of dna - pnas · proc. nati. acad. sci. usa vol. 89, pp. 6492-6496, july 1992 biochemistry...
TRANSCRIPT

Proc. Nati. Acad. Sci. USAVol. 89, pp. 6492-6496, July 1992Biochemistry
The synapsis event in the homologous pairing of DNAs: RecArecognizes and pairs less than one helical repeat ofDNA
(homologous rmombinatlon/gene mapping/triple-stranded DNA)
PEGGY HSIEH, CAROL S. CAMERINIOTERO*, AND R. DANIEL CAMERINI-OTEROGenetics and Biochemistry Branch, National Institute of Diabetes and Digestive and Kidney Diseases, National Institutes of Health, Bethesda, MD 20892
Communicated by Howard A. Nash, April 9, 1992 (received for review March S, 1992)
ABSTRACT A key step in homologous recombination isthe alignment and pairing of homologous DNAs. The Esche-rnchia coli RecA protein initiates iring by binding to single-strand DNA, forming a helical nucleoprotein filament. Wedemonstrate that in the presence of the nonhydrolyzable ATPanalogue adenosine 5'-[vthioJtrlphosphate and ADP, RecAcan pair a homologous oligonucleotide 15 bases long with aduplex DNA to yield synaptic complexes consisting of theoligonucleotide and duplex DNA stabilized by RecA. RecA canpair as few as eight bases of homology to form such synapticcomplexes. The homologous DNAs remain paired to each otherupon removal of RecA provided that the length of sharedhomology is at least 26 base pairs. Based on our findings andthe work of others, we propose that in vitro, one helical turn ofa RecA nucleoprotein filament containing approximately sixRecA monomers and 15 bases of single-strand DNA is thefunctional unit sufficient to carry out the homology search.
The essential problem in genetic recombination is how ho-mologousDNA sequences find each other and align. Definingthe basic pairing structure that carries out the homologysearch is critical for unraveling the mechanism of homolo-gous pairing by recombinase proteins like the Escherichiacoli RecA protein (reviewed in ref. 1). We reasoned thatdetermination of the structure of strand-exchange interme-diates might be facilitated by limiting pairing to very smallregions. Recently, we described the stable pairing at the endof a linear duplex of very short regions of homology (<60base pairs (bp)] by RecA or recombinases from human cellsor Drosophila embryos (2). The deproteinized product ofthese reactions appears to be a DNA triple helix; Rao et al.(3) have also obtained evidence for such a triple helix. Whilethis structure is interesting in its own right, it remains to beseen how it relates to bona fide strand-exchange intermedi-ates, since it is unlikely that RecA dissociates from strand-exchange intermediates (reviewed in ref. 4). In addition,pairing or synapsis of homologous DNAs during conjuga-tional recombination in E. coli (5) and in meiotic recombi-nation in yeast (6) is probably initiated by invasion of asingle-strand end into a region ofuninterrupted duplex DNA.
In this paper, we show that in the presence of adenosine5'-y-[thio]triphosphate (ATP[,yS]) and ADP cofactors, RecAcan pair an oligonucleotide and a homologous supercoiledduplexDNA to yield a synaptic complex product stabilized bya small number of RecA protomers. We observe that in vitroRecA can pair as few as eight bases, corresponding to abouthalf a helical turn of the RecA nucleoprotein filament (1).
MATERIALS AND METHODSMaterials. Purified E. coli RecA was generously provided
by Stephen C. Kowalczykowski, University of California at
Davis. pBR322 and pUC18 plasmid DNAs and M13mpl8replicative-form DNA were from Pharmacia. Oligonucleo-tides were synthesized and purified by passage over a MonoQ column (Pharmacia) (7). Oligonucleotides were 5'-end-labeled with [y-32P]ATP (New England Nuclear) and T4polynucleotide kinase (Pharmacia) (7).
Oligonucleotides homologous to pUC18 (see Fig. 2)spanned positions 230-285, 230-267, 230-255, and 230-249(8). Oligonucleotides completely homologous to pBR322 (seeFig. 3) spanned positions 15-29, 10-29, and 4359-29 (9). The20L series of oligonucleotides (see Fig. 4) had homology topBR322 at positions 10-29, 20-29, 22-29, 24-29, and 26-29.The 20R series (see Fig. 4) had homology to positions 20-39,20-29, 20-27, 20-25, and 20-23. DNA concentrations areexpressed as moles of nucleotide or by weight.
Synaptic-Complex Formation. Synaptic complexes wereformed by incubating 1.8 1xM oligonucleotide (15 ng), 18 ILMduplex DNA (150 ng), and 1.5 ,uM RecA (1.5 pug) in 25 ,4 of20 mM Tris-HCI, pH 7.5/0.4 mM dithiothreitol/12.5 mMMgCl2/0.3 mM ATP[IyS] (Fluka)/1.1 mM ADP (Sigma) for 15min at 37C. Following synaptic complex formation, 10-20units of the appropriate restriction endonuclease (New En-gland Biolabs) was added and incubation continued for anadditional 5 min. The reaction was quenched by the additionof SDS (1%) and EDTA (10 mM). Reaction mixtures wereelectrophoresed at 0.6 V/cm in 1% agarose gels in 40mM Trisacetate, pH 8.0/1 mM EDTA with ethidium bromide (1,ug/ml) for 14-16 hr at room temperature. Quantitation wasdetermined by densitometer scanning of Polaroid 665 nega-tives. One hundred percent protection of the duplex corre-sponds to the duplex signal from 150 ng of duplex DNAincubated without restriction endonuclease minus the duplexsignal from a control reaction containing duplex DNA, RecA,and the appropriate restriction endonuclease. In some cases,synaptic complex assays were quenched by the addition of1% SDS and electrophoresed at 0.6 V/cm in 1% agarose gelsin 89 mM Tris borate, pH 8.3/5 mM MgCl2 for 14-16 hr atroom temperature.
Deproteinized Joint Molecules. Synaptic complexes wereformed as described above except that 5'-32P-labeled oligo-nucleotides were used. The reactions were quenched by theaddition of SDS and EDTA. After electrophoresis and auto-radiography, joint molecules were quantified by densitomet-ric scanning.
RESULTSFormation of Synaptic Complexes and Joint Molecules.
Formation of synaptic complexes is accomplished by incu-bating a duplex DNA, such as a supercoiled plasmid DNA,with a homologous oligonucleotide and RecA (Fig. 1). The
Abbreviation: ATP[yS], adenosine 5'-[y-thio]triphosphate.*Present address: Diabetes Branch, National Institute of Diabetesand Digestive and Kidney Diseases, National Institutes of Health,Bethesda, MD 20892.
6492
The publication costs of this article were defrayed in part by page chargepayment. This article must therefore be hereby marked "advertisement"in accordance with 18 U.S.C. §1734 solely to indicate this fact.

Proc. Natl. Acad. Sci. USA 89 (1992) 6493
SDS
FIG. 1. Scheme for the formation of synaptic complexes andstable joint molecules. A duplex DNA and a homologous oligonu-cleotide are incubated in the presence of E. coli RecA to form acomplex we call a synaptic complex, in which the two DNAs arepaired within a RecA nucleoprotein filament. The formation ofsynaptic complexes is monitored by the inability of a restrictionendonuclease (R.E.) to cleave the duplex within the region of pairing.IfRecA is removed from synaptic complexes by the addition of SDS,stable deproteinized joint molecules result.
oligonucleotide spans a restriction endonuclease recognitionsite in the duplex DNA. Formation of a synaptic complexinvolving RecA, oligonucleotide, and the duplex DNA ren-
ders the duplex resistant to cleavage by the restrictionendonuclease. The restriction endonuclease footprint corre-
sponding to a synaptic complex can be visualized in ethidiumbromide-stained agarose gels as supercoiled plasmid DNAremaining after incubation of complexes with the appropriaterestriction endonuclease. RecA can be dissociated from thesesynaptic complexes by adding EDTA and SDS, and depro-teinized joint molecules result in which the 32P-labeled oli-gonucleotide migrates on agarose gels at the position ofduplex DNA. Since our primary aim was to isolate synapticcomplexes or a closely related structure that might representan intermediate in the strand exchange reaction, a usefulmeasure for such synaptic complexes was that they yieldedstable joint molecules consisting solely of the two pairedDNAs when RecA was removed. We began by definingparameters that optimize the formation of such stable, de-proteinized joint molecules. Subsequently, we characterizedthe synaptic complexes that gave rise to suchjoint molecules.When RecA and the DNA substrates were coincubated in
the presence of ATP, deproteinized joint molecules were
recovered after 1 min at 37TC. However, the yield of jointmolecules diminished with increasing incubation times. After3 min, recovery of joints was reduced by half; after 15 min,no joints were recovered (data not shown). Since in allexperiments the samples were deproteinized and electropho-resed under identical conditions after incubation with RecA,we postulate that the decreasing yield ofjoint molecules was
not due to instability of the deproteinized products them-selves but was due to a RecA-promoted dissociation in thepresence of ATP (10). The short half-life of these jointmolecules formed in the presence of ATP made it infeasibleto isolate synaptic complexes under these reaction condi-tions.
After some experimentation, we observed that in theabsence of ATP, but in the presence of 0.3 mM ATP[yS] and1.1 mM ADP, RecA formed joint molecules between a
homologous 32P-labeled oligonucleotide and pUC18 DNAthat were stable upon removal of protein (Fig. 2B). As few as26 bases ofhomology shared between the oligonucleotide andthe duplex DNA were sufficient for the formation of jointmolecules in this assay, whereas 20 bases of homology werenot sufficient. Quantitation of the recovery of stable jointmolecules in three experiments indicates that 20-50% of thepUC18 duplex was paired with a homologous oligonucleotide56 bases long when the complexes were electrophoresed inTris/acetate/EDTA buffer. As was observed for synapticcomplexes (see below), the efficiency ofjoint-molecule for-mation by RecA increased as a function of the length ofhomology available for pairing. When the superhelical strainwas relieved upon linearization of the duplex at a restriction
AHind III Sac
5 ..AAGCTTGCATGCCTGCAGGTCGACTCTAGAGGATCCCCGGGTACCGAGcTcGAATTc....3' ...TTCGAACGTACGGACGTCCAGCTGAGATCTCCTAGGGGCCCATGGCTCGAGCTTAAG...
t tHind III Sac _
31
B
5-
TC GAACGTACGGACGTC CAGCTGAGATCTCCTAGGGGCCCATGGCTCG AGCTTAAG 56
38
26
20
1 2 3 4 5 6 7 8 9 10oligo 20 -56- -38- -26- -20- 38RecA + - + - + - + - + +Sad - + + + + + + + + Hindill
...ugly.*$4 v * *...* *m~~~~~~~~~.:**
sC -.-
c1 2 3
oligo 20 -56-RecA + - +Sad - + +
4 5-38-
_+
+ +
6 7-26-_ +
+ +
8 9-20-
_+
+ +
1038+
Hindill
SC --
FIG. 2. Joint molecules and synaptic complexes formed by RecA.(A) Oligonucleotides having 56, 38, 26, or 20 bases of homology topUC18 duplexDNA that span a Sac I site. (B) Joint molecules formedby RecA. 32P-labeled oligonucleotide (oligo), pUC18 supercoiledplasmid DNA, and RecA protein were coincubated as described inMaterials and Methods. After incubation with the appropriate re-striction endonuclease, the reaction mixtures were brought to 1%SDS and electrophoresed in a 1% agarose gel containing ethidiumbromide. Autoradiography reveals the formation ofjoint moleculesas indicated by the presence of 32p label migrating at the position ofsupercoiled (sc) plasmid DNA. (C) Synaptic complexes formed byRecA. Visualization of the ethidium bromide-stained gel prior toautoradiography reveals the footprint of synaptic complexes repre-sented by the presence of supercoiled plasmid DNA remaining afterincubation with a restriction endonuclease. 1, Linear duplex.
endonuclease site located outside the region of pairing, thedeproteinized joint molecules dissociated (Fig. 2B, lane 10).The formation of joint molecules exhibited a sharp opti-
mum at an oligonucleotide concentration of 1.2 uM, with 0.9juM oligonucleotide yielding no joint molecules; increasingthe oligonucleotide concentration to 1.8 ,uM resulted in no
R.E.
RecAooO. o.
Biochemistry: Hsieh et al.

Proc. Natl. Acad. Sci. USA 89 (1992)
further increase in the yield of joint molecules (data notshown). The amount of RecA used in these assays (1.5 AuM)is saturating with respect to the concentration of bases in thesingle-strand oligonucleotides. The ready detection of syn-aptic complexes and joint molecules was dependent on thepresence of both ATP['yS] and ADP in a 1:4 molar ratio.Alteration of the ratio of ATP[yS] to ADP or the concentra-tion of these two cofactors reduced the efficiency of bothsynaptic-complex and joint-molecule formation (L. Ferrinand R.D.C.-O., unpublished observations). For example, at0.1 mM ATP[yS] and 1.0 mM ADP, the yield of jointmolecules was reduced by half. Joint-molecule assays per-formed in the absence of ADP but with 0.25, 0.5, 1, 5, or 10mM ATP[yS] yielded at most 25% (at 10mM ATP[,yS]) of thejoint molecules obtained under optimal conditions. Finally,no joint molecules were observed when ADP was the solenucleotide cofactor.Mg2+ stabilizes deproteinized joint molecules containing
superhelical DNA. Synaptic complexes were deproteinizedby the addition of SDS alone and the reaction products wereelectrophoresed in agarose gels containing 5 mM MgCl2. Therecovery ofjoint molecules in the presence of Mg2+ was 2- to4-fold higher than when Mg2+ was omitted from the electro-phoresis step. However, no qualitative differences wereobserved; i.e., stable joint molecules were formed witholigonucleotides containing 26 bases but not 20 bases ofhomology (data not shown).Having defined reaction conditions that yielded stable joint
molecules, we turned to the formation of synaptic complexescontaining a RecA nucleoprotein filament. Synaptic com-plexes were formed with as few as 20 bases of homologyshared between the oligonucleotide and the duplex DNA(Fig. 2C, lane 9). Quantitation of the supercoiled DNAprotected from digestion by Sac I indicated that 70-75% ofthe duplexDNA was present as synaptic complexes when theoligonucleotide contained 56 or 38 bases of homology (lanes3 and 5). Fifty-five percent and 20% of the duplex wereconverted to synaptic complexes with 26 and 20 bases,respectively, ofhomology. Formation of synaptic complexesrequired the presence of both RecA and the homologousoligonucleotide. As expected, when the duplex was incu-bated with oligonucleotide and RecA, but without restrictionenzyme, the supercoiled DNA remained intact (lane 1). Thefootprint of the synaptic complex did not extend appreciablybeyond the region of the duplex that was colinear with theoligonucleotide; for example, HindIII cleaved at a site 14 bpfrom the region spanned by the 38-base oligonucleotide (lane10). If, however, synaptic complexes were formed with the56-base oligonucleotide, protection from HindIII cleavagewas observed (data not shown).We mapped the extent of protection from restriction en-
donuclease cleavage conferred by a RecA synaptic complex(data not shown). When we formed three separate synapticcomplexes with one of three different 20-base oligonucleo-tides spanning different regions of the polylinker of pUC18,we found that the footprint of the synaptic complex extends11-14 bases beyond the 5' and 3' ends of the paired oligo-nucleotide. We do not know whether the footprint resultedfrom steric hindrance by the synaptic complex or from a localdeformation of the duplex DNA that prevented bindingand/or cleavage by a restriction endonuclease.
Superhelical strain is not essential for the formation ofsynaptic complexes involving very short regions of homol-ogy. RecA converted 78% of a linear pBR322 duplex tosynaptic complexes when reacted with a homologous oligo-nucleotide 33 bases long (data not shown).The formation of synaptic complexes involving 56 bp of
homology does not exhibit directionality. Synaptic com-plexes were formed with about equal efficiencies when weused a 56-base oligonucleotide homologous to the polylinker
region of pUC18 or one of several other oligonucleotideshaving the same 56-base sequence plus 20 bases of nonho-mologous sequence at the 5' end, at the 3' end, or at both the5' and 3' ends of the oligonucleotide (data not shown).A Minimum Structure That Carries Out the Homology
Search. Three oligonucleotides, 33, 15, or 13 bases long andhomologous to a pBR322 sequence (Fig. 3A), were incubatedwith pBR322 supercoiled plasmid DNA in the presence ofRecA. The 15-base oligomer was of sufficient length to formsynaptic complexes with a 50-70%o yield as evidenced byresistance to cleavage by HindIII and Cla I (Fig. 3C, lanes 4and 7). In this assay, use ofa 33-base oligomer resulted in theconversion of 80-90%o of the duplex to synaptic complexes(i.e., protection from cleavage by EcoRI, Cla I, and HindIII),but a 13-base oligomer yielded no synaptic complexes (datanot shown). The formation of synaptic complexes requiredthe presence ofboth oligonucleotide and RecA. As expected,the footprint of the synaptic complex did not extend appre-ciably beyond the region of pairing (see Fig. 3C, lane 10).
Since the formation of a presynaptic filament composed ofRecA and single-strand DNA is a prerequisite for pairing (1),it is likely that the failure to form synaptic complexes with a13-base oligonucleotide is attributable to the previously de-scribed inability of RecA to bind to single-strand DNAsshorter than 15 bases in the presence of ATP[yS] (11). We
A
B
cLANESOLIGO 15b -
REC AREST ENZ -
5' AGCTTATCATCGATA 3'
5' ...AGCTTATCATCGATA...3' ...TCGAATAGTAGCTAT...
Cla Hind III
Hind Cia Eco RI
FIG. 3. Minimal searching unit for homologous pairing. (A)Oligonucleotides 33, 15, or 13 bases long that are homologous topBR322. Positions of EcoRI (E), Cla I (C), and HindIII (H) restric-tion endonuclease sites in the duplex are shown. (B) Nucleotidesequence of the 15-base oligonucleotide and corresponding duplexsequence showing positions of Cla I and HindIII cleavage. Bases inthe duplex that comprise all or part of the recognition sequence forCla I and HindIII are indicated in bold. (C) Formation of synapticcomplexes with the 15-base oligonucleotide (oligo 15b). Lane 1,incubation without oligonucleotide, RecA, or restriction enzyme;lanes 2-4, incubation with HindIll; lanes 5-7, incubation with Cla I;lanes 8-10, incubation with EcoRI.
6494 Biochemistry: Hsieh et al.
Proc. Natl. Acad. Sci. USA 89 (1992) 6495
Table 1. Oligonucleotide series with decreasing homologyto pBR322
Homology,Series Sequence (5' to 3') bases
20L TTGACAGCTTATCAZTCGAT- 20GAATATATGCATCATCGATA 10GAATATATGCCACATCGATA 8GAATATATGCCATGTCGATA 6GAATATATGCCATGGAGATA 4GAATATATGCCATGGATCGT 0
20R ATCATCGATALGCTTTAATG 20ATCATCGATAGAATATATGC 10ATCATCGAGCGAATATATGC 8ATCATCTCGCGAATATATGC 6ATCAGATCGCGAATATATGC 4CGACGATCGCGAATATATGC 0
Bold type corresponds to homologous pBR322 sequence. Under-lined sequence corresponds to the Cla I site on the duplex.
confirm below that this is the case under our experimentalconditions.
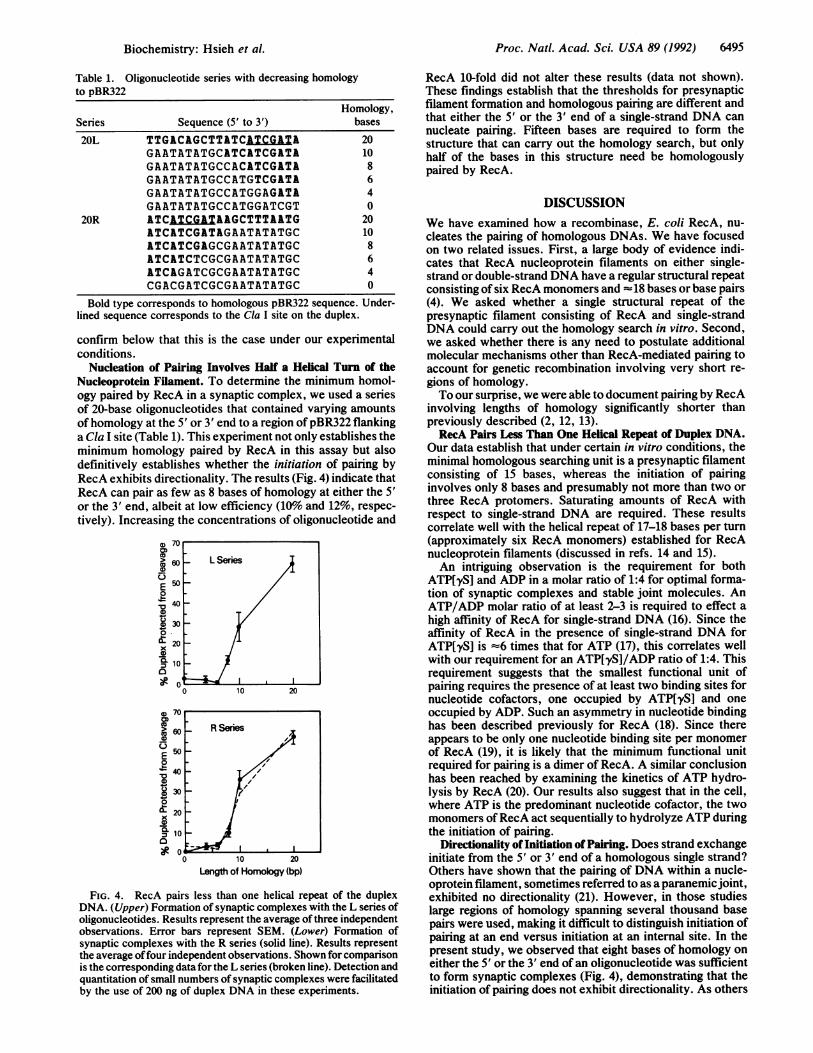
Nucleation of Pairing Involves Half a Helical Turn of theNucleoprotein Filament. To determine the minimum homol-ogy paired by RecA in a synaptic complex, we used a seriesof 20-base oligonucleotides that contained varying amountsofhomology at the 5' or 3' end to a region ofpBR322 flankinga Cla I site (Table 1). This experiment not only establishes theminimum homology paired by RecA in this assay but alsodefinitively establishes whether the initiation of pairing byRecA exhibits directionality. The results (Fig. 4) indicate thatRecA can pair as few as 8 bases of homology at either the 5'or the 3' end, albeit at low efficiency (10%o and 12%, respec-tively). Increasing the concentrations of oligonucleotide and
CD0,
Cu
E!0
4V'a,0
01x
a,
'U0,7cm
~60
E 500
z40
~30
X20
'C=
C]Or
10 20
Length of Homology (bp)
FIG. 4. RecA pairs less than one helical repeat of the duplexDNA. (Upper) Formation of synaptic complexes with the L series ofoligonucleotides. Results represent the average of three independentobservations. Error bars represent SEM. (Lower) Formation ofsynaptic complexes with the R series (solid line). Results representthe average offour independent observations. Shown for comparisonis the corresponding data for the L series (broken line). Detection andquantitation of small numbers of synaptic complexes were facilitatedby the use of 200 ng of duplex DNA in these experiments.
RecA 10-fold did not alter these results (data not shown).These findings establish that the thresholds for presynapticfilament formation and homologous pairing are different andthat either the 5' or the 3' end of a single-strand DNA cannucleate pairing. Fifteen bases are required to form thestructure that can carry out the homology search, but onlyhalf of the bases in this structure need be homologouslypaired by RecA.
DISCUSSIONWe have examined how a recombinase, E. coli RecA, nu-cleates the pairing of homologous DNAs. We have focusedon two related issues. First, a large body of evidence indi-cates that RecA nucleoprotein filaments on either single-strand or double-strand DNA have a regular structural repeatconsisting of six RecA monomers and -18 bases or base pairs(4). We asked whether a single structural repeat of thepresynaptic filament consisting of RecA and single-strandDNA could carry out the homology search in vitro. Second,we asked whether there is any need to postulate additionalmolecular mechanisms other than RecA-mediated pairing toaccount for genetic recombination involving very short re-gions of homology.To our surprise, we were able to document pairing by RecA
involving lengths of homology significantly shorter thanpreviously described (2, 12, 13).RecA Pairs Less Than One Helical Repeat of Duplex DNA.
Our data establish that under certain in vitro conditions, theminimal homologous searching unit is a presynaptic filamentconsisting of 15 bases, whereas the initiation of pairinginvolves only 8 bases and presumably not more than two orthree RecA protomers. Saturating amounts of RecA withrespect to single-strand DNA are required. These resultscorrelate well with the helical repeat of 17-18 bases per turn(approximately six RecA monomers) established for RecAnucleoprotein filaments (discussed in refs. 14 and 15).An intriguing observation is the requirement for both
ATP[yS] and ADP in a molar ratio of 1:4 for optimal forma-tion of synaptic complexes and stable joint molecules. AnATP/ADP molar ratio of at least 2-3 is required to effect ahigh affinity of RecA for single-strand DNA (16). Since theaffinity of RecA in the presence of single-strand DNA forATP[yS] is =6 times that for ATP (17), this correlates wellwith our requirement for an ATP[yS]/ADP ratio of 1:4. Thisrequirement suggests that the smallest functional unit ofpairing requires the presence of at least two binding sites fornucleotide cofactors, one occupied by ATP[yS] and oneoccupied by ADP. Such an asymmetry in nucleotide bindinghas been described previously for RecA (18). Since thereappears to be only one nucleotide binding site per monomerof RecA (19), it is likely that the minimum functional unitrequired for pairing is a dimer of RecA. A similar conclusionhas been reached by examining the kinetics of ATP hydro-lysis by RecA (20). Our results also suggest that in the cell,where ATP is the predominant nucleotide cofactor, the twomonomers ofRecA act sequentially to hydrolyze ATP duringthe initiation of pairing.
Directionality of Initiation of Pairing. Does strand exchangeinitiate from the 5' or 3' end of a homologous single strand?Others have shown that the pairing of DNA within a nucle-oprotein filament, sometimes referred to as a paranemicjoint,exhibited no directionality (21). However, in those studieslarge regions of homology spanning several thousand basepairs were used, making it difficult to distinguish initiation ofpairing at an end versus initiation at an internal site. In thepresent study, we observed that eight bases of homology oneither the 5' or the 3' end of an oligonucleotide was sufficientto form synaptic complexes (Fig. 4), demonstrating that theinitiation of pairing does not exhibit directionality. As others
Biochemistry: Hsieh et al.

Proc. Natl. Acad. Sci. USA 89 (1992)
have observed (reviewed in ref. 1), subsequent steps, includ-ing stable joint-molecule formation, have a directionality.
Conversion of Synaptic Complexes to Joint Molecules. WhenRecA was removed from the synaptic complexes, we ob-served stable joint molecules consisting of an oligonucleotidepaired to a supercoiled duplex over lengths of homologysimilar to those previously reported (2). Our data are con-sistent with a mechanistic pathway in which the formation ofsynaptic complexes gives rise to stable joint molecules. Theformation of both deproteinized joint molecules and synapticcomplexes containing RecA is dependent on the length ofhomology available for pairing. Also, the formation of bothsynaptic complexes and joint molecules occurs with rela-tively high efficiency for 56 bases of homology; that is, upondeproteinization of these synaptic complexes, most of theduplex DNA is still paired with the oligonucleotide in stablejoint molecules in the presence of Mg2+ (see above). Inaddition, synaptic complexes are formed sufficiently rapidlyto be on the reaction pathway (22). Under no conditionsexamined did we observe joint molecules in the absence ofsynaptic-complex formation. In other systems, the use ofnonhydrolyzable nucleoside triphosphate analogues such asATP[yS] has been successfully exploited to characterizestructures that presumably bear a close relationship to a truereaction intermediate. In addition, RecA can carry out strandexchange in the presence of ATP[yS] (23, 24). We proposethat the synaptic complexes described here are structurallyrelated to a strand-exchange intermediate; however, demon-stration that the putative intermediate reacts sufficientlyrapidly to yield product to be on the pathway (see ref. 22) willrequire further work.Although there is evidence for the formation of a triple
helix when a single-strand circular DNA is paired at the endofa linear duplex (2, 3), we have no evidence that the synapticcomplexes or joint molecules formed between a supercoiledduplex DNA and an oligonucleotide have such a triple-helicalstructure. The restriction endonuclease protection assay forsynaptic complexes does not possess sufficient resolution toreveal the disposition of the three DNA strands in the regionof pairing. That the joint molecules in this study are stabilizedby Mg2+ and superhelicity whereas the ones formed fromlinear duplexes are not suggests that these two sets of jointmolecules are different. Thus, although the structure of thepaired region in joint molecules containing supercoiled du-plexes could be a triplex or closely related structure, it isequally likely that strand exchange may have occurred,creating a displacement loop (D-loop).
Biological Implications. Our observations on the pairing ofexceedingly short regions of homology by RecA are consis-tent with genetic data from both prokaryotes and eukaryotesindicating that short homologies (<50 bp) are substrates forrecombination in vivo, albeit at low frequencies (referencescited in ref. 2). In addition, recent evidence points to recom-bination involving <10 bp in yeast (25) as well as in the humanmitochondrial genome (refs. 26 and 27 and references there-in). Our data establish a plausible explanation for suchresults-that RecA-like proteins can mediate such recombi-nation events involving short homologies in vivo-and thatthere is no need to invoke other mechanisms (e.g., copychoice) to explain such phenomena.Based on the synaptic complexes described here, it was
recently shown that RecA possesses a high degree of dis-crimination for sequence homology, allowing targeting of anoligonucleotide to a specific site on duplex DNA of largecomplexity, and it is now possible to achieve site-specificexcision of large DNA fragments from human genomic DNA,a useful step in mapping this and other complex genomes (28).In that study it was demonstrated that synaptic complexesinhibit the action of other DNA-modifying enzymes, such asDNA methyltransferases. If the targeting by RecA of an
oligonucleotide to a duplex sequence in a complex genome isa paradigm for the pairing of homologous chromosomes ineukaryotes, then a study of the parameters of the in vitroreaction, such as the effects of sequence context and sub-strate concentrations on the kinetics ofpairing, might providesome insight into the process of chromosome pairing duringmeiosis.
Finally, one can now employ an extensive array of meth-ods used to study sequence-specific DNA-binding proteins toexamine the alignment of an oligonucleotide with a targetduplex DNA in a RecA synaptic complex. The alignment byRecA of an oligonucleotide to its "target" sequence on aDNA duplex is very specific, involves a limited number ofRecA molecules and DNA bases, and represents the thresh-old event for homologous pairing. We hope to be able torelate this structure to our previously described triple-helicalstructure (2).
We thank Lance Ferrin, Moon Kim, Richard Proia, and TammyTobin of the National Institutes of Health for comments on themanuscript. We thank Stephen Kowalczykowski for providing E.coli RecA protein and George Poy for oligonucleotide syntheses. Weare grateful to Linda Robinson for her assistance.
1. Cox, M. M. & Lehman, I. R. (1987) Annu. Rev. Biochem. 56,229-262.
2. Hsieh, P., Camerini-Otero, C. S. & Camerini-Otero, R. D.(1990) Genes Dev. 4, 1951-1963.
3. Rao, B. J., Dutreix, M. & Radding, C. M. (1991) Proc. Nati.Acad. Sci. USA 88, 2984-2988.
4. Kowalczykowski, S. C. (1991) Annu. Rev. Biophys. Biophys.Chem. 20, 539-575.
5. Smith, G. R. (1991) Cell 64, 19-27.6. Sun, H., Treco, D. & Szostak, J. W. (1991) Cell 64, 1155-1161.7. Hsieh, P. & Camerini-Otero, R. D. (1989) J. Biol. Chem. 264,
5089-5097.8. Norrander, J., Kempe, T. & Messing, J. (1983) Gene 26,
101-106.9. Sutcliffe, J. G. (1978) Cold Spring Harbor Symp. Quant. Biol.
49, 561-570.10. Shibata, T., Ohtani, T., Iwabuchi, M. & Ando, T. (1982) J. Biol.
Chem. 257, 13981-13986.11. Leahy, M. C. & Radding, C. M. (1986) J. Biol. Chem. 261,
6954-6960.12. Gonda, D. K. & Radding, C. M. (1983) Cell 34, 647-654.13. Cheng, S., Van Housten, B., Gamper, H. B., Sancar, A. &
Hearst, J. E. (1988) J. Biol. Chem. 263, 15110-15117.14. Egelman, E. H. & Stasiak, A. (1986) J. Mol. Biol. 191, 677-697.15. Heuser, J. & Griffith, J. (1989) J. Mol. Biol. 210, 473-484.16. Menetski, J. P., Varghese, A. & Kowalczykowski, S. C. (1988)
Biochemistry 27, 1205-1212.17. Kowalczykowski, S. C. (1986) Biochemistry 25, 5872-5881.18. Lauder, S. D. & Kowalczykowski, S. C. (1991) J. Biol. Chem.
266, 5450-5458.19. Cotterill, S. M., Satterthwait, A. C. & Fersht, A. R. (1982)
Biochemistry 21, 4332-4337.20. Menge, K. L. & Bryant, F. R. (1988) Biochemistry 27, 2635-
2640.21. Wu, A. M., Kahn, R., DasGupta, C. & Radding, C. M. (1982)
Cell 30, 37-44.22. Fersht, A. (1985) Enzyme Structure and Mechanism (Freeman,
New York), pp. 193-219.23. Menetski, J. P., Bear, D. G. & Kowalczykowski, S. C. (1990)
Proc. Natl. Acad. Sci. USA 87, 21-25.24. Rosselli, W. & Stasiak, A. (1990) J. Mol. Biol. 216, 335-352.25. Schiestl, R. H. & Petes, T. D. (1991) Proc. Natl. Acad. Sci.
USA 88, 7585-7589.26. Rotig, A., Cormier, V., Blanche, S., Bonnefont, J.-P., Ledeist,
F., Romero, N., Schmitz, J., Rustin, P., Fischer, A., Saudu-bray, J.-M. & Munnich, A. (1990) J. Clin. Invest. 86, 1601-1608.
27. Zeviani, M., Bresolin, N., Gellera, C., Bordoni, A., Pannacci,M., Amati, P., Moggio, M., Servidei, S., Scarlato, G. &DiDonato, S. (1990) Am. J. Hum. Genet. 47, 904-914.
28. Ferrin, L. J. & Camerini-Otero, R. D. (1991) Science 254,1494-1497.
64% Biochemistry: Hsieh et al.