op vol. 268, no. 33, issue of november 25, pp. the and molecular … · 2001-06-25 · the journal...
TRANSCRIPT

THE JOURNAL OP BIOLOGICAL CHEMISTRY 0 1993 by The American Society for Biochemistry and Molecular Biology, Inc.
Vol. 268, No. 33, Issue of November 25, pp. 2496744975,1993 Printed in U.S.A.
Post-translational Processing of Proopiomelanocortin (POMC) in Mouse Pituitary Melanotroph Tumors Induced by a POMC-Simian Virus 40 Large T Antigen Transgene*
(Received for publication, May 20, 1993)
Malcolm J. LowSgT, Bin Liuf, Gary D. HarnmerSlI, Marcel0 Rubinsteinf, and Richard G. Allen#** From the $ Vollum Institute for Advanced Biomedical Research, **Center for Research on Occupational and Environmental Toxicology, and the §Department of Biochemistry and Molecular Biology, Oregon Health Sciences University, Portland, Oregon 97201 ~
Mice harboring a transgene composed of proopiome- lanocortin (POMC) gene promoter sequences (nucleo- tides -706 to +64) ligated to the simian virus (SV) 40 early gene encoding large T antigen developed large POMC-expressing pituitary tumors. Histologically the tumors arose from the intermediate lobe, contained nuclear SV40 T antigen and POMC peptides, but did not express other pituitary hormones. POMC process- ing in the pituitary tumors was indistinguishable from normal mouse intermediate lobe melanotropbs and was characterized by high proportions of acetylated and carboxyl-terminal shortened &endorphins, and amino- terminal acetylated a-melanocyte-stimulating hor- mone, and virtually no adrenocorticotropic hormone (ACTH)(1-39), 8-lipotropin, or POMC. The tumors contained abundant levels of mRNA for the prohor- mone convertase PC2 and undetectable levels of PC1. Normal mouse neurointermediate lobe also has a high ratio of PC2/PC1 expression that is distinct from the relative abundance of PC1 in anterior lobe and AtT- 20 corticotroph cells. In contrast, extracts from tumors transplanted subcutaneously in nude mice contained predominantly nonacetylated forms of D-endorphin( 1- 31) and -(l-27), very little ACTH(1-39), almost no corticotropin-like intermediate peptide or a-melano- cyte-stimulating hormone, and higher proportions of intact POMC. Surprisingly, despite the less efficient proteolytic cleavage, a transplanted tumor expressed both PC 1 and PC2. These studies are the first biochem- ical documentation of a melanotroph pituitary tumor in a rodent species and provide a new model for the investigation of pituitary oncogenesis and the molecu- lar basis of tissue-specific prohormone post-transla- tional processing.
The post-translational processing of proopiomelanocortin
* This work was supported by National Institutes of Health Grant DK 40457 (to M. J. L.), National Science Foundation Grant 91- 08426 (to R. G . A,), and Fundacibn Autorchas (to M. R.). The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “adver- tisement’’ in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
7l To whom all correspondence should be addressed Vollum Insti- tute for Advanced Biomedical Research, L474, Oregon Health Sci- ences University, 3181 S.W. Sam Jackson Park Rd., Portland, OR 97201. Tel.: 503-494-4672; Fax: 503-494-4976.
I( Current address: School of Medicine, Dept. of Internal Medicine, University of California, San Francisco, CA 94143-0114.
(POMC)’ is essential for the production of bioactive peptides including adrenocorticotropic hormone (ACTH), a-melano- cyte stimulating hormone (MSH), and @-endorphin (1). These processing steps include a series of endoproteolytic cleavages, carboxyl-terminal amidation, and amino-terminal acetylation reactions, many of which have been characterized at the biochemical level (2-4). Cell populations residing in the an- terior (corticotrophs) and intermediate (melanotrophs) lobes of the pituitary gland, as well as POMC secreting neurons have characteristic processing phenotypes that appear to be regulated during development and possibly in response to environmental factors (5, 6). Mature corticotrophs and the AtT-20 cell line derived from a mouse corticotroph tumor secrete predominantly ACTH(1-39), @-lipotropin, and small amounts of nonacetylated @-endorphin(1-31). Melanotrophs have more complete proteolytic processing of POMC and secrete acetylated a-MSH, corticotropin-like intermediate peptide, y-lipotropin, and predominantly acetylated and car- boxyl-terminal truncated forms of @-endorphin. Overpro- duction of POMC and the secretion of POMC-derived pep- tides causing Cushing’s syndrome are hallmarks of cortico- troph- and melanotroph-derived adenomas (7-9) and certain neuroendocrine tumors arising in extrapituitary tissues (10, 11). There is wide variation in the proportions of POMC- derived peptides in individual pituitary and extrapituitary tumors from all species suggesting that the oncogenic process affects the post-translational processing steps in a stochastic pattern (12-15). The processing patterns found in tumor extracts and the identification of large molecular weight POMC products secreted from these tumors suggest that incomplete proteolytic processing may contribute residual biological activity involved in the pathogenesis of Cushing’s syndrome (14, 15, 65).
A considerable body of evidence implicates the subtilisn- like proteases prohormone convertase (PC) PC1 (identical to PC3) and PC2 in the endoproteolytic processing of POMC (2, 16-20). Consistent with its endogenous expression in anterior lobe and AtT2O corticotrophs, PC1 is capable of cleaving POMC to ACTH and @-lipotropin in heterologous expression systems (21,22). Antisense RNA to PC1 has also been shown to block the processing of POMC in AtT2O cells (23). The combination of PC1 and PC2 cleaves POMC in at least 5 basic cleavage sites producing a spectrum of peptides closely resembling the normal intermediate lobe (21, 22). PC2 alone
The abbreviations used are: POMC, proopiomelanocortin; ACTH, adrenocorticotropic hormone; MSH, melanocyte-stimulating hor- mone; SV40, Simian virus 40; Tag, large T antigen; RP-HPLC, reversed-phase high performance liquid chromatography; PC, pro- hormone convertase; LPH, lipotropin.
24967

24968 POMC Processing in Mouse Melanotroph Tumors
appears to be capable of high specific activity cleavage at the paired basic residues at the ACTH amino terminus and at the y-LPH/@-endorphin junction as well as at the KKRR residues within ACTH, although there is controversy about its activity at the KR site at the carboxyl terminus of ACTH (21, 22). PC2 is expressed abundantly in the intermediate lobe (18,22, 23) consistent with its putative role as a major POMC proc- essing enzyme. However, the proteolytic preferences of the processing enzymes identified to date are insufficient to ac- count for all of the known processing steps in the generation of the cell-specific bioactive peptides derived from the pro- hormone POMC, in particular the KK cleavage at the @- endorphin carboxyl terminus required for generation of @- endorphin(1-27) and -(1-26) (21).
Targeted tumorigenesis utilizing a tissue-specific gene pro- moter and the Simian virus (SV) 40 large T antigen has been a frequently used strategy to induce neoplasms in endocrine glands in transgenic mice. The resulting tumors have been used to characterize the multistep processes involved in tumor evolution (24), as a sensitive reporter of gene expression from promoter fragments (25), and as a source of tissue for the propagation of novel immortalized cell lines (26). A particular advantage of the transgenic approach is the transformation of cells that rarely produce spontaneous tumors. For example, both gonadotrophs and a precursor cell type for somatomam- motrophs in the mouse pituitary have been immortalized and provided an enriched source of cell-type specific transcription factors for the analysis of pituitary hormone gene expression (27, 28).
The purpose of our study was to characterize the effects of pituitary cell transformation on the post-translational proc- essing of POMC by producing transgenic mice with a fusion gene encoding SV40 large T antigen under the transcriptional control of a rat POMC promoter. Three independent founder mice developed pituitary tumors and a single transgenic ped- igree has been maintained for multiple generations. HPLC and radioimmunoassay for POMC-derived peptides showed that the processing patterns in the tumors were virtually identical to those characteristic of normal intermediate lobe melanotrophs. After serial subcutaneous transplantation in nude mice, extracts of the pituitary tumors showed processing patterns unlike those found in either corticotrophs or melan- otrophs, indicating a loss of specific steps in the biosynthetic pathways of POMC-derived peptide maturation. The higher proportions of unprocessed POMC and P-endorphin(1-31) and -(l-27), and the absence of acetylated a-MSH and acet- ylated @-endorphins from the transplanted tumors most closely resembles the POMC processing patterns found in the developing pituitary gland, however, the changes in processing phenotype apparently bore no direct relationship to the tu- mors' expression of PC1 and PC2. These observations are the first biochemical documentation of melanotroph pituitary tumors in mice and provide a new system to characterize the post-translational processing of POMC.
EXPERIMENTAL PROCEDURES
Transgenic Mice-The pronuclei of (B6D2FI X B6D2Fl)Fz hybrid fertilized mouse oocytes were microinjected with approximately 500 copies of a linearized fusion gene containing rat POMC gene regula- tory sequences -706 to +64 ligated to a 2700-base pair StuI to BamHI fragment of the wild type SV40 early gene encoding large T antigen essentially as described previously (29). Transgenic mice were iden- tified by dot-blot hybridization of tail DNA using an SV40 T antigen- specific radiolabeled probe (30) and the males were outbred to female Swiss Webster mice. All animal studies were conducted in accordance with the Public Health Service Guide for the Care and Use of Laboratory Animals.
Tumor Transplantation-Freshly excised primary pituitary tumors
or secondary transplanted tumors were minced under sterile condi- tions and enzymatically dissociated with trypsin and DNase as de- scribed (31). 6 X lo6 cells were resuspended in 0.5 ml of sterile, 0.9% NaCl and injected subcutaneously over the left shoulder blade of female NIH nude mice under asceptic conditions.
Histology and Immunohistochemistry-Mice were anesthetized with 2% Avertin 0.15 m1/10 g body weight intraperitoneally and perfused transcardially with 10% formalin. 10-20-pm sections of pituitary, brain, thymus, gonads, and adrenal glands were prepared with a cryostat and collected onto gelatin-coated slides for staining with hematoxylin and eosin or for immunohistochemistry. Primary antisera were detected either by the avidin-biotin-peroxidase complex method using a Vectastain kit (Vector Labs, Burlingame, CA) and diaminobenzidine or by indirect immunofluorescence as described previously (32). The primary antisera were directed against the fol- lowing antigens and were used at the specified dilutions: SV40 large T antigen (Tag) (donated by D. Hanahan) 1:lOOO; cytokeratin (DAKO, Carpenteria, CA) 1:200; rat @-luteinizing hormone, ACTH IC-1, rat &thyroid-stimulating hormone, human growth hormone S2, and rat prolactin rPrl S9 (donated by the National Hormone and Pituitary Program, National Institute of Diabetes and Digestive and Kidney Diseases, Bethesda, MD) 15000, 1:1000, 1:5000, 1:1000, and 1:25, respectively.
Northern Blot Analysis of mRNA-Normal Swiss Webster mouse pituitary glands, transgenic mouse pituitary tumors, and transplanted nude mouse tumors were snap frozen on dry ice. Total RNA was extracted by the guanidinium thiocyanate method followed by pellet- ing through a CsCl cushion (33). Northern blots were performed using standard methods (34). POMC mRNA and Tag mRNA were detected simultaneously using random primer 32P-labeled mouse POMC cDNA (kindly provided by M. Uhler (35)) and SV40 Tag probes and washed to a final stringency of 0.2 X SSC, 0.1% SDS at 60 "C (1 X SSC is 0.15 M NaCl, 0.015 M sodium citrate, pH 7.2). Dopamine D, receptor mRNA was detected using a human D, cDNA probe (kindly provided by 0. Civelli (36)) and washed to a final stringency of 0.2 X SSC, 0.1% SDS at 52 "C. Mouse PC1 and PC2 mRNA were detected using mouse cDNA probes (kindly provided by N. G. Seidah (18)) and washed to a final stringency of 0.2 X SSC, 0.1% SDS st 60 'C.
Fractionation of POMC Peptides by RP-HPLC and Radioimmuno- assays-Normal pituitaries and tumor tissue (50-100 mg) were ho- mogenized in 1 ml of 30% glacial acetic acid containing 0.3 mg/ml phenylmethylsufonyl fluoride, 0.5 mg/ml bovine serum albumin, fro- zen and thawed three times, diluted 3 X with water, centrifuged, and the supernatant divided into aliquots, lyophilized, and stored at -20 "C. The extracts were resuspended in 13.6% acetonitrile (AcN), 0.1% trifluoroacetic acid and fractionated by reversed-phase high performance liquid chromatography (RP-HPLC) using a Vydac Pro- tein C4 column with a linear gradient of AcN in 0.1% trifluoroacetic acid and a flow rate of 1 ml/min. Fractions were assayed for p- endorphin immunoactivity with a midportion specific p-endorphin antiserum as described previously (37). The antiserum recognizes POMC and all molecules containing @endorphin on an equimolar basis (38). The sensitivity of the immunoassay was 20-40 pg/tube. The a-MSH, y-LPH (p-MSH), and the ACTH immunoassays have also been described in detail (37, 39). The ACTH assay recognizes the carboxyl terminus of ACTH(1-39), the a-MSH assay is 85% acetyl-specific, and the y-LPH assay recognizes 0-LPH as well as y- LPH (39). Synthetic peptides were obtained from Peninsula Labo- ratories, Belmont, CA, and BaChem, Torrance, CA. Corticosterone was measured in unextracted plasma using a radioimmunoassay kit (Ventrex, Portland, ME) and the manufacturer's instructions.
Further Characterization of POMC Peptides by Ion Exchange RP- HPLC-Ion exchange chromatography of pooled fractions was per- formed using a polysulfoethyl aspartamide HPLC cation exchange column (Nest Group, Southborough, MA). 100-200 pl of the fractions indicated in Fig. 6D were pooled, carrier bovine serum albumin (100 pg) was added, and the samples frozen and lyophilized. The samples were resuspended in 250 pl of buffer A for injection onto the column. Synthetic @-endorphin peptides were used to characterize the column and eluted with a gradient of 0-400 mM KC1 in 80 min at a flow rate of 1 ml/min. These peptides eluted at times directly related to their net charge at pH 3.0. Thus, 8-endorphins eluted in the order: (1-26), (1-27), Ac-(1-31), and (1-31). Buffer A was 5 mM KPO, and buffer B was 400 mM KC1 in buffer A. Fractions were assayed directly (20- 50 pl) for @-endorphin immunoreactivity. The recovery of immuno- reactive material was 85% or greater for each sample analyzed.

POMC Processing in Mouse Melanotroph Tumors 24969
RESULTS
Phenotype and Morphology-Three founder transgenic mice from a total of nine identified by dot-blot analysis of tail DNA for SV40 large T antigen sequences developed obvious pituitary tumors. Founder mouse No. 427 was bred success- fully prior to succumbing to its pituitary tumor and estab- lished a pedigree that has maintained the pituitary tumor phenotype unchanged for 15 generations through successive crosses to outbred Swiss Webster females. Microscopic ab- normalities of the pituitary intermediate lobes were evident as early as 3 weeks of age and consisted of multifocal nodules of SV40 Tag immunoreactive melanotrophs. Tag immunopo- sitive cells were also demonstrated occasionally in the anterior lobes (Fig. 1 and 2). The neoplastic melanotrophs had a reduced cytoplasmic to nuclear ratio and pleomorphic nuclei with prominent nucleoli. Colocalization of SV40 Tag and ACTH immunoreactivity was found in variable proportions of both the melanotrophs and corticotrophs, although the most intensely immunofluorescent Tag positive cells were devoid of ACTH staining.
At 6 weeks of age many of the mice had a diffuse hyperplasia of the intermediate lobe easily visible to the unaided eye, however, the general architecture of the pituitary remained intact. Serial examination of transgenic mice a t 3-week inter- vals clearly revealed a progressive growth of the intermediate lobes culminating in tumor masses overflowing the sella tur- cica and compressing the overlying brain. Any incipient an- terior lobe nodules in addition to the normal anterior lobes were engulfed and destroyed by the intermediate lobe tumors, presumably by competition for nutrient supply by the more rapidly dividing transformed melanotrophs. Mice succumbed to massive tumors of the pituitary gland (ranging to 150 mg
n
FIG. 1. ACTH-like immunoreactivity ( A and C) and SV40 T antigen ( E and D ) in a pituitary gland from a 3-week-old transgenic mouse. The intermediate lobe ( I L ) is enlarged and both the intermediate and anterior lobes (AL) have multifocal nodules of T antigen positive cells. An arrou points at a nodule in the anterior lobe. The intermediate lobe is a patchwork of cells demonstrating all combinations of strong and weak expression of both T antigen and POMC (simultaneous dual immunofluorescence technique; NL, neural lobe; the outlined areas in A and B are enlarged in C and D, respectively; scale bar represents 300 pm).
FIG. 2. At 3 weeks of age ( le f t ) multifocal nodules of abnor- mal cells are present in a normal sized intermediate lobe (arrowheads). Pituitary tumor cells from an advanced neoplasm contain widely variable cytoplasmic immunopositivity for ACTH (center) and nuclear localization of SV40 T antigen (right). Left, hematoxylin and eosin stains, scale bar represents 100 pm; center and right, avidin-biotin-peroxidase complex technique, differential inter- ference contrast, scale bar represents 50 pm.
I
FIG. 3. Adrenal gland from 12-week-old control (left) and transgenic littermate bearing a large pituitary tumor (right). The cortex is hyperplastic in the transgenic mouse while the centrally located medulla is unaffected. Hematoxylin and eosin stains, scale bar represents 500 pm.
wet weight compared to a normal gland weight of 2 mg) by 12-20 weeks. Larger tumors sometimes contained tiny resid- ual islands of immunohistochemically normal anterior lobe cells buried within them but there was no evidence of hormone expression other than POMC peptides in the neoplastic cells (data not shown). Transformed cells from the large tumors varied widely in their content of Tag, ACTH-like and p- endorphin-like peptides by immunohistochemical staining (Fig. 2).
Two founder mice and approximately 20% of the 427 ped- igree mice developed massive thymic hyperplasia in addition to or preceding their pituitary tumors. No POMC mRNA or P-endorphin peptides were detected in the hyperplastic thy- mus glands. No tumors of the brain, reproductive tract, or any other extrapituitary site were found in the transgenic mice.
Transgenic mice with pituitary tumors characteristically had a 50% increase in adrenal weight compared to sibling controls (7.4 & 0.5 uersw 4.7 & 0.2 mg, X & S.E., n = 9). Histologically the enlarged adrenals had cortical hyperplasia in the zonae fasiculata and reticularis (Fig. 3). The tumor bearing mice invariably rapidly lost weight in their final week and never developed any increased adipose tissue. Tissue content of &endorphin and plasma P-endorphin and corticos- terone levels in normal and transgenic mice are compared in Table I.
Dissociated pituitary tumor cells readily formed subcuta- neous tumors when implanted into nude mice but not outbred Swiss Webster or hybrid B6D2 F1 mice. Tumors were serially transplanted up to three times for a total growth period of 8 months following dissociation of the original primary tumors.

24970 POMC Processing in Mouse Melanotroph Tumors TABLE I
Tissue and plasma content of &endorphin immunoreactivity and plasma corticosterone in normal mice, transgenic mice, and nude mice implanted subcutaneously with pituitary tumor cells
&Endorphin immunoreactivity in
NIL (normal mice)” Tumors (transgenic or nude) Plasma &endorphin corticosterone Plasma
pmolltissw pmollmg wet weight m l m l nglml Normal mice NILb 696 f 59 1393 f 117 <55 41 f 7 Transgenic mice primary tumors 1088 f 169 12 f 2 1007 f 49 679 f 100 Nude mice with transplanted tumors 1052 f 353 2 f 0.6 ND‘ >loo0
Mean f S.E., n = 3-5.
ND, not determined. * NIL, neurointermediate lobe.
Tag -
POMC -
FIG. 4. A, Northern blot analysis of POMC and T antigen mRNA transcripts. Lane 1, primary pituitary tumor; lanes 2 and 3, trans- planted tumors; lane 4, tumor cells maintained in vitro. B, Northern blot analysis of dopamine DZ receptor mRNA transcripts. Lane 1 , mormal mouse pituitary; lane 2, primary pituitary tumor; lane 3, transplanted tumor; lane 4, tumor cells maintained in vitro. 20 pg of total RNA was loaded per lane in each Northern blot.
Northern blot analysis of mRNA extracted from normal pi- tuitary glands, primary pituitary tumors, transplanted tu- mors, and cultured tumor cells showed a pattern of increasing abundance of Tag RNA and decreasing abundance of both POMC RNA and dopamine DP receptor RNA (Fig. 4). The relative abundance of POMC and Tag mRNA was confirmed in three separate experiments. Despite less efficient expres- sion of the POMC gene, the transplanted tumors continued to store @-endorphin immunoreactive material and stimulate corticosterone release (Table I).
Molecular Forms of @-Endorphin and ACTH-related POMC Peptides in Normal Pituitary, Primary Pituitary Tumors, and Transplanted Tumor Tissue-The molecular forms of POMC- derived peptides were characterized in extracts of pituitary tumor tissue from descendants of transgenic founder mouse No. 427. All primary tumors examined (total of 5) showed essentially identical results and representative data are pre- sented in the figures. Fig. 5 compares the @-endorphin-related peptides extracted from normal Swiss Webster mouse neu- rointermediate lobe (A), normal mouse anterior lobe (B), and two individual pituitary tumors (C and D). Qualitatively, the relative proportions of tumor @-endorphin(l-31), acetylated @-endorphin(l-31), acetylated @-endorphin( 1-27), and acety-
lated @-endorphin(1-26) were very similar to those found in the normal mouse intermediate lobe and distinct from normal mouse anterior lobe. Peak 6 in normal anterior lobe (Fig. 5B) was identified as @-LPH by the coelution of @-endorphin and y L P H immunoreactivity (data not shown). A minor, but possibly significant difference, was that tumor extracts con- tained a slightly higher percentage of unprocessed POMC than the normal tissues.
The pattern of post-translational processing was strikingly different in the six transplanted tumors compared to all the primary tumors that were analyzed. Fig. 6, A-D, shows the @- endorphin processing profile of four independent transplanted tumors. Notably, the transplanted tumor extracts contained a high percentage of @-endorphin(l-31) (peak 1) and variable quantities of two peaks (7 and 8) that eluted close to, but distinct from, acetylated @-endorphin(1-31). To further char- acterize these peptides, we pooled material from the three major peaks shown in Fig. 6D and analyzed them by ion exchange HPLC and immunoassay. Peak 1 eluting in frac- tions 43-45 was confirmed to be nonacetylated @-endor- phin(1-31) (Fig. 7A). Peak 7 eluting in fractions 48-50 con- tained mostly nonacetylated P-endorphin( 1-27) and minor amounts of acetylated @-endorphin( 1-31) and nonacetylated @-endorphin(1-26) (Fig. 7B). Peak 8 eluting in fractions 52- 53 was nonacetylated @-endorphin(1-26) (data not shown). Additionally, as in the primary pituitary tumors, there was relatively more unprocessed POMC (peak 5 in Fig. 6, A-D). These results indicate that relatively little acetylation of @- endorphin(1-31) was taking place and that carboxyl-short- ening of @-endorphin was occurring, but with reduced effi- ciency.
Because acetylation of @-endorphins was markedly reduced we reassayed the HPLC column fractions to determine if there was a similar reduction in acetylated forms of a-MSH. Further analyses of the HPLC fractions were performed using a relatively acetyl-specific a-MSH assay and a carboxyl- terminal specific ACTH assay. Fig. 8A shows that the primary pituitary tumors processing closely resembled normal mouse intermediate lobe a-MSH profiles. The primary tumors con- tained large relative amounts of diacetyl-a-MSH (peak 11) and there was a minor amount of intact ACTH(1-39) (data not shown). In contrast, the transplanted tumors from nude mice (Fig. 8B) contained no diacetyl-a-MSH and very little des- and monoacetyl-a-MSH (peak 9 and 10). The profile of ACTH-related peptides in the transplanted tumors (Fig. 8C) showed no corticotropin-like intermediate peptide, almost no ACTH( 1-39), unidentified peaks of later eluting material, and unprocessed POMC (peak 5). In other studies (40) we have demonstrated that the phosphorylated and glycosylated ACTH(1-39) contained in normal mouse anterior pituitary extracts elutes as four major peaks at 23-36 min on the Cq system used here. Partial cleavage at the KR site between

POMC Processing in Mouse Melanotroph Tumors 24971
FIG. 5. Molecular forms of B-en- dorphin containing molecules in normal mouse intermediate and an- terior lobes and primary pituitary tumors. Mouse pituitary tissue was ana- lyzed by RP-HPLC and @-endorphin im- munoassay as described under “Experi- mental Procedures.” The elution times of the peptides were as follows: 1, @- endorphin(1-31) (45-47 rnin); 2, acetyl- @-endorphin(l-31) (53-54 rnin); 3, ace- tyl-@-endorphin(l-27) (59 rnin); 4, ace- tyl-@-endorphin(1-26) (62 min); and 5, POMC (73-75 min). The elution time of peak 6 (@-LPH) is shown by the arrow and was confirmed using an immunoas- say for y L P H (not shown). A, interme- diate lobe, 1.5 neurointermediate lobes were fractionated and analyzed. B , an- terior lobe, a small portion of one ante- rior lobe dissected completely free of me- lanotrophs was fractionated and ana- lyzed. C, POMC-tag pituitary tumor No. 1; and D, POMC-Tag pituitary tumor No. 2. 50 mg wet weight equivalent of the tumor extracts were fractionated.
so - . A. Intermediate
3
40-
20-
10 -
, B. Anterior 6
4.
3- 1
Tumor c. Pituitary
1
0
ACTH and @-LPH sequences in the transplanted tumors was confirmed by the presence of y-LPH detected with the y- LPH radioimmunoassay (data not shown). The later eluting material (peaks between 50 and 65 min) was not characterized further, however, the other major ACTH containing material in normal pituitary tissues is the 20-kDa ACTH biosynthetic intermediate. Therefore these peaks most likely represent a mixture of the 20-kDa biosynthetic intermediate cleaved at the KR paired basic pair between ACTH and @-LPH and the intermediate carboxyl-terminally extended by y-LPH se- quences. The processing profiles of primary and transplanted melanotroph tumors are depicted schematically in Fig. 9 to illustrate the sites of efficient, partial, or absent endoproteo- lytic cleavage and the loss of acetylation function by the transplanted tumors.
Expression of PC1 and PC2 in Normal Pituitary Tissues and Mehnotroph Tumors-To determine whether the change in POMC processing accompanying transplantation of the tumors was due to altered expression of PC1 or PC2, we analyzed total RNA from AtT-20 cells, mouse anterior lobes, neurointermediate lobes, primary pituitary tumors, and a transplanted tumor by Northern blots (Fig. 10). PC1 expres- sion (Fig. 1OA) was highest in AtT-20 cells, moderately abun- dant in normal anterior lobe and a transplanted tumor, low in normal neurointermediate lobes, and undetectable in three independent primary tumors. The identical pattern of PC1 expression was confirmed by an RNase protection assay (data not shown). PC2 expression (Fig. 10B) was highest in normal
24l 40
5
D. Pituitary Tumor
3
1
5
60 80 0 20 40 60 EO
time (rnin)
neurointermediate lobe, moderately but variably abundant in the primary and transplanted tumors, and barely detectable in AtT-20 cells and normal anterior lobes. Equivalent amounts of total RNA were loaded on the gels based on the pattern of ribosomal RNA staining (Fig. lOC).
DISCUSSION
The localization of tumors induced by the POMC-Tag fusion gene was highly tissue-specific for the pituitary gland of several independent lines of transgenic mice. Pituitary expression of the transgene was expected based on previous observations in transgenic mice produced by the microinjec- tion of reporter genes containing identical regulatory se- quences from the rat POMC gene (41,42). Because the SV40 large T antigen is capable of transforming a wide range of murine cell types, including neurons (43-45), it was conceiv- able that the transgene used in these studies would be a more sensitive marker for expression of the POMC 5”flanking sequences in the arcuate nucleus of the hypothalamus or reproductive tract (46,47). The absence of expression in these tissues as characterized by immunohistochemistry and the lack of tumor formation is further evidence that the rat POMC gene sequences between -706 and +64 are not generally sufficient for productive transcriptional activity in extrapi- tuitary locations.
Several transgenes encoding SV40 Tag under the transcrip- tional control of neuroendocrine gene promoters have induced thymic neoplasia with identical characteristics to the mice in
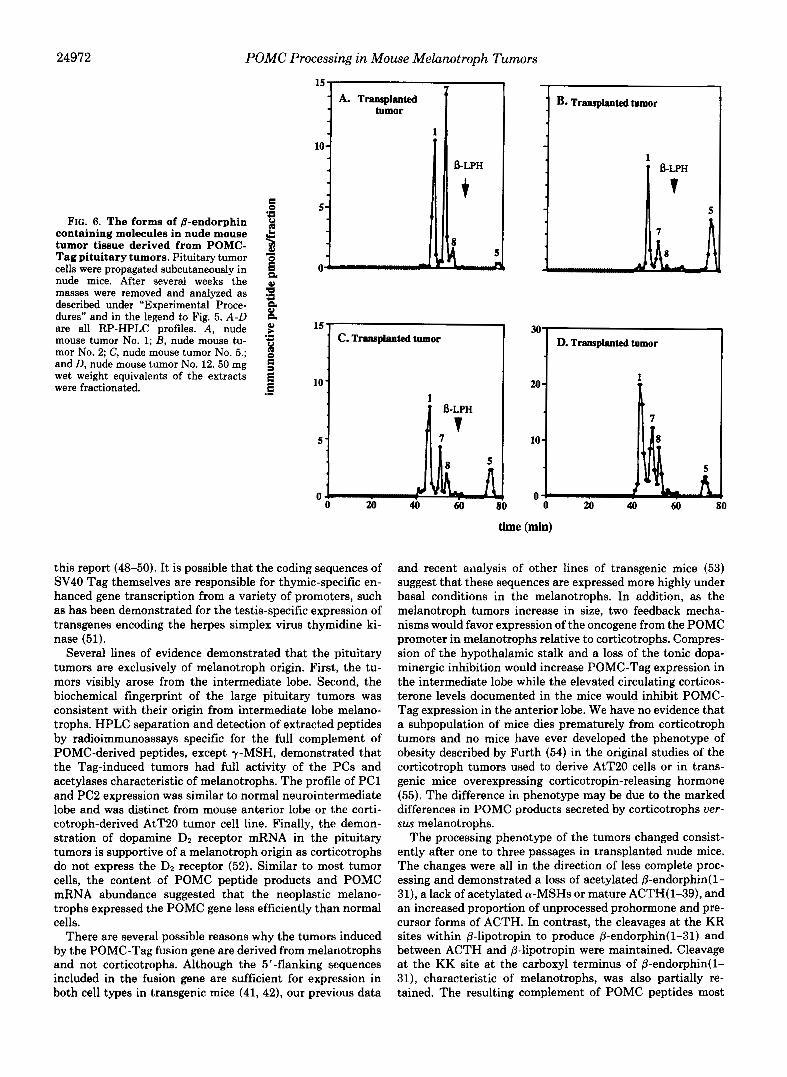
24972 POMC Processing in Mouse Melanotroph Tumors
.I
FIG. 6. The forms of &endorphin 2 containing molecules in nude mouse tumor tissue derived from POMC- $ Tag pituitary tumors. Pituitary tumor o cells were propagated subcutaneously in nude mice. After several weeks the masses were removed and analyzed as 3 described under “Experimental Proce- g dures” and in the legend to Fig. 5. A-D p f . are all RP-HPLC profiles. A, nude $! mouse tumor No. 1; B, nude mouse tu- mor No. 2; C, nude mouse tumor No. 5.; 3 and D, nude mouse tumor No. 12.50 mg 3 wet weight equivalents of the extracts were fractionated. .I 3
Y
c. Transpianted tumor
10 -
5‘
I ?
1 6-LPH I r I I
10 -
lu 0 . 40 80 0 20 40 60 - 0 u, BI
this report (48-50). It is possible that the coding sequences of SV40 Tag themselves are responsible for thymic-specific en- hanced gene transcription from a variety of promoters, such as has been demonstrated for the testis-specific expression of transgenes encoding the herpes simplex virus thymidine ki- nase (51).
Several lines of evidence demonstrated that the pituitary tumors are exclusively of melanotroph origin. First, the tu- mors visibly arose from the intermediate lobe. Second, the biochemical fingerprint of the large pituitary tumors was consistent with their origin from intermediate lobe melano- trophs. HPLC separation and detection of extracted peptides by radioimmunoassays specific for the full complement of POMC-derived peptides, except yMSH, demonstrated that the Tag-induced tumors had full activity of the PCs and acetylases characteristic of melanotrophs. The profile of PC1 and PC2 expression was similar to normal neurointermediate lobe and was distinct from mouse anterior lobe or the corti- cotroph-derived AtTZO tumor cell line. Finally, the demon- stration of dopamine DP receptor mRNA in the pituitary tumors is supportive of a melanotroph origin as corticotrophs do not express the Dz receptor (52). Similar to most tumor cells, the content of POMC peptide products and POMC mRNA abundance suggested that the neoplastic melano- trophs expressed the POMC gene less efficiently than normal cells.
There are several possible reasons why the tumors induced by the POMC-Tag fusion gene are derived from melanotrophs and not corticotrophs. Although the 5”flanking sequences included in the fusion gene are sufficient for expression in both cell types in transgenic mice (41, 42), our previous data
time (min)
D
and recent analysis of other lines of transgenic mice (53) suggest that these sequences are expressed more highly under basal conditions in the melanotrophs. In addition, as the melanotroph tumors increase in size, two feedback mecha- nisms would favor expression of the oncogene from the POMC promoter in melanotrophs relative to corticotrophs. Compres- sion of the hypothalamic stalk and a loss of the tonic dopa- minergic inhibition would increase POMC-Tag expression in the intermediate lobe while the elevated circulating corticos- terone levels documented in the mice would inhibit POMC- Tag expression in the anterior lobe. We have no evidence that a subpopulation of mice dies prematurely from corticotroph tumors and no mice have ever developed the phenotype of obesity described by Furth (54) in the original studies of the corticotroph tumors used to derive AtT2O cells or in trans- genic mice overexpressing corticotropin-releasing hormone (55). The difference in phenotype may be due to the marked differences in POMC products secreted by corticotrophs uer- sus melanotrophs.
The processing phenotype of the tumors changed consist- ently after one to three passages in transplanted nude mice. The changes were all in the direction of less complete proc- essing and demonstrated a loss of acetylated @-endorphin(1- 31), a lack of acetylated a-MSHs or mature ACTH(1-39), and an increased proportion of unprocessed prohormone and pre- cursor forms of ACTH. In contrast, the cleavages at the KR sites within @-lipotropin to produce @-endorphin( 1-31) and between ACTH and @-lipotropin were maintained. Cleavage at the KK site at the carboxyl terminus of @-endorphin(1- 31), characteristic of melanotrophs, was also partially re- tained. The resulting complement of POMC peptides most

POMC Processing in Mouse Melanotroph Tumors 24973
10 I 1
30 40 50 time (min)
7. Ion exchange HPLC of &endorphin containing mol- ecules in nude mouse tumor tissue derived from POMC-Tag pituitary tumors. Peaks from the RP-HPLC column shown in Fig. 6D were analyzed as described under “Experimental Procedures.” A, pooled fractions 43-45 (peak 1) from Fig. 6Q and B, pooled fractions 48-50 (peak 7) from Fig. 6D. The elution positions of synthetic 8- endorphin peptides are indicated.
closely resembles the processing profile found in the devel- oping mouse or rat pituitary at late embryonic stages or postnatal day 1 (56, 57). During development, the carboxyl- terminal shortening of P-endorphin(1-31) and the amino- terminal acetylation of a-MSH and @-endorphins appear after establishment of the endoproteolytic cleavages at KR sites (57). A recent report found that proglucagon processing in SV40 Tag-induced intestinal carcinomas in transgenic mice also changed markedly with progressive tumor growth sug- gesting that processing plasticity may be a common feature of neoplastic endocrine cells (58).
PC2 gene expression was relatively abundant in the primary pituitary tumors consistent with its presumptive role in nor- mal melanotrophs in the post-translational processing of POMC (21,22). Interestingly, despite the loss or reduction in processing at several paired basic amino acid sites of POMC in the transplanted tumors, both PC2 and PC1 were expressed at the mRNA level. These data suggest that the active en- zymes may not be efficiently produced in the transplanted tumors either because of a block in translation or in activation of the proenzymes. Alternatively, other changes in the cell biology of the transplanted tumor cells involving the intra-
C. ~rrnsplrntedtumor 4:
3:
2: ACTH (1-39)
1:
0
ACTE RIA
t
.I. ,
0 20 40 60 80 0 40 80
time FIG. 8. The forms of a-MSH- ( A and B ) and ACTH-(C)
related molecules in a primary pituitary tumor extract ( A ) and in a subcutaneously transplanted tumor extract ( B and C). The RP-HPLC fractions were reassayed from those depicted in Figs. 5 0 and 6B. The elution times of the peptides were as follows: (9) desacetyl-a-MSH (18 rnin); (10) monoacetyl-a-MSH (20 rnin); (11) diacetyl-a-MSH (22 min); and 5, POMC (73-75 min). The elution positions of corticotropin-like intermediate peptide and syn- thetic ACTH(1-39) are indicated.
cellular transport of the prohormone may alter either the accessibility or residence time in an appropriate convertase- rich compartment. A further possibility is that an enzyme distinct from PC1 and PC2 is physiologically relevant for POMC processing in melanotrophs and that this novel enzy- matic activity is lost after tumor transplantation. Direct stud- ies of the enzymatic activities in addition to PC gene expres- sion are necessary to distinguish between these possibilities.
Our data does not allow us to make definitive conclusions concerning the molecular events responsible for the change in POMC processing. Transplantation of the tumors is nec- essarily associated with an increase in the total number of cell divisions and age of the neoplasm. The altered processing

24974 POMC Processing in Mouse Melanotroph Tumors A NORMAL MOUSE INTERMEDIATE LOBE AND PRIMARY TUMORS
NI RK RR KR KKRR KR KR KK IC
N.D. N.D. c c c c c c c "
"
I p-ENDORPHIN (1-27) 10 WNDORPHIN (1-28)
B SUBCUTANEOUSLY TRANSPLANTED PITUITARY TUMORS
41 RK RR KR KKRR KR KR KK IC N.D. N.D. -
LNDoAPHlN (141) I KK I
FIG. 9. Summary and comparison of POMC processing in normal mouse melanotrophs, primary pituitary tumors, and subcutaneously transplanted pituitary tumors. A, in normal mouse melanotrophs and primary pituitary tumors five paired basic amino acids are cleaved efficiently, giving rise to the peptides de- picted. The majority of a-MSH and 0-endorphin peptides are amino- terminally acetylated. B, in the transplanted tumors specific endo- proteolytic cleavages are markedly reduced in these melanotroph- derived cells. Amino-terminal acetylation of &endorphin is also re- duced and diacetyl-a-MSH is absent. Thus, specific post-transla- tional processing steps are lost after transplantation. N , amino ter- minus; C, carboxyl terminus; K , lysine; R, arginine; solid arrow, efficient cleavage; open arrow, partial cleavage; -, absent cleavage; ND, not determined.
may be due to general effects of transformation induced by SV40 Tag and would also occur in the primary tumors in situ after a sufficient length of time if the tumors were not fatal as a result of mass effect. Arguing against a simple loss of all differentiated function, however, was the emergence of PC1 expression a t a level comparable to normal anterior lobe. Alternatively, the extracellular environment of the subcuta- neously transplanted tumors may directly alter the processing phenotype either because of a loss of factors present in the tumors' normal location or exposure to novel factors. Previous studies have suggested that POMC processing patterns can be altered in vivo by glucocorticoids (59), vasopressin (60), and interleukin-2 (61). Furthermore, in the neurointermediate lobe both PC1 and PC2 are regulated in response to dopami- nergic agents and in AtT-20 cells PC1 is regulated in response to glucocorticoids and corticotropin-releasing hormone (23).
This study is the first biochemical documentation of a melanotroph pituitary tumor in mice. It was recently reported that vasopressin-SV40 large T antigen transgenic mice occa- sionally developed intermediate lobe tumors that stained with an antibody to ACTH, in addition to somatotroph tumors, but the forms of POMC peptides have not been characterized (62). Intermediate lobe pituitary adenomas are relatively com- mon, however, in canine and equine species (8,9). In the dog, two types of melanotrophs have been described. Type A cells produce a-MSH and 8-endorphin similarly to mouse melan- otrophs while type B cells are more like corticotrophs in their
A
. .
' .. ?.
2.7 kb - r)l B
C
28 S -
18s -
PC 1
PC2
FIG. 10. Northern blot analysis of PC1 ( A ) and PC2 (B) expression in AtT20 cells, normal mouse pituitary lobes, and POMC-SV40 Tag-induced transgenic mouse tumors. 20 pg of total RNA were loaded per lane and duplicate blots were probed for the indicated mRNAs. The positions of the 18 S and 28 S ribosomal bands are indicated. C, methylene blue stain of the RNA from one of the blots showing that equivalent total amounts were present in each lane.
complement of POMC peptides (63,64). Tumors of either cell type are capable of producing Cushing's syndrome in dogs, however, with elevated glucocorticoid levels. The melanotroph tumors induced in transgenic mice are most like the canine type A. The processing pattern is identical to normal mouse intermediate lobe cells and the mice have striking elevations in plasma 8-endorphin and corticosterone with histological evidence of adrenocortical hyperplasia. Although plasma ACTH levels were not measured in this study, it is likely that the melanotroph tumors secrete enough ACTH to cause the adrenocortical hyperplasia as has been found in canine Cush- ing's syndrome. Large molecular weight products of POMC in combination with ACTH could also be responsible for stimulation of the adrenal gland (65).
The establishment of a line of transgenic mice that repro- ducibly develops melanotroph pituitary tumors provides a new tool for the study of pituitary oncogenesis and the biochemical processes regulating POMC post-translational processing. It should also be possible to derive immortalized cell lines rep- resenting the melanotroph cell lineage that will complement the existing AtT20 mouse corticotroph cell line in studies of

POMC Processing in Mouse Melanotroph Tumors 24975
the cell-specific regulation of POMC gene expression and processing.
Acknowledgments-We thank V. Fairchild-Huntress and M. Lev- ine for excellent technical assistance, Drs. M. Uhler, J. Roberts, N. Seidah, and 0. Civelli for DNA clones, Dr. G. Thomas for helpful discussions, Dr. A. Heckendorfer of the Nest group (Southborough, MA) for prompt and attentive service, J. Shigi, I. Schoffstoll, and N. Kurkinen for the artwork, and Dr. D. Hanahan and the National Hormone and Pituitary Program for gifts of antisera.
REFERENCES
2. Barr, P. J. (1991) Cell 6 6 , 1-3 1. Eipper, B. A,, and Mains, R. E. (1980) Endocr. Reu. 1 , 1-27
3. Mains, R. E., and Eipper, B. A. (1984) Endocrinology 115 , 1683-1690 4. Eipper, B. A., and Mains, R. E. (1981) J. Biol. Chem. 266,5689-5695 5. Smith, A. I., and Funder, J. W. (1988) Endocr. Reu. 9 , 159-179 6. Akil, H., Young, E. A., Watson, S. J., and Coy, D. H. (1981) Peptides 2 ,
8. Peterson, M. E., Krieger, D. T., Drucker, W. D., and Halmi, N. S. (1982) 7. Krieger, D. T. (1983) Endocr. Reu. 4 , 22-43
9. Wilson, M. G., Nicholson, W. E., Holscher, M. A., Sherrell, B. J., Mount,
10. Jex. R. K.. van Heerden. J. A.. Camenter. P. C.. and Grant. C. S. (1985)
289-292
Acta. Endocrinol. (Copenh.) 1 0 1 , 15-24
C. D., and Orth, D. N. (1982) Endocrinology 110,941-954
Am. J . Surg. 149,276-282 ' . G. M. (1986) Clin. Endocrinol. 24,699-713
11. Howlett, T. A., Drury, P. L., Perry, L., Doniach, I., Rees, L. H., and Besser,
12. Vieau, D., Massias, J.-F., Girard, F., Luton, J.-P., and Bertagna, X. (1989)
13. de Keyzer, Y., Bertagna, X., Lenne, F., Girard, F., Luton, J.-P., and Kahn, Clin. Endocrinol. 3 1 , 691-700
14. Hale, A. C., Besser, G. M., and Rees, L. H. (1986) J. Endocrinol. 1 0 8 , 49- A. (1985) J . Clin. Inuest. 76,1892-1898
15. Ratter, S. J., Gillies, G., Hope, J., Hale, A. C., Grossman, A., Gaillard, R., 56
Cook, D., Edwards, C. R. W., and Rees, L. H. (1983) Clin. Endocrinol. 18,211-218
16. Smeekens, S. P., and Steiner, D. F. (1990) J. Biol. Chem. 266,2997-3000 17. Seidah, N. G., Gaspar, L., Mion, P., Marcinkiewicz, M., Mbikay, M.,
18. Seidah, N. G., Marcinkiewicz, M., Benjannet, S., Gaspar, L., Beaubien, G., Chrbtien, M. (1990) DNA ( N . Y . ) 9,415-424
fitt,ejLM. G., Mbikay, M., and Chrbtien, M. (1991) Mol. Endocrinol. 5 ,
19. Smeekens, S. P., Avruch, A. S., LaMendola, J., Chan, S. J., and Steiner, D.
20. Nakayama, K., Hosaka, M., Hatsuzawa, K., and Murakami, K. (1991) J . F. (1991) Proc. Natl. Acad. Sci. U. S . A. 88, 340-344
21. Thomas, L., Leduc, R., Thorne, B. A., Smeekens, S. P., Steiner, D. F., and Biochem. (Tokyo) 109,803-806
22. Benjannet, S., Rondeau, N., Day, R., Chrbtien, M., and Seidah, N. G. (1991) Thomas, G. (1991) Proc. Natl. Acad. Sci. U. S. A. 88,5297-5301
23. Bloomquist, B. T., Eipper, B. A., and Mains, R. E. (1991) Mol. Endocrinol. Proc. Natl. Acad. Sci. U. S. A. 88,3564-3568
24. Folkman, J., Watson, K., Ingber, D., and Hanahan, D. (1989) Nature 3 3 9 , 6,2014-2024
25. Efrat, S., Teitelman, G., Anwar, M., Ruggiero, D., and Hanahan, D. (1988) 58-61
Neuron 1.605-613
111-122
26. Gilligan, A.:Jewett,-L., Simon, D., Damjanov, I., Matschinsky, F. M., Weik, H., Pinkec, C., and Knowles, B. B. (1989) Diabetes 38,1056-1062
27. Horn. F.. Wmdle. J. J.. Barnhart. K. M.. and Mellon. P. L. (1992) Mol. ceil. sbl. 12,2143-2153
. ,
28. Lew, D., Brady, H., Klausing, K., Yaginuma, K., Theill, L. E., Stauber, C.,
29. Low, M. J. (1992) in Methods in Molecular Biology (Longstaff, A,, and
30. Hogan, B. N., Costantini, N., and Lacy, E. (1986) Manipulating the Mouse
Karin, M., and Mellon, P. L. (1993) Genes & Deu. 7,683-693
Reuest, P., eds) Vol. 13, Humana Press, Totowa, NJ
Embryo: A Laboratory Manual, Cold Spring Harbor Laboratory, Cold Spring Harbor, NY
31. Hammer, G. D., Mueller, G., Liu, B., Petrides, J. S., Roos, B. A., and Low, M. J. (1992) Endocrinology 130 , 1975-1985
32. Low, M. J., Goodman, R. H., and Ebert, K. M. (1989) Mol. Endocrinol. 3 , 21-12~~20.1.1
33. Cathala, G., Savouret, J. F., Mendez, B., West, B. L., Karin, M., Martial,
34. Sambrook, J., Fritsch, E. F., and Maniatis, T. (1989) Molecular Cloni A J. A., and Baxter, J. D. (1983) DNA (N. Y.) 2,329-335
e r a t o r y Manual, 2nd Ed., Cold Spring Harbor Laboratory, ?!old Sprmg Harbor, NY
35. Uhler, M., Herbert, E., D'Eustachio, P., and Ruddle, F. D. (1983) J. Biol. Chem. 258.9444-9453
36. Grandy, D., Marchionni, M., Makam, H., Stofko, R., Alfono, M., Froth- in ham, L., Fischer, J., Burke, K., Bunzow, J., Server, A., and Civelli, 0.
37. Hatfield, J. M., Allen, R. G., Stack, J., and Ronnekleiv, 0. (1988) Deu. Biol. (1889) Proc. Natl. Acad. Sci. U. S. A. 86,9762-9766
- - - -. - -
38. Thomas, G., Thorne, B. A,, Thomas, L., Allen, R. G., Hruhy, D. E., Fuller,
39. Allen, R. G., Hatfield, J. M., and Stack, J. (1988) Deu. Biol. 126,156-163 40. Allen, R. G., Herbert, G., Hinman, M., Shihuya, H., and Pert, C. B. (1978)
41. Hammer, G. D., Fairchild-Huntress, V., and Low, M. J. (1990) Mol. Endo-
126,164-172
R., and Thorner, J. (1988) Science 241,226-230
Proc. Natl. Acad. Sci. U. S . A. 75,49724976
42. Liu, B., Hammer, G. D., Rubinstein, M., Mortrud, M., and Low, M. J.
43. Lee, Y. C., Asa, S. L., and Drucker D. J. (1992) J. Biol. Chem. 267,10705-
crinol. 4 , 1689-1697
(1992) Mol. Cell. Biol. 1 2 , 3978-3990
44. Mellon, P. L., Windle J. J., Goldsmith, P. C., Padula, C. A., Roberts, J. L.,
45. Hammang, J. P., Baefge, E. E., Behringer, R. R., Brinster, R. L., Palmiter,
46. Gee, C. E., Ching-ling, C. C., Roberts J. L., Thompson, R., and Watson, S.
47. Kiloatrick. D. L.. Borland. K.. and Jin. D. F. (1987) Proc. Natl. Acad. Sci.
10708
and Weiner, R. I. (1990) Neuron 6, 1-10
R. D., and Messing, A. (1990) Neuron 4,775-782
J. (1983) Nature 306,374-376
0. S. A. '84,5695-5699 ' '
48. Reynolds, R. K., Hoekzema, G. S., Vogel, J., Hinrichs, S. H., and Jay, G .
49. Botteri, F. B., van der Putten H, Wong, D. F., Sauvage, C. A,, and Evans, (1988) Proc. Natl. Acad. Sci. U. S. A. 85,3135-3139
50. Jat, P. S., Noble, M. D., Ataliotis, P., Tanaka, Y., Yannoutsos, N., Larsen, R. M. (1987) Mol. Cell. Biol. 7,3178-3184
51. AI-Shawi, R., Burke, J., Jones, C. T., Simons, Harrison, S., Buxton, D., L., and Kioussis, D. (1991) Proc. Natl. Acad. Sci. U. S . A. 88,5096-5100
Maley, S., Chandley, A,, and Bishop, J. 0. (1991) Mol. Cell. Biol. 11,
. ,
52. De Souza, E. B. (1986) Endocrinology 119 , 1534-1542 53. Rubinstein, M., Mortrud, M., Liu, B., and Low, M. J. (1993) Neuroendo-
54. Furth, J. (1955) Rec. Prog. Horm. Res. 11, 221-255 55. Stenzel-Poore, M. P., Cameron, V. A., Vaughan, J., Sawchenko, P. E., and
56. Rius, R. A,, Chikuma, T., and Loh, Y. P. (1991) Deu. Brain Res. 6 0 , 179-
57. Allen, R. G., Pintar, J. E., Stack, J., and Kendall, J. W. (1984) Dew Biol.
4207-4216
crinology, in press
Vale, W. (1992) Endocrinology 130,3378-3386
185
58. Brubaker, P. L., Lee, Y. C., and Drucker, D. J. (1992) J. Biol. Chem. 2 6 7 ,
59. Noel, G., and Mains, R. E. (1991) Endocrinology 129 , 1317-1325 60. Canny, B. J., Smith, I. A,, Clements, J. A., and Funder, J. W. (1988)
Neuroendocrinology 48,603-610 61. Zakarian, S., Eleazar, M. S., and Silvers, W. K. (1989) Nature 339 , 553-
556 62. Stefaneanu, L., Rindi, G., Horvath, E., Murphy, D., Polak, J. M., and
Kovacs, K. (1992) Endocrinology 130 , 1789-1795 63. Peterson, M. E. (1987) Front. Horm. Res. 17 , 37-47 64. Peterson, M. E., Orth, D. N., Halmi, N. S., Zielinski, A. C., Davis, D. R.,
65. London, D. R., Gibson, S., and White, A. (1992) ACTH and its recursors Chavez, F. T., and Drucker, W. D. (1986) Endocrinology 119 , 720-730
sor syndrome? Proceedin s of the 74th Annual Meetin of the Endocrine m Cushing's syndrome: ectopic ACTH syndrome or ectopic AC$Hprecur-
Society, The Endocrine 8ociety Press, San Antonio, 'fX (Ahstr. 590)
102,43-50
2072a20733