observations on the efficiency of dystrophic muscle in vitro · mice with a form of hereditary...
TRANSCRIPT

Journal of Neurology, Neurosurgery, and Psychiatry, 1973, 36, 565-573
Observations on the efficiency of dystrophicmuscle in vitro
WILLIAM W. HOFMANN AND ERNST 0. RUPRECHT
From the Department of Neurology, Technical University of Munich, Munich, Germany
SUMMARY A study of muscles of the dystrophic mouse has failed to substantiate earlier claims thatthese muscles were especially resistant to fatigue in vitro or that fast muscles are preferentiallydamaged. It has been found that the fast muscle selected for previous studies is very often unable towithstand isolation in an organ bath if it is working, and both the difficulty in removing the normalgastrocnemius muscle intact and the need to trim it surgically contribute independently toward itsdeterioration in vitro. The smaller dystrophic gastrocnemius muscle is less liable to excision damage,is able to satisfy its resting metabolic needs in nutrient solution, and requires no damaging dissection,but is nevertheless unable to recover normally from fatigue. Using EDL and soleus muscles whichare small enough to withstand isolation in vitro, no differences are found between fatigue patterns ofnormal and dystrophic specimens. Responses to rest, KCI, and 2 mM caffeine are also quite similar,and the only distinguishing biomechanical characteristic we have found in dystrophic mouse muscleis a weaker contraction and a longer total twitch time.
It has been reported, and apparently not widelydisputed, that the gastrocnemius muscle frommice with a form of hereditary muscular dys-trophy is much more resistant to fatigue thanthat from a normal mouse (Sandow and Brust,1958, 1962; Eberstein and Sandow, 1963,Hinterbuchner, Angyan, and Hirsch, 1966).There is nothing in the well-known morpho-logical or biochemical abnormalities of the dys-trophic muscle cell which could be expected toconfer such remarkable durability, and it is verydifficult to reconcile the findings in vitro witheither clinical experience or the motor perform-ance of the afflicted mice. During the course ofexperiments with the same type of dystrophicmice and controls we have observed that widelydivergent fatigue patterns may be recorded indifferent muscles from the same animal, depend-ing upon the conditions of the experiment, andwe have therefore re-examined in both slow andfast muscles the question of relative preservationof strength in muscular dystrophy.
METHODS
All experiments were performed in vitro on musclesremoved from the hind limbs of mice. Twelve dys-
trophic mice of the strain ddy 1291 and 30 normalcontrols of similar age and weight were used. All theanimals were from 8 to 12 weeks of age. The muscleswere removed during ether anaesthesia with theassistance of a watch-maker's loupe and were testedin a specially constructed organ bath having twoseparate force transducers attached. The batharrangement permitted a pair of muscles to bestudied simultaneously, usually a slow and a fastmuscle from the same animal. Muscles from bothhind limbs were often tested in the same experiment,and then either both slow muscles or both fastmuscles were examined together. The temperature ofthe bath was controlled by means of a water jacketand was maintained within 1.50 C during an experi-ment. The range of temperatures selected in allexperiments was 29 ± 1-50 C.The muscles were removed as quickly as possible
and were equilibrated at room temperature for atleast 20 minutes before testing in vigorously oxy-genated solution of the composition described byElmqvist and Quastel (1965). Before mounting, thesoleus muscles were carefully teased away from theunder surface of the gastrocnemius muscle under adissecting microscope, and, in the early experiments,the latter muscle was then gently separated into twoor more separate bundles. Though great care was1 Jackson Laboratory, Bar Harbor, Maine, U.S.A.
565
Protected by copyright.
on February 22, 2020 by guest.
http://jnnp.bmj.com
/J N
eurol Neurosurg P
sychiatry: first published as 10.1136/jnnp.36.4.565 on 1 August 1973. D
ownloaded from

William W. Hofmann and Ernst 0. Ruprecht
taken in the original muscle excision, microscopicexamination of the smaller bundles of the gastro-cnemius muscle usually showed that there werelarge numbers of fibres that had been transected justbelow the very short tendon of origin. In a few casesit was felt that the muscle had been removed withthe heads of origin intact, but then it was alwaysfound that many fibres had been torn or cut in theprocess of separating the whole muscle into smallerbundles. The smaller, flatter, dystrophic gastrocnemiiwere much easier to remove, as the heads of origintended to be flat bands easily accessible deep in thepopliteal fossa, and it was even possible in some ofthese muscles to excise a clearly identifiable piece ofproximal tendon. Further, because of their reducedweight and volume, the dystrophic muscles neededno trimming to remain viable in the organ bath.
Because of the difficulties in obtaining truly com-parable specimens, the gastrocnemius preparationwas then rejected after the initial experiments, exceptas a vehicle for the soleus, and the extensor digitorumlongus (EDL) was used as the fast muscle. The EDLwas very nearly the same weight and shape as thesoleus muscle from the same animal and so lent itselfnicely to an additional comparison between slow andfast muscles. In several experiments the fatiguepatterns of EDL and a relatively small bundle ofgastrocnemius muscle from the same normal animalwere compared and, in two dystrophic animals, asubdissected gastrocnemius bundle from one leg wastested in the same bath with the whole gastrocnemiusmuscle from the other.Recording was by means of individual transducer
units; each containing a strain gauge (Phillips PR9810F) which was part of a separate bridge circuit.The output of these instruments was linear throughthe tension range of the test muscle (up to 80 g) andwas calibrated with known weights during everyexperiment. Since the transducers allowed a dis-placement of about 2 mm/100 g of force, the muscleswere not tested under truly isometric conditions.However, exactly the same recording technique wasused for all the 85 muscles examined. The tensionrecords were obtained with a direct-writing, multi-channel oscillograph with a flat frequency responseup to 500 Hz.2 This instrument gave very satisfactoryand continuous records of serial twitch and tetanusresponses but did not permit detailed analysis oftension rise times. Total twitch times were thereforemeasured from onset to base line return. Bothmuscles lay across large silver electrode bars ofabout 1 mm diameter and were stimulated by meansof a constant current/constant voltage stimulator.3
2 Elema-S^honander Mingograf 8, Siemenis, Germany.3 Multistim, Disa Elektronik, Denmark.
The preparations were not curarized. Rectangularelectrical pulses of 0-5 msec duration and 40 Vamplitude were delivered at either 1 or 100 Hz.Higher tetanic stimulation frequencies did not giveconsistent increases in force at 290 C. After settingthe transducers so that the twitch responses weremaximal, the muscles were carried through a stimula-tion programme designed to test certain aspects offatigue. Sample twitches and tetani were first ob-tained while the muscles were fresh, and then stimuliat 1 Hz were continued, interrupted every four tofive minutes by tetani of from 400 to 1,000 impulseseach, until the responses fatigued and became toosmall to measure accurately. With the dystrophicmuscles the measurement end-point was oftenreached more quickly, as the responses tended to beof lower amplitude to start with. The total stimula-tion programme was much more prolonged thanthat used by earlier workers, extending in severalcases to over 10,000 impulses. After the fatigue pro-gramme all muscles were then tested sequentially forrecovery with rest, ability to recover after a KCl-induced contracture, and for the ability to respond to2mM caffeine.
Measurements were made of the initial twitch andtetanus responses and thereafter at various pointsfrom 100 to 500 impulses apart in the 'run-down'.Tetanus: twitch ratios were calculated from thehighest values for each, and the first measurementsof twitch parameters were always made with freshmuscles before the first tetanus had been delivered.Total twitch time measurements were then repeatedat from 2,000-3,000 impulses. The twitch fatiguecurves of the muscles were all plotted simply as afraction of the first and displayed as a separategroup for each type of preparation. In order to beable to compare results with those of other workers,however, relative twitch values were specificallytabulated at 1,200-2,000 impulses. Relative tetanusfatigue was plotted at 5,000-5,200 impulses. To testthe effects of rest, KC1, and caffeine, the maximumtwitch and tetanus tensions (as well as the fractionaldecay of tension within each tetanus) of the lastresponses before each change were compared withthose some minutes later. Since the rest periods, KCI,or caffeine were given at various times after thebeginning of an experiment, the responses recordedwere always those of a partly fatigued muscle, butthe aim of these experiments was to compare therecuperative powers in the control and diseasedfibres. The muscles selected for comparison werethose in which the test of interest had been made atnearly the same point on the fatigue curve, or afternearly the same total number of impulses. At the endof the experiment the tendons and all traces of fatwere removed from each muscle under a dissecting
566
Protected by copyright.
on February 22, 2020 by guest.
http://jnnp.bmj.com
/J N
eurol Neurosurg P
sychiatry: first published as 10.1136/jnnp.36.4.565 on 1 August 1973. D
ownloaded from
Observations on the efficiency of dystrophic muscle in vitro
microscope, and, after a single blotting, the muscleswere weighed.
RESULTS
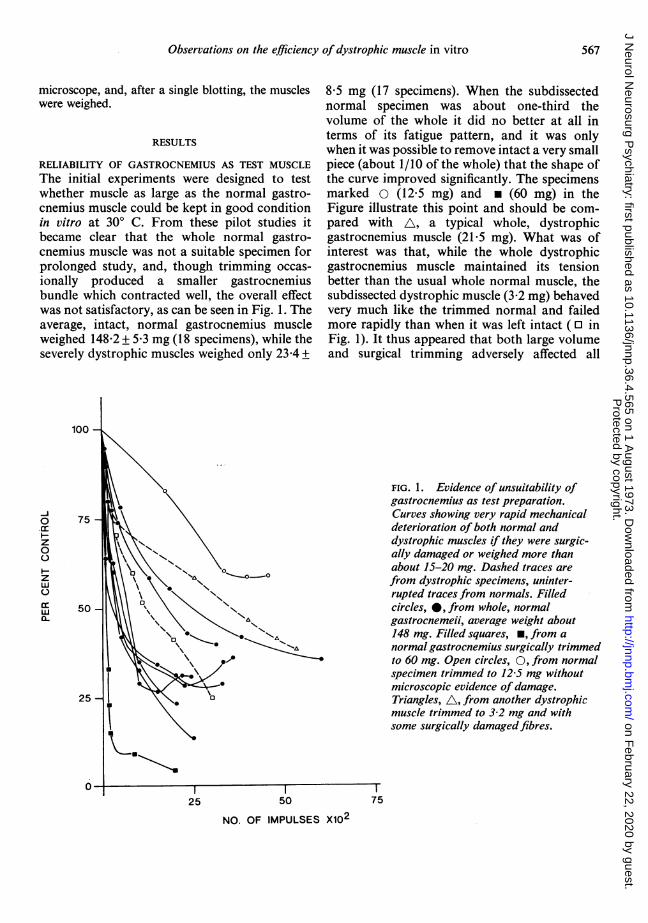
RELIABILITY OF GASTROCNEMIUS AS TEST MUSCLEThe initial experiments were designed to testwhether muscle as large as the normal gastro-cnemius muscle could be kept in good conditionin vitro at 300 C. From these pilot studies itbecame clear that the whole normal gastro-cnemius muscle was not a suitable specimen forprolonged study, and, though trimming occas-ionally produced a smaller gastrocnemiusbundle which contracted well, the overall effectwas not satisfactory, as can be seen in Fig. 1. Theaverage, intact, normal gastrocnemius muscleweighed 148-2 + 5-3 mg (18 specimens), while theseverely dystrophic muscles weighed only 23-4 +
-J0I-Cz0
I-z
crUJa-
100 -
75
50
25
8 5 mg (17 specimens). When the subdissectednormal specimen was about one-third thevolume of the whole it did no better at all interms of its fatigue pattern, and it was onlywhen it was possible to remove intact a very smallpiece (about 1/10 of the whole) that the shape ofthe curve improved significantly. The specimensmarked 0 (12-5 mg) and * (60 mg) in theFigure illustrate this point and should be com-pared with A, a typical whole, dystrophicgastrocnemius muscle (21-5 mg). What was ofinterest was that, while the whole dystrophicgastrocnemius muscle maintained its tensionbetter than the usual whole normal muscle, thesubdissected dystrophic muscle (3-2 mg) behavedvery much like the trimmed normal and failedmore rapidly than when it was left intact (0 inFig. 1). It thus appeared that both large volumeand surgical trimming adversely affected all
FIG. 1. Evidence of unsuitability ofgastrocnemius as test preparation.Curves showing very rapid mechanicaldeterioration of both normal anddystrophic muscles if they were surgic-ally damaged or weighed more thanabout 15-20 mg. Dashed traces arefrom dystrophic specimens, uninter-rupted traces from normals. Filledcircles, 0, from whole, normalgastrocnemeii, average weight about148 mg. Filled squares, m, from anormal gastrocnemius surgically trimmedto 60 mg. Open circles, 0, from normalspecimen trimmed to 12-5 mg withoutmicroscopic evidence of damage.Triangles, A, from another dystrophicmuscle trimmed to 3-2 mg and withsome surgically damagedfibres.
25 50 75
NO. OF IMPULSES X102
567
Protected by copyright.
on February 22, 2020 by guest.
http://jnnp.bmj.com
/J N
eurol Neurosurg P
sychiatry: first published as 10.1136/jnnp.36.4.565 on 1 August 1973. D
ownloaded from

William W. Hofmann and Ernst 0. Ruprecht
0
z0
zU
50
NO. OF IMPULSES X102
(a)
75 25 50 75
NO. OF IMPULSES X102
(b)
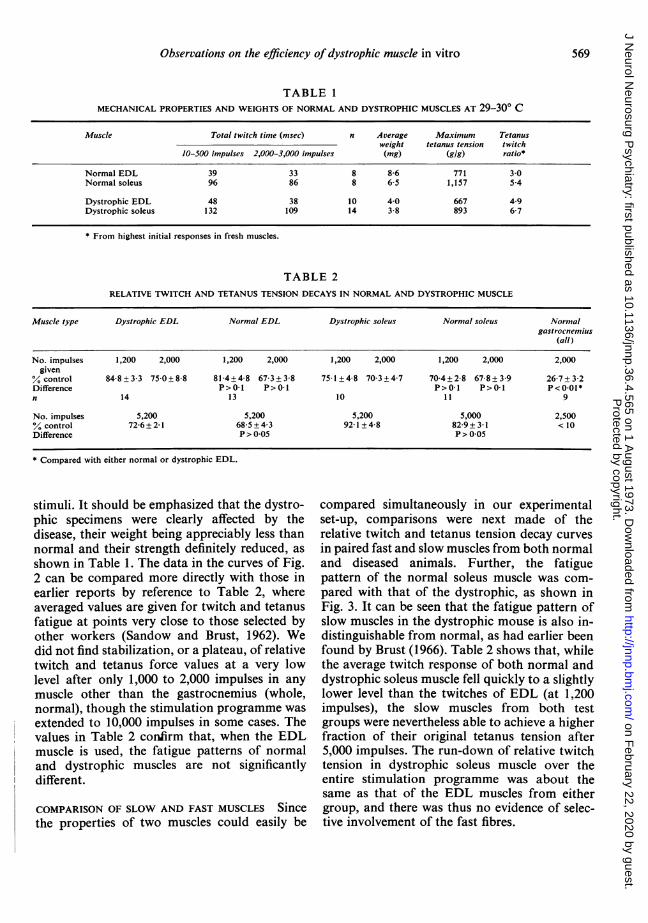
FIG. 2. Relative fatiguability of intact, fast muscles in normal and dystrophic mice. (a) Fatigue curves ofseveral normal EDL muscles; (b) same for dystrophic EDL muscles. Note occasional positive treppe phenom-enon in both groups. Stimulation at I Hz. Temperature 28.5-30O0° C. Each curve fitted by eye to plots of valuesfrom a single muscle.
muscles under the conditions of our experi-ments.
Additional evidence of unsuitability of thegastrocnemius was obtained after the muscles'twitch tensions had dropped to about one halfof their original values. In neither a normalwhole gastrocnemius muscle nor a trimmedbundle was there the slightest recovery with 10-25 minutes' rest, and there was either a very tinyor no response to both isotonic KCl and to2 mM caffeine. Further, with repeated stimula-tion there was no shortening of contraction andrelaxation times in any gastrocnemius specimen.In fact, the tetanus rise and fall time in thesepreparations increased slowly until the musclebecame exhausted. These results are to be con-
trasted with the partly reversible post-fatigueeffects in the smaller extensor digitorum longusand soleus muscles (see below) and are identicalwith responses of muscles intentionally damaged
before testing. Comparisons between the EDLfrom one leg of a normal mouse with the partlydissected or whole gastrocnemius muscle fromthe other revealed that the smaller musclefatigued much less rapidly and that it couldclearly recover after a partial run-down. Thedivergence of fatigue curves in these two normalmuscles was identical with that reported bySandow and Brust (1962) for dystrophic versusnormal gastrocnemii.
COMPARISON OF FAST MUSCLES IN GOOD CONDITIONSince the gastrocnemius muscle would not pro-vide reliable data, the comparison between fastmuscles was made with the extensor digitorumlongus muscle (EDL) in the remaining animals.In the groups of curves of Fig. 2a and Fig. 2b itcan be seen that the EDL muscles from the con-
trol and dystrophic animals did not show signifi-cantly different behaviour during long runs of
0c:.
z00
z
wa-
25
568P
rotected by copyright. on F
ebruary 22, 2020 by guest.http://jnnp.bm
j.com/
J Neurol N
eurosurg Psychiatry: first published as 10.1136/jnnp.36.4.565 on 1 A
ugust 1973. Dow
nloaded from
Observations on the efficiency of dystrophic muscle in vitro
TABLE 1MECHANICAL PROPERTIES AND WEIGHTS OF NORMAL AND DYSTROPHIC MUSCLES AT 29-30 C
Muscle Total twitch time (msec) n Average Maximum Tetanusweight tetanus tension twitch
10-500 impulses 2,000-3,000 impulses (mg) (g/g) ratio*
Normal EDL 39 33 8 8-6 771 3-0Normal soleus 96 86 8 6 5 1,157 5-4
Dystrophic EDL 48 38 10 4-0 667 4-9Dystrophic soleus 132 109 14 3-8 893 6-7
* From highest initial responses in fresh muscles.
TABLE 2RELATIVE TWITCH AND TETANUS TENSION DECAYS IN NORMAL AND DYSTROPHIC MUSCLE
Muscle type Dystrophic EDL Normal EDL Dystrophic soleus Normal soleus Normalgastrocnemius
(all)
No. impulses 1,200 2,000 1,200 2,000 1,200 2,000 1,200 2,000 2,000given
Y. control 848±33 75-0±8-8 81-4+4-8 67-3±3-8 75-1±4-8 703±47 70-4±2-8 67-8±3-9 26-7±3-2Difference P>0-1 P>0-1 P>0-1 P>0-1 P<0-01*n 14 13 10 11 9
No. impulses 5,200 5,200 5,200 5,000 2,500%/ control 72-6±2-1 68-5 ± 4-3 92-1 ±4-8 82 9± 3 1 < 10Difference P > 0-05 P> 0 05
* Compared with either normal or dystrophic EDL.
stimuli. It should be emphasized that the dystro-phic specimens were clearly affected by thedisease, their weight being appreciably less thannormal and their strength definitely reduced, asshown in Table 1. The data in the curves of Fig.2 can be compared more directly with those inearlier reports by reference to Table 2, whereaveraged values are given for twitch and tetanusfatigue at points very close to those selected byother workers (Sandow and Brust, 1962). Wedid not find stabilization, or a plateau, of relativetwitch and tetanus force values at a very lowlevel after only 1,000 to 2,000 impulses in anymuscle other than the gastrocnemius (whole,normal), though the stimulation programme wasextended to 10,000 impulses in some cases. Thevalues in Table 2 confirm that, when the EDLmuscle is used, the fatigue patterns of normaland dystrophic muscles are not significantlydifferent.
COMPARISON OF SLOW AND FAST MUSCLES Since
the properties of two muscles could easily be
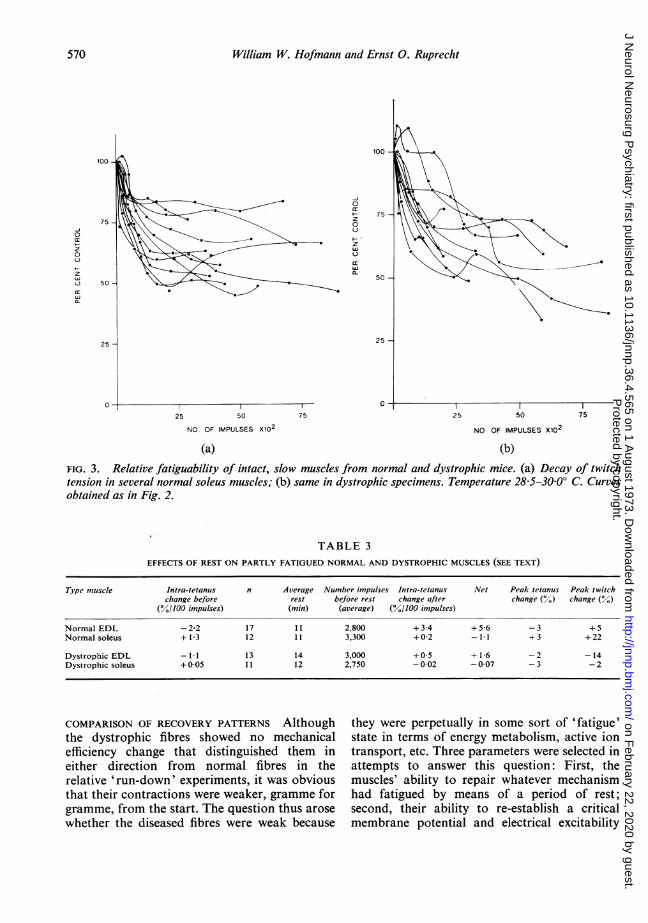
compared simultaneously in our experimentalset-up, comparisons were next made of therelative twitch and tetanus tension decay curvesin paired fast and slow muscles from both normaland diseased animals. Further, the fatiguepattern of the normal soleus muscle was com-pared with that of the dystrophic, as shown inFig. 3. It can be seen that the fatigue pattern ofslow muscles in the dystrophic mouse is also in-distinguishable from normal, as had earlier beenfound by Brust (1966). Table 2 shows that, whilethe average twitch response of both normal anddystrophic soleus muscle fell quickly to a slightlylower level than the twitches of EDL (at 1,200impulses), the slow muscles from both testgroups were nevertheless able to achieve a higherfraction of their original tetanus tension after5,000 impulses. The run-down of relative twitchtension in dystrophic soleus muscle over theentire stimulation programme was about thesame as that of the EDL muscles from eithergroup, and there was thus no evidence of selec-tive involvement of the fast fibres.
569
Protected by copyright.
on February 22, 2020 by guest.
http://jnnp.bmj.com
/J N
eurol Neurosurg P
sychiatry: first published as 10.1136/jnnp.36.4.565 on 1 August 1973. D
ownloaded from
William W. Hofmann and Ernst 0. Ruprecht
0cc
z0
z
ccLLJCL
0Crz0
z
0
a.
25 50
NO. OF IMPULSES X102
(a)
NO OF IMPULSES X102
(b)
FIG. 3. Relative fatiguability of intact, slow muscles from normal and dystrophic mice. (a) Decay of twitchtension in several normal soleus muscles; (b) same in dystrophic specimens. Temperature 28&5-30O° C. Curvesobtained as in Fig. 2.
TABLE 3EFFECTS OF REST ON PARTLY FATIGUED NORMAL AND DYSTROPHIC MUSCLES (SEE TEXT)
Type mluscle Intra-tetanus n Average Number impulses Intra-tetanus Net Peak tetanus Peak twitchchange bejore rest before rest change after change (%) change (%)
(%/100 impulses) (min) (average) (%/100 impulses)
Normal EDL -2-2 17 11 2,800 +3-4 +5-6 -3 +5Normalsoleus +1-3 12 11 3,300 +0-2 -1.1 +3 +22
Dystrophic EDL -1 1 13 14 3,000 +0 5 + 1-6 -2 -14Dystrophic soleus +0 05 11 12 2,750 -0-02 -0-07 -3 -2
COMPARISON OF RECOVERY PATTERNS Althoughthe dystrophic fibres showed no mechanicalefficiency change that distinguished them ineither direction from normal fibres in therelative 'run-down' experiments, it was obviousthat their contractions were weaker, gramme forgramme, from the start. The question thus arosewhether the diseased fibres were weak because
they were perpetually in some sort of 'fatigue'state in terms of energy metabolism, active iontransport, etc. Three parameters were selected inattempts to answer this question: First, themuscles' ability to repair whatever mechanismhad fatigued by means of a period of rest;second, their ability to re-establish a criticalmembrane potential and electrical excitability
570
Protected by copyright.
on February 22, 2020 by guest.
http://jnnp.bmj.com
/J N
eurol Neurosurg P
sychiatry: first published as 10.1136/jnnp.36.4.565 on 1 August 1973. D
ownloaded from

Observations on the efficiency of dystrophic muscle in vitro
TABLE 4POTASSIUM CONTRACTURES IN NORMAL AND DYSTROPHIC MUSCLE
Muscle type Initial response Recovery
Average no. Average time Average half Reactive con- Percent last Percent last Interval nimpulses and to peak and relaxation tracture twitch and tetanus and (min)
range range (sec) time and strength (Y. range rangerange (sec) last tetanus)
and range
Normal EDL 6,200 6 14 27 76 45 5 7(2,500-11,600) (4-8) (10-22) (11-36) (32-114) (23-77)
Normal soleus 6,200 8 21 55 48 51 5 8(2,500-11,600) (4-18) (10-41) (35-90) (46-92) (41-87)
Dystrophic EDL 5,600 10 12 32 60 61 5 12(2,200-11,000) (5-18) (5-18) (15-60) (30-92) (35-100)
Dystrophic soleus 5,600 19 20 43 69 45 5 12(2,200-11,000) (12-51) (11-40) (17-66) (23-85) (19-80)
after a sustained depolarization, and, thirdly,their ability to respond to a potentiating or 'de-fatiguing' drug.As shown in Table 3, recovery with rest alone
was compared in normal and dystrophic EDL aswell as in normal and dystrophic soleus muscle.The very complex changes that followed the restperiod can be considered in terms of three basicfactors; a change in the peak tension availableduring the tetanus, a changed ability to sustainthat tetanus tension, or a change in the maximumtwitch tension. These measurements cannot giveany direct information about the links betweenexcitation and contraction or about the intensityof the active state in the contractile proteinsthemselves because, in a partly fatigued, multi-fibre preparation, the changes after rest may beentirely the result of recruitment. In any case,the normal muscle seems to do slightly betterthan the dystrophic (certainly not the reverse)and may have some type of greater reserve whichis not defined by the test.The KCI effects and recovery are illustrated in
Table 4, the data on the left indicating thefatigued muscles' ability to respond, those onthe right, their ability to recover excitabilityafter five minutes again in normal nutrientsolution. In each of the partly fatigued musclesthe strength of the contracture after KCI was buta fraction of that generated by the last tetanus.Such a finding is not unexpected in view of thefact that, even in the smallest specimens (2-3 mg), diffusion lag precluded simultaneousdepolarization of all the fibres, and it is clear
that, in the multifibre preparation, the KCI con-tracture cannot be used to distinguish failureof excitation-contraction coupling, or surfacecauses of fatigue, from disturbances in thecontractile proteins themselves (Eberstein andSandow, 1963). The data show that the KCIcontracture was weakest in the normal fastmuscles, the normal EDL achieving only aboutone-quarter the force of a tetanus given electric-ally at the same point on the run-down curve.The slow muscles of both dystrophic and normalanimals gave proportionally stronger responses.With regard to the time course of the contrac-tures in all the muscles, it must be emphasizedthat the KCI exposure was never more than 60seconds, after which the bath was very thorough-ly washed. All the muscles had, moreover,already begun to relax by the time the KCI wasremoved, and so the withdrawal of the ionpresumably had little to do with the relaxation.It can be seen that slow muscles give more pro-longed contractures with slower rise times thanthose of fast muscles in both normal and dys-trophic animals, and that the development ofcontracture tension in dystrophic soleus muscleis slowest of all.. The fast muscles of normalmice gave a quicker rise to peak contracturetension than dystrophic muscles, though therelaxation times were about the same.The ability of the various muscles to repolarize
to some critical value was estimated by recording,at a standard interval, the relative return oftwitch and tetanus tension. The rate at which themembranes would approach normal excitability
571
Protected by copyright.
on February 22, 2020 by guest.
http://jnnp.bmj.com
/J N
eurol Neurosurg P
sychiatry: first published as 10.1136/jnnp.36.4.565 on 1 August 1973. D
ownloaded from

William W. Hofmann and Ernst 0. Ruprecht
would, of course, be influenced greatly by thelength of their exposure to the high [K +]solution (Milligan and Edwards, 1965), and inthis connection it is important to emphasize thatall muscles were treated in exactly the same way.Taking exposure time as equal in all experiments,probably the most significant factor in theelectrophysiological recovery would be thedegree to which actual concentration changesacross the membrane had progressed during thedepolarization. The aim of this part of theexperiments was to try to learn if a brief periodin pure, isotonic KCl would cause major 'leaks'in a diseased fibre, after which energy-dependention pumping would be needed to restoreexcitability. If such were the case, the dystrophicmuscles would require more time to recover orwould not recover at all.The data in Table 4 indicate that the slow and
fast muscles from both dystrophic and normalmice can recover from one half to three-quartersof their contracture strength shortly after KCIdepolarization and that the normal seems tohave no particular advantage. Statistical methodscould not be applied to either twitch or tetanusrecovery data as the KCl tests were not all madeat the same points on the fatigue curves of thevarious muscles. It was concluded that thediseased muscle had not suffered either a greaterdisturbance of its internal ionic milieu or anydifficulty with active ion transport and restora-tion of membrane polarization after KClexposure.The caffeine experiments can be summarized
by saying that every muscle was exposed to a2mM concentration of the alkaloid at some pointin the 'run-down' programme. Sometimes thedrug was applied before KCl and sometimesafter, and, in all cases, there were impressivechanges. Within a few seconds both twitch andtetanus tensions had begun to increase, and,where the tension of the previously fatigued fastmuscles had shown a tendency to decreasewithin a tetanus, or to rise very slowly to peak,the caffeine record usually showed a return tothe original, stable pattern. The caffeine poten-tiation was about the same in normal and dys-trophic fibres, and in neither group did a 2 mMconcentration produce a measurable contracture.On the other hand, twitch relaxation times wereclearly prolonged in slow and fast muscles of
both animal groups, and a tetanus after caffeinewas followed by a post-tetanic contracture, mostmarked in the soleus muscle.
DISCUSSION
Since we have not been able to substantiate theclaim that dystrophy confers on either fast orslow skeletal muscle an unusual resistance tofatigue it is essential to describe any technical orexperimental variations which might accountfor the divergent findings. First, we have used adifferent muscle, because in a series of earlyexperiments it became clear that neither a whole,nor a partially subdissected, normal gastrocnem-ius muscle could be kept in good condition invitro. The smaller dystrophic gastrocnemiusmuscle, which could more easily be removedintact and which required no trimming to reduceits volume, performed better in terms of itsrelative fatiguability, but if a normal gastrocnem-ius specimen could be reduced to the same weight,approximately, as the soleus muscle withoutinjury, then that small bundle fatigued no morerapidly than the dystrophic muscle. The import-ance of injury in muscle performance was clearlyshown for the dystrophic gastrocnemius muscle,which failed after subdissection just as quickly asthe trimmed normal muscle. Even the intact,whole dystrophic gastrocnemius muscle showeda very limited ability to recover after 'run-down',and so it appeared that this relatively largemuscle, too, was damaged irreversibly by longstimulation in vitro. Taken together, all thesefindings led us to conclude that no reliable datacould be obtained from the gastrocnemiusmuscle and that what had previously been calledrapid fatigue in relatively large pieces of normalmuscle was in fact cell death.
All significant differences in relative fatigu-ability disappeared when smaller fast musclesfrom the two groups of animals were compared,and it was further observed that the fatiguepatterns of the slow soleus muscles were in-distinguishable.Another point of experimental difference was
that we used a higher temperature. There is noreason to suppose that this change biased resultsin the direction of better survival of muscle, as itis well known that higher temperatures causeearlier deterioration of muscles in vitro. Thirdly,
572
Protected by copyright.
on February 22, 2020 by guest.
http://jnnp.bmj.com
/J N
eurol Neurosurg P
sychiatry: first published as 10.1136/jnnp.36.4.565 on 1 August 1973. D
ownloaded from

Observations on the efficiency of dystrophic muscle in vitro
we did not curarize our preparations. While it istheoretically possible to argue that, after eachdirect shock to the muscles there could have beenrepetitive, indirect activation of some fibres,which would then fatigue earlier, there is noobvious mechanism by which this phenomenonwould have equalized the performance of themuscles. Furthermore, it is clear that indirectactivation would very quickly have failed en-tirely under the conditions of our experiments.No arguments can be made that the EDL
muscle is not affected by the disease or that itdoes not represent the group of pale, fastmuscles of the extremities. Its twitch time was, ifanything, shorter than that of the gastrocnemiusmuscle, suggesting that it may be even freer ofthe admixture of type I fibres known to exist inthe larger muscle (Wuerker, McPhedran, andHenneman, 1965; Dubowitz, 1968). A higherproportion of type IL fibres in the EDL couldhardly be expected to make it more resistant tofatigue during long trains of stimulation in vitro.
In certain other aspects of muscle behaviourour results also have not confirmed the findingsof previous investigators. For example, we didnot find that dystrophic muscle, though slowerthan normal at the beginning, responded toactivity with a greater than normal increase ofcontraction and relaxation rates during repeti-tive stimulation (Sandow and Brust, 1962;Hinterbuchner et al., 1966). The diseased EDLdid show longer total twitch times than thenormal, but both responded with the same rela-tive increases in the velocity of shortening andrelaxing during the first 1,000 to 2,000 impulses.On the other hand, the whole or subdissectednormal gastrocnemius muscle characteristicallydeveloped a progressive delay in both rise topeak tetanic tension and relaxation very soonafter stimulation was begun and then quicklywent on to irreversible exhaustion. This latterbehaviour is typical of a damaged or anoxicmuscle and does not suggest that the dystrophicfibres, by comparison, are especially resistant tostress.
In these experiments we have found far moresimilarity between the diseased and normalfibres than differences with the tests we haveused, and we believe that the previously reportedresiliency of dystrophic muscle, as well as theapparent greater vulnerability of fast fibres to
the disease process (Brust, 1966) were both theresult of an experimental artefact. Laying asidethe paradoxical durability phenomenon, onestill has the question how it can be that tissues sogrossly deranged histologically as dystrophicmuscle fibres can still function so much like thenormal. One possibility is that the finding resultsfrom a major sampling error, the fibres respond-ing to the present series of test being largelyspared by the disease process, while the othershave either not achieved development pastrudimentary stages (Goldspink and Rowe, 1968)or have been replaced with fat or connectivetissue.
The authors acknowledge with thanks the generoussupport of the Deutsche Forschungs Gemeinschaftand the Technical University of Munich and thefacilities provided by Professor A. Struppler, Direc-tor of the Department of Neurology.
REFERENCES
Brust, M. (1966). Relative resistance to dystrophy of slowskeletal muscle of the mouse. American Journal of Physi-ology, 210, 445-451.
Dubowitz, V. (1968). Further studies on fibre types in skeletalmuscle. In Research in Muscular Dystrophy, pp. 100-115.Proceedings of the Fourth Symposium on CurrentResearch, 1968. Edited by members of the MuscularDystrophy Group. Pitman: London.
Eberstein, A., and Sandow, A. (1963). Fatigue mechanismsin muscle fibres. In The Effects of Use and Disuse onNeuromuscular Functions, pp. 515-526. Edited by E.Gutmann and P. Hnik. Elsevier: Amsterdam.
Elmqvist, D., and Quastel, D. M. J. (1965). A quantitativestudy of end-plate potentials in isolated human muscle.Journal of Physiology, 178, 505-529.
Goldspink, G., and Rowe, R. W. D. (1968). The growth anddevelopment of muscle fibres in normal and dystrophicmice. In Research in Muscular Dystrophy, pp. 116-131.Proceedings of the Fourth Symposium on CurrentResearch, 1968. Edited by members of the MuscularDystrophy Group. Pitman: London.
Hinterbuchner, L. P., Angyan, A., and Hirsch, M. (1966).Effect of series of tetani on dystrophic and normal musclesof mouse. American Journal ofPhysiology, 211, 915-918.
Milligan, J. V., and Edwards, C. (1965). Some factorsaffecting the time course of the recovery of contractureability following a potassium contracture in frog striatedmuscle. Journal of General Physiology, 48, 975-983.
Sandow, A., and Brust, M. (1958). Contractility of dystrophicmouse muscle. American Journal of Physiology, 194, 557-563.
Sandow, A., and Brust, M. (1962). Effects of activity oncontractions of normal and dystrophic mouse muscles.American Journal ofPhysiology, 202, 815-820.
Wuerker, R. B., McPhedran, A. M., and Henneman, E.(1965). Properties of motor units in a heterogeneous palemuscle (M. gastrocnemius) of the cat. Journal of Neuro-physiology, 28, 85-99.
573
Protected by copyright.
on February 22, 2020 by guest.
http://jnnp.bmj.com
/J N
eurol Neurosurg P
sychiatry: first published as 10.1136/jnnp.36.4.565 on 1 August 1973. D
ownloaded from