off-target function of the sonic hedgehog inhibitor cyclopamine in
TRANSCRIPT

Therapeutic Discovery
Off-Target Function of the Sonic Hedgehog InhibitorCyclopamine in Mediating Apoptosis via Nitric Oxide–Dependent Neutral Sphingomyelinase 2/Ceramide Induction
Marisa Meyers-Needham1,2, Jocelyn A. Lewis2,3, Salih Gencer1,2, R. David Sentelle1,2, Sahar A. Saddoughi1,2,Christopher J. Clarke1,2, Yusuf A. Hannun1,2, Haakan Norell2,4, TelmaMartins da Palma2,4, Michael Nishimura2,4,Jacqueline M. Kraveka2,3, Zohreh Khavandgar5, Monzur Murshed5, M. Ozgur Cevik1,2, and Besim Ogretmen1,2
AbstractSonic hedgehog (SHh) signaling is important in the pathogenesis of various human cancers, such as
medulloblastomas, and it has been identified as a valid target for anticancer therapeutics. The SHh inhibitor
cyclopamine induces apoptosis. The bioactive sphingolipid ceramide mediates cell death in response to
various chemotherapeutic agents; however, ceramide’s roles/mechanisms in cyclopamine-induced apoptosis
are unknown. Here, we report that cyclopamine mediates ceramide generation selectively via induction of
neutral sphingomyelin phosphodiesterase 3, SMPD3 (nSMase2) in Daoy human medulloblastoma cells.
Importantly, short interfering RNA-mediated knockdown of nSMase2 prevented cyclopamine-induced
ceramide generation and protected Daoy cells from drug-induced apoptosis. Accordingly, ectopic wild-type
N-SMase2 caused cell death, compared with controls, which express the catalytically inactive N-SMase2
mutant. Interestingly, knockdown of smoothened (Smo), a target protein for cyclopamine, or Gli1, a down-
stream signaling transcription factor of Smo, did not affect nSMase2. Mechanistically, our data showed that
cyclopamine induced nSMase2 and cell death selectively via increased nitric oxide (NO) generation by
neuronal-nitric oxide synthase (n-NOS) induction, in Daoy medulloblastoma, and multiple other human
cancer cell lines. Knockdown of n-NOS prevented nSMase2 induction and cell death in response to cyclo-
pamine. Accordingly, N-SMase2 activity-deficient skin fibroblasts isolated from homozygous fro/fro (fragi-
litas ossium) mice exhibited resistance to NO-induced cell death. Thus, our data suggest a novel off-target
function of cyclopamine in inducing apoptosis, at least in part, by n-NOS/NO-dependent induction of N-
SMase2/ceramide axis, independent of Smo/Gli inhibition. Mol Cancer Ther; 11(5); 1092–102. �2012 AACR.
IntroductionThe Sonic hedgehog (SHh) signaling pathway plays a
critical role in normal cerebellar development and hasbeen implicated in the pathogenesis of medulloblasto-ma and other cancers of the brain, prostate, lung, breast,
and colon (1–3). SHh is regulated by the transmembranereceptor patched (Ptch), which when altered or mutat-ed, results in SHh pathway activation and cell growthdysregulation (4–6). The SHh ligand binds Ptch, whichthen alleviates Ptch-mediated suppression of Smo,which activates Gli, a family of transcription factorsinvolved in the regulation of numerous genes control-ling cell division, growth, and/or apoptosis, leading toproliferation and/or inhibition of cell death (7, 8). Thus,SHh is a novel therapeutic target for the treatment ofcancers, including brain tumors (9, 10). Cyclopamine(Fig. 1A, top) is a Smo antagonist, which inhibitsgrowth, and induces apoptosis in various cancer cells,including medulloblastoma (11–15). However, off-tar-get functions of cyclopamine in inducing apoptosis,independent of Smo/SHh inhibition, have not beenclearly defined previously.
The bioactive ceramide is a precursor for the synthesisof more complex sphingolipids via multiple pathways(16–18). Stress-induced ceramide generation in responseto various stimuli, such as anticancer agents, mediatescell-cycle arrest, growth inhibition, and/or apoptosis (19).Ceramide is generated mainly via de novo synthesis by
Authors'Affiliations: 1Department of Biochemistry andMolecular Biology,2Hollings Cancer Center, 3Department of Pediatrics, Division of PediatricOncology, and 4Department of Surgery, Medical University of SouthCarolina, Charleston, South Carolina; and 5Faculty of Dentistry, McGillUniversity, Montreal, Quebec, Canada
Note: Supplementary material for this article is available at MolecularCancer Therapeutics Online (http://mct.aacrjournals.org/).
Current address for J.A. Lewis: St. Jude Children's Research Hospital,Pediatrics, Pediatric Hematology and Oncology, 262 Danny Thomas Pl,Memphis, TN 38105.
Current address for M.O. Cevik: Adiyaman University School of Medicine,Department of Medical Genetics, Adiyaman, 02040, Turkey.
Corresponding Author: Besim Ogretmen, Medical University of SouthCarolina, 86 Jonathan Lucas Street, Charleston, SC 29425. Phone: 843-792-0940; Fax: 843-792-2556; E-mail: [email protected]
doi: 10.1158/1535-7163.MCT-11-0705
�2012 American Association for Cancer Research.
MolecularCancer
Therapeutics
Mol Cancer Ther; 11(5) May 20121092
on November 21, 2018. © 2012 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst March 27, 2012; DOI: 10.1158/1535-7163.MCT-11-0705

ceramide synthases 1–6 (CerS1–6; ref. 20) or via hydrolysisof sphingomyelin (SM) by SMases with pH optima inacidic, neutral, or alkaline conditions (21). Activation ofneutral (N)-SMases1–2 in response to chemotherapy hasbeen reported to generate ceramide, thereby inducing celldeath (22–24). However, whether cyclopamine inducesceramide generation has not been described previously.Therefore, our focus was to define roles and mechan-
isms of cyclopamine-induced apoptosis and determinewhether this occurs, at least in part, via induction ofceramide generation by inhibition of SHh/Smo-depen-dent or SHh/Smo-independent manner in Daoy humanmedulloblastoma and UM-SCC-14A or UM-SCC-1human head neck squamous cell carcinoma (HNSCC)cells.
Materials and MethodsCell cultureThe Daoy medulloblastoma line (American Type Cul-
ture Collection) was grown in minimum essential medi-um with 10% FBS and 1% penicillin/streptomycin. WT,
þ/fro, and activity-deficient fro/fro skin fibroblastsisolated from new born mice (25) were cultured inDulbecco’s Modified Eagle’s Medium with 10% FBSand 1% penicillin/streptomycin. UM-SCC-1 and UM-SCC-14A cells were obtained from Dr. Thomas Carey(University of Michigan). Cell lines used in this studywere not authenticated. Cells were treated at final con-centrations of 5 to 20 mg/mL from cyclopamine stocksolution (20 mg/mL; LC Laboratories) dissolved in100% ethanol. Cyclopamine aliquots were dissolved at55�C.
Measurement of ceramideEndogenous ceramides were measured by liquid chro-
matography/tandemmass spectrometry (LC/MS-MS) asdescribed (26).
Short interfering RNA and plasmidsShort interfering RNAs (siRNA) for nSMase1 and
nSMase2 were obtained from Ambion (Applied Biosys-tems). Gli1 and SMO siRNAs were custom designed by
Figure 1. Cyclopamine induces cell death via caspase-3 activation and loss of mitochondrial membrane potential in Daoy cells. A, effects of increasingconcentrations of cyclopamine (top) at 0, 2.5, 5, 10, 25, and 50 mg/mL (24–48 hours) on cell viability were measured by MTT assay (bottom). Activationof caspase-3 (B) or lossofmitochondrialmembranepotential (C) in response tocyclopamine (10mg/mL, 24hours), as comparedwith controls,weremeasured.D, effects of cyclopamine on total ceramide accumulation were measured by LC/MS-MS, normalized to inorganic phosphate (Pi).
Cyclopamine Induces N-SMase2/Ceramide-Dependent Apoptosis
www.aacrjournals.org Mol Cancer Ther; 11(5) May 2012 1093
on November 21, 2018. © 2012 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst March 27, 2012; DOI: 10.1158/1535-7163.MCT-11-0705

QIAGEN and Invitrogen, respectively. Nontargeting(scrambled, SCR) siRNA #2 was obtained from Dhar-macon. Transfections (40 nmol/l, 48 hours) were carriedout with DharmaFECT as described by the manufac-turer. Knockdown of inducible NO synthase (iNOS) andneuronal NOS (nNOS) was carried out with siRNAsobtained from Dharmacon. Plasmids for wild-type (wt)and mutant N-SMase2, containing V5 tags were used asdescribed (27).
Detection of cell deathCell death was measured by various techniques: (i)
activation of caspase-3, measured by fluorometry usinga caspase-3 activity assay kit (R&D Systems); (ii) loss ofmitochondrial membrane potential, detected by flowcytometry using the JC-1 mitochondrial membranepotential detection kit (Cell Technology); (iii) depletionof cellular ATP, measured by the ATP colorimetric/fluo-rometric assay kit (Abcam); and (iv) detection of AnnexinV–aminoactinomycin D (7-AAD) staining, measured byflow cytometry using the BD Pharmingen PE Annexin VApoptosis Detection Kit, as described by the manufac-turers. Growth inhibition was measured by MTT andtrypan blue assay.
Measurement of SMase activityN-SMase and A-SMase activities were assayed in vitro
as described previously, using 14C-[methyl]-SM as sub-strate (27).
Detection of NO by flow cytometryTreated and untreated cells were incubated with
fluorescent dyes (1 hour). Cells were trypsinsized,washed with PBS, and stained with 7-AAD, before flowcytometry analysis on BD FACScan (BD Biosciences).For hCatalase-transduced cells, before addition ofCD34 antibody, cells were blocked in 30% humanAB-serum (Gemini) for 30 minutes, preventing back-ground staining. Fluorochrome-labeled antibodieswere added, and cells were stained with 7-AAD, andflow cytometry analysis was conducted with the BDFACScan (BD Biosciences). Data for viable (7-AAD)cells were analyzed and visualized by the FlowJo soft-ware (Tree Star).
Statistical analysisData are represented as mean� SEM, unless otherwise
indicated. Data represent at least 2 independent trialscarried out as duplicates. Error bars on graphs representSDs. An unpaired Student t test was carried out withPrism/GraphPad software; P < 0.05 was considered sig-nificant (28).
Details of chemicals, RNA isolation, quantitative PCR(qPCR), Western blotting, target sequences of nSMase1and nSMase2 siRNAs, catalase expression, and activityassays can be found in Supplementary Materials andMethods.
ResultsCyclopamine induces cell death and increasesceramide generation/accumulation
Cyclopamine (Fig. 1A, top) has shown some efficacyagainst desmoplasticmedulloblastomas inpreclinical andclinical studies (29–31). To confirm cyclopamine inducescell death, we treated Daoy human desmoplastic cerebel-lar medulloblastoma cells with increasing concentrationsof cyclopamine (0–50 mg/mL) and examined its effectson cell growth and cell death; measuring survival, cas-pase-3 activity, and loss of mitochondrial membranepotential. Cyclopamine inhibited growth in a dose-dependent manner (IC50 �5 mg/mL, 48 hours, and�10 mg/mL, 24 hours) compared with vehicle-treatedcontrols (Fig. 1A, bottom). Accordingly, cyclopamineincreased caspase-3 activity around 2-fold, which wasconsistent with a loss of mitochondrial membrane poten-tial, as measured by increased accumulation of cyto-plasmic JC-1 (�8-fold), compared with controls (Fig.1B and C, respectively). Pretreatment with z-VAD (10mg/mL) almost completely prevented caspase-3 activa-tion and loss of mitochondrial membrane potential inresponse to cyclopamine (Fig. 1B and C, respectively).Thus, these data are consistent with previous studies,showing that cyclopamine induces caspase-3–dependentmitochondrial apoptosis.
Next, we investigated whether cyclopamine-inducedapoptosis is mediated by induction of ceramide via Smo-signaling inhibition, or via off-target functions of cyclo-pamine, independent of SHh/Smo inhibition. Cyclopa-mine (5 or 10 mg/mL, 24 hours) increased total ceramideapproximately 2.5- or 3-fold, respectively, increasing totalceramide from 20 (in controls) to 50 to 60 pmol/nmol Picyclopamine-treated cells, respectively (Fig. 1D). Therewere no significant changes in sphingosine or S1P inresponse to cyclopamine (data not shown). Moreover,cyclopamine (10 mg/mL, 24 hours) induced C14-, C16-,C18-, C20-, and C22-ceramide generation approximately2.5-, 3.5-, 15-, 6-, or 6.5-fold, respectively, compared withvehicle-treated controls (Supplementary Fig. S1A). Com-parable increases were also observed with dihydro-C14-C22-ceramides (Supplementary Fig. S1B). Similar datawere also obtained when cells were treated with 5 mg/mLcyclopamine (24 hours),which increased endogenousC14-C22-ceramide generation comparedwith controls (Supple-mentary Fig. S1A). Thus, these data suggest that cyclopa-mine induces endogenousceramidegeneration, consistentwith its proapoptotic effects in Daoy cells.
Cyclopamine-mediated ceramide generation isdependent on nSMase2 induction
To assess whether de novo generation of ceramideplays a role in cyclopamine-induced apoptosis, wepretreated cells with fumonisin B1 (FB1) and myriocin(MYR) at 50 mmol/L and 50 nmol/L, respectively, andexamined cyclopamine-induced caspase-3 activation inDaoy cells (10 mg/mL, 24 hours). FB1 or MYR did not
Meyers-Needham et al.
Mol Cancer Ther; 11(5) May 2012 Molecular Cancer Therapeutics1094
on November 21, 2018. © 2012 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst March 27, 2012; DOI: 10.1158/1535-7163.MCT-11-0705
prevent cyclopamine-induced caspase-3 activation(Supplementary Fig. S2A), indicating de novo generationof ceramide might not be involved in cyclopamine-induced apoptosis. We also examined the effects ofsiRNA-mediated knockdown of CerS1 on growth inhi-bition in response to cyclopamine. CerS1 is known tomainly generate C18-ceramide, and cyclopamineinduced C18-ceramide generation approximately 15-fold compared with controls in Daoy cells (see Supple-mentary Fig. S1A). Downregulation of CerS1 with siR-NAs did not protect cells from cyclopamine-inducedgrowth inhibition compared with controls (Supplemen-tary Fig. S2B). Effectiveness of siRNAs for knockdownof CerS1 in Daoy cells was confirmed with qPCR. Anapproximately 40% to 60% decrease in CerS1 wasobserved compared with controls in the absence/pres-
ence of cyclopamine (Supplementary Fig. S2C). Simi-larly, ectopic expression of wt CerS1, or its catalyticallyinactive form with the H122A mutation (32), did notenhance or prevent the growth inhibitory effects ofcyclopamine (Supplementary Fig. S2D). Expression ofwt and mutant CerS1-FLAG proteins was confirmed byWestern blotting using anti-FLAG antibody comparedwith vector-transfected controls (Supplementary Fig.S1E, lanes 2–3 and 1, respectively). Actin was used asa loading control (Supplementary Fig. S2E). Thus, thesedata suggest that cyclopamine-induced cell death isindependent of CerS1 activation in these cells.
Because activation of N-SMase2 is known to mediateapoptosis (33, 34), we determined whether cyclopamineaffects nSMase2 with qPCR. Cyclopamine increasednSMase2 approximately 3- and 6-fold (12–24 hours,
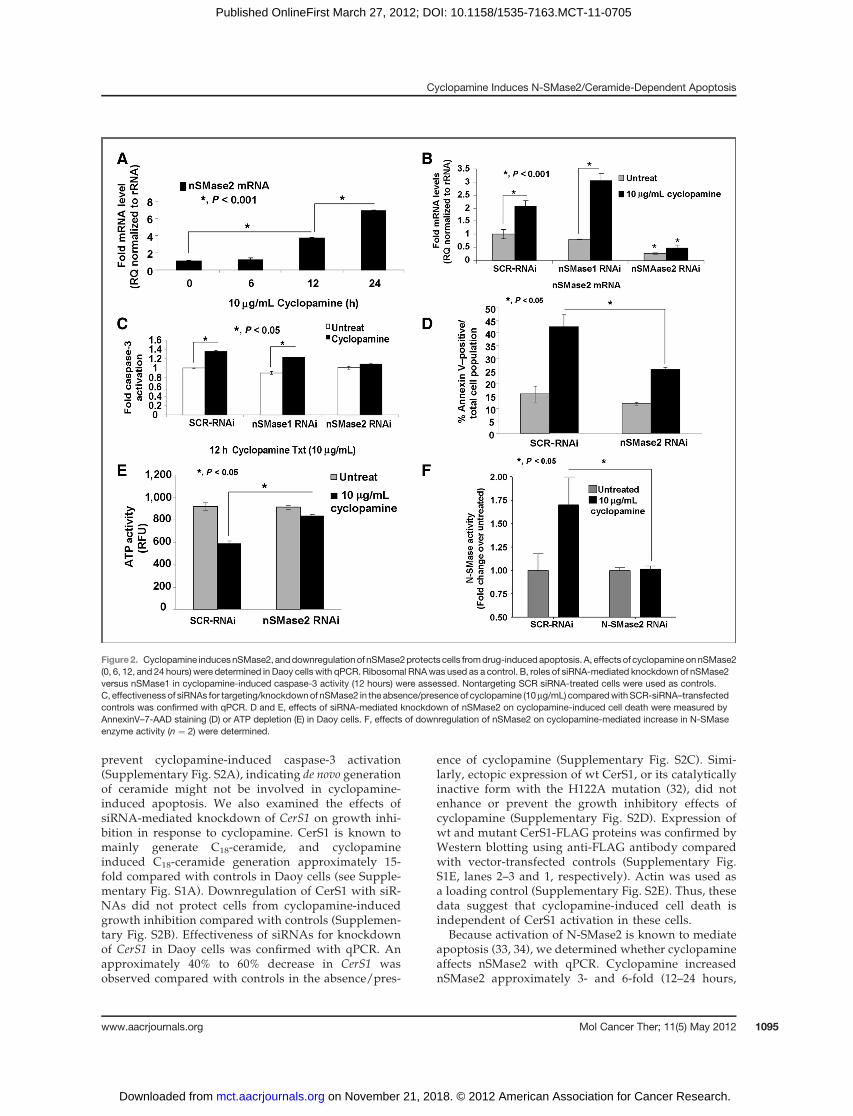
Figure2. Cyclopamine inducesnSMase2, anddownregulationof nSMase2protects cells fromdrug-induced apoptosis. A, effects of cyclopamineonnSMase2(0, 6, 12, and 24 hours) were determined in Daoy cells with qPCR. Ribosomal RNAwas used as a control. B, roles of siRNA-mediated knockdown of nSMase2versus nSMase1 in cyclopamine-induced caspase-3 activity (12 hours) were assessed. Nontargeting SCR siRNA-treated cells were used as controls.C, effectivenessof siRNAs for targeting/knockdownof nSMase2 in the absence/presenceof cyclopamine (10mg/mL) comparedwithSCR-siRNA–transfectedcontrols was confirmed with qPCR. D and E, effects of siRNA-mediated knockdown of nSMase2 on cyclopamine-induced cell death were measured byAnnexinV–7-AAD staining (D) or ATP depletion (E) in Daoy cells. F, effects of downregulation of nSMase2 on cyclopamine-mediated increase in N-SMaseenzyme activity (n ¼ 2) were determined.
Cyclopamine Induces N-SMase2/Ceramide-Dependent Apoptosis
www.aacrjournals.org Mol Cancer Ther; 11(5) May 2012 1095
on November 21, 2018. © 2012 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst March 27, 2012; DOI: 10.1158/1535-7163.MCT-11-0705

respectively), whereas treatment at 6 hours had noeffect (Fig. 2A). Cyclopamine did not induce caspase-3 activity at 6 hours (Supplementary Fig. S3A), treat-ment at 12 hours slightly, but significantly, increasedcaspase-3 activity (�30%, P < 0.05). Knockdown ofnSMase2, but not nSMase1 (Supplementary Fig. S3B),(�75% compared with SCR controls, as determined byqPCR, Fig. 2B), prevented cyclopamine-induced cas-pase-3 activation (Fig. 2C). These data were also con-sistent with studies in which siRNA-mediated knock-down of nSMase2 abrogated cyclopamine-induced celldeath, as measured by increased Annexin V staining or
depletion of cellular ATP (Fig. 2D and E, respectively).Importantly, increased nSMase2 was also consistentwith increased enzyme activity of N-SMase2 (�1.7-fold)in response to cyclopamine (10 mg/mL, 12 hours), whichwas prevented by knockdown of nSMase2 using siR-NAs (Fig. 2F). Cyclopamine had no effect on A-SMaseactivity in these cells (data not shown). Increase innSMase2 by cyclopamine (10 mg/mL) was also observedin UM-SCC-14A or UM-SCC-1 human HNSCC cells, inwhich it was increased approximately 4- and 8-fold, or1.5- and 5-fold at 12 and 24 hours, respectively (Sup-plementary Fig. S3C and S3D, respectively).
Figure 3. N-SMase2–generated ceramide is important in cyclopamine-induced cell death in Daoy cells. Ceramide (A) and SM (B) in response to cyclopamine(10 mg/mL), in the absence/presence of siRNAs targeting nSMase1 or nSMase2, were measured in Daoy cells by LC/MS-MS. SCR-siRNA–treated cells wereused as controls. C, effects of downregulation of nSMase2 versus nSMase1 with siRNAs on cell viability were determined with trypan blue exclusion assay.D, roles of nSMase2/ceramide in the induction of apoptosis were examined with trypan blue exclusion assay after transfection of Daoy cells with vectorscontaining cDNAs of the wt or catalytically inactive mutant of N-SMase2, compared with vector only transfected cells. Ectopic expression of wt or mutantN-SMase2-V5 was confirmed by measurement of SMase activity (E) andWestern blotting (F) with anti-V5 antibody (as duplicates) in Daoy cells as comparedwith vector-transfected controls. Actin was used as a loading control in F (bottom).
Meyers-Needham et al.
Mol Cancer Ther; 11(5) May 2012 Molecular Cancer Therapeutics1096
on November 21, 2018. © 2012 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst March 27, 2012; DOI: 10.1158/1535-7163.MCT-11-0705

Overall, these data suggest that cyclopamine inducesnSMase2 in multiple human cancer cell lines, and knock-down of nSMase2 prevents cell death in response tocyclopamine, indicating an important role for N-SMase2in this process.
Downregulation of nSMase-2 prevents cyclopamine-induced ceramide generation and cell deathTo determine whether increased nSMase2 plays a role
in cyclopamine-mediated ceramide generation (10 mg/mL,24 hours), we measured endogenous ceramide and SMby LC/MS-MS in the absence/presence of siRNAs againstnSMase1 or nSMase2 in Daoy cells. Data revealed thatcyclopamine increased total ceramide approximately 2- to3.5-fold in the presence of SCR or N-SMase1 siRNAs, andknockdown of nSMase2 prevented cyclopamine-mediatedceramide generation (Fig. 3A). Consistent with these data,total SM was significantly decreased (�70%, P < 0.05) inresponse to cyclopamine in the absence/presence of SCRor nSMase1 siRNAs (Fig. 3B). Concentrations of ceramidesand SM in response to cyclopamine in the absence/pres-ence of nSMase1 or nSMase2 siRNAs were depicted inSupplementary Fig. S4A and S4B. As observed earlier, C18-ceramide was the main species generated by cyclopamine(�20-fold), and accordingly, a significant decrease in C18-SMwas also observed (Supplementary Fig. S4C and S4D).Interestingly, although knockdown of nSMase2 prevented
ceramidegeneration (Fig. 3A), it did not attenuate decreasein SM (Fig. 3B), suggesting the hydrolysis of SM withoutincreased ceramide generation by an unknown mecha-nism, or inhibition of SM synthases. These data suggestthat cyclopamine induces ceramide generation by induc-tion of nSMase2.
We then examined whether downregulation ofnSMase2, which prevented ceramide generation, alsoattenuated cyclopamine-mediated cell death (10 mg/mL,24hours).Data showed that siRNA-mediatedknockdownof nSMase2 significantly protected growth inhibition ofDaoy cells (�50%, P < 0.05) compared with SCR-siRNA–transfected controls in response to cyclopamine (Fig. 3C),consistent with the protective effects of downregulationof nSMase2 on cyclopamine-mediated apoptosis (Fig. 2C–E). Taken together, these data suggest that increasedceramide generation by elevation of nSMase2 mRNA andactivity is involved, at least in part, in caspase-3 activation,and cell death in response to cyclopamine.
Accordingly, wt N-SMase2 expression significantlydecreased cell growth, and increased apoptosis (�30%,P< 0.05) inDaoy cells comparedwith cells transfectedwiththe catalytically inactive form of N-SMase2 (Fig. 3D), orvector-transfectedcontrols (datanot shown).Expressionofwt andmutant N-SMase2-V5 (27) was confirmed byWest-ern blotting using anti-V5 antibody, and activity of wtcomparedwith catalytically inactivemutantN-SMase2-V5
Figure4. Upregulationof nSMase2andceramidegeneration in response to cyclopamine is independent of the inhibitionofSmo/Gli of theSHhpathway inDaoycells. Effects of downregulation of nSMase2 or nSMase1 using siRNAs on the inhibition of Smo (A) or Gli1 (B) in the absence/presence of cyclopamine(10mg/mL, 24 hours) were determined byqPCR.SCR-siRNA–transfected cells were usedas controls. Potential roles of SmoorGli1 inhibition on the regulationof nSMase2were examined after siRNA-mediated knockdown ofSmo (C) orGli1 (D) using qPCR. Effectiveness of siRNAs on knockdown ofSmo orGli1werealso confirmed using qPCR (C and D, respectively). SCR-siRNA–transfected cells were used as controls.
Cyclopamine Induces N-SMase2/Ceramide-Dependent Apoptosis
www.aacrjournals.org Mol Cancer Ther; 11(5) May 2012 1097
on November 21, 2018. © 2012 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst March 27, 2012; DOI: 10.1158/1535-7163.MCT-11-0705

or vector-transfected controls were confirmed (Fig. 3Eand 3F), supporting that the N-SMase2/ceramide axisplays an important role for inducing cell death in thesecells.
Cyclopamine-induced nSMase-2 is independent ofSmo inhibition
Because cyclopamine is an antagonist of Smo, it wasimportant to determine whether cyclopamine-inducednSMase2 is dependent or independent of Smo inhibition.First, we examined the effects of downregulation ofnSMase2 or nSMase1 using siRNAs on Smo and GlimRNAs in the absence/presence of cyclopamine (10mg/mL, 24 hours) by qPCR in Daoy cells. Cyclopaminereduced Smo and Gli by approximately 60%, but down-regulation of nSMase2 or nSMase1 had no effect on inhi-bition of the Smo/Gli axis by cyclopamine (Fig. 4A and B,respectively). Thus, these data confirmed that inhibition
of the Smo/Gli axis by cyclopamine is regulated indepen-dently of N-SMase2.
Then, we examined the effects of downregulation ofSmo or Gli1 using siRNAs on nSMase2. We reasonedwhether cyclopamine-mediated nSMase2 elevation isregulated downstream of Smo/Gli1 inhibition, knock-down of Smo or Gli1 should also increase nSMase2.Interestingly, data showed while siRNAs successfullyreduced Smo or Gli1 by approximately 90% or 80%, theydid not increase N-SMase2 (Fig. 4C and D). Similarly,pharmacologic inhibitors of Gli or Smo, GANT-61 orSANT-1 (5–10 mmol/L, 24–48 hours) had no effect onnSMase2 (data not shown), despite successfully reduc-ing expression of Gli1, and its downstream target Bcl2(Supplementary Fig. S5A and S5B). Taken together,these data indicate that induction of nSMase2 inresponse to cyclopamine is independent of the Smo/Gli axis of SHh signaling in Daoy cells.
Figure 5. Cyclopamine-mediated increased nSMase2 is regulated by ROS/RNS generation. Effects of oxidative stress via ROS/RNS generation oncyclopamine-mediated overexpression of nSMase2 were examined in the absence/presence of antioxidant NAC (500 mmol/L) using qPCR in Daoy (A) andUM-SCC-14A (B) cells. C, ROS/RNS induction in response to cyclopamine (10 mg/mL) at 0, 3, 6, 9, 12, and 15 hours was measured by DCFDA by flowcytometry in Daoy cells. D, effects of cyclopamine (10 mg/mL, 4 hours) on ROS/RNS generation (left) and potential mitochondrial colocalization (middle andright) in Daoy cells were visualized by confocal microscopy using DCFDA (green) and mitotracker (red) as compared with vehicle-treated controls (lower andupper, respectively). E, effects of NAC (500 mmol/L) on prevention of ROS/RNS generation in response to cyclopamine (10 mg/mL, 6 hours) in UM-SCC-14Acells were measured by DCFDA by flow cytometry.
Meyers-Needham et al.
Mol Cancer Ther; 11(5) May 2012 Molecular Cancer Therapeutics1098
on November 21, 2018. © 2012 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst March 27, 2012; DOI: 10.1158/1535-7163.MCT-11-0705

Cyclopamine-induced nSMase-2 is regulated byoxidative stressBecause oxidative stress is known to induce N-SMase
activity, and ceramide-mediated apoptosis (35–37), wethen examined whether cyclopamine enhances nSMase2via induction of reactive oxygen species (ROS)/nitrogenspecies (RNS). First, we determined whether pretreat-ment with NAC (0.5 mmol/L), an antioxidant, alterednSMase2 in the absence/presence of cyclopamine. Inter-estingly, while treatment with cyclopamine inducednSMase2 around 8-fold (10 mg/mL, 24 hours), pretreat-ment with NAC almost completely prevented cyclopa-mine-mediated nSMase2 in Daoy (Fig. 5A) and also inUM-SCC-14A cells (Fig. 5B), suggesting a role for cyclo-pamine-induced ROS/RNS in increased nSMase2, whichwas observed not only in Daoy cells, but was alsoobserved in other human cancer cells, such as HNSCCs.Induction of ROS/RNS in response to cyclopamine(10 mg/mL) at various time points was also confirmed bystaining the cells with dichlorofluorescin diacetate(DCFDA) and flow cytometry analysis. Data showedcyclopamine treatment induced ROS/RNS generationwithin 3 to 6 hours of treatment, increasing DCFDAfluorescence approximately 2- to 3-fold, respectively (Fig.5C). Cyclopamine-mediated ROS/RNS generation wasalso detected by confocal microscopy, in which greenfluorescence of DCFDA, a probe for ROS/RNS generation(38, 39), was visualized in Daoy cells compared withcontrols (Fig. 5D, bottom and top right, respectively).Interestingly, there was no significant colocalization ofthe greenfluorescence ofDCFDAwith the redmitotrackerin these cells in response to cyclopamine (Fig. 5D). Thus,these data suggest cyclopamine induces ROS/RNS gen-eration, which is not selectively induced in mitochondria.Protective effects of NAC on cyclopamine-induced ROS/RNS generation were confirmed by DCFDA and flowcytometry. Pretreatment with NAC prevented the gener-ation of ROS/RNS, shifting the DCFDA fluorescencesignal to the left when compared with cyclopamine-trea-tedUM-SCC-14AorUM-SCC-1 cells, respectively (Fig. 5Eand Supplementary Fig. S6A).Thus, these data suggest that cyclopamine induces
early ROS/RNS generation (within 3–6 hours), whichprecedes increased nSMase2 detected at 12 hours, andthat cyclopamine-mediated ROS/RNS is important ininduction of nSMase2.
Cyclopamine increases nSMase-2 via induction ofNO generationWe then determined the mechanism by which cyclo-
pamine induces ROS/RNS generation, which subse-quently leads to increasednSMase2. First, to assesswheth-er H2O2 generation was involved, we overexpressedhuman catalase, which increased its enzyme activity sig-nificantly (by >20-fold, P < 0.001) compared with controls(Supplementary Fig. S6B–S6D), and then determined itseffects onnSMase2 and cell death in the absence/presenceof cyclopamine (10 mg/mL, 24 hours). Catalase overex-
pression did not prevent increased nSMase2 or Daoy celldeath in response to cyclopamine (Supplementary Fig.S6B–S6D), suggesting they were independent of H2O2
generation. In addition, buthionine sulfoximine (2.5mmol/L, 16 hours), a known inhibitor of glutathione(GSH) synthesis, inducing oxidative stress via accumula-tion of intracellular ROS, had no significant effect onnSMase2 or apoptosis (Supplementary Fig. S6E–S6F).
Wenext examinedwhether RNS generation plays a rolein this process. To determine RNS generation, effects ofcyclopamine on DAF fluorescence, a probe for peroxyni-trite [ONOO(-)] and NO generation, was examined inDaoy cells using flow cytometry. As shown in Fig. 6A,cyclopamine (10 mg/mL, 4 hours) enhanced DAF andDCFDA, but not DHE (which detects superoxide anion),fluorescence compared with controls, suggesting cyclo-pamine induces RNS, but not ROS, via generation ofONOO(-) and/or NO. Pretreatment with MnTBAP, aONOO(-) scavenger, did not prevent nSMase2 upregula-tion (Supplementary Fig. S7A), whereas L-NAME pre-treatment, an inhibitor ofNOS, almost completely bluntedcyclopamine-induced nSMase2 (Fig. 6B).
In reciprocal experiments, DETA, a well-known NOdonor, significantly increased nSMase2 (�15-fold, P <0.05; Fig. 6C). Thus, these novel data indicate that cyclo-pamine-induced nSMase2 is selectively regulated viaNO,but not via ONOO(-), generation in Daoy cells. To definethemechanismbywhichNOgeneration inducesnSMase2and cell death in response to cyclopamine, we examinedthe roles of i-NOS, endothelial NOS (e-NOS), or n-NOS.Cyclopamine (10 mg/mL, 24 hours) increased (�2-fold,measured by qPCR) n-NOS (Supplementary Fig. S7B), butit had no significant effect on e-NOS or i-NOS (data notshown). Importantly, siRNA-mediated knockdown ofn-NOS (�70%, measured by qPCR; Supplementary Fig.S7C) prevented nSMase2 induction and cell death (Fig. 6Dand E, respectively). Similar data were also obtained inUM-SCC-22A cells, in which knockdown of n-NOS usingsiRNAs significantly blocked cyclopamine-induced celldeath, compared with SCR-siRNA–transfected controls(Fig. 6F), suggesting a role for n-NOS in cyclopamine-mediated nSMase2 induction and cell death.
These results were also consistent when wt versusactivity-deficient skin fibroblasts isolated from new bornhomozygous fro/fro (fragilitas ossium) mice (25) weretreated with DETA. Treatment of wt cells with increasingconcentrations of NO-donor DETA elevated N-SMase2(data not shown), and inhibited cell viability (Fig. 6G),whereas fro/fro skin fibroblasts with inactive N-SMase2,exhibited resistance to DETA-mediated cell death (Fig.6G), supporting the role of N-SMase2 in DETA/NO-mediated cell death. Interestingly, in contrast to humancancer cells, these noncancerous wt and fro/fro skinfibroblasts were equally sensitive to cyclopamine-medi-ated cell death, suggesting that SHh/Smo plays animportant role for the growth and/or proliferation ofthese cells, regardless of their N-SMase activity (data notshown).
Cyclopamine Induces N-SMase2/Ceramide-Dependent Apoptosis
www.aacrjournals.org Mol Cancer Ther; 11(5) May 2012 1099
on November 21, 2018. © 2012 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst March 27, 2012; DOI: 10.1158/1535-7163.MCT-11-0705

Overall, these data suggest that increased nSMase2/ceramide and cell death in response to cyclopamine isregulated, at least in part, by the n-NOS/NO axis, in Daoymedulloblastoma and HNSCC cells.
DiscussionHere, possible roles and mechanisms of interaction
between the SHh inhibitor/anticancer drug cyclopamine,and the bioactive sphingolipid ceramide, in apoptosis ofDaoy and HNSCC cells were examined. Unexpected andnovel data revealed that cyclopamine induces apoptosisin part via selective induction of nSMase2/increasedceramide. Mechanistically, induction of nSMase2 wasregulated selectively by n-NOS/NO in response to cyclo-pamine, which was independent of Smo/Gli1 inhibition.
Cyclopamine was shown to induce apoptosis (1, 9, 14).Another Smo antagonist GDC-0449 (40) is currently inclinical trials againstmedulloblastomas and various othercancers. In fact, amino acid substitution at a conservedaspartic acid residue of SMO, which interrupted GDC-0449 binding, was reported to cause drug resistance in theclinic (41). Ceramide mediates drug-induced apoptosis.However, any involvement of ceramide in cyclopamine-induced apoptosis has not previously been reported. Ournovel data revealed that cyclopamine results in robustceramide generation via induction of nSMase2,which is atleast partly critical in cyclopamine-induced cell death inDaoy and HNSCC cells. These data are somewhat inagreement with a previous study, in which cyclopa-mine-induced apoptosis was partially rescued by Gli
Figure 6. Cyclopamine-mediated induction of nSMase2 is regulated selectively by NO generation in Daoy cells. A, generation of NO/ONOO(-) in response tocyclopamine (10 mg/mL for 4 hours) was measured by DAF, DCFDA, or DHE by flow cytometry in Daoy cells. Effects of L-NAME (50 mmol/L; B) versus DETA(3 mmol/L, 12 hours; C) on nSMase2 in the absence/presence of cyclopamine (10 mg/mL, 24 hours) were measured by qPCR. Effects of siRNA-mediatedknockdown of n-NOS on nSMase2 (D), or cell death in Daoy (E) and UM-SCC-14A (F) cells were measured by qPCR, or trypan blue exclusion assay,respectively. G, cells isolated fromwt or fro/fromicewere treatedwith increasing concentrations of cyclopamine, and effects on cell growthweremeasuredbytrypan blue exclusion assay. Vehicle-treated cells were used as controls.
Meyers-Needham et al.
Mol Cancer Ther; 11(5) May 2012 Molecular Cancer Therapeutics1100
on November 21, 2018. © 2012 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst March 27, 2012; DOI: 10.1158/1535-7163.MCT-11-0705

overexpression in Daoy cells, indicating an involvementof othermechanisms in this process (42). Unexpected dataadditionally suggested cyclopamine-inducedN-SMase2/ceramide generation is independent of SHh/Smo/Gliinhibition, but is regulated by NO stress/signaling inthese cells.To our knowledge, no previous data involving cyclo-
pamine-induced NO generation in cancer cells exist.Although ceramide was reported to play roles in sodiumnitroprusside (an NO donor)-induced apoptosis (43), anyrole of NO in inducing nSMase2 has not been previouslyreported. However, many excellent previous reports exist(35–37), which showed the involvement of ROS/RNS inSMase regulation and cell death in various cell types. Forexample, H2O2was shown to activate N-SMase2, which isprevented byGSH in human airway epithelial cells (HAE;ref. 44). In aging rat hepatocytes, however, decreasedGSHinduced N-SMase2, whereas in young hepatocytes, inhi-bition of GSH synthesis activated N-SMase (45), suggest-ing that ROS/N-SMase2 regulation might be contextdependent. This was also consistent with the roles of RNSin SMase regulation. It was shown that ONOO(-) inducedA-SMase, but not N-SMase, in HAE cells (35). However,our data indicate a role for n-NOS/NO in the activation ofN-SMase but not A-SMase in Daoy and HNSCC cells.Thesedata together suggest that SMase regulation byRNSis also context dependent, and it might be differentiallyregulated in noncancerous HAE versus some cancercell types. The cell type/context-dependent regulationof ceramide metabolism by NO is also consistent withearlier studies, in which NO donor induced apoptosis incultured fibroblasts but not in keratinocytes (46). More-over, L-NAME, inhibitor of NOS, increased ceramideformation and apoptosis in keratinocytes, but not infibroblasts (46).NOgeneration is regulatedmainly by e-NOS, i-NOS, or
n-NOS (47). Interestingly, our novel data suggest thatcyclopamine increased n-NOS mRNA, and knockdownof n-NOS (48) prevented cyclopamine-inducedN-SMase2and cell death, indicating its involvement in this process.
However, specific mechanisms involved in n-NOS/NOgeneration in response to cyclopamine remain unknown.
Moreover, mechanisms by which cyclopamine-induced n-NOS/NO results in increased nSMase2 areunknown. It was reported previously that daunorubicinactivated the nSMase2 promoter via Sp1/Sp3 transcrip-tion factors in MCF-7 human breast cancer cells, increas-ing ceramide accumulation and cell death (49). Recently,all-trans retionic acid was also shown to induce nSMase2,resulting inMCF-7 growth arrest (50). These studies are inagreement with our data, suggesting that various anti-cancer drugs, including cyclopamine, induce nSMase2,leading to increased ceramide generation and apoptosis.
In summary, our data show a novel off-target functionof cyclopamine by inducing n-NOS/NO-dependentnSMase2 expression, and ceramide generation, which, inpart, was necessary for drug-induced apoptosis. Thesedata may have important implications for the Smo-inde-pendent apoptotic roles of cyclopamine in the treatment ofvarious human cancers, in which the n-NOS/NO/N-SMase2/ceramide axis is intact, but SHh activation mightbe partly dispensable.
Disclosure of Potential Conflicts of InterestNo potential conflicts of interest were disclosed.
AcknowledgmentsThe authors thank Dr. J. Schnellmann for her editorial review and Dr. J.
Houghton and her group (Cleveland Clinic) for helpful discussions andsharing SHh pathway molecular tools.
Grant SupportThisworkwas supported in part by research funding obtained from the
NIH (CA088932, DE016572, and CA097165), and from the Angel Walk,Rally Foundation for Childhood Cancer Research, Chase after a Cure, andMonica Kreber Golf Tournament. Lipid measurements and flow cytome-try were conducted in facilities constructed with support from NIH (C06RR015455).
The costs of publication of this article were defrayed in part by thepayment of page charges. This article must therefore be hereby markedadvertisement in accordance with 18 U.S.C. Section 1734 solely to indicatethis fact.
Received September 8, 2011; revised February 23, 2012; accepted March14, 2012; published OnlineFirst March 27, 2012.
References1. Taipale J, Beachy PA. The Hedgehog and Wnt signalling pathways in
cancer. Nature 2001;411:349–54.2. TakebeN,Harris PJ,WarrenRQ, IvySP. Targeting cancer stemcells by
inhibiting Wnt, Notch, and Hedgehog pathways. Nat Rev Clin Oncol2011;8:97–106.
3. Ng JM, Curran T. The Hedgehog's tale: developing strategies fortargeting cancer. Nat Rev Cancer 2011;11:493–501.
4. Mazumdar T, DeVecchio J, Shi T, Jones J, Agyeman A, Houghton JA.Hedgehog signaling drives cellular survival in human colon carcinomacells. Cancer Res 2011;71:1092–102.
5. O'Toole SA, Machalek DA, Shearer RF, Millar EK, Nair R, Schofield P,et al. Hedgehogoverexpression is associatedwith stromal interactionsand predicts for poor outcome in breast cancer. Cancer Res2011;71:4002–14.
6. Shi T, Mazumdar T, Devecchio J, Duan ZH, Agyeman A, Aziz M, et al.cDNAmicroarray gene expression profiling of hedgehog signaling path-way inhibition in human colon cancer cells. PLoS One 2010;5:e13054.
7. Bar EE, Chaudhry A, Farah MH, Eberhart CG. Hedgehog signalingpromotes medulloblastoma survival via Bc/II. Am J Pathol2007;170:347–55.
8. Karhadkar SS, Bova GS, Abdallah N, Dhara S, Gardner D, Maitra A,et al. Hedgehog signalling in prostate regeneration, neoplasia andmetastasis. Nature 2004;431:707–12.
9. Berman DM, Karhadkar SS, Hallahan AR, Pritchard JI, Eberhart CG,Watkins DN, et al. Medulloblastoma growth inhibition by hedgehogpathway blockade. Science 2002;297:1559–61.
10. Sanchez P, Ruiz i Altaba A. In vivo inhibition of endogenous braintumors through systemic interference of Hedgehog signaling in mice.Mech Dev 2005;122:223–30.
11. Taipale J, Chen JK, CooperMK,Wang B,Mann RK,Milenkovic L, et al.Effects of oncogenic mutations in Smoothened and Patched can bereversed by cyclopamine. Nature 2000;406:1005–9.
12. Kiselyov AS. Targeting the hedgehog signaling pathway with smallmolecules. Anticancer Agents Med Chem 2006;6:445–9.
Cyclopamine Induces N-SMase2/Ceramide-Dependent Apoptosis
www.aacrjournals.org Mol Cancer Ther; 11(5) May 2012 1101
on November 21, 2018. © 2012 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst March 27, 2012; DOI: 10.1158/1535-7163.MCT-11-0705

13. Bar EE, Stearns D. New developments in medulloblastoma treatment:the potential of a cyclopamine-lovastatin combination. Expert OpinInvestig Drugs 2008;17:185–95.
14. Enguita-German M, Schiapparelli P, Rey JA, Castresana JS. CD133þ
cells from medulloblastoma and PNET cell lines are more resistant tocyclopamine inhibition of the sonic hedgehog signaling pathway thanCD133� cells. Tumour Biol 2010;31:381–90.
15. Katoh Y, Katoh M. Hedgehog target genes: mechanisms of carcino-genesis induced by aberrant hedgehog signaling activation. Curr MolMed 2009;9:873–86.
16. Pruett ST, Bushnev A, Hagedorn K, AdigaM, Haynes CA, SullardsMC,et al. Biodiversity of sphingoid bases ("sphingosines") and relatedamino alcohols. J Lipid Res 2008;49:1621–39.
17. Stiban J, Tidhar R, Futerman AH. Ceramide synthases: roles in cellphysiology and signaling. Adv Exp Med Biol 2010;688:60–71.
18. Ogretmen B, Hannun YA. Biologically active sphingolipids in cancerpathogenesis and treatment. Nat Rev Cancer 2004;4:604–16.
19. DengX, Yin X, AllanR, LuDD,Maurer CW,Haimovitz-FriedmanA, et al.Ceramide biogenesis is required for radiation-induced apoptosis in thegerm line of C. elegans. Science 2008;322:110–5.
20. Pewzner-Jung Y, Ben-Dor S, Futerman AH. When do Lasses (longev-ity assurance genes) become CerS (ceramide synthases)?: insightsinto the regulation of ceramide synthesis. J Biol Chem 2006;281:25001–5.
21. Wu BX, Clarke CJ, Hannun YA. Mammalian neutral sphingomyeli-nases: regulation and roles in cell signaling responses. Neuromole-cular Med 2010;12:320–30.
22. Edelmann B, Bertsch U, Tchikov V, Winoto-Morbach S, Perrotta C,Jakob M, et al. Caspase-8 and caspase-7 sequentially mediate pro-teolytic activation of acid sphingomyelinase in TNF-R1 receptosomes.Embo J 2011;30:379–94.
23. Truman JP, Garcia-Barros M, Kaag M, Hambardzumyan D, StancevicB, ChanM, et al. Endothelial membrane remodeling is obligate for anti-angiogenic radiosensitization during tumor radiosurgery. PLoS One2010;5:e12310.
24. Filosto S, Castillo S, Danielson A, Franzi L, Khan E, Kenyon N, et al.Neutral sphingomyelinase 2: a novel target in cigarette smoke-inducedapoptosis and lung injury. Am J Respir Cell Mol Biol 2011;44:350–60.
25. Aubin I, Adams CP, Opsahl S, Septier D, Bishop CE, Auge N, et al. Adeletion in the gene encoding sphingomyelin phosphodiesterase 3(Smpd3) results in osteogenesis and dentinogenesis imperfecta in themouse. Nat Genet 2005;37:803–5.
26. Bielawski J, Pierce JS, Snider J, Rembiesa B, Szulc ZM, Bielawska A.Sphingolipid analysis by high performance liquid chromatography-tandem mass spectrometry (HPLC-MS/MS). Adv Exp Med Biol2010;688:46–59.
27. Clarke CJ, Cloessner EA, Roddy PL, Hannun YA. Neutral sphingo-myelinase-2 (nSMase2) is the primary neutral sphingomyelinase iso-formactivatedby tumor necrosis factor-alpha inMCF-7 cells. BiochemJ 2011;435:381–90.
28. Mukhopadhyay A, Saddoughi SA, Song P, Sultan I, Ponnusamy S,Senkal CE, et al. Direct interaction between the inhibitor 2 and cer-amide via sphingolipid-protein binding is involved in the regulation ofprotein phosphatase 2A activity and signaling. FASEB J 2009;23:751–63.
29. Coon V, Laukert T, Pedone CA, Laterra J, Kim KJ, Fults DW. Moleculartherapy targeting Sonic hedgehog and hepatocyte growth factorsignaling in a mouse model of medulloblastoma. Mol Cancer Ther2010;9:2627–36.
30. NishimakiH,Kasai K, Kozaki K, TakeoT, IkedaH,SagaS, et al. A role ofactivated Sonic hedgehog signaling for the cellular proliferation of oralsquamous cell carcinoma cell line. Biochem Biophys Res Commun2004;314:313–20.
31. Kelleher FC. Hedgehog signaling and therapeutics in pancreatic can-cer. Carcinogenesis 2011;32:445–51.
32. Spassieva S, Seo JG, Jiang JC, Bielawski J, Alvarez-Vasquez F,Jazwinski SM, et al. Necessary role for the Lag1p motif in (dihydro)ceramide synthase activity. J Biol Chem 2006;281:33931–8.
33. Canals D, Perry DM, Jenkins RW, Hannun YA. Drug targeting ofsphingolipid metabolism: sphingomyelinases and ceramidases. Br JPharmacol 2011;163:694–712.
34. Devillard R, Galvani S, Thiers JC, Guenet JL, Hannun Y, Bielawski J,et al. Stress-induced sphingolipid signaling: role of type-2 neutralsphingomyelinase in murine cell apoptosis and proliferation. PLoSOne 2010;5:e9826.
35. Castillo SS, Levy M, Thaikoottathil JV, Goldkorn T. Reactive nitrogenand oxygen species activate different sphingomyelinases to induceapoptosis in airway epithelial cells. Exp Cell Res 2007;313:2680–6.
36. Jana A, Pahan K. Oxidative stress kills human primary oligodendro-cytes via neutral sphingomyelinase: implications for multiple sclerosis.J Neuroimmune Pharmacol 2007;2:184–93.
37. Ichi I, Kamikawa C, Nakagawa T, Kobayashi K, Kataoka R, Nagata E,et al. Neutral sphingomyelinase-induced ceramide accumulation byoxidative stress during carbon tetrachloride intoxication. Toxicology2009;261:33–40.
38. Ischiropoulos H, Gow A, Thom SR, Kooy NW, Royall JA, Crow JP.Detection of reactive nitrogen species using 2,7-dichlorodihydrofluor-escein and dihydrorhodamine 123. Methods Enzymol 1999;301:367–73.
39. Bilski P, Belanger AG, Chignell CF. Photosensitized oxidation of 20,70-dichlorofluorescin: singlet oxygen does not contribute to the formationof fluorescent oxidation product 20,70-dichlorofluorescein. Free RadicBiol Med 2002;33:938–46.
40. Tremblay MR, Nesler M, Weatherhead R, Castro AC. Recent patentsfor Hedgehog pathway inhibitors for the treatment of malignancy.Expert Opin Ther Pat 2009;19:1039–56.
41. YauchRL, DijkgraafGJ, Alicke B, Januario T, AhnCP,Holcomb T, et al.Smoothened mutation confers resistance to a Hedgehog pathwayinhibitor in medulloblastoma. Science 2009;326:572–4.
42. Bar EE, Chaudhry A, Farah MH, Eberhart CG. Hedgehog signalingpromotes medulloblastoma survival via Bc/II. Am J Pathol2007;170:347–55.
43. Takeda Y, TashimaM, Takahashi A, Uchiyama T, Okazaki T. Ceramidegeneration in nitric oxide-induced apoptosis. Activation of magne-sium-dependent neutral sphingomyelinase via caspase-3. J BiolChem 1999;274:10654–60.
44. Levy M, Castillo SS, Goldkorn T. nSMase2 activation and traffickingare modulated by oxidative stress to induce apoptosis. BiochemBiophys Res Commun 2006;344:900–5.
45. Rutkute K, Asmis RH,Nikolova-KarakashianMN. Regulation of neutralsphingomyelinase-2 by GSH: a new insight to the role of oxidativestress in aging-associated inflammation. J Lipid Res 2007;48:2443–52.
46. Gallala H, Macheleidt O, Doering T, Schreiner V, Sandhoff K. Nitricoxide regulates synthesis of gene products involved in keratinocytedifferentiation and ceramide metabolism. Eur J Cell Biol 2004;83:667–79.
47. Mocellin S. Nitric oxide: cancer target or anticancer agent? CurrCancer Drug Targets 2009;9:214–36.
48. Hirst DG, Robson T. Nitric oxide physiology and pathology. MethodsMol Biol 2011;704:1–13.
49. Ito H, Murakami M, Furuhata A, Gao S, Yoshida K, Sobue S, et al.Transcriptional regulation of neutral sphingomyelinase 2 geneexpression of a human breast cancer cell line, MCF-7, induced bythe anti-cancer drug, daunorubicin. Biochim Biophys Acta2009;1789:681–90.
50. Clarke CJ, Mediwala K, Jenkins RW, Sutton CA, Tholanikunnel BG,Hannun YA. Neutral sphingomyelinase-2 mediates growth arrest byretinoic acid through modulation of ribosomal S6 kinase. J Biol Chem2011;286:21565–76.
Meyers-Needham et al.
Mol Cancer Ther; 11(5) May 2012 Molecular Cancer Therapeutics1102
on November 21, 2018. © 2012 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst March 27, 2012; DOI: 10.1158/1535-7163.MCT-11-0705

2012;11:1092-1102. Published OnlineFirst March 27, 2012.Mol Cancer Ther Marisa Meyers-Needham, Jocelyn A. Lewis, Salih Gencer, et al. Sphingomyelinase 2/Ceramide Induction
Dependent Neutral−in Mediating Apoptosis via Nitric Oxide Off-Target Function of the Sonic Hedgehog Inhibitor Cyclopamine
Updated version
10.1158/1535-7163.MCT-11-0705doi:
Access the most recent version of this article at:
Material
Supplementary
http://mct.aacrjournals.org/content/suppl/2012/03/27/1535-7163.MCT-11-0705.DC1
Access the most recent supplemental material at:
Cited articles
http://mct.aacrjournals.org/content/11/5/1092.full#ref-list-1
This article cites 50 articles, 13 of which you can access for free at:
Citing articles
http://mct.aacrjournals.org/content/11/5/1092.full#related-urls
This article has been cited by 9 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications Department at
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://mct.aacrjournals.org/content/11/5/1092To request permission to re-use all or part of this article, use this link
on November 21, 2018. © 2012 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst March 27, 2012; DOI: 10.1158/1535-7163.MCT-11-0705