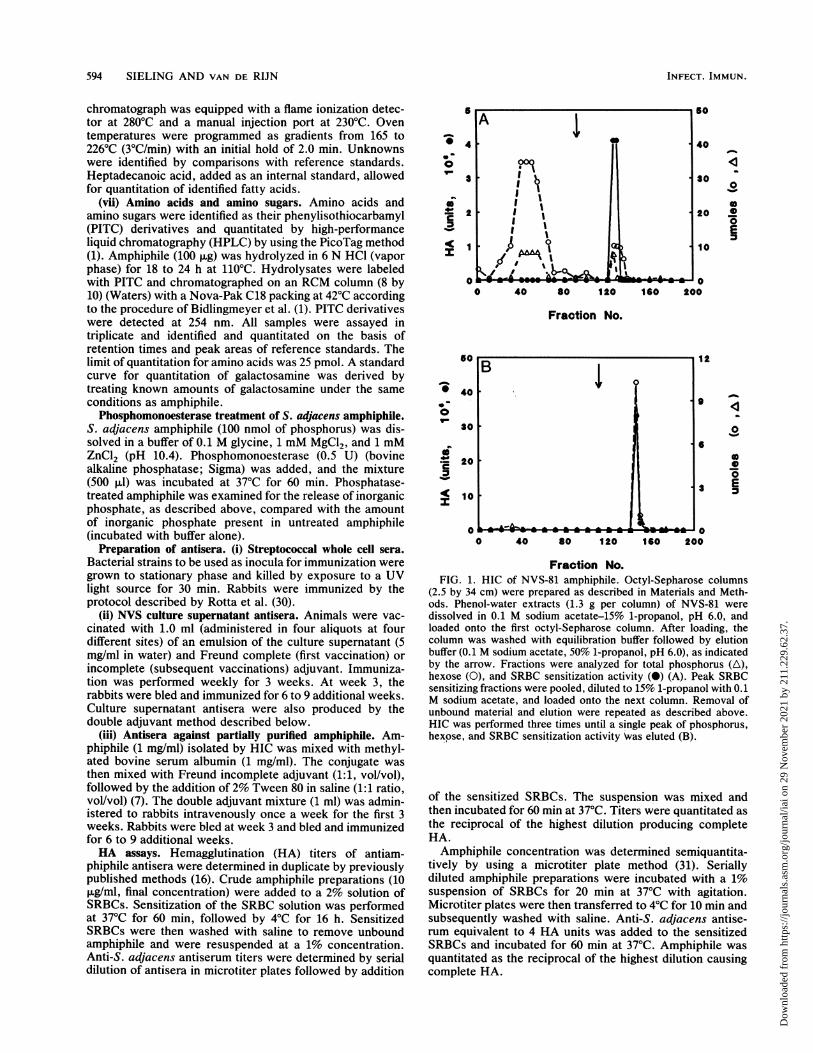
Fraction No.FIG. 1. HIC of NVS-81 amphiphile. Octyl-Sepharose columns
(2.5 by 34 cm) were prepared as described in Materials and Meth-ods. Phenol-water extracts (1.3 g per column) of NVS-81 weredissolved in 0.1 M sodium acetate-15% 1-propanol, pH 6.0, andloaded onto the first octyl-Sepharose column. After loading, thecolumn was washed with equilibration buffer followed by elutionbuffer (0.1 M sodium acetate, 50% 1-propanol, pH 6.0), as indicatedby the arrow. Fractions were analyzed for total phosphorus (A),hexose (0), and SRBC sensitization activity (0) (A). Peak SRBCsensitizing fractions were pooled, diluted to 15% 1-propanol with 0.1M sodium acetate, and loaded onto the next column. Removal ofunbound material and elution were repeated as described above.HIC was performed three times until a single peak of phosphorus,hexose, and SRBC sensitization activity Was eluted (B).
of the sensitized SRBCs. The suspension was mixed andthen incubated for 60 min at 37°C. Titers were quantitated asthe reciprocal of the highest dilution producing completeHA.Amphiphile concentration was determined semiquantita-
tively by using a microtiter plate method (31). Seriallydiluted amphiphile preparations were incubated with a 1%suspension of SRBCs for 20 min at 37°C with agitation.Microtiter pla-tes were then transferred to 4°C for 10 min andsubsequently washed with saline. Anti-S. adjacens antise-rum equivalent to 4 HA units was added to the sensitizedSRBCs and incubated for 60 min at 37°C. Amphiphile wasquantitated as the reciprocal of the highest dilution causingcomplete HA.
INFECT. IMMUN.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 29
Nov
embe
r 20
21 b
y 21
1.22
9.62
.37.
S. ADJACENS GROUP ANTIGEN 595
0-%4
0
0
.0
0~
30
20
10
30
0X X'_* ,_- a*
0 8 16 24 32 40
Fraction No.FIG. 2. Gel filtration chromatography (Sephadex G-75) of
NVS-81 amphiphile. HIC-isolated NVS-81 amphiphile (4 mg) wasrehydrated in 0.1 M sodium acetate-0.5% Chapso (pH 6.5) andloaded onto a column of Sephadex G-75 (2.5 by 39 cm) equilibratedin the same buffer. Amphiphile was eluted from the column, andfractions (5 ml) were collected and analyzed for antigenic activity byCIE (O and A), expressed as peak height in millimeters, and SRBCsensitization activity (0). Fractions containing both SRBC sensitiz-ing activity and antigenic activity (peak A, fractions 14 to 17) werepooled as amphiphile, whereas fractions 18 to 20 were pooled andrechromatographed to recover additional amphiphile. Peak B con-tained less than 1% of the SRBC sensitization activity of peak A andwas not analyzed further. VO, Voided volume; VT, total volume.
A
C
RESULTS
Purification of S. adjacens amphiphile. Preliminary exper-
iments were designed to ascertain the S. adjacens strainwhich produced the most amphiphile, the growth phase foroptimal amphiphile production, and the quantity of cell-associated versus extracellular amphiphile. On the basis ofresults of these preliminary experiments, S. adjacens am-
phiphile was purified from 360 liters of strain NVS-81stationary-phase culture supernatant. Enzyme-treated phe-nol-water extracts were subjected to HIC on octyl-Sepha-rose. Figure 1A (first HIC column) illustrates that fall-through fractions contained contaminating hexose andphosphorus. The SRBC-sensitizing fractions were pooledand rerun on two successive HIC columns. By the third HICcolumn (Fig. 1B), amphiphile migrated as a single peakcomposed of SRBC sensitization activity, hexose, and phos-phorus.Though HIC yielded a single peak, gel filtration chroma-
tography on Sephadex G-75 revealed two distinct peaks (Fig.2) when examined by the SRBC sensitization assay and CIE.Peak A possessed SRBC sensitization activity, indicating thepresence of amphiphile, whereas peak B had less than 1% ofthe SRBC sensitization activity of peak A. Amphiphilefractions (14 to 17, peak A) were pooled and examinedfurther to determine the homogeneity of the purified am-
phiphile. Fractions containing both amphiphile and contam-inating antigen (fractions 18 to 20) were pooled and rechro-matographed to isolate additional amphiphile.
B
D
Jo K..
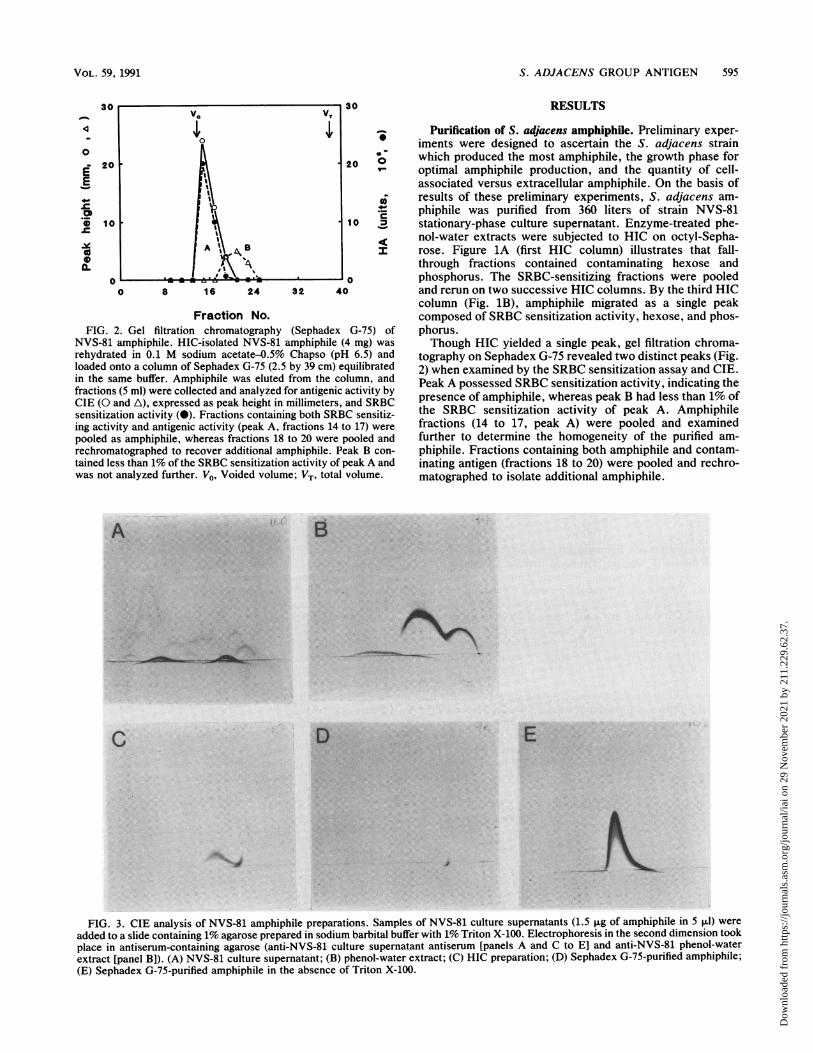
FIG. 3. CIE analysis of NVS-81 amphiphile preparations. Samples of NVS-81 culture supernatants (1.5 ,ug of amphiphile in 5 ,ul) wereadded to a slide containing 1% agarose prepared in sodium barbital buffer with 1% Triton X-100. Electrophoresis in the second dimension tookplace in antiserum-containing agarose (anti-NVS-81 culture supernatant antiserum [panels A and C to El and anti-NVS-81 phenol-waterextract [panel B]). (A) NVS-81 culture supernatant; (B) phenol-water extract; (C) HIC preparation; (D) Sephadex G-75-purified amphiphile;(E) Sephadex G-75-purified amphiphile in the absence of Triton X-100.
VOL. 59, 1991
*__- L _ I
-.ddmbb-
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 29
Nov
embe
r 20
21 b
y 21
1.22
9.62
.37.

596 SIELING AND VAN DE RIJN
FIG. 4. SDS-PAGE of NVS-81 amphiphile. Purified NVS-81
amphiphile (1.5 p.g; right lane) was loaded onto a 12.5% polyacryl-
amide gel. Electrophoresis was performed at 40 mA, after which the
gel was silver stained by the method of Morrissey (26). LPS (1.5 p.g;
left lane) and LTA (0.25 jig; center lane) were electrophoresed
adjacent to the amphiphile for comparison.
Demonstration of a homogeneous preparation of S. adjacens
amphiphile. CIE was performed to detect antigenic species
of the amphiphile preparation throughout the purification
process. Amphiphile preparations were electrophoresed in
agarose containing anti-S. adjacens antiserum. The CIE
slides (Fig. 3) demonstrate that more than 15 antigens were
precipitated in crude culture supernatant (Fig. 3A) and at
least 3 were precipitated in the phenol-water extract (Fig.
3B). CIE of HIC-isolated amphiphile in the presence of
Triton X-100 (Fig. 3C) revealed two antigens in the am-
phiphile preparation. The use of Triton X-100 in CIE of
HIC-purified amphiphile was necessary, since in the absence
of detergent the contaminating antigen was undetectable. A
similar phenomenon was observed in the purification of the
S. defectivus amphiphile (11). Analysis of G-75-purified
amphiphile by GTE revealed a single antigen in the presence
or absence (Fig. 3D and E, respectively) of Triton X-100.
CIE analysis with four additional anti-NVS-81 antisera also
precipitated a single antigen from the G-75-purified material,confirming the antigenically homogeneous nature of theamphiphile (data not shown). RIE analysis also detected a
single antigen when purified amphiphile was titrated to a
quantity at which 1% protein contamination would be de-tectable.To further analyze the homogeneity of the purified am-
phiphile, SDS-PAGE was performed (Fig. 4). Amphiphile(1.5 ,ug; Fig. 4, right lane) was run on a 12.5% polyacryl-amide gel, and the gel was silver stained by the method ofMorrissey (26). SDS-PAGE of LPS (left lane) produced a
characteristic stepladder-like staining pattern, whereas LTA(center lane) produced only a single, diffuse band. S. ad-jacens amphiphile produced a staining pattern which resem-
bled that of LPS.Purification summary. S. adjacens amphiphile was quan-
titated at each purification step for its HA activity (SRBCsensitization) and RIE peak height. Additionally, total phos-phorus and hexose were examined at each purification step.The results from these analyses from one of two independentpreparations are presented in Table 1. From 360 liters ofNVS-81 stationary-phase culture supernatant, 1.8 x 108 HAunits of the S. adjacens amphiphile was present in crudeform.
Nuclease and protease treatments of the culture superna-tant, followed by a phenol-water extraction, reduced theprotein level to less than 0.2% of the amphiphile. Theamount of amphiphile recovered, 138 mg, represented 23%of the amphiphile present in the starting material. HIC andgel filtration chromatography were used to remove phospho-rus- and hexose-containing contaminants. These chromato-graphic methods also reduced nucleic acid below quantifi-able levels (<0.3%, dry weight). Purified amphiphile (22 mg;8.5 x 106 HA units) corresponded to a recovery of 3.6% ofthe amphiphile present in the culture supernatant.
Characterization of S. adjacens group antigen. (i) Molecularsize determination. The molecular size of S. adjacens am-
phiphile was determined by gel filtration chromatography on
a Sephacryl S-200 column. The fractions collected were
examined for SRBC sensitization activity and RIE peakheight (Fig. 5). Dextran molecular weight standards (66,300,40,200, and 10,000 Da) were loaded on the Sephacryl S-200column and eluted in the same manner as amphiphile. Themolecular size of the S. adjacens group antigen was deter-mined to be approximately 23,000 to 24,000 Da from multiplecolumn analyses.
(ii) Chemical characterization. Table 2 shows the compo-
nents detected in the S. adjacens amphiphile, as determinedby the methods outlined above. Phosphorus (580 and 641nmol/mg of amphiphile) was designated as a molar ratio of1.00 in order to compare the relative abundance of each
TABLE 1. Purification of S. adjacens amphiphilea
Purification step Amphiphile HA units Phosphorus Hexose(mg) (106) (pmol) (pmol)b % Recovery
Culture supernatantc 610 180.0 15916.3 22071.4 100Phenol-water extract 138 70.0 15733.5d 4209.3 22.6HIC 69 44.0 51.4 49.8 11.3Gel filtration chromatographye 22 8.5 12.7 11.0 3.6
a Values are the means of duplicate analyses.b Quantitated by using glucose as the standard.c From 360 liters of NVS-81 stationary-phase culture fluid (preparation A).d Value similar to previous step because dialysis did not completely remove phosphate buffer from phenol-water extract.e Sephadex G-75.
INFECT. IMMUN.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 29
Nov
embe
r 20
21 b
y 21
1.22
9.62
.37.

S. ADJACENS GROUP ANTIGEN 597
50
40
30
20
10
0
75
60 °
45
._
30 0
0
15 0
0.
4 6 8 10 12
Elution volume (ml)
0
FIG. 5. Molecular size determination of purified NVS-81 am-phiphile. Sephadex G-75-purified amphiphile (53 ,ug) was dissolvedin 0.1 M sodium acetate-0.5% Chapso (pH 6.5) and loaded onto acolumn of Sephacryl S-200 (0.7 by 28.5 cm) equilibrated with thesame buffer. Amphiphile was eluted from the column, and fractions(0.5 ml) were collected and analyzed for antigenic activity by RIE(0) and SRBC sensitization activity (0). Void (V0) and total (VT)volumes were determined by loading blue dextran and 14C aminoacids, respectively, on the same S-200 column. For molecular sizedetermination, dextrans (average molecular sizes, 66,300, 40,200,and 10,000) were chromatographed, and peak values are indicatedby arrows.
amphiphile component. Two independent preparations (Aand B) of amphiphile yielded equivalent relative chemicalcompositions. Ribitol was present at 467 and 611 nmol/mg inthe two samples, respectively. For these studies, ribitol wasquantitated as the sum of anhydroribitol and ribitol. Galac-tose was detected as the primary sugar component of theS. adjacens group antigen (796 and 897 nmol/mg of am-phiphile). Galactosamine, detected at 586 and 758 nmol/mg,constitutes a second abundant carbohydrate in the S. ad-jacens amphiphile. Amino acid analysis of the amphiphileidentified alanine as the only amino acid detectable atquantifiable levels (64 and 40 nmol/mg of amphiphile).
Fatty acid analysis of the S. adjacens amphiphile (Table 3)revealed an average total of 146 nmol of fatty acid per mg ofamphiphile. The major fatty acid detected was palmitic acid(55 nmol), followed in order of relative abundance by palm-itoleic acid (34 nmol), oleic acid (27 nmol), stearic acid (19
TABLE 2. Chemical composition of S. adjacens amphiphileaAmt
Molar ratioComponent ,ug/mg nmol/mg
A B A B A B
Phosphorusb 18.0 19.9 580 641 1.00 1.00Ribitolc 71.0 92.9 467 611 0.80 0.95Galactosed 143.3 161.5 796 897 1.37 1.40Galactosaminee 104.9 135.7 586 758 1.01 1.18Alaninee 5.6 3.6 64 40 0.11 0.06Fatty acidsf 34.9 39.4 138 155 0.24 0.24
a Values are expressed as the means of triplicate analyses.b Quantitated as total phosphorus by the method of Lowry et al. (23).c Quantitated as anhydroribitol and ribitol released from ribitol phosphate.d Quantitated as alditol acetates by gas chromatography.e Quantitated as PITC derivatives by HPLC.f Quantitated as methyl esters by gas chromatography.
TABLE 3. Fatty acid composition of S. adjacens amphiphilea
AmtFatty acid Molar ratioCig/mg nmol/mg
C14:0 1.8 8 0.23C16:0 14.0 55 1.59C16:1 8.8 34 1.00C18:0 5.5 19 0.56C18:1 7.7 27 0.80C20:1 0.6 2 0.06C20:2 0.3 1 0.03
a Values are expressed as the means of triplicate analyses from twoindependent preparations of amphiphile.
nmol), myristic acid (8 nmol), and two C20 unsaturated fattyacids (C20:1, 2 nmol; C20:2, 1 nmol).
Since phosphodiester bonds are commonly found in gram-positive bacterial amphiphiles (8), experiments were per-formed to determine the nature of phosphate linkages in theS. adjacens amphiphile. Treatment of S. adjacens am-phiphile with phosphomonoesterase did not result in therelease of inorganic phosphate. These results indicated thepresence of phosphodiester bonds within the amphiphilestructure. To summarize, the chemical composition of the S.adjacens amphiphile was as follows. Phosphorus, ribitol,galactose, galactosamine, alanine, and fatty acids werepresent in a molar ratio of 1.00:0.88:1.39:1.10:0.08:0.24.
DISCUSSIONPurification of the S. adjacens group antigen was carried
out to provide a sufficient quantity for its chemical charac-terization. The abundance of extracellular amphiphile andthe ease of its isolation relative to cell-associated amphiphileled to the use of stationary-phase culture supernatants as thesource for the purification of S. adjacens amphiphile. Thequantity recovered (22 mg) corresponded to a 3.6% yield ofamphiphile from the initial culture supernatant. Amphiphilesof other gram-positive bacteria are also released from thecell (up to 92% of total amphiphile recovered) during station-ary phase in either the acylated or deacylated form (17).Since release of amphiphiles is not dependent upon cell lysis,functions for secreted amphiphiles have been postulated(25). Weinreb et al. (39) demonstrated that LTAs of severalgram-positive bacteria were capable of sensitizing humanerythrocytes for destruction by autologous complement.These investigators suggest that extracellular amphiphilemight play a role in bacterial infection by rendering hosttissue susceptible to destruction by complement. Extracel-lular LTA could also act as a mediator for host cell destruc-tion, due to its function as a carrier for the oxygen-stablehemolysin streptolysin S (32).The S. adjacens amphiphile was more readily observed by
CIE analysis in the absence rather than the presence ofdetergent (Triton X-100), a phenomenon similar to the anti-genic behavior of the NVS serotype I amphiphile (11), nowdesignated as the S. defectivus group antigen. Detergent-induced dissociation of amphiphile micelles might be oneexplanation for the lack of antibody precipitation. Alterna-tively, the S. adjacens amphiphile may contain antigenicsites requiring a specific conformation which is disrupted inthe presence of detergent. Wessels and Kasper (40) sug-gested that conformational epitopes exist for bacterialpolysaccharides. By using pneumococcal oligosaccharidesof increasing lengths, they demonstrated that large polysac-
i00
0-
ci
VOL. 59, 1991
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 29
Nov
embe
r 20
21 b
y 21
1.22
9.62
.37.

598 SIELING AND VAN DE RIJN
charides (22 repeating units) inhibited antibody recognitionof fixed pneumococcal polysaccharide in an enzyme-linkedimmunosorbent assay (ELISA), whereas double or singlerepeating units were 34 and 36,000 times less effective atELISA inhibition, respectively. Knox et al. (20) foundevidence for potential conformational epitopes in LTA bydemonstrating that a deacylated amphiphile precipitatedonly 30% of the antibody precipitated by intact LTA. Ifconformational epitopes exist for the S. adjacens am-phiphile, the predominant antigenic conformation isolatedfrom culture supernatants is probably the aggregated ormicellar form, since in this form it more readily precipitatesanti-amphiphile antisera.The S. adjacens group antigen was characterized as a
lipopolyribitol teichoic acid similar to that of the S. defecti-vus amphiphile (10). Earlier studies on the serotype or groupantigens of NVS demonstrated that they were not LTAs (3).In contrast to the structure of LTA, the S. adjacens am-phiphile did not contain a glycerolphosphate backbone, asneither paper chromatography nor gas chromatography de-signed for phosphorylated sugars detected glycerol phos-phate (14). Possessing ribitol, galactose, and galactosamine(467, 796, and 586 nmollmg, respectively) as the primarycarbohydrate components detected, the composition of theS. adjacens amphiphile is more reminiscent of lipopolyribitolteichoic acids than the poly(glycerol phosphate) structure ofLTA. Similarity to LPS and nonpoly(glycerol phosphate)polysaccharides was supported when the amphiphile wassubjected to SDS-PAGE. The silver-stained banding patternof S. adjacens amphiphile resembled those of LPS and the S.defectivus group antigen more than the diffuse pattern seenfor LTA. However, SDS-PAGE (Fig. 4) demonstrated alarger apparent size for the S. adjacens amphiphile thaneither LPS (approximate molecular size, 5,000 Da) or LTA(7,500 Da). The molecular size (23,000-24,000 Da) deter-mined for the S. adjacens amphiphile by gel filtration con-firmed its larger size relative to that of either LPS or LTA.Again this size more closely resembles that obtained for theS. defectivus group antigen (10).
Preliminary information regarding the structure of theamphiphile was provided by evidence that the phosphatepresent was stable to phosphomonoesterase treatment, indi-cating the presence of phosphodiester bonds. Phosphodi-ester linkages are commonly found among the pneumococcalcapsular polysaccharides bridging sugars within the polysac-charide backbone or linking side chain components (22).Alanine (0.06 to 0.11 nmollmg) is presumably present as aside chain component in the S. adjacens amphiphile, analo-gous to its position in LTA and other gram-positive am-phiphiles (8). The observed difference in alanine quantitybetween the two preparations of amphiphile was probablydue to the lability of alanine esters in these molecules (9).Compositional analysis of the S. adjacens amphiphile doesnot provide a basis for a structural model, therefore compar-isons to other amphiphile structures with similar composi-tions are speculative. Current experiments examining thepolysaccharide linkages and positions of phosphodiesterbonds will provide more detailed structural information.
Fatty acids comprised approximately 11% of the S. ad-jacens amphiphile's components. The fatty acid residuesmay be bound directly to galactose in the amphiphile viaester linkages or by N acylation through galactosamine.Direct ester linkage of fatty acids to glucose is commonamong LTAs (8), whereas N acylation of amphiphile fattyacids would be analogous to the fatty acid substitution ofLPS through glucosamine. An alternative mode of acylation
of the S. adjacens amphiphile would be via a phosphogly-colipid complex similar to the LTA of S. faecalis (33). Thepresence of glycerol- or glycerol phosphate-linked fattyacids is unlikely due to the absence of detectable glycerolphosphate. However, a terminal diacylglycerol or phosphati-dyldiacylglycerol cannot be ruled out.
Quantitation of components within the S. adjacens am-phiphile revealed that only 38 to 45% of the purified am-phiphile weight was described. The lower quantity of dryweight accounted for may be due to a number of reasons.Neither an elemental analysis nor ash content for inorganiccontaminants was performed due to the limited quantity ofamphiphile available. This we expect would account for themajority of the remainder of the material. Two unidentifiedminor phosphate-containing compounds were detected bygas chromatography, but only at very low concentrations.Glycosidic linkages involving amino sugars are known to berelatively resistant to hydrolysis (15), but there were nocompounds detected by paper chromatography that wereunidentifiable. However, neither of these latter two possibil-ities fully explains the low percentage of analysis.The purified and chemically characterized S. adjacens
amphiphile may now be examined for its predominant anti-genic epitope, which might provide clues as to the involve-ment of NVS amphiphiles in bacterial endocarditis. By usinga rabbit model for bacterial endocarditis, van de Rijn (34)demonstrated that animals immunized with killed NVS wereprotected against endocarditis when challenged with NVS.Protection was serotype specific (now group specific), how-ever, which suggests a role for this group antigen in elicitinga protective immune response against NVS (35). Finally,analysis of the biological activities of the S. adjacens am-phiphile can now be performed with the purified material,which may provide evidence for a pathogenic function of theamphiphile in bacterial endocarditis.
ACKNOWLEDGMENTS
This work was supported by Public Health Service grant HL-30069 from the National Institutes of Health and was supported inpart by the Protein Analysis and Membrane Lipid Core Laboratoriesof the Cancer Center of Wake Forest University (CA12197).
REFERENCES1. Bidlingmeyer, B. A., S. A. Cohen, and T. L. Tarvin. 1984. Rapid
analysis of amino acids using pre-column derivatization. J.Chromatogr. 336:93-104.
2. Bouvet, A., F. Grimont, and P. A. D. Grimont. 1989. Strepto-coccus defectivus sp. nov. and Streptococcus adjacens sp.nov., nutritionally variant streptococci from human clinicalspecimens. Int. J. Syst. Bacteriol. 39:290-294.
3. Bouvet, A., I. van de Rijn, and M. McCarty. 1981. Nutritionallyvariant streptococci from patients with endocarditis: growthparameters in a semisynthetic medium and demonstration of achromophore. J. Bacteriol. 146:1075-1082.
4. Bouvet, A., F. Villeroy, F. Cheng, C. Lamesch, R. Wiliamson,and L. Gutman. 1985. Characterization of nutritionally variantstreptococci by biochemical tests and penicillin-binding pro-teins. J. Clin. Microbiol. 22:1030-1034.
5. Dubois, M., K. A. Gilles, J. K. Hamilton, P. A. Rebers, and F.Smith. 1956. Colorimetric method for determination of sugarsand related substances. Anal. Chem. 28:350-356.
6. Elliott, S. D. 1963. Teichoic acid and the group antigen of lacticstreptococci (group N). Nature (London) 200:1184-1185.
7. Fiedel, B. A., and R. W. Jackson. 1976. Immunogenicity of apurified and carrier-complexed streptococcal lipoteichoic acid.Infect. Immun. 13:1585-1590.
8. Fischer, W. 1988. Physiology of lipoteichoic acids in bacteria.Adv. Microb. Physiol. 29:233-302.
INFECT. IMMUN.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 29
Nov
embe
r 20
21 b
y 21
1.22
9.62
.37.

S. ADJACENS GROUP ANTIGEN 599
9. Fischer, W., H. U. Koch, P. Rosel, and F. Fiedler. 1980. Alanineester-containing native lipoteichoic acids do not act as lipotei-choic acid carrier. Isolation, structural and functional charac-terization. J. Biol. Chem. 255:4557-4562.
10. George, M., and I. van de Ron. 1988. Nutritionally variantstreptococcal serotype I antigen. Characterization as a lipid-substituted poly(ribitol phosphate). J. Immunol. 140:2008-2015.
11. George, M., and I. van de Rijn. 1988. Purification of serotype Iantigen from nutritionally variant streptococci. Infect. Immun.56:1222-1231.
12. Gephart, J. F., and J. A. Washington II. 1982. Antimicrobialsusceptibilities of nutritionally variant streptococci. J. Infect.Dis. 146:536-539.
13. Grimes, W. J., and S. Greegor. 1976. Carbohydrate composi-tions of normal, spontaneously transformed, and virally trans-formed cells derived from BALB/c mice. Cancer Res. 36:3905-3910.
14. Harvey, D. J., and M. G. Horning. 1973. Characterization of thetrimethylsilyl derivatives of sugar phosphates and related com-pounds by gas chromatography and gas chromatography-massspectrometry. J. Chromatogr. 76:51-62.
15. Hellerqvist, C. G., and B. J. Sweetman. 1990. Mass spectrome-try of carbohydrates. Methods Biochem. Anal. 34:91-143.
16. Hewett, M. J., K. W. Knox, and A. J. Wicken. 1970. Studies onthe group F antigen of lactobacilli: detection of antibodies byhemagglutination. J. Gen. Microbiol. 60:315-322.
17. Joseph, R., and G. D. Shockman. 1975. Synthesis and excretionof glycerol teichoic acid during growth of two streptococcalspecies. Infect. Immun. 12:333-338.
18. Kessler, R. E., and G. D. Shockman. 1979. Precursor-productrelationship of intracellular and extracellular lipoteichoic acid ofStreptococcus faecium. J. Bacteriol. 137:869-877.
19. Kessler, R. E., and I. van de Rijn. 1979. Quantitative immuno-electrophoretic analysis of Streptococcus pyogenes membrane.Infect. Immun. 26:892-902.
20. Knox, K. W., M. J. Hewett, and A. J. Wicken. 1970. Studies onthe group F antigen of lactobacilli: antigenicity and serologicalspecificity of teichoic acid preparations. J. Gen. Microbiol.60:303-313.
21. Laemmli, U. K. 1970. Cleavage of structural proteins during theassembly of the head of bacteriophage T4. Nature (London)227:680-685.
22. Larm, O., and B. Lindberg. 1976. The pneumococcal polysac-charides: a re-examination. Adv. Carbohydr. Chem. 33:295-322.
23. Lowry, 0. H., N. R. Roberts, K. Y. Leiner, M. Wu, and A. L.Farr. 1954. The quantitative histochemistry of the brain. J. Biol.Chem. 207:1-17.
24. Lowry, 0. H., W. J. Rosebrough, A. L. Farr, and R. J. Randall.1951. Protein measurement with the Folin-phenol reagent. J.Biol. Chem. 193:265-275.
25. Markham, J. L., K. W. Knox, A. J. Wicken, and M. J. Hewett.1975. Formation of extracellular lipoteichoic acid by oral strep-tococci and lactobacilli. Infect. Immun. 12:378-386.
26. Morrissey, J. H. 1981. Silver stain for proteins in polyacryl-amide gels: a modified procedure with enhanced uniform sensi-
tivity. Anal. Biochem. 117:307-310.27. Roberts, R. B., A. G. Krieger, N. L. Schiller, and K. C. Gross.
1979. Viridans streptococcal endocarditis: the role of variousspecies, including pyridoxal-dependent streptococci. Rev. In-fect. Dis. 1:955-966.
28. Rogozinski, M. 1964. The methanol-sulfuric acid esterificationmethods. II. An improved extraction procedure. J. Gas Chro-matogr. 2:328-329.
29. Rosan, B. 1978. Absence of glycerol teichoic acids in certainoral streptococci. Science 201:918-920.
30. Rotta, J., R. M. Krause, R. C. Lancefield, W. Everly, and H.Lackland. 1971. New approaches for the laboratory recognitionof M types of group A streptococci. J. Exp. Med. 134:1298-1315.
31. Silvestri, L. J., R. A. Craig, E. M. Ingram, E. M. Hoffman, andA. S. Bleiweis. 1978. Purification of lipoteichoic acids by usingphosphatidyl choline vesicles. Infect. Immun. 22:107-118.
32. Theodore, T. S., and G. B. Calandra. Streptolysin S carrieractivity of lipoteichoic acid, p. 271-278. In G. D. Shockman andA. J. Wicken (ed.), Chemistry and biological activities ofbacterial surface amphiphiles. Academic Press, New York.
33. Toon, P., P. E. Brown, and J. Baddiley. 1972. The lipid-teichoicacid complex in the cytoplasmic membrane of Streptococcusfaecalis NCIB 8191. Biochem. J. 127:399-409.
34. van de Rijn, I. 1985. Role of culture conditions and immuniza-tion in experimental nutritionally variant streptococcal en-docarditis. Infect. Immun. 50:641-646.
35. van de Run, I. 1988. Analysis of cross-protection betweenserotypes and passively transferred immune globulin in experi-mental nutritionally variant streptococcal endocarditis. Infect.Immun. 56:117-121.
36. van de Rijn, I., and A. Bouvet. 1984. Characterization of apH-dependent chromophore from nutritionally variant strepto-cocci. Infect. Immun. 43:28-31.
37. van de Ron, I., and M. George. 1984. Immunochemical study ofnutritionally variant streptococci. J. Immunol. 133:2220-2225.
38. van Driel, D., A. J. Wicken, M. R. Dickson, and K. W. Knox.1973. Cellular location of the lipoteichoic acids of Lactobacillusfermenti NCTC 6991 and Lactobacillus casei NCTC 6375. J.Ultrastruct. Res. 43:483-497.
39. Weinreb, B. D., G. D. Shockman, E. H. Beachey, A. J. Swift, andJ. A. Winkelstein. 1986. The ability to sensitize host cells fordestruction by autologous complement is a general property oflipoteichoic acid. Infect. Immun. 54:494-499.
40. Wessels, M. R., and D. L. Kasper. 1989. Antibody recognition ofthe type 14 pneumococcal capsule. Evidence for a conforma-tional epitope in a neutral polysaccharide. J. Exp. Med. 169:2121-2131.
41. Wicken, A. J., S. D. Elliott, and J. Baddiley. 1963. The identityof streptococcal group D antigen and teichoic acid. J. Gen.Microbiol. 31:231-239.
42. Wicken, A. J., and K. W. Knox. 1975. Lipoteichoic acids: a newclass of bacterial antigen. Science 187:1161-1167.
43. Wicken, A. J., and K. W. Knox. 1980. Bacterial cell surfaceamphiphiles. Biochim. Biophys. Acta 604:1-26.
VOL. 59, 1991
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 29
Nov
embe
r 20
21 b
y 21
1.22
9.62
.37.